УРОВЕНЬ ТЕХНИКИ
Область изобретения
[0001] Настоящее изобретение относится к способам и материалам для доставки нуклеиновой кислоты, вакцинации и/или лечения злокачественных новообразований. Например, настоящее изобретение относится к аденовирусам (Ad) и способам применения аденовирусов для лечения таких заболеваний, как злокачественные новообразования. В одном из аспектов изобретения аденовирус, представленный в настоящем описании, можно использовать в качестве онколитического средства.
[0002] Несмотря на огромные усилия, злокачественные новообразования остаются основной проблемой здравоохранения в США с более 1,6 миллионами новых случаев только за 2017 год (National Cancer Institute, "Cancer Stat Facts: Cancer of Any Site," seer.cancer.gov/statfacts/html/all.html). Обычные способы терапии, такие как химиотерапия, лучевая терапия и хирургическое вмешательство, зачастую не приводят к успеху, особенно в случае злокачественных новообразований на поздних стадиях. Одной из причин этого является то, что злокачественные клетки могут устранять или модифицировать компоненты, на которые нацелены эти способы терапии, и эффективно избегать уничтожения.
[0003] Онколитическая виротерапия может представлять собой альтернативный подход для лечения злокачественных новообразований с использованием селективно реплицирующихся вирусов для уничтожения опухолей, активации адаптивных иммунных ответов и обеспечения пожизненного иммунитета против опухолей (Russell et al., 2017 Molecular Therapy 25:1107-1116).
[0004] Настоящее изобретение относится к способам и материалам для доставки нуклеиновой кислоты, вакцинации и/или лечения злокачественных новообразований. Например, настоящее изобретение относится к способам и материалам для лечения злокачественных новообразований посредством введения одного или более рекомбинантных Ad (например, одного или более из Ad657 и его вариантов) в качестве онколитического средства. В варианте осуществления рекомбинантный Ad можно получать из первого Ad (например, может включать геном первого Ad, такого как Ad6, также обозначаемого как рекомбинантный остов Ad), и он может включать HVR гексона из второго Ad, такого как Ad57. Если рекомбинантный Ad включает геном Ad6 и HVR гексона Ad57, рекомбинантный Ad может являться химерным Ad, обозначаемым как Ad657 (см. Nguyen, et al. Oncolytic Virotherapy 7:43-51, 2018).
[0005] В одном из аспектов настоящее изобретение относится к способам вакцинации против инфекционного заболевания с использованием одного или более рекомбинантных Ad (например, одного или более Ad657 и его вариантов). В одном из аспектов настоящее изобретение относится к способам лечения злокачественных новообразований с использованием одного или более рекомбинантных Ad (например, одного или более из Ad657 и его вариантов) в качестве онколитического средства. В некоторых случаях один или более рекомбинантных Ad (например, один или более Ad657) можно использовать для снижения количества злокачественных клеток (например, посредством инфицирования и уничтожения злокачественных клеток) у млекопитающего. В некоторых случаях один или более рекомбинантных Ad (например, один или более Ad657 и его вариантов) можно использовать для стимуляции противоопухолевых иммунных ответов у млекопитающего. В некоторых случаях один или более рекомбинантных Ad (например, один или более Ad657 и его вариантов) можно использовать для стимуляции иммунных ответов против инфекционных заболеваний у млекопитающего.
[0006] Как показано в настоящем описании, если Ad657 вводят посредством внутривенной инъекции мышам, имеющим подкожные опухоли рака предстательной железы человека DU145, Ad657 сначала инфицирует печень, а затем достигает отдаленных опухолей. И Ad6, и Ad657 опосредовали значительную задержку роста опухоли и пролонгирование выживаемости, при этом Ad6 опосредует более высокую эффективность.
[0007] Настоящее изобретение относится к способам и материалам для доставки нуклеиновой кислоты, вакцинации и/или лечения злокачественных новообразований. Например, настоящее изобретение относится к способам и материалам для лечения злокачественных новообразований посредством введения одного или более рекомбинантных Ad (например, одного или более Ad657 и его вариантов) в качестве онколитического средства. В варианте осуществления рекомбинантный Ad можно получать из первого Ad (например, он может включать геном первого Ad, такого как Ad6, также обозначаемого как рекомбинантный остов Ad) и может включать HVR гексона из второго Ad, такого как Ad57. В случаях, когда рекомбинантный Ad включает геном Ad6 и HVR гексона Ad57, рекомбинантный Ad может являться химерным Ad, обозначаемым как Ad657. В одном из аспектов настоящее изобретение относится к способам вакцинации против инфекционного заболевания с использованием одного или более рекомбинантных Ad (например, одного или более Ad657 и его вариантов). В одном из аспектов настоящее изобретение относится к способам лечения злокачественных новообразований с использованием одного или более рекомбинантных Ad (например, одного или более Ad657 и его вариантов) в качестве онколитического средства. В некоторых случаях один или более рекомбинантных Ad (например, один или более Ad657) можно использовать для снижения количества злокачественных клеток (например, посредством инфицирования и уничтожения злокачественных клеток) у млекопитающего. В некоторых случаях один или более рекомбинантных Ad (например, один или более Ad657 и его вариантов) можно использовать для стимуляции противоопухолевых иммунных ответов у млекопитающего. В некоторых случаях один или более рекомбинантных Ad(например, один или более Ad657 и его вариантов) можно использовать для стимуляции иммунных ответов против инфекционных заболеваний у млекопитающего.
[0008] Как показано в настоящем описании, если Ad657 вводят посредством внутривенной инъекции мышам, имеющим подкожные опухоли рака предстательной железы человека DU145, Ad657 сначала инфицирует печень, а затем достигает отдаленных опухолей. И Ad6, и Ad657 опосредовали значительную задержку роста опухоли и пролонгирование выживаемости, при этом Ad6 опосредует более высокую эффективность.
[0009] Кроме того, секвестрация печени является значительной проблемой, по существу, для любого онколитического вируса, если его используют в качестве внутривенной системной терапии. Если вирус инфицирует гепатоциты и уничтожает их, это будет приводить к повреждению печени при низких дозах и гибели при более высоких дозах. Примечательно, что введение Ad по изобретению, т.е. химерного вектора Ad657 и его вариантов, опосредовало неожиданное более низкое повреждение печени, чем в случае Ad5 или Ad6. Таким образом, уникальная комбинация платформы Ad6 с HVR 1-7 из Ad657 опосредовала изменения биораспределения и терапии, ненаблюдаемые в случае природных вирусов.
[00010] Кроме того, как показано в настоящем описании, иммунизацию макак-резусов с использованием вакцин реплицирующегося одноциклового аденовируса (SC-Ad657), экспрессирующих только gp160 ВИЧ-1 клады B, интраназальным (IN) и внутримышечным (IM) путями сравнивали со слизистым и системным путями вакцинации. Вакцины SC-Ad сами по себе приводят к значительным титрам циркулирующих антител против Env всего лишь после однократной иммунизации. Животные, иммунизированные только путем IM, имели высокий уровень периферических фолликулярных T-хелперов (pTfh) в крови, но низкий уровень Tfh в лимфоузлах, и имели более низкую активность антителозависимой клеточной цитотоксичности (ADCC) антитела. Животные, иммунизированные путем IN, имели высокий уровень Tfh в лимфоузлах, но низкий уровень pTfh в крови, и имели более высокую ADCC антител. Если иммунизированных животных подвергали заражению ректально с помощью SHIVSF162P3, все они становились инфицированными, но животные, примированные через слизистые оболочки, имели значимо более низкую вирусную нагрузку в своем желудочно-кишечном тракте. Аналогично, Ad657, несущий гены антигенов гепатита C, может приводить к ответам цитотоксических T-лимфоцитов (CTL) против гепатита и цитомегаловируса. С помощью Ad657 можно доставлять и экспрессировать терапевтические гены, включая цитокины, подобные 4-1BBL, гранулоцитарно-макрофагальному колониестимулирующему фактору (ГМ-КСФ) и ИЛ-21. Результаты, представленные в настоящем описании, свидетельствуют о том, что рекомбинантные Ad можно использовать в качестве носителя для локальной или системной доставки нуклеиновой кислоты, вакцин и/или онколитической виротерапии злокачественных новообразований.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[00011] В целом, один из аспектов настоящего изобретения относится к рекомбинантному Ad, содержащему (a) геном Ad из первого штамма Ad и (b) нуклеиновую кислоту, кодирующую гексоновый полипептид из второго штамма Ad, где одна или более гипервариабельных областей (HVR) гексонового полипептида отличаются от HVR, кодируемых геномом Ad. Первый штамм Ad может являться первым штаммом Ad человека, и второй штамм Ad может являться вторым штаммом Ad человека, отличающимся от первого штамма Ad человека. Первый штамм Ad и второй штамм Ad могут серотипически отличаться. Первый штамм Ad может являться штаммом Ad6 человека, и второй штамм Ad может являться штаммом Ad57 человека. Рекомбинантный Ad также может включать один или более нацеливающих полипептидов, антигенных полипептидов, ферментов, замен аминокислот, пегилированных участков, лигандов, меток и т.п.
[00012] Рекомбинантный Ad можно использовать в качестве вектора для генной вакцинации, введения/доставки генотерапевтического средства или онколитической виротерапии.
[00013] В дополнительном варианте осуществления рекомбинантный Ad содержит (a) геном Ad из первого штамма Ad и (b) нуклеиновую кислоту, кодирующую по меньшей мере один гексоновый полипептид из одного или более штаммов Ad, где гипервариабельные области (HVR) гексонового полипептида из одного или более штаммов Ad отличаются от HVR, кодируемых первым геномом Ad.
[00014] В дополнительном варианте осуществления рекомбинантный Ad может являться репликационно-компетентным или условно-репликативным Ad (например, CRAd).
[00015] В другом аспекте настоящее изобретение относится к рекомбинантному и/или химерному Ad, содержащему (a) нуклеиновую кислоту, кодирующую первый гексоновый полипептид, и (b) второй гексоновый полипептид, где аминокислотная последовательность первого гексонового полипептида отличается от аминокислотной последовательности второго гексонового полипептида. Аминокислотная последовательность гипервариабельной области (HVR) первого гексонового полипептида может отличаться от аминокислотной последовательности гипервариабельной области второго гексонового полипептида. Нуклеиновая кислота может быть из первого штамма Ad, и второй гексоновый полипептид может быть из второго штамма Ad. Первый штамм Ad может являться первым штаммом Ad человека, и второй штамм Ad может являться вторым штаммом Ad человека, отличающимся от первого штамма Ad человека. Первый штамм Ad и второй штамм Ad могут серотипически отличаться. Штамм Ad может являться штаммом Ad6 человека, и второй штамм Ad может являться штаммом Ad57 человека. Рекомбинантный Ad также может включать нацеливающий полипептид. Нацеливающий полипептид может включать аминокислотную последовательность TARGEHKEEELI (SEQ ID NO: 1).
[00016] В дополнительном варианте осуществления рекомбинантный Ad содержит a) нуклеиновую кислоту, кодирующую первый гексоновый полипептид, и (b) второй гексоновый полипептид из одного или более штаммов Ad, где аминокислотная последовательность первого гексонового полипептида отличается от аминокислотной последовательности второго гексонового полипептида из одного или более штаммов Ad.
[00017] В дополнительном варианте осуществления рекомбинантный Ad может являться репликационно-компетентным Ad или условно-репликативным Ad (например, CRAd).
[00018] В другом аспекте изобретение относится к материалам и способам для лечения млекопитающего, имеющего злокачественное новообразование. Способы могут включать или состоять, по существу, из введения млекопитающему, имеющему злокачественное новообразование, рекомбинантного Ad, содержащего (a) геном Ad из первого штамма Ad и (b) по меньшей мере один гексоновый полипептид из одного или более штаммов Ad, где одна или более гипервариабельных областей (HVR) гексонового полипептида отличаются от HVR, кодируемых геномом Ad, и/или Ad, содержащего (a) нуклеиновую кислоту, кодирующую первый гексоновый полипептид и (b) второй гексоновый полипептид, где аминокислотная последовательность первого гексонового полипептида отличается от аминокислотной последовательности второго гексонового полипептида. Млекопитающее может являться человеком. Злокачественное новообразование может являться раком предстательной железы, раком яичников, раком легких, печеночноклеточной карциномы, раком поджелудочной железы, раком почки, меланомой, злокачественным новообразованием головного мозга, раком толстого кишечника, лимфомой, миеломой, лимфоцитарным лейкозом или миелогенным лейкозом. Введение может включать системное или локальное введение (например, внутривенное, внутриопухолевое, внутримышечное, внутриорганное введение и введение в лимфоузлы).
[00019] В настоящем описании показано, что с помощью Ad657 и его вариантов можно доставлять терапевтические гены в клетки для экспрессии терапевтических полипептидов. Таким образом, рекомбинантные Ad, включая химерные Ad, можно использовать в качестве носителя для локальной или системной доставки нуклеиновой кислоты, вакцин и/или онколитической виротерапии злокачественных новообразований.
[00020] Подробности одного или более вариантов осуществления изобретения приведены в примерах и на сопутствующих чертежах, а также в описании ниже. Другие признаки, цели и преимущества изобретения будут очевидны из описания и чертежей и из формулы изобретения. Рекомбинантный аденовирус (Ad), содержащий (a) геном Ad, кодирующий гексоновый полипептиды из первого штамма Ad, и (b) нуклеиновую кислоту, кодирующую по меньшей мере один гексоновый полипептид из одного или более других штаммов Ad, где по меньшей мере одна гипервариабельная область (HVR) гексонового полипептида отличается от HVR, кодируемых геномом Ad первого штамма Ad.
[00021] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где первый штамм Ad и один или более других штаммов Ad серотипически отличаются.
[00022] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где первый штамм Ad является штаммом Ad6 человека, и где второй штамм Ad является штаммом Ad57 человека.
[00023] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, дополнительно содержащему нуклеиновую кислоту, кодирующую нацеливающий полипептид, антиген, фермент, рецептор, лиганд или метку.
[00024] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где нацеливающий полипептид содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-41 и SEQ ID NO: 46-47.
[00025] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный рекомбинантный Ad является репликационно-компетентным Ad.
[00026] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где репликационно-компетентный Ad является одноцикловым Ad или условно-репликативным Ad (CRAd).
[00027] Дополнительный аспект изобретения относится к такому рекомбинантному аденовирусу (Ad), содержащему (a) Ad капсидные полипептиды из первого штамма Ad и (b) по меньшей мере один гексоновый полипептид из одного или более других штаммов Ad, где гипервариабельные области (HVR) гексонового полипептида или капсидные полипептиды отличаются от HVR или капсидных полипептидов первого штамма Ad.
[00028] Дополнительный аспект изобретения относится к такому рекомбинантному аденовирусу (Ad), содержащему (a) нуклеиновую кислоту, кодирующую первый гексоновый полипептид, и (b) нуклеиновую кислоту, кодирующую второй гексоновый полипептид, где аминокислотная последовательность первого гексонового полипептида отличается от аминокислотной последовательности второго гексонового полипептида.
[00029] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где нуклеиновая кислота, кодирующая по меньшей мере один гексоновый полипептид, отличается от аминокислотной последовательности гипервариабельной области указанного второго гексонового полипептида.
[00030] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанная нуклеиновая кислота получена из первого штамма Ad, и где указанный второй гексоновый полипептид получен из второго штамма Ad.
[00031] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный первый штамм Ad является первым штаммом Ad человека, и где указанный второй штамм Ad является вторым штаммом Ad человека, отличающимся от указанного первого штамма Ad человека.
[00032] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный первый штамм Ad и указанный второй штамм Ad серотипически отличаются.
[00033] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный первый штамм Ad является штаммом Ad6 человека, и где указанный второй штамм Ad является штаммом Ad57 человека.
[00034] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, дополнительно содержащему нацеливающий полипептид.
[00035] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный нацеливающий полипептид содержит аминокислотную последовательность TARGEHKEEELI (SEQ ID NO: 1).
[00036] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный рекомбинантный Ad является репликационно-компетентным Ad.
[00037] Дополнительный аспект изобретения относится к такому рекомбинантному Ad, где указанный репликационно-компетентный Ad является одноцикловым Ad или условно-репликативным Ad (CRAd).
[00038] Дополнительный аспект изобретения относится к способу лечения млекопитающего, имеющего злокачественное новообразование, где указанный способ включает введение указанному млекопитающему рекомбинантного аденовируса (Ad), как представлено в настоящем описании.
[00039] Дополнительный аспект изобретения относится к такому способу, где указанное злокачественное новообразование выбрано из группы, состоящей из рака предстательной железы, рака яичников, рака легких, печеночноклеточной карциномы, рака поджелудочной железы, рака почки, меланомы, злокачественного новообразования головного мозга, рака толстого кишечника, лимфомы, миеломы, лимфоцитарного лейкоза и миелогенного лейкоза.
[00040] Дополнительный аспект изобретения относится к такому способу, где указанное введение включает системное введение.
[00041] Дополнительный аспект изобретения относится к такому способу, где системное введение включает внутримышечное, интраназальное или внутривенное введение.
[00042] Дополнительный аспект изобретения относится к такому способу, где указанное введение включает локальное введение.
[00043] Дополнительный аспект изобретения относится к такому способу, где указанное локальное введение включает внутриопухолевую инъекцию.
[00044] Если не определено иначе, все технические и научные термины, используемые в настоящем описании, обладают значением, общепринято понимаемым специалистами в области, к которой принадлежит настоящее изобретение. Хотя для практического осуществления настоящего изобретения можно использовать способы и материалы, схожие или эквивалентные представленным в настоящем описании, ниже описаны подходящие способы и материалы. Все публикации, патентные заявки, патенты и другие ссылки, упомянутые в настоящем описании, включены в него в качестве ссылки в полном объеме. В случае противоречия настоящее описание, включая определения, будет обладать приоритетом. Кроме того, материалы, способы и примеры являются исключительно иллюстративными, и не предназначены для ограничения изобретения.
[00045] Подробности одного или более вариантов осуществления изобретения приведены на сопутствующих чертежах и в описании ниже. Другие признаки, цели и преимущества изобретения будут очевидны из описания, чертежей и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00046] На фигуре 1 показана трансляция контекст-специфических пептидов из фага в аденовирус. A) Диаграмма библиотеки фагового дисплея, содержащей β-листы HI волокна Ad5, которые структурно затрудняют библиотеку случайных 12-мерных пептидов. Ниже приведено изображение структурно схожего участка между β7- и β8-листами в HVR5 гексона Ad5. B) Первичные выравнивания аминокислот 12.51 и 12.52 в библиотеке HI и их локализация при инсерции в HVR5 гексона. C) Представление GFP-Luc-экспрессирующихся вирусов Ad5, модифицированных с помощью пептидов.
[00047] На фигуре 2 показана трансдукция in vitro. А) Экспрессия GFP по результатам флуоресцентной микроскопии клеток С2С12, инфицированных 104 vp/клетку указанных векторов через 2 дня после инфекции. В) Люциферазная активность в клетках С2С12 через 2 дня после инфекции с различными MOI указанных Ad.
[00048] На фигуре 3 показана трансдукция in vivo у мышей. А) Люциферазная визуализация мышей через 1 день после инъекции внутривенным (IV) или внутримышечным (IM) путем. Мышам, подвергнутым инъекции IM, вводили 109 vp в каждую четырехглавую мышцу. Мышам, подвергнутым инъекции IV, вводили 1010 vp через хвостовую вену. В) Количественный анализ люциферазной активности посредством визуализации в указанные дни после инъекции. *р<0,05 по результатам одностороннего ANOVA. ***р<0,001 по результатам одностороннего ANOVA.
[00049] На фигуре 4 показана трансдукция in vivo на хомяках. Люциферазная активность в мышцах хомяков через 1 день после инъекции 1010 vp IM-путем. **р<0,01 при использовании t-критерия Стьюдента.
[00050] На фигуре 5 показаны иммунные ответы на генную иммунизацию через 16 недель после однократной IM иммунизации. Мышей, показанных на фиг. 3, обескровливали через 16 недель после IM инъекции и анализировали их сыворотки в серийных разведениях посредством ELISA для детекции антител против белка GFP. *р<0,05, **р<0,01, ***р<0,001 по результатам одностороннего ANOVA. ****р<0,0001 по результатам одностороннего ANOVA. Все мыли, которым инъецировали Ad, продуцировали значительное количество антител против GFP по сравнению с группой PBS при разведениях сыворотки от 1:10000 до 1:1000. Сравнение Ad5-GL-HVR-12.51 и 12.52 с Ad5-GL показано черными звездочками. Сравнение Ad5-GL-HVR-12.51 и Ad5-GL-HVR-12.52 показано серой звездочкой. Серой штриховой и точечной пунктирной линией при OD 0,06 показан 95%-ный доверительный интервал, позволяющий различать антитела, отличающиеся от группы PBS.
[00051] На фигуре 6 показано филогенетическое древо полногеномных последовательностей серотипов аденовируса человека.
[00052] На фигуре 7 показано выравнивание Ad5, 6 и 57, где показаны вариации областей гексона и ЕЗ. (А) Выравнивание Pustell ДНК геномов Ad6 и Ad57. Рамками указаны области гексона и ЕЗ, где вариация является наиболее высокой между двумя вирусами. (В) Выравнивание ClustalW аминокислот гипервариабельной области в гексоновых белках из Ad5, Ad6 и Ad57. Выравнивания осуществляли с помощью MacVector.
[00053] На фигуре 8 показано изображение конструкции Ad657 при замене HVR Ad6 на HVR Ad57. Сокращения: HVR, гипервариабельные области.
[00054] На фигуре 9 показана онколитическая активность in vitro. Клетки LNCaP и DU145 обрабатывали указанными вирусами с указанными vp/клетку в течение 5 дней. Клетки окрашивали кристалл-виолетом и измеряли жизнеспособность клеток по OD595. Жизнеспособность клеток (%) вычисляли, разделяя OD образцов на среднее OD необработанных контрольных клеток на том же 96-луночном планшете и умножая это количество на 100. (A) Уничтожение клеток LNCaP. (B) Уничтожение клеток DU145. Сокращения: vp, вирусная частица.
[00055] На фигуре 10 показаны эффекты онколитических Ad в отношении повреждения печени. Мышам C57BL/6 (n=6 на группу) инъецировали 1011 vp каждого вируса через хвостовую вену. (A) Выживаемость по результатам анализа Каплана-Мейер. (B) Кровь отбирали для измерений АЛТ через 3 дня после инъекции (****p<0,001 по результатам ANOVA). Сокращения: АЛТ, аланинаминотрансфераза; vp, вирусная частица.
[00056] На фигуре 11 показан противоопухолевая активность Ad6 и Ad657 в ксенотрансплантатах опухоли DU145 у "голых" мышей после однократного i.v. введения. "Голым" мышам (n=9 на группу), несущим развившиеся опухоли DU145, инъецировали i.v. однократную дозу 3×1010 vp указанных вирусов или PBS. (A) Эффект однократной i.v. инъекции в отношении роста опухоли. Размеры опухоли измеряли калипером и вычисляли объем опухоли как ширина2×длина×1/2. Данные представлены в виде среднего значения±SE. *p<0,05, ****p=0,0001 при анализе с помощью ANOVA или t-критерия Стьюдента, как описано. Черными звездочками с черной стрелкой, указывающей вверх, показаны статистические различия между группой Ad6 и группой PBS в выбранный день, описанный в тексте. Серыми звездочками и стрелкой, указывающей вверх, показаны различия между группой Ad657 и группой PBS в указанный день. Затененными белыми звездочками с серой стрелкой, указывающей вниз, показаны статистические различия между группами Ad6 и Ad657 в указанный день. (B) Эффект однократной i.v. инъекции в отношении выживаемости. Животных умерщвляли, когда объем опухоли достигал 2000 мкл, или когда достигали соответствия другим критериям умерщвления (например, изъязвления), и строили кривые выживаемости Каплана-Мейера (*p<0,05, **p<0,01 при использовании логарифмического рангового критерия). Сокращения: i.v., внутривенный.
[00057] На фигуре 12 показана люциферазная визуализация "голых" мышей. Через четыре дня после однократной i.v. инъекции 3×1010 vp Ad6 и Ad657-GFP-Luc с делециями части 12,5K, 6,7K, 19K, 11,6K (ADP), 10,4K (RIDα), 14,5K (RIDβ) и 14,7K и частичной делецией E4 34K. Сокращения: i.v., внутривенный; vp, вирусная частица.
[00058] На фигуре 13 показана люциферазная визуализация "голых" мышей. Примечания: Через четырнадцать дней после однократной i.v. инъекции 3×1010 vp Ad657-GFP-Luc. Сокращения: i.v., внутривенный; vp, вирусная частица.
[00059] На фигуре 14 показаны титры связывания Env ВИЧ в плазме. Иммунизации разными SC-Ad и белками gp140 указаны крупными стрелками над графиком. Титры связывания gp140 F8 в средней точке по результатам ELISA показаны для каждого животного до и после каждой иммунизации. Штриховой пунктирной линией указан нижний предел чувствительности для антител в этом анализе. Символы распределены по оси x в каждый момент времени, чтобы была возможность наблюдать отдельные измерения. SC-Ad6-Ebov представляет собой вакцину Ad отрицательного контроля. Этой группе животных не вводили бустерную дозу gp140. *p<0,05, **p<0,01, ***p< 0,001, ****p<0,0001 по результатам одностороннего ANOVA при сравнении с группой SC-Ad6-Ebov.
[00060] На фигуре 15 показаны титры нейтрализации ВИЧ в плазме. Нейтрализацию указанных вирусов осуществляли с использованием анализа нейтрализации TZM-bl. Все значения вычисляли по сравнению с лунками, содержащими только вирус. Каждая точка соответствует среднему значению для каждого животного.
[00061] На фигуре 16 показана активность ADCC в плазме. Образцы плазмы тестировали с использованием эффекторных клеток CD16-KHYG-1 для уничтожения клеток-мишеней CEM.NKR.CCR5.CD4+-Luc, инфицированных SHIVSF162P3. Каждая точка соответствует среднему значению для каждого животного. *p<0,05, ***p< 0,001, ****p<0,0001 по результатам одностороннего ANOVA при сравнении с группой SC-Ad6-Ebov.
[00062] На фигуре 17 показана активность ADCC в слизистой оболочке. Вагинальные смывы и образцы слюны тестировали с использованием эффекторных клеток CD16-KHYG-1 для уничтожения клеток-мишеней CEM.NKR.CCR5.CD4+-Luc, инфицированных SHIVSF162P3. Каждая точка соответствует среднему значению для каждого животного. *p<0,05 по результатам одностороннего ANOVA при сравнении с группой SC-Ad6-Ebov.
[00063] На фигуре 18 показаны ИФНγ-секретирующие клетки из PBMC и лимфоузлов. Клетки PBMC и лимфоузлов анализировали посредством ELISPOT с помощью окрашивания на ИФНγ. Обозначение "против Env" относится к клеткам, которые стимулировали консервативными пептидами Env ВИЧ и SC-Ad. Подсчитывали общее количество образующих пятна клеток (SFC) для каждого из типов стимулированных клеток и корректировали его по контрольной среде в качестве фона. Каждая точка соответствует среднему значению для каждого животного. *p<0,05 по результатам одностороннего ANOVA.
[00064] На фигуре 19 показан транспорт и активация T-клеток в слизистых оболочках. T-клетки собирали из биоптатов прямой кишки, полученных после второго бустерного введения белка, и анализировали посредством проточной цитометрии на CD4, CD8, интегрин α4β7, CD69 и FoxP3. Каждая точка соответствует среднему значению для каждого животного.
[00065] На фигуре 20 показан ответ Tfh-клеток в крови и лимфоузлах. Клетки PBMC и лимфоузлов, собранные на неделе 40, стимулировали белком Env ВИЧ-1, а затем исследовали на коэкспрессию CD3+, CD4+, CXCR5+ и ИЛ-21. Каждая точка соответствует среднему значению для каждого животного. *p<0,05 по результатам одностороннего ANOVA.
[00066] На фигуре 21 показана защита от повторной ректальной провокационной пробы SHIVSF162P3. Указанные группы ректально заражали с использованием 4,3 TCID50 (на PBMC макак-резусов) SHIVSF162P3 на еженедельной основе. Образцы плазмы анализировали на копии вирусной РНК SHIV. Животных с копиями РНК более 10 считали инфицированными, и количество заражений, необходимых для инфицирования этого животного, использовали в качестве событий для анализа выживаемости способом Каплана-Мейера.
[00067] На фигуре 22 показано заражение и вирусная нагрузка SHIVSF162P3. A) Животных, показанных на фигуре 8, группировали по их исходному пути примирования SC-Ad (IM или IN), получая группы по 8, и осуществляли анализ способом Каплана-Мейера. B) Уровни вирусной РНК SHIVSF162P3 в плазме в течение исследования заражения.
[00068] На фигуре 23 показана вирусная нагрузка SHIV в тканях. РНК из PBMC и трупных тканей собирали и осуществляли qPCR для детекции вирусной РНК SHIV.
[00069] На фигуре 24 показаны вакцины одноциклового аденовируса, используемые в примере 7. A) Изображение серотипов 6 и 657 SC-Ad, несущих гены оболочки F8 и G4 ВИЧ клады B. B) Выравнивание гексонов Ad клады C, включая гексоны Ad6 и 57, экспонируемые на вакцинах.
[00070] На фигуре 25 показано титрование связывания env ВИЧ в слюне и смывах из влагалища. Уровни OD450 по результатам ELISA показаны для указанных образцов в указанных разведениях при тестировании против F8. Низкий уровень антител в этих образцах слизистых предотвращает достижение насыщения анализа. По этой причине значения EC50 нельзя легко вычислить для большинства животных. Макак-резус Rh13-091 в группе IN-IM-IM являлся единственным животным, у которого можно было вычислить EC50 (EC50=4580). Схожие результаты наблюдали при ELISA с использованием gp140 SF162.
[00071] На фигуре 26 показан Ad657, экспрессирующий гены антигенов gB гепатита C и цитомегаловируса (CMV), приводящие к активности in vivo цитотоксических T-лимфоцитов (CTL). Показано уничтожение нагруженных пептидом гепатита C клеток-мишеней у мышей, вакцинированных Ad657-HCV, а не gB CMV.
[00072] На фигуре 27 показана экспрессия ГМ-КСФ человека с Ad657, индуцирующая пролиферацию ГМ-КСФ-зависимых эритробластов человека TF-1.
[00073] Фигура 28 является графиком, на котором показана однократная IV инъекция Ad6 относительно опухолей легких A549.
[00074] Фигура 29 является графиком, на котором показана однократная IV или IT инъекция Ad6 относительно опухолей поджелудочной железы Panc1.
[00075] Фигура 30 является графиком, на котором показана однократная IV инъекция Ad6 относительно рака почки у иммунокомпетентных хомяков.
[00076] Фигура 31 является графиком, на котором показана люциферазная активность в клетках меланомы B16 и клетках рака легких A549 при воздействии Ad, экспонирующих связывающие клетки пептиды 12.51 в HVR5 гексона.
[00077] Фигура 32 является графиком, на котором показана люциферазная активность в печеночноклеточной карциноме и раке почки при воздействии Ad, экспонирующих связывающие клетки пептиды 12.51 в HVR5 гексона.
[00078] На фигуре 33 показано изображение, на котором скомбинирована инсерция отдельных HVR из разных серотипов Ad с инсерцией клеточно-нацелевающих/ненацеливающих пептидов или новых аминокислот, таких как цистеин, в гексон для направленной химической модификации и экранирования. Показаны химерные конструкции HVR, в которых комбинируют разные HVR из разных серотипов Ad для модуляции природных взаимодействий с клетками и факторами крови, улучшающие фармакологию, при комбинировании с инсерцией связывающихся с клетками и клеточно-ненацеливающих пептидов в разных HVR для изменения проникновения в клетку и клеточного избегания. Если заменяют одну HVR из 100 Ad, то получают 100 разных гексоновых химер. Если в каждую из всех 7 HVR вносят разные HVR Ad, эта комбинаторная библиотека будет эквивалентна 7100 вариантам. Если 1 пептид встраивают в 7 HVR, это будет эквивалентно 7×7100 вариантам. Если 10 разных пептидов встраивают в 7 HVR, это будет равно 10×7×7100 вариантам и т.д.
[00079] На фигуре 34 показаны карты плазмид для типичных комбинаторных гексонов и комбинаций пептидов. Показаны гексоны с HVR1 из Ad6 и HVR 2-7 из Ad57, а также инсерции клеточно-нацеливающих пептидов в отдельные HVR.
[00080] На фигуре 35 показаны химерные конструкции HVR, в которых комбинируют разные HVR из разных серотипов Ad для модуляции природных взаимодействий с клетками и факторами крови, улучшающих фармакологию, при комбинировании с инсерцией связывающихся с клетками и клеточно-ненацеливающих пептидов в разных HVR для изменения проникновения в клетку и клеточного избегания. В этом примере единичную аминокислоту цистеин встраивают в HVR1 и HVR5 Ad657 для модуляции фармакологии и делают возможной направленную конъюгацию полимеров, подобных полиэтиленгликолю или другим остаткам, подобным средствам визуализации, подобным флуорофорам.
[00081] На фигуре 36 показана конъюгация полиэтиленгликоля (PEG) с Ad657-HVR1-C. A) Электрофорез в ПААГ с SDS белков Ad с пегилированием или без него. Стрелкой показано повышение размера из-за химического присоединения PEG к гексону. B) Эффекты нацеленного пегилирования посредством малеимид-PEG и ненацеливающего NHS-PEG в отношении инфекции вируса.
[00082] На фигуре 37 показана конъюгация полиэтиленгликоля (PEG) с Ad657-HVR5-C. A) Электрофорез в ПААГ с SDS белков Ad с пегилированием или без него. B) Визуализация в ближней ИК-области ПААГ с SDS белков Ad с пегилированием или без него и с меткой для визуализации в ближней ИК-области IR800 и без нее. C) Трансдукция in vivo после интраперитонеальной инъекции малеимид-пегилированого Ad657-HVR5-C посредством люциферазной визуализации.
[00083] На фигуре 38 показана люциферазная визуализация "голых" мышей. A) Через 1, 4, 7, 14, 28 и 42 дня после однократной I.V. инъекции средства Ad6 относительно отдаленных опухолей предстательной железы DU145. B) Через 3, 7 и 19 дней после I.V. инъекции реплицирующегося Ad5-GFPLUC мышам, несущим опухоли предстательной железы LNCaP.
[00084] На фигуре 39 показана схема опухолеспецифических условно-репликативных Ad (CRAd) dl1101+dl1107, имеющих модификацию в гене E1A.
[00085] Фигура 40 является графиком, на котором показано, что и Ad6, и Ad657 можно использовать в качестве CRAd для таргетированной терапии злокачественных новообразований.
[00086] Фигура 41 является схемой, на которой показаны Ad-терапевтические циклы. A) Схема переключения серотипа с Ad. B) Схема примера терапевтического цикла, где Ad6 и Ad657 можно использовать для многочисленных раундов лечения посредством переключения серотипа в комбинации с ковалентной конъюгацией с полимером.
[00087] На фигуре 42 показано переключение серотипа и целевая люциферазная активность в опухолях предстательной железы DU145 после однократной IV инъекции Ad6 и Ad6-F35 с делециями в генах E3A (12,5K, 6,7K, 19K, 11,6K), но сохранением генов E3B (10,4K, 14,5K и 14,7K) и сохранением E4 34K. Мышам, опухоли которых были резистентны к предшествующей однократной IV инъекции Ad657 и CRAd657, оба с интактными генами E3, инъецировали указанные векторы посредством однократной IV инъекции.
[00088] Фигура 43 является схемой репликационно-компетентного Ad (RC-Ad), где экспрессию E1 контролируют с помощью нативного промотора E1; варианта CRAd-пробазин-E1A (Ad-PB), где экспрессию E1 контролируют с помощью специфического для предстательной железы промотора пробазина; CRAd-dl1101, где устранено связывание пути p300, восприимчивое к пути ИФН в нормальных клетках; CRAd-dl1107, где устранение связывания pRB позволяет вирусу уничтожать злокачественные клетки с нарушениями пути RB, но он репрессирован в RB+ нормальных клетках; CRAd-dl1101/07, где устранено связывание пути p300, восприимчивое к пути ИФН, устранение связывания pRB позволяет вирусу уничтожать злокачественные клетки с нарушениями пути RB, но он репрессирован в RB+ нормальных клетках.
[00089] На фигуре 44 (A и B) показан эффект инфекции репликационно-компетентного Ad5, Ad6, Ad657 в отношении незлокачественных клеток и модификация Ad6 и Ad657 для их превращения в условно-репликативные Ad (CRAd).
[00090] На фигуре 45 показано уничтожение злокачественных клеток репликационно-компетентными Ad5, Ad6, Ad657 и указанными CRAd.
[00091] На фигуре 46 показана модификация Ad6 и Ad657 для их превращения в условно-репликативные Ad (CRAd).
[00092] На фигуре 47 показана модификация Ad6 и Ad657 для их превращения в условно-репликативные Ad (CRAd).
[00093] На фигуре 48 показаны эффекты in vivo репликационно-компетентного Ad6 или указанных CRAd в отношении роста опухолей DU145 у мышей.
[00094] На фигуре 49 показаны эффекты in vivo репликационно-компетентного Ad657 и условно-репликативного Ad657-dl1101/07 (оба с интактными областями E3) in vivo после однократной внутривенной инъекции у мышей, несущих опухоли предстательной железы человека.
[00095] На фигуре 50 показано, что пегилирование приводит к детаргетингу аденовируса в печень in vivo.
[00096] На фигуре 51 показана модификация Ad657 с более коротким волокном из AdC68 шимпанзе и добавление гена E4 34K с "вобблом" кодона изменяет эффективность in vitro.
[00097] На фигуре 52 показано уничтожение вирусом 6/56/6 клетки рака предстательной железы человека с модификациями CRAd и без них.
[00098] На фигуре 53 показана продукция антитела в ответ на антиген злокачественной опухоли человека рецептор фолиевой кислоты альфа после однократной внутримышечной иммунизации мышей BALB/c с помощью CRAd657-dl1101/07-FOLR с интактной областью E3.
[00099] На фигуре 54 показаны участки на HVR Ad, которые могут быть модифицированы, например, посредством пегилирования или "бапилирования" с использованием пептиды-акцепторы биотина (BAP).
[000100] Фигура 55 является схемой вариантов Ad, имеющих мутации в белке E1 для превращения вируса в условно-репликативный Ad (CRAd).
[000101] На фигуре 56 показаны аминокислотные последовательности N-концевой части полипептида E1A дикого типа и N-конца E1A вариантов CRAd, dl1101, dl1107 и dl1101/1107.
[000102] На фигуре 57 показана схема разных генов уклонения от иммунологического надзора E3 в видах Ad клады C экземпляра Ad6 и видах Ad клады D экземпляра Ad26. Оба Ad экспрессируют варианты с разным размером и последовательностью генов E3 12,5K, 6,7K, 19K, 10,4K (RIDα), 14,5K (RIDβ) и 14,7K, а также изображение функций этих кодируемых E3 белков.
[000103] На фигуре 58 показаны эффекты пегилирования и делеции E3 в отношении онколитической вирусной противоопухолевой активности вирусов Ad6-Luc у иммунокомпетентных хомяков. И Ad6-Luc, и Ad6-Luc-20K PEG имеют все гены E3 и E4 34K интактные. Ad6-deltaE3-Luc имеет частичную делецию E3 12,5K и E4 34K и полную делецию генов E3 6,7K, 19K, 11,6K (ADP), 10,4K (RIDα), 14,5K (RIDβ) и 14,7K. В этой модели на иммунокомпетентных животных онколитическая эффективность утрачивается, если эти гены уклонения от иммунологического надзора отсутствуют в онколитическом аденовирусе.
[000104] Фигура 59 является картой плазмиды Ad657 с частичной делецией E3 12,5K и E4 34K и полной делецией генов E3 6,7K, 19K, 11,6K (ADP), 10,4K (RIDα), 14,5K (RIDβ) и 14,7K.
[000105] На фигуре 60 показаны 657 конструкций CRAd с модификациями CRAd dl1101/1107 и без них и с делециями выбранных генов уклонения от иммунологического надзора E3 и без них.
[000106] На фигуре 61 показан CRAd657 с участком инсерции E3. Он является CRAd657 с модификациями CRAd dl1101/1107, представленными в настоящем описании, и без них и с модификациями генов уклонения от иммунологического надзора E3 и без них.
[000107] На фигуре 62 показаны CRAd657 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7. Он являются CRAd657 с модификациями CRAd dl1101/1107, представленными на предшествующих изображениях, и без них и с модификациями генов уклонения от иммунологического надзора E3 и без них. В некоторых случаях ген E4 34K с "вобблом" кодона включают после E4 и до волокна для компенсации частичной делеции E4 34K, если делетированы гены E3B.
[000108] На фигуре 63 показаны CRAd657 +/- волокно Ad35 или нить C68 шимпанзе +/- пептид K7. Они являются CRAd657 с модификациями CRAd dl1101/1107, представленными на предшествующих изображениях, и без них и с модификациями генов уклонения от иммунологического надзора или без них E3.
[000109] На фигуре 64 показаны CRAd657 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7, экспрессирующие рецептор фолиевой кислоты альфа.
[000110] На фигуре 65 показаны CRAd657 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7, экспрессирующие гранулоцитарно-макрофагальный колониестимулирующий фактор (ГМ-КСФ).
[000111] На фигуре 66 показаны CRAd657 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7, экспрессирующие 4-1BBL, или ГМ-КСФ, или ИЛ-21, или CD40L и комбинации в одном вирусе.
[000112] На фигуре 67 показаны Ad6/57 с HVR1 Ad6 и HVR 2-7 Ad57 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7.
[000113] На фигуре 68 показаны Ad6/57/6 с HVR1Ad6, HVR 2-6 Ad57, HVR7 Ad6 +/- волокно Ad35 или нить C68 шимпанзе +/- пептид K7.
[000114] На фигуре 69 показаны Ad6/57/6 с HVR1Ad6, HVR 2-6 Ad57, HVR7 Ad6 +/- волокно Ad35 или волокно C68 шимпанзе +/- пептид K7, экспрессирующие GFP-люциферазу.
[000115] На фигуре 70 показана люциферазная визуализация после переключения серотипа. Мышей, несущим опухоли предстательной железы LNCaP в своих боках, лечили посредством однократной IV инъекции Ad657 или CRAd657. Мышей с остаточными опухолями через 5 месяцев после однократной IV инъекции лечили посредством переключения серотипа указанных вариантов Ad6/57/6, экспрессирующих GFP-люциферазу с вариантами волокон и кодон-оптимизированным геном E4 34K и без них. Указанные варианты Ad6/57/6 включают вирус Ad6/57/6, имеющий разные модификации волокна, включая добавленные 7 лизинов на волокне (K7), волокно C68 шимпанзе, пересаженное на волокно Ad6 после его придающего гибкость домена KKTK, и вирус Ad6/57/6 с волокном Ad35. Через 7 дней мышей визуализировали на люциферазную активность.
[000116] На фигуре 71 показаны клетки рака легких человека A549, обработанные указанными вирусными частицами (vp) на клетку Ad6/57/6 с вариантами волокон и кодон-оптимизированным геном E4 34K и без них. Через 7 дней клетки окрашивали кристалл-виолетом и анализировали лунки с помощью спектрофотометра для чтения планшетов. Высокая OD свидетельствует о наличии жизнеспособных клеток. Низкая OD свидетельствует о гибели и утрате адгезивных клеток.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[000117] Настоящее изобретение относится к способам и материалам для доставки нуклеиновой кислоты, вакцинации и/или лечения злокачественных новообразований. Например, настоящее изобретение относится к способам и материалам для доставки нуклеиновой кислоты белков/полипептидов, вакцинации и/или лечения злокачественных новообразований с использованием одного или более рекомбинантных Ad (например, Ad657 и его вариантов) в качестве онколитического средства.
[000118] Икосаэдр аденовируса состоит из до 720 копий его гексонового белка. Вирус не использует этот белок для связывания рецепторов, но эта нано-решетка из повторяющихся белков представляет собой матрицу для взаимодействий (например, природных взаимодействий и неприродных взаимодействий) с белками, клетками и лекарственными средствами. Антитела, которые могут нейтрализовать Ad, могут быть нацелены на гипервариабельные области (HVR) гексонового полипептида на Ad.
[000119] В некоторых случаях настоящее изобретение относится к рекомбинантным Ad, имеющим онколитическую противоопухолевую активность. Например, рекомбинантный Ad можно получать из первого Ad, и он может включать HVR гексона из одного или более разных Ad. HVR можно получать из любых биологических видов Ad клады C, например, Ad1, Ad2, Ad5, Ad6 и Ad57. В варианте осуществления рекомбинантный Ad можно получать из первого Ad, и он может включать одну или более HVR гексона из по меньшей мере одного другого Ad, где по меньшей мере один HVR гексона отличается от HVR первого Ad. Первый штамм Ad может являться штаммом Ad6 человека, и второй штамм Ad может являться штаммом Ad57 человека. На карте челночной плазмиды гексона (фигура 34) показана комбинация инсерции отдельных HVR из разных серотипов Ad с инсерцией клеточно-нацеливающих/ненацеливающих пептидов или новых аминокислот, таких как цистеин, в гексон для направленной химической модификации и экранирования. В варианте осуществления рекомбинантные Ad содержат замены аминокислот, например, замену цистеинов в полипептидах и модификации, такие как пегилирование и бапилирование. В настоящем описании показана возможность нацеливать модификации полимеров и другие химические модификации на цистеины, встроенные в гексон Ad657.
[000120] Продемонстрирован Ad657 в качестве онколитического средства против рака предстательной железы человека. HVR Ad6 заменяли HVR из Ad57 для получения химерного онколитического вируса клады C Ad657. Ad657 и Ad6 тестировали в качестве системных онколитических терапевтических средств посредством однократной i.v. инъекции "голым" мышам, несущим злокачественные опухоли человека. Ad657 можно использовать в качестве локальной или системной онколитической виротерапии злокачественных новообразований. Эти данные также свидетельствуют о неожиданных эффектах переключения серотипа с онколитическими Ad клады C.
[000121] В некоторых случаях настоящее изобретение относится к способам применения одного или более рекомбинантных Ad, представленных в настоящем описании, для лечения млекопитающего имеющего злокачественное новообразование, инфекционное заболевание и/или генетическое заболевание или имеющего риск их развития. Например, один или более рекомбинантных Ad можно вводить млекопитающему, имеющему злокачественное новообразование или имеющему риск его развития, для снижения количества злокачественных клеток (например, посредством инфицирования и уничтожения злокачественных клеток) у млекопитающего (например, человека). Например, один или более рекомбинантных Ad можно вводить млекопитающему, имеющему злокачественное новообразование или имеющему риск его развития, для стимуляции противоопухолевых иммунных ответов у млекопитающего (например, человека).
[000122] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантный Ad657 и его варианты), не разрушаются иммунной системой млекопитающего. Например, рекомбинантный Ad не разрушается антигенпрезентирующими клетками (APC), макрофагами и/или другими иммунными клетками млекопитающего, которому вводят рекомбинантный Ad.
[000123] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантный Ad657 и его варианты), можно вводить в виде многих (например, двух или более) раундов лечения. Например, первый рекомбинантный Ad, представленный в настоящем описании, может избегать антител, которые могут нейтрализовать второй рекомбинантный Ad, представленный в настоящем описании, и наоборот. В случаях, когда млекопитающее, имеющее злокачественное новообразование, лечат с использованием одного или более рекомбинантных Ad, представленных в настоящем описании, млекопитающего можно подвергать первому раунду лечения с помощью первого рекомбинантного Ad, а затем можно подвергать второму раунду лечения с помощью второго рекомбинантного Ad.
[000124] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad имеющие онколитическую противоопухолевую активность, такие как рекомбинантный Ad657 и его варианты), могут являться репликационно-компетентными Ad (RC-Ad). Например, RC-Ad может являться RC-Ad, включающим нуклеиновую кислоту, кодирующую полипептид E1 (например, E1+ RC-Ad). Например, RC-Ad может являться одноцикловым Ad (SC-Ad), включающим делецию одной или более нуклеиновых кислот, кодирующих один или более полипептидов, ассоциированных с продукцией инфекционного вирусного потомства (например, pIIIa и E3). Например, RC-Ad может являться условно-репликативным Ad (CRAd). Примеры одноцикловых Ad и того, как их получать и использовать, приведены в других источниках (публикации международной патентной заявки № WO2009/111738).
[000125] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантный Ad657 и его варианты), могут являться репликационно-дефектными Ad (RD-Ad). Например, RD-Ad может являться RD-Ad, включающим делецию нуклеиновой кислоты, кодирующей полипептид E1 (например, E1-делетированным RD-Ad).
[000126] В примерах в настоящем описании показано, что CRAd 657 и его варианты являются условно-репликативными Ad (CRAd) в злокачественных клетках, и что инфекция клеток CRAd 657 и его варианты снижает жизнеспособность клеток и объем опухоли. Таким образом, CRAd 657 и его варианты можно использовать в качестве локальной или системной онколитической виротерапии для индивидуумов со злокачественными новообразованиями.
[000127] Кроме того, показано, что CRAd можно использовать для экспрессии антигенов и в качестве вакцины для вакцинации против вирусов, например, вируса иммунодефицита человека (ВИЧ), вируса папилломы человека (HPV) и вируса гепатита C (HCV).
[000128] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантный Ad657 и его варианты), могут связываться с рецептором поверхности клетки (например, для облегчения проникновение вируса в клетку). Например, рекомбинантный Ad, представленный в настоящем описании, может связываться с коксаки-аденовирусными рецепторами (CAR) и/или Fc-рецепторами (например, FcμR и FcγR), рецепторами комплемента (например, CR3 и/или C2qR).
[000129] В одном из аспектов изобретения CRAd могут содержать нуклеиновые кислоты, кодирующие полипептиды, гетерологичные в отношении Ad, например, антигенов, рецепторов поверхности клетки, клеточно-нацеливающих полипептидов и т.п. Например, CRAd-657-dl1101/1107-FolR является рекомбинантным Ad, содержащим интактный E3 и экспрессирующим рецептор фолиевой кислоты альфа человека. В настоящем описании показано, что CRAd можно использовать для получения антител против известных антигенов злокачественных новообразований, например, рецептора фолиевой кислоты альфа.
[000130] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантные Ad657), могут избегать связывания (например, не связываются) со скэвенджер-рецептором (например, для облегчения проникновения вируса в клетку). Например, рекомбинантный Ad, представленный в настоящем описании, избегает связывания с рецепторами SREC и/или рецепторами SR-A.
[000131] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантные Ad657), могут избегать фагоцитоза.
[000132] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантные Ad657), являются непатогенными (например, для млекопитающего, подвергаемого лечению, как представлено в настоящем описании).
[000133] В некоторых случаях рекомбинантные Ad, представленные в настоящем описании (например, рекомбинантные Ad, имеющие онколитическую противоопухолевую активность, такие как рекомбинантные Ad657), могут инфицировать делящиеся клетки (например, могут инфицировать только делящиеся клетки).
[000134] Рекомбинантный Ad, представленный в настоящем описании, может являться любым подходящим рекомбинантным Ad (например, рекомбинантным Ad, имеющим онколитическую противоопухолевую активность), получаемым посредством технологии рекомбинантных ДНК и способами, известными специалистам в этой области. Рекомбинантный Ad может являться любым Ad, получаемым посредством рекомбинации материала (например, нуклеиновой кислоты и/или полипептида), из любого организма, иного, чем Ad, из которого получают рекомбинантный Ad. Например, рекомбинантный Ad может включать один или более материалов, не встречающихся в природе в этом Ad (например, не встречающихся в природе в этом Ad до рекомбинации). В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, может являться химерным Ad (например, может включать вирусные элементы из двух или более (например, двух, трех, четырех, пяти или более) разных геномов Ad).
[000135] Эти варианты осуществления также относятся к Ad, в которых комбинируют разные HVR из разных Ad (т.е. осуществляют перестановку HVR). Например, HVR1 из Ad6 с HVR 2-7 из Ad57 или HVR1 и 7 из Ad6 с HVR 2-6 из Ad57, или HVR 1 и 7 из Ad6 и HVR 2-6 из Ad657.
[000136] Нуклеиновая кислота и/или полипептиды, не встречающиеся в природе в Ad, могут быть из любого подходящего источника. В некоторых случаях нуклеиновая кислота и/или полипептид, не встречающийся в природе в этом Ad, может быть из невирусного организма. В некоторых случаях нуклеиновая кислота и/или полипептид, не встречающийся в природе в этом Ad, может быть из иного вируса, чем Ad. В некоторых случаях нуклеиновая кислота и/или полипептид, не встречающийся в природе в этом Ad, может быть из Ad, полученного из другого биологического вида. В некоторых случаях нуклеиновая кислота и/или полипептид, не встречающийся в природе в этом Ad, может быть из другого штамма Ad (например, серотипически отличающихся штаммов). В некоторых случаях нуклеиновая кислота и/или полипептид, не встречающийся в природе в этом Ad, может являться синтетической нуклеиновой кислотой и/или синтетическим полипептидом.
[000137] Рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657), можно получать (например, он может включать геномный остов) из любого подходящего Ad. В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно получать из Ad, имеющего низкую серопревалентность. Например, 50% или менее (например, 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 5% или менее) млекопитающих (например, людей) могут подвергаться воздействию Ad, из которого получают рекомбинантный Ad, представленный в настоящем описании. Что касается серопревалентности, аденовирусы клады C Ad6 и Ad657 имеют более низкую превалентность, чем вирус архетипа Ad5. В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно получать из Ad, имеющего сниженные или устраненные побочные эффекты (например, фагоцитоз и повреждение печени). Рекомбинантный Ad можно получать из Ad, выделенного из любого подходящего вида животного. Например, Ad можно выделять из людей, не являющихся человеком приматов (например, обезьян, таких как виды Мартышковых, подобных макаку-резусу), рыб, лягушек и змей. В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно получать из Ad человека (HAd или HAdV). Рекомбинантный Ad можно получать из любого вида Ad (например, A, B, C, D, E, F или G). В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно получать из вида Ad клады C (например, Ad человека клады C (HAd-C)). Рекомбинантный Ad можно получать из любого подходящего серотипа Ad (например, 2, 5, 6 или 57). В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно получать из Ad серотипа 6 (Ad6; например, Ad6 человека).
[000138] В некоторых случаях рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657 и его варианты), может включать геном Ad, содержащий одну или более модификаций в одной или более нуклеиновых кислот, кодирующих полипептид (или его фрагменты), и/или один или более вирусных элементов генома Ad. Одна или более модификаций могут являться любой подходящей модификацией. В некоторых случаях модификация может являться эффективной в ингибировании способности модифицированного полипептида связываться с другим полипептидом, таким как p300 и/или pRB. В некоторых случаях модификация может являться эффективной в нейтрализации одного или более интерфероновых путей. Примеры модификаций, которые можно осуществлять в отношении нуклеиновой кислоты, кодирующей полипептид, или вирусного элемента, включают, в качестве неограничивающих примеров, замены, делеции, инсерции и мутации.
[000139] Ad, например, Ad657 и его варианты, могут быть модифицированы и сохранять все гены E1A или модифицированы для делеции выбранных областей и функций кодируемых белков.
[000140] На фигуре 57 показана схема разных генов уклонения от иммунологического надзора E3 в экземпляре вида Ad клады C Ad6 и экземпляре вида Ad клады D Ad26, а также изображение функций этих кодируемых E3 белков. Оба Ad экспрессируют варианты разного размера и последовательности генов E3 12,5K, 6,7K, 19K, 10,4K (RIDα), 14,5K (RIDβ) и 14,7K. 19K снижает экспонирование белков MHC I и MIC на поверхности клетки для защиты инфицированных клеток от T-клеток и NK-клеток. Белки RID защищают инфицированные клетки от индуцирующих гибель лигандов (FAS, TRAIL, TNFR и EGFR). 14,7K ингибирует внутреннюю активацию апоптоза в инфицированных клетках. Виды Ad клады C также экспрессируют 11,6K, известный как белок гибели клеток, инфицированных аденовирусом (ADP). Гиперэкспрессия ADP ускоряет гибель клеток, но, в целом, гибель клеток остается той же. Вирусы клады D также экспрессируют два новых варианта, названные 49K и 31K. Секретируемая форма 49K связывается с CD46 на T-клетках и NK-клетках, что приводит к отрицательной регуляции этих клеток и менее эффективному уничтожению клеток, имеющих дефицит MHC I класса, NK-клетками. Плазмиды Ad657 модифицированы для сохранения всех нативных генов уклонения от иммунологического надзора E3 (12,5K, 6,7K, 19K, 11,6K (ADP), 10,4K (RIDα), 14,5K (RIDβ) и 14,7K) и E4 34K или для делеции выбранных областей. Ad657 и его варианты также модифицированы с помощью добавления 49K и 31K для обеспечения этих вирусных платформ на основе видов клады C этими дополнительными функциями.
[000141] Ad, например, Ad657 и его варианты, можно модифицировать для сохранения всех генов уклонения от иммунологического надзора E3 или для делеции выбранных областей и функций кодируемых белков. Что касается мутаций E3: 19k отрицательно регулирует белки MHCI и MIC на инфицированных клетках; гиперэкспрессия ADP ускоряет гибель клеток, но не повышает количество уничтожаемых клеток; белки 10k и 14k (RIDα и RIDβ) комбинируют для блокирования уничтожения клеток под действием внешних белков апоптоза, подобных FAS, TRAIL, ФНО, TNFR и EGFR; белок 14,7k ингибирует внутреннюю передачу сигнала апоптоза.
[000142] Сохранение этих белков E3 может позволить онколитическому средству персистировать дольше, и их делеция может повышать стимуляцию иммунной системы.
[000143] Данные тестирования онколитической эффективности позволяют предполагать, что интактный E3 опосредует лучшую эффективность.
[000144] Конструкции DE3 имеют делетированную часть 12,5k до 14,7k включительно, конструкции DE3A имеют делетированную часть E3 12,5k до 19k включительно, и конструкции DE3ADP имеют делетированную часть E3 12,5k до ADP включительно.
[000145] Неожиданно, делеция всех генов E3 делает онколитический вирус менее эффективным в репрессии роста опухоли.
[000146] В варианте осуществления настоящее изобретение относится к одноцикловому аденовирусу, например, SC-Ad657 и его вариантам. Рекомбинантные вирусы SC-Ad с гетерологичными нуклеиновыми кислотами, кодирующими полипептиды, оценивали для использования в качестве вакцины. Вакцины SC-Ad657 сами по себе приводили к значительным титрам циркулирующих антител против белка оболочки ВИЧ после всего лишь однократной иммунизации.
[000147] Аналогично, Ad657, несущие гены антигенов гепатита B и C, могут приводить к ответам цитотоксических T-лимфоцитов (CTL) против гепатита и цитомегаловируса.
[000148] Ad657 модифицировали посредством инсерции синтетических пептидов из вируса папилломы человека в HVR5. В варианте осуществления аминокислотная последовательность варианта гексона Ad657-HVR5-HPV определена в SEQ ID NO: 57. Модификация делает возможным экспонирование этого антигена в целях вакцинации, а также перенаправление посредством связывания с белками, взаимодействующими с пептидами HPV.
[000149] В дополнительном варианте осуществления в настоящем описании показана экспрессия Ad657 гранулоцитарно-макрофагального колониестимулирующего фактора человека (ГМ-КСФ).
[000150] Таким образом, в примерах в настоящем описании показано, что рекомбинантные Ad, например, Ad657 и его варианты, можно использовать для экспрессии гетерологичных белков, например, полипептидных антигенов и клеточно-нацеливающих полипептидов.
[000151] В некоторых случаях рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657), может включать геном Ad, содержащий одну или более замен. Например, можно заменять одну или более нуклеиновых кислот, кодирующих полипептид (или его фрагменты) и/или один или более вирусных элементов, кодируемых геномом Ad. Замена может являться любой подходящей заменой. В некоторых случаях одну или более нуклеиновых кислот, кодирующих полипептид капсида, генома первого Ad можно заменять одной или более нуклеиновыми кислотами, кодирующими полипептид капсида второго Ad, для получения химерного Ad. Например, если рекомбинантный Ad включает геном из первого Ad, где нуклеиновую кислоту, кодирующую полипептид капсида в геноме, заменяют нуклеиновой кислотой, кодирующей полипептид капсида из второго Ad (например, иного Ad, чем остов Ad), нуклеиновая кислота, кодирующая полипептид капсида из второго Ad может экспрессировать один или более капсидных полипептидов, и экспрессируемые полипептиды капсида могут встраиваться в капсид рекомбинантного Ad. Неограничивающие примеры капсидных полипептидов включают гексоновые полипептиды, полипептиды волокна, полипептиды основания пентона, полипептиды IIIa, полипептиды IX и полипептиды pVI.
[000152] Белок волокна Ad является комплексом из трех, по-видимому, идентичных субъединиц, опосредующих исходную стадию прикрепления клеток. Нативный белок волокна Ad6 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 60, и связывается с CAR.
[000153] В одном из аспектов изобретения получают волокно-модифицированные рекомбинантные и химерные Ad, имеющие белки волокна, не являющиеся нативными для родительского Ad или Ad "остова".
[000154] Химерный Ad, химера волокна AdF35, имеет аминокислотную последовательность SEQ ID NO: 61 и является более коротким, чем белки волокна Ad5 и Ad6, и перенаправляет вирус на CD46.
[000155] Волокно-модифицированный рекомбинантный Ad, содержащий волокно K7, имеющее последовательность SEQ ID NO: 62, нацеливает вирус на гепаринсульфат протеогликаны и отрицательные заряды на клетках.
[000156] Рекомбинантный, химерный Ad, волокно 6/FC68, содержащее последовательность SEQ ID NO: 63, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6, и связывается с CAR.
[000157] Рекомбинантный, химерный Ad, волокно 6/FC68-K7, содержащее последовательность SEQ ID NO: 64, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6. Волокно 6/FC68-K7 связывается с CAR и перенаправляет на гепаринсульфат и отрицательные заряды.
[000158] Рекомбинантный, химерный Ad, волокно 6/FC68-HI-K7, содержащее последовательность SEQ ID NO: 65, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6. Волокно 6/FC68-HI-K7 связывается с CAR и перенаправляет на гепаринсульфат и отрицательные заряды.
[000159] В некоторых случаях рекомбинантный Ad может включать геном из первого Ad, где нуклеиновую кислоту, кодирующую гексоновый полипептид (например, HVR нуклеиновой кислоты, кодирующей гексоновый полипептид), в геноме заменяют нуклеиновой кислотой, кодирующей гексоновый полипептид (например, HVR нуклеиновой кислоты, кодирующей гексоновый полипептид) из второго Ad. В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, может включать геном из первого Ad, имеющий одну или более HVR, замененных одной или более HVR из второго Ad. Например, рекомбинантный Ad может являться химерой, в частности, Ad657 (например, может включать геном Ad6, где HVR гексона заменяют на HVR гексона Ad57). В случаях, когда рекомбинантный Ad включает геном из первого Ad, где нуклеиновую кислоту, кодирующую гексоновый полипептид в геноме заменяют нуклеиновой кислотой, кодирующей гексоновый полипептид из второго Ad, рекомбинантный Ad может включать от приблизительно 1 до приблизительно 720 гексоновых полипептидов из второго Ad. Например, если рекомбинантный Ad является Ad657, Ad657 может включать геном Ad6 и 720 гексоновых полипептидов, включая HVR гексона Ad57.
[000160] В некоторых случаях рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657), может включать геном Ad, содержащий одну или более делеций нуклеиновых кислот. Делеция нуклеиновой кислоты может являться любой подходящей делецией нуклеиновой кислоты. Делеция нуклеиновой кислоты может являться полной делецией (например, делецией нуклеиновой кислоты, кодирующей полипептид) или частичной делецией (например, делецией одного или более нуклеотидов в нуклеиновой кислоте, кодирующей полипептид). Делеция нуклеиновой кислоты может снижать или устранять транскрипцию и трансляцию полипептида, кодируемого делетированной нуклеиновой кислотой. Делеции можно подвергать любую подходящую нуклеиновую кислоту. В некоторых случаях делеции можно подвергать нуклеиновую кислоту, кодирующую полипептид, ассоциированный с продукцией инфекционного потомства. Примеры нуклеиновых кислот, которые можно подвергать делеции и/или модифицировать в рекомбинантном Ad, представленном в настоящем описании, могут кодировать E1 (например, E1A и E1B), E2, E3, E4, pIIIA, волокно, E1B и включают вирусные энхансеры и промоторы. Например, рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657), может включать геном Ad, содержащий делецию одного или более нуклеотидов в нуклеиновой кислоте, кодирующей полипептид E1. В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, может включать одну или более замен в нуклеиновой кислоте, кодирующей полипептид E1.
[000161] В конкретных вариантах осуществления рекомбинантный Ad, представленный в настоящем описании, модифицируют так, чтобы он содержал промотор пробазина, содержащий, например, нуклеиновую кислоту SEQ ID NO: 48; рекомбинантный Ad, представленный в настоящем описании, модифицируют так, чтобы он содержал делецию dl1101 в нуклеиновой кислоте, кодирующей полипептид E1; рекомбинантный Ad, представленный в настоящем описании, модифицируют так, чтобы он содержал делецию dl1107 в нуклеиновой кислоте, кодирующей полипептид E1; рекомбинантный Ad, представленный в настоящем описании модифицируют так, чтобы он содержал делецию dl1101 и делецию dl1107. См. примеры в настоящем описании и фигуру 56 на предмет N-концевых аминокислотных последовательностей полипептида E1A, например, E1A Ad дикого типа, и варианты CRAd-657-dl1101, CRAd-657-dl1107 и CRAd-657-dl1101/1107.
[000162] В варианте осуществления вариант CRAd-657-dl1101/1107-FolR содержит интактный E3 и экспрессирует рецептор фолиевой кислоты альфа человека, обнаруживаемый на злокачественных клетках.
[000163] В основном, Ad можно модифицировать так, чтобы они включали модификации CRAd, представленные в настоящем описании.
[000164] В некоторых случаях рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657), может включать геном Ad, содержащий одну или более инсерций нуклеиновых кислот. Например, инсерция нуклеиновой кислоты может включать нуклеиновую кислоту, кодирующую полипептид. Нуклеиновую кислоту можно встраивать в любое подходящее место в геноме рекомбинантного Ad, представленного в настоящем описании. В некоторых случаях нуклеиновую кислоту, кодирующую полипептид, можно встраивать в HVR (например, петлю HVR 5) генома рекомбинантного Ad, представленного в настоящем описании. Например, если нуклеиновую кислоту, кодирующую полипептид, встраивают в HVR генома рекомбинантного Ad, представленного в настоящем описании, с нуклеиновой кислоты, кодирующей полипептид, могут экспрессироваться один или более полипептидов, и экспрессируемые полипептиды могут встраиваться в капсид рекомбинантного Ad. В случаях, когда нуклеиновую кислоту, кодирующую полипептид, встраивают в HVR генома рекомбинантного Ad, представленного в настоящем описании, рекомбинантный Ad может презентировать от приблизительно 1 до приблизительно 720 полипептидов, кодируемых встроенной нуклеиновой кислотой, на своей поверхности. Инсерция нуклеиновой кислоты может представлять собой нуклеиновую кислоту, кодирующую любой подходящий полипептид. В некоторых случаях инсерция нуклеиновой кислоты может кодировать полипептидный антиген.
[000165] В некоторых случаях инсерция нуклеиновой кислоты может кодировать нацеливающий полипептид. Неограничивающие примеры нацеливающих полипептидов, которые можно включать в рекомбинантный Ad, представленный в настоящем описании, включают пептид 12.51 (TARGEHKEEELI; SEQ ID NO: 1), пептид 12.52 (LRQTGAASAVWG; SEQ ID NO: 2), 12.53 (ARRADTQWRGLE; SEQ ID NO: 3), VSV (GTWLNPGFPPQSCGYATVT; SEQ ID NO: 4), RGD (CDCRGDCFC; SEQ ID NO: 5), альфа-4-интегрин-связывающий пептид (NMSLDVNRKA; SEQ ID NO: 6), Met 3-4 (ISLSSHRATWVV; SEQ ID NO: 7), L10.1F (WTMGLDQLRDSSWAHGGFSA; SEQ ID NO: 8), L10.1RGDF (WTMGLDQLRGDSSWAHGGFS; SEQ ID NO: 9), L10.2F (RSVSGTEWVPMNEQHRGAIW; SEQ ID NO: 10), L10.5F (TELRTHTSKELTIRTAASSD; SEQ ID NO: 11), S5,1 (DRAIGWQDKLYKLPLGSIHN; SEQ ID NO: 12), DU9C.1 (MGSWEKAALWNRVSASSGGA; SEQ ID NO: 13), DU9C.2 (MAMGGKPERPADSDNVQVRG; SEQ ID NO: 14), DU9A.7 (MASRGDAGEGSTQSNTNVPS; SEQ ID NO: 15), XS.1 (GPEDTSRAPENQQKTFHRRW; SEQ ID NO: 16), REDVmyc (MGREDVGEQKLISEEDLGGS; SEQ ID NO: 17), RGD-4C (ACDCRGDCFCG; SEQ ID NO: 18), REDV-4C (ACDCREDVCFCG; SEQ ID NO: 19), SKBR5C1 (GQIPITEPELCCVPWTEAFY; SEQ ID NO: 20), 231R10,1 (PQPPNSTAHPNPHKAPPNTT; SEQ ID NO: 21), HepaCD8 (VRWFPGGEWGVTHPESLPPP; SEQ ID NO: 22), K20 (KKKKKKKKKKKKKKKKKKK; SEQ ID NO: 23), BAP (GLNDIFEAQKIEWH; SEQ ID NO: 24), CALM BP (CAAARWKKAFIAVSAANRFKKIS; SEQ ID NO: 25), EBV (EDPGFFNVEIPEFP; SEQ ID NO: 26), #1-5 (GGHGRVLWPDGWFSLVGISP; SEQ ID NO: 27), ##4*-5 (MARTVTANVPGMGEGMVVVPC; SEQ ID NO: 28), 1-1 (GVSKRGLQCHDFISCSGVPW; SEQ ID NO: 29), 1-2 (NQSIPKVAGDSKVFCWWCAL; SEQ ID NO: 30), 1-3 (QSTPPTKHLTIPRHLRNTLI; SEQ ID NO: 31), 1-4 (DMSFQLVTPFLKALPTGWRG; SEQ ID NO: 32), 1-5 (GGHGRVLWPDGWFSLVGISP; SEQ ID NO: 33), 1-5con (FSLVGISP; SEQ ID NO: 34), 1-6 (QIMMGPSLGYYMPSESIFAY; SEQ ID NO: 35), 2-11 (ISWDIWRWWYTSEDRDAGSA; SEQ ID NO: 36), 2-14 (VWGMTTSDHQRKTERLDSPE; SEQ ID NO: 37), 2-20 (MTSAQTSEKLKAETDRHTAE; SEQ ID NO: 38), 2-9 (MGSRSAVGDFESAEGSRRP; SEQ ID NO: 39), 3b-6 (MGRTVQSGDGTPAQTQPSVN; SEQ ID NO: 40), 4*-5 (MARTVTANVPGMGEGMVVVP; SEQ ID NO: 41), пептиды CLL, PD-1, полипептиды GLA (например, фактор X), гены антигенов, слитые белки, фузогенные гликопротеины, одноцепочечные антитела и белки капсида других вирусов. Нацеливающий полипептид может нацеливать любой подходящий тип клетки. Неограничивающие примеры типов клеток, которые можно подвергать таргетингу посредством нацеливающего полипептида, включенного в рекомбинантный Ad, представленный в настоящем описании, включают мышечные клетки (например, клетки скелетных мышц), опухоли, злокачественные клетки, клетки почки, клетки печени, клетки слизистых оболочек, углеводы и мембраны клеток.
[000166] В этом примере показано, что пептиды, выбранные в совместимом структурном контексте на фаговых библиотеках, можно подвергать трансляции в гексоновый белок Ad. Например, в случае пептида 12.51, этот участок инсерции повышает трансдукцию в мышцах, при этом снижая неспецифическую инфекцию в печени. Таким образом, такой рекомбинантный Ad, нацеленный на мышечную ткань, можно использовать в качестве вектора для генетической мышечной вакцинации или генной терапии/доставки в мышцу.
[000167] В некоторых случаях инсерция нуклеиновой кислоты может приводить к ненацеливанию вируса (например, посредством нарушения взаимодействий клетки и белка, происходящей в указанной HVR). В некоторых случаях инсерция нуклеиновой кислоты может кодировать детектируемую метку. Неограничивающие примеры детектируемых меток включают флуорофоры (например, зеленый флуоресцентный белок (GFP), mCherry и mBFP) и ферменты (например, люциферазу, ДНКазы, протеазы, транспортеры и полимеразы).
[000168] Настоящее изобретение также относится к экспрессирующим векторам, содержащим рекомбинантный Ad, представленный в настоящем описании (например, рекомбинантный Ad, имеющий онколитическую противоопухолевую активность, такой как рекомбинантный Ad657 и его варианты). Экспрессирующие векторы могут нести рекомбинантный Ad, представленный в настоящем описании, в другую клетку (например, злокачественную клетку), где он может реплицироваться и/или экспрессироваться. Экспрессирующий вектор, также часто обозначаемый как экспрессирующая конструкция, как правило, является плазмидой или вектором, имеющим энхансерную/промоторную область, контролирующую экспрессию конкретной нуклеиновой кислоты. При встраивании в клетку экспрессирующий вектор может использовать клеточный аппарат синтеза белка для продукции вируса в клетке. В некоторых случаях экспрессирующие векторы, содержащие рекомбинантные Ad, представленные в настоящем описании, могут являться вирусными векторами. Например, экспрессирующий вектор, содержащий рекомбинантный Ad, представленный в настоящем описании, может являться ретровирусным вектором. В некоторых случаях экспрессирующие векторы, включающие рекомбинантный Ad, представленный в настоящем описании, также можно конструировать так, чтобы сделать возможной инсерцию одного или более трансгенов (например, в участке множественного клонирования). Например, экспрессирующие векторы, включающие рекомбинантный Ad, представленный в настоящем описании, также могут включать нуклеиновую кислоту, кодирующую детектируемую метку. Неограничивающие примеры детектируемых меток включают флуорофоры (например, зеленый флуоресцентный белок (GFP), mCherry и mBFP), и ферменты (например, люциферазу, рекомбиназы, нуклеазы и факторы транскрипции).
[000169] Настоящее изобретение также относится к способам и материалам для применения одного или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657). В некоторых случаях рекомбинантный Ad, представленный в настоящем описании, можно использовать для лечения млекопитающего, имеющего злокачественное новообразование или имеющего риск его развития. Например, способы лечения млекопитающего, имеющего злокачественное новообразование или имеющего риск его развития, могут включать введение одного или более рекомбинантных Ad, представленных в настоящем описании, млекопитающему. В некоторых случаях способы лечения млекопитающее, имеющего злокачественное новообразование или имеющего риск развития, могут включать введение одного или более экспрессирующих векторов, кодирующих рекомбинантный Ad, представленный в настоящем описании, или нуклеиновую кислоту, кодирующую рекомбинантный Ad, представленный в настоящем описании, млекопитающему. В некоторых случаях один или более рекомбинантных Ad, представленных в настоящем описании, можно вводить млекопитающему для снижения количества злокачественных клеток у млекопитающего (например, супрессии и/или задержки роста опухоли) и/или повышения выживаемости млекопитающего.
[000170] Показан таргетинг злокачественной опухоли посредством онколитических аденовирусов с переключением серотипа. Мышам, несущим опухоли предстательной железы DU145 или LNCaP в их боках, лечили однократно посредством внутривенной (IV) инъекции Ad657. Этим мышам второй раз вводили альтернативные варианты онколитических вирусов Ad6 или Ad6/57/6, имеющие модификации волокна и экспрессирующие GFP-люциферазу, и измеряли люциферазную активность посредством визуализации. Ad6 имеет гексон Ad6 и волокно Ad6, нацеленное на CAR. Ad6-F35 имеет гексон Ad6 и волокно Ad35, нацеленное на CD46. Ad6/57/6 имеет HVR1 и 7 из Ad6 и HVR 2-6 из Ad57. Вирусы Ad6/57/6 имеют волокно Ad6, волокно AdC68 или волокно Ad35. Эти данные, представленные на фигуре 9, демонстрируют неожиданную возможность переключения серотипа онколитических средств с использованием вирусов, нацеленных на опухоль с более низким нецелевым инфицированием печени.
[000171] В другом примере переключения серотипа мышей, несущих опухоли предстательной железы LNCaP на своих боках, подвергали однократной внутривенной (IV) инъекции Ad657 или CRAd657. Этим мышам второй раз, через 5 месяцев, вводили альтернативный онколитический вирус Ad6/57/6, экспрессирующий GFP-люциферазу и варианты волокна K7 (с добавлением 7 лизинов), F35 (с волокном Ad35) или KKTK-C68 (волокно C68 шимпанзе, слитое после придающего гибкость домена KKTK Ad6). Вирус KKTK-C68 также имеет добавленный кодон-оптимизированный ген E4 34K для повышения вирусной продуктивности. Люциферазную активность измеряли посредством визуализации. Все Ad6/57/6 имеют гексон с HVR1 и 7 из Ad6 и HVR 2-6 из Ad57. Ad6/57/6 и KKTK-C68 имеют волокна, нацеленные на CAR. Ad6/57/6-F35 имеет волокно Ad35, нацеленное на CD46. K7 повышает связывание с отрицательными зарядами на клетках, включая связывание с гепаринсульфат протеогликанами. На фигуре 70 показана возможность переключения серотипа онколитических средств с использованием вирусов, нацеленных на опухоль с более низким нецелевым инфицированием печени.
[000172] Любое подходящее млекопитающее, имеющее злокачественное новообразование или имеющее риск его развития, инфекционное заболевание и/или генетическое заболевание можно лечить, как представлено в настоящем описании. Например, людей, не являющихся человеком приматов (например, обезьян), лошадей, крупный рогатый скот, свиней, собак, кошек, мышей, крыс и кормовых животных, имеющих злокачественное новообразование, инфекционное заболевание и/или генетическое заболевание, можно лечить от злокачественного новообразования, как представлено в настоящем описании. В некоторых случаях можно лечить человека, имеющего злокачественное новообразование. В некоторых случаях млекопитающее (например, человек), которого лечат, как представлено в настоящем описании, не является природным хозяином Ad, используемого для получения рекомбинантного Ad, представленного в настоящем описании (например, рекомбинантного Ad, имеющего онколитическую противоопухолевую активность, такого как рекомбинантный Ad657). Например, у человека, которого лечат с помощью рекомбинантного Ad657, представленного в настоящем описании, может отсутствовать какой-либо уже существующий адаптивный иммунитет против Ad6.
[000173] Млекопитающего, имеющего какой-либо тип злокачественного новообразования, можно лечить, как представлено в настоящем описании. В некоторых случаях злокачественное новообразование может включать одну или более солидных опухолей. В некоторых случаях злокачественное новообразование может являться гемобластозом. Неограничивающие примеры злокачественных новообразований можно лечить, как представлено в настоящем описании, включают рак предстательной железы, рак яичников, рак легких, печеночноклеточную карциному, рак поджелудочной железы, рак почки, меланому, злокачественное новообразование головного мозга, рак толстого кишечника, лимфому, миелому и лейкозы (например, лимфоцитарные лейкозы и миелогенные лейкозы).
[000174] В некоторых случаях способы, представленные в настоящем описании, также могут включать идентификацию млекопитающего как имеющего злокачественное новообразование. Неограничивающие примеры способов идентификации млекопитающего как имеющего злокачественное новообразование включают физикальный осмотр, лабораторные анализы (например, крови и/или мочи), биопсию, способы визуализации (например, рентгенографию, ПЭТ/КТ, МРТ и/или УЗИ), сцинтиграфию (например, остеосцинтиграфию), эндоскопию и/или генетические анализы. После идентификации в качестве имеющего злокачественное новообразование, инфекционное заболевание и/или генетическое заболевание, млекопитающему можно вводить или инструктировать его по самостоятельному введению одного или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657), или нуклеиновой кислоты (например, экспрессирующего вектора), кодирующей один или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657).
[000175] Один или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657), можно вводить любым подходящим путем. В некоторых случаях введение может являться локальным введением. В некоторых случаях введение может являться системным введением. Неограничивающие примеры путей введения включают внутривенное, внутримышечное, подкожное, пероральное, интраназальное, ингаляционное, трансдермальное, парентеральное, внутриопухолевое, прямокишечно-уретральное, субкапсулярное, вагинальное и ректальное введение. В случаях, если проводят множество раундов лечения, первый раунд лечения может включать введение одного или более рекомбинантных Ad, представленных в настоящем описании, млекопитающему (например, человеку) первым путем (например, внутривенно), и второй раунд лечения может включать введение один или более рекомбинантных Ad, представленных в настоящем описании, млекопитающему (например, человеку) вторым путем (например, внутримышечно).
[000176] В рамках изобретения термин "фармацевтическая композиция" относится к комбинации одного или более рекомбинантных и/или химерных Ad по настоящему изобретению с носителем, инертным или активным, делающим композицию особенно подходящей для терапевтического применения. Один или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657), можно составлять в композиции (например, фармацевтической композиции) для введения млекопитающему (например, млекопитающему, имеющему злокачественное новообразование или имеющему риск его развития). Например, один или более рекомбинантных Ad можно составлять в фармацевтически приемлемой композиции для введения млекопитающему, имеющему злокачественное новообразование или имеющему риск его развития. В некоторых случаях один или более рекомбинантных Ad можно составлять вместе с одним или более фармацевтически приемлемыми носителями (добавками) и/или дилюентами. Фармацевтическую композицию можно составлять для введения в твердой или жидкой форме, включая, в качестве неограничивающих примеров, стерильные растворы, суспензии, составы с замедленным высвобождением, таблетки, капсулы, пилюли, порошки, крахмальные капсулы и гранулы. Фармацевтически приемлемые носители, наполнители и носители, которые можно использовать в фармацевтической композиции, представленной в настоящем описании, включают, в качестве неограничивающих примеров, физиологический раствор (например, фосфатно-солевой буфер, ионообменный материал, оксид алюминия, стеарат алюминия, лецитин, белки сыворотки, такие как сывороточный альбумин человека, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, смеси неполных глицеридов насыщенных растительных жирных кислот, воду, соли или электролиты, такие как протамин сульфат, гидрофосфат натрия, гидроортофосфат калия, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, вещества на основе целлюлозы, натрий-карбоксиметилцеллюлоза, полиакрилаты, воски, полиэтилен-полиоксипропилен-блок-сополимеры, полиэтиленгликоль и ланолин). Подходящие фармацевтические носители описаны в "Remington's Pharmaceutical Sciences", E.W. Martin, 18th Edition. Выбор и использование подходящих эксципиентов описаны в Gennaro, ed., Remington: The Science and Practice of Pharmacy, 20th Ed. (Lippincott Williams & Wilkins, 2003).
[000177] Композицию (например, фармацевтическую композицию), включающую один или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантные Ad657), можно вводить млекопитающему (например, млекопитающему имеющему злокачественное новообразование или имеющему риск его развития) в качестве вакцины. Вакцина может являться профилактической или терапевтической.
[000178] В некоторых случаях способы, представленные в настоящем описании, также могут включать введение млекопитающему (например, млекопитающему, имеющему злокачественное новообразование) одного или более дополнительных средств, используемых для лечения злокачественных новообразований. Одно или более дополнительных средств, используемых для лечения злокачественных новообразований, могут включать любое подходяще противоопухолевое средство. В некоторых случаях лечение злокачественного новообразования может включать хирургическое вмешательство. В некоторых случаях лечение злокачественного новообразования может включать лучевую терапию. В некоторых случаях лечение злокачественного новообразования может включать проведение медикаментозной терапии, такой как химиотерапия, гормональная терапия, направленная терапия и/или цитотоксическая терапия. Например, млекопитающему, имеющему злокачественное новообразование, можно вводить один или более рекомбинантных Ad, представленных в настоящем описании (например, рекомбинантных Ad, имеющих онколитическую противоопухолевую активность, таких как рекомбинантный Ad657 и его варианты), и вводить одно или более дополнительных средств, используемых для лечения злокачественных новообразований. В случаях, когда млекопитающее, имеющее злокачественное новообразование, лечат с помощью одного или более рекомбинантных Ad, представленных в настоящем описании, и лечат с помощью одного или более дополнительных средств, используемых для лечения злокачественных новообразований, инфекционного заболевания и/или генетического заболевания, дополнительные средства, используемые для лечения злокачественных новообразований, инфекционного заболевания и/или генетического заболевания, можно вводить одновременно или независимо. Например, один или более рекомбинантных Ad, представленных в настоящем описании, и одно или более дополнительных средств, используемых для лечения злокачественных новообразований, инфекционного заболевания и/или генетического заболевания, можно составлять совместно для получения единой композиции. В некоторых случаях один или более рекомбинантные Ad, представленные в настоящем описании, можно вводить первыми, а одно или более дополнительных средств, используемых для лечения злокачественных новообразований, инфекционного заболевания и/или генетического заболевания, можно вводить вторыми, или наоборот.
ПРИМЕРЫ
Пример 1. Аденовирусы.
[000179] В течение 50 лет было известно всего четыре вида Ad человека клады C, включая Ad1, Ad2, Ad5 и Ad6 (см., например, Weaver et al., 2011 Virology. 412:19-27). В 2001 годы идентифицировали пятый вид аденовируса клады C как штамм полевого изолята #16700, и анализ нейтрализации вируса с использованием антисывороток против Ad1, 2, 5 и 6 показал высокие уровни нейтрализации (реципрокные титры 500-16000), если каждую антисыворотку использовали против когнатного вируса (Lukashev et al., 2008 J Gen Virol. 89:380-388). В отличие от этого, антитела против Ad1, 2 и 5 имеют слабую перекрестную реактивность против #16700 (реципрокные титры 32-64). Сыворотка против Ad6 демонстрировала более высокую перекрестную реактивность против #16700, но нейтрализация требовала в 10 раз более высоких концентраций сывороток для нейтрализации #16700 по сравнению с самим Ad6. При последующих сравнениях последовательностей подтверждали, что #16700 является новым видом аденовируса клады C, и его переименовывали в Ad57 (см., например, Walsh et al., 2011 J. Clin Microbiol. 49:3482-3490).
[000180] 15 Ad человека оценивали на онколитическую активность в отношении злокачественных новообразований молочной железы, яичника, печени, предстательной железы, почки и B-клеточных злокачественных новообразований. В тестах против опухолей предстательной железы человека DU145 вид Ad6 клады C являлся более активным после однократной внутриопухолевой или внутривенной (i.v.) инъекции, чем виды Ad5 клады C или виды вирусов Ad11 и Ad35 клады B. Ad6 также являлся более эффективным, чем Ad5 и Ad11, у иммунокомпетентных сирийских хомячков.
Конструирование рекомбинантных аденовирусов.
[000181] Рекомбинантные аденовирусы конструировали посредством технологии рекомбинантных ДНК способами, известными специалистам в этой области. Рекомбинантный Ad получают из первого Ad (например, он может включать геном первого Ad, такого как Ad6), и он может включать HVR гексона из второго Ad, такого как Ad57. В случаях, когда рекомбинантный Ad включает геном Ad6 и HVR гексона Ad57, рекомбинантный Ad может являться химерным Ad, обозначаемый как Ad657.
[000182] Для получения Ad657 синтезировали последовательность HVR Ad57 и встраивали ее в гексон Ad6 в плазмиду в кассете FRT-Zeocin®-FRT между pVI и гексоном. Их рекомбинировали в различные плазмиды pAd6 для получения Ad657 и его вариантов. Аминокислотная последовательность гексона Ad657 приведена в SEQ ID NO: 49. См. карты плазмид вариантов Ad657 на фигурах 34 и 59-65.
[000183] Что касается вариантов Ad657, последовательность HVR Ad57 синтезировали с использованием HVR1, модифицированной с помощью цистеина, аминокислот, придающих гибкость, и участков рестрикции, чтобы сделать возможными инсерции других пептидов. Ее встраивали в гексон Ad6 в плазмиду с кассетой FRT-Zeocin®-FRT между pVI и гексоном. Их рекомбинировали в различные плазмиды pAd6 для получения вариантов Ad657 с цистеином в HVR1, вариант, обозначаемый как Ad657-HVR1-XXA, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 50.
[000184] Что касается вариантов Ad657, последовательность HVR Ad57 синтезировали с использованием HVR5, модифицированной с помощью цистеина, аминокислот, придающих гибкость, и участков рестрикции, чтобы сделать возможными инсерции других пептидов. Ее встраивали в гексон Ad6 в плазмиду с кассетой FRT-Zeocin®-FRT между pVI и гексоном. Их рекомбинировали в различные плазмиды pAd6 для получения вариантов Ad657 с цистеином в HVR5, вариант, обозначаемый как Ad657-HVR5-XXA, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 51.
[000185] Что касается вариантов Ad657, последовательность HVR Ad57 синтезировали с использованием HVR1, модифицированной с помощью цистеина, аминокислот, придающих гибкость, и участков рестрикции, чтобы сделать возможными инсерции других пептидов. Ее встраивали в гексон Ad6 в плазмиду с кассетой FRT-Zeocin®-FRT между pVI и гексоном. Их рекомбинировали в различные плазмиды pAd6 для получения Ad657 варианты без цистеина в HVR1, но с участками рестрикции, делающими возможными инсерции пептидов в HVR1, вариант, обозначаемый как Ad657-HVR1-XA, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 52.
[000186] Что касается вариантов Ad657, последовательность HVR Ad57 синтезировали с использованием HVR5, модифицированной с помощью цистеина, аминокислот, придающих гибкость, и участков рестрикции, чтобы сделать возможными инсерции других пептидов. Ее встраивали в гексон Ad6 в плазмиду с кассетой FRT-Zeocin®-FRT между pVI и гексоном. Их рекомбинировали в различные плазмиды pAd6 для получения Ad657 варианты без цистеина в HVR5, но с участками рестрикции, делающими возможными инсерции пептидов в HVR5, вариант, обозначаемый как Ad657-HVR5-XA, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 53.
[000187] Что касается вариантов Ad657, Ad657 HVR1-XA последовательность модифицировали посредством инсерции пептида-акцептора биотина в HVR1. Их рекомбинировали в различные плазмиды pAd6 для получения вариантов Ad657 с BAP в HVR1, вариант, обозначаемый как Ad657-HVR1-PSTCD, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 54.
[000188] Инсерция пептида-акцептора биотина приводит к ненацеливанию вариантов вируса в отношении печени, позволяет перенаправлять вирус с использованием авидина или стрептавидина и биотинилированных лигандов и позволяет очищать вирус на колонках с мономерным авидином или стрептавидином.
[000189] Что касается вариантов Ad657, последовательность HVR1-XA Ad657 модифицировали посредством инсерции пептида-акцептора биотина в HVR1. Их рекомбинировали в различные плазмиды pAd6 для получения вариантов Ad657 с BAP в HVR1, вариант, обозначаемый как Ad657-HVR5-PSTCD, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 55.
[000190] Что касается вариантов Ad657, последовательность HVR5-XA Ad657 модифицировали посредством инсерции синтетической петли V1/V2 из оболочки ВИЧ в HVR5, вариант, обозначаемый как Ad657-HVR5-V1/V2, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 56.
[000191] Инсерция синтетической петли V1/V2 из оболочки ВИЧ делает возможным экспонирование этого антигена так, чтобы он служил в качестве вакцины, а также перенаправление посредством связывания с белками, взаимодействующими с оболочкой ВИЧ.
[000192] Что касается вариантов Ad657, последовательность HVR5-XA Ad657 модифицировали посредством инсерции синтетических пептидов из вируса папилломы человека в HVR5, вариант, обозначаемый как Ad657-HVR5-HPV, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 57.
[000193] Инсерция синтетических пептидов из вируса папилломы человека делает возможным экспонирование пептидов HPV в качестве антигенов в целях вакцинирования, а также перенаправление посредством связывания с белками, взаимодействующими с пептидами HPV.
[000194] В другом аспекте изобретения получали химерные Ad, имеющие HVR1 Ad6 и HVR 2-7 Ad57, химера, обозначаемая как химера HVR Ad6/57, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 58.
[000195] В еще одном аспекте изобретения получали химерные Ad, имеющие HVR1 и 7 Ad6 и HVR 2-6Ad57, химера, обозначаемая как химера HVR Ad6/57/6, содержит гексон, имеющий аминокислотную последовательность SEQ ID NO: 59.
Пример 2. Перенаправленный и ненацеленный рекомбинантный аденовирус для доставки генов.
[000196] Аденовирусы являются надежными векторами для доставки генов и генной иммунизации. Архетип аденовируса, используемый в большинстве случаев, является серотипом 5 вида аденовируса человека клады C (HAdV-C5 или Ad5). In vitro Ad5 связывается и проникает в клетки посредством комбинированных взаимодействий его белков волокна и основания пентона с рецепторами поверхности клетки. Тримерное волокно связывается с коксаки-аденовирусным рецептором (CAR), и клетки, в которых отсутствует CAR, являются относительно резистентными к инфекции, если они также не экспрессируют αv-интегрины, которые могут связываться мотивом RGD на основании пентона.
[000197] In vivo эти взаимодействия также используют, но их важность варьируются в зависимости от пути инъекции. При непосредственной инъекции в солидную ткань или опухоль, взаимодействия CAR и интегринов доминируют. При внутривенной (IV) инъекции эти взаимодействия становятся вторичными из-за эффектов связывания Ad5 с витамин-K-зависимыми факторами свертывания крови. Фактор свертывания X (FX) связывается с субнаномолярной аффинностью с гексонами Ad5 и, таким образом, позволяет Ad5 эффективно трансдуцировать гепатоциты после IV инъекции. В отсутствие FX трансдукция печени значительно снижается.
[000198] Аденовирусные векторы являются в некоторой степени уникальными из-за их способности нести очень крупные последовательности кДНК до 36 тысяч пар оснований (т.п.н.) по сравнению с другими векторами, подобными векторам аденоассоциированного вируса (AAV) с последовательностью ДНК всего 4,5 т.п.н. Эта нагрузочная емкость обосновывает ранние исследования векторов Ad для мышечной генной терапии при доставке очень крупных трансгенов, подобных кДНК дистрофина размером 14 т.п.н. Введение IV новорожденным мышам может опосредовать мышечную генную терапию, но эта возможность утрачивается в случае взрослых мышей. Снижение трансфекции с возрастом частично является результатом очень крупного размера вирионов Ad (т.е. 100 нм), а также утраты рецептора CAR на мышечных клетках с возрастом. В настоящее время внутримышечный (IM) путь является чаще всего используемым путем для генных вакцин при использовании Ad5 и других серотипов, несмотря на то, что CAR отсутствует на клетках скелетных мышц.
[000199] Таким образом, трансдукции мышц с помощью Ad5 и другого серотипа может быть достаточно для генной терапии или генной вакцинации. Однако отсутствие первичного рецептора вируса в мышцах снижает эффективность вируса и требует доставки большего количества вектора для достижения желаемых эффектов.
Конструирование пептид-модифицированных гексонов в аденовирусе.
[000200] Пептиды из 12 аминокислот (12-мерные) на мышечных клетках мыши C2C12 подвергали селекции из библиотеки случайных пептидов, экспонируемых между β-листами H и I из области "выступа" Ad5 (фигура 1A). Пептиды 12.51 (TARGEHKEEELI; SEQ ID NO: 1) и 12.52 (LRQTGAASAVWG; SEQ ID NO: 2) подвергали селекции против миобластов с предварительной очисткой относительно нецелевых клеток для получения пептидов, которые будут специфическим для связывания мышечных клеток. В большинстве случаев небольшие пептиды имеют относительно низкую аффинность. Таким образом, предполагают, что экспонирование этих пептидов, подвергнутых селекции в отношении мышц, на 720 копиях гексона Ad5 может сделать возможной лучшую доставку генов в мышцы. Этот участок инсерции также может обладать преимуществом инактивации связывания FX с гексоном для "ненацеливания" вектора в отношении печени, если любой вектор попадет в кровоток после IM инъекции.
[000201] Инсерция этих пептидов, подвергнутых селекции в отношении мышц, между двумя другими β-листами, что также затрудняет гипервариабельную петлю на вирус, оценивали способность модифицированных Ad модулировать тропизм. Пептиды 12.51 и 12.52 встраивали в петлю гипервариабельной области (HVR) 5, затрудненной β7- и β8-листами в гексоне Ad5. Оценивали способность этих вирусов in vivo трансдуцировать ткани после внутривенных и внутримышечных инъекций мышам и хомякам.
Аденовирусы
[000202] E1-делетированный Ad5-GL (RD-Ad5-GL) экспрессирует слитый белок зеленый флуоресцентный белок-люциферазу (GFP-люциферазу, GL), как описано в других источниках (см., например, Crosby et al., 2002 J. Virol., 76:6893-6899; Khare et al., 2011 Mol. Ther., 19:1254-1262; и Khare et al., 2012 J. Virol., 86:2293-2301). Пептиды 12.51 и 12.52, подвергнутые селекции в отношении мышц, встраивали вместо HVR5 на гексоне Ad5 между β7- и β8-листами, структурно схожими с β-листами H и I из области "выступа" Ad5 (фигура 1A), способами, известными специалистам в этой области. Пептидами с лидерной последовательностью, придающей гибкость, заменяют целую петлю HVR5 в Ad5 (фигура 1B). Эти модифицированные последовательности гексона встраивали в репликационно-дефектный Ad5, экспрессирующий слитый белок зеленый флуоресцентный белок-люциферазу (RD-Ad5-GL) посредством Red-зависимой рекомбинации у бактерий (см., например, Campos et al., 2004 Hum. Gene Ther., 15:1125-1130). Пептид-модифицированные Ad конструировали посредством инсерции отжигаемых олигонуклеотидов, кодирующих пептиды 12.51 и 12.52 (фигура 1B), в плазмиду pHVR5 дисплея (фигура 1A внизу) для получения рекомбинантных Ad, Ad5-HVR5-12.51 и Ad5-HVR5-12.52.
[000203] Получаемые плазмиды расщепляли и использовали для Red-зависимой рекомбинации в pAd5-GL. Эти векторы подвергали "спасению" в клетках 293, очищенных на двух последовательных градиентах CsCl, обессоливали на хроматографических колонках Econopac 10-DG (Bio-Rad) в 50 мМ Трис, pH 8, с 0,5 M сахарозой и хранили при -80°C.
Тестирование вируса in vitro
[000204] Миобласты мыши C2C12 приобретали в American Type Tissue Culture (Manassas, VA). Клетки 293 получали из Microbix, Toronto, Ontario, Canada. Клетки поддерживали в DMEM с 10% FBS, Invitrogen.
[000205] Мышечные клетки C2C12 высевали в 6-луночные планшеты (Corning) за день до инфекции. Вирусы использовали для инфицирования клеток в DMEM с 5% FBS. Клетки инкубировали в течение 2 дней до наблюдения зеленой флуоресценции.
[000206] Эксперименты на животных осуществляли с одобрения Институционального комитета по содержанию и использованию животных Клиники Мэйо в соответствии с положениями Закона о благополучии животных, Политики благополучия животных PHS и принципами Руководства по содержанию и использованию лабораторных животных NIH. Самок мышей CD-1 (Charles River) анестезировали и инъецировали им внутримышечно (IM) или внутривенно (IV) 1010 vp указанных вирусов в указанные моменты времени. Животных анестезировали, инъецировали им 3 мг D-люциферин (Molecular Imaging Products) и визуализировали с помощью системы Xenogen. Позднее животных анестезировали и собирали кровь в пробирки для отделения сыворотки для ELISA.
[000207] ELISA осуществляли следующим образом. Планшеты Immulon 4 HBX (Thermo) инкубировали с 100 нг на лунку белка GFP в 1-кратном фосфатно-солевом буфере (PBS) при 4°C в течение ночи, промывали и блокировали 5% молоком в Трис-забуференном физиологическом растворе с 0,1% Tween 20 (TBST) при комнатной температуре в течение 2 часов. Подготавливали разведения от 1:100000 до 1:11000 каждого образца сыворотки в блокирующем буфере. Лунки промывали и добавляли по 100 мкл каждого образца в покрытые GFP планшеты в трех параллелях и инкубировали в течение 3 часов при комнатной температуре. Лунки промывали и в каждую лунку добавляли разведение 1/10000 HRP-конъюгированного вторичного антитела козы против мыши (Pierce Chemical). Планшеты инкубировали в течение 2 часов при комнатной температуре, промывали и добавляли 50 мкл 1 Step Ultra TMB ELISA (Thermo Fisher Scientific Inc.) для детекции HRP, а затем 50 мкл 2 M H2SO4. Оптическую плотность при 450 нм определяли с помощью Beckman Coulter DTX 880.
[000208] Статистические анализы осуществляли с использованием Prism (Graphpad). Статистическую значимость вычисляли посредством одностороннего ANOVA, а затем HSD Тьюки.
Трансдукция in vitro в мышечных клетках мыши C2C12
[000209] RD-Ad5-GL, RD-Ad5-GL-HVR5-12.51 и RD-Ad5-HVR5-12.52 использовали для инфицирования миобластов мышь C2C12 при различной множественности заражения (MOI) в терминах вирусных частиц (vp)/клетку. Если зеленую флуоресценцию слитого белка GFP наблюдали посредством флуоресцентной микроскопии, оба из пептид-модифицированных векторов опосредовали значимо лучшую трансдукцию, чем RD-Ad5-GL (фигура 2A). Когда измеряли люциферазную активность, наблюдали значимые повышения в RD-Ad5-GL-12.51- и 12.52-инфицированных клетках (фигура 2B). Когда один из пептидов, 12.51, встраивали обратно в область "выступа" Ad5, пептид повышал трансдукцию миобластов C2C12 in vitro в 14 раз.
Трансдукция in vivo после внутримышечной инъекции у мышей
[000210] 109 vp Ad5-GL, Ad5-GL-HVR5-12.51 и Ad5-GL-HVR5-12.52 инъецировали мышам IM путем в обе четырехглавые мышцы и осуществляли люциферазную визуализацию в разные моменты времени (фигура 3A, сверху). Ad5-GL-HVR5-12.51 приводил к в 2-3 раза более высокой люциферазной активности, чем Ad5-GL во все временные точки (p<0,05 в день 1 по результатам одностороннего ANOVA) (фигура 3B слева). Активность Ad5-GL-HVR5-12.52 в мышце была более низкой, чем Ad5-GL и HVR5-12.51, в отличие от его более высокой активности in vitro.
Трансдукция in vivo после внутривенной инъекции у мышей
[000211] 3×1010 vp Ad5-GL, Ad5-GL-HVR5-12.51 и Ad5-GL-HVR5-12.52 инъецировали мышам IV путем и осуществляли люциферазную визуализацию (фигура 3A внизу). В отличие от результатов в случае мышц, только немодифицированный Ad5-GL опосредовал сильную трансдукцию печени. Трансдукция печени Ad5-GL была в 60 раз более высокой, чем в случае Ad5-GL-HVR5-12.51 и Ad5-GL-HVR5-12.52 (p<0,001 в день 1 по результатам одностороннего ANOVA) (фигура 3B справа), что свидетельствует о том, что рекомбинантные Ad нацелены на мышечную ткань, при этом снижая нецелевое инфицирование в печени.
Трансдукция in vivo после внутримышечной инъекции у хомяков
[000212] Для тестирования того, работает ли 12.51-модифицированн вектор на иных биологических видах, чем мыши, 1010 vp Ad5-GL и Ad5-GL-HVR5-12.51 инъецировали IM путем в обе четырехглавые мышцы более крупных сирийских хомячков и осуществляли люциферазную визуализацию через 24 часа (фигура 4). В этом случае Ad5-GL-HVR5-12.51 опосредовал в 7 раз более высокую люциферазную активность, чем Ad5-GL (p<0,01 в день 1 по результатам одностороннего ANOVA).
Генная иммунизация после внутримышечной инъекции у мышей
[000213] Через 16 недель после IM инъекции собирали сыворотки мышей, подвергнутых лечению, как описано выше и как показано на фигуре 3, и анализировали в серийных разведениях посредством ELISA на антитела против кодируемого трансгеном GFP (фигура 5). Все мыши, которым инъецировали Ad, генерировали значимые уровни антител против GFP по сравнению с группой PBS при разведениях сыворотки от 1:10000 до 1:1000 (p<0,0001 по результатам одностороннего ANOVA). Однако Ad5-GL-HVR-12.51 приводил к более высоким уровням антител, чем Ad5 или Ad5-HVR5-12.52. При разведениях сыворотки от 1:1000 до 1:10000 Ad5-GL-HVR-12.51 был значимо выше, чем Ad5-GL (p<0,01-0,0001 по результатам одностороннего ANOVA). При разведении сыворотки 1:1000 12.51 был значимо выше, чем 12.52 (p<0,05). При разведениях сыворотки 1:1000 и 1:10000 Ad5-GL-12.52 был значимо выше, чем Ad5-GL (p<0,05-0,001).
[000214] В этом примере показано, что пептиды, выбранные в совместимом структурном контексте на фаговых библиотеках, могут транслироваться в белок гексона Ad. Например, в случае пептида 12.51 этот участок инсерции повышает трансдукцию мышц, при этом снижая нецелевое инфицирование в печени. Таким образом, такой рекомбинантный Ad, нацеленный на мышечную ткань, можно использовать в качестве вектора для генной вакцинации в мышцы или генной терапии/доставки в мышцы.
[000215] Дополнительный аспект изобретения относится к рекомбинантным и/или химерным Ad, содержащим другие клеточно-нацеливающие пептиды, встроенные в HVR Ad657 и петли HI Ad6 и C68, описанные в таблице 1.
Таблица 1
Другие клеточно-нацеливающие пептиды, встроенные в HVR Ad657 и петли HI Ad6 и C68
[000216] Синтезировали ДНК, кодирующую указанные пептиды, и комплементарную ей ДНК, фланкированные липкими концами для лигирования в плазмиды Ad, например, плазмиды гексона XA или челночные плазмиды волокна pAd6-NdePfl. Эти отжигаемые олигонуклеотиды лигировали в HVR или петлю HI Ad. Эти плазмиды использовали для рекомбинации в различные плазмиды остова Ad.
[000217] Некоторые пептиды предназначены для таргетинга новых рецепторов на клетках. Другие пептиды, подобные небольшому BAP, можно использовать для таргетинга авидина и очистки, если вирус выращивают в клетках, экспрессирующих бактериальную биотинлигазу BirA. Пептид кальмодулина позволяет вирусу связываться с кальмодулином или слитыми белками кальмодулина для перенаправления или очистки вируса.
[000218] Таким образом, такие рекомбинантные Ad, нацеленные на специфические ткани/клеточные рецепторы, можно использовать в качестве вектора для генной вакцинации или генной терапии в целевых клетках и/или тканях.
Пример 3. Инсерция отдельных HVR из разных серотипов Ad с инсерцией клеточно-нацеливающих/ненацеливающих пептидов или новых аминокислот.
[000219] На карте челночной плазмиды гексона (фигура 34) показана комбинация инсерции отдельных HVR из разных серотипов Ad с инсерцией клеточно-нацеливающих/ненацеливающих пептидов или новых аминокислот, таких как цистеин, в гексон для направленной химической модификации и экранирования.
[000220] В определенных вариантах осуществления связывающие клетки пептиды 12.51, VSV, RGD (см. таблицу 1) встраивают в HVR 1 или HVR 5, где варианты осуществления служат в качестве примеров инсерции этих и других пептидов в любых из HVR Ad (фигура 34). В другом примере показано, что в эти HVR встраивают инсерцию пептида-акцептора биотина (BAP), делающую возможным перенаправление вектора с помощью авидина или стрептавидина и биотинилированных лигандов или с помощью слитых белков авидина или стрептавидина. Инсерция BAP также позволяет очищать вирусы на колонки с мономерным авидином или стрептавидином для получения вектора. Аналогично, Ad57-HVR1-XXA и XA представляют собой пример инсерции цистеина в этот участок, чтобы сделать возможной направленную химическую модификацию с помощью малеимида или других цистеин-реактивных средств (фигура 34).
[000221] Эти варианты осуществления также относятся к Ad, в которых комбинируют разные HVR из разных Ad (т.е. перестановка HVR). Например, HVR1 из Ad6 м HVR 2-7 из Ad57 или HVR1 и 7 из Ad6 с HVR 2-6 из Ad57. В дополнительном варианте осуществления вирус 6/56/6 имеет HVR 1 и 7 из Ad6 и HVR 2-6 из Ad657.
Пример 4. Направленная химическая конъюгация цистеин-модифицированного гексон-модифицированного Ad657-HVR5C.
[000222] Фигура 35 является изображением вариантов Ad, на котором показана комбинация инсерции отдельных HVR из разных серотипов Ad с инсерцией новых аминокислот, таких как цистеин, в гексон для направленной химической модификации и экранирования.
[000223] Сравнение эффектов ненаправленной химической конъюгации и направленной химической конъюгации в отношении экранирования и функции цистеин-модифицированного гексон-модифицированного Ad657-HVR1C (фигура 36). В этом примере показана возможность нацеливания полимера и других химических модификаций на цистеины, встроенные в гексон Ad. Ненацеленный PEG инактивирует вирусную инфекцию, в то время как цистеин-направленное пегилирование позволяет сохранить функции вируса.
[000224] В одном из аспектов изобретения предусмотрено применение полимеров или встроенных пептидов/белков для ненацеливания, перенацеливания и экранирования от антител, белков, клеток. На фигуре 54 показаны участки HVR Ad, которые можно модифицировать, например, посредством пегилирования или "бапилирования".
[000225] В варианте осуществления разные серотипы и/или варианты Ad включают экранирование полимером, чтобы сделать возможной многократное введение вариантов Ad6 и Ad657. Приведен пример терапевтического цикла, где Ad6 и Ad657 можно использовать для множества раундов лечения благодаря переключению серотипа в комбинации с ковалентной конъюгации полимера (фигура 41B).
[000226] Ad657-HVR1C, экспрессирующий GFP-люциферазу, получали из клеток и очищали с помощью градиентов CsCl. Вирус ковалентно модифицировали с помощью полиэтиленгликоля 5 кДа (PEG). Вирус обрабатывали NHS-PEG, случайным образом реагирующим с аминами/лизинами на вирусных белках, или малеимид-PEG, реагирующим специфически с цистеином, встраиваемым в HVR1 с использованием челночной плазмиды XXA. Затем эти немодифицированные или модифицированные вирусы очищали посредством конечного центрифугирования в CsCl с последующим обессоливанием. Указанный вирус разделяли с помощью ПААГ в SDS, окрашивали SyproRuby и визуализировали (фигура 36A). Показано, что NHS-пегилирование позволяет случайным образом модифицировать многие вирусные белки, о чем свидетельствуют повышения кажущейся массы белков (показаны стрелками). В отличие от этого, направленная реакция малеимид-PEG с цистеином в HVR1 позволяет модифицировать только гексон и не повреждать другие вирусные капсомерные белки. Оценивали эффекты пегилирования в отношении функции вируса.
[000227] Указанные вирусы инкубировали с клетками A549 и измеряли их способность инфицировать клетки посредством анализа люциферазы. Это свидетельствует о том, что случайное NHS-пегилирование снижает активность вируса более чем на 90%, в то время как малеимид-PEG не снижает (фигура 36B).
[000228] Иммунокомпетентным сирийским хомячкам приживляли подкожные опухоли рака почки HaK. Когда они достигали объема 200 мкл, хомякам однократно внутривенно инъецировали указанные вирусы Ad6, сконструированные с E3 (DE3) и без него и со случайным NHS-пегилированием или без него. Измеряли размер опухоли с течением времени. Данные свидетельствуют о том, что делеция всех генов E3 в онколитическом вирусе Ad6-deltaE3-Luc делает вирус менее эффективным в снижении объема опухоли, чем родительский онколитический вирус Ad6-Luc. Данные также свидетельствуют о том, что Ad6 можно пегилировать и сохранять их эффективность (см. Ad6-Luc vs. Ad6-Luc-20 кДа PEG) (фигура 58).
[000229] Направленная химическая конъюгация цистеин-модифицированных гексон-модифицированных Ad, например, Ad657-HVR5C. Ad657-HVR5C, экспрессирующий GFP-люциферазу, получали из клеток и очищали с помощью градиентов CsCl. Вирус ковалентно модифицировали с использованием флуорофора ближней ИК-области малеимид-IR800, малеимид-биотина или 5 кДа малеимид-PEG, реагирующего специфически с цистеином, встраиваемым в HVR5 с использованием челночной плазмиды XXA. Указанные Ad и модифицированные Ad разделяли с помощью ПААГ с SDS, окрашивали SyproRuby и визуализировали (фигура 37A). Электрофорез в ПААГ с SDS вирусных белков с последующей визуализацией в ближней ИК-области показал, что HVR-C можно метить средством для визуализации (фигура 37B). Эффекты пегилирования в отношении функции вируса Ad in vivo демонстрировали посредством интраперитонеальной инъекции пегилированного вируса Ad. Способность инфицировать клетки мышей, несущих опухоли, демонстрировали с помощью детектируемой люциферазной активности посредством визуализации. На фигуре 37C показана возможность нацеливания полимера и других химических модификаций на цистеины, встроенные в область гексона Ad657. Кроме того, показано, что пегилирование вызывает ненацеливание аденовируса на печень in vivo (фигура 50).
Пример 5. Экспрессия Ad657 гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ) человека.
[000230] Ad657, несущий кДНК ГМ-КСФ человека, использовали для инфицирования клеток A549 и к ГМ-КСФ-зависимым клеткам TF-1 добавляли различные количества супернатанта. Повышение количества клеток свидетельствует об экспрессии функционального цитокина человека (фигура 27). Данные свидетельствуют о том, что рекомбинантные Ad можно использовать для экспрессии гетерологичных белков.
Пример 6. Онколитический аденовирус Ad657 для системной виротерапии против злокачественных клеток.
[000231] Выравнивание выбранных полных геномов Ad, с помощью которого получали филогенетическое дерево, в котором Ad57 кластеризуют с другими видами вирусов клады C с наибольшей гомологией с Ad6, показано на фигуре 6.
[000232] Ad57 выглядит почти идентичным Ad6 с дивергенцией последовательностей в гипервариабельных областях гексона (HVR) и генах уклонения от иммунологического надзора E3 (фигура 7). Другие экспонируемые вирусные белки капсида, включая волокно, основание пентона, IIIa и IX, являются, по существу, идентичными между Ad6 и Ad57. Данные анализа нейтрализации соответствуют тому, что большинство аденовирус-нейтрализующих антител нацелены на HVR Ad. Считают, что причиной низкой перекрестной реактивности между Ad6-антисывороткой и Ad57 являются антитела, которые могут быть нацелены на распространенный белок волокна (Lukashev et al., 2008 J Gen Virol. 89:380-388).
[000233] В этом примере показана применимость Ad657 в качестве онколитического средства против рака предстательной железы человека. HVR Ad6 заменяли HVR из Ad57 для получения химерного онколитического вируса клады C, названного Ad657. Ad657 и Ad6 тестировали в качестве системной онколитической терапии посредством однократной i.v. инъекции "голым" мышам, несущим опухоли рака предстательной железы человека. Тропизм этого вируса к печени и опухоли оценивали на мышиных моделях рака предстательной железы следующим образом.
[000234] Клетки карциномы предстательной железы человека DU145 приобретали в American Type Culture Collection (ATCC; Manassas, VA, USA) и проверяли отсутствие в них специфических патогенов посредством тестирования IMPACT RADIL. Клетки 293 получали в Microbix, Toronto, Ontario, Canada. Клетки поддерживали в DMEM с 10% FBS (Invitrogen, Grand Island, NY, USA).
[000235] Геном Ad6 штамма Tonsil 99 (ATCC VR-1083) клонировали, как описано в других источниках (см., например, Weaver et al., 2013 PLoS One. 8:e73313). Кассету, соответствующую гексону Ad57 между природными участками ApaI и SacI, синтезировали в Genscript. Этот фрагмент клонировали в челночную плазмиду pUC57-Гексон Ad6-FZF, содержащую гены pVI и гексона Ad6 с кассетой гена резистентности FRT FRT-Zeocin® между ними для гомологичной рекомбинации у бактерий, как описано в других источниках (см., например, Campos et al., 2004 Hum Gene Ther. 15:1125-1130 и Khare et al., 2012 J Virol. 86:2293-2301). Фрагмент Ad6 ApaI-SacI заменяли фрагментом Ad57, получая плазмиду pUC57-Гексон Ad6/57-FZF. Ее рекомбинировали в геном Ad6 посредством Red-зависимой рекомбинации (Campos et al., 2004 Hum Gene Ther. 15:1125-1130). На фигуре 59 показана карта плазмиды Ad657 с делецией E3. Вирусы подвергали "спасению" посредством трансфекции в клетках 293 и продуцировали в камерах для культивирования CellStack 10 (Corning Life Sciences, Lowell, MA, USA). Вирусы, экспрессирующие слитый белок зеленый флуоресцентный белок-люциферазу (GFP-Luc), содержат экспрессирующую кассету CMV-GFP-Luc, встроенную между волокном Ad и E4, и делецию E3, чтобы освободить место для этой инсерции. Вирусы очищали с помощью двух градиентов CsCl и вычисляли количество вирусных частиц (vp) по OD260.
[000236] Для исследования онколитической активности in vitro клетки обрабатывали при указанной множественности заражения (MOI) в терминах vp/клетку в DMEM с 5% FBS и антибиотиком-противогрибковым средством (Invitrogen, Grand Island, NY, USA). Через пять дней удаляли среду и клетки обрабатывали кристалл-виолетом (0,05% кристалл-виолетом, 3,7% формальдегидом в фосфатно-солевом буфере; Invitrogen, Grand Island, NY, USA) в течение 10 минут. Клетки дважды промывали PBS, а затем инкубировали в течение ночи при 37°C в 0,1% додецилсульфате натрия в PBS для солюбилизации кристалл-виолета. Оптическую плотность кристалл-виолета измеряли при OD595 с помощью спектрофотометра для чтения планшетов Beckman Coulter DTX 880. Жизнеспособность клеток (%) вычисляли, разделяя OD образцов на среднюю OD необработанных контрольных клеток на том же 96-луночном планшете и умножая это число на 100.
[000237] Животных держали в виварии Клиники Мэйо по руководствам Международной ассоциации оценки и аккредитации лабораторных исследований на животных. Исследования были одобрены Институциональным комитетом по содержанию и использованию животных Клиники Мэйо в соответствии с положениями Закона о благополучии животных, Политики благополучия животных PHS. Развитие подкожных опухолей у "голых" мышей возрастом 4 недели (Harlan Sprague Dawley, Indianapolis, IN, USA) инициировали посредством подкожной инъекции (s.c.) 1×107 клеток DU145 в 100 мкл DMEM/50% матригеля (BD Biosciences, San Jose, CA, USA). Объемы опухолей вычисляли с использованием уравнения ширина2×длина×1/2. Когда опухоли достигали объема ~200 мкл, мышей распределяли по разным группам и повергали их однократной i.v. инъекции в хвостовую вену. Животных умерщвляли, когда объем опухоли достигал 2000 мкл, или если животные находились в агонии, состоянии дистресса, или если над опухолью произошел разрыв кожи.
[000238] Для измерений аланинаминотрансферазы (АЛТ) в крови группам по шесть мышей C57BL/6 инъецировали i.v. 1011 vp Ad5, Ad6 или Ad657 в хвостовую вену и собирали кровь через 3 дня для измерения АЛТ с использованием анализа активности АЛТ (Sigma-Aldrich, St. Louis, MO, USA).
[000239] Статистический анализ осуществляли с помощью Prism (Graphpad) посредством ANOVA повторных измерений или одностороннего ANOVA с последующим тестом Тьюки HSD. Строили кривые выживаемости Каплана-Мейера и сравнивали их с помощью логарифмического рангового критерия.
[000240] Гены капсомерных белков Ad57 являются почти идентичными Ad6, за исключением HVR гексона (фигуры 6 и 7). Для получения химерного вируса Ad57 и Ad6 кассету, соответствующую HVR гексона Ad57, рекомбинировали в геном Ad6 дикого типа (фигура 8). Этот вирус подвергали "спасению", продуцировали в клетках 293 и очищали на градиентах CsCl. Учитывая, что основой вирусного генома являлся Ad6, эти гексон-химерные вирусы обозначают как Ad657 (фигура 59).
[000241] Онколитическую активность in vitro оценивали посредством инфицирования клеток LNCap и DU145 с использованием 10, 100 или 1000 vp/клетку. Для сравнения повреждения печени Ad5, Ad6 и Ad657 высокую дозу 1011 vp каждого вируса инъецировали в хвостовую вену иммунокомпетентным мышам C57BL/6. У животных, которым инъецировали Ad5, наступала агония в течение 2 дней, и их приходилось умерщвлять (фигура 10A). Выживаемость Ad5 и Ad6 была значимо ниже по сравнению с PBS (p=0,0001 и 0,0009, соответственно, по результатам анализа логарифмического рангового критерия). Выживаемость Ad657 также была снижена по сравнению с PBS (p=0,0578). Выживаемость после воздействия Ad6 или Ad657 была значимо лучше, чем у мышей, которым вводили Ad5 (p=0,0001 и 0,0001, соответственно). Выживаемость в случае Ad6 и Ad657 значимо не отличалась (p=0,248).
[000242] АЛТ измеряли в крови через 3 дня после инъекции у животных, выживших после Ad6 и Ad657. Животных, которым вводили Ad5, не тестировали, т.к. основную часть группы приходилось умерщвлять. Этот анализ показал, что Ad6 провоцировал относительно низкие уровни повреждения печени в терминах высвобождения фермента АЛТ печени в кровоток (фигура 10B). Обе группы Ad6 и Ad657 имели низкие, но значимые, уровни АЛТ по сравнению с мышами, которым вводили PBS (p<0,001 по результатам одностороннего ANOVA с критерием множественного сравнения Тьюки для обоих вирусов). Ad657 приводил к более низким уровням АЛТ, чем Ad6 (p<0,001 по результатам ANOVA). Это соответствовало более высоким уровням инфекции Ad6 в печени, чем Ad657 после i.v. инъекции экспрессирующих люциферазу вирусов (фигура 12). Одно животное в группе Ad657 было утрачено после забора крови в день 3. Через 6 дней у большинства животных в группе Ad6 началась агония (фигура 10A). В отличие от этого, 50% животных в группе Ad657 выживали в течение более 2 недель после введения.
[000243] Для сравнения онколитической активности Ad6 и Ad657 против опухолей предстательной железы DU145 человека, "голым" мышам приживляли s.c. клетки DU145. Животных распределяли по группам по схожим размерам опухолей в среднем 200 мкл, и группам по девять мышей однократно вводят i.v. дозу 3×1010 vp Ad6 или Ad657 (фигура 11).
[000244] Эта однократная i.v. инъекция Ad6 и Ad657 приводила к снижению размеров опухолей по сравнению с контрольными животными, которым инъецировали PBS. Опухоли были значимо меньше в группе Ad6 в течение 7 дней по сравнению с группой PBS (p<0,05 по результатам двухстороннего ANOVA с использованием критерия множественного сравнения Тьюки). Опухоли в группе Ad657 значимо отличались от опухолей в группе PBS ко дню 14 (p<0,01 по результатам ANOVA). И Ad6, и Ad657 поддерживали значимые различия с PBS до дня 38 (p<0,0001 по результатам двухстороннего ANOVA). Это сравнение заканчивали в день 38, когда первое животное в группе PBS приходилось умерщвлять, и последующее сравнение будет искажено из-за изменения количества животных. Размеры опухолей в группах Ad6 и Ad657 не отличались значимо до дня 38, когда в группе Ad657 был значимо большим объем опухоли (p<0,05) по результатам двухстороннего ANOVA (фигура 11A). Это различие между размерами опухолей в группах Ad6 и Ad657 сохранялось до дня 52 (p=0,04, t-критерий Стьюдента), а затем опухоли переставали значимо отличаться.
[000245] Когда оценивали общую выживаемость, и Ad6, и Ad657 значимо продлевали выживаемость по сравнению с животными, которым вводили PBS (фигура 11B, p<0,01 и 0,05, соответственно, по результатам анализа логарифмического рангового критерия). Общая выживаемость в группе Ad6 была значимо лучше, чем в группе Ad657 (p<0,05). Однако это являлось артефактом выживаемости, приписываемым полностью тому, что трех из животных в группе Ad657 приходилось умерщвлять по руководствам Институционального комитета по содержанию и использованию животных (IACUC) из-за образования язв на коже над опухолью, а не избыточному размеру опухоли. В некоторых случаях изъязвление конкретно ассоциировано с эффективным контролем опухоли. Подобно Ad6, Ad657, экспрессирующий GFP-люциферазу, приводил к значимой активности люциферазы в отдаленных подкожных опухолях DU145 после однократной i.v. инъекции (фигура 12 и фигура 13). Это позволяет предполагать, что и Ad6, и Ad657 могут опосредовать онколитические эффекты в опухолях предстательной железы после однократного системного введения.
[000246] В этом примере показано, что Ad657 можно использовать в качестве локальной или системной онколитической виротерапии рака предстательной железы. Эти данные также свидетельствуют о неожиданных эффектах переключения серотипа онколитических Ad клады C.
[000247] Онколитическую активность Ad оценивали в опухолевых клетках и/или злокачественных опухолях. Оценивали однократную IV инъекцию Ad6 в отношении опухолевых клеток легких A549 (фигура 28); оценивали однократную IV или внутриопухолевую (IT) инъекцию Ad6 в отношении опухолей поджелудочной железы Panc1 (фигура 29), и оценивали однократную IV инъекцию Ad6 в отношении рака почки у иммунокомпетентных хомяков (фигура 30).
[000248] На фигуре 38 показана люциферазная визуализация "голых" мышей. A) через 1, 4, 7, 14, 28 и 42 дня после однократной I.V. инъекции Ad6 в отношении отдаленных опухолей предстательной железы DU145. B) через 3, 7 и 19 дней после I.V. инъекции реплицирующегося Ad5-GFPLUC мышам, несущим опухоли предстательной железы LNCaP.
[000249] Можно сделать вывод, что после IV инъекции Ad6 достигает отдаленных клеток-мишеней.
[000250] В другом варианте осуществления Ad, экспрессирующие люциферазу, с пептидами 12.51 и 12.52, полученными с помощью библиотеки пептидов, или без них, встроенными в HVR5 гексона, инкубировали на указанных линиях клеток, клетках меланомы B16 и клетках карциномы легких A549, с указанными количествами вирусных частиц (vp) и измеряли люциферазную активность. Показана улучшенная инфекция злокачественных клеток Ad, несущими пептид-модифицированные гексоны (фигура 31).
Улучшенная инфекция злокачественных клеток Ad, несущими пептид-модифицированные гексоны.
[000251] Ad, экспрессирующие люциферазу с пептидами 12.51 и 12.52, полученными с помощью библиотеки пептидов, встроенными в HVR5 гексона, инкубировали на указанных линиях клеток печеночноклеточной карциномы с использованием 104 vp каждого вируса и измеряли люциферазную активность. Показана улучшенная инфекция злокачественных клеток Ad, несущими пептид-модифицированные гексоны (фигура 32).
Пример 7. Направляемые дивергентным ВИЧ-1 иммунные ответы, достигаемые посредством системной и слизистой иммунизации реплицирующимися одноцикловыми аденовирусами на макаках-резусах.
[000252] Большинство генных аденовирусных вакцин представляют собой векторы репликационно-дефектного Ad (RD-Ad), у которых ген E1 делетирован для предотвращения репликации и инициации инфекций Ad. У хелпер-зависимых аденовирусов (HD-Ad) все гены Ad делетированы, и они также являются репликационно-дефектными. Вакцина Ad с делетированным E1 может инфицировать клетку, доставлять одну копию гена антигена и экспрессировать одну копию (например, "1X") этого антигена. Они являются безопасными, но не реплицируют или экспрессируют трансгены.
[000253] В отличие от этого, вакцина E1+ репликационно-компетентного Ad (RC-Ad) может инфицировать ту же клетку, реплицировать ДНК гена антигена 10000 раз, продуцировать значительно больше антигена и провоцировать более сильные иммунные ответы, чем векторы с делетированным E1. Хотя RC-Ad является более активным, чем RD-Ad, репликационно-компетентные Ad могут представлять значительный риск выраженных аденовирусных инфекций у людей.
[000254] Чтобы воспользоваться преимуществом репликации ДНК трансгена, но избежать риска аденовирусных инфекций, разрабатывали векторы одноциклового Ad (SC-Ad) с делецией гена ключевого вирусного позднего белка, pIIIa (Crosby et al., 2014. Virology 462-463:158-165; Crosby et al., 2015 J Virol 89:669-675; Anguiano-Zarate et al., 2018 J Infectious Dis 218:1883-1889; и Crosby et al., 2017 Genes (Basel) 8:E79). SC-Ad сохраняют свои гены E1, что позволяет реплицировать их геном, но отсутствие pIIIa блокирует продукцию инфекционного потомства вирусов. SC-Ad реплицируют свои геномы и трансгены, а также RC-Ad (до 10000 раз; Crosby et al., 2014. Virology 462-463:158-165). RC- и SC-Ad продуцируют больше трансгенного белка, чем векторы RD-Ad (Crosby et al., 2014. Virology 462-463:158-165). SC-Ad генерируют более устойчивые и более персистирующие иммунные ответы, чем RD-Ad или RC-Ad (Crosby et al., 2015 J Virol 89:669-675). При параллельном сравнении SC-Ad приводит к образованию значимо большего количества антител и лучшей защите от вируса гриппа (Crosby et al., 2017 J Virol 91:e00720-16).
[000255] В этом исследовании макаков-резусов иммунизировали SC-Ad, экспрессирующими последовательности оболочки клады B, полученными от пациента с ВИЧ-1 до и после того, как их гуморальный ответ подвергся увеличению широты нейтрализации. Макаков иммунизировали посредством однократной системной IM иммунизации или однократной слизистой интраназальной (IN) иммунизации. Затем животных подвергали бустерной иммунизации теми же или альтернативными путями SC-6 Ad с последующим бустерным введением белка. В этом примере описано, как эти разные стратегии иммунизации SC-Ad влияли на образование ВИЧ-связывающих, вызывающих антителозависимую клеточную цитотоксичность (ADCC) и нейтрализующих антител, а также их эффекты в отношении клеточных иммунных ответов, включая фолликулярные T-хелперы (pTfh) в крови и лимфоузлах.
Одноцикловой аденовирус, экспрессирующий белок оболочки ВИЧ-1 gp140.
[000256] Последовательности белка оболочки gp160 клады B, возникающего до (G4) и непосредственно перед пиком увеличения широты нейтрализации антител (F8), от пациента с ВИЧ VC10014 (Malherbe et al., 2014 J Virol 88:12949-12967) использовали в качестве иммуногенов. Оптимизированные в отношении мотивов последовательности gp160 G4 и F8 рекомбинировали в SC-Ad на основе серотипов 6 и 57 Ad человека (см., например, Crosby et al., 2014. Virology 462-463:158-165; Crosby et al., 2015 J Virol 89:669-675; Anguiano-Zarate et al., 2018 J Infectious Dis 218:1883-1889; и Nguyen et al., 2018 Oncolytic Therapy 7:43-51). Также использовали контрольный SC-Ad, экспрессирующий гликопротеин вируса Эбола. Вирусы подвергали "спасению" и очищали, как описано в других источниках (см., например, Crosby et al., 2014. Virology 462-463:158-165; Crosby et al., 2015 J Virol 89:669-675; и Anguiano-Zarate et al., 2018 J Infectious Dis 218:1883-1889).
[000257] Ген оболочки F8, клонированный из инфицированного ВИЧ клады B индивидуума, использованный в векторе SC-Ad, подвергали оптимизации мотивов и модифицировали посредством сайт-специфического мутагенеза для экспрессии нерасщепленного, тримерного gp140. Подробности экспрессии, очистки и антигенных характеристик описаны в других источниках (см., например, Malherbe et al., 2014 J Virol 88:12949-12967).
[000258] Взрослых самок макак-резусов (Macaca mulatta) индийского происхождения содержали в Michael Keeling Center for Comparative Medicine and Research в University of Texas MD Anderson Cancer Center, Bastrop TX в колонии молодняка без специфических патогенов. Все содержание животных осуществляли в соответствии с политикой и процедурами Клиники Мэйо и University of Texas MD Anderson Cancer Center, положениями с положениями Закона о благополучии животных, Политики благополучия животных PHS и принципами Руководства по содержанию и использованию лабораторных животных NIH.
[000259] Макак анестезировали кетамином и иммунизировали интраназальным (IN) или внутримышечным (IM) путем с использованием 2×1010 вирусных частиц (vp) указанной вакцины SC-Ad. Животных подвергали бустерной иммунизации 50 мкг очищенного, тримерного рекомбинантного gp140 F8, комбинированного с адъювантом Adjuplex™, IM путем, как описано в других источниках (см., например, Malherbe et al., 2014 J Virol 88:12949-12967; и Hessell et al., 2016 J Immunol 196:3064-3078).
[000260] Образцы периферической венозной крови собирали в ЭДТА. Перед изоляцией мононуклеарных клеток периферической крови (PBMC) плазму разделяли и хранили при -80°C. PBMC получали из крови в градиенте плотности фиколл-гипака. Слюну и вагинальные смывы, собранные с помощью тупферов Weck-Cel в 1 мл PBS, содержащего ингибиторы протеаз, перемешивали на центрифуге типа "вортекс" и собирали супернатанты после центрифугирования при 2000 об/мин, как описано в других источниках (см., например, Kozlowski et al., 1997 Infect Immun 65:1387-1394). Образцы хранили замороженными при -80°C до дальнейшего использования.
Анализ ELISPOT для детекции антиген-специфических, ИФНγ-продуцирующих клеток
[000261] Свежевыделенные PBMC стимулировали белком gp140 F8 (1 мкг/мл) или термически инактивированным Ad6 (7,0×108 vp/лунку) для определения количества ИФНγ-продуцирующих клеток способом иммуноферментных пятен (ELISPOT) с использованием методологии, описанной в других источниках (см., например, Nehete et al., 2017 Comp Med 67:67-78; Nehete et al., 2013 PLoS One 8:e79836; и Nehete et al., 2017 J Am Assoc Lab Anim Sci 56:509-519). В кратком изложении, аликвоты PBMC (105/лунку) высевали в двух параллелях в лунки 96-луночных планшетов (планшеты из поливинилидендифторида с противоореольным слоем, MAIP S45, Millipore, Bedford, MA), предварительно покрытых первичным антителом против ИФНγ, и лимфоциты стимулировали Con A, белком gp140 F8 или термически инактивированным Ad6. После инкубации в течение 30-36 часов при 37°C клетки удаляли, лунки тщательно промывали PBS и проявляли по инструкциям производителя. Результаты выражали как ИФНγ образующих пятна клеток (SFC) на 105 PBMC после вычитания параллельных лунок только со средой (отрицательный контроль) и считали положительными при двухкратном превышении фона и превышении 5 SFC/105 PBMC.
ELISA антител
[000262] Титры антител, связывающихся с оболочкой ВИЧ-1, измеряли в образцах плазмы, собранных через регулярные интервалы, против gp140 F8 или gp140 SF162, как описано в других источниках (Malherbe et al., 2014 J Virol 88:12949-12967; и Hessell et al., 2016 J Immunol 196:3064-3078).
Анализ нейтрализации
[000263] Нейтрализацию ВИЧ осуществляли с использованием анализы нейтрализации TZM-bl, как описано в других источниках (Malherbe et al., 2014 J Virol 88:12949-12967; и Hessell et al., 2016 J Immunol 196:3064-3078). Все значения вычисляли по сравнению с лунками только с вирусом.
Антителозависимая клеточная цитотоксичность (ADCC)
[000264] Клетки-мишени CEM.NKR.CCR5.CD4+-Luc инфицировали 50 нг SHIVSF162P3 и культивировали в течение 4 дней, как описано в других источниках (см., например, Alpert et al., 2012 PLoS Pathog 8:e1002890). К инфицированным мишеням добавляли двухкратные серийные разведения каждого образца в течение 20 минут при комнатной температуре. Эффекторные клетки CD16-KHYG-1 добавляли при соотношении эффектор-мишень 10:1 и их инкубировали еще в течение 8 часов. Клетки лизировали и измеряли люциферазную активность с помощью спектрофотометра для чтения планшетов Bio-Tek.
Проточная цитометрия
[000265] Клетки, собранные из ректальных биоптатов и биоптатов лимфоузлов, инкубировали в течение ночи с 0,2 мкг gp140 или только средой в присутствие GolgiPlug™ (BD Biosciences, San Jose, CA, USA) в течение последних 4 часов. После культивирования клетки собирали и инкубировали на льду в течение 45 минут с панелью антител человека, перекрестно реагирующих с образцами макак-резусов. Панели включали следующие меченые флуорохромом антитела: против CD8 (Qdot655), против α4β7 (PE) и против CXCR5 (PE), все их которых получены из Nonhuman Primate Reagent Resource; против CD69 (BV737, клон FN50) и FoxP3 (PECy5, клон: PCH101), полученное из eBioscience, против ИЛ-21 (BV421, клон: 3A3-N2.1), против CD45 (BV786, D058-1283) и против CD3 (клон SP34-2, меченое PE-Cy7), все из BD Bioscience (San Jose, CA); против CD4 (Pacific Blue, клон OKT4) из ThermoFisher Scientific (Waltham, MA). Разведения антител определяли по инструкциям производителя. Погибшие клетки исключали с использованием набора для окрашивания фиксируемых погибших клеток LIVE/DEAD®, полученного из Invitrogen (Carlsbad, CA). Затем клетки дважды промывали PBS, содержащим 2% FBS и 2 мМ ЭДТА, а затем фиксировали и пермеабилизовали с помощью набора FoxP3 Fix/Perm (ThermoFisher Scientific, Waltham, MA). Внутриклеточные маркеры FoxP3 и ИЛ-21 окрашивали в буфере для пермеабилизации. Использовали и компенсационные контроли (OneComp eBeads, (ThermoFisher Scientific, Waltham, MA), и контроли флуоресценции с комбинацией детектируемых меток без одной (FMO). Все образцы собирали с помощью анализатора LSR Fortessa X-20 (BD Biosciences, San Jose, CA) и анализировали с использованием программного обеспечения FlowJo (FlowJo, LLC, Ashland, Oregon). Собирали приблизительно от 2×105 до 1×106 событий на образец.
Ректально заражение SHIVSF162P3
[000266] Вирус SHIVSF162P3 получали из сбора R157 3 (3.16.12). Этот сток имел содержание P27 66 нг/мл, содержание РНК Log ~9,35, TCID50 в PBMC макак-резусов индийского происхождения: 1288/мл, и TCID50 в клетках TZM-bl: 4,1×104/мл. Использовали 1 мл разведения 1:300 стока. Это эквивалентно 4,3 TCID50 в PBMC макак-резусов и 137 TCID50 в клетках TZM-bl. Эту дозу использовали для еженедельного интраректального (IR) заражения. Образцы плазмы анализировали на количество копий РНК вируса SHIV в Leidos Biomedical Research, Inc., Frederick National Laboratory. Животных с количеством копий РНК выше 10 считали инфицированными и количество заражений, необходимых для инфицирования этого животного, использовали в качестве событий для анализа выживаемости Каплана-Мейера. После инфицирования животное больше не заражали. Вирусную нагрузку в плазме подвергали периодическому мониторингу тем же способом до конца исследования.
Вирусная нагрузка SHIVSF162P3 в тканях
[000267] В конце исследования собирали PBMC и трупные ткани. PBMC и образцы кишечника анализировали на РНК вируса SHIVSF162P3 посредством qPCR. Для всего статистического анализа использовали программное обеспечение Graph Pad Prism 7.
SC-Ad, экспрессирующий gp160 ВИЧ-1
[000268] Последовательности белка оболочки вируса клады B (gp160G4) идентифицировали до и непосредственно перед пиком увеличения широты нейтрализации антител (F8 gp160) от пациента с ВИЧ VC10014. Эти последовательности gp160 встраивали в SC-Ad6 и SC-Ad657 под контролем сильного промотора цитомегаловируса (фигура 24A). Ad57 является Ad человека клады C, почти идентичным Ad6, с изменением в гипервариабельных областях гексона (HVR) и в генах уклонения от иммунологического надзора E3 (фигура 24B). Большинство Ad-нейтрализующих антител нацелены против HVR гексона Ad (Pichla-Gollon et al., 2007 J Virol 81:1680-1689; и Sumida et al., 2005 J Immunol 174:7179-7185). Учитывая это, HVR Ad6 заменяли HVR из Ad57 для получения химерного вектора Ad клады C, обозначаемого как Ad657. И SC-Ad6, и SC-Ad657 сохраняют все гены Ad, включая E1, и не содержат функциональные гены pIIIA и E3 (фигура 24A). Таким образом, оба SC-Ad могут реплицировать свои геномы для амплификации экспрессии gp160, но не приводят к образованию потомства вирусов Ad. Оба вируса подвергали "спасению", продуцировали в клетках 293-IIIA и очищали с помощью градиентов CsCl. При использовании для инфекции клеток A549 оба вектора приводили к продукции gp160, что определяли с помощью вестерн-блоттинга.
[000269] Различные векторы Ad ранее тестировали на макаках-резусах посредством системного внутримышечного (IM) введения и введения различными слизистыми путями, включая желудочный зонд, капсулы с кишечнорастворимым покрытием, интраназальный (IN) и интравагинальный (IVAG) путь. Тестирование SC-Ad-G4 путями IM, IN и IVAG на небольших животных показало, что примирование путем IVAG приводило к незначительным гуморальным ответам. В отличие от этого, иммунизация IN путем у мышей и хомяков приводила к сильным гуморальным ответам. Учитывая эти данные и потенциальные затруднения в осуществлении иммунизации IVAG людей, выбирали IN путь для слизистого пути иммунизации в последующих исследованиях на макаках.
Однократная слизистая и системная иммунизация макак-резусов
[000270] Использовали 2×1010 vp SC-Ad6-G4 Env для вакцинации групп по 8 самок макак-резусов посредством однократной IM или IN иммунизации (фигура 14). Эта доза является относительно низкой, являясь в приблизительно 7,5 раз более низкой, чем при использовании вакцин RC-Ad с оболочкой ВИЧ, доставляемых с помощью смешанной IN и IM иммунизации. Группу вектора отрицательного контроля иммунизировали IN с помощью SC-Ad6, экспрессирующего гликопротеин вируса Эбола (gp). Через четыре недели образцы плазмы анализировали на Env-связывающие антитела против gp140 F8 (фигура 14). Наблюдали значимо более высокие титры связывания в средней точке в группе иммунизированных IM путем после однократной иммунизации (p<0,01 по результатам ANOVA). Титры SF162-нейтрализующих антител (NAb) также повышались в этот момент времени, но не достигали значимости по результатам ANOVA для групп отдельных путей введения.
Бустерное введение IM по сравнению с IN при использовании SC-Ad6 на неделе 4
[000271] Показано, что нейтрализующих антител против аденовируса, получаемых при однократной IM иммунизации Ad, можно избегать посредством бустерного введения другим путем (Xiang et al., 2003 J Virol 77:10780-10789). Для тестирования концепции этого пути введения, чтобы сделать возможным повторное использование того же серотипа Ad на макаках, каждую SC-Ad6-примированную группу разделяли на 2 группы по 4. Каждое животное подвергали бустерному введению SC-Ad6, экспрессирующего альтернативный Env F8, на неделе 4 IM или IN путем. Образцы плазмы, собранные через 3 недели после бустерного введения, демонстрировали повышение титров связывания в средней точке у животных, которых примировали и подвергали бустерному введению в группах IM-IM, IM-IN и IN-IM. Не наблюдали детектируемых антител в группе IN-IN (фигура 14).
Бустерное введение SC-Ad657 на неделе 13
[000272] Затем животных подвергали бустерной иммунизации посредством переключения серотипа с использованием SC-Ad657, экспрессирующего Env G4, на неделе 13. Тот же путь использовали в предшествующей бустерной иммунизации. Титры на неделе 15 свидетельствовали о том, что IM-примированные животные имели повышенные титры связывания Env приблизительно 350, но эти уровни не отличались значимо от контролей (фигура 14). В отличие от этого, антитела в группе IN-IM-IM были значимо выше, чем в группе контрольного вектора и группе IN-IN-IN (p<0,01). В группе IN-IN-IN снова не наблюдали антител против Env даже после 3 иммунизаций.
Бустерное введение рекомбинантного тримерного белка Env на неделе 24
[000273] В большинстве исследований вакцин против ВИЧ иммунизацию Ad расширяют с использований бустерных введений белков для амплификации гуморальных ответов. Например, в недавнем исследовании векторы RD-Ad26 использовали дважды и осуществляли бустерное введение три раза с адъювантным белком gp140 (Barouch et al., 2015 Science 349(6245):320-4). Для определения того, будет ли эта стратегия усиливать вакцины SC-Ad, все группы SC-Ad-Env подвергали бустерному введению 50 мкг рекомбинантного тримерного белка gp140 F8, смешанного с адъювантом ADJUPLEX™, IM путем. Тримерный белок F8 повышал титры связывания в средней точке на два порядка во всех группах (фигура 14). Эта иммунизация белком также повышала IN-IN-IN до уровней, сравнимых с другими группами, даже если Env-связывающие антитела не определяли после более ранних иммунизаций SC-Ad.
Связывающие и нейтрализующие антитела в плазме после второго бустерного введения белка.
[000274] Животных второй раз подвергали бустерному введению белка на неделе 38. Эти повышало титры F8-связывающих антител в плазме до приблизительно 105 к неделе 40, и все группы значимо отличались от контролей (фигура 14). Титры нейтрализующих антител (NAb) против вируса SF162 группы 1A повышались с 100 до 10000 на неделе 40 (фигура 15). NAb против вируса SS1196 группы 1B и вируса JRCSF группы 2 повышались до 100 у большинства животных, за исключением двух животных в группе IN-IN-IN, титры которых находились на фоновых уровнях (фигура 15).
Активность ADCC после второго бустерного введения белка
[000275] Активность антителозависимой клеточной цитотоксичности (ADCC) на неделе 40 в плазме тестировали в отношении инфицированных клеток SHIVSF162P3. Активность ADCC, как правило, была более высокой у животных, подвергнутых по меньшей мере одной слизистой IN иммунизации SC-Ad (фигура 16). Все животные, подвергнутые слизистой иммунизации, имели значимо более высокий максимальный % ADCC, чем контрольные животные, которым вводили SC-Ad-Ebola (p<0,05, 0,0001, 0,0001 для IM-IN-IN, IN-IM-IM и IN-IN-IN, соответственно). При сравнении по 50% титров ADCC только группы, IN-примированные SC-Ad, имели значимо более высокую активность ADCC, чем контроли (p<0,05 и 0,001 по результатам ANOVA для группы IN-IM-IM и IN-IN-IN).
Гуморальные ответы в слюне и вагинальных смывах после второго бустерного введения белка
[000276] С помощью указанных выше данных осуществляли мониторинг системных гуморальных ответов в плазме. Образцы слюны и вагинальных смывов также собирали на неделе 40 и измеряли антитела в этих участках слизистых оболочек. Когда слюну и вагинальные смывы анализировали на связывание env F8 и SF162 посредством ELISA, эти ответы наблюдали в большинстве групп, за исключением контрольной группы SC-Ad-Ebola (фигура 25).
[000277] По-видимому, в этом случае имел место регионарный эффект в отношении этих антител. У животных, иммунизированных с помощью SC-Ad, главным образом, IN путем (IM-IN-IN и IN-IN-IN), связывающие антитела были более высокими в слюне вблизи этого места иммунизации, но более низкими в более отдаленном участке во влагалище (фигура 25). Когда активность ADCC измеряли в этих слизистых образцах, эти ответы сильно варьировались (фигура 17). Несмотря на это, наблюдали более высокую активность ADCC в группе IN-IN-IN по сравнению с контрольными животными (p<0,05 по результатам ANOVA).
Системные клеточные иммунные ответы после однократного бустерного введения белка
[000278] На неделе 38 PBMC анализировали посредством ELISPOT на T-клетки против Env и против аденовируса на образцах, собранных непосредственно перед вторым бустерным введением белка Env F8. Все Env-иммунизированные животные имели Env-специфические ИФНγ-секретирующие клетки в PBMC (фигура 18A). Уровень Env-специфических ИФНγ-секретирующих SFC, как правило, был повышен у животных, подвергнутых по меньшей мере одной слизистой иммунизации. Однако ИФНγ-секретирующие SFC были значимо более высокими только в группе введения SC-Ad IN-IN-IN, когда их сравнивали с контрольными животными, иммунизированными SC-Ad Ebola (p<0,05 по результатам ANOVA). SFC против Ad являлись относительно низкими во всех группах по сравнению с SFC против Env в этот момент времени.
Системные клеточные иммунные ответы после второго бустерного введения белка
[000279] На неделе 40 PBMC и клетки паховых лимфоузлов анализировали на Env-специфические ИФНγ-секретирующих SFC посредством ELISPOT (фигура 18B). Это бустерное введение белка повышало Env-специфические SFC в PBMC и лимфоузлах до уровней, схожих с уровнями у всех Env-иммунизированных животных.
Клеточный транспорт в слизистых оболочках
[000280] При проточной цитометрии биоптатов прямой кишки на неделе 40 определяли схожие количества α4β7 CD4+ и CD8+ клеток в участках прямой кишки (фигура 19A). Наблюдали тенденцию в сторону повышения количеств в IN-примированных группах, но она не достигала значимости. Количества активированных CD69+ CD4+ клеток в тканях прямой кишки были схожими между группами (фигура 19B). Аналогично, FoxP3+ CD4+ клетки в этом участке слизистой оболочки не различались значимо (фигура 19B).
Распределение антиген-специфических Tfh-клеток
[000281] CXCR5+ ИЛ-21+ CD4+ фолликулярные T-хелперы (Tfh) измеряли в PBMC и образцах лимфоузлов на неделе 40 (фигура 20). Животные, иммунизированные Ad и белком только IM путем, имели значимо более высокие уровне периферических Tfh-клеток (pTfh) в PBMC, чем в других группах (фигура 20). В лимфоузлах Tfh-клетки имели наименьший уровень в контроле и группе только IM введения. В отличие от этого, приблизительно половина животных, подвергнутых по меньшей мере одной IN слизистой иммунизации, имели детектируемые Tfh-клетки в своих лимфоузлах после последнего бустерного введения белка (фигура 20).
Ректальное заражение SHIVSF162P3
[000282] Иммунизированных макак ректально заражали изолятом ВИЧ SHIVSF162P3. В исследование добавляли четырех неиммунизированных контрольных животных и каждую группу еженедельно заражали посредством ректальной инокуляции 1 мл разведения 1:300 стока SHIVSF162P3, предоставленного NIH. Это заражение эквивалентно 4,3 TCID50 на PBMC макак-резусов и 137 TCID50 на клетках TZM-bl. После первого заражения заражались 2 животные в неиммунизированной контрольной группе и 2 животных в группе IM-IM-IM (фигура 21). Одно животное в каждой из групп смешанного введения (IM-IN-IN и IN-IM-IM) заражались после первого заражения. Ни одно из животных в группе IN-IN-IN не заражалось после первого заражения. Вирусная нагрузка в плазме свидетельствовала о том, что все животные, за исключением животного в группе Ebola, достигали высокой вирусной нагрузки после 3 заражений (фигура 22B). Животные в группе IN-IN-IN имели задержку в достижении этой высокой вирусной нагрузки.
[000283] При продолжении заражений заражали животных во всех группах, за исключением одного животного в группе Ebola, оставшегося неинфицированным после 7 заражений. Аллели Trim5α и MHC анализировали ретроспективно (таблица 2). При этом анализе не наблюдали очевидно протективных генов у резистентного животного в группе Ebola. Большинство животных нельзя классифицировать по аллелям, которые могут делать их умеренно защищенными, но большинство групп включало по меньшей мере одно животное с более высокой вероятностью защиты благодаря этим аллелям. Следует отметить, что 2/4 животных в группах IM-IM-IM и IN-IN-IN имели аллели Trim5a и MHC, с помощью которых можно прогнозировать более высокую вероятность врожденной защиты против SIVsmm и, возможно, SHIVSF162P3 (таблица 2).
[000284] Таблица 2. Ретроспективный скрининг на протективные аллели генов SIV.
A08, A11, B01
[000285] Когда животных классифицировали на основе того, примировали ли их путем IM или IN SC-Ad, и анализировали выживаемость способом Каплана-Мейера, инфекция восьми IM-примированных животных соответствовала таковой у контрольных животных (фигура 22A). В отличие от этого, наблюдали задержку инфекций у восьми животных, примированных SC-Ad-Env слизистым IN путем.
Вирусная нагрузка в трупных тканях
[000286] Исследование заражения прекращали через 9 недель после первого заражения. Выделяли PBMC и ткани кишечника, очищали РНК и оценивали на вирусные геномы SHIV (фигура 22B). Трупные PBMC имели различные уровни вирусной РНК SHIV с, в некоторой степени, более низкими уровнями в группе IN-IN-IN, чем в группе IM-IM-IM. Средние уровни вирусной РНК в толстом кишечнике был в 15 раз более низким в группе IN-IN-IN, чем в группе IM-IM-IM (фигура 23). Это различие не достигало значимости по результатам ANOVA, но с помощью двухстороннего t-критерия Стьюдента получали значение p 0,0079.
[000287] В этом примере показано, что реплицирующиеся векторы SC-Ad можно использовать в качестве надежной и безопасной платформы для вакцинации против ВИЧ-1 и других инфекционных заболеваний. С помощью SC-Ad можно амплифицировать гены антигена и цитокинов в до 10000 раз в инфицированных клетках человека. Иммунный ответ амплифицируют до уровней, сильно превышающих уровни, опосредованные векторами RD-Ad, в настоящее время тестируемыми в качестве вакцин против ВИЧ-1 на людях. Вакцины против ВИЧ можно преобразовывать в вакцинные платформы, где амплифицируют гены антигенов ВИЧ с использованием векторов SC-Ad, например, векторов SC-Ad на основе рекомбинантных Ad, имеющих низкую серопревалентность.
Пример 8. Анализ in vivo цитотоксических T-лимфоцитов (CTL) на иммунные ответы против антигена вируса гепатита C (HCV).
[000288] Мышей иммунизировали Ad657, экспрессирующим кДНК гликопротеина B (gB) цитомегаловируса (CMV) или антиген 2.4 HCV. В сингенные клетки вводили пептид HCV и метили сукцинимидиловым сложным эфиром карбоксифлуоресцеина (CFSE) перед инъекцией иммунизированным мышам. Наблюдали активность когнатных CTL против HCV по утрате меченых клеток у животных, иммунизированных HCV, но не CMV (фигура 26).
Пример 9. Условно-репликативные Ad (CRAd).
[000289] Схема мутаций в Ad6, Ad657 и его вариантах, включая мутации в белке E1 для преобразования вируса в условно-репликативный Ad (CRAd), показана на фигуре 39 и фигуре 43. Они включают dl1101 и/или dl1107, блокирующие связывание с p300 и pRB, соответственно.
[000290] На фигуре 56 показаны N-концевые аминокислотные последовательности E1A в Ad дикого типа, а также варианты E1A dl1101, E1A dl1107 и E1A dl1101/1107 Ad.
[000291] Также показана замена промотора E1 Ad последовательностью ДНК простат-специфического промотора пробазина-E1 SEQ ID NO: 44 для получения CRAd, Ad-PB (фигура 55). Промотор пробазина является андроген-зависимым, таким образом, он будет действовать в андроген-чувствительных опухолях, подобных LNCaP, но не в андроген-резистентных опухолях, подобных DU145.
[000292] Клетки A549 инфицировали указанными вариантами Ad6 или Ad657 в указанных концентрациях вируса (vp/клетку) и измеряли жизнеспособность клеток посредством окрашивания кристалл-виолетом через 5 дней (фигура 40).
[000293] Уничтожение незлокачественных клеток репликационно-дефектным Ad (RD-Ad), Ad6, CRAd6-dl1101/dl1107 или CRAd6-PB. Показана модификация Ad6 и Ad657 в условно-репликативные Ad (CRAd) (фигура 44).
[000294] На фигуре 45 показано уничтожение злокачественных клеток репликационно-компетентным Ad5, Ad6, Ad657 и указанными CRAd. Показана модификация Ad6 и Ad657 в условно-репликативные Ad (CRAd).
[000295] Результаты, показанные на фигуре 46, свидетельствуют о модификации Ad6 и Ad657 в условно-репликативные Ad (CRAd) в клетках рака молочной железы.
[000296] Результаты, показанные на фигуре 47, свидетельствуют о модификации Ad6 и Ad657 в условно-репликативные Ad (CRAd) в клетках рака предстательной железы и клетках рака легких.
[000297] Эффекты in vivo репликационно-компетентного Ad6 или указанных CRAd в отношении роста опухолей DU145 у мышей. На фигуре 48 показана модификация Ad6 и Ad657 в условно-репликативные Ad (CRAd) in vivo после однократной внутривенной инъекции мышам, несущим опухоли предстательной железы человека.
[000298] Эффекты in vivo репликационно-компетентного Ad6 или указанных CRAd в отношении выживаемости мышей с опухолями DU145. На фигуре 49 показана модификация Ad6 и Ad657 в условно-репликативные Ad (CRAd) in vivo после однократной внутривенной инъекции мышам, несущим опухоли предстательной железы человека.
[000299] На фигуре 51 показано, что модификация Ad657 с более коротким волокном из AdC68 шимпанзе снижает эффективность.
[000300] В варианте осуществления вирус Ad 6/56/6 имеет HVR 1 и 7 из Ad6 и HVR 2-6 из Ad657. На фигуре 52 показан вирус Ad 6/56/6, уничтожающий клетки рака легких человека, с модификациями CRAd и без них.
[000301] Уничтожение опухолевых клеток вариантами Ad, включающими мутации в белке E3. Иммунокомпетентным сирийским хомячкам приживляли подкожные опухоли рака почки HaK. Когда они достигали объема 200 мкл, хомякам однократно внутривенно инъецировали указанные вирусы Ad6, сконструированные с E3 (DE3) и без него и со случайным NHS-пегилированием или без него. Измеряли размеры опухолей с течением времени. Данные свидетельствовали о том, что делеция всех генов E3 делает онколитический вирус менее эффективным (Ad6-deltaE3-Luc по сравнению с Ad6-Luc) (фигура 58).
[000302] Белок волокна Ad является комплексом трех, по-видимому, идентичных субъединиц, опосредующих исходную стадию прикрепления. Нативный белок волокна Ad6 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 60, и связывается с CAR.
[000303] В дополнительном аспекте изобретения получали волокно-модифицированные рекомбинантные Ad, имеющие разные белки волокна, не являющиеся нативными для родительского Ad. Получали рекомбинантные Ad, включая CRAd, содержащие белки капсида из разных штаммов Ad, например, рекомбинантные Ad, содержащие гетерологичный полипептид волокна Ad35 или полипептид волокна C68 шимпанзе +/- пептид K7 (фигуры 62-69).
[000304] Химерный Ad, химера волокна AdF35, имеет аминокислотную последовательность SEQ ID NO: 61 и является более коротким, чем белки волокна Ad5 и Ad6, и перенаправляет вирус на CD46.
[000305] Волокно-модифицированный рекомбинантный Ad, содержащий волокно K7, имеющее последовательность SEQ ID NO: 62, нацеливает вирус на гепаринсульфат протеогликаны и отрицательные заряды на клетках.
[000306] Рекомбинантный, химерный Ad, волокно 6/FC68, содержащее последовательность SEQ ID NO: 63, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6, и связывается с CAR.
[000307] Рекомбинантный, химерный Ad, волокно 6/FC68-K7, содержащее последовательность SEQ ID NO: 64, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6. Волокно 6/FC68-K7 связывается с CAR и перенаправляет на гепаринсульфат и отрицательные заряды.
[000308] Рекомбинантный, химерный Ad, волокно 6/FC68-HI-K7, содержащее последовательность SEQ ID NO: 65, является химерным Ad, имеющим белок волокна из аденовируса C68 шимпанзе. Белок волокна является более коротким, чем белки волокна Ad5 или Ad6. Волокно 6/FC68-HI-K7 связывается с CAR и перенаправляет на гепаринсульфат и отрицательные заряды.
Пример 10. Переключение серотипа аденовирусов.
[000309] Фигура 41 является схемой, на которой показаны Ad-терапевтические циклы. В варианте осуществления приведен пример переключения серотипа с использованием разных Ad с течением курса лечения (фигура 41A).
Таргетинг опухоли предстательной железы после переключения серотипа онколитических аденовирусов.
[000310] Мышей, несущих опухоли предстательной железы DU145 на своих боках, подвергали однократной внутривенной (IV) инъекции Ad657 или CRAd657. Этим мышам во второй раз вводили альтернативный онколитический вирус Ad6 или Ad6-F35, экспрессирующий GFP-люциферазу, и измеряли люциферазную активность посредством визуализации. Ad6 имеет гексон Ad6 и волокно Ad6, нацеленное на CAR. Ad6-F35 имеет гексон Ad6 и волокно Ad35, нацеленное на CD46. На фигуре 42 показана возможность переключения серотипа онколитических средств с использованием вирусов, нацеленных на опухоль, с более низким нецелевым инфицированием печени.
[000311] В другом примере переключения серотипа мышей, несущих опухоли предстательной железы LNCaP на своих быках, подвергали однократной внутривенной (IV) инъекции с использованием 3e10 вирусных частиц (vp) Ad657 или CRAd657. Этим мышам во второй раз через 5 месяцев вводили 3e10 vp альтернативного онколитического вируса Ad6/57/6, экспрессирующего GFP-люциферазу и варианты волокна K7 (с добавлением 7 лизинов), F35 (с волокном Ad35) или KKTK-C68 (волокно C68 шимпанзе, слитое после придающего гибкость домена KKTK Ad6). Вирус KKTK-C68 также содержит добавленный кодон-оптимизированный ген E4 34K для повышения вирусной продуктивности. Люциферазную активность измеряли через 7 дней посредством визуализации. Все Ad6/57/6 имели гексон с HVR1 и 7 из Ad6 и HVR 2-6 из Ad57. Ad6/57/6 и KKTK-C68 имели волокна, нацеленные на CAR. Ad6/57/6-F35 имел волокно Ad35, нацеленное на CD46. K7 повышает связывание с отрицательными зарядами на клетках, включая связывание гепаринсульфат протеогликанов. На фигуре 70 показана возможность переключения серотипа онколитических средств с использованием вирусов, нацеленных на опухоль, с более низким нецелевым инфицированием печени.
Переключение серотипа во время вакцинации не являющихся человеком приматов.
[000312] На фигурах 14-25 показано, что макак-резусов иммунизировали с использованием реплицирующегося одноциклового Ad6, экспрессирующего оболочку ВИЧ, а затем осуществляли бустерную иммунизацию посредством переключения серотипа с использованием одноциклового Ad657, экспрессирующего оболочку ВИЧ. После этих иммунизаций каждое животное подвергали бустерному введению белка оболочки. На каждой фигуре показано образование адаптивных гуморальных или клеточных иммунных ответов и то, как животные преодолевали ректальное заражение вирусом SHIV SF162P3. На фигуре 14 показано значение переключения серотипа, где замена на Ad657 приводит к значительному повышению гуморальных ответов.
Пример 11. Онколитические противоопухолевые вакцины.
[000313] Мышей BALB/c внутримышечно иммунизировали 1010 вирусными частицами CRAd-657-dl1101/1107-FolR с интактным E3, экспрессирующими рецептор фолиевой кислоты альфа человека, или PBS. Сыворотки собирали через 2 недели после однократной иммунизации и анализировали на антитела против рецептора фолиевой кислоты альфа посредством ELISA с использованием антитела против IgM для детекции (все антитела являются IgM при этом типе ранней временной точки после иммунизации). Данные свидетельствуют об образовании антител против известного антигена злокачественной опухоли - рецептора фолиевой кислоты альфа - при использовании этого CRAd. p - 0,07 при использовании t-критерия Стьюдента (фигура 53).
Пример 12. Эффекты генов уклонения от иммунологического надзора E3 в отношении онколитической активности.
[000314] На фигуре 57 показана схема разных генов уклонения от иммунологического надзора E3 в Ad. E3 19K защищает инфицированные клетки от T-клеток и NK-клеток. Белки RID защищают инфицированные клетки от индуцирующих гибель лигандов (FAS, TRAIL, TNFR и EGFR). 14,7K ингибирует внутреннюю активацию апоптоза в инфицированных клетках. Ad клады C также экспрессируют 11,6K, известный в качестве белка гибели клеток, инфицированных аденовирусом (ADP). Гиперэкспрессия ADP ускоряет гибель клеток, но общая гибель клеток является той же. Виды 49K связываются с CD46 на T-клетках и NK-клетках, что приводит к отрицательной регуляции этих клеток и менее эффективному уничтожению клеток, имеющих дефицит MHC I класса, NK-клетками.
[000315] На фигуре 58 показано, что частичная делеция E3 12,5K и полная делеция генов E3 6,7K, 19K, 11,6K (ADP), 10,4K (RIDα), 14,5K (RIDβ) и 14,7K снижает онколитическую эффективность в модели рака почки на иммунокомпетентных хомяках, когда эти гены уклонения от иммунологического надзора отсутствуют в онколитическом аденовирусе.
Другие варианты осуществления
[000316] Следует понимать, что хотя настоящее изобретение описано в комбинации с подробным описанием, изложенное выше описание представлено в иллюстративных целях, а не для ограничения объема изобретения, определяемого объемом формулы изобретения. Другие аспекты, преимущества и модификации входят в объем следующей формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MAYO FOUNDATION FOR MEDICAL EDUCATION AND RESEARCH
<120> АДЕНОВИРУСЫ И СПОСОБЫ ПРИМЕНЕНИЯ АДЕНОВИРУСОВ
<130> ADZE 1 US SEQ
<150> 62/770,631
<151> 2018-11-21
<160> 78
<170> PatentIn version 3.5
<210> 1
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 1
Thr Ala Arg Gly Glu His Lys Glu Glu Glu Leu Ile
1. 5 10
<210> 2
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 2
Leu Arg Gln Thr Gly Ala Ala Ser Ala Val Trp Gly
1. 5 10
<210> 3
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 3
Ala Arg Arg Ala Asp Thr Gln Trp Arg Gly Leu Glu
1. 5 10
<210> 4
<211> 19
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 4
Gly Thr Trp Leu Asn Pro Gly Phe Pro Pro Gln Ser Cys Gly Tyr Ala
1. 5 10 15
Thr Val Thr
<210> 5
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 5
Cys Asp Cys Arg Gly Asp Cys Phe Cys
1. 5
<210> 6
<211> 10
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 6
Asn Met Ser Leu Asp Val Asn Arg Lys Ala
1. 5 10
<210> 7
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 7
Ile Ser Leu Ser Ser His Arg Ala Thr Trp Val Val
1. 5 10
<210> 8
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 8
Trp Thr Met Gly Leu Asp Gln Leu Arg Asp Ser Ser Trp Ala His Gly
1. 5 10 15
Gly Phe Ser Ala
20
<210> 9
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 9
Trp Thr Met Gly Leu Asp Gln Leu Arg Gly Asp Ser Ser Trp Ala His
1. 5 10 15
Gly Gly Phe Ser
20
<210> 10
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 10
Arg Ser Val Ser Gly Thr Glu Trp Val Pro Met Asn Glu Gln His Arg
1. 5 10 15
Gly Ala Ile Trp
20
<210> 11
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 11
Thr Glu Leu Arg Thr His Thr Ser Lys Glu Leu Thr Ile Arg Thr Ala
1. 5 10 15
Ala Ser Ser Asp
20
<210> 12
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 12
Asp Arg Ala Ile Gly Trp Gln Asp Lys Leu Tyr Lys Leu Pro Leu Gly
1. 5 10 15
Ser Ile His Asn
20
<210> 13
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 13
Met Gly Ser Trp Glu Lys Ala Ala Leu Trp Asn Arg Val Ser Ala Ser
1. 5 10 15
Ser Gly Gly Ala
20
<210> 14
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 14
Met Ala Met Gly Gly Lys Pro Glu Arg Pro Ala Asp Ser Asp Asn Val
1. 5 10 15
Gln Val Arg Gly
20
<210> 15
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 15
Met Ala Ser Arg Gly Asp Ala Gly Glu Gly Ser Thr Gln Ser Asn Thr
1. 5 10 15
Asn Val Pro Ser
20
<210> 16
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 16
Gly Pro Glu Asp Thr Ser Arg Ala Pro Glu Asn Gln Gln Lys Thr Phe
1. 5 10 15
His Arg Arg Trp
20
<210> 17
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 17
Met Gly Arg Glu Asp Val Gly Glu Gln Lys Leu Ile Ser Glu Glu Asp
1. 5 10 15
Leu Gly Gly Ser
20
<210> 18
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 18
Ala Cys Asp Cys Arg Gly Asp Cys Phe Cys Gly
1. 5 10
<210> 19
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 19
Ala Cys Asp Cys Arg Glu Asp Val Cys Phe Cys Gly
1. 5 10
<210> 20
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 20
Gly Gln Ile Pro Ile Thr Glu Pro Glu Leu Cys Cys Val Pro Trp Thr
1. 5 10 15
Glu Ala Phe Tyr
20
<210> 21
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 21
Pro Gln Pro Pro Asn Ser Thr Ala His Pro Asn Pro His Lys Ala Pro
1. 5 10 15
Pro Asn Thr Thr
20
<210> 22
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 22
Val Arg Trp Phe Pro Gly Gly Glu Trp Gly Val Thr His Pro Glu Ser
1. 5 10 15
Leu Pro Pro Pro
20
<210> 23
<211> 19
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 23
Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys
1. 5 10 15
Lys Lys Lys
<210> 24
<211> 14
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 24
Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His
1. 5 10
<210> 25
<211> 23
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 25
Cys Ala Ala Ala Arg Trp Lys Lys Ala Phe Ile Ala Val Ser Ala Ala
1. 5 10 15
Asn Arg Phe Lys Lys Ile Ser
20
<210> 26
<211> 14
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 26
Glu Asp Pro Gly Phe Phe Asn Val Glu Ile Pro Glu Phe Pro
1. 5 10
<210> 27
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 27
Gly Gly His Gly Arg Val Leu Trp Pro Asp Gly Trp Phe Ser Leu Val
1. 5 10 15
Gly Ile Ser Pro
20
<210> 28
<211> 21
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 28
Met Ala Arg Thr Val Thr Ala Asn Val Pro Gly Met Gly Glu Gly Met
1. 5 10 15
Val Val Val Pro Cys
20
<210> 29
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 29
Gly Val Ser Lys Arg Gly Leu Gln Cys His Asp Phe Ile Ser Cys Ser
1. 5 10 15
Gly Val Pro Trp
20
<210> 30
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 30
Asn Gln Ser Ile Pro Lys Val Ala Gly Asp Ser Lys Val Phe Cys Trp
1. 5 10 15
Trp Cys Ala Leu
20
<210> 31
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 31
Gln Ser Thr Pro Pro Thr Lys His Leu Thr Ile Pro Arg His Leu Arg
1. 5 10 15
Asn Thr Leu Ile
20
<210> 32
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 32
Asp Met Ser Phe Gln Leu Val Thr Pro Phe Leu Lys Ala Leu Pro Thr
1. 5 10 15
Gly Trp Arg Gly
20
<210> 33
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 33
Gly Gly His Gly Arg Val Leu Trp Pro Asp Gly Trp Phe Ser Leu Val
1. 5 10 15
Gly Ile Ser Pro
20
<210> 34
<211> 8
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 34
Phe Ser Leu Val Gly Ile Ser Pro
1. 5
<210> 35
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 35
Gln Ile Met Met Gly Pro Ser Leu Gly Tyr Tyr Met Pro Ser Glu Ser
1. 5 10 15
Ile Phe Ala Tyr
20
<210> 36
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 36
Ile Ser Trp Asp Ile Trp Arg Trp Trp Tyr Thr Ser Glu Asp Arg Asp
1. 5 10 15
Ala Gly Ser Ala
20
<210> 37
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 37
Val Trp Gly Met Thr Thr Ser Asp His Gln Arg Lys Thr Glu Arg Leu
1. 5 10 15
Asp Ser Pro Glu
20
<210> 38
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 38
Met Thr Ser Ala Gln Thr Ser Glu Lys Leu Lys Ala Glu Thr Asp Arg
1. 5 10 15
His Thr Ala Glu
20
<210> 39
<211> 19
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 39
Met Gly Ser Arg Ser Ala Val Gly Asp Phe Glu Ser Ala Glu Gly Ser
1. 5 10 15
Arg Arg Pro
<210> 40
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 40
Met Gly Arg Thr Val Gln Ser Gly Asp Gly Thr Pro Ala Gln Thr Gln
1. 5 10 15
Pro Ser Val Asn
20
<210> 41
<211> 20
<212> белок
<213> Искусственная последовательность
<220>
<223> Нацеливающий полипептид
<400> 41
Met Ala Arg Thr Val Thr Ala Asn Val Pro Gly Met Gly Glu Gly Met
1. 5 10 15
Val Val Val Pro
20
<210> 42
<211> 132
<212> белок
<213> Искусственная последовательность
<220>
<223> N-концевой полипептид E1A аденовируса
<400> 42
Met Arg His Ile Ile Cys His Gly Gly Val Ile Thr Glu Glu Met Ala
1. 5 10 15
Ala Ser Leu Leu Asp Gln Leu Ile Glu Glu Val Leu Ala Asp Asn Leu
20 25 30
Pro Pro Pro Ser His Phe Glu Pro Pro Thr Leu His Glu Leu Tyr Asp
35 40 45
Leu Asp Val Thr Ala Pro Glu Asp Pro Asn Glu Glu Ala Val Ser Gln
50 55 60
Ile Phe Pro Glu Ser Val Met Leu Ala Val Gln Glu Gly Ile Asp Leu
65 70 75 80
Phe Thr Phe Pro Pro Ala Pro Gly Ser Pro Glu Pro Pro His Leu Ser
85 90 95
Arg Gln Pro Glu Gln Pro Glu Gln Arg Ala Leu Gly Pro Val Ser Met
100 105 110
Pro Asn Leu Val Pro Glu Val Ile Asp Leu Thr Cys His Glu Ala Gly
115 120 125
Phe Pro Pro Ser
130
<210> 43
<211> 112
<212> белок
<213> Искусственная последовательность
<220>
<223> N-концевой полипептид E1A
<400> 43
Met Arg His Ile Glu Glu Val Leu Ala Asp Asn Leu Pro Pro Pro Ser
1. 5 10 15
His Phe Glu Pro Pro Thr Leu His Glu Leu Tyr Asp Leu Asp Val Thr
20 25 30
Ala Pro Glu Asp Pro Asn Glu Glu Ala Val Ser Gln Ile Phe Pro Glu
35 40 45
Ser Val Met Leu Ala Val Gln Glu Gly Ile Asp Leu Phe Thr Phe Pro
50 55 60
Pro Ala Pro Gly Ser Pro Glu Pro Pro His Leu Ser Arg Gln Pro Glu
65 70 75 80
Gln Pro Glu Gln Arg Ala Leu Gly Pro Val Ser Met Pro Asn Leu Val
85 90 95
Pro Glu Val Ile Asp Leu Thr Cys His Glu Ala Gly Phe Pro Pro Ser
100 105 110
<210> 44
<211> 119
<212> белок
<213> Искусственная последовательность
<220>
<223> N-концевой полипептид E1A
<400> 44
Met Arg His Ile Ile Cys His Gly Gly Val Ile Thr Glu Glu Met Ala
1. 5 10 15
Ala Ser Leu Leu Asp Gln Leu Ile Glu Glu Val Leu Ala Asp Asn Leu
20 25 30
Pro Pro Pro Ser His Phe Glu Pro Pro Thr Leu His Glu Leu Tyr Asp
35 40 45
Leu Asp Val Thr Ala Pro Glu Asp Pro Asn Glu Glu Ala Val Ser Gln
50 55 60
Ile Phe Pro Glu Ser Val Met Leu Ala Val Gln Glu Gly Ile Asp Leu
65 70 75 80
Phe Thr Phe Pro Pro Ala Pro Gly Ser Pro Glu Pro Pro His Leu Ser
85 90 95
Arg Gln Pro Glu Gln Pro Glu Gln Arg Ala Leu Gly Pro Val Cys His
100 105 110
Glu Ala Gly Phe Pro Pro Ser
115
<210> 45
<211> 99
<212> белок
<213> Искусственная последовательность
<220>
<223> N-концевой полипептид E1A
<400> 45
Met Arg His Ile Glu Glu Val Leu Ala Asp Asn Leu Pro Pro Pro Ser
1. 5 10 15
His Phe Glu Pro Pro Thr Leu His Glu Leu Tyr Asp Leu Asp Val Thr
20 25 30
Ala Pro Glu Asp Pro Asn Glu Glu Ala Val Ser Gln Ile Phe Pro Glu
35 40 45
Ser Val Met Leu Ala Val Gln Glu Gly Ile Asp Leu Phe Thr Phe Pro
50 55 60
Pro Ala Pro Gly Ser Pro Glu Pro Pro His Leu Ser Arg Gln Pro Glu
65 70 75 80
Gln Pro Glu Gln Arg Ala Leu Gly Pro Val Cys His Glu Ala Gly Phe
85 90 95
Pro Pro Ser
<210> 46
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> Связывающий клетки пептид
<400> 46
Arg Glu Asp Val Tyr
1. 5
<210> 47
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Связывающий пептид рака молочной железы
<400> 47
Ile Ser Leu Ser Ser His Arg Ala Thr Trp Val Val
1. 5 10
<210> 48
<211> 1608
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота пробазина-E1
<400> 48
tcgagcgacg gtatcgataa gcttggagct tatgatagca tcttgttctt agtctttttc 60
ttaataggga cataaagccc acaaataaaa atatgcctga agaatgggac aggcattggg 120
cattgtccat gcctagtaaa gtactccaag aacctatttg tatactagat gacacaatgt 180
tctagccaag cttggtagtc atcatgttta aacatctacc attccagtta agaaaatatg 240
atagcatctt gttcttagtc tttttcttaa tagggacata aagcccacaa ataaaaatat 300
gcctgaagaa tgggacaggc attgggcatt gtccatgcct agtaaagtac tccaagaacc 360
tatttgtata ctagatgaca caatgtcaat gtctgtgtac aactgccaac tgggatgcaa 420
gacactgccc atgccaatca tcctgaaaag cagctataaa aagcaggaag ctactctgca 480
ccttgtcagt gaggtccaga tacctccctc gagcggccgc gacgcgcagt gtatttatac 540
ccggtgagtt cctcaagagg ccactcttga gtgccagcga gtagagtttt ctcctccgag 600
ccgctccgac accgggactg aaaatgagac atattatctg ccacggaggt gttattaccg 660
aagaaatggc cgccagtctt ttggaccagc tgatcgaaga ggtactggct gataatcttc 720
cacctcctag ccattttgaa ccacctaccc ttcacgaact gtatgattta gacgtgacgg 780
cccccgaaga tcccaacgag gaggcggttt cgcagatttt tcccgagtct gtaatgttgg 840
cggtgcagga agggattgac ttattcactt ttccgccggc gcccggttct ccggagccgc 900
ctcacctttc ccggcagccc gagcagccgg agcagagagc cttgggtccg gtttctatgc 960
caaaccttgt gccggaggtg atcgatctta cctgccacga ggctggcttt ccacccagtg 1020
acgacgagga tgaagagggt gaggagtttg tgttagatta tgtggagcac cccgggcacg 1080
gttgcaggtc ttgtcattat caccggagga atacggggga cccagatatt atgtgttcgc 1140
tttgctatat gaggacctgt ggcatgtttg tctacagtaa gtgaaaatta tgggcagtcg 1200
gtgatagagt ggtgggtttg gtgtggtaat ttttttttaa tttttacagt tttgtggttt 1260
aaagaatttt gtattgtgat tttttaaaag gtcctgtgtc tgaacctgag cctgagcccg 1320
agccagaacc ggagcctgca agacctaccc ggcgtcctaa attggtgcct gctatcctga 1380
gacgcccgac atcacctgtg tctagagaat gcaatagtag tacggatagc tgtgactccg 1440
gtccttctaa cacacctcct gagatacacc cggtggtccc gctgtgcccc attaaaccag 1500
ttgccgtgag agttggtggg cgtcgccagg ctgtggaatg tatcgaggac ttgcttaacg 1560
agtctgggca acctttggac ttgagctgta aacgccccag gccataag 1608
<210> 49
<211> 959
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 49
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Val Asn Ala Met Asn Glu Ala Asn Ala Ile Gln Pro
275 280 285
Lys Leu Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr
290 295 300
His Leu Ser Tyr Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala Met
305 310 315 320
Leu Gly Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg
325 330 335
Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly
340 345 350
Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln
355 360 365
Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly
370 375 380
Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr
385 390 395 400
Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu
405 410 415
Pro Asn Tyr Cys Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr
420 425 430
Gln Ala Ile Lys Ala Thr Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala
435 440 445
Gln Asp Asn Thr Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn
450 455 460
Phe Ala Met Glu Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu
465 470 475 480
Tyr Ser Asn Ile Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro
485 490 495
Thr Asn Val Glu Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn
500 505 510
Lys Arg Val Val Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly
515 520 525
Ala Arg Trp Ser Leu Asp Tyr Met Asp Asn Val Asn Pro Phe Asn His
530 535 540
His Arg Asn Ala Gly Leu Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly
545 550 555 560
Arg Tyr Val Pro Phe His Ile Gln Val Pro Gln Lys Phe Phe Ala Ile
565 570 575
Lys Asn Leu Leu Leu Leu Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe
580 585 590
Arg Lys Asp Val Asn Met Val Leu Gln Ser Ser Leu Gly Asn Asp Leu
595 600 605
Arg Val Asp Gly Ala Ser Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala
610 615 620
Thr Phe Phe Pro Met Ala His Asn Thr Ala Ser Thr Leu Glu Ala Met
625 630 635 640
Leu Arg Asn Asp Thr Asn Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala
645 650 655
Ala Asn Met Leu Tyr Pro Ile Pro Ala Asn Ala Thr Asn Val Pro Ile
660 665 670
Ser Ile Pro Ser Arg Asn Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr
675 680 685
Arg Leu Lys Thr Lys Glu Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro
690 695 700
Tyr Tyr Thr Tyr Ser Gly Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr
705 710 715 720
Leu Asn His Thr Phe Lys Lys Val Ala Ile Thr Phe Asp Ser Ser Val
725 730 735
Ser Trp Pro Gly Asn Asp Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile
740 745 750
Lys Arg Ser Val Asp Gly Glu Gly Tyr Asn Val Ala Gln Cys Asn Met
755 760 765
Thr Lys Asp Trp Phe Leu Val Gln Met Leu Ala Asn Tyr Asn Ile Gly
770 775 780
Tyr Gln Gly Phe Tyr Ile Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser
785 790 795 800
Phe Phe Arg Asn Phe Gln Pro Met Ser Arg Gln Val Val Asp Asp Thr
805 810 815
Lys Tyr Lys Asp Tyr Gln Gln Val Gly Ile Ile His Gln His Asn Asn
820 825 830
Ser Gly Phe Val Gly Tyr Leu Ala Pro Thr Met Arg Glu Gly Gln Ala
835 840 845
Tyr Pro Ala Asn Val Pro Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp
850 855 860
Ser Ile Thr Gln Lys Lys Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile
865 870 875 880
Pro Phe Ser Ser Asn Phe Met Ser Met Gly Ala Leu Thr Asp Leu Gly
885 890 895
Gln Asn Leu Leu Tyr Ala Asn Ser Ala His Ala Leu Asp Met Thr Phe
900 905 910
Glu Val Asp Pro Met Asp Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu
915 920 925
Val Phe Asp Val Val Arg Val His Gln Pro His Arg Gly Val Ile Glu
930 935 940
Thr Val Tyr Leu Arg Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
945 950 955
<210> 50
<211> 967
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 50
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Ser Ser Cys Ser Ser Gly Gly
145 150 155 160
Thr Glu Glu Glu Glu Gln Leu Pro Gln Gln Arg Asn Gly Lys Lys Thr
165 170 175
His Val Tyr Ala Gln Ala Pro Phe Ala Gly Glu Ala Ile Asn Lys Asn
180 185 190
Gly Leu Gln Ile Gly Thr Asn Gly Ala Ala Thr Glu Gly Asn Lys Glu
195 200 205
Ile Tyr Ala Asp Lys Thr Tyr Gln Pro Glu Pro Gln Ile Gly Glu Ser
210 215 220
Gln Trp Asn Glu Ala Glu Ser Ser Val Ala Gly Gly Arg Val Leu Lys
225 230 235 240
Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg Pro Thr
245 250 255
Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly Lys Leu
260 265 270
Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Val Asn Ala Met
275 280 285
Asn Glu Ala Asn Ala Ile Gln Pro Lys Leu Val Leu Tyr Ser Glu Asp
290 295 300
Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly Lys
305 310 315 320
Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro Asn
325 330 335
Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr
340 345 350
Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser Gln
355 360 365
Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr
370 375 380
Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met
385 390 395 400
Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu
405 410 415
Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly
420 425 430
Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn Gly
435 440 445
Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu Arg
450 455 460
Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn Leu Asn
465 470 475 480
Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr Leu
485 490 495
Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp Asn
500 505 510
Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro Gly Leu
515 520 525
Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp Tyr Met
530 535 540
Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala Gly Leu Arg Tyr
545 550 555 560
Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro Phe His Ile Gln
565 570 575
Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu Leu Leu Pro Gly
580 585 590
Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val Asn Met Val Leu
595 600 605
Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly Ala Ser Ile Lys
610 615 620
Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro Met Ala His Asn
625 630 635 640
Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp Thr Asn Asp Gln
645 650 655
Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu Tyr Pro Ile Pro
660 665 670
Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser Arg Asn Trp Ala
675 680 685
Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr Lys Glu Thr Pro
690 695 700
Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly Ser Ile
705 710 715 720
Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr Phe Lys Lys Val
725 730 735
Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly Asn Asp Arg Leu
740 745 750
Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val Asp Gly Glu Gly
755 760 765
Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp Phe Leu Val Gln
770 775 780
Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe Tyr Ile Pro Glu
785 790 795 800
Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn Phe Gln Pro Met
805 810 815
Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp Tyr Gln Gln Val
820 825 830
Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val Gly Tyr Leu Ala
835 840 845
Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn Val Pro Tyr Pro
850 855 860
Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln Lys Lys Phe Leu
865 870 875 880
Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser Asn Phe Met Ser
885 890 895
Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu Tyr Ala Asn Ser
900 905 910
Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro Met Asp Glu Pro
915 920 925
Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val Val Arg Val His
930 935 940
Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu Arg Thr Pro Phe
945 950 955 960
Ser Ala Gly Asn Ala Thr Thr
965
<210> 51
<211> 955
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 51
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Ser Cys Ser Ser Gly Gly Thr Pro Lys Leu Val Leu
275 280 285
Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr
290 295 300
Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln
305 310 315 320
Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile
325 330 335
Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly
340 345 350
Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr
355 360 365
Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg
370 375 380
Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val
385 390 395 400
Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys
405 410 415
Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys
420 425 430
Ala Thr Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr
435 440 445
Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu
450 455 460
Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile
465 470 475 480
Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu
485 490 495
Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val
500 505 510
Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser
515 520 525
Leu Asp Tyr Met Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala
530 535 540
Gly Leu Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro
545 550 555 560
Phe His Ile Gln Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu
565 570 575
Leu Leu Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val
580 585 590
Asn Met Val Leu Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly
595 600 605
Ala Ser Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro
610 615 620
Met Ala His Asn Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp
625 630 635 640
Thr Asn Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu
645 650 655
Tyr Pro Ile Pro Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser
660 665 670
Arg Asn Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr
675 680 685
Lys Glu Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr
690 695 700
Ser Gly Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr
705 710 715 720
Phe Lys Lys Val Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly
725 730 735
Asn Asp Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val
740 745 750
Asp Gly Glu Gly Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp
755 760 765
Phe Leu Val Gln Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe
770 775 780
Tyr Ile Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn
785 790 795 800
Phe Gln Pro Met Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp
805 810 815
Tyr Gln Gln Val Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val
820 825 830
Gly Tyr Leu Ala Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn
835 840 845
Val Pro Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln
850 855 860
Lys Lys Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser
865 870 875 880
Asn Phe Met Ser Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu
885 890 895
Tyr Ala Asn Ser Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro
900 905 910
Met Asp Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val
915 920 925
Val Arg Val His Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu
930 935 940
Arg Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
945 950 955
<210> 52
<211> 964
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 52
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Ser Ser Gly Gly Thr Glu Glu
145 150 155 160
Glu Glu Gln Leu Pro Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr
165 170 175
Ala Gln Ala Pro Phe Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln
180 185 190
Ile Gly Thr Asn Gly Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala
195 200 205
Asp Lys Thr Tyr Gln Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn
210 215 220
Glu Ala Glu Ser Ser Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr
225 230 235 240
Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn
245 250 255
Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln
260 265 270
Val Glu Met Gln Phe Phe Ser Thr Ser Val Asn Ala Met Asn Glu Ala
275 280 285
Asn Ala Ile Gln Pro Lys Leu Val Leu Tyr Ser Glu Asp Val Asn Met
290 295 300
Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly Lys Ser Asp Asp
305 310 315 320
Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro Asn Arg Pro Asn
325 330 335
Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser
340 345 350
Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala
355 360 365
Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu
370 375 380
Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln
385 390 395 400
Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly
405 410 415
Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly Gly Ile Gly
420 425 430
Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn Gly Asn Gly Gly
435 440 445
Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu Arg Asn Glu Ile
450 455 460
Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn Leu Asn Ala Asn Leu
465 470 475 480
Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr Leu Pro Asp Lys
485 490 495
Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp Asn Pro Asn Thr
500 505 510
Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro Gly Leu Val Asp Cys
515 520 525
Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp Tyr Met Asp Asn Val
530 535 540
Asn Pro Phe Asn His His Arg Asn Ala Gly Leu Arg Tyr Arg Ser Met
545 550 555 560
Leu Leu Gly Asn Gly Arg Tyr Val Pro Phe His Ile Gln Val Pro Gln
565 570 575
Lys Phe Phe Ala Ile Lys Asn Leu Leu Leu Leu Pro Gly Ser Tyr Thr
580 585 590
Tyr Glu Trp Asn Phe Arg Lys Asp Val Asn Met Val Leu Gln Ser Ser
595 600 605
Leu Gly Asn Asp Leu Arg Val Asp Gly Ala Ser Ile Lys Phe Asp Ser
610 615 620
Ile Cys Leu Tyr Ala Thr Phe Phe Pro Met Ala His Asn Thr Ala Ser
625 630 635 640
Thr Leu Glu Ala Met Leu Arg Asn Asp Thr Asn Asp Gln Ser Phe Asn
645 650 655
Asp Tyr Leu Ser Ala Ala Asn Met Leu Tyr Pro Ile Pro Ala Asn Ala
660 665 670
Thr Asn Val Pro Ile Ser Ile Pro Ser Arg Asn Trp Ala Ala Phe Arg
675 680 685
Gly Trp Ala Phe Thr Arg Leu Lys Thr Lys Glu Thr Pro Ser Leu Gly
690 695 700
Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly Ser Ile Pro Tyr Leu
705 710 715 720
Asp Gly Thr Phe Tyr Leu Asn His Thr Phe Lys Lys Val Ala Ile Thr
725 730 735
Phe Asp Ser Ser Val Ser Trp Pro Gly Asn Asp Arg Leu Leu Thr Pro
740 745 750
Asn Glu Phe Glu Ile Lys Arg Ser Val Asp Gly Glu Gly Tyr Asn Val
755 760 765
Ala Gln Cys Asn Met Thr Lys Asp Trp Phe Leu Val Gln Met Leu Ala
770 775 780
Asn Tyr Asn Ile Gly Tyr Gln Gly Phe Tyr Ile Pro Glu Ser Tyr Lys
785 790 795 800
Asp Arg Met Tyr Ser Phe Phe Arg Asn Phe Gln Pro Met Ser Arg Gln
805 810 815
Val Val Asp Asp Thr Lys Tyr Lys Asp Tyr Gln Gln Val Gly Ile Ile
820 825 830
His Gln His Asn Asn Ser Gly Phe Val Gly Tyr Leu Ala Pro Thr Met
835 840 845
Arg Glu Gly Gln Ala Tyr Pro Ala Asn Val Pro Tyr Pro Leu Ile Gly
850 855 860
Lys Thr Ala Val Asp Ser Ile Thr Gln Lys Lys Phe Leu Cys Asp Arg
865 870 875 880
Thr Leu Trp Arg Ile Pro Phe Ser Ser Asn Phe Met Ser Met Gly Ala
885 890 895
Leu Thr Asp Leu Gly Gln Asn Leu Leu Tyr Ala Asn Ser Ala His Ala
900 905 910
Leu Asp Met Thr Phe Glu Val Asp Pro Met Asp Glu Pro Thr Leu Leu
915 920 925
Tyr Val Leu Phe Glu Val Phe Asp Val Val Arg Val His Gln Pro His
930 935 940
Arg Gly Val Ile Glu Thr Val Tyr Leu Arg Thr Pro Phe Ser Ala Gly
945 950 955 960
Asn Ala Thr Thr
<210> 53
<211> 952
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 53
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Ser Gly Gly Thr Pro Lys Leu Val Leu Tyr Ser Glu
275 280 285
Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly
290 295 300
Lys Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro
305 310 315 320
Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met
325 330 335
Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser
340 345 350
Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser
355 360 365
Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser
370 375 380
Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile
385 390 395 400
Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu
405 410 415
Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn
420 425 430
Gly Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu
435 440 445
Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn Leu
450 455 460
Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr
465 470 475 480
Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp
485 490 495
Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro Gly
500 505 510
Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp Tyr
515 520 525
Met Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala Gly Leu Arg
530 535 540
Tyr Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro Phe His Ile
545 550 555 560
Gln Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu Leu Leu Pro
565 570 575
Gly Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val Asn Met Val
580 585 590
Leu Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly Ala Ser Ile
595 600 605
Lys Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro Met Ala His
610 615 620
Asn Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp Thr Asn Asp
625 630 635 640
Gln Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu Tyr Pro Ile
645 650 655
Pro Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser Arg Asn Trp
660 665 670
Ala Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr Lys Glu Thr
675 680 685
Pro Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly Ser
690 695 700
Ile Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr Phe Lys Lys
705 710 715 720
Val Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly Asn Asp Arg
725 730 735
Leu Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val Asp Gly Glu
740 745 750
Gly Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp Phe Leu Val
755 760 765
Gln Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe Tyr Ile Pro
770 775 780
Glu Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn Phe Gln Pro
785 790 795 800
Met Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp Tyr Gln Gln
805 810 815
Val Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val Gly Tyr Leu
820 825 830
Ala Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn Val Pro Tyr
835 840 845
Pro Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln Lys Lys Phe
850 855 860
Leu Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser Asn Phe Met
865 870 875 880
Ser Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu Tyr Ala Asn
885 890 895
Ser Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro Met Asp Glu
900 905 910
Pro Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val Val Arg Val
915 920 925
His Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu Arg Thr Pro
930 935 940
Phe Ser Ala Gly Asn Ala Thr Thr
945 950
<210> 54
<211> 1033
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 54
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Ser Thr Gly Glu Ile Pro Ala
145 150 155 160
Pro Leu Ala Gly Thr Val Ser Lys Ile Leu Val Lys Glu Gly Asp Thr
165 170 175
Val Lys Ala Gly Gln Thr Val Leu Val Leu Glu Ala Met Lys Met Glu
180 185 190
Thr Glu Ile Asn Ala Pro Thr Asp Gly Lys Val Glu Lys Val Leu Val
195 200 205
Lys Glu Arg Asp Ala Val Gln Gly Gly Gln Gly Leu Ile Lys Ile Gly
210 215 220
Gly Gly Thr Glu Glu Glu Glu Gln Leu Pro Gln Gln Arg Asn Gly Lys
225 230 235 240
Lys Thr His Val Tyr Ala Gln Ala Pro Phe Ala Gly Glu Ala Ile Asn
245 250 255
Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly Ala Ala Thr Glu Gly Asn
260 265 270
Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln Pro Glu Pro Gln Ile Gly
275 280 285
Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser Val Ala Gly Gly Arg Val
290 295 300
Leu Lys Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg
305 310 315 320
Pro Thr Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly
325 330 335
Lys Leu Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Val Asn
340 345 350
Ala Met Asn Glu Ala Asn Ala Ile Gln Pro Lys Leu Val Leu Tyr Ser
355 360 365
Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro
370 375 380
Gly Lys Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met
385 390 395 400
Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu
405 410 415
Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala
420 425 430
Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu
435 440 445
Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe
450 455 460
Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile
465 470 475 480
Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro
485 490 495
Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr
500 505 510
Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala
515 520 525
Glu Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn
530 535 540
Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu
545 550 555 560
Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser
565 570 575
Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro
580 585 590
Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp
595 600 605
Tyr Met Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala Gly Leu
610 615 620
Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro Phe His
625 630 635 640
Ile Gln Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu Leu Leu
645 650 655
Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val Asn Met
660 665 670
Val Leu Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly Ala Ser
675 680 685
Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro Met Ala
690 695 700
His Asn Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp Thr Asn
705 710 715 720
Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu Tyr Pro
725 730 735
Ile Pro Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser Arg Asn
740 745 750
Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr Lys Glu
755 760 765
Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly
770 775 780
Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr Phe Lys
785 790 795 800
Lys Val Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly Asn Asp
805 810 815
Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val Asp Gly
820 825 830
Glu Gly Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp Phe Leu
835 840 845
Val Gln Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe Tyr Ile
850 855 860
Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn Phe Gln
865 870 875 880
Pro Met Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp Tyr Gln
885 890 895
Gln Val Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val Gly Tyr
900 905 910
Leu Ala Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn Val Pro
915 920 925
Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln Lys Lys
930 935 940
Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser Asn Phe
945 950 955 960
Met Ser Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu Tyr Ala
965 970 975
Asn Ser Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro Met Asp
980 985 990
Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val Val Arg
995 1000 1005
Val His Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu Arg
1010 1015 1020
Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
1025 1030
<210> 55
<211> 1021
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 55
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Thr Gly Glu Ile Pro Ala Pro Leu Ala Gly Thr Val
275 280 285
Ser Lys Ile Leu Val Lys Glu Gly Asp Thr Val Lys Ala Gly Gln Thr
290 295 300
Val Leu Val Leu Glu Ala Met Lys Met Glu Thr Glu Ile Asn Ala Pro
305 310 315 320
Thr Asp Gly Lys Val Glu Lys Val Leu Val Lys Glu Arg Asp Ala Val
325 330 335
Gln Gly Gly Gln Gly Leu Ile Lys Ile Gly Gly Gly Thr Pro Lys Leu
340 345 350
Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu
355 360 365
Ser Tyr Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala Met Leu Gly
370 375 380
Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn
385 390 395 400
Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu
405 410 415
Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg
420 425 430
Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg
435 440 445
Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro
450 455 460
Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn
465 470 475 480
Tyr Cys Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala
485 490 495
Ile Lys Ala Thr Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp
500 505 510
Asn Thr Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala
515 520 525
Met Glu Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser
530 535 540
Asn Ile Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn
545 550 555 560
Val Glu Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg
565 570 575
Val Val Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg
580 585 590
Trp Ser Leu Asp Tyr Met Asp Asn Val Asn Pro Phe Asn His His Arg
595 600 605
Asn Ala Gly Leu Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr
610 615 620
Val Pro Phe His Ile Gln Val Pro Gln Lys Phe Phe Ala Ile Lys Asn
625 630 635 640
Leu Leu Leu Leu Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys
645 650 655
Asp Val Asn Met Val Leu Gln Ser Ser Leu Gly Asn Asp Leu Arg Val
660 665 670
Asp Gly Ala Ser Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe
675 680 685
Phe Pro Met Ala His Asn Thr Ala Ser Thr Leu Glu Ala Met Leu Arg
690 695 700
Asn Asp Thr Asn Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn
705 710 715 720
Met Leu Tyr Pro Ile Pro Ala Asn Ala Thr Asn Val Pro Ile Ser Ile
725 730 735
Pro Ser Arg Asn Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu
740 745 750
Lys Thr Lys Glu Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr
755 760 765
Thr Tyr Ser Gly Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn
770 775 780
His Thr Phe Lys Lys Val Ala Ile Thr Phe Asp Ser Ser Val Ser Trp
785 790 795 800
Pro Gly Asn Asp Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg
805 810 815
Ser Val Asp Gly Glu Gly Tyr Asn Val Ala Gln Cys Asn Met Thr Lys
820 825 830
Asp Trp Phe Leu Val Gln Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln
835 840 845
Gly Phe Tyr Ile Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe
850 855 860
Arg Asn Phe Gln Pro Met Ser Arg Gln Val Val Asp Asp Thr Lys Tyr
865 870 875 880
Lys Asp Tyr Gln Gln Val Gly Ile Ile His Gln His Asn Asn Ser Gly
885 890 895
Phe Val Gly Tyr Leu Ala Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro
900 905 910
Ala Asn Val Pro Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp Ser Ile
915 920 925
Thr Gln Lys Lys Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe
930 935 940
Ser Ser Asn Phe Met Ser Met Gly Ala Leu Thr Asp Leu Gly Gln Asn
945 950 955 960
Leu Leu Tyr Ala Asn Ser Ala His Ala Leu Asp Met Thr Phe Glu Val
965 970 975
Asp Pro Met Asp Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu Val Phe
980 985 990
Asp Val Val Arg Val His Gln Pro His Arg Gly Val Ile Glu Thr Val
995 1000 1005
Tyr Leu Arg Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
1010 1015 1020
<210> 56
<211> 1015
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 56
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Ser Ser Asn Phe Thr Arg Glu Gly Asn Val Thr Tyr
275 280 285
Lys Glu Glu Met Asp Lys Val Lys Asn Cys Ser Phe Asn Val Thr Thr
290 295 300
Gly Ile Arg Asp Lys Lys Gln Lys Val Asn Ala Leu Phe Tyr Arg Leu
305 310 315 320
Asp Ile Thr Pro Leu Asp Glu Asn Asn Asn Asn Ser Ser Glu Tyr Arg
325 330 335
Leu Ile Asn Ser Gly Gly Thr Pro Lys Leu Val Leu Tyr Ser Glu Asp
340 345 350
Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly Lys
355 360 365
Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro Asn
370 375 380
Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr
385 390 395 400
Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser Gln
405 410 415
Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr
420 425 430
Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met
435 440 445
Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu
450 455 460
Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly
465 470 475 480
Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn Gly
485 490 495
Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu Arg
500 505 510
Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn Leu Asn
515 520 525
Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr Leu
530 535 540
Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp Asn
545 550 555 560
Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro Gly Leu
565 570 575
Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp Tyr Met
580 585 590
Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala Gly Leu Arg Tyr
595 600 605
Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro Phe His Ile Gln
610 615 620
Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu Leu Leu Pro Gly
625 630 635 640
Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val Asn Met Val Leu
645 650 655
Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly Ala Ser Ile Lys
660 665 670
Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro Met Ala His Asn
675 680 685
Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp Thr Asn Asp Gln
690 695 700
Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu Tyr Pro Ile Pro
705 710 715 720
Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser Arg Asn Trp Ala
725 730 735
Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr Lys Glu Thr Pro
740 745 750
Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly Ser Ile
755 760 765
Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr Phe Lys Lys Val
770 775 780
Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly Asn Asp Arg Leu
785 790 795 800
Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val Asp Gly Glu Gly
805 810 815
Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp Phe Leu Val Gln
820 825 830
Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe Tyr Ile Pro Glu
835 840 845
Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn Phe Gln Pro Met
850 855 860
Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp Tyr Gln Gln Val
865 870 875 880
Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val Gly Tyr Leu Ala
885 890 895
Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn Val Pro Tyr Pro
900 905 910
Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln Lys Lys Phe Leu
915 920 925
Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser Asn Phe Met Ser
930 935 940
Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu Tyr Ala Asn Ser
945 950 955 960
Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro Met Asp Glu Pro
965 970 975
Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val Val Arg Val His
980 985 990
Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu Arg Thr Pro Phe
995 1000 1005
Ser Ala Gly Asn Ala Thr Thr
1010 1015
<210> 57
<211> 987
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 57
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Ser Gln Ala Glu Pro Asp Arg Ala His Tyr Asn Ile
275 280 285
Val Thr Phe Cys Cys Lys Cys Asp Gln Leu Leu Arg Arg Glu Val Tyr
290 295 300
Asp Phe Ala Phe Arg Asp Leu Ser Gly Gly Thr Pro Lys Leu Val Leu
305 310 315 320
Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr
325 330 335
Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln
340 345 350
Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile
355 360 365
Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly
370 375 380
Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr
385 390 395 400
Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg
405 410 415
Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val
420 425 430
Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys
435 440 445
Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys
450 455 460
Ala Thr Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr
465 470 475 480
Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu
485 490 495
Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile
500 505 510
Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu
515 520 525
Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val
530 535 540
Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser
545 550 555 560
Leu Asp Tyr Met Asp Asn Val Asn Pro Phe Asn His His Arg Asn Ala
565 570 575
Gly Leu Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly Arg Tyr Val Pro
580 585 590
Phe His Ile Gln Val Pro Gln Lys Phe Phe Ala Ile Lys Asn Leu Leu
595 600 605
Leu Leu Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe Arg Lys Asp Val
610 615 620
Asn Met Val Leu Gln Ser Ser Leu Gly Asn Asp Leu Arg Val Asp Gly
625 630 635 640
Ala Ser Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala Thr Phe Phe Pro
645 650 655
Met Ala His Asn Thr Ala Ser Thr Leu Glu Ala Met Leu Arg Asn Asp
660 665 670
Thr Asn Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala Ala Asn Met Leu
675 680 685
Tyr Pro Ile Pro Ala Asn Ala Thr Asn Val Pro Ile Ser Ile Pro Ser
690 695 700
Arg Asn Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr Arg Leu Lys Thr
705 710 715 720
Lys Glu Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro Tyr Tyr Thr Tyr
725 730 735
Ser Gly Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr Leu Asn His Thr
740 745 750
Phe Lys Lys Val Ala Ile Thr Phe Asp Ser Ser Val Ser Trp Pro Gly
755 760 765
Asn Asp Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile Lys Arg Ser Val
770 775 780
Asp Gly Glu Gly Tyr Asn Val Ala Gln Cys Asn Met Thr Lys Asp Trp
785 790 795 800
Phe Leu Val Gln Met Leu Ala Asn Tyr Asn Ile Gly Tyr Gln Gly Phe
805 810 815
Tyr Ile Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser Phe Phe Arg Asn
820 825 830
Phe Gln Pro Met Ser Arg Gln Val Val Asp Asp Thr Lys Tyr Lys Asp
835 840 845
Tyr Gln Gln Val Gly Ile Ile His Gln His Asn Asn Ser Gly Phe Val
850 855 860
Gly Tyr Leu Ala Pro Thr Met Arg Glu Gly Gln Ala Tyr Pro Ala Asn
865 870 875 880
Val Pro Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp Ser Ile Thr Gln
885 890 895
Lys Lys Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile Pro Phe Ser Ser
900 905 910
Asn Phe Met Ser Met Gly Ala Leu Thr Asp Leu Gly Gln Asn Leu Leu
915 920 925
Tyr Ala Asn Ser Ala His Ala Leu Asp Met Thr Phe Glu Val Asp Pro
930 935 940
Met Asp Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu Val Phe Asp Val
945 950 955 960
Val Arg Val His Gln Pro His Arg Gly Val Ile Glu Thr Val Tyr Leu
965 970 975
Arg Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
980 985
<210> 58
<211> 959
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 58
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Asp Glu Asp Asp Thr Gln Val Gln Val
130 135 140
Ala Ala Glu Asp Asp Gln Asp Asp Asp Glu Glu Glu Glu Gln Leu Pro
145 150 155 160
Gln Gln Arg Asn Gly Lys Lys Thr His Val Tyr Ala Gln Ala Pro Phe
165 170 175
Ala Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn Gly
180 185 190
Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr Gln
195 200 205
Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser Ser
210 215 220
Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys
225 230 235 240
Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly Val
245 250 255
Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln Phe
260 265 270
Phe Ser Thr Ser Val Asn Ala Met Asn Glu Ala Asn Ala Ile Gln Pro
275 280 285
Lys Leu Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr
290 295 300
His Leu Ser Tyr Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala Met
305 310 315 320
Leu Gly Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg
325 330 335
Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly
340 345 350
Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln
355 360 365
Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly
370 375 380
Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr
385 390 395 400
Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu
405 410 415
Pro Asn Tyr Cys Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr
420 425 430
Gln Ala Ile Lys Ala Thr Asn Gly Asn Gly Gly Ala Thr Thr Trp Ala
435 440 445
Gln Asp Asn Thr Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn
450 455 460
Phe Ala Met Glu Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu
465 470 475 480
Tyr Ser Asn Ile Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro
485 490 495
Thr Asn Val Glu Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn
500 505 510
Lys Arg Val Val Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly
515 520 525
Ala Arg Trp Ser Leu Asp Tyr Met Asp Asn Val Asn Pro Phe Asn His
530 535 540
His Arg Asn Ala Gly Leu Arg Tyr Arg Ser Met Leu Leu Gly Asn Gly
545 550 555 560
Arg Tyr Val Pro Phe His Ile Gln Val Pro Gln Lys Phe Phe Ala Ile
565 570 575
Lys Asn Leu Leu Leu Leu Pro Gly Ser Tyr Thr Tyr Glu Trp Asn Phe
580 585 590
Arg Lys Asp Val Asn Met Val Leu Gln Ser Ser Leu Gly Asn Asp Leu
595 600 605
Arg Val Asp Gly Ala Ser Ile Lys Phe Asp Ser Ile Cys Leu Tyr Ala
610 615 620
Thr Phe Phe Pro Met Ala His Asn Thr Ala Ser Thr Leu Glu Ala Met
625 630 635 640
Leu Arg Asn Asp Thr Asn Asp Gln Ser Phe Asn Asp Tyr Leu Ser Ala
645 650 655
Ala Asn Met Leu Tyr Pro Ile Pro Ala Asn Ala Thr Asn Val Pro Ile
660 665 670
Ser Ile Pro Ser Arg Asn Trp Ala Ala Phe Arg Gly Trp Ala Phe Thr
675 680 685
Arg Leu Lys Thr Lys Glu Thr Pro Ser Leu Gly Ser Gly Tyr Asp Pro
690 695 700
Tyr Tyr Thr Tyr Ser Gly Ser Ile Pro Tyr Leu Asp Gly Thr Phe Tyr
705 710 715 720
Leu Asn His Thr Phe Lys Lys Val Ala Ile Thr Phe Asp Ser Ser Val
725 730 735
Ser Trp Pro Gly Asn Asp Arg Leu Leu Thr Pro Asn Glu Phe Glu Ile
740 745 750
Lys Arg Ser Val Asp Gly Glu Gly Tyr Asn Val Ala Gln Cys Asn Met
755 760 765
Thr Lys Asp Trp Phe Leu Val Gln Met Leu Ala Asn Tyr Asn Ile Gly
770 775 780
Tyr Gln Gly Phe Tyr Ile Pro Glu Ser Tyr Lys Asp Arg Met Tyr Ser
785 790 795 800
Phe Phe Arg Asn Phe Gln Pro Met Ser Arg Gln Val Val Asp Asp Thr
805 810 815
Lys Tyr Lys Asp Tyr Gln Gln Val Gly Ile Ile His Gln His Asn Asn
820 825 830
Ser Gly Phe Val Gly Tyr Leu Ala Pro Thr Met Arg Glu Gly Gln Ala
835 840 845
Tyr Pro Ala Asn Val Pro Tyr Pro Leu Ile Gly Lys Thr Ala Val Asp
850 855 860
Ser Ile Thr Gln Lys Lys Phe Leu Cys Asp Arg Thr Leu Trp Arg Ile
865 870 875 880
Pro Phe Ser Ser Asn Phe Met Ser Met Gly Ala Leu Thr Asp Leu Gly
885 890 895
Gln Asn Leu Leu Tyr Ala Asn Ser Ala His Ala Leu Asp Met Thr Phe
900 905 910
Glu Val Asp Pro Met Asp Glu Pro Thr Leu Leu Tyr Val Leu Phe Glu
915 920 925
Val Phe Asp Val Val Arg Val His Gln Pro His Arg Gly Val Ile Glu
930 935 940
Thr Val Tyr Leu Arg Thr Pro Phe Ser Ala Gly Asn Ala Thr Thr
945 950 955
<210> 59
<211> 962
<212> белок
<213> Искусственная последовательность
<220>
<223> Гексоновый полипептид
<400> 59
Met Ala Thr Pro Ser Met Met Pro Gln Trp Ser Tyr Met His Ile Ser
1. 5 10 15
Gly Gln Asp Ala Ser Glu Tyr Leu Ser Pro Gly Leu Val Gln Phe Ala
20 25 30
Arg Ala Thr Glu Thr Tyr Phe Ser Leu Asn Asn Lys Phe Arg Asn Pro
35 40 45
Thr Val Ala Pro Thr His Asp Val Thr Thr Asp Arg Ser Gln Arg Leu
50 55 60
Thr Leu Arg Phe Ile Pro Val Asp Arg Glu Asp Thr Ala Tyr Ser Tyr
65 70 75 80
Lys Ala Arg Phe Thr Leu Ala Val Gly Asp Asn Arg Val Leu Asp Met
85 90 95
Ala Ser Thr Tyr Phe Asp Ile Arg Gly Val Leu Asp Arg Gly Pro Thr
100 105 110
Phe Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu Ala Pro Lys Gly
115 120 125
Ala Pro Asn Ser Cys Glu Trp Glu Gln Asn Glu Thr Ala Gln Val Asp
130 135 140
Ala Gln Glu Leu Asp Glu Glu Glu Asn Glu Ala Asn Glu Ala Gln Ala
145 150 155 160
Arg Glu Gln Glu Gln Ala Lys Lys Thr His Val Tyr Ala Gln Ala Pro
165 170 175
Leu Ser Gly Glu Ala Ile Asn Lys Asn Gly Leu Gln Ile Gly Thr Asn
180 185 190
Gly Ala Ala Thr Glu Gly Asn Lys Glu Ile Tyr Ala Asp Lys Thr Tyr
195 200 205
Gln Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Asn Glu Ala Glu Ser
210 215 220
Ser Val Ala Gly Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro
225 230 235 240
Cys Tyr Gly Ser Tyr Ala Arg Pro Thr Asn Ser Asn Gly Gly Gln Gly
245 250 255
Val Met Val Glu Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met Gln
260 265 270
Phe Phe Ser Thr Ser Val Asn Ala Met Asn Glu Ala Asn Ala Ile Gln
275 280 285
Pro Lys Leu Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp
290 295 300
Thr His Leu Ser Tyr Lys Pro Gly Lys Ser Asp Asp Asn Ser Lys Ala
305 310 315 320
Met Leu Gly Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe
325 330 335
Arg Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met
340 345 350
Gly Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu
355 360 365
Gln Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile
370 375 380
Gly Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser
385 390 395 400
Tyr Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu
405 410 415
Leu Pro Asn Tyr Cys Phe Pro Leu Gly Gly Ile Gly Ile Thr Asp Thr
420 425 430
Phe Gln Ala Val Lys Thr Thr Ala Ala Asn Gly Asp Gln Gly Asn Thr
435 440 445
Thr Trp Gln Lys Asp Ser Thr Phe Ala Glu Arg Asn Glu Ile Gly Val
450 455 460
Gly Asn Asn Phe Ala Met Glu Ile Asn Leu Asn Ala Asn Leu Trp Arg
465 470 475 480
Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr Leu Pro Asp Lys Leu Lys
485 490 495
Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp Asn Pro Asn Thr Tyr Asp
500 505 510
Tyr Met Asn Lys Arg Val Val Ala Pro Gly Leu Val Asp Cys Tyr Ile
515 520 525
Asn Leu Gly Ala Arg Trp Ser Leu Asp Tyr Met Asp Asn Val Asn Pro
530 535 540
Phe Asn His Pro Arg His Ala Gly Leu Arg Tyr Arg Ser Met Leu Leu
545 550 555 560
Gly Asn Gly Arg Tyr Val Pro Phe His Ile Gln Val Pro Gln Lys Phe
565 570 575
Phe Ala Ile Lys Asn Leu Leu Leu Leu Pro Gly Ser Tyr Thr Tyr Glu
580 585 590
Trp Asn Phe Arg Lys Asp Val Asn Met Val Leu Gln Ser Ser Leu Gly
595 600 605
Asn Asp Leu Arg Val Asp Gly Ala Ser Ile Lys Phe Asp Ser Ile Cys
610 615 620
Leu Tyr Ala Thr Phe Phe Pro Met Ala His Asn Thr Ala Ser Thr Leu
625 630 635 640
Glu Ala Met Leu Arg Asn Asp Thr Asn Asp Gln Ser Phe Asn Asp Tyr
645 650 655
Leu Ser Ala Ala Asn Met Leu Tyr Pro Ile Pro Ala Asn Ala Thr Asn
660 665 670
Val Pro Ile Ser Ile Pro Ser Arg Asn Trp Ala Ala Phe Arg Gly Trp
675 680 685
Ala Phe Thr Arg Leu Lys Thr Lys Glu Thr Pro Ser Leu Gly Ser Gly
690 695 700
Tyr Asp Pro Tyr Tyr Thr Tyr Ser Gly Ser Ile Pro Tyr Leu Asp Gly
705 710 715 720
Thr Phe Tyr Leu Asn His Thr Phe Lys Lys Val Ala Ile Thr Phe Asp
725 730 735
Ser Ser Val Ser Trp Pro Gly Asn Asp Arg Leu Leu Thr Pro Asn Glu
740 745 750
Phe Glu Ile Lys Arg Ser Val Asp Gly Glu Gly Tyr Asn Val Ala Gln
755 760 765
Cys Asn Met Thr Lys Asp Trp Phe Leu Val Gln Met Leu Ala Asn Tyr
770 775 780
Asn Ile Gly Tyr Gln Gly Phe Tyr Ile Pro Glu Ser Tyr Lys Asp Arg
785 790 795 800
Met Tyr Ser Phe Phe Arg Asn Phe Gln Pro Met Ser Arg Gln Val Val
805 810 815
Asp Asp Thr Lys Tyr Lys Asp Tyr Gln Gln Val Gly Ile Ile His Gln
820 825 830
His Asn Asn Ser Gly Phe Val Gly Tyr Leu Ala Pro Thr Met Arg Glu
835 840 845
Gly Gln Ala Tyr Pro Ala Asn Val Pro Tyr Pro Leu Ile Gly Lys Thr
850 855 860
Ala Val Asp Ser Ile Thr Gln Lys Lys Phe Leu Cys Asp Arg Thr Leu
865 870 875 880
Trp Arg Ile Pro Phe Ser Ser Asn Phe Met Ser Met Gly Ala Leu Thr
885 890 895
Asp Leu Gly Gln Asn Leu Leu Tyr Ala Asn Ser Ala His Ala Leu Asp
900 905 910
Met Thr Phe Glu Val Asp Pro Met Asp Glu Pro Thr Leu Leu Tyr Val
915 920 925
Leu Phe Glu Val Phe Asp Val Val Arg Val His Gln Pro His Arg Gly
930 935 940
Val Ile Glu Thr Val Tyr Leu Arg Thr Pro Phe Ser Ala Gly Asn Ala
945 950 955 960
Thr Thr
<210> 60
<211> 528
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 60
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Ser
35 40 45
Leu Arg Leu Ser Glu Pro Leu Val Thr Ser His Gly Met Leu Ala Leu
50 55 60
Lys Met Gly Ser Gly Leu Ser Leu Asp Gln Ala Gly Asn Leu Thr Ser
65 70 75 80
Asn Thr Ile Thr Val Ser Gln Pro Leu Lys Lys Thr Lys Ser Asn Ile
85 90 95
Thr Leu Glu Thr Ser Ala Pro Leu Thr Val Ser Ser Gly Ala Leu Thr
100 105 110
Met Ala Thr Thr Ser Pro Leu Val Val Ser Asp Asn Thr Leu Thr Met
115 120 125
Gln Ser Gln Ala Pro Leu Thr Val Gln Asp Ser Lys Leu Ser Ile Ala
130 135 140
Thr Lys Glu Pro Leu Thr Val Leu Asp Gly Lys Leu Ala Leu Gln Thr
145 150 155 160
Ser Ala Pro Leu Ser Ala Thr Asp Asn Asn Ala Leu Thr Ile Thr Ala
165 170 175
Ser Pro Pro Leu Thr Thr Ala Asn Gly Ser Leu Ala Val Thr Met Glu
180 185 190
Asn Pro Leu Tyr Asn Asn Asn Gly Lys Leu Gly Leu Lys Ile Gly Gly
195 200 205
Pro Leu Gln Val Ala Thr Asp Ser His Ala Leu Thr Leu Gly Thr Gly
210 215 220
Gln Gly Val Ala Val His Asn Asn Leu Leu His Thr Lys Val Thr Gly
225 230 235 240
Ala Ile Gly Phe Asp Thr Ser Gly Asn Met Glu Leu Lys Thr Gly Asp
245 250 255
Gly Leu Tyr Val Asp Ser Ala Gly Pro Asn Gln Lys Leu His Ile Asn
260 265 270
Leu Asn Thr Thr Lys Gly Leu Ala Phe Asp Asn Thr Ala Ile Thr Ile
275 280 285
Asn Ala Gly Lys Gly Leu Glu Phe Glu Thr Asp Ser Ser Asn Gly Asn
290 295 300
Pro Ile Lys Thr Lys Ile Gly Ser Gly Ile Gln Tyr Asn Thr Asn Gly
305 310 315 320
Ala Met Val Ala Lys Leu Gly Thr Gly Leu Ser Phe Asp Ser Ser Gly
325 330 335
Ala Ile Thr Met Gly Ser Ile Asn Asn Asp Arg Leu Thr Leu Trp Thr
340 345 350
Thr Pro Asp Pro Ser Pro Asn Cys Arg Ile Ala Ser Asp Lys Asp Cys
355 360 365
Lys Leu Thr Leu Ala Leu Thr Lys Cys Gly Ser Gln Ile Leu Gly Thr
370 375 380
Val Ser Ala Leu Ala Val Ser Gly Asn Met Ala Ser Ile Asn Gly Thr
385 390 395 400
Leu Ser Ser Val Asn Leu Val Leu Arg Phe Asp Asp Asn Gly Val Leu
405 410 415
Met Ser Asn Ser Ser Leu Asp Lys Gln Tyr Trp Asn Phe Arg Asn Gly
420 425 430
Asp Ser Thr Asn Gly Gln Pro Tyr Thr Tyr Ala Val Gly Phe Met Pro
435 440 445
Asn Leu Lys Ala Tyr Pro Lys Thr Gln Ser Lys Thr Ala Lys Ser Asn
450 455 460
Ile Val Ser Gln Val Tyr Leu Asn Gly Asp Lys Ser Lys Pro Leu His
465 470 475 480
Phe Thr Ile Thr Leu Asn Gly Thr Asp Glu Thr Asn Gln Val Ser Lys
485 490 495
Tyr Ser Ile Ser Phe Ser Trp Ser Trp Asn Ser Gly Gln Tyr Thr Asn
500 505 510
Asp Lys Phe Ala Thr Asn Ser Tyr Thr Phe Ser Tyr Ile Ala Gln Glu
515 520 525
<210> 61
<211> 324
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 61
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Thr
35 40 45
Leu Lys Cys Leu Thr Pro Leu Thr Thr Thr Gly Gly Ser Leu Gln Leu
50 55 60
Lys Val Gly Gly Gly Leu Thr Val Asp Asp Thr Asp Gly Thr Leu Gln
65 70 75 80
Glu Asn Ile Arg Ala Thr Ala Pro Ile Thr Lys Asn Asn His Ser Val
85 90 95
Glu Leu Ser Ile Gly Asn Gly Leu Glu Thr Gln Asn Asn Lys Leu Cys
100 105 110
Ala Lys Leu Gly Asn Gly Leu Lys Phe Asn Asn Gly Asp Ile Cys Ile
115 120 125
Lys Asp Ser Ile Asn Thr Leu Trp Thr Gly Ile Asn Pro Pro Pro Asn
130 135 140
Cys Gln Ile Val Glu Asn Thr Asn Thr Asn Asp Gly Lys Leu Thr Leu
145 150 155 160
Val Leu Val Lys Asn Gly Gly Leu Val Asn Gly Tyr Val Ser Leu Val
165 170 175
Gly Val Ser Asp Thr Val Asn Gln Met Phe Thr Gln Lys Thr Ala Asn
180 185 190
Ile Gln Leu Arg Leu Tyr Phe Asp Ser Ser Gly Asn Leu Leu Thr Asp
195 200 205
Glu Ser Asp Leu Lys Ile Pro Leu Lys Asn Lys Ser Ser Thr Ala Thr
210 215 220
Ser Glu Thr Val Ala Ser Ser Lys Ala Phe Met Pro Ser Thr Thr Ala
225 230 235 240
Tyr Pro Phe Asn Thr Thr Thr Arg Asp Ser Glu Asn Tyr Ile His Gly
245 250 255
Ile Cys Tyr Tyr Met Thr Ser Tyr Asp Arg Ser Leu Phe Pro Leu Asn
260 265 270
Ile Ser Ile Met Leu Asn Ser Arg Met Ile Ser Ser Asn Val Ala Tyr
275 280 285
Ala Ile Gln Phe Glu Trp Asn Leu Asn Ala Ser Glu Ser Pro Glu Ser
290 295 300
Asn Ile Ala Thr Leu Thr Thr Ser Pro Phe Phe Phe Ser Tyr Ile Thr
305 310 315 320
Glu Asp Asp Asn
<210> 62
<211> 534
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 62
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Ser
35 40 45
Leu Arg Leu Ser Glu Pro Leu Val Thr Ser His Gly Met Leu Ala Leu
50 55 60
Lys Met Gly Ser Gly Leu Ser Leu Asp Gln Ala Gly Asn Leu Thr Ser
65 70 75 80
Asn Thr Ile Thr Val Ser Gln Pro Leu Lys Lys Thr Lys Ser Asn Ile
85 90 95
Thr Leu Glu Thr Ser Ala Pro Leu Thr Val Ser Ser Gly Ala Leu Thr
100 105 110
Met Ala Thr Thr Ser Pro Leu Val Val Ser Asp Asn Thr Leu Thr Met
115 120 125
Gln Ser Gln Ala Pro Leu Thr Val Gln Asp Ser Lys Leu Ser Ile Ala
130 135 140
Thr Lys Glu Pro Leu Thr Val Leu Asp Gly Lys Leu Ala Leu Gln Thr
145 150 155 160
Ser Ala Pro Leu Ser Ala Thr Asp Asn Asn Ala Leu Thr Ile Thr Ala
165 170 175
Ser Pro Pro Leu Thr Thr Ala Asn Gly Ser Leu Ala Val Thr Met Glu
180 185 190
Asn Pro Leu Tyr Asn Asn Asn Gly Lys Leu Gly Leu Lys Ile Gly Gly
195 200 205
Pro Leu Gln Val Ala Thr Asp Ser His Ala Leu Thr Leu Gly Thr Gly
210 215 220
Gln Gly Val Ala Val His Asn Asn Leu Leu His Thr Lys Val Thr Gly
225 230 235 240
Ala Ile Gly Phe Asp Thr Ser Gly Asn Met Glu Leu Lys Thr Gly Asp
245 250 255
Gly Leu Tyr Val Asp Ser Ala Gly Pro Asn Gln Lys Leu His Ile Asn
260 265 270
Leu Asn Thr Thr Lys Gly Leu Ala Phe Asp Asn Thr Ala Ile Thr Ile
275 280 285
Asn Ala Gly Lys Gly Leu Glu Phe Glu Thr Asp Ser Ser Asn Gly Asn
290 295 300
Pro Ile Lys Thr Lys Ile Gly Ser Gly Ile Gln Tyr Asn Thr Asn Gly
305 310 315 320
Ala Met Val Ala Lys Leu Gly Thr Gly Leu Ser Phe Asp Ser Ser Gly
325 330 335
Ala Ile Thr Met Gly Ser Ile Asn Asn Asp Arg Leu Thr Leu Trp Thr
340 345 350
Thr Pro Asp Pro Ser Pro Asn Cys Arg Ile Ala Ser Asp Lys Asp Cys
355 360 365
Lys Leu Thr Leu Ala Leu Thr Lys Cys Gly Ser Gln Ile Leu Gly Thr
370 375 380
Val Ser Ala Leu Ala Val Ser Gly Asn Met Ala Ser Ile Asn Gly Thr
385 390 395 400
Leu Ser Ser Val Asn Leu Val Leu Arg Phe Asp Asp Asn Gly Val Leu
405 410 415
Met Ser Asn Ser Ser Leu Asp Lys Gln Tyr Trp Asn Phe Arg Asn Gly
420 425 430
Asp Ser Thr Asn Gly Gln Pro Tyr Thr Tyr Ala Val Gly Phe Met Pro
435 440 445
Asn Leu Lys Ala Tyr Pro Lys Thr Gln Ser Lys Thr Ala Lys Ser Asn
450 455 460
Ile Val Ser Gln Val Tyr Leu Asn Gly Asp Lys Ser Lys Pro Leu His
465 470 475 480
Phe Thr Ile Thr Leu Asn Gly Thr Asp Glu Thr Asn Gln Val Ser Lys
485 490 495
Tyr Ser Ile Ser Phe Ser Trp Ser Trp Asn Ser Gly Gln Tyr Thr Asn
500 505 510
Asp Lys Phe Ala Thr Asn Ser Tyr Thr Phe Ser Tyr Ile Ala Gln Glu
515 520 525
Lys Lys Lys Lys Lys Lys
530
<210> 63
<211> 425
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 63
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Ser
35 40 45
Leu Arg Leu Ser Glu Pro Leu Val Thr Lys Asn Gly Glu Ile Thr Leu
50 55 60
Lys Leu Gly Glu Gly Val Asp Leu Asp Ser Ser Gly Lys Leu Ile Ser
65 70 75 80
Asn Thr Ala Thr Lys Ala Ala Ala Pro Leu Ser Phe Ser Asn Asn Thr
85 90 95
Ile Ser Leu Asn Met Asp His Pro Phe Tyr Thr Lys Asp Gly Lys Leu
100 105 110
Ser Leu Gln Val Ser Pro Pro Leu Asn Ile Leu Arg Thr Ser Ile Leu
115 120 125
Asn Thr Leu Ala Leu Gly Phe Gly Ser Gly Leu Gly Leu Arg Gly Ser
130 135 140
Ala Leu Ala Val Gln Leu Val Ser Pro Leu Thr Phe Asp Thr Asp Gly
145 150 155 160
Asn Ile Lys Leu Thr Leu Asp Arg Gly Leu His Val Thr Thr Gly Asp
165 170 175
Ala Ile Glu Ser Asn Ile Ser Trp Ala Lys Gly Leu Lys Phe Glu Asp
180 185 190
Gly Ala Ile Ala Thr Asn Ile Gly Asn Gly Leu Glu Phe Gly Ser Ser
195 200 205
Ser Thr Glu Thr Gly Val Asp Asp Ala Tyr Pro Ile Gln Val Lys Leu
210 215 220
Gly Ser Gly Leu Ser Phe Asp Ser Thr Gly Ala Ile Met Ala Gly Asn
225 230 235 240
Lys Glu Asp Asp Lys Leu Thr Leu Trp Thr Thr Pro Asp Pro Ser Pro
245 250 255
Asn Cys Gln Ile Leu Ala Glu Asn Asp Ala Lys Leu Thr Leu Cys Leu
260 265 270
Thr Lys Cys Gly Ser Gln Ile Leu Ala Thr Val Ser Val Leu Val Val
275 280 285
Gly Ser Gly Asn Leu Asn Pro Ile Thr Gly Thr Val Ser Ser Ala Gln
290 295 300
Val Phe Leu Arg Phe Asp Ala Asn Gly Val Leu Leu Thr Glu His Ser
305 310 315 320
Thr Leu Lys Lys Tyr Trp Gly Tyr Arg Gln Gly Asp Ser Ile Asp Gly
325 330 335
Thr Pro Tyr Thr Asn Ala Val Gly Phe Met Pro Asn Leu Lys Ala Tyr
340 345 350
Pro Lys Ser Gln Ser Ser Thr Thr Lys Asn Asn Ile Val Gly Gln Val
355 360 365
Tyr Met Asn Gly Asp Val Ser Lys Pro Met Leu Leu Thr Ile Thr Leu
370 375 380
Asn Gly Thr Asp Asp Ser Asn Ser Thr Tyr Ser Met Ser Phe Ser Tyr
385 390 395 400
Thr Trp Thr Asn Gly Ser Tyr Val Gly Ala Thr Phe Gly Ala Asn Ser
405 410 415
Tyr Thr Phe Ser Tyr Ile Ala Gln Glu
420 425
<210> 64
<211> 432
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 64
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Ser
35 40 45
Leu Arg Leu Ser Glu Pro Leu Val Thr Lys Asn Gly Glu Ile Thr Leu
50 55 60
Lys Leu Gly Glu Gly Val Asp Leu Asp Ser Ser Gly Lys Leu Ile Ser
65 70 75 80
Asn Thr Ala Thr Lys Ala Ala Ala Pro Leu Ser Phe Ser Asn Asn Thr
85 90 95
Ile Ser Leu Asn Met Asp His Pro Phe Tyr Thr Lys Asp Gly Lys Leu
100 105 110
Ser Leu Gln Val Ser Pro Pro Leu Asn Ile Leu Arg Thr Ser Ile Leu
115 120 125
Asn Thr Leu Ala Leu Gly Phe Gly Ser Gly Leu Gly Leu Arg Gly Ser
130 135 140
Ala Leu Ala Val Gln Leu Val Ser Pro Leu Thr Phe Asp Thr Asp Gly
145 150 155 160
Asn Ile Lys Leu Thr Leu Asp Arg Gly Leu His Val Thr Thr Gly Asp
165 170 175
Ala Ile Glu Ser Asn Ile Ser Trp Ala Lys Gly Leu Lys Phe Glu Asp
180 185 190
Gly Ala Ile Ala Thr Asn Ile Gly Asn Gly Leu Glu Phe Gly Ser Ser
195 200 205
Ser Thr Glu Thr Gly Val Asp Asp Ala Tyr Pro Ile Gln Val Lys Leu
210 215 220
Gly Ser Gly Leu Ser Phe Asp Ser Thr Gly Ala Ile Met Ala Gly Asn
225 230 235 240
Lys Glu Asp Asp Lys Leu Thr Leu Trp Thr Thr Pro Asp Pro Ser Pro
245 250 255
Asn Cys Gln Ile Leu Ala Glu Asn Asp Ala Lys Leu Thr Leu Cys Leu
260 265 270
Thr Lys Cys Gly Ser Gln Ile Leu Ala Thr Val Ser Val Leu Val Val
275 280 285
Gly Ser Gly Asn Leu Asn Pro Ile Thr Gly Thr Val Ser Ser Ala Gln
290 295 300
Val Phe Leu Arg Phe Asp Ala Asn Gly Val Leu Leu Thr Glu His Ser
305 310 315 320
Thr Leu Lys Lys Tyr Trp Gly Tyr Arg Gln Gly Asp Ser Ile Asp Gly
325 330 335
Thr Pro Tyr Thr Asn Ala Val Gly Phe Met Pro Asn Leu Lys Ala Tyr
340 345 350
Pro Lys Ser Gln Ser Ser Thr Thr Lys Asn Asn Ile Val Gly Gln Val
355 360 365
Tyr Met Asn Gly Asp Val Ser Lys Pro Met Leu Leu Thr Ile Thr Leu
370 375 380
Asn Gly Thr Asp Asp Ser Asn Ser Thr Tyr Ser Met Ser Phe Ser Tyr
385 390 395 400
Thr Trp Thr Asn Gly Ser Tyr Val Gly Ala Thr Phe Gly Ala Asn Ser
405 410 415
Tyr Thr Phe Ser Tyr Ile Ala Gln Glu Lys Lys Lys Lys Lys Lys Lys
420 425 430
<210> 65
<211> 441
<212> белок
<213> Искусственная последовательность
<220>
<223> Полипептид волокна
<400> 65
Met Lys Arg Ala Arg Pro Ser Glu Asp Thr Phe Asn Pro Val Tyr Pro
1. 5 10 15
Tyr Asp Thr Glu Thr Gly Pro Pro Thr Val Pro Phe Leu Thr Pro Pro
20 25 30
Phe Val Ser Pro Asn Gly Phe Gln Glu Ser Pro Pro Gly Val Leu Ser
35 40 45
Leu Arg Leu Ser Glu Pro Leu Val Thr Lys Asn Gly Glu Ile Thr Leu
50 55 60
Lys Leu Gly Glu Gly Val Asp Leu Asp Ser Ser Gly Lys Leu Ile Ser
65 70 75 80
Asn Thr Ala Thr Lys Ala Ala Ala Pro Leu Ser Phe Ser Asn Asn Thr
85 90 95
Ile Ser Leu Asn Met Asp His Pro Phe Tyr Thr Lys Asp Gly Lys Leu
100 105 110
Ser Leu Gln Val Ser Pro Pro Leu Asn Ile Leu Arg Thr Ser Ile Leu
115 120 125
Asn Thr Leu Ala Leu Gly Phe Gly Ser Gly Leu Gly Leu Arg Gly Ser
130 135 140
Ala Leu Ala Val Gln Leu Val Ser Pro Leu Thr Phe Asp Thr Asp Gly
145 150 155 160
Asn Ile Lys Leu Thr Leu Asp Arg Gly Leu His Val Thr Thr Gly Asp
165 170 175
Ala Ile Glu Ser Asn Ile Ser Trp Ala Lys Gly Leu Lys Phe Glu Asp
180 185 190
Gly Ala Ile Ala Thr Asn Ile Gly Asn Gly Leu Glu Phe Gly Ser Ser
195 200 205
Ser Thr Glu Thr Gly Val Asp Asp Ala Tyr Pro Ile Gln Val Lys Leu
210 215 220
Gly Ser Gly Leu Ser Phe Asp Ser Thr Gly Ala Ile Met Ala Gly Asn
225 230 235 240
Lys Glu Asp Asp Lys Leu Thr Leu Trp Thr Thr Pro Asp Pro Ser Pro
245 250 255
Asn Cys Gln Ile Leu Ala Glu Asn Asp Ala Lys Leu Thr Leu Cys Leu
260 265 270
Thr Lys Cys Gly Ser Gln Ile Leu Ala Thr Val Ser Val Leu Val Val
275 280 285
Gly Ser Gly Asn Leu Asn Pro Ile Thr Gly Thr Val Ser Ser Ala Gln
290 295 300
Val Phe Leu Arg Phe Asp Ala Asn Gly Val Leu Leu Thr Glu His Ser
305 310 315 320
Thr Leu Lys Lys Tyr Trp Gly Tyr Arg Gln Gly Asp Ser Ile Asp Gly
325 330 335
Thr Pro Tyr Thr Asn Ala Val Gly Phe Met Pro Asn Leu Lys Ala Tyr
340 345 350
Pro Lys Ser Gln Ser Ser Thr Thr Lys Asn Asn Ile Val Gly Gln Val
355 360 365
Tyr Met Asn Gly Asp Val Ser Lys Pro Met Leu Leu Thr Ile Thr Leu
370 375 380
Asn Gly Thr Asp Asp Ser Gly Gly Ser Ser Gly Lys Lys Lys Lys Lys
385 390 395 400
Lys Lys Ala Ser Gly Gly Ser Ser Thr Tyr Ser Met Ser Phe Ser Tyr
405 410 415
Thr Trp Thr Asn Gly Ser Tyr Val Gly Ala Thr Phe Gly Ala Asn Ser
420 425 430
Tyr Thr Phe Ser Tyr Ile Ala Gln Glu
435 440
<210> 66
<211> 27
<212> белок
<213> Mus musculus
<400> 66
Pro Val Thr Leu Thr Ile Thr Leu Thr Ala Arg Gly Glu His Lys Glu
1. 5 10 15
Glu Glu Leu Ile Gly Ala Tyr Tyr Ser Met Ser
20 25
<210> 67
<211> 27
<212> белок
<213> Mus musculus
<400> 67
Pro Val Thr Leu Thr Ile Thr Leu Leu Arg Gln Thr Gly Ala Ala Ser
1. 5 10 15
Ala Val Trp Gly Gly Ala Tyr Tyr Ser Met Ser
20 25
<210> 68
<211> 23
<212> белок
<213> Аденовирус типа 5
<400> 68
Glu Met Gln Phe Phe Ser Thr Thr Glu Ala Thr Ala Gly Asn Gly Asp
1. 5 10 15
Asn Leu Thr Pro Lys Val Val
20
<210> 69
<211> 26
<212> белок
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 69
Glu Met Gln Phe Phe Ser Gly Ser Thr Ala Arg Gly Glu His Lys Glu
1. 5 10 15
Glu Glu Leu Ile Gly Thr Pro Lys Val Val
20 25
<210> 70
<211> 27
<212> белок
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 70
Glu Met Gln Phe Phe Ser Gly Ser Met Leu Arg Gln Thr Gly Ala Ala
1. 5 10 15
Ser Ala Val Trp Gly Gly Thr Pro Lys Val Val
20 25
<210> 71
<211> 407
<212> белок
<213> Аденовирус типа 5
<400> 71
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Pro Cys Glu Trp Asp
1. 5 10 15
Glu Ala Ala Thr Ala Leu Glu Ile Asn Leu Glu Glu Glu Asp Asp Asp
20 25 30
Asn Glu Asp Glu Val Asp Glu Gln Ala Glu Gln Gln Lys Thr His Val
35 40 45
Phe Ser Gln Ala Pro Tyr Ser Gly Ile Asn Ile Thr Lys Glu Gly Ile
50 55 60
Gln Ile Gly Val Glu Gly Gln Thr Pro Lys Tyr Ala Asp Lys Thr Phe
65 70 75 80
Gln Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Tyr Glu Thr Glu Ile
85 90 95
Asn His Ala Ala Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro
100 105 110
Cys Tyr Gly Ser Tyr Ala Lys Pro Thr Asn Glu Asn Gly Gly Gln Gly
115 120 125
Ile Leu Val Lys Gln Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met
130 135 140
Gln Phe Phe Ser Thr Thr Glu Ala Thr Ala Gly Asn Gly Asp Asn Leu
145 150 155 160
Thr Pro Lys Val Val Leu Tyr Ser Glu Asp Val Asp Ile Glu Thr Pro
165 170 175
Asp Thr His Ile Ser Tyr Met Pro Thr Ile Lys Glu Gly Asn Ser Arg
180 185 190
Glu Leu Met Gly Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala
195 200 205
Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn
210 215 220
Met Gly Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp
225 230 235 240
Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser
245 250 255
Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp
260 265 270
Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp
275 280 285
Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly Gly Val Ile Asn Thr Glu
290 295 300
Thr Leu Thr Lys Val Lys Pro Lys Thr Gly Gln Glu Asn Gly Trp Glu
305 310 315 320
Lys Asp Ala Thr Glu Phe Ser Asp Lys Asn Glu Ile Arg Val Gly Asn
325 330 335
Asn Phe Ala Met Glu Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe
340 345 350
Leu Tyr Ser Asn Ile Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Ser
355 360 365
Pro Ser Asn Val Lys Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met
370 375 380
Asn Lys Arg Val Val Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu
385 390 395 400
Gly Ala Arg Trp Ser Leu Asp
405
<210> 72
<211> 418
<212> белок
<213> Аденовирус типа 6
<400> 72
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Glu
1. 5 10 15
Gln Asn Glu Thr Ala Gln Val Asp Ala Gln Glu Leu Asp Glu Glu Glu
20 25 30
Asn Glu Ala Asn Glu Ala Gln Ala Arg Glu Gln Glu Gln Ala Lys Lys
35 40 45
Thr His Val Tyr Ala Gln Ala Pro Leu Ser Gly Ile Lys Ile Thr Lys
50 55 60
Glu Gly Leu Gln Ile Gly Thr Ala Asp Ala Thr Val Ala Gly Ala Gly
65 70 75 80
Lys Glu Ile Phe Ala Asp Lys Thr Phe Gln Pro Glu Pro Gln Val Gly
85 90 95
Glu Ser Gln Trp Asn Glu Ala Asp Ala Thr Ala Ala Gly Gly Arg Val
100 105 110
Leu Lys Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg
115 120 125
Pro Thr Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly
130 135 140
Lys Leu Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Thr Asn
145 150 155 160
Ala Thr Asn Glu Val Asn Asn Ile Gln Pro Thr Val Val Leu Tyr Ser
165 170 175
Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro
180 185 190
Lys Met Gly Asp Lys Asn Ala Lys Val Met Leu Gly Gln Gln Ala Met
195 200 205
Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu
210 215 220
Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala
225 230 235 240
Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu
245 250 255
Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe
260 265 270
Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile
275 280 285
Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro
290 295 300
Leu Gly Gly Ile Gly Ile Thr Asp Thr Phe Gln Ala Val Lys Thr Thr
305 310 315 320
Ala Ala Asn Gly Asp Gln Gly Asn Thr Thr Trp Gln Lys Asp Ser Thr
325 330 335
Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu
340 345 350
Ile Asn Leu Asn Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile
355 360 365
Ala Leu Tyr Leu Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu
370 375 380
Ile Ser Asp Asn Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val
385 390 395 400
Ala Pro Gly Leu Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser
405 410 415
Leu Glu
<210> 73
<211> 414
<212> белок
<213> Аденовирус типа 57
<400> 73
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Asp
1. 5 10 15
Glu Asp Asp Thr Gln Val Gln Val Ala Ala Glu Asp Asp Gln Asp Asp
20 25 30
Asp Glu Glu Glu Glu Gln Leu Pro Gln Gln Arg Asn Gly Lys Lys Thr
35 40 45
His Val Tyr Ala Gln Ala Pro Phe Ala Gly Glu Ala Ile Asn Lys Asn
50 55 60
Gly Leu Gln Ile Gly Thr Asn Gly Ala Ala Thr Glu Gly Asn Lys Glu
65 70 75 80
Ile Tyr Ala Asp Lys Thr Tyr Gln Pro Glu Pro Gln Ile Gly Glu Ser
85 90 95
Gln Trp Asn Glu Ala Glu Ser Ser Val Ala Gly Gly Arg Val Leu Lys
100 105 110
Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg Pro Thr
115 120 125
Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly Lys Leu
130 135 140
Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Val Asn Ala Met
145 150 155 160
Asn Glu Ala Asn Ala Ile Gln Pro Lys Leu Val Leu Tyr Ser Glu Asp
165 170 175
Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly Lys
180 185 190
Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro Asn
195 200 205
Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr
210 215 220
Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser Gln
225 230 235 240
Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr
245 250 255
Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met
260 265 270
Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu
275 280 285
Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly
290 295 300
Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn Gly
305 310 315 320
Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu Arg
325 330 335
Asn Glu Ile Gly Val Gly Asn Asn Phe Ala Met Glu Ile Asn Leu Asn
340 345 350
Ala Asn Leu Trp Arg Asn Phe Leu Tyr Ser Asn Ile Ala Leu Tyr Leu
355 360 365
Pro Asp Lys Leu Lys Tyr Asn Pro Thr Asn Val Glu Ile Ser Asp Asn
370 375 380
Pro Asn Thr Tyr Asp Tyr Met Asn Lys Arg Val Val Ala Pro Gly Leu
385 390 395 400
Val Asp Cys Tyr Ile Asn Leu Gly Ala Arg Trp Ser Leu Asp
405 410
<210> 74
<211> 348
<212> белок
<213> Аденовирус типа 1
<400> 74
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Glu
1. 5 10 15
Gln Glu Glu Pro Thr Gln Glu Met Ala Glu Glu Leu Glu Asp Glu Glu
20 25 30
Glu Ala Glu Glu Glu Glu Ala Glu Glu Glu Ala Glu Ala Pro Gln Ala
35 40 45
Asp Gln Lys Val Lys Lys Thr His Val Tyr Ala Gln Ala Pro Leu Ala
50 55 60
Gly Glu Lys Ile Thr Ala Asn Gly Leu Gln Ile Val Ser Asp Thr Gln
65 70 75 80
Thr Glu Gly Asn Pro Val Phe Ala Asp Pro Thr Tyr Gln Pro Glu Pro
85 90 95
Gln Val Gly Glu Ser Gln Trp Asn Glu Ala Glu Ala Thr Ala Ser Gly
100 105 110
Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser
115 120 125
Tyr Ala Arg Pro Thr Asn Lys Asn Gly Gly Gln Gly Ile Leu Val Ala
130 135 140
Asn Asn Gln Gly Ala Leu Glu Ser Lys Val Glu Met Gln Phe Phe Ala
145 150 155 160
Pro Ser Gly Thr Ala Met Asn Glu Arg Asn Ala Val Gln Pro Ser Ile
165 170 175
Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Ile
180 185 190
Ser Tyr Lys Pro Ser Lys Thr Asp Glu Asn Ser Lys Ala Met Leu Gly
195 200 205
Gln Gln Ala Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn
210 215 220
Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu
225 230 235 240
Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg
245 250 255
Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg
260 265 270
Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro
275 280 285
Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn
290 295 300
Tyr Cys Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Gly
305 310 315 320
Ile Lys Ser Asn Gly Asn Gly Asn Pro Gln Asn Trp Thr Lys Asn Asp
325 330 335
Asp Phe Ala Ala Arg Asn Glu Ile Gly Val Gly Asn
340 345
<210> 75
<211> 352
<212> белок
<213> Аденовирус типа 2
<400> 75
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Glu
1. 5 10 15
Gln Thr Glu Asp Ser Gly Arg Ala Val Ala Glu Asp Glu Glu Glu Glu
20 25 30
Asp Glu Asp Glu Glu Glu Glu Glu Glu Glu Gln Asn Ala Arg Asp Gln
35 40 45
Ala Thr Lys Lys Thr His Val Tyr Ala Gln Ala Pro Leu Ser Gly Glu
50 55 60
Thr Ile Thr Lys Ser Gly Leu Gln Ile Gly Ser Asp Asn Ala Glu Thr
65 70 75 80
Gln Ala Lys Pro Val Tyr Ala Asp Pro Ser Tyr Gln Pro Glu Pro Gln
85 90 95
Ile Gly Glu Ser Gln Trp Asn Glu Ala Asp Ala Asn Ala Ala Gly Gly
100 105 110
Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr
115 120 125
Ala Arg Pro Thr Asn Pro Phe Gly Gly Gln Ser Val Leu Val Pro Asp
130 135 140
Glu Lys Gly Val Pro Leu Pro Lys Val Asp Leu Gln Phe Phe Ser Asn
145 150 155 160
Thr Thr Ser Leu Asn Asp Arg Gln Gly Asn Ala Thr Lys Pro Lys Val
165 170 175
Val Leu Tyr Ser Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu
180 185 190
Ser Tyr Lys Pro Gly Lys Gly Asp Glu Asn Ser Lys Ala Met Leu Gly
195 200 205
Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn
210 215 220
Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu
225 230 235 240
Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg
245 250 255
Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg
260 265 270
Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro
275 280 285
Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn
290 295 300
Tyr Cys Phe Pro Leu Gly Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala
305 310 315 320
Ile Lys Ala Asn Gly Asn Gly Ser Gly Asp Asn Gly Asp Thr Thr Trp
325 330 335
Thr Lys Asp Glu Thr Phe Ala Thr Arg Asn Glu Ile Gly Val Gly Asn
340 345 350
<210> 76
<211> 336
<212> белок
<213> Аденовирус типа 5
<400> 76
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Pro Cys Glu Trp Asp
1. 5 10 15
Glu Ala Ala Thr Ala Leu Glu Ile Asn Leu Glu Glu Glu Asp Asp Asp
20 25 30
Asn Glu Asp Glu Val Asp Glu Gln Ala Glu Gln Gln Lys Thr His Val
35 40 45
Phe Ser Gln Ala Pro Tyr Ser Gly Ile Asn Ile Thr Lys Glu Gly Ile
50 55 60
Gln Ile Gly Val Glu Gly Gln Thr Pro Lys Tyr Ala Asp Lys Thr Phe
65 70 75 80
Gln Pro Glu Pro Gln Ile Gly Glu Ser Gln Trp Tyr Glu Thr Glu Ile
85 90 95
Asn His Ala Ala Gly Arg Val Leu Lys Lys Thr Thr Pro Met Lys Pro
100 105 110
Cys Tyr Gly Ser Tyr Ala Lys Pro Thr Asn Glu Asn Gly Gly Gln Gly
115 120 125
Ile Leu Val Lys Gln Gln Asn Gly Lys Leu Glu Ser Gln Val Glu Met
130 135 140
Gln Phe Phe Ser Thr Thr Glu Ala Thr Ala Gly Asn Gly Asp Asn Leu
145 150 155 160
Thr Pro Lys Val Val Leu Tyr Ser Glu Asp Val Asp Ile Glu Thr Pro
165 170 175
Asp Thr His Ile Ser Tyr Met Pro Thr Ile Lys Glu Gly Asn Ser Arg
180 185 190
Glu Leu Met Gly Gln Gln Ser Met Pro Asn Arg Pro Asn Tyr Ile Ala
195 200 205
Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr Tyr Asn Ser Thr Gly Asn
210 215 220
Met Gly Val Leu Ala Gly Gln Ala Ser Gln Leu Asn Ala Val Val Asp
225 230 235 240
Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr Gln Leu Leu Leu Asp Ser
245 250 255
Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met Trp Asn Gln Ala Val Asp
260 265 270
Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu Asn His Gly Thr Glu Asp
275 280 285
Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly Gly Val Ile Asn Thr Glu
290 295 300
Thr Leu Thr Lys Val Lys Pro Lys Thr Gly Gln Glu Asn Gly Trp Glu
305 310 315 320
Lys Asp Ala Thr Glu Phe Ser Asp Lys Asn Glu Ile Arg Val Gly Asn
325 330 335
<210> 77
<211> 347
<212> белок
<213> Аденовирус типа 6
<400> 77
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Glu
1. 5 10 15
Gln Asn Glu Thr Ala Gln Val Asp Ala Gln Glu Leu Asp Glu Glu Glu
20 25 30
Asn Glu Ala Asn Glu Ala Gln Ala Arg Glu Gln Glu Gln Ala Lys Lys
35 40 45
Thr His Val Tyr Ala Gln Ala Pro Leu Ser Gly Ile Lys Ile Thr Lys
50 55 60
Glu Gly Leu Gln Ile Gly Thr Ala Asp Ala Thr Val Ala Gly Ala Gly
65 70 75 80
Lys Glu Ile Phe Ala Asp Lys Thr Phe Gln Pro Glu Pro Gln Val Gly
85 90 95
Glu Ser Gln Trp Asn Glu Ala Asp Ala Thr Ala Ala Gly Gly Arg Val
100 105 110
Leu Lys Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg
115 120 125
Pro Thr Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly
130 135 140
Lys Leu Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Thr Asn
145 150 155 160
Ala Thr Asn Glu Val Asn Asn Ile Gln Pro Thr Val Val Leu Tyr Ser
165 170 175
Glu Asp Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro
180 185 190
Lys Met Gly Asp Lys Asn Ala Lys Val Met Leu Gly Gln Gln Ala Met
195 200 205
Pro Asn Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu
210 215 220
Met Tyr Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala
225 230 235 240
Ser Gln Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu
245 250 255
Ser Tyr Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe
260 265 270
Ser Met Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile
275 280 285
Ile Glu Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro
290 295 300
Leu Gly Gly Ile Gly Ile Thr Asp Thr Phe Gln Ala Val Lys Thr Thr
305 310 315 320
Ala Ala Asn Gly Asp Gln Gly Asn Thr Thr Trp Gln Lys Asp Ser Thr
325 330 335
Phe Ala Glu Arg Asn Glu Ile Gly Val Gly Asn
340 345
<210> 78
<211> 343
<212> белок
<213> Аденовирус типа 57
<400> 78
Tyr Asn Ala Leu Ala Pro Lys Gly Ala Pro Asn Ser Cys Glu Trp Asp
1. 5 10 15
Glu Asp Asp Thr Gln Val Gln Val Ala Ala Glu Asp Asp Gln Asp Asp
20 25 30
Asp Glu Glu Glu Glu Gln Leu Pro Gln Gln Arg Asn Gly Lys Lys Thr
35 40 45
His Val Tyr Ala Gln Ala Pro Phe Ala Gly Glu Ala Ile Asn Lys Asn
50 55 60
Gly Leu Gln Ile Gly Thr Asn Gly Ala Ala Thr Glu Gly Asn Lys Glu
65 70 75 80
Ile Tyr Ala Asp Lys Thr Tyr Gln Pro Glu Pro Gln Ile Gly Glu Ser
85 90 95
Gln Trp Asn Glu Ala Glu Ser Ser Val Ala Gly Gly Arg Val Leu Lys
100 105 110
Lys Thr Thr Pro Met Lys Pro Cys Tyr Gly Ser Tyr Ala Arg Pro Thr
115 120 125
Asn Ser Asn Gly Gly Gln Gly Val Met Val Glu Gln Asn Gly Lys Leu
130 135 140
Glu Ser Gln Val Glu Met Gln Phe Phe Ser Thr Ser Val Asn Ala Met
145 150 155 160
Asn Glu Ala Asn Ala Ile Gln Pro Lys Leu Val Leu Tyr Ser Glu Asp
165 170 175
Val Asn Met Glu Thr Pro Asp Thr His Leu Ser Tyr Lys Pro Gly Lys
180 185 190
Ser Asp Asp Asn Ser Lys Ala Met Leu Gly Gln Gln Ser Met Pro Asn
195 200 205
Arg Pro Asn Tyr Ile Ala Phe Arg Asp Asn Phe Ile Gly Leu Met Tyr
210 215 220
Tyr Asn Ser Thr Gly Asn Met Gly Val Leu Ala Gly Gln Ala Ser Gln
225 230 235 240
Leu Asn Ala Val Val Asp Leu Gln Asp Arg Asn Thr Glu Leu Ser Tyr
245 250 255
Gln Leu Leu Leu Asp Ser Ile Gly Asp Arg Thr Arg Tyr Phe Ser Met
260 265 270
Trp Asn Gln Ala Val Asp Ser Tyr Asp Pro Asp Val Arg Ile Ile Glu
275 280 285
Asn His Gly Thr Glu Asp Glu Leu Pro Asn Tyr Cys Phe Pro Leu Gly
290 295 300
Gly Ile Gly Val Thr Asp Thr Tyr Gln Ala Ile Lys Ala Thr Asn Gly
305 310 315 320
Asn Gly Gly Ala Thr Thr Trp Ala Gln Asp Asn Thr Phe Ala Glu Arg
325 330 335
Asn Glu Ile Gly Val Gly Asn
340
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИВАЛЕНТНЫЕ PD-L1-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2020 |
|
RU2816646C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2758596C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ОБЛАДАЮЩИЕ ПЕРЕКРЕСТНОЙ РЕАКТИВНОСТЬЮ С ВИРУСОМ ЗИКА | 2019 |
|
RU2811697C2 |
| АНТИТЕЛА К МИОСТАТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2778945C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2799522C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| АНТИТЕЛА К С5 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746356C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
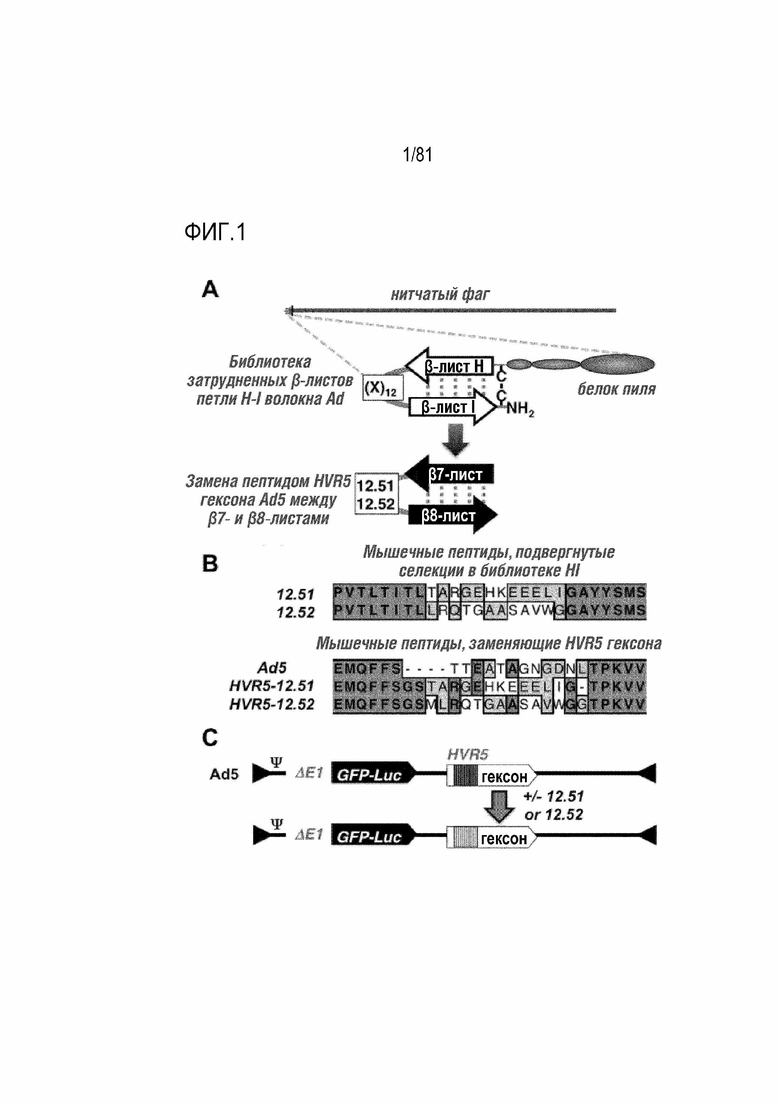
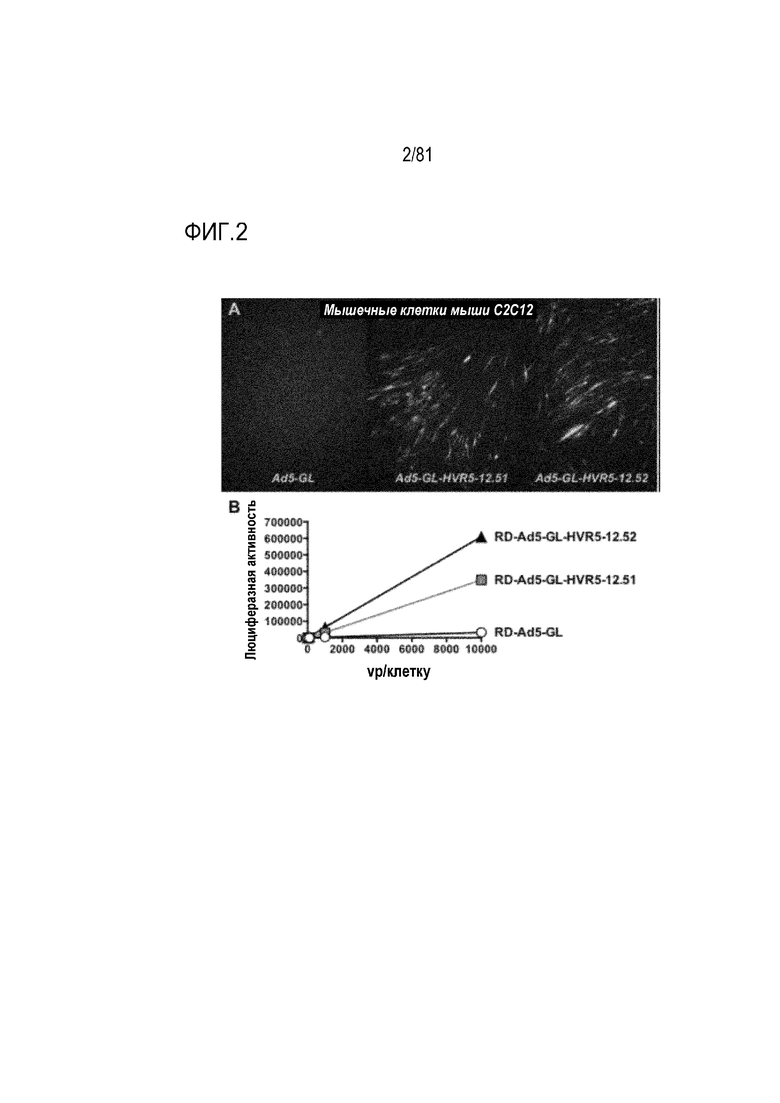
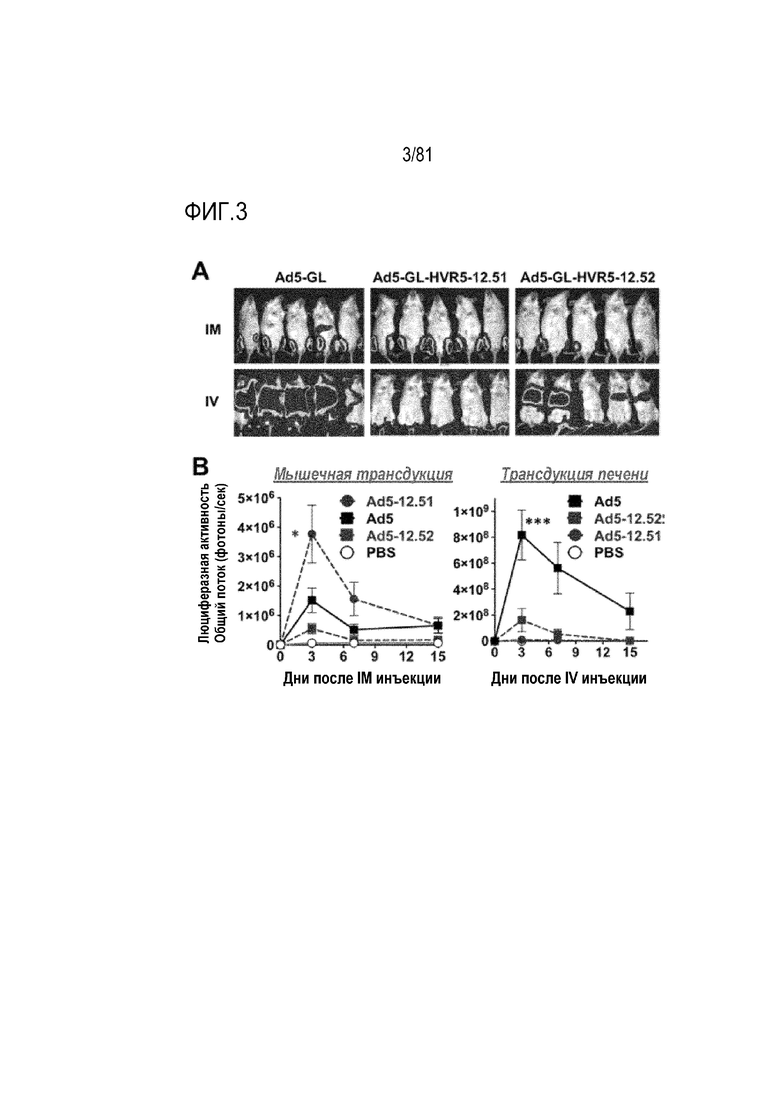
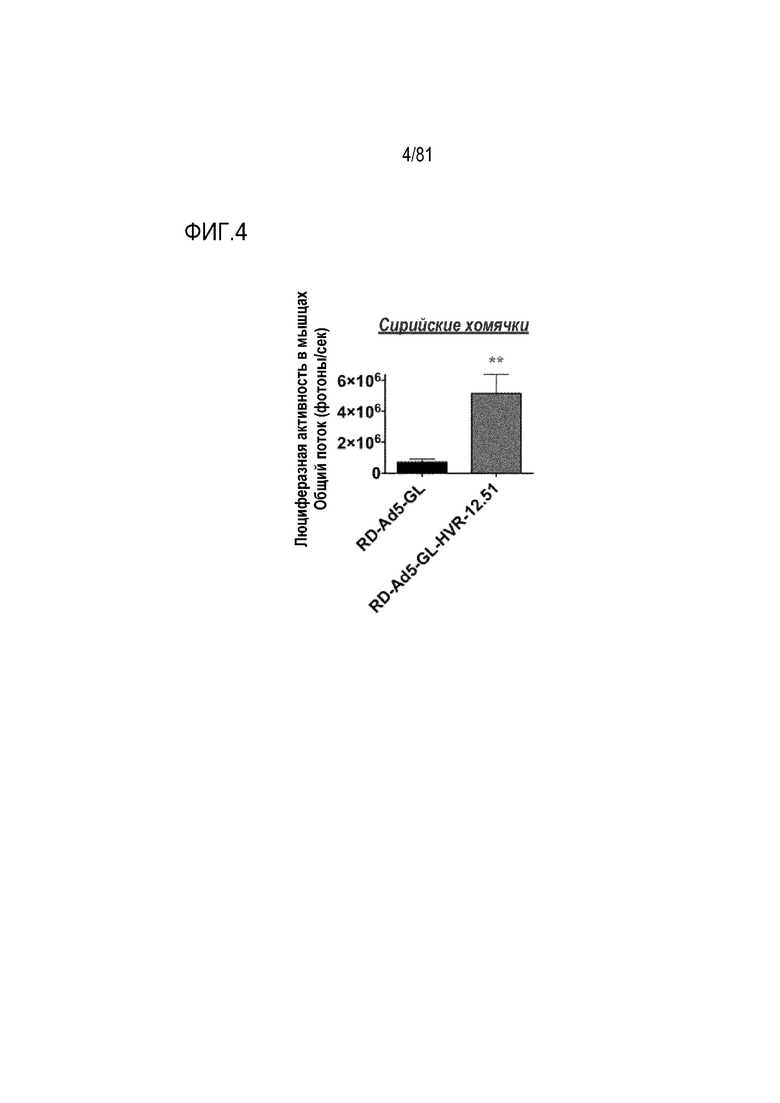
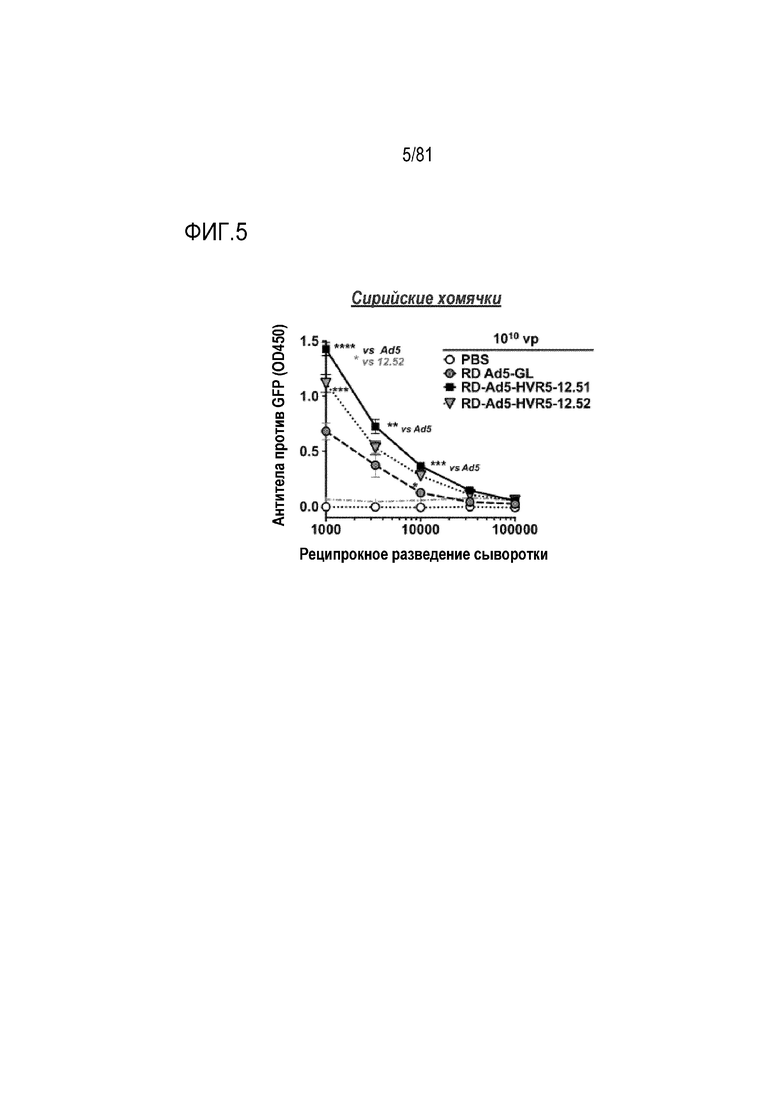
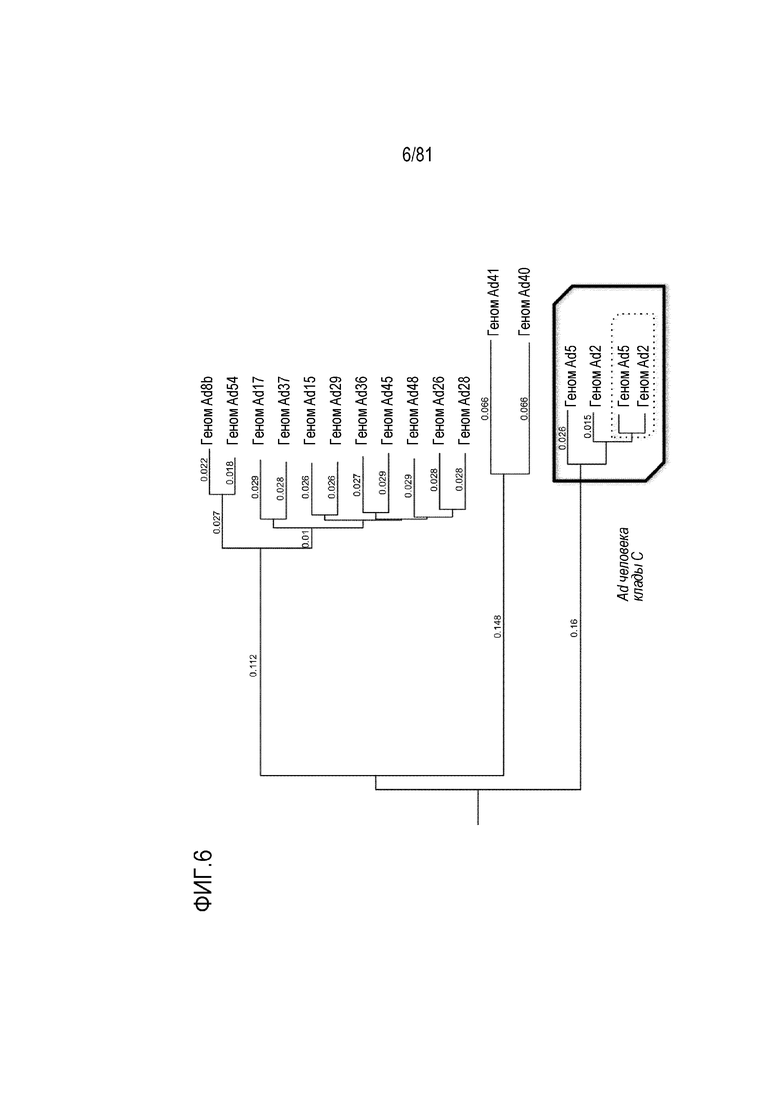

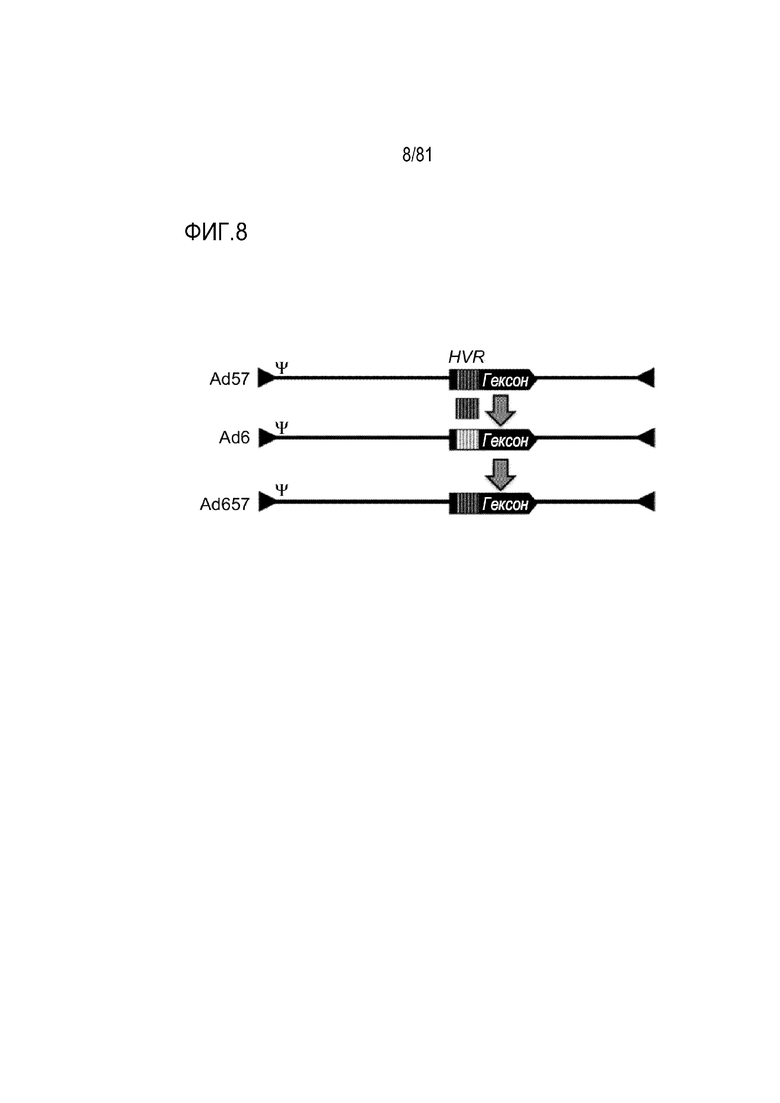
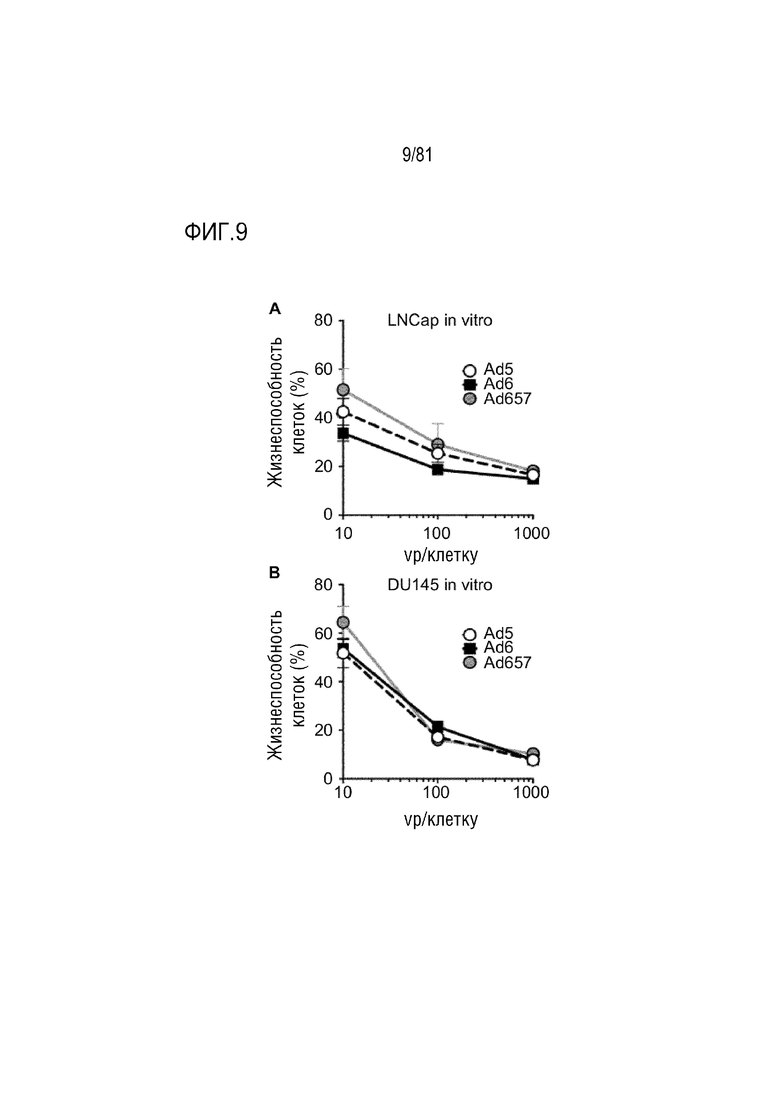
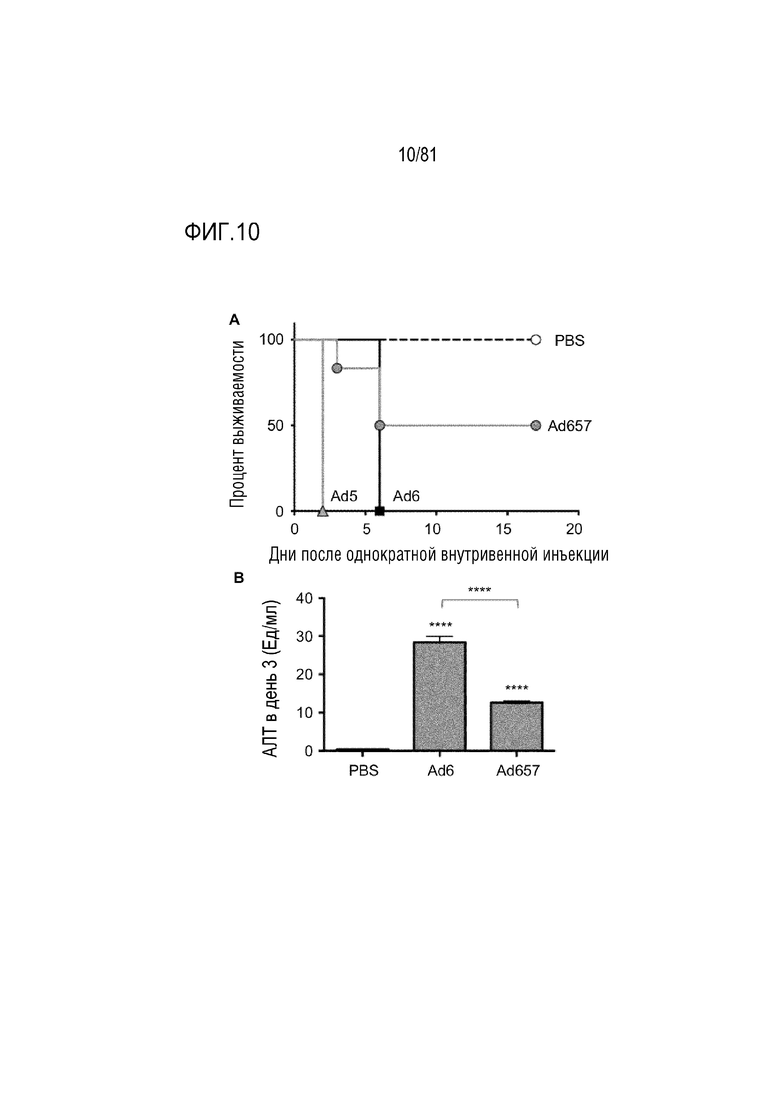
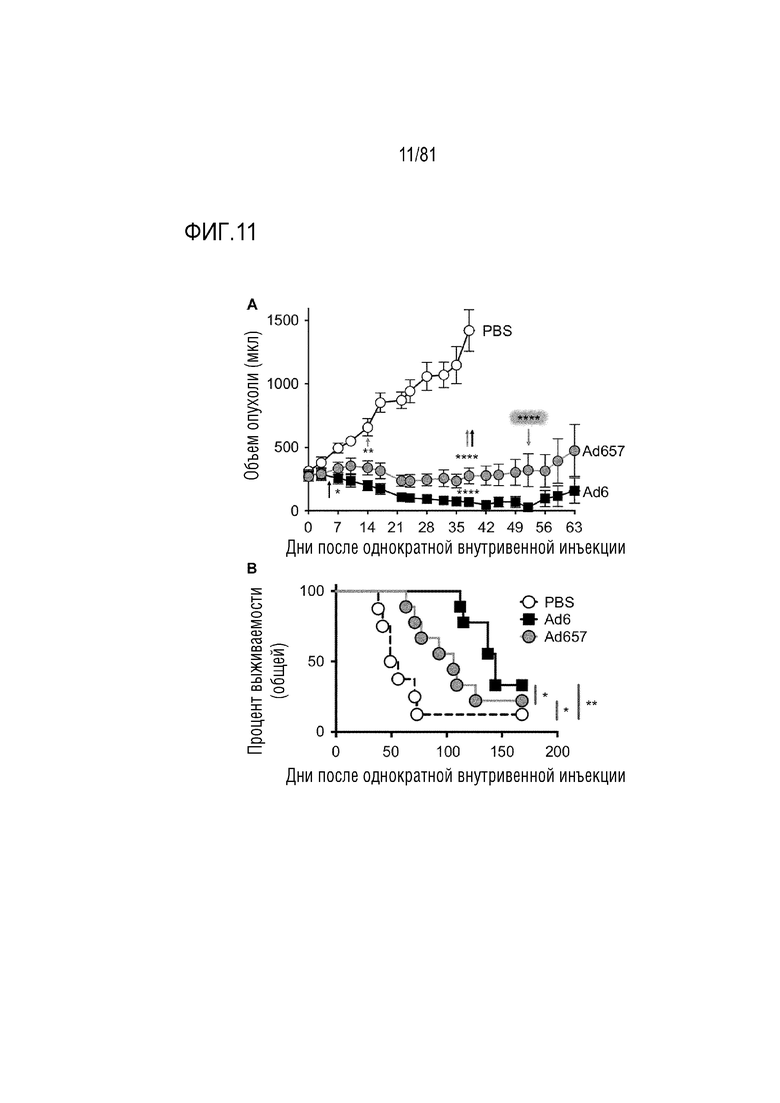
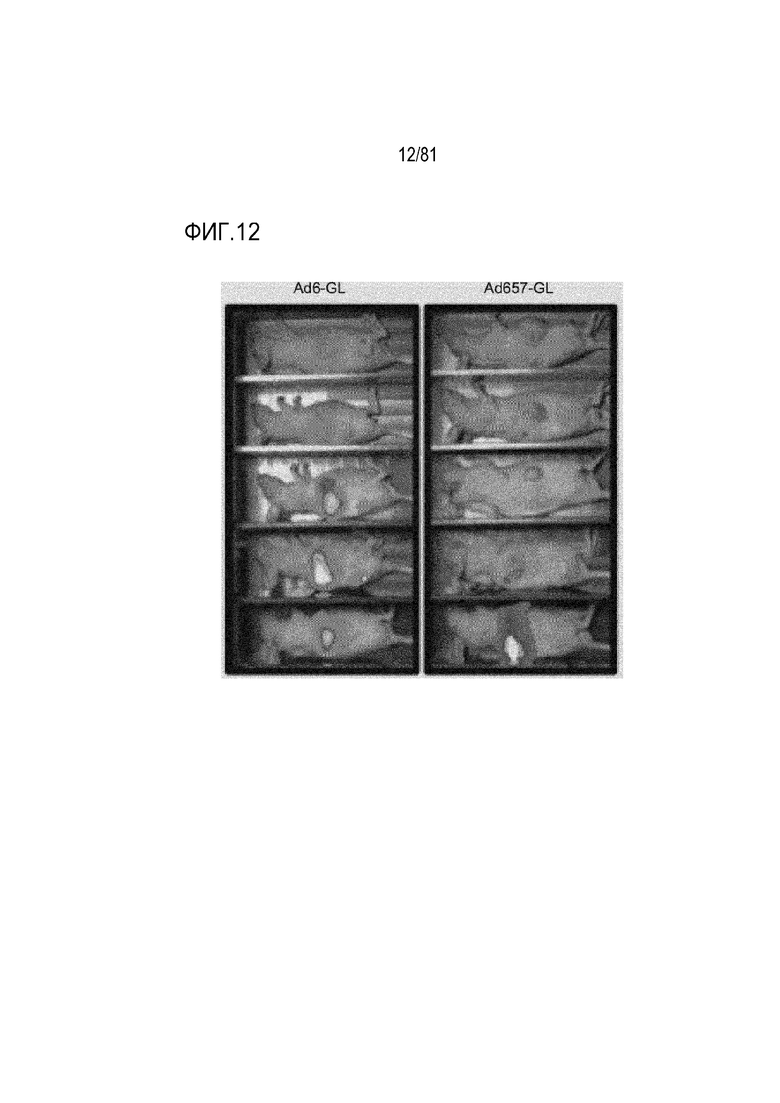

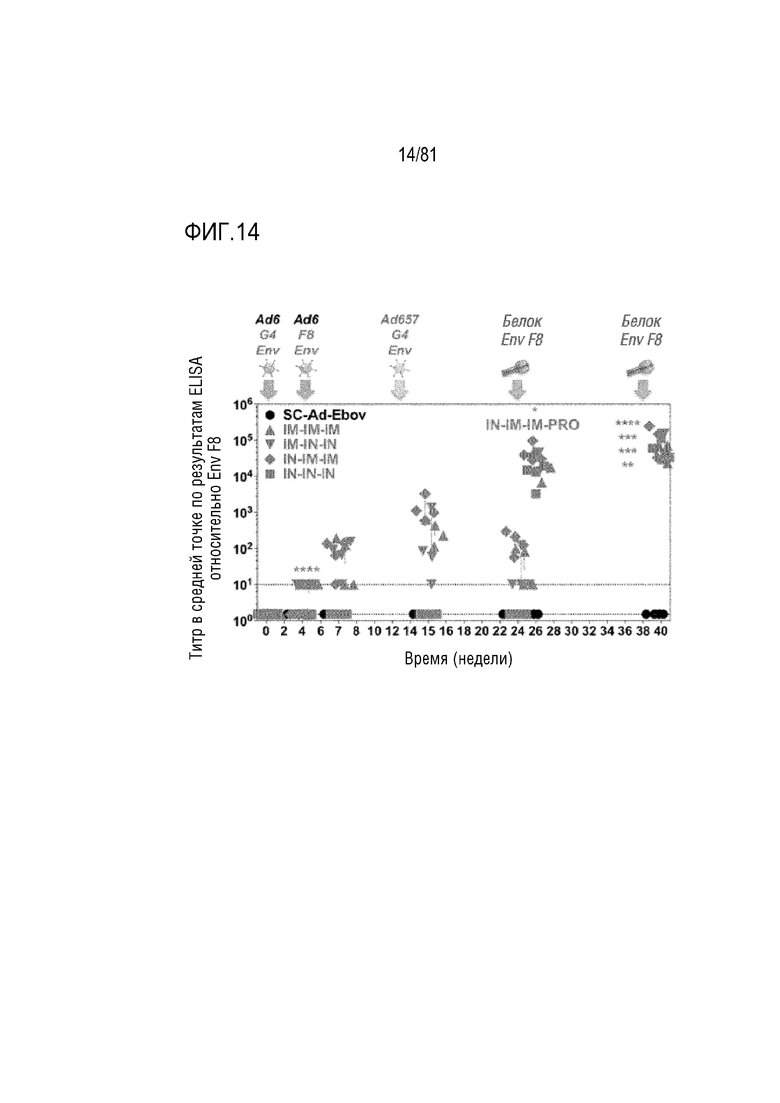
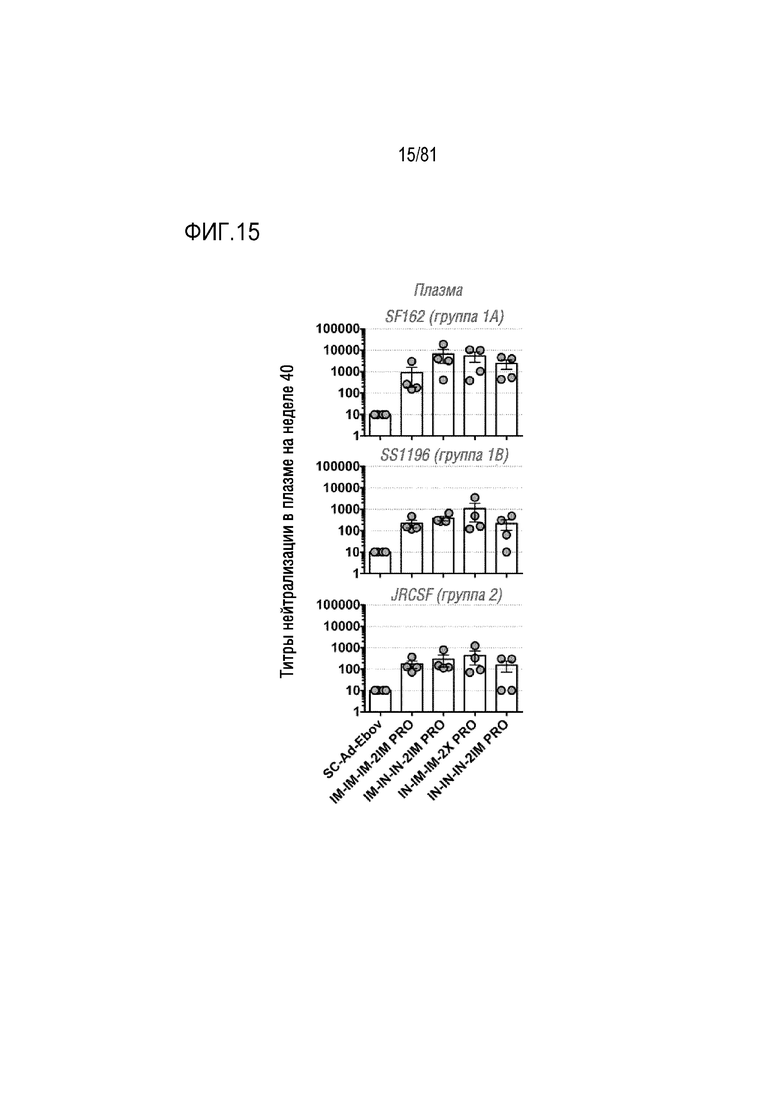
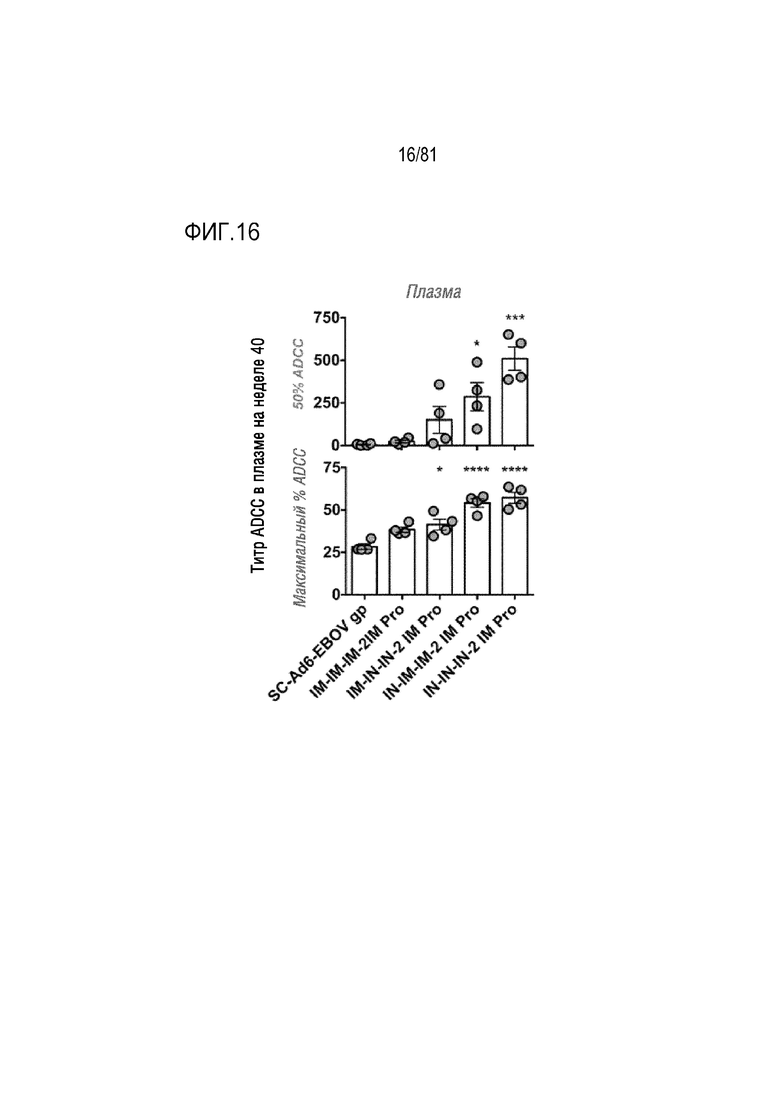
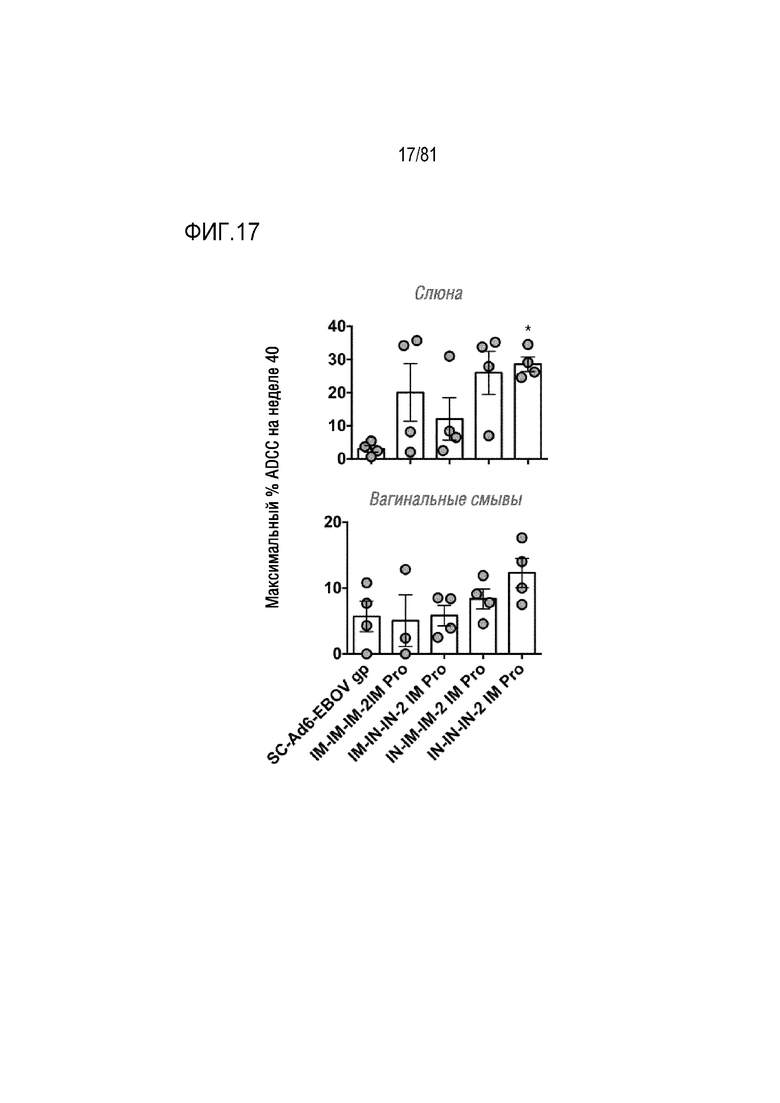
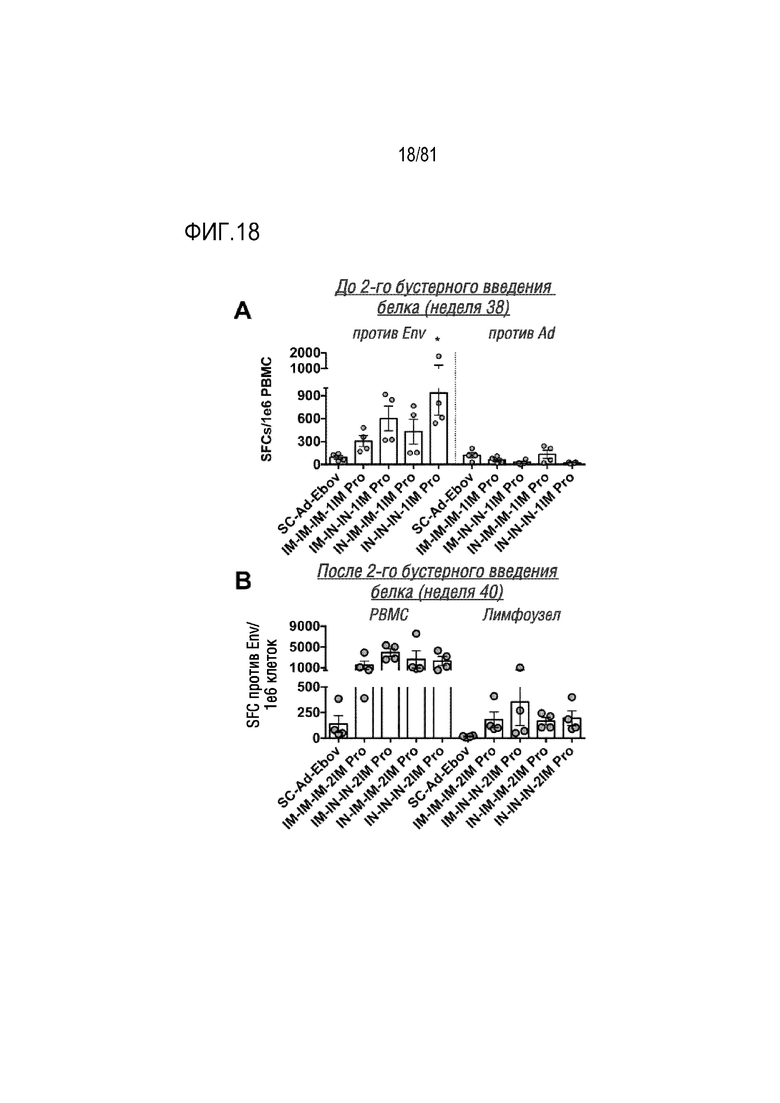
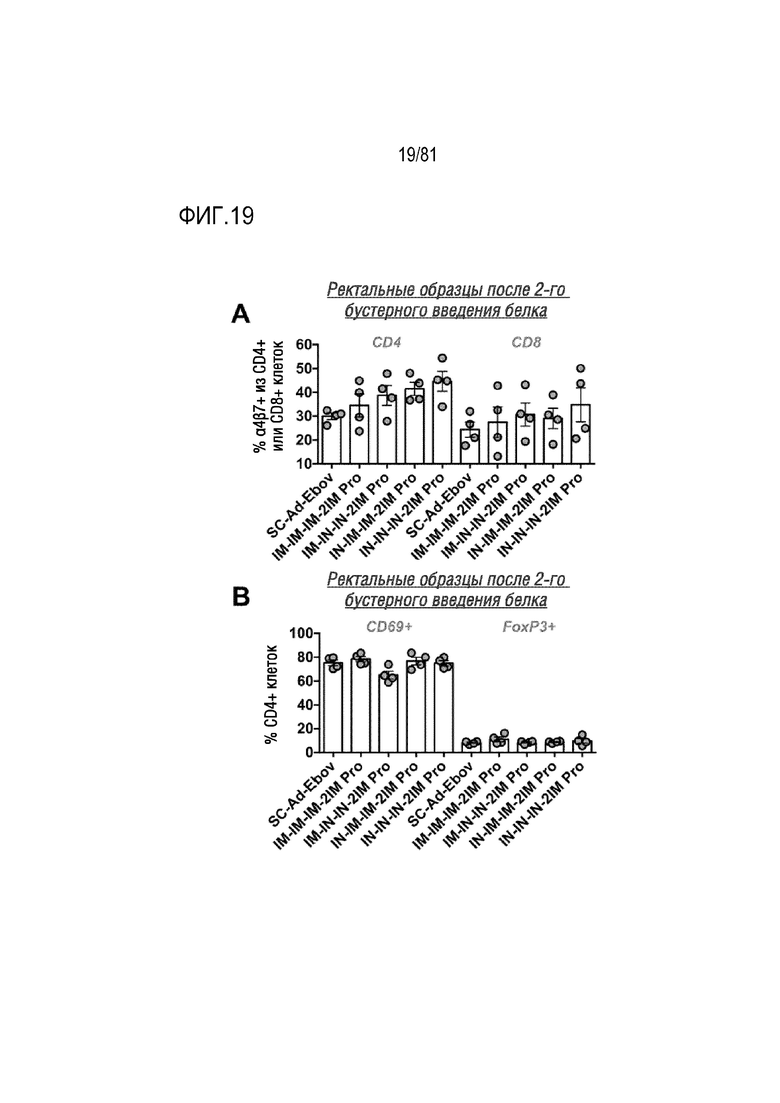
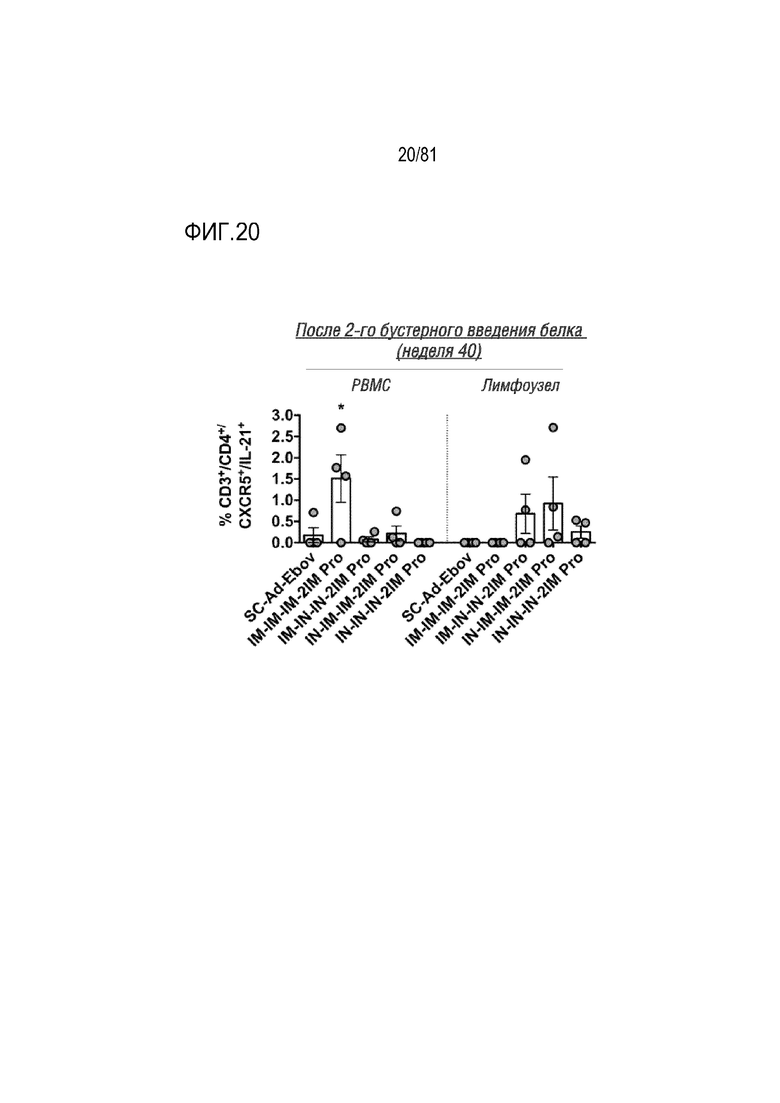
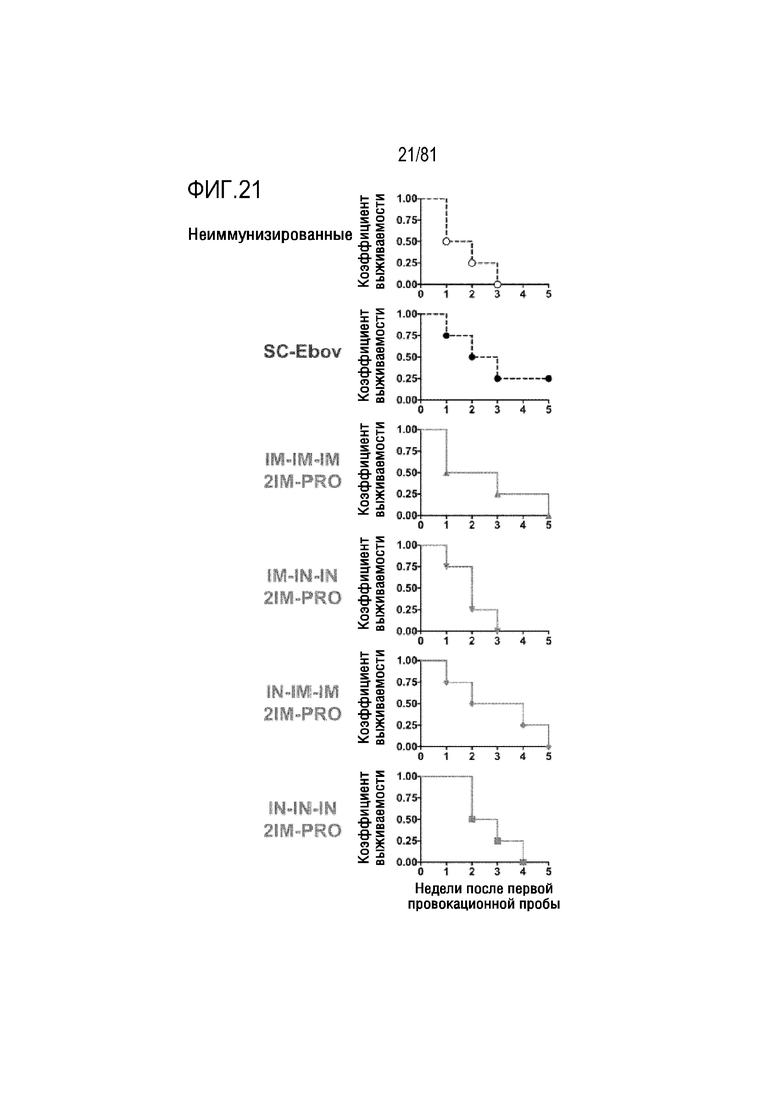
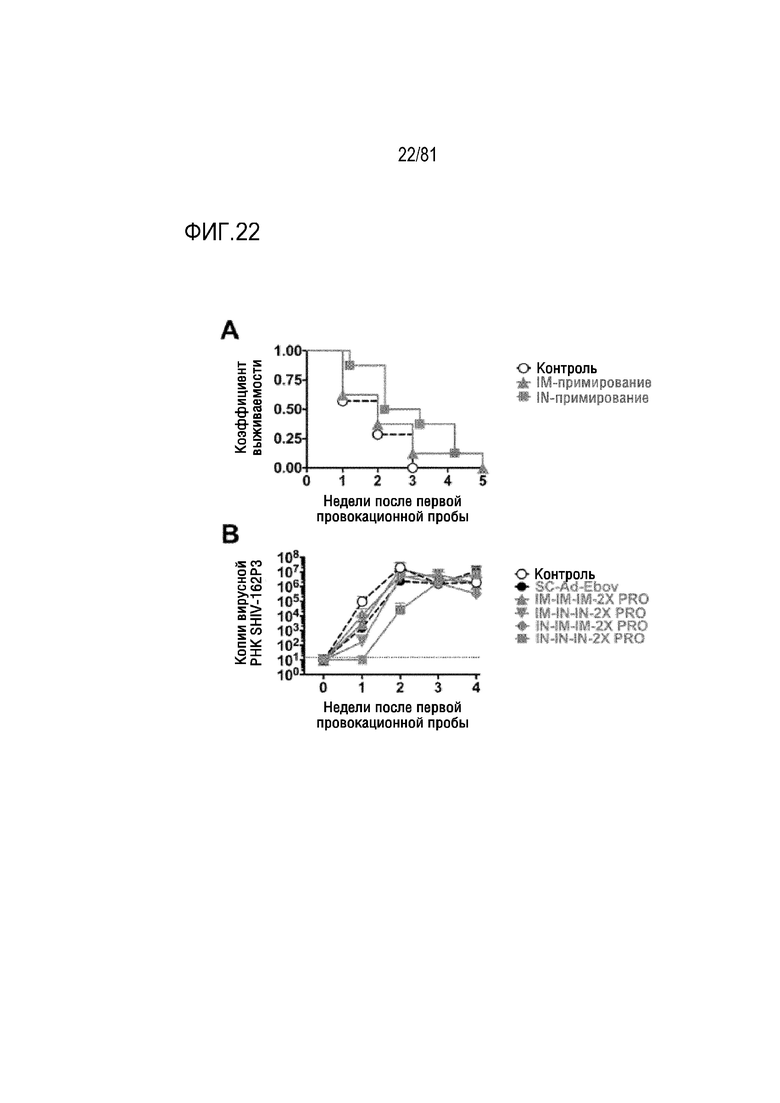
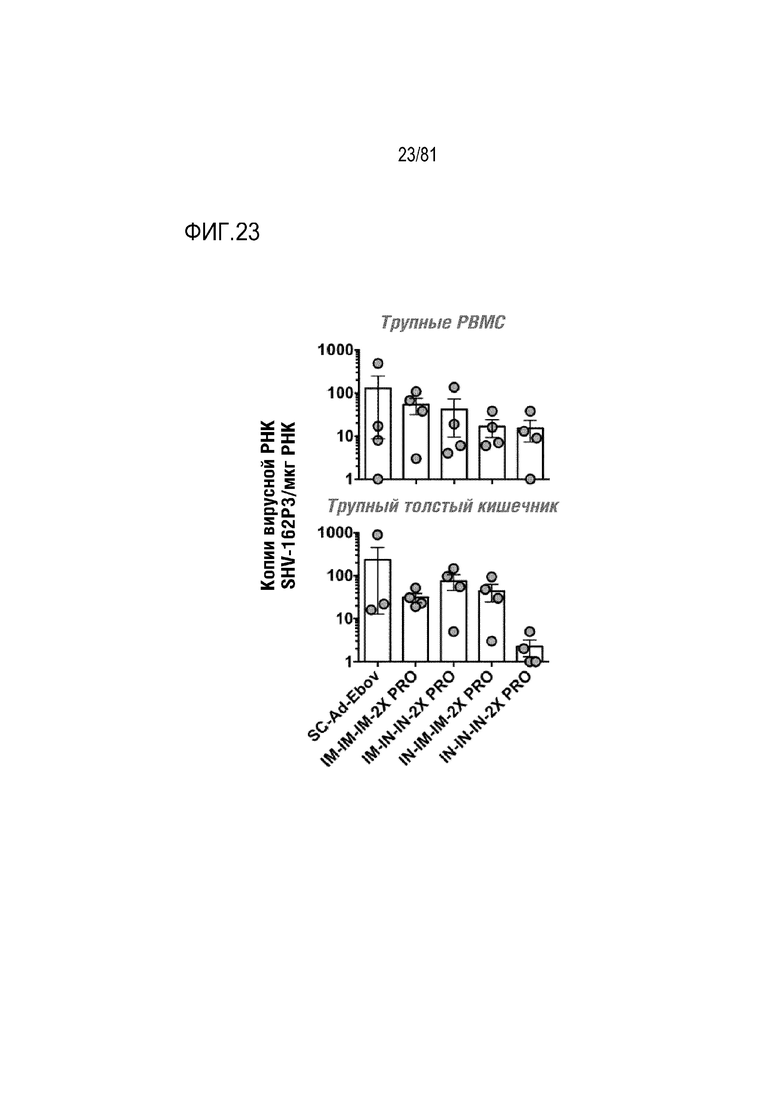
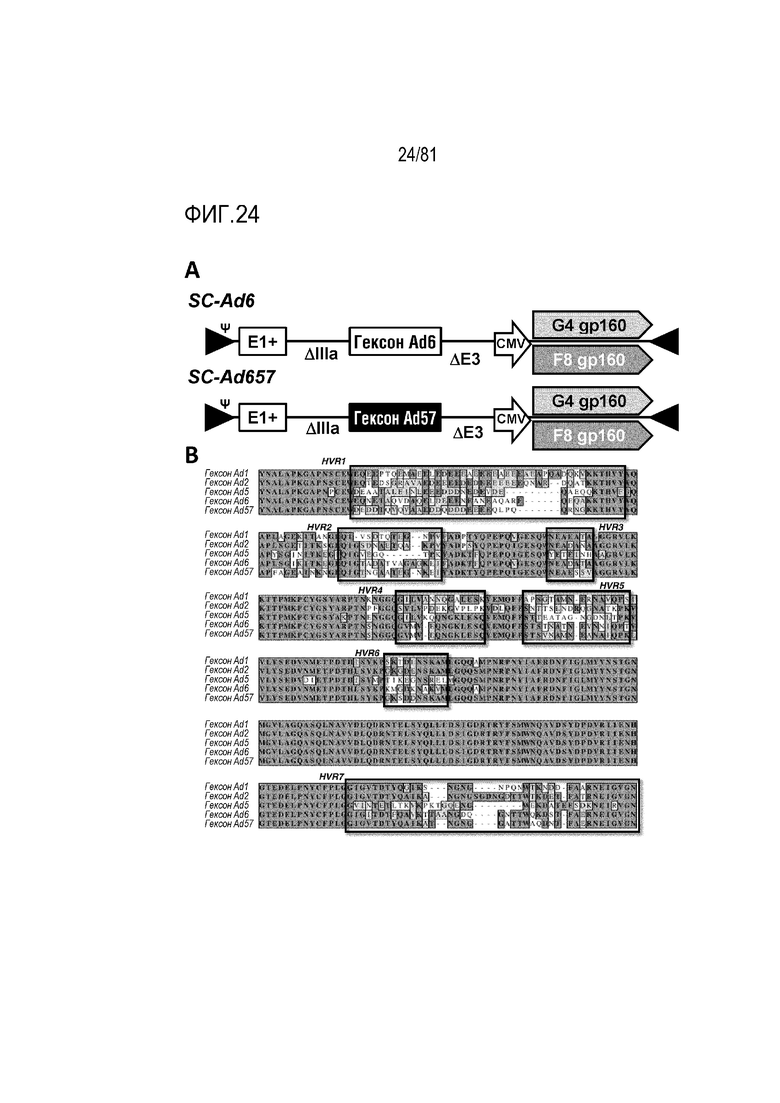
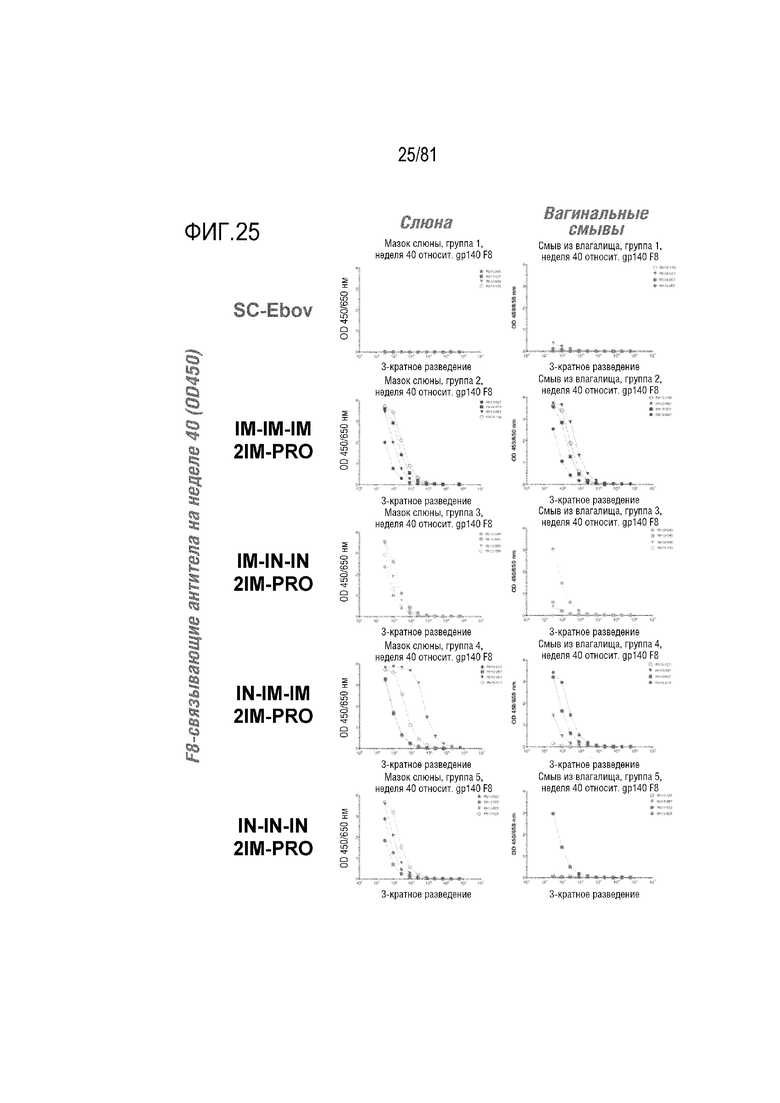
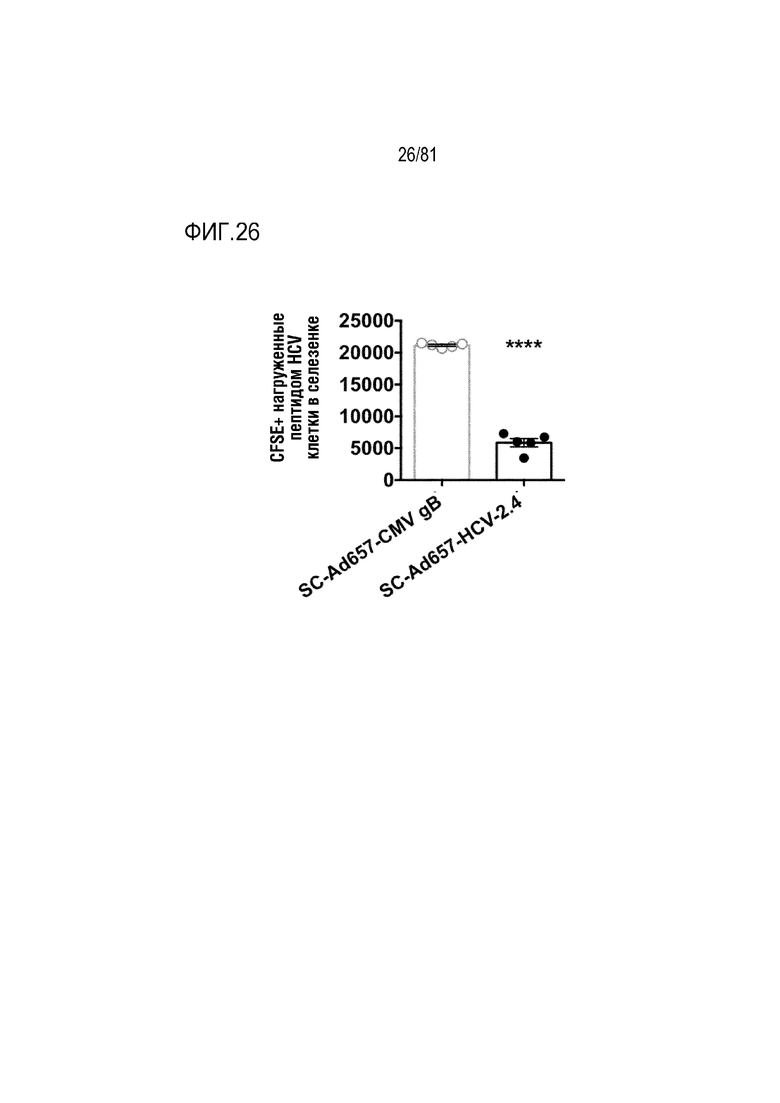
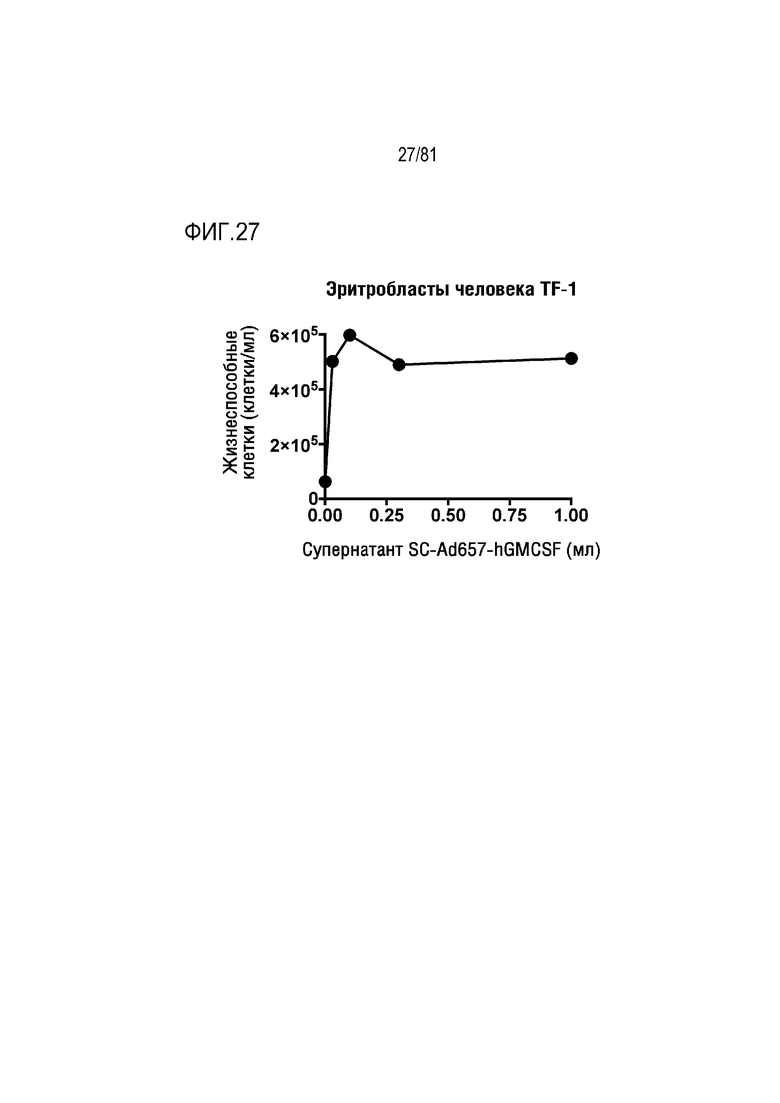
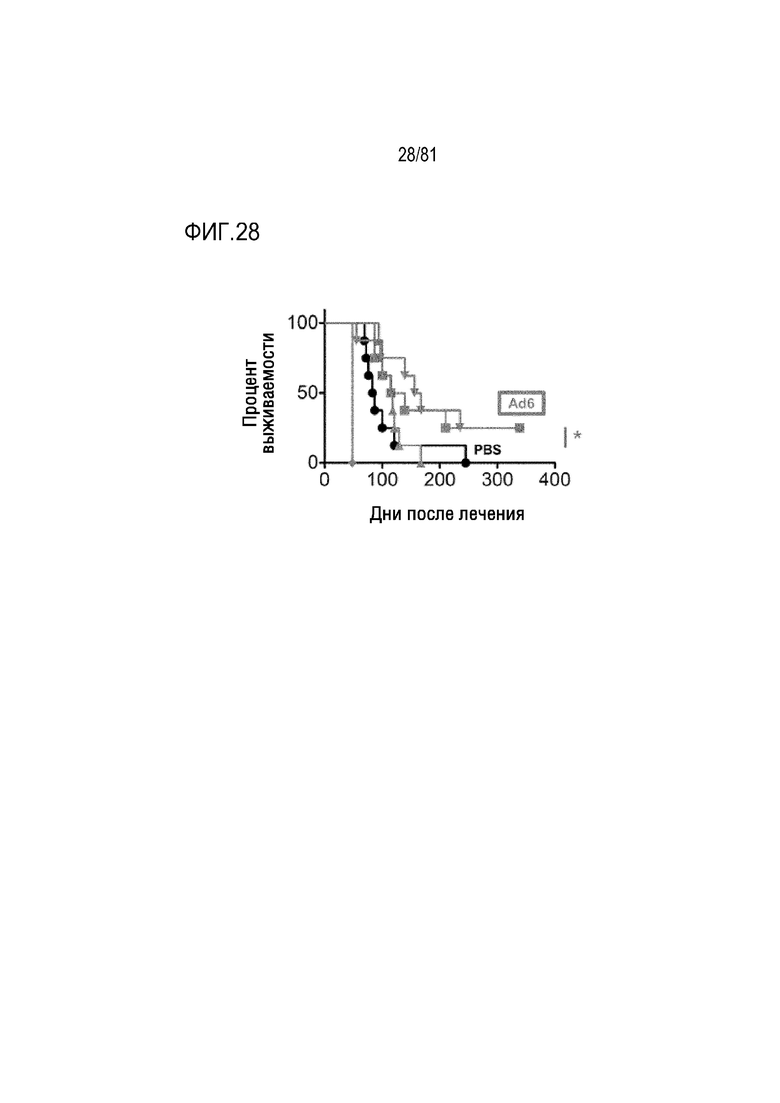
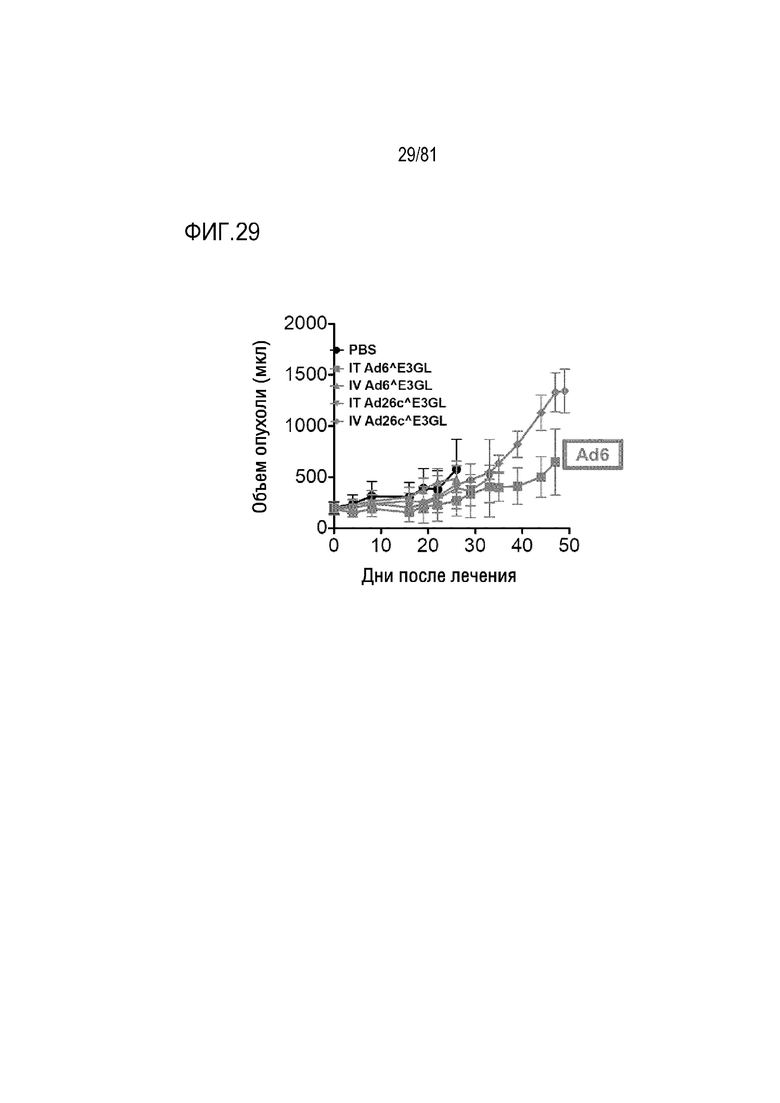
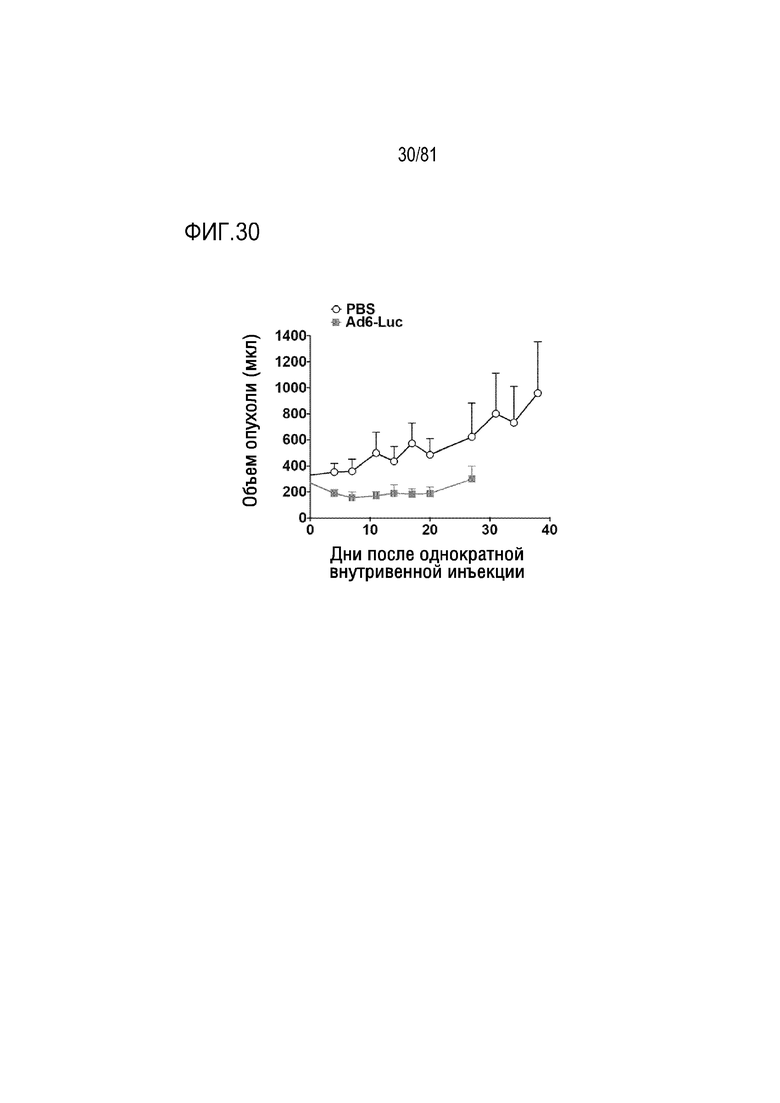
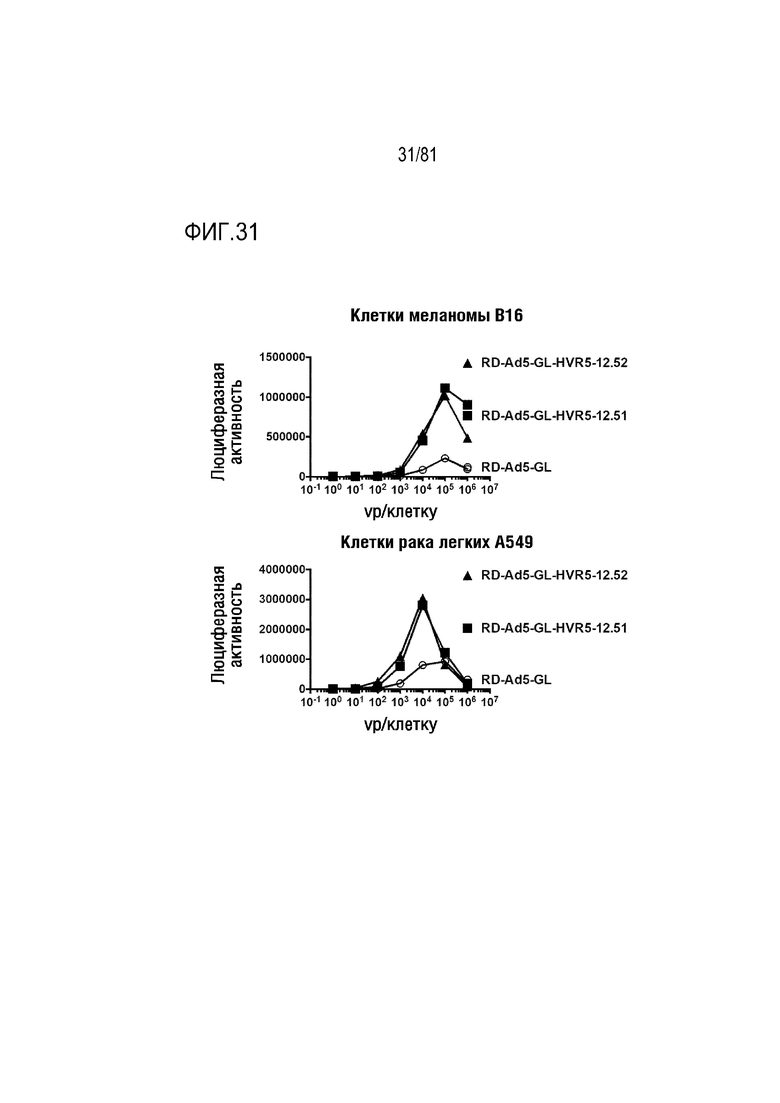
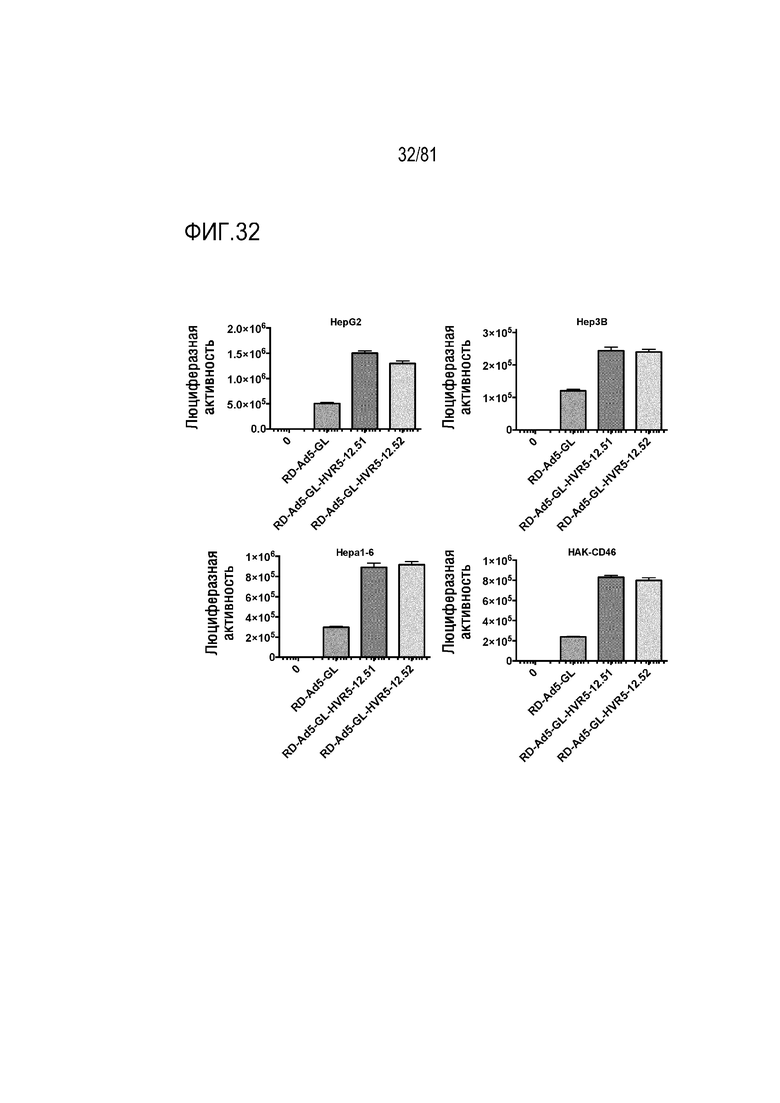

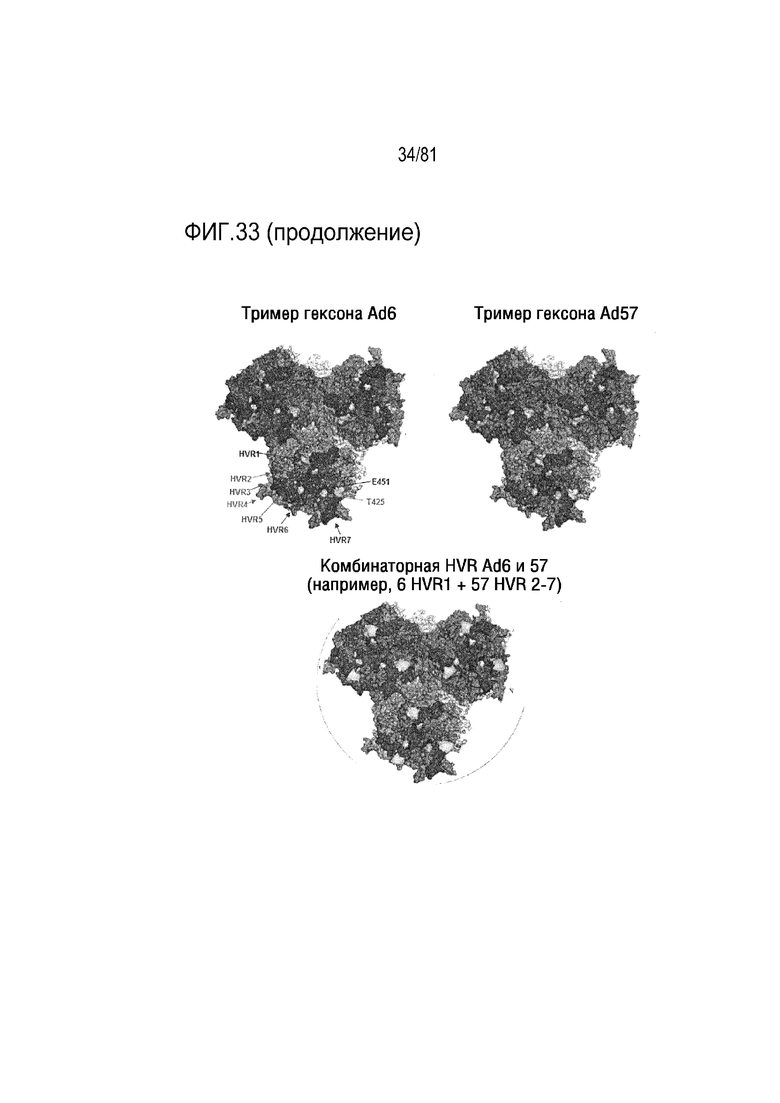

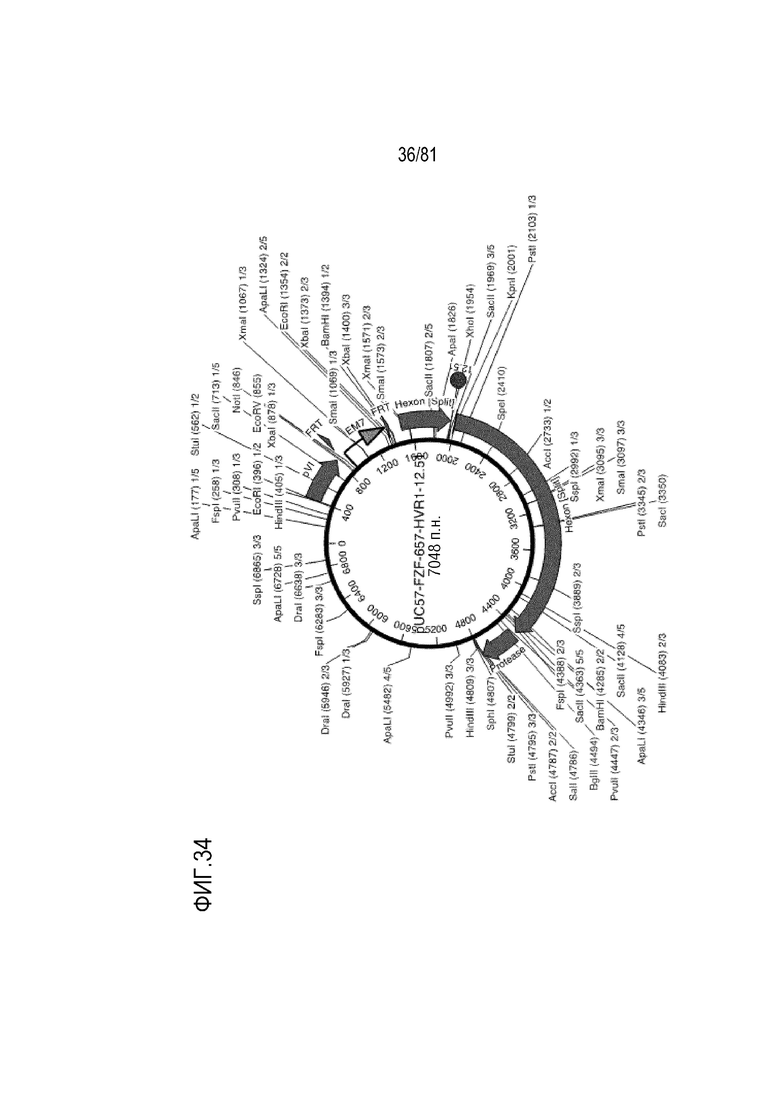

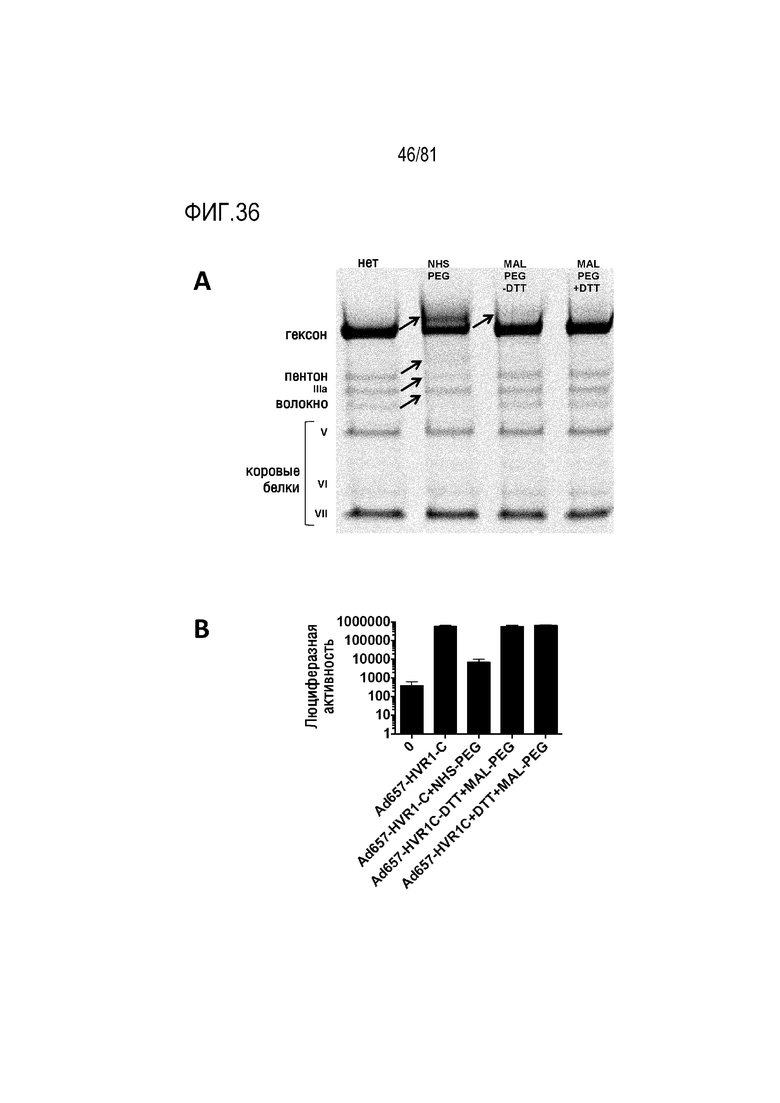
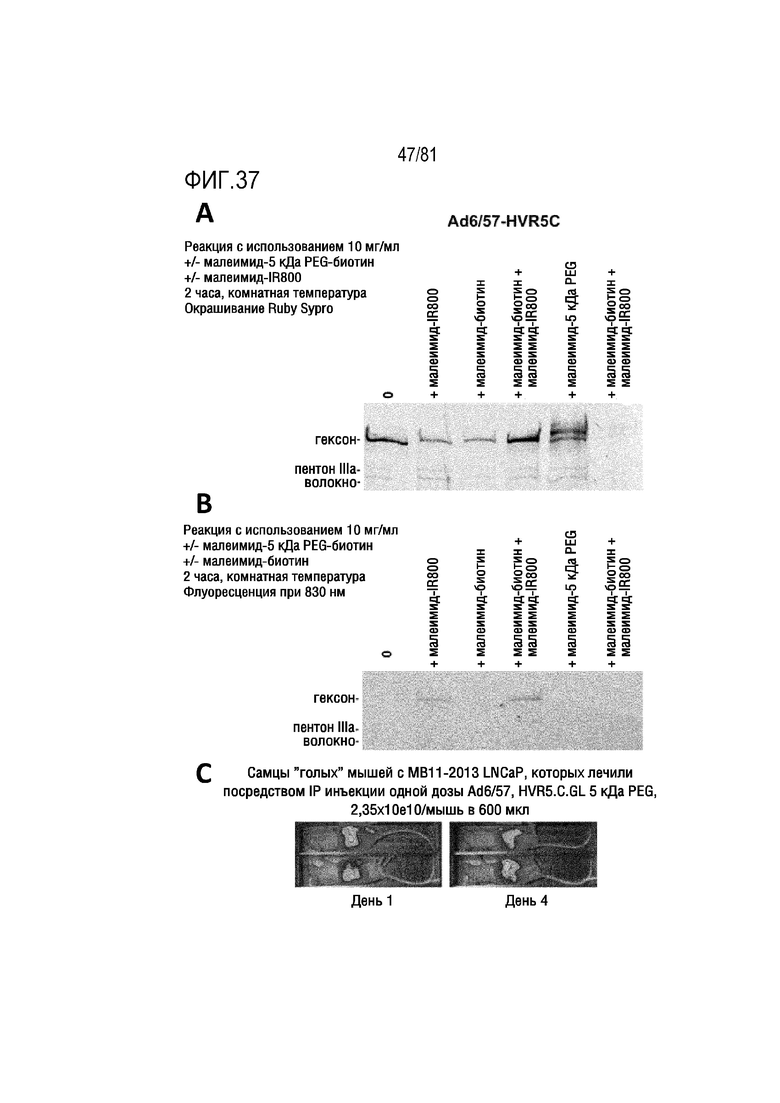
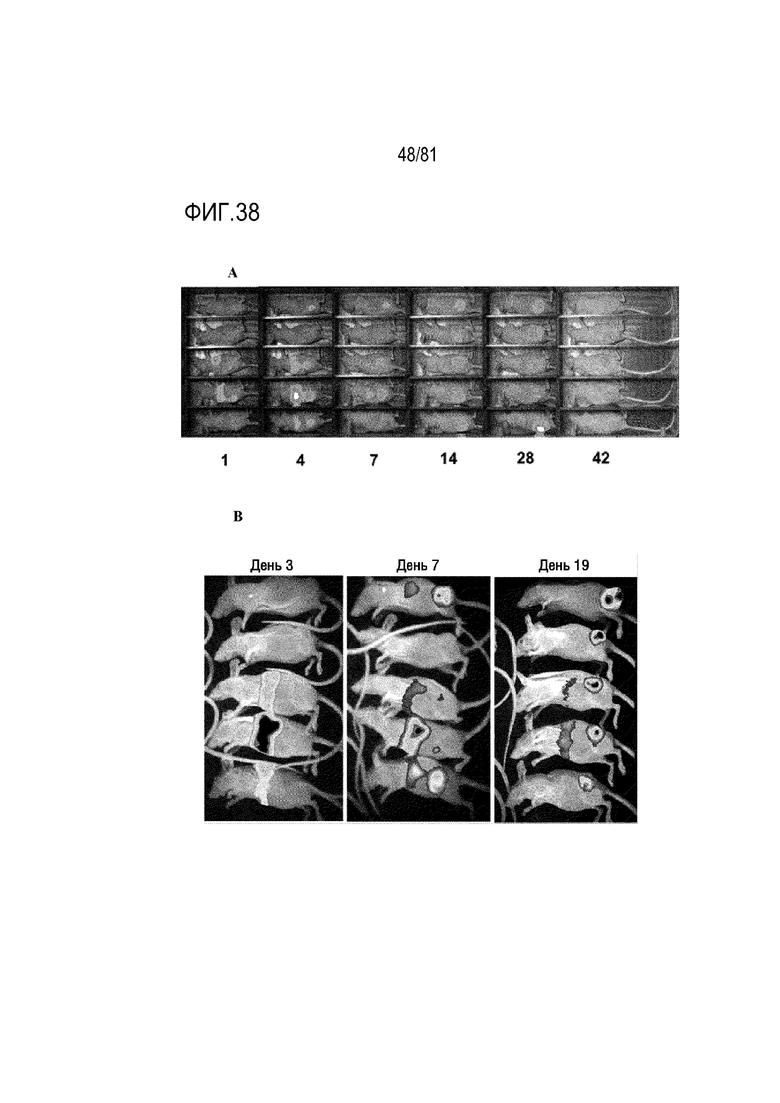
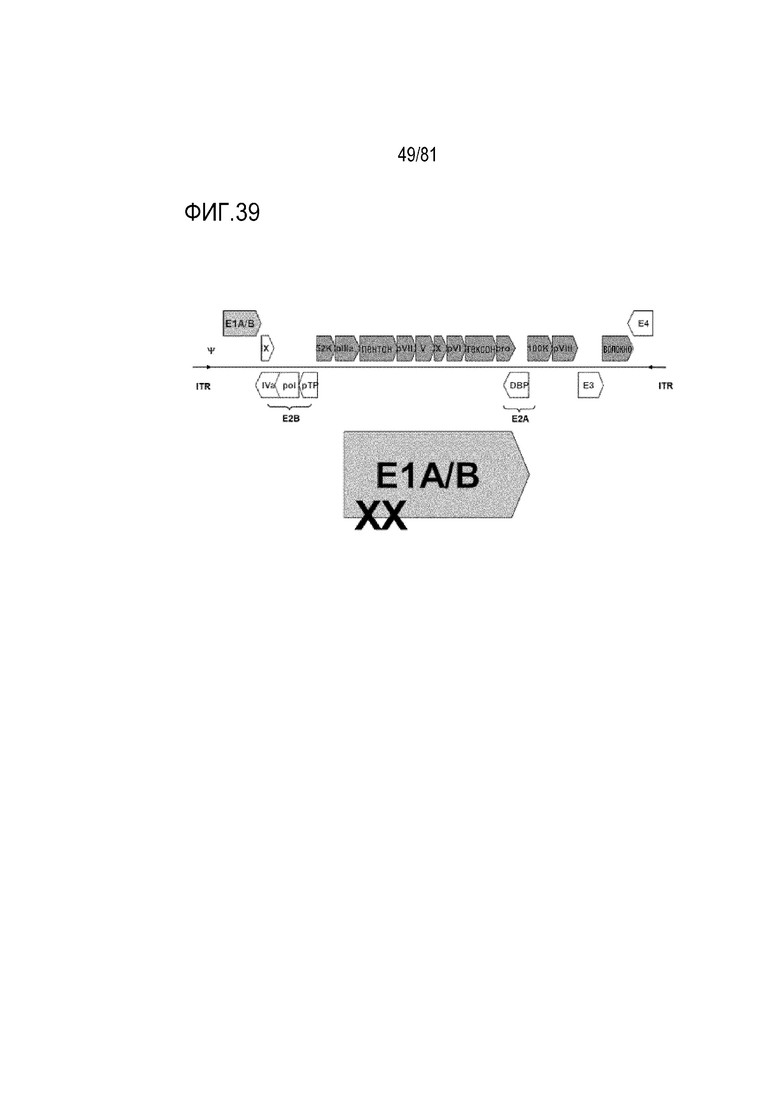
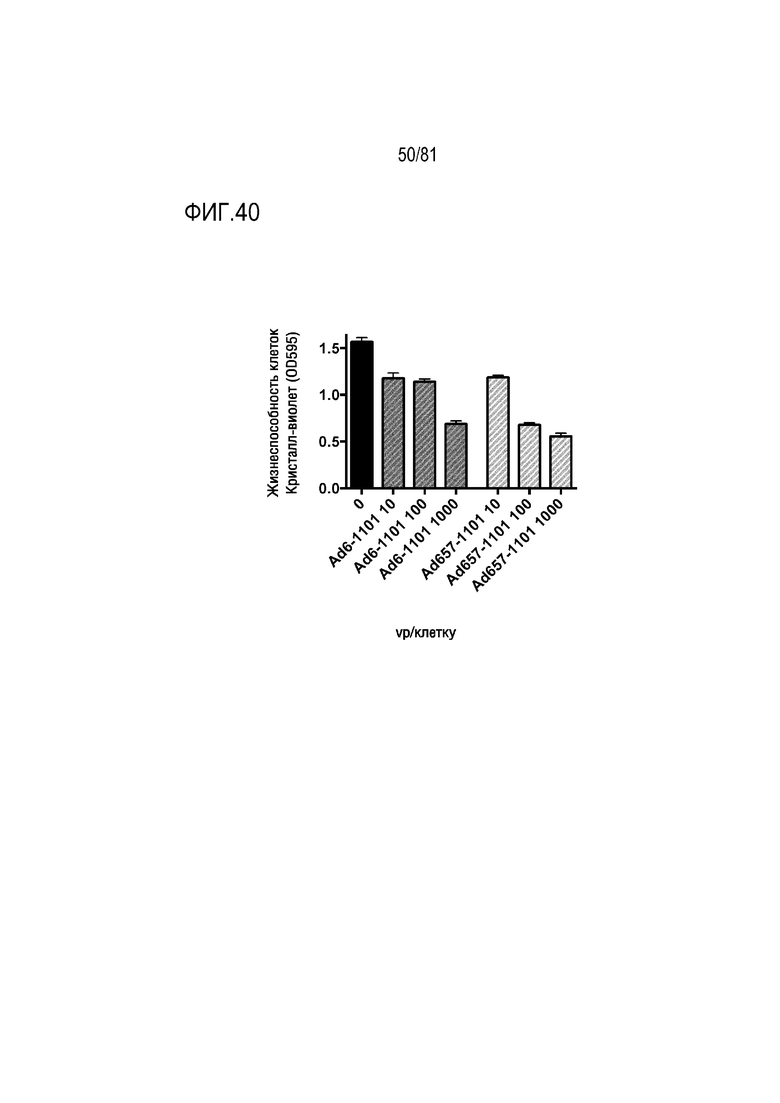
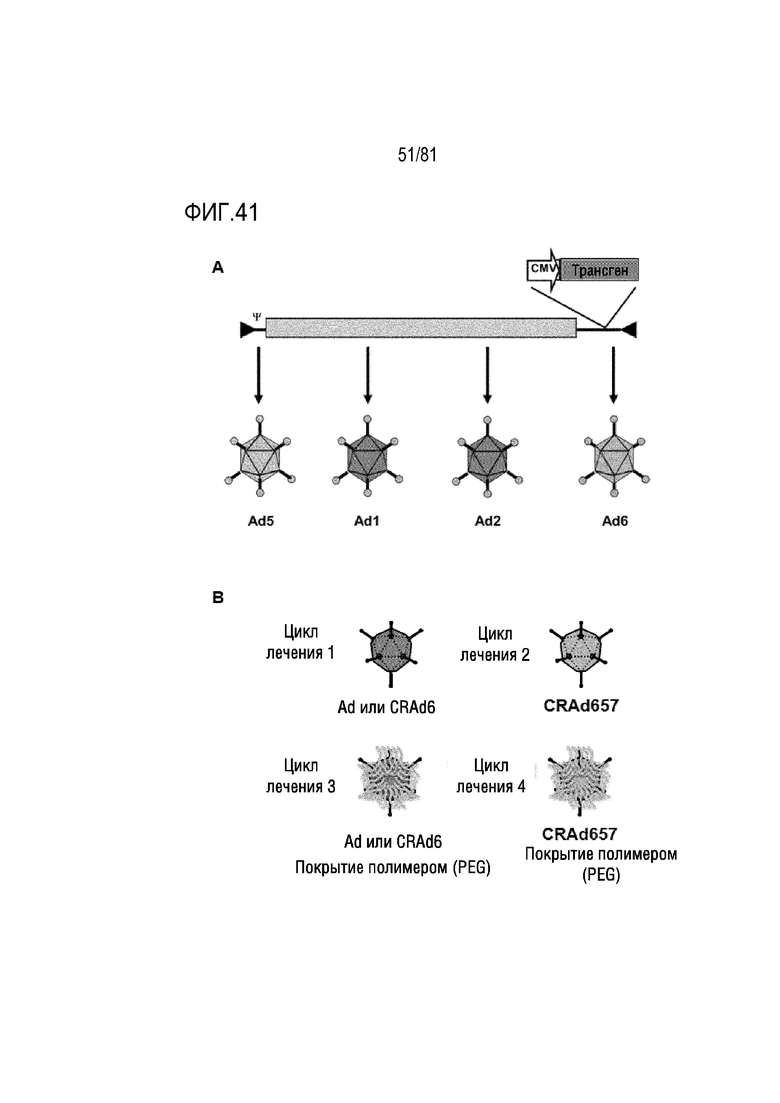
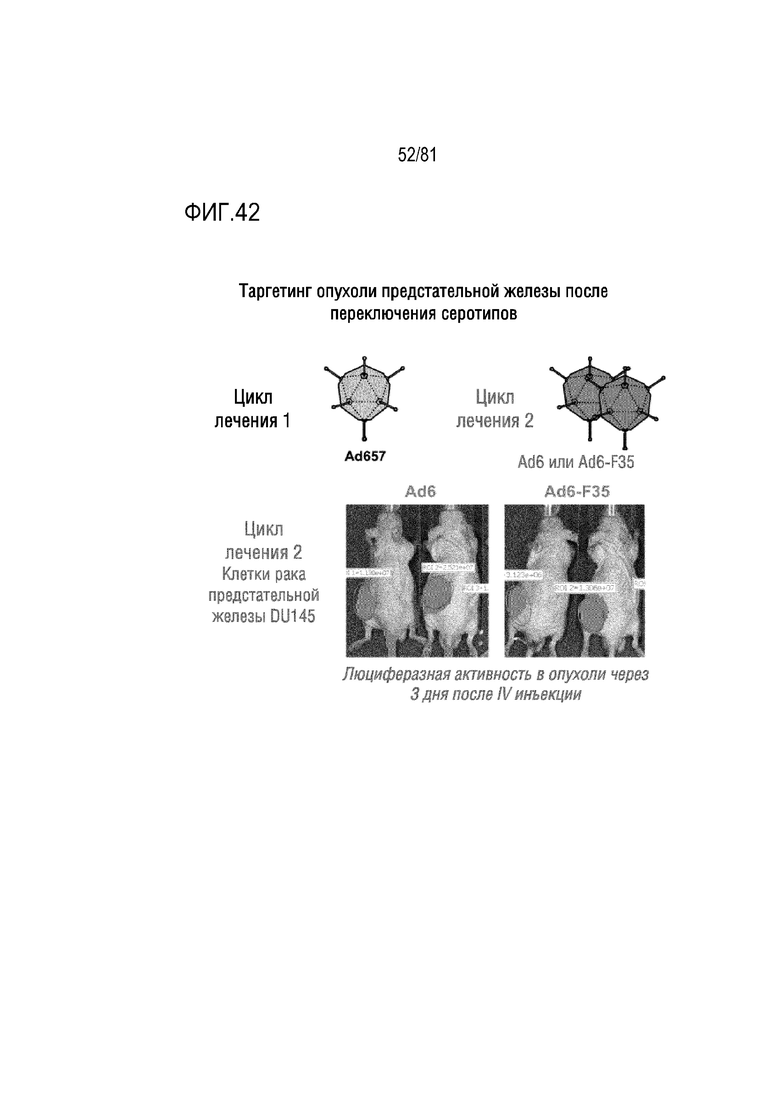
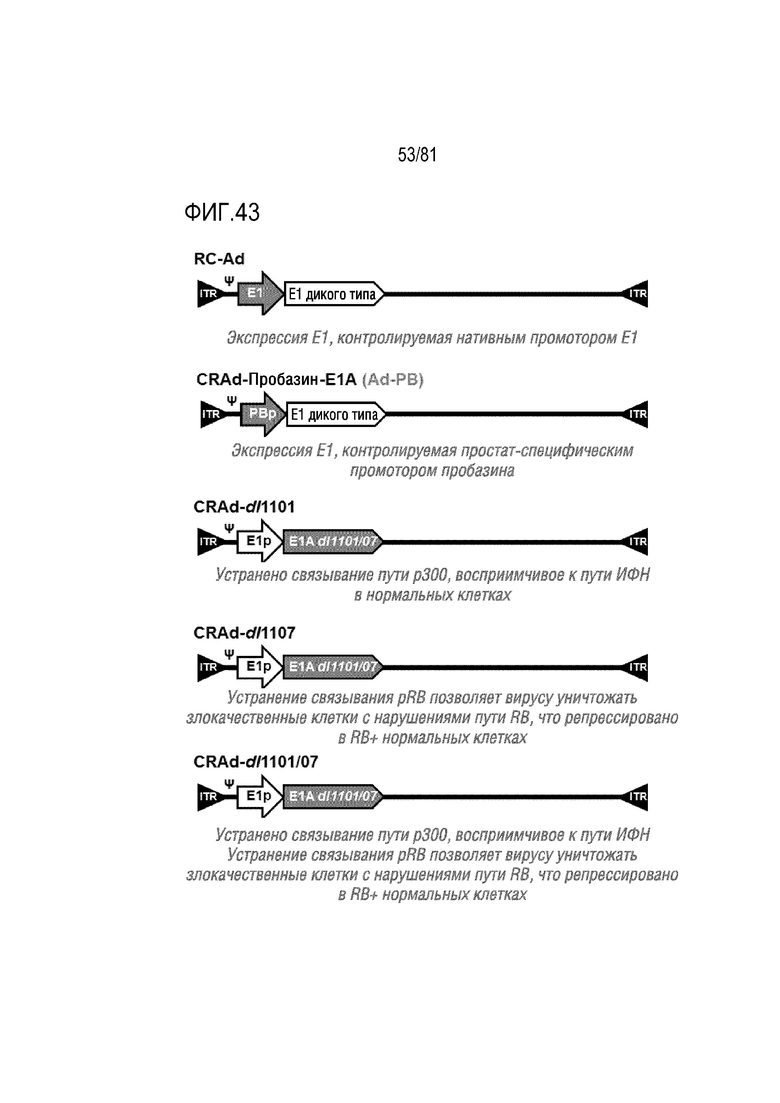
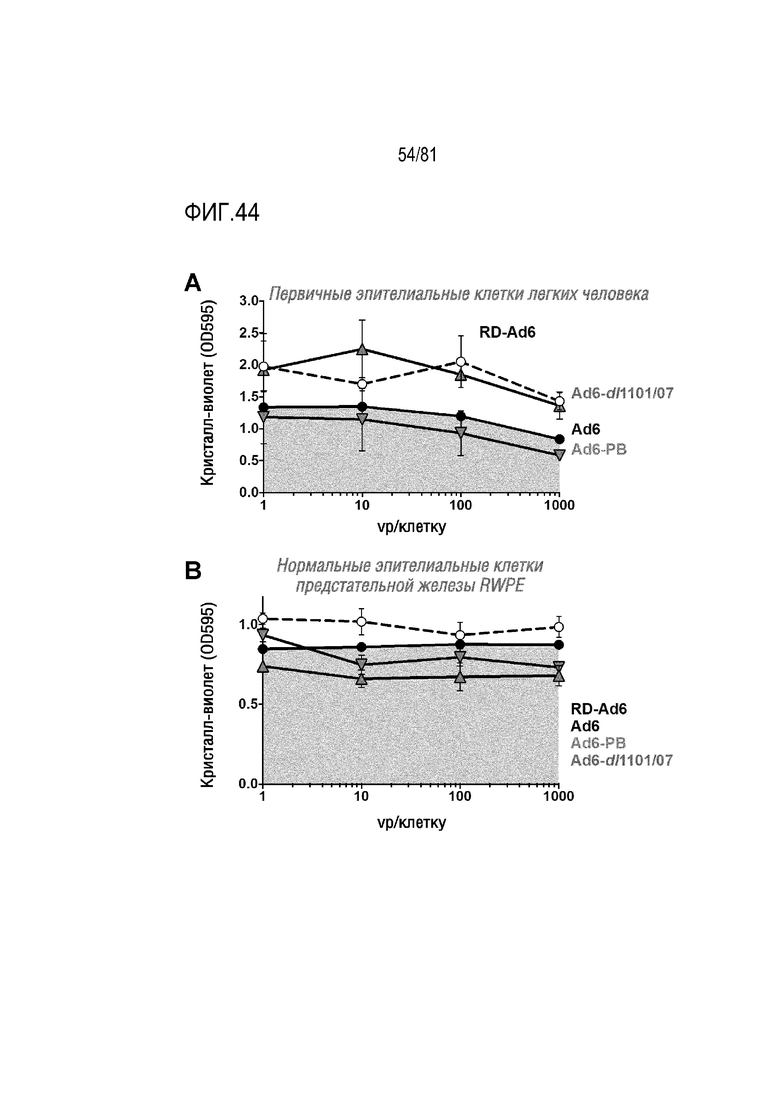
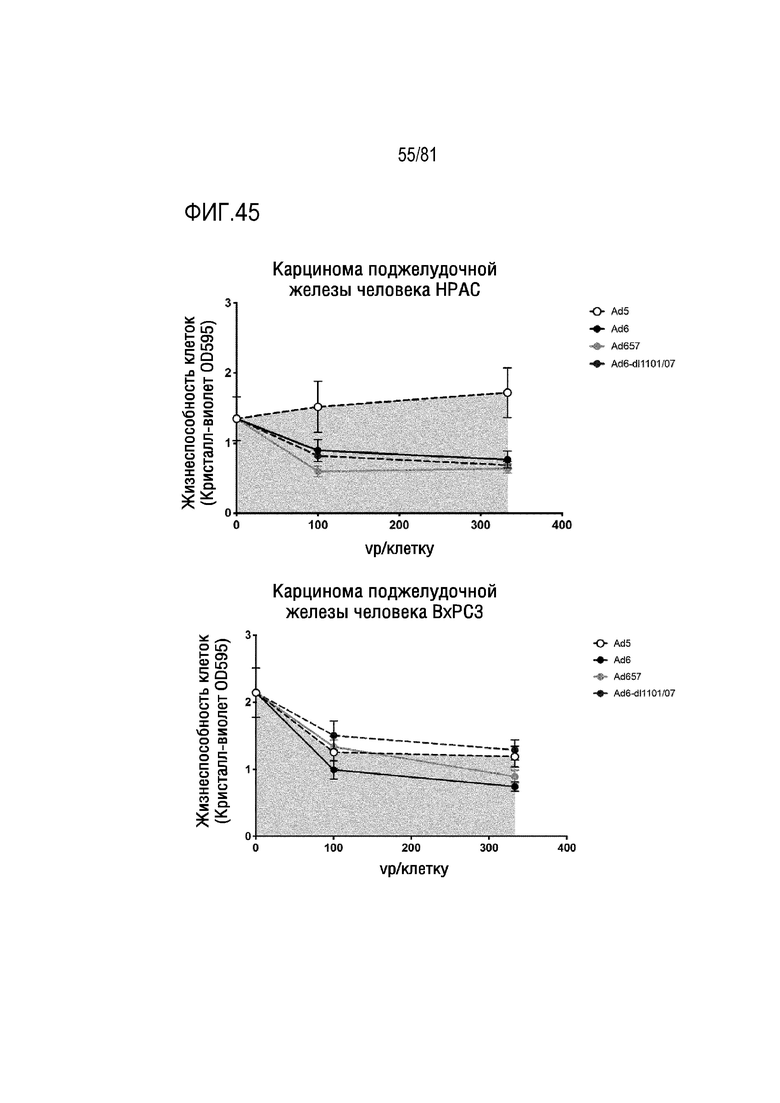
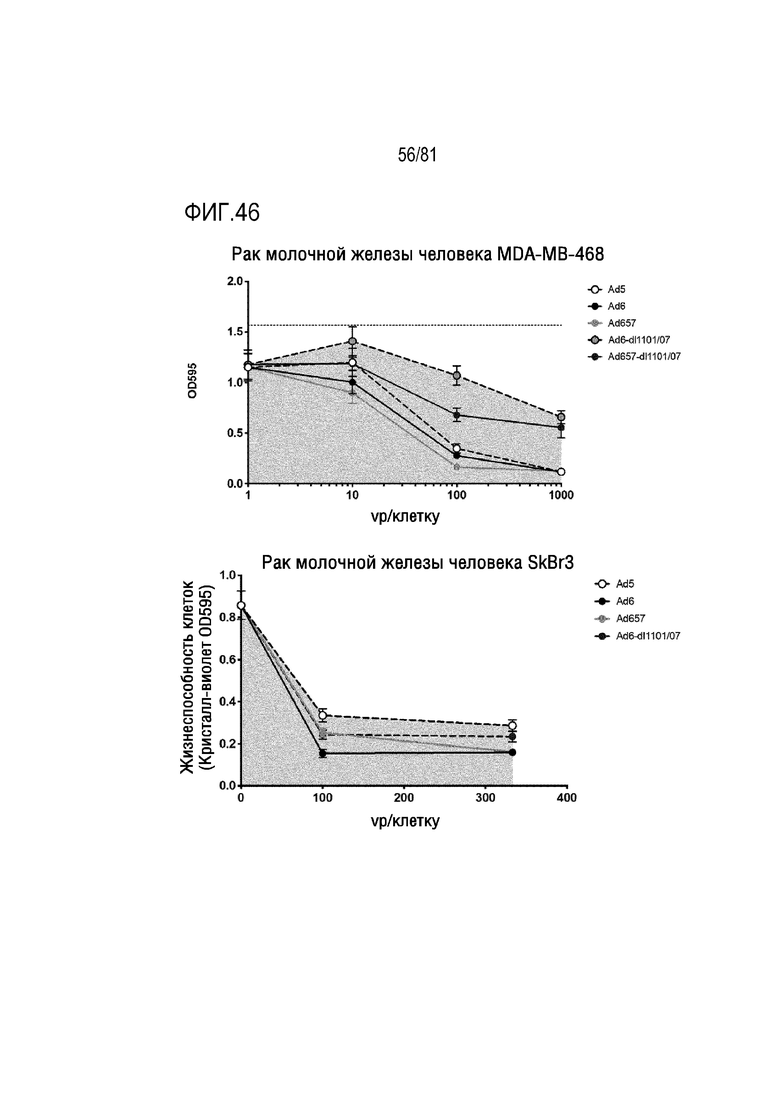
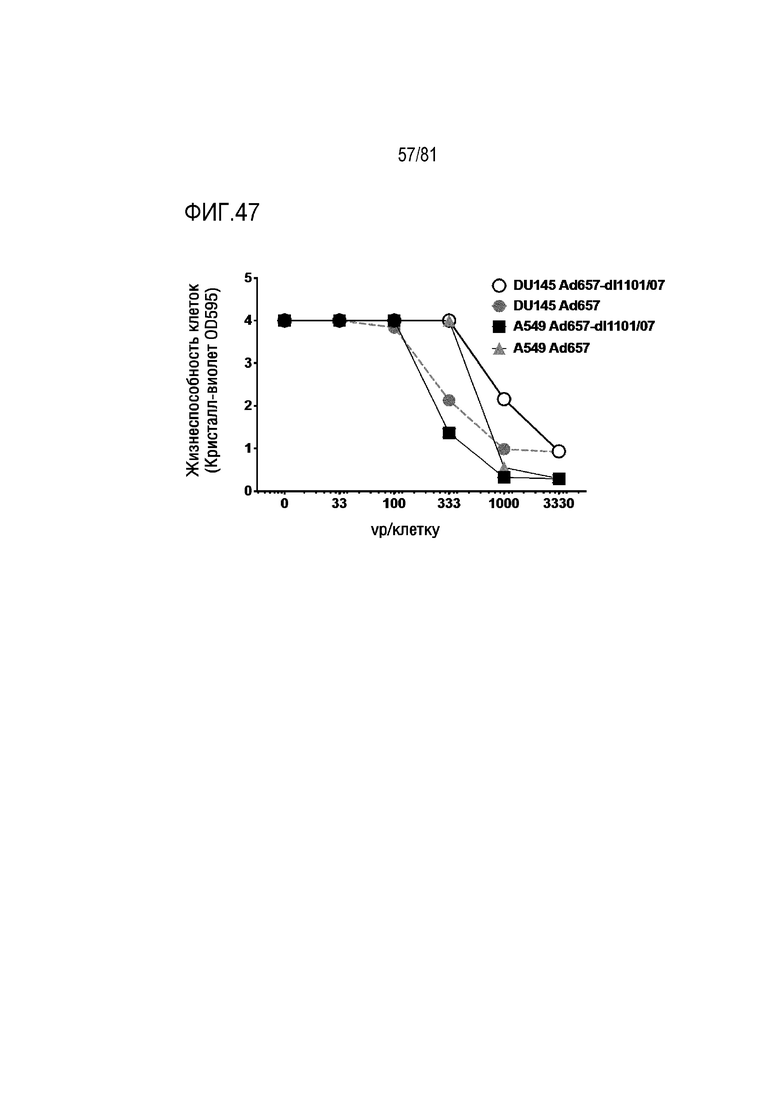
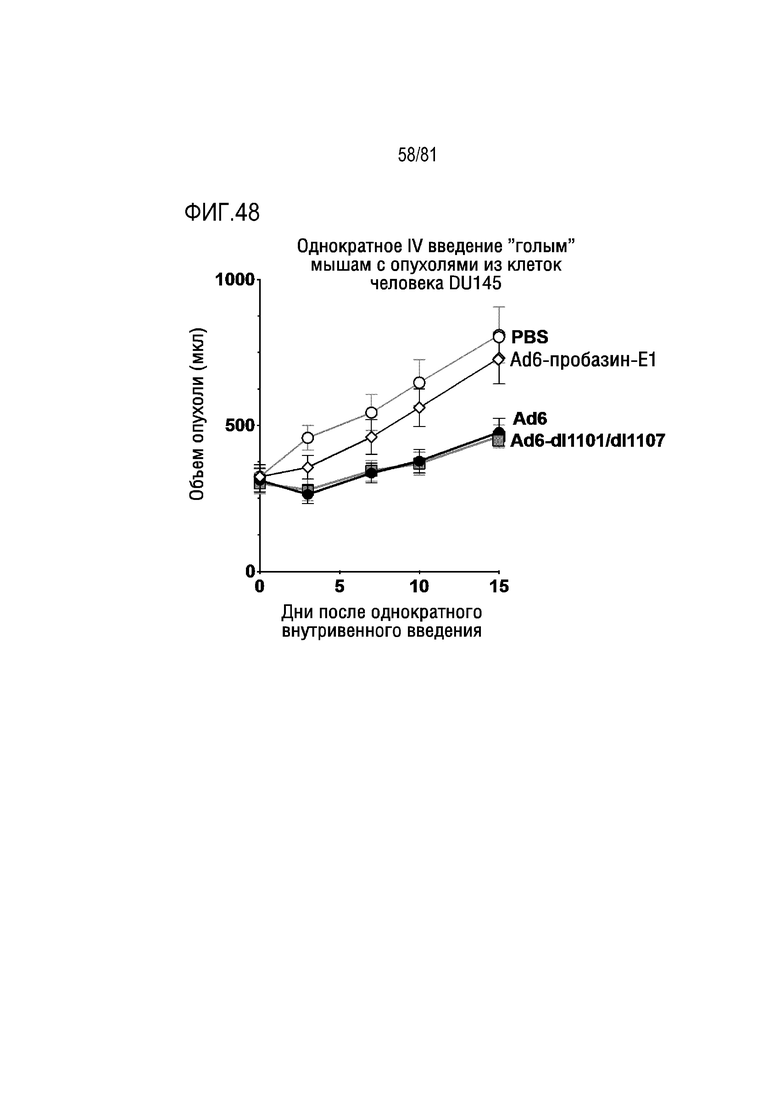
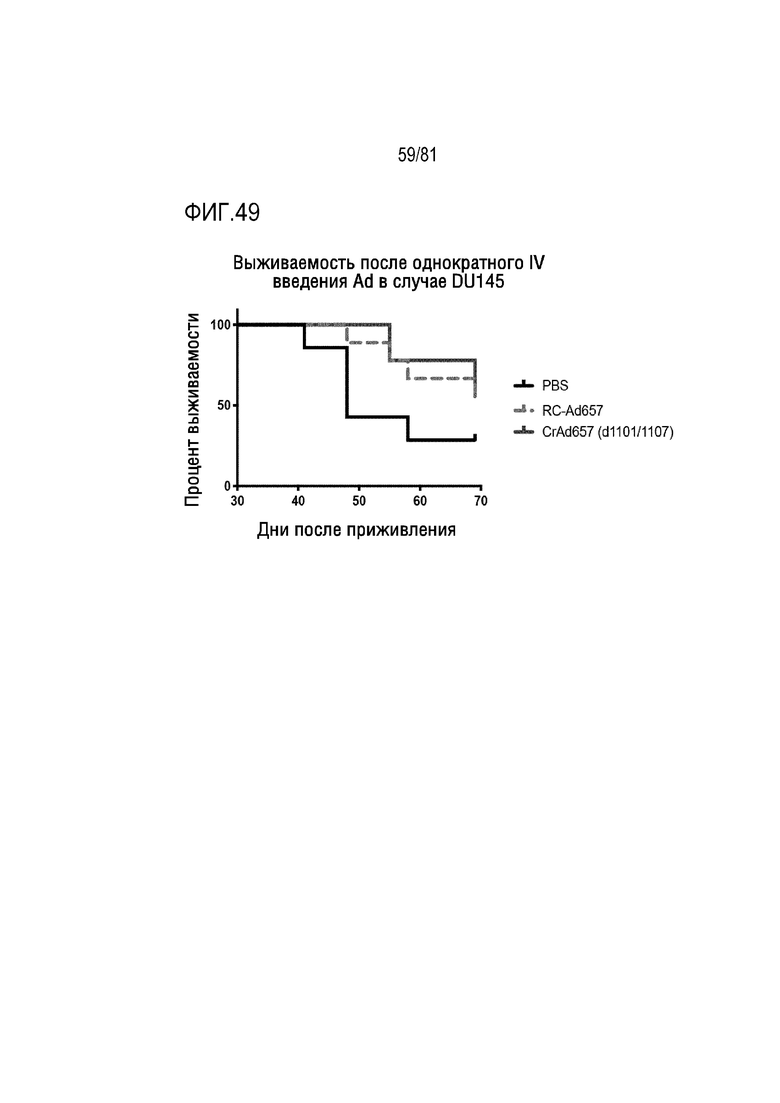
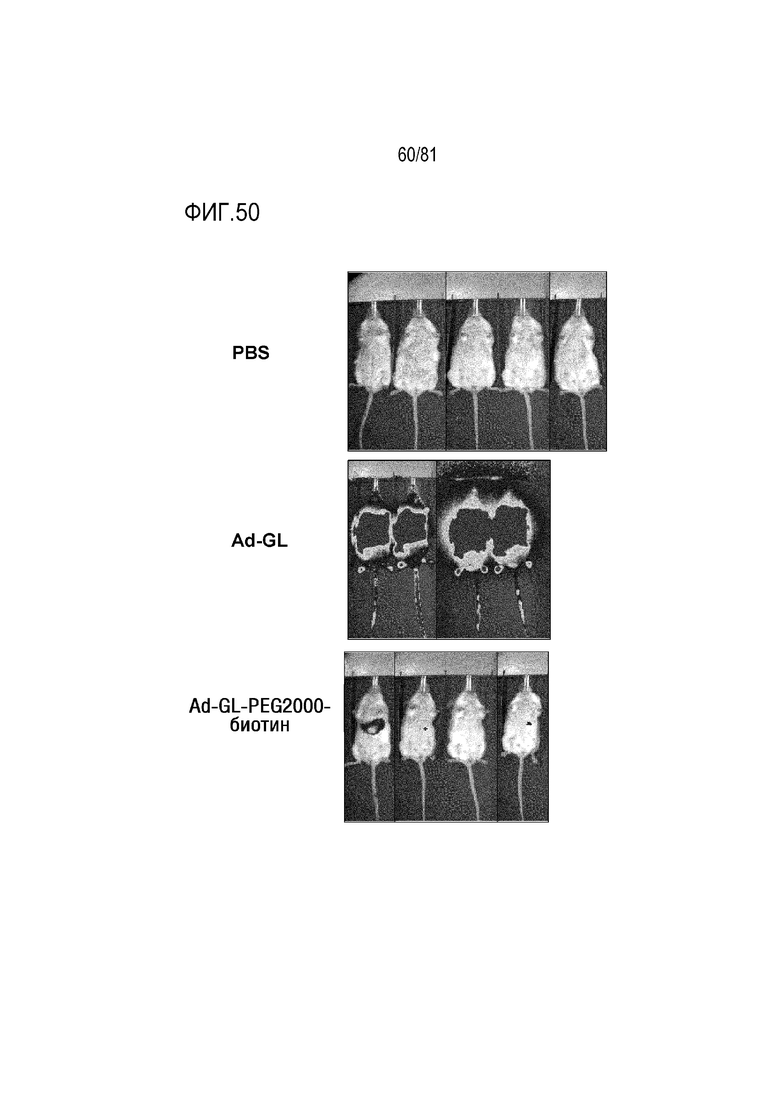
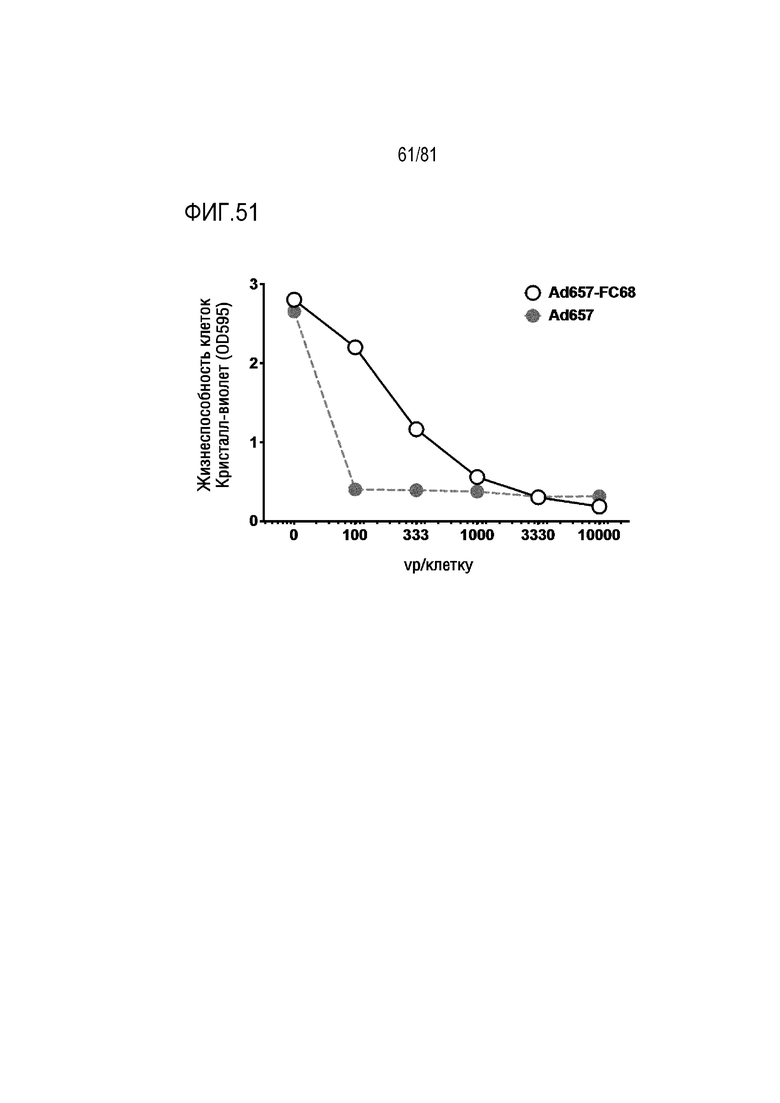
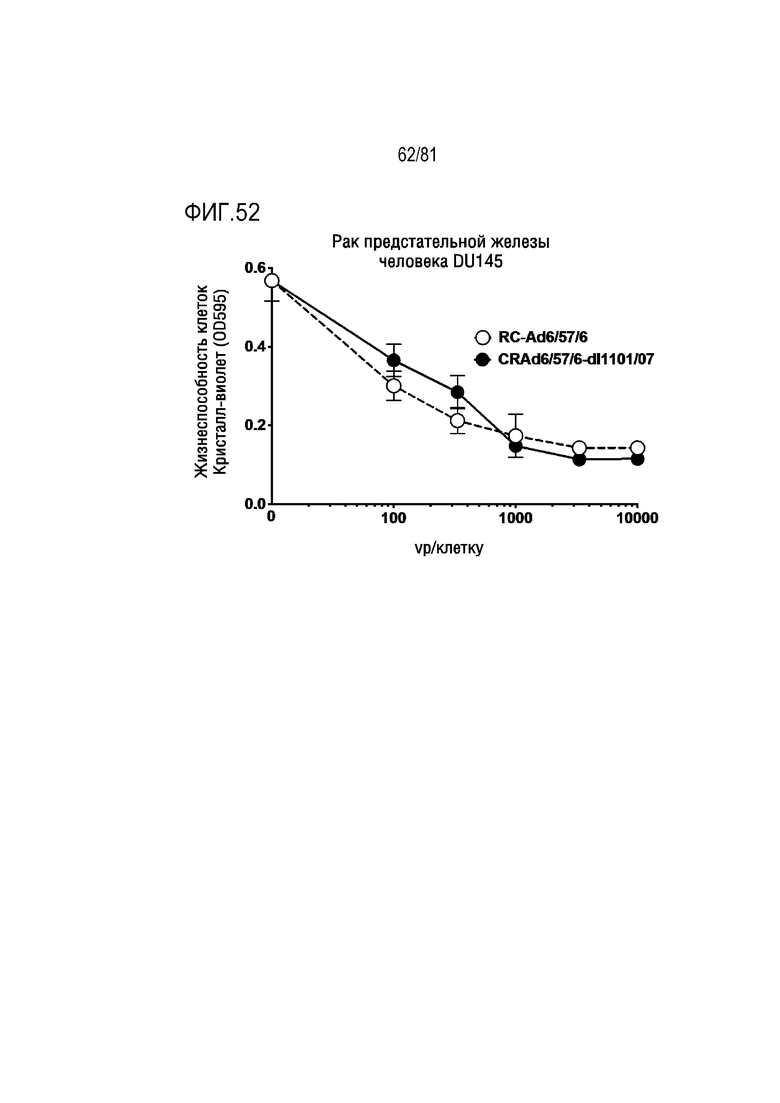
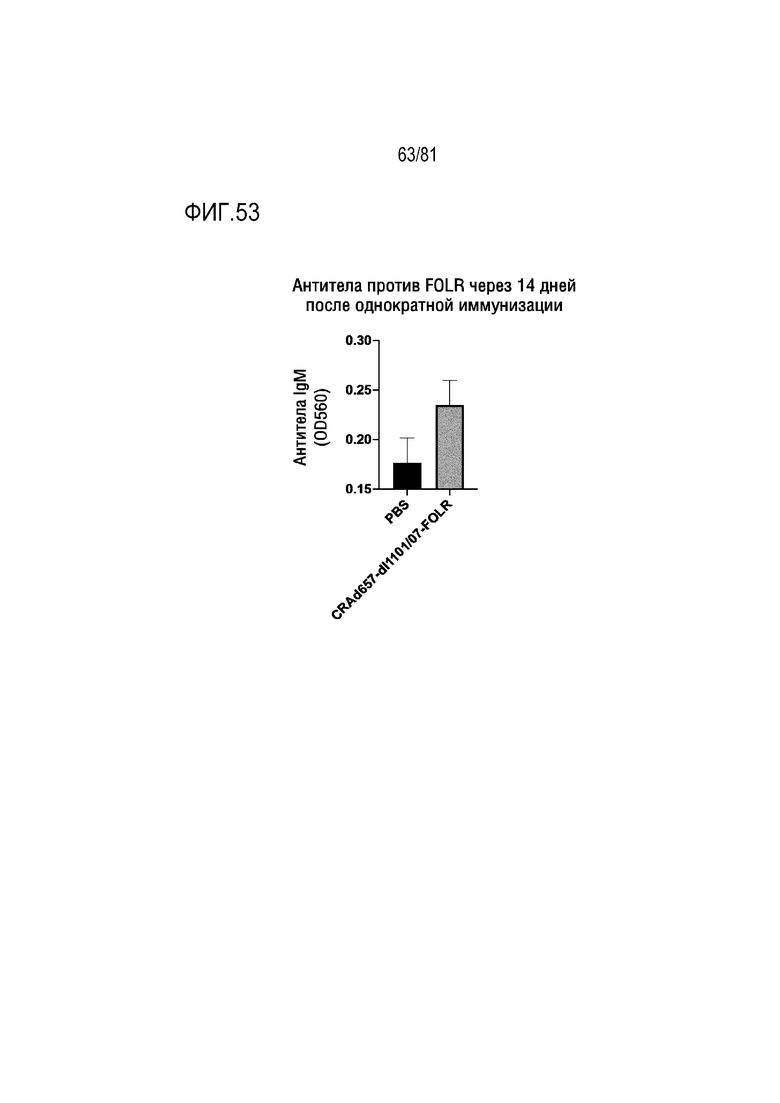
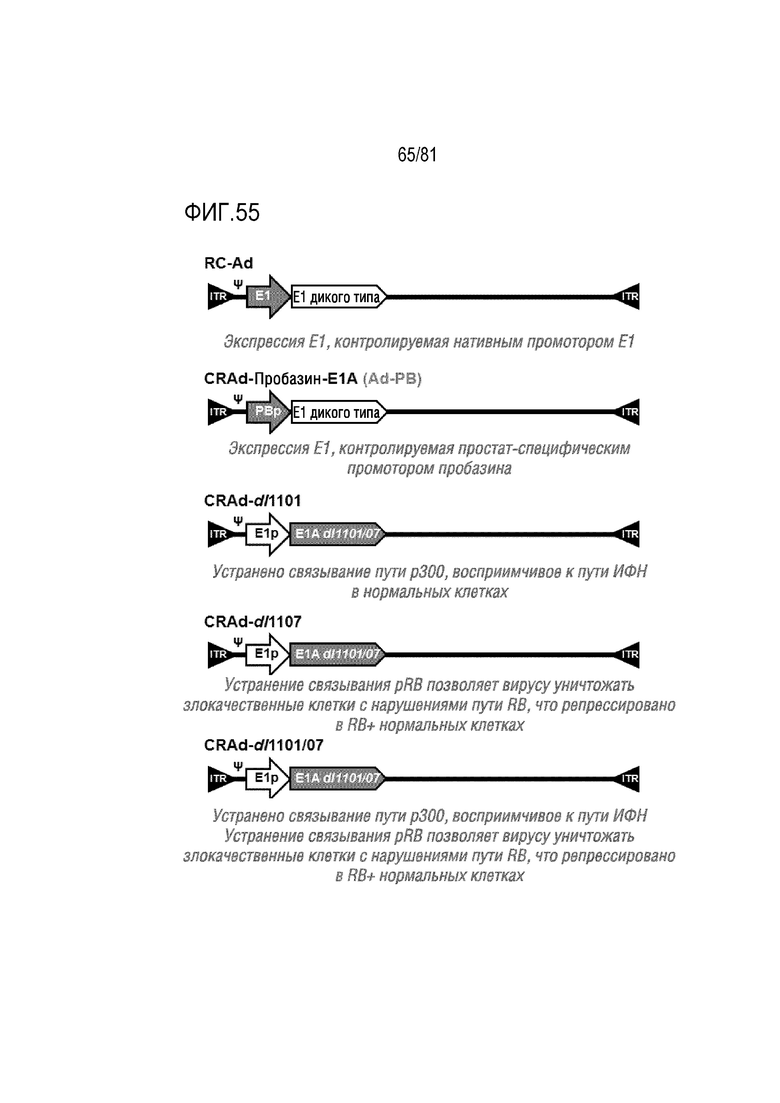

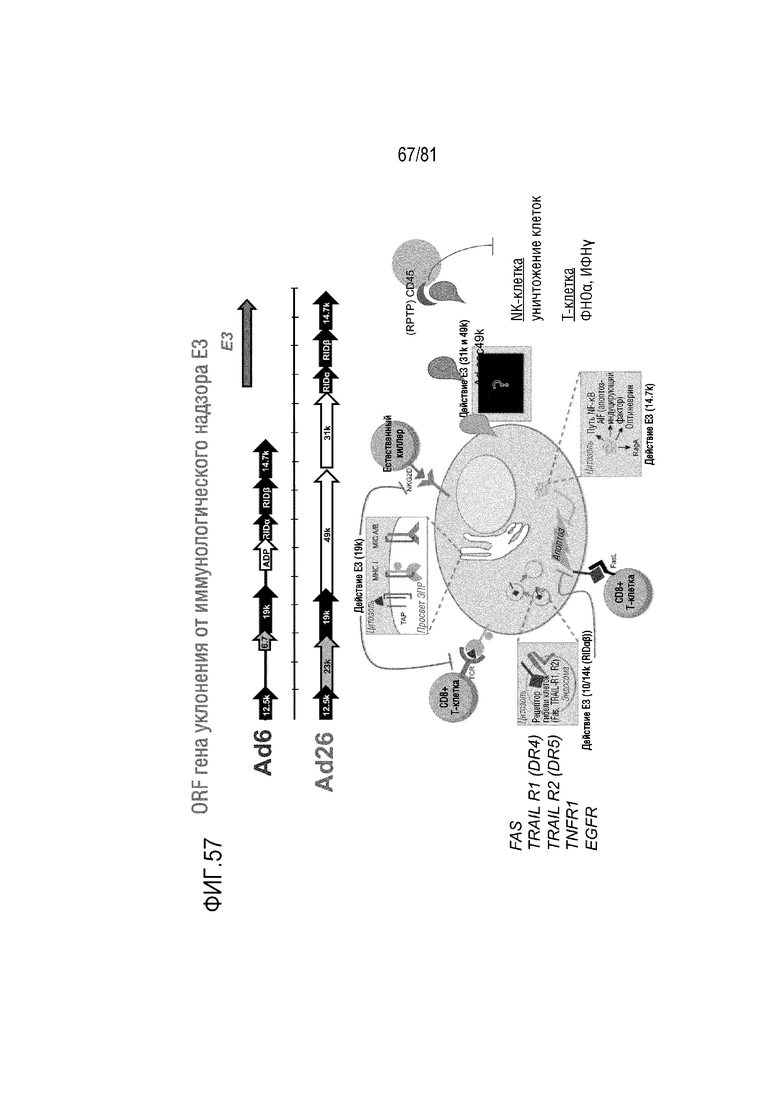
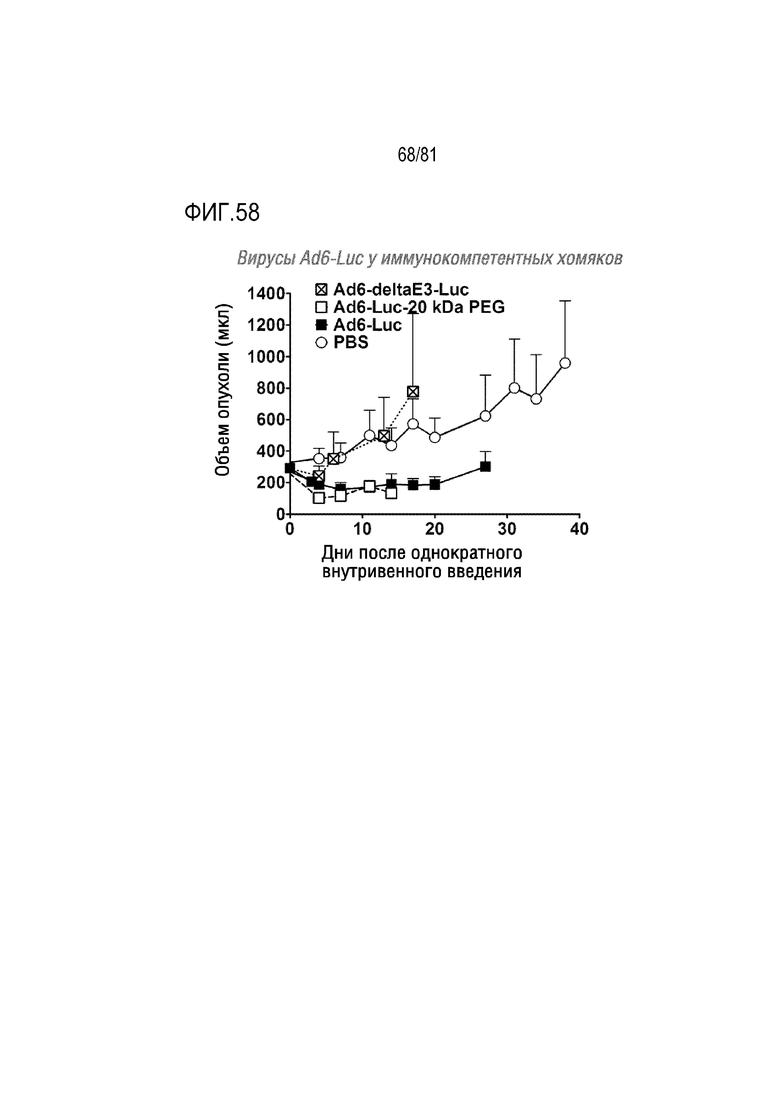
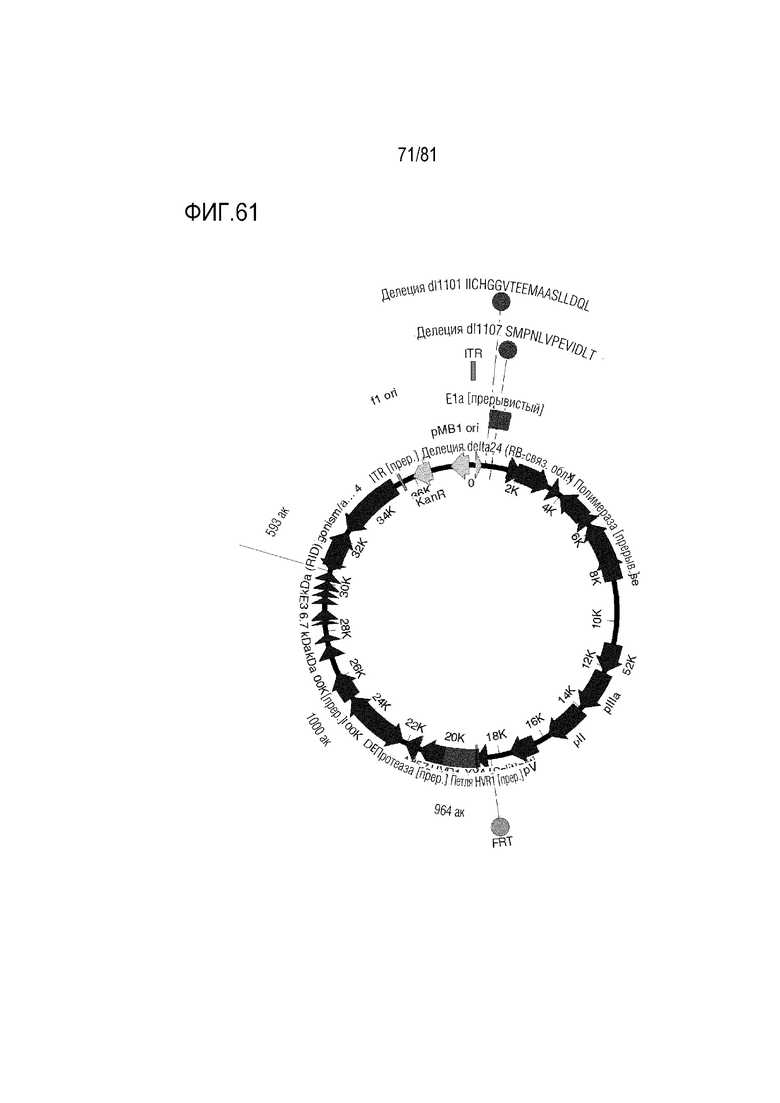
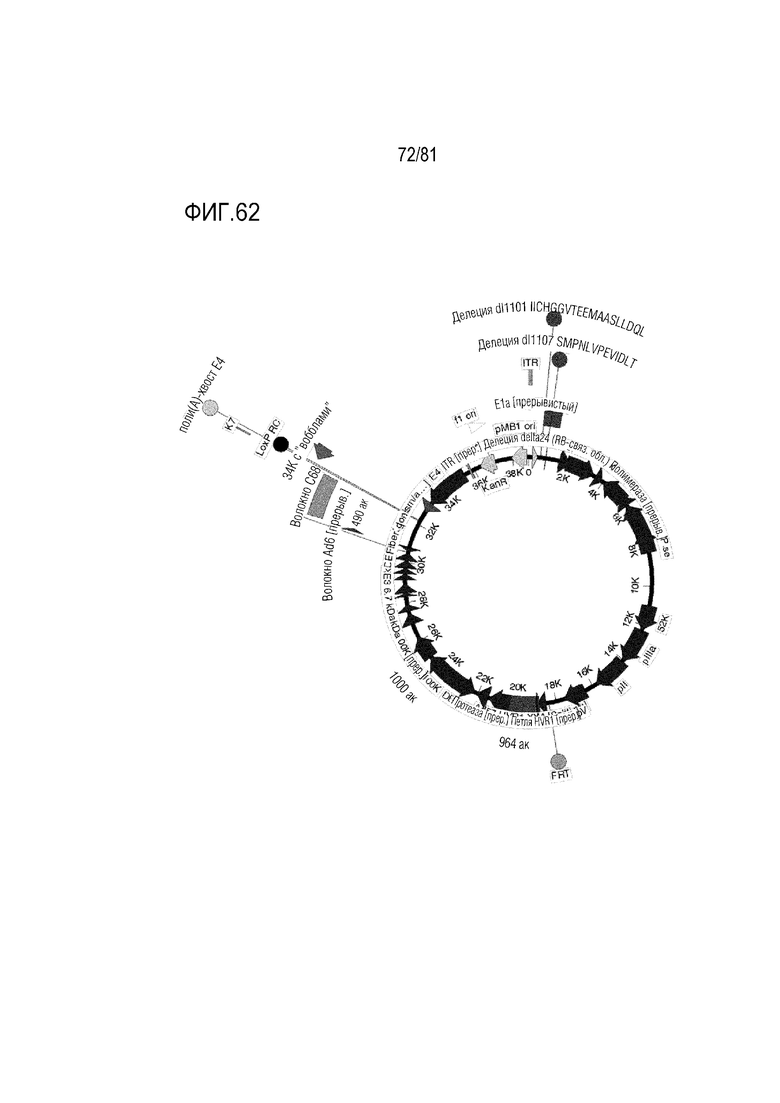
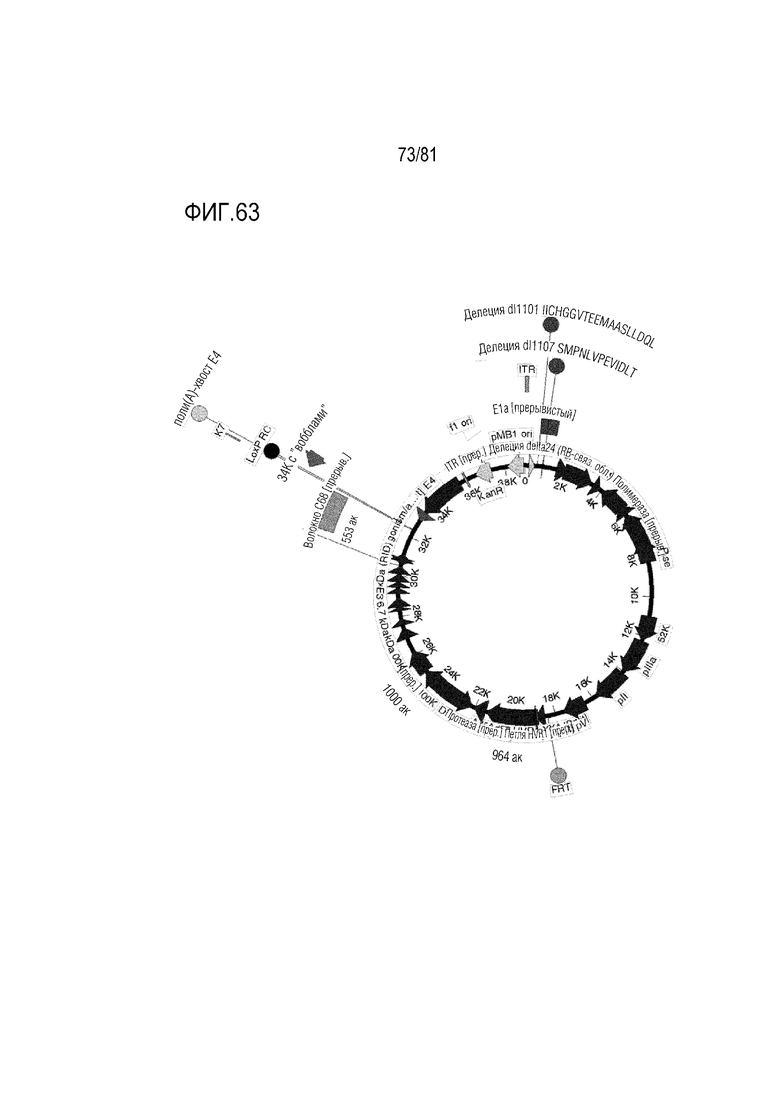
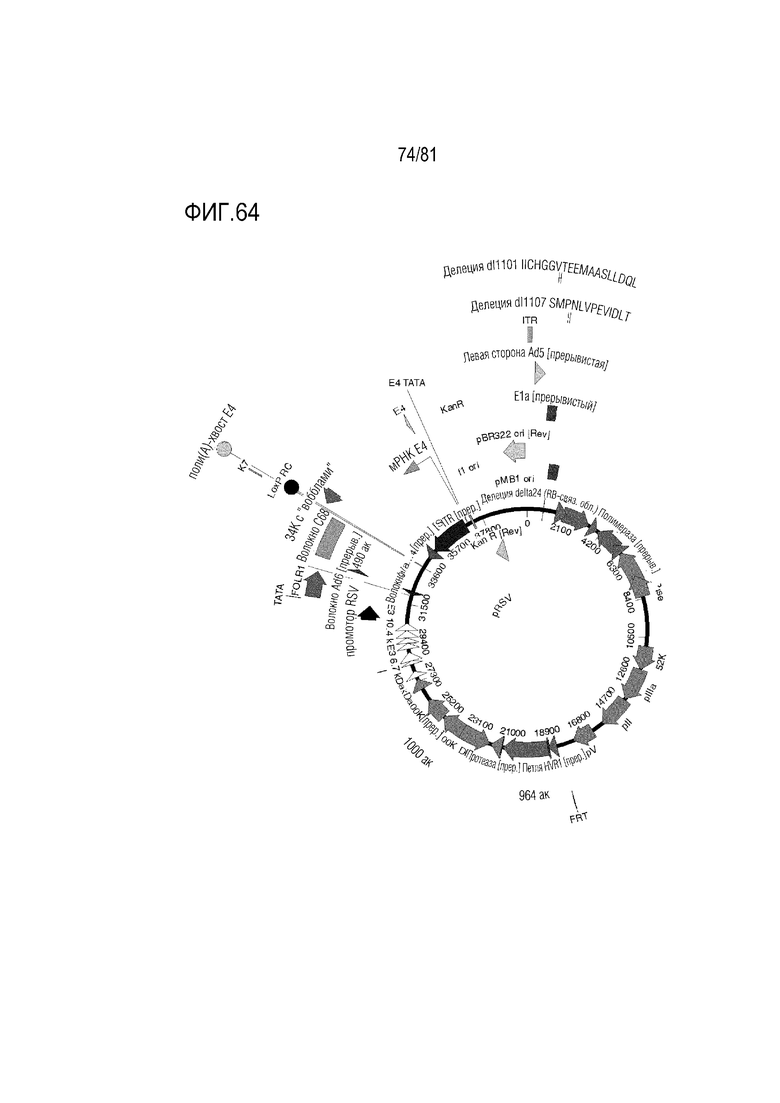
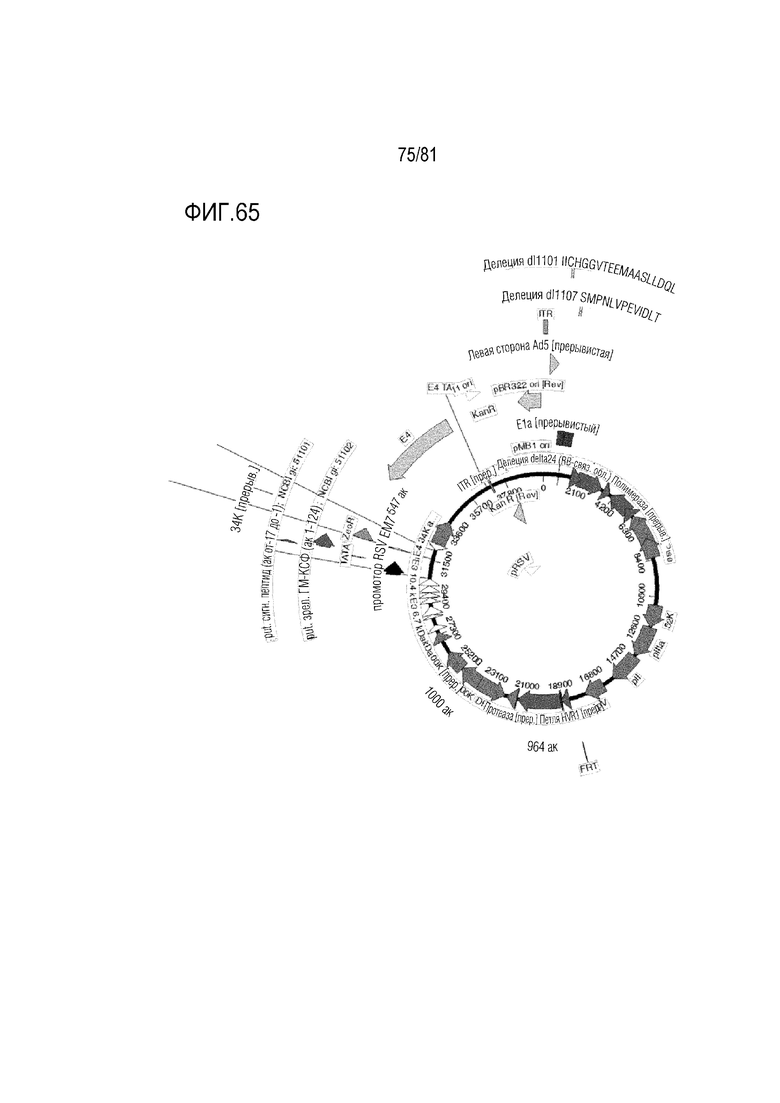
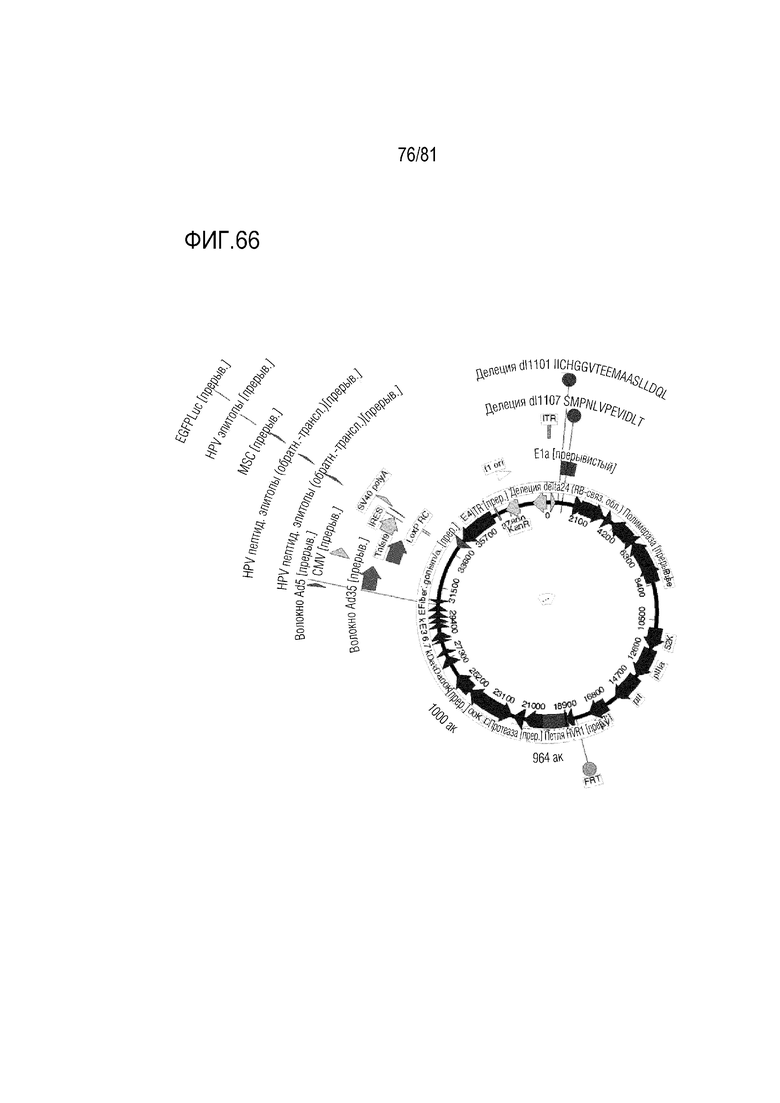
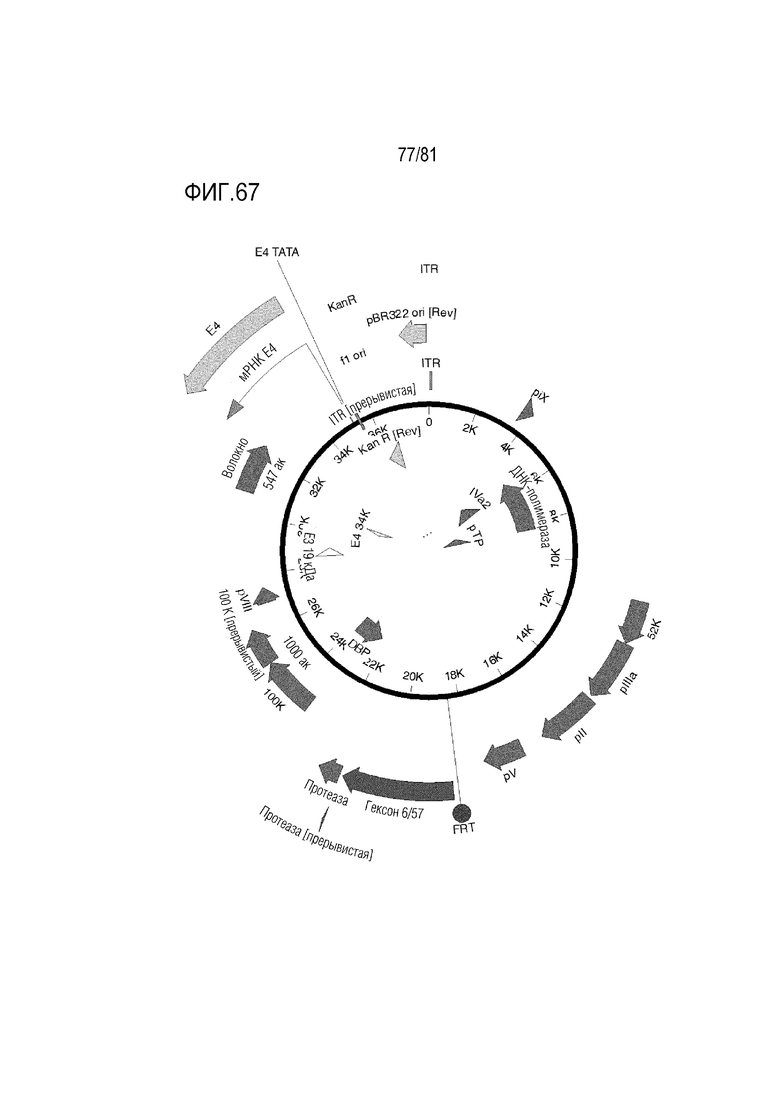
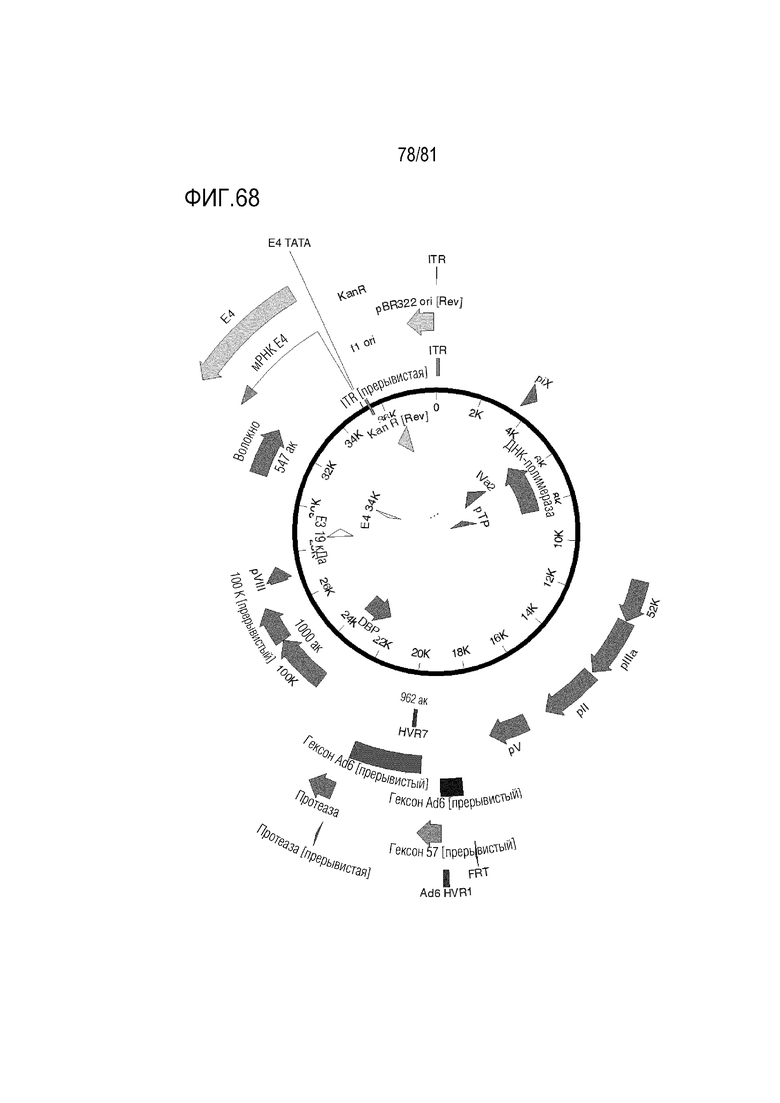
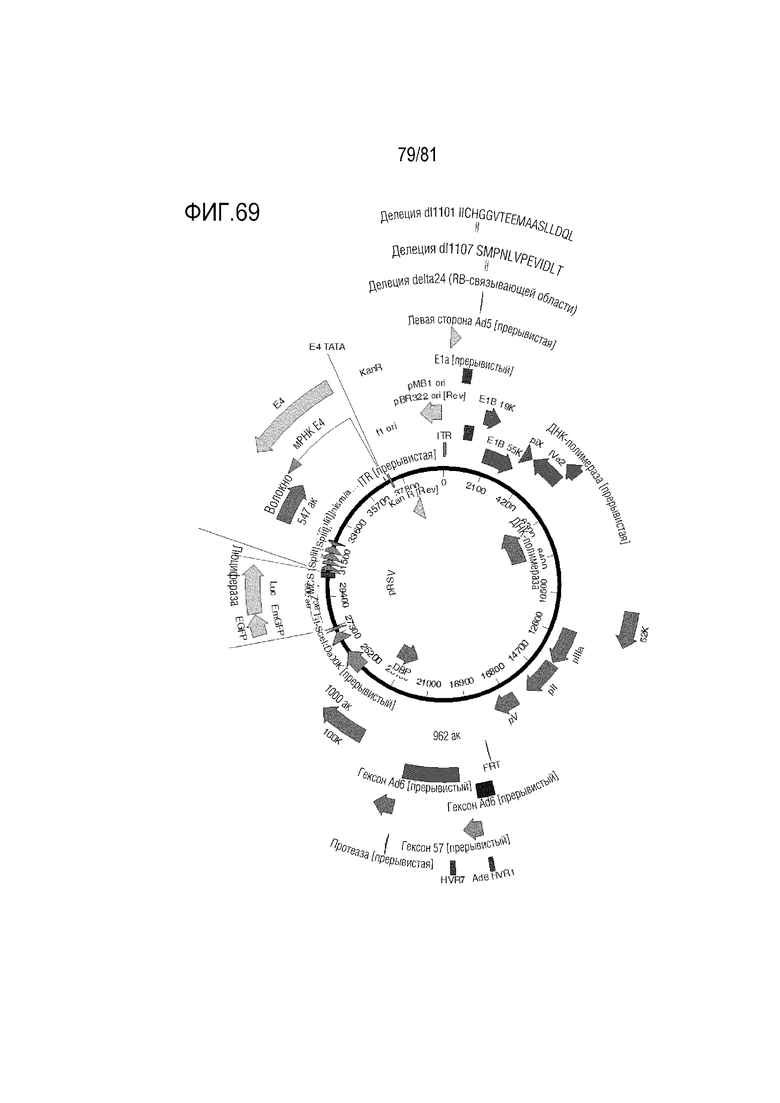
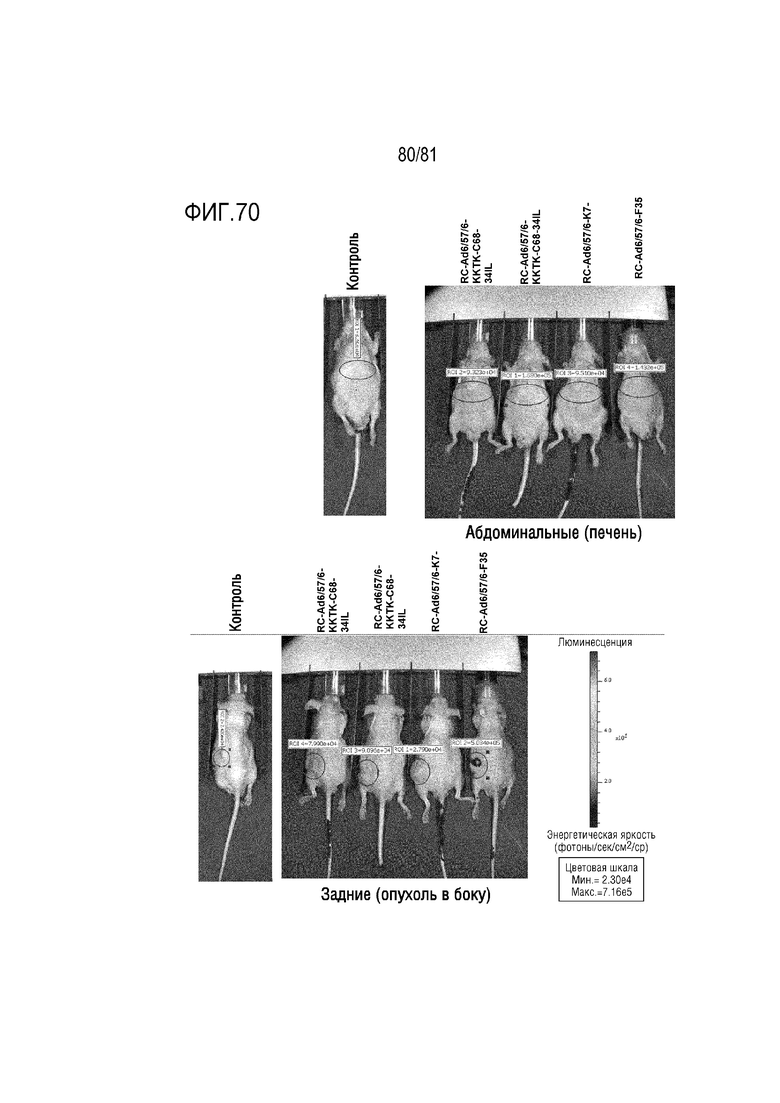
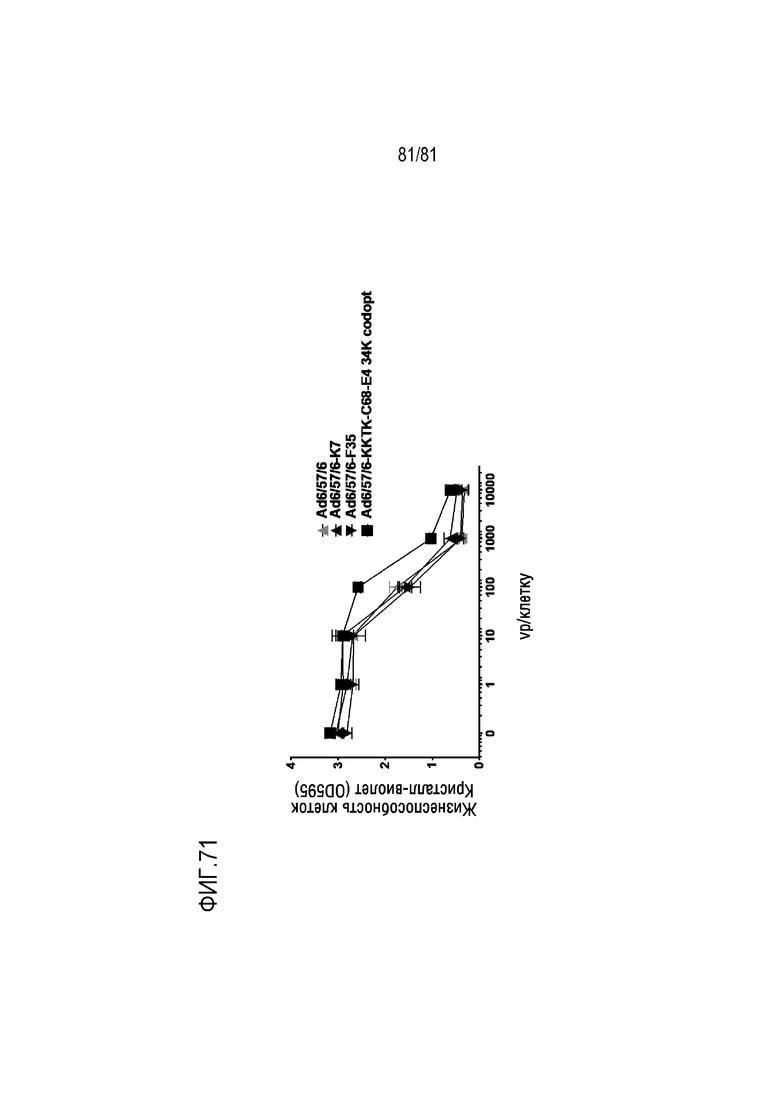
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая рекомбинантный онколитический аденовирус (Ad) на основе Ad штамма Ad6 человека, способ лечения млекопитающего, имеющего злокачественное новообразование, применение рекомбинантного онколитического аденовируса для лечения злокачественных новообразований и применение рекомбинантного онколитического аденовируса в качестве лекарственного средства для лечения злокачественных новообразований. В одном из вариантов реализации рекомбинантный онколитический аденовирус содержит полипептиды капсидного гексона, где по меньшей мере два полипептида гипервариабельной области гексона (HVR) полипептида капсидного гексона Ad6 человека заменены по меньшей мере двумя полипептидами HVR из Ad штамма Ad57 человека. Изобретение расширяет арсенал рекомбинантных онколитечических аденовирусов. 5 н. и 22 з.п. ф-лы, 71 ил., 2 табл., 12 пр.
1. Рекомбинантный онколитический аденовирус (Ad) на основе Ad штамма Ad6 человека, который содержит полипептиды капсидного гексона, где по меньшей мере два полипептида гипервариабельной области гексона (HVR) полипептида капсидного гексона Ad6 человека заменены по меньшей мере двумя полипептидами HVR из Ad штамма Ad57 человека.
2. Рекомбинантный околитический Ad по п.1, где полипептид HVR1 Ad штамма Ad6, содержащий аминокислоты 16-52 SEQ ID NO:77, заменен полипептидом HVR1 Ad штамма Ad57, содержащим аминокислоты 16-51 SEQ ID NO:78,
полипептид HVR 2 Ad штамма Ad6, содержащий аминокислоты 68-83 SEQ ID NO: 77, заменен полипептидом HVR 2 Ad штамма Ad57, содержащим аминокислоты 67-81 SEQ ID NO: 78,
полипептид HVR 3 Ad штамма Ad6, содержащий аминокислоты 101-107 SEQ ID NO: 77, заменен полипептидом HVR 3 Ad штамма Ad57, содержащим аминокислоты 99-105 SEQ ID NO: 78,
полипептид HVR 4 Ad штамма Ad6, содержащий аминокислоты 137-149 SEQ ID NO: 77, заменен полипептидом HVR 4 Ad штамма Ad57, содержащим аминокислоты 135-147 SEQ ID NO: 78,
полипептид HVR 5 Ad штамма Ad6, содержащий аминокислоты 156-171 SEQ ID NO: 77, заменен полипептидом HVR 5 Ad штамма Ad57, содержащим аминокислоты 154-169 SEQ ID NO: 78,
полипептид HVR 6 Ad штамма Ad6, содержащий аминокислоты 193-202 SEQ ID NO: 77, заменен полипептидом HVR 6 Ad штамма Ad57, содержащим аминокислоты 191-200 SEQ ID NO: 78, и
полипептид HVR 7 Ad штамма Ad6, содержащий аминокислоты 307-347 SEQ ID NO: 77, заменен полипептидом HVR 7 Ad штамма Ad57, содержащим аминокислоты 305-343 SEQ ID NO: 78.
3. Рекомбинантный околитический Ad по п.1, содержащий полипептиды капсидного гексона, содержащие аминокислотную последовательность, представленную в SEQ ID NO:49.
4. Рекомбинантный околитический Ad по п.2 или 3, который представляет собой Ad657.
5. Рекомбинантный околитический Ad по п.1, где полипептид HVR 2 Ad штамма Ad6, содержащий аминокислоты 68-83 SEQ ID NO: 77, заменен полипептидом HVR 2 Ad штамма Ad57, содержащим аминокислоты 67-81 SEQ ID NO: 78,
полипептид HVR 3 Ad штамма Ad6, содержащий аминокислоты 101-107 SEQ ID NO: 77, заменен полипептидом HVR 3 Ad штамма Ad57, содержащим аминокислоты 99-105 SEQ ID NO: 78,
полипептид HVR 4 Ad штамма Ad6, содержащий аминокислоты 137-149 SEQ ID NO: 77, заменен полипептидом HVR 4 Ad штамма Ad57, содержащим аминокислоты 135-147 SEQ ID NO: 78,
полипептид HVR 5 Ad штамма Ad6, содержащий аминокислоты 156-171 SEQ ID NO: 77, заменен полипептидом HVR 5 Ad штамма Ad57, содержащим аминокислоты 154-169 SEQ ID NO: 78, и
полипептид HVR 6 Ad штамма Ad6, содержащий аминокислоты 193-202 SEQ ID NO: 77, заменен полипептидом HVR 6 Ad штамма Ad57, содержащим аминокислоты 191-200 SEQ ID NO: 78.
6. Рекомбинантный околитический Ad по п.1, содержащий полипептиды капсидного гексона, содержащие аминокислотную последовательность, представленную в SEQ ID NO:59.
7. Рекомбинантный околитический Ad по п.5 или 6, который представляет собой Ad6/57/6.
8. Рекомбинантный околитический Ad по любому из пп. 1-7, содержащий один или более нацеливающих полипептидов, антигенных полипептидов, ферментов, рецепторов или лигандов.
9. Рекомбинантный околитический Ad по п.8, где нацеливающий полипептид содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-41 и SEQ ID NO: 46-47.
10. Рекомбинантный околитический Ad по любому из пп. 1-9, где замена аминокислоты является заменой цистеином, и модификация аминокислоты выбрана из пегилирования и бапилирования.
11. Рекомбинантный околитический Ad по любому из пп. 1-10, где рекомбинантный Ad является условно-репликативным Ad (CRAd).
12. Рекомбинантный околитический Ad по любому из пп. 1-10, содержащий нуклеиновые кислоты, кодирующие аминокислотную последовательность SEQ ID NO:43, SEQ ID NO:44 или SEQ ID NO:45.
13. Рекомбинантный околитический Ad по любому из пп. 1-11, содержащий один или более полипептидов модифицированных волокон.
14. Рекомбинантный онколитический аденовирус (Ad) по п.13, где модифицированные полипептиды волокна содержат гетерологичный полипептид волокна Ad35, необязательно содержащий 7 добавленных лизинов (пептид K7), или полипептид волокна C68 шимпанзе, необязательно содержащий 7 добавленных лизинов (пептид K7), или их комбинации.
15. Рекомбинантный онколитический аденовирус (Ad) по п.14, где полипептиды модифицированного волокна содержат аминокислотную последовательность SEQ ID NO:61, SEQ ID NO:62, SEQ ID NO:63, SEQ ID NO:64, SEQ ID NO:65 или их комбинации.
16. Рекомбинантный околитический Ad по любому из пп. 1-15, содержащий одну или более нуклеиновых кислот, кодирующих один или более гетерологичных белков.
17. Рекомбинантный околитический Ad по п.16, где гетерологичные белки выбраны из терапевтических полипептидов, нацеливающих полипептидов, антигенных полипептидов, антител, ферментов, рецепторов и лигандов.
18. Рекомбинантный онколитический аденовирус (Ad) по п.17, где антигенный полипептид представляет собой альфа-полипептид человеческого фолатного рецептора.
19. Рекомбинантный околитический Ad по п.17, где гетерологичные белки выбраны из антител, лиганда 4-1BB (4-1BBL), гранулоцитарно-макрофагального колониестимулирующего фактора (hGMCSF), лиганда CD40 (CD40L), белка запрограммированной гибели 1 (PD-1), интерлейкина-21 (IL-21) и пептидов хронического лимфоцитарного лейкоза (CLL).
20. Способ лечения млекопитающего, имеющего злокачественное новообразование, где указанный способ включает введение указанному млекопитающему рекомбинантного онколитического аденовируса (Ad) по любому из пп. 1-19.
21. Способ по п.20, где указанное злокачественное новообразование выбрано из группы, состоящей из рака предстательной железы, рака яичников, рака легких, печеночноклеточной карциномы, рака поджелудочной железы, рака почки, меланомы, злокачественного новообразования головного мозга, рака толстого кишечника, лимфомы, миеломы, лимфоцитарного лейкоза и миелогенного лейкоза.
22. Способ по любому из пп. 20-21, где указанное введение включает системное введение.
23. Способ по п.22, где системное введение включает внутримышечное, интраназальное или внутривенное введение.
24. Способ по любому из пп. 20-21, где указанное введение включает местное введение.
25. Способ по п.24, где указанное местное введение включает внутриопухолевую инъекцию.
26. Применение рекомбинантного онколитического Ad по любому из пп. 1-19 для лечения злокачественных новообразований.
27. Применение рекомбинантного онколитического аденовируса (Ad) на основе человеческого Ad штамма Ad6, который содержит полипептиды капсидного гексона, где по меньшей мере два полипептида гипервариабельной области гексона (HVR) полипептида капсидного гексона Ad6 человека заменены по меньшей мере двумя полипептидами HVR из человеческого Ad штамма Ad57, в качестве лекарственного средства для лечения злокачественных новообразований.
| US 7741099 B2 22.06.2010 | |||
| Устройство для зажима заготовок при их резке | 1990 |
|
SU1785488A3 |
| КОНСТРУИРОВАНИЕ РЕКОМБИНАНТА ОНКОЛИТИЧЕСКОГО АДЕНОВИРУСА, СПЕЦИФИЧЕСКИ ЭКСПРЕССИРУЮЩЕГО ИММУНОМОДУЛЯТОРНЫЙ ФАКТОР GM-CSF В ОПУХОЛЕВЫХ КЛЕТКАХ, И ЕГО ПРИМЕНЕНИЕ | 2004 |
|
RU2361611C2 |
| US 9683025 B2 20.06.2017 | |||
| Davison, A | |||
| J | |||
| Genetic content and evolution of adenoviruses | |||
| Journal of General Virology, 2003, 84(11), 2895-2908 | |||
| Weaver, E | |||
| A., Hillestad, M | |||
| L., Khare, R., Palmer, D., Ng, P., & Barry, M | |||
| A | |||
| Characterization of species C human adenovirus | |||

