Перекрестная ссылка на родственные заявки
[0001] Данная заявка испрашивает приоритет и преимущество по предварительной заявке на патент США № 62/418,468, поданной 7 ноября 2016 г., содержание которой включено в данный документ во всей своей полноте.
Область техники
[0002] Настоящее изобретение относится к композиции, содержащей последовательность рекомбинантной нуклеиновой кислоты, для создания in vivo одного или более синтетических антител к OspA и их функциональных фрагментов, а также к способу предотвращения и/или лечения бактериальной инфекции у субъекта посредством введения указанной композиции.
Уровень техники
[0003] Болезнь Лайма вызывается бактерией Borrelia burgdorferi и передается человеку через укус зараженного Ixodes scapularis (также называемого «клещ черноногий»). В настоящее время для лечения множества заболеваний одобрены терапевтические антитела. К сожалению, производство и доставка очищенных антител являются дорогостоящими. Кроме того, терапия антителами должна проводиться повторно переходя с еженедельного на ежемесячный прием, что является сложной задачей в обеспечении эффективного лечения для предотвращения или снижения риска развития хронической болезни Лайма у пациента.
[0004] Таким образом, в данной области существует потребность в улучшенных терапевтических средствах для предотвращения и/или лечения инфекции Borrelia burgdorferi и сопутствующей болезни Лайма. Настоящее изобретение удовлетворяет эту потребность.
Сущность изобретения
[0005] В одном варианте осуществления настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей одно или более синтетических антител, причем молекула нуклеиновой кислоты содержит по меньшей мере одно из: а) нуклеотидной последовательности, кодирующей синтетическое антитело к OspA, и b) нуклеотидной последовательности, кодирующей фрагмент синтетического антитела к OspA.
[0006] В одном варианте осуществления молекула нуклеиновой кислоты дополнительно содержит нуклеотидную последовательность, кодирующую домен расщепления.
[0007] В одном варианте осуществления молекула нуклеиновой кислоты кодирует по меньшей мере одну аминокислотную последовательность, выбранную из: а) аминокислотной последовательности по меньшей мере на около 95% идентичной по всей длине аминокислотной последовательности аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27; b) аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27; и c) фрагмента аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27.
[0008] В одном варианте осуществления молекула нуклеиновой кислоты содержит по меньшей мере одно из: a) нуклеотидной последовательности по меньшей мере на около 95% идентичной по всей длине последовательности нуклеиновой кислоты последовательности нуклеиновой кислоты SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 или SEQ ID NO:23; b) нуклеотидной последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 или SEQ ID NO:23; и c) фрагмента нуклеотидной последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 или SEQ ID NO:23.
В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более вариабельных областей тяжелой цепи и вариабельных областей легкой цепи. В одном варианте осуществления последовательность, кодирующая вариабельную область тяжелой цепи, выбрана из: a) нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO:4, SEQ ID NO:10, SEQ ID NO:16 или SEQ ID NO:22; b) нуклеотидной последовательности, кодирующей аминокислотную последовательность по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO:4, SEQ ID NO:10, SEQ ID NO:16 или SEQ ID NO:22, c) нуклеотидной последовательности SEQ ID NO:3, SEQ ID NO:9, SEQ ID NO:15 или SEQ ID NO:21, и d) нуклеотидной последовательности по меньшей мере на 95% идентичной нуклеотидной последовательности SEQ ID NO:3, SEQ ID NO:9, SEQ ID NO:15 или SEQ ID NO:21. В одном варианте осуществления последовательность, кодирующая вариабельную область легкой цепи, выбрана из: e) нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18 или SEQ ID NO:24; f) нуклеотидной последовательности, кодирующей аминокислотную последовательность по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18 или SEQ ID NO:24, g) нуклеотидной последовательности SEQ ID NO:5, SEQ ID NO:11, SEQ ID NO:17 или SEQ ID NO:23, и h) нуклеотидной последовательности по меньшей мере на 95% идентичной нуклеотидной последовательности SEQ ID NO:5, SEQ ID NO:11, SEQ ID NO:17 или SEQ ID NO:23.
[0009] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:2, SEQ ID NO:8, SEQ ID NO:14 и SEQ ID NO:20.
[0010] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более из: вариабельной области тяжелой цепи, содержащей SEQ ID NO: 4 и вариабельной области легкой цепи, содержащей SEQ ID NO: 6. В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO:2.
[0011] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 10 и вариабельной области легкой цепи, содержащей SEQ ID NO: 12. В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO:8.
[0012] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 16 и вариабельной области легкой цепи, содержащей SEQ ID NO: 18. В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO:14.
[0013] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 22 и вариабельной области легкой цепи, содержащей SEQ ID NO: 24. В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO:20.
[0014] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую одну или более из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 26 и вариабельной области легкой цепи, содержащей SEQ ID NO: 27. В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO:25.
[0015] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность по меньшей мере на около 95% идентичную по всей длине последовательности нуклеиновой кислоты одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 и SEQ ID NO:23.
[0016] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 и SEQ ID NO:23.
[0017] В одном варианте осуществления нуклеотидная последовательность кодирует лидерную последовательность.
[0018] В одном варианте осуществления молекула нуклеиновой кислоты представляет собой вектор экспрессии.
[0019] В одном варианте осуществления настоящее изобретение относится к молекуле аминокислоты, включающей одно или более синтетических антител, причем аминокислотная молекула содержит по меньшей мере одно из выбранного из группы, состоящей из аминокислотной последовательности, содержащей синтетическое антитело к OspA, и аминокислотной последовательности, содержащей фрагмент синтетического антитела к OspA.
[0020] В одном варианте осуществления аминокислотная молекула дополнительно содержит домен расщепления.
[0021] В одном варианте осуществления аминокислотная молекула содержит по меньшей мере одну аминокислотную последовательность, выбранную из: а) аминокислотной последовательности по меньшей мере на около 95% идентичную по всей длине аминокислотной последовательности аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27; b) аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27; и c) фрагмента аминокислотной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 или SEQ ID NO:27.
[0022] В одном варианте осуществления аминокислотная молекула содержит одну или более вариабельных областей тяжелой цепи и вариабельных областей легкой цепи. В одном варианте осуществления последовательность, содержащая вариабельную область тяжелой цепи, выбрана из a) аминокислотной последовательности SEQ ID NO:4, SEQ ID NO:10, SEQ ID NO:16 или SEQ ID NO:22; и b) аминокислотной последовательности по меньшей мере на 95% идентичной аминокислотной последовательности SEQ ID NO:4, SEQ ID NO:10, SEQ ID NO:16 или SEQ ID NO:22. В одном варианте осуществления последовательность, содержащая вариабельную область легкой цепи, выбрана из: c) аминокислотной последовательности SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18 или SEQ ID NO:24; и d) аминокислотной последовательности по меньшей мере на 95% идентичной аминокислотной последовательности SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18 или SEQ ID NO:24.
[0023] В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность SEQ ID NO:2, SEQ ID NO:8, SEQ ID NO:14 и SEQ ID NO:20.
[0024] В одном варианте осуществления аминокислотная молекула содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 4 и вариабельной области легкой цепи, содержащей SEQ ID NO: 6. В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность, как определено в SEQ ID NO:2.
[0025] В одном варианте осуществления аминокислотная молекула содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 10 и вариабельной области легкой цепи, содержащей SEQ ID NO: 12. В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность, как определено в SEQ ID NO:8.
[0026] В одном варианте осуществления аминокислотная молекула содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 16 и вариабельной области легкой цепи, содержащей SEQ ID NO: 18. В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность, как определено в SEQ ID NO:14.
[0027] В одном варианте осуществления аминокислотная молекула содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 22 и вариабельной области легкой цепи, содержащей SEQ ID NO: 24. В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность, как определено в SEQ ID NO:20.
[0028] В одном варианте осуществления аминокислотная молекула содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 26 и вариабельной области легкой цепи, содержащей SEQ ID NO: 27. В одном варианте осуществления аминокислотная молекула содержит аминокислотную последовательность, как определено в SEQ ID NO:25.
[0029] В одном варианте осуществления аминокислотная последовательность содержит лидерную последовательность.
[0030] В одном варианте осуществления настоящее изобретение относится к композиции, содержащей нуклеиновую кислоту, кодирующую одно или более синтетических антител, причем нуклеиновая кислота содержит по меньшей мере одно из: а) нуклеотидной последовательности, кодирующей синтетическое антитело к OspA, и b) нуклеотидной последовательности, кодирующей фрагмент синтетического антитела к OspA.
[0031] В одном варианте осуществления композиция дополнительно содержит фармацевтически приемлемое вспомогательное вещество.
[0032] В одном варианте осуществления настоящее изобретение относится к композиции, содержащей молекулу аминокислоты, включающую одно или более синтетических антител, причем аминокислотная молекула содержит по меньшей мере одно, выбранное из группы, состоящей из аминокислотной последовательности, содержащей синтетическое антитело к OspA, и аминокислотной последовательности, содержащей фрагмент синтетического антитела к OspA.
[0033] В одном варианте осуществления настоящее изобретение относится к способу предотвращения или лечения заболевания у субъекта, причем способ включает введение субъекту нуклеиновой кислоты, кодирующей одно или более синтетических антител, причем нуклеиновая кислота содержит по меньшей мере одно из: а) нуклеотидной последовательности, кодирующей синтетическое антитело к OspA, и b) нуклеотидной последовательности, кодирующей фрагмент синтетического антитела к OspA. В одном варианте осуществления способ включает введение субъекту композиции, содержащей нуклеиновую кислоту.
[0034] В одном варианте осуществления настоящее изобретение относится к способу предотвращения или лечения заболевания у субъекта, включающему введение субъекту аминокислотной молекулы, включающей одно или более синтетических антител, причем аминокислотная молекула содержит по меньшей мере одно, выбранное из группы, состоящей из аминокислотной последовательности, содержащей синтетическое антитело к OspA, и аминокислотной последовательности, содержащей фрагмент синтетического антитела к OspA.
[0035] В одном варианте осуществления заболевание представляет собой инфекцию, вызванную Borrelia. В одном варианте осуществления заболевание представляет собой болезнь Лайма.
[0036] В одном варианте осуществления способ дополнительно включает введение субъекту антибиотика.
Краткое описание графических материалов
[0037] На фиг. 1 проиллюстрирована схема, используемая для проведения испытания на иммуногенность при приведении в контакт с клещом. Пять мышей C3H на группу иммунизировали контрольным или тестируемым ДНК-моноклональным антителом (DMAb) за пять дней до приведения в контакт с клещом. Сыворотку собирали во время иммунизации или через 21 день после приведения в контакт с клещом.
[0038] На фиг. 2 проиллюстрированы результаты экспериментов, демонстрирующие иммунизацию мышей DMAb 319-44 wt, 319-44 mod1 или 221-7 wt, обеспечиваемых обнаруживаемых уровней антител в сыворотке на 3-й день, и что DMAb 319-44 wt обеспечивает 60% защиту от болезни Лайма, тогда как 319-44 mod1 DMAb обеспечивает 80% защиту.
[0039] На фиг. 3 проиллюстрированы результаты из испытания DMAb Лайма. Мыши, которым вводили DMAb 319-44 wt, 319-44 mod1 или 221-7 wt показали устойчивую реакцию анти-человеческого-IgG на 21-й день.
[0040] На фиг. 4 проиллюстрированы результаты экспериментов, демонстрирующие боррелиацидную активность DMAb против B. burgdorferi. Все четыре DMab (319-44 mod1, 319-44 wt, 221-7 mod9 и 221-7 wt) были боррелиацидными против B. burgdorferi.
[0041] На фиг. 5 проиллюстрированы результаты экспериментов, демонстрирующие то, что форма дозировки 319-44 DMAb приводит к повышению уровня антител in vivo. Результаты показывают уровни человеческого IgG у мышей C3H/HeNCrl, n=5/группу. Составленное 319-44mod1 (доза 300 мкг)=~ 7 мкг/мл в день 7.
[0042] На фиг. 6, включающей фигуры 6А-6С, проиллюстрированы результаты экспериментов, демонстрирующие тот факт, что трехуровневая стратегия оптимизации приводит к увеличению экспрессии in vivo 221-7 mod 9. На фиг. 6А проиллюстрировано время ввода пробы и анализа, использованное для экспериментов. На фиг. 6В проиллюстрировано результаты экспериментов, демонстрирующие то, что составленное 221-7 mod 9 DMAb вызывало более устойчивую реакцию анти-человеческого-IgG, чем не составленное 221-7 mod 9 DMAb. На фиг. 6C проиллюстрировано результаты экспериментов, демонстрирующие то, что составленное DMAb 221-7 mod 9 имело более высокий уровень связывания hisOspA, чем не составленное DMAb 221-7 mod 9.
[0043] На фиг. 7, включающей фигуры 7А-7В, проиллюстрированы результаты экспериментов, демонстрирующие то, что инъекция DMAb привела к образованию антитела Лайма in vivo. На фиг. 7А проиллюстрированы результаты экспериментов, демонстрирующие то, что инъекция DMAb 319-44 и, в меньшей степени, не составленного DMAb 221-7 wt, давала более устойчивую реакцию человеческого IgG, чем один вектор (pVax), начиная по меньшей мере через два дня после инъекции. На фиг. 7B проиллюстрированы результаты экспериментов, демонстрирующие то, что инъекция DMAb 319-44 и, в меньшей степени, не составленного 221-7 wt DMAb, имела более высокий уровень связывания hisOspA, чем один pVax.
[0044] На фиг. 8 изображен график процента защиты от последствий приведения в контакт с клещами, предоставленной мышам C3H, иммунизированным различными DMAb. Последнее представляет собой графическое изображение данных на фиг. 2.
Подробное описание изобретения
[0045] Настоящее изобретение относится к композициям, содержащим последовательность рекомбинантной нуклеиновой кислоты, кодирующую антитело, ее фрагмент, ее вариант или их комбинацию. Композиция может быть введена нуждающемуся в этом субъекту для облегчения in vivo экспрессии и образования синтетического антитела.
[0046] В частности, полипептиды тяжелой цепи и легкой цепи, экспрессируемые с последовательностей рекомбинантных нуклеиновых кислот, могут быть собраны в синтетическое антитело. Полипептид тяжелой цепи и полипептид легкой цепи могут взаимодействовать друг с другом так, что сборка приводит к получению синтетического антитела, способного связывать антиген, являющийся более иммуногенным, по сравнению с антителом, не прошедшим описанную в данном документе сборку, и способного вызывать или индуцировать иммунный ответ против антигена.
[0047] Кроме того, данные синтетические антитела более быстро генерируются в организме субъекта, чем антитела, которые вырабатываются в ответ на индуцированный антигеном иммунный ответ. Синтетические антитела способны эффективно связывать и нейтрализовать ряд антигенов. Синтетические антитела также способны эффективно защищать от и/или стимулировать выживаемость при заболевании.
Определения
[0048] Если не указано иное, все технические и научные термины, используемые в данном документе, имеют такое же значение, которое обычно подразумевается специалистом в данной области техники. В случае противоречия, приоритет имеет настоящий документ, включая определения. Предпочтительные способы и материалы описаны ниже, хотя способы и материалы, аналогичные или эквивалентные тем, которые описаны в данном документе, могут быть использованы на практике или при проверке настоящего изобретения. Все публикации, патентные заявки, патенты и другие ссылки, упоминаемые в данном документе, в полном объеме включены посредством ссылки. Описанные в данном документе материалы, способы и примеры являются исключительно иллюстративными и не подразумевают ограничения.
[0049] В контексте данного документа подразумевается, что термины «содержит (-ат)», «включает (-ют)», «имеющий», «имеет», «может» и их варианты являются открытыми переходными фразами, терминами или словами, которые не исключают возможности наличия дополнительных действий или структур. Формы единственного числа включают отсылки к множественному числу, если иное четко не следует из контекста. В настоящем изобретении также предусмотрены другие варианты осуществления, «содержащие», «состоящие из» и «состоящие преимущественно из» представленных в данном документе вариантов осуществления или элементов, приведенных явным образом или нет.
[0050] «Антитело» может означать антитело классов IgG, IgM, IgA, IgD или IgE или их фрагменты или производные, включая Fab, F(ab')2, Fd и одноцепочечные антитела и их фрагменты. Антитело может представлять собой антитело, выделенное из образца сыворотки млекопитающего, поликлональное антитело, прошедшее аффинную очистку антитело или их смеси, которые проявляют достаточную специфичность связывания с необходимым эпитопом или полученной из него последовательностью.
[0051] В контексте данного документа «фрагмент антитела» относится к части интактного антитела, содержащей антигенсвязывающий участок или вариабельную область. Эта часть не включает константные домены тяжелой цепи (т. е. CH2, CH3 или CH4 в зависимости от изотипа антитела) Fc-области интактного антитела. Примеры фрагментов антител включают, но не ограничиваются этим, фрагменты Fab, фрагменты Fab', фрагменты Fab'-SH, фрагменты F(ab')2, фрагменты Fd, фрагменты Fv, диатела, одноцепочечные Fv (scFv), одноцепочечные полипептиды, содержащие только один вариабельный домен легкой цепи, одноцепочечные полипептиды, содержащие три CDR вариабельного домена легкой цепи, одноцепочечные полипептиды, содержащие только одну вариабельную область тяжелой цепи, одноцепочечные полипептиды, содержащие три CDR вариабельной области тяжелой цепи.
[0052] «Антиген» относится к белкам, которые способны генерировать иммунный ответ в организме-хозяине. Антиген может распознаваться и связываться антителом. Антиген может иметь происхождение, связанное с организмом или внешней средой.
[0053] В контексте данного документа «кодирующая последовательность» или «кодирующая нуклеиновая кислота» относится к нуклеиновой кислоте (РНК или ДНК), которая содержит нуклеотидную последовательность, кодирующую приведенное в данном документе антитело. Кодирующая последовательность может дополнительно содержать сигналы инициации и терминации, функционально связанные с регуляторными элементами, включая промотор и сигнал полиаденилирования, способными управлять экспрессией в клетках индивида или млекопитающего, которому вводят нуклеиновую кислоту. Кодирующая последовательность может дополнительно содержать последовательности, которые кодируют сигнальные пептиды.
[0054] В контексте данного документа термин «комплементарная» может означать нуклеиновую кислоту, характеризующуюся Уотсон-Криковским (например, A-T/U и C-G) или Хугстиновским спариванием оснований между нуклеотидами или нуклеотидными аналогами нуклеиновых кислот.
[0055] В контексте данного документа термин «постоянный ток» используется для определения тока, проходящего или воздействующего на ткань или клетки, определяющие указанную ткань, в течение длительности электрического импульса, подаваемого на ту же самую ткань. Электрический импульс подается из описанных в данном документе устройств для электропорации. Ток остается постоянным в указанной ткани в течение действия электрического импульса, так как предложенное в данном документе устройство для электропорации имеет цепь обратной связи, предпочтительно характеризующуюся мгновенной обратной связью. Цепь обратной связи может измерять сопротивление ткани (или клеток) во время длительности импульса и давать команду устройству для электропорации для изменения вырабатываемой электрической энергии (например, повышения напряжения) так, чтобы ток в одной ткани оставался постоянным в течение электрического импульса (порядка микросекунд) и от импульса к импульсу. В некоторых вариантах осуществления цепь обратной связи содержит контроллер.
[0056] В контексте данного документа термины «обратная связь по току» или «обратная связь» могут использоваться взаимозаменяемо и могут означать активный ответ предложенных устройств для электропорации, который включает измерение тока в ткани между электродами и соответствующее изменение вырабатываемой устройством ЭП энергии, чтобы поддерживать ток на постоянном уровне. Этот постоянный уровень задается пользователем до инициации последовательности импульсов или электрической обработки. Обратная связь может осуществляться электропорационным компонентом, например, контроллером, устройства для электропорации, так как электрический контур в нем может непрерывно отслеживать ток в ткани между электродами и сравнивать этот отслеживаемый ток (или ток в ткани) с заданным током и непрерывно проводить корректировку вырабатываемой энергии для поддержания отслеживаемого тока на заданном уровне. Контур обратной связи может характеризоваться мгновенным действием, так как он является аналоговым замкнутым контуром обратной связи.
[0057] В контексте данного документа термин «децентрализованный ток» может означать профиль электрического тока, подаваемого из различных матриц с игольчатыми электродами описанных в данном документе устройств для электропорации, при этом профили минимизируют или, предпочтительно, устраняют появление связанного с электропорацией теплового стресса в любой области электропорируемой ткани.
[0058] В контексте данного документа взаимозаменяемые термины «электропорация», «электропермеабилизация» или «электрокинетическое усиление» («ЭП») могут относиться к применению трансмембранных электрических импульсов для индукции микроскопических путей (пор) в биомембране; их наличие позволяет биомолекулам, таким как плазмиды, олигонуклеотиды, миРНК, лекарства, ионы и вода, проходить с одной стороны клеточной мембраны на другую.
[0059] В контексте данного документа термин «эндогенное антитело» может относиться к антителу, генерируемому у субъекта, которому вводят эффективную дозу антигена для индукции гуморального иммунного ответа.
[0060] В контексте данного документа термин «механизм обратной связи» может относиться к процессу, осуществляемому программным обеспечением или аппаратным обеспечение (или техническим обеспечением), который заключается в получении и сравнении импеданса необходимой ткани (до, во время и/или после подачи импульса энергии) с заданным значением, предпочтительно током, и коррекции подаваемого импульса энергии для достижения заданного значения. Механизм обратной связи может осуществляться аналоговым замкнутым контуром.
[0061] «Фрагмент» может обозначать полипептидный фрагмент антитела, который является функциональным, т. е. может связываться с необходимой мишенью и имеет такое же предполагаемое действие, что и полноразмерное антитело. Фрагмент антитела может быть на 100% идентичным полноразмерному антителу, за исключением отсутствия по меньшей мере одной аминокислоты в N- и/или С-конце, в каждом случае с сигнальными пептидами и/или метионином в позиции 1 или без. Фрагменты могут содержать 20% или более, 25% или более, 30% или более, 35% или более, 40% или более, 45% или более, 50% или более, 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более процентов длины конкретного полноразмерного антитела, за исключением добавления любого гетерологичного сигнального пептида. Фрагмент может содержать фрагмент полипептида, который на 95% или более, 96% или более, 97% или более, 98% или более или 99% или более идентичен антителу, и дополнительно содержать N-концевой метионин или гетерологичный сигнальный пептид, который не был включен при расчете процента идентичности. Фрагменты могут дополнительно содержать N-концевой метионин и/или сигнальный пептид, такой как сигнальный пептид иммуноглобулина, например, сигнальный пептид IgE или IgG. N-концевой метионин и/или сигнальный пептид могут быть связаны с фрагментом антитела.
[0062] Фрагмент последовательности нуклеиновой кислоты, которая кодирует антитело, может быть на 100% идентичным полноразмерной последовательности, за исключением отсутствия по меньшей мере одного нуклеотида в 5' и/или 3' конце, в каждом случае с последовательностями, кодирующими сигнальные пептиды и/или метионин в позиции 1, или без. Фрагменты могут содержать 20% или более, 25% или более, 30% или более, 35% или более, 40% или более, 45% или более, 50% или более, 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более процентов длины конкретной полноразмерной кодирующей последовательности, за исключением добавления любого гетерологичного сигнального пептида. Фрагмент может содержать фрагмент, который кодирует полипептид, который на 95% или более, 96% или более, 97% или более, 98% или более или 99% или более идентичен антителу, и дополнительно, необязательно, содержать последовательность, кодирующую N-концевой метионин или гетерологичный сигнальный пептид, который не был включен при расчете процента идентичности. Фрагменты могут дополнительно содержать последовательности для N-концевого метионина и/или сигнального пептида, такого как сигнальный пептид иммуноглобулина, например, сигнальный пептид IgE или IgG. Кодирующая последовательность, кодирующая N-концевой метионин и/или сигнальный пептид, может быть связана с фрагментом кодирующей последовательности.
[0063] В контексте данного документа термин «генетическая конструкция» относится к молекулам ДНК или РНК, которые содержат нуклеотидную последовательность, которая кодирует белок, такой как антитело. Кодирующая последовательность содержит сигналы инициации и терминации, функционально связанные с регуляторными элементами, включая промотор и сигнал полиаденилирования, способными управлять экспрессией в клетках индивида, которому вводят нуклеиновую кислоту. В контексте данного документа термин «форма с возможностью экспрессии» относится к генным конструкциям, которые содержат необходимые регуляторные элементы, функционально связанные с кодирующей последовательностью, которая кодирует белок так, что в случае присутствия в клетке индивида будет происходить экспрессия кодирующей последовательности.
[0064] Употребляемые в данном документе термины «идентичная» или «идентичность» в контексте двух или более последовательностей нуклеиновых кислот или полипептидов могут означать, что последовательности имеют заданный процент остатков, являющихся одинаковыми на протяжении заданной области. Процент можно рассчитывать путем оптимального выравнивания двух последовательностей, сравнения двух последовательностей на протяжении заданной области, определения числа позиций, в которых в обеих последовательностях находится идентичный остаток, для получения числа совпадающих позиций, деления числа совпадающих позиций на общее число позиций в заданной области и умножения результата на 100 для получения процента идентичности последовательностей. В случае, когда две последовательности имеют разную длину или выравнивание приводит к получению одного или более ступенчатых концов и заданная область сравнения включает только одну последовательность, остатки этой одной последовательности включаются в знаменатель, но не в числитель при расчете. При сравнении ДНК и РНК тимин (T) и урацил (U) могут считаться эквивалентными. Идентичность можно оценивать вручную, или используя компьютерный алгоритм выравнивания последовательностей, такой как BLAST или BLAST 2.0.
[0065] В контексте данного документа термин «импеданс» может использоваться при обсуждении механизма обратной связи и может быть преобразован в значение силы тока по закону Ома, делая возможным сравнение с текущим значением силы тока.
[0066] В контексте данного документа термин «иммунный ответ» может означать активацию иммунной системы организма-хозяина, например, млекопитающего, в ответ на внесение одной или более нуклеиновых кислот и/или пептидов. Иммунный ответ может иметь форму клеточного или гуморального ответа или их обоих.
[0067] В контексте данного документа термины «нуклеиновая кислота» или «олигонуклеотид», или «полинуклеотид» могут означать по меньшей мере два нуклеотида, ковалентно связанных вместе. Описание одной цепи также определяет последовательность комплементарной цепи. Таким образом, нуклеиновая кислота также включает комплементарную цепь описанной одной цепи. Много вариантов нуклеиновой кислоты можно использовать в тех же целях, что и заданную нуклеиновую кислоту. Таким образом, нуклеиновая кислота также включает по существу идентичные нуклеиновые кислоты и их комплементарные последовательности. Одна цепь обеспечивает зонд, который может гибридизироваться с последовательностью-мишенью в жестких условиях гибридизации. Таким образом, нуклеиновая кислота также включает зонд, который гибридизируется в жестких условиях гибридизации.
[0068] Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными или могут содержать части как двухцепочечной, так и одноцепочечной последовательности. Нуклеиновая кислота может представлять собой ДНК, как геномную, так и кДНК, РНК или гибрид, когда нуклеиновая кислота может содержать комбинации дезоксирибо- и рибонуклеотидов, и комбинации оснований, включая урацил, аденин, тимин, цитозин, гуанин, инозин, ксантин, гипоксантин, изоцитозин и изогуанин. Нуклеиновые кислоты можно получать методами химического синтеза или рекомбинантными методами.
[0069] В контексте данного документа выражение «функционально связанный» может означать, что экспрессия гена находится под управлением промотора, с которым он пространственно соединен. Промотор может располагаться 5' (выше) или 3' (ниже) гена, находящегося под его управлением. Расстояние между промотором и геном может быть приблизительно таким же самым, что и расстояние между этим промотором и геном, которым он управляет, в гене, из которого получен промотор. Как известно в данной области техники это расстояние можно варьировать без потери функциональности промотора.
[0070] В контексте данного документа термины «пептид», «белок» или «полипептид» могут означать связанную последовательность аминокислот и могут быть природными, синтетическими или представлять собой модификацию или комбинацию природных и синтетических.
[0071] В контексте данного документа термин «промотор» может означать синтетическую или полученную из природного источника молекулу, которая способна обеспечивать, активировать или усиливать экспрессию нуклеиновой кислоты в клетке. Промотор может содержать одну или более специфических транскрипционных регуляторных последовательностей для дополнительного усиления экспрессии и/или для изменения пространственной экспрессии и/или временной экспрессии. Промотор также может содержать удаленные энхансерные или репрессорные элементы, которые могут быть расположены на расстоянии до нескольких тысяч пар оснований от начала участка транскрипции. Промотор может быть получен из источников, включая вирусы, бактерии, грибы, растения, насекомых и животных. Промотор может регулировать экспрессию генного компонента конститутивным или дифференциальным образом по отношению к клетке, ткани или органу, в которых происходит экспрессия, или по отношению к стадии развития, на которой происходит экспрессия, или в ответ на внешние стимулы, такие как физиологический стресс, патогены, ионы металлов или индуцирующие агенты. Типовые примеры промоторов включают промотор бактериофага T7, промотор бактериофага T3, промотор SP6, оператор-промотор lac, промотор tac, поздний промотор SV40, ранний промотор SV40, промотор RSV-LTR, промотор CMV IE, ранний промотор SV40 или поздний промотор SV40 и промотор CMV IE.
[0072] Термины «сигнальный пептид» и «лидерная последовательность» взаимозаменяемо используются в данном документе и относятся к аминокислотной последовательности, которая может быть связана с амино-концом приведенного в данном документе белка. Сигнальные пептиды/лидерные последовательности, как правило, управляют локализацией белка. Используемые в данном документе сигнальные пептиды/лидерные последовательности предпочтительно облегчают секрецию белка из клетки, в которой он вырабатывается. После секреции из клетки сигнальные пептиды/лидерные последовательности часто отщепляются от остатка белка, часто называемого зрелым белком. Сигнальные пептиды/лидерные последовательности связаны с N-концом белка.
[0073] В контексте данного документа «жесткие условия гибридизации» могут означать условия, в которых первая последовательность нуклеиновой кислоты (например, зонд) будет гибридизироваться со второй последовательностью нуклеиновой кислоты (например, мишенью), например, в комплексной смеси нуклеиновых кислот. Жесткие условия зависят от последовательности и будут разными в разных обстоятельствах. Жесткие условия можно выбрать так, что они были на около 5-10°C ниже температуры плавления (Tп) для конкретной последовательности при определенных ионной силе и pH. Tп может представлять собой температуру (при определенных ионной силе, pH и концентрации нуклеиновой кислоты), при которой 50% зондов, комплементарных мишени, гибридизируются с последовательностью-мишенью в равновесном состоянии (так как последовательности-мишени присутствуют в избытке, при Tп, 50% зондов оказываются занятыми в равновесном состоянии). Жесткие условия могут быть такими, при которых концентрация соли составляет менее чем около 1,0 M ионов натрия, например, около 0,01-1,0 M концентрация ионов натрия (или других солей) при pH 7,0-8,3, а температура составляет по меньшей мере около 30°C для коротких зондов (например, около 10-50 нуклеотидов) и по меньшей мере около 60°C для длинных зондов (например, более чем около 50 нуклеотидов). Жесткие условия также можно обеспечить путем добавления дестабилизирующих агентов, таких как формамид. Для избирательной или специфической гибридизации положительный сигнал может по меньшей мере в 2-10 раз превышать фоновую гибридизацию. Примеры жестких условий гибридизации включает следующие: 50% формамид, 5x SSC и 1% ДСН, инкубация при 42°C, или 5x SSC, 1% ДСН, инкубация при 65°C с промывкой в 0,2x SSC и 0,1% ДСН при 65°C.
[0074] В контексте данного документа взаимозаменяемо используемые термины «субъект» и «пациент» относятся к любому позвоночному, включая, но не ограничиваясь этим, млекопитающее (например, корову, свинью, верблюда, ламу, лошадь, козу, кролика, овцу, хомяков, морскую свинку, кошку, собаку, крысу и мышь, отличного от человека примата (например, обезьяну, такую как яванский или резус-макак, шимпанзе и т. д.) и человека). В некоторых вариантах осуществления субъект может представлять собой человека или не человека. Субъект или пациент может проходить другие формы лечения.
[0075] В контексте данного документа выражение «по существу комплементарная» может означать, что первая последовательность по меньшей мере на 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична комплементарной цепи второй последовательности на протяжении 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или более нуклеотидов или аминокислот, или что две последовательности гибридизируются в жестких условиях гибридизации.
[0076] В контексте данного документа выражение «по существу идентичные» может означать, что первая и вторая последовательность идентичны по меньшей мере на 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% на протяжении области из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100 или более нуклеотидов или аминокислот, или, в случае нуклеиновых кислот, что первая последовательность является по существу комплементарной с комплементарной цепью второй последовательности.
[0077] В контексте данного документа термин «синтетическое антитело» относится к антителу, которое кодируется последовательностью рекомбинантной нуклеиновой кислоты, описанной в данном документе, и вырабатывается в организме субъекта.
[0078] В контексте данного документа термин «лечение» может означать защиту субъекта от заболевания посредством предотвращения, супрессии, подавления или полного устранения заболевания. Предотвращение заболевания включает введение субъекту вакцины согласно настоящему изобретению до начала заболевания. Супрессия заболевания включает введение субъекту вакцины согласно настоящему изобретению после индукции заболевания, но до его клинического проявления. Подавление заболевания включает введение субъекту вакцины согласно настоящему изобретению после клинического проявления заболевания.
[0079] Термин «вариант», используемый в данном документе в отношении нуклеиновой кислоты, может означать (i) часть или фрагмент указанной нуклеотидной последовательности; (ii) комплементарную цепь указанной нуклеотидной последовательности или ее часть; (iii) нуклеиновую кислоту, которая является по существу идентичной указанной нуклеиновой кислоте или ее комплементарной цепи; или (iv) нуклеиновую кислоту, которая гибридизируется в жестких условиях с указанной нуклеиновой кислотой, ее комплементарной цепью или последовательностью, по существу идентичной ей.
[0080] Термин «вариант» в отношении пептида или полипептида, который отличается по аминокислотной последовательности вследствие вставки, делеции или консервативной замены аминокислот, но сохраняет по меньшей мере один вид биологической активности. Вариант также может означать белок с аминокислотной последовательностью, которая является по существу идентичной указанному белку с аминокислотной последовательностью, который сохраняет по меньшей мере один вид биологической активности. Консервативные замены аминокислот, т. е. замещение аминокислоты другой аминокислотой с аналогичными свойствами (например, гидрофильностью, степенью и распределением заряженных областей), известны в данной области техники как включающие, как правило, незначительные изменения. Эти незначительные изменения можно частично определить на основании индекса гидропатичности аминокислот, как известно в данной области техники. Kyte с соавт., J. Mol. Biol. 157:105-132 (1982). Индекс гидропатичности аминокислоты основан на оценке ее гидрофобности и заряда. В данной области техники известно, что аминокислоты с аналогичными индексами гидропатичности можно взаимно заменять с сохранением функции белка. В одном аспекте проводят замену аминокислот, имеющих индекс гидропатичности ± 2. Гидрофильность аминокислот также можно использовать для определения замен, которые бы привели к получению белков, сохраняющих биологическую функцию. Учет гидрофильности аминокислот в контексте пептида позволяет рассчитывать наибольшую локальную среднюю гидрофильность этого пептида, что является полезным показателем, который, как сообщалось, хорошо коррелирует с антигенностью и иммуногенностью. Патент США № 4554101, в полном объеме включенный в данный документ посредством ссылки. Замена аминокислот, имеющих аналогичные значения гидрофильности, может привести к получению пептидов, сохраняющих биологическую активность, например, иммуногенность, как известно в данной области техники. Замены можно проводить с аминокислотами, имеющими значения гидрофильности в пределах ± 2 относительно друг друга. Как на индекс гидрофобности, так и на значение гидрофильности аминокислот влияет конкретный тип боковой цепи этой аминокислоты. В соответствии с этим наблюдением понятно, что аминокислотные замены, совместимые с биологической функцией, зависят от относительного сходства аминокислот и, в частности, боковых цепей этих аминокислот, проявляемого в гидрофобности, гидрофильности, заряде, размере и других свойствах.
[0081] Вариант может представлять собой последовательность нуклеиновой кислоты, которая является по существу идентичной на протяжении всей длины последовательности всего гена или ее части. Последовательность нуклеиновой кислоты может быть на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной на протяжении всей длины последовательности гена или ее части. Вариант может представлять собой аминокислотную последовательность, которая является по существу идентичной на протяжении всей длины аминокислотной последовательности или ее части. Аминокислотная последовательность может быть на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной на протяжении всей длины аминокислотной последовательности или ее части.
[0082] В контексте данного документа термин «вектор» может означать последовательность нуклеиновой кислоты, содержащую точку начала репликации. Вектор может представлять собой плазмиду, бактериофаг, бактериальную искусственную хромосому или дрожжевую искусственную хромосому. Вектор может представлять собой ДНК- или РНК-вектор. Вектор может представлять собой как самореплицирующийся внехромосомный вектор, так и вектор, который интегрируется в геном организма-хозяина.
[0083] В случае перечисления в данном документе числовых диапазонов явным образом и с той же степенью точности подразумевается и каждое промежуточное число. Например, в случае диапазона 6-9, кроме чисел 6 и 9 подразумеваются числа 7 и 8, а в случае диапазона 6,0-7,0, явным образом подразумеваются числа 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 и 7,0.
Композиция
[0084] Настоящее изобретение относится к композиции, содержащей последовательность рекомбинантной нуклеиновой кислоты, кодирующую антитело, его фрагмент, его вариант или их комбинацию. При введении нуждающемуся в этом субъекту композиция может приводить к генерации синтетического антитела в организме субъекта. Синтетическое антитело может связывать молекулу-мишень (т. е. антиген), присутствующую у субъекта. Такое связывание может нейтрализовать антиген, блокировать распознавание антигена другой молекулой, например, белком или нуклеиновой кислотой, и вызывать или индуцировать иммунный ответ на антиген.
[0085] В одном варианте осуществления композиция содержит нуклеотидную последовательность, кодирующую синтетическое антитело. В одном варианте осуществления композиция содержит нуклеиновую кислоту, содержащую первую нуклеотидную последовательность, кодирующую первое синтетическое антитело, и вторую нуклеотидную последовательность, кодирующую второе синтетическое антитело. В одном варианте осуществления нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую домен расщепления.
[0086] В одном варианте осуществления предложена композиция одной или более нуклеиновых кислот, кодирующих одну или более тяжелых цепей и легких цепи синтетического антитела. В одном варианте осуществления молекула нуклеиновой кислоты содержит последовательность, кодирующую лидерный пептид тяжелой цепи или легкой цепи. В одном варианте осуществления композиция содержит первую нуклеиновую кислоту, кодирующую тяжелую цепь синтетического антитела, и вторую нуклеиновую кислоту, кодирующую легкую цепь синтетического антитела. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая тяжелую цепь синтетического антитела, содержит последовательность, кодирующую сигнальный пептид тяжелой цепи IgG человека, вариабельную тяжелую область и константную тяжелую область. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая легкую цепь синтетического антитела, содержит последовательность, кодирующую сигнальный пептид человеческой каппа легкой цепи, вариабельную легкую область и константную легкую область.
[0087] В одном варианте осуществления композиция содержит одну нуклеиновую кислоту, кодирующую как тяжелую цепь, так и легкую цепь синтетического антитела. В одном варианте осуществления молекула нуклеиновой кислоты содержит последовательность, кодирующую лидерный пептид как тяжелой цепи, так и легкой цепи. В одном варианте осуществления композиция содержит одну молекулу нуклеиновой кислоты, кодирующую: сигнальный пептид тяжелой цепи IgG человека, вариабельную область тяжелой цепи, константную область тяжелой цепи, участок расщепления фурином, линкер GSG, пептид P2A, сигнальный пептид человеческой каппа легкой цепи, вариабельную легкую область и константную легкую область.
[0088] В одном варианте осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую антитело к OspA. В одном варианте осуществления антитело к OspA представляет собой DMAb-319-44 mod1. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-319-44 mod1, содержит одну или более кодон-оптимизированные нуклеотидные последовательности, кодирующие вариабельные области VH и VL SEQ ID NO:4 и SEQ ID NO:6 соответственно. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-319-44 mod1, кодирует аминокислотную последовательность, приведенную в SEQ ID NO:2.
[0089] В одном варианте осуществления антитело к OspA представляет собой DMAb-319-44 wt. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-319-44 wt, содержит одну или более кодон-оптимизированные нуклеотидные последовательности, кодирующие вариабельные области VH и VL SEQ ID NO:10 и SEQ ID NO:12 соответственно. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-319-44 wt, кодирует аминокислотную последовательность, приведенную в SEQ ID NO:8.
[0090] В одном варианте осуществления антитело к OspA представляет собой DMAb-221-7 mod9. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-221-7 mod9, содержит одну или более кодон-оптимизированные нуклеотидные последовательности, кодирующие вариабельные области VH и VL SEQ ID NO:16 и SEQ ID NO:18 соответственно. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-221-7 mod9, кодирует аминокислотную последовательность, приведенную в SEQ ID NO:14.
[0091] В одном варианте осуществления антитело к OspA представляет собой DMAb-221-7 wt. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-221-7 wt, содержит одну или более кодон-оптимизированные нуклеотидные последовательности, кодирующие вариабельные области VH и VL SEQ ID NO:22 и SEQ ID NO:24 соответственно. В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело DMAb-221-7, кодирует аминокислотную последовательность, приведенную в SEQ ID NO:20.
[0092] В одном варианте осуществления нуклеотидная последовательность, кодирующая мышиное антитело DMAb-221-7 mod9, содержит одну или более кодон-оптимизированные нуклеотидные последовательности, кодирующие вариабельные области VH и VL SEQ ID NO:26 и SEQ ID NO:27 соответственно. В одном варианте осуществления нуклеотидная последовательность, кодирующая мышиное антитело DMAb-221-7 mod9, кодирует аминокислотную последовательность, приведенную в SEQ ID NO:25.
[0093] В одном варианте осуществления нуклеотидная последовательность, кодирующая антитело к OspA, содержит последовательность, выбранную из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 и SEQ ID NO:23.
[0094] Композиция согласно настоящего изобретения может обеспечивать лечение, предотвращение и/или защиту от заболевания, расстройства или состояния, связанного с бактериальной активностью бактерии, экспрессирующей белок OspA. В некоторых вариантах реализации композиция может обеспечивать лечение, предотвращение и/или защиту от бактериальной инфекции. В некоторых вариантах реализации композиция может обеспечивать лечение, предотвращение и/или защиту от инфекции, вызванной Borrelia spp. В некоторых вариантах реализации композиция может обеспечивать лечение, предотвращение и/или защиту от болезни Лайма.
[0095] Синтетическое антитело может обеспечивать лечение, предотвращение и/или защиту от заболевания у субъекта, которому вводят композицию. Синтетическое антитело посредством связывания антигена может обеспечивать лечение, предотвращение и/или защиту от заболевания у субъекта, которому вводят композицию. Синтетическое антитело может способствовать выживаемости при заболевании у субъекта, которому вводят композицию. Синтетическое антитело может обеспечивать по меньшей мере около 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% выживаемости при заболевании у субъекта, которому вводят композицию.
[0096] Композиция может приводить к генерации синтетического антитела в организме субъекта в течение по меньшей мере около 1 часа, 2 часов, 3 часов, 4 часов, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов, 10 часов, 11 часов, 12 часов, 13 часов, 14 часов, 15 часов, 20 часов, 25 часов, 30 часов, 35 часов, 40 часов, 45 часов, 50 часов или 60 часов после введения композиции субъекту. Композиция может приводить к генерации синтетического антитела в организме субъекта в течение по меньшей мере около 1 суток, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 7 суток, 8 суток, 9 суток или 10 суток после введения композиции субъекту. Композиция может приводить к генерации синтетического антитела в организме субъекта в течение от около 1 часа до около 6 суток, от около 1 часа до около 5 суток, от около 1 часа до около 4 суток, от около 1 часа до около 3 суток, от около 1 часа до около 2 суток, от около 1 часа до около 1 суток, от около 1 часа до около 72 часов, от около 1 часа до около 60 часов, от около 1 часа до около 48 часов, от около 1 часа до около 36 часов, от около 1 часа до около 24 часов, от около 1 часа до около 12 часов или от около 1 часа до около 6 часов после введения композиции субъекту.
[0097] При введении нуждающемуся в этом субъекту композиция может приводить к генерации синтетического антитела в организме субъекта быстрее, чем происходит генерация эндогенного антитела у субъекта, которому вводят антиген, чтобы индуцировать гуморальный иммунный ответ. Композиция может приводить к генерации синтетического антитела по меньше мере на около 1 сутки, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 7 суток, 8 суток, 9 суток или 10 суток раньше генерации эндогенного антитела у субъекта, которому вводят антиген, чтобы индуцировать гуморальный иммунный ответ.
[0098] Композиция согласно настоящему изобретению может иметь характеристики, необходимые для эффективных композиций, такие как безопасность в том смысле, что композиция не приводит к болезни или смерти; защита от болезни; обеспечение легкости введения, небольшое количество побочных явлений, биологическая стабильность и низкая стоимость, приходящаяся на дозу.
Последовательность рекомбинантной нуклеиновой кислоты
[0099] Как описано выше, композиция может содержать последовательность рекомбинантной нуклеиновой кислоты. Последовательность рекомбинантной нуклеиновой кислоты может кодировать антитело, его фрагмент, его вариант или их комбинацию. Антитело более подробно описано ниже.
[00100] Последовательность рекомбинантной нуклеиновой кислоты может представлять собой гетерологичную последовательность нуклеиновой кислоты. Последовательность рекомбинантной нуклеиновой кислоты может содержать одну или более гетерологичных последовательностей нуклеиновой кислоты.
[00101] Последовательность рекомбинантной нуклеиновой кислоты может представлять собой оптимизированную последовательность нуклеиновой кислоты. Такая оптимизация может повышать или изменять иммуногенность антитела. Оптимизация также может улучшать транскрипцию и/или трансляцию. Оптимизация может включать в себя одно или более из следующего: лидерную последовательность с низким содержанием GC для повышения транскрипции; стабильность мРНК и оптимизацию кодонов; добавление последовательности Козака (например, GCC ACC) для повышения трансляции; добавление лидерной последовательности иммуноглобулина (Ig), кодирующей сигнальный пептид; добавление внутренней последовательности IRES и удаление по мере возможности цис-действующих мотивов последовательностей (т. е. внутренних TATA-боксов).
Конструкция последовательности рекомбинантной нуклеиновой кислоты
[00102] Последовательность рекомбинантной нуклеиновой кислоты может содержать одну или более конструкций последовательности рекомбинантной нуклеиновой кислоты. Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более компонентов, которые более подробно описаны ниже.
[00103] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, которая кодирует полипептид тяжелой цепи, его фрагмент, его вариант или их комбинацию. Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, которая кодирует полипептид легкой цепи, его фрагмент, его вариант или их комбинацию. Конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать гетерологичную последовательность нуклеиновой кислоты, которая кодирует участок расщепления протеазой или пептидазой. Конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать гетерологичную последовательность нуклеиновой кислоты, которая кодирует участок внутренней посадки рибосомы (IRES). IRES может представлять собой IRES вируса или IRES эукариотической клетки. Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать одну или более лидерных последовательностей, причем лидерная последовательность кодирует сигнальный пептид. Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более промоторов, один или более интронов, одну или более областей терминации транскрипции, один или более инициирующих кодонов, один или более терминирующих или стоп-кодонов и/или один или более сигналов полиаденилирования. Последовательность рекомбинантной нуклеиновой кислоты также может содержать одну или более линкерных или метящих последовательностей. Метящая последовательность может кодировать гемагглютининовую (ГА) метку.
Полипептид тяжелой цепи
[00104] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную нуклеиновую кислоту, кодирующую полипептид тяжелой цепи, его фрагмент, его вариант или их комбинацию. Полипептид тяжелой цепи может содержать вариабельную область тяжелой цепи (VH) и/или по меньшей мере одну константную область тяжелой цепи (CH). По меньшей мере одна константная область тяжелой цепи может содержать константную область тяжелой цепи 1 (CH1), константную область тяжелой цепи 2 (CH2) и константную область тяжелой цепи 3 (CH3) и/или шарнирную область.
[00105] В некоторых вариантах осуществления полипептид тяжелой цепи может содержать область VH и область CH1. В других вариантах осуществления полипептид тяжелой цепи может содержать область VH, область CH1, шарнирную область, область CH2 и область CH3.
[00106] Полипептид тяжелой цепи может содержать набор определяющих комплементарность областей («CDR»). Набор CDR может содержать три гипервариабельные области из области VH. Начиная с N-конца полипептида тяжелой цепи, эти CDR обозначаются «CDR1», «CDR2» и «CDR3» соответственно. CDR1, CDR2 и CDR3 полипептида тяжелой цепи могут вносить свой вклад в связывание или распознавание антигена.
Полипептид легкой цепи
[00107] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную нуклеиновую кислоту, кодирующую полипептид легкой цепи, его фрагмент, его вариант или их комбинацию. Полипептид легкой цепи может содержать вариабельную область легкой цепи (VL) и/или константную область легкой цепи (CL).
[00108] Полипептид легкой цепи может содержать набор определяющих комплементарность областей («CDR»). Набор CDR может содержать три гипервариабельные области из области VL. Начиная с N-конца полипептида легкой цепи, эти CDR обозначаются «CDR1», «CDR2» и «CDR3» соответственно. CDR1, CDR2 и CDR3 полипептида легкой цепи могут вносить свой вклад в связывание или распознавание антигена.
Участок расщепления протеазой
[00109] Конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать гетерологичную нуклеиновую кислоту, кодирующую участок расщепления протеазой. Участок расщепления протеазой может распознаваться протеазой или пептидазой. Протеаза может представлять собой эндопептидазу или эндопротеазу, например, но не ограничиваясь этим, фурин, эластазу, HtrA, кальпаин, трипсин, химотрипсин, трипсин и пепсин. Протеаза может представлять собой фурин. В других вариантах осуществления протеаза может представлять собой сериновую протеазу, треониновую протеазу, цистеиновую протеазу, аспартатную протеазу, металлопротеазу, протеазу глутаминовой кислоты или любую протеазу, которая расщепляет внутреннюю пептидную связь (т. е. не расщепляет N-концевую или С-концевую пептидную связь).
[00110] Участок расщепления протеазой может содержать одну или более аминокислотных последовательностей, которые стимулируют расщепление или повышают его эффективность. Одна или более аминокислотных последовательностей могут стимулировать или повышать эффективность образования или генерации дискретных полипептидов. Одна или более аминокислотных последовательностей могут содержать 2А пептидную последовательность.
Линкерная последовательность
[00111] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать одну или более линкерных последовательностей. Линкерная последовательность может пространственно разделять или связывать один или более описанных в данном документе компонентов. В других вариантах осуществления линкерная последовательность может кодировать аминокислотную последовательность, которая пространственно разделяет или связывает два или более полипептидов.
Промотор
[00112] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более промоторов. Один или более промоторов могут представлять собой любой промотор, способный управлять генной экспрессией и регулировать генную экспрессию. Такой промотор представляет собой цис-действующий элемент последовательности, необходимый для транскрипции с помощью ДНК-зависимой РНК-полимеразы. Выбор промотора, применяемого для управления генной экспрессией, зависит от конкретного применения. Промотор может располагаться приблизительно на таком же расстоянии от точки начала транскрипции в конструкции последовательности рекомбинантной нуклеиновой кислоты, что и от точки начала транскрипции в своем природном окружении. При этом это расстояние можно варьировать без потери функциональности промотора.
[00113] Промотор может быть функционально связан с гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи и/или полипептид легкой цепи. Промотор может представлять собой промотор, эффективный для экспрессии в эукариотических клетках. Промотор, функционально связанный с кодирующей последовательностью, может представлять собой промотор ЦМВ, промотор вируса обезьян 40 (SV40), такой как ранний промотор SV40 и поздний промотор SV40, промотор вируса опухоли молочной железы мышей (MMTV), промотор вируса иммунодефицита человека (ВИЧ), такой как промотор длинных концевых повторов (ДКП) бычьего вируса иммунодефицита (БВИ), промотор вируса Молони, промотор вируса лейкоза птиц (ВЛП), промотор цитомегаловируса (ЦМВ), такой как немедленно-ранний промотор ЦМВ, промотор вируса Эпштейна-Барр (ВЭБ) или промотор вируса саркомы Рауса (ВСР). Промотор также может представлять собой промотор из человеческого гена, такого как человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатинин, человеческий полиэдрин или человеческий металлотионеин.
[00114] Промотор может представлять собой конститутивный промотор или индуцибельный промотор, который инициирует транскрипцию, только когда клетка-хозяин подвергается воздействию некоторого внешнего стимула. В случае многоклеточного организма промотор также может быть специфическим в отношении конкретной ткани или органа, или стадии развития. Промотор также может быть тканеспецифическим промотором, таким как промотор, специфический в отношении мышц или кожи, природный или синтетический. Примеры таких промоторов описаны в публикации заявки на патент США № US20040175727, содержание которой в полном объеме включено в данный документ посредством ссылки.
[00115] Промотор может быть связан с энхансером. Энхансер может располагаться выше кодирующей последовательности. Энхансер может представлять собой энхансер человеческого актина, человеческого миозина, человеческого гемоглобина, человеческого мышечного креатинина или вирусный энхансер, например, из ЦМВ, FMDV, РСВ или ВЭБ. Энхансеры полинуклеотидной функции описаны в патентах США № 5593972, 5962428 и W094/016737, содержание которых в полном объеме включено посредством ссылки.
Интрон
[00116] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более интронов. Каждый интрон может содержать функциональные донорные и акцепторные участки сплайсинга. Интрон может содержать энхансер сплайсинга. Интрон может содержать один или более сигналов, необходимых для эффективного сплайсинга.
Область терминации транскрипции
[00117] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать одну или более областей терминации транскрипции. Область терминации транскрипции может располагаться ниже кодирующей последовательности, чтобы обеспечивать эффективную терминацию. Область терминации транскрипции может быть получена из того же гена, что и вышеописанный промотор, или может быть получена из одного или более других генов.
Инициирующий кодон
[00118] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более инициирующих кодонов. Инициирующий кодон может располагаться выше кодирующей последовательности. Инициирующий кодон может находиться в рамке с кодирующей последовательностью. Инициирующий кодон может быть связан с одним или более сигналами, необходимыми для эффективной инициации трансляции, например, но не ограничиваясь этим, участком связывания рибосомы.
Терминирующий кодон
[00119] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более терминирующих или стоп-кодонов. Терминирующий кодон может располагаться ниже кодирующей последовательности. Терминирующий кодон может находиться в рамке с кодирующей последовательностью. Терминирующий кодон может быть связан с одним или более сигналами, необходимыми для эффективной терминации трансляции.
Сигнал полиаденилирования
[00120] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более сигналов полиаденилирования. Сигнал полиаденилирования может содержать один или более сигналов, необходимых для эффективного полиаденилирования транскрипта. Сигнал полиаденилирования может располагаться ниже кодирующей последовательности. Сигнал полиаденилирования может представлять собой сигнал полиаденилирования SV40, сигнал полиаденилирования ДКП, сигнал полиаденилирования бычьего гормона роста (bGH), сигнал полиаденилирования человеческого гормона роста (hGH) или сигнал полиаденилирования человеческого β-глобина. Сигнал полиаденилирования SV40 может представлять собой сигнал полиаденилирования из плазмиды pCEP4 (Invitrogen, San Diego, CA).
Лидерная последовательность
[00121] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать одну или более лидерных последовательностей. Лидерная последовательность может кодировать сигнальный пептид. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина (Ig), например, но не ограничиваясь этим, сигнальный пептид IgG и сигнальный пептид IgE.
Компоновка конструкции последовательности рекомбинантной нуклеиновой кислоты
[00122] Как описано выше, последовательность рекомбинантной нуклеиновой кислоты может содержать одну или более конструкций последовательности рекомбинантной нуклеиновой кислоты, при этом каждая конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более компонентов. Один или более компонентов подробно описаны выше. Один или более компонентов, когда они включены в конструкцию последовательности рекомбинантной нуклеиновой кислоты, могут располагаться в любом порядке по отношению друг к другу. В некоторых вариантах осуществления один или более компонентов могут располагаться в конструкции последовательности рекомбинантной нуклеиновой кислоты так, как описано ниже.
Компоновка 1
[00123] В одной компоновке первая конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид тяжелой цепи, а вторая конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид легкой цепи. Например, в одном варианте осуществления первая последовательность рекомбинантной нуклеиновой кислоты кодирует полипептид тяжелой цепи, имеющий аминокислотную последовательность по меньшей мере на 95% гомологичную одной из SEQ ID NO:4, SEQ ID NO:10, SEQ ID NO:16 и SEQ ID NO:22. В одном варианте осуществления первая последовательность рекомбинантной нуклеиновой кислоты содержит последовательность нуклеиновой кислоты по меньшей мере на 95% гомологичную SEQ ID NO:3, SEQ ID NO:9, SEQ ID NO:15, SEQ ID NO:21. В одном варианте осуществления вторая последовательность рекомбинантной нуклеиновой кислоты кодирует полипептид легкой цепи, имеющий аминокислотную последовательность по меньшей мере на 95% гомологичную одной из SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18 и SEQ ID NO:24. В одном варианте осуществления вторая последовательность рекомбинантной нуклеиновой кислоты содержит последовательность нуклеиновой кислоты по меньшей мере на 95% гомологичную SEQ ID NO:5, SEQ ID NO:11, SEQ ID NO:17, SEQ ID NO:23.
[00124] Первая конструкция последовательности рекомбинантной нуклеиновой кислоты может быть помещена в вектор. Вторая конструкция последовательности рекомбинантной нуклеиновой кислоты может быть помещена во второй или отдельный вектор. Размещение конструкции последовательности рекомбинантной нуклеиновой кислоты в векторе более подробно описано ниже.
[00125] Первая конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать промотор, интрон, область терминации транскрипции, инициирующий кодон, терминирующий кодон и/или сигнал полиаденилирования. Первая конструкция последовательности рекомбинантной нуклеиновой кислоты может дополнительно содержать лидерную последовательность, причем лидерная последовательность расположена выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид тяжелой цепи. Соответственно, сигнальный пептид, кодируемый лидерной последовательностью, может быть связан пептидной связью с полипептидом тяжелой цепи.
[00126] Вторая конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать промотор, инициирующий кодон, терминирующий кодон и сигнал полиаденилирования. Вторая конструкция последовательности рекомбинантной нуклеиновой кислоты может дополнительно содержать лидерную последовательность, причем лидерная последовательность расположена выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид легкой цепи. Соответственно, сигнальный пептид, кодируемый лидерной последовательностью, может быть связан пептидной связью с полипептидом легкой цепи.
[00127] Соответственно, один пример компоновки 1 может включать первый вектор (и, таким образом, первую конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH и CH1, и второй вектор (и, таким образом, вторую конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид легкой цепи, который содержит VL и CL. Второй пример компоновки 1 может включать первый вектор (и, таким образом, первую конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH, CH1, шарнирную область, CH2 и CH3, и второй вектор (и, таким образом, вторую конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид легкой цепи, который содержит VL и CL.
Компоновка 2
[00128] Во второй компоновке конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид тяжелой цепи, и гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид легкой цепи. Гетерологичная последовательность нуклеиновой кислоты, кодирующая полипептид тяжелой цепи, может располагаться выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид легкой цепи. В альтернативном варианте гетерологичная последовательность нуклеиновой кислоты, кодирующая полипептид легкой цепи, может располагаться выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид тяжелой цепи.
[00129] Конструкцию последовательности рекомбинантной нуклеиновой кислоты можно помещать в вектор, как более подробно описано ниже.
[00130] Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать гетерологичную последовательность нуклеиновой кислоты, кодирующую участок расщепления протеазой и/или линкерную последовательность. При включении в конструкцию последовательности рекомбинантной нуклеиновой кислоты гетерологичная последовательность нуклеиновой кислоты, кодирующая участок расщепления протеазой, может располагаться между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи. Соответственно, участок расщепления протеазой позволяет разделять полипептид тяжелой цепи и полипептид легкой цепи на разные полипептиды после экспрессии. В других вариантах осуществления при включении в конструкцию последовательности рекомбинантной нуклеиновой кислоты линкерной последовательности она может располагаться между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
[00131] Конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать промотор, интрон, область терминации транскрипции, инициирующий кодон, терминирующий кодон и/или сигнал полиаденилирования. Конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один или более промоторов. Конструкция последовательности рекомбинантной нуклеиновой кислоты также может содержать два промотора так, что один промотор может быть связан с гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, а второй промотор может быть связан с гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи. В других вариантах осуществления конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать один промотор, связанный с гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
[00132] Конструкция последовательности рекомбинантной нуклеиновой кислоты может дополнительно содержать две лидерные последовательности, причем первая лидерная последовательность расположена выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, а вторая лидерная последовательность расположена выше (или на 5' конце) гетерологичной последовательности нуклеиновой кислоты, кодирующей полипептид легкой цепи. Соответственно, сигнальный пептид, кодируемый первой лидерной последовательностью, может быть связан пептидной связью с полипептидом тяжелой цепи, а второй сигнальный пептид, кодируемый второй лидерной последовательностью, может быть связан пептидной связью с полипептидом легкой цепи.
[00133] Соответственно, один пример компоновки 2 может включать вектор (и, таким образом, конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH и CH1, и полипептид легкой цепи, который содержит VL и CL, в котором линкерная последовательность расположена между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
[00134] Второй пример компоновки 2 может включать вектор (и, таким образом, конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH и CH1, и полипептид легкой цепи, который содержит VL и CL, в котором гетерологичная последовательность нуклеиновой кислоты, кодирующая участок расщепления протеазой, расположена между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
[00135] Третий пример компоновки 2 может включать вектор (и, таким образом, конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH, CH1, шарнирную область, CH2 и CH3, и полипептид легкой цепи, который содержит VL и CL, в котором линкерная последовательность расположена между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
[00136] Четвертый пример компоновки 2 может включать вектор (и, таким образом, конструкцию последовательности рекомбинантной нуклеиновой кислоты), кодирующий полипептид тяжелой цепи, который содержит VH, CH1, шарнирную область, CH2 и CH3, и полипептид легкой цепи, который содержит VL и CL, в котором гетерологичная последовательность нуклеиновой кислоты, кодирующая участок расщепления протеазой, расположена между гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид тяжелой цепи, и гетерологичной последовательностью нуклеиновой кислоты, кодирующей полипептид легкой цепи.
Экспрессия из конструкции последовательности рекомбинантной нуклеиновой кислоты
[00137] В других вариантах осуществления конструкция последовательности рекомбинантной нуклеиновой кислоты может содержать, в числе одного или более компонентов, гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид тяжелой цепи, и/или гетерологичную последовательность нуклеиновой кислоты, кодирующую полипептид легкой цепи. Соответственно, конструкция последовательности рекомбинантной нуклеиновой кислоты может облегчать экспрессию полипептида тяжелой цепи и/или полипептида легкой цепи.
[00138] В случае применения описанной выше компоновки 1 первая конструкция последовательности рекомбинантной нуклеиновой кислоты может облегчать экспрессию полипептида тяжелой цепи, а вторая конструкция последовательности рекомбинантной нуклеиновой кислоты может облегчать экспрессию полипептида легкой цепи. В случае применения описанной выше компоновки 2 конструкция последовательности рекомбинантной нуклеиновой кислоты может облегчать экспрессию полипептида тяжелой цепи и полипептида легкой цепи.
[00139] После экспрессии, например, но не ограничиваясь этим, в клетке, организме или млекопитающем, полипептид тяжелой цепи и полипептид легкой цепи могут формировать синтетическое антитело. В частности, полипептид тяжелой цепи и полипептид легкой цепи могут взаимодействовать друг с другом так, что сборка приводит к получению синтетического антитела, способного связывать антиген. В других вариантах осуществления полипептид тяжелой цепи и полипептид легкой цепи могут взаимодействовать друг с другом так, что сборка приводит к получению синтетического антитела, являющегося более иммуногенным по сравнению с антителом, не прошедшим описанную в данном документе сборку. В других вариантах осуществления полипептид тяжелой цепи и полипептид легкой цепи могут взаимодействовать друг с другом так, что сборка приводит к получению синтетического антитела, способного вызывать или индуцировать иммунный ответ против антигена.
Вектор
[00140] Описанную выше конструкцию последовательности рекомбинантной нуклеиновой кислоты можно помещать в один или более векторов. Один или более векторов могут содержать точку начала репликации. Один или более векторов могут представлять собой плазмиду, бактериофаг, бактериальную искусственную хромосому или дрожжевую искусственную хромосому. Один или более векторов могут представлять собой как самореплицирующийся внехромосомный вектор, так и вектор, который интегрируется в геном организма-хозяина.
[00141] Векторы включают, но не ограничиваются этим, плазмиды, векторы экспрессии, рекомбинантные вирусы, любую форму рекомбинантного вектора «голой ДНК» и т. п. «Вектор» содержит нуклеиновую кислоту, которая может инфицировать, трансфицировать, временно или постоянно трансдуцировать клетку. Следует понимать, что вектор может представлять собой «голую» нуклеиновую кислоту или нуклеиновую кислоту в комплексе с белком или липидом. Вектор необязательно содержит вирусные или бактериальные нуклеиновые кислоты, и/или белки, и/или мембраны (например, клеточную мембрану, вирусную липидную оболочку и т. д.). Векторы включают, но не ограничиваются этим, репликоны (например, репликоны РНК, бактериофаги), к которым могут быть прикреплены и становиться реплицированными фрагменты ДНК. Таким образом, векторы включают, но не ограничиваются этим, РНК, автономную самореплицирующуюся циркулярную или линейную ДНК или РНК (например, плазмиды, вирусы и т. п., см., например, патент США. № 5,217,879), и включают как экспрессионные, так и не экспрессионные плазмиды. В некоторых вариантах осуществления вектор включает линейную ДНК, ферментативную ДНК или синтетическую ДНК. Когда рекомбинантный микроорганизм или клеточная культура описывается как хозяин «вектора экспрессии», это означает, что они включают как внехромосомную циркулярную, так и линейную ДНК, и ДНК, которые были включены в хромосому (-ы) хозяина. Когда вектор поддерживается в клетке-хозяине, вектор может либо стабильно реплицироваться клетками во время митоза как автономная структура, либо может быть включен в геном хозяина.
[00142] Один или более векторов могут представлять собой гетерологичную экспрессионную конструкцию, которая в общем случае является плазмидой, которую используют для внесения конкретного гена в клетку-мишень. После того, как вектор экспрессии попадает в клетку, полипептид тяжелой цепи и/или полипептид легкой цепи, которые кодируются конструкцией последовательности рекомбинантной нуклеиновой кислоты, начинают вырабатываться рибосомальными комплексами клеточного аппарата транскрипции и трансляции. Один или более векторов могут экспрессировать большие количества стабильной матричной РНК и, следовательно, белков.
Вектор экспрессии
[00143] Один или более векторов могут представлять собой кольцевую плазмиду или линейную нуклеиновую кислоту. Кольцевая плазмида и линейная нуклеиновая кислота способны управлять экспрессией конкретной нуклеотидной последовательности в соответствующей клетке субъекта. Один или более векторов, содержащих конструкцию последовательности рекомбинантной нуклеиновой кислоты, могут быть химерными, что означает, что по меньшей мере один из их компонентов является гетерологичным по отношению к по меньшей мере одному из других его компонентов.
Плазмида
[00144] Один или более векторов могут представлять собой плазмиду. Плазмиду можно применять для трансфекции клеток конструкцией последовательности рекомбинантной нуклеиновой кислоты. Плазмиду можно применять для внесения конструкции последовательности рекомбинантной нуклеиновой кислоты в организм субъекта. Плазмида также может содержать регуляторную последовательность, которая может хорошо подходить для генной экспрессии в клетке, в которую внесена плазмида.
[00145] Плазмида также может содержать точку начала репликации млекопитающего для внехромосомной поддержки плазмиды и выработки множества копий плазмиды в клетке. Плазмида может представлять собой pVAX1, pCEP4 или pREP4 от Invitrogen (г. Сан-Диего, штат Калифорния, США), которые могут содержать точку начала репликации вируса Эпштейна-Барр и область, кодирующую ядерный антиген EBNA-1, которая может обеспечивать многокопийную эписомальную репликацию без интеграции. Скелет плазмиды может представлять собой pAV0242. Плазмида может представлять собой дефективную по репликации аденовирусную плазмиду типа 5 (Ad5).
[00146] Плазмида может представлять собой pSE420 (Invitrogen, San Diego, Calif.), которую можно использовать для выработки белка в Escherichia coli (E. coli). Плазмида также может представлять собой pYES2 (Invitrogen, г. Сан-Диего, штат Калифорния, США), которую можно использовать для выработки белка в штаммах дрожжей Saccharomyces cerevisiae. Плазмида также может принадлежать полной бакуловирусной системе экспрессии MAXBAC™ (Invitrogen, San Diego, Calif.), которую можно использовать для выработки белка в клетках насекомых. Плазмида также может представлять собой pcDNA I или pcDNA3 (Invitrogen, San Diego, Calif.), которые можно использовать для выработки белка в клетках млекопитающих, таких как клетки яичника китайского хомяка (CHO).
РНК
[00147] В одном варианте осуществления нуклеиновая кислота представляет собой РНК. В одном варианте осуществления РНК транскрибирована из описанной в настоящем документе последовательности ДНК. Например, в некоторых вариантах осуществления, РНК кодируется последовательностью ДНК по меньшей мере на 90% гомологичной одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 и SEQ ID NO:23. В другом варианте осуществления, нуклеотидная последовательность содержит последовательностью РНК, транскрибированную последовательностью ДНК, кодирующую полипептидную последовательность по меньшей мере одну из SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26 и SEQ ID NO:27, или ее вариант, или ее фрагмент. Соответственно, в одном варианте осуществления в изобретении предложена РНК, кодирующая одно или более DMAb. РНК может иметь дополнительную цепь. Соответственно, в некоторых вариантах осуществления РНК может быть транслирована клетками без необходимости каких-либо промежуточных этапов репликации, таких как обратная транскрипция. Молекула РНК, подходящая для использования в изобретении, может иметь 5′-кэп (например, 7-метилгуанозин). Этот кэп может усиливать трансляцию РНК in vivo. 5′-нуклеотид РНК, используемой в изобретении, может иметь группу трифосфата на 5′-конце. В кэппированной РНК она может быть связана с 7-метилгуанозином через мостик 5′-к-5′. РНК может иметь поли-A хвост на 3′-конце. Около своего 3′-конца она может также включать последовательность, отвечающую за распознавание поли-A полимеразы (например, AAUAAA). РНК, используемая в изобретении, обычно является одноцепочечной. РНК, используемая в изобретении, может содержать синтетическую РНК. В некоторых вариантах осуществления молекула РНК представляет собой «голую РНК» молекулу. В одном варианте осуществления РНК содержится в пределах вектора.
[00148] В одном варианте осуществления РНК имеет 5' и 3' НТО. В одном варианте осуществления 5'-НТО имеет длину от нуля до 3000 нуклеотидов. Длину последовательностей 5'- и 3'-НТО, которые будут добавлены к кодирующему участку, можно изменить различными способами, включая, помимо прочего, разработку праймеров для ПЦР, которые ренатурируются с различными участками НТО. Используя такой подход, специалист в данной области техники сможет модифицировать длину 5'- и 3'-НТО, необходимую для достижения оптимальной эффективности трансляции после трансфекции транскрибированной РНК.
[00149] 5'- и 3'-НТО могут быть природными, эндогенными 5'- и 3'- НТО для представляющего интерес гена. Альтернативно, последовательности НТО, которые не являются эндогенными по отношению к представляющему интерес гену, могут быть добавлены путем введения последовательностей НТО в прямые и обратные праймеры, либо с помощью любых других модификаций матрицы. Использование последовательностей НТО, которые не являются эндогенными по отношению к представляющему интерес гену, может быть полезным при модификации стабильности и/или эффективности трансляции РНК. Например, известно, что богатые AU элементы в последовательностях 3'-НТО могут снижать стабильность РНК. Таким образом, 3' НТО могут выбираться или разрабатываться таким образом, чтобы повышать стабильность транскрибированной РНК, исходя из свойств НТО, которые хорошо известны в данной области техники.
[00150] В одном варианте осуществления 5'-НТО может содержать последовательность Козак эндогенного гена. Альтернативно, при добавлении 5' НТО, которая не является эндогенной по отношению к представляющей интерес нуклеиновой кислоте, путем ПЦР, как было описано выше, консенсусная последовательность Козак может быть переделана путем добавления последовательности 5' НТО. Последовательность Козак может повышать эффективность трансляции некоторых транскриптов РНК, однако похоже, что она не требуется для всех РНК, чтобы обеспечить эффективную трансляцию. Необходимость последовательностей Козак для многих РНК известна в данной области техники. В других вариантах осуществления 5'-НТО может быть получена из РНК-вируса, чей геном РНК является стабильным в клетках. В других вариантах осуществления различные нуклеотидные аналоги могут использоваться в 3'- или 5'-НТО, чтобы воспрепятствовать деградации РНК экзонуклеазами.
[00151] В одном варианте осуществления РНК содержит кэп на 5'-конце и 3'-поли(A)-хвост, которые определяют связывание рибосомы, инициацию трансляции и стабильность РНК в клетке.
[00152] В одном варианте осуществления РНК представляет собой нуклеозид-модифицированную РНК. Нуклеозид-модифицированная РНК обладает особыми преимуществами по сравнению с немодифицированной РНК, включая, например, повышенную стабильность, низкую или отсутствующую врожденную иммуногенность и улучшенную трансляцию.
Кольцевой и линейный вектор
[00153] Один или более векторов могут представлять собой кольцевую плазмиду, которая может трансформировать клетку-мишень путем интеграции в клеточный геном или существовать внехромосомно (например, автономно-реплицирующаяся плазмида с точкой начала репликации). Вектор может представлять собой pVAX, pcDNA3.0 или provax, или любой другой вектор эскпрессии, способный экспрессировать полипептид тяжелой цепи и/или полипептид легкой цепи, кодируемые конструкцией последовательности рекомбинантной нуклеиновой кислоты.
[00154] Также в данном документе предложена линейная нуклеиновая кислота или линейная экспрессионная кассета («ЛЭК»), которую возможно эффективно доставлять субъекту путем электропорации и которая способна экспрессировать полипептид тяжелой цепи и/или полипептид легкой цепи, кодируемые конструкцией последовательности рекомбинантной нуклеиновой кислоты. ЛЭК может представлять собой любую линейную ДНК, в которой отсутствует какой-либо фосфатный остов. ЛЭК может не содержать каких-либо генов устойчивости к антибиотикам и/или фосфатный остов. ЛЭК может не содержать другие последовательности нуклеиновых кислот, не связанные с экспрессией необходимого гена.
[00155] ЛЭК может быть получена из любой плазмиды, которую возможно линеаризовать. Плазмида может быть способна экспрессировать полипептид тяжелой цепи и/или полипептид легкой цепи, кодируемые конструкцией последовательности рекомбинантной нуклеиновой кислоты. Плазмида может представлять собой pNP (Пуэрто-Рико/34) или pM2 (Новая Каледония/99). Плазмида может представлять собой WLV009, pVAX, pcDNA3.0 или provax, или любой другой вектор экспрессии, способный экспрессировать полипептид тяжелой цепи и/или полипептид легкой цепи, кодируемые конструкцией последовательности рекомбинантной нуклеиновой кислоты.
[00156] ЛЭК может представлять собой pcrM2. ЛЭК может представлять собой pcrNP. pcrNP и pcrMR могут быть получены из pNP (Пуэрто-Рико/34) и pM2 (Новая Каледония/99) соответственно.
Вирусные векторы
[00157] В одном варианте осуществления предложены вирусные векторы, способные доставлять нуклеиновую кислоту настоящего изобретения в клетку. Вектор экспрессии можно доставлять в клетку в форме вирусного вектора. Технологии вирусных векторов хорошо известны в данной области и описаны, например, у Sambrook et al. (2001), и у Ausubel et al. (1997), и в других справочниках по вирусологии и молекулярной биологии. Вирусы, используемые в качестве векторов, включают, но не ограничиваются этим, ретровирусы, аденовирусы, аденоассоциированные вирусы, вирусы герпеса и лентивирусы. В основном, подходящий вектор содержит участок начала репликации, функциональный по меньшей мере в одном организме, промоторную последовательность, подходящие участки узнавания рестрикционных эндонуклеаз и один или более селектируемых маркеров. (См., например, WO 01/96584; WO 01/29058; и патент США № 6,326,193). Вирусные векторы и особенно ретровирусные векторы стали наиболее широко используемым способом переноса генов в клетки млекопитающих, например клетки человека. Другие вирусные векторы могут быть получены из лентивируса, поксвируса, вируса простого герпеса I типа, аденовирусов и аденоассоциированных вирусов и т. п. См., например, патент США №№ 5,350,674 и 5,585,362.
Способ получения вектора
[00158] В данном документе предложен способ получения одного или более векторов, в которые была помещена последовательность рекомбинантной нуклеиновой кислоты. После конечного этапа субклонирования вектор можно использовать для инокуляции клеточной культуры в крупномасштабном ферментационном чане с помощью известных в данной области техники способов.
[00159] В других вариантах осуществления после конечного этапа субклонирования вектор можно использовать с одним или более устройствами для электропорации (ЭП). Устройства для ЭП более подробно описаны ниже.
[00160] Один или более векторов можно готовить или получать, используя комбинацию известных устройств и методик, но предпочтительно их получают с помощью методики получения плазмид, описанной в лицензированной, одновременно находящейся на рассмотрении предварительной заявке США № 60/939,792, поданной 23 мая 2007 г. В некоторых примерах описанные в данном документе ДНК-плазмиды можно готовить в концентрациях, превышающих или равных 10 мг/мл. Методики получения также включают применение различных устройств и протоколов, которые являются общеизвестными для специалистов в данной области техники, в дополнение описанным в заявке на патент США, серийный № 60/939792, включая описанные в лицензированном патенте США № 7,238,522, изданном 3 июля 2007 г. Вышеуказанные заявка и патент, серийный № 60/939,792 и патент США № 7,238,522, соответственно, в полном объеме включены в данный документ.
Антитело
[00161] Как описано выше, последовательность рекомбинантной нуклеиновой кислоты может кодировать антитело, его фрагмент, его вариант или их комбинацию. Антитело может связываться или вступать в реакцию с антигеном, который более подробно описан ниже.
[00162] Антитело может содержать набор определяющих комплементарность областей («CDR») тяжелой цепи и легкой цепи, перемежающих соответственно набор каркасных областей («FR») тяжелой цепи и легкой цепи, которые обеспечивают поддержку CDR и определяют пространственную взаимосвязь CDR относительно друг друга. Набор CDR может содержать три гипервариабельные области из V-области тяжелой или легкой цепи. Начиная с N-конца тяжелой или легкой цепи, эти CDR обозначаются «CDR1», «CDR2» и «CDR3» соответственно. Следовательно, антигенсвязывающий участок может включать шесть CDR, составляющих набор CDR из каждой V-области тяжелой и легкой цепи.
[00163] Протеолитический фермент папаин предпочтительно расщепляет молекулы IgG с получением нескольких фрагментов, два из которых (фрагменты F(ab)) содержат ковалентный гетеродимер, который содержит интактный антигенсвязывающий участок. Фермент пепсин способен расщеплять молекулы IgG с получением нескольких фрагментов, включая фрагмент F(ab')2, который содержит оба антигенсвязывающих участка. Соответственно, антитело может представлять собой Fab или F(ab')2. Fab может содержать полипептид тяжелой цепи и полипептид легкой цепи. Полипептид тяжелой цепи Fab может содержать область VH и область CH1. Полипептид легкой цепи Fab может содержать область VL и область CL.
[00164] Антитело может представлять собой иммуноглобулин (Ig). Ig может представлять собой, например, IgA, IgM, IgD, IgE и IgG. Иммуноглобулин может содержать полипептид тяжелой цепи и полипептид легкой цепи. Полипептид тяжелой цепи иммуноглобулина может содержать область VH, область CH1, шарнирную область, область CH2 и область CH3. Полипептид легкой цепи иммуноглобулина может содержать область VL и область CL.
[00165] Антитело может представлять собой поликлональное или моноклональное антитело. Антитело может представлять собой химерное антитело, одноцепочечное антитело, антитело с созревшей аффинностью, человеческое антитело, гуманизированное антитело или полностью человеческое антитело. Гуманизированное антитело может представлять собой антитело от отличного от человека вида, которое связывает необходимый антиген, имеющее одну или более определяющих комплементарность областей (CDR) от отличного от человека вида и каркасные области из молекулы человеческого иммуноглобулина.
[00166] Антитело может представлять собой биспецифическое антитело, более подробно описанное ниже. Антитело может представлять собой бифункциональное антитело, также более подробно описанное ниже.
[00167] Как описано выше, антитело может генерироваться у субъекта после введения субъекту композиции. Антитело может характеризоваться определенным временем полужизни в организме субъекта. В некоторых вариантах осуществления антитело можно модифицировать, чтобы продлить или укоротить его время полужизни в организме субъекта. Такие модификации более подробно описаны ниже.
[00168] Антитело может быть дефукозилированным, как более подробно описано ниже.
[00169] В одном варианте осуществления антитело связывается с антигеном Borrelia spp. В одном варианте осуществления антитело связывается с OspA.
[00170] Антитело можно модифицировать, чтобы снизить или предотвратить антителозависимое усиление (АЗУ) заболевания, связанного с антигеном, как более подробно описано ниже.
Биспецифическое антитело
[00171] Последовательность рекомбинантной нуклеиновой кислоты может кодировать биспецифическое антитело, его фрагмент, его вариант или их комбинацию. Биспецифическое антитело может связываться или вступать в реакцию с двумя антигенами, например, двумя антигенами, более подробно описанными ниже. Биспецифическое антитело может состоять из фрагментов двух описанных в данном документе антител, что, таким образом, позволяет биспецифическому антителу связываться или вступать в реакцию с двумя необходимыми молекулами-мишенями, которые могут включать антиген, более подробно описанный ниже, лиганд, включая лиганд для рецептора, рецептор, включая лиганд-связывающий участок на рецепторе, комплекс лиганд-рецептор и маркер.
[00172] В изобретении предложены новые биспецифические антитела, содержащие первый антигенсвязывающий участок, который специфически связывается с первой мишенью, и второй антигенсвязывающий участок, который специфически связывается со второй мишенью, с особенно предпочтительными свойствами, такими как воспроизводимость, стабильность, аффинность связывания, биологическая активность, специфическое нацеливание на определенные Т-клетки, эффективность нацеливания и сниженная токсичность. В некоторых случаях существуют биспецифические антитела, которые связываются с первой мишенью с высокой аффинностью, а со второй мишенью - с низкой аффинностью. В других случаях существуют биспецифические антитела, которые связываются с первой мишенью с низкой аффинностью, а со второй мишенью - с высокой аффинностью. В других случаях существуют биспецифические антитела, которые связываются с первой мишенью с желаемой аффинностью и со второй мишенью с желаемой аффинностью.
[00173] В одном варианте осуществления биспецифическое антитело представляет собой двухвалентное антитело, которое содержит a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и b) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном.
[00174] Молекула биспецифического антитела в соответствии с настоящим изобретением может иметь два участка связывания любой желаемой специфичности. В некоторых вариантах осуществления один из участков связывания способен связываться с опухолеассоциированным антигеном. В некоторых вариантах осуществления, участок связывания, включенный в Fab-фрагмент, представляет собой участок связывания, специфичный антигена Borrelia spp. В некоторых вариантах осуществления участок связывания, включенный в одноцепочечный Fv-фрагмент, представляет собой участок связывания, специфичный для антигена Borrelia spp, такого как антиген OspA.
[00175] В некоторых вариантах осуществления один из участков связывания молекулы антитела в соответствии с настоящим изобретением способен связываться со специфической рецепторной молекулой T-клетки и/или со специфической рецепторной молекулой естественной клетки-киллера (NK-клетки). Специфический рецептор Т-клетки также называют «Т-клеточным рецептором» (TCR), который позволяет Т-клетке связываться и, при наличии дополнительных сигналов, активироваться и реагировать на эпитоп/антиген, представленный другой клеткой, называемой антигенпрезентирующей клеткой (APC). Известно, что Т-клеточный рецептор напоминает Fab-фрагмент встречающегося в природе иммуноглобулина. Он, как правило, является одновалентным и охватывает α- и β-цепи, в некоторых вариантах осуществления он охватывает γ-цепи и δ-цепи (выше). Соответственно, в некоторых вариантах осуществления TCR представляет собой TCR (альфа/бета), а в некоторых вариантах осуществления он представляет собой TCR (гамма/дельта). Т-клеточный рецептор образует комплекс с корецептором Т-клеток CD3. CD3 представляет собой белковый комплекс и состоит из четырех разных цепей. У млекопитающих комплекс содержит цепь CD3γ, цепь CD36 и две цепи CD3E. Эти цепи связываются с молекулой, известной как Т-клеточный рецептор (TCR), и ζ-цепью, чтобы генерировать сигнал активации в Т-лимфоцитах. Поэтому в некоторых вариантах осуществления специфический Т-клеточный рецептор представляет собой корецептор Т-клеток CD3. В некоторых вариантах осуществления специфический Т-клеточный рецептор представляет собой CD28, белок, который также экспрессируется на Т-клетках. CD28 может обеспечивать костимулирующие сигналы, необходимые для активации Т-клеток. CD28 играет важную роль в пролиферации и выживаемости Т-клеток, производстве цитокинов и развитии Т-хелперов типа-2. Дополнительным примером специфического Т-клеточного рецептора является CD134, также называемый Ox40. Экспрессия CD134/OX40 происходит через 24-72 часа после активации и может быть использована для определения вторичной костимулирующей молекулы. Другим примером Т-клеточного рецептора является 4-1 BB, способный связываться с лигандом 4-1 BB на антигенпрезентирующих клетках (APC), в результате чего генерируется костимулирующий сигнал для Т-клетки. Другим примером рецептора, преимущественно обнаруживаемого на Т-клетках, является CD5, который на низких уровнях также встречается на В-клетках. Другим примером рецептора, модифицирующего функции Т-клеток, является CD95, также известный как Fas-рецептор, который опосредует передачу апоптического сигнала Fas-лигандом, экспрессируемым на поверхности других клеток. Сообщается, что CD95 модулирует сигнальные пути, управляемые TCR/CD3, в находящихся в состоянии покоя Т-лимфоцитах.
[00176] Примером специфической рецепторной молекулы NK-клеток является CD16, низкоаффинный Fc-рецептор и NKG2D. Пример рецепторной молекулы, присутствующей на поверхности как Т-клеток, так и естественных клеток-киллеров (NK), является CD2 и другие представители суперсемейства CD2. CD2 на T- и NK-клетках способен действовать как костимулирующая молекула.
[00177] В некоторых вариантах осуществления первый участок связывания молекулы антитела связывается с антигеном Borrelia spp, и второй участок связывания связывается со специфической рецепторной молекулой T-клетки и/или специфической рецепторной молекулой естественной клетки-киллера (NK).
[00178] В некоторых вариантах осуществления первый участок связывания молекулы антитела связывается с антигеном Borrelia spp, и второй участок связывания связывается со специфической рецепторной молекулой T-клетки и/или специфической рецепторной молекулой естественной клетки-киллера (NK). В некоторых вариантах осуществления первый участок связывания молекулы антитела связывается с антигеном Borrelia spp, и второй участок связывания связывается с одним из перечисленного: CD3, Т-клеточный рецептор (TCR), CD28, CD16, NKG2D, Ox40, 4-1BB, CD2, CD5 и CD95.
[00179] В некоторых вариантах осуществления первый участок связывания молекулы антитела связывается со специфической рецепторной молекулой T-клетки и/или специфической рецепторной молекулой естественной клетки-киллера (NK), и второй участок связывания связывается с антигеном Borrelia spp. В некоторых вариантах осуществления первый участок связывания антитела связывается со специфической рецепторной молекулой T-клетки и/или специфической рецепторной молекулой естественной клетки-киллера (NK), и второй участок связывания связывается с антигеном Borrelia spp. В некоторых вариантах осуществления первый участок связывания антитела связывается с одним из перечисленного: CD3, Т-клеточный рецептор (TCR), CD28, CD16, NKG2D, Ox40, 4-1BB, CD2, CD5 и CD95, и второй участок связывания связывается с антигеном Borrelia spp. В одном варианте осуществления антиген Borrelia spp представляет собой OspA.
Бифункциональное антитело
[00180] Последовательность рекомбинантной нуклеиновой кислоты может кодировать бифункциональное антитело, его фрагмент, его вариант или их комбинацию. Бифункциональное антитело может связываться или вступать в реакцию с описанным ниже антигеном. Бифункциональное антитело также можно модифицировать, чтобы придать антителу дополнительную функциональность помимо распознавания и связывания антигена. Такая модификация может включать, но не ограничивается, сопряжение с фактором H или его фрагментом. Фактор H представляет собой растворимый регулятор активации комплемента и, таким образом, может вносить свой вклад в иммунный ответ посредством комплемент-опосредованного лизиса (КОЛ).
Продление времени полужизни антитела
[00181] Как описано выше, антитело можно модифицировать, чтобы продлить или укоротить время полужизни антитела в организме субъекта. Модификация может продлевать или укорачивать время полужизни антитела в сыворотке субъекта.
[00182] Модификация может присутствовать в константной области антитела. Модификация может представлять собой одну или более аминокислотных замен в константной области антитела, которые продлевают время полужизни антитела по сравнению со временем полужизни антитела, не содержащего одну или более аминокислотных замен. Модификация может представлять собой одну или более аминокислотных замен в домене CH2 антитела, которые продлевают время полужизни антитела по сравнению со временем полужизни антитела, не содержащего одну или более аминокислотных замен.
[00183] В некоторых вариантах осуществления одна или более аминокислотных замен в константной области могут включать замещение остатка метионина в константной области остатком тирозина, остатка серина в константной области остатком треонина, остатка треонина в константной области остатком глутамата или любую их комбинацию, продлевая, таким образом, время полужизни антитела.
[00184] В других вариантах осуществления одна или более аминокислотных замен в константной области могут включать замещение остатка метионина в домене CH2 остатком тирозина, остатка серина в домене CH2 остатком треонина, остатка треонина в домене CH2 остатком глутамата или любую их комбинацию, продлевая, таким образом, время полужизни антитела.
Дефукозилирование
[00185] Последовательность рекомбинантной нуклеиновой кислоты может кодировать антитело, которое не является фукозилированным (т. е. дефукозилированное антитело или нефукозилированное антитело), его фрагмент, его вариант или их комбинацию. Фукозилирование включает добавление к молекуле сахара фукозы, например, присоединение фукозы к N-гликанам, О-гликанам и гликолипидам. Соответственно, в дефукозилированном антителе фукоза не присоединена к углеводным цепям константной области. В свою очередь, это отсутствие фукозилирования может улучшать связывание FcγRIIIa и антителозависимую клеточную цитотоксическую (АЗКЦ) активность антитела по сравнению с фукозилированным антителом. Следовательно, в некоторых вариантах осуществления нефукозилированное антитело может проявлять повышенную активность АЗКЦ по сравнению с фукозилированным антителом.
[00186] Антитело можно модифицировать так, чтобы предотвратить или ингибировать фукозилирование антитела. В некоторых вариантах осуществления такое модифицированное антитело может проявлять повышенную активность АЗКЦ по сравнению с немодифицированным антителом. Модификация может присутствовать в тяжелой цепи, легкой цепи или их комбинации. Модификация может представлять собой одну или более аминокислотных замен в тяжелой цепи, одну или более аминокислотных замен в легкой цепи или их комбинацию.
Сниженный ответ АЗУ
[00187] Антитело можно модифицировать, чтобы снизить или предотвратить антителозависимое усиление (АЗУ) заболевания, связанного с антигеном, но при этом нейтрализовать антиген.
[00188] В некоторых вариантах осуществления антитело можно модифицировать так, чтобы оно содержало одну или более аминокислотных замен, которые снижают или предотвращают связывание антитела с FcγR1a. Одна или более аминокислотных замен могут присутствовать в константной области антитела. Одна или более аминокислотных замен могут включать замещение остатка лейцина остатком аланина в константной области антитела, т. е. также называемое в данном документе LA, мутацией LA или заменой LA. Одна или более аминокислотных замен могут включать замещение каждого из двух остатков лейцина остатком аланина в константной области антитела, которое также называется в данном документе LALA, мутацией LALA или заменой LALA. Присутствие замен LALA может предотвращать или блокировать связывание антитела с FcγR1a и, таким образом, модифицированное антитело не усиливает заболевание или не вызывает АЗУ заболевания, связанного с антигеном, но при этом нейтрализует антиген.
Антиген
[00189] Синтетическое антитело направлено на данный антиген или его фрагмент или вариант. Антиген может представлять собой последовательность нуклеиновой кислоты, аминокислотную последовательность, полисахарид или их комбинацию. Последовательность нуклеиновой кислоты может представлять собой ДНК, РНК, кДНК, их вариант, их фрагмент или их комбинацию. Аминокислотная последовательность может представлять собой белок, пептид, его вариант, его фрагмент или их комбинацию. Полисахарид может представлять собой полисахарид, кодируемый нуклеиновой кислотой.
[00190] Антиген может быть получен из бактерии. Антиген может быть связан с бактериальной инфекцией. В одном варианте осуществления антиген может быть бактериальным фактором вирулентности.
[00191] В одном варианте осуществления синтетическое антитело настоящего изобретения нацелено на два или более антигенов. В одном варианте осуществления по меньшей мере один антиген для биспецифического антитела выбран из антигенов, описанных в настоящем документе. В одном варианте осуществления два или более антигенов выбраны из антигенов, описанных в настоящем документе.
Бактериальные антигены
[00192] Бактериальный антиген может представлять собой бактериальный антиген, его фрагмент или его вариант. Бактерия может представлять собой Spirochaetae phylum. Бактерия может представлять собой бактерию вызывающую заболевание. Бактерия может представлять собой бактерию Borrelia spp. Бактерия может представлять собой одну или более бактерий Borrelia spp включая, но не ограничиваясь, Borrelia burgdorferi, Borrelia lusitaniae, Borrelia afzelii, Borrelia bissettii, Borreliella bavariensis, Borrelia chilensis, Borrelia garinii, Borrelia valaisiana, Borrelia spielmanii и Borrelia finlandensis.
[00193] Бактериальный антиген может представлять собой антиген Borrelia spp, его фрагмент или его вариант. Антиген Borrelia spp может быть получен из бактериального продукта, который позволяет Borrelia spp реплицироваться или выживать. Бактериальный продукт, который позволяет Borrelia spp реплицироваться или выживать включает, но не ограничивается, структурные компоненты, ферменты и токсины. Такой продукт может представлять собой липопротеин, протеин внешней поверхности клетки, продукт необходимый для инфекционности или персистенции у позвоночных хозяев, и продукт вовлеченный в подвижность и хемотаксис.
[00194] В одном варианте осуществления антиген представляет собой липопротеин (например, BptA). В одном варианте осуществления антиген представляет собой протеин внешней поверхности клетки (e.g. OspA, OspB и OspC). В одном варианте осуществления антиген представляет собой продукт необходимый для инфекционности или персистенции у позвоночных хозяев(например, PncA, DbpA, DbpB, Bgp, P66 и VlsE).
Вспомогательные вещества и другие компоненты композиции
[00195] Композиция может дополнительно содержать фармацевтически приемлемое вспомогательное вещество. Фармацевтически приемлемое вспомогательное вещество может представлять собой функциональные молекулы, такие как молекулы базовых растворов, носителей или разбавителей. Фармацевтически приемлемое вспомогательное вещество может представлять собой облегчающий трансфекцию агент, который может включать поверхностно-активные агенты, такие как иммуностимулирующие комплексы (ИМКОМ), неполный адъювант Фрейнда, аналог ЛПС, включая монофосфориллипид A, мурамилпептиды, аналоги хинона, везикулы, такие как сквален и сквален, гиалуроновая кислота, липиды, липосомы, ионы кальция, вирусные белки, полианионы, поликатионы или наночастицы, или другие известные облегчающие трансфекцию агенты.
[00196] Облегчающий трансфекцию агент представляет собой полианион, поликатион, включая поли-L-глутамат (LGS), или липид. Облегчающий трансфекцию агент представляет собой поли-L-глутамат, а поли-L-глутамат может присутствовать в композиции в концентрации менее 6 мг/мл. Облегчающий трансфекцию агент, который также может включать поверхностно-активные агенты, такие как иммуностимулирующие комплексы (ИМКОМ), неполный адъювант Фрейнда, аналог ЛПС, включая монофосфориллипид A, мурамилпептиды, аналоги хинона и везикулы, такие как сквален и сквален, и гиалуроновую кислоту, также можно использовать для введения в сочетании с композицией. Композиция также может содержать облегчающий трансфекцию агент, такой как липиды, липосомы, включая лецитиновые липосомы или другие липосомы, известные в данной области техники, в виде смеси ДНК-липосом (смотрите, например, W09324640), ионы кальция, вирусные белки, полианионы, поликатионы или наночастицы, или другие известные облегчающие трансфекцию агенты. Облегчающий трансфекцию агент представляет собой полианион, поликатион, включая поли-L-глутамат (LGS), или липид. Концентрация трансфекционного агента в вакцине составляет менее 4 мг/мл, менее 2 мг/мл, менее 1 мг/мл, менее 0,750 мг/мл, менее 0,500 мг/мл, менее 0,250 мг/мл, менее 0,100 мг/мл, менее 0,050 мг/мл или менее 0,010 мг/мл.
[00197] Композиция может дополнительно содержать генетический вспомогательный агент, как описано в заявке на патент США № 021,579, поданной 1 апреля 1994 г., содержание которой в полном объеме включено путем ссылки.
[00198] Композиция может содержать ДНК в количестве от около 1 нанограмма до 100 миллиграммов; от около 1 микрограмма до около 10 миллиграммов; или предпочтительно от около 0,1 микрограмма до около 10 миллиграммов; или более предпочтительно от около 1 миллиграмма до около 2 миллиграммов. В некоторых предпочтительных вариантах осуществления композиция в соответствии с настоящим изобретением содержит от около 5 нанограмм до около 1000 микрограмм ДНК. В некоторых предпочтительных вариантах осуществления композиция может содержать от около 10 нанограмм до около 800 микрограмм ДНК. В некоторых предпочтительных вариантах осуществления композиция может содержать от около 0,1 до около 500 микрограмм ДНК. В некоторых предпочтительных вариантах осуществления композиция может содержать от около 1 до около 350 микрограмм ДНК. В некоторых предпочтительных вариантах осуществления композиция может содержать от около 25 до около 250 микрограмм, от около 100 до около 200 микрограмм, от около 1 нанограмма до 100 миллиграмм; от около 1 микрограмма до около 10 миллиграмм; от около 0,1 микрограмма до около 10 миллиграмм; от около 1 миллиграмма до около 2 миллиграмм, от около 5 нанограмм до около 1000 микрограмм, от около 10 нанограмм до около 800 микрограмм, от около 0,1 до около 500 микрограмм, от около 1 до около 350 микрограмм, от около 25 до около 250 микрограмм, от около 100 до около 200 микрограмм ДНК.
[00199] Композиция может быть приготовлена в соответствии с применяемым режимом введения. Инъекционная фармацевтическая композиция может быть стерильной, апирогенной и не содержать частиц. Можно использовать изотонический состав или раствор. Добавки для придания изотоничности могут включать хлорид натрия, декстрозу, маннит, сорбит и лактозу. Композиция может содержать сосудосуживающий агент. Изотонические растворы могут включать фосфатно-солевой буфер. Композиция может дополнительно содержать стабилизаторы, включая желатин и альбумин. Стабилизаторы могут обеспечивать стабильность состава при комнатной температуре или температуре окружающей среды в течение длительных периодов времени, включая LGS или поликатионы или полианионы.
Способ получения синтетического антитела
[00200] Настоящее изобретение также относится к способу генерации синтетического антитела. Этот способ может включать введение нуждающемуся в этом субъекту композиции, применяя способ доставки, более подробно описанный ниже. Соответственно, синтетическое антитело генерируется в организме субъекта или in vivo после введения субъекту композиции.
[00201] Этот способ также может включать внесение композиции в одну или более клеток и, следовательно, синтетическое антитело может генерироваться или вырабатываться в одной или более клетках. Этот способ также может включать внесение композиции в одну или более тканей, например, но без ограничений, кожу и мышцы, и, следовательно, синтетическое антитело может генерироваться или вырабатываться в одной или более тканях.
Способ определения или скрининга антитела
[00202] Настоящее изобретение дополнительно относится к способу определения или скрининга описанного выше антитела, которое вступает в реакцию или связывается с описанным выше антигеном. В способе определения или скрининга антитела можно использовать антиген с применением методик, известных специалистам в данной области техники, для определения или скрининга антитела. Такие методики включают, но не ограничиваются этим, выбор антитела из библиотеки (например, фагового дисплея) и иммунизацию животного с последующим выделением и/или очисткой антитела.
Способ доставки композиции
[00203] Настоящее изобретение также относится к способу доставки композиции нуждающемуся в этом субъекту. Способ доставки может включать введение композиции субъекту. Введение может включать, но не ограничивается этим, инъекцию ДНК с in vivo электропорацией или без нее, опосредованную липосомами доставку и облегчаемую наночастицами доставку.
[00204] Млекопитающее, получающее доставку композиции, может представлять собой человека, примата, отличного от человека примата, корову, крупный рогатый скот, овцу, козу, антилопу, бизона, буйвола, бизона, быков, оленя, ежей, слонов, ламу, альпака, мышей, крыс и кур.
[00205] Композицию можно вводить разными путями, включая пероральный, парентеральный, сублингвальный, трансдермальный, ректальный, трансмукозальный, местный, ингаляционный, буккальный, интраплевральный, внутривенный, внутриартериальный, внутрибрюшинный, подкожный, внутримышечный, интраназальный, интратекальный и внутрисуставной или их комбинацию. В случае ветеринарного применения композицию можно вводить в подходящем приемлемом составе в соответствии с общепринятой ветеринарной практикой. Ветеринар может без труда определить режим дозирования и путь введения, которые наиболее подходит для конкретного животного. Композицию можно вводить с помощью традиционных шприцов, безыгольных устройств для инъекций, «пушек для бомбардировки микрочастицами» или других физических способов, таких как электропорация («ЭП»), «гидродинамический способ» или ультразвук.
Электропорация
[00206] Введение композиции посредством электропорации можно осуществлять, используя устройства для электропорации, которые могут быть выполнены с возможностью подачи в необходимые ткани млекопитающего импульса энергии, эффективного для стимуляции образования обратимых пор в клеточных мембранах, и предпочтительно импульс энергии представляет собой постоянный ток, аналогичный предварительно заданному пользователем входному току. Устройство для электропорации может содержать электропорационный компонент и комплект электродов или комплект рукояток. Электропорационный компонент может содержать и включать один или более различных элементов устройств для электропорации, включая: контроллер, генератор формы тока, тестер импеданса, устройство регистрации формы сигнала, элемент ввода, элемент отчета о состоянии, порт связи, компонент памяти, источник питания и переключатель питания. Электропорацию можно осуществлять, используя устройство для in vivo электропорации, например, систему CELLECTRA EP (Inovio Pharmaceuticals, Plymouth Meeting, PA) или электропоратор Elgen (Inovio Pharmaceuticals, Plymouth Meeting, PA), чтобы облегчить трансфекцию клеток плазмидой.
[00207] Электропорационный компонент может функционировать как один элемент устройств для электропорации, а другие элементы являются отдельными элементами (или компонентами), связанными с электропорационным компонентом. Электропорационный компонент может функционировать как более чем один элемент устройств для электропорации, который может быть связан с другими элементами устройств для электропорации, отдельными от электропорационного компонента. Элементы и устройства для электропорации, являющиеся частями одного электромеханического или механического устройства, могут не ограничиваться, так как эти элементы могут функционировать как одно устройство или как отдельные элементы, связанные друг с другом. Электропорационный компонент может подавать импульс энергии, который обеспечивает постоянный ток в необходимой ткани, и содержит механизм обратной связи. Комплект электродов может включать матрицу из электродов, содержащую некоторое количество пространственно упорядоченных электродов, при этом комплект электродов получает импульс энергии от электропорационного компонента и доставляет его в необходимую ткань через электроды. По меньшей мере один из некоторого количества электродов является нейтральным во время подачи импульса энергии и измеряет импеданс в необходимой ткани и передает значение импеданса на электропорационный компонент. Механизм обратной связи может получать измеренное значение импеданса и может корректировать импульс энергии, подаваемый электропорационным компонентом, для поддержания постоянного тока.
[00208] Некоторое количество электродов могут подавать импульс энергии на децентрализованную схему. Некоторое количество электродов могут подавать импульс энергии на децентрализованную схему посредством управления электродами согласно программируемой последовательности, а программируемая последовательность вводится пользователем в электропорационный компонент. Программируемая последовательность может включать некоторое количество импульсов, подаваемых в последовательности, причем каждый импульс из некоторого количества импульсов подается по меньшей мере двумя активными электродами с одним нейтральным электродом, который измеряет импеданс, а следующий импульс из некоторого количества импульсов подается другим из по меньшей мере двух активных электродов с одним нейтральным электродом, который измеряет импеданс.
[00209] Механизм обратной связи может осуществляться аппаратным или программным обеспечением. Механизм обратной связи может осуществляться аналоговым замкнутым контуром. Обратная связь происходит каждые 50 мкс, 20 мкс, 10 мкс или 1 мкс, но предпочтительно представляет собой обратную связь в режиме реального времени или мгновенную (т. е. по существу мгновенную по определению доступными методами определения времени ответа). Нейтральный электрод может измерять значение импеданса в необходимой ткани и передавать значение импеданса на механизм обратной связи, а механизм обратной связи в ответ на значение импеданса корректирует импульс энергии для поддержания значения постоянного тока, аналогичного предварительно заданному току. Механизм обратной связи может поддерживать постоянный ток непрерывно и мгновенно во время подачи импульса энергии.
[00210] Примеры устройств для электропорации и способов электропорации, которые могут облегчить доставку композиции согласно настоящему изобретению, включают описанные в патенте США № 7245963 авторства Draghia-Akli, с соавт., публикации патента США 2005/0052630, поданной Smith, с соавт., содержание которых в полном объеме включено в данный документ посредством ссылки. Другие устройства для электропорации и способы электропорации, которые можно использовать для облегчения доставки композиции, включают предложенные в одновременно находящейся на рассмотрении заявке одних заявителей на патент США, серийный № 11/874072, поданной 17 октября 2007 г., которая заявляет приоритет согласно 35 USC 119(e) предварительных заявок США № 60/852,149, поданной 17 октября 2006 г., и 60/978,982, поданной 10 октября 2007 г., которые в полном объеме включены в данный документ посредством ссылки.
[00211] В патенте США № 7245963 авторства Draghia-Akli, с соавт. описаны модульные системы электродов и их применение для облегчения внесения биомолекул в клетки выбранной ткани в организме или растении. Модульные системы электродов могут содержать некоторое количество игольчатых электродов; гиподермическую иглу; электрический соединитель, который обеспечивает проводящее соединение программируемого импульсного контроллера постоянного тока с некоторым количеством игольчатых электродов; и источник питания. Оператор может брать некоторое количество игольчатых электродов, которые находятся на опорной конструкции и плотно вставлять их в выбранную ткань в организме или растении. Затем происходит доставка биомолекул в выбранную ткань через гиподермическую иглу. Происходит активация программируемого импульсного контроллера постоянного тока и применение электрического импульса постоянного тока к некоторому количеству игольчатых электродов. Применяемый электрический импульс постоянного тока облегчает внесение биомолекулы в клетку между некоторым количеством электродов. Полное содержание патента США № 7245963 включено в данный документ посредством ссылки.
[00212] В публикации патента США 2005/0052630, поданной Smith, с соавт., описано устройство для электропорации, которое можно использовать, чтобы эффективным образом облегчать внесение биомолекул в клетки выбранной ткани в организме или растении. Устройство для электропорации содержит электрокинетическое устройство («ЭКУ устройство»), чья работа определяется программным или встроенным программным обеспечением. ЭКУ устройство создает ряд программируемых профилей импульсов постоянного тока между электродами в матрице под управлением пользователем и на основании введенных параметров импульсов и позволяет хранить и получать данные по форме тока. Устройство для электропорации также содержит сменный электродный диск, содержащий матрицу из игольчатых электродов, центральный инъекционный канал для инъекционной иглы и съемный направляющий диск. Полное содержание публикации патента США 2005/0052630 включено в данный документ посредством ссылки.
[00213] Электродные матрицы и способы, описанные в патенте США № 7245963 и публикации патента США 2005/0052630, можно адаптировать для глубокого проникновения не только в ткани, такие как мышцы, но также в другие ткани или органы. Вследствие конфигурации электродной матрицы инъекционная игла (для доставки биомолекулы по выбору) также полностью вставляется в целевой орган, а инъекция проводится перпендикулярно целевой ткани в области, которая заранее намечена электродами. Электроды, описанные в патенте США № 7245963 и публикации патента США 2005/005263, предпочтительно имеют длину 20 мм и калибр 21.
[00214] Кроме того, подразумевается, что в некоторых вариантах осуществления, которые включают устройства для электропорации и их применение, используются устройства для электропорации, описанные в следующих патентах: патент США 5273525, выданный 28 декабря 1993 г., патенты США 6,110,161, выданный 29 августа 2000 г., 6261281, выданный 17 июля 2001 г., и 6958060, выданный 25 октября 2005 г., и патент США 6939862, выданный 6 сентября 2005 г. Кроме того, в данном документе предусмотрено применение материалов из патентов, относящихся к предмету обсуждения, таких как патент США 6697669, выданный 24 февраля 2004 г., который относится к доставке ДНК с помощью различных устройств, и патент США 7328064, выданный 5 февраля 2008, относящийся к способу инъекции ДНК. Вышеуказанные патенты в полном объеме включены посредством ссылки.
Способ лечения
[00215] Также в данном документе предложен способ лечения, защиты от и/или предотвращения заболевания у нуждающегося в этом субъекта путем генерации синтетического антитела в организме субъекта. Способ может включать введение композиции субъекту. Введение композиции субъекту можно осуществлять, используя описанный выше способ доставки.
[00216] В некоторых вариантах осуществления в изобретении предложен способ лечения, защиты от и/или предотвращения инфекции Borrelia spp. В одном варианте осуществления способ обеспечивает лечение, защиту от и/или предотвращение болезни Лайма.
[00217] После генерации синтетического антитела в организме субъекта синтетическое антитело может связываться или вступать в реакцию с антигеном. Такое связывание может нейтрализовать антиген, блокировать распознавание антигена другой молекулой, например, белком или нуклеиновой кислотой, и вызывать или индуцировать иммунный ответ на антиген, осуществляя, таким образом, лечение, защиту от и/или предотвращение заболевания, связанного с антигеном, у субъекта.
[00218] Синтетическое антитело может обеспечивать лечение, предотвращение и/или защиту от заболевания у субъекта, которому вводят композицию. Синтетическое антитело посредством связывания антигена может обеспечивать лечение, предотвращение и/или защиту от заболевания у субъекта, которому вводят композицию. Синтетическое антитело может способствовать выживаемости при заболевании у субъекта, которому вводят композицию. В одном варианте осуществления синтетическое антитело может обеспечить повышенную выживаемость субъекта с заболеванием по сравнению с ожидаемой выживаемостью у имеющего заболевание субъекта, которому не вводили синтетическое антитело. В различных вариантах осуществления синтетическое антитело может обеспечивать повышение выживаемости у субъекта с заболеванием, которому вводили композицию, по меньшей мере на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% больше по сравнению с ожидаемой выживаемостью при отсутствии введения композиции. В одном варианте осуществления синтетическое антитело может обеспечить повышенную защиту от заболевания у субъекта по сравнению с ожидаемой защитой у субъекта, которому не вводили синтетическое антитело. В различных вариантах осуществления синтетическое антитело может обеспечивать защиту от заболевания у субъекта, которому вводили композицию, по меньшей мере на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% больше по сравнению с ожидаемой защитой при отсутствии введения композиции.
Использование в комбинации с антибиотиками
[00219] В настоящем изобретении также предложен способ лечения, защиты от и/или предотвращения заболевания у нуждающегося в этом субъекта посредством введения комбинации синтетического антитела и терапевтического антибиотика.
[00220] Синтетическое антитело и антибиотик можно вводить, используя любой подходящий способ, чтобы обе составляющие комбинации синтетического антитела и антибиотика присутствовали в организме субъекта. В одном варианте осуществления способ может включать введение первой композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, и введение второй композиции, содержащей антибиотик, менее чем через 1, менее чем через 2, менее чем через 3, менее чем через 4, менее чем через 5, менее чем через 6, менее чем через 7, менее чем через 8, менее чем через 9 или менее чем через 10 суток после введения синтетического антитела. В одном варианте осуществления способ может включать введение первой композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, и введение второй композиции, содержащей антибиотик, более чем через 1, более чем через 2, более чем через 3, более чем через 4, более чем через 5, более чем через 6, более чем через 7, более чем через 8, более чем через 9 или более чем через 10 суток после введения синтетического антитела. В одном варианте осуществления способ может включать введение первой композиции, содержащей антибиотик, и введение второй композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, менее чем через 1, менее чем через 2, менее чем через 3, менее чем через 4, менее чем через 5, менее чем через 6, менее чем через 7, менее чем через 8, менее чем через 9 или менее чем через 10 суток после введения антибиотика. В одном варианте осуществления способ может включать введение первой композиции, содержащей антибиотик, и введение второй композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, более чем через 1, более чем через 2, более чем через 3, более чем через 4, более чем через 5, более чем через 6, более чем через 7, более чем через 8, более чем через 9 или более чем через 10 суток после введения антибиотика. В одном варианте осуществления способ может включать введение первой композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, и введение второй композиции, содержащей антибиотик, одновременно. В одном варианте осуществления способ может включать введение первой композиции, содержащей синтетическое антитело по изобретению, любым из способов, подробно описанным выше, и введение второй композиции, содержащей антибиотик, одновременно. В одном варианте осуществления способ может включать введение одной композиции, содержащей синтетическое антитело по изобретению и антибиотик.
[00221] Неограничивающие примеры антибиотиков, которые можно использовать в комбинации с синтетическим антителом согласно изобретению, включают аминогликозиды (например, гентамицин, амикацин, тобрамицин), хинолоны (например, ципрофлоксацин, левофлоксацин), цефалоспорины (например, цефтазидим, цефепим, цефоперазон, цефпиром, цефтобипрол), антипсевдомонадные пенициллины: карбоксипенициллины (например, карбенициллин и тикарциллин) и уреидопенициллины (например, мезлоциллин, азлоциллин и пиперациллин), карбапенемы (например, меропенем, имипенем, дорипенем), полимиксины (например, полимиксин B и колистин) и монобактамы (например, азтреонам).
Получение синтетических антител In Vitro и Ex Vivo
[00222] В одно варианте осуществления синтетическое антитело получают in vitro или ex vivo. Например, В одном варианте осуществления молекула нуклеиновой кислоты кодирующая синтетическое антитело может быть введена и экспресирована in vitro или ex vivo клетки. Способы введения и экспрессии генов в клетке известны в данной области техники. В контексте вектора экспрессии вектор может быть легко введен в клетку-хозяин, например, клетку млекопитающего, клетку бактерии, дрожжевую клетку или клетку насекомого с помощью любого способа в данной области техники. Например, вектор экспрессии может быть перенесен в клетку-хозяин физическим, химическим или биологическим путем.
[00223] К физическим способам введения полинуклеотида в клетку-хозяин относятся осаждение фосфатом кальция, липофекция, бомбардировка частицами, микроинъекция, электропорация и т. п. Способы получения клеток содержащих векторы и/или экзогенные нуклеиновые кислоты хорошо известны в данной области техники. См., например, Sambrook et al. (2012, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York). Предпочтительным способом введения полинуклеотида в клетку-хозяин является трансфекция фосфатом кальция.
[00224] К биологическим способам введения представляющего интерес полинуклеотида в клетку-хозяин относится использование векторов ДНК и РНК. Вирусные векторы и особенно ретровирусные векторы стали наиболее широко используемым способом переноса генов в клетки млекопитающих, например клетки человека. Другие вирусные векторы могут быть получены из лентивируса, поксвируса, вируса простого герпеса I типа, аденовирусов и аденоассоциированных вирусов и т. п. См., например, патент США №№ 5,350,674 и 5,585,362.
[00225] К химическим способам введения полинуклеотида в клетку-хозяина относятся системы коллоидной дисперсии, такие как макромолекулярные комплексы, нанокапсулы, микросферы, гранулы, и системы на основе липидов, включая эмульсии масло-в-воде, мицеллы, смешанные мицеллы и липосомы. Примером коллоидной системы, используемой в качестве средства доставки in vitro и in vivo, является липосома (например, искусственная мембранная везикула).
[00226] В случае использования невирусной системы доставки примером средства доставки является липосома. Предусмотрено использование липидных составов для введения нуклеиновых кислот в клетку-хозяин (in vitro, ex vivo или in vivo). В другом аспекте изобретения нуклеиновая кислота может быть связана с липидом. Нуклеиновая кислота, связанная с липидом, может быть инкапсулирована в водном внутреннем пространстве липосомы, рассеяна по липидному бислою липосомы, присоединена к липосоме посредством линкерной молекулы, которая связана как с липосомой, так и с олигонуклеотидом, заключена в липосоме, связана в комплекс с липосомой, диспергирована в растворе, содержащем липид, смешана с липидом, объединена с липидом, может содержаться в виде суспензии в липиде, содержатся или быть связана в комплекс с мицеллой или любым другим образом связана с липидом. Композиции, связанные с липидом, липидом/ДНК или липидом/вектором экспрессии, не ограничиваются какой-либо конкретной структурой в растворе. Например, они могут присутствовать в бислойной структуре, такой как мицеллы, или иметь «разрушенную» структуру. Также они могут быть просто рассеяны в растворе, предположительно образуя агрегаты, не имеющие одинаковых размера или формы. Липиды представляют собой жиры, которые могут быть природными или синтетическими липидами. Например, липиды включают в себя жировые капли, которые встречаются естественным образом в цитоплазме, и также класс соединений, которые содержат длинноцепочечные алифатические углеводороды и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
[00227] Настоящее изобретение имеет множество аспектов, проиллюстрированных следующими неограничивающими примерами.
Примеры
[00228] Настоящее изобретение дополнительно проиллюстрировано в следующих примерах. Следует понимать, что эти примеры, хотя в них и указаны предпочтительные варианты осуществления изобретения, приведены исключительно в иллюстративных целях. Из приведенного выше обсуждения и этих примеров специалист в данной области техники может установить важные характеристики этого изобретения и, не выходя за рамки его сущности и объема, может осуществлять различные изменения и модификации изобретения, чтобы адаптировать его под различные потребности и условия. Таким образом, различные модификации изобретения в дополнение к показанным и описанным в данном документе станут очевидны для специалистов в данной области техники из вышеприведенного описания. Также подразумевается, что такие модификации входят в объем прилагаемой формулы изобретения.
Example 1
[00229] Представленные в данном документе исследования демонстрируют получение функциональных «ДНК-моноклональных антител» (DMAb) к OspA посредством внутримышечной электропорации плазмидной ДНК. Кодон-оптимизированные последовательности ДНК вариабельной области из моноклональных антител к OspA синтезировали в константном домене IgG1 человека. Плазмидную ДНК, кодирующую антитело, доставляли мышам C3H. Данное исследование подтверждает, что DMAb является альтернативой существующим вариантам биологической терапии.
[00230] Далее будут описаны материалы и способы.
[00231] Животные и белки, и ведение плазмид, и доставка DMAb дикого типа или модифицированные консенсусного типа DMAb специфичные к OspA вводили мышам C3H (5 мышей на группу). Для данных введений 300 мкг плазмой ДНК вводили внутримышечно (ВМ) с ЕР опосредованной доставкой посредством системы MID-EP (CELLECTRA®; Inovio Pharmaceuticals, Блю Белл, Пенсильвания). Параметры пульсов для доставки были следующими; 3 пульса 0,5 А постоянного тока, с интервалом в 1 с и продолжительностью 52 мс. Каждое животное получало одну дозу либо экспериментального, либо контрольного составов плазмиды за пять дней до приведения в контакт с клещом. Сыворотки от мышей отбирали через 21 день после приведения в контакт с клещом для анализа (Фиг. 1).
[00232] Конструирование плазмид ДНК DMAb-319-44 wt и DMAb-221-7 wt ДНК плазмиды p319-44wt и p221-7wt, кодирующие DMAb-319-44wt и DMAb-221-1wt соответственно, полностью кодируют mAb IgG1 человека, вариабельные области которых были получены из анти-hisOspA-антител 319-44 и 221-7 соответственно. Каждый трансген состоял из генов тяжелой и легкой цепей, разделенных сайтом расщепления фурином в сочетании с последовательностью самообработки P2A. Трансгены были кодонами и РНК были оптимизированы для экспрессии у человека, синтезированы GenScript и клонированы в модифицированные векторы экспрессии у млекопитающих pVax1 (Invitrogen) под контролем промотора гена немедленного раннего ответа цитомегаловируса человека.
[00233] Конструирование плазмид ДНК DMAb-319-44 mod1 и DMAb-221-7 mod9 Чтобы улучшить экспрессию DMAb p319-44wt и p221-7wt, последовательности вариабельной области легкой и тяжелой цепей, DMAb 'wt' были дополнительно оптимизированы с помощью целевого подхода, который фокусировался на повышении стабильности антитела посредством значительных модификаций каркасной области, чтобы улучшить продуцирование антитела в условиях in vitro и in vivo. Во-первых, было обнаружено DMAb с высокой экспрессией, которое действовало в качестве акцепторного каркаса для прививки. Данное акцепторное DMAb, нацеленное на антиген HER2 и, как показано, экспрессирует > 5 мкг/мл человеческого IgG у мышей с иммунодефицитом после однократной дозы 100 мкг ДНК, состоит из тяжелых и легких цепей из высокостабильных семейств зародышевых линий hVH3 и hVκ1, соответственно. Чтобы создать оптимизированный 319-44mod1, мы привили три CDR и дополнительные 13 критических каркасных остатков тяжелой и легкой цепи 319-44wt на гены DMAb с высокой экспрессией. Для 221-7mod9, мы привили три CDR и дополнительные 11 критических каркасных остатков тяжелой и легкой цепи 221-7wt на гены DMAb с высокой экспрессией.
[00234] Анализ приведения в контакт с клещами Мышам вводили 300 мкг pDMAb-319-44 wt, pDMAb-319-44 mod1, pDMAb-221-7 wt, pDMAb-221-7 mod9 или отрицательный контроль (pDVSF-3 LSLS) за пять дней до приведения в контакт с клещом с Borrelia. Сыворотку собирали через двадцать один день после приведения в контакт с клещами и анализировали на наличие Borrelia и на IgG ответ. Для анализа инфекции в тканях мышей умерщвляли через 21 день после приведения в контакт с клещами, и клетки собирали и затем культивировали и наблюдали в течение дополнительного периода времени, до 50 дней после приведения в контакт с клещами, на наличие Borrelia burgdorferi (Фиг. 1).
[00235] Ниже описаны результаты экспериментов
[00236] Характеристика DMAb-319-44 mod1 и DMAb-221-7 mod9 in vivo - Мышам вводили pDMAb-319-44 wt, pDMAb-319-44 mod1, pDMAb-221-7 wt или pDMAb-221-7 mod9 внутримышечно с последующей усиленной доставкой посредством EP. Однократную инъекцию плазмиды ДНК проводили за пять дней до приведения в контакт с клещами, и сыворотка была собрана через 21 день после приведения в контакт с клещами (Фиг. 1). Данные на Фиг. 2 представлены (от отдельных мышей из pDMAb-319-44 wt, PDMAb-319-44 mod1, pDMAb-221-7 wt и контрольной группы) как OD 450 нм, что пропорционально уровню Ig/Fab. Данные показали, что относительные уровни Fab после однократного введения pDMAb-319-44 wt, pDMAb-319-44 mod1 и pDMAb-221-7 wt были обнаружены через 3 дня после введения DMAb. Уровни pDMAb-319-44 wt и pDMAb-319-44 mod1 были выше на 3-й день и привели к увеличению числа мышей, «защищенных» от последствий приведения в контакт с клещами. Серийное разведение pDMAb показало, что мыши, которым вводили разведение pDMAb в соотношении от 1: 100 до 1: 24000, показали значительно более высокие уровни IgG на 21 день, чем мыши, которым вводили отрицательный контроль (Фиг. 3).
[00237] Оценивали боррелиацидную активность pDMAb-319-44 wt, pDMAb-319-44 mod1, pDMAb-221-7 wt и pDMAb-221-7 mod9. Мыши, которым вводили pDMAb-319-44 wt, pDMAb-319-44 mod1, pDMAb-221-7 wt и pDMAb-221-7 mod9, показали снижение процента живой Borrelia по сравнению с мышами, которым вводили контрольное антитело (Фиг. 4). pDMAb-319-44 wt, pDMAb-319-44 mod1 и pDMAb-221-7 wt были выбраны для эксперимента по заражению на основании их сильной боррелиацидной активности.
[00238] Мыши, которым вводили pDMAb-319-44 mod1, показали аналогичный или слегка повышенный ответ IgG по сравнению с мышами, которым вводили pDMAb-319-44 wt (Фиг. 5). Состав увеличивает дозу 300 мкг до уровня приблизительно 7 мкг/мл на день 7. Составленная дозировка (например, добавление стабилизаторов и/или адъювантов) с гиалуронидазой приводила к большей экспрессии in vivo как WT, так и mod1 DMAb (Фиг. 5).
[00239] Трехуровневая стратегия оптимизации привела к увеличению экспрессии in vivo pDMAb-221-7 mod 9 (Фиг. 6). Состав увеличивает дозу 300 мкг до уровня > 10 мкг/мл на день 7.
[00240] Исследования in vivo демонстрируют увеличение антител Лайма (антител к OspA) и связанное с этим увеличение IgG человека от 2 до по меньшей мере 7 дней после инъекции (Фиг. 7). Следует отметить, что 221-7 отражает введение wt DMAb без состава.
[00241] Введение антитела pDMAb-319-44 mod1 обеспечивает 80% защиту от инфекции Borrelia. Что является увеличением по сравнению с защитой, обеспечиваемой антителом pDMAb-319-44 wt (Фиг. 8).
[00242] ДНК является гибкой платформой для доставки генов и нуклеотидных последовательностей in vivo. ДНК-кодируемые mAb защищают от бактерий и имеют сравнимую эффективность с белком IgG mAb.
[00243] ДНК-технология облегчает доставку терапии mAb, делая ее рутинной и расширяет доступ к мировому рынку. DMAb можно комбинировать с технологией ДНК-вакцины для обеспечения немедленного и стойкого иммунитета.
Последовательности:
SEQ ID NO:1 нуклеотидная последовательность DMAb 319-44 mod1 CDR: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа)- функционально связанный с двумя стоп-кодонами.
SEQ ID NO:2 аминокислотная последовательность DMAb 319-44 mod1 CDR: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа).
SEQ ID NO:3 нуклеотидная последовательность DMAb 319-44 mod1 тяжелой цепи (анти-HER2 DMAb оптимизированный CDR графт): Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO:4 аминокислотная последовательность DMAb 319-44 mod1 тяжелой цепи (анти-HER2 DMAb оптимизированный CDR графт): Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 5 нуклеотидная последовательность легкой цепи DMAb 319-44 mod1: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO: 6 аминокислотная последовательность легкой цепи DMAb 319-44 mod1: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO:7 нуклеотидная последовательность DMAb 319-44 wt: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа)- функционально связанный с двумя стоп-кодонами.
SEQ ID NO:8 аминокислотная последовательность DMAb 319-44 wt: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа).
SEQ ID NO:9 нуклеотидная последовательность тяжелой цепи DMAb 319-44 wt: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO:10 аминокислотная последовательность тяжелой цепи DMAb 319-44 wt: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 11 нуклеотидная последовательность легкой цепи DMAb 319-44 wt: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO: 12 аминокислотная последовательность легкой цепи DMAb 319-44 wt: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO: 13 нуклеотидная последовательность DMAb 221-7 mod 9 полноразмерной одиночной плазмиды IgG1 человека: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа)- функционально связанный с двумя стоп-кодонами.
SEQ ID NO: 14 аминокислотная последовательность DMAb 221-7 mod 9 полноразмерной одиночной плазмиды IgG1 человека: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа).
SEQ ID NO: 15 нуклеотидная последовательность DMAb 221-7 mod 9 тяжелой цепи IgG1 человека: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 16 аминокислотная последовательность DMAb 221-7 mod 9 тяжелой цепи IgG1 человека: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 17 нуклеотидная последовательность DMAb 221-7 mod 9 легкой цепи IgG1 человека: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO: 18 аминокислотная последовательность DMAb 221-7 mod 9 легкой цепи IgG1 человека: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO:19 нуклеотидная последовательность DMAb 221-7 wt: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа)- функционально связанный с двумя стоп-кодонами.
SEQ ID NO:20 аминокислотная последовательность DMAb 221-7 wt: Тяжелый сигнальный пептид IgG человека-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа человека-VL-CL (каппа).
SEQ ID NO: 21 нуклеотидная последовательность тяжелой цепи DMAb 221-7 wt: тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO:22 аминокислотная последовательность тяжелой цепи DMAb 221-7 wt: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 23 нуклеотидная последовательность легкой цепи DMAb 221-7 wt: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO: 24 аминокислотная последовательность легкой цепи DMAb 221-7 wt: сигнальный пептид легкой цепи каппа человека, вариабельная легкая область и константная легкая область.
SEQ ID NO:25 аминокислотная последовательность DMAb 221-7 mod 9 CDR мыши: Тяжелый сигнальный пептид IgG2а-VH-CH1-шарнирная область-CH2-CH3-нестандартный участок расщепления фурином -'GSG' линкер и P2A пептид-сигнальный пептид легкой цепи каппа-VL-CL (каппа).
SEQ ID NO:26 аминокислотная последовательность тяжелой цепи DMAb 221-7 mod 9 IgG2a мыши: Тяжелый сигнальный пептид IgG человека, вариабельная тяжелая область, константная тяжелая область 1, шарнирная область, константная тяжелая область 2 и константная тяжелая область 3.
SEQ ID NO: 27 аминокислотная последовательность легкой цепи DMAb 221-7 mod 9 IgG2a мыши: сигнальный пептид легкой цепи каппа, вариабельная легкая область и константная легкая область.
[00244] Понятно, что вышеприведенное подробное описание и сопроводительные примеры являются всего лишь иллюстративными, и их не следует воспринимать как ограничение объема изобретения, который определяется исключительно прилагаемой формулой изобретения и ее эквивалентами.
[00245] Для специалистов в данной области техники очевидно существование различных изменений и модификаций описанных вариантов осуществления. Такие изменения и модификации, включая, без ограничения, относящиеся к химическим структурам, заместителям, производным, промежуточным соединениям, синтезу, композициям, составам или способам применения изобретения, можно осуществлять, не отступая от его сущности и объема.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Вейнер, Дэвид Б.
Силике, Флингей
<120> КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА
<130> 206108-0071-00-WO.606915
<150> США 62/418468
<151> 07.11.2016
<160> 27
<170> PatentIn версия 3.5
<210> 1
<211> 2193
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность DMAb 319-44 mod1 CDR
<400> 1
atggactgga cttggagaat cctgttcctg gtcgccgccg ctactgggac tcacgccgag
60
atgcagctgg tcgaatccgg gggaggcctg gtgcagcctg gcggaagcct gcgactgtcc
120
tgcgctggct ctggatacat cttcgcaact tattggattg gatgggtccg ccaggcacca
180
gggaagggac tggaatgggt gggaatcatc tacccaaacg actctgatac aagatatagt
240
ccccggttca aaggccgctt taccatcagt gccgacaagt caattaacac agcttacctg
300
cagatgaatt ccctgcgagc agaggacacc gccgtgtact attgcgcccg gacacgctgg
360
tatttcgatc tgtggggaca ggggaccctg gtcacagtga gctccgcctc aaccaaaggg
420
cctagcgtgt ttcccctggc tccttctagt aagtcaacta gcgggggcac cgccgctctg
480
ggatgtctgg tgaaggatta cttccctgag ccagtcacag tgagctggaa ctccggcgct
540
ctgaccagcg gagtccacac atttccagca gtgctgcagt caagcggact gtacagcctg
600
tcctctgtgg tcacagtccc cagttcaagc ctggggactc agacctatat ctgcaacgtg
660
aatcacaaac cctccaatac taaggtcgac aagaaagtgg aacctaaatc ttgtgataag
720
acacatactt gccctccctg tccagcacct gagctgctgg gagggcctag cgtgttcctg
780
tttccaccca agccaaaaga caccctgatg atcagcagaa cacctgaagt cacttgcgtg
840
gtcgtggacg tgagccacga ggaccccgaa gtcaagttta actggtacgt ggatggcgtc
900
gaggtgcata atgctaagac caaaccccgc gaggaacagt acaactccac ttatcgagtc
960
gtgtctgtcc tgaccgtgct gcaccaggac tggctgaacg gaaaggagta taagtgcaaa
1020
gtgtccaaca aggccctgcc agcacccatc gagaagacaa tttctaaggc taaagggcag
1080
ccaagggaac cccaggtgta cactctgcct ccaagcagag acgagctgac aaaaaaccag
1140
gtcagcctga cttgtctggt gaaggggttc tatccatccg atatcgcagt ggagtgggaa
1200
tctaatggcc agcccgaaaa caattacaag accacacccc ctgtgctgga ctctgatggc
1260
agtttctttc tgtatagcaa actgaccgtg gacaagtccc ggtggcagca gggaaacgtc
1320
ttttcctgct ctgtgatgca tgaggccctg cacaatcatt acacccagaa aagtctgtca
1380
ctgagcccag ggaaacgagg caggaagagg agatccggct ctggagcaac aaacttctcc
1440
ctgctgaagc aggccgggga tgtggaggaa aatcctggcc caatggtcct gcagacccag
1500
gtgtttatca gtctgctgct gtggatttca ggagcctacg gggacatcca gctgacacag
1560
tctccctcct ctctgagtgc atcacctggc gatcgagtca ccattacatg tagggccagc
1620
cagtccgtga gttcaagcta cctggcttgg tatcagcaga agcctggaaa agcaccaaag
1680
ctgctgatct acggagcatc ctctagagcc actggagtgc ccagccggtt ctctgggagt
1740
ggctcaggaa ccgactttac tctgaccatt agttcactgc agcccgagga tttcgccacc
1800
tactattgcc agcagtatgg cagctcccct ctgacttttg gcggagggac caaagtggaa
1860
atcaagcgaa ctgtcgcagc ccccagcgtg ttcatctttc cacccagtga cgagcagctg
1920
aagagcggca ccgcttccgt ggtgtgcctg ctgaacaatt tctaccctag ggaagccaaa
1980
gtccagtgga aggtggataa cgctctgcag tcaggcaata gccaggagtc cgtgacagaa
2040
caggactcta aagatagtac ttattcactg tctagtacac tgactctgag caaggcagac
2100
tacgagaagc ataaagtgta tgcctgcgaa gtcactcacc aggggctgcg gtcacccgtc
2160
acaaaatctt tcaacagagg ggaatgttga taa
2193
<210> 2
<211> 729
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность DMAb 319-44 mod1 CDR
<400> 2
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Gly Ser Gly Tyr Ile Phe
35 40 45
Ala Thr Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Ile Ile Tyr Pro Asn Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Arg Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Asn
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Thr Arg Trp Tyr Phe Asp Leu Trp Gly Gln Gly
115 120 125
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys Arg Gly Arg Lys Arg Arg Ser Gly Ser Gly Ala Thr Asn Phe Ser
465 470 475 480
Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn Pro Gly Pro Met Val
485 490 495
Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser Gly Ala
500 505 510
Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
515 520 525
Pro Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser
530 535 540
Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
545 550 555 560
Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Val Pro Ser Arg
565 570 575
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
580 585 590
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ser
595 600 605
Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
610 615 620
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
625 630 635 640
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
645 650 655
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
660 665 670
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
675 680 685
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
690 695 700
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg Ser Pro Val
705 710 715 720
Thr Lys Ser Phe Asn Arg Gly Glu Cys
725
<210> 3
<211> 1395
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность тяжелой цепи DMAb 319-44 mod1
(анти-HER2
DMAb оптимизированный CDR графт)
<400> 3
atggactgga cttggagaat cctgttcctg gtcgccgccg ctactgggac tcacgccgag
60
atgcagctgg tcgaatccgg gggaggcctg gtgcagcctg gcggaagcct gcgactgtcc
120
tgcgctggct ctggatacat cttcgcaact tattggattg gatgggtccg ccaggcacca
180
gggaagggac tggaatgggt gggaatcatc tacccaaacg actctgatac aagatatagt
240
ccccggttca aaggccgctt taccatcagt gccgacaagt caattaacac agcttacctg
300
cagatgaatt ccctgcgagc agaggacacc gccgtgtact attgcgcccg gacacgctgg
360
tatttcgatc tgtggggaca ggggaccctg gtcacagtga gctccgcctc aaccaaaggg
420
cctagcgtgt ttcccctggc tccttctagt aagtcaacta gcgggggcac cgccgctctg
480
ggatgtctgg tgaaggatta cttccctgag ccagtcacag tgagctggaa ctccggcgct
540
ctgaccagcg gagtccacac atttccagca gtgctgcagt caagcggact gtacagcctg
600
tcctctgtgg tcacagtccc cagttcaagc ctggggactc agacctatat ctgcaacgtg
660
aatcacaaac cctccaatac taaggtcgac aagaaagtgg aacctaaatc ttgtgataag
720
acacatactt gccctccctg tccagcacct gagctgctgg gagggcctag cgtgttcctg
780
tttccaccca agccaaaaga caccctgatg atcagcagaa cacctgaagt cacttgcgtg
840
gtcgtggacg tgagccacga ggaccccgaa gtcaagttta actggtacgt ggatggcgtc
900
gaggtgcata atgctaagac caaaccccgc gaggaacagt acaactccac ttatcgagtc
960
gtgtctgtcc tgaccgtgct gcaccaggac tggctgaacg gaaaggagta taagtgcaaa
1020
gtgtccaaca aggccctgcc agcacccatc gagaagacaa tttctaaggc taaagggcag
1080
ccaagggaac cccaggtgta cactctgcct ccaagcagag acgagctgac aaaaaaccag
1140
gtcagcctga cttgtctggt gaaggggttc tatccatccg atatcgcagt ggagtgggaa
1200
tctaatggcc agcccgaaaa caattacaag accacacccc ctgtgctgga ctctgatggc
1260
agtttctttc tgtatagcaa actgaccgtg gacaagtccc ggtggcagca gggaaacgtc
1320
ttttcctgct ctgtgatgca tgaggccctg cacaatcatt acacccagaa aagtctgtca
1380
ctgagcccag ggaaa
1395
<210> 4
<211> 465
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи DMAb 319-44
mod1 (анти-HER2
DMAb оптимизированный CDR графт)
<400> 4
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Gly Ser Gly Tyr Ile Phe
35 40 45
Ala Thr Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Ile Ile Tyr Pro Asn Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Arg Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Asn
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Thr Arg Trp Tyr Phe Asp Leu Trp Gly Gln Gly
115 120 125
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys
465
<210> 5
<211> 705
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность легкой цепи DMAb 319-44 mod1
<400> 5
atggtcctgc agacccaggt gtttatcagt ctgctgctgt ggatttcagg agcctacggg
60
gacatccagc tgacacagtc tccctcctct ctgagtgcat cacctggcga tcgagtcacc
120
attacatgta gggccagcca gtccgtgagt tcaagctacc tggcttggta tcagcagaag
180
cctggaaaag caccaaagct gctgatctac ggagcatcct ctagagccac tggagtgccc
240
agccggttct ctgggagtgg ctcaggaacc gactttactc tgaccattag ttcactgcag
300
cccgaggatt tcgccaccta ctattgccag cagtatggca gctcccctct gacttttggc
360
ggagggacca aagtggaaat caagcgaact gtcgcagccc ccagcgtgtt catctttcca
420
cccagtgacg agcagctgaa gagcggcacc gcttccgtgg tgtgcctgct gaacaatttc
480
taccctaggg aagccaaagt ccagtggaag gtggataacg ctctgcagtc aggcaatagc
540
caggagtccg tgacagaaca ggactctaaa gatagtactt attcactgtc tagtacactg
600
actctgagca aggcagacta cgagaagcat aaagtgtatg cctgcgaagt cactcaccag
660
gggctgcggt cacccgtcac aaaatctttc aacagagggg aatgt
705
<210> 6
<211> 235
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи DMAb 319-44 mod1
<400> 6
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Pro Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr
100 105 110
Gly Ser Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
115 120 125
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
130 135 140
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
145 150 155 160
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
165 170 175
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
180 185 190
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
195 200 205
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg Ser
210 215 220
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 7
<211> 2190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность DMAb 319-44 wt
<400> 7
atggattgga cctggaggat tctgtttctg gtcgccgccg ctactggaac tcacgccgag
60
atgcagctgg tgcagtctgg agccgaagtg aagaaacctg gcgaaagcct gaagatctcc
120
tgcaaaggat ctgggtacat cttcgccaca tattggattg gctgggtgcg acagatgcca
180
ggcaagggac tggagtggat gggaatcatc tacccaaacg actccgatac tcgctattct
240
cccagttttc agggccaggt gaccatcagt gccgacaagt caattaatac agcttacctg
300
cagtggagct ccctgaaagc ctctgacacc gctatgtact attgcgcccg gacacgctgg
360
tatttcgatc tgtggggacg aggcacactg gtcaccgtga gcagcgcctc tacaaagggg
420
cctagcgtgt tccccctggc tccttcaagc aaatcaacca gcggcggaac agccgctctg
480
ggatgtctgg tgaaggatta cttccctgag ccagtcaccg tgagctggaa ctccggagct
540
ctgacaagcg gggtgcacac ttttccagca gtcctgcagt cctctggact gtacagcctg
600
agttcagtgg tcactgtgcc cagctcctct ctggggactc agacctatat ctgcaacgtc
660
aatcacaagc ccagcaatac caaagtcgac aagaaagtgg aacctaagag ctgtgataaa
720
acacatactt gccctccctg tccagcacct gagctgctgg gcggcccttc cgtgttcctg
780
tttccaccca agccaaaaga cacactgatg atttcccgca ctcctgaagt gacctgcgtg
840
gtcgtggacg tgagccacga ggaccccgaa gtgaagttca actggtacgt ggatggcgtc
900
gaggtgcata atgctaagac caaacccagg gaggaacagt acaactcaac ttatagagtc
960
gtgagcgtcc tgaccgtgct gcaccaggac tggctgaacg gaaaggagta taagtgcaaa
1020
gtgagcaata aggctctgcc agcacccatc gagaaaacaa ttagcaaggc aaaagggcag
1080
ccacgggaac cccaggtgta cactctgcct ccatcccgcg acgagctgac aaagaaccag
1140
gtgtctctga cttgtctggt caaagggttc tatccatctg atatcgccgt ggagtgggaa
1200
agtaatggcc agcccgaaaa caattacaag accacacccc ctgtgctgga ctctgatggc
1260
agtttctttc tgtatagtaa gctgaccgtg gacaaatcac ggtggcagca gggaaacgtc
1320
ttttcctgct ctgtgatgca tgaggccctg cacaatcatt acacacagaa gagtctgtca
1380
ctgagcccag gcaagcgagg aaggaaaagg agatccggat ctggggcaac taacttcagc
1440
ctgctgaaac aggccggcga tgtggaggaa aatcctggac caatggtcct gcagacccag
1500
gtgtttatct ccctgctgct gtggatttct ggggcttacg gcgaaatcgt gctgacccag
1560
tcccccggga cactgagtct gtcacctggc gagagagcta ccctgtcttg tcgggcaagc
1620
cagtccgtga gttcaagcta cctggcttgg tatcagcaga agcctggcca ggcaccaagg
1680
ctgctgatct acggagcatc ctctcgcgcc actgggattc ccgaccgatt ctctggcagt
1740
ggatcaggga ccgacttcac cctgacaatt agcagactgg agcccgaaga cttcgccgtg
1800
tactattgcc agcagtatgg cagttcacct ctgacttttg gagggggcac caaggtcgaa
1860
atcaaaaccg tggcagcccc cagcgtcttc atttttccac cctccgacga gcagctgaag
1920
agcggcacag catccgtggt gtgcctgctg aacaatttct accctagaga ggccaaggtc
1980
cagtggaaag tggataacgc tctgcagtca ggaaatagcc aggagtccgt gacagaacag
2040
gactctaagg atagtactta ttcactgagc tccactctga ccctgagcaa agcagattac
2100
gagaagcata aagtgtatgc ctgcgaggtc acccaccagg ggctgaggtc tccagtcaca
2160
aaatccttca atagaggcga atgttgataa
2190
<210> 8
<211> 728
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 319-44 wt
<400> 8
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ile Phe
35 40 45
Ala Thr Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu
50 55 60
Glu Trp Met Gly Ile Ile Tyr Pro Asn Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Asn
85 90 95
Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Thr Arg Trp Tyr Phe Asp Leu Trp Gly Arg Gly
115 120 125
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys Arg Gly Arg Lys Arg Arg Ser Gly Ser Gly Ala Thr Asn Phe Ser
465 470 475 480
Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn Pro Gly Pro Met Val
485 490 495
Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser Gly Ala
500 505 510
Tyr Gly Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser
515 520 525
Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser
530 535 540
Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
545 550 555 560
Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg
565 570 575
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg
580 585 590
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser
595 600 605
Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr Val
610 615 620
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
625 630 635 640
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
645 650 655
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
660 665 670
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
675 680 685
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
690 695 700
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg Ser Pro Val Thr
705 710 715 720
Lys Ser Phe Asn Arg Gly Glu Cys
725
<210> 9
<211> 1395
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность тяжелой цепи DMAb 319-44 wt
<400> 9
atggattgga cctggaggat tctgtttctg gtcgccgccg ctactggaac tcacgccgag
60
atgcagctgg tgcagtctgg agccgaagtg aagaaacctg gcgaaagcct gaagatctcc
120
tgcaaaggat ctgggtacat cttcgccaca tattggattg gctgggtgcg acagatgcca
180
ggcaagggac tggagtggat gggaatcatc tacccaaacg actccgatac tcgctattct
240
cccagttttc agggccaggt gaccatcagt gccgacaagt caattaatac agcttacctg
300
cagtggagct ccctgaaagc ctctgacacc gctatgtact attgcgcccg gacacgctgg
360
tatttcgatc tgtggggacg aggcacactg gtcaccgtga gcagcgcctc tacaaagggg
420
cctagcgtgt tccccctggc tccttcaagc aaatcaacca gcggcggaac agccgctctg
480
ggatgtctgg tgaaggatta cttccctgag ccagtcaccg tgagctggaa ctccggagct
540
ctgacaagcg gggtgcacac ttttccagca gtcctgcagt cctctggact gtacagcctg
600
agttcagtgg tcactgtgcc cagctcctct ctggggactc agacctatat ctgcaacgtc
660
aatcacaagc ccagcaatac caaagtcgac aagaaagtgg aacctaagag ctgtgataaa
720
acacatactt gccctccctg tccagcacct gagctgctgg gcggcccttc cgtgttcctg
780
tttccaccca agccaaaaga cacactgatg atttcccgca ctcctgaagt gacctgcgtg
840
gtcgtggacg tgagccacga ggaccccgaa gtgaagttca actggtacgt ggatggcgtc
900
gaggtgcata atgctaagac caaacccagg gaggaacagt acaactcaac ttatagagtc
960
gtgagcgtcc tgaccgtgct gcaccaggac tggctgaacg gaaaggagta taagtgcaaa
1020
gtgagcaata aggctctgcc agcacccatc gagaaaacaa ttagcaaggc aaaagggcag
1080
ccacgggaac cccaggtgta cactctgcct ccatcccgcg acgagctgac aaagaaccag
1140
gtgtctctga cttgtctggt caaagggttc tatccatctg atatcgccgt ggagtgggaa
1200
agtaatggcc agcccgaaaa caattacaag accacacccc ctgtgctgga ctctgatggc
1260
agtttctttc tgtatagtaa gctgaccgtg gacaaatcac ggtggcagca gggaaacgtc
1320
ttttcctgct ctgtgatgca tgaggccctg cacaatcatt acacacagaa gagtctgtca
1380
ctgagcccag gcaag
1395
<210> 10
<211> 465
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи DMAb 319-44 wt
<400> 10
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ile Phe
35 40 45
Ala Thr Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu
50 55 60
Glu Trp Met Gly Ile Ile Tyr Pro Asn Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Asn
85 90 95
Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Thr Arg Trp Tyr Phe Asp Leu Trp Gly Arg Gly
115 120 125
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys
465
<210> 11
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность легкой цепи DMAb 319-44 wt
<400> 11
atggtcctgc agacccaggt gtttatctcc ctgctgctgt ggatttctgg ggcttacggc
60
gaaatcgtgc tgacccagtc ccccgggaca ctgagtctgt cacctggcga gagagctacc
120
ctgtcttgtc gggcaagcca gtccgtgagt tcaagctacc tggcttggta tcagcagaag
180
cctggccagg caccaaggct gctgatctac ggagcatcct ctcgcgccac tgggattccc
240
gaccgattct ctggcagtgg atcagggacc gacttcaccc tgacaattag cagactggag
300
cccgaagact tcgccgtgta ctattgccag cagtatggca gttcacctct gacttttgga
360
gggggcacca aggtcgaaat caaaaccgtg gcagccccca gcgtcttcat ttttccaccc
420
tccgacgagc agctgaagag cggcacagca tccgtggtgt gcctgctgaa caatttctac
480
cctagagagg ccaaggtcca gtggaaagtg gataacgctc tgcagtcagg aaatagccag
540
gagtccgtga cagaacagga ctctaaggat agtacttatt cactgagctc cactctgacc
600
ctgagcaaag cagattacga gaagcataaa gtgtatgcct gcgaggtcac ccaccagggg
660
ctgaggtctc cagtcacaaa atccttcaat agaggcgaat gt
702
<210> 12
<211> 234
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи DMAb 319-44 wt
<400> 12
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
35 40 45
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
50 55 60
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
65 70 75 80
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
100 105 110
Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 13
<211> 2205
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность DMAb 221-7 mod 9
полноразмерного IgG1 человека:
Единичная плазмида
<400> 13
atggattgga catggaggat tctgtttctg gtcgccgccg ccacaggaac ccacgccgaa
60
gtgcagctgg tggaatcagg cggagggctg gtgcagccag gcggaagtct gcgactgtca
120
tgcgccgtga gtgggtactc attcactagc tattggattg gatgggtgcg gcaggcacca
180
ggcaagggac tggagtgggt gggattcatc taccccgggg actccgatac acgctatagt
240
ccttctttca aaggcaggtt cacaatctct gccgacaaaa gcatttccac tgcttatctg
300
cagatgaact ccctgcgggc tgaggatacc gcagtgtact attgcgccag gggcattctg
360
aggtacttcg actggtttct ggactattgg gggcagggca ccctggtcac agtgtcaagc
420
gcctctacca aaggaccaag cgtgttccca ctggctcctt cctctaagtc tactagtggc
480
ggaaccgccg ctctgggatg tctggtgaag gattacttcc ctgagccagt cacagtgtcc
540
tggaactctg gcgctctgac cagcggagtc cacacatttc ccgcagtgct gcagagttca
600
ggcctgtact ccctgagctc cgtggtcaca gtcccttcta gttcactggg aactcagacc
660
tatatctgca acgtgaatca caaaccttcc aatactaagg tcgacaagaa agtggaacca
720
aaatcttgtg ataagacaca tacttgccct ccctgtccag cacctgagct gctgggcggc
780
ccaagcgtgt tcctgtttcc acccaagccc aaagataccc tgatgattag caggacacca
840
gaagtcactt gcgtggtcgt ggacgtgtcc cacgaggacc ccgaagtcaa gttcaactgg
900
tacgtggacg gcgtcgaggt gcataatgct aagaccaaac caagagagga acagtacaac
960
tcaacctatc gggtcgtgag cgtcctgaca gtgctgcacc aggactggct gaacggaaag
1020
gagtataagt gcaaagtgtc taacaaggcc ctgccagctc ccatcgagaa gactattagc
1080
aaggctaaag ggcagccacg cgaaccccag gtgtacaccc tgcctccatc acgagatgag
1140
ctgacaaaaa accaggtctc tctgacttgt ctggtgaagg gattctatcc ctctgacatc
1200
gcagtggagt gggaaagtaa tgggcagcct gaaaacaatt acaagaccac accccctgtg
1260
ctggacagtg atggatcatt ctttctgtat agtaaactga ccgtggataa gtcaagatgg
1320
cagcagggga acgtcttttc atgcagcgtg atgcatgagg ccctgcacaa tcattacacc
1380
cagaagtccc tgtctctgag tcctggcaaa cggggacgca agaggagatc aggaagcggg
1440
gctacaaact tctccctgct gaagcaggca ggggacgtgg aggaaaatcc tggcccaatg
1500
gtcctgcaga cccaggtgtt tatctccctg ctgctgtgga tttctggggc atacggcgac
1560
atccagctga cacagtctcc cagctccctg tccgcatctg tcggcgaccg agtgaccatc
1620
acatgtaggg ccagccaggg gatttctagt ggctcagcat ggtaccagca gaagcctggg
1680
aaagcaccaa agctgctgat ctatgacgtg tctagcctgg aatccggagt gcctagccgg
1740
ttctccggat caggaagtgg gacagacttt actctgacca tttcaagcct gcagcctgag
1800
gatttcgcca cttactattg ccagcagttc aatagctatc tgctgacttt tggacagggc
1860
accaaagtgg aaatcaagag gactgtcgca gcccctagcg tgttcatttt tccaccctcc
1920
gatgagcagc tgaagagcgg caccgcttcc gtggtgtgcc tgctgaacaa cttctaccca
1980
cgcgaggcca aagtccagtg gaaggtggac aacgctctgc agtctggaaa tagtcaggag
2040
tcagtgactg aacaggacag caaagattcc acctattctc tgtcctctac actgactctg
2100
agcaaggcag actacgagaa gcataaagtg tatgcctgcg aagtcaccca ccaggggctg
2160
tcctcaccag tcactaaatc tttcaatcgg ggagaatgtt gataa
2205
<210> 14
<211> 733
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 221-7 mod 9
полноразмерного IgG1 человека
Единичная плазмида
<400> 14
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys Arg Gly Arg Lys Arg Arg Ser Gly Ser Gly
465 470 475 480
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
485 490 495
Pro Gly Pro Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu
500 505 510
Trp Ile Ser Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser
515 520 525
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
530 535 540
Ser Gln Gly Ile Ser Ser Gly Ser Ala Trp Tyr Gln Gln Lys Pro Gly
545 550 555 560
Lys Ala Pro Lys Leu Leu Ile Tyr Asp Val Ser Ser Leu Glu Ser Gly
565 570 575
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
580 585 590
Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln
595 600 605
Gln Phe Asn Ser Tyr Leu Leu Thr Phe Gly Gln Gly Thr Lys Val Glu
610 615 620
Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
625 630 635 640
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
645 650 655
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
660 665 670
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
675 680 685
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
690 695 700
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
705 710 715 720
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
725 730
<210> 15
<211> 1409
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность тяжелой цепи DMAb 221-7 mod 9
IgG1 человека
<400> 15
atggattgga catggaggat tctgtttctg gtcgccgccg ccacaggaac ccacgccgaa
60
gtgcagctgg tggaatcagg cggagggctg gtgcagccag gcggaagtct gcgactgtca
120
tgcgccgtga gtgggtactc attcactagc tattggattg gatgggtgcg gcaggcacca
180
ggcaagggac tggagtgggt gggattcatc taccccgggg actccgatac acgctatagt
240
ccttctttca aaggcaggtt cacaatctct gccgacaaaa gcatttccac tgcttatctg
300
cagatgaact ccctgcgggc tgaggatacc gcagtgtact attgcgccag gggcattctg
360
aggtacttcg actggtttct ggactattgg gggcagggca ccctggtcac agtgtcaagc
420
gcctctacca aaggaccaag cgtgttccca ctggctcctt cctctaagtc tactagtggc
480
ggaaccgccg ctctgggatg tctggtgaag gattacttcc ctgagccagt cacagtgtcc
540
tggaactctg gcgctctgac cagcggagtc cacacatttc ccgcagtgct gcagagttca
600
ggcctgtact ccctgagctc cgtggtcaca gtcccttcta gttcactggg aactcagacc
660
tatatctgca acgtgaatca caaaccttcc aatactaagg tcgacaagaa agtggaacca
720
aaatcttgtg ataagacaca tacttgccct ccctgtccag cacctgagct gctgggcggc
780
ccaagcgtgt tcctgtttcc acccaagccc aaagataccc tgatgattag caggacacca
840
gaagtcactt gcgtggtcgt ggacgtgtcc cacgaggacc ccgaagtcaa gttcaactgg
900
tacgtggacg gcgtcgaggt gcataatgct aagaccaaac caagagagga acagtacaac
960
tcaacctatc gggtcgtgag cgtcctgaca gtgctgcacc aggactggct gaacggaaag
1020
gagtataagt gcaaagtgtc taacaaggcc ctgccagctc ccatcgagaa gactattagc
1080
aaggctaaag ggcagccacg cgaaccccag gtgtacaccc tgcctccatc acgagatgag
1140
ctgacaaaaa accaggtctc tctgacttgt ctggtgaagg gattctatcc ctctgacatc
1200
gcagtggagt gggaaagtaa tgggcagcct gaaaacaatt acaagaccac accccctgtg
1260
ctggacagtg atggatcatt ctttctgtat agtaaactga ccgtggataa gtcaagatgg
1320
cagcagggga acgtcttttc atgcagcgtg atgcatgagg ccctgcacaa tcattacacc
1380
cagaagtccc tgtctctgag tcctggcaa
1409
<210> 16
<211> 470
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи DMAb 221-7 mod
9 IgG1 человека
<400> 16
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 17
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность легкой цепи DMAb 221-7 mod 9
IgG1 человека
<400> 17
atggtcctgc agacccaggt gtttatctcc ctgctgctgt ggatttctgg ggcatacggc
60
gacatccagc tgacacagtc tcccagctcc ctgtccgcat ctgtcggcga ccgagtgacc
120
atcacatgta gggccagcca ggggatttct agtggctcag catggtacca gcagaagcct
180
gggaaagcac caaagctgct gatctatgac gtgtctagcc tggaatccgg agtgcctagc
240
cggttctccg gatcaggaag tgggacagac tttactctga ccatttcaag cctgcagcct
300
gaggatttcg ccacttacta ttgccagcag ttcaatagct atctgctgac ttttggacag
360
ggcaccaaag tggaaatcaa gaggactgtc gcagccccta gcgtgttcat ttttccaccc
420
tccgatgagc agctgaagag cggcaccgct tccgtggtgt gcctgctgaa caacttctac
480
ccacgcgagg ccaaagtcca gtggaaggtg gacaacgctc tgcagtctgg aaatagtcag
540
gagtcagtga ctgaacagga cagcaaagat tccacctatt ctctgtcctc tacactgact
600
ctgagcaagg cagactacga gaagcataaa gtgtatgcct gcgaagtcac ccaccagggg
660
ctgtcctcac cagtcactaa atctttcaat cggggagaat gt
702
<210> 18
<211> 234
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи DMAb 221-7 mod 9
IgG1 человека
<400> 18
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Ser Gly Ser Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Asp Val Ser Ser Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn
100 105 110
Ser Tyr Leu Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 19
<211> 2202
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность DMAb 221-7 wt
<400> 19
atggattgga catggaggat tctgtttctg gtcgccgccg ccacaggaac ccacgccgaa
60
gtgcagctgg tgcagtcagg ctccgaggtg aagaaaccag gcgaaagtct gaaaatctca
120
tgccaggtga gtgggtactc attcactagc tattggattg gatgggtgcg gcagatgcca
180
ggcaagggac tggagcgagt gggattcatc taccccgggg actccgatac acgctatagt
240
ccttcatttc agggccaggt gacaatctct gccgacaaaa gcatttccac tgcttatctg
300
cagtggagct ccctgaaggc ttccgatacc gcaatgtact attgcgccag gggcattctg
360
aggtacttcg actggtttct ggactattgg gggcagggca ccctggtcac agtgtcaagc
420
gcctctacca aaggaccaag cgtgttccca ctggctcctt cctctaagtc tactagtggc
480
ggaaccgccg ctctgggatg tctggtgaag gattacttcc ctgagccagt cacagtgtcc
540
tggaactctg gcgctctgac cagcggagtc cacacatttc ccgcagtgct gcagagttca
600
ggcctgtact ccctgagctc cgtggtcaca gtcccttcta gttcactggg aactcagacc
660
tatatctgca acgtgaatca caaaccttcc aatactaagg tcgacaagaa agtggaacca
720
aaatcttgtg ataagacaca tacttgccct ccctgtccag cacctgagct gctgggcggc
780
ccaagcgtgt tcctgtttcc acccaagccc aaagataccc tgatgattag caggacacca
840
gaagtcactt gcgtggtcgt ggacgtgtcc cacgaggacc ccgaagtcaa gttcaactgg
900
tacgtggacg gcgtcgaggt gcataatgct aagaccaaac caagagagga acagtacaac
960
tcaacctatc gggtcgtgag cgtcctgaca gtgctgcacc aggactggct gaacggaaag
1020
gagtataagt gcaaagtgtc taacaaggcc ctgccagctc ccatcgagaa gactattagc
1080
aaggctaaag ggcagccacg cgaaccccag gtgtacaccc tgcctccatc acgagatgag
1140
ctgacaaaaa accaggtctc tctgacttgt ctggtgaagg gattctatcc ctctgacatc
1200
gcagtggagt gggaaagtaa tgggcagcct gaaaacaatt acaagaccac accccctgtg
1260
ctggacagtg atggatcatt ctttctgtat agtaaactga ccgtggataa gtcaagatgg
1320
cagcagggga acgtcttttc atgcagcgtg atgcatgagg ccctgcacaa tcattacacc
1380
cagaagtccc tgtctctgag tcctggcaaa cggggacgca agaggagatc aggaagcggg
1440
gctacaaact tctccctgct gaagcaggca ggggacgtgg aggaaaatcc tggcccaatg
1500
gtcctgcaga cccaggtgtt tatctccctg ctgctgtgga tttctggggc atacggcgcc
1560
atccagctga cacagtctcc cagctccctg tccgcatctg tcggcgaccg agtgaccatc
1620
acatgtaggg ccagccaggg gatttctagt ggctcagcat ggtaccagca gaagcctggg
1680
aaagcaccaa agctgctgat ctatgacgtg tctagcctgg aatccggagt gcctagccgg
1740
ttctccggat caggaagtgg gacagagttt actctgacca tttcaagcct gcagcctgag
1800
gatttcgcca cttactattg ccagcagttc aatagctatc tgctgacttt tggagggggc
1860
accaaagtgg aaatcaagac tgtcgcagcc cctagcgtgt tcatttttcc accctccgat
1920
gagcagctga agagcggcac cgcttccgtg gtgtgcctgc tgaacaactt ctacccacgc
1980
gaggccaaag tccagtggaa ggtggacaac gctctgcagt ctggaaatag tcaggagtca
2040
gtgactgaac aggacagcaa agattccacc tattctctgt cctctacact gactctgagc
2100
aaggcagact acgagaagca taaagtgtat gcctgcgaag tcacccacca ggggctgagg
2160
tcaccagtca ctaaatcttt caatcgggga gaatgttgat aa
2202
<210> 20
<211> 732
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 221-7 wt
<400> 20
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Gln Ser Gly Ser Glu Val Lys Lys
20 25 30
Pro Gly Glu Ser Leu Lys Ile Ser Cys Gln Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu
50 55 60
Glu Arg Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys Arg Gly Arg Lys Arg Arg Ser Gly Ser Gly
465 470 475 480
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
485 490 495
Pro Gly Pro Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu
500 505 510
Trp Ile Ser Gly Ala Tyr Gly Ala Ile Gln Leu Thr Gln Ser Pro Ser
515 520 525
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
530 535 540
Ser Gln Gly Ile Ser Ser Gly Ser Ala Trp Tyr Gln Gln Lys Pro Gly
545 550 555 560
Lys Ala Pro Lys Leu Leu Ile Tyr Asp Val Ser Ser Leu Glu Ser Gly
565 570 575
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu
580 585 590
Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln
595 600 605
Gln Phe Asn Ser Tyr Leu Leu Thr Phe Gly Gly Gly Thr Lys Val Glu
610 615 620
Ile Lys Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
625 630 635 640
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
645 650 655
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
660 665 670
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
675 680 685
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
690 695 700
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg
705 710 715 720
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
725 730
<210> 21
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность тяжелой цепи DMAb 221-7 wt
<400> 21
atggattgga catggaggat tctgtttctg gtcgccgccg ccacaggaac ccacgccgaa
60
gtgcagctgg tgcagtcagg ctccgaggtg aagaaaccag gcgaaagtct gaaaatctca
120
tgccaggtga gtgggtactc attcactagc tattggattg gatgggtgcg gcagatgcca
180
ggcaagggac tggagcgagt gggattcatc taccccgggg actccgatac acgctatagt
240
ccttcatttc agggccaggt gacaatctct gccgacaaaa gcatttccac tgcttatctg
300
cagtggagct ccctgaaggc ttccgatacc gcaatgtact attgcgccag gggcattctg
360
aggtacttcg actggtttct ggactattgg gggcagggca ccctggtcac agtgtcaagc
420
gcctctacca aaggaccaag cgtgttccca ctggctcctt cctctaagtc tactagtggc
480
ggaaccgccg ctctgggatg tctggtgaag gattacttcc ctgagccagt cacagtgtcc
540
tggaactctg gcgctctgac cagcggagtc cacacatttc ccgcagtgct gcagagttca
600
ggcctgtact ccctgagctc cgtggtcaca gtcccttcta gttcactggg aactcagacc
660
tatatctgca acgtgaatca caaaccttcc aatactaagg tcgacaagaa agtggaacca
720
aaatcttgtg ataagacaca tacttgccct ccctgtccag cacctgagct gctgggcggc
780
ccaagcgtgt tcctgtttcc acccaagccc aaagataccc tgatgattag caggacacca
840
gaagtcactt gcgtggtcgt ggacgtgtcc cacgaggacc ccgaagtcaa gttcaactgg
900
tacgtggacg gcgtcgaggt gcataatgct aagaccaaac caagagagga acagtacaac
960
tcaacctatc gggtcgtgag cgtcctgaca gtgctgcacc aggactggct gaacggaaag
1020
gagtataagt gcaaagtgtc taacaaggcc ctgccagctc ccatcgagaa gactattagc
1080
aaggctaaag ggcagccacg cgaaccccag gtgtacaccc tgcctccatc acgagatgag
1140
ctgacaaaaa accaggtctc tctgacttgt ctggtgaagg gattctatcc ctctgacatc
1200
gcagtggagt gggaaagtaa tgggcagcct gaaaacaatt acaagaccac accccctgtg
1260
ctggacagtg atggatcatt ctttctgtat agtaaactga ccgtggataa gtcaagatgg
1320
cagcagggga acgtcttttc atgcagcgtg atgcatgagg ccctgcacaa tcattacacc
1380
cagaagtccc tgtctctgag tcctggcaaa
1410
<210> 22
<211> 470
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи DMAb 221-7 wt
<400> 22
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Gln Ser Gly Ser Glu Val Lys Lys
20 25 30
Pro Gly Glu Ser Leu Lys Ile Ser Cys Gln Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu
50 55 60
Glu Arg Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 23
<211> 699
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность легкой цепи DMAb 221-7 wt
<400> 23
atggtcctgc agacccaggt gtttatctcc ctgctgctgt ggatttctgg ggcatacggc
60
gccatccagc tgacacagtc tcccagctcc ctgtccgcat ctgtcggcga ccgagtgacc
120
atcacatgta gggccagcca ggggatttct agtggctcag catggtacca gcagaagcct
180
gggaaagcac caaagctgct gatctatgac gtgtctagcc tggaatccgg agtgcctagc
240
cggttctccg gatcaggaag tgggacagag tttactctga ccatttcaag cctgcagcct
300
gaggatttcg ccacttacta ttgccagcag ttcaatagct atctgctgac ttttggaggg
360
ggcaccaaag tggaaatcaa gactgtcgca gcccctagcg tgttcatttt tccaccctcc
420
gatgagcagc tgaagagcgg caccgcttcc gtggtgtgcc tgctgaacaa cttctaccca
480
cgcgaggcca aagtccagtg gaaggtggac aacgctctgc agtctggaaa tagtcaggag
540
tcagtgactg aacaggacag caaagattcc acctattctc tgtcctctac actgactctg
600
agcaaggcag actacgagaa gcataaagtg tatgcctgcg aagtcaccca ccaggggctg
660
aggtcaccag tcactaaatc tttcaatcgg ggagaatgt
699
<210> 24
<211> 233
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи DMAb 221-7 wt
<400> 24
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Ser Gly Ser Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Asp Val Ser Ser Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn
100 105 110
Ser Tyr Leu Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Arg Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 25
<211> 738
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 221-7 mod 9 CDR мыши
<400> 25
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
130 135 140
Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly Gly Thr Thr Gly
145 150 155 160
Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
165 170 175
Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
180 185 190
Phe Pro Ala Leu Leu Gln Ser Gly Leu Tyr Thr Leu Ser Ser Ser Val
195 200 205
Thr Val Thr Ser Asn Thr Trp Pro Ser Gln Thr Ile Thr Cys Asn Val
210 215 220
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg
225 230 235 240
Val Pro Ile Thr Gln Asn Pro Cys Pro Pro Leu Lys Glu Cys Pro Pro
245 250 255
Cys Ala Ala Pro Asp Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
260 265 270
Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Met Val Thr
275 280 285
Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser
290 295 300
Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His
305 310 315 320
Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile
325 330 335
Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn
340 345 350
Asn Arg Ala Leu Pro Ser Pro Ile Glu Lys Thr Ile Ser Lys Pro Arg
355 360 365
Gly Pro Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Ala Glu
370 375 380
Glu Met Thr Lys Lys Glu Phe Ser Leu Thr Cys Met Ile Thr Gly Phe
385 390 395 400
Leu Pro Ala Glu Ile Ala Val Asp Trp Thr Ser Asn Gly Arg Thr Glu
405 410 415
Gln Asn Tyr Lys Asn Thr Ala Thr Val Leu Asp Ser Asp Gly Ser Tyr
420 425 430
Phe Met Tyr Ser Lys Leu Arg Val Gln Lys Ser Thr Trp Glu Arg Gly
435 440 445
Ser Leu Phe Ala Cys Ser Val Val His Glu Val Leu His Asn His Leu
450 455 460
Thr Thr Lys Thr Ile Ser Arg Ser Leu Gly Lys Arg Gly Arg Lys Arg
465 470 475 480
Arg Ser Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly
485 490 495
Asp Val Glu Glu Asn Pro Gly Pro Met Val Leu Gln Thr Gln Val Phe
500 505 510
Ile Ser Leu Leu Leu Trp Ile Ser Gly Ala Tyr Gly Asp Ile Gln Leu
515 520 525
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
530 535 540
Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Gly Ser Ala Trp Tyr
545 550 555 560
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Val Ser
565 570 575
Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
580 585 590
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
595 600 605
Thr Tyr Tyr Cys Gln Gln Phe Asn Ser Tyr Leu Leu Thr Phe Gly Gln
610 615 620
Gly Thr Lys Val Glu Ile Lys Arg Ala Asp Ala Ala Pro Thr Val Ser
625 630 635 640
Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala Ser Val
645 650 655
Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp
660 665 670
Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser Trp Thr
675 680 685
Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr Leu Thr
690 695 700
Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys Glu Ala
705 710 715 720
Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn Arg Asn
725 730 735
Glu Cys
<210> 26
<211> 475
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 221-7 mod 9 CDR мыши
<400> 26
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Thr His Ala Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Phe
35 40 45
Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Phe Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser
65 70 75 80
Pro Ser Phe Lys Gly Arg Phe Thr Ile Ser Ala Asp Lys Ser Ile Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Ile Leu Arg Tyr Phe Asp Trp Phe Leu Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
130 135 140
Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly Gly Thr Thr Gly
145 150 155 160
Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
165 170 175
Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
180 185 190
Phe Pro Ala Leu Leu Gln Ser Gly Leu Tyr Thr Leu Ser Ser Ser Val
195 200 205
Thr Val Thr Ser Asn Thr Trp Pro Ser Gln Thr Ile Thr Cys Asn Val
210 215 220
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg
225 230 235 240
Val Pro Ile Thr Gln Asn Pro Cys Pro Pro Leu Lys Glu Cys Pro Pro
245 250 255
Cys Ala Ala Pro Asp Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
260 265 270
Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Met Val Thr
275 280 285
Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser
290 295 300
Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His
305 310 315 320
Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile
325 330 335
Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn
340 345 350
Asn Arg Ala Leu Pro Ser Pro Ile Glu Lys Thr Ile Ser Lys Pro Arg
355 360 365
Gly Pro Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Ala Glu
370 375 380
Glu Met Thr Lys Lys Glu Phe Ser Leu Thr Cys Met Ile Thr Gly Phe
385 390 395 400
Leu Pro Ala Glu Ile Ala Val Asp Trp Thr Ser Asn Gly Arg Thr Glu
405 410 415
Gln Asn Tyr Lys Asn Thr Ala Thr Val Leu Asp Ser Asp Gly Ser Tyr
420 425 430
Phe Met Tyr Ser Lys Leu Arg Val Gln Lys Ser Thr Trp Glu Arg Gly
435 440 445
Ser Leu Phe Ala Cys Ser Val Val His Glu Val Leu His Asn His Leu
450 455 460
Thr Thr Lys Thr Ile Ser Arg Ser Leu Gly Lys
465 470 475
<210> 27
<211> 234
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DMAb 221-7 mod 9 CDR мыши
<400> 27
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Ser Gly Ser Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Asp Val Ser Ser Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn
100 105 110
Ser Tyr Leu Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
130 135 140
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
145 150 155 160
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
165 170 175
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
195 200 205
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
210 215 220
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
225 230
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| КОМПОЗИЦИИ И СПОСОБЫ РЕДАКТИРОВАНИЯ ГЕНОВ | 2019 |
|
RU2804665C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2019 |
|
RU2791022C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |

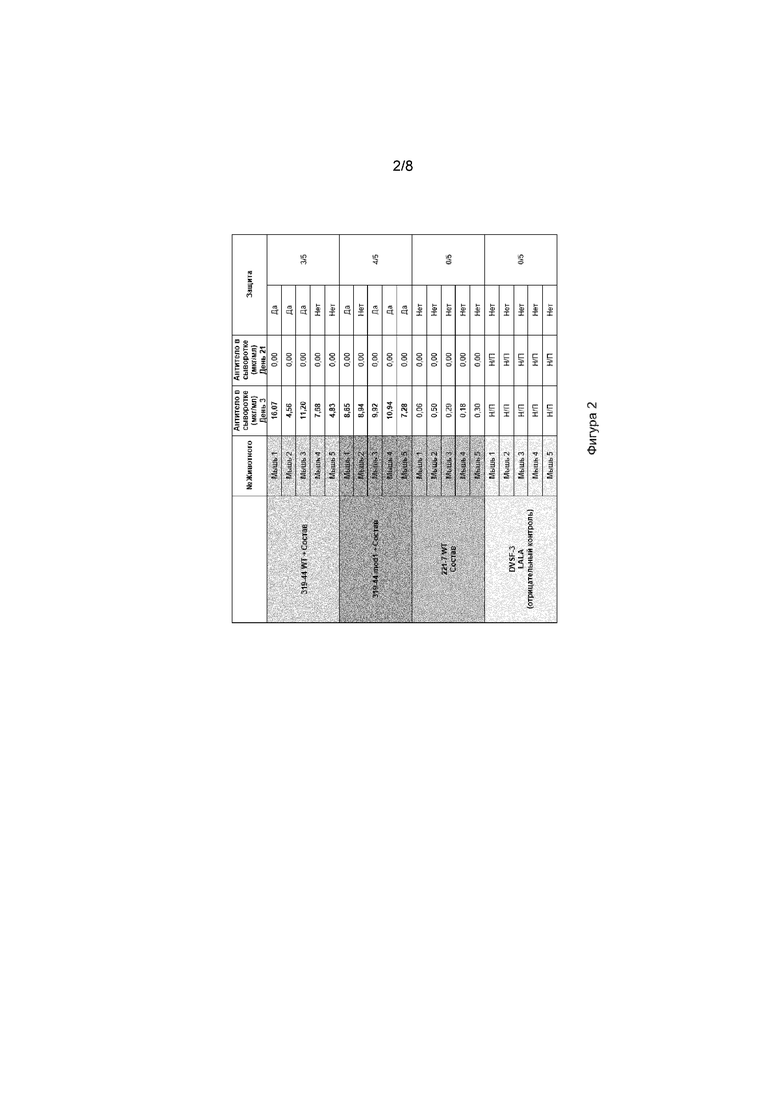





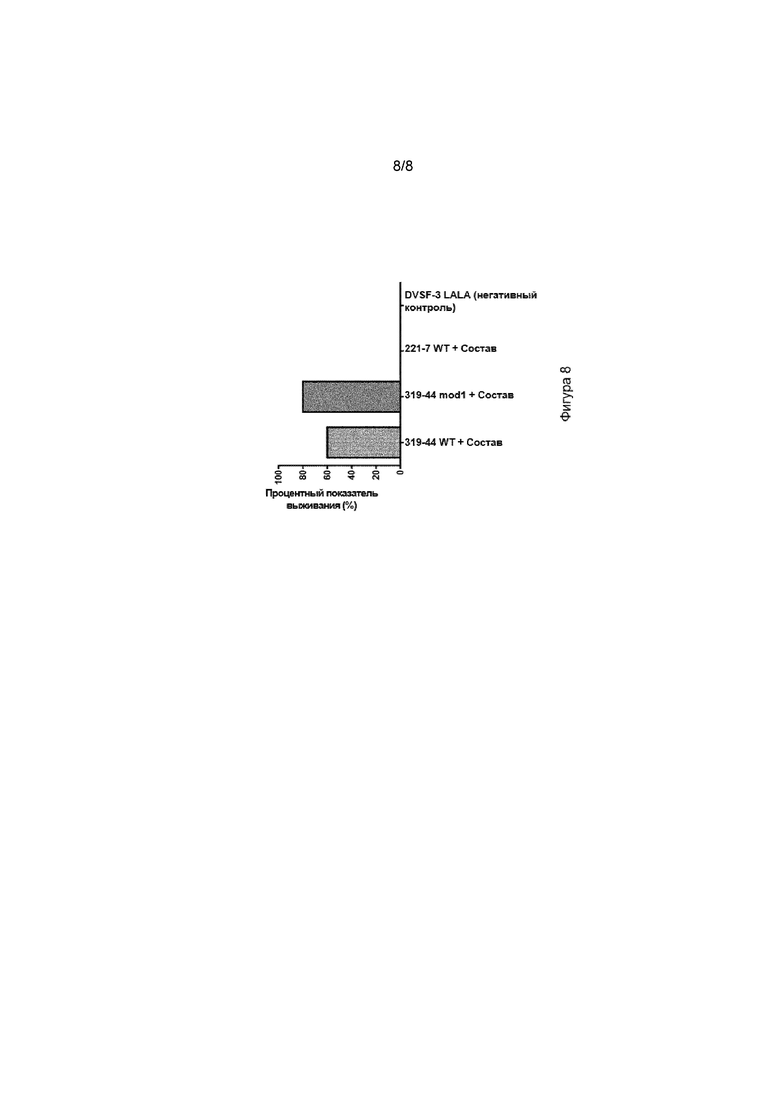
Изобретение относится к области биотехнологии, конкретно к рекомбинантным нуклеиновым кислотам, кодирующим антитела к антигену Borrelia, и может быть использовано в медицине для предотвращения или лечения болезни Лайма. Предложена молекула нуклеиновой кислоты, кодирующая одно или более синтетических антител к OspA или их функциональных фрагментов, которая в составе вектора экспрессии при введении пациенту обеспечивает генерацию специфических антител против возбудителя болезни Лайма. 3 н. и 8 з.п. ф-лы, 8 ил., 1 пр.
1. Молекула нуклеиновой кислоты, кодирующая одно или более синтетических антител, где молекула нуклеиновой кислоты содержит по меньшей мере одно выбранное из группы, состоящей из
а) нуклеотидной последовательности, кодирующей синтетическое антитело к OspA, и
b) нуклеотидной последовательности, кодирующей функциональный фрагмент синтетического антитела к OspA, где указанный функциональный фрагмент связывается с белком OspA или его участком сходным образом с указанным антителом к OspA; и
где указанная молекула нуклеиновой кислоты кодирует по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из
i) аминокислотной последовательности, представленной в SEQ ID NO: 2, SEQ ID NO: 4 или SEQ ID NO: 6; и
ii) функционального фрагмента аминокислотной последовательности, представленной в SEQ ID NO: 2, SEQ ID NO: 4 или SEQ ID NO: 6; и/или
где указанная молекула нуклеиновой кислоты содержит по меньшей мере одно выбранное из группы, состоящей из
iii) нуклеотидной последовательности, представленной в SEQ ID NO: 1, SEQ ID NO: 3 или SEQ ID NO: 5; и
iv) функционального фрагмента нуклеотидной последовательности, представленной в SEQ ID NO: 1, SEQ ID NO: 3 или SEQ ID NO: 5.
2. Молекула нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты дополнительно содержит домен расщепления.
3. Молекула нуклеиновой кислоты по п. 1, где молекула нуклеиновой кислоты кодирует по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из
i) аминокислотной последовательности, представленной в SEQ ID NO: 2, SEQ ID NO: 4 или SEQ ID NO: 6; и
ii) функционального фрагмента аминокислотной последовательности, представленной в SEQ ID NO: 2, SEQ ID NO: 4 или SEQ ID NO: 6.
4. Молекула нуклеиновой кислоты по п. 1, где молекула нуклеиновой кислоты содержит по меньшей мере одно выбранное из группы, состоящей из
i) нуклеотидной последовательности, представленной в SEQ ID NO: 1, SEQ ID NO: 3 или SEQ ID NO: 5; и
ii) функционального фрагмента нуклеотидной последовательности, представленной в SEQ ID NO: 1, SEQ ID NO: 3 или SEQ ID NO: 5.
5. Молекула нуклеиновой кислоты по п. 1, содержащая нуклеотидную последовательность, кодирующую одну или более из вариабельной области тяжелой цепи и вариабельной области легкой цепи, причем последовательность, кодирующая вариабельную область тяжелой цепи, выбрана из группы, состоящей из:
i) нуклеотидной последовательности, кодирующей аминокислотную последовательность, представленную в SEQ ID NO: 4;
ii) нуклеотидной последовательности, представленную в SEQ ID NO: 3, и
причем последовательность, кодирующая вариабельную область легкой цепи, выбрана из группы, состоящей из:
iii) нуклеотидной последовательности, кодирующей аминокислотную последовательность, представленную в SEQ ID NO: 6; и
iv) нуклеотидной последовательности, представленной в SEQ ID NO: 5.
6. Молекула нуклеиновой кислоты по п. 5, содержащая нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную в SEQ ID NO: 2.
7. Молекула нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты дополнительно содержит лидерную последовательность.
8. Молекула нуклеиновой кислоты по любому из пп. 1-7, где молекула нуклеиновой кислоты дополнительно содержит вектор экспрессии.
9. Композиция для предотвращения или лечения болезни Лайма, содержащая вектор экспрессии для экспрессии эффективного количества молекулы нуклеиновой кислоты по любому из пп. 1-8.
10. Композиция по п. 9, дополнительно содержащая фармацевтически приемлемое вспомогательное вещество.
11. Способ предотвращения или лечения болезни Лайма у субъекта, где способ включает введение субъекту композиции по любому из пп. 9, 10.
| WO 2016025331 А1, 18.02.2016, WO 2014093894 А2, 19.06.2014, БАРСКОВА В | |||
| Г | |||
| и др | |||
| Критерии позитивности для IgG антител в методе иммунного блоттинга при болезни Лайма, Научно-практическая ревматология, 2001, н | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| M | |||
| Effects of amino acid sequence changes on antibody-antigen interactions, Research in Immunology, 1994, v | |||
| Заслонка для русской печи | 1919 |
|
SU145A1 |

