В данном документе сообщается о способах отбора и получения мультиспецифических терапевтических молекул, изготовленных из полипептид-полинуклеотидного комплекса, где специфичности терапевтической молекулы выбраны в зависимости от фенотипа терапевтических мишеней.
Уровень техники
За последние годы было разработано и подвергнуто клиническим испытаниям широкое разнообразие опухолеспецифических терапевтических белков, включая антитела, фрагменты антител и лиганды рецепторов клеточной поверхности. Эти терапевтические белки были конъюгированы с несколькими классами терапевтических токсинов, такими как низкомолекулярные лекарственные вещества, ферменты, радиоизотопы, белковые токсины и другие токсины для специфической доставки пациентам.
Эффективная доставка к месту заболевания является необходимым условием для высокой эффективности и низкой токсичности любой терапевтической молекулы. Например, в данном контексте могут использоваться антитела. Если антитело само по себе не является терапевтическим, то конъюгация лекарственного препарата с антителом позволяет достичь высокого уровня локализации лекарственного средства в нужном месте в организме человека. Это увеличивает эффективную концентрацию лекарственного средства в его целевой области, тем самым оптимизируя терапевтический эффект агента. Кроме того, с целевой доставкой врач может иметь возможность понизить дозу терапевтического агента, что особенно актуально, если полезная нагрузка препарата имеет ассоциированные токсичности, или если он будет использоваться для лечения хронических заболеваний (см., например, McCarron, Р.А., et al., Mol. Interventions 5 (2005) 368-380).
О создании биспецифических антител, например, сообщается в WO 2004/081051. Был разработан и создан широкий спектр форматов биспецифических антител (см., например, Fischer, N. and Leger, О., Pathobiology 74 (2007) 3-14). О хелатирующих рекомбинантных антителах (CRAb) первоначально сообщалось в Neri, D., et al. (Neri, D., et al., J. Mol. Biol. 246 (1995) 367-373). Wright, M.J. and Deonarain, MP. (Molecular Immunology 44 (2007) 2860-2869) сообщали о библиотеке фагового дисплея для создания хелатизированных рекомбинантных антител.
О молекулярных носителях для целевой доставки лекарственных препаратов сообщается в Backer, M.V., et al., Bioconjugate Chem. 13 (2002) 462-467. В WO 2010/118169 сообщается о человеческих белковых матрицах с контролируемой фармакокинетикой в сыворотке крови. Способы и композиции, связанные с пептидами и белками с C-концевыми элементами, имеют перекрестную ссылку на родственные заявки и описаны в WO 2009/105671. В WO 2007/038658 сообщается о конъюгатах «антитело-лекарственный препарат» и способах их применения. Композиции и способы для целевой биологической доставки молекулярных носителей приведены в WO 2004/062602. В WO 2002/072141 сообщается о целевых лигандах.
В WO 2009/037659 сообщается о магнитном обнаружении малых группировок. Об обнаружении однородного анализируемого вещества сообщается в WO 2006/137932. В US 2008/0044834 сообщается о трехкомпонентном биосенсоре для обнаружения макромолекул и других анализируемых веществ. О разработке и синтезе биспецифических реагентов сообщается в WO 95/05399.
В US 2002/051986 сообщается о способах обнаружения анализируемого вещества с помощью нуклеиновокислотного репортера. В WO 95/053990 сообщается о разработке и синтезе биспецифичных реагентов с использованием двухцепочечных ДНК в качестве химически и пространственно сшивающих линкеров.
Gosuke, Н., et al. сообщают о применении L-ДНК в качестве молекулярной метки (Nucl. Acids Symp. Ser. 49 (2005) 261-262). Применение амфипатических спиралей для продукции функциональных гибко связанных димерных Fv-фрагментов с высокой авидностью в E. coli описано в Pack, P., et al. (Biochem. 31 (1992) 1579-1584). Kostelny, S.A., et al. сообщают о формировании биспецифического антитела с использованием лейциновых молний (J. Immunol. 148 (1992) 1547-1553). Димерное биспецифическое миниантитело, объединяющее две специфичности с авидностью, описано в Muller, К.М., et al. (FEBS Lett. 432 (1998) 45-49). Goldenberg, D.M., et al. сообщают о продукции многофункциональных антител способом «dock-and-lock» для улучшения визуализации и лечения рака за счет предварительного нацеливания (J. Nuc. Med. 49(2008) 158-163).
Сущность изобретения
В данном документе сообщается о способе получения высокоспецифической мультиспецифической терапевтической молекулы с заданными свойствами для лечения заболевания, такого как рак, у пациента, нуждающегося в лечении, где терапевтическая молекула адаптирована к характеристикам заболевания пациента и/или к генотипу/фенотипу пациента.
Такая адаптация достигается путем изготовления молекулы с заданными свойствами с учетом генотипа/фенотипа вызывающих заболевание/пораженных клеток пациента.
На первом этапе определяется генотип/фенотип клеток (например, наличие и число/количество специфических для заболевания антигенов клеточной поверхности), которые станут мишенями терапевтической молекулы. Это может быть достигнуто, например, с помощью методик клеточной визуализации, таких как иммуногистохимическое окрашивание (IHC, иммуногистохимия) клеток пациента, полученных, например, из крови и/или биопсийного материала, с использованием флуоресцентно меченных моноспецифических (терапевтических или диагностических) антител. Альтернативно генотип/фенотип клеток может быть проанализирован после окрашивания мечеными терапевтическими или диагностическими антителами с использованием способов на основе FACS. Методики визуализации in vivo, включая оптическую визуализацию, молекулярную визуализацию, флуоресцентную визуализацию, биолюминесцентную визуализацию, ЯМР, ПЭТ, ОФЭКТ, КТ и прижизненную микроскопию, также могут быть использованы для определения генотипа/фенотипа связанных с заболеванием клеток пациента. В зависимости от определенного генотипа/фенотипа клеток пациента, связанных с заболеванием, может быть выбрана/выбирается комбинация нацеливающих/связывающих группировок с заданными свойствами, и они объединяются в терапевтическую молекулу. Такая терапевтическая молекула может быть, например, биспецифическим антителом.
Такие терапевтические молекулы с заданными свойствами i) будут высокоспецифичными, ii) будут иметь хорошую эффективность и iii) будут вызывать меньше побочных эффектов по сравнению с обычными выбранными терапевтическими средствами. Это может быть достигнуто путем придания терапевтической молекуле улучшенной адресности и/или улучшенных свойств доставки с заданными свойствами, например, для полезной нагрузки терапевтического препарата в месте его предполагаемого действия.
Улучшенная доставка терапевтической молекулы к месту его действия, такому как, например, раковая клетка, может быть достигнута за счет более высокой/повышенной селективности и/или специфичности целевой терапевтической молекулы по сравнению с обычными выбранными терапевтическими молекулами. Терапевтическая молекула содержит по меньшей мере две группировки, которые специфически связываются с различными антигенами (например, с двумя различными поверхностными маркерами) или с различными эпитопами на одном и том же антигене (например, с двумя различными эпитопами на одном и том же поверхностном маркере).
Повышенная селективность и/или специфичность терапевтической молекулы с заданными свойствами может быть достигнута за счет одновременного связывания обеих нацеливающих группировок с их соответствующими мишенями/эпитопами, т.е. она достигается за счет эффекта авидности. Особенно подходящим является сочетание двух связывающих группировок, имеющих аффинность от низкой до средней к их соответствующим мишеням/эпитопам. Кроме того, нецелевое связывание значительно уменьшается или даже может быть полностью устранено.
Специфичности связывания предоставляются по отдельности исходными компонентами, из которых формируется мультиспецифическая терапевтическая молекула. Таким образом, можно сделать мультиспецифическую терапевтическую молекулу с заданными свойствами, такую как биспецифическое антитело, просто путем определения поверхностных маркеров, присутствующих на клетке, например на раковой клетке, и конъюгации соответствующих связывающих группировок, таких как фрагменты антител, которые специфически связываются с этими поверхностными маркерами, с нуклеиновой кислотой и соединения их с помощью линкерного нуклеотида.
Было обнаружено, что для направленной доставки эффекторной группировки особенно полезным является комплекс, содержащий полипептидный и полинуклеотидный компоненты. Эффекторная группировка, полипептидный компонент и полинуклеотидный линкер комплекса нековалентно связаны друг с другом. Это делает возможным модульное производство отдельных компонентов комплекса. Благодаря модульной архитектуре комплекса отдельные компоненты могут быть изменены без необходимости изменения других компонентов комплекса. Это позволяет легко и эффективно собирать множество вариантов комплекса, например, для получения библиотеки, на основе которой может быть выбрана высокоспецифическая мультиспецифическая терапевтическая молекула с заданными свойствами.
Одним из аспектов, описанном в данном документе, является способ выбора по меньшей мере двух связывающих группировок из коллекции/библиотеки связывающих группировок, которые собирают в одну мультиспецифическую связывающую молекулу путем инкубации (a) Fab-фрагмента антитела или scFv-фрагмент антитела, каждый из которых содержит или конъюгирован с первым элементом или элементом первой связывающей пары, где этот Fab-фрагмент или scFv специфически связывается с первым маркером клеточной поверхности или с первым эпитопом первого маркера клеточной поверхности, (b) Fab-фрагмента антитела или scFv-фрагмента антитела, каждый из которых содержит или конъюгирован с первым элементом или элементом второй связывающей пары, где этот Fab-фрагмент или scFv-фрагмент антитела специфически связывается со вторым маркером клеточной поверхности или со вторым эпитопом первого маркера клеточной поверхности, и (с) линкера, содержащего на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары, для использования в качестве терапевтического агента. Такой агент имеет улучшенные свойства нацеливания/доставки.
Одним из аспектов, описанных в данном документе, является способ получения мультиспецифической связывающей молекулы, включающий следующие этапы:
(i) определение маркеров клеточной поверхности, присутствующих в образце, содержащем клетку, и i) выбор по меньшей мере первого маркера клеточной поверхности и, возможно, второго маркера клеточной поверхности, или ii) выбор множества маркеров клеточной поверхности, соответствующего числу специфичностей связывания мультиспецифической связывающей молекулы,
(ii) инкубация (а) множества связывающих группировок, каждая из которых содержит первый элемент или участника связывающей пары, где каждая из связывающих группировок специфически связывается с другим маркером клеточной поверхности или его лигандом или эпитопом того же маркера клеточной поверхностности, где каждый первый элемент или участник связывающей пары связывается только с соответствующим вторым элементом или элементом и не связывается с любым другим вторым элементом или участником связывающей пары, и (b) линкера, содержащего соответствующие вторые элементы связывающих пар,
и, таким образом получение мультиспецифической связывающей молекулы.
Одним из аспектов, описанных в данном документе, является способ получения биспецифического антитела, включающий следующие этапы:
(i) определение маркеров клеточной поверхности, присутствующих в образце, содержащем клетку, и выбор первого поверхностного маркера и второго поверхностного маркера,
(ii) инкубация (а) Fab-фрагмента антитела или scFv-фрагмент антитела, содержащего или конъюгированного с первым элементом или участником первой связывающей пары, где этот Fab-фрагмент или scFv специфически связывается с первым маркером клеточной поверхности, (b) Fab-фрагмента антитела или scFv-фрагмента антитела, содержащего или конъюгированного с первым элементом или участником второй связывающей пары, где этот Fab-фрагмент или scFv-фрагмент антитела специфически связывается со вторым маркером клеточной поверхности, и (с) линкера, содержащего на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары.
и, таким образом получение биспецифического антитела.
Одним из аспектов, описанных в данном документе, является способ определения комбинации связывающих группировок для мультиспецифической связывающей молекулы, включающий следующие этапы:
(i) определение специфичности связывания и/или селективности и/или аффинности и/или эффекторной функции и/или периода полужизни in vivo множества мультиспецифических связывающих молекул, где во множестве мультиспецифических связывающих молекул содержится каждая (возможная) комбинация связывающих группировок,
и
(ii) выбор мультиспецифической связывающей молекулы с подходящей специфичностью связывания и/или селективностью и/или аффинностью и/или эффекторной функцией и/или периодом полужизни in vivo, и тем самым определение комбинации антигенсвязывающих группировок.
Одним из аспектов, описанных в данном документе, является способ определения комбинации антигенсвязывающих сайтов, включающий следующие этапы:
(i) определение специфичности связывания и/или селективности и/или аффинности и/или эффекторной функции и/или периода полужизни in vivo множества биспецифических антител, полученных путем объединения каждого элемента первого множества Fab-фрагментов антител или scFv-фрагментов антител, содержащего или конъюгированного с первым элементом первой связывающей пары, с каждым элементом второго множества Fab-фрагментов антител или scFv-фрагментов антител, содержащим или конъюгированным с первым элементом второй связывающей пары, и линкером, содержащим на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары,
где первое множество специфически связывается с первым маркером клеточной поверхности, а второе множество специфически связывается со вторым маркером клеточной поверхности,
и
(ii) выбор биспецифического антитела с подходящей специфичностью связывания и/или селективностью и/или аффинностью и/или эффекторной функцией и/или периодом полужизни in vivo, и тем самым определение комбинации антигенсвязывающих сайтов.
Одним из аспектов, описанных в данном документе, является биспецифическое антитело, содержащее
a) первый Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается с первым поверхностным маркером и
ii) который конъюгирован с первым элементом первой связывающей пары,
b) второй Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается со вторым поверхностным маркером и
ii) который конъюгирован с первым элементом второй связывающей пары, и
c) энантиомерный ДНК-полинуклеотидный линкер,
i) который конъюгирован со вторым элементом первой связывающей пары и
ii) который конъюгирован со вторым элементом второй связывающей пары,
при этом первый и второй Fab-фрагмент или scFv-фрагмент антитела образуют нековалентный комплекс.
Ниже приведены воплощения всех аспектов, описании в данном документе. При этом нужно отметить, что каждое воплощение может быть объединено с каждым из аспектов, а также со всеми другими отдельными воплощениями, приведенными в данном документе.
В одном воплощении связывающие группировки независимо друг от друга выбраны среди связывающей группировки на основании дарпинового домена, связывающей группировки на основании антикалинового домена, связывающей группировки на основании фрагмента Т-клеточного рецептора, такого как домен scTCR, связывающей группировки на основании верблюжьего VH-домена, связывающей группировки на основании десятого домена фибронектина III типа, связывающей группировки на основании тенасцинового домена, связывающей группировки на основании кадгеринового домена, связывающей группировки на основании ICAM-домена, связывающей группировки на основании домена титина, связывающей группировки на основании GCSF-R-домена, связывающей группировки на основании домена цитокинового рецептора, связывающей группировки на основании домена ингибитора гликозидазы, связывающей группировки на основании домена супероксидсупероксиддисмутазы или фрагментов антител (Fab- или scFv-фрагментов).
В одном воплощении всех аспектов мультиспецифическая связывающая молекула является биспецифическим антителом, или первая и вторая связывающие группировки независимо друг от друга являются фрагментами антитела.
В одном воплощении фрагмент антитела выбран из группы, содержащей Fv, Fab, Fab', Fab'-SH, F(ab')2, димерное антитело, линейное антитело, scFv, scFab и dsFv.
В одном воплощении по меньшей мере два компонента биспецифического антитела, содержащего эффекторную группировку, специфичности связывания и полинуклеотидный линкер, нековалентно связаны друг с другом.
В одном воплощении связывающая группировка выбрана среди антител, фрагментов антител, рецепторов, лигандов рецепторов и связывающих мишень каркасов, при условии, что лиганд рецептора не является полипептидным лигандом рецептора инкретина.
В одном воплощении фрагмент антитела выбран из группы, содержащей Fv, Fab, Fab', Fab'-SH, F(ab')2, димерное антитело, линейное антитело, scFv, scFab и dsFv.
В одном воплощении связывающий мишень каркас выбран среди дарпиновг гемопексин-подобных молекул и антикалинов.
В одном воплощении рецептор выбран среди фрагментов Т-клеточного рецептора и scTCR.
В одном воплощении мультиспецифическая связывающая молекула является комплексом, включающим
а) первую связывающую группировку,
i) которая специфически связывается с первым маркером клеточной поверхности или его лигандом и
ii) которая конъюгирована с первым элементом первой связывающей пары,
b) вторую связывающую группировку,
i) которая специфически связывается со вторым маркером клеточной поверхности или его лигандом и
ii) которая конъюгирована с первым элементом второй связывающей пары, и
c) полинуклеотидный линкер,
i) который конъюгирован со вторым элементом первой связывающей пары и
ii) который конъюгирован со вторым элементом второй связывающей пары.
В одном воплощении биспецифическое антитело представляет собой комплекс, содержащий
a) первый Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается с первым маркером клеточной поверхности и
ii) который конъюгирован с первым элементом первой связывающей пары,
b) второй Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается со вторым маркером клеточной поверхности и
ii) который конъюгирован с первым элементом второй связывающей пары и
c) полинуклеотидный линкер,
i) который конъюгирован со вторым элементом первой связывающей пары и
ii) который конъюгирован со вторым элементом второй связывающей пары.
В одном воплощении комплекс является нековалентным комплексом.
В одном воплощении комплекс также содержит другой полипептид, i) который специфически связывается со второй мишенью, и ii) который конъюгирован с первым элементом второй связывающей пары, а полинуклеотидный линкер конъюгирован со вторым элементом второй связывающей пары.
В одном воплощении комплекс также содержит эффекторную группировку, которая конъюгирована с полинуклеотидом, который комплементарен по меньшей мере части полинуклеотидного линкера.
В одном воплощении комплекс также содержит эффекторную группировку, конъюгированную с полинуклеотидом, который i) комплементарен по меньшей мере части полинуклеотида, который конъюгирован с первой или второй связывающей группировкой или Fab-фрагментом или scFv-фрагментом антитела, и ii) не комплементарен полинуклеотидному линкеру.
В одном воплощении первая и вторая связывающие группировки или Fab-фрагмент или scFv-фрагмент антитела связываются с одной и той же мишенью и с неперекрывающимися эпитопами на ней.
В одном воплощении полинуклеотидный линкер содержит от 8, 10, 15, 20, 25, 50, 100 нуклеотидов. В одном воплощении полинуклеотидный линкер содержит до 500, 750, 1000 или 2000 нуклеотидов. В одном воплощении полинуклеотидный линкер содержит от 10 до 500 нуклеотидов.
В одном воплощении полинуклеотидный линкер является энантиомерной ДНК. В одном воплощении энантиомерная ДНК является L-ДНК. В одном воплощении L-ДНК является одноцепочечной L-ДНК (оц-L-ДНК, ss-L-DNA).
В одном воплощении эффекторная группировка выбрана из группы, состоящей из связывающей группировки, меченой группировки и биологически активной группировки.
В одном воплощении полинуклеотидный линкер конъюгирован со связывающей группировкой или Fab-фрагментом или scFv-фрагментом антитела на его первом или втором конце.
В одном воплощении полинуклеотидный линкер конъюгирован с двумя вторыми элементами двух связывающих пар, где второй элемент первой связывающей пары конъюгирован с первым концом полинуклеотидного линкера, а второй элемент второй связывающей пары конъюгирован со вторым концом полинуклеотидного линкера.
В одном воплощении первый и второй элементы первой связывающей пары содержат нуклеиновокислотные последовательности SEQ ID № 05 и SEQ ID № 08, соответственно.
В одном воплощении первый и второй элементы второй связывающей пары содержат нуклеиновокислотные последовательности SEQ ID № 06 и SEQ ID № 07, соответственно.
В одном воплощении способ включает следующие этапы:
a) синтез первой связывающей группировки или Fab-фрагмента или scFv-фрагмента антитела, который специфически связывается с первым маркером клеточной поверхности или его лигандом, и который конъюгирован с первым элементом первой связывающей пары,
b) синтез второй связывающей группировки или Fab-фрагмента или scFv-фрагмента антитела, который специфически связывается со вторым маркером клеточной поверхности или его лигандом, и который конъюгирован с первым элементом второй связывающей пары,
c) синтез полинуклеотидного линкера, который конъюгирован с вторым элементом первой связывающей пары, и который конъюгирован со вторым элементом второй связывающей пары, и
d) формирование комплекса путем объединения синтезированных компонентов.
Другим аспектом, описанным в данном документе, является фармацевтическая композиция, содержащая мультиспецифическую связывающую молекулу или биспецифическое антитело, описанном в данном документе, и, возможно, фармацевтически приемлемый носитель.
Другим аспектом, описанным в данном документе, является мультиспецифическая связывающая молекула или биспецифическое антитело, описанное в данном документе, для применения в качестве лекарственного средства.
Также аспектом, описанным в данном документе, является мультиспецифическая связывающая молекула или биспецифическое антитело, описанное в данном документе, для применения в лечении рака.
Другим аспектом, описанным в данном документе, является применение мультиспецифической связывающей молекулы или биспецифического антитела, описанного в данном документе, в производстве лекарственного средства.
В одном воплощении лекарственное средство предназначено для лечения рака.
Одним из аспектов, описанных в данном документе, является способ лечения индивидуума, страдающего от рака, который включает введение индивидууму эффективного количества мультиспецифической связывающей молекулы или биспецифического антитела, описанного в данном документе.
Подробное описание воплощений изобретения
I. Определения
Объекты в единственном числе используются в данном документе для обозначения одного или более чем одного (т.е. по меньшей мере одного) из грамматических объектов пункта. В качестве примера, "антитело" означает одно антитело или более чем одно антитело.
«Акцепторная человеческая каркасная область» является каркасной областью, включающей аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученной из каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, определенной ниже. Акцепторная человеческая каркасная область, «полученная из» каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, может содержать одну и ту же аминокислотную последовательность, или она может содержать аминокислотные замены. В некоторых воплощениях число аминокислотных замен составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее, или 2 или менее. В некоторых воплощениях акцепторная человеческая каркасная область VL по своей последовательности идентична VL каркасной последовательности человеческого иммуноглобулина или консенсусной человеческой каркасной последовательности.
Понятие «аффинность» относится к силе суммарных общих нековалентных взаимодействий между одним сайтом связывания молекулы (например, полипептида или антитела) и его партнером по связыванию (например, мишенью или антигеном). Если не указано иное, то термин «аффинность связывания», используемый в данном документе, относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между элементами пары связывания (например, в полипептид-полинуклеотидном комплексе или между полипептидом и его мишенью, или между антителом и его антигеном). Аффинность молекулы X в отношении ее партнера Y может быть выражена, в основном, константой диссоциации (kD). Аффинность может быть измерена обычными способами, известными в данной области, такими как поверхностный плазменный резонанс, а также включая те, которые описаны в данном документе.
Понятие «антитела со зрелой аффинностью» относится к антителу с одним или более чем одним изменением в одной или более чем одной гипервариабельной области (HVR) по сравнению с родительским антителом, которое не содержит такие изменения, причем такие изменения приводят к повышению аффинности антитела к антигену.
Термин "запертый" означает, что эффектор защищен защитной группой, имеющей контролируемое время полужизни в сыворотке крови и жидкостях организма. Защитная группа может быть ферментативно отщеплена эндогенными ферментами. Защитная группа может быть удалена, расщеплена, разложена, ферментативно расщеплена или метаболизирована вторым эффектором, который вводят снаружи путем инъекции или перорально, таким как аскорбиновая кислота. Запертые эффекторные молекулы могут быть активированы с помощью ферментов, которые в природе находятся в жидкостях организма. Запертые эффекторные группировки могут быть активированы восстанавливающими агентами, также находящимися в жидкостях организма, такими как аскорбиновая кислота.
Термин "эффекторная группировка" означает любую молекулу или комбинацию молекул, чья активность, желательно, должна быть доставлена (в) и/или локализована в клетке. Эффекторные группировки включают, но не ограничиваясь ими, метки, цитотоксины (например, экзотоксин Pseudomonas, рицин, абрин, дифтерийный токсин и т.п.), ферменты, факторы роста, факторы транскрипции, лекарственные вещества, радионуклиды, лиганды, антитела, Fc-области антител, липосомы, наночастицы, вирусные частицы, цитокины и т.п.
Термин «антитело» в данном документе используется в самом широком его смысле и охватывает различные структуры антитела, в том числе, но не ограничиваясь ими, моноклональные антитела и фрагменты антител, до тех пор пока они проявляют нужную антигенсвязывающую активность.
Термин «фрагмент антитела» относится к фрагменту полного или полноразмерного антитела, который сохраняет способность связываться с антигеном. Примеры фрагментов антитела включают, но не ограничиваясь ими, Fv, FAB, FAB', FAB'-SH, F(ab')2; димерные антитела; линейные антитела; одноцепочечные молекулы антител (например, scFv). Обзор определенных фрагментов антител см. в Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134. Более детально, термин «фрагмент антитела» охватывает (i) FAB-фрагмент, т.е. одновалентный фрагмент антитела, состоящий из доменов VL, VH, CL и СН1 (обсуждение FAB- и Р(ab')2-фрагментов, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное время полужизни in vivo, см. в 5869046), (ii) F(ab')2-фрагмент, т.е. двухвалентный фрагмент, содержащий два FAB-фрагмента, связанных дисульфидным мостиком в шарнирной области, (iii) Fd-фрагмент, состоящий из доменов VH и СН1, (iv) Fv-фрагмент, состоящий из VL- и VH-доменов одной ветви антитела (см., например, Plueckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore (eds.), (Springer-Verlag, New York), (1994) pp. 269-315, WO 93/16185, US 5,571,894, US 5,587,458), (v) dAb-фрагмент (см., например, Ward, E.S., et al., Nature 341 (1989) 544-546), который состоит из VH-домена, и (vi) выделенная область, определяющая комплементарность (CDR). Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с использованием рекомбинантных способов синтетическим линкером, который позволяет им быть сделанными в виде одной белковой цепи, в которой VL- и VH-области объединяются и образуют одновалентные молекулы (известные как одноцепочечный Fv (scFv), см., например, Bird, R.E., et al., Science 242 (1988) 423-426; Huston, J.S., et al., Proc. Natl. Acad. Sci. USA 85 (1988) 5879-5883). Эти фрагменты антител могут быть получены с использованием обычных способов, известных специалистам в данной области, и могут быть подвергнуты скринингу на предмет их связывающих свойств таким же образом, как и интактные антитела.
Понятие "антитело, которое связывается с тем же эпитопом", что и референсное антитело, относится к антителу, которое блокирует связывание референсного антитела с его антигеном в конкурентном анализе на 50% или более, и, наоборот, референсное антитело блокирует связывание антитела с его антигеном в конкурентном анализе на 50% или более.
Термин "химерное" антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида.
"Класс" антитела относится к типу константного домена или константной области, которой обладает его тяжелая цепь. Есть пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
«Химиотерапевтический агент» представляет собой химическое соединение, используемое в лечении рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN™); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбохон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфарамид и триметиломеламин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорофосфамид, эстрамустин, ифосфамид, мехлоретамин, оксигидрохлорид мехлоретамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урацил мустард; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики, такие как аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин, эпирубицин, эсорубицин, идарубицин, марцелломицин, митомицины, микофенольная кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флюдарабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин, 5-FU; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; анти-адреналиновые средства, такие как аминоглутетимид, митотан, трилостан; вещество, пополняющее запас фолиевой кислоты, такое как фролиновая кислота; ацеглатон; альдофосфамидный гликозид; аминолевулиновая кислота; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демекольцин; диазиквон; элфорнитин; эллиптиний ацетат; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидамин; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; подофиллиновая кислота; 2-этилгидразид; прокарбазин; PSK®; разоксан; сизофиран; спирогерманий; тенуазоновая кислота; триазиквуон; 2,2',2''-трихлортриэтиламин; уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-С»); циклофосфамид; тиотепа; таксаны, например паклитаксел (TAXCOL®, Bristol-Myers Squibb Oncology, Принстон, Нью-Джерси) и доксетаксел (TAXOTERE®, Rhône-Poulenc Rorer, Антони, Франция); хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митомицин С; митоксантрон; винкристин; винорелбин; навелбин; новантрон; тенипозид; дауномицин; аминоптерин; кселода; ибандронат; СРТ-Н; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноевая кислота; эсперамицины; капецитабин; и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных препаратов. В это определение также включены антигормональные агенты, которые регулируют или ингибируют воздействие гормонов на опухоли, такие как антиэстрогены, включая, например, тамоксифен, ралоксифен, ингибирующие ароматазу 4(5)-имидазолы, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен (Fareston); и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; и их фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных препаратов.
Понятие "антиангиогенный агент" относится к соединению, которое блокирует или препятствует в некоторой степени развитию кровеносных сосудов. Антиангиогенный агент, например, может быть малой молекулой или антителом, которое связывается с фактором роста или рецептором фактора роста, вовлеченным в стимуляцию ангиогенеза. Антиангиогенный фактор в одном воплощении представляет собой антитело, которое связывается с сосудистым эндотелиальным фактором роста (VEGF).
Термин "цитокин" является общим термином для белков, высвобождаемых одной популяцией клеток, которые действуют на другие клетки как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и традиционные полипептидные гормоны. В число цитокинов входит гормон роста, такой как человеческий гормон роста, человеческий N-метионин-гормон роста и бычий гормон роста; паратгормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреотропный гормон (TSH) и лютеинизирующий гормон (LH); фактор роста гепатоцитов; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухоли α и β; ингибирующее вещество Мюллера; мышиный гонадотропин-ассоциированный пептид; ингибин; активин; сосудистый эндотелиальный фактор роста; интегрин; тромбопоэтин (ТРО); факторы роста нервов, такие как NGF-β; фактор роста тромбоцитов; трансформирующие факторы роста (TGF), такие как TGF-α и TGF-β; инсулиноподобный фактор роста I и II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, такие как интерферонт α, β и γ; колониестимулирующие факторы (CSF), такие как макрофагальный CSF (M-CSF); гранулоцитарно-макрофагальный CSF (GM-CSF); и гранулоцитарный CSF (GCSF); интерлейкины (IL), такие как IL-I, IL-Ia, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-IO, IL-II, IL-12; фактор некроза опухоли, такой как TNF-α или TNF-β; и другие полипептидные факторы, включая LIF и kit-лиганд (KL). Используемый в данном документе термин «цитокин» включает белки из природных источников или из культуры рекомбинантных клеток, а также биологически активные эквиваленты цитокинов с нативной последовательностью.
Термин "fMLP" обозначает трипептид, состоящий из N-формилметионина, лейцина и фенилаланина. В одном воплощении эффекторная группировка представляет собой fMLP или его производное.
Термин "фенотип пациента" обозначает состав рецепторов клеточной поверхности типа клеток от пациента. Состав может быть как качественным, так и количественным. Клетка, для которой определяется/дан генотип, может быть отдельной клеткой или образцом, содержащим клетки.
Термин "пролекарство" относится к предшественнику или производной форме фармацевтически активного вещества, которая является менее цитотоксической в отношении опухолевых клеток по сравнению с исходным лекарственным средством и способна ферментативно активироваться или превращаться в более активную исходную форму. См., например, Wilman, "Prodrugs in Cancer Chemotherapy" Biochemical Society Transactions, Vol.14, 615th Meeting Belfast (1986) pp. 375-382, и Stella, et al., "Prodrugs: A Chemical Approach to Targeted Drug Delivery", Directed Drug Delivery, Borchardt, et al., (eds.), pp. 247-267, Humana Press (1985). Пролекарства, которые могут быть использованы в качестве эффекторной группировки, включают, но не ограничиваясь ими, фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, пролекарства, модифицированные D-аминокислотами, гликозилированные пролекарства, b-лактамсодержащие пролекарства, пролекарства, содержащие возможно замещенный феноксиацетамид или пролекарства, содержащие возможно замещенный фенилацетамид, 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые могут быть преобразованы в более активное лекарственное средство без цитотоксичности. Примеры цитотоксических лекарственных средств, которые могут быть получены в форме пролекарства для применения в данном изобретении, включают, но не ограничиваясь ими, те химиотерапевтические агенты, которые описаны в данном документе.
Термин "цитотоксическая группировка" относится к веществу, которое ингибирует или предотвращает клеточное функционирование и/или вызывает гибель или разрушение клеток. Цитотоксические агенты включают, но не ограничиваясь ими, радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты-ингибиторы роста; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или их варианты; и различные противоопухолевые или противораковые агенты, описанные в данном документе.
Понятие "эффективное количество" агента, например фармацевтической композиции, относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата.
Термин «Fc-область» в данном документе используется для определения C-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Этот термин включает Fc-области нативной последовательности и вариантные Fc-области. В одном воплощении Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбокси-конца тяжелой цепи. Тем не менее, C-концевой лизин (Lys447) Fc-области может присутствовать или отсутствовать. Если не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области осуществляется в соответствии с системой нумерации ЕС, также называемой индексом ЕС, как описано в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD(1991).
Понятие «каркасного участка», или «FR», относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно появляются в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактное антитело» и «целое антитело» используются в данном документе взаимозаменяемо для обозначения антитела, имеющего структуру, по существу аналогичную структуре нативного антитела, или содержащего тяжелые цепи с Fc-областью, определенной в данном документе. Такое антитело, как правило, содержит две тяжелые цепи и две легкие цепи.
«Человеческое антитело» является таким антителом, которое имеет аминокислотную последовательность, соответствующую антителу, полученному от человека или из клетки человека или полученному из нечеловеческого источника, который использует репертуар человеческих антител или другие последовательности, кодирующие человеческие антитела. Это определение человеческого антитела специально исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
Понятие «гуманизированного» антитела относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых воплощениях гуманизированное антитело будет включать по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют таковым в нечеловеческом антителе, и все или по существу все FR соответствуют таковым в человеческом антителе. Гуманизированное антитело, возможно, может содержать по меньшей мере часть константной области антитела, полученной из человеческого антитела. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подверглось гуманизации.
Термин «гипервариабельная область», или «HVR», используемый в данном документе, относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными в последовательности и/или образуют структурно определенные петли («гипервариабельные петли»). Как правило, нативные четырехцепочечные антитела содержат шесть HVR, три в VH (Н1, Н2, Н3) и три в VL (L1, L2, L3). HVR обычно содержат аминокислотные остатки из гипервариабельных петель и/или из «областей, определяющих комплементарность» (CDR), причем последние имеют наивысшую изменчивость последовательности и/или участвуют в распознавании антигена. Иллюстративные гипервариабельные петли возникают на аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) (см. Chothia, С. and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917). Иллюстративные CDR (CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3) возникают на аминокислотных остатках 24-34 в L1, 50-56 в L2, 89-97 в L3, 31-35B в Н1, 50-65 в Н2 и 95-102 в Н3 (см. Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)). 3a исключением CDR1 в VH, CDR, как правило, содержат аминокислотные остатки, которые образуют гипервариабельные петли. CDR также включают «остатки, определяющие специфичность», или «SDR», которые являются остатками, контактирующими с антигеном. SDR содержатся в областях CDR, называемых сокращенно -CDR, или a-CDR. Иллюстративные a-CDR (a-CDR-L1, a-CDR-L2, а-CDR-L3, a-CDR-H1, a-CDR-H2 и a-CDR-H3) возникают на аминокислотных остатках 31-34 в L1, 50-55 в L2, 89-96 в L3, 31-35В в Н1, 50-58 в Н2 и 95-102 в Н3 (см. Almagro, J.С. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633). Если не указано иное, то HVR-остатки и другие остатки в вариабельном домене (например, FR-остатки) пронумерованы в данном документе в соответствии с Kabat et al., см. выше.
«Иммуноконъюгат» представляет собой антитело или фрагмент антитела, конъюгированный с одной или более чем одной молекулой, полученной не из антитела, в том числе, но не ограничиваясь ими, с элементом связывающей пары, нуклеиновой кислотой или эффекторной группировкой.
«Индивидуум» или «субъект» является млекопитающим. Млекопитающие включают, но не ограничиваясь ими, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых воплощениях индивидуум или субъект является человеком.
Термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих природные мутации или возникающих в процессе производства препарата моноклональных антител, при этом такие варианты, как правило, присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, понятие «моноклональное» указывает на характер антитела, полученного по существу из гомогенной популяции антител, и не должно быть истолковано как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела или фрагменты моноклональных антител, которые будут использоваться в комплексе, описанном в данном документе, могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, гибридомную методику, методики рекомбинантной ДНК, методики фагового дисплея, а также методики, использующие трансгенных животных, содержащих все или часть локусов человеческого иммуноглобулина, при этом такие методики и другие иллюстративные методики получения моноклональных антител описаны в данном документе.
Термин «одновалентный связывающий полипептид» или «одновалентный связывающий фрагмент антитела» означает молекулу, содержащую только один сайт или область для связывания с его мишенью или антигеном. Примерами одновалентных связывающих полипептидов являются пептиды, пептидные миметики, аптамеры, малые органические молекулы (ингибиторы, способные специфически связываться с целевым полипептидом), дарпины, белки, содержащие анкириновые повторы, домен типа Кунитц, однодоменные антитела (см: Hey, T., et al., Trends Biotechnol. 23 (2005) 514-522), (природные) лиганды рецептора клеточной поверхности, одновалентные фрагменты полноразмерных антител и т.п. Например, полноразмерное антитело имеет два связывающих сайта для своей мишени и, таким образом, является двухвалентным, в то время как scFv- или FAB'-фрагмент антитела имеет только один сайт связывания для своей мишени и, таким образом, является одновалентным. В случае одновалентных антител или фрагментов антител, используемых в качестве полипептида, этот сайт называется паратопом.
Понятие «голое антитело» или «фрагмент голого антитела» относится к антителу или группировку антитела, который не конъюгирован с группировкой, полученной не из антитела (например, нуклеиновой кислотой или цитотоксической группировкой или радиоактивной меткой).
Понятие «нативные антитела» относится к природным молекулам иммуноглобулинов с различными структурами. Например, нативные IgG-антитела являются гетеротетрамерными гликопротеинами весом примерно 150000 дальтон, состоящими из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидной связью. От N-конца к С-концу каждая тяжелая цепь содержит вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, а затем три константных домена (СН1, СН2 и СН3). Аналогичным образом от N-конца к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, с последующим константным легким (CL) доменом. Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
Термин «фармацевтическая композиция» относится к композиции, которая находится в такой форме, чтобы обеспечить биологическую активность активного ингредиента, содержащегося в нем, чтобы он был эффективным, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому будет осуществляться введение состава.
Понятие «фармацевтически приемлемого носителя» относится к ингредиенту в фармацевтической композиции, помимо активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемые носители включают, но не ограничиваясь ими, буфер, эксципиент, стабилизатор или консервант.
Термин "полинуклеотид" или "нуклеиновокислотная последовательность" обозначает короткие, как правило, одноцепочечные полинуклеотиды, которые содержат по меньшей мере 8 нуклеотидов и не более чем примерно 1000 нуклеотидов. В одном воплощении полинуклеотид имеет в длину по меньшей мере 9, или 10, или 11, или 12, или 15, или 18, или 21, или 24, или 27, или 30 нуклеотидов. В одном воплощении полинуклеотид имеет в длину не более 200, или 150, или 100, или 90, или 80, или 70, или 60, или 50, или 45, или 40, или 35, или 30 нуклеотидов. В другом воплощении полинуклеотид имеет в длину по меньшей мере 9, или 10, или 11, или 12, или 15, или 18, или 21, или 24, или 27, или 30 нуклеотидов, и не более 200, или 150, или 100, или 90, или 80, или 70, или 60, или 50, или 45, или 40, или 35, или 30 нуклеотидов.
Термин "L-полинуклеотид" означает нуклеиновую кислоту, которая содержит более 50% L-нуклеотидов в виде мономерных структурных блоков, таких как L-ДНК. В одном воплощении L-полинуклеотид содержит только L-нуклеотиды. Число нуклеотидов в таких L-полинуклеотидах следует понимать в диапазоне от одного L-нуклеотида до любого числа. Тем не менее, в одном воплощении число L-нуклеотидов составляет по меньшей мере 10, или 15, или 20, или 25, или 30, или 35, или 40, или 45, или 50, или 55, или 60, или 70, или 80, или 90, или 100 нуклеотидов. L-полинуклеотиды изготовлены из L-A, L-G, L-C, L-U, L-T и их комбинаций, где L-A обозначает L-рибозу-аденин и т.д. L-полидезоксинуклеотиды изготовлены из L-dA, L-dG, L-dC, L-dU, L-dT и их комбинаций, где L-dA означает L-дезоксирибозу-аденин и т.д.
Термин "полинуклеотидный линкер" обозначает группировку, связывающую две нуклеотидные последовательности вместе. В одном воплощении полинуклеотидный линкер представляет собой полинуклеотид. В одном воплощении полинуклеотидный линкер содержит по меньшей мере один полинуклеотид и по меньшей мере один неполинуклеотид. Неполинуклеотид может представлять собой полипептид, полимер или полисахарид. В одном воплощении полинуклеотидный линкер содержит полинуклеотид длиной от 10 до 30 нуклеотидов и линейный полиэтиленгликоль.
"Полипептид" представляет собой полимер, состоящий из аминокислот, соединенных пептидными связями, природных или полученных синтетически. Полипептиды, содержащие менее примерно 20 аминокислотных остатков, могут упоминаться как "пептиды", тогда как молекулы, состоящие из двух или более полипептидов или содержащие один полипептид длиной более 100 аминокислотных остатков, могут упоминаться как "белки". Полипептид может также содержать неаминокислотные компоненты, такие как углеводные группы, ионы металлов или сложные эфиры карбоновых кислот. Неаминокислотные компоненты могут быть добавлены клеткой, в которой экспрессируется полипептид, и могут варьировать в зависимости от типа клетки. В данном документе полипептиды определены в терминах их основной аминокислотной структуры или нуклеиновой кислоты, кодирующей ее. Дополнения, такие как углеводные группы, как правило, не указаны, тем не менее, они могут присутствовать.
Термин "полипептидный эпитоп" обозначает сайт связывания на полипептидной мишени, связанной соответствующим одновалентным связывающим полипептидом. Как правило, он состоит из аминокислот. Связывающий полипептид либо связывается с линейным эпитопом, т.е. эпитопом, состоящим из участка от 5 до 12 последовательных аминокислот, либо связывающий полипептид связывается с трехмерной структурой, образованной пространственным расположением нескольких коротких участков полипептидной мишени. Трехмерные эпитопы, распознаваемые связывающим полипептидом, например, сайтом распознавания антигена или паратопом антитела или фрагментом антитела, можно рассматривать как трехмерные объекты на поверхности молекулы антигена. Эти особенности подходят именно соответствующему сайту связывания связывающего полипептида, и, таким образом, связывание между связывающим полипептидом и его мишенью облегчается.
Термин «специфически связывается» означает, что полипептид или антитело или фрагмент антитела связывается с его мишенью с константой диссоциации (KD) 10-8 М или менее, в одном воплощении от 10-5 М до 10-13 М, в одном воплощении от 10-5 М до 10-10 М, в одном воплощении от 10-5 М до 10-7 М, в одном воплощении от 10-8 М до 10-13 М, или в одном воплощении от 10-9 М до 10-13 М. Этот термин также используется для указания того, что полипептид не связывается специфически с другими присутствующими биомолекулами, т.е. он связывается с другими биомолекулами с константой диссоциации (KD) 10-4 М или более, в одном воплощении от 10-4 М до 1 М.
Используемый в данном документе термин «лечение» (и его грамматические варианты, такие как «лечить») относится к клиническому вмешательству в попытке изменить естественное развитие заболевания у индивидуума, которого лечат, и которое может быть выполнено либо для профилактики, либо в ходе Клинической патологии. Желательные эффекты лечения включают, но не ограничиваясь ими, предотвращение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любого прямого или косвенного патологического последствия заболевания, предотвращение метастазирования, уменьшение скорости прогрессирования заболевания, улучшение или временное облегчение болезненного состояния, достижение ремиссии или улучшение прогноза. В некоторых воплощениях комплексы, которые описаны в данном документе, используются для задержки развития заболевания или замедления прогрессии заболевания.
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела, как правило, имеют аналогичную структуру, при этом каждый домен содержит четыре консервативные каркасные области (FR, framework region) и три гипервариабельные области (HVR, hypervariable region) (см., например, Kindt, et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007)). Один VH- или VL-домен может быть достаточным для придания специфичности связывания с антигеном. Кроме того, антитела, которые связывают конкретный антиген, могут быть выделены с помощью VH- или VL-домена из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов, соответственно (см., например, Portolano, S. et al., J. Immunol. 150 (1993) 880-887; Clackson, T. et al., Nature 352 (1991) 624-628).
Термин «вектор», используемый в данном документе, относится к нуклеиновокислотной молекуле, способной размножать другую нуклеиновую кислоту, с которой она связана. Этот термин включает вектор как самореплицирующуюся нуклеиновокислотную структуру, а также вектор, включенный в геном клетки-хозяина, в которую он был введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называют в данном документе «экспрессионными векторами».
II. Мультиспецифические связывающие молекулы с заданными свойствами
В большинстве заболеваний с клеточной основой одним из перспективных подходов является нацеливание на клетки, связанные с болезнью, через связывание рецепторных молекул на основе антитела. Тем не менее, уровень экспрессии клинически значимых поверхностных рецепторов (= мишеней) варьирует от пациента к пациенту, и эффективность стандартных лекарственных препаратов на основе антител, таким образом, является очень разной. Это относится особенно к би- и мультиспецифическим связывающим молекулам, чей режим действия заключается в нацеливании на два различных эпитопа/рецептора одновременно.
Один перспективный подход состоит в разработке лекарственного средства (здесь би- или мультиспецифической связывающей молекулы) специально для конкретной/индивидуальной ситуации соответствующего пациента.
Основываясь на данных о профиле экспрессии клинически значимых поверхностных рецепторов на ассоциированных с заболеваниями клетках пациента, из библиотеки были специально выбраны серии связывающих группировок (например, Fab-фрагментов) и объединены с мультиспецифической связывающей молекулой в качестве специфического для пациента лекарственного препарата. Эти выбранные связывающие молекулы специально выбраны по отношению к соответствующим ассоциированным с заболеванием клеткам, таким как, например, опухолевые клетки, на основании, например, уровня экспрессии поверхностных рецепторов и, таким образом, нужд и фенотипа конкретного пациента.
Вариации в длине линкера, который связывает/соединяет связывающие группировки, позволяют выбирать правильную гибкость и расстояние, которые могут потребоваться для одновременного связывания обеих связывающих группировок и, таким образом, для селективности и/или специфичности и/или эффективности.
Кроме того, могут быть добавлены полезные нагрузки, такие как эффекторные функции или токсины, путем специфической гибридизации полезной нагрузки с линкером; Эта возможность дополнительно повышает существование терапевтических применений.
Мультиспецифические связывающие молекулы, специфические для выбранного пациента, могут быть протестированы в различных клеточных in vitro анализах/на образцах клеток в отношении соответствующих критериев (например, оптимальное связывание/партнеры связывания, оптимальная длина линкера и т.д.):
- определение статуса фосфорилирования фосфотирозинкиназ
- определение ингибирования JNK
- определение молекулярно индуцированного апоптоза
- анализ связывания, осуществляемый с моноспецифической против мультиспецифической связывающей молекулы
- определение ингибирования пролиферации.
При таком подходе возможно создание терапевтических молекул с заданными свойствами и, таким образом, высокоэффективных терапевтических молекул. Эти молекулы будут иметь сниженные побочные эффекты за счет улучшенного нацеливания/доставки (например, полезной нагрузки для опухолевых клеток), и улучшенное нацеливание на клетку-мишень основано на более высокой селективности и специфичности нацеливающего компонента (содержащего по меньшей мере две связывающие молекулы).
Более высокая селективность и специфичность мультиспецифической связывающей молекулы происходит из-за одновременного связывания (авидности) за счет комбинации двух «низкоаффинных» связывающих агентов, что снижает возможность связывания «мимо мишени».
Каждая клетка от индивидуума отличается в отношении экспрессированных молекул клеточной поверхности, таких как рецепторы, по количеству и виду. Это особенно верно для раковых клеток и нераковых клеток. Таким образом, клетка может быть охарактеризована представленными на клеточной поверхности молекулами.
Такая характеризация может быть осуществлена с помощью методик на основе клеточной визуализации in vivo. Методики визуализации in vivo включают, например, оптическую визуализацию, молекулярную визуализацию, флуоресцентную визуализацию, биолюминесценцию визуализацию, ЯМР, ПЭТ, ОФЭКТ, КТ и прижизненную микроскопию. Методики визуализации in vitro включают, например, иммуногистохимическое окрашивание клеток пациента, например, флуоресцентно меченными антителами, распознающими специфические маркеры клеточной поверхности, и анализ сигналов флуоресценции путем микроскопии. Альтернативно, генотип/фенотип клеток может быть проанализирован после окрашивания мечеными терапевтическими или диагностическими антителами с использованием способов на основе FACS.
В одном воплощении генотип/фенотип клеток, полученных от пациента, определяется способом на основе FACS. В одном воплощении маркеры клеточной поверхности определяют с помощью флуоресцентно меченных диагностических или терапевтических антител. В одном воплощении используются флуоресцентно меченные терапевтические антитела.
Некоторые заболевания могут коррелировать с изменением количества специфических молекул клеточной поверхности или с возникновением новой молекулы клеточной поверхности.
Лица, страдающие от такого заболевания, будут демонстрировать в определенных диапазонах паттерны маркеров клеточной поверхности, специфические для заболевания и/или индивидуума.
Это нужно принимать во внимание в целях получения для такого человека терапевтического агента с заданными свойствами.
Известно большое число терапевтических антител, направленных против молекул клеточной поверхности и их лигандов, которые могут быть использованы для выбора и построения мультиспецифических нацеливающих группировок с заданными свойствами, таких как Ритуксан/Мабтера/Ритуксимаб, 2Н7/Окрелизумаб, Зевалин/Ибризумомаб, Арзерра/Офатумумаб (CD20), HLL2/Эпратузумаб, Инотузомаб (CD22), Зенапакс/Даклизумаб, Симулект/Базиликсимаб (CD25), Герцептин/Трастузумаб, Пертузумаб (HER2/ERBB2), Милотарг/Гемтузумаб (CD33), Раптива/Эфализумаб (CD11a), Эрбитукс/Цетуксимаб (EGFR, рецептор эпидермального фактора роста), IMC-1121В (рецептор VEGF-2), Тизабри/Натализумаб (α4-субъединица интегринов α4β1 и α4β7), РеоПро/Абциксимаб (gpIIb-gpIIa и αvβ3-интегрин), Ортоклон ОКТ3/Муромонаб-CD3 (CD3), Бенлиста/Белимумаб (BAFF), Толеркс/Отеликсизумаб (CD3), Солирис/Экулизумаб (белок комплемента С5), Актемра/Тоцилизумаб (IL-6R), Панорекс/Эдреколомаб (ЕрСАМ, молекула адгезии эпителиальных клеток), СЕА-САМ5/Лабетузумаб (CD66/CEA, карциноэмбриональный антиген), СТ-11 (PD-1, ингибитор T-клеточного рецептора запрограммированной гибели 1, CD-d279), H224G11 (рецептор c-Met), SAR3419 (CD19), IMC-А12/Циксутумумаб (IGF-1R, рецептор инсулиноподобного фактора роста 1), MEDI-575 (PDGF-R, рецептор фактора роста тромбоцитов), СР-675, 206/Тремелимумаб (антиген 4 цитотоксических Т-лимфоцитов), RO5323441 (фактор роста плаценты или PGF), HGS1012/Мапатумумаб (TRAIL-R1), SGN-70 (CD70), Ведотин (SGN-35)/Брентуксимаб (CD30) и ARH460-16-2 (CD44).
Известны различные способы определения маркеров клеточной поверхности, присутствующих в образце, например, от пациента. Один иллюстративный способ основан на флуоресцентно активированной сортировке клеток (FACS), в частности, на анализе специфически окрашенных и отсортированных клеточных популяций. В этом способе фенотипирование образца (клеточной популяции) достигается путем анализа отдельных клеток в отношении представленных маркеров клеточной поверхности с использованием флуоресцентно меченных антител, направленных против этих маркеров, возможно, включая статистическое распределение поверхностных маркеров в клеточной популяции. Это особенно подходит для применения терапевтических антител, которые были помечены флуоресцентной меткой для этой цели, вместе с тем это обеспечивается тем, что позднее мультиспецифическая связывающая молекула с заданными свойствами будет связываться с тем же эпитопом, что и диагностическое антитело. Мультиспецифические связывающие молекулы/биспецифические антитела, о которых сообщается в данном документе, могут быть использованы при получении лекарственных средств для лечения, например, онкологического заболевания, сердечно-сосудистого заболевания, инфекционного заболевания, воспалительного заболевания, аутоиммунного заболевания, метаболического (например, эндокринного) заболевания или неврологического (например, нейродегенеративного) заболевания. Иллюстративными не ограничивающими примерами этих заболеваний являются болезнь Альцгеймера, неходжкинскйе лимфомы, B-клеточные острые и хронические лимфолейкозы, лимфома Беркитта, лимфома Ходжкина, волосатоклеточный лейкоз, острые и хронические миелолейкозы, Т-клеточные лимфомы и лейкемии, множественная миелома, глиома, макроглобулинемия Вальденстрема, карциномы (например, карциномы ротовой полости, желудочно-кишечного тракта, толстой кишки, желудка, дыхательного тракта, легких, молочной железы, яичника, предстательной железы, матки, эндометрия, шейки матки, мочевого пузыря, поджелудочной железы, костей, печени, желчного пузыря, почек, кожи и яичек), меланомы, саркомы, глиомы и раки кожи, острая идиопатическая тромбоцитопеническая пурпура, хроническая идиопатическая тромбоцитопеническая пурпура, дерматомиозит, хорея Сиденгама, миастения, системная красная волчанка, волчаночный нефрит, ревматизм, полиэндокринные синдромы, буллезный пемфигоид, сахарный диабет, пурпура Шенлейна-Геноха, постстрептококковой нефрит, узловатая эритема, синдром Такаясу, болезнь Аддисона, ревматоидный артрит, рассеянный склероз, саркоидоз, язвенный колит, эритема, IgA-нефропатия, узелковый полиартрит, болезнь Бехтерева, синдром Гудпасчера, обЛитерирующий тромбангиит, синдром Шегрена, первичный билиарный цирроз печени, аутоиммунный тиреоидит, тиреотоксикоз, склеродермия, хронический активный гепатит, полимиозит/дерматомиозит, полихондрит, пузырчатка обыкновенная, гранулематоз Вегенера, мембранозная нефропатия, боковой амиотрофический склероз, сухотка спинного мозга, гигантоклеточный артериит/полимиалгия, злокачественная анемия, быстро прогрессирующий гломерулонефрит, псориаз или фиброзирующий альвеолит.
Известно большое число маркеров клеточной поверхности и их лигандов. Например, сообщалось, что раковые клетки экспрессируют по меньшей мере один из следующих маркеров клеточной поверхности и/или лигандов, в том числе, но не ограничиваясь ими, карбоангидразу IX, альфа-фетопротеин, альфа-актинин-4, A3 (антиген, специфичный для антитела А33) АРТ-4, В7, Ва-733, BAGE, BrE3-антиген, СА125, CAMEL, САР-1, CASP-8/m, CCCL19, CCCL21, CD1, CD1a, CD2, CD3, CD4, CDS, CD8, CD1-1A, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD29, CD30, CD32b, CD33, CD37, CD38, CD40, CD40L, CD45, CD46, CD54, CD55, CD59, CD64, CD66a-e, CD67, CD70, CD74, CD79a, CD80, CD83, CD95, CD126, CD133, CD138, CD147, CD154, CDC27, CDK-4/m, CDKN2A, CXCR4, CXCR7, CXCL12, HIF-I-альфа, специфический антиген колоректального рака p (CSAp), СЕА (СЕАСАМ5), СЕАСАМ6, c-met, DAM, EGFR, EGFRvIII, EGP-1, EGP-2, ELF2-M, Ep-CAM, FIt-1, FIt-3, рецептор фолиевой кислоты, антиген G250, GAGE, GROB, HLA-DR, HM1.24, человеческий хорионический гонадотропин (HCG) и его субъединицы, HER2/neu, HMGB-1, индуцируемый гипоксией фактор (HIF-1), HSP70-2M, HST-2 или 1а, IGF-1R, IFN-гамма, IFN-альфа, IFN-бета, IL-2, IL-4R, IL-6R, IL-13R, IL-15R, IL-17R, IL-18R, IL-6, IL-8; IL-12, IL-15, IL-17, IL-18, IL- 25, инсулиноподобный фактор роста 1 (IGF-1), антиген KC4, антиген KS-1, KS1-4, Le-Y, LDR/FUT, фактор, ингибирующий миграцию макрофагов (MIF), MAGE, MAGE-3, MART-1, MART-2, NY-ESO-1, TRAG-3, mCRP, МСР-1, MIP-1A, MIP-1B, MIF, MUC1, MUC2, MUC3, MUC4, MUC5, MUM-1/2, MUM-3, NCA66, NCA95, NCA90, муцин рака поджелудочной железы, плацентарный фактор роста, р53, PLAGL2, простатическую кислую фосфатазу, PSA, PRAME, PSMA, P1GF, ILGF, ILGF-1R, IL-6, IL-25, RS5, RANTES, Т101, SAGE, S100, сурвивин, сурвивин-2В, ТАС, TAG-72, тенасцин, рецепторы TRAIL, TNF-альфа, антиген Tn, антигены Томсена-Фриденрейха, антигены некроза опухоли, VEGFR, ED-B фибронектина, WT-1, антиген 17-1А, факторы комплемента С3, C3a, C3b, С5а, С5, маркер ангиогенеза, bcl-2, bcl-6, Kras, сМЕТ, онкогенный маркер и онкогенный продукт (см., например, Sensi et al., Clin. Cancer Res. 12 (2006) 5023-5032; Parmiani et al, J. Immunol. 178 (2007) 1975-1979; Novellino et al., Cancer Immunol. Immunother. 54 (2005) 187-207).
Таким образом, антитела, распознающие специфические рецепторы клеточной поверхности, в том числе их лиганды, могут быть использованы для специфического и селективного нацеливания и связывания с большим числом/множеством маркеров клеточной поверхности, ассоциированных с заболеванием. Маркер клеточной поверхности представляет собой полипептид, расположенный на поверхности клетки (например, на связанной с заболеванием клетке), который, например, связан с событием сигналинга или со связыванием лиганда.
В одном воплощении для лечения рака/опухоли используются мультиспецифические связывающие молекулы/биспецифические антитела, которые нацелены на связанные с опухолью антигены, такие как те, о которых сообщается в Herberman, "Immunodiagnosis of Cancer", in Fleisher ed., "The Clinical Biochemistry of Cancer", page 347 (American Association of Clinical Chemists, 1979) и в US 4150149; US 4361544; и US 4444744.
Отчеты об ассоциированных с опухолью антигенах (ТАА, tumor associated antigens) включают Mizukami et al., Nature Med. 11 (2005) 992-997; Hatfield et al., Curr. Cancer Drug Targets 5 (2005) 229-248; Vallbohmer et al., J. Clin. Oncol. 23 (2005) 3536-3544; и Ren et al., Ann. Surg. 242 (2005) 55-63), каждый из которых включен в данный документ посредством ссылки в связи с идентифицированными ТАА.
Там, где это заболевание включает лимфому, лейкемию или аутоиммунное нарушение, целевые антигены могут быть выбраны из группы, состоящей из CD4, CD5, CD8, CD14, CD15, CD19, CD20, CD21, CD22, CD23, CD25, CD33, CD37, CD38, CD40, CD40L, CD46, CD54, CD67, CD74, CD79a, CD80, CD126, CD138, CD154, CXCR4, В7, MUC1 или Ia, НМ1.24, HLA-DR, тенасцина, VEGF, P1GF, ED-B фибронектина, онкогена, онкогенного продукта (например, c-met или PLAGL2), CD66a-d, антигенов некроза, IL-2, Т101, TAG, IL-6, MIF, TRAIL-R1 (DR4) и TRAIL-R2 (DR5).
Известно большое число биспецифических антител, направленных против двух различных мишеней, таких как BCMA/CD3, различные антигены семейства HER в комбинации (EGFR, HER2, HER3), CD19/CD3, IL17RA/IL7R, IL-6/IL-23, IL-1-бета/IL-8, IL-6 или IL-6R/ IL-21 или IL-21R, когда первая специфичность направлена на гликоэпитоп антигена, выбранного из группы, состоящей из структур Lewis х, Lewis b и Lewis y, структур Globo Н, KH1, антигена Tn, антигена TF и углеводных структуры муцинов, CD44, гликолипидов и гликосфинголипидов, таких как Gg3, Gb3, GD3, GD2, Gb5, Gm1, Gm2, сиалилтетраозилцерамид, а вторая специфичность направлена на тирозинкиназный ErbB-рецептор, выбранный из группы, состоящей из EGFR, HER2, HER3 и HER4, GD2 в комбинации со вторым сайтом связывания антигена связан с иммунной клеткой, выбранной из группы, состоящей из Т-лимфоцитов, NK клеток, В-лимфоцитов, дендритных клеток, моноцитов, макрофагов, нейтрофилов, мезенхимальных стволовых клеток, нервных стволовых клеток, ANG2/VEGF, VEGF/PDGFR-бета, фактора роста эндотелия сосудов (VEGF) акцептора 2/CD3, PSMA/CD3, EPCAM/CD3, комбинации антигенов, выбранных из группы, состоящей из VEGFR-1, VEGFR-2, VEGFR-3, FLT3, C-FMS/CSF1R, RET, c-Met, EGFR, Her2/neu, HER3, HER4,, IGFR, PDGFR, c-KIT, BCR, интегрина и MMP с водорастворимым лигандом, выбранным из группы, состоящей из VEGF, EGF, PIGF, PDGF, HGF и ангиопоэтина, ERBB-3/C-MET, ERBB-2/C-MET, рецептора EGF 1/CD3, EGFR/HER3, PSCA/CD3, C-MET/CD3, ENDOSIALIN/CD3, EPCAM/CD3, IGF-1R/CD3, FAPALPHA/CD3, EGFR/IGF-1R, IL 17A/F, EGF-рецептора 1/CD3 и CD19/CD16.
Таким образом, обнаружили, что при использовании модульного подхода, о котором сообщается в данном документе, могут быть получены биспецифические терапевтические антитела с заданными свойствами. Эти антитела имеют заданные свойства в отношении молекул клеточной поверхности, фактически присутствующих на клетках индивидуума, нуждающегося в лечении, или в отношении лигандов, взаимодействующих с такой молекулой клеточной поверхности. При определении молекулярного статуса клеточной поверхности индивидуума может быть выбрана комбинация терапевтических мишеней с заданными свойствами.
При таком образовании биспецифических терапевтических агентов с заданными свойствами путем объединения двух отдельных терапевтических молекул для одновременного нацеливания и связывания двух различных эпитопов можно ожидать совокупный/синергический эффект по сравнению с отдельными терапевтическими молекулами.
При использовании уже имеющихся моноспецифических терапевтических связывающих группировок, таких как производные терапевтических антител, может быть быстро и легко получена продукция нужной мультиспецифической связывающей молекулы.
Эти связывающие молекулы/антитела с разработанной авидностью могут связываться с двумя или более маркерами клеточной поверхности, присутствующими на одной клетке. Это связывание является авидным, только если все/обе связывающие группировки одновременно связываются с клеткой. Для этой цели особенно подходят антитела с аффинностью от средней до высокой. С другой стороны, это также позволяет в процессе скрининга исключить менее специфические комбинации специфичностей связывания.
Подход "Combimatrix"
Желательно совместить первую связывающую группировку, такую как Fab-фрагмент антитела, с другой специфически связывающейся группировкой, такой как Fab-фрагмент второго антитела. Кроме того, можно провести скрининг того, показывает ли первая связывающая группировка лучшие свойства при соединении ее с множеством различных других связывающих группировок. С помощью так называемого подхода Combimatrix простым способом можно по-разному нацелить множество комбинаций связывающих группировок. Следует отметить, что вторые связывающие группировки либо могут связываться с различными мишенями/эпитопами/антигенами, либо могут связываться с тем же антигеном, но с другими эпитопами, либо могут связываться с тем же эпитопом, но быть другими вариантами одной связывающей группировки (например, гуманизированными кандидатами).
В этом случае автоматизированная платформа может выполнять задачи пипетирования, очистки и объединения связывающих группировок и их реакций или производных. Подходящей является любая платформа, которая использует, например, 96-луночные планшеты или другие форматы с высокой пропускной способностью, например, пипетирующий робот Eppendorf epMotion 5075vac.
Вначале выполняют клонирование конструкций, кодирующих связывающую группировку (например, Fab-фрагмент антитела). Плазмиду с нуклеиновой кислотой, кодирующей связывающую группировку, обычно получают путем синтеза гена, где C-концевая область закодированной связывающей группировки содержит сортазный мотив и His-метку. Плазмиды по отдельности переносят в отдельные лунки многолуночного планшета (может быть загружен целый планшет). После этого плазмиды расщепляют смесью рестрикционных ферментов, чтобы вырезать область, кодирующую связывающую группировку. Желательно разработать весь генный синтез таким образом, чтобы для всех плазмид была необходима одинаковая смесь рестрикционных ферментов. Впоследствии, этап очистки дает очищенные фрагменты ДНК. Эти фрагменты лигируют в плазмидный остов, который был вырезан из акцепторного вектора той же смесью рестрикционных ферментов, что указана выше. Альтернативно, процедура клонирования может быть выполнена с помощью SLIC-опосредованного этапа клонирования. После лигирования автоматизированная платформа переносит все лигазные смеси в другой многолуночный планшет с компетентными клетками Е. coli (например, Top10 Multi Shot, Invitrogen) и выполняет реакцию трансформации. Клетки культивируют до нужной плотности. Из аликвоты культивационной смеси могут быть сделаны глицериновые стоки. Из культуры выделяют плазмиду (например, с помощью мини-набора для выделения плазмид (например, NucleoSpin 96 Plasmid, Macherey& Nagel)). Идентичность плазмиды проверяют путем расщепления аликвоты подходящей рестрикционной смесью и с помощью электрофореза в SDS-геле (например, Е-Gel 48, Invitrogen). После этого в новый планшет может быть загружена аликвота плазмиды для проведения контрольной реакции секвенирования.
На следующем этапе экспрессируют связывающие группировки. Для этого клетки НЕК высевают в многолуночный планшет с (например, 48-луночный планшет) и трансфицируют выделенными плазмидами (содержащими область, кодирующую связывающую группировку, в соответствующем базовом векторе). Трансфицированные клетки НЕК культивируют в течение нескольких дней и собирают (например, фильтрацией через фильтровальный планшет 1,2 мкм и 0,22 мкм с помощью вакуумной станции). Титры можно контролировать путем выполнения, например, ELISA.
Связывающие группировки могут быть ковалентно связаны с соответствующими элементами олигонуклеотидных связывающих пар с помощью опосредованной сортазой реакции транспептидации. Связывающую группировку и реакционную смесь сортазы объединяют в многолуночном формате. После инкубации при 37°С в течение 4-16 ч собирают конъюгаты «связывающая группировка - олигонуклеотид» с помощью процедуры отрицательной His-Tag-селекции (смесь наносят на His MultiTrap HP plates (GE Healthcare) и фильтруют, в результате чего все молекулы, которые все еще имеют His-метку, связываются с хроматографической колонкой, в то время как все другие молекулы, такие как олигонуклеотидные конъюгаты, находятся в фильтрате); буферный обмен с фильтратом должен быть проведен, например, путем нанесения конъюгата «связывающая группировка - олигонуклеотид» на ультрафильтрационную мембрану или с помощью планшета, содержащего аффинную среду, которая специфична для связывающей группировки; после замены буфера, который также удаляет избыток свободных олигонуклеотидов, конъюгаты «связывающая группировка - олигонуклеотид» могут быть связаны с образованием мультиспецифической связывающей молекулы.
Мультиспецифические связывающие молекулы сделаны с использованием подхода Combimatrix, см. таблицу ниже.
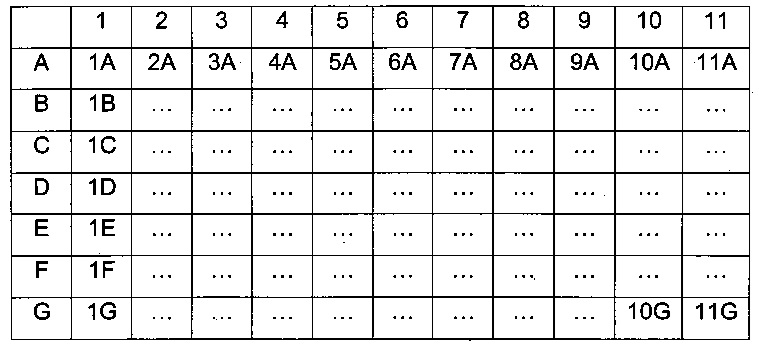
В первом ряду многолуночного планшета пипетировали различные конъюгаты «связывающая группировка - олигонуклеотид» в эквимолярных концентрациях в каждой лунке (за исключением первой лунки первого ряда), обозначенной арабскими цифрами (например, от 1 до 11). В первом столбце того же планшета пипетировали различные конъюгаты «связывающая группировка - олигонуклеотид» в эквимолярных концентрациях в каждой лунке (за исключением первой лунки первого столца), обозначенной буквами (например, от А до G). После этого все конъюгаты «связывающая группировка - олигонуклеотид» в первом ряду объединяли со всеми конъюгатами «связывающая группировка - олигонуклеотид» в первом столбце (например, получая 77 комбинаций в 96-луночном планшете), обозначая их комбинацией цифры и буквы (например, 1А-11G). Ко всем комбинациям добавляли линкерную молекулу в эквимолярных соотношениях с конъюгатами «связывающая группировка - олигонуклеотид» и соответствующий буфер (например, PBS с 150 мМ NaCl, 1.5 мМ MgCl2). Реакцию связывая можно проводить при комнатной температуре или денатурируя смесь при 60°С, а затем медленно ее охлаждая. Затем может быть выполнен дополнительный этап очистки, например, путем эксклюзионной хроматографии. После этого мультиспецифические связывающие молекулы готовы для оценки в клеточных анализах.
Способы, описанные в данном документе
Одним из аспектов, о которых сообщается в данном документе, является способ получения биспецифического антитела, включающий следующие этапы:
(i) определение поверхностных маркеров, присутствующих на поверхности клетки в образце, и выбор первого поверхностного маркера и второго поверхностного маркера,
(ii) инкубация (a) Fab-фрагмента антитела или scFv-фрагмент антитела, конъюгированного с первым партнером или элементом первой связывающей пары, где этот Fab-фрагмент или scFv специфически связывается с первым поверхностным маркером, (b) Fab-фрагмента антитела или scFv-фрагмента антитела, конъюгированного с первым элементом второй связывающей пары, где этот Fab-фрагмент или scFv-фрагмент антитела специфически связывается со вторым поверхностным маркером, и (с) линкера, содержащего на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары.
и, таким образом, получение биспецифического антитела.
Одним из аспектов, описанных в данном документе, является способ определения комбинации антигенсвязывающих сайтов, включающий следующие этапы:
(i) определение специфичности связывания и/или аффинности и/или эффекторной функции и/или периода полужизни in vivo множества биспецифических антител, полученных путем объединения каждого элемента первого множества Fab-фрагментов антител или scFv-фрагментов антител с каждым элементом второго множества Fab-фрагментов антител или scFv-фрагментов антител, и линкером, содержащим на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары,
где первое множество специфически связывается с первым маркером клеточной поверхности, а второе множество специфически связывается со вторым маркером клеточной поверхности,
и
(ii) выбор биспецифического антитела с подходящей специфичностью связывания и/или аффинностью и/или эффекторной функцией и/или периодом полужизни in vivo, и тем самым определение комбинации антигенсвязывающих сайтов.
В одном воплощении биспецифическое антитело представляет собой комплекс, содержащий
a) первый Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается с первым маркером клеточной поверхности, и
ii) который конъюгирован с первым элементом первой связывающей пары,
b) второй Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается со вторым маркером клеточной поверхности, и
ii) который конъюгирован с первым элементом второй связывающей пары, и
с) полинуклеотидный линкер,
i) который конъюгирован со вторым элементом первой связывающей пары, и
ii) который конъюгирован со вторым элементом второй связывающей пары.
Ниже приведены воплощения всех аспектов, описанных в данном документе.
В одном воплощении комплекс является нековалентным комплексом.
В одном воплощении комплекс также содержит эффекторную группировку, которая конъюгирована с полинуклеотидом, который комплементарен по меньшей мере части полинуклеотидного линкера.
В одном воплощении комплекс также содержит другой полипептид, i) который специфически связывается со второй мишенью, и ii) который конъюгирован с первым элементом второй связывающей пары, а полинуклеотидный линкер конъюгирован со вторым элементом второй связывающей пары.
В одном воплощении комплекс также содержит эффекторную группировку, конъюгированную с полинуклеотидом, который i) комплементарен по меньшей мере части полинуклеотида, который конъюгирован с первой эффекторной группировкой, и ii) не комплементарен полинуклеотидному линкеру.
В одном воплощении первый и второй Fab-фрагмент или scFv-фрагмент антитела связываются с одной и той же мишенью и с неперекрывающимися эпитопами на ней.
В одном воплощении полинуклеотидный линкер содержит от 8 до 1000 нуклеотидов. В одном воплощении полинуклеотидный линкер содержит от 10 до 500 нуклеотидов.
В одном воплощении полинуклеотидный линкер является энантиомерной ДНК. В одном воплощении энантиомерная ДНК является L-ДНК. В одном воплощении L-ДНК является одноцепочечной L-ДНК (оц-L-ДНК, ss-L-DNA).
В одном воплощении эффекторная группировка выбрана из группы, состоящей из связывающей группировки, меченой группировки и биологически активной группировки.
В одном воплощении полинуклеотидный линкер конъюгирован с Fab-фрагментом или scFv-фрагментом антитела на его первом или втором конце.
В одном воплощении полинуклеотидный линкер конъюгирован с двумя вторыми элементами двух связывающих пар, где второй элемент первой связывающей пары конъюгирован с первым концом полинуклеотидного линкера, а второй элемент второй связывающей пары конъюгирован со вторым концом полинуклеотидного линкера.
В одном воплощении первый и второй элементы первой связывающей пары содержат нуклеиновокислотные последовательности SEQ ID №05 и SEQ ID №08, соответственно.
В одном воплощении первый и второй элементы второй связывающей пары содержат нуклеиновокислотные последовательности SEQ ID №06 и SEQ ID №07, соответственно.
В одном воплощении способ включает следующие этапы:
a) синтез первого Fab-фрагмента или scFv-фрагмента антитела, который специфически связывается с первым маркером клеточной поверхности, и который конъюгирован с первым элементом первой связывающей пары,
b) синтез второго Fab-фрагмента или scFv-фрагмента антитела, который специфически связывается со вторым маркером клеточной поверхности, и который конъюгирован с первым элементом второй связывающей пары,
c) синтез полинуклеотидного линкера, который конъюгирован со вторым элементом первой связывающей пары и со вторым элементом второй связывающей пары, и
d) формирование комплекса путем объединения синтезированных компонентов.
Полипептид-полинуклеотидный комплекс
В данном документе сообщается о мультиспецифической связывающей молекуле, например, биспецифическом антителе, которая является комплексом, содержащим по меньшей мере два компонента, которые соединены путем нековалентного взаимодействия, в результате чего компоненты более устойчивы к протеолитической и ферментативной деградации in vivo, чем выделенная РНК или ДНК, особенно D-ДНК. Комплекс имеет высокую аффинность к своей мишени, используя авидность связывания, и имеет хорошую растворимость. Комплекс может быть использован для доставки к мишени одной или более чем одной эффекторной группировки.
Было обнаружено, что комплекс, содержащий смесь полипептидных и полинуклеотидных частей, особенно L-полинуклеотидных частей, отвечает этим требованиям и особенно подходит для доставки эффекторной группировки in vivo.
Если клетка-мишень имеет по меньшей мере две молекулы клеточной поверхности, то мультиспецифическая связывающая молекула (например, биспецифическое антитело), описанная в данном документе, содержит линкерный полинуклеотид и два или более двух полипептидов (связывающих группировок), которые специфически связываются с неперекрывающимися эпитопами, и она построена таким образом, что линкерный полинуклеотид имеет оптимальную длину для синергического связывания полипептидов, которые специфически связываются с этими молекулами клеточной поверхности.
Одним из аспектов, описанных в данном документе, является полипептид-полинуклеотидный комплекс следующей формулы:
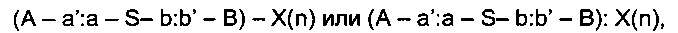
где А, а также В, являются связывающими группировками, которые специфически связываются с мишенью,
где а':а, а также b:b', являются связывающими парами, где а' и а не мешают связыванию b с b', и наоборот,
где S представляет собой линкерный полинуклеотид,
где (: X) обозначает эффекторную группировку, связанную ковалентно или через связывающую пару по меньшей мере с одним из a', a, b, b' или S,
где (n) представляет собой целое число,
где (-) представляет собой ковалентную связь, и
где (:) представляет собой нековалентную связь.
Также аспектом, описанным в данном документе, является способ получения полипептид-полинуклеотидного комплекса формулы:
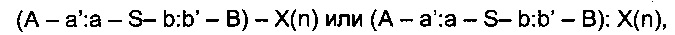
описанного выше, включающий следующие этапы:
а) синтез А-а' и b'-В, соответственно,
б) синтез линкера a-S-b, и
в) формирование комплекса с данной формулой,
где эффекторная группировка X связана по меньшей мере с одним из а', а, b, b' или S на этапе a), b) или c).
На основании отдельных компонентов может быть получен комплекс, описанный в данном документе, в соответствии со стандартными процедурами путем гибридизации между элементами связывающей пары, конъюгированными с отдельными компонентами комплекса.
Для того чтобы получить комплекс, например, со стехиометрией 1:1:1, комплекс может быть отделен с помощью хроматографии от других побочных продуктов конъюгации. Эта процедура может быть облегчена с помощью меченного красителем элемента связывающей пары и/или заряженного линкера. С помощью такого рода меченого и сильно отрицательного элемента связывающей пары моноконъюгированные связывающие группировки/полипептиды легко отделяются от немеченых связывающих группировок/полипептидов и связывающих группировок/полипептидов, которые несут более одного линкера, поскольку разница в заряде и молекулярном весе может быть использована для разделения. Флуоресцентная метка может быть использована для очистки комплекса от несвязанных компонентов, таких как меченый одновалентный связывающий элемент.
Одним из аспектов, описанных в данном документе, является способ получения комплекса «связывающая группировка - полинуклеотид», включающего следующие компоненты:
a) связывающую группировку, такую как полипептид, которая специфически связывается с мишенью и конъюгирована с первым элементом связывающей пары,
b) полинуклеотидный линкер, конъюгированный на своем первом конце со вторым элементом связывающей пары, и
c) эффекторную группировку, конъюгированную с полинуклеотидом, который комплементарен по меньшей мере части полинуклеотидного линкера,
при этом способ включает следующие этапы: а) синтез i) связывающей группировки, специфически связывающейся с мишенью и конъюгированной с первым элементом связывающей пары, и ii) эффекторной группировки, конъюгированной с полинуклеотидом, который комплементарен по меньшей мере части полинуклеотидного линкера, соответственно, b) синтез полинуклеотидного линкера, конъюгированного на своем первом конце со вторым элементом связывающей пары, и с) формирование комплекса «связывающая группировка - полинуклеотид» путем гибридизации синтезированных компонентов.
Другой аспект, описанный в данном документе, представляет собой способ получения комплекса «связывающая группировка - полинуклеотид», включающего следующие компоненты:
a) первую связывающую группировку, такую как полипептид, которая специфически связывается с первой мишенью и конъюгирована с первым элементом первой связывающей пары,
b) вторую связывающую группировку, такую как полипептид, которая специфически связывается с второй мишенью и конъюгирована с первым элементом второй связывающей пары, и
c) полинуклеотидный линкер, конъюгированный на своем первом конце со вторым элементом первой связывающей пары и на своем втором конце со вторым элементом второй связывающей пары,
при этом способ включает следующие этапы: а) синтез первой связывающей группировки, специфически связывающейся с первой мишенью и конъюгированной с первым элементом первой связывающей пары, и второй эффекторной группировки, специфически связывающейся со второй мишенью и конъюгированной с первым элементом второй связывающей пары, соответственно, и b) синтез полинуклеотидного линкера, конъюгированного на своем первом конце со вторым элементом первой связывающей пары, и на своем втором конце со вторым элементом второй связывающей пары, и с) формирование комплекса «связывающая группировка - полинуклеотид» путем гибридизации синтезированных компонентов.
Комплекс может дополнительно содержать один или несколько противоионов Y, чтобы уравнять заряд. Примерами подходящих отрицательно заряженных противоионов являются галогениды, OH-, карбонатная, алкилкарбоксилатная, например, трифторацетатная, сульфатная, гексафторфосфатная и тетрафторборатная группы. Особенно подходящими являются гексафторфосфатная, трифторацетатная и тетрафторборатная группы. Другими подходящими положительно заряженными противоионами являются одновалентные катионы, такие как ионы щелочных металлов и/или ионов аммония.
Полная библиотека комплексов, представленных в данном документе, может быть легко получена и проанализирована, и подходящий связывающий агент из такой библиотеки может быть получен в большом объеме, если требуется.
Понятие библиотеки относится к набору комплексов, представленных в данном документе, где связывающая группировка, длина полинуклеотидного линкера регулируются таким образом, чтобы наилучшим образом соответствовать требованиям, установленным для связывающего агента.
Например, можно легко использовать для первого применения набор полинуклеотидных линкеров, охватывающий весь спектр от 1 нм до 100 нм с шагом примерно 10 нм. Затем в районе самой подходящей длины, полученной в первом раунде, длину линкера снова легко доработать.
В данном документе также сообщается о способе выбора комплекса «связывающая группировка - полинуклеотид» из библиотеки, содержащей множество комплексов с полинуклеотидным линкером различной длиной. В одном воплощении этого способа несколько линкерных молекул с полинуклеотидными линкерами различной длины синтезируют и используют для формирования комплекса, описанного в данном документе, который содержит полинуклеотидные линкеры различной длины, и выбирают те комплексы, которые имеют улучшение Kдисс по меньшей мере в 5 раз по сравнению с лучшей из двух одновалентных полипептидных связующих молекул. В одном воплощении выбор двухвалентного связывающего агента с нужной Kдисс осуществляют с помощью анализа BIAcore, описанного в примерах.
Одним из аспектов, описанных в данном документе, является комплекс, содержащий
а) связывающую группировку (например, полипептид), которая специфически связывается с первой мишенью и которая конъюгирована с первой одноцепочечной L-ДНК-группировкой,
б) вторую связывающую группировку (например, полипептид), которая специфически связывается со второй мишенью и которая конъюгирована со второй одноцепочечной L-ДНК-группировкой, и
с) линкер, который содержит на его первом (или 3') конце первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке, и который содержит на его втором (или 5') конце вторую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
Одним из аспектов, описанных в данном документе, является комплекс, включающий
a) FAB-фрагмент антитела или scFv, который специфически связывается с первой мишенью и который конъюгирован с первой одноцепочечной L-ДНК-группировкой
b) FAB-фрагмент антитела или scFv, который специфически связывается со второй мишенью и который конъюгирован со второй одноцепочечной L-ДНК-группировкой, и
с) линкер, который содержит на его первом (или 3') конце первую одноцепочечную L-ДНК-линкерную группировку, комплементарную первой одноцепочечной L-ДНК-группировке, и который содержит на его втором (или 5') конце вторую одноцепочечную L-ДНК-линкерную группировку, комплементарную второй одноцепочечной L-ДНК-группировке.
Первая одноцепочечная L-ДНК-группировка не гибридизуется со второй одноцепочечной L-ДНК-группировкой и не гибридизуется со второй одноцепочечной L-ДНК-линкерной группировкой. В свою очередь, вторая одноцепочечная L-ДНК-группировка не гибридизуется с первой одноцепочечной L-ДНК-группировкой и не гибридизуется с первой одноцепочечной L-ДНК-линкерной группировкой.
Далее приведены воплощения всех аспектов, представленных в данном документе:
В одном воплощении связывающая группировка, которая специфически связывается с мишенью, является антителом или фрагментом антитела. В одном воплощении фрагмент антитела представляет собой Fab.
В одном воплощении первая и/или вторая одноцепочечная L-ДНК-группировка имеет длину от 10 до 50 нуклеотидов. В одном воплощении длина составляет от 15 до 35 нуклеотидов. В одном воплощении длина составляет от 20 до 30 нуклеотидов.
В одном воплощении линкер содержит первую одноцепочечную L-ДНК-линкерную группировку, вторую одноцепочечную L-ДНК-линкерную группировку и одноцепочечную стыковочную группировку. В одном воплощении линкер также содержит линейную ненуклеотидную группировку. В одном воплощении линейная ненуклеотидная группировка представляет собой полипептид или неионный полимер. В одном воплощении неионный полимер является линейным полиэтиленгликолем. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 100 единиц этиленгликоля. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 50 единиц этиленгликоля. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 25 единиц этиленгликоля.
В одном воплощении комплекс включает
a) полипептид, который специфически связывается с первой мишенью и который конъюгирован с первой одноцепочечной L-ДНК-группировкой,
b) полипептид, который специфически связывается со второй мишенью и который конъюгирован со второй одноцепочечной L-ДНК-группировкой, и
с) линкер, который содержит на его первом (или 3') конце первую одноцепочечную L-ДНК-линкерную группировку, комплементарную первой одноцепочечной L-ДНК-группировке, и который содержит на его втором (или 5') конце вторую одноцепочечную L-ДНК-линкерную группировку, комплементарную второй одноцепочечной L-ДНК-группировке, и который содержит третью одноцепочечную L-ДНК-линкерную группировку между первой и второй одноцепочечными L-ДНК-группировками.
В одном воплощении линкер содержит в направлении от 3' к 5':
- первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке,
- стыковочную одноцепочечную L-ДНК-группировку, и
- вторую одноцепочечную L L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
Стыковочная одноцепочечная L-ДНК-группировка не гибридизуется с первой одноцепочечной L-ДНК-группировкой или комплементарной ей первой одноцепочечной линкерной группировкой, и не гибридизуется со второй одноцепочечной L-ДНК-группировкой или комплементарной ей второй одноцепочечной L-ДНК-линкерной группировкой.
В одном воплощении линкер содержит в направлении от 3' к 5':
- первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке,
- линейную ненуклеотидную группировку,
- стыковочную одноцепочечную L-ДНК-группировку и
- вторую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
В одном воплощении линкер содержит в направлении от 3' к 5':
- первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке,
- стыковочную одноцепочечную L-ДНК-группировку,
- ненуклеотидную группировку и
- вторую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
В одном воплощении линкер содержит в направлении от 3' к 5':
- первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке,
- ненуклеотидную группировку,
- стыковочную одноцепочечную L-ДНК-группировку и
- вторую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
В одном воплощении линкер содержит в направлении от 3' к 5':
- первую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна первой одноцепочечной L-ДНК-группировке,
- первую ненуклеотидную группировку,
- стыковочную одноцепочечную L-ДНК-группировку,
- вторую ненуклеотидную группировку,
- вторую одноцепочечную L-ДНК-линкерную группировку, которая комплементарна второй одноцепочечной L-ДНК-группировке.
В одном воплощении первая ненуклеотидная группировка и вторая ненуклеотидная группировка являются одинаковыми или различными. В одном воплощении линейная ненуклеотидная группировка представляет собой полипептид или неионный полимер. В одном воплощении неионный полимер является линейным полиэтиленгликолем. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 100 единиц этиленгликоля. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 50 единиц этиленгликоля. В одном воплощении линейный полиэтиленгликоль содержит от 1 до 25 единиц этиленгликоля.
Компонент связывающей группировки
Методики моноклональных антител делают возможной продукцию специфически связывающих агентов в форме специфически связывающихся моноклональных антител или их фрагментов. Для создания моноклональных антител или их фрагментов могут быть использованы такие методики как иммунизация мышей, кроликов, хомячков или любых других млекопитающих полипептидом, т.е. мишенью антитела, и/или нуклеиновой кислотой, кодирующей полипептид. Альтернативно, моноклональные антитела или их фрагменты могут быть получены с использованием фаговых библиотек scFv (одноцепочечная вариабельная область), в частности человеческих scFv (см., например, US 5885793, WO 92/01047, WO 99/06587).
В одном воплощении связывающая группировка, которая специфически связывается с мишенью, является фрагментом одновалентного антитела. В одном воплощении фрагмент одновалентного антитела получен из моноклонального антитела.
Фрагменты одновалентных антител включают, но не ограничиваясь ими, Fab, Fab'-SH, однодоменное антитело, F(ab')2) Fv- и scFv-фрагменты. Таким образом, в одном воплощении фрагмент одновалентного антитела выбран из группы, включающей Fab, Fab'-SH, однодоменное антитело, F(ab')2, Fv- и scFv-фрагменты.
В одном воплощении по меньшей мере одна из связывающих группировок комплекса, описанного в данном документе, представляет собой однодоменное антитело, или Fab-фрагмент, или Fab'-фрагмент моноклонального антитела.
В одном воплощении обе связывающие группировки комплекса, описанного в данном документе, независимо друг от друга представляют собой однодоменное антитело, или Fab-фрагмент, или Fab'-фрагмент моноклонального антитела.
В одном воплощении обе связывающие группировки комплекса, описанного в данном документе, являются однодоменными антителами, или Fab-фрагментами, или Fab'-фрагментами.
В одном воплощении мишени или эпитопы, которые специфически связываются связывающими группировками, не перекрываются.
Димерные антитела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими (см., например, ЕР 0404097, WO 93/01161, Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134, и Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448). Тримерные и тетрамерные антитела также описаны в Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134.
Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В некоторых воплощениях однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Уолтем, Массачусетс; US 6248516).
Fv представляет собой минимальный фрагмент антитела, который содержит полный антигенсвязывающий сайт и лишен константной области. Обзор scFv см., например, в Plueckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore (eds.), (Springer-Verlag, New York, 1994), pp. 269-315, WO 93/16185, US 5571894, US 5587458. Как правило, шесть гипервариабельных областей (HVR) придают антигенсвязывающую специфичность антителу. Тем не менее, даже один вариабельный домен (или половина Fv, содержащая только три HVR, специфичных к антигену) обладает способностью распознавать и связывать его антиген.
В одном воплощении фрагменты одновалентных антител являются двухцепочечными Fv-видами, состоящими из димера вариабельных доменов одной тяжелой и одной легкой цепи в тесной нековалентной ассоциации.
В одном воплощении фрагменты одновалентных антител являются одноцепочечными Fv (scFv)-видами, состоящими из вариабельных доменов одной тяжелой цепи и одной легкой цепи, ковалентно связанных посредством гибкого пептидного линкера.
Fab-фрагмент антитела содержит вариабельные домены тяжелой цепи и легкой цепи, а также константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи.
Fab'-фрагменты отличаются от Fab-фрагмента добавлением нескольких остатков на карбокси-конце СН1-домена тяжелой цепи, включая один или более чем один цистеин из шарнирной области антитела.
Fab'-SH обозначает Fab', в котором цистеиновый остаток (остатки) константных доменов несут свободную тиольную группу.
Различные методики были разработаны для получения фрагментов антител. Традиционно фрагменты антител могут быть получены путем протеолитического расщепления полноразмерных антител (см., например, Morimoto, K., et al., J. Biochem. Biophys. Meth. 24(1992) 107-117, Brennan, M., et al., Science 229 (1985) 81-83). Например, расщепление полноразмерных антител папаином дает два одинаковых антигенсвязывающих фрагмента, называемых «Fab»-фрагментами, каждый с одним антигенсвязывающим сайтом, и остаточный "Fc"-фрагмент, название которого отражает его способность легко кристаллизоваться. Обзор некоторых фрагментов антител см. в Hudson, P.J., et al., Nat. Med. 9 (2003) 129-134.
Фрагменты антител также могут быть получены напрямую рекомбинантными способами. Фрагменты антител Fab, Fv и scFv могут быть экспрессированы и секретированы, например, из Е. coli, таким образом, позволяя легко получить большие количества этих фрагментов. Фрагменты антител могут быть выделены из фаговых библиотек антител в соответствии со стандартными процедурами. Альтернативно, Fab'-SH-фрагменты могут быть непосредственно выделены из E. coli. (Carter, P., et al., Bio/Technology 10 (1992) 163-167). Системы клеток млекопитающих также можно использовать для экспрессии и, при желании, для секреции фрагментов антител.
В одном воплощении связывающая группировка, которая специфически связывается с антигеном, представляет собой однодоменное антитело. В некоторых воплощениях однодоменное антитело представляет собой человеческое однодоменное антитело (см., например, US 6248516). В одном воплощении однодоменное антитело состоит из всей или части вариабельного домена тяжелой цепи антитела.
Однодоменное антитело представляет собой одноцепочечную полипептидную цепь, содержащую весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела.
В некоторых воплощениях связывающая группировка связывается с ее мишенью с константой диссоциации (KD) ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ или ≤ 0,001 нМ (например 10-8 М или менее, например от 10-8 М до 10-13 М, например от 10-9 М до 10-13 М).
В некоторых воплощениях связывающая группировка связывается с ее мишенью с константой диссоциации (KD) от 10-5 М до 10-13 М, или от 10-5 М до 10-10 М, или от 10-5 М до 10-8 М.
В одном воплощении, в котором связывающая группировка представляет собой антитело или фрагмент антитела, константа диссоциации определяется в анализе связывания антигена, меченного радиоактивным изотопом (RIA), проводимом с Fab-фрагментом антитела и его антигеном, как описано далее.
Аффинность связывания Fab с антигеном в растворе измеряют, уравновешивая Fab с минимальной концентрацией (125I)-меченого антигена в присутствии серии титров немеченого антигена с последующим захватом связанного антигена на планшете, покрытом анти-Fab-антителом (см., например, Chen, Y., et al., J. Mol. Biol. 293 (1999) 865-881). Для создания условий анализа многолуночные планшеты MICROTITER® (Thermo Scientific) покрывают в течение ночи 5 мкг/мл захватывающего анти-FAB антитела (Cappel Labs) в 50 мМ растворе карбоната натрия (pH 9,6), а затем блокируют 2% (вес/объем) раствором бычьего сывороточного альбумина в PBS в течение 2-5 часов при комнатной температуре (примерно 23°С). В неадсорбирующем планшете (Nunc №269620) смешивают 100 пМ или 26 пМ [125I]-антигена с серийными разведениями Fab, представляющего интерес (например, совместимого с оценкой анти-VEGF-антитела, Fab-12, в Presta, L.G., et al., Cancer Res. 57 (1997) 4593-4599). Затем FAB, представляющий интерес, инкубируют в течение ночи; тем не менее, инкубацию можно продолжать в течение более долгого периода (например, примерно 65 часов) для гарантии того, что равновесие достигнуто. После этого смеси переносят в планшет для захвата и инкубируют при комнатной температуре (например, в течение одного часа). Затем раствор удаляют и планшет промывают восемь раз 0,1% раствором полисорбата 20 (TWEEN-20®) в PBS. После высушивания планшетов добавляют 150 мкл/лунка сцинтиллянта (MICROSCINT-20™; Packard) и подсчитывают на гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Выбирают концентрации каждого FAB, которые дают 20% или меньше от максимального связывания, для использования в анализах на конкурентное связывание.
Согласно другому воплощению константа диссоциации определяется с помощью анализов поверхностного плазмонного резонанса с использованием BIACORE®-2000 или BIACORE®-3000 или BIACORE® А-100 (BIAcore, Inc., Пискатавэй, Нью-Джерси) при 25°C с иммобилизованным антигеном на чипе СМ5 примерно на уровне 10 единиц ответа (RU).
Вкратце, биосенсорные чипы с карбоксиметилированным декстраном (СМ5, BIACORE, Inc) активируют N-этил-N'-(3-диметиламинопропил)карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят в 10 мМ растворе ацетата натрия, pH 4,8, до концентрации 5 мкг/мл (примерно 0,2 мкМ) и вводят со скоростью потока 5 мкл/мин до достижения примерно 10 единиц ответа (RU) связанного белка. После введения антигена вводят 1 М этаноламин для блокировки непрореагировавших групп. Для измерений кинетики двукратные серийные разведения FAB (от 0,78 нМ до 500 нМ) вводят в PBS с 0,05% поверхностно-активного вещества полисорбата 20 (TWEEN-20™) (PBST) при 25°C со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывают с помощью простой модели связывания Ленгмюра 1:1 (программное обеспечение BIACORE® Evaluation Software версии 3.2), одновременно выстраивая сенсограммы ассоциации и диссоциации. Константа равновесия диссоциации (KD) рассчитывается как отношение koff/kon (см., например, Chen, Y. et al., J. Mol. Biol. 293 (1999) 865-881). Если on-скорость превышает 106 M-1 с-1 в анализе поверхностного плазмонного резонанса, описанном выше, то on-скорость может быть определена с помощью методики тушения флуоресценции, которая измеряет увеличение или уменьшение интенсивности флуоресцентного излучения (возбуждение = 295 нм; излучение = 340 нм; 16 нм полосовой фильтр) при 25°C с 20 нМ антитела против антигена (FAB-форма) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена, измеряемых спектрометром, таком как спектрофотометр, оборудованный системой быстрого смешивания с последующей остановкой (stop-flow) (Aviv Instruments) или спектрофотометр SLM-AMINCO™ серии 8000 (ThermoSpectronic) со встряхивающейся кюветой.
В случае, когда две связывающие молекулы распознают два независимых сайта связывания, может иметь место явление кооперативного связывания, которое может находиться в зависимости от длины полинуклеотидного линкера.
Эффект кооперативного связывания физически характеризуется тем, что свободные энергии связывания ΔG°1 и ΔG°2 суммируются, образуя ΔG°кооп: ΔG°1+ΔG°2=ΔG°кооп.
В соответствии с уравнением Гиббса ΔG°кооп = -RTInKDкооп, ΔG°кооп формирует продукт из аффинностей KD1 и KD2.
Повышение свободной энергии связывания Гиббса за счет кооперативности резко увеличивает аффинность связывания (KDкооп) и специфичность связывания.
Специфичность связывания дополнительно увеличивается, когда адресные сайты связывания независимо расположены на двух разных молекулах-мишенях, которые, например, могут быть совместно локализованы на поверхности опухолевой клетки.
Связывающая группировка, специфически связывающаяся с мишенью, вероятно, несет одну или более чем одну свободную ОН-, СООН-, NH2- и/или SH-группу, которая потенциально может вступать в реакцию с определенными реагентами сочетания. Чтобы избежать (побочных) реакций во время реакции конъюгации, можно выбрать одну из реакций сочетания, приведенных в следующей таблице 1.
В таблице 1 представлен обзор реакционноспособных групп для ковалентного связывания полипептидов с соответствующим элементом связывающей пары, а также для ковалентного связывания линкера с соответствующими элементами связывающей пары.
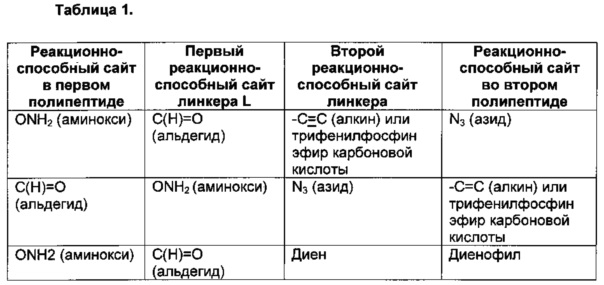
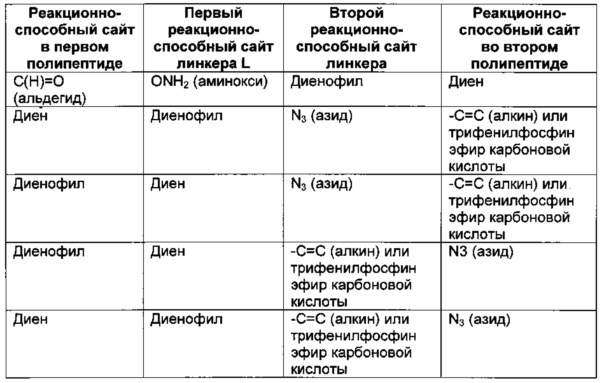
Приведенные выше биортогональные реакции сочетания особенно подходят для конъюгации одновалентных связывающих полипептидов. Если два связывающихся элемента не несут определенных реакционноспособных функциональных групп, например, в случае комбинации двух аптамеров, то есть больше свободы в выборе реакционноспособных сайтов. Поэтому в дополнение или в комбинации с парами соответствующих реакционноспособных сайтов, указанных в приведенной выше таблице, может быть использован амино/активный сложный эфир (например, сложный эфир NHS) и SH/SH или SH/малеинимидо для ортогонального сочетания.
Одновалентный связывающий полипептид также может быть синтетическим пептидом или пептидомиметиком. В случае если полипептид химически синтезирован, то аминокислоты с ортогональной химической реактивностью могут быть включены в ходе такого синтеза (см., например, de Graaf, A.J., et al., Bioconjug. Chem. 20 (2009) 1281-1295). Поскольку большое разнообразие ортогональных функциональных групп имеется и может быть введено в синтетический пептид, конъюгация такого пептида с линкером является стандартной химической реакцией.
Полинуклеотидный компонент
Комплекс, описанный в данном документе, включает (полинуклеотидный) линкер. Линкер либо может быть ковалентно связан с полипептидом(ами), либо (полинуклеотидный) линкер и полипептид(ы) могут быть связаны друг с другом с помощью специфической связывающей пары.
Когда используются (полинуклеотидные) линкеры различной длины, в результате могут быть получены сложные конструкции с различными расстояниями между первым и вторым полипептидом, специфически связывающиеся с мишенью. Это обеспечивает оптимальное расстояние и/или гибкость.
Термин «полинуклеотид» понимается в широком смысле и включает ДНК и РНК, а также их аналоги и модификации.
В одном воплощении полинуклеотидный линкер состоит из смеси различных типов мономеров до тех пор, пока более 20% мономеров являются нуклеозидами. В одном воплощении полинуклеотидный линкер состоит из смеси различных типов мономеров до тех пор, пока более 30% мономеров являются нуклеозидами. В одном воплощении полинуклеотидный линкер состоит из смеси различных типов мономеров до тех пор, пока более 40% мономеров являются нуклеозидами. В одном воплощении полинуклеотидный линкер состоит из смеси различных типов мономеров до тех пор, пока более 50% мономеров являются нуклеозидами.
Например, линкер может состоять исключительно из нуклеозидов, или он может быть смесью нуклеозидов и аминокислот, и/или сахарных остатков, и/или диолов, и/или фосфосахарных звеньев, и/или неионных полимерных структурных блоков.
Олигонуклеотид может содержать, например, замещенный нуклеотид, несущий заместитель в стандартных основаниях дезоксиаденозине (dA), дезоксигуанозине (dG), дезоксицитозине (dC), дезокситимидине (dT), дезоксиурациле (dU). Примерами таких замещенных нуклеотидов являются 5-замещенные пиримидины (такие как 5-метил-dC, аминоаллил-dU или -dC, 5-(аминоэтил-3-акрилимидо)-dU, 5-пропинил-dU или -dC), 5-галогензамещенный dU или -dC, N-замещенные пиримидины (например, N4-этил-dC), N-замещенные пурины (например, N6-этил-dA, N2-этил-dG), 8-замещенные пурины (например, 8-[6-амино)-гекс-1-ил]-8-амино-dG или -dA), 8-галогензамещенный dA или -dG, 8-алкил-dG или -dA и 2-замещенный dA (например, 2-амино-dA).
Олигонуклеотид может содержать нуклеотидный или нуклеозидный аналог. Т.е. природные нуклеотиды можно заменить на нуклеотидные аналоги, такие как 5-нитроиндол-D-рибозид, 3-нитро-пиррол-D-рибозид, дезоксиинозин (dI), дезоксиксантоксин (dX), 7-деаза-dG, -dA, -dI или -dX, 7-деаза-8-аза-dG, -dA, -dI или -dX, 8-аза-dA, -dG, -dI или -dX, D-формицин, псевдо-dU, псевдо-изо-dC, 4-тио-dT, 6-тио-dG, 2-тио-dT, изо-dG, 6-метил-изо-dC, N8-связанный 8-аза-7-деаза-dA, 5,6-дигидро-5-аза-dC, этено-dA или пиролло-dC. Как очевидно специалисту в данной области, олигонуклеотид в комплементарной нити должен быть выбран таким образом, чтобы образование дуплекса было специфичным. Если, например, в одной нити используется 5-метил-изо-dC (например, (а)), то в комплементарной цепи должен находиться изо-dG (например (а')).
В одном воплощении олигонуклеотидный остов линкера модифицирован так, что он содержит замещенные остатки сахаров, аналоги сахаров, модификации в межнуклеозидной фосфатной группировке и/или ПНК (с основной цепью без фосфата и d-рибозы).
Олигонуклеотид, например, может содержать нуклеотид с замещенной дезоксирибозой, такой как 2'-метокси-, 2'-фтор-, 2'-метилселено-, 2'-аллилокси-, 4'-метил-dN (где N представляет собой нуклеотид, например, A, G, С, Т или U).
Аналоги Сахаров представляют собой, например, ксилозу, рибозу с 2',4'-мостиком, таким как (2'-O, 4'-С метилен) (олигомер, известный как LNA) или (2'-O, 4'-С-этилена) (олигомер, известный также ENA), L-рибозу, L-D-рибозу, гекситол (олигомер, известный как HNA), циклогексенил (олигомер, известный как CeNA), алтритол (олигомер, известный как ANA), аналог трициклической рибозы, в котором атомы С3' и С5' соединены этиленовым мостиком, который конденсирован с циклопропановым кольцом (олигомер, известный как трицикло-ДНК), глицерин (олигомер, известный как GNA), глюкопиранозу (олигомер, известный как Homo DNA), карбарибозу (с циклопентаном вместо тетрагидрофурановой субъединицы), гидроксиметилморфолин (олигомеры, известные как морфолино-ДНК).
Также известно большое число модификаций межнуклеозидной фосфатной группировки, чтобы не мешать гибридизационным свойствам, и такие модификации остова также могут быть объединены с замещенными нуклеотидами или нуклеотидными аналогами. Примерами являются фосфортиоатные, фосфордитиоатные, фосфорамидатные и метилфосфонатные олигонуклеотиды.
Вышеуказанные модифицированные нуклеотиды, нуклеотидные аналоги, а также полинуклеотидные модификации остова могут быть при желании объединены в полинуклеотиде, содержащемся в комплексе, описанном в данном документе.
(Полинуклеотидный) линкер имеет длину от 1 нм до 100 нм. В одном воплощении (полинуклеотидный) линкер имеет длину от 4 нм до 80 нм. В одном воплощении (полинуклеотидный) линкер имеет длину от 5 нм до 50 нм или от 6 нм до 40 нм. В одном воплощении (полинуклеотидный) линкер имеет длину 10 нм или более или 15 нм или более. В одном воплощении (полинуклеотидный) линкер имеет длину от 10 нм до 50 нм.
В одном воплощении участники связывающей пары, конъюгированные с (полинуклеотидным) линкером, имеют длину по меньшей мере 2,5 нм каждый.
Длина (полинуклеотидного) линкера может быть рассчитана с использованием известных расстояний связей и валентных углов компонентов, которые химически подобны группировкам. Такие длины связей приведены для некоторых молекул в стандартных руководствах (см., например, CRC Handbook of Chemistry and Physics, 91st edition (2010-2011), раздел 9).
При расчете длины спейсера или линкера применяются следующие аппроксимации: а) для вычисления длины ненуклеозидных группировок используется средняя длина связи 130 пм с углом связи 180° независимо от характера связанных атомов, b) один нуклеотид в одной цепи считается 500 пм, и в) один нуклеотид в двойной цепи считается 330 пм.
Значение 130 пм основано на вычислении расстояния между двумя концевыми атомами углерода в цепи C(sp3)-C(sp3)-C(sp3) с углом связей 109°28' и расстоянием 153 пм между двумя C(sp3), которое составляет примерно 250 пм, что переводится с предполагаемым углом связей 180° и длиной связи между двумя C(Sp3) 125 пм. Принимая во внимание, что гетероатомы, такие как Р и S, и С-атомы sp2 и sp1 также могут быть частью линкера, берется значение 130 пм. Если линкер содержит циклическую структуру, такую как циклоалкил или арил, то расстояние вычисляется аналогичным образом путем подсчета числа связей циклической структуры, которые являются частью общей цепи атомов, которые определяют расстояние.
Длина (полинуклеотидного) линкера в комплексе, описанном в данном документе, при желании может быть изменена. Для того чтобы легко сделать доступными линкеры различной длины, т.е. библиотеку, можно иметь простой синтетический доступ к различным линкерам такой библиотеки. Подходящим является комбинаторный твердофазный синтез линкера. Поскольку линкеры должны быть синтезированы с длиной примерно до 100 нм, стратегию синтеза выбирают таким образом, что мономерные синтетические структурные блоки собирают в ходе твердофазного синтеза с высокой эффективностью. Синтез дезоксиолигонуклеотидов на основании сборки фосфорамидита в качестве мономерных структурных блоков отвечает этому требованию. В таком линкере мономерные единицы в пределах линкера соединены в каждом случае через фосфатную группировку или аналог фосфата.
(Полинуклеотидный) линкер может содержать, в качестве одного воплощения, свободные положительно и/или отрицательно заряженные группы полифункциональных аминокарбоновых кислот, например, амино-, карбоксилатные или фосфатные группы. Например, носители заряда могут быть получены из трифункциональных аминокарбоновых кислот, которые содержат а) аминогруппу и две карбоксилатные группы, или b) две аминогруппы и одну карбоксилатную группу. Примерами таких трифункциональных аминокарбоновых кислот являются лизин, орнитин, гидроксилизин, а,В-диаминопропионовая кислота, аргинин, аспарагиновая кислота и глутаминовая кислота, карбоксиглутаминовая кислота и симметричные трифункциональные карбоновые кислоты, такие как описанные в ЕР 0618192 или US 5519142. Альтернативно, одну из карбоксилатных групп в трифункциональных аминокарбоновых кислотах можно заменить фосфатной, сульфонатной или сульфатной группой. Примером такой трифункциональной аминокислоты является фосфосерин.
(Полинуклеотидный) линкер также может содержать, в качестве одного воплощения, незаряженные гидрофильные группы. Подходящими примерами незаряженных гидрофильных групп являются этиленоксидные или полиэтиленоксидные группы, содержащие, в частности, по меньшей мере три структурных блока, таких как этиленоксид, сульфоксид, сульфон, амид карбоновой кислоты, эфир карбоновой кислоты, амид фосфоновой кислоты, сложный эфир фосфоновой кислоты, амид фосфорной кислоты, сложный эфир фосфорной кислоты, амид сульфоновой кислоты, сложный эфир сульфоновой кислоты, амид серной кислоты и сложный эфир серной кислоты. Амидные группы в одном воплощении являются первичными амидными группами, особенно остатками амидов карбоновых кислот в боковых группах аминокислот, например, аминокислот аспарагина и глутамина. Сложные эфиры, в частности, получают из гидрофильных спиртов, в частности С1-С3-спиртов, или диолов, или триолов.
Энантиомерная L-ДНК известна своей ортогональной гибридизацией, устойчивостью к нуклеазам и простотой синтеза полинуклеотидов переменной длины.
В одном воплощении все полинуклеотиды в комплексе представляют собой энантиомерную L-ДНК или L-PHK. В одном воплощении все полинуклеотиды в комплексе представляют собой энантиомерную L-ДНК.
Энантиомерная одноцепочечная L-ДНК (оц-L-ДНК) сочетает в себе высокую молекулярную гибкость и стабильность в жидкостях организма. При использовании одноцепочечной L-ДНК в качестве линкера между двумя или более независимыми связывающими молекулами эти связывающие молекулы могут привыкнуть к практически любому углу связывания и расстоянию связывания, которые зависят только от длины оц-L-ДНК-линкера.
В одном воплощении (полинуклеотидный) линкер синтезируют по сегментам, которые могут гибридизоваться друг с другом.
В этом случае линкер может быть образован путем гибридизации сегментов друг с другом. Полученная линкерная конструкция включает олигонуклеотидные дуплексные части. В случае если линкер построен таким образом, последовательность гибридизуемой полинуклеотидной группировки, формирующей дуплекс, выбирается таким образом, чтобы не произошла гибридизация или влияние на нуклеиновые кислоты связывающей пары.
В одном воплощении полинуклеотидный линкер синтезируют по сегментам оц-L-ДНК, например, А и В, которые могут гибридизоваться друг с другом.
В этом случае полинуклеотидный линкер может быть построен путем гибридизации сегментов друг с другом. Таким образом, длина линкера может самостоятельно регулироваться согласно расстоянию между двумя сайтами связывания просто путем последовательного применения структурных блоков, образующих конкатемер, т.е. А и В в качестве примера. Линкер характеризуется тем, что нуклеиновокислотные концы установленного линкера гибридизуются с оц-L-ДНК-мечеными связывающими молекулами при температуре ниже точки плавления (т.е. ТМ1), чем межконкатемерная температура плавления (т.е. ТМ2, таким образом ТМ2>ТМ1). Для анализа окончательной длины полноразмерного линкера полученный комплекс инкубировали при третьей температуре (т.е. ТМ3), которая выше первой температуры плавления, но ниже второй температуры плавления (т.е. ТМ3>ТМ1 и ТМ3<ТМ2). Температурно элюированный линкер может быть проанализирован с помощью стандартных способов, например, с помощью агарозного геля, окрашенного этидия бромидом. Длина линкера также может быть рассчитана, так как известна длина каждого конкатемера. В одном воплощении отдельные конкатемеры могут быть помечены.
Части дуплекса могут повышать ригидность олигонуклеотидного линкера. Это может быть использовано для уменьшения подвижности и гибкости линкера.
В одном воплощении один или более L-ДНК-олигонуклеотид гибридизуют с олигонуклеотидным L-ДНК-линкером.
В этом воплощении олигонуклеотидный линкер имеет повышенную ригидность за счет образования L-ДНК-дуплекса.
В одном воплощении гибрид L-ДНК/полиэтиленгликоль используется в качестве (олигонуклеотидного) линкера.
В одном воплощении гибрид L-ДНК/D-ДНК/полиэтиленгликоль используется в качестве (олигонуклеотидного) линкера.
В одном воплощении гибрид L-ДНК/D-ДНК/полиэтиленгликоль/полипептид используется в качестве (олигонуклеотидного) линкера.
В одном воплощении один или более L-ДНК-олигонуклеотид гибридизован с L-ДНК/полиэтиленгликоль-гибридным (олигонуклеотидным) линкером.
В одном воплощении один или более L-ДНК-олигонуклеотид, который ковалентно связан с молекулой полиэтиленгликоля переменной длины, гибридизован с олигонуклеотидом поли-L-ДНК-этиленгликоль-гибридным (олигонуклеотидным) линкером.
В одном воплощении гибрид L-ДНК/D-ДНК используется в качестве (олигонуклеотидного) линкера.
В одном воплощении гибрид L-ДНК/D-ДНК используется в качестве (олигонуклеотидного) линкера, где один или более D-ДНК-олигонуклеотид гибридизован с олигонуклеотидой D-ДНК-частью (олигонуклеотидного) линкера с образованием двухцепочечной D-ДНК.
В одном воплощении гибрид L-ДНК/D-ДНК используется в качестве линкера, где один или более L-ДНК-олигонуклеотид гибридизован с олигонуклеотидной L-ДНК-частью (олигонуклеотидного) линкера с образованием двухцепочечной L-ДНК.
Образование двухцепочечных, т.е. спиральных, ДНК-дуплексов можно использовать для модификации или регулировки периода полужизни комплекса in vivo, делая его доступным ферментативной активности нуклеаз.
Простой способ построить (полинуклеотидный) линкер заключается в использовании стандартных D- или L-нуклеозид-фосфорамидитных структурных блоков.
В одном воплощении используется одноцепочечный участок dT.
Это выгодно, потому что dT не требует введения в основание защитной группы.
Гибридизация может быть использована для того, чтобы варьировать длину (полинуклеотидного) линкера (расстояние между элементами связывающей пары на концах полинуклеотидного линкера) и гибкость спейсера, так как длина двухцепочечной нити уменьшается по сравнению с одиночной нитью, и двухцепочечная нить является более жесткой, чем одиночная нить.
Для гибридизации в одном воплощении используются олигонуклеотиды, модифицированные функциональной группировкой.
Олигонуклеотид, используемый для гибридизации, может иметь один или два концевых удлинения, не гибридизующихся с линкером, и/или может быть разветвленным внутри. Такие концевые удлинения, которые не гибридизуются с линкером (и не мешают элементам связывающих пар), могут быть использованы для дальнейших событий гибридизации.
В одном воплощении олигонуклеотид, гибридизующийся с концевым удлинением, является олигонуклеотидом, содержащим эффекторной группировку.
Этот меченый олигонуклеотид опять может содержать концевые удлинения или быть разветвленным, чтобы обеспечить дальнейшую гибридизацию, таким образом может быть получен полинуклеотидный агрегат или дендример. Поли-олигонуклеиновокислотный дендример, в частности, используется для того, чтобы получить полиметку, либо для того, чтобы получить высокую локальную концентрацию эффекторной группировки.
Модифицированные нуклеотиды, которые не мешают гибридизации полинуклеотидов, могут быть включены в этих полинуклеотиды. Подходящими модифицированными нуклеотидами являются С5-замещенные пиримидины или С7-замещенные 7-деаза-пурины. Полинуклеотиды могут быть модифицированы внутри или на 5'- или 3'-конце ненуклеотидными группировками, которые используются для введения эффекторной группировки.
В одном воплощении такие ненуклеотидные группировки находятся в пределах (полинуклеотидного) линкера между двумя элементами связывающей пары, конъюгированными с его концами.
В литературе известно множество различных ненуклеотидных структурных блоков для конструирования полинуклеотида, и большое их разнообразие является коммерчески доступным. Для введения эффекторной группировки могут использоваться либо ненуклеозидные бифункциональные структурные блоки, либо ненуклеозидные трифункциональные структурные блоки в качестве CPG для концевого мечения или в качестве фосфорамидита для внутреннего мечения (см., например, Wojczewski, С., et al., Synlett 10 (1999) 1667-1678).
Бифункциональные спейсерные структурные блоки в одном воплощении являются ненуклеозидными компонентами. Например, такими линкерами являются С2-С18-алкил-, алкенил-, алкинильные углеродные цепи, в то время как алкил-, алкенил-, алкинильные цепи могут быть прерваны дополнительными этиленокси- и/или амидными группировками или квартернизированными катионными аминогруппировками в целях повышения гидрофильности линкера. В качестве ненуклеозидных бифункциональных линкеров также могут быть использованы циклические группировки, такие как С5-С6-циклоалкил, C4N-, C5N-, C4O-, C5O-гетероциклоалкил, фенил, который, возможно, может быть замещен одним или двумя С1-С6-алкильными группами. Подходящие бифункциональные структурные блоки содержат С3-С6-алкильные группировки и от три- до гексаэтиленгликольные цепи. В таблицах 2а и 2b показаны некоторые примеры нуклеотидных бифункциональных спейсерных структурных блоков с различной гидрофильностью, различной жесткостью и различными зарядами. Один атом кислорода соединен с кислотно-лабильной защитной группой, предпочтительно диметокситритилом, а другой является частью фосфорамидита.
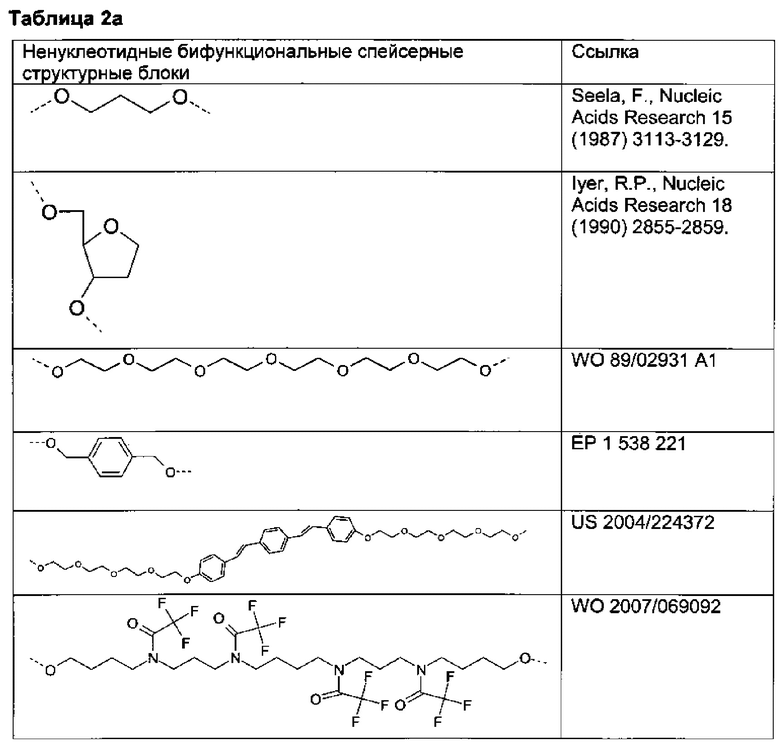
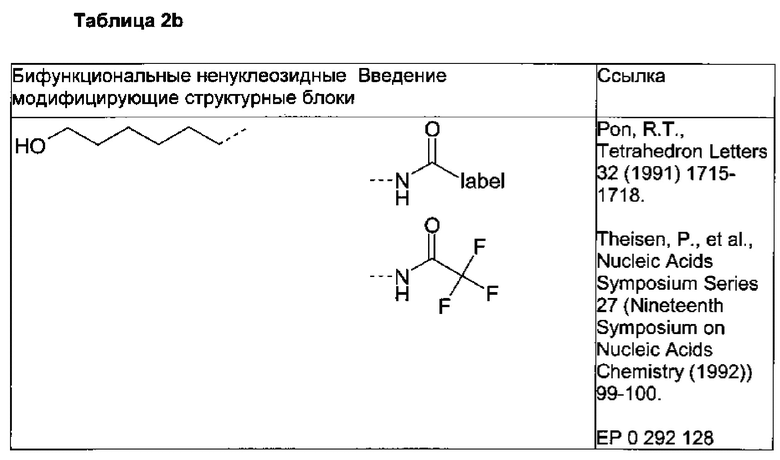
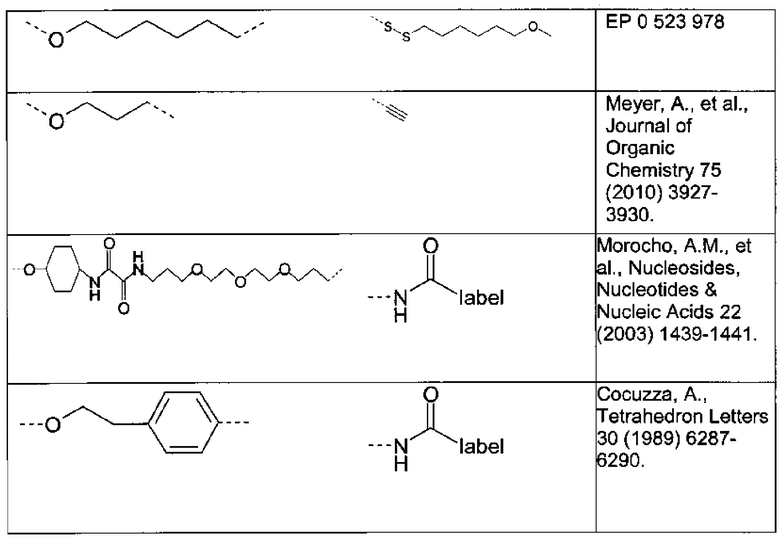
Таким образом, трифункциональные структурные блоки обеспечивают позиционирование функциональной группировки в любом месте в пределах полинуклеотида. Трифункциональные структурные блоки также являются предварительным условием для синтеза с использованием твердых носителей, например, стекла с контролируемым размером пор (CPG), которые используются для 3'-концевого мечения полинуклеотидов. В этом случае трифункциональные линкеры соединены с функциональной группировкой или - при необходимости - с защищенной функциональной группировкой через С2-С18-алкил-, -алкенил-, -алкинильные углеродные цепи, в то время как ал кил-, алкенил-, алкинильные цепи могут быть прерваны дополнительными этиленокси- и/или амидными группировками для повышения гидрофильности линкера, и включают гидроксильную группу, которая присоединена через способный к расщеплению спейсер к твердой фазе, и гидроксильную группу, которая защищена кислотно-лабильной защитной группой. После удаления этой защитной группы высвобождается гидроксильная группа, которая затем может реагировать с фосфорамидитом.
Трифункциональные структурные блоки могут быть ненуклеозидными или нуклеозидными.
Ненуклеозидные трифункциональные структурные блоки представляют собой С2-С18-алкил-, -алкенил-, -алкинильные углеродные цепи, в то время как алкил, алкенил, алкинил, возможно, могут быть прерваны дополнительными этиленокси- и/или амидными группировками для повышения гидрофильности линкера. Другие трифункциональные структурные блоки представляют собой циклические группы, такие как С5-С6-циклоалкил, C4N-, C5N-, C4O-, C5O-гетероциклоалкил, фенил, который, возможно, может быть замещен одной или двумя С1-С6-алкильными группами. Циклические и ациклические группы могут быть замещены одной (С1-С18)алкил-O-РС-группой, в то время как С1-С18-алкил содержит группировки (этиленокси)n, (амид)m, где n и m независимо друг от друга равны от 0 до 6, a PG представляет собой кислотно-лабильную защитную группу. Предпочтительными трифункциональными структурными блоками являются группировки С3-С6-алкил, циклоалкил, C5O-гетероциклоалкил, возможно, содержащие одну амидную связь и замещенные С1-С6-алкил-O-PG-группой, где PG представляет собой кислотно-лабильную защитную группу, предпочтительно монометокситритил, диметокситритил, пиксил, ксантил, где наиболее предпочтительным является диметокситритил.
Неограничивающие, но подходящие примеры ненуклеозидных трифункциональных структурных блоков приведены, например, в таблице 3.
Нуклеозидные трифункциональные структурные блоки используются для внутреннего мечения, когда это необходимо, чтобы не влиять на гибридизационные свойства полинуклеотида по сравнению с немодифицированным полинуклеотидом. Поэтому нуклеозидные структурные блоки содержат основание или аналог основания, который все еще способен к гибридизации с комплементарным основанием. Общая формула соединения для мечения нуклеиновокислотной последовательности одного или более из a, a', b, b' или S, содержащихся в комплексе, описанном в данном документе, приведена в формуле II.

где PG представляет собой кислотно-лабильную защитную группу, в частности монометокситритил, диметокситритил, пиксил, ксантил, особенно диметокситритил, где Y представляет собой С2-С18-алкил, -алкенил, -алкинил, где алкил, алкенил, алкинил может содержать этиленокси- и/или амидные группировки, где Y предпочтительно представляет собой С4-С18-алкил, -алкенил или -алкинил и содержит одну амидную группировку, и где X представляет собой функциональную группировку.
Для такой замены могут быть выбраны конкретные позиции основания, чтобы свести к минимуму влияние на гибридизационные свойства. Поэтому следующие позиции особенно подходят для замещения: а) природными основаниями: урацил, замещенный в С5, цитозин, замещенный в С5 или N4, аденин, замещенный в С8 или N6, и гуанин, замещенный в С8 или N2, и b) аналогами оснований: 7-деаза-А- и 7-деаза-G, замещенный в С7, 7-деаза-8-аза- и 7-деаза-8-аза-С замещенный в С7, 7-деаза-аза-2-амино-А, замещенный в С7, псевдоуридин, замещенный в N1, и формицин, замещенный в N2.
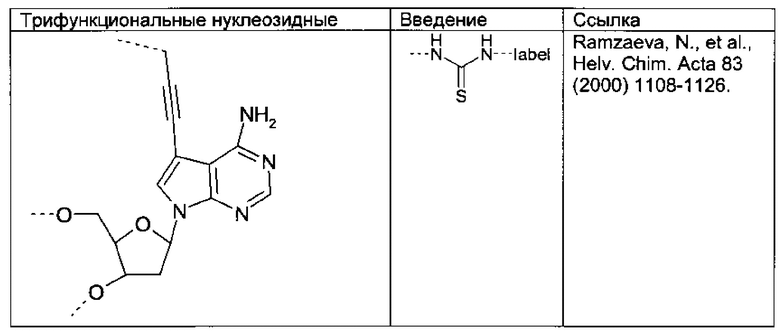
В таблице 4 концевой атом кислорода бифункциональной группировки или один из концевых атомов кислорода трифункциональной группировки являются частью фосфорамидита, который не показан подробно во всех деталях, но очевиден специалисту в данной области. Второй концевой атом кислорода трифункционального структурного блока защищен кислотно-лабильной защитной группой PG, определенной выше для формулы II.
Постсинтетическая модификация является другой стратегией введения ковалентно связанной функциональной группировки в линкер. При таком подходе аминогруппу вводят с помощью бифункциональных или трифункциональных структурных блоков в ходе твердофазного синтеза. После отщепления от субстрата и очистки аминомодифицированного линкера этот линкер подвергают взаимодействию с активированным сложным эфиром функциональной группировки или с бифункциональным реагентом, где одна функциональная группа является активным сложным эфиром. Особенно подходящими активными сложными эфирами являются NHS-эфир или пентафторфениловый эфир.
Постсинтетическая модификация особенно полезна для введения функциональной группировки, которая не стабильна в ходе твердофазного синтеза и удаления защиты. Примерами являются модификация трифенилфосфинкарбоксиметиловым эфиром для лигирования по Штаудингеру (Wang, Charles C.-Y., et al., Bioconjugate Chemistry 14 (2003) 697-701), модификация дигоксигенином или введение малеинимидо-группы с помощью коммерчески доступного сульфо-SMCC.
Компонент связывающей пары
В одном воплощении каждый элемент связывающей пары имеет молекулярную массу 10 кДа или менее. В одном воплощении молекулярная масса каждого элемента связывающей пары составляет 8 кДа, или 7 кДа, или 6 кДа, или 5 кДа, или 4 кДа, или менее.
Константа диссоциации, т.е. аффинность связывания, для связывающей пары составляет по меньшей мере 10-8 М (=10-8 моль/л = 108 л/моль). Элементы обеих связывающих пар в комплексе, описанном в данном документе, являются различными. Разница между связывающими парами а:а' и b:b' является признанной, если, например, константа диссоциации противоположного связывания, например, связывания а и а' с b или b', больше константы диссоциации пары а:а' в 10 раз или более.
В одном воплощении константа диссоциации противоположного связывания, т.е. связывания а и а' с b или b', соответственно, больше константы диссоциации пары а:а' в 20 раз или более. В одном воплощении константа диссоциации больше константы диссоциации пары а:а' в 50 раз или более. В одном воплощении константа диссоциации противоположного (перекрестно-реактивного) связывания больше константы диссоциации связывающей пары в 100 раз или более.
В одном воплощении элементы связывающих пар выбраны из группы, состоящей из димеров домена лейциновой молнии и гибридизирующихся нуклеиновокислотных последовательностей. В одном воплощении обе связывающие пары являются димерами домена лейциновой молнии.
В одном воплощении обе связывающие пары являются гибридизующимися нуклеиновокислотными последовательностями. В одном воплощении все элементы связывающей пары являются L-ДНК-последовательностями. В одном воплощении обе связывающие пары являются гибридизующимися L-ДНК.
В одном воплощении, оба элемента связывающих пар являются доменами лейциновой молнии.
Термин "домен лейциновой молнии" обозначает домен димеризации, характеризующийся наличием остатка лейцина на каждом седьмом остатке на участке приблизительно 35 остатков. Домены лейциновой молнии представляют собой пептиды, которые способствуют олигомеризации белков, в которых они находятся. Лейциновые молнии первоначально были найдены в нескольких ДНК-связывающих белках (Landschulz, W.H., et al., Science 240 (1988) 1759-1764). Среди известных лейциновых молний есть природные пептиды и их производные, которые димеризуются или тримеризуются. Примерами доменов лейциновой молнии, пригодных для получения растворимых многомерных белков, являются такие, о которых сообщается в WO 94/10308, и домен лейциновой молнии, полученный из белка D сурфактанта легких (SPD), как сообщалось в Норре, H.J., et al., FEBS Lett. 344 (1994) 191-195.
Домены лейциновой молнии образуют димеры (связывающие пары), удерживаемые вместе биспиральной альфа-спиральной катушкой. Спиральная катушка имеет 3,5 остатка на виток, что означает, что каждый седьмой остаток занимает эквивалентное положение по отношению к оси спирали. Стандартный набор лейцинов внутри гибкой катушки стабилизирует структуру за счет гидрофобных и взаимодействий Ван-дер-Ваальса.
Если домены лейциновой молнии образуют первую связывающую пару и вторую связывающую пару, то обе последовательности лейциновой молнии различны, т.е. элементы первой связывающей пары не связываются с элементами второй связывающей пары. Домены лейциновой молнии могут быть выделены из природных белков, известных тем, что они содержат такие домены как факторы транскрипции. Один домен лейциновой молнии, например, может происходить из фактора транскрипционных fos, а второй из фактора транскрипции jun. Домены лейциновой молнии также могут быть разработаны и синтезированы искусственно с использованием стандартных методик синтеза и конструирования, известных в данной области.
В одном воплощении обе связывающие пары являются гибридизующимися нуклеиновокислотными последовательностями.
Таким образом, элементы каждой связывающей пары, т.е. а и а', а также b и b', гибридизуются друг с другом, соответственно. Нуклеиновокислотные последовательности, содержащиеся в первой связывающей паре с одной стороны и во второй связывающей паре с другой стороны, являются разными, т.е. не гибридизуются друг с другом.
В одном воплощении обе связывающие пары являются гибридизующимися нуклеиновокислотными парами, где гибридизующиеся нуклеиновокислотные последовательности различных связывающих пар не гибридизуются друг с другом.
Другими словами, нуклеиновые кислоты первой связывающей пары гибридизуются друг с другом, но не связываются с любой из нуклеиновых кислот второй связывающей пары или не мешают их гибридизации, и наоборот. Кинетику гибридизации и специфичность гибридизации можно легко контролировать с помощью анализа температуры плавления. Специфическая гибридизация связывающей пары и отсутствие воздействия считаются признанными, если температура плавления связывающей пары по меньшей мере на 20°С выше по сравнению с любой возможной комбинацией с другими связывающими парами или комбинацией элементов связывающей пары.
Нуклеиновокислотные последовательности, образующие связывающую пару, могут содержать, в принципе, любой природный нуклеотид или его аналог, и могут иметь, в принципе, модифицированный или не модифицированный остов, как описано выше, при условии, что он способен образовывать стабильный дуплекс за счет многократного спаривания оснований. «Стабильный» означает, что температура плавления дуплекса выше 30°С, особенно выше 37°С.
Двойная цепь в одном воплощении состоит из двух полностью комплементарных одноцепочечных полинуклеотидов.
Тем не менее, несоответствия или вставки возможны до тех пор, пока сохраняется стабильность при 37°С.
Нуклеиновокислотный дуплекс может быть дополнительно стабилизирован путем межцепочечного сшивания. Известно несколько соответствующих способов поперечного сшивания, например, способы с использованием псоралена или на основании тионуклеозидов.
Нуклеиновокислотные последовательности, представляющие элементы связывающей пары в одном воплощении, состоят из 12-50 нуклеотидов. В одном воплощении такие нуклеиновокислотные последовательности состоят из 15-35 нуклеотидов.
РНКазы являются вездесущими, и необходимо принять специальные меры, чтобы избежать нежелательного расщепления связывающих пар и/или линкерных последовательностей на основе РНК. Хотя связывающие пары и/или линкеры на основе РНК могут использоваться, особенно подходящими являются связывающие пары и/или линкеры на основе ДНК.
Соответствующие гибридизующиеся нуклеиновокислотные последовательности могут быть легко разработаны для обеспечения более чем двух пар ортогонально комплементарных полинуклеотидов, что позволяет легко разрабатывать и использовать более двух связывающих пар. Еще одно преимущество использования гибридизирующихся нуклеиновокислотных последовательностей в комплексе, описанном в данном документе, заключается в том, что легко могут быть введены изменения. Коммерчески доступны модифицированные структурные блоки, которые, например, позволяют легко синтезировать полинуклеотид, содержащий функциональную группировку. Такая функциональная группировка может быть легко введена в любую желаемую позицию или в любой из элементов первой и/или второй связывающей пары и/или полинуклеотидного линкера, при условии, что все они представляют собой полинуклеотид.
(Полинуклеотидный) линкер, содержащий элементы связывающих пар на своих концах, может быть предусмотрен и синтезирован в виде одного полинуклеотида. Каждый из полипептидов, специфически связывающихся с мишенью, может быть соединен с гибридизующимися нуклеиновокислотными последовательностями, т.е. элементами связывающих пар. Длина (полинуклеотидного) линкера может быть легко изменена любым нужным образом.
В зависимости от биохимической природы полипептида, который специфически связывается с мишенью, используются различные стратегии для конъюгации с элементом связывающей пары. В случае если полипептид является природным или получен рекомбинантным способом и содержит от 50 до 500 аминокислотных остатков в длину, специалист в данной области может легко следовать стандартным процедурам, описанным в руководствах (см., например, Hackenberger, C.P.R., and Schwarzer, D., Angew. Chem. Int. Ed. 47 (2008) 10030-10074).
В одном воплощении для конъюгации используется реакция малеинимидо-группировки с остатком цистеина в полипептиде.
Эта реакция сочетания является особенно подходящей в случае, например, когда FAB- или FAB'-фрагмент антитела используется как одновалентный связывающий полипептид.
В одном воплощении выполняется реакция сочетания элемента связывающей пары с С-концом полипептида.
С-концевая модификация белка, например, FAB-фрагмента, может быть выполнена, например, как описано в Sunbul, М. and Yin, J., Org. Biomol. Chem. 7 (2009) 3361-3371.
В целом, сайт-специфическая реакция и ковалентное связывание элемента связывающей пары с одновалентным связывающим полипептидом основаны на преобразовании природной аминокислоты в аминокислоту с реакционной группой, которая ортогональна реактивности других функциональных групп, присутствующих в полипептиде.
Например, конкретный цистеин в контексте редкой последовательности может быть ферментативно преобразован в альдегид (см., например, Frese, М-А. and Dierks, Т., ChemBioChem 10 (2009) 425-427). Также можно получить нужную аминокислотную модификацию, используя способность некоторых ферментов специфически реагировать с природной аминокислотой в данном контексте последовательности (см., например: Taki, М., et al., Prot. Eng. Des. Sel. 17 (2004) 119-126, Gautier, A., et al., Chem. Biol. 15 (2008) 128-136; Bordusa, F., in Highlights in Bioorganic Chemistry (2004), Schmuck, C. and Wennemers, H., (eds.), Wiley VCH, Weinheim, pp. 389-403).
Сайт-специфическое взаимодействие и ковалентное сочетание элемента связывающей пары с одновалентным связывающим полипептидом также может быть достигнуто путем селективной реакции концевых аминокислот с соответствующими модифицирующими реагентами.
Реакционная способность N-концевого цистеина с бензонитрилами (см. Ren, Н., et al., Angew. Chem. Int. Ed. 48 (2009) 9658-9662) может быть использована для достижения сайт-специфического ковалентного сочетания.
Нативное химическое лигирование также может быть основано на С-концевых остатках цистеина (Taylor, Е., et al., Nucl. Acids Mol. Biol. 22 (2009) 65-96).
EP 1074563 сообщает о способе конъюгации, который основан на более быстрой реакции цистеина, расположенного в участке отрицательно заряженных аминокислот, с цистеином, расположенным в участке положительно заряженных аминокислот.
Эффекторный компонент
Эффекторная группировка может быть выбрана из группы, состоящей из связывающей группировки, группировки для мечения, биологически активной группировки и реакционноспособной группировки. Если в комплексе присутствует более чем одна эффекторная группировка, то каждая такая эффекторная группировка в каждом случае независимо может быть связывающей группировкой, группировкой для мечения, биологически активной группировкой или реакционноспособной группировкой. Связывающая группировка будет выбрана так, чтобы она не влияла на каждую из связывающих пар.
В одном воплощении эффекторная группировка выбрана из группы, состоящей из связывающей группировки, группировки для мечения и биологически активной группировки.
В одном воплощении эффекторная группировка является связывающей группировкой.
Примерами связывающих группировок являются элементы пары биоаффинного связывания, которые могут специфически взаимодействовать друг с другом. Подходящими парами биоаффинного связывания являются гаптен или антиген и антитело; биотин или аналоги биотина, такие как аминобиотин, иминобиотин или дестиобиотин, и авидин или стрептавидин; сахар и лектин, полинуклеотид и комплементарный полинуклеотид, рецептор и лиганд, например, рецептор стероидного гормона и стероидный гормон; и пара из 104-аминокислотного фрагмента бычьей рибонуклеазы А (известного как S-белок) и 15-аминокислотного фрагмента бычьей рибонуклеазы А (известного как S-пептид).
В одном воплощении эффекторная группировка представляет собой связывающую группировку и ковалентно связана по меньшей мере с одним из компонентов комплекса.
В одном воплощении меньший элемент пары биоаффинного связывания, например, биотин или его аналог, лиганд рецептора, гаптен или полинуклеотид, ковалентно связан по меньшей мере с одним из полинуклеотидов, входящих в комплекс, описанный в данном документе.
В одном воплощении эффекторая группировка является связывающей группировкой, выбранной среди гаптена, биотина или аналогов биотин, таких как аминобиотин, иминобиотин или дестиобиотин; полинуклеотида и стероидного гормона.
В одном воплощении эффекторная группировка является группой для мечения.
Группа для мечения может быть выбрана среди любых известных обнаруживаемых групп.
В одном воплощении группа для мечения выбрана среди таких как меток как группы люминесцентного мечения, например хемилюминесцентные группы, например акридиния сложные эфиры или диоксетаны, или флуоресцентные метки, например, флуоресцеин, кумарин, родамин, оксазин, ресоруфин, цианин и их производные, люминесцентные комплексы металлов, такие как комплексы рутения или европия, ферменты, которые используются для CEDIA (иммуноанализ с имитированным донором энзима, например, ЕР 0061888), микрочастицы или наночастицы, например латексные частицы или золи металлов, и радиоизотопы.
В одном воплощении группа для мечения представляет собой люминесцентный комплекс металла и соединение, имеющее структуру с общей формулой (I):

где М представляет собой двухвалентный или трехвалентный катион металла, выбранный среди ионов редкоземельных или переходных металлов, L1, L2 и L3 являются одинаковыми или различными и означают лиганды по меньшей мере с двумя азотсодержащими гетероциклами, где L1, L2 и L3 связаны с катионом металла через атомы азота, X представляет собой реакционноспособную функциональную группу, которая ковалентно связана по меньшей мере с одним из лигандов L1, L2 и L3 через линкер Y, n представляет собой целое число от 1 до 10, особенно от 1 до 4, m представляет собой 1 или 2, или в особенности 1, и А обозначает противоион, который может потребоваться для выравнивания заряда.
Комплекс металла в одном воплощении представляет собой люминесцентный комплекс металла, т.е. комплекс металла, который подвергается обнаруживаемой реакции люминесценции после соответствующего возбуждения.
Реакция люминесценции, например, может быть обнаружена с помощью измерения флуоресценции или электрохемилюминесценции. Катион металла в этом комплексе, например, является переходным металлом или редкоземельным металлом.
Металл в одном воплощении представляет собой рутений, осмий, рений, иридий, родий, платину, индий, палладий, молибден, технеций, медь, хром или вольфрам. Особенно подходящими являются рутений, иридий, рений, осмий и хром. Наиболее подходящим является рутений.
Золотые наностержни (GNR, gold nanorod) также могут быть использованы в качестве группировки для мечения в комплексах, описанных в данном документе. Наностержни могут иметь длину от 10 до 100 нм включительно, в том числе и все целые числа между ними.
В одном воплощении GNR имеют среднюю длину 70-75 нм.
GNR могут иметь диаметр от 5 до 45 нм включительно, в том числе и все целые числа между ними.
В одном воплощении GNR имеют средний диаметр 25-30 нм. GNR могут быть сделаны из чистого золота или могут быть золотыми на 90-99% включительно, включая все целые числа между ними.
В различных воплощениях GNR могут содержать до 1% серебра на их поверхности и могут содержать бромид цетилтриметиламмония (СТАВ).
В связи с этим GNR могут быть изготовлены любым подходящим способом. Например, электрохимический синтез в растворе, шаблонный синтез с использованием мембран, фотохимический синтез, микроволновой синтез и обеспечиваемое затравками выращивание являются подходящими и не ограничивающими примерами способов изготовления GNR.
В одном воплощении золотые наностержни изготовлены с использованием способа обеспечиваемого затравками выращивания в бромиде цетилтриметиламмония (СТАВ).
Для того, чтобы сформировать комплексы из золотых наностержней и РНК-полинуклеотидов, поверхности золотых наностержней могут быть функционализированы таким образом, что они придают положительный дзета-потенциал, подходящий для электростатического комплексообразования GNR с ДНК- или РНК-полинуклеотидами. Может быть использован любой подходящий способ создания положительного дзета-потенциал на золотых наностержнях. Например, поверхности золотых наностержней могут быть функционализированы бифункциональными молекулами, такими как тиолированный ПЭГ-NH2 или тиолированный ПЭГ-COOH.
В одном воплощении функционализация поверхности достигается путем покрытия СТАВ-покрытых золотых наностержней вначале анионным полиэлектролитом поли(3,4-этилендиоксити-6-фен)/полистиролсульфатом (PEDT/PSS), а затем катионным полиэлектролитом полидиаллилдиметиламмония хлоридом (PDDAC).
Так получают золотые наностержни с катионным поверхностным зарядом (положительным дзета-потенциалом), а также маскируют СТАВ-слой (см., например, Ding, Н., et al., J. Phys. Chem. С 111 (2007) 12552-12557).
Положительно заряженные золотые наностержни образуют электростатические комплексы с ДНК-полинуклеотидами за счет электростатических взаимодействий.
Образование наноплексов может быть подтверждено наблюдаемым красным смещением пика в локализованном продольном плазмоном резонансе золотых наностержней, а также ограниченной электрофоретической подвижностью наноплексов при гель-электрофорезе.
В одном воплощении эффекторная группировка X представляет собой терапевтически активное вещество.
Терапевтически активные вещества могут быть эффективными в различных способах, например, в ингибировании рака за счет повреждения ДНК-матрицы путем алкилирования, путем сшивания или двухцепочечного расщепления ДНК. Другие терапевтически активные вещества могут блокировать синтез РНК путем интеркаляции. Некоторые агенты являются ядами, блокирующими митотическое веретено, такими как алкалоиды барвинка, или антиметаболитами, которые ингибируют активность ферментов, или гормональными и противогормональными препаратами. Эффекторная группировка может быть выбрана среди алкилирующих агентов, антиметаболитов, противоопухолевых антибиотиков, алкалоидов барвинка, эпиподофиллотоксинов, нитрозомочевин, гормональных и противогормональных агентов и токсинов.
Подходящими алкилирующими агентами являются циклофосфамид, хлорамбуцил, бусульфан, мелфалан, ифосфамид, тиотепа или азотистый иприт.
Подходящими антиметаболиами являются метотрексат, 5-фторурацил, цитозинарабинозид, 6-тиогуанин, 6-меркаптопутин.
Подходящими противоопухолевыми антибиотиками являются доксорубицин, даунорубицин, идорубицин, нимитоксантрон, дактиномицин, блеомицин, митомицин и пликамицин.
Подходящими ядами, блокирующими митотическое веретено, являются майтанзин и майтанзиноиды, алкалоиды барвинка и эпиподофиллотоксины, которые могут быть представлены винкристином, винбластином, виндестином, этопозидом, тенипозидом.
Кроме того, подходящие таксановые агенты могут быть представлены паклитакселом, доцетакселом, SB-T-1214.
Подходящими нитрозомочевинами являются кармустин, ломустином семустин, стрептозоцин.
Подходящими гормональными и противогормональными агентами являются адренокортикоиды, эстрогены, антиэстрогены, прогестины, ингибиторы ароматазы, андрогены, антиандрогены.
Подходящими случайными синтетическими агентами являются дакарбазин, гексаметилмеламин, гидроксимочевина, митотан, прокарбазид, цисплатин, карбоплатин.
Подходящими факторами хемотаксиса моноцитов являются f-Met-Leu-Phe (fMLP), f-Met-Leu-Phe-o-метиловый эфир, формил-норлейцин-фенилаланин, формил-метионил-фенилаланин.
Подходящими факторами привлечения NK-клеток являются IL-12, IL-15, IL-18, IL-2 и CCL5, FC-часть антитела.
В одном воплощении эффекторная группировка X представляет собой Fc-область антитела или ее фрагмент.
В одном воплощении Fc-область человеческого антитела относится или к человеческому IgG1-подклассу, или к человеческому IgG2-подклассу, или к человеческому IgG3-подклассу, или к человеческому IgG4-подклассу.
В одном воплощении антитело Fc-область человеческого антитела представляет собой Fc-область человеческого IgG1-подкласса или человеческого IgG4-подкласса.
В одном воплощении Fc-область человеческого антитела содержит мутацию природного аминокислотного остатка по меньшей мере в одной из следующих аминокислотных позиций: 228, 233, 234, 235, 236, 237, 297, 318, 320, 322, 329 и/или 331, с заменой другим остатком, где остатки в Fc-области антитела пронумерованы в соответствии с ЕС-индексом по Кабату.
В одном воплощении Fc-область человеческого антитела содержит мутацию природного аминокислотного остатка в позиции 329 и по меньшей мере одну мутацию по меньшей мере одного аминокислотного остатка, выбранного из группы, включающей аминокислотные остатки в позиции 228, 233, 234, 235, 236, 237, 297, 318, 320, 322 и 331, с заменой на другой остаток, где остатки в Fc-области пронумерованы в соответствии с ЕС-индексом по Кабату. Замена этих конкретных аминокислотных остатков приводит к изменению эффекторной функции Fc-области по сравнению с немодифицированной Fc-областью (дикого типа).
В одном воплощении Fc-область человеческого антитела имеет сниженную аффинность к FcγRIIIA и/или FcγRIIA и/или FcγRI по сравнению с конъюгатом, содержащим соответствующую Fc-область IgG дикого типа.
В одном воплощении аминокислотный остаток в позиции 329 Fc-области человеческого антитела замещен на глицин, или аргинина, или аминокислотный остаток, достаточно большой, чтобы разрушить «пролиновый сэндвич» в Fc-области.
В одном воплощении мутация природного аминокислотного остатка в Fc-области человеческого антитела является по меньшей мере одной из S228P, Е233Р, L234A, L235A, L235E, N297A, N297D, P329G и/или P331S.
В одном воплощении мутация представляет собой L234A и L235A, если Fc-область антитела относится к человеческому IgG1-подклассу, или S228P и L235E, если Fc-область антитела относится к человеческому IgG4-подклассу.
В одном воплощении Fc-область антитела содержит мутацию P329G.
Эффекторная группировка может быть связана либо ковалентно, либо с помощью дополнительной связывающей пары по меньшей мере с одним из компонентов комплекса. Эффекторная группировка может содержаться от одного до нескольких (n) раз в комплексе, описанном в данном документе, где (n) представляет собой целое число, 0 или 1 или более чем 1. В одном воплощении (n) составляет от 1 до 1000000. В одном воплощении (n) составляет от 1000 до 300000. В одном воплощении (n) составляет от 1 до 50. В одном воплощении (n) составляет от 1 до 10 или от 1 до 5. В одном воплощении (n) составляет 1 или 2.
Для ковалентного связывания эффекторной группировки по меньшей мере с одним из компонентов в комплексе может быть использована любая подходящая реакция сочетания. Также можно включить функциональную группу с помощью соответствующих структурных блоков при синтезе элементов первой и/или второй связывающей пары и/или (полинуклеотидного) линкера, особенно элементов связывающих пар, конъюгированных с полипептидом или (полинуклеотидным) линкером.
Особенно подходящими являются способы конъюгации, образующие связи, которые по существу (или почти) неиммуногенны. Таким образом, особенно подходящими являются пептидные (т.е. амидные), сульфидные (пространственно затрудненные), дисульфидные, гидразоновые или эфирные связи. Эти связи почти неиммуногенны и демонстрируют разумную стабильность в сыворотке крови (см., например, Senter, P.D., Curr. Opin. Chem. Biol. 13 (2009) 235-244; WO 2009/059278; WO 95/17886).
В одном воплощении эффекторная группировка связана с (полинуклеотидным) линкером комплекса, описанного в данном документе.
В одном воплощении эффекторная группировка ковалентно связана с элементом связывающей пары, конъюгированным с полипептидом или (полинуклеотидным) линкером комплекса, описанного в данном документе.
Если эффекторная группировка расположена в гибридизующемся полинуклеотиде, то особенно подходящим является связывание ее с модифицированным нуклеотидом или прикрепление к межнуклеозидному атому фосфора (см., например, WO 2007/059816).
Бифункциональные структурные блоки (описанные выше) могут быть использованы для соединения эффекторной группировки или - при необходимости - защищенной эффекторной группировки с фосфорамидитной группой для прикрепления структурного блока на 5'-конце (обычный синтез) или на 3'-конце (обратный синтез) к концевой гидроксильной группе растущей полинуклеотидной цепи.
Трифункциональные структурные блоки (описанные выше) могут быть использованы для соединения (i) эффекторной группировки или - при необходимости - защищенной эффекторной группировки, (ii) фосфорамидитной группы для реакции сочетания с репортером или эффекторной группировкой или - при необходимости - защищенной эффекторной группировкой в ходе полинуклеотидного синтеза с гидроксильной группой растущей полинуклеотидной цепи и (iii) гидроксильной группы, которая защищена кислото-лабильной защитной группой, особенно защитной диметокситритил-группой. После удаления этой кислотно-лабильной защитной группы высвобождается гидроксильная группа, которая может вступать в реакцию с другими фосфорамидитами.
Эффекторная группировка связана в одном воплощении по меньшей мере с одним из элементов первой и/или второй связывающей пары или с полинуклеотидным линкером с помощью дополнительной третьей связывающей пары. В одном воплощении третья связывающая пара представляет собой пару гибридизующихся нуклеиновокислотных последовательностей. Элементы третьей связывающей пары не мешают связыванию элементов других связывающих пар друг с другом.
Дополнительная связывающая пара, с которой может быть связана эффекторная группировка, в частности, может представлять собой домен лейциновой молнии или гибридизующуюся нуклеиновую кислоту. В случае если эффекторная группировка связана по меньшей мере с одним из элементов первой и/или второй связывающей пары или (полинуклеотидным) линкером с помощью дополнительного элемента пары связывания, элемент связывающей пары, с которой связана эффекторная группировка, и элементы первой и второй связывающих пар, соответственно, выбраны так, чтобы они имели различную специфичность. Каждый из элементов первой и второй связывающих пар и связывающей пары, с которой связана эффекторная группировка, связывается (например, гибридизуется) с его соответствующим элементом, не мешая связыванию всех других связывающих пар.
В одном воплощении комплементарные нуклеиновые кислоты связывающих пар и/или (полинуклеотидный) линкер по меньшей мере частично состоят из L-ДНК, или L-PHK, или ЗНК (закрытой нуклеиновой кислоты), или изо-С-нуклеиновой кислоты, или изо-G-нуклеиновой кислоты, или любой их комбинации. В одном воплощении (полинуклеотидный) линкер состоит по меньшей мере на 50% из L-ДНК, или L-PHK, или ЗНК, или изо-С-нуклеиновой кислоты, или изо-G-нуклеиновой кислоты, или любой их комбинации. В одном воплощении (полинуклеотидный) линкер представляет собой L-полинуклеотид (шпигельмер). В одном воплощении L-полинуклеотид представляет собой L-ДНК.
В одном воплощении (полинуклеотидный) линкер представляет собой ДНК. В одном воплощении (полинуклеотидный) линкер представляет собой L-стереоизомер ДНК, также известный как бета-L-ДНК, или L-ДНК, или зеркально отраженную ДНК.
У этой стереоизомерной ДНК есть преимущества, такие как ортогональный характер гибридизации, что означает, что дуплекс образуется только между двумя комплементарными одиночными нитями L-ДНК, но не образуется между одиночными нитями L-ДНК и комплементарной нитью D-ДНК, устойчивость к нуклеазам и легкий синтез даже длинного линкера. Простота синтеза и изменчивость длины спейсера являются важными условиями для получения библиотеки линкеров. (Полинуклеотидные) линкеры различной длины используются для идентификации комплексов, описанных в данном документе, которые имеют полинуклеотидный линкер оптимальной длины, таким образом, для получения оптимального расстояния между двумя полипептидами, специфически связывающими мишень.
В одном воплощении комплекс является нековалентным комплексом. В одном воплощении нековалентный комплекс образуется с помощью связывающих пар.
В некоторых воплощениях эффекторная группировка является терапевтическим лекарственным средством.
Например, эффекторная группировка может быть терапевтическим радионуклидом, гормоном, цитокином, интерфероном, антителом или фрагментом антитела, нуклеиновокислотным аптамером, ферментом, полипептидом, токсином, цитотоксином, химиотерапевтическим агентом или сенсибилизатором радиации.
Одним из аспектов, описанных в данном документе, является способ применения комплекса, описанного в данном документе.
Например, в данном документе описан способ уничтожения клетки, в котором комплекс, описанный в данном документе, вводят в клетку в количестве, достаточном, чтобы уничтожить эту клетку.
В одном воплощении клетка представляет собой раковую клетку.
В данном описании также сообщается о способе замедления или остановки роста раковой клетки у млекопитающего, в котором комплекс, описанный в данном документе, вводят млекопитающему в количестве, достаточном, чтобы замедлить или остановить рост раковой клетки.
В одном воплощении способ представляет собой способ ингибирования роста или пролиферации раковой клетки.
В одном воплощении полипептид, специфически связывающийся с мишенью, специфически связывается с молекулой клеточной поверхности клетки. В одном воплощении молекула клеточной поверхности специфически присутствует на раковых клетках.
В одном воплощении первый и второй полипептид, специфически связывающиеся с мишенью, независимо друг от друга выбраны из группы, состоящей из антитела, фрагмента антитела, одноцепочечной вариабельной области антитела, малой пептидной молекулы, циклического полипептида, пептидомиметика и аптамера.
В одном воплощении первый и второй полипептиды, специфически связывающиеся с мишенью, являются одновалентными связывающими полипептидами.
В одном воплощении полипептид представляет собой фрагмент антитела. В одном воплощении фрагмент антитела получен из интернализованного антитела, которое специфически связывается с молекулой клеточной поверхности.
Конъюгация эффекторной группировки с комплексом, описанным в данном документе, позволяет конкретно локализовать эффекторную группировку в нужном месте на клетке. Локализация увеличивает эффективную концентрацию эффекторной группировки на клетке-мишени и, таким образом, оптимизирует влияние эффекторной группировки. Кроме того, комплекс может быть введен в более низкой дозе по сравнению с ненацеливающей эффекторной группировкой. Это может быть особенно актуально, если эффекторная группировка имеет ассоциированную токсичность или если ее необходимо использовать для лечения хронических заболеваний.
L-ДНК является нуклеотидом, используемым в образовании комплексов, описанных в данном документе. L-ДНК сама по себе не гибридизуется с природной формой ДНК (D-ДНК) или РНК. Так как L-ДНК не является природным субстратом для многих ферментов, стабильность L-ДНК in vivo может быть больше, чем у D-ДНК. Дуплексы L-ДНК имеют такие же физические характеристики в плане растворимости, стабильности дуплекса и селективности, как у D-ДНК, но образуют левоспиральную двойную спираль. Следует понимать, что L-полинуклеотид, упоминаемый в данном документе, может также включать некоторые D-полинуклеотиды.
Из-за химической природы L-полинуклеотидов они не метаболизируются, так что фармакокинетика, лежащая в основе использования L-нуклеотидов, не зависит или по меньшей мере зависит не до такой степени от процессов специфической деградации ДНК. Таким образом, в связи с повышенной стабильностью L-полинуклеотидов время полужизни in vivo комплекса, описанного в данном документе, у млекопитающего фактически неограниченно. Особое значение имеет тот факт, что L-полинуклеотиды не являются нефротоксичными.
В одном воплощении млекопитающее выбрано среди человека, обезьяны, собаки, кошки, лошади, крысы или мыши. В одном воплощении полинуклеотидный линкер содержит D-ДНК- и L-ДНК-нуклеотиды, т.е. полинуклеотидный линкер представляет собой смесь D-ДНК и L-ДНК.
С помощью этого линкера можно спроектировать время полужизни полинуклеотидного линкера, т.е. время полужизни in vivo олигонуклеотидного линкера может быть подобрано и доведено до предполагаемого применения комплекса.
Каждый из полинуклеотидов, присутствующих в комплексе, описанном в данном документе, может содержать одну или более эффекторную группировку. Эффекторные группировки позволяют применять комплекс, описанный в данном документе, для лечения заболевания. Эффекторные группировки могут быть использованы, например, для транспортных целей, т.е. для доставки эффекторной функции, и/или для модуляции фармакокинетического характера, и/или для модуляция физико-химических свойств.
В одном воплощении эффекторная группировка выбрана среди липофильных группировок, пептидов, белков, углеводов и липосом.
В одном воплощении полинуклеотид представляет собой L-полинуклеотид.
L-поли(дезокси)нуклеотиды могут присутствовать в виде одно- или двухцепочечного полинуклеотида. Как правило, L-поли(дезокси)нуклеотид присутствует в виде одноцепочечной нуклеиновой кислоты, которая может формировать (определенные) вторичные структуры, а также третичные структуры. В таких вторичных структурах также могут присутствовать двухцепочечные участки. Тем не менее, L-поли(дезокси)нуклеотид также может присутствовать, по меньшей мере частично, в виде двухцепочечной молекулы в том смысле, что две нити, которые комплементарны друг другу, гибридизуются. L-полинуклеотиды(ы) также может быть модифицирован. Модификация может быть связана с отдельными нуклеотидами в полинуклеотиде.
Для того чтобы избежать образования вторичной структуры, в одном воплощении может быть использован 2,4-дигидрокси-5-метилпиримидин (Т) в качестве нуклеотида.
L-полинуклеотиды в комплексе, описанном в данном документе, в одном воплощении восприимчивы к "самогибридизации".
Таким образом, L-полинуклеотиды способны к более легкой гибридизации с комплементарными L-полинуклеотидными последовательностями, но не образуют стабильный дуплекс с природной ДНК или РНК.
В одном воплощении нуклеотиды в L-ДНК-сегменте имеют конформацию 1'S, 3'R и 4'S.
В одном воплощении L-ДНК-полинуклеотидный линкер конъюгирован через гибридизацию элементов связывающей пары на его концах с полипептидом(ами) комплекса.
В одном воплощении полинуклеотидный линкер имеет длину по меньшей мере 1 нм. В одном воплощении полинуклеотидный линкер имеет длину от 6 нм до 100 нм. В одном воплощении полинуклеотидный линкер имеет длину по меньшей мере 70 нуклеотидов.
Полинуклеотидный линкер также может содержать последовательность метки. Последовательность метки может быть выбрана из обычно используемых меток для распознавания белка, таких как YPYDVPDYA (HA-Tag, SEQ ID № 64) или GLNDIFEAQKIEWHE (Avi-Tag, SEQ ID № 65).
Таким образом, в одном воплощении способов, описанных в данном документе, комплекс, описанный в данном документе, не содержащий эффекторной группировки, вначале вводят и позволяют ему связаться с его мишенью (мишенями), а затем вводят эффекторную группировку, конъюгированную с полинуклеотидом, комплементарным по меньшей мере части (полинуклеотидного) линкера. Таким образом, эффекторная группировка in situ локализуется вместе с комплексом, связанным с его мишенью, за счет гибридизации с комплексом, описанным в данном документе.
Следует понимать, что комплекс, описанный в данном документе, не ограничен какой-либо конкретной нуклеиновокислотной последовательностью, или какой-либо связывающей группировкой (полипептидом), специфически связывающейся с мишенью, или конкретными типами клеток, или конкретными условиями, или конкретными способами и т.п., и они могут варьировать, и многочисленные модификации и вариации будут очевидны специалистам в данной области.
В одном воплощении способов, описанных в данном документе, комплекс связывается с клеточной поверхностью опухолевой клетки и локально обогащает ее эффекторной группировкой до высокой плотности или высокой локальной концентрации.
В одном воплощении эффекторная группировка является меченой оц-L-ДНК, которую вводят одновременно или последовательно для исходно целевой ассоциации комплекса.
Меченая оц-L-ДНК-эффекторная группировка гибридизуется с оц-L-ДНК (олигонуклеотидным) линкером комплекса.
Связанный с мишенью комплекс используется для активации врожденного иммунного ответа, а именно, чтобы привлечь цитотоксические лимфоциты, также называемые натуральными киллерами (NK-клетками). NK-клетки играют важную роль в отторжении опухолей и клеток, инфицированных вирусами. Они уничтожают клетки, высвобождая небольшие цитоплазматические гранулы с белками, называемыми перфорином и гранзимом, которые вызывают гибель клетки-мишени путем апоптоза.
В одном воплощении комплекс, описанный в данном документе, используется для привлечения NK-клеток в непосредственную близость от связанного комплекса. В одном воплощении оц-L-ДНК, конъюгированную с цитокином, используют в качестве эффекторной группировки.
Эта меченная цитокином эффекторная группировка может быть использована для привлечения NK-клеток. Цитокины, участвующие в NK-активации, включают IL-12, IL-15, IL-18, IL-2 и CCL5.
В одном воплощении оц-L-ДНК, конъюгированную с Fc-частью антитела, используют в качестве эффекторной группировки.
NK-клетки, наряду с макрофагами и некоторыми другими типами клеток, экспрессируют молекулу Fc-рецептора (FcR) (FC-гамма-RIII = CD16), активирующий биохимический рецептор, который связывается с Fc-частью антител. Это позволяет NK-клеткам нацеливаться на клетки, против которых был мобилизован гуморальный ответ, и лизировать клетки за счет антителозависимой клеточной цитотоксичности (ADCC).
В одном воплощении одна или более оц-L-ДНК или комбинация оц-L-ДНК, конъюгированных с одной или более Fc-частью, используется в качестве эффекторных группировок.
В этом воплощении комплекс может быть использован для модуляции ADCC и/или активации комплемента (CDC).
В одном воплощении этот комплекс используют в способе скрининга инженерных Fc-компартментов на их эффективность в привлечении ADCC и CDC.
В одном воплощении комплекс используют для ингибирования фосфатазы семенной жидкости.
В этом воплощении комплекс может быть использован для предотвращения инактивации NK-клеток.
В одном воплощении полипептид, специфически связывающийся с мишенью, например, антитело или фрагмент антитела, специфически связывающийся с молекулой клеточной поверхности, конъюгирован с лигандом целевого рецептора или большой молекулой, которая более просто захватывается эндоцитозными механизмами клетки, для того чтобы увеличить поглощение комплекса клеткой, представляющей мишень.
Комплекс, связанный с мишенью, может быть интернализован за счет эндоцитоза и эффекторной группировки, высвобождаемой внутри клетки.
Связывающая группировка (полипептид), специфически связывающаяся с мишенью, в одном воплощении представляет собой фрагмент антитела.
Термин "одноцепочечный фрагмент вариабельной области" или "scFv" обозначает вариабельную антигенсвязывающую область одной легкой цепи антитела и одной тяжелой цепи антитела, связанные между собой ковалентной связью, имеющей длину, достаточную, чтобы позволить частям легкой и тяжелой цепей формировать антигенсвязывающий сайт.Такой линкер может быть таким коротким, как ковалентная связь. Особенно подходящие линкеры содержат от 2 до 50 аминокислотных остатков, в особенности от 5 до 25 аминокислотных остатков.
Другие фрагменты антител могут представлять собой димерные антитела, впервые описанные Holliger, P., et al. (PNAS (USA) 90 (1993) 6444-6448). Они могут быть построены с использованием тяжелых и легких цепей антитела, а также с использованием отдельных CDR-областей антитела. Как правило, димеры антител содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким, чтобы позволить спаривание двух доменов одной и той же цепи. Соответственно, домены VH и VL одного фрагмента вынуждены спариваться с комплементарными доменами VH и VL другого фрагмента, тем самым образуя два антигенсвязывающих участка. Тримерные антитела могут быть построены аналогичным образом с тремя антигенсвязывающими сайтами.
Fv-фрагмент антитела содержит полный антигенсвязывающий сайт, который включает VL-домен и VH-домен, удерживаемые вместе за счет нековалентных взаимодействий. Fv-фрагменты также включают конструкции, в которых домены VH и VL сшиты с помощью глутаральдегида, межмолекулярных дисульфидных связей или других линкеров. Вариабельные домены тяжелой и легкой цепей могут быть слиты вместе с образованием одноцепочечного вариабельного фрагмента (scFv), который сохраняет исходную специфичность родительского антитела. Димеры одноцепочечного Fv (scFv), впервые описанные Gruber, М., et al., J. Immunol. 152 (1994) 5368-5374, могут быть построены с использованием тяжелых и легких цепей антитела, а также с помощью отдельных CDR-областей антитела. Многие методики, известные в данной области, могут быть использованы для получения специфически связывающихся конструкций, подходящих для комплекса, описанного в данном документе (см., например, US 2007/0196274, US 2005/0163782).
Биспецифические антитела могут быть получены путем химической сшивки или по методике межвидовой гибридомы. Альтернативно, молекулы биспецифического антитела могут быть получены с помощью рекомбинантных методик. Димеризация может усиливаться при уменьшении длины линкера, соединяющего VH- и VL-домены, до длины примерно от 15 аминокислот, что обычно используется для получения scFv-фрагментов, примерно до 5 аминокислот.Эти линкеры благоприятны для внутрицепочечной сборки доменов VH и VL. Подходящим коротким линкером является SGGGS (SEQ ID № 66), но могут быть использованы и другие линкеры. Таким образом, два фрагмента собираются в димерную молекулу. Дальнейшее уменьшение длины линкера до 0-2 аминокислотных остатков может дать молекулы тримерных или тетрамерных антител.
В одном воплощении связывающая группировка (полипептид), специфически связывающаяся с мишенью, например, антитело, специфически связывающееся с рецептором клеточной поверхности, может быть связана с лигандом целевого рецептора или большой молекулой, которая более просто захватывается эндоцитозными механизмами клетки.
В этом воплощении комплекс может быть использован для увеличения поглощения комплекса клеткой, представляющей мишень. Комплекс, связанный с мишенью, может быть интернализован за счет эндоцитоза и эффекторной группировки, высвобождаемой за счет кислого гидролиза или ферментативной активности, когда эндоцитозный пузырек сливается с лизосомами.
Комплексы, описанные в данном документе, могут быть использованы для доставки эффекторной группировки внутриклеточно и внеклеточно. Комплекс может быть использован для распознавания раковых клеток in situ, что делает их привлекательными кандидатами для разработки нацеливающих лекарственных веществ.
Когда желательно нековалентное связывание компонента с другим компонентом (или частицей или капсулой), то могут быть использованы соответствующие ассоциативные взаимодействия, которые включают, но не ограничиваясь ими, антитело-антиген, рецептор-гормон, пары авидин-биотин, стрептавидин-биотин, металл-хелат, малая молекула/полинуклеотид (см., например, Dervan, Р.В., Bioorg. Med. Chem. 9 (2001) 2215-2235; Zahn, Z.Y. and Dervan, P.B., Bioorg. Med. Chem. 8 (2000) 2467-2474); полинуклеотид/комплементарный полинуклеотид (например, димерные и тримерные спирали), аптамер/малая молекула, аптамер/полипептид, суперспираль и полинуклеотид/полипептид (например, мотивы «цинковый палец», «спираль-поворот-спираль», «лейциновая молния» и «спираль-петля-спираль», которые связываются с ДНК-последовательностями).
Комплекс, описанный в данном документе, может быть использован для доставки в клетку различных эффекторных группировок, таких как цитотоксические лекарственные препараты, в том числе лекарственные препараты, компоненты, испускающих излучение, молекулы растительного, грибного или бактериального происхождения, биологические белки и их смеси. Цитотоксический лекарственный препарат, например, может быть внутриклеточно действующим цитотоксическим лекарственным препаратом, например, испускающим слабопроникающее излучение, в том числе, например, слабопроникающее α-излучение большой энергии.
В одном воплощении эффекторная группировка представляет собой инкапсулированный в липосому лекарственный препарат (например, противоопухолевый препарат, такой как арбаксан, доксорубицин, памидронат динатрия, анастрозол, экземестан, циклофосфамид, эпирубицин, торемифен, летрозол, трастузумаб, мегестролтамоксифен, паклитаксел, доцетаксел, капецитабин, гозерелина ацетат, золедроновая кислота, винбластин и т.д.), антиген, который стимулирует распознавание связанной клетки компонентами иммунной системы, антитело, которое специфически связывается с компонентами иммунной системы и направляет их в клетки, и т.п.
В одном воплощении эффекторная группировка может содержать радиосенсибилизатор, который усиливает цитотоксическое действие ионизирующего излучения (например, такого, какое может быть получено с 60Co или источником рентгеновского излучения) на клетку.
В одном воплощении эффекторная группировка выбрана среди хемотаксических факторов моноцитов, или f-Met-Leu-Phe (fMLP), или f-Met-Leu-Phe-o-метилового эфира, или формил-норлейцин-фенилаланина, или формил-метионил-фенилаланина, или их производных.
В одном воплощении эффекторная группировка является реакционноспособной группой.
Реакционноспособная группа может быть выбрана среди любых известных реакционноспособных групп, таких как амино-, сульфгидрил-, карбоксилат-, гидроксил-, азидо-, алкинил- или алкенильных групп.
В одном воплощении реакционноспособная группа выбрана среди малеинимидо-, сукцинимидил-, дитиопиридил-, нитрофенилэфир-, гексафторфенилэфирных групп.
Если формат действия зависит от создания на мишени высокой локальной концентрации эффектора, как в случае fMLP в качестве эффекторной группировки, то L-ДНК-природа линкерных группировок позволяет специфическую гибридизацию со вторым L-ДНК-олигонуклеотидом, модифицированным одинаковыми или разными эффекторными группировками.
Число эффекторных группировок, которые связаны со второй L-ДНК, должно быть ограничено, чтобы не было ответа, индуцированного одной эффекторной модифицированной L-ДНК. При желании вторая L-ДНК ставит под угрозу другой сайт, который способен к специфической гибридизации с третьим L-ДНК-олигонуклеотидом, модифицированным с той же самой или другой эффекторной группировкой. Поскольку легко выбрать много различных последовательностей, которые специфически образуют дуплекс в присутствии других дуплексов, то легко может быть построен мультимерный комплекс.
Мультимерные комплексы могут быть построены с использованием олигонуклеотидов с перекрывающимися последовательностями, образующими линейный многомерный комплекс, или с использованием разветвленных олигонуклеотидов, в которых ветви способны к гибридизации с третьим олигонуклеотидом, что приводит к образованию разветвленных многомерных комплексов.
В одном воплощении эффекторная группировка является альфа-излучателем, т.е. радиоактивным изотопом, который испускает альфа-частицы. Подходящие альфа-излучатели включают, но не ограничиваясь ими, Bi, 213Bi, 211At и т.п.
Эффекторная группировка также может содержать лиганд, эпитопную метку, Fc-область антитела или антитело.
Ферментативно активные токсины и их фрагменты могут быть выбраны среди фрагмента дифтерийного токсина А, несвязывающих активных фрагментов дифтерийного токсина, экзотоксина А (из Pseudomonas aeruginosa), цепи А рицина, цепи А абрина, цепи А модецина, альфа-сарцина, некоторых белков Aleurites fordii, некоторых белков Dianthin, белков Phytolacca americana (PAP, PAPII и PAP-S), ингибитора Morodica charantia, курцина, кротина, ингибитора Saponaria officinalis, гелонина, митогиллина, рестриктоцина, феномицина и эномицина.
В одном воплощении один или более L-ДНК-олигонуклеотид, модифицированный высокой плотностью запертых эффекторных группировок, гибридизован с L-ДНК-линкером.
Раковые клетки отличаются от нормальных клеток различным образом, одним из которых является молекулярный состав клеточной поверхности. Измененные химические реакции на поверхности позволяют раковым клеткам эффективно реагировать на внешние сигналы для роста и выживания и непосредственно взаимодействовать с различными элементами ткани хозяина для миграции, входа в кровоток, экстравазации и колонизации в удаленном месте. Кроме того, что они выступают в качестве маркеров злокачественных клеток, поверхностные молекулы опухолевых клеток являются ценными мишенями для терапии благодаря их относительно легкой доступности для нацеливающих молекул, вводимых в кровоток или во внеклеточное пространство (Feng, A., et al., Mol. Cancer Ther. 7 (2008) 569-578).
Предполагаемые опухолеспецифические антигены включают, но не ограничиваясь ими, СЕА, CD20, HER1, HER2/neu, HER4, PSCA, PSMA, СА-125, СА-19-9, c-Met, MUC1, RCAS1, Ер-САМ, Melan-A/MART1, RHA-MM, VEGF, EGFR, интегрины, ED-B фибронектина, ChL6, Lym-1, CD1b, CD3, CD5, CD14, CD20, CD22, CD33, CD56, TAO-72, рецептор интерлейкина-2 (IL-2R), ферритин, нейронную молекулу клеточной адгезии (NCAM), ассоциированный с меланомой антиген, ганглиозид Gm, рецептор EOF, тенасцин, c-Met (HGFR).
В одном воплощении антитело специфически связывается с посттрансляционно модифицированной мишенью на рецепторе клеточной поверхности. В одном воплощении мишень посттрансляционно модифицирована путем фосфорилирования или гликозилирования.
В одном воплощении первый полипептид и второй полипептид связываются с одним и тем же или с перекрывающимися эпитопами.
Было обнаружено, что посттрансляционно модифицированный целевой полипептид может быть обнаружен в комплексе, состоящем из двух одновалентных полипептидов, специфически связывающихся с мишенью, которые связаны друг с другом с помощью полинуклеотидной линкера, где первый полипептид связывается с полипептидным эпитопом мишени, второй полипептид связывается с посттрансляционной модификацией полипептида, где каждый одновалентный связывающий элемент имеет Kдисс в диапазоне от 10-2/с до 10-3/с, и где комплекс имеет Kдисс 10-4/с или менее.
Известны различные типы ковалентных модификаций аминокислот. Посттрансляционные модификации описаны, например, в Mann and Jensen (2003) и в Seo and Lee (2004), которые включены в данный документ посредством ссылки (Mann, М. and Jensen, O.N., Biochemistry 21 (2003) 255-261; Seo, J. and Lee, K.-J., Biochemistry and Molecular Biology 37/1 (2004) 35-44).
В одном воплощении посттрансляционная модификация выбрана из группы, состоящей из ацетилирования, фосфорилирования, ацилирования, метилирования, гликозилирования, убиквитинилирования, сумоилирования, сульфатизации и нитрования.
Ацетилирование (изменение молекулярной массы +42 Да) является достаточно стабильной вторичной модификацией. Примерами являются ацетилирование, которое находится на N-концах многих белков, или ацетилирование на остатках лизина или серина. Обычно ацетилирование остатка лизина находится на одной или более четко определенной позиции (позициях) в полипептидной цепи, в то время как другие остатки лизина ацетилируются реже или вовсе не ацетилируются.
Фосфорилирование и дефосфорилирование белка (конечный баланс, который может упоминаться как статус фосфорилирования), как известно, является одним из ключевых элементов в регуляции биологической активности белка. Низкий процент фосфорилированных аминокислотных остатков уже может быть достаточным, чтобы вызвать определенную биологическую активность. Фосфорилирование приводит к увеличению массы на 80 Да (увеличение молекулярной массы). Аминокислоты тирозин (Y), серии (S), треонин (Т), гистидин (Н) и аспарагиновая кислота (D) могут быть фосфорилированы. Чем сложнее биологическая функция полипептида, тем более сложным является соответствующий паттерн возможных сайтов фосфорилирования. Особенно это относится и хорошо известно для мембраносвязанных рецепторов, особенно так называемых рецепторных тирозинкиназ (RTK). Как уже предполагает номенклатура, по меньшей мере часть внутриклеточного сигналинга RTK опосредуется статусом фосфорилирования определенного тирозина во внутриклеточном домене таких RTK.
Полипептиды могут быть ацилированы фарнезил-, миристоил- или пальмитоил-группами. Ацилирование обычно происходит на боковой цепи остатка цистеина.
Метилирование в качестве вторичной модификации происходит через боковую цепь остатка лизина. Было показано, что связывающие свойства регуляторных белков, которые способны связываться с нуклеиновой кислотой, например, могут быть модулированы за счет метилирования.
Гликозилирование является обычной вторичной модификацией. Оно имеет большое влияние на белок-белковые взаимодействия, на солюбилизацию белков, а также их стабильность. Известно два различных типа гликозилирования: N-связанное (через боковые цепи аминокислоты N (аспарагина)) и О-связанное (через боковые цепи серина (S) или треонина (Т)). Было выявлено много различных полисахаридов (линейных или с разветвленными боковыми цепями), некоторые из них содержат производные сахара, такие как O-Glc-NAc.
Как известно, убиквитинилирование и сумоилирование, соответственно, влияют на период полужизни белков в кровотоке. Убиквитинилирование может служить сигналом уничтожения, что приводит к расщеплению и/или удалению убиквитинилированных полипептидов.
Сульфатация через остаток тирозина (Y), по-видимому, играет важную роль в модуляции белок-белковых (клетка-клетка) взаимодействий, а также взаимодействий белок-лиганд.
Нитрование остатков тирозина (Y), по-видимому, является критерием окислительного повреждения, например, такого как при воспалительных процессах.
L-дезоксинуклеозид-фосфорамидитные группировки L-dT, L-dC, L-dA и L-dG могут быть получены в соответствии с литературными данными (см., например, Urata, Н., et al., Nucl. Acids Res. 20 (1992) 3325-3332; Shi, Z.D., et al., Tetrahed. 58 (2002) 3287-3296). Производное L-дезоксирибозы может быть синтезировано из L-арабинозы за 8 этапов. L-дезоксинуклеозиды могут быть получены путем гликозилирования соответствующих нуклеотидных производных L-дезоксирибозным производным. После получения продуктов с нуклеозидными фосфорамидитами они могут быть встроены в олигодезоксинуклеозиды с помощью способа твердофазного синтеза ДНК. Олигомер может быть очищен с помощью ВЭЖХ с обращенной фазой и электрофореза в полиакриламидном геле (PAGE).
L-ДНК может быть синтезирована как ДНК в больших масштабах с помощью стандартных протоколов синтеза.
Для экспрессии и очистки scFv-фрагментов антитела нуклеиновая кислота, кодирующая scFv, может быть клонирована в вектор для экспрессии и/или секреции, такой как pUC119mycHis, что приводит к добавлению эпитопной метки c-myc и гексагистидиновой метки на С-конец scFv. Для создания (scFv')2-димера для иммуногистохимии (Adams, G.P., et al., Cancer Res. 53 (1993) 4026-4034) эпитопная метка c-myc может быть генетически удалена из pUC119mycHis, и свободный цистеин может быть введен на С-конец scFv перед гексагистидиновой меткой. scFv или димер белка (scFv')2 может быть собран из бактериальной периплазмы и очищен с помощью аффинной хроматографии с иммобилизованными ионами металла и с помощью гель-фильтрации (Nielsen, U.B., et al., Biochim. Biophys. Acta 1591 (2002) 109-118).
Альтернативно, scFv могут быть получены путем введения структурных генов, кодирующих scFv, в вектор экспрессии, добавляющий с-myc и гексагистидиновую метку на С-конец (Liu, В., et al., Cancer Res. 64 (2004) 704-710). Чтобы получить растворимый (scFv)2, можно использовать второй вектор для добавления цистеина и гексагистидиновой метки на С-конец. После индукции с IPTG фрагменты антител могут быть выделены из периплазматического пространства бактерий на шариках из никеля и нитрилотриуксусной кислоты. Для FACS и иммуногистохимических исследований scFv могут быть биотинилированы с помощью EZ-Link Sulfo-NHS-LC-Biotin (Pierce) в соответствии с инструкциями производителя.
Для определения константы диссоциации (KD) клеточную линию, экспрессирующую соответствующую поверхностную молекулу-мишень, можно выращивать до 90% конфлюэнтности в подходящей среде, такой как RPMI 1640 с добавлением 10% FCS. Клетки могут быть собраны путем короткого расщепления трипсином (0,2%) в 2 ммоль/л растворе EDTA/PBS. Биотинилированные scFv можно инкубировать с 105 клеток в течение 4 часов при 4°С в PBS/0,25% бычьего сывороточного альбумина. Связанные scFv могут быть обнаружены с помощью стрептавидин-фикоэритрина и проанализированы с помощью FACS. Данные могут быть представлены в виде кривой, и значения KD могут быть рассчитаны с использованием GraphPad Prism (программное обеспечение Graph-Pad).
Для иммуногистохимии могут быть использованы срезы ткани из замороженных и/или залитых в парафин блоков. Для иммуногистохимического анализа срезы ткани могут быть инкубированы с очищенным димерным scFv (например, 50 мкг/мл в 2% растворе молока/PBS) при 4°С в течение четырех часов, промыты PBS, инкубированы с анти-(His)6-антителом, разведенным 1:400 (Santa Cruz Biotechnology), а затем с биотинилированным антителом против анти(His)6-антитела, разведенным 1:400 (Vector Lab), и со стрептавидином, конъюгированным с пероксидазой хрена, разведенным 1:400 (Sigma). Связывание может быть обнаружено с использованием диаминобензидина в качестве субстрата (Sigma).
Альтернативно, замороженные срезы исследуемых и контрольных тканей могут быть окрашены биотинилированными scFv (250 нмоль/л) при комнатной температуре в течение одного часа. scFv, которые не связываются с анализируемыми клеточными линиями при FACS, могут быть использованы в качестве контроля для всех экспериментов. Связанные scFv могут быть обнаружены с помощью конъюгата стрептавидина и пероксидазы хрена с использованием субстрата 3,3'-диаминобензидина. Окрашенные ткани могут быть контрастно окрашены гематоксилином, высушены в 70%, 95% и 100% этаноле, залиты и проанализированы.
В частности, см. US 2003/0152987, касающийся иммуногистохимии (IHC) и флуоресцентной гибридизации in situ (FISH) для выявления сверхэкспрессии и амплификации HER2 (включен в данный документ посредством ссылки).
Для определения интернализации может быть проведена следующая процедура. Для экспериментов с флуоресцентной микроскопией клетки могут быть выращены примерно до 80% конфлюэнтности в 24-луночных планшетах и инкубированы совместно с нецелевыми или целевыми комплексами, меченными 1,1-диоктадецил-3,3,3',3'-тетраметилиндокарбоцианин-5,5'-цисульфоновой кислотой, в течение четырех часов при 37°С. Клетки могут быть промыты PBS и исследованы с помощью флуоресцентного микроскопа Nikon Eclipse ТЕ300. Для FACS-анализа клетки могут быть инкубированы с комплексами, меченными 1,1-диоктадецил-3,3,3',3'-тетраметилиндокарбоцианин-5,5'-дисульфоновой кислотой, при 37°С в течение двух часов, удалены путем расщепления трипсином, помещены в глициновый буфер (pH 2,8; 150 ммоль/л NaCl) при комнатной температуре на 5 минут для удаления связанных с поверхностью липосом и проанализированы с помощью FACS (LSRII; BD Biosciences). Средние значения интенсивности флуоресценции могут быть использованы для расчета процента интернализированных липосом (устойчивых к обработке глицином) среди всех связанных с клеткой липосом (до обработки глицином).
Для анализов ингибирования роста и интернализации раковые клетки при конфлюэнтности примерно 30-80% могут быть инкубированы с различными концентрациями аффинно очищенного комплекса при 37°С в течение 72 ч в среде, содержащей 1% FCS. Статус роста может быть оценен с помощью анализа с использованием соли тетразолия 3-(4,5-диметилтизаол-2-ил)-2,5-дифенилтетразоля бромида (Promega), и IC50 может быть рассчитана с использованием KaleidaGraph 3.5 (Synergy Software). Для анализа интернализации комплекс может быть биотинилирован сульфо-NHS-LC-биотином (Pierce) и инкубирован с клетками-мишенями при 37°С в течение различных периодов времени. Клетки могут быть промыты 100 мМ глициновым буфером (рН 2,8), фиксированы 2% формальдегидом, пермеабилизованы 100% ледяным метанолом и инкубированы со стрептавидин-FITC. Окрашенные клетки сначала могут быть оценены с помощью флуоресцентного микроскопа Axiophot (Zeiss), а затем изучены с помощью конфокального лазерного флуоресцентного микроскопа Leica TCS NT (Leica).
Для определения токсичности клетки могут быть внесены в 96-луночные планшеты по 6000 на лунку и инкубированы с комплексом, описанном в данном документе, в различных концентрациях (0-10 мкг/мл) в течение двух часов при 37°С. После удаления комплекса клетки могут быть промыты один раз средой RPMI 1640 с добавлением 10% FCS и инкубированы еще в течение 70 ч при 37°С. Жизнеспособность клеток может быть проанализирована с помощью набора Cell Counting Kit-8 (Dojindo) в соответствии с инструкциями изготовителя. Эти данные могут быть выражены в виде процента жизнеспособных клеток по сравнению с необработанными контрольными клетками.
Для определения цитотоксичности in vitro раковые клетки могут быть высеяны в 96-луночные планшеты (6000 клеток на лунку, например, для клеток РС3 и Du-145, или 10000 клеток на лунку, например, для клеток LNCaP) и инкубированы с комплексом, описанном в данном документе (0-10 мкг/мл) в течение 4 ч при 37°С. Клетки могут быть дважды промыты дополненной RPMI 1640 для удаления лекарственных препаратов и инкубированы в свежей среде еще в течение 72 ч при 37°С. Жизнеспособность клеток может быть проанализирована путем окрашивания 3-(4,5-диметилтизаол-2-ил)-2,5-дифенилтетразолия бромидом (Carmichael, J., et al., Cancer Res. 47 (1987) 936-942), и результаты могут быть считаны при 570 нм с использованием ридера для микротитровальных планшетов (SpectraMax 190, Molecular Devices). Жизнеспособность клеток может быть определена с использованием набора Cell Counting Kit-8 (Dojindo) в соответствии с инструкциями изготовителя.
Для анализа внутриклеточной доставки может быть использован следующий способ. Для оценки внутриклеточной доставки комплекса этот комплекс может быть добавлен к клеткам вместе с 1 мкг/мл очищенного (His)6-меченого scFv, инкубирован при 37°С в течение 30 мин и промыт три раза солевым раствором, содержащим 1 мМ EDTA, для удаления комплексов, связанных с поверхностью клеток, которые не были интернализованы. Поглощение комплекса может быть определено путем микрофлуориметрии с помощью микрофлуориметра Gemini (Molecular Devices) и с помощью перевернутого флуоресцентного микроскопа (Nikon).
Рекомбинантные способы
Как правило, связывающие группировки, такие как фрагменты антител или элементы связывающей пары, могут быть получены с использованием рекомбинантных способов и композиций, например, описанных в US 4816567.
В одном воплощении предложена выделенная нуклеиновая кислота, кодирующая каждую связывающую группировку комплекса, описанного в данном документе.
Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела, (например, легкую и/или тяжелую цепи антитела) и/или аминокислотную последовательность элемента связывающей пары.
В одном воплощении предложена клетка-хозяйка, содержащая такую нуклеиновую кислоту. В одном воплощении предложена клетка-хозяйка, которая содержит (например, была трансформирована ими): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном воплощении клетка является эукариотической, например, клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, Y0, NS0, Sp2/0). Для экспрессии фрагментов антитела и полипептидов в бактериях см., например, US 5648237, US 5789199 и US 5840523; Charlton, K.А., In: Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp. 245-254. После экспрессии антитело может быть выделено из бактериальной клеточной массы в растворимой фракции и может быть далее очищено.
Кроме прокариотов, для векторов, кодирующих антитела, подходящими клонирующими или экспрессирующими хозяевами являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи (см. Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414; и Li, Н. et al., Nat. Biotech. 24 (2006) 210-215).
Культуры растительных клеток также могут быть использованы в качестве хозяев (см., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429).
Клетки позвоночных животных также могут быть использованы в качестве хозяев. Например, могут быть использованы клеточные линии млекопитающих, приспособленные к росту в суспензии. Другими примерами используемых клеточных линий от млекопитающих являются линия почки обезьяны CV1, трансформированная SV40 (COS-7); человеческая эмбриональная почечная линия (HEK 293), клетки почек детенышей хомяка (BHK); мышиные клетки Сертоли (ТМ4-клетки, описанные, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зеленой обезьяны (VERO-76); клетки шейки матки человеческой карциномы (HELA); клетки почки собаки (MDCK); клетки печени крысы buffalo (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); TRI-клетки, описанные, например, в Mather, J.P. et al., Annals N.Y. Acad. Sci. 383 (1982) 44-68; клетки MRC 5 и клетки FS4. Другие используемые линии клеток от млекопитающих включают клетки яичника китайского хомячка (СНО), в том числе DHFR-CHO-клетки (Urlaub, G. et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев от млекопитающих, подходящих для продукции антител, см., например, в Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp. 255-268 (2003).
Клетки-хозяева, используемые для получения полипептидов комплекса, описанного в данном документе, могут быть культивированы в различных средах. Коммерчески доступные среды, такие как F10 Хэма (Sigma), минимальная поддерживающая среда ((MEM), (Sigma), RPMI-1640 (Sigma) и среда Игла в модификации Дульбекко ((DMEM), Sigma), подходят для культивирования клеток-хозяев. Кроме того, любая из сред, описанных в Ham, R.G., et al., Meth. Enzymol. 58 (1979) 44-93, Barnes, D., et al., Anal. Biochem. 102 (1980) 255-270, US 4767704, US 4657866, US 4927762, US 4560655, US 5122469, WO 90/03430, WO 87/00195 и US Re 30985, может быть использована в качестве культуральной среды для клеток-хозяев. Любая из этих сред может быть дополнена при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальций, магний и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как препарат GENTAMYCIN™), микроэлементами (определяются как неорганические компоненты, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки также могут быть включены в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, рН и т.п., являются такими, которые ранее использовались для клетки-хозяйки, выбранной для экспрессии, и будут очевидны специалисту в данной области.
При использовании рекомбинантных методик связывающая группировка/полипептид могут быть получены внутриклеточно, в периплазматическом пространстве или непосредственно секретированы в среду. Если полипептид продуцируется внутриклеточно, то на первом этапе удаляют дисперсный дебрис, клетки-хозяева или лизированные фрагменты, например, путем центрифугирования или ультрафильтрации. Например, в Carter, P., et al., Bio/Technology 10 (1992) 163-167 описана процедура выделения антител, которые секретируются в периплазматическое пространство E. coli. Вкратце, клеточную массу размораживают в присутствии ацетата натрия (рН 3,5), EDTA и фенилметилсульфонилфторида (PMSF) в течение примерно 30 минут. Клеточный дебрис можно удалить центрифугированием. Если полипептид секретируется в среду, то супернатанты таких экспрессионных систем, как правило, сначала концентрируют с помощью коммерчески доступного фильтра для концентрирования белков, например, ультрафильтрационного аппарата Amicon или Millipore Pellicon. Ингибитор протеазы, такой как PMSF, может быть включен в любой из предыдущих этапов для ингибирования протеолиза, а антибиотики могут быть включены для предотвращения роста случайных контаминантов. Композиция с полипептидом, полученная из клеток, может быть очищена, например, с помощью гидроксилапатитной хроматографии, гель-электрофореза, диализа и аффинной хроматографии, при этом аффинная хроматография является предпочтительной методикой для очистки антител. Пригодность белка А в качестве аффинного лиганда зависит от вида и подкласса любого иммуноглобулинового Fc-домена, который присутствует в антителе. Белок А может быть использован для очистки антител, которые основаны на человеческих тяжелых цепях γ1; γ2 или γ4 (Lindmark, R., et al., J. Immunol. Meth. 62 (1983) 1-13). Белок G рекомендуется для всех мышиных подклассов и человеческого γ3 (Guss, В., et al., EMBO J. 5 (1986) 1567-1575). Матрица, к которой прикрепляется аффинный лиганд, наиболее часто является агарозой, но также доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокую скорость потока и более короткое время обработки, чем можно достичь с агарозой. Если антитело содержит СН3-домен, то для очистки используется Bakerbond ABXTMresin (Bakerbond ABX™resin (J.Т. Baker, Филипсбург, Нью-Джерси, США). Другие методики очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, ВЭЖХ с обращенной фазой, хроматография на диоксиде кремния, хроматография на Heparin-SEPHAROSE™, хроматографии на анионообменной или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония, также доступны в зависимости от антитела, которое должно быть получено.
После каждого этапа предварительной очистки смесь, содержащую представляющий интерес полипептид и загрязняющие вещества, можно подвергать хроматографии гидрофобных взаимодействий при низком рН с использованием элюирующего буфера при рН примерно 2,5-4,5, особенно выполняемой при низких концентрациях соли (например, примерно 0-0,25 М соль).
Комплекс и его отдельные компоненты, описанные в данном документе, при желании могут быть выделены и очищены. Нежелательные компоненты реакционной смеси, в которой формируется комплекс, например, представляют собой полипептиды и полинуклеотиды, которые не завершают желаемый комплекс, но составляют его структурные блоки. В одном воплощении комплекс очищают до чистоты более 80%, определяемой с помощью аналитической эксклюзионной хроматографии. В некоторых воплощениях комплекс очищают до чистоты более 90%, 95%, 98% или 99% по весу, определяемой с помощью аналитической эксклюзионной хроматографии, соответственно. Альтернативно, чистоту можно легко определить, например, с помощью SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием, например, красителя Кумасси синего или серебряного окрашивания для обнаружения белка. В случае если чистоту оценивают на уровне комплекса, то для отделения комплекса от побочных продуктов можно применять эксклюзионную хроматографию и контролировать OD при 260 нм для оценки его чистоты.
Иммуноконъюгаты
Изобретение также предусматривает комплексы, в которых по меньшей мере одна из связывающих группировок, которая специфически связывается с мишенью или линкером, также конъюгирована с одной или более эффекторной группировкой, например, цитотоксическим агентом, таким как химиотерапевтические агенты или лекарства, ингибирующие рост агенты, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном воплощении эффекторная группировка является лекарственным средством, включая, но не ограничиваясь ими, мейтансиноид (см. патенты US 5208020, US 5416064, ЕР 0425235); ауристатин, такой как монометилауристатиновые лекарственные группировки DE и DF (ММАЕ и MMAF) (см. патенты US 5635483, US 5780588, US 7498298); доластатин; калихеамицин или его производное (см. патенты US 5712374, US 5714586, US 5739116, US 5767285, US 5770701, US 5770710, US 5773001, US 5877296; Hinman, L.M., et al., Cancer Res. 53 (1993) 3336-3342, Lode, H.N., et al., Cancer Res. 58 (1998) 2925-2928), антрациклины, такие как дауномицин или доксорубицин (см. Kratz, F., et al., Current Med. Chem. 13 (2006) 477-523, Jeffrey, S.C., et al., Bioorg. Med. Chem. Letters 16 (2006) 358-362, Torgov, M.Y., et al., Bioconjug. Chem. 16 (2005) 717-721, Nagy, A., et al., Proc. Natl. Acad. Sci. USA 97 (2000) 829-834, Dubowchik, G.M., et al., Bioorg. Med. Chem. Lett. 12 (2002) 1529-1532, King, H.D., et al., J. Med. Chem. 45 (2002) 4336-4343, и US 6,630,579), метотрексат; виндезин; таксаны, такие как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и СС1065.
В одном воплощении эффекторная группировка является ферментативно активным токсином или его фрагментом, включая, но не ограничиваясь ими, дифтерийную А-цепь, несвязывающие активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolacca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Saponaria officinalis, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены.
В одном воплощении эффекторная группировка является радиоактивным атомом. Множество радиоактивных изотопов доступно для получения радиоконъюгатов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Когда радиоконъюгат используется для обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, Tc99m или I123, или спиновую метку для визуализации путем ядерного магнитного резонанса (NMR) (также известного как магнитно-резонансная томография, MRI), такую как снова I123, I131, In111, F19, С13, N15, О17 гадолиний, марганец или железо.
Эффекторная группировка может быть конъюгирована с любым компонентом комплекса, описанного в данном документе, с использованием множества бифункциональных связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(параазидобензоил)гександиамин), производные бис-диазония (такие как бис-(парадиазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен, как описано в Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен-триамин-пентауксксная кислота (МХ-DTPA) является примером хелатирующего агента для конъюгации радионуклеотида с комплексом (см. WO94/11026). Линкер для конъюгации токсической группировки с комплексом, описанным в данном документе, может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного агента в клетке. Например, может быть использован кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); US №5208020).
Эффекторная группировка может быть конъюгирована с соединением комплекса, описанного в данном документе, но не ограниченного такими конъюгатами, полученными с помощью сшивающих реагентов, включая, но не ограничиваясь ими, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые коммерчески доступны (например, Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Фармацевтические композиции
Фармацевтические композиции мультиспецифической связывающей молекулы (такой как биспецифическое антитело), описанной в данном документе, получают путем смешивания такой мультиспецифической связывающей молекулы, имеющей требуемую степень чистоты, с одним или более чем одним возможным фармацевтически приемлемым носителем (Osol, A. (ed.), Remington's Pharmaceutical Sciences, 16th edition, Mack Publishing Company (1980)), в виде лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители, как правило, нетоксичны для реципиентов в используемых дозах и концентрациях и включают, но не ограничиваясь ими: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетониума хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол, 3-пентанол и m-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Типичные фармацевтически приемлемые носители в данном документе также включают дисперсные агенты промежуточных лекарственных средств, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, человеческие растворимые РН-20 гликопротеины гиалуронидазы, такие как rhuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые иллюстративные sHASEGP и способы их применения, в том числе rhuPH20, описаны в патентных публикациях US №№2005/0260186 и 2006/0104968. В одном аспекте sHASEGP комбинирован с одной или более чем одной дополнительной гликозамингликаназой, такой как хондроитиназа.
Иллюстративные лиофилизированные составы антител описаны в патенте US 6267958. Водные составы антител включают те, которые описаны в US 6171586 и WO 2006/044908, последние составы включают гистидин-ацетатный буфер.
Состав в данном документе может также содержать более одного активного ингредиента, что нужно при лечении по конкретным показаниям, особенно с дополняющими активностями, которые не влияют друг на друга. Такие активные ингредиенты соответствующим образом присутствуют в комбинации в количествах, которые эффективны для данной цели.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, с помощью методик коацервации или путем межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методики раскрыты в Remington's Pharmaceutical Sciences 16th edition, Osol, A. (ed.), (1980).
Могут быть изготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, при этом матрицы находятся в виде формованных изделий, например пленок или микрокапсул.
Составы, используемые для введения in vivo, обычно являются стерильными. Стерильность может быть легко достигнута, например, путем фильтрации через стерилизующие фильтрационные мембраны.
Терапевтические способы и композиции
Любая из мультиспецифических связывающих молекул (например, биспецифических антител), описанных в данном документе, может быть использована в терапевтических способах.
В одном аспекте предложена мультиспецифическая связывающая молекула или биспецифическое антитело, описанное в данном документе, для применения в качестве лекарственного средства. В другом аспекте предложена мультиспецифическая связывающая молекула или биспецифическое антитело для применения в лечении рака. В некоторых воплощениях предложена мультиспецифическая связывающая молекула или биспецифическое антитело для применения в способе лечения. В некоторых воплощениях данное изобретение предусматривает мультиспецифическую связывающую молекулу или биспецифическое антитело для применения в способе лечения индивидуума с раком, включающем введение индивидууму эффективного количества мультиспецифической связывающей молекулы или биспецифического антитела. В одном таком воплощении способ также включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента. «Индивидуум» в соответствии с любым из вышеописанных воплощений предпочтительно представляет собой человека.
В другом аспекте изобретение предусматривает применение мультиспецифической связывающей молекулы или биспецифического антитела, описанного в данном документе, в производстве или изготовлении лекарственного средства. В одном воплощении лекарственное средство предназначено для лечения рака. В другом воплощении лекарственное средство предназначено для применения в способе лечения рака, включающем введение индивидууму с раком эффективного количества лекарственного средства. «Индивидуум» в соответствии с любым из вышеописанных воплощений может быть человеком.
В другом аспекте, описанном в данном документе, предусмотрен способ лечения рака. В одном воплощении способ включает введение индивидууму с таким раком эффективного количества мультиспецифической связывающей молекулы или биспецифического антитела, описанного в данном документе. «Индивидуум» в соответствии с любым из вышеописанных воплощений может быть человеком.
В другом аспекте, описанном в данном документе, предусмотрены фармацевтические композиции, содержащие любую из мультиспецифических связывающих молекул или биспецифических антител, предложенных в данном документе, например, для применения в любом из указанных выше терапевтических способов. В одном воплощении фармацевтическая композиция содержит любую из мультиспецифических связывающих молекул или биспецифических антител, предложенных в данном документе, и фармацевтически приемлемый носитель. В другом воплощении фармацевтическая композиция содержит любую из мультиспецифических связывающих молекул или биспецифических антител, предложенных в данном документе, и по меньшей мере один дополнительный терапевтический агент.
Мультиспецифические связывающие молекулы или биспецифические антитела, описанные в данном документе, могут быть использованы в терапии по отдельности или в комбинации с другими агентами. Например, мультиспецифическая связывающая молекула или биспецифическое антитело, описанное в данном документе, может быть введено совместно по меньшей мере с одним дополнительным терапевтическим агентом.
Такие комбинированные терапии, отмеченные выше, охватывают комбинированное введение (где два или более двух терапевтических агентов включены в одни и те же или отдельные составы), а также раздельное введение, и в этом случае введение мультиспецифической связывающей молекулы или биспецифического антитела изобретения может происходить до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта. Мультиспецифические связывающие молекулы или биспецифические антитела, описанные в данном документе, также могут быть использованы в сочетании с лучевой терапией.
Мультиспецифическая связывающая молекула или биспецифическое антитело, описанное в данном документе, может быть введено с помощью любых подходящих средств, в том числе путем парентерального, внутрилегочного и интраназального, и, если желательно местное лечение, внутриочагового введения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Дозирование может быть осуществлено любым подходящим способом, например с помощью инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или хроническим. В данном описании рассматриваются различные схемы дозировки, включая, но не ограничиваясь ими, одно или множество введений в различные временные точки, болюсное введение и импульсную инфузию.
Мультиспецифические связывающие молекулы и биспецифические антитела, описанные в данном документе, будут собраны в состав, дозированы и введены в соответствии с надлежащей медицинской практикой. Факторы, подлежащие рассмотрению в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки агента, способ введения, схему введения и другие факторы, известные практикующим врачам. Мультиспецифическая связывающая молекула или биспецифическое антитело не обязательно, но может быть собрано в состав с одним или более чем одним агентом, используемым в настоящее время для профилактики или лечения рассматриваемого нарушения. Эффективное количество таких других агентов зависит от количества мультиспецифической связывающей молекулы или биспецифического антитела, присутствующего в составе, от типа заболевания или лечения и от других факторов, описанных выше. Как правило, они используются в тех же дозах и с теми же путями введения, как описано в данном документе, или в дозировках от 1 до 99% от описанных в данном документе, или в любой дозировке и любым путем, который является эмпирически/клинически подходящим.
Для профилактики или лечения заболевания подходящая доза мультиспецифической связывающей молекулы или биспецифического антитела изобретения, описанного в данном документе, (при использовании его отдельно или в сочетании с одним или более чем одним другим дополнительным терапевтическим агентом) будет зависеть от типа заболевания, подлежащего лечению, от типа мультиспецифической связывающей молекулы или биспецифического антитела, от тяжести и течения заболевания, от того, вводят ли мультиспецифическую связывающую молекулу или биспецифическое антитело с профилактической или терапевтической целью, от предыдущей терапии, истории болезни пациента и ответа на антитела и по усмотрению лечащего врача. Мультиспецифическая связывающая молекула или биспецифическое антитело вводят пациенту подходящим образом одномоментно или серийно. В зависимости от типа и тяжести заболевания изначальной кандидатной дозировкой мультиспецифической связывающей молекулы или биспецифического антитела для введения пациенту может быть дозировка примерно от 1 мкг/кг до 15 мг/кг (например, от 0,1 мг/кг до 10 мг/кг) независимо от того, например, вводится ли оно в ходе одного или нескольких отдельных введений или путем непрерывной инфузии. Одна типичная суточная доза может варьировать от примерно 1 мкг/кг до 100 мг/кг или более в зависимости от указанных выше факторов. В случае повторных введений в течение нескольких дней или дольше в зависимости от состояния лечение, как правило, может быть продолжительным до требуемого подавления симптомов заболевания. Один из примеров дозировки мультиспецифической связывающей молекулы или биспецифического антитела будет находиться в диапазоне от примерно 0,05 мг/кг до примерно 10 мг/кг.Таким образом, пациенту может быть введена одна или более чем одна доза примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любая их комбинация). Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, так, чтобы пациент получал от примерно двух до примерно двадцати или, например, примерно шесть доз мультиспецифической связывающей молекулы или биспецифического антитела). Может быть введена первоначальная более высокая нагрузочная доза с последующей одной или более чем одной более низкой дозой. Тем не менее, могут использоваться и другие режимы дозирования. Прогресс этой терапии легко контролировать с помощью обычных методик и анализов.
Изделия
В одном аспекте, описанном в данном документе, предусматривается изделие, содержащее материалы, используемые для лечения, профилактики и/или диагностики нарушений, описанных выше. Изделие содержит контейнер и этикетку или листок-вкладыш, прикрепленный или связанный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для внутривенно вводимых растворов и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит композицию саму по себе или в сочетании с другой композицией, эффективной для лечения, профилактики и/или диагностики состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенно вводимого раствора или флакон с пробкой, прокалываемой иглой для подкожной инъекции). По меньшей мере один активный агент в композиции представляет собой комплекс, описанный в данном документе. На этикетке или вкладыше в упаковку указывается, что композиция используется для лечения выбранного состояния. Кроме того, изделие может включать: (а) первый контейнер с композицией, содержащейся в нем, где композиция содержит комплекс, описанный в данном документе; и (б) второй контейнер с композицией, содержащейся в нем, где композиция также содержит цитотоксический или другой терапевтический агент. Изделие в этом воплощении изобретения может также содержать вкладыш в упаковку, указывающий, что композиции могут быть использованы для лечения конкретного состояния. Альтернативно или дополнительно изделие может также включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, например, бактериостатическую воду для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Он также может включать другие материалы, желательные с коммерческой и потребительской точки зрения, в том числе другие буферы, разбавители, фильтры, иглы и шприцы.
Следующие примеры, графические материалы и последовательности приведены для облегчения понимания данного изобретения, истинный объем которого изложен в прилагаемой формуле изобретения. Понятно, что в изложенных процедурах могут быть сделаны модификации без отхода от сущности данного изобретения.
Последовательности
SEQ ID № 01 VH (mAb 1.4.168)
SEQ ID № 02 VL (mAb 1.4.168)
SEQ ID № 03 VH (mAb 8.1.2)
SEQ ID № 04 VL (mAb 8.1.2)
SEQ ID № 05 17-членная оцДНК (ковалентно связанная 5'-концом с FAB' МКА а против тропонина Т и FAB' 1.4.168 с IGF-1R, соответственно)
SEQ ID № 06 19-членная оцДНК (ковалентно связанная 3'-концом с FAB' МКА b против тропонина Т и FAB' 8.1.2 с фосфорилированным IGF-1R, соответственно)
SEQ ID № 07 комплементарная 19-членная оцДНК (используемая в качестве части линкера)
SEQ ID № 08 комплементарная 17-членная оцДНК (используемая в качестве части линкера)
SEQ ID № 09 Эпитоп "А" для антитропонинового антитела а.
SEQ ID № 10 Эпитоп "В" для антитропонинового антитела b.
SEQ ID № 11 IGF-1R (1340-1366)
SEQ ID № 12 hlnsR (1355-1382)
SEQ ID № 13 35-членный L-ДНК-полинуклеотидный линкер
SEQ ID № 14 75-членный L-ДНК-полинуклеотидный линкер
SEQ ID № 15 95-членный L-ДНК-полинуклеотидный линкер
SEQ ID № 16 аминокислотная последовательность тяжелой цепи 4D5 FAB'
SEQ ID № 17 аминокислотная последовательность легкой цепи 4D5 FAB'
SEQ ID № 18 аминокислотная последовательность тяжелой цепи 2С4 FAB'
SEQ ID № 19 аминокислотная последовательность легкой цепи 2С4 FAB'
SEQ ID № 20 Остатки 22-645 во внеклеточном домене (ECD) ErbB2
SEQ ID № 21 5'-AGT СТА ТТА ATG СТТ CTG C-XXX-Y-Z-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 5'-аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
SEQ ID № 22 5'-Y-Z-XXX-AGT TCT АТС GTC GTC CA-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 5'-Аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси, введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
SEQ ID № 23 5`-GCA GAA GCA TTA ATA GAC T (биотин-dT)-GG ACG ACG ATA GAA CT-3'
SEQ ID № 24 5'-GCA GAA GCA TTA ATA GAC T ТТТТТ-(биотин-dT)-TTTTT GG ACG ACG ATA GAA CT-3'
SEQ ID № 25 5`-GCA GAA GCA TTA ATA GAC T TTTTTTTTTTTTTTT-(биотин-dT)-TTTTTTTTTTTTTTT GG ACG ACG ATA GAA CT-3'.
SEQ ID № 26 вариабельный домен тяжелой цепи анти-HER2-антитела 4D5
SEQ ID № 27 VH CDR1
SEQ ID № 28 VH CDR2
SEQ ID № 29 VH CDR3
SEQ ID № 30 вариабельный домен легкой цепи анти-HER2-антитела 4D5
SEQ ID № 31 VL CDR1
SEQ ID № 32 VL CDR2
SEQ ID № 33 VL CDR3
SEQ ID № 34 вариабельный домен тяжелой цепи анти-HER2-антитела 2С4
SEQ ID № 35 VH CDR1
SEQ ID № 36 VH CDR2
SEQ ID № 37 VH CDR3
SEQ ID № 38 вариабельный домен легкой цепи анти-HER2-антитела 2С4
SEQ ID № 39 VL CDR1
SEQ ID № 40 VL CDR2
SEQ ID № 41 VL CDR3
SEQ ID № 42 5`-X-AGT СТА TTA ATG CTT CTG C-ZZZ-Y-; X = флуоресцеин, Y = С7-аминолинкер, Z = С3-спейсер
SEQ ID № 43 5`-X AGT СТА TTA ATG CTT CTG C-ZZZ-Y-; X=Cy5, Y=С7-аминолинкер, Z=C3-спейсер
SEQ ID № 44 5`-X-ZZZ-AGT TCT АТС GTC GTC CA-Y-3`; Х = Аминолинкер, Y = Флуоресцеин, Z=С3-спейсер
SEQ ID № 45 5`-X-(AGT СТА TTA ATG CTT CTG C)-(ZZZ)-y-; Х = Флуоресцеин, Y = С7-аминолинкер, Z = С3-спейсер
SEQ ID № 46 5`-X-(ZZZ-(AGT TCT АТС GTC GTC CAJ-Y-3`; Х = Аминолинкер, Y = Флуоресцеин, Z = С3-спейсер
SEQ ID № 47 5`-G CAG AAG CAT TAA TAG ACT-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 48 5`-G CAG AAG CAT TAA TAG ACT-(T40)-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 49 5`-[B-L]G CAG AAG CAT TAA TAG АСТ-(биотин-dT)-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 50 5`-[B-L]G CAG AAG CAT TAA TAG АСТ-Т5-(биотин-dT)-T5-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 51 5`-[B-L]G CAG AAG CAT TAA TAG АСТ-Т20-(биотин-dT)-T20-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 52 5`-[B-L] G CAG AAG CAT TAA TAG АСТ-Т30-(биотин-dT)-T30-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 53 S`-GCA GAA GCA TTA ATA GAC T T5-(биотин-dT)-T5 TG GAC GAC GAT AGA ACT-3'
SEQ ID № 54 5`-GCA GAA GCA TTA ATA GAC T T10-(биотин-dT)-T10 TGG ACG ACG ATA GAA CT-3'
SEQ ID № 55 5`-GCA GAA GCA TTA ATA GAC T T15-(биотин-dT)-Т15 TGG ACG ACG ATA GAA CT-3'
SEQ ID № 56 5`-GCA GAA GCA TTA ATA GAC T Т20-(биотин-dT)-Т20 TGG ACG ACG ATA GAA CT-3'
SEQ ID № 57 5`-G CAG AAG CAT TAA TAG АСТ-спейсер C18-(биотин-dT)-спейсер C18-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 58 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер C18)2-(биотин-dT)-(спейсер C18)2-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 59 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер C18)3-(биотин-dT)-(спейсер C18)3-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 60 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер C18)4-(биотин-dT)-(спейсер C18)4-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 61 5`-G CAG AAG CAT TAA TAG ACT-T20-(Dig-dT)-T20-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 62 5`-G CAG AAG CAT TAA TAG ACT-(Dig-dT)-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 63 5`-G CAG AAG CAT TAA TAG ACT-(биотин-dT)-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 64 YPYDVPDYA
SEQ ID № 65 GLNDIFEAQKIEWHE
SEQ ID № 66 SGGGS
SEQ ID № 67 f-Met-Leu-Phe (fMLP)
SEQ ID № 68 f-Met-Leu-Phe-o-метиловый эфир
SEQ ID № 69 IgG1 константный домен
SEQ ID № 70 IgG2 константный домен
SEQ ID № 71 IgG4 константный домен
SEQ ID № 72 Cy5-Y-ATG CGA-GTA CCT TAG AGT С-Z-Cy5
SEQ ID № 73 5`-G CAG AAG CAT TAA TAG ACT-T20-GAC TCT AAG GTA CTC GCA T-T20-TGG ACG ACG ATA GAA CT-3'
SEQ ID № 74 Сортазная метка
SEQ ID № 75 Олигонуклеотидный элемент связывающей пары
SEQ ID № 76 L-ДНК-линкер.
Графические материалы
Фиг. 1 Схема структуры анализа BIAcore. оц-L-ДНК-билинкеры были представлены на сенсоре BIAcore SA. Проточная ячейка 1 служила контролем (не показана).
Фиг. 2 Сенсограммы BIAcore для взаимодействия HER2 ECD с оц-D-ДНК-меченными FAB-фрагментами.
Фиг. 3 Сенсограммы BIAcore, показывающие зависимое от концентрации взаимодействие комплекса, описанного в данном документе, содержащего 35-член (= 35 нуклеотидов в длину) в качестве линкерного полинуклеотида, с HER2 ECD.
Фиг. 4 Сенсограммы BIAcore, показывающие зависимое от концентрации взаимодействие комплекса, описанного в данном документе, содержащего 75-член (= 75 нуклеотидов в длину) в качестве линкерного полинуклеотида, с HER2 ECD.
Фиг. 5 Сенсограммы BIAcore, показывающие зависимое от концентрации взаимодействие комплекса, описанного в данном документе, содержащего 95-член (= 95 нуклеотидов в длину) в качестве линкерного полинуклеотида, с HER2 ECD.
Фиг. 6 Схема структуры анализа BIAcore: поликлональное козлиное антитело против человеческого IgG-Fc-гамма было иммобилизовано на сенсоре BIAcore SA. Проточная ячейка 1 служила контролем (не показана).
Фиг. 7 Сенсограммы BIAcore показывают наложение графиков сигналов взаимодействия от введений 50 нМ «анти-HER2-антитело 2С4-FAB'-оц-L-ДНК (2С4)», «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК (4D5)» и полностью собранного комплекса (2С4-75-член-405), соединенного с помощью 75-членного оц-L-ДНК-линкера.
Фиг. 8 Сенсограмма BIAcore показывает наложение графиков измерений в зависимости от концентрации полностью собранного 75-членного комплекса в качестве анализируемого вещества в растворе, взаимодействующем с представленной на поверхностью huFc-химерой HER2 ECD.
Фиг. 9 График уровней ответа из фиг. 8 в зависимости от анализируемой концентрации полностью собранного комплекса.
Фиг. 10 Эксперименты аналитической гель-фильтрации по оценке эффективности сборки анти-pIGF1-R-комплекса. Диаграммы a, b и с показывают профиль элюции отдельных компонентов комплекса (флуоресцеин-оц-FAB' 1.4.168, Cy5-оц-Fab' 8.1.2 и линкерная ДНК (Т=0)). Диаграмма d показывает профиль элюции после того, как три компонента, необходимые для формирования двухвалентного связывающего агента, были смешаны в молярном соотношении 1:1:1. Толстая (нижняя) кривая представляет поглощение, измеренное при 280 нм, указывающее на присутствие оц-FAB'-белков или линкерной ДНК, соответственно. Тонкая (верхняя) кривая в b) и d) (поглощение при 495 нм) указывает на присутствие Cy5, а тонкая (верхняя) кривая в а) и средняя кривая в d) (поглощение при 635 нм) указывает на присутствие флуоресцеина. Сравнение объемов элюции отдельных компонентов комплекса (VEssFab' 1.4.168 ~ 15 мл; VEssFab' 8.1.2 ~ 15 мл; VEлинкер ~ 16 мл) с объемом элюции реакционной смеси (VEmix ~ 12 мл) показывает, что реакция сборки комплекса прошла успешно (величина выхода: ~ 90%). Основной пик при 280 нм, который представляет элюированный комплекс, хорошо совпадает с основными пиками в каналах 495 нм и 695 нм, что доказывает присутствие и оц-FAB' 8.1.2, и оц-Fab'1.4.168 в пике, представляющем двухвалентный связывающий агент.
Фиг. 11 Схема эксперимента BIAcore: схематично и иллюстративно показаны две связывающие молекулы в растворе. T=0-Dig, двухвалентный связывающий агент, и T=40-Dig, двухвалентный связывающий агент. Эти двухвалентные связывающие агенты отличаются только длиной их линкера (без дополнительных Т против 40 дополнительных Т, отделяющих две гибридизующиеся нуклеиновокислотные последовательности). Кроме того, были использованы оц-FAB'-фрагменты 8.1.2 и 1.4.168.
Фиг. 12 Сенсограмма BIAcore с наложением трех кинетических графиков, показывающая взаимодействие 100 нМ двухвалентного связывающего агента (состоящего из оц-FAB 8.1.2 и оц-FAB 1.4.168, гибридизованных на T40-Dig оцДНК-линкере) с иммобилизованным пептидом pIGF-1R по сравнению с характеристиками связывания 100 нМ оц-FAB 1.4.168 или 100 нМ оц-FAB 8.1.2 с тем же пептидом. Высокая эффективность связывания достигается только с комплексной конструкцией, что ясно показывает, что кооперативный связывающий эффект комплекса повышает аффинность по сравнению с пептидом-мишенью pIGF-1R.
Фиг. 13 Сенсограмма BIAcore с наложением трех кинетических графиков, показывающая взаимодействие двухвалентного связывающего агента, состоящего из оц-FAB 8.1.2 и оц-FAB 1.4.168, гибридизованных на T40-Dig оцДНК-линкере с иммобилизованным пептидом pIGF-1R (фосфорилированный IGF-1R), IGF-1R или IR (фосфорилированный рецептора инсулина). Высокая эффективность связывания достигается с пептидом pIGF-1R, что ясно показывает, что кооперативный связывающий эффект комплекса повышает специфичность против пептида-мишени pIGF-1R по сравнению, например, с пептидом фосфорилированного рецептора инсулина (IR).
Фиг. 14 Сенсограмма BIAcore с наложением двух кинетических графиков, показывающая взаимодействие 100 нМ двухвалентного связывающего агента, состоящего из оц-FAB' 8.1.2 и оц-FAB' 1.4.168, гибридизованных на Т40-Dig оцДНК-линкере), и смеси 100 нМ оц-FAB' 8.1.2 и 100 нМ оц-FAB' 1.4.168 без линкерной ДНК. Высокая эффективность связывания достигается только с двухвалентным связывающим агентом, в то время как смесь оц-FAB' без линкера не показывает заметного кооперативного связывающего эффекта, несмотря на то, что общая концентрация этих оц-FAB' составляла 200 нМ.
Фиг. 15 Схематическое изображение сэндвич-анализа BIAcore. Этот анализ был использован для изучения доступности эпитопов для обоих антител на фосфорилированном IGF-1R-пептиде. <MIgGFcy>R представляет собой кроличье противомышиное антитело, используемое для захвата мышиного антитела М-1.4.168. Затем М-1.4.168 используется для захвата пептида plGF-1R. М-8.1.2, наконец, формирует сэндвич, состоящий из М-1.4.168, пептида и М-8.1.2.
Фиг. 16 Сенсограмма BIAcore показывает сигнал связывания (жирная линия) вторичного антитела 8.1.2. с пептидом pIGF-1R после того, как он был захвачен антителом 1.4.168 на чипе BIAcore. Другие сигналы (тонкие линии) являются контрольными сигналами: даны линии для гомологичного контроля с 500 нМ 1.4.168, 500 нМ целевого несвязанного антитела <CKMM>М- 33-IgG; и 500 нМ целевого несвязанного контрольного антитела <TSH>M-1.20-IgG, соответственно. Ни в одном из этих контролей не обнаружены события связывания.
Фиг. 17 Схематическое изображение анализа BIAcore, представляющее комплексы на сенсорной поверхности. На проточной ячейке 1 (=FC1) (не показана) захвачен амино-ПЭО-биотин. На FC2, FC3 и Fc4 иммобилизованы двухвалентные связывающие агенты с увеличивающейся длиной линкера. Анализируемое вещество 1: пептид IGF-1R, содержащий эпитоп оц-FAB М-1.4.168 (тонкая линия) - фосфо-эпитоп оц-FAB М-8.1.2 не представлен, потому что этот пептид не является фосфорилированным; анализируемое вещество 2: пептид pIGF-1R, содержащий фосфо-эпитоп оц-FAB М-8.1.2 (Р) и эпитоп оц-FAB М-1.4.168 (тонкая линия). Анализируемое вещество 3: pINR-пептид, содержащий перекрестно взаимодействующий фосфо-эпитоп оц-FAB М-8.1.2, но не содержащий эпитоп для М-1.4.168.
Фиг. 18 Кинетические данные из эксперимента с комплексом. Т40-би-комплекс с оц-FAB 8.1.2 и оц-FAB 1.4.168 показывает 1300-кратное уменьшение скорости диссоциации KD=2,79Е-05/с) против pIGF-1R при сравнении с pINR (KD=3.70Е -02).
Фиг. 19 Сенсограмма BIAcore, показывающая зависимое от концентрации измерение Т40-би-комплекса против пептида pIGF-1R (фосфорилированного пептида IGF-1R).
Фиг. 20 Сенсограмма BIAcore, показывающая зависимое от концентрации измерение Т40-би-комплекса против пептида IGF-1R (нефосфорилированного пептида pIGF-1R).
Фиг. 21 Сенсограмма BIAcore, показывающая зависимое от концентрации измерение Т40-би-комплекса против пептида pINR (фосфорилированного пептида рецептора инсулина).
Фиг. 22 Окрашивание опухолевых клеток Cy5-меченным ксоларом (Xolair®) и герцептином (Herceptin®).
Фиг. 23 NIRF-визуализации клеток KPL-4.
Фиг. 24 Окрашивание ксенотрансплантатов KPL-4 ex vivo.
Фиг. 25 Эксклюзионный профиль свежеприготовленного комплекса «4D5-95-член-2С4». Верхний сигнал: сигнал 260 нм, нижний сигнал: сигнал 280 нм. Между началом на 0,0 мин и пиком элюции на 5,64 мин агрегаты не обнаружены.
Фиг. 26 Эксклюзионный профиль комплекса «4D5-95-член-2С4» после цикла замораживания и оттаивания. Верхний сигнал: сигнал 260 нм, нижний сигнал: сигнал 280 нм. Между началом на 0,0 мин и пиком элюции на 5,71 мин агрегаты не обнаружены.
Фиг. 27 Иллюстративная хроматограмма (аналитическая SEC) биспецифической связывающей молекулы.
Фиг. 28 Электрофореграммы из системы LabChip (Perkin Elmer) мониторинга конъюгата «Fab-фрагмент антитела - олигонуклеотид» на различных этапах процесса.
Пример 1
Формирование FAB-оцДНК-конъюгатов
Были использованы два моноклональных антитела, связывающихся с человеческим сердечным тропонином Т на различных непересекающихся эпитопах, эпитопе a и эпитопе b, соответственно. Оба эти антитела используются в текущем анализе Roche Elecsys™ Troponin Т, в котором тропонин Т обнаруживают в формате сэндвич-иммуноанализа.
Очистку моноклональных антител из культурального супернатанта проводили с использованием способов из области химии белка в соответствии с уровнем техники.
Очищенные моноклональные антитела подвергали протеазному расщеплению либо предварительно активированным папаином (МКА против эпитопа a), либо пепсином (МКА против эпитопа b) с получением F(ab')2-фрагментов, которые затем восстанавливали до FAB'-фрагментов низкой концентрацией цистеамина при 37°С. Реакцию останавливали путем отделения цистеамина на колонке Sephadex G-25 (GE Healthcare) от части образца, содержащей полипептид.
FAB'-фрагменты конъюгировали с описанными ниже активированными оцДНКа- и оц-ДНКb-олигонуклеотидами.
а) конъюгат А оцДНК-FAB антитела против тропонина Т (эпитопа А)
Для получения конъюгата А «оцДНКа-FAB антитела против эпитопа а тропонина Т» использовали производное SEQ ID № 05, т.е. 5'-AGT СТА TTA ATG CTT CTG C(=SEQ ID № 5)-XXX-Y-Z-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 3'-аминомодификатор С6, введенный через 3'-аминомодификатор TFA Amino С-6 Icaa CPG (ChemGenes), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
б) конъюгат В оцДНК-FAB антитела против тропонина Т (эпитопа В)
Для получения конъюгата В «оцДНКb-FAB антитела против эпитопа b тропонина Т» использовали производное SEQ ID № 06, т.е. 5'-Y-Z-XXX-AGT ТСТ АТС GTC GTC СА-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 5'-аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси, введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
Олигонуклеотиды SEQ ID № 05 или 06, соответственно, были синтезированы с использованием способов олигонуклеотидного синтеза в соответствии с уровнем техники. Введение мелеинимидогруппы проводили с помощью реакции аминогруппы в Y с сукцинимидил-группой в Z, которая была встроена в процессе твердофазного олигонуклеотидного синтеза.
Одноцепочечные ДНК-конструкции, показанные выше, несут тиольную реакционноспособную малеимидогруппу, которая реагирует с цистеином из шарнирной области FAB', образованной при обработке цистеамином. Для того чтобы получить высокий процент однократно меченного FAB'-фрагмента, относительное молярное отношение оцДНК-фрагмента к FAB'-фрагменту поддерживают на низком уровне. Очистка однократно меченных FAB'-фрагментов (оцДНК:FAB' = 1:1) происходит с помощью анионообменной хроматографии (колонка: MonoQ, GE Healthcare). Проверка эффективного мечения и очистки проводится путем аналитической эксклюзионной хроматографии и SDS-PAGE.
Пример 2
Формирование биотинилированных линкерных молекул
Олигонуклеотиды, используемые в оцДНК-линкерах L1, L2 и L3, соответственно, были синтезированы с использованием способов синтеза олигонуклеотидов в соответствии с уровнем техники и с использованием биотинилированного фосфорамидитного реагента для биотинилирования.
Линкер 1 (=L1), биотинилированный оцДНК-линкер 1 без спейсера, имеет следующий состав:
5'-GCA GAA GCA TTA ATA GAC T (биотин-dT)-GG ACG ACG ATA GAA CT-3'. Он включает оцДНК-олигонуклеотиды SEQ ID №№ 7 и 8, соответственно, и биотинирован с использованием биотин-dT(5'-диметокситритилокси-5-[N-((4-t-бутилбензоил)-биотинил)-аминогексил)-3-акрилимидо]-2'-дезоксиуридин-3'-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита (Glen Research).
Линкер 2 (= L2), биотинилированный оцДНК-линкер 2 с 10-членным спейсером, имеет следующий состав:
5'-GCA GAA GCA TTA ATA GAC T T5-(биотин-dT)-T5 GG ACG ACG ATA GAA CT-3'. Он включает оцДНК-олигонуклеотиды SEQ ID №№ 7 и 8, соответственно, двойные олигонуклеотидные участки из пяти тимидинов каждый, и биотинилирован с использованием биотин-dT(5'-диметокситритилокси-5-[N-((4-t-бутилбензоил)-биотинил)-аминогексил)-3-акрилимидо]-2'-дезоксиуридин-3'-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита (Glen Research).
Линкер 3 (L3 =), биотинилированный оцДНК-линкер 3 с 30-членным спейсером, имеет следующий состав:
5'-GCA GAA GCA TTA ATA GAC T Т15-(биотин-dT)-Т15 GG ACG ACG ATA GAA CT-3'. Он включает оцДНК-олигонуклеотиды SEQ ID №№ 7 и 8, соответственно, двойные олигонуклеотидные участки из пятнадцати тимидинов каждый, и биотинилирован с использованием биотин-dT(5'-диметокситритилокси-5-[N-((4-t-бутилбензоил)-биотинил)-аминогексил)-3-акрилимидо]-2'-дезоксиуридин-3'-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита (Glen Research).
Пример 3
Эпитопы а и b, соответственно, для одновалентных элементов, связывающих тропонин Т
Синтетические пептиды были сконструированы так, что по отдельности имели только умеренную аффинность к соответствующему FAB'-фрагменту, полученному из антител a и b, соответственно, против тропонина Т.
а) эпитоп "А" для антитела а содержится в:
SEQ ID № 9 = ERAEQQRIRAEREKEUUSLKDRIEKRRRAERAEамид, где U представляет собой β-аланин.
b) эпитоп "В" для антитела b содержится в:
SEQ ID № 10=SLKDRIERRRAERAEOOERAEQQRIRAEREKEамид, где О представляет собой аминотриоксаоктановую кислоту.
Специалисту в данной области будет понятно, что можно совместить эти два содержащих эпитопы пептида двумя способами, и оба варианта были разработаны и получены путем линейного объединения эпитопов "А" и "В". Последовательности обоих вариантов, линейные последовательности эпитопов «А"-"В" (=TnT 1) и "В"-"А" (=TnT 2), соответственно, были получены с использованием способов синтеза пептидов в соответствии с уровнем техники.
Последовательности эпитопов "А" и "В", соответственно, были изменены по сравнению с исходными эпитопами на человеческой последовательности сердечного тропонина Т (P45379/UniProtKB) в целях снижения аффинности связывания с каждым из FAB. В этих условиях динамика влияния гетеро-двухвалентного связывания видна лучше, например, при анализе аффинности связывания по методике BIAcore.
Пример 4
Анализ биомолекулярного взаимодействия
Для этого эксперимента использовали инструмент BIAcore 3000 (GE Healthcare) с сенсором BIAcore SA, установленным в системе на Т=25°С Предварительная подготовка была проведена в 100 мкл/мин с 3-кратным введением 1 М NaCl в 50 мМ NaOH в течение 1 мин и 10 мМ HCl в течение 1 мин.
HBS-ET (10 мМ HEPES pH 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Tween® 20) использовали в качестве системного буфера. Буфер для образцов был идентичен системному буферу.
Система BIAcore 3000 находилась под управлением программного обеспечения V1.1.1. Проточная ячейка 1 была насыщена 7 RU D-биотина. На проточной ячейке 2 было иммобилизовано 1063 RU биотинилированного оцДНК-линкера L1. На проточной ячейке 3 было иммобилизовано 879 RU биотинилированного оцДНК-линкера L2. На проточной ячейке 4 было иммобилизовано 674 RU биотинилированного оцДНК-линкера L3.
После этого конъюгат А «FAB-фрагмент-ДНК» вводили при 600 нМ. Конъюгат В «FAB-фрагмент-ДНК» вводили в систему при 900 нМ. Конъюгаты вводили в течение 3 мин со скоростью потока 2 мкл/мин. Конъюгаты последовательно вводили для контроля соответствующего сигнала насыщения для каждого конъюгата «FAB-фрагмент-ДНК» на соответствующем линкере. Комбинации FAB запускали с одним конъюгатом А «FAB-фрагмент-ДНК», с одним конъюгатом В «FAB-фрагмент-ДНК», и с обоими конъюгатами А и В «FAB-фрагмент-ДНК», присутствующими на соответствующем линкере. Стабильные исходные уровни были получены после того, как линкеры были насыщены конъюгатами «FAB-фрагмент-ДНК», что было предпосылкой для дальнейших кинетических измерений.
Пептидные анализируемые вещества TnT1 и TnT2 вводили в качестве анализируемых веществ в растворе в систему для взаимодействия с представленными на поверхности FAB-фрагментами.
TnT1 вводили при концентрации анализируемого вещества 500 нМ, TnT2 вводили при 900 нМ. Оба пептида вводили со скоростью 50 мкл/мин в течение времени ассоциации 4 минуты. Диссоциацию наблюдали в течение 5 минут. Регенерацию проводили путем введения 50 мМ NaOH в течение 1 минуты со скоростью 50 мкл/мин во все проточные ячейки.
Кинетические данные определяли с помощью программного обеспечения BIAevaluation (v.4.1). Скорость диссоциации KD (1/с) пептидов TnT1 и TnT2 из соответствующей комбинации представленных на поверхности FAB-фрагментов определяли в соответствии с линейной Моделью подбора Ленгмюра 1:1. Время полужизни комплекса в минутах было рассчитано в соответствии с решением кинетического уравнения первого порядка: In(2)/(60*кДа).
Результаты:
Экспериментальные данные, приведенные в таблицах 5 и 6, соответственно, показывают увеличение стабильности комплекса между анализируемым веществом (TnT1 или TnT2), соответственно, и различными гетеродвухвалентными FAB-FAB-конъюгатами А-В по сравнению с одновалентным конъюгатом дцДНК-FAB А или В, соответственно. Этот эффект показан в каждой таблице в строке 1 в сравнении со строками 2 и 3.
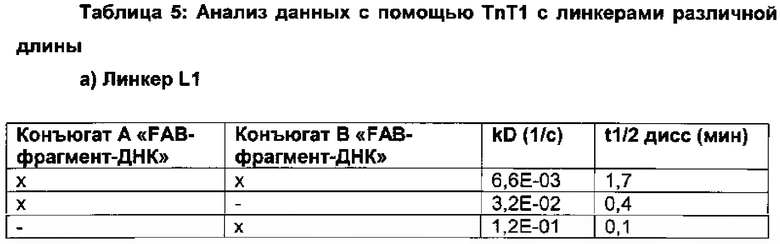

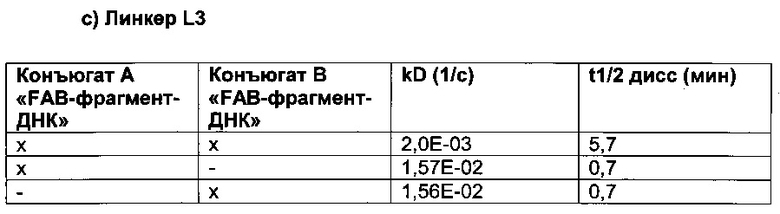
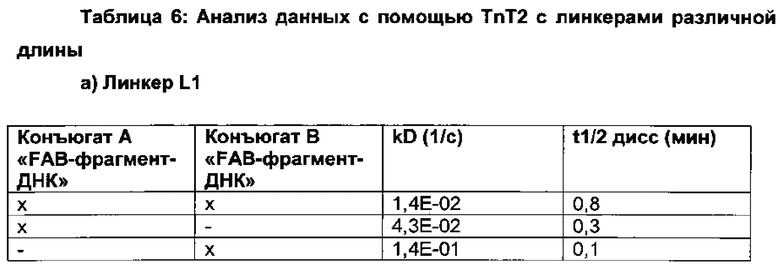


Эффект авидности также зависит от длины линкера. В подтаблицах, приведенных после таблицы 1, 30-членный линкер L3 демонстрирует самый низкий уровень диссоциации или самую высокую стабильность комплекса, в подтаблицах, приведенных после таблицы 2, 10-членный L2 линкер демонстрирует самый низкий уровень диссоциации или самую высокую стабильность комплекса. Эти данные, взятые вместе, показывают, что изменчивость длины линкера, присущая подходу данного изобретения, представляет большую пользу и преимущества.
Пример 5
Формирование конъюгатов «FAB'-оцДНК»
Было использовано два моноклональных антитела, связывающихся с человеческим HER2 (ErbB2 или p185neu) на различных неперекрывающихся эпитопах А и В. Первое антитело представляет собой анти-HER2-антитело 4D5 (huMAb4D5-8, rhuMab HER2, трастузумаб или HERCEPTIN®; см. US 5821337, включенный в данный документ в полном объеме посредством ссылки). Второе антитело представляет собой анти-HER2-антитело 2С4 (пертузумаб).
Очистку моноклональных антител из культурального супернатанта проводили с использованием способов из области химии белка в соответствии с уровнем техники.
Очищенные моноклональные антитела подвергали протеазному расщеплению либо предварительно активированным папаином, либо пепсином с получением F(ab')2-фрагментов. Затем их восстанавливали до FAB'-фрагментов низкой концентрацией цистеамина при 37°С. Реакцию останавливали путем отделения цистеамина на колонке Sephadex G-25 (GE Healthcare) от части образца, содержащей полипептид.
Полученные FAB'-фрагменты конъюгировали с активированными оцДНК-полинуклеотидами.
а) конъюгат «анти-HER2-антитело 4D5 FAB'-оцДНК»
Для получения конъюгата «анти-HER2-антитело 4D5 FAB'-оцДНК» использовали производное SEQ ID № 05, т.е. 5-AGT СТА TTA ATG CTT CTG С(=SEQ ID № 05)-XXX-Y-Z-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 5'-аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
b) анти-HER2-антитело 2С4 FAB'-оцДНК-конъюгат
Для получения анти-HER2-антитело 2С4 FAB'-оцДНК-конъюгата использовали производное SEQ ID № 06, т.е. 5'-Y-Z-XXX-AGT TCT АТС GTC GTC СА-3', где X = пропилен-фосфат, введенный через фосфорамидит С3 (3-(4,4'-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research), где Y = 5'-аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), и где Z = 4[N-мелеинимидометил]циклогексан-1-карбокси, введенный через сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (ThermoFischer).
Полинуклеотиды SEQ ID № 05 или 06, соответственно, были синтезированы с использованием способов полинуклеотидного синтеза в соответствии с уровнем техники. Введение мелеинимидогруппы проводили с помощью реакции аминогруппы в Y, которая была встроена в процессе твердофазного полинуклеотидного синтеза с сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилатом (ThermoFischer).
Одноцепочечные ДНК-конструкции несут тиольную реакционноспособную малеимидогруппу, которая реагирует с цистеином из шарнирной области FAB', образованной при обработке цистеамином. Для того чтобы получить высокий процент однократно меченного FAB'-фрагмента, относительное молярное отношение оцДНК-фрагмента к FAB'-фрагменту поддерживают на низком уровне,. Очистка однократно меченных Fab'-фрагментов (оцДНК:FAB'=1:1) происходит с помощью анионообменной хроматографии (колонка: MonoQ, GE Healthcare). Проверка эффективного мечения и очистки проводится путем аналитической эксклюзионной хроматографии и SDS-PAGE.
Пример 6
Анализ биомолекулярного взаимодействия
Для этого эксперимента использовали инструмент BIAcore Т100 (GE Healthcare) с сенсором BIAcore SA, установленным в системе при Т=25°С Предварительная подготовка была проведена путем трехкратного введения 1 М NaCl в 50 мМ NaOH, рН 8,0, в течение 1 мин со скоростью 100 мкл/мин, и затем введения 10 мМ HCl в течение 1 мин. Системным буфером был HBS-EP (10 мМ HEPES рН 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Р20). Буфером для образцов был системный буфер, дополненный 1 мг/мл CMD (карбоксиметилдекстрана).
Биотинилированные оц-L-ДНК-линкеры захватывали на SA-поверхности в соответствующих проточных ячейках. Проточная ячейка 1 была насыщена амино-ПЭО-биотином (Pierce).
40 RU биотинилированного 35-членного олигонуклеотидного линкера было захвачено на проточной ячейке 2. 55 RU биотинилированного 75-членного олигонуклеотидного линкера было захвачено на проточной ячейке 3. 60 RU биотинилированного 95-членного олигонуклеотидного линкера было захвачено на проточной ячейке 4.
250 нМ конъюгата «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК» вводили в систему в течение 3 мин. 300 нМ конъюгата «анти-HER2-антитело 2C4-FAB'-оц-L-ДНК» вводили в систему со скоростью 2 мкл/мин в течение 5 мин. ДНК-меченые FAB-фрагменты вводили отдельно или в комбинации.
В качестве контроля в систему вводили только 250 нМ конъюгата «анти-HER2-антитело 4D5-FAB'-оц-D-ДНК» и 300 нМ конъюгата «анти-HER2-антитело 2C4-FAB'-оц-D-ДНК». В качестве дополнительного контроля вместо ДНК-меченых FAB-фрагментов вводили буфер. После гибридизации оц-L-ДНК-меченых FAB-фрагментов на соответствующих оц-L-ДНК-билинкерах анализируемое вещество в растворе hHER2 ECD вводили в различных сериях концентраций от 24 нМ, 8 нМ, 3 нМ, 1 нМ, 0,3 нМ, 0 нМ в систему в течение фазы ассоциации 3,5 мин со скоростью 100 мкл/мин. Фазу диссоциации контролировали при 100 мкл/мин в течение 15 мин. Регенерацию системы проводили путем введения 100 мМ глицинового буфера (глицин рН 11, 150 мМ NaCl) в течение 30 сек со скоростью 20 мкл/мин с последующим вторым введением воды в течение 1 мин со скоростью 30 мкл/мин.
Сигналы измеряли в виде сенсограмм с разрешением во времени и с зависимостью от концентрации анализируемого вещества. Данные оценивали с помощью программного обеспечения BIAcore BIAevaluation 4.1. В качестве модели для подгонки использовали стандартную модель бинарного связывания Ленгмюра.
Результаты:
Взаимодействие HER2 ECD не наблюдалось, когда в систему вводили оц-D-ДНК-меченые FAB-фрагменты, так как оц-D-ДНК-меченые FAB-фрагменты не гибридизовались со шпигельмерными оц-L-ДНК-линкерами, представленными на сенсорной поверхности (фиг. 2).
Таблица 7: Кинетические результаты эксперимента комплексообразования. Линкер: представленный на поверхности биотинилированный оц-L-ДНК-полинуклеотидный линкер, Oligo_35mer-Bi, Oligo_75mer-Bi и Oligo_95mer-Bi, отличающиеся длиной линкера. оц-L-ДНК-FAB: 2С4-оц-L-ДНК: анти-HER2-антитело 2C4-FAB'-оц-L-ДНК, помеченный 19-членным флуоресцеином. 4D5-оц-L-ДНК: анти-HER2-антитело 4D5-FAB'-оц-L-ДНК, помеченный 17-членным флуоресцеином. 4D5-+2C4-оц-L-ДНК: представленное на поверхности сочетание обоих фрагментов. LRU: масса в единицах ответа, которая гибридизована на серсорной поверхности. Антиген: 87 кДа HER2 ECD использовали в качестве анализируемого вещества в растворе. ka: скорость ассоциации в (1/Мс). kd: скорость диссоциации (1/с): t1/2 дисс: время полужизни комплекса антигена, рассчитанное в часах согласно решению In(2)/kD*3600 кинетического уравнения первого порядка. kD: аффинность в молях. kD: аффинность, рассчитанная в пикомолях. Rmax: максимальный сигнал ответа анализируемого вещества при насыщении в единицах ответа (RU). MR: молярное соотношение, показывающее стехиометрию взаимодействия. Chi2, U-значение: показатель качества измерений.
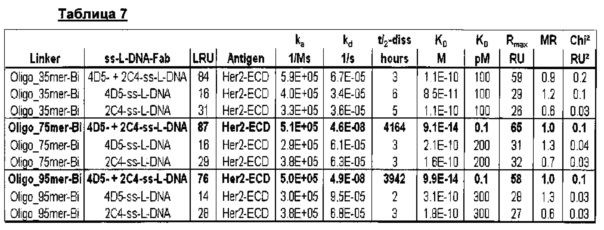
Сенсограммы BiaCore показывают зависимые от концентрации измерения взаимодействия 35-членного комплекса HER2 ECD (фиг. 3). Этот линкер состоит исключительно из последовательностей гибридизационных мотивов ДНК-меток. Кинетические данные указывают, что полностью собранный комплекс не показывает улучшений в кинетической эффективности. Это происходит из-за недостаточной длины линкера и отсутствия гибкости 35-членного комплекса.
Сенсограммы BiaCore показывают зависимые от концентрации измерения взаимодействия 75-членного комплекса HER2 ECD (фиг. 4). 75-членный линкер несет поли-T" для увеличения длины линкера по сравнению с 35-членным линкером. Кинетические данные указывают, что полностью собранный комплекс показывает значительное улучшение кинетической эффективности. Это связано с оптимальной длиной линкера и гибкостью 75-члена.
Сенсограммы BiaCore показывают зависимые от концентрации измерения взаимодействия 95-членного комплекса HER2 ECD (фиг. 5). 95-членный линкер несет поли-Т для увеличения длины линкера по сравнению с 35-членным линкером. Кинетические данные указывают, что полностью собранный комплекс показывает значительное улучшение кинетической эффективности. Это связано с увеличенной длиной линкера и гибкостью 95-членного комплекса.
Настройки анализа BIAcore включают следующие (см. также фиг. 1): оц-L-ДНК-билинкеры находились на сенсоре BIAcore SA. Проточная ячейка 1 служила контролем. В качестве анализируемого вещества в растворе был использован HER2 ECD. Конъюгаты «анти-HER2-антитело 2С4-FAB'-оц-L-ДНК» и «анти-HER2-антитело 2С4-FAB'-оц-L-ДНК» гибридизовали с линкерами, представленными на поверхности.
Здесь в первый раз показано событие полностью функционального кооперативного связывания между герцептин-FAB и пертузумаб-FAB, связанными друг с другом с помощью оц-L-ДНК-линкера с высокой гибкостью. Данные в таблице 3 представляют доказательство наличия события кооперативного связывания. Несмотря на то, что значения Rmax полностью собранного комплекса были примерно в два раза больше высоты сигнала конструкций с одиночным FAB, значения молярного соотношения точно равны 1 (MR=1). Это ясно указывает на наличие одновременного кооперативного связывания обоих FAB-фрагментов. Комплекс считается одной молекулой со стехиометрией связывания Ленгмюра 1:1. Несмотря на наличие двух независимо связанных поверхностей раздела HER2, между одним комплексом и двумя HER2-доменами нельзя обнаружить межмолекулярного связывания.
Константы авидности для синергетических пар моноклональных антител или для химически сшитых биспецифических F(ab')2, как правило, только до 15 раз превышают константы аффинности для отдельных моноклональных антител, что значительно меньше, чем теоретическая авидность, ожидаемая для идеального сочетания между реагентами (Cheong, H.S., et al., Biochem. Biophys. Res. Commun. 173 (1990) 795-800). He вдаваясь глубоко в эту теорию, одной из причин этого может быть то, что отдельные взаимодействия эпитопа/паратопа, участвующие в синергетическом связывании (которое приводит к высокой авидности), должны быть ориентированы определенным образом относительно друг друга для оптимальной синергии.
Кроме того, данные, представленные в таблице 7, свидетельствуют о том, что короткий 35-членный линкер, который состоит только из гибридизационных оц-L-ДНК-мотивов, не показывает достаточной гибкости или/и длины линкера для получения эффекта кооперативного связывания. 35-членный линкер является жесткой L-ДНК-конструкцией в виде двойной спирали. Гибридизация формирует двойную спираль L-ДНК, которая короче и менее гибкая, чем оц-L-ДНК-последовательность. Спираль демонстрирует уменьшенные степени свободы и может рассматриваться как жесткая линкерная конструкция. Таблица 7 показывает, что 35-членный линкер не способен участвовать в кооперативном связывании.
Увеличение длины линкера за счет очень гибкой поли-Т-оц-L-ДНК с образованием 75-члена и 95-члена, соответственно, предусматривает увеличение аффинности и особенно стабильности kD (1/с) комплекса антигена.
Значения chi2 показывают высокое качество измерений. Все измерения показывают крайне небольшие ошибки. Данные могут быть установлены по модели Ленгмюра 1:1 с остаточным отклонением +/- 1 RU, малые значения Chi2, и только 10 итерационных расчетов были необходимы для получения данных.
Эффект кооперативного связывания работает в соответствии с физическими законами, т.е. свободные энергии связывания ΔG1 и ΔG2 суммируются. Аффинности умножаются: KDкооп=KD1×KD2. Кроме того, скорости диссоциации также умножаются: KDкооп=kd1×KD2. Это именно то, что наблюдается в эксперименте с 75-членным и 95-членным линкером. Это приводит к очень долгому периоду полужизни комплекса: 4146 часов (173 дня) и 3942 часов (164 дня), соответственно. Значения аффинностей находятся в области 100 фмоль/л. Очевидно, что происходит явление кооперативного связывания.
Скорости ассоциации всех полностью собранных комплексов являются более высокими по сравнению с отдельно гибридизующимися конструкциями. Несмотря на более высокую молекулярную массу увеличивается скорость ассоциации.
Здесь мы смогли показать, что трастузумаб и пертузумаб, связанные между собой в комплексе, описанном в данном документе, одновременно связываются с внеклеточным доменом HER-2 (ECD). Оба FAB-фрагмента связываются с истинными эпитопами на HER2 ECD (PDB 1S78 и PDB 1N82). Кроме того, оба FAB-фрагмента сильно отличаются по углам их связывания. При использовании 75-членного оц-L-ДНК-линкера оптимальной длины (30 нм) и его полезных свойств в плане гибкости и длины может быть показано событие кооперативного связывания.
Сигналы измеряли в виде сенсограмм с разрешением во времени и с зависимостью от концентрации анализируемого вещества. Данные оценивали с помощью программного обеспечения BIAcore BIAevaluation 4.1. В качестве модели для подгонки использовали стандартную модель бинарного связывания Ленгмюра.
Пример 7
Дополнительный анализ биомолекулярного взаимодействия
На инструмент BIAcore 3000 был установлен сенсорный чип СМ-5. Предварительная подготовка сенсора была проведена в соответствии с рекомендациями производителя (GE Healthcare, Упсала, Швеция). Системным буфером был (10 мМ HEPES рН 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Tween® 20). Системный буфер также использовался в качестве буфера для образцов. Система работала при 25°С в соответствии с управляющим программным обеспечением 4.1.
30 мкг/мл поликлонального козлиного антитела против человеческого IgG-Fc гамма (Jackson Laboratories, США) в 10 мМ ацетатном буфере, pH 4,5, иммобилизовали путем стандартной химической реакции NHS/EDC при 13952 RU на проточной ячейке 1 и 15047 RU на проточной ячейке 2. Регенерацию системы проводили путем введения со скоростью 20 мкл/мин 10 мМ глицинового буфера, рН 1,5, в течение 20 сек, затем 10 мМ глицинового буфера, рН 1,7, в течение 1 мин, и 10 мМ глицинового буфера, рН 1,5, в течение 30 сек. В. проточную ячейку 1 в качестве референса вводили 5 нМ huIgG (Bayer Healthcare) в течение 1 мин со скоростью 10 мкл/мин.
В проточную ячейку 2 вводили 10 нМ химеры «человеческий HER2 внеклеточный рецептор FC» (hHER2 ECDpresSFc) в течение 1 мин со скоростью 10 мкл/мин. Как правило, 100 единиц ответа предварительно подготовленного гомодимерного hHER2 ECDpresSFc было захвачено через человеческую FC-часть на проточной ячейке 2 с помощью козлиного антитела против человеческого IgG-Fc гамма. Как правило, 130 единиц ответа huIgG было захвачено через человеческую FC-часть на проточной ячейке 1.
Сигнал на проточной ячейке 2 был представлен в сравнении с проточной ячейкой 1.
оц-L-ДНК-меченые FAB-фрагменты «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК» и «анти-HER2 антитело 2C4-FAB'-оц-L-ДНК» гибридизовали с 75-членным оц-L-ДНК-линкером в молярном соотношении 1:1:1. Полностью установленный комплекс 2С4-75-член-4D5 вводили в систему при 50 нМ в течение трех минут.В качестве контроля в систему вводили отдельные FAB-фрагменты при 50 нМ.
Сразу же по окончании введения в систему вводили 250 нМ стрептавидина или системный буфер в течение 3 мин со скоростью 10 мкл/мин. Поскольку 75-членный линкер содержит отдельную биотиновую группировку в центре его последовательности, SA должен работать в качестве зонда для распознавания биотина в линкере, но не наличия FAB-фрагментов.
В другом эксперименте полностью собранный комплекс 4D5-75-член-2С4 вводили в систему в различных концентрациях 0 нМ, 0,6 нМ, 1,9 нМ, 2х 5,6 нМ, 16,7 нМ, 50 нМ со скоростью 10 мкл/мин в течение 3 мин. Проводили мониторинг уровней ответа анализируемого вещества hHER2 ECDpresSFc в зависимости от концентрации. Уровни ответа наносили в зависимости от концентрации hHER2 ECDpresSFc. Данные визуализировали с помощью программного обеспечения Origin 7. Данные подгоняли с помощью уравнения Хилла y=Vmax*xn/(kn+xn), как предусмотрено программой Origin 7.
Структура анализа BIAcore была следующей (см. также фиг. 6): Поликлональное козлиное антитело против человеческого IgG-Fc-гамма было иммобилизовано на сенсоре BIAcore СМ5 и служило в качестве системы захвата для huFc-химеры HER2 ECD. Вводили «анти-HER2-антитело 2С4-FAB'-оц-L-ДНК» (2С4 FAB), «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК» (4D5 FAB) и полностью собранные комплексы с последующим введением стрептавидина (SA). Целью эксперимента было продемонстрировать наличие и доступность биотиновой группировки в 75-членном оц-L-ДНК-линкере.
Результаты эксперимента приведены на фиг. 7. Сенсограмма BIAcore показывает наложение графиков сигналов взаимодействия от введений 50 нМ «анти-HER2-антитело 2С4-FAB'-оц-L-ДНК» (2С4), «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК» (4D5) и полностью собранного комплекса (2С4-75-член-4D5), соединенного с помощью 75-членного оц-L-ДНК-линкера. Наложение графиков показывает, что из-за более высокой массы 137 «Да полностью собранный связывающий комплекс (2С4-75-член-4D5 + буфер) создает высокий уровень сигнала ответа по сравнению с введениями FAB-фрагмента (2С4 + буфер, 4D5 + буфер). Каждый из FAB-фрагментов имеет рассчитанную молекулярную массу 57 кДа. Сразу же после окончания введения на 420 секунде вводили 250 нМ стрептавидин или системный буфер. Двусторонняя стрелка показывает сдвиг сигнала 14 RU (ΔRU), индуцированный введением 250 нМ стрептавидина (2С4-75-член-4D5 + SA), по сравнению с введением буфера (2С4-75-член-4D5 + буфер). FAB-фрагменты показывают отсутствие сдвига сигнала после введения SA и сохранение сигнала на уровне буфера ((2С4 + SA), (2С4 + буфер), (4D5 + SA), (4D5 + буфер)). Стрептавидин является эффекторной группировкой. Это показывает доступность оц-L-ДНК-линкера.
На фиг. 8 приведена сенсограмма BIAcore, показывающая наложение графиков измерений в зависимости от концентрации полностью собранного 75-членного комплекса в качестве анализируемого вещества в растворе, взаимодействующем с представленной на поверхности huFc-химерой HER2 ECD. Черные линии представляют подгонку данных по Ленгмюру 1:1. Кинетические данные, скорость ассоциации ka=1,25*105 1/Мс, скорость диссоциации KD=3,39*10-5 1/с, константа аффинности 0,3 нМ.
Уровни ответа на фиг. 8 откладывали в зависимости от анализируемой концентрации полностью собранного комплекса (фиг. 9). Данные были подогнаны под уравнение Хилла, и был определен фактор Хилла (Origin 7). Уравнение: y=Vmax*xn(kn+xn), Chi2/DoF=0,6653, R2=0,99973; n=1,00201 +/- 0,06143.
В таблице 8 показаны кинетические данные в формате анализа BIAcore, приведенного на фиг. 6. Эффект кооперативного связывания может быть получен с комплексом в растворе. Молярные соотношения показывают, что именно отдельный комплекс распознает отдельную химеру HER2 ECD. Кинетические данные, скорость ассоциации ка 1/Мс, скорость диссоциации kd 1/с, константа аффинности KD (М) и в (нМ), максимальный сигнал ответа связывания (Rmax), количество захваченного лиганда huFc Chim Her2ECD (RU), время полужизни комплекса в соответствии с t1/2дисс Ленгмюра. Молярное отношение MR, показывающее стехиометрию событий связывания. Ошибка chi2. 4D5-2C4-75-член является полностью собранным комплексом. 4D5-75-член и 2С4-75-член являются FAB-фрагментами, но гибридизуются с 75-членным оц-L-ДНК-линкером.
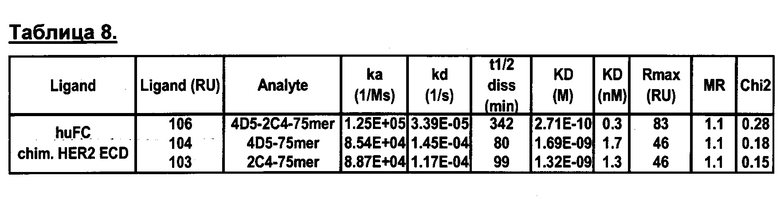
Данные, представленные в таблице 8, показывают, что полностью собранный комплекс, соединенный через 75-членный оц-L-ДНК-линкер, демонстрирует кооперативное связывание. Отдельные FAB-фрагменты показывают более низкую аффинность по сравнению с полностью собранным комплексом. Уровни сигналов на Rmax показывают повышенную молекулярную массу комплекса по сравнению с отдельными FAB-фрагментами. Несмотря на более высокий уровень сигнала молярные соотношения составляют точно 1:1. Это показывает, что статистически каждый комплекс связывается с отдельной huFc-химерной молекулой HER2 ECD.
Фактор усиления за счет кооперативности не является столь высоким по сравнению с предыдущим форматом анализа, в котором комплекс был собран на сенсорной поверхности. KDкооп увеличивается до 6 раз. Без связи с какой-либо теорией, это может быть следствием природы гомодимерного huFc химерного HER2 ECD. Потенциально двойное связующее вещество распознает два отдельных HER2 ECD в huFc-химере HER2 и не может в полной мере обеспечить кооперативность.
Эффективная доставка эффекторной группировки в виде метки может быть показана с помощью анализа FACS (см. следующий пример) с использованием фикоэритрин-меченого стрептавидинового зонда на живых клетках. Меченный стрептавидином зонд может легко получить доступ к биотиновой группировке в 75-членной оц-L-ДНК-линкерной конструкции.
Данные измерений, описанных выше, используются для создания графика Хилла (фиг. 9). Анализ Хилла комплексов показывает, что явления связывания FAB-фрагментов являются независимыми друг от друга и не мешают друг другу. Кооперативное связывание с точки зрения структурного нарушения молекулы HER2 обнаружено не было, коэффициент Хилла (n=1,00201 +/- 0,06143) равен точно 1. Таким образом, химический состав линкера, природа оц-L-ДНК-линкера и олиго-меченого FAB-фрагмента не влияют отрицательно на распознавание молекулы-мишени.
Пример 8
Другие комплексы - синтез и характеризация
Синтез способных к гибридизации олигонуклеотидов
Следующие аминомодифицированные предшественники, содержащие последовательности, приведенные в SEQ ID №№ 05 и 06, соответственно, были синтезированы в соответствии со стандартными способами. Приведенные ниже олигонуклеотиды содержат не только так называемый аминолинкер, но также флуоресцентный краситель. Специалист в данной области легко поймет, что этот флуоресцентный краситель очень удобен для облегчения очистки олигонуклеотидов как таковых, а также компонентов, включающих их.
a) 5'-флуоресцеин-AGT СТА TTA ATG CTT CTG С-(спейсер С3)3-С7аминолинкер-;
b) 5'-Cy5 AGT СТА TTA ATG CTT CTG С-(спейсер С3)3-С7аминолинкер-;
c) 5'-аминолинкер-(спейсер C3)3-AGT TCT АТС GTC GTC СА-флуоресцеин-3';
d) 5'-флуоресцеин-(бета L AGT СТА TTA ATG CTT CTG С)-(спейсер С3)3-С7аминолинкер-; (бета-L указывает, что это L-ДНК-олигонуклеотид); и
e) 5'-аминолинкер-(спейсер С3)3-(бета L-AGT ТСТ АТС GTC GTC СА)-флуоресцеин-3 (бета-L указывает, что это L-ДНК-олигонуклеотид).
Синтез проводили на синтезаторе ABI 394 в масштабе 10 мкмоль в режиме с тритилом (для 5'-аминомодификации) или без тритила (для 3-аминомодификации) с использованием коммерчески доступных CpG в качестве твердой подложки и стандартных фосфорамидитов dA(bz), dT, dG (iBu) и dC(Bz) (Sigma Aldrich).
В процессе синтеза олигонуклеотидов для введения С3-спейсера, метки и аминокислотных группировок, соответственно, были использованы следующие амидиты, аминомодификаторы и CPG-подложки:
- спейсер фосфорамидит С3(3-(4,4-диметокситритилокси)пропил-1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research);
- 5'-аминомодификатор вводится с помощью 5'-аминомодификатора С6 (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research);
- 5'-флуоресцеин фосфорамидит-6-(3',6'-дипивалоилфлуоресцеинил-6-карбоксамидо)-гексил-1-O-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research);
- Cy5™ фосфорамидит 1-[3-(4-монометокситритилокси)пропил]-1'-[3-[(2-цианоэтил)-(N,N-диизопропилфосфорамидитил]пропил]-3,3,3',3'-хлорид-тетраметилиндодикарбоцианин (Glen Research);
- LightCycler Fluorescein CPG 500 (Roche Applied Science); и
- 3'-аминомодификатор TFA Amino C-6 Icaa CPG 500 A (ChemGenes).
Для Cy5-меченых олигонуклеотидов были использованы фосфорамидиты dA(tac), dT, dG(tac), dC(tac) (Sigma Aldrich), и снятие защитной группировки проводили с помощью 33% аммиака в течение 2 ч при комнатной температуре.
L-ДНК-олигонуклеотиды были синтезированы с помощью бета-L-dA(bz)-, dT-, dG(iBu)- и dC(Bz)- фосфорамидитов (ChemGenes)
Очистку флуоресцеин-модифицированных способных к гибридизации олигонуклеотидов проводили в два этапа: сначала олигонуклеотиды очищали путем обращенно-фазовой ВЭЖХ (Merck-Hitachi-HPLC; колонка RP-18; градиентная система [А: 0,1 М (Et3NH)OAc (рН 7,0)/MeCN 95:5; В: MeCN]: 3 мин, 20% В в А, 12 мин, 20-50% В в А, и 25 мин, 20% В в А со скоростью потока 1,0 мл/мин, обнаружение при 260 нм. Фракции (мониторинг с помощью аналитической ОФ-ВЭЖХ), содержащие требуемый продукт, объединяли и выпаривали досуха. (Олигонуклеотиды, модифицированные на 5'-конце монометокситритилом, защищенным алкиламино-группой, детритилировали путем инкубации с 20% уксусной кислотой в течение 20 мин). Олигомеры, содержащие флуоресцеин в качестве метки, снова очищали путем ионообменной хроматографии на ВЭЖХ [колонка Mono Q: буфер А: гидроксид натрия (10 ммоль/л; рН примерно 12); буфер В: 1 М хлорид натрия, растворенный в гидроксиде натрия (10 ммоль/л; рН примерно 12); градиент: 30 минут от 100% буфера А до 100% буфера В, скорость потока 1 мл/мин; обнаружение при 260 нм]. Продукт обессоливали путем диализа.
Су5-меченые олигомеры использовали после первой очистки на обращенно-фазовой ВЭЖХ (Merck-Hitachi-HPLC; колонка RP-18; градиентная система [А: 0,1 М (Et3NH)OAc (зН 7,0)/MeCN 95:5; В: MeCN]: 3 мин, 20% В в А, 12 мин, 20-50% В в А, и 25 мин, 20% В в А со скоростью потока 1,0 мл/мин, обнаружение при 260 нм. Олигомеры обессоливали путем диализа и лиофилизировали на эвапораторе Speed-Vac, получая твердые вещества, которые замораживали при -24°С.
Активация способных к гибридизации олигонуклеотидов
Аминомодифицированные олигонуклеотиды из примера 2 растворяли в 0,1 М натрий-боратном буфере, зН 8,5 (с=600 мкмоль) и подвергали взаимодействию с 18-кратным мольным избытком сульфо-SMCC (сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилата), растворенного в DMF (с=3 мг/100 мкл) от Thermo Scientific, продукт реакции тщательно диализовали против воды, чтобы удалить продукт гидролиза сульфо-SMCC 4-[N-малеимидометил]циклогексан-1-карбоксилат.
Диализат концентрировали путем выпаривания и непосредственно использовали для конъюгации с одновалентным связывающим веществом, содержащим тиольную группу.
Синтез линкерных олигонуклеотидов, содержащих способные к гибридизации олигонуклеотиды на обоих концах
Олигонуклеотиды синтезировали с помощью стандартных способов на синтезаторе ABI 394 в масштабе 10 мкмоль в режиме с тритилом с использованием коммерчески доступных dT-CPG в качестве твердой подложки и стандартных фосфорамидитов dA(bz), dT, dG(iBu) и dC(Bz) (Sigma Aldrich).
L-ДНК-олигонуклеотиды синтезировали с использованием коммерчески доступного бета-L-dT-CPG в качестве твердой подложки и beta-L-dA(Bz)-, dT-, dG(iBu)- и dC(Bz)-фосфорамидитов (ChemGenes).
Очистку олигонуклеотидов проводили, как описано в примере 3, путем обращенно-фазовой ВЭЖХ. Фракции (мониторинг с помощью аналитической ОФ-ВЭЖХ), содержащие требуемый продукт, объединяли и выпаривали досуха. Детритилирование проводили путем инкубации с 80% уксусной кислотой в течение 15 мин. Уксусную кислоту удаляли путем выпаривания. Остаток растворяли в воде и лиофилизировали.
В процессе синтеза олигонуклеотидов для введения С18-спейсера, дигоксигенина и биотиновой группы были использованы следующие амидиты и CPG-подложки:
- спейсер фосфорамидит 18 (18-O-диметокситритилгексаэтиленгликоль, 1-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research);
- биотин-dT(5'-диметокситритилокси-5-[N-((4-t-бутилбензоил)-биотинил)-аминогексил)-3-акрилимидо]-2'-дезоксиуридин-3'-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит (Glen Research);
- биотин фосфорамидит-1-диметокситритилокси-2-(N-биотинил-4-аминобутил)-пропил-3-O-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит и
- 5'-диметокситритил-5- [N-(трифторацетиламиногексил) -3-акрилимидо] 2'-дезоксиуридин, 3'-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидит для аминомодификации и последующего мечения дигоксигенин-N-гидроксил-сукцинимидила эфиром.
Были синтезированы следующие мостовые конструкции или линкеры:
Линкер 1: 5'-G CAG AAG CAT ТАА TAG ACT-TGG ACG ACG ATA GAA CT-3'
Линкер 2: 5`-G CAG AAG CAT TAA TAG ACT-(T40)-TGG ACG ACG ATA GAA CT-3'
Линкер 3: 5`-[B-L]G CAG AAG CAT TAA TAG ACT-(биотин-dT)-TGG ACG ACG ATA GAA CT-3'
Линкер 4: 5`-[B-L]G CAG AAG CAT TAA TAG ACT-T5-(биотин-dT)-T5-TGG ACG ACG ATA GAA CT-3'
Линкер 5: 5`-[B-L]G CAG AAG CAT TAA TAG АСТ-Т20-(биотин-dT)-Т20-TGG ACG ACG ATA GAA CT-3'
Линкер 6: 5`-[B-L] G CAG AAG CAT TAA TAG ACT-T30-(биотин-dT)-T30-TGG ACG ACG ATA GAA CT-3'
Линкер 7: 5`-GCA GAA GCA TTA ATA GAC T T5-(биотин-dT)-T5 TG GAC GAC GAT AGA ACT-3'
Линкер 8: 5`-GCA GAA GCA TTA ATA GAC T Т10-(биотин-dT)-Т10 TGG ACG ACG ATA GAA CT-3'
Линкер 9: 5`-GCA GAA GCA TTA ATA GAC T T15-(биотин-dT)-T15 TGG ACG ACG ATA GAA CT-3'
Линкер 10: 5`-GCA GAA GCA TTA ATA GAC T Т20-(биотин-dT)-Т20 TGG ACG ACG ATA GAA CT-3'
Линкер 11: 5`-G CAG AAG CAT TAA TAG ACT-Spacer C18-(биотин-dT)-спейсер C18-TGG ACG ACG ATA GAA CT-3'
Линкер 12: 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер С18)2-(биотин-dT)-(спейсер C18)2-TGG ACG ACG ATA GAA CT-3'
Линкер 13: 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер С18)3-(биотин-dT)-(спейсер C18)3-TGG ACG ACG ATA GAA CT-3'
Линкер 14: 5`-G CAG AAG CAT TAA TAG АСТ-(спейсер C18)4-(биотин-dT)-(спейсер C18)4-TGG ACG ACG ATA GAA CT-3'
Линкер 15. 5`-G CAG AAG CAT TAA TAG ACT-T20-(Dig-dT)-T20-TGG ACG ACG ATA GAA CT-3'
Линкер 16: 5`-G CAG AAG CAT TAA TAG ACT-(Dig-dT)-TGG ACG ACG ATA GAA CT-3'
Линкер 17: 5`-G CAG AAG CAT TAA TAG ACT-(биотин-dT)-TGG ACG ACG ATA GAA CT-3'
Приведенные выше примеры мостовых конструкций включают по меньшей мере первый способный к гибридизации олигонуклеотид и второй способный к гибридизации олигонуклеотид. Линкеры с 3 по 18, в дополнение к способным к гибридизации нуклеиновокислотным участкам, содержат центральный биотинилированный или дигоксигенилированный тимидин, соответственно, или спейсер, состоящий из тимидиновых блоков приведенной выше длины.
5'-гибридизующийся олигонуклеотид соответствует SEQ ID № 07, а 3'-гибридизующийся олигонуклеотид соответствует SEQ ID № 08, соответственно. Олигонуклеотид SEQ ID № 07 будет легко гибридизоваться с олигонуклеотидом SED ID № 06. Олигонуклеотид SEQ ID № 08 будет легко гибридизоваться с олигонуклеотидом SED ID № 05.
В приведенных выше примерах мостовых конструкций [B-L] указано, что дана последовательность L-ДНК-олигонуклеотидов; спейсер С 18, биотин и биотин dT, соответственно, относятся к С18-спейсеру, биотину и биотину-dT как производным приведенных выше структурных блоков; а Т с числом указывает на число тимидиновых остатков, входящих в линкер в данной позиции.
Сборка комплекса
A) Расщепление IgG и мечение FAB'-фрагментов посредством оцДНК
Очищенные моноклональные антитела расщепляли с помощью протеазы пепсина, получая F(ab')2-фрагменты, которые затем восстанавливали до FAB'-фрагментов путем обработки низкой концентрацией цистеамина при 37°С. Реакцию останавливали путем отделения цистеамина на колонке PD 10. FAB'-фрагменты метили активированным олигонуклеотидом, полученным в соответствии с примером 3. Эта одноцепочечная ДНК (=оцДНК) несет тиол-реактивную малеимидогруппу, которая реагирует с цистеинами FAB'-шарнирной области. Для того чтобы получить высокий процент однократно меченных FAB'-фрагментов, относительное молярное отношение оцДНК к FAB'-фрагменту поддерживают на низком уровне. Очистка однократно меченных FAB'-фрагментов (оцДНК:FAB'=1:1) происходит с помощью ионообменной хроматографии (колонка: Source 15 Q РЕ 4.6/100, Pharmacia/GE). Проверка эффективной очистки проводится путем аналитической эксклюзионной хроматографии и SDS-PAGE.
B) Сборка комплекса, включающего два полипептида, специфически связывающихся с мишенью
Анти-pIGF-1R-комплекс основан на двух FAB'-фрагментах, которые нацелены на различные эпитопы внутриклеточного домена IGF-1R: FAB' 8.1.2, обнаруживает сайт фосфорилирования (pTyr 1346), a FAB' 1.4.168 обнаруживает нефосфорилированный сайт белка-мишени. FAB'-фрагменты были ковалентно связаны с одноцепочечной ДНК (оцДНК): FAB' 1.4.168 с 17-членной оцДНК, содержащей SEQ ID № 05 и флуоресцеин в качестве флуоресцентного маркера, a FAB' 8.1.2 с 19-членной оцДНК, содержащей SEQ ID № 06 и Cy5 в качестве флуоресцентного маркера. Далее эти FAB' с ковалентно связанной 17-членной или 19-членной оцДНК называются оц-FAB' 1.4.168 и оц-FAB' 8.1.2, соответственно. Сборка комплекса опосредуется линкером (т.е. мостовой конструкцией, состоящей из двух комплементарных оцДНК-олигонуклеотидов (SEQ ID №. 7 и 8 соответственно), которые гибридизуются с соответствующими оцДНК этих оц-FAB'-фрагментов. Расстояние между двумя оц-FAB'-фрагментами комплекса может быть изменено с помощью спейсеров, например С18-спейсера или ДНК разной длины, соответственно.
Для оценки сборки компонентов комплекса оц-FAB' 8.1.2, оц-FAB' 1.4.168 и линкерные конструкции (1) (= линкер 17 из примера 2.4) 5'-G CAG AAG CAT TAA TAG ACT T(-Bi)-TGG ACG ACG ATA GAA CT-3' и (2) (= линкер 10 из примера 2.4) 5'-G CAG AAG CAT TAA TAG ACT-T(20)-T(-Bi)-(T20)-TGG ACG ACG ATA GAA CT-3') смешивали в эквимолярных соотношениях при комнатной температуре. После этапа инкубации в течение 1 минуты реакционную смесь анализировали на аналитической колонке для гель-фильтрации (Superdex™ 200, 10/300 GL, GE Healthcare). Сравнение объемов элюирования (VE) отдельных компонентов комплекса с VE реакционной смеси показывает, что комплекс был успешно сформирован (фиг. 10).
Эксперимент BIAcore по оценке связывания анти-pIGF-1R-комплекса с иммобилизованными IGF-1R- и IR-пептидами
Для этого эксперимента использовали инструмент BIAcore 2000 (GE Healthcare) с сенсором BIAcore SA, установленным в системе при Т=25°С. Предварительная подготовка была проведена на скорости 100 мкл/мин 3-кратным введением 1 М NaCl в 50 мМ NaOH в течение 1 мин и 10 мМ HCl в течение 1 мин.
HBS-ET (10 мМ HEPES рН 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Tween® 20) использовали в качестве системного буфера. Буфер для образцов был идентичен системному буферу. Система BIAcore 2000 находилась под управлением программного обеспечения V1.1.1.
Затем биотинилированные пептиды захватывали на поверхности SA в соответствующих проточных ячейках. 16 RU IGF-1R(1340-1366)[1346-pTyr; Glu(Bi-РЕС-1340]амида (т.е. - тирозин 1346 фосфорилирован - пептид SEQ ID № 11, содержащий ПЭГ-линкер, связанный через глутаминовую кислоту, соответствующую позиции 1340, и биотинилированный на другом конце линкера) было захвачено на проточной ячейке 2. 18 RU IGF-1R(1340-1366); Glu(Bi-PEG-1340]амида (т.е. - тирозин 1346 не фосфорилирован - пептид SEQ ID № 11, содержащий ПЭГ-линкер, связанный через глутаминовую кислоту, соответствующую позиции 1340, и биотинилированный на другом конце линкера) было захвачено на проточной ячейке 3. 20 RU hlnsR(1355-1382)[1361-pTyr; Glu(Bi-PEG-1355]амида (т.е. - тирозин 1361 фосфорилирован - пептид SEQ ID № 12, содержащий ПЭГ-линкер, связанный через глутаминовую кислоту, соответствующую позиции 1355 человеческого рецептора инсулина, и биотинилированный на другом конце линкера) было захвачено на проточной ячейке 4. Наконец, все проточные ячейки насыщались d-биотином.
Для образования комплекса был использован протокол сборки, описанный выше. Когда выполнялись отдельные циклы только с одним из двух оц-FAB', отсутствие или наличие линкерной ДНК не влияло на кривые ассоциации или диссоциации.
100 нМ анализируемого вещества (т.е. в этих экспериментах двухвалентного двойного связывающего агента) в растворе вводили со скоростью 50 мкл/мин в течение времени ассоциации 240 сек и наблюдали диссоциацию в течение 500 сек. Эффективного восстановления достигали с помощью этапа введения 80 мМ NaOH в течение 1 мин со скоростью 50 мкл/мин. Проточная ячейка 1 использовалась в качестве референса. Введение «пустого» буфера использовали вместо введения антигена для получения данных двойного референса, вычитая сигнал буфера.
В каждом цикле измерений во все четыре проточные ячейки вводили одно из следующих анализируемых веществ в растворе: 100 нМ оц-FAB' 8.1.2, 100 нМ оц-FAB' 1.4.168, смесь 100 нМ оц-FAB' 8.1.2 и 100 нМ оц-FAB', 100 нМ двухвалентный связывающий агент, состоящий из оц-FAB' 8.1.2 и оц-FAB' 1.4.168, гибридизованный с линкером (3) (5`-G CAG AAG CAT TAA TAG ACT-T(20)-T(-Dig)-(T20)-TGG ACG ACG ATA GAA СТ-3'(=линкер 15)), и 100 нМ двухвалентный связующий агент, состоящий из оц-FAB' 8.1.2 и оц-FAB' 1.4.168, гибридизованный с линкером (1) (5'-G CAG AAG CAT TAA TAG ACT-T(-Dig) -TGG ACG ACG ATA GAA СТ-3'(=линкер 16)), соответственно.
Сигналы наблюдали в форме сенсограмм BIAcore в зависимости от времени.
Точки отчетности были установлены на конец фазы ассоциации анализируемого вещества (Binding Late, BL) и на конец фазы диссоциации анализируемого вещества (Stability Late, SL) для контроля высоты сигнала ответа для каждого взаимодействия. Скорости диссоциации kd (1/с) были рассчитаны в соответствии с линейной подгонкой Ленгмюра 1:1 с помощью программного обеспечения для оценки BIAcore 4.1. Время полужизни комплекса в минутах рассчитывали по формуле In(2)/(60*kD).
Сенсограммы (фиг. 11-14) показывают усиление специфичности и стабильности комплекса в связывании pIGF-1R при использовании оц-FAB' 1.4.168 и оц-FAB' 1.4.168 в виде комплекса (= двухвалентный связывающий агент), вероятно, благодаря лежащему в основе кооперативному связывающему эффекту. FAB' 1.4.168 в одиночку не демонстрирует перекрестную реактивность с пептидом pIR, но не различает фосфорилированную и нефосфорилированную формы IGF-1R (Т1/2 дисс = 3 мин в обоих случаях). Тем не менее, FAB' 8.1.2 связывается только с фосфорилированной версией пептида pIGF-1R, но проявляет некоторую нежелательную перекрестную реактивность с фосфорилированным рецептором инсулина. Комплекс хорошо различает пептид pIGF-1R и оба других пептида (см. фиг. 13) и, таким образом, помогает преодолеть проблемы неспецифического связывания. Обратите внимание, что выигрыш в специфичности теряется, когда оба FAB' применяются без линкерной ДНК (фиг. 14). Усиление аффинности комплекса по отношению к пептиду pIGF-1R проявляется в повышенном времени полудиссоциации по сравнению с отдельными FAB' и смесью FAB' без линкерной ДНК (фиг. 12 и фиг. 14). Хотя тестируемые комплексы с двумя различными ДНК-линкерами в целом разделяют положительное влияние на специфичность и аффинность связывания мишени, более длинный линкер (с T40-Dig в качестве спейсера) (т.е. линкер 15) кажется быть предпочтительным с точки зрения обоих критериев.
Анализ BIAcore сэндвич-типа M-1.4.168-IgG и М-8.1.2
Использовали инструмент BIAcore Т100 (GE Healthcare) с установленным в системе сенсором BIAcore СМ5. Предварительная подготовка сенсора была проведена введением 0,1% SDS, 50 мМ NaOH, 10 мМ HCl и 100 мМ H3PO4 в течение 1 мин со скоростью 100 мкл/мин.
Системным буфером был HBS-ET (10 мМ HEPES pH 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Tween® 20). Буфер для образцов был идентичен системному буферу.
Система BIAcore Т100 находилась под управлением программного обеспечения V1.1.1. Поликлональное кроличье IgG-антитело <IgGFCγM>R (Jackson ImmunoResearch Laboratories Inc.) в концентрации 30 мкг/мл в 10 мМ Na-ацетате с pH 4,5 иммобилизовали при 10000 RU на проточных ячейках 1, 2, 3 и 4, соответственно, с помощью химической реакции EDC/NHS в соответствии с инструкциями изготовителя. Наконец, сенсорную поверхность блокировали с помощью 1 М этаноламина. Весь эксперимент проводили при 13°С.
500 нМ первичного МКА M-1.004.168-IgG захватывали в течение 1 мин при 10 мкл/мин на поверхности <IgGFCγM>R. 3 мкМ смеси IgG-фрагментов (IgG классов IgG1, IgG2a, IgG2b, IgG3), содержащей блокирующий раствор, вводили со скоростью 30 мкл/мин в течение 5 мин. Пептид IGF-1R(1340-1366)[1346-pTyr; Glu(Bi-PEG-1340]амид вводили в концентрации 300 нМ в течение 3 мин со скоростью 30 мкл/мин. Вторичное антитело M-8.1.2-IgG в концентрации 300 нМ вводили со скоростью 30 мкл/мин. Сенсор регенерировали с использованием 10 мМ глицин-HCl, рН 1,7, при 50 мкл/мин в течение 3 мин.
На фиг. 15 представлены условия анализа. На фиг. 18 приведены результаты измерений. Измерения четко показывают, что оба моноклональных антитела способны одновременно связываться с двумя различными не родственными эпитопами на соответствующем целевом пептиде. Это является необходимым условием для любых последующих экспериментов с целью получения явлений кооперативного связывания.
Анализ BIAcore комплекса на сенсорной поверхности
Использовали инструмент BIAcore 3000 (GE Healthcare) с сенсором BIAcore SA, установленным в системе на Т=25°С. Предварительная подготовка системы была проведена на скорости 100 мкл/мин с трехкратным введением 1 М NaCl в 50 мМ NaOH в течение 1 мин и 10 мМ HCl в течение 1 мин.
Системным буфером был HBS-ET (10 мМ HEPES рН 7,4, 150 мМ NaCl, 1 мМ EDTA, 0,05% Tween® 20). Буфер для образцов был идентичен системному буферу.
Система BIAcore 3000 находилась под управлением программного обеспечения V4.1.
124 RU амино-ПЭО-биотина захватывали на референсной проточной ячейке 1. 1595 RU биотинилированного 14,6 кДа Т0-Bi 37-членного оцДНК-линкера (1) (5'-G CAG AAG CAT ТАА TAG ACT-T(-Bi)-TGG ACG ACG ATA GAA CT-3') (= линкер 17 из примера 2.4) и 1042 RU биотинилированного 23,7 кДа T40-Bi 77-членного оцДНК-линкера (2) (5`-G CAG AAG CAT TAA TAG АСТ-Т(20)-(биотин-dT)-(T20)-TGG ACG ACG ATA GAA CT-3' = линкер 10) захватывали на разных проточных ячейках.
300 нМ оц-FAB 8.1.2 и 300 нМ оц-FAB 1.004.168 вводили в систему со скоростью 50 мкл/мин в течение 3 мин. В качестве контроля вводили только 300 нМ оц-FAB 8.1.2 или 300 нМ оц-FAB 1.004.168, чтобы проверить кинетический вклад каждого оц-FAB. В качестве контроля вместо оц-FAB вводили буфер. Пептиды IGF-1R(1340-1366)[1346-pTyr]амид, INR(1355-1382)[1361-pTyr]амид и IGF-1R(1340-1366)амид вводили в систему со скоростью 50 мкл/мин в течение 4 мин, свободно в растворе, в концентрации 0 нМ, 4 нМ, 11 нМ, 33 нМ (дважды), 100 нМ и 300 нМ. В другом воплощении для измерения аффинности в сравнении с пептидами IGF-1R(1340-1366)[1346-pTyr]амид использовали концентрации 0 нМ, 0,4 нМ, 1,1 нМ, 3,3 нМ (дважды), 10 нМ и 30 нМ.
Диссоциацию контролировали на скорости 50 мкл/мин в течение 5,3 мин. Систему регенерировали после каждой концентрировании 12-секундным введением 250 мМ NaOH и перезагружали оц-FAB-лигандом.
Фиг. 17 схематически описывает условия анализа на приборе BIAcore. Таблицы, приведенные на фиг. 18, показывают количественные результаты этого подхода. Фиг. 19, фиг. 20 и фиг. 21 изображая иллюстративные результаты BIAcore в этих условиях анализа.
Таблица на фиг. 18 демонстрирует преимущества концепции комплекса. Двойной связывающий агент Т40 (двойной связывающий агент с линкером 10 из примера 2.4, т.е. линкер со спейсером из Т20-биотин-dT-Т20) дает двукратное увеличение времени полужизни комплекса антигена (414 мин) и трехкратное увеличение аффинности (10 пМ) по сравнению с двойным связывающим агентом Т0 (т.е. двойным связывающим агентом с линкером 16) со значениями 192 мин и 30 пМ, соответственно. Это подчеркивает необходимость оптимизации длины линкера для получения оптимального эффекта кооперативного связывания.
Двойной связывающий агент Т40 (т.е. двойной связывающий агент, содержащий линкер T40-Bi (линкер 10)) демонстрирует аффинность 10 пМ по сравнению с фосфорилированным пептидом IGF-1R (таблица на фиг. 18, фиг. 19). Это является 2400-кратным повышением аффинности по сравнению с фосфорилированным пептидом рецептора инсулина (24 нМ) и 100-кратным повышением по сравнению с нефосфорилированным пептидом IGF-1R.
Таким образом, достигается цель в повышении специфичности и аффинности за счет комбинации двух отдельных и разделенных событий связывания.
Эффект кооперативного связывания становится особенно очевидным для скоростей диссоциации по сравнению с фосфорилированным пептидом IGF-1R, где комплекс антигена демонстрирует время полужизни 414 мин по сравнению с 0,5 мин у одновалентного связывающего агента 8.1.2 и по сравнению с 3 мин у одновалентного связывающего агента 1.4. 168 в одиночку, соответственно.
Кроме того, полностью собранная конструкция примерно умножает свои скорости диссоциации kd (1/с) по сравнению с одиночными FAB-гибридизующимися конструкциями (фиг. 21, фиг. 20, фиг. 21 и таблица на фиг. 18). Интересно, что скорость ассоциации ka (1/Мс) также слегка увеличивается по сравнению с событиями взаимодействия одиночных FAB, это может быть из-за увеличения молекулярной гибкости конструкции.
Пример 9
Анализы связывания - in vitro и ex vivo
Олигонуклеотид для обнаружения зонд-Cy5
оц-L-ДНК-олигонуклеотид для обнаружения, зонд-Cy5 5' Cy5-Y- ATG CGA-GTA CCT TAG AGT С -Z-Cy5 3' (SEQ ID № 72), был синтезирован с использованием стандартных способов олигонуклеотидного синтеза. Метка Cy5 была введена за счет реакции аминогрупп с Cy5-монореактивным NHS-эфиром. (GE Healthcare Lifescience, STADT, LAND). Для нуклеотидов были использованы L-ДНК-амидиты (ChemGenes, STADT, LAND). 5'- и 3'-аминогруппы были введены в процессе твердофазного олигонуклеотидного синтеза, где Y = 5'-аминомодификатор С6, введенный через (6-(4-монометокситритиламино)гексил-(2-цианоэтил)-(N,N-диизопропил)-фосфорамидит (Glen Research), a Z = 3'-аминомодификатор С6, введенный через 3'-аминомодификатор TFA Amino С6 long chain aminoalkyl Controlled Pore Glass 1000 A (ChemGenes).
Линкерный олигонуклеотид двойного связывания
оц-L-ДНК-олигонуклеотидный линкер SEQ ID № 73 5'-G CAG AAG CAT TAA TAG ACT-T20-GAC TCT AAG GTA CTC GCA T-T20-TGG ACG ACG ATA GAA CT-3' был синтезирован с использованием стандартных способов олигонуклеотидного синтеза.
Сборка комплекса
Комплекс был собран путем гибридизации «анти-HER2-антитело 2C4-FAB'-оц-L-ДНК», меченного FITC, и «анти-HER2-антитело 4D5-FAB'-оц-L-ДНК», меченного FITC, в эквимолярном соотношении с оц-L-ДНК-линкером SEQ ID № 73. Для проверки правильной сборки комплекса комплекс подвергали этапу хроматографии SEC и фильтровали через стерилизующий фильтр.
Анализ связывания in vitro
Клетки человеческого рака молочной железы KPL-4 высевали в концентрации 2×106 клеток/мл в объеме 30 мкл в μ-слайды VI (ibidi, Германия). Через три часа после этого добавляли 70 мкл среды (RPMI 1640, 2 мМ L-глутамина, 10% FCS) для адгезии клеток.
После инкубации в течение 24 часов при 37°С и в атмосфере с 5% CO2 и насыщенной паром (эффективной для всех следующих инкубаций), супернатант удаляли и клетки промывали один раз 100 мкл PBS для удаления остатков среды.
Для последующего применения 50 мкл комплекса 4D5-2C4, полученного выше, метили FITC-раствором (С=2,5 мкг/мл) и инкубировали в течение 45 минут, после чего один раз промывали 100 мкл PBS и далее инкубировали с 50 мкл ДНК-зонда (SEQ ID № 72) в эквимолярном количестве (0,13 мкг/мл).
Процедуру предварительного смешивания выполняли, сначала смешивая комплекс и зонд для обнаружения. После этого смесь добавляли к клеткам (концентрации см. выше) и инкубировали в течение 45 минут.
Xolair®, гуманизированное моноклональное IgG1-антитело, нацеленное на человеческий IgE-иммуноглобулин, использовали в качестве отрицательного контроля, a Herceptin®, помеченное Cy5, нацеленное на человеческий HER-2-рецептор, использовали в качестве положительного контроля. Оба антитела использовали в одной и той же концентрации (2,5 мкг/мл).
Затем супернатант удаляли и клетки промывали один раз 100 мкл PBS. Затем ядра клеток окрашивали DAPI, добавляя 50 мкл раствора HOECHST33342 (С = 10 мкг/мл) и инкубировали в течение 15 минут. Чтобы удалить краситель для окрашивания клеток, клетки дважды промывали 100 мкл PBS после удаления супернатанта. Еще 120 мкл PBS добавляли, чтобы сохранить клетки влажными, чтобы обеспечить их жизнеспособность. Все разведения проводили с использованием среды (без L-глутамина и FCS), чтобы обеспечить жизнеспособность клеток и избежать отделения клеток. После этой процедуры слайды обследовали путем флуоресцентного мультиспектрального анализа с использованием системы NUANCE (CRi, Кембридж, США). Изображения нормировали для сопоставимости интенсивности флуоресценции.
Анализ ex vivo
Иммунодефицитным мышам SCID beige с установленными опухолями KPL-4 (ортотопически имплантированными) внутривенно вводили 50 мкг комплекса в 100 мкл PBS, и через 18 часов вводили Cy5-меченый ДНК-зонд в эквимолярной концентрации (2,63 мкг на мышь). Опухоли эксплантировали через 48 часов и обследовали путем флуоресцентного мультиспектрального анализа с использованием системы MAESTRO (CRi, Кембридж, США).
Результаты
Анализ связывания in vivo
Комплекс был дважды мечен FITC за счет каждого из его Fab'-оц-L-ДНК-компонентов. Зонд для обнаружения представляет собой дважды Cy5-меченый оц-L-ДНК 20-членный олигонуклеотидный зонд, который может быть гибридизован с 95-членным оц-L-ДНК-линкером комплекса.
В отличие от Xolair-Cy5 (без сигнала флуоресценции, отрицательный контроль) Herceptin-Cy5 специфически окрашивал опухолевые клетки (фиг. 22). FITC-меченый комплекс 4D5-2С4-95-член специфически связывается с опухолевыми клетками KPL-4, как можно видеть при последовательной инкубации с зондом для обнаружения (измеренного в канале флуоресценции Cy5), который локализован вместе с комплексом на опухолевых клетках, что указывает на гибридизацию олигонуклеотида для обнаружения зонда-Cy5 с комплексом. В режиме последовательной инкубации, а также при предварительном смешивании может быть продемонстрировано специфическое окрашивание опухолевых клеток на Her-2-антиген (фиг. 2).
На фиг. 22 показано в ближнем инфракрасном излучении изображение раковых клеток, инкубированных с комплексом и зондом для обнаружения (визуализация NIRF). В правом верхнем углу фигуры показана схема полностью собранного 4D5-2С4-95-членного комплекса, гибридизованного с Cy5-меченым олигонуклеотидом для обнаружения. В средней правой части фигуры показан эскиз Cy5-меченого герцептина. В правом нижнем углу фигуры показан столбец интенсивности сигнала.
В верхней левой части фиг. 22 показано связывание Cy5-меченого герцептина (Herceptin®) с раковыми клетками (положительный контроль). Мембраны клеток KPL-4 выглядят как яркие освещенные кольца, окружающие DAPI-окрашенные клеточные ядра. В левой нижней части показана инкубация Cy5-меченого ксолара (Xolair®) (отрицательный контроль). Окрашивания мембраны обнаружено не было, но имело место DAPI-окрашивание клеточных ядер. В нижней средней части фигуры показано связывание комплекса 4D5-2C4-FITC. Флуоресцеиновый сигнал мембранно-связанного комплекса выглядит как освещенные кольца, окружающие DAPI-окрашенные клеточные ядра. В верхней средней части фигуры показано связывание комплекса 4D5-2C4-FITC и Cy5-меченого зонда для обнаружения. Можно видеть обнаружение комплекса через Cy5-меченный оц-L-ДНК-зонд обнаружения, который был последовательно гибридизован. Cy5-сигнал олигонуклеотида обнаружения выглядит как окрашивание мембраны, дающее яркие освещенные кольца, окружающие DAPI-окрашенные клеточные ядра.
На фиг. 23 показано в ближнем инфракрасном излучении (NIRF) изображение клеток KPL-4. На фиг. 23А показаны результаты последовательного применения FITC-меченного комплекса 4D5-2C4 и Cy5-меченого зонда обнаружения. На фиг 23В показаны результаты инкубации клеток KPL-4 с предварительно перемешанным FITC-меченым комплексом 4D5-2C4 и Cy5-меченым зондом обнаружения. Оба изображения показывают локализованные в мембране сигналы. В качестве контроля клетки окрашивали DAPI.
Эксперимент показывает, что комплекс, описанный в данном документе, вначале может быть применен для специфического нацеливания HER-2-положительных клеток. На втором этапе меченый зонд обнаружения может быть применен для гибридизации связанного комплекса с мишенью. Флуоресцентно меченый зонд обнаружения, таким образом, является доказательством концепции отложенного времени, последовательного применения и специфического нацеливания эффекторной группировки на основе олигонуклеотида. В этом случае полезная нагрузка представляет собой флуоресцентную метку для визуализации клетки in vitro.
Анализ связывания ex vivo
Как показано на фиг. 24 (левое изображение), сильный флуоресцентный сигнал можно обнаружить в экспериментальных условиях, когда образец сначала инкубировали с комплексом, а затем с Cy5-меченым зондом обнаружения. Напротив, флуоресцентный сигнал не обнаруживается в опухолях, где в ксенотрансплантат KPL-4 ранее был введен только Cy5-меченый зонд обнаружения (правое изображение).
На фиг. 24 показана эксплантированная опухоль KPL-4, которую подвергли визуализации NIRF. На первом изображении показаны Cy5-флуоресцентные сигналы, полученные из трех опухолей KPL-4, эксплантированных у мышей, которым последовательно вводили сначала комплекс 4D5-2C4, а затем зонд обнаружения. На правом изображении показано, что ни один флуоресцентный сигнал не был получен из трех опухолей KPL-4, когда трем мышам вводили только зонд обнаружения, без комплекса 4D5-2C4.
Пример 10
Ингибирование клеточной пролиферации клеточной линии рака молочной железы MDA-MB-175
2×104 клеток рака молочной железы MDA-MB-175, культивируемых в среде DMEM/F12, дополненной 10% фетальной телячьей сывороткой, 2 мМ глутамином и пенициллином/стрептомицином, высевали в 96-луночные планшеты. Антитела и комплекс, соответственно, добавляли в указанных концентрациях на следующий день (от 40 до 0,0063 мкг/мл). На 6 дней инкубации добавляли аламаровый синий и инкубировали планшеты в течение 3-4 ч в инкубаторе для тканевых культур. Измеряли флуоресценцию (возбуждение 530 нм/испускание 590) и рассчитывали процент ингибирования, используя необработанные клетки в качестве референса.
Результаты
Анти-HER2-антитело 2С4 (пертузумаб) показало максимальное ингибирование 44%. Анти-HER2-антитело герцептин показало максимальное ингибирование 9%. Комплекс, описанный в данном документе, содержащий FAB-фрагменты пертузумаба и герцептина, показал максимальное ингибирование 46%.
Нужно отметить, что пертузумаб был протестирован как полноразмерное IgG-антитело с двумя HER2-связывающими сайтами, в то время как комплекс содержит один Fab-фрагмент пертузумаба с одним HER2-связывающим сайтом.
Пример 11
Стабильность комплекса при замораживании-оттаивании
Комплекс собирали путем гибридизации «анти-HER2-антитело-2С4-FAB'-оц-L-ДНК», меченного FITC, и «анти-HER2-антитело-4D5-Fab'-оц-L-ДНК», меченного FITC, в эквимолярном соотношении с оц-L-ДНК-линкером SEQ ID № 73. Для того чтобы проверить правильную сборку комплекса, комплекс подвергали этапу хроматографии SEC и фильтровали через стерилизующий фильтр.
50 мкл комплекса (1,5 мг/мл) анализировали с помощью аналитической SEC, используя колонку TSK3000 (GE). Рабочим буфером был 0,1 М KH2PO4 pH 6,8. Скорость потока составляла 1 мл/мин. Хроматограмма показана на фиг. 25.
После замораживания и оттаивания комплекс подвергали повторной хроматографии. 50 мкл комплекса (1,5 мг/мл) анализировали с помощью аналитической SEC, используя колонку TSK3000 (GE). Рабочим буфером был 0,1 М KH2PO4, pH 6,8. Скорость потока составляла 1 мл/мин. Хроматограмма показана на фиг. 26.
Пример 12
Клонирование и экспрессия связывающих группировок
Описание основной/стандартной плазмиды для экспрессии у млекопитающих
Нужные белки экспрессировали путем временной трансфекции эмбриональных клеток почек человека (HEK 293). Для экспрессии нужного гена/белка использовали транскрипционный блок, включающий следующие функциональные элементы:
- немедленно-ранний энхансер и промотор цитомегаловируса человека (P-CMV), включая интрон А,
- 5'-нетранслируемую область гена человеческого зародышевого антитела(5'-UTR),
- сигнальную последовательность тяжелой цепи мышиного иммуноглобулина (SS),
- ген/белок, который нужно экспрессировать (например, тяжелую цепь полноразмерного антитела), и
- сигнальную последовательность полиаденилирования («полиА») бычьего гормона роста (BGH рА).
Помимо экспрессионного блока/кассеты, содержащей нужный ген, который должен быть экспрессирован, основная/стандартная плазмида включает:
- сайт начала репликации из вектора pUC18, который разрешает репликацию этой плазмиды в Е. coli, и
- ген β-лактамазы, который придает устойчивость к ампициллину в Е. coli.
Клонирование
Во-первых, выполняется клонирование конструкции, кодирующей связывающую группировку (например, Fab-фрагмент антитела). Плазмиду с нуклеиновой кислотой, кодирующей связывающую группировку, обычно получают путем синтеза гена, где С-концевая область закодированной связывающей группировки содержит сортазный повтор и His-метку. Плазмиды по отдельности переносят в отдельные лунки многолуночного планшета (может быть загружен целый планшет). После этого плазмиды расщепляют смесью рестрикционных ферментов, чтобы вырезать область, кодирующую связывающую группировку. Желательно разработать весь генный синтез таким образом, чтобы для всех плазмид была необходима одна смесь рестрикционных ферментов. Впоследствии, этап очистки дает очищенные фрагменты ДНК. Эти фрагменты лигируют в плазмидный остов, который был вырезан из акцепторного вектора той же смесью рестрикционных ферментов, что указана выше. Альтернативно, процедура клонирования может быть выполнена с помощью SLIC-опосредованного этапа клонирования. После лигирования автоматизированная платформа переносит все лигазные смеси в другой многолуночный планшет с компетентными клетками E. coli (например, Top10 Multi Shot, Invitrogen) и выполняет реакцию трансформации. Клетки культивируют до нужной плотности. Из аликвоты культивационной смеси могут быть сделаны глицериновые стоки. Из культуры выделяют плазмиду (например, с помощью мини-набора для выделения плазмид (например, NucleoSpin 96 Plasmid, Macherey& Nagel)). Идентичность плазмиды проверяют путем расщепления аликвоты подходящей рестрикционной смесью и с помощью электрофореза в SDS-геле (например, Е-Gel 48, Invitrogen). После этого в новый планшет может быть загружена аликвота плазмиды для проведения контрольной реакции секвенирования.
Экспрессия
Fab-фрагменты антител получали путем временной трансфекции клеток HEK293 (клеток, полученных из линии эмбриональных клеток почек человека 293), культивируемых в среде 17 (Invitrogen Corp.). Для трансфекции использовали реагент для трансфекции "293-Fectin" (Invitrogen). Fab-фрагменты антител экспрессировали из двух различных плазмид, кодирующих полноразмерную легкую цепь и соответствующую усеченную тяжелую цепь, содержащую С-концевую последовательность LPXTG (SEQ ID № 74). При трансфекции две плазмиды использовали в эквимолярном соотношении. Трансфекцию выполняли в соответствии с инструкцией производителя. Клеточные культуральные супернатанты, содержащие Fab-фрагменты антител, собирали через семь дней после трансфекции. Супернатанты хранили замороженными до очистки.
Клеточные культуральные супернатанты, содержащие Fab-фрагменты антител, фильтровали и очищали путем двух хроматографических этапов. Fab-фрагменты антител захватывали с помощью аффинной хроматографии с использованием колонки HisTrap HP Ni-NTA (GE Healthcare), уравновешенной PBS, содержащим 20 мМ имидазола (1 мМ KH2PO4, 10 мМ Na2HPO4, 137 мМ NaCl, 2,7 мМ KCl, 20 мМ имидазола), pH 7,4. Несвязанные белки удаляли путем промывания уравновешивающим буфером. Меченный гистидином белок элюировали линейным градиентом имидазола от 20 мМ до 400 мМ в PBS (1 мМ KH2PO4, 10 мМ Na2HPO, 137 мМ NaCl, 2,7 мМ KCl, 400 мМ имидазола) в объеме десяти колонок. Эксклюзионную хроматографию на Superdex 200™ (GE Healthcare) использовали в качестве второго этапа очистки. Эксклюзионную хроматографию проводили в 40 мМ Tris-HCl буфере, 0,15 М NaCl, рН 7,5. Fab-фрагменты антител концентрировали с помощью центрифужного фильтрационного блока Ultrafree-CL, снабженного мембраной Biomax-SK (Millipore, Биллерика, Массачусетс) и хранили при -80°С.
Белковую концентрацию Fab-фрагментов антител определяли путем измерения оптической плотности (OD) при 280 нм с использованием молярного коэффициента экстинкции, рассчитанного на основании аминокислотной последовательности. Чистоту и правильное образование Fab антител анализировали с помощью SDS-PAGE в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотреитола) и окрашивания Кумасси.
Пример 13
Создание биспецифических связывающих молекул за счет связывания конъюгатов «Fab-фрагмент антитела - олигонуклеотид»
Связывание Fab-фрагментов антител с олигонуклеотидами проводили с использованием фермента сортазы А. Таким образом, молекула с группировкой, содержащей пептид LPXTG (SEQ ID № 74), была ковалентно присоединена к другой молекуле, обладающей GG-группировкой. Таким образом, олигонуклеотид имел одну из группировок, а Fab-фрагмент антитела имел соответствующую другую группировку. Ферментативная реакция может быть более выгодной, чем химическая реакция, из-за более высоких оборотов, более высокой специфичности, меньших побочных продуктов и менее опасных отходов.
Реакцию Fab-фрагмента антитела с олигонуклеотидом проводили в профильтрованной среде HEK после экспрессии Fab-фрагмента антитела путем добавления 10х буфера для сортазы (1х: 50 мМ Tris-HCl, 150 мМ NaCl, 5 мМ CaCl2, pH 7,5), фермента сортазы (0,15 мкг фермента на 1 мкг Fab) и олигонуклеотида в 4-кратном молярном избытке по отношению к Fab-фрагменту антитела. Олигонуклеотиды состояли из 18-членной L-ДНК, в которой два остатка глицина были добавлены через аминолинкер и спейсер (5'-(Gly)4-аминолинкер-(спейсер C3)3-AG ТТС TAT CGT CGT ССА-флуоресеин-3') (SEQ ID № 75). Инкубацию проводили при 37°С в течение 4-16 ч.
Для очистки применяли (полу)автоматизированной подход. Поскольку не все Fab-фрагменты антител превращаются в конъюгаты, эти две популяции должны быть разделены. Это было достигнуто путем отрицательного His-Tag-отбора при помощи 96-луночного фильтрационного планшета His MultiTrap HP (GE Healthcare), который был загружен никель-сефарозой. Планшет промывали дважды 400 мкл воды на лунку и фильтровали под вакуумом, после чего уравновешивали дважды 400 мкл на лунку связывающего буфера (25 мМ Tris-HCl, 200 мМ NaCl, 10 мМ имидазола, рН 8,0) и фильтровали под вакуумом. К сортазной реакционной смеси добавляли равный объем связывающего буфера. Смесь наносили на колонку и инкубировали в течение 5 минут. При применении вакуума на колонке конъюгаты фильтровались через колонку, в то время как неконъюгированные Fab-фрагменты антитела оставались связанными с колонкой через ее His-Tag-группировку. Поскольку используемая сортаза была изменена путем генной инженерии, благодаря чему содержала His-Tag-группировку, она также связывалась с колонкой, так что полученный фильтрат не содержал фермент.
На следующем этапе свободные олигонуклеотиды удаляли из образца, так как они помешали бы последующей реакции связывания. Есть два возможных решения этой задачи: либо ультрафильтрация, либо подход, основанный на аффинности. Для ультрафильтрации подходящими являются такие устройства, которые задерживают конъюгат, при этом позволяя свободным олигонуклеотидам проходить через мембрану, например, 96-луночный планшет Zeba 96-well Spin Desalting Plates, 40K MWCO (Thermo Scientific, кат. №89807) или AcroPrep 96, 30K (Pall, cat.no. 5035), или любые другие устройства, подходящие для ультрафильтрации. После нанесения образца и этапа фильтрации образцы трижды промывают связывающим буфером (PBS). Раствор переносят в новый планшет и доводят объем до 200 мкл. Отбирают аликвоту для количественной оценки. В качестве альтернативы ультрафильтрации может быть применен подход на основе аффинности, для которого необходима матрица, которая связывает конъюгат, при этом оставляя несвязанными свободные олигонуклеотиды. Примерами такой матрицы являются KappaSelect (GE Healthcare, кат. №17-5458-01), CaptoL (GE Healthcare, кат. №17-5478-99) или CaptureSelect IgG-CH1 Affinity Matrix (ВАС, кат. №191.3120.05). Матрица может быть доступна в виде колонок в 96-луночном фильтровальном планшете, или она может быть куплена в виде суспензии. В последнем случае она может быть аликвотирована в 96-луночный фильтровальный планшет, такой как MSGVN2250 (MultiScreen HTS Millipore, кат. № MSGVN2250), который служит установочным планшетом/планшетом-носителем. Аффинность матрицы может быть специфичной для легкой цепи (как в случае KappaSelect и CaptoL), или она может быть специфичной для тяжелой цепи (как в случае Capture Select IgG-CH1). Поскольку для каждой матрицы могут быть применены различные протоколы, в качестве примера следующий план основан на протоколе для KappaSelect. Планшеты, содержащие KappaSelect, промывают водой, затем три раза 400 мкл PBS с рН 4,0. После этого наносят образец (разведенный в PBS, pH 4,0) и оставляют для связывания с матрицей в течение 90 мин в условиях перемешивания. Выполняют три этапа промывания с помощью 400 мкл PBS, рН 4,0, а затем два этапа элюирования с помощью 200 мкл буфера для элюции (0,1 М глицин, 250 мМ NaCl, 5% ПЭГ, pH 2,5). После элюирования на каждые 200 мкл буфера добавляют 30 мкл нейтрализующего буфера (1 М Tris-HCl, pH 8,0). При необходимости выполняют этап ультрафильтрации, чтобы перевести образец в другой буфер, такой как PBS.
Для создания биспецифичных связывающих молекул два конъюгата «Fab-фрагмент антитела - олигонуклеотид» пипетируют вместе, включая линкер L-ДНК ((5`-G CAG AAG CAT TAA TAG ACT- T10-GAC TCT AAG GTA CTC GCA T -T10-TGG ACG ACG ATA GAA CT-3', SEQ ID № 76), в эквимолярных соотношениях. Линкерная ДНК гибридизуется с 5'-концом конъюгата «Fab-фрагмент первого антитела - олигонуклеотид», в то время как его 3'-конец комплементарен конъюгату «Fab-фрагмент второго антитела - олигонуклеотид», тем самым создавая физическую связь между двумя различными конъюгатами «Fab-фрагмент антитела - олигонуклеотид». Для правильной гибридизации раствор нагревают до 60°С и затем медленно охлаждают до комнатной температуры, а затем хранят при 4°С. Для быстрых протоколов также достаточна инкубация при комнатной температуре в течение нескольких минут.
Биспецифическую связывающую молекулу очищают с помощью препаративной эксклюзионной хроматографии. На колонке Superdex200 с двухкратным PBS в качестве рабочего буфера белковые фракции разделяют в соответствии с их размером. Типичная хроматограмма (аналитическая SEC) показана на фиг. 27. В реакции связывания формируются высокомолекулярные виды, которые явно отличаются от чистого Fab, конъюгата и промежуточного продукта, состоящего из конъюгата «Fab-фрагмент одного антитела -олигонуклеотид», связанного с линкером. Фракция, содержащая биспецифическую связывающую молекулу, может быть дополнительно проанализирована в клеточных анализах.
Препаративный подход
Если необходимы большие количества биспецифической связывающей молекулы или если существуют другие ограничения, то может быть применен так называемый препаративной подход. При этом реакционную смесь с сортазой наносят на колонку Superdex200 с однократным PBS в качестве рабочего буфера. Фракции собирают в объеме 0,4 мл. Затем аликвоты всех релевантных фракций анализируют с помощью системы LabChip (Perkin Elmer) для определения тех фракций, которые содержат конъюгат «Fab-фрагмент антитела - олигонуклеотид». Кроме того, аликвоты всех релевантных фракций загружают в агарозный гель, при этом к образцу перед его внесением в гель добавляют так называемый «олигонуклеотид-ловушку». Это олигонуклеотид-ловушка комплементарен олигонуклеотиду из конъюгата «Fab-фрагмент антитела - олигонуклеотид», в результате чего образуется дцДНК-группировка, которую можно легче визуализировать на агарозном геле, чем оцДНК конъюгата «Fab-фрагмент антитела - олигонуклеотид» в одиночку. Те фракции, которые содержат конъюгат «Fab-фрагмент антитела - олигонуклеотид» (как видно на системе LabChip) и которые не содержат свободный олигонуклеотид (как видно на агарозном геле), объединяют и, при необходимости, концентрируют путем ультрафильтрации, например, с помощью Amicon Ultra 0.5ML.10K (Millipore). Для реакции связывания две молекулы конъюгатов «Fab-фрагмент антитела - олигонуклеотид» пипетируют вместе, включая линкерную L-ДНК, в эквимолярных соотношениях и обрабатывают, как описано в предыдущем разделе.
Эффективность сортазной реакции и процесса очистки можно контролировать с помощью белковых гелей или с помощью системы LabChip (Perkin Elmer). Последняя предоставляет электросферограммы с определением размеров и концентраций. Пример такого цикла, в котором были проанализированы образцы на различных этапах технологического процесса, показан на фиг. 28. Обратите внимание, что пики ниже 10 кДа являются так называемыми системными пиками, которые свойственны устройству LabChip и не относятся к образцам. Fab-фрагмент антитела имеет размер 55-56 кДа в зависимости от буфера. Фиг. 28-1 показывает исходный материал (чистый Fab-фрагмент антитела), фиг. 28-2 показывает начало сортазной реакции (обратите внимание на появление сортазы с массой 27 кДа). В конце сортазной реакции (фиг. 28-3) на 64 кДа после пика Fab-фрагмента антитела появляется заметный пик, представляющий конъюгат «Fab-фрагмент антитела - олигонуклеотид» (скорость связывания/эффективность примерно 60%). После очистки путем отрицательной His-Tag-селекции и ультрафильтрации можно видеть почти чистый пик конъюгата «Fab-фрагмент антитела - олигонуклеотид» (фиг. 28-4).
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АНТИГЕНАМ, АКТИВИРУЮЩИМ Т-КЛЕТКИ, И ОПУХОЛЕВОМУ АНТИГЕНУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2605390C2 |
| СПОСОБ ПОЛУЧЕНИЯ И ОТБОРА МОЛЕКУЛ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2646159C2 |
| АНТИТЕЛА, НЕ СОДЕРЖАЩИЕ FC-ФРАГМЕНТА, ВКЛЮЧАЮЩИЕ ДВА FAB-ФРАГМЕНТА, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2617970C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| Вирус-кодируемые регулируемые агенты, связывающие T-клетки (CATVERT) или NK-клетки (CATVERN), нацеленные на рак | 2020 |
|
RU2827779C2 |
| КОНЪЮГАТЫ ПОЛИПЕПТИДНОГО ТОКСИНА И АНТИТЕЛА, СОЕДИНЕННЫХ КОВАЛЕНТНОЙ СВЯЗЬЮ | 2014 |
|
RU2682754C2 |
| ХИМЕРНЫЙ БЕЛОК, СОСТАВЛЕННЫЙ ИЗ ДОМЕНА АНТАГОНИСТА NGF И ДОМЕНА АНТАГОНИСТА TNFα | 2015 |
|
RU2678810C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2663123C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2774414C2 |










































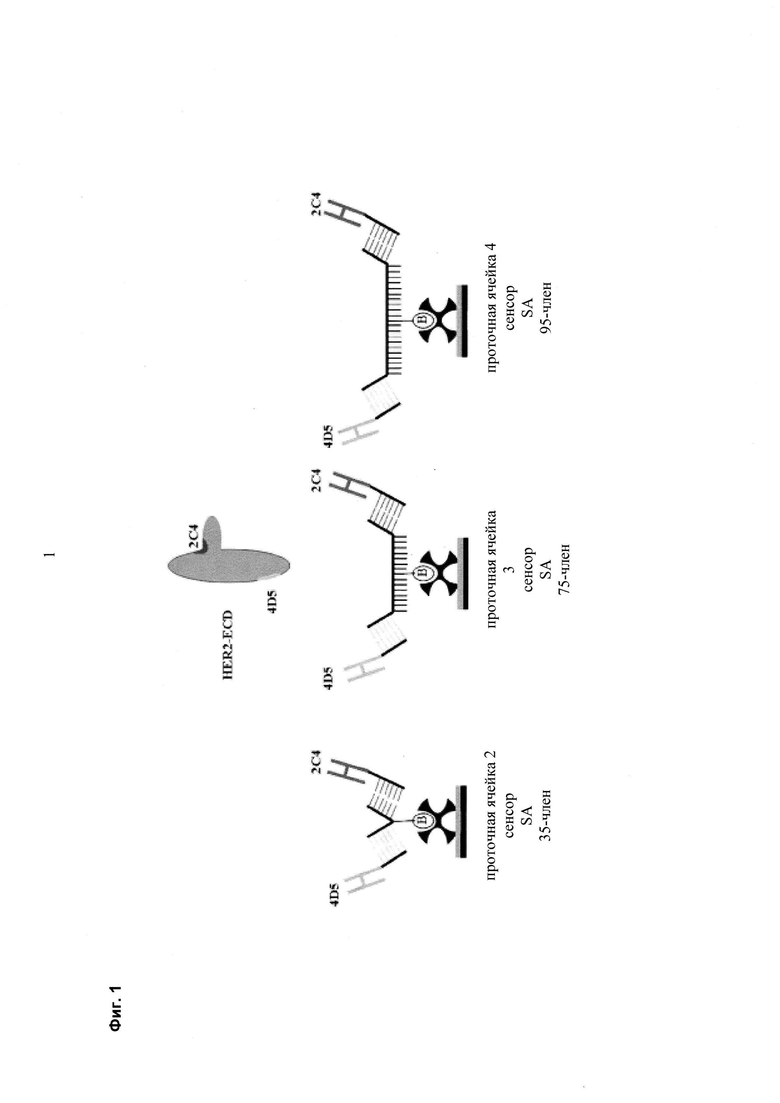
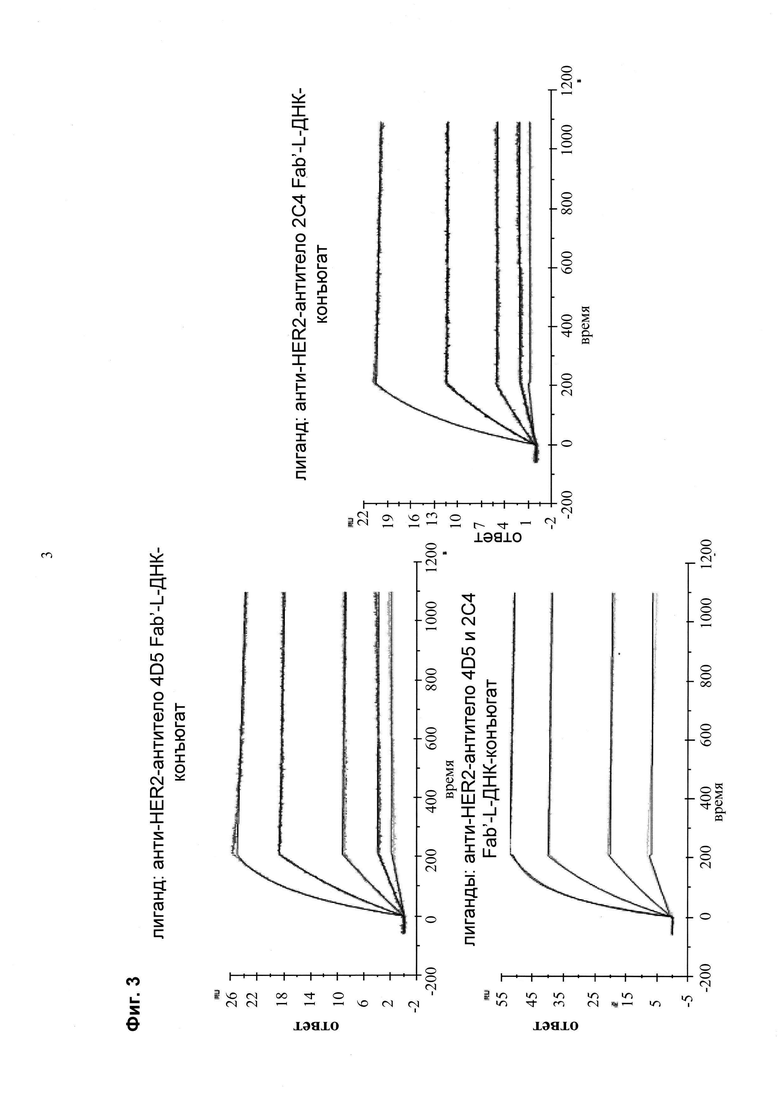
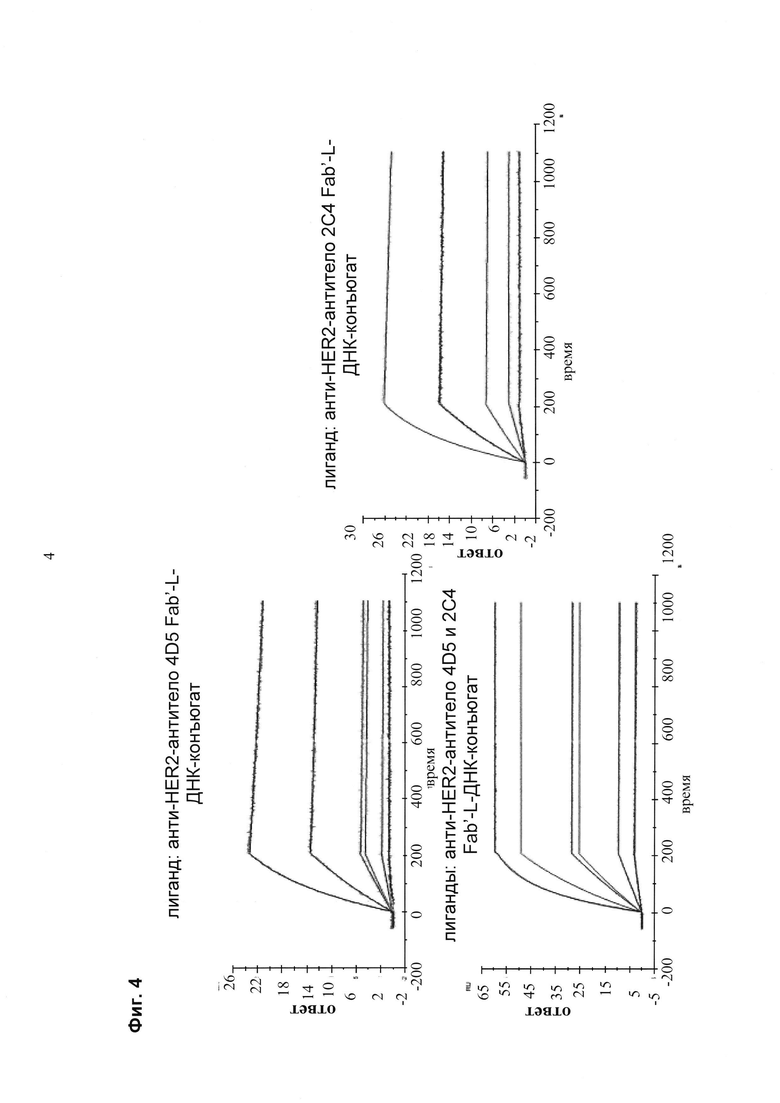
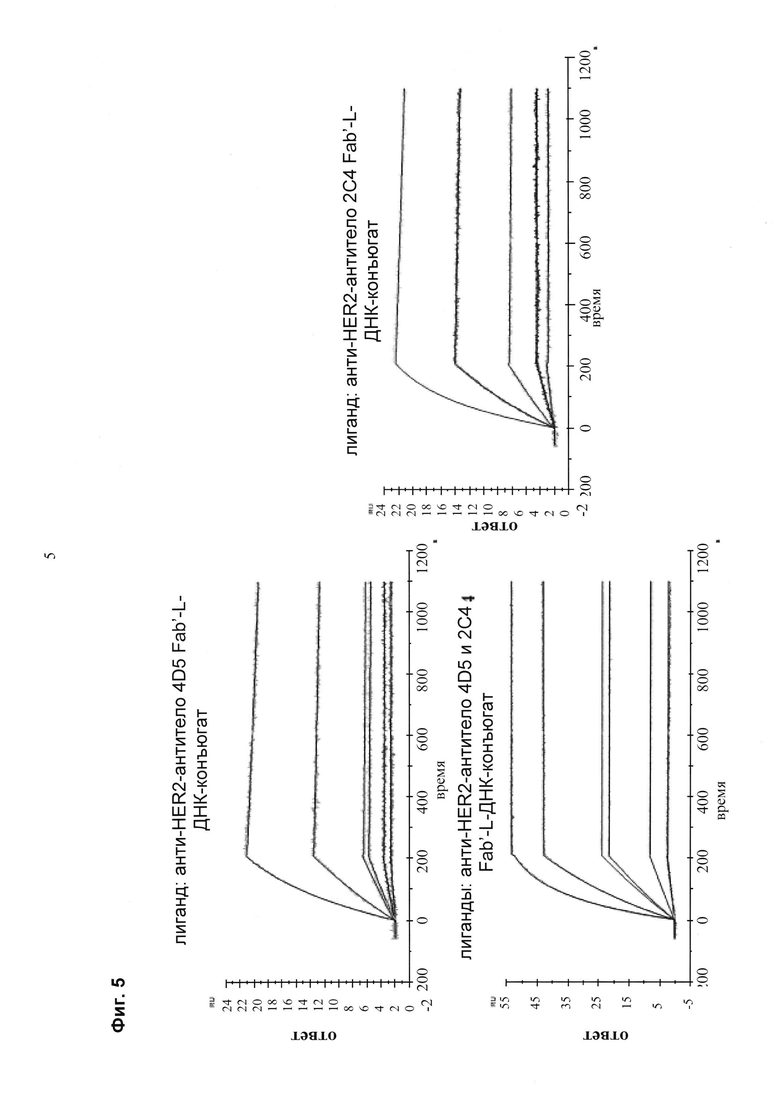
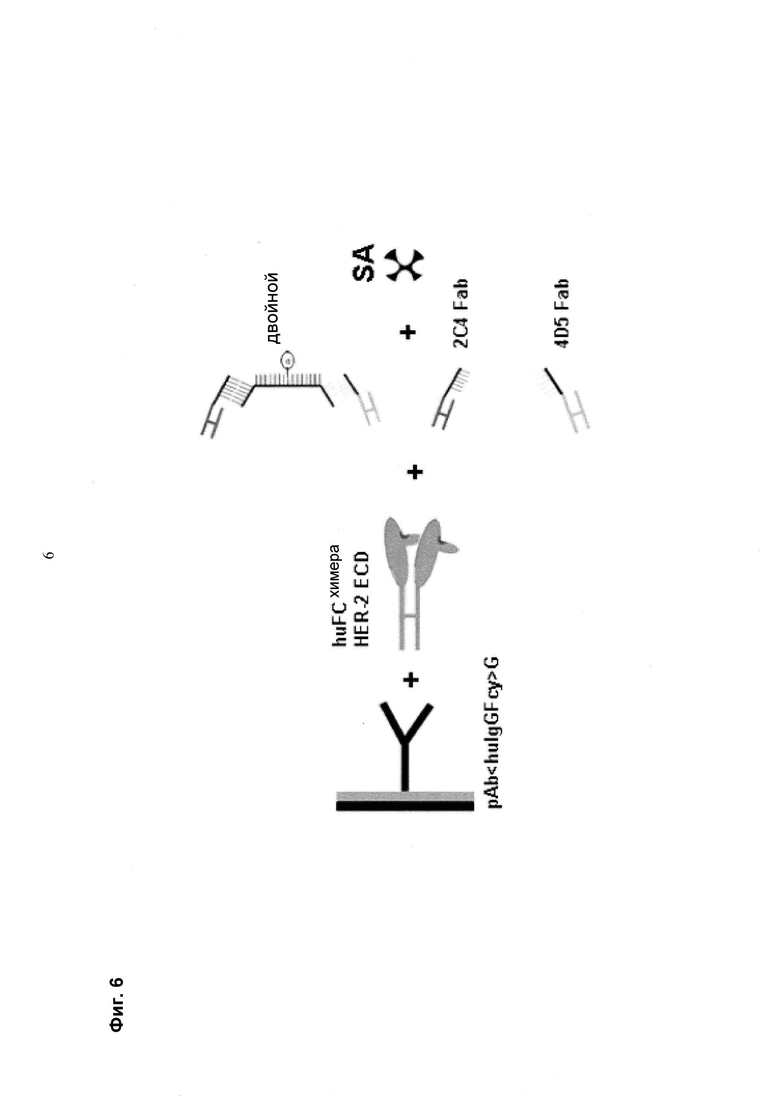
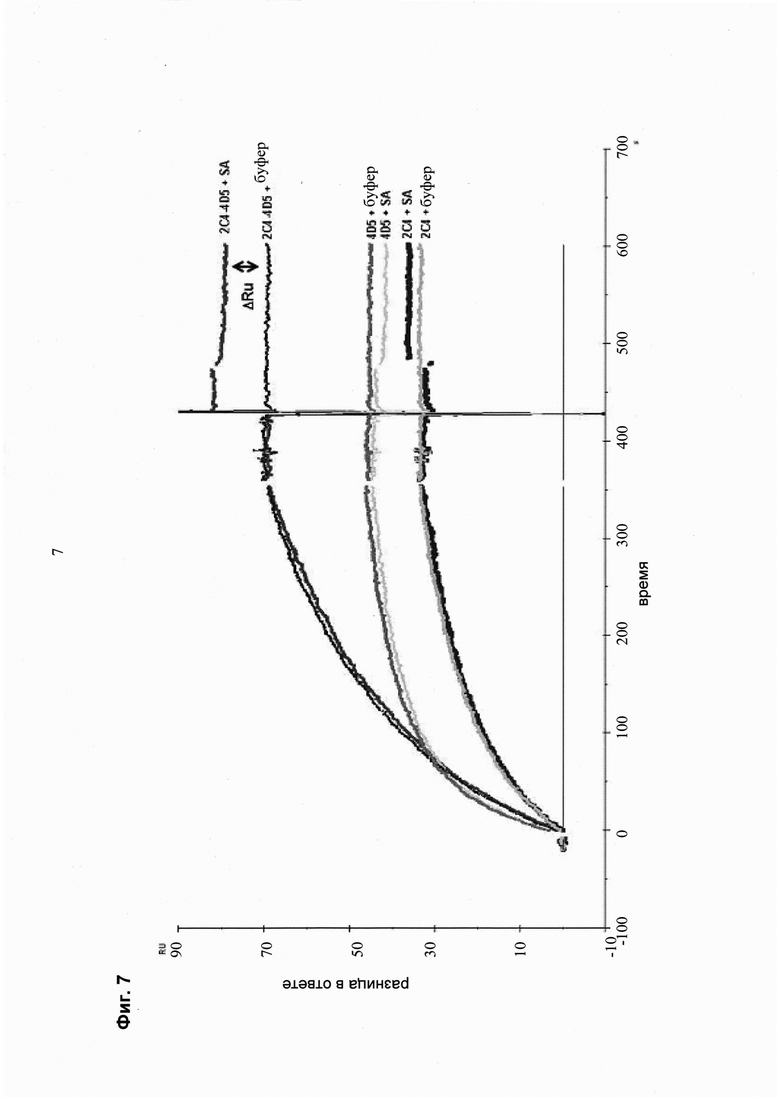
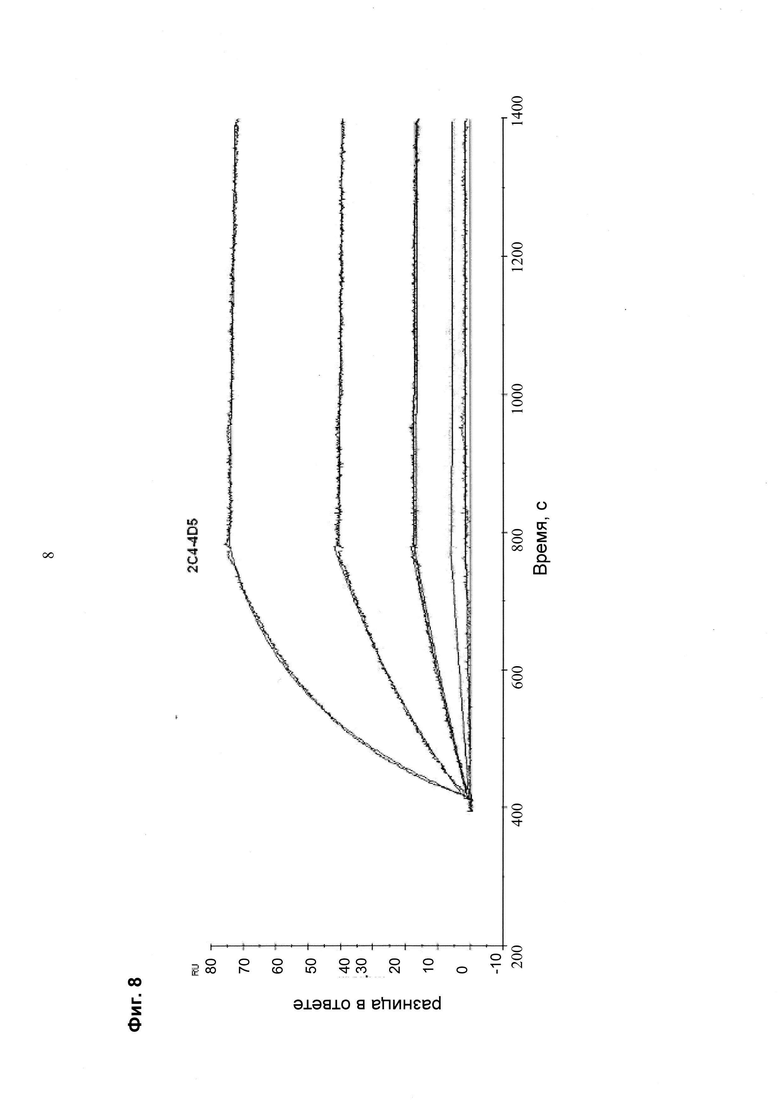
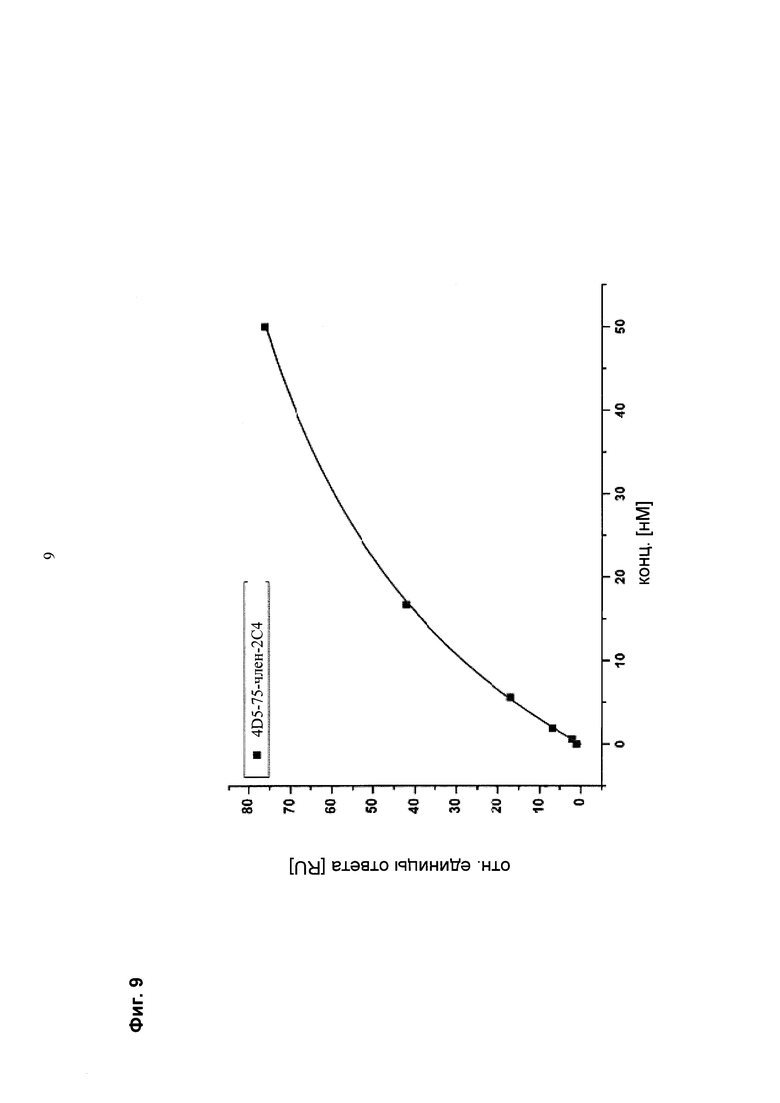
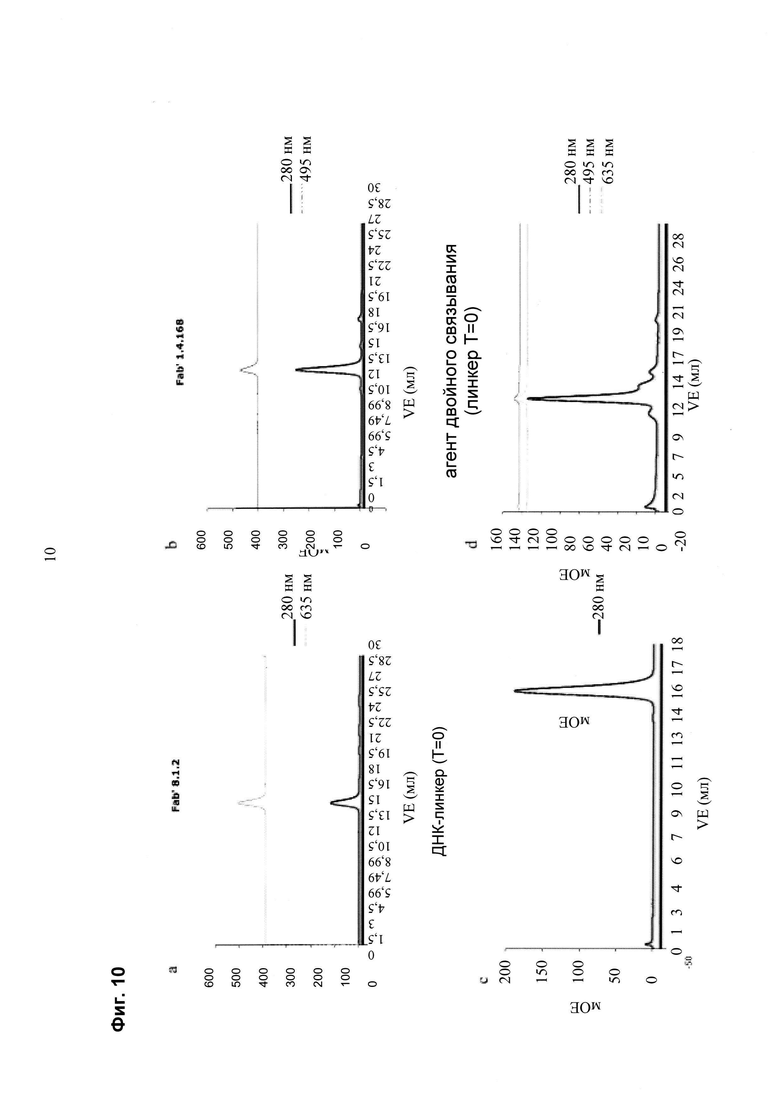
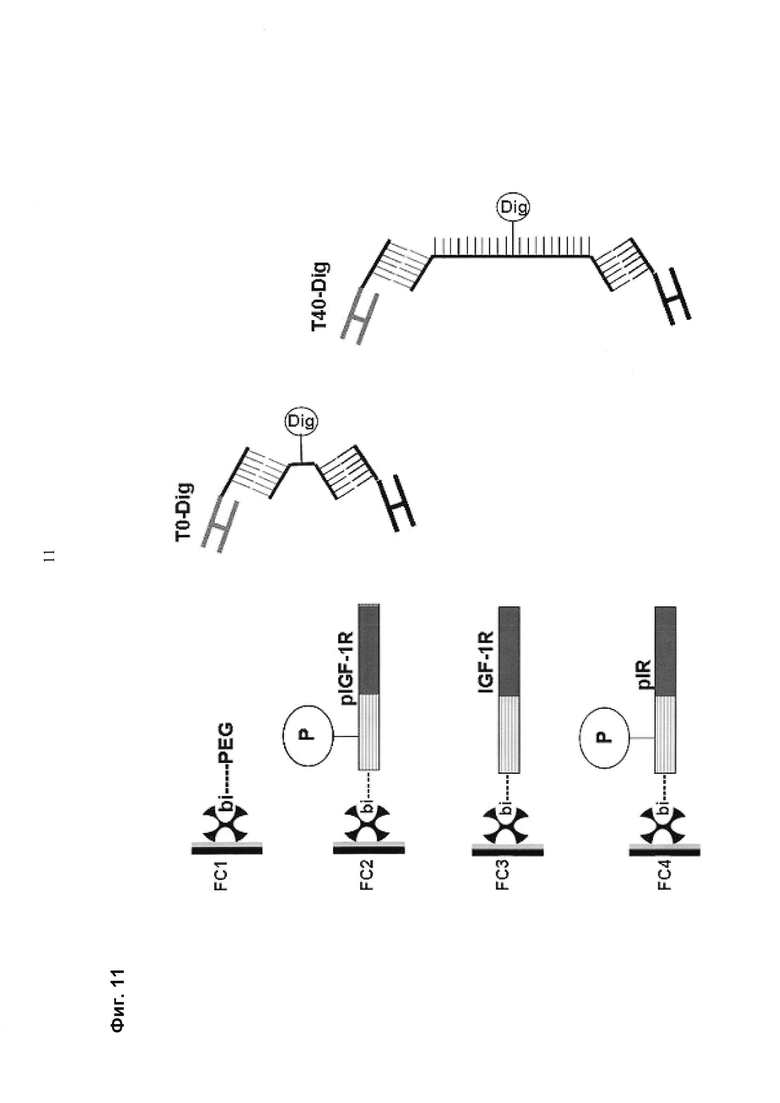
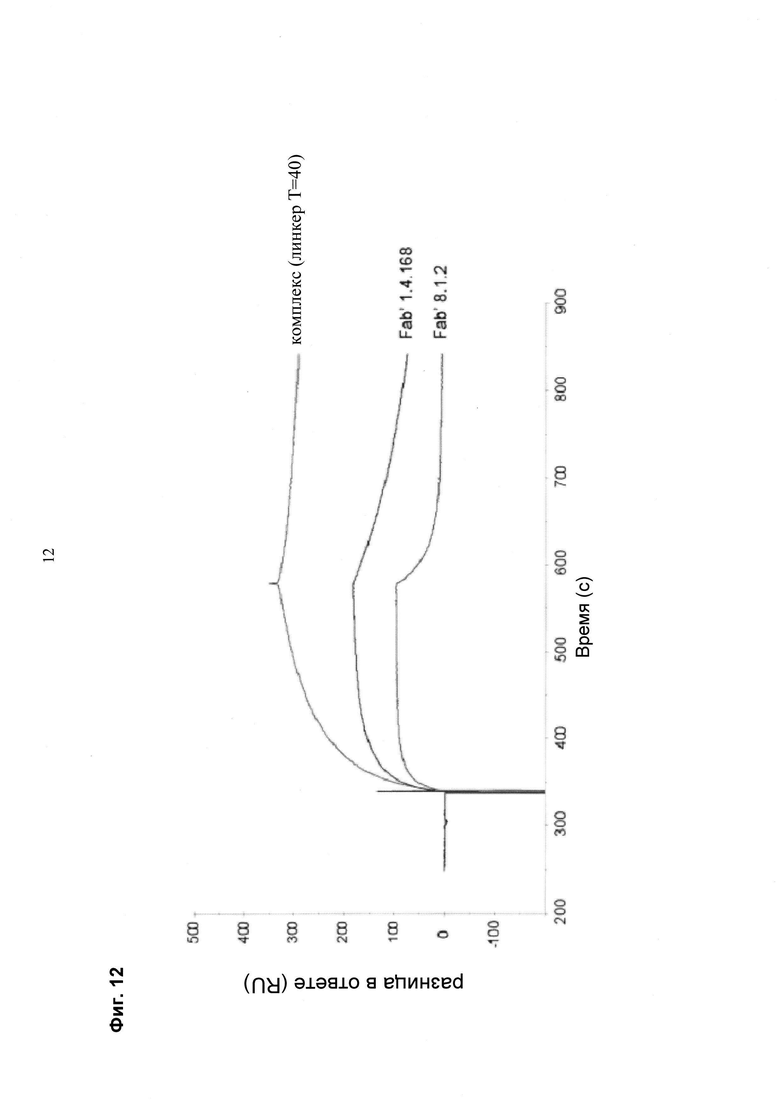
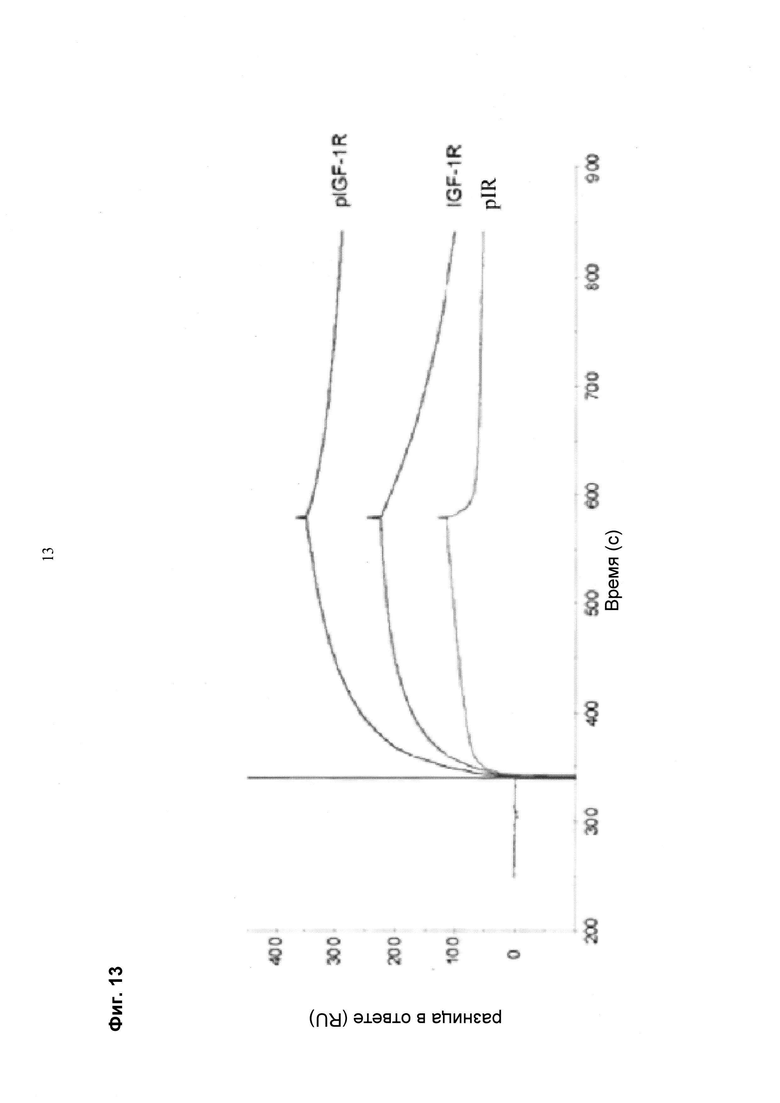
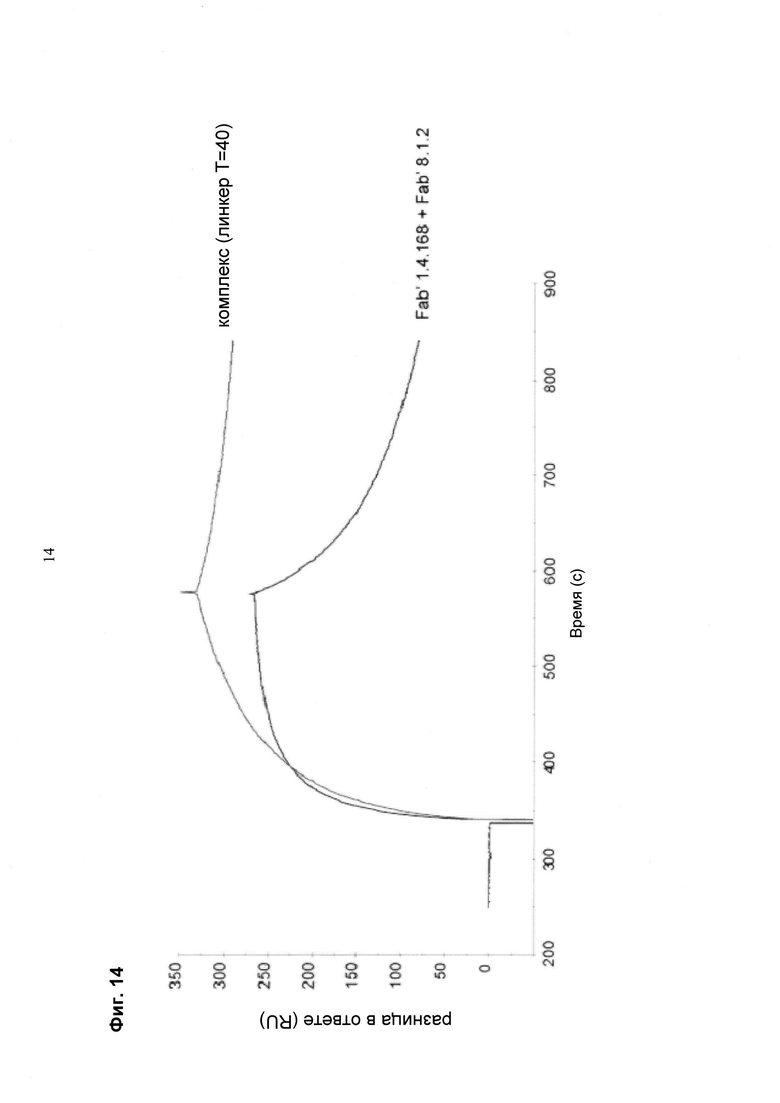
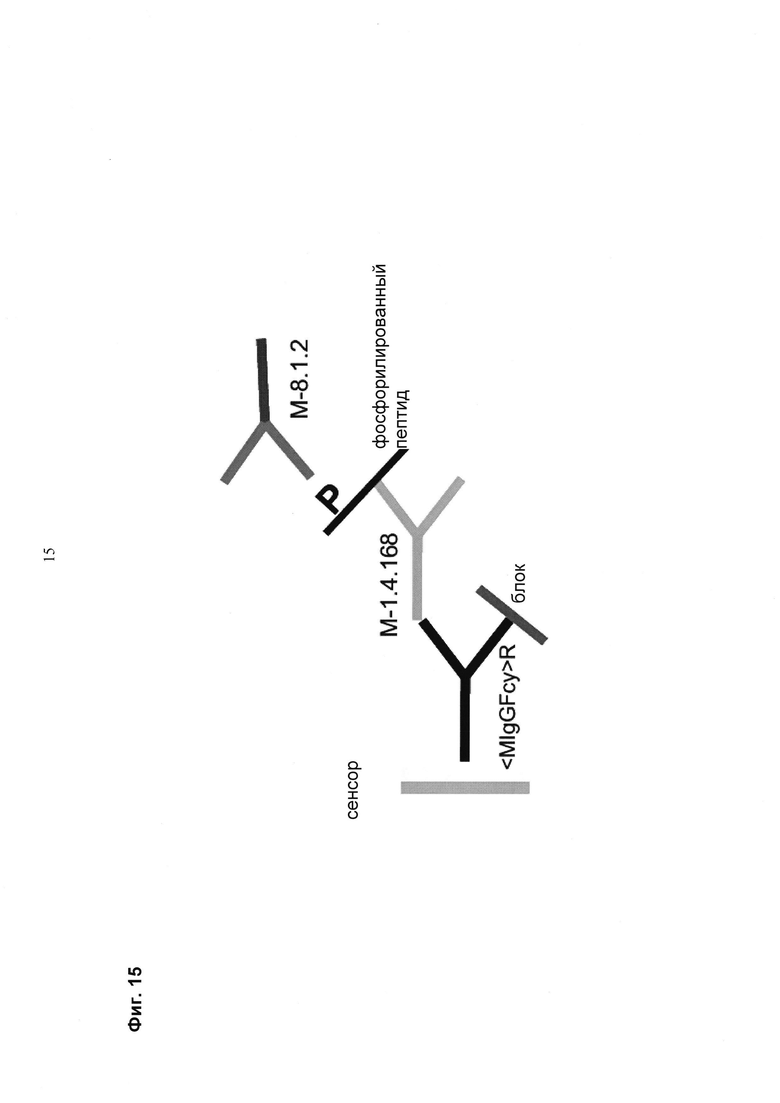
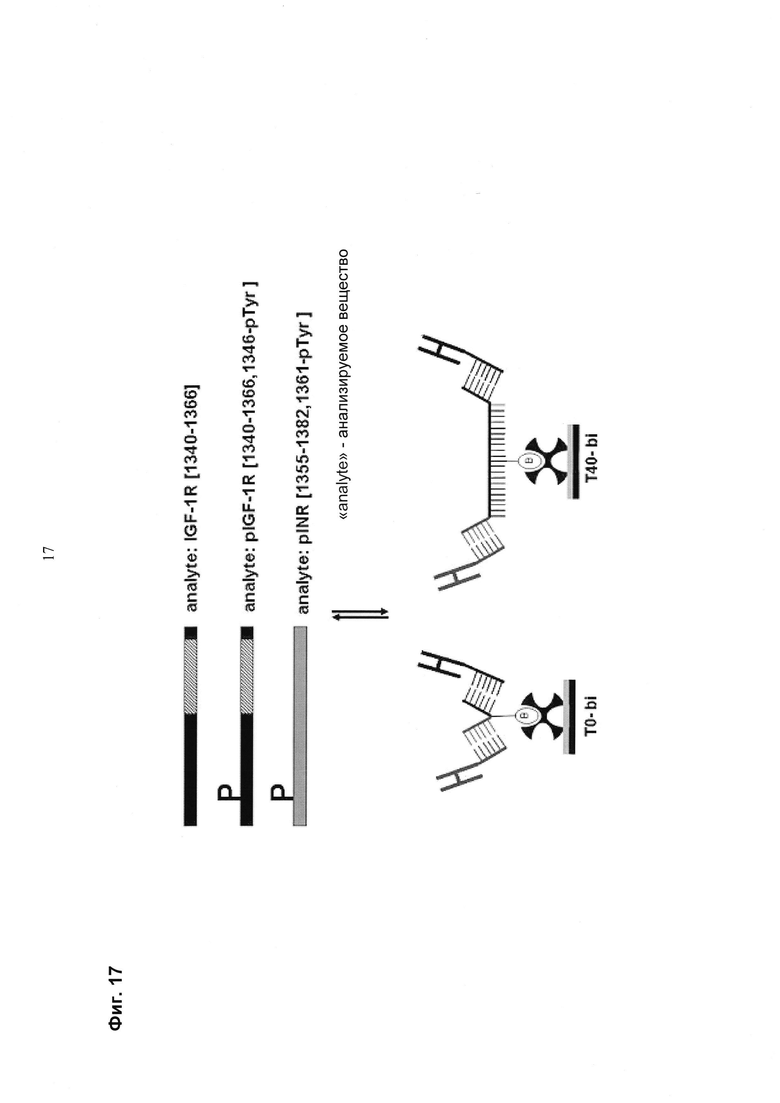
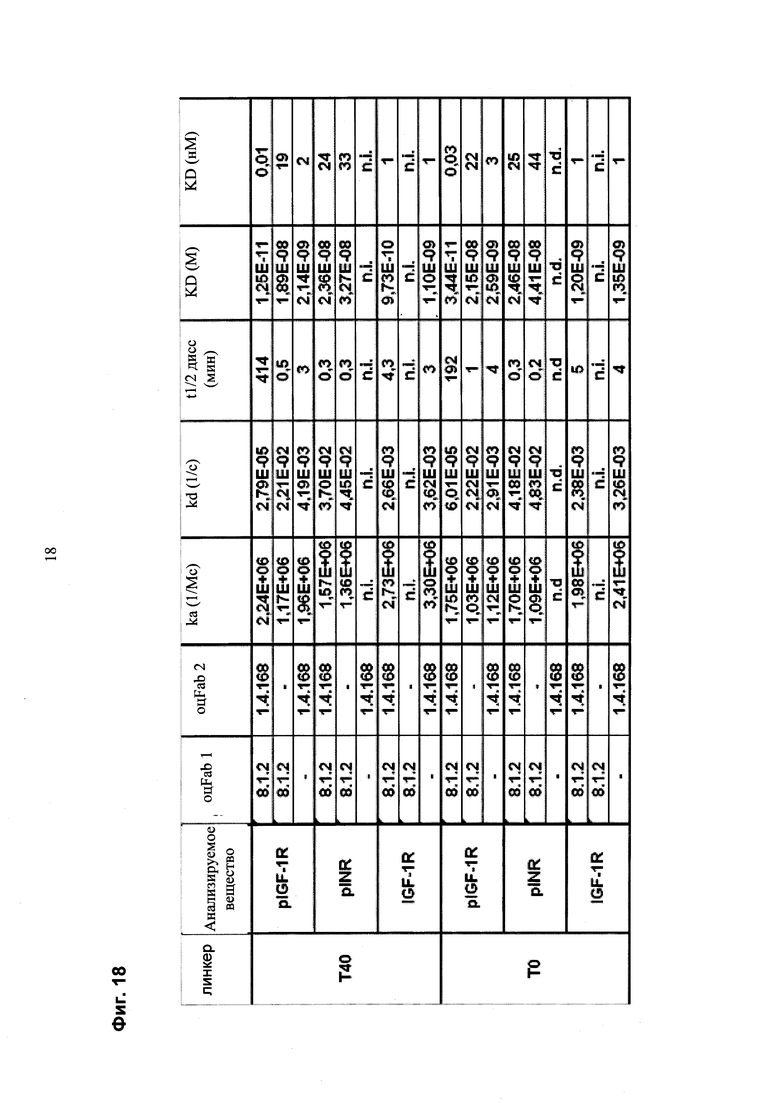
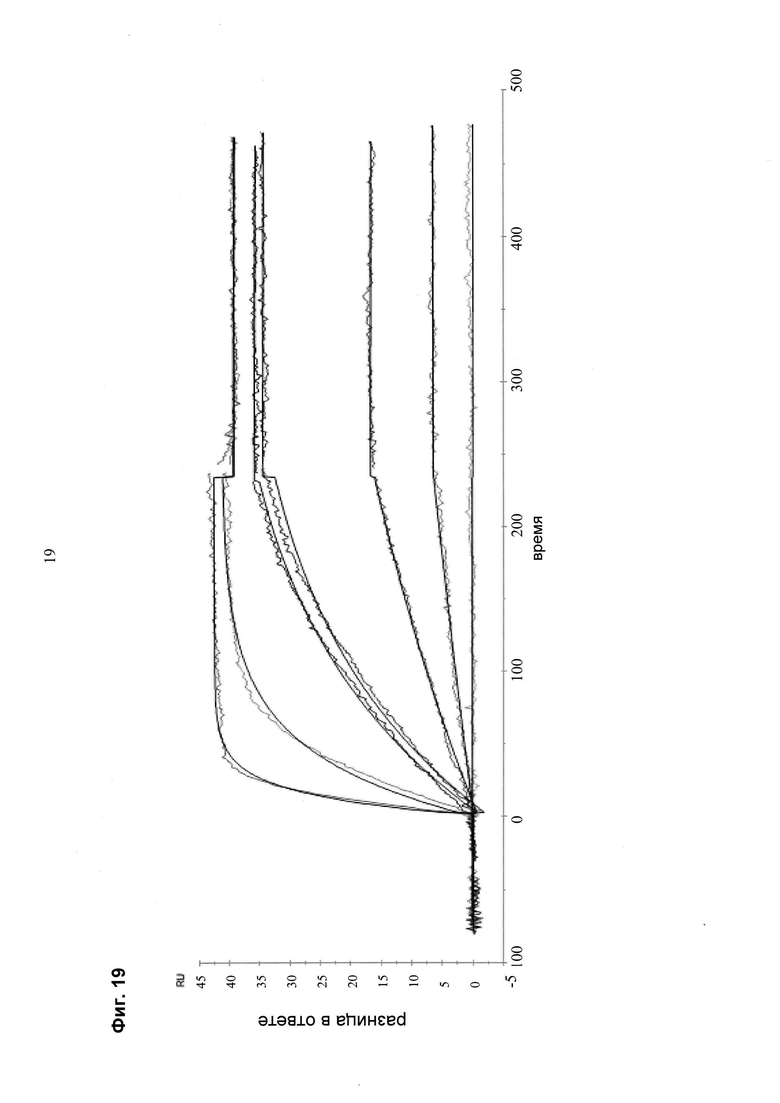
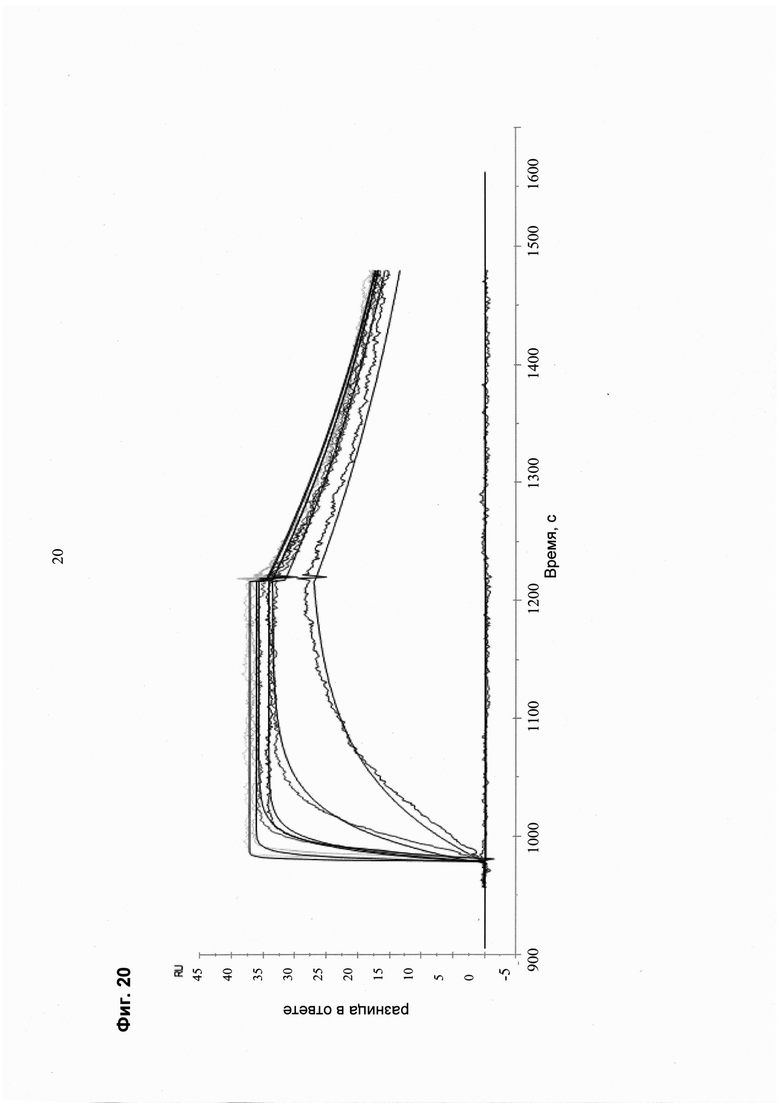
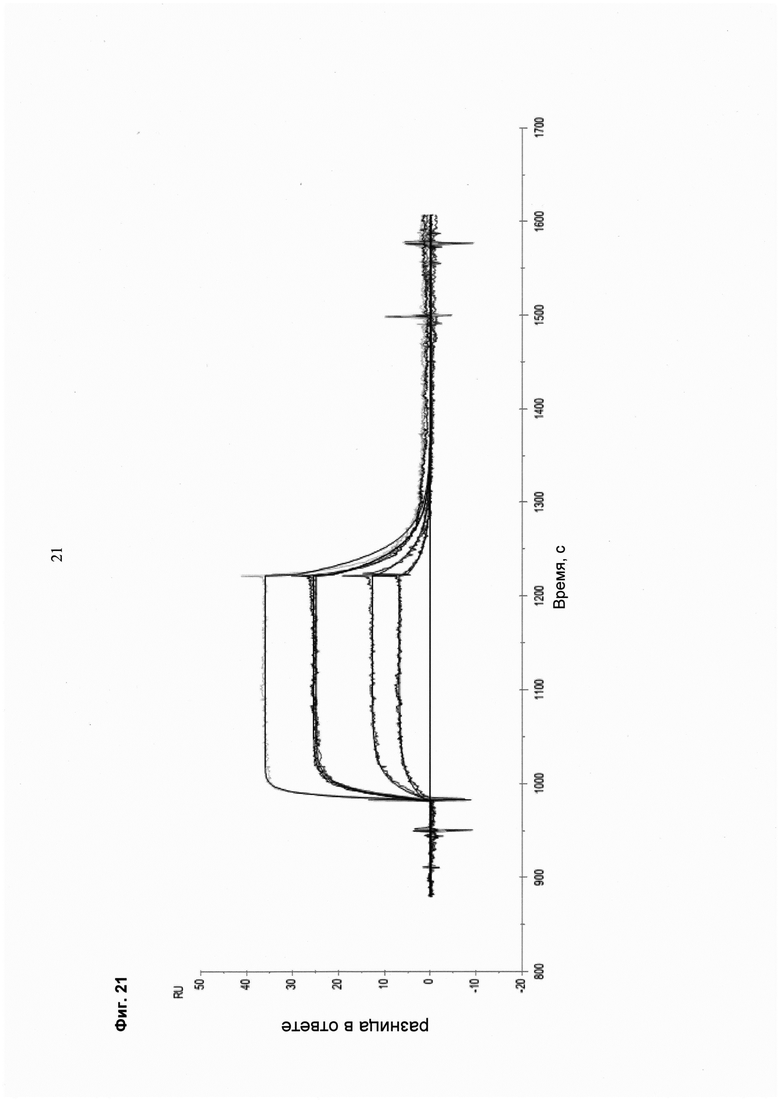
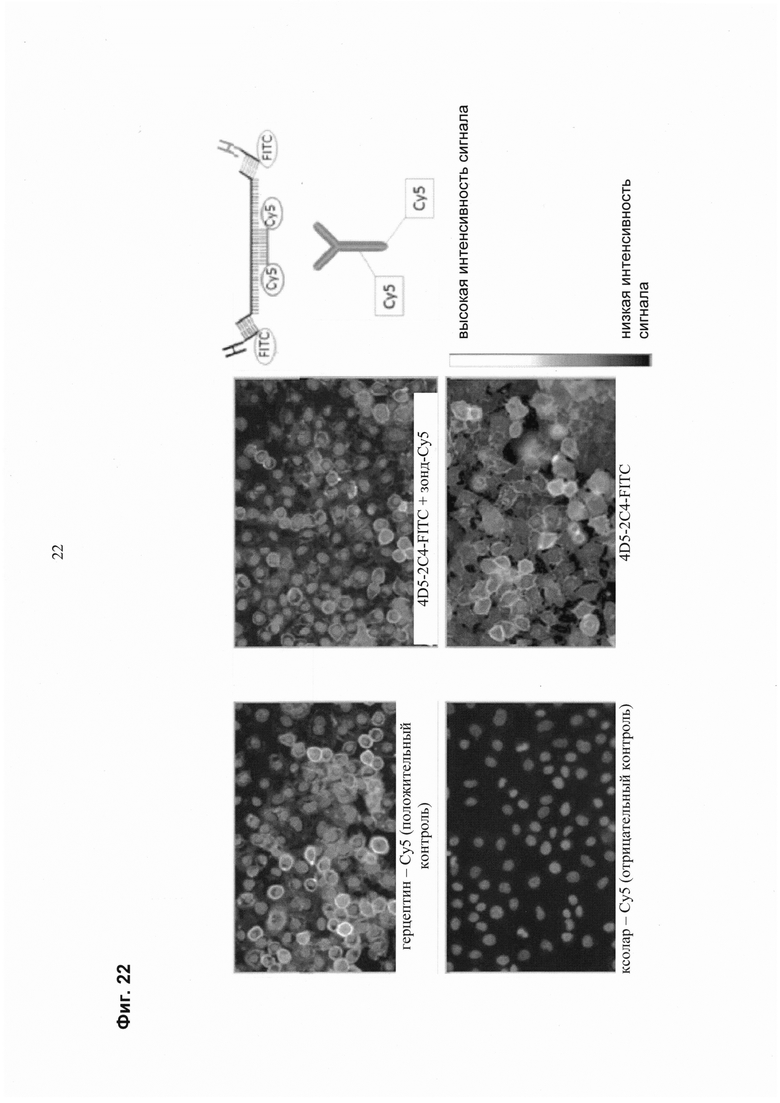
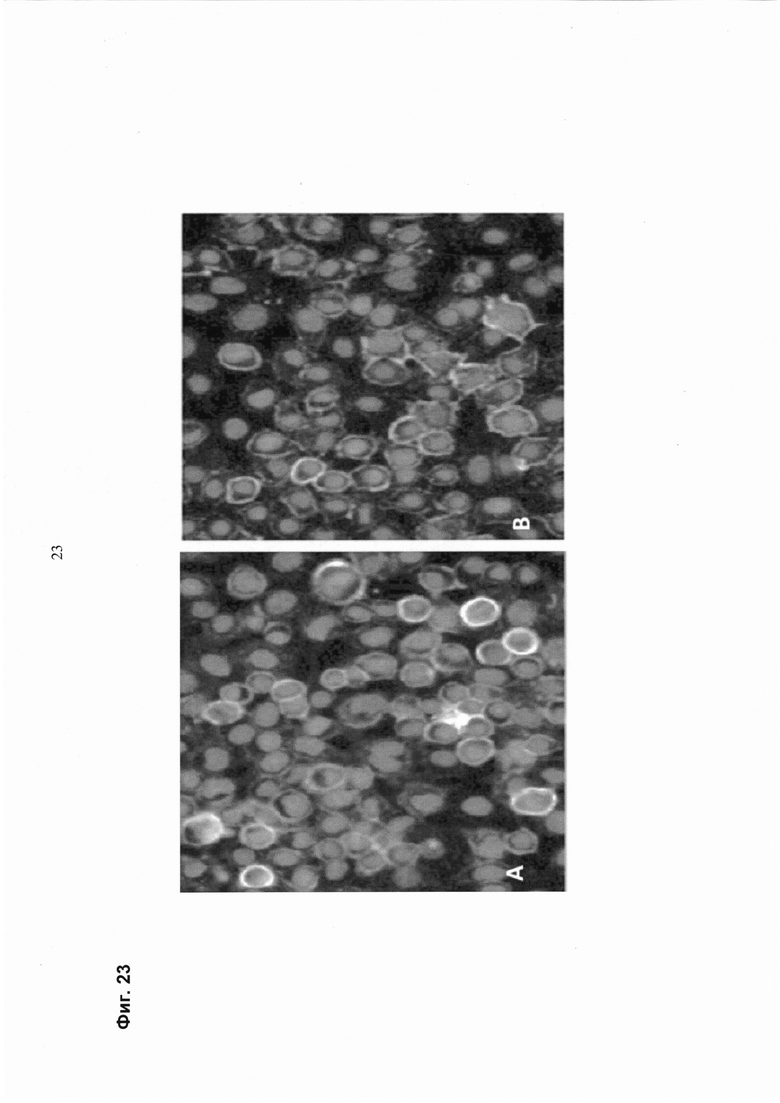

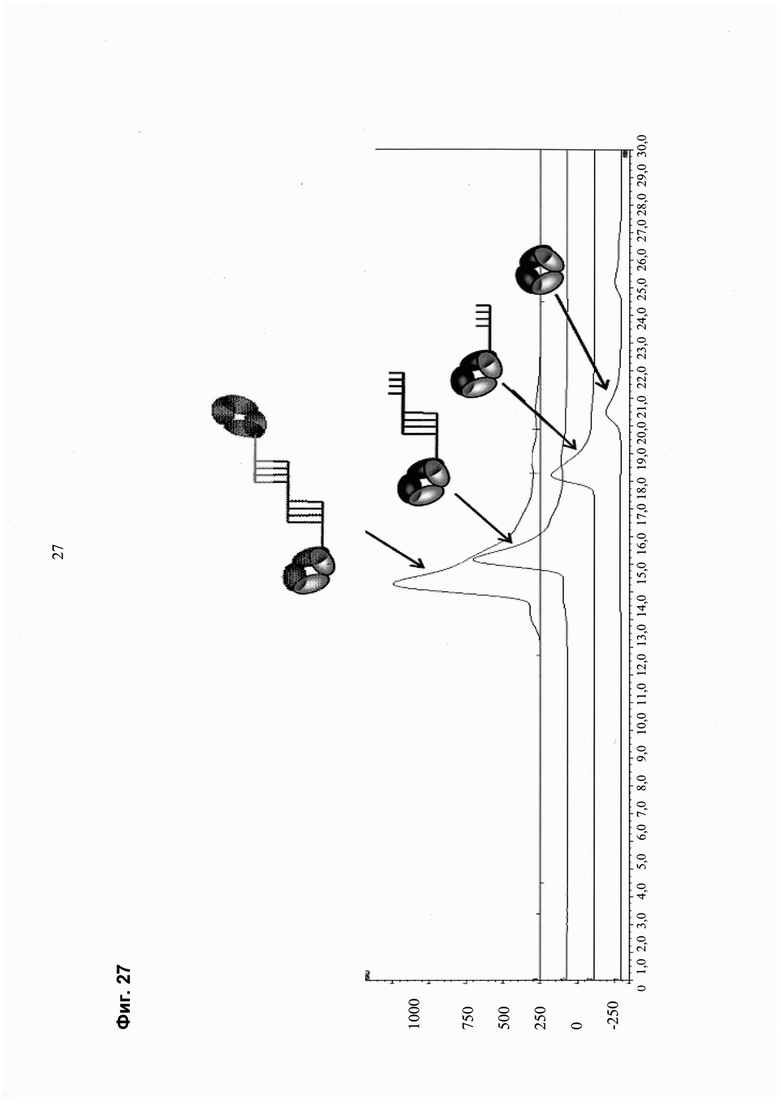
Изобретение относится к области биотехнологии, конкретно к способам определения комбинации антигенсвязывающих сайтов и получения биспецифического антитела, что может быть использовано в медицине. Заявленные способы включают следующий этап: определение специфичности связывания и/или аффинности множества биспецифических антител, полученных путем объединения каждого элемента первого множества Fab-фрагментов антитела или scFv-фрагментов антитела с каждым элементом второго множества Fab-фрагментов антитела или scFv-фрагментов антитела и с линкером, содержащим на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары. Изобретение позволяет эффективно получать мультиспецифические терапевтические молекулы, такие как биспецифические антитела, с заданными свойствами в отношении клеточных поверхностных молекул, присутствующих на клетке у пациента, нуждающегося в лечении, или в отношении лигандов, взаимодействующих с такой клеточной поверхностной молекулой. 2 н. и 9 з.п. ф-лы, 28 ил., 8 табл., 13 пр.
1. Способ получения биспецифического антитела, включающий следующие этапы:
(i) определение поверхностных маркеров, присутствующих на поверхности клетки в образце, и выбор первого поверхностного маркера и второго поверхностного маркера,
(ii) инкубация в течение 4-16 ч (а) Fab-фрагмента антитела или scFv-фрагмента антитела, конъюгированного с первым элементом первой связывающей пары, где этот Fab-фрагмент или scFv-фрагмент антитела специфически связывается с первым поверхностным маркером, (b) Fab-фрагмента антитела или scFv-фрагмента антитела, конъюгированного с первым элементом второй связывающей пары, где этот Fab-фрагмент или scFv-фрагмент антитела специфически связывается со вторым поверхностным маркером, и (с) энантиомерного полинуклеотидного ДНК-линкера, содержащего на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары,
и, таким образом, получение биспецифического антитела.
2. Способ определения комбинации антигенсвязывающих сайтов, включающий следующие этапы:
(i) определение специфичности связывания и/или аффинности множества биспецифических антител, полученных путем объединения каждого элемента первого множества Fab-фрагментов антител или scFv-фрагментов антител, каждый из которых связан с тем же первым элементом первой связывающей пары, с каждым элементом второго множества Fab-фрагментов антител или scFv-фрагментов антител, каждый из которых связан с тем же первым элементом второй связывающей пары, и энантиомерным полинуклеотидным ДНК-линкером, содержащим на одном его конце второй элемент первой связывающей пары и на соответствующем другом конце второй элемент второй связывающей пары,
где первое множество специфически связывается с первым маркером клеточной поверхности, а второе множество специфически связывается со вторым маркером клеточной поверхности, и
(ii) выбор биспецифического антитела с подходящей специфичностью связывания и/или аффинностью и тем самым определение комбинации антигенсвязывающих сайтов.
3. Способ по любому из пп. 1, 2, где биспецифическое антитело представляет собой комплекс, включающий
a) первый Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается с первым поверхностным маркером и
ii) который конъюгирован с первым элементом первой связывающей пары,
b) второй Fab-фрагмент или scFv-фрагмент антитела,
i) который специфически связывается со вторым поверхностным маркером и
ii) который конъюгирован с первым элементом второй связывающей пары, и
c) энантиомерный полинуклеотидный ДНК-линкер,
i) который конъюгирован со вторым элементом первой связывающей пары и
ii) который конъюгирован со вторым элементом второй связывающей пары.
4. Способ по п. 3, где комплекс является нековалентным комплексом.
5. Способ по п. 1 или 2, где связывающая пара является комплементарной парой полинуклеотидов.
6. Способ по п. 3, где комплекс также содержит эффекторную группировку, конъюгированную с полинуклеотидом, который комплементарен по меньшей мере части линкера.
7. Способ по п. 6, где комплекс содержит вторую эффекторную группировку, конъюгированную с полинуклеотидом, который комплементарен по меньшей мере части полинуклеотида, который конъюгирован с первой эффекторной группировкой, и не комплементарен полинуклеотидному линкеру.
8. Способ по п. 1 или 2, где первый и второй Fab-фрагмент или scFv-фрагмент антитела связываются с одной и той же мишенью и с неперекрывающимися эпитопами на ней.
9. Способ по п. 1 или 2, где полинуклеотидный линкер имеет последовательность, выбранную из SEQ ID № 05, SEQ ID № 06, SEQ ID № 07 и SEQ ID № 08.
10. Способ по п. 1 или 2, где энантиомерная ДНК представляет собой L-ДНК.
11. Способ по п. 10, где L-ДНК представляет собой одноцепочечную L-ДНК (оц-L-ДНК).
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| HAYASHI G | |||
| et al., Application of L-DNA as a molecular tag, Nucleic Acids Sym | |||
| Ser., 2005, v.49, p.261-262 | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ ДЛЯ МОДУЛИРОВАНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ | 2005 |
|
RU2433831C2 |

