Изобретение относится к медицине, терапии наследственных заболеваний и молекулярной биологии и может, в комплексе с направляющей РНК, быть использовано для редактирования точковых патогенных мутаций митохондриальной ДНК (мтДНК), ассоциированных с различными наследственными митохондриальными патологиями.
Известен «Способ лечения атрофии зрительного нерва различной этиологии» (патент РФ №2375019 2003 г. A61F 9/007, А61K 35/28, А61Р 27/02), обеспечивающий улучшение или стойкую стабилизацию зрительных функций на небольшом временном промежутке, поскольку не направлен на коррекцию наследственного материала митохондрий.
Известно изобретение «Митохондриальные таргетные антиоксиданты» (Mitochondrially targeted antioxidants) (патент США № US 6331532 1998 г. C07F 9/54, C07F 9/572, C07F 9/655, А61K 31/665, А61K 31/66), которое является технологией импорта функциональных молекул в митохондрии, с использованием липофильных агентов с иммобилизованными молекулами антиоксидантов, но не дает возможности осуществлять редактирование патогенных мутаций в митохондриальной ДНК.
Известно изобретение «Система доставки нуклеиновых кислот в митохондрии» (Mitochondrial nucleic acid delivery systems) (патент Канады №CA2678572 2008 г. A61K 48/00D2, C12N 15/864A, C12N 15/90B4). Недостатками данной технологии является то, что она не подразумевает доставку в митохондрии белков, поэтому не может быть использована для импорта нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10 в митохондрии, следовательно, не подходит для работы с системами RGEN/Cas9-BE4-Gam и RGEN/Cas9-АВЕ7.10.
Известно изобретение «Генетическая конструкция на основе системы редактирования генома CRISPR/Cas9, кодирующая нуклеазу Cas9, специфически импортируемую в митохондрии клеток человека» (патент РФ №2634395, 2015 год). Технология позволяет направлять в митохондрии клеток человека нуклеазу SpCas9, и при совместном использовании с соответствующей направляющей РНК может обеспечивать сдвиг уровня гетероплазмии по определенным патогенным мутациям мтДНК.
Недостатком данной системы является неспособность редактировать точечные мутации в мтДНК в состоянии гомоплазмии. Кроме того, система вносит двухцепочечные разрывы в мтДНК, что губительно сказывается на функционировании клеток при непрерывном использовании.
Из данных отечественной и зарубежной литературы, патентов и патентных заявок авторам не известно использование генетических конструкций на базе систем RGEN/Cas9-BE4-Gam и RGEN/Cas9-ABE7.10 для редактирования точечных патогенных мутаций мтДНК путем доставки нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10 в митохондрии; генетические конструкции отличаются модификацией последовательности кодирующей Cas9-BE4-Gam и Cas9-ABE7.10, обеспечивающей доставку данных нуклеаз в митохондрии.
Отличительным признаком изобретения является модификация нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10, обеспечивающая их доставку в митохондрии для проведения редактирования оснований мтДНК, без стадии внесения в структуру мтДНК двухцепочечных разрывов. Новизну представляют нуклеотидные последовательности конструкций, которые кодируют модифицированные Cas9-BE4-Gam и Cas9-ABE7.10.
Задачей заявляемого изобретения, является исправление точковых патогенных мутаций митохондриальной ДНК, которые ассоциированы с наследственными митохондриальными патологиями.
Поставленная задача решается тем, что генетические конструкции заявляемого изобретения, после попадания в цитоплазму клеток, обеспечивает экспрессию кодируемых им молекул нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10, которые используя собственный аппарат клетки транспортируются в митохондрии, где обеспечивают исправление точковых мутаций в мтДНК путем распознавания специфического участка последовательности посредством взаимодействия со специфической направляющей РНК и последующим дезаминированием нуклеотидного основания. Далее, в ходе репликации молекулы мтДНК, напротив дезаминированного основания используется комплементарный ему нуклеотид, являющийся «исправленным вариантом» мутации.
Техническим результатом изобретения является обеспечение системы редактирования точечных мутаций в мтДНК и способа доставки нуклеаз в митохондрии клеток человека.
Принцип функционирования предлагаемых веществ базируется на особенностях работы систем RGEN/Cas9-BE4-Gam и RGEN/Cas9-ABE7.10. Отметим сразу, что системы работают независимо и не подразумевают их одновременной доставки в клетки человека. Системы решают разные задачи. Так, система RGEN/Cas9-BE4-Gam позволяет исправить мутацию C:G на Т:А, тогда как система RGEN/Cas9-ABE7.10 позволяет исправлять более частные замены А:Т на G:C. Выбор системы определяется типом точковых мутаций, требующих редактирования. После доставки в митохондрии клеток человека, нуклеаза взаимодействует с направляющей РНК (импортируется совместно с нуклеазой), образуя комплекс, который специфически взаимодействует с участком двухцепочечной ДНК комплементарным участку направляющей РНК. В результате такого взаимодействия, функциональные домены в составе нуклеаз Cas9-BE4-Gam или Cas9-ABE7.10 осуществляют дезаминирования основания в цепи мтДНК. В частности, нуклеаза Cas9-BE4-Gam дезаминирует цитозин, который становится комплементарен аденину. В ходе репликации мтДНК напротив аденина митохондриальная ДНК-полимераза интегрирует тимин. Нуклеаза Cas9-ABE7.10 дезаминирует аденин, превразая его в инозин, который воспринимается митохондриальной ДНК-полимераза как гуанин. В дальнейших раундах репликации напротив инозина появляется цитозин.
Генетические конструкции, кодирующие нуклеазы Cas9-BE4-Gam и Cas9-ABE7.10 с детерминантной импорта в митохондрии клеток человека представляет собой плазмидные векторы разработанные на базе ранее созданного нами вектора pMitoCas9 (патент РФ №2634395, 2015 год). Каждый из плазмидных векторов обеспечивает возможность трансформации компетентных клеток E.coli с последующей наработкой большого количества копий. Позволяют отбирать трансформированные колонии на селективной среде содержащей антибиотик. Обеспечивают экспрессию нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10 в клетках млекопитающих и человека, а так же доставку продуктов трансляции в митохондрии. Проникая в митохондрии нуклеазы Cas9-BE4-Gam и Cas9-АВЕ7.10 могут быть использованы для редактирования точечных мутаций в мтДНК, при условии ко-трансфекции с определенной направляющей РНК. Выбор мутаций однако ограничен наличием РАМ последовательности в генетическом окружении мутации. С другой стороны, известны технологии изменения узнавания РАМ сайта. Таким образом, в дальнейшем методами генной инженерии можно расширить спектр редактируемых заявленными нуклеазами мутаций в мтДНК.
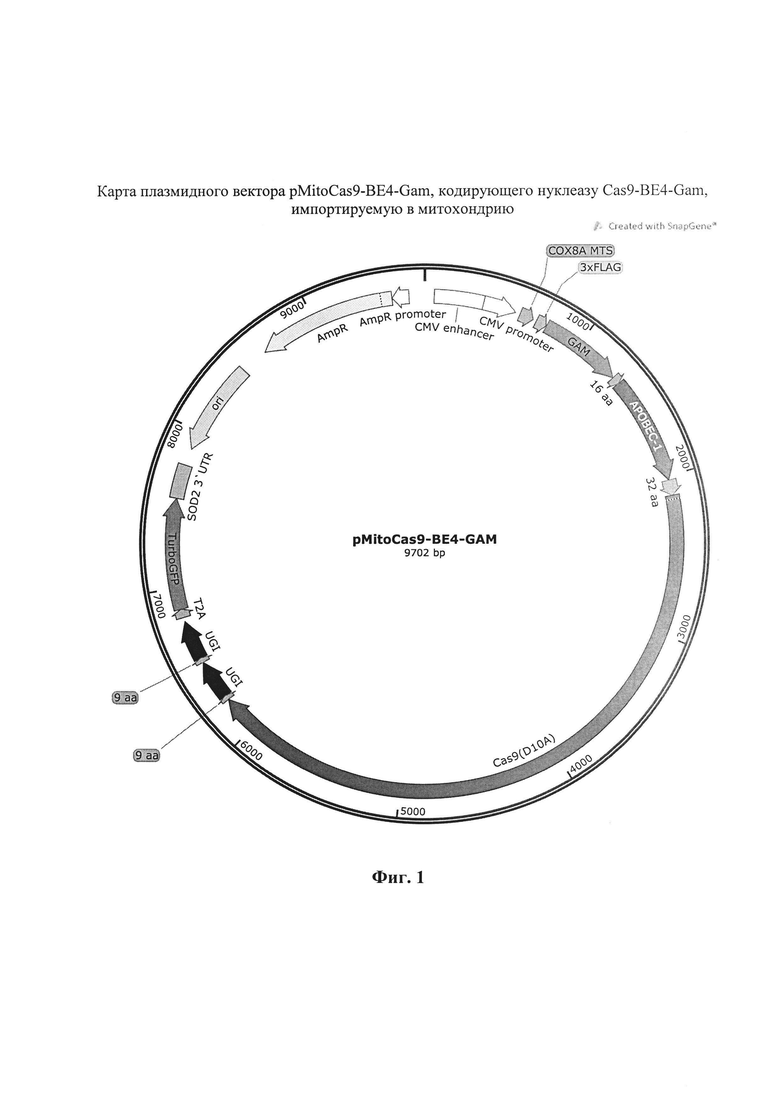
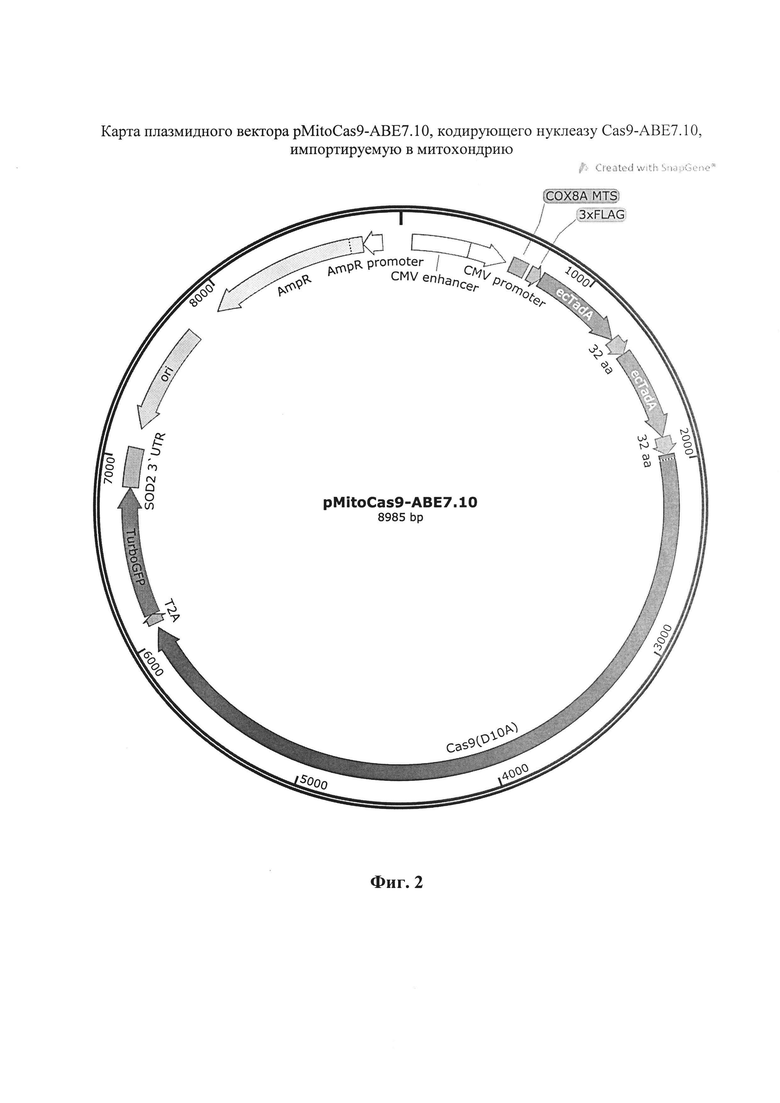
Генетическая конструкция pMitoCas9-BE4-Gam содержит ориджин репликации, ген устойчивости к ампициллину, промотор цитомегаловируса митохондриальную лидерную последовательность гена СОХ8А, 3×FLAG, последовательность кодирующую модифицированную нуклеазу Cas9-BE4-Gam, Т2А, ген TurboGFP, 3' UTR гена SOD2. Отличие генетической конструкции pMitoCas9-ABE7.10 заключается только в замене последовательности, кодирующей Cas9-BE4-Gam на последовательность Cas9-ABE7.10.
Описание способа получения генетической конструкции заявляемого изобретения.
Карты генетических конструкций pMitoCas9-BE4-Gam и pMitoCas9-ABE7.10 были построены при помощи программного обеспечения SnapGene. Для сборки был использован каркас разработанной ранее генетической конструкции pMitoCas9. Используя протокол бесшовного клонирования Гибсона, мы заменили нуклеотидную последовательность, кодирующую Cas9 на Cas9-BE4-Gam и Cas9-ABE7.10, соответственно. Амплификацию фрагментов ДНК с конструкций проводили при помощи полимеразы Q5 (NEB, США) на амплификаторе С1000 Touch (Bio-Rad, США). Олигонуклеотидные праймеры синтезировали с помощью фосфорамидитного метода на AMS-2000 («Биоссет», Россия), очищали методом обращено-фазовой хроматографии на OPS-1000 («Биоссет»), с применением реагентов компании «Glen Research)) (США).
На фиг. 1 представлена карта генетической конструкции pMitoCas9-BE4-Gam, кодирующей нуклеазу Cas9-BE4-Gam, импортируемую в митохондрию. На фиг. 2 представлена карта генетической конструкции pMitoCas9-ABE7.10, кодирующей нуклеазу Cas9-ABE7.10, импортируемую в митохондрию.
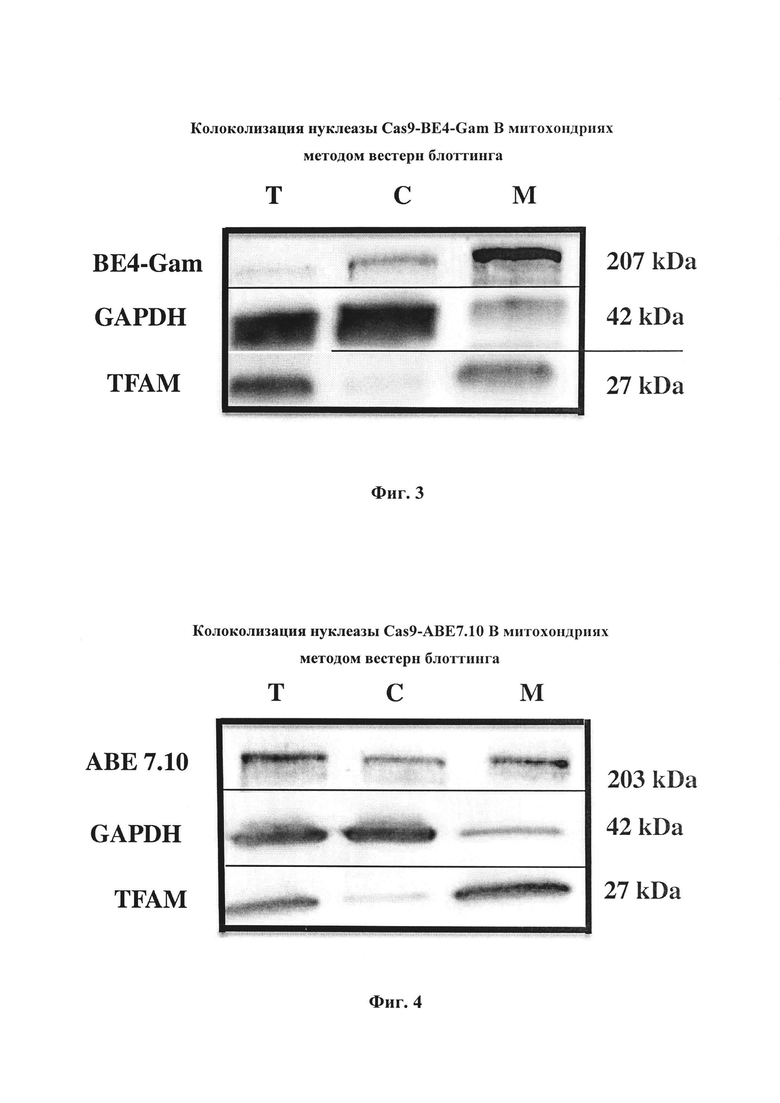
Оценка эффективности импорта нуклеаз Cas9-BE4-Gam и Cas9-ABE7.10 в митохондрии проводилась с использованием метода вестерн блоттинга.
Для приготовления экстрактов митохондриальных и тотальных белков клетки на 100 мм культуральных чашках дважды промывали 2 мл PBS, инкубировали 10 мин в 1,5 мл PBS при температуре тающего льда, отделяли от пластика специальным пластиковым лифтером и переносили в 15 мл центрифужные пробирки. Клетки осаждали центрифугированием 3 мин при 1000 об/мин. Для получения тотальных клеточных экстрактов клеточные осадки лизировали в 3-х объемах (по отношению к плотному осадку клеток) буфера, содержащего 20 мМ Трис-HCl (рН 7,4), 250 мМ NaCl, 0.25% детергента NP-40, коктейль ингибиторов протеаз и фосфатаз (Sigma, США) в разведении 1:100. Экстракты центрифугировали 15 мин при 4оС и 14000 g и супернатанты использовали для последующих опытов. Для получения митохондриальных и цитоплазматических белковых экстрактов использовался метод дифференциального центрифугирования и набор реактивов Qproteome Mitochondria Isolation Kit (Qiagen, Германия) в соответствии с протоколом производителя. Для лизиса митохондрий использовался буфер, содержащий 25 мМ Трис НСl рН 7.5, 150 мМ NaCl, 1% Тритон X100, 1 мМ ЭДТА.
Концентрацию белка определяли с помощью набора реактивов ВСА Protein Assay (Pierce) в соответствии с протоколом производителя. Для получения рабочего раствора смешивали реагент А смешивали с реагентом В в соотношении 50:1. К 2 мкл экстракта добавляли 100 мкл рабочего раствора и инкубировали полученный раствор 30 мин при 37°С. Оптическую плотность измеряли на спектрофотометре NanoPhotometer Р360 (Implen). Концентрацию белка рассчитывали по калибровочной кривой, построенной по результатам измерений оптических плотностей образцов БСА с известной концентрацией.
Электрофорез белков в денатурирующем полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и иммуноблотинг. Электрофорез проводили в геле Any kD™ Mini-PROTEAN® TGX Stain-Free™ Protein Gel (Bio-Rad). Объем пробы брали из расчета 20 мкг общего белка на дорожку. Пробы переносили в пробирки, содержащие равный объем буфера для нанесения проб на гель (4% SDS, 20% глицерина, 200 мМ дитиотреитола, 120 мМ Трис-НСl (рН 6.8), 0.002%) бромфенолового синего), инкубировали в термостате 5 мин при 95°С и наносили на ПААГ (Laemmli, 1970). Электрофоретическое разделение белков проводили при постоянном напряжении, составляющем 250 В на гель. Электрофоретически разделенные белки переносили с геля на мембрану PVDF (Bio-Rad) с помощью полусухого электропереноса в камере Trans-Blot® Turbo™ Transfer System (Bio-Rad) при постоянном напряжении, составляющем 25 В на гель. Буфер для переноса содержал 47,9 мМ Трис-HCl, 38,6 мМ глицина, 10% метанола. Для блокировки сайтов неспецифического связывания антител мембрану инкубировали 1 ч в блокирующем растворе (5% non-fat dry milk в TBST (150 мМ NaCl, 10 мМ Трис-HCl (рН 8.0), 0.05% Tween 20). Антитела для гибридизации разводили в блокирующем растворе в соответствии с рекомендациями производителей. В качестве вторых антител использовали видоспецифичные антитела, конъюгированные с пероксидазой хрена (HRP). HRP на мембране визуализировали с помощь реактива Novex™ ECL Chemiluminescent Substrate Reagent Kit (Invitrogen). Для этого компоненты набора смешали в соотношении 1:1, из расчета 0.1 мл раствора на 1 см2 мембраны, наносили на мембрану и инкубировали в течение 5 мин. Изображения препаратов, активированных с помощью хемилюминесценции, получали на приборе ChemiDoc Touch Imaging System (BioRad, США) (фиг. 3 и 4).
Использование заявляемых генетических конструкций позволяет осуществлять терапию наследственных митохондриальных патологий.












Изобретение относится к медицине, терапии наследственных заболеваний и молекулярной биологии и может быть использовано для редактирования точечных патогенных мутаций, ассоциированных с наследственными митохондриальными патологиями. Генетические конструкции pMitoCas9-BE4-Gam и pMitoCas9-ABE7.10, кодирующие нуклеазы Cas9-BE4-Gam и Cas9-ABE7.10 соответственно, импортируются в митохондрии, где обеспечивают исправление точковых мутаций в мтДНК путем распознавания специфического участка последовательности посредством взаимодействия со специфической направляющей РНК и последующим дезаминированием нуклеотидного основания. Далее, в ходе репликации молекулы мтДНК, напротив дезаминированного основания используется комплементарный ему нуклеотид, являющийся «исправленным вариантом» мутации. Так, нуклеаза Cas9-BE4-Gam позволяет исправить мутацию C:G на Т:А, тогда как Cas9-ABE7.10 позволяет исправлять более частные замены А:Т на G:C. Выбор системы определяется типом точковых мутаций, требующих редактирования. Изобретение позволяет обеспечить систему редактирования точечных мутаций в мтДНК и способ доставки нуклеаз в митохондрии. 2 н.п. ф-лы, 4 ил.
1. Генетическая конструкция pMitoCas9-BE4-Gam, кодирующая нуклеазу Cas9-BE4-Gam SEQ ID N1, импортируется в митохондрии благодаря наличию детерминанты импорта белков гена СОХ8А и последовательности 3' UTR гена SOD2 в структуре мРНК, структура конструкции pMitoCas9-BE4-Gam показана на фиг. 1.
2. Генетическая конструкция pMitoCas9-ABE7.10, кодирующая нуклеазу Cas9-АВЕ7.10 SEQ ID N2, импортируется в митохондрии благодаря наличию детерминанты импорта белков гена СОХ8А и последовательности 3' UTR гена SOD2 в структуре мРНК, структура конструкции pMitoCas9-ABE7.10 показана на фиг. 2.
| Генетическая конструкция pMitoAsCpf1, кодирующая нуклеазу AsCpf1 с детерминантной импорта в митохондрии клеток человека | 2016 |
|
RU2662994C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ НА ОСНОВЕ СИСТЕМЫ РЕДАКТИРОВАНИЯ ГЕНОМА CRISPR/CAS9, КОДИРУЮЩАЯ НУКЛЕАЗУ CAS9, СПЕЦИФИЧЕСКИ ИМПОРТИРУЕМУЮ В МИТОХОНДРИИ КЛЕТОК ЧЕЛОВЕКА | 2015 |
|
RU2634395C1 |
| ОИЩЕНКО К.Е.и др | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |

