Изобретение относится к области микробиологии, молекулярной генетики и может быть использовано в медицинской промышленности, в частности, по выделению, очистке и определению концентрации IgG человека и антител. Изобретение также может применяться в биотехнологии в качестве доступного реагента для выделения иммуноглобулина G (IgG) из крови человека и млекопитающих аффинной хроматографией, а также для освобождения сыворотки крови от IgG, что позволяет определять другие белки, присутствующие в сыворотке в более низких концентрациях.
Иммуноглобулины или антитела - белки соединения плазмы крови, образующиеся в ответ на введение в организм человека или теплокровных животных бактерий, вирусов, белковых токсинов и других антигенов. Связываясь активными участками (центрами) с бактериями или вирусами, антитела препятствуют их размножению или нейтрализуют выделяемые ими токсические вещества.
Антитела являются особым классом гликопротеинов, имеющихся на поверхности В-лимфоцитов в виде мембран связанных рецепторов и в сыворотке крови. Антитела являются важнейшим фактором специфического гуморального иммунитета и используются иммунной системой для идентификации и нейтрализации чужеродных объектов - например, бактерий и вирусов. Антитела выполняют две функции: антигенсвязывающую и эффекторную (вызывают тот или иной иммунный ответ, например, запускают классическую схему активации комплемента). Антитела состоят из двух легких и двух тяжелых цепей, которые связаны дисульфидными мостиками. У млекопитающих выделяют пять классов антител (иммуноглобулинов) - IgG, IgA, IgM, IgD, IgE, различающихся между собой по строению и аминокислотному составу тяжелых цепей и по выполняемым эффекторным функциям. IgG - основной класс иммуноглобулинов, содержащихся в сыворотке крови (70-75% от всех антител). IgG представлен четырьмя подклассами (IgG1, IgG2, IgG3, IgG4), каждый из которых выполняет свои уникальные функции. IgG, в основном, обеспечивает вторичный иммунный ответ, начиная вырабатываться спустя несколько дней после иммуноглобулинов класса М. Он сохраняется в организме длительно - таким образом, не дает повторно заболеть перенесенной инфекцией (например, ветряной оспой), обеспечивает иммунитет, направленный на нейтрализацию вредных токсических веществ микроорганизмов, имеет малые размеры, что позволяет ему беспрепятственно проникать во время беременности через плаценту к плоду, защищая его от инфекций. Благодаря высокому содержанию в сыворотке крови, IgG имеет наибольшее значение в противоинфекционном иммунитете. Поэтому об эффективности вакцинации судят по наличию его в сыворотке крови.
Мировой рынок антител значительно вырос за счет увеличения применения их в научных исследованиях, в диагностике и терапии. Антитела имеют важное значение для иммунодиагностики, для протеомики и для хроматографии. Антитела обычно выделяют из плазмы, сыворотки, асцитической жидкости, клеточной питательной среды, яичного желтка, растительных экстрактов, бактериальных и дрожжевых культур. Все эти источники в дополнение к антителам содержат различные белки. Следовательно, становится необходимой эффективная очистка антител, которая также необходима при их химической модификации, такой как мечение флуоресцентным, радиоактивным или ферментным зондом.
Очистка антител может быть достигнута с помощью различных методологий на основе конкретных физических или химических свойств антител, таких, как размер, растворимость, заряд, гидрофобность и аффинность связывания. Препараты IgG, полученные из нормальной или иммунной сыворотки и плазмы крови, широко применяются в медицинской практике.
Метод Кона по-прежнему наиболее часто используемый основной метод фракционирования плазмы человека. Он основан на разнице в растворимости основных белков плазмы благодаря взаимодействию пяти параметров: рН, температура ионная сила, концентрация этанола и концентрацию белка [Cohn EJ, et.al, 1946]. На протяжении многих лет этот метод изменялся для улучшения качества, чистоты и стоимости продукции. Эти модификации, в основном, основывались на замене или сочетании одного, или более стадий фракционирования [Oncley JL, et. al, 1949, Kistler Р, et. al, 1962]. Однако, процесс последовательного осаждения по методу Кона вызывает существенные потери белков, в результате и низкий выход продукта. Были введены и другие изменения, такие как введение других агентов для осаждения белков, в том числе каприловой кислоты [Steinbuch М, et. al., 1969], полиэтиленгликоля [Poison A, et. al., 1972, Нао Y, et. al., 1980], сульфата аммония и риванола Steinbuch М., et. al., 1980]. Но эти изменения в настоящее время не используются в масштабах производства.
Нынешний высокий спрос на крови-производные продукты, особенно на иммуноглобулин G для внутривенного введения, на который резко вырос спрос в последнее десятилетие, делает приоритетной проблему выделения IgG из плазмы крови. За последние годы накоплены данные о том, что одним из недостатков существующих препаратов является наличие измененных молекулярных форм IgG, оказывающие существенное влияние на иммунологические свойства препаратов. Эта причина, а также соосаждение вместе с основным продуктом посторонних белковых примесей, обусловили переход зарубежных фирм на более современную хроматографическую технологию. Традиционно процесс очистки антител включает стадию осаждения и фильтрации, затем несколько стадий ионообменной хроматографии, используя либо анионит, либо катионит. На последнем этапе, чтобы достичь высокой степени очистки антител, часто используют гельфильтрацию. [G. Li. R. Stewart et. al, 2002, D.K. Follmann, et. al., 2004, Ana C.A. Roque et. al, 2007, K. Tanaka, et. al, M. Vargas et. al, 2012]. При этом получение IgG включает в себя 5-6 стадий хроматографии, которые в данном изобретении предлагается заменить аффинной хроматографией с использованием в качестве лиганда рекомбинантного G белка стрептококка, специфически связывающего IgG. Полученный в данном изобретении новый штамм-продуцент позволяет нарабатывать достаточное количество G белка для аффинных колонок.
Аффинная хроматография является одним из наиболее широко используемых методов для очистки антител. Использование аффинной очистки уменьшает неспецифические взаимодействия, повышает выход и устраняет нежелательные примеси. Аффинная хроматография представляет собой метод разделения биологических молекул, который основан на специфических взаимодействиях между белком и специфическим лигандом, связанным с матрицей. Техника обладает высокой селективностью и, следовательно, дает высокое разрешение, а само вещество может быть очищено в несколько тысяч раз. Аффинная хроматография является уникальной технологией очистки, так, как только она позволяет очистить биомолекулы на основе их биологической функции или химической структуры. Один шаг аффинной очистки дает огромную экономию времени по сравнению с менее селективными многоступенчатыми процедурами. Успешная аффинная очистка требует биоспецифического лиганда, который может быть ковалентно связан с хроматографической матрицей. Матрица является инертным носителем и должна быть однородной, макропористой, обеспечивающей высокую связывающую способность даже для больших биомолекул, химически и механически стабильной, обладать чрезвычайно низкой неспецифической адсорбцией. Кроме того, матрица должна способствовать химической активации, тем самым, обеспечивая присоединение необходимых лигандов. Наиболее изученным и широко используемым реагентом активации является бромистый циан - CNBr. Выпускаются предварительно активированные матрицы, что позволяет избежать проблемы химической активации матрицы. Всем перечисленным выше свойствам отвечает сефароза - агароза в виде шариков. В основном, все виды агарозы отличаются диаметром частиц и количеством внутренних их сщивок. Одной из лучших матриц считается коммерческая CNBr-activated Sepharose 4 Fast Flow.
Выбор лиганда для аффинной хроматографии зависит от двух факторов: лиганд должен обладать обратимым сродством к целевому белку (в данном случае, к антителу) и должен содержать химические группы, через которые он может быть иммобилизован на матрице без нарушения связывающей активности. За последнее десятилетие было разработано множество лигандов для улучшения очистки белков.
Появление и использование бактериальных рецепторов в качестве биоспецифического лиганда для очистки антител явилось мощным инструментом для их обнаружения и очистки. Использование технологии рекомбинантных ДНК дало возможность манипулировать их свойствами для решения многих задач.
Выделение IgG и его фрагментов основано на высоком сродстве к белку А и белку G. Белки А и G - это бактериальные протеины, выделенные из клеточных структур Staphylococcus aureus и Streptococcus, соответственно, которые на сефарозной матрице образуют чрезвычайно полезный и простой сорбент. Белок G обладает способностью связывать Fc часть иммуноглобулина G большинства видов млекопитающих. В отличие от хорошо изученного IgG-связывающего рецептора - стафилококкового белка А, белок G связывает все подклассы иммуноглобулина G человека (белок А не связывает третий подкласс человеческого IgG), а также имеет более широкий спектр связывания IgG различных млекопитающих. Однако белок G связывает также альбумин, α2-макроглобулин и киноген. Для получения, только IgG-связывающего G белка были разработаны генно-инженерные формы G белка, поскольку сайты связывания для этих белков находятся на разных концевых участках молекулы белка G (Guss В., et. al, 1996; Goward C.R., et. al, 1990).
Благодаря своей способности связывать Fc фрагмент IgG белок G может быть использован в иммунологических исследованиях, в ELISA диагностических тест-системах для создания наборов для идентификации инфекционных заболеваний, выявления антигенов возбудителей и антител к ним. Кроме того, G белок может быть эффективным лигандом для аффинной хроматографии.
Описано использование рекомбинантного бифункционального белка G для освобождения сыворотки человека от IgG и альбумина и для получения очищенных препаратов IgG и альбумина из той же сыворотки аффинной хроматографией, в которой в качестве лигандов использованы бифункциональный и рекомбинантный монофункциональный IgG-связывающий белок, полученный из штамма стрептококка группы G, G148 [Гупалова Т.В. с соавт., 1996].
Данное техническое решение принято в качестве прототипа заявляемой группы изобретений.
Этот метод имеет недостатки из-за использования бифункционального G белка, который связывает ЧСА с меньшей эффективностью, чем IgG. 90% -ное удаление ЧСА происходило, когда на колонку наносили только 10 мкл сыворотки, что явно недостаточно. Кроме того, G белок, который связывает только IgG, имеет недостатки из-за очень сложного пути клонирования гена G белка, начиная с конструкции банка генов штамма в фаговом EMBL3a лямбде векторе и дальнейшем клонировании в плазмидных векторах через получение гена, кодирующего двух рецепторный белок, имеющий как IgG-, так и альбумин-связывающую активности. В результате такого пути клонирования была получена последовательность, имеющая только второй и третий IgG-связывающие домены, области W и М, которые отвечают за связывания белка с клеточной стенкой и мембраной (патент Гупалова Т.В. с соавт., 1996).
Современные технологии, такие, как полимеразная цепная реакция, позволяют производить клонирование только нужного фрагмента гена.
Задачей изобретения является решение актуальной проблемы - создание нового отечественного штамма-продуцента G белка с использованием более активного штамма стрептококка группы G, выделенного от человека, обладающего большей способностью связывать IgG человека и млекопитающих. Показано, что обнаружен штамм стрептококка группы G - G4223, выделенный со слизистой оболочки верхних дыхательных путей человека (Гупалова Т.В. с соавт., 1999). Оказалось, что G белок, экспрессируемый этим штаммом, содержит, как и штамм G148 стрептококка группы G, три IgG-связывающих домена, но активность связывания им IgG выше.
Получение рекомбинантного штамма с высокой экспрессией IgG-связывающего белка для выделения и очистки человеческого IgG, а также любых антител, имеющих сродство к G белку, - актуальная проблема.
Сущность заявляемого изобретения состоит в том, что для создания сорбента для выделения или удаления человеческого IgG из сыворотки крови, выделения и очистки антител, имеющих сродство к G белку, предлагается использовать новый рекомбинантный IgG-связывающий полипептид.
Заявлена группа изобретений, объединенных единым изобретательским замыслом, которая включает следующие объекты:
1. Рекомбинантная ДНК (обозначенная как pG4223), полученная в результате полимеразной цепной реакции (ПЦР) с использованием хромосомной ДНК штамма G4223 CГG, уникальных праймеров PG1 и PG2, и последующего клонирования с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США).
2. Рекомбинантная плазмидная ДНК pQE 30-pG4223, несущая рекомбинантную ДНК pG4223.
3. Штамм-продуцент E.coli M15-G4223, позволяющий при определенных условиях экспрессировать рекомбинантный полипептид G4223.
4. Рекомбинантный полипептид G4223, содержащий последовательность трех IgG-связывающих доменов, обладающий высокой способностью связывать IgG и имеющий большую IgG-связывающую активность, чем рекомбинантный IgG-связывающий G-белок стрептококка группы G, полученный клонированием гена G, белка из стрептококка G148.
5. Аффинный сорбент, в котором к матрице - активированной цианбромом сефарозе 4 В присоединен, в качестве лиганда, новый рекомбинантный IgG-связывающий полипептид G4223.
Техническая суть и достигаемый при использовании заявленной группы изобретений технический результат состоят в следующем.
IgG - один из наиболее распространенных белков плазмы крови человека и млекопитающих. Он нашел широкое применение в медицине, диагностике и научных исследованиях.
В последние десятилетия выявлен и изучен белок бактерий, обладающий рецепторной активностью в отношении IgG, так называемый белок G, выделенный из стрептококков групп С и G. Изучение структуры IgG-связывающего белка имеет большое значение для технологии создания белковых реагентов, актуальных в иммунохимии, протеомике, биотехнологии и клинической диагностике.
Использование рекомбинантных полипептидов позволяет: исключить трудоемкий процесс приготовления специфических антител к IgG (для которых используются лабораторные животные), избежать неспецифических "фоновых" реакций, часто встречающихся в иммунологии, стандартизовать используемый IgG- связывающий полипептид, выделять IgG из сыворотки крови в одну стадию очистки.
Белок G может быть выделен из стрептококка энзиматическим расщеплением папаином (нативный белок) либо получен генно-инженерным путем (рекомбинантный белок). Выделение G белка из стрептококка энзиматическим путем характеризуется низким выходом, гетерогенностью получаемого продукта и трудоемкостью технологического процесса, связанного с использованием патогенного микроба. Ввиду этого предпочтительным является получение рекомбинатного белка.
Известен рекомбинатный белок G, получаемый рядом лабораторий путем клонирования гена G белка в непатогенном хозяине [Guss В., et. al., 1986, Fahhnestock. S.R., et. al, 1986, Goward С.R., 1990]. Описанные препараты G белка были либо гетерогенны, либо при получении белка в высокоочищенном состоянии выход его довольно низкий.
Получение рекомбинантного G белка, описано Гуссом (Guss В., et. al, 1986), который сконструировал банк генов стрептококка G148 в фаговом EMBL За лямбда векторе. Был изолирован положительный фаговый клон SPG, дающий экспрессию IgG связывающей активности. При дальнейшем клонировании с целью получения ДНК меньшего размера с экспрессией функционального G белка был проклонирован фрагмент гена G белка в векторе pUC18. Из субклона, продуцирующего G белок, белок был выделен, очищен аффинной хроматографией и исследован электрофоретически в SDS-PAGE. В SDS-полиакриламидном геле было получено два бенда белка с молекулярным весом 32 и 38 kD. Описанный автором G белок оказался гетерогенным, и выход его был низок, кроме того он связывал и альбумин человека.
Причиной гетерогенности полученного Гуссом препарата G белка явилось наличие двух внутренних дополнительных промоторов в структурной части гена G белка. Выход G белка был низкий, так как экспрессия G белка обеспечивалась функцией двух внутренних дополнительных промоторов, которые обычно не функционируют в условиях нормального развития стрептококка.
Укороченный рекомбинантный стрептококковый G белок был получен Говардом [Goward С.R., 1990], который проклонировал ген G белка как Hind III фрагмент геномной ДНК G 148. Очищенные фрагменты геномной ДНК были лигированы с HindIII фрагментом плазмидного вектора pUC 8 и трансформированы в E.coli TG2. Из полученных клонов с использованием меченых олигонуклеотидов, методом гибридизации, были обнаружены положительные клоны. Затем, используя сложный путь сайт-направленного мутагенеза, был найден фрагмент, кодирующий укороченный G белок [Goward С.R., 1990]. В результате был получен белок, утративший половину первого IgG-связывающего домена и обладающий за счет этого неполноценной IgG-связывающей частью G белка.
Для клонирования гена G белка также была получена библиотека генов штамма G 148 в фаговом EMBL3a лямбде векторе (Гупалова Т.В. с соавт., 1996). Из 50000 рекомбинантных фаговых клонов был отобран клон, дающий наибольшую экспрессию IgG-связывающей активности G белка. Для нахождения минимального фрагмента ДНК, содержащего ген G белка, проводили субклонирование в E.coli JM109. 1,5 kb EcoRI/HindIII фрагмент был клонирован в pUC8.
Из клона, дающего наибольшую экспрессию, была выделена рекомбинантная плазмида p3G. Многие стрептококковые штаммы, экспрессирующие G белок, также связывают и человеческий сывороточный альбумин. Полученный клон, экспрессирующий G белок, обладал как IgG-, так и альбумин-связывающей активностью. Для получения только IgG связывающего G белка необходимо было проклонировать фрагмент гена G белка, кодирующего синтез конкретного рецепторного участка. В подходе, использованном Гуссом, рамка считывания лактозного оперона, в котором не осуществлялось клонирование гена G белка, была нарушена. Для сохранения данной рамки считывания клонирование осуществлялось в векторе pUC8 в область сайта рестриктазы HindIII, после чего ДНК обрабатывалась ферментом Кленова. Фрагмент гена G белка рестрицировали рестриктазами Sau3A и Hindlll, после чего он также обрабатывался ферментом Кленова. Лигирование происходило в данном случае по тупым концам. При этом сохранилась нормальная рамка считывания лактозного оперона, и структурная часть гена G белка транскрибировалась без нарушения рамки считывания; образовывался рекомбинатный продукт, обладающий IgG связывающими свойствами G белка с высоким уровнем экспрессии. Альбумин связывающей активности обнаружено при этом не было. Отобранный в данном случае рекомбинантный клон содержал плазмиду p7G.
Таким образом, был получен штамм-продуцент, который был назван E.coli JM109.7G. Однако, в результате такого пути клонирования была получена последовательность, имеющая только второй и третий IgG-связывающие домены, области W и М, которые отвечают за связывания белка с клеточной стенкой и мембраной. Поэтому, необходимо создать новый штамм-продуцент, способный экспрессировать большое количество рекомбинантного IgG-связывающего полипептида, обладающего высокой способностью связывать IgG.
Появление современной технологии, такой, как полимеразная цепная реакция, позволяет намного быстрее производить клонирование только нужного фрагмента гена. В данном изобретении клонирование гена G белка проводили с помощью полимеразной цепной реакции.
Согласно заявленному изобретению получен рекомбинантный полипептид G4223 и его штамм-продуцент. Полипептид G4223 содержит последовательность трех IgG-связывающих доменов. Полипептид обладает высокой способностью связывать IgG. Его использование возможно для выделения и очистки человеческого IgG, любых антител, имеющих сродство к G белку, а также для создания сорбентов для эффективного удаления IgG из сыворотки крови.
Поставленная задача решалась конструированием уникальных праймеров, направленных к участкам гена, кодирующих полипептид, состоящий из трех IgG-связывающих доменов. С целью быстрого и простого получения рекомбинантного полипептида G4223 для клонирования фрагмента ДНК была использована система экспрессионных плазмид pQE. Для создания штамма-продуцента рекомбинантного полипептида G4223 был использован штамм Е. coli M15. В результате экспериментальной работы этот штамм приобрел новые свойства: способность экспрессировать рекомбинантный полипептид G4223. Новый штамм получил название Е. coli M15-G4223. Штамм задепонирован в коллекции ФГБНУ «ИЭМ» под №725.
В процессе работы была создана уникальная рекомбинантная ДНК р G4223, полученная в результате ПЦР, с использованием хромосомной ДНК штамма G4223 стрептококка группы G, уникальных праймеров PG1 и PG2, и последующего клонирования с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США). Была также создана рекомбинантная плазмидная ДНК pQE 30-pG4223, несущая рекомбинантную ДНК pG4223, и штамм-продуцент Е. coli M15-G4223, позволяющий при определенных условиях экспрессировать рекомбинантный полипептид G4223.
В процессе работы получен фрагмент гена, кодирующий область с тремя IgG-связывающими доменами штамма G4223 стрептококка группы G, размером 630 п. н. с помощью ПЦР с использованием праймеров PG1 и PG2 и хромосомной ДНК штамма G4223 стрептококка группы G. Также авторами осуществлено клонирование pG4223 с использованием экспрессионной плазмиды pQE-30 и последующей трансформацией полученной рекомбинантной плазмиды (обозначенной как pQE 30-pG4223) в гетерологическую систему E.coli M15. Авторами получены штамм-продуцент рекомбинантного полипептида G4223, обозначенный как Е. coli M15-G4223. Также авторами реализована экспрессия рекомбинантного полипептида G4223 клетками штамма Е. coli M15-G4223 с последующей одноступенчатой аффинной очисткой с использованием Ni-сефарозы (GE Healthcare, Sweden).
Получение рекомбинантного полипептида.
Источником хромосомной ДНК послужил штамм G4223 стрептококка группы G (выделенный от человека). ДНК, выделенная фенольно-хлороформной экстракцией, была использована в качестве матрицы в ПЦР с целью получения фрагмента гена pG4223 (630 п. н.). Олигонуклеотидные праймеры представлены в таблице 1.

Подчеркнутые звенья нуклеотидной последовательности указывают на сайты рестрикции.
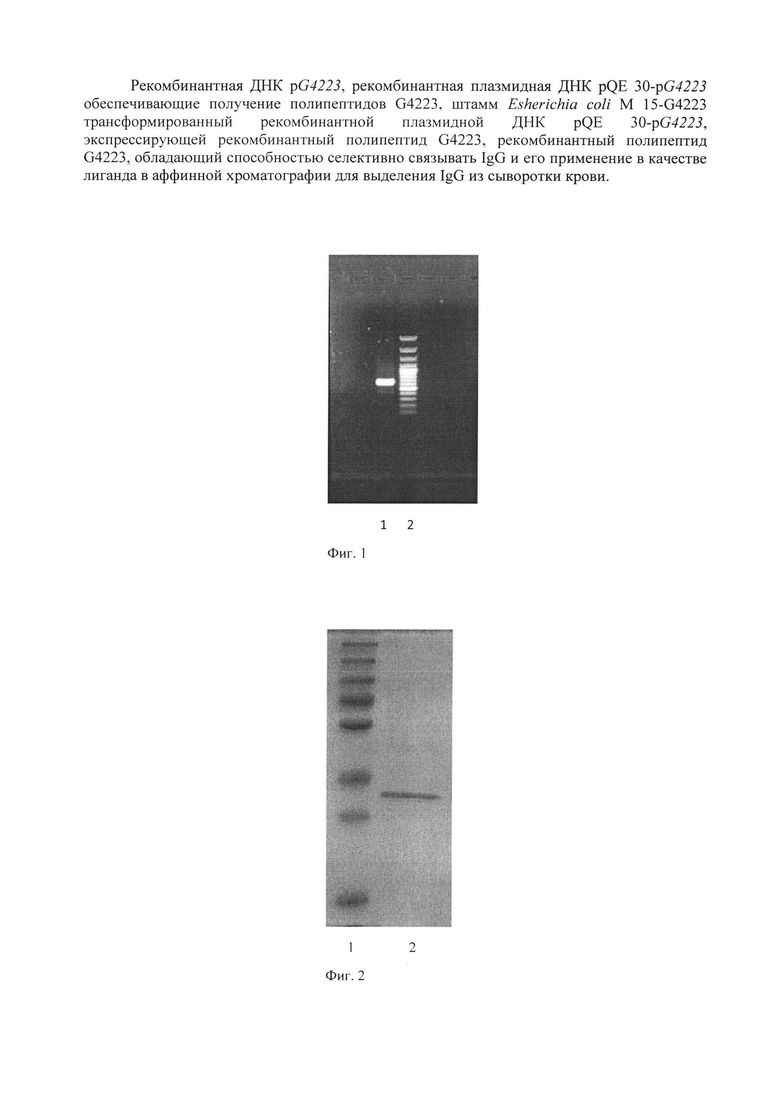
Анализ результатов ПЦР осуществлен путем разделения фрагментов ДНК в 1% агарозном геле при помощи горизонтального электрофореза (фиг. 1).
Выделение искомого амплифицированного участка ДНК проведено с использованием набора «QIAquick Gel Extraction Kit» (Qiagen, США).
Полученный фрагмент клонировали с использованием экспрессионной плазмиды pQE 30. При подготовке к клонированию была проведена рестрикция выделенного из агарозы фрагментов pG4223 (630 н.п.) и плазмиды pQE 30 ферментами BamHI и KpnI с образованием липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле, а затем лигировали.
Продуктом лигирования проводили кальциевую трансформацию штамма E.coli М 15. Процесс трансформации осуществляли по методике трансформации кишечной палочки, используя плотные селективные среды, содержащие 100 мкг/мл ампициллина, 25 мкг/мл канамицина. [Maniatis Т, Fritsch E.F., Sambrook J. Molecular cloning: a laboratory manual. - Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 867, 1982].
Антибиотикоустойчивые трансформанты откалывали на две параллельные чашки Петри с антибиотиками. Колонии с одной чашки переносили на нитроцеллюлозную мембрану и вскрывали раствором, содержащим 0,2 N NaOH, 0,1% SDS и 0,5% β-меркаптоэтанола. Мембрану инкубировали 1 час при комнатной температуре в блокирующем растворе (2 части 3% молока и 1 часть фосфатно-солевого буфера (ФСБ)) для удаления неспецифического связывания и затем в блокирующем растворе, содержащем IgG, меченный пероксидазой хрена (Sigma, USA).
Конъюгирование пероксидазы хрена с IgG проводили перйодатным методом [Фримель Г. Иммунологические методы: М. Медицина: 438-439, 1987]. Затем мембрану последовательно отмывали блокирующим раствором и ФСБ. Пероксидазную активность проявляли раствором ТМБ для мембран (Sigma, USA). Из трансформантов с наиболее выраженной экспрессией IgG-связывающей активности выделяли рекомбинантные плазмиды с помощью Mini-prep, plasmid DNA purification kit (Qiagen, USA). Наличие в них рекомбинантных ДНК подтверждали в ПЦР с исходными праймерами PG1 и PG2. Плазмидную ДНК pQE 30-pG4223 использовали как матрицу в ПЦР с праймерами pQE1 и pQE2. Амплификат выделяли после электрофореза в 1% агарозном геле и секвенировали.
Итоговые данные совпали с известными нуклеотидными последовательностями, кодирующими фрагмент с тремя IgG-связывающими доменами штамма G148 стрептококка группы G. [Guss В., et. al., 1986].
Культуру штамма Е. coli М 15, содержащую рекомбинантную плазмиду pQE 30-pG4223, обозначенную как Е. coli М 15-G4223, культивировали в жидкой среде LB с добавлением антибиотиков (ампициллин (100 мкг/мл) и канамицин (25 мкг/мл)) до поздней логарифмической фазы роста (OD600=0,9). Затем экспрессию рекомбинантного белка индуцировали добавлением изопропил-бета-D-тиогалактопиранозида (IPTG), и клетки культивировали еще 4 часа. После этого клетки осаждали центрифугированием, отмывали лизирующим буфером А (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 20 мМ имидазола, рН 8,0) и суспендировали в том же буфере, добавляя ингибитор протеаз, фенилметилсульфонилфторид (ФМСФ), до концентрации 1 мМ. Суспензию клеток вскрывали ультразвуком и центрифугировали. Надосадочную жидкость наносили на колонку, заполненную Ni-сефарозой. После того, как белок связывался с Ni-сефарозой, колонку промывали буфером А для удаления не связавшихся белков. Рекомбинантный белок элюировали буфером Б (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 250 мМ имидазола, рН 8,0). После аффинной хроматографии белок диализовали 18 часов против дистиллированной воды.
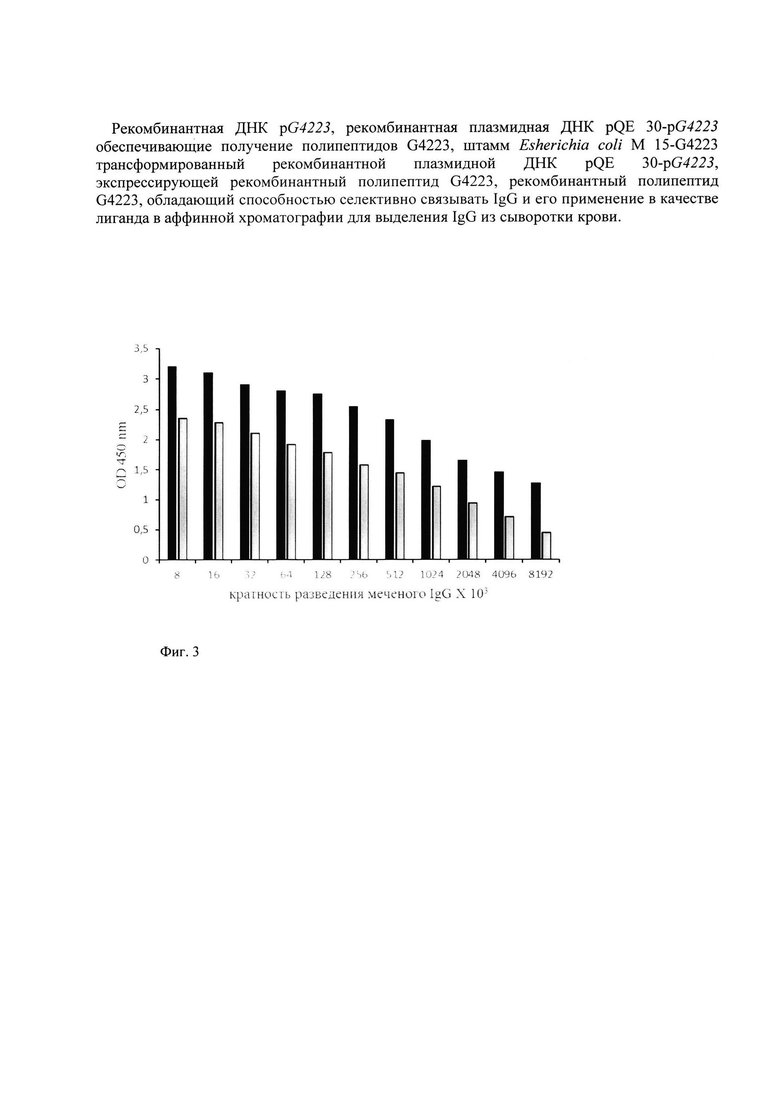
Выделенный рекомбинантный полипептид анализировали методом электрофореза в полиакриламидном геле, который позволил сделать заключение об удовлетворительном качестве очистки полипептида и также об его молекулярной массе, сравнивая пробег полипептида G4223 с пробегом белков известной молекулярной массы (Precision Plus Protein standards (161-0373), Bio-Rad, США). Молекулярная масса полипептида G4223 оказалась равной (33,0±0,5) кДа (фиг. 2).
Таким образом, рекомбинантный полипептид G4223 содержит аминокислотную последовательность IgG-связывающего полипептида стрептококка группы G штамма G4223, содержащую 210 аминокислот, ковалентно связанную с 22 аминокислотными остатками, кодируемыми pQE 30.
Изучение взаимодействия рекомбинантного полипептида G4223 с IgG при сравнении с аналогичной реакцией с IgG-связывающим полипептидом штамма G148, полученным ранее (Гупалова Т.В. с соавт., 1996).
Способность полипептида G4223 и IgG-связывающего полипептида штамма G148 связывать IgG, представленного IgG - поликлональным иммуноглобулином G человека, меченного пероксидазой хрена, и препаратами очищенных полипептидов G4223 и IgG-связывающего полипептида штамма G148, оценивали с помощью прямого метода иммуноферментного анализа (ИФА). На планшет сорбировали полипептиды в концентрации 1 мкг/мл. При этом оценивали количество связавшегося с ними меченого IgG. На фиг. 3 показана гистограмма, отражающая сопоставление IgG-связывающей активности полипептидов G4223 и IgG-связывающего полипептида штамма G148. Исследуемый полипептид G4223 и IgG-связывающий полипептид штамма G148 обладают IgG-связывающей активностью в отношении меченого IgG, причем количество IgG, связанного полипептидом G4223 больше, чем в случае IgG-связывающего полипептида штамма G148.
Средние значения оптической плотности по данным нескольких опытов приведены на гистограмме (фиг. 3), где черный столбик - полипептид G4223, серый столбик - IgG-связывающий полипептид штамма G148, по оси абсцисс -кратность разведений меченого IgG, по оси ординат - оптическая плотность (OD450).
Создание аффинного носителя: сефарозы 4В-IgG-связывающего G4223 полипептида. Выделение очищенного препарата IgG.
В качестве матрицы аффинного носителя использовали сефарозу 4 В, активированную бромцианом, ("Sigma", США). Лигандом, иммобилизованным на матрице, служил рекомбинантный IgG-связывающий G4223 полипептид.
Предварительно сефарозу 4 В, активированную бромцианом, промывали несколько раз холодной 1 мM HCl, каждый раз центрифугировали, надосадок выбрасывали, добавляя свежую порцию холодной 1 мM HCl. Затем суспензию сефарозы промывали 0,1 М бикарбонатным буфером, рН 8,3.
Рекомбинантный IgG-связывающий полипептид G4223, 35 мг, иммобилизовали на 5 мл сефарозы 4В, активированной бромцианом в 0,1 М бикарбонатном буфере, рН 8,3, содержащем 0,5 М NaCl и заполняли полученным сорбентом колонку. Количество IgG-связывающего G4223 полипептида, присоединившегося к матрице, составило 30 мг. Сорбент промывали в колонке (1.5 × 5 см) последовательно 0,1 М ацетатным буфером, рН 4,0, и 0,1 М бикарбонатным буфером, рН 8,3, содержащим 0,5 М NaCl, затем уравновешивали 0,01 М фосфатным буфером, рН 7.4, содержащим 0.85% NaCl, (ФСБ) и хранили при 4°С.
Для проверки емкости сорбента на сорбент, уравновешенный ФСБ, рН 7,4, последовательно наносили 5, 10 и 15 мл сыворотки человека. Полноту освобождения сыворотки от IgG проверяли методом блоттинга. Для этого на нитроцеллюлозную мембрану (НЦ) наносили пробу исходной сыворотки, пробу сыворотки, не связавшуюся с сорбентом, и элюированную с сорбента пробу. НЦ мембрану переносили в блокирующий раствор, содержащий IgG-связывающий полипептид, меченный пероксидазой, и выдерживали их 15 минут при перемешивании. Мембрану последовательно отмывали блокирующим раствором и ФСБ. Отмытую мембрану подсушивали на воздухе и помещали в раствор 3,3/,5,5/-Тетраметилбензидин (ТМБ, жидкий субстрат для мембран, Sigma). Окрашивание происходило в течение нескольких секунд.
В пробе сыворотки, не связавшейся с сорбентом при нанесении 5, 10 и 15 мл человеческой сыворотки, практически полностью отсутствовал IgG. Связавшийся с сорбентом IgG элюировали 0,1 М глициновым буфером, рН 2,2. Во фракциях объемом 1 мл определяли оптическую плотность (ОП) при 280 нм. Фракции с высокими значениями ОП объединяли и значение рН доводили 2.0 М NaOH до значения 7.5 и диализовали 18 часов против ФСБ. Наличие и качество полученного IgG определяли методом электрофореза по методу Лэммли [Laemmli U.K. Nature (London), 1970 p.227] в 12% полиакриламидном геле (ПААГ) в присутствии 0,1% додецилсульфата натрия (ДДС Na) на приборе "Bio_Rad" (США), а его концентрацию методом Лоури. Перед нанесением образцы белков в растворе, содержащем 2% ДДС Na и 0,5% β-меркаптоэтанола, погружали на 2 мин в кипящую водяную баню. Электрофорез проводили при силе тока 50-60 мА в течение 1-2 часов. Затем гель три раза промывали дистиллированной водой и в течение 30 минут окрашивали в растворе Кумасси синий R 250 «Bio-Rad» (США), после чего промывали в 10% ледяной уксусной кислоты с 10% изопропанола.
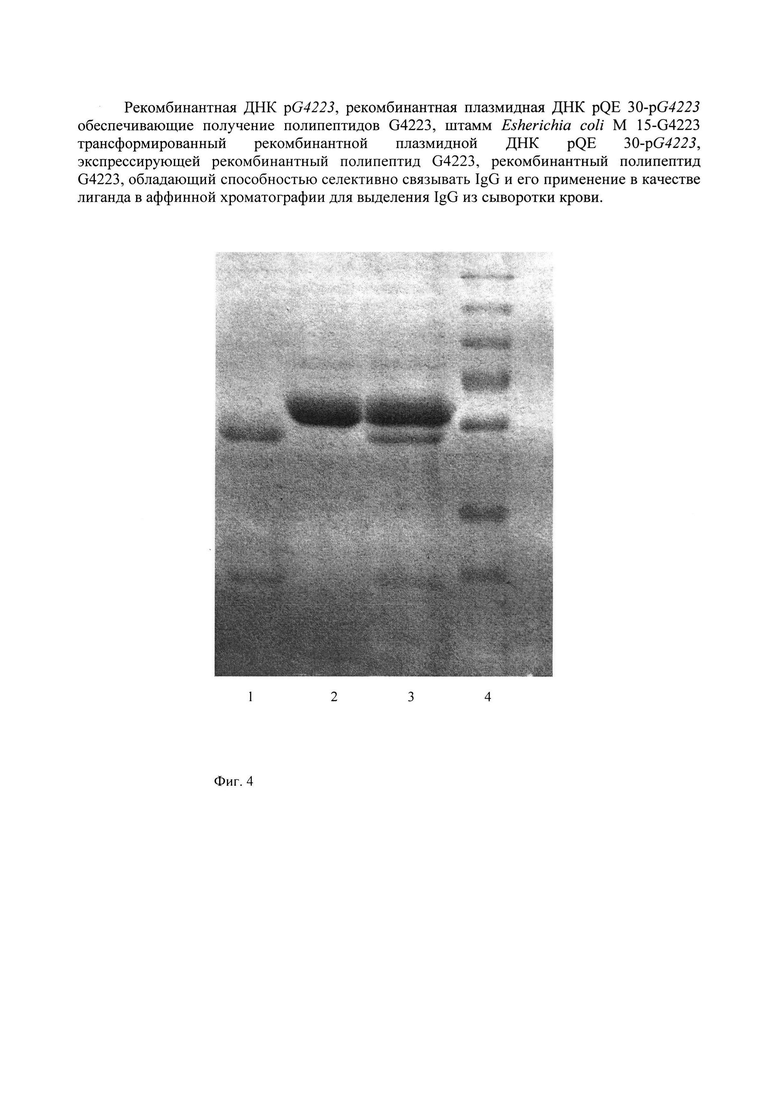
Проба IgG, очищенная аффинной хроматографией, элюированная с колонки, представлена на фиг. 4. Из рисунка следует, что препарат IgG получен в высокоочищенном состоянии.
Таким образом, создан сорбент сефароза 4В-IgG-связывающий G4223 полипептид, позволяющий получать очищенные препараты IgG.
На полученном сорбенте с таким же успехом можно получать очищенные препараты IgG любых антител, полученных от млекопитающих, таких, как кролик, мышь или морская свинка, так как белок G вообще, а в данном случае IgG-связывающий G4223 полипептид обладает способностью связывать Fc часть иммуноглобулина G большинства видов млекопитающих
Пример 1. Получение фрагментов ДНК pG4223 методом ПЦР.
К 0,25 мкг геномной ДНК, выделенной фенольно-хромосомной экстракцией из штамма G4223 CГG добавляли по 10 микромолей каждого из специфических праймеров, фланкирующих исследуемую последовательность, по 0,2 мМ четырех дезоксирибонуклеотидтрифосфатов и 12,5 мкл смеси (ThermoScientific DreamTag Green PCR Master Mix (2x), объем доводили дистиллированной водой (Water, nuclease-free) до 25 мкл. Пробирку помещали в амплификатор и инкубировали при 94°С 2 мин. Программа ПЦР состояла из: денатурации при 94°С - 30 сек, отжига праймеров - 60°С - 1 мин и синтеза - 72°С - 1 мин. Этот цикл повторялся 30 раз, после чего смесь инкубировалась при 72°С 10 мин. В работе использовали олигонуклеотидные праймеры, приведенные в Таблице 1. ПЦР продукты разделяли в 1% агарозном геле в горизонтальном электрофорезе (фиг. 1). Выделение амплифицированного участка ДНК проводили с использованием набора «QIAquick Gel Extraction Kit» (Qiagen, США). Анализ размера полученного фрагмента ДНК проводили, исходя из сравнения его электрофоретической подвижности с электрофоретической подвижностью маркера молекулярных весов (100 п. н. ДНК-маркер, Хеликон).
На фиг. 1 показана электрофореграмма амплифицированного ДНК-фрагмента pG4223, где 1 - 100 п. н. ДНК - маркер, (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар), 2 - продукт ПЦР.
В результате ПЦР с праймерами РА1 и РА2 был получен фрагмент 630 п. н., что продемонстрировано на фиг. 1.
Пример 2. Клонирование ДНК-фрагмента pG4223 с использованием экспрессионной плазмиды pQE 30.
Плазмида pQE 30 и фрагмент, полученный в результате ПЦР были обработаны двумя рестрикционными ферментами BamHI и KpnI, что привело к образованию липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле, а затем выделяли из агарозы с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, США). В ходе клонирования к 5 мкл фрагмента ДНК pG4223 добавляли 1 мкл фрагмента ДНК pQE 30, полученных после рестрикции и вырезанных из агарозы, 2 мкл десятикратного лигазного буфера и 1 мкл лигазы фага Т4. Объем доводили дистиллированной водой до 20 мкл. Смесь инкубировали при 10° в течение 12 часов. В результате проведенного клонирования была получена рекомбинантная плазмида, обозначенная как pQE 30-pG4223, несущая рекомбинантную ДНК (обозначенную как pG4223), состоящую из 630 п. н. и 66 п. н. фрагмента плазмиды pQE-30.
Пример 3. Трансформация культуры Е. coli М 15 плазмидной ДНК pQE 30-pG4223.
Рекомбинантную плазмиду трансформировали в гетерологичную систему Е. coli М 15. В качестве положительного контроля параллельно проводили трансформацию Е. coli М 15 исходной плазмидной ДНК pQE 30.
Генетическим маркером плазмидной ДНК pQE 30 является маркер amp, кодирующий бета-лактомазу, что обеспечивает устойчивость к ампициллину клеток, несущих эту плазмиду. Культуру клеток Е. coli М 15 культивировали в бульоне LB (США) с канамицином (25 мкг/мл) в течение 12 часов при 37° и интенсивном перемешивании. Затем инокулят пересевали на новый объем той же среды (на 10 мл среды - 0,1 мл инокулята) и инкубировали при 37° в течение 2-3 часов при перемешивании до ОД600=0,3. Выращенные клетки в объеме 1,5 мл центрифугировали в течение 1 мин. при 12000 об/мин., и полученный осадок суспендировали в 200 мкл раствора 0,1 М CaCl2. Далее смесь выдерживали во льду 1 час. После центрифугирования осадок ресуспендировали в 187, 5 мкл 0,1 М CaCl2 и 64,5 мкл дистиллированной воды. Затем в пробирку вносили 0,2 мкг плазмидной ДНК и инкубировали смесь во льду 1 час. Далее смесь выдерживали 2 мин. при 42°С. После этого к смеси добавляли 1 мл бульона LB и инкубировали 1 час при 37°С. После осаждения центрифугированием (8000 об/мин. в течение 1 мин) клетки высевали на чашки с 1% L-агаром (Difco, США), содержащим ампициллин (100 мкг/мл) и канамицин (25 мкг/мл). Через 18 часов роста клеток при 37°С проводили отбор клонов-трансформантов. Антибиотик устойчивые трансформанты переносили на две параллельные чашки Петри с антибиотиками. Колонии с одной чашки переносили на нитроцеллюлозную мембрану и вскрывали раствором, содержащим 0,2 N NaOH, 0,1% SDS и 0,5% β-меркаптоэтанола. Мембрану инкубировали 1 час в блокирующем растворе, содержащем IgG, меченный пероксидазой хрена (Sigma, США). Затем ее последовательно отмывали блокирующим раствором и раствором ФСБ. Пероксидазную активность проявляли раствором ТМБ для мембран
Таким образом, полученные данные позволяют сделать вывод о том, что в результате проведенного клонирования был получен рекомбинантный клон, несущий плазмиду pQE 30-pG4223 с рекомбинантной ДНК pG4223.
Пример 4. Очистка рекомбинантного полипептида G4223.
Культуру штамма Е. coli М 15-G4223 выращивали в бульоне LB с добавлением ампициллина в концентрации 100 мкг/мл и канамицина в концентрации 25 мкг/мл в течение ночи при интенсивном перемешивании. Затем клетки пересевали на 1000 мл той же среды и инкубировали при 37°С в течение 2-3 часов при интенсивном перемешивании до ОД600=0,9. Экспрессию рекомбинантного белка индуцировали добавлением раствора изопропил-бета-D-тиогалактопиранозида до конечной концентрации 2 мМ, после чего клетки инкубировали при тех же условиях еще 4 часа. Полученную суспензию клеток центрифугировали при 4000 об/мин. 20 мин. Надосадочную жидкость сливали, а клетки суспендировали в буфере А ((20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 20 мМ имидазола, рН 8,0), добавляя ингибитор протеаз фенилметилсульфонилфторид (ФМСФ) до конечной концентрации 1 мМ. Для лизирования клеток была использована трехкратная ультразвуковая обработка при 4°С в течение 20 сек. с перерывом в 40 сек. в ультразвуковом дезинтеграторе (УЗДН-1У4.2, Англия). Лизат клеток центрифугировали при 20000 об/мин. и 4°С в течение 20 мин. Надосадочную жидкость пропускали через 0,45 мкм фильтр (Millipore, США) и затем ее наносили на колонку с Ni-сефарозой, предварительно уравновешенную буфером А. Далее колонку промывали тем же буфером до тех пор, пока значение ОД280 выходящего раствора не был больше, чем 0,01. Белок элюировали раствором буфера Б (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 250 мМ имидазола, рН 8,0). Элюат собирали по фракциям по 500 мкл и измеряли в них значения ОД280. Фракции с наибольшими значениями ОД280, объединяли и диализовали против дистиллированной воды в течение 12 часов. На фиг. 2 представлена электрофореграмма рекомбинантного полипептида G4223.
На фиг. 2 показана электрофореграмма рекомбинантного полипептида G4223, где 1 - маркер молекулярной массы, (сверху вниз: 250, 130, 95, 72, 55, 36, 28, и 17 кДа), 2 - препарат очищенного белка G4223.
Молекулярную массу полипептида G4223 определяли, сравнивая пробег белка G4223 с пробегом белков известной молекулярной массы (Precision Plus Protein standarts (161-0373), Bio-Rad, США). Молекулярная масса белка G4223 оказалась равной (33±0,5) кДа.
Пример 5. Связывание сывороточного меченого IgG адсорбированным полипептидом G4223.
Анализ IgG-связывающей активности проведен методом прямого ИФА. Для проведения ИФА использовали планшеты NUNC MaxiSorb (Denmark).
Приготовление разведенных препаратов меченого IgG осуществляли с применением раствора ФСБ. Удаление не связавшихся реагентов проводили трехкратным промыванием планшетов раствором ФСБ с 0,05% твин 20 (ФСБТ).
Полипептид G4223 и IgG-связывающий полипептид штамма G148, полученный ранее, разведенные в ФСБ, рН 8,0 до 1 мкг/мл, адсорбировали на планшет в течение 18 часов при 4°С. После трехкратного отмывания в планшеты вносили разведенный меченый IgG. Инкубацию проводили при 37°С в течение 1 часа. Далее планшет 3 раза отмывали от несвязавшихся реагентов. Активность фермента-метки пероксидазы хрена определяли с использованием хромогена - тетраметилбензидина (ТМБ), растворенного в 0,1 М цитрат-фосфатном буфере, рН 5,0 с добавлением перекиси водорода. Реакцию останавливали добавлением 2N серной кислоты. Оптическую плотность определяли при длине волны 450 нм на мультискане.
Установлено, что полипептид G4223 имеет высокую IgG-связывающую активность, которая выше, чем у рекомбинантного IgG-связывающего полипептида штамма G148, что продемонстрировано на фиг. 3.
На фиг. 3 представлено сравнение эффективности связывания полипептида G4223 и IgG-связывающего полипептида штамма G148, с IgG, где черный столбик - полипептид G4223, серый столбик - IgG-связывающий полипептид штамма G148, по оси абсцисс - кратность разведений меченого IgG, по оси ординат - оптическая плотность (OD450).
Пример 6. Способ получения IgG из сыворотки крови на аффинном носителе: сефарозе 4B-G4223-IgG-связывающий полипептид.
На аффинный сорбент в колонке (1.5 × 5 см) сефароза 4B-G4223-IgG-связывающий полипептид было нанесено 5, 10 или 15 мл сыворотки крови. Не связавшиеся белки элюировали ФСБ, связавшийся с сорбентом IgG элюировали 0,1 М глициновым буфером, рН 2.2. Пробу исходной сыворотки и пробу элюированного IgG оценивали методом блоттинга. Результаты блоттинга показали во всех пробах сыворотки, не связавшейся с сорбентами, отсутсвие IgG. Проба IgG, элюированного с сорбента, была проверена методом СДС-ПААГ электрофореза (фиг. 4).
На фиг. 4 показана электрофореграмма:
1 - проба IgG, элюированного и очищенного аффинной хроматографией на сорбенте сефароза 4B-G4223-IgG-связывающий полипептид,
2 - проба исходной сыворотки,
3 - проба сыворотки, не связанная с сорбентом на колонке и освобожденная от IgG,
4 - маркер молекулярных весов (сверху вниз: 250, 130, 95, 72, 55, 36, 28, и 17 кДа).
Таким образом, создан сорбент сефароза 4В-IgG-связывающий G4223 полипептид, позволяющий получать очищенные препараты IgG. На полученном сорбенте с таким же успехом можно получать очищенные препараты IgG любых антител, полученных от млекопитающих, таких, как кролик, мышь или морская свинка, так как белок G вообще, а в данном случае и IgG-связывающий G4223 полипептид, обладает способностью связывать Fc часть иммуноглобулина G большинства видов млекопитающих.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ФГБНУ «ИЭМ» (FSBSI “IEM”)
<120> Рекомбинантная ДНК pG4223, рекомбинантная плазмидная ДНК pQE 30-рG4223 обеспечивающие получение полипептидов G4223, штамм Esсherichia coli M 15-G4223 трансформированный рекомбинантной плазмидной ДНК pQE 30-рG4223, экспрессирующей рекомбинантный полипептид G4223, рекомбинантный полипептид G4223, обладающий способностью селективно связывать IgG и его применение в качестве лиганда в аффинной хроматографии для выделения IgG из сыворотки крови.
<140> 2017146174
<141>26.12.2017
<150>
<151>
<160> 2
<210> 1
<211> 696
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность фрагмента гена IgG-связывающего полипептида G4223 Streptococcus группы G, экспрессированного в системе Escherichia coli
<400> 1
аtgagaggat cgcatcacca tcaccatcac ggatcccgag ctgcattacc taagactgac 60 acttacactt acaaattaat ccttaatggt aaaacattga aaggcgaaac aactactgaa 120 gctgttgatg ctgctactgc agaaaaagtc ttcaaacaat acgctaacga caacggtgtt 180 gacggtgaat ggacttacga cgatgcgaсt aagaccttta cagttactga aaaaccagaa 240 gtgatcgatg cgtctgaatt aacaccagcc gtgacaactt acaaacttgt tattaatggt 300 aaaacattga aaggcgaaac aactactgaa gctgttgatg ctgctactgc agaaaaagtc 360 ttcaaacaat acgctaacga caacggtgtt gacggtgaat ggacttacga cgatgcgact 420 aagaccttta cagttactga aaaaccagaa gtgatcgatg cgtctgaatt aacaccagcc 480 gtgacaactt acaaacttgt tattaatggt aaaacattga aaggcgaaac aactactaaa 540 gcagtagacg cagaaactgc agaaaaagcc ttcaaacaat acgctaacga caacggtgtt 600 gatggtgttt ggacttatga tgatgcgact aagaccttta cggtaactga aatggttagg 660 ggtaccccgg tcgacctgca gccaagctta attagc
<210> 2
<211> 232
<212> Аминокислотная последовательность
<213> Искусственная последовательность
<223> Аминокислотная последовательность IgG-связывающего полипептида G4223, содержащего область с тремя IgG-связывающими доменами.
<400> 2
Met Arg Gly Ser His His His His His His Gly Ser Arg Ala Ala Leu
5 10 15
Pro Lys Thr Asp Thr Tyr Thr Tyr Lys Leu Ile Leu Asn Gly Lys Thr
20 25 30
Leu Lys Gly Glu Thr Thr Thr Glu Ala Val Asp Ala Ala Thr Ala Glu
35 40 45
Lys Val Phe Lys Gln Tyr Ala Asn Asp Asn Gly Val Asp Gly Glu Trp
50 55 60
Thr Tyr Asp Asp Ala Thr Lys Thr Phe Thr Val Thr Glu Lys Pro Glu
65 70 75 80
Val Ile Asp Ala Ser Glu Leu Thr Pro Ala Val Thr Thr Tyr Lys Leu
85 90 95
Val Ile Asn Gly Lys Thr Leu Lys Gly Glu Thr Thr Thr Glu Ala Val
100 105 110
Asp Ala Ala Thr Ala Glu Lys Val Phe Lys Gln Tyr Ala Asn Asp Asn
115 120 125
Gly Val Asp Gly Glu Trp Thr Tyr Asp Asp Ala Thr Lys Thr Phe Thr
130 135 140
Val Thr Glu Lys Pro Glu Val Ile Asp Ala Ser Glu Leu Thr Pro Ala
145 150 155 160
Val Thr Thr Tyr Lys Leu Val Ile Asn Gly Lys Thr Leu Lys Gly Glu
165 170 175
Thr Thr Thr Lys Ala Val Asp Ala Glu Thr Ala Glu Lys Ala Phe Lys
180 185 190
Gln Tyr Ala Asn Asp Asn Gly Val Asp Gly Val Trp Thr Tyr Asp Asp
195 200 205
Ala Thr Lys Thr Phe Thr Val Thr Glu Met Val Arg Gly Thr Pro Val
210 215 220
Asp Leu Gln Pro Ser Leu Ile Ser
225 230
<---
Группа изобретений относится к биотехнологии и касается получения генетической конструкции, обеспечивающей синтез в клетках Escherichia coli рекомбинантного полипептида G4223 (рG4223). Предложены рекомбинантная ДНК pG4223, имеющая нуклеотидную последовательность SEQ ID NO: 1 и кодирующая рG4223, характеризующийся аминокислотной последовательностью SEQ ID NO: 2 и обладающий способностью селективно связывать человеческий иммуноглобулин G (IgG); рекомбинантная плазмидная ДНК pQE 30-pG4223, представляющая собой плазмиду pQE 30, несущую рекомбинантную ДНК pG4223, обеспечивающая получение рG4223; штамм E.coli M15-G4223, продуцирующий рG4223, полученный на основе штамма Е. coli М 15 и содержащий рекомбинантную плазмиду pQE30-pG4223; полипептид рG4223 с аминокислотной последовательностью SEQ ID NO: 2, обладающий способностью селективно связывать IgG, и его применение в качестве лиганда в аффинном сорбенте для удаления IgG из сыворотки крови и одновременно для получения очищенного препарата IgG. Изобретения позволяют получать IgG-связывающий рекомбинантный полипептид G4223, обладающий способностью селективно связывать Fc часть IgG большинства видов млекопитающих. 6 н.п. ф-лы, 1 табл., 4 ил., 6 пр.
1. Рекомбинантная ДНК pG4223, кодирующая рекомбинантный полипептид G4223, характеризующийся аминокислотной последовательностью SEQ ID NO: 2 и обладающий способностью селективно связывать человеческий иммуноглобулин G (IgG), имеющая нуклеотидную последовательность SEQ ID NO: 1.
2. Рекомбинантная плазмидная ДНК pQE 30-pG4223, представляющая собой плазмиду pQE 30, несущую рекомбинантную ДНК pG4223 по п. 1, обеспечивающая получение рекомбинантного полипептида G4223.
3. Штамм E.coli M15-G4223, продуцирующий рекомбинантный полипептид G4223, характеризующийся аминокислотной последовательностью SEQ ID NO: 2, причем штамм E.coli M15-G4223 получен на основе штамма Escherichia coli М15 и содержит рекомбинантную плазмидную ДНК pQE 30-pG4223 по п. 2.
4. Рекомбинантный полипептид G4223, экспрессируемый штаммом E.coli M15-G4223 по п. 3, обладающий способностью селективно связывать IgG и характеризующийся аминокислотной последовательностью SEQ ID NO: 2.
5. Аффинный сорбент для удаления IgG из сыворотки крови и одновременно для получения очищенного препарата IgG, состоящий из смеси матрицы, представляющей собой активированную цианобромом сефарозу 4В, предварительно промытую холодной 1мM HCl и 0,1 М бикарбонатным буфером, рН 8,3, и добавленного к ней лиганда - IgG-связывающего полипептида G4223 по п.4, которая проинкубирована ночь при 4°С с постоянным перемешиванием.
6. Применение рекомбинантного полипептида G4223 по п. 4 для удаления IgG из сыворотки крови и одновременно для получения очищенного препарата IgG человека с использованием аффинного сорбента по п. 5.

