ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
Настоящая заявка заявляет преимущество предварительной заявки на патент США с серийным номером 61/790907, поданной 15 марта 2013 года, содержание которой включено в данный документ в полном объеме посредством ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДОСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
Копия перечня последовательностей предоставлена в электронном виде с помощью EFS-Web в виде перечня последовательностей в формате ASCII с названием файла “2912939- 20179W001_Sequence_Listing.txt”, созданного 10 марта 2014 года, и имеющего размер 4,14 килобайт, и поданного одновременно с описанием. Перечень последовательностей, содержащийся в этом документе в формате ASCII, является частью описания и включен в данный документ в полном объеме посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области молекулярной биологии растений, более конкретно к идентификации и применению регуляторных элементов у растений.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
В настоящее время существует сильная потребность в трансгенных растениях, которые экспрессируют биотехнологически важные белковые продукты на высоком или индуцируемом уровне. Манипуляция с культурными растениями для изменения и/или улучшения фенотипических характеристик (таких как продуктивность или качество) требует экспрессии гетерологичных генов в растительных тканях. Такие генетические манипуляции стали возможными благодаря двум открытиям: способности трансформировать гетерологичный генетический материал в растительную клетку и существованию промоторов, которые способны управлять экспрессией гетерологичного генетического материала.
В число наиболее часто используемых промоторов входят промотор гена нопалин-синтазы (NOS) (Ebert et al., Proc. Natl. Acad. Sci. U.S.A. 84:5745-5749 (1987)); промотор гена октопин-синтазы (OCS), каулимовирусные промоторы, такие как 19S промотор вируса мозаики цветной капусты (CaMV) (Lawton et al., Plant Mol. Biol. 9:315-324 (1987)); 35S промотор CaMV (Odell et al., Nature 313:810-812 (1985)) и 35S промотор вируса мозаики норичника (Sanger et al., Plant Mol. Biol. 14:433-43 (1990)); светоиндуцируемый промотор из гена малой субъединицы RUBISCO (рибулозо-1,5-бифосфаткарбоксилаза/оксигеназа) (Pellegrineschi et al., Biochem. Soc. Trans. 23(2):247-250 (1995)); промотор гена Adh (Walker et al., Proc. Natl. Acad. Sci. U.S.A. 84:6624-66280 (1987)); промотор гена сахароза-синтазы (Yang et al., Proc. Natl. Acad. Sci. U.S.A. 87:4144-4148 (1990)); промотор комплекса R-генов (Chandler et al., Plant Cell 1:1175-1183 (1989)); промотор гена хлорофилл a/b-связывающего белка и тому подобное.
Идентификация и выделение регуляторных элементов, пригодных для сильной или индуцируемой экспрессии генов в микроорганизмах и растениях, будут полезны для разработки коммерческих сортов трансгенных растений.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Предполагаются композиции и способы для регуляции экспрессии генов в растении. Композиции содержат нуклеотидные последовательности из Glycine max и их варианты, которые инициируют транскрипцию в растении. В частности, предполагается участок инициации транскрипции, выделенный из гена гамма-белка тонопласта и белка плазматической мембраны Glycine max. Дополнительные композиции по настоящему изобретению содержат нуклеотидные последовательности, изложенные в SEQ ID NO: 1 и 2, а также их варианты и фрагменты. Композиции по настоящему изобретению также включают экспрессионные кассеты, содержащие промотор по настоящему изобретению, функционально связанный с гетерологичной нуклеотидной последовательностью, представляющей интерес. Настоящее изобретение дополнительно предусматривает векторы, содержащие экспрессионные кассеты, а также растения и растительные клетки, содержащие стабильно встроенную в их геном экспрессионную кассету, описанную выше. Кроме того, композиции включают трансгенные семена таких растений.
Представляющая интерес последовательность, которая может модифицировать фенотип растения, является функционально связанной с промотором. Такая модификация может включать, например, модуляцию продуцирования эндогенного продукта, или она может включать продуцирование экзогенного продукта экспрессии для обеспечения новой функции или продукта в растении. Например, данным описанием охватывается гетерологичная нуклеотидная последовательность, кодирующая генный продукт, который придает устойчивость к гербицидам или вредителям.
ОПИСАНИЕ ФИГУР
На фигуре 1 показан высокий уровень экспрессии люциферазы в случае, когда ее ген находится под контролем промоторов Pbdc6 (SEQ ID NO: 1) и Pbdc7 (SEQ ID NO: 2).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам для регуляции экспрессии генов в растениях или растительных клетках. Композиции по настоящему изобретению содержат новые нуклеотидные последовательности для промоторов сои. В частности, настоящее изобретение предусматривает выделенные молекулы нуклеиновой кислоты промотора, содержащие нуклеотидную последовательность, изложенную в SEQ ID NO: 1 или 2, а также ее фрагменты и варианты. Кроме того, предполагаются трансформированные растения, растительные клетки и семена.
Промоторные последовательности по настоящему изобретению, когда они собраны в ДНК-конструкции таким образом, что промотор функционально связан с нуклеотидной последовательностью, представляющей интерес, управляют экспрессией нуклеотидной последовательности в клетках организма, стабильно трансформированного этой ДНК-конструкцией, в частности, в растительных клетках. Промоторные последовательности также могут быть пригодны в качестве зондов для выделения других промоторных последовательностей или генов сои, в качестве молекулярных маркеров и т.п.
Способы экспрессии нуклеотидной последовательности в растении предусматривают введение в растительные клетки экспрессионной кассеты, содержащей промотор по настоящему изобретению, функционально связанный с нуклеотидной последовательностью, представляющей интерес, и регенерацию трансформированного растения из растительной клетки.
Используемые в данном документе формы единственного числа обозначают один или более одного (т.е. по меньшей мере один) грамматического объекта, приведенного в такой форме. Например, "элемент" означает один или несколько элементов.
Подразумевается, что используемый в данном документе термин "молекула нуклеиновой кислоты" включает молекулы ДНК (например, кДНК или геномной ДНК) и молекулы РНК (например, мРНК), а также аналоги ДНК или РНК, полученные с использованием нуклеотидных аналогов. Молекула нуклеиновой кислоты может быть однонитевой или двунитевой, но предпочтительно представляет собой двунитевую ДНК.
"Выделенная" или "очищенная" молекула нуклеиновой кислоты или ее биологически активная часть практически не содержит другого клеточного материала или культуральной среды, если она получена с помощью рекомбинантных технологий, или практически не содержит химических предшественников или других химических продуктов, если она синтезирована химическим путем. Предпочтительно, "выделенная" нуклеиновая кислота не содержит последовательностей (предпочтительно, кодирующих белок последовательностей), которые в естественных условиях фланкируют нуклеиновую кислоту (т.е. последовательности, расположенные на 5'- и 3'-концах нуклеиновой кислоты) в геномной ДНК организма, из которого получена нуклеиновая кислота. В контексте настоящего изобретения "выделенный" при использовании в отношении молекул нуклеиновой кислоты исключает выделенные хромосомы. Например, в различных вариантах осуществления настоящего изобретения молекула промотора может содержать менее приблизительно 5 т.н., 4 т.н., 3 т.н., 2 т.н., 1 т.н., 0,5 т.н. или 0,1 т.н. нуклеотидной последовательности, которая в естественных условиях фланкирует молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой получена нуклеиновая кислота. Различные аспекты настоящего изобретения описаны более подробно в следующих подразделах.
Выделенные молекулы нуклеиновой кислоты и их варианты и фрагменты
Нуклеотидные последовательности по настоящему изобретению включают промоторные последовательности, изложенные в SEQ ID NO: 1 и 2 и их варианты. Под "промотором" подразумевают последовательность нуклеиновой кислоты, которая функционирует для управления транскрипцией нижележащей кодирующей последовательности. Промотор, как правило, включает содержит последовательность ДНК, гомологичную консенсусной последовательности 5'-TATAAT-3' (ТАТА-боксу), расположенной приблизительно за 10-30 пар оснований в направлении 5' относительно сайта начала транскрипции ("кэп"), которая обладает способностью направлять РНК-полимеразу для инициации синтеза РНК. Промоторы могут дополнительно включать другие последовательности распознавания, как правило, расположенные выше или в направлении 5' относительно ТАТА-бокса, называемые вышележащими промоторными элементами, которые влияют на уровень инициации транскрипции. Они включают CAAT-бокс, который часто находится приблизительно в 30-70 парах оснований в направлении 5' относительно ТАТА-бокса и характеризуется гомологией с канонической формой 5'-CCAAT-3' (Breathnach and Chambon (1981) Ann. Rev. Biochem. 50:349-383). В растениях, CAAT-бокс иногда заменен последовательностью, известной как AGGA-бокс, участком, в котором адениновые остатки симметрично фланкируют триплет G(или T)NG (Messing et al. (1983), в Genetic Engineering of Plants, T. Kosuge, C. Meredith and A. Hollaender (eds.), Plenum Press, New York, pp. 211-227). Эти элементы вместе с другими транскрипционными и трансляционными регуляторными последовательностями нуклеиновой кислоты (также называемые "управляющими последовательностями") необходимы для экспрессии последовательности ДНК, представляющей интерес. Не описанные в данном документе способы выделения и идентификации регуляторных элементов, такие как энхансеры и элементы, ответственные за экспрессию кодирующего участка в определенных тканях или в определенное время, хорошо известны в уровне техники. См., например, патенты США №№ 5635618; 6218140; 6303370; 6310197 и 6355864.
Под "коровым промотором" подразумевается промотор без промоторных элементов. Коровый промотор содержит необходимые для функционирования промотора нуклеотидные последовательности, в том числе ТАТА-бокс и сайт инициации транскрипции. Такой участок обычно присутствует с некоторым изменениями в большинстве промоторов. Участок корового промотора часто называется минимальным промоторным участком, она сама по себе является функциональной для обеспечения базального уровня транскрипции. В различных вариантах осуществления настоящего изобретения последовательность корового промотора для Pbdc6 примерно соответствует нуклеотидам с 29 по 318 в SEQ ID NO: 1; ТАТА примерно соответствует нуклеотидам с 288 по 296 в SEQ ID NO: 1; и сайт инициации трансляции соответствует положению нуклеотида 318 в SEQ ID NO: 1. В других вариантах осуществления настоящего изобретения, последовательность корового промотора для Pbdc7 примерно соответствует нуклеотидам с 1341 по 1643 в SEQ ID NO: 2; ТАТА примерно соответствует нуклеотидам с 1603 по 1608 в SEQ ID NO: 2; и сайт инициации трансляции соответствует положению нуклеотида 1643 в SEQ ID NO: 2. Специалисту в данной области техники будет понятно, что участок корового промотора может отличаться от приведенных выше положений на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более нуклеотидов в направлении выше или ниже относительно сайта инициации транскрипции, и эти изменения в последовательности корового промотора могут допускаться.
Молекулы нуклеиновой кислоты, которые представляют собой фрагменты раскрытых промоторных последовательностей, также охватываются настоящим изобретением. Под "фрагментом" подразумевается часть промоторной последовательности. Фрагмент нуклеотидной последовательности может быть биологически активным и, следовательно, способным инициировать транскрипцию функционально связанной нуклеотидной последовательности в растении, или он может представлять собой фрагмент, который можно применять в качестве гибридизационного зонда или ПЦР праймера с использованием способов, описанных ниже. Анализы для определения того, способны ли такие фрагменты снижать уровни экспрессии или изменять основные свойства экспрессии, т.е. конститутивную или индуцируемую экспрессию, хорошо известны в уровне техники.
Молекулы нуклеиновых кислот, которые представляют собой фрагменты промоторной последовательности могут содержать по меньшей мере приблизительно 20, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600 смежных нуклеотидов или нуклеотиды в количестве вплоть до числа нуклеотидов, присутствующих в промоторной последовательности полной длины, раскрытой в данном документе (например, 1230 нуклеотидов для SEQ ID NO: 1 или 1688 нуклеотидов для SEQ ID NO: 2), в зависимости от предполагаемого применения. Под "смежными" нуклеотидами подразумевается остатки нуклеиновой кислоты, которые непосредственно примыкают друг к другу. Биологически активные фрагменты промоторов по настоящему изобретению будут сохранять промоторную активность (т.е. инициировать транскрипцию). Под "сохранением промоторной активности" подразумевается, что фрагмент будет характеризоваться по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, по меньшей мере, приблизительно 70% или по меньшей мере приблизительно 80% от промоторной активности промотора полной длины. Биологически активную часть промотора можно получить путем выделения части одной из промоторных нуклеотидных последовательностей по настоящему изобретению и оценки активности этой части промотора. Способы измерения активности промотора, хорошо известны в уровне техники. См. раздел, озаглавленный "Оценка активности промотора" для примеров подходящих способов.
Такие фрагменты, как правило, будут содержать последовательность распознавания ТАТА в конкретной промоторной последовательности. Эти фрагменты можно получить путем расщепления встречающейся в естественных условиях промоторной нуклеотидной последовательности, описанной в данном документе, ферментами рестрикции, путем синтеза нуклеотидной последовательности из встречающейся в естественных условиях последовательности ДНК промотора или посредством применения технологии ПЦР. См., в частности, Mullis et al. (1987) Methods Enzymol. 155:335-350, и Erlich, ed. (1989) PCR Technology (Stockton Press, New York). Варианты этих фрагментов промотора, таких как полученные в результате сайт-направленного мутагенеза, также охватываются композициями по настоящему изобретению.
Также охватываются варианты промоторных последовательностей, раскрытых в данном документе. Под "вариантом" подразумевается достаточно идентичная последовательность. Промоторные последовательности, охватываемые настоящим изобретением, являются достаточно идентичными нуклеотидной последовательности SEQ ID NO: 1 или 2. Под "достаточно идентичной" подразумевается нуклеотидная последовательность, которая по меньшей мере приблизительно на 70% или 75%, приблизительно на 80% или 85% идентична последовательности, приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична последовательности по сравнению с эталонной последовательностью при использовании одной из программ выравнивания, как описано в данном документе.
Встречающиеся в естественных условиях варианты можно быть идентифицировать с применением хорошо известных методик молекулярной биологии, таких как полимеразная цепная реакция (ПЦР) и методики гибридизации, как указано ниже. Вариантные нуклеотидные последовательности также включают полученные синтетическим путем нуклеотидные последовательности, которые были созданы, например, с использованием сайт-направленного мутагенеза, но которые все еще имеют промоторную активность, как определено в данном документе.
Варианты, охватываемые настоящим изобретением, являются биологически активными, то есть они продолжают обладать необходимой биологической активностью нативной последовательности, то есть сохраняют промоторную активность (т.е. инициируют транскрипцию). Под "сохранением промоторной активности" подразумевается, что вариант будет характеризоваться активностью, составляющей по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 80% или более от промоторной активности у нативной последовательности. Способы измерения промоторной активности хорошо известны в уровне техники. См. раздел, озаглавленный "Оценка промоторной активности" для примеров подходящих способов.
Специалисту в данной области техники также будет понятно, что изменения могут быть внесены путем мутации в нуклеотидных последовательностях по настоящему изобретению без изменения способности промотора управлять экспрессией в растительной клетке. Таким образом, вариантные выделенные молекулы нуклеиновой кислоты можно создать путем введения одной или нескольких нуклеотидных замен, добавлений или делеций в соответствующую нуклеотидную последовательность, раскрытую в данном документе. Мутации могут быть введены с помощью стандартных методик, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Такие вариантные нуклеотидные последовательности также охватываются настоящим изобретением.
В качестве альтернативы, вариантные нуклеотидные последовательности можно получить путем введения мутаций случайным образом вдоль всей последовательности или ее части, например, путем насыщающего мутагенеза, и полученных мутантов можно подвергнуть скринингу в отношении способности к управлению экспрессией функционально связанной нуклеотидной последовательности в растительной клетке.
Под "функционально связанным" подразумевается функциональная связь между промотором и второй последовательностью, где промоторная последовательность инициирует и опосредует транскрипцию последовательности ДНК, соответствующей второй последовательности. Как правило, но не всегда, функционально связанный означает, что последовательности нуклеиновой кислоты являются смежными, а в случае, если необходимо объединить два белок-кодирующих участка, они являются смежными и находятся в одной и той же рамке считывания.
Для определения процентной идентичности двух нуклеиновых кислот, последовательности выравнивают с целью получения оптимального сравнения. Процентная идентичность у двух последовательностей зависит от числа идентичных положений, общих для последовательностей (т.е. процентная идентичность = число идентичных положений/общее число положений (например, перекрывающиеся положения) × 100). В одном варианте осуществления настоящего изобретения две последовательности имеют одинаковую длину. Процентная идентичность у двух последовательностей можно определить с использованием методик, аналогичных описанным ниже, с или без обеспечения возможности гэпов. При расчете процентной идентичности, как правило, подсчитывают точные совпадения.
Определение процентной идентичности у двух последовательностей можно выполнить с использованием математического алгоритма. Неограничивающим примером математического алгоритма, применяемого для сравнения двух последовательностей, является алгоритм по Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264, модифицированный как в Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877. Такой алгоритм включен в программу BLASTN по Altschul et al. (1990) J. Mol. Biol. 215:403. Поиск нуклеотидов BLAST может быть выполнен с помощью программы BLASTN, оценка = 100, длина слова = 12, чтобы получить нуклеотидные последовательности, гомологичные промоторам по настоящему изобретению. Чтобы получить выравнивания с гэпами с целью сравнения, можно применять Gapped BLAST, как описано в Altschul et al. (1997) Nucleic Acids Res. 25:3389. В качестве альтернативы, PSI-BLAST может быть использован для выполнения итеративного поиска, который обнаруживает дальнее родство между молекулами. См., Altschul et al. (1997), выше. При применении программ BLAST, Gapped BLAST и PSI-BLAST можно быть использовать параметры по умолчанию у соответствующих программ (например, BLASTN). См., www.ncbi.nlm.nih.gov. Другой неограничивающий пример математического алгоритма, применяемого для сравнения последовательностей, представляет собой алгоритм ClustalW (Higgins et al. (1994) Nucleic Acids Res. 22:4673-4680). ClustalW сравнивает последовательности, и производит выравнивание по всей длине последовательности ДНК, и, следовательно, может предоставить данные о консервативности последовательности нуклеотидной последовательности полной длины. Алгоритм ClustalW используется в нескольких коммерчески доступных пакетах программного обеспечения для анализа ДНК, таких как модуль ALIGNX в наборе программного обеспечения vector NTi Program Suite (Informax, Inc). Неограничивающим примером программного обеспечения, пригодного для анализа выравниваний с использованием ClustalW, является GeneDoc™. Genedoc™ (Karl Nicholas) позволяет оценить сходство ДНК и идентичность у нескольких генов. Другим предпочтительным, но неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм по Myers and Miller (1988) CABIOS 4:11-17. Такой алгоритм включен в программу ALIGN (версия 2.0), которая является частью пакета программного обеспечения для выравнивания последовательностей GCG (доступного от Accelrys, Inc., 9865 Scranton Rd., Сан-Диего, Калифорния, США).
Если не указано иное, GAP версия 10, в которой используется алгоритм по Needleman and Wunsch (1970) J. Mol. Biol. 48(3):443-453, будет применяться для определения идентичности или сходства последовательности с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа за открытие гэпа 50 и штрафа за продолжение гэпа 3 и оценочной матрицы nwsgapdna.cmp; % идентичности или % сходства для аминокислотной последовательности с использованием штрафа за открытие гэпа 8 и штрафа за продолжение гэпа 2 и оценочной программы BLOSUM62. Также могут быть использованы эквивалентные программы. Под "эквивалентной программой" подразумевается любая программа для сравнения последовательностей, которая для любых двух последовательностей в запросе создает выравнивание, имеющее идентичные совпадения нуклеотидных остатков и идентичную процентную идентичность по сравнению с соответствующим выравниванием, создаваемым GAP версии 10.
С помощью таких способов, как ПЦР, гибридизация и т.п., можно идентифицировать соответствующие последовательности из других организмов, в частности, других растений, такие последовательности характеризуются существенной идентичностью с последовательностями по настоящему изобретению. См., например, Sambrook J., and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY); и Innis, et al. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, NY). Последовательности, которые идентифицированы по их идентичности с промоторными последовательностями, изложенными в данном документе, охватываются настоящим изобретением.
Олигонуклеотидные праймеры можно сконструировать для применения в реакциях ПЦР для амплификации соответствующих последовательностей ДНК из кДНК или геномной ДНК из растения, представляющего интерес. Способы конструирования ПЦР праймеров и ПЦР-клонирования, как правило, известны в уровне техники и описаны в Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). См., также, Innis et al., eds. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, New York); Innis and Gelfand, eds. (1995) PCR Strategies (Academic Press, New York); и Innis and Gelfand, eds. (1999) PCR Methods Manual (Academic Press, New York). Известные способы ПЦР включают способы с использованием спаренных праймеров, гнездовых праймеров, одиночных специфических праймеров, вырожденных праймеров, ген-специфических праймеров, вектор-специфических праймеров и частично несоответствующих праймеров.
В способе гибридизации всю известную нуклеотидную последовательность или ее часть можно использовать для скрининга кДНК или геномных библиотек. Способы создания таких кДНК и геномных библиотек, как правило, известны в уровне техники и описаны в Sambrook and Russell, 2001, выше. Зонды для гибридизации могут быть фрагментами геномной ДНК, фрагментами кДНК, фрагментами РНК или другими олигонуклеотидами и могут быть помечены детектируемой группой, такой как 32P, или любым другим детектируемым маркером, таким как другие радиоизотопы, флуоресцентное соединение, фермент или кофактор фермента. Зонды для гибридизации можно получить путем мечения синтетических олигонуклеотидов, исходя из известной последовательности промотора, описанной в данном документе. Также можно применять вырожденные праймеры, созданные на основе консервативных нуклеотидов в нуклеотидной последовательности. Зонд, как правило, содержит участок нуклеотидной последовательности, который гибридизуется в жестких условиях по меньшей мере приблизительно с 12, по меньшей мере приблизительно с 25, по меньшей мере приблизительно с 50, 75, 100, 125, 150, 175, 200, 250, 300, 350 или 400 последовательными нуклеотидами в промоторной последовательности по настоящему изобретению или ее фрагменте или варианте. Получение зондов для гибридизации, как правило, известно в уровне техники и описано в Sambrook and Russell, 2001, выше, включенной в данный документ посредством ссылки.
Например, полная промоторная последовательность, описанная в данном документе, или одна или несколько ее частей, можно применять в качестве зонда, способного к специфической гибридизации с соответствующей промотор-подобной последовательностью. Для достижения специфической гибридизации в различных условиях, такие зонды имеют в своем составе последовательности, которые являются уникальными и имеют длину по меньшей мере приблизительно 10 нуклеотидов или по меньшей мере приблизительно 20 нуклеотидов. Такие зонды могут быть использованы для амплификации соответствующих промоторных последовательностей из выбранного организма посредством ПЦР. Эта технология может быть использована для выделения дополнительных кодирующих последовательностей из необходимого организма или в качестве диагностического анализа для определения наличия кодирующих последовательностей в организме. Методики гибридизации включают гибридизационный скрининг высеянных ДНК библиотек (либо бляшек, либо колоний; см, например, Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY).
Гибридизацию таких последовательностей можно проводить в жестких условиях. Под "жесткими условиями" или "жесткими условиями гибридизации" подразумеваются условия, при которых зонд гибридизуется со своей целевой последовательностью в детектируемо большей степени, нежели с другими последовательностями (например, по меньшей мере в 2 раза больше по сравнению с фоном). Жесткие условия зависят от последовательности и будут отличаться при разных обстоятельствах. Контролируя жесткость условий гибридизации и/или условия отмывки, можно идентифицировать целевые последовательности, которые комплементарны зонду на 100% (гомологичное зондирование). В качестве альтернативы, жесткость условий можно регулировать для обеспечения возможности некоторого несовпадения в последовательностях для того, чтобы выявить более низкие степени сходства (гетерологичное зондирование). Как правило, зонд имеет длину менее приблизительно 1000 нуклеотидов или менее 500 нуклеотидов.
Как правило, жесткими условиями будут считаться условия, в которых концентрация соли составляет менее приблизительно 1,5 М ионов Na, обычно при концентрации ионов Na (или других солей) приблизительно 0,01-1,0 М при рН 7,0-8,3, и температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°С для длинных зондов (например, более 50 нуклеотидов). Жесткие условия можно также получить добавлением дестабилизирующих средств, таких как формамид. Иллюстративные условия низкой жесткости включают гибридизацию в буферном растворе с 30-35% формамида, 1 М NaCl, 1% SDS (додецилсульфат натрия) при 37°C и отмывку в 1X-2X SSC (20X SSC = 3,0 М NaCl/0,3 М цитрат тринатрия) при 50-55°С. Иллюстративные условия умеренной жесткости включают гибридизацию в 40-45% формамиде, 1,0 М NaCl, 1% SDS при 37°С и отмывку в 0,5-1X SSC при 55-60°С. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамиде, 1 М NaCl, 1% SDS при 37°С и отмывку в 0,1X SSC при 60-65°C. Необязательно, буферы для отмывки могут содержать от приблизительно 0,1% до приблизительно 1% SDS. Продолжительность гибридизации, как правило, составляет менее приблизительно 24 часов, обычно от приблизительно 4 до приблизительно 12 часов. Необязательно, буферы для отмывки могут содержать от приблизительно 0,1% до приблизительно 1% SDS.
Специфичность, как правило, зависит от послегибридизационных отмывок, причем решающими факторами являются ионная сила и температура конечного раствора для отмывки. Для гибридов ДНК-ДНК Тm можно аппроксимировать из уравнения по Meinkoth and Wahl (1984) Anal. Biochem. 138:267-284: Тm = 81,5°C + 16,6 (log M) + 0,41 (%GC) - 0,61 (% формамида) - 500/L; где М представляет собой молярность одновалентных катионов, %GC представляет собой процентное содержание гуанозиновых и цитозиновых нуклеотидов в ДНК, % формамида представляет собой процентное содержание формамида в растворе для гибридизации; и L представляет собой длину гибрида в парах оснований. Тm представляет собой температуру (при определенных ионной силе и рН), при которой 50% комплементарной целевой последовательности гибридизуются с идеально совпадающим зондом. Тm снижается примерно на 1°С для каждого 1% несовпадения; следовательно, Тm, условия гибридизации и/или отмывки можно регулировать для гибридизации с последовательностями необходимой идентичности. Например, если производят поиск последовательностей с идентичностью >90%, Тm можно уменьшить на 10°C. Как правило, жесткие условия выбирают так, чтобы температура при них была приблизительно на 5°С ниже температуры плавления (Тm) для конкретной последовательности и ее комплементарной последовательности при определенных ионной силе и pH. Тем не менее, при очень жестких условиях гибридизацию и/или отмывку можно проводить при температуре, которая на 1, 2, 3 или 4°C ниже температуры плавления (Тm); при умеренно жестких условиях гибридизацию и/или отмывку можно проводить при температуре, которая на 6, 7, 8, 9 или 10°C ниже температуры плавления (Тm); при условиях низкой жесткости гибридизацию и/или отмывку можно проводить при температуре, которая на 11, 12, 13, 14, 15 или 20°С ниже температуры плавления (Тm). С использованием уравнения, композиций для гибридизации и отмывки и необходимой Тm специалист в данной области техники поймет, что изменения жесткости условий гибридизации и/или растворах для отмывки, по сути, уже описаны. Если необходимый уровень несовпадений приводит к Тm менее 45°C (водный раствор) или 32°C (раствор с формамидом), концентрацию SSC можно повысить для того, чтобы можно было использовать более высокую температуру. Исчерпывающее руководство по гибридизации нуклеиновых кислот приведено в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology- Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, New York); и Ausubel et al., eds. (1995) Current Protocols in Molecular Biology, Chapter 2 (Greene Publishing and Wiley-Interscience, New York). См., Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY).
Выделенные последовательности, которые характеризуются промоторной активностью и гибридизуются в жестких условиях с промоторными последовательностями, раскрытыми в данном документе, или их фрагментами, охватываются настоящим изобретением.
Способы применения
Способы по настоящему изобретению направлены на экспрессию гетерологичных нуклеотидных последовательностей в растениях и растительных клетках под контролем промоторной последовательности по настоящему изобретению. Трансгенные растения могут иметь изменения в фенотипе, в том числе без ограничения измененный механизм защиты от патогенов или насекомых, повышенную устойчивость к одному или нескольким гербицидам, повышенную способность выдерживать стрессовые условия окружающей среды, модифицированную способность вырабатывать крахмал, модифицированный уровень выработки крахмала, модифицированное содержание масла и/или его состав, модифицированную способность использовать, распределять и/или запасать азот и т.п. Эти результаты могут быть достигнуты путем экспрессии гетерологичных генов или повышенной экспрессии эндогенных продуктов в растениях. В качестве альтернативы, результаты могут быть достигнуты за счет снижения экспрессии одного или нескольких эндогенных продуктов, особенно ферментов, транспортеров или кофакторов, или путем влияния на поглощение питательных веществ у растения.
Как правило, нуклеотидная последовательность промотора по настоящему изобретению обеспечивается в экспрессионной кассете с нуклеотидной последовательностью, представляющей интерес, обычно гетерологичной нуклеотидной последовательностью, для экспрессии в растении, представляющем интерес. Под "гетерологичной нуклеотидной последовательностью" подразумевается последовательность, которая в естественных условиях не является функционально связанной с промоторной последовательностью, в том числе не встречающиеся в естественных условиях множественные копии встречающихся в естественных условиях последовательностей ДНК. Хотя эта нуклеотидная последовательность является гетерологичной по отношению к промоторной последовательности, она может быть гомологичной, или нативной, или гетерологичной, или чужеродной по отношению к растению-хозяину. Понятно, что промотор может также управлять экспрессией гомологичной или нативной нуклеотидной последовательности. В некоторых случаях трансформированное растение может иметь изменение в фенотипе. Гетерологичные последовательности нуклеиновой кислоты включают те, которые являются экзогенными или не присутствующими в нетрансформированных растительных клетках, а также те, которые могут являться эндогенными или присутствующими в нетрансформированных растительных клетках. "Гетерологичный", как правило, относится к последовательностям нуклеиновой кислоты, которые не являются эндогенными по отношению к клетке или части нативного генома, в котором они находятся, а были добавлены в клетку посредством инфекции, трансфекции, микроинъекции, электропорации, посредством бомбардировки микрочастицами или т.п.
Любую последовательность, представляющую интерес, можно экспрессировать при помощи промоторных последовательностей по настоящему изобретению. Такие гетерологичные нуклеотидные последовательности включают кодирующие последовательности, обеспечивающие выносливость к гербицидам, кодирующие последовательности, обеспечивающие инсектицидный эффект, кодирующие последовательности, обеспечивающие нематоцидный эффект, кодирующие последовательности, обеспечивающие антимикробный эффект, кодирующие последовательности, обеспечивающие противогрибковый эффект, кодирующие последовательности, обеспечивающие противовирусный эффект, кодирующие последовательности, обеспечивающие выносливость к биотическим и абиотическим стрессам, или последовательности, модифицирующие признаки растений, такие как урожайность, качество зерна, содержание питательных веществ, качество и количество крахмала, фиксация и/или утилизация азота и содержание и/или состав масла, но не ограничиваются ими.
Более конкретные гены, представляющие интерес, согласно настоящему изобретению включают без ограничения гены, которые улучшают урожайность, гены, которые улучшают привлекательность культур, гены, кодирующие белки, придающие устойчивость к абиотическому стрессу, такому как засуха, температура, засоление, токсичные металлы или микроэлементы, или гены, которые придают устойчивость к токсину, такому как пестициды и гербициды, или к биотическому стрессу, такому как заражение грибками, вирусами, бактериями, насекомыми и нематодами, а также развитие болезней, связанных с этими организмами. Понятно, что любой ген, представляющий интерес, может быть функционально связан с промоторными последовательностями по настоящему изобретению и может экспрессироваться в растении.
Эти гетерологичные нуклеотидные последовательности могут кодировать белки, вовлеченные в обеспечение устойчивости к болезням или вредителям. Под "устойчивостью к болезням" или "устойчивостью к вредителям" подразумевается, что растения избегают вредных симптомов, которые являются последствием взаимодействий растение-патоген. Все из генов белков, обеспечивающих устойчивость к болезням и вредителям, таких как лизоцимы или цекропины, для антибактериальной защиты, или белков, таких как дефензины, глюканазы или хитиназы, для противогрибковой защиты, или эндотоксинов Bacillus thuringiensis, ингибиторов протеаз, коллагеназ, лектинов или гликозидаз для борьбы с нематодами или насекомыми, представляют собой примеры пригодных генных продуктов. Примеры генов, представляющих интерес, можно найти, например, на сайте www.nbiap.vt.edu/cfdocs/fieldtests2.cffn.
"Вредитель" включает насекомых, грибы, бактерии, вирусы, нематод, клещей, зудней и т.п., но не ограничивается ими. Насекомые-вредители включают насекомых, выбранных из отрядов Coleoptera, Diptera, Hymenoptera, Lepidoptera, Mallophaga, Homoptera, Hemiptera, Orthroptera, Thysanoptera, Dermaptera, Isoptera, Anoplura, Siphonaptera, Trichoptera и т.д., в особенности, Coleoptera, Lepidoptera и Diptera. Вирусы включают вирус мозаики табака или огурца, вирус кольцевой пятнистости, вирус некроза, вирус карликовой мозаики маиса т.д., но не ограничиваются ими. Нематоды включают без ограничения паразитических нематод, таких как яванская галловая нематода, цистообразующая нематода и ранящие нематоды, в том числе нематод Heterodera spp., Meloidogyne spp. и Globodera spp.; в особенности, представителей цистообразующих нематод, в том числе без ограничения Heterodera glycines (соевую цистообразующую нематоду); Heterodera schachtii (свекловичную нематоду); Heterodera avenae (злаковую цистообразующую нематоду) и Globodera rostochiensis и Globodera pailida (картофельную нематоду). Ранящие нематоды включают без ограничения Pratylenchus spp. Грибковые вредители включают вызывающих листовую, желтую, полосчатую и стеблевую ржавчины.
Термин "белок, обеспечивающий устойчивость к гербицидам" или белок, полученный в результате экспрессии "молекулы нуклеиновой кислоты, кодирующей устойчивость к гербицидам", включает белки, которые на клеточном уровне придают способность переносить более высокую концентрацию гербицида, чем клетки, которые не экспрессируют белок, или переносить определенную концентрацию гербицида в течение более длительного периода времени, чем клетки, которые не экспрессируют белок. Свойства устойчивости к гербицидам могут быть введены в растения с генами, кодирующими устойчивость к гербицидам, которые действуют посредством ингибирования активности ацетолактат-синтазы (ALS), в особенности, гербицидам сульфонилмочевинного типа, с генами, кодирующими устойчивость к гербицидам, которые действуют посредством ингибирования активности глутамин-синтазы, таким как фосфинотрицин или баста (например, ген bar), глифосат (например, ген EPSP-синтазы и ген GAT) или другие подобные гены, известные в уровне техники.
Гены, которые улучшают урожайность культуры, включают гены карликовости, такие как Rhtl и Rht2 (Peng et al. (1999) Nature 400:256-261), и гены, которые повышают рост растений, такие как ген аммоний-индуцируемой глутамат-дегидрогеназы. Гены, улучшающие привлекательность культур, включают, например, гены, которые позволяют растениям иметь пониженное содержание насыщенных жиров, гены, которые повышают питательную ценность растений, и гены, которые повышают содержание белка в зерне. Генами, которые улучшают выносливость к засолению, являются гены, позволяющие повысить рост или обеспечивают возможность роста растения в среде с более высокой соленостью, чем естественная окружающая среда растения, в которое были введены ген(ы) выносливости к засолению.
Также предполагаются способы идентификации регуляторных элементов (например, промоторов, терминаторов и энхансеров). Под "регуляторным элементом" или "регуляторным участком" подразумевается часть нуклеиновой кислоты, находящаяся выше или ниже гена, которая может состоять из ДНК или РНК или ДНК и РНК, и которая участвует в экспрессии гена. Регуляторные элементы могут быть способны опосредовать органоспецифичность или контролировать активацию генов в зависимости от периода развития или временного периода и включают промоторные элементы, коровые промоторные элементы, элементы, индукция которых происходит в ответ на внешние стимулы, элементы, которые активируются конститутивно, терминаторы транскрипции, сигналы полиаденилирования и элементы, которые снижают или повышают активность промотора, такие как отрицательные регуляторные элементы или транскрипционные энхансеры, соответственно. Под "цис-действующей" подразумевается последовательность, которая является физически смежной с транскрибируемой последовательностью. Цис-действующие последовательности, как правило, взаимодействуют с белками или другими молекулами для осуществления (включения/выключения, регулирования, модуляции и т.д.) транскрипции. Под "транскрипционным энхансером" подразумевается последовательность нуклеиновой кислоты, которая, располагаясь поблизости к промотору и присутствуя в транскрипционной среде, способна поддерживать транскрипцию, обеспечивая повышенную транскрипционную активность по сравнению с активностью, получаемой в результате от промотора в отсутствие энхансера. Энхансеры могут функционировать выше, в пределах или ниже относительно гена даже на расстоянии 50 килобаз от сайта инициации транскрипции. Энхансеры также могут функционировать независимо от их ориентации. Под "терминатором транскрипции" подразумевают последовательность ДНК, которая включает последовательность пар нуклеотидных оснований, необходимую для снижения или устранения транскрипции.
Под "сигналом полиаденилирования" подразумевается последовательность, которая управляет терминацией транскрипции и трансляции.
Регуляторные последовательности для применения в растениях можно клонировать из сои путем конструирования одного или нескольких ПЦР праймеров на основе последовательности гена растения или регуляторного элемента. Способ может предусматривать конструирование по меньшей мере одного праймера, способного к гибридизации с нуклеотидной последовательностью из растения, применение праймера для амплификации ДНК из растения сои с получением амплифицированной ДНК и тестирование амплифицированной ДНК на предмет регуляторной активности последовательности. Под "регуляторной активностью последовательности" подразумевается способность к осуществлению транскрипции или трансляции гена. Она включает промоторную активность, активность транскрипционного энхансера, активность в отношении терминации транскрипции и активность в отношении полиаденилирования. Способы измерения или определения промоторной активности хорошо известны в уровне техники (см. раздел, озаглавленный “Оценка промоторной активности"). Способы измерения или тестирования энхансерной активности хорошо известны в уровне техники (см., например, патенты США №№ 6806064, 6818757 и 6784289). Способы измерения или тестирования терминаторной активности хорошо известны в уровне техники (см., например, патент США № 5093252). Способы измерения или тестирования активности в отношении полиаденилирования хорошо известны в уровне техники (см., например, патент США № 6632637).
В качестве альтернативы, регуляторные элементы можно идентифицировать и клонировать с помощью других подходов. Например, геномные или субгеномные библиотеки сои можно сконструировать с использованием векторов на основе BAC, космид или фага лямбда. Библиотеки можно подвергнуть зондированию с использованием промоторных элементов из растений. В качестве альтернативы, библиотеки можно подвергнуть зондированию с использованием кодирующих участков гена из растения. Полученные клоны можно секвенировать и определить цис-действующие элементы, окружающие кодирующие участки сои. В качестве альтернативы, фрагменты из кодирующих участков различных генов сои можно амплифицировать из геномной ДНК с помощью ПЦР с использованием праймеров, сконструированных из консервативных участков генов растений, таких как консервативные участки из маиса. Фрагменты кодирующих участков сои можно использовать для исследования геномных библиотек, как описано.
Цис-действующие элементы можно клонировать с помощью ПЦР с обратной транскрипцией. Последовательность кодирующих участков генов сои можно получить, как описано выше, затем сконструировать праймеры для ПЦР и использовать ПЦР с обратной транскрипцией для клонирования ДНК, фланкирующей кодирующие участки, с использованием методик, хорошо известных в уровне техники.
Антисмысловая последовательность
Гетерологичная нуклеотидная последовательность, которая функционально связана с промотором сои, раскрытым в данном документе, может представлять собой антисмысловую нуклеотидную последовательность для целевого гена. Под "антисмысловой нуклеотидной последовательностью" подразумевается последовательность, которая находится в обратной ориентации по отношению к нормальной ориентации 5'-3' данной нуклеотидной последовательности. Экспрессия антисмысловой последовательности ДНК в растительной клетке препятствует нормальной экспрессии целевого гена. Антисмысловая нуклеотидная последовательность кодирует РНК транскрипт, который является комплементарным и способным к гибридизации с эндогенными матричными РНК (мРНК), вырабатываемыми при транскрипции целевого гена. Таким образом, продуцирование нативного белка, кодируемого целевым геном, подавляется и достигается желаемый фенотипический ответ. Модификации с помощью антисмысловых последовательностей можно осуществить, поскольку последовательности гибридизуются с соответствующей мРНК и препятствуют ее экспрессии. Можно быть использовать антисмысловые конструкции, последовательность которых приблизительно на 70%, 80%, 85%, 90% или 95% идентична соответствующим антисмысловым последовательностям.
Более того, участки антисмысловых нуклеотидов можно использовать для нарушения экспрессии целевого гена. Как правило, можно использовать последовательности по меньшей мере из 50 смежных нуклеотидов, 100 смежных нуклеотидов, 200 смежных нуклеотидов или более. Таким образом, промоторные последовательности, раскрытые в данном документе, могут быть функционально связаны с антисмысловыми последовательностями ДНК с целью снижения или подавления экспрессии нативного белка в растении.
Экспрессионные кассеты и векторы трансформации растений
Трансформацию растительных клеток можно осуществлять одним из нескольких способов, известных в уровне техники. Под "растением" подразумеваются целые растения, органы растений (например, листья, стебли, корни и т.д.), семена, растительные клетки, побеги, зародыши и их потомство. Растительные клетки могут быть дифференцированными или недифференцированными (например, каллюсом, суспензионной культурой клеток, протопластами, клетками листа, клетками корня, клетками флоэмы, пыльцой). "Трансгенные растения", или "трансформированные растения", или "стабильно трансформированные" растения, или клетки, или ткани относятся к растениям, которые имеют экзогенные последовательности нуклеиновой кислоты или фрагменты ДНК, встроенные или интегрированные в растительную клетку. Под "стабильной трансформацией" подразумевается, что нуклеотидная конструкция, введенная в растение, интегрируется в геном растения и способна наследоваться его потомством.
Промоторная последовательность по настоящему изобретению может обеспечиваться в виде экспрессионной кассеты, что позволяет ей управлять экспрессией гетерологичной последовательности, представляющей интерес, в растительных клетках. Под "экспрессионной кассетой" подразумевается конструкция ДНК, которая способна приводить к экспрессии белка из открытой рамки считывания в клетке. Кассета будет включать в 5'-3' направлении транскрипции участок инициации транскрипции, включающий одну из промоторных нуклеотидных последовательностей, раскрытых в данном документе, или их вариантов или фрагментов, функционально-связанных с гетерологичной последовательностью, представляющей интерес, и участок терминации трансляции и транскрипции (т.е. участок терминации), функциональный в растениях. Кассета может дополнительно содержать по меньшей мере один дополнительный ген, который нужно котрансформировать в организм, такой как селектируемый маркерный ген. В качестве альтернативы, дополнительный ген(ы) можно обеспечить на нескольких экспрессионных кассетах. Такую экспрессионная кассета снабжена несколькими сайтами рестрикции для вставки гетерологичной последовательности, представляющей интерес, под транскрипционным контролем регуляторных участков.
Часто такие конструкции также будут содержать 5 'и 3' нетранслируемые участки. Такие конструкции могут также содержать транслируемую "сигнальную последовательность" или "лидерную последовательность" для облегчения котрансляционного или посттрансляционного транспорта пептида, представляющего интерес, к определенным внутриклеточным структурам, таким как хлоропласт (или другой пластид), эндоплазматический ретикулум или аппарат Гольджи, или для дальнейшей секреции. Например, ген может быть сконструирован таким образом, чтобы содержать сигнальный пептид для облегчения транспорта пептида в эндоплазматический ретикулум. Также может быть предпочтительным конструирование экспрессионной кассеты для растений, содержащей интрон так, чтобы процессинг мРНК интрона был необходим для экспрессии. Под "сигнальной последовательностью" подразумевают последовательность, о которой известно или предполагается, что она приводит к котрансляционному или посттрансляционному транспорту пептида через клеточную мембрану. У эукариот это обычно включает секрецию в аппарат Гольджи с получением в результате некоторого гликозилирования. Под "лидерной последовательностью" подразумевают любую последовательность, которая при трансляции приводит к образованию аминокислотной последовательности, достаточной для запуска котрансляционного транспорта пептидной цепи к внутриклеточной органелле. Таким образом, это понятие включает в себя направленный лидерными последовательностями транспорт и/или гликозилирование по прохождении в эндоплазматический ретикулум, прохождение в вакуоли, пластиды, в том числе хлоропласты, митохондрии и т.п.
Под "3' нетранслируемым участком" подразумевается нуклеотидная последовательность, расположенная ниже по отношению к кодирующей последовательности. Сигнальные последовательности полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные воздействовать на добавление трактов полиадениловой кислоты к 3' концу предшественника мРНК, представляют собой 3' нетранслируемые участки. Под "5' нетранслируемым участком" подразумевается нуклеотидная последовательность, расположенная выше по отношению к кодирующей последовательности. Другие нетранслируемые элементы, расположенные выше или ниже, включают энхансеры. Энхансеры представляют собой нуклеотидные последовательности посредством повышения экспрессии промоторного участка. Энхансеры хорошо известны в уровне техники и включают без ограничения энхансерный участок SV40 и энхансерный элемент 35S.
Участок терминации транскрипции может быть нативным по отношению к участку инициации транскрипции, содержащему промоторную нуклеотидную последовательность по настоящему изобретению, может быть нативным по отношению к последовательности ДНК, представляющей интерес, или может быть получен из другого источника. Подходящие участки терминации доступны из Ti-плазмиды A. tumefaciens, такие как участки терминации транскрипции генов октопин-синтазы и нопалин-синтазы. См. также, Guerineau et al. (1991) Mol. Gen. Genet. 262:141-144; Proudfoot (1991) Cell 64:671- 674; Sanfacon et al. (1991) Genes Dev. 5:141-149; Mogen et al. (1990) Plant Cell 2:1261-1272; Munroe et al. (1990) Gene 91:151-158; Balias et al. (1989) Nucleic Acids Res. 17:7891-7903; и Joshi et al. (1987) Nucleic Acid Res. 15:9627-9639.
Если это целесообразно, ген(ы), представляющий(ие) интерес, можно оптимизировать для повышенной экспрессии в трансформированной клетке-хозяине. То есть гены можно синтезировать с использованием предпочтительных для клетки-хозяина кодонов для улучшенной экспрессии, или их можно синтезировать с использованием кодонов в соответствии с предпочтительной для хозяина частотой использования кодонов. Как правило, содержание GC в составе гена будет повышено. См., например, Campbell and Gowri (1990) Plant Physiol. 92:1-11, для обсуждения предпочтительного для хозяина использования кодонов. Способы синтеза предпочтительных для растений генов известны в уровне техники. См., например, патенты США №№ 6320100; 6075185; 5380831 и 5436391; опубликованные заявки на патент США № 20040005600 и № 20010003849; и Murray et al. (1989) Nucleic Acids Res. 17:477-498, включенные в данный документ посредством ссылки.
В одном варианте осуществления настоящего изобретения нуклеиновые кислоты, представляющие интерес, направляются в хлоропласт для экспрессии. Таким образом, в том случае, если нуклеиновая кислота, представляющая интерес, не вводится непосредственно в хлоропласт, экспрессионная кассета будет дополнительно содержать нуклеиновую кислоту, кодирующую транзитный пептид или сигнальную последовательность для направления продукта гена, представляющего интерес, в хлоропласты. Такие транзитные пептиды известны в уровне техники. См., например, Von Heijne et al. (1991) Plant Mol. Biol. Rep. 9:104-126; Clark et al. (1989) J. Biol. Chem. 264:17544-17550; Della-Cioppa et al. (1987) Plant Physiol. 84:965-968; Romer et al. (1993) Biochem. Biophys. Res. Commun. 196:1414-1421; и Shah et al. (1986) Science 233:478-481.
Представляющие интерес нуклеиновые кислоты, которые нужно направить в хлоропласт, могут быть оптимизированы для экспрессии в хлоропластах с учетом различий в использовании кодонов между ядром растения и этой органеллой. В этом случае представляющие нуклеиновые кислоты могут быть синтезированы с использованием кодонов, предпочтительных для хлоропластов. См., например, патент США № 5380831, включенный в данный документ посредством ссылки.
Как правило, эта "экспрессионная кассета для растений" будет вставлена в "вектор для трансформации растений". Под "вектором для трансформации" подразумевается молекула ДНК, которая необходима для эффективной трансформации клетки. Такая молекула может состоять из одной или нескольких экспрессионных кассет и может быть организована в виде более чем одной "векторной" молекулы ДНК. Например, бинарные векторы представляют собой векторы для трансформации растений, в которых используются два несмежных ДНК вектора для кодирования всех необходимых цис- и транс-действующих функций для трансформации растительных клеток (Hellens and Mullineaux (2000) Trends in Plant Science 5:446-451). "Вектор" относится к конструкции нуклеиновой кислоты, предназначенной для передачи между различными клетками-хозяевами. "Экспрессионный вектор" относится к вектору, который характеризуется способностью к встраиванию, интеграции и экспрессии гетерологичных последовательностей или фрагментов ДНК в чужеродной клетке. Под "введением" подразумевается предоставление организму, который нужно трансформировать, нуклеотидной конструкции таким образом, чтобы конструкция получала доступ к внутреннему пространству по меньшей мере одной клетки организма.
Этот вектор для трансформации растений может состоять из одного или нескольких ДНК векторов, необходимых для достижения трансформации растения. Например, обычной практикой в данной области техники является применение векторов для трансформации растений, которые состоят из более чем одного смежного сегмента ДНК. В данной области техники эти векторы часто называют "бинарными векторами". Бинарные векторы, а также векторы с хелперными плазмидами наиболее часто используется для Agrobacterium-опосредованной трансформации, где размер и сложность сегментов ДНК, необходимых для достижения эффективной трансформации, являются довольно большими, и выгодно разделить функции между отдельными молекулами ДНК. Бинарные векторы обычно содержат плазмидный вектор, содержащий цис-действующие последовательности, необходимые для переноса Т-ДНК (например, левую граничную область и правую граничную область), селектируемый маркер, который сконструирован с возможностью его экспрессии в растительной клетке, и "представляющий интерес ген" (ген, сконструированный с возможностью экспрессии в растительной клетке, из которой желательно получение трансгенных растений). Также на данном плазмидном векторе присутствуют последовательности, необходимые для репликации у бактерий.
Цис-действующие последовательности расположены в таким образом, чтобы способствовать эффективному переносу в растительные клетки и экспрессии в них. Например, селектируемый маркерный ген и ген, представляющий интерес, расположены между левой и правой граничными областями. Часто, второй плазмидный вектор содержит транс-действующие факторы, которые опосредуют перенос Т-ДНК из Agrobacterium в растительные клетки. Эта плазмида часто содержит функциональные элементы вирулентности (Vir гены), которые обеспечивают возможность инфицирования растительных клеток Agrobacterium и переноса ДНК путем расщепления последовательностей граничных областей и vir-опосредованного переноса ДНК, как этот процесс понимается в данной области техники (Hellens and Mullineaux (2000) Trends in Plant Science, 5:446-451). Несколько типов штаммов Agrobacterium (например, LBA4404, GV3101, EHA101, EHA105 и т.д.) можно использовать для трансформации растений. Второй плазмидный вектор не является необходимым для трансформации растений другими способами, такими как бомбардировка микрочастицами, микроинъекция, электропорация, трансформация под воздействием полиэтиленгликоля и т.д.
Трансформация растений
Способы по настоящему изобретению предусматривают введение нуклеотидной конструкции в растение. Под "введением" подразумевается предоставление растению нуклеотидной конструкции таким образом, что конструкция получает доступ к внутреннему пространству клетки растения. Способы по настоящему изобретению не требуют использования конкретного способа введения нуклеотидной конструкции в растение, главное, чтобы нуклеотидная конструкция получала доступ к внутреннему пространству по меньшей мере одной клетки растения. Способы введения нуклеотидных конструкций в растения известны в уровне техники, в том числе без ограничения способы стабильной трансформации, способы временной трансформации и вирус-опосредованные способы.
Под "растением" подразумеваются целые растения, органы растений (например, листья, стебли, корни и т.д.), семена, растительные клетки, побеги, зародыши и их потомство. Растительные клетки могут быть дифференцированными или недифференцированными (например, каллюс, суспензионная культура клеток, протопласты, клетки листьев, клетки корней, клетки флоэмы, пыльца).
"Трансгенные растения", или "трансформированные растения", или "стабильно трансформированные" растения, или клетки, или ткани относятся к растениям, которые имеют встроенные или интегрированные в клетку растения экзогенные последовательности нуклеиновой кислоты или фрагменты ДНК. Эти последовательности нуклеиновой кислоты включают последовательности, которые являются экзогенными или не присутствуют в нетрансформированной растительной клетке, а также последовательности, которые могут быть эндогенными или присутствовать в нетрансформированной растительной клетке. "Гетерологичный", как правило, относится к последовательностям нуклеиновых кислот, которые не являются эндогенными для клетки или части нативного генома, в котором они находятся, а были добавлены в клетку посредством инфекции, трансфекции, микроинъекции, электропорации, бомбардировки микрочастицами или т.п.
Трансгенные растения по настоящему изобретению экспрессируют одну или несколько новых последовательностей токсина, описанных в данном документе. В различных вариантах осуществления настоящего изобретения трансгенное растение дополнительно содержит один или несколько дополнительных генов для устойчивости к насекомым (например, Cryl, такие как члены семейств CrylA, CrylB, CrylC, CrylD, CrylE и CrylF; Cry2, такие как члены семейства Cry2A; Cry9, такие как члены семейств Cry9A, Cry9B, Cry9C, Cry9D, Cry9E и Cry9F; и т.д.). Специалисту в данной области техники будет понятно, что трансгенное растение может содержать любой ген, придающий агрономическое свойство, представляющее интерес. В различных вариантах осуществления настоящего изобретения промотор по настоящему изобретению можно применять для управления экспрессией одного или нескольких генов, описанных в патентных публикациях, перечисленных в Таблице 1, содержание которых включено в данный документ в полном объеме посредством ссылки.
Трансформацию растительных клеток можно осуществлять с помощью одного из нескольких способов, известных в уровне техники. Пестицидный ген по настоящему изобретению можно модифицировать для получения или усиления экспрессии в растительных клетках. Как правило, конструкция, которая экспрессирует такой белок, будет содержать промотор для управления транскрипцией гена, а также 3-нетранслируемый участок для обеспечения возможности терминации транскрипции и полиаденилирования. Строение таких конструкций хорошо известно в уровне техники. В некоторых случаях полезным может быть конструирование гена таким образом, чтобы полученный пептид секретировался или каким-либо иным образом направлялся в растительной клетке. Например, ген можно сконструировать таким образом, чтобы он содержал сигнальный пептид для обеспечения транспорта пептида в эндоплазматический ретикулум. Предпочтительным также может быть конструирование экспрессионной кассеты для растений, содержащей интрон, так, чтобы для экспрессии требовался процессинг мРНК интрона.
Как правило, эта "экспрессионная кассета" будет вставлена в "вектор для трансформации растений". Этот вектор для трансформации растений может состоять из одного или нескольких ДНК векторов, необходимых для достижения трансформации растения. Например, обычной практикой в данной области техники является применение векторов для трансформации растений, которые состоят из более чем одного смежного сегмента ДНК. Эти векторы часто упоминаются в данной области техники как "бинарные векторы". Бинарные векторы, а также векторы с хелперными плазмидами наиболее часто используется для Agrobacterium-опосредованной трансформации, где размер и сложность сегментов ДНК, необходимых для достижения эффективной трансформации, являются довольно большими, и выгодно разделить функции между отдельными молекулами ДНК. Бинарные векторы обычно содержат плазмидный вектор, содержащий цис-действующие последовательности, необходимые для переноса Т-ДНК (например, левую и правую граничную область), селектируемый маркер, который разработан с возможностью его экспрессии в растительной клетке, и "представляющий интерес ген" (ген, сконструированный с возможностью экспрессии в растительной клетке, из которой желательно получение трансгенных растений). Также на данном плазмидном векторе присутствуют последовательности, необходимые для репликации у бактерий. Цис-действующие последовательности расположены в таким образом, чтобы обеспечивать возможность эффективного переноса в растительные клетки и экспрессии в них. Например, селектируемый маркерный ген и пестицидный ген расположены между левой и правой граничными областями. Часто, второй плазмидный вектор содержит транс-действующие факторы, которые опосредуют перенос Т-ДНК из Agrobacterium в растительные клетки. Эта плазмида часто содержит функциональные элементы вирулентности (Vir гены), обеспечивают возможность инфицирования растительных клеток Agrobacterium и переноса ДНК путем расщепления последовательностей граничных областей и vir-опосредованного переноса ДНК, как этот процесс понимается в данной области техники (Hellens and Mullineaux (2000) Trends in Plant Science, 5:446-451). Несколько типов штаммов Agrobacterium (например, LBA4404, GV3101, EHA101, EHA105 и т.д.) могут быть использованы для трансформации растений. Второй плазмидный вектор не является необходимым для трансформации растений с помощью других способов, таких как бомбардировка микрочастицами, микроинъекция, электропорация, трансформация под воздействием полиэтиленгликоля и т.д.
В целом способы трансформации растений включают перенос гетерологичной ДНК в растительные клетки-мишени (например, незрелые или зрелые зародыши, суспензионные культуры, недифференцированный каллюс, протопласты и т.д.) с последующим применением максимального порогового уровня соответствующего селектирующего средства (в зависимости от селектируемого маркерного гена) с целью выделения трансформированных растительных клеток из группы нетрансформированной клеточной массы. Экспланты, как правило, переносят на свежую порцию той же питательной среды и культивируют с применением стандартных подходов. Впоследствии, после помещения в регенерационную среду, дополненную максимальным пороговым уровнем селектирующего средства, трансформированные клетки дифференцируются в побеги. Затем побеги переносят на селективную среду для укоренения с целью выделения укорененных побегов или проростков. Затем трансгенное растение вырастает в зрелое растение и производит фертильные семена (например, Hiei et al. (1994) The Plant Journal 6:271-282; Ishida et al. (1996) Nature Biotechnology 14:745-750). Экспланты, как правило, переносят на свежую порцию той же питательной среды и культивируют с применением стандартных подходов. Общее описание методик и способов получения трансгенных растений представлено в Ayres and Park (1994) Critical Reviews in Plant Science 13:219-239; и Bommineni and Jauhar (1997) Maydica 42:107-120. Поскольку трансформированный материал содержит много клеток, как трансформированные, так и нетрансформированные клетки присутствуют в любой части подвергнутого воздействию целевого каллюса, или ткани, или группы клеток. Возможность уничтожения нетрансформированных клеток и обеспечения возможности пролиферации трансформированных клеток приводит к получению трансформированных растительных культур. Часто возможность удаления нетрансформированных клеток является ограничением для быстрого выделения трансформированных растительных клеток и успешного получения трансгенных растений.
Протоколы трансформации, как и протоколы для введения нуклеотидных последовательностей в растения, могут изменяться в зависимости от типа растения или растительной клетки, т.е. однодольных или двудольных, выбранных для трансформации. Получение трансгенных растений можно осуществлять с помощью одного из нескольких способов, в том числе без ограничения с помощью микроинъекции, электропорации, прямого переноса генов, введения гетерологичной ДНК в растительные клетки при помощи Agrobacterium (Agrobacterium-опосредованной трансформации), бомбардировки растительных клеток гетерологичной чужеродной ДНК, прикрепленной на частицы, баллистического ускорения частиц, трансформации аэрозольным потоком (опубликованная заявка на патент США № 20010026941; патент США № 4945050; публикация международной заявки № WO 91/00915; опубликованная заявка на патент США № 2002015066), трансформации с использованием Lec1, а также различных других способов переноса ДНК, не опосредованного использованием частиц.
Способы трансформации хлоропластов хорошо известны в уровне техники. См., например, Svab et al. (1990) Proc. Natl. Acad. Sci. USA 87:8526-8530; Svab and Maliga (1993) Proc. Natl. Acad. Sci. USA 90:913-917; Svab and Maliga (1993) EMBO J. 12:601-606. Способ основан на доставке с использованием пушки частиц с ДНК, содержащей селективный маркер и целенаправленной доставке ДНК в геном пластиды посредством гомологичной рекомбинации. Кроме того, трансформацию пластид можно осуществлять путем трансактивации молчащего пластидного трансгена с помощью тканеспецифичной экспрессии кодируемой ядерным геномом и направляемой в пластиды РНК-полимеразы. О такой системе сообщалось в McBride et al. (1994) Proc. Natl. Acad. Sci. USA 91:7301-7305.
После интеграции гетерологичной чужеродной ДНК в клетки растений применяют максимально допустимый пороговый уровень соответствующего селектирующего средства в составе среды для уничтожения нетрансформированных клеток, а также отделения и обеспечения пролиферации предположительно трансформированных клеток, которые выживают в результате данной селекционной обработки, путем регулярного переноса на свежую среду. Путем непрерывного пассирования и воздействия соответствующей селекции идентифицируют и обеспечивают пролиферацию клеток, которые трансформированы плазмидным вектором. Для подтверждения наличия интегрированного гетерологичного гена, представляющего интерес, в геном трансгенного растения можно применять молекулярные и биохимические способы.
Клетки, которые были трансформированы, можно вырастить в растения в соответствии с традиционными способами. См., например, McCormick et al. (1986) Plant Cell Reports 5:81-84. Эти растения можно затем вырастить и переопылить либо с той же трансформированной линией, либо с другими линиями, и идентифицировать полученный в результате гибрид с конститутивной экспрессией необходимой фенотипической характеристики. Два или больше поколений можно вырастить для подтверждения того, что экспрессия необходимой фенотипической характеристики стабильно сохраняется и наследуется, а затем семена можно собрать для подтверждения того, что экспрессия необходимой фенотипической характеристики была достигнута. В этом случае настоящее изобретение предполагает получение трансформированных семян (также упоминаемых как "трансгенные семена"), содержащих нуклеотидную конструкцию по настоящему изобретению, например, экспрессионную кассету по настоящему изобретению, стабильно встроенную в их геном.
Растения
Настоящее изобретение можно применять для трансформации любых видов растений, в том числе без ограничения двудольных и однодольных. Примеры растений, представляющих интерес, включают без ограничения кукурузу (маис), сорго, пшеницу, подсолнечник, томат, крестоцветные, перцы, картофель, хлопчатник, рис, сою, сахарную свеклу, сахарный тростник, табак, ячмень, масличный рапс, Brassica sp., люцерну, рожь, просо, сафлор, арахис, сладкий картофель, маниок, кофе, кокос, ананас, цитрусовые деревья, какао, чай, банан, авокадо, инжир, гуаву, манго, оливу, папайю, кешью, макадамию, миндаль, овес, овощи, декоративные и хвойные растения.
Овощи включают без ограничения томаты, латук, зеленые бобы, лимскую фасоль, горох и членов рода Curcumis, таких как огурец, дыня и мускусная дыня. Декоративные растения включают без ограничения азалию, гортензию, гибискус, розы, тюльпаны, нарциссы, петунии, гвоздики, пуансеттию и хризантему. Предпочтительно, растения по настоящему изобретению являются сельскохозяйственными культурами (например, маис, сорго, пшеница, подсолнечник, томат, крестоцветные, перцы, картофель, хлопчатник, рис, соя, сахарная свекла, сахарный тростник, табак, ячмень, масличный рапс и т.д.).
Данное изобретение является особенно подходящим для любого члена класса однодольных растений, в том числе без ограничения для маиса, риса, ячменя, овса, пшеницы, сорго, ржи, сахарного тростника, ананаса, ямса, лука, банана, кокоса и финика.
Оценка трансформации растений
После введения гетерологичной чужеродной ДНК в растительные клетки трансформацию или интеграцию гетерологичной ДНК в геном растения подтверждают с помощью различных способов, таких как анализ нуклеиновых кислот или белков и метаболитов, связанных с интегрированной ДНК.
ПЦР-анализ является быстрым способом для скрининга трансформированных клеток, тканей или побегов на наличие встроенной ДНК на ранней стадии до пересадки в почву (Sambrook and Russell, 2001. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). ПЦР проводят с использованием специфичных олигонуклеотидных праймеров для гена, представляющего интерес, или фонового элемента из вектора Agrobacterium (без учета гена, представляющего интерес) и т.д.
Трансформацию растения можно подтвердить с помощью анализом геномной ДНК методом Саузерн-блоттинга (Sambrook and Russell, 2001, выше). В целом, общую ДНК экстрагируют из трансформанта, расщепляют подходящими ферментами рестрикции, фракционируют в агарозном геле и переносят на нитроцеллюлозную или нейлоновую мембрану. Затем проводят зондирование мембраны или "блота", например, целевым фрагментом ДНК, меченным радиоактивным 32Р, для подтверждения интеграции введенной ДНК в геном растения согласно стандартным методикам (Sambrook and Russell, 2001, выше).
В анализе методом Нозерн-блоттинга РНК выделяют из конкретных тканей трансформанта, фракционируют в агарозном геле с формальдегидом, переносят промоканием на нейлоновый фильтр согласно стандартным методикам, которые обычно используются в данной области техники (Sambrook and Russell, 2001, выше). Экспрессию РНК, кодируемой гетерологичным геном, функционально связанным с промотором TripPro5, затем проверяют при помощи гибридизации фильтра с радиоактивным зондом, полученным из гетерологичного гена, с помощью способов, известных в уровне техники (Sambrook and Russell, 2001, выше).
Оценка промоторной активности
Доступны многочисленные способы оценки промоторной активности в растениях.
Функционирование промотора в ходе экспрессии гена, представляющего интерес, под его регуляторным контролем можно исследовать либо на стадии транскрипции, либо трансляции. На стадии транскрипции уровни РНК можно исследовать при помощи гибридизации ДНК-РНК (т.е. анализа методом Нозерн-блоттинг), конкурентной ПЦР с обратной транскрипцией и анализа с защитой от РНКазы.
На стадии транскрипции промоторную активность можно определить с использованием специфических функциональных анализов для синтезированного белка (например, ферментативной активности или с помощью иммунологического анализа белка). Например, активность репортерного гена, как например, β-глюкуронидазная активность, люциферазная активность или флуоресценция GFP, можно отслеживать в разные моменты времени после трансформации. Активность репортерного гена можно отслеживать по ферментативной активности, путем окрашивания клеток или тканей субстратом для фермента, кодируемого репортерным геном или путем прямой визуализации при соответствующей длине волны света (см., например, Wang et al. (2000) Plant Science 156:201-211). Вестерн-блоттинг можно проводить на трансгенных растениях для подтверждения наличия белка, кодируемого представляющим интерес геном, функционально связанным с промотором TripPro5, в соответствии со стандартными процедурами (Sambrook and Russell, 2001, выше) с использованием антител, которые связываются с одним или несколькими эпитопами, присутствующими на белке. Можно определить промоторные последовательности полной длины, делеции и мутации в промоторной последовательности, а уровни их экспрессии можно сравнить. См., например, патент США № 6072050; и Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York), включенные в данные документ посредством ссылки.
Следующие примеры предлагаются для иллюстрации, а не в качестве ограничения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Пример 1. Идентификация конститутивных промоторов сои
Общедоступные базы данных транскриптома сои использовали для идентификации генов, которые экспрессируются на высоком уровне в различных тканях (листья, стручок, цветы, корни и т.д.). Промоторные участки (выше относительно первого ATG) этих генов амплифицировали при помощи ПЦР из геномной ДНК сои (Jack) и сцепляли с кодирующим участком гена люциферазы и терминатором Pinll. Эти промотор-содержащие векторы трансформировали в Agrobacterium. Трансформированные Agrobacterium использовали для пропитывания листовых дисков молодой сои или фасоли. После 2 дней инкубации при 25°C при 16-часовом освещении листовые диски гомогенизировали в PBS буфере для экстракции белка. Затем, растворимые белки анализировали на предмет люциферазной активности с помощью системы анализа люциферазной активности STEADY-GLO® от Promega. Люциферазная активность в виде среднего значения от трех независимых наборов пропитанных листовых дисков сои для каждого вектора показана на Фигуре 1. Pbdc6 и Pbdc7 показали активность, сравнимую с Pubi3 из Arabidopsis. Pbdc6 (SEQ ID NO: 1) получали из Glyma03g34310, который кодирует гамма-внутренний белок тонопласта. Pbdc7 (SEQ ID NO: 2) получали из Glyma23g42220, который кодирует внутренний белок плазматической мембраны.
Пример 2. Анализ in planta
Последовательности ДНК, несущие промоторы Pbdc6 и Pbdc7, соответственно, клонировали в pSZ8133 для сцепления этих промоторов с grg23Ace5. Полученные бинарные векторы, pSZ8806 и pSZ8807, трансформировали в Agrobacterium LBA4404 и использовали для получения трансгенных растений сои. Приблизительно 150 трансгенных трансформантов для каждого вектора анализировали с использованием опрыскивания 4X глифосатом. Устойчивость к 4X глифосату оценивали через неделю после опрыскивания (таблица 2, 0 означает отсутствие устойчивости, и 4 представляет наиболее сильную устойчивость). UBQ3 использовали в качестве контроля. Pbdc6 и Pbdc7 вновь показали уровень устойчивости, сравнимый с Pubi3At.
Устойчивость к 4x глифосату (представлена в процентах растений, оцененных в каждой из категорий)
Все публикации и заявки на патенты, упомянутые в описании, указывают на уровень квалификации специалистов в данной области техники, к которой относится настоящее изобретение. Все публикации и заявки на патенты включены в данный документ посредством ссылки в той же степени, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно указана как включенная посредством ссылки.
Несмотря на то, что настоящее изобретение было описано в некоторых деталях для иллюстрации и примера в целях ясности понимания, будет очевидным, что определенные изменения и модификации могут быть осуществлены в пределах объема приложенной формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Байер Кроп-Сайенс ЭлПи
Чжан, Шижун
<120> КОНСТИТУТИВНЫЕ ПРОМОТОРЫ СОИ
<130> 2912939-20179WO01
<160> 2
<170> PatentIn версия 3.5
<210> 1
<211> 1230
<212> ДНК
<213> Glycine max
<400> 1
ctcgaaacag aatatagcag tgattccagg tgcacggtgc cacctgtttg ctaaaaaaca 60
aaaataattg agggggttaa ctaagctaat ccaaaccaga gatcagagcg ggagattaat 120
tttaataatt ctcttagtct cttcattaag atcctttttt tcatatcaag ttaaacttta 180
tgcaaaatat catattttca tttcagaaaa acaaaaaagc tactgtacat acaacattaa 240
gtttaaaata aacaagtaca gtttacccac gtgtcagtcg cggtcttgta ataatacctg 300
cttttttccg gtgggtcacc agagagtcac agtgcttatc cccttatctc attcattata 360
ttatgatgaa ttatgccctt ataattatag attaactact actttgcagt tataaactta 420
aaattcacaa ttctctctat tatttctttg cacacgcgat ataatataaa actctagagc 480
tatcgtcatt cacaacttta aaaaaattaa aaaagttaat tatattagag tgcaatagct 540
cactctcttt ctctcacgga accatccaga tatacaaaaa agactagaca atgtccactt 600
atcagcgccc tacatggctc ttatccatct cggacacgtg tcagtcctgg gacggctctg 660
gcccaaacga ttcgctgtct taagagcaac caacagtgat gtggggatat actccttgtt 720
ttttttttct tcaaataatt atgtataatg tatttataat ataatttttt tacttttaaa 780
ataattattc tgtatagttt cgatgataat ttatgctaaa tgtttatgta aaattattta 840
cactttttaa tacatagaca ttaacttttt ttctctcata aatgtttaaa ataaaaggaa 900
aaacaattaa taatttgaaa tgaagaacct gtcccaggct gtcaccaaat aggattcttt 960
cttttccggc ttggtgtttt tgaaaaactt gattcctgac atttttggaa gcattgcgca 1020
tattttccaa ataggaaaat aaaaataacc gggaaccggg tacagccggt ggaatcctag 1080
gcaggctcgt cttcaatcac ggtatataag aagctacgaa gcgaaaacga atagcatagc 1140
gaaacagaga gtggtttcag tgagtgatcg gtgttcactt gatccttcat aagtcgtcat 1200
catcatcatc atcatcaccg ttgattgaag 1230
<210> 2
<211> 1688
<212> ДНК
<213> Glycine max
<400> 2
ctaacgtttt tggctcacaa ctcaaactga tcaagaacct ttcattttat taacgttatc 60
cttacaagtc actcagacta atcaaggcac taaactaatg catattaggt aatgcaaata 120
aataatgacg ctcccatgaa tattcaaatg gtttcttttg ctttttgctt aacgactttt 180
gtatctctac gaattacttg agaaaaagct gctattatta ttatccaact atataaacaa 240
atgaaagcta cagttaagga catggcctat taacaatata tgtagacttg atcattgtct 300
catccacgag atagaaacaa aatatataaa agggctcatt atgcttattt agttcaagga 360
agagaggaaa atgagttatg catgtagcca gaatgaacat ttgatcatgg acgtgagata 420
agttaatcgc tgacagccat gtgccgacca tgtttttata aatgaaaaga aagaaagaaa 480
tgttcgtata taataattta cgggcacaag aaccttgtta ataattatca ttatcttttt 540
tttttttctg aaaaccttgt ttctctaatc attgatgtat atttgtaatc tctctccaac 600
tccttaccat gtagtgagga gtgaatttca tattaacatt ggtcattaca atattatcaa 660
ctttcgcgct aaatcagaat atatataaac tttcgcgcta aatattctta aaatattggt 720
attttggtag gtaaagattt acaatcacga ataagtaata aagaattttt catacgcatt 780
caatgattcc gaccatgtgt tatttgtttt gaaataccta tatgagagac tgagagcatc 840
ttgttattta tgactgctta ttaattttcc cttcatgcct tatctaatta gtttaaatat 900
attatttctc cttgtataaa aaaaaattat gatttctcca accatacata ttagagaata 960
acttgaaatt atattcaacg tattaattgc attaccttta acgtgccaaa ataataaata 1020
aaactaaaaa ctactacaat cataaatcgc gtgtggttga attgagacaa attctattct 1080
aaaaaagaaa aacattaaca aaaagagaaa gaaaaaaaaa attgttgaca cctgacagcg 1140
gtaacaggga agtagcggta ggagattggc gtgtcggttt ccaactctgg aatccaacgt 1200
gccaaactga gaatgcagga gaaagagaca cgtgtccaat tgcaggcgcg agttcaacgt 1260
gacaattcga aagccttgac aatcgcaccg cccagcatcg aacgcagaca aggaccacgt 1320
ggaattcggt ccctgtatcc gtcaaaacgt tttttaccct cttcttcttc agttcttcca 1380
tttttatttt tttttcaaac cacagtaatc cacgttccag tgctgcgcgg aacatggtcg 1440
gtctttctag gagtggttgg aatcccgcca gctaggacaa accccatcaa tcattggtcc 1500
ccatcaaaca aaaacatttt taaaaattca acatattacg ccacgggacc cacctcccac 1560
cacccctcac cctcacttct attaactcaa acctattccg gttataaatc cgcaaccctc 1620
gttcttacta actcactcac tcacaactca gtgaaagaga atccgaggcg aagagaagag 1680
aaaaatta 1688
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕГУЛЯТОРНЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УСИЛЕНИЯ СЕМЯ-СПЕЦИФИЧНОЙ И/ИЛИ СЕМЯ-ПРЕДПОЧТИТЕЛЬНОЙ ГЕННОЙ ЭКСПРЕССИИ В РАСТЕНИЯХ | 2010 |
|
RU2692924C2 |
| РЕГУЛЯТОРНЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УСИЛЕНИЯ СЕМЯ-СПЕЦИФИЧНОЙ И/ИЛИ СЕМЯ-ПРЕДПОЧТИТЕЛЬНОЙ ГЕННОЙ ЭКСПРЕССИИ В РАСТЕНИЯХ | 2010 |
|
RU2559534C2 |
| СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV | 2012 |
|
RU2627595C2 |
| АНТИГЕННЫЕ ПЕПТИДЫ NEISSERIA | 2000 |
|
RU2281956C2 |
| ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ХИМЕРНЫЙ ГЕН, РЕКОМБИНАНТНЫЙ ВЕКТОР, ШТАММ БАКТЕРИЙ, ТОКСИН, ИНСЕКТИЦИДНАЯ КОМПОЗИЦИЯ, СПОСОБ ПОЛУЧЕНИЯ ТОКСИНА, СПОСОБ ПОЛУЧЕНИЯ УСТОЙЧИВОГО К НАСЕКОМЫМ РАСТЕНИЯ И СПОСОБ БОРЬБЫ С НАСЕКОМЫМИ | 1999 |
|
RU2240003C2 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |
| ЭНХАНСЕРНЫЕ ЭЛЕМЕНТЫ CPMV | 2015 |
|
RU2699982C2 |
| ГЕН АХМ1554, КОДИРУЮЩИЙ ДЕЛЬТА-ЭНДОТОКСИН, И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2016 |
|
RU2755282C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2774709C2 |
| НОВЫЕ ГЕНЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2000 |
|
RU2241749C2 |
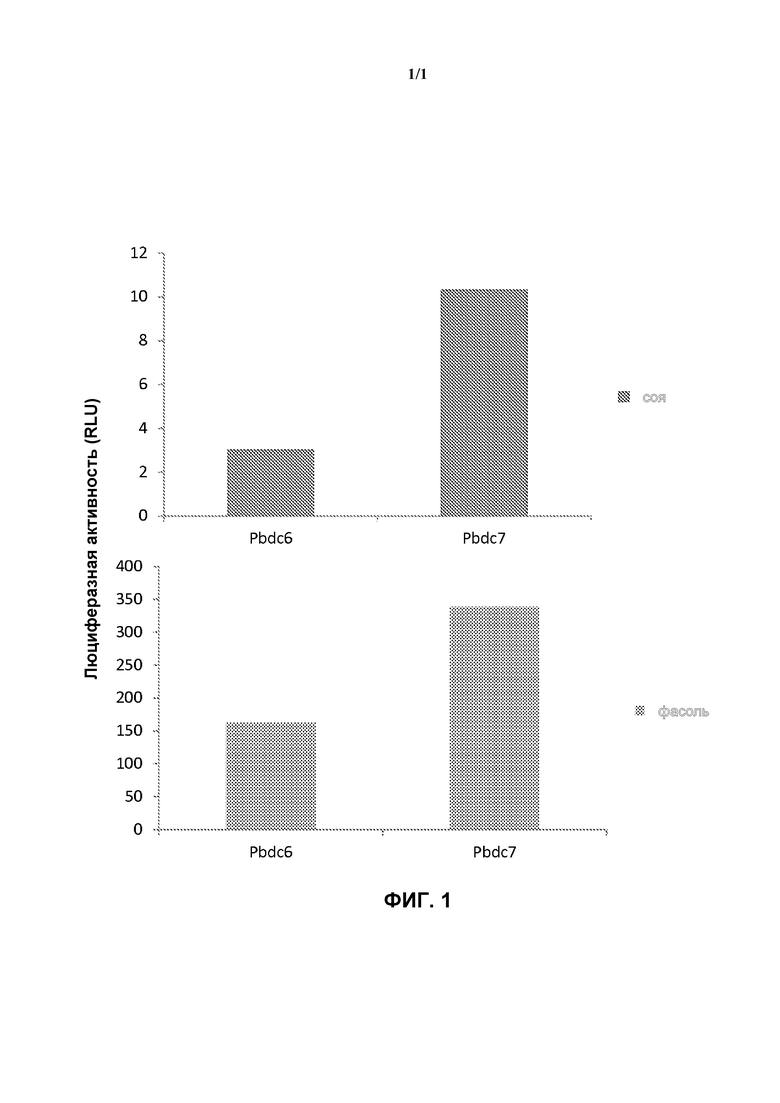
Изобретение относится к области биотехнологии. Предложена экспрессионная кассета, содержащая промоторную нуклеотидную последовательность гена, кодирующего внутренний белок плазматической мембраны сои, функционально связанную с гетерологичной нуклеиновой кислотой. Также представлены трансгенные растения и растительные клетки, содержащие в своем геноме стабильно встроенную экспрессионную кассету по изобретению. Кроме того, изобретение относится к способам экспрессии гетерологичной нуклеотидной последовательности в растении с помощью промоторных последовательностей, раскрытых в данном документе. Способы предусматривают стабильное встраивание в геном растительной клетки гетерологичной нуклеотидной последовательности, функционально связанной с промотором по настоящему изобретению, и регенерацию стабильно трансформированного растения, которое экспрессирует нуклеотидную последовательность. Изобретение обеспечивает повышенный уровень экспрессии генов в трансгенных растениях. 12 н. и 19 з.п. ф-лы, 1 ил., 2 табл., 2 пр. 
1. Экспрессионная кассета, содержащая молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, функционально связанную с гетерологичной нуклеиновой кислотой, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, изложенной в SEQ ID NO: 2; и
(b) нуклеотидной последовательности, которая по меньшей мере на 99% идентична последовательности, изложенной в SEQ ID NO: 2, где указанная последовательность инициирует транскрипцию в растительной клетке.
2. Экспрессионная кассета по п. 1, где указанная нуклеотидная последовательность функционально связана с гетерологичной нуклеотидной последовательностью, представляющей интерес.
3. Растительная клетка для придания растению устойчивости к вредителям, предназначенная для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, содержащая в своем геноме стабильно встроенную экспрессионную кассету по п. 2, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к вредителям.
4. Растительная клетка по п. 3, где указанная растительная клетка происходит из двудольного растения.
5. Растительная клетка по п. 4, где указанное двудольное растение представляет собой сою.
6. Растение, устойчивое к вредителям растения, содержащее в своем геноме стабильно встроенную экспрессионную кассету по п. 1, предназначенное для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, где указанная нуклеотидная последовательность функционально связана с гетерологичной нуклеиновой кислотой, представляющей интерес, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к вредителям.
7. Растение по п. 6, где указанное растение представляет собой двудольное растение.
8. Растение по п. 7, где указанное двудольное представляет собой сою.
9. Трансгенное семя, устойчивое к вредителям растения, предназначенное для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, содержащее экспрессионную кассету по п. 1, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к вредителям.
10. Способ экспрессии нуклеотидной последовательности в растении, причем указанный способ предусматривает введение в растительную клетку экспрессионной кассеты, содержащей промотор, функционально связанный с гетерологичной нуклеиновой кислотой, представляющей интерес, где указанный промотор содержит нуклеотидную последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, изложенной в SEQ ID NO: 2; и
(b) нуклеотидной последовательности, которая по меньшей мере на 99% идентична последовательности, изложенной в SEQ ID NO: 2, где указанная последовательность инициирует транскрипцию в растительной клетке; и
регенерацию трансформированного растения из указанной растительной клетки, где указанное растение содержит в своем геноме стабильно встроенную указанную экспрессионную кассету.
11. Способ по п. 10, где указанное растение представляет собой однодольное растение.
12. Способ по п. 10, где указанное растение представляет собой двудольное растение.
13. Способ по п. 11, где указанное однодольное представляет собой маис.
14. Способ по п. 10, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к вредителям.
15. Способ экспрессии нуклеотидной последовательности в растительной клетке, причем указанный способ предусматривает введение в растительную клетку экспрессионной кассеты, содержащей промотор, функционально связанный с гетерологичной нуклеиновой кислотой, представляющей интерес, где указанный промотор содержит нуклеотидную последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, изложенной в SEQ ID NO: 2; и
(b) нуклеотидной последовательности, которая по меньшей мере на 99% идентична последовательности, изложенной в SEQ ID NO: 2, где указанная последовательность инициирует транскрипцию в растительной клетке.
16. Способ по п. 15, где растительная клетка представляет собой клетку однодольного растения.
17. Способ по п. 15, где растительная клетка представляет собой клетку двудольного растения.
18. Способ по п. 16, где указанное однодольное растение представляет собой маис.
19. Способ по п. 15, где гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к вредителям.
20. Растение, устойчивое к гербицидам, содержащее в своем геноме стабильно встроенную экспрессионную кассету по п. 2, предназначенную для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, где указанная гетерологичная нуклеотидная последовательность, представляющая интерес, придает указанному растению устойчивость к гербицидам.
21. Способ по п. 10, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к гербицидам.
22. Способ по п. 15, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к гербицидам.
23. Растение, устойчивое к засолению, содержащее в своем геноме стабильно встроенную экспрессионную кассету по п. 2, предназначенную для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, где указанная гетерологичная нуклеотидная последовательность, представляющая интерес, придает указанному растению устойчивость к засолению.
24. Способ по п. 10, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к засолению.
25. Способ по п. 15, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к засолению.
26. Растение, устойчивое к патогенам, содержащее в своем геноме стабильно встроенную экспрессионную кассету по п. 2, предназначенную для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, где указанная гетерологичная нуклеотидная последовательность, представляющая интерес, придает указанному растению устойчивость к патогенам.
27. Способ по п. 10, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к патогенам.
28. Способ по п. 15, где указанная гетерологичная нуклеиновая кислота кодирует генный продукт, который придает устойчивость к патогенам.
29. Растительная клетка для придания растению устойчивости к гербицидам, предназначенная для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, содержащая в своем геноме стабильно встроенную экспрессионную кассету по п. 2, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к гербицидам.
30. Растительная клетка для придания растению устойчивости к засолению, предназначенная для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, содержащая в своем геноме стабильно встроенную экспрессионную кассету по п. 2, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к засолению.
31. Растительная клетка для придания растению устойчивости к патогенам, предназначенная для экспрессии гетерологичной нуклеотидной последовательности, представляющей интерес, содержащая в своем геноме стабильно встроенную экспрессионную кассету по п. 2, где указанная гетерологичная нуклеиновая кислота, представляющая интерес, придает указанному растению устойчивость к патогенам.
| WO 2012127373 A1, 27.09.2012 | |||
| US 2010186127 A1, 22.07.2010 | |||
| US 2009133159 A1, 21.05.2009 | |||
| US 8026412 B2, 27.09.2011 | |||
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |
| HERNANDEZ-GARCIA C.M | |||
| et al., A soybean (Glycine max) polyubiquitin promoter gives strong constitutive expression in transgenic soybean, Plant Cell Rep., 2009, Vol.28, Issue 5, pp.837-849 | |||
| SAEED H.A | |||
| et | |||

