Область техники, к которой относится изобретение
Настоящее изобретение относится к константным областям антитела, аминокислотную последовательность которых модифицируют на основании константной области природного антитела, антителам, содержащим такие константные области, фармацевтическим композициям, содержащим такие антитела, и способам их получения.
Предшествующий уровень техники
Антитела привлекают внимание в качестве фармацевтических средств вследствие того, что они являются высокостабильными в крови и обладают немногими побочными эффектами (непатентные документы 1 и 2). Практически все фармацевтические средства на основе антител, представленные в настоящее время на рынке, представляют собой антитела подкласса IgG1 человека. К настоящему времени проведено много исследований в отношении антителозависимой клеточной цитотоксичности (далее в настоящем описании обозначаемой как ADCC) и обусловленной комплементом цитотоксичности (далее в настоящем описании обозначаемой как CDC), которые представляют собой эффекторные функции антител класса IgG, и для класса IgG человека опубликовано, что антитела подкласса IgG1 обладают наибольшей активностью ADCC и активностью CDC (непатентный документ 3). Кроме того, антителозависимый клеточный фагоцитоз (ADCP), который представляет собой фагоцитоз клеток-мишеней, опосредованный
антителами класса IgG, также показан в качестве одной из эффекторных функций антитела (непатентные документы 4 и 5).
Для того чтобы антитело IgG проявляло ADCC, CDC или ADCP, необходимо, что бы Fc-область антитела связывалась с рецептором антитела, который находится на поверхности эффекторных клеток, таких как клетки-киллеры, естественные клетки-киллеры и активированные макрофаги (далее в настоящем описании обозначаемые как FcγR), и различных компонентах комплемента. У людей изоформы FcγRIa, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb были опубликованы в качестве семейства белков FcγR, а также были опубликованы соответствующие аллотипы (непатентный документ 6).
Повышение цитотоксических эффекторных функций, таких как ADCC, ADCP и CDC, привлекает внимание в качестве перспективных способов повышения противоопухолевых эффектов антител. С использованием моделей на мышах описали важную роль FcγR-опосредованных эффекторных функций антител для их противоопухолевых эффектов (непатентные документы 7 и 8). Кроме того, наблюдали корреляцию между клиническими эффектами у людей и высокоаффинным полиморфным аллотипом (V158) и низкоаффинным полиморфным аллотипом (F158) FcγRIIIa (непатентный документ 9). Эти опубликованные данные продемонстрировали, что антитела, содержащие Fc-область, которую оптимизировали для специфического связывания с FcγR, опосредуют более выраженную эффекторную функцию, вследствие этого демонстрируют более выраженные противоопухолевые эффекты.
Баланс активностей связывания антитела в отношении
активирующего рецептора, состоящего из FcγRIa, FcγRIIa, FcγRIIIa и FcγRIIIb, и ингибирующего рецептора, состоящего из FcγRIIb, является важным элементом при оптимизации эффекторной функции антитела. Использование Fc-области, которая повышает активность связывания с активирующими рецепторами и снижает активность связывания с ингибирующими рецепторами, может обеспечить возможность придания антителам оптимальных эффекторных функций (непатентный документ 10). Наоборот, использование Fc-области, которая обладает пролонгированной или сниженной активностью связывания с активирующими рецепторами и повышенной активностью связывания с ингибирующими рецепторами, может обеспечивать возможность придания антителам иммуносупрессивного эффекта (непатентный документ 11). В отношении связывания Fc-области и FcγR продемонстрировано, что несколько аминокислотных остатков в шарнирной области антитела и домене СН2 и цепь сахаров, добавляемые к Asn в положении 297 (нумерация EU), который связан с доменом СН2, являются важными для связывания Fc-области и FcγR (непатентные документы 12, 13 и 14). Это исследование проводили в отношении вариантов Fc-области, которые обладают различными свойствами связывания с FcγR в основном на этом участке связывания, и получали варианты Fc-области, которые обладают более высокими активностями связывания с активацией FcγR (патентные документы 1 и 2). Например, Lazar et al. успешно увеличивали связывание FcγRIIIa (V158) человека приблизительно в 370 раз заменой Ser в положении 239, Ala в положении 330 и Ile в положении 332 (нумерация EU) IgG1 человека на Asp, Leu и Glu,
соответственно (непатентный документ 15 и патентный документ 2). Отношение связывания с FcγRIIIa к связыванию с FcγIIb (отношение А/I) этого варианта является повышенным приблизительно в 9 раз по сравнению с таким отношением для дикого типа. Кроме того, Lazar et al. успешно увеличивали связывание с FcγIIb приблизительно в 430 раз (непатентный документ 16). Shinkawa et al. успешно увеличивали связывание FcγRIIIa приблизительно в 100 раз удалением фукозы из цепи сахаров, добавляемой к Asn в положении 297 (нумерация EU) (непатентный документ 17).
Однако опубликованные в настоящее время функциональные модификации Fc-областей имеют ограничения, и существует необходимость в лучших функциональных модификациях Fc-областей.
Документы известного уровня техники
[Патентные документы]
[Патентный документ 1] WO 2000/042072
[Патентный документ 2] WO 2006/019447
[Патентный документ 3] WO 2009/041062
[Патентный документ 4] WO 2006/106905
[Непатентные документы]
[Непатентный документ 1] Nature Biotechnology, 23, 1073-1078 (2005)
[Непатентный документ 2] Eur. J. Pharm. Biopharm, 59(3), 389-96 (2005)
[Непатентный документ 3] Chemical Immunology, 65, 88 (1997)
[Непатентный документ 4] Cancer Res., 68, 8049-8057 (2008)
[Непатентный документ 5] Blood, 113, 3735-3743 (2009)
[Непатентный документ 6] Immunol. Lett., 82, 57-65 (2002)
[Непатентный документ 7] Pro. Nat. Acad. Sci., 95: 652-656 (1998)
[Непатентный документ 8] Nature Medicine, 6: 443-446 (2000)
[Непатентный документ 9] Blood, 99: 754-758 (2002)
[Непатентный документ 10] Science, 310, 1510-1512 (2005)
[Непатентный документ 11] Science, 291, 484-486 (2001)
[Непатентный документ 12] Chemical Immunology, 65, 88 (1997)
[Непатентный документ 13] Eur. J. Immunol., 23, 1098 (1993)
[Непатентный документ 14] Immunology, 86, 319 (1995)
[Непатентный документ 15] Pro. Nat. Acad. Sci., 103, 4005-4010 (2006)
[Непатентный документ 16] Mol. Immun., 45, 3926-3933 (2008)
[Непатентный документ 17] J. Biol. Chem., 278, 3466-3473 (2003)
Сущность изобретения
Задачи, подлежащие решению посредством изобретения Настоящее изобретение выполнено с учетом указанных выше обстоятельств. Задачей настоящего изобретения является предоставление полипептидов с улучшенной функцией Fc-области по сравнению с общепринятыми гомодимеризованными полипептидами, содержащими Fc-область; фармацевтических композиций, содержащих полипептиды; терапевтических средств или профилактических средств для иммуновоспалительных заболеваний, содержащих фармацевтические композиции; терапевтических средств или профилактических средств для различных типов злокачественных опухолей и способов их получения. Кроме того, целью настоящего
изобретения является предоставление способов улучшения функции Fc-области по сравнению с общепринятыми гомодимеризованными полипептидами, содержащими Fc-область.
Средства решения задач
Авторы настоящего изобретения провели исследование, посвященное решению указанных выше проблем. В результате авторы настоящего изобретения получили гетеродимеризованные полипептиды, содержащие Fc-область, состоящую из двух полипептидов с различными аминокислотными последовательностями (первого полипептида и второго полипептида) и, таким образом, успешно получили гетеродимеризованный полипептид, содержащий Fc-область с улучшенной функцией Fc-области по сравнению с общепринятыми гомодимерами, в которых Fc-область состоит только из первого полипептида или только из второго полипептида.
Более конкретно, настоящее изобретение относится к [1]-[78], указанным ниже:
[1] полипептиду, содержащему Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, и где полипептид отличается тем, что функция Fc-области является измененной по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, и о сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид;
[2] полипептиду [1], где в Fc-область вводят по меньшей мере одну или более мутаций аминокислот;
[3] полипептиду [1] или [2], где мутация аминокислоты включает по меньшей мере одну мутацию аминокислоты, которая улучшает функцию Fc-области, когда мутацию вводят только в одну из Fc-областей по сравнению с тем, когда мутацию вводят Fc-области первого полипептида и второго полипептида, и когда мутацию не вводят;
[4] полипептиду [2] или [3], где по меньшей мере одну или более мутаций аминокислот вводят в домен СН2 Fc-области;
[5] полипептиду [4], где по меньшей мере одну или более мутаций аминокислот вводят в аминокислотный участок, выбранный из группы, состоящей из:
Ala в положении 231 (нумерация EU);
Pro в положении 232 (нумерация EU);
Glu в положении 233 (нумерация EU);
Leu в положении 234 (нумерация EU);
Leu в положении 235 (нумерация EU);
Gly в положении 236 (нумерация EU);
Gly в положении 237 (нумерация EU);
Pro в положении 238 (нумерация EU);
Ser в положении 239 (нумерация EU);
Val в положении 240 (нумерация EU);
Asp в положении 265 (нумерация EU);
Val в положении 266 (нумерация EU);
Ser в положении 267 (нумерация EU);
His в положении 268 (нумерация EU);
Glu в положении 269 (нумерация EU);
Asp в положении 270 (нумерация EU);
Pro в положении 271 (нумерация EU);
Gln в положении 295 (нумерация EU);
Tyr в положении 296 (нумерация EU);
Ser в положении 298 (нумерация EU);
Tyr в положении 300 (нумерация EU);
Ser в положении 324 (нумерация EU);
Asn в положении 325 (нумерация EU);
Lys в положении 326 (нумерация EU);
Ala в положении 327 (нумерация EU);
Leu в положении 328 (нумерация EU);
Pro в положении 329 (нумерация EU);
Ala в положении 330 (нумерация EU);
Pro в положении 331 (нумерация EU);
Ile в положении 332 (нумерация EU);
Glu в положении 333 (нумерация EU);
Lys в положении 334 (нумерация EU);
Thr в положении 335 (нумерация EU);
Ile в положении 336 (нумерация EU) и
Ser в положении 337 (нумерация EU) в домене СН2 Fc-области;
[6] полипептиду по любому из [1]-[5], где изменение функции Fc-области представляет собой по меньшей мере одно или более изменений, выбранных из группы, состоящей из повышения активности связывания и улучшения селективности связывания с рецептором Fcγ;
[7] полипептиду [6], где изменение функции Fc-области представляет собой повышение активности связывания с рецептором Fcγ;
[8] полипептиду [7], где рецептор Fcγ представляет собой по меньшей мере один или более рецепторов, выбранных из группы, состоящей из FcγRIa, FcγRIIa R, FcγRIIa H, FcγRIIb и FcγRIIIa;
[9] полипептиду [8], где рецептор Fcγ представляет собой FcγRIa;
[10] полипептиду [9], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблиц 2-1 и 2-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[11] полипептиду [9], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблиц 2-1, 2-2 и 2-3 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[12] полипептиду [8], где рецептор Fcγ представляет собой FcγRIIa R;
[13] полипептиду [12], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблиц 3-1 и 3-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[14] полипептиду [12], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблиц 3-1 и 3-2 настоящего
описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[15] полипептиду [8], где рецептор Fcγ представляет собой FcγRIIa Н;
[16] полипептиду [15], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 4 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[17] полипептиду [15], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 4 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[18] полипептиду [8], где рецептор Fcγ представляет собой FcγRIIb;
[19] полипептиду [18], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 5 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[20] полипептиду [18], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 5 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[21] полипептиду [8], где рецептор Fcγ представляет собой FcγRIIIa;
[22] полипептиду [21], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 6 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[23] полипептиду [21], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 6 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[24] полипептиду [6], где изменение функции Fc-области представляет собой улучшение селективности активности связывания с рецептором Fcγ;
[25] полипептиду [24], где улучшение селективности активности связывания с рецептором Fcγ относится к селективности в отношении активирующего рецептора Fcγ и ингибирующего рецептора Fcγ;
[26] полипептиду [25], где в числе рецептором Fcγ активирующий рецептор Fcγ представляет собой по меньшей мере один или более рецепторов, выбранных из группы, состоящей из FcγRIa, FcγRIIa R, FcγRIIa H и FcγRIIIa, и ингибирующий рецептор Fcγ представляет собой FcγRIIb;
[27] полипептиду [26], где активирующий рецептор Fcγ
представляет собой FcγRIa, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIa является селективно повышенной по сравнению с активностью связывания с FcγRIIb;
[28] полипептиду [27], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблиц 19-1, 19-2, 19-3 и 19-4 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[29] полипептиду [27], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 19-1, 19-2, 19-3, 19-4 и 19-5 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[30] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIa, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIa является селективно сниженной по сравнению с активностью связывания с FcγRIIb;
[31] полипептиду [30], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблиц 23-1 и 23-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[32] полипептиду [30], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 23-1 и 23-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[33] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIa R, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIa R является селективно повышенной по сравнению с активностью связывания с FcγRIIb;
[34] полипептиду [33], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 20-1 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[35] полипептиду [33], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 20-1, 20-2 и 20-3 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[36] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIa R, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIa R является селективно сниженной по сравнению с активностью связывания с FcγRIIb;
[37] полипептиду [36], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблицы 24-1 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[38] полипептиду [36], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 24-1 и 24-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[39] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIa Н, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIa Н является селективно повышенной по сравнению с активностью связывания с FcγRIIb;
[40] полипептиду [39], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 21-1 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[41] полипептиду [39], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 21-1, 21-2 и 21-3 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[42] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIa Н, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIa Н является селективно сниженной по сравнению с активностью связывания с FcγRIIb;
[43] полипептиду [42], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблиц 25-1 и 25-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[44] полипептиду [42], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 25-1, 25-2 и 25-3 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[45] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIIa, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIIa является селективно повышенной по сравнению с активностью связывания с FcγRIIb;
[46] полипептиду [45], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 22-1 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[47] полипептиду [45], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 22-1, 22-2 и 22-3 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[48] полипептиду [26], где активирующий рецептор Fcγ представляет собой FcγRIIIa, и ингибирующий рецептор Fcγ представляет собой FcγRIIb, который отличается тем, что активность связывания с FcγRIIIa является селективно сниженной по сравнению с активностью связывания с FcγRIIb;
[49] полипептиду [48], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблиц 26-1 и 26-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[50] полипептиду [48], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 26-1, 26-2, 26-3 и 26-4 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[51] полипептиду [1], [21] или [45], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из:
замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y или Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на W;
замены аминокислоты А в положении 330 (нумерация EU) на М или K, и
замены аминокислоты K в положении 334 (нумерация EU) на Е или L, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[52] полипептиду [1], [21] или [45], где мутацию вводят по меньшей мере в одну аминокислоту, выбранную из Leu в положении 234, Leu в положении 235, Gly в положении 236, Ser в положении 239, His в положении 268, Asp в положении 270, Ser в положении 298, Lys в положении 326, Ala в положении 327, Leu в положении 328, Ala в положении 330 и Lys в положении 334 (нумерация EU), в аминокислотной последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[53] полипептиду [1], [21] или [45], где мутацию вводят по меньшей мере в одну аминокислоту, выбранную из Leu в положении 234, Leu в положении 235, Gly в положении 236, Ser в положении 239, His в положении 268, Asp в положении 270, Ser в положении
298, Ala в положении 327, Leu в положении 328 и Lys в положении 334 (нумерация EU), в аминокислотной последовательности любого полипептида из первого полипептида и второго полипептида, составляющих Fc-область; и мутацию вводят по меньшей мере в одну аминокислоту, выбранную из Asp в положении 270, Lys в положении 326, Ala в положении 330 и Lys в положении 334 (нумерация EU) в аминокислотную последовательность другого полипептида;
[54] полипептиду [1], [21] или [45], где по меньшей мере одну мутацию аминокислоты, выбранную из группы, состоящей из:
замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y или Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты А в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на W и
замены аминокислоты K в положении 334 (нумерация EU) на L,
вводят в аминокислотную последовательность любого полипептида из первого полипептида и второго полипептида, составляющих Fc-область, и по меньшей мере одну мутацию аминокислот, выбранную из группы, состоящей из:
замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М
или K и
замены аминокислоты K в положении 334 (нумерация EU) на Е,
вводят в аминокислотную последовательность другого полипептида;
[55] полипептиду [1], [21] или [45], где любую одну группу мутаций от (i) до (vi) вводят в аминокислотную последовательность любого полипептида из первого полипептида и второго полипептида, составляющих Fc-область, и любую одну группу мутаций от (vii) до (ix) вводят в аминокислотную последовательность другого полипептида:
(i) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(ii) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iii) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iv) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(v) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(vi) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты A в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на W и
замены аминокислоты K в положении 334 (нумерация EU) на L;
(vii) замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты K в положении 334 (нумерация EU) на Е;
(viii) замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты K в положении 334 (нумерация EU) на Е;
(ix) замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на K и
замены аминокислоты K в положении 334 (нумерация EU) на Е;
[56] полипептиду, содержащему Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, и где полипептид отличается тем, что функция Fc-области является измененной по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, или по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид;
[57] полипептиду [56], где изменение функции Fc-области
представляет собой по меньшей мере одно или более изменений, выбранных из группы, состоящей из повышения активности связывания, снижения связывания и улучшения селективности связывания полипептида с рецептором Fcγ;
[58] полипептиду [57], где изменение функции Fc-области дополнительно представляет собой изменение, которое улучшает физико-химическую стабильность;
[59] полипептиду [58], где изменение, которое улучшает физико-химическую стабильность, означает, что Tm полипептида, отличающегося тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, является выше, чем Tm полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, или Tm полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид;
[60] полипептиду любого из [57]-[59], где изменение функции Fc-области представляет собой повышение активности связывания с рецептором Fcγ, и рецептор Fcγ представляет собой по меньшей мере один или более рецепторов, выбранных из группы, состоящей из FcγRIa, FcγRIIa R, FcγRIIa H, FcγRIIb и FcγRIIIa;
[61] полипептиду [60], где рецептор Fcγ представляет собой FcγRIa;
[62] полипептиду [61], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 31-1, 31-2 и 31-3 настоящего описания, вводят в аминокислотные последовательности первого
полипептида и/или второго полипептида, составляющих Fc-область;
[63] полипептиду [60], где рецептор Fcγ представляет собой FcγRIIa R;
[64] полипептиду [63], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 32-1 и 32-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[65] полипептиду [60], где рецептор Fcγ представляет собой FcγRIIa Н;
[66] полипептиду [65], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 33-1 и 33-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[67] полипептиду [60], где рецептор Fcγ представляет собой FcγRIIb;
[68] полипептиду [67], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 34-1 и 34-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[69] полипептиду [60], где рецептор Fcγ представляет собой FcγRIIIa;
[70] полипептиду [69], где по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций
аминокислот, описанных в таблицах 35-1 и 35-2 настоящего описания, вводят в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область;
[71] полипептиду любого из [1]-[70], где дополнительно вводят изменение аминокислоты для обеспечения различий изоэлектрических точек первого полипептида и второго полипептида;
[72] полипептиду [71], где изменение аминокислоты для обеспечения различия изоэлектрических точек, отличается тем, что введение по меньшей мере одной мутации аминокислоты на участке аминокислот, выбранных из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Val в положении 263, Glu в положении 272, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 364, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Pro в положении 387, Asn в положении 390, Val в положении 397 и Val в положении 422 (нумерация EU), в аминокислотной последовательности первого полипептида и/или второго полипептида;
[73] полипептиду [71], где изменение аминокислоты для обеспечения различия изоэлектрических точек, отличается
введением мутации по меньшей мере в одной аминокислоте, выбранной из группы, состоящей из Gln в положении 196, Ile в положении 199, Val в положении 263, Glu в положении 272, Gly в положении 316, Leu в положении 358, Ser в положении 364, Ser в положении 383, Pro в положении 387 и Val в положении 397 (нумерация EU), в аминокислотной последовательности любого полипептида из первого полипептида и второго полипептида, и введением мутации по меньшей мере в одной аминокислоте, выбранной из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390 и Val в положении 422 (нумерация EU), в аминокислотной последовательности другого полипептида;
[74] полипептиду любого из [1]-[73], где полипептид представляет собой антигенсвязывающую молекулу;
[75] полипептиду [74], где антигенсвязывающая молекула представляет собой антитело, биспецифическое антитело или слитую с Fc молекулу, такую как слитый с пептидом Fc белок или слитый с каркасной областью Fc белок;
[76] фармацевтической композиции, содержащей полипептид [74]
или [75] и приемлемому носителю с медицинской точки зрения;
[77] способу изменения функции полипептида, содержащего Fc-область, который включает стадии гетеродимеризации Fc-области введением мутации аминокислот в первом полипептиде и/или втором полипептиде, из которого состоит Fc-область, и введением мутации аминокислот для изменения функции Fc-области по сравнению со случаем, когда Fc-область образует гомодимер, и
[78] способу получения полипептида, содержащего Fc-область, который включает стадии гетеродимеризации Fc-области введением мутации аминокислоты в первом полипептиде и/или втором полипептиде, из которого состоит Fc-область, и введением мутации аминокислоты для изменения функции Fc-области по сравнению со случаем, когда Fc-область образует гомодимер.
Эффекты изобретения
Настоящее изобретение относится к полипептидам, которые являются подходящими в качестве фармацевтических средств, где их активности связывания и физические свойства (например, стабильность и однородность) улучшили изменением аминокислотной последовательности константной области антитела.
Краткое описание чертежей
На фиг. 1 представлена структура комплекса Fc-области и FcRn. FcRn связывается с СН2 и СН3 каждой Н-цепи антитела и связан со всем антителом симметричным образом.
На фиг. 2 представлена структура комплекса IgA и FcαR, который представляет собой рецептор IgA. FcαR связывается с Сα2 и Сα3 каждой Н-цепи IgA и связан со всем антителом симметричным
образом.
На фиг. 3 представлена структура комплекса Fc-области и FcγRIII. Н-цепь, СН2 и СН3, которые находятся с левой стороны фигуры, обозначают как НА-цепь, СНА2 и СНА3 и те, которые находятся с правой стороны, обозначают как HB-цепь, CHB2 и CHB3, соответственно.
На фиг. 4 представлены подробные взаимодействия А327 в каждой Н-цепи и FcγRIII. На (А) представлено взаимодействие А327 в CHA2 и FcγRIII. На (В) представлено взаимодействие А327 в СНВ2НВ и FcγRIII. В FcγRIII цвета черный, серый и белый означают щелочной участок, нейтральный участок и кислотный участок, соответственно.
На фиг. 5 представлено сравнение активности связывания антител с FcγR, в которые вводили D356K, H435R и/или K439E. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5) и GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5).
На фиг. 6 представлено сравнение активности связывания антител с FcγR, в которые вводили G237A. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ
ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A26/GpH7-B3/GpL16-k0 (SEQ ID NO: 6, 4 и 5) и GpH7-A26/GpL16-k0 (SEQ ID NO: 6 и 5).
На фиг. 7 представлено сравнение активности связывания с FcγR гомодимеризованных антител и гетеродимеризованных антител, в которые вводили G237L. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A29/GpH7-B3/GpL16-k0 (SEQ ID NO: 7, 4 и 5) и GpH7-A29/GpL16-k0 (SEQ ID NO: 7 и 5).
На фиг. 8 представлено сравнение активности связывания с FcγR гомодимеризованных антител и гетеродимеризованных антител, в которые вводили L328E. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A42/GpH7-B3/GpL16-k0 (SEQ ID NO: 8, 4 и 5) и GpH7-A42/GpL16-k0 (SEQ ID NO: 8 и 5).
На фиг. 9 представлено сравнение активности связывания с FcγR гомодимеризованных антител и гетеродимеризованных антител, в которые вводили L328D. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой
GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A43/GpH7-B3/GpL16-k0 (SEQ ID NO: 9, 4 и 5) и GpH7-A43/GpL16-k0 (SEQ ID NO: 9 и 5).
На фиг. 10 представлено сравнение активности связывания гомодимеризованных антител и гетеродимеризованных антител с FcγR, в которые вводили L234E. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A5/GpH7-B16/GpL16-k0 (SEQ ID NO: 3, 10 и 5) и GpH7-B16/GpL16-k0 (SEQ ID NO: 10 и 5).
На фиг. 11 представлено сравнение активности связывания с FcγR гомодимеризованных антител и гетеродимеризованных антител, в которые вводили L234D. Активность связывания GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5), GpH7-A5/GpH7-B17/GpL16-k0 (SEQ ID NO: 3, 11 и 5) и GpH7-B17/GpL16-k0 (SEQ ID NO: 11 и 5).
На фиг. 12 представлены взаимодействия P329 в Fc-области с FcγRIII. Н-цепь, СН2 и СН3, которые находятся с левой стороны фигуры, обозначают как НА-цепь, СНА2 и СНА3 и те, которые находятся с правой стороны обозначают как HB-цепь, СНВ2 и СНВ3,
соответственно. На этой фигуре продемонстрировано, что Pro в положении 329 (нумерация EU) в Fc-области взаимодействует с FcγRIII в основном через СНА2, который представляет собой домен СН2. Н-цепь, СН2 и СН3, которые находятся с левой стороны фигуры, обозначают как НА-цепь, CHA2 и СНА3 и те, которые находятся с правой стороны обозначают как HB-цепь, СНВ2 и СНВ3, соответственно.
На фиг. 13 представлено сравнение эффектов в отношении активности связывания с FcγR гомодимеризованных антител и гетеродимеризованных антител, в которые вводили P329R, P329K, P329D или Р329Е. Активность связывания GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-A5/GpH7-B12/GpL16-k0 (SEQ ID NO: 3, 12 и 5), GpH7-A5/GpH7-B13/GpL16-k0 (SEQ ID NO: 3, 13 и 5), GpH7-A5/GpH7-B14/GpL16-k0 (SEQ ID NO: 3, 14 и 5), GpH7-A5/GpH7-B15/GpL16-k0 (SEQ ID NO: 3, 15 и 5), GpH7-Bl2/GpL16-k0 (SEQ ID NO: 12 и 5), GpH7-B13/GpL16-k0 (SEQ ID NO: 13 и 5), GpH7-B14/GpL16-k0 (SEQ ID NO: 14 и 5) и GpH7-B15/GpL16-k0 (SEQ ID NO: 15 и 5).
На фиг. 14 представлено сравнение активности связывания с каждым FcγR гетеродимеризованного антитела с G237A, введенным в одну из Н-цепей, когда P329R вводят в одну и ту же цепь Н-цепь или в другую Н-цепь. Активность связывания GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности
представляли собой GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-A5/GpH7-B12/GpL16-k0 (SEQ ID NO: 3, 12 и 5), GpH7-A48/GpH7-B3/GpL16-k0 (SEQ ID NO: 16, 4 и 5), GpH7-A26/GpH7-B3/GpL16-k0 (SEQ ID NO: 6, 4 и 5), GpH7-A26/GpH7-B12/GpL16-k0 (SEQ ID NO: 6, 12 и 5), и GpH7-A45/GpH7-B3/GpL16-k0 (SEQ ID NO: 17, 4 и 5).
На фиг. 15 представлено сравнение активности связывания с каждым FcγR гетеродимеризованного антитела с L234D, вводимым в одну из Н-цепей, когда P329R вводят в одну и ту же Н-цепь или в другую Н-цепь. Активность связывания GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5) с каждым FcγR определяли как 100. Используемые для оценки образцы и их последовательности представляли собой GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-A5/GpH7-B12/GpL16-k0 (SEQ ID NO: 3, 12 и 5), GpH7-A48/GpH7-B3/GpL16-k0 (SEQ ID NO: 16, 4 и 5), GpH7-A5/GpH7-B17/GpL16-k0 (SEQ ID NO: 3, 11 и 5), GpH7-A48/GpH7-B17/GpL16-k0 (SEQ ID NO: 16, 11 и 5) и GpH7-A5/GpH7-B41/GpL16-k0 (SEQ ID NO: 3, 18 и 5).
На фиг. 16 представлено сравнение активности связывания с FcγRIa гомодимеризованных антител и гетеродимеризованных антител, в которые вводили одинаковые изменения. По горизонтальной оси представлены значения Ho/Con и по вертикальной ось представлены значения Не/Со. He/Con представляет собой значение, получаемое делением активности связывания с FcγRIa гетеродимеризованного антитела СрН7-А5/вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для одной из Н-цепей, на активность
связывания с FcγRIa гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100. Ho/Con представляет собой значение, получаемое делением активности связывания с FcγRIa варианта гомодимеризованного антитела GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для обеих Н-цепей, на активность связывания с FcγRIa гомодимеризованного антитела GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100.
На фиг. 17 представлено сравнение активности связывания с FcγRIIa R гомодимеризованных антител и гетеродимеризованных антител, в которых вводили указанные изменения. По горизонтальной оси представлены значения Ho/Con и по вертикальной оси представлены значения Не/Со. He/Con представляет собой значение, получаемое делением активности связывания с FcγRIIa R гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для одной из Н-цепей, на активность связывания с FcγRIIa R гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в котором используют немутантный GpH7-B3. Ho/Con представляет собой значение, получаемое делением активности связывания с FcγRIIa R варианта гомодимеризованного антитела GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 в обеих Н-цепях, на активность связывания с FcγRIIa R гомодимеризованного антитела GpH7-B3/GpL16-k0 (SEQ ID
NO: 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100.
На фиг. 18 представлено сравнение активности связывания с FcγRIIa Н гомодимеризованных антител и гетеродимеризованных антител, в которые вводили одинаковые изменения. По горизонтальной оси представлены значения Ho/Con и по вертикальной оси представлены значения Не/Со. He/Con представляет собой значение, получаемое делением активность связывания с FcγRIIa Н гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для одной из Н-цепей, на активность связывания с FcγRIIa Н гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в котором используют немутантный GpH7-B3. Ho/Con представляет собой значение, получаемое делением активности связывания с FcγRIIa Н гомодимеризованного антитела варианта GpH7-B3/GpL16-k0, в котором используют мутантый вариант GpH7-B3 для обеих Н-цепей, на активность связывания с FcγRIIa Н гомодимеризованного антитела GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100.
На фиг. 19 представлено сравнение активности связывания с FcγRIIb гомодимеризованных антител и гетеродимеризованных антител, в которые вводят одинаковые изменения. По горизонтальной оси представлены значения Ho/Con и по вертикальной оси представлены значения Не/Со. He/Con представляет собой значение, получаемое делением активность
связывания с FcγRIIb гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для одной из Н-цепей, на активность связывания с FcγRIIb гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в котором используют немутантный GpH7-B3. Ho/Con представляет собой значение, получаемое делением активности связывания с FcγRIIb гомодимеризованного антитела вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для обеих Н-цепей, на -активность связывания с FcγRIIb гомодимеризованного антитела GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100.
На фиг. 20 представлено сравнение активности связывания с FcγRIIIa гомодимеризованных антител и гетеродимеризованных антител, в которые вводят одинаковые изменения. По горизонтальной оси представлены значения Ho/Con и по вертикальной оси представлены значения Не/Со. He/Con представляет собой значение, получаемое делением активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, в котором используют мутантный вариант GpH7-B3 для одной из один Н-цепей, на активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в котором используют немутантный GpH7-B3. Ho/Con представляет собой значение, получаемое делением активности связывания с FcγRIIIa гомодимеризованного антитела вариант GpH7-B3/GpL16-k0, в котором
используют немутантный вариант GpH7-B3 для обеих Н-цепей, на активность связывания с FcγRIIIa гомодимеризованного антитела GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5), в котором используют немутантный GpH7-B3, и умножением результата на 100.
На фиг. 21 представлена концептуальная диаграмма, в которой сравнивают связывание с FcγR каждого из гомодимеризованных антител и гетеродимеризованных антител, в которых используют Н-цепи, в которые вводили изменения. В случае, когда нанесенные на график точки находятся в области i, это означает, что изменение, вводимое в Fc-область, оказывает эффект, приводящий к He/Con>100, Ho/Con<100, Не/Con>Но/Con. В случае, когда нанесенные на график точки находятся в области ii, это означает, что изменение, вводимое в Fc-область, оказывает эффект, приводящий к He/Con>100, Ho/Con>100, Не/Con>Но/Con. В случае, когда нанесенные на график точки находятся в области iii, это означает, что изменение, вводимое в Fc-область, вызывает эффект, приводящий к He/Con>100, Ho/Con>100, Не/Con<Но/Con.
На фиг. 22 представлено число комбинаций, получаемых, когда вводят три типа изменений в любую из Н-цепей гетеродимеризованного антитела. Каждая из закрашенной окружности ( ), закрашенного треугольника (
), закрашенного треугольника ( ) и закрашенного квадрата (
) и закрашенного квадрата ( ) относится к различным изменениям.
) относится к различным изменениям.
На фиг. 23 представлено взаимодействие FcγRIII и каждого из остатков S239, А330 и I332 в Fc-области антитела. Цитировано из Proc. Natl. Acad. Sci. USA, 103, 4005-4010, 2006.
На фиг. 24 представлено сравнение активностей связывания с
активирующим FcγR и ингибирующим FcγR. Оно представляет собой концептуальную диаграмму, в которой сравнивают активности связывания каждого варианта с активирующим FcγR и ингибирующим FcγR. По вертикальной оси представлена активность каждого варианта по отношению к активирующему FcγR (активирующему рецептору) и по горизонтальной оси представлена активность связывания каждого варианта с ингибирующим FcγR (ингибирующим рецептором). Каждые активности связывания природного антитела с активирующим FcγR и ингибирующим FcγR обозначали как 100. Когда вариант variant повышал активность связывания с активирующим FcγR выше природного антитела и снижал активность связывания с ингибирующим FcγR, антитело наносят на график в области а (закрашенная область). Когда вариант повышал активность связывания с ингибирующим FcγR выше природного антитела и снижал активность связывания с активирующим FcγR, антитело наносят на график в области с (заштрихованная область).
На фиг. 25 представлено сравнение активностей связывания с активирующим FcγR и ингибирующим FcγR. Оно представляет собой концептуальную диаграмму, в которой сравнивают активности связывания каждого варианта с активирующим FcγR и ингибирующим FcγR. По вертикальной оси представлена активность связывания природного антитела с активирующим FcγR (активирующим рецептором) и по горизонтальной оси представлена активность связывания с ингибирующим FcγR (ингибирующий рецептор). Каждые активности связывания природного антитела с активирующим FcγR и ингибирующим
FcγR обозначали как 100. Когда значение, получаемое делением активности связывания варианта с активирующим FcγR на активность связывания с ингибирующим FcγR, равно 1,2 или более, антитело наносят на график в области b (закрашенная область). Когда значение, получаемое делением активности связывания варианта с активирующим FcγR на активность связывания с ингибирующим FcγR, равно 0,8 или менее, антитело наносят на график в области d (заштрихованная область).
На фиг. 26 представлено сравнение активности связывания гетеродимеризованного антитела с FcγRIa и FcγRIIb. Оно представляет собой диаграмму, в которой сравнивают активности связывания измененного гетеродимеризованного антитела с FcγRIa, который представляет собой активирующий FcγR, и с FcγRIIb, который представляет собой ингибирующий FcγR. По горизонтальной оси представлены значения Ho/Con для ингибирующего FcγR и по вертикальной оси представлены значения He/Con для активирующего FcγR. He/Con представляет собой значение, получаемое делением FcγR-активность связывания гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, которое содержит мутантный Fc, на активность связывания с FcγR гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), в которое не содержит введения изменений, и умножением результата на 100.
На фиг. 27 представлено сравнение активностей связывания гетеродимеризованного антитела с FcγRIIa R и FcγRIIb. Оно представляет собой диаграмму, в которой сравнивают активности
связывания измененного гетеродимеризованного антитела с FcγRIIa R, который представляет собой активирующий FcγR, и с FcγRIIb, который представляет собой ингибирующий FcγR. По горизонтальной оси представлены значения He/Con для ингибирующего FcγR и по вертикальной оси представлены значения He/Con для активирующего FcγR. He/Con представляет собой значение, получаемое делением активность связывания с FcγR гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, которое содержит мутантный Fc, на активность связывания с FcγR гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), которое не содержит введения изменений, и умножением результата на 100.
На фиг. 28 представлено сравнение активностей связывания гетеродимеризованного антитела с FcγRIIa Н и FcγRIIb. Оно представляет собой диаграмму, в которой сравнивают активности связывания измененного гетеродимеризованного антитела с FcγRIIa Н, который представляет собой активирующий FcγR, и с FcγRIIb, который представляет собой ингибирующий FcγR. По горизонтальной оси представлены значения He/Con для ингибирующего FcγR и по вертикальной оси представлены значения He/Con для активирующего FcγR. He/Con представляет собой значение, получаемое делением активности связывания с FcγR гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, которое содержит мутантный Fc, на активность связывания с FcγR гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), которое не содержит введения изменений, и умножением результата на 100.
На фиг. 29 представлено сравнение активностей связывания гетеродимеризованного антитела с FcγRIIIa и FcγRIIb. Оно представляет собой диаграмму, в которой сравнивают активности связывания измененного гетеродимеризованного антитела с FcγRIIIa, который представляет собой активирующий FcγR, и с FcγRIIb, который представляет собой ингибирующий FcγR. По горизонтальной оси представлены значения He/Con для ингибирующего FcγR и по вертикальной оси представлены значения He/Con для активирующего FcγR. He/Con представляет собой значение, получаемое делением активности связывания с FcγR гетеродимеризованного антитела GpH7-А5/вариант GpH7-B3/GpL16-k0, которое содержит мутантный Fc, на активность связывания с FcγR гетеродимеризованного антитела GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), которое не содержит введения изменений, и умножением результата на 100.
На фиг. 30 представлено сравнение эффектов комбинаций L234Y, G236W и S298A с S239D, A330L и I332E в отношении термостабильности антител. Оно представляет собой график, в котором сравнивают изменения содержания мономеров после того, как проводили ускоренное исследование стабильности при нагревании (40°С в течение двух недель или четырех недель) гомодимеризованных антител и гетеродимеризованных антител с L234Y, G236W и S298A, гомодимеризованных антител и гетеродимеризованных антител с S239D, A330L и I332E, и гетеродимеризованных антител, в которые вводят L234Y, G236W и S298A в одну из Н-цепей и вводят S239D, A330L и I332E в другую Н-цепь. Используемые для оценки образцы и их последовательности
представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-TA7/GpH7-B3/GpL16-k0 (SEQ ID NO: 31, 4 и 5), GpH7-A5/GpH7-B78/GpL16-k0 (SEQ ID NO: 3, 41 и 5), GpH7-TA7/GpH7-TA45/GpL16-k0 (SEQ ID NO: 31, 32 и 5), GpH7-A57/GpH7-B78/GpL16-k0 (SEQ ID NO: 40, 41 и 5) и GpH7-TA7/GpH7-B78/GpL16-k0 (SEQ ID NO: 31, 41 и 5).
На фиг. 31 представлен результат анализа активности ADCC гетеродимеризованных антител. Используемая для оценки линия клеток представляла собой SK-pca13a, и отношение Е/Т составляло 50. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5), GpH7-A5/GpH7-B78/GpL16-k0 (SEQ ID NO: 3, 41 и 5), GpH7-TA7/GpH7-B3/GpL16-k0 (SEQ ID NO: 31, 4 и 5), GpH7-A57/GpH7-B78/GpL16-k0 (SEQ ID NO: 40, 41 и 5), GpH7-TA7/GpH7-TA45/GpL16-k0 (SEQ ID NO: 31, 32 и 5) и GpH7-TA7/GpH7-B78/GpL16-k0 (SEQ ID NO: 31, 41 и 5). По вертикальной оси представлена цитотоксическая активность антитела и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 32 представлены аминокислотные остатки, из которых состоят Fc-области IgG1 (SEQ ID NO: 76), IgG2 (SEQ ID NO: 77), IgG3 (SEQ ID NO: 78) и IgG4 (SEQ ID NO: 79), и их отношение к нумерации EU по Kabat (в настоящем описании, также обозначаемой как индекс EU).
На фиг. 33 представлен результат анализа активностей ADCC гетеродимеризованных антител Fc, описанных в примере 12.
Используемая для оценки линия клеток представляла собой SK-рса13а, и отношение Е/Т составляло 50. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их последовательности представляли собой GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5), GpH7-Kn033/GpH7-HI033/GpL16-k0 (SEQ ID NO: 51, 56 и 5), GpH7-Kn032/GpH7-HI032/GpL16-k0 (SEQ ID NO: 53, 58 и 5), GpH7-Kn045/GpH7-HI048/GpL16-k0 (SEQ ID NO: 54, 59 и 5), GpH7-Kn056/GpH7-HI055/GpL16-k0 (SEQ ID NO: 55, 60 и 5) и GpH7-Kn037/GpH7-HI036/GpL16-k0 (SEQ ID NO: 52, 57 и 5). По вертикальной оси представлена цитотоксическая активность антитела и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 34 представлен результат анализа активности ADCC гетеродимеризованного антитела H240-Kn061/H240-Hl071/L73-k0. Используемая для оценки линия клеток представляла собой SKE18, и отношение Е/Т составляло 50. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их последовательности представляли собой H240-Kn033/H240-Hl033/L73-k0 (SEQ ID NO: 84, 85 и 106), H240-Kn032 /H240-Hl032/L73-k0 (SEQ ID NO: 86, 87 и 106), H240-Kn061 /H240-Hl071/L73-k0 (SEQ ID NO: 81, 82 и 106) и Н240-дефукозил_G1d (аминокислотная последовательность Н240-дефукозил_G1d является аналогичной аминокислотной последовательности H240-G1d (SEQ ID NO: 83), но фукозу удаляют)/L73-k0 (SEQ ID NO: 83 и 106). По вертикальной оси представлена цитотоксическая активность антитела и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 35 представлена активность связывания с FcγRI
точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы. Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRI на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 36 представлена активность связывания с FcγRIIa R точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы. Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRIIa R на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 37 представлена активность связывания с FcγRIIa Н точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы. Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRIIa Н на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 38 представлена активность связывания с FcγRIIb точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы.
Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRIIb на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 39 представлена активность связывания с FcγRIIIa F точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы. Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRIIIa F на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 40 представлена активность связывания с FcγRIIIa V точечного мутанта, в котором используют гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы. Относительная KD по вертикальной оси отражает значение, получаемое делением KD (моль/л) H240-Kn061/H240-Hl071/L73-k0 для FcγRIIIa V на KD каждого варианта. Числа по горизонтальной оси отражают ранги, где относительные KD располагают в возрастающем порядке.
На фиг. 41 представлен результат анализа активности ADCC гетеродимеризованного антитела H240-Kn072/H240-Hl076/L73-k0. Используемая для оценки линия клеток представляла собой MIAPaCa-2, и отношение Е/Т составляло 25. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их
последовательности представляли собой H240-Kn033/H240-Hl033/L73-k0 (SEQ ID NO: 84, 85 и 106), H240-Kn061/H240-Hl071/L73-k0 (SEQ ID NO: 81, 82 и 106), Н240-дефукозил_C1d/L73-k0 (SEQ ID NO: 83 и 106) и H240-Kn072/H240-Hl076/L73-k0 (SEQ ID NO: 90, 91 и 106). По вертикальной оси представлена цитотоксическая активность антитела, и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 42 представлен результат анализа активности ADCC улучшенного антитела, получаемого из гетеродимеризованного антитела H240-Kn072/H240-Hl076/L73-k0. Используемая для оценки линия клеток представляла собой DLD-1, и отношение Е/Т составляло 50. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их последовательности представляли собой H240-Kn033/H240-Hl033/L73-k0 (SEQ ID NO: 84, 85 и 106), Н240-дефукозил_G1d/L73-k0 (SEQ ID NO: 83 и 106), H240-Kn113/H240-Hl076/L73-k0 (SEQ ID NO: 92, 91 и 106), H240-Kn115/H240-Hl076/L73-k0 (SEQ ID NO: 93, 91 и 106) и H240-Kn125/H240-Hl076/L73-k0 (SEQ ID NO: 94, 91 и 106). По вертикальной оси представлена цитотоксическая активность антитела и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 43 представлен результат анализа активности ADCC гетеродимеризованного антитела H240-Kn067/H240-Hl068/L73-k0 и т.д. Используемая для оценки линия клеток представляла собой DLD-1, и отношение Е/Т составляло 50. В качестве эффекторной клетки использовали РВМС человека. Используемые для оценки образцы и их последовательности представляли собой Н240-
Kn033/H240-Hl033/L73-k0 (SEQ ID NO: 84, 85 и 106), H240-дефукозил_G1d/L73-k0 (SEQ ID NO: 83 и 106), H240-Kn067/H240-Hl068/L73-k0 (SEQ ID NO: 95, 96 и 106), H240-Kn120/H240-Hl068/L73-k0 (SEQ ID NO: 99, 96 и 106), и H240-Kn126/H240-Hl068/L73-k0 (SEQ ID NO: 100, 96 и 106). По вертикальной оси представлена цитотоксическая активность антитела и по горизонтальной оси представлена концентрация антитела (мкг/мл).
На фиг. 44 представлена кристаллическая структура Fc (WT) /FcγR2a R типа (PDB ID=3RY6, J. Imunol., 2011, 187, 3208-321). На фигуре проиллюстрированы боковые цепи Gln127, Leu132 и Phe160, которые представляют собой три остатка, которые различаются в FcγRIIa типе R и FcγRIIb в области вблизи поверхности взаимодействия FcγRIIa R типа и Fc в этой структуре. Соответствующие аминокислотные остатки в FcγRIIb показаны с использованием однобуквенных кодов в скобках.
На фиг. 45 представлен комплекс внеклеточного домена FcKn (120Hl068)/FcγRIIb, структуру которого определяли рентгеновским кристаллографическим анализом. Домен СН2 и домен СН3 те, которые находятся с левой стороны, обозначают как домен А и, те которые находятся справа, обозначают как домен В, соответственно.
На фиг. 46 представлена структура вокруг Lys127 (Gln в FcγRIIa R типа) внеклеточной области FcγRIIb в комплексе Fc (Kn120/Hl068)/внеклеточной области FcγRIIb, структуру которого определяли рентгеновским кристаллографическим анализом. Вследствие того, что не наблюдали электронной плотности боковой цепи для Tyr296 Fc (Kn120/Hl068), для боковой цепи не
конструировали модели, отличные от атома Сα.
На фиг. 47 представлена структура вокруг Ser132 (Leu в FcγRIIa R типа) внеклеточной области FcγRIIb в комплексе Fc (Kn120/Hl068)/внеклеточного домена FcγRIIb, структуру которого определяли рентгеновским кристаллографическим анализом. Вследствие того, что не наблюдали электронной плотности боковой цепи для D327 Fc (Kn120/Hl068), для боковой цепи не конструировали модели, отличные от атома Сα.
На фиг. 48 представлена структура вокруг Tyr160 (Phe в FcγRIIa R типа) внеклеточной области FcγRIIb в комплексе Fc (Kn120/Hl068)/внеклеточной области FcγRIIb, структуру которого определяли рентгеновским кристаллографическим анализом.
На фиг. 49 представлен комплекс Fc (ВР208)/внеклеточной области FcγRIIb, структуру которого определяли рентгеновским кристаллографическим анализом. Домен СН2 и домен СН3 те, которые находятся с левой стороны, обозначают как домен А и те, которые находятся справа, обозначают как домен В, соответственно.
На фиг. 50 представлено сравнение доменов СН2 А структуры комплекса Fc (ВР208)/внеклеточной области FcγRIIb, определяемой рентгеновским кристаллографическим анализом, и структуры комплекса Fc (WT)/внеклеточной области FcγRIIa (код PDB: 3RY6), определяемой рентгеновским кристаллографическим анализом. На фигуре структуру комплекса Fc (ВР208)/внеклеточной области FcγRIIb изображают жирной линией и структуру комплекса Fc (WT)/внеклеточной области FcγRIIa изображают тонкой линией. Домен СН2 А отдельно проиллюстрирован в структуре комплекса Fc
(WT)/внеклеточной области FcγRIIa.
На фиг. 51 представлена структура вокруг Ser239 домена СН2 В Fc (ВР208) в комплексе Fc (ВР208)/внеклеточной области FcγRIIb, которую определяют рентгеновским кристаллографическим анализом.
На фиг. 52 представлена структура вокруг Ser239 домена СН2 А Fc (ВР208) в комплексе Fc (ВР208)/внеклеточной области FcγRIIb, которую определяют рентгеновским кристаллографическим анализом.
На фиг. 53 представлены результаты аналитической катионообменной хроматографии характерного измененного антитела H240-AK072/H240-BH076/L73-k0. A: H240-AK072/H240-BH076/L73-k0, В: H240-FA021/H240-FB084/L73-k0.
На фиг. 54 представлены результаты катионообменной хроматографии фракционирования (А) и аналитической катионообменной повторной хроматографии собираемых фракций (В) H240-FA021/H240-FB084/L73-k0, которое представляет собой характерное измененное антитело H240-AK072/H240-BH076/L73-k0.
Способ осуществления изобретения
Следующие ниже определения предоставлены для облегчения понимания настоящего изобретения, описываемого в настоящем описании.
Настоящее изобретение относится к полипептидам, содержащим Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, где полипептид отличается тем, что функция Fc-области является измененной по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из
гомодимера, содержащего только первый полипептид, и по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид. Кроме того, настоящее изобретение относится к способам получения полипептида и способам изменения функций содержащего Fc-область полипептида.
В настоящем изобретении "полипептид, содержащий Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид" может представлять собой полипептидный комплекс, состоящий из первого полипептида и второго полипептида, а также многих других полипептидов.
В настоящем описании "первый полипептид" и "второй полипептид" означают полипептиды, из которых состоит Fc-область антитела. Термины "первый полипептид" и "второй полипептид" означают, что их последовательности отличаются друг от друга, и предпочтительно по меньшей мере последовательности домена СН2 являются различными. Полипептиды могут представлять собой, например, полипептиды, из которых состоит Fc-область природного IgG, или полипептиды, получаемые изменением полипептидов, из которых состоит Fc-область природного IgG.
"Природные IgG" относится к полипептидам, которые принадлежат к классу антител, фактически кодируемых генами иммуноглобулина гамма, и содержат аминокислотную последовательность идентичную аминокислотной последовательности IgG, встречающихся в природе. Например, природный IgG человека относится к природному IgG1 человека, природному IgG2 человека,
природному IgG3 человека, природному IgG4 человека или т.п. Природные IgG также включают спонтанно получаемых из них мутантов.
"Полипептиды" по настоящему изобретению, как правило, относится к пептидам или белкам длиной приблизительно десять аминокислот или более. Кроме того, как правило, они представляют собой полипептиды, получаемые из организмов, но конкретно не ограничены, и, например, они могут представлять собой полипептиды, содержащие искусственным образом сконструированную последовательность. Кроме того, они могут представлять собой любой из природных полипептидов, синтетических полипептидов, рекомбинантных полипептидов или т.п. Белковые молекулы по настоящему изобретению относятся к молекулам, содержащим полипептид.
Предпочтительные примеры полипептидов по настоящему изобретению включают антитела. Более предпочтительные примеры включают природные IgG, в частности природные IgG человека. "Природные IgG" относятся к полипептидам, принадлежащим к классу антител, фактически кодируемых генами иммуноглобулина гамма, и содержат аминокислотную последовательность, идентичную аминокислотной последовательности встречающихся в природе IgG. Например, природный IgG человека означает природный IgG1 человека, природный IgG2 человека, природный IgG3 человека, природный IgG4 человека или т.п. Природные IgG также включают спонтанно получаемых из них мутантов.
Несмотря на то, что константная область типа IgK (каппа, k-цепь), IgL1, IgL2, IgL3, IgL6 и IgL7 (лямбда, l-цепь) содержится
в константной области легкой цепи антитела, она может представлять собой любую константную область легкой цепи. Для константной области IgK (каппа) человека и константной области IgL7 (лямбда) человека в "Sequences of proteins of immunological interest", NIH Publication No. 91-3242 описан ряд последовательностей аллотипа, являющихся результатом генетического полиморфизма, и любую из них можно использовать в настоящем изобретении. Кроме того, в настоящем изобретении константная область легкой цепи может представлять собой константную область легкой цепи, которую изменили путем замены, добавлений, делеций, вставок и/или модификаций аминокислот или т.п. Для Fc-области антитела, например, существуют Fc-области типов IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 и IgM. Например, Fc-область антитела IgG человека можно использовать в качестве Fc-области антитела по настоящему изобретению, и Fc-области антитела IgG1 человека являются предпочтительными. Fc-области, которые можно использовать в качестве Fc-области по настоящему изобретению, представляют собой, например, такие, которые выделяют из константных областей природного IgG, или конкретно константную область, выделяемую из природного IgG1 человека (SEQ ID NO: 76), константную область, выделяемую из природного IgG2 человека (SEQ ID NO: 77), константную область, выделяемую из природного IgG3 человека (SEQ ID NO: 78), и константную область, выделяемую из природного IgG4 человека (SEQ ID NO: 79). На фиг. 32 представлены последовательности константных областей природных IgG1, IgG2, IgG3 и IgG4. Константные области природных IgG также включают спонтанно
получаемых из них мутантов. Для константных областей антител IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека в "Sequences of proteins of immunological interest", NIH Publication No. 91-3242 описан ряд последовательностей аллотипов, являющихся результатом генетического полиморфизма, и любую из них можно использовать в настоящем изобретении. В частности, для последовательности IgG1 человека аминокислотная последовательность в положениях от 356 до 358 (нумерация EU) может представлять собой DEL или ЕЕМ.
Кроме того, опубликовано, что сила взаимодействия Fc-области антитела и FcγR зависит от концентрации ионов Zn2+ (Immunology Letters, 143, (2012) 60-69). Для антитела продемонстрировано сильное взаимодействие Fc-области и FcγR при более высокой концентрации ионов Zn2+ Fc-области. Хелатирование Zn2+ His310 и His435, содержащихся в СН3 Fc-области антитела, открывает каждый домен СН2 Fc-области в дистальном положении. Это облегчает взаимодействие домена СН2 и FcγR и усиливает взаимодействие Fc-области и FcγR. Неограничивающий вариант осуществления Fc-области по настоящему изобретению относится к Fc-области, в которой His в положении 310, His в положении 435, His в положении 433 и/или Asn в положении 434 (нумерация EU) являются хелатированными ионами Zn2+.
В настоящем изобретении "Fc-область" относится к области, состоящей из шарнирной области или ее части, домена СН2 и домена СН3, в молекуле антитела. Согласно нумерации EU (в настоящем описании также называемой индекс EU), Fc-область класса IgG
относится, например, к области от цистеина в положении 226 до С-конца или от пролина в положении 230 до С-конца, но не ограничена этим.
Fc-область можно получать предпочтительно повторным элюированием фракций, адсорбированных на колонке с белком А или колонке с белком G после частичного расщепления моноклональных антител IgG1, IgG2, IgG3, IgG4 или т.п.с использованием протеазы, такой как пепсин. Протеаза является конкретно не ограниченной при условии, что она может расщеплять полноразмерное антитело таким образом, чтобы получать Fab и F(ab')2 ограниченным образом, соответствующим способом подбирая условия реакции фермента, такие как рН, и примеры включают пепсин и папаин.
В настоящем описании "гетеродимеризация" означает составление одного полипептида из двух полипептидов с различными аминокислотными последовательностями, и "гетеродимер" означает полипептид, состоящий из двух полипептидов с различными аминокислотными последовательностями. Кроме того, "гомодимеризация" означает составление одного полипептида из двух полипептидов, содержащих одинаковые аминокислотные последовательности, и "гомодимер" означает полипептид, состоящий из двух полипептидов, содержащих одинаковые аминокислотные последовательности, или полипептид, состоящий из полипептидов, содержащих одинаковые аминокислотные последовательности, за исключением изменений, проводимых для эффективной гетеродимеризации, или изменений, проводимых для эффективной очистки гетеродимеров, или полипептид, состоящий из
полипептидов, содержащих одинаковые аминокислоты, за исключением изменений, которые не проводили с целью улучшения функции Fc. В настоящем изобретении "гетеродимер" или "гомодимер" предпочтительно означает "гетеродимеризацию" или "гомодимеризацию" для Fc-области или предпочтительно означает "гетеродимеризацию" или "гомодимеризацию" для СН2 в Fc-области. Кроме того, "родительский полипептид" означает полипептид до введения изменений, таких как мутации аминокислот.
Мутацию аминокислоты по настоящему изобретению можно использовать отдельно или в комбинации с множественными мутациями аминокислот.
Когда используют множественные мутации аминокислот в комбинации, число комбинируемых мутаций является конкретно не ограниченным, и его можно устанавливать соответствующим образом в диапазоне, в котором можно достигать целей изобретения, и примеры включают две или более 30 или менее и предпочтительно две или более 15 или менее.
Когда комбинируют множественные мутации аминокислот, мутации аминокислот можно вводить только в один из двух полипептидов, из которых состоит Fc-область, или их можно соответствующим образом распределять в двух полипептидах.
Кроме того, в настоящем изобретении для получения большего модифицирующего функцию эффекта в Fc-области предпочтительно вводить по меньшей мере одну мутацию аминокислоты, которая улучшает функцию Fc-области, когда мутацию вводят только в один из полипептидов, по сравнению с тем, когда мутацию не вводят, и когда мутацию вводят в обе Fc-области двух полипептидов.
Подлежащий изменению участок не является конкретно ограниченным при условии, что он находится в Fc-области, и его можно соответствующим образом устанавливать в диапазоне, в котором можно достигать целей настоящего изобретения, и примеры включают шарнирную область, область СН2 и область СН3.
Более предпочтительно подлежащий изменению участок представляет собой домен СН2. Домен СН2 относится к положениям от 231 до 340 (нумерация EU), и домен СН3 относится к положениям от 341 до 447 (нумерация EU). Например, при введении мутаций в аминокислотную последовательность константной области, выделяемой из IgG1 человека, можно изменять аминокислотные остатки в одном или нескольких положениях, выбранных из числа 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312,
313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, и 477 (нумерация EU).
Более конкретно при внесении изменений в аминокислотную последовательность константной области IgG1 человека можно изменять аминокислотные остатки в одном или нескольких положениях, выбранных из числа 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387,
388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, и 477 (нумерация EU).
Более конкретно при внесении изменений в аминокислотную последовательность константной области IgG1 человека можно изменять аминокислотные остатки в одном или нескольких положения, выбранных из числа 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339 и 340 (нумерация EU).
Более конкретно при введении изменений в аминокислотную последовательность константной области IgG1 человека можно изменять аминокислотные остатки в одном или нескольких положениях, выбранных из числа 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 265, 266, 267, 268, 269, 270, 271, 295, 296, 298, 300, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336 и 337 (нумерация EU).
В настоящем изобретении изменение аминокислот означает любую замену, делецию, добавление, вставку и модификацию или их
сочетание. В настоящем изобретении изменение аминокислоты можно перефразировать как мутация аминокислот и их используют синонимично.
Замена
При замене аминокислотных остатков замену на отличный аминокислотный остаток проводят с целью изменения аспектов, таких как (а)-(с), описанных ниже:
a) структуры основной цепи полипептида в области листовой или спиральной структуры;
b) электрического заряда или гидрофобности в участке-мишени или
c) размера боковой цепи.
Аминокислотные остатки классифицируют на следующие ниже группы в зависимости от основных свойств их боковых цепей:
1) гидрофобные: норлейцин, met, ala, val, leu и ile;
2) нейтральные гидрофильные: cys, ser, thr, asn и gln;
3) кислые: asp и glu;
4) основные: his, lys и arg;
5) остатки, которые влияют на ориентацию цепи: gly и pro, и
6) ароматические: trp, tyr и phe.
Замену аминокислотных остатков в каждой из этих групп аминокислот обозначают как консервативную замену и замену аминокислотного остатка различных групп обозначают как неконсервативную замену.
Замены в настоящем изобретении могут представлять собой консервативные замены или неконсервативные замены, или комбинацию консервативных замен и неконсервативных замен.
В дополнение к мутациям аминокислот, вводимым по настоящему изобретению, полипептиды по настоящему изобретению могут дополнительно содержат дополнительные изменения. Например, дополнительные изменения можно выбирать из любой из замен аминокислот, делеций и модификаций или их сочетаний.
Например, полипептиды по настоящему изобретению можно произвольно изменять в диапазоне, который конкретно не изменяет предполагаемую функцию полипептида. Когда полипептид по настоящему изобретению представляет собой антитело, можно изменять его тяжелую цепь и легкую цепь. Например, такие мутации можно проводить посредством консервативной замены аминокислотных остатков. Кроме того, даже если изменение меняет предполагаемую функцию полипептида по настоящему изобретению, изменение можно проводить при условии, что изменение функции находится в диапазоне задач настоящего изобретения.
Изменение аминокислотной последовательности по настоящему изобретению относится к посттрансляционной модификации. Конкретная посттрансляционная модификация может представлять собой добавление или делецию цепи сахаров. Например, в константной области IgG1 аминокислотный остаток в положении 297 (нумерация EU) может представлять собой модифицированную цепь сахаров. Структура цепи сахаров для модификации не является ограниченной. Как правило, антитела, экспрессируемые в эукариотических клетках, содержат гликозилирование в константной области. Таким образом, антитела, экспрессируемые в клетках, таких как указанные ниже, являются обычно модифицированными по некоторому типу цепи сахаров:
- продуцирующие антитело клетки млекопитающих,
- эукариотические клетки, трансформированные экспрессирующим вектором, содержащим ДНК, кодирующую антитело.
Представленные в настоящем описании эукариотические клетки включают дрожжевые клетки и клетки животных. Например, клетки СНО и клетки HEK293H представляют собой характерные клетки животных, используемы в трансформации экспрессирующим вектором, содержащим кодирующую антитело ДНК. С другой стороны, антитело по настоящему изобретению также включает антитела без гликозилирования на этом участке. Антитела, константная область которых не является гликозилированной, можно получать посредством экспрессии кодирующего антитело гена в прокариотические клетки, так как Escherichia coli.
В настоящем изобретении дополнительные изменения конкретно включают, например, добавление сиаловой кислоты к цепи сахаров Fc-области (MAbs., 2010 Sep-Oct; 2(5): 519-27).
Когда полипептид по настоящему изобретению представляет собой антитело, в участок константной области антитела можно вводить, например, замены аминокислот, которые улучшают активность связывания с FcRn (J. Immunol., 2006 Jan 1; 176(1): 346-56; J. Bio.l Chem.,., 2006 Aug 18; 281(33):23514-24; Int. Immunol., 2006 Dec; 18(12): 1759-69; Nat Biotechnol., 2010 Feb; 28(2):157-9; WO/2006/019447; WO/2006/053301 и WO/2009/086320), и замены аминокислот для улучшения гетерогенности или стабильности антитела (WO/2009/041613).
Для получения гетеродимеризованного полипептида по настоящему изобретению необходимо, чтобы полипептиды, содержащие
аминокислоты, которые отличаются друг от друга, являлись ассоциированными, или представляющий интерес гетеродимеризованный полипептид выделяют из других гомодимеризованных полипептидов.
Для ассоциации полипептидов, содержащих различные аминокислоты друг от друга и содержащих Fc-область, можно применять технологию подавления непредусмотренной ассоциации между Н-цепями посредством введения электростатического отталкивания на поверхности второй константной области Н-цепи антитела (СН2) или третьей константной области Н-цепи (СН3) (WO 2006/106905).
В технологии подавления непредусмотренной ассоциации между Н-цепями введением электростатического отталкивания на поверхности СН2 или СН3 примеры аминокислотных остатков при контактировании на поверхности других константных областей Н-цепи включают остаток в положении 356 (нумерация EU), остаток в положении 439 (нумерация EU), область, находящуюся перед остатком в положении 357 (нумерация EU), остаток в положении 370 (нумерация EU), остаток в положении 399 (нумерация EU) и остаток в положении 409 (нумерация EU) в домене СН3.
Более конкретно, например, для антитела, содержащего два типа доменов СН3 Н-цепи, можно получать антитело, в котором от одной до трех пар аминокислотных остатков, выбранных из аминокислотных остатков, представленных ниже в (1)-(3), в домене СН3 первой Н-цепи, имеют одинаковый тип заряда:
1) аминокислотные остатки в положениях 356 и 439 (нумерация EU), которые представляют собой аминокислотные остатки,
содержащиеся в домене СН3 Н-цепи;
2) аминокислотные остатки в положениях 357 и 370 (нумерация EU), которые представляют собой аминокислотные остатки, содержащиеся в домене СН3 Н-цепи, и
3) аминокислотные остатки в положениях 399 и 409 (нумерация EU), которые представляют собой аминокислотные остатки, содержащиеся в домене СН3 Н-цепи.
Кроме того, можно получать антитело, в котором от одной до трех пар аминокислотных остатков, соответствующих парам аминокислотных остатков, указанных выше в (1)-(3), обладающих аналогичным типом заряда в домене СН3 первой Н-цепь, обладают зарядом, противоположным соответствующим аминокислотным остаткам в указанном выше домене СН3 первой Н-цепи, где пары аминокислотных остатков в домене СН3 второй Н-цепи, который отличается от домена СН3 первой Н-цепи, выбраны из пар аминокислотных остатков, указанных выше в (1)-(3).
Соответствующие указанные выше аминокислотные остатки (1)-(3) при ассоциации располагают вблизи друг от друга. Специалисты в данной области могут выявлять участки, которые соответствуют указанным выше аминокислотным остаткам (1)-(3), посредством гомологичного моделирования и т.п. с использованием коммерчески доступного программного обеспечения для желаемого домена СН3 Н-цепи или константной области Н-цепи, и при необходимости можно изменять аминокислотные остатки этих участков.
В указанных выше антителах, например, "несущие заряд аминокислотные остатки" предпочтительно выбрать из аминокислотных остатков, входящих в любую из групп (X) или (Y)
ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D), и (Y) лизин (К), аргинин (R) и гистидин (Н).
Для указанных выше антител фраза "обладающий одинаковым типом заряда" означает, что, например, все два или более аминокислотных остатка представляют собой аминокислотные остатки, входящие в указанные выше группы (X) и (Y). Фраза "обладающий противоположным зарядом" означает, например, когда по меньшей мере один из двух или более аминокислотных остатков представляет собой аминокислотный остаток, входящий в любую из указанных выше групп (X) и (Y), оставшиеся аминокислотные остатки представляют собой аминокислотные остатки, входящие в другую группу.
В предпочтительном варианте осуществления указанного выше антитела домен СН3 первой Н-цепи и домен СН3 второй Н-цепи можно поперечно сшивать посредством дисульфидных связей.
В настоящем изобретении подлежащие изменению аминокислотные остатки не ограничены аминокислотными остатками описанной выше константной области антитела или вариабельной области антитела. Специалисты в данной области могут находить аминокислотные остатки, которые образуют область контакта в мутантах полипептидов или гетеромультимерах, посредством гомологичного моделирования и т.п. с использованием коммерчески доступного программного обеспечения и могут изменять аминокислотные остатки на этих участках для регуляции ассоциации.
Также можно использовать другие известные технологии для ассоциации полипептидов по настоящему изобретению, содержащих
различные аминокислоты и содержащих Fc-область. Содержащие различные аминокислоты и содержащие Fc-область полипептиды можно эффективно ассоциировать друг с другом, заменяя аминокислотную боковую цепь, содержащуюся в одной из вариабельных областей Н-цепи антитела, на более крупную боковую цепь ("выступ") и заменяя аминокислотную боковую цепь, содержащуюся в противоположной вариабельной области, на другую Н-цепь с меньшей боковой цепью ("впадина") для обеспечения возможности расположения "выступа" во "впадине" (WO 1996/027011 и Ridgway J.B. et al.. Protein Engineering, (1996) 9, 617-621; Merchant A.M. et al.. Nature Biotechnology, (1998) 16, 677-681).
Кроме того, также можно использовать другие известные технологии для ассоциации полипептидов, содержащих различные аминокислоты и содержащих Fc-область. Ассоциацию полипептидов, содержащих различные последовательности, можно эффективно индуцировать посредством комплементарной ассоциации СН3 с использованием сконструированного домена СН3 с заменой цепей, получаемого заменой участка СН3 в одной из Н-цепей антитела на получаемую из IgA последовательность, соответствующую этому участку, и введением соответствующей получаемой из IgA последовательности в комплементарный участок СН3 на другой Н-цепи (Protein Engineering Design & Selection, 23; 195-202, 2010). Эту известную технологию также можно использовать для эффективной индукции ассоциации полипептидов, содержащих различные аминокислоты и содержащих Fc-область.
Кроме того, также можно использовать технологии получения гетеродимеризованных антител, в которых используют ассоциации
СН1 и CL антитела и ассоциации VH и VL, которые описаны в WO 2011/028952.
Кроме того, даже в случаях, когда гетеродимеризованные полипептиды невозможно эффективно получать, гетеродимеризованные полипептиды можно получать путем их выделения и очистки из гомодимеризованных полипептидов. При получении гетеродимеризованного полипептида, состоящего из первого полипептида и второго полипептида, которые содержат отличные друг от друга последовательности, гомодимеризованные полипептиды, состоящие только из двух первых полипептидов, и гомодимеризованный полипептид, состоящий только из двух вторых полипептидов, смешивают в качестве примесей. В качестве способа эффективного удаления этих двух типов гомодимеризованных полипептидов можно использовать известные технологии. Опубликовано, что способом можно очищать два типа гомодимеров и представляющее интерес гетеродимеризованное антитело посредством ионообменной хроматографии путем создания различия изоэлектрических точек введением замены аминокислот в вариабельные области двух типов Н-цепей (WO 2007114325). К настоящему времени в качестве способа очистки гетеродимеризованных антител опубликовано, что способом с использованием белка А очищают гетеродимеризованное антитело, содержащее Н-цепь IgG2a мыши, которая связывается с белком А, и Н-цепь IgG2b крысы, которая не связывается с белком А (WO 98050431 и WO 95033844).
Кроме того, можно эффективно очищать одно гетеродимеризованное антитело с использованием Н-цепей, в
которых аминокислотные остатки на участке связывания IgG-белок А в положениях 435 и 436 (нумерация EU) заменяют аминокислотами, приводящими к другим аффинностям к белку А, такими как Tyr или His, для изменения взаимодействия каждой из Н-цепей с белком А и с использованием колонки с белком А.
Многие такие замены и технологии, например, две или более из них, можно использовать в комбинации. Кроме того, при необходимости эти изменения можно проводить отдельно для первого полипептида и второго полипептида. Полипептиды по настоящему изобретению также могут представлять собой полипептиды, получаемые на основе продуктов, указанных выше изменений.
Аминокислотную последовательность можно изменять различными способами, известными специалистам в данной области. Такие способы включают способ сайт-специфического мутагенеза (Hashimoto-Gotoh Т., Mizuno Т., Ogasahara Y. and Nakagawa M., (1995) An oligodeoxyribonucleotide-directed dual amber method for site-directed mutagenesis. Gene, 152:271-275; Zoller M.J. and Smith M., (1983) Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors. Methods Enzymol., 100: 468-500; Kramer W., Drutsa V., Jansen H.W., Kramer В., Pflugfelder M. and Fritz H.J., (1984) The gapped duplex DNA approach to oligonucleotide-directed mutation construction. Nucleic Acids Res., 12:9441-9456; Kramer W. and Fritz H.J., (1987) Oligonucleotide-directed construction of mutations via gapped duplex DNA Methods. Enzymol., 154, 350-367, и Kunkel Т.А. (1985) Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA, 82: 488-492), способ
мутации путем ПЦР и способ кассетной мутации, но не являются ограниченными ими.
В настоящем изобретении фраза "функция Fc-области" относится, например, к активности связывания Fc-области с рецептором Fcγ (повышению активности связывания или снижению активности связывания), селективности Fc-области в отношении изоформы рецептора Fcγ (улучшению селективности связывания), физико-химической стабильности Fc-области, активности ADCC и активности ADCP. В настоящем описании селективность Fc-области в отношении изоформы рецептора Fcγ означает селективное связывание с конкретными изоформами рецептора Fcγ. Физико-химическая стабильность Fc-области означает, например, термостабильность Fc-области, стабильность к протеазе, стабильность к химической обработке, стабильность к замораживанию-оттаиванию, стабильность при хранении, стабильность в кислотных условиях, фотостабильность, стабильность к ассоциированному с встряхиванием или ассоциированному с концентрацией стрессу, или сохранение растворимости в широком диапазоне состояний раствора. Кроме того, функция Fc-области может относиться к функции, объединяющей две или более функции из активности связывания Fc-области с рецептором Fcγ, селективности Fc-области в отношении изоформ рецептора Fcγ и физико-химической стабильности Fc-области; и это означает, например, функцию, объединяющую активность связывания Fc-области с рецептором Fcγ и селективность Fc-области в отношении изоформы рецептора Fcγ, функцию, объединяющую активность связывания Fc-области с рецептором Fcγ и
физико-химическую стабильность Fc-области, функцию, объединяющую селективность Fc-области в отношении изоформы рецептора Fcγ и физико-химическую стабильность Fc-области, и функцию, объединяющую активность связывания Fc-области с рецептором Fcγ, селективность Fc-области в отношении изоформы рецептора Fcγ и физико-химическую стабильность Fc-области.
В настоящем изобретении "изменение функции Fc-области" означает, например, повышение, снижение или т.п. активности связывания Fc-области с рецептором Fcγ, когда функция Fc-области относится к активности связывания Fc-области с рецептором Fcγ. Улучшение селективности означает, например, повышение активности связывания с рецептором Fcγ и одновременно сохранение или снижение активности связывания с другими рецепторами Fcγ. Альтернативно, улучшение селективности означает, например, снижение активности связывания с определенными рецепторами Fcγ при сохранении или повышении активности связывания с другими рецепторами Fcγ. Кроме того, например, когда функция Fc-области относится к селективности Fc-области в отношении подтипа рецептора Fcγ, "изменение функции Fc-области" означает улучшение или снижение селективности Fc-области в отношении подтипа рецептора Fcγ. Альтернативно, например, когда функция Fc-области относится к физико-химической стабильности Fc-области, "изменение функции Fc-области" означает улучшение или снижение физико-химической стабильности Fc-области, подавления снижения стабильности или т.п., и более конкретно это означает, например, улучшение или снижение значения Tm домена СН2, подавления
снижения значения Tm или т.п.
Например, улучшение объединенных функций активности связывания Fc-области с рецептором Fcγ, селективности Fc-области в отношении изоформы рецептора Fcγ и физико-химической стабильности Fc-области не обязательно должно представлять собой улучшение в каждой из активности связывания Fc-области с рецептором Fcγ, селективности Fc-области в отношении изоформы рецептора Fcγ и физико-химической стабильности Fc-области по сравнению с контролем, и улучшение является приемлемым при условии, что функция Fc-области улучшается в целом. В противоположность этому, например, снижение объединенных функций активности связывания Fc-области с рецептором Fcγ, селективности Fc-области в отношении изоформы рецептора Fcγ и физико-химической стабильности Fc-области необязательно представляет собой снижение каждой из активности связывания Fc-области с рецептором Fcγ, селективности Fc-области в отношении изоформы рецептора Fcγ и физико-химической стабильности Fc-области по сравнению с контролем, и снижение является приемлемым при условии, что функция Fc-области снижается в целом.
В настоящем изобретении "рецепторы Fcγ" (в настоящем описании обозначаемые как рецепторы Fcγ, FcγR или FcgR) относятся к рецепторам, которые могут связываться с Fc-областью IgG1, IgG2, IgG3 и IgG4, и фактически означают любой член семейства белков, кодируемых генами рецептора Fcγ. У людей это семейство включает FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc;
FcγRII (CD32), включая изоформы FcγRIIa (в том числе аллотипы Н131 (тип Н) и R131 (тип R)), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), включая изоформы FcγRIIIa (в том числе аллотипы V158 и F158) и FcγRIIIb (в том числе аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2), и любые FcγR человека, изоформы или аллотипы FcγR, которые еще откроют, но не ограничиваются ими. FcγR включают получаемые у человека, мыши, крысы, кролика и обезьяны FcγR, но не ограничиваются ими, и их можно получать из любого организма. FcγR мыши включают FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD16-2 или FcγRIV), и любые FcγR мыши или изоформы или аллотипы FcγR, которые еще откроют, но не ограничиваются ими. Предпочтительные примеры таких рецепторов Fcγ включают FcyRI (CD64), FcyRIIa (CD32), FcγRIIb (CD32), FcγRIIIa (CD16) и/или FcγRIIIb (CD16) человека.
Для FcγR существуют активирующие рецепторы, несущие иммунорецепторный активирующий мотив на основе тирозина (ITAM), и ингибирующий рецептор, несущий иммунорецепторный ингибирующий мотив на основе тирозина (ITIM). FcγR классифицируют на активирующие FcγR: FcγRI, FcγRIIa R, FcγRII H, FcγRIIIa, и FcγRIIIb, и ингибирующий FcγR: FcγRIIb.
Полинуклеотидная последовательность и аминокислотная последовательность FcγRI указаны в NM_000566.3 и NP_000557.1, соответственно;
полинуклеотидная последовательность и аминокислотная
последовательность FcγRIIa указаны в ВС020823.1 и ААН20823.1, соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIb указаны в ВС146678.1 и AAI46679.1, соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIa указаны в ВС033678.1 и 7ААН33678.1, соответственно; и
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIb указаны в ВС128562.1 и AAI28563.1, соответственно (регистрационный номер RefSeq).
В FcγRIIa существует два аллотипа: один, где аминокислота в положении 131 FcγRIIa представляет собой гистидин (тип Н), и другой, где эта аминокислота замещена аргинином (тип R) (J. Ехр. Med, 172:19-25, 1990). В FcγRIIb существует два аллотипа один, где аминокислота в положении 232 FcγRIIb представляет собой изолейцин (тип I), и другой, где эта аминокислота замещена треонином (тип Т) (Arthritis. Rheum., 46:1242-1254 (2002)). В FcγRIIIa существует два аллотипа: один, где аминокислота в положении 158 FcyRIIIa представляет собой валин (типа V), и другой эта аминокислота замещена фенилаланином (тип F) (J. Clin. Invest., 100(5):1059-1070 (1997)). Кроме того, в FcγRIIIb существует два аллотипа: типа NA1 и типа NA2 (J. Clin. Invest., 85: 1287-1295 (1990)).
Одно из веществ (лиганд) при наблюдении взаимодействия иммобилизуют на золотой тонкой пленке на сенсорном чипе, и
посредством пропускания света с обратной стороны сенсорного чипа, таким образом, чтобы происходило полное отражение на поверхности между золотой тонкой пленкой и стеклом, в части отраженного света получают участок сниженной интенсивности отражения (сигнал SPR). Когда другое одно из веществ (аналит) при наблюдении взаимодействия получают с возможностью растекаться на поверхности сенсорного чипа, и лиганд связывается с аналитом, масса молекулы иммобилизованного лиганда увеличивается, и показатель преломления растворителя на поверхности сенсорного чипа изменяется. В результате этого изменения показателя преломления смещается положение сигнала SPR (с другой стороны, положение сигнала возвращается, когда это связывание диссоциирует). Система Biacore указывает на количество указанного выше смещения или более конкретно на переменную времени по массе, нанося на график изменение массы на поверхности сенсорного чипа по ординате в виде данных измерения (сенсограммы). Количество аналита, связанного с лигандом, захваченным на поверхности сенсорного чипа, определяют из сенсограммы (количество изменения ответа на сенсограмме до и после взаимодействия с аналитом). Однако вследствие того, что количественная мера связывания также зависит от количества лиганда, необходимо проводить сравнение в условиях, где предполагают, что количество лиганда является практически одинаковым. Параметры кинетики, такие как константы скорости ассоциации (ka) и константы скорости диссоциации (kd) определяют на основании кривой сенсограммы и из отношения этих констант определяют аффинности (KD). В способе BIACORE предпочтительно
используют способ измерения ингибирования. Пример способа измерения ингибирования описан в Proc. Natl. Acad. Sci. USA, (2006) 103 (11), 4005-4010.
В настоящем изобретении повышается или не повышается, снижается или не снижается активность связывания полипептида или в Fc-области с каждым типом рецептора Fcγ по настоящему изобретению можно определять, например, с использованием Biacore (GE Healthcare), который представляет собой анализатор взаимодействия, в котором используют явление поверхностного плазмонного резонанса (SPR), как показано в примерах. Biacore включает любые модели, такие как Biacore Т100, Т200, Х100, А100, 4000, 3000, 2000, 1000 и С. Для сенсорного чипа можно использовать любые сенсорные чипы для Biacore, такие как чип СМ7, СМ5, СМ4, СМ3, C1, SA, NTA, L1, НРА и Au. Для подвижного буфера наряду с НВЕ-ЕР+ можно использовать буфер, получаемый с использованием  , фосфорной кислоты, ACES, Tris, лимонной кислоты и т.п. для подведения рН до нейтрального рН, такого как 7,4. Измерения можно проводить в диапазоне от 4°С до 37°С. Белок для захвата антител, такой как белок А, белок G или белок L, который захватывает антитело, антитело против IgG человека, Fab против IgG человека, антитело против L-цепи человека, антитело против Fc человека, антигенный белок или антигенный пептид иммобилизуют на сенсорном чипе способами связывания, такими как аминное связывание, дисульфидное связывание или альдегидное связывание. В качестве аналитов применяют различные типы рецепторов Fcγ, такие как рецептор Fcγ I, IIа R типа, IIа Н типа,
, фосфорной кислоты, ACES, Tris, лимонной кислоты и т.п. для подведения рН до нейтрального рН, такого как 7,4. Измерения можно проводить в диапазоне от 4°С до 37°С. Белок для захвата антител, такой как белок А, белок G или белок L, который захватывает антитело, антитело против IgG человека, Fab против IgG человека, антитело против L-цепи человека, антитело против Fc человека, антигенный белок или антигенный пептид иммобилизуют на сенсорном чипе способами связывания, такими как аминное связывание, дисульфидное связывание или альдегидное связывание. В качестве аналитов применяют различные типы рецепторов Fcγ, такие как рецептор Fcγ I, IIа R типа, IIа Н типа,
IIb, IIIa F типа, V типа и IIIb, и измеряли взаимодействия с получением сенсорограмм. На данной стадии измерения можно проводить, регулируя концентрацию рецептора Fcγ до уровня в диапазоне от нескольких мкМ до нескольких пМ в зависимости от силы взаимодействия, такой как KD образца, подлежащего измерению. Константу диссоциации (KD) получают на основании измерения и посредством наблюдения снижается или повышается значение KD, можно определять повышается или понижается активность связывания Fc-области или полипептида по настоящему изобретению к различным рецепторам Fcγ. Когда захват проводят посредством захватывающего антитело белка, иммобилизованного на сенсорном чипе, в качестве показателя используют уровень заряда в сенсограмме до и после нанесения различных рецепторов Fcγ в качестве аналитов для антител на сенсорный чип; и в зависимости от степени увеличения значения, можно определять повышается или понижается активность связывания полипептида или Fc-области по настоящему изобретению к различным рецепторам Fcγ Альтернативно, вместо антител также можно иммобилизировать различные рецепторы Fcγ на сенсорные чипы, и приводить их во взаимодействие с образцом антитела, подлежащим оценке. Повышается или снижается активность связывания Fc-области или полипептида по настоящему изобретению с различными рецепторами Fcγ можно определять на основании снижения или увеличения значений KD, рассчитываемых из сенсорограмм взаимодействия или на основании степени увеличения сенсорограмм до и после обеспечения взаимодействия образца антитела.
В частности, активность связывания Fc-области с рецептором Fcγ можно измерять путем скринигна гомогенным анализом усиленной за счет эффекта близости люминесценции (ALPHA) способом BIACORE, в котором используют явление поверхностного плазмонного резонанса (SPR) или т.п.в дополнение к ELISA или активируемой флуоресценцией сортировки клеток (FACS) (Proc. Natl. Acad. Sci. USA, (2006) 103 (11): 4005-4010).
Скрининг ALPHA проводят технологией ALPHA, в которой используют два типа гранул, донор и акцептор, в основе которой лежат следующие ниже принципы. Люминесцентные сигналы детектируют только, когда молекулы, связанные с донорными гранулами, физически взаимодействуют с молекулами, связанными с акцепторными гранулами, и два типа гранул находятся в непосредственной близости друг к другу. Возбуждаемый лазером фотосенсибилизатор в донорных гранулах трансформирует кислород в окружающей среде в возбужденное состояние синглетного кислорода. Синглетный кислород рассеивается вокруг донорных гранул, и когда он попадает на соседние акцепторные гранулы, в гранулах индуцируется хемилюминесцентная реакция, и в итоге излучается свет. Когда связанные с донорными гранулами молекулы не взаимодействуют с молекулами, связанными с акцепторными гранулами, хемилюминесцентной реакции не происходит, т.к. синглетный кислород, продуцируемый донорными гранулами, не попадает на акцепторные гранулы.
Например, биотинилированный тестовый полипептид является связанным со стрептавидином на донорных гранулах, и рецептор Fcγ, меченый глутатион-S-трансферазой, (GST) связан с акцепторными
гранулами. При отсутствии конкурентного полипептида тестовый полипептид взаимодействует с рецептором Fcγ и продуцирует сигналы 520-620 нм. Немеченый полипептид конкурирует с тестовым полипептидом за взаимодействие с рецептором Fcγ. Относительные активности связывания можно определять, количественно определяя снижение флуоресценции, наблюдаемой в результате конкурентного связывания. Биотинилирование полипептида с использованием Sulfo-NHS-биотина и т.п.является хорошо известным. Способ экспрессии рецептора Fcγ и GST в клетке, несущей слитый ген, получаемый слиянием слитого полинуклеотида, кодирующего рецептор Fcγ в рамке считывания с полинуклеотидом, кодирующим GST в экспрессирующем векторе, и проведение очистки с использованием колонки с глутатионом соответствующим образом адаптируют в качестве способа мечения рецептора Fcγ GST. Получаемые сигналы предпочтительно анализируют, например, их подгонкой к модели конкурентного связывания с одним участком, в которой используют нелинейный регрессионный анализ, с использованием программного обеспечения, такого как GRAPHPAD PRISM (GraphPad, San Diego).
Мечение не ограничено использованием GST и можно использовать любую метку, такую как гистидиновая метка, МВР, СВР, метка Flag, метка НА, метка V5 и метка с-myc. Кроме того, связывание тестового полипептида с донорными гранулами не ограничено использованием реакции биотина-стрептавидина. В частности, когда тестовый полипептид представляет собой антитело или пептид, несущий Fc, такой как слитый с Fc полипептид, можно рассматривать способ связывания тестового полипептида через
распознающий Fc белок, такой как белок А или белок G, на донорных гранулах.
Снижение связывания FcγR или активности связывания с FcγR относится к связыванию с FcγR по существу с меньшей активностью связывания, чем активность связывания родительского полипептида, когда анализы проводят с использованием фактически одинакового количества подлежащих сравнению полипептидов.
Гетеродимеризованные полипептиды с ослабленным, сниженным или уменьшенным связыванием с FcγR или активностью связывания с FcγR относятся к таким, которые связываются с FcγR по существу с меньшей активностью связывания, чем другие гомодимеризованные полипептиды, когда анализы проводят с использованием фактически одинакового количества подлежащих сравнению полипептидов.
Например, в значениях KD, определяемых указанные выше способами измерения, отношение значений KD (значение KD родительского полипептида/значение KD мутантного полипептида) составляет предпочтительно 0,99 или менее, 0,95 или менее, 0,9 или менее, 0,8 или менее, 0,7 или менее, 0,5 или менее, 0,3 или менее, и 0,1 или менее и более предпочтительно 0,08 или менее, 0,05 или менее, 0,02 или менее, 0,01 или менее и 0,001 или менее. В настоящем описании, отношение значений KD также называют отношением KD.
Значения KD, определяемые указанными выше способами измерения, предпочтительно являются повышенными на 1 пМ или более, и более предпочтительно повышенными на 10 пМ, 100 пМ, 1 нМ или более, 2 нМ или более, 3 нМ или более, 5 нМ или более, 10
нМ или более, 20 нМ или более, 50 нМ или более, 100 нМ или более, или 1 мМ или более. Значения KD, определяемые указанными выше способами измерения, предпочтительно составляют 1 пМ или более и предпочтительно 10 пМ или более, 100 пМ или более, 1 нМ или более, 10 нМ или более, 100 нМ или более, 500 нМ или более, 1 мМ или более, 3 мМ или более, или 5 мМ или более.
Усиление, повышение или улучшение связывания FcγR или активности связывания с FcγR относится к связыванию FcγR по существу с большей активностью связывания по сравнению с активностью связывания родительского полипептида, когда анализы проводят с использованием фактически одинакового количества подлежащих сравнению полипептидов.
Гетеродимеризованные полипептиды с усиленным, повышенным или улучшенным связыванием с FcγR или активностью связывания с FcγR относятся к таким, которые связываются с FcγR по существу с большей активность связывания, чем активность связывания гомодимеризованных полипептидов, когда анализы проводят с использованием фактически одинакового количества подлежащих сравнению полипептидов.
Например, в значениях KD, определяемых указанными выше способами измерения, отношение значений KD (значение KD родительского полипептида/значение KD мутантного полипептида) предпочтительно составляет 1,1 или более, 1,2 или более, 1,3 или более, 1,5 или более, 1,8 или более, 2 или более, или 3 или более и более предпочтительно 5 или более, 10 или более, 100 или более, 250 или более, или 1000 или более. В настоящем описании
отношение значений KD также называют отношением KD.
Значения KD, определяемые указанными выше способами измерения, предпочтительно явялются сниженными на 1 пМ или более и более предпочтительно сниженными на 10 пМ, 100 пМ, 1 нМ или более, 2 нМ или более, 3 нМ или более, 5 нМ или более, 10 нМ или более, 20 нМ или более, 50 нМ или более, 100 нМ или более, или 1 мМ или более.
Значения KD, определяемые указанными выше способами измерения, предпочтительно составляют 5 мМ или менее и более предпочтительно 3 мМ или менее, 1 мМ или менее, 0,5 мМ или менее, 0,1 мМ или менее, 0,01 мМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,001 нМ или менее, или 1 пМ или менее.
В настоящем изобретении, когда изменение функции Fc-области полипептида повышает активность связывания с рецептором Fcγ, можно вводить мутации аминокислот в аминокислотные последовательности первого полипептида и/или второго полипептида, из которого состоит указанная выше Fc-область. Тип и диапазон мутации аминокислот, которые можно вводить, не являются конкретно ограниченными.
Опубликовано, что сила взаимодействия Fc-области антитела и FcγR зависит от концентрации ионов Zn2+ (Immunology Letters, 143 (2012) 60-69). Для антитела показано более сильное взаимодействие между Fc-областью и FcγR, когда концентрация ионов Zn2+ Fc-области является выше. Хелатирование Zn2+ His310 и His435, содержащихся в СН3 Fc-области антитела, открывает домен СН2 Fc-области в дистальном положении. Это облегчает взаимодействие
домена СН2 и FcγR и усиливает взаимодействие Fc-область и FcγR. Неограничивающий вариант осуществления антитела по настоящему изобретению относится к Fc-области, в котором His в положении 310, His в положении 435, His в положении 433 и/или Asn в положении 434 (нумерация EU) хелатируют Zn2+.
Когда рецептор Fсγ представляет собой FcγRIa по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблиц 2-1 и 2-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fcγ представляет собой FcγRIa, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблиц 2-1, 2-2 и 2-3 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
Когда рецептор Fcγ представляет собой FcγRIIa R, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблиц 3-1 и 3-2 в настоящем описании, можно водить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fсγ представляет собой FcγRIIa R, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблиц 3-1 и 3-2 в настоящем
описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
Когда рецептор Fcγ представляет собой FcγRIIa Н, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 4 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fcγ представляет собой FcγRIIa Н, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 4 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
Когда рецептор Fcγ представляет собой FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 5 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fсγ представляет собой FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 5 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-
область.
Когда рецептор Fcγ представляет собой FcγRIIIa, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области i таблицы 6 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fcγ представляет собой FcγRIIIa, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области ii таблицы 6 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда рецептор Fcγ представляет собой FcγRIIIa, более конкретно по меньшей мере одну или более (например, две или три) мутаций аминокислот, выбранных из группы, состоящей из замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y или Q; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты S в положении 239 (нумерация EU) на М; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты D в положении 270 (нумерация EU) на Е; замены аминокислоты S в положении 298 (нумерация EU) на А; замены аминокислоты К в положении 326 (нумерация EU) на D; замены аминокислоты А в положении 327 (нумерация EU) на D; замены аминокислоты L в положении 328 (нумерация EU) на W; замены аминокислоты А в положении 330
(нумерация EU) на М или К и замены аминокислоты К в положении 334 (нумерация EU) на Е или L, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Даже более конкретно, когда рецептор Fcγ представляет собой FcγRIIIa, по меньшей мере одну или более (например, две или три) мутаций аминокислот, выбранных из группы, состоящей из замены аминокислоты S в положении 239 (нумерация EU) на D; замены аминокислоты А в положении 330 (нумерация EU) на L и замены аминокислоты I в положении 332 (нумерация EU) на Е, можно вводить в аминокислотную последовательность любого одного из полипептидов, из которых состоит Fc-область, первого полипептида или второго полипептида и по меньшей мере одну или более (например, две или три) мутаций аминокислот, выбранных из группы, состоящей из замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W и замены аминокислоты S в положении 298 (нумерация EU) на А, можно вводить в аминокислотную последовательность другого полипептида.
Даже более конкретно, когда рецептор Fcγ представляет собой FcγRIIIa, мутацию можно вводить по меньшей мере в одну или более (например, две или три) аминокислот, выбранных из числа Leu в положении 234, Leu в положении 235, Gly в положении 236, Ser в положении 239, His в положении 268, Asp в положении 270, Ser в положении 298, Ala в положении 327, Leu в положении 328 и Lys в положении 334 (нумерация EU), в аминокислотной последовательности любого из полипептидов, из которых состоит
Fc-область, первый полипептид или второй полипептид, и можно вводить мутацию по меньшей мере в одну или более (например, две или три) аминокислот, выбранных из числа Asp в положении 270, Lys в положении 326, Ala в положении 330 и Lys в положении 334 (нумерация EU), в аминокислотной последовательности другого полипептида.
Аминокислоту, подлежащую изменению, можно выбирать соответствующим образом, и предпочтительно по меньшей мере одна или более (например, две или три) мутаций аминокислот выбраны из группы, состоящей из:
замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y или Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты А в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на W; и
замены аминокислоты К в положении 334 (нумерация EU) на L, можно вводить в аминокислотную последовательность любого из полипептидов, из которого состоит Fc-область, первый полипептид или второй полипептид, и по меньшей мере одну или более (например, две или три) мутаций аминокислот, выбранных из группы, состоящей из:
замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты К в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М или К и
замены аминокислоты К в положении 334 (нумерация EU) на Е, можно вводить в аминокислотную последовательность другого полипептида.
Более предпочтительно любую серию мутаций от (i) до (vi) можно вводить в аминокислотную последовательность любого из полипептидов, из которых состоит Fc-область, первый полипептид или второй полипептид, и любую серию из мутаций от (vii) до (ix) можно вводить в аминокислотную последовательность другого полипептида:
(i) замены аминокислоты L в положении 234 (нумерация EU) на
Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D; и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(ii) замены аминокислоты L в положении 234 (нумерация EU) на
Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iii) замены аминокислоты L в положении 234 (нумерация EU)
на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е; и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iv) замены аминокислоты L в положении 234 (нумерация EU) на
Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(v) замены аминокислоты L в положении 234 (нумерация EU) на
Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(vi) замены аминокислоты L в положении 234 (нумерация EU) на
Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты А в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на W и
замены аминокислоты К в положении 334 (нумерация EU) на L;
(vii) замены аминокислоты К в положении 326 (нумерация EU)
на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты К в положении 334 (нумерация EU) на Е;
(viii) замены аминокислоты D в положении 270 (нумерация EU)
на Е;
замены аминокислоты К в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты К в положении 334 (нумерация EU) на Е;
(ix) замены аминокислоты D в положении 270 (нумерация EU) на
Е;
замены аминокислоты К в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на К и
замены аминокислоты К в положении 334 (нумерация EU) на Е.
Селективность активности связывания можно определять измерением активности связывания полипептида с соответствующей изоформой рецептора Fcγ, а затем определением их отношений. Например, количество связывания и значение KD с FcγR можно использовать в качестве показателя активности связывания.
В настоящем описании "улучшение селективности активности связывания" означает, что, например, отношение активности связывания тестируемого полипептида с изоформой рецептора Fcγ
(активность связывания родительского полипептида тестируемого полипептида с первой изоформой рецептора Fcγ/активность связывания тестируемого полипептида со второй изоформой рецептора Fcγ) повышается на 0,1 или более или предпочтительно на 0,2 или более, 0,5 или более, 1 или более, 2 или более, 3 или более, 5 или более, 7 или более, 8 или более, 9 или более, 10 или более, 15 или более, 20 или более, 30 или более, 50 или более, 70 или более, 100 или более, 150 или более, 200 или более, 500 или более, или 1000 или более, по сравнению с отношением активностей связывания тестируемого полипептида с изоформами рецептора Fcγ (активность связывания родительского полипептида тестируемого полипептида с первой изоформой рецептора Fcγ/активность связывания родительского полипептида тестируемого полипептида со второй изоформой рецептора Fcγ), определяемым указанным выше способа измерения. Кроме того, уменьшение селективности изоформы рецептора Fcγ означает, что, например, отношение активностей связывания тестируемого полипептида с изоформами рецептора Fcγ уменьшается на 0,1 или более или предпочтительно на 0,2 или более, 0,5 или более, 1 или более, 2 или более, 3 или более, 5 или более, 7 или более, 8 или более, 9 или более, 10 или более, 15 или более, 20 или более, 30 или более, 50 или более, 70 или более, 100 или более, 150 или более, 200 или более, 500 или более, или 1000 или более, по сравнению с отношением активности связывания родительского пептида тестируемого полипептида с изоформами рецептора Fcγ, определяемым указанным выше способом измерения.
В настоящем описании, в качестве показателя селективности также можно использовать, например, отношение А/I, которое показывает отношение активности связывания с активирующим FcγR и ингибирующим FcγR. Значения, получаемые делением KD тестируемого полипептида для FcγRIIb на KD тестируемого полипептида для FcγRIIa Н типа или R типа использовали в качестве соответствующих отношений А/I. Отношение А/I предпочтительно составляет 1,1 или более, 1,5 или более, 2 или более, 3 или более, или 5 или более и более предпочтительно 6 или более, 8 или более, или 9 или более.
В настоящем описании в качестве показателя селективности можно использовать, например, отношение FcγRIIIa F/FcγRIIb, которое представляет собой значение, получаемое делением KD для FcγRIIb на KD для FcγRIIIa F. Значения, получаемые делением KD тестируемого полипептида для FcγRIIb на KD тестируемого полипептида для FcγRIIIa определяли как соответствующие отношения FcγRIIIa F/FcγRIIb. Отношение FcγRIIIa F/FcγRIIb предпочтительно составляет 1,1 или более, 1,5 или более, 2 или более, 3 или более, или 5 или более и более предпочтительно 10 или более, 20 или более, 30 или более, 40 или более, 50 или более, 60 или более, 70 или более, 80 или более, 90 или более, 100 или более, 110 или более, 120 или более, 130 или более, 140 или более, 150 или более, 200 или более, 210 или более, 220 или более, 230 или более, или 240 или более.
В настоящем изобретении, когда изменение функции Fc-области полипептида представляет собой улучшение селективности
активности связывания с рецептором Fcγ, мутацию аминокислот можно вводить в аминокислотную последовательность первого полипептида и/или второго полипептида, составляющих Fc-область. Тип и диапазон мутаций аминокислот, которые можно вводить, не является конкретно ограниченным.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIa, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное повышение активности связывания с FcγRIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблиц 19-1, 19-2, 19-3 и 19-4 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное повышение активности связывания с FcγRIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 19-1, 19-2, 19-3, 19-4 и 19-5 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае когда активирующий рецептор Fcγ представляет собой FcγRIa, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное снижение активности связывания с FcγRIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы,
состоящей из мутаций аминокислот, описанных в области с таблиц 23-1 и 23-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное снижение активности связывания с FcγRIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 23-1 и 23-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIa R, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное повышение активности связывания с FcγRIIa R по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 20-1 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное повышение активности связывания с FcγRIIa R по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 20-1, 20-2 и 20-3 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или
второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIa R, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIa R по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблицы 24-1 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIa R по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 24-1 и 24-2 в настоящем описании можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIa Н, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное повышение активности связывания с FcγRIIa Н по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 21-1 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда
улучшение селективности представляет собой селективное повышение активности связывания с FcγRIIa Н по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 21-1, 21-2 и 21-3 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIa Н, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIa Н по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблиц 25-1 и 25-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIa Н по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d таблиц 25-1, 25-2 и 25-3 в настоящем описании можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIIa, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное повышение
активности связывания с FcγRIIIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области а таблицы 22-1 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное повышение активности связывания с FcγRIIIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области b таблиц 22-1, 22-2 и 22-3 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда активирующий рецептор Fcγ представляет собой FcγRIIIa, ингибирующий рецептор Fcγ представляет собой FcγRIIb, и улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области с таблиц 26-1 и 26-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область. Кроме того, когда улучшение селективности представляет собой селективное снижение активности связывания с FcγRIIIa по сравнению с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в области d
таблиц 26-1, 26-2, 26-3 и 26-4 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В настоящем описании селективное повышение активности связывания с желаемым рецептором Fcγ означает любой из перечисленных ниже случаев:
i) активность связывания с желаемым рецептором Fcγ повышают, и активности связывания с рецепторами, отличными от желаемого рецептора Fcγ не изменяют или уменьшают;
ii) активность связывания с желаемым рецептором Fcγ повышают, и активности связывания с рецепторами, отличными от желаемого рецептора Fcγ также повышают, но степень повышения активности связывания с рецепторами, отличными от желаемого рецептора Fcγ является меньше чем степень повышения активности связывания с желаемым рецептором Fcγ, или
iii) активность связывания с желаемым рецептором Fcγ снижают, но степень снижения активности связывания является меньше чем степень снижения активностей связывания с рецепторами Fcγ, отличными от желаемого рецептора Fcγ.
Кроме того, селективное снижение активности связывания с желаемым рецептором Fcγ означает любой из следующих случаев:
i) активность связывания с желаемым рецептором Fcγ снижают, и активности связывания с рецепторами, отличными от желаемого рецептора Fcγ не изменяют или повышают;
ii) активность связывания с желаемым рецептором Fcγ снижают,
и активности связывания с рецепторами, отличными от желаемого рецептора Fcγ также снижают, но степень снижения активности связывания с рецепторами, отличными от желаемого рецептора Fcγ является меньше чем степень снижения активности связывания с желаемым рецептором Fcγ, или
iii) активность связывания с желаемым рецептором Fcγ повышают, но степень повышения активности связывания является меньше чем степень повышения активности связывания с рецепторами Fcγ, отличными от желаемого рецептора Fcγ.
В настоящем описании физико-химическая стабильность полипептида означает, например, термодинамическую стабильность полипептида, которую можно определять с использованием, например, значения Tm домена СН2 в качестве показателя. Значения Tm можно измерять посредством кругового дихроизма (CD), дифференциального сканирующего калориметра (DSC) или дифференциальной сканирующей флуориметрии (DSF).
Для вычисления значения Tm измеряют изменение средней остаточной молярной эллиптичности (θ), которая сопровождает повышение температуры, посредством CD. Измерительное устройство включает, например, устройство измерения дисперсии кругового дихроизма (JASCO Corporation). Когда измеряют спектры CD при подходящей длине волны (например, 208 нм или 222 нм) при повышении температуры, θ повышается при определенной температуре и становится постоянным значением при более высоких температурах. Температура, при которой вычисляют среднюю точку между низкой температурой θ и высокой температурой θ, принимают
как Tm. Для измерения можно использовать, например, раствор белка, получаемый с использованием лимонной кислоты, Tris, фосфатного раствора или т.п., в концентрации нескольких сотен мкг/мл.
Для вычисления значения Tm DSC измеряют изменение калорий, которое сопровождает повышение температуры. Измерительное устройство включает MicroCal VP-DSC и Micro Cal Capillary DSC (оба от DKSH Japan). Белковый раствор и буфер помещают в измерительные ячейки, и когда измеряют разницы температур между ячейками при повышении температуры, наблюдают, что при определенных температурах начинается изменение в сторону эндотермической реакции. Эту температуру принимают за равную Tm. Для измерения можно использовать, например, раствор белка, получаемый с использованием цитратного буфера, TBS, PBS, гистидинового буфера или т.п.в концентрации от нескольких десятков мкг/мл до нескольких сотен мкг/мл.
Для вычисления значения Tm DSF детектируют экспозицию гидрофобных остатков, которая сопровождает повышение температуры, с использованием флуоресцентного реагента (например, SYPRO Orange), который специфически связывается с гидрофобными остатками. Раствор белка и флуоресцентный реагент смешивают в соответствующих отношениях, и когда измеряют интенсивности флуоресценции при повышении температуры с использованием устройства ПЦР в режиме реального времени, при определенной температуре наблюдают повышение интенсивности флуоресценции. Эту температуру принимают за равную Tm. Примеры измерительного устройства включают Rotor-Gene Q (QIAGEN) и
систему анализа CFX96 ПЦР в режиме реального времени (Bio-Rad). Для измерения можно использовать, например, раствор белка, получаемый с использованием PBS, гистидинового буфера или т.п.в концентрации от нескольких десятков мкг/мл до нескольких сотен мкг/мл.
В настоящем описании улучшенная физико-химическая стабильность полипептида означает, например, что значение Tm домена СН2 в Fc-области тестируемого полипептида, определяемое указанным выше способом измерения, повышается на 0,1 градуса или более, предпочтительно 0,2 градуса или более, 0,3 градуса или более, 0,4 градуса или более, 0,5 градусов или более, 1 градус или более, 2 градуса или более, 3 градуса или более, 4 градуса или более, 5 градусов или более, или 10 градусов или более по сравнению со значением Tm домена СН2 в Fc-области контрольного полипептида. Кроме того, улучшенная физическая стабильность полипептида относится к уменьшенному снижению физической стабильности полипептида, и например, снижение значения Tm домена СН2 в Fc-области тестируемого полипептида уменьшают относительно значения Tm домена СН2 в Fc-области контрольного полипептида на 0,1 градуса или более, предпочтительно 0,2 градуса или более, 0,3 градуса или более, 0,4 градуса или более, 0,5 градусов или более, 1 градус или более, 2 градуса или более, 3 градуса или более, 4 градуса или более, 5 градусов или более, или 10 градусов или более, как указанным выше способом измерения.
В настоящем описании "снижение физической стабильности полипептида" означает, что значение Tm домена СН2 в Fc-области
тестируемого полипептида, определяемое указанным выше способом измерения, снижается на 0,1 градуса или более, предпочтительно 0,2 градуса или более, 0,3 градуса или более, 0,4 градуса или более, 0,5 градусов или более, 1 градус или более, 2 градуса или более, 3 градуса или более, 4 градуса или более, 5 градусов или более, или 10 градусов или более по сравнению со значением Tm домена СН2 в Fc-области контрольного пептида.
Настоящее изобретение также включает полипептид, содержащий Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, и где функция Fc-области является измененной относительно функции Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, или по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид.
В полипептиде изменение функции Fc-области может представлять собой изменение, которое дополнительно улучшает физико-химическую стабильность в дополнение по меньшей мере к одному или нескольким изменениям, выбранным из группы, состоящей из повышения активности связывания, снижения связывания и улучшения селективности активности связывания полипептида с рецептором Fcγ, и при условии, что любую из этих функций изменяют, можно сказать, что изменяют функцию Fc-области по настоящему изобретению.
В настоящем изобретении фраза "когда мутации аминокислот вводят в Fc-область в первый полипептид и второй полипептид,
функция Fc-области не изменяется" означает, что, когда вводят одинаковые мутации аминокислот в первый полипептид и второй полипептид, желаемую функцию не улучшают. Например, это означает, что когда предусматривают повышение активности связывания полипептида с рецептором Fcγ, активность связывания не изменяется или снижается; когда предусматривают снижение активности связывания, активность связывания не изменяется или повышается; когда предусматривают улучшение селективности активности связывания, селективность не улучшается, и когда предусматривают улучшение физико-химической стабильности полипептида, стабильность не изменяется или снижается. В отношении мутации аминокислоты фраза "когда ее вводят только в одну из Fc-областей, функцию Fc-области изменяют" означает, что желаемую функцию улучшают, когда мутацию аминокислот вводят только в любой из первого полипептида или второго полипептида. Например, это означает, что когда предусматривают повышение активности связывания полипептида с рецептором Fcγ, активность связывания повышается; когда предусматривают снижение активности связывания, активность связывания снижается; когда предусматривают улучшение селективности активности связывания, селективность улучшается, и когда предусматривают улучшение физико-химической стабильности полипептида, стабильность улучшается.
Настоящее изобретение также относится к полипептиду, который отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, и который отличается тем, что имеет более высокую Tm по сравнению с Tm
полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, или полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид. Наряду с изменением, которое улучшает физико-химическую стабильность, т.е. наличие высокой Tm, полипептид также может содержать дополнительные изменения функции Fc-области.
В случае, когда дополнительное изменение функции Fc-области представляет собой повышение активности связывания с FcγRIa, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 31-1, 31-2 и 31-3 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда дополнительное изменение функции Fc-области представляет собой повышение активности связывания с FcγRIIa R, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 32-1 и 32-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда дополнительное изменение функции Fc-области представляет собой повышение активности связывания с FcγRIIa Н, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 33-1 и 33-2 в настоящем описании, можно вводить в аминокислотные
последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда дополнительное изменение функции Fc-области представляет собой повышение активность связывания с FcγRIIb, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 34-1 и 34-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В случае, когда дополнительное изменение функции Fc-области представляет собой повышение активность связывания с FcγRIIIa, по меньшей мере одну или более мутаций аминокислот, выбранных из группы, состоящей из мутаций аминокислот, описанных в таблицах 35-1 и 35-2 в настоящем описании, можно вводить в аминокислотные последовательности первого полипептида и/или второго полипептида, составляющих Fc-область.
В настоящем изобретении комбинация первого полипептида и второго полипептида, в которые вводят мутации аминокислот являются конкретно не ограниченными, и примеры включают комбинации различных типов/или одинакового типа полипептидов, выбранных из полипептидов, описанных в SEQ ID NO:2-4 и 6-60. Кроме того, предпочтительные примеры включают комбинацию полипептидов, содержащих первый полипептид и второй полипептид, описанные в примерах в настоящем описании (комбинация Н-цепей двух антител и L-цепи одиночного антитела).
Полипептид по настоящему изобретению может представлять собой антигенсвязывающую молекулу. В настоящем изобретении
вследствие того, что связывающая антитело молекула является конкретно не ограниченной по типу, предпочтительные примеры включают антитело, биспецифическое антитело или слитую с Fc молекулу, такую как слитый белок с пептидом Fc или слитый с каркасной областью Fc белок.
Антитело
Кроме того, предоставлено антитело в качестве полипептида по настоящему изобретению.
Термин "антитело/антитела" в настоящем изобретении используют в самом широком смысле и при условии, что обладает желаемой биологической активностью, оно включает любое антитело, такое как моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, варианты антител, фрагменты антител, полиспецифические антитела (например, биспецифические антитела), химерные антитела и гуманизированные антитела.
В отношении антител по настоящему изобретению типа антигена и происхождение антитела не являются ограниченными, и они могут представлять собой любой тип антител. Происхождение антител является конкретно не ограниченным, и примеры включают антитела человека, антитела мыши, антитела крысы и антитела кролика.
Способы получения антител хорошо известны специалистам в данной области, и, например, моноклональные антитела можно получать гибридомным способом (Kohler and Milstein, Nature, 256:495 (1975)), или способом рекомбинации (патент США №4816567). Альтернативно, их можно выделять из фаговой библиотеки антител (Clackson et al., Nature, 352:624-628 (1991); Marks et
al., J. Mol. Biol., 222:581-597 (1991)). Альтернативно, их можно выделять из одного клона В-клетки (N. Biotechnol., 28 (5):253-457 (2011)).
Гуманизированное антитело также называют реконструированное антитело человека. Конкретно, известны гуманизированные антитела, получаемые пересадкой CDR антитела, не являющегося человеком животного, такого как антитело мыши, в антитело человека и т.п. Также известны общепринятые технологии генетической инженерии получения гуманизированных антител. Конкретно, например, известна ПЦР с достройкой перекрывающихся участков в качестве способа пересадки CDR антитела мыши в FR человека.
Вектор для экспрессии гуманизированного антитела можно получать встраиванием ДНК, кодирующей вариабельную область антитела, в которой лигируют три CDR и четыре FR и ДНК, кодирующую константную область антитела человека в экспрессирующий вектор, таким образом, что эти ДНК подвергают слиянию в рамке считывания. После такой интеграции вектор трансфицируют в хозяина с получением рекомбинантных клеток, эти клетки культивируют и экспрессируют ДНК, кодирующую гуманизированное антитело, с получением гуманизированного антитела в культуре клеток (см., европейскую патентную публикацию №ЕР 239400 и международную патентную публикацию №WO 1996/002576).
По мере необходимости аминокислотный остаток в FR можно замещать таким образом, что CDR реконструированного антитела человека образует соответствующий антигенсвязывающий участок.
Например, мутацию можно вводить в аминокислотную
последовательность FR способом ПЦР, который используют для пересадки CDR мыши в FR человека.
Желаемое антитело человека можно получать иммунизацией ДНК с использованием трансгенного животного, содержащего полный репертуар генов антитела человека (см. международную публикацию №№WO 1993/012227, WO 1992/003918, WO 1994/002602, WO 1994/025585, WO 1996/034096 и WO 1996/033735) в качестве животного для иммунизации.
Кроме того, известны технологии получения антитела человека посредством пэннинга с использованием библиотеки антител человека. Например, способом фагового дисплея обеспечивают экспрессию V-области антитела человека на поверхности фага в виде одноцепочечного антитела (scFv). Можно выбирать экспрессирующий scF фаг, который связывается с антигеном. ДНК последовательность, кодирующую V-область связывающего антиген антитела человека можно определять, анализируя гены выбранного фага. После определения ДНК последовательности scFv, которое связывается с антигеном, можно получать экспрессирующий вектор слиянием V-области последовательности в рамке считывания с последовательностью С-области желаемого антитела человека, а затем встраивать это в подходящий экспрессирующий вектор. Экспрессирующий вектор вводят в подходящие клетки для экспрессии, такие как описанные выше клетки, и можно получать антитело человека посредством экспрессии кодирующего антитело человека гена. Такие способы уже известны (см. международную публикацию №№WO 1992/001047, WO 1992/020791, WO 1993/006213, WO
1993/011236, WO 1993/019172, WO 1995/001438 и WO 1995/15388).
Вариабельные области, из которых состоят антитела по настоящему изобретению, могут представлять собой вариабельные области, который распознают любой антиген.
В настоящем описании не существует конкретного ограничения для антигена, и он может представлять собой любые антигены. Примеры антигена включают 17-IA, 4-1 ВВ, 4Dc, 6-кето-PGFla, 8-изо-PGF2a, 8-оксо-dG, рецептор аденозина Al, А33, АСЕ, АСЕ-2, активин, активин А, активин АВ, активин В, активин С, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RUB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17 /ТАСЕ, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессины, адипонектин, АДФ-рибозилциклазу-1, aFGF, AGE, ALCAM, ALK, ALK-1, ALK-7, аллерген, альфа-1-антихимотрипсин, альфа-1-антитрипсин, альфа-синуклеин, антагонист альфа-V/бета-1, аминин, амилин, бета амилоид, вариабельную область тяжелой цепи иммуноглобулина амилоида, вариабельную область легкой цепи иммуноглобулина амилоида, андроген, ANG, ангиотензиноген, лиганд ангиопоэтина-2, анти-Id, антитромбин III, антракс, APAF-1, АРЕ, APJ, аро А1, сывороточный амилоид аро A, Apo-SAA, АРР, APRIL, AR, ARC, ART, артемин, аспаргиновую кислоту, атриальный натрийуретический фактор, атриальный натрийуретический пептид, атриальные натрийуретические пептиды А, атриальные натрийуретический пептиды В, атриальные натрийуретические пептиды С, интегрин av/b3, Axl, В7-1, В7-2, В7-Н, ВАСЕ, ВАСЕ-1, защитный антиген Bacillus anthracis, Bad, BAFF, BAFF-R, Bag-1, ВАК, Bax, BCA-1, BCAM, BeI, BCMA, BDNF, b-ECGF, бета-2-микроглобулин,
беталактамазу, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, стимулятор В-лимфоцитов (BIyS), BMP, BMP-2 (BMP-2a), BMP-3 (остеогенин), BMP-4 (BMP-2b), BMP-5, BMP-6 (Vgr-1), BMP-7 (OP-1), BMP-8 (BMP-8a), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BMPR-II (BRK-3), BMP, BOK, бомбезин, костный нейротрофический фактор, бычий гормон роста, BPDE, BPDE-ДНК, BRK-2, ВТС, молекулу клеточной адгезии В-лимфоцитов, С10, С1-ингибитор, C1q, С3, С3а, С4, С5, С5а (комплемент 5а), СА125, CAD-8, кадгерин-3, кальцитонин, цАМФ, карбоангидразу-IX, раково-эмбриональный антиген (СЕА), ассоциированный с карциномой антиген, кардиотрофин-1, катепсин А, катепсин В, катепсин C/DPPI, катепсин D, катепсин Е, катепсин Н, катепсин L, катепсин О, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, ССК2, CCL, CCL1/I-309, CCL11/эотаксин, CCL12/MCP-5, CCL13/MCP-4, CCL14/HCC-1, CCL15/HCC-2, CCL16/HCC-4, CCL17/TARC, CCL18/PARC, CCL19/ELC, CCL2/MCP-1, CCL20/MIP-3-альфа, CCL21/SLC, CCL22/MDC, CCL23/MPIF-1, CCL24/эотаксин-2, CCL25/TECK, CCL26/эотаксин-3, CCL27/СТАСК, CCL28/MEC, CCL3/M1P-1-альфа, CCL3L1/LD-78-бета, CCL4/MIP-1-бета, CCL5/RANTES, CCL6/C10, CCL7/MCP-3, CCL8/MCP-2, CCL9/10/МТР-1-гамма, CCR, CCR1, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD10, CD105, CD11a, CD11b, CD11c, CD123, CD13, CD137, CD138, CD14, CD140a, CD146, CD147, CD148, CD15, CD152, CD16, CD164, CD18, CD19, CD2, CD20, CD21, CD22, CD23, CD25, CD26, CD27L, CD28, CD29, CD3, CD30, CD30L, CD32, CD33 (белки р67), CD34, CD37, CD38, CD3E, CD4, CD40, CD40L, CD44, CD45, CD46, CD49a, CD49b, CD5, CD51, CD52, CD54, CD55, CD56, CD6, CD61, CD64, CD66e, CD7, CD70, CD74, CD8, CD80 (B7-1), CD89, CD95,
CD105, CD158a, CEA, CEACAM5, CFTR, cGMP, рецептор CGRP, CINC, CKb8-1, клауидн 18, CLC, токсин Clostridium botulinum, токсин Clostridium difficile, токсин Clostridium perfringens, c-Met, CMV, CMV UL, CNTF, CNTN-1, фактор комплемента 3 (С3), фактор комплемента D, связывающий кортикостероид глобулин, рецептор колониестимулирующего фактора 1, СОХ, C-Ret, CRG-2, CRTH2, СТ-1, СТАСК, CTGF, CTLA-4, CX3CL1/фракталкин, CX3CR1, CXCL, CXCLl/Gro-альфа, CXCL10, CXCL11/I-TAC, CXCL12/SDF-1-альфа/бета, CXCL13/BCA-1, CXCL14/BRAK, CXCL15/Lungkine, CXCL16, CXCL16, CXCL2/Gro-бетa CXCL3/Gro-гаммa, CXCL3, CXCL4/PF4, CXCL5/ENA-78, CXCL6/GCP-2, CXCL7/NAP-2, CXCL8/IL-8, CXCL9/Mig, CXCLlO/IP-10, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цистатина С, цитокератин опухолеассоциированный антиген, DAN, DCC, DcR3, DC-SIGN, комплементзависимый стимулятор гемолиза, лиганд дельта-подобного белка 4, des(1-3)-IGF-1 (IGF-1 головного мозга), Dhh, оксидазу DHICA, Dickkopf-1, дигоксин, дипептидилпептидазу IV, DK1, DNAM-1, ДНКазу, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), содержащий EGF-подобный домен белок 7, эластазу, эластин, EMA, EMMPRIN, ENA, ENA-78, эндосиалин, рецептор эндотелина, эндотоксин, энкефалиназу, eNOS, Eot, эотаксин, эотаксин-2, эотаксин, ЕрСАМ, эфрин B2/EphB4, рецептор тирозинкиназы Epha2, рецептор эпидермального фактора роста (EGFR), рецептор ErbB2, рецептор тирозинкиназы ЕrbB3, ERCC, EREG, эритропоэтин (ЕРО), рецептор эритропоэтина, Е-селектин, ЕТ-1, Exodus-2, белок F RSV, F10, F11, F12, F13, F5, F9, фактор Iа, фактор IX, фактор Ха, фактор VII, фактор VIII, фактор VIIIc, Fas, Fc-альфа-R, Fc-эпсилон-RI, Fc-гамма-IIb, Fc-
гамма-RI, Fc-гамма-RIIa, Fc-гамма-RIIIa, Fc-гамма-RIIIb, FcRn, FEN-1, ферритин, FGF, FGF-19, FGF-2, рецептор FGF-2, FGF-3, FGF-8, FGF-кислый, FGF-основной, FGFR, FGFR-3, фибрин, белок активатор фибробластлв (FAP), фактор роста фибробластов, фактор роста фибробластов 10, фибронектин, FL, FLIP, Flt-З, лиганд FLT3, рецептор фолата, фолликулостимулирующий гормон (FSH), фракталкин (СХ3С), свободную тяжелую цепь, свободную легкую цепь, FZD1, FZD10, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, G250, Gas 6, GCP-2, GCSF, G-CSF, рецептор G-CSF, GD2, GD3, GDF, GDF-1, GDF-15 (MIC-1), GDF-3 (Vgr-2), GDF-5 (BMP-14/CDMP-1), GDF-6 (BMP-13/CDMP-2), GDF-7 (BMP-12/CDMP-3), GDF-8 (миостатин), GDF-9, GDNF, гельсолин, GFAP, GF-CSF, GFR-альфа-1, GFR-aльфa12, GER-альфа13, GF-b1, гликопротеин оболочки gH, GITR, глюкагон, рецептор глюкагона, рецептор глюкагоноподобного пептида 1, Glut 4, глутаминаткарбоксипептидазу II, рецепторы гликопротеинового гормона, гликопротеин IIb/IIIa (GP IIb/IIIa), глипикан-3, GM-CSF, рецептор GM-CSF, gp130, gp140, gp72, гранулоцит-CSF (G-CSF), GRO/MGSA, рилизинг-фактор гормона роста, GRO-β, GRO-γ, H. pylori, гаптен (NP-cap или NIP-cap), HB-EGF, НСС, НСС 1, гликопротеин оболочки HCMV gВ, HCMV UL, гемопоэтический фактор роста (HGF), Hep В gp120, гепараназу, кофактор гепарина II, фактор роста гепатоцитов, защитный антиген Bacillus anthracis, гликопротеин Е2 вируса гепатита С, гепатит Е, гепсидин, Her1, Her2/neu (ErbB-2), Неr3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), HGF, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMw-MAA), белки оболочки ВИЧ, такие как GP120, петля V3 gp 120 MIB ВИЧ,
HLA, HLA-DR, HM1,24, HMFG РЕМ, HMGB-1, HRG, Hrk, HSP47, Hsp90, гликопротеин gD HSV, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (hGH), сывороточный альбумин человека, активатор плазминогена типа ткани человека (t-PA), хантингтин, HVEM, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFN-альфа, IFN-бета, IFN-гамма, IgA, рецептор IgA, IgE, IGF, связывающие белки IGF, IGF-1, IGF-1 R, IGF-2, IGFBP, IGFR, IL, IL-1, IL-10, рецепторы IL-10, IL-11, рецепторы IL-11, IL-12, рецепторы IL-12, IL-13, рецепторы IL-13, IL-15, рецепторы IL-15, IL-16, рецепторы IL-16, IL-17, рецепторы IL-17, IL-18 (IGIF), рецепторы IL-18, IL-1-альфа, IL-1-бета, рецепторы IL-1, IL-2, рецепторы IL-2, IL-20, рецепторы IL-20, IL-21, рецепторы IL-21, IL-23, рецепторы IL-23, рецепторы IL-2, IL-3, рецепторы IL-3, IL-31, рецепторы IL-31, рецепторы IL-3, IL-4, рецепторы IL-4, IL-5, рецепторы IL-5, IL-6, рецепторы IL-6, IL-7, рецепторы IL-7, IL-8, рецепторы IL-8, IL-9, рецепторы IL-9, иммунный комплекс с иммуноглобулином, иммуноглобулины, INF-альфа, рецепторы INF-альфа, INF-бета, рецепторы INF-бета, INF-гамма, рецепторы INF-гамма, IFN I тип, рецептор IFN I типа, грипп, ингибин, ингибин α, Ингибин β, iNOS, инсулин, А-цепь инсулина, В-цепь инсулина, инсулиноподобный фактор роста 1, инсулиноподобный фактор роста 2, связывающие белки инсулиноподобного фактора роста, интегрин, интегрин альфа-2, интегрин альфа-3, интегрин альфа-4, интегрин альфа-4/бета-1, интегрин альфа-V/бета-3, интегрин альфа-V/бета-6, интегрин альфа-4/бета-7, интегрин альфа-5/бета-1, интегрин альфа-5/бета-3, интегрин альфа-5/бета-6, интегрин альфа-σ
(альфа-V), интегрин альфа-θ, интегрин бета-1, интегрин бета-2, интегрин бета-3 (GPIIb-IIIa), IP-10, I-TAC, JE, калликреин, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин 2, калликреин 5, калликреин 6, калликреин L1, калликреин L2, калликреин L3, калликреин L4, каллистатин, КС, KDR, фактор роста кератиноцитов (KGF), фактор роста кератиноцитов 2 (KGF-2), KGF, иммуноглобулинподобный рецептор киллерных клеток, лиганд Kit (KL), тирозинкиназы Kit, ламинин 5, LAMP, LAPP (амилин, островковый амилоидный полипептид), LAP (TGF-1), связанный с латентностью пептид, латентный TGF-1, латентный TGF-1 bpl, LBP, LDGF, LDL, рецептор LDL, LECT2, Lefty, лептин, лютеинизирующий гормон (LH), антиген Lewis-Y, связанный с Lewis-Y антиген, LFA-1, LFA-3, рецепторы LFA-3, Lfo, LIF, LIGHT, липопротеины, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, легкое поверхностно-активное вещество, лютеинизирующий гормон, лимфотактин, рецептор лимфотоксина-бета, рецептор лизосфинголипидов, Мас-1, макрофаг-CSF (M-CSF), MAdCAM, MAG, МАР2, MARC, маспин, МСАМ, МСК-2, МСР, МСР-1, МСР-2, МСР-3, МСР-4, МСР-I (MCAF), M-CSF, MDC, MDC (67 а.к.), MDC (69 а.к.), мегзин, Меr, семейство рецепторов тирозинкиназы МЕТ, металлопротеазы, мембранный гликопротеин OX2, мезотелин, рецептор MGDF, MGMT, МНС (HLA-DR), микробный белок, MIF, MIG, MIP, MIP-1α, MIP-1β, MIP-3α, MIP-3β, MIP-4, МК, ММАС1, ММР, ММР-1, ММР-10, ММР-11, ММР-12, ММР-13, ММР-14, ММР-15, ММР-2, ММР-24, ММР-3, ММР-7, ММР-8, ММР-9, моноцитарный хемотаксический белок, фактор, подавляющий колониеобразование моноцитов, гонадотропин-ассоциированный пептид мыши, MPIF, Мро, MSK, MSP,
MUC-16, MUC18, муцин (Mud), мюллерову ингибирующую субстанцию, Mug, MuSK, миелин-ассоциированный гликопротеин, ингибирующий предшественники миелоида фактор 1 (MPIF-I), NAIP, нанотело, NAP, NAP-2, NCA 90, NCAD, N-кадгерин, NCAM, неприлизин, молекулу адгезии нейрональных клеток, неросерпин, фактор роста нейронов (NGF), нейротрофин-3, нейротрофин-4, нейротрофин-6, нейропилин 1, нейротурин, NGF-бета, NGFR, NKG20, N-метионил гормон роста человека, nNOS, NO, Nogo-A, рецептор Nogo, неструктурный белок 3 типа (NS3) из вируса гепатита С, NOS, Npn, NRG-3, NT, NT-3, NT-4, NTN, OB, OGG1, онкостатин M, OP-2, OPG, OPN, OSM, рецепторы OSM, остеоиндуктивные факторы, остеопонтин, OX40L, OX40R, окисленный LDL, р150, р95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, Р-кадгерин, PCNA, PCSK9, PDGF, рецептор PDGF, PDGF-AA, PDGF-AB, PDGF-BB, PDGF-D, PDK-1, РЕСАМ, PEDF, РЕМ, PF-4, PGE, PGF, PGI2, PGJ2, PIGF, PIN, PLA2, фактор роста плаценты, плацентарная щелочная фосфатаза (PLAP), плацентарный лактоген, ингибитор активатора плазминогена 1, фактор роста тромбоцитов, plgR, PLP, полигликолиеевые цепи различного размера (например, PEG-20, PEG-30, PEG40), РР14, прекалликреин, прионный белок, прокальцитонин, белок программируемой гибели клеток 1, проинсулин, пролактин, пропротеинконвертазу РС9, прорелаксин, специфический мембранный антиген предстательной железы (PSMA), белок А, белок С, белок D, белок S, белок Z, PS, PSA, PSCA, PsmAr, PTEN, PTHrp, Ptk, PTN, гликопротеиновый лиганд Р-селектина 1, R51, RAGE, RANK, RANKL, RANTES, релаксин, А-цепь релаксина, В-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, Ret, ретикулон 4, ревматоидные факторы, RLI Р76,
RPA2, RPK-1, RSK, RSV Fgp, S100, RON-8, SCF/KL, SCGF, склеростин, SDF-1, SDF1a, SDF1b, серин, сыворотный амилоид P, сывороточный альбумин, sFRP-3, Shh, шигаподобный токсин II, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, рецептор 1 сфингозин-1-фосфата, стафилококковую липотейхоевую кислоту, Stat, STEAP, STEAP-II, фактор стволовых клеток (SCF), стерптокиназу, супероксиддисмутазу, стиндекан-1, ТАСЕ, TACI, TAG-72 (опухолеассоциированный гликопротеин-72), TARC, ТВ, ТСА-3, Т-клеточный рецептор альфа/бета, TdT, ТЕСК, TEM1, ТЕМ5, ТЕМ7, ТЕМ8, тенасцин, TERT, тестикулярная PLAP-подобная щелочная фосфатаза, TfR, TGF, TGF-альфа, TGF-бета, Pan-специфический TGF-бета, TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета R1 (ALK-5), TGF-бета-1, TGF-бета-2, TGF-бета-3, TGF-бета-4, TGF-бета-5, TGF-I, тромбин, тромбопоэтин (ТРО), рецептор тимусного стромального лимфопоэтина, тимус Ck-l, тиреотропновый гормон (TSH), тироксин, тироксинсвязывающий глобулин, Tie, TIMP, TIQ, тканевой фактор, протеазный ингибитор тканевого фактора, белок тканевого фактора, TMEFF2, Tmpo, TMPRSS2, рецептор TNF I, рецептор TNF II, TNF-альфа, TNF-бета, TNF-бета-2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2/DR4), TNFRSF10B (TRAIL R2 DR5/KILLER/TRICK-2A/TRICK-B), TNFRSF10C (TRAIL R3 DcR1/LIT/TRID), TNFRSF10D (TRAIL R4 DcR2/TRUNDD), TNFRSF11A (RANK ODF R/TRANCE R), TNFRSF11B (OPG OCIF/TR1), TNFRSF12 (TWEAK R FN14), TNFRSF12A, TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR/HveA/LIGHT R/TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ/TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF R1 CD120a/p55-60), TNFRSF1B (TNF RII
CD120b/p75-80), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRSF25 (DR3 Apo-3/LARD/TR-3/TRAMP/WSL-l), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII/TNFC R), TNFRSF4 (OX40 ACT35/TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1/APT1/CD95), TNFRSF6B (DcR3 M68/TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1 BB CD137/ILA), TNFRST23 (DcTRAIL R1 TNFRH1), TNFSF10 (лиганд TRAIL Apo-2/TL2), TNFSF11 (TRANCE/RANK лиганд ODF/лиганд OPG), TNFSF12 (лиганд TWEAK Аро-3/лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS/TALL1/THANK/TNFSF20), TNFSF14 (лиганд LIGHT HVEM/LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд GITR лиганд AITR/TL6), TNFSF1A (TNF-a конектин/DIF/TNFSF2), TNFSF1B (TNF-b LTa/TNFSF1), TNFSF3 (LTb TNFC/р33), TNFSF4 (лиганд OX40 gp34/TXGP1), TNFSF5 (CD40 лиганд CD154/gp39/HIGM1/IMD3/TRAP), TNFSF6 (лиганд Fas лиганд Аро-1/лиганд APT1), TNFSF7 (CD27 лиганд CD70), TNFSF8 (CD30 лиганд CD153), TNFSF9 (лиганд 4-1 BB лиганд CD137), TNF-α, TNF-β, TNIL-I, метаболит токсина, TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, трансферриновый рецептор, трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-бета, трансмембранный гликопротеин NMB, транстиретин, TRF, Trk, TROP-2, трофобластный гликопротеин, TSG, TSLP, фактор некроза опухоли (TNF), опухолеассоциированный антиген СА 125, опухолеассоциированный антиген, экспрессирующий родственный Lewis Y углевод, TWEAK, ТХВ2, Ung, uPAR, uPAR-1, урокиназу, VAP-1, фактор роста эндотелия сосудов (VEGF), васпин, VCAM, VCAM-1, VECAD, VE-кадгерин, УЕ-кадгерин-2, VEFGR-1 (flt-1), VEFGR-2, рецептор VEGF (VEGFR), VEGFR-3 (flt-4), VEGI, VIM, вирусные антигены, рецептор VitB12, рецептор витронектина, VLA,
VLA-1, VLA-4, интегрин VNR, фактор фон Виллебранда (vWF), WIF-1, WNT1, WNT10A, WNT10B, WNT11, WNT16, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, XCL1, XCL2/SCM-1-бета, ХСL1/лимфотактин, XCR1, XEDAR, XIAP, XPD.
Одно или более изменений аминокислотных остатков можно вводить в аминокислотные последовательности, из которых состоят вариабельные области, при условии, что сохраняются их активности связывания с антигеном. При изменении аминокислотной последовательности вариабельной области не существует конкретного ограничения в отношении участка изменения и числа изменяемых аминокислот. Например, соответствующим образом можно изменять аминокислоты, содержащиеся в CDR и/или FR. При изменении аминокислот в вариабельной области, предпочтительно сохраняется активность связывания без конкретного ограничения, и, например, по сравнению с состоянием до изменения, активность связывания составляет 50% или более, предпочтительно 80% или более и более предпочтительно 100% или более. Кроме того, активность связывания можно повышать изменениями аминокислот. Например, активность связывания может являться в 2, 5, 10 раз выше или т.п.по сравнению с активностью связывания до изменения. В антителах по настоящему изобретению изменение аминокислотной последовательности может представлять собой по меньшей мере одну замену, добавление, делецию, вставку и модификацию аминокислотных остатков.
Например, модификация N-концевого глутамина вариабельной области в пироглутаминовой кислоте посредством
пироглутамилированием является хорошо известной специалистам в данной области модификацией. Таким образом, когда тяжелая цепь N-конца представляет собой глутамин, антитела по настоящему изобретению содержат вариабельные области, в которых глутамин модифицируют до пироглутаминовой кислоты.
Вариабельные области антитела по настоящему изобретению могут содержать любые последовательности, и они могут представлять собой вариабельные области антител любой природы, таких как антитела мыши, антитела крысы, антитела кролика, антитела козы, антитела верблюда, гуманизированные антитела, получаемые гуманизацией таких не принадлежащих человеку антител, и антитела человека. "Гуманизированные антитела", также обозначаемые как "реконструированные антитела человека", представляют собой антитела, в которых определяющие комплементарность области (CDR) антитела, получаемого от не являющегося человеком млекопитающего, например, антитела мыши, трансплантируют в CDR антитела человека. Способы определения CDR являются известными (Kabat et al., Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989), 342:877). Также известны общепринятые технологии их генетической рекомбинации (см. публикацию европейской патентной заявки №ЕР 125023 и WO 96/02576). Кроме того, эти антитела могут содержать различные замены аминокислот, вводимые в их вариабельные области для улучшения их связывания с антигеном, фармакокинетики, стабильности и антигенности. Вариабельные области антител по настоящему изобретению могут повторно связываться с антигенами
вследствие их зависимости от рН при связывании антигена (WO/2009/125825).
В константных областях легкой цепи антител содержатся константные области типа k-цепей и L-цепей, а также любая из константных областей легкой цепи является приемлемой. Кроме того, константные области легкой цепи по настоящему изобретению могут представлять собой константные области легкой цепи с изменениями аминокислот, такими как замены, добавки, делеции, вставки и/или модификации.
Например, для константных областей тяжелой цепи антитела по настоящему изобретению можно использовать константные области тяжелой цепи антител IgG человека, и предпочтительными являются константные области тяжелой цепи антител IgG1 человека.
Вариабельные области, из которых состоит антитело по настоящему изобретению, могут представлять собой вариабельные области, которые распознают любой антиген. Один или более аминокислотных остатков в аминокислотной последовательности, из которой состоит вариабельная область тяжелой цепи, можно изменять при условии, что сохраняется активность связывания с антигеном.
Кроме того, изменение вариабельной области проводят с целью повышения активности связывания, улучшения специфичности, снижения pI, обеспечения свойства зависимости от рН при связывании антигена, улучшения термостабильности связывания, улучшения растворимости, улучшения стабильности химической модификации, улучшения гетерогенности, получаемой для цепей сахаров, устранения Т-клеточного эпитопа, который снижает
иммуногенность, определяемую компьютерным моделированием или анализом in vitro с использованием Т-клеток, вводя Т-клеточный эпитоп, который активирует регуляторные Т-клетки, или т.п. (mAbs 3:243-247, 2011).
Кроме того, полипептид по настоящему изобретению может представлять собой молекулу слитого с Fc белка, получаемую связыванием Fc-области с другим белком, биологически активным пептидом или т.п.(слитый с пептидом Fc белок), или молекулу слитого с Fc белка, получаемую связыванием Fc-область с внеклеточным матриксом, состоящим из полимеров, таких как коллаген или полимолочная кислота (слитый с каркасной областью Fc белок).
Примеры другого белка или биологически активного пептида включают рецепторы, молекулы адгезии, лиганды и ферменты, но не ограничены ими.
Предпочтительные примеры молекул слитого с Fc белка по настоящему изобретению включают белки с доменом Fc, слитые с рецепторным белком, который связывается с мишенью, и такие примеры включают слитый белок TNFR-Fc, слитый белок IL1R-Fc, слитый белок VEGFR-Fc и слитый белок CTLA4-Fc (Nat Med., 2003 Jan; 9(1):47-52; BioDrugs. 2006; 20(3):151-60). Кроме того, белок для слияния с полипептидом по настоящему изобретению может представлять собой любую молекулу при условии, что она связывается с молекулой-мишенью, и примеры включают молекулы scFv (WO 2005/037989), молекулы однодоменных антител (WO 2004/058821; WO 2003/002609), подобные антителам молекулы (Current Opinion in Biotechnology, 2006, 17:653-658; Current
Opinion in Biotechnology, 2007, 18:1-10; Current Opinion in Structural Biology, 1997, 7:463-469, и Protein Science, 2006, 15:14-27), такие как DARPin (WO 2002/020565), Affibody (WO 1995/001937), Avimer (WO 2004/044011; WO 2005/040229) и Adnectin (WO 2002/032925). Кроме того, антитела и молекулы слитого с Fc белка могут представлять собой полиспецифические антитела, которые связываются со многими типами молекул-мишеней или эпитопов, такие как биспецифические антитела.
Кроме того, антитела по настоящему изобретению включают продукты модификации антител. Такие продукты модификации антител включают, например, антитела, связанные с различными молекулами, такими как полиэтиленгликоль (PEG) и цитотоксические вещества. Такие продукты модификации антител можно получать, химически модифицируя антитела по настоящему изобретению. В данной области уже установлены способы модификации антител.
Антитела по настоящему изобретению также могут представлять собой биспецифические антитела. "Биспецифическое антитело" относится к антителу, которое содержит в молекуле одиночного антитела вариабельные области, которые распознают различные эпитопы. Эпитопы могут содержаться в одной молекуле или в различных молекулах.
Полипептиды по настоящему изобретению можно получать известными специалистам в данной области способами. Например, антитела можно получать описанными ниже способами, но способы не являются ограниченными ими.
Существуют различные известные комбинации клетки-хозяина /экспрессирующего вектора для получения антител
посредством введения выделенных генов, кодирующих полипептид, в соответствующих хозяев. Все эти экспрессирующие системы можно применять для выделения антигенсвязывающих молекул по настоящему изобретению. Подходящие эукариотические клетки, используемые в качестве клеток-хозяев, включают клетки животных, растительные клетки и клетки грибков. В частности, клетки животных включают, например, следующие ниже клетки.
1) Клетки млекопитающих: СНО (линия клеток яичника китайского хомяка), COS (линия клеток почки обезьяны), миелома (Sp2/O, NS0 и т.п.), ВНК (линия клеток почки новорожденного хомяка), НЕК293 (линия эмбриональных клеток почки человека с расщепленной ДНК аденовируса (Ad)5), клетка PER. С6 (линия эмбриональных клеток сетчатки человека, трансформированных генами (Ad5) Е1А и Е1В аденовируса 5 типа), Hela, Vero или т.п. (Current Protocols in Protein Science (May, 2001, Unit 5, 9, Table 5.9.1)).
2) Клетки амфибий: ооциты Xenopus или т.п. и
3) Клетки насекомых: sf9, sf21, Тn5 или т.п.
Экспрессируют ДНК, кодирующую тяжелую цепь антитела, в которой один или более аминокислотных остатков в Fc-области заменяли другими представляющими интерес аминокислотами, и ДНК, кодирующую легкую цепь антитела. ДНК, кодирующую тяжелую цепь, в которой заменяют один или несколькими аминокислотными остатками в Fc-области на другие представляющие интерес аминокислоты, можно получать, например, получая ДНК, кодирующую Fc-область природной тяжелой цепи, и вводя соответствующую замену, таким образом, чтобы кодон, кодирующий конкретную аминокислоту в Fc-
области, кодировал другую представляющую интерес аминокислоту.
Альтернативно, ДНК, кодирующую тяжелую цепь, в которой один или более аминокислотных остатков в Fc-области заменяют другими представляющими интерес аминокислотами, также можно получать, конструируя, а затем химически синтезируя ДНК, кодирующую белок, в котором один или более аминокислотных остатков в Fc-области природной тяжелой цепи заменяют другими представляющими интерес аминокислотами. Положение и тип замены аминокислот не являются конкретно ограниченными. Кроме того, изменение не ограничено заменой, и изменение может представлять собой любую из делеции, добавления или вставки или их сочетания.
Альтернативно, ДНК, кодирующую тяжелую цепь, в которой один или более аминокислотных остатков в Fc-области заменяют другими представляющими интерес аминокислотами, можно получать как комбинацию неполных ДНК. Такие комбинации неполных ДНК включают, например, комбинацию ДНК, кодирующей вариабельную область, и ДНК, кодирующей константную область, и комбинацию ДНК, кодирующей Fab-область и ДНК, кодирующей Fc-область, но не являются ограниченными ими. Кроме того, ДНК, кодирующую легкую цепь, можно аналогичным образом получать в виде комбинации неполных ДНК.
Способы экспрессии описанных выше ДНК включают описанные ниже способы. Например, экспрессирующий тяжелую цепь вектор конструируют посредством введения ДНК, кодирующей вариабельную область тяжелой цепи, в экспрессирующий вектор совместно с ДНК, кодирующей константную область тяжелой цепи. Аналогично, экспрессирующий легкую цепь вектор конструируют посредством
введения ДНК, кодирующей вариабельную область легкой цепи, в экспрессирующий вектор совместно с ДНК, кодирующей константную область легкой цепи. Альтернативно, эти гены тяжелой и легкой цепей можно вводить в один вектор.
При встраивании ДНК, кодирующей представляющее интерес антитело в экспрессирующий вектор, ДНК вводят таким образом, чтобы антитело экспрессировались под контролем регулирующей экспрессию области, такой как энхансер или промотор. Затем клетки-хозяева трансформируют таким экспрессирующим вектором для экспрессии антитела. В таких случаях можно использовать подходящую комбинацию хозяина и экспрессирующего вектора.
Примеры векторов включают векторы М13, векторы pUC, pBR322, pBluescript и pCR-Script. Альтернативно, с целью субклонирования и удаления кДНК в дополнение к описанным выше векторам можно использовать pGEM-T, pDIRECT, рТ7 и т.п.
Экспрессирующие векторы являются особенно пригодными при использовании векторов для получения антител по настоящему изобретению. Например, когда клетка-хозяин представляет собой Е. coli, такую как JM109, DH5a, НВ101 и XL1-Blue, экспрессирующие векторы должны содержать промотор, который обеспечивает эффективную экспрессию в Е. coli, например, промотор lacZ (Ward et al., Nature, (1989) 341:544-546; FASЕВ J., (1992) 6:2422-2427; полностью включенные в настоящее описание посредством ссылки), промотор araB (Better et al., Science, (1988) 240:1041-1043; полностью включенную в настоящее описание посредством ссылки), промотор Т7 или т.п. В дополнение к описанным выше векторам такие векторы включают pGEX-5X-1 (Pharmacia),
"QIAexpress system" (QIAGEN), pEGFP, или рЕТ (в этом случае хозяин предпочтительно представляет собой BL21, который экспрессирует РНК-полимеразу Т7).
Векторы могут содержать сигнальные последовательности для секреции полипептида. Если полипептид секретируется в периплазме Е. coli, в качестве сигнальных последовательностей для секреции полипептида, можно использовать сигнальную последовательность pellB (Lei S.P. et al., J. Bacteriol., (1987) 169:4379; полностью включенную в настоящее описание посредством ссылки). Вектор можно вводить в клетки-хозяева, например, способом с использованием липофектина, способом с использованием фосфата кальция и способом с использованием DEAE-декстрана.
В дополнение к экспрессирующим векторам на основе Е. coil векторы для получения полипептидов по настоящему изобретению включают, например, экспрессирующие векторы млекопитающих (например, pcDNA3 (Invitrogen), pEGF-BOS (Nucleic Acids. Res., 1990, 18(17):p5322; полностью включенную в настоящее описание посредством ссылки), pEF и pCDM8), получаемые их клеток насекомого экспрессирующие векторы (например, "бакуловирусную экспрессирующую систему Bac-to-BAC" (GIBCO BRL) и pBacPAK8), получаемые из растений экспрессирующие векторы (например, рМН1 и рМН2), получаемые из вирусов животных экспрессирующие векторы (например, pHSV, pMV и pAdexLcw), ретровирусные экспрессирующие векторы (например, pZIPneo), дрожжевые экспрессирующие векторы (например, "Pichia Expression Kit" (Invitrogen), pNV11 и SP-Q01) и экспрессирующие векторы Bacillus subtilis (например, pPL608 и pKTHSO).
Для экспрессии в клетках животного, таких как клетки СНО, COS, NIH3T3 и HEK293, векторы должны содержать промотор, необходимый для экспрессии в клетках, например, промотор SV40 (Mulligan et al., Nature, (1979) 277:108; полностью включенной в настоящее описание посредством ссылки), промотор MMTV-LTR, промотор EF1a (Mizushima et al., Nucleic Acids Res., (1990) 18:5322; полностью включенной в настоящее описание посредством ссылки), промотор CAG (Gene, (1990) 18:5322; полностью включенной в настоящее описание посредством ссылки) и промотор CMV, и более предпочтительно они содержат ген для отбора трансформированных клеток (например, ген устойчивости к лекарственным средствам, что позволяет проводить оценку с использованием средства (неомицина, G418 или т.п.)). Векторы с такими характеристиками включают, например, рМАМ, pDR2, pBK-RSV, pBK-CMV, pOPRSV и рОР13. Кроме того, в некоторых случаях для увеличения числа копий гена можно дополнительно совместно экспрессировать белок EBNA1, и в этом случае используют вектор, содержащий точку начала репликации OriP (Biotechnol. Bioeng., 2001, Oct 20; 75 (2):197-203; и Biotechnol. Bioeng., 2005 Sep 20; 91(6):670-7).
Кроме того, для стабильной экспрессии гена и увеличения числа копий гена в клетках можно использовать следующий способ: в клетки СНО с дефицитом в пути синтеза нуклеиновых кислот вводят вектор, который содержит ген DHFR, который компенсирует дефицит (например, pCHOI), и амплифицируют вектор с использованием метотрексата (МТХ). Альтернативно, для временной экспрессии гена можно использовать следующий способ: клетки COS,
содержащие на хромосоме ген, обеспечивающий экспрессию антигена SV40 Т, трансформируют вектором с участком начала репликации SV40 (pcD и т.п.). Также можно использовать участки начала репликации, выделяемые из вируса полиомы, аденовируса, вируса папилломы крупнорогатого скота (BPV) и т.п. Для увеличения числа копий гена в клетках-хозяевах экспрессирующие векторы могут дополнительно содержать селективные маркеры, такие как ген аминогликозидтрансферазы (АРН), ген тимидинкиназы (ТK), ген ксантингуанинфосфорибозилтрансферазы Е. coli (Ecogpt) и ген дигидрофолатредуктазы (dhfr).
Антитела можно получать, например, культивированием трансформированных клеток, а затем выделением антител из внутреннего содержимого трансформированных клеток или из среды для культивирования. Антитела можно выделять и очищать соответствующей комбинацией способов, таких как центрифугирование, фракционирование сульфатом аммония, высаливание, ультрафильтрация, 1q, FcRn, колонка с белком А, белком G, аффинная хроматография, ионообменная хроматография и гельфильтрационная хроматография.
В качестве эффективного способа получения биспецифических антител можно использовать технологию "выступы во впадины". В частности, для получения гетеродимеризованного полипептида по настоящему изобретению необходимо наличие ассоциации между полипептидами, содержащими аминокислоты, которые отличаются друг от друга, или необходимо отделять представляющий интерес гетеродимеризованный полипептид от других гомодимеризованных полипептидов.
Для ассоциации полипептидов, содержащих отличающиеся друг от друга аминокислоты и содержащих Fc-область, можно применять технологию подавления непредусмотренной ассоциации между Н-цепями посредством введения электростатического отталкивания на поверхности второй константной области Н-цепи антитела (СН2) или третьей константной области Н-цепи (СН3) (WO 2006/106905).
В технологии подавления непредусмотренной ассоциации между Н-цепями посредством введения электростатического отталкивания на поверхности СН2 или СН3 примеры аминокислотных остатков в области контактирования на поверхности других константных областей Н-цепи включают остаток в положении 356 (нумерация EU), остаток в положении 439 (нумерация EU), область, находящуюся перед остатком в положении 357 (нумерация EU), остаток в положении 370 (нумерация EU), остаток в положении 399 (нумерация EU) и остаток в положении 409 (нумерация EU) в домене СН3.
Более конкретно, например, для антитела, содержащего два типа доменов СН3 Н-цепи, можно получать антитело, в котором от одной до трех пар аминокислотных остатков, выбранных из аминокислотных остатков, представленных ниже в (1)-(3), в домене СН3 первой Н-цепи, имеют одинаковый тип заряда:
1) аминокислотные остатки в положениях 356 и 439 (нумерация EU), которые представляют собой аминокислотные остатки, содержащиеся в домене СН3 Н-цепи;
2) аминокислотные остатки в положениях 357 и 370 (нумерация EU), которые представляют собой аминокислотные остатки, содержащиеся в домене СН3 Н-цепи, и
3) аминокислотные остатки в положениях 399 и 409 (нумерация
EU), которые представляют собой аминокислотные остатки, содержащиеся в домене СН3 Н-цепи.
Кроме того, можно получать антитело, в котором от одной до трех пар аминокислотных остатков, соответствующих парам аминокислотных остатков, указанных выше в (1)-(3), обладающих аналогичным типом заряда в домене СН3 первой Н-цепь, обладают зарядом, противоположным соответствующим аминокислотным остаткам в указанном выше домене СН3 первой Н-цепи, где пары аминокислотных остатков в домене СН3 второй Н-цепи, который отличается от домена СН3 первой Н-цепи, выбраны из пар аминокислотных остатков, указанных выше в (1)-(3).
Соответствующие указанные выше аминокислотные остатки (1)-(3) при ассоциации располагают вблизи друг от друга. Специалисты в данной области могут выявлять участки, которые соответствуют указанным выше аминокислотным остаткам (1)-(3), посредством гомологичного моделирования и т.п. с использованием коммерчески доступного программного обеспечения для желаемого домена СН3 Н-цепи или константной области Н-цепи, и при необходимости можно изменять аминокислотные остатки этих участков.
В указанных выше антителах, например, "несущие заряд аминокислотные остатки" предпочтительно выбрать из аминокислотных остатков, входящих в любую из групп (X) или (Y) ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D), и
(Y) лизин (K), аргинин (R) и гистидин (Н). Для указанных выше антител фраза "обладающий одинаковым типом заряда" означает, что, например, все два или более
аминокислотных остатка представляют собой аминокислотные остатки, входящие в указанные выше группы (X) и (Y). Фраза "обладающий противоположным зарядом" означает, например, когда по меньшей мере один из двух или более аминокислотных остатков представляет собой аминокислотный остаток, входящий в любую из указанных выше групп (X) и (Y), оставшиеся аминокислотные остатки представляют собой аминокислотные остатки, входящие в другую группу.
В предпочтительном варианте осуществления указанного выше антитела домен СН3 первой Н-цепи и домен СН3 второй Н-цепи можно поперечно сшивать посредством дисульфидных связей.
В настоящем изобретении подлежащие изменению аминокислотные остатки не ограничены аминокислотными остатками описанной выше константной области антитела или вариабельной области антитела. Специалисты в данной области могут находить аминокислотные остатки, которые образуют область контакта в мутантах полипептидов или гетеромультимерах, посредством гомологичного моделирования и т.п. с использованием коммерчески доступного программного обеспечения и могут изменять аминокислотные остатки на этих участках для регуляции ассоциации.
Также можно использовать другие известные технологии для ассоциации полипептидов по настоящему изобретению, содержащих различные аминокислоты и содержащих Fc-область. Содержащие различные аминокислоты и содержащие Fc-область полипептиды можно эффективно ассоциировать друг с другом, заменяя аминокислотную боковую цепь, содержащуюся в одной из вариабельных областей Н-цепи антитела, на более крупную боковую цепь ("выступ") и
заменяя аминокислотную боковую цепь, содержащуюся в противоположной вариабельной области, на другую Н-цепь с меньшей боковой цепью ("впадина") для обеспечения возможности расположения "выступа" во "впадине" (WO 1996/027011 и Ridgway J.B. et al., Protein Engineering, (1996) 9, 617-621; Merchant A.M. et al., Nature Biotechnology, (1998) 16, 677-681).
Кроме того, для ассоциации полипептидов, содержащих различные аминокислоты и содержащих Fc-область, также можно использовать другие известные технологии. Ассоциацию полипептидов, содержащих различные последовательности, можно эффективно индуцировать посредством комплементарной ассоциации СН3 с использованием сконструированного домена СН3 с заменой цепей, получаемого заменой участка СН3 в одной из Н-цепей антитела на получаемую из IgA последовательность, соответствующую этому участку, и введением соответствующей получаемой из IgA последовательности в комплементарный участок СН3 на другой Н-цепи (Protein Engineering Design & Selection, 23; 195-202, 2010). Эту известную технологию также можно использовать для эффективной индукции ассоциации полипептидов, содержащих различные аминокислоты и содержащих Fc-область.
Кроме того, также можно использовать технологии получения гетеродимеризованных антител, в которых используют ассоциации СН1 и CL антитела и ассоциации VH и VL, которые описаны в WO 2011/028952.
Кроме того, даже в случаях, когда гетеродимеризованные полипептиды невозможно эффективно получать, гетеродимеризованные полипептиды можно получать путем их выделения и очистки из
гомодимеризованных полипептидов. При получении гетеродимеризованного полипептида, состоящего из первого полипептида и второго полипептида, которые содержат отличные друг от друга последовательности, гомодимеризованные полипептиды, состоящие только из двух первых полипептидов, и гомодимеризованный полипептид, состоящий только из двух вторых полипептидов, смешивают в качестве примесей. В качестве способа эффективного удаления этих двух типов гомодимеризованных полипептидов можно использовать известные технологии. Опубликовано, что способом можно очищать два типа гомодимеров и представляющее интерес гетеродимеризованное антитело посредством ионообменной хроматографии путем создания различия изоэлектрических точек введением замены аминокислот в вариабельные области двух типов Н-цепей (WO 2007114325).
В отношении изменения аминокислот для обеспечения различия изоэлектрических точек изменение аминокислот, которые необходимо вводить, не являются конкретно ограничеными при условии, что получают различие между изоэлектрическими точками двух образующих ассоциацию полипептидов, и оно также может включать изменения аминокислот, проводимые с целями, такими как снижение иммуногенности. Изменяемые аминокислоты предпочтительно представляют собой аминокислоты в положениях с небольшим влиянием на активность связывания с рецептором Fcγ. Кроме того, оно может представлять собой изменение аминокислот, которое повышает активность связывания с желаемым рецептором Fcγ. Для таких изменений предпочтительно вводить по меньшей мере одну мутацию аминокислот в положение аминокислот, выбранное из
конкретно из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Val в положении 263, Glu в положении 272, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 364, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Pro в положении 387, Asn в положении 390, Val в положении 397 и Val в положении 422 (нумерация EU), в аминокислотной последовательности первого полипептида и/или второго полипептида. Кроме того, предпочтительно вводить мутацию по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Ile в положении 199, Asn в положении 203, Lys в положении 214, Glu в положении 272, Lys в положении 274, Lys в положении 288, Lys в положении 290, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390, Val в положении 397 и Val в положении 422 (нумерация EU). Кроме того, более предпочтительно вводить мутацию по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении
138, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Ile в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Lys в положении 288, Leu в положении 358, Asn в положении 384 и Val в положении 397 (нумерация EU).
Более конкретно предпочтительно, чтобы мутацию вводили по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gln в положении 196, Ile в положении 199, Val в положении 263, Glu в положении 272, Gly в положении 316, Leu в положении 358, Ser в положении 364, Ser в положении 383, Pro в положении 387 и Val в положении 397 (нумерация EU), в аминокислотной последовательности любого полипептида из первого полипептида и второго полипептида; и предпочтительно мутацию вводят по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390 и Val в положении 422 (нумерация EU), в аминокислотной последовательности другого полипептида. Кроме того, предпочтительно, чтобы мутацию вводили по меньшей мере в одну аминокислоту, выбранную из группы,
состоящей из Gln в положении 196, Ile в положении 199, Glu в положении 272, Leu в положении 358, Ser в положении 383 и Val в положении 397 (нумерация EU), в аминокислотной последовательности одного из полипептидов; и предпочтительно мутацию вводят по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Ile в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Lys в положении 288, Lys в положении 290, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390 и Val в положении 422 (нумерация EU), в аминокислотной последовательности другого полипептида. Кроме того, более предпочтительно, чтобы мутацию вводили по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gln в положении 196, Ile в положении 199, Leu в положении 358 и Val в положении 397 (нумерация EU), в аминокислотной последовательности одного из полипептидов; и чтобы мутацию вводили по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении 138, Lys в положении 147, Ser в положении 192, Leu в положении 193, Ile в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Lys в положении 288 и Asn в положении 384 (нумерация EU), в аминокислотной последовательности другого полипептида.
Изменения аминокислот конкретно не ограничены при условии, что изменение проводят для получения различия изоэлектрических
точек между двумя образующими ассоциацию полипептидами после изменения.
Примеры предпочтительного изменения для повышения изоэлектрической точки включают замену аминокислоты в положении 196 на Lys, замену аминокислоты в положении 263 на Lys, замену аминокислоты в положении 272 на Lys, замену аминокислоты в положении 316 на Lys, замену аминокислоты в положении 364 на Lys, замену аминокислоты в положении 358 на Lys, замену аминокислоты в положении 383 на Lys, замену аминокислоты в положении 387 на Lys и замену аминокислоты в положении 397 на Lys (нумерация EU). Примеры предпочтительного изменения для снижения изоэлектрической точки включают замену аминокислоты в положении 137 на Glu, замену аминокислоты в положении 138 на Glu, замену аминокислоты в положении 139 на Glu, замену аминокислоты в положении 147 на Glu, замену аминокислоты в положении 198 на Glu, замену аминокислоты в положении 203 на Asp, замену аминокислоты в положении 214 на Thr, замену аминокислоты в положении 274 на Gln, замену аминокислоты в положении 278 на Glu, замену аминокислоты в положении 288 на Glu, замену аминокислоты в положении 290 на Glu, замену аминокислоты в положении 316 на Glu, замену аминокислоты в положении 317 на Glu, замену аминокислоты в положении 320 на Glu, замену аминокислоты в положении 324 на Glu, замену аминокислоты в положении 335 на Glu, замену аминокислоты в положении 337 на Asp, замену аминокислоты в положении 340 на Glu, замену аминокислоты в положении 358 на Glu, замену аминокислоты в положении 360 на Glu, замену аминокислоты в
положении 362 на Glu, замену аминокислоты в положении 383 на Glu, замену аминокислоты в положении 384 на Glu, замену аминокислоты в положении 385 на Glu, замену аминокислоты в положении 386 на Glu, замену аминокислоты в положении 390 на Glu и замену аминокислоты в положении 422 на Glu (нумерация EU).
При комбинации изменений аминокислот, которые проводят с целью отличной от получения различия изоэлектрических точек, например, для снижения антигенности, можно комбинировать замену аминокислоты в положении 138 на Ser, замену аминокислоты в положении 192 на Asn, замену аминокислоты в положении 193 на Phe и замену аминокислоты в положении 199 на Thr (нумерация EU).
В настоящее время в качестве способа очистки гетеродимеризованного антитела опубликован способ, в котором используют белок А для очистки гетеродимеризованного антитела, содержащего Н-цепь IgG2a мыши, которая связывается с белком А, и Н-цепь IgG2b крысы, которая не связывается с белком A (WO 98050431 и WO 95033844).
Кроме того, можно эффективно очищать одно гетеродимеризованное антитело с использованием Н-цепей, в которых аминокислотные остатки в положениях 435 и 436 (нумерация EU), которые представляют собой участок связывания IgG и белка А, заменяют на аминокислоты, такие как Tyr и His, которые обладают различной аффинностью к белку А, для изменения взаимодействия между каждой из Н-цепей и белком А, и с использованием колонки с белком А. Ряд, например, две или более, таких замен и технологий можно использовать в комбинации. Кроме того, при необходимости эти изменения можно применять отдельно
для первого полипептида и второго полипептида. Полипептиды по настоящему изобретению могут представлять собой полипептиды, получаемые на основе полипептидов, в которых применяли указанные выше изменения.
Настоящее изобретение также относится к способу получения полипептида, содержащего Fc-область, который включает стадии гетеродимеризации Fc-области введением мутации аминокислот в первый полипептид и/или второй полипептид, из которых состоит Fc-область, и введением мутации аминокислот для изменения функции Fc-области по сравнению с тем, когда Fc-область образует гомодимер.
Примеры включают способ получения, включающий следующие ниже стадии:
a) в полипептиде, содержащем Fc-область, введения мутации аминокислоты в первый полипептид и/или второй полипептид, из которых состоит Fc-область;
b) определения функции Fc-области гетеродимера, состоящего из первого полипептида и второго полипептида, в которые вводят мутацию на стадии а), и
c) отбора полипептида с измененной функцией Fc-области по сравнению с родительским полипептидом или по сравнению с тем, когда Fc-область гомодимеризуют введением мутации аминокислоты.
В этом способе получения после стадии а) можно проводить еле дующую стадию:
d) экспонирования содержащего Fc-область гетеродимеризованного полипептида, состоящего из первого полипептида и второго полипептида, на предоставленных рибосомах,
фагах или дрожжах.
Предпочтительный вариант осуществления представляет собой способ получения полипептида, содержащего Fc-область, который включает стадии:
a) изменения нуклеиновой кислоты, кодирующей полипептид, таким образом, чтобы функция Fc-области являлась измененной по сравнению с родительским полипептидом или по сравнению с тем, когда Fc-область образует гомодимер, посредством введения мутации аминокислот;
b) введения нуклеиновой кислоты в клетку-хозяина и культивирование клетки для экспрессии полипептида, и
c) выделения полипептида из культуры клеток-хозяина.
В настоящее изобретение также включены антитела и молекулы слитого с Fc белка, получаемые способом получения.
Тип и диапазон мутаций аминокислот, вводимых настоящим способом, конкретно не ограничены, и примеры включают мутации аминокислот, участвующие в изменение каждой функции Fc-области, описываемой в настоящем описании (более конкретно, мутации аминокислот, конкретно описанные в таблицах в примерах).
Настоящее изобретение также относится к способу изменения функции полипептида, содержащего Fc-область, который включает стадии гетеродимеризации Fc-области введением мутации аминокислоты в первый полипептид и/или второй полипептид, из которых состоит Fc-область, для изменения функции Fc-области по сравнению с тем, когда Fc-область образует гомодимер, посредством введения мутации аминокислот.
Примеры включают способы изменения, включающие следующие
ниже стадии:
a) в полипептиде, содержащем Fc-область, введения мутации аминокислоты в первый полипептид и/или второй полипептид, из которых состоит Fc-область;
b) определения функции Fc-области гетеродимера, состоящего из первого полипептида и второго полипептида, в которые вводят мутацию на стадии а), и
c) отбора полипептида с измененной функцией Fc-области по сравнению с родительским полипептидом или по сравнению с тем, когда Fc-область гомодимеризуют введением мутации аминокислоты.
В этом способе изменения после стадии а) можно проводить следующую стадию:
d) экспонирования содержащего Fc-область гетеродимеризованного полипептида, состоящего из первого полипептида и второго полипептида, на предоставленных рибосомах, фагах или дрожжах.
Предпочтительный вариант осуществления представляет собой способ изменения полипептида, содержащего Fc-область, который включает стадии:
a) изменения нуклеиновой кислоты, кодирующей полипептид, таким образом, чтобы функция Fc-области являлась измененной по сравнению с родительским полипептидом или по сравнению с тем, когда Fc-область образует гомодимер, посредством введения мутации аминокислот;
b) введения нуклеиновой кислоты в клетку-хозяина и культивирование клетки для экспрессии полипептида, и
c) выделения полипептида из культуры клеток-хозяина.
В настоящее изобретение также включены антитела и молекулы слитого с Fc белка, получаемые способом изменения.
Тип и диапазон мутаций аминокислот, вводимых настоящим способом, конкретно не ограничены, и примеры включают мутации аминокислот, участвующие в изменение каждой функции Fc-области, описываемой в настоящем описании (более конкретно мутации аминокислот, конкретно описанные в таблицах в примерах).
Кроме того, настоящее изобретение относится к нуклеиновой кислоте, кодирующей полипептид, содержащий Fc-область, где полипептид отличается тем, что Fc-область состоит из гетеродимера, содержащего первый полипептид и второй полипептид, и где полипептид отличается тем, что функция Fc-области является измененной по сравнению с функций Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только первый полипептид, и/или по сравнению с функцией Fc-области полипептида, отличающегося тем, что Fc-область состоит из гомодимера, содержащего только второй полипептид. Нуклеиновая кислота по настоящему изобретению может находиться в любой форме, такой как ДНК или РНК.
Настоящее изобретение также относится к векторам, содержащим описанные выше нуклеиновые кислоты по настоящему изобретению. Специалисты в данной области могут соответствующим образом выбирать тип вектора можно в зависимости от клеток-хозяев, в которые необходимо вводить вектор. Векторы включают, например, такие векторы, как описанные выше.
Кроме того, настоящее изобретение относится к клеткам-хозяевам, трансформированным описанными выше векторами по
настоящему изобретению. Специалисты в данной области может выбирать подходящие клетки-хозяева. Клетки-хозяева включают, например, такие клетки-хозяева, как описанные выше.
Фармацевтические композиции
Настоящее изобретение относится к фармацевтическим композициям, содержащим полипептид по настоящему изобретению.
В дополнение к антителам или молекулам слитого с Fc белка, которые представляют собой полипептиды по настоящему изобретению, можно формулировать фармацевтические композиции по настоящему изобретению с фармацевтически приемлемыми носителями известными способами. Например, композиции можно использовать парентерально, когда антитела или молекулы слитого с Fc белка формулируют в стерильном растворе или суспензии для инъекций с использованием воды или любой другой фармацевтически приемлемой жидкости. Например, композиции можно формулировать, соответствующим образом комбинируя антитела или молекулы слитого с Fc белка с фармакологически приемлемыми носителями или средами, в частности со стерильной водой или физиологическим раствором, растительными маслами, эмульгаторами, суспендирующими средствами, поверхностно-активными веществами, стабилизаторами, ароматизаторами, эксципиентами, носителями, консервантами, связывающими средствами и т.п., смешивая их в стандартной дозе и форме, согласно требованиям общепринятой фармацевтической реализации. Конкретные примеры носителей включают легкую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кармеллозу кальция, кармеллозу натрия, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу,
поливинилацетальдиэтиламиноацетат, поливинилпирролидон, желатин, среднецепочечные триглицериды, отвержденное полиоксиэтиленом касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганическую соль и т.п. Содержание активного ингредиента в таком составе регулируют таким образом, чтобы можно было получать подходящую дозу в необходимом диапазоне.
Стерильные композиции для инъекций можно формулировать с использованием носителей, таких как дистиллированная вода для инъекций, в соответствии со стандартными протоколами.
Водные растворы, используемые для инъекций, включают, например, физиологический раствор и изотонические растворы, содержащие глюкозу или другие адъюванты, такие как D-сорбит, D-манноза, D-маннит, и хлорид натрия. Их можно использовать в сочетании с подходящими солюбилизаторами, такими как спирт, в частности этанол, многоатомными спиртами, такими как пропиленгликоль и полиэтиленгликоль, и неионными поверхностно-активными веществами, такими как полисорбат 80™ и НСО-50.
Масла включают сезамовые масла и соевые масла, и их можно комбинировать с солюбилизаторами, такими как бензилбензоат или бензиловый спирт.Их можно также формулировать с буферами, например, фосфатными буферами или буферами на основе ацетата натрия; анальгетиками, например, прокаином гидрохлорида; стабилизаторами, например, бензиловым спиртом или фенолом; или антиоксидантами. Получаемые инъекционные растворы, как правило, разливают по аликвотам в подходящие ампулы.
Введение предпочтительно проводят парентерально, и, в
частности, оно включает инъекцию, интраназальное введение, внутрилегочное введение и чрескожное введение. Например, инъекции можно вводить системно или местно посредством внутривенной инъекции, внутримышечной инъекции, интраперитонеальной инъекции или подкожной инъекции.
Кроме того, способ введения можно соответствующим образом выбирать в зависимости от возраста и симптомов пациента. Стандартную лекарственную форму фармацевтической композиции, содержащей полипептид или полинуклеотид, кодирующий полипептид, можно выбирать, например, из диапазона от 0,0001 до 1,000 мг на кг массы тела. Альтернативно, дозирование может находиться, например, в диапазоне от 0,001 до 100000 мг/пациенту. Однако дозирование не является ограниченным этими значениями. Дозирование и способ введения изменяются в зависимости от массы тела, возраста пациента и симптомов, и специалисты в данной области могут соответствующим образом их выбирать.
В настоящем изобретении фармацевтические композиции, содержащие полипептиды по настоящему изобретению, описанные выше, являются пригодными в качестве активных ингредиентов для терапевтических средств или профилактических средств для злокачественной опухоли, иммуновоспалительных заболеваний и т.п.
Как используют в настоящем описании, трехбуквенные и однобуквенные коды для соответствующих аминокислот представляют собой такие, как указано ниже:
Аланин: Ala (А)
Аргинин: Arg (R)
Аспарагин: Asn (N)
Аспарагиновая кислота: Asp (D)
Цистеин: Cys (С)
Глутамин: Gln (Q)
Глутаминовая кислота: Glu (E)
Глицин: Gly (G)
Гистидин: His (H)
Изолейцин: Ile (I)
Лейцин: Leu (L)
Лизин: Lys (K)
Метионин: Met (М)
Фенилаланин: Phe (F)
Пролин: Pro (Р)
Серин: Ser (S)
Треонин: Thr (Т)
Триптофан: Trp (W)
Тирозин: Tyr (Y)
Валин: Val (V).
Все цитируемые в настоящем описании документы известного уровня техники полностью включены посредством ссылки.
ПРИМЕРЫ
В настоящем описании ниже настоящее изобретение дополнительно конкретно описано со ссылкой на примеры, но не следует это интерпретировать как ограничивающее его.
[Пример 1] Объяснение принципа улучшения распознавания FcγR гетеродимеризованными антителами
Антитело взаимодействует через свою Fc-область с различными молекулами, такими как FcRn, FcγR и комплементы. Одна молекула
FcRn, которая представляет собой лиганд Fc, связывается с каждой из тяжелых цепей (Н-цепи) антитела. Таким образом, две молекулы FcRn связываются с одной молекулой антитела (фиг. 1). In vivo FcRn экспрессируется на клеточной мембране. Таким образом, антитело распознает две молекулы FcRn симметричным образом через идентичные участки на своих соответствующих Н-цепях in vivo (Nature, 372:379-383, 1994). Кроме того, аналогично взаимоотношению IgG и FcRn одна молекула IgA, которая принадлежит к тому же семейству иммуноглобулинов, как IgG, распознает симметричным образом две молекулы FcaR, который представляет собой рецептор IgA (фиг. 2) (Nature, 423:614-620, 2003).
Однако в отличие от FcRn и других только одна молекула FcγR связывается с одной молекулой антитела (фиг. 3) (JBC, 276:16469-16477, 2001). IgG распознает FcγR через домены СН2 двух Н-цепей; однако участки взаимодействия с FcγR различаются в двух Н-цепях. Например, когда Н-цепь, представленную с левой стороны фиг. 3, определяют как HA-цепь и Н-цепь с правой стороны как HB-цепь, Ala в положении 327 (нумерация EU) в каждой из HA-цепи и HB-цепи взаимодействует с FcγR. Однако существует различие свойств парных остатков, с которыми взаимодействуют соответствующие Н-цепи (фиг. 4). HA-цепь взаимодействует с FcγRIII гидрофобным образом при Trp в положениях 87 и 110 (нумерация EU), тогда как HB-цепь взаимодействует с FcγRIII при His в положении 131 (нумерация EU). Таким образом, когда Ala в положении 327 (нумерация EU) заменяют высокогидрофобной аминокислотой, такой как Trp, можно снижать
активность связывания с FcγR HB-цепи, даже если это обладает эффектом улучшения активности связывания с FcγR HA-цепи. Таким образом, при оптимизации взаимодействия Fc-области IgG с FcγR посредством изменения аминокислот необходимо учитывать асимметричный эффект двух Н-цепей в отношении FcγR. Однако на известном уровне техники проводили аналогичное изменение в двух Н-цепях для оптимизации взаимодействия Fc-области IgG с FcγR (WO 2006/019447 и WO 2000/042072). Таким образом, учитывая асимметричное взаимодействие Fc-области IgG с FcγR, можно точнее оптимизировать взаимодействие IgG и FcγR посредством введения различных изменений в Н-цепи. Таким образом, взаимодействие с FcγR можно точнее оптимизировать посредством применения гетеродимеризованного антитела, получаемого в результате введения различных изменений в две Н-цепи для оптимизации взаимодействия Fc-области с FcγR по сравнению с гомодимеризованным антителом, получаемом в результате введения аналогичного изменения в Н-цепи, которую проводят в известном уровне техники.
[Пример 2] Доказательство принципа улучшения распознавания FcγR гетеродимеризованными антителами
Оценивали можно ли точнее оптимизировать активность связывания с FcγR антитела с использованием гетеродимеризованного антитела, в котором вводили различные изменения в две Н-цепи, по сравнению с гомодимеризованным антителом известного уровня техники.
Общепринято поиск изменений, которые повышают связывание
FcγR, проводили с использованием гомодимеризованного антитела, получаемого в результате введения аналогичного изменения в обе Н-цепи антитела. Однако, как описано в примере 1, антитело взаимодействует с FcγR асимметричным образом, и когда аналогичное изменение вводят в две Н-цепи, изменение в одной Н-цепи может повышать активность связывания с FcγR, тогда как изменение в другой Н-цепи может наоборот ингибировать связывание. Активность связывания с FcγR необязательно является повышенной в гомодимеризованном антителе, получаемом в результате введения такого изменения в обе Н-цепи. Однако гетеродимеризованное антитело, получаемое в результате введения изменения только в одну из двух Н-цепей, может обладать повышенной активностью связывания с FcγR.
Для тестирования этой гипотезы в отношении связывания FcγR гетеродимеризованное антитело, содержащее первый полипептид, в котором только в одну Н-цепь вводили изменение, которое, как предполагали, изменяет активность связывания с FcγR, и второй полипептид без указанного выше изменения, сравнивали с гомодимеризованным антителом, содержащим первый полипептид, в котором только в одну Н-цепь вводили изменение, которое как предполагали, изменяет активность связывания с FcγR. Основываясь на предыдущем принципе, когда активность связывания с FcγR повышают посредством изменения, гомодимеризованное антитело всегда превосходит гетеродимеризованное антитело. Однако если Fc антитела распознает FcγR асимметричным образом, предполагают, что гетеродимеризованное антитело проявляет большую активность
связывания с FcγR по сравнению с гомодимеризованным антителом в зависимости от типа изменения.
Вариабельная область Н-цепи используемого антитела представляла собой вариабельную область антитела против глипикана-3, которое содержит CDR рН7 антитела против глипикана-3 с улучшенной кинетикой в плазме, описанной в WO 2009/041062. Вариабельная область называется GpH7 (SEQ ID NO: 1). Константные области описанных ниже Н-цепей антитела использовали в комбинации с GpH7. Когда константную область Н-цепи антитела называют H1, последовательность Н-цепи антитела, содержащую вариабельную область GpH7, обозначают как GpH7-H1. Изменения аминокислот обозначают таким образом, как D356K. Первое буквенное обозначение (например, "D" D356K) представляет собой однобуквенный код, представляющий аминокислотный остаток перед изменением, и следующее число (например, "356" D356K) указывает положение изменения (нумерация EU). Последнее буквенное обозначение (например, "K" D356K) представляет собой однобуквенный код, представляющий аминокислотный остаток после изменения. GpH7-G1d (SEQ ID NO: 2), получаемую в результате удаления С-концевого Gly и Lys из IgG1, содержащего вариабельную область GpH7, GpH7-A5 (SEQ ID NO: 3), получаемую в результате введения мутаций D356K и H435R в GpH7-G1d, и GpH7-B3 (SEQ ID NO: 4), получаемую в результате введения К439Е в GpH7-G1d, получали способом, описанный в справочном примере 1. Мутации D356K и K439E вводили в каждую Н-цепь для обеспечения эффективного образования гетеродимера характерных Н-цепей при получении гетеродимеризованного антитела, содержащего два типа
Н-цепей (WO 2006/106905). Для обеспечения эффективного разделения гетеродимеризованных и гомодимеризованных форм вводили H435R, которое представляет собой изменение, которое ингибирует связывание с белком А (см. справочные примеры 3, 4 и 5). При этом используемая L-цепь антитела представляла собой GpL16-k0 (SEQ ID NO: 5), которая представляет собой L-цепь антитела против глипикан-3 с улучшенной кинетикой в плазме, описанной в WO 2009/041062.
Для доказательства принципа гетеродимеризованного антитела в родительские полипептиды GpH7-A5 и GpH7-B3 вводили мутации для конструкции модифицированных вариантов и оценивали варианты. Конструируемые экспрессирующие векторы использовали для трансфекции клеток FreeStyle293 (Invitrogen) способом, описанным в справочном примере 1. Экспрессируемые антитела очищали способом, описанным в справочном примере 1. При экспрессии гомодимеризованного антитела экспрессирующий вектор, вводимый с L-цепыо GpL16-k0 антитела, использовали совместно с экспрессирующим вектором, вводимый в один тип последовательности Н-цепи антитела. При экспрессии гетеродимеризованного антитела экспрессирующий вектор, вводимый в L-цепь GpL16-k0 антитела, как используют для указанного выше гомодимеризованного антитела, и экспрессирующий вектор, вводимый в последовательность, получаемую в результате введения дополнительного изменения в GpH7-A5, содержащую изменение D356K, в качестве одной Н-цепь антитела, и экспрессирующий вектор, вводимый в последовательность, получаемую в результате введения дополнительного изменения, в GpH7-B3, содержащую изменение
K439E, в качестве другой Н-цепи антитела, использовали для получения эффективной экспрессии гетеродимеризованных антител. Экспрессируемое и очищенное антитело обозначают, например, как GpH7-H1/GpH7-H2/GpL16-k0, когда экспрессирующий вектор, используемый для экспрессии одной Н-цепи антитела гетеродимеризованного антитела, представляет собой GpH7-H1, экспрессирующий вектор для другой Н-цепи антитела представляет собой GpH7-H2, и экспрессирующий вектор для L-цепи антитела представляет собой GpL16-k0. В этой системе последовательность, в которую вводят изменения D356K и H435R, соответствует H1, и последовательность, в которую вводят изменения К439Е, соответствует Н2. Например, когда экспрессирующие векторы, используемые для экспрессии Н-цепи и L-цепи антитела гомодимеризованного антитела, представляют собой соответственно GpH7-H1 и GpL16-k0, гомодимеризованное антитело обозначают как GpH7-H1/GpL16-k0. Получаемые антитела использовали для измерения активности связывания с FcγR способом, описанным в справочном примере 2.
Сначала оценивали, оказывает ли эффект изменения D356K и H435R, вводимые в GpH7-A5, и изменение К439Е, вводимое в GpH7-В3, для получения и очистки гетеродимеров, на активность связывания с FcγR по сравнению с нативным IgG. В качестве контроля экспрессировали GpH7-G1d/GpL16-k0 (SEQ ID NO: 2 и 5, соответственно) с использованием плазмид, вводимых в GpH7-G1d и GpL16-k0 в качестве Н-цепи и L-цепи антитела, соответственно, и очищали способом справочного примера 1. Аналогично, получали гомодимеризованное антитело GpH7-A5/GpL16-k0 (SEQ ID NO: 3 и 5,
соответственно), в две Н-цепи которого вводили D356K и H435R, гомодимеризованное антитело GpH7-B3/GpL16-k0 (SEQ ID NO: 4 и 5, соответственно), в две Н-цепи вводили K439Е, и гетеродимеризованное антитело GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5, соответственно), в одну из Н-цепей которого вводили D356K и H435R и в другие вводили K439E. Эти антитела и их активности связывания с каждым FcγR сравнивали способом, описанным в справочном примере 2, и результат представлен на фиг. 5.
Результат измерения демонстрировал, что не существовало значимого отличия активности связывания с каждым FcγR для GpH7-G1d/GpL16-k0 и GpH7-A5/GpH7-B3/GpL16-k0. Кроме того, по отношению к каждому FcγR GpH7-A5/GpL16-k0 и GpH7-B3/GpL16-k0 сохраняли по меньшей мере приблизительно 80% активности связывания GpH7-G1d/GpL16-k0. На основании указанного выше результата определяли, что связывание с FcγR GpH7-A5/GpH7-B3/GpL16-k0, GpH7-A5/GpL16-k0 и GpH7-B3/GpL16-k0 не снижалось значительно по сравнению с GpH7-G1d/GpL16-k0, и, таким образом, можно сравнивать варианты, получаемые в результате введения мутаций в каждую Н-цепь этих антител, в отношении активности связывания с каждым FcγR.
Затем конструировали GpH7-A26 (SEQ ID NO: 6), получаемую в результате введения мутации G237A в GpH7-A5, способом, описанным в справочном примере 1. С использованием GpL16-k0 в качестве L-цепи и GpH7-A26 и GpH7-B3 в качестве Н-цепи способом, описанным в справочном примере 1, экспрессировали гетеродимеризованное
антитело GpH7-A26/GpH7-B3/GpL16-k0 (SEQ ID NO: 6, 4 и 5, соответственно), в котором только в одну из Н-цепей вводили G237A. Аналогично, с использованием GpH7-A26 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-A26/GpL16-k0 (SEQ ID NO: 6 и 5, соответственно), в две Н-цепи которого вводили G237A. Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 6). Результат демонстрировали, что гетеродимеризованное антитело GpH7-A26/GpH7-B3/GpL16-k0 обладало повышенными активностями связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом гомодимеризованное антитело GpH7-A26/GpL16-k0, в котором вводили аналогичное изменение в обе Н-цепи, обладало сниженными активностями связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Эти результаты демонстрируют, что G237A представляет собой изменение, которое повышает активности связывания с FcγRIIa R и FcγRIIb, при введении только в одну Н-цепь, тогда как, при введении в обе Н-цепи снижает активности связывания с FcγRIIa R и FcγRIIb.
Затем способом, описанным в справочном примере 1, конструировали GpH7-A29 (SEQ ID NO: 7), получаемую в результате введения мутации G237L в GpH7-A5. Способом, описанным в справочном примере 1, экспрессировали гетеродимеризованное антитело GpH7-A29/GpH7-B3/GpL16-k0 (SEQ ID NO: 7, 4 и 5, соответственно), в котором только в одну из Н-цепей вводили
G237L, с использованием GpL16-k0 в качестве L-цепи и GpH7-A29 и GpH7-B3 в качестве Н-цепи. Аналогично, с использованием GpH7-A29 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-A29/GpL16-k0 (SEQ ID NO: 7 и 5, соответственно), в две Н-цепи которого вводили G237L. Эти антитела и их активности связывания с каждым FcγR оценивали способом, описанным в справочном примере 2 (фиг. 7). Гетеродимеризованное антитело GpH7-A29/GpH7-B3/GpL16-k0 обладало повышенными активностями связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом гомодимеризованное антитело GpH7-A29/GpL16-k0, две Н-цепи которого содержали аналогичное изменение, обладало сниженной активностью связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Эти результаты демонстрируют, что G237L представляет собой изменение, которое обладает эффектом повышения активностей связывания с FcγRIIa R и FcγRIIb при введении только в одну Н-цепь, тогда как, при введении в обе Н-цепи снижает активности связывания с FcγRIIa R и FcγRIIb.
Затем способом, описанным в справочном примере 1, конструировали GpH7-A42 (SEQ ID NO: 8), получаемую в результате введения мутации L328E в GpH7-A5. С использованием GpL16-k0 в качестве L-цепь и GpH7-A42 и GpH7-B3 в качестве Н-цепи способом, описанным в справочном примере 1, экспрессировали гетеродимеризованное антитело GpH7-A42/GpH7-B3/GpL16-k0 (SEQ ID NO: 8, 4 и 5, соответственно), в котором только в одну из Н-цепей
вводили L328E. Аналогично, с использованием GpH7-A42 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-A42/GpL16-k0 (SEQ ID NO: 8 и 5, соответственно), в две Н-цепи которого вводили L328E. Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 8). Гетеродимеризованное антитело GpH7-A42/GpH7-B3/GpL16-k0 обладало повышенной активностью связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом, гомодимеризованное антитело GpH7-A42/GpL16-k0, в две Н-цепи которого вводили аналогичное изменение, обладало сниженной активность связывания с FcγRIIa R и повышенной активностью связывания с FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0; однако степень повышения являлась больше для GpH7-A42/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили L328E. Эти результаты демонстрируют, что L328E представляет собой изменение, которое является более эффективным в отношении повышения активностей связывания с FcγRIIa R и FcγRIIb при введении только в одну Н-цепь, чем при введении в обе Н-цепи.
Затем способом, описанным в справочном примере 1, конструировали GpH7-A43 (SEQ ID NO: 9), получаемую в результате введения мутации L328D в GpH7-A5. С использованием GpL16-k0 в качестве L-цепи и GpH7-A43 и GpH7-B3 в качестве Н-цепи способом, описанным в справочном примере 1, экспрессировали Гетеродимеризованное антитело GpH7-A43/GpH7-B3/GpL16-k0 (SEQ ID
NO: 9, 4 и 5, соответственно), в которое только в одну из Н-цепей вводили L328D. Аналогично, с использованием GpH7-A43 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-A43/GpL16-k0 (SEQ ID NO: 9 и 5, соответственно), в две Н-цепи которого вводили L328D. Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 9). Гетеродимеризованное антитело GpH7-A43/GpH7-B3/GpL16-k0 обладало повышенной активностью связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом гомодимеризованное антитело GpH7-A43/GpL16-k0, в две Н-цепи которого вводили аналогичное изменение, обладало сниженной активность связывания с FcγRIIa R и повышенной активностью связывания с FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0; однако степень повышения являлась больше для GpH7-A43/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили L328D. Эти результаты демонстрируют, что L328D представляет собой изменение, которое является более эффективным в отношении повышения активностей связывания с FcγRIIa R и FcγRIIb при введении только в одну Н-цепь, чем при введении в обе Н-цепи.
Затем способом, описанным в справочном примере 1, конструировали GpH7-B16 (SEQ ID NO: 10), получаемую в результате введения мутации L234E в GpH7-B3. С использованием GpL16-k0 в качестве L-цепи и GpH7-A5 и GpH7-B16 в качестве Н-цепи способом, описанным в справочном примере 1, экспрессировали
гетеродимеризованное антитело GpH7-A5/GpH7-B16/GpL16-k0 (SEQ ID NO: 3, 10 и 5, соответственно), в котором только в одну из Н-цепей вводили L234E. Аналогично, с использованием GpH7-B16 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-B16/GpL16-k0 (SEQ ID NO: 10 и 5, соответственно), в две Н-цепи которого вводили L234E. Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 10). Гетеродимеризованное антитело GpH7-A5/GpH7-B16/GpL16-k0 обладало повышенными активностями связывания с FcγRIIIa F и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом гомодимеризованное антитело GpH7-B16/GpL16-k0, в две Н-цепи которого вводили аналогичное изменение, обладало сниженными активностями связывания с FcγRIIIa F и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Эти результаты демонстрируют, что L234E представляет собой изменение, которое является эффективным в отношении повышения активностей связывания с FcγRIIIa F и FcγRIIb при введении только в одну Н-цепь, тогда как оно снижает активности связывания с FcγRIIIa F и FcγRIIb при введении в обе Н-цепи.
Затем способом, описанным в справочном примере 1, конструировали GpH7-B17 (SEQ ID NO: 11), получаемую в результате введения мутации L234D в GpH7-B3. С использованием GGpL16-k0 в качестве L-цепи и GpH7-A5 и GpH7-B17 в качестве Н-цепи способом, описанным в справочном примере 1, экспрессировали гетеродимеризованное антитело GpH7-A5/GpH7-B17/GpL16-k0 (SEQ ID
NO: 3, 11 и 5, соответственно), в две Н-цепи которого вводили L234D. Аналогично, с использованием GpH7-B17 в качестве Н-цепи и GpL16-k0 в качестве L-цепи способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело GpH7-B17/GpL16-k0 (SEQ ID NO: 11 и 5, соответственно), в две Н-цепи которого вводили L234D. Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 11). Гетеродимеризованное антитело GpH7-A5/GpH7-B17/GpL16-k0 обладало повышенной активностью связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом гомодимеризованное антитело GpH7-B17/GpL16-k0, в две Н-цепи которого вводили аналогичное изменение, обладало сниженными активностями связывания с FcγRIIa R и FcγRIIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Эти результаты демонстрируют, что L234D представляет собой изменение, которое эффективно повышает активности связывания с FcγRIIa R и FcγRIIb при введении только в одну Н-цепь, тогда как, оно снижает активности связывания с FcγRIIa R и FcγRIIb при введении в обе Н-цепи.
Указанные выше открытия демонстрируют, что даже если гомодимеризованное антитело, в две Н-цепи которого вводили аналогичное изменение, обладает сниженной активностью связывания с FcγR, возможно повышать активность связывания с FcγR, конструируя Гетеродимеризованное антитело, в котором только в одну из Н-цепей вводят изменение.
Таким образом, указанные выше открытия демонстрируют, можно
обеспечивать превосходные свойства связывания с FcγR домена Fc антитела применением гетеродимеризованного антитела, в две Н-цепи которого вводят различные изменения, по сравнению с гомодимеризованным антителом, получаемым общепринятым способом введения аналогичного изменения в обе Н-цепи.
[Пример 3] Подтверждение направления распознавания FcγR гетеродимеризованных антител
Как показано в примере 2, было продемонстрировано, что с использованием гетеродимеризованных антител можно получать большую активность связывания с FcγR по сравнению с гомодимеризованными антителами. Когда изменения, описанные в примере 2, вводили в обе Н-цепи антитела, активность связывания с FcγR даже снижалась по сравнению с природным антителом. Это открытие позволяет предположить, что такое изменение приводит к повышению активности связывания с FcγR при введении в одну Н-цепь гетеродимеризованного антитела; однако когда мутацию вводят в обе Н-цепи при связывании с FcγR остаток после изменения повышает связывание в одной цепи, при этом ингибирует взаимодействие с FcγR в другой цепи. В настоящем описании состояние, когда FcγR связывается в направлении с внутренней стороны, на фиг. 3 определяют как "связывание в направлении X", тогда как в противоположном случае связывание с внешней стороны обозначают как "связывание в направлении Y". Предполагали, что в гетеродимеризованном антителе изменяется любое из направления Х и направления Y активности связывания с FcγR, тогда как в гомодимеризованном антителе аналогичным образом изменяются
активности связывания с FcγR в направлении Х и направлении Y.
Для экспериментального подтверждения гипотезы авторы настоящего изобретения выявили изменения, которые в основном ингибируют связывание с FcγR только в одном из направления Х и направления Y, и вводили такое изменение в одну из Н-цепей и вводили в аналогичную цепь или другую Н-цепь изменение, которое повышает активность связывания с FcγR, для подтверждения ингибирования активности связывания с FcγR. Целью авторов настоящего изобретения являлось открытие способа, который можно применять в комбинации с указанным выше изменением. Изучали данные о конформационном состоянии в отношении изменений, которые участвуют в связывании с FcγR антитела, но участвуют только в связывании только в одной из Н-цепей. В результате в качестве кандидата выявили Р329. Р329 в HA-цепи образует гидрофобное ядро с Trp в положениях 87 и 110 в FcγRIII, при этом Р329 в HB-цепи непосредственно не взаимодействует с FcγRIII (Nature, 372:379-383, 1994) (фиг. 12). Например, предполагали, что замена Р329 HA-цепи на электрически заряженный остаток вызывает коллапс гидрофобного ядра, и приводит к ингибированию связывания в направлении X, представленном на фиг. 12; однако теоретически было рассчитано, что не существует значительного влияния на связывание в направлении Y, вследствие того, что Р329 HB-цепи, который не участвует в связывании в Y направлении на другой стороне, остается незамещенным.
Мутации, электрически заряженные R, K, D и Е вводили в GpH7-В3 при Р329 соответственно для получения последовательностей
GpH7-B12, GpH7-B13, GpH7-B14 и GpH7-B15 (SEQ ID NO: 12-15) и конструировали экспрессирующие векторы, в которые вводили эти последовательности, способом, описанным в справочном примере 1. Каждый из конструируемых экспрессирующих векторов комбинировали с GpH7-A5 и GpL16-k0 для экспрессии гетеродимеризованных антител или комбинировали их с одним GpL16-k0, но не с другими Н-цепями для экспрессии гомодимеризованных антител. Эти антитела экспрессировали и очищали способом, описанным в справочном примере 1. Получаемые очищенные антитела представляют собой следующие: гетеродимеризованные антитела GpH7-A5/GpH7-B12/GpL16-k0, GpH7-A5/GpH7-B13/GpL16-k0, GpH7-A5/GpH7-B14/GpL16-k0 и GpH7-A5/GpH7-B15/GpL16-k0 и гомодимеризованные антитела GpH7-B12/GpL16-k0, GpH7-B13/GpL16-k0, GpH7-B14/GpL16-k0 и GpH7-B15/GpL16-k0. Получаемые антитела использовали для оценки активности связывания с каждым FcγR способом, описанным в справочном примере 2. Результат представлен на фиг. 13.
Активность связывания с FcγR GpH7-A5/GpH7-B12/GpL16-k0 и GpH7-A5/GpH7-B13/GpL16-k0, которые представляют собой гетеродимеризованные антитела, в которые вводили P329R и Р329К соответственно, составляла приблизительно от 1/5 до 1/4 активности связывания с каждым FcγR GpH7-A5/GpH7-B3/GpL16-k0. При этом для гомодимеризованных антител GpH7-B12/GpL16-k0 и GpH7-B13/GpL16-k0 не наблюдали связывание с FcγR. С другой стороны, в отношении FcγRIa активность связывания с FcγR GpH7-A5/GpH7-B14/GpL16-k0 и GpH7-A5/GpH7-B15/GpL16-k0, которые представляют собой гетеродимеризованные антитела, в которые вводили P329D и
Р329Е соответственно, составляла приблизительно от 1/5 до 1/4 активности связывания с каждым FcγR GpH7-A5/GpH7-B3/GpL16-k0; в отношении FcγR, отличного от FcγRIa, они сохраняли 50% или более активности связывания. Гомодимеризованные антитела GpH7-B14/GpL16-k0 и GpH7-B15/GpL16-k0 сохраняли только активность связывания с FcγRIa, и активность составляла 1/5 или менее активность связывания с FcγRIa GpH7-A5/GpH7-B3/GpL16-k0. GpH7-B12 и GpH7-B13 вводят с основными остатками, тогда как GpH7-B14 и GpH7-B15 вводят с кислыми остатками. Таким образом, предполагали, что активность связывания с FcγR сильнее ингибируется заменой Р329 основным остатком, таким как Аrg и Lys. Результат, демонстрирующий, что активность связывания с FcγR сохраняется, когда изменение вводят только в одну Н-цепь, тогда как связывание являлось почти не детектируемым, когда аналогичное изменение вводили в обе Н-цепи, подтверждает гипотезу, что замена гидрофильного остатка на Р329 в одной Н-цепи ингибирует связывание в одном направлении, хотя связывание в другом направлении сохраняется.
Затем оценивали, усиливают ли мутации G237A и L234D, которые повышают активность связывания с FcγR посредством гетеродимеризации, которые были выявлены, как описано в примере 2, взаимодействие с FcγR исключительно в одном направлении, сравнивая активность связывания с каждым Fc каждого варианта, который получали таким образом, чтобы он содержал изменение P329R в комбинации с указанными выше изменениями. Способом, описанным в справочном примере 1, конструировали экспрессирующие
векторы, в которые вводили GpH7-B12 (SEQ ID NO: 12), получаемую в результате введения мутации P329R в GpH7-B3, таким образом, что P329R вводят в аналогичную Н-цепь, как вводят К439Е; GpH7-A48 (SEQ ID NO: 16), получаемую в результате введения мутации P329R в GpH7-A5, таким образом, что P329R вводят в аналогичную Н-цепь, как вводят D356K и H435R; GpH7-A45 (SEQ ID NO: 17), получаемую в результате введения мутаций G237A и P329R в GpH7-A5, и GpH7-B41 (SEQ ID NO: 18), получаемую в результате введения мутаций L234D и P329R в GpH7-B5. С использованием этих экспрессирующих векторов и экспрессирующего вектора GpL16-k0, соответствующего L-цепи антитела, антитела экспрессировали, таким образом, что P329R и G237A или L234D вводят только в одну из Н-цепей, способом, описанным в справочном примере 1 (таблица 1).

В столбце "образец" указаны названия антител, в столбцах "HI" и "Н2" указано название константных областей Н-цепи соответствующих антител, и в столбце "участок мутации" указаны мутации, которые являются отличными по сравнению GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела также показана SEQ ID NO.
Комбинацию G237A и P329R оценивали, применяя GpH7-A5/GpH7-B3/GpL16-k0 в качестве полипептида для введения изменений; GpH7-A5/GpH7-B12/GpL16-k0 и GpH7-A48/GpH7-B3/GpL16-k0 в качестве
вариантов, в одну из Н-цепей которых вводили P329R; GpH7-A45/GpH7-B3/GpL16-k0 в качестве варианта, в котором G237A вводили в аналогичную цепь, как вводили P329R, и GpH7-A26/GpH7-B12/GpL16-k0 в качестве варианта, в котором G237A вводили в другую цепь по сравнению с P329R. Их сравнивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2 (фиг. 14).
Когда GpH7-A5/GpH7-B12/GpL16-k0, в одну из Н-цепей которого вводили P329R, сравнивали с GpH7-A48/GpH7-B3/GpL16-k0, не наблюдали значимого отличия в характере связывания с FcγR. Таким образом, предполагали, что P329R не оказывает влияния на связывание с FcγR даже при введении в Н-цепь, в которую вводили D356K и H435R, или при введении в Н-цепь, в которую вводили K439Е.
С другой стороны, GpH7-A26/GpH7-B3/GpL16-k0, которое представляет собой гетеродимеризованное антитело, в котором только в одну из Н-цепей вводили G237A, демонстрировало повышенное связывание с FcγRIIa R и IIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Однако по сравнению с GpH7-A5/GpH7-B12/GpL16-k0, в котором только в одну из Н-цепей вводили P329R, GpH7-A26/GpH7-B12/GpL16-k0, в котором в другую Н-цепь вводили G237A, демонстрировало сниженное связывание с FcγR, и, таким образом, не наблюдали эффект G237A в отношении повышения связывания FcγRIIa R и IIb. При этом по сравнению с GpH7-A48/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили P329R, GpH7-A45/GpH7-B3/GpL16-k0, в котором G237A вводили в
аналогичную Н-цепь, как вводили P329R, демонстрировало повышенное связыванием с FcγRIIa R и IIb, и, таким образом, наблюдали эффект G237A в отношении повышения связывания с FcγRIIa R и IIb. Вследствие того, что связывание антитела и FcγR значительно ингибировалось, когда G237A вводили в цепь, отличную от цепи, в которую вводили P329R, указанные выше результаты демонстрируют, что, когда G237A, вводимую в GpH7-A5, комбинируют с P329R, вводимой в GpH7-B3, они распознают связывание антитела и FcγR в одинаковом направлении.
Аналогичным образом также оценивали изменение L234D (фиг. 15). GpH7-A5/GpH7-B17/GpL16-k0, которое представляет собой гетеродимеризованное антитело, в котором только в одну из Н-цепей вводили L234D, демонстрировало повышенное связывание с FcγRIIa R и IIb по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Однако по сравнению с GpH7-A48/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили P329R, GpH7-A48/GpH7-B17/GpL16-k0, в котором L234D вводили в другую Н-цепь, демонстрировало сниженное связывание с FcγR, отличным от FcγRIa, и, таким образом, не наблюдали эффекта L234D в отношении повышения связывания с FcγRIIa R и IIb. При этом по сравнению с GpH7-A5/GpH7-B12/GpL16-k0, в котором только в одну из Н-цепей вводили P329R, GpH7-A5/GpH7-B41/GpL16-k0, в котором L234D вводили в аналогичную Н-цепь, как вводили P329R, демонстрировало повышенное связывание с FcγRIIa R и IIb, и, таким образом, наблюдали эффект L234D в отношении повышения связывания с FcγRIIa R и IIb. Вследствие того, что связывание антитела и FcγR значительно ингибировалось,
когда L234D вводили в цепь, отличную от цепи, в которую вводили P329R, указанные выше результаты демонстрируют, что, когда L234D, вводимую в GpH7-B3, комбинируют с P329R, вводимой в GpH7-А5, они распознают связывание антитела и FcγR в одинаковом направлении.
Таким образом, обе G237A и L234D повышают (/снижают) активность связывания с FcγR антитела в аналогичном направлении как P329R.
Таким образом, гетеро-изменение повышало связывание с FcγR исключительно в одном из направления Х и направления Y. Это открытие демонстрирует, что взаимодействие с FcγR усиливается асимметричным образом в гетеродимеризованных антителах.
[Пример 4] Сравнение связывания с FcγR гетеродимеризованного антитела со связыванием гомодимеризованного антитела
Как описано в примере 2, авторы настоящего изобретения выявили, что связывание с FcγR антитела через его домен Fc можно повышать посредством введения различных изменений в две Н-цепи антитела, в отличие от введения одинакового изменения в обе Н-цепи. Таким образом, для выявления изменений с таким свойством авторы настоящего изобретения провели описанный ниже эксперимент.
Для конструкции вариантов GpH7-B3 аминокислоты, для которых предполагали, что они участвуют в связывании с FcγR, и аминокислоты вокруг них в GpH7-B3 (SEQ ID NO: 4), конструируемой как описано в примере 2, в частности Leu в положении 234 (нумерация EU), Leu в положении 235 (нумерация EU), Gly в
положении 236 (нумерация EU), Gly в положении 237 (нумерация EU), Pro в положении 238 (нумерация EU), Ser в положении 329 (нумерация EU), Asp в положении 265 (нумерация EU), Val в положении 266 (нумерация EU), Ser в положении 267 (нумерация EU), His в положении 268 (нумерация EU), Glu в положении 269 (нумерация EU), Asp в положении 270 (нумерация EU), Pro в положении 271 (нумерация EU), Gln в положении 295 (нумерация EU), Tyr в положении 296 (нумерация EU), Ser в положении 298 (нумерация EU), Tyr в положении 300 (нумерация EU), Ser в положении 324 (нумерация EU), Asn в положении 325 (нумерация EU), Lys в положении 326 (нумерация EU), Ala в положении 327 (нумерация EU), Leu в положении 328 (нумерация EU), Pro в положении 329 (нумерация EU), Ala в положении 330 (нумерация EU), Pro в положении 331 (нумерация EU), Ile в положении 332 (нумерация EU), Glu в положении 333 (нумерация EU), Lys в положении 334 (нумерация EU), Thr в положении 335 (нумерация EU), Ile в положении 336 (нумерация EU) и Ser в положении 337 (нумерация EU) каждые заменяли 18 типами аминокислот, за исключением исходной аминокислоты и цистеина. Каждый вариант GpH7-B3 называли "А_В", где А представляет собой информацию о типе аминокислоты, обозначаемым однобуквенным кодом, и нумерацию EU изменяемого остатка, и В означает информацию об аминокислоте после замены. Например, когда Gly заменяют на Leu в положении 234 (нумерация EU), В3_вариант называют L234_01G. С целью описания в информации об аминокислоте после замены число, характерное для аминокислоты, добавляют перед каждым однобуквенным кодом. В частности, используемые символы
представляют собой следующие: 01G для Gly, 02A для А1а, 03V для Val, 04F для Phe, 05P для Pro, 06M для Met, 07I для Ilе, 08L для Leu, 09D для Asp, 10E для Glu, 11K для Lys, 12R для Arg, 13S для Ser, 14T для Thr, 15Y для Туг, 16Н для His, 18N для Asn, 19Q для Gln и 20W для Trp.
Гомодимеризованные антитела, в обе Н-цепи которых вводили мутация, получали следующим ниже способом. Антитела экспрессировали с использованием варианта GpH7-B3 в качестве Н-цепи и GpL16-k0 (SEQ ID NO: 5) в качестве L-цепи. Антитела получали способом, описанным в справочном примере 1. Получаемые таким образом гомодимеризованные антитела, в обе Н-цепи которых вводили мутацию, обозначают как Но Ab.
Гетеродимеризованные антитела, в которых только в одну из Н-цепей вводили мутацию, получали следующим ниже способом. Антитела экспрессировали с использованием варианта GpH7-B3 и GpH7-A5 (SEQ ID NO: 3) в качестве Н-цепи и GpL16-k0 (SEQ ID NO: 5) в качестве L-цепи. Антитела получали способом, описанным в справочном примере 1. Получаемые таким образом Гетеродимеризованные антитела, в которых только в одну из Н-цепей вводили мутацию, обозначают как Не Ab.
В качестве контрольного гомодимеризованного антитела получали GpH7-B3/GpL16-k0, которое получали с использованием GpH7-B3 (SEQ ID NO: 4) в качестве Н-цепи и GpL16-k0 (SEQ ID NO: 5) в качестве L-цепи, способом, описанным в справочном примере 1. Это контрольное гомодимеризованное антитело обозначают как HoCon Ab. Как определено в примере 2, активность связывания HoCon Ab с каждым FcγR значительно не изменяется по сравнению с нативным
IgG1.
В качестве контрольного гетеродимеризованного антитела получали GpH7-A5/GpH7-B3/GpL16-k0, которое получали с использованием GpH7-A5 (SEQ ID NO: 3) и GpH7-B3 (SEQ ID NO: 4) в качестве Н-цепи и GpL16-k0 (SEQ ID NO: 5) в качестве L-цепи, способом, описанным в справочном примере 1. Это контрольное гетеродимеризованное антитело обозначают как HeCon Ab. Как определено в примере 2, активность связывания HeCon Ab с каждым FcγR значительно не изменялась по сравнению с нативным IgG1.
Получаемые Но Ab, He Ab, HeCon Ab и HoCon Ab анализировали в отношении активность связывания с FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb и FcγRIIIa способом, описанным в справочном примере 2. В отношении результата анализа для каждого FcγR строили график следующим ниже способом. Активность связывания Не Ab с каждым FcγR делили на активность связывания HeCon Ab, а затем умножали на 100, это обозначают как He/Con. Активность связывания Но Ab с каждым FcγR делили на активность связывания HoCon Ab, а затем умножали на 100, это обозначают как Но/Con. Для гомодимеризованных и гетеродимеризованных антител, получаемых с использованием варианта GpH7-B3, содержащего представляющее интерес изменение, значения Но/Con и Не/Con наносили на график на горизонтальных и вертикальных осях соответственно. Результаты для соответсвующих FcγR, т.е. FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb и FcγRIIIa, приведены на фиг. 16-20. На основании значений Не/Con и Но/Con каждое изменение можно интерпретировать, как указано ниже.
1. Когда значения He/Con и Ho/Con равны 100: это означает, что активность связывания с FcγR гетеродимеризованного антитела Не Ab и гомодимеризованного антитела Но Ab, которые содержат вариант GpH7-B3, в который вводили представляющую интерес мутацию, является эвивалентной активности связывания с FcγR контрольного гетеродимеризованного антитела и контрольного гомодимеризованного антитела, соответственно.
2. Когда значения He/Con и Ho/Con равны 100 или менее: это означает, что активность связывания с FcγR гетеродимеризованного антитела He Ab и гомодимеризованного антитела Но Ab, которые содержат вариант GpH7-B3, в который вводили представляющую интерес мутацию, является слабее, чем активность связывания с FcγR контрольного гетеродимеризованного антитела и контрольного гомодимеризованного антитела, соответственно.
3. Когда значения Не/Con и Но/Con равны 100 или более: это означает, что активность связывания с FcγR гетеродимеризованного антитела Не Ab и гомодимеризованного антитела Но Ab, которые содержат вариант GpH7-B3, в который вводили представляющую интерес мутацию, является больше активности связывания с FcγR контрольного гетеродимеризованного антитела и контрольного гомодимеризованного антитела, соответственно.
4. Когда значение Не/Con является больше значения Но/Con: это означает, что активность связывания с FcγR гетеродимеризованного антитела Не Ab, содержащего вариант GpH7-B3, в который вводили представляющую интерес мутацию, является сильнее активности гомодимеризованного антитела Но Ab.
5. Когда значение He/Con является меньше значения Ho/Con: это означает, что активность связывания с FcγR гетеродимеризованного антитела Не Ab, содержащего вариант GpH7-В3, в который вводили представляющую интерес мутацию, является слабее активности гомодимеризованного антитела Но Ab.
Основываясь на интерпретации указанных выше пунктов от 1 до 5, экспериментальные точки на фиг. 16-20 можно классифицировать так, как представлено на фиг. 21.
Когда изменение находится в области i на фиг. 21, это означает, что изменение при введении в обе Н-цепи гомодимеризованного антитела снижает его связывание с FcγR, тогда как аналогичное изменение при его введении только в одну Н-цепь гетеродимеризованного антитела обладает эффектом повышения его связывания с FcγR. Таким образом, изменение повышает связывание с FcγR только гетеродимеризованного антитела. В отношении каждого FcγR изменения, содержащиеся в области i, приведены в таблице 2 (таблицы от 2-1 до 2-3), таблице 3 (таблицы 3-1 и 3-2), таблице 4, таблице 5 и таблице 6.
Когда изменение находится в области ii на фиг. 21, это означает, что изменение соответствует точке, повышающей связывание с FcγR при введении в обе Н-цепи гомодимеризованного антитела или при введении только в одну Н-цепь гетеродимеризованного антитела, и эффект повышения связывания является больше в гетеродимеризованном антителе. В частности, изменения в этой области обладают большим эффектов повышения связывания с FcγR в гетеродимеризованном антителе, чем в
гомодимеризованном антителе. В отношении каждого FcγR изменения, содержащиеся в области ii, представлены в таблице 2 (таблицах от 2-1 до 2-3), таблице 3 (таблицах 3-1 и 3-2), таблице 4, таблице 5 и таблице 6.
В каждой таблице He/Con, относящееся к активности связывания с FcγRIa, FcγRIIa H, FcγRIIa R, FcγRIIb и FcγRIIIa, обозначают как He/Con_1a, He/Con_2aH, He/Con_2aR, He/Con_2b и He/Con_3a, соответственно; Ho/Con, относящееся к активности связывания с FcγRIa, FcγRIIa H, FcγRIIa R, FcγRIIb и FcγRIIIa, обозначают как Ho/Con_1a, Ho/Con_2aH, Ho/Con_2aR, Ho/Con_2b и Ho/Con_3a, соответственно.
Когда изменение находится в области iii на фиг. 21, это означает, что изменение, соответствующее точке, при введении в обе Н-цепи гомодимеризованного антитела или при введении только в одну Н-цепь гетеродимеризованного антитела повышает связывание с FcγR, и эффект повышения связывания является сильнее в гомодимеризованном антителе. В частности, изменения в этой области обладают большим эффектом повышения связывания с FcγR в гомодимеризованном антителе, чем в гетеродимеризованном антителе.
В таблицах ниже изменения аминокислот указаны таким образом, как А327_03V. Первое буквенное обозначение (например, "А" A327_03V) представляет собой однобуквенный код, соответствующий аминокислотному остатку перед изменением, и следующее число (например, "327" A327_03V) указывает на положение изменения (нумерация EU). Последнее число и буквенное обозначение
(например, "03V" A327_03V) указывает на буквенное обозначение, где аминокислотный остаток после изменения показан однобуквенным кодом (число, соответствующее типу аминокислоты, + буквенное обозначение). Это обозначают следующим образом: 01G (Gly), 02А (Ala), 03V (Val), 04F (Phe), 05P (Pro), 06M (Met), 07I (Ile), 08L (Leu), 09D (Asp), 10E (Glu), 11K (Lys), 12R (Arg), 13S (Ser), 14T (Thr), 15Y (Tyr), 16H (His), 17C (Cys), 18N (Asn), 19Q (Gln) и 20W (Trp). Таким образом, например, "A327_03V" означает "замену V на А в положении аминокислоты 327 (нумерация EU)".
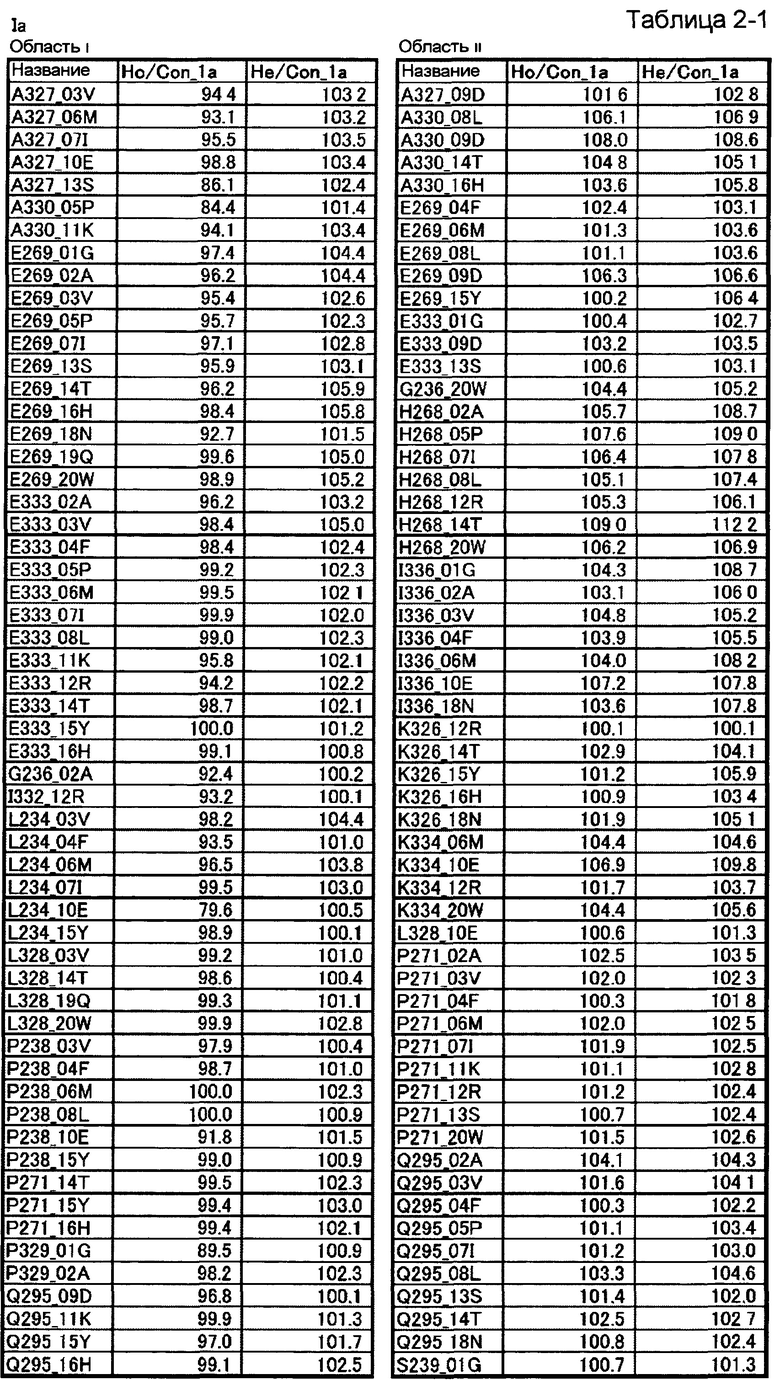
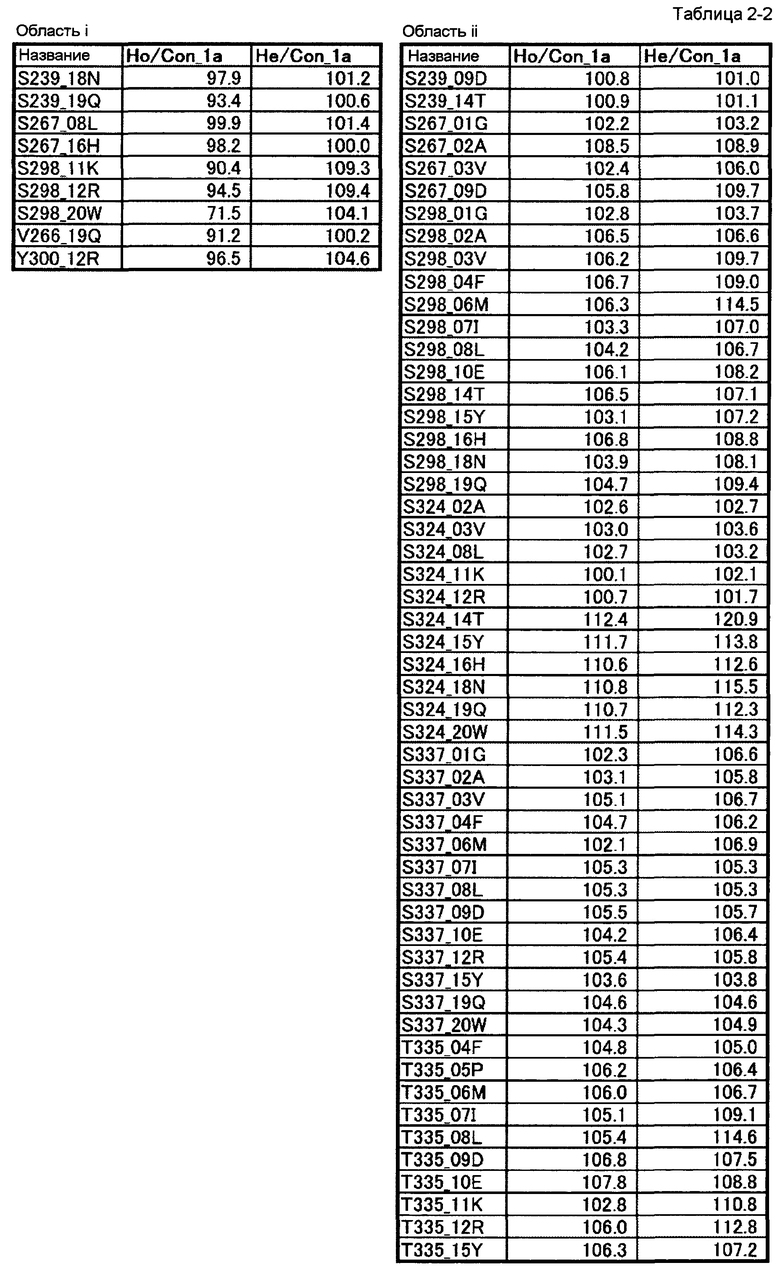

По отношению к FcγRIa представлены изменения, находящиеся в областях i и ii (фиг. 21).


По отношению к FcγRIIa R представлены изменения, находящиеся в областях i и ii (фиг. 21).
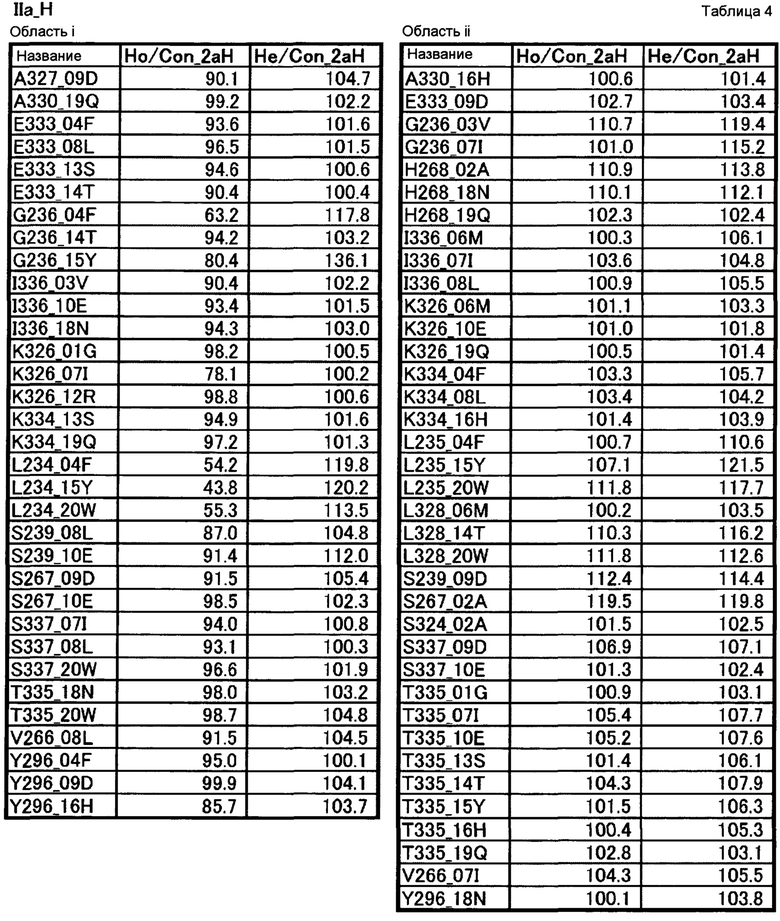
По отношению к FcγRIIa H представлены изменения, находящиеся в областях i и ii (фиг. 21).
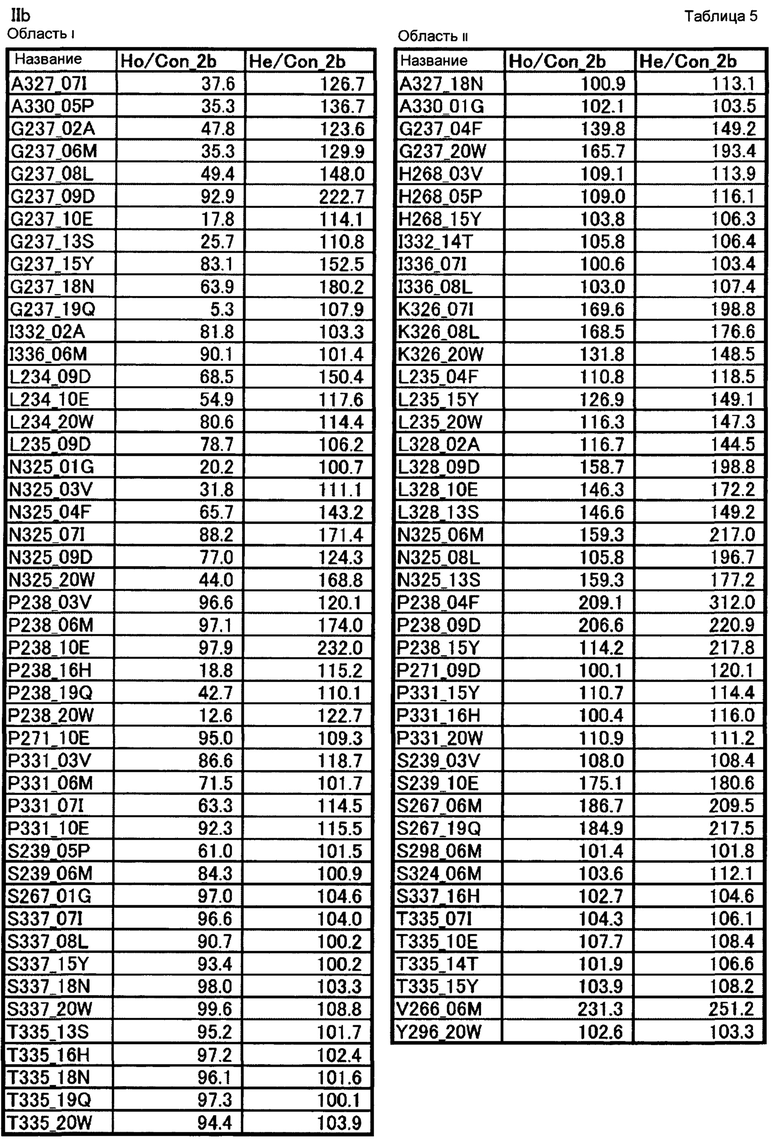
По отношению к FcγRIIb представлены изменения, находящиеся в
областях i и ii (фиг. 21).
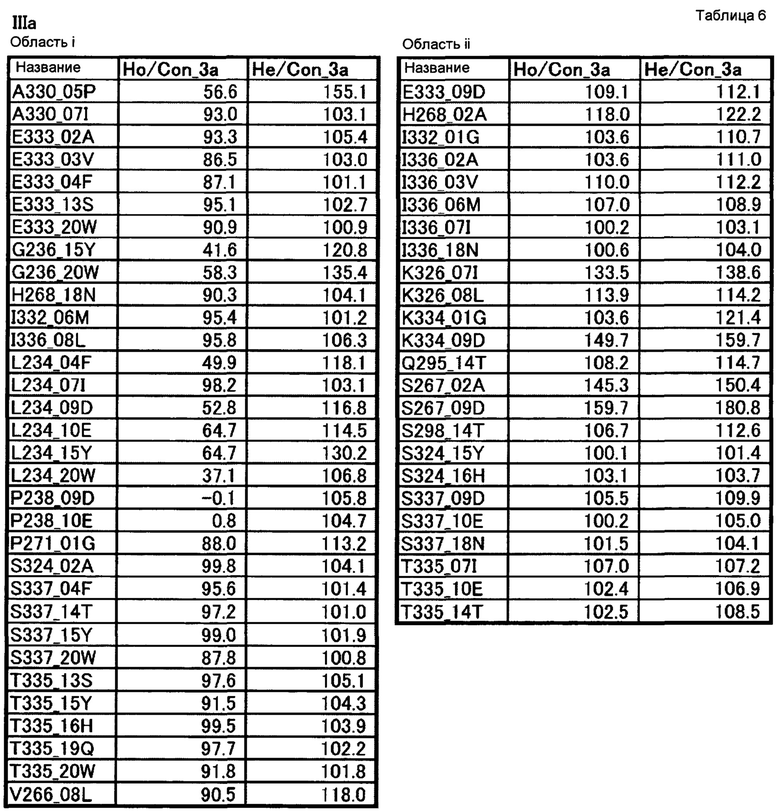
По отношению к FcγRIIIa представлены изменения, находящиеся в областях i и ii (фиг. 21).
Предполагают, что изменения, выявленные в этом примере наряду с примером 2, подтверждают принцип улучшения способности распознавания FcγR гетеродимеризованным антителом, описанным в примере 1. Считают, что изменения в области i на фиг. 21 снижают активность связывания с FcγR при введении в обе Н-цепи
общепринятым способом, и их не считали изменениями, которые повышают связывание. Однако авторам настоящего изобретения удалось продемонстрировать, что такие изменения также улучшают активность связывания с FcγR посредством их введения только в одну из двух Н-цепей.
[Пример 5] Способ определения комбинаций изменений гетеродимеризованных антител
Как описано в примере 3 эффект изменения в отношении связывания с FcγR гетеродимеризованного антитела различается по направлению в зависимости от изменения. По этой причине, когда несколько изменений комбинируют для дополнительного повышения или снижения связывания FcγR гетеродимеризованного антитела, необходимо выравнивать направление эффекта каждого изменения в отношении связывания с FcγR. Если два изменения вводят таким образом, что их эффекты в отношении повышения связывания FcγR антитела различаются по направлению, эффекты изменений взаимно устраняются, и не наблюдают эффект повышения связывания несмотря на комбинацию изменений для повышения связывания с FcγR. Однако не существует способа предварительного теоретического расчета, в какую Н-цепь необходимо вводить изменение. Таким образом, для выявления подходящего способа комбинации, как правило, необходимо получать антитела, в которых представляющие интерес изменения вводят в одну и ту же цепь или различные Н-цепи, и их сравнивают друг с другом в отношении активности связывания с FcγR. Например, когда все три различных типа изменений вводят в гомодимеризованное антитело, можно получать один типа антитела,
в котором каждое изменение вводят в обе Н-цепи. Однако в случае гетеродимеризованного антитела необходимо определять, в какую Н-цепь необходимо вводить каждое изменение. В этом случае, как показано на фиг. 22, максимально существует четыре комбинации, которые необходимо испытывать, т.е. число вариантов, которые необходимо получать для оценки комбинаций изменений является очень большим, и это является неэффективным. Таким образом, авторы настоящего изобретения тестировали способ определения направления, в котором представляющее интерес изменение обладает эффектом в отношении связывания с FcγR антитела, посредством комбинации изменений с P329R, которая ингибирует связывание с FcγR антитела только в одном направлении.
Авторы настоящего изобретения исследовали, как изменяется связывание с FcγRIIIa, когда каждое из изменений L234Y, G236W и S298A, которые повышают связывание с FcγRIIIa, вводят в аналогичную Н-цепь, как вводят P329R, или в другую Н-цепь гетеродимеризованного антитела, выявленного в примере 4. Сначала представляющие интерес варианты экспрессировали и получали способом, описанным в справочном примере 1, с использованием GpH7-A5 в качестве Н-цепи и GpL16-k0 в качестве L-цепи, и в качестве другой Н-цепи GpH7-HA5 (SEQ ID NO: 19), GpH7-HA6 (SEQ ID NO: 20) и GpH7-HA11 (SEQ ID NO: 21), получаемые в результате введения L234Y, G236W и S298A, соответственно, в GpH7-B12, в которую вводили изменение P329R. Кроме того, представляющие интерес антитела экспрессировали и получали способом, описанным в справочном примере 1, с использованием в качестве Н-цепи GpH7-
А48 (SEQ ID NO: 16), получаемую в результате введения P329R в GpH7-A5, и GpL16-k0 в качестве L-цепи, и в качестве другой Н-цепи GpH7-B3-01-15Y (SEQ ID NO: 22), GpH7-B3-03-20W (SEQ ID NO: 23) и GpH7-B3-15-02A (SEQ ID NO: 24), получаемые в результате введения L234Y, G236W и S298A, соответственно, в GpH7-B3. Получаемые таким образом антитела называли GpH7-A5/GpH7-HA5/GpL16-k0 (SEQ ID NO: 3, 19 и 5, соответственно), GpH7-A5/GpH7-HA6/GpL16-k0 (SEQ ID NO: 3, 20 и 5, соответственно), GpH7-A5/GpH7-HA11/GpL16-k0 (SEQ ID NO: 3, 21 и 5, соответственно), GpH7-A48/GpH7-B3-01-15Y/GpL16-k0 (SEQ ID NO: 16, 22 и 5, соответственно), GpH7-A48/GpH7-B3-03-20W/GpL16-k0 (SEQ ID NO: 16, 23 и 5, соответственно) и GpH7-A48/GpH7-B3-15-02A/GpL16-k0 (SEQ ID NO: 16, 24 и 5, соответственно). Варианты сравнивали друг с другом в отношении активности связывания с FcγRIIIa способом, описанным в справочном примере 2. Эффекты комбинации L234Y, G236W и S298A с P329R приведены в таблице 7.

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 (SEQ ID NO: 3, 4 и 5, соответственно) за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Основываясь на результате, сравнивали изменения в отношении активности связывания с FcγRIIIa, когда каждое изменение вводили в аналогичную Н-цепь, как вводили P329R или Н-цепь, отличную от той, в которую вводили P329R. В случае L234Y активность связывания GpH7-A5/GpH7-HA5/GpL16-k0, соответствующего первому случаю, составляла 60, тогда как активность связывания GpH7-A48/GpH7-B3-01-15Y/GpL16-k0, соответствующего последнему случаю, составляла 11; таким образом, связывание с FcγR ингибировали, когда изменение вводили в Н-цепь, отличную от той, в которую вводили P329R. В случае G236W активность связывания GpH7-A5/GpH7-HA6/GpL16-k0, соответствующего первому случаю, составляла 56, тогда как активность связывания GpH7-A48/GpH7-B3-03-20W/GpL16-k0, соответствующего последнему случаю составляла 13; таким образом, связывание с FcγR ингибировали, когда изменение вводили в Н-цепь, отличную от той, в которую вводили P329R. В случае S298A активность связывания GpH7-A5/GpH7-HA11/GpL16-k0,
соответствующего первому случаю, составляла 84, тогда как активность связывания GpH7-A48/GpH7-B3-15-02A/GpL16-k0, соответствующего последнему случаю, составляла 47; таким образом, активность связывания с FcγR ингибировали, когда изменение вводили в Н-цепь, отличную от той, в которую вводили P329R. Для всех изменений связывание с FcγRIIIa ингибировали, когда изменение вводили в Н-цепь, отличную от той, в которую вводили P329R, как описано в последнем случае. Если Н-цепь, в которую вводили P329R, соответствовала HA-цепи фиг. 3, считают, что P329R ингибирует связывание в направлении X. Вследствие того, что комбинации, которые приводили к значительному ингибированию связывания, представляют собой такие как, когда L234Y, G236W или S298A вводили в Н-цепь, отличную от той, в которую вводили P329R, в этом случае изменения необходимо вводить в HB-цепь. Вследствие того, что эффект повышения связывания с FcγRIIIa заметно ингибировали, когда любое из изменений L234Y, G236W и S298A вводили в Н-цепь, отличную от той в которую вводили P329R, все изменения при введении в HB-цепь повышали связывание с FcγRIIIa в направлении X, которое ингибируют P329R. Таким образом, предполагали, что связывание FcγRIIIa можно дополнительно усиливать введением этих изменений в аналогичную Н-цепь.
Описанную выше гипотезу тестировали, определяя, повышают ли связывание с FcγR введением двух из L234Y, G236W и S298A в аналогичную Н-цепь или в различные Н-цепи. Экспрессирующие векторы, в которые вводили GpH7-TA1 (SEQ ID NO: 25), GpH7-TA2
(SEQ ID NO: 26) и GpH7-TA3 (SEQ ID NO: 27), получаемые в результате введения L234Y, G236W и S298A в GpH7-A5, соответственно, и экспрессирующие векторы, в которые вводили GpH7-B3-01-15Y (SEQ ID NO: 22), GpH7-B3-03-20W (SEQ ID NO: 23) и GpH7-B3-15-02A (SEQ ID NO: 24), получаемые в результате введения L234Y, G236W и S298A в GpH7-B3, соответственно, конструировали способом, описанным в справочном примере 1. Кроме того, конструировали экспрессирующие векторы, в которые вводили, как указано ниже: GpH7-TA4 (SEQ ID NO: 28), получаемую в результате введения L234Y и G236W в GpH7-A5; GpH7-TA5 (SEQ ID NO: 29), получаемую в результате введения L234Y и S298A в GpH7-A5, и GpH7-TA6 (SEQ ID NO: 30), получаемую в результате введения G236W и S298A в GpH7-A5. Их комбинировали таким образом, как показано в таблице 8, и к каждой комбинации добавляли GpL16-k0 в качестве L-цепи. Представляющие интерес антитела экспрессировали и получали способом, описанным в справочном примере 1. В отношении экспрессируемых образцов информация о Н-цепях и участках мутации и анализ результатов в отношении связывания с FcγRIIIa антитела приведены в таблице 8.
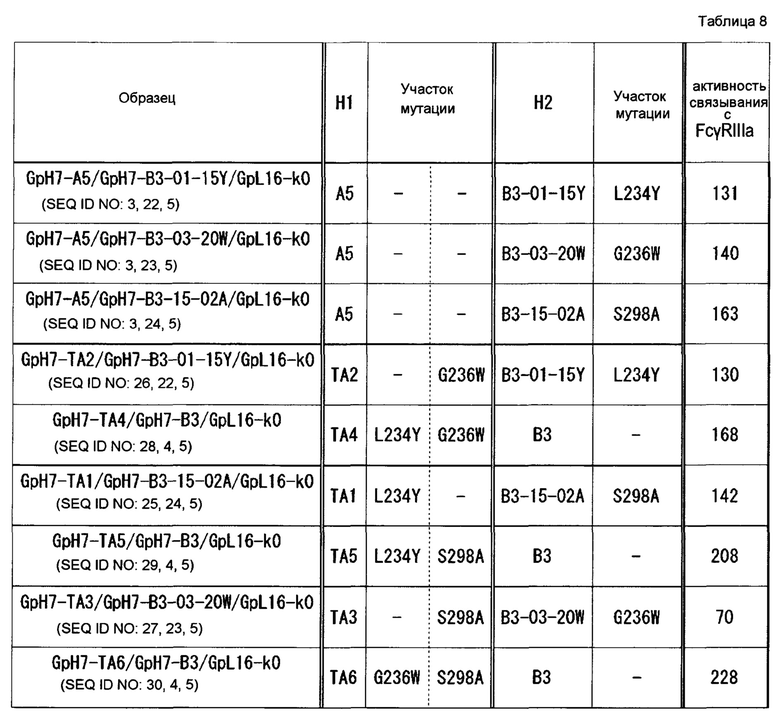
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Эффект комбинации L234Y и G236W оценивали на основании результата, представленного в таблице 8. Активность связывания с FcγRIIIa GpH7-TA2/GpH7-B3-01-15Y/GpL16-k0, в котором L234Y и G236W вводили в различные Н-цепи, составляла 130 и не повышалась по сравнению с активностью связывания 131 GpH7-A5/GpH7-B3-01-15Y/GpL16-k0, в котором вводили только L234Y, при этом она снижалась по сравнению с активностью связывания 140 GpH7-A5/GpH7-B3-03-20W/GpL16-k0, в котором вводили только G236W. При этом активность связывания GpH7-TA4/GpH7-B3/GpL16-k0, в котором L234Y и G236W вводили в аналогичные Н-цепи, составляла 168, и повышалась по сравнению с активностью связывания GpH7-A5/GpH7-B3-01-15Y/GpL16-k0, в котором вводили только L234Y, и GpH7-A5/GpH7-B3-03-20W/GpL16-k0, в котором вводили только G236W. Как теоретически рассчитано, этот результат демонстрирует, что L234Y и G236W при введении в аналогичную Н-цепь дополнительно повышает активность связывания с FcγRIIIa.
Затем оценивали эффект комбинации L234Y и S298A на основании таблицы 8. Активность связывания с FcγRIIIa GpH7-TA1/GpH7-B3-15-02A/GpL16-k0, в котором L234Y и S298A вводили в различные Н-цепи, составляла 142 и повышалась по сравнению с активностью связывания 131 GpH7-A5/GpH7-B3-01-15Y/GpL16-k0, в котором вводили только L234Y, при этом она снижалась по сравнению с активностью связывания 163 GpH7-A5/GpH7-B3-15-02A/GpL16-k0, в котором вводили только S298A. Таким образом, вследствие того, что активность связывания GpH7-TA1/GpH7-B3-15-02A/GpL16-k0 не повышалась по сравнению с той, когда водили только S298A, можно
сказать, что, когда S298A и L234Y вводили в различные Н-цепи, не получали эффекта дополнительного повышения активности связывания с FcγRIIIa. При этом активность связывания с FcγRIIIa GpH7-TA5/GpH7-B3/GpL16-k0, в котором L234Y и S298A вводили в аналогичную Н-цепь, составляла 208 и повышалась по сравнению с GpH7-A5/GpH7-B3-01-15Y/GpL16-k0, в котором вводили только L234Y, и GpH7-A5/GpH7-B3-15-02A/GpL16-k0, в котором вводили только S298A. Как теоретически рассчитано, этот результат демонстрирует, что L234Y и S298A при введении в аналогичную Н-цепь дополнительно повышают активность связывания с FcγRIIIa.
Затем оценивали эффект комбинации G236W и S298A на основании таблицы 8. Активность связывания с FcγRIIIa GpH7-TA3/GpH7-B3-03-20W/GpL16-k0, в котором G236W и S298A вводили в различные Н-цепи, составляла 70 и снижалась по сравнению с активностью связывания 140 GpH7-A5/GpH7-B3-03-20W/GpL16-k0, в котором вводили только G236W, и активностью связывания 163 GpH7-A5/GpH7-B3-15-02A/GpL16-k0, в котором вводили только S298A. При этом активность связывания GpH7-TA6/GpH7-B3/GpL16-k0, в котором G236W и S298A вводили в аналогичную Н-цепь, составляла 228 и повышалась по сравнению с GpH7-A5/GpH7-B3-03-20W/GpL16-k0, в котором вводили только G236W, и GpH7-A5/GpH7-B3-15-02A/GpL16-k0, в котором вводили только S298A. Как теоретически рассчитано, этот результат демонстрирует, что G236W и S298A при введении в аналогичную Н-цепь дополнительно повышают активность связывания с FcγRIIIa.
Как изначально было теоретически рассчитано, эти результаты
демонстрируют, что L234Y, G236W и S298A повышают связывание, только когда каждое из них вводят в аналогичную Н-цепь. Эти данные подтверждают, что L234Y, G236W и S298A, когда содержатся в аналогичной цепи, повышают связывание с FcγR в аналогичном направлении. Таким образом, это демонстрирует, что можно определять способ соответствующей комбинации двух изменений на основании результата, теоретически рассчитанного из результата сравнения активности связывания с FcγRIIIa, комбинируя P329R с каждым из изменений. Другими словами, комбинация с P329R представляет собой пригодный способ теоретического расчета способа комбинации изменений в гетеродимеризованных антителах. Этот способ можно использовать для выявления других пригодных комбинаций изменений.
Комбинации двух или более изменений рассматривали на основании результата сравнения активностей связывания с FcγRIIIa, комбинируя P329R с каждым из изменений. Демонстрировали, что L234Y и G236W, G236W и S298A, и S298A и L234Y при введении в аналогичную Н-цепь, соответственно, усиливают взаимодействие FcγRIIIa в аналогичном направлении. Таким образом, на основании этого результата предполагали, что все из L234Y, G236W и S298A повышают активность связывания с FcγRIIIa в аналогичном направлении, и, таким образом, предполагали, что эти изменения при введении в аналогичную Н-цепь максимально повышают активность связывания с FcγRIIIa. Для проверки этой гипотезы конструировали GpH7-TA4, GpH7-TA5 и GpH7-TA6 введением в GpH7-A5 каждого из групп изменений из L234Y и G236W, L234Y и S298A и
G236W и S298A; GpH7-B3-01-15Y, GpH7-B3-03-20W и GpH7-B3-15-02A конструировали введением L234Y, G236W и S298A, соответственно, в GpH7-B3; экспрессирующие векторы, в которые вводили указанные выше конструкции, получали в соответствии со справочным примером 1. Векторы комбинировали таким образом, чтобы вводить три изменения L234Y, G236W и S298A в любую Н-цепь, и добавляли GpL16-k0 в качестве L-цепи, и экспрессировали и получали способом, описанным в справочном примере 1, представляющие интерес антитела. Кроме того, для экспрессии и очистки представляющего интерес антитела конструировали GpH7-TA7 (SEQ ID NO: 31), получаемую в результате введения трех изменений, L234Y, G236W и S298A в GpH7-A5, и комбинировали с GpH7-B3 и GpL16-k0 способом, описанным в справочном примере 1. Список антител, получаемых, как описано в настоящем описании, и результат сравнения активности связывания с FcγRIIIa антител приведен в таблице 9.
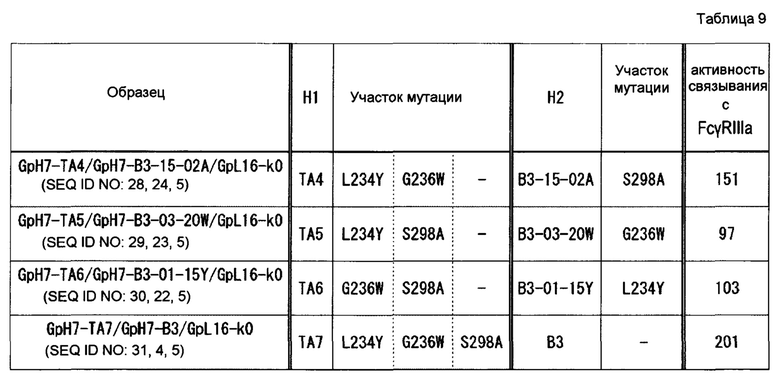
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи
соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Как было теоретически рассчитано на основании результата сравнения активности связывания с FcγRIIIa, комбинируя P329R с каждым изменением, для GpH7-TA7/GpH7-B3/GpL16-k0, в котором L234Y, G236W и S298A вводили в аналогичную Н-цепь, демонстрировали наибольшее повышение связывания с FcγRIIIa. Таким образом, результат демонстрирует, что можно теоретически рассчитывать способ, соответствующей комбинации двух или более изменений, сравнивая активность связывания с FcγRIIIa, комбинируя P329R с каждым изменением.
[Пример 6] Сравнение общепринятого гомодимеризованного антитела и нового гетеродимеризованного антитела на основе гетеродимеризованного антитела
Результаты, представленные в таблицах 7, 8 и 9 примера 5, демонстрируют, что гетеро-изменения, каждое из которых отдельно повышает активность связывания с FcγRIIIa при комбинации соответствующим образом друг с другом, могут дополнительно повышать связывание с FcγRIIIa. В частности, демонстрировали, что изменения L234Y, G236W и S298A при введении в аналогичную Н-цепь
дополнительно повышают активность связывания с FcγR.
Затем авторы настоящего изобретения исследовали даже при комбинации многих изменений, обладает ли еще получаемое гетеродимеризованное антитело характеристикой гетеродимеризованного антитела, которое при введении в него многих изменений, демонстрирует в большей степени повышенное связывание с FcγR, чем гомодимеризованное антитело, в котором вводили соответствующие многие изменения. В частности, GpH7-TA7 и GpH7-TA45 (SEQ ID NO: 32), получаемые в результате введения L234Y, G236W и S298A в GpH7-A5 и GpH7-B3, соответственно, получали способом, описанным в справочном примере 1. Как показано в таблице 10, гетеродимеризованные антитела GpH7-TA7/GpH7-B3/GpL16-k0 и GpH7-A5/GpH7-TA45/GpL16-k0, в которых L234Y, G236W и S298A вводили только в одну Н-цепь, и гомодимеризованное антитело GpH7-TA7/GpH7-TA45/GpL16-k0, в котором L234Y, G236W и S298A вводили в обе Н-цепи, экспрессировали и очищали способом, описанным в справочном примере 1. Эти антитела сравнивали в отношении связывания с FcγRIIIa способом, описанным в справочном примере 2 (таблица 10).

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Представленный в таблице 10 результат демонстрирует, что гомодимеризованное антитело GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили L234Y, G236W и S298A, демонстрировало сниженное связывание с FcγRIIIa по сравнению с гетеродимеризованными антителами GpH7-TA7/GpH7-B3/GpL16-k0 и GpH7-A5/GpH7-TA45/GpL16-k0, в которых только в одну из Н-цепей вводили L234Y, G236W и S298A. Это демонстрирует, что характеристика изменений L234Y, G236W и S298A, которые повышают активность связывания с FcγR гетеродимеризованного антитела, но снижают активность связывания с FcγR гомодимеризованного антитела, сохраняется, даже когда многие изменения комбинируют друг с другом.
Затем авторы настоящего изобретения исследовали, сохраняет ли гетеродимеризованное антитело, в котором только в одну из Н-цепей вводили L234Y, G236W и S298A, направленность связывания FcγRIIIa, обсуждаемую в примере 3.
GpH7-TA8 (SEQ ID NO: 33) и GpH7-B12 (SEQ ID NO: 12),
получаемые в результате введения P329R в GpH7-TA7 и GpH7-B3, соответственно, получали способом, описанным в справочном примере 1. Как представлено в таблице 11, гетеродимеризованное антитело GpH7-TA8/GpH7-B3/GpL16-k0, в котором L234Y, G236W и S298A в аналогичную Н-цепь, как вводили P329R, и гетеродимеризованное антитело GpH7-TA7/GpH7-B12/GpL16-k0, в котором L234Y, G236W и S298A вводили в Н-цепь, отличную от той, в которую вводили P329R, получали способом, описанным в справочном примере 1. Эти антитела сравнивали с гетеродимеризованным антителом GpH7-TA7/GpH7-B3/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, в отношении активности связывания с FcγRIII способом, описанным в справочном примере 2 (таблица 11).
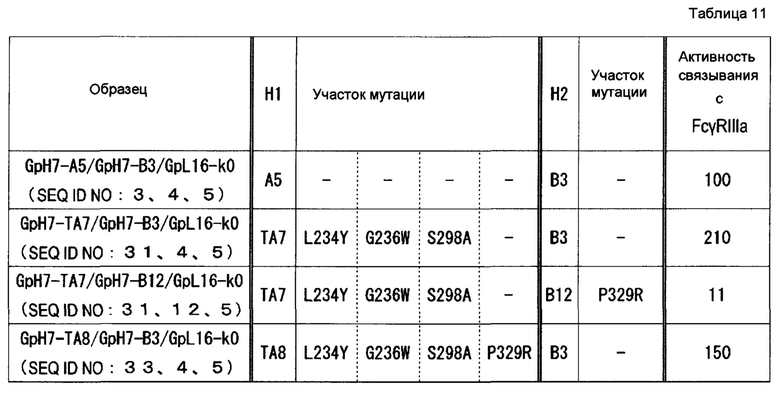
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0
("-": когда не существует конкретной мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Представленный в таблице 11 результат демонстрирует, что гетеродимеризованное антитело GpH7-TA8/GpH7-B3/GpL16-k0, в котором группу изменений L234Y, G236W и S298A в аналогичную Н-цепь, как вводили P329R, и гетеродимеризованное антитело GpH7-TA7/GpH7-B12/GpL16-k0, в котором группу изменений L234Y, G236W и S298A вводили в Н-цепь, отличную от той, в которую вводили P329R, демонстрировали сниженные активности связывания с FcγRIIIa по сравнению с GpH7-TA7/GpH7-B3/GpL16-k0. Активность связывания с FcγRIIIa GpH7-TA7/GpH7-B12/GpL16-k0 составляла 11 и заметно снижалась по сравнению с активностью 150 GpH7-TA8/GpH7-B3/GpL16-k0. Этот результат демонстрирует характеристику, заключающуюся в том, что активность связывания с FcγR заметно снижается, если многие изменения L234Y, G236W и S298A вводят в Н-цепь, когда их вводят в Н-цепь, отличную от той, в которую вводят P329R, что наблюдали в примере 3 для изменений L234Y, G236W и S298A.
Указанные выше открытия демонстрируют, что соответствующая комбинация изменений, которая повышает связывание с FcγR, может дополнительно повышать активность связывания с FcγR, при этом сохраняя характеристику гетеродимеризованного антитела.
[Пример 7] Сравнение с известным уровнем техники: сравнение
гетеродимеризованных вариантов и антител с измененными аминокислотами, которые повышают связывание с FcγRIIIa
Уже известны изменения, которые повышают активность ADCC путем повышение связывания с FcγRIIIa. Например, известны изменения S239D, I332E и A330L в качестве мутаций, которые повышают связывание с FcγRIIIa в большей степени при введении в обе Н-цепи антитела (Proc. Natl. Acad. Sci. USA, 103, 4005-4010, 2006). Демонстрировали, что противоопухолевую активность антитела повышают посредством повышения антителозависимой клеточной цитотоксической (ADCC) активности. Повышение связывания с FcγRIIIa антитела представляет собой эффективный способ повышения полезности антител в качестве фармацевтических средств. Однако, как продемонстрировано в примерах выше, полагают, что существуют ограничения в отношении повышения активности связывания с FcγR с использованием гомодимеризованных антител. Таким образом, предполагали, что активность связывания с FcγR можно дополнительно повышать посредством гетеро-изменения.
Как указано в примере 1, домен Fc антитела взаимодействует с FcγR асимметричным образом. В случае антитела, в котором вводили изменения S239D, I332E и A330L, учитывая трехмерную структуру, полагают, что в HA-цепи все измененные остатки S239D, I332E и A330L участвуют в усилении взаимодействия с FcγR, тогда как в Нв-цепи остатки, отличные от S239D, не участвуют в контактировании с FcγR и не способствуют повышению активности связывания с FcγR (фиг. 23). Таким образом, вследствие асимметричности взаимодействия домена Fc и FcγR каждое изменение, вводимое
общепринятой технологией изменения антитела, в достаточной степени не способно взаимодействовать с FcγR, и полагают, что это является достаточным для оптимизации взаимодействия антитела/FcγR. Например, в случае описанных выше изменений S239D, I332E и A330L, полагают, что связывание с FcγRIIIa можно дополнительно повышать введением изменений, которые усиливают взаимодействие с FcγRIIIa со стороны Нв-цепи, вместо введения указанных выше изменений в Нв-цепь. Таким образом, связывание с FcγR можно дополнительно повышать, применяя технологию получения гетеродимеризованных антител по настоящему изобретению посредством введения различных изменений в каждую Н-цепь антитела (далее в настоящем описании, обозначаемую как "технология гетеродимеризованного антитела"), вместо технологии введения аналогичного изменения в обе Н-цепи антитела (далее в настоящем описании, обозначаемой как "известный уровень техники" или "технология гомодимеризованного антитела").
Учитывая трехмерную структуру комплекса Fc/FcγRIIIa, полагают, что S298 в отличие от S239D, I332E и A330L взаимодействует с FcγR только в Нв-цепи, как представлено на фиг. 23 (JBC, 276:16469-16477, 2001). Таким образом, полагают, что, когда изменение вводят в S298, остаток после мутации замены также взаимодействует с FcγRIIIa со стороны Нв-цепи. Как видно в примере 5, полагают, что L234Y и G236W повышают взаимодействие с FcγR в аналогичном направлении как S298A. Таким образом, полагают, что, когда S239D, A330L и I332E вводят в аналогичную Н-цепь, а L234Y, G236W и S298A вводят в другую Н-цепь, все
вводимые изменения могут взаимодействовать с FcγR одновременно, приводя к дополнительному повышению взаимодействия с FcγR.
Для оценки указанной выше гипотезы авторы настоящего изобретения проводили следующие ниже эксперименты. Направление распознавания FcγR каждым изменением определяли способом, описанным в примере 5, с использованием антител, содержащих Н-цепь, в которую вводили изменение L234Y, G236W, S298A, S239D, A330L или I332E и вводили P329R в аналогичную или другую Н-цепь. Способом, описанным в справочном примере 1, конструировали экспрессирующие векторы, в которые вводили, как указано ниже: GpH7-A5; GpH7-A48 (SEQ ID NO: 16), получаемую в результате введения P329R в GpH7-A5; GpH7-HA7 (SEQ ID NO: 34), получаемую в результате введения S239D и P329R в GpH7-B3; GpH7-HA15 (SEQ ID NO: 35), получаемую в результате введения A330L и P329R в GpH7-B3; GpH7-HA18 (SEQ ID NO: 36), получаемую в результате введения I332E и P329R в GpH7-B3; GpH7-HA5 (SEQ ID NO: 19), получаемую в результате введения L234Y и P329R в GpH7-B3; GpH7-HA6 (SEQ ID NO: 20), получаемую в результате введения G236W и P329R в GpH7-B3; GpH7-HA11 (SEQ ID NO: 21), получаемую в результате введения S298A и P329R в GpH7-B3; GpH7-B3-06-09D (SEQ ID NO: 37), получаемую в результате введения S239D в GpH7-B3; GpH7-B3-20-08L (SEQ ID NO: 38), получаемую в результате введения A330L в GpH7-B3; GpH7-B3-22-10E (SEQ ID NO: 39), получаемую в результате введения I332E в GpH7-B3; GpH7-B3-01-15Y (SEQ ID NO: 22), получаемую в результате введения L234Y в GpH7-B3; GpH7-B3-03-20W (SEQ ID NO: 23), получаемую в результате введения G236W в GpH7-В3,
и GpH7-B3-15-02A (SEQ ID NO: 24), получаемую в результате введения S298A в GpH7-B3. Экспрессирующие векторы для характерных Н-цепей комбинировали таким способом, что каждое из изменений L234Y, G236W, S298A, S239D, A330L и I332E содержится в аналогичной Н-цепи как P329R или в Н-цепи, отличной от той, в которой содержится P329R, и комбинировали с ними экспрессирующий вектор GpL16-k0 для L-цепи. Представляющие интерес антитела экспрессировали и получали способом, описанным в справочном примере 1. Получаемые антитела использовали для анализа активности связывания с FcγRIIIa. Результат оценки с использованием P329R в отношении направления распознавания FcγRIIIa L234Y, G236W, S298A, S239D, A330L и I332E приведен в таблице 12.

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Активность связывания с FcγRIIIa указывают в виде относительной активности связывания при принятии связывания с FcγRIIIa GpH7-A5/GpH7-B3/GpL16-k0 за 100. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого
антитела.
На основании результата в отношении каждого изменения сравнивают активность связывания с FcγRIIIa, когда каждое изменение вводили в аналогичную Н-цепь, как вводят P329R, и в Н-цепь, отличную от той, в которую вводят P329R. В случае S239D активность связывания GpH7-A5/GpH7-HA7/GpL16-k0, соответствующего первому случаю, составляла 3, тогда как активность связывания GpH7-A48/GpH7-B3-06-09D/GpL16-k0, соответствующего последнему случаю, составляла 123; таким образом, связывание с FcγRIIIa ингибировали, когда изменение вводили в аналогичную Н-цепь, как вводили P329R. В случае A330L активность связывания GpH7-A5/GpH7-HA15/GpL16-k0, соответствующего первому случаю, составляла 32, тогда как активность связывания GpH7-A48/GpH7-B3-20-08L/GpL16-k0, соответствующего последнему случаю, составляла 60; таким образом, связывание FcγRIIIa ингибировали, когда изменение вводили в аналогичную Н-цепь, как вводили P329R. В случае I332E активность связывания GpH7-A5/GpH7-HA18/GpL16-k0, соответствующего первому случаю, составляла 35, тогда как активность связывания GpH7-A48/GpH7-B3-22-10E/GpL16-k0, соответствующего второму случаю, составляла 189; таким образом, связывания с FcγRIIIa ингибировали, когда изменение вводили в аналогичную Н-цепь, как вводили P329R. В отношении всех изменений связывание с FcγRIIIa ингибировали, когда изменение вводили в аналогичную Н-цепь, как вводили P329R, т.е. как в первом случае. Если Н-цепь, в которую вводили P329R,
соответствовала HA-цепи фиг. 23, полагают, что P329R ингибирует связывание в направлении X. Вследствие того, что комбинации, которые значительно ингибируют связывание, представляют собой такие, когда S239D, A330L или I332E водили в аналогичную Н-цепь, как вводили P329R, таким образом, в этом случае, изменения вводили в HA-цепь. Вследствие того, что эффект повышения связывания с FcγRIIIa заметно ингибировали, когда любое из изменений S239D, A330L и I332E вводили в аналогичную Н-цепь, как вводили P329R, все изменения при введении в HA-цепь повышают связывание с FcγRIIIa в направлении X, которое ингибируют P329R. Таким образом, полагают, что связывание с FcγRIIIa можно дополнительно повышать введением этих изменений в аналогичную Н-цепь. На основании примера 5 предполагали, что все L234Y, G236W и S298A при введении в Нв-цепь повышают связывание в направлении X. Как обсуждается в примерах 5 и 6, можно выявить способ соответствующей комбинации соответствующих изменений, комбинируя их с P329R. На основании указанных выше результатов, для повышения связывания с FcγRIIIa в направлении X, как представлено на фиг. 3, S239D, A330L и I332E необходимо вводить в HA-цепь, тогда как L234Y, G236W и S298A необходимо вводить в Нв-цепь. Таким образом, полагали, что связывание с FcγRIIIa в направлении Х можно дополнительно повышать введением соответствующих групп изменений в различные Н-цепи.
Для оценки указанной выше гипотезы способом, описанным в справочном примере 1, конструировали экспрессирующие векторы, в которые вводили, как указано ниже:
GpH7-A57 (SEQ ID NO: 40), получаемую в результате введения всех из S239D, A330L и I332E в GpH7-A5;
GpH7-B78 (SEQ ID NO: 41), получаемую в результате введения всех из S239D, A330L и I332E в GpH7-B3;
GpH7-TA7 (SEQ ID NO: 31), получаемую в результате введения всех из L234Y, G236W и S298A в GpH7-A5, и
GpH7-TA45 (SEQ ID NO: 32), получаемую в результате введения всех из L234Y, G236W и S298A в GpH7-B3. С использованием экспрессирующих векторов и GpH7-A5, GpH7-B3 и GpL16-k0 следующие ниже антитела экспрессировали и получали способом, описанным в справочном примере 1:
гетеро GpH7-TA7/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, а в другую вводили S239D, A330L и I332E;
GpH7-TA7/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили L234Y, G236W и S298A;
GpH7-TA7/GpH7-TA45/GpL16-k0, в котором в обе Н-цепи вводили L234Y, G236W и S298A;
GpH7-A5/GpH7-B78/GpL16-k0, в котором только в одну из Н-цепей вводили S239D, A330L и I332E, и
GpH7-A57/GpH7-B78/GpL16-k0, в котором в обе Н-цепи вводили S239D, A330L и I332E. Получаемые антитела сравнивали в отношении активности связывания с FcγRIIIa с использованием в качестве показателя KD для FcγRIIIa, которую определяли способом, описанным в справочном примере 2. Оценка результата эффекта комбинации L234Y, G236W и S298A с S239D, A330L и I332E приведена в таблице 13.
.
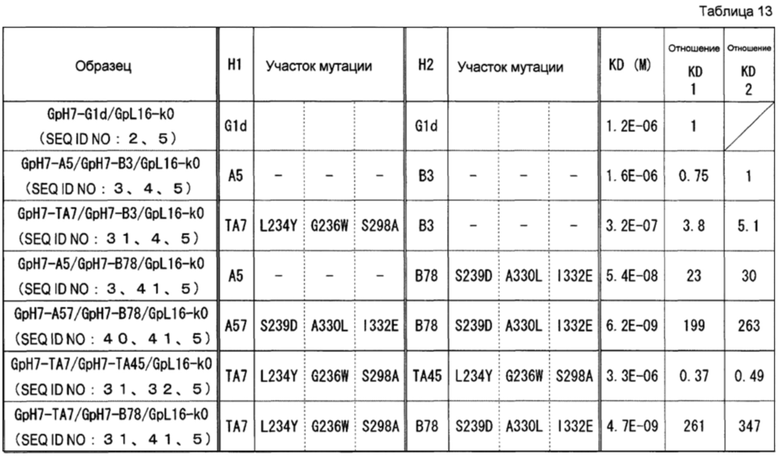
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-G1d/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначают как "отношение KD 1", при этом значение, получаемое делением KD GpH7-A5/GpH7-B3/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD 2". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Принимая во внимание результат таблицы 13, когда GpH7-A5/GpH7-B3/GpL16-k0, в котором каждый из D356K, H435R и K439Е вводили в одну Н-цепь, сравнивают с GpH7-G1d/GpL16-k0, которой представляет собой нативный IgG1, разница в активности связывания с FcγRIIIa составляет 0,75 раз, и не наблюдали значимого отличия. Таким образом, полагали, что изменения D356K, H435R и K439Е не влияют на активность связывания с FcγRIIIa.
Каждое изменение оценивали в отношении его эффекта на гомодимеризованные антитела, которые используют в известном уровне техники. Активность связывания с FcγRIIIa гомодимеризованного антитела GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E, повышалась приблизительно в 260 раз по сравнению с активность связывания GpH7-A5/GpH7-B3/GpL16-k0. В противоположность этому активность связывания с FcγRIIIa гомодимеризованного антитела GpH7-TA7/GpH7-TA45/GpL16-k0,
в обе Н-цепи которого вводили L234Y, G236W и S298A, снижалась в 0,49 раз. Это демонстрирует, что только группа изменений S239D, A330L и I332E обладает эффектом повышения активности связывания с FcγRIIIa гомодимеризованных антител.
Гетеродимеризованные антитела, в которых только в одну из Н-цепей вводили каждую группу изменений, оценивали в отношении эффекта каждой группы изменений. Активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-A5/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили S239D, A330L и I332E, повышалась приблизительно в 30 раз по сравнению с активностью связывания GpH7-A5/GpH7-B3/GpL16-k0, тогда как активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-TA7/GpH7-B3/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, повышалась в 5,1 раз. Этот результат демонстрирует, что группа изменений S239D, A330L и I332E обладает большим эффектом в отношении повышения активности связывания с FcγRIIIa.
Каждую группу изменений оценивали в отношении отличия их эффекта в гомодимеризованном антителе и гетеродимеризованном антителе. В отношении S239D, A330L и I332E активность связывания с FcγRIIIa гетеродимеризованного антитела повышалась в 30 раз по сравнению с активностью связывания GpH7-A5/GpH7-B3/GpL16-k0, тогда как активность связывания гомодимеризованного антитела повышалась приблизительно в 260 раз, демонстрируя, что изменения при введении в гомодимеризованные антитела дополнительно повышают активность связывания с FcγRIIIa. При этом в отношении
L234Y, G236W и S298A, активность связывания с FcγRIIIa гетеродимеризованного антитела повышалась в 5,1 раз по сравнению с активностью связывания GpH7-A5/GpH7-B3/GpL16-k0, однако активность гомодимеризованного антитела снижалась в 0,49 раз. Этот результат демонстрирует, что группа изменений L234Y, G236W и S298A обладает эффектом повышения связывания с FcγRIIIa только в гетеродимеризованных антителах, как обсуждается в примере 5.
Демонстрировали, что в гомодимеризованных антителах, только группа изменений S239D, A330L и I332E обладает эффектом повышения связывания с FcγRIIIa, а также в гетеродимеризованных антителах группа изменений S239D, A330L и I332E в большей степени повышает связывание с FcγRIIIa. Принимая во внимание общепринятый принцип, если рассматривают комбинацию группы изменений S239D, A330L и I332E с группой изменений L234Y, G236W, теоретически можно рассчитывать, что эффект повышения связывания с FcγRIIIa является больше в гомодимеризованном антителе GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводят только группу изменений S239D, A330L и I332E, которая значительно повышает связывание с FcγRIIIa гетеродимеризованного антитела и гомодимеризованного антитела. Однако активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-TA7/GpH7-B78/GpL16-k0, в котором S239D, A330L и I332E вводили в одну Н-цепь и L234Y, G236W и S298A вводили в другую Н-цепь, повышалась приблизительно в 350 раз по сравнению с активностью связывания GpH7-A5/GpH7-B3/GpL16-k0; эффект повышения связывания являлся сильнее, чем для гомодимеризованного антитела, в обе Н-цепи
которого вводили S239D, A330L и I332E. Это подтверждает гипотезу, что, когда группу изменений S239D, A330L и I332E и группу изменений L234Y, G236W и S298A вводят в различные Н-цепи, все вводимые изменения повышают активность связывания с FcγRIIIa в HA-цепи и HB-цепи, и эффект является больше, чем в случае, когда группу изменений S239D, A330L и I332E вводят в обе Н-цепи.
Таким образом, демонстрируют, что применение гетеродимеризованных антител вместо общепринятых гомодимеризованных антител позволяет точнее оптимизировать асимметричное взаимодействие домена Fc и FcγRIIIa для конструкции доменов Fc, обладающих большей активностью связывания.
Принимая во внимание фиг. 23, предполагали, что A330L и I332E взаимодействуют с FcγR только в HA-цепи, тогда как S239D взаимодействует с FcγR в HA-цепи и HB-цепи. Фактически, KD для FcγRIIIa гетеродимеризованного антитела GpH7-A5/GpH7-B78/GpL16-k0, в котором только в одну из Н-цепей, вводили S239D, A330L и I332E, составляла 5,4Е-8, тогда как KD гомодимеризованного антитела GpH7-A57/GpH7-B78/GpL16-k0 составляла 6,2Е-9; таким образом, активность связывания с FcγRIIIa повышалась в 8,7 раз. Если предполагать, что различие активности связывания обусловлено участием S239D в повышении связывания в обеих Н-цепях, полагают, что это различие устраняют введением S239D в обе Н-цепи. Для проверки этой гипотезы конструировали GpH7-A53 (SEQ ID NO: 42), получаемую в результате введения S239D в GpH7-А5, и комбинировали ее с GpH7-B78, в которую вводили S239D, A330L и I332E, и экспрессию и получение проводили способом,
описанным в справочном примере 1. Эффект комбинации S239D и S239D, A330L и I332E оценивали, сравнивая связывание с FcγRIIIa гетеродимеризованных и гомодимеризованных антител с S239D, A330L и I332E способом, описанным в справочном примере 2 (таблица 14).

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-A5/GpH7-B78/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
В таблице 14 продемонстрировано, что активность связывания с FcγRIIIa GpH7-A53/GpH7-B78/GpL16-k0, в котором в одну из Н-цепей вводят S239D, A330L и I332E, а в другую вводят S239D, повышалась в 4,9 раза, по сравнению с активностью связывания GpH7-A5/GpH7-B78/GpL16-k0, в котором только в одну из Н-цепей вводили S239D, A330L и I332E; при этом активность снижалась только в 1,8 раз по сравнению с активностью GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи
которого вводили S239D, A330L и I332E. Этот результат демонстрирует, что S239D взаимодействует с FcγRIIIa в обеих Н-цепях, как указано выше для гипотезы. Полагали, что взаимодействие с FcγRIIIa можно дополнительно повышать введением указанного выше изменения.
Авторы настоящего изобретения тестировали можно ли дополнительно повышать активность связывания с FcγRIIIa введением S239D в гетеродимеризованное антитело GpH7-TA7/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, а в другую вводили S239D, A330L и I332E. GpH7-TA22 (SEQ ID NO: 43) конструировали посредством введения S239D в GpH7-TA7, а затем встраивали в экспрессирующий вектор и комбинировали получаемый экспрессирующий вектор с GpL16-k0 и GpH7-B78, получаемой введением S239D, A330L и I332E в GpH7-B3, и представляющее интерес антитело экспрессировали и получали способом, описанным в справочном примере 1. Кроме того, гетеродимеризованное антитело GpH7-TA22/GpH7-B78/GpL16-k0 получали введением L234Y, G236W, S239D и S298A в одну Н-цепь и S239D, A330L и I332E в другую Н-цепь. Для оценки эффекта комбинации S239D с S239D, A330L и I332E антитела сравнивали в отношении активности связывания с FcγRIIIa способом, описанным в справочном примере 2 (таблица 15).
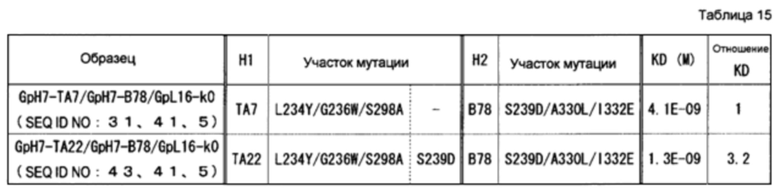
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-TA7/GpH7-B78/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
В таблице 15 продемонстрировано, что активность связывания GpH7-TA22/GpH7-B78/GpL16-k0, получаемого в результате введения S239D в Н-цепь, которая не содержит S239D, в гетеродимеризованном антителе GpH7-TA7/GpH7-B78/GpL16-k0, в котором в одну из Н-цепей вводили L234Y, G236W и S298A, а в другую Н-цепь S239D, A330L и I332E, повышалась в 3,2 раза по сравнению с активностью связывания GpH7-TA7/GpH7-B78/GpL16-k0. Этот результат демонстрирует, что связывание с FcγRIIIa можно дополнительно повышать применением S239D.
Затем авторы настоящего изобретения рассматривали дополнительное введение изменений Y296W и K334G, которые повышают связывание с FcγRIIIa, которые выявили, как описано в примере 4.
Сначала изучали, в какую Н-цепь необходимо вводить Y296W. Конструировали GpH7-TA52 (SEQ ID NO: 44), получаемую в результате введения мутация Y296W в GpH7-TA7, и экспрессировали и получали в комбинации с GpH7-B78 способом, описанным в справочном примере 1. Кроме того, конструировали GpH7-TA58 (SEQ ID NO: 45), получаемую в результате введения Y296W в GpH7-B78, и экспрессировали и получали в комбинации с GpH7-TA22 способом, описанным в справочном примере 1. Для оценки эффектов комбинаций с Y296W, получаемые антитела сравнивали в отношении активности связывания с FcγRIIIa способом, описанным в справочном примере 2 (таблица 16).

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-TA7/GpH7-B78/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей
Н-цепи и L-цепи каждого антитела.
Результат демонстрировал, что Y296W при введении в Н-цепь, отличную от той, в которую вводили L234Y, G236W и S298A, повышал активность связывания с FcγRIIIa только в 1,2 раз после введения по сравнению с активностью до введения, при этом Y296W при введении в аналогичную Н-цепь повышала активность связывания с FcγRIIIa в 1,8 раза по сравнению с активностью связывания GpH7-TA7/GpH7-B78/GpL16-k0. На основании этого результата полагают, что Y296W при введении в аналогичную Н-цепь, как вводят L234Y, G236W и S298A, обладает эффектом повышения связывания с FcγRIIIa.
Затем конструировали GpH7-TA54 (SEQ ID NO: 46), получаемую в результате введения Y296W в GpH7-TA22, и экспрессировали и получали в комбинации с GpH7-B78 способом, описанным в справочном примере 1. Кроме того, конструировали GpH7-TA58 (SEQ ID NO: 45), получаемую в результате введения Y296W в GpH7-B78, и экспрессировали и получали в комбинации с GpH7-TA22 способом, описанным в справочном примере 1. Для оценки эффекта комбинации с Y296W получаемые антитела сравнивали в отношении активности связывания с FcγRIIIa способом, описанным в справочном примере 2 (таблица 17).
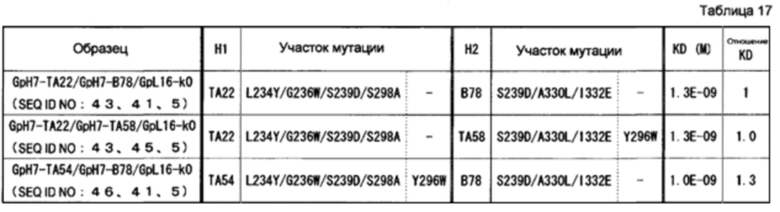
В столбце "образец" указаны названия антител, в столбцах
"H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-TA22/GpH7-B78/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Результат, представленный в таблице 17, демонстрирует, что, когда Y296W вводили в Н-цепь, которая отличается от той, в которую вводили L234Y, G236W и S298A, не существовало различия активности связывания с FcγRIIIa после введения по сравнению с активностью до введения. Когда Y296W вводили в аналогичную Н-цепь, как вводили L234Y, G236W и S298A, активность связывания с FcγRIIIa повышалась в 1,3 раз по сравнению с активностью связывания GpH7-TA22/GpH7-B78/GpL16-k0. На основании этого результата предполагали, что Y296W при введении в аналогичную Н-цепь, как вводят L234Y, G236W и S298A, обладает эффектом повышения активности связывания с FcγRIIIa.
Затем аналогичным образом также оценивали K334G. Конструировали GpH7-TA40 (SEQ ID NO: 47), получаемую в результате введения K334G в GpH7-TA7, и экспрессировали и получали в комбинации с GpH7-B78 способом, описанным в справочном примере 1. Кроме того, конструировали GpH7-TA50 (SEQ ID NO: 48), получаемую в результате введения K334G в GpH7-B78, и экспрессировали и получали в комбинации с GpH7-TA7 способом,
описанным в справочном примере 1. Для оценки эффекта комбинаций с K334G получаемые антитела сравнивали в отношении связывания с FcγRIIIa способом, описанным в справочном примере 2 (таблица 18).
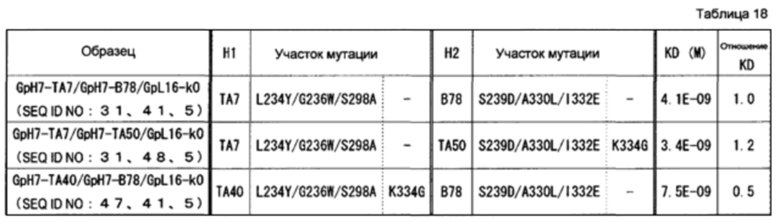
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-TA7/GpH7-B78/GpL16-k0 для FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Результат, представленный в таблице 18, демонстрирует, что, когда K334G вводили в аналогичную Н-цепь, как вводили L234Y, G236W и S298A, активность связывания с FcγRIIIa снижалась на половину после введения по сравнению с активностью до введения. Когда K334G вводили в другую Н-цепь, активность связывания с FcγRIIIa повышалась в 1,2 раз по сравнению с активностью GpH7-TA7/GpH7-B78/GpL16-k0. На основании этого результата полагали, что K334G при введении в Н-цепь, отличную от той, в которую вводят L234Y, G236W и S298A, обладает эффектом повышения
активности связывания с FcγRIIIa.
[Пример 8] Улучшение селективности активирующего FcγR или ингибирующего FcγR
Существуют активирующий FcγR, который содержит ITAM, и ингибирующий FcγR, который содержит ITIM. Характерные активирующие FcγR (активирующий рецептор) включают FcγRIa, FcγRIIa и FcγRIIIa, тогда как характерные ингибирующие FcγR (ингибирующий рецептор) включают FcγRIIb. В отношении антител, направленных против злокачественной опухоли, полагают, что важную роль играет отношении активности связывания с активирующим FcγR, механизм действия которого основан на активности ADCC или антителозависимой клеточной фагоцитарной (ADCP) активности, к активности связывания с ингибирующим FcγR (Nature Medicine, 6:443-446, 2000).
Для антител, направленных против злокачественной опухоли, желательно повышать их активность связывания с активирующим FcγR наряду со снижением их активности связывания с ингибирующим FcγR. В частности, желаемые изменения, такие как изменения, содержащиеся в области а, представленной на фиг. 24, представляют собой изменения, которые способствуют тому, что антитело сильнее связывается с активирующим FcγR по сравнению с нативным антителом и слабее связывается с ингибирующим FcγR по сравнению с нативным антителом, т.е. изменения, которые селективно повышают связывание с активирующим FcγR. Кроме того, желаемые изменения, такие как изменения в области b,
представленной на фиг. 25, представляют собой изменения, которые обеспечивают то, что отношение активности связывания с активирующим FcγR к активности связывания с ингибирующим FcγR является больше чем для нативного антитела. Можно сказать, что такие изменения селективно повышают активность связывания с активирующим FcγR по сравнению с ингибирующий FcγR.
Гетеродимеризованные антитела Не Abs, в которых одну из Н-цепей вводили изменение, анализировали в отношении их активностей связывания с каждым активирующим FcγR и ингибирующим FcγR способом, описанным в примере 4. Результат приведен на фиг. 26, 27, 28 и 29 с оценкой отношения активностей связывания с каждым активирующим FcγR и ингибирующим FcγR для каждого гетеродимеризованного антитела. Активирующие FcγR на фиг. 26, 27, 28 и 29 представляют собой FcγRIa, FcγRIIa (R), FcγRIIa (H) и FcγRIIIa, соответственно.
Изменения, находящиеся в области на фиг. 26, соответствующей а и b на фиг. 24 и 25, перечислены в таблице 19 (таблице от 19-1 до 19-5). Аналогично, в отношении FcγRIIa (R) (фиг. 27), FcγRIIa (H) (фиг. 28), FcγRIIIa (фиг. 29) изменения, находящиеся в области, соответствующей а и b, перечислены в таблице 20 (таблицах от 20-1 до 20-3), таблице 21 (таблицах от 21-1 до 21-3) и таблице 22 (таблицах от 22-1 до 22-3).




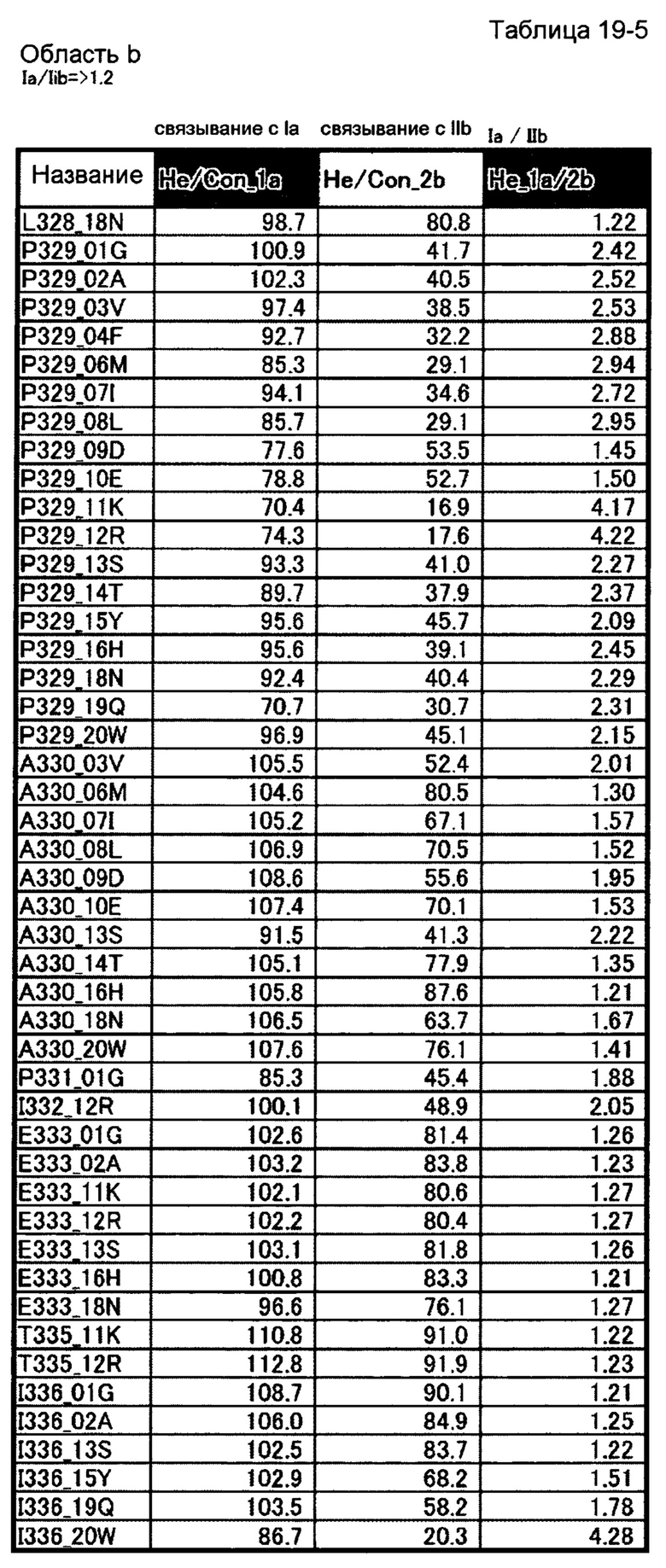
В таблице приведен список изменений, которые селективно повышают связывание с FcγRIa по сравнению с FcγRIIb.
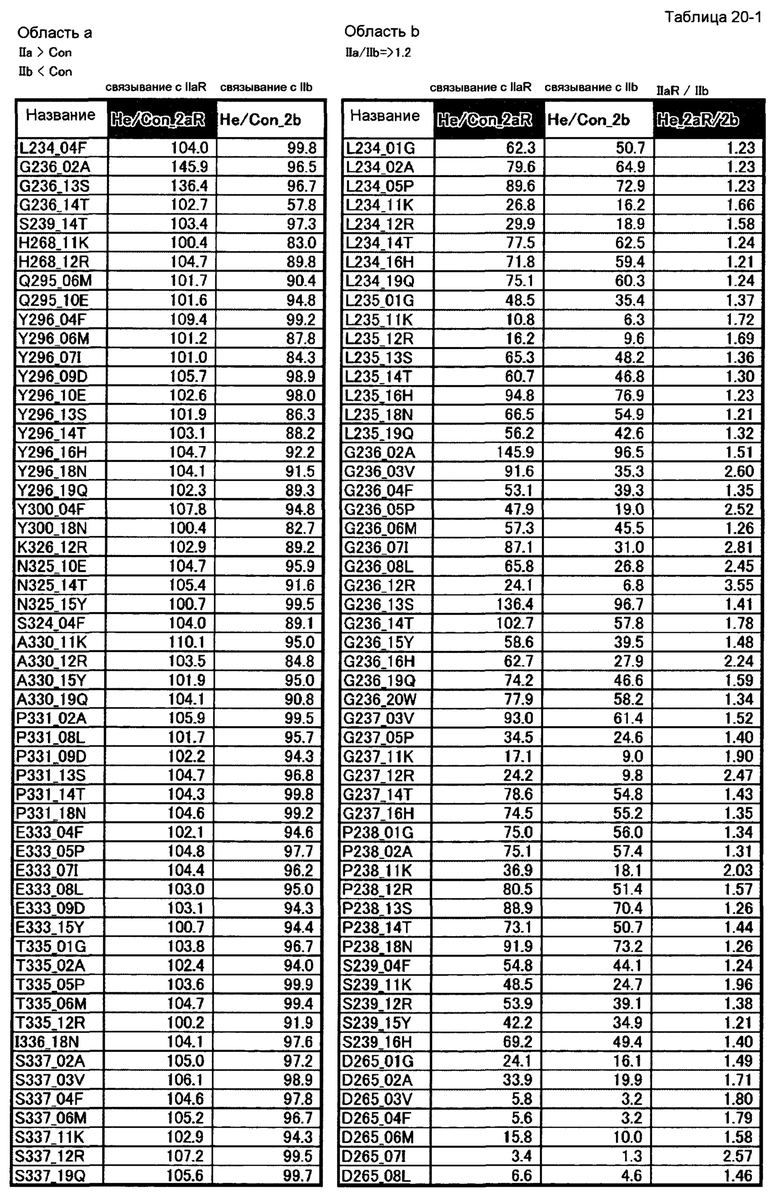

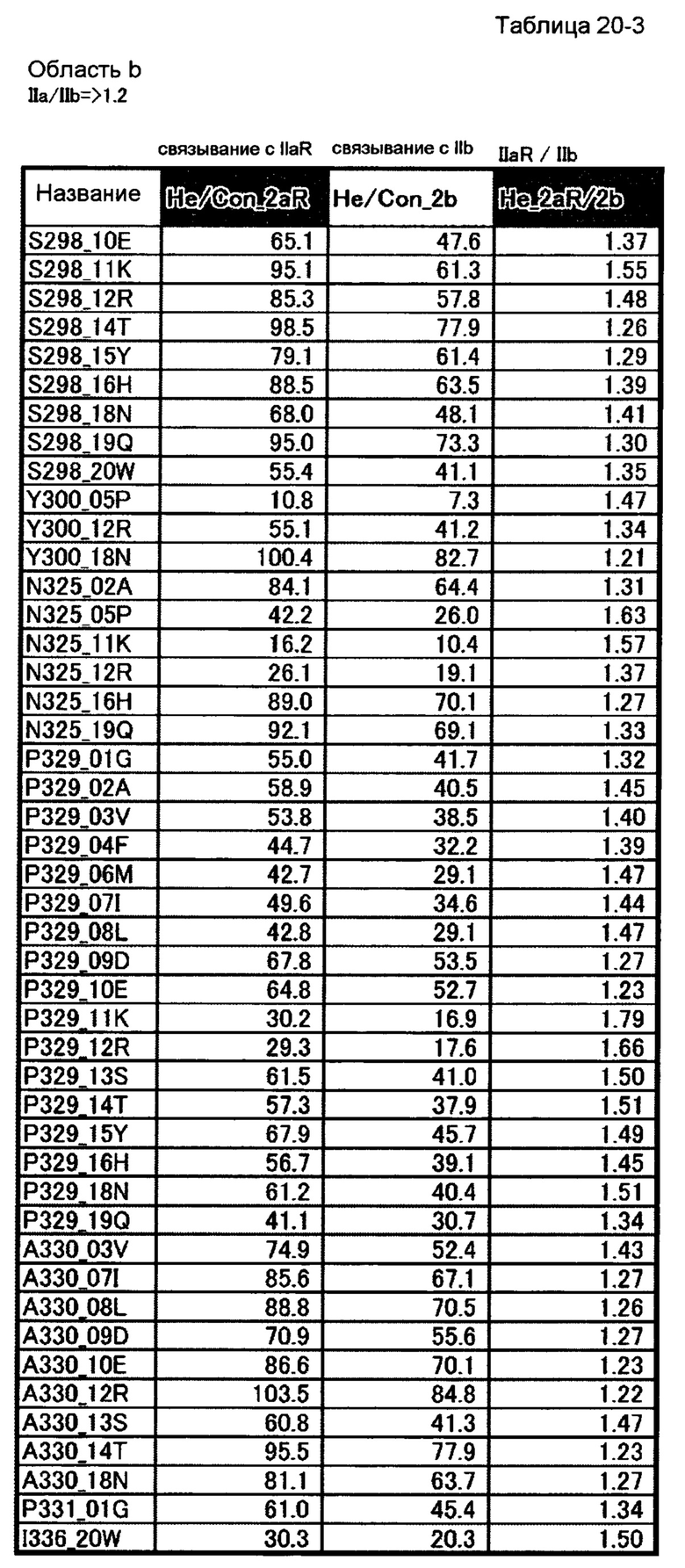
В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIa (R) по сравнению с FcγRIIb.



В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIa (H) по сравнению с FcγRII.
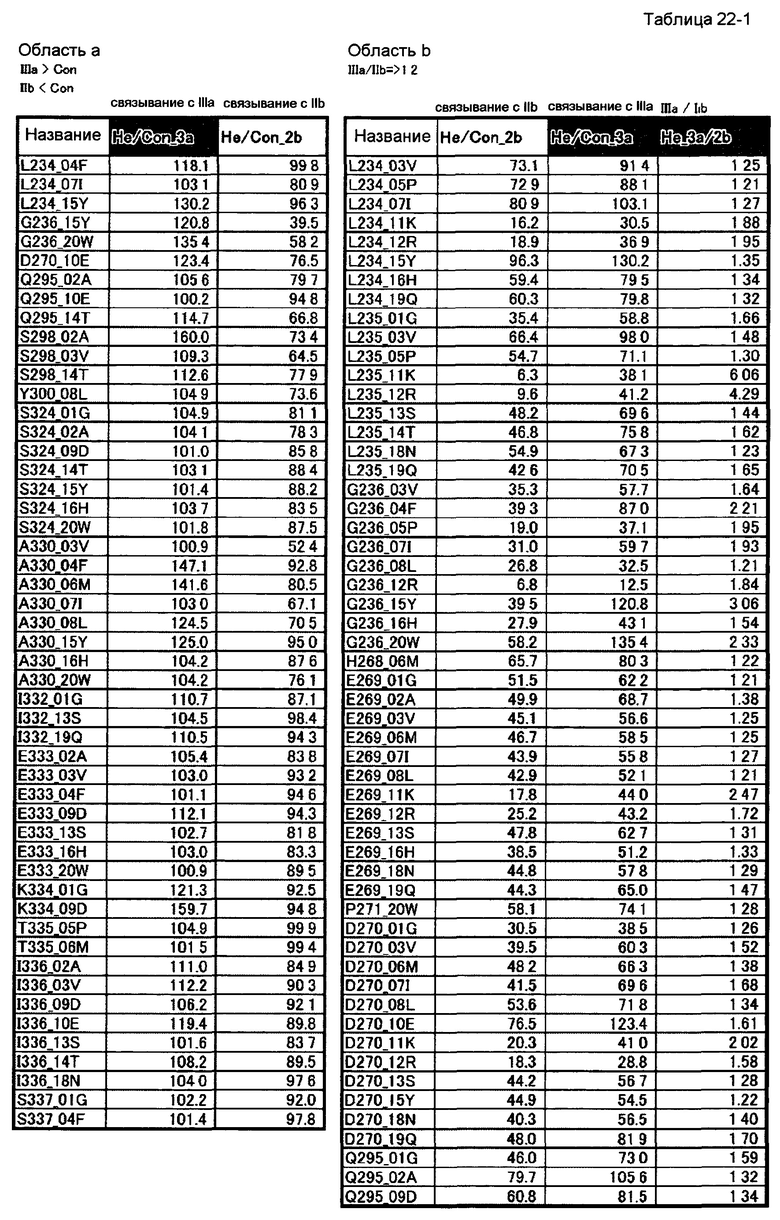
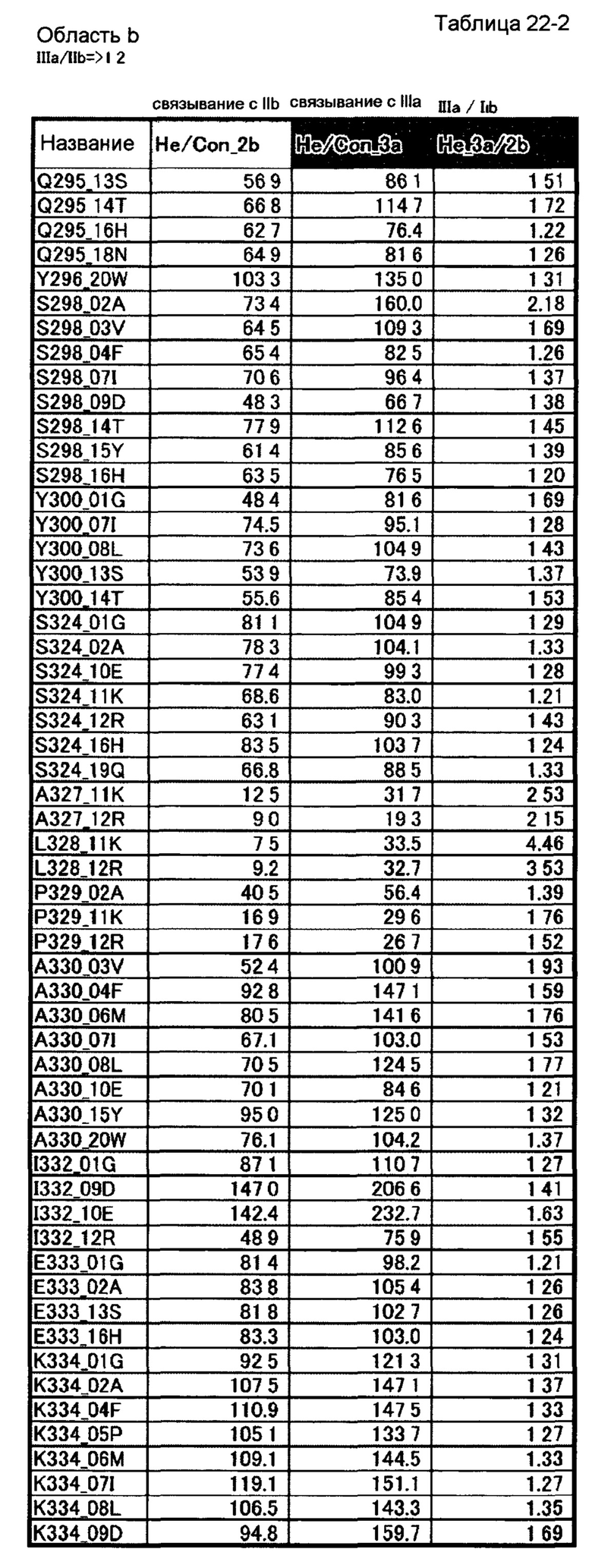

В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIIa по сравнению с FcγRIIb.
С другой стороны, FcγRIIb единственный ингибирующий FcγR играет важную роль в аутоиммунных заболеваниях и воспалительных заболеваниях (J. Exp. Med., 203, 2157-2164, 2006; J. immunol., 178, 3272-3280, 2007). Также показано, что антитела, содержащие домен Fc с повышенной активностью связывания с FcγRIIb, могут являться эффективными при лечении аутоиммунных заболеваний, обусловленных В-клетками (Mol. Immunology, 45, 3926-3933, 2008). В случае антител, которые направлены на лечение аутоиммунных заболеваний и воспалительных заболеваний, активность ADCC и активность ADCP может усугублять патологические состояния через активирующий FcγR. Таким образом, желательно повышать активность связывания с ингибирующим FcγR, при этом как можно больше снижая активность связывания с активирующим FcγR. В частности, как и изменения, находящиеся в области с на фиг. 24, желательно изменения обладают эффектом повышения активности связывания с
ингибирующим FcγR по сравнению с нативным антителом и эффектом снижения активности связывания с активирующим FcγR. Можно сказать, что такие изменения представляют собой изменения, которые селективно повышают активность связывания с ингибирующим FcγR по сравнению с активирующим FcγR. Кроме того, как и изменения, находящиеся в области d на фиг. 25, желательно изменения обладают эффектом повышения отношения активности связывания с ингибирующим FcγR к активности связывания с активирующим FcγR по сравнению с нативным антителом. Можно сказать, что такие изменения представляют собой изменения, которые селективно повышают активность связывания с ингибирующим FcγR по сравнению с активирующим FcγR.
На основании фиг. 26, 27, 28 и 29 на основании оценки отношения активности связывания с ингибирующим FcγR и активирующим FcγR для каждого из указанных выше гетеродимеризованных антител для изменений, представленных на каждой фигуре, изменения, находящиеся в областях, соответствующих с и d на фиг. 24 и 25, приведены в виде списка в таблице 23 (таблицах 23-1 и 23-2), таблице 24 (таблицах 24-1 и 24-2), таблице 25 (таблицах 25-1 до 25-3), таблице 26 (таблицах 26-1 до 26-4).


В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIb по сравнению с FcγRIa.
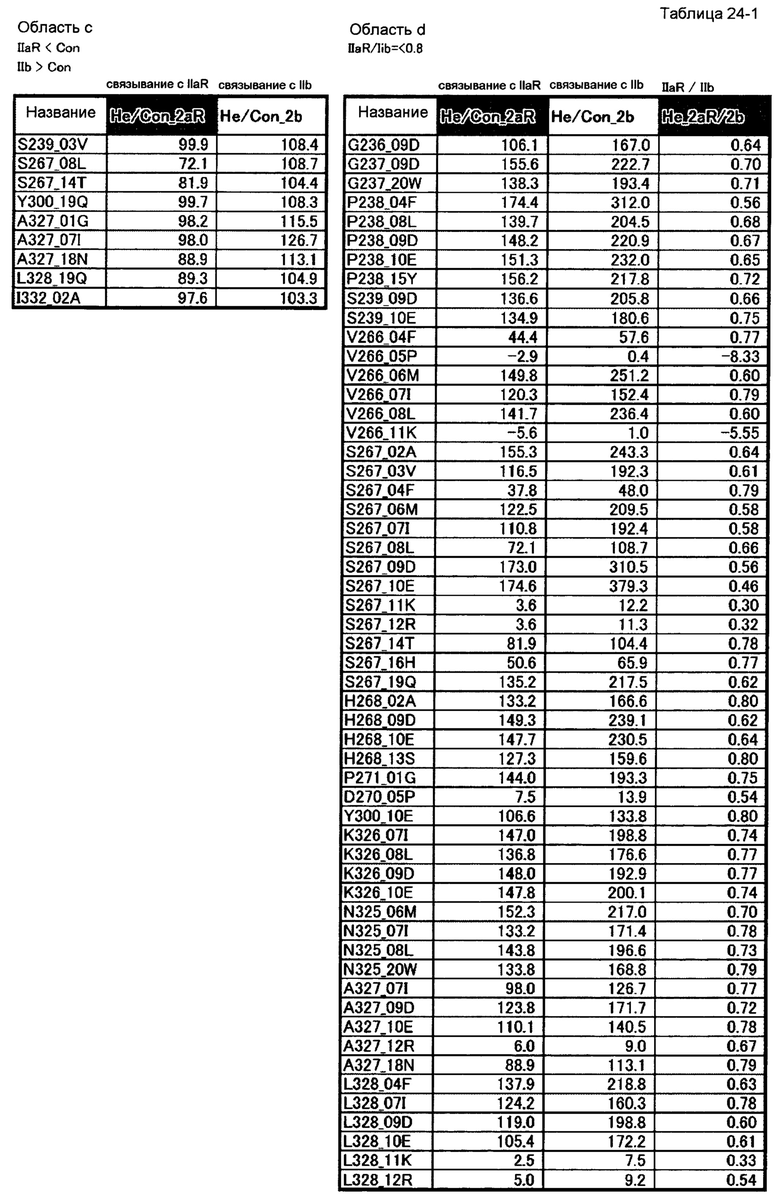
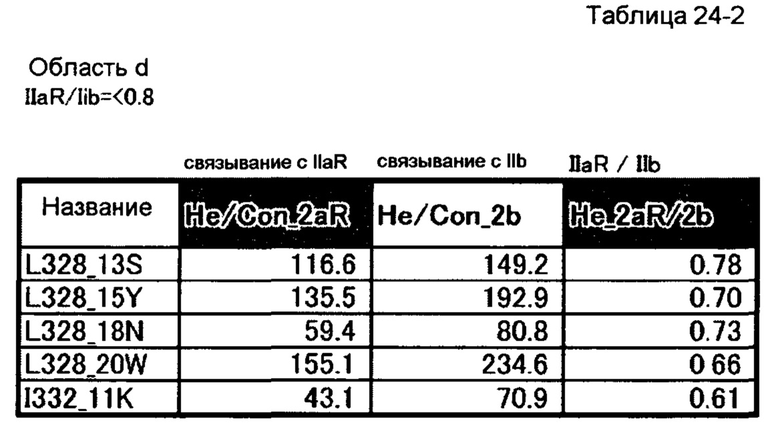
В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIb по сравнению с FcγRIIa (R).
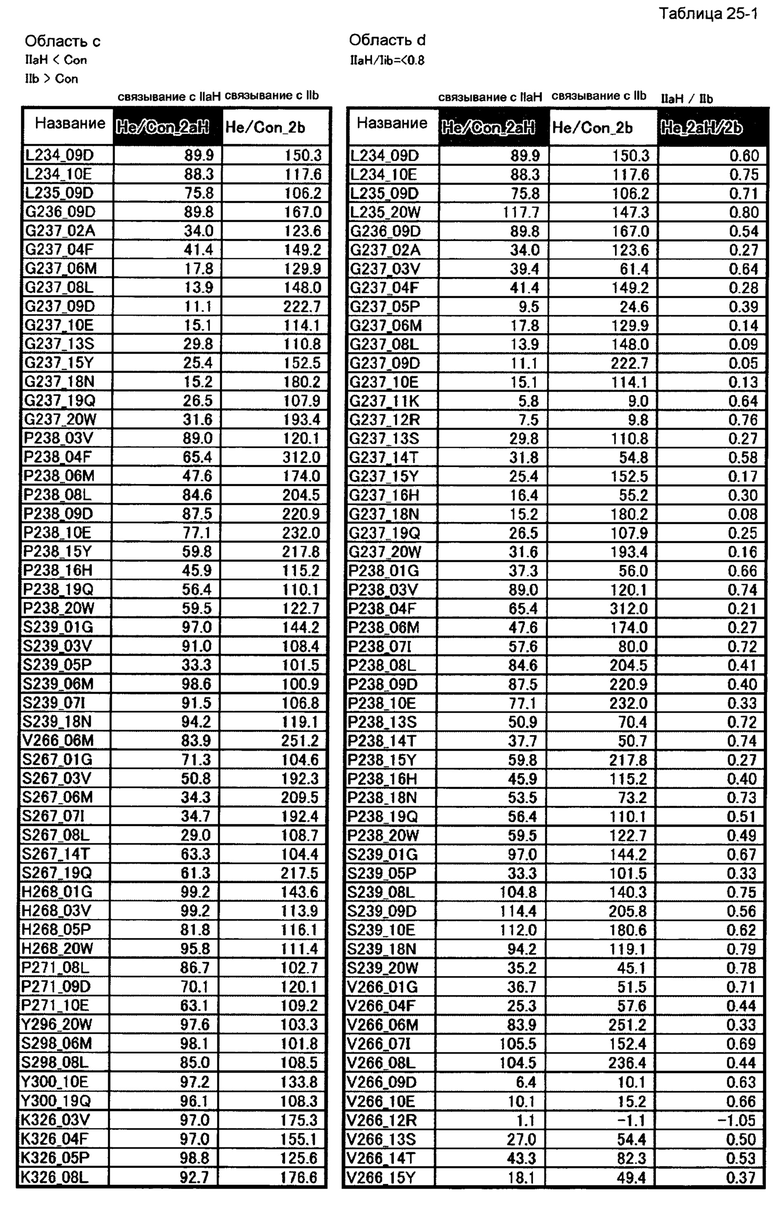


В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIb по сравнению с FcγRIIa (H).

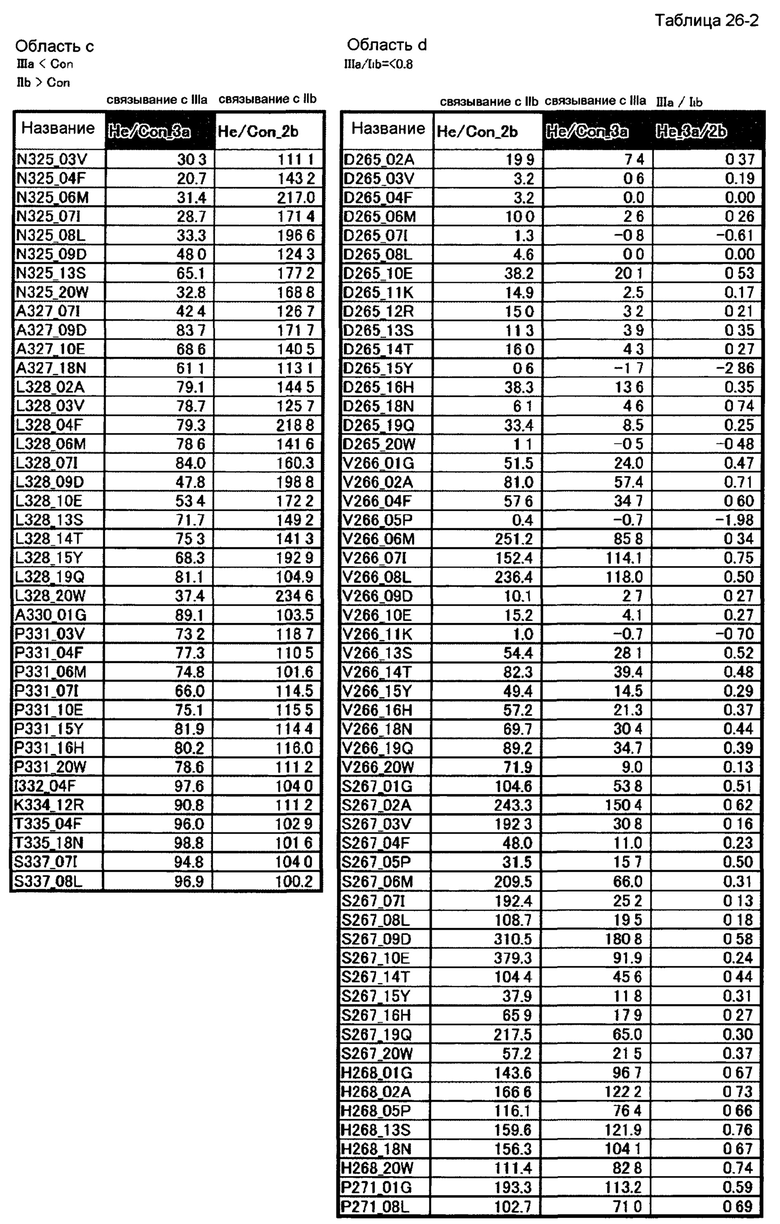

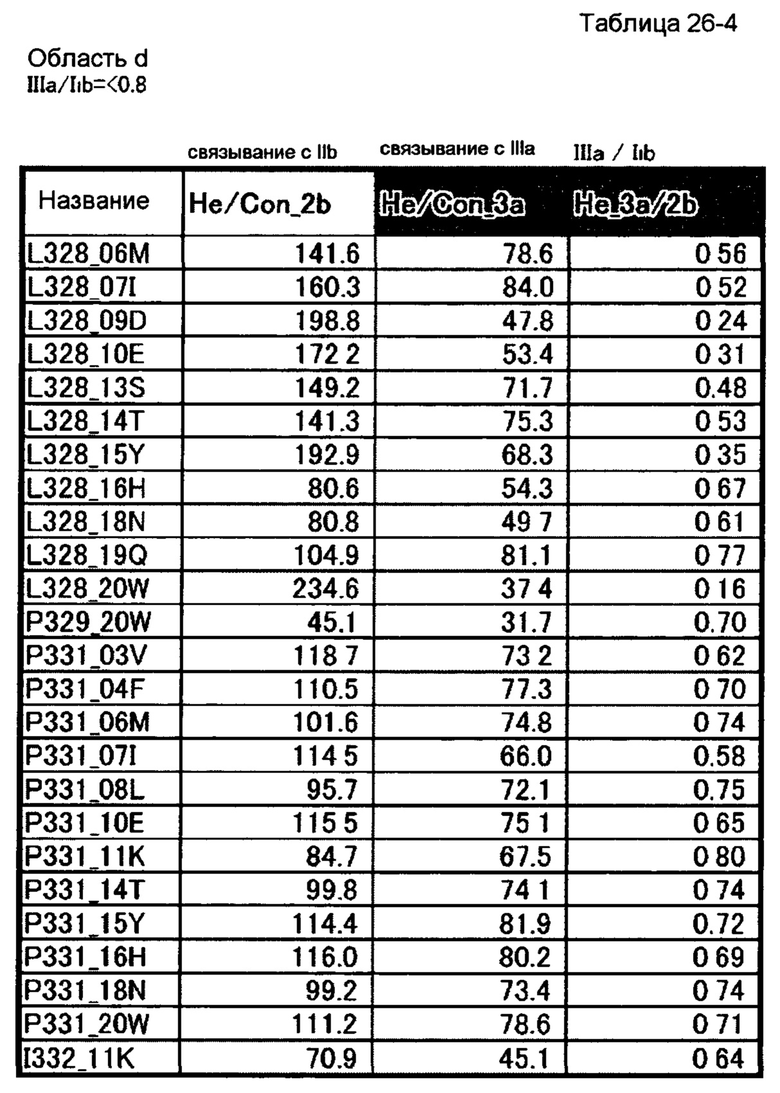
В таблице приведен список изменений, которые селективно повышают связывание с FcγRIIb по сравнению с FcγRIIIa.
[Пример 9] Оценка гетеродимеризованных антител в отношении физико-химической стабильности
Когда антитела разрабатывают в качестве фармацевтических средств, предполагают, что антитела обладают высокой физико-химической стабильностью. Например, в отношении изменения антитела, где описанные выше S239D, A330L и I332E вводят в обе цепи антитела, опубликовано, что домен Fc антитела становится термодинамически нестабильным вследствие вводимого изменения (Molecular Immunology, 45, 1872-1882, 2008), и такая сниженная термостабильность затрудняет разработку антител в качестве фармацевтических средств. Для повышения пригодности антител в качестве фармацевтических средств и облегчения разработки, также важно повышать активность связывания с FcγR и сохранять физико-химическую стабильность. В случае гомодимеризованных антител изменения вводят в обе Н-цепи, и это означает, что молекула антитела, в которую ввели один тип изменения, содержит изменения на двух участках. С другой стороны, в случае гетеродимеризованных антител можно выбирать, в какую Н-цепь вводить изменение, и таким образом, даже при введении одного типа изменения его вводят только на одном участке в молекуле антитела. Как обсуждается в примере 7, в зависимости от типа изменения в отношении эффект повышения активности связывания с FcγRIIIa иногда достаточно вводить изменение в одну Н-цепь. Если изменение обладает эффектом снижения физико-химической стабильности антитела, антителу можно придавать эффект повышения активности связывания с FcγRIIIa введением изменения только в одну Н-цепь, при этом минимизируя физико-химическую дестабилизацию антитела.
Для проверки этой гипотезы авторы настоящего изобретения оценивали, какой остаток из S239D, A330L и I332E по существу способствует дестабилизации СН2 домена. Способом, описанным в справочном примере 1, получали экспрессирующие векторы, в которые вводили, как указано ниже: GpH7-B3-06-09D (SEQ ID NO: 37), GpH7-B3-20-08L (SEQ ID NO: 38) и GpH7-B3-22-10E (SEQ ID NO: 39), получаемые в результате введения изменений S239D, A330L
и I332E, соответственно, в GpH7-B3. С использованием GpL16-k0 в качестве L-цепи представляющие интерес антитела экспрессировали и получали способом, описанным в справочном примере 1. Кроме того, в качестве контроля экспрессировали и получали с использованием GpL16-k0 и GpH7-B3, которые не содержали изменения, представляющее интерес антитело. Соответствующие антитела сравнивали в отношении Tm домена СН2 анализом теплового сдвига способом, описанным в справочном примере 5 (таблица 27). Далее в настоящем описании, если не указано иное, Tm означает Tm домена СН2.

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации), в столбце "Tm" указаны Tm каждого антитела, и в столбце "ΔTm" указана разница Tm для каждого антитела и GpH7-B3/GpL16-k0. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи
каждого антитела.
Когда в гомодимеризованное антитело GpH7-B3-06-09D/GpL16-k0, в которое вводили S239D, гомодимеризованное антитело GpH7-B3-20-08L/GpL16-k0, в которое вводили A330L и гомодимеризованное антитело GpH7-B3-22-10E/GpL16-k0, в которое вводили I332E, сравнивали с GpH7-B3/GpL16-k0, их Tm являлись сниженными на 3°С, 1°С и 8°С, соответственно. Этот результат демонстрирует, что из трех изменений I332E обладал большим эффектом снижения Tm СН2, таким образом, полагают, что I332E также способствует снижению Tm антитела, в которое вводят группу изменений S239D, A330L и I332E.
Боковая цепь I332E окружена гидрофобными аминокислотами, такими как V240, V323 и L328. В антителе, в которое вводят I332E, гидрофобное взаимодействие с окружающими остатками устранено вследствие замены гидрофобного Ile на гидрофильный Glu, и полагают, что это способствует дестабилизации домена Fc. При этом, как обсуждается в примере 7, взаимодействие I332E с FcγRIIIa происходит исключительно в одной из Н-цепи. По этой причине полагают, что, когда I332 сохраняют незамещенным в другой Н-цепи, которая не участвует во взаимодействии с FcγRIIIa, можно сохранять термодинамическую стабильность, при этом обеспечивая эффект повышения связывания с FcγRIIIa. Затем авторы настоящего изобретения вводили I332E только в одну Н-цепь и оценивали, повышается ли Tm по сравнению с тем, когда I332E вводят в обе Н-цепи. Конструировали экспрессирующие векторы, в которые вводили GpH7-A44 (SEQ ID NO: 49) и GpH7-B80 (SEQ ID
NO: 50), получаемые в результате введения I332E в GpH7-A5 и GpH7-В3, соответственно. Их комбинировали с GpH7-B3, GpH7-A5 и GpL16-k0 для экспрессии и получения следующих ниже антител способом, описанным в справочном примере 1: гетеродимеризованных антител GpH7-A5/GpH7-B80/GpL16-k0 и GpH7-A44/GpH7-B3/GpL16-k0, в которых только в одну из Н-цепей вводили I332E, и гомодимеризованного антитела GpH7-A44/GpH7-B80/GpL16-k0, в обе Н-цепи которого вводили I332E. В качестве контроля получали GpH7-A5/GpH7-B3/GpL16-k0. Каждое антитело оценивали в отношении активности связывания с FcγRIIIa способом, описанным в справочном примере 2. Кроме того, антитела сравнивали в отношении Tm домена СН2 анализом теплового сдвига способом, описанным в справочном примере 5 (таблицы 28 и 29).
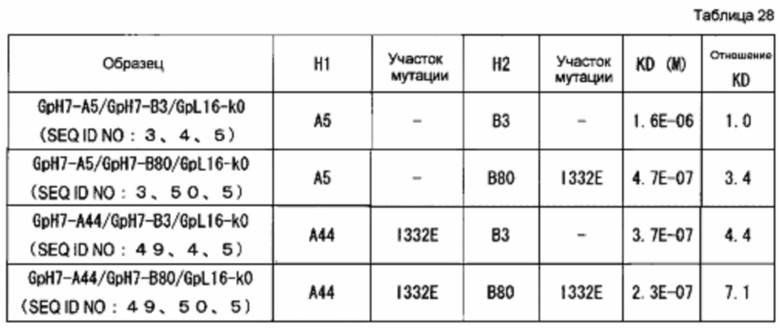
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-A5/GpH7-B3/GpL16-k0 для
FcγRIIIa на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.

В таблице представлены Tm СН2 антител с заменой I332E в одной Н-цепи и антител с заменой I332E в обеих Н-цепях.
В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи соответствующих антител и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": когда не существует конкретной мутации), в столбце "Tm" указаны Tm каждого антитела, и в столбце "ΔTm" указана разница Tm для каждого антитела и GpH7-B3/GpL16-k0. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Результат, представленный в таблице 28, демонстрирует, что активность связывания с FcγRIIIa гетеродимеризованного антитела GpH7-A5/GpH7-B80/GpL16-k0, в котором только в одну из Н-цепей вводили I332E, повышалась приблизительно в 3 раза по сравнению с активностью GpH7-A5/GpH7-B3/GpL16-k0, при этом активность GpH7-A44/GpH7-B3/GpL16-k0
повышалась приблизительно в 4 раза по сравнению с активностью GpH7-A5/GpH7-B3/GpL16-k0. На основании этого полагали, что эффект I332E в отношении повышения активности связывания с FcγRIIIa значительно не изменяется, когда I332E вводят в GpH7-A5 или вводят его в GpH7-B3. При этом активность связывания с FcγRIIIa GpH7-A44/GpH7-B80/GpL16-k0, в обе Н-цепи которого вводили I332E, повышалась приблизительно в 7 раз по сравнению с активностью GpH7-A5/GpH7-B3/GpL16-k0. Эти открытия указывают на то, что I332E обладает эффектом значительного повышения активности связывания с FcγRIIIa даже в случае введения только в одну Н-цепь, если не вводят в обе Н-цепи, как рассматривают на основании трехмерной структуры.
Кроме того, результат, представленный в таблице 29, указывает на то, что Tm гетеродимеризованных антител GpH7-A5/GpH7-B80/GpL16-k0 и GpH7-A44/GpH7-B3/GpL16-k0, в которые I332E вводили только в одну Н-цепь, повышались на 4°С по сравнению с Tm GpH7-A5/GpH7-B3/GpL16-k0, которое представляет собой их исходную молекулу Fc. Таким образом, полагали, что не существует разницы во влиянии I332E на Tm антитела независимо от того, когда I332E вводят в GpH7-A5 или GpH7-B3. При этом Tm гомодимеризованного антитела GpH7-A44/GpH7-B80/GpL16-k0, в обе Н-цепи которого вводили I332E, снижалась на 10°С по сравнению с Tm GpH7-A5/GpH7-B3/GpL16-k0. Tm гетеродимеризованного антитела, в котором только в одну из Н-цепей вводили I332E, сохранялась на 6°С выше, чем Tm гомодимеризованного антитела, в обе Н-цепи которого вводили I332E. Этот результат демонстрирует, что
снижение Tm можно подавлять применением гетеродимеризованного антитела, в котором только в одну, а не в обе Н-цепи вводят I332E. Это демонстрирует, что технология гетеродимеризованного антитела является пригодной для сохранения физико-химической стабильности антитела.
Изменение I332E является превосходным в отношении эффекта повышения связывания с FcγRIIIa, однако, если с ним используют общепринятые гомодимеризованные антитела, их термодинамическая стабильность значительно снижается, и это является проблемой, когда антитела используют в качестве фармацевтических средств. Однако результаты, представленные в таблицах 28 и 29, демонстрируют, что применение гетеродимеризованного антитела обеспечивает возможность использовать эффект I332E в отношении повышения связывания с FcγRIIIa при сохранении физико-химической стабильности антитела. На основании этого открытия полагают, что гетеродимеризованные антитела представляют собой превосходную технологию более точной регуляции активности связывания с FcγR и физико-химической стабильности антител.
GpH7-TA7/GpH7-B78/GpL16-k0 содержит S239D, A330L и I332E только в одной Н-цепи и таким образом, вероятно, сохраняет высокую Tm по сравнению с GpH7-A57/GpH7-B78/GpL16-k0, в котором обе Н-цепи содержат S239D, A330L и I332E. Таким образом, гетеродимеризованные и гомодимеризованные антитела, получаемые в результате комбинации групп изменений L234Y, G236W и S298A с группой изменений S239D, A330L и I332E, описанной в примере 7, анализировали в отношении Tm способом, описанным в справочном
примере 5.
Для оценки этого способом, описанным в справочном примере 1, получали гетеродимеризованное антитело GpH7-TA7/GpH7-B78/GpL16-k0, в котором в одну из Н-цепей вводили группу изменений L234Y, G236W и S298A, а в другую Н-цепь вводили группу изменений S239D, A330L и I332E; гетеродимеризованное антитело GpH7-TA7/GpH7-B3/GpL16-k0, в котором только в одну из Н-цепей вводили группу изменений L234Y, G236W и S298A; гомодимеризованное антитело GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили группу изменений L234Y, G236W и S298A; гетеродимеризованное антитело GpH7-A5/GpH7-B78/GpL16-k0, в котором только в одну из Н-цепей вводили S239D, A330L и I332E, и гомодимеризованное антитело GpH7-A57/GpH7-B78/GpL16-k0, в котором в обе Н-цепи вводили S239D, A330L и I332E. Влияние комбинации L234Y, G236W и S298A с S239D, A330L и I332E на Tm оценивали, сравнивая Tm домена СН2 соответствующих антител анализом теплового сдвига способом, описанным в справочном примере 5 (таблица 30).

В столбце "образец" указаны названия антител, в столбцах "H1" и "Н2" указаны названия константной области Н-цепи каждого антитела, в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-Gld/GpL16-k0 ("-": когда не существует конкретной мутации), в столбце "Tm" указаны Tm каждого антитела, и в столбце "ΔTm" указана разница Tm для каждого антитела и GpH7-A5/GpH7-B3/GpL16-k0. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
GpH7-A5/GpH7-B3/GpL16-k0, в которое вводили изменения D356K/H435R и K439Е для повышения эффективности образования гетеродимеризованного антитела, сравнивали с GpH7-G1d/GpL16-k0, которое представляет собой нативный IgG1. Tm СН2 понижалась на 1°С.
Гомодимеризованные антитела по технологии известного уровня техники оценивали в отношении эффекта каждой группы изменений. Tm гомодимеризованного антитела GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E, снижалась на 20°С по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом снижения Tm не наблюдали для гомодимеризованного антитела GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили группу изменений L234Y, G236W и S298A. Таким образом, полагали, что группа изменений L234Y, G236W и S298A не обладает эффектом снижения Tm гомодимеризованных антител.
Гетеродимеризованные антитела, в которых только в одну из Н-цепей вводили каждую группу изменений, оценивали в отношении
эффекта каждой группы изменений. Tm гетеродимеризованного антитела GpH7-A5/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили S239D, A330L и I332E, снижалась на 8°С по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом снижение Tm не наблюдали для гетеродимеризованного антитела GpH7-TA7/GpH7-B3/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A. На основании этих открытий полагали, что группа изменений L234Y, G236W и S298A также не обладает эффектом снижения Tm гетеродимеризованных антител.
Tm GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E, снижалась на 21°С по сравнению с нативным IgG1. Tm GpH7-A5/GpH7-B78/GpL16-k0, в котором только одна из Н-цепей содержит изменения S239D, A330L и I332E, составляла 60°С, и это поддерживало Tm на 10°С или более выше по сравнению с Tm гомодимеризованного антитела. Как представлено в таблице 13 примера 7, связывание с FcγRIIIa гомодимеризованного антитела с S239D, A330L и I332E повышалось приблизительно в 9 раз по сравнению с гетеродимеризованным антителом с S239D, A330L и I332E. S239D, A330L и I332E при введении в обе Н-цепи в значительной степени повышают связывание с FcγRIIIa, но значительно снижают Tm.
Кроме того, Tm GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили L234Y, G236W и S298A, снижалась только на 1°С по сравнению с нативным антителом. Полагают, что снижение Tm является следствием влияния D356K/H435R и K439Е, используемых для конструкции гетеродимеризованного антитела, как указано
выше, а не вследствие L234Y, G236W и S298A. Это также продемонстрировано тем фактом, что Tm GpH7-TA7/GpH7-B3/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, также снижалась только на 1°С.
Наконец, Tm GpH7-TA7/GpH7-B78/GpL16-k0, одна из Н-цепей которого содержит L234Y, G236W и S298A, а другая Н-цепь содержит S239D, A330L и I332E, снижалась на 10°С по сравнению с нативным антителом и являлась практически эквивалентной Tm GpH7-A5/GpH7-B78/GpL16-k0, в котором в одну из Н-цепей вводили S239D, A330L и I332E. Однако, как представлено в таблице 13 примера 7, связывание с FcγRIIIa GpH7-TA7/GpH7-B78/GpL16-k0 повышается в 10 раз или более по сравнению со связыванием GpH7-A5/GpH7-B78/GpL16-k0.
Таким образом, демонстрировали что, с использованием гетеродимеризованного антитела GpH7-TA7/GpH7-B78/GpL16-k0, в котором в одну из Н-цепей вводили L234Y, G236W и S298A и в другую вводили S239D, A330L и I332E, можно повышать связывание с FcγRIIIa, а также можно повышать Tm на 10°С или более по сравнению с гомодимеризованным антителом GpH7-A57/GpH7-B78/GpL16-k0 с S239D, A330L и I332E.
Затем описанные выше образцы, для которых проводили измерение Tm, дополнительно оценивали в отношении термодинамической стабильности ускоренным испытанием стабильности под воздействием тепла, описанным в справочном примере 6 (при 40°С в течение двух или четырех недель) (фиг. 30).
Когда GpH7-A5/GpH7-B3/GpL16-k0 сравнивали с нативным
антителом GpH7-G1d/GpH7-G1d/GpL16-k0 содержания мономеров первого и последнего снижались на 1,27% и 1,86% после четырех недель, соответственно, и не наблюдали значимого отличия. Таким образом, полагали, что изменения D356K/H435R и K439Е, используемые для конструкции гетеродимеризованного антитела практически не оказывают влияния на изменение содержания мономеров в ускоренном испытании стабильности под воздействием тепла.
В отношении GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E, содержание мономеров снижалось приблизительно на 16% после четырех недель. При этом содержание мономеров GpH7-A5/GpH7-B78/GpL16-k0, в котором только одна из Н-цепей содержит изменения S239D, A330L и I332E, снижалось на 9,63% после четырех недель. Таким образом, демонстрировали, что эффект более стабильного сохранения содержания мономеров получали с использованием гетеродимеризованных антител, в которых только в одну из Н-цепей вводили S239D, A330L и I332E.
Кроме того, содержание мономеров GpH7-TA7/GpH7-TA45/GpL16-k0, в котором в обе Н-цепи вводили L234Y, G236W и S298A, и GpH7-TA7/GpH7-B3/GpL16-k0, в котором в одну из Н-цепей вводили L234Y, G236W и S298A, снижалось только на 1,78% и 1,42% после четырех недель, соответственно. Не наблюдали четкого различия в изменении содержания мономеров по сравнению с нативным антителом. Таким образом, полагали, что L234Y, G236W и S298A не оказывают влияния на содержание мономеров в ускоренном испытании стабильности под воздействием тепла, даже когда их вводят в одну
Н-цепь или обе Н-цепи.
Наконец, содержание мономеров GpH7-TA7/GpH7-B78/GpLl6-k0, в котором одна из Н-цепей содержит L234Y, G236W и S298A, а другая Н-цепь содержит S239D, A330L и I332E, снижалось на 2,47% после четырех недель. Содержание мономеров только незначительно снижалось по сравнению с 1,8 6% для нативного антитела. Таким образом, демонстрировали, что с использованием гетеродимеризованного антитела GpH7-TA7/GpH7-B78/GpL16-k0, в котором одна из Н-цепей содержит L234Y, G236W и S298A, а другая Н-цепь содержит S239D, A330L и I332E, можно повышать связывание с FcγRIIIa, а также можно получать эффект сохранения содержания мономеров на высоком уровне в ускоренном испытании стабильности под воздействием тепла по сравнению с гомодимеризованным антителом GpH7-A57/GpH7-B78/GpL16-k0 с S239D, A330L и I332E.
Таким образом, демонстрировали, что по сравнению с общепринятыми гомодимеризованными антителами технология гетеродимеризованных антител может не только повышать связывание с FcγR, а также улучшать стабильность, и таким образом повышать ценность антител в качестве фармацевтических средств в большей степени, чем гомодимеризованных антител.
[Пример 10] Поиск изменений, которые улучшают связывание с FcγR, но не снижают стабильность
Как описано в примере 9, активность связывания с FcγR повышают введением изменений в Н-цепь. Однако такие изменения могут снижать физико-химическую стабильность СН2, т.е. снижать Tm. Однако, как описано в примере 9, такие свойства являются
неблагоприятными, в частности, когда антитела используют в качестве фармацевтических средств. Как описано в примере 9, пригодным является использование гетеродимеризованного антитела, в котором только в одну из Н-цепей вводили изменения для повышения активности связывания с FcγR наряду с подавлением дестабилизации СН2. В частности, в отношении изменений, которые снижают Tm гомодимеризованных антител, даже если наблюдают, что они повышают активность связывания с FcγR общепринятых гомодимеризованных антител, таких как антител соответствующие областям ii и iii, представленным на фиг. 21, их гетеродимеризация обеспечивает то, что Tm является выше Tm гомодимеризованных антител при повышении активности связывания с FcγR по сравнению с нативным антителом.
Для выявления таких изменений гомодимеризованные антитела в областях ii и iii, представленных на фиг. 21, анализировали в отношении Tm способом, описанным в справочном примере 5. Список изменений, которые снижают Tm по сравнению с нативным антителом, приведен в таблицах 31-35.
В таблице 31 (таблицах от 31-1 до 31-3) представлены данные для Ia, где Tm составляет 68°С или менее для области ii или iii, в таблице 32 (таблицах 32-1 и 32-2) представлены данные для IIaR, где Tm составляет 68°С или менее для области ii или iii, в таблице 33 (таблицах 33-1 и 33-2) представлены данные для IIaH, где Tm составляет 68°С или менее для области ii или iii, в таблице 34 (таблицах 34-1 и 34-2) представлены данные для IIb, где Tm составляет 68°С или менее для области ii или iii, в
таблице 35 (таблицах 35-1 и 35-2) представлены данные для IIIa, где Tm составляет 68°С или менее для области ii или iii.
Антитела, которые обладают улучшенным связыванием с FcγR по сравнению с нативным антителом, и стабильность которых значительно не снижается, можно получать с использованием гетеродимеризованного антитела, в котором только в одну из Н-цепей вводят эти изменения.



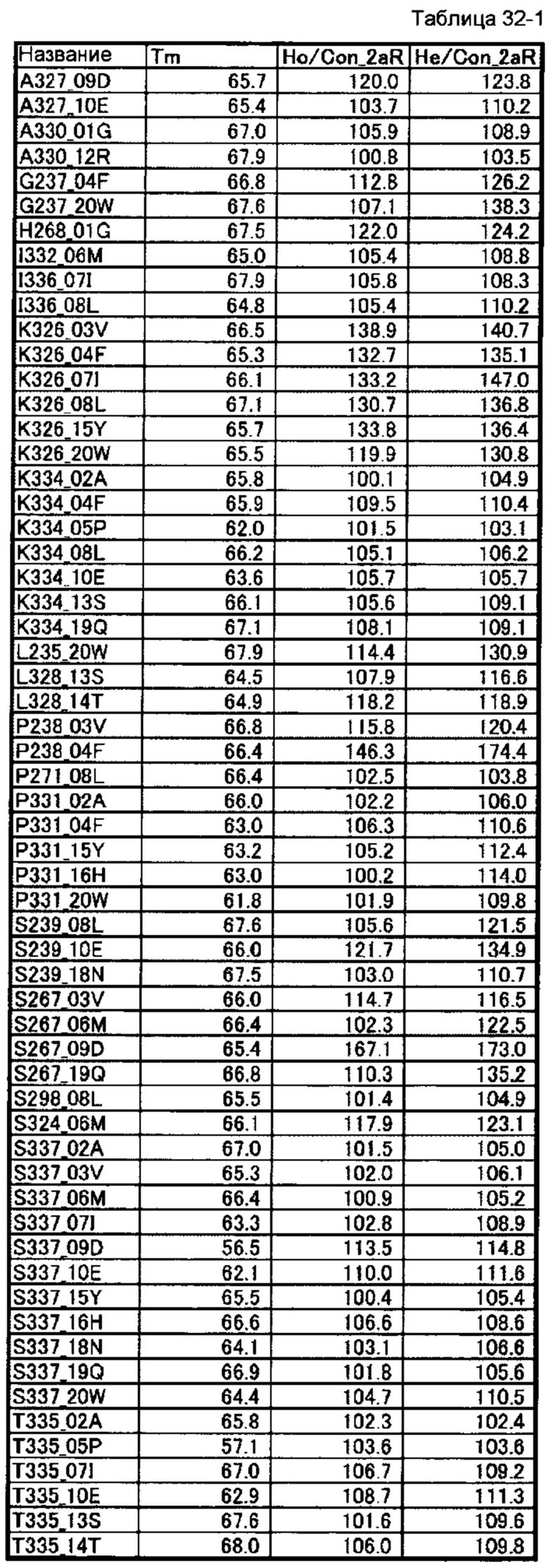
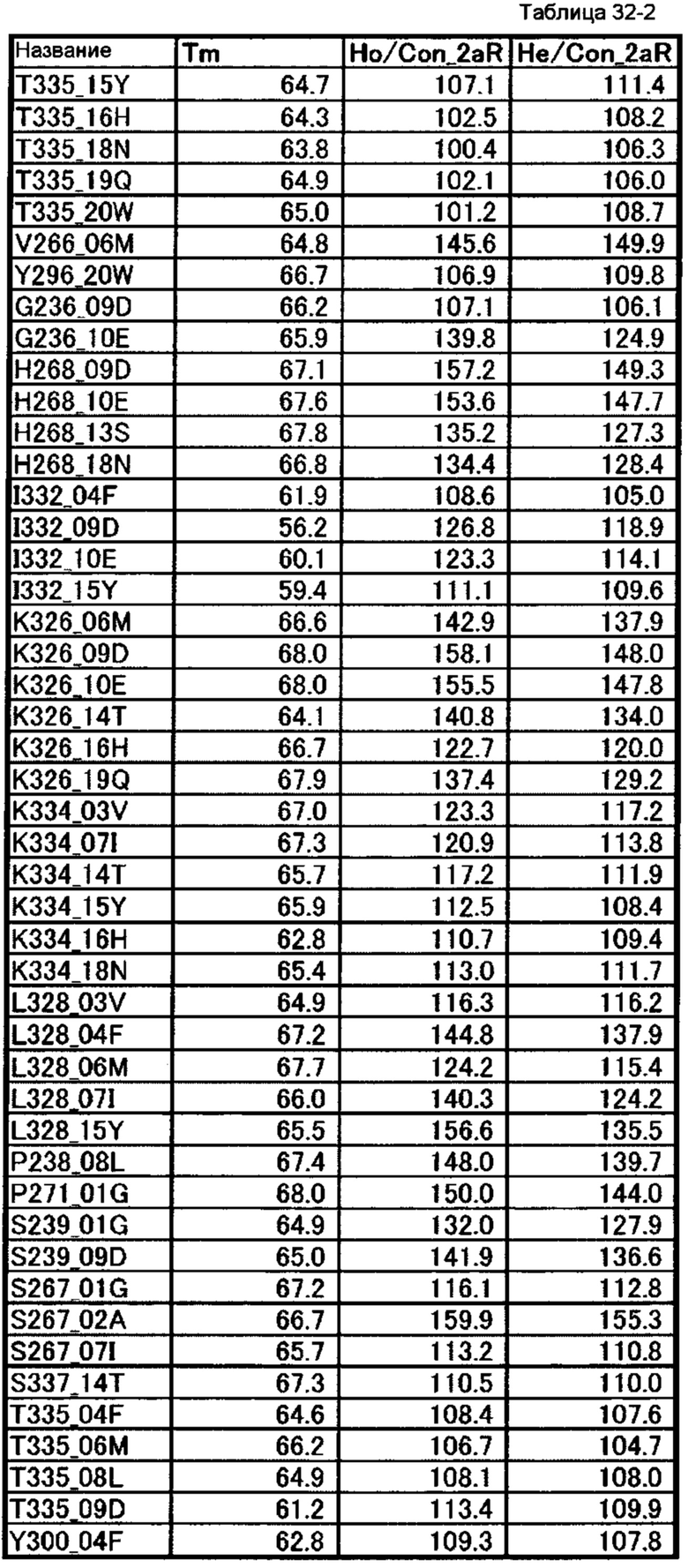
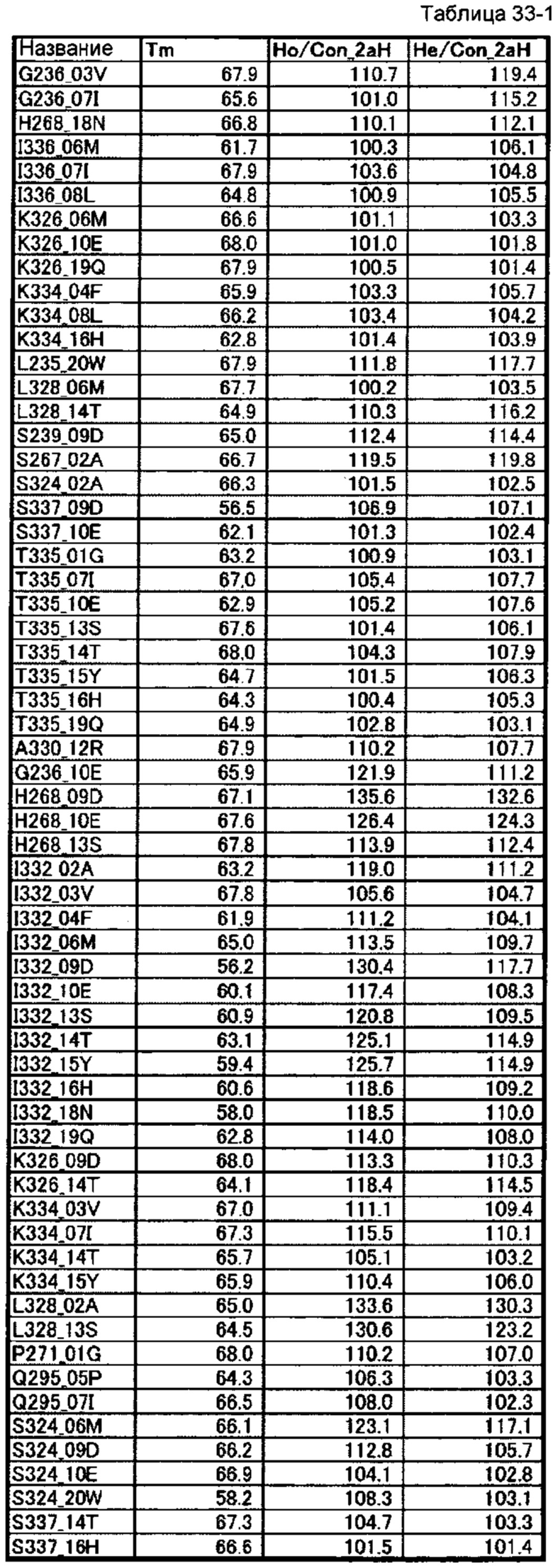
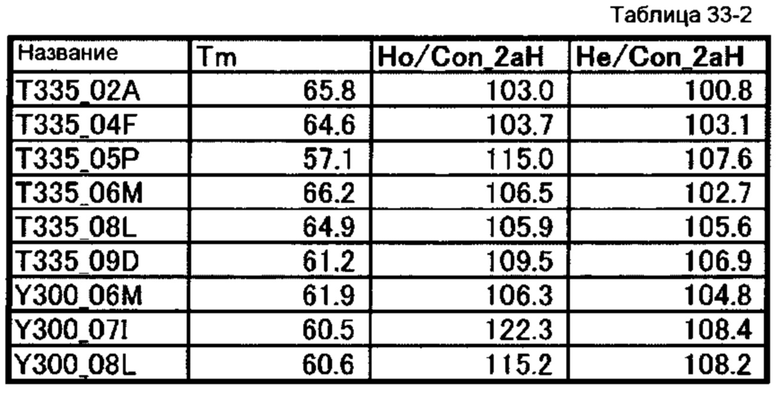

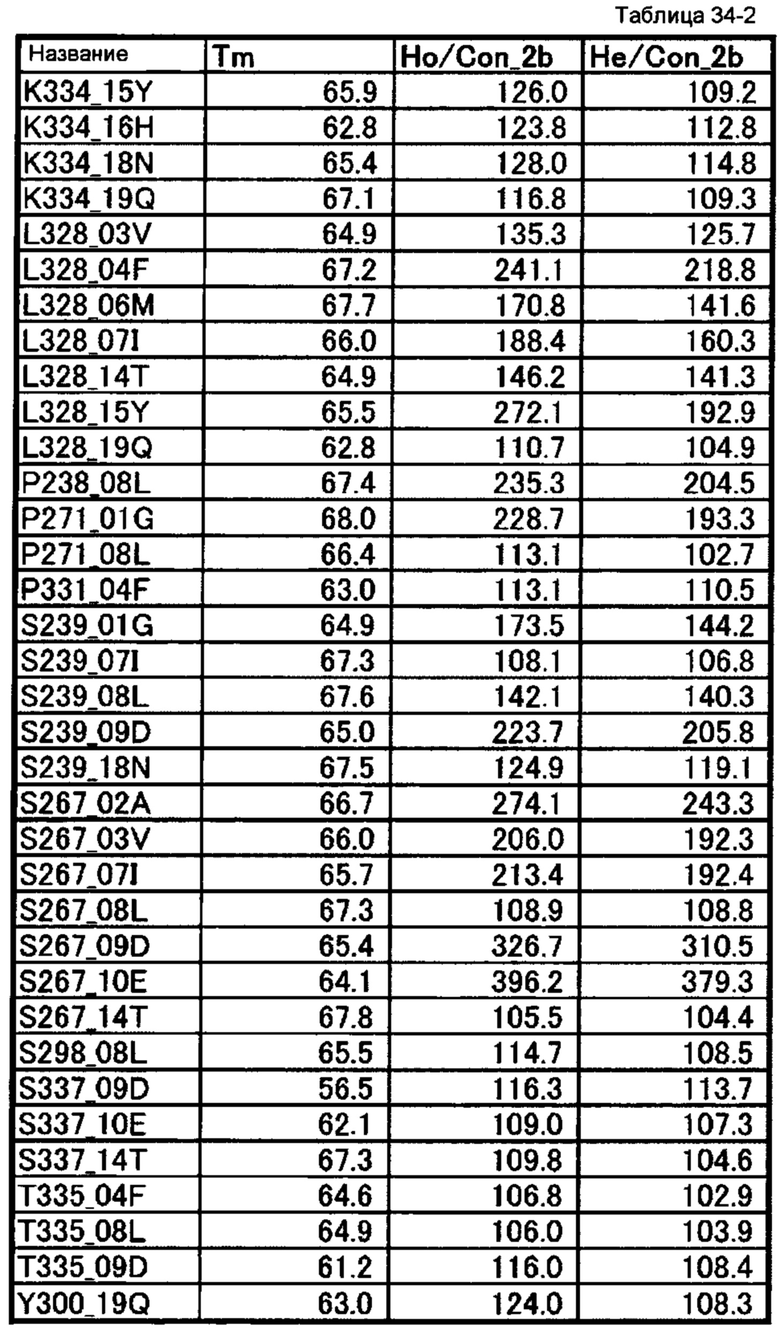


[Пример 11] Анализ активности ADCC гетеродимеризованных антител с улучшенной способностью распознавать FcγRIIIa
Как обсуждается в примере 7, с использованием гетеродимеризованного антитела активность связывания с FcγRIIIa эффективно повышали больше, чем активность вариантов, получаемых общепринятой технологией гомодимеризованного антитела. Антитела индуцируют NK-клетки через FcγRIIIa проявлять антителозависимую клеточную цитотоксичность против клеток, экспрессирующих антиген-мишень. Для оценки, на сколько гетеродимеризованные антитела обладают не только повышенной активностью связывания с FcγRIIIa, но и повышенной активностью ADCC, гетеродимеризованные антитела с повышенной активностью связывания с FcγRIIIa, представленные в таблице 13 примера 7, гомодимеризованные антитела и нативный IgG1 анализировали в отношении активности ADCC способом, описанным в справочном примере 7. Результат
представлен на фиг. 31.
На основании результата, приведенного на фиг. 31, когда GpH7-G1d/GpL16-k0, которое представляет собой нативный IgG1, сравнивают с GpH7-A5/GpH7-B3/GpL16-k0, в котором каждое из D356K, H435R и K439E вводили в одну Н-цепь, не наблюдают значимого отличия в активности ADCC. Таким образом, полагали, что изменения D356K, H435R и K439E не влияют на активность ADCC.
Затем для оценки, наблюдают ли также аналогичную тенденцию, как наблюдают для повышающего связывание эффекта, для активности ADCC, анализировали гомодимеризованные антитела, в которые изменение, которое повышает активность связывания с FcγRIIIa, вводили в обе Н-цепи общепринятым способом. GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили L234Y, G236W и S298A, сравнивали с GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E. В отношении активности связывания с FcγRIIIa связывание GpH7-A57/GpH7-B78/GpL16-k0 заметно повышалось по сравнению с GpH7-A5/GpH7-B3/GpL16-k0, тогда как связывание GpH7-TA7/GpH7-TA45/GpL16-k0 снижалось. Аналогично, активность ADCC GpH7-A57/GpH7-B78/GpL16-k0 повышалась больше, чем активность ADCC GpH7-A5/GpH7-B3/GpL16-k0, тогда как активность GpH7-TA7/GpH7-TA45/GpL16-k0 снижалась по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Таким образом, в отношении гомодимеризованных антител наблюдали корреляцию между уровнем активности связывания с FcγRIIIa и уровнем активности ADCC.
Затем анализировали гетеродимеризованные антитела, в которых только в одну из Н-цепей вводили изменение, которое повышает
активность связывания с FcγRIIIa, для оценки наблюдают ли аналогичную тенденцию, как наблюдают для повышающего связывание эффекта, для активности ADCC. GpH7-TA7/GpH7-B3/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, сравнивали с GpH7-A5/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили S239D, A330L и I332E. Активности связывания с FcγRIIIa GpH7-A5/GpH7-B78/GpL16-k0 и GpH7-TA7/GpH7-B3/GpL16-k0 повышались по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Аналогичную тенденцию наблюдали для активности ADCC. Кроме того, активность связывания с FcyRIIIa GpH7-A5/GpH7-B78/GpL16-k0 повышалась больше, чем активность связывания GpH7-TA7/GpH7-B3/GpL16-k0, однако аналогичную тенденцию получали для активности ADCC. Таким образом, не только гомодимеризованные антитела, а также гетеродимеризованные антитела обладают корреляцией между уровнем активности связывания с FcγRIIIa и активностью ADCC.
Затем относительно каждой из групп изменений L234Y, G236W и S298A и изменений S239D, A330L и I332E оценивали, существует ли корреляция между эффектами повышения активности связывания с FcγRIIIa и активностью ADCC для гетеродимеризованных антител и гомодимеризованных антител. Сначала GpH7-A5/GpH7-B78/GpL16-k0, которое представляет собой гетеродимеризованное антитело, в котором в одну из Н-цепей вводили группу изменений S239D, A330L и I332E, сравнивали с GpH7-A57/GpH7-B78/GpL16-k0, которое представляет собой гомодимеризованное антитело, в обе Н-цепи которого вводили группу, эффект повышения активности связывания с FcγRIIIa являлся выше в гомодимеризованном антителе по
сравнению с гетеродимеризованным антителом, однако различия в активности ADCC не наблюдали. Кроме того, когда GpH7-TA7/GpH7-B3/GpL16-k0, которое представляет собой гетеродимеризованное антитело, в котором только в одну из Н-цепей вводили группу изменений L234Y, G236W и S298A, сравнивали с GpH7-TA7/GpH7-TA45/GpL16-k0, которое представляет собой гомодимеризованное антитело, в обе Н-цепи которого вводили группу, в отношении активности связывания с FcγRIIIa, связывание гетеродимеризованного антитела повышалось в большей степени, чем связывание GpH7-A5/GpH7-B3/GpL16-k0, тогда как связывание гомодимеризованного антитела снижалось. Аналогичную тенденцию наблюдали для активности ADCC. Таким образом, полагали, что эффект группы изменений L234Y, G236W и S298A в отношении повышения активности связывания с FcγRIIIa только в одном направлении отражается в активности ADCC. На основании этих результатов полагали, что в гетеродимеризованных антителах, в которых только в одну из Н-цепей вводили группу изменений, и гомодимеризованных антителах, в которых в обе Н-цепи вводили группу, существует корреляция между уровнем активности связывания с FcγRIIIa и активностью ADCC.
Затем гетеродимеризованное антитело GpH7-TA7/GpH7-B78/GpL16-k0, в одну из Н-цепей которого вводили L234Y, G236W и S298A, а в другую Н-цепь вводили S239D, A330L и I332E, сравнивали с гомодимеризованным антителом GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E. В отношении активности связывания с FcγRIIIa оба обладали заметно повышенными
активностями связывания по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. Аналогичную тенденцию наблюдали для активности ADCC. Кроме того, активность связывания с FcyRIIIa GpH7-TA7/GpH7-B78/GpL16-k0 повышалась больше, чем активность GpH7-A57/GpH7-B78/GpL16-k0, и GpH7-TA7/GpH7-B78/GpL16-k0 также обладало большей активностью ADCC.
Как описано выше, в отношении групп изменений L234Y, G236W и S298A и изменений S239D, A330L и I332E наблюдали, что последняя группа изменений S239D, A330L и I332E обладает большим эффектом повышения активности ADCC независимо от введения в одну Н-цепь или обе Н-цепи. С другой стороны, демонстрировали, что, когда группу изменений L234Y, G236W и S298A и группу изменений S239D, A330L и I332E соответственно вводят в различные Н-цепи, эффект повышения активности ADCC является больше, чем когда S239D, A330L и I332E, которые обладают сильным эффектом повышения активности ADCC в гетеродимеризованных и гомодимеризованных антителах, вводят в обе Н-цепи.
Таким образом, демонстрировали, что корреляцию между уровнем активности связывания с FcγRIIIa и уровнем ADCC, таким как уровень, наблюдаемый в гомодимеризованных антителах технологии известного уровня техники, также наблюдали, когда сравнивали гетеродимеризованные антитела друг с другом и когда сравнивали гетеродимеризованные и гомодимеризованные антитела. Это указывает на то, что применение технологии гетеродимеризованного антитела обеспечивает получение антител, обладающих активностью ADCC, превосходящей антитела общепринятых технологий.
[Пример 12] Сравнение общепринятых гомодимеризованных
антител и новых гетеродимеризованных антител по отношению к FcγRIIa
Как описано в примере 1, полагают, что FcγRIIIa играет важную роль в лекарственной эффективности фармацевтических средств на основе антител. Кроме того, уделяли внимание роли FcγRIIa, которую он играет в лекарственной эффективности получаемых из IgG1 фармацевтических средств на основе антител в дополнение к FcγRIIIa.
Для FcγRIIa существуют аллотипы, называемые типами R и Н, которые содержат Arg и His в положении 131 аминокислоты, соответственно, и известно, что они отличаются активностью связывания с IgG2 человека (Tissue Antigens, 2003, 61, 189-202). Известно, что восприимчивость к инфекции изменяется в зависимости от различия в аллотипе FcγRIIa (Tissue Antigens, 2003:61:189-202). Полагают, что это обусловлено тем, что активность связывания с IgG2 FcγRIIa изменяется в зависимости от различия в аллотипе, и в результате механизм устойчивости к патогенам посредством IgG2 различается (Infection and Immunity, 1995, 63, 73-81). При этом известно, что клетки, экспрессирующие FcγRIV мыши, соответствуют клеткам, экспрессирующим FcγRIIa человека, и опубликовано, что FcγRIV играет важную роль в лекарственной эффективности антитела против CD20 на модели на мышах. Эти открытия позволяют предположить, что у человека FcγRIIa играет аналогичную роль (The Journal of Experimental Medicine, 2004:199:1659-1669; The Journal of Experimental Medicine, 2006, 203, 743-753; Immunity, 2005, 23, 41-51).
Фактически, опубликовано, что антитело, в котором активность связывания с FcγRIIa Fc-области является повышенной по сравнению с IgG1, повышает активность опосредованного макрофагами антителозависимого клеточного фагоцитоза (ADCP) по сравнению с IgG1 (Molecular Cancer Therapeutics, 2008, 7, 2517-2527). Кроме того, на модели ксенотрансплантата на мышах антитело против CD19, содержащее домен Fc с повышенной ADCP, демонстрирует более выраженный противоопухолевый эффект по сравнению с IgG1 (Nature Medicine, 2000, 6, 443-446). Домен Fc такого антитела обладает повышенной активностью связывания с FcγRIIa обезьяны. CD19 экспрессируется на поверхности В-клеток. Опубликовано, что такое антитело при введении обезьянам повышает элиминацию В-клеток по сравнению с антителом против CD19, содержащим Fc-область IgG1 (Science, 2005, 310, 1510-1512).
На основании этих опубликованных данных предполагают, что эффективность фармацевтических средств на основе антител, в частности противоопухолевый эффект, можно дополнительно улучшать, повышая активность связывания с FcγRIIa наряду с повышением активности связывания с FcγRIIIa. Антитела с такими характеристиками получали общепринятыми технологиями (Molecular Cancer Therapeutics, 2008, 7, 2517-2527). Однако поскольку полагают, что FcγRIIa связывается с доменом Fc антитела асимметричным образом, предполагают, что активность связывания с FcγRIIa можно дополнительно повышать технологией гетеродимеризованного антитела, как описано в примере 11. Для проверки этого на основании результата, представленного в
примере 4, выбирали изменения, которые повышают активность связывания со всеми FcγRIIIa, FcγRIIa R типа и FcγRIIa Н типа по сравнению с нативным IgG, и комбинировали их для введения мутаций, которые повышают активность связывания со всеми FcγRIIIa, FcγRIIa R типа и FcγRIIa Н типа, и получали гетеродимеризованные антитела таким образом, чтобы они содержали комбинацию Н-цепей с различным связыванием с FcγR. Антитела оценивали в отношении активности связывания с каждым FcγR.
При этом в отличие от этих активирующих FcγR, FcγRIIb, который представляет собой ингибирующий FcγR, индуцирует внутриклеточный сигнал, который подавляет иммунный ответ. Опубликовано, что у мышей с нокаутом по FcyRIIb противоопухолевый эффект антител повышается (Nature Medicine, 2000, 6, 443-446) и активируется антителоопосредованная элиминация В-клеток (The Journal of Experimental Medicine, 2006, 203, 743-753), демонстрируя, что FcγRIIb играет важную роль в лекарственной эффективности антител in vivo. При этом наблюдали корреляцию между противоопухолевым эффектом подкласса IgG мыши и отношением активности связывания с активирующим FcγR к активности связывания с ингибирующим FcγR (отношением А/I) каждого подкласса IgG (Science, 2005, 310, 1510-1512). Эти опубликованные данные позволяют предположить, что отношения А/I являются важными для эффекторной функции антитела посредством иммунитета. В частности, когда антитела получают, чтобы они обладали более высоким отношением А/I, их эффекторная функция является повышенной, и такие антитела являются пригодными. Однако было
теоретически рассчитано, что крайне трудно повышать отношение А/I путем повышения активности связывания с FcγRIIa без повышения активности связывания с FcγIIb, вследствие того, что гомология последовательностей FcγRIIa, который представляет собой активирующий FcγR, и FcγRIIb, который представляет собой ингибирующий FcγR, составляет 93% в их внеклеточных доменах, что является крайне высокой. Полагают, что подобно FcγRIIIa и FcγRIIa FcγRIIb связывается с доменом Fc антитела асимметричным образом. Общепринятыми технологиями взаимодействие с FcγR можно регулировать только введением аналогичного изменения в обе Н-цепи антитела. В противоположность этому применение технологии гетеродимеризованного антитела обеспечивает более точную регуляцию и, таким образом, обеспечивает улучшение отношения А/I даже для FcγRIIa и FcγRIIb, последовательности которых являются очень похожими. Таким образом, авторы настоящего изобретения также оценивали с этой точки зрения, превосходит ли технология гетеродимеризованного антитела общепринятые технологии.
В этой оценке для эффективного получения гетеродимеризованных антител для константной области Н-цепи антитела использовали технологию "выступы во впадины". Технология "выступы во впадины" представляет собой технологию, которая обеспечивает активацию гетеродимеризации Н-цепей и эффективное получение представляющих интерес гетеродимеризованных антител путем замены аминокислотной боковой цепи на большую боковую цепь (выступ) в домене СН3 одной Н-цепи и замены аминокислотной боковой цепь на меньшую боковую цепь
(впадина) в домене СН3 другой Н-цепи, таким образом, что выступ помещается во впадину (Nature, 372, 379-383 (1994)). Н-цепь, в константную область которой вводили изменения Y349C и T366W, для которых предполагают, что они содержат большие аминокислотные боковые цепи в домене СН3, обозначают как "цепь выступа". Когда в нее вводят дополнительное изменение, название константной области Н-цепи начинается с символа "Kn", за которым следует трехзначное число, такое как Kn001. Н-цепь, в константную область которой вводят изменения D356C, T336S, L368A и Y407V, для которых предполагают, что они содержат меньшие аминокислотные боковые цепи в домене СН3, обозначают "цепь впадины". Когда в нее вводят дополнительное изменение, название константной области Н-цепи начинается с символа "Hl", за которым следует трехзначное число, и, таким образом, ее обозначают как Нl001. Кроме того, когда константные области Н-цепи антитела представляют собой Kn001 и Нl001, последовательности Н-цепей антитела, вариабельные области которого содержат GpH7, обозначают как GpH7-Kn001 и GpH7-Hl001, соответственно. Антитело, очищаемое после экспрессии, обозначают, например, как GpH7-Kn001/GpH7-Hl001/GpL16-k0, когда последовательность, соответствующая Н-цепи антитела, используемой для экспрессии гетеродимеризованного антитела, представляет собой GpH7-Kn001, и последовательность, соответствующая другой Н-цепи антитела, представляет собой GpH7-Hl001, и последовательность, соответствующая L-цепи антитела, представляет собой GpL16-k0.
Сначала GpH7-Kn033 (SEQ ID NO: 51), получаемую в результате введения изменений Y349C и T336W в константную область, для
GpH7-G1d и GpH7-Hl033 (SEQ ID NO: 56), получаемую в результате введения изменений D356C, T336S, L368A и Y407V в константную область, для GpH7-G1d конструировали способом, описанным в справочном примере 1. При экспрессии гетеродимеризованных антител для их эффективной экспрессии использовали следующие ниже экспрессирующие векторы:
экспрессирующий вектор, в который вводили GpL16-k0, для L-цепи антитела; для одной Н-цепи антитела экспрессирующий вектор, в который вводили последовательность, в которую вводили дополнительное изменение в GpH7-Kn033 (SEQ ID NO: 51), в которую вводили изменения Y349C и T336W; и для другой Н-цепи антитела экспрессирующий вектор, в который вводили последовательность, в которую вводили дополнительное изменение в GpH7-Hl033 (SEQ ID NO: 56), в которую вводили изменения D356C, T336S, L368A и Y407V.
Антитела, для которых необходимо повышать связывание со всеми FcγRIIIa, FcγRIIa R типа и FcγRIIa Н типа, получали, как описано ниже на основании информации, получаемой в примере 4, касательно влияния каждого изменения на связывание антитела с каждым FcγR. При введении различных изменений в константные области соответствующих Н-цепей антитела GpH7-Kn033 и GpH7-Hl033 использовали в качестве родительских полипептидов. Способом, описанным в справочном примере 1, конструировали GpH7-Kn045 (SEQ ID NO: 54), получаемую в результате введения L234Y, L235Y, G236A, H268D и S298A в GpH7-Kn033, GpH7-Kn056 (SEQ ID NO: 55), получаемую в результате введения L234Y, L235Y, G236A, H268D, Q295L и S298A в GpH7-Kn033, GpH7-Hl048 (SEQ ID NO: 59), получаемую в результате введения G236A, S239D, А330K и I332E в
GpH7-Hl033, и GpH7-Hl055 (SEQ ID NO: 60), получаемую в результате введения G236A, S239D, Q295L, А330М и I332E в GpH7-Hl033.
Затем соответствующие Н-цепи комбинировали описанным ниже способом и экспрессировали антитела в соответствии со справочным примером 1. GpH7-Kn033/GpH7-Hl033/GpL16-k0, получаемое в результате применения одной технологии "выступы во впадины" к G1d, экспрессировали с использованием GpH7-Kn033 и GpH7-Hl033 в качестве Н-цепи и GpL16-k0 в качестве L-цепи. Гетеродимеризованное антитело GpH7-Kn045/GpH7-Hl048/GpL16-k0 экспрессировали с использованием GpH7-Kn045 и GpH7-Hl048 в качестве Н-цепи и GpL16-k0 в качестве L-цепи. Гетеродимеризованное антитело GpH7-Kn045/GpH7-Hl055/GpL16-k0 экспрессировали с использованием GpH7-Kn045 и GpH7-Hl055 в качестве Н-цепи и GpL16-k0 в качестве L-цепи. Гетеродимеризованное антитело GpH7-Kn056/GpH7-Hl055/GpL16-k0 экспрессировали с использованием GpH7-Kn056 и GpH7-Hl055 в качестве Н-цепи и GpL16-k0 в качестве L-цепи.
Антитела, основанные на применение общепринятых технологий, для сравнения получали согласно Pro. Nat. Acad. Sci., 103, 4005-4010 (2006), как описано ниже. Изменение G236A/S239D/I332E, для которого опубликовано, что оно повышает связывание со всеми FcγRIIIa, FcγRIIa R типа и FcγRIIa Н типа, вводили в GpH7-Kn033 и GpH7-Hl033 для конструкции GpH7-Kn037 (SEQ ID NO: 52) и GpH7-Hl036 (SEQ ID NO: 57), соответственно. Кроме того, изменение S239D/A330L/I332E, для которого опубликовано, что оно повышает связывание с FcγRIIIa, вводили в GpH7-Kn033 и GpH7-Hl033 для конструкции GpH7-Kn032 (SEQ ID NO: 53) и GpH7-Hl032 (SEQ ID
NO: 58), соответственно. Эти Н-цепи комбинировали и экспрессировали антитела в соответствии со справочным примером 1. Гомодимеризованное антитело GpH7-Kn037/GpH7-Hl036/GpL16-k0, получаемое в результате введения G236A/S239D/I332E в обе Н-цепи GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d, экспрессировали с использованием GpH7-Kn037 и GpH7-Hl036 в качестве Н-цепи и GpL16-k0 в качестве L-цепи. Гомодимеризованное антитело GpH7-Kn032/GpH7-Hl032/GpL16-k0, получаемое в результате введения S239D/A330L/I332E в обе Н-цепи GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d, экспрессировали с использованием GpH7-Kn032 и GpH7-Hl032 в качестве Н-цепи и GpL16-k0 в качестве L-цепи.
Эти антитела оценивали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 2. Результат приведен таблице 36. При этом отношение KD каждого антитела приведено в таблице 37, и отношение А/I, которое представляет собой отношение KD для FcγRIIIa, приведено в таблице 38.

В таблице представлена активность связывания с каждым FcγR гетеродимеризованных антител с повышенным связыванием с FcγRIIa и IIIa.
В столбце "образец" указаны названия антител, в столбце "Kn" и "Hl" указаны названия константных областей цепи выступ и цепи впадина каждого антитела, и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0 ("-": когда не существует конкретной мутации).

В таблице представлена активность связывания с каждым FcγR гетеродимеризованных антител с повышенным связыванием с FcγRIIa и IIIa.
В столбце "образец" указаны названия антител, в столбце "Kn" и "Hl" указаны названия константных областей цепи выступ и цепи впадина каждого антитела, и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-Kn033/GpH7-Hl033/GpL16-k0 для FcγR на KD каждого антитела, обозначали как "отношение KD". Также приведены SEQ ID NO для аминокислотных последовательностей
Н-цепи и L-цепи каждого антитела.

В таблице представлено отношение активности связывания с активирующим FcγR к активности связывания с ингибирующим FcγR.
В столбце "образец" указаны названия антител, в столбце "Kn" и "Hl" указаны названия константных областей цепи выступ и цепи впадина каждого антитела, и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0 ("-": когда не существует конкретной мутации). Значение, получаемое делением KD GpH7-Kn033/GpH7-Hl033/GpL16-k0 для FcγRIIb на KD каждого антитело для FcγRIIa Н типа и R типа, обозначали как "отношение А/I" для каждого типа. Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Результаты, представленные в таблицах 36 и 37, демонстрируют, что по отношению к GpH7-Kn037/GpH7-Hl036/GpL16-k0, в обе Н-цепи которого вводили G236A/S239D/I332E, его связывание с FcγRIIa типа Н повышалось в 22 раза, его связывание
с FcγRIIa типа R повышалось в 43 раза, и его связывание с FcγRIIIa F повышалось в 161 раз по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d. При этом результат, представленный в таблице 38, демонстрирует, что отношение A/I GpH7-Kn037/GpH7-Hl036/GpL16-k0 составляло 8,6 для FcγRIIa типа Н и 13 для FcγRIIa типа R, которые повышались по сравнению соответственно с 6,2 и 4,9 для GpH7-Kn033/GpH7-Hl033/GpL16-k0.
Результаты, представленные в таблицах 36 и 37, демонстрируют, что по отношению к GpH7-Kn032/GpH7-Hl032/GpL16-k0, в обе Н-цепи которого вводили S239D/A330L/I332E, его связывание с FcyRIIa типа Н повышалось в 1,2 раза, его связывание с FcγRIIa типа R повышалось в 3,0 раза, и его связывание с FcγRIIIa F повышалось в 381 раз по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d. При этом результат, представленный в таблице 38, демонстрирует, что отношение A/I GpH7-Kn032/GpH7-Hl032/GpL16-k0 составляло 0,93 для FcγRIIa типа H и 1,8 для FcγRIIa типа R, которые снижались по сравнению с GpH7-Kn033/GpH7-Hl033/GpLl6-k0.
Результаты, представленные в таблицах 36 и 37, демонстрируют, что по отношению к GpH7-Kn045/GpH7-Hl048/GpL16-k0, в одну из Н-цепей которого вводили L234Y/L235Y/G236A/H268D/S298A, а в другую Н-цепь G236A/S239D/A330K/I332E, его связывание с FcγRIIa типа Н
повышалось в 52 раза, его связывание с FcγRIIa типа R повышалось в 154 раза, и его связывание с FcγRIIIa F повышалось в 419 раза по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу получаемую в результате применения одной технологии "выступы во впадины" к G1d. Результат также демонстрируют, что для Н и R типов активность связывания с FcγRIIa повышалась по сравнению с гомодимеризованным антителом GpH7-Kn037/GpH7-Hl036/GpL16-k0, в обе Н-цепи которого вводили G236A/S239D/I332E по технологии известного уровня техники. Кроме того, активность связывания с FcγRIIIa F незначительно повышалась по сравнению с гомодимеризованным антителом GpH7-Kn032/GpH7-Hl032/GpL16-k0, в обе Н-цепи которого вводили S239D/A330L/I332E по технологии известного уровня техники. Результат, представленный в таблице 38, демонстрирует, что отношение А/I GpH7-Kn045/GpH7-Hl048/GpL16-k0 составлял 9,5 для FcγRIIa типа Н и 22 для FcγRIIa типа R, которое повышалось по сравнению с любым из GpH7-Kn033/GpH7-Hl033/GpL16-k0, GpH7-Kn037/GpH7-Hl036/GpL16-k0 и GpH7-Kn032/GpH7-Hl032/GpL16-k0. Результат демонстрирует, что с применением технологии гетеродимеризованного антитела повышают активности связывания с FcγRIIa и FcγRIIIa F GpH7-Kn045/GpH7-Hl048/GpL16-k0, и антитело связывается более селективно с активирующим FcγR по сравнению с тем, когда применяют общепринятую технологию.
Результаты, представленные в таблицах 36 и 37 демонстрируют, что по отношению к GpH7-Kn045/GpH7-Hl055/GpL16-k0, в одну из Н-цепей которого вводили L234Y/L235Y/G236A/H268D/S298A, а в другую
Н-цепь вводили G236A/S239D/Q295L/A330M/I332E, его связывание с FcγRIIa типа Н повышалось в 21 раз, его связывание с FcγRIIa типа R повышалось в 56 раз, и его связывание с FcγRIIIa F повышалось в 985 раз по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d. Результат также демонстрирует, что для Н и R типов активность связывания с FcγRIIa являлась практически эквивалентной активности гомодимеризованного антитела GpH7-Kn037/GpH7-Hl036/GpL16-k0, в обе Н-цепи которого вводили G236A/S239D/I332E по технологии известного уровня техники. Активность связывания с FcγRIIIa F повышалась по сравнению с гомодимеризованным антителом GpH7-Kn032/GpH7-Hl032/GpL16-k0, в обе Н-цепи которого вводили S239D/A330L/I332E по технологии известного уровня техники. Результат, представленный в таблице 38, демонстрирует, что отношение A/I GpH7-Kn045/GpH7-Hl055/GpL16-k0 составляло 8,3 для FcγRIIa типа Н и 18 для FcγRIIa типа R, которое являлось повышенным по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0 и GpH7-Kn032/GpH7-Hl032/GpL16-k0, и по сравнению с GpH7-Kn037/GpH7-Hl036/GpL16-k0 оно являлось практически эквивалентным для FcγRIIa типа Н и повышалось для FcγRIIa типа R. Этот результат демонстрирует, что с использованием технологии гетеродимеризованного антитела в GpH7-Kn045/GpH7-Hl055/GpL16-k0 дополнительно повышают активность связывания с FcγRIIIa, при этом в сравнимой степени повышают его активность связывания с FcγRIIa, и антитело связывается более селективно с активирующим FcγR по
сравнению с тем, когда используют общепринятую технологию.
Результаты, представленные в таблицах 36 и 37 демонстрируют, что по отношению к GpH7-Kn056/GpH7-Hl055/GpL16-k0, в одну из Н-цепей которого вводили L234Y/L235Y/G236A/H268D/Q295L/S298A, а в другую Н-цепь вводили G236A/S239D/Q295L/A330M/I332E, его связывание с FcγRIIa типа Н повышалось в 20 раз, его связывание с FcγRIIa типа R повышалось в 44 раза, и его связывание с FcγRIIIa F повышалось в 1114 раза по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0, которое представляет собой молекулу, получаемую в результате применения одной технологии "выступы во впадины" к G1d. Этот результат также демонстрирует, что для Н и R типов активность связывания с FcγRIIa являлась эквивалентной активности гомодимеризованного антитела GpH7-Kn037/GpH7-Hl036/GpL16-k0, в обе Н-цепи которого вводили G236A/S239D/I332E по технологии известного уровня техники. Активность связывания с FcγRIIIa F повышалась по сравнению с гомодимеризованным антителом GpH7-Kn032/GpH7-Hl032/GpL16-k0, в обе Н-цепи которого вводили S239D/A330L/I332E по технологии известного уровня техники. Результат, представленный в таблице 38, демонстрирует, что отношение A/I GpH7-Kn056/GpH7-Hl055/GpL16-k0 составляло 8,7 для FcγRIIa типа Н и 16 для FcγRIIa типа R, которое являлось повышенным по сравнению с GpH7-Kn033/GpH7-Hl033/GpL16-k0 и GpH7-Kn032/GpH7-Hl032/GpL16-k0, и по сравнению с GpH7-Kn037/GpH7-Hl036/GpL16-k0 оно являлось практически эквивалентным для FcγRIIa типа Н и повышалось для FcγRIIa типа R. Результат демонстрирует, что с использованием технологии гетеродимеризованного антитела в
GpH7-Kn056/GpH7-Hl055/GpL16-k0 дополнительно повышают активность связывания с FcγRIIIa, при этом его активность связывания с FcγRIIa повышается в сравнимой степени, и антитело связывается более селективно с активирующим FcγR по сравнению с тем, когда применяют общепринятую технологию.
[Пример 13] Сравнение с общепринятой технологией: оценка термостабильности гетеродимеризованных антител с повышенной активностью связывания с FcγRIIa и FcγRIIIa
Как описано в примере 9, полагают, что получаемые общепринятой технологией гомодимеризованные антитела с повышенной активностью связывания с FcγR, являются физико-химически нестабильными, и это снижает их ценность в качестве фармацевтических средств. С другой стороны, выявлено, что технология гетеродимеризованного антитела являлась подходящей для регуляции эффекта каждого изменения в отношении повышения активности связывания с FcγR и влияния физико-химического аспекта, и что возможным являлось повышение активности связывания с FcγR без снижения физико-химической стабильности. В этом примере тестируют, не обладают ли антитела с повышенными активностями связывания с FcγRIIa и FcγRIIIa, которые представляют собой активирующий FcγR, аналогично сниженной физико-химической стабильностью, в частности термодинамической стабильностью. Каждое антитело, оцениваемое в отношении его активности связывания с FcγR в примере 11, анализировали в отношении Tm области СН2 способом, описанным в справочном примере 5. Результат представлен в таблице 39.

В таблице приведены Tm антител с повышенными активностями связывания с FcγRIIa и FcγRIIIa.
Результат, представленный в таблице 39, демонстрирует, что все гетеродимеризованные антитела GpH7-Kn045/GpH7-Hl048/GpL16-k0, GpH7-Kn045/GpH7-Hl055/GpL16-k0 и GpH7-Kn056/GpH7-Hl055/GpL16-k0 сохраняли высокую Tm по сравнению с гомодимеризованными антителами GpH7-Kn037/GpH7-Hl036/GpL16-k0 и GpH7-Kn032/GpH7-Hl032/GpL16-k0 известного уровня техники. Как описано в примере 11, гетеродимеризованные антитела обладают свойствами более подходящими для получения опосредованной FcγR эффекторной функции по сравнению с общепринятыми гомодимеризованными антителами. В частности, описанное выше открытие демонстрирует, что связывание с FcγR можно точно регулировать без снижения физико-химической стабильности антител технологией гетеродимеризованного антитела.
[Пример 14] Эффект комбинации изменений, которые улучшают
селективность в отношении FcγRIIIa F активирующего FcγR.
Как описано в примере 8, пригодными являются технологии улучшения селективности в отношении активирующего FcγR и ингибирующего FcγR. В этом примере тестируют, является ли гетеродимеризация эффективной для повышения отношения связывания с FcγRIIIa F, активирующим Fcγ, к связыванию с FcγRIIb, ингибирующим FcγR, т.е. эффективной для улучшения селективности, как описано в примере 8. В частности, L234Y, G236W и S298A (область а, представленная в таблице 22-1), которые представляют собой изменения, которые повышают отношение связывания с FcγRIIIa F, активирующим Fcγ, к связыванию с FcγRIIb, ингибирующим FcγR, комбинировали с S239D, A330L и I332E, оцениваемыми в примере 7, для тестирования, можно ли получать улучшение селективности в гетеродимеризованных антителах по сравнению с гомодимеризованными антителами.
Для проверки этого способом, описанным в справочном примере 1, конструировали экспрессирующие векторы, в которые вводили GpH7-A57 (SEQ ID NO: 40), получаемую в результате введения всех S239D, A330L и I332E в GpH7-A5; GpH7-B78 (SEQ ID NO: 41), получаемую в результате введения всех S239D, A330L и I332E в GpH7-B3; GpH7-TA7 (SEQ ID NO: 31), получаемую в результате введения всех L234Y, G236W и S298A в GpH7-A5, и GpH7-TA45 (SEQ ID NO: 32), получаемую в результате введения всех L234Y, G236W и S298A в GpH7-B3. С использованием этих экспрессирующих векторов и GpH7-A5, GpH7-B3 и GpL16-k0 экспрессировали и получали способом, описанным в справочном примере 1, гетеро-GpH7-TA7/GpH7-B78/GpL16-k0,
в одну из Н-цепей которого вводили L234Y, G236W и S298A, а в другую вводили S239D, A330L и I332E; GpH7-TA7/GpH7-B3/GpL16-k0, только в одну из Н-цепей которого вводили L234Y, G236W и S298A; GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили L234Y, G236W и S298A; GpH7-A5/GpH7-B78/GpL16-k0, только в одну из Н-цепей которого вводили S239D, A330L и I332E, и GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E. Получаемые антитела анализировали в отношении KD для FcγRIIIa и FcγRIIb способом, описанным в справочном примере 2. Являлось ли каждое антитело улучшенным в отношении селективности связывания с FcγRIIIa, оценивали с использованием в качестве показателя отношения FcγRIIIa/FcγRIIb, которое получают делением KD каждого антитела для FcγRIIb на KD каждого антитела для FcγRIIIa. Результат оценки приведен в таблице 40.

В столбце "образец" указаны названия антител, в столбцах
"H1" и "H2" указаны названия константной области Н-цепи в каждом антителе, и в столбце "участок мутации" указаны мутации, которые отличаются по сравнению с GpH7-A5/GpH7-B3/GpL16-k0 ("-": используемый, когда не существует конкретной мутации). Значения, получаемые делением KD каждого антитела для FcγRIIb на KD каждого антитела для FcγRIIIa F обозначают как "отношение FcγRIIIa F/FcγRIIb". Также приведены SEQ ID NO для аминокислотных последовательностей Н-цепи и L-цепи каждого антитела.
Когда на основании результатов, представленных в таблице 40, нативный IgG1 GpH7-G1d/GpL16-k0 сравнивают с GpH7-A5/GpH7-B3/GpL16-k0, в котором каждое D356K, H435R и K439Е вводят в одну Н-цепь, отношение FcγRIIIa/FcγRIIb составляет 2,5 и 1,9, соответственно, и не наблюдают большого отличия между ними. Это позволяет предположить, что изменения D356K, H435R и K439Е не влияют на селективность связывания с FcγRIIIa.
Проверяли эффект каждого изменения на гомодимеризованные антитела современного уровня техники. Отношение FcγRIIIa/FcγRIIb гомодимеризованного антитела GpH7-A57/GpH7-B78/GpL16-k0, в обе Н-цепи которого вводили S239D, A330L и I332E, составляло 100 и повышалось по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом отношение FcγRIIIa/FcγRIIb гомодимеризованного антитела GpH7-TA7/GpH7-TA45/GpL16-k0, в обе Н-цепи которого вводили L234Y, G236W и S298A, составляло 5,3. По отношению к гомодимеризованным антителам демонстрировали, что комбинации S239D, A330L и I332E обладают большим эффектом улучшения селективности связывания с FcγRIIIa.
Затем гетеродимеризованные антитела, только в одну из Н-цепей которых вводили каждую группу изменений, оценивали в отношении эффекта каждой группы изменений. Отношение FcγRIIIa/FcγRIIb гетеродимеризованного антитела GpH7-A5/GpH7-B78/GpL16-k0, в одну Н-цепь которого вводили S239D, A330L и I332E, составляло 7,9 и повышалось по сравнению с GpH7-A5/GpH7-B3/GpL16-k0. При этом отношение FcγRIIIa/FcγRIIb гетеродимеризованного антитела GpH7-TA7/GpH7-B3/GpL16-k0, в одну Н-цепь которого вводили L234Y, G236W и S298A, составляло 14. Эти результаты демонстрируют, что по отношению к гетеродимеризованным антителам группа изменений L234Y, G236W и S298A обладает большим эффектом повышения селективности связывания с FcγRIIIa.
Оценивали различие эффекта каждой группы изменений в гомодимеризованных антителах и гетеродимеризованных антителах. Касательно S239D, A330L и I332E отношение FcγRIIIa/FcγRIIb гетеродимеризованного антитела с ними составляло 7,9, тогда как аналогичное отношение гомодимеризованного антитела с ними составляло 100. Эти результаты демонстрируют, что S239D, A330L и I332E обладают эффектом улучшения селективности связывания с FcγRIIIa при гетеродимеризации, и эффект дополнительно повышается при гомодимеризации. С другой стороны, касательно L234Y, G236A и S298A отношение FcγRIIIa/FcγRIIb гетеродимеризованного антитела с ними составляло 14, тогда как аналогичное отношение гомодимеризованного антитела с ними составляло 5,3. Эти результаты демонстрируют, что L234Y, G236A и S298A обладают
эффектом улучшения селективности связывания с FcγRIIIa при гетеродимеризации, но эффект снижается при гомодимеризации. Результаты демонстрируют, что группа изменений S239D, A330L и I332E в гомодимеризованных антителах обладает большим эффектом улучшения селективности связывания с FcγRIIIa, чем группа изменений L234Y, G236A и S298A, при этом группа изменений L234Y, G236A и S298A в гетеродимеризованных антителах обладает большим эффектом улучшения селективности связывания с FcγRIIIa, чем группа изменений S239D, A330L и I332E.
Демонстрировали, что отношение FcγRIIIa/FcγRIIb гетеродимеризованного антитела GpH7-TA7/GpH7-B78/GpL16-k0 с группой изменений L234Y, G236A и S298A в комбинации с группой изменений S239D, A330L и I332E составляло 244, и, таким образом, их эффект улучшения селективности связывания с FcγRIIIa являлся больше по сравнению с гетеродимеризованным антителом GpH7-TA7/GpH7-B3/GpL16-k0, только одна из Н-цепей которого содержала группа изменений L234Y, G236A и S298A, гомодимеризованным антителом GpH7-TA7/GpH7-TA45/GpL16-k0, обе Н-цепи которого содержат группу изменений L234Y, G236A и S298A, гетеродимеризованным антителом GpH7-A5/GpH7-B78/GpL16-k0, только одна из Н-цепей которого содержит группу изменений S239D, A330L и I332E, и гомодимеризованным антителом GpH7-A57/GpH7-B78/GpL16-k0, обе Н-цепи которого содержат группу изменений S239D, A330L и I332E. Полагают, что эти результаты представляют собой сумму эффекта группы изменений L234Y, G236A и S298A в гетеродимеризованном антителе в отношении улучшения
селективности связывания с FcγRIIIa и эффекта группы изменений S239D, A330L и I332E в гетеродимеризованном антителе. В частности, выявлено, что гетеродимеризованные антитела демонстрируют превосходный эффект улучшения селективности связывания с FcγRIIIa по сравнению с гомодимеризованными антителами.
В частности, показано, что использование гетеродимеризованных антител вместо общепринятых гомодимеризованных антител обеспечивает более точную оптимизацию асимметричного взаимодействия Fc-области и FcγRIIIa и конструкцию Fc-областей, обладающих большей селективностью связывания с FcγRIIIa.
[Пример 15] Измерение активности ADCC гетеродимеризованных антител, которые демонстрируют повышенное связывание с FcγRIIa
Активность ADCC GpH7-G1d/GpL16-k0, GpH7-Kn033/GpH7-Hl033/GpL16-k0, GpH7-Kn037/GpH7-Hl036/GpL16-k0, GpH7-Kn032/GpH7-Hl032/GpL16-k0, GpH7-Kn045/GpH7-Hl048/GpL16-k0 и GpH7-Kn056/GpH7-Hl055/GpL16-k0, получаемых в примере 12, оценивали способом, описанным в справочном примере 7. Результаты приведены на фиг. 33.
Когда GpH7-G1d/GpL16-k0 и GpH7-Kn033/GpH7-Hl033/GpL16-k0 сравнивали в отношении активности ADCC на основании фиг. 33, они обладали сравнимой активностью ADCC. Этот результат демонстрирует, что выступы во впадины даже при введении в Fc-область антитела не влияют на связывание с FcγR или активность ADCC.
Оба гетеродимеризованных антитела GpH7-Kn045/GpH7-Hl048/GpL16-k0 и GpH7-Kn056/GpH7-Hl055/GpL16-k0, описанных в примере 12, демонстрировали большую активность ADCC по сравнению с антителом GpH7-Kn033/GpH7-Hl033/GpL16-k0 до введения изменения. При этом гетеродимеризованные антитела GpH7-Kn045/GpH7-Hl048/GpL16-k0 и GpH7-Kn056/GpH7-Hl055/GpL16-k0 демонстрировали активность ADCC, сравнимую с активностью гомодимеризованного антитело GpH7-Kn037/GpH7-Hl036/GpL16-k0, обе Н-цепи которого содержат изменение G236A/S239D/I332E, для которого опубликовано, что оно повышает связывание с FcγRIIa R и FcγRIIa Н и активность ADCP, и с активностью антитела GpH7-Kn032/GpH7-Hl032/GpL16-k0, получаемого в результате применения существующего повышения активности ADCC.
В частности, как продемонстрировано в примере 12, GpH7-Kn045/GpH7-Hl048/GpL16-k0 и GpH7-Kn056/GpH7-Hl055/GpL16-k0 не только демонстрируют дополнительно повышенное связывание с FcγRIIa R и FcγRIIa Н по сравнению с существующей техникой, а также обладают повышающим активность ADCC эффектом, сравнимым с существующей техникой повышения активности ADCC. В частности, гетеродимеризованные антитела, оцениваемые в настоящем описании, превосходят существующую технику не только в том, что они обладают повышающим активность ADCC эффектом, сравнимым с тем, который получают существующей техникой, а также демонстрируют повышенное связывание с FcγRIIa Н и FcγRIIa R.
[Пример 16] Получение гетеродимеризованного антитела Н240-Kn061/H240-Hl071/L73-k0, которое демонстрирует повышенное
связывание с FcγRIIIa.
Как описано в примере 11, демонстрировали, что гетеродимеризованные антитела с повышенной активностью связывания с FcγRIIIa обладают улучшенной активностью ADCC. В примере 11 эффект демонстрировали с антителами против GPC3. Проводя аналогичный эксперимент с использованием антитела против эпирегулина (EREG), оценивали, можно ли наблюдать аналогичный эффект для других антигенов. В настоящем описании вариабельной области последовательность Н-цепи антитела против EREG обозначают как Н240 (SEQ ID NO: 80), и его последовательность L-цепи, включая вариабельную и константную области, обозначают как L73-k0 (SEQ ID NO: 106).
На основании результатов примера 4 получали новые варианты, содержащие Н-цепи, которые обладают повышенным связыванием с FcγRIIIa. В настоящем описании описанную в примере 12 технику "выступы во впадины" использовали в качестве техники гетеродимеризации. В частности, H240-Kn033 (SEQ ID NO: 84), получаемую в результате введения изменений Y349C и T336W в константную область H240-G1d (SEQ ID NO: 83), и Н240-Нl033 (SEQ ID NO: 85), получаемую в результате введения изменений D356C, T336S, L368A и Y407V в константную область H240-G1d, получали способом, описанным в справочном примере 1. Затем получали Н240-Kn061 (SEQ ID NO: 81) введением L234Y, L235Y, G236W, H268D и S298A в H240-Kn033 (SEQ ID NO: 84) способом, описанным в справочном примере 1. Н240-Hl071 (SEQ ID NO: 82) конструировали введением K326D, А330М и K334Е в Н240-Hl033 (SEQ ID NO: 85)
способом, описанным в справочном примере 1. Гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 экспрессировали, комбинируя H240-Kn061, Н240-Hl071 и L73-k0 способом, описанным в справочном примере 1.
Варианты, в которые вводили S239D, A330L и I332E, для которых опубликовано, что они повышают связывание FcγRIIIa, конструировали аналогичным образом, как описано в примере 12, чтобы использовать для сравнения. В частности, H240-Kn032 (SEQ ID NO: 86) и Н240-Hl032 (SEQ ID NO: 87), получаемые в результате введения S239D, A330L и I332E в H240-Kn033 (SEQ ID NO: 84) и Н240-Hl033 (SEQ ID NO: 85), соответственно, получали способом, описанным в справочном примере 1. Гомодимеризованное антитело H240-Kn032/H240-Hl032/L73-k0 экспрессировали, комбинируя Н240-Kn032, Н240-Hl032 и L73-k0 способом, описанным в справочном примере 1.
Затем для сравнения получали дефукозилированное антитело, для которого опубликовано, что оно повышает связывание с FcγRIIIa (Glycobiol., Vol. 17 no. 1, pp. 104-118 (2006)). Функция транспортера фукозы ингибируется в клетках, где экспрессию гена транспортера фукозы искусственно подавляли в обеих гомологичных хромосомах. Антитела, не содержащие фукозу, можно получать с использованием таких клеток (WO 2006/067913 и т.д.). Альтернативно, антитела, не содержащие фукозу, также можно получать, продуцируя антитела в клетках с усиленной экспрессией бета-1,4-N-ацетилглукозаминилтрансферазы III и Гольджи альфа-маннозидазы II (Ferrara et al., Biotechnol. Bioeng., (2006) 93 (5), 851-861). H240-G1d (SEQ ID NO: 83) и L73-k0 (SEQ ID NO: 106)
экспрессировали в комбинации и получали антитело Н240-дефукозил_G1d/L73-k0 (SEQ ID NO: 83 и 106) дефукозилированием H240-G1d/L73-k0 с применением описанных выше технологий, известных специалистам в данной области.
Кроме того, H240-Kn033/H240-Hl033/L73-k0 экспрессировали в качестве контроля, комбинируя H240-Kn033 (SEQ ID NO: 84), Н240-Hl033 (SEQ ID NO: 85) и L73-k0 (SEQ ID NO: 106) способом, описанным в справочном примере 1.
Активность связывания антител с каждым FcγR оценивали способом, описанным в справочном примере 8, и результаты приведены в таблице 41.
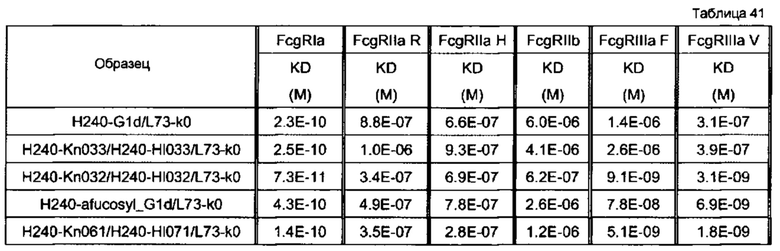
Результаты, представленные в таблице 41, демонстрируют, что связывание гетеродимеризованного антитела Н240-Kn061/Н240-Hl071/L73-k0, в частности с FcγRIIIa F и FcγRIIIa V повышалось по сравнению с H240-Kn033/H240-Hl033/L73-k0. Вследствие того, что гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 представляет собой вариант, получаемый в результате введения L234Y/L235Y/G236W/H268D/S298A и K326D/A330M/K334E в Н240-Kn033/H240-Hl033/L73-k0, можно говорить, что связывание с FcγR вводимых изменений повышалось.
Результаты, представленные в таблице 41, демонстрируют, что связывание с FcγRIIIa F и FcγRIIIa V гетеродимеризованного антитела H240-Kn061/H240-Hl071/L73-k0 повышалось по сравнению с Н240-дефукозил_C1d/L73-k0 и H240-Kn032/H240-Hl032/L73-k0, получаемых в результате применения существующей техники повышения активности ADCC. Этот результат демонстрирует, что гетеродимеризованное антитело обладает значительным эффектом в отношении повышения связывания с FcγRIIIa по сравнению с общепринятой техникой повышения активности ADCC на основе гомодимеризованного антитела и техники повышения активности ADCC на основе дефукозилирования.
Кроме того, в отношении связывания с FcγRIIa, которое, как полагают, является важным для повышения активности ADCP, связывание с FcγRIIa Н гетеродимеризованного антитела повышалось по сравнению с двумя антителами, и его связывание с FcγRIIa R повышалось относительно Н240-дефукозил_C1d/L73-k0 и по сравнению со связыванием H240-Kn032/H240-Hl032/L73-k0.
Оценивали, обладает ли гетеродимеризованное антитело Н240-Kn061/H240-Hl071/L73-k0 свойством гетеродимеризованного антитела, которое заключается в том, что оно обладает повышенной активностью связывания с FcγR по сравнению с гомодимеризованными антитела, содержащими каждую его Н-цепь. В гетеродимеризованное антитело вводили H240-Kn061/H240-Hl071/L73-k0, L234Y/L235Y/G236W/H268D/S298A в Н240-Kn061, которая представляет собой одну из Н-цепей, и вводили K326D/A330M/K334E в Н240-Hl071, которая представляет собой другую Н-цепь. Гетеродимеризованное
антитело сравнивали с гомодимеризованными антителами, содержащими каждую его Н-цепь, для оценки обладает ли гетеродимеризованное антитело большей активностью связывания с каждым FcγR. В частности, Н240-Hl134 (SEQ ID NO: 88), получаемую в результате введения L234Y/L235Y/G236W/H268D/S298A в Н240-Hl033 и H240-Kn132 (SEQ ID NO: 89), получаемую в результате введения K326D/A330M/K334E в Н240-Kn033, конструировали способом, описанным в справочном примере 1. С использованием этих экспрессирующих векторов гомодимеризованное антитело Н240-Kn061/H240-Hl134/L73-k0, обе Н-цепи которого содержат L234Y/L235Y/G236W/H268D/S298A, и гомодимеризованное антитело H240-Kn132/H240-Hl071/L73-k0, обе Н-цепи которого содержат K326D/A330M/K334E, экспрессировали способом, описанным в справочном примере 1. Связывание с FcγRIIIaF и FcγRIIIa V гомодимеризованных антител и гетеродимеризованного антитела H240-Kn061/H240-Hl071/L73-k0, каждая Н-цепь которого содержит L234Y/L235Y/G236W/H268D/S298A или K326D/A330M/K334E, измеряли способом, описанным в справочном примере 8. Результаты приведены в таблице 42.

Результаты, представленные в таблице 42, демонстрируют, что активность связывания с FcγRIIIa F и FcγRIIIa V гетеродимеризованного антитела H240-Kn061/H240-Hl071/L73-k0, одна из Н-цепей которого содержит L234Y/L235Y/G236W/H268D/S298A, а другая Н-цепь содержит K326D/A330M/K334E, является выше, чем активность гомодимеризованного антитела Н240-Kn061/Н240-Hl134/L73-k0, обе Н-цепи которого содержат L234Y/L235Y/G236W/H268D/S298A, и гомодимеризованного антитела H240-Kn132/H240-Hl071/L73-k0, обе Н-цепи которого содержат K326D/A330M/K334E. А именно, демонстрировали, что Н240-Kn061/H240-Hl071/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно обладает повышенной активностью связывания с FcγR по сравнению с гомодимеризованными антителами, содержащими каждую его Н-цепь.
Затем сравнивали активности ADCC H240-Kn033/H240-Hl033/L73-k0, H240-Kn032/H240-Hl032/L73-k0, Н240-дефукозил_C1d/L73-k0 и H240-Kn061/H240-Hl071/L73-k0 способом, описанным в справочном примере 9. Результаты представлены на фиг. 34.
Результаты, представленные на фиг. 34, демонстрируют, что H240-Kn061/H240-Hl071/L73-k0 обладает значительно большей активностью ADCC по сравнению с H240-Kn033/H240-Hl033/L73-k0. Кроме того, активность ADCC являлась больше чем активность Н240-Kn032/H240-Hl032/L73-k0 и Н240-дефукозил_C1d/L73-k0, получаемых в результате применения существующей техники повышения активности ADCC. А именно, демонстрировали, Н240-Kn061/Н240-Hl071/L73-k0 обладает большей активностью ADCC, чем активность, получаемая существующей техникой повышения активности ADCC.
[Пример 17] Получение дополнительных вариантов с использованием гетеродимеризованного антитела Н240-Kn061/Н240-Hl071/L73-k0 в качестве матрицы и их оценка
H240-Kn061/H240-Hl071/L73-k0, демонстрирующее превосходную активность ADCC, открыли в примере 16 выше. Для выявления дополнительных вариантов с превосходной активностью, всего приблизительно 420 типов вариантов, в которых аминокислоты в положениях от 231 до 242 (нумерация EU) замещали 18 типами аминокислот, за исключением Cγs и исходной аминокислоты, в каждой из Н-цепей Н240-Kn061 и Н240-Hl071, получали с использованием H240-Kn061/H240-Hl071/L73-k0 в качестве матрицы способом, описанным в справочном примере 1. Антитела оценивали в отношении связывания с каждым FcγR. В частности, рассчитывали значение KD каждого варианта для каждого из FcγRI, FcγRIIa R, FcγRIIa H, FcγRIIb, FcγRIIIa F и FcγRIIIa V способом, описанным в справочном примере 8, и значение KD H240-Kn061/H240-Hl071/L73-k0 для каждого из FcγRI, FcγRIIa R, FcγRIIa H, FcγRIIb, FcγRIIIa F и FcγRIIIa V делили на значение KD, получаемое как указано выше. Получаемое значение определяли как относительную KD и использовали в качестве показателя оценки. В частности, когда значение KD H240-Kn061/H240-Hl071/L73-k0 принимали за 1, в качестве показателя оценки использовали изменение степени кратности значения KD каждого варианта для каждого FcγR. Когда относительная KD является больше, это значит, что связывание варианта с каждым FcγR значительно больше повышается по сравнению с H240-Kn061/H240-Hl071/L73-k0.
Графики, в которых относительные KD соответствующих вариантов для FcγRI, FcγRIIa R, FcγRIIa H, FcγRIIb, FcγRIIIa F или FcγRIIIa V показаны по вертикальной оси и ранговые числа при ранжировании относительных KD в возрастающем порядке показаны на горизонтальной оси, продемонстрированы на фиг. 35, 36, 37, 38, 39 и 40, соответственно.
На основании результата анализа выявляли варианты, которые повышают связывание с любым одним или несколькими из FcγRIIa R, FcγRIIIa F и FcγRIIIa V без повышения связывания с FcγRIIb по отношению к H240-Kn061/H240-Hl071/L73-k0. Изменения и H-цепи, в которые вводили изменения, приведены в таблице 43 (изменения, которые повышают связывание с FcγRIIa R и FcγRIIIa без повышения связывания с FcγRIIb). Выбирали изменения, в которых относительная KD равна 1 или менее для FcγRIIb и значение относительной KD равно 1,3 или более для любого одного или нескольких из FcγRIIa R, FcγRIIIa F и FcγRIIIa V.

Значения, представленные в таблице 43 выше, обозначают относительную KD каждого варианта для каждого FcγR.
Изменения обладают эффектом повышения связывания с FcγRIIa, который играет важную роль в активности ADCP, и связывании с FcγRIIIa, который играет важную роль в активности ADCC, без повышения связывания с FcγRIIb, ингибирующим FcγR. Таким образом, можно предполагать большую противоопухолевую активность, т.к. введение этих изменений повышает активности ADCC и ADCP без повышения иммуносупрессивного действия антитела.
Кроме того, выявляли варианты, которые повышают связывание с FcγRIIa H и FcγRIIa R без снижения связывания с FcγRIIIa по отношению к H240-Kn061/H240-Hl071/L73-k0. Изменения и H-цепи, в которые вводили изменения, приведены в таблице 44 (изменения, которые повышают связывание с FcγRIIa без снижения связывания с FcγRIIIa). Выбирали изменения, в которых относительная KD равна 0,7 или более для FcγRIIIa F и FcγRIIIa V, и относительная KD равна 1,5 или более для FcγRIIa H и FcγRIIa R.

Значения, представленные в таблице 44 выше, обозначают
относительную KD каждого варианта для каждого FcγR.
Изменения повышают связывание с FcγRIIa, который играет важную роль в активности ADCP, без снижения связывания с FcγRIIIa который играет важную роль в активности ADCC, и некоторые из изменений повышают связывание с FcγRIIIa. Таким образом, посредством введением изменений можно предполагать большую противоопухолевую активность и, таким образом, повышать активности ADCC и ADCP или любую из активности ADCC или ADCP.
Кроме того, выявляли варианты, которые снижают связывание с FcγRIIb без снижения связывания с FcγRIIIa по отношению к H240-Kn061/H240-Hl071/L73-k0. Изменения и H-цепи, в которые вводили изменения, приведены в таблице 45 (изменения, которые снижают связывание с FcγRIIb при сохранении связывания с FcγRIIIa). Выбирали изменения, в которых относительная KD равна 1 или более для FcγRIIIa F и FcγRIIIa V, и относительная KD равна 0,5 или менее для FcγRIIb.

Значения, представленные в таблице 45 выше, означают относительную KD каждого варианта для каждого FcγR.
Изменения снижают связывание с FcγRIIb, ингибирующим FcγR, без снижения связывания с FcγRIIIa, который играет важную роль в активности ADCC. Таким образом, можно предполагаю большую противоопухолевую активность, т.к. введение изменений снижает иммуносупрессивное действие антитела без снижения активности ADCC.
Кроме того, вследствие того, что гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно связывается с большей силой с FcγR по сравнению с гомодимеризованными антителами, содержащими каждую его H-цепь, полагают, что варианты, получаемые введением изменений в H240-Kn061/H240-Hl071/L73-k0, обладают аналогичным свойством гетеродимеризованного антитела.
[Пример 18] Изменения, которые могут замещать изменения, вводимые в гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0
На основании результатов, представленных на фиг. 35, 36, 37, 38, 39 и 40, получаемых в примере 17, оценивали, являлись ли изменения в гетеродимеризованном антителе H240-Kn061/H240-Hl071/L73-k0 заменяемыми другими изменениями. В настоящем описании "заменяемое изменение" относится к изменению, которое при введении приводит к связыванию с FcγRIIIa F и FcγRIIIa V, составляющему 0,7 или более, и связыванию с FcγRIIb, составляющему 1,3 или менее от величины до введения изменения.
В отношении вариантов, получаемых в примере 17, изменения
вводили в антитела в положениях аминокислот от 231 до 242 (нумерация EU). В гетеродимеризованном антителе H240-Kn061/H240-Hl071/L73-k0 вводили L234Y/L235Y/G236W/H268D/S298A в H-цепь H240-Kn061 и вводили K326D/A330M/K334E в H-цепь H240-Hl071. Из изменений, вводимых в H-цепь H240-Kn061, на основании результатов, приведенных в примере 17, можно оценивать являются ли заменяемыми L234Y, L235Y и G236W другими аминокислотами.
Участки изменения включают положения 234, 235 и 236 (нумерация EU) в H-цепи H240-Kn061, которая представляет собой одну из H-цепей H240-Kn061/H240-Hl071/L73-k0, но не включают участки изменений, вводимых в H-цепь H240-Hl071, которая представляет собой другую H-цепь. В числе вариантов, получаемых в этом эксперименте, полагают, что варианты, в которые вводили изменения в положениях 234, 235 и 236 (нумерация EU) в H-цепи H240-Kn061, варианты, обладающие связыванием с FcγRIIIa F и FcγRIIIa V, которое составляет 0,7 или более, и связыванием с FcγRIIb, которое составляет 1,3 или менее от связывания по сравнению с H240-Kn061/H240-Hl071/L73-k0, обладают сравнимой или превосходящей активностью по сравнению с H240-Kn061/H240-Hl071/L73-k0. Таким образом, полагали, что возможными являются замены без снижения превосходных свойств гетеродимеризованного антитела H240-Kn061/H240-Hl071/L73-k0. В отношении изменений, которые соответствуют описанным выше условиям, изменения и H-цепи, в которые их вводили, приведены в таблице 46 (изменения, которые обеспечивают сохранение сравнимой активности или придают активность, превосходящую активность H240-Kn061/H240-Hl071/L73-k0).
В таблице 46, указанной выше, "H-цепь, в которую вводили изменение" означает какую H-цепь H240-Kn061/H240-Hl071/L73-k0 она заменяют, и в "изменении" числовое обозначение представляет собой номер остатка в соответствии с нумерацией EU, первое буквенное обозначение представляет собой аминокислоту, соответствующую указанному номеру остатка в H240-Kn061/H240-Hl071/L73-k0, и последнее буквенное обозначение представляет собой заменяемую аминокислоту.
На основании результатов в числе изменений, вводимых в
константную область H-цепи H240-Kn061, приведены заменяемые участки и аминокислоты, как представлено в таблице 47 (участки изменений в H240-Kn061, в которых изменение можно заменять при сохранении активности, сравнимой с активностью H240-Kn061/H240-Hl071/L73-k0, и заменяемые аминокислоты).

В таблице 47, представленной выше, "изменяемое положение" относится к номеру остатка в соответствии с нумерацией EU в H240-Kn061, и "заменяемая аминокислота" относится к аминокислоте, которая даже при замене в этом участке аминокислотой, представленной в этой таблице, обеспечивает активность, сравнимую с активностью H240-Kn061/H240-Hl071/L73-k0, т.е. к аминокислоте, которую можно заменять.
Из изменений, вводимых в H240-Kn061/H240-Hl071/L73-k0, в отношении H268D и S298A в H240-Kn061 и K326D, A330M и K334E в H240-Hl071 изменения не вводят в соответствующие участки в эксперименте, описанном в примере 17. Таким образом, наличие или отсутствие заменяемых изменений в описанных выше участках также обсуждается ниже на основании результатов, приведенных в примере 4. В частности, на основании результатов, приведенных в примере 4, три изменения, которые приводили к значению He/Con_3aF 130 или более и которые демонстрировали наиболее сильный эффект в участках, т.е. такие, которое демонстрировали повышение в 1,3
раз или более по сравнению с величиной до введения изменения в He/Con_3aF, выбирали в качестве показателя связывания с FcγRIIIa F в гетеродимеризованном антителе, в котором только в одну из H-цепей вводили изменение. Результаты приведены в таблице 48 (участки изменений, заменяемые изменениями H268D, S298A, K326D, A330M или K334E H240-Kn061/H240-Hl071/L73-k0, заменяемые аминокислоты и получаемая активность связывания с FcγRIIIa F).

В таблице 48, представленной выше, "изменяемое положение" относится к номеру остатка в соответствии с нумерацией EU. "Заменяемая аминокислота" относится к трем заменам, которые приводят к 1,3-кратному или более повышению по сравнению с He/Con_3aF до введения изменения в качестве показателя связывания с FcγRIIIa F в гетеродимеризованном антителе, в
котором изменение вводили только в одну H-цепь, как описано в примере 4, и которое демонстрирует наиболее сильный эффект в участках. "He/Con_3aF" представляет собой значение, определяемое в примере 4.
На основании результатов, представленных в таблице 48, заменяемые аминокислоты на каждом участке изменения приведены в таблице 49 (заменяемы участки изменения H268D, S298A, K326D, A330M или K334E a H240-Kn061/H240-Hl071/L73-k0 и заменяемые аминокислоты).

На основании таблицы 49, даже если D H268D заменяют Е или А в H240-Kn061/H240-Hl071/L73-k0, полагают, что получают сравнимую активность. Аналогично, даже когда D K326D представляет собой Т или I, или даже когда М A330M представляет собой Р или F, или даже когда D K334D представляет собой Е или I, полагают, что получают сравнимую активность. При этом в отношении S298A никаких изменений, с которыми, как полагают, можно получать сравнимую активность, ну удалось найти.
Кроме того, т.к. гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно повышает активность связывания с FcγR по сравнению с гомодимеризованными антителами,
содержащими каждую его H-цепь, полагают, что варианты, получаемые введением изменение в H240-Kn061/H240-Hl071/L73-k0, обладают аналогичным свойством гетеродимеризованного антитела.
[Пример 19] Введение D270E в гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 и его оценка
Затем для дополнительного улучшения H240-Kn061/H240-Hl071 авторы настоящего изобретения пытались дополнительно повысить связывание с FcγRIIIa, которое, как полагали, повышает активность ADCC и дополнительно снижает связывание с FcγRIIb, который, как полагают, снижает противоопухолевую активность антитела через иммуносупрессивные сигналы. В частности, D270E, изменение, выявленное в примере 4, которое повышает связывание с FcγRIIIa и снижает связывание с FcγRIIb, вводили в обе Н-цепе H240-Kn061/H240-Hl071/L73-k0. Последовательности, получаемые введением D270E в H240-Kn061 и H240-Hl071, называли H240-Kn072 (SEQ ID NO: 90) и H240-Hl076 (SEQ ID NO: 91) и комбинировали с L73-k0 для экспрессии и получения гетеродимеризованного антитела H240-Kn072/H240-Hl076/L73-k0 способом, описанным в примере 1. Наряду с этим антитело H240-Kn033/H240-Hl033/L73-k0, H240-Kn032/H240-Hl032/L73-k0, H240-дефукозил_G1d/L73-k0 и H240-Kn061/H240-Hl071/L73-k0 анализировали в отношении активности связывания с каждым FcγR способом, описанным в справочном примере 8. Результаты приведены в таблице 50.
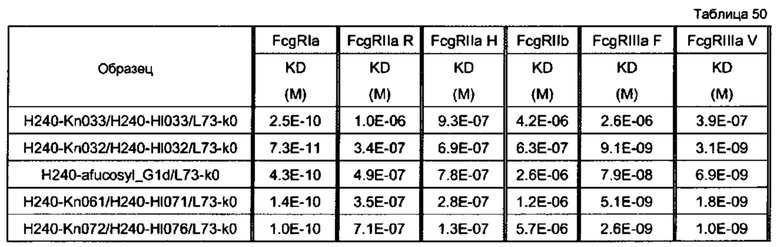
В таблице 50 продемонстрировано, что аналогично H240-Kn061/H240-Hl071/L73-k0 H240-Kn072/H240-Hl076/L73-k0 более сильно связывается с FcγRIIIa F и FcγRIIIa V по сравнению с H240-Kn032/H240-Hl032/L73-k0 и H240-дефукозил_C1d/L73-k0, получаемыми в результате применения существующей техники повышения активности ADCC, и кроме того он более сильно связывается по сравнению с H240-Kn061/H240-Hl071/L73-k0. Связывание с FcγRIIb H240-Kn072/H240-Hl076/L73-k0 снижалось относительно связывания H240-Kn032/H240-Hl032/L73-k0 и H240-дефукозил_C1d/L73-k0, получаемых существующей техникой повышения активности ADCC, и кроме того оно снижалось относительно связывания H240-Kn061/H240-Hl071/L73-k0.
В частности, вследствие того, что введение D270E повышает связывание с FcγRIIIa, которое повышает активность ADCC, предполагают более сильную активность ADCC, и вследствие того, что связывание с FcγRIIb, которое преобразует иммуносупрессивные сигналы, ослабевает, предполагают снижение иммуносупрессивного действия антитела. Таким образом, полагают, что H240-Kn072/H240-Hl076/L73-k0 обладает противоопухолевым эффектом, который превосходит аналогичный эффект H240-Kn061/H240-Hl071/L73-k0.
Затем активность ADCC H240-Kn072/H240-Hl076/L73-k0 сравнивали с активностью H240-Kn061/H240-Hl071/L73-k0, H240-Kn033/H240-Hl033/L73-k0 и H240-дефукозил_C1d/L73-k0. Результаты представлены на фиг. 41.
Результаты, представленные на фиг. 41, демонстрируют, что H240-Kn072/H240-Hl076/L73-k0 обладает активностью ADCC, которая является значительно больше активности H240-Kn033/H240-Hl033/L73-k0. Кроме того, активность ADCC H240-Kn072/H240-Hl076/L73-k0 являлась больше активности афукозилованного антитела H240-дефукозил_C1d/L73-k0, получаемого в результате применения существующей техники повышения активности ADCC, и являлась сравнимой с активностью H240-Kn061/H240-Hl071/L73-k0.
Эти результаты демонстрируют, что гетеродимеризованное антитело H240-Kn072/H240-Hl076/L73-k0 не только обладает более сильной активностью ADCC, чем активность, получаемая существующей техникой повышения активности ADCC, а также демонстрирует сниженное связывание с FcγRIIb, и, таким образом, представляет собой в большей степени превосходное антитело, чем антитела, получаемые существующей техникой.
Кроме того, вследствие того, что гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно связывается более сильно с FcγR, чем гомодимеризованные антитела, содержащие каждую его H-цепь, полагают, что H240-Kn072/H240-Hl076/L73-k0, получаемый введением D270E в обе H-цепи H240-Kn061/H240-Hl071/L73-k0, обладает аналогичным свойством гетеродимеризованного антитела.
[Пример 20] Дополнительное улучшение гетеродимеризованного антитела H240-Kn072/H240-Hl076/L73-k0
Авторы настоящего изобретения принимали попытки дополнительно улучшать H240-Kn072/H240-Hl076/L73-k0, которое открыли в примере 19. В частности, H240-Kn072/H240-Hl076/L73-k0 комбинировали с изменениями Y234E, Y235N, Y235Q и S239M, которые при введении в H240-Kn061, дополнительно обеспечивают превосходные свойства H240-Kn061/H240-Hl071/L73-k0, открытому в примере 18.
H240-Kn113 (SEQ ID NO: 92), получаемую в результате введения Y234E и Y235N в H240-Kn072, H240-Kn115 (SEQ ID NO: 93), получаемую в результате введения S239M в H240-Kn072, и H240-Kn125 (SEQ ID NO: 94), получаемую в результате введения в Y235Q и S239M в H240-Kn072, получали способом, описанным в справочном примере 1. H240-Kn113/H240-Hl076/L73-k0, H240-Kn115/H240-Hl076/L73-k0 и H240-Kn125/H240-Hl076/L73-k0 получали, комбинируя H240-Hl076 в качестве H-цепи, L73-k0 в качестве L-цепи и H240-Kn113, H240-Kn115, и H240-Kn125 в качестве другой H-цепи способом, описанным в справочном примере 1. Описанные выше антитела оценивали в отношении связывания с каждым FcγR способом, описанным в справочном примере 8, наряду с нативным IgG1 Н240-G1d/L73-k0, H240-Kn033/H240-Hl033/L73-k0, модифицированным на основании использования "выступов во впадины", дефукозилорованного антитела H240-дефукозил_G1d/L73-k0, получаемого существующей техникой повышения активности ADCC, и гомодимеризованного антитела H240-Kn032/H240-Hl032/L73-k0, в обе H-цепи которого вводили повышающее активность ADCC изменение
S239D/A330L/I332E. Результаты приведены в таблице 51.
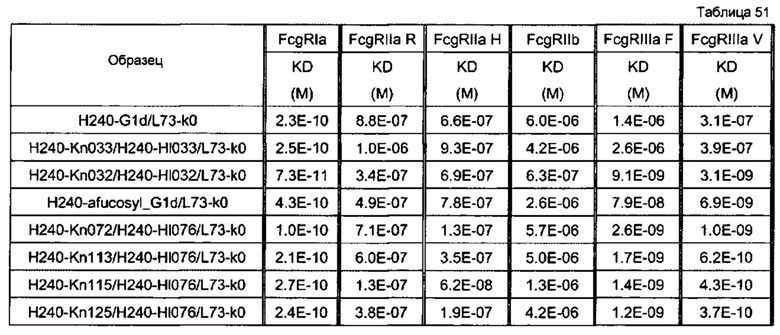
По сравнению с H240-Kn072/H240-Hl076/L73-k0 H240-Kn113/H240-Hl076/L73-k0 демонстрировало сравнимое связывание с FcγRIIb, ингибирующим FcγR, и повышенное связывание с FcγRIIIa F и FcγRIIIa V, которые играют важную роль в активности ADCC. В отношении связывания с FcγRIIb, ингибирующим FcγR, степень связывания являлась сравнимой даже при сравнении с нативным антителом IgG1. В отношении FcγRIIIa связывание с FcγRIIIa F и FcγRIIIa V являлось повышенным по сравнению с дефукозилированным антителом H240-дефукозил_C1d/L73-k0, получаемым существующей технологией повышения активности ADCC, и гомодимеризованным антителом H240-Kn032/H240-Hl032/L73-k0, в обе H-цепи которого вводили повышающие активность ADCC изменения S239D/A330L/I332E. Описанные выше открытия подтверждают возможность того, что Н240-Kn113/H240-Hl076/L73-k0 не повышает иммуносупрессивные действия по сравнению с нативным IgG1 и демонстрирует более сильный противоопухолевый эффект по сравнению с эффектом дефукозилированного антитела, получаемого в результате
применения существующего повышающего активность ADCC изменения, и эффектом гомодимеризованного антитела.
По сравнению с H240-Kn113/H240-Hl076/L73-k0 H240-Kn115/H240-Hl076/L73-k0 демонстрировало повышенное связывание с FcγRIIIa F и FcγRIIIa V, которые играют важную роль в активности ADCC. Кроме того, связывание с FcγRIIa R и FcγRIIa H, которые являются важными для активности ADCP, являлось более повышенным по сравнению со связыванием нативного IgG1 H240-G1d/L73-k0, H240-Kn033/H240-Hl033/L73-k0, получаемого в результате применения к нему "выступов во впадины", дефукозилированного антитела Н240-дефукозил_G1d/L73-k0, получаемого существующей техникой повышения активности ADCC, и гомодимеризованного антитела H240-Kn032/H240-Hl032/L73-k0, в обе H-цепи которого вводили повышающие активность ADCC изменения S239D/A330L/I332E.
Как и в случае H240-Kn113/H240-Hl076/L73-k0 H240-Kn125/H240-Hl076/L73-k0 демонстрировал более повышенное связывание с FcγRIIIa F и FcγRIIIa V, которые играют важную роль в активности ADCC, чем H240-Kn115/H240-Hl076/L73-k0 при сохранении связывания с FcγRIIb, ингибирующим FcγR, на сравнимом с IgG1 уровне. H240-Kn125/H240-Hl076/L73-k0 демонстрировал повышенное связывание с FcγRIIa H, аллотипом FcγRIIa, а также сниженное связывание с FcγRIIb и повышенное связывание с обоими аллотипами FcγRIIIa по сравнению с дефукозилированным антителом H240-дефукозил_G1d/L73-k0, получаемым существующей техникой повышения активности ADCC, и гомодимеризованным антителом H240-Kn032/H240-Hl032/L73-k0, в обе H-цепи которого вводили повышающее активность ADCC изменение
S239D/A330L/I332E. Таким образом, можно предполагать, что H240-Kn125/H240-Hl076/L73-k0 повышает активности ADCP и ADCC, а также больше снижает иммуносупрессивное действие, чем гомодимеризованное антитело и дефукозилированное антитело, получаемые в результате применения существующего повышающего активность ADCC изменения.
Затем активности ADCC H240-Kn113/H240-Hl076/L73-k0, H240-Kn115/H240-Hl076/L73-k0 и H240-Kn125/H240-Hl076/L73-k0 сравнивали с активностями H240-Kn033/H240-Hl033/L73-k0 и дефукозилированного антитела H240-дефукозил_G1d/L73-k0, получаемого существующей техникой повышения активности ADCC. Результаты представлены на фиг. 42.
Как продемонстрировано на фиг. 42, все гетеродимеризованные антитела обладали превосходной активностью ADCC по сравнению с дефукозилированным антителом, получаемым существующей техникой повышения активности ADCC.
Профили связывания с каждым FcγR и результат сравнения активности ADCC с существующими технологиями демонстрируют, что Y234E, Y235N, Y235Q и S239M, которые вводили в H240-Kn072 H240-Kn072/H240-Hl076/L73-k0, представляют собой изменения, которые придают H240-Kn072/H240-Hl076/L73-k0 дополнительные превосходные характеристики.
Вследствие того, что гетеродимеризованное антитело H240-Kn061/H240-Hl071/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно связывается сильнее с FcγR, чем гомодимеризованные антитела, содержащие каждую его H-цепь, предполагают, что H240-Kn072/H240-Hl076/L73-k0, получаемое
введением D270Eв обе H-цепи H240-Kn061/H240-Hl071/L73-k0, обладает аналогичным свойством гетеродимеризованного антитела. Аналогично, полагают, что H240-Kn113/H240-Hl076/L73-k0, H240-Kn115/H240-Hl076/L73-k0 и H240-Kn125/H240-Hl076/L73-k0, получаемые введением дополнительных изменений в H240-Kn072/H240-Hl076/L73-k0, также обладают аналогичным признаком гетеродимеризованного антитела.
[Пример 21] Получение гетеродимеризованных антител, которые обладают повышенным связыванием с FcγRIIa и FcγRIIIa
На основании результата, представленного в примере 4, получали варианты с Н-цепями, которые демонстрируют повышенное связывание с FcγRIIa и FcγRIIIa. В частности, H240-Kn067 (SEQ ID NO: 95) получали введением L234Y, L235Y, G236W, H268D, S298A, A327D в H240-Kn033 (SEQ ID NO: 84) и H240-Hl068 (SEQ ID NO: 96) получали введением D270E, K326D, A330K и K334E в H240-Hl033 (SEQ ID NO: 85) способом, описанным в справочном примере 1. Гетеродимеризованное антитело H240-Kn067/H240-Hl068/L73-k0 экспрессировали, комбинируя H240-Kn067, H240-Hl068 и L73-k0, способом, описанным в справочном примере 1.
Сначала оценивали, обладает ли гетеродимеризованное антитело H240-Kn067/H240-Hl068/L73-k0 свойством гетеродимеризованного антитела, которое заключается в том, что оно повышает активность связывания с FcγR по сравнению с гомодимеризованными антителами, содержащими каждую его H-цепь. В гетеродимеризованное антитело H240-Kn067/H240-Hl068/L73-k0 вводили L234Y/L235Y/G236W/H268D/S298A/A327D в H240-Kn067 в качестве Н-цепи
и вводили D270E/K326D/A330K/K334E в H240-Hl068 в качестве другой H-цепь. Затем H240-Hl136 (SEQ ID NO: 97), получаемую в результате введения L234Y/L235Y/G236W/H268D/S298A/A327D в H240-Hl033, и H240-Kn133 (SEQ ID NO: 98), получаемую в результате введения D270E/K326D/A330K/K334E в H240-Kn033, конструировали способом, описанным в справочном примере 1. С использованием этих экспрессирующих векторов способом, описанным в справочном примере 1, экспрессировали гомодимеризованное антитело Н240-Kn067/H240-Hl136/L73-k0, обе H-цепи которого содержат L234Y/L235Y/G236W/H268D/S298A/A327D, и гомодимеризованное антитело H240-Kn113/H240-Hl068/L73-k0, обе H-цепи которого содержат D270E/K326D/A330K/K334E. Гомодимеризованные антитела и гетеродимеризованное антитело H240-Kn067/H240-Hl068/L73-k0, одна из H-цепей которого содержит L234Y/L235Y/G236W/H268D/S298A/A327D, а другая H-цепь содержит D270E/K326D/A330K/K334E, анализировали в отношении связывания с FcγRIIa H, FcγRIIa R, FcγRIIIaF и FcγRIIIa V способом, описанным в справочном примере 8. Результаты представлены в таблице 52.

Кроме того, на основании этого варианта H240-Kn067/H240-Hl068/L73-k0 комбинировали с изменениями Y235Q и S239M, которые при введении в H240-Kn061 дополнительно придают H240-Kn061/H240-Hl071/L73-k0
превосходные характеристики, как обсуждается в примере 18. В частности, H240-Kn120 (SEQ ID NO: 99), получаемую в результате введения S239M в H240-Kn067, и H240-Kn126 (SEQ ID NO: 100), получаемую в результате введения Y235Q в H240-Kn120, получали способом, описанным в справочном примере 1. H240-Kn067/H240-Hl068/L73-k0, H240-Kn120/H240-Hl068/L73-k0 и H240-Kn126/H240-Hl068/L73-k0 получали комбинацией H240-Hl068 в качестве H-цепи и L73-k0 в качестве L-цепи с H240-Kn067, H240-Kn120, и H240-Kn126 в качестве другой H-цепи способом, описанным в справочном примере 1. Описанные выше антитела анализировали в отношении связывания с каждым FcγR способом, описанным в справочном примере 8. Результаты представлены в таблице 53.

Этот результат демонстрирует, что все H240-Kn067/H240-Hl068/L73-k0, H240-Kn120/H240-Hl068/L73-k0 и H240-Kn126/H240-Hl068/L73-k0 демонстрируют сравнимое или повышенное связывание с FcγRIIIa по сравнению с существующими антителами с повышенной активностью: H240-дефукозил_C1d/L73-k0 и H240-Kn032/H240-Hl032/L73-k0. Кроме того, связывание с FcγRIIa R и FcγRIIa H, которые играют важную роль в активности ADCP, являлось
повышенным по сравнению с каждой из существующих технологий. Описанное выше открытие позволяет предположить, что все из гетеродимеризованных антител H240-Kn067/H240-Hl068/L73-k0, H240-Kn120/H240-Hl068/L73-k0 и H240-Kn126/H240-Hl068/L73-k0, получаемые в настоящем описании, обладают сравнимой или большей активностью ADCC и превосходной активностью ADCP относительно активностей, получаемых существующими технологиями.
В частности, H240-Kn120/H240-Hl068/L73-k0 демонстрировал в большей степени повышенное связывание с FcγRIIa R и FcγRIIa H даже по сравнению с гомодимеризованным антителом Н240-Kn037/H240-Hl036/L73-k0, обе H-цепи которого содержат изменения G236A/S239D/I332E, для которых опубликовано, что они повышают связывание с FcγRIIa R и FcγRIIa H, а также повышают активность ADCP. В частности, H240-Kn120/H240-Hl068/L73-k0 демонстрирует в большей степени повышенное связывание с FcγRIIIa F и FcγRIIIa V, чем антитела, получаемые существующей техникой повышения активности ADCC, и демонстрирует в большей степени повышенное связывание с FcγRIIa R и FcγRIIa H, чем антитела, получаемые существующей техникой повышения активности ADCP. Таким образом, H240-Kn120/H240-Hl068/L73-k0 представляет собой антитело, которое может обладать более сильными активностями ADCC и ADCP, чем антитела, получаемые существующими технологиями.
Затем в соответствии со справочным примером 9 сравнивали активность ADCC каждого варианта с активностью дефукозилированного антитела H240-дефукозил_G1d/L73-k0, получаемого существующей техникой повышения активности ADCC.
Результат представлен на фиг. 43.
Результат, представленный на фиг. 43, демонстрирует, что все из гетеродимеризованных антител, получаемые на этот раз, обладают активностью ADCC, сравнимой или превосходящей активность дефукозилированного антитела, получаемого существующей техникой повышения активности ADCC.
Профили связывания с каждым FcγR и результат сравнения активности ADCC с существующими технологиями демонстрируют, что все H240-Kn067/H240-Hl068/L73-k0, H240-Kn120/H240-Hl068/L73-k0 и H240-Kn126/H240-Hl068/L73-k0, получаемые на этот раз, представляют собой гетеродимеризованные антитела, которые с высокой вероятностью обладают активностью ADCC, сравнимой или превосходящей активность, получаемую существующей техникой повышения активности ADCC, а также обладают повышенной активностью ADCP посредством связывания с FcγRIIa.
[Пример 22] Оценка гетеродимеризованного антитела H240-AK072/H240-BH076/L73-k0 в отношении активности связывания с каждым FcγR и активности ADCC
Как указано выше, H240-Kn072/H240-Hl076/L73-k0, которое представляет собой гетеродимеризованное антитело, получаемое в результате применения "выступов во впадины", оценивали в отношении его связывания с каждым FcγR и активности ADCC. В настоящем описании оценивали, наблюдают ли также свойство гетеродимеризованного антитела, которое заключается в том, что гетеродимеризованное антитело, содержащее две различные H-цепи, обладает повышенной активностью активность связывания с FcγR по
сравнению с гомодимеризованными антителами, содержащими каждую его H-цепь, в случае использования D356K, H435R и K439E в качестве альтернативной техники получения гетеродимеризованного антитела.
Сначала конструировали H240-AK072 (SEQ ID NO: 104) введением L234Y/L235Y/G236W/H268D/D270E/S298A, изменения, вводимого в H240-Kn072, которая представляет собой H-цепь H240-Kn072/H240-Hl076/L73-k0, в H240-A5E (SEQ ID NO: 102), получаемую в результате введения D356K и H435R в H240-G1dE (SEQ ID NO: 101), которая представляет собой аллотип H240-G1d (SEQ ID NO: 83). Затем конструировали H240-BH076 (SEQ ID NO: 105) введением D270E/K326D/A330M/K334E, которое вводили в H240-Hl076 в качестве другой H-цепи, в H240-B3E (SEQ ID NO: 103), получаемую в результате введения K439E в H240-G1DE. Гетеродимеризованное антитело H240-AK072/H240-BH076/L73-k0 экспрессировали и получали комбинацией H240-AK072, H240-BH076 и L73-k0 способом, описанным в примере 1. Аналогично, H240-AK072 и L73-k0, и H240-BH076 и L73-k0 каждые комбинировали для экспрессии и получения гомодимеризованных антител H240-AK072/L73-k0 и H240-BH076/L73-k0. Антитела сравнивали в отношении связывания с каждым FcγR способом, описанным в справочном примере 8. Результаты представлены в таблице 54.

Описанные выше результаты подтверждают, что гетеродимеризованное антитело H240-AK072/H240-BH076/L73-k0 обладает свойством гетеродимеризованного антитела, которое заключается в том, что оно связывается с FcγR более сильно, чем гомодимерные антитела H240-AK072/L73-k0 и H240-BH076/L73-k0, каждое из которых содержит любую его H-цепь.
Затем H240-A5E, H240-B3E и L73-k0 комбинировали и экспрессировали для получения H240-A5E/H240-B3E/L73-k0 способом, описанным в справочном примере 1. H240-G1d/L73-k0, H240-A5E/H240-B3E/L73-k0, H240-дефукозил_G1d/L73-k0 и гетеродимеризованное антитело H240-AK072/H240-BH076/L73-k0 оценивали в отношении связывания с каждым FcγR способом, описанным в справочном примере 8. Результаты приведены в таблице 55.

Результаты демонстрируют, что связывание H240-AK072/H240-BH076/L73-k0 с FcγRIIIa F и FcγRIIIa V повышалось по сравнению с H240-G1d/L73-k0 и дефукузилированным антителом H240-дефукозил_G1d/L73-k0, получаемым существующей техникой повышения активности ADCC. При этом вследствие того, что активность связывания с FcγR являлась сравнимой для H240-G1d/L73-k0 и H240-A5E/H240-B3E/L73-k0, полагали, что повышенная активность
связывания H240-AK072/H240-BH076/L73-k0 обусловлена изменениями L234Y/L235Y/G236W/H268D/D270E/S298A и D270E/K326D/A330M/K334E, вводимыми в каждую из H-цепей.
[Пример 23] Рентгеновский кристаллографический анализ комплекса между Fc (Kn120/Hl068) и внеклеточной областью FcγRIIb
H240-Kn120/H240-Hl068/L73-k0, получаемое как описано в примере 21, не только обладало повышенной активностью связывания с FcγRIIIa и FcγRIIa типа Н и с аллотипом FcγRIIa R, а также обладало повышенной активностью связывания с FcγRIIb, ингибирующим рецептором. Полагают, что повышенное связывание с FcγRIIb вызывает иммуносупрессивные эффекты. Таким образом, снижением связывания с FcγRIIb можно получать дополнительное повышение активности ADCC, что является целью настоящего изобретения.
Однако 93% аминокислотной последовательности внеклеточной области FcγRIIa и FcγRIIb совпадает и является высоко гомологичными друг другу. Кроме того, согласно анализу на основе кристаллографической структуры (J. Imunol., 2011, 187, 3208-3217) комплекса Fc нативного IgG1 (далее в настоящем описании сокращенно обозначаемого как Fc (WT)) и внеклеточной области FcγRIIa типа R, около взаимодействующих поверхностей обоих белков только три аминокислоты (Gln127, Leu132 и Phe160) в FcγRIIa типа R отличались по сравнению с FcγRIIb (фиг. 44). По этой причине было теоретически рассчитано, что снижение только активности связывания с FcγRIIb при сохранении активности связывания с FcγRIIa типа R является крайне затруднительным. Затем авторы
настоящего изобретения предполагали, что если они получат информацию о приблизительной кристаллографической структуре комплекса между участком Fc (Fc (Kn120/Hl068)) H240-Kn120/H240-Hl068/L73-k0 и внеклеточной областью FcγRIIb и более подробно исследуют мутации аминокислот, которые необходимо вводить то, можно повысить возможность выявления изменения, которое селективно снизит активность связывания с FcγRIIb. Таким образом, комплекс между Fc (Kn120/Hl068) и внеклеточной областью FcγRIIb анализировали рентгеновским кристаллографическим анализом.
В результате авторам настоящего изобретения удалось определить трехмерную структуру комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb при разрешении 2,78  рентгеновским кристаллографическим анализом. Структура, получаемая в результате анализа, представлена на фиг. 45. Выявлено, что внеклеточная область FcγRIIb является связанной и располагается между двумя доменами CH2 Fc, что является сходным с трехмерными структурами ранее анализируемых комплексов между Fc (WT) и соответствующих внеклеточных областей FcγRIIIa (Proc. Natl. Acad. Sci. USA, 2011, 108, 12669-126674), FcγRIIIb (Nature, 2000, 400, 267-273; J. Biol. Chem., 2011, 276, 16469-16477) и FcγRIIa.
рентгеновским кристаллографическим анализом. Структура, получаемая в результате анализа, представлена на фиг. 45. Выявлено, что внеклеточная область FcγRIIb является связанной и располагается между двумя доменами CH2 Fc, что является сходным с трехмерными структурами ранее анализируемых комплексов между Fc (WT) и соответствующих внеклеточных областей FcγRIIIa (Proc. Natl. Acad. Sci. USA, 2011, 108, 12669-126674), FcγRIIIb (Nature, 2000, 400, 267-273; J. Biol. Chem., 2011, 276, 16469-16477) и FcγRIIa.
Затем описывали структуры вблизи трех остатков аминокислот, которые располагаются около поверхности связывания с Fc и являются различными в FcγRIIa типа R и FcγRIIb. На фиг. 46 представлена структура вблизи Lys127 (Gln в FcγRIIa типа R). Ближайший остаток в FcγRIIb представляет собой Ala в положении
298 (нумерация EU), располагаемый в домене CH2 В Fc, представленном на фиг. 46. Этот остаток находится в непосредственном контакте с FcγRIIb на поверхности связывания. Таким образом, полагали, что введение крупного остатка, способного взаимодействовать с Lys127, являлось затруднительным. Другие соседние аминокислотные остатки слишком удалены от Lys127, и невозможно выявить мутаций, способных к непосредственному взаимодействию. На фиг. 47 представлена структура вблизи Ser132 (Leu в FcγRIIa типа R). Это остаток также является слишком удаленным от Fc, и невозможно выявить мутаций, способных к непосредственному взаимодействию с этим Ser. Наконец, на фиг. 48 представлена структура вблизи Tyr160 (Phe в FcγRIIa типа R). Этот Tyr образует водородную связь с карбонильным кислородом основной цепи Gly в положении 236 (нумерация EU) в домене CH2 A Fc. В этом случае введением мутации при Gly236 в положении 236 (нумерация EU) можно снижать только активность связывания с FcγRIIb, если мутация вызывает изменение структуры петли и, таким образом, устраняет водородную связь. Кроме того, когда вводят мутацию с любой большой боковой цепью в положении Gly в положении 236 (нумерация EU), теоретически рассчитано, что боковая цепь непосредственно взаимодействует с боковой цепью в положении 160 в FcγRIIa или FcγRIIb, и активность связывания с FcγRIIb можно селективно снижать с использованием различия между Phe160 в FcγR2a типа R и Tyr160 в FcγRIIb.
Экспериментальный способ
[Экспрессия и очистка Fc (Kn0120/Hl068)]
Fc (Kn0120/Hl068) получали, как указано ниже. Сначала Cys в положении 220 (нумерация EU) H240-Kn120 (SEQ ID NO: 99) и H240-Hl068 (SEQ ID NO: 96) заменяли на Ser. Затем генетическую последовательность Fc (Kn0120) и Fc (Hl068) от Glu в положении 236 (нумерация EU) до его C-конца клонировали ПЦР. С использованием этой клонированной генетической последовательности способом справочного примера 1 проводили получение экспрессирующих векторов и экспрессию и очистку Fc (Kn0120) и Fc (Hl068). Cys в положении 220 (нумерация EU) образует дисульфидную связь с Cys L-цепи в общем IgG1. Когда получают только Fc, L-цепь совместно не экспрессируют, и, таким образом, этот остаток заменяли Ser, предотвращая образование ненужной дисульфидной связи.
[Экспрессия и очистка внеклеточной области FcγRIIb]
Ее проводили способом справочного примера 8.
[Очистка комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb]
К 1,5 мг образца внеклеточной области FcγRIIb, получаемого для кристаллизации, добавляли 0,15 мг Endo F1 (Protein Science, 1996, 5:2617-2622), экспрессируемого и очищаемого из Escherichia coli в виде слитого белка глутатион-S-трансферазы. Его можно было хранить при комнатной температуре в течение трех суток в 0,1 М Bis-Tris буфере при pH 6,5, и расщепляли N-связанный олигосахарид, оставляя N-ацетилглюкозамин, непосредственно
связанный с Asn. Затем этот образец внеклеточной области FcγRIIb, который подвергали обработке с отщеплением углеводов, концентрировали ультрафильтрацией с 10000 MWCO, и очищали гельфильтрационной хроматографией (Superdex 200 10/300) с использованием колонки, уравновешенной в 20 мМ HEPES при pH 7,5, содержащей 0,1 М NaCl. Кроме того, к получаемой фракции внеклеточной области FcγRIIb с отщепленными углеводами добавляли Fc(Kn0120/Hl068) таким образом, чтобы молярное отношение внеклеточной области FcγRIIb находилось в небольшом избытке, и после концентрации ультрафильтрацией с 10000 MWCO образец комплекса Fc (Kn0120/Hl068)/внеклеточной области FcγRIIb получали посредством очистки гельфильтрационной хроматографией (Superdex 200 10/300) с использованием колонки, уравновешенной в 25 мМ HEPES при pH 7,5, содержащей 0,1 М NaCl.
[Кристаллизация комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb]
Образец комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb концентрировали до приблизительно 10 мг/мл ультрафильтрацией с 10000 MWCO, и кристаллизацию проводили способом диффузии паров из висячей капли в комбинации с затравочным способом. Для кристаллизации использовали планшет VDXm (Hampton Research). С использованием резервуарного раствора, содержащего 0,1 М Bis-Tris (pH 6,0), 14,4% PEG3350 и 0,2 М сульфата аммония, получали кристаллизационные капли при коэффициенте смешения резервуарного раствора : кристаллизуемого образца = 0,85 мл : 0,85 мл. Затем проводили затравку нанесением
полосок с использованием Seeding Tool (Hampton Research) для переноса затравочных кристаллов из кристаллов комплекса, получаемого при аналогичном условии. Капли выдерживали при 20°C. Это эффективно приводило к образованию столбчатых кристаллов.
[Измерение рентгенодифракционных данных кристалла комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb]
Один из получаемых одиночных кристаллов комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb выдерживали в растворе 0,1 М Bis-Tris pH 6,0, 17,5% PEG3350, 0,2 М сульфата аммония, 16% глицерина (об./об.). Кристалл извлекали из раствора с использованием иглы с прикрепленной небольшой нейлоновой петлей и замораживали в жидком азоте, а затем измеряли рентгенодифракционные данные в лаборатории синхротронного излучения Photon Factory BL-17A в High Energy Accelerator Research Organization. Во время измерения кристалл постоянно располагали в струе азота при -178°C для поддержания замороженного состояния и получали всего 360 рентгенографических дифракционных изображений с использованием детектора Quantum 315r CCD (ADSC), прикрепленного к каналу пучка с вращением кристалла 0,5° за один раз. Определение параметров ячейки, разметки дифракционных пятен и обработку дифракционных данных из получаемых дифракционных изображений проводили с использованием программы Xia2 (J. Appl. Cryst., 2010, 43:186-190), пакета программ XDS (Acta. Cryst., 2010, D66: 125-132) и Scala (Acta. Cryst., 2006, D62:72-82), и в итоге получали данные дифракционной интенсивности с разрешением до 2,78 . Кристалл
принадлежит к пространственной группе P41212 и имеет следующие параметры ячейки а=152,94 , b=152,94 , с=82,24 , α=90°, β=90°, γ=90°.
[Рентгеновский кристаллографический анализ комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb]
Кристаллическую структуру комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb определяли способом молекулярного замещения с использованием программы Phaser (J. Appl. Cryst., 2007, 40:658-674). На основании размера получаемой кристаллической решетки и молекулярной массы комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb теоретически рассчитывали, что число комплексов в асимметричной единице равно одному. На основании структурных координат кода PDB: 3SGJ, который представляет собой кристаллическую структуру комплекса Fc(WT)/внеклеточной области FcγRIIIa, участки аминокислотных остатков положений А-цепи 239-340 и положений В-цепи 239-340 получали в виде отдельных координат и использовали их соответственно в качестве моделей для исследования доменов CH2 Fc. Участки аминокислотных остатков положений А-цепи 341-444 и положений В-цепи 341-443 получали в виде одного набора координат из аналогичных структурных координат кода PDB: 3SGJ, и использовали его в качестве модели для исследования доменов CH3 Fc. Наконец, на основании структурных координат кода PDB: 2FCB, который представляет собой кристаллическую структуру внеклеточной области FcγRIIb, получали участки аминокислотных остатков положений А-цепи 6-178 и использовали в качестве модели
для исследования FcγRIIb. Ориентацию и положение каждой модели исследования в кристаллической решетке определяли для домена CH3 Fc, внеклеточной области FcγRIIb и домена CH2 Fc на основании функции вращения и функции трансляции с получением исходной модели для кристаллической структуры комплекса Fc (Kn120/Hl068)/внеклеточной области FcγRIIb. Когда на получаемой исходной модели проводили моделирование жесткого тела, в котором двигают два домена CH2 Fc, два домена CH3 Fc и внеклеточную область FcγRIIb, кристаллографический фактор достоверности, величина R становилась 41,4%, и величина Free R становилась 42,6% для данных дифракционной интенсивности от 25 до 3,0 в этой точке. Кроме того, структурное уточнение с использованием программы REFMAC5 (Acta Cryst., 2011, D67, 355-367) и редактирование модели для наблюдения карт электронной плотности, коэффициент которых имеет 2Fo-Fc или Fo-Fc, которые рассчитывают на основании экспериментально установленного структурного фактора Fo, рассчитываемого структурного фактора Fc и рассчитываемой фазы с использованием модели, проводили программой Coot (Acta Cryst., 2010, D66:486-501), и уточнение модели проводили повторением этих стадий. В итоге, в результате включения молекул воды в модель на основе карт электронной плотности, в которых используют 2Fo-Fc или Fo-Fc в качестве коэффициента, и последующего уточнения, кристаллографический фактор достоверности, значения R и значения Free R модели, содержащей 4964 неводородных атома, становились 22,8% и 27,7% для 24274 данных дифракционной интенсивности с разрешением от 25
до 2,78 , соответственно.
[Пример 24] Получение антител, сохраняющих активность связывания с FcγRIIaR или обладающих повышенной активностью связывания с FcγRIIaR и обладающих сниженной активностью связывания с FcγRIIb
Вариант H240-Kn120/H240-Hl068/L73-k0, открытый в примере 21, обладал повышенной активностью связывания с FcγRIIaR и FcγRIIaH, которые являются важными для активности ADCP, и кроме того, его активность связывания с ингибирующим FcγRIIb являлась сниженной приблизительно в 50 раз по сравнению с активностью нативного IgG1. Для получения высокой активности ADCP предпочтительно как можно больше снижать активность связывания с ингибирующим FcγRIIb. Таким образом, авторы настоящего изобретения проводили поиск изменений, которые обеспечивают снижение активности связывания с FcγRIIb при сохранении активности связывания с активирующими FcγRIIaR и FcγRIIaH. Как описано в примере 23, результат кристаллографического анализа комплекса между Fc H240-Kn120/H240-Hl068/L73-k0 и внеклеточной областью FcγRIIb демонстрировал, что Tyr160 в FcγRIIb образовывал водородную связь с карбонильным кислородом основной цепи Gly236, содержащегося в домене CH2 A Fc (Kn120/Hl068). В FcγRIIaR и FcγRIIaH соответствующее положение занято Phe160, и маловероятно, что существует описанное выше взаимодействие. Это подтверждает возможность того, что если взаимодействие с Tyr160 в FcγRIIb можно устранять введением изменения при Gly236, активность
связывания с FcγRIIb можно снижать при сохранении активности связывания с FcγRIIa типа R, т.е. активность связывания с FcγRIIb можно селективно снижать. В частности, авторы настоящего изобретения предполагали, что взаимодействие с Y160 можно устранять, если вводить реверсию G236 и изменение структуры петли заменой Ser, Val, Ile или Thr на G236 в H-цепи, получаемой из H240-Hl068, которая взаимодействует с Y160 FcγRIIb в связывании между FcγRIIb и H240-Kn120/H240-Hl068/L73-k0. При этом, тем не менее, на растоянии Lys127 FcγRIIb может находиться в электростатическом взаимодействии с Е294 домена CH2 A Fc (Kn120/Hl068). Таким образом, авторы настоящего изобретения рассматривали возможность того, что взаимодействие с FcγRIIb можно снижать введением электростатического отталкивания заменой Е294 на положительно заряженный Lys или Arg.
H240-Kn179 (SEQ ID NO: 107) и H240-Kn180 (SEQ ID NO: 108), получаемые в результате введения каждого из E294R и E294K в H240-Kn120 (SEQ ID NO: 99) и H240-Hl073 (SEQ ID NO: 109), H240-Hl085 (SEQ ID NO: 110), H240-Hl086 (SEQ ID NO: 111) и H240-Hl089 (SEQ ID NO: 112), получаемые в результате введения каждого из G236S, G236V, G236I и G236T в H240-Hl068 (SEQ ID NO: 96), получали способом, описанным в справочном примере 1. H240-Kn120/H240-Hl073/L73-k0, H240-Kn120/H240-Hl085/L73-k0, H240-Kn120/H240-Hl086/L73-k0, H240-Kn120/H240-Hl089/L73-k0, H240-Kn179/H240-Hl068/L73-k0 и H240-Kn180/H240-Hl068/L73-k0 получали комбинацией каждой из H240-Kn120 и H240-Kn180 в качестве H-цепи, L73-k0 в качестве L-цепи и H240-Hl073, H240-Hl085, H240-Hl086 и
H240-Hl089 в качестве дургой H-цепи способом, описанным в справочном примере 1. Описанные выше варианты оценивали в отношении их связывания с FcγR способом, описанным в справочном примере 8. Результаты представлены в таблице 56. Закрашенную серым KD H240-Kn120/H240-Hl073/L73-k0 в таблице рассчитывали при условии, что kd составляет 5×10-5 сек-1 или менее, вследствие того, что значение kd для FcγRIa являлось меньше, чем предел измерения 5×10-5 сек-1 вне измеряемого диапазона от 5×10-5 сек-1 до 1 сек-1 для константы диссоциации (kd) в Biacore4000, используемом в анализе.
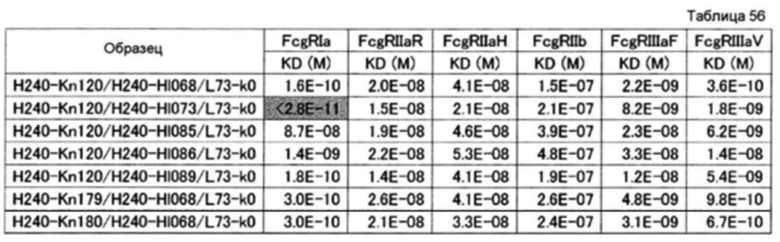
Кроме того, в таблице 57 представлены значения, получаемые делением KD H240-Kn120/H240-Hl068/L73-k0 для каждого из FcγRIIaR, FcγRIIaH и FcγRIIb на KD каждого варианта, относительная KD, определяемая при принятии KD H240-Kn120/H240-Hl068/L73-k0 для FcγRIIaR, FcγRIIaH или FcγRIIb за 1.

Результаты демонстрируют, что все из шести типов изменений, получаемых для этой оценки, сохраняли или повышали активности связывания с FcγRIIaR и FcγRIIaH и снижали активность связывания с FcγRIIb по сравнению с H240-Kn120/H240-Hl068/L73-k0.
Затем изменения, анализируемые в таблице 57, комбинировали для дополнительного снижения активности связывания с FcγRIIb. В этом эксперименте целью авторов настоящего изобретения являлось дополнительно подавление активности связывания с FcγRIIb посредством комбинированного применения введения E294K или E294R в H240-Kn120 и введения G236T в H240-Hl068. Как продемонстрировано в таблице 57, все изменения снижают активности связывания с FcγRIIIa F и FcγRIIIa V. Затем в дополнение к этим изменениям вводили I332E, для которого было опубликовано, что оно повышает активность связывания с FcγRIIIa, и изменение Y235N, которое повышает активность связывания с FcγRIIIa, как описано в примере 18, для дополнительного подавления активности связывания с FcγRIIb и повышения активности связывания с FcγRIIIa.
H240-Kn192 (SEQ ID NO: 113), получаемую в результате введения
Y235N и E294K в H240-Kn120 (SEQ ID NO: 99), H240-Kn193 (SEQ ID NO: 114), получаемую в результате введения Y235N и E294R в H240-Kn120 (SEQ ID NO: 99), и H240-Hl204 (SEQ ID NO: 115), получаемую в результате введения G236T и I332E в H240-Hl068 (SEQ ID NO: 96), получали способом, описанным в справочном примере 1. H240-Kn179/H240-Hl089/L73-k0, H240-Kn180/H240-Hl089/L73-k0, H240-Kn192/H240-Hl204/L73-k0 и H240-Kn193/H240-Hl204/L73-k0 получали комбинацией каждой из H240-Kn192 и H240-Kn193 в качестве H-цепи, L73-k0 в качестве L-цепи и H240-Hl089 и H240-Hl204 в качестве другой H-цепи способом, описанным в справочном примере 1. Описанные выше варианты оценивали в отношении их связывания с FcγR способом, описанным в справочном примере 8. Результат представлен в таблице 58. Закрашенную серым KD H240-Kn120/H240-Hl073/L73-k0 в таблице рассчитывали при условии, что kd составляет 5×10-5 сек-1 или менее, вследствие того, что значение kd для FcγRIa являлось меньше, чем предел измерения 5×10-5 сек-1 вне измеряемого диапазона от 5×10-5 сек-1 до 1 сек-1 для константы диссоциации (kd) в Biacore4000, используемом в анализе.

Кроме того, значения, получаемые делением KD Н240-Kn120/H240-Hl068/L73-k0 для каждого FcγRIIaR, FcγRIIaH и FcγRIIb на KD каждого варианта, относительная KD, определяемая при принятии KD H240-Kn120/H240-Hl068/L73-k0 для FcγRIIaR, FcγRIIaH или FcγRIIb за 1, представлены в таблице 59.

H240-Kn179/H240-Hl089/L73-k0 и H240-Kn180/H240-Hl089/L73-k0, получаемые в результате введения E294K или E294R в H240-Kn120 и введения G236T в H240-Hl068, демонстрировали повышенное связывание с FcγRIIaR и FcγRIIaH по сравнению с H240-Kn120/H240-Hl068/L73-k0, при этом их связывание с FcγRIIb снижалось в 0,4 раза по сравнению со связыванием H240-Kn120/H240-Hl068/L73-k0. Кроме того, эффект снижения связывания с FcγRIIb составлял от 1,5 до двух раз по сравнению с H240-Kn120/H240-Hl089/L73-k0, H240-Kn179/H240-Hl068/L73-k0 и H240-Kn180/H240-Hl068/L73-k0, только одна из цепей которого содержала каждое изменение.
H240-Kn192/H240-Hl204/L73-k0 и H240-Kn193/H240-Hl204/L73-k0, получаемые в результате введения I332E и Y235N в указанные выше
варианты, сохраняли связывание с FcγRIIaR, FcγRIIaH и FcγRIIb и повышали связывание с FcγRIIIaF и FcγRIIIaV в два и 8 раз, соответственно, по сравнению с H240-Kn179/H240-Hl089/L73-k0 и H240-Kn180/H240-Hl089/L73-k0 до введения изменений.
[Пример 25] Повышение активности связывания гетеродимеризованного антитела H240-Kn120/H240-Hl068/L73-k0 с активирующим FcγR
В примере 24 варианты, обладающие сниженной активностью связывания с ингибирующим FcγRIIb при сохранении или повышении связывания с FcγRIIaR и FcγRIIaH, получали введением изменений в H240-Kn120/H240-Hl068/L73-k0. В этом анализе авторы настоящего изобретения проводили попытки повысить активность связывания с активирующими FcγR: FcγRIIaR, FcγRIIaH, FcγRIIIaF и FcγRIIIaV.
H240-Kn138 (SEQ ID NO: 116), получаемую в результате введения L328W в H240-Kn120; H240-Kn173 (SEQ ID NO: 117), получаемую в результате введения I332Q в H240-Kn120; H240-Kn178 (SEQ ID NO: 118), получаемую в результате введения K334Y в H240-Kn120; H240-Kn166 (SEQ ID NO: 119), получаемую в результате введения L328A в H240-Kn120; H240-Kn172 (SEQ ID NO: 120), получаемую в результате введения I332M в H240-Kn120, и H240-Kn149 (SEQ ID NO: 121), получаемую в результате введения L328W и K334L в H240-Kn120, получали способом, описанным в справочном примере 1. Кроме того, H240-Hl147 (SEQ ID NO: 122), получаемую в результате введения L328W в H240-Hl068; H240-Hl170 (SEQ ID NO: 123), получаемую в результате введения L328A в H240-Hl068; H240-Hl174 (SEQ ID NO: 124), получаемую в результате введения I332E в H240-Hl068;
H240-Hl150 (SEQ ID NO: 125), получаемую в результате введения I332T в H240-Hl068; H240-Hl182 (SEQ ID NO: 126), получаемую в результате введения А231Н в H240-Hl068, и H240-Hl177 (SEQ ID NO: 127), получаемую в результате введения I332Q в H240-Hl068, получали в качестве другой H-цепи. H240-Kn120/H240-Hl170/L73-k0, H240-Kn120/H240-Hl150/L73-k0, H240-Kn138/H240-Hl068/L73-k0, H240-Kn120/H240-Hl174/L73-k0, H240-Kn173/H240-Hl068/L73-k0, H240-Kn178/H240-Hl068/L73-k0, H240-Kn120/H240-Hl182/L73-k0, H240-Kn138/H240-Hl147/L73-k0, H240-Kn166/H240-Hl170/L73-k0, H240-Kn172/H240-Hl177/L73-k0 и H240-Kn149/H240-Hl068/L73-k0 получали способом, описанным в справочном примере 1 с использованием L73-k0 (SEQ ID NO: 106) в качестве L-цепи, H240-Kn138 (SEQ ID NO: 116), H240-Kn173 (SEQ ID NO: 117), H240-Kn178 (SEQ ID NO: 118), H240-Kn149 (SEQ ID NO: 121), H240-Kn166 (SEQ ID NO: 119) и H240-Kn172 (SEQ ID NO: 120) в качестве H-цепи и H240-Hl170 (SEQ ID NO: 123), H240-Hl150 (SEQ ID NO: 125), H240-Hl174 (SEQ ID NO: 124), H240-Hl182 (SEQ ID NO: 126), H240-Hl147 (SEQ ID NO: 122) и H240-Hl177 (SEQ ID NO: 127) в качестве другой H-цепи. Эти варианты оценивали в отношении связывания с FcγR способом, описанным в справочном примере 8. Результаты представлены в таблице 60.

Кроме того, значения, получаемые делением KD Н240-Kn120/H240-Hl068/L73-k0 для каждого FcγRIa FcγRIIaR, FcγRIIaH, FcγRIIb, FcγRIIa F и FcγRIIIa V на KD каждого варианта, относительная KD, определяемая при принятии KD H240-Kn120/H240-Hl068/L73-k0 для каждого из FcγRIIaR, FcγRIIaH и FcγRIIb за 1, приведены в таблице 61.

Варианты, представленные в этой таблице, демонстрируют повышенное связывание по меньшей мере с одним FcγR из FcγRIIaR, FcγRIIaH, FcγRIIIaF и FcγRIIIaV по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn120/H240-Hl170/L73-k0, получаемое в результате введения L328A в H240-Hl068, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активностями связывания с FcγRIIaR и FcγRIIaH, повышенными в 2,3 раза по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn120/H240-Hl150/L73-k0, получаемое в результате введения I332T в H240-Hl068, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активность связывания с FcγRIIaH, повышенной в 1,2 раза по сравнению с H240-Kn120/H240-Hl068/L73-k0, при сохранении активности связывания с FcγRIIaR.
H240-Kn138/H240-Hl068/L73-k0, получаемое в результате введения L328W в H240-Kn120, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, has активность связывания с FcγRIIaH, повышенной в 1,6 раз по сравнению с H240-Kn120/H240-Hl068/L73-k0, при сохранении активности связывания с FcγRIIaR.
H240-Kn120/H240-Hl174/L73-k0, получаемое в результате введения I332E в H240-Hl068, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания с FcγRIIIaF, повышенной в 4,3 раза, и активностью связывания с FcγRIIIaV, повышенной в 10 раз по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn173/H240-Hl068/L73-k0, получаемое в результате
введения I332Q в H240-Kn120, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания с FcγRIIIaF, повышенной в 1,2 раза оп сравнению с активностью H240-Kn120/H240-Hl068/L73-k0, при сохранении активности связывания с FcγRIIIaV.
H240-Kn178/H240-Hl068/L73-k0, получаемое в результате введения K334Y в H240-Kn120, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания cFcγRIIIaF, повышенной в 1,6 раза, и активностью связывания с FcγRIIIaV, повышенной в 1,9 раза по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn120/H240-Hl182/L73-k0, получаемое в результате введения А231Н в H240-Hl068, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0 обладает активностью связывания с FcγRIIIaV, повышенной в 1,2 раза по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn138/H240-Hl147/L73-k0, получаемое в результате введения L328W в обе H-цепи H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания с FcγRIIaR, повышенной в 1,8 раз по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn166/H240-Hl170/L73-k0, получаемое в результате введения L328A в обе H-цепи H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания с FcγRIIaH, повышенной в 1,9 раза по сравнению с H240-Kn120/H240-Hl068/L73-k0.
H240-Kn172/H240-Hl177/L73-k0, получаемое в результате введения I332M в H240-Kn120, которая представляет собой H-цепь
H240-Kn120/H240-Hl068/L73-k0, и введения I332Q в H240-Hl068, которая представляет собой другую H-цепь, обладает активностью связывания с FcγRIIIaV, повышенной в 1,6 раза по сравнению с активностью H240-Kn120/H240-Hl068/L73-k0, при сохранении активности связывания с FcγRIIIaF.
H240-Kn149/H240-Hl068/L73-k0, получаемое в результате введения L328W и K334L в H240-Kn120, которая представляет собой H-цепь H240-Kn120/H240-Hl068/L73-k0, обладает активностью связывания с FcγRIIaH, повышенной в 1,6 раза по сравнению с активностью H240-Kn120/H240-Hl068/L73-k0, при сохранении активностей связывания с FcγRIIaR и FcγRIIIaV.
Описанные выше результаты позволяют предположить, что варианты обладают сильной активностью ADCP или ADCC по сравнению с H240-Kn120/H240-Hl068/L73-k0.
[Пример 26] Получение гетеродимеризованного антитела с повышенной активностью связывания с FcγRIIb
Опубликовано, что у людей FcγRIa (CD64A), FcγRIIa (CD32A), FcγRIIb (CD32B), FcγRIIIa (CD16A) и FcγRIIIb (CD16B) являются изоформами семейства белков FcγR человека, и также опубликованы их аллотипы (Immunol. Lett., 82(1-2), 57-65, 2002). FcγRIa, FcγRIIa, и FcγRIIIa обладают иммуноактивирующими функциями, и называются активирующими FcγR, тогда как FcγRIIb обладает иммуносупрессивными функциями, и называется ингибирующим FcγR (Nat. Rev. Immunol., 10, 328-343, 2010).
FcγRIIb представляет собой единственные FcγR,
экспрессируемый на В-клетках (Eur. J. Immunol., 19 1379-1385, 1989). Опубликовано, что взаимодействие Fc-области антитела с FcγRIIb подавляет первичный иммунный ответ В-клеток (J. Ехр. Med., 129, 1183-1201, 1969). Кроме того, опубликовано, что когда FcγRIIb на В-клетках и B-клеточном рецепторе (BCR) являются поперечно связанными посредством иммунного комплекса в крови, активация В-клеток подавляется, и продукция антител В-клетками подавляется (Immunol. Lett., 88, 157-161, 2003). В этой иммуносупрессивной передаче сигнала, опосредованной BCR и FcγRIIb, необходим иммунорецепторный тирозиновый ингибирующий мотив (ITIM), содержащийся во внутриклеточном домене FcγRIIb (Science, 256, 1808-1812, 1992; Nature, 368, 70-73, 1994). Когда ITIM фосфорилируется при передаче сигнала, вовлекается содержащая SH2 инозитолполифосфат-5-фосфатаза (SHIP), трансдукция сигнального каскада других активирующих FcγR ингибируется, и воспалительный иммунный ответ подавляется (Science, 290, 84-89, 2000). Кроме того, опубликовано, что агрегация одного FcγRIIb временно подавляет приток кальция вследствие поперечного связывания BCR и B-клеточной пролиферации BCR-зависимым путем без индукции апоптоза продуцирующих IgM В-клеток (J. Imunol., 181, 5350-5359, 2008).
Кроме того, FcγRIIb также экспрессируется на дендритных клетках, макрофагах, активированных нейтрофилах, тучных клетках и базофилах. FcγRIIb ингибирует функцию активирующего FcγR, такую как фагоцитоз и выделение воспалительных цитокинов в этих клетках, и подавляет воспалительные иммунные ответы (Nat. Rev.
Immunol., 10, 328-343, 2010).
В настоящее время важность иммуносупрессивных функций FcγRIIb установили посредством исследований с использованием мышей с нокаутом по FcγRIIb. Опубликовано, что у мышей с нокаутом по FcγRIIb гуморальный иммунитет не регулируется соответствующим образом (J. Immunol., 163, 618-622, 1999), восприимчивость к индуцируемому коллагеном артриту (CIA) является повышенной (J. Exp. Med., 189, 187-194, 1999), присутствуют волчаночноподобные симптомы, и присутствуют симптомы подобные синдрому Гудпасчера (J. Exp. Med., 191, 899-906, 2000).
Кроме того, опубликовано, что регуляторная недостаточность FcγRIIb связана с аутоиммунными заболеваниями человека. Например, опубликовано отношение генетического полиморфизма в трансмембранной области и промоторной областью FcγRIIb к частоте развития системной красной волчанке (SLE) (Hum. Genet., 117, 220-227, 2005; J. Bio.l Chem., 282, 1738-1746, 2007; Arthritis Rheum., 54, 3908-3917, 2006; Nat. Med., 11, 1056-1058, 2005; J. Immunol., 176, 5321-5328, 2006) и снижение экспрессии FcγRIIb на поверхности В-клеток у пациентов с SLE (J. Exp. Med., 203, 2157-2164, 2006; J. Immunol., 178, 3272-3280, 2007).
На основании моделей на мышах и клинических данных соответственно считают, что FcγRIIb играет роль контроля развития аутоиммунных заболеваний и воспалительных заболеваний в основном через вовлечение В-клеток, и он представляет собой перспективную молекулу-мишень для контроля аутоиммунных заболеваний и воспалительных заболеваний.
Известно, что IgG1, в основном используемый в качестве коммерчески доступного фармацевтического средства на основе антител, связывается не только с FcγRIIb, а также в значительной степени с активирующим FcγR (Blood, 113, 3716-3725, 2009). Существует возможность разработки фармацевтических средств на основе антител, обладающих большими иммуносупрессивными свойствами по сравнению со свойствами IgG1, с использованием Fc-области с повышенным связывание с FcγRIIb или селективно улучшенной активностью связывания с FcγRIIb по сравнению с активирующим FcγR. Например, предполагали, что применением антитела, содержащего вариабельную область, которая связывается с BCR, и Fc с повышенным связыванием с FcγRIIb можно ингибировать активацию B-клеток (Mol. Immunol., 45, 3926-3933, 2008). Опубликовано, что поперечное связывание FcγRIIb на В-клетках и IgE, связанного с B-клеточным рецептором, подавляет дифференцировку В-клеток в плазматических клетках, что в результате приводит к подавлению продукции IgE, и у мышей с трансплантированными РВМС человека сохраняются концентрации IgG и IgM человека, тогда как концентрация IgE человека снижается (Acad. News., doi: 10, 1016, jaci. 2011, 11, 029). Наряду с IgE опубликовано, что, когда FcγRIIB и CD79b, образующие комплекс B-клеточного рецептора поперечно связываются антителом, подавляется B-клеточная пролиферация in vitro, и ослабевают симптомы на модели индуцируемого коллагеном артрита (Arthritis Rheum., 62, 1933-1943, 2010).
Наряду с В-клетками опубликовано, что поперечное связывание
FcεRI и FcγRIIb на тучных клетках с использованием молекул, в которых участок Fc IgG с повышенным связыванием с FcγRIIb является слитым с участком Fc IgE, который связывается с рецептором IgE FcεRI, вызывает фосфорилирование FcγRIIb, таким образом, подавляя FcsRI-зависимый приток кальция. Это позволяет предположить, что возможно ингибировать дегрануляцию через стимуляцию FcγRIIb посредством повышения связывания с FcγRIIb (Immunol. Let., doi: 10, 1016/j. imlet. 2012.01.008).
Таким образом, предполагают, что антитело, содержащее Fc с улучшенной активностью связывания с FcγRIIb, является перспективным в качестве терапевтического средства для воспалительных заболеваний, таких как аутоиммунные заболевания.
Кроме того, предполагали, что мутанты с повышенным связыванием с FcγRIIb представляют собой перспективные терапевтические средства для злокачественной опухоли, а также терапевтические средства для воспалительных заболеваний, таких как аутоиммунные заболевания. В настоящее время выявлено, что FcγRIIb играет важную роль в агонистической активности агонистических антител против антитела к семейству рецепторов TNF. В частности, предполагали, что взаимодействие с FcγRIIb является необходимым для агонистической активности антител против CD40, DR4, DR5, CD30 и CD137, которые входят в семейство рецепторов TNF (Science, 333, 1030-1034, 2011; Cancer Cell, 19, 101-1113, 2011; J. Clin. Invest., 2012, doi: 10, 1172/JCI61226; J. Immunol., 171, 562-, 2003; Blood, 108, 705-, 2006; J. Immunol., 166, 4891, 2001). В непатентном документе (Science, 333, 1030-1034,
2011) продемонстрировано, что использование антител с повышенным связыванием с FcγRIIb повышает противоопухолевый эффект антител к CD40. Таким образом, предполагают, что антитела с повышенным связыванием с FcγRIIb обладают эффектом повышения агонистической активности агонистических антител, включая антитела против антитела к семейству рецепторов TNF.
Опубликованы антитела, содержащие Fc с улучшенной активностью связывания с FcγRIIb (Mol. Immunol., 45, 3926-3933, 2008). В этом документе активность связывания с FcγRIIb улучшали добавлением изменений, таких как S267E/L328F, G236D/S267E и S239D/S267E, к Fc-области антитела. В документе антитело, в которое вводили мутацию S267E/L328F, более сильно связывается с FcγRIIb. Таким образом, дополнительно повышая связывание с FcγRIIb, предполагают, что можно повышать описанную выше функцию, опосредованную FcγRIIb.
Кроме того, антитела с введенной мутацией S267E/L328E сохраняют аналогичный уровень связывания с FcγRIa и FcγRIIa типа Н как связывание природного IgG1. Однако в другой публикации продемонстрировано, что это изменение повышает связывание с типом R FcγRIIa в несколько сотен раз до аналогичного уровня связывания с FcγRIIb, что означает, что селективность связывания с FcγRIIb не улучшается оп сравнению с типом R FcγRIIa (Eur. J. Immunol., 23, 1098-1104, 1993).
Даже если связывание с FcγRIIb повышают по сравнению со связыванием IgG1, предполагают, что только эффект повышения
связывания с FcγRIIa, а не повышение связывания с FcγRIIb оказывает влияние на клетки, такие как тромбоциты, которые экспрессируют FcγRIIa, но не экспрессируют FcγRIIb (Nat. Rev. Immunol., 10, 328-343, 2010). Например, известно, что группа пациентов, которой вводили бевацизумаб, антитело против VEGF, подвергается повышенному риску тромбоэмболии (J. Natl. Cancer Inst., 99, 1232-1239, 2007). Кроме того, тромбоэмболию, аналогично, наблюдали в клиническом испытании, в котором тестировали антитело против лиганда CD40, и клиническое исследование прекращали (Arthritis Rheum., 48, 719-727, 2003). В обоих случаях для этих антител в дальнейших исследованиях с использованием моделей на животных и т.п. предполагали, что вводимые антитела вызывают агрегацию тромбоцитов через связывание с FcγRIIa на тромбоцитах и образуют тромбы (J. Thromb. Haemost., 7, 171-181, 2008; J. Immunol., 185, 1577-1583, 2010). При системной красной волчанке, которая является аутоиммунным заболеванием, тромбоциты активируются через FcγRIIa-зависимый механизм, и опубликовано, что активация тромбоцитов коррелирует с тяжестью симптомов (Sci. Transl. Med., vol 2, issue 47, 47-63, 2010). Даже если связывание с FcγRIIb является повышенным, введение антитела с повышенным связыванием с FcγRIIa таким пациентам, которые уже подвергаются высокому риску развития тромбоэмболии, повысит риск развития тромбоэмболии, таким образом, является крайне опасным.
Кроме того, опубликовано, что антитела с повышенным связыванием с FcγRIIa повышают опосредованный макрофагами
антителозависимый клеточный фагоцитоз (ADCP) (Mol. Cancer. Ther., 7, 2517-2527, 2008). Когда антигены антитела подвергаются фагоцитозу макрофагами, одновременно сами антитела также подвергаются фагоцитозу. В этом случае пептидные фрагменты, получаемые из этих антител, также презентируются в качестве антигена, и антигенность может становиться более высокой, таким образом, повышая риск продукции антител против антител (антител к антителам). Более конкретно, повышение связывания с FcγRIIa повысит риск продукции антител против антител, и это заметно снизит их ценность в качестве фармацевтических средств.
Более конкретно, ценность в качестве фармацевтических средств значительно снижается, когда повышается связывание с FcγRIIa, которое приводит к повышенному риску образования тромбов посредством агрегации тромбоцитов, высокой антигенности и повышенного риска продукции антител к антителам.
С этой точки зрения указанный выше Fc с повышенным связыванием с FcγRIIb демонстрирует заметно повышенное связывание с типом R FcγRIIa по сравнению со связыванием природного IgG1. Таким образом, его ценность в качестве фармацевтического средства для пациентов, несущих тип R FcγRIIa, значительно снижается. Типы Н и R FcγRIIa наблюдают у кавказцев и афроамериканцев приблизительно с аналогичной частотой (J. Clin. Invest., 97, 1348-1354, 1996; Arthritis Rheum., 41, 1181-1189, 1998). Таким образом, когда этот Fc использовали для лечения аутоиммунных заболеваний, количество пациентов, которые могут безопасно использовать его, при этом получая от него эффекты, в
качестве фармацевтического средства является ограниченным.
Кроме того, для дендритных клеток с дефицитом FcγRIIb или дендритных клеток, в которых взаимодействие FcγRIIb и участка Fc антитела ингибируют антителом к FcγRIIb, опубликовано, что дендритные клетки созревают спонтанно (J. Clin. Invest., 115, 2914-2923, 2005; Proc. Natl. Acad. Sci. USA, 102, 2910-2915, 2005). Эта публикация позволяет предположить, что FcγRIIb активно подавляет созревание дендритных клеток в равновесном состоянии, при котором не возникает воспаления и т.п. Наряду с FcγRIIb на поверхности дендритных клеток экспрессируется FcγRIIa, таким образом, даже если повышают связывание с ингибирующим FcγRIIb, и если также повышают связывание с активирующим FcγR, таким как FcγRIIa, в результате можно способствовать созреванию дендритных клеток. Более конкретно, считают, что улучшение не только активности связывания с FcγRIIb, а также отношения активности связывания с FcγRIIb к активности связывания с FcγRIIa является важным для получения антител с иммуносупрессивным действием.
Таким образом, при рассмотрении поколения фармацевтических средств, в которых используют опосредованное связыванием с FcγRIIb иммуносупрессивное действие, существует необходимость в Fc, который не только обладает повышенной активностью связывания с FcγRIIb, а также обладает связыванием с обоими FcγRIIa, аллотипами типов Н и R, которое сохраняется на аналогичном уровне или является сниженным до уровня, ниже уровня природного IgG1.
При этом в настоящее время опубликованы случаи, когда изменения аминокислоты вводили в Fc-области для повышения селективности связывания с FcγRIIb (Mol. Immunol., 40, 585-593, 2003). Однако все варианты, для которых указано, что они обладают улучшенной селективностью в отношении FcγRIIb, как опубликовано в этом документе, демонстрировали сниженное связывание с FcγRIIb по сравнению со связыванием природного IgG1. Таким образом, предполагают, что этим вариантам затруднительно фактически индуцировать опосредованную FcγRIIb иммуносупрессивную реакцию более сильную по сравнению с IgG1.
Кроме того, вследствие того, что FcγRIIb играет важную роль в указанных выше агонистических антителах, предполагают, что повышение их активности связывания повышает агонистическую активность. Однако когда связывание с FcγRIIa аналогично повышают, проявляются непредусмотренные активности, такие как активность ADCC и активность ADCP, и это может вызывать побочные эффекты. Также с этой точки зрения предпочтительно иметь возможность селективно повышать активность связывания с FcγRIIb.
На основании этих результатов для получения фармацевтических средств на основе антител, предназначенных для лечения аутоиммунных заболеваний и злокачественной опухоли с использованием FcγRIIb, важно, чтобы по сравнению со средствами на основе природного IgG, активности связывания с обоими аллотипами FcγRIIa сохранялись или снижались, и связывание с FcγRIIb повышалось. Однако FcγRIIb обладает 93% идентичностью последовательности во внеклеточной области с аналогичной
последовательностью FcγRIIa, который представляет собой один из активирующих FcγR, и они являются очень структурно схожими. Существуют аллотипы FcγRIIa H типа и R типа, в которых аминокислота в положении 131 представляет собой His (тип Н) или Arg (тип R), но каждый из них по-разному реагирует с антителами (J. Exp. Med., 172, 19-25, 1990). Таким образом, для получения Fc-области, которая селективно связывается с FcγRIIb, наиболее трудной задачей может быть придание Fc-области антитела свойства селективно улучшенной активности связывания с FcγRIIb, что включает различие этих гомологичных последовательностей и снижение или неповышение активности связывания с каждым аллотипом FcγRIIa, при повышении активности связывания с FcγRIIb. К настоящему моменту не получены варианты, обладающие достаточной селективностью в отношении FcγRIIb. В US 2009/0136485 опубликованы варианты с повышенной активностью связывания с FcγRIIb, однако степень повышения является низкой, и существует необходимость в получении вариантов, обладающих свойствами, аналогичными описанным выше свойствам.
В этой оценке авторы настоящего изобретения исследовали получение вариантов, активность связывания с FcγRIIb которых селективно повышали с использованием гетеродимеризованных антител. Вариабельную область IL6R (SEQ ID NO: 128), которая представляет собой вариабельную область антитела против рецептора интерлейкин 6 человека, описанную в WO 2009/125825, использовали в качестве вариабельной области H-цепи антитела. Кроме того, получали IL6R-G1d (SEQ ID NO: 129), содержащую G1d, в
которой отсутствует C-концевой Gly и Lys IgG1 человека, а затем вводили K439E в IL6R-G1d для конструкции IL6R-B3 (SEQ ID NO: 130). При этом E233D, G237D, P238D, H268D, P271G и A330R вводили в IL6R-B3 для конструкции IL6R-BP208 (SEQ ID NO: 131). Кроме того, S267E и L328F вводили в IL6R-B3 для конструкции IL6R-BP253 (SEQ ID NO: 132), содержащей существующий Fc с повышенной активностью связывания с FcγRIIb. L-цепь тоцилизумаба, IL6R-L (SEQ ID NO: 133), использовали в качестве общей L-цепи антитела и использовали в комбинации с каждой H-цепью способом, описанным в справочном примере 1, для получения гомодимеров IL6R-B3/IL6R-L, IL6R-BP208/IL6R-L и IL6R-BP253/IL6R-L. Эти варианты оценивали в отношении активности связывания с FcγR Ia, FcγR IIaR, FcγR IIaH, FcγR IIb и FcγR IIIaV способом, описанным в справочном примере 8. Результаты представлены в таблице 62. Когда в таблице 62 клетка закрашена серым, это означает, что делали заключение, что связывание FcγR с IgG является слишком слабым, чтобы надлежащим образом анализировать кинетическим анализом. Таким образом, когда взаимодействие каждого измененного антитела и FcγR являлось слабым, и устанавливали, что невозможно провести корректный анализ указанным выше кинетическим анализом, KD для таких взаимодействий рассчитывали с использованием следующего уравнения модели связывания 1:1, описанного в программном обеспечении Handbook BR1006-48 Edition АЕ Biacore Т100.
Поведение взаимодействующих молекул согласно модели связывания 1:1 на Biacore можно описать уравнением 1,
приведенным ниже.
Уравнение 1
Req=C⋅Rmax/(KD+C)+RI
Req: график уровней связывания в равновесном состоянии в зависимости от концентрации аналита
С: концентрация
RI: вклад объемного коэффициента преломления в образце
Rmax: связывающая способность аналита с поверхностью
После преобразования этого уравнения KD можно выразить в виде уравнения 2, приведенного ниже.
Уравнение 2
KD=C⋅Rmax/(Req-RI)-C
KD можно рассчитывать путем замены значений Rmax, RI и С в этом уравнении. Значения RI и С можно определять из сенсограммы результатов измерения и условий измерения. Rmax рассчитывали следующим ниже способом. В качестве цели сравнения для антител, которые обладали достаточно сильными взаимодействиями, как оценивали одновременно в аналогичном цикле измерения, значение Rmax получали посредством общей подгонки с использованием модели связывания Ленгмюра 1:1, а затем его делили на количество антитела сравнения, захваченное на сенсорном чипе, и умножали на захваченное количество измененного антитела, подлежащего оценке.

Значения, получаемые делением KD IL6R-B3/IL6R-L для каждого из FcγRIa FcγRIIaR, FcγRIIaH и FcγRIIb на соответствующую KD каждого модифицированного варианта, относительная KD, определяемая при принятии KD IL6R-B3/IL6R-L для FcγRIIaR, FcγRIIaH или FcγRIIb за 1, представлены в таблице 63.

Как представлено в таблице 63, IL6R-BP253/IL6R-L, которое представляет собой существующее антитело с повышенной активностью связывания с FcγRIIb, обладает активностью связывания с FcγRIIb, повышенной приблизительно в 350 раз, и активностью связывания с FcγRIIaR, повышенной приблизительно в 500 раз по сравнению с антителом IgG1 человека (IL6R-B3/IL6R-L) без введения изменения. При этом активность связывания с FcγRIIb IL6R-BP208/IL6R-L составляет приблизительно в 100 раз большую и таким образом превосходит существующее антитело с повышенной
активностью связывания с FcγRIIb, однако его активность связывания с FcγRIIaR составляет в 1,3 раз большую и является сравнимой с активностью типа IgG1. Таким образом, IL6R-BP208/IL6R-L представляет собой антитело превосходное в отношении селективности к FcγRIIb.
Затем авторы настоящего изобретения рассматривали, что с целью улучшения активности связывания с FcγR2b и селективности, необходимо получить информацию о кристаллографической структуре комплекса внеклеточной области FcγRIIb и Fc (ВР208), который представляет собой Fc IL6R-BP208/IL6R-L, и более точно оценить мутации аминокислот, подлежащие введению. Таким образом, комплекс Fc (ВР208) и внеклеточной области FcγRIIb анализировали рентгеноструктурной кристаллографией экспериментальным способом, описанным ниже.
[Экспрессия и очистка Fc (ВР208)]
Fc (ВР208) получали, как указано ниже. Сначала Cys в положении 220 (нумерация EU) IL6R-BP208 заменяли на Ser. Затем генетическую последовательность Fc (ВР208) от Glu в положении 236 (нумерация EU) до его C-конца клонировали ПЦР. С использованием клонируемой генетической последовательности способом проводили справочного примера 1 получение экспрессирующих векторов и экспрессию и очистку Fc (ВР208). Cys в положении 220 (нумерация EU) образует дисульфидную связь с Cys L-цепи в общем IgG1. Когда получают только Fc, L-цепь совместно не экспрессируют, и таким образом, этот остаток заменяли Ser для предотвращения образования ненужной дисульфидной связи.
[Экспрессия и очистка внеклеточной области FcγRIIb]
Ее проводили способом справочного примера 8.
[Очистка комплекса Fc (ВР208)/внеклеточной области FcγRIIb]
К 1,5 мг образца внеклеточной области FcγRIIb, получаемого для кристаллизации, добавляли 0,15 мг Endo F1 (Protein Science, 1996, 5:2617-2622), экспрессируемого и очищаемого из Escherichia coli в виде слитого белка глутатион-S-трансферазы. Его можно было хранить при комнатной температуре в течение трех суток в 0,1 М Bis-Tris буфере при pH 6,5, и расщепляли N-связанный олигосахарид, оставляя N-ацетилглюкозамин, непосредственно связанный с Asn. Затем этот образец внеклеточной области FcγRIIb, который подвергали обработке с отщеплением углеводов, концентрировали ультрафильтрацией с 50000 MWCO, и очищали гельфильтрационной хроматографией (Superdex 200 10/300) с использованием колонки, уравновешенной в 20 мМ HEPES при pH 7,5, содержащей 0,1 М NaCl. Кроме того, к получаемой фракции внеклеточной области FcγRIIb с отщепленными углеводами добавляли Fc(Kn0120/Hl068) таким образом, чтобы молярное отношение внеклеточной области FcγRIIb находилось в небольшом избытке, и после концентрации ультрафильтрацией с 10000 MWCO образец комплекса Fc (ВР208)/внеклеточной области FcγRIIb получали посредством очистки гельфильтрационной хроматографией (Superdex 200 10/300) с использованием колонки, уравновешенной в 25 мМ HEPES при pH 7,5, содержащей 0,1 М NaCl.
[Кристаллизация комплекса Fc (ВР208)/внеклеточной области FcγRIIb]
Образец комплекса Fc (ВР208) / внеклеточной области FcγRIIb концентрировали до приблизительно 10 мг/мл с использованием фильтра для ультрафильтрации 10000 MWCO, и кристаллизовали способом диффузии паров из висячей капли в комбинации с затравочным способом. Для кристаллизации использовали планшет VDXm (Hampton Research). С использованием резервуарного раствора, содержащего 0,1M Bis-Tris (pH 6,0), 19% PEG3350 и 0,2М двухосновного фосфата калия, получали кристаллизационные капли при коэффициенте смешения резервуарного раствора : кристализуемого образца = 0,85 мл : 0,85 мл. Для получения раствора затравочного кристалла получаемые при аналогичном условии кристаллы комплекса измельчали с использованием затравочных гранул (Hampton Research). 0,15 мкл разбавленного раствора, получаемого из раствора затравочного кристалла, добавляли к кристаллизационным каплям, которые герметизировали в лунках, содержащих резервуары, и выдерживали при 20°C. Это эффективно приводило к образованию плоских кристаллов.
[Измерение рентгенодифракционных данных кристалла комплекса Fc (ВР208)/внеклеточной области FcγRIIb]
Один из получаемых одиночных кристаллов комплекса Fc (ВР208)/внеклеточной области FcγRIIb выдерживали в растворе 0,1М Bis-Tris pH 6,5, 24% PEG3350, 0,2М двухосновного фосфата калия, 10% этиленгликоля (об./об.). Кристалл извлекали из раствора с использованием иглы с прикрепленной небольшой нейлоновой петлей и замораживали в жидком азоте, а затем измеряли рентгенодифракционные данные посредством BL32XUin Spring-8. Во
время измерения кристалл постоянно располагали в струе азота при -178°C для поддержания замороженного состояния и получали всего 300 рентгенографических дифракционных изображений с использованием детектора МХ-225НЕ CCD (RAYONIX), прикрепленного к каналу пучка с вращением кристалла 0,6° за один раз. Определение параметров ячейки, разметки дифракционных пятен и обработку дифракционных данных из получаемых дифракционных изображений проводили с использованием программы Xia2 (J. Appl. Cryst., 2010, 43: 186-190), пакета программ XDS (Acta. Cryst., 2010, D66: 125-132) и Scala (Acta. Cryst., 2006, D62: 72-82), и в итоге получали данные дифракционной интенсивности с разрешением до 2,81  . Кристалл принадлежит к пространственной группе C2221 и имеет следующие параметры ячейки а=15б,69 , b=260,17 , с=56,85 , α=90°, β=90°, γ=90°.
. Кристалл принадлежит к пространственной группе C2221 и имеет следующие параметры ячейки а=15б,69 , b=260,17 , с=56,85 , α=90°, β=90°, γ=90°.
[Рентгеновский кристаллографический анализ комплекса Fc (ВР208) / внеклеточной области FcγRIIb]
Кристаллическую структуру комплекса Fc (ВР208) / внеклеточной области FcγRIIb определяли способом молекулярного замещения с использованием программы Phaser (J. Appl. Cryst., 2007, 40: 658-674). На основании размера получаемой кристаллической решетки и молекулярной массы комплекса Fc (ВР208) / внеклеточной области FcγRIIb теоретически рассчитывали, что число комплексов в асимметричной единице равно одному. На основании структурных координат кода PDB: 3SGJ, который представляет собой кристаллическую структуру комплекса Fc(WT) / внеклеточной области FcyRIIIa, участки аминокислотных остатков положений А-цепи 239-340
и положений В-цепи 239-340 получали в виде отдельных координат и использовали их соответственно в качестве моделей для исследования доменов СН2 Fc. Участки аминокислотных остатков положений А-цепи 341-444 и положений В-цепи 341-443 получали в виде одного набора координат из аналогичных структурных координат кода PDB: 3SGJ, и использовали его в качестве модели для исследования доменов СН3 Fc. Наконец, на основании структурных координат кода PDB: 2FCB, который представляет собой кристаллическую структуру внеклеточной области FcγRIIb, получали участки аминокислотных остатков положений А-цепи 6-178 и использовали в качестве модели для исследования Fc(BP208). Авторы настоящего изобретения пытались определить ориентации и положения каждой модели поиска доменов СН 3Fc, внеклеточной области FcγRIIb и домена СН2 Fc в кристаллических решетках с использованием функции вращения и функции трансляции, но им не удалось определить положение одного из доменов СН2. Затем на основании кристаллической структуры комплекса Fc (WT) / внеклеточной области FcγRIIIa определяли положение последнего домена СН2 А на основании карты электронной плотности, которую рассчитывали на основании фазы, определяемой из оставшихся терх частей. Таким образом, авторы настоящего изобретения получали исходную модель кристаллической структуры комплекса Fc (ВР208) / внеклеточной области FcγRIIb. Когда на получаемой исходной модели проводили моделирование жесткого тела, в котором двигают два домена СН2 Fc, два домена СН3 Fc и внеклеточную область FcγRIIb, кристаллографический фактор
достоверности величина R становилась 42,6%, и величина Free R становилась 43,7% для данных дифракционной интенсивности от 25 до 3,0 в этой точке. Кроме того, структурное уточнение с использованием программы REFMAC5 (Acta Cryst., 2011, D67, 355-367) и редктирование модели для наблюдения карт электронной плотности, коэффициент которых имеет 2Fo-Fc или Fo-Fc, которые рассчитывают на основании экспериментально установленного структурного фактора Fo, рассчитываемого структурного фактора Fc и рассчитываемой фазы с использованием модели, проводили программой Coot (Acta Cryst., 2010, D66: 486-501), и уточнение модели проводили повторением этих стадий. В итоге, в результате включения молекул воды в модель на основе карт электронной плотности, в которых используют 2Fo-Fc или Fo-Fc в качестве коэффициента, и последующего уточнения, кристаллографический фактор достоверности, значения R и значения Free R модели, содержащей 4794 неводородных атома, становились 24,4% и 27,9% для 27259 данных дифракционной интенсивности с разрешением от 25 до 2,81 , соответственно.
Трехмерную структуру комплекса Fc (ВР208) / внеклеточной области FcγRIIb определяли при разрешении 2,81 структурным анализом. Структура, получаемая в результате анализа, представлена на фиг. 49. Выявляли, что внеклеточная область FcγRIIb является связанной и располагается между двумя доменами СН2 Fc, что является сходным с трехмерными структурами ранее анализируемых комплексов между Fc (WT), которые представляют собой Fc нативного IgG, и соответствующих внеклеточных областей
FcγRIIIa (Proc. Natl. Acad. Sci. USA, 2011, 108, 12669-126674), FcγRIIIb (Nature, 2000, 400, 267-273; J. Biol. Chem., 2011, 276, 16469-16477) и FcγRIIa.
Однако при более подробном изучении Fc (ВР208) выявляли изменение в структуре петли при 233-239 после шарнирной области в домене СН2 А вследствие влияния вводимых мутаций G237D и P238D по сравнению с Fc (WT), связанного с FcyRIIa (фиг. 50). Результат демонстрировал, что амид основной цепи G237 в Fc (ВР208) образовывал сильную водородную связь с боковой цепью Tyr160 в FcγRIIb. В FcγRIIa Phe содержится вместо этого Tyr160 и он не способен образовывать такую водородную связь. Это позволяет предположить, что описанная выше водородная связь оказывает важный вклад в приобретение селективности, т.е. улучшение активности связывания с FcγRIIb и снижение связывания с FcγRIIa.
На основании результатов структурного анализа авторы настоящего изобретения провели точную оценку возможных изменений для дополнительного повышения активности и выявили S239 в качестве кандидата участка введения изменения. Как продемонстрировано на фиг. 51, Ser239 домена СН2 В располагается в положении, в направлении которого выступает Lys117 FcγRIIb естественным образом для структуры. Однако вследствие того, что описанным выше анализом не наблюдали электронную плотность для Lys117 FcγRIIb, полагают, что Lys117 не принимает определенной структуры и оказывает только ограниченный эффект на взаимодействие с Fc (ВР208) в данной ситуации. Кода S239 домена СН2 В заменяют на отрицательно заряженный D или Е, можно
предполагать электростатическое взаимодействие с положительно заряженным Lys117 FcγRIIb, таким образом, приводящее к улучшению активности связывания с FcγRII.
С другой стороны, наблюдение структуры S239 в домене СН2 А позволяет предположить, что боковая цепь этой аминокислоты посредством формирования водородной связи с основной цепью G236 стабилизирует структуру петли в положениях от 233 до 239, включая D237, который образует водородная связь с боковой цепью Tyr160 FcγRIIb, следующим за шарнирной областью (фиг. 52). Стабилизация структуры петли в конформации связывания подавляет снижение энтропии при связывании и приводит к повышению свободной энергии связывания, приводя к улучшению активности связывания. При этом, когда S239 домена СН2 А заменяют на D или Е, структура петли становится нестабильной вследствие утраты водородной связи с основной цепью G236. Кроме того, в непосредственной близости может возникать электростатическое отталкивание с D265, приводя к дополнительной дестабилизации структуры петли. Энергия для дестабилизации направлена на снижение энергии взаимодействия с FcγRIIb, приводя к снижению активности связывания. В частности, в случае гомодимеризованного антитела, в обе Н-цепи которого вводили S239D или S239E, повышающий активность связывания эффект вследствие электростатического взаимодействия домена СН2 В с Lys117 FcγRIIb нейтрализуется снижающим активность связывания эффектом вследствие дестабилизации структуры петли в домене СН2 А, и, таким образом, может не приводить к повышению активности
связывания. С другой стороны, в случае гетеродимеризованного антитела, только в одну из Н-цепей которого вводили S239D или S239E, вследствие того, что поддерживается стабилизация структуры петли S239 домена СН2 А, полагали, что активность связывания повышается, что соответствует новому образованному электростатическому взаимодействию с Lys117 FcγRIIb вследствие введения S239D или S239E в домен СН2 В.
Для проверки описанной выше гипотезы авторы настоящего изобретения проводили попытки получения антител с дополнительно повышенной активностью связывания с FcγRIIb введением мутации S239D или S239E в Fc-область только одной Н-цепи с использованием IL6R-BP208/IL6R-L в качестве матрицы. В качестве Н-цепи антитела получали IL6R-BP256 (SEQ ID NO: 134), получаемую в результате введения S239D в IL6R-BP208, и IL6R-BP257 (SEQ ID NO: 135), получаемую в результате введения S239E в IL6R-BP208. Кроме того, в качестве другой Н-цепи получали IL6R-A5 (SEQ ID NO: 136), получаемую в результате введения мутации D356K и H435R в IL6R-G1d, и IL6R-AP002 (SEQ ID NO: 137), получаемую в результате введения E233D, G237D, P238D, H268D, P271G и A330R а IL6R-A5 (SEQ ID NO: 136). IL6R-L (SEQ ID NO: 133), которая представляет собой L-цепь из тоцилизумаба, использовали в качестве L-цепи общего антитела и получали гомодимеризованные антитела IL6R-B3/IL6R-L, IL6R-BP208/IL6R-L, IL6R-BP253/IL6R-L, IL6R-BP256/IL6R-L и IL6R-BP257/IL6R-L и гетеродимеризованные антитела IL6R-AP002/IL6R-BP256/IL6R-L и IL6R-AP002/IL6R-BP257/IL6R-L совместно с соответствующими Н-цепями способом, описанным в справочном примере 1. Варианты оценивали в отношении
активности связывания с FcγR Ia, FcγR IIaR, FcγR IIaH и FcγRIIb способом, описанным в справочном примере 8. Результаты представлены в таблице 64.

Когда клетка закрашена серым в таблице 64, это означает, что делали вывод, что связывание FcγR с IgG являлось очень слабым, чтобы надлежащим образом анализировать кинетическим анализом. Таким образом, когда взаимодействие каждого измененного антитела и FcγR являлось слабым, и устанавливали, что невозможно провести корректный анализ указанным выше кинетическим анализом, KD для таких взаимодействий рассчитывали с использованием следующего уравнения модели связывания 1:1, описанного в программном обеспечении Handbook BR1006-48 Edition АЕ Biacore Т100.
Поведение взаимодействующих молекул согласно модели связывания 1:1 на Biacore можно описать уравнением 1, приведенным ниже.
Уравнение 1

Req: график уровней связывания в равновесном состоянии в зависимости от концентрации аналита
С: концентрация
RI: вклад объемного коэффициента преломления в образце
Rmax: связывающая способность аналита с поверхностью
После преобразования этого уравнения KD можно выразить в виде уравнения 2, приведенного ниже.
Уравнение 2

KD можно рассчитывать путем замены значений Rmax, RI и С в этом уравнении. Значения RI и С можно определять из сенсограммы результатов измерения и условий измерения. Rmax рассчитывали следующим ниже способом. В качестве цели сравнения для антител, которые обладали достаточно сильными взаимодействиями, как оценивали одновременно в аналогичном цикле измерения, значение Rmax получали посредством общей подгонки с использованием модели связывания Ленгмюра 1:1, а затем его делили на количество антитела сравнения, захваченное на сенсорном чипе, и умножали на захваченное количество измененного антитела, подлежащего оценке.
При этом значения, получаемые делением KD IL6R-B3/IL6R-L для каждого из FcγRIa FcγRIIaR, FcγRIIaH и FcγRIIb на KD каждого варианта, относительная KD, определяемая при принятии KD IL6R-B3/IL6R-L для каждого из FcγRIIaR, FcγRIIaH и FcγRIIb за 1, и значения, получаемые делением KD каждого варианта для FcγRIIaR на KD для FcγRIIb приведены в таблице 65.
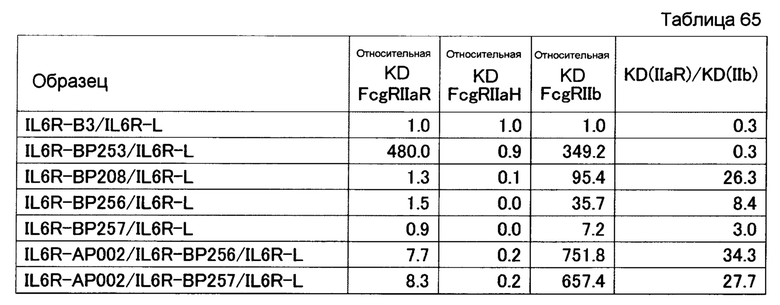
Как продемонстрировано в таблице 65, IL6R-BP256/IL6R-L и IL6R-BP257/IL6R-L, получаемые в результате введения S239D или S239E в обе Н-цепи IL6R-BP208/IL6R-L, демонстрировали сниженную активность связывания с FcγRIIb, а также сниженную активность связывания с FcγRIIaR по сравнению с вариантом IL6R-BP208/IL6R-L до введения. При этом связывание с FcγRIIb IL6R-AP002/IL6R-BP256/IL6R-L и IL6R-AP002/IL6R-BP257/IL6R-L, получаемых в результате введения S239D или S239E в одну Н-цепь IL6R-BP208/IL6R-L, повышалось в 752 раз и 657 раз, соответственно, и, таким образом, их активности связывания с FcγRIIb являлись выше, чем активность, получаемая существующей техникой. Кроме того, активности связывания с FcγRIIaR повышалась в 7,7 раз и 8,3 раз, соответственно, тогда как активность IL6R-BP208/IL6R-L повышалась в 1,3 раза. В таблице KD(IIaR)/KD(IIb) представляет собой значение, получаемое делением KD каждого варианта для FcγRIIaR на KD для FcγRIIb. Когда значение является больше, это означает, что селективность в отношении FcγRIIb является больше. Значение для IL6R-BP253/IL6R-L, которое представляет собой существующее антитело с повышенной активностью связывания с
FcγRIIb, составляет 0,3, подтверждая, что селективность не улучшали по сравнению с типом IgG1, тогда как значение для IL6R-BP208/IL6R-L составляет 26,3, означая, что оно обладает высокой селективностью в отношении FcγRIIb. В этом эксперименте для IL6R-AP002/IL6R-BP256/IL6R-L и IL6R-AP002/IL6R-BP257/IL6R-L, получаемых в результате введения S239D или S239E в одну Н-цепь IL6R-BP208/IL6R-L, демонстрировали значения KD(IIaR)/KD(IIb) 34,3 и 27,7, соответственно, и они являлись более улучшенными по сравнению с IL6R-BP208/IL6R-L.
Результат сравнительной оценки вариантов с использованием типа IgG1 (GpH7-B3/GpL16-k0) в качестве матрицы, которую проводили, как описано в примере 4, демонстрирует, что S239D и S239E обладают эффектом, как показано в таблице 66. В таблице Ho/Con_2aR и Но/Con_2b представляют собой уровни активности связывания с FcγRIIaR и с FcγRIIb при принятии за 100 уровней активности контрольного гомодимеризованного антитела. При этом He/Con_2aR и Не/Con_2b представляют собой уровни связывания с FcγRIIaR и FcγRIIb, принятии за 100 уровней связывания контрольного гетеродимеризованного антитела.

В таблице 66 продемонстрировано, что изменения S239D и S239E, вводимые в этом эксперименте, при введении в нативный IgG1 независимо от введения в одну или обе цепи, повышали
связывание с FcRIIaR и FcγRIIb. Однако изменения при введении в обе Н-цепи IL6R-BP208/IL6R-L снижали активности связывания с FcγRIIaR и FcγRIIb и только при введении в одну Н-цепь повышали активности связывания с FcγRIIaR и FcγRIIb. Результат демонстрирует, что эффект изменений различается от эффекта, получаемого с использованием типа IgG1 в качестве матрицы. Таким образом, для этих изменений демонстрировали получение описанного выше эффекта только при введении в IL6R-BP208/IL6R-L.
[Пример 27] Конструирование константной области аминокислотной последовательности для улучшения возможности разделения и очищения гомодимеров и гетеродимеров
[Выбор участка замены остатка]
Совместная экспрессия двух типов Н-цепи (обозначаемых как А-цепь и В-цепь) для получения гетеродимеризованных антител приводит к образованию: гомодимеризованного антитела, получаемого в результате димеризиции Н-цепей, которые обе представляют собой А-цепи, гомодимеризованного антитела, получаемого в результате димеризация Н-цепей, которые обе представляют собой В-цепи, и гетеродимеризованного антитела, получаемого в результате димеризации Н-цепей, одна из которых представляет собой А-цепь, и другая представляет собой В-цепь. Известный способ эффективного разделения и очистки представляющих интерес гетеродимеризованных антител заключается в регуляции изоэлектрической точки антитела и возможности удержания и разделения на ионообменной колонке посредством замены аминокислотных остатков в каждой вариабельной области (WO
2007/114325). Однако вследствие того, что каждое антитело содержит различную последовательность в своей вариабельной области, в частности в области CDR, описанный выше способ обладает ограниченной возможностью применения. Затем в качестве более универсального способа замены остатков в антителах для очистки гетеродимеризованных антител авторы настоящего изобретения исследовали способ регуляции изоэлектрической точки и возможности удержания и разделения на ионообменной колонке посредством замены остатков только в константной области антитела.
В основном полагают, что разделение на ионообменной колонке зависит от электрических зарядов на поверхности молекул, и во многих случаях анализируют условия разделения путем изучения изоэлектрической точки молекулы-мишени. Таким образом, в этом примере предполагают, что разделение на ионообменной колонке также улучают посредством замены аминокислотных остатков, из которых состоит константная область антитела, таким способом, который приводит к разнице электрических точек гомодимеризованных антител и гетеродимеризованных антител, которые необходимо разделять.
При этом предполагали, что не только чистый ионообменный способ, а также гидрофобное взаимодействие участвуют в разделении ионообменной хроматографией (Peng Liu et al., J. Chromatogr. A. 2009 Oct 30; 1216 (44): 7497-504). По этой причине в разделении и очистке на основе описанного выше способа, в дополнение к ионообменной хроматографии можно использовать гидрофобную хроматографию.
Способы замены остатков для изменения изоэлектрической точки включают способы замены нейтрального остатка на основной или кислый остаток и способы замены основного или кислого остатка на нейтральный остаток. Более эффективные подходы включают способы замены положительно заряженного остатка на отрицательно заряженный остаток и способы замены отрицательно заряженного остатка на положительно заряженный остаток.
В описанных выше способах каждая часть последовательности антитела может представлять собой кандидата для участка замены остатка, что приводит к изменению изоэлектрической точки. Однако случайная замена, которая приводит к ненативной последовательности, может повышать риск иммуногенности и не представляет собой подходящий способ с точки зрения фармацевтического применения.
Остатки можно заменять так, чтобы минимизировать число Т-клеточных эпитопов, вовлеченных в иммуногенность, с целью минимизации повышения риска иммуногенности. Один возможный способ включает использование последовательностей подкласса IgG. Подклассы IgG человека включают IgG1, IgG2, IgG3 и IgG4. Заменой участков последовательности антитела на последовательности отличных подклассов способом, описанным в WO 2007/114325, можно изменять изоэлектрическую точку при подавлении повышения числа Т-клеточных эпитопов.
Доступные альтернативные способы включают средства теоретического расчета Т-клеточных эпитопов компьютерным моделированием, такие как Epibase.
Epibase Light (Lonza) представляет собой средство
компьютерного моделирования для теоретического расчета связывающей способности между 9-мономерным пептидом и основным аллелем DRB1 с использованием алгоритма FASTER (Expert. Opin. Biol. Ther., 2007 Mar; 7 (3): 405-18). Это средство обеспечивает идентификацию Т-клеточных эпитопов, которые сильно или умеренно связываются с МНС II класса.
Расчет отражает относительную распространенность аллотипов DRB1. Для этой цели можно использовать следующую ниже относительную распространенность в популяциях кавказцев.
DRB1*1501 (24,5%), DRB1*0301 (23,7%), DRB1*0701 (23,3%), DRB1*0101 (15,0%), DRB1*1101 (11,6%), DRB1*1302 (8,2%), DRB1*1401/1454 (4,9%), DRB1*0901 (2,3%), DRB1*1502 (0,5%), DRB1*1202 (0,1%).
Все эпитопы в каждой модифицированной последовательности антитела, для которых демонстрируют сильное или умеренное связывание, определяют посредством алгоритма FASTER, а затем выделяют критические эпитопы после исключения зародышевых последовательностей человека и последовательности соединения вариабельной области и константной области. Число Т-клеточных эпитопов, увеличиваемое заменой любого аминокислотного остатка для каждого остатка в любой последовательности, рассчитывают с использованием функции рандомизации этого средства. Это обеспечивает выбор участков замены остатка, которая приводит к изменению изоэлектрической точки без увеличения числа Т-клеточных эпитопов.
Н240-AK072 (SEQ ID NO: 104) и Н240-ВН076 (SEQ ID NO: 105) анализировали посредством Epibase. В таблице 67
продемонстрировано по отношению к Н240-AK072 число Т-клеточных эпитопов, которые можно изменять заменой любого остатка, тогда как в таблице 68 продемонстрировано число для Н240-ВН076. На основании результатов можно выбирать замену остатка, которая приводит к изменению изоэлектрической точки без увеличения Т-клеточных эпитопов.











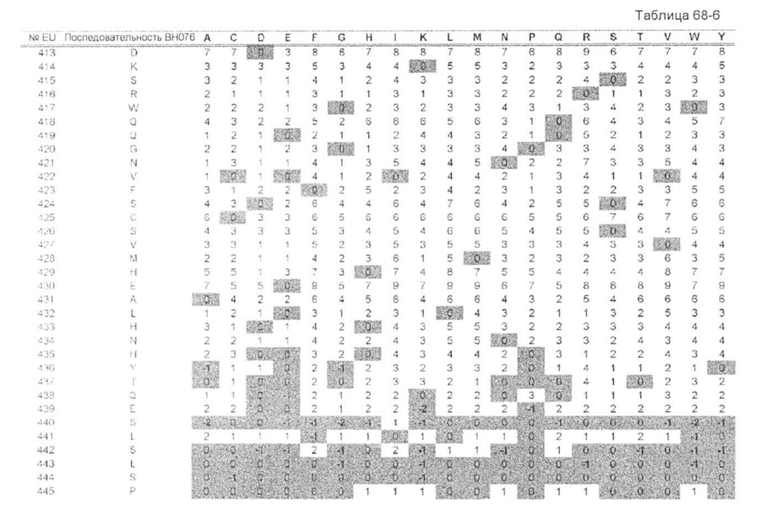
Описанными выше способами и их сочетанием описанные ниже варианты Н240-AK072 и Н240-ВН076 (таблицы 69 и 70) конструировали с учетом участков замены остатка для изменения изоэлектрической точки.


| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕТЕРОДИМЕРИЗОВАННЫЙ ПОЛИПЕПТИД | 2012 |
|
RU2641256C2 |
| FcγRIIb-СПЕЦИФИЧНОЕ Fc-АНТИТЕЛО | 2012 |
|
RU2608504C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАРИАНТОВ FcyRIIB-СПЕЦИФИЧЕСКОЙ Fc-ОБЛАСТИ | 2013 |
|
RU2820161C1 |
| СПОСОБ ИЗМЕНЕНИЯ УДЕРЖАНИЯ В ПЛАЗМЕ И ИММУНОГЕННОСТИ АНТИГЕНСВЯЗЫВАЮЩЕЙ МОЛЕКУЛЫ | 2012 |
|
RU2799423C1 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ЭЛИМИНАЦИИ АНТИГЕНОВ | 2012 |
|
RU2772771C1 |
| ВАРИАНТЫ FcγRIIB-СПЕЦИФИЧЕСКОЙ Fc-ОБЛАСТИ | 2013 |
|
RU2729831C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| ОПТИМИЗИРОВАННЫЕ Fc-ВАРИАНТЫ, ИМЕЮЩИЕ ИЗМЕНЕННОЕ СВЯЗЫВАНИЕ С FcγR, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2337107C2 |
| FC ВАРИАНТЫ АНТИТЕЛА | 2012 |
|
RU2607014C2 |


















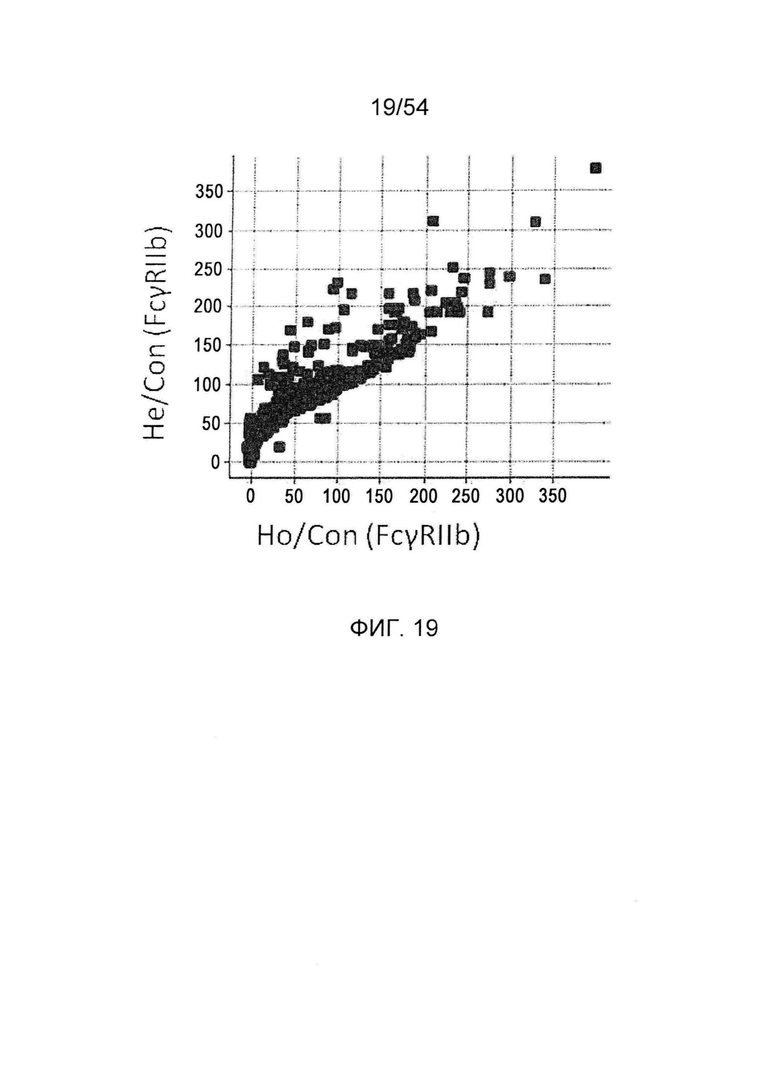
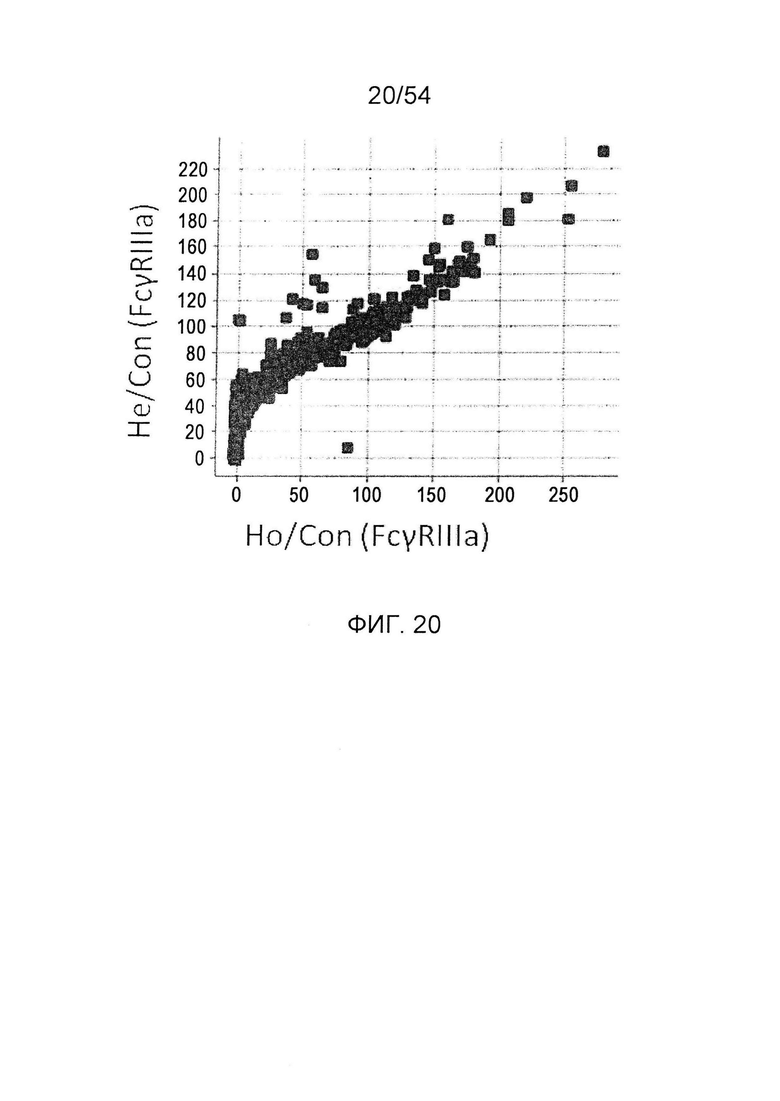






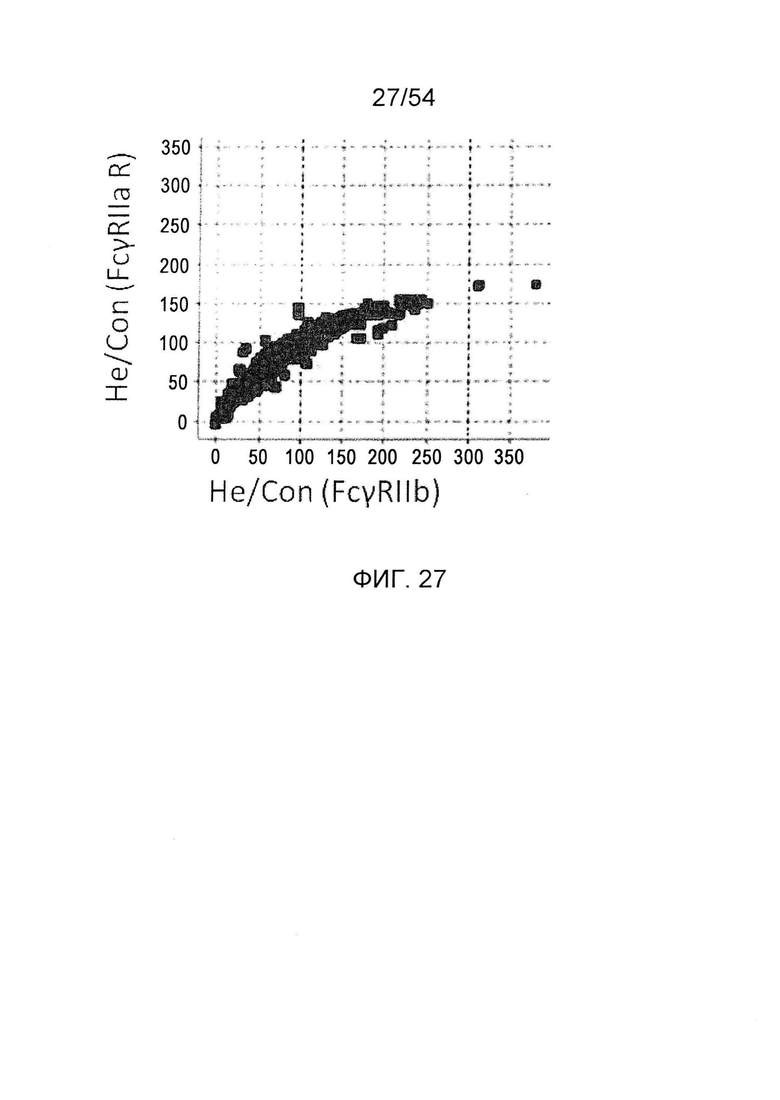
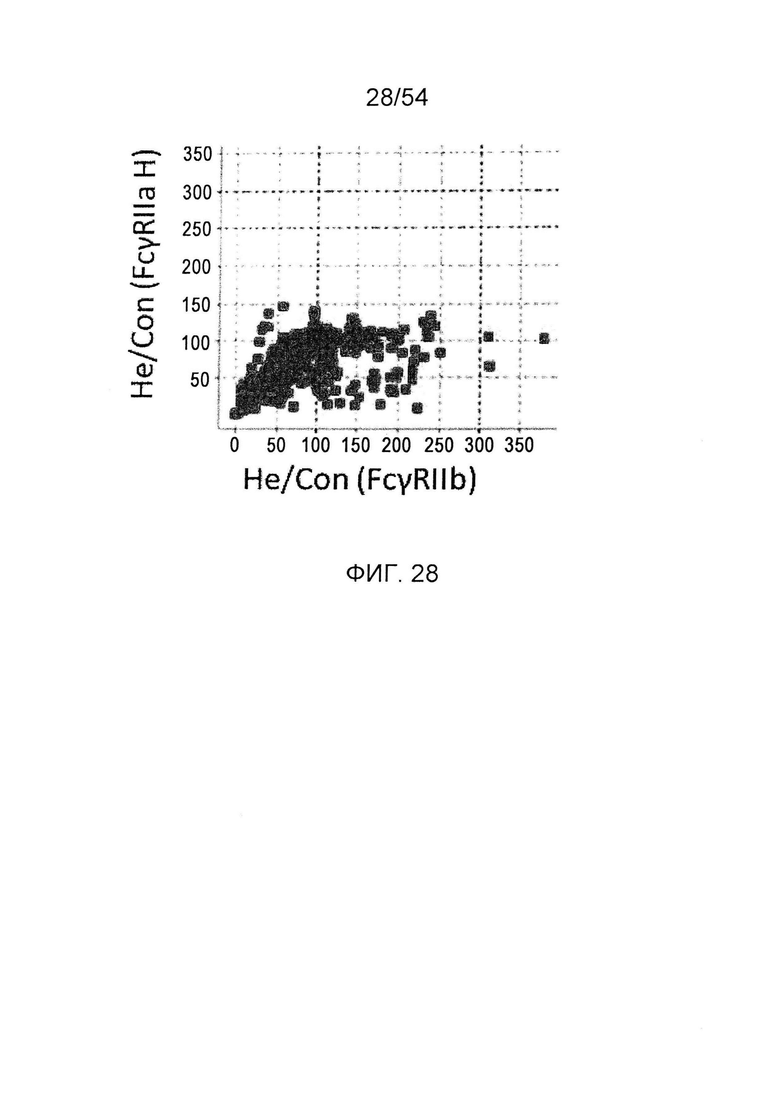

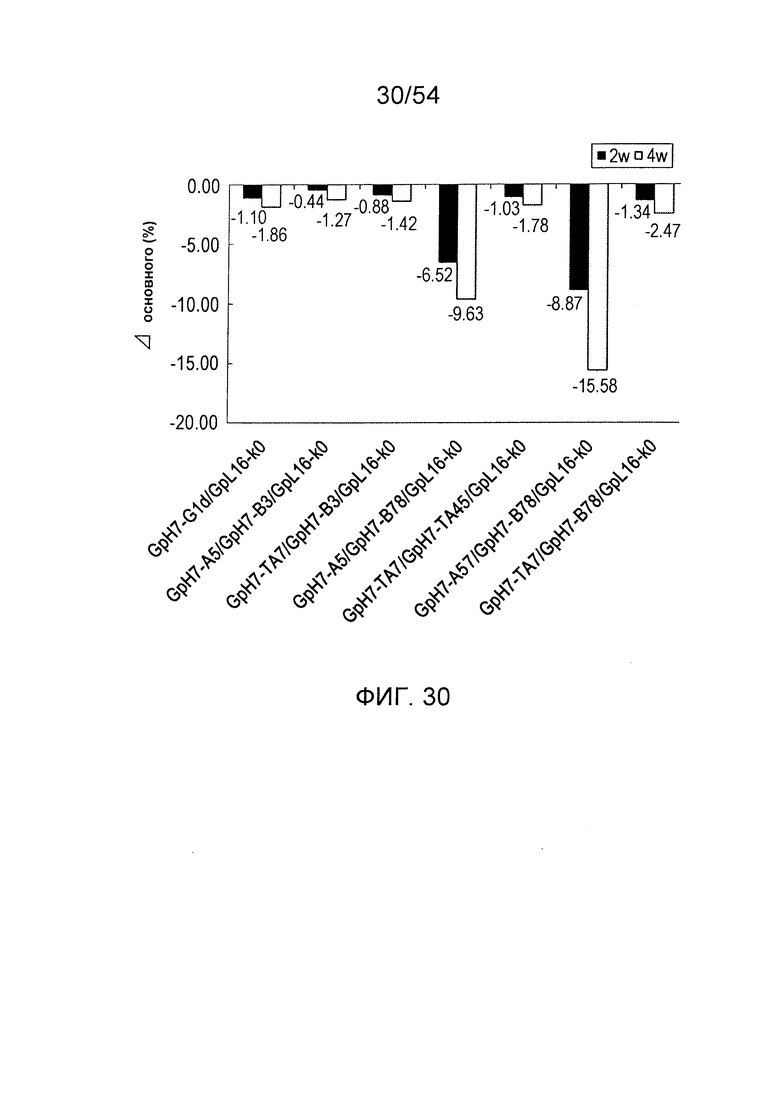
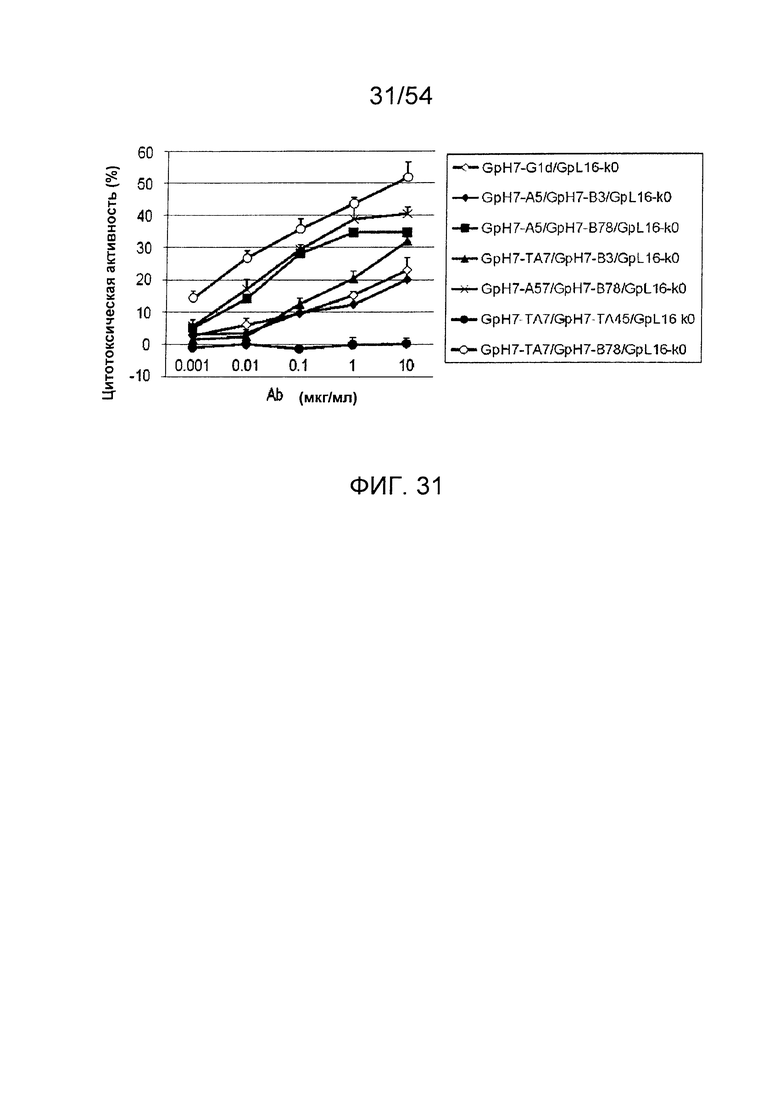


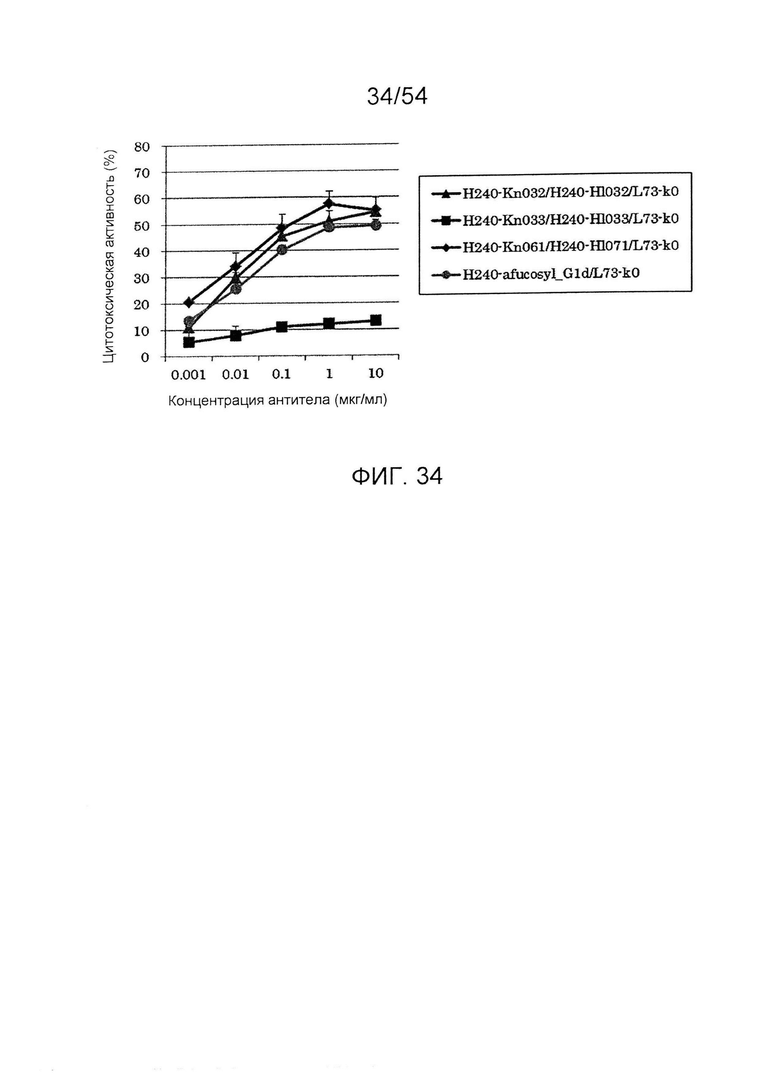




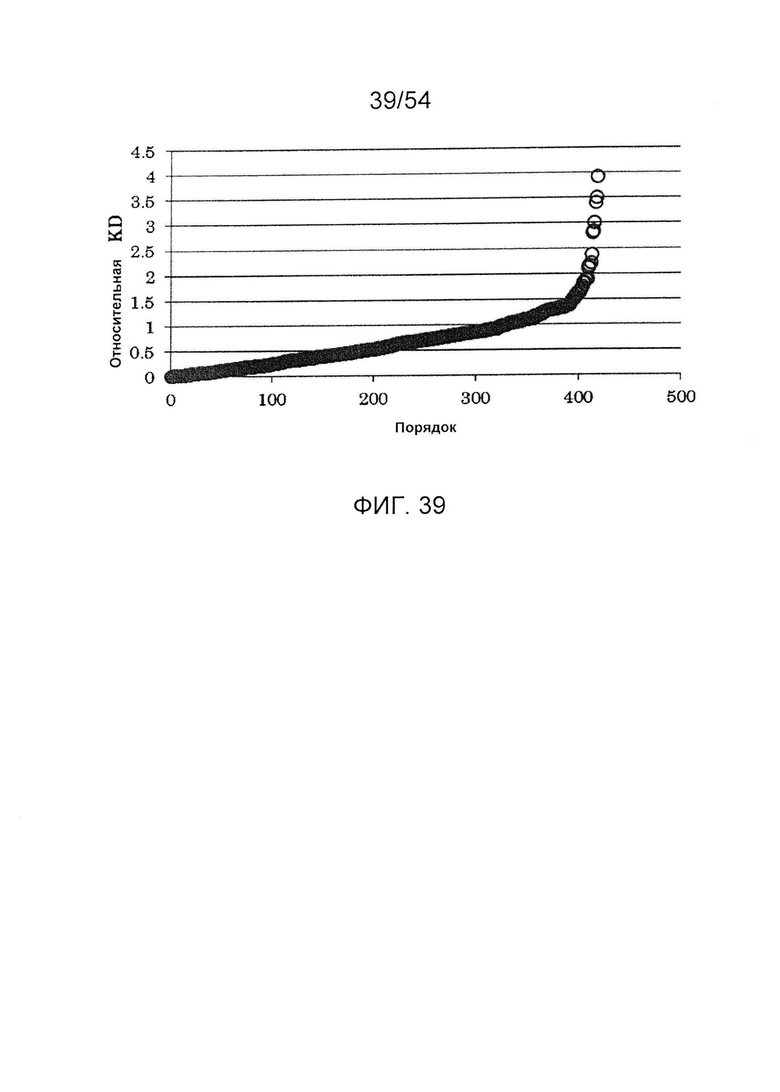
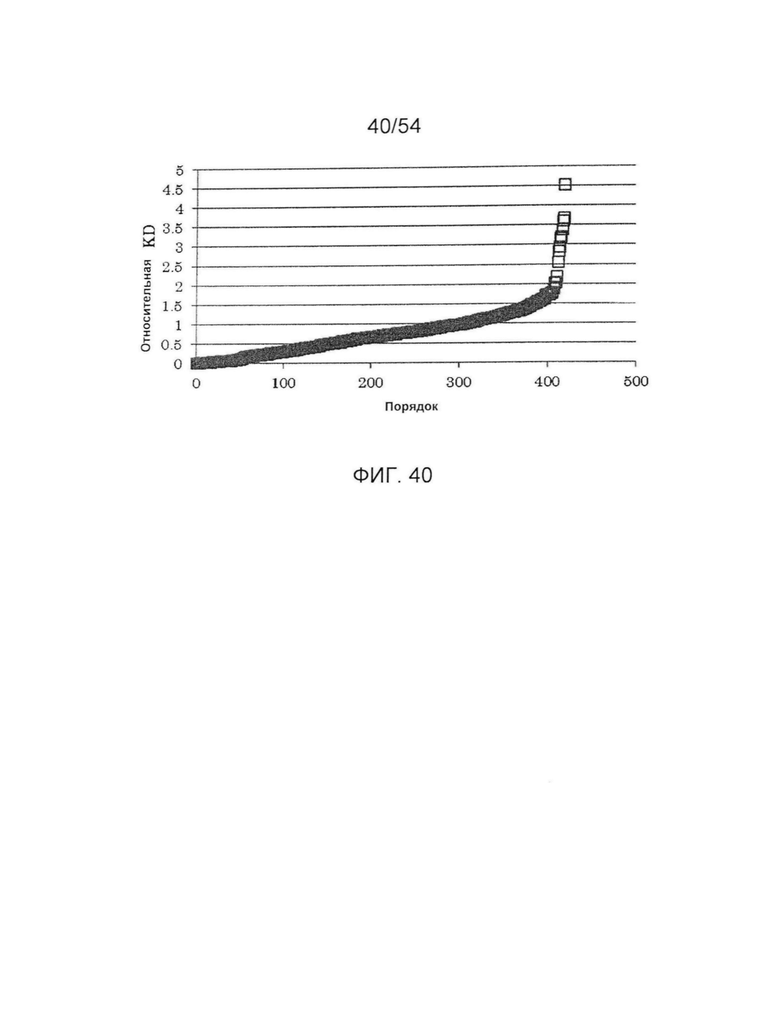

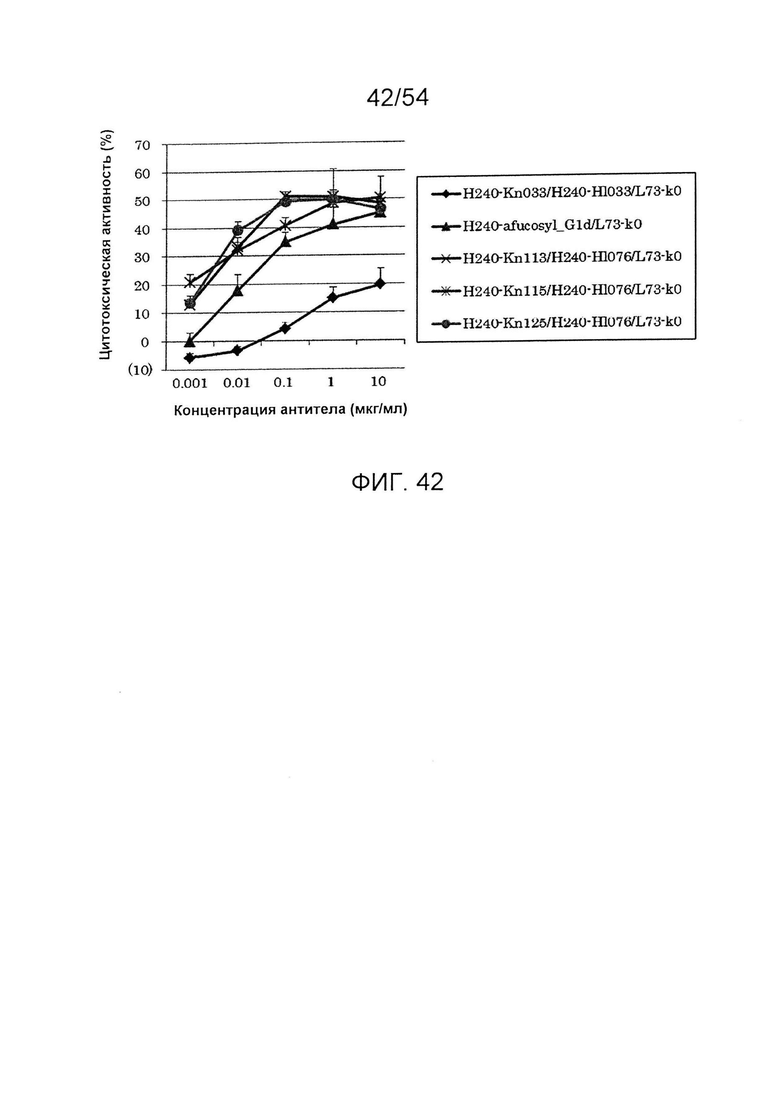










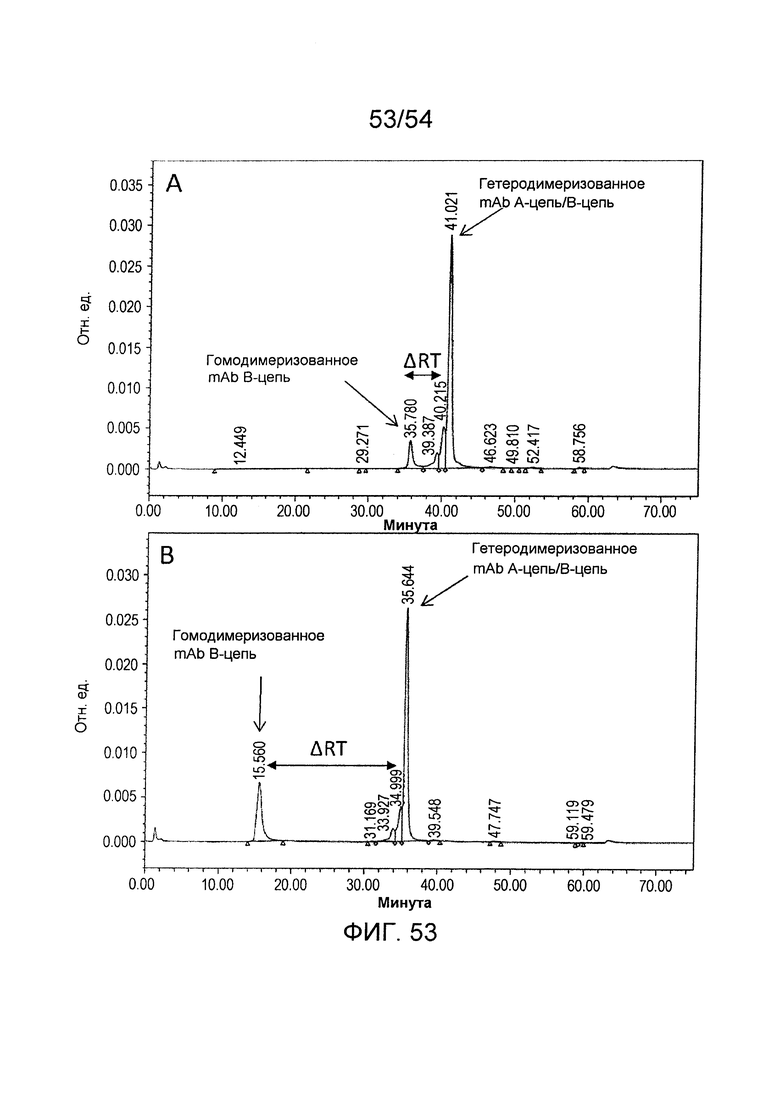

Изобретение относится к биотехнологии. Описан способ изменения функции полипептида, содержащего Fc-область IgG, который включает стадии гетеродимеризации Fc-области введением аминокислотной мутации(й) в первый полипептид и/или второй полипептид, образующий Fc-область, и введением аминокислотной мутации(й), чтобы изменить функцию Fc-области по сравнению с тем, когда Fc-область образует гомодимер, где любая одна серия мутаций от (i) до (vi) введена в аминокислотную последовательность любого полипептида из первого полипептида или второго полипептида, составляющих Fc-область IgG, и любая одна серия мутаций от (vii) до (ix) введена в аминокислотную последовательность другого полипептида. Также описан способ гетеродимеризации Fc-области посредством введения мутации(й) аминокислот в первый полипептид и/или второй полипептид, составляющих Fc-область, и введения мутации(й) аминокислот для изменения функции Fc-области по сравнению с тем, когда Fc-область образует гомодимер, в котором один из наборов мутаций (i)-(vi) вводили в аминокислотную последовательность первого полипептида или второго полипептида, составляющего Fc-область IgG, любой набор мутаций из (vii)-(ix) введен в аминокислотную последовательность другого полипептида. Изобретение позволяет улучшить функции Fc-области по сравнению с общепринятыми гомодимеризованными полипептидами, содержащими Fc-область. 2 н. и 10 з.п. ф-лы, 54 ил., 80 табл., 38 пр.
1. Способ изменения функции полипептида, содержащего Fc-область IgG, который включает стадии гетеродимеризации Fc-области введением аминокислотной мутации(й) в первый полипептид и/или второй полипептид, образующий Fc-область, и введением аминокислотной мутации(й), чтобы изменить функцию Fc-области по сравнению с тем, когда Fc-область образует гомодимер, где любая одна серия мутаций от (i) до (vi) введена в аминокислотную последовательность любого полипептида из первого полипептида или второго полипептида, составляющих Fc-область IgG, и любая одна серия мутаций от (vii) до (ix) введена в аминокислотную последовательность другого полипептида:
(i) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(ii) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iii) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Q;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты D в положении 270 (нумерация EU) на Е и
замены аминокислоты S в положении 298 (нумерация EU) на А;
(iv) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(v) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А и
замены аминокислоты А в положении 327 (нумерация EU) на D;
(vi) замены аминокислоты L в положении 234 (нумерация EU) на Y;
замены аминокислоты L в положении 235 (нумерация EU) на Y;
замены аминокислоты G в положении 236 (нумерация EU) на W;
замены аминокислоты S в положении 239 (нумерация EU) на М;
замены аминокислоты Н в положении 268 (нумерация EU) на D;
замены аминокислоты S в положении 298 (нумерация EU) на А;
замены аминокислоты А в положении 327 (нумерация EU) на D;
замены аминокислоты L в положении 328 (нумерация EU) на N и
замены аминокислоты К в положении 334 (нумерация EU) на L;
(vii) замены аминокислоты К в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты К в положении 334 (нумерация EU) на Е;
(viii) замены аминокислоты D в положении 20 (нумерация EU) на Е;
замены аминокислоты К в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на М и
замены аминокислоты К в положении 334 (нумерация EU) на Е;
(ix) замены аминокислоты D в положении 270 (нумерация EU) на Е;
замены аминокислоты K в положении 326 (нумерация EU) на D;
замены аминокислоты А в положении 330 (нумерация EU) на K и
замены аминокислоты K в положении 334 (нумерация EU) на Е.
2. Способ по п. 1, дополнительно содержащий стадию введения аминокислоты для обеспечения разницы изоэлектрических точек первого полипептида и второго полипептида.
3. Способ по п. 2, где изменение аминокислоты для обеспечения разницы изоэлектрических точек отличается введением по меньшей мере одной мутации аминокислоты в аминокислотный участок, выбранный из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Val в положении 263, Glu в положении 272, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 364, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Pro в положении 387, Asn в положении 390, Val в положении 397 и Val в положении 422 (нумерация EU), в аминокислотной последовательности первого полипептида и/или второго полипептида.
4. Способ по п. 2, где изменение аминокислоты для обеспечения разницы изоэлектрических точек отличается введением мутации по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gln в положении 196, Ile в положении 199, Val в положении 263, Glu в положении 272, Gly в положении 316, Leu в положении 358, Ser в положении 364, Ser в положении 383, Pro в положении 387 и Val в положении 397 (нумерация EU), в аминокислотной последовательности любого полипептида из первого полипептида или второго полипептида; и введением мутации по меньшей мере в одну аминокислоту, выбранную из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Tyr в положении 198, Не в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390 и Val в положении 422 (нумерация EU), в аминокислотной последовательности другого полипептида.
5. Способ по п. 1, где полипептид представляет собой антигенсвязывающую молекулу.
6. Способ по п. 5, где антигенсвязывающая молекула представляет собой антитело, биспецифическое антитело или слитую с Fc молекулу, такую как слитый с пептидом Fc белок или слитый с каркасной областью белок.
7. Способ получения полипептида, содержащего Fc-область IgG, который включает стадии гетеродимеризации Fc-области посредством введения мутации(й) аминокислот в первый полипептид и/или второй полипептид, составляющих Fc-область, и введения мутации(й) аминокислот для изменения функции Fc-области по сравнению с тем, когда Fc-область образует гомодимер, в котором один из наборов мутаций (i)-(vi) вводили в аминокислотную последовательность первого полипептида или второго полипептида, составляющего Fc-область IgG, любой набор мутаций из (vii)-(ix) введен в аминокислотную последовательность другого полипептида:
i. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты Н в положении 268 (нумерация EU) на D и замены аминокислоты S в положении 298 (нумерация EU) на А;
ii. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты D в положении 270 (нумерация EU) на Е и замены аминокислоты S в положении 298 (нумерация EU) на А;
iii. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Q; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты S в положении 239 (нумерация EU) на М; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты D в положении 270 (нумерация EU) на Е и замены аминокислоты S в положении 298 (нумерация EU) на А;
iv. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты S в положении 298 (нумерация EU) на А и замены аминокислоты А в положении 327 (нумерация EU) на D;
v. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты S в положении 239 (нумерация EU) на М; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты S в положении 298 (нумерация EU) на А и замены аминокислоты А в положении 327 (нумерация EU) на D;
vi. замены аминокислоты L в положении 234 (нумерация EU) на Y; замены аминокислоты L в положении 235 (нумерация EU) на Y; замены аминокислоты G в положении 236 (нумерация EU) на W; замены аминокислоты S в положении 239 (нумерация EU) на М; замены аминокислоты Н в положении 268 (нумерация EU) на D; замены аминокислоты S в положении 298 (нумерация EU) на А; замены аминокислоты А в положении 327 (нумерация EU) на D; замены аминокислоты L в положении 328 (нумерация EU) на W и замены аминокислоты K в положении 334 (нумерация EU) на L;
vii. замены аминокислоты K в положении 326 (нумерация EU) на D; замены аминокислоты А в положении 330 (нумерация EU) на М и замены аминокислоты K в положении 334 (нумерация EU) на Е;
viii. замены аминокислоты D в положении 270 (нумерация EU) на Е; замены аминокислоты К в положении 326 (нумерация EU) на D; замены аминокислоты А в положении 330 (нумерация EU) на М и замены аминокислоты K в положении 334 (нумерация EU) на Е;
ix. замены аминокислоты D в положении 270 (нумерация EU) на Е; замены аминокислоты К в положении 326 (нумерация EU) на D; замены аминокислоты А в положении 330 (нумерация EU) на K и замены аминокислоты К в положении 334 (нумерация EU) на Е.
8. Способ по п. 7, где дополнительно вводят изменение аминокислоты для обеспечения различий изоэлектрических точек первого полипептида и второго полипептида.
9. Способ по п. 8, где изменение аминокислоты для обеспечения различия изоэлектрических точек характеризуется введением по меньшей мере одной мутации аминокислоты на аминокислотном участке, выбранном из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Gln в положении 196, Tyr в положении 198, Ile в положении 199, Asn в положении 203, Lys в положении 214, Val в положении 263, Glu в положении 272, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 364, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Pro в положении 387, Asn в положении 390, Val в положении 397 и Val в положении 422 (нумерация EU), в аминокислотной последовательности первого полипептида и/или второго полипептида.
10. Способ по п. 8, где изменение аминокислоты для обеспечения различия изоэлектрических точек характеризуется введением мутации по меньшей мере в одной аминокислоте, выбранной из группы, состоящей из Gln в положении 196, Ile в положении 199, Val в положении 263, Glu в положении 272, Gly в положении 316, Leu в положении 358, Ser в положении 364, Ser в положении 383, Pro в положении 387 и Val в положении 397 (нумерация EU), в аминокислотной последовательности любого полипептида из первого полипептида или второго полипептида; и введением мутации по меньшей мере в одной аминокислоте, выбранной из группы, состоящей из Gly в положении 137, Gly в положении 138, Thr в положении 139, Lys в положении 147, Ser в положении 192, Leu в положении 193, Tyr в положении 198, Не в положении 199, Asn в положении 203, Lys в положении 214, Lys в положении 274, Tyr в положении 278, Lys в положении 288, Lys в положении 290, Gly в положении 316, Lys в положении 317, Lys в положении 320, Lys в положении 324, Thr в положении 335, Ser в положении 337, Lys в положении 340, Leu в положении 358, Lys в положении 360, Gln в положении 362, Ser в положении 383, Asn в положении 384, Gly в положении 385, Gln в положении 386, Asn в положении 390 и Val в положении 422 (нумерация EU), в аминокислотной последовательности другого полипептида.
11. Способ по п. 7, где полипептид представляет собой антигенсвязывающую молекулу.
12. Способ по п. 11, где антигенсвязывающая молекула представляет собой антитело, биспецифическое антитело или слитую с Fc молекулу, такую как слитый с пептидом Fc белок или слитый с каркасной областью Fc белок.
| WO 2011107989 A1, 09.09.2011 | |||
| RU 2006142852 А, 10.06.2008 | |||
| US 20080051563 А1, 28.02.2008 | |||
| WO 2006133486 А, 21.12.2006. |

