Изобретение относится к области молекулярной биотехнологии и касается Т-лимфоцитов, имеющих новый рецептор.
В процессе иммунного ответа на антигены бактерий, вирусов и опухолей в организме формируется популяция клеток памяти - лимфоцитов, способных к ускоренному и усиленному ответу на тот же антиген, введенный повторно. Встреча организма с антигеном ведет к дифференцировке наивных Т-лимфоцитов в эффекторные клетки, а затем в клетки памяти. Это пул клеток, составляющий около 5% от общего количества лимфоцитов, способных длительно персистировать и осуществлять эффективный иммунный ответ при повторной встрече организма с антигеном (Rocha В., 1997; Jacob J. et al., 1997, Rocha В., 1999, Murali K.K. et al., 1998). Все известные методы вакцинации основаны на возникновении клеток памяти.
Специфическое узнавание антигена Т-лимфоцитами происходит благодаря наличию Т-клеточного рецептора (ТКР). ТКР состоит из α- и β-цепей, которые являются уникальными для каждого клона Т-лимфоцитов. Пул Т-лимфоцитов организма представляют собой смесь клеток, на поверхности каждой из которых присутствует своя уникальная комбинация α- и β-цепей ТКР, которая передается ее потомкам, образующимся в результате деления (клонам).
Получение Т-лимфоцитов с ТКР нужной специфичности происходит в несколько этапов:
1. Проведение Т-клеточного клонирования. В пуле Т-лимфоцитов индуцируют размножение отдельных клеток с нужной специфичностью ТКР путем добавления к ним специфического антигена. Чтобы получить достаточное количество таких клеток, проводят несколько раундов рестимуляции их пролиферации специфическим антигеном. В результате получают необходимое количество Т-лимфоцитов, несущих ту же комбинацию α- и β-цепей ТКР, что и исходная клетка (клон). Данная процедура требует значительных временных и технических затрат, связанных с длительностью роста индивидуальных клонов и необходимостью подбора и обеспечения достаточно сложных (иногда индивидуальных) условий их поддержания и дальнейшего тестирования.
2. Проведение генного клонирования, идентификация и определение нуклеотидных последовательностей α- и β-цепей каждого клона Т-лимфоцитов. Полученные генетические последовательности при помощи экспрессионных векторов вводят в активированные Т-лимфоциты, в результате чего их потомки приобретают способность к экспрессии ТКР с новой заданной специфичностью.
Этот подход был использован для адоптивной иммунотерапии опухолей в онкологии [Robbins PF, Morgan RA, Feldman SA., Yang JC, Sherry RM, Dudley ME, Wunderlich JR, Nahvi AV, Helman LJ, Mackall CL, Kammula US, Hughes MS, Restifo NP, Raffeld M, Lee CC, Levy CL, Li YF, El-Gamil M, Schwarz SL, Laurencot C, Rosenberg SA.Tumor regression in patients with metastatic synovial cell sarcoma and melanoma using genetically engineered lymphocytes reactive with NY-ESO-1.J Clin Oncol. 2011 Mar 1; 29(7):917-24.; Blankenstein T, Leisegang M, Uckert W, Schreiber H. Targeting cancer-specific mutations by T cell receptor gene therapy. Curr Opin Immunol. 2015; 33:112-9].
Недостатки данного подхода: необходимость проведения процедуры клеточного клонирования. Использована полная конфигурация ТКР, состоящая из α- и β-цепей.
Ранее мы обнаружили, что среди Т-лимфоцитов присутствуют Т-лимфоциты с ТКР, специфичность которых к заданному антигену определяет только α-цепь [Казанский Д.Б. Трансгенные технологии создания иммунологической защиты организма. В сборнике докладов семинара Фонда перспективных исследований "Проблемные вопросы иммунологии", 03 октября 2014 г., Москва, ООО "Б-принт", 2015, С. 17-25]. Такие ТКР распознают антиген вне зависимости от того, в комбинации с какой β-цепью он присутствует на мембране Т-лимфоцита. Это дает возможность избежать процедуры клеточного клонирования и отказаться от трансгенного переноса α-цепей при получении ТКР с нужной специфичностью. В данном способе технология идентификации нужного рецептора сводится к получению библиотек кДНК из поликлональных клеток памяти и определению и секвенированию генов биологически активных α-цепей методом секвенирования нового поколения (NGS-секвенирования). Данный подход позволяет значительно сократить процедуру поиска и идентификации нужных рецепторов и облегчает техническое выполнение переноса в клеточный геном генетических конструкций благодаря уменьшению их размеров. Этот способ предполагает использование индивидуальных трансгенных α-цепей, которые при взаимодействии с эндогенными β-цепями, присутствующими в клетке, формируют ТКР для защиты от онкологических заболеваний.
В качестве одного из подходов для лечения некоторых раковых заболеваний (меланомы, лейкозов) используют лимфоциты с генетически модифицированным химерным рецептором (CAR) (С.Slaney, B.Scheidt, А. Davenport, P.Beavis, J.Westwood, S.Mardiana, D.Tscharke, S.Ellis, H. M. Prince, J.Trapani, R. Johnstone, M. Smyth, M.Teng, A. Ali, Z.Yu, S. Rosenberg, N. Restifo, P. Neeson, P. Darcy, M.Kershaw. Dual-specific Chimeric Antigen Receptor T Cells and an Indirect Vaccine Eradicate a Variety of Large Solid Tumors in an Immunocompetent, Self-antigen Setting. Clin Cancer Res; 23(10) May 15, 2017; Kershaw MH, Westwood JA, Darcy PK. Gene-engineered T cells for cancertherapy. Nat Rev Cancer 2013; 13:525-41.).
Недостаток данного подхода: CAR - это полностью искусственный рецептор, сочетающий элементы Т- и В-клеточных рецепторов. Для изготовления химерного рецептора приходится использовать фрагменты нескольких различных молекул и результирующий белок часто становится иммуногенным [Maus MV, Haas AR, Beatty GL, Albelda SM, Levine BL, Liu X, Zhao Y, Kalos M, June CH. T cells expressing chimeric antigen receptors can cause anaphylaxis in humans. Cancer Immunol Res. 2013 Jul; 1(1):26-31. doi: 10.1158/2326-6066.CIR-13-0006].
Известен подход, направленный на усиление иммунного ответа при помощи получения высокоаффинных ТКР путем отбора и мутагенеза цепей ТКР [Schendel D, Wilde S, Frankenberger В, Uckert W. High affinity T cell receptor and use thereof. Patent US 8697854 B2. Priority date 2008-11-24; Jakobsen BK, Harwood N, Liddy NR T cell receptors. Patent WO 2011001152 A1; Priority date 2009-07-03].
Отличие данного подхода от заявляемого состоит в использовании полной конфигурации ТКР, состоящей из α- и β-цепей.
Еще один способ терапии опухолей трансгенными ТКР - ретровирусная трансдукция Т-лимфоцитов в культуре in vitro с их последующей активацией и получением лимфокинактивированных киллеров [Schendel DJ. Expression of transgenic t cell receptors in lak-t cells. Patent US 20110020308 Al. Priority date: 2006-12-12].
Отличие данного подхода от предлагаемого в использовании полной конфигурации ТКР, состоящей из α- и β-цепей.
Известен метод усиления иммунного ответа на раковые опухоли и патогены, где описаны модифицированные Т-лимфоциты, одновременно экспрессирующие ТКР или химерный антигенный рецептор и антитело (Патент WO 2017147383A1 (2017-08-31)).
Недостатком данного метода в необходимости экспрессии трансгенной β-цепи ТКР и процедуры Т-клеточного клонирования для идентификации α- и β-цепей таких ТКР.
Предлагается метод усиления иммунного ответа путем модификации различных ТКР, приводящей к увеличению уровня их экспрессии на поверхности Т-лимфоцита (Патент WO 2016170320 A1 (2016-10-27)).
Недостатком данного метода является необходимость присутствия в генетической конструкции ТКР β-цепи и необходимость процедуры Т-клеточного клонирования для идентификации α- и β-цепей таких ТКР.
Задачей заявляемого изобретения является: разработка нового способа создания противоопухолевой иммунологической защиты с помощью трансгенеза Т-лимфоцитов в системе in vitro и in vivo.
Задача решается получением Т-лимфоцитов с экспрессией α-цепи ТКР. Трансдукцию Т-лимфоцитов мыши проводили ретровирусными частицами, содержащими ген α-цепи ТКР 1D1 (SEQ ID NO 1, GenBank: DQ983579.1) с последующим адоптивным переносом модифицированных клеток мышам -реципиентам с целью оценки функциональной активности трансдуцированных клеток in vitro и in vivo.
Технический результат изобретения: разработан способ создания противоопухолевой иммунологической защиты с помощью трансгенеза Т-лимфоцитов.
Изобретение иллюстрируется фиг. 1, табл. 1-3 и перечнем использованных последовательностей.
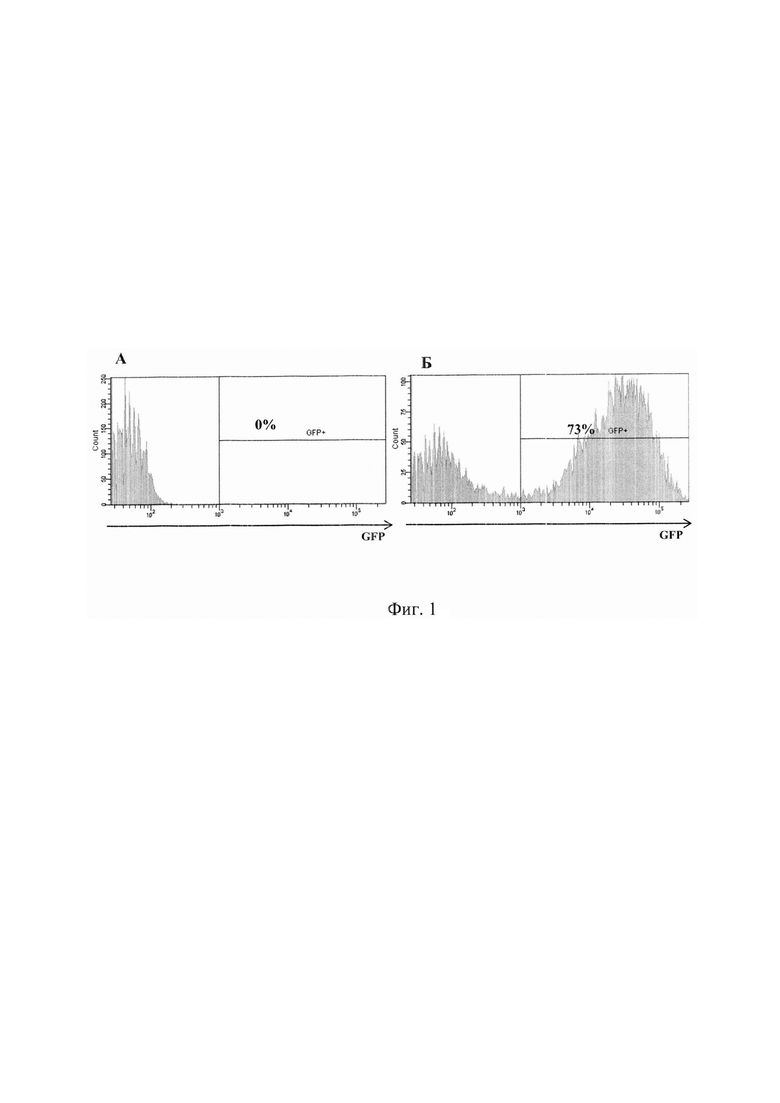
На фиг. 1 представлена оценка уровня трансдукции клеток вирусом Migr1-PGK-1-GFP методом проточной цитофлуориметрии. А- нетрансдуцированные клетки (контроль); Б- Т-клетки, трансдуцированные вирусом.
На табл. 1 представлена оценка функциональной активности трансдуцированных Т-лимфоцитов в системе in vitro. Соотношение киллер-мишень 10:1.
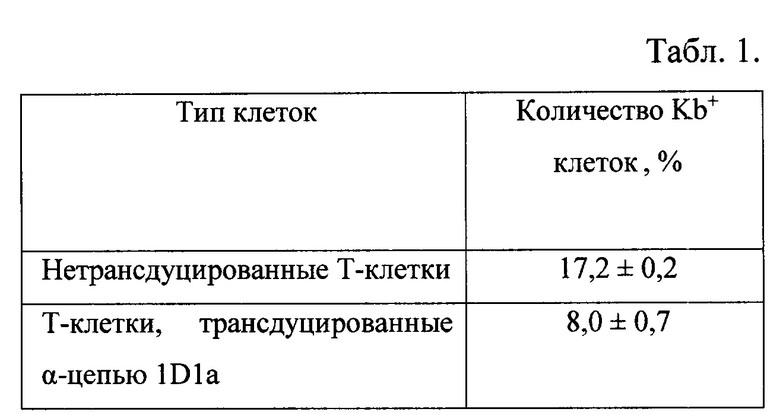
На табл. 2 представлена оценка функциональной активности трансдуцированных Т-лимфоцитов в системе in vivo. Абсолютное количество клеток лимфомы EL-4 на 6 день после адоптивного переноса Т-лимфоцитов. Соотношение киллер-мишень 10:1.
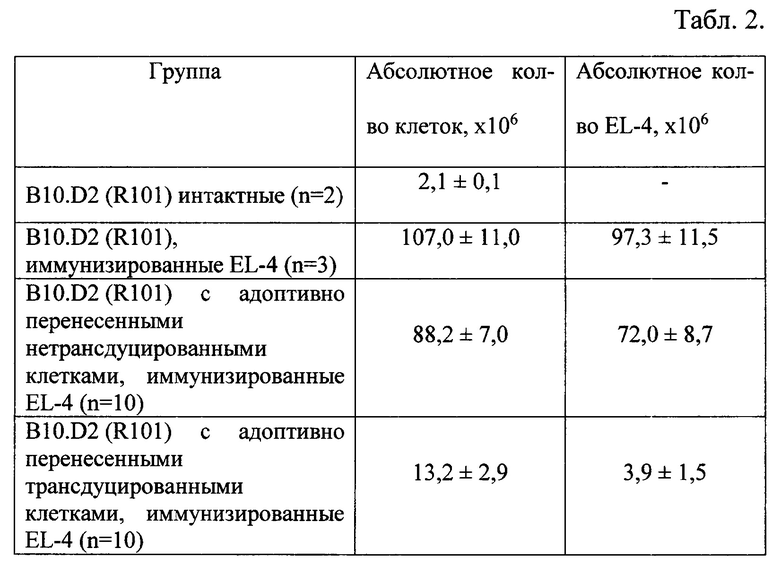
На табл. 3 представлена оценка защитной активности трансдуцированных Т-лимфоцитов в системе in vivo. Анализ клеток EL-4 в перитонеальной полости мышей B10.D2(R101) с адоптивно перенесенными трансдуцированными Т-лимфоцитами за 7, 14 и 28 дней до введения клеток опухоли. Соотношение киллер-мишень 10:1.
Способ осуществляется следующим образом.
1. Создание генетических конструкций.
В работе для сборки частиц ретровируса использовали два ДНК-вектора с генетическими конструкциями. Для получения первого ДНК-вектора коммерческую генетическую конструкцию MigR1 (Addgene, США), содержащую ген зеленого флуоресцентного белка (green fluorescent protein, GFP), модифицировали путем удаления из нее последовательности гена GFP и введения промотора PGK и гена α-цепи ТКР 1D1 (SEQ ID NO 1). Созданный вектор получил название MigR1-PGK-1D1a. Так как к цепи 1D1a не существует коммерческих антител, то для оценки уровня трансдукции была создана вторая генетическая конструкция, которая была получена также на основе MigR1 (Addgene, США) путем вставки промотора PGK в область сайта для множественного клонирования этой плазмиды. Полученный таким образом вектор MigR1-PGK-GFP, содержащий ген GFP, использовали в дальнейшей работе для контроля эффективности процедуры введения генетических конструкций в целевые клетки.
В качестве второго ДНК-вектора использовали коммерческую генетическую конструкцию pCL-Eco (Addgene, США). Данная плазмида содержит все гены белков, необходимых для сборки вирусной частицы с экотропной оболочкой (белок оболочки (env), полимераза (pol), ревертаза (rev), интеграза).
2. Получение частиц ретровирусов.
2.1. Трансфекция клеток линии HEK293T и получение среды, содержащей вирусные частицы.
2.1.1 Подготовка клеток HEK293T, пакующей вирусные частицы.
Клетки пакующей линии HEK293T выращивали в культуральном флаконе площадью 75 см2(Costar, США) в 12 мл стандартной ростовой среды DMEM (Панэко, Россия), содержащей с 4,5 г/л глюкозы и 10% эмбриональной телячьей сыворотки (Панэко, Россия) до достижения конфлюэнтности 60-80%.
2.1.2. Трансфекция HEK293T, получение вирус-содержащей среды.
В 1,5 мл стерильной воды смешивали плазмиды MigR1-PGK-1D1a, содержащую ген α-цепи ТКР 1D1(SEQ ID NO 1), и pCl-Есо в соотношении 1:1 по 20 мкг каждой плазмиды. К смеси добавляли 75 мкл 2М CaCl2 и перемешивали путем пипетирования. Затем по каплям при помешивании вносили 750 мкл двукратного HBS буфера (50 мМ HEPES, рН 7.05; 10 мМ KCl, 12 мМ декстроза, 280 мМ NaCl, 1,5 мМ Na2HPO4). Смесь инкубировали 5 мин при комнатной температуре и по каплям вносили в среду роста, содержащуюся в культуральных флаконах с клетками пакующей линии. Трансфекцию клеток HEK293T проводили в течение 24 часов, после чего удаляли модифицированную ростовую среду и заменяли на стандартную среду роста DMEM.
Контроль эффективности трансфекции осуществляли при помощи белка GFP. Для этого параллельно при тех же условиях проводили трансфекцию клеток HEK293T плазмидами MigR1-PGK-GFP, содержащей ген GFP, и pCL-Eco. Наличие экспрессии белка GFP в клетках HEK293T оценивали при помощи флюоресцентного микроскопа (Nikon, Япония). Уровень трансфекции экстраполировали на клетки HEK293T с 1D1 геном (SEQ ID NO 1). Трансфекцию считают успешной, если не менее, чем у 60% клеток пакующей линии наблюдают флюоресценцию GFP. Сбор ростовой среды клеток HEK293T проводили через 48 и 72 часа после заражения клеток HEK293T плазмидами MigR1-PGK-1D1a и pCl-Есо. Полученную таким образом вирус-содержащую среду в свежем виде использовали для трансдукции Т-клеток мыши. Возможно замораживание вирус-содержащей среды при температуре -70°С. Допускается однократное размораживание.
3. Получение лимфоцитов из селезенки и лимфатических узлов мыши.
Мышей линии B10.D2(R101) умерщвляли путем цервикальной дислокации и извлекали селезенку и лимфоузлы (мезентериальный, 2 подмышечных и 2 паховых). Селезенку помещали в гомогенизатор Поттера с 3 мл питательной среды RPMI-1640 (Панэко, Россия), содержащей 10 мкг/мл ципрофлоксацина (KRK, Словения). После гомогенизации селезенки суспензию клеток переносили в центрифужную пробирку и осаждали центрифугированием при 1500 об/мин в течение 5 мин при 4°С. Надосадочную жидкость удаляли, осадок клеток ресуспендировали и проводили гипотонический лизис эритроцитов. Для этого к осадку спленоцитов добавляли 360 мкл стерильной дистиллированной воды, пробирку встряхивали в течение 10-20 сек и добавляли 40 мкл 10-кратного фосфатно-солевого буфера и 6-7 мл 1-кратного фосфатно-солевого буфера. Затем клетки осаждали центрифугированием при 1500 об/мин в течение 5 мин при 4°С, удаляли надосадочную жидкость и ресуспендировали осадок в 3 мл питательной среды RPMI-1640 (Панэко, Россия), содержащей 10 мкг/мл ципрофлоксацина (KRK, Словения).
Лимфатические узлы гомогенизировали описанным выше образом, осаждали центрифугированием при 1500 об/мин в течение 5 мин при 4°С. Надосадочную жидкость удаляли, осадок клеток ресуспендировали в 2 мл питательной среды RPMI-1640 (Панэко, Россия) и вносили в суспензию спленоцитов. Полученную смесь клеток осаждали центрифугированием при 1500 об/мин в течение 5 мин при 4°С, удаляли надосадочную жидкость и ресуспендировали в 3 мл полной среды (ПС) для культивирования лимфоцитов, состоящей из питательной среды RPMI 1640 (Панэко, Россия) с добавлением 10% сыворотки эмбрионов крупного рогатого скота (Панэко, Россия), 5×10-5М 2-меркаптоэтанола (Merck, США), 2 мМ L-глютамина (Sigma, США), 20 мМ HEPES (Панэко, Россия) и 10 мкг/мл ципрофлоксацина (KRK, Словения). Количество спленоцитов в суспензии подсчитывали в камере Горяева в присутствии смеси красителей 1% трипанового синего и 1% эозина в соотношении 1:1 и доводили до концентрации 107кл/мл.
3.1. Активация лимфоцитов мыши.
Смесь клеток селезенки и лимфатических узлов мыши в количестве 5×107 клеток помещали в 5 мл ПС в культуральные флаконы с площадью роста 25 см2 (Costar, США). В среду дополнительно вносили митоген Т-лимфоцитов конканавалин А (КонА, Sigma, США) до конечной концентрации 3 мкг/мл и интерлейкин-2 (ИЛ-2) до конечной концентрации 10 МЕ/мл (Neto Е.Н., Coelho A.L., Sampaio A.L. и др. Activation of human T lymphocytes via integrin signaling induced by RGD-disintegrins. Biochim.Biophys. Acta.2007, 1773(2): 176-84). Лимфоциты культивировали в СО2-инкубаторе при 37°С, 5% СО2 в течение 24 часов. Данный подход обеспечивал активацию и пролиферацию только Т-лимфоцитов селезенки и лимфатических узлов мыши (Neveu P.J., Perdoux D. Polyclonal activation of guinea pig spleen lymphocytes- Int. Arch. Allergy. Appl. Immunol. 1985; 78(4):401-405).
4. Трансдукция Т-лимфоцитов мыши вирусными частицами.
Трансдукцию активированных Т-лимфоцитов мыши вирусными частицами, содержащими ген α-цепи ТКР 1D1 (SEQ ID NO 1), проводили в два этапа.
1) Через 24 часа после активации подсчитывали количество спленоцитов с помощью камеры Горяева и переносили в 6-луночный планшет (Costar, США) из расчета 12×106 клеток на лунку. Затем среду отбирали и добавляли вирус-содержащую среду роста клеток HEK293T из расчета 6 мл среды на одну лунку и 3 мл ПС. Конечный объем среды в каждой лунке составлял 9 мл. В лунки планшета также вносили КонА (3 мкг/мл), ИЛ-2 (10МЕ/мл) и полибрен до конечной концентрации 8 мкг/мл. Клетки центрифугировали (спинокулировали) при 2000 g в течение 90 мин при комнатной температуре. Затем лимфоциты помещали в СО2-инкубатор.
2) По истечении 16-20 часов культивирования проводили еще одну смену среды роста спленоцитов на вирус содержащую среду и повторно выполняли трансдукцию спленоцитов при тех же параметрах спинокуляции. По окончании второй спинокуляции спленоциты культивировали в СО2-инкубаторе при 37°С, 5% СО2 в течение 2 часов, после чего проводили замену вирус-содержащей среды на свежую ПС, содержащую ИЛ-2 и КонА в указанных выше концентрациях, и продолжали культивирование в течение 48 часов при тех же условиях.
5. Оценку уровня трансдукции Т- лимфоцитов проводили путем анализа методом проточной цитометрии (FacsCantoII, Beckton Dickenson, США) с использованием флуоресцентно меченных антител к молекуле CD3, которая является специфическим маркером Т- лимфоцитов. Уровень трансдукции определяли по экспрессии белка GFP.
Анализ клеток на проточном цитофлуориметре. При анализе на проточном цитофлуориметре исключали слипшиеся клетки, а также мертвые клетки, окрашенные пропидий йодидом. Среди одиночных живых лимфоцитов определяли Т-лимфоциты по наличию маркера CD3, для которых оценивают уровень экспрессии белка GFP по флуоресценции в области спектра 520 нм.
5.1. Обработка результатов цитофлюориметрического анализа.
Результаты анализа клеток на проточном цитофлуориметре обрабатывали при помощи программного обеспечения FlowJo (Beckton Dickenson, США). Функциональную эффективность трансдуцированных Т-лимфоцитов определяли при уровне трансдукции не менее 50% (Фиг.).
6. Оценка функциональной активности трансдуцированных Т-лимфоцитов в системе in vitro.
Т-лимфоциты, трансдуцированные геном α-цепи 1D1 (SEQ ID NO 1), в количестве 500×103 клеток помещали в культуру с клетками лимфомы EL-4 в соотношении трансдуцированные Т-лимфоциты: EL-4 = 10:1. Культивирование проводили в 96 луночных пластиковых планшетах (Costar, США) в ПС во влажной атмосфере в СО2-инкубаторе при 37°С, 5% СО2 в течение 48 часов. В качестве контроля использовали активированные нетрансдуцированные клетки, а также Т-клетки, трансдуцированные GFP, в аналогичном соотношении.
По истечении времени инкубации методом проточной цитофлуориметрии проводили анализ относительного количества опухолевых клеток, которые определяли при помощи флюоресцентно меченых антител к маркеру Кb (Табл. 1).
Было показано, что Т-лимфоциты, трансдуцированные геном α-цепи 1D1a (SEQ ID NO 1), значительно быстрее устраняют опухолевые клетки лимфомы EL-4 in vitro по сравнению с нетрансдуцированными клетками (контроль). При этом данный эффект являлся специфичным, т.к. в культуре с Т- лимфоцитами, трансдуцированными GFP, обнаруженное количество EL-4 было сопоставимо с контролем (14.1% и 17.2%, соответственно, Табл. 1).
7. Оценка функциональной активности трансдуцированных Т-лимфоцитов в системе in vivo.
7.1. Адоптивный перенос трансдуцированных Т-лимфоцитов. После оценки уровня трансдукции Т-лимфоциты, трансдуцированные ретровирусными частицами, содержащими ген α-цепи ТКР 1D1 (SEQ ID NO 1), вводили внутрибрюшинно мышам линии B10.D2(R101) из расчета 5,0×106 трансдуцированных клеток в 1 мл фосфатно-солевого буфера. В качестве контроля использовали нетрансдуцированные Т-лимфоциты, которые активировали и культивировали аналогично клеткам, подвергшимся трансдукции. Контрольные Т-лимфоциты вводили в количестве, равном количеству адоптивно перенесенных трансдуцированных Т- лимфоцитов.
7.2. Приготовление лаважа брюшной полости.
Через 6 дней после введения клеток опухоли и адоптивного переноса Т-лимфоцитов, мышей умерщвляли методом цервикальной дислокации и извлекали клетки из брюшной полости. Для этого в перитонеальную полость мышей вводили 2 мл охлажденной среды RPMI-1640 и отбирали жидкость. Полученную клеточную суспензию отмывали центрифугированием (200×g, 5 мин) и подсчитывали количество клеток при помощи камеры Горяева в присутствии смеси красителей трипанового синего и эозина. Часть клеток (1×106) использовали для анализа на проточном цитофлуориметре, окрашивая флюоресцентно меченными антителами к молекуле Кb, для определения опухолевых клеток EL-4. Статистическую обработку данных проводили по статистическому критерию для малых выборок Манна-Уитни.
При адоптивном переносе трансдуцированных лимфоцитов в соотношении киллер: мишень 10:1 было обнаружено в 25 раз меньшее количество опухолевых клеток в лаваже мышей - реципиентов на 6 день после иммунизации EL-4 по сравнению с животными, которым вводили нетрансдуцированные лимфоциты (Табл. 2).
8. Определение продолжительности сохранения состояния иммунологической памяти у животных с перенесенными трансгенными Т-лимфоцитами.
Для оценки защитного потенциала трансдуцированных Т-лимфоцитов проводили адоптивный перенос трансдуцированных или нетрансдуцированных (контрольных) Т-лимфоцитов в количестве 5,0×106 клеток/мышь. Через 7, 14 или 28 дней мышей иммунизировали клетками лимфомы EL-4 в дозе 5,0×105 клеток/мышь. Таким образом, соотношение введенных Т-клеток и клеток EL-4 составило 10:1. Группы были составлены из 5 животных.
Защитный эффект трансдуцированных Т-лимфоцитов проявлялся следующим образом: введение лимфоцитов за 7 и 14 дней до иммунизации EL-4 способствовало полному либо ускоренному отторжению клеток опухоли к 6 дню и не проявлялся в случае адоптивного переноса за 28 дней до иммунизации (Табл. 3). Аналогичный защитный эффект наблюдали при соотношении киллер: мишень = 20:1.
Таким образом, при внутрибрюшинном адоптивном переносе трансдуцированных Т-лимфоцитов в соотношении 10:1 или 20:1 к клеткам лимфомы EL-4 формируется иммунологическая защита мышей дикого типа B10.D2 (R101) сроком на 14 дней.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ создания противоинфекционной иммунологической защиты к Salmonella typhimurium и Listeria monocytogenes с помощью трансгенеза Т-лимфоцитов | 2018 |
|
RU2706554C1 |
| Линия мышей, трансгенных по альфа-цепи Т-клеточного рецептора клеток памяти, для изучения их функциональной активности | 2017 |
|
RU2691484C2 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2012 |
|
RU2688185C2 |
| Способ терапии болезни Тея-Сакса и болезни Сандхоффа с помощью генетически модифицированных мезенхимных стволовых клеток человека со сверхэкспрессией β-гексозаминидазы A | 2020 |
|
RU2748383C1 |
| ЛЕНТИВИРУСНЫЕ ВЕКТОРЫ, ПСЕВДОТИПИРОВАННЫЕ МУТАНТНЫМИ BaEV ГЛИКОПРОТЕИНАМИ | 2012 |
|
RU2618864C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР | 2013 |
|
RU2665548C2 |
| Генетические конструкции и их смеси для антиВИЧ терапии | 2017 |
|
RU2666991C1 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2013 |
|
RU2700765C2 |
| МОДИФИЦИРОВАННЫЕ МОНОЦИТЫ/МАКРОФАГИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2766690C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2015 |
|
RU2685479C2 |

Изобретение относится к области биохимии, в частности к способу создания противоопухолевой иммунной защиты организма к клеткам лимфомы EL-4. Изобретение позволяет эффективно противостоять клеткам лимфомы EL-4. 1 ил., 3 табл., 1 пр.
Способ создания противоопухолевой иммунологической защиты организма к клеткам лимфомы EL-4 при помощи трансдукции Т-лимфоцитов ретровирусными частицами, содержащими ген ТКР, представленный последовательностью SEQ ID NO:1, характеризующийся тем, что трансгенез Т-лимфоцитов мыши in vitro проводится путем введения гена только α-цепи ТКР, специфичного к клеткам опухоли, а затем производится адоптивный перенос животным Т-лимфоцитов, модифицированных ретровирусными частицами, содержащими ген ТКР, представленный последовательностью SEQ ID NO:1, с последующей функциональной оценкой активности Т-лимфоцитов in vivo.
| ЗВЕЗДОВА Е.С | |||
| и др., КОРЕЦЕПТОРНАЯ ФУНКЦИЯ CD4 В ОТВЕТЕ НА МОЛЕКУЛУ MHC КЛАССА I, Молекулярная биология, том 42, N.4, с.662-672 | |||
| АНТИТЕЛА ПРОТИВ αβTCR | 2012 |
|
RU2630656C2 |
| МНОГОКОМПОНЕНТНАЯ ИММУНОГЕННАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ ЗАБОЛЕВАНИЯ, ВЫЗВАННОГО β-ГЕМОЛИТИЧЕСКИМИ СТРЕПТОКОККАМИ (БГС) | 2009 |
|
RU2478396C2 |
| WO 2011020783 A2, 24.02.2011.. | |||

