Область изобретения
Настоящее изобретение относится к мутанту фосфолипазы С с высокой ферментативной активностью.
Предшествующий уровень техники
Дегуммирование является важной стадией в очистке масел и жиров. Традиционный способ дегуммирования путем гидратации подразумевает большие финансовые затраты, большой расход материалов и энергии и серьезное загрязнение окружающей среды. Поэтому в последние годы много усилий было приложено для применения ферментативного дегуммирования в процессе дегуммирования при очистке масел и жиров, и был достигнут большой прогресс. По сравнению с традиционными способами ферментативное дегуммирование может увеличивать экономическую выгоду, уменьшать энергетические затраты, снижать выбросы, уменьшать экологическое загрязнение окружающей среды, то есть имеет огромные преимущества в отношении защиты окружающей среды, экономической целесообразности и качества. Ферментом, используемым в дегуммировании масел и жиров, является фосфолипаза. По сравнению с другими ферментами для дегуммирования, фосфолипаза С (PLC) обладает существенными преимуществами, такими как увеличение выхода диацилглицерина (DAG) и уменьшение потерь выхода масел.
PC-PLC из Bacillus cereus (BC-PC-PLC) представляет собой фосфолипазу С, которую изучали ранее. Полная длина BC-PC-PLC составляет 283 аминокислоты, включая сигнальный пептид из 24 аминокислот и лидерный пептид из 14 аминокислот. Зрелый пептид BC-PC-PLC имеет 245 аминокислот (Johansen et al. 1988). Известно, что кристаллическая структура BC-PC-PLC состоит из множественных спиральных доменов с каталитическим сайтом D55 и по меньшей мере тремя сайтами связывания Zn2+ (Hough et al. 1989). Менее изучена гетерологичная экспрессия BC-PC-PLC за исключением таковой в Bacillus subtilis и Pichia pastoris (Durban et al.2007; Seo et al. 2004).
В предыдущих исследованиях авторы настоящего изобретения получили мутант со значительно повышенной по сравнению с диким типом специфической ферментативной активностью путем введения мутаций в N56, N63, N131 и N134 BC-PC-PLC с заменой на Н, D, S и D, соответственно. Для дополнительного повышения специфической ферментативной активности фосфолипазы С и эффективности дегуммирования с использованием фосфолипазы С авторы настоящего изобретения выбрали аминокислоты по положениям 6, 8, 10, 104 и 205 в BC-PC-PLC для случайного насыщающего мутагенеза. Полагают, что путем направленного развития существующей мутантной фосфолипазы С специфическая ферментативная активность может быть дополнительно повышена, с тем чтобы получить более эффективную фосфолипазу С, улучшить эффективность дегуммирования и увеличить выход диацилглицерина (DAG) в процессе дегуммирования.
Краткое изложение сущности изобретения
В настоящем изобретении предложен выделенный полипептид, выбранный из группы, состоящей из:
(1) полипептида, представленного в SEQ ID NO: 7; и
(2) полипептида, имеющего идентичность последовательности SEQ ID NO: 7 по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, где аминокислотные остатки полипептида, соответствующие по меньшей мере одной из аминокислот 6, 8, 10 и 104 в SEQ ID NO: 7, являются такими же как аминокислоты 6, 8, 10 и/или 104 в SEQ ID NO: 7, соответственно, и полипептид обладает активностью фосфолипазы С с SEQ ID NO: 7.
В одном или более чем одном воплощении аминокислота 6 в SEQ ID NO: 7 представляет собой пролин или триптофан; аминокислота 8 представляет собой аланин, лейцин или изолейцин; каждая из аминокислот 10 и 104 независимо представляет собой серии или треонин.
В одном или более чем одном воплощении выделенный полипептид является таким, как представлено в SEQ ID NO: 4.
В данном изобретении дополнительно предложен выделенный полипептид, имеющий идентичность последовательности SEQ ID NO: 4 по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, где аминокислотные остатки выделенного полипептида, соответствующие аминокислотам 6, 8, 10 и 104 в SEQ ID NO: 4, представляют собой пролин, валин, серии и серии, соответственно; предпочтительно, полипептид получен из Bacillus subtilis.
Также в настоящем изобретении предложена полинуклеотидная последовательность, выбранная из группы, состоящей из:
(1) полинуклеотидной последовательности, кодирующей полипептид согласно любому из воплощений, изложенных здесь; и
(2) последовательности, комплементарной полинуклеотидной последовательности (1); и
(3) фрагмента от 15 до 30 п. о. (пар оснований) последовательности (1) или (2); предпочтительно, полинуклеотидная последовательность является такой, как
представлено в SEQ ID NO: 3.
Также в настоящем изобретении предложена конструкция нуклеиновой кислоты, содержащая полинуклеотидную последовательность согласно любому из воплощений, изложенных здесь, и одну или более чем одну регуляторную последовательность, функционально связанную с данной полинуклеотидной последовательностью.
В одном или более чем одном воплощении конструкция нуклеиновой кислоты представляет собой вектор.
В одном или более чем одном воплощении вектор представляет собой экспрессионный вектор или клонирующий вектор.
Также в настоящем изобретении предложена генетически сконструированная клетка-хозяин, содержащая полинуклеотидную последовательность или конструкцию нуклеиновой кислоты согласно любому из воплощений, изложенных здесь.
Также в настоящем изобретении предложена композиция, содержащая полипептид согласно любому из раскрытых здесь воплощений и возможно адъюванты, предпочтительно адъюванты представляют собой адсорбирующие вещества, выбранные из группы, состоящей из: активированного угля, оксида алюминия, диатомовой земли, пористых керамических веществ и пористого стекла.
Также в настоящем изобретении предложено применение полипептида, полинуклеотидной последовательности, конструкции нуклеиновой кислоты, клетки-хозяина или композиции согласно любому из раскрытых здесь воплощений, в очистке масел и жиров, модификации фосфолипидов, модификации пищевых продуктов, пищевой промышленности и фармацевтической промышленности.
Также в настоящем изобретении предложен способ ферментативного дегуммирования, включающий дегуммирование с использованием полипептида согласно любому из раскрытых здесь воплощений; предпочтительно способ включает стадию приведения в контакт полипептида с неочищенным маслом; предпочтительно эта стадия включает приведение в контакт полипептида с неочищенным маслом при 50-70°С.
В одном или более чем одном воплощении способ включает один или более чем один из следующих признаков:
(1) исходя из массы неочищенного масла, полипептид добавляют в количестве от 10 до 1000 миллионных долей (м.д.), предпочтительно от 50 до 500 м.д., более предпочтительно от 100 до 300 м.д.;
(2) дегуммирование включает перемешивание при 50-60°С в течение 1-3 часов и затем повышение температуры до 80-90°С и выдерживание в течение 1-10 минут; и
(3) неочищенное масло выбрано из группы, состоящей из соевого масла, подсолнечного масла, арахисового масла, рапсового масла, масла из рисовых отрубей, кукурузного масла, оливкового масла, пальмового масла, пальмоядрового масла, пальмового олеина, масла канола, касторового масла, кокосового масла, кориандрового масла, хлопкового масла, масла лесного ореха, конопляного масла, льняного масла, масла из косточек манго, масла пенника лугового, копытного масла, сафлорового масла, масла камелии (camellia), талового масла и масла камелии (tsubaki).
Краткое описание графических материалов
Фиг. 1: Специфическая ферментативная активность мутантов.
Фиг. 2: Результаты теста дегуммирования в небольшом объеме.
Фиг. 3: Стандартная кривая для определения активности фосфолипазы.
Подробное описание изобретения
Следует понимать, что в объеме настоящего изобретения вышеизложенные технические признаки изобретения и технические признаки, специально описанные далее (например, в Примерах), можно комбинировать друг с другом, тем самым образуя предпочтительное(ые) техническое(ие) решение(я).
В настоящем изобретении предложен выделенный полипептид с аминокислотной последовательностью, представленной в SEQ ID NO: 7, или выделенный полипептид, имеющий идентичность последовательности SEQ ID NO: 7 по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, где аминокислотные остатки полипептида, соответствующие по меньшей мере одной из аминокислот 6, 8, 10 и 104 в SEQ ID NO: 7, являются такими же как аминокислоты 6, 8, 10 и/или в 104 SEQ ID NO: 7, соответственно, и полипептид обладает активностью фосфолипазы С с SEQ ID NO: 7.
Здесь термин «выделенный» означает форму или вещество, которые не существуют в природе. Неограничивающие примеры выделенных веществ включают любое не встречающееся в природе вещество и любое вещество, которое по меньшей мере частично отделено от одного или более чем одного или всех из встречающихся в природе компонентов, ассоциированных с ним в природе, включая любой фермент, вариант, нуклеиновую кислоту, белок, пептид или кофактор, но не ограничиваясь ими. Для SEQ ID NO: 7 аминокислотные остатки (Хаа) в положениях 6 и 8 могут представлять собой аминокислотные остатки, имеющие неполярную боковую цепь, включая аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин и триптофан, но не ограничиваясь ими; аминокислотные остатки (Хаа) в положениях 10 и 104 могут представлять собой аминокислоты, имеющие незаряженную полярную боковую цепь, включая глицин, аспарагин, глутамин, серии, треонин, тирозин или цистеин, но не ограничиваясь ими. Предпочтительно аминокислота 6 в SEQ ID NO: 7 представляет собой пролин или триптофан; аминокислота 8 представляет собой аланин, лейцин или изолейцин; каждая из аминокислот 10 и 104 независимо представляет собой серии или треонин. Аминокислотная последовательность репрезентативного полипептида показана в SEQ ID NO: 4.
Также настоящее изобретение включает полипептиды, имеющие одну или более чем одну (обычно от 1 до 10, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) аминокислотную мутацию (делецию, вставку и/или замену) на основе SEQ ID NO: 7, в то же время сохраняя активность фосфолипазы С, присущей аминокислотной последовательности, представленной в SEQ ID NO: 4. В некоторых воплощениях аминокислотная мутация представляет собой добавление одной или нескольких (обычно в пределах 20, предпочтительно в пределах 10, более предпочтительно в пределах 8) аминокислот на С-конце и/или N-конце SEQ ID NO: 7. Следует понимать, что одна или более чем одна аминокислотная мутация обычно не включает случай, где все аминокислоты (Хаа) в положениях 6, 8, 10 и 104 в SEQ ID NO: 7 мутированы.
Предпочтительно мутация представляет собой консервативную замену. Например, в данной области техники консервативная замена с использованием аминокислот с близкими или похожими свойствами обычно не изменяет функцию данного полипептида или белка. Выражение «аминокислоты с близкими или похожими свойствами» включает, например, семейство аминокислот, имеющих похожие боковые цепи. Эти семейства включают аминокислоты с основной боковой цепью (например, лизин, аргинин, гистидин), аминокислоты с кислотной боковой цепью (например, аспартат, глутамат), аминокислоты с незаряженной полярной боковой цепью (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), аминокислоты с неполярной боковой цепью (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), аминокислоты с β-разветвленной боковой цепью (например, треонин, валин, изолейцин) и аминокислоты с ароматической боковой цепью (например, тирозин, фенилаланин, триптофан, гистидин). Следовательно, замена одного или нескольких аминокислотных остатков в полипептиде по настоящему изобретению на другой аминокислотный остаток из семейства аминокислот того же класса боковых цепей не будет существенно изменять активность полипептида.
В настоящем изобретении термин «случайный насыщающий мутагенез» относится к процессу использования NNK вырожденных кодонов в ПЦР-праймерах для введения мутаций в сайт, который может охватывать все 20 аминокислот для достижения насыщающего мутагенеза. Между тем, поскольку множественные сайты выбраны для мутагенеза, комбинация является случайной. В настоящем описании этот способ обозначают как случайный насыщающий мутагенез.
Дополнительно, насколько известно специалистам в данной области техники, процесс генетического клонирования часто требует конструирования подходящих сайтов для эндонуклеаз, которые в итоге вводят один или более чем один нерелевантный остаток на конце белка, подлежащего экспрессии, но это не влияет на активность исследуемого белка. В другом примере для конструирования слитого белка, для стимуляции экспрессии рекомбинантного белка, для получения рекомбинантного белка, который автоматически секретируется из клетки-хозяина, или для облегчения очистки рекомбинантного белка часто желательно иметь добавленные на N-конце, С-конце или других подходящих участках рекомбинантного белка некоторые аминокислотные последовательности, например, включая линкерные пептиды, сигнальные пептиды, лидерные пептиды, концевые удлинения, глутатион-S-трансферазу (GST), связывающий мальтозу белок Е, Белок А, метки, такие как 6His или Flag, или подходящие сайты расщепления протеазами, но не ограничиваясь ими. Следует понимать, что присутствие этих аминокислотных последовательностей не влияет на активность получаемого полипептида. Следовательно, настоящее изобретение также включает полипептид, имеющий одну или более чем одну аминокислоту на С-конце и/или N-конце полипептида по настоящему изобретению или в подходящем участке белка по изобретению, которая облегчает конструирование вектора, экспрессирующего полипептид, экспрессию и/или очистку полипептида. При этом эти полипептиды по-прежнему обладают активностью фосфолипазы С, как описано здесь.
Следовательно, в определенных воплощениях настоящего изобретения предложена аминокислотная последовательность, имеющая идентичность последовательности SEQ ID NO: 7 по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%. Кроме того, в настоящем изобретении предложена аминокислотная последовательность, имеющая идентичность последовательности SEQ ID NO: 4 по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%. В аминокислотных последовательностях, имеющих такую идентичность с последовательностями SEQ ID NO: 4 и 7, не все аминокислотные остатки в положениях, соответствующих аминокислотам 6, 8, 10 и 104 в SEQ ID NO: 4 и 7, мутированы; предпочтительно аминокислотные последовательности имеют такие же аминокислотные остатки как в SEQ ID NO: 4 и 7 в положениях, соответствующих аминокислотам 6, 8, 10 и 104 в SEQ ID NO: 4 и 7, например, имеют пролин, валин, серии и серии в положениях, соответствующих аминокислотам 6, 8, 10 и 104 в SEQ ID NO: 4. Более предпочтительно аминокислотная последовательность, имеющая такую идентичность последовательности, получена из Bacillus subtilis. Здесь понятие «идентичность последовательности» используют для описания соответствия между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями. Методы, известные в данной области техники, можно использовать для расчета идентичности последовательностей. Например, идентичность последовательности между двумя аминокислотными последовательностями можно определить с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, Journal of Molecular Biology, 48:443-453), входящего в программу Needle пакета EMBOSS (EMBOSS: European Molecular Biology Open Software Suite, Rice et al., 2000, Trends in Genetics, 16:276-277). Альтернативно, можно использовать BLASTP на сайте NCBI для расчета идентичности последовательности между двумя аминокислотными последовательностями.
В зависимости от хозяина, используемого для рекомбинантной продукции, полипептид по настоящему изобретению может быть гликозилированным или негликозилированным.
Полинуклеотид
В настоящем изобретении предложена нуклеотидная последовательность, кодирующая полипептид по настоящему изобретению, или комплементарная ей последовательность. SEQ ID NO: 3 представляет собой пример кодирующей последовательности полипептида по настоящему изобретению. Термин «кодирующая последовательность» включает последовательность нуклеиновой кислоты, кодирующую полипептид по настоящему изобретению (особенно SEQ ID NO: 7). Последовательность, кодирующая полипептид по настоящему изобретению, может быть идентична, например, последовательности кодирующей области как показано в SEQ ID NO: 3 или ее вырожденному варианту. Как его используют здесь, термин «вырожденный вариант» в настоящем описании относится к нуклеотидной последовательности, которая кодирует такую же аминокислотную последовательность, но отличается по нуклеотидной последовательности.
Последовательность, кодирующая полипептид по настоящему изобретению, включает кодирующую последовательность, кодирующую только зрелый полипептид; кодирующую последовательность зрелого полипептида и различные дополнительные кодирующие последовательности; кодирующую последовательность зрелого полипептида (и возможно дополнительную кодирующую последовательность) и некодирующую последовательность.
Также настоящее изобретение относится к варианту вышеописанных полинуклеотидов, который кодирует фрагменты, аналоги, производные и варианты той же самой аминокислотной последовательности по настоящему изобретению. Эти варианты нуклеотидов включают варианты с заменами, варианты с делециями и варианты со вставками. Как известно в данной области техники, аллельный вариант представляет собой альтернативную форму полинуклеотида, которая может быть обусловлена одной или более чем одной нуклеотидной заменой, делецией или вставкой, но существенно не изменяет функцию кодируемого белка.
Также в настоящее изобретение включен фрагмент последовательности нуклеиновой кислоты (такой как SEQ ID NO: 3 или комплементарная ей последовательность), кодирующей полипептид по настоящему изобретению. Длина «фрагмента нуклеиновой кислоты», как используют здесь, составляет по меньшей мере 15 нуклеотидов, предпочтительно по меньшей мере 30 нуклеотидов, более предпочтительно по меньшей мере 50 нуклеотидов и наиболее предпочтительно по меньшей мере 100 или более чем 100 нуклеотидов. Фрагмент нуклеиновой кислоты можно использовать в методах амплификации нуклеиновой кислоты (например, ПЦР) для определения и/или выделения полинуклеотидов, кодирующих полипептиды по настоящему изобретению. Следовательно, в некоторых воплощениях фрагмент нуклеиновой кислоты имеет длину от 15 до 30 оснований. Известные методы можно использовать для выбора подходящих фрагментов нуклеиновых кислот из последовательности нуклеиновой кислоты по изобретению, которые используют в качестве праймеров или зондов.
Обычно кодирующие последовательности полипептидов по настоящему изобретению или их фрагменты можно получить с помощью методов ПЦР-амплификации, рекомбинации или искусственного синтеза. Для ПЦР-амплификации праймеры могут быть сконструированы в зависимости от соответствующей нуклеотидной последовательности, раскрытой в настоящем изобретении, в частности, последовательности открытой рамки считывания. Доступную в продаже библиотеку кДНК или библиотеку кДНК, полученную с использованием рутинных методов специалистом в данной области техники, можно использовать в качестве матрицы для амплификации соответствующей последовательности. Для более длинных последовательностей обычно требуются две или более индивидуальных ПЦР-амплификации, после которых следует лигирование амплифицированных по отдельности фрагментов друг с другом в соответствующем порядке.
Конструкция нуклеиновой кислоты
Также настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей выделенный полинуклеотид по настоящему изобретению, функционально связанный с одной или более чем одной регуляторной последовательностью, которая управляет экспрессией кодирующей последовательности в подходящей клетке-хозяине в условиях, подходящих для регуляторных последовательностей. Термин «функционально связанный» означает, что регуляторные последовательности расположены в соответствующих положениях, так чтобы контролировать и направлять экспрессию данной полинуклеотидной последовательности. С полинуклеотидом, кодирующим полипептид по настоящему изобретению, можно проводить различные манипуляции для обеспечения экспрессии полипептида.
Регуляторная последовательность может представлять собой подходящую промоторную последовательность, нуклеотидную последовательность, распознаваемую клеткой-хозяином для экспрессии полинуклеотида, кодирующего полипептид по настоящему изобретению. Промоторная последовательность содержит последовательности для регуляции транскрипции, которые опосредуют экспрессию полипептида. Промотор может представлять собой любую нуклеотидную последовательность, обладающую транскрипционной активностью в выбранной клетке-хозяине, включая мутантные, укороченные и гибридные промоторы, и может быть получен из гена, кодирующего внеклеточный или внутриклеточный полипептид, который является гомологичным или гетерологичным для данной клетки-хозяина.
Примеры подходящих промоторов для управления транскрипцией конструкций нуклеиновой кислоты по настоящему изобретению, особенно в бактериальных клетках-хозяевах, представляют собой промоторные последовательности, имеющие происхождение от промотора фага Т7, lac-оперона Е. coli, гена агаразы Streptomyces coelicolor, гена левансахаразы Bacillus subtilis, гена альфа-амилазы Bacillus licheniformis, гена альфа-амилазы Bacillus amyloliquefaciens, гена пенициллиназы Bacillus licheniformis и так далее.
Примеры подходящих промоторов для управления транскрипцией конструкции нуклеиновой кислоты по настоящему изобретению в хозяевах-клетках мицелиального гриба представляют собой промоторы, имеющие происхождение из генов: ТАКА-амилазы Aspergillus oryzae, аспарагиновой протеазы Rhizomucor miehei, нейтральной альфа-амилазы Aspergillus niger, кислотно-стабилизированной альфа-амилазы Aspergillus niger, глюкоамилазы Aspergillus niger или Aspergillus awamori (glaA), целлобиогидролазы I Trichoderma reesei, целлобиогидролазы II Trichoderma reesei, щелочной протеазы Aspergillus oryzae, триозофосфатизомеразы Aspergillus oryzae, эндоглюканазы Trichoderma reesei и так далее, или мутантные, укороченные и гибридные формы этих промоторов.
В хозяевах-дрожжах полезные промоторы могут быть получены из генов енолазы Saccharomyces cerevisiae (ENO-1), галактокиназы Saccharomyces cerevisiae (GAL1), алкогольдегидрогеназы Saccharomyces cerevisiae, глицеральдегид-3-фосфат-дегидрогеназы, триозофосфатизомеразы Saccharomyces cerevisiae, 3-фосфоглицераткиназы Saccharomyces cerevisiae, алкогольоксидазы Pichia pastoris. Другие подходящие промоторы для дрожжевых клеток-хозяев описаны в Romanos et al., 1992, Yeast 8:423-488.
Также регуляторная последовательность может представлять собой подходящий терминатор транскрипции, последовательность, распознаваемую клеткой-хозяином для остановки транскрипции. Терминатор функционально связан с 3'-концом нуклеотидной последовательности, кодирующей полипептид. Любой терминатор, который является функциональным в выбранной клетке-хозяине, может быть использован в настоящем изобретении.
Предпочтительный терминатор для бактериальных хозяев может представлять собой терминатор из фага Т7.
Предпочтительный терминатор для клеток-хозяев мицелиального гриба получен из генов ТАКА-амилазы Aspergillus oryzae, глюкоамилазы Aspergillus niger, антранилатсинтазы Aspergillus nidulans, альфа-глюкозидазы Aspergillus niger.
Предпочтительный терминатор для дрожжевых клеток-хозяев получен из генов енолазы Saccharomyces cerevisiae, цитохрома С Saccharomyces cerevisiae, глицеральдегид-3-фосфат-дегидрогеназы Saccharomyces cerevisiae, алкоголь оксидазы Pichia pastoris и им подобных.
Также регуляторная последовательность может представлять собой подходящую лидерную последовательность, нетранслируемый участок мРНК, который важен для трансляции в клетках-хозяевах. Лидерная последовательность функционально связана с 5'-концом нуклеотидной последовательностьи, кодирующей полипептид. Любой терминатор, который является функциональным в выбранной клетке-хозяине, можно использовать в настоящем изобретении.
Также регуляторная последовательность может представлять собой участок, кодирующий сигнальный пептид, который кодирует аминокислотную последовательность, связанную с концом полипептида и направляет кодируемый полипептид по секреторному пути клетки. 5'-Конец кодирующей нуклеотидной последовательности может неотъемлемо содержать участок, кодирующий сигнальный пептид, естественным путем связанный с трансляционной рамкой считывания, содержащей сегмент кодирующей области, кодирущий секретируемый полипептид. Альтернативно, 5'-конец кодирующей последовательности может содержать участок, кодирующий сигнальный пептид, который является экзогенным для кодирующей области. Когда кодирующая последовательность от природы не содержит участок, кодирующий сигнальный пептид, может потребоваться экзогенный участок, кодирующий сигнальный пептид. Альтернативно, экзогенный участок, кодирующий сигнальный пептид, может просто заменять природный участок, кодирующий сигнальный пептид, для усиления секреции полипептида. Однако, любой участок, кодирующий сигнальный пептид, который направляет экспрессированный полипептид по пути секреции в выбранной клетке-хозяине (то есть секреции в культуральную среду), можно использовать в настоящем изобретении.
В определенных воплощениях конструкция нуклеиновой кислоты по настоящему изобретению представляет собой экспрессионную кассету. Термин «экспрессионная кассета» относится к полному набору элементов, необходимых для экспрессии гена, включающему промоторы, кодирующие последовательности и последовательности хвостов поли-А.
Вектор
Также настоящее изобретение относится к векторам, содержащим полинуклеотиды или конструкции нуклеиновых кислот по настоящему изобретению, включая экспрессионные векторы и клонирующие векторы. Экспрессионный вектор может представлять собой любой вектор (такой как плазмида или вирус), к которым удобно применять технологии рекомбинантной ДНК, и может приводить к экспрессии интересуемой нуклеотидной последовательности. Клонирующий вектор обычно обладает способностью размножаться в клетке-хозяине после введения в данную клетку-хозяина.
Выбор вектора обычно зависит от совместимости вектора с клеткой-хозяином, в которую вводят вектор. Вектор может быть линейным или представлять собой замкнутую кольцевую плазмиду.
Вектор может представлять собой самореплицирующийся вектор, то есть вектор, который существует в экстрахромосомной форме, репликация которой не зависит от репликации хромосомы, такой как плазмида, экстрахромосомный элемент, мини-хромосомы или искусственная хромосома. Такой вектор может содержать любые средства для обеспечения саморепликации. Альтернативно, вектор может представлять собой вектор, который при введении в клетку-хозяина встраивается в геном и реплицируется с хромосомой, в которую он встроен. Кроме того, можно использовать один вектор или одну плазмиду, или два или более чем два вектора, или две или более чем две плазмиды, или два или более чем два транспозона, которые вместе содержат общую ДНК для введения в геном клетки-хозяина.
Предпочтительно вектор по настоящему изобретению содержит один или более чем один селектируемый маркер, позволяющий легко отбирать трансформированные, трансфицированные, трансдуцированные клетки и тому подобное. Селектируемые маркеры представляют собой гены, продукты экспрессии которых обеспечивают устойчивость к антибиотикам или вирусам, устойчивость к тяжелым металлам, прототрофию для ауксотрофов и так далее.
Предпочтительно вектор по настоящему изобретению содержит элементы, которые позволяют вектору встраиваться в геном клетки-хозяина или которые дают вектору способность к саморепликации в клетках независимо от генома.
Более чем одна копия полинуклеотидов по настоящему изобретению может быть введена в клетку-хозяина для увеличения выхода продукта гена. Множество копий полинуклеотидов можно получить путем встраивания по меньшей мере одной дополнительной копии последовательности в геном клетки-хозяина или путем включения полинуклеотида и селектируемого гена-маркера, способных к амплификации. Клетки, содержащие амплифицированную копию селектируемого гена-маркера и вследствие этого содержащие дополнительную копию полинуклеотида, могут быть отобраны путем культивирования клетки в присутствии подходящего агента селекции.
Вектор по настоящему изобретению предпочтительно содержит искусственно синтезированную последовательность, содержащую множественные сайты распознавания экдонуклеазами рестрикции, которая может обеспечивать разнообразие положений вставки или схем вставки для экзогенных ДНК.
Более предпочтительно вектор экспрессии по настоящему изобретению представляет собой вектор, который можно использовать для экспрессии в Pichia pastoris. Предпочтительно вектор по настоящему изобретению представляет собой таковой, используемый в доступных в продаже Pichia pastoris, таких как pPIC, pPICZ, pAO, pGAP или pGAPZ, или вектор из этих же видов.
Клетка-хозяин
Также настоящее изобретение относится к рекомбинантной клетке-хозяину, содержащей полинуклеотид или конструкцию нуклеиновой кислоты по настоящему изобретению, которую используют для рекомбинантной продукции полипептидов. Вектор, содержащий полинуклеотид по настоящему изобретению, вводят в клетку-хозяина, так что этот вектор поддерживается как часть хромосомы или как экстрахромосомный самореплицирующийся вектор, как описано ранее. Выбор клетки-хозяина во многом зависит от гена, кодирующего полипептид, и его источника.
Клетка-хозяин может представлять собой одноклеточный микроорганизм или не являющийся одноклеточным микроорганизм. Одноклеточные микроорганизмы, такие как грамположительные бактерии, включают клетку Bacillus, например, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus megaterium, Bacillus subtilis, Bacillus licheniformis, Bacillus coagulans, Bacillus stearothermophilus, Bacillus thuringiensis и им подобные; или клетку Streptomyces, например, Streptomyces lividans; или грамотрицательные бактерии, такие как Е. coli и Pseudomonas sp., но не ограничиваясь ими. В предпочтительном аспекте бактериальный хозяин представляет собой Bacillus subtilis, Е. coli, Bacillus licheniformis, Bacillus stearothermophilus и клетки E.coli.
Клетка-хозяин может представлять собой эукариотическую клетку, такую как клетка млекопитающего, насекомого, растения, дрожжей или гриба. В предпочтительном аспекте клетка-хозяин представляет собой эукариотическую клетку. Как используют здесь, «эукариотический» включает Ascomycota, Basidiomycota, Chytridiomycota, Zygomycota, Oomycota и тому подобное.
В более предпочтительном аспекте клетка-хозяин представляет собой клетку Ascomycota, такую как Saccharomyces sp., Pichia sp., Yarrowia sp., Candida sp.и Komagataella sp.
В наиболее предпочтительном аспекте клетка-хозяин представляет собой Pichia pastoris, Saccharomyces cerevisiae, Yarrowia lipolytica и им подобные. В другом наиболее предпочтительном аспекте клетка-хозяин представляет собой клетку Pichia pastoris.
Способ получения
После получения кодирующей последовательности полипептида можно использовать способ получения полипептида по настоящему изобретению, включающий: (а) культивирование клетки-хозяина, содержащей вектор для экспрессии полипептида, в условиях, подходящих для продуцирования полипептида; и (б) выделение полипептида.
В способе получения по настоящему изобретению клетки можно культивировать в среде, подходящей для продуцирования полипептида, с помощью методов, известных в данной области техники. Например, клетки можно культивировать во встряхиваемых колбах в лабораторных или промышленных ферментерах и в небольших объемах ферментации или в промышленных объемах ферментации (включая непрерывную, периодическую, периодическую с подпиткой или твердофазную ферментацию) и культивировать в подходящей среде и условиях, обеспечивающих экспрессию и/или выделение полипептида. Культивирование выполняют в подходящей среде, содержащей источники углерода и азота и неорганические соли, с использованием методов, известных в данной области техники. Подходящие среды могут быть приобретены у коммерческих поставщиков или могут быть изготовлены в соответствии с опубликованным составом. Если полипептид секретируется в среду, этот полипептид может быть выделен непосредственно из среды. Если полипептид не секретируется в среду, он может быть выделен из клеточных лизатов.
Альтернативно, полипептид по настоящему изобретению можно синтезировать методом химического синтеза, известным в данной области техники. Методы химического синтеза полипептида включают методы твердофазного синтеза и жидкофазного синтеза, где твердофазный синтез является наиболее широко применяемым. Методы твердофазного синтеза включают два общепринятых метода, Fmoc и tBoc, но не ограничиваясь ими. Обычно в качестве нерастворимого твердого носителя используют смолу, аминокислоты обычно соединяют друг с другом от С-конца (карбоксильный конец) к N-концу (амино-конец) в пептидную цепь, и цикл присоединения каждой аминокислоты состоит из следующих трех стадий: 1) снятие защиты: в защищенной аминокислоте защитную группу аминокислоты необходимо удалить с помощью растворителя для снятия защиты; 2) активация: карбоксильную группу присоединяемой аминокислоты активируют с помощью активатора; и 3) сочетание: активированный карбоксил подвергают взаимодействию с доступной аминогруппой предыдущей аминокислоты с образованием пептидной связи. Этот цикл повторяют до тех пор, пока пептидная цепь не удлинится до желаемой длины. Наконец, соединение между твердым носителем и пептидной цепью расщепляют раствором для расщепления, и желаемая аминокислотная последовательность может быть получена. Вышеописанный химический синтез можно проводить в автоматическом синтезаторе пептидов с программным управлением, и такие приборы включают двухканальный синтезатор пептидов Tribute от Protein Technologies, UV Online Monitor System from CS Bio Company, трехканальный синтезатор Focus XC от Aapptec и им подобные, но не ограничиваясь ими.
Описанный здесь полипептид может быть выделен способом, известным в данной области техники. Например, полипептид может быть выделен из среды традиционными способами, включая центрифугирование, фильтрование, ультрафильтрацию, экстракцию, хроматографию, сушку распылением, лиофильную сушку, упаривание, осаждение или им подобное, но не ограничиваясь этим.
Полипептид по настоящему изобретению можно очищать разнообразными методами, известными в данной области техники, включая метод хроматографии (например ионообменной, аффинной, гидрофобной, хроматофокусирующей, эксклюзионной), электрофореза (например изоэлектрофокусирования), метод на основе различия в растворимости (такой как осаждение высаливанием), SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) или экстракции, для получения по существу чистого полипептида, но не ограничиваясь ими. Свойства и применение полипептида
Полипептид по настоящему изобретению обладает активностью фосфолипазы С, которую можно использовать для очистки масел и жиров, модификации фосфолипидов, модификации пищевых продуктов и в различных аспектах пищевой промышленности и фармацевтической промышленности, включая выпечку, детергенты, улучшение фильтрования водных растворов или сиропа и тому подобное. При использовании для дегуммирования полипептид по настоящему изобретению может улучшать эффективность дегуммирования и увеличивать выход DAG во время процесса дегуммирования.
Полипептид по настоящему изобретению может быть представлен в форме чистого ферментативного препарата или в форме композиции. Эта композиция может представлять собой порошкообразную композицию, жидкую композицию и пастообразную композицию. Если полипептид представлен в форме композиции, такая композиция может содержать различные эксципиенты в соответствии с различным применением композиции, содержащей фермент. Эксципиенты, известные в данной области техники, можно добавлять в композиции по настоящему изобретению, и такие эксципиенты включают сорбит, сорбат калия, метилбензоат, этилбензоат, сахарозу, маннит, трегалозу, крахмал, хлорид натрия, хлорид кальция, другие стабилизаторы, но не ограничиваясь ими, или одно или более других веществ.
Количество полипептида по настоящему изобретению, используемого в способе по настоящему изобретению, может быть практически определено.
Ферментативное дегуммирование
Также в настоящем изобретении предложен способ ферментативного дегуммирования, включающий добавление полипептида по настоящему изобретению к неочищенному маслу для дегуммирования. Обычно неочищенное масло нагревают до 50-70°С, предпочтительно до 50-60°С, и затем добавляют полипептид по настоящему изобретению, а именно фосфолипазу С.
Фосфолипазу С по настоящему изобретению обычно добавляют в форме водного раствора. Исходя из массы неочищенного масла фермент добавляют в количестве от 10 до 1000 м.д., предпочтительно от 50 до 500 м.д., более предпочтительно от 100 до 300 м.д.
Условия дегуммирования обычно включают следующее: перемешивание при 50-60°С в течение 1-3 часов и затем нагревание до 80-90°С и выдерживание в течение 1-10 минут.
Неочищенное масло, подходящее для способа дегуммирования по настоящему изобретению, включает соевое масло, подсолнечное масло, арахисовое масло, рапсовое масло, масло из рисовых отрубей, кукурузное масло, оливковое масло, пальмовое масло, пальмоядровое масло, пальмовый олеин, масло канола, касторовое масло, кокосовое масло, кориандровое масло, хлопковое масло, масло лесного ореха, конопляное масло, льняное масло, масло из косточек манго, масло пенника лугового, копытное масло, сафлоровое масло, масло камелии (camellia), талловое масло, масло камелии (tsubaki) и другие растительные масла, но не ограничиваясь ими.
Далее настоящее изобретение проиллюстрировано конкретными примерами. Экспериментальные способы без уточнения конкретных условий в примерах ниже выполнены в общепринятых условиях, таких как изложено в Sambrook et al., "Molecular Cloning: A Laboratory Manual" (New York: Cold Spring Harbor Laboratory Press (Cold Spring Harbor Laboratory Press), 1989), или в условиях, рекомендованных производителем. Использование и дозировка реагентов, если специально не оговорено, находятся в соответствии с традиционным использованием и дозировкой.
Экспериментальные материалы
1. Экспериментальные штаммы и плазмиды
Штаммы: Pichia pastoris SMD1168 (Invitrogen, номер по каталогу No. С17500), Е. coli DH5a (TAKARA, номер по каталогу No. D9057A).
2. Среды и растворы
Жидкая среда LB: 0,5% дрожжевого экстракта, 1% триптона, 1% NaCl, рН 7,0.
Твердая среда LB: 1,5% агара, добавленного в жидкую среду LB.
Жидкая среда YPD: 1% дрожжевого экстракта, 2% пептона, 2% глюкозы.
Твердая среда YPD: 2% агара, добавленного в жидкую среду LB.
Твердая среда MGYS: 1,34% основы азотного агара для дрожжей (YNB), содержащей сульфат аммония и не содержащей аминокислоты; 1% глицерина; 1 М сорбит; 4×10-5% D-биотина; 2% агара.
Среда ВММ для скрининга на основе фосфолипидов соевых бобов: 1,34% основы азотного агара для дрожжей (YNB), содержащей сульфат аммония, не содержащей аминокислоты; 4×10-5% D-биотина; 0,5% метанола (стерилизованного); 2% эмульсии фосфолипидов соевых бобов; 0,1М буфер лимонная кислота-цитрат натрия с рН 6,6; 2% агара; 10 мкМ ZnSO4×7 H2O.
2%-ная эмульсия фосфолипидов соевых бобов: 2 г фосфолипидов соевых бобов, 100 мл H2O, гомогенизировали в высокоскоростном гомогенизаторе при 8000 об/мин в течение 1 мин.
Жидкая среда BMGY: 1% дрожжевого экстракта; 2% пептона; 1,34% основы азотного агара для дрожжей (YNB), содержащей сульфат аммония и не содержащей аминокислоты; 1% глицерина; 4×10-5% D-биотина; 0,1 М буфер дигидрофосфат калия -гидроортофосфат калия с рН 6,0.
Жидкая среда BMMY: 1% дрожжевого экстракта; 2% пептона; 1,34% основы азотного агара для дрожжей (YNB), содержащей сульфат аммония и не содержащей аминокислоты; 0,3% ZnSO4⋅7H2O; 0,5% метанола (стерилизованного); 4×10-5% D-биотина (стерилизованного); 0,1 М буфер лимонная кислота-цитрат натрия с рН 6,6.
3. Определение ферментативной активности методом pNPPC Построение стандартной кривой для определения активности фосфолипазы: 0,01391 г пара-нитрофенола растворяли в 50 мл стерильной воды для получения 2 ммоль/л рабочего раствора. Количество каждого реагента показано в таблице ниже. Объем реакции и условия реакции для построения стандартной кривой согласуются с условиями для измерения ферментативной активности образца в эксперименте.
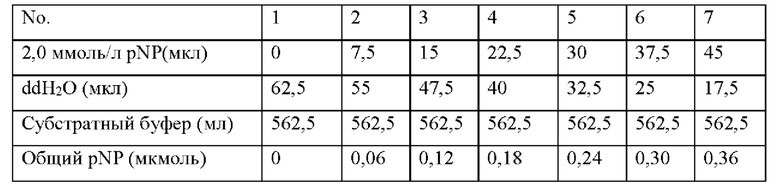
Вышеуказанные растворы смешивали и проводили их обработку при 37°С в течение 15 минут. Затем добавляли 500 мкл 0,5 н. NaOH и измеряли поглощение при 410 нм. Полученная в результате стандартная кривая представлена на фиг. 3.
4. Реакционный буфер
0,1 М буфер борная кислота-борат натрия (рН 7,6), 20 мМ pNPPC.
5. Расчет ферментативной активности
25 мкл ферментативного раствора для тестирования добавляли в 600 мкл вышеописанного реакционного буфера и проводили реакцию при 37°С в течение 15 мин. 500 мкл 0,5 н. NaOH добавляли для остановки реакции и измеряли поглощение при 410 нм.
Ферментативная активность образца (Ед/мл)=А (поглощение при 410 нм)×0,1935×фактор разведения 10/15.
Улучшенный набор реактивов для определения концентрации белка по методу Брэдфорда получали от Shanghai Sangon Bioengineering Co., Ltd.; фермент для ПЦР -ДНК-полимеразу PrimeSTAR®HS получали от TaKaRa (Dalian) Co., Ltd; ДНК-лигазу T4 получали от Fermentas Co., Ltd.
Пример 1: Конструирование и скрининг библиотеки насыщающих мутаций
Библиотека случайных насыщающих мутаций для аминокислот 6, 8, 10, 104 и 205 была сконструирована Synbio Technologies (Suzhou) Co., Ltd. с использованием вектора pmAO-PLC-N63DN131SN134D-Y56H (CN 201680072289.5, последовательность ДНК PLC-N63DN131SN134D-Y56H представлена в SEQ ID NO: 1, и его аминокислотная последовательность представлена в SEQ ID NO: 2) в качестве матрицы. Этой плазмидной библиотекой трансформировали штамм DH5a Е. coli и все клоны Е. coli промывали жидкой средой LB (содержащей 100 мкг/мл ампициллина) и культивировали при 37°С в течение 4 ч. Плазмиды экстрагировали и линеаризовали SalI, и выделяли фрагмент примерно 8,5 т.п.о. Компетентные клетки Pichia pastoris SMD1168 трансформировали 500 нг вектора путем электро-трансформации. Трансформанты высевали на чашки MGYS и культивировали при 30°С в течение 3 суток для получения библиотеки мутантов Pichia pastoris PLC-N63DN131SN134D-Y56H. Моноклональные штаммы отбирали и переносили на чашку со средой ВММ для скрининга на основе фосфолипидов соевых бобов. Отбирали один из клонов с относительно большим белым кольцом преципитации и обозначали его как 31#.
Пример 2: Анализ последовательности мутанта 31#
Штамм 31# инокулировали в 3 мл жидкой среды YPD и культивировали в течение ночи при 30°С, затем из клеток экстрагировали геномную ДНК. Последовательность ДНК PLC штамма 31# амплифицировали методом ПЦР с использованием ДНК-полимеразы PrimeSTAR®HS, пары праймеров АОХ1-5/АОХ1-3 и геномной ДНК штамма 31# в качестве матрицы. Полученную последовательность секвенировали в Shanghai Sangon Biotech Co., Ltd. с использованием пары праймеров АОХ1-5/АОХ1-3:
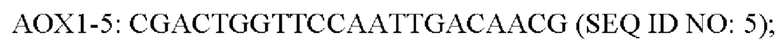
Результат секвенирования ДНК PLC 31# представлен в SEQ ID NO: 3. Согласно этому результату 5 оснований последовательности ДНК PLC 31# мутированы. Аминокислотная последовательность представлена в SEQ ID NO: 4, где аминокислоты в положениях 6, 8, 10 и 104 были мутированы с заменой лизина, лизина, глицина и лизина на пролин, валин, серии и серии, соответственно.
Пример 3: Ферментирование во встряхиваемой колбе мутанта 31# и определение его ферментативной активности
Штаммы 31# и исходные штаммы (то есть, SMD1168, трансфицированные вектором pmAO-PLC-N63DN131SN134D-Y56H) активировали в жидкой YPD и затем инокулировали в среду BMGY и подвергали встряхиванию при 220 об/мин при 30°С в течение ночи. Культуру переносили в среду BMMY с начальной OD600 равной 6.
Сначала проводили индукцию 2% метанолом с добавлением 1% метанола через 24 ч, 32 ч, 48 ч и 56 ч, соответственно, и отбирали образцы через 72 ч. Полученные образцы концентрировали в 40 раз методом обессоливания ультрафильтрацией с помощью ультрафильтрационных трубок с порогом отсечения молекулярной массы 40 кДа. Обработанные образцы добавляли в буфер (20 мМ буфер лимонная кислота-цитрат натрия (рН 6,6), 10 мкМ ZnSO4).
0,5 мкл концентрата ферментативного бульона добавляли в 600 мкл реакционного буфера pNPPC и проводили реакцию при 37°С в течение 15 мин. Добавляли 500 мкл 0,5 и. NaOH для остановки реакции и измеряли поглощение при 410 нм. Активность PLC для каждого образца ферментативного бульона рассчитывали по стандартной кривой. Концентрацию белка в ферментативном бульоне в колбе после встряхивания определяли с помощью реактива Брэдфорда для получения специфической ферментативной активности. Результаты представлены на фиг. 1. Специфическая ферментативная активность образца ферментативного бульона мутанта 31# была в 1,88 раз выше, чем таковая у образца ферментативного бульона исходного штамма.
Пример 4: Тест дегуммирования 31#
100 г неочищенного соевого масла нагревали до 55°С и добавляли в него 50 м.д. и 100 м.д. образца ферментативного бульона мутанта 31# и исходного штамма, полученных в Примере 3, соответственно. Водная фаза в полученной системе составляла 3%. Систему подвергали воздействию больших сдвиговых усилий (10000 об/мин) в течение 1 мин с помощью высокоскоростного миксера, перемешивали (750 об/мин) при 55°С в течение 2 ч, нагревали до 85°С и выдерживали в течение 5 мин и центрифугировали при 12000 об/мин в течение 10 мин. Примерно 10 г образца масла верхней фазы использовали для определения содержания DAG методом HPLC (высокоэффективная жидкостная хроматография). Увеличение количества DAG для образца ферментативного бульона мутанта 31# и образца ферментативного бульона исходного штамма по сравнению с неочищенным маслом показано на фиг. 2. Увеличение количества DAG в образце ферментативного бульона мутанта 31# при 50 м.д. фермента и в образце ферментативного бульона исходного штамма при 100 м.д. фермента является одинаковым. Следовательно, требуемое количество фермента образца ферментативного бульона мутанта 31# можно наполовину уменьшить по сравнению с образцом ферментативного бульона исходного штамма.
--->
Sequence Listing
<110> WILMAR (SHANGHAI) BIOTECHNOLOGY RESEARCH & DEVELOPMENT CENTER CO., LTD
<120> PHOSPHOLIPASE C MUTANT WITH HIGH ENZYME ACTIVITY
<130> 188686
<150> CN 201811620055.7
<151> 2018-12-28
<160> 7
<170> SIPOSequenceListing 1.0
<210> 1
<211> 738
<212> DNA
<213> Artificial Sequence
<220>
<223> Последовательность ДНК PLC-N63DN131SN134D-Y56H
<400> 1
tggtcagctg aggacaagca taaggaaggt gtgaatagtc acttatggat cgtgaaccgt 60
gccattgata taatgtctag gaatacaact ctggttaagc aagatagagt tgctcaattg 120
aatgaatggc gtacagagct agagaatggc atctacgctg ctgatcatga aaacccctat 180
tacgatgaca gtaccttcgc ttctcacttt tacgatccag acaacggaaa gacatatatc 240
ccattcgcca agcaagctaa ggagactgga gctaagtact tcaagttggc tggagagtca 300
tacaagaata aagacatgaa gcaggccttc ttttatcttg ggttgtcatt gcattatttg 360
ggcgatgtca accaacctat gcatgccgca tcctttacgg acctgtccta tccacagggt 420
tttcactcca agtacgagaa ctttgtcgat actattaaag acaactacaa agttaccgat 480
gggaacggat attggaattg gaaaggcacc aaccctgaag aatggattca cggtgcagca 540
gtagttgcaa aacaggacta ctctggaatt gtcaatgaca ataccaaaga ttggtttgtg 600
aaagccgcag tctcccagga atatgcagat aaatggagag ctgaagttac acctatgact 660
ggtaaacgac taatggatgc ccaaagagtt actgctggtt acattcaatt atggttcgac 720
acttacggtg acaggtaa 738
<210> 2
<211> 245
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность PLC-N63DN131SN134D-Y56H
<400> 2
Trp Ser Ala Glu Asp Lys His Lys Glu Gly Val Asn Ser His Leu Trp
1 5 10 15
Ile Val Asn Arg Ala Ile Asp Ile Met Ser Arg Asn Thr Thr Leu Val
20 25 30
Lys Gln Asp Arg Val Ala Gln Leu Asn Glu Trp Arg Thr Glu Leu Glu
35 40 45
Asn Gly Ile Tyr Ala Ala Asp His Glu Asn Pro Tyr Tyr Asp Asp Ser
50 55 60
Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Asn Gly Lys Thr Tyr Ile
65 70 75 80
Pro Phe Ala Lys Gln Ala Lys Glu Thr Gly Ala Lys Tyr Phe Lys Leu
85 90 95
Ala Gly Glu Ser Tyr Lys Asn Lys Asp Met Lys Gln Ala Phe Phe Tyr
100 105 110
Leu Gly Leu Ser Leu His Tyr Leu Gly Asp Val Asn Gln Pro Met His
115 120 125
Ala Ala Ser Phe Thr Asp Leu Ser Tyr Pro Gln Gly Phe His Ser Lys
130 135 140
Tyr Glu Asn Phe Val Asp Thr Ile Lys Asp Asn Tyr Lys Val Thr Asp
145 150 155 160
Gly Asn Gly Tyr Trp Asn Trp Lys Gly Thr Asn Pro Glu Glu Trp Ile
165 170 175
His Gly Ala Ala Val Val Ala Lys Gln Asp Tyr Ser Gly Ile Val Asn
180 185 190
Asp Asn Thr Lys Asp Trp Phe Val Lys Ala Ala Val Ser Gln Glu Tyr
195 200 205
Ala Asp Lys Trp Arg Ala Glu Val Thr Pro Met Thr Gly Lys Arg Leu
210 215 220
Met Asp Ala Gln Arg Val Thr Ala Gly Tyr Ile Gln Leu Trp Phe Asp
225 230 235 240
Thr Tyr Gly Asp Arg
245
<210> 3
<211> 738
<212> DNA
<213> Artificial Sequence
<220>
<223> Кодирующая последовательность мутанта
<400> 3
tggtcagctg aggacccgca tgttgaatcg gtgaatagtc acttatggat cgtgaaccgt 60
gccattgata taatgtctag gaatacaact ctggttaagc aagatagagt tgctcaattg 120
aatgaatggc gtacagagct agagaatggc atctacgctg ctgatcatga aaacccctat 180
tacgatgaca gtaccttcgc ttctcacttt tacgatccag acaacggaaa gacatatatc 240
ccattcgcca agcaagctaa ggagactgga gctaagtact tcaagttggc tggagagtca 300
tacaagaatt ctgacatgaa gcaggccttc ttttatcttg ggttgtcatt gcattatttg 360
ggcgatgtca accaacctat gcatgccgca tcctttacgg acctgtccta tccacagggt 420
tttcactcca agtacgagaa ctttgtcgat actattaaag acaactacaa agttaccgat 480
gggaacggat attggaattg gaaaggcacc aaccctgaag aatggattca cggtgcagca 540
gtagttgcaa aacaggacta ctctggaatt gtcaatgaca ataccaaaga ttggtttgtg 600
aaagccgcag tctcccagga atatgcagat aaatggagag ctgaagttac acctatgact 660
ggtaaacgac taatggatgc ccaaagagtt actgctggtt acattcaatt atggttcgac 720
acttacggtg acaggtaa 738
<210> 4
<211> 245
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность мутанта
<400> 4
Trp Ser Ala Glu Asp Pro His Val Glu Ser Val Asn Ser His Leu Trp
1 5 10 15
Ile Val Asn Arg Ala Ile Asp Ile Met Ser Arg Asn Thr Thr Leu Val
20 25 30
Lys Gln Asp Arg Val Ala Gln Leu Asn Glu Trp Arg Thr Glu Leu Glu
35 40 45
Asn Gly Ile Tyr Ala Ala Asp His Glu Asn Pro Tyr Tyr Asp Asp Ser
50 55 60
Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Asn Gly Lys Thr Tyr Ile
65 70 75 80
Pro Phe Ala Lys Gln Ala Lys Glu Thr Gly Ala Lys Tyr Phe Lys Leu
85 90 95
Ala Gly Glu Ser Tyr Lys Asn Ser Asp Met Lys Gln Ala Phe Phe Tyr
100 105 110
Leu Gly Leu Ser Leu His Tyr Leu Gly Asp Val Asn Gln Pro Met His
115 120 125
Ala Ala Ser Phe Thr Asp Leu Ser Tyr Pro Gln Gly Phe His Ser Lys
130 135 140
Tyr Glu Asn Phe Val Asp Thr Ile Lys Asp Asn Tyr Lys Val Thr Asp
145 150 155 160
Gly Asn Gly Tyr Trp Asn Trp Lys Gly Thr Asn Pro Glu Glu Trp Ile
165 170 175
His Gly Ala Ala Val Val Ala Lys Gln Asp Tyr Ser Gly Ile Val Asn
180 185 190
Asp Asn Thr Lys Asp Trp Phe Val Lys Ala Ala Val Ser Gln Glu Tyr
195 200 205
Ala Asp Lys Trp Arg Ala Glu Val Thr Pro Met Thr Gly Lys Arg Leu
210 215 220
Met Asp Ala Gln Arg Val Thr Ala Gly Tyr Ile Gln Leu Trp Phe Asp
225 230 235 240
Thr Tyr Gly Asp Arg
245
<210> 5
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Праймер
<400> 5
cgactggttc caattgacaa cg 22
<210> 6
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Праймер
<400> 6
ggcaaatggc attctgacat cctc 24
<210> 7
<211> 245
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность мутанта
<220>
<221> MUTAGEN
<222> (6)..(6)
<223> Xaa представляет собой аланин, валин, лейцин,
изолейцин, пролин,фенилаланин, метионин или триптофан
<220>
<221> MUTAGEN
<222> (8)..(8)
<223> Xaa представляет собой аланин, валин, лейцин,
изолейцин, пролин,фенилаланин, метионин или триптофан
<220>
<221> MUTAGEN
<222> (10)..(10)
<223> Xaa представляет собой глицин, аспарагин, глутамин,
серин, треонин,тирозин или цистеин
<220>
<221> MUTAGEN
<222> (104)..(104)
<223> Xaa представляет собой глицин, аспарагин, глутамин,
серин, треонин,тирозин или цистеин
<400> 7
Trp Ser Ala Glu Asp Xaa His Xaa Glu Xaa Val Asn Ser His Leu Trp
1 5 10 15
Ile Val Asn Arg Ala Ile Asp Ile Met Ser Arg Asn Thr Thr Leu Val
20 25 30
Lys Gln Asp Arg Val Ala Gln Leu Asn Glu Trp Arg Thr Glu Leu Glu
35 40 45
Asn Gly Ile Tyr Ala Ala Asp His Glu Asn Pro Tyr Tyr Asp Asp Ser
50 55 60
Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Asn Gly Lys Thr Tyr Ile
65 70 75 80
Pro Phe Ala Lys Gln Ala Lys Glu Thr Gly Ala Lys Tyr Phe Lys Leu
85 90 95
Ala Gly Glu Ser Tyr Lys Asn Xaa Asp Met Lys Gln Ala Phe Phe Tyr
100 105 110
Leu Gly Leu Ser Leu His Tyr Leu Gly Asp Val Asn Gln Pro Met His
115 120 125
Ala Ala Ser Phe Thr Asp Leu Ser Tyr Pro Gln Gly Phe His Ser Lys
130 135 140
Tyr Glu Asn Phe Val Asp Thr Ile Lys Asp Asn Tyr Lys Val Thr Asp
145 150 155 160
Gly Asn Gly Tyr Trp Asn Trp Lys Gly Thr Asn Pro Glu Glu Trp Ile
165 170 175
His Gly Ala Ala Val Val Ala Lys Gln Asp Tyr Ser Gly Ile Val Asn
180 185 190
Asp Asn Thr Lys Asp Trp Phe Val Lys Ala Ala Val Ser Gln Glu Tyr
195 200 205
Ala Asp Lys Trp Arg Ala Glu Val Thr Pro Met Thr Gly Lys Arg Leu
210 215 220
Met Asp Ala Gln Arg Val Thr Ala Gly Tyr Ile Gln Leu Trp Phe Asp
225 230 235 240
Thr Tyr Gly Asp Arg
245
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Полипептид, обладающий активностью фосфолипазы С, и его применение | 2019 |
|
RU2819269C2 |
| НОВЫЙ ПОЛИПЕПТИД И СПОСОБ ПОЛУЧЕНИЯ L-ЛЕЙЦИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2811433C1 |
| СПОСОБ СНИЖЕНИЯ ОШИБОЧНОГО ВКЛЮЧЕНИЯ НЕКАНОНИЧЕСКИХ АМИНОКИСЛОТ С РАЗВЕТВЛЕННОЙ ЦЕПЬЮ | 2019 |
|
RU2816654C2 |
| ВАРИАНТЫ СКВАЛЕНГОПЕНЦИКЛАЗЫ (SHC) | 2020 |
|
RU2829857C1 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| Модифицированный полипептид с пониженной активностью цитратсинтазы и способ получения L-аминокислоты с использованием этого полипептида | 2019 |
|
RU2732815C1 |
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| Новые гибридные ActRIIB белки-ловушки лигандов для лечения заболеваний, связанных с мышечной атрофией | 2016 |
|
RU2834707C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| АНТИТЕЛА, БЛОКИРУЮЩИЕ CD73 | 2020 |
|
RU2819204C2 |
Изобретение относится к области биотехнологии, конкретно к рекомбинантным полипептидам с активностью фосфолипазы С, и может быть использовано для дегуммирования масел и жиров. Предложен мутантный полипептид фосфолипазы С, аминокислотная последовательность которого по меньшей мере на 95% идентична SEQ ID NO: 4 и характеризуется при этом наличием пролина, валина, серина и серина в положениях аминокислотных остатков, соответствующих положениям 6, 8, 10 и 104 в SEQ ID NO: 4 соответственно. Изобретение обеспечивает получение мутантной фосфолипазы C с высокой ферментативной активностью, что позволяет улучшить эффективность дегуммирования и увеличить выход диацилглицерина (DAG) во время дегуммирования. 7 н. и 5 з.п. ф-лы, 3 ил., 1 табл., 4 пр.
1. Выделенный полипептид, характеризующийся повышенной специфической ферментативной активностью фосфолипазы С, имеющий идентичность последовательности SEQ ID NO: 4 по меньшей мере на 95%, где аминокислотные остатки выделенного полипептида, соответствующие аминокислотам 6, 8, 10 и 104 в SEQ ID NO: 4, представляют собой пролин, валин, серин и серин соответственно.
2. Выделенный полипептид по п. 1, где аминокислотная последовательность выделенного полипептида является такой, как представлено в SEQ ID NO: 4, или он получен из Bacillus subtilis.
3. Нуклеиновая кислота, кодирующая мутантную фосфолипазу С с повышенной специфической ферментативной активностью и содержащая полинуклеотидную последовательность, выбранную из группы, состоящей из:
(1) полинуклеотидной последовательности, кодирующей полипептид по любому из пп. 1, 2; и
(2) последовательности, комплементарной полинуклеотидной последовательности (1); и
(3) полинуклеотидной последовательности, представленной в SEQ ID NO: 3.
4. Конструкция нуклеиновой кислоты для экспрессии мутантной фосфолипазы С с повышенной специфической ферментативной активностью, содержащая нуклеиновую кислоту по п. 3 и одну или более чем одну регуляторную последовательность, функционально связанную с полинуклеотидной последовательностью.
5. Конструкция нуклеиновой кислоты по п. 4, представляющая собой экспрессионный вектор или клонирующий вектор.
6. Генетически сконструированная клетка-хозяин для экспрессии мутантной фосфолипазы С с повышенной специфической ферментативной активностью, содержащая нуклеиновую кислоту по п. 3 или конструкцию нуклеиновой кислоты по п. 4 или 5.
7. Композиция, характеризующаяся повышенной специфической ферментативной активностью фосфолипазы С, содержащая полипептид по любому из пп. 1, 2 и адъювант.
8. Композиция по п. 7, где адъювант представляет собой адсорбирующее вещество, выбранное из группы, состоящей из активированного угля, оксида алюминия, диатомовой земли, пористых керамических веществ и пористого стекла.
9. Применение полипептида по любому из пп. 1, 2, нуклеиновой кислоты по п. 3, конструкции нуклеиновой кислоты по п. 4 или 5, клетки-хозяина по п. 6 или композиции по п. 7 или 8 для ферментативного дегуммирования.
10. Применение по п. 9, где указанное дегуммирование используют в очистке масел и жиров, модификации фосфолипидов или модификации пищевых продуктов.
11. Способ ферментативного дегуммирования, включающий дегуммирование с использованием полипептида по любому из пп. 1, 2, где указанный способ включает стадию приведения в контакт полипептида по любому из пп. 1, 2 с неочищенным маслом.
12. Способ по п. 11, где указанный способ включает один или более чем один из следующих признаков:
(1) полипептид добавляют в количестве от 10 до 1000 миллионных долей (м.д.) исходя из массы неочищенного масла;
(2) дегуммирование включает перемешивание при 50-60°C в течение 1-3 часов с последующим повышением температуры до 80-90°C и выдерживанием в течение 1-10 минут; и
(3) неочищенное масло выбрано из группы, состоящей из соевого масла, подсолнечного масла, арахисового масла, рапсового масла, масла из рисовых отрубей, кукурузного масла, оливкового масла, пальмового масла, пальмоядрового масла, пальмового олеина, масла канола, касторового масла, кокосового масла, кориандрового масла, хлопкового масла, масла лесного ореха, конопляного масла, льняного масла, масла из косточек манго, масла пенника лугового, копытного масла, сафлорового масла, масла камелии (camellia), таллового масла и масла камелии (tsubaki);
(4) указанная стадия включает приведение в контакт полипептида с неочищенным маслом при 50-70°C.
| US 2017058235 A1, 02.03.2017 | |||
| WO 2015140275 A1, 24.09.2015 | |||
| US 2013011887 A1, 10.01.2013 | |||
| CN 106884009 A, 23.06.2017 | |||
| EA 200401407 A1, 24.02.2006 | |||
| CN 106459934 A, 22.02.2017 | |||
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ ФОСФОЛИПАЗЫ С | 2012 |
|
RU2500811C1 |
| YAMPOLSKY L.Y | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Коловратный двигатель | 1924 |
|
SU1459A1 |
| KESKIN O | |||
| et al | |||
| A new, | |||

