Область изобретения
Настоящее изобретение относится к нуклеиново-кислотным конструкциям, к клеткам-хозяевам, содержащим такие конструкции, и к их применению в нуклеиново-кислотных вакцинах. Кроме того, изобретение относится к вакцинным препаратам, содержащим такие конструкции, и к применению таких препаратов в медицине. В частности, изобретение относится к ДНК вакцинам, которые полезны при профилактике и лечении ВИЧ-инфекций, более конкретно при введении путем опосредованной частицами доставки.
Предпосылки изобретения
ВИЧ-1 (HIV-1) является главной причиной синдрома приобретенного иммунодефицита (СПИД), который рассматривают как одну из основных проблем мирового здравоохранения. Хотя во всем мире были предприняты широкомасштабные исследования по получению вакцины, такие усилия, тем не менее, не увенчались успехом.
Описаны необолочечные белки ВИЧ-1, которые включают, например, белки внутренней структуры, такие как продукты генов gag и pol, и другие неструктурные белки, такие как Rev, Nef, Vif и Tat (Green et al., New England J. Med., 324, 5, 308 et seq (1991) и Bryant et al. (Ed. Pizzo), Pediatr. Infect. Dis. J., 11, 5, 390 et seq (1992)).
Ген Gag транслируется с полноразмерной РНК с образованием полипротеина-предшественника, который потом расщепляется на 3-5 капсидных белков; белок матрикса, капсидный белок и белок, связывающий нуклеиновые кислоты, а также протеазу (1. Fundamental Virology, Fields BN, Knipe D.M. and Howley M., 1996; 2. Fields Virology, vol. 2, 1996).
Ген gag кодирует белок-предшественник Gag 55 килодальтон (кД), также называемый р55, который экспрессируется с несплайсированной вирусной мРНК. В процессе трансляции N-конец р55 подвергается миристоилированию, которое запускает его связывание с цитоплазматической частью клеточных мембран. Связанный с мембраной полипротеин Gag рекрутирует две копии вирусной геномной ДНК параллельно с другими вирусными и клеточными белками, которые запускают почкование вирусной частицы с поверхности инфицированной клетки. После почкования р55 расщепляется кодируемой вирусом протеазой (продукт гена pol) в процессе созревания вируса на четыре белка меньшего размера, обозначенных МА (матрикс [р17]), СА (капсид [р24]), NC (нуклеокапсид [р9]) и р6 (4).
Кроме трех основных белков Gag, все предшественники Gag содержат несколько других районов, которые отщепляются и остаются в вирионе в виде пептидов различных размеров. Эти белки играют различные роли, например белок р2 предположительно играет роль в регуляции активности протеазы и вносит вклад в правильное определение времени протеолитического процессинга.
Полипептид МА образуется из N-терминального миристоилированного конца р55. Большая часть молекул МА остается присоединенной к внутренней поверхности липидного бислоя вириона, стабилизируя частицу. Подгруппа МА рекрутируется внутри более глубоких слоев вириона, где она становится частью комплекса, который доставляет вирусную ДНК в ядро (5). Эти молекулы МА способствуют ядерному транспорту вирусного генома, поскольку кариофильный сигнал на МА распознается клеточным механизмом ядерного импорта. Это явление дает возможность ВИЧ инфицировать неделящиеся клетки, что является необычным свойством для ретровируса.
Белок р24 (СА) образует коническую сердцевину вирусных частиц. Продемонстрировано, что циклофилин А взаимодействует с районом р24 белка р55, приводя к его включению в частицы ВИЧ. Взаимодействие между Gag и циклофилином А существенно, поскольку прерывание этого взаимодействия циклоспорином А ингибирует вирусную репликацию.
Район NC Gag ответственен за специфичность распознавания так называемого сигнала упаковки ВИЧ. Сигнал упаковки состоит из четырех шпилечных структур, локализованных вблизи 5' конца вирусной РНК, и достаточен в качестве посредника включения гетерологичной РНК в вирионы ВИЧ-1. NC связывается с сигналом упаковки с помощью взаимодействий, опосредованных двумя мотивами «цинковые пальцы». NC также способствует обратной транскрипции.
Район полипептида р6 является посредником взаимодействий между р55 Gag и вспомогательным белком Vpr, приводящих к включению Vpr в собирающиеся вирионы. Район р6 также содержит так называемый поздний домен, который необходим для эффективного высвобождения почкующихся вирионов из инфицированной клетки.
Ген pol кодирует два белка, содержащих две активности, необходимые вирусу при ранней инфекции, белок RT (обратную транскриптазу) и белок интегразу, необходимый для интеграции вирусной ДНК в клеточную ДНК. Первичный продукт Pol расщепляется протеазой вириона с образованием амино-концевого пептида RT, который содержит активности, необходимые для синтеза ДНК (РНК и ДНК-зависимую ДНК-полимеразу, рибонуклеазу Н) и карбокси-концевого белка интегразы. RT ВИЧ представляет собой гетеродимер полноразмерной RT (р66) и продукта расщепления (р51), в котором отсутствует карбокси-концевой домен РНКазы и интегразы.
RT является одним из наиболее высококонсервативных белков, кодируемых ретровирусным геномом. Двумя основными активностями RT являются ДНК-полимераза и рибонуклеаза Н. Активность ДНК-полимеразы RT использует РНК и ДНК в качестве матриц взаимозаменяемо и, подобно всем известным ДНК-полимеразам, не способна инициировать синтез ДНК de novo, но ей необходима ранее существующая молекула, которая служит в качестве праймера (РНК).
Активность РНКазы Н, свойственная всем белкам RT, играет существенную роль удаления РНК генома по мере продолжения синтеза ДНК при ранней репликации. Она избирательно разрушает РНК из всех гибридных молекул РНК-ДНК. Структурно полимераза и РНКаза Н занимают отдельные неперекрывающиеся домены в пределах Pol, покрывая две трети аминокислотной последовательности Pol.
Каталитическая субъединица р66 упакована в 5 различных поддоменов. Ее амино-конец 23 обладает частью активности RT. На ее карбокси-конце находится домен РНКазы Н.
После инфекции клетки-хозяина ретровирусный РНК геном копируется обратной транскриптазой, которая присутствует в инфекционной частице, в линейную двунитевую (ds, double strand) ДНК. Интеграза (обзор приведен в Skalka AM'99 Adv in Virus Res, 52: 271-273) распознает концы вирусной ДНК, укорачивает их и сопровождает вирусную ДНК к сайту хромосомы хозяина, катализируя интеграцию. Мишенями для интеграции могут быть многие сайты ДНК хозяина. Хотя интеграза достаточна для катализа интеграции in vitro, она является не единственным белком, связанным с комплексом вирусная ДНК in vivo - большой белок - вирусная ДНК, выделенным из инфицированных клеток, был назван комплексом преинтеграции. Этот комплекс способствует приобретению генов клетки-хозяина вирусными геномами-потомками.
Интеграза состоит из 3 отдельных доменов, N-концевого домена, каталитической сердцевины и С-концевого домена. Каталитический сердцевинный домен содержит все необходимое для химии полинуклеотидильного переноса.
Известно, что белок Nef вызывает удаление CD4, рецептора ВИЧ, с клеточной поверхности, но биологическая значимость этой функции является предметом споров. Кроме того, Nef взаимодействует с биохимическим путем передачи сигнала Т-клеток и индуцирует активное состояние, которое, в свою очередь, может способствовать более эффективной генной экспрессии. Некоторые изоляты ВИЧ имеют мутации в этом районе, которые являются причиной отсутствия кодирования ими функционального белка, и у них несколько ослаблена репликация и патогенез in vivo.
ДНК вакцины обычно состоят из бактериального плазмидного вектора, в который встроены сильный промотор, целевой ген, который кодирует антигенный пептид, и последовательности полиаденилирования/терминации транскрипции. Целевой ген может кодировать полноразмерный белок или просто антигенную пептидную последовательность, родственную патогену, опухоли или другому агенту, против которого необходима защита. Эту плазмиду можно выращивать в бактериях, таких как, например, Е.coli, а затем выделять и получать ее препарат в подходящей среде в зависимости от предназначенного пути введения, после чего ее вводят хозяину. После введения эта плазмида захватывается клетками хозяина, где продуцируется кодируемый белок. Плазмидный вектор предпочтительно следует конструировать без точки начала репликации, которая является функциональной в эукариотических клетках, чтобы предотвратить репликацию плазмиды в млекопитающем-хозяине для ее участия в интеграции в хромосомную ДНК.
Существует ряд преимуществ ДНК вакцинации перед традиционными методиками вакцинации. Во-первых, предсказывают, что в связи с тем, что белки, которые кодируются последовательностью ДНК, синтезируются в хозяине, структура или конформация этого белка будет подобна нативному белку, связанному с болезненным состоянием. Вероятно также, что ДНК вакцинация будет предоставлять защиту против различных штаммов вируса посредством генерации ответа цитотоксических Т-лимфоцитов, которые распознают эпитопы из консервативных белков. Кроме того, поскольку плазмиды захватываются клетками хозяина, где может продуцироваться антигенный белок, можно вызвать продолжительный иммунный ответ. Эта методика также предоставляет возможность комбинирования различных иммуногенов в одном препарате, что способствует одновременной иммунизации в отношении ряда болезненных состояний.
Полезная информация по предпосылкам изобретения в отношении ДНК вакцинации представлена в Donnelly et al. "DNA vaccines". Ann. Rev. Immunol. 1997, 15: 617-648, описание которой включено здесь в полном объеме путем ссылки.
Краткое изложение сущности изобретения
В настоящем изобретении предложены новые конструкции для применения в нуклеиново-кислотных вакцинах для профилактики и лечения ВИЧ-инфекций и СПИДа.
Соответственно, в первом аспекте предложена молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую белок gag ВИЧ или его фрагмент, сцепленную с нуклеотидной последовательностью, кодирующей дополнительный антиген ВИЧ или его фрагмент, и оперативно сцепленную с гетерологичным промотором. Фрагмент указанной нуклеотидной последовательности будет кодировать эпитоп ВИЧ и типично кодирует пептид из по меньшей мере 8 аминокислот. Эта нуклеотидная последовательность предпочтительно представляет собой последовательность ДНК и предпочтительно содержится внутри плазмиды без точки начала репликации. Препарат таких молекул нуклеиновой кислоты готовят с фармацевтически приемлемыми эксципиентом, носителями, разбавителями или адъювантами для получения фармацевтической композиции, подходящей для лечения и/или профилактики ВИЧ-инфекции и СПИД.
В предпочтительном воплощении препарат последовательности ДНК готовят на поверхности инертных частиц или гранул, подходящих для опосредованной частицами доставки лекарства. Предпочтительно эти гранулы являются золотыми.
В предпочтительном воплощении изобретения предложена последовательность ДНК, которая с высокой экспрессией кодирует белок gag, причем эта последовательность оптимизирована таким образом, что использование кодонов подобно таковому для генов в клетках млекопитающих. В частности, белок gag оптимизирован таким образом, что оно подобно таковому для генов человека с высокой экспрессией.
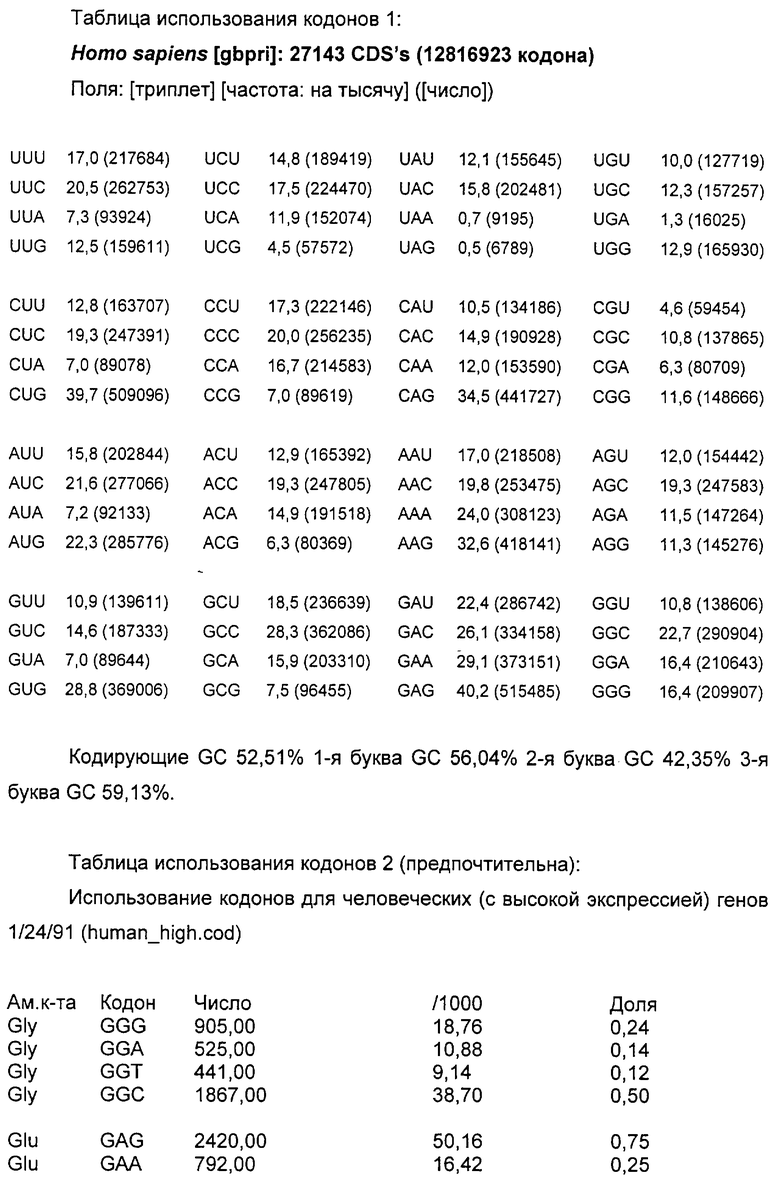
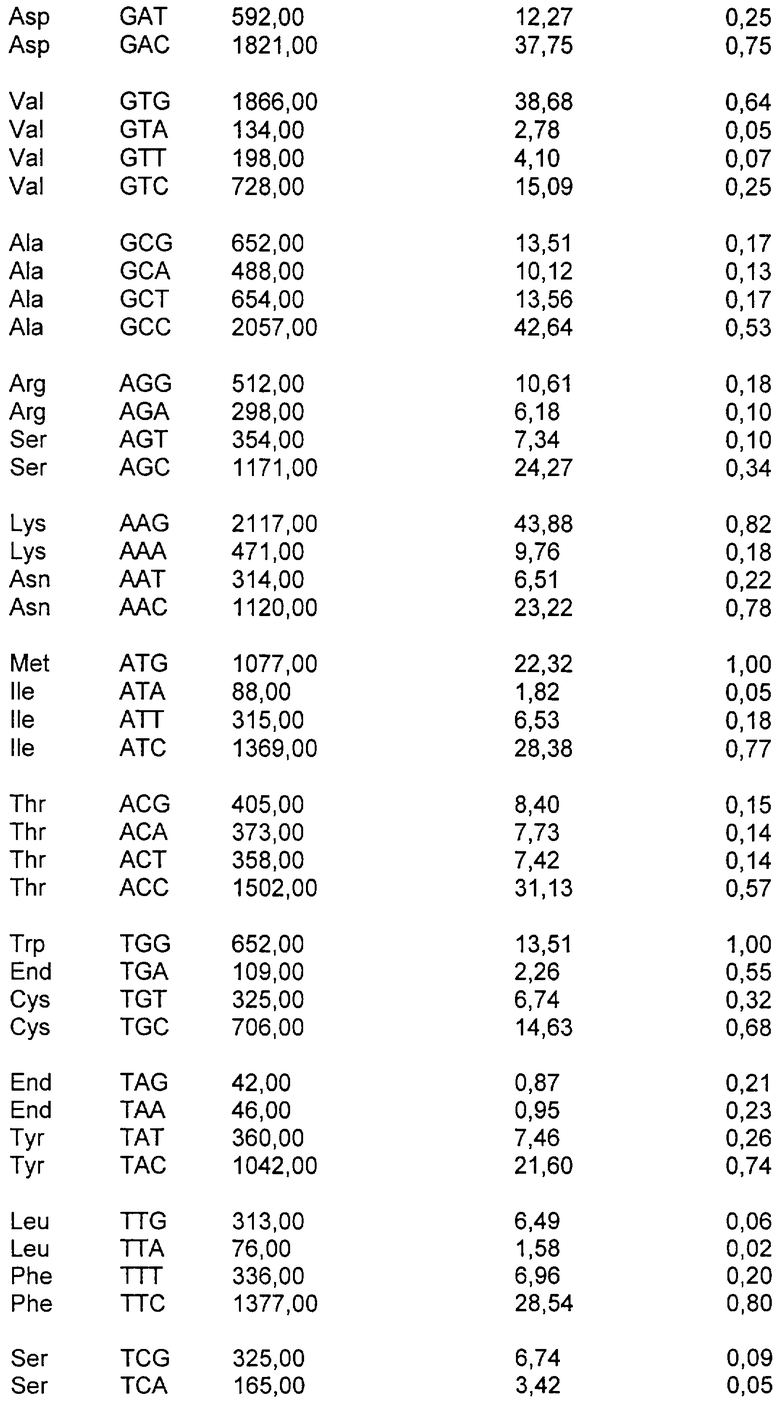
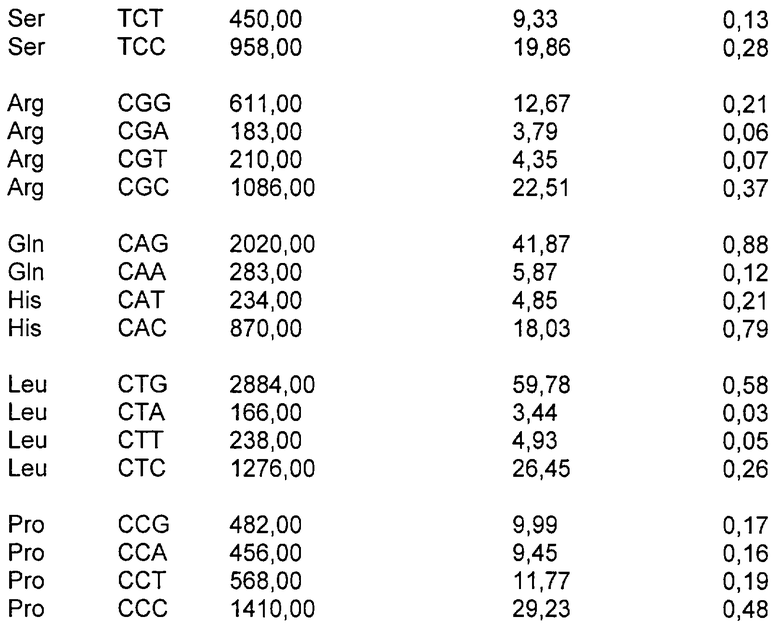
Код ДНК имеет 4 буквы (А, Т, С и G) и использует их для записи трехбуквенных «кодонов», которые представляют аминокислоты белков, кодируемых в генах организма. Линейная последовательность кодонов на протяжении молекулы ДНК транслируется в линейную последовательность аминокислот в белке (белках), кодируемом этими генами. Код является высоковырожденным, поскольку 61 кодон кодирует 20 природных аминокислот и 3 кодона представляют собой «стоп»-сигналы. Таким образом, большинство аминокислот кодируется более чем одним кодоном - в действительности, некоторые из них кодируются четырьмя различными кодонами или большим их числом.
Если более чем один кодон доступен для кодирования данной аминокислоты, наблюдали, что паттерны использования кодонов организмов в высокой степени неслучайны. Различные виды проявляют различные тенденции в отборе кодонов, и, кроме того, использование кодонов может быть в значительной степени различным у одного вида между генами, которые экспрессируются на высоких и низких уровнях. Эта тенденция различается у вирусов, растений, бактерий и клеток млекопитающих, и некоторые виды проявляют более сильную тенденцию отличия от случайного выбора кодонов, чем другие. Например, люди и другие млекопитающие имеют менее сильную тенденцию, чем некоторые бактерии или вирусы. По этим причинам существует значительная вероятность того, что ген млекопитающего, экспрессирующийся в Е.coli, либо чужеродный или рекомбинантный ген, экспрессирующийся в клетках млекопитающих, будет иметь несоответствующее распределение кодонов для эффективной экспрессии. Считают, что присутствие гетерологичной последовательности кластеров кодонов ДНК или обилие кодонов, которые редко наблюдают у хозяина, в котором должна осуществляться экспрессия, позволяет предсказать низкие уровни гетерологичной экспрессии в этом хозяине.
В воплощении настоящего изобретения предложена полинуклеотидная последовательность gag, которая кодирует аминокислотную последовательность, где паттерн использования кодонов этой полинуклеотидной последовательности подобен таковому для генов млекопитающих с высокой экспрессией. Предпочтительно эта полинуклеотидная последовательность представляет собой последовательность ДНК. Желательно, чтобы паттерн использования кодонов этой полинуклеотидной последовательности был типичен для генов человека с высокой экспрессией.
В полинуклеотидах по настоящему изобретению паттерн использования кодонов изменен по сравнению с таковым, типичным для вирусов иммунодефицита человека, в сторону смещения кодонов, более близкого для целевого организма, например, млекопитающего, в частности человека. «Коэффициент использования кодонов» является мерой того, насколько близко паттерн кодонов данной полинуклеотидной последовательности подобен таковому для целевого вида. Частоты кодонов можно узнать из литературных источников для генов многих видов с высокой экспрессией (см., например, Nakamura et al., Nucleic Acids Research, 1996, 24: 214-215). Частоты кодонов для каждого из 61 кодона (выраженные в виде числа кодонов, встречающихся на 1000 кодонов выбранного класса генов) нормализованы по каждой из двадцати природных аминокислот, так что значение для наиболее часто используемого кодона для каждой аминокислоты приведено к 1, и частоты для менее распространенных кодонов нанесены на шкалу между 0 и 1. Таким образом, для каждого из 61 кодонов определено значение 1 или ниже для генов целевого вида с высокой экспрессией. Чтобы вычислить коэффициент использования кодонов для конкретного полинуклеотида относительно генов данного вида с высокой экспрессией, отмечают значение на шкале для каждого кодона конкретного полинуклеотида и определяют геометрическое среднее всех этих значений (путем деления суммы натуральных логарифмов этих значений на суммарное число кодонов и взятия антилогарифма). Этот коэффициент будет иметь значение между 0 и 1, и, чем выше этот коэффициент, тем больше кодонов в полинуклеотиде являются часто используемыми кодонами. Если полинуклеотидная последовательность имеет коэффициент использования кодонов 1, то все кодоны являются «наиболее частыми» кодонами для генов целевого вида с высокой экспрессией.
В соответствии с настоящим изобретением паттерн использования кодонов полинуклеотида предпочтительно должен исключать кодоны со значением RSCU менее чем 0,2 по сравнению с генами целевого организма с высокой экспрессией. Альтернативно паттерн использования кодонов должен исключать кодоны, составляющие <10% кодонов, используемых для конкретной аминокислоты. Значение относительного использования кодонов-синонимов (RSCU) представляет собой наблюдаемое число кодонов, разделенное на ожидаемое число, если все кодоны для данной аминокислоты были использованы одинаково часто. Полинуклеотид по настоящему изобретению будет, как правило, иметь коэффициент использования кодонов (или RSCU) для генов человека с высокой экспрессией выше чем 0,3, предпочтительно выше чем 0,4, наиболее предпочтительно выше чем 0,5. Таблицы использования кодонов для человека можно также найти в Genebank.
Для сравнения высокоэкспрессирующийся ген бета-актина имеет RSCU 0,747. Таблица использования кодонов для Homo sapiens представлена ниже:
В соответствии со следующим аспектом изобретения предложен вектор экспрессии, который содержит и способен направлять экспрессию полинуклеотидной последовательности согласно первому аспекту изобретения, в частности паттерн использования кодонов полинуклеотидной последовательности gag типичен для генов млекопитающих с высокой экспрессией, предпочтительно для генов человека с высокой экспрессией. Этот вектор может быть подходящим для направления экспрессии гегерологичной ДНК в клетках бактерий, насекомых или млекопитающих, в частности в клетках человека. В одном воплощении этот вектор экспрессии представляет собой р7313 (см. фиг.1).
В третьем воплощении предложен ген gag под контролем гетерологичного промотора, слитого с последовательностью ДНК, кодирующей Nef или его фрагмент или обратную транскриптазу (RT) ВИЧ или ее фрагмент. Участок, представляющий собой ген gag, может составлять либо N, либо С-концевую часть слитой конструкции.
В предпочтительном воплощении ген gag не кодирует пептид gag p6. Предпочтительно ген NEF укорочен с удалением последовательности, кодирующей N-концевой район, то есть с удалением 30-85, предпочтительно 60-85, типично примерно 81, предпочтительно 65 N-концевых аминокислот.
В следующем воплощении ген RT также оптимизирован таким образом, что подобен гену человека с высокой экспрессией. RT предпочтительно кодирует мутацию, которая по существу инактивирует какую-либо активность обратной транскриптазы. Предпочтительная мутация инактивации включает замену W (триптофана) 229 на К (лизин).
Согласно следующему аспекту изобретения предложена клетка-хозяин, содержащая нуклеотидную последовательность согласно изобретению или вектор экспрессии согласно изобретению. Эта клетка-хозяин может представлять собой бактериальную клетку, например E.coli, клетку млекопитающего, например человеческую, либо она может представлять собой клетку насекомого. Клетки млекопитающих, содержащие вектор согласно настоящему изобретению, могут представлять собой культивируемые клетки, трансфицированные in vitro, либо они могут быть трансфицированы in vivo путем введения вектора млекопитающему.
Далее в настоящем изобретении предложена фармацевтическая композиция, содержащая полинуклеотидную последовательность согласно изобретению. Предпочтительно эта композиция содержит ДНК-вектор. В предпочтительных воплощениях эта композиция содержит множество частиц, предпочтительно золотых частиц, покрытых ДНК, содержащей вектор, кодирующий полинуклеотидную последовательность по изобретению. Предпочтительно эта последовательность кодирует аминокислотную последовательность gag ВИЧ, где паттерн использования кодонов этой полинуклеотидной последовательности типичен для генов млекопитающих с высокой экспрессией, особенно для генов человека. В альтернативных воплощениях эта композиция содержит фармацевтически приемлемый эксципиент и ДНК-вектор согласно второму аспекту настоящего изобретения. Эта композиция может также включать адъювант.
Таким образом, одно из воплощений изобретения является таким, что векторы по изобретению используют с иммуностимуляторными агентами. Предпочтительно иммуностимуляторный агент вводят одновременно с нуклеиново-кислотным вектором по изобретению и в предпочтительных воплощениях их включают в один препарат вместе. Такие иммуностимуляторные агенты включают (хотя этот перечень никоим образом не является исчерпывающим и не устраняет другие агенты) синтетические имидазохинолины, такие как имихимод [imiquimod, S-26308, R-837] (Harrison et al. 'Reduction of recurrent HSV disease using imiquimod alone or combined with a glycoprotein vaccine', Vaccine 19: 1820-1826, (2001)) и резихимод [resiquimod, S-28463, R-848] (Vasilakos et al. 'Adjuvant activities of immune response modifier R-848: Comparison with CpG ODN', Cellular Immunology 204: 64-74 (2000)), Шиффовы основания карбонилов и аминов, которые конститутивно экспрессируются на поверхностях антигенпрезентирующих клеток и Т-клеток (Rhodes, J. et al. Therapeutic potentiation of the immune system by costimulatory Schiff-base-forming drugs', Nature 377: 71-75 (1995)), молекулы цитокинов, хемокинов и кости муляторные молекулы либо в виде белка, либо в виде пептида, которые включают провоспалительные цитокины, такие как GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор), ИЛ-1 (интерлейкин-1)-альфа, ИЛ-1-бета, ФНО (фактор некроза опухоли)-альфа и ФНО-бета, индукторы Th1, такие как интерферон-гамма, ИЛ-2, ИЛ-12, ИЛ-15 и ИЛ-18, индукторы Th2, такие как ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13, а также другие гены хемокинов и костимуляторов, такие как МСР-1, MIP-1-альфа, MIP-1-бета, RANTES, TCA-3, CD80, CD86 и CD40L, другие лиганды-мишени иммуностимуляции, такие как CTLA-4 и L-селектин, белки, стимулирующие апоптоз, и пептиды, такие как Fas (49), синтетические адъюванты на основе липидов, такие как ваксфектин [vaxfectin] (Reyes et al., 'Vaxfectin enhances antigen specific antibody titres and maintains Th1 type immune responses to plasmid DNA immunization', Vaccine 19: 3778-3786), сквален, альфа-токоферол, полисорбат 80, DOPC и холестерин, эндотоксин [LPS] (Beutler, В. 'Endotoxin, Toll-like receptor 4, and the afferent limb of innate immunity', Current Opinion in Microbiology 3: 23-30 (2000)); олиго- и динуклебтиды CpG, Sato, Y. et al., 'Immunostimulatory DNA sequences necessary for effective intradermal gene immunization', Science 273 (5273): 352-354 (1996), Hemmi, H. et al., 'A Toll-like receptor recognizes bacterial DNA', Nature 408: 740-745, (2000) и другие потенциальные лиганды, которые запускают рецепторы Toll для продуцирования Th-1-индуцирующих цитокинов, такие как синтетические микобактериальные липопротеины, микобактериальный протеин p19, пептидогликан, тейхоевая кислота и липид А.
Некоторые предпочтительные адъюванты, вызывающие ответ преимущественно Th-1-типа, включают, например, производное липида А, такое как монофосфориллипид А или предпочтительно 3-де-О-ацилированный монофосфориллипид А. Адъюванты MPL® можно получить от Corixa Corporation (Seattle, WA; см., например, патенты США №№4436727, 4877611, 4866034 и 4912094). CpG-содержащие олигонуклеотиды (в которых динуклеотид CpG не метилирован) также вызывают ответ преимущественно Th-1-типа. Такие олигонуклеотиды хорошо известны и описаны, например, в WO 96/02555, WO 99/33488 и в патентах США №№6008200 и 5856462. Иммуностимуляторные последовательности ДНК также описаны, например, Sato et al., Science 273: 352, 1996. Другой предпочтительный адъювант содержит сапонин, такой как Quil А, или его производные, включая QS21 и QS7 (Aquila Biopharmaceuticals Inc., Framingham, MA), эсцин, дигитонин или сапонины Gypsophila или Chenopodium quinoa.
Предложено также применение полинуклеотида согласно изобретению или вектора согласно изобретению при лечении или профилактике ВИЧ-инфекции.
В настоящем изобретении также предложены способы лечения или предупреждения ВИЧ-инфекций, любых симптомов или заболеваний, связанных с ними, при которых вводят эффективное количество полинуклеотида, вектора или фармацевтической композиции согласно изобретению. Введение фармацевтической композиции может принимать форму одной или более чем одной индивидуальной дозы, например, в виде повторных доз одной и той же ДНК-плазмиды, или в виде режима «первичная вакцинация - ревакцинация» терапевтической вакцинации. В некоторых случаях «первичную» вакцинацию можно осуществлять путем опосредованной частицами доставки ДНК полинуклеотида согласно настоящему изобретению, предпочтительно встроенного в вектор плазмидного происхождения, а ревакцинацию можно осуществлять путем введения рекомбинантного вирусного вектора, содержащего ту же полинуклеотидную последовательность, или путем ревакцинации белком в адъюванте. Наоборот, первичную вакцинацию можно осуществлять вирусным вектором или белковым препаратом, типично белковым препаратом в адъюванте, а ревакцинацию осуществлять ДНК-вакциной по настоящему изобретению. Можно использовать многократные дозы первичной вакцинации и/или ревакцинации.
В воплощениях изобретения рассматривают фрагменты белков gag, nef или RT. Например, полинуклеотид по изобретению может кодировать фрагмент белка gag, nef или RT ВИЧ. Полинуклеотид, который кодирует фрагмент по меньшей мере из 8, например из 8-10 аминокислот, либо вплоть до 20, 50, 60, 70, 80, 100, 150 или 200 аминокислот в длину, считают находящимся в пределах объема изобретения, если кодируемый олиго- или полипептид проявляет антигенные свойства ВИЧ. В частности, но не исключительно, данный аспект изобретения охватывает ситуацию, когда полинуклеотид кодирует фрагмент или полноразмерную последовательность белка ВИЧ и может представлять один или более чем один дискретный эпитоп данного белка. Такие фрагменты могут быть оптимизированными по кодонам, так что этот фрагмент имеет паттерн использования кодонов, который подобен таковому для гена млекопитающего с высокой экспрессией.
Предпочтительные конструкции согласно настоящему изобретению включают:
1. р17, р24, слитые с укороченным NEF (без нуклеотидов, кодирующих концевые аминокислоты 1-65);
2. р17, р24, RT, укороченный NEF (без нуклеотидов, кодирующих концевые аминокислоты 1-65);
3. р17, р24 (оптимизированный gag), укороченный NEF (без нуклеотидов, кодирующих концевые аминокислоты 1-65);
4. р17, р24 (оптимизированный gag), RT (оптимизированный), укороченный NEF (без нуклеотидов, кодирующих концевые аминокислоты 1-85);
5. р17, р24, RT (оптимизированный), укороченный NEF (без нуклеотидов, кодирующих концевые аминокислоты 1-65);
6. Укороченный NEF (без нуклеотидов 1-65), слитый с оптимизированным р17, p24gag;
7. Особенно предпочтительные конструкции по изобретению включают тройные слияния RT-NEF-Gag и RT-Gag-Nef, в частности:
8. Оптимизированный RT, укороченный NEF и оптимизированный р17, р24 (gag) (RNG);
9. Оптимизированный RT, оптимизированный р17, 24 (gag), укороченный Nef (без ак 1-65)(RGN).
Предпочтительно, чтобы конструкции ВИЧ имели происхождение от ВИЧ-изолята В или изолята С, особенно изолята В.
Как обсуждено выше, настоящее изобретение включает векторы экспрессии, которые содержат нуклеотидные последовательности по изобретению. Конструирование таких векторов экспрессии является рутинной методикой молекулярной биологии и включает использование плазмидной ДНК и соответствующих инициаторов, промоторов, энхансеров и других элементов, таких как, например, сигналы полиаденилирования, которые могут быть необходимы и которые расположены в правильной ориентации, чтобы дать возможность экспрессии белка. Другие подходящие векторы должны быть очевидны специалистам в данной области техники. В качестве дополнительного примера в этом отношении авторы изобретения ссылаются на Sambrook et al. Molecular Cloning: a Laboratory Manual, 2nd Edition, CSH Laboratory Press (1989).
Предпочтительно полинуклеотид по изобретению или для использования по изобретению в векторе оперативно сцеплен с регуляторной последовательностью, которая способна обеспечить экспрессию кодирующей последовательности клеткой-хозяином, то есть этот вектор представляет собой вектор экспрессии. Термин «оперативно сцеплен» относится к соседнему положению, где описанные компоненты находятся во взаимоотношениях, позволяющих им функционировать надлежащим образом. Регуляторная последовательность, такая как промотор, «оперативно сцепленная» с кодирующей последовательностью, расположена таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с этой регуляторной последовательностью.
Векторы могут представлять собой, например, плазмиды, искусственные хромосомы (например, ВАС, РАС, YAC), вирусные или фаговые векторы, снабженные точкой начала репликации, возможно, промотором для экспрессии полинуклеотида и, возможно, регулятором этого промотора. Векторы могут содержать один или более чем один селективный ген-маркер, например, ген устойчивости к ампициллину или канамицину в случае бактериальной плазмиды или ген устойчивости для вектора грибов. Векторы можно использовать in vitro, например, для продуцирования ДНК или РНК, либо использовать для трансфекции или трансформации клетки-хозяина, например клетки-хозяина млекопитающего, например, для продуцирования белка, кодируемого вектором. Векторы могут быть также адаптированы для использования in vivo, например, при способе ДНК-вакцинации или генотерапии.
Промотры и другие сигналы регуляции экспрессии могут быть выбраны таким образом, чтобы быть совместимыми с клеткой-хозяином, для которой предназначена экспрессия. Например, промоторы млекопитающих включают металлотионеиновый промотор, который можно индуцировать в ответ на тяжелые металлы, такие как кадмий, и промотор β-актина. Можно также использовать вирусные промоторы, такие как промотор большого Т-антигена SV40, быстрый ранний (IE, immediate early) промотор цитомегаловируса человека (CMV), промотор LTR вируса саркомы Рауса, аденовирусный промотор или промотор HPV, в частности, регуляторный район вверх по течению (URR, upstream regulatory region) HPV. Все эти промоторы хорошо описаны и легко доступны в данной области техники.
Предпочтительным промоторным элементом является быстрый ранний промотор CMV за исключением интрона А, но включая экзон 1. Промоторный элемент может представлять собой минимальный промоторный элемент или усиленный промотор, причем усиленный промотор предпочтителен. Соответственно, предложен вектор, содержащий полинуклеотид по изобретению под контролем раннего промотора IE HCMV.
Примеры подходящих вирусных векторов включают векторы вируса простого герпеса, векторы вируса осповакцины или альфа-вируса и ретровирусов, включая лентивирусы, аденовирусы и аденоассоциированные вирусы. Методики переноса генов, использующие эти вирусы, известны специалистам в данной области техники. Ретровирусные векторы, например, можно использовать для стабильной интеграции полинуклеотида по изобретению в геном хозяина, хотя такая рекомбинация не является предпочтительной. Аденовирусные векторы, дефектные по репликации, напротив, поддерживаются в эписомном состоянии и, следовательно, дают возможность транзитной экспрессии. Векторы, способные направлять экспрессию в клетках насекомых (например, бакуловирусные векторы), в клетках человека, в дрожжах или в бактериях, можно использовать для продуцирования количеств белка ВИЧ, кодируемого полинуклеотидами по настоящему изобретению, например, для использования в качестве субъединичных вакцин или в иммунологических анализах.
Полинуклеотиды согласно изобретению обладают пользой при продуцировании путем экспрессии кодируемых белков, причем эта экспрессия может иметь место in vitro, in vivo или ex vivo. Нуклеотиды, следовательно, могут быть вовлечены в синтез рекомбинантного белка, например, для повышения выходов, либо действительно могут находить применение в качестве терапевтических агентов как таковых, применяемых при методиках ДНК-вакцинации. Если полинуклеотиды по настоящему изобретению используют при продуцировании кодируемых белков in vitro или ex vivo, клетки, например, в клеточной культуре следует модифицировать таким образом, чтобы включить в них полинуклеотид, экспрессия которого необходима. Такие клетки включают транзитные или, предпочтительно, стабильные клеточные линии млекопитающих. Конкретные примеры клеток, которые можно модифицировать путем вставки векторов, кодирующих полипептид согласно изобретению, включают клетки млекопитающих НЕК293Т, СНО, HeLa, 293 и клетки COS. Предпочтительно выбранная клеточная линия должна быть не только стабильной, но такой, чтобы давать возможность гликозилирования и экспрессии зрелого полипептида на клеточной поверхности. Экспрессия может быть достигнута в трансформированных ооцитах. Полипептид можно экспрессировать с полинуклеотида по настоящему изобретению в клетках трансгенного животного, представляющего собой не человека, предпочтительно мыши. Трансгенное животное, представляющее собой не человека, экспрессирующее полипептид с полинуклеотида по изобретению, включено в объем изобретения.
Далее в изобретении предложен способ вакцинации субъекта млекопитающего, при котором ему вводят эффективное количество такой вакцины или вакцинной композиции. Наиболее предпочтительно векторы экспрессии для применения в ДНК-вакцинах, вакцинных композициях и иммунотерапевтических агентах будут представлять собой плазмидные векторы.
ДНК-вакцины можно вводить в форме «голой ДНК», например, в жидком препарате, который вводят, используя шприц или струю высокого давления, либо готовят препарат ДНК с липосомами или раздражающим усилителем трансфекции, либо вводят путем опосредованной частицами доставки ДНК (PMDD, particle mediated DNA delivery). Все эти системы доставки хорошо известны в данной области техники. Вектор можно вводить млекопитающему, например, с помощью системы доставки вирусных векторов.
Композиции по настоящему изобретению можно доставлять с помощью ряда путей, таких как внутримышечный, подкожный, внутрибрюшинный, внутривенный или введение в слизистую оболочку.
В предпочтительном воплощении композицию доставляют внутрикожно. В частности, композицию доставляют с помощью «генного ружья», в частности методик введения путем бомбардировки частицами, которые включают нанесение вектора в виде покрытия на гранулу (например, золотую), которую затем вводят под высоким давлением в эпидермис, например, так, как описано в Haynes et al., J. Biotechnology 44: 37-42 (1996).
В одном иллюстративном примере управляемое газом ускорение частиц может быть достигнуто с помощью таких устройств, которые выпускаются PowderJect Pharmaceuticals PLC (Oxford, UK) и PowderJect Vaccines Inc. (Madison, WI), и некоторые примеры таких устройств описаны в патентах США №№5846796, 6010478, 5865796, 5584807 и в Европейском патенте №0500799. Данный подход дает возможность безыгольной доставки, где сухой порошкообразный препарат микроскопических частиц, таких как полинуклеотид, ускоряют до высокой скорости внутри струи газа гелия, генерируемой ручным устройством, движущей частицы в целевую ткань-мишень, обычно в кожу.
Частицы предпочтительно представляют собой золотые гранулы 0,4-4,0 мкм, более предпочтительно 0,6-2,0 мкм, покрытые конъюгатом ДНК, а затем упакованные в картридж или кассету для помещения в «генное ружье».
В родственном воплощении другие устройства и способы, которые могут быть полезны для управляемой газом безыгольной инъекции композиций по настоящему изобретению, включают устройства, выпускаемые Bioject, Inc. (Portland, OR), и некоторые примеры таких устройств описаны в патентах США №№4790824, 5064413, 5312335, 5383851, 5399163, 5520639 и 5993412.
Векторы, которые содержат нуклеотидные последовательности, кодирующие антигенные пептиды, вводят в таком количестве, которое будет профилактически или терапевтически эффективным. Количество, которое следует вводить, как правило, находится в интервале от одного пикограмма до 1 миллиграмма, предпочтительно от 1 пикограмма до 10 микрограммов нуклеотида на дозу для опосредованной частицами доставки и от 100 нанограммов до 1 миллиграмма, предпочтительно от 10 микрограммов до 1 миллиграмма нуклеотида на дозу для других путей. Точное количество может значительно варьироваться в зависимости от массы пациента, подлежащего иммунизации, и от пути введения.
Для иммуногенного компонента, содержащего нуклеотидную последовательность, кодирующую антигенный пептид, возможно введение на однократной основе или повторное введение, например, от 1 до 7 раз, предпочтительно от 1 до 4 раз, через интервалы между примерно 1 сутками и примерно 18 месяцами. Однако данный режим будет значительно варьироваться в зависимости от размера подлежащего лечению пациента, от количества вводимой нуклеотидной последовательности, от пути введения и от других факторов, которые будут очевидны специалистам в области ветеринарии или практикующим врачам. Пациент может получать одно или более чем одно из других противовирусных лекарств против ВИЧ в виде части их общего режима введения. Кроме того, нуклеиново-кислотный иммуноген можно вводить с адъювантом.
Адъювантный компонент, описанный здесь, можно подобным образом вводить с помощью ряда различных путей введения, как, например, с помощью перорального, интраназального, легочного, внутримышечного, подкожного, внутрикожного или местного путей. Предпочтительно адъювантный компонент вводят с помощью внутрикожного или местного путей. Наиболее предпочтителен местный путь. Это введение может иметь место между примерно 14 сутками перед введением и примерно 14 сутками после введения нуклеотидной последовательности, предпочтительно между примерно 1 сутками перед введением и примерно 3 сутками после введения нуклеотидной последовательности. Адъювантный компонент в воплощении вводят по существу одновременно с введением нуклеотидной последовательности. Под «по существу одновременным» подразумевают, что введение адъювантного компонента предпочтительно в то же время, что и введение нуклеотидной последовательности, или, если не в то же время, то, по меньшей мере, в пределах нескольких часов в любую сторону от введения нуклеотидной последовательности. В наиболее предпочтительном протоколе введения адъювантный компонент будут вводить по существу одновременно с введением нуклеотидной последовательности. Очевидно, данный протокол можно варьировать, если необходимо, в соответствии с типом вариаций, описанным выше. Предпочтительно, чтобы адъювант представлял собой производное 1Н-имидазо[4,5-с]хинолин-4-амина, такое как имихимод. Типично имихимод будет представлен в виде местного препарата в виде крема, и его будут вводить в соответствии с вышеописанным протоколом.
Снова в зависимости от таких вариаций доза введения этого производного будет также варьировать, но может, например, находиться в интервале между примерно 0,1 мг на кг до примерно 100 мг на кг, где «на кг» относится к массе тела подлежащего лечению млекопитающего. Это введение производного 1Н-имидазо[4,5-с]хинолин-4-амина предпочтительно следует повторять с каждым последующим или повторным введением нуклеотидной последовательности. Наиболее предпочтительно доза введения будет находиться между примерно 1 мг на кг и примерно 50 мг на кг. В случае схемы «первичная вакцинация - ревакцинация», как описано выше, имихимод или другое производное 1Н-имидазо[4,5-с]хинолин-4-амина можно вводить либо с первичной вакцинацией, либо с ревакцинацией, либо и с первичной вакцинацией, и с ревакцинацией.
Хотя возможно, чтобы адъювантный компонент содержал только производные 1Н-имидазо[4,5-с]хинолин-4-амина в виде сырого химического вещества, для введения предпочтительно, чтобы он находился в форме фармацевтического препарата. Таким образом, адъювантный компонент будет предпочтительно содержать 1Н-имидазо[4,5-с]хинолин-4-амин, комбинированный с одним или более чем одним фармацевтически приемлемым носителем и, возможно, с другими терапевтическими ингредиентами. Носитель (носители) должен быть «приемлемым» в том смысле, что он должен быть совместимым с другими ингредиентами в препарате и не быть вредным для его реципиента. Природа этих препаратов будет естественно варьироваться в соответствии с предназначенным путем введения, и они могут быть получены способами, хорошо известными в области фармацевтики. Все способы включают стадию приведения в контакт производного 1Н-имидазо[4,5-с]хинолин-4-амина с подходящими носителем или носителями. Как правило, эти препараты готовят путем однородного и тесного приведения в контакт этого производного с жидкими носителями или тонко измельченными твердыми носителями, либо и с теми, и с другими, а затем, если необходимо, придают этому продукту форму желаемого препарата. Препараты по настоящему изобретению, подходящие для перорального введения, могут быть представлены в виде дискретных единиц, таких как капсулы, крахмальные облатки или таблетки, каждая из которых содержит предопределенное количество активного ингредиента; в виде порошка или гранул; в виде раствора или суспензии в водной жидкости или в неводной жидкости; либо в виде жидкой эмульсии масло-в-воде или эмульсии вода-в-масле. Активный ингредиент может также быть представлен в виде болюса, электуария или пасты.
Таблетку можно изготовить путем прессования или формования, возможно, с одним или более чем одним вспомогательным ингредиентом. Прессованные таблетки можно изготовить путем прессования в подходящем аппарате активного ингредиента в свободно-текучей форме, такой как порошок или гранулы, возможно, смешанного со связующим агентом, смазкой, инертным разбавителем, смазывающим, поверхностно-активным или диспергирующим агентом. Формованные таблетки можно изготовить путем формования в подходящем аппарате смеси порошкообразного соединения, увлажненного инертным жидким разбавителем.
Таблетки могут быть, возможно, покрыты оболочкой или помечены, и их можно изготовить таким образом, чтобы обеспечить медленное или регулируемое высвобождение активного ингредиента.
Препараты для инъекции с помощью, например, внутримышечного, внутрибрюшинного или подкожного пути введения включают водные или неводные стерильные инъекционные растворы, которые могут содержать антиоксиданты, буферы, бактериостатики и растворенные вещества, которые делают этот препарат изотоническим с кровью предназначенного реципиента; и водные или неводные стерильные суспензии, которые могут включать суспендирующие агенты и загустители. Эти препараты могут быть представлены в контейнерах, содержащих однократную дозу или многократные дозы, например, в запаянных ампулах и флаконах, и их можно хранить в сухом замороженном (лиофилизированном) состоянии, требующем только добавления жидкого стерильного носителя, например воды для инъекций, непосредственно перед применением. Инъекционные растворы и суспензии, которые готовят непосредственно перед применением, можно изготовить из стерильных порошков, гранул и таблеток вышеописанного типа. Препараты, подходящие для легочного введения через трансбуккальную или назальную полость, представлены таким образом, что частицы, содержащие активный ингредиент, желательно имеющие диаметр в интервале от 0,5 до 7 микрон, доставляют в бронхиальное дерево реципиента. Возможности для таких препаратов таковы, что они находятся в форме тонкоизмельченных порошков, которые могут быть удобно представлены либо в виде прокалываемой капсулы, подходящим материалом для которой является, например, желатин, для применения в ингаляционном устройстве, либо альтернативно в виде самостоятельно распыляемого препарата, содержащего активный ингредиент, подходящий жидкий пропеллент и, возможно, другие ингредиенты, такие как сурфактант и/или твердый разбавитель. Самостоятельно распыляемые препараты можно также применять, если активный ингредиент распределен в форме капелек раствора или суспензии. Такие самостоятельно распыляемые препараты аналогичны известным в данной области техники и могут быть изготовлены с помощью разработанных методик. Они соответствующим образом снабжены либо ручным, либо автоматически функционирующим клапаном, имеющим желаемые характеристики распыления; предпочтительно этот клапан представляет собой клапан мерного типа, поставляющий фиксированный объем, например, от 50 до 100 мкл при каждой операции с ним.
При следующей возможности адъювантный компонент может находиться в форме раствора для применения в аэрозольном ингаляторе или небулайзере, с помощью которого ускоренный поток воздуха или ультразвуковое возбуждение используют для получения аэрозоля из мелких капелек для ингаляции.
Препараты, подходящие для интраназального введения, обычно включают формы, подобные описанным выше для легочного введения, хотя для таких препаратов предпочтительно иметь диаметр частиц в интервале от примерно 10 до примерно 200 микрон, чтобы обеспечить удерживание в носовой полости. Этого можно достичь, соответственно, путем использования порошка из частиц подходящего размера или выбора подходящего клапана. Такие подходящие препараты включают крупные порошки, имеющие диаметр частиц в интервале от примерно 20 до примерно 500 микрон, для введения путем быстрой ингаляции через носовой ход из контейнера, удерживаемого вблизи носа, и носовые капли, содержащие примерно от 0,2 до 5% мас./мас. активного ингредиента в водных или масляных растворах. В одном воплощении изобретения возможно, чтобы вектор, который содержит нуклеотидную последовательность, кодирующую антигенный пептид, вводили в том же препарате, что и производное 1Н-имидазо[4,5-с]хинолин-4-амина. Следовательно, в данном воплощении иммуногенный и адъювантный компонент находятся в одном препарате.
В воплощении адъювантный компонент готовят в форме, подходящей для введения с помощью «генного ружья», и его вводят этим путем по существу одновременно с введением нуклеотидной последовательности. Для изготовления препаратов, подходящих для введения данным способом, может быть необходимо, чтобы производное 1Н-имидазо[4,5-с]хинолин-4-амина было лиофилизировано и прикреплено на поверхности, например, золотых гранул, которые подходят для введения с помощью «генного ружья». В альтернативном воплощении адъювантный компонент можно вводить в виде сухого порошка с помощью проталкивания высоким давлением газа.
Даже если их не включают в препарат вместе, может быть подходящим, чтобы адъювантный компонент вводили в тот же или примерно в тот же сайт введения, что и нуклеотидную последовательность.
Другие подробности о фармацевтических препаратах можно найти в Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvania (1985), причем данное описание включено здесь в полном объеме путем ссылки.
Подходящие методики введения «голого» полинуклеотида или вектора пациенту также включают местное введение с подходящим носителем. Нуклеиновую кислоту можно вводить местным путем на кожу или на поверхности слизистых оболочек, например, путем интраназального, перорального, интравагинального или интраректального введения. «Голый» нуклеотид или вектор может быть представлен вместе с фармацевтически приемлемым эксципиентом, таким как фосфатно-солевой буферный раствор (ФСБ). Захват ДНК можно дополнительно усилить путем использования вспомогательных агентов, таких как бупивакаин (bupivacaine), либо отдельно, либо включенных в препарат ДНК. Другие способы введения нуклеиновой кислоты непосредственно реципиенту включают ультразвуковую, электрическую стимуляцию, электропорацию и микропосев, которые описаны в патенте США №5697901.
Захват нуклеиново-кислотных конструкций можно усилить с помощью некоторых известных методик трансфекции, например тех, которые включают использование трансфекционных агентов. Примеры этих агентов включают катионные агенты, например фосфат кальция и ДЭАЭ-декстран, и липофектанты, например липофектам и трансфертам. Дозировку нуклеиновой кислоты, которую нужно вводить, можно менять.
Нуклеиново-кислотную последовательность по настоящему изобретению можно также вводить посредством специализированных векторов доставки, применяемых в генотерапии. Подходы генотерапии обсуждены, например, Verme et al., Nature 1997, 389: 239-242. Можно использовать как вирусные, так и невирусные векторные системы. Системы на вирусной основе включают системы на основе ретровирусов, лентивирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, Canarypox и вируса осповакцины. Системы на невирусной основе включают прямое введение нуклеиновых кислот, методику инкапсуляции в микросферы (поли(лактид-ко-гликозид)) и системы на основе липосом. Вирусные и невирусные системы доставки можно комбинировать, где это желательно для обеспечения инъекций ревакцинации после исходной вакцинации, например исходной «первичной» ДНК-вакцинации с использованием невирусного вектора, такого как плазмиды, с последующей одной или более чем одной ревакцинацией с использованием вирусного вектора или системы на невирусной основе. Подобным образом в изобретении рассмотрены системы первичной вакцинации - ревакцинации полинуклеотидом по изобретению с последующей ревакцинацией белком в адъюванте, либо наоборот.
Нуклеиново-кислотную последовательность по настоящему изобретению можно также вводить с помощью трансформированных клеток. Такие клетки включают клетки, собранные от субъекта. «Голый» полинуклеотид или вектор по настоящему изобретению можно вводить в такие клетки in vitro, после чего возвращать эти трансформированные клетки субъекту. Полинуклеотид по изобретению может интегрировать в нуклеиновую кислоту, уже находящуюся в клетке, посредством событий гомологичной рекомбинации. Трансформированную клетку можно, если желательно, выращивать in vitro, и одну или более чем одну из полученных в результате клеток использовать по настоящему изобретению. Клетки можно вводить в подходящем сайте пациента с помощью известных хирургических или микрохирургических методик (например, трансплантации, микроинъекции и т.д.).
Фармацевтические композиции по настоящему изобретению могут включать адъювантные соединения, как подробно описано выше, или другие вещества, которые могут служить для усиления иммунного ответа, индуцированного белком, который кодируется ДНК. Эти белки могут кодироваться ДНК либо по отдельности, либо в виде слияния с антигеном, либо могут быть включены в препарат в качестве не-ДНК-элементов. Примеры веществ адъювантного типа, которые могут быть включены в препараты по настоящему изобретению, включают убихитин, связанный с липосомами мембранный белок (LAMP, lyposomal associated membrane protein), коровый антиген вируса гепатита В, лиганд FLT-3 (цитокин, важный при генерировании профессиональных антигенпрезентирующих клеток, в частности дендритных клеток) и другие цитокины, такие как ИФН-γ и GM-CSF. Другие предпочтительные адъюванты включают имихимод и резимхимод, а также тукарасол (Tucarasol). Особенно предпочтителен имихимод.
В настоящем изобретении в предпочтительных воплощениях изобретения предложено применение молекулы нуклеиновой кислоты, как описано здесь, для лечения или профилактики ВИЧ-инфекции. Эту молекулу нуклеиновой кислоты предпочтительно вводят с имихимодом. Имихимод предпочтительно вводят местным путем, тогда как молекулу нуклеиновой кислоты предпочтительно вводят с помощью опосредованной частицами доставки.
Соответственно, в настоящем изобретении предложен способ лечения субъекта, страдающего ВИЧ-инфекцией или восприимчивого к ВИЧ-инфекции, при котором вводят молекулу нуклеиновой кислоты, как описано здесь, и имихимод.
Теперь настоящее изобретение будет проиллюстрировано ссылкой на приведенные ниже примеры.
ПРИМЕРЫ
Пример 1: Оптимизация р55 gag (p17, р24, р13), так чтобы использование кодонов было подобно генам человека с высокой экспрессией
Целевой ген
Сборку синтетического гена, кодирующего антиген р55 gag ВИЧ-1 изолят В штамм НХВ2 (GenBank entry K03455), оптимизированного для экспрессии в клетках млекопитающих, осуществляли с помощью перекрывающихся олигонуклеотидов путем ПЦР.
Оптимизация включала изменение паттерна использования кодонов вирусного гена с получением частоты кодонов, более близкой к таковой, обнаруживаемой в генах человека с высокой экспрессией. Кодоны оценивали, используя статистическую программу Visual Basic, названную Syngene (современная версия Calcgene, написанная R.S. Hale and G. Thompson, Protein Expression and Purification, Vol.12, pp.185-188, 1998).
Клонирование:
Продукт ПЦР gag 1528 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами NotI и BamHI и лигировали в вектор WRG7077, рестрицированный Notl/BamHI. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста.
Клоны секвенировали и проверяли на ошибки. Ни один клон не был на 100% правильным. Районы правильной последовательности из двух клонов, следовательно, объединяли с помощью перекрывающейся ПЦР, используя соответствующие комбинации серии олигонуклеотидов оптимизации, с получением полноразмерного гена gag, оптимизированного по кодонам. Затем определили, что этот конечный клон содержит делецию единственного нуклеотида, которая приводит в результате к сдвигу рамки считывания и преждевременной терминации трансляции. Эту делецию репарировали путем вырезания района гена, содержащего неправильную последовательность, и клонирования в правильную последовательность эквивалентного района другого клона. Таким образом, получили конечный клон р55 gag, оптимизированный по кодонам: Gagoptrpr2 (см. фиг.2).
Пример 2: Получение слитого гена р17/р24/укороченный Nef
Целевой ген
Участки р17 и р24 гена р55 gag происхождения ВИЧ-1 изолят В штамм НХВ2 амплифицировали с помощью ПЦР с плазмиды рНХВ/Pr (В. Maschera, E. Furfine and E.D. Blair, 1995, J. Virol. 69, 5431-5436). PHXB/Pr 426 п.о. от 3'конца гена nef HXB2 амплифицировали с той же плазмиды. Поскольку ген nef НХВ2 содержит кодон преждевременной терминации, для репарации этого кодона использовали две перекрывающиеся ПЦР (TGA [стоп-кодон] на TGG [Trp]).
Продукты ПЦР линкер р17/р24 и линкер trNEF соединяли с образованием слитого гена (фиг.3) в реакции ПЦР (антисмысловой).
Продукт ПЦР 1542 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами NotI и BamHI и клонировали в сайты NotI и BamHI вектора WRG7077. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста.
Пример 3: Получение слитого гена Gag p17/24opt/trNef1 ('Gagopt/Nef)
Целевой ген
Участок р17/р24 оптимизированного по кодонам гена р55 gag происхождения ВИЧ-1 изолят В штамм НХВ2 амплифицировали с помощью ПЦР с плазмиды pGagoptTrpr2. Укороченный ген Nef HXB2 с репарированным кодоном преждевременной терминации (TGA [стоп-кодон] на TGG [Trp]) амплифицировали с помощью ПЦР с плазмиды 7077trNef20. Эти два продукта ПЦР должны были иметь перекрывающиеся концы, так чтобы два гена можно было соединить во второй ПЦР.
Продукт ПЦР 1544 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами NotI и BamHI и клонировали (см. фиг.3-4) в сайты NotI и BamHI вектора WRG7077. Таким образом ген помещали между промотором/интроном А CMV и сигналом полиаденилирования бычьего гормона роста.
Пример 4: Плазмида p7077-RT3 клон #А
Синтетический ген, кодирующий участок RT гена pol ВИЧ-1 изолят В штамм HXB2, оптимизировали для экспрессии в клетках млекопитающих путем сборки с помощью перекрывающихся олигонуклеотидов путем ПЦР. Эта клонированная последовательность эквивалентна положениям 2550-4222 последовательности сравнения HXB2 (GenBank entry K03455). Для обеспечения экспрессии эта клонированная последовательность имеет два дополнительных кодона на 5'конце, отсутствующих в исходном гене - AUG GGC (Met Gly).
Оптимизация включала изменение паттерна использования кодонов вирусного гена с получением частоты кодонов, более близкой к обнаруживаемой в человеческих генах с высокой экспрессией, исключая при этом редко используемые кодоны. Кодоны оценивали, используя статистическую программу Visual Basic, названную Syngene (современная версия Calcgene, написанная R.S. Hale and G. Thompson, Protein Expression and Purification, Vol. 12, pp.185-188, 1998).
Конечный клон конструировали из двух промежуточных клонов, #16 и #21.
Клонирование
Продукты ПЦР 1,7 т.п.н. очищали в геле, подвергали рестрикции NotI и BamHI и очищали с помощью ПЦР, после чего лигировали с pWRG7077, рестрицированным Notl/BamHI. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста. Клоны секвенировали. Ни один клон не был на 100% правильным, но клон #16 корректировали путем замены фрагмента KpnI-BamHI 403 п.о., содержащего 3 ошибки, правильным фрагментом KpnI-BamHI из клона #21. Конечный клон проверяли секвенированием (см. фиг.5).
Пример 5: Оптимизированный RT
Целевой ген
Синтетический ген, кодирующий участок RT гена pol ВИЧ-1 изолят В штамм НХВ2, оптимизированный для экспрессии в клетках млекопитающих, вырезали из плазмиды p7077-RT3 в виде фрагмента Notl/BamHI 1697 п.о., очищали в геле и клонировали в сайты NotI и BamHI p7313-ie (полученного из pspC31), чтобы поместить этот ген вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика (R7004 р27) (фиг.6).
Пример 6
Плазмида: 7077 trNef20
Целевой ген
Вставка содержит участок гена Nef из ВИЧ-1 изолят В штамм НХВ2. 195 п.о. делетировали с 5'конца этого гена, удаляя кодоны для первых 65 аминокислот Nef. Кроме того, кодон преждевременной терминации в опубликованной последовательности nef HXB2 репарировали (TAG на TGG [Trp]), как было описано для плазмиды p17/24trNEF1. Эту укороченную последовательность nef амплифицировали с помощью ПЦР с плазмиды p17/24trNef1. Эта клонированная последовательность эквивалентна положениям 8992-9417 последовательности сравнения НХВ2 (GenBank entry К03455). Для обеспечения экспрессии эта клонированная последовательность имеет дополнительный кодон на 5'конце, отсутствующий в исходном гене - AUG (Met).
Праймеры:
StrNef (смысловой)
ATAAGAATGCGGCCGCCATGGTGGGTTTTCCAGTCACACCTT [SEQ ID NO:1]
AstrNef (антисмысловой)
CGCGGATCCTCAGCAGTTCTTGAAGTACTCC [SEQ ID NO:2]
ПЦР: 94°C 2 мин, затем 25 циклов: 94°С 30 сек, 50°С 30 сек, 72°С 2 мин, окончание 72°С 5 мин.
Клонирование:
Продукт ПЦР RT 455 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами Notl и BamHI и лигировали в рестрицированный Notl/BamHI вектор WRG7077. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста.
Пример 7
Плазмида: 7077RT 8
Целевой ген
RT-участок гена pol имеет происхождение от ВИЧ-1 изолят В штамм НХВ2. Этот участок амплифицировали с помощью ПЦР с плазмиды р7077pol14. Клонированная последовательность эквивалентна положениям 2550-4234 последовательности сравнения НХВ2 (GenBank entry K03455). Для обеспечения экспрессии эта клонированная последовательность имеет два дополнительных кодона на 5'конце, отсутствующих в исходном гене - AUG GGC (Met Gly).
Праймеры:
SRT (смысловой)
ATAAGAATGCGGCCGCCATGGGCCCCATTAGCCCTATTGAGACT [SEQ ID NO:3]
ASRT (антисмысловой)
CGCGGATCCTTAATCTAAAAATAGTACTTTCCTGATT [SEQ ID NO:4]
ПЦР: 94°C 2 мин, затем 25 циклов: 94°С 30 сек, 50°С 30 сек, 72°С 4 мин, окончание 72°С 5 мин.
Клонирование:
Продукт ПЦР RT 1720 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами NotI и BamHI и лигировали в рестрицированный Notl/BamHI вектор WRG7077. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста.
Пример 8
p17/24opt/RT/trNef1 ('Gagopt/RT/Nef')
В эту конструкцию включена ПЦР, которая вызывает аминокислотную замену R на Н.
Целевой ген
Участок р17/р24 оптимизированного по кодонам гена р55 gag происхождения ВИЧ-1 изолят В штамм НХВ2 амплифицировали с помощью ПЦР с плазмиды pGagoptTrpr2. Кодирующую последовательность RT амплифицировали с плазмиды 7077RT 8. Укороченный ген Nef HXB2 с репарированным кодоном преждевременной терминации (TGA [стоп-кодон] на TGG [Trp]) амплифицировали с помощью ПЦР с плазмиды 7077trNef20. Эти три продукта ПЦР должны были иметь перекрывающиеся концы, так чтобы три гена можно было соединить во второй ПЦР.
Праймеры:
(P17/24)
Sp17p24opt (смысловой)
ATAAGAATGCGGCCGCCATGGGTGCCCGAGCTTCGGT [SEQ ID NO:5]
Asp14p24optRT линкер (антисмысловой)
TGGGGCCCATCAACACTCTGGCTTTGTGTC [SEQ ID NO:6]
ПЦР: 94°С 1 мин, затем 20 циклов: 94°С 30 сек, 50°С 30 сек, 72°С 2 мин, окончание 72°С 4 мин.
Продукт ПЦР p17/24opt 1114 п.о. очищали в геле.
(RT)
Sp17p24optRT линкер (смысловой)
CAGAGTGTTGATGGGCCCCATTAGCCCTAT [SEQ ID NO:7]
ASRTtrNef линкер (антисмысловой)
AACCCACCATATCTAAAAATAGTACTTTCC [SEQ ID NO:8]
ПЦР: как описано выше.
Продукт ПЦР RT 1711 п.о. очищали в геле.
(5' укороченный nef)
SRTtrNef линкер (смысловой)
CTATTTTTAGATATGGTGGGTTTTCCAGTCAC [SEQ ID NO:9]
AstrNef (антисмысловой)
CGCGGATCCTCAGCAGTTCTTGAAGTACTCC [SEQ ID NO:10]
ПЦР: как описано выше.
Продукт ПЦР 448 п.о. очищали в геле.
Затем эти три продукта ПЦР сшивали во второй ПЦР с праймерами Sp17/24opt и AstrNef.
ПЦР: 94°С 1 мин, затем 30 циклов: 94°С 30 сек, 50°С 30 сек, 72°С 4 мин, окончание 72°С 4 мин.
Продукт ПЦР 3253 п.о. очищали в геле, подвергали рестрикции рестрикционными эндонуклеазами NotI и BamHI и клонировали в сайты NotI и BamHI вектора WRG7077. Таким образом ген помещали между промотором/интроном A CMV и сигналом полиаденилирования бычьего гормона роста.
Пример 9
Плазмида: pGRN#16 (p17/p24opt corr/RT/trNef)
Целевой ген:
Наблюдали, что полипротеин, генерируемый p17/24opt/RT/Nef13 ('Gagopt/RT/Nef'), экспрессирует укороченный продукт ˜30 кДа благодаря кластеру невыгодных кодонов внутри р24 вокруг аминокислоты 270. Эти кодоны заменяли оптимальными кодонами с помощью сшивающего ПЦР-мутагенеза. В качестве матрицы использовали p17/24opt/RT/trNef13, чтобы амплифицировать участок Gag 5' до мутации с праймерами Sp17/p24opt и GTR-A и участок Gag 3' до мутации с праймерами GTR-S и Asp17/p24optRT линкер. Перекрывание продуктов, содержащих замены кодонов, и очищенные в геле продукты сшивали вместе, используя праймеры Sp17/p24opt и Asp17/p24optRT линкер. Этот продукт подвергали рестрикции NotI и Agel и вставляли в подобным образом ресрицированный p17/24opt/RT/trNef13 с образованием pGRN. Клон #16 проверяли и размножали.
Праймеры:
5' ПЦР:
Sp17p24opt (смысловой)
ATAAGAATGCGGCCGCCATGGGTGCCCGAGCTTCGGT [SEQ ID NO:11]
GTR-A (антисмысловой)
GCGCACGATCTTGTTCAGGCCCAGGATGATCCACCGTTTATAGATTTCTCC [SEQ ID NO:12]
3' ПЦР:
Смысловой: GTR-S (смысловой)
ATCCTGGGCCTGAACAAGATCGTGCGCATGTACTCTCCGACATCCATCC [SEQ ID NO:13]
Asp17/p24optRT линкер (антисмысловой)
TGGGGCCCATCAACACTCTGGCTTTGTGTC [SEQ ID NO:14]
Условия ПЦР для индивидуальных продуктов и сшивки с использованием ДНК полимеразы PWO (Roche):
95°С 1 мин, затем 20 циклов 95°С 30 с, 55°С 30 с, 72°С 180 с, окончание 72°С 120 с и хранение 4°С.
Продукт 1114 п.о. очищали в геле и подвергали рестрикции NotI и Agel с высвобождением фрагмента 6647 п.о., который очищали в геле и лигировали в рестрицированный Notl/Agel и очищенный в геле p17/24opt/RT/trNef13 с образованием pGRN#16.
Пример 10
Плазмида: p73i-GRN2 клон #19 (p17/p24(opt)/RT(opt)trNef) - репарированный
Целевой ген:
Участок р17/р24 оптимизированного по кодонам gag, оптимизированный по кодонам RT и укороченный ген nef из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Плазмиды, содержащие ген trNef, имеющие происхождение от плазмиды p17/24trNef1, содержат ошибку ПЦР, которая приводит к аминокислотной замене R на Н 19-й аминокислоты от конца nef. Эту ошибку корректировали с помощью ПЦР-мутагенеза, корректированный продукт ПЦР nef сшивали с оптимизированным по кодонам RT из p7077-RT3, и сшитый фрагмент подвергали рестрикции Apal и BamHI и клонировали в рестрицированный Apal/BamHI p73i-GRN.
Праймеры:
ПЦР coRT из p7077-RT3 с использованием праймеров:
(полимераза = PWO (Roche) везде).
Смысловой: U1
GAATTCGCGGCCCGCGATGGGCCCCATCAGTCCCATCGAGACCGTGCCGGTGAAGCTGAAACCCGGGAT [SEQ ID NO:15]
AscoRT-Nef
GGTGTGACTGGAAAACCCACCATCAGCACCTTTCTAATCCCCGC [SEQ ID NO:16]
Цикл: 95°C (30 с), затем 20 циклов 95°С (30 с), 55°С (30 с), 72°С (180 с), затем 72°С (120 с) и хранение при 4°С.
Продукт ПЦР 1,7 т.п.о. очищали в геле.
ПЦР 5' Nef из p17/24trNef1 с использованием праймеров:
Смысловой: S-Nef
ATGGTGGGTTTTCCAGTCACACC [SEQ ID NO:17]
Антисмысловой: ASNef-G:
GATGAAATGCTAGGCGGCTGTCAAACCTC [SEQ ID NO:18]
Цикл: 95°С (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (120 с) и хранение при 4°С.
ПЦР 3' Nef из p17/24trNef1 с использованием праймеров:
Смысловой: SNEF-G
GAGGTTTGACAGCCGCCTAGCATTTCATC [SEQ ID NO:19]
Антисмысловой:
AStrNef (антисмысловой)
CGCGGATCCTCAGCAGTTCTTGAAGTACTCC [SEQ ID NO:20]
Цикл: 95°C (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (120 с) и хранение при 4°С.
Продукты ПЦР очищали в геле. Сначала два продукта Nef сшивали, используя праймеры 5' (S-Nef) и 3' (AstrNef).
Цикл: 95°С (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (180 с) и хранение при 4°С.
Продукт ПЦР очищали с помощью ПЦР и сшивали с продуктом RT, используя праймеры U1 и AstrNef:
Цикл: 95°С (30'с), затем 20 циклов 95°С (30 с), 55°С (30 с), 72°С (180 с), затем 72°С (180 с) и хранение при 4°С.
Продукт 2,1 т.п.о. очищали в геле и подвергали рестрикции Apal и BamHI. Плазмиду p73i-GRN также подвергали рестрикции Apal и BamHI, очищали в геле и лигировали с Apal-BamHI RT3trNef с восстановлением гена р17/p24(opt)/RT(opt)trNef.
Пример 11
p73i-GN2 клон #2 (p17/p24opt/trNef) - репарированный
Целевой ген:
Участок р17/р24 оптимизированного по кодонам гена gag и укороченный ген Nef из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Плазмиды, содержащие ген trNef, имеющие происхождение от плазмиды p17/24trNef1, содержат ошибку ПЦР, которая приводит к аминокислотной замене R на Н 19-й аминокислоты от конца nef. Эту ошибку корректировали с помощью ПЦР мутагенеза, и корректированный фрагмент подвергали рестрикции BglII и BamHI и клонировали в рестрицированный BglII/ BamHI p73i-GN (фиг.12). Восстанавливали корректированный слитый ген p17/p24opt/trNef вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
ПЦР 5' Nef из p17/24trNef1 с использованием праймеров:
(полимераза = PWO (Roche) везде)
Смысловой: S-Nef
ATGGTGGGTTTTTCCAGTCACACC [SEQ ID NO:21]
Антисмысловой: ASNef-G:
GATGAAATGCTAGGCGGCTGTCAAACCTC [SEQ ID NO:22]
Цикл: 95°C (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (120 с) и хранение при 4°С.
ПЦР 5' Nef из p17/24trNef1 с использованием праймеров:
Смысловой: SNEF-G
GAGGTTTGACAGCCGCCTAGCATTTCATC [SEQ ID NO:23]
Антисмысловой: AStrNef
CGCGGATCCTCAGCAGTTCTTGAAGTACTCC [SEQ ID NO.-24]
Цикл: 95°С (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (120 с) и хранение при 4°С.
Продукты ПЦР очищали в геле и сшивали, используя праймеры 5' (S-Nef) и 3' (AstrNef).
Цикл: 95°С (30 с), затем 15 циклов 95°С (30 с), 55°С (30 с), 72°С (60 с), затем 72°С (180 с) и хранение при 4°С.
Продукт ПЦР очищали с помощью ПЦР, подвергали рестрикции BglII/BamHI и фрагмент 367 п.о. очищали в геле и клонировали в рестрицированный BglII/BamHI очищенный в геле p73i-GN.
Пример 12
Плазмида: p73i-RT w229k (инактивированный RT)
Целевой ген:
Конструирование инактивированного гена RT вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
В связи с беспокойством по поводу использования активных видов RT ВИЧ в терапевтической вакцине была желательна инактивация этого гена. Эта инактивация была достигнута с помощью ПЦР-мутагенеза RT (имеющего происхождение от p73i-GRN2) положения аминокислоты 229 с Trp на Lys (R7271 р1-28).
Праймеры:
ПЦР 5' RT + мутация с использованием праймеров:
(полимераза = PWO (Roche) везде)
Смысловой: RT3-u:1
GAATTCGCGGCCGCGATGGGCCCCATCAGTCCCATCGAGACCGTGCCGGTGAAGCTGAAACCCGGGAT [SEQ ID NO:25]
Антисмысловой: AscoRT-Trp229Lys
GGAGCTCGTAGCCCATCTTCAGGAATGGCGGCTCCTTCT [SEQ ID NO:26]
Цикл:
1×[94°C (30 c)]
15×[94°C (30 c)/55°C (30 c)/72°C (60 с)]
1×[72°C (180 c)]
ПЦР очистка в геле
ПЦР 3' RT + мутация с использованием праймеров:
Антисмысловой: RT3-L:1
GAATTCGGATCCTTACAGCACCTTTCTAATCCCCGCACTCACCAGCTTGTCGACCTGCTCGTTGCCGC [SEQ ID NO:27]
Смысловой: ScoRT-Trp229Lys
CCTGAAGATGGGCTACGAGCTCCATG [SEQ ID NO:28]
Цикл:
1×[94°C (30 c)]
15×[94°C (30 c)/55°C (30 c)/72°C (60 с)]
1×[72°С (180 с)]
ПЦР очистка в геле
Продукты ПЦР очищали в геле и 5' и 3' концы RT сшивали, используя праймеры 5' (RT3-U1) и 3' (RT3-L1).
Цикл:
1×[94°С (30 с)]
15×[94°С (30 с)/55°С (30 с)/72°С (120 с)]
1×[72°С (180 с)]
Продукт ПЦР очищали в геле и клонировали в p7313ie, используя сайты рестрикции NotI и BamHI, с образованием p73i-RT w229k [см. фиг.13].
Пример 13
Плазмида: p73i-Tgrn (#3)
Целевой ген:
Участок р17/р24 оптимизированного по кодонам gag, оптимизированный по кодонам RT и укороченный ген nef из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Тройные слитые конструкции, которые содержат активную форму RT, могут быть неприемлемы законом для использования у людей, поэтому инактивация RT была достигнута путем вставки фрагмента, вырезанного Nhel и Apal, из p73i-RT w229k в Nhel/Apal рестрикт p73i-GRN2#19 (фиг.14). Результатом этого была замена W→К в положении 229 в RT.
Пример 14
P73i-Tnrg (#16)
Целевой ген:
Укороченный Nef, инактивированный оптимизированный по кодонам RT и участок р17/р24 оптимизированного по кодонам гена gag из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Порядок генов в полипротеине, кодируемом p73i-Tgrn, перестраивали с помощью ПЦР и ПЦР-сшивки с образованием p73i-Tnrg (фиг.15). Каждый ген амплифицировали с помощью ПЦР и очищали в геле, после чего сшивали эти гены с помощью ПЦР с образованием единого полипротеина. Этот продукт очищали в геле, подвергали ферментативному гидролизу Notl/BamHI и лигировали в Notl/BamHI рестрикт p7313ie.
Праймеры:
TrNef ПЦР
S-Nef (NotI)
CATTAGAGCGGCCGCGATGGTGGGTTTTCCAC [SEQ ID NO:29]
AS-Nef-coRT линкер
GATGGGACTGATGGGGCCCATGCAGTTCTTGAACTACTCCGG [SEQ ID NO:30]
RTw229k ПЦР
S-coRT
ATGGGCCCCATCAGTCCCATCGAG [SEQ ID NO:31]
AS-coRT-p17p24 линкер
CAGTACCGAAGCTCGGGCACCCATCAGCACCTTTCTAATCCCCGC [SEQ ID NO:32]
P17p24opt ПЦР
S-p17p24opt
ATGGGTGCCCGAGCTTCGGTACTG [SEQ ID NO:33]
AS-p17p24opt (BamHI)
GATGGGGGATCCTCACAACACTCTGGCTTTGTGTCC [SEQ ID NO:34]
Условия ПЦР для индивидуальных продуктов и сшивок с использованием ДНК полимеразы VENT (NEB):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (120 с [р17р24 или RT] или 60 с [trNef])]
1×[72°С (240 с)]
Продукты ПЦР очищали в геле и использовали в ПЦР сшивке с использованием праймеров S-trNef (NotI) и AS-p17p24opt (BamHI):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (210 с)]
1×[72°С (240 с)]
Продукт 3000 п.о. очищали в геле и подвергали рестрикции NotI и BamHI и этот фрагмент очищали с помощью ПЦР и лигировали в гидролизованный Notl/BamHI и очищенный в геле p7313ie с образованием p73i-Tnrg.
Пример 15:
1. Плазмида: p73i-Tngr (#3)
Целевой ген
Укороченный Nef, участок р17/р24 оптимизированного по кодонам гена gag и инактивированный оптимизированный по кодонам RT из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Порядок генов в полипротеине, кодируемом p73i-Tgrn, перестраивали с помощью ПЦР и ПЦР-сшивки с образованием p73i-Tngr (фиг.16). Оптимизированный по кодонам р17/р24 и RT конструировали в виде единого продукта и сшивали с помощью ПЦР с амплифицированным trNef. Этот продукт очищали в геле, подвергали ферментативному гидролизу Notl/BamHI и лигировали в Notl/BamHI рестрикт p7313ie.
Праймеры:
P17/p24-RT3' ПЦР
Sp17p24opt (смысловой)
ATGGGTGCCCGAGCTTCGGTACTG [SEQ ID NO:35]
RT3 L:1 (антисмысловой)
GAATTCGGATCCTTACAGCACCTTTCTAATCCCCGCACTCACCAGCTTGTCGACCTGCTCGTTGCCGC [SEQ ID NO:36]
TrNef5' ПЦР
S-Nef (NotI)
CATTAGAGCGGCCGCGATGGTGGGTTTTCCAC [SEQ ID NO:37]
AS-Nef-p17p24
CAGTACCGAAGCTCGGGCACCCATGCAGTTCTTGAACTACTCCGG [SEQ ID NO:38]
Условия ПЦР для индивидуальных продуктов и сшивки с использованием ДНК полимеразы VENT (NEB):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (180 с [р17р24 + RT], или 60 с [trNef], или 210 с [сшивка])]
1×[72°С (240 с)]
Продукт ПЦР 3000 п.о. очищали в геле и подвергали рестрикции NotI и BamHI, и этот фрагмент очищали с помощью ПЦР и лигировали в гидролизованный Notl/BamHl и очищенный в геле p7313ie с образованием p73i-Tngr.
Пример 16:
Плазмида: p73i-Trgn (#6)
Целевой ген
Инактивированный оптимизированный по кодонам RT, участок р17/р24 оптимизированного по кодонам гена gag и укороченный ген Nef из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV+экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Порядок генов в этой конструкции был достигнут с помощью ПЦР амплификации p17p24-trNef и RTw229k из плазмид p73i-GN2 и p73i-RTw229k соответственно. Осуществляли ПЦР-сшивку и продукт очищали в геле и рестрицировали Notl/BamHl, после чего лигировали с гидролизованным Notl/BamHl p7313ie. Секвенирование выявило, что р17р24 не был полностью оптимизирован. Затем фрагмент 700 п.о. Agel/Munl вырезали из кодирующего района и заменяли фрагментом MunI/Agel из p73i-Tgrn#3, содержащего правильную кодирующую последовательность (см. фиг.17).
Праймеры:
P17p24-trNef ПЦР
S-p17p24opt
ATGGGTGCCCGAGCTTCGGTACTG [SEQ ID No:39]
AstrNef (BamHI)
RTw229k
RT3-U:1
GAATTCGCGGCCGCGATGGGCCCCATCAGTCCCATCGAGACCGTGCCGGTGAAGCTGAAACCCGGGAT [SEQ ID NO:40]
AS-coRT-p17p24opt линкер
CAGTACCGAAGCTCGGGCACCCATCAGCACCTTTCTAATCCCCGC [SEQ ID NO:41]
Условия ПЦР для индивидуальных продуктов и сшивки с использованием ДНК полимеразы VENT (NEB):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (120 с (ПЦР) или 180 с (сшивка)]
1×[72°С (240 с)]
Продукт ПЦР-сшивки 3000 п.о. очищали в геле и подвергали рестрикции NotI и BamHI и этот фрагмент очищали с помощью ПЦР и лигировали в гидролизованный NotI/BamHI и очищенный в геле p7313ie с образованием p73i-Tngr. Анализ последовательности показал, что последовательность р17р24, полученная из p73i-GN2, не была полностью оптимизирована по кодонам, и что эта последовательность была перенесена в новую плазмиду. Эту последовательность очищали путем вырезания фрагмента 700 п.о. из p73i-Tngr Munl и Agel и замены его 700 п.о. продуктом ферментативного гидролиза Munl/Agel из p73i-Tgrn с образованием конструкции p73i-Tngr#6.
Пример 17:
Плазмида: p73i-Trng (#11)
Целевой ген
Инактивированный оптимизированный по кодонам RT, укороченный Nef и участок р17/р24 оптимизированного по кодонам гена gag и из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Порядок генов в этой конструкции был достигнут с помощью ПЦР-амплификации генов RT-trNef и р17р24 из p73i-Tgrn. Осуществляли ПЦР-сшивку двух фрагментов ДНК и продукт 3 т.п.о. очищали в геле и рестрицировали NotI/BamHI, после чего лигировали с гидролизованным NotI/BamHI p7313ie и получили p73i-Trng (#11).
Праймеры:
RTw229k-trNef
RT3-U:1
GAATTCGCGGCCGCGATGGGCCCCATCAGTCCCATCGAGACCGTGCCGGTGAAGCTGAAACCCGGGAT [SEQ ID NO:42]
AS-Nef-p17p24opt линкер
CAGTACCGAAGCTCGGGCACCCATGCAGTTCTTGAACTACTCCGG [SEQ ID NO:43]
P17p24
S-p17p24opt
ATGGGTGCCCGAGCTTCGGTACTG [SEQ ID NO:44]
AS-p17p24opt (BamHI)
GATGGGGGATCCTCACAACACTCTGGCTTTGTGTCC [SEQ ID NO:45]
Условия ПЦР для индивидуальных продуктов и сшивки с использованием ДНК полимеразы VENT (NEB):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (120 с (ПЦР генов) или 180 с (сшивка)]
1×[72°С (240 с)]
Продукт ПЦР сшивки 3000 п.о. очищали в геле и подвергали рестрикции NotI и BamHI и этот фрагмент очищали с помощью ПЦР и лигировали в гидролизованный Notl/BamHI и очищенный в геле p7313ie с образованием p73i-Tngr.
Пример 18:
p73i-Tgnr (#f1)
Целевой ген
Участок р17/р24 оптимизированного по кодонам gag, укороченный Nef и инактивированный оптимизированный по кодонам ген RT и из ВИЧ-1 изолят В штамм НХВ2 вниз по течению от короткого промотора HCMV + экзон 1 и вверх по течению от сигнала полиаденилирования β-глобина кролика.
Порядок генов в этой конструкции был достигнут с помощью ПЦР-амплификации p17p24-trNef и RTw229k из плазмид p73i-GN2 и p73i-RTw229k соответственно. Осуществляли ПЦР-сшивку и продукт очищали в геле и рестрицировали Notl/BamHI, после чего лигировали с гидролизованным Notl/BamHI p7313ie. Две ошибки последовательности, локализованные в последовательности (р17р24 и RT), последовательно репарировали путем замены правильными участками этих генов, используя сайты рестрикции внутри полипротеина (см. фиг.19).
Праймеры:
P17p24-trNef ПЦР
S-p17p24opt
ATGGGTGCCCGAGCTTCGGTACTG [SEQ ID NO:46]
AS-Nef-coRT линкер
GATGGGACTGATGGGGCCCATGCAGTTCTTGAACTACTCCGG [SEQ ID NO:47]
RTw229k
S-coRT
ATGGGCCCCATCAGTCCCATCGAG [SEQ ID NO:48]
RT3-L:1
GAATTCGGATCCTTACAGCACCTTTCTAATCCCCGCACTCACCAGCTTGTCGACCTGCTCGTTGCCGC [SEQ ID NO:49]
Условия ПЦР для индивидуальных продуктов и сшивки с использованием ДНК полимеразы VENT (NEB):
1×[94°C (30 c)]
25×[94°С (30 с)/55°С (30 с)/72°С (120 с (ПЦР) или 180 с (сшивка)]
1×[72°С (240 с)]
Продукт ПЦР-сшивки 3000 п.о. очищали в геле и подвергали рестрикции NotI и BamHI и этот фрагмент очищали с помощью ПЦР и лигировали в гидролизованный Notl/BamHI и очищенный в геле p7313ie с образованием p73i-Tngr. Секвенирование выявило, что р17р24 не был полностью оптимизирован, и поэтому фрагмент 700 п.о. вырезали Agel/Munl из кодирующего района и заменяли его фрагментом Munl/Agel из p73i-Tgrn#3, содержащей правильную кодирующую последовательность. Полипротеин также содержал одиночную точечную мутацию (G2609A), приводящую в результате к аминокислотной замене Th2 на Ala в участке RT полипротеина. Эту мутацию корректировали ферментативным гидролизом конструкции Apal/BamHI и очисткой с помощью ПЦР для удаления мутированной последовательности, которую заменяли путем лигирования с гидролизованным Apal/BamHI участком RT из p73i-Tgnr.
Пример 19:
Получение покрытой плазмидой «золотой суспензии» для ДНК-картриджей «генного ружья»
Плазмидную ДНК (примерно 1 мкг/мкл), примерно 100 мкг, и 2 мкм золотых частиц, примерно 50 мг (PowderJect), суспендировали в 0,05 М спермидине, примерно 100 мкл (Sigma). ДНК осаждали на золотых частицах путем добавления 1 М CaCl2, примерно 100 мкл (American Pharmaceutical Partners, Inc., USA). Комплекс ДНК/золото инкубировали в течение 10 минут при комнатной температуре, промывали 3 раза в абсолютном этаноле, примерно 3×1 мл (предварительно высушенном на молекулярном сите 3Å (BDH)). Образцы ресуспендировали в абсолютном этаноле, содержащем 0,05 мг/мл поливинилпирролидона (PVP, Sigma), и разделяли на три равные аликвоты в 1,5 мл микроцентрифужных пробирках (Eppendorf). Эти аликвоты предназначены для анализа (а) «золотой суспензии», (б) элюата - плазмиды, элюированной из (а) и (б) для получения покрытых плазмидой/золотом картриджей Tefzel для «генного ружья» (см. пример 3 ниже). Для получения образцов (а) и (б) пробирки, содержащие плазмидную ДНК/»золотую суспензию» в этаноле/PVP, центрифугировали в течение 2 минут при максимальной скорости в микроцентрифуге Eppendorf 5418, супернатант удаляли, и «золотую суспензию» высушивали в течение 10 минут при комнатной температуре. Образец (а) ресуспендировали до 0,5-1,0 мкг/мкл плазмидной ДНК в ТЕ рН 8,0, допуская примерно 50% покрытия. Для элюции образец (б) ресуспендировали до 0,5-1,0 мкг/мкл плазмидной ДНК в ТЕ рН 8,0 и инкубировали при 37°С в течение 30 минут, энергично встряхивая, а затем центрифугировали в течение 2 минут при максимальной скорости в микроцентрифуге Eppendorf 5418, и супернатант, элюат, отделяли и хранили при -20°С. Точную концентрацию элюированной ДНК определяли с помощью спектрофотометрического количественного определения, используя Genequant II (Pharmacia Biotech).
Пример 20:
Препараты картриджей для ДНК-иммунизации
Изготовление препаратов картриджей для устройства переноса генов Accell осуществляли, как описано ранее (Eisenbraun et al DNA and Cell Biology, 1993, Vol 12, No 9, pp.791-797; Pertner et al). Коротко, плазмидную ДНК наносили в виде покрытия на 2 мкм золотые частицы (DeGussa Corp., South Plainfield, N. J., USA) и загружали в трубки Tefzel, которые после этого резали на отрезки длиной 1,27 см, которые служили в качестве картриджей, и хранили их в эксикаторе при 4°С до использования. При типичной вакцинации каждый картридж содержал 0,5 мг золотого покрытия с 0,5 мкг суммарной ДНК/картридж.
Пример 21:
Иммунный ответ на антигены ВИЧ после ДНК-вакцинации с использованием генного ружья
Мышей (n=3/группа) вакцинировали антигенами, кодируемыми нуклеиновой кислотой и локализованными в двух векторах. В р7077 использован промотор IE HCMV, включающий интрон А и экзон 1 (промотор fcmv). P73i доставляет тот же антиген, но содержит промотор HCMV IE (промотор icmv), в котором отсутствует интрон А, но включен экзон 1.
Плазмиду доставляли в выбритый участок кожи живота, являющийся мишенью, мышей F1 (C3H×Balb/c). Мышам давали первичную иммунизацию 2×0,5 мкг ДНК на сутки 0, ревакцинировали 2×0,5 мкг ДНК на сутки 35 и определяли клеточный ответ на сутки 40, используя анализ ELIspot ИФН-гамма.
- P73I - пустой вектор
- Р7077 - пустой вектор
- p7077 GRN - (промотор f CMV) Gag, RT, Net
- P73I GRN - (промотор i CMV) Gag, RT, Net
- P73I GR3N - (промотор CMV) оптимизированный Gag, оптимизированный RT, Nef
- P7077 GN - (промотор f CMV) Gag, Nef
- P73I GN - (промотор i CMV) Gag, Nef
Цитотоксические Т-клеточные ответы
Цитотоксический Т-клеточный ответ оценивали на основании ограниченного Т-клеточным CD8+ анализа ELIspot IFN-γ спленоцитов, собранных через 5 суток. Мышей забивали путем цервикальной дислокации и собирали селезенки в охлажденном во льду ФСБ. Спленоциты препарировали в фосфатно-солевом буферном растворе (ФСБ), после чего осуществляли лизис красных кровяных клеток (1 минута в буфере, состоящем из 155 мМ NH4Cl, 10 мМ КНСО3, 0,1 мМ ЭДТА). После двух отмывок в ФСБ для удаления твердых частиц однородную клеточную суспензию разделяли на аликвоты в планшеты ELISPOT, предварительно покрытые антителом захвата IFN-γ, и стимулировали соответствующим пептидом, ограниченным CD8 (Gag, Nef или RT). После культивирования в течение ночи клетки, продуцирующие IFN-γ, визуализировали путем нанесения антитела против мышиного IFN-γ, меченного биотином (Pharmingen), а затем конъюгированной со стрептавидином щелочной фосфатазы, и проводили количественное определение, используя анализ изображения.
Результаты данного эксперимента представлены на фиг.20, 21 и 22.
Пример 22:
Иммуногенность вакцинных конструкций
1. Клеточные анализы
Клеточный иммунный ответ включает цитотоксические клетки CD8 и хелперные клетки CD4. Чувствительным способом обнаружения специфичных клеток CD8 и CD4 является ELISPOT-анализ, который можно использовать для количественного определения числа клеток, способных секретировать интерферон-γ или ИЛ-2. ELISPOT-анализ основан на захвате цитокинов, секретируемых из индивидуальных клеток. Коротко, специализированные микротитровальные планшеты покрывают антителами против цитокина. Спленоциты, выделенные из иммунизированных животных, инкубируют в течение ночи в присутствии специфичных пептидов, представляющих известные эпитопы (CD8), или белков (CD4). Если клетки стимулированы на высвобождение цитокинов, они будут связываться с антителами на поверхности планшета, окруженные локально индивидуальными продуцирующими клетками. Цитокины остаются прикрепленными к антителу покрытия после лизиса клеток, и после этого планшеты промывают. Этот анализ проявляют тем же способом, что и ELISA-анализ (твердофазный иммуноферментный анализ), с использованием системы амплификации биотин/авидин. Число пятен равно числу клеток, продуцирующих цитокины.
Ответы CD8 на следующие K2d-ограниченные мышиные эпитопы: Gag (AMQMLKETI), Nef (MTYKAAVDL) и RT (YYDPSKDLI), и ответы CD4 на белки Gag и RT регистрировали для всех 6 конструкций. Результаты этих анализов анализировали статистически, и конструкции оценивали в соответствии с их иммуногенностью. Результат показан на фиг.23.
2. Гуморальные анализы
Образцы крови собирали для анализа на антитела на 7 и 14 сутки после ревакцинации от двух экспериментов. Сыворотку отделяли и хранили в замороженном состоянии до измерения титра антител с использованием специфичных ELISA-анализов. Все образцы тестировали на антитела к Gag, Nef и RT. Коротко, планшеты для ELISA покрывали соответствующим белком. Избыток белка отмывали, после чего разводили образцы сыворотки и инкубировали в планшетах. Образцы сыворотки отмывали, и антисыворотку против мыши конъюгировали с подходящей меткой. Планшет проявляли и считывали на считывающем устройстве для планшетов. Результаты представлены на фиг.24.
3. Данные по антителам
Титры антител измеряли для всех шести конструкций в четырех экспериментах. Конструкция p73i-GNR постоянно не давала ответов антител на Gag и давала ограниченные ответы антител на Nef. Причина этого неясна, поскольку Т-клеточные ответы наблюдали от спленоцитов, выделенных из тех же мышей, что указывало на экспрессию белка Gag in vivo.
Ранжирование образования специфичных антител Gag представляло собой:
RHG>GRN>NRG>RGN>NGR>GNR
Анализ данных клеточной иммунологии
Задачей было ранжирование 6 конструкций на основании данных счета пятен в результате 3 иммунологических экспериментов. Оценивали три серии ответов:
Ответы CD8 на Gag, Nef и RT на сутки 7 (7 суток после первичной иммунизации)
Ответы CD4 на Gag и RT на сутки 35 (7 суток после ревакцинации)
Ответы CD8 на Gag, Nef и RT на сутки 35 (7 суток после ревакцинации)
Каждый ответ (например, ответ CD8 на Gag) моделировали, используя модель линейного смешанного эффекта, в программе SAS версия 8. Модель включала фиксированные эффекты конструкции, присутствует или отсутствует конкретный антиген (Gag, Nef или RT) и присутствует или отсутствует ИЛ-2. Кроме того, для ответов CD8, где данные были доступны от каждой индивидуальной мыши, субъект включали в качестве случайного эффекта в модель. Модель включала взаимосвязанные параметры, позволяющие дифференцировать эффект конструкции для каждой комбинации антигенов (присутствие/отсутствие) и ИЛ-2 (присутствие/отсутствие).
На основании этой модели разность в скорректированном среднем ответе между каждой конструкцией и р7313 (контрольная группа) оценивали отдельно для каждой комбинации антигенов (присутствие/отсутствие) и ИЛ-2 (присутствие/отсутствие) вместе с р-значением, указывающим на статистическую значимость этой разности. На основании разностей и р-значений в присутствии антигена и в отсутствие ИЛ-2 конструкции ранжировали, обозначая баллом 6 конструкцию с самой большой разностью, 5 со следующей самой большой и т.д., но 0 для любых конструкций, где разность не была статистически значимой при 5% уровне.
Допущения данной модели - что остатки были распределены в соответствии с нормальным распределением с постоянной вариансой, оценивали, используя графические методы и анализы чувствительности, где моделировали, во-первых, логарифмическое преобразование и, во-вторых, квадратный корень ответа. Ранжирование конструкций не было чувствительным к отклонениям от допущений модели.
Вычислив ранжирования для каждого ответа в каждом эксперименте отдельно, суммарное ранжирование для 3 серий ответов вычисляли по всем трем экспериментам. В приведенной ниже таблице показано суммарное ранжирование по всем трем экспериментам.
Суммарное ранжирование конструкций для каждой из 3 серий ответов, объединенное по всем 3 иммунологическим экспериментам
RNG имеет самый высокий показатель для обеих серий ответов на сутки 35 и второй самый высокий показатель после RGN на сутки 7. RGN также проявляет высокие показатели для обеих серий ответов на сутки 35.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| ВАКЦИНА ПРОТИВ ВИЧ/СПИД | 2010 |
|
RU2475264C2 |
| ХИМЕРНЫЙ ГЕН CR3 И КОДИРУЕМЫЙ ИМ ХИМЕРНЫЙ БЕЛОК CR3 (ВАРИАНТЫ), ИНДУЦИРУЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ-1 | 2002 |
|
RU2302461C2 |
| ВИЧ ВАКЦИНА, ОСНОВАННАЯ НА НАПРАВЛЕННОСТИ МАКСИМИЗИРОВАННЫХ Gag И Nef НА ДЕНДРИТНЫЕ КЛЕТКИ | 2009 |
|
RU2539765C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| СПОСОБ ПОДАВЛЕНИЯ ВИЧ-ИНФИЦИРОВАННЫХ КЛЕТОК МЛЕКОПИТАЮЩЕГО И БЕЛКОВЫЙ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2165703C2 |
| ИСКУССТВЕННЫЙ БЕЛОК-ИММУНОГЕН TCI, СОДЕРЖАЩИЙ МНОЖЕСТВЕННЫЕ CTL-ЭПИТОПЫ ОСНОВНЫХ АНТИГЕНОВ ВИЧ-1, ИСКУССТВЕННЫЙ ГЕН TCI, КОДИРУЮЩИЙ ПОЛИЭПИТОПНЫЙ БЕЛОК-ИММУНОГЕН TCI | 2002 |
|
RU2238946C2 |
| Антитела против белка р17 ВИЧ-1 субтипа А | 2019 |
|
RU2727673C1 |
| СИНТЕТИЧЕСКИЙ ГЕН, КОДИРУЮЩИЙ ЗЕЛЕНЫЙ ФЛУОРЕСЦЕНТНЫЙ БЕЛОК, СПОСОБ ПОЛУЧЕНИЯ ГЕНА, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУРЫ КЛЕТОК | 1997 |
|
RU2233329C2 |



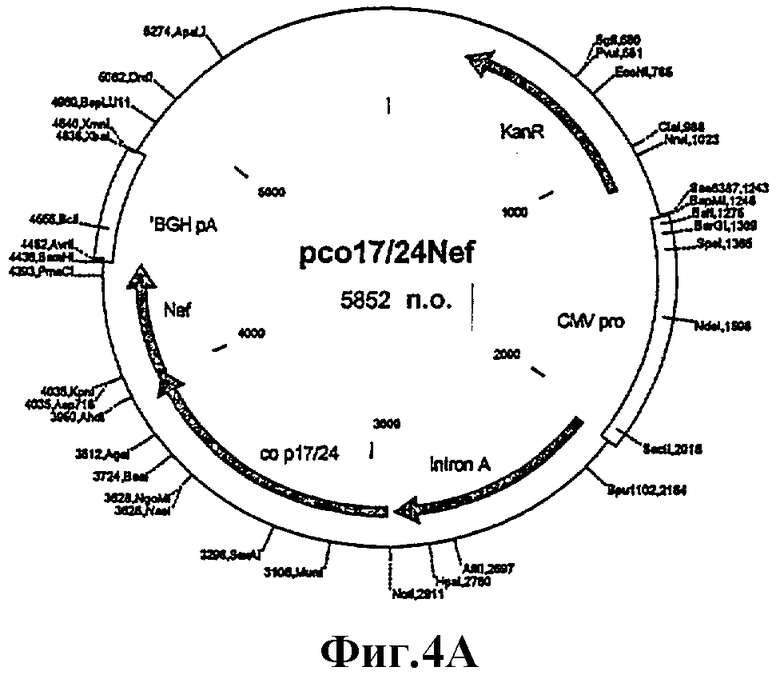















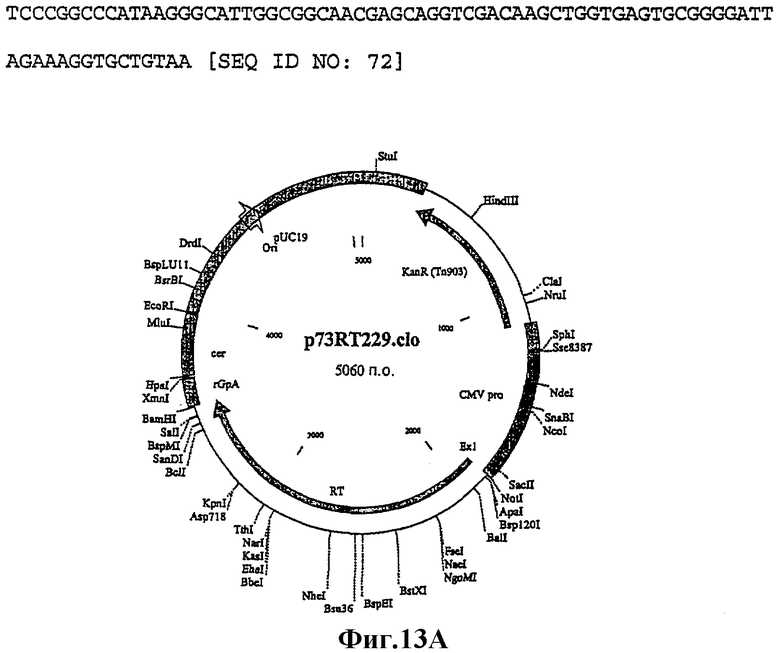
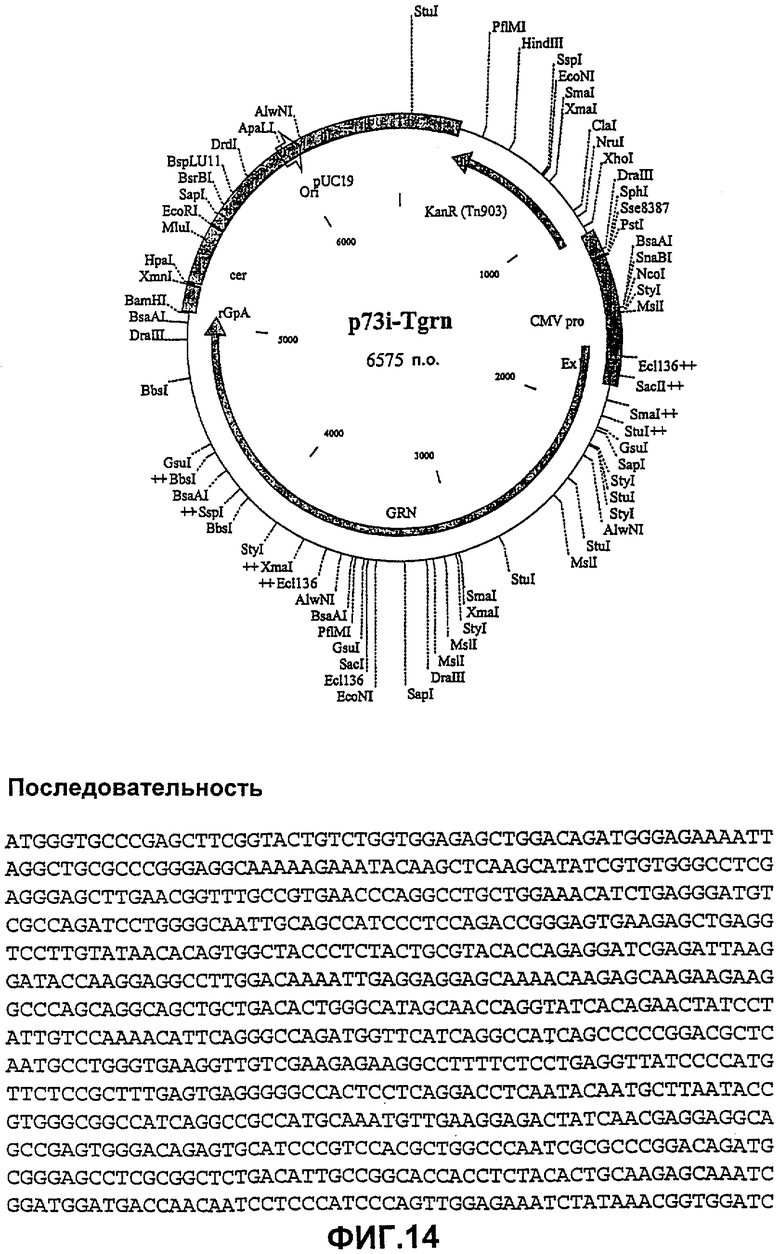




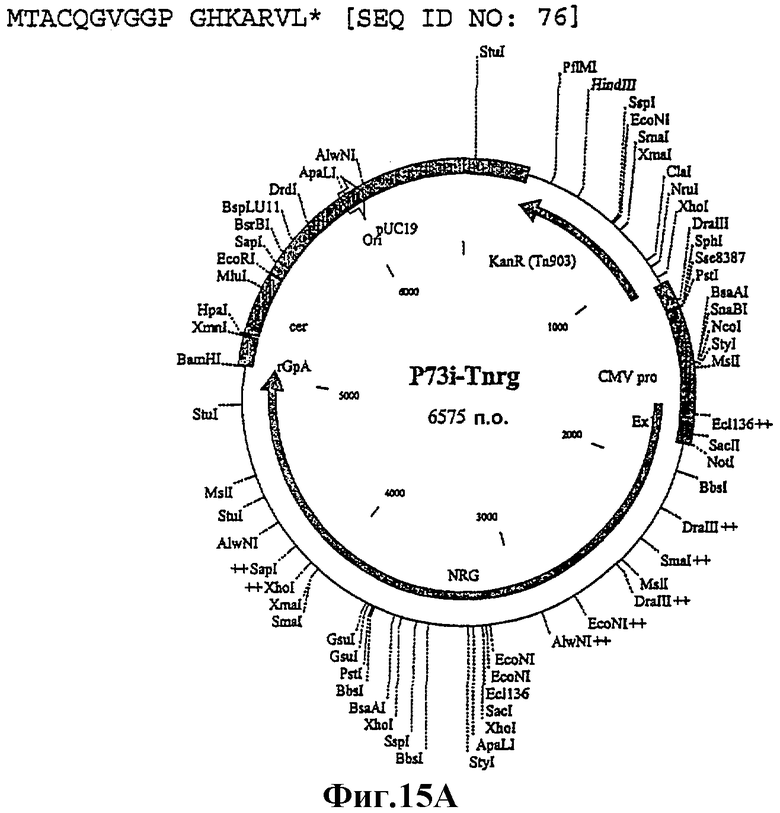








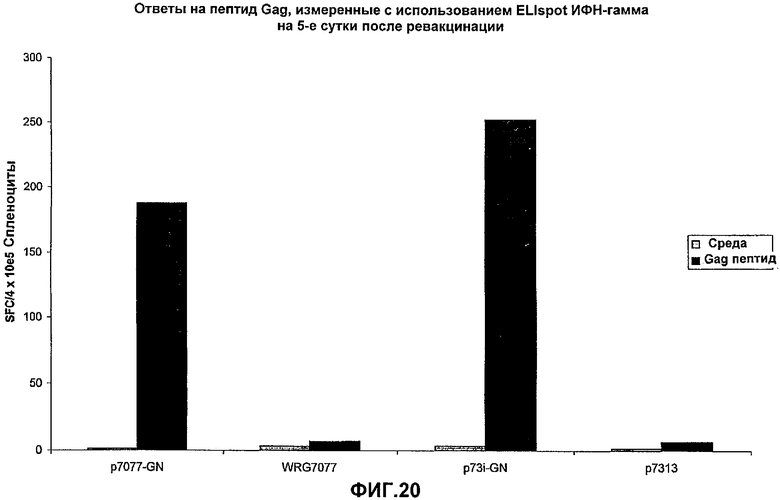

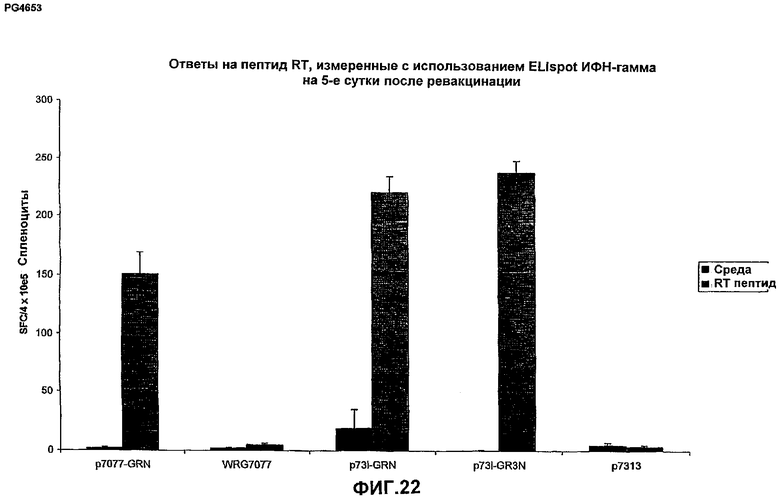
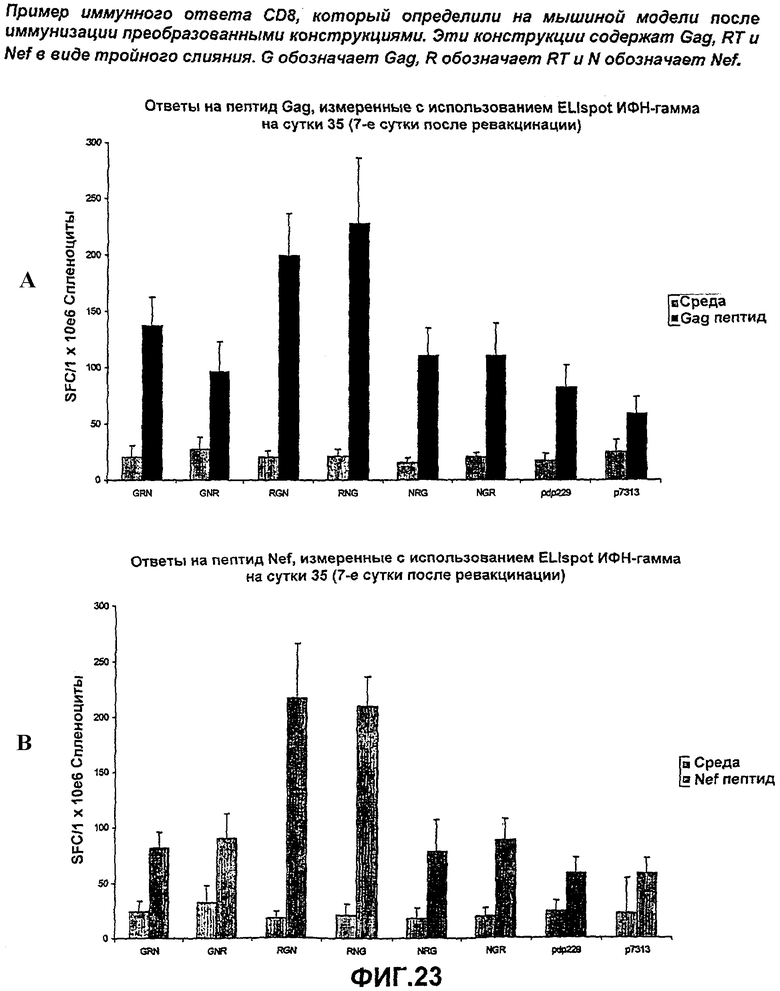
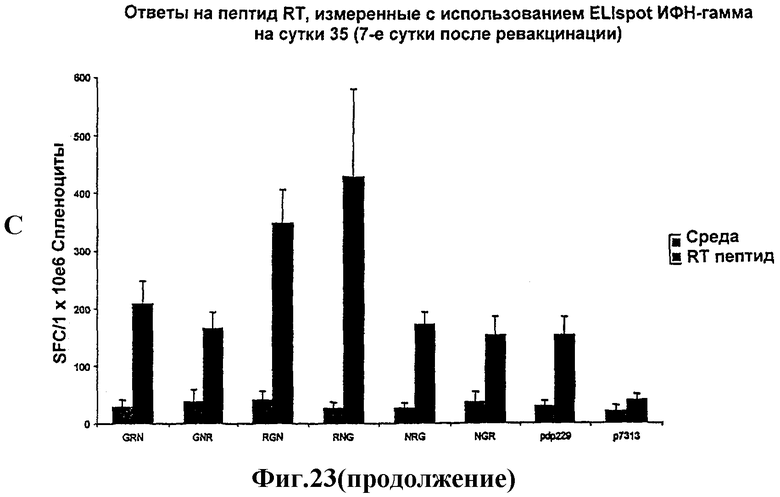
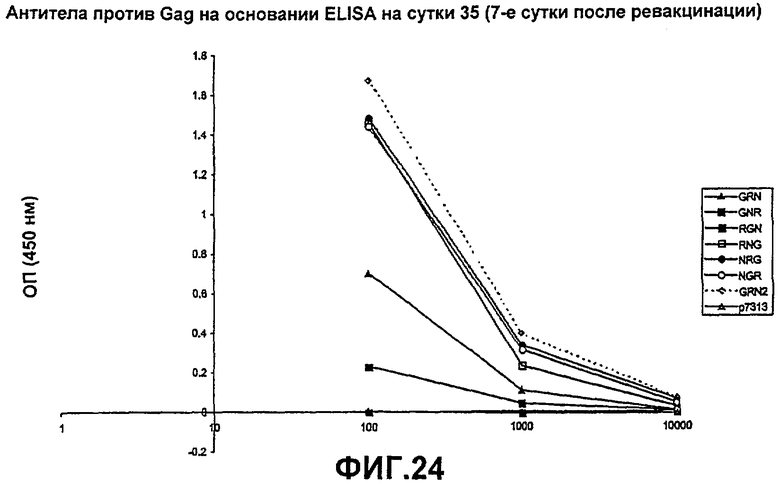
Изобретение относится к нуклеиново-кислотным конструкциям, а также к вакцинным препаратам, содержащим такие конструкции, и к применению таких препаратов в медицине. Нуклеотидная последовательность, которая кодирует белок gag ВИЧ-1 (HIV-1) или его фрагмент, содержащий эпитоп gag, и белок Nef ВИЧ-1 или его фрагмент, содержащий эпитоп nef, и белок RT или его фрагмент, содержащий эпитоп RT, оперативно сцеплена с гетерологичным промотором, причем последовательность gag оптимизирована по кодонам. Данная последовательность входит в состав фармацевтической композиции и используется для лечения и/или профилактики ВИЧ-инфекции и СПИДа. 7 н. и 9 з.п. ф-лы, 3 табл., 24 ил.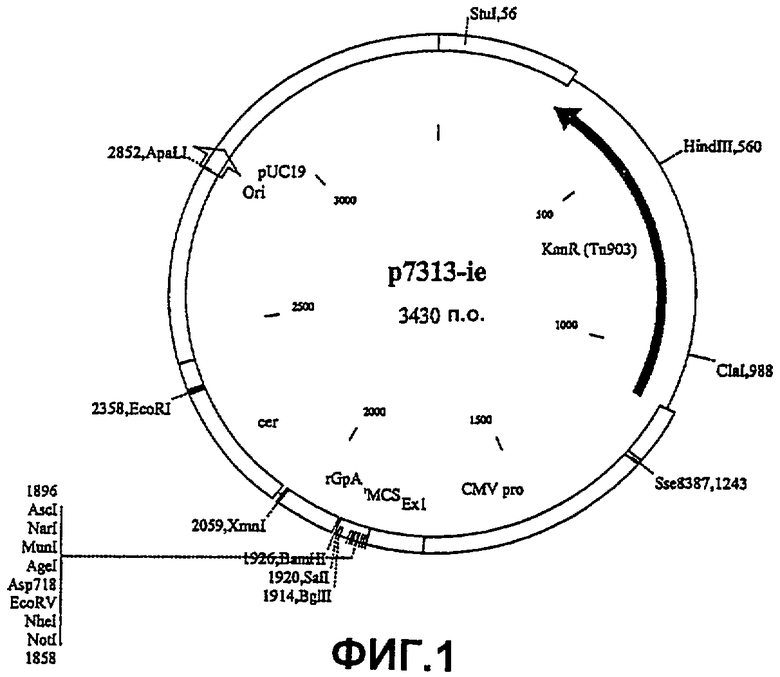
| Приспособление для шуровки топлива в топках с движущейся решеткой | 1934 |
|
SU39302A1 |
| HUANG Y | |||
| et al., "Human immunodeficiency virus type 1-specific immunity after genetic immunization is enhanced by modification of Gag and Pol expression", J | |||
| Virol | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| KOTSOPOULOU E | |||
| et al., "A Rev-independent human immunodeficiency virus type 1 (HIV-1)-based vector that exploits a codon-optimized HIV-1 gag-pol gene", J Virol | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |

