Изобретение относится к области иммунологии, микробиологии и молекулярной генетики и может быть использовано в медицинской промышленности для создания вакцин против вируса SARS-CoV-2.
Коронавирусы (Coronaviridae) - семейство вирусов, содержащее более 60 видов РНК-содержащих патогенных вирусов, объединенных в четыре подсемейства, которые поражают человека и животных. Коронавирусы имеют сходную организацию генома, представленного цепочкой из положительной цепи однонитевой РНК из 25-30 тысяч нуклеотидов.
Коронавирус человека впервые был выделен в 1965 году от больных ОРВИ Д. Тиррелом из носоглотки при остром рините. В 2002-2003 годах в Китае распространение вируса SARS-CoV, бета-коронавируса В привело к вспышке атипичной пневмонии или тяжелого острого респираторного синдрома (ТОРС, SARS). В 2015 году в Южной Корее была зафиксирована вспышка ближневосточного респираторного синдрома (MERS), причиной которой стал вирус MERS-CoV, бета-коронавирус С. В декабре 2019 года начавшаяся эпидемия атипичной пневмонии, вызванная ранее неизвестным бета-коронавирусом, обозначенным как SARS-CoV-2, быстро переросла в пандемию COVID-19, охватив практически все страны и континенты, и население более 164 миллионов человек. Несмотря на приятые меры по борьбе с короновирусом, распространение SARS-CoV-2 привело к гибели более 3,4 миллиона человек. К настоящему времени население планеты пережило две-три вспышки заболевания COVID-19. Усилия по борьбе с COVID-19 привели к закрытию границ между странами, строгому карантину и ограничениям на перевозки, что серьезно подорвало экономику многих стран.
Коронавирусы имеют РНК около 26-30 тысяч пар оснований. Это означает, что коронавирусы обладают крупнейшей несегментированной РНК среди всех известных вирусов, то есть являются сложнейшими по структуре среди известных вирусов. Геном вируса состоит из 30000 нуклеотидов и кодирует два репликативных полипротеина pp1a и pp1ab, из которых в следующий проход репликации/трансляции формируется копия РНК вируса, а также 8 отдельных мРНК-шаблонов для белков вирусов, которые бесконечно их генерируют.[Thiel V. Coronaviruses: Molecular and Cellular Biology. Caister Academic Press (2007); King, Andrew M.Q.; Adams, Michael J.; Carstens, Eric В.; Lefkowitz, Elliot J, Virus Taxonomy, 2012].
При адгезии клеток-мишеней вирусом SARS-CoV-2 основная роль принадлежит поверхностному белку, обозначенному как S-белок, который продуцируется на поверхности шиловидных отростков белковой оболочки вируса и связывает трансмембранный рецептор эукариотических клеток (АСЕ2) [Xintian Xu, Ping Chen, Jingfang Wang, Jiannan Feng, Hui Zhou. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission (англ.) // SCIENCE CHINA Life Sciences, 2020].
Благодаря появлению в январе 2002 года в международной базе данных генома SARS-CoV-2 у международного научного сообщества появилась возможность активизировать усилия по созданию вакцинных препаратов против данного вида вируса. Разработки вакцин ведутся по нескольким направлениям: использование аттенуированных или инактивированных вирусов, создание рекомбинантных полипептидов на основе вирусных белков, создание живых вакцин с использованием аденовирусов и лентивирусов содержащих вирусные белки, создание мРНК-вакцин. Большинство вакцинных препаратов содержат поверхностный S-белок или ДНК, кодирующую S-белок, и вызывают продукцию специфических анти-IgG.
Интерес представляет непосредственное получение поверхностного S-белка, изучение его антигенных свойств, перспективы применения в тест-системах для обнаружения специфических антител, а также возможности для применения при производстве вакцинных препаратов, в состав которых входит S-белок.
Прототипом данного изобретения является создание живой вакцины на основе биологически активного штамма Enterococcus faecium L3 за счет включения в структуру его пилей фрагмента гена, кодирующего S-белок коронавируса SARS-CoV-2 [Патент на изобретение № RU 2745626 С1 «Создание живой вакцины против коронавирусной инфекции COVID-19 на основе пробиотического штамма Enterococcus faecium L3 и живая вакцина Enterococcus faecium L3-pentF-covid-19»].
Особенностью прототипа является встраивание фрагмента гена, кодирующего S-белок коронавируса SARS-CoV-2, неспосредственно в геном клеток штамма Enterococcus faecium L3.
Задачей данного изобретения стало получение рекомбинантной ДНК, обеспечивающей получение рекомбинантного белка Cov1, обладающего иммуногенными свойствами, причем получение его на основе поверхностного S-белка вируса SARS-Cov-2 с целью использования при получении вакцин против вируса SARS-CoV-2 и в диагностике специфических антител против вируса SARS-Cov-2.
Задачей данного изобретения стало создание штамма-продуцента рекомбинантного белка Cov1.
Поставленная задача решалась конструированием уникальных праймеров, с помощью которых в полимеразной цепной реакции (ПЦР) получали фрагмент нуклеотидной последовательности, кодирующий часть аминокислотной последовательности S-белка, которая при образовании третичной структуры белка Cov1 приводила к формированию антигенных детерминант, доступных для выработки специфических антител. С целью быстрого и простого получения рекомбинантного белка Cov1 для клонирования фрагмента ДНК была использована система экспрессионных плазмид pQE. Для создания штамма-продуцента рекомбинантного белка Cov1 был использован штамм Е.coli M15 (The QIAexpress System, Qiagen, США). В результате экспериментальной работы этот штамм приобрел новые свойства: способность продуцировать рекомбинантный белок Cov1. Новый штамм получил название Е.coli M15- Cov1.
Сущностью предлагаемого изобретения является создание уникальной рекомбинантной ДНК (обозначенной как cov1), полученной в результате полимеразной цепной реакции (ПЦР) с использованием синтезированной ДНК, кодирующей S-белок коронавируса SARS-CoV-2 (синтез ДНК был проведен компанией Евроген), уникальных праймеров SEQ ID NO: 1 и SEQ ID NO: 2 и последующего клонирования с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США) в E.coli M15. Полученная рекомбинантная ДНК cov1, имеющая нуклеотидную последовательность SEQ ID NO: 5, кодирует уникальный рекомбинантный белок Cov1, имеющий аминокислотную последовательность SEQ ID NO: 6.
Также сущностью предлагаемого изобретения является создание рекомбинантной плазмидной ДНК pQE-cov1, несущей рекомбинантную ДНК cov1, и штамма-продуцента Е. coli M15-Cov1, позволяющего при определенных условиях продуцировать рекомбинантный белок Cov1.
Авторами получен фрагмент гена, кодирующего S-белок вируса SARS-CoV-2 (обозначенный как cov, размером 526 п.н.) с помощью полимеразной цепной реакции с использованием праймеров SEQ ID NO: 1 и SEQ ID NO: 2 и синтезированной ДНК, кодирующей S-белок коронавируса SARS-CoV-2 (синтез ДНК был проведен компанией Евроген). Также авторами осуществлено клонирование cov с использованием экспрессионной плазмиды pQE-30 и последующей трансформацией полученной рекомбинантной плазмиды (обозначенной как pQE-cov1) в гетерологическую систему Е.coli M15. Авторами получен штамм-продуцент рекомбинантного белка Cov1, обозначенный как Е. coli M15-Cov1. Также авторами реализована продукция рекомбинантного белка Cov1 клетками штамма Е. coli M15-Cov1 с последующей одноступенчатой аффинной очисткой с использованием Ni-сефарозы (Amersham, США).
Авторами получены анти-Cov1 антитела (IgG) при подкожном введении рекомбинантного белка Cov1 млекопитающим (мышам и кроликам).
Авторы также продемонстрировали применение Cov1 для тестирования специфических антител в сыворотках от больных, перенесших инфекционное заражение вирусом SARs-CoV-2.
Представленный новый рекомбинантный белок Cov1, имеет желаемую физиологическую стабильность и вызывает синтез анти-Cov1 антител. Рекомбинантный белок Cov1 может быть использован при получении вакцин, содержащих S-белок. Рекомбинантный белок Cov1 полезен для тестирования специфических антител в сыворотках от больных, перенесших инфекционное заражение вирусом SARs-CoV-2. Получение рекомбинантного белка Cov1
Фрагмент ДНК, кодирующий S-белок вируса SARS-CoV-2, был синтезирован фирмой Евроген на основе известной нуклеотидной последовательности, полученной из базы данных. Для получения фрагмента ДНК размером 526 п.н., обозначенного как cov, использовали метод полимеразной цепной реакции. Полученный в результате ПЦР фрагмент ДНК cov выделяли из реакционной смеси с помощью с помощью набора «QIAquick PCR purification Kit» (Qiagen, США).
Полученный фрагмент cov клонировали с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США). При подготовке к клонированию была проведена двойная рестрикция выделенного фрагмента cov (526 п.н.) и плазмиды pQE-30 (3462 п.н.) ферментами BamHI и SacI с образованием липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле. Рестрицированный фрагмент cov (размером 514 п.н., обозначенный как cov') и рестрицированную плазмиду pQE-30 (обозначенную как pQE-30', размером 3446 п.н.) выделяли из агарозного геля с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, США). В результате проведенного клонирования была получена рекомбинантная плазмидная ДНК (обозначенная как pQE-cov1), несущая рекомбинантную ДНК (обозначенную как cov1, имеющую нуклеотидную последовательность SEQ ID NO: 5 и состоящую из 514 п.н. фрагмента cov' и 55 п.н. фрагмента плазмиды pQE-30).
Рекомбинантная плазмидная ДНК pQE-cov1 была трансформирована в Е. coli M15 (The QIAexpress System, Qiagen, США) с помощью кальциевого метода.
Клоны-трансформанты отбирали после роста на плотных селективных средах, учитывая маркер устойчивости к ампициллину, который несла плазмида pQE-30. Из клонов-трансформантов были отобраны клоны, №13, 15, 17 содержащие рекомбинантную плазмиду pQE-cov1, наличие которой подтверждали ПЦР с использованием исходных праймеров SEQ ID NO: 3 и SEQ ID NO: 4 и последующим электрофоретическим анализом полученных фрагментов ДНК в 1,0% агарозном геле.
Клоны Е. col M15, содержащие рекомбинантную плазмиду pQE-cov1, (обозначенные как Е. col M15-Cov1) проверяли на способность экспрессировать белок Cov1. Для получения очищенного препарата рекомбинантного белка клетки Е.coli M15-Cov1 культивировали на бульоне Terrific Broth (ТВ) (Helicon, Росссия) с добавлением антибиотиков (ампициллин (100 мкг/мл) и канамицин (25 мкг/мл)) до поздней логарифмической фазы роста (OD600=0,7+0,9). Затем, продукцию рекомбинантного белка индуцировали добавлением изопропил-бета-D-тиогалактопиранозида (IPTG), и клетки культивировали еще 4,5 часа. После этого клетки осаждали центрифугированием, отмывали забуференным физиологическим раствором (PBS), рН=7.4). После повторного центрифугирования надосадочную жидкость сливали, а клетки осадка замораживали при -70°С. Для дальнейшего выделения белка осадок размораживали при комнатной температуре, далее осадок ресуспендировали в 10 мл буфера А (8 М мочевина (Panreac, Испания), 0,1 М Na2HPO4, 0,1 М NaH2PO4, рН=8,0). Полученную суспензию осторожно перемешивали в течение часа при комнатной температуре для полного вскрытия клеток. Суспензию центрифугировали при 13400 об/мин и 4°С в течение 20 мин. Надосадочную жидкость пропускали через 0,45 мкм фильтры (Millipore, США). Далее профильтрованную надосадочную жидкость наносили на колонку с Ni-сефарозой (Qiagen, США), предварительно уравновешенной буфером А. После 30 мин. инкубации при комнатной температуре колонку промывали при 4°С тем же буфером до тех пор, пока значения OD280 выходящего раствора не были меньше, чем 0,01. Белок элюировали раствором буфера Б (8 М мочевина (Panreac, Испания), 0,1 М Na2HPO4, 0,1 М NaH2PO4, 0,4 М имидазола рН=8,0), создавая градиент имидазола от 0 до 0,4 М. Элюат собирали фракциями по 0,5-1,0 мл и анализировали в 12% SDS-PAGE. Фракции, в которых содержался белок объединяли и диализовали сначала относительно буфера В (3М мочевина, 0,1 М Na2HPO4, 0,1 М NaH2PO4, рН=9,1) при 4°С в течение 1,5 часов без перемешивания, затем относительно буфера Г (0,02 М Na2HPO4, 0,02 М NaH2PO4, 0,4 М NaCl, рН=9,1) без перемешивания при 4°С в течение 12 часов. После диализа белок Cov1 стерилизовали фильтрованием с помощью одноразовых фильтров Millipore (0,45 мкм) и хранили при 6°С.
Молекулярную массу белка Cov1 определяли с помощью 12% SDS-PAGE, сравнивая пробег белка Cov1 с пробегом белков известной молекулярной массы (Precision Plus Protein™ Standards Unstained (161-0363, Bio-Rad, США). Молекулярная масса рекомбинантного белка Cov1 оказалась равной (24,5±0,5) кДа.
Полученный рекомбинантный белок Cov1 содержит 168 аминокислотных остатков белка S вируса SARS-CoV-2, ковалентно связанных с 21 аминокислотным остатком, кодируемым плазмидой pQE-30 (The QIAexpress System, Qiagen, США).
Таким образом, результатом изобретения являются следующие объекты, связанные единым изобретательским замыслом:
• рекомбинантная ДНК cov1, имеющая нуклеотидную последовательность SEQ ID NO: 5;
• рекомбинантная плазмидная ДНК pQE-cov1, представляющая собой плазмиду pQE-30, несущую рекомбинантную ДНК cov1',
• штамм Е. coli M15, содержащий рекомбинантную плазмидную ДНК pQE-cov1, и продуцирующий рекомбинантный белок Cov1;
• рекомбинантный белок Cov1, имеющий аминокислотную последовательность SEQ ID NO: 6, состоящую из 189 аминокислотных остатков.
Рекомбинантный белок Cov1 обладает следующими свойствами:
а) при введении в организм млекопитающим (мышам и кроликам) вызывает синтез анти-Cov1 антител (IgG);
б) В иммуноферментном анализе (ИФА) связывает анти-Cov1 антитела из сывороток от больных COVID-19
Ниже приводятся конкретные примеры, иллюстрирующие некоторые варианты изобретения, но не ограничивающие его.
Пример 1. Получение ДНК-фрагмента cov методом полимеразной цепной реакции
К 0,25 мкг синтезированной ДНК (фрагмент ДНК синтезирован фирмой Евроген), добавляли 10 мкМ каждого из специфических праймеров (SEQ ID NO: 1 и SEQ ID NO: 2), фланкирующих исследуемую последовательность, буфер с магнием для полимеразы, по 0,2 мМ каждого из 4 дезоксирибонуклеотидтрифосфатов, объем доводили водой до 25 мкл. Добавляли 0,2 мкл термостабильной ДНК полимеразы. На поверхность жидкости наслаивали 40 мкл минерального масла. Пробирки помещали в амплификатор («Терцик», Россия). Смесь инкубировали при 94°С в течение 3 минут. Прибор программировали на цикл денатурации 94°С на 15 секунд, цикл отжига праймеров при 56°С на 15 секунд, цикл синтеза ДНК при 72°С на 30 секунд. Последовательность таких циклов повторялась 35 раз. После чего смесь инкубировали при 72°С в течение 1 минуты.
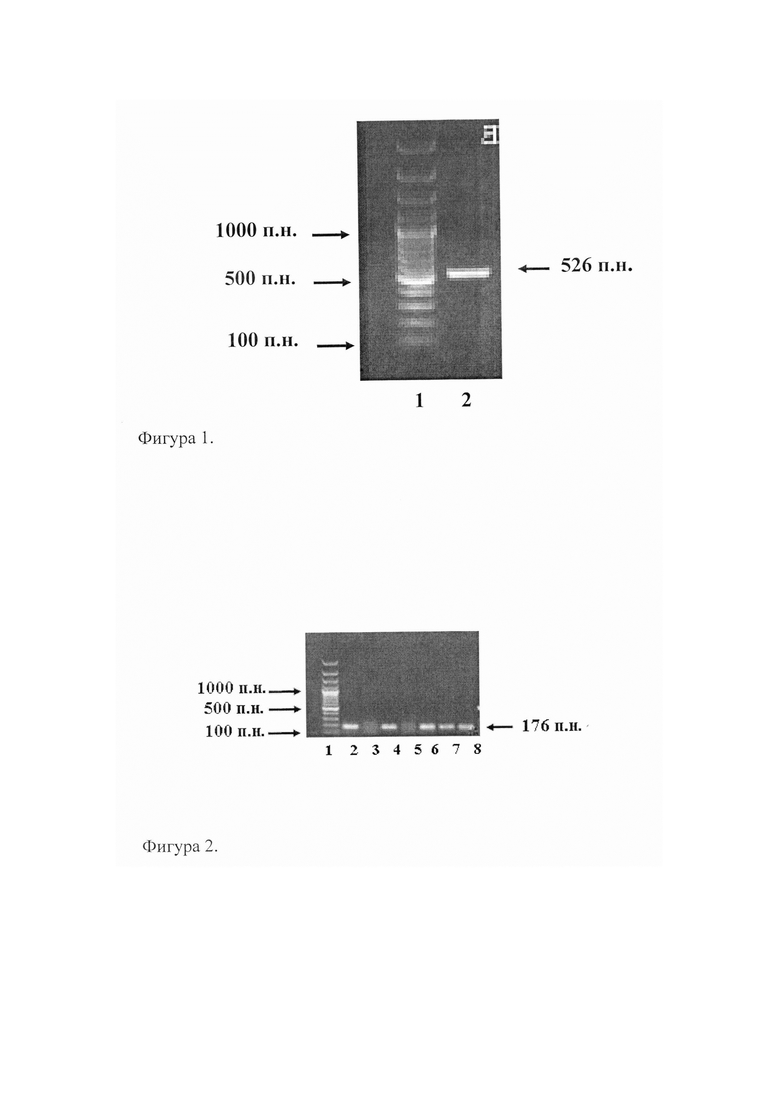
В результате полимеразной цепной реакции с праймерами SEQ ID NO: 1 и SEQ ID NO: 2, был получен ДНК-фрагмент 526 п.н., обозначенный как cov. Кроме того, на 5'-концах праймеров находились сайты распознавания эндонуклеазами BamHI и SacI с целью последующего клонирования ДНК-фрагмента cov. Результат ПЦР оценивали с помощью горизонтального электрофореза в 1% агарозном геле (фиг. 1). Размеры полученных фрагментов ДНК рассчитывали с помощью компьютерной программы SEQAID, исходя из сравнения их электрофоретической подвижности с электрофоретической подвижностью маркера молекулярных весов (100 п.н. ДНК-маркер, Сибэнзим, Россия). На фиг. 1 представлена электрофореграмма получения ДНК-фрагмента cov методом ПЦР: 1-100 п.н. ДНК-маркер (GeneRular 100 bp Plus DNA Ladder); 2 - продукт ПЦР, полученный с праймерами SEQ ID NO: 1 и SEQ ID NO: 2.
Пример 2. Клонирование ДНК-фрагмента cov с использованием экспрессионной плазмиды pQE-30
Клонирование ДНК-фрагмента cov осуществляли с использованием экспрессионной плазмиды pQE-30 («The QIAexpress System», Qiagen, США). При подготовке к клонированию была проведена двойная рестрикция выделенного из ПЦР смеси cov (526 п.н.) и плазмиды pQE-30 (3462 п.н.) ферментами BamHI и SacI с образованием липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле. Размеры полученных фрагментов ДНК рассчитывали, как описано в примере 1. Рестрицированный фрагмент cov (размером 514 п.н., обозначенный как cov') и рестрицированную плазмиду pQE-30 (обозначенную как pQE-30', размером 3446 п.н.) выделяли из агарозного геля с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, США).
В ходе клонирования к 30 мкл фрагмента ДНК cov' добавляли 2 мкл плазмиды pQE-30' (The QIAexpress System, Qiagen, США) в соотношении концентраций 4:1, 5 мкл десятикратного лигазного буфера и 1 мкл лигазы, выделенной из фага Т4. Объем доводили водой до 50 мкл. Смесь инкубировали в течение 1 часа при комнатной температуре и при температуре 4°С в течение 12 часов. В результате проведенного клонирования была получена рекомбинантная плазмидная ДНК (обозначенная как pQE-cov1), несущая рекомбинантную ДНК (обозначенную как cov1, имеющую нуклеотидную последовательность SEQ ID NO: 5 и состоящую из 514 п.н. фрагмента гена, кодирующего S-белок, и 55 п.н. фрагмента плазмиды pQE-30).
Пример 3. Трансформация культуры Е. coli M15 плазмидной ДНК pQE-cov1
Рекомбинантную плазмидную ДНК pQE-cov1 трансформировали в гетерологичную систему Е. coli M15. В качестве положительного контроля параллельно проводили трансформацию Е. coli M15 исходной плазмидной ДНК pQE-30. Генетическим маркером плазмидной ДНК pQE-30 был ген amp, кодирующий бета-лактамазу, что обеспечивало устойчивость к ампициллину клеток, несущих данную плазмиду. Культуру клеток Е. coli M15 культивировали на бульоне LB (Luria-Bertani Broth, Amresco, США) с канамицином (25 мкг/мл) в течение 12 часов при 37°С и интенсивном перемешивании, пересевали на новый объем той же среды (1/50V) и инкубировали при 37°С в течение 2 часов при интенсивном перемешивании до ОД600=0,35. Выращенные клетки в объеме 1,5 мл центрифугировали в течение 1 мин при 12000 об/мин и полученный осадок ресуспендировали в 200 мкл 0,1 М CaCl2. Далее смесь инкубировали во льду 1 час. После центрифугирования, осадок ресуспендировали в 250 мкл 0,075 М CaCl2. Затем в пробирку вносили 0,2-0,3 мкг плазмидной ДНК и инкубировали смесь во льду 0,5 час. Далее смесь выдерживали 2 мин при 42°С и снова переносили в лед на 5 мин. После этого к смеси добавляли 1 мл бульона LB с канамицином (25 мкг/мл) и инкубировали 0,5 часа при 37°С и интенсивном перемешивании. После осаждения центрифугированием (8000 об/мин, в течение 2 мин) клетки высевали на чашки с 1% L-агаром (Difco, США), содержащим ампициллин (100 мкг/мл) и канамицин (25 мкг/мл). Отбор клонов-трансформантов проводили через 18 часов роста клеток при 37°С. В качестве отрицательного контроля высевали чистую культуру клеток Е. coli М15.
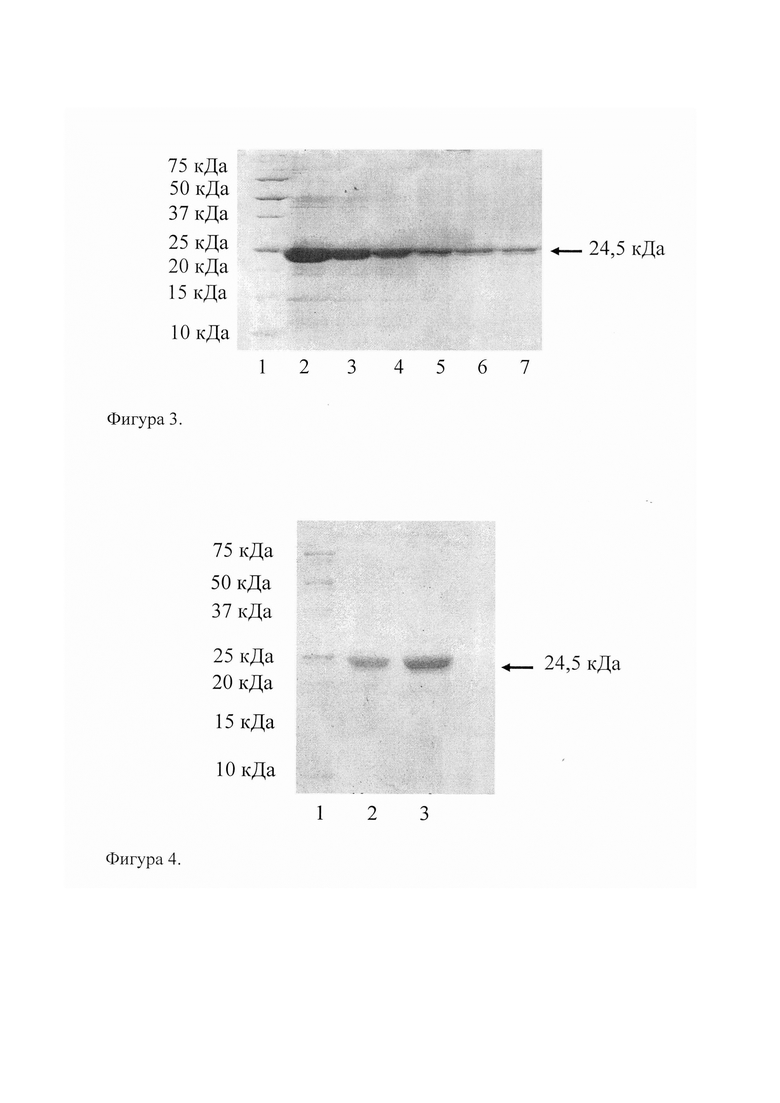
Наличие рекомбинантной ДНК cov1 в плазмиде pQE-cov1 определяли методом ПЦР с использованием праймеров SEQ ID NO: 3 и SEQ ID NO: 4 и последующим электрофоретическим анализом полученных фрагментов ДНК в 1,0% агарозном геле. Размеры полученных фрагментов ДНК рассчитывали, как описано в примере 1. На фиг. 2 представлен электрофорез продуктов ПЦР, полученных в результате амплификации ДНК, выделенной из различных клонов-трансформантов: 1-100 п.н. ДНК-маркер (GeneRular 100 bp Plus DNA Ladder); 2 - продукт ПЦР исходной ДНК; 3 - клон, несущий исходную плазмиду pQE-30; 4-8 - рекомбинантные клоны, несущие плазмиду pQE-cov1.
Полученные данные позволяют сделать вывод о том, что в результате проведенного клонирования были получены рекомбинантные клоны, несущие плазмиду pQE-cov1 с рекомбинантной ДНК cov1, которые получили название Е. coli M15-Cov1.
Пример 4. Очистка рекомбинантного белка Cov1
Культуру штамма Е. coli M15-Cov1 выращивали на бульоне Terrific Broth с добавлением ампициллина в концентрации 100 мкг/мл и канамицина в концентрации 25 мкг/мл в течение ночи при интенсивном перемешивании. Клетки пересевали на 150 мл той же среды (1/50V) и инкубировали при 37°С в течение 2-3 часов при интенсивном перемешивании до OD600=0,7 0,9. Экспрессию рекомбинантного белка индуцировали добавлением раствора изопропил-бета-D-тиогалактопиранозида до конечной концентрации 0,001 М, после чего инкубировали клетки при тех же условиях еще 4,5 часа. Полученную суспензию клеток центрифугировали при 6000 об/мин 5 мин или при 4700 об/мин 25 мин. Однократно отмывали клеточный осадок забуференным физиологическим раствором (PBS), рН=7.4. После повторного центрифугирования надосадочную жидкость сливали, а клетки осадка замораживали при -70°С. Осадок в замороженном состоянии можно хранить достаточно долго, как минимум 6 месяцев. Для дальнейшего выделения белка осадок размораживали при комнатной температуре, далее осадок ресуспендировали в 10 мл буфера А (8 М мочевина (Panreac, Испания), 0,1 М Na2HPO4, 0,1 М NaH2PO4; рН=8,0). Полученную суспензию осторожно перемешивали в течение часа при комнатной температуре для полного вскрытия клеток.
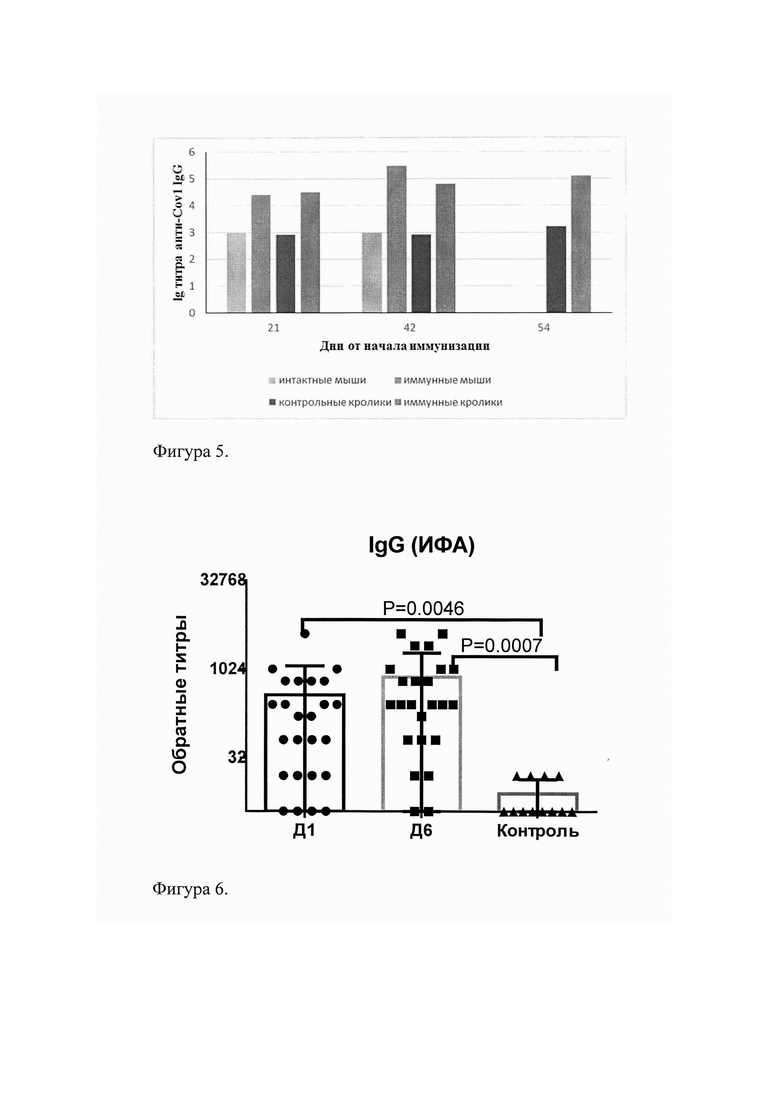
Суспензию центрифугировали при 13400 об/мин и 4°С в течение 20 мин. Надосадочную жидкость пропускали через 0,45 мкм фильтры (Millipore, США). Далее профильтрованную надосадочную жидкость наносили на колонку с Ni-сефарозой (Qiagen, США), предварительно уравновешенной буфером А. После 30 мин. инкубации при комнатной температуре колонку промывали при 4°С тем же буфером до тех пор, пока значения OD280 выходящего раствора не были меньше, чем 0,01. Белок элюировали раствором буфера Б (8 М мочевина (Panreac, Испания), 0,1 М Na2HPO4, 0,1 М NaH2PO4, 0,4 М имидазола рН=8,0), создавая градиент имидазола от 0 до 0,4 М. Элюат собирали фракциями по 0,5-1,0 мл и анализировали в 12% SDS-PAGE. Фракции, в которых содержался белок объединяли и диализовали сначала относительно буфера В (3М мочевина, 0,1 М Na2HPO4, 0,1 М NaH2PO4, рН=9,1) при 4°С в течение 1,5 часов без перемешивания, затем относительно буфера Г (0,02 М Na2HPO4, 0,02 М NaH2PO4, 0,4 М NaCl, рН=9,1) без перемешивания при 4°С в течение 12 часов. На фиг. 3 представлена электрофореграмма рекомбинантного белка Cov1 на разных этапах очистки с помощью Ni-сефарозы: 1 - маркер молекулярной массы (Precision Plus Protein™ Standards Unstained, BioRad, CIIIA); 2-7 фракции препарата чистого белка Cov1 (К=1,0 мг/мл) при элюции в градиенте имидазола от 0 до 0,4 М.
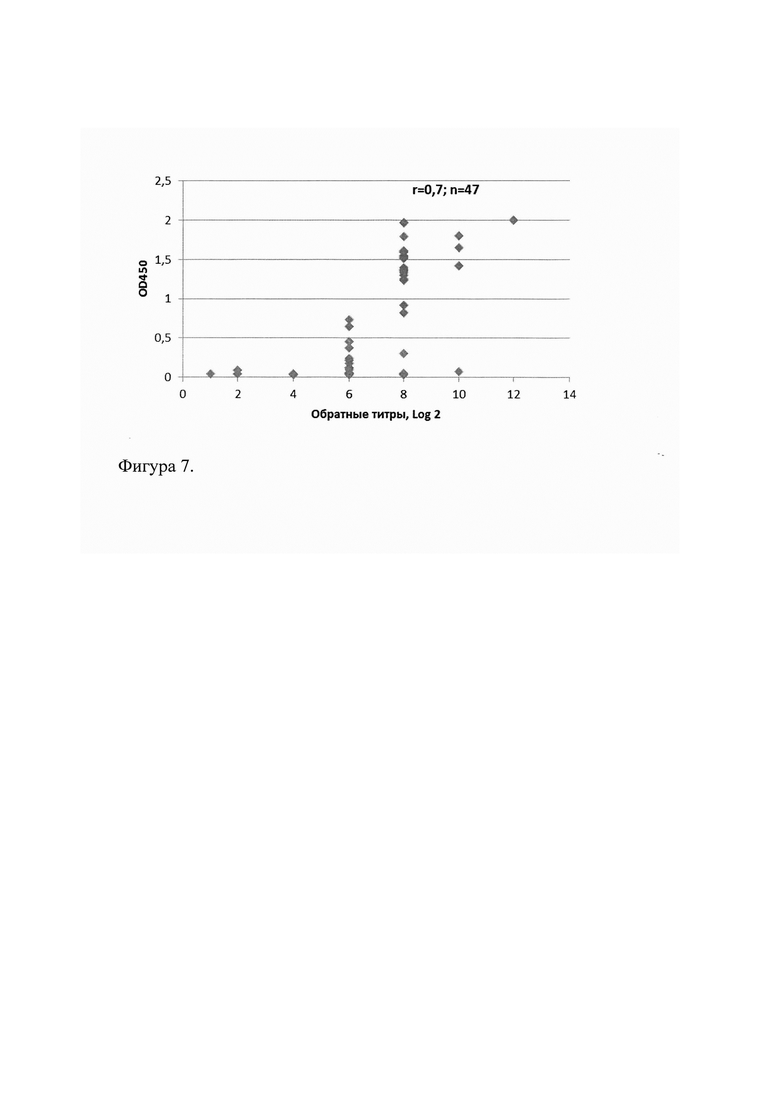
Молекулярную массу Cov1 определяли с помощью 12% SDS-PAGE, сравнивая пробег белка Cov1 с пробегом белков известной молекулярной массы (Precision Plus Protein™ Standards Unstained (161-0363), Bio-Rad, США). Молекулярная масса рекомбинантного белка Cov1 оказалась равной (24,5±0,5) кДа. На фиг. 4 представлена электрофореграмма рекомбинантного белка Cov1 после диализа: 1 - маркер молекулярной массы (Precision Plus Protein™Standards Unstained, BioRad, CIIIA); 2 - чистый белок Cov1 (5 мкл, К=0,5 мг/мл); 3 - чистый белок Cov1 (5 мкл, К=1,0 мг/мл).
Пример 5. Динамика изменения титра антител к рекомбинантному белку Cov1 после иммунизации млекопитающих
Гуморальный иммунный ответ к рекомбинантному белку Cov1 изучали на беспородных белых мышах (самках, массой 16÷18 г), и кроликах (самках, массой 2,5 кг), полученных из питомника «Рапполово».
Иммунизацию проводили двукратно и подкожно с адъювантом гидроокиси алюминия (Thermo Scientific, США). В эксперименте использовали по 10 мышей в контрольной и иммунной группах и 4 кролика (1 контрольный и 3 иммунных), которым вводили 0,2 мл и 0,5 мл Cov1 с адъювантом в соотношении объемов 1:1, соответственно. Для мышей при первой инъекции вводили 0,2 мл Cov1 с адъювантом в соотношении объемов 1:1 и на 22 день проводили бустирование 0,2 мл белка Cov1 с адъювантом. Мышам дважды вводили по 20 мкг белка Cov1 на одну мышь. Кроликам дважды вводили подкожно по 0,5 мл белка Cov1 с адъювантом: 120 мкг при первой инъекции и 70 мкг при второй инъекции на одного кролика. Контрольному кролику дважды вводили физиологический раствор с адъювантом в соотношении объемов 1:1.
Мышиные антитела к белку Cov1 определяли с 21-го по 42-й день от начала иммунизации, используя сыворотки от каждого животного. Кроличьи антитела к белку Cov1 определяли с 21-го по 54-й день от начала иммунизации, используя сыворотки от каждого животного.
Анти-Cov1 антитела определяли с помощью двухслойного метода иммуноферментного анализа (ИФА). В лунки полистиролового планшета с высокой сорбционной емкостью (Costar, США) вносили по 100 мкл антигена (рекомбинантный белок Cov1) в концентрации 2 мкг/мл. Сорбцию проводили в 0,1 М бикарбонатном буфере (рН=9,5) в течение 16 20 часов при 4°С. После этого содержимое из планшета удаляли и добавляли в лунки по 150 мкл забуференного физиологического раствора, рН=7,4 (PBS), содержащего 0,05% Твин-20 (PBST). Инкубацию проводили при 37°С в течение 30 мин. Содержимое планшета удаляли и трижды промывали PBST. Затем в лунки планшета вносили иммунную или нормальную (в качестве отрицательного контроля) сыворотку (с начальным разведением 1:100) с последующим шагом разведения равным двум. Все разведения осуществляли в PBST, и каждую пробу дублировали. Инкубацию проводили в течение 1 часа при 37°С. Далее содержимое планшета удаляли, и дважды промывали планшет PBST. После этого в лунки добавляли по 100 мкл анти IgG мыши-ПХ конъюгата или анти IgG кролика-ПХ конъюгата (Sigma, США). После часовой инкубации при 37°С содержимое планшета удаляли и трижды отмывали планшет PBST и один раз PBS, чтобы избежать ингибирующего действия Твин-20 на ферментативную активность пероксидазы. Далее для визуализации реакции в лунки планшета вносили по 100 мкл субстратной смеси (ТМВ (3,3',5,5'-Tetramethylbenzidine dihydrochloride Monohydrate, Sigma, США) в 0,1 M фосфатно-цитратном буфере, рН=5,0, 2 мкл 30% перекиси водорода), которую готовили непосредственно перед применением. Планшет инкубировали в течение 30 мин в темноте при комнатной температуре, и реакцию останавливали внесением в лунки по 30 мкл 50% концентрированной серной кислоты. Реакцию регистрировали при длине волны 450 нм с помощью прибора Titertek Multiskan (США).
Титр специфических антител определяли на разных сроках от начала иммунизации. Через три недели от начала эксперимента по иммунизации лабораторных животных было отмечено появление в крови специфических антител к белку Cov1. На фиг. 5 представлены средние величины титров, полученные после тестирования сывороток от каждого животного. Результаты, выраженные графически на фиг. 5, показали наличие гуморального иммунного ответа на введение рекомбинантного белка Cov1. При этом максимальное значение титра антител к белку Cov1 наблюдалось на 42 день (со средним титром 4,0×10-5) и на 54 день (со средним титром 1,3×10-5) от начала иммунизации у мышей и кроликов, соответственно. Пример 6. Определение анти-Cov1 антител в сыворотках от больных
В тестировании использовали архивные сыворотки после текущих лабораторных исследований крови пациентов с разной степенью тяжести COVID-19. Исследование одобрено Локальным этическим комитетом ФГБНУ «ИЭМ» (протокол 3/20 от 06.05.2020). Было изучено 28 парных проб, полученных при поступлении пациентов в стационар и через 5-7 дней пребывания в стационаре. В качестве контрольных образцов использовались сыворотки крови пациентов, полученные до распространения COVID-19 в начале 2019 года. Сыворотки исследовали с помощью иммуноферментного анализа (ИФА) на наличие антител IgG, специфичных к белкам SARS-CoV-2. Для этого рекомбинантным белком Cov1 в концентрации 0,5 мкг/мл сенсибилизировали 96-луночные панели для ИФА. Далее реакцию проводили как описано ранее [Rowe Т., Abernathy R.A., Hu-Primmer J., Thompson W.W., Lu X., Lim W., Fukuda K., Cox N.J., Katz J.M. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. Jclin microbiol. 1999; 37(4), 937-43.].
Оценку статистической значимости различий проводили с помощью непараметрического критерия Манна-Уитни. Различия считались значимыми при Р=0,05. На фиг. 6 представлены значения обратных титров анти-Cov1 антител в сыворотках от пациентов с подтвержденной инфекцией SARS-CoV-2 на двух сроках пребывания в стационаре: Д1 - 1-й день пребывания в стационаре; Д6 - 6-й день пребывания в стационаре; Контроль - сыворотки пациентов МНЦ ФГБНУ «ИЭМ», обследованных в 2019 г.
Дополнительно 47 сывороток пациентов с подтвержденной инфекцией COVID-19 были изучены с помощью коммерческого набора для выявления IgG антител к коронавирусу SARS-CoV-2 «SARS-Cov-2 IgG Screen» (Imbian, Россия). На фиг. 7 продемонстрирован корреляционный анализ уровней сывороточных антител среди больных COVID-19 при тестировании с помощью двух тест-систем: по оси абсцисс представлены уровни антител, определенные с использованием рекомбинантного белка; по оси ординат представлены уровни антител, определенные с помощью коммерческого набора для выявления IgG антител к коронавирусу «SARS-Cov-2 IgG Screen».
Сравнение двух тест-систем для выявления сывороточных антител человека к SARS-CoV-2 с использованием рекомбинантного белка Cov1 и коммерческого набора показало высокий уровень корреляции (коэффициент ранговой корреляции Спирмена=0,7, n=47). Полученный результат свидетельствует о возможности использования рекомбинантного белка Cov1 для создания тест-систем при детекции антител к SARS-CoV-2 в сыворотках пациентов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ФГБНУ «ИЭМ»
<120> Рекомбинантная ДНК, обеспечивающая получение рекомбинантного
белка Cov1, обладающего иммуногенными свойствами в отношении
вируса SARS-CoV-2
<140>
<141>
<150>
<151>
<160> 6
<210> 1
<211> 24
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность прямого праймера,
позволяющая получить 526 п.н. фрагмент ДНК, обозначенный как сov
<400> 1
Aaggatccat acatatgggt ttcc
<210> 2
<211> 19
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность обратного праймера,
позволяющая получить 526 п.н. фрагмент ДНК, обозначенный как сov
<400> 2
Tgtcgacgga gctcgaatt
<210> 3
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность прямого праймера,
позволяющая получить 176 п.н. фрагмент сov
<400> 3
Ttggcagaga cattgctgac
<210> 4
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность обратного праймера,
позволяющая получить 176 п.н. фрагмент сov
<400> 4
Agcaacaggg acttctgtgc
<210> 5
<211> 569
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<223> Нуклеотидная последовательность рекомбинантной ДНК сov1,
кодирующая аминокислотную последовательность
рекомбинантного белка Cov1
<400> 5
Catcaccatc accatcacgg atccatacat atgggtttcc aacccactaa tggtgttggt 60
taccaaccat acagagtagt agtactttct tttgaacttc tacatgcacc agcaactgtt 120
tgtggaccta aaaagtctac taatttggtt aaaaacaaat gtgtcaattt caacttcaat 180
ggtttaacag gcacaggtgt tcttactgag tctaacaaaa agtttctgcc tttccaacaa 240
tttggcagag acattgctga cactactgat gctgtccgtg atccacagac acttgagatt 300
cttgacatta caccatgttc ttttggtggt gtcagtgtta taacaccagg aacaaatact 360
tctaaccagg ttgctgttct ttatcaggat gttaactgca cagaagtccc tgttgctatt 420
catgcagatc aacttactcc tacttggcgt gtttattcta caggttctaa tgtttttcaa 480
acacgtgcag gctgtttaat aggggctgaa catgtcaaca acgaattcga gctcggtacc 540
ccgggtcgac ctgcagccaa gcttaatta
<210> 6
<211> 189
<212> Аминокислотная последовательность
<213> Искусственная последовательность
<223> Аминокислотная последовательность рекомбинантного белка Cov1
<400> 6
His His His His His His Gly Ser Ile His Met Gly Phe Gln Pro Thr
5 10 15
Asn Gly Val Gly Tyr Gln Pro Tyr Arg Val Val Val Leu Ser Phe Glu
20 25 30
Leu Leu His Ala Pro Ala Thr Val Cys Gly Pro Lys Lys Ser Thr Asn
35 40 45
Leu Val Lys Asn Lys Cys Val Asn Phe Asn Phe Asn Gly Leu Thr Gly
50 55 60
Thr Gly Val Leu Thr Glu Ser Asn Lys Lys Phe Leu Pro Phe Gln Gln
65 70 75 80
Phe Gly Arg Asp Ile Ala Asp Thr Thr Asp Ala Val Arg Asp Pro Gln
85 90 95
Thr Leu Glu Ile Leu Asp Ile Thr Pro Cys Ser Phe Gly Gly Val Ser
100 105 110
Val Ile Thr Pro Gly Thr Asn Thr Ser Asn Gln Val Ala Val Leu Tyr
115 120 125
Gln Asp Val Asn Cys Thr Glu Val Pro Val Ala Ile His Ala Asp Gln
130 135 140
Leu Thr Pro Thr Trp Arg Val Tyr Ser Thr Gly Ser Asn Val Phe Gln
145 150 155 160
Thr Arg Ala Gly Cys Leu Ile Gly Ala Glu His Val Asn Asn Glu Phe
165 170 175
Glu Leu Gly Thr Pro Gly Arg Pro Ala Ala Lys Leu Asn
180 185
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Выделенный рекомбинантный вирус на основе вируса гриппа для индукции специфического иммунитета к вирусу гриппа и/или профилактики заболеваний, вызванных вирусом гриппа | 2021 |
|
RU2813150C2 |
| Способ создания живого штамма энтерококка L3-SARS на основе биологически активного штамма Е. faecium L3 | 2021 |
|
RU2782529C1 |
| Рекомбинантный вакцинный штамм для живой интраназальной вакцины, обеспечивающей сочетанную профилактику гриппозной и коронавирусной инфекций | 2022 |
|
RU2782531C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2783313C1 |
| Рекомбинантный белок GM, обладающий способностью связывать α2-макроглобулин и его применение в качестве лиганда в аффинной хроматографии для выделения α2-макроглобулина из сыворотки крови человека | 2019 |
|
RU2758604C2 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2021 |
|
RU2761879C1 |
| Вакцина для профилактики или лечения коронавирусной инфекции на основе генетической конструкции | 2020 |
|
RU2747762C1 |
| Рекомбинантные ДНК pG4223 и плазмидная ДНК pQE 30-pG4223, штамм Escherichia coli M 15-G4223, обеспечивающие получение полипептида G4223, селективно связывающего IgG, и его применение в аффинной хроматографии для выделения IgG | 2017 |
|
RU2715672C2 |
Изобретение относится к области иммунологии, микробиологии и молекулярной генетики и может быть использовано в медицинской промышленности при производстве вакцин против вируса SARS-COV-2 и диагностикумов для его идентификации. Сущностью предлагаемого изобретения является создание рекомбинантной ДНК (обозначенной как cov1), полученной в результате полимеразной цепной реакции (ПЦР) с использованием синтезированной ДНК, кодирующей S-белок коронавируса SARS-CoV-2, уникальных праймеров SEQ ID NO: 1 и SEQ ID NO: 2 и рекомбинантной плазмиды pQE-cov1. Полученная с использованием экспрессионной плазмиды pQE-30 в клетках Е.coli M15 рекомбинантная ДНК cov1 кодирует уникальный рекомбинантный белок Cov1 молекулярной массы 24,5±0,5 кДа, который вызывает образование антител к белку S коронавируса SARS-CoV-2. В результате применения предлагаемого изобретения создана рекомбинантная плазмидная ДНК pQE-cov1, представляющая собой плазмидную ДНК pQE-30, несущую рекомбинантную ДНК cov1, и штамм-продуцент Е.coli M15-Cov1, позволяющий продуцировать рекомбинантный белок Cov1. 4 н.п. ф-лы, 7 ил., 5 пр.
1. Рекомбинантная ДНК cov1, кодирующая рекомбинантный белок Cov1, обладающий иммуногенными свойствами в отношении вируса SARS-CoV-2, и имеющая нуклеотидную последовательность SEQ ID NO:5.
2. Рекомбинантная плазмидная ДНК pQE-cov1 для продукции рекомбинантного белка Cov1, представляющая собой плазмидную ДНК pQE-30, несущую рекомбинантную ДНК по п.1.
3. Штамм E.coli М15-Cov1, трансформированный рекомбинантной плазмидой по п.2 и продуцирующий рекомбинантный белок Cov1.
4. Рекомбинантный белок Cov1 для иммунизации млекопитающих, состоящий из 168 аминокислотных остатков S-белка вируса SARS-CoV-2, ковалентно связанных с 21 аминокислотным остатком, кодируемым плазмидой pQE-30, имеющий аминокислотную последовательность SEQ ID NO: 6.

