Ссылка на родственные заявки
Согласно настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой на выдачу патента США №61/988581, поданной 5 мая 2014 года, и в соответствии с предварительной заявкой на выдачу патента США №62/067836, поданной 23 октября 2014 года, описания каждой из которых полностью включены в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
В настоящем документе описываются отличные от человека животные, содержащие последовательности нуклеиновых кислот, кодирующие белок С3 и/или белок С5, которые содержат человеческую последовательность, а также трансгенные отличные от человека животные, содержащие ген С3 и/или С5, который полностью или частично является человеческим. Также в настоящем документе описываются отличные от человека животные, которые экспрессируют человеческие или гуманизированные белки С3 и/или С5, и способы применения отличных от человека животных, содержащих человеческие или гуманизированные последовательности нуклеиновых кислот С3 и/или С5.
Предшествующий уровень техники
Белки комплемента С5 и С3 являются мишенями для терапевтического воздействия при лечении множества заболеваний, нарушений и патологических состояний человека, которые ассоциированы с активацией комплемента, например, глазные воспалительные заболевания и дегенеративные заболевания сетчатки. Оценку фармакокинетики (PK) и фармакодинамики (PD) терапевтических молекул, которые специфически направлены на белки С5 человека или С3 человека, обычно проводят на отличных от человека животных, например, грызунах, например, мышах или крысах. Тем не менее, PD таких терапевтических молекул невозможно правильно определить в определенных отличных от человека животных, поскольку данные терапевтические молекулы не направлены на эндогенные белки С5 или С3.
Кроме того, оценка терапевтической эффективности специфичных для человека антагонистов белков С5 или С3 в различных моделях заболеваний, связанных с активированным комплементом, на отличных от человека животных проблематична на отличных от человека животных, у которых такие видоспецифичные антагонисты не взаимодействуют с эндогенными белками С5 или С3.
В соответствии с этим существует необходимость в отличных от человека животных, например, грызунах, например, относящихся к мышам или крысам животных, например, мышах или крысах, у которых гены С5 и/или С3 отличного от человека животного полностью или частично гуманизированы или заменены (например, в эндогенных не являющихся человеческими локусах) на гены С5 и/или С3 человека, содержащие последовательности, кодирующие человеческие или гуманизированные белки С5 и/или С3, соответственно. Существует потребность в таких гуманизированных отличных от человека животных, которые экспрессируют человеческие или гуманизированные белки С5 и/или С3 в сыворотке при концентрациях, схожих с концентрациями белков С5 и/или С3, соответственно, присутствующих в сыворотке отличного от человека животного соответствующей возрастной группы, которые экспрессируют функциональные белки С5 и/или С3, но не содержат человеческие или гуманизированные гены С5 и/или С3, соответственно.
На протяжении данного описания изобретения упоминаются различные патентные заявки и другие типы публикаций (например, журнальные статьи, записи электронных баз данных и так далее. Описание всех патентов, патентных заявок и других публикаций, цитируемых в настоящем документе, таким образом полностью включены в качестве ссылки во всех отношениях.
Краткое раскрытие
Предложены отличные от человека животные, содержащие последовательности нуклеиновых кислот, кодирующие С3 и/или С5, которые содержат последовательность человека.
Предложены трансгенные отличные от человека животные, содержащие ген С3 и/или С5, который полностью или частично является человеческим.
Предложены отличные от человека животные, которые экспрессируют человеческие или гуманизированные белки С3 и/или С5.
Предложены отличные от человека животные, несущие замену (полностью или частично) эндогенных генов С5 и/или С3 отличного от человека животного.
Предложены отличные от человека животные, содержащие гуманизацию С5 и/или С3 (полностью или частично) в эндогенном не являющимся человеческим локусе С5 и/или С3.
Предложены отличные от человека животные, которые обладают человеческими или гуманизированными генами С5 и/или С3, где отличные от человека животные не экспрессируют эндогенный белок С5 и/или С3 и экспрессируют человеческий или гуманизированный белок С5 и/или С3 в сыворотке при концентрациях, схожих с концентрациями белков С5 и/или С3, соответственно, присутствующих в сыворотке отличного от человека животного соответствующей возрастной группы, которые экспрессируют функциональные эндогенные белки С5 и/или С3, но не содержат замены.
Согласно одному аспекту предложены отличные от человека животные, содержащие человеческую или гуманизированную последовательность нуклеиновой кислоты С3 и/или С5.
Согласно одному аспекту предложены генетически модифицированные отличные от человека животные, которые содержат замену в эндогенном локусе С5 и/или С3 гена, кодирующего эндогенный С5 и/или С3, на ген, кодирующий человеческий или гуманизированный белок С5 и/или С3. Предложены грызуны, например, мыши или крысы, которые содержат замену эндогенного гена С5 в эндогенном локусе С5 крысы на ген С5 человека и/или содержат замену эндогенного гена С3 в эндогенном локусе С3 крысы на ген С3 человека. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мыши или крысы, содержащие замену в эндогенном крысином локусе С5 гена крысы, кодирующего белок С5, на ген С5 человека, кодирующий человеческий или гуманизированный белок С5, где экспрессия гена С5 человека, кодирующего человеческий или гуманизированный белок С5, находится под контролем регуляторных элементов грызуна в эндогенном локусе С5 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления ген С5 человека, кодирующего человеческий или гуманизированный белок С5, содержит с 2экзона по 41экзон гена C5 человека.
Согласно одному варианту осуществления грызун представляет собой мышь, которая неспособна экспрессировать белок С5 мыши.
Согласно одному варианту осуществления грызун представляет собой мышь, которая экспрессирует белок С3 мыши, кодируемый эндогенным геном С3 мыши.
Согласно одному варианту осуществления грызун представляет собой мышь, которая экспрессирует человеческий или гуманизированный белок С3.
Согласно одному варианту осуществления грызун представляет собой мышь, которая содержит замену в эндогенном локусе С3 мыши мышиного гена, кодирующего белок С3 на ген С3 человека, кодирующего человеческий или гуманизированный белок С3.
Согласно одному варианту осуществления экспрессия гена С3 человека, кодирующего человеческий или гуманизированный белок С5, находится под контролем регуляторных элементов в эндогенном локусе С3 мыши.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мыши или крысы, содержащие замену в эндогенном локусе С3 грызуна гена грызуна, кодирующего белок С3, на ген С3 человека, кодирующий человеческий или гуманизированный белок С3, где экспрессия гена С3 человека, кодирующего человеческий или гуманизированный белок С3 находится под контролем регуляторных элементов грызуна в эндогенном локусе С3 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления ген С3 человека, кодирующий человеческий или гуманизированный белок С3, содержит с 1 экзона по 41 экзон гена С3 человека.
Согласно одному варианту осуществления ген С3 человека, кодирующий человеческий или гуманизированный белок С3, содержит со 2 экзона по 41 экзон гена С3 человека.
Согласно одному варианту осуществления грызун представляет собой мышь, которая неспособна экспрессировать белок С3 мыши.
Согласно одному варианту осуществления грызун представляет собой мышь, которая экспрессирует мышиный белок С5, кодируемый эндогенным геном С5 мыши.
Согласно одному варианту осуществления грызун представляет собой мышь, которая экспрессирует человеческий или гуманизированный белок С5.
Согласно одному варианту осуществления грызун представляет собой мышь, которая содержит замену в эндогенном мышином локусе С5 гена мыши, кодирующего белок С5, на ген С5 человека, кодирующий человеческий или гуманизированный белок С5.
Согласно одному варианту осуществления экспрессия гена С5 человека, кодирующего человеческий или гуманизированный белок С5, находится под контролем мышиных регуляторных элементов в эндогенном локусе С5 мыши.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мышь или крыса, которые экспрессируют человеческий или гуманизированный белок С5, где грызун, который экспрессирует человеческий или гуманизированный белок С5, содержит нормальную систему комплемента, например, уровни белков комплемента в крови, плазме или сыворотки крови грызуна, экспрессирующего человеческий или гуманизированный белок С5, схожи с уровнями белков комплемента в крови, плазме или сыворотки крови грызуна, который экспрессирует функциональный эндогенный белок С5. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мышь или крыса, которые экспрессируют белок С5 из гена С5 человека, где грызун экспрессирует человеческий или гуманизированный белок С5 в его сыворотку. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления в сыворотке крови грызуна, который экспрессирует человеческий или гуманизированный белок С5, содержится приблизительно такой же уровень белка С5, как и у грызуна, который экспрессирует функциональный эндогенный белок С5, например, мышь или крыса дикого типа. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С5 в сыворотке при концентрации, по меньшей мере, приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190% или 200% от уровня белка С5, присутствующего в сыворотке крови мыши соответствующей возрастной группы, которая экспрессирует функциональный эндогенный белок С5, но не содержит замены эндогенного гена С5 в эндогенном мышином локусе С5 на ген С5 человека.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С5 в сыворотке при концентрации в диапазоне от приблизительно 10% до приблизительно 200%, в диапазоне от приблизительно 20% до приблизительно 150% или в диапазоне от приблизительно 30% до приблизительно 100% от уровня мышиного белка С5, присутствующего в сыворотке крови мыши соответствующей возрастной группы, которая экспрессирует функциональный эндогенный белок С5, но не содержит замены эндогенного гена С5 в эндогенном мышином локусе С5 на ген С5 человека.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С5 в сыворотке при концентрации в диапазоне от приблизительно 10 мкг/мл до приблизительно 150 мкг/мл, в диапазоне от приблизительно 10 мкг/мл до приблизительно 125 мкг/мл или в диапазоне от приблизительно 15 мкг/мл до приблизительно 100 мкг/мл.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С5 в сыворотке при концентрации, по меньшей мере, приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 125 или 150 мкг/мл.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мышь или крыса, которые экспрессируют человеческий или гуманизированный белок С3, где грызун, который экспрессирует человеческий или гуманизированный белок С3 содержит нормальную систему комплемента, то есть, уровни белков комплемента в крови, плазме или сыворотки крови грызуна, экспрессирующего человеческий или гуманизированный белок С3, схожи с уровнями белков комплемента в крови, плазме или сыворотки крови грызуна, который экспрессирует функциональный эндогенный белок С3. Согласно одному варианту осуществления, грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному аспекту предложены генетически модифицированные грызуны, например, мышь или крыса, которые экспрессируют белок С3 из гена С3 человека, где грызун экспрессирует человеческий или гуманизированный белок С3 в его сыворотку. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления в сыворотке грызуна, который экспрессирует человеческий или гуманизированный белок С3, содержится приблизительно такой же уровень белка С3, как у грызуна, который экспрессирует функциональный эндогенный белок С3, например, у мыши или крысы дикого типа. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С3 (hC3) в сыворотке при концентрации, по меньшей мере, приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190% или 200% от уровня белка С3, присутствующего в сыворотке крови мыши соответствующей возрастной группы, которая экспрессирует функциональный эндогенный белок С3, но не содержит замены эндогенного гена С3 в эндогенном мышином локусе С3 на ген С3 человека.
Согласно одному варианту осуществления мышь экспрессирует человеческий белок С3 в сыворотке при концентрации в диапазоне от приблизительно 10% до приблизительно 200%, в диапазоне от приблизительно 20% до приблизительно 150% или в диапазоне от приблизительно 30% до приблизительно 100% от уровня мышиного белка С3, присутствующего в сыворотке крови мыши соответствующей возрастной группы, которая экспрессирует функциональный эндогенный белок С3, но не содержит замены эндогенного гена С3 в эндогенном мышином локусе С3 на ген С3 человека.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С3 в сыворотке при концентрации в диапазоне от приблизительно 100 мкг/мл до приблизительно 1500 мкг/мл, в диапазоне от приблизительно 200 мкг/мл до приблизительно 1250 мкг/мл или в диапазоне от приблизительно 300 мкг/мл до приблизительно 1000 мкг/мл.
Согласно одному варианту осуществления мышь экспрессирует человеческий или гуманизированный белок С3 в сыворотке при концентрации, по меньшей мере, приблизительно 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1250 или 1500 мкг/мл.
Согласно одному аспекту предложены генетически модифицированные грызуны, содержащие ген С5 человека, содержащие замену в эндогенном локусе С5 грызуна гена грызуна, кодирующего белок С5, на ген С5 человека, кодирующий человеческий или гуманизированный белок С5, где экспрессия гена С5 человека, кодирующего человеческий или гуманизированный белок С5, находится под контролем регуляторных элементов или последовательностей грызуна в эндогенном локусе С5 грызуна, и где грызун дополнительно содержит ген С3 человека, включающий замену в эндогенном локусе С3 грызуна гена грызуна, кодирующего белок С3, на ген С3 человека, кодирующего человеческий или гуманизированный белок С3, где экспрессия гена С3 человека, кодирующего человеческий или гуманизированный белок С3, находится под контролем регуляторных элементов или последовательностей грызуна в эндогенном локусе С3 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления мышь неспособна экспрессировать мышиный белок С5 и неспособна экспрессировать мышиный белок С3.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна в локусе С5 грызуна и/или локусе С3 грызуна происходят из мыши или крысы.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна представляют собой эндогенные регуляторные элементы или последовательности грызуна в локусе С5 грызуна и/или локусе С3 грызуна происходят из мыши или крысы.
Согласно одному аспекту предложено отличное от человека животное, например, грызун, например, мышь или крыса, которое экспрессирует человеческие или гуманизированные белки С5 и/или С3, где отличное от человека животное экспрессирует человеческие или гуманизированные белки С5 и/или С3 из эндогенного не относящегося к человеческому локуса С5 и/или C3 эндогенного не относящегося к человеческому локуса С3. Согласно варианту осуществления отличное от человека животное представляет собой грызуна. Согласно варианту осуществления, грызун представляет собой мышь. Согласно варианту осуществления грызун представляет собой крысу.
Согласно одному аспекту предложена генетически модифицированная мышь, которая экспрессирует человеческий или гуманизированный белок С5 из эндогенного мышиного локуса С5, где эндогенный мышиный локус С5 был заменен полностью или частично на ген С5 человека.
Согласно одному варианту осуществления приблизительно 75,8 т.п.н. в эндогенном локусе С5 мыши, включающем в себя экзоны со 2 по 42 и 3'-нетранслируемую последовательность, удаляют и заменяют на приблизительно 97 т.п.н. последовательности гена С5 человека, содержащего экзоны со 2 по 41 гена С5 человека. Согласно особому варианту осуществления ген С5 человека содержит экзоны со 2 по 42 гена С5 человека из ВАС CTD-2559I19 человека. Согласно особому варианту осуществления ген С5 содержит экзон 1 С5 мыши и экзоны со 2 по 42 С5 человека.
Согласно одному аспекту предложена генетически модифицированная мышь, которая содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный белок С5, где нуклеотидная последовательность, кодирующая человеческий или гуманизированный белок С5, полностью или частично заменяет эндогенную нуклеотидную последовательность, кодирующую эндогенный белок С5 мыши.
Согласно одному аспекту предложена генетически модифицированная мышь, которая экспрессирует человеческий или гуманизированный белок С3 из эндогенного локуса С3 мыши, где эндогенный ген С3 мыши заменен полностью или частично на ген С3 человека.
Согласно одному варианту осуществления часть эндогенного локуса С3 мыши, включающую в себя 5'-регуляторные элементы против хода транскрипции от 1 экзона по 41 экзон, удаляют и заменяют на последовательность гена С3 человека, содержащую 5'-регуляторные элементы против хода транскрипции от 1 экзона по 41 экзон гена С3 человека. Согласно особому варианту осуществления, ген С3 человека содержит целую кодирующую область С3 человека.
Согласно одному варианту осуществления часть эндогенного локуса С3 мыши, включающую в себя часть интрона 1 и экзоны с 1 по 41, удаляют и заменяют последовательностью гена С3 человека, содержащую часть интрона 1 и с 2 экзона по 41 экзон гена С3 человека. Согласно особому варианту осуществления ген С3 содержит экзон 1 С3 мыши и экзоны со 2 по 41 С3 человека.
Согласно одному аспекту предложен способ получения гуманизированного по С5 грызуна, включающий замену последовательности гена С5 грызуна, кодирующую белок С5 грызуна, на последовательность гена С5 человека, содержащую один или несколько экзонов последовательности гена С5 человека, кодирующей человеческий или гуманизированный белок С5. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С5 грызуна, и последовательность гена С5 человека, содержащая один или несколько экзонов последовательности гена С5 человека, кодирующей человеческий или гуманизированный белок С5, функционально связана с регуляторными элементами или последовательностями грызуна в эндогенном локусе С5 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна происходят из мыши. Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна происходят из крысы.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна представляют собой эндогенные регуляторные элементы или последовательности грызуна в локусе С5 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления последовательность гена С5 человека, заменяющая последовательность гена С5 грызуна, содержит, по меньшей мере, один экзон последовательности гена С5 человека. Согласно другим вариантам осуществления последовательность гена С5 человека, заменяющая последовательность гена С5 грызуна, содержит, по меньшей мере, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 экзонов последовательности гена С5 человека. Согласно одному варианту осуществления последовательность гена С5 человека, заменяющая последовательность гена С5 грызуна, содержит все из 41 экзона последовательности гена С5 человека. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С5 грызуна, и последовательность гена С5 человека, содержащая один или несколько экзонов последовательности гена С5 человека, кодирующей человеческий или гуманизированный белок С5, функционально связана с эндогенными регуляторными элементами или последовательностями грызуна в эндогенном локусе С5 грызуна.
Согласно одному аспекту предложен способ получения гуманизированных по С5 мышей, включающий замену последовательности гена С5 мыши, кодирующей белок С5 мыши, на последовательность гена С5 человека, кодирующую человеческий или гуманизированный белок С5.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С5 мыши, и ген С5 человека, кодирующий человеческий или гуманизированный белок С5, функционально связан с регуляторными элементами или последовательностями мыши в эндогенном локусе С5 мыши.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С5 мыши, и ген С5 человека, кодирующий человеческий или гуманизированный белок С5, функционально связан с эндогенными регуляторными элементами или последовательностями мыши в эндогенном локусе С5 мыши.
Согласно одному аспекту предложен способ получения гуманизированного по С3 грызуна, включающий замену последовательности гена С3 грызуна, кодирующей белок С3 грызуна, на последовательность гена С3 человека, содержащую один или несколько экзонов последовательности гена С3 человека, кодирующей человеческий или гуманизированный белок С3. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С3 грызуна, и последовательность гена С3 человека, содержащая один или несколько экзонов последовательности гена С3 человека, кодирующей человеческий или гуманизированный белок С3, функционально связана с регуляторными элементами или последовательностями грызуна в эндогенном локусе С3 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна происходят из мыши. Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна происходят из крысы.
Согласно одному варианту осуществления регуляторные элементы или последовательности грызуна представляют собой эндогенные регуляторные элементы или последовательности грызуна в локусе С3 грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления последовательность гена С3 человека, заменяющая последовательность гена С3 грызуна, содержит, по меньшей мере, одни экзон последовательности гена С3 человека. Согласно другим вариантам осуществления последовательность гена С3 человека, заменяющая последовательность гена С3 грызуна, содержит, по меньшей мере, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 экзонов последовательности гена С3 человека. Согласно одному варианту осуществления последовательность гена С3 человека, заменяющая последовательность гена С5 грызуна содержит все из 41 экзона последовательности гена С3 человека. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С3 грызуна, и последовательность гена С3 человека, содержащая один или несколько экзонов последовательности гена С3 человека, кодирующей человеческий или гуманизированный белок С3, функционально связана с эндогенными регуляторными элементами или последовательностями грызуна в эндогенном локусе С3 грызуна.
Согласно одному аспекту предложен способ получения гуманизированной по С3 мыши, включающий замену последовательности гена С3 мыши, кодирующей белок С3 мыши, на последовательность гена С3 человека, кодирующую человеческий или гуманизированный белок С3.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С3 мыши, и ген С3 человека, кодирующий человеческий или гуманизированный белок С3, функционально связан с эндогенными регуляторными элементами или последовательностями мыши в эндогенном локусе С3 мыши.
Согласно одному варианту осуществления замена происходит в эндогенном локусе С3 мыши, и ген С3 человека, кодирующий человеческий или гуманизированный белок С3, функционально связан с регуляторными элементами или последовательностями мыши в эндогенном локусе С3 мыши.
Согласно различным аспектам генетически модифицированные отличные от человека животные, например, грызуны, например, мыши или крысы, описываемые в настоящем документе, содержат генетические модификации в их зародышевой линии.
Согласно одному аспекту предложен эмбрион отличного от человека животного, например, грызуна, например, мыши или крысы, содержащий генетическую модификацию, как описано в настоящем документе.
Согласно одному аспекту предложен эмбрион-хозяин отличного от человека животного, например, грызуна, например, мыши или крысы, который содержит донорную клетку, которая содержит генетическую модификацию, как описано в настоящем документе.
Согласно одному аспекту предложена плюрипотентная или тотипотентная клетка отличного от человека животного, например, грызуна, например, мыши или крысы, содержащая генетическую модификацию, как описано в настоящем документе. Согласно одному варианту осуществления клетка представляет собой клетку грызуна. Согласно одному варианту осуществления клетка представляет собой клетку мыши. Согласно одному варианту осуществления клетка представляет собой ES клетку грызуна. Согласно одному варианту осуществления клетка представляет собой ES клетку мыши.
Согласно одному аспекту предложена яйцеклетка отличного от человека животного, например, грызуна, например, мыши или крысы, где яйцеклетка отличного от человека животного содержит эктопическую хромосому отличного от человека животного, где эктопическая хромосома отличного от человека животного содержит генетическую мутацию, как описано в настоящем документе. Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна. Согласно одному варианту осуществления грызун представляет собой мышь. Согласно одному варианту осуществления грызун представляет собой крысу.
Согласно одному аспекту эмбрион, яйцеклетка или клетка мыши, которая генетически модифицирована, чтобы содержать ген С5 человека или ген С3 человека, представляет собой мышь, которая принадлежит к линии C57BL, выбранную из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Согласно другому варианту осуществления мышь представляет собой линию 129, выбранную из группы, состоящей из линии, которая представляет собой 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (смотри, например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10: 836, смотри также Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). Согласно особому варианту осуществления генетически модифицированная мышь представляет собой смешанный тип упомянутой выше линии 129 и упомянутой выше линии C57BL/6. Согласно другому особому варианту осуществления мышь представляет собой смешанный тип упомянутых выше линий 129 или смешанный тип упомянутых выше линий BL/6. Согласно особому варианту осуществления линия 129 смешанного типа представляет собой линию 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь представляет собой линию BALB, например, линию BALB/c. Согласно еще одному варианту осуществления мышь представляет собой смешанный тип линии BALB и другой упомянутой выше линии. Согласно одному варианту осуществления мышь представляет собой швейцарскую мышь (Swiss) или швейцарскую мышь Вебстер (Swiss Webster).
Согласно дополнительному аспекту в настоящем документе предложены способы идентификации соединения, способного модулировать активацию системы комплемента, включающие введение соединения любому из отличных от человека животных, например, грызуну, например, мыши или крысе, раскрываемому и описываемому в настоящем документе; и проведения анализа того, модулируется ли активация системы комплемента у грызуна, тем самым идентифицируя соединение, способное модулировать активацию системы комплемента. Согласно одному варианту осуществления соединение представляет собой низкомолекулярное химическое соединение, антитело, белок, ингибирующую нуклеиновую кислоту или любую их комбинацию. Согласно другому варианту осуществления любого из вариантов осуществления, описываемых в настоящем документе, соединение модулирует активацию системы комплемента, увеличивая активность комплемента. Согласно другому варианту осуществления любого из вариантов осуществления, описываемых в настоящем документе, соединение модулирует активацию системы комплемента, снижая активность комплемента. Согласно другому варианту осуществления любого из вариантов осуществления, описываемых в настоящем документе, анализируют сыворотку грызуна для того, чтобы определить модулируется ли активация системы комплемента у грызуна. В дополнительном варианте осуществления анализ представляет собой скрининговое исследование системы комплемента СН50.
Каждый из аспектов и вариантов осуществления, описываемых в настоящем документе, может применяться совместно, если только это не исключено либо однозначно, либо явно из контекста варианта осуществления или аспекта.
Краткое описание чертежей
На фигуре 1 приведена иллюстрация без соблюдения масштаба геномных локусов С5 мыши (сверху) и человека (снизу). Показаны области генов С5 мыши и человека, которые удаляют и заменяют, соответственно, чтобы получить гуманизированных по С5 мышей.
На фигуре 2 приведена иллюстрация без соблюдения масштаба мышиного (mC3) и гуманизированного (hC3) геномных локусов С3. (А) Ген С3 мыши, охватывающий 5'-регуляторные элементы и кодирующую область от экзона 1 до экзона 41, удаляют и заменяют на 5'-регуляторные элементы и кодирующую область от экзона 1 до экзона 41 гена С3 человека и участок loxP, как указано жирной линией и стрелкой, соответственно. (В) Ген С3 мыши, охватывающий часть интрона 1 и кодирующую область от экзона 2 до экзона 41, удаляют и заменяют на часть интрона 1 и кодирующую область от экзона 2 до экзона 41 гена С5 человека и участок loxP, как указано жирной линией и стрелкой, соответственно.
На фигуре 3 показан комплемент мыши, который обладает более низкой активностью при тесте на гемолиз, чем комплемент человека.
На фигуре 4 показано, что гемолитическая активность комплемента блокируется видоспецифичными моноклональными антителами против С5 видоспецифичным образом.
На фигуре 5 показано, что гуманизированные по С5 мыши представляют собой функциональную нокаут-мышь по С5. (А) Добавление белка С5 или С3 человека восстанавливает гемолитическую активность в сыворотке человека с удаленным С5 или С3, соответственно. (В) Белок С5 мыши, а не человека, восстанавливает гемолитическую активность в сыворотках С5-/- и гуманизированных по С5 мышей. (С) Уровни белка С5 человека в сыворотке гуманизированной по С5 мыши в три раза ниже, чем в сыворотке человека. (D) Уровни С5 комплемента до введения препарата у гуманизированных по С5 мышей отличались у самцов и самок. В сыворотке гуманизированной по С5 мыши присутствует С5 человека в диапазоне от 20 до 100 мкг/мл.
На фигуре 6 показано, что добавление белков С3 и С5 человека, а не одного только белка С5 человека, восстанавливает гемолитическую активность в сыворотке С5-/- мыши.
На фигуре 7 показано, что для достижения уровней гемолитической активности у мышей, характерных для дикого типа, необходимо добавление в сыворотку гуманизированной по С5 мыши белка С3 человека.
На фигуре 8 показаны результаты теста на гемолиз, проведенного на образцах сыворотки гуманизированных по С5 мышей, подвергнутых обработке антителом против С5 человека (C5Ab), используя 400 мкг/мл белка С3 человека (А) и 800 мкг/мл белка С3 человека (В).
На фигуре 9 показаны результаты фармакокинетического анализа, определяющего уровни антител против С5 человека в зависимости от времени после инъекции для всех животных в исследовании (А) и для самок (сверху) и самцов (снизу) животных (В).
На фигуре 10А показано, что добавление белка С3 человека к сыворотке мыши дикого типа может усиливать гемолитическую функцию, но не до такой же степени, как наблюдается в сыворотке человека в норме (для каждой точки на графике по мышам, смешивали сыворотку от 3 животных). На фигуре 10В показано, что гемолитическую функцию можно восстановить в гуманизированных мышах C5hu/hu, когда добавлен белок С3 человека.
На фигуре 11 показано, что добавление белка С3 человека к сыворотке, полученной из нокаут-мышей по С3, может восстановить гемолитическую функцию (для каждой точки на графике по мышам, смешивали сыворотку от 3 животных).
На фигуре 12 показано, что добавление одного только белка С3 человека к сыворотке нокаут-мыши по С5 не может восстановить гемолитическую функцию (для каждой точки на графике по мышам, смешивали сыворотку от 3 животных).
Подробное раскрытие
Система комплемента является важнейшим компонентом врожденная иммунной системы и играет важную роль в качестве защитного механизма против инвазивных патогенов, подготавливает адаптивная иммунную реакцию и способствует удалению иммунных комплексов и апоптотических клеток. В то время как система комплемента играет ключевую роль во множестве защитных иммунных функций, активация системы комплемента является важным медиатором для повреждения ткани при развитии широкого диапазона аутоиммунных и воспалительных заболеваний.
В изобретении, описываемом в настоящем документе, предложены, помимо всего прочего, отличные от человека животные, которые экспрессируют человеческие или гуманизированные белки С5 и/или С3 в сыворотке при концентрациях, схожих с экспрессией эндогенных белков С5 и/или С3 у отличных от человека животных дикого типа. Успешная инженерия отличных от человека животных, способных экспрессировать человеческие или гуманизированные белки С5 и/или С3, обеспечивает необходимое и применимое средство для воспроизведения системы комплемента человека на лабораторных животных, пригодных для широкомасштабных и высокопроизводительных скрининговых исследований лекарственных веществ. Также в настоящем документе предложены способы применения отличных от человека животных (таких как грызуны), описываемых в настоящем документе, для идентификации соединений, способных модулировать активацию системы комплемента. Способы основаны, отчасти, на находке авторов изобретения, что в то время как активность белка С5 системы комплемента является видоспецифичной (то есть, белок С5 человека не способен заменить белок С5 мыши), активность белка С3 системы комплемента человека способна заменить белок С3 мыши для активности комплемента сыворотки мыши.
Общие методики
Практика настоящего изобретения будет применять, если не указано иное, общепринятые методики молекулярной биологии, микробиологии, клеточной биологии, биохимии, химии нуклеиновых кислот и иммунологии, которые хорошо известны специалистам в данной области. Такие методики полностью объясняются в литературе, такой как Molecular Cloning: A Laboratory Manual, четвертое издание (Sambrook et al., 2012) и Molecular Cloning: A Laboratory Manual, третье издание (Sambrook and Russel, 2001), (вместе обозначаемые в настоящем документе как «Sambrook»); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987, включая дополнения вплоть до 2014); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Antibodies: A Laboratory Manual, второе издание, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (Greenfield, ed., 2014), Beaucage et al. eds., Current Protocols in Нуклеиновая кислота Chemistry, John Wiley & Sons, Inc., New York, 2000, (включая дополнения вплоть до 2014) и Gene Transfer and Expression in Mammalian Cells (Makrides, ed., Elsevier Sciences B.V., Amsterdam, 2003).
Определения
Как используется в настоящем документе, термин «белок» включает в себя полипептиды, пептиды, фрагменты полипептидов и слитые полипептиды.
Как используется в настоящем документе, «нуклеиновая кислота» обозначает два или более дезоксирибонуклеотида и/или рибонуклеотида, ковалентно связанных вместе либо в одно-, либо в двухцепочечной форме.
Под фразой «функционально связанный» обозначают функциональную связь между последовательностью, контролирующей экспрессию нуклеиновой кислоты (такой как промотор), и второй последовательностью нуклеиновой кислоты, где последовательность, контролирующая экспрессию, управляет транскрипцией нуклеиновой кислоты, соответствующей второй последовательности.
Термин «замена» в отношении замены гена означает внесение экзогенного генетического материала в эндогенный генетический локус, тем самым заменяя весь или часть эндогенного гена на ортологическую или гомологичную последовательность нуклеиновой кислоты. В одном случае эндогенный не являющийся человеческим ген или его фрагмент заменяют на соответствующий ген человека или его фрагмент. Соответствующий ген человека или его фрагмент представляет собой ген человека или фрагмент, который является ортологом, гомологом, или является по существу идентичным или таким же по структуре и/или функции, как и эндогенный не являющийся человеческим ген или его фрагмент, который заменяют. Как продемонстрировано ниже в примерах, нуклеотидные последовательности эндогенных не являющихся человеческими (например, грызуна, такого как мышь) локусов гена С3 и/или С5 заменяли нуклеотидными последовательностями, соответствующими локусам генов С3 и/или С5 человека. Согласно другому варианту осуществления замена гена может происходить, когда эндогенный ген удаляют или приводят в нефункциональное состояние (такое, как при вставке миссенс-мутации или преждевременного стоп-кодона) и соответствующий ген человека или его фрагмент встраивают в зародышевую линию в отдельное местоположение.
Термин «гуманизированный», как применяют в фразах «гуманизированный аллель С3» или «гуманизированный аллель С5» или «гуманизированный ген С3» или «гуманизированный ген С5» включает в себя, но не ограничиваясь этим, варианты осуществления, где весь или часть эндогенного не являющегося человеческим гена или аллеля С3 и/или С5 заменяют соответствующей частью гена или аллеля С3 и/или С5 человека. Например, согласно некоторым вариантам осуществления термин «гуманизированный» относится к полной замене кодирующей области (например, экзонов) эндогенного не являющегося человеческим гена или аллеля С3 и/или С5 на соответствующую кодирующая область гена или аллеля С3 и/или С5 человека, в то время как эндогенная некодирующая область(и) (такая как, но не ограничиваясь этим, промотор, 5'- и/или 3'-нетранслируемая область(и), энхансерные элементы и так далее) отличного от человека животного не заменяется. Согласно некоторым вариантам осуществления отличное от человека животное представляет собой грызуна, такого как крыса или мышь.
Фраза «гуманизированный белок» включает в себя, но, не ограничиваясь этим, варианты осуществления, в которых весь или часть кодируемого эндогенного не являющегося человеческим белка С3 и/или С5 заменяют соответствующей частью белка С3 и/или С5 человека. Согласно некоторым вариантам осуществления «гуманизированный белок» может кодироваться гуманизированным геном или аллелем С3 и/или С5, и тем не менее кодировать целый белок С3 и/или С5 человека (такой как, но, не ограничиваясь этим, в ситуации, где все из кодирующих областей (например, экзоны) эндогенного не являющегося человеческим гена или аллеля С3 и/или С5 заменяют соответствующими кодирующими областями гена или аллеля С3 и/или С5 человека, но эндогенная некодирующая область(и) (такая как, но не ограничиваясь этим, промотор, 5'- и/или 3'-нетранслируемая область(и), энхансерные элементы и так далее) отличного от человека животного не заменяется). Согласно некоторым вариантам осуществления отличное от человека животное представляет собой грызуна, такого как крыса или мышь.
Используемый в настоящем документе термин «модулирует» означает способность соединения изменять активность системы комплемента. Согласно одному варианту осуществления модулирование системы комплемента соединением означает снижение активации системы комплемента у субъекта (например, грызуна, такого как мышь). Согласно другому варианту осуществления модулирование системы комплемента соединением означает увеличение опосредованной комплементом активности у субъекта.
Используемый в настоящем документе термин «грызун» означает любого члена отряда Rodentia. Примеры грызунов включают в себя без ограничения мышей, крыс, белок, луговых собачек, дикобразов, бобров, морских свинок и хомячков. Согласно одному варианту осуществления грызун представляет собой крысу. Согласно другому варианту осуществления грызун представляет собой мышь.
Если не указано иное в настоящем документе, все технические и научные термины имеют такое же значение, как обычно понимает специалист в данной области, к которой принадлежит данное изобретение.
Используемые в настоящем документе термины единственного числа включают в себя множественное число, если контекст явно не указывает на иное.
Предполагается, что каждое максимальное количественное ограничение, приводимое на протяжении данного описания, включает в себя меньшее количественное ограничение, как если бы такие меньшие количественные ограничения были бы в явном виде написаны в настоящем документе. Каждое минимальное количественное ограничение, приводимое на протяжении данного описания, будет включать в себя каждое более высокое количественное ограничение, как если бы такие более высокие количественные ограничения были бы в явном виде написаны в настоящем документе. Каждый диапазон числовых значений, приводимый на протяжении данного описания, будет включать в себя каждый более узкий диапазон числовых значений, который попадает в такой долее широкий диапазон числовых значений, как если бы такие более узкие диапазоны числовых значений были бы в явном виде написаны в настоящем документе.
Система комплемента
Комплемент, важный компонент иммунной системы, состоит из более чем 30 сывороточных и клеточных белков, которые вовлечены в два связанных биохимических каскада, классический и альтернативный пути. Комплемент функционирует, чтобы помогать иммунной системе разрушать инвазивные микроорганизмы и поддерживать гомеостаз ткани.
Тем не менее, чрезмерная или нерегулируемая активация системы комплемента вызывает повреждение ткани и ассоциирована со множеством заболеваний, нарушений и патологических состояний человека, которые связаны с активацией комплемента, например, глазные воспалительные заболевания и дегенеративные заболевания сетчатки (смотри Makrides (1998) Therapeutic inhibition of the complement system, Pharmacological Reviews 50(1): 59-87; и Mollnes et al. (2006) Strategies of therapeutic complement inhibition, Molecular Immunology 43; 107-121). Другие заболевания или состояния, которые, как известно, ассоциированы с нарушенной активацией комплемента, включают в себя без ограничения аллергическую астму и сопутствующие воспаление дыхательных путей и гиперреактивность дыхательных путей («AHR»), хроническое обструктивное заболевание легких («COPD»), аллергический бронхолегочный аспергиллез, аллергический альвеолит, эозинофильную пневмонию, эмфизему, бронхит, аллергический бронхит, бронхоэктазию, муковисцидоз, туберкулез, пневмонит гиперчувствительности, профессиональную бронхиальную астму, саркоид, реактивные заболевания дыхательных путей, интерстициальное заболевание легких, гиперэозинофильный синдром, ринит, синусит, бронхиальную астму физического напряжения, вызванную загрязнением среды астму, кашлевую астму, паразитарное заболевание легких, респираторно-синцитиальную вирусную («RSV») инфекцию, парагриппозную вирусную («PIV») инфекцию, риновирусную («RV») инфекцию и аденовирусную инфекцию и ишемически-реперфузионное повреждение (see, например, публикацию патентной заявки США №2005/0260198, которая включена в настоящий документ в качестве ссылки).
Ген и белок С5 системы комплемента
Ген С5 кодирует сывороточный белок С5 системы комплемента, который играет важную роль при развитии воспаления и уничтожения клетки. Зрелый белок С5 состоит из α- и β--цепей, которые связаны дисульфидной связью. Белок С5 расщепляется конвертазой, приводя к активации пептида С5а, анафилатоксина, происходящего из α-полипептида, который в большей степени является провоспалительным, и C5b, макромолекулярного продукта расщепления, происходящего из β-полипептида, который образует комплекс с другими компонентами комплемента для образования мембраноатакующего комплекса (MAC), который вовлечен в осмотический лизис клеток-мишеней.
С5 человека. Официальное сокращение: С5; ID гена в NCBI: 727; первичный источник: HGNC:1331; ID мРНК в RefSeq: NM_001735.2; ID в UniProt: P01031; Genomic assembly: GRCh38; локализация: chr9:120,952,335-121,050,275 - цепь.
Ген C5 человека локализован на 9 хромосоме, в 9q33-q34. Ген С5 человека содержит 41 экзон и кодирует полипептид-предшественник длиной 1676 аминокислот, включающий в себя сигнальный пептид из 18 аминокислот, β-цепь из 655 аминокислот и α-цепь из 999 аминокислот. Во время активации системы комплемента α-цепь расщепляется, тем самым образуя С5а из 74 аминокислот, который является сильным провоспалительным анафилатоксином.
Недостаточность С5 у людей связана с повышенной предрасположенностью к тяжелым рецидивирующим инфекциям.
Ген С5 является консервативным среди некоторых видов, включая приматов, например, шимпанзе, макака-резус, других млекопитающих, например, собака, корова, грызун, например, мышь, курицу, данио и лягушку.
С5 мыши. Официальное сокращение: Hc; ID гена в NCBI: 15139; первичный источник: MGI:96031; ID мРНК в RefSeq: NM_010406.2; ID в UniProt: Р06684; Genomic assembly: GRCm38; локализация: chr2:34,983,331-35,061,449 - цепь.
Ген C5 мыши локализован на 2хромосоме, в 2 23.22 сМ. Ген С5 мыши содержит 42 экзона и кодирует полипептид-предшественник длиной 1680 аминокислот, включающий в себя сигнальный пептид из 18 аминокислот, β-цепь из 656 аминокислот и α-цепь из 1002 аминокислот. Во время активации системы комплемента α-цепь расщепляется, тем самым образуя С5а из 77 аминокислот, который является сильным провоспалительным анафилатоксином.
Недостаточность С5 наблюдается у нескольких обычных линий лабораторных мышей, которые содержат делецию 2 пар нуклеотидов около 5'-конца кДНК, которая приводит в неспособности С5 секретироваться; таким образом, данные мыши являются лишенными функционального С5, то есть, нокаутами по С5 (6 таких линий представляют собой A/HeJ, AKR/J, DBA/2J, NZB/B1NJ, SWR/J и B10.D2/oSnJ) (смотри, например, Wetzel et al. (1990) Deficiency of the murine fifth complement component (C5): a 2-base pair deletion in a 5' exon, J Biol Chem 265: 2435-2440). Нокаут-мыши по C5 можно также получить, следуя стандартным способам, известным в данной области.
Ген и белок С3 системы комплемента
Ген С3 кодирует сывороточный белок С3 системы комплемента, который играет центральную роль в активации классического и альтернативного путей активации системы комплемента.
Ген С3 человека локализован на 19 хромосоме на 19р13.3-р13.2. Ген С3 человека содержит 41 экзон и кодирует полипептид-предшественник из 1663 аминокислот, включающий в себя сигнальный пептид из 22 аминокислот, β-цепь из 645 аминокислот и α-цепь из 992 аминокислот. Во время активации системы комплемента α-цепь расщепляется, тем самым образуя 9 различных пептидов, включая С5а из 77 аминокислот, который является сильным провоспалительным анафилатоксином.
Недостаточность С3 у людей ассоциирована с повышенной предрасположенностью к бактериальным инфекциям.
Ген С3 является консервативным среди нескольких видов, включая приматов, например, шимпанзе, макака-резус, других млекопитающих, например, собака, корова, грызун, например, мышь, курицу, данио и лягушку.
Ген С3 мыши локализован на 17 хромосоме в 17 29.72 сМ19. Ген С3 мыши содержит 41 экзон и кодирует полипептид-предшественник из 1663 аминокислот, включающий в себя сигнальный пептид из 24 аминокислот, β-цепь из 642 аминокислот и α-цепь из 993 аминокислот. Во время активации системы комплемента α-цепь расщепляется, тем самым образуя 9 различных пептидов, включая С3а из 78 аминокислот, который является сильным провоспалительным анафилатоксином.
Нокаут-мыши по С3 были получены, следуя стандартным способам, известным в данной области (see, например, Drouin et al. (2001) Cutting edge: the absence of 3 demonstrates a role for complement in Th2 effector functions in a murine model of pulmonary allergy, J Immunology 167: 4141-4144).
Видоспецифичность белков комплемента C5 и С3
Показанные в настоящем документе гуманизированные по С5 мыши являются функционально дефицитными по С5, поскольку, по меньшей мере, некоторые из эндогенных белков комплемента проявляют видоспецифичность, то есть, эндогенные белки комплемента мыши не способны функционально взаимодействовать с белком С5 человека. Сыворотку, полученную из гуманизированных по С5 мышей или нокаут-мышей по С5, проверяли в отношении активности комплемента; не наблюдали никакой гемолитической активности комплемента, пока не добавляли оба белка С3 и С5 человека.
Также показанное в настоящем документе добавление С3 человека в сыворотки нокаут-мышей по С3 было способно восстанавливать гемолитическую функцию; активность комплемента наблюдали у нокаут-мышей по С3, когда к сыворотке добавляли белок С3 человека.
Активность белка С5 комплемента является видоспецифичной, то есть, белок С5 человека не может заменить белок С5 мыши в анализе активности (то есть, гемолиза) сывороточного комплемента мыши.
Активность белка С3 комплемента, напротив, очевидно, не является схожим образом видоспецифичной, то есть, белок С3 человека способен заменить белок С3 мыши в анализе активности (то есть, гемолиза) сывороточного комплемента мыши.
Терапевтическое ингибирование системы комплемента
Две мишени для терапевтического ингибирования комплемента представляют собой пептиды активации системы комплемента С5а и С3а, которые получаются при активации системы комплемента посредством ферментативного расщепления как при классическом, так и альтернативном путях из белков комплемента С5 и С3, соответственно (смотри, Markides (1998); Mollnes et al. (2006)).
Например, блокировка расщепления С5 антителами, например, антителами, описанными в патенте США №6355245, является возможной стратегией для ингибирования комплемента при заболеваниях, ассоциированных с активацией комплемента.
Видоспецифичность ингибиторов С5 и С3 человека
Терапевтические молекулы-кандидаты, которые направлены на белки С5 или С3 комплемента, типично оценивают в отношении фармакокинетики (PK) и фармакодинамики (PK) на отличных от человека животных, например, грызунах, например, мышах или крысах. Такие терапевтические молекулы также проверяют в отношении терапевтической эффективности in vivo на отличном от человека животном, например, грызуне, например, мыши или крысе, в моделях заболеваний, нарушений или патологических состояний человека, ассоциированных с активацией комплемента.
Тем не менее, терапевтические молекулы, которые специфичны для белков С5 или С3 комплемента человека, то есть, специфичные для человека ингибиторы С5 или С3, невозможно адекватно оценить в отношении PD или терапевтической эффективности in vivo на грызунах, в частности на мышах, поскольку мишени для данных терапевтических молекул отсутствуют. Данная проблема не решается с применением трансгенных отличных от человека животных, например, грызунов, например, мышей или крыс, экспрессирующих белки С5 или С3 комплемента человека, вследствие упомянутой выше видоспецифичности данных белков.
В соответствии с этим, для оценки PD и терапевтической эффективности in vivo специфичного для человека антагониста или ингибитора белка С5 или С3 на отличных от человека животных, например, грызунах, например, мышах или крысах, требуется заменить эндогенные белки С5 и/или С3 на белки С5 и/или С3 человека. Кроме того, для того, чтобы избежать возможных проблем повышенной или пониженной экспрессией белков С5 и/или С3 человека, требуется встраивать гены С5 и/или С3 человека в геном отличных от человека животных, например, грызунов, например, мышей или крыс, в эндогенные локусы генов С5 и/или С3 и экспрессировать белки С5 и/или С3 человека у отличных от человека животных, например, грызунов, например, мышей или крыс, под контролем, по меньшей мере, частично, эндогенных регуляторных элементов для С5 и/или С3.
Получение гуманизированных по С3 и/или С5 отличных от человека животных
Способы получения гуманизированных по С3 и/или С5 животных по настоящему изобретению хорошо известны в данной области (смотри, в общем случае, Gene Targeting: A Practical Approach, Joyner, ed., Oxford University Press, Inc. (2000)). Согласно одному варианту осуществления получение мышей может необязательно включать разрушение мышиных генов С3 и/или С5 и введение гена, кодирующего человеческий или гуманизированный С3 и/или С5 в мышиный геном. Согласно одному варианту осуществления введение гена, кодирующего человеческий или гуманизированный С3 и/или С5, происходит в то же самое месторасположение, как и эндогенные мышиные гены С3 и/или С5.
Трансгенных отличных от человека животных по изобретению можно получить, вводя трансгены в зародышевую линию животного. Эмбриональные клетки-мишени на различных стадиях развития можно использовать для введения трансгенов. Применяют различные способы в зависимости от стадии развития эмбриональной клетки-мишени. Определенную линию(ии) любого животного, используемого в практике данного изобретения, отбирают в отношении хорошего общего состояния здоровья, хорошего выхода эмбрионов, хорошей пронуклеарной визуальной доступности в эмбрионе и хорошего полового соответствия. Когда необходимо получить трансгенных мышей, часто используют линии, такие как линии C57BL/6 или C57BL/6 × DBA/2 F1, или FVB (коммерчески приобретаемые в Charles River Labs, Boston, Mass., The Jackson Laboratory, Bar Harbor, ME, или Taconic Labs.).
Введение трансгена в эмбрион можно проводить любыми способами, известными в данной области, такими как, например, микроинъекция, электропорация или липофекция. Например, трансген(ы) можно вводить в млекопитающее посредством микроинъекции конструкции в пронуклеусы оплодотворенной яйцеклетки(яйцеклеток) млекопитающих, чтобы заставить одну или несколько копий конструкции остаться в клетках развивающегося млекопитающего(их). После введения трансгенной конструкции в оплодотворенную яйцеклетку, яйцеклетку можно инкубировать in vitro в течение различного времени или реплантировать в суррогатный организм-хозяина или и то, и другое. Инкубирование in vitro до зрелости находится в рамках данного изобретения. Один общепринятый способ заключается в инкубировании эмбрионов in vitro в течение приблизительно 1-7 дней в зависимости от вида и затем в реплантации их в суррогатный организм-хозяина.
Реплантацию проводят, используя стандартные способы. Обычно суррогатный организм-хозяина анестезируют и эмбрионы вводят в фаллопиеву трубу. Количество эмбрионов, имплантируемых в определенный организм-хозяина будет меняться от вида, но обычно будет совместим с количеством потомства вида, приносимого в природе.
Ретровирусную инфекцию также можно использовать для введения трансгена в отличное от человека животное. Развитие не являющегося человеческим эмбриона можно культивировать in vitro до стадии бластоцисты. Во время данного периода бластомеры могут быть мишенями для ретровирусной инфекции (Jaenich, R. (1976) PNAS 73: 1260-1264). Эффективное инфицирование бластомеров получают при ферментативной обработке для удаления вителлинового слоя (Manipulating the Mouse Embryo, Hogan eds. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1986). Вирусная векторная система, используемая для введения трансгена, типично представляет собой неспособный к репликации ретровирус, несущий трансген (Jahner et al. (1985) PNAS 82: 6927-6931; Van der Putten et al. (1985) PNAS 82: 6148-6152).
Третий тип клетки-мишени для введения трансгена представляет собой эмбриональную стволовую (ES) клетку. Трансгены можно эффективно вводить в ES клетки при помощи трансфекции ДНК или опосредованной ретровирусом трансдукции. Такие трансформированные ES клетки можно вслед за этим объединять с бластоцитами из отличного от человека животного. ES клетки затем заселяют в эмбрион и они участвуют в развитии зародышевой линии полученного химерного животного.
Трансгенные животные, содержащие гуманизированный С3 и/или С5, можно скрещивать с другими животными. Способ получения заключается в том, чтобы сконструировать ряд млекопитающих, причем каждый из которых содержит одну из требуемых конструкций или трансгенов. Такие млекопитающие размножаются совместно путем ряда скрещиваний, обратных скрещиваний и отборов, в конечном итоге получая одно млекопитающее, содержащее все необходимые конструкции и/или трансгены, где млекопитающее является в остальное части конгенным (генетически идентичным) дикому типу, за исключением наличия требуемых конструкций и/или траснгена(ов). Согласно одному варианту осуществления мышь, содержащую человеческий или гуманизированный ген С3 и/или С5 получают таким образом.
Типично, скрещивание и обратное скрещивание осуществляют спариванием братьев и сестер или родительской линии с потомством в зависимости от цели каждой определенной стадии в процессе размножения. В определенных случаях может быть необходимым получить большое количество потомства для того, чтобы получить одного потомка, который содержит каждую из нокаутных конструкций и/или трансгенов в соответствующем хромосомном месторасположении. В дополнение, может быть необходимо скрещивать или обратно скрещивать несколько поколений, чтобы в конечном итоге получить требуемый генотип.
Применение гуманизированных по С3 и/или С5 отличных от человека животных для идентификации соединений, способных модулировать систему комплемента
В некоторых аспектах в настоящем документе предложены способы идентификации терапевтической молекулы-кандидата (то есть, соединения) способного модулировать активацию системы комплемента. В способе используют любых из гуманизированных по С3 и/или С5 грызунов (например, мышей или крыс), описываемых в настоящем документе. Согласно некоторым вариантам осуществления соединения-кандидаты вводят непосредственно грызунам после экспериментальной индукции активации системы комплемента (например, в модели почечной ишемии/реперфузии) и оценивают эффект указанных соединений в отношении их способности модулировать систему комплемента. Согласно другим вариантам осуществления соединения-кандидаты контактируют с сывороткой, полученной из данных животных и активность комплемента оценивают, применяя любую общепринятую методику оценки in vitro (такие как, в качестве неограничивающих примеров, исследования СН50).
Согласно некоторым вариантам осуществления соединение-кандидат может модулировать активацию системы комплемента снижая, уменьшая или ингибируя активацию системы комплемента. Соединение может снижать активацию системы комплемента у любого грызуны, описываемого в настоящем документе на любой процент из 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 21%, 22%, 23%, 24%, 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% по сравнению с контрольными грызунами, которых не обрабатывали соединением-кандидатом.
Согласно другому варианту осуществления соединение-кандидат может снижать, уменьшать или ингибировать активацию системы комплемента в течение вплоть до 1 часа, 2 часов, 3 часов, 4 часов, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов, 10 часов, 11 часов, 12 часов, 18 часов, 24 часов, 30 часов, 36 часов, 42 часов, 48 часов, 54 часов, 60 часов, 66 часов, 72 часов, 4 дней, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней, 12 дней, 13 дней, 14 дней или трех недель, включительно (включая любые периоды времени среди данных значений).
Соединение-кандидат может дополнительно модулировать активацию системы комплемента согласно другим вариантам осуществления, увеличивая, усиливая или активизируя активность комплемента. Соединение может увеличивать активацию системы комплемента у любого из грызунов, описываемых в настоящем документе, на любой процент из 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 21%, 22%, 23%, 24%, 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% по сравнению с контрольными грызунами, которых не обрабатывали соединением-кандидатом.
Согласно другому варианту осуществления соединение-кандидат может увеличивать, усиливать или активизировать активность комплемента в течение вплоть до 1 часа, 2 часов, 3 часов, 4 часов, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов, 10, часов, 11, часов, 12 часов, 18 часов, 24 часов, 30 часов, 36 часов, 42 часов, 48 часов, 54 часов, 60 часов, 66 часов, 72 часов, 4 дней, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней, 12 дней, 13 дней, 14 дней или трех недель, включительно (включая любые периоды времени среди данных значений).
Соединения-кандидаты могут без ограничения представлять собой низкомолекулярные химические соединения, антитела, белки, ингибирующие нуклеиновые кислоты или любую их комбинацию.
1. Антитела
В некоторых аспектах соединение-кандидат связывается (такое, которое предпочтительно связывается) с белком комплемента (таким как, но не ограничиваясь этим, С5 или С3 (например, С3а)) и представляет собой антитело. Согласно некоторым вариантам осуществления, антитела являются антагонистами С5 и/или С3 и могут снижать активацию системы комплемента. Согласно другим вариантам осуществления антитела являются агонистами С5 и/или С3 и могут увеличивать активацию системы комплемента.
Варианты антител можно также получить на основе информации, известной в данной области, по существу не влияя на активность антитела. Например, варианты антител могут содержать, по меньшей мере, один аминокислотный остаток в молекуле антител, замененный на другой остаток. Для антител сайты, представляющие наибольший интерес для замещающего мутагенеза, в большинстве случаев включают в себя гипервариабельные области, но изменения каркасной области (FR) также рассматриваются.
Для антител, один тип замещающего варианта охватывает замещение одного или нескольких остатков гипервариабельной области исходного антитела (например, гуманизированного антитела или антитела человека). В большинстве случаев, полученный вариант(ы), выбранные для дальнейшей разработки, будут обладать улучшенными биологическими свойствами по сравнению с исходным антителом, из которого их получают. Пригодный способ получения таких замещающих вариантов включает в себя созревание аффинности с использованием фагового дисплея. Вкратце, несколько сайтов гипервариабельной области (например, 6-7 сайтов) мутируют, чтобы получить все возможные замещения аминокислот в каждом сайте. Получаемые таким образом антитела проявляются из нитеобразных частиц фагов, как слияние с продуктом гена III М13, упакованного внутри каждой частицы. Проявляемые с помощью фагов варианты затем подвергаются скринингу в отношении их биологической активности (например, аффинность связывания), как описано в настоящем документе. Для того чтобы идентифицировать кандидатов на модификацию среди сайтов гипервариабельной области, можно проводить мутагенез с аланиновым сканированием для идентификации остатков гипервариабельной области, вносящих значительный вклад в связывание антигена.
Молекулы нуклеиновых кислот, кодирующие варианты аминокислотных последовательностей антитела, можно получить при помощи множества способов, известных в данной области. Данные способы включают в себя, но не ограничиваясь этим, выделение из природного источника (в случае существующих в природе вариантов последовательностей аминокислот) или получение с помощью опосредуемого олигонуклеотидами (или сайт-направленного) мутагенеза, PCR мутагенеза и кассетного мутагенеза полученного ранее варианта или невариантной версии антитела.
Может потребоваться введение одной или нескольких модификаций аминокислот в области Fc полипептидов иммуноглобулина по изобретению, тем самым приводя к получению варианта области Fc. Вариант области Fc может содержать последовательность области Fc человека (например, область Fc IgG1, IgG2, IgG3 или IgG4 человека), содержащую модификацию аминокислоты (например, замещения) в одном или нескольких положениях аминокислот, включая положения шарнирного цистеина.
Варианты области Fc с измененными (то есть, улучшенными или ослабленными) связыванием Clq и/или комплементзависимой цитотоксичностью (CDC) описываются в публикации международной заявки на патент №: W099/51642 (включенной в настоящий документ в качестве ссылки). Такие варианты могут содержать замену аминокислоты в одном или нескольких положениях аминокислот области Fc. В отношении вариантов области Fc, смотри также Duncan & Winter, Nature 322: 738-40 (1988); патент США №5648260; патент США №5624821; и публикации международной заявки на патент №: W094/29351, описания каждой из которых включены в настоящий документ в качестве ссылки.
2. Не являющиеся антителом связывающие полипептиды
В некоторых аспектах соединение-кандидат связывается (такое, которое предпочтительно связывается) с белком комплемента (таким как, но не ограничиваясь этим, С5 или С3 (например, С3а)) и представляет собой, не являющийся антителом связывающий полипептид. Согласно некоторым вариантам осуществления не являющийся антителом связывающий полипептид является антагонистом С5 и/или С3 и может снижать активацию системы комплемента. Согласно другим вариантам осуществления не являющийся антителом связывающий полипептид являются агонистом С5 и/или С3 и может увеличивать активацию системы комплемента.
Связывающие полипептиды можно химически синтезировать, применяя известную методологию синтеза полипептидов, или можно получить и очистить, применяя рекомбинантную технологию. Связывающие полипептиды обычно составляют, по меньшей мере, приблизительно 5 аминокислот в длину, альтернативно, по меньшей мере, приблизительно 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100 аминокислот в длину или более, где такие связывающие полипептиды, которые способны связываться с мишенью, такой как любой из белков комплемента {например, С5 или С3), обсуждаются в настоящем документе.
Связывающие полипептиды можно идентифицировать без излишнего экспериментирования, применяя хорошо известные методики. В этом отношении отмечено, что методики для скрининга библиотек полипептидов для связывающих полипептидов, которые способны связываться с мишенью, хорошо известны в данной области (смотри, например, патенты США №№5556762, 5750373, 4708871, 4833092, 5223409, 5403484, 5571689, 5663143; публикации заявок РСТ №№WO 84/03506 и WO 84/03564; Geysen et al, Proc. Natl. Acad. Sci. U.S.A., 81: 3998-4002 (1984); Geysen et al, Proc. Natl. Acad. Sci. U.S.A., 82: 178-182 (1985); Geysen et al., J. Immunol. Meth,, 102: 259-274 (1987); Clackson, T. et al., (1991) Nature, 352: 624; Kang, A.S. et al., (1991) Proc. Natl. Acad. Sci. USA, 88: 8363 и Smith, G.P. (1991) Current Opin. Biotechnol, 2: 668, описания каждого из которых включены в настоящий документ в качестве ссылки.
Способы получения библиотек пептидов и скрининга данных библиотек также описаны в патентах США №№5723286, 5432018, 5580717, 5427908, 5498530, 5770434, 5734018, 5698426, 5763192 и 5723323, описания каждого из которых включены в настоящий документ в качестве ссылки.
Связывающие полипептиды можно модифицировать, чтобы увеличить их ингибирующий и/или терапевтический эффект (включая, например, повышенную аффинность, улучшенные фармакокинетические свойства, такие как время полужизни, стабильность и скорость клиренса, пониженная токсичность и так далее). Такие модификации включают в себя без ограничения гликозилирование, пегилирование, замену на не встречающуюся в природе, но функционально эквивалентную аминокислоту, связывающие группы и так далее.
3. Малые молекулы
В некоторых аспектах соединение-кандидат (такое, которое предпочтительно связывается) с белком комплемента (таким как С5 или С3 (например, С3а)) и представляет собой малую молекулу. Согласно некоторым вариантам осуществления малая молекула является антагонистом С5 и/или С3 и может снижать активацию системы комплемента. Согласно другим вариантам осуществления малая молекула является агонистом С5 и/или С3 и может увеличивать активацию системы комплемента.
Малые молекулы предпочтительно представляют собой органические молекулы, отличные от связывающих полипептидов или антител, как определено в настоящем документе. Органические малые молекулы можно идентифицировать и химически синтезировать, применяя известную методологию (смотри, например, публикации заявок РСТ №№. WO 00/00823 и WO 00/39585). Органические малые молекулы размером обычно меньше, чем приблизительно 2000 Дальтон, альтернативно, размером меньше, чем приблизительно 1500, 750, 500, 250 или 200 Дальтон, где такие органические малые молекулы, которые способны связываться с полипептидом, как описано в настоящем документе, можно идентифицировать без излишнего экспериментирования, применяя хорошо известные методики. В этом отношении отмечено, что методики для скрининга библиотек малых молекул для молекул, которые способны связываться с полипептидной мишенью, хорошо известны в данной области (смотри, например, публикации заявок РСТ №№WO 00/00823 и WO 00/39585).
Органические малые молекулы могут представлять собой, например, аальдегиды, кетоны, оксимы, гидразоны, семикарбазоны, карбазиды, первичные амины, вторичные амины, третичные амины, N-замещенные гидразины, гидразиды, спирты, простые эфиры, тиолы, простые тиоэфиры, дисульфиды, карбоксильные кислоты, сложные эфиры, амиды, мочевины, карбаматы, карбонаты, кетали, тиокетали, ацетали, тиоацетали, арилгалогениды, арилсульфонаты, алкилгалогениды, алкилсульфонаты, ароматические соединения, гетероциклические комплексы, анилины, алкены, алкины, диолы, аминоспирты, оксазолидины, оксазолины, триазолидины, тиазолины, енамины, мульфонамиды, эпоксиды, азиридины, изоцианаты, сульфонилхлориды, диазосоединения, кислотные хлориды, хлорангидриды или тому подобное.
В некоторых аспектах низкомолекулярное химическое соединение является компонентом комбинаторной библиотеки химических веществ. Комбинаторная библиотека химических веществ представляет собой коллекцию множества видов химических соединений, содержащий меньшие субъединицы или мономеры. Комбинаторные библиотеки существуют во множестве размеров, в диапазоне от нескольких сотен до многих сотен из тысячи различных видов химических соединений. Существует также множество типов библиотек, включающих в себя олигомерные и полимерные библиотеки, составленные из соединений, таких как карбогидраты, олигонуклеотиды и малые органические молекулы и так далее. Такие библиотеки обладают множеством применений, такими как при иммобилизации и хроматографическом разделении химических соединений, а также идентификации и описании характеристик лигандов, способных связываться с молекулой-мишенью (например, С5 и/или С3) или опосредующих интересующую биологическую активность (такую как, но не ограничиваясь этим, ингибирование или активизирование активности комплемента).
Различные методики для синтеза библиотек соединений на твердофазной подложке известны в данной области. Твердофазные подложки типично представляют собой полимерные объекты с поверхностью, которая функционализирована для связывания субъединиц или мономеров с образованием соединений библиотеки. Синтез одной библиотеки, как правило, включает большое количество твердофазных подложек. Для того чтобы получить комбинаторную библиотеку, твердофазные подложки подвергают реакции с одной или несколькими субъединицами соединений и с одним или некоторым количеством реагентов в тщательно контролируемых с предварительно определенной последовательностью химических реакциях. Другими словами, субъединицы библиотеки «растут» на твердофазных подложках. Чем больше библиотека, тем больше количество реакций требуется, усложняя задачу отслеживания химического состава множества видов соединений, которые составляют библиотеку. Согласно некоторым вариантам осуществления малые молекулы обладают размером меньше, чем приблизительно 2000 Дальтон, альтернативно, размером меньше, чем приблизительно 1500, 750, 500, 250 или 200 Дальтон.
Низкомолекулярные средства, описываемые в любом из аспектов в настоящем документе, можно получить в любом типе химической реакции, которая химической реакции, которая может быть выполнена на твердой подложке. Такие химические реакции включают в себя, но не ограничиваются этим, 2+2-циклоприсоединение, включая захват бутадиена; [2+3]-циклоприсоединение, включая синтез изоксазолинов, фуранов и модифицированных пептидов; образование ацеталя, включая иммобилизацию диолов, альдегидов и кетонов; альдольную конденсацию, включая получение производных альдегидов, синтез пропандиолов; бензоиновую конденсацию, включая получение производных альдегидов; циклоконденсации, включая бензодиазепины и гидантоины, тиазолидины, обратимые миметики (turn mimetics), порфирины, фталоцианины; циклизацию Дикмана, включая циклизацию сложных диэфиров; реакцию Дильса-Альдера, включая получение производных акриловой кислоты; электрофильное присоединение, включая присоединение спиртов к алкенам; реакцию Гриньяра, включая получение производных альдегидов; реакцию Хека, включая синтез двузамещенных алкенов; реакцию Генри, включая синтез нитрилоксидов in situ (смотри, 2+3-циклоприсоединение); каталитическую гидрогенизацию, включая синтез феромонов и пептидов (гидрогенизацию алкенов); реакцию Михаэля, включая синтез сульфанилкетонов, бицикло[2.2.2]октанов; реакцию Мицунобу, включая синтез ариловых эфиров, пептидилфосфонатов и простых тиоэфиров; нуклеофильные ароматические замещения, включая синтез хинолонов; окисление, включая синтез альдегидов и кетонов; циклоприсоединение Паузена-Ханда, включая циклизацию норборнадиена с фентинолом; фотохимическую циклизацию, включая синтез гелиценов; реакции с металлоорганическими соединениями, включая получение производных альдегидов и хлорангидридов; восстановление с комплексными гидридами и соединениями олова, включая восстановление карбонилов, карбоновых кислот, сложных эфиров и нитрогрупп; реакцию Соаи, включая восстановление карбоксильных групп; реакцию Стилла, включая синтез бифенильный производных; реакцию Сторка, включая синтез замещенных циклогексанонов; восстановительное аминирование, включая синтез хинолонов; реакцию Сузуки, включая синтез производных фенилуксусной кислоты и реакцию Виттиг-Хорнера включая реакции альдегидов, феромонов и сульфонилкетонов.
Ссылки, описывающие синтез химических библиотек, а также и деконволюцию отдельных соединений из данных библиотек на отдельных твердофазных подложках, можно найти в заявке на патент США №2009/0032592; Needels et al., (1993), Proc. Natl. Acad. Sci. USA 90: 10700-10704; и публикации заявки РСТ №WO 97/15390, описание которых включены в настоящий документ в качестве ссылки.
4. Ингибирующие нуклеиновые кислоты
Согласно одному аспекту данного изобретения соединение-кандидат на модулирование комплемента представляет собой один или несколько олигонуклеотидов, направленных на компонент (такой как мРНК) системы комплемента. Ингибирующая нуклеиновая кислота может без ограничения представлять собой любое из антисмыслового олигонуклеотида, малой ингибирующей РНК (ми РНК) или рибозима.
Олигонуклеотид по изобретению может представлять собой мРНК, кодирующую белковый компонент системы комплемента. Олигонуклеотиды будут связываться с мРНК и препятствовать их функциям, либо опосредуя их разрушение, либо предотвращая трансляцию белков. Абсолютной комплементарности, хотя она и предпочтительно, не требуется. «Комплементарность» олигонуклеотидной последовательности к части РНК, как обозначается в настоящем документе, означает последовательность, обладающую достаточной комплементарностью, чтобы иметь способность гибридизоваться с РНК, образуя стабильных дуплекс. Способность к гибридизации будет зависеть как от степени комплементарности, так и длины олигонуклеотида. В большинстве случаев, чем длиннее гибридизующаяся нуклеиновая кислота, тем больше несоответствия оснований с РНК она может содержать и все-таки образовать стабильный дуплекс. Специалисты в данной области могут установить допустимую степень несоответствия при помощи стандартных способов, чтобы определить точку плавления гибридизованного комплекса.
В основном, комплементарные Олигонуклеотиды, произведенные для гибридизации с мРНК для белков системы комплемента, направлены на любой участок мРНК, включая 5'-нетранслируемую область мРНК, комплемент инициирующего кодона AUG или 3'-нетранслируемую область.
Олигонуклеотиды могут содержать альтернативные межнуклеозидные связи, такие как, но не ограничиваясь этим, фосфотиоат (Mag at al., Nucleic Acids Res. 19: 1437-1441, 1991; и патент США №5644048), пептидная нуклеиновая кислота или PNA (Egholm, Nature, 3685: 566-568, 1993; и патент США №6656687), фосфорамид (Beaucage, Methods Mol. Biol. 20:33-61, 1993), фосфородитиоат (Capaldi et al., Nucleic Acids Res., 28: E40, 2000). Другие аналоги олигонуклеотидов включают в себя, но не ограничиваясь этим, морфолиновый олигонуклеотид (Summerton, Biochim. Biophys. Acta, 1489: 141-158, 1999), запирающие олигонуклеотиды (Wahlestedt wt al., Proc. Natl. Acad. Sci. USA, 97: 5633-5638, 2000), пептидную нуклеиновую кислоту или PNA (Nielsen et al., 1993; Hyrup and Nielsen, 1996) или модифицированные 2-о-(2-метокси)этилом 5'- и 3'-концевые олигонуклеотиды (McKay et al., J. Biol. Chem., 274: 1715-1722, 1999). Все из данных ссылок тем самым явно включены в качестве ссылок. Нуклеиновые кислоты могут содержать любую комбинацию дезоксирибо- и рибо- нуклеотидов и любую комбинацию оснований, включающих в себя урацил, тимин, цитозин, гуанин, инозин, ксантин, гипоксантин, изоцитозин, изогуанин и так далее
Комплементарные олигонуклеотиды по изобретению могут содержать, по меньшей мере, одну часть модифицированного основания, которое выбрано из группы, включающей в качестве неограничивающих примеров 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксилметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеуозин, инозин. N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеуозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусная кислота (v), вибутоксозин, псевдоурацил, квеуозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусная кислота (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N2-карбоксипропил)урацил, (acp3)w, и 2,6-диаминопурин.
Комплементарные олигонуклеотиды могут также содержать, по меньшей мере, одну модифицированную сахарную часть, выбранную из группы, включающей в качестве неограничивающих примеров арабинозу, 2-фторарабинозу, ксилулозу и гексозу.
Комплементарные олигонуклеотиды должны быть длиной, по меньшей мере, десять нуклеотидов и длина может находить в диапазоне от 10 до приблизительно 50 нуклеотидов, такая как длина 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 нуклеотидов.
Оценка эффективности соединений-кандидатов
Терапевтическая эффективность модулирующих комплемент соединений-кандидатов, подходящие дозировки и способы по настоящему изобретению в данном отличном от человека животном (таком как грызун, например, мышь или крыса), можно определить в соответствии с анализами системы комплемента in vitro, хорошо известными специалистам в данной области. Каскад комплемента приводит к появлению большого количества определенных продуктов во время нормального хода активации, и были разработаны чувствительные и специфичные анализы и они коммерчески доступны для большинства данных продуктов активации, включая небольшие фрагменты активации С3а, С4а и С5а и более крупные фрагменты активации Bb и sC5b-9. В большинстве из данных анализов используют моноклональные антитела, которые вступают в реакцию с новыми антигенами, обученными на фрагментах комплемента, а не на нативных белках, из которых они образуются, делая такие анализы очень простыми и специфичными. Технология ИФА является особенно распространенной, хотя радиоиммунологический анализ (РИА) также используется для определения С3а и С5а. Данные последние анализы определяют и непроцессированные фрагменты и их процессированные фрагменты (то есть, основные формы, обнаруживаемые в кровотоке). Определение С3а обеспечивает присутствие чувствительного, независимого от каскада показателя активации системы комплемента. Обнаружение продукта жидкой фазы при формировании мембраноатакующего комплекса, sC5b-9, предоставляет доказательство, что комплемент активируется до конца. дополнительно, при классическом пути также образуется С4а, определение которого предоставляет информацию об активности ингибитора С3b, и активирует ли такой ингибитор классический путь.
В дополнение, несколько моделей активация системы комплемента in vivo также доступны специалистам для оценки способности соединения-кандидата модулировать систему комплемента. Например, система комплемента непосредственно связана с ишемически-реперфузионным повреждением, которое продолжается после ишемического события и последующего восстановления тока крови. Примеры повреждений, которые могут вызывать ишемически-реперфузионное повреждение включают в себя без ограничения инфаркт миокарда, церебральные ишемические события, желудочно-кишечную ишемию и множество аспектов сосудистой хирургии, операции на сердце, травмы и трансплантации. Активация система комплемента играет роль при воспалительных событиях ишемически-реперфузионного повреждения. Ишемическое повреждение приводит к изменениям клеточной мембраны, влияя на липиды, углеводы или белки внешней поверхности, так что данные выставленные на поверхность эпитопы изменяются и могут действовать как неоантигены (модифицированные аутоантигены). Участие классического пути комплемента в ишемически-реперфузионном повреждении доказано при помощи мышей, генетически лишенных либо С3, либо С4, которые демонстрируют равную защиту от местного повреждения задней конечности и в модели повреждения на животном (Austen et al. (2003) Int J Immunopath Pharm 16: 1). Дополнительно, активация системы комплемента, как также было показано, вовлечена в модель ишемического повреждения почек (Guo et al. (2005) Ann Rev Immunol 23: 821).
Изобретение можно дополнительно понять при ссылке на следующие ниже примеры, которые предложены в качестве иллюстрации и не предназначены для ограничения.
Примеры
Пример 1
Замена эндогенного гена С5 мыши на ген С5 человека
Ген С5 человека из 97 т.п.н., содержащий кодирующие экзоны со 2 по 42 гена С5 человека заменял 75,8 т.п.н. локуса мышиного гена С5, охватывающего кодирующие экзоны со 2 по 41 и включающий часть 3'-нетранслируемой области. Смотри фигуру 1.
Направленную конструкцию для замены гена С5 мыши на ген С5 человека за одну направленную стадию конструировали с использованием технологии генной инженерии VelociGene® (смотри Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech, 21(6): 652-659). ДНК С5 мыши и человека получали из клонов искусственной бактериальной хромосомы (ВАС) bMQ-2277L16 и CTD-2559I19, соответственно. Вкратце, линейную направленную конструкцию, полученную при клонировании репарация гэпов, содержащую гомологичные плечи против хода транскрипции и по ходу транскрипции С5 мыши, фланкирующие 97 т.п.н. последовательности С5 человека, протяженностью от интрона 1 только против хода транскрипции кодирующего экзона 2 до кодирующего экзона 41 и 3'-нетранслируемой области (геномные координаты: GRCh38: chr9:120,952,335-121,050,276 (- цепь)) и фланкированную loxP кассету отбора neo электропорировали в эмбриональные стволовые (ES) клетки мыши F1H4 (гибрид C57BL/6 × 129 F1). Корректно направленные ES клетки (MAID 7140) далее электропорировали с транзиторным экспрессирующим Cre вектором, чтобы удалить кассету отбора по чувствительности к лекарственному препарату. Направленные клоны ES клеток без кассеты отбора по чувствительности к лекарственному препарату (MAID 7141) вводили эмбриону мыши SW на 8-клеточной стадии развития при помощи способа VelociMouse® (смотри, патенты США №№7294754, 7576259, 7659442 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1): 91-99). VelociMice ® (мыши F0, целиком происходящие из донорной ES клетки), несущие гуманизированный ген С5, идентифицировали при генотипировании в отношении потери аллели мыши и приобретения аллели человека, применяя модификацию аллельного анализа (смотри, Valenzuela et al. (2003)).
Клоны корректно направленных ES клеток идентифицировали по анализу на потерю нативного аллеля (LONA) (Valenzuela et al. 2003), в котором количество копий нативного немодифицированного гена С5 определяли по двум количественным полимеразным цепным реакциям (qPCR) TaqMan™, специфичным для последовательно в гене С5 мыши, которую предназначали для удаления. Анализы qPCR включали в себя следующие ниже наборы праймеров-зондов (записанные от 5'- к 3'-концу): прямой праймер против хода транскрипции, CCTCCGGTTA ACTGGTTTGT GAT (SEQ ID №: 1); обратный праймер против хода транскрипции, GCAGTGAATG GTAGACTTCC СА (SEQ ID №: 2); зонд против хода транскрипции, FAM-AGTGACTTTA CTTTGGTTGT TCTGCTCACA-BHQ (SEQ ID №: 3); прямой праймер по ходу транскрипции, CCGGGAAAGG AAACCAAGAC (SEQ ID №: 4); обратный праймер по ходу транскрипции, CAGCACAGAC TGAGGATCCA A (SEQ ID №: 5); зонд по ходу транскрипции, FAM-AGACTGTCAT TCTGGCCCGG ACCT-BHQ (SEQ ID №: 6); в котором FAM означает флуоресцентный зонд 5-карбоксифлуоресцеин и BHQ означает гаситель флуоресценции типа гаситель флуоресценции «черная дыра» (Biosearch Technologies). ДНК, очищенную из клонов ES клетки, которые взяли направленный вектор и встроили в свои геномы, объединяли с набором TaqMan™ Gene Expression Master Mix (Life Technologies) согласно указаниям производителя в 384-луночном планшете для PCR (MicroAmp™ Optical 384-Well Reaction Plate, Life Technologies) и запускали циклы в Applied Biosystems Prism 7900HT, который собирал данные по флуоресценции по время прохождения PCR и определял пороговый цикл (Ct), неполный цикл PCR, при котором накопленная флуоресценция достигает предварительно установленного порога. Специфичные для С5 qPCR против хода транскрипции и по ходу транскрипции и две qPCR для ненаправленных генов сравнения запускали для каждого образца ДНК. Вычисляли различия в величинах Ct (ΔCt) среди каждого специфичного для С5 qPCR и каждой qPCR гена сравнения и затем исследованную разницу между каждым ΔCt и медианой ΔCt для всех образцов вычисляли, чтобы получить величины ΔΔCt для каждого образца. Количество копий гена С5 в каждом образце вычисляли по следующей ниже формуле: количество копий = 2×2-ΔΔCt. Корректно направленные клоны, содержащие потерю одной из нативных копий, будут обладать количеством копий гена С5 равным единице. Подтверждение, что последовательность гена С5 человека заменяла удаленную последовательность гена С5 мыши в гуманизированном аллели, проводили анализом qPCR TaqMan™, который содержал следующие ниже наборы праймер-зонд (записанные от 5'- к 3'-концу): человеческий прямой праймер против хода транскрипции, TGCTCTTAAA GGACCATCAT САС (SEQ ID №: 7); человеческий обратный праймер против хода транскрипции, GTAAACGGAT AACGGAAAGG ATAGA (SEQ ID №: 8); человеческий зонд против хода транскрипции, FAM-CCATTTCCAA ATGCACTTCC CAGC-BHQ (SEQ ID №: 9); человеческий прямой праймер по ходу транскрипции, CCCACTGAAC TATCACAGGA AACA (SEQ ID №: 10); человеческий обратный праймер по ходу транскрипции, GGAGATGCAT TCCAGTCTCT GTTA (SEQ ID №: 11); человеческий зонд по ходу транскрипции, FAM-TGAAGATGAC CTCCGGATGT AACGG-BHQ (SEQ ID №: 12).
Такой же анализ LONA применяли для анализа ДНК, очищенной из хвостовых биопсий мышей, полученных из направленных ES клеток, чтобы определить их генотипы по С5 и подтвердить, что гуманизированный аллель С5 был передан через зародышевую линию. Двух потомков-гетерозигот в отношении замены скрещивали, чтобы получить мышь, которая является гомозиготной в отношении замены эндогенного гена С5 мыши на ген С5 человека. Детенышей, которые являются гомозиготными в отношении замены, используют для фенотипирования.
Против хода транскрипции место соединение мышиного локуса и последовательности, содержащей ген С5 человека, как конструируют, находится в пределах 5'-TGAAAAACCT ССАТТАСТАС TTCTACAGAT GCCCACAGTG GTCTTGCATT CTATCCTTGT /(GCGATCGC)/ TCTGCCATCT CCTACTAGGC ATGTGGGGAA GGGGAATTCA GATGATGGTT GGAAATCTGG АААТТСТТТС CTCTCTTTTG TAATTTGCCT (SEQ ID №: 13), где последний нуклеотид С5 мыши перед первым нуклеотидом гена С5 человека является последним Т в CTTGT, и первый нуклеотид последовательности С5 человека является первым Т в TCTGC, который охватывает сайт AsiSI (внутри скобок), где точные места соединений показаны наклонными чертами вправо. По ходу транскрипции место соединения последовательности, содержащей ген С5 человека и фланкированную loxP кассету отбора по neo, как конструируют, находится внутри 5'-GCAAATAGGG CAGGCCACAG TGGCCTAATT AACCCACAAT GCAGCTGTCA AATATCAAAG AAGGTTTTGT GAATTAGCCT/CTCGAGATAA CTTCGTATAA TGTATGCTAT ACGAAGTTAT ATGCATGGCC TCCGCGCCGG GTTTTGGCGC CTCCCGCGGG (SEQ ID №: 14), где последний нуклеотид последовательности С5 человека представляет собой Т в AGCCT, и первый нуклеотид фланкированной loxP кассеты отбора по neo представляет собой первый С в CTCGA, причем точное место соединения указано наклонной чертой вправо. Место соединения по ходу транскрипции последовательности мышиного локуса С5, как конструируют, находится в 5'-ATGTCTGGAA TAACTTCGTA TAATGTATGC TATACGAAGT TATGCTAGTA ACTATAACGG TCCTAAGGTA GCGAGCTAGC/CAGGCTATTA GGCTTCTGTC CCCAACTGTG GTGGCAAATA GGCCAGACCA CAGTCACCAG ATGAAGCCAT AAATGCAGCT (SEQ ID №: 15), где последний нуклеотид фланкированной loxP кассеты отбора по neo представляет собой второй С в CTAGC, и первый нуклеотид мышиного локуса С5 представляет собой первый С в CAGGC, причем точное место соединения указано наклонной чертой вправо. По ходу транскрипции место соединения последовательности, содержащей ген С5 человека и мышиный локус, как конструируют, находится в 5'-GCAAATAGGG CAGGCCACAG TGGCCTAATT AACCCACAAT GCAGCTGTCA AATATCAAAG AAGGTTTTGT GAATTAGCCT/CTCGAG(ATAA CTTCGTATAA TGTATGCTAT ACGAAGTTAT) GCTAGTAACT ATAACGGTCC TAAGGTAGCG AGCTAGC/CA GGCTATTAGG CTTCTGTCCC CAACTGTGGT GGCAAATAGG CCAGACCACA GTCACCAGAT GAAGCCATAA ATGCAGCTGA (SEQ ID №: 16), где последний нуклеотид последовательности С5 человека представляет собой «Т» в AGCCT, и первый нуклеотид последовательности С5 мыши представляет собой «С» в CAGGC; который охватывает сайт loxP (внутри скобок), который остается после удаления фланкированной loxP кассеты отбора по neo, где точные места соединений указаны наклонными чертами вправо.
Пример 2
Замена эндогенного гена С3 мыши на ген С3 человека
Версия 1 - Замена с использованием промотора и кодирующих экзонов с 1 по 41 С3 человека
Ген С3 человека, содержащий 5'-регуляторные элементы и все кодирующие экзоны с 1 по 41 гена С3 человека заменял мышиный локус гена С5, охватывающего 5'-регуляторные элементы и все кодирующие экзоны с 2 по 41. Способы, описываемые выше в примере 1, в основном использовали для замены последовательностей гена С3 мыши на последовательности гена С3 человека. Смотри фигуру 2А.
Предварительно, направленную делецию 25 т.п.н. гена С3 мыши получали в ES клетках мыши при помощи замены кодирующих экзонов с 2 по 41 и введения 900 п. н. 3' в сайт полиаденилирования с фланкированной loxP кассетой отбора по neo. Полученные в результате ES клетки мыши, ES клетки 12132, представляют собой гетерозиготных нокаутов по С3. Клетки 12132 использовали, чтобы получить нокаут-мышей по С3 согласно процедурам, известным в данной области.
Получали направленную конструкцию, содержащую гомологичные плечи против хода транскрипции и по ходу транскрипции получали, фланкирующие последовательность С3 человека, протянувшуюся с 5'-регуляторных элементов против хода транскрипции от кодирующего экзона 1 до кодирующего экзона 41 и 3'-нетранслируемой области, и фланкированную loxP кассету отбора по hygro, и электропорировали в ES клетки 12132. Корректно направленные ES клетки (MAID 6148) далее электропорировали с транзиторным экспрессирующим Cre вектором, чтобы удалить кассету отбора по чувствительности к лекарственному препарату. Направленные клоны ES клеток без кассеты отбора по чувствительности к лекарственному препарату (MAID 6149) вводили эмбриону мыши на 8-клеточной стадии развития. Мышей F0, полностью полученных из донорной ES клетки, несущей гуманизированный ген С3, идентифицировали при генотипировании в отношении потери аллели мыши и приобретения аллели человека, как описано в примере 1.
Подтверждение, что фланкированная loxP кассета отбора по neo заменяла удаленную последовательность гена С3 мыши в нокаутных по С3 ES клетках 12132, проводили при помощи анализа qPCR TaqMan™, который содержал следующие ниже наборы праймер-зонд (записанные от 5'- к 3'-концу): 12132TU, интрон 1 С3 мыши: прямой праймер, GGCCTGATTA CATGGACCTG ТС (SEQ ID №: 17); обратный праймер, CCCAGGCTTG GCTGGAATG (SEQ ID №: 18); зонд, FAM-TGTCCACTCT GGAAGCCCAG GC-BHQ (SEQ ID №: 19); 12132TD, 3' экзона 21 С3 мыши: прямой праймер, GCCAGGAGAG GAAGCTGGAG (SEQ ID №: 20); обратный праймер, TGGCTCAGCA GTAAAGAACA С (SEQ ID №: 21); зонд, FAM-ACAGATTGCT GTGAGCTGCC CAAA-BHQ (SEQ ID №: 22); кассета neo: прямой праймер, GGTGGAGAGG CTATTCGGC (SEQ ID №: 23); обратный праймер, GAACACGGCG GCATCAG (SEQ ID №: 24); зонд, FAM-TGGGCACAAC AGACAATCGG CTG-BHQ (SEQ ID №: 25).
Подтверждение, что последовательность гена С3 человека заменяла удаленную последовательность гена С3 мыши в гуманизированном аллеле, проводили при помощи анализа qPCR TaqMan™, который содержал следующие ниже наборы праймер-зонд (записанные от 5'- к 3'-концу): mC3-1, промотор С3 мыши: прямой праймер, GCCAGCCTAG ССТАСТТСА (SEQ ID №: 26); обратный праймер, GCCACCCATC CCAGTTCT (SEQ ID №: 27); зонд, FAM- CAGCCCAGGC CCTTTAGATT GCA-BHQ(SEQ ID №: 28); mC3-2, 3'-нетранслируемая область С3 мыши, прямой праймер, TACGGTGTTA GGTTCACTAT TGGT (SEQ ID №: 29); обратный праймер, GTCGCCAGCA GTCTCATACA G (SEQ ID №: 30); зонд, CAL оранжевый-AGTGGGCATC CCTTGCCAGG C-BHQ(SEQ ID №: 31); кассета hygro: прямой праймер, TGCGGCCGAT CTTAGCC (SEQ ID №: 32); обратный праймер, TTGACCGATT CCTTGCGG (SEQ ID №: 33); зонд, FAM-ACGAGCGGGT TCGGCCCATT C-BHQ (SEQ ID №: 34); hC3-1, промотор С3 человека: прямой праймер, GGGCCTCCTA AGTTTGTTGA GTATC (SEQ ID №: 35); обратный праймер, CAGGGCTGGT TCCCTAGAAA ТС (SEQ ID №: 36); зонд, FAM-TACAATAGCA GGCACAGCAC ССА-BHQ (SEQ ID №: 37); hC3-2, интрон 1 С3 человека: прямой праймер, GGCTGAGAGT GGGAGTCATG (SEQ ID №: 38); обратный праймер, GCACTTGCCA ATGCCATTAT С (SEQ ID №: 39); зонд, FAM-CTGCTGTCCT GCCCATGTGG TTG-BHQ (SEQ ID №: 40); hC3-3, экзон 41 С3 человека: прямой праймер, CGAATGCCAA GACGAAGAGA АС (SEQ ID №: 41); обратный праймер, GGGCACCCAA AGACAACCAT (SEQ ID №: 42); зонд, CAL оранжевый-CAGAAACAAT GCCAGGACCT CGGC-BHQ (SEQ ID №: 43).
Такой же анализ LONA применяли для анализа ДНК, очищенной из хвостовых биопсий мышей, полученных из целевых ES клеток, чтобы определить их генотипы по С3 и подтвердить, что гуманизированный аллель С3 был передан через зародышевую линию. Двух потомков-гетерозигот в отношении замены скрещивали, чтобы получить мышь, которая является гомозиготной в отношении замены эндогенного гена С3 мыши на ген С3 человека. Детенышей, которые являются гомозиготными в отношении замены, используют для фенотипирования.
Место соединения последовательности мышиного локуса С3 и последовательности, содержащей ген С3 человека, место соединения последовательности, содержащей ген С3 человека и фланкированную loxP кассету отбора по hygro и место соединения последовательности фланкированной loxP кассеты отбора по hygro и мышиного локуса С3, определяли как описано выше в примере 1.
Версия 2 - Замена с использованием кодирующих экзонов с по 41 С3 человека
Ген С3 человека, содержащий кодирующие экзоны с 2 по 41 гена С3 человека заменял мышиный локус гена С5, охватывающий кодирующие экзоны с 2 по 41. Способы, описываемые выше в примере 1, в основном применялись для замены последовательностей гена С3 мыши на последовательности гена С3 человека. Смотри фигуру 2В.
Предварительно, получали целевую делецию из 25 т.п.н. гена С3 мыши в ES клетках мыши посредством замены кодирующих экзонов со 2 по 41 и внесения 900 п. н. 3' в сайт полиаденилирования с фланкированной loxP кассетой отбора по neo. Полученные в результате ES клетки мыши, ES клетки 12132, представляют собой гетерозиготных нокаутов по С3.
Получали направленную конструкцию, содержащую гомологичные плечи против хода транскрипции и по ходу транскрипции С3 мыши, фланкирующие последовательность С3 человека, продолжающуюся против хода транскрипции от кодирующего экзона 2 до кодирующего экзона 41 и 3'-нетранслируемой области и фланкированной loxP кассетой отбора по hygro, и электропорировали ее в ES клетки 12132. Корректно направленные ES клетки (MAID 6155) далее электропорировали с транзиторным экспрессирующим Cre вектором, чтобы удалить кассету отбора по чувствительности к лекарственному препарату. Направленные клоны ES клеток без кассеты отбора по чувствительности к лекарственному препарату (MAID 6156) вводили эмбриону мыши на 8-клеточной стадии развития. Мышей F0, полностью происходящих от донорной ES клетки, несущей гуманизированный ген С3, идентифицировали при генотипировании в отношении потери аллели мыши и приобретения аллели человека, как описано в примере 1.
Подтверждение, что фланкированная loxP кассета отбора по neo заменяла удаленную последовательность гена С3 мыши в нокаутных по С3 ES клетках 12132 и что последовательность гена С3 человека заменяла удаленную последовательность гена С3 мыши в гуманизированном аллеле, проводили при помощи анализа qPCR TaqMan™, который содержал такие же наборы праймер-зонд (записанные от 5'- к 3'-концу), ка описано выше в версии.
Такой же анализ LONA применяли для анализа ДНК, очищенной из хвостовых биопсий мышей, полученных из направленных ES клеток, чтобы определить их генотипы по С3 и подтвердить, что гуманизированный аллель С3 был передан через зародышевую линию. Двух потомков-гетерозигот в отношении замены скрещивали, чтобы получить мышь, которая является гомозиготной в отношении замены эндогенного гена С3 мыши на ген С3 человека. Детенышей, которые являются гомозиготными в отношении замены, используют для фенотипирования.
Место соединения последовательности мышиного локуса С3 и последовательности, содержащей ген С3 человека, место соединения последовательности, содержащей ген С3 человека и фланкированную loxP кассету отбора по hygro и место соединения последовательности фланкированной loxP кассеты отбора по hygro и мышиного локуса С3 определяли, как описано выше в примере 1.
Пример 3
Анализ гемолитической активности комплемента в сыворотке мыши
Комплемент был непосредственно связан с глазными воспалительными заболеваниями и дегенеративными заболеваниями сетчатки (смотри Mullins et al. (2000) Drusen associated with aging and age-related macular degeneration contain proteins common to extracellular deposits associated with atherosclerosis, elastosis, amyloidosis, and dense deposit disease, FASEB J, 14(7): 835-846). Блокировка расщепления C5 моноклональным антителом (mAb) является возможной терапевтической стратегией для данных нарушений (смотри Copland et al. (2009) Systemic and local anti-C5 therapy reduces the disease severity in экспериментальный аутоиммунный увеоретинит, Clinical and Experimental Immunology, 159: 303-314). Для оценки способов терапии, направленных на систему комплемента, таких как антагонисты С5 и/или С3, необходимы анализы, для того чтобы оценить функцию кандидатных терапевтических средств in vitro и в подходящих in vivo моделях заболеваний, нарушений и патологических состояний человека, ассоциированных с активацией комплемента.
Способы
Гемолитическую активность комплемента оценивали, используя классический анализ на комплемент СН50 (Gilcas et al. (2001) Classical pathway evaluation, Current Protocols in Immunology, Unit 13.1). кровь мыши собирали из сердца и коагулировали в пробирках эппендорф на льду в течение 1 часа; сыворотку разделяли центрифугированием и хранили при at -70°С. Эритроциты (Е) овцы сенсибилизировали посредством инкубации с кроличьим гемолизином против иммуноглобулинов овцы в течение 20 мин при температуре 37°С. Сенсибилизированные клетки (Еа) инкубировали в течение 1 часа при температуре 37°С с двойным разведением обработанной или контрольной сыворотки от мыши или человека. Интактные клетки Еа осаждали центрифугированием и надосадочную жидкость удаляли для измерения поглощения при 541 нм, индекса гемолиза. Клетки Еа, инкубированные только с буфером (0% лизиса) или с ddH2O (100% лизиса) служили в качестве отрицательного и положительного контролей, соответственно. Процент гемолиза и СН50 в каждой лунке вычисляли относительно контроля с ddH2O (Holt et al. (2001) Targeted deletion of the CD59 gene causes spontaneous intravascular hemolysis and hemoglobinuria, Blood, 98(2): 442-449). Мышей, включая линии C5hu/hu, C5-/-, C3hu/hu, С3-/- и соответствующие контрольные линии, получали с использованием технологии Velocigene®, как описано в примерах 1 и 2. Нормальные, лишенные С5 и лишенные С3-сыворотки человека получали от Quidel Corp. и использовали их в качестве контролей или для сравнения. Компоненты комплемента добавляли для проверки, может ли восстановиться функция гемолиза. Тестируемые сыворотки инкубировали в течение 40 мин при температуре 4°С с компонентами комплемента; С5 человека (Sigma-Aldrich или Quidel Corp., 50 мкг/мл), С3 человека (Sigma-Aldrich или Quidel Corp., 10, 400, 1200 мкг/мл) и мышиным С5 (Regeneron Pharmaceuticals, 50 мкг/мл). mAb против С5 человека (100 мкг/мл) и mAb против С5 мыши (Hycult Biotech, 10 мкг/мл) использовали для подтверждения специфичности и применимости данной системы для дальнейшего поиска лекарственных средств.
Результаты
Сыворотки мышей дикого типа обладали приблизительно в 10 меньшей гемолитической активностью комплемента по сравнению с нормальными сыворотками человека. Как показано на фигуре 3, гемолитическая активность комплемента, как определено с использованием значения СН50, была, по меньшей мере, в 10 раз ниже в трех сериях сывороток нормальных мышей или мышей дикого типа по сравнению с двумя сериями сывороток человека.
Моноклональные антитела против С5 блокируют гемолиз видоспецифичным образом. Как показано на фигуре 4, моноклональное антитело против С5 мыши (msC5Ab) блокировало гемолитическую активность комплемента в нормальной сыворотке мыши, и mAb против С5 человека (C5Ab) блокировало гемолитическую активность комплемента в нормальной сыворотке человека и в лишенной С5 сыворотке человека, дополненной белком С5 человека, но не нормальной сывороткой мыши.
Белки С5 и С3 человека проявляют видоспецифичную гемолитическую активность. Как показано на фигуре 5А, белки С5 и С3 человека способны восстановить гемолитическую активность комплемента в сыворотке человека с удаленным С5 и С3, соответственно.
Как показано на фигуре 5В, для сывороток от мышей С5-/- и С3-/- показана потеря активности комплемента, и в сыворотке гуманизированной мыши C5hu/hu не показали никакой гемолитической активности, по существу равную для сыворотки от мышей С5-/-и С3-/-. Добавление белка С5 мыши к сыворотке гуманизированной по C5hu/hu мыши восстанавливало гемолитическую активность комплемента, но добавление белка С5 человека в сыворотку гуманизированной по C5hu/hu мыши не восстанавливало активность комплемента.
Как показано на фигуре 5С, потеря гемолитической активности комплемента в сыворотке гуманизированной по C5hu/hu мыши, по-видимому, происходит не из-за отсутствия белка С5 человека, поскольку количество белка С5 человека, присутствующего в сыворотке данных мышей составляло приблизительно одну треть от количества белка С5, присутствующего в нормальной сыворотке человека. На фигуре 5D показано изменение среди самцов и самок гуманизированных по С5 мышей в отношении уровней С5 комплемента перед использованием препарата. Количество белка С5 человека, присутствующего в сыворотке таких мышей, находился в диапазоне от 20 до 100 мкг/мл, которого было достаточно, чтобы проявить гемолитическую активность в сыворотке. Концентрация С5 человека, в норме добавляемого в данных экспериментах, составляла 50 мкг/мл.
Оба белка С5 и С3 человека необходимы для восстановления гемолитической активности в сыворотках нокаут-мышей по С5. Как показано на фигуре 6, гемолитическая активность комплемента не могла быть восстановлена добавлением белка С5 человека в сыворотку мыши С5-/-, тогда как добавление белков С3 и С5 человека вместе восстанавливало гемолиз в сыворотке мыши С5-/-.
Добавление белка С3 человека восстанавливает гемолитическую активность в сыворотке мыши C5hu/hu. Как показано на фигуре 7, белок С3 человека является достаточным для восстановления гемолитической активности комплемента в сыворотке мышей C5hu/hu.
Основываясь на указанных выше результатах, гуманизированные мыши C5hu/hu и/или C3hu/hu являются подходящими отличными от человека животными для оценки PK и PD специфичных для человека антагонистов С5 и/или С3. Основываясь на указанных выше результатах, сыворотка дважды гуманизированной мыши C5hu/hu и C3hu/hu может проявлять гемолитическую активность комплемента без необходимости добавления белков С5 и/или С3 человека, тем самым образуя подходящую линию мышей для проверки in vivo терапевтической эффективности специфичных для человека антагонистов С5 и/или С3.
Заключение
Сыворотка мыши дикого типа обладает существенно более низкой гемолитической активностью комплемента по сравнению с сывороткой человека. Генетически модифицированные мыши, экспрессирующие С5 человека, являются функциональными мышиными нокаутами по С5, возможно вследствие того, что конвертаза мыши не может расщепить белок С5 человека. В сыворотке гуманизированной С5 мыши необходимо добавление белка С3 человека, чтобы достичь уровней гемолитическая активность комплемента дикого типа.
Пример 4
Применения гуманизированных по С5 и/или С3 мышей
Гуманизированные по С5 и/или С3 мыши применимы для оценки фармакодинамики (PD) специфичных для человека соединений С5 и/или С3 (таких как, антагонисты, например, нейтрализующие антитела против С5 или против С3).
Анализы фармакокинетики (PK) и PD на гуманизированных по С5 и/или С3 мышах проводят согласно стандартным процедурам, известным в данной области, например, как описано в патенте США №6355245 в отношении антитела против С5.
Гуманизированные по С5 и/или С3 мыши применимы для тестирования in vivo терапевтической эффективности специфичных для человека антагонистов С5 или С3, например, нейтрализующих антител против С5 или против С3, во множестве моделей заболеваний, например, и без ограничения: возрастная макулярная дегенерация (AMD) в сухой форме; влажная форма AMD; экспериментальный аутоиммунный увеоретинит (EAU); внутриглазная гипертензия; и повреждение зрительного нерва. (Смотри, например, Makrides (1998) и Mollnes et al. (2006) в отношении перечня заболеваний, нарушений и патологических состояний, ассоциированных с активацией комплемента).
Сухую форму AMD можно вызвать у гуманизированных по С5 и/или С3 мышей при помощи рациона с высоким содержанием жира и повреждения ультрафиолетом (смотри, например, Exp Eye Res. 2002, 75(5): 543-553), сигаретного дыма (смотри, например, Invest Ophthalmol Vis Sci. 2006, 47(2): 729-737), карбоксиэтилпиррола (СЕР) (смотри, например, Invest. Ophthalmol. Vis. Sci. 2010, 51: 1276-1281; Nat Med. 2012, 18(5): 791-798).
Гуманизированных по C5 и/или С3 мышей можно также скрещивать с другими трансгенными моделями AMD, такими как, например, АроЕ(-/-) (смотри, например, Invest. Ophthalmol. Vis. Sci. 2000, 41: 2035-2042), Mdm1 (смотри, например, Hum. Mol. Genet. 2008, 17: 3929-3941), Sod1(-/-) (смотри, например, PNAS 2006; 103: 11282-11287), LDLR(-/-) (смотри, например, Br J Ophthalmol 2005, 89: 1627-1630) и DICER1 (-/-) (смотри, например, Nature. 2011; 471(7338): 325-330).
Влажную форму AMD можно вызвать у гуманизированных по С5 и/или С3 мышей посредством индуцированной лазерным излучением хориоидальной неоваскуляризации (CNV) (смотри Nat Protoc. 2013, 8(11): 2197-2211).
EAU можно вызвать у гуманизированных по С5 и/или С3 мышей (смотри, например, Clin Exp Immunol. 2010, 159(3): 303-314).
Внутриглазную гипертензию можно вызвать у гуманизированных по С5 и/или С3 мышей (смотри, например, Experimental Eye Research 2006, 83: 620-628).
Повреждение зрительного нерва можно вызвать у гуманизированных по С5 и/или С3 мышей (смотри, например, J Neurotrauma 2003, 20(9): 895-904.
Гуманизированных по С5 мышей можно использовать для оценки специфичных для человека антагонистов С5 во множестве моделей заболеваний, например, и без ограничения: пароксизмальная ночная гематурия; нейромиелит зрительного нерва; пересадка почки; и повреждение почек. (Смотри, например, патент США №6355245.)
Анализы терапевтической эффективности с использованием гуманизированных по С5 и/или С3 мышей проводят согласно стандартным процедурам, известным в данной области, например, как описано в патенте США№6355245 в отношении антитела против С5.
Пример 5
Фармакокинетическое/фармакодинамическое исследование антитела против С5 на гуманизированных по С5 гомозиготных мышах
В данном примере используют самцов и самок гуманизированных по С5 гомозиготных мышей, полученных согласно предшествующим примерам, для проверки способности антитела против С5 снижать активацию системы комплемента.
Способы
Всего 25 мышей (самцов и самок) C5hu/hu использовали в данном исследовании. Сыворотку (20 мкл) собирали от двух мышей в каждой определенной временной точке, которая включала временную точку «до введения препарата», а также и временные точки через 3 ч, 6 ч, 1 д, 2 д, 4 д, 1 Н, 2 Н, 4 Н после подкожного введения 15 мг/кг моноклонального антитела против С5 человека. Образцы хранили при температуре -20°С до анализа изменений фармакокинетики (PK) и фармакодинамики (PD) при активации системы комплемента.
Гемолитическую активность комплемента оценивали, как описано выше в примере 3. Как указано выше, С3 человека (Sigma-Aldrich или Quidel Corp., 400 и 800 мкг/мл) добавляли к сыворотке для воспроизведения системы комплемента человека.
Результаты
Как показано на фигуре 8, подкожная инъекция 15 мг/кг антитела против С5 человека снижает активацию системы комплемента в течение, по меньшей мере, одной недели после инъекции. Данный эффект наблюдали независимо от того, добавляли ли белок С3 человека в концентрации 400 мкг/мл (фигура 8А) или 800 мкг/мл (фигура 8В) в гемолитических анализах.
Как показано на фигуре 9, анализ PK моноклонального антитела к С5 выявил клиренс с течением времени для данного антитела с 2 из 5 мышей, обладающих недетектируемыми уровнями на 28 день.
Пример 6
Перекрестная активность белков комплемента человека и мыши
В данном примере исследуется перекрестная активность белков С3 и С5 человека и мыши с использованием сконструированных мышей, описанных в предыдущих примерах. Как обсуждается в примере 3 и как показано на фигуре 6, белки С5 человека и С3 мыши не являются достаточными для восстановления гемолитической активности у мышей, с отсутствием эндогенного С5. В данном исследовании наблюдали способность С5 мыши и С3 человека восстанавливать гемолитическую активность в нокаут-мышах по С3.
Способы
Гемолитическую активность комплемента оценивали, как описано в примере 3.
Мышей, включая С5-/-, С3-/- и соответствующие контрольные линии, получали с использованием технологии Velocigene®, как описано в примерах 1 и 2. Нормальные, лишенные С3сыворотки человека получали от Quidel Corp. и использовали их в качестве контролей или для сравнения. Компоненты комплемента добавляли для проверки, может ли восстановиться функция гемолиза. Тестируемые сыворотки инкубировали в течение 40 мин при температуре 4°С с компонентами комплемента; С3 человека (Sigma-Aldrich или Quidel Corp., 50, 100, 500, 1200 мкг/мл); и С5 человека (Sigma-Aldrich или Quidel Corp., 50 мкг/мл),). mAb против С5 человека (100 мкг/мл) и mAb против С5 мыши (Hycult Biotech, 10 мкг/мл) использовали, чтобы подтвердить специфичность и применимость данной системы для дальнейшего поиска лекарственных средств.
Результаты
Как показано на фигуре 10А, добавление белка С3 человека к сыворотке мышей дикого типа может усилить гемолитическую функцию в отношении гемолитической функции, наблюдаемой в контролях дикого типа. Тем не менее, когда сравнивали с добавлением белка С3 человека обратно в лишенную С3 сыворотку человека, наблюдаемое увеличение является меньшим, чем усиление, показанное у людей, дополнительно, как иллюстрируется на фигуре 10В, процентное соотношение гемолиза восстанавливается при использовании сыворотки гуманизированной мыши 5hu/hu; дополненной белком С3 человека.
Когда данный эксперимент повторяли с использованием сывороток, полученных из нокаутированных по С3 животных, было показано, что добавление белка С3 человека к сыворотке С3-/- восстанавливает гемолитическую функцию, особенно при увеличении концентраций добавляемого С3 человека (фигура 11).
В отличие от этого, на фигуре 12 показано, что добавление С3 человека к сывороткам, полученным от животного без С5, не восстанавливает гемолитическую функцию.
Заключения
В заключение отметим, что данные полученные из настоящего примера демонстрируют, что в то время как С3 человека способе перекрестно реагировать с мышиным С5, С3 мыши схожим образом не способен перекрестно реагировать с С5 человека для восстановления активности комплемента in vitro. Краткое изложение данных результатов представлено ниже в таблице 1
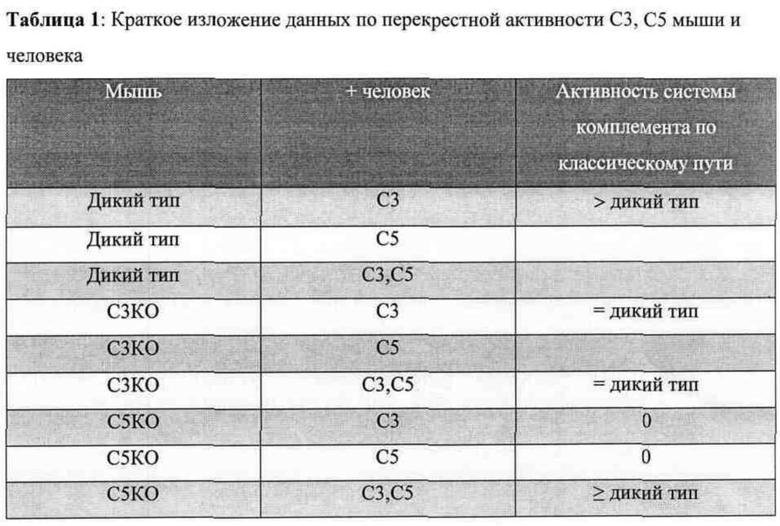
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
Hu, Ying
Latuszek, Adriana
Cao, Jingtai
Mujica, Alexander
Wiegand, Stanley
McWhirter, John
Murphy, Andrew
Macdonald, Lynn
<120> Humanized C5 and C3 Animals
<130> 47206-502001WO
<140> PCT/US2015/029111
<141> 2015-05-04
<150> US 61/988,581
<151> 2014-05-05
<150> US 62/067,836
<151> 2014-10-23
<160> 43
<170> PatentIn version 3.5
<210> 1
<211> 23
<212> DNA
<213> Artificial Construct
<220>
<223> Synthetic Construct
<400> 1
cctccggtta actggtttgt gat 23
<210> 2
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 2
gcagtgaatg gtagacttcc ca 22
<210> 3
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 3
agtgacttta ctttggttgt tctgctcaca 30
<210> 4
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 4
ccgggaaagg aaaccaagac 20
<210> 5
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 5
cagcacagac tgaggatcca a 21
<210> 6
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 6
agactgtcat tctggcccgg acct 24
<210> 7
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 7
tgctcttaaa ggaccatcat cac 23
<210> 8
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 8
gtaaacggat aacggaaagg ataga 25
<210> 9
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 9
ccatttccaa atgcacttcc cagc 24
<210> 10
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 10
cccactgaac tatcacagga aaca 24
<210> 11
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 11
ggagatgcat tccagtctct gtta 24
<210> 12
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 12
tgaagatgac ctccggatgt aacgg 25
<210> 13
<211> 158
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 13
tgaaaaacct ccattactac ttctacagat gcccacagtg gtcttgcatt ctatccttgt 60
gcgatcgctc tgccatctcc tactaggcat gtggggaagg ggaattcaga tgatggttgg 120
aaatctggaa attctttcct ctcttttgta atttgcct 158
<210> 14
<211> 160
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 14
gcaaataggg caggccacag tggcctaatt aacccacaat gcagctgtca aatatcaaag 60
aaggttttgt gaattagcct ctcgagataa cttcgtataa tgtatgctat acgaagttat 120
atgcatggcc tccgcgccgg gttttggcgc ctcccgcggg 160
<210> 15
<211> 160
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 15
atgtctggaa taacttcgta taatgtatgc tatacgaagt tatgctagta actataacgg 60
tcctaaggta gcgagctagc caggctatta ggcttctgtc cccaactgtg gtggcaaata 120
ggccagacca cagtcaccag atgaagccat aaatgcagct 160
<210> 16
<211> 239
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 16
gcaaataggg caggccacag tggcctaatt aacccacaat gcagctgtca aatatcaaag 60
aaggttttgt gaattagcct ctcgagataa cttcgtataa tgtatgctat acgaagttat 120
gctagtaact ataacggtcc taaggtagcg agctagccag gctattaggc ttctgtcccc 180
aactgtggtg gcaaataggc cagaccacag tcaccagatg aagccataaa tgcagctga 239
<210> 17
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 17
ggcctgatta catggacctg tc 22
<210> 18
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 18
cccaggcttg gctggaatg 19
<210> 19
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 19
tgtccactct ggaagcccag gc 22
<210> 20
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 20
gccaggagag gaagctggag 20
<210> 21
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 21
tggctcagca gtaaagaaca c 21
<210> 22
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 22
acagattgct gtgagctgcc caaa 24
<210> 23
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 23
ggtggagagg ctattcggc 19
<210> 24
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 24
gaacacggcg gcatcag 17
<210> 25
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 25
tgggcacaac agacaatcgg ctg 23
<210> 26
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 26
gccagcctag cctacttca 19
<210> 27
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 27
gccacccatc ccagttct 18
<210> 28
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 28
cagcccaggc cctttagatt gca 23
<210> 29
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 29
tacggtgtta ggttcactat tggt 24
<210> 30
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 30
gtcgccagca gtctcataca g 21
<210> 31
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 31
agtgggcatc ccttgccagg c 21
<210> 32
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 32
tgcggccgat cttagcc 17
<210> 33
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 33
ttgaccgatt ccttgcgg 18
<210> 34
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 34
acgagcgggt tcggcccatt c 21
<210> 35
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 35
gggcctccta agtttgttga gtatc 25
<210> 36
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 36
cagggctggt tccctagaaa tc 22
<210> 37
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 37
tacaatagca ggcacagcac cca 23
<210> 38
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 38
ggctgagagt gggagtcatg 20
<210> 39
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 39
gcacttgcca atgccattat c 21
<210> 40
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 40
ctgctgtcct gcccatgtgg ttg 23
<210> 41
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 41
cgaatgccaa gacgaagaga ac 22
<210> 42
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 42
gggcacccaa agacaaccat 20
<210> 43
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Construct
<400> 43
cagaaacaat gccaggacct cggc 24
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННАЯ МОДЕЛЬ НАРУШЕНИЙ СО СТОРОНЫ ПОЧЕК И ПЕЧЕНИ | 2018 |
|
RU2772583C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2799086C2 |
| ГРЫЗУНЫ С ГУМАНИЗИРОВАННЫМ IL-7 | 2013 |
|
RU2654565C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2703139C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2020 |
|
RU2819731C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2788523C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2012 |
|
RU2730643C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН ФАКТОРА АКТИВАЦИИ В-КЛЕТОК | 2014 |
|
RU2670016C1 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ГЕНОМ IL-15 | 2014 |
|
RU2674910C2 |
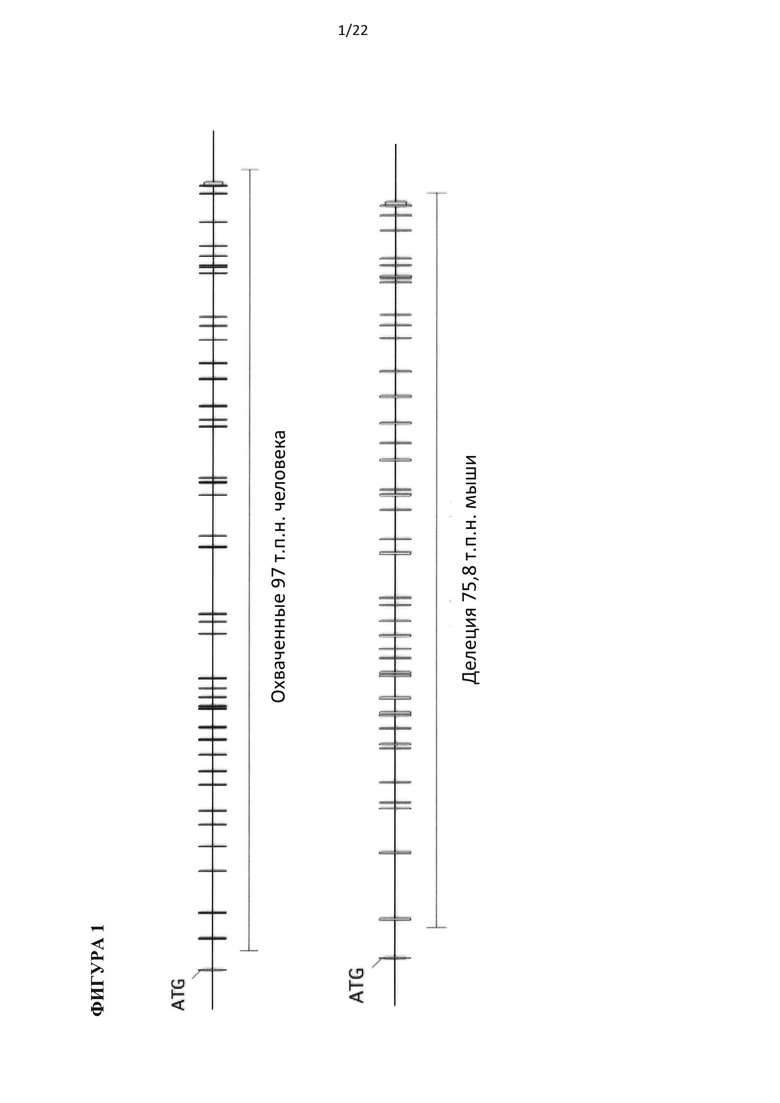
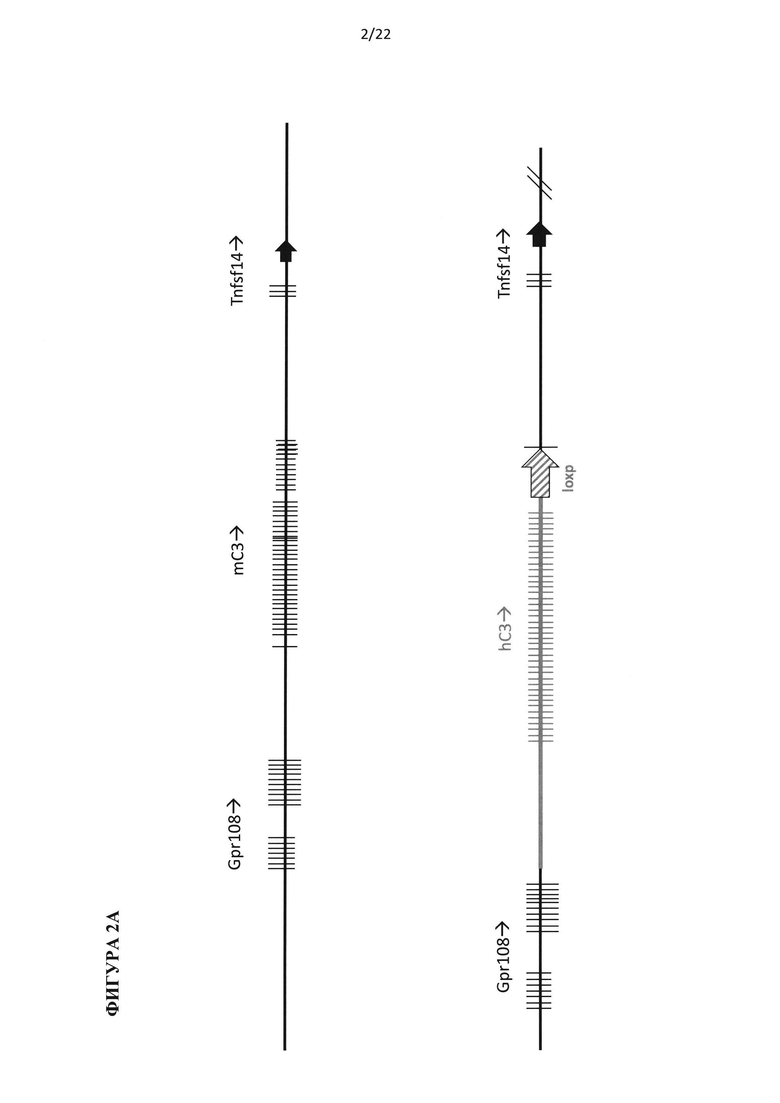
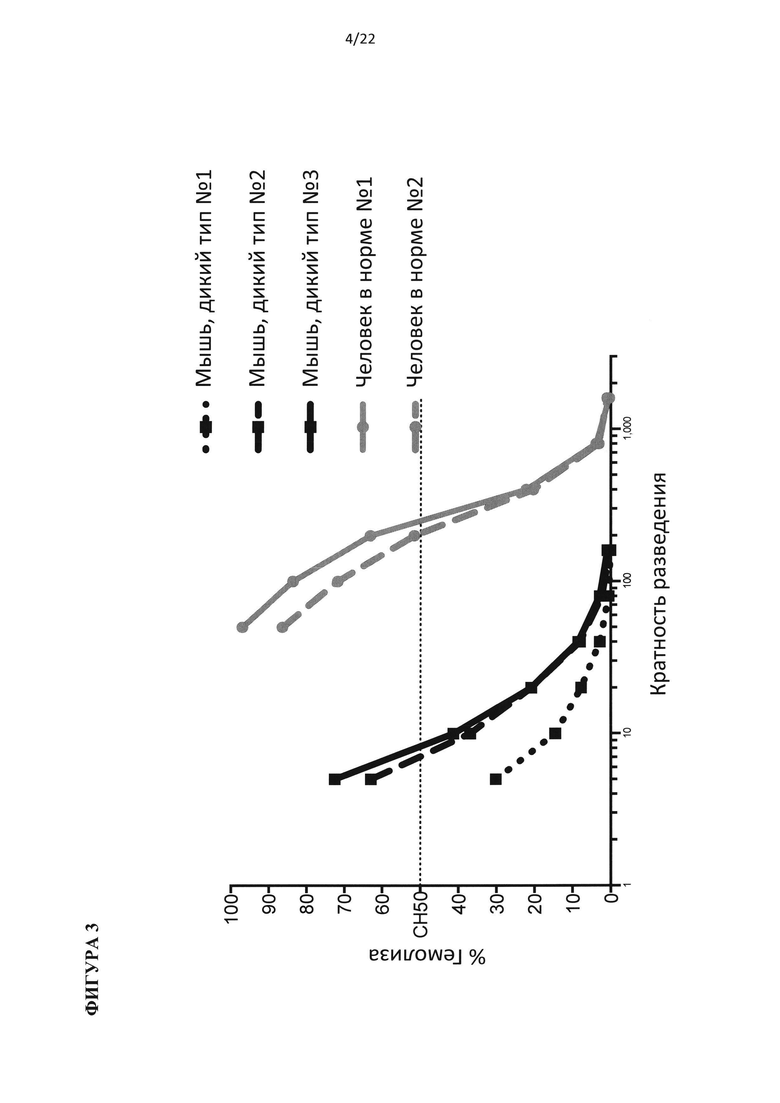
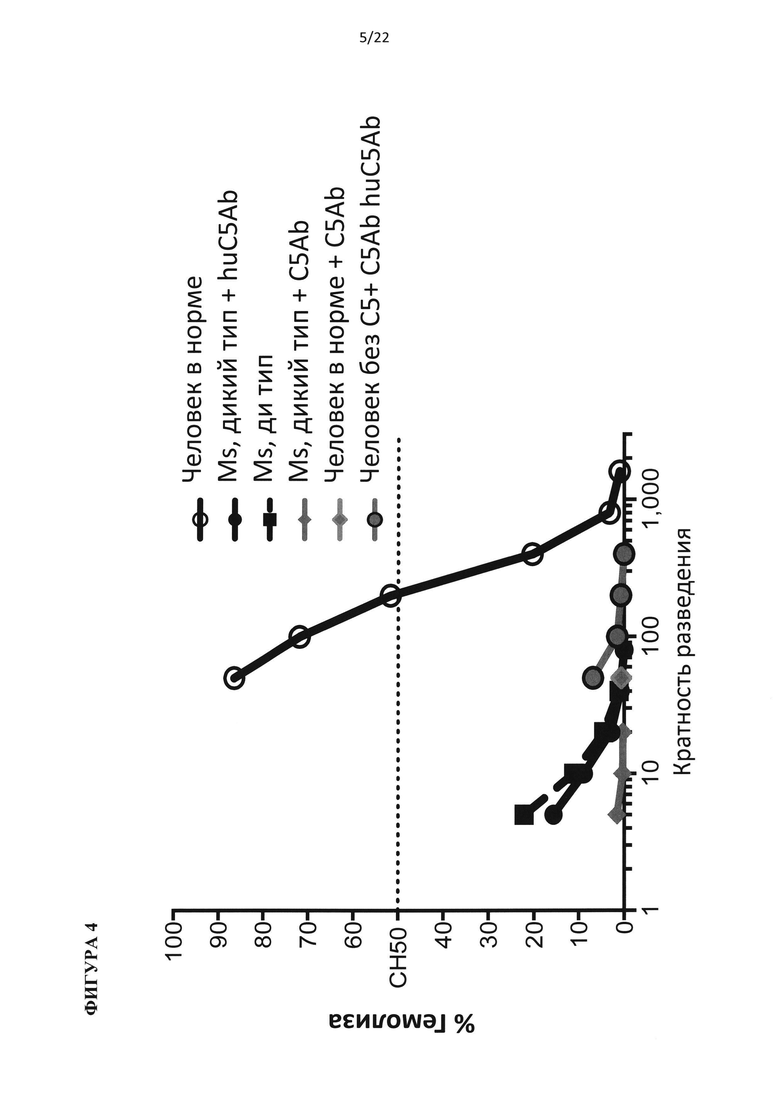
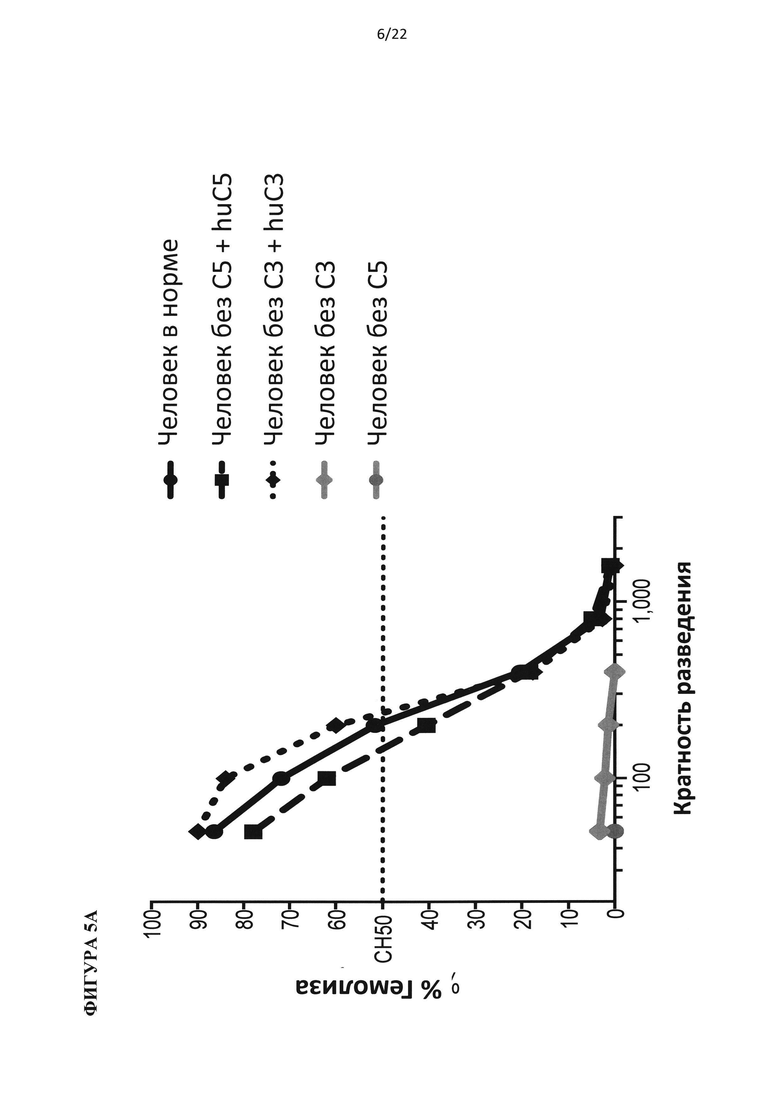
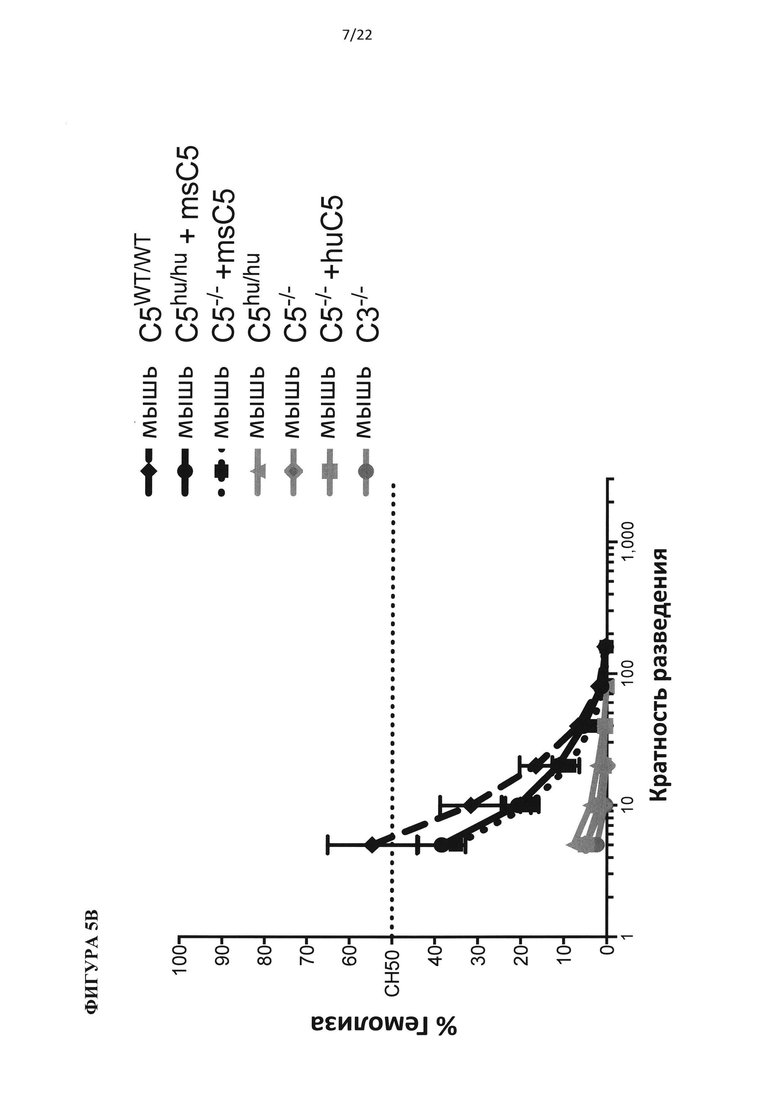
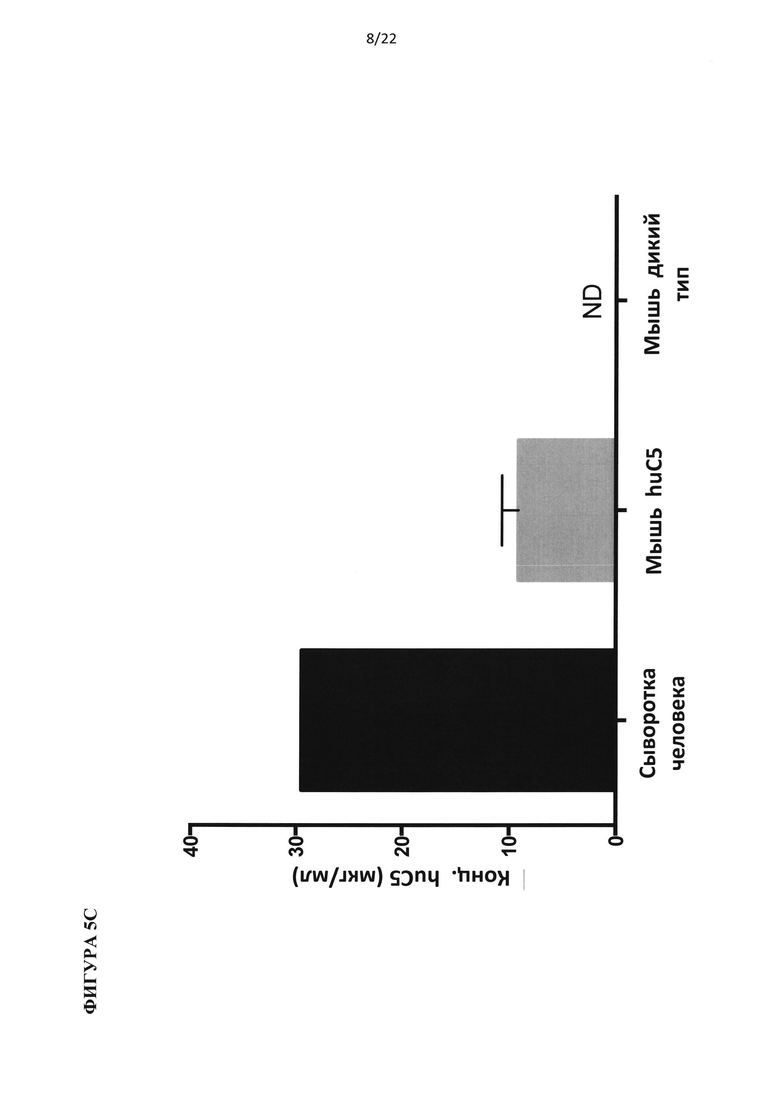
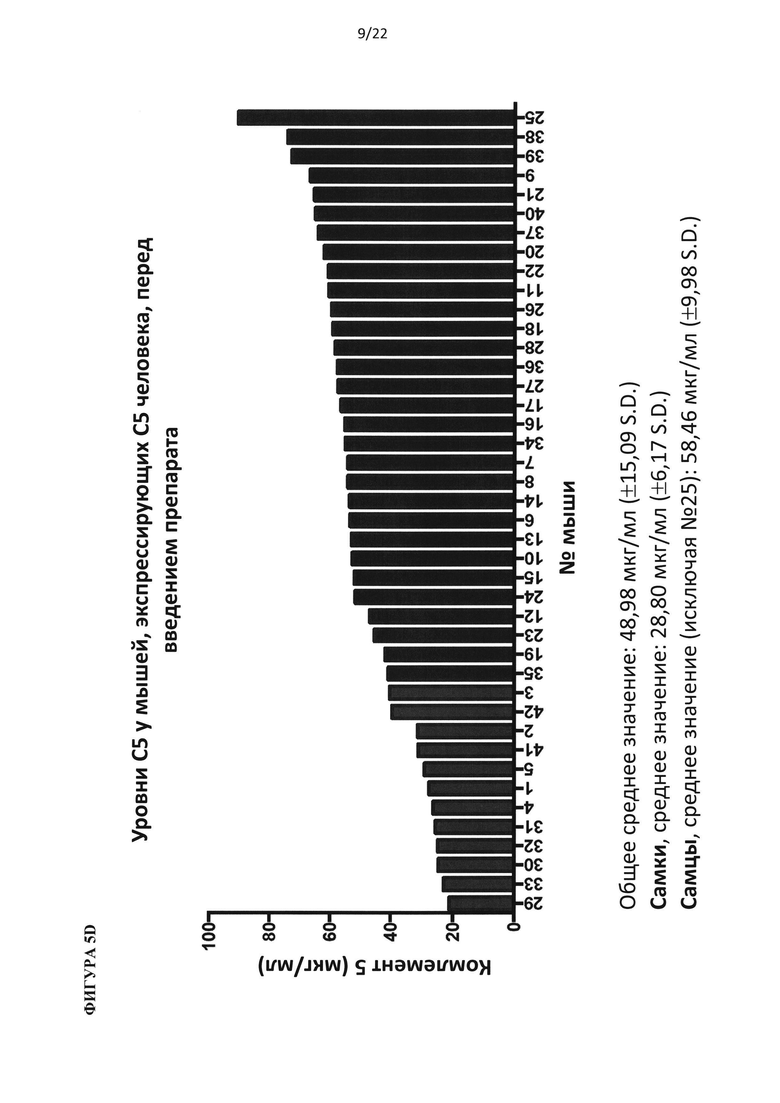

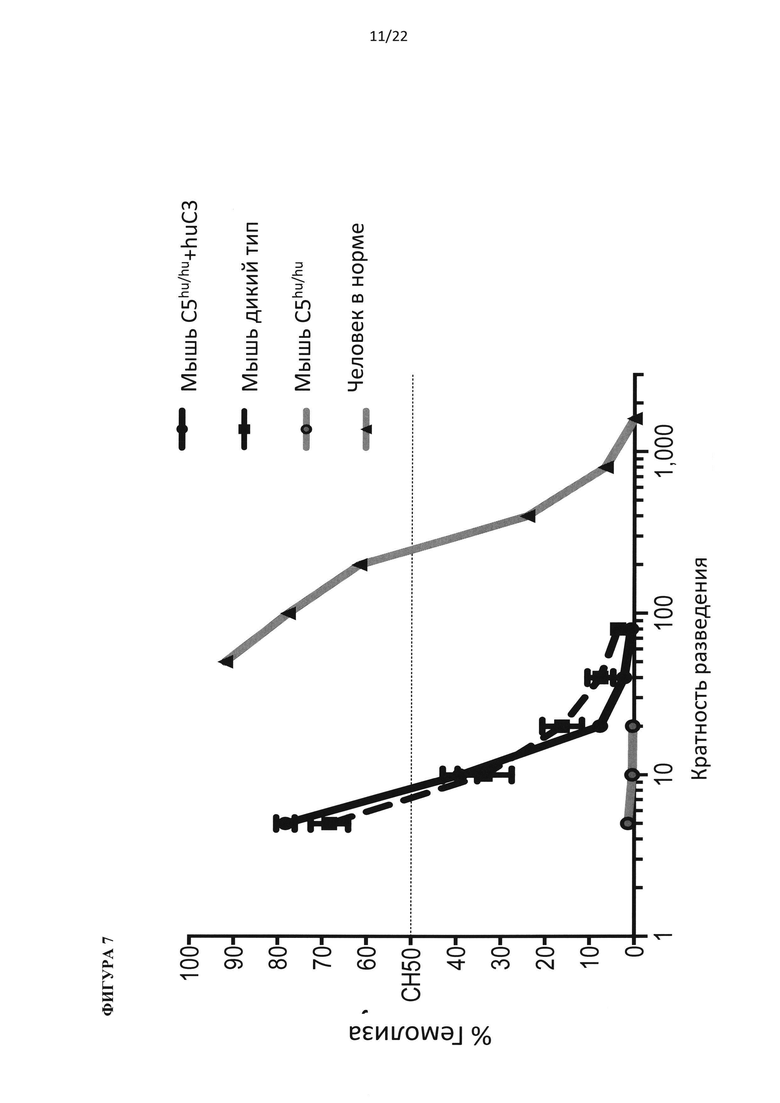
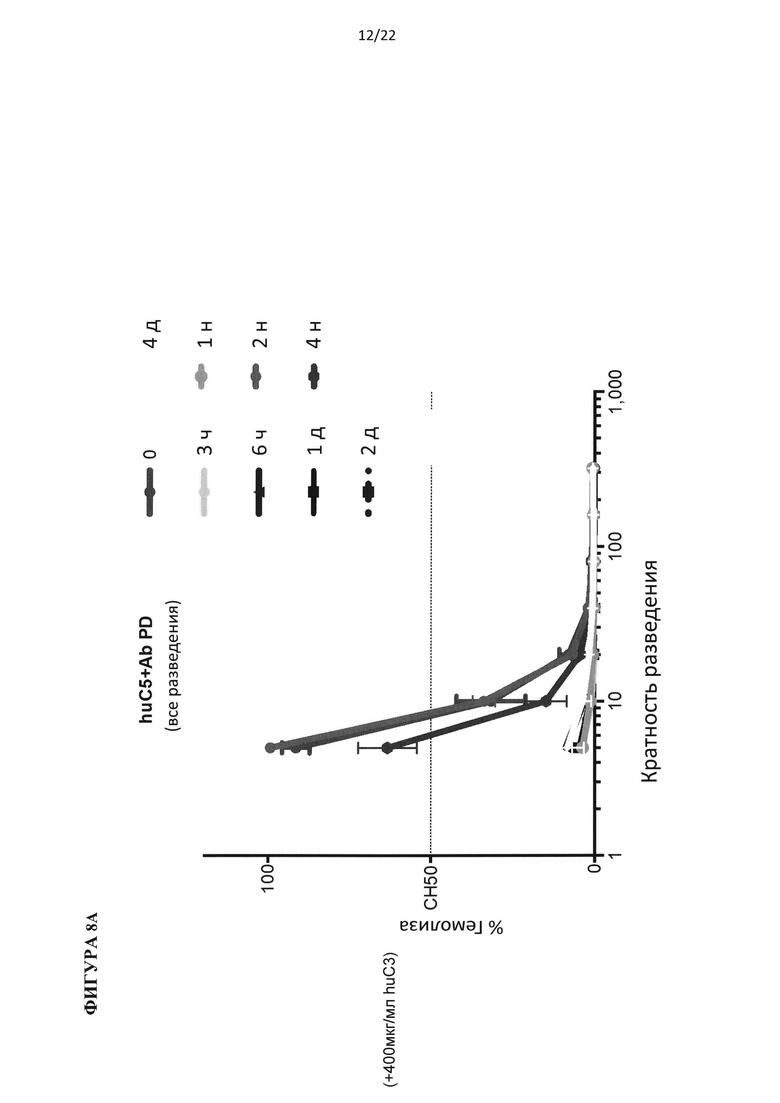
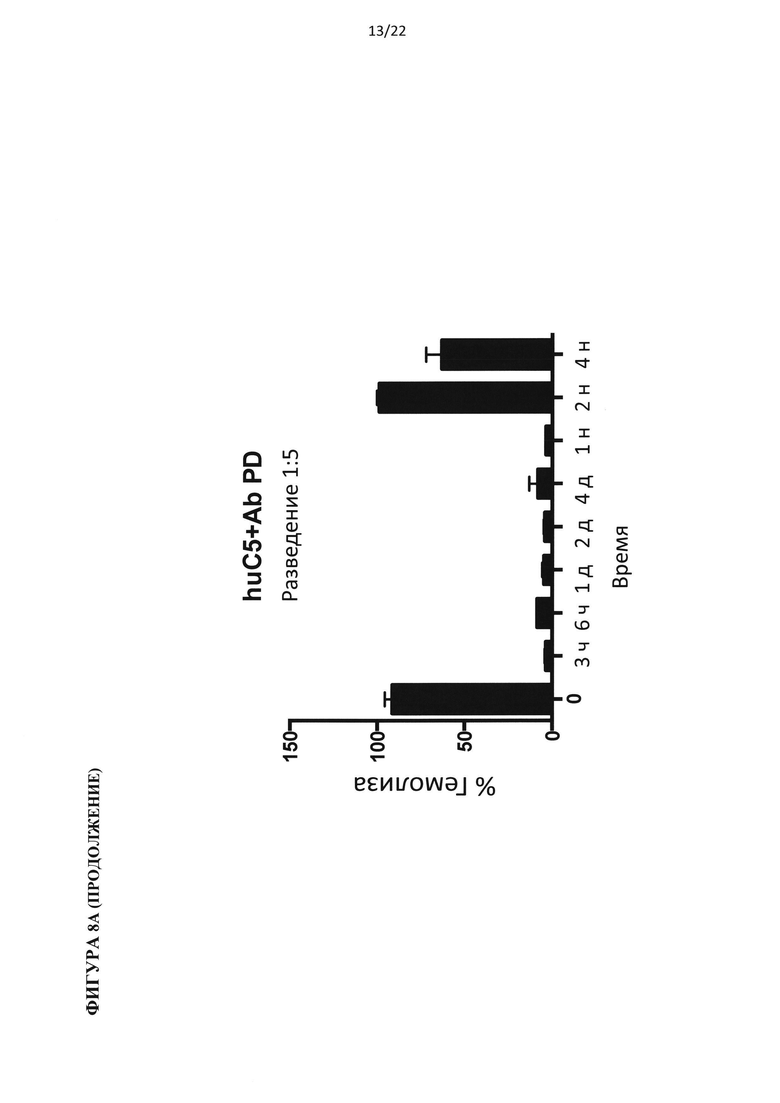
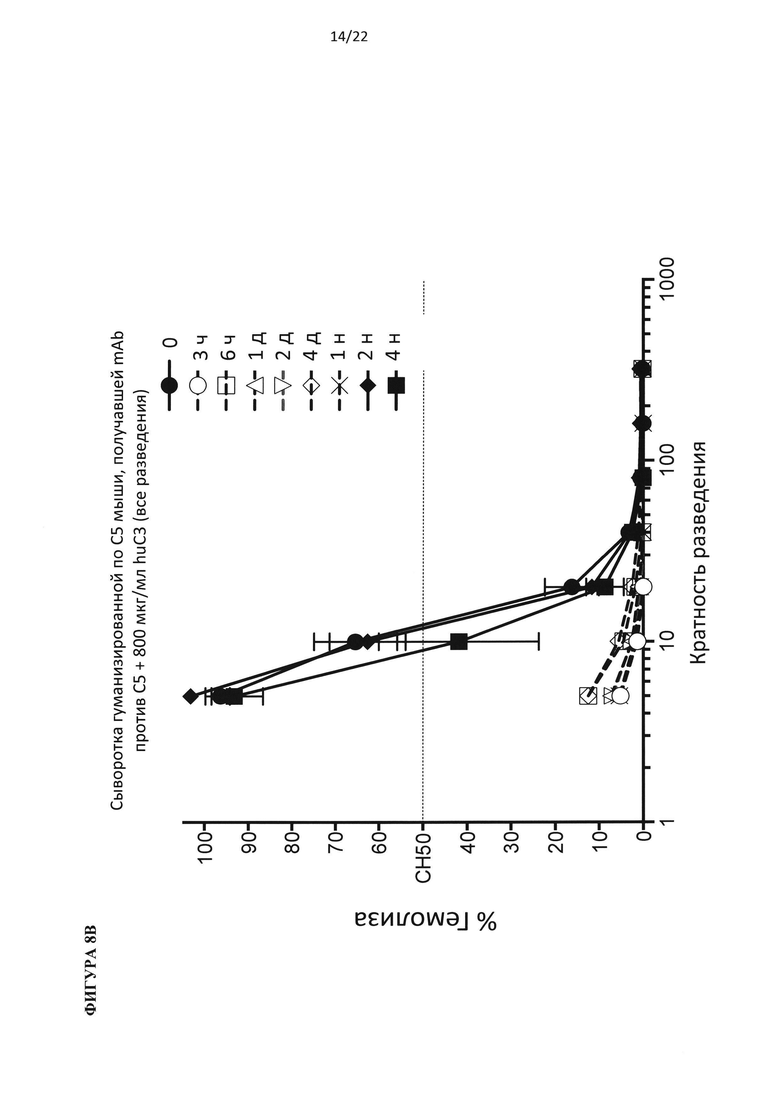
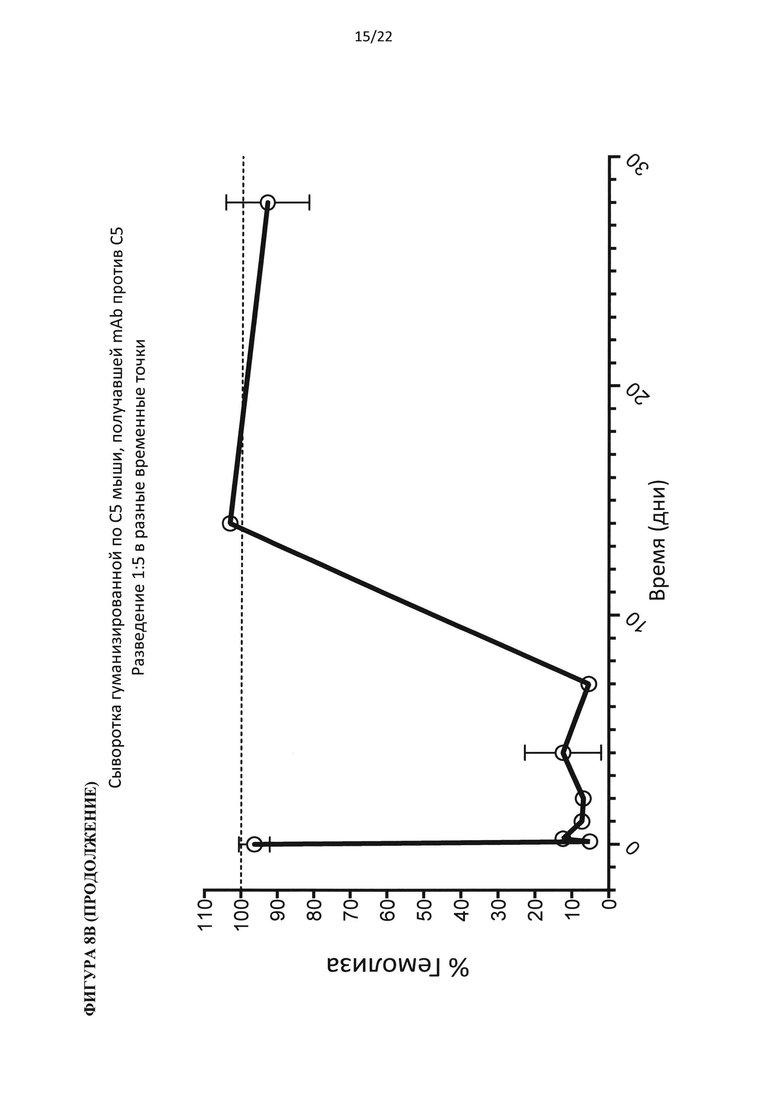
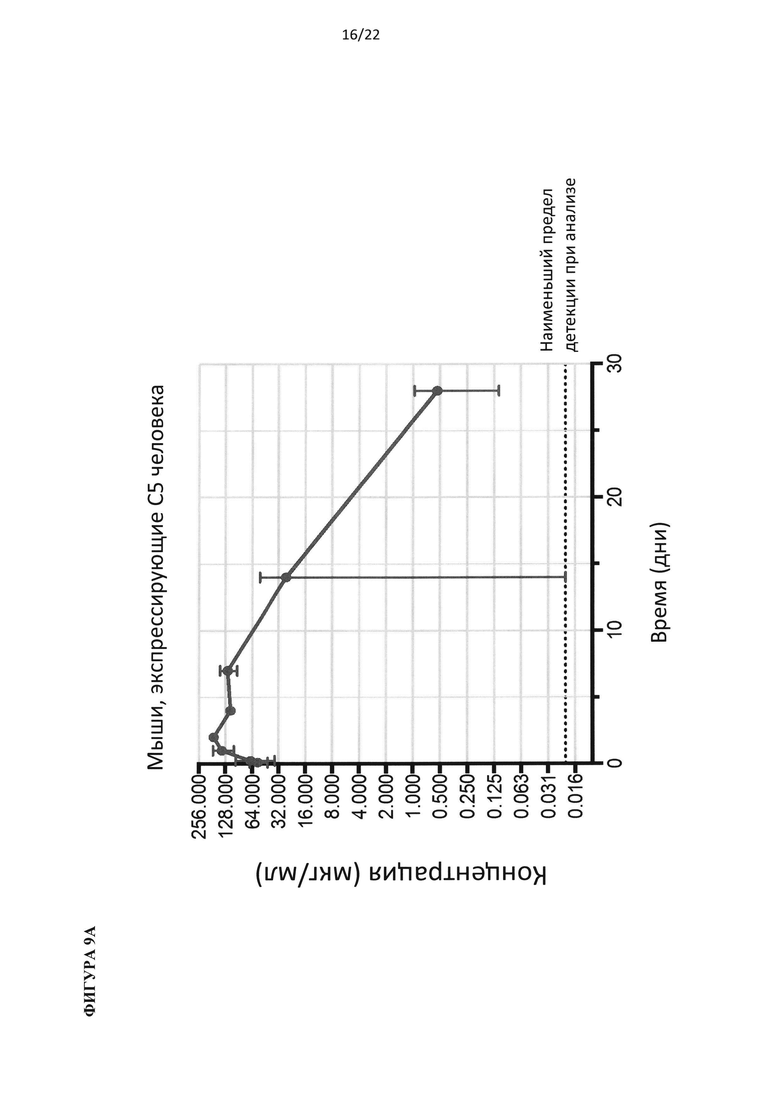
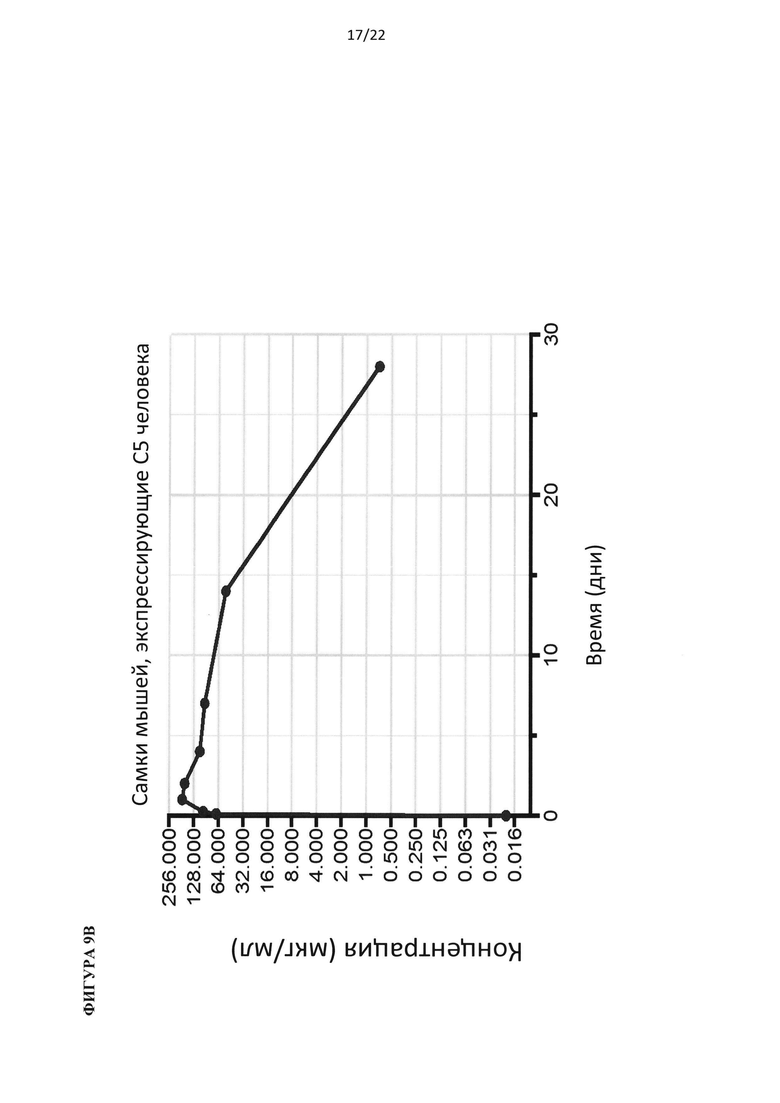
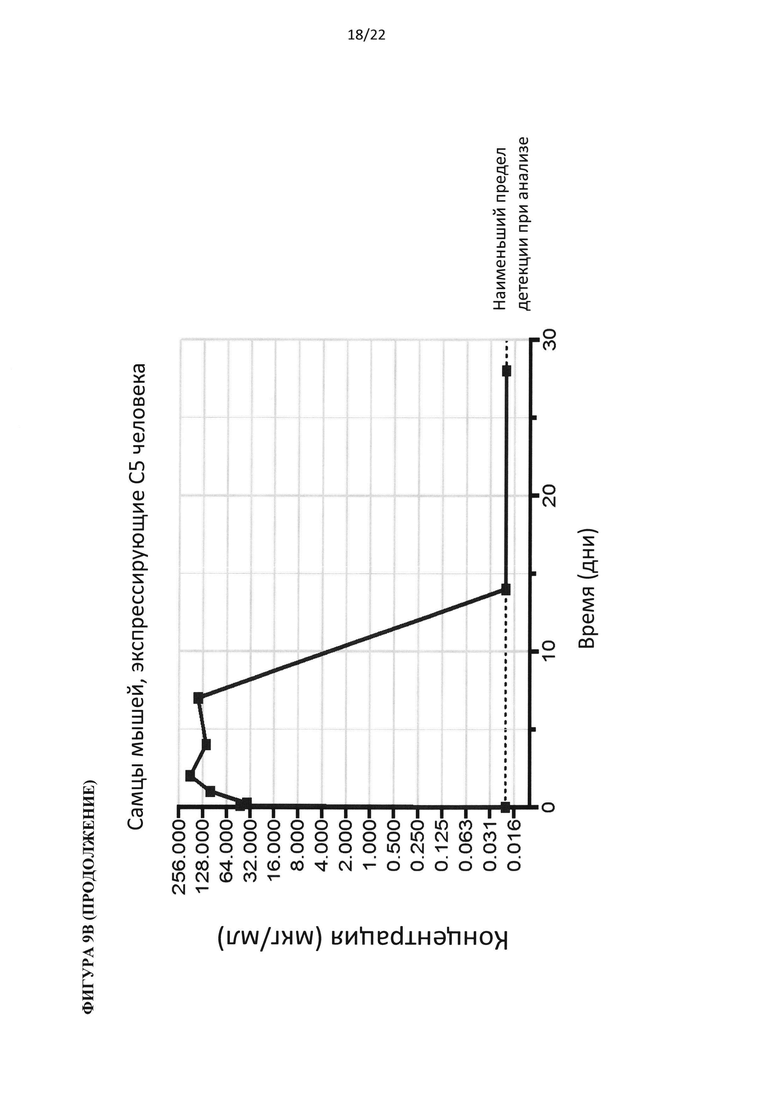

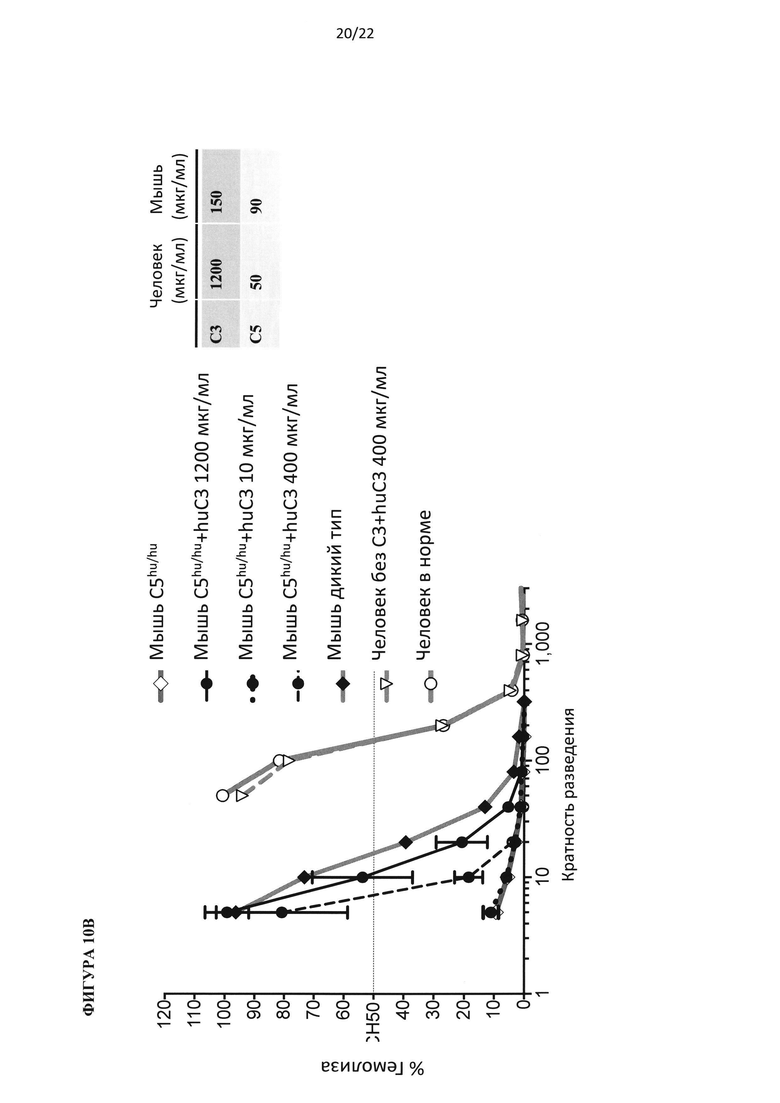
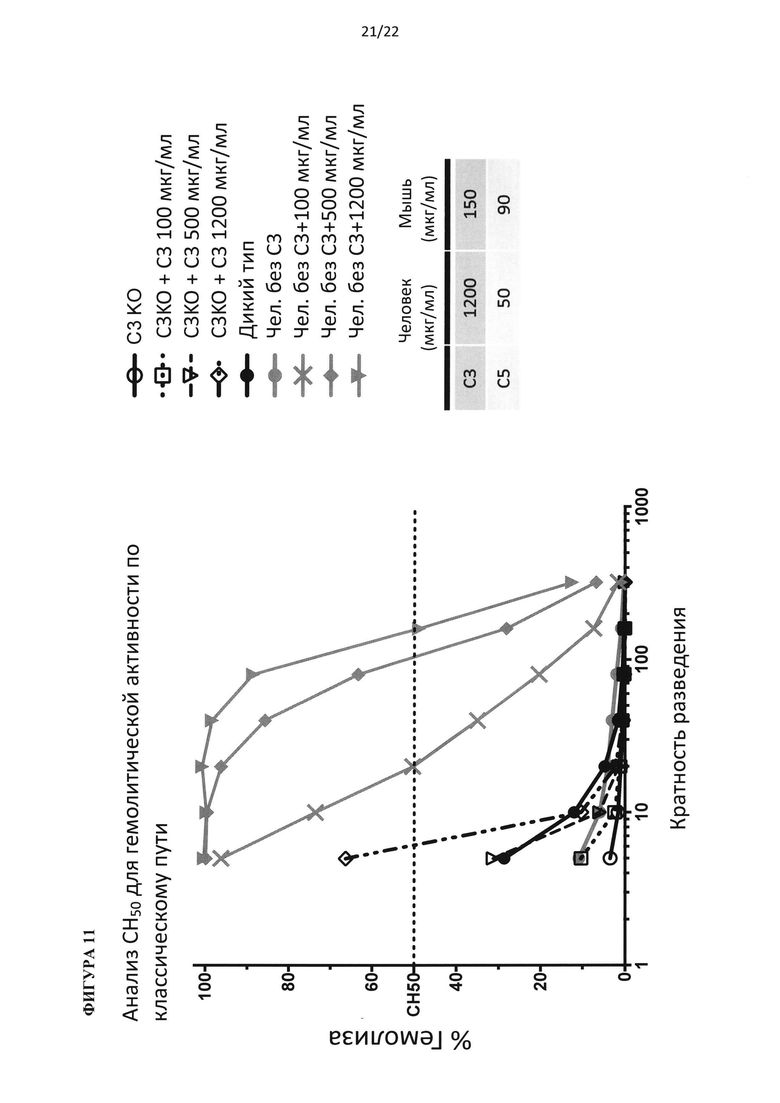
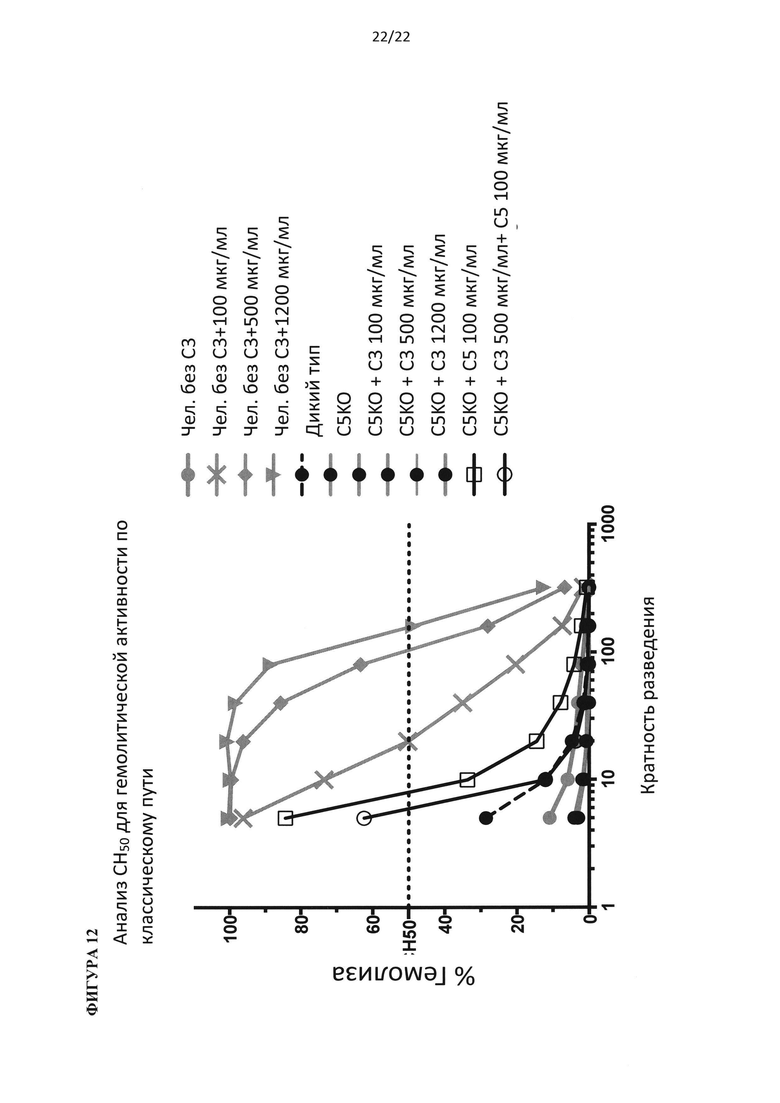
Изобретение относится к области биохимии, в частности к грызуну для экспрессии модифицированного гена С3, геном которого содержит замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3, а также к способу его получения, его эмбриону и клетке. Также раскрыт грызун для экспрессии модифицированного гена С3 и модифицированного гена С5. Геном вышеуказанного грызуна содержит замену последовательности гена С5 грызуна на последовательность гена С5 человека с образованием модифицированного гена С5 и замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3. Изобретение также относится к способу идентификации соединения, способного модулировать активацию системы комплемента, предусматривающему использование вышеуказанного грызуна. Изобретение позволяет эффективно осуществлять идентификацию соединения, способного модулировать активацию системы комплемента. 8 н. и 18 з.п. ф-лы, 12 ил., 1 табл., 6 пр.
1. Грызун для экспрессии модифицированного гена С3, где геном грызуна содержит замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3,
где модифицированный ген С3 содержит экзон 1 эндогенного гена С3 грызуна и со 2 экзона по 41 экзон гена С3 человека,
где экспрессия модифицированного гена С3 находится под контролем регуляторных элементов грызуна в эндогенном локусе С3 грызуна,
и где грызун представляет собой мышь или крысу.
2. Грызун для экспрессии модифицированного гена С3,
где геном грызуна содержит замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3,
где модифицированный ген С3 содержит с 1 экзона по 41 экзон гена С3 человека, и где грызун представляет собой мышь или крысу.
3. Грызун по п. 1 или 2, в котором грызун представляет собой мышь.
4. Грызун по п. 1 или 2, в котором грызун представляет собой крысу.
5. Грызун по п. 3, в котором грызун представляет собой мышь, которая неспособна экспрессировать белок С3 мыши.
6. Грызун по п. 3, в котором грызун представляет собой мышь, которая экспрессирует белок С5 мыши, кодируемый эндогенным геном С5 мыши.
7. Грызун по п. 3, в котором грызун представляет собой мышь, которая экспрессирует человеческий или гуманизированный белок С5.
8. Грызун по п. 7, в котором мышь содержит замену последовательности гена С5 мыши в эндогенном локусе С5 мыши на последовательность гена С5 человека с образованием модифицированного гена С5.
9. Грызун по п. 8, в котором экспрессия модифицированного гена С5 находится под контролем регуляторных элементов мыши в эндогенном локусе С5 мыши.
10. Способ получения С3 гуманизированного грызуна, включающий замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3, где модифицированный ген С3 содержит экзон 1 эндогенного гена С3 грызуна и 2-41 экзоны гена С3 человека,
где экспрессия модифицированного гена С3 находится под контролем регуляторных элементов грызуна в эндогенном локусе С3 грызуна,
и где грызун представляет собой мышь или крысу.
11. Способ получения С3 гуманизированного грызуна, включающий замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3, в котором модифицированный ген С3 содержит с 1 экзона по 41 экзон гена С3 человека, и
где грызун представляет собой мышь или крысу.
12. Способ по п. 10 или 11, в котором грызун представляет собой мышь.
13. Способ по п. 10 или 11, в котором грызун представляет собой крысу.
14. Грызун для экспрессии модифицированного гена С3 и модифицированного гена С5,
в котором геном грызуна содержит замену последовательности гена С5 грызуна на последовательность гена С5 человека с образованием модифицированного гена С5, где последовательность гена С5 человека содержит со 2 экзона по 41 экзон гена С5 человека, а экспрессия модифицированного гена С5 находится под контролем регуляторных элементов или последовательностей грызуна в эндогенном локусе С5 грызуна,
где геном грызуна дополнительно содержит замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3, где (i) модифицированный ген С3 содержит экзон 1 эндогенного гена С3 грызуна и со 2 экзона по 41 экзон гена С3 человека, и экспрессия модифицированного гена С3 находится под контролем регуляторных элементов грызуна в эндогенном локусе С3 грызуна или (ii) модифицированный ген С3 содержит с 1 экзона по 41 экзон гена С3 человека; и
где грызун представляет собой мышь или крысу.
15. Грызун по п. 14, в котором грызун представляет собой мышь.
16. Грызун по п. 14, в котором грызун представляет собой крысу.
17. Грызун по п. 14, в котором грызун представляет собой мышь, которая неспособна экспрессировать белок С5 мыши и неспособна экспрессировать белок С3 мыши.
18. Способ идентификации соединения, способного модулировать активацию системы комплемента, включающий:
a. введение соединения грызуну по любому из пп. 1-9 или 14-17; и
b. анализ того, модулируется ли активация системы комплемента у грызуна, тем самым идентифицируя соединение, способное модулировать активацию системы комплемента.
19. Способ по п. 18, в котором соединение представляет собой низкомолекулярное химическое соединение.
20. Способ по п. 18, в котором соединение представляет собой белок.
21. Способ по п. 18, в котором белок представляет собой антитело.
22. Способ по п. 18, в котором соединение представляет собой ингибирующую нуклеиновую кислоту.
23. Способ по любому из пп. 18-22, в котором сыворотку грызуна анализируют для того, чтобы определить, модулируется ли активация системы комплемента у грызуна.
24. Способ по п. 23, в котором анализ представляет собой скрининговое исследование системы комплемента CH50.
25. Эмбриональная стволовая (ES) клетка грызуна для получения С3 гуманизированного грызуна, содержащая замену последовательности гена С3 грызуна в эндогенном локусе С3 грызуна на последовательность гена С3 человека с образованием модифицированного гена С3, где (i) модифицированный ген С3 содержит экзон 1 эндогенного гена С3 грызуна и со 2 экзона по 41 экзон гена С3 человека, и экспрессия модифицированного гена С3 находится под контролем регуляторных элементов грызуна в эндогенном локусе С3 грызуна или (ii) модифицированный ген С3 содержит с 1 экзона по 41 экзон гена С3 человека; и где ES клетка грызуна представляет собой ES клетку мыши или ES клетку крысы.
26. Эмбрион грызуна для получения С3 гуманизированного грызуна, содержащий ES клетку грызуна по п. 25, где эмбрион грызуна представляет собой эмбрион мыши или эмбрион крысы.
| ADRIANNA LATUSZEK et al., Evaluation of complement activity in wild type, C5 knockout and humanized mice for drug discovery, ARVO Annual Meeting Abstract, April 2014, Volume 55, Issue 13 | |||
| MICHAEL A | |||
| FLIERL et al., Functions of the complement components C3 and C5 during sepsis, FASEB J., 2008, Vol | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| DENNIS P | |||
| VIK et al., | |||

