Изобретение относится к области молекулярной биологии и протеомики и может быть использовано в медицинской практике в качестве доступного реагента для освобождения сыворотки крови от двух белков, альбумина (ЧСА) и иммуноглобулина G (IgG), находящихся в ней в высоких концентрациях. Удаление двух мажорных белков из сыворотки крови позволит определить другие белки, присутствующие в сыворотке в более низких концентрациях.
В последние годы одним из перспективных направлений при создании методов диагностики различных заболеваний является использование протеомных технологий, область применения которых тесно связана с поиском диагностических маркеров заболеваний. Основной идеей такой целевой «маркерной» диагностики является идентификация определенных молекул или их комплексов, которые присутствуют только в пораженных тканях или клетках и/или выделяются во внешнюю среду или внутренние среды организма.
Объектами исследования могут являться биологические жидкости (плазма или сыворотка крови, спинно-мозговая, синовиальная, амниотическая, бронхоальвеолярная жидкости, моча), клетки и биоптаты тканей человека, а также протеомы микроорганизмов - возбудителей заболеваний [Arevalo-Ferro С. et al., Environmental Microbiology 5 (12): 1350-1369, 2003].
Сыворотка (плазма) крови легкодоступна и наиболее удобна для использования в диагностических целях. Сыворотка является богатым источником биохимических продуктов, которые могут выступать в качестве показателей, характеризующих физиологическое и клиническое состояние пациента. Например, уровень гормонов, холестерина, ферментов или других белков может предоставить информацию о сердечно-сосудистых заболеваниях или пищевом статусе, вирусной болезни или раке. Поэтому много усилий прилагается для поиска новых биомаркеров для диагностики различных заболеваний, все больше применяют аналитических методов для изучения профиля содержания протеинов сыворотки крови человека. Методами протеомного анализа возможно проанализировать до 10000 индивидуальных белков в одном образце и зафиксировать изменения их концентраций, что позволяет проводить диагностику и мониторинг течения заболевания.
Главной задачей протеомики является выявление механизма взаимодействия огромного числа белков и пептидов в одном организме. Очень часто прослеживается тесная связь между изменениями в белковом составе и болезненным состоянием человека. Поэтому новые данные в протеомике используются для быстрой разработки новых лекарственных свойств и новейших методов лечения болезней, с которыми медицина боролась веками. На сегодняшний день большинство фармакологических средств воздействует на белки. Протеомика со своим системным подходом может помочь идентифицировать и оценить важность появления новых белков гораздо эффективнее, что, в свою очередь, ускорит разработку новых диагностических тестов и терапевтических средств. Протеом сыворотки можно анализировать различными методами, среди которых одним из наиболее эффективных является метод двумерного электрофореза (2-DE). Разрешение этого метода настолько велико, что позволяет разделить два практически идентичных белка, отличающихся одной заряженной аминокислотой. В результате проведения 2-DE получают двумерную «белковую карту». Для выявления биомаркеров сравнивают «протеомные карты» образцов при патологическом состоянии и в норме, обнаруживая белки, уровень экспрессии которых изменяется при развитии патологии Далее эти белки вырезают из геля и проводят гидролиз какой-либо эндопептидазой, например трипсином (трипсинолиз) для получения коротких (в среднем 10-15 аминокислотных остатков) пептидов, которые идентифицируют на следующем этапе анализа. MALDI-TOF-масс-спектрометры позволяют проводить прямой масс-спектрометрический анализ белков сыворотки крови (прямое белковое профилирование) и получать уникальные масс-спектры с высокой точностью и разрешением, характеризующие исследуемый объект по типу «отпечатков пальцев» [М.А. Stowers et. al., Rapid Commun. Mass Spectrom. 14: 829-833, 2000]. Они позволяют очень быстро анализировать большое количество образцов, требуют сравнительно небольшого количества биологического материала, обладают высокой чувствительностью и хорошим разрешением для низкомолекулярных белков и пептидов, что делает особенно перспективным их использование для поиска биомаркеров. Следующим этапом по фрагментам пептида возможно установить их аминокислотную последовательность. Существуют базы данных, содержащие аминокислотные последовательности белков, и, осуществляя запрос по базе данных по известной пептидной последовательности, идентифицируется белок [Говорун В.М., Арчаков А.И., Биохимия, 67, 10: 1341-1359, 2002].
Однако применение технологий протеомики для поиска потенциальных диагностико-прогностических биомаркеров в крови пациентов ограничено присутствием ЧСА и IgG в больших концентрациях. Их удаление из сыворотки крови должно облегчить обнаружение других белков, находящихся, как в области высокомолекулярных значений (70 кДа), экранированных ЧСА, так и в области 30-50 кДа, где широкая полоса IgG скрывает белки, мигрирующие в этой области. Именно соотношение минорных компонентов в плазме представляет наибольший интерес для протеомных исследований.
В большинстве случаев для удаления ЧСА используют лиганд Cibacron blue, который, обладая сильным неспецифическим связыванием, препятствует проведению анализа двумерным электрофорезом [Laura F. Steel et. al., Proteomics 3: 601-609, 2003]. Помимо ЧСА с этим лигандом связываются также и другие сывороточные белки. Поэтому для увеличения специфичности реакции стали использовать сорбенты с присоединенными к ним антителами к ЧСА, что оказалось невыгодным из-за низкой емкости сорбента и высокой стоимости [V. Polaskova, Electrophoresis, 31: 471-482, 2010]. Для удаления ЧСА из сыворотки иногда пользуются антителами к ЧСА, а IgG - монофункциональным G белком. В последнее время появились коммерческие наборы Aurum™ Affi-Gel Blue kit (Bio-Rad), the ProteoPrep immunoaffinity albumin and IgG depletion kit (Sigma), the Multiple Affinity Removal System (MARS column Agilent Technologies). Эти наборы основаны на использовании аффинных колонок, к которым присоединены антитела к ЧСА и иммуноглобулину IgG. Работа с этими наборами показала, что помимо ЧСА и IgG удаляются и многие другие сывороточные белки, что свидетельствует о недостаточной специфичности этих наборов [Elisa Bellei, Stefania Bergamini, Amino Acids, 40: 145-156, 2011]. Все существующие приемы основаны на реакции «антиген-антитело», которая может давать неспецифические реакции. Поэтому перечисленные приемы недостаточно эффективны. Некоторые зарубежные авторы пытались создать вариант рекомбинантного ЧСА-связывающего полипептида из стрептококка группы G, выделенного от человека, который бы имел высокую аффинность по отношению к ЧСА [A. Jonsson et al, Engeneering, Design Selection, 21, 8: 515-527, 2008]. Был получен вариант рекомбинантного ЧСА-связывающего полипептида, который обладал очень высокой аффинностью для ЧСА с Ко, порядка 10-15. Однако эти работы не увенчались успехом, так как присоединение данного полипептида к сорбенту для аффинной хроматографии не увеличило емкости этого сорбента, что сделало невозможным его использование для полного освобождения сыворотки от ЧСА [A. Jonsson et al, Engeneering, Design Selection, 21, 8: 515-527, 2008].
Описано использование рекомбинантного бифункционального белка G, (одновременно связывающего ЧСА и IgG), для удаления из сыворотки крови ЧСА и IgG [Е.А. Бормотова, Т.В. Гупалова. Биотехнология, №2: 49-55, 2009].
Данное техническое решение принято в качестве прототипа заявляемой группы изобретений.
Этот метод имеет недостатки из-за использования бифункционального G белка, который связывает ЧСА с меньшей эффективностью, чем IgG. 90%-ное удаление ЧСА происходило, когда на колонку наносили только 10 мкл сыворотки, что явно недостаточно.
Задачей изобретения является решение актуальной проблемы - создание нового способа эффективного удаления ЧСА и IgG из сыворотки крови, основанного на взаимодействии «белок-белок» с использованием рекомбинантных рецепторных ЧСА- и IgG- связывающих полипептидов, полученных из стрептококков, качество очистки которых обеспечивает практически 100% специфичность сорбента.
Заявлена группа изобретений, объединенных единым изобретательским замыслом, которая включает следующие объекты:
1. Рекомбинантная ДНК (обозначенная как рА4), полученная в результате полимеразной цепной реакции (ПНР) с использованием хромосомной ДНК штамма DG13 стрептококка группы G, уникальных праймеров РА1 и РА2 и последующего клонирования с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США).
2. Рекомбинантная плазмидная ДНК pQE 30-pA4, несущая рекомбинантную ДНК рА4.
3. Штамм-продуцент Е. coli M15-A4, позволяющий при определенных условиях экспрессировать рекомбинантный полипептид А4.
4. Рекомбинантный полипептид А4, содержащий последовательность двух ЧСА-связывающих GA модулей и области W, обладающий высокой способностью связывать ЧСА и имеющий меньший молекулярный вес, чем полипептид А2 [Т.В. Гупалова и др., Биотехнология, №3: 75-84, 2012].
5. Аффинный сорбент, в котором к матрице, представляющей собой активированную цианбромом сефарозу 4 В, присоединен ЧСА-связывающий полипептид.
6. Аффинный комбинированный сорбент, состоящий одновременно из двух сорбентов: сефарозы 4В-IgG-связывающего полипептида и сорбента по пункту 5.
7. Способ последовательного удаления ЧСА и IgG из сыворотки крови методом аффинной хроматографии, отличающийся тем, что в качестве аффинных носителей в последовательно соединенных колонках используются сефароза 4В-IgG-связывающий полипептид и сорбент по п.5.
8. Способ одновременного удаления ЧСА и IgG из сыворотки крови, отличающийся тем, что в качестве сорбента используют аффинный комбинированный сорбент по п.6.
Техническая суть и достигаемый при использовании заявленной группы изобретений технический результат состоят в следующем.
ЧСА и IgG - наиболее распространенные белки плазмы крови человека и млекопитающих.
В последние десятилетия выявлены и изучены белки бактерий, обладающие рецепторной активностью в отношении ЧСА и IgG.
Стрептококки (Streptococcus) групп А, С и G экспрессируют поверхностные белки, которые связывают ЧСА [Myhre E. and Kronvall G. Infect. Immun. 27: 6-14, 1980; Wideback K. et. al., Acta Pathol. Microbiol. Immunol. Scand. Sect. B. 91: 373-382, 1983].
Среди таких поверхностных белков стрептококков группы G интерес представляет белок G, полноразмерная молекула которого обладает способностью связывать IgG человека и различных млекопитающих, а также ЧСА человека и животных [Bjorck L. et. al. Mol.ImmunoL 24: 1113-1122, 1987].
Из штамма стрептококков группы G, G148, выделенного от человека, были получены рекомбинантный G белок, который одновременно связывал ЧСА и IgG, бифункциональный G белок и монофункциональный G белок, который связывал только IgG, то есть IgG-связывающий полипептид [Гупалова Т.В., Вестник Академии Медицинских наук, 3: 44-49, 1996].
Из штамма Finegoldia magnus выделен РАВ белок, связывающий ЧСА [de Chateau М., Bjorck L., J. of Biol. Chem. 269: 12147-12151, 1994]. Анализ его аминокислотной последовательности выявил домен в 45 аминокислотных остатков, отвечающий за связывание с ЧСА.
Было высказано предположение, что этот домен перемещен из стрептококков групп С и G в ЧСА-связывающий домен штамма Finegoldia magna. [Johansson M.U., Nilsson H. et al, J. Mol. BioL, 2002; 316: 1083-1099]. Такой межвидовой обмен структурно был определен как подвижный модуль и обозначен как GA модуль (G related albumin-binding domain).
Последовательность GA модуля имеет гомологию с ЧСА-связывающей областью белка G [Akerstrom В. et al., J of Biol. Chem. 262: 13388-13391, 1987;
Sjobring U. et. al., J. Immun. 140: 1595-1599, 1988; Nygren P. et. al., J. Mol. Recognit. 1: 69-74, 1988].
В отличие от РАВ белка, который не имеет гомологичных повторов, белок G, выделенный из штамма G 148, содержит три гомологичные повторяющиеся последовательности, т.е. три GA модуля.
Из штамма Streptococcus canis DG12, выделенного из коровьего молока, получен ЧСА-связывающий белок, который содержит два GA модуля. Он не связывает IgG, но по способности связывать ЧСА превосходит белок G [Sjobring U., Infection and Immun. 60: 3601-3608, 1992]. Это свойство делает его привлекательным для практического применения.
Изучение структуры IgG и ЧСА-связывающих белков имеет большое значение для технологии создания белковых реагентов, актуальных в иммунохимии, протеомике, биотехнологии и клинической диагностике.
Использование рекомбинантных полипептидов позволяет: исключить трудоемкий процесс приготовления специфических антител к ЧСА или IgG (для которых используются лабораторные животные), избежать неспецифических "фоновых" реакций, часто встречающихся в иммунологии, стандартизовать используемые ЧСА- или IgG- связывающие полипептиды.
Впервые рекомбинантный ЧСА рецептор был получен шведскими авторами путем клонирования фрагмента гена ЧСА рецептора в непатогенном хозяине [Nygren P.-A. et al., J. Mol. Recognition. 1: 69-74, 1988]. В данной работе для клонирования АВ области (области, ответственной за связывание ЧСА) рестрицировали плазмиду pSPG2 ферментом EcoRI с последующей обработкой фрагментом Кленова, добавлением синтетического линкера SalI и лигированием. При рестрикции SalI и PstI был получен фрагмент в 640 н.п., который изолировали из агарозы и вставляли между теми же сайтами в вектор pEML8. Фрагмент гена вырезали из полученной плазмиды, используя EcoRI и HindIII сайты и вставляли в плазмиду pEG, конструкция которой описана в работе Eliasson [Eliasson M. et al.; J. Biol. Chem. 263: 4323-4327, 1988].
В результате такой модификации была получена плазмида pB1B2, ответственная за ЧСА-связывающую активность. Из субклона, продуцирующего ЧСА-связывающий белок, был выделен белок. Он был очищен аффинной хроматографией. Этот белок оказался гетерогенным с деградацией очищенного продукта. Причиной гетерогенности, очевидно, являлся довольно сложный путь его генетического конструирования. Практического применения из-за своих недостатков этот белок не находит.
В 1996 году был проклонирован фрагмент гена, кодирующий третий GA модуль белка G штамма G148. Полученный белок G148-GA3 был использован для изучения структуры ЧСА-связывающего модуля [Kraulis P. et al., FEBS Lett. 378: 190-194, 1996], т.е. только в исследовательских целях.
В 1992 году из стрептококка группы G животного происхождения, штамма DG 12, был проклонирован фрагмент гена, кодирующий ЧСА-связывающий фрагмент, с использованием плазмидного вектора pKK233-2 и определена его нуклеотидная последовательность [Sjobring U. Infect, and Immun. 60: 3601-3608, 1992].
Получены рекомбинантные ЧСА-связывающие рецепторные полипептиды A1 и А2. Полипептид A1 был получен из стрептококка группы G, штамма G148, выделенного от человека, в Отделе молекулярной микробиологии в 1996 году [Орлова С.Н. и др. Биотехнология. 8: 13-21, 1996]. Штамм, продуцирующий полипептид A1, задепонирован в коллекции НИИЭМ СЗО РАМП, №357.
Особенностью этого полипептида является то, что он содержит ЧСА-связывающую область, охватывающую три GA модуля. Был изучен фрагмент гена, который кодирует полипептид А1. Изучение этого рецепторного полипептида доказало его способность связываться исключительно с основным белком плазмы человека - ЧСА. Данное свойство полипептида было использовано в диагностических тест-системах, позволяющих определять концентрацию ЧСА в различных биологических жидкостях, в частности в моче. Однако при выделении и очистке рекомбинантного полипептида А1 выход активного реагента в достаточном количестве был затруднен, что привело к необходимости создания нового ЧСА-связывающего препарата, обладающего большей активностью, чем полипептид А1 белка G.
Полипептид А2 был получен из штамма DG 13 стрептококка группы G, выделенного из коровьего молока [Гупалова Т.В. и др. Биотехнология, №3: 75-84, 2012]. Штамм, продуцирующий полипептид А2, задепонирован в коллекции НИИЭМ СЗО РАМН, №593. Показано, что полипептид А2 обладает большей ЧСА-связывающей активностью по сравнению с полипептидом А1 из штамма стрептококка группы G человеческого происхождения. Создан штамм-продуцент E. coli M 15-A2, позволяющий получить полипептид А2, обладающий ЧСА-связывающей активностью, с высоким выходом. Полипептид А2 был использован в методе РФА для определения микроальбуминурии. Он характеризуется последовательностью из 346 аминокислот, в которой первые 13 аминокислот, кодирующиеся последовательностью плазмиды pQE 32, ковалентно связаны с последующими 333 аминокислотами, кодируемыми последовательностью фрагмента хромосомной ДНК штамма DG 13 стрептококка группы G и идентичными последовательности ЧСА-связывающего белка из штамма DG 12 стрептококка группы G [Sjobring U. Infect, and Immun. 60: 3601-3608, 1992]. Эта последовательность состоит из 11 аминокислот, которые совместимы с последней частью лидерного пептида, то есть она содержит ряд гидрофобных остатков с небольшой незаряженной аминокислотой в конце. Затем идет уникальная последовательность из 55 аминокислот, обозначенная N, за которой идут две области, обозначенные Е1 (28 аминокислот) и Е2 (29 аминокислот). За ними идут последовательности двух GA модулей. В следующей последовательности, обозначенной W, обнаружена последовательность из четырех аминокислот (LysProGluVal), повторяющаяся 12 раз.
Молекулярный вес ЧСА-связывающего белка, последовательность которого состоит из 346 аминокислот, составляет 48 кДа. Отсутствие у такого большого белка оптимальной конформации может вызвать стерические затруднения при его взаимодействии с сорбентом. Поэтому необходимо создать новый рекомбинантный полипептид, связывающий ЧСА с такой же высокой активностью, как и полипептид А2, но имеющий меньшую последовательность и вследствие этого меньший молекулярный вес.
Согласно заявленному изобретению получен рекомбинантный полипептид А4 и его штамм-продуцент. Рекомбинантный полипептид А4 содержит последовательность двух ЧСА-связывающих GA модулей и W область и обладает высокой способностью связывать ЧСА и меньшим молекулярным весом. Его использование возможно при создании сорбента для эффективного удаления ЧСА из сыворотки крови.
Поставленная задача решалась конструированием уникальных праймеров, направленных к участкам гена, кодирующего полипептид, состоящий из двух ЧСА-связывающих GA модулей и W области. С целью быстрого и простого получения рекомбинантного полипептида А4 для клонирования фрагмента ДНК была использована система экспрессионных плазмид pQE. Для создания штамма-продуцента рекомбинантного полипептида А4 был использован штамм Е. coli M15. В результате экспериментальной работы этот штамм приобрел новые свойства: способность экспрессировать рекомбинантный полипептид А4. Новый штамм получил название Е. coli M15-A4. Штамм задепонирован в коллекции НИИЭМ СЗО РАМН, №715.
В процессе работы была создана уникальная рекомбинантная ДНК рА4, полученная в результате ПЦР, с использованием хромосомной ДНК штамма DG13 стрептококка группы G, уникальных праймеров РА1 и РА2 и последующего клонирования с использованием экспрессионной плазмиды pQE-30 (The QIAexpress System, Qiagen, США).
Была также создана рекомбинантная плазмидная ДНК pQE 30-pA4, несущая рекомбинантную ДНК рА4, и штамм-продуцент Е. coli M15-A4, позволяющий при определенных условиях экспрессировать рекомбинантный полипептид А4.
В процессе работы получен фрагмент гена, кодирующий область с двумя ЧСА-связывающими GA модулями и областью W штамма DG13 стрептококка группы G, обозначенный как рА4, размером 612 п.н. с помощью ПЦР с использованием праймеров РА1 и РА2 и хромосомной ДНК штамма DG13 стрептококка группы G. Также авторами осуществлено клонирование рА4 с использованием экспрессионной плазмиды pQE-30 и последующей трансформацией полученной рекомбинантной плазмиды (обозначенной как pQE 30-pA4) в гетерологическую систему Е. coli M15. Авторами получен штамм-продуцент рекомбинантного полипептида А4, обозначенный как Е. coli M15-A4. Также авторами реализована экспрессия рекомбинантного полипептида А4 клетками штамма Е. coli M15-A4 с последующей одноступенчатой аффинной очисткой с использованием Ni-сефарозы (GE Healthcare, Sweden).
Получение рекомбинантного полипептида А4.
Источником хромосомной ДНК послужил штамм DG 13 стрептококка группы G (выделенный из коровьего молока). ДНК, выделенная фенольно-хлороформной экстракцией, была использована в качестве матрицы в ПЦР с целью получения фрагмента гена рА4 (612 п.н.) Олигонуклеотидные праймеры представлены в таблице 1.

Подчеркнутые звенья нуклеотидной последовательности указывают на сайты рестрикции.
Анализ результатов ПЦР осуществлен путем разделения фрагментов ДНК в 1% агарозном геле при помощи горизонтального электрофореза (фиг.1).
Выделение искомого амплифицированного участка ДНК проведено с использованием набора «QIAquick Gel Extraction Kit» (Qiagen, США).
Полученный фрагмент клонировали с использованием экспрессионной плазмиды pQE 30. При подготовке к клонированию была проведена рестрикция выделенного из агарозы фрагмента рА4 (612 н.п.) и плазмиды pQE 30 ферментами BamHI и Kpnl с образованием липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле, а затем лигировали.
Продуктами лигирования проводили кальциевую трансформацию штамма E.coli M 15. Процесс трансформации осуществляли по методике трансформации кишечной палочки, используя плотные селективные среды, содержащие 100 мкг/мл ампициллина, 25 мкг/мл канамицина [Maniatis Т, Fritsch E.F., Sambrook J. Molecular cloning: a laboratory manual. - Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 867, 1982].
Антибиотикоустойчивые трансформанты откалывали на две параллельные чашки Петри с антибиотиками. Колонии с одной чашки переносили на нитроцеллюлозную мембрану и вскрывали раствором, содержащим 0,2 N NaOH, 0,1% SDS и 0,5% β-меркаптоэтанола. Мембрану инкубировали 1 час при комнатной температуре в блокирующем растворе (2 части 3% молока и 1 часть фосфатно-солевого буфера (ФСБ)) для удаления неспецифического связывания и затем в блокирующем растворе, содержащем ЧСА, меченный пероксидазой хрена (Sigma, USA).
Конъюгирование пероксидазы хрена с ЧСА проводили перйодатным методом [Фримель Г. Иммунологические методы. М.: Медицина: 438-439, 1987]. Затем мембрану последовательно отмывали блокирующим раствором и ФСБ. Пероксидазную активность проявляли раствором парафенилендиамина в концентрации 1 мг/мл с 0,15% перекисью водорода. Из трансформантов с наиболее выраженной экспрессией ЧСА-связывающей активности выделяли рекомбинантные плазмиды с помощью Mini-prep, plasmid DNA purification kit (Qiagen, USA). Наличие в них рекомбинантной ДНК подтверждали в ПЦР с исходными праймерами РА1 и РА2. Плазмидную ДНК pQE 30-рА4 использовали как матрицу в ПЦР с праймерами pQEl и pQE2. Амплификат выделяли после электрофореза в 1% агарозном геле и секвенировали.
Итоговые данные совпали с известной нуклеотидной последовательностью, кодирующей область с двумя ЧСА-связывающими GA модулями [Sjobring U. Infect, and Immun. 60: 3601-3608, 1992].
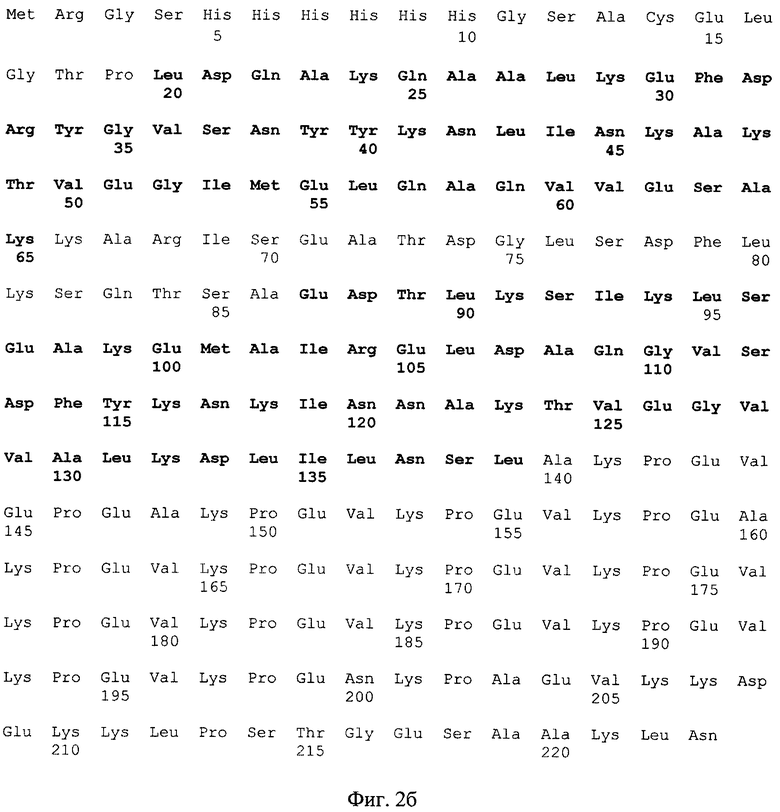
На фиг. 2а представлена нуклеотидная последовательность, состоящая из 612 п. н. рекомбинантной ДНК рА4 и 57 п. н. плазмиды pQE 30, и на фиг. 2б - аминокислотная последовательность, состоящая из 223 аминокислотных остатков, в которой 204 аминокислоты полипептида А4 кодируются рекомбинантной ДНК рА4 и первые 19 аминокислот - плазмидой pQE 30. Жирным шрифтом выделены последовательности ЧСА-связывающих GA модулей.
Культуру штамма Е. coli М 15, содержащую рекомбинантную плазмиду pQE 30-рА4, обозначенную как Е. coli М 15-А4, культивировали в жидкой среде BHI (Brain Heart Infusion Broth, Gibco, CIIIA) с добавлением антибиотиков (ампициллин (100 мкг/мл) и канамицин (25 мкг/мл)) до поздней логарифмической фазы роста (OD600=0,7-0,9). Затем экспрессию рекомбинантного белка индуцировали добавлением изопропил-бета-D-тиогалактопиранозида (IPTG), и клетки культивировали еще 4 часа. После этого клетки осаждали центрифугированием, отмывали лизирующим буфером А (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 20 мМ имидазола, pH 8,0) и суспендировали в том же буфере, добавляя ингибитор протеаз, фенилметилсульфонилфторид (ФМСФ), до концентрации 1 мМ. Суспензию клеток вскрывали ультразвуком и центрифугировали. Надосадочную жидкость наносили на колонку, заполненную Ni-сефарозой. После того, как белок связывался с Ni-сефарозой, колонку промывали буфером А для удаления не связавшихся белков. Рекомбинантный белок элюировали буфером Б (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 250 мМ имидазола, pH 8,0). После аффинной хроматографии белок диализовали 18 часов против дистиллированной воды.
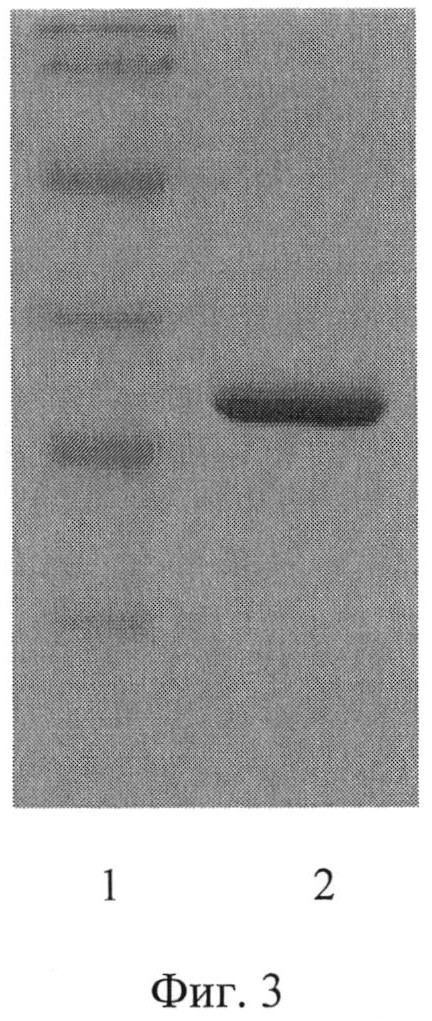
Выделенный рекомбинантный полипептид анализировали методом электрофореза в полиакриламидном геле, который позволил сделать заключение об удовлетворительном качестве очистки полипептида и также об его молекулярной массе, сравнивая пробег полипептида А4 с пробегом белков известной молекулярной массы (Precision Plus Protein standards (161-0373), Bio-Rad, США). Молекулярная масса полипептида А4 оказалась равной (27,0±0,5) кДа (фиг.3).
Таким образом, рекомбинантный полипептид А4 содержит аминокислотную последовательность ЧСА-связывающего полипептида стрептококка группы G штамма DG 13, содержащую 204 аминокислоты, ковалентно связанную с 19 аминокислотными остатками, кодируемыми pQE 30.
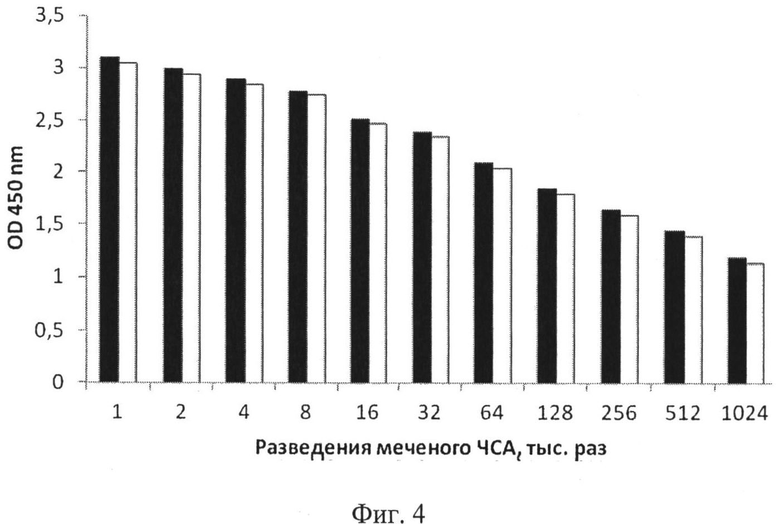
Изучение взаимодействия рекомбинантного полипептида А4 с ЧСА при сравнении с аналогичной реакцией с полипептидом А2
Способность полипептидов А4 и А2 связывать ЧСА, представленного следующими образцами: 1) ЧСА - поликлональный сывороточный альбумин человека, меченный пероксидазой хрена, 2) поликлональный немеченый ЧСА, оценивали с помощью прямого метода рецепторно-ферментного анализа (РФА) [Гупалова и др. Биотехнология, №3: 75-84, 2012]. На планшет сорбировали полипептиды в концентрации 1 мкг/мл. При этом оценивали количество связавшегося с ними меченого ЧСА. На фиг.4 показана гистограмма, отражающая сопоставление ЧСА-связывающей активности полипептидов А4 и А2. Исследуемые полипептиды А4 и А2 обладают ЧСА-связывающей активностью в отношении меченого ЧСА, причем количество ЧСА, связанного полипептидом А4, примерно такое же, что и в случае полипептида А2.
Способ удаления ЧСА и IgG из сыворотки крови, т.е. создание аффинных носителей: сефарозы 4В-ЧСА-связывающего полипептида и сефарозы 4В-IgG-связывающего полипептида.
Получение рекомбинантного полипептида, используемого для создания одного из аффинных носителей, IgG-связывающего полипептида, описано в статье [Гупалова и др. Вестник Академии Медицинских наук, 3: 44-49, 1996] и в патенте №2056859 «Рекомбинантный IgG-связывающий G-белок стрептококка группы G» [Гупалова Т.В., 1996].
В качестве матрицы аффинных носителей использовали цианбромированную сефарозу 4 В (Sigma). Лигандами, иммобилизованными на матрице, служили рекомбинантные IgG- и ЧСА-связывающие полипептиды.
Ранее такой подход для удаления ЧСА и IgG из сыворотки не применялся.
Предварительно циан-бромированную сефарозу 4 В промывали несколько раз холодной 1 mM HCl, каждый раз центрифугировали, надосадок выбрасывали, добавляя свежую порцию холодной 1 mM HCl. Затем суспензию сефарозы промывали 0,1 М бикарбонатным буфером, pH 8,3.
Затем к трем порциям приготовленной, как описано выше, циан-бромированной сефарозы 4 В добавляли ЧСА-связывающие полипептиды А2 [Гупалова Т.В., Биотехнология, №3: 75-84, 2012], А4 и IgG-связывающий полипептид соответственно и инкубировали при 4°C при перемешивании. После инкубации заполняли три колонки сефарозой 4В-ЧСА-связывающим полипептидом А2, сефарозой 4В-ЧСА-связывающим полипептидом А4 и сефарозой 4В-IgG-связывающим полипептидом. Колонки промывали сначала бикарбонатным буфером с 0,5% NaCl, pH 8,0, затем 0,1 М глициновым буфером, pH 2,2 и снова бикарбонатным буфером с 0,5% NaCl. Колонки уравновешивали раствором ФСБ, pH 7,4 и хранили при 4°C.
При приготовлении аффинных сорбентов было обнаружено, что при иммобилизации сефарозы 4 В ЧСА-связывающим полипептидом А4 его связалось больше за счет меньшего молекулярного веса. Поэтому при практически одинаковой ЧСА-связывающей активности у полипептидов А2 и А4 (фиг.3) предпочтительнее использовать аффинный сорбент на основе полипептида А4.
Для проверки емкости сорбентов на две приготовленные колонки с сефарозой 4В-ЧСА-связывающим полипептидом А4 и сефарозой 4B-IgG-связывающим полипептидом, уравновешенных ФСБ, pH 7,4, последовательно наносили 100, 200, 400 и 500 мкл сыворотки. Полноту освобождения сыворотки от ЧСА и IgG проверяли методом блоттинга. Для этого на нитроцеллюлозные мембраны (НЦ) наносили пробы исходной сыворотки, пробы сыворотки, не связавшиеся с сорбентом и элюированные с колонок пробы. Одни НЦ мембраны переносили в блокирующий раствор, содержащий ЧСА, меченный пероксидазой, а другие - содержащий IgG, меченный пероксидазой, и выдерживали их 40 минут. Мембраны последовательно отмывали блокирующим раствором и ФСБ. Отмытые мембраны подсушивали на воздухе и помещали в раствор 3,3/,5,5/-Тетраметилбензидин (ТМБ, жидкий субстрат для мембран, Sigma). Окрашивание происходило в течение нескольких секунд.
В пробах сывороток, не связавшихся с сорбентами и прошедших через аффинные колонки при нанесении 100, 200, 400 и даже 500 мкл сыворотки, практически полностью отсутствовали как ЧСА, так и IgG.
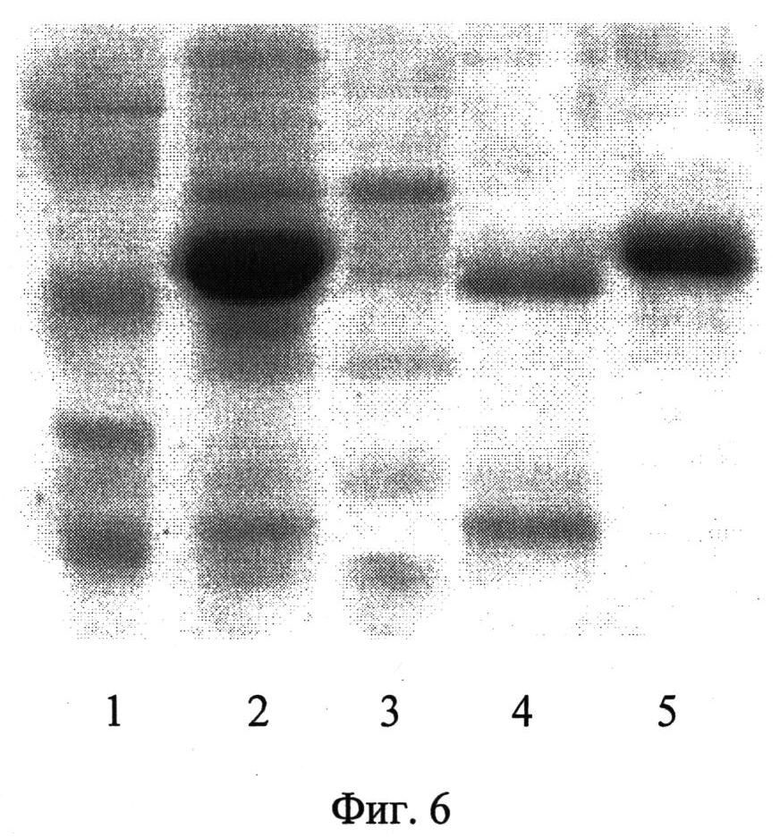
Две аффинные колонки с сефарозой 4В-IgG-связывающим и с сефарозой 4В-ЧСА-связывающим А4 полипептидами были соединены между собой. На первую из них нанесли 400 мкл сыворотки. Проба, не связавшаяся с первой колонкой, перетекала на вторую колонку. Аффинную хроматографию проводили, как описано выше. Пробы исходной сыворотки, сыворотки, не связавшейся с сорбентами обеих колонок и пробы элюированных ЧСА и IgG, оценивали методом блоттинга. Результаты блоттинга показали, как и в первом эксперименте, отсутствие в пробе сыворотки, не связавшейся с сорбентами, как ЧСА, так и IgG (фиг.5а и 5б). Удаление ЧСА и IgG из сыворотки было проверено также методом СДС-ПААГ электрофореза (фиг.6). Как следует из фиг.6, проба сыворотки после пропускания ее последовательно через две аффинные колонки, освобождалась практически полностью от ЧСА и IgG (фиг.6, (3)). Проба IgG, очищенная аффинной хроматографией, элюированная с первой колонки, представлена на фиг.6, (4), проба очищенного ЧСА представлена на фиг.6, (5).
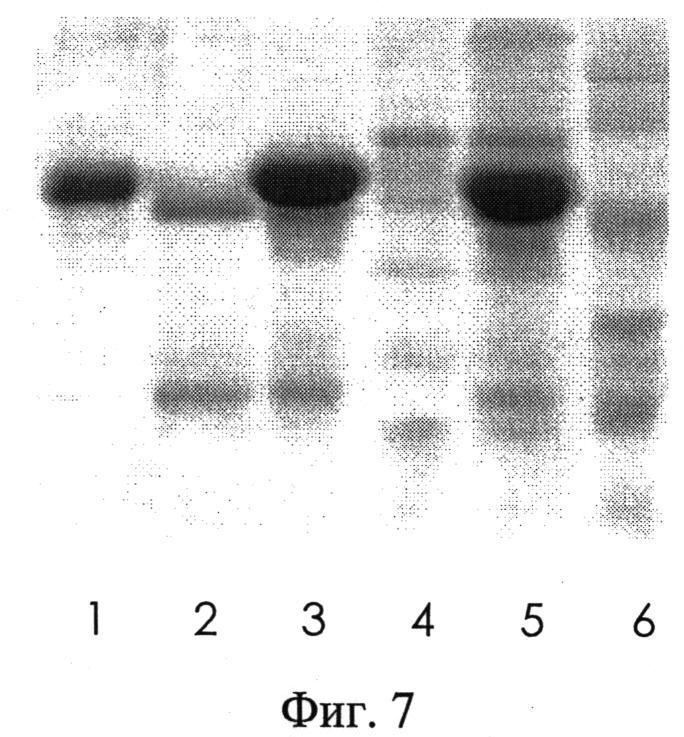
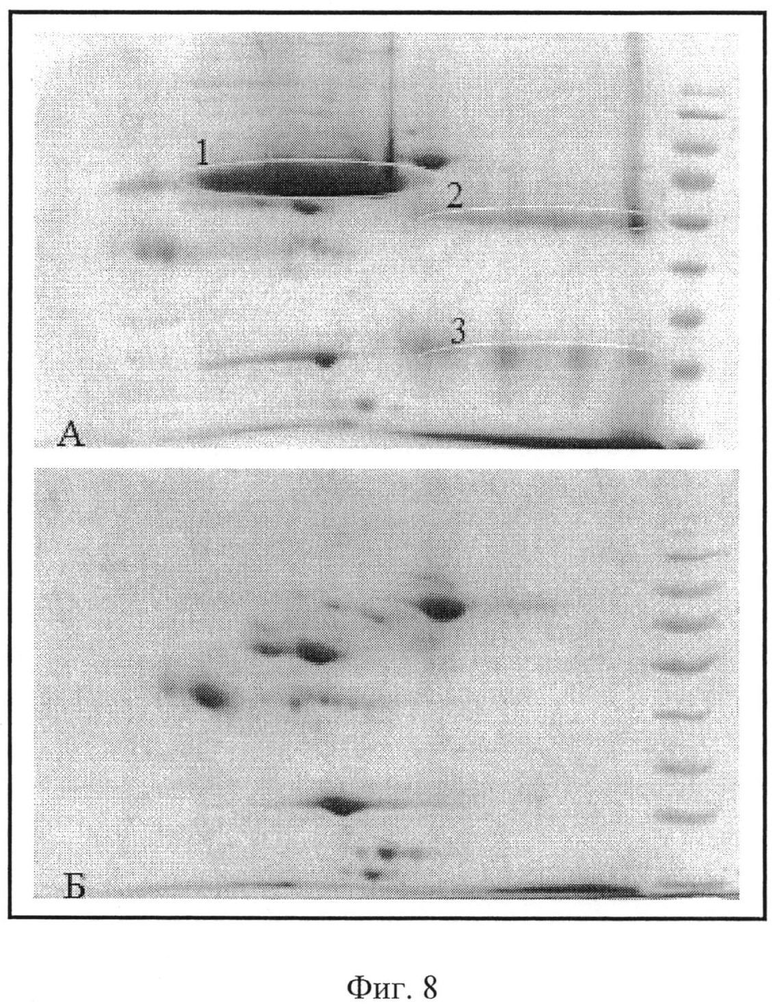
Сорбент с присоединенным IgG-связывающим полипептидом и сорбент с присоединенным ЧСА-связывающим А4 полипептидом смешали и получили комбинированный сорбент. На колонку с комбинированным сорбентом было нанесено 400 мкл сыворотки. Аффинную хроматографию проводили, как описано ранее. Пробы исходной сыворотки, сыворотки, не связавшейся с сорбентом комбинированной колонки и элюированной пробы, содержащей ЧСА и IgG, были проанализированы в СДС-ПААГ электрофорезе (фиг.7). Пробы сыворотки до и после пропускания ее через колонку с комбинированным сорбентом проанализировали методом 2-DE (фиг.8). Как видно из фиг.7, проба сыворотки после пропускания ее через аффинную колонку освободилась от ЧСА и IgG (фиг.7, (4)). Фиг.7, (3) показывает наличие ЧСА и IgG в пробе сыворотки, элюированной с колонки. Затем эта проба была пропущена через колонку с IgG-связывающим полипептидом, чтобы разделить ЧСА и IgG друг от друга и получить их в очищенном состоянии. Проба IgG была элюирована с колонки, а проба ЧСА прошла через колонку. Как показано на фиг.7, ЧСА и IgG получены в высокоочищенном состоянии (фиг.7, (1 и 2)). Пробы исходной сыворотки и сыворотки, пропущенной через колонку и не связавшейся с сорбентом, были проанализированы методом 2-DE, который показал освобождение сыворотки от ЧСА и IgG (фиг.8 Б). Освобождение сыворотки от ЧСА IgG позволяет обнаружить белки, находящиеся как в области высокомолекулярных значений (70 kDa), экранированных ЧСА, так и в области 30-50 kDa, где широкая полоса IgG скрывает белки, мигрирующие в этой области.
Таким образом, созданные сорбенты: сефароза 4В-IgG-связывающий полипептид, сефароза 4В-ЧСА-связывающий А4 полипептид и комбинированный сорбент, состоящий из обоих сорбентов, позволяют практически полностью удалять из сыворотки ЧСА и IgG и выявлять белки, находящиеся в сыворотке в низких концентрациях, и тем самым перейти к поиску и характеристике новых биомаркеров.
Ниже приводятся конкретные примеры, иллюстрирующие некоторые варианты изобретения, но не ограничивающие его.
Пример 1. Получение фрагмента ДНК рА4 методом ПЦР.
К 0,25 мкг геномной ДНК, выделенной фенольно-хромосомной экстракцией из штамма DG 13 стрептококка группы G, добавляли по 10 микромолей каждого из специфических праймеров, фланкирующих исследуемую последовательность, буфер с магнием для полимеразы, по 0,2 мМ четырех дезоксирибонуклеотидтрифосфатов, объем доводили водой до 25 мкл и добавляли 0,5 мкл термостабильной ДНК полимеразы. На поверхность жидкости наслаивали 40 мкл минерального масла. Пробирки помещали в амплификатор и инкубировали при 94°C 2 мин. Программа ПЦР состояла из: денатурации при 94°C - 30 сек, отжига праймеров - 60°C - 1 мин и синтеза - 72°C - 1 мин. Этот цикл повторялся 30 раз, после чего смесь инкубировалась при 72°C 10 мин. В работе использовали олигонуклеотидные праймеры, приведенные в Таблице 1. ПЦР продукты разделяли в 1% агарозном геле в горизонтальном электрофорезе (фиг.1). Выделение амплифицированного участка ДНК проводили с использованием набора «QIAquick Gel Extraction Kit» (Qiagen, США). Анализ размера полученного фрагмента ДНК проводили, исходя из сравнения его электрофоретической подвижности с электрофоретической подвижностью маркера молекулярных весов (100 п.н. ДНК-маркер, Хеликон).
На фиг.1 показана электрофореграмма амплифицированного ДНК-фрагмента рА4, где 1-100 п.н. ДНК - маркер, (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар), 2 - продукт ПЦР.
В результате ПЦР с праймерами РА1 и РА2 был получен фрагмент 612 п.н., что продемонстрировано на фиг.1.
Пример 2. Клонирование ДНК-фрагмента рА4 с использованием экспрессионной плазмиды pQE 30.
Плазмида pQE 30 и фрагмент, полученный в результате ПЦР, были обработаны двумя рестрикционными ферментами BamHI и KpnI, что привело к образованию липких концов. Продукты рестрикции разделяли с помощью горизонтального электрофореза в 1% агарозном геле, а затем выделяли из агарозы с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, США). В ходе клонирования к 5 мкл фрагмента ДНК рА4, добавляли 1 мкл фрагмента ДНК pQE 30, полученных после рестрикции и вырезанных из агарозы, 2 мкл десятикратного лигазного буфера и 1 мкл лигазы фага Т4.
Объем доводили дистиллированной водой до 20 мкл. Смесь инкубировали при 10°C в течение 12 часов. В результате проведенного клонирования была получена рекомбинантная плазмида, обозначенная как pQE 30-рА4, несущая рекомбинантную ДНК (обозначенную как рА4), состоящую из 612 п.н. и 57 п.н. фрагмента плазмиды pQE-30.
Пример 3. Трансформация культуры Е. coli M 15 плазмидной ДНК pQE 30-pA4.
Рекомбинантную плазмиду трансформировали в гетерологичную систему Е. coli M 15. В качестве положительного контроля параллельно проводили трансформацию Е. coli M 15 исходной плазмидной ДНК pQE 30.
Генетическим маркером плазмидной ДНК pQE 30 является маркер amp, кодирующий бета-лактомазу, что обеспечивает устойчивость к ампициллину клеток, несущих эту плазмиду. Культуру клеток Е. coli M 15 культивировали в бульоне BHI (Brain Heart Infusion Broth, Gibco, США) с канамицином (25 мкг/мл) в течение 12 часов при 37°C и интенсивном перемешивании. Затем инокулят пересевали на новый объем той же среды (на 10 мл среды - 0,1 мл инокулята) и инкубировали при 37°C в течение 2-3 часов при перемешивании до OD600=0,3. Выращенные клетки в объеме 1,5 мл центрифугировали в течение 1 мин при 12000 об/мин и полученный осадок суспендировали в 200 мкл раствора 0,1 M CaCl2. Далее смесь выдерживали во льду 1 час. После центрифугирования осадок ресуспендировали в 187, 5 мкл 0,1 M CaCl2 и 64,5 мкл дистиллированной воды. Затем в пробирку вносили 0,2 мкг плазмидной ДНК и инкубировали смесь во льду 1 час. Далее смесь выдерживали 2 мин при 42°C и снова переносили смесь в лед на 10 мин. После этого к смеси добавляли 1 мл бульона BHI и инкубировали 1 час при 37°C. После осаждения центрифугированием (8000 об/мин, в течение 1 мин) клетки высевали на чашки с 1% L-агаром (Difco, США), содержащим ампициллин (100 мкг/мл) и канамицин (25 мкг/мл). Через 18 часов роста клеток при 37°C проводили отбор клонов-трансформантов. Антибиотикоустойчивые трансформанты переносили на две параллельные чашки Петри с антибиотиками. Колонии с одной чашки переносили на нитроцеллюлозную мембрану и вскрывали раствором, содержащим 0,2 N NaOH, 0,1% СДС и 0,5% β-меркаптоэтанола. Мембрану инкубировали 1 час в блокирующем растворе, содержащем ЧСА, меченный пероксидазой хрена (Sigma, США). Затем ее последовательно отмывали блокирующим раствором и раствором ФСБ. Пероксидазную активность проявляли раствором парафенилендиамина в концентрации 1 мкг/мл с 0,15 М перекисью водорода.
Таким образом, полученные данные позволяют сделать вывод о том, что в результате проведенного клонирования были получены рекомбинантные клоны, несущие плазмиды pQE 30-pA4 с рекомбинантной ДНК pA4.
Пример 4. Очистка рекомбинантного полипептида А4. Культуру штамма Е. coli M 15-A4 выращивали в бульоне BHI с добавлением ампициллина в концентрации 100 мкг/мл и канамицина в концентрации 25 мкг/мл в течение ночи при интенсивном перемешивании. Затем клетки пересевали на 800 мл той же среды и инкубировали при 37°C в течение 2-3 часов при интенсивном перемешивании до OD600=0,7-0,9. Экспрессию рекомбинантного белка индуцировали добавлением раствора изопропил-бета-D-тиогалактопиранозида до конечной концентрации 2 мМ, после чего клетки инкубировали при тех же условиях еще 4 часа. Полученную суспензию клеток центрифугировали при 4000 об/мин 20 мин. Надосадочную жидкость сливали, а клетки суспендировали в буфере А (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 20 мМ имидазола, pH 8,0), добавляя ингибитор протеаз фенилметилсульфонилфторид (ФМСФ) до конечной концентрации 1 мМ. Для лизирования клеток была использована трехкратная ультразвуковая обработка при 4°C в течение 20 сек с перерывом в 40 сек в ультразвуковом дезинтеграторе (УЗДН-1 У4.2, Англия). Лизат клеток центрифугировали при 20000 об/мин и 4°C в течение 20 мин. Надосадочную жидкость пропускали через 0,45 мкм фильтр (Millipore, США) и затем ее наносили на колонку с Ni-сефарозой, предварительно уравновешенную буфером А. Далее колонку промывали тем же буфером до тех пор, пока значения ОД280 выходящего раствора не были больше, чем 0,01. Белок элюировали раствором буфера Б (20 мМ Na2HPO4, 20 мМ NaH2PO4, 500 мМ NaCl, 250 мМ имидазола, pH 8,0). Элюат собирали по фракциям по 500 мкл и измеряли в них значения ОД280. Фракции с наибольшими значениями ОД280 объединяли и диализовали против дистиллированной воды в течение 12 часов. На фиг.3 представлена электрофореграмма рекомбинантного полипептида А4.
На фиг.3 показана электрофореграмма рекомбинантного полипептида А4, где 1 - маркер молекулярной массы (сверху вниз: 113, 92, 52, 35, 28 и 21 кДа), 2 - препарат очищенного белка А4.
Молекулярную массу полипептида А4 определяли, сравнивая пробег белка А4 с пробегом белков известной молекулярной массы (Precision Plus Protein standards (161-0373), Bio-Rad, США). Молекулярная масса белка А4 оказалась равной (27±0,5) кДа.
Пример 5. Связывание сывороточного меченого ЧСА адсорбированными полипептидами А2 и А4.
Анализ ЧСА-связывающей активности проведен методом прямого РФА. Для проведения РФА использовали планшеты NUNC MaxiSorp (Danmark).
Приготовление разведенных препаратов меченого ЧСА осуществляли с применением раствора ФСБ. Удаление несвязавшихся реагентов проводили трехкратным промыванием планшетов раствором ФСБ с 0,05% твин 20 (ФСБТ).
Полипептиды А2 и А4, разведенные в ФСБ, pH 8,0 до 1 мкг/мл, адсорбировали на планшет в течение 18 часов при 4°C. После трехкратного отмывания в планшеты вносили разведенный меченый ЧСА. Инкубацию проводили при 37°C в течение 1 часа. Далее планшеты 3 раза отмывали от несвязавшихся реагентов. Активность фермента-метки пероксидазы хрена определяли с использованием хромогена - тетраметилбензидина (ТМБ), растворенного в 0,1 М цитрат-фосфатном буфере, pH 5,0 с добавлением перекиси водорода. Реакцию останавливали добавлением 2N серной кислоты. Оптическую плотность определяли при длине волны 450 нм на мультискане.
Установлено, что полипептид А4, подобно полипептиду А2, имеет примерно такую же высокую ЧСА-связывающую активность, что продемонстрировано на фиг.4.
На фиг.4 представлено сравнение эффективности связывания полипептидов А2 и А4 с ЧСА, где черный столбик - полипептид А2, белый столбик - полипептид А4, по оси абсцисс - разведения меченого ЧСА, по оси ординат - оптическая плотность (OD45o).
Пример 6. Способ удаления ЧСА и IgG из сыворотки крови на аффинных носителях: сефарозе 4В-IgG-связывающем полипептиде и на сефарозе 4В-ЧСА-связывающем полипептиде.
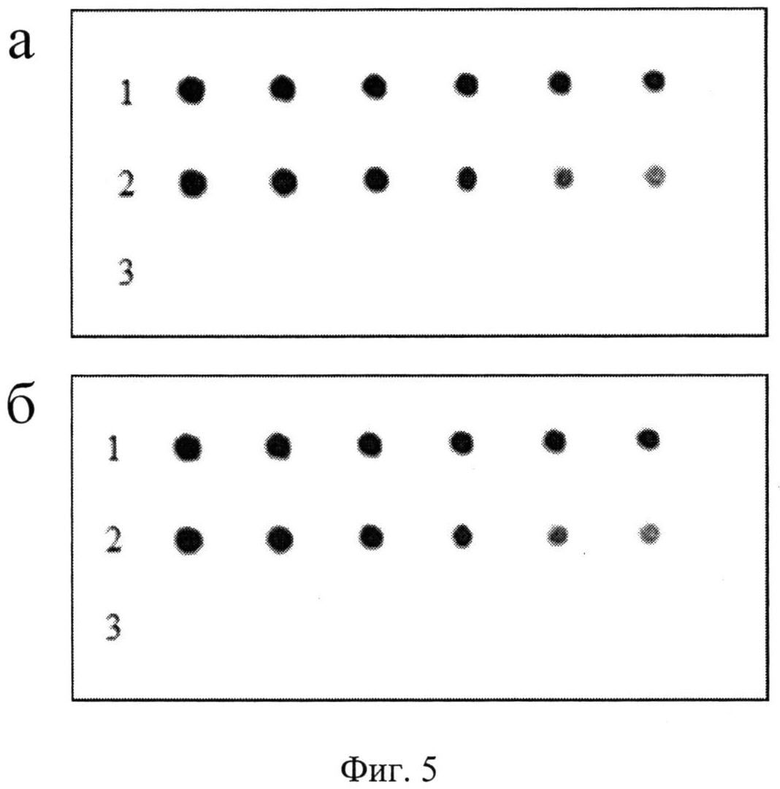
Две аффинные колонки с сефарозой 4В-IgG-связывающим и с сефарозой 4В-ЧСА-связывающим А4 полипептидами были соединены между собой и на первую из них нанесли 400 мкл сыворотки. Проба, не связавшаяся с первой колонкой, перетекала на вторую колонку. Аффинную хроматографию проводили, как описано выше. Пробы исходной сыворотки, сыворотки, не связавшейся с сорбентами обеих колонок и пробы элюированных ЧСА и IgG, оценивали методом блоттинга. Результаты блоттинга показали, как и в первом эксперименте, отсутствие в пробе сыворотки, не связавшейся с сорбентами, как ЧСА, так и IgG (фиг.5а и 5б).
На фигуре 5 представлены две НЦ, одна из которых, проинкубирована с ЧСА-связывающим полипептидом, меченным пероксидазой (фиг.5а), а вторая - с IgG-связывающим полипептидом, меченным пероксидазой (фиг.5б).
На фиг.5а (нитроцеллюлозная мембрана с проявленными пятнами):
1 - пятна, соответствующие двукратным разведениям исходной сыворотки,
2 - пятна, соответствующие разведениям элюированного ЧСА,
3 - отсутствие пятен, соответствующих разведениям сыворотки, которая протекла через аффинную колонку сорбентом и освободилась от ЧСА.
На фиг.5б (нитроцеллюлозная мембрана с проявленными пятнами):
1 - пятна, соответствующие двукратным разведениям исходной сыворотки,
2 - пятна, соответствующие разведениям элюированного IgG,
3 - отсутствие пятен, соответствующих разведениям сыворотки, которая протекла через аффинную колонку сорбентом и освободилась от IgG.
Удаление ЧСА и IgG из сыворотки было проверено также методом СДС-ПААГ электрофореза (фиг.6).
На фигуре 6 показана электрофореграмма:
1 - маркер молекулярных весов (сверху вниз: 113, 92, 52, 35, 28 кДа),
2 - проба исходной сыворотки,
3 - проба сыворотки, не связанной с сорбентами на колонках и освобожденная от ЧСА и IgG,
4 - проба IgG, элюированного и очищенного аффинной хроматографией,
5 - проба ЧСА, элюированного и очищенного аффинной хроматографией.
Пример 7. Способ удаления ЧСА и IgG из сыворотки крови на комбинированном аффинном носителе.
Сорбент с присоединенным IgG-связывающим полипептидом и сорбент с присоединенным ЧСА-связывающим А4 полипептидом смешали и получили комбинированный сорбент, на котором можно удалять из сыворотки одновременно и IgG, и ЧСА. На колонку с комбинированным сорбентом было нанесено 400 мкл сыворотки. Аффинную хроматографию проводили, как описано ранее. Пробы исходной сыворотки, сыворотки, не связавшейся с сорбентом комбинированной колонки и элюированной пробы, содержащей ЧСА и IgG, были проанализированы в СДС-ПААГ электрофорезе (фиг.7). Пробы сыворотки до и после пропускания ее через колонку с комбинированным сорбентом проанализировали методом 2-DE (фиг.8).
На фиг.7 представлена электрофореграмма пробы исходной сыворотки, пробы сыворотки, которая не связалась с комбинированным сорбентом и освободилась от ЧСА и IgG, пробы, содержащей ЧСА и IgG, элюированной с комбинированного сорбента и проб ЧСА и IgG, элюированных с сорбентов, содержащих ЧСА-связывающий полипептид или IgG-связывающий полипептид. Проба, элюированная с комбинированной колонки и содержащая ЧСА и IgG, была пропущена через колонку с IgG-связывающим полипептидом, чтобы разделить ЧСА и IgG друг от друга и получить их в очищенном состоянии. Проба IgG была элюирована с колонки, а проба ЧСА прошла через колонку.
На фиг.7 СДС-ПААГ электрофорез:
1 - проба IgG, элюированного и очищенного аффинной хроматографией;
2 - проба ЧСА, элюированного и очищенного аффинной хроматографией;
3 - проба, элюированная с комбинированного сорбента, содержащая ЧСА и IgG;
4 - проба сыворотки, не связанной с сорбентами на колонках и освобожденной от ЧСА и IgG;
5 - проба исходной сыворотки;
6 - маркер молекулярных весов (сверху вниз: 113, 92, 52, 35, 28 и 21 кДа). На фиг.8 показаны электрофореграммы после проведения 2DE. На фиг.8А-2DE исходной сыворотки эллипсами обозначены белки:
1 - ЧСА;
2 - IgG, тяжелые цепи;
3 - IgG, легкие цепи.
На фиг.8Б-2DE пробы сыворотки, освобожденной от ЧСА и IgG. Освобождение сыворотки от ЧСА и IgG позволяет обнаружить белки, находящиеся как в области высокомолекулярных значений (70 kDa), экранированных ЧСА, так и в области 30-50 kDa, где широкая полоса IgG скрывает белки, мигрирующие в этой области.




Изобретения относятся к области молекулярной биологии и касаются рекомбинантной ДНК pA4, рекомбинантной плазмидной ДНК pQE 30-А4, штамма Esherichia coli M15-A4, рекомбинантного полипептида А4, обладающего способностью селективно связывать человеческий сывороточный альбумин (ЧСА), аффинного сорбента, содержащего такой полипептид, аффинного комбинированного сорбента и способов последовательного удаления ЧСА и IgG из сыворотки крови. Охарактеризованный аффинный комбинированный сорбент получен смешением аффинного сорбента, содержащего указанный рекомбинантный полипептид А4, и аналогичного аффинного сорбента, в котором в качестве лиганда использован известный рекомбинантный IgG-связывающий полипептид. Представленные изобретения могут быть использованы в медицинской практике для освобождения сыворотки крови от двух белков, альбумина (ЧСА) и иммуноглобулина G (IgG), находящихся в ней в высоких концентрациях. Удаление двух мажорных белков из сыворотки крови позволит определить другие белки, присутствующие в сыворотке в более низких концентрациях. 8 н.п. ф-лы, 9 ил., 7 пр., 1 табл.
1. Рекомбинантная ДНК рА4, имеющая нуклеотидную последовательность, представленную на фиг. 2а, кодирующая рекомбинантный полипептид А4, обладающий способностью селективно связывать человеческий сывороточный альбумин - ЧСА.
2. Рекомбинантная плазмидная ДНК pQE 30-рА4, представляющая собой плазмиду pQE 30, несущую рекомбинантную ДНК рА4 по п.1, и обеспечивающая получение рекомбинантного полипептида А4.
3. Штамм Esherichia coli М 15-А4, продуцирующий рекомбинантный полипептид А4, полученный на основе штамма Esherichia coli М15 и содержащий рекомбинантную плазмидную ДНК pQE 30-рА4 по п. 2.
4. Рекомбинантный полипептид А4, обладающий способностью селективно связывать ЧСА и характеризующийся аминокислотной последовательностью, представленной на фиг. 26, в которой первые 19 аминокислот кодируются последовательностью плазмиды pQE 30, ковалентно связанные с последующими 204 аминокислотами, кодируемыми последовательностью ЧСА-связывающего фрагмента хромосомной ДНК штамма DG 13 стрептококка группы G-CTG.
5. Аффинный сорбент, состоящий из смеси матрицы, представляющей собой активированную цианбромом сефарозу 4 В, предварительно промытую холодной ImM HCl и 0,1 М бикарбонатным буфером, pH 8,3, и добавленного к ней раствора лиганда - ЧСА-связывающего полипептида А4 по п. 4, которая проинкубирована при постоянном перемешивании в течение 18 ч при 4°C.
6. Аффинный комбинированный сорбент, полученный смешением равных количеств двух сорбентов: аффинного сорбента по п. 5 и аналогичного аффинного сорбента, в котором в качестве лиганда использован известный рекомбинантный IgG-связывающий полипептид.
7. Способ последовательного удаления ЧСА и IgG из сыворотки крови методом аффинной хроматографии, отличающийся тем, что пробу сыворотки пропускают через первую колонку, содержащую аффинный сорбент, в котором в качестве лиганда использован рекомбинантный IgG-связывающий полипептид, а затем пропускают через соединенную с ней вторую колонку, содержащую аффинный сорбент по п. 5.
8. Способ одновременного удаления ЧСА и IgG из сыворотки крови, отличающийся тем, что пробу сыворотки пропускают через колонку, содержащую аффинный комбинированный сорбент по п. 6.
| РЕКОМБИНАНТНЫЙ IGG-СВЯЗЫВАЮЩИЙ G-БЕЛОК СТРЕПТОКОККА ГРУППЫ G | 1993 |
|
RU2056859C1 |
| Способ получения сорбентов для аффинной хроматографии сериновых протеаз | 1977 |
|
SU734215A1 |
| Е.А.БОРМОТОВА и др., Использование рекомбинантного бекла, связывающего альбумин и иммуноглобулин G, для целей протеомики, Биотехнология, 2009, N2, c.49-54 | |||
| ULF SJOBRING, Isolation and Molecular Characterization of a Novel Albumin-Binding Protein from Group G Streptococci, Infection and Immunity, Sept.1992, Vol.60, No.9, pp.3601-3608. | |||

