Настоящее изобретение относится к способу идентификации соединения, которое связывается с одним или несколькими полипептидами или модулирует активность одного или нескольких полипептидов, кодирующих один или несколько рецепторов, которые вовлекаются в детекцию и перцепцию жирных кислот.
Хорошо известно, что животные-компаньоны, кошки и собаки, очень разборчивы в пище. Животные зачастую отказываются принимать пищу, которую ели в течение некоторого времени, или отказываются есть больше какого минимального количества. Частично этот феномен можно объяснить тонкими различиями в сенсорном профиле сырьевого материала. Эти различия могут быть неощутимыми для человека, принимающего пищу, но за счет особенностей в обонянии и пищеварительной системе, характерных для животных-компаньонов, кошки и собаки могут ощущать эти различия. Такие отличительные сенсорные особенности могут быть связаны с естественными вариациями в используемой для кормления животных пище, или возникать в том случае, когда определенный продукт применяется редко или в незначительном количестве, и, в этой связи, заменяется альтернативным продуктом. Это может расстраивать хозяина такого животного, который думает, что животное не получает удовольствия от пищи и страдает от этого. При этом, животное может недополучать требуемое количество необходимых питательных компонентов, если оно не потребляет соответствующее количество доступной ему пищи. В этой связи, очевидно, что имеется потребность в способе активации животных-компаньонов к приему пищи, которую ему дают. Было предложено много подходов для решения этой проблемы. Большая часть коммерчески доступного питания для животных сделано в рамках определенных вкусовых качеств и/или текстуры. Однако, владельцы животных-компаньонов хорошо знают, что их питомец может внезапно, без какой-либо видимой причины, отказаться от того вкуса, который, по мнению хозяина животного, должен быть наилучшим. Было проведено множество исследований для изучения вкусовых предпочтений животных, в которых им давалась возможность сделать выбор из различных продуктов.
Вкусовая перцепция у животных-млекопитающих управляется вкусовыми рецепторами, обнаруженными во вкусовых бугорках на языке, которые, как считается, воспринимают пять вкусов: соленый, сладкий, горький, кислый и умами. Вкус пищи определяется тем, какой рецептор она стимулирует. Хотя некоторые рецепторы характеризуется межвидовой гомологией, превалирование, частота и активность каждого рецептора зависит от вида, и, соответственно, как это можно ожидать, в случае травоядного животного будет требоваться иной вкусовой стимул, чем в случае плотоядного животного. Вкусовые рецепторы кошек и собак имеют некоторую гомологию с вкусовыми рецепторами людей, хотя, как это известно, различные рецепторы у кошек и собак характеризуются иным уровнем активации, и/или иной предпочтительностью, чем у человека.
В основном, считается, что перцепция жиров в пище определяется ощущениями в ротовой полости и, в некоторой степени, зависит и от запаха. При этом, и у человека, и у полевых грызунов недавно были идентифицированы вкусовые рецепторы, реагирующие на жирные кислоты (Cartoni et al., 2010; Galindo et al., 2012; Martin et al., 2011), и это указывает на то, что реакция на вкус также вовлекается в детекцию и перцепцию жировых компонентов.
Было высказано предположение о том, что GPR120 (известный также как GPR129, O3FAR1, PGR4, FFAR4) представляет собой связанный с G-белком рецептор на клеточной поверхности, содержащий семь трансмембранных доменов (а также внеклеточную часть), вовлекаемый в детекцию определенных жирных кислот, а также ассоциированную с G-белком внутриклеточную часть, вовлекаемую в сигнальную трансдукцию. Было высказано предположение, что GPR120 связывает жирные кислоты со средней длиной цепи и длинноцепочечные жирные кислоты, такие как олеиновая кислоты и линолевая кислота, в их свободной форме. Предполагается, что обе изоформы (сплайс-варианты) GPR120 рецептора присутствуют у человека, GPR120L и GPR120S, на эндокринных клетках толстой кишки. Было высказано предположение, что длинная изоформа не обладает сигнальной функцией в процессе вкусовой перцепции.
GPR120 экспресируется в различных тканях у млекопитающих и, как известно, вовлекается в стимуляцию секреции холецистокинина (ССК) из линии секреторных клеток кишечника STC-1, вдобавок к тому, что, как было показано, GPR120 оказывает стимулирующий эффект на секрецию глюкагон-подобного пептида (GLP-1). GPR120 также экспресируется в гипофизе, в связи с чем исследовалась его возможная роль в регуляции стресса. GPR120 рассматривается как рецептор для ненасыщенных длинноцепочечных жирных кислот, который вовлекается в секрецию GLP-1, который также действует как сенсибилизатор инсулина, как противовоспалительное средство и средство против ожирения. Были, в этой связи, предложения рассматривать агонисты или антагонисты GPR120 в качестве потенциальных лекарственных средств, применяемых при лечении различных метаболических заболеваний, таких как диабет. Однако, GPR120 надо еще исследовать на наличие у него способности улучшать вкусовую привлекательность пищи.
В заявке на Европейский патент ЕР 1688138 (Takeda Pharmaceutical Company Limited) описан специфический агент, регулирующий функцию рецептора человека 14273 (рецептор GRP120). В этом документе приводятся низкомолекулярные синтетические агонисты или антагонисты, способные стимулировать GRP120. Указывается, что данные вещества могут применяться для лечения переедания, диабета и ожирения. Описаны также альтернативные агенты, способные подавлять GRP120, и их применение для лечения анорексии. Область указанной заявки ограничена рецепторами GRP120 у человека и мышей и относится к идентификации и применению соединений с терапевтической целью.
В публикации по патентной заявке WO 2007/134613 (Rheo-Sciеnce A/S) описывается экспрессия рецептора GRP120 в различных тканях млекопитающих. В этой заявке предлагается использование соединения для модуляции экспрессии GRP120 с целью лечения, ослабления, профилактики и/или диагностики диабета и/или ожирения, а именно: для терапевтического применения у людей.
В заявке на Европейский патент ЕР1932920А1 (Eisai R&D Management Co.Ltd.) раскрывается способ определения, изменяет ли соединение стимулирующую клетки активность, опосредованную человеческим GRP120, для целей его терапевтического применения.
В публикации по патентной заявке WO 2011/159297A1 (Metabolex Inc.) описаны агонисты человеческого и крысиного GRP120 и их применение при лечении метаболических заболеваний, включающих диабет, и заболеваний, связанных со слабым гликемическим контролем. В указанной заявке отмечалось, что агонисты GRP120 вводились мышам для выявления их эффектов на секрецию инсулина, глюкагон-подобного пептида 1 и различных других гормонов. Было показано, что агонисты GRP120 могут снижать уровень глюкозы в крови после внутрибрюшинной глюкозной провокации, проведенной у мышей.
Bharat Shimpukade et al. (Journal of Medicinal Chemistry, Discovery of a Potent and Selective Agonist of GRP120, 2012 May; 59(9): 4511-4515) описывает агонист человеческого GRP120 для его терапевтического применения.
Qi Sun et al (Molecular Pharmacology, Structure-Activity Relationships of GRP agonists based on a Docking Stimulation, 2010 Nov; 78(5): 804-810) описывает агонист человеческого GRP120 для его терапевтического применения.
Takafumi Hara et al (Naunyn-Schmied Arch Pharmacol, Novel Selective Ligands for Free Acid Receptors GRP120 and GRP40, 2009 Sep; 380(3): 247-255) сделали попытку идентифицировать новые терапевтические лиганды для рецептора человеческого GRP120. Однако, указанные авторы смогли идентифицировать лишь частичные агонисты.
Takayoshi Suzuki et al (Journal of Medicinal Chemistry, Identification of G Protein-Coupled Receptor 12-Selective Agonists Derived from PPARγ Agonists, 2008 Dec 11; 51(23):7640-7644) указывают на необходимость выявления GRP120-селективных агонистов, поскольку они могут использоваться в качестве терапевтических средств.
CD36 (также известный как FAT, GP3B, GP4, GPIV, SCARB3, рецептор тромбоспондина) не относится к семейству рецепторов, связанных с G-белком (он относится к классу В семейства фагоцитарных рецепторов), который отличается от других вкусовых рецепторов жирных кислот, известных у людей, и является необычным в этом плане.
Известно, что домашние кошки разборчивы в питании, и многие владельцы кошек считают, что их питомец ест определенную пищу в определенные дни. В этой связи, возможность быть уверенным, что кошка хорошо среагирует на конкретный пищевой продукт будет гарантировать неизменный прием этого пищевого продукта данным животным, а также создаст уверенность в том, что хозяин такого животного будет считать, что его питомец счастлив и здоров.
Собаки также могут быть привередливы в отношении пищи или, в случае некоторых животных, неразборчивы в выборе пищи. За счет улучшения вкусовой перцепции пищевого продукта, можно с большей уверенностью полагать, что собаки будут съедать определенный пищевой продукт, причем с высокой степенью надежности, гарантированно и неизменно.
В настоящее время предпочтительность для кошек и собак тех или иных вкусовых стимулов оценивают в рамках пищевых тестов, которые могут быть неэффективны с точки зрения их стоимости, временных затрат и получаемых результатов. Кроме того, затруднительна в этом случае идентификация нового вкусового стимула, так как нужно исследовать большое число соединений с использованием для этого тестов по оценке вкусовых преференций у животных, для определения того вкуса, который может быть действительно привлекательным для кошек и собак. И для таких методов тестирования требуется относительно большие количества каждого из исследуемых соединений.
Исходя из вышесказанного, имеется потребность в надежных, высокоэффективных способах скрининга, направленного на идентификацию тех вкусовых соединений, которые могут связывать и стимулировать (или даже модулировать) некоторые вкусовые рецепторы у животных, в частности, у кошек и собак.
В этой связи, в одном своем аспекте настоящее изобретение относится к идентификации соединения, которое связывается с полипептидом и/или модулирует активность полипептида, где указанный полипептид включает:
(i) последовательность рецептора GPR120 кошки или собаки или CD36 кошки или собаки;
(ii) аминокислотную последовательность, приведенную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 и SEQ ID NO:7;
(iii) аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:1 или SEQ ID NO:3;
(iv) аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:5 или SEQ ID NO:7;
(v) аминокислотную последовательность, включающую аминокислоты на участке 127-279 в SEQ ID NO:3 или SEQ ID NO:7;
(vi) функциональный фрагмент по пп. (i), (ii), (iii), (iv) или (v),
где указанный способ включает определение, может ли исследуемое соединение связываться с данным полипептидом и/или модулировать активность этого полипептида.
Авторы настоящего изобретения показали, что полипептиды, включающие последовательность SEQ ID NO:1 и SEQ ID NO:3, представляют собой аминокислотные последовательности кошачьих гомологов рецепторов жирных кислот GPR120 и CD36 у человека, соответственно. SEQ ID NO:5 и SEQ ID NO:7 представляют собой аминокислотные последовательности рецепторов GPR120 и CD36 у собаки, соответственно. Аналогичные последовательности у человека показаны в виде SEQ ID NO:9 и SEQ ID NO:11.
Указанные последовательности были получены из геномной ДНК кошки и собаки после ресеквенирования и затем было проведено их сравнение с соответствующими последовательностями человека и предсказанными последовательностями кошки и собаки. Были обнаружены некоторые различия в генной последовательности кошачьего CD36, при сравнении соответствующих опубликованных данных и результатов, полученных авторами при его выделении и анализе. Настоящее изобретение также относится к использованию функциональных и аллельных вариантов, которые могут отличаться по последовательности, но которые тем не менее сохраняют способность к стимуляции под действием жирных кислот, что ведет к перцепции жирных кислот животным и что фактически представляет собой метод скрининга.
Предполагается, что последовательности, выделенные от кошек и собак, будут активными, функциональными рецепторами, благодаря их близости, на уровне последовательности, к доступным последовательностям GPR120 и СD36 у человека, крысы и мыши (по меньшей мере 82% сходство). Нет каких-либо оснований полагать, что такие рецепторы не будут функционировать in vivo, особенно, ввиду того факта, что, как показали авторы настоящего изобретения, наблюдается определенная, явная реакция у исследованных животных, кошек и собак, на олеиновую и линолевую кислоты, которые, как известно, связываются с эквивалентными рецепторами человека, а, кроме того, в тестах in vitro было показано, что в присутствии известных лигандов, эти рецепторы связываются с ними и соответствующим образом реагируют.
Таким образом, в одном своем аспекте настоящего изобретения, предлагаемый способ представляет собой in vitro способ. Указанный in vitro способ может включать:
- определение биологической активности полипептида в отсутствие и при наличии исследуемого соединения;
- идентификацию такого агента, который связывается с полипептидом или модулирует биологическую активность полипептида, когда имеется различие в биологической активности, в том случае, когда исследуемый агент отсутствует, и в том варианте, когда он присутствует.
Указанный in vitro способ может также включать приведение во взаимодействие полипептида с исследуемым соединением.
Методы детекции, используемые в настоящем изобретении, могут включать использование меченого соединения/агента и затем определение, после промывки, какие соединения, из числа исследуемых, продолжают оставаться в связанном состоянии с рецепторами. Детектирование активности, индуцированной связыванием соединения с рецептором, может представлять собой мониторинг концентрации свободного кальция внутри клетки, которая повышается в результате активации, получившей название кальциевый флюс, которая хорошо известна специалистам в данной области. Указанный мониторинг может проводиться путем детекции флуоресценции, такой как, например, детекция в присутствии чувствительного на кальций флуоресцентного красителя, или по методу люминесцентной детекции, с использованием люминесцирующего белка. Альтернативный способ включает мониторинг активности цГМФ, который также известен специалистам в данной области.
Участок между аминокислотными остатками 127 и 279 в CD36 вовлекается в связывание длинноцепочечных жирных кислот у людей, и, таким образом, полипептид, включающий эту часть в SEQ ID NO:3 или SEQ ID NO:5, может использоваться в способе настоящего изобретения. Полипептид, включающий участок между аминокислотными остатками 155 и 183, в SEQ ID NO:3 или SEQ ID NO:5, может использоваться в способе скрининга по настоящему изобретению.
Рассматриваемый способ может также включать одновременное использование двух полипептидов по настоящему изобретению, поскольку белок CD36 может действовать как шаперон, позволяя агенту или соединению взаимодействовать с белком GPR120 или усиливать взаимодействие между GPR120 и жирной кислотой или другим активирующим соединением. Таким образом, такой in vitro способ, включающий использование обоих полипептидов, также входит в настоящее изобретение, как еще один его аспект. Соответственно, настоящее изобретение относится к способу, включающему определение биологической активности полипептида GR120 (SEQ ID NO:1 или SEQ ID NO:5) или его фрагмента в присутствии полипептида CD36 (SEQ ID NO:3 или SEQ ID NO:7, соответственно), в отсутствие и при наличии исследуемого соединения, и затем идентификацию такого соединения, которое вызывает изменение активности, в сравнении с активностью в отсутствие агента.
Способы скрининга агентов, которые могут модулировать биологическую активность полипептида, хорошо известны в данной области и могут включать использование твердых носителей, на которых иммобилизованы полипептиды по настоящему изобретению.
Агенты, идентифицированные с помощью таких методов скрининга, могут ингибировать/действовать как антагонисты или активировать/действовать как агонисты биологическую активность/в отношении биологической активности пептида по настоящему изобретению. Таким образом, указанные соединения могут использоваться в качестве антагонистов или агонистов рецепторов.
Соединения, идентифицированные с помощью таких методов in vitro скрининга, далее могут быть исследованы in vivo, например, в ходе экспериментов по скармливанию этих соединений животным.
Настоящее изобретение также относится к способу идентификации вкусового вещества, которое связывает и/или модулирует активность рецептора GPR120 и/или рецептора CD36, где указанный рецептор GPR120 или рецептор CD36 представляет собой рецептор кошки, собаки или человека (SEQ ID NОNО: 1, 3, 5, 7, 9,11), или где рецептор GPR120 по меньшей мере на 87% идентичен SEQ ID NO:1 и где рецептор CD36 по меньшей мере на 82% идентичен SEQ ID NO:3.
Рецептор GPR120 может быть на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичен любой из SEQ ID NОNО: 1, 5 или 9.
Рецептор CD36 может быть на 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичен любой из SEQ ID NО NО: 3, 7 или 11.
Идентификация таких вкусовых веществ может позволить получать пищевые добавки с более приятным вкусом для их включения в пищу кошек, собак или человека.
Желательно идентифицировать такие соединения, которые обладали бы более полезными свойствами, чем уже известные соединения, которые связываются с GPR120 и/или CD36; соответствующие примеры включают соединения, которые легче/дешевле производить; соединения, которые можно применять в меньших количествах, с достижением тех же эффектов, которые демонстрируют уже известные соединения; соединения, которые взаимодействуют синергически с другими соединениями.
Настоящее изобретение также относится к рецепторам, известным как FFAR1 (GPR40), FFAR2 (GPR43) и/или FFAR3 (GPR41). Эти полипептиды были описаны ранее как соединения, связывающиеся с жирными кислотами, но их никогда не рассматривали в контексте вкусовых веществ, а также применительно к использованию в пище для животных, кошек и собак.
Соответственно, описанные в настоящем изобретении способы идентификации соединений, которые связывают и/или модулируют биологическую активность рецепторов FFAR1, 2 или 3, также включены в область настоящего изобретения.
Все свойства приведенных здесь аспектов применимы к каждому из них, с соответствующими поправками, mutatis mutandis.
Аминокислотные последовательности описаны здесь с использованием стандартного однобуквенного кода. Указанные последовательности описаны в направлении от N-конца к С-концу, слева направо. Аминокислоты, которые могут быть введены в состав данных пептидов, включают любую аминокислоту из числа известных природных аминокислот.
Кроме того, пептиды по настоящему изобретению могут включать модифицированные аминокислоты, то есть аминокислоты, которые в естественном состоянии в природе не встречаются. Например, пептиды по настоящему изобретению могут включать норлейцин или другие, известные в данной области модифицированные аминокислоты.
Пептиды по настоящему изобретению могут состоять только из тех аминокислотных последовательностей, которые описаны здесь, но могут включать и другие аминокислоты, в дополнение к этим последовательностям. Описанные здесь полипептидные последовательности могут содержать дополнительные аминокислоты на N-конце (амино-конце) и/или С-конце последовательностей, особенно в случае их использования в способе скрининга по настоящему изобретению. Такие дополнительные аминокислоты могут способствовать иммобилизации полипептиды для целей скрининга или могут позволить ввести рассматриваемый полипептид в состав белка слияния, в качестве его части, для целей облегчения детекции биологической активности.
Полипептиды по настоящему изобретению включают гомологи или производные указанных выше последовательностей, которые сохранили способность связывать средне- и длинноцепочечные жирные кислоты. Так, в пептид может быть введено большое число консервативных аминокислотных замещений, без существенных структурных или функциональных изменений. Соответственно, возможно провести замену одной аминокислоты другой, «близкого» типа, например, замену одной гидрофобной аминокислоты другой гидрофобной аминокислотой. В данной области известны приемлемые варианты аминокислотного замещения. В случае таких гомологов и производных, степень идентичности с конкретными, описанными здесь последовательностями менее важна, чем тот факт, что такой гомолог или такое производное должны сохранять способность связывать жирную кислоту, запуская сигнал далее по цепи. Однако, в подходящем варианте, указанные гомологи или производные обладают по меньшей мере 90% идентичностью к описанным здесь последовательностям. Наиболее предпочтительно, указанные гомологи или производные обладают по меньшей мере 91%, 92%, 93%, 94%, 95%, 95%, 96%, 97%, 98%, 99% идентичностью.
Процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот определяют при выравнивании последовательностей для целей оптимального сравнения (например, в первую последовательность могут быть введены гэпы для наилучшего сопоставления с данной последовательностью) и при последующем сравнении аминокислотных остатков и нуклеотидов в соответствующих положениях. «Наилучшее выравнивание» представляет собой такой вариант выравнивания двух последовательностей, который приводит к наивысшему проценту идентичности. Процент идентичности определяется по числу идентичных аминокислотных остатков и нуклеотидов в сравниваемых последовательностях (т.е. процент идентичности=число идентичных положений/общее число позиций х 100).
Определение процента идентичности между двумя последовательностями может быть выполнено с использованием математического алгоритма, известного специалистам в данной области. Один пример математического алгоритма, используемого для сравнения двух последовательностей, включает алгоритм Karlin and Schul (1990), описанный в работе Proc. Natl. Acad. Sci. USA, 87:2264-2268, модифицированный в алгоритм Karlin and Schul (1993), Proc. Natl. Acad. Sci. USA, 90: 5873-5877. Программы NBLAST и XBLAST, Altschul, et al. (1990), описанные в работе J. Mol. Biol.215: 403-410, включают такой алгоритм. BLAST-анализ нуклеотидов может быть проведен по программе NBLAST, где количество очков установлено на уровне 100, длина слова=12, для получения нуклеотидных последовательностей, гомологичных молекулам нуклеиновых кислот по настоящему изобретению. BLAST-анализ белков может быть проведен по программе XBLAST, где количество очков установлено на уровне 50, длина слова=3, для получения аминокислотных последовательностей, гомологичных молекулам белка по настоящему изобретению. Для проведения локального выравнивания последовательностей, содержащих бреши, может быть использована соответствующая BLAST-программа для анализа последовательностей, содержащих бреши, Gapped BLAST, описанная в работе Altschul et al.(1997), Nucleic Acids Research, 25:3389-3402. Альтернативно, может использоваться программа PSI-BLAST для проведения итеративного поиска, который позволяет детектировать отдаленное сходство между молекулами (Id). При использовании для анализа программ BLAST, Gapped BLAST и PSI- BLAST, могут использоваться заданные по умолчанию параметры соответствующих программ (например, XBLAST и NBLAST). См. сайт http://www.ncbi.nlm.nih.gov. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Myers and Miller, CABIOS (1989). Программа ALIGN (версия 2.0), которая входит в состав программного обеспечения, используемого для проведения локального выравнивания последовательностей CGC, включает такой алгоритм. Другие, известные в данной области алгоритмы, используемые для анализа последовательностей, включают ADVANCE и ADAM, описанные в работе Torellis and Robotti (1994), Comput. Appl. Biosci., 10:3-5; и алгоритм FASTA, описанный в работе Pearson and Lipman (1988), Proc. Natl. Acad. Sci, 85: 2444-8. В алгоритме FASTA, параметр ktup представляет собой контрольную опцию, которая устанавливает чувствительность и скорость поиска.
Эксперименты по функциональной оценке GPR120 и CD36 белков человека и грызунов были проведены с использованием линолевой и олеиновой кислот, и многие из них включали эксперименты с использованием в качестве моделей подопытных грызунов с выключенным/нокаутированным геном. Хотя указанные эксперименты продемонстрировали эффект отсутствия рецептора, но конкретно они не показали, что данная молекула активирует рецептор. Один пример, указывающий на наличие реакций на линолевую и олеиновую кислоту, выявленных в экспериментах in vitro c использованием человеческого GPR120, описан в работе Galindo et al. (2011), Chem. Sen. Эксперименты с CD36 человека, в варианте использования линолевой кислоты, описаны в работе Kuda et al. (2013, J.Niol.Chem). Cartoni et al. (2010, J. Neurosci) показали, что мыши с выключенным геном по GPR120 демонстрировали измененные реакции на линолевую и олеиновую кислоты. В экспериментах других авторов (Gaillard et al., 2008, FASEB) было показано, что клетки, содержащие вкусовой рецептор СD36 (СD36+) были чувствительными на линолевую кислоту, тогда как СD36- клетки, без соответствующего вкусового рецептора, таким свойством не обладали. Как было показано, человеческие гомологи и гомологи от грызунов описанных здесь полипептидов связываются с такого рода специфическими длинноцепочечными жирными кислотами. По всей видимости, эквивалентные последовательности от кошки и собаки также связывают такие молекулы, что следует из реакции in vivo на жирные кислоты, используемые в возрастающих концентрациях, как это описано здесь. Кроме того, приведенные в настоящем описании тесты in vitro, в которых были использованы кошачьи рецепторы, демонстрируют наличие позитивной активации под действием линолевой и олеиновой кислоты.
Пептиды, подходящие для использования в способах скрининга по настоящему изобретению, могут быть получены методами химического синтеза, которые хорошо известны в данной области. Так, например, указанные пептиды могут быть синтезированы химически, с использованием метода твердофазного синтеза пептидов. Используемые для этой цели способы основаны на методах синтеза в твердой или жидкой фазе (см., например, J.M. Steward and J.D. Young, Solid Phase Peptide Synthesis, 2nd Ed, Pierce Chemical Co., Rockford III (1984), и G. Barany and R.B. Merrifield, The Peptides: Analysis Synthesis, Biology editors E. Gross and J. Meienhofer, Vol 2, Academic Press, New York, 1980, pp.3-254, в том, что касается технологии твердофазного синтеза; и M. Bodansky, Principles of Peptide Synthesis, Springer-Verlag, Berlin, 1984, и E.Gross and J. Meienhofer, Eds., The Peptides: Analysis, Synthesis, Biology, supra, Vol.1, в том, что касается классического синтеза в фазе раствора). В данной области известны также другие методы пептидного синтеза.
Альтернативно, пептиды могут быть получены путем экспрессии молекул нуклеиновой кислоты, кодирующих предшественники пептидов. В другом аспекте, настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей предшественника пептидов по настоящему изобретению. Такие нуклеиновые кислоты могут быть синтезированы с использованием для этого методов, хорошо известных в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Edition, Sambrook and Russell, 2001, Cold Spring Harbor Laboratory Press).
Кроме того, такие кодирующие пептиды нуклеиновые кислоты, могут быть включены в состав соответствующего вектора на основе нуклеиновой кислоты. В еще одном своем аспекте, настоящее изобретение относится к вектору, включающему нуклеиновую кислоту по настоящему изобретению. Указанный вектор может содержать промоторный элемент, оперативно связанный с последовательностью нуклеиновой кислоты, кодирующей пептид Подходящие векторы и способы получения таких векторов известны в данной области.
Нуклеиновая кислота по настоящему изобретению или вектор по настоящему изобретению могут быть введены в клетку-хозяина. Соответственно, в дополнительном аспекте настоящее изобретение относится к клетке, включающей нуклеиновую кислоту или вектор по настоящему изобретению. Указанная клетка может представлять собой изолированную клетку, такую как СНО К1 клетка, или другую, подходящую из числа известных, стабильную клеточную линию.
В другом аспекте, настоящее изобретение относится к слитым/гибридным белкам, включающим полипептиды, приведенные в настоящем описании. Такие слитые белки могут содержать детектируемый маркер, функциональную группу, такую, как носитель, метку, стабилизирующую последовательность, или включать в своем составе механизм, с помощью которого может быть детектирован факт связывания жирной кислоты. Подходящие метки включают FLAG маркер, His маркер, MYC маркер, белок, связывающий мальтозу, и другие известные в данной области метки. Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим такие слитые белки, к векторам, содержащим нуклеиновые кислоты, кодирующие слитые белки, а также к клеткам организма-хозяина, включающим такие нуклеиновые кислоты или векторы.
Способы синтеза таких слитых белков хорошо известны в данной области.
Способ по настоящему изобретению может представлять собой виртуальный способ, или способ in silico. Такой способ может включать:
(i) прогнозирование (3-х-мерной) 3D-структуры полипептида;
(ii) скрининг предсказанной 3D-структуры полипептида in silico, применительно к 3D-структуре исследуемого соединения;
(iii) прогнозирование, будет ли данное исследуемое соединение взаимодействовать с сайтом связывания полипептида; и
(iv) идентификацию соединения, которое связывает полипептид и модулирует биологическую активность полипептида, в том случае, когда 3D структура такого соединения соответствует сайту связывания в 3D структуре полипептида.
Такие технологии и методы хорошо известны специалистам в данной области. Модели GPR120 были созданы с использованием, для целей моделирования гомологий, кристаллических структур других GPR из группы А, в качестве матриц, которые были получены из банка белков (Protein Data Bank), и такая процедура может быть выполнена любым специалистом со средним уровнем знаний в данной области. В этом случае использовали пакет прикладных программ Modeler. Такие процедуры, как имитация и минимизация, применительно к отдельным жирным кислотам (например, линолевой кислоты) проводили по методам, известным специалистам в данной области. При этом можно использовать любой подходящий для целей моделирования пакет прикладных программ, как и в случае программ для имитации.
Соединение, идентифицированное в ходе скрининга in silico по настоящему изобретению, как соединение способное связываться с GPR120 или CD36, может быть далее протестировано c использованием in vitro способа по настоящему изобретению. Дополнительно или альтернативно, такое соединение может быть исследовано in vivo, например, в рамках тестов, проводимых при кормлении животных.
В еще одном аспекте, настоящее изобретение относится к соединениям, которые модулируют биологическую активность GPR120 или CD35, согласно первому аспекту настоящего изобретения.
В этой связи, способ по первому аспекту настоящего изобретения представляет собой подходящий способ скрининга, направленный на идентификацию вкусовых соединений, которые могут использоваться в корме для кошек и собак, так чтобы их применение гарантировало в течение длительного времени принятие и потребление таких кормов животными.
Таким образом, в дополнительном аспекте настоящее изобретение относится к пищевому/кормовому продукту, включающему агент или соединение, идентифицированный/идентифицированное по способу настоящего изобретения.
Рассматриваемый здесь корм может представлять собой любой корм из числа известных в данной области. Соединение, идентифицированное по способу настоящего изобретения, может быть включено в состав любого продукта, который кошка или собака может потреблять в качестве пищи. Соответственно, настоящее изобретение охватывает любые стандартные пищевые продукты, добавки, корм для домашних животных, напитки, закусочные пищевые продукты и лакомства. Указанный пищевой продукт представляет собой предпочтительно продукт, подвергнутый кулинарной обработке. Он может представлять собой мясо или продукт животного происхождения (такой как говядина, курица, индейка, ягненок, плазма крови, костный мозг и т.п., или сочетания двух или более таких продуктов). Альтернативно, указанный пищевой продукт может представлять собой не содержащий мяса продукт (предпочтительно включающий заменители мяса, такие как соя, кукурузный глютен или соевый продукт) для обеспечения в продукте источника белка. Указанный продукт может содержать дополнительные белковые источники, такие как концентрат соевого белка, молочные белки, глютен и т.п. Указанный продукт может также содержать источник крахмала, такой как желатинизированный крахмал, такой как один или несколько видов зернового продукта (например, пшеница, кукуруза, рис, ячмень, овес и т.п.) или может не содержать крахмал. Типичный сухой коммерчески доступный корм для кошек содержит примерно 10-70% неочищенного белка, примерно 10-60% жира, тогда как оставшаяся часть представляет собой углеводы, включающие пищевые волокна и зольные вещества. Настоящее изобретение, в частности, относится к корму для домашних животных, согласно настоящему описанию, который представляет собой твердую пищу, пищевой продукт или пищевую добавку для кошки или собаки. В контексте настоящего описания, термин «домашний», применительно к кошке, обозначает кошек, в частности, Felis domesticus (Felis catus), и термин «домашний», применительно к собаке, обозначает собак, в частности, Canis lupus familiaris. Предпочтительно, указанный корм для домашних животных соответствует потребностям животного в макроэлементах.
Предпочтительные особенности каждого аспекта настоящего изобретения соответствуют тем, которые были указаны для каждого из других его аспектов, включая соответствующие изменения, mutatis mutandis.
Все цитированные здесь документы приведены в настоящем описании во всей полноте, допустимой действующим законодательством.
Примеры
Далее настоящее изобретение будет описано со ссылкой на соответствующие, приведенные ниже неограничивающие примеры. В качестве ссылочного материала приведены сопроводительные чертежи, где:
На фиг.1 показана аминокислотная последовательность GPR120 кошки;
На фиг.2 показана нуклеотидная последовательность GPR120 кошки;
На фиг.3 показана аминокислотная последовательность CD36 кошки;
На фиг.4 показана нуклеотидная последовательность CD36 кошки;
На фиг.5 показана аминокислотная последовательность GPR120 собаки;
На фиг.6 показана нуклеотидная последовательность GPR120 собаки;
На фиг.7 показана аминокислотная последовательность CD36 собаки;
На фиг.8 показана нуклеотидная последовательность CD36 собаки;
На фиг.9 показана аминокислотная последовательность человеческого GPR120;
На фиг.10 показана нуклеотидная последовательность человеческого GPR120;
На фиг.11 показана аминокислотная последовательность человеческого CD36;
На фиг.12 показана нуклеотидная последовательность человеческого CD36;
На фиг.13 показано различие по последовательности между SEQ ID NO:3 и опубликованными данными по последовательности CD36 кошки;
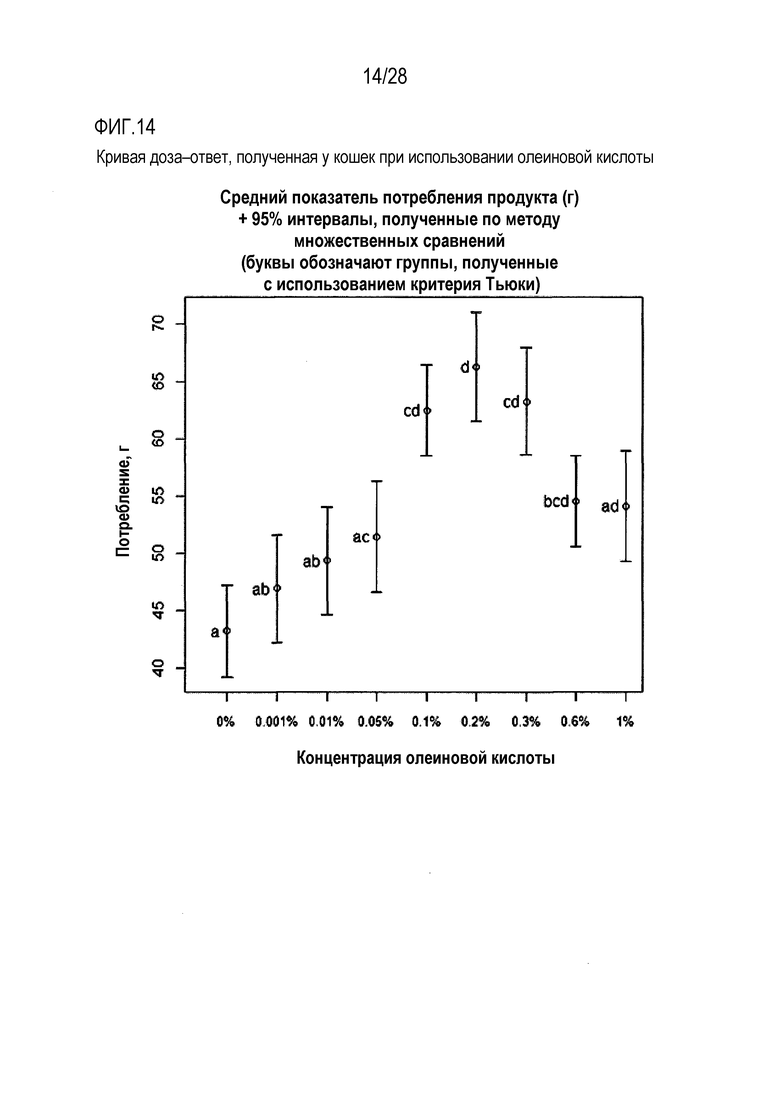
На фиг.14 приведена кривая доза-ответ для олеиновой кислоты у кошки;
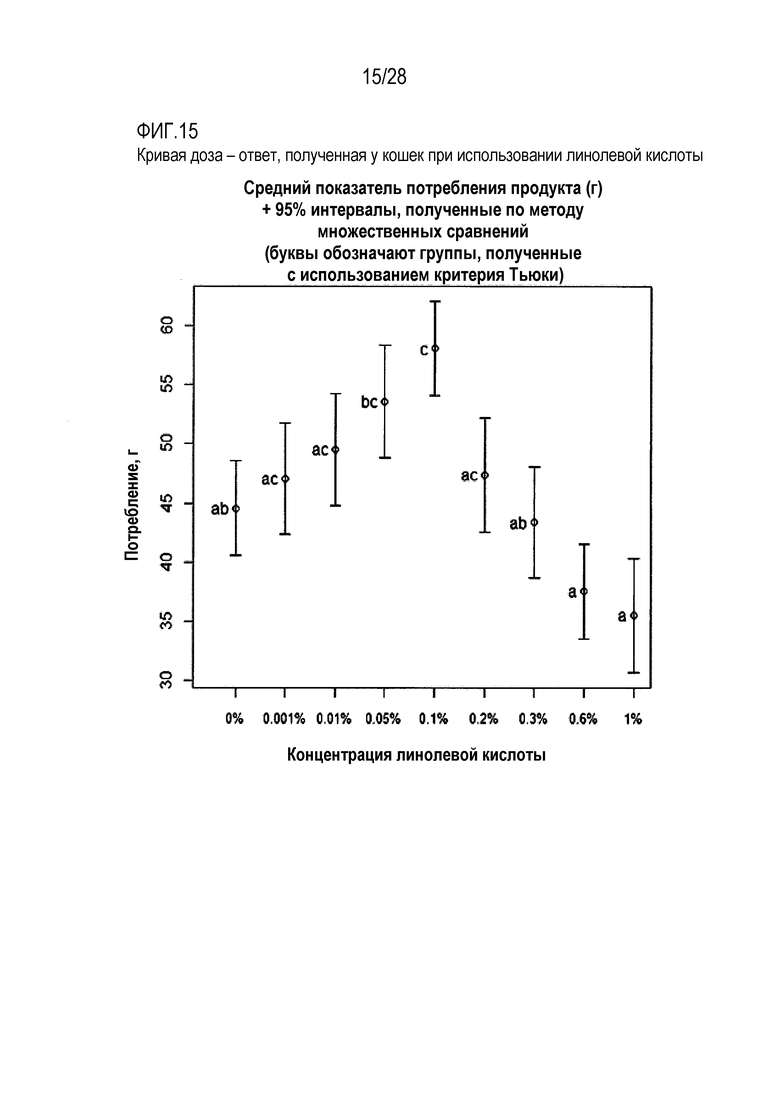
На фиг.15 приведена кривая доза-ответ для линолевой кислоты у кошки;
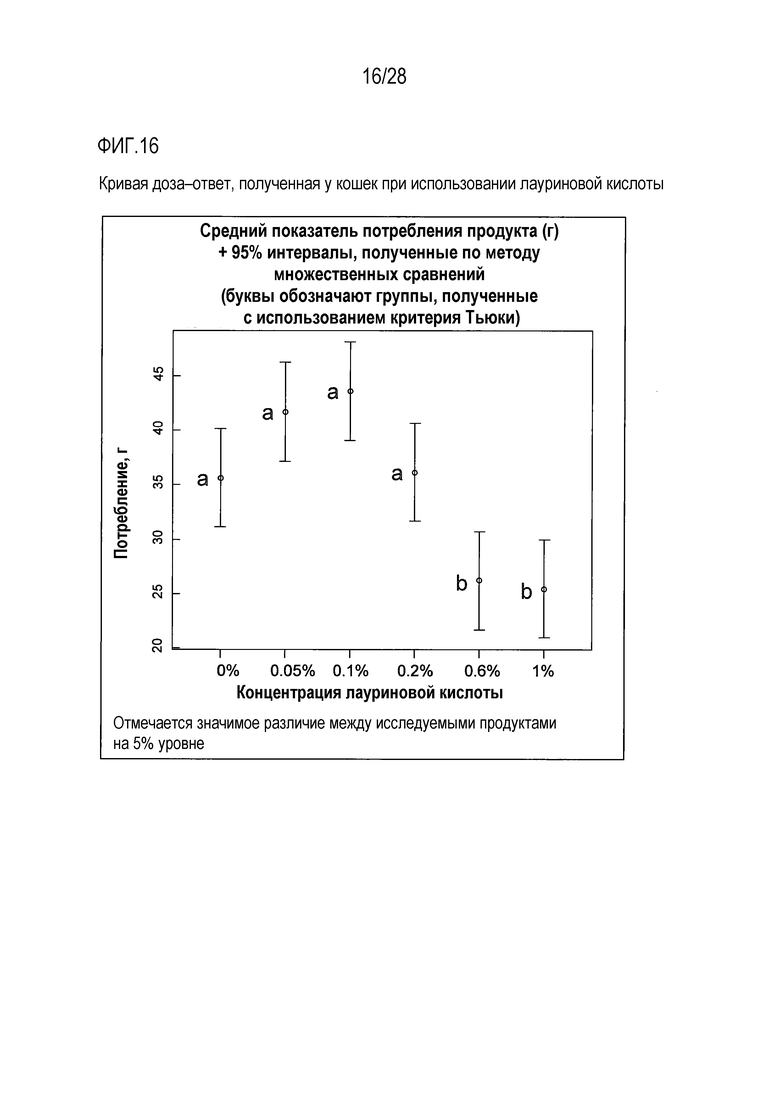
На фиг.16 приведена кривая доза-ответ для лауриновой кислоты у кошки;
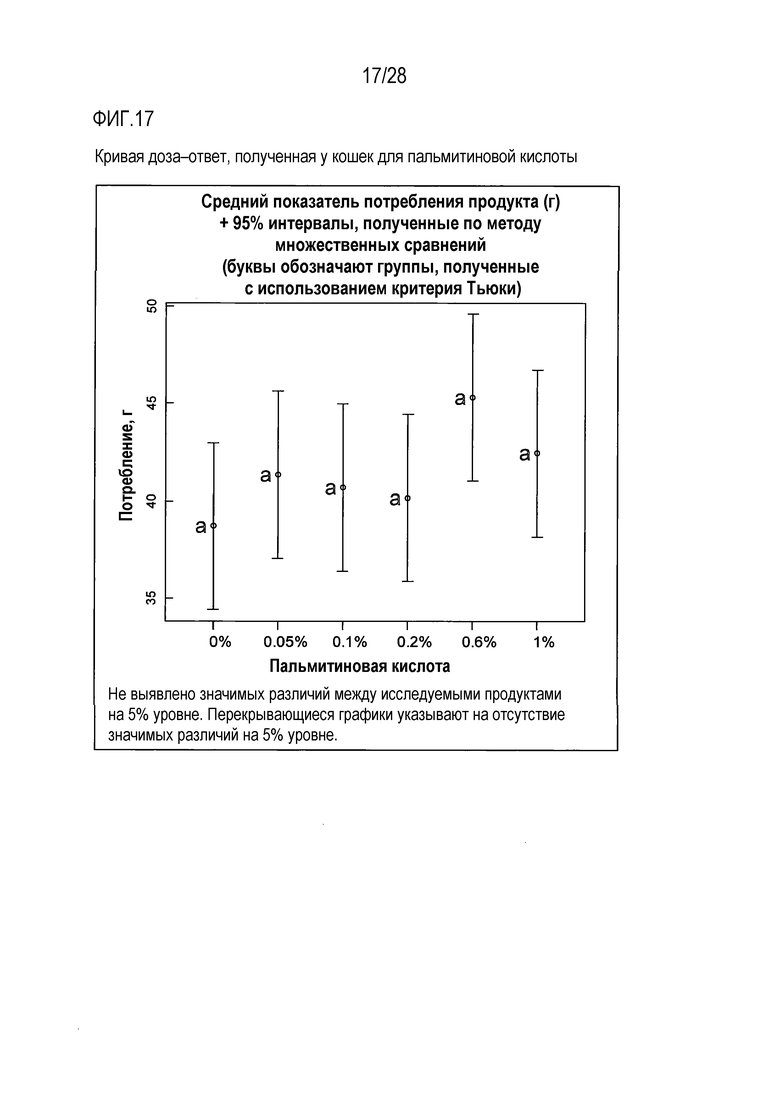
На фиг.17 приведена кривая доза-ответ для пальмитиновой кислоты у кошки;
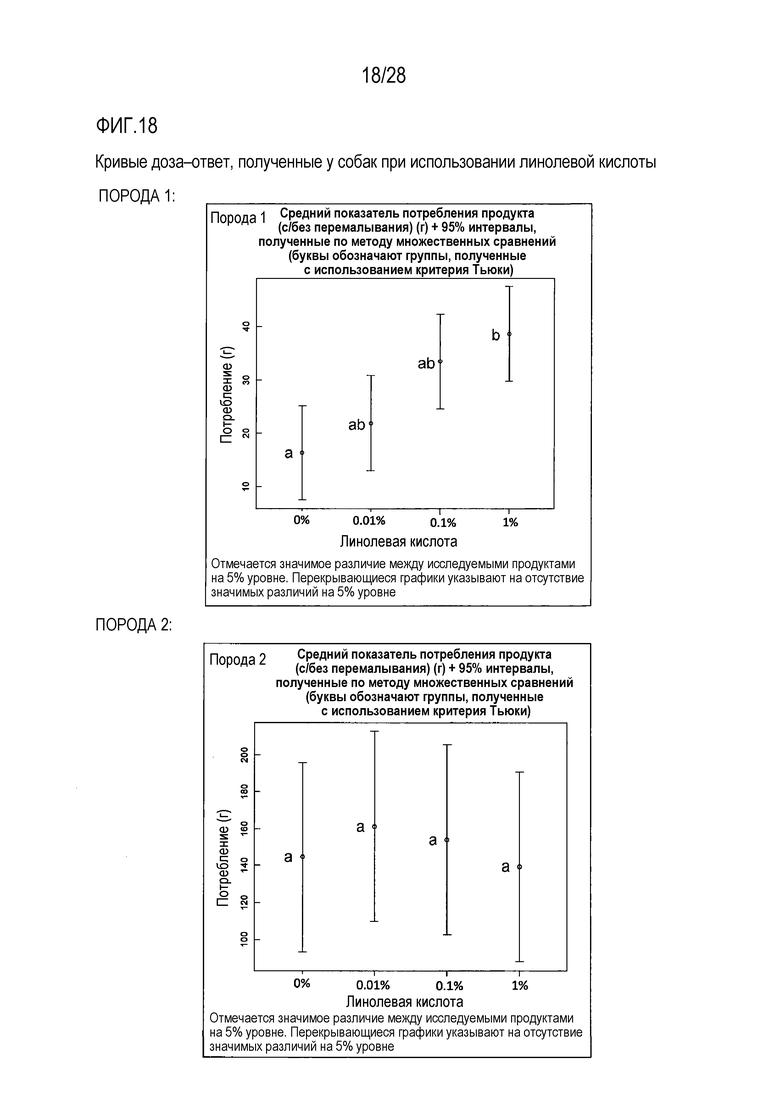
На фиг.18 приведены кривые доза-ответ для линолевой кислоты у собаки;
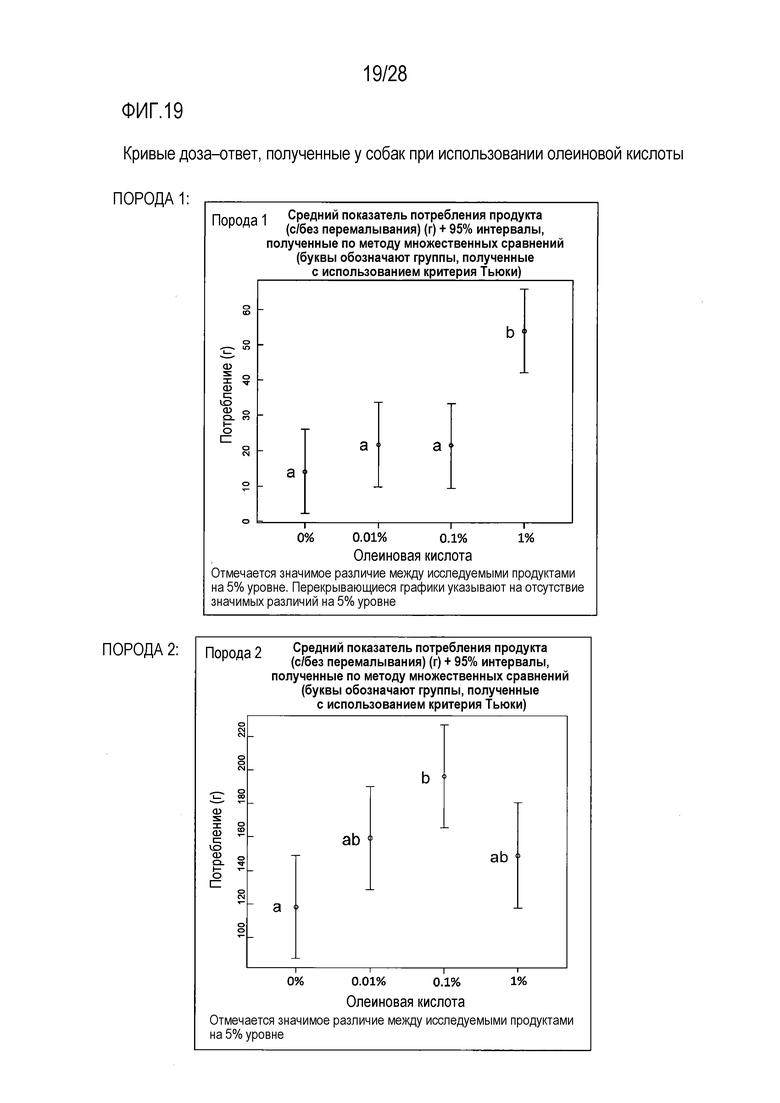
На фиг.19 приведены кривые доза-ответ для олеиновой кислоты у собаки;
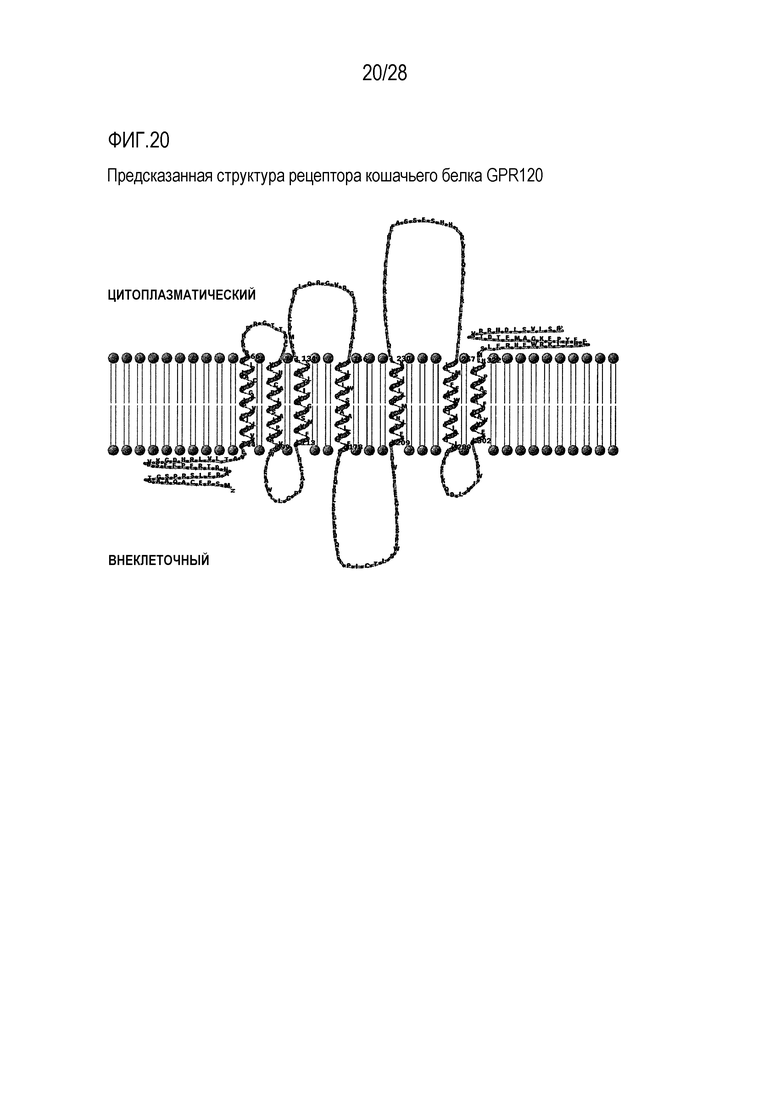
На фиг.20 показана предсказанная структура GPR120 у кошки;
На фиг.21 показана предсказанная структура человеческого CD36;
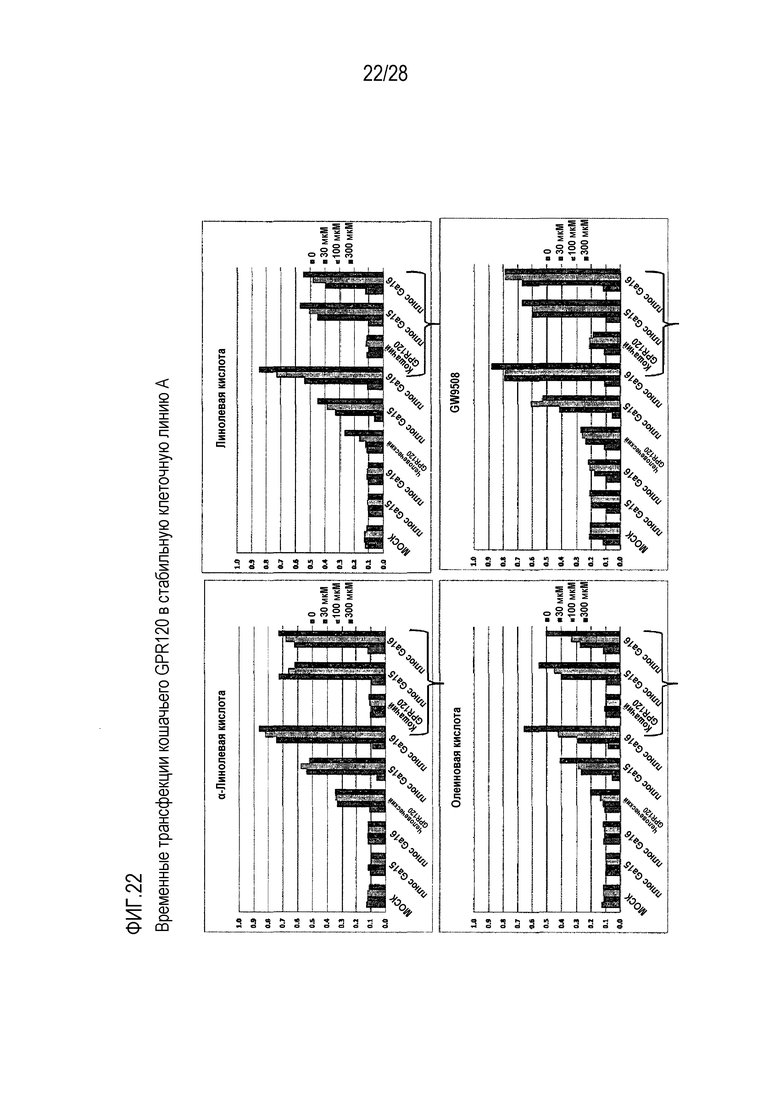
На фиг.22 проиллюстрированы временные трансфекции GPR120 кошки в стабильную клеточную линию;
На фиг.23 проиллюстрированы временные трансфекции GPR120 кошки в СНОК1 клетки;
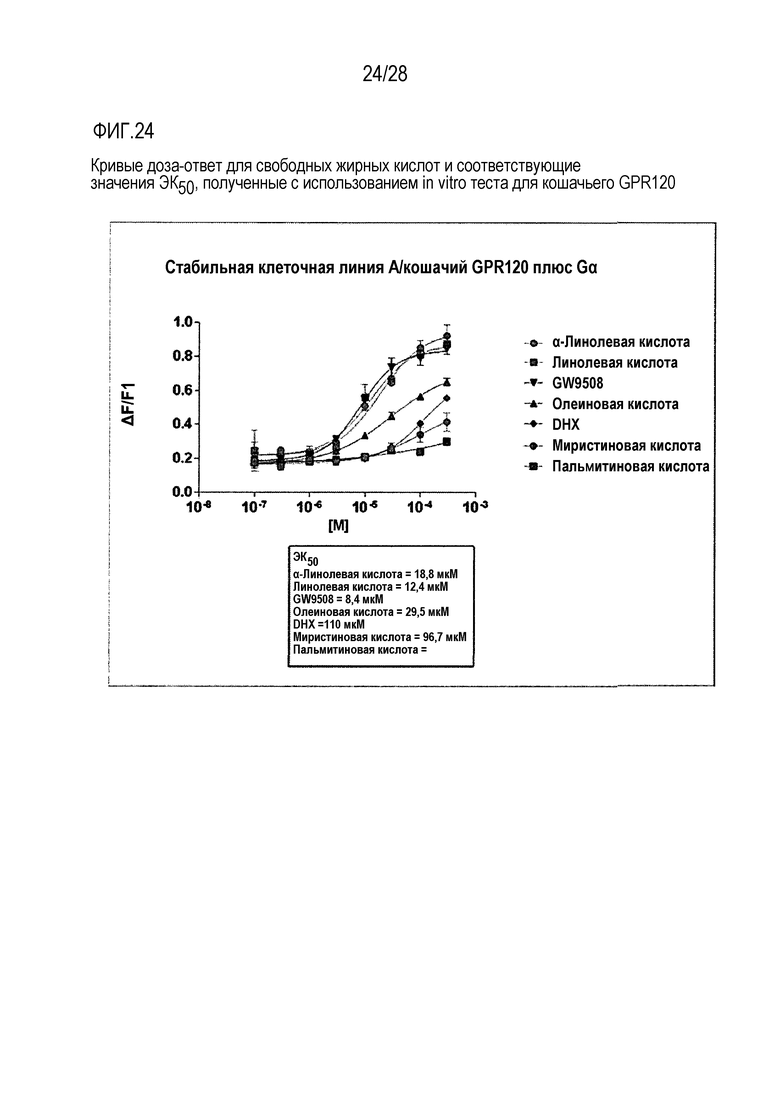
На фиг.24 приведены кривые доза-ответ для свободных жирных кислот и значение ЕК50, полученные в тесте in vitro с использованием GPR120 кошки;
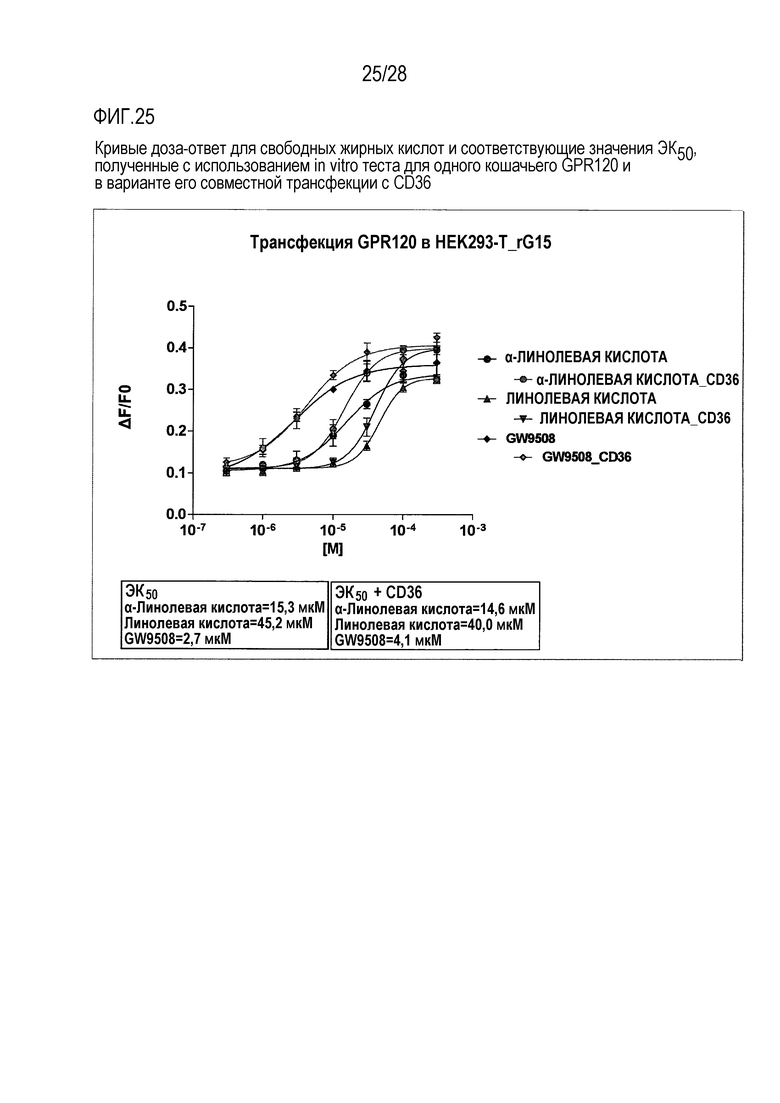
На фиг.25 приведены кривые доза-ответ для свободных жирных кислот и значение ЕК50, полученные в тесте in vitro с использованием только GPR120 кошки и при его совместной трансфекции с CD36;
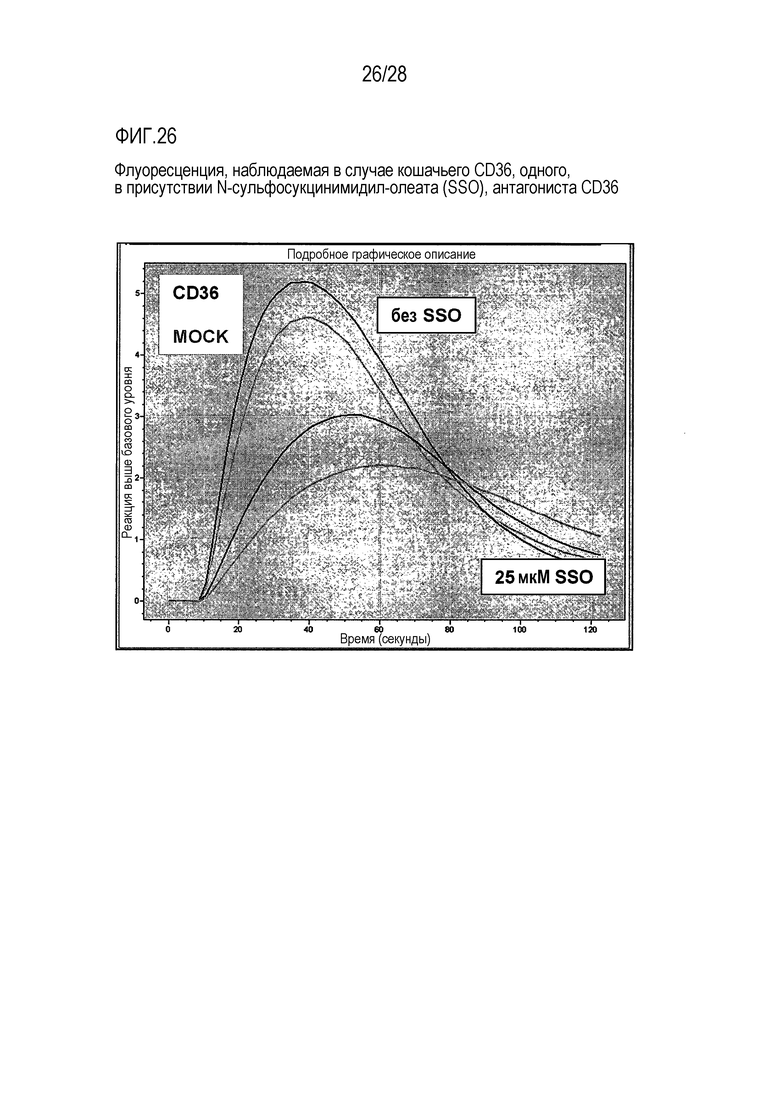
На фиг. 26 проиллюстрирована флуоресценция, наблюдаемая при использовании только CD36 кошки, в присутствии SSO, антагониста CD36;
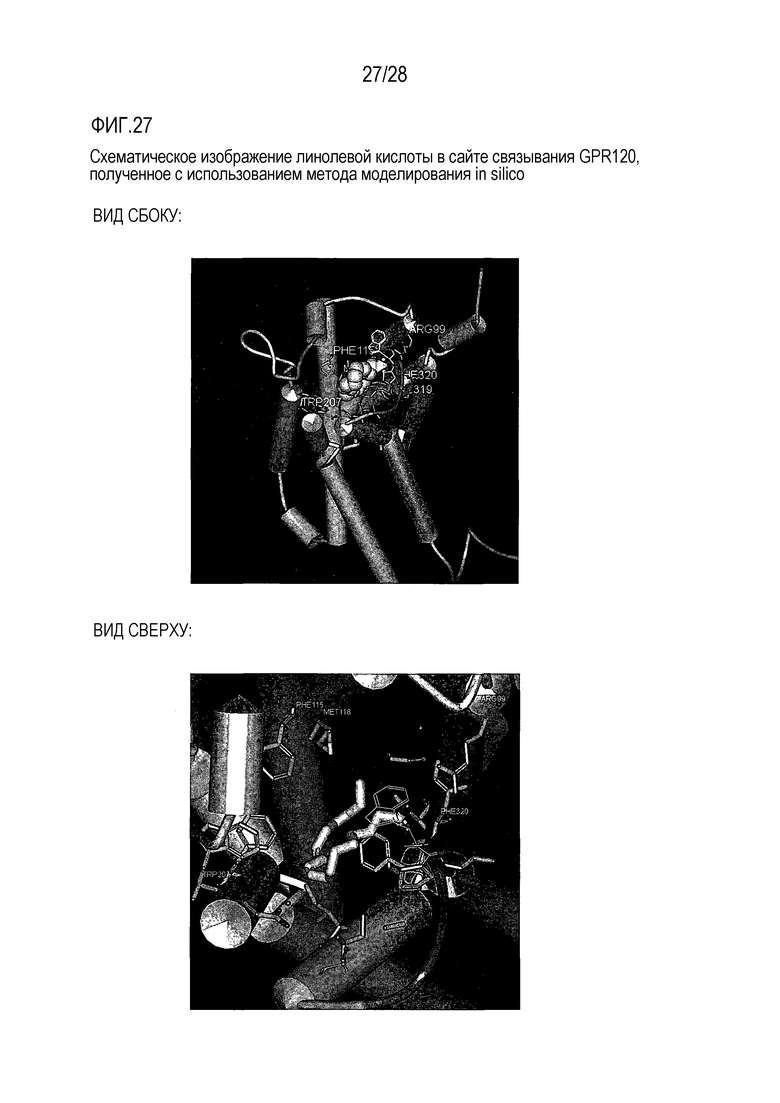
На фиг. 27 схематически, в рамках метода in silico, показана линолевая кислота в сайте связывания GPR120;
На фиг. 28 схематически, в рамках метода in silico, показана олеиновая кислота в сайте связывания GPR120.
Пример 1
Определение корректной последовательности GPR120 кошки
Выделяли ДНК от 26 кошек с использованием буккального мазка. Отбирали по 2 мазка от каждой кошки. ДНК экстрагировали с использованием набора для выделения ДНК (Qiagen DNeasy Blood and Tissue) и далее проводили ее секвенирование. Использованные при этом праймеры были сконструированы, на основании доступных данных по кошачьей геномной последовательности, таким образом, чтобы фланкировать экзонные участки. Там, где это было возможно, все экзонные участки были секвенированы в обоих направлениях. Далее, последовательности были проанализированы с использованием Sequencher v51 (Gene Codes, USA). Сравнивали консенсусные последовательности от 26 кошек с доступными данными по последовательностям и далее конструировали окончательную консенсусную последовательность для всех экзонов.
Эти последовательности построены на основании данных по ресеквенированию WCPN кошек, с использованием данных по секвенированию РНК и доступных данных по последовательностям для кошек и человека.
Подтвержденная кодирующая последовательность fGPR120 соответствует последовательности, приведенной в Ensembl. Данный вариант представляет собой корректную изоформу, тогда как другая, длинная изоформа, идентифицированная у человека, не способна к сигнальной функции.
Пример 2
Определение корректной последовательности GPR120 кошки
Выделяли ДНК от 26 кошек с использованием буккального мазка. Отбирали по 2 мазка от каждой кошки. ДНК экстрагировали с использованием набора для выделения ДНК (Qiagen DNeasy Blood and Tissue) и далее проводили ее секвенирование. Использованные при этом праймеры были сконструированы, на основании доступных данных по кошачьей геномной последовательности, таким образом, чтобы фланкировать экзонные участки. Там, где это было возможно, все экзонные участки были секвенированы в обоих направлениях. Далее, последовательности были проанализированы с использованием Sequencher v51 (Gene Codes, USA). Сравнивали консенсусные последовательности от 26 кошек с доступными данными по последовательностям и далее конструировали итоговую консенсусную последовательность для всех экзонов.
Эти последовательности построены на основании данных по ресеквенированию WCPN для кошек, с использованием данных по секвенированию РНК, данных по секвенированию кДНК из вкусовых рецепторов кошек и доступных данных по последовательностям для кошек и человека.
Доступные на Ensembl, последовательности транскриптов и в случае человека, и в случае кошки, содержат секции после первого стоп-кодона. Вполне возможно, что у кошки, как и в случае человека, первая часть последовательности транскрипта до первого стоп-кодона, представляет собой первичную кодирующую последовательность. В положении 300 находится фрагмент из 8 остатков аденина. Это отличается от предсказанной в Ensembl последовательности транскрипта, но дает в результате белок из 427 аминокислот, который соответствует другой, найденной у кошек изоформе, а также соответствует по длине человеческому белку. Таким образом, ни один из транскриптов, предсказанных на Ensemble, точно не соответствует этой последовательности, но секвенирование кДНК из вкусовых сосочков кошек показывает, что эта конфигурация транскрипта является корректной.
Пример 3
Определение корректной последовательности GPR120 собаки
Выделяли ДНК от 84 собак с использованием образца крови небольшого объема. Секвенирование полного генома с использованием платформы illuminа проводили для всех образцов с достижением среднего показателя охвата 15х. Полученные данные картировали на стандартном геноме с использованием Bowtie2. Участки, которые представляли интерес, экстрагировали внутрилабораторных Perl скриптов. Далее идентифицировали экзонные участки, с последующим конструированием консенсусной последовательности.
Указанные последовательности построены на основании данных по секвенированию генома собаки, с включением данных по секвенированию РНК и доступных данных по последовательностям для собаки и человека.
Подтвержденная кодирующая последовательность GPR120 собаки соответствует последовательности, приведенной в Ensembl.
Пример 4
Определение корректной последовательности CD36 собаки
Выделяли ДНК от 84 собак с использованием образца крови небольшого объема. Секвенирование полного генома с использованием платформы illuminа проводили для всех образцов с достижением среднего показателя охвата 15х. Полученные данные картировали на стандартном геноме с использованием Bowtie2. Участки, которые представляли интерес, экстрагировали внутрилабораторных Perl скриптов. Далее идентифицировали экзонные участки, с последующим конструированием консенсусной последовательности.
Указанные последовательности построены на основании данных по секвенированию генома собаки, с включением данных по секвенированию РНК и доступных данных по последовательностям для собаки и человека.
Подтвержденная кодирующая последовательность CD36 собаки соответствует последовательности, приведенной в Ensembl.
Пример 5
Тест на кормление кошек с целью определения их реакции на олеиновую кислоту
Была использована панель гелевых кормов для кошек с целью определения вкусовых качеств варьирующих концентраций олеиновой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 8 концентраций олеиновой кислоты, в диапазоне значений от 0,001% олеиновой кислоты до 1% олеиновой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием олеиновой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации олеиновой кислоты.
Продукты, включавшие олеиновую кислоту в концентрациях 0,1%, 0,2%, 0,3% и 0,6% (вес/объем) характеризовались более высоким уровнем потребления их в ходе тестирования, чем контрольная, слепая проба, указывая на то, что подопытные кошки были способны оценивать вкус олеиновой кислоты.
Пример 6
Тест на кормление кошек с целью определения их реакции на линолевую кислоту
Была использована панель гелевых кормов для кошек с целью определения вкусовых качеств варьирующих концентраций линолевой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 8 концентраций линолевой кислоты, в диапазоне значений от 0,001% линолевой кислоты до 1% линолевой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием линолевой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации линолевой кислоты.
Было показано, что продукт с концентрацией линолевой кислоты 0,1%, потреблялся в ходе тестирования в значительной большей мере, чем контрольная, слепая проба (с 0% содержанием линолевой кислоты). В этом эксперименте отмечалось негативное, вызывающее неприятие продукта влияние более высоких концентраций, со снижением потребления, в сравнении с контрольной, слепой пробой (с 0% содержанием линолевой кислоты), указывая на то, что подопытные кошки были способны оценивать вкус линолевой кислоты.
Пример 7
Тест на кормление кошек с целью определения их реакции на лауриновую кислоту
Была использована панель гелевых кормов для кошек с целью определения вкусовых качеств варьирующих концентраций лауриновой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 5 концентраций лауриновой кислоты, в диапазоне значений от 0,05% лауриновой кислоты до 1% лауриновой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием лауриновой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации лауриновой кислоты.
Лауриновая кислота в концентрации 0,1% отличалась наивысшим уровнем потребления в ходе тестирования, в сравнении с контрольной, слепой пробой (с 0% содержанием лауриновой кислоты). Тогда как самые высокие исследованные концентрации, 0,6% и 1% лауриновой кислоты оказывали негативное, вызывающее неприятие продукта влияние, в сравнении с контрольной, слепой пробой (с 0% содержанием лауриновой кислоты), указывая на то, что подопытные кошки были способны оценивать вкус лауриновой кислоты.
Пример 8
Тест на кормление кошек с целью определения их реакции на пальмитиновую кислоту
Была использована панель гелевых кормов для кошек с целью определения вкусовых качеств варьирующих концентраций пальмитиновой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 5 концентраций пальмитиновой кислоты, в диапазоне значений от 0,05% пальмитиновой кислоты до 1% пальмитиновой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием пальмитиновой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации пальмитиновой кислоты.
В ходе тестировано не было выявлено существенных различий в потреблении продуктов с разными исследованными концентрациями пальмитиновой кислоты, в связи с тем, что она представляет собой твердое вещество при комнатной температуре (точка плавления 63°С). В этой связи, пальмитиновая кислота не могла взаимодействовать с рецепторами жирной кислоты и связываться с этими рецепторами, вызывая при этом вкусовую реакцию у кошек.
Пример 9
Тест на кормление собак с целью определения их реакции на линолевую кислоту
Были исследованы две разные группы собак с целью оценки вкусовых качеств разных концентраций линолевой кислоты. Каждая группа представляла собой одну породу собак; каждая другая группа представляла собой другую породу.
Была использована панель гелевых кормов для собак с целью определения вкусовых качеств варьирующих концентраций линолевой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 3 концентраций линолевой кислоты, в диапазоне значений от 0,01% линолевой кислоты до 1% линолевой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием линолевой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации линолевой кислоты.
Разные породы собак демонстрировали разную реакцию на линолевую кислоту. Порода 1 характеризовалась выраженной позитивной, в плане усиления потребления, реакцией на линолевую кислоту в ее наивысшей исследованной концентрации 1%, в сравнении с контрольной, слепой пробой (с 0% содержанием линолевой кислоты), тогда как в случае породы 2 наблюдалось менее выраженные различия между исследованными в ходе тестирования концентрациями линолевой кислоты.
Пример 10
Тест на кормление собак с целью определения их реакции на олеиновую кислоту
Были исследованы две разные группы собак с целью оценки вкусовых качеств разных концентраций олеиновой кислоты. Каждая группа представляла собой одну породу собак; каждая другая группа представляла собой другую породу.
Была использована панель гелевых кормов для собак с целью определения вкусовых качеств варьирующих концентраций олеиновой кислоты в монадической среде. Характер доза-ответ оценивали с использованием 3 концентраций олеиновой кислоты, в диапазоне значений от 0,01% олеиновой кислоты до 1% олеиновой кислоты. Все продукты (включая слепую пробу, представлявшую собой вариант с 0% содержанием олеиновой кислоты) содержали 25 мМ L-гистидина, в качестве потребляемого/положительного вкусового вещества, для повышения базового потребления геля, что позволяло идентифицировать потенциальное негативное влияние той или иной концентрации олеиновой кислоты.
Разные породы собак демонстрировали разную реакцию на олеиновую кислоту. Порода 1 характеризовалась выраженной позитивной, в плане усиления потребления, реакцией на олеиновую кислоту в ее наивысшей исследованной концентрации 1%, в сравнении с контрольной, слепой пробой (с 0% содержанием олеиновой кислоты), тогда как в случае породы 2 наблюдалось более выраженная позитивная, в плане усиления потребления, реакция на олеиновую кислоту в концентрации 0,1%, в сравнении с контрольной, слепой пробой (с 0% содержанием олеиновой кислоты).
Пример 11
Разработка способа тестирования in vitro рецепторов GPR120 и GPR120+CD36 и его применение
Вначале, генные последовательности для целевых рецепторов GPR120+CD36 подтверждали путем ресеквенирования генов от кошек и собак.
В случае GPR120 (FFAR4, O3FAR1), полученные последовательности сравнивали с доступными стандартными последовательностями кошки и собаки, а также со стандартной человеческой последовательностью. Использовали последовательности, относящиеся к короткой изоформе.
В случае CD36, полученные последовательности сравнивали с доступными стандартными последовательностями кошки и собаки, а также со стандартной человеческой последовательностью.
После установления целевых последовательностей, проводили их синтез и клонирование в векторах экспрессии pcDNA3.1Hydro и pcDNA3.1G418. Далее, эти конструкции подвергали временной трансфекции в иммортализованную клеточную линию СНО К1 и другие обычно применяемые клеточные линии с использованием Липофектамина 2000 (Lipofectamine 2000) и проводили дальнейшее тестирование с целью достижения удачной экспрессии целевого белка. Указанное тестирование проводили с использованием кальций-чувствительного флуоресцентного красителя (Fluo8). Трансфицированные клетки высевали в 384-ячеечные аналитические планшеты. При внесении исследуемых клеток с красителем, с последующей провокацией клеток агонистом для рецептора, ответную реакцию клеток можно зафиксировать на приборе FLIPRTETRA путем измерения усиления флуоресценции, ассоциированной с внутриклеточным высвобождением кальция, что, таким образом, будет подтверждать функциональную экспрессию рецептора. Применение в этом случае соответствующих экспериментальных контролей позволяет оценивать наблюдаемую реакцию как специфическую, поскольку устранена любая возможность того, что эта реакция является неспецифической или того, что усиление флуоресценции определяется иными факторами, нежели высвобождение внутриклеточного кальция клетками.
Как для человеческого, так и для кошачьего GPR120 были продемонстрированы специфические реакции на жирные кислоты в микромолярном диапазоне, в случае их временной экспрессии в клеточной линии А или клеточной линии СНОК1 (фиг.24 и фиг.25, соответственно). При этом, в случае человеческого рецептора не требовалось наличие экзогенного G-белка, тогда как в случае экспрессии кошачьего рецептора в стабильной клеточной линии А он был необходим. Для всех исследованных соединений были построены кривые доза-ответ (фиг.26) и вычислены значения показателя ЕК50. На фиг.27 проиллюстрирован эффект совместной трансфекции GPR120 и CD36.
Пример 12
Разработка способа тестирования in vitro рецептора CD36 и его применение
Вначале, генную последовательность для целевого рецептора CD36 подтверждали путем ресеквенирования генов от кошек и собак.
Полученные CD36 последовательности сравнивали с доступными стандартными последовательностями кошки и собаки, а также со стандартной человеческой последовательностью.
После установления целевой последовательности, проводили ее синтез и клонирование в векторе экспрессии pcDNA3.1Hydro. Далее, эту конструкцию подвергали временной трансфекции в иммортализованную клеточную линию СНО К1 и другие обычно применяемые клеточные линии с использованием Липофектамина 2000 (Lipofectamine 2000) и проводили дальнейшее тестирование с целью достижения удачной экспрессии целевого белка. Указанное тестирование проводили с использованием кальций-чувствительного флуоресцентного красителя (Fluo8). Трансфицированные клетки высевали в 384-ячеечные аналитические планшеты. При внесении исследуемых клеток с красителем, с последующей провокацией клеток агонистом для рецептора, ответную реакцию клеток можно зафиксировать на приборе FLIPRTETRA путем измерения усиления флуоресценции, ассоциированной с внутриклеточным высвобождением кальция, что, таким образом, будет подтверждать функциональную экспрессию рецептора. Применение в этом случае соответствующих экспериментальных контролей позволяет оценивать наблюдаемую реакцию как специфическую, поскольку устранена любая возможность того, что эта реакция является неспецифической или того, что усиление флуоресценции определяется иными факторами, нежели высвобождение внутриклеточного кальция клетками.
Дальнейшие эксперименты с использованием CD36 проводили с целью выяснения, будет ли сульфо-N-сукцинимидил-олеат (SSO), как предположительный антагонист CD36, ингибировать CD36-опосредованный инфлюкс кальция после предварительной обработки Тапсигаргином (Thapsigargin). Реакция клеток, в которые был трансфицирован CD36, или клеток, взятых в качестве ложного контроля, показана на фиг.28.
Пример 13
Разработка модели для оценки рецептора GPR120 in silico и ее применение
Модели GPR120 были разработаны с использованием, в качестве матриц для целей моделирования гомологий, кристаллических структур GPCR из группы А, которые доступны от банка белков (Protein Data Bank). Использовали пакет прикладных программ Modeler.
Проводили также процедуры имитации и минимизации, применительно к индивидуальным свободным жирным кислотам (например, линолевой кислоте). Для этой цели использовали программу Sharmm из Discovery Studio.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ИДЕНТИФИКАЦИИ МОДУЛЯТОРОВ GPR92 | 2017 |
|
RU2742608C2 |
| СОЕДИНЕНИЯ, МОДУЛИРУЮЩИЕ АКТИВНОСТЬ РЕЦЕПТОРОВ GPR92, И КОРМОВЫЕ ПРОДУКТЫ ДЛЯ ДОМАШНИХ ЖИВОТНЫХ, СОДЕРЖАЩИЕ ИХ | 2019 |
|
RU2807217C2 |
| СОЕДИНЕНИЯ, МОДУЛИРУЮЩИЕ АКТИВНОСТЬ КАЛЬЦИЙЧУВСТВИТЕЛЬНЫХ РЕЦЕПТОРОВ, ДЛЯ МОДУЛЯЦИИ ВКУСА КОКУМИ И КОРМОВЫЕ ПРОДУКТЫ ДЛЯ ДОМАШНИХ ЖИВОТНЫХ, СОДЕРЖАЩИЕ ИХ | 2017 |
|
RU2759563C2 |
| МОДИФИЦИРОВАННЫЙ ЭРИТРОПОЭТИН, К КОТОРОМУ ПРИСОЕДИНЕНА ВОДОРАСТВОРИМАЯ ДЛИННОЦЕПОЧЕЧНАЯ МОЛЕКУЛА | 2010 |
|
RU2549986C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640254C2 |
| ИНДУЦИРУЮЩИЙ ИММУНИТЕТ АГЕНТ | 2009 |
|
RU2511039C2 |
| ПЕПТИДНЫЕ ВАКЦИНЫ ПРОТИВ ИНТЕРЛЕЙКИНА-31 | 2019 |
|
RU2786441C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2761663C2 |
| КАНИНИЗИРОВАННЫЕ МЫШИНЫЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ PD-1 | 2014 |
|
RU2676158C1 |
| АНТИТЕЛА ПРОТИВ NGF И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2019 |
|
RU2838175C2 |


































Изобретение относится к биотехнологии и представляет собой способ идентификации соединения, которое модулирует активность полипептида, включающего GPR120 кошки, который содержит аминокислотную последовательность, приведенную в SEQ ID NO:1, или ее функциональный фрагмент, где способ включает: контактирование полипептида с соединением; измерение биологической активности полипептида в отсутствие и в присутствии соединения и идентификацию соединения в качестве соединения, которое модулирует биологическую активность полипептида, в том случае, когда имеется разница по биологической активности в отсутствие соединения и в присутствии соединения. Изобретение позволяет модулировать вкусоароматику пищевых продуктов. 4 н. и 9 з.п. ф-лы, 28 ил., 3 пр.
1. Способ идентификации соединения, которое модулирует активность полипептида, включающего GPR120 кошки, который содержит аминокислотную последовательность, приведенную в SEQ ID NO:1, или ее функциональный фрагмент,
где способ включает:
(а) контактирование полипептида с соединением;
(b) измерение биологической активности полипептида в отсутсвие и в присутствии соединения и
(с) идентификацию соединения в качестве соединения, которое модулирует биологическую активность полипептида, в том случае, когда имеется разница по биологической активности в отсутствие соединения и в присутствии соединения.
2. Способ идентификации вкусообразующего соединения, содержащий:
(а) контактирование полипептида, содержащего GPR120 кошки, с соединением;
(b) измерение биологической активности полипептида в отсутствие и в присутствии соединения и
(с) идентификацию соединения в качестве вкусообразующего соединения в том случае, когда имеется разница по биологической активности в отсутствие соединения и в присутствии соединения,
где GPR120 кошки содержит аминокислотную последовательность, приведенную в SEQ ID NO:1, или ее функциональный фрагмент.
3. Способ по п. 1 или 2, дополнительно включающий:
(a) прогнозирование 3-мерной (3D)структуры полипептида;
(b) скрининг предсказанной 3D структуры полипептида in silico на основе 3D структуры исследуемого соединения;
(c) выявление, будет ли данное исследуемое соединение соответствовать с сайтом связывания полипептида; и
(d) идентификацию исследуемого соединения в качестве соединения, которое связывает полипептид и модулирует биологическую активность полипептида, в том случае, когда 3D структура такого соединения соответствует сайту связывания в 3D структуре полипептида.
4. Способ по любому из пп. 1, 2, где биологическую активность полипептида измеряют в присутствии второго полипептида, содержащего CD36.
5. Способ по любому из пп. 1, 2, где биологическую активность полипептида измеряют в присутствии второго полипептида, содержащего экзогенный G-белок.
6. Способ по любому из пп. 1, 2, где полипептид экспрессируется в клетке.
7. Способ по любому из пп. 1, 2, где полипептид экспрессируется вектором.
8. Способ по любому из пп. 1, 2, где биологическую активность полипептида измеряют путем мониторинга концентрации кальция или активности cGMP в клетке.
9. Способ по любому из пп. 1, 2, где концентрацию кальция мониторят с помощью флуоресцентной детекции или люминесцентной детекции.
10. Способ по п. 9, где флуоресцентная детекция включает флуоресцирующую краску, чувствительную к кальцию.
11. Способ по любому из пп. 1, 2, дополнительно включающий тестирование соединения при исследовании питания животных.
12. Набор для определения, происходит ли активация соединением полипептида, включающего GPR120, содержащий аминокислотную последовательность, приведенную в SEQ ID NO:1, или ее функциональный фрагмент, где набор содержит указанный полипептид и одно или несколько исследуемых соединений.
13. Слитый белок для идентификации соединения, которое модулирует активность полипептида, включающий полипептид, который включает GPR120 кошки, содержащий аминокислотную последовательность, приведенную в SEQ ID NO:1, или ее функциональный фрагмент, и дополнительный компонент, выбранный из группы, содержащей детектируемый маркер, функциональную группу или стабилизирующую последовательность.
| NCBI DATABASE ХР_003982755.1, 15.02.2013, найдено в Интернет по адресу https://www.ncbi.nlm.nih.gov/protein/410952168?sat=18&satkey=3322721; | |||
| WO 2013050523 A1, 11.04.2013 | |||
| УЛУЧШИТЕЛЬ ОСНОВНОГО ВКУСА, СОДЕРЖАЩИЙ ВЫСШУЮ ДЛИННОЦЕПОЧЕЧНУЮ НЕНАСЫЩЕННУЮ ЖИРНУЮ КИСЛОТУ И/ИЛИ ЕЕ ЭФИР | 2004 |
|
RU2340213C2 |

