Область, к которой относится изобретение
Настоящее изобретение относится к вариантам Fc-области антитела, в которые была(и) введена(ы) аминокислотная(ые) модификация(и), и которые, по сравнению с Fc-областью, не имеющей такой(их) аминокислотной(ых) модификации(й), обладают более высокой FcγRIIb-связывающей активностью и/или более высокой селективностью связывания с FcγRIIb, но не с FcγRIIа (типа R); и к полипептидам, содержащим указанные варианты Fc-области, а также к фармацевтическим композициям, содержащим указанные полипептиды.
Предшествующий уровень техники
Антитела представляют особый интерес как фармацевтические средства, поскольку они обладают высокой стабильностью в крови и имеют мало побочных эффектов (непатентные документы 1 и 2). В настоящее время почти все терапевтические антитела, имеющиеся в продаже, представляют собой человеческие антитела подкласса IgG1. Одной из известных функций антител класса IgG является антитело-зависимая клеточно-опосредуемая цитотоксичность (далее называемая ADCC-активностью) (непатентный документ 3). В антителе, обладающим ADCC-активностью, Fc-область должна связываться с Fcγ-рецептором (далее обозначаемым FcγR), который представляет собой антитело-связывающий рецептор, присутствующий на поверхности эффекторных клеток, таких как клетки-киллеры, природные клетки-киллеры и активированные макрофаги.
Сообщалось, что человеческие изоформы FcγRIa (CD64A), FcγRIIa (CD32A), FcγRIIb (CD32B), FcγRIIIa (CD16A), FcγRIIIb (CD16B) представляют собой белки семейства FcγR, и были также описаны их соответствующие аллотипы (непатентный документ 7). FcγRIa, FcγRIIa и FcγRIIIa называются активирующими FcγR, поскольку они обладают иммунологически активными функциями, а FcγRIIb называется ингибирующим FcγR, поскольку он обладает иммуносупрессорными функциями (непатентный документ 8).
Было показано, что при связывании Fc-области с FcγR важную роль играют некоторые аминокислотные остатки, присутствующие в шарнирной области и в CH2-домене антитела, а также сахарная цепь, присоединенная к аминокислоте Asn в положении 297 (в соответствии с Европейской нумерацией), которая связана с CH2-доменом (непатентные документы 4, 5 и 6). Были проведены исследования различных вариантов, обладающих FcγR-связывающими свойствами, главным образом, антител с мутациями, введенными в их сайты, и были получены варианты Fc-области, обладающие более высокой активностью связывания с активирующим FcγR (патентные документы 1, 2, 3 и 4).
Если активирующий FcγR перекрестно связан с иммунным комплексом, то он фосфорилирует тирозиновые активирующие мотивы иммунорецептора (ITAM), содержащиеся во внутриклеточном домене, или общую γ-цепь FcR (партнера по связыванию); активирует SYK, осуществляющий передачу сигнала, и запускает воспалительный иммунный ответ посредством инициации каскада сигналов активации (непатентный документ 9).
FcγRIIb представляет собой FcγR, экспрессирующийся только на В-клетках (непатентный документ 10). Сообщалось, что взаимодействие Fc-области антитела с FcγRIIb приводит к подавлению первичного иммунного ответа В-клеток (непатентный документ 11). Кроме того, сообщалось, что если FcγRIIb, присутствующий на В-клетках, и В-клеточный рецептор (BCR) перекрестно связываются посредством иммунного комплекса в кровотоке, то наблюдается подавление активации В-клеток и подавление продуцирования антитела В-клетками (непатентный документ 12). Для такой передачи иммуносупрессорного сигнала, опосредуемого BCR и FcγRIIb, необходимо присутствие тирозинового ингибирующего мотива иммунорецептора (ITIM), содержащегося во внутриклеточном домене FcγRIIb (непатентные документы 13 и 14). При фосфорилировании ITIM после передачи сигнала происходит рекрутинг SH2-содержащей инозитполифосфат-5-фосфатазы (SHIP), ингибирование передачи других каскадов сигналов активирующего FcγR и подавление воспалительного иммунного ответа (непатентный документ 15). Кроме того, сообщалось, что агрегация лишь одного FcγRIIb приводит к временному подавлению притока кальция, обусловленного перекрестным связыванием BCR, и к пролиферации В-клеток по BCR-независимому механизму без индуцирования апоптоза IgМ-продуцирующих В-клеток (непатентный документ 16).
Кроме того, FcγRIIb также экспрессируется на дендритных клетках, макрофагах, активированных нейтрофилах, тучных клетках и базофилах. FcγRIIb ингибирует функции активирующего FcγR, такие как фагоцитоз и высвобождение воспалительных цитокинов в этих клетках, и подавляет воспалительные иммунные ответы (непатентный документ 8).
Важная роль иммуносупрессорных функций FcγRIIb была выявлена в исследованиях, проводимых на FcγRIIb-дефицитных мышах (нокаут-мышах). Сообщалось, что у FcγRIIb-дефицитных мышей не наблюдается соответствующей регуляции гуморального иммунного ответа (непатентный документ 17), а также наблюдается повышенная чувствительность к индуцированному коллагеном артриту (CIA) (непатентный документ 18), наличие симптомов, напоминающих волчанку, и симптомов, напоминающих синдром Гудпасчера (непатентный документ 19).
Кроме того, сообщалось, что такая неадекватная регуляция FcγRIIb ассоциируется с развитием аутоиммунных заболеваний у человека. Так, например, сообщалось о наличии взаимосвязи между генетическим полиморфизмом в трансмембранной области и в промоторной области FcγRIIb и частотой развития системной красной волчанки (СКВ) (непатентные документы 20, 21, 22, 23 и 24), а также о снижении уровня экспрессии FcγRIIb на поверхности В-клеток у пациентов с СКВ (непатентные документы 25 и 26).
Исследования, проводимые на мышиных моделях, и клинические исследования показали, что FcγRIIb играет определенную роль в подавлении аутоиммунных заболеваний и воспалительных заболеваний посредством связывания с конкретными В-клетками, а поэтому такая молекула представляет особый интерес как мишень для подавления аутоиммунных заболеваний и воспалительных заболеваний.
Известно, что IgG1, используемый, главным образом, в качестве коммерчески доступного терапевтического антитела, связывается не только с FcγRIIb, но также обладает высокой активностью связывания с активирующим FcγR (непатентный документ 27). Поэтому, с использованием Fc-области, обладающей более высокой активностью связывания с FcγRIIb или более высокой селективностью связывания с FcγRIIb по сравнению с активирующим FcγR, могут быть разработаны терапевтические антитела, обладающие более эффективными иммуносупрессорными свойствами по сравнению с IgG1. Так, например, было высказано предположение, что использование антитела, имеющего вариабельную область, которая связывается с BCR, и Fc-область, которая обладает повышенной способностью связываться с FcγRIIb, дает возможность ингибировать активацию В-клеток (непатентный документ 28). Сообщалось, что перекрестное связывание FcγRIIb на В-клетках и IgE, связанного с B-клеточным рецептором, приводило к подавлению дифференцировки В-клеток в клетки плазмы, что, в свою очередь, приводило к подавлению продуцирования IgЕ, а у мышей, которым были трансплантированы человеческие МКПК, концентрации человеческого IgG и IgМ сохранялись, тогда как концентрация человеческого IgЕ снижалась (непатентный документ 29). Сообщалось, что если FcγRIIb и CD79b, которые представляют собой молекулы-компоненты комплекса В-клеточного рецептора, перекрестно связываются не только с IgЕ, но с другим антителом, то пролиферация В-клеток подавляется in vitro, и при этом может наблюдаться ослабление симптомов артрита у модели с артритом, индуцированным коллагеном (непатентный документ 30).
Сообщалось, что перекрестное связывание FcεRI и FcγRIIb не только на В-клетках, но и на тучных клетках, посредством молекул, в которых Fc-часть IgG, обладающего повышенной активностью связывания с FcγRIIb, присоединена к Fc-части IgЕ, связывающейся с рецептором FcεRI IgE, приводит к фосфорилированию FcγRIIb и, тем самым, к подавлению FcεRI-зависимого притока кальция. Это дает основание предположить, что ингибирование дегрануляции посредством стимуляции FcγRIIb может быть обусловлено повышением активности связывания с FcγRIIb (непатентный документ 31).
В соответствии с этим было высказано предположение, что антитело, имеющие Fc с повышенной активностью связывания с FcγRIIb, может представлять особый интерес как терапевтическое средство для лечения воспалительных заболеваний, таких как аутоиммунные заболевания.
Кроме того, сообщалось, что активация макрофагов и дендритных клеток посредством Toll-подобного рецептора 4, обусловленная стимуляцией ЛПС, подавляется в присутствии иммунного комплекса «антитело-антиген», и, предполагается, что такой эффект также обусловлен действием иммунного комплекса, опосредуемым FcγRIIb (непатентные документы 32 и 33). Поэтому ожидается, что использование антител с повышенной активностью связывания с FcγRIIb позволит усиливать подавление активации сигнала, опосредуемого TLR, и, таким образом, можно предположить, что такие антитела представляют особый интерес как терапевтические средства для лечения воспалительных заболеваний, таких как аутоиммунные заболевания.
Кроме того, было высказано предположение, что мутанты с повышенной активностью связывания с FcγRIIb представляют особый интерес как терапевтические средства для лечения рака, а также как терапевтические средства для лечения воспалительных заболеваний, таких как аутоиммунные заболевания. Было обнаружено, что FcγRIIb играет важную роль в агонистической активности антител-агонистов против суперсемейства рецепторов TNF. В частности, было высказано предположение, что взаимодействие с FcγRIIb необходимо для агонистической активности антител против CD40, DR4, DR5, CD30 и CD137, которые принадлежат к семейству рецепторов TNF (непатентные документы 34, 35, 36, 37, 38, 39 и 40). В непатентном документе 34 описано использование антител с повышенной активностью связывания с FcγRIIb в целях усиления противоопухолевого эффекта анти-CD40 антител. В соответствии с этим, можно предположить, что антитела с повышенной активностью связывания с FcγRIIb обладают действием, направленным на повышение агонистической активности антител-агонистов, включая антитела против суперсемейства рецепторов TNF.
Кроме того, было показано, что пролиферация клеток может быть ингибирована с использованием антитела, которое распознает Kit, то есть тирозинкиназный рецептор (RTK), и перекрестно связывается с FcγRIIb на Kit-экспрессирующих клетках. Аналогичные эффекты наблюдались даже в тех случаях, когда этот рецептор Kit является конститутивно активированным и имеет мутации, приводящие к онкогенезу (непатентный документ 41). Поэтому предполагается, что использование антител с повышенной активностью связывания с FcγRIIb будет приводить к усилению ингибирующего действия на клетки, экспрессирующие RTK, имеющий конститутивно активированные мутации.
Имеются также данные об антителах, содержащих Fc с повышенной активностью связывания с FcγRIIb (непатентный документ 28). В этом документе FcγRIIb-связывающая активность была усилена путем введения модификаций, таких как S267E/L328F, G236D/S267E и S239D/S267E, в Fc-область антитела. Из этих антител, антитело, в которое была введена мутация S267E/L328F, исключительно сильно связывалось с FcγRIIb и сохраняло на том же уровне свою способность связываться с FcγRIа и FcγRIIа типа H, где остаток в положении 131 FcγRIIа представляет собой His, соответствующий остатку природного IgG1. Однако в другой работе показано, что при введении такой модификации уровень связывания антитела с FcγRIIа типа R, где остатком в положении 131 FcγRIIа является Arg, в несколько сотен раз выше, чем уровень связывания этого антитела с FcγRIIb, что означает, что его селективность связывания с FcγRIIb не отличается от селективности связывания с FcγRIIа типа R (патентный документ 5).
Считается, что на клетки, такие как тромбоциты, которые экспрессируют FcγRIIа, но не экспрессируют FcγRIIb, влияет только эффект усиления связывания с FcγRIIа, но не эффект усиления связывания с FcγRIIb (непатентный документ 8). Так, например, известно, что у группы пациентов, которым вводили бевацизумаб, то есть антитело против VEGF, наблюдался повышенный риск развития тромбоэмболии (непатентный документ 42). Кроме того, аналогичным образом, тромбоэмболия наблюдалась в клинических испытаниях на образование антител против лиганда CD40, а поэтому такое клиническое испытание было прервано (непатентный документ 43). В результате последних исследований, проводимых на животных-моделях с использованием этих антител, было высказано предположение, что вводимые антитела способствуют агрегации тромбоцитов посредством связывания с FcγRIIа на тромбоцитах и образуют кровяные сгустки (непатентные документы 44 и 45). В случае системной красной волчанки, которая является аутоиммунным заболеванием, тромбоциты активируются по FcγRIIа-зависимому механизму, и сообщалось, что такая активация тромбоцитов коррелирует с тяжестью симптомов (непатентный документ 46). Введение антитела с повышенной активностью связывания с FcγRIIа таким пациентам с высоким риском развития тромбоэмболии приводит к повышению риска развития тромбоэмболии и, таким образом, представляет исключительную опасность для пациента.
Кроме того, сообщалось, что антитела с повышенной активностью связывания с FcγRIIа усиливают опосредуемый макрофагами антитело-зависимый клеточный фагоцитоз (ADCP) (непатентный документ 47). Если антигены, связанные с антителами, подвергаются фагоцитозу под действием макрофагов, то считается, что сами эти антитела также одновременно подвергаются фагоцитозу. Если антитела вводят в качестве фармацевтических средств, то предполагается, что пептидные фрагменты, происходящие от вводимых антител, также присутствуют в качестве антигена, что тем самым повышает риск продуцирования антител против терапевтических антител (антитерапевтические антитела). Более конкретно, при повышении активности связывания с FcγRIIа повышается риск продуцирования антител против терапевтических антител, что значительно снижает ценность этих антител как фармацевтических средств. Кроме того, было высказано предположение, что FcγRIIb на дендритных клетках вносит свой вклад в периферическую толерантность посредством ингибирования активации дендритных клеток, вызываемой иммунными комплексами, образованными между антигенами и антителами, или посредством супрессии презентации антигена Т-клеткам посредством активирующих Fcγ-рецепторов (непатентный документ 48). Поскольку FcγRIIа также экспрессируется на дендритных клетках, то в случае использования антител, имеющих Fc с повышенной селективностью связывания с FcγRIIb, в качестве фармацевтических средств презентация антигенов этими дендритными клетками становится маловероятной, и вследствие повышения селективности связывания с FcγRIIb риск продуцирования антитела против лекарственного средства может быть относительно снижен. А поэтому, такие антитела также могут представлять определенную ценность.
Более конкретно, при повышении активности связывания с FcγRIIа, ценность антител как фармацевтических средств значительно снижается, что приводит к повышению риска образования тромбов в результате агрегации тромбоцитов и к повышению риска продуцирования антител против терапевтических средств из-за повышенной иммуногенности.
С этой точки зрения, вышеупомянутый Fc-вариант с повышенной активностью связывания с FcγRIIb обладает значительно более высокой активностью связывания с FcγRIIа типа R по сравнению с природным IgG1. Поэтому его ценность как фармацевтического средства для пациентов, имеющих FcγRIIа типа R, значительно снижается. FcγRIIа типа Н и R встречается у людей индоевропейской расы и у афроамериканцев приблизительно с одинаковой частотой (непатентные документы 49 и 50). Поэтому, если такой Fc-вариант используется для лечения аутоиммунных заболеваний, то число пациентов, у которых может наблюдаться безопасный и благоприятный эффект от введения этого фармацевтического средства, ограничено.
Кроме того, сообщалось, что дендритные клетки, дефицитные по FcγRIIb, или дендритные клетки, в которых взаимодействие между FcγRIIb и Fc-частью антитела ингибируются анти-FcγRIIb антителом, способны к созреванию (непатентные документы 51 и 52). Эти данные позволяют предположить, в дендритных клетках в стабильном состоянии не наблюдаются воспаление и активация, а поэтому предполагается, что FcγRIIb активно подавляет созревание дендритных клеток. FcγRIIа, помимо FcγRIIb, экспрессируются на поверхности дендритных клеток, поэтому, даже при повышении активности связывания с ингибирующим FcγRIIb, а также при повышении активности связывания с активирующим FcγR, таким как FcγRIIа, может происходить стимуляция созревания дендритных клеток. Более конкретно, повышение активности связывания не только FcγRIIb, но также и отношение активности связывания с FcγRIIb к активности связывания с FcγRIIа, рассматривается как важный фактор при получении антител, обладающих иммуносупрессорным действием.
Поэтому, при получении фармацевтических средств, обладающих иммуносупрессорным действием, опосредуемым связыванием с FcγRIIb, необходимо создать Fc-вариант, который будет обладать не только повышенной активностью связывания с FcγRIIb, но также и способностью связываться с FcγRIIа аллотипов типа Н и R, где такое связывание будет сохраняться на уровне, аналогичном уровню связывания природного IgG1, или на более низком уровне.
С другой стороны, сообщалось, что в некоторых случаях аминокислотные модификации могут быть введены в Fc-область для повышения селективности связывания с FcγRIIb (непатентный документ 53). Однако все варианты, которые якобы имеют повышенную селективность к FcγRIIb, как упоминается в этом документе, обладают более низкой способностью связываться с FcγRIIb по сравнению с природным IgG1. Поэтому считается, что фактически индуцирование этими вариантами иммуносупрессорной FcγRIIb-опосредуемой реакции, которая была бы более сильной, чем реакция под действием IgG1, представляет определенные трудности.
Кроме того, поскольку FcγRIIb играет важную роль в продуцировании вышеупомянутых антител-агонистов, то предполагается, что усиление их активности связывания будет приводить к усилению агонистической активности. Однако, при аналогичном усилении активности связывания с FcγRIIа, может наблюдаться непредусмотренная активность, такая как ADCC-активность и ADCP-активность, а поэтому могут возникать побочные эффекты. С этой точки зрения также предпочтительно, чтобы такая повышенная активность связывания с FcγRIIb была селективной.
Исходя из этих результатов, важно отметить, что при продуцировании терапевтических антител, используемых для лечения аутоиммунных заболеваний и рака с применением FcγRIIb, активность связывания с FcγRIIа обоих аллотипов сохранялась на том же уровне как и для природного IgG, либо такая активность была более низкой, а активность связывания с FcγRIIb была более высокой. Однако, последовательность рецептора FcγRIIb во внеклеточной области на 93% идентична последовательности рецептора FcγRIIа, который является одним из активирующих FcγR, и такие последовательности по своей структуре являются почти аналогичными. Существуют аллотипы FcγRIIа типа Н и R, в которых аминокислотой в положении 131 является His (типа Н) или Arg (типа R), и реакции каждого из них с антителами отличаются друг от друга (непатентный документ 54). Поэтому проблема продуцирования варианта Fc-области, обладающего более высокой селективностью связывания с FcγRIIb по сравнению с селективностью связывания с FcγRIIа каждого аллотипа, заключается в трудности идентификации в высокой степени гомологичных последовательностей FcγRIIа и FcγRIIb. До сих пор, фактически, еще не получены варианты, обладающие достаточной активностью и селективностью связывания с FcγRIIb. В патентном документе 5 описаны варианты, обладающие повышенной активностью связывания с FcγRIIb, однако, степень такого повышения является низкой, а поэтому необходимо разработать варианты, которые обладали бы свойствами, аналогичными свойствам, описанным выше.
Документы предшествующего уровня техники
[Патентные документы]
[Патентный документ 1] WO 2000/42072
[Патентный документ 2] WO 2006/019447
[Патентный документ 3] WO 2004/99249
[Патентный документ 4] WO 2004/29207
[Патентный документ 5] US 2009/0136485
[Непатентные документы]
[Непатентный документ 1] Nat Biotechnol, 23, 1073-1078, 2005
[Непатентный документ 2] Eur J Pharm Biopharm, 59 (3), 389-96, 2005
[Непатентный документ 3] Chem Immunol, 65, 88-110, 1997
[Непатентный документ 4] J Biol Chem, 276, 16478-16483, 2001
[Непатентный документ 5] Eur J Immunol 23, 1098-1104, 1993
[Непатентный документ 6] Immunology, 86, 319-324, 1995
[Непатентный документ 7] Immunol Lett, 82, 57-65, 2002
[Непатентный документ 8] Nat Rev Immunol, 10, 328-343, 2010
[Непатентный документ 9] Nat Rev Immunol, 8, 34-47, 2008
[Непатентный документ 10] Eur J Immunol 19, 1379-1385, 1989
[Непатентный документ 11] J Exp Med 129, 1183-1201, 1969
[Непатентный документ 12] Immunol Lett 88, 157-161, 2003
[Непатентный документ 13] Science, 256, 1808-1812, 1992
[Непатентный документ 14] Nature, 368, 70-73, 1994
[Непатентный документ 15] Science, 290, 84-89, 2000
[Непатентный документ 16] J Imunol, 181, 5350-5359, 2008
[Непатентный документ 17] J Immunol, 163, 618-622, 1999
[Непатентный документ 18] J Exp Med, 189, 187-194, 1999
[Непатентный документ 19] J Exp Med, 191, 899-906, 2000
[Непатентный документ 20] Hum, Genet, 117, 220-227, 2005
[Непатентный документ 21] J Biol Chem, 282, 1738-1746, 2007
[Непатентный документ 22] Arthritis Rheum, 54, 3908-3917, 2006
[Непатентный документ 23] Nat Med, 11, 1056-1058, 2005
[Непатентный документ 24] J Immunol, 176, 5321-5328, 2006
[Непатентный документ 25] J. Exp Med, 203, 2157-2164, 2006
[Непатентный документ 26] J Immunol 178, 3272-3280, 2007
[Непатентный документ 27] Blood, 113, 3716-3725, 2009
[Непатентный документ 28] Mol Immunol, 45, 3926-3933, 2008
[Непатентный документ 29] J Allergy Clin Immunol, 2012, 129: 1102-1115
[Непатентный документ 30] Arthritis Rheum, 62, 1933-1943, 2010
[Непатентный документ 31] Immunol let, doi: 10.1016/j.imlet.2012.01.008)
[Непатентный документ 32] J. Immunol, 2009, 183: 4509-4520
[Непатентный документ 33] J. Immunol, 2009, 182: 554-562
[Непатентный документ 34] Science, 333, 1030-1034, 2011
[Непатентный документ 35] Cancer Cell 19, 101-113, 2011
[Непатентный документ 36] J Clin Invest, 122 (3), 1066-1075, 2012
[Непатентный документ 37] J Immunol 171, 562-, 2003
[Непатентный документ 38] Blood, 108, 705-, 2006
[Непатентный документ 39] J Immunol 166, 4891, 2001
[Непатентный документ 40] doi: 10.1073/pnas.1208698109
[Непатентный документ 41] Immunol let, 2002, 143: 28-33
[Непатентный документ 42] J Natl Cancer Inst, 99, 1232-1239, 2007
[Непатентный документ 43] Arthritis Rheum, 48, 719-727, 2003
[Непатентный документ 44] J Thromb Haemost, 7, 171-181, 2008
[Непатентный документ 45] J Immunol, 185, 1577-1583, 2010
[Непатентный документ 46] Sci Transl Med, vol 2, issue 47, 47-63, 2010
[Непатентный документ 47] Mol Cancer Ther 7, 2517-2527, 2008
[Непатентный документ 48] J. Immunol, 2007, 178: 6217-6226
[Непатентный документ 49] J Clin Invest, 97, 1348-1354, 1996
[Непатентный документ 50] Arthritis Rheum, 41, 1181-1189, 1998
[Непатентный документ 51] J Clin Invest 115, 2914-2923, 2005
[Непатентный документ 52] Proc Natl Acad Sci USA, 102, 2910-2915, 2005
[Непатентный документ 53] Mol Immunol, 40, 585-593, 2003
[Непатентный документ 54] J Exp Med, 172, 19-25, 1990
Описание сущности изобретения
[Проблемы, которые могут быть решены с помощью настоящего изобретения]
Настоящее изобретение было разработано с учетом вышеупомянутых факторов. Целью настоящего изобретения является получение варианта Fc-области путем введения аминокислотной(ых) модификации(й) в Fc-область антитела, где указанный вариант обладает повышенной активностью связывания с FcγRIIb и/или повышенной селективностью связывания с FcγRIIb по сравнению с активностью и/или селективностью с FcγRIIа (R-типа), в отличие от Fc-области, в которую не была введена аминокислотная модификация; получение полипептида, содержащего вариант Fc-области; и получение фармацевтической композиции, содержащей указанный полипептид.
[Способ решения указанных проблем]
Авторами настоящего изобретения были проведены исследования, посвященные получению варианта Fc-области, обладающей повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа (типа R), в отличие от немодифицированной Fc-области, путем введения аминокислотной(ых) модификации(й) в Fc-область, и получению полипептида, содержащего указанный вариант Fc-области. В результате, авторами настоящего изобретения было обнаружено, что активность связывания с FcγRIIb и/или селективность связывания с FcγRIIb по сравнению с активностью и/или селективностью связывания с FcγRIIа (R-типа) может быть повышена путем объединения варианта Fc-области, в котором аминокислота в положении 238 (в соответствии с Европейской нумерацией) в Fc-области была заменена другой(ими) аминокислотой(ами).
Более конкретно, настоящее изобретение относится:
[1] к варианту Fc-области, в котором аминокислота в положении 238 в соответствии с Европейской нумерацией и по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 233, 234, 235, 237, 264, 265, 266, 267, 268, 269, 271, 272, 274, 296, 326, 327, 330, 331, 332, 333, 334, 355, 356, 358, 396, 409 и 419 в соответствии с Европейской нумерацией, заменены другими аминокислотами, где активность связывания варианта с Fcγ-рецепторами [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 15,0 или более;
[2] к варианту Fc-области [1], где аминокислоты в положениях 238, 268 и 271 в соответствии с Европейской нумерацией заменены другими аминокислотами, и где, кроме того, по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 233, 237, 264, 267, 272, 296, 327, 330, 332 и 396 в соответствии с Европейской нумерацией, заменена другими аминокислотами;
[3] к варианту Fc-области, в котором аминокислота в положении 238 в соответствии с Европейской нумерацией представляет собой Asp, и который содержит по меньшей мере одну аминокислоту, выбранную из группы аминокислот, состоящей из аминокислоты Asp в положении 233, аминокислоты Tyr в положении 234, аминокислоты Phe в положении 235, аминокислоты Asp в положении 237, аминокислоты Ile в положении 264, аминокислоты Glu в положении 265, аминокислот Phe, Leu или Met в положении 266, аминокислот Ala, Glu, Gly или Gln в положении 267, аминокислот Asp, Gln или Glu в положении 268, аминокислоты Asp в положении 269, аминокислоты Gly в положении 271, аминокислот Asp, Phe, Ile, Met, Asn, Pro или Gln в положении 272, аминокислоты Gln в положении 274, аминокислот Asp или Phe в положении 296, аминокислот Ala или Asp в положении 326, аминокислоты Gly в положении 327, аминокислот Lys, Arg или Ser в положении 330, аминокислоты Ser в положении 331, аминокислот Lys, Arg, Ser или Thr в положении 332, аминокислот Lys, Arg, Ser или Thr в положении 333, аминокислот Arg, Ser или Thr в положении 334, аминокислот Ala или Gln в положении 355, аминокислоты Glu в положении 356, аминокислоты Met в положении 358, аминокислот Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в положении 396, аминокислоты Arg в положении 409 и аминокислоты Glu в положении 419, где активность связывания варианта с Fcγ-рецепторами [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 15,0 или более;
[4] к варианту Fc-области [3], где аминокислотой в положении 238 является Asp, аминокислотой в положении 268 является Asp или Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией, и где указанный вариант Fc-области также содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из аминокислоты Asp в положении 233, аминокислоты Asp в положении 237, аминокислоты Ile в положении 264, аминокислоты Ala или Gly в положении 267, аминокислоты Asp или Pro в положении 272, аминокислоты Asp в положении 296, аминокислоты Gly в положении 327, аминокислоты Arg в положении 330, аминокислоты Thr в положении 332 и аминокислоты Leu или Met в положении 396;
[5] к варианту Fc-области по любому из п.п. [1]-[4], где величина [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 50,0 или более;
[6] к варианту Fc-области по любому из п.п. [1]-[4], где величина [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 100,0 или более;
[7] к варианту Fc-области по любому из п.п. [1]-[6], где величина [величина KD для FcγRIIа (R-типа) полипептида, содержащего вариант Fc-области]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 10,0 или более;
[8] к варианту Fc-области по любому из п.п. [1]-[6], где величина [величина KD для FcγRIIа (R-типа) полипептида, содержащего вариант Fc-области]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 20,0 или более;
[9] к варианту Fc-области по любому из п.п. [1]-[8], где указанный вариант Fc-области содержит любую из нижеследующих групп аминокислотных модификаций (а)-(х), таких как:
(а) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(b) аминокислотные модификации в положениях 238, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(c) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(d) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(е) аминокислотные модификации в положениях 238, 233, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(f) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(g) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 327 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(h) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(i) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(j) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(k) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(l) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(m) аминокислотные модификации в положениях 238, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(n) аминокислотные модификации в положениях 238, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(o) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(p) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(q) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 327, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(r) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(s) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(t) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(u) аминокислотные модификации в положениях 238, 233, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(v) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(w) аминокислотные модификации в положениях 238, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области; и
(x) аминокислотные модификации в положениях 238, 233, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области;
[10] к варианту Fc-области по любому из п.п. [1]-[8], где указанный вариант Fc-области содержит любую из нижеследующих аминокислотных последовательностей (а)-(х), таких как:
(a) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(b) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(c) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(d) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Gly или Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(e) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(f) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(g) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(h) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(i) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(j) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(k) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(l) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(m) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(n) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(o) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(p) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(q) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met в соответствии с Европейской нумерацией;
(r) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(s) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(t) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(u) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(v) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Gly, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(w) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией; и
(x) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
[11] к варианту Fc-области, состоящему из любой аминокислотной последовательности, выбранной из SEQ ID NО: 43-68, SEQ ID NО:70, SEQ ID NО: 71 и SEQ ID NО: 75-77;
[12] к полипептиду, содержащему по меньшей мере два варианта Fc-области по любому из п.п. [1]-[11], где указанные два варианта Fc-области являются связанными;
[13] к полипептиду по п. [12], где аминокислотные последовательности двух связанных вариантов Fc-области в полипептиде являются одинаковыми;
[14] к полипептиду по п. [12], где аминокислотные последовательности двух связанных вариантов Fc-области в полипептиде являются различными;
[15] к полипептиду по п. [14], где аминокислотные последовательности двух связанных вариантов Fc-области имеют отличающуюся(иеся) аминокислоту(ы) по меньшей мере в одном положении, выбранном из положений 235, 236, 237, 238 и 239 в соответствии с Европейской нумерацией в указанном варианте Fc-области;
[16] к полипептиду по п. [15], где одной из аминокислотных последовательностей двух связанных вариантов Fc-области является аминокислотная последовательность, содержащая по меньшей мере одну аминокислоту, выбранную из аминокислот Asp, Gln, Glu или Thr в положении 235, аминокислоты Asn в положении 236, аминокислот Phe или Trр в положении 237, аминокислот Glu, Gly или Asn в положении 238 и аминокислот Asp или Glu в положении 239 в соответствии с Европейской нумерацией;
[17] к полипептиду по любому из п.п. [12]-[16], где полипептидом, содержащим вариант Fc-области, является антитело IgG;
[18] к полипептиду по любому из п.п. [12]-[16], где полипептидом, содержащим вариант Fc-области, является молекула слитого Fc-белка; и
[19] к фармацевтической композиции, содержащей полипептид по любому из п.п. [12]-[18].
Кроме того, настоящее изобретение относится к способу повышения активности связывания Fc-области с FcγRIIb и к повышению селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа) путем введения в Fc-область согласно изобретению аминокислотной(ых) модификации(й). Настоящее изобретение также относится к способу подавления продуцирования антител против полипептида, содержащего Fc-область, путем введения в Fc-область согласно изобретению аминокислотной(ых) модификации(й).
Настоящее изобретение также относится к терапевтическому или профилактическому средству для лечения иммунных воспалительных заболеваний, где указанное средство содержит полипептид согласно изобретению. Кроме того, настоящее изобретение относится к способу лечения или предупреждения иммунных воспалительных заболеваний, где указанный способ включает стадию введения индивидууму полипептида согласно изобретению. Кроме того, настоящее изобретение относится к набору, применяемому в способе согласно изобретению для лечения или предупреждения иммунных воспалительных заболеваний, где указанный набор содержит полипептид согласно изобретению. Настоящее изобретение также относится к применению полипептида согласно изобретению в целях получения терапевтического или профилактического средства для лечения иммунных воспалительных заболеваний. Кроме того, настоящее изобретение относится к полипептиду согласно изобретению, применяемому в способе лечения или предупреждения иммунных воспалительных заболеваний согласно изобретению.
Настоящее изобретение относится к ингибитору активации В-клеток, тучных клеток, дендритных клеток и/или базофилов, где указанный ингибитор содержит полипептид согласно изобретению. Кроме того, настоящее изобретение относится к способу ингибирования активации В-клеток, тучных клеток, дендритных клеток и/или базофилов, где указанный способ включает введение индивидууму полипептида согласно изобретению. Настоящее изобретение также относится к набору для его применения в способе ингибирования активации В-клеток, тучных клеток, дендритных клеток и/или базофилов, где указанный набор содержит полипептид согласно изобретению. Настоящее изобретение относится к применению полипептида согласно изобретению для продуцирования ингибиторов активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. Настоящее изобретение также относится к полипептиду согласно изобретению, применяемому в способе ингибирования активации В-клеток, тучных клеток, дендритных клеток и/или базофилов согласно изобретению.
Кроме того, настоящее изобретение относится к терапевтическому средству для лечения заболеваний, ассоциированных с дефицитом белка, необходимого для организма, где указанное средство содержит полипептид согласно изобретению. Настоящее изобретение также относится к способу лечения заболеваний, ассоциированных с дефицитом белка, необходимого для организма, где указанный способ включает введение индивидууму полипептида согласно изобретению. Кроме того, настоящее изобретение относится к набору, применяемому в способе согласно изобретению для лечения заболеваний, ассоциированных с дефицитом белка, необходимого для организма, где указанный набор содержит полипептид согласно изобретению. Настоящее изобретение относится к применению полипептида согласно изобретению в целях получения терапевтического средства для лечения заболеваний, ассоциированных с дефицитом белка, необходимого для организма. Настоящее изобретение также относится к полипептиду согласно изобретению, применяемому в способе согласно изобретению для лечения заболеваний, ассоциированных с дефицитом белка, необходимого для организма.
Кроме того, настоящее изобретение относится к средству для подавления пролиферации вируса, где указанное средство содержит полипептид согласно изобретению. Настоящее изобретение также относится к способу подавления пролиферации вируса, где указанный способ включает введение индивидууму полипептида согласно изобретению. Кроме того, настоящее изобретение относится к набору, применяемому в способе подавления пролиферации вируса, где указанный набор содержит полипептид согласно изобретению. Настоящее изобретение относится к применению полипептида согласно изобретению в целях получения средства для подавления пролиферации вируса. Настоящее изобретение также относится к полипептиду согласно изобретению, применяемому в способе согласно изобретению для подавления пролиферации вируса.
[Эффекты настоящего изобретения]
Настоящее изобретение относится к вариантам Fc-области, которые, в отличие от немодифицированной Fc-области, обладают повышенной активностью связывания с FcγRIIb и/или повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа (R-типа). С использованием полипептидов, содержащих варианты Fc-области, могут быть усилены сигналы, ингибирующие воспалительные иммунные ответы, опосредуемые фосфорилированием ITIM FcγRIIb. Кроме того, благодаря сообщению Fc-области такого свойства, как селективное связывание с FcγRIIb, может быть ингибировано продуцирование антитела против лекарственного средства. Кроме того, с использованием вариантов Fc-области согласно изобретению в виде полипептида, обладающего активностью связывания с человеческим FcRn при кислотных значениях рН и содержащего антигенсвязывающий домен, в котором антигенсвязывающая активность антигенсвязывающей молекулы варьируется в зависимости от концентрации ионов, может быть стимулировано удаление полипептид-связывающих антигенов, которые присутствуют в плазме.
Краткое описание графического материала
На фиг. 1 представлен график, на котором показаны результаты оценки агрегации тромбоцитов, вызываемой иммунным комплексом «омализумаб-G1d-3/IgE» в анализе на агрегацию тромбоцитов, проводимом с использованием тромбоцитов, выделенных у донора, имеющего полиморфизм FcγRIIа (R/H).
На фиг. 2 представлен график, на котором показаны результаты оценки агрегации тромбоцитов, вызываемой иммунным комплексом «омализумаб-G1d-3/IgE» в анализе на агрегацию тромбоцитов, проводимом с использованием тромбоцитов, выделенных у донора, имеющего полиморфизм FcγRIIа (Н/H).
На фиг. 3 представлен график, на котором показаны результаты оценки экспрессии CD62p на промытой мембранной поверхности тромбоцитов. На залитой черным кривой показан результат, полученный при проведении реакции с PBS с последующей стимуляцией путем добавления ADP, а на незаштрихованной кривой показан результат, полученный при проведении реакции с иммунным комплексом с последующей ADP-стимуляцией.
На фиг. 4 представлен график, на котором показаны результаты оценки экспрессии активного интегрина на промытой мембранной поверхности тромбоцитов. На залитой черным кривой показан результат, полученный при проведении реакции с PBS с последующей стимуляцией путем добавления ADP, а на незаштрихованной кривой показан результат, полученный при проведении реакции с иммунным комплексом с последующей ADP-стимуляцией.
На фиг. 5 показан комплекс Fc(P208)/внеклеточная область FcγRIIb, определенный с помощью рентгеноструктурного анализа. Для CH2-домена и CH3-домена Fc-области, часть, показанная слева, определена как домен A, а часть, показанная справа, определена как домен В.
На фиг. 6 проиллюстрировано сравнение, сделанное путем суперпозиции кристаллической структуры комплекса Fc(P208)/внеклеточная область FcγRIIb и кристаллической структуры комплекса Fc(WT)/внеклеточная область FcγRIIа, определенных с помощью рентгеноструктурного анализа, (код PDB: 3RY6) для CH2-домена А Fc-области, где такое сравнение проводили методом наименьших квадратов исходя из расстояния между парами атомов Cα. На этой фигуре, структура комплекса Fc(P208)/внеклеточная область FcγRIIb показана жирными линиями, а структура комплекса Fc(WT)/внеклеточная область FcγRIIа показана тонкими линиями. Следует отметить, что структура комплекса Fc(WT)/внеклеточная область FcγRIIа показана только для СН2-домена А Fc-части.
На фиг. 7 детально представлена структура вокруг Asp в положении 237 (в соответствии с Европейской нумерацией) CH2-домена A Fc-части, главная цепь которой образует водородную связь с Tyr в положении 160 FcγRIIb, как показал рентгеноструктурный кристаллографический анализ комплекса Fc(P208)/внеклеточная область FcγRIIb.
На фиг. 8 представлена структура аминокислотных остатков вокруг боковой цепи Asp в положении 237 (в соответствии с Европейской нумерацией) CH2-домена A Fc-части, главная цепь которой образует водородную связь с Tyr в положении 160 FcγRIIb, как показал рентгеноструктурный анализ комплекса Fc(P208)/внеклеточная область FcγRIIb.
На фиг. 9 графически проиллюстрирована суперпозиция определенных с помощью рентгеноструктурного анализа кристаллических структур комплекса Fc(P238D)/внеклеточная область FcγRIIb, описанного в сравнительном примере 7, и комплекса Fc(P208)/внеклеточная область FcγRIIb для CH2-домена В Fc-части, где такую суперпозицию осуществляли для сравнения области вокруг петли в положениях 266-271, соответствующих Европейской нумерации, с применением метода наименьших квадратов исходя из расстояния между парами атомов Cα. В этой петле вариант Fc(P208) имеет модификацию Н268D в положении 268 (в соответствии с Европейской нумерацией) и модификацию Р271G в положении 271 (в соответствии с Европейской нумерацией) по сравнению с вариантом Fc(P238D).
На фиг. 10 представлена структура вокруг Ser239 CH2-домена B Fc-части вместе с картой электронной плотности, полученной с помощью рентгеноструктурного анализа комплекса Fc(P208)/внеклеточная область FcγRIIb, в котором в качестве коэффициента был использован 2Fo-Fc.
На фиг. 11 проиллюстрировано сравнение, сделанное путем суперпозиции трехмерной структуры комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа и трехмерной структуры комплекса Fc(P208)/внеклеточная область FcγRIIb, определенных с помощью рентгеноструктурного анализа, где такое сравнение было осуществлено методом наименьших квадратов, исходя из расстояния между парами атомов Cα.
На фиг. 12 проиллюстрировано сравнение кристаллической структуры комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа, определенной с помощью рентгеноструктурного анализа, и кристаллической структуры комплекса Fc(P208)/внеклеточная область FcγRIIb, определенной с помощью рентгеноструктурного анализа, вокруг Asp в положении 237 (в соответствии с Европейской нумерацией) в CH2-домене А Fc-части, вместе с картой электронной плотности, полученной с помощью рентгеноструктурного анализа, в котором в качестве коэффициента был использован 2Fo-Fc.
На фиг. 13 проиллюстрировано сравнение кристаллической структуры комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа, определенной с помощью рентгеноструктурного анализа, и кристаллической структуры комплекса Fc(P208)/внеклеточная область FcγRIIb, определенной с помощью рентгеноструктурного анализа, вокруг Asp в положении 237 (в соответствии с Европейской нумерацией) в CH2-домене В Fc-части, вместе с картой электронной плотности, полученной с помощью рентгеноструктурного анализа, в котором в качестве коэффициента был использован 2Fo-Fc.
На фиг. 14 проиллюстрировано сравнение последовательностей константных областей G1d и G4d. На этой фигуре обведенные рамкой аминокислоты означают части, в которых аминокислотные остатки G1d и G4d отличаются друг от друга.
На фиг. 15 представлены величины активности связывания каждого варианта с FcγRIIb на горизонтальной оси и величины активности связывания каждого варианта с FcγRIIаR на вертикальной оси. Модификации, показанные на этой фигуре, а именно, G237W, G237F, G236N, P238G, P238N, P238E и P238D, означают модификации, введенные в GpH7-B3. A5/B3 означает GpH7-A5/GpH7-B3/GpL16-k0, цепи которых не содержат каких-либо модификаций, а вариант, содержащий P238D только в одной цепи, обозначен GpH7-A5/GpH7-BF648/GpL16-k0.
На фиг. 16 представлены величины KD для связывания каждого варианта с FcγRIIb на горизонтальной оси и величины KD для связывания каждого варианта с FcγRIIаR на вертикальной оси. На этой фигуре IL6R-B3/IL6R-L и IL6R-G1d/IL6R-L означают антитела, имеющие нативные человеческие последовательности IgG, которые служат в качестве сравнительного контроля при оценке каждого из этих вариантов. IL6R-BP264/IL6R-L представляет собой исходный вариант, используемый для продуцирования каждого из вариантов. IL6R-BP404/IL6R-L представляет собой вариант, полученный путем введения модификации L234Y в обе цепи IL6R-BP264/IL6R-L и обладающий повышенной активностью связывания с FcγRIIb по сравнению с вариантом IL6R-BP264/IL6R-L до введения в него указанной модификации.
На фиг. 17 проиллюстрировано сравнение активности связывания с FcγRIа и с FcγRIIb. На этой фигуре отмечено связывание антитела, в котором остаток Pro в положении 238 (в соответствии с Европейской нумерацией) был заменен остатком Asp, и связывание антитела, в котором остаток Leu в положении 328 (в соответствии с Европейской нумерацией) был заменен остатком Glu. Термин «мутация А» означает мутацию, полученную путем замены остатка Pro в положении 238 (в соответствии с Европейской нумерацией) остатком Asp, а термин «мутация В» означает мутацию, полученную путем замены остатка Leu в положении 328 (в соответствии с Европейской нумерацией) остатком Glu.
На фиг. 18 проиллюстрировано сравнение активности связывания с FcγRIIа Н-типа и с FcγRIIb. На этой фигуре отмечено связывание антитела, в котором остаток Pro в положении 238 (в соответствии с Европейской нумерацией) был заменен остатком Asp, и связывание антитела, в котором остаток Leu в положении 328 (в соответствии с Европейской нумерацией) был заменен остатком Glu. Термин «мутация А» означает мутацию, полученную путем замены остатка Pro в положении 238 (в соответствии с Европейской нумерацией) остатком Asp, а термин «мутация В» означает мутацию, полученную путем замены остатка Leu в положении 328 (в соответствии с Европейской нумерацией) остатком Glu.
На фиг. 19 проиллюстрировано сравнение активности связывания с FcγRIIа R-типа и с FcγRIIb. На этой фигуре отмечено связывание антитела, в котором остаток Pro в положении 238 (в соответствии с Европейской нумерацией) был заменен остатком Asp, и связывание антитела, в котором остаток Leu в положении 328 (в соответствии с Европейской нумерацией) был заменен остатком Glu. Термин «мутация А» означает мутацию, полученную путем замены остатка Pro в положении 238 (в соответствии с Европейской нумерацией) остатком Asp, а термин «мутация В» означает мутацию, полученную путем замены остатка Leu в положении 328 (в соответствии с Европейской нумерацией) остатком Glu.
На фиг. 20 проиллюстрировано сравнение активности связывания с FcγRIIIа и с FcγRIIb. На этой фигуре отмечено связывание антитела, в котором остаток Pro в положении 238 (в соответствии с Европейской нумерацией) был заменен остатком Asp, и связывание антитела, в котором остаток Leu в положении 328 (в соответствии с Европейской нумерацией) был заменен остатком Glu. Термин «мутация А» означает мутацию, полученную путем замены остатка Pro в положении 238 (в соответствии с Европейской нумерацией) остатком Asp, а термин «мутация В» означает мутацию, полученную путем замены остатка Leu в положении 328 (в соответствии с Европейской нумерацией) остатком Glu.
На фиг. 21 показано сходство между аминокислотными остатками, составляющими константные области IgG1, IgG2, IgG3 и IgG4 и пронумерованными в соответствии с Европейской нумерацией (соответствующей здесь индексу EU).
На фиг. 22 представлен график, где на горизонтальной оси отложены относительные величины активности связывания каждого PD-варианта с FcγRIIb, а на вертикальной оси отложены относительные величины активности связывания каждого PD-варианта с FcγRIIа R-типа. Величину активности связывания каждого PD-варианта с каждым из FcγR делят на величину активности связывания IL6R-F652/IL6R-L, которое представляет собой контрольное антитело до введения модификации (модификации Fc с заменой Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp) с каждым FcγR, и полученную величину умножают на 100, в результате чего получают величину относительной активности связывания каждого PD-варианта с каждым FcγR. На графике F652 представлена величина для IL6R-F652/IL6R-L.
На фиг. 23 представлен график, где на вертикальной оси отложены относительные величины FcγRIIb-связывающей активности вариантов, полученных путем введения модификации в вариант GpH7-B3, который не имеет модификации P238D, а на горизонтальной оси отложены относительные величины FcγRIIb-связывающей активности вариантов, полученных путем введения модификации в вариант IL6R-F652, который имеет модификацию P238D. Величину активности связывания каждого варианта с FcγRIIb делят на величину активности связывания FcγRIIb с антителом до его модификации, а затем полученную величину умножают на 100, в результате чего получают величину относительной активности связывания. В данном случае область А содержит модификации, повышающие активность связывания с FcγRIIb в обоих случаях, где модификацию вводят в вариант GpH7-B3, который не имеет модификации P238D, и в IL6R-F652/IL6R-L, который имеет модификацию P238D. Область В содержит модификации, повышающие активность связывания с FcγRIIb, если данная модификации была введена в вариант GpH7-B3, который не имеет модификации P238D, но не повышающие активности связывания с FcγRIIb, если данная модификации была введена в вариант IL6R-F652, который имеет модификацию P238D.
На фиг. 24 представлена кристаллическая структура комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг. 25 графически проиллюстрирована суперпозиция кристаллической структуры комплекса внеклеточной области Fc(P238D)/внеклеточная область FcγRIIb и структуры-модели комплекса Fc(WT)/внеклеточная область FcγRIIb на внеклеточную область FcγRIIb и CH2-домен А Fc, где указанную суперпозицию осуществляют методом наименьших квадратов исходя из расстояния между парами атомов Cα.
На фиг. 26 проиллюстрировано детальное сравнение структуры вокруг замены P238D после суперпозиции кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и структуры-модели комплекса Fc(WT)/внеклеточная область FcγRIIb только на СН2-домен А Fc или только на СН2-домен В Fc, где указанное сравнение проводят методом наименьших квадратов, исходя из расстояния между парами атомов Cα.
На фиг. 27 показано, что водородная связь присутствует между главной цепью Gly в положении 237 (в соответствии с Европейской нумерацией) в СН2-домене А Fc и Tyr в положении 160 FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг. 28 показано, что электростатическое взаимодействие наблюдается между Asp в положении 270 (в соответствии с Европейской нумерацией) в СН2-домене В Fc и Arg в положении 131 FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточной области FcγRIIb.
На фиг. 29 представлен график, где на горизонтальной оси отложены величины относительной активности связывания каждого варианта 2В с FcγRIIb, а на вертикальной оси отложены относительные величины активности связывания каждого варианта 2В с FcγRIIа R-типа. Величину активности связывания каждого варианта 2В с каждым FcγR делят на величину активности связывания с контрольным антителом до модификации (модификации Fc с заменой Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp) в каждом FcγR, и полученную величину умножают на 100, в результате чего получают величину относительной активности связывания каждого варианта 2В с каждым FcγR.
На фиг. 30 показаны Glu в положении 233 (в соответствии с Европейской нумерацией) в цепи А Fc и окружающие остатки во внеклеточной области FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг. 31 показаны Ala в положении 330 (в соответствии с Европейской нумерацией) в цепи А Fc и окружающие остатки во внеклеточной области FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг. 32 проиллюстрировано сравнение структуры с Pro в положении 271 (в соответствии с Европейской нумерацией) в цепи В Fc после суперпозиции кристаллических структур комплекса Fc(P238D)/внеклеточная область FcγRIIb и комплекса Fc(WT)/внеклеточная область FcγRIIb со структурой цепи В Fc, где указанное сравнение проводили методом наименьших квадратов исходя из расстояния между парами атомов Cα.
На фиг. 33 проиллюстрировано изменение концентраций антигенсвязывающих молекул человеческого FcγRIIb в плазме трансгенных мышей после введения этим мышам Fv4-IgG1 или Fv4-P587.
На фиг. 34 проиллюстрировано изменение концентраций человеческого IL-6R или человеческого FcγRIIb в плазме трансгенных мышей после введения этим мышам Fv4-IgG1 или Fv4-P587.
На фиг. 35 проиллюстрирован неограничивающий механизм выведения растворимых антигенов из плазмы при введении антител, которые связываются с антигенами в зависимости от концентрации ионов, где указанные антитела обладают повышенной активностью связывания с FcγR при нейтральном рН по сравнению с активностью связывания известных нейтрализующих антител.
На фиг. 36 проиллюстрировано изменение концентраций антигенсвязывающих молекул человеческого FcγRIIb и человеческого FcRn в плазме трансгенных мышей после введения этим мышам Fv4-IgG1, Fv4-P587 или Fv4-P587_LS.
На фиг. 37 проиллюстрировано изменение концентраций человеческого IL-6R, человеческого FcγRIIb и человеческого FcRn в плазме трансгенных мышей после введения этим мышам Fv4-IgG1, Fv4-P587 или Fv4-P587_LS.
Способ осуществления изобретения
Настоящее изобретение относится к варианту Fc-области, обладающему повышенной активностью связывания с FcγRIIb и/или повышенной селективностью связывания с FcγRIIb по сравнению с активностью и/или селективностью связывания с FcγRIIа (R-типа), в отличие от Fc-области, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и); и к полипептиду, содержащему вариант Fc-области.
Более конкретно, настоящее изобретение относится к варианту Fc-области, содержащему аминокислотную последовательность, которая имеет комбинацию аминокислотной замены в положении 238 (в соответствии с Европейской нумерацией) и другой(их) специфической(их) аминокислотной(ых) модификации(й); и к полипептиду, содержащему такой вариант Fc-области. Кроме того, настоящее изобретение относится к способу повышения активности связывания с FcγRIIb и/или селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от Fc-области, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и), где указанный способ включает введение аминокислотной(ых) модификации(й) в Fc-область. Настоящее изобретение также относится к способу ингибирования продуцирования антител против Fc-области путем введения аминокислотной(ых) модификации(й) в Fc-область и последующего введения такого варианта Fc-области в организм, причем такого ингибирования не происходит, если в Fc-область не была(и) введена(ы) аминокислотная(ые) модификация(и).
Термин «полипептиды» согласно изобретению в общих чертах означает пептиды или белки, длина которых составляет приблизительно десять или более аминокислот. Кроме того, такие полипептиды могут быть получены от любых организмов без каких-либо ограничений, и такие полипептиды могут представлять собой, например, полипептиды, содержащие искусственно сконструированную последовательность. Кроме того, такими полипептидами могут быть природные полипептиды, синтетические полипептиды, рекомбинантные полипептиды или т.п.
Предпочтительными примерами полипептидов согласно изобретению являются антитела. Более предпочтительными примерами являются природные IgG, а в частности, природные человеческие IgG. Термин «природные (нативные) IgG» означает полипептиды, принадлежащие к классу антител, обычно кодируемых генами гамма-иммуноглобулина и содержащих аминокислотную последовательность, идентичную аминокислотной последовательности природных IgG. Так, например, термин «природный человеческий IgG» означает природный человеческий IgG1, природный человеческий IgG2, природный человеческий IgG3, природный человеческий IgG4 или т.п. Природными IgG также являются мутанты, спонтанно продуцируемые из этих IgG.
Если константная область типа IgK (каппа-цепь, k-цепь), IgL1, IgL2, IgL3, IgL6 и IgL7 (лямбда-цепь, λ-цепь) присутствует в константной области легкой цепи антитела, то она может представлять собой любую константную область легкой цепи. Для константной области человеческого IgK (каппа) и константной области человеческого IgL7 (лямбда), множество последовательностей аллотипов, образующихся в результате генетического полиморфизма, описаны в публикации NIH No.91-3242 «Sequences of proteins of immunological interest», и любая из этих последовательностей может быть использована в настоящем изобретении. Кроме того, в настоящем изобретении константной областью легкой цепи может быть константная область легкой цепи, в которую были введены аминокислотные замены, добавления, делеции, инсерции и/или модификации или т.п. В антителах, например, в антителах IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 и IgM имеются Fc-области. Так, например, Fc-областью антитела согласно изобретению может служить Fc-область человеческого антитела IgG, а предпочтительными являются Fc-области человеческого антитела IgG1. Fc-областями, которые могут быть использованы в настоящем изобретении, являются, например, Fc-области, происходящие от константных областей природного IgG, или, в частности, константных областей природного человеческого IgG1 (SEQ ID NO:11), константных областей природного человеческого IgG2 (SEQ ID NО:12), константных областей природного человеческого IgG3 (SEQ ID NО:13) и константных областей природного человеческого IgG4 (SEQ ID NО:14). На фиг. 21 представлены последовательности константной области природных IgG1, IgG2, IgG3 и IgG4. Константными областями природных IgG также являются мутанты, спонтанно продуцируемые из этих IgG. Для константных областей человеческих антител IgG1, IgG2, IgG3 и IgG4, множество последовательностей аллотипов, образующихся в результате генетического полиморфизма, описаны в публикации NIH No.91-3242 «Sequences of proteins of immunological interest», и любая из этих последовательностей может быть использована в настоящем изобретении. В частности, аминокислотная последовательность человеческого IgG1 в положениях 356-358 (в соответствии с Европейской нумерацией) может представлять собой DEL или EEM.
Термин «Fcγ-рецепторы» (называемые здесь Fcγ-рецепторами, FcγR или FcgR) означает рецепторы, которые могут связываться с Fc-областью моноклональных антител IgG1, IgG2, IgG3 и IgG4, и, практически, означает любой член семейства белков, кодируемых генами Fcγ-рецептора. У человека это семейство включает FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIа (включая аллотипы H131 (H-типа) и R131 (R-типа)), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIb-с; а также FcγRIII (CD16), включая изоформы FcγRIIIа (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2) и любые неограничивающие примеры человеческих изоформ или аллотипов FcγR, которые пока еще не были обнаружены. Сообщалось, что FcγRIIb1 и FcγRIIb2 представляют собой варианты сплайсинга человеческого FcγRIIb. Кроме того, в литературе также описан вариант сплайсинга, обозначаемый FcγRIIb3 (J. Exp Med, 1989, 170: 1369). Человеческий FcγRIIb, помимо этих вариантов сплайсинга, включает все варианты сплайсинга, зарегистрированные в NCBI под номерами NP_001002273.1, NP_001002274.1, NP_001002275.1, NP_001177757.1 и NP_003992.3. Кроме того, человеческий FcγRIIb включает любой ранее описанный генетический полиморфизм, а также FcγRIIb (Arthritis Rheum., 2003, 48: 3242-52, Hum. Mol. Genet, 2005, 14: 2881-92, и Arthritis Rheum, 2002 May; 46(5): 1242-54) и каждый генетический полиморфизм, который будет описан в будущем.
Неограничивающими примерами FcγR являются человеческие, мышиные, крысиные, кроличьи и обезьяньи FcγR, а также FcγR, которые могут происходить от любого организма. Мышиными FcγR являются FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD16-2), и любые неограничивающие примеры мышиных изоформ или аллотипов FcγR, которые пока еще не были обнаружены. Предпочтительными примерами таких Fcγ-рецепторов являются человеческие FcγRI (CD64), FcγRIIА (CD32), FcγRIIВ (CD32), FcγRIIIА (CD16) и/или FcγRIIIВ (CD16).
Полинуклеотидная последовательность и аминокислотная последовательность FcγRI представлены в SEQ ID NO:1 (NM_000566.3) и 2 (NP_000557.1), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIА представлены в SEQ ID NO:3 (BC020823.3) и 4 (AAH20823.4), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIB представлены в SEQ ID NO:5 (BC146678.1) и 6 (AAI46679.1), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIА представлены в SEQ ID NO:7 (BC033678.1) и 8 (AAH33678.1), соответственно; и
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIВ представлены в SEQ ID NO:9 (BC128562.1) и 10 (AAI28563.1), соответственно (регистрационный номер RefSeq указан в скобках).
FcγRIIа имеет два аллотипа, в одном из которых аминокислотой в положении 131 является гистидин (Н-типа), а в другом эта аминокислота заменена аргинином (R-типа)(J. Exp. Med, 172, 19-25, 1990).
В настоящем изобретении термин «Fc-область, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и)» или аналогичный термин означает Fc-область до введения в нее аминокислотной(ых) модификации(й) согласно изобретению. В настоящем изобретении этот термин может означать, например, Fc-область нативного IgG или Fc-область IgG, полученную путем введения в нативный IgG модификации, не являющейся аминокислотной(ыми) модификацией(ями) согласно изобретению. Кроме того, в настоящем изобретении термин «вариант Fc-области» означает Fc-область, в которой по меньшей мере одна аминокислота была заменена другой аминокислотой согласно изобретению без введения аминокислотной(ых) модификации(й) согласно изобретению. В настоящем изобретении Fc-область, в которой «по меньшей мере одна аминокислота была заменена другой аминокислотой», включает Fc-область, в которую была введена указанная аминокислотная модификация, и Fc-область, состоящую из той же самой аминокислотной последовательности.
Термин «природные IgG» означает полипептиды, принадлежащие к классу антител, обычно кодируемых генами гамма-иммуноглобулина и содержащих аминокислотную последовательность, идентичную аминокислотной последовательности природных IgG. Так, например, термин «природный человеческий IgG» означает нативный человеческий IgG1, нативный человеческий IgG2, нативный человеческий IgG3, нативный человеческий IgG4 или т.п. Природными IgG также являются мутанты, спонтанно продуцируемые из этих IgG.
Термин «Fc-область нативного IgG» означает Fc-область, содержащую аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, происходящей от природного IgG. Константная область тяжелой цепи нативного IgG представлена на фиг. 21 (SEQ ID NO:11-14), и на этой фиг. 21, например, показаны Fc-области в константной области тяжелой цепи, происходящей от нативного человеческого IgG1, Fc-области в константной области тяжелой цепи, происходящей от нативного человеческого IgG2, Fc-области в константной области тяжелой цепи, происходящей от нативного человеческого IgG3, и Fc-области в константной области тяжелой цепи, происходящей от нативного человеческого IgG4. Fc-областями нативных IgG также являются мутанты, спонтанно продуцированные из этих IgG.
В соответствии с настоящим изобретением, активность связывания полипептида, содержащего вариант Fc-области или вариант Fc-области согласно изобретению, с FcγR каждого типа, независимо от того является ли она повышенной, постоянной или пониженной, может быть определена, например, путем мониторинга снижения или увеличения величины константы диссоциации (KD), вычисленной по результатам анализа сенсограммы, где различные FcγR, используемые в качестве аналита, подвергают взаимодействию с антителами, иммобилизованными на сенсорных чипах или захваченными на сенсорных чипах посредством белка А, белка L, белка А/G, G-белка, антител против цепи лямбда, антител против цепи каппа, антигенных пептидов, антигенных белков или т.п. с применением оборудования BIACORE, которое представляет собой анализатор взаимодействия, работающий на основе поверхностного плазмонного резонанса (ППР), как описано в примерах. Альтернативно, такая активность может быть определена путем мониторинга увеличения или уменьшения величины, полученной путем деления величины изменения резонансной единицы (RU) на сенсограмме до и после взаимодействия FcγR различных типов, используемых в качестве аналита, с антителами, иммобилизованными на сенсорных чипах или захваченными на сенсорных чипах посредством белка А, белка L, белка А/G, G-белка, антител против цепи лямбда, антител против цепи каппа, антигенных пептидов, антигенных белков или т.п., на величину изменения резонансных единиц (RU) до и после иммобилизации или захвата антител на сенсорном чипе. Кроме того, активность может быть определена, например, путем мониторинга увеличения или уменьшения величины константы диссоциации (KD), вычисленной по результатам анализа сенсограммы, где оцениваемый образец, такой как антитело, используемое в качестве аналита, подвергают взаимодействию на сенсорном чипе, на котором FcγR иммобилизован непосредственно или посредством антитела против метки. Альтернативно, активность может быть определена, например, путем мониторинга увеличения или уменьшения величин на сенсограмме до и после взаимодействия оцениваемого образца, такого как антитело, используемое в качестве аналита, с сенсорным чипом, на котором FcγR иммобилизован непосредственно или посредством антитела против метки.
В частности, активность связывания варианта Fc-области с Fcγ-рецептором может быть определена путем скрининга, проводимого посредством проксимального гомогенного анализа на активацию люминесценции (ALPHA), с применением метода BIACORE на основе поверхностного плазмонного резонанса (ППР) или т.п., а также с помощью ELISA или клеточного сортинга с активацией флуоресценции (FACS) (Proc. Natl. Acad. Sci. USA (2006) 103 (11), 4005-4010).
ALPHA-скрининг осуществляют с применением ALPHA-технологии, в которой используются две сферы, «донор» и «акцептор», и которая основана на нижеследующих принципах. Люминесцентные сигналы детектируются только в том случае, когда молекулы, связанные с донорными сферами, физически взаимодействуют с молекулами, связанными с акцепторными сферами, и эти две сферы расположены в непосредственной близости друг к другу. Фотосенсибилизатор с лазерным излучением, присутствующий в донорных сферах, превращает кислород окружающей среды в синглетный кислород в возбужденном состоянии. Синглетный кислород диспергируется вокруг донорных сфер, и когда он достигает соседних акцепторных сфер, в этих сферах индуцируется хемилюминесцентная реакция, приводящая, в конечном счете, к излучению света. Если молекулы, связанные с донорными сферами, не взаимодействуют с молекулами, связанными с акцепторными сферами, то хемилюминесцентной реакции не происходит, поскольку синглетный кислород, продуцируемый донорными сферами, не достигает акцепторных сфер.
Так, например, биотинилированный полипептидный комплекс связывается с донорными сферами, а Fcγ-рецептор, меченный глутатион-S-трансферазой (GST), связывается с акцепторными сферами. В отсутствии конкурирующего полипептидного комплекса, содержащего вариант Fc-области, полипептидный комплекс, содержащий Fc-область дикого типа, взаимодействует с Fcγ-рецептором и продуцирует сигналы на длине волны 520-620 нм. Полипептидный комплекс, содержащий немеченую мутантную Fc-область, конкурирует с полипептидным комплексом, содержащим Fc-область дикого типа, за связывание с Fcγ-рецептором. Относительная активность связывания может быть определена путем количественной оценки снижения интенсивности флуоресценции, наблюдаемого после такого конкурентного связывания. Биотинилирование полипептидных комплексов, таких как антитела, с использованием сульфо-NHS-биотина и т.п. хорошо известно специалистам. Экспрессию Fcγ-рецептора и GST в клетке, несущей слитый ген, продуцируемый путем присоединения полинуклеотида, кодирующего Fcγ-рецептор, с сохранением рамки считывания, к полинуклеотиду, кодирующему GST в экспрессионном векторе, и проведение очистки на колонке с глутатионом осуществляют методом мечения Fcγ-рецептора ферментом GST. Полученные сигналы, предпочтительно, анализируют, например, путем приведения их в соответствие с односайтовой моделью конкуренции посредством нелинейного регрессионного анализа с помощью компьютерной программы, такой как GRAPHPAD PRISM (GraphPad Inc., San Diego).
При наблюдении взаимодействия, одно из веществ (лиганд) иммобилизуют на тонкой золотой пленке на сенсорном чипе, после чего, при облучении светом с обратной стороны сенсорного чипа так, чтобы на границе между тонкой золотой пленкой и стеклом наблюдалось полное отражение, в части отраженного света образуется участок с пониженной интенсивностью отражения (ППР-сигнал). При наблюдении взаимодействия, в случае, если другое вещество (аналит) наносят на поверхность сенсорного чипа, и если лиганд связывается с аналитом, то масса иммобилизованной лигандной молекулы повышается, что приводит к изменению показателя преломления растворителя на поверхности сенсорного чипа. В результате такого изменения показателя преломления происходит сдвиг положения ППР-сигнала (с другой стороны, при диссоциации такого связывания, сигнал возвращается в свое первоначальное положение). Система Biacore регистрирует величину вышеупомянутого изменения или, более конкретно, изменение массы в зависимости от времени путем построения кривой изменения массы на поверхности сенсорного чипа, отложенной на ординате по данным измерения (сенсограммы). По этой сенсограмме определяют количество аналита, связанного с лигандом, захваченным на поверхности сенсорного чипа. Кинетические параметры, такие как константы скорости ассоциации (ka) и константы скорости диссоциации (kd), определяют на кривых сенсограммы, а константы диссоциации (KD) определяют как отношение этих констант. В анализе BIACORE, предпочтительно, применяется метод оценки ингибирования. Пример такого метода оценки ингибирования описан в Proc. Natl. Acad. Sci. USA (2006) 103 (11): 4005-4010.
Fc-область с пониженной активностью связывания с FcγR или полипептид, содержащий такую Fc-область, означают вариант Fc-области или полипептид, содержащий такой вариант Fc-области, который связывается с FcγR со значительно более низкой активностью связывания, чем полипептид, содержащий родительскую Fc-область; при этом анализы на такое снижение активности связывания осуществляют с использованием, в основном, одинаковых количеств полипептида, содержащего Fc-область, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и) (где указанный полипептид также называется полипептидом, содержащим родительские Fc-области, или родительским полипептидом), и полипептида, содержащего по меньшей мере одну аминокислотную модификацию в Fc-области (где указанный полипептид также называется полипептидом, содержащим вариант Fc-области, или модифицированным полипептидом).
Кроме того, Fc-область с повышенной активностью связывания с FcγR или полипептид, содержащий такую Fc-область, означают вариант Fc-области или полипептид, содержащий такой вариант Fc-области, который связывается с FcγR со значительно более высокой активностью связывания, чем полипептид, содержащий родительскую Fc-область; при этом анализы на такое повышение активности связывания осуществляют с использованием, в основном, одинаковых количеств полипептида, содержащего родительскую Fc-область, и полипептида, содержащего вариант Fc-области.
Полипептид с постоянной активностью связывания с FcγR означает полипептид, который связывается с FcγR с активностью связывания, эквивалентной активности связывания родительского полипептида или по существу не отличающейся от активности связывания родительского полипептида; при этом анализы на такую постоянную активность осуществляют с использованием, в основном, одинаковых количеств полипептида, содержащего родительскую Fc-область, и полипептида, содержащего вариант Fc-области.
В настоящем изобретении повышенная активность связывания с FcγRIIb, предпочтительно, означает, например, активность, при которой отношение величин KD [величина KD для связывания полипептида, содержащего родительскую Fc-область, с FcγRIIb]/[величина KD для связывания полипептида, содержащего вариант Fc-области, с FcγRIIb], измеренное с применением вышеупомянутого метода оценки, предпочтительно имеет значение, равное 15,0 или выше; 20,0 или выше; 25,0 или выше; 30,0 или выше; 35,0 или выше; 40,0 или выше; 45,0 или выше; 50,0 или выше; 55,0 или выше; 60,0 или выше; 65,0 или выше; 70,0 или выше; 75,0 или выше; 80,0 или выше; 85,0 или выше; 90,0 или выше; 95,0 или выше; или 100,0 или выше.
Кроме того, выражение «вариант Fc-области согласно изобретению обладает повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа» означает, что:
(i) активность связывания с FcγRIIb является повышенной, и активность связывания с FcγRIIа является постоянной или пониженной;
(ii) активность связывания с FcγRIIb является повышенной, и активность связывания с FcγRIIа также является повышенной, однако, степень повышения активности связывания с FcγRIIа ниже степени повышения активности связывания с FcγRIIb; или
(iii) активность связывания с FcγRIIb является пониженной, но степень снижения такой активности связывания ниже, чем степень снижения активности связывания с FcγRIIа. Независимо от того, имеет или не имеет вариант Fc-области согласно изобретению повышенную селективность связывания с FcγRIIb, но не с FcγRIIа, такая селективность связывания может быть определена, например, путем сравнения отношения величины KD для связывания полипептида, содержащего вариант Fc-области согласно изобретению, с FcγRIIа к величине KD для связывания полипептида, содержащего вариант Fc-области согласно изобретению, с FcγRIIb (величина KD для FcγRIIа/величина KD для FcγRIIb), где такое отношение величины KD для связывания полипептида, содержащего вариант Fc-области согласно изобретению, с FcγRIIа к величине KD для связывания полипептида, содержащего вариант Fc-области согласно изобретению, с FcγRIIb (величина KD для FcγRIIа/величина KD для FcγRIIb), определяют как описано в вышеупомянутых примерах. В частности, если величина отношения KD для полипептида, содержащего вариант Fc-области согласно изобретению, превышает величину отношения KD для полипептида, содержащего родительскую Fc-область, то полипептид, содержащий вариант Fc-области согласно изобретению, может быть определен как полипептид, обладающий повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа, по сравнению с полипептидом, содержащим родительскую Fc-область. В частности, поскольку активность связывания с FcγRIIа (R-типа), вероятно, коррелирует с активностью связывания с FcγRIIb, по сравнению с FcγRIIа (Н-типа), то выбор аминокислотной(ых) модификации(й), которая(ые) может (могут) способствовать повышению селективности связывания с FcγRIIb, но не с FcγRIIа (R-типа), будет играть важную роль в повышении селективности связывания с FcγRIIb по сравнению с селективностью связывания с другими FcγR, не являющимися FcγRIIb.
Селективность связывания FcγRIIа (R-типа) и FcγRIIb выражают, например, как отношение величин KD [величина KD для связывания полипептида, содержащего вариант Fc-области, с FcγRIIа (R-типа)]/[величина KD для связывания полипептида, содержащего вариант Fc-области, с FcγRIIb], составляющее предпочтительно 10,0 или более, а более предпочтительно 20,0 или более, где указанную величину KD измеряют методом, описанном выше.
Селективность связывания FcγRIIа (Н-типа) и FcγRIIb выражают, например, как отношение величин KD [величина KD для связывания полипептида, содержащего вариант Fc-области, с FcγRIIа (Н-типа)]/[величина KD для связывания полипептида, содержащего вариант Fc-области, с FcγRIIb], составляющее предпочтительно 100 или более, 200 или более, 300 или более, 400 или более, 500 или более, а более предпочтительно 600 или более, 700 или более, 800 или более или 900 или более, где указанную величину KD измеряют методом, описанным выше.
Кроме того, активность связывания полипептидов согласно изобретению с различными FcγR, независимо от того, является ли она повышенной, постоянной или пониженной, может быть определена по увеличению или снижению активности связывания полипептидов согласно изобретению с различными FcγR, как описано выше в примерах. В настоящем изобретении активность связывания полипептидов с различными FcγR означает величину, полученную путем определения разности относительных величин (RU) на сенсограммах до и после взаимодействия различных FcγR, используемых в качестве аналита, с каждым полипептидом, и деления этой разности на разность величин RU на сенсограммах до и после захвата полипептидов на сенсорных чипах.
Неограничивающие варианты Fc-области согласно изобретению имеют величину KD (моль/л) для FcγRIIb, составляющую, например, 9×10-7 или менее, предпочтительно 5×10-7 или менее, более предпочтительно 3×10-7 или менее, еще более предпочтительно 1×10-7 или менее, а наиболее предпочтительно 5×10-8 или менее.
Термин «Fc-область» означает фрагмент, состоящий из шарнирной области или ее части, СН2-домена и СН3-домена, присутствующих в молекуле антитела. В соответствии с Европейской нумерацией (также называемой здесь EU-индексом) (см. фиг. 21), например, Fc-область класса IgG, означает, например, область, которая простирается от цистеина в положении 226 до С-конца или от пролина в положении 230 до С-конца, но не ограничивается этими областями.
Fc-область может быть получена, предпочтительно, путем повторного элюирования фракции, адсорбированной на колонке с белком А, после частичного гидролиза моноклональных антител IgG1, IgG2, IgG3, IgG4 или т.п. с использованием протеазы, такой как пепсин. Такой протеазой может быть любая протеаза, при условии, что она может гидролизовать полноразмерное антитело так, чтобы фрагменты Fab и F(ab')2 продуцировались посредством рестрикционного гидролиза в ферментативной реакции, осуществляемой при соответствующем рН, и посредством соответствующих ферментов, и примерами таких ферментов являются пепсин и папаин.
Настоящее изобретение относится к вариантам Fc-области, имеющим комбинацию модификаций, которые включают замену аминокислоты в положении 238 (в соответствии с Европейской нумерацией) другой аминокислотой и замену по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 233, 234, 235, 237, 264, 265, 266, 267, 268, 269, 271, 272, 274, 296, 326, 327, 330, 331, 332, 333, 334, 355, 356, 358, 396, 409 и 419 (в соответствии с Европейской нумерацией), другой аминокислотой в Fc-области человеческого IgG (IgG1, IgG2, IgG3, IgG4). Путем объединения замены аминокислоты в положении 238 (в соответствии с Европейской нумерацией) с заменой по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 233, 234, 237, 264, 265, 266, 267, 268, 269, 271, 272, 274, 296, 326, 327, 330, 331, 332, 333, 334, 355, 356, 358, 396, 409 и 419 (в соответствии с Европейской нумерацией), другой аминокислотой в Fc-области человеческого IgG, можно получить полипептид, содержащий вариант Fc-области, обладающий повышенной активностью связывания с FcγRIIb и/или повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа, а в частности, с FcγRIIа (R-типа), в отличие от полипептида, содержащего Fc-область, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и). Другими аминокислотными модификациями, которые могут быть объединены с аминокислотной модификацией в положении 238 (в соответствии с Европейской нумерацией), предпочтительно являются модификации в положениях 233, 237, 264, 267, 268, 271, 272, 296, 327, 330, 332, 333 и 396 (в соответствии с Европейской нумерацией), а в частности, модификации в положениях 233, 237, 264, 267, 268, 271, 296, 330 и 396 (в соответствии с Европейской нумерацией). В частности, для повышения активности связывания с FcγRIIb или повышения селективности связывания с FcγRIIb по сравнению с FcγRIIа, предпочтительной комбинацией аминокислотных модификаций является комбинация модификаций в положениях аминокислот 238, 268 и 271 (в соответствии с Европейской нумерацией) с модификацией по меньшей мере одной аминокислоты в положениях, выбранных из положений 233, 237, 264, 267, 272, 296, 327, 330, 332 и 396 (в соответствии с Европейской нумерацией).
Модифицированные аминокислоты не имеют конкретных ограничений, при условии, что эти аминокислоты будут способствовать повышению активности связывания с FcγRIIb или повышению селективности связывания с FcγRIIb, а не с FcγRIIа, по сравнению с активностью до их модификации, при этом предпочтительной аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 234 является Tyr, аминокислотой в положении 235 является Phe, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 265 является Glu, аминокислотой в положении 266 является Phe, Leu или Met, аминокислотой в положении 267 является Ala, Glu, Gly или Gln, аминокислотой в положении 268 является Asp, Gln или Glu, аминокислотой в положении 269 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, Phe, Ile, Met, Asn, Pro или Gln, аминокислотой в положении 274 является Gln, аминокислотой в положении 296 является Asp или Phe, аминокислотой в положении 326 является Ala или Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Lys, Arg или Ser, аминокислотой в положении 331 является Ser, аминокислотой в положении 332 является Lys, Arg, Ser или Thr, аминокислотой в положении 333 является Lys, Arg, Ser или Thr, аминокислотой в положении 334 является Arg, Ser или Thr, аминокислотой в положении 355 является Ala или Gln, аминокислотой в положении 356 является Glu, аминокислотой в положении 358 является Met, аминокислотой в положении 396 является Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr, аминокислотой в положении 409 является Arg, а аминокислотой в положении 419 является Glu в соответствии с Европейской нумерацией. В частности, при объединении модификаций аминокислот в положениях 238, 268 и 271 (в соответствии с Европейской нумерацией) с модификациями по меньшей мере одной аминокислоты в положениях, выбранных из положений 233, 264, 267, 272, 296, 327, 330, 332 и 396 (в соответствии с Европейской нумерацией), предпочтительной аминокислотой в положении 238 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 272 является Asp или Pro, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, аминокислотой в положении 332 является Thr, а аминокислотой в положении 396 является Leu или Met в соответствии с Европейской нумерацией.
В соответствии с настоящим изобретением, помимо этих модификаций, в Fc-область может быть добавлена по меньшей мере одна другая модификация. Такая добавленная модификация не имеет конкретных ограничений, при условии, что она будет приводить к повышению активности связывания с FcγRIIb и/или селективности связывания с FcγRIIb, а не с FcγRIIа. Кроме того, модификации могут быть осуществлены путем введения совокупности модификаций, где часть Fc-области заменена соответствующей частью Fc-области другого изотипа. Так, например, активность связывания с FcγRIIb и/или селективность связывания с FcγRIIb может быть повышена путем объединения вышеупомянутых аминокислотных модификаций с заменой аминокислотной последовательности, простирающейся от аминокислоты Ala в положении 118 до Thr в положении 225 (в соответствии с Европейской нумерацией) в Fc-области, происходящей от IgG1, аминокислотной последовательности, простирающейся от аминокислоты Ala в положении 118 до Pro в положении 222 (в соответствии с Европейской нумерацией) в Fc-области, происходящей от IgG4. Конкретный пример включает комбинацию аминокислотных модификаций, вводимых в IL6R-BP230, и модификации, такой как замена аминокислотной последовательности, простирающейся от аминокислоты Ala в положении 118 до Thr в положении 225 (в соответствии с Европейской нумерацией) в G1d, аминокислотной последовательности, простирающейся от аминокислоты Ala в положении 118 до Pro в положении 222 (в соответствии с Европейской нумерацией) в G4d, как описано в примере 7 для IL6R-BP478/IL6R-L.
Из этих модификаций предпочтительными являются модификации, которые приводят к значительному повышению активности связывания с FcγRIIb или к значительному повышению селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа). Примерами таких предпочтительных комбинаций аминокислотных модификаций являются нижеследующие модификации (a)-(x):
(а) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(b) аминокислотные модификации в положениях 238, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(c) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(d) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(е) аминокислотные модификации в положениях 238, 233, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(f) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(g) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 327 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(h) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(i) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(j) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(k) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(l) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(m) аминокислотные модификации в положениях 238, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(n) аминокислотные модификации в положениях 238, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(o) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(p) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(q) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 327, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(r) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(s) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(t) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(u) аминокислотные модификации в положениях 238, 233, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(v) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(w) аминокислотные модификации в положениях 238, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области; и
(x) аминокислотные модификации в положениях 238, 233, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области.
Кроме того, из этих комбинаций более предпочтительными комбинациями аминокислотных модификаций являются нижеследующие комбинации модификаций аминокислотных последовательностей (a)-(x), таких как:
(a) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(b) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(c) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(d) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Gly или Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(e) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(f) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(g) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(h) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(i) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(j) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является аминокислотой Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(k) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(l) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(m) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(n) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(o) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(p) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(q) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met в соответствии с Европейской нумерацией;
(r) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, и аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(s) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(t) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(u) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(v) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Gly, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(w) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, и аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией; и
(x) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией.
Кроме того, аминокислотные модификации, введенные для другой(их) цели(ей), могут быть объединены в полипептидах, содержащих вариант Fc-области согласно изобретению. Так, например, могут быть добавлены аминокислотные замены, повышающие активность связывания с FcRn (J. Immunol. 2006 Jan 1; 176(1): 346-56; J. Biol. Chem. 2006 Aug 18; 281(33): 23514-24; Int. Immunol. 2006 Dec., 18(12): 1759-69; Nat. Biotechnol. 2010 Feb. 28(2): 157-9; WO/2006/019447; WO/2006/053301 и WO/2009/086320), и аминокислотные замены, повышающие гетерогенность или стабильность антитела (WO/2009/041613). Альтернативно, полипептиды, продуцируемые путем сообщения полипептидам, содержащим вариант Fc-области согласно изобретению, свойств, стимулирующих элиминацию антигенов, как описано в WO 2011/122011 или PCT/JP2011/072550, и полипептиды, сообщающие способность повторно связываться с множеством антигенных молекул, как описано в WO 2009/125825, WO 2012/073992 или WO 2013/047752, также входят в объем настоящего изобретения. Альтернативно, для повышения способности полипептидов сохраняться в плазме, аминокислотные модификации, способствующие снижению рI константной области (WO/2012/016227), могут быть объединены в полипептиде, содержащем вариант Fc-области согласно изобретению. Альтернативно, для сообщения способности связываться с другими антигенами, аминокислотные модификации, описанные в EP 1752471 и EP 1772465, могут быть объединены в CH3 полипептида, содержащего вариант Fc-области согласно изобретению.
Если полипептид, содержащий вариант Fc-области согласно изобретению, представляет собой антигенсвязывающую молекулу, такую как антитело, то аминокислотные модификации, способствующие повышению активности связывания с человеческим FcRn при кислотных значениях рН, могут быть объединены в целях усиления действия антигенсвязывающей молекулы, направленного на элиминацию антигенов из плазмы. Более конкретно, модификации, используемые для повышения активности связывания с человеческим FcRn при кислотных значениях рН, могут быть введены в антитело IgG, например, путем замены Met на Leu в положении 428 и замены Asn на Ser в положении 434 в соответствии с Европейской нумерацией (Nat. Biotechnol., 2010 28: 157-159); путем замены Asn на Ala в положении 434 (Drug Metab Dispos 2010 Apr.; 38(4): 600-5); путем замены Met на Tyr в положении 252, замены Ser на Thr в положении 254 и замены Thr на Glu в положении 256 (J. Biol. Chem., 2006, 281: 23514-23524); путем замены Thr на Gln в положении 250 и замены Met на Leu в положении 428 (J. Immunol. 2006, 176(1): 346-56); путем замены Asn на His в положении 434 (Clinical Pharmacology & Therapeutics (2011) 89(2): 283-290) или замен, описанных в WO 2010106180, WO 2010045193, WO 2009058492, WO 2008022152, WO 2006050166, WO 2006053301, WO 2006031370, WO 2005123780, WO 2005047327, WO 2005037867, WO 2004035752, WO 2002060919 или т.п.
Кроме того, недавно сообщалось, что молекула антитела, полученная путем замены Asn на His в положении 434 (в соответствии с Европейской нумерацией) в гуманизированном анти-CD4 антителе в целях повышения активности связывания с человеческим FcRn при кислотных значениях рН и улучшения свойств, таких как пролонгированное время присутствия в плазме, связывается с ревматоидными факторами (РФ) (Clin. Pharmacol. Ther. 2011 Feb.; 89(2): 283-90). Было показано, что это антитело имеет Fc-область человеческого IgG1, которая, после замены Asn на His в положении 434 в сайте связывания с FcRn, связывается с ревматоидными факторами, распознающими сайт с указанными заменами.
Как описано выше, имеются сообщения о различных модификациях, способствующих повышению активности связывания с человеческим FcRn при кислотных значениях рН, но при этом введение этих модификаций в сайт связывания с FcRn в Fc-области приводит к повышению аффинности с ревматоидными факторами, которые распознают этот сайт.
Однако, при введении модификаций, которые не снижают активности связывания с FcRn, а снижают только активность связывания с ревматоидными факторами в этом сайте Fc-области, могут быть получены антигенсвязывающие молекулы, обладающие повышенной активностью связывания с человеческим FcRn при кислотных значениях рН, но не обладающие аффинностью по отношению к ревматоидным факторам.
Для введения модификаций, снижающих активность связывания с ревматоидными факторами, были сделаны замены в положениях 248-257, 305-314, 342-352, 380-386, 388, 414-421, 423, 425-437, 439 и 441-444 в соответствии с Европейской нумерацией. Предпочтительными являются модификации в положениях 387, 422, 424, 426, 433, 436, 438 и 440. Особенно предпочтительными являются замена Val на Glu или Ser в положении 422, замена Ser на Arg положении 424, замена His на Asp в положении 433, замена Tyr на Thr в положении 436, замена Gln на Arg или Lys в положении 438 и замена Ser на Glu или Asp в положении 440. Эти модификации могут быть использованы отдельно или в комбинации друг с другом во множестве положений.
Альтернативно, для снижения активности связывания с ревматоидными факторами, в этот сайт может быть введена последовательность гликозилирования N-типа. В частности, последовательность Asn-Xxx-Ser/Thr (Xxx представляет собой любую аминокислоту кроме Pro) известна как последовательность гликозилирования N-типа. Присоединение сахарной цепи N-типа путем введения этой последовательности в данный сайт Fc-области приводит к ингибированию связывания с РФ благодаря стерическому затруднению, вызываемому сахарной цепью N-типа. Модификациями, используемыми для присоединения сахарной цепи N-типа, предпочтительно являются замена Lys на Asn в положении 248, замена Ser на Asn в положении 424, замена Tyr на Asn в положении 436, замена Gln на Thr в положении 438 и замена Gln на Asn в положении 438. Особенно предпочтительной модификацией является замена Ser на Asn в положении 424.
Предпочтительным примером полипептида, содержащего вариант Fc-области согласно изобретению, является полипептид, содержащий по меньшей мере два варианта Fc-области, где указанные два варианта Fc-области, по всей вероятности, связаны друг с другом в антителе IgG. При использовании антитела IgG в качестве полипептида согласно изобретению, тип константной области не имеет конкретных ограничений, и могут быть использованы IgG различных изотипов (подклассов), таких как IgG1, IgG2, IgG3 и IgG4. Антителами IgG согласно изобретению предпочтительно являются человеческий IgG, а более предпочтительно человеческий IgG1 и человеческий IgG4. Аминокислотные последовательности константных областей тяжелой цепи человеческого IgG1 и человеческого IgG4 являются известными. Множество последовательностей аллотипов со свойственным им генетическим полиморфизмом описаны в публикации «Sequences of proteins of immunological interest» NIH Publication No.91-3242 для константной области человеческого IgG1, и любая из этих последовательностей может быть использована в настоящем изобретении.
Два ассоциированных варианта Fc-области, включенных в вышеупомянутый полипептид, могут представлять собой варианты Fc-области, в которые была(и) введена(ы) одна и та же или одни и те же аминокислотные модификации (далее, этот полипептид будет называться полипептидом, содержащим гомологичные варианты Fc-области), или варианты Fc-области, содержащие различные аминокислотные последовательности, в каждую из которых была(и) введена(ы) другая(ие) аминокислотная(ые) модификация(и), или, альтернативно, варианты Fc-области, содержащие различные аминокислотные последовательности, где только в одну из Fc-областей была(и) введена(ы) аминокислотная(ые) модификация(и) (далее, этот полипептид будет называться полипептидом, содержащим гетерологичные варианты Fc-области). Из аминокислотных модификаций, вводимых только в одну из Fc-областей, предпочтительной модификацией в части петлевой структуры является модификация в положениях 233-239 (в соответствии с Европейской нумерацией) в СН2-домене Fc-области, где указанная модификация способствует связыванию с FcγRIIb и FcγRIIа; а более предпочтительной модификацией является модификация, способствующая повышению активности связывания с FcγRIIb и/или селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа) в СН2-области структуры петли одной из Fc-областей, и аминокислотная модификация, способствующая дестабилизации петлевой структуры СН2-области в другой Fc-области. Примерами аминокислотных модификаций, которые могут дестабилизировать петлевую структуру СН2-области, может быть замена по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 235, 236, 237, 238 и 239, другой аминокислотой. В частности, структура петли СН2-области может быть дестабилизирована, например, путем замены аминокислоты в положении 235 на Asp, Gln, Glu или Thr, замены аминокислоты в положении 236 на Asn, замены аминокислоты в положении 237 на Phe или Trp, замены аминокислоты в положении 238 на Glu, Gly или Asn, и замены аминокислоты в положении 239 на Asp или Glu в соответствии с Европейской нумерацией.
Для получения полипептида, содержащего гетерологичные варианты Fc-области согласно изобретению, необходимо, чтобы варианты Fc-области, имеющие аминокислоты, отличающиеся друг от друга, были связанными, либо чтобы полипептид, содержащий представляющие интерес гетерологичные варианты Fc-области, был отделен от других полипептидов, содержащих гомологичные варианты Fc-области.
Для связывания полипептидов, имеющих различные аминокислоты, может быть применен метод ингибирования непредусмотренного связывания Н-цепей, заключающийся в сообщении электростатического отталкивания на границе второй константной области Н-цепи антитела (CH2) или третьей константной области Н-цепи антитела (CH3) (WO 2006/106905).
В методе ингибирования непредусмотренного связывания Н-цепей, заключающемся в сообщении электростатического отталкивания на границе CH2 или CH3, примерами аминокислотных остатков, контактирующих на границе других константных областей Н-цепи, являются остаток в положении 356 (в соответствии с Европейской нумерацией), остаток в положении 439 (в соответствии с Европейской нумерацией), область, расположенная перед остатком в положении 357 (в соответствии с Европейской нумерацией), остаток в положении 370 (в соответствии с Европейской нумерацией), остаток в положении 399 (в соответствии с Европейской нумерацией) и остаток в положении 409 (в соответствии с Европейской нумерацией) СН3-домена.
Более конкретно, например, может быть получено антитело, содержащее СН3-домены Н-цепи двух типов, то есть антитело, в котором 1-3 пары аминокислотных остатков, выбранных из аминокислотных остатков, представленных ниже в пунктах (1)-(3), в первом СН3-домене Н-типа, имеют одинаковый заряд, то есть:
(1) аминокислотные остатки в положениях 356 и 439 (в соответствии с Европейской нумерацией), которые представляют собой аминокислотные остатки, присутствующие в СН3-домене Н-цепи;
(2) аминокислотные остатки в положениях 357 и 370 (в соответствии с Европейской нумерацией), которые представляют собой аминокислотные остатки, присутствующие в СН3-домене Н-цепи; и
(3) аминокислотные остатки в положениях 399 и 409 (в соответствии с Европейской нумерацией), которые представляют собой аминокислотные остатки, присутствующие в СН3-домене Н-цепи.
Кроме того, может быть продуцировано антитело, в котором 1-3 пары аминокислотных остатков, соответствующих парам аминокислотных остатков, указанных выше в (1)-(3) и имеющих одинаковый заряд в СН3-домене первой Н-цепи, имеют заряды, противоположные зарядам соответствующих аминокислотных остатков в вышеупомянутом СН3-домене первой Н-цепи, где пары аминокислотных остатков выбраны из пар аминокислотных остатков, указанных выше в (1)-(3), в СН3-домене второй Н-цепи, который отличается от СН3-домена первой Н-цепи.
Соответствующие аминокислотные остатки (1)-(3), упомянутые выше, расположены близко друг от друга, если они являются связанными. Специалист в данной области может самостоятельно найти сайты, соответствующие вышеупомянутым аминокислотным остаткам (1)-(3), путем моделирования гомологии и т.п. с использованием коммерчески доступной компьютерной программы для нужного СН3-домена Н-цепи или константной области Н-цепи, и, при необходимости, аминокислотные остатки этих сайтов могут быть модифицированы.
В вышеупомянутых антителах, например, «заряженные аминокислотные остатки» предпочтительно выбраны из аминокислотных остатков, входящих в любую из представленных ниже групп (X) или (Y):
(X) глутаминовая кислота (E) и аспарагиновая кислота (D), и
(Y) лизин (K), аргинин (R) и гистидин (H).
В вышеупомянутых антителах выражение «имеющий один и тот же заряд» означает, что, например, все два или более аминокислотных остатков представляют собой аминокислотные остатки, входящие в любую из вышеупомянутых групп (X) и (Y). Выражение «имеющий противоположный заряд» означает, что, например, если по меньшей мере один из двух или более аминокислотных остатков представляет собой аминокислотный остаток, входящий в любую из вышеупомянутых групп (X) и (Y), то остальные аминокислотные остатки представляют собой аминокислотные остатки, входящие в другую группу.
В предпочтительном варианте вышеупомянутого антитела, СН3-домен первой Н-цепи и СН3-домен второй Н-цепи могут быть перекрестно связаны дисульфидными связями.
В настоящем изобретении модифицируемые аминокислотные остатки не ограничиваются аминокислотными остатками константной области антитела или вариабельной области антитела, описанных выше. Специалист в данной области может самостоятельно найти аминокислотные остатки, которые образуют границу в полипептидных мутантах или гетеромультимерах, путем моделирования гомологии и т.п. с использованием коммерчески доступной компьютерной программы, и аминокислотные остатки в этих сайтах могут быть модифицированы для регуляции связывания.
Кроме того, для связывания гетерологичных вариантов Fc-области могут быть применены и другие известные методы. В частности, такой метод осуществляют путем замены аминокислотной боковой цепи, присутствующей в вариабельной области одной из Н-цепей антитела, более крупной боковой цепью (узел, который означает «утолщение»), и путем замены аминокислотной боковой цепи, присутствующей в вариабельной области другой Н-цепи, более короткой боковой цепью (дырка, которая означает «пустоту»), в результате чего такой узел может быть помещен в дырку. Это может стимулировать эффективное связывание полипептидов, содержащих Fc-область и имеющих различные аминокислоты (WO 1996/027011; Ridgway JB et al., Protein Engineering (1996) 9, 617-621; Merchant AM et al. Nature Biotechnology (1998) 16, 677-681).
Кроме того, для гетерологичного связывания вариантов Fc-области могут быть также применены и другие известные методы. Связывание полипептидов, имеющих различные последовательности, может быть эффективно индуцировано посредством комплементарного связывания СН3 с использованием СН3-домена, сконструированного путем замены цепи, то есть путем замены части СН3 одной из Н-цепей антитела на происходящую от IgА последовательность, соответствующую этой части, и введения в комплементарную часть СН3 другой Н-цепи последовательности, происходящей от IgА и соответствующей этой части (Protein Engineering Design & Selection, 23; 195-202, 2010). Этот известный метод может быть также применен для эффективного индуцирования связывания полипептидов, содержащих Fc-область и имеющих различные аминокислоты.
Кроме того, могут быть также применены методы продуцирования гетеродимеризованных антител путем связывания СН1 и CL антитела и связывания VH и VL антитела, как описано в WO 2011/028952.
Как описано в WO 2008/119353 и WO 2011/131746, может быть также применен метод продуцирования гетеродимеризованных антител, где сначала получают гомодимеризованные антитела двух типов с последующим их инкубированием в восстанавливающих условиях для диссоциации, а затем их снова подвергают ассоциации.
Как описано в публикации J. Mol. (2012) 420, 204-219, может быть также применен метод продуцирования гетеродимеризованных антител путем введения заряженных остатков Lys, Arg, Glu и Asp так, чтобы в СН3 IgG1 и IgG2 наблюдалось электростатическое отталкивание.
Кроме того, как описано в WO 2012/058768, может быть также применен метод продуцирования гетеродимеризованных антител путем введения модификаций в CH2- и CH3-области.
Кроме того, даже в тех случаях, если полипептиды, содержащие гетерологичные варианты Fc-области, не могут быть эффективно продуцированы, то полипептиды, содержащие гетерологичные варианты Fc-области, могут быть получены путем их отделения и очистки от полипептидов, содержащих гетерологичные варианты Fc-области. При продуцировании полипептида, содержащего гетерологичные варианты Fc-области, состоящие из первого полипептида и второго полипептида, имеющих различные последовательности, полипептиды, содержащие гомологичную Fc-область, состоящую только из первого из двух полипептидов, смешивают с полипептидом, содержащим гомологичную Fc-область, состоящую только из второго из двух полипептидов, в виде примеси. Эти известные технологии могут быть использованы как метод эффективного удаления указанных полипептидов двух типов, содержащих гомологичную Fc-область. Сообщалось, что этот метод позволяет проводить очистку гомодимеров двух типов и представляющего интерес гетеродимеризованного антитела с помощью ионообменной хроматографии благодаря созданию различий изоэлектрических точек путем введения аминокислотных замен в вариабельные области Н-цепей двух типов (WO 2007114325). В настоящее время, для очистки гетеродимеризованных антител применяется метод с использованием белка А, позволяющий проводить очистку гетеродимеризованного антитела, содержащего Н-цепь мышиного IgG2а, которая связывается с белком А, и Н-цепь крысиного IgG2b, которая не связывается с белком А (WO 98050431, WO 95033844).
Кроме того, одно гетеродимеризованное антитело может быть эффективно очищено с использованием Н-цепей, в которых аминокислотные остатки в сайте связывания IgG-белок А в положениях 435 и 436 (в соответствии с Европейской нумерацией) были заменены аминокислотами Tyr или His с получением антитела, обладающего другой аффинностью к белку А, то есть другой аффинностью связывания каждой из Н-цепей с белком А, и такое антитело может быть очищено на колонке с белком А.
Множество таких замен и методов, например, два или более из них, могут быть применены в комбинации друг с другом. Кроме того, при необходимости, эти модификации могут быть введены по отдельности в первый полипептид и во второй полипептид. Полипептиды согласно изобретению могут быть также получены из продуктов с вышеупомянутыми модификациями.
В настоящем изобретении термин «аминокислотная модификация» означает любую замену, любую делецию, любое добавление, любую инсерцию и любую другую модификацию или их комбинации. Другими словами, в настоящем изобретении аминокислотная модификация может называться аминокислотной мутацией, и эти термины могут быть использованы как синонимы.
При замене аминокислотных остатков, замена одного аминокислотного остатка другим аминокислотным остатком может быть осуществлена в зависимости от различных факторов, таких как факторы, описанные ниже (a)-(c):
(а) структура полипептидного остова в области, имеющей складчатую или спиральную структуру;
(b) электрический заряд или гидрофобность в сайте-мишени; или
(с) размер боковой цепи.
Аминокислотные остатки могут быть подразделены на нижеследующие группы по их общим свойствам боковых цепей:
(1) гидрофобные остатки: норлейцин, met, ala, val, leu и ile;
(2) нейтральные гидрофильные остатки: cys, ser, thr, asn и gln;
(3) кислотные остатки: asp и glu;
(4) основные остатки: his, lys и arg;
(5) остатки, влияющие на ориентацию цепи: gly и pro; и
(6) ароматические остатки: trp, tyr и phe.
Замена аминокислотных остатков, входящих в каждую из этих групп аминокислот, называется консервативной заменой, а замена аминокислотных остатков, входящих в другие группы, называется неконсервативной заменой. В настоящем изобретении такими заменами могут быть консервативные замены или неконсервативные замены или комбинации консервативных замен и неконсервативных замен.
Модификации аминокислотной последовательности получают различными методами, известными специалистам. Такими методами являются, но не ограничиваются ими, метод сайт-направленного мутагенеза (Hashimoto-Gotoh, T, Mizuno, T, Ogasahara, Y, and Nakagawa, M. (1995) An oligodeoxyribonucleotide-directed dual amber method for site-directed mutagenesis. Gene 152, 271-275; Zoller, MJ and Smith, M. (1983) Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors. Methods Enzymol. 100, 468-500; Kramer, W, Drutsa, V, Jansen, HW, Kramer, B, Pflugfelder, M and Fritz, HJ (1984) The gapped duplex DNA approach to oligonucleotide-directed mutation construction. Nucleic Acids Res. 12, 9441-9456; Kramer W and Fritz HJ (1987) Oligonucleotide-directed construction of mutations via gapped duplex DNA Methods. Enzymol. 154, 350-367; и Kunkel, TA (1985) Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA. 82, 488-492), ПЦР-мутагенез и кластерный мутагенез.
Модификация аминокислот согласно изобретению включает посттрансляционную модификацию. Специфическая посттрансляционная модификация может представлять собой присоединение или делецию сахарной цепи. Так, например, в константной области IgG1, состоящей из аминокислотной последовательности SEQ ID NO:11, аминокислотный остаток в положении 297 (в соответствии с Европейской нумерацией) может быть модифицирован сахарной цепью. Структура сахарной цепи, используемая для модификации, не имеет конкретных ограничений. Вообще говоря, антитела, экспрессирующиеся в эукариотических клетках, содержат в своей константной области сайт гликозилирования. Поэтому антитела, экспрессирующиеся в клетках, таких как клетки, указанные ниже, обычно модифицируют сахарной цепью нескольких типов, где указанными клетками являются:
- антитело-продуцирующие клетки млекопитающих
- эукариотические клетки, трансформированные экспрессионным вектором, содержащим ДНК, кодирующую антитело.
Описанными здесь эукариотическими клетками являются дрожжевые клетки и клетки животных. Так, например, репрезентативными клетками животных, используемыми для трансформации экспрессионным вектором, содержащим ДНК, кодирующую антитело, являются клетки CHO и клетки НЕК293Н. С другой стороны, в константную область согласно изобретению также включены последовательности, не гликозилированные в этом положении. Антитела, константная область которых не является гликозилированной, могут быть получены посредством экспрессии антитело-кодирующего гена в прокариотических клетках, таких как Escherichia coli.
В частности, например, к сахарной цепи Fc-области может быть присоединена сиаловая кислота (MAbs. 2010 Sep-Oct; 2(5): 519-27).
Кроме того, настоящее изобретение относится к антителам, содержащим любой описанный выше вариант Fc-области.
В настоящем изобретении термин «антитело/антитела» употребляется в самом широком смысле, при условии, что такое антитело обладает нужной биологической активностью, и этот термин включает любое антитело, такое как моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, варианты антител, фрагменты антител, полиспецифические антитела (мультиспецифические антитела)(например, биспецифические антитела (диантитела)), химерные антитела и гуманизированные антитела.
Что касается антител согласно изобретению, то тип и происхождение антител не имеет конкретных ограничений, и такими антителами могут быть антитела любого типа. Происхождение антител также не имеет конкретных ограничений, например, такими антителами являются человеческие антитела, мышиные антитела, крысиные антитела и кроличьи антитела.
Методы продуцирования антител хорошо известны специалистам, например, моноклональные антитела могут быть получены гибридомным методом (Kohler and Milstein, Nature 256:495 (1975) или рекомбинантным методом (патент США № 4816567). Альтернативно, эти антитела могут быть выделены из фаговой библиотеки антител (Clackson et al., Nature 352:624-628 (1991); Marks et al., J. Mol. Biol. 222:5 81-597 (1991)).
Гуманизированное антитело также называется реконструированным человеческим антителом. В частности, гуманизированные антитела получают путем присоединения CDR антитела животного, не являющегося человеком, такого как мышиное антитело, к человеческому антителу, и такие методы также известны специалистам. Также известны стандартные методы генной инженерии, применяемые для получения гуманизированных антител. В частности, например, метод ПЦР с перекрывающимся удлинением известен как метод присоединения CDR мышиного антитела к человеческим FR.
Вектор для экспрессии гуманизированного антитела может быть получен путем встраивания ДНК, кодирующей вариабельную область антитела, в которой три CDR и четыре FR лигированы друг с другом, и ДНК, кодирующей константную область человеческого антитела, в экспрессионный вектор так, чтобы эти ДНК были присоединены друг к другу с сохранением рамки считывания. После переноса этого интегрирующего вектора хозяину для получения рекомбинантных клеток, эти клетки культивируют, и ДНК, кодирующую гуманизированное антитело, подвергают экспрессии для продуцирования гуманизированного антитела в клеточной культуре (см. публикацию Европейской патентной заявки EP 239400 и публикацию Международной патентной заявки WO 1996/002576).
При необходимости, аминокислотный остаток в FR может быть заменен так, чтобы CDR реконструированного человеческого антитела образовывали соответствующий антигенсвязывающий сайт. Так, например, мутация может быть введена в аминокислотную последовательность FR ПЦР-методом, применяемым для присоединения мышиных CDR к человеческим FR.
Нужное человеческое антитело может быть получено путем ДНК-иммунизации трансгенного животного, имеющего полный репертуар генов человеческого антитела (см. публикации Международных заявок WO 1993/012227, WO 1992/003918, WO 1994/002602, WO 1994/025585, WO 1996/034096 и WO 1996/033735), и это антитело может быть использовано для иммунизации.
Кроме того, методы получения человеческого антитела посредством пэннинга с использованием библиотеки человеческих антител известны специалистам. Так, например, V-область человеческого антитела экспрессируют на поверхности фага в виде одноцепочечного антитела (scFv) методом фагового представления. При этом может быть отобран scFv-экспрессирующий фаг, который связывается с антигеном. Последовательность ДНК, кодирующая V-область связанного с антигеном человеческого антитела, может быть определена путем анализа генов выбранного фага. После определения последовательности ДНК scFv, которое связывается с антигеном, экспрессионный вектор может быть получен путем присоединения, в той же рамке считывания, последовательности V-области к последовательности нужной С-области человеческого антитела и последующего встраивания полученной последовательности в подходящий экспрессионный вектор. Экспрессионный вектор вводят в клетки, подходящие для экспрессии, как описано выше, и человеческое антитело может быть получено посредством экспрессии гена, кодирующего человеческое антитело. Эти методы уже известны специалистам (см. публикации Международных заявок WO 1992/001047, WO 1992/020791, WO 1993/006213, WO 1993/011236, WO 1993/019172, WO 1995/001438 и WO 1995/015388).
Вариабельными областями, входящими в состав антител согласно изобретению, могут быть вариабельные области, распознающие любой антиген.
В настоящем изобретении антигенами могут быть любые антигены без каких-либо конкретных ограничений. Примерами таких антигенов предпочтительно являются лиганды (цитокины, хемокины и т.п.), рецепторы, раковые антигены, антигены МНС, антигены дифференцировки, иммуноглобулины и иммунные комплексы, частично содержащие иммуноглобулины.
Примерами цитокинов являются интерлейкины 1-18, колониестимулирующие факторы (G-CSF, M-CSF, GM-CSF и т.п.), интерфероны (IFN-α, IFN-β, IFN-γ и т.п.), факторы роста (EGF, FGF, IGF, NGF, PDGF, TGF, HGF и т.п.), факторы некроза опухоли (TNF-α и TNF-β), лимфотоксин, эритропоэтин, лептин, SCF, TPO, MCAF и BMP.
Примерами хемокинов являются хемокины CC, такие как CCL1-CCL28, хемокины СХС, такие как CXCL1-CXCL17, хемокины С, такие как XCL1-XCL2, и хемокины СХ3С, такие как CX3CL1.
Примерами рецепторов являются рецепторы, принадлежащие к таким семействам, как семейство рецепторов гемопоэтического фактора роста, семейство рецепторов цитокинов, семейство рецепторов тирозинкиназного типа, семейство рецепторов серин/треонин-киназного типа, семейство рецепторов TNF, семейство рецепторов, сопряженных с G-белком, семейство рецепторов GPI якорного типа, семейство рецепторов тирозин-фосфатазного типа, семейство рецепторов факторов адгезии и семейство рецепторов гормонов. Рецепторы, принадлежащие к этим семействам, и их свойства описаны во многих документах, таких как Cooke BA., King RJB., Van der Molen HJ. Ed. New Comprehesive Biochemistry Vol.18B "Hormones and their Actions Part II" pp.1-46 (1988) Elsevier Science Publishers BV., Patthy (1990) 61(1), 13-14); Ullrich et al. (Cell (1990) 61(2), 203-212), Massagué (Сеll (1992) 69(6): 1067-1070); Miyajima, et al (Annu. Rev. Immunol. (1992) 10, 295-331), Taga et al. (FASEB J. (1992) 6, 3387-3396); Fantl et al. (Annu. Rev. Biochem. (1993), 62: 453-481), Smith et al. (Cell (1994) 76(6): 959-962) и Flower DR. Flower (Biochim. Biophys. Acta (1999) 1422 (3): 207-234).
Примерами специфических рецепторов, принадлежащих к вышеупомянутым семействам рецепторов, предпочтительно являются рецепторы человеческого или мышиного эритропоэтина (EPO) (Blood (1990) 76(1), 31-35 и Cell (1989) 57(2): 277-285); рецепторы человеческого или мышиного гранулоцитарного колониестимулирующего фактора (G-CSF) (Proc. Natl. Acad. Sci. USA. (1990), 87(22), 8702-8706; mG-CSFR (Cell (1990) 61(2): 341-350); рецепторы человеческого или мышиного тромбопоэтина (TPO) (Proc. Natl. Acad. Sci. USA. (1992) 89(12): 5640-5644, EMBO J. (1993) 12(7): 2645-53); рецепторы человеческого или мышиного инсулина (Nature (1985) 313 (6005): 756-761); рецепторы человеческого или мышиного лиганда Flt-3 (Proc. Natl. Acad. Sci. USA. (1994) 91(2): 459-463); рецепторы человеческого или мышиного тромбоцитарного фактора роста (PDGF) (Proc. Natl. Acad. Sci. USA. (1988) 85(10): 3435-3439); рецепторы человеческого или мышиного интерферона (IFN-α и β) (Cell (1990) 60(2): 225-234) и Cell (1994) 77(3): 391-400); рецепторы человеческого или мышиного лептина, рецепторы человеческого или мышиного гормона роста (GH), рецепторы человеческого или мышиного интерлейкина (IL)-10, рецепторы человеческого или мышиного инсулиноподобного фактора роста (IGF)-I, рецепторы ингибирующего фактора человеческого или мышиного лейкоза (LIF) и рецепторы человеческого или мышиного цилиарного нейтротропного фактора (CNTF).
Раковые антигены представляют собой антигены, которые экспрессируются в процессе озлокачествования клеток, и такие антигены также называются опухолеспецифическими антигенами. Аномальные сахарные цепи, образующиеся на клеточных поверхностях или белковых молекулах раковых клеток, также называются раковыми антигенами, и такие антигены также являются раковыми антигенами с сахарной цепью. Примерами раковых антигенов предпочтительно является антиген GPC3, который представляет собой рецептор, принадлежащий к вышеупомянутому семейству рецепторов GPI якорного типа, и который также экспрессируется в некоторых раковых опухолях, включая рак печени (Int. J. Cancer (2003) 103(4): 455-65); EpCAM, который экспрессируется в некоторых раковых опухолях, включая рак легких (Proc. Natl. Acad. Sci. USA. (1989) 86(1): 27-31); CA19-9, CA15-3 и сиалил-SSEA-1 (SLX).
Антигены MHC, грубо говоря, подразделяются на антигены MHC класса I и антигены MHC класса II. Антигенами MHC класса I являются HLA-A, -B, -C, -E, -F, -G и -H, а антигенами MHC класса II являются HLA-DR, -DQ и -DP.
Антигенами дифференцировки могут быть CD1, CD2, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15s, CD16, CD18, CD19, CD20, CD21, CD23, CD25, CD28, CD29, CD30, CD32, CD33, CD34, CD35, CD38, CD40, CD41a, CD41b, CD42a, CD42b, CD43, CD44, CD45, CD45RO, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD51, CD54, CD55, CD56, CD57, CD58, CD61, CD62E, CD62L, CD62P, CD64, CD69, CD71, CD73, CD95, CD102, CD106, CD122, CD126 и CDw130.
Иммуноглобулины включают IgA, IgM, IgD, IgG и IgE. Иммунные комплексы включают компонент по меньшей мере любого из этих иммуноглобулинов.
Другими антигенами являются, например, нижеследующие молекулы: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, аденозиновый рецептор A1, A33, ACE, ACE-2, активин, активин А, активин AB, активин B, активин C, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, APE, APJ, APP, APRIL, AR, ARC, ART, артемин, анти-Id антитело, аспарагиновая кислота, атриальный натриуретический пептид, интегрин av/b3, Axl, b2M, B7-1, B7-2, B7-H, B-лимфоцитарный стимулирующий фактор (BlyS), BACE, BACE-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bcl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, остеогенин BMP-3, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, остеогенный нейротропный фактор, BPDE, BPDE-DNA, BTC, фактор комплемента 3 (C3), C3a, C4, C5, C5a, C10, CA125, CAD-8, кальцитонин, cAMP, канцероэмбриональный антиген (CEA), антиген, ассоциированный с раковой опухолью, катепсин A, катепсин B, катепсин C/DPPI, катепсин D, катепсин E, катепсин H, катепсин L, катепсин O, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок p67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (B7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, ботулинический токсин, токсины Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, антиген, ассоциированный с опухолевым цитокератином, DAN, DCC, DcR3, DC-SIGN, фактор регуляции комплемента (фактор, ускоряющий разрушение опухоли), des(1-3)-IGF-I (IGF-I головного мозга), Dhh, дигоксин, DNAM-1, ДНКаза, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, белок активации фибробластов (FAP), Fas, FcR1, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа 1, GFR-альфа 2, GFR-альфа 3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, рилизинг-фактор гормона роста, гаптен (NP-кэп или NIP-кэп), HB-EGF, HCC, гликопротеин оболочки gB HCMV, гликопротеин оболочки gH HCMV, HCMV UL, гемопоэтический фактор роста (HGF), gp120 Hep B, гепараназа, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGFA, высокомолекулярный антиген, ассоциированный с меланомой (HMW-MAA), gp120 ВИЧ, V3-петля gp 120 ВИЧ IIIB, HLA, HLA-DR, HM1.24, HMFG PEM, HRG, Hrk, человеческий кардиомиозин, человеческий цитомегаловирус (HCMV), человеческий гормон роста (HGH), HVEM, I-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, рецептор IgA, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL- 1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-23, интерферон (INF)-альфа, INF-β, INF-гамма, ингибин, iNOS, A-цепь инсулина, В-цепь инсулина, инсулиноподобный фактор роста 1, интегрин-альфа 2, интегрин-альфа 3, интегрин-альфа 4, интегрин-α4/β1, интегрин-альфа 4/бета 7, интегрин-альфа 5 (альфа V), интегрин-α5/β1, интегрин-альфа 5/бета 3, интегрин-альфа 6, интегрин-бета 1, интегрин-бета 2, интерферон-гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин L1, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bp1, LBP, LDGF, LECT2, Lefty, антиген Lewis-Y, Lewis-Y-ассоциированный антиген, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, поверхностный антиген легких, лютеинизирующий гормон, рецептор лимфотоксина-бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCAM, MCK-2, MCP, M-CSF, MDC, Mer, металлопротеазы, рецептор MGDF, MGMT, MHC (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, мюллеровский ингибирующий фактор, Mug, MuSK, NAIP, NAP, NCAD, адгерин N-C, NCA 90, NCAM, NCAM, неприлизин, нейротропин-3, -4 или -6, нейротурин, фактор роста нервных клеток (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGG1, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, щелочная фосфатаза плаценты (PLAP), PlGF, PLP, PP14, проинсулин, прорелаксин, белок C, PS, PSA, PSCA, мембранный антиген, специфичный для предстательной железы (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, A-цепь релаксина, В-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, серин, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (опухолеассоциированный гликопротеин-72), TARC, TCA-3, T-клеточный рецептор (например, T-клеточный рецептор альфа/бета), TdT, TECK, TEM1, TEM5, TEM7, TEM8, TERT, PLAP-подобная щелочная фосфатаза яичек, TfR, TGF, TGF-альфа, TGF-бета, антиген, специфичный к TGF-бета всех типов, TGF-бета RI (ALK-5), TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета 1, TGF-бета 2, TGF-бета 3, TGF-бета 4, TGF-бета 5, тромбин, Ck-1 тимуса, тиреотропный стимулирующий гормон, Tie, TIMP, TIQ, тканевый фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа,бета, TNF-бета 2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD120a, p55-60), TNFRSF1B (TNF RII CD120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (OX40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL R1 TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (лиганд TRAIL Apo-2, TL2), TNFSF11 (лиганд TRANCE/RANK ODF, лиганд OPG), TNFSF12 (лиганд TWEAK Apo-3, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд GITR AITR, лиганд TL6), TNFSF1A (коннектин TNF-α, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд OX40 gp34, TXGP1), TNFSF5 (лиганд CD40, CD154, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Fas, лиганд Apo-1, лиганд APT1), TNFSF7 (лиганд CD27, CD70), TNFSF8 (лиганд CD30, CD153), TNFSF9 (лиганд 4-1BB, лиганд CD137), TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TSG, TSLP, опухолеассоциированный антиген CA125, опухолеассоциированный антиген, экспрессирующий Lewis Y-ассоциированные углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный антиген, VLA, VLA-1, VLA-4, интегрин VNR, фактор фон Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, Aβ, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный ЛНП, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин A, хромогранин B, tau, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор B, фактор D, фактор H, пропердин, склеростин, фибриноген, фибрин, протромбин, тромбин, тканевый фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Xa, фактор XI, фактор XIa, фактор XII, фактор XIIa, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и S1P, а также рецепторы гормонов и факторов роста.
В аминокислотные последовательности, составляющие вариабельные области, могут быть введены одна или более модификаций аминокислотных остатков, при условии, что они будут сохранять антигенсвязывающую активность. При модификации аминокислотной последовательности вариабельной области, сайты модификации и число модифицируемых аминокислот не имеют конкретных ограничений. Так, например, аминокислоты, присутствующие в CDR и/или FR, могут быть соответствующим образом модифицированы. При модификации аминокислот в вариабельной области, активность связывания предпочтительно должна быть сохранена, но без каких-либо конкретных ограничений; например, она должна составлять 50% или более, предпочтительно 80% или более, а более предпочтительно 100% или более от активности до модификации. Кроме того, активность связывания может быть повышена путем модификации аминокислот. Так, например, активность связывания может в 2, 5, 10 или более раз превышать активность до модификации. В антителах согласно изобретению, модификацией аминокислотной последовательности могут быть по меньшей мере одна замена, по меньшей мере одно добавление, по меньшей мере одна делеция и по меньшей мере одна модификация аминокислоты.
Так, например, замена N-концевого глутамина вариабельной области на пироглутаминовую кислоту, осуществляемая посредством пироглутамилирования, представляет собой модификацию, хорошо известную специалистам. Таким образом, если у N-конца тяжелой цепи присутствует глутамин, то антитела согласно изобретению содержат вариабельные области, в которых глутамин заменен пироглутаминовой кислотой.
Вариабельные области антител согласно изобретению могут иметь любые последовательности, и такие вариабельные области антител могут происходить от любого источника, например, такими антителами могут быть мышиные антитела, крысиные антитела, кроличьи антитела, козьи антитела, верблюжьи антитела, гуманизированные антитела, полученные путем гуманизации нечеловеческих антител, и человеческие антитела. «Гуманизированные антитела», также называемые «реконструированными человеческими антителами», представляют собой антитела, в которых гипервариабельные области (CDR) антитела, происходящего от млекопитающего, не являющегося человеком, например, от мышиного антитела, были заменены на CDR человеческого антитела. Методы идентификации CDR известны специалистам (Kabat et al, Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md; Chothia et al., Nature (1989) 342: 877). Стандартные методы рекомбинации генов также известны специалистам (см. публикацию Европейской патентной заявки № EP 125023 и WO 96/02576). Кроме того, эти антитела могут иметь различные аминокислотные замены, введенные в их вариабельные области для повышения уровня связывания с антигеном, для улучшения фармакокинетических свойств, для повышения стабильности и для снижения иммуногенности. Вариабельные области антител согласно изобретению могут обладать способностью связываться с антигенами повторно, что обусловлено зависимостью связывания антигена от рН (WO 2009/125825).
Константные области легкой цепи антитела представляют собой константные области цепи каппа-типа и лямбда-типа, однако, приемлемой может быть любая из константных областей легкой цепи. Кроме того, константные области легкой цепи согласно изобретению могут представлять собой константные области легкой цепи с аминокислотными модификациями, такими как замены, делеции, добавления и/или инсерции.
Так, например, для конструирования константных областей тяжелой цепи антитела согласно изобретению могут быть использованы константные области тяжелой цепи человеческих антител IgG, при этом предпочтительными являются константные области тяжелой цепи человеческих антител IgG1 и человеческих антител IgG4.
Кроме того, варианты Fc-области согласно изобретению могут быть получены в виде молекул слитых белков Fc путем присоединения к другим белкам, физиологически активным пептидам и т.п. В настоящем изобретении термин «слитый белок» означает химерный полипептид, содержащий по меньшей мере два различных полипептида, которые в природе спонтанно не связываются друг с другом. Примерами таких других белков и биологически активных пептидов являются, но не ограничиваются ими, рецепторы, адгезивные молекулы, лиганды и ферменты.
Предпочтительными примерами молекул слитых белков Fc согласно изобретению являются белки, содержащие Fc-область, присоединенную к рецепторному белку, который связывается с мишенью, и примерами таких белков являются слитый белок TNFR-Fc, слитый белок IL1R-Fc, слитый белок VEGFR-Fc и слитый белок CTLA4-Fc (Nat. Med. 2003 Jan; 9(1): 47-52, BioDrugs 2006; 20(3):151-60). Кроме того, белком, присоединенным к полипептиду согласно изобретению, может быть любая молекула, при условии, что она будет связываться с молекулой-мишенью, и примерами таких молекул являются молекулы scFv (WO 2005/037989), молекулы однодоменных антител (WO 2004/058821; WO 2003/002609), антитело-подобные молекулы (Current Opinion in Biotechnology 2006, 17: 653-658; Current Opinion in Biotechnology 2007, 18: 1-10; Current Opinion in Structural Biology 1997, 7: 463-469 и Protein Science 2006, 15: 14-27), такие как DARP-ины (WO 2002/020565), аффинное антитело (WO 1995/001937), авимер (WO 2004/044011, WO 2005/040229) и аднектин (WO 2002/032925). Кроме того, молекулы антител и Fc-слитых белков могут представлять собой мультиспецифические антитела, которые связываются с молекулами-мишенями или эпитопами многих типов.
Кроме того, антителами согласно изобретению являются модификации антител. Такими модификациями антител являются, например, антитела, связанные с различными молекулами, такими как полиэтиленгликоль (ПЭГ) и цитотоксические вещества. Такие модифицированные антитела могут быть получены путем химической модификации антител согласно изобретению. Методы модификации антител хорошо известны специалистам в данной области.
Антителами согласно изобретению могут быть также биспецифические антитела. Термин «биспецифическое антитело» означает антитело, которое, в одной своей молекуле, имеет вариабельные области, распознающие различные эпитопы. Эпитопы могут присутствовать в одной молекуле или в различных молекулах.
Полипептиды согласно изобретению могут быть получены методами, известными специалистам. Так, например, антитела могут быть получены методами, описанными ниже, но не ограничивающимися ими.
Была экспрессирована ДНК, кодирующая тяжелую цепь антитела, в которой один или более аминокислотных остатков в Fc-области были заменены другими представляющими интерес аминокислотами, а также была экспрессирована ДНК, кодирующая легкую цепь антитела. ДНК, кодирующая тяжелую цепь, в которой один или более аминокислотных остатков в Fc-области были заменены другими представляющими интерес аминокислотами, может быть получена, например, путем конструирования ДНК, кодирующей природную тяжелую цепь Fc-области, и введения соответствующей замены так, чтобы кодон, кодирующий конкретную аминокислоту в Fc-области, кодировал другую представляющую интерес аминокислоту.
Альтернативно, ДНК, кодирующая тяжелую цепь, в которой один или более аминокислотных остатков в Fc-области были заменены другими представляющими интерес аминокислотами, может быть также получена путем конструирования и химического синтеза ДНК, кодирующей белок, в котором один или более аминокислотных остатков в природной тяжелой цепи Fc-области были заменены другими представляющими интерес аминокислотами. Положение и тип аминокислотной замены не имеют конкретных ограничений. Кроме того, модификация не ограничивается только заменой, то есть такой модификацией может быть любая делеция, любое добавление или любая инсерция или их комбинации.
Альтернативно, ДНК, кодирующая тяжелую цепь, в которой один или более аминокислотных остатков в Fc-области были заменены другими представляющими интерес аминокислотами, может быть получена в виде комбинации неполных ДНК. Такими комбинациями неполных ДНК являются, например, но не ограничиваются ими, комбинация ДНК, кодирующей вариабельную область, и ДНК, кодирующей константную область, и комбинация ДНК, кодирующей Fab-область, и ДНК, кодирующей Fc-область. Кроме того, ДНК, кодирующая легкую цепь, может быть аналогичным образом получена в виде комбинации неполных ДНК.
Методы экспрессии вышеописанных ДНК включают методы, описанные ниже. Так, например, экспрессионный вектор для тяжелой цепи конструируют путем встраивания ДНК, кодирующей вариабельную область тяжелой цепи, в экспрессионный вектор вместе с ДНК, кодирующей константную область тяжелой цепи. Аналогичным образом, экспрессионный вектор для легкой цепи конструируют путем встраивания ДНК, кодирующей вариабельную область легкой цепи, в экспрессионный вектор вместе с ДНК, кодирующей константную область легкой цепи. Альтернативно, эти гены тяжелой и легкой цепей могут быть встроены в один вектор.
ДНК, кодирующую представляющее интерес антитело, встраивают в экспрессионный вектор так, чтобы антитело экспрессировалось под контролем области регуляции экспрессии, такой как энхансер или промотор. Затем клетки-хозяева трансформируют этим экспрессионным вектором для экспрессии антитела. В таких случаях может быть использована соответствующая комбинация хозяина и экспрессионного вектора.
Примерами векторов являются векторы M13, векторы pUC, pBR322, pBluescript и pCR-Script. Альтернативно, для субклонирования и вырезания кДНК, помимо вышеописанных векторов, могут быть использованы pGEM-T, pDIRECT, pT7 и т.п.
Экспрессионные векторы являются особенно подходящими для продуцирования полипептидов согласно изобретению. Так, например, если клеткой-хозяином является E. coli, такая как JM109, DH5α, HB101 и XL1-Blue, то экспрессионные векторы должны содержать промотор, обеспечивающий эффективную экспрессию в E. coli, например, промотор lacZ (см. публикации Ward et al., Nature (1989) 341: 544-546; FASEB J. (1992) 6, 2422-2427, которые во всей своей полноте вводятся в настоящее описание посредством ссылки), промотор araB (см. публикацию Better et al., Science (1988) 240: 1041-1043, которая во всей своей полноте вводится в настоящее описание посредством ссылки), промотор T7 или т.п. Такими векторами, помимо вышеописанных векторов, являются pGEX-5X-1 (Pharmacia), «QIAexpress system» (QIAGEN), pEGFP или pET (в этом случае хозяином предпочтительно является BL21, который экспрессирует РНК-полимеразу Т7).
Векторы могут содержать сигнальные последовательности для секреции полипептида. В случае секреции полипептида в периплазму E. coli, в качестве сигнальной последовательности для такой секреции может служить сигнальная последовательность pelB (см. публикацию Lei, SP et al. J. Bacteriol. (1987) 169, 4397, которая во всей своей полноте вводится в настоящее описание посредством ссылки). Вектор может быть встроен в клетки-хозяева методом с использованием липофектина, фосфата кальция и DEAE-дектрана.
Помимо экспрессионных векторов E. coli, векторами для продуцирования полипептидов согласно изобретению являются, например, экспрессионные векторы млекопитающих (например, pcDNA3 (Invitrogen), pEGF-BOS (см. публикацию Nucleic Acids. Res. 1990, 18(17), p. 5322, которая во всей своей полноте вводится в настоящее описание посредством ссылки), pEF и pCDM8); экспрессионные векторы, происходящие от клеток насекомых (например, «бакуловирусная экспрессионная система Bac-to-BAC» (GIBCO BRL) и pBacPAK8); экспрессионные векторы, происходящие от растений (например, pMH1 и pMH2); экспрессионные векторы, происходящие от вируса животного (например, pHSV, pMV и pAdexLcw); ретровирусные экспрессионные векторы (например, pZIPneo); дрожжевые экспрессионные векторы (например, «Набор для экспрессии в дрожжах Pichia» («Pichia Expression Kit» Invitrogen), pNV11 и SP-Q01); и экспрессионные векторы Bacillus subtilis (например, pPL608 и pKTH50).
Векторы для экспрессии в клетках животных, таких как клетки CHO, COS и NIH3T3, должны иметь промоторы, необходимые для экспрессии в таких клетках, например, промотор SV40 (см. публикацию Mulligan et al., Nature (1979) 277, 108, которая во всей своей полноте вводится в настоящее описание посредством ссылки); промотор MMTV-LTR, промотор EF1α (см. публикацию Mizushima et al., Nucleic Acids Res. (1990) 18, 5322, которая во всей своей полноте вводится в настоящее описание посредством ссылки); промотор CAG (см. публикацию Gene. (1991) 108, 193, которая во всей своей полноте вводится в настоящее описание посредством ссылки) и промотор CMV, а более предпочтительно, такие векторы должны содержать ген для отбора трансформированных клеток (например, ген резистентности к лекарственному средству, который позволяет проводить такой отбор с использованием агента (неомицина, G418 или т.п.)). Векторами, обладающими такими свойствами, являются, например, pMAM, pDR2, pBK-RSV, pBK-CMV, pOPRSV и pOP13.
Кроме того, для стабильной экспрессии гена и амплификации различных копий генов в клетках может быть применен описанный ниже метод, в котором клетки CHO, дефицитные по механизму синтеза нуклеиновой кислоты, вводят вместе с вектором, который несет ген DHFR, компенсирующий такой дефицит (например, pCHOI), а вектор амплифицируют с использованием метотрексата (MTX). Альтернативно, для временной экспрессии гена может быть применен описанный ниже метод, в котором клетки COS, имеющие ген, экспрессирующий Т-антиген SV40 на хромосоме этих клеток, трансформируют вектором, имеющим ориджин репликации SV40 (pcD и т.п.). Могут быть также использованы ориджины репликации, происходящие от полиомавируса, аденовируса, вируса бычьей папилломы (BPV) и т.п. Для амплификации ряда копий генов в клетках-хозяевах могут быть также использованы экспрессионные векторы, содержащие селективные маркеры, такие как ген аминогликозид-трансферазы (APH), ген тимидин-киназы (TK), ген ксантин-гуанин-фосфорибозилтрансферазы E. coli (Ecogpt) и ген дигидрофолат-редуктазы (dhfr).
Антитела могут быть собраны, например, путем культивирования трансформированных клеток с последующим выделением антител из трансформированных клеток или из культуральной среды. Антитела могут быть выделены и очищены с применением соответствующих комбинированных методов, таких как центрифугирование, фракционирование сульфатом аммония, высаливание, ультрафильтрация, метод с использованием 1q, FcRn и колонки с белком A и G-белком, аффинная хроматография, ионообменная хроматография и гель-фильтрация.
Кроме того, настоящее изобретение относится к способам продуцирования полипептида, содержащего вариант Fc-области антитела, обладающего повышенной активностью связывания с FcγRIIb, по сравнению с полипептидом, содержащим родительскую Fc-область, где указанный способ включает введение в вариант Fc-области по меньшей мере одной аминокислотной модификации.
Примерами способов продуцирования являются способы, включающие следующие стадии:
(a) добавление по меньшей мере одной аминокислотной модификации в Fc-область полипептидов, содержащих Fc-область;
(b) определение активности связывания полипептидов, модифицированных в стадии (а) с FcγRIIb; и
(c) отбор полипептидов, содержащих вариант Fc-области, обладающий повышенной активностью связывания с FcγRIIb, по сравнению с полипептидом, содержащим родительскую Fc-область.
Предпочтительным вариантом осуществления изобретения является способ продуцирования полипептида, содержащего вариант Fc-области, где указанный включает стадии:
(a) модификации полипептид-кодирующей нуклеиновой кислоты для повышения активности связывания с FcγRIIb по сравнению с активностью полипептида, содержащего родительскую Fc-область;
(b) введения нуклеиновой кислоты в клетки-хозяева и их культивирования для индуцирования экспрессии; и
(c) сбора полипептида из культуры клеток-хозяев.
Кроме того, антитела и молекулы Fc-слитого белка, полученные этим способом, также входят в объем настоящего изобретения.
Настоящее изобретение также относится к способу продуцирования полипептида, содержащего вариант Fc-области, обладающий повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область, где указанный способ включает введение в вариант Fc-области полипептида антитела, содержащего вариант Fc-области, по меньшей мере одной аминокислотной модификации.
Примером способа продуцирования является способ, включающий стадии:
(a) добавления по меньшей мере одной аминокислотной модификации в Fc-область полипептидов, содержащих Fc-область;
(b) определения активности связывания полипептидов, модифицированных в стадии (а), с FcγRIIа и с FcγRIIb; и
(c) отбора полипептида, содержащего вариант Fc-области, обладающий повышенной селективностью связывания с FcγRIIb, по сравнению с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область.
Предпочтительным вариантом осуществления изобретения является способ продуцирования полипептидов, содержащих вариант Fc-области, где указанный способ включает стадии:
(a) модификации нуклеиновой кислоты, кодирующей полипептид, содержащий родительскую Fc-область, для повышения селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от указанного полипептида;
(b) введения нуклеиновой кислоты в клетку-хозяина и культивирования этой клетки в условиях, подходящих для экспрессии такой нуклеиновой кислоты; и
(c) сбора полипептида из культуры клеток-хозяев.
Антитела и молекулы слитого белка Fc, полученные этим способом, также входят в объем настоящего изобретения.
Кроме того, настоящее изобретение относится к способу продуцирования полипептида, содержащего вариант Fc-области, обладающей повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область, где указанный способ включает добавление по меньшей мере одной аминокислотной модификации в вариант Fc-области полипептида антитела, содержащего вариант Fc-области антитела.
Примером способа продуцирования является способ, включающий стадии:
(a) добавления по меньшей мере одной аминокислотной модификации в Fc-область полипептида, содержащего Fc-область;
(b) определения активности связывания полипептида, модифицированного в стадии (а), с FcγRIIа и с FcγRIIb; и
(c) отбора полипептида, содержащего вариант Fc-области, обладающий повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область.
Предпочтительным вариантом осуществления изобретения является способ продуцирования полипептидов, содержащих вариант Fc-области, где указанный способ включает стадии:
(a) модификации нуклеиновой кислоты, кодирующей полипептид, содержащий родительскую Fc-область, для повышения активности связывания с FcγRIIb и повышения селективности связывания с FcγRIIb по сравнению с FcγRIIа (R-типа), в отличие от указанного полипептида;
(b) введения нуклеиновой кислоты в клетку-хозяина и культивирования этой клетки в условиях, подходящих для экспрессии такой нуклеиновой кислоты; и
(c) сбора полипептида из культуры клеток-хозяев.
Антитела и молекулы Fc-слитого белка, полученные этим способом, также входят в объем настоящего изобретения.
Настоящее изобретение также относится к способам продуцирования полипептида, в которых уровень продуцирования антитела против полипептида ниже, чем уровень продуцирования антитела против полипептида, содержащего родительскую Fc-область, при его введении in vivo, где указанные способы включают добавление по меньшей мере одной аминокислотной модификации в Fc-область полипептида, содержащего Fc-область антитела.
Примером способа продуцирования является способ, включающий следующие стадии:
(a) добавление по меньшей мере одной аминокислотной модификации в Fc-область полипептида, содержащего Fc-область;
(b) подтверждение подавления продуцирования антитела при введении in vivo полипептида, содержащего Fc-область, модифицированную в стадии (а), по сравнению с полипептидом, содержащим родительскую Fc-область.
Подавление продуцирования антител против полипептида может быть подтверждено способами введения полипептида животному и т.п. Альтернативно, подавление продуцирования антитела может быть определено путем измерения активности связывания с FcγRIIа и с FcγRIIb и мониторинга повышения величины, полученной путем деления величины KD для FcγRIIа на величину KD для FcγRIIb. Считается, что такие полипептиды могут быть использованы в качестве фармацевтических средств, поскольку они могут подавлять продуцирование антитела без активации активирующего FcγR.
В вышеупомянутых способах продуцирования предпочтительно, чтобы активность связывания с FcγRIIb и селективность связывания с FcγRIIb была выше, чем активность и селективность связывания с FcγRIIа (R-типа).
Примером предпочтительного варианта вышеупомянутого способа продуцирования является модификация Fc-области человеческого IgG, а именно, замена аминокислоты в положении 238 (в соответствии с Европейской нумерацией) другой аминокислотой и замены по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 233, 234, 235, 237, 264, 265, 266, 267, 268, 269, 271, 272, 274, 296, 326, 327, 330, 331, 332, 333, 334, 355, 356, 358, 396, 409 и 419 в соответствии с Европейской нумерацией, другой аминокислотой, введенной в Fc-область. Другими аминокислотными модификациями, объединенными с аминокислотной модификацией в положении 238 (в соответствии с Европейской нумерацией), предпочтительно являются модификации аминокислот в положениях 233, 237, 264, 267, 268, 271, 272, 296, 327, 330, 332, 333 и 396 в соответствии с Европейской нумерацией, а в частности, модификации аминокислот в положениях 233, 237, 264, 267, 268, 271, 296, 330 и 396. В частности, предпочтительная комбинация аминокислотных модификаций, вводимых для повышения активности связывания с FcγRIIb или повышения селективности связывания с FcγRIIb по сравнению с FcγRIIа, включает, например, комбинацию аминокислотных модификаций в положениях 238, 268 и 271 (в соответствии с Европейской нумерацией) вместе модификацией по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 233, 237, 264, 267, 272, 296, 327, 330, 332 и 396 (в соответствии с Европейской нумерацией).
Модифицированные аминокислоты не имеют конкретных ограничений, при условии, что эти аминокислоты будут способствовать повышению активности связывания с FcγRIIb или повышению селективности связывания с FcγRIIb, а не с FcγRIIа, по сравнению с активностью до их модификации, при этом предпочтительной аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 234 является Tyr, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 265 является Glu, аминокислотой в положении 266 является Phe, Leu или Met, аминокислотой в положении 267 является Ala, Glu, Gly или Gln, аминокислотой в положении 268 является Asp, Gln или Glu, аминокислотой в положении 269 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, Phe, Ile, Met, Asn, Pro или Gln, аминокислотой в положении 274 является Gln, аминокислотой в положении 296 является Asp или Phe, аминокислотой в положении 326 является Ala или Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Lys, Arg или Ser, аминокислотой в положении 331 является Ser, аминокислотой в положении 332 является Lys, Arg, Ser или Thr, аминокислотой в положении 333 является Lys, Arg, Ser или Thr, аминокислотой в положении 334 является Arg, Ser или Thr, аминокислотой в положении 355 является Ala или Gln, аминокислотой в положении 356 является Glu, аминокислотой в положении 358 является Met, аминокислотой в положении 396 является Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr, аминокислотой в положении 409 является Arg, а аминокислотой в положении 419 является Glu. Более предпочтительными модифицированными аминокислотами являются аминокислоты, способствующие повышению активности связывания с FcγRIIb, а также повышению селективности связывания с FcγRIIb, а не с FcγRIIа, по сравнению с немодифицированными аминокислотами. В частности, при объединении модификаций аминокислот в положениях 238, 268 и 271 (в соответствии с Европейской нумерацией) с модификациями по меньшей мере одной аминокислоты в положениях, выбранных из положений 233, 264, 267, 272, 296, 327, 330, 332 и 396 (в соответствии с Европейской нумерацией), предпочтительной аминокислотой в положении 238 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 272 является Asp или Pro, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, аминокислотой в положении 332 является Thr, а аминокислотой в положении 396 является Leu или Met в соответствии с Европейской нумерацией.
Из этих модификаций предпочтительными являются модификации, которые приводят к значительному повышению активности связывания с FcγRIIb или к значительному повышению селективности связывания с FcγRIIb, а не с FcγRIIа (R-типа). Примерами таких предпочтительных комбинаций аминокислотных замен, используемых в качестве модификаций, являются нижеследующие комбинации модификаций (a)-(x):
(а) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(b) аминокислотные модификации в положениях 238, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(c) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(d) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(е) аминокислотные модификации в положениях 238, 233, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(f) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(g) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 327 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(h) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(i) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(j) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(k) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(l) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(m) аминокислотные модификации в положениях 238, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(n) аминокислотные модификации в положениях 238, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(o) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(p) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(q) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 327, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(r) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(s) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(t) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(u) аминокислотные модификации в положениях 238, 233, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(v) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(w) аминокислотные модификации в положениях 238, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области; и
(x) аминокислотные модификации в положениях 238, 233, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области.
Кроме того, из этих комбинаций более предпочтительными комбинациями аминокислотных модификаций являются нижеследующие комбинации модификаций аминокислотных последовательностей (a)-(x), таких как:
(a) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(b) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(c) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(d) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Gly или Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(e) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(f) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(g) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(h) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(i) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(j) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(k) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(l) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(m) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(n) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(o) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(p) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(q) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met в соответствии с Европейской нумерацией;
(r) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, и аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(s) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(t) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(u) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(v) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Gly, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(w) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией; и
(x) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией.
Настоящее изобретение также относится к способу модификации полипептида для продуцирования полипептида, обладающего повышенной активностью связывания с FcγRIIb или селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа), по сравнению с полипептидом, содержащим родительскую Fc-область. Настоящее изобретение также относится к способу модификации полипептида для продуцирования полипептида, обладающего повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа), по сравнению с полипептидом, содержащим родительскую Fc-область.
Настоящее изобретение также относится к способам модификации полипептида для продуцирования полипептида, в которых подавляется продуцирование антитела против этого полипептида, вводимого in vivo, по сравнению с продуцированием антитела против полипептида, содержащего родительскую Fc-область.
Пример предпочтительного варианта осуществления изобретения включает комбинацию аминокислотных модификаций, описанных в способе продуцирования полипептидов, содержащих варианты Fc-области, обладающие повышенной активностью связывания с FcγRIIb или повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа). Пример более предпочтительного варианта осуществления изобретения включает комбинацию аминокислотных модификаций, описанных в способе продуцирования полипептидов, содержащих варианты Fc-области, обладающие повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа).
Кроме того, настоящее изобретение относится к нуклеиновой кислоте, кодирующей полипептид, содержащий Fc-область, имеющую по меньшей мере одну аминокислотную модификацию, где указанный полипептид включает вариант Fc-области, обладающий повышенной активностью связывания с FcγRIIb или повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область. Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей полипептид, содержащий Fc-область, имеющую по меньшей мере одну аминокислотную модификацию, где указанный полипептид включает вариант Fc-области, обладающий повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа), в отличие от полипептида, содержащего родительскую Fc-область. Нуклеиновой кислотой согласно изобретению может быть любая форма нуклеиновой кислоты, например, ДНК или РНК.
Настоящее изобретение также относится к векторам, содержащим описанные выше нуклеиновые кислоты согласно изобретению. Тип вектора может быть соответствующим образом выбран специалистом в зависимости от клеток-хозяев, в которые вводят этот вектор. Такими векторами являются, например, векторы, описанные выше.
Кроме того, настоящее изобретение относится к клеткам-хозяевам, трансформированным вышеописанными векторами согласно изобретению. Соответствующие клетки-хозяева могут быть выбраны самим специалистом. Клетками-хозяевами являются, например, клетки-хозяева, описанные выше. Конкретными примерами являются нижеследующие клетки-хозяева.
Если в качестве клеток-хозяев используются эукариотические клетки, то такими клетками могут быть клетки животных, клетки растений или клетки грибов. В частности, клетками животных являются, например, следующие клетки:
(1) клетки млекопитающих: СНО (клеточная линия яичника китайского хомячка); COS (клеточная линия почек обезьян), клетки миеломы (Sp2/O, NS0 и т.п.), BHK (клеточная линия почек детеныша хомячка), клетки Hela, клетки Vero, клетки HEK293 (клеточная линия почек человеческого эмбриона, содержащая фрагментированную ДНК аденовируса (Ad5), клетки Freestyle 293, клетки PER.C6 (клеточная линия сетчатки человеческого эмбриона, трансформированная генами E1A и E1B аденовируса типа 5 (Ad5) и т.п. (Current Protocols in Protein Science (May, 2001, Unit 5.9, Table 5.9.1));
(2) клетки амфибий: ооциты Xenopus или т.п.; и
(3) клетки насекомых: sf9, sf21, Tn5 или т.п.
Кроме того, известно, что в качестве системы для экспрессии гена антитела могут быть использованы клетки растений, происходящие от растений рода Nicotiana, таких как Nicotiana tabacum. Для трансформации клеток растений могут быть соответствующим образом использованы культивированные клетки каллуса.
Кроме того, в качестве клеток грибов могут быть использованы следующие клетки:
- дрожжи рода Saccharomyces, такие как Saccharomyces serevisiae, и рода Pichia, такие как Pichia pastoris; и
- нитчатые грибы рода Aspergillus, такие как Aspergillus niger.
Кроме того, настоящее изобретение относится к способу повышения активности связывания с FcγRIIb и/или повышения селективности связывания с FcγRIIb, а не с FcγRIIа (R-типа), по сравнению с активностью полипептида, содержащего родительскую Fc-область, где указанный способ включает добавление в Fc-область полипептида, содержащего Fc-область, по меньшей мере одной аминокислотной модификации.
Настоящее изобретение также относится к способу подавления продуцирования антител против полипептида, содержащего Fc-область, по сравнению с продуцированием антител против полипептида, содержащего родительскую Fc-область, при введении этого полипептида in vivo, где указанный способ включает добавление в Fc-область указанного полипептида по меньшей мере одной аминокислотной модификации.
Примером предпочтительного варианта осуществления изобретения является комбинация аминокислотных модификаций, описанных в способе продуцирования полипептидов, содержащих варианты Fc-области, обладающие повышенной активностью связывания с FcγRIIb и/или повышенной селективностью связывания с FcγRIIb, а не с FcγRIIа (R-типа).
Полипептиды, полученные с применением любого из вышеупомянутых способов, также входят в объем настоящего изобретения.
Настоящее изобретение относится к фармацевтическим композициям, содержащим полипептид, включающий вариант Fc-области согласно изобретению.
Фармацевтические композиции согласно изобретению могут быть приготовлены известными методами и, помимо вышеописанных молекул антитела или слитого белка Fc согласно изобретению, содержат фармацевтически приемлемые носители. Так, например, если антитела приготовлены в виде стерильного раствора или суспензии для инъекции с использованием воды или любой другой фармацевтически приемлемой жидкости, то такие композиции могут быть введены парентерально. Так, например, такие композиции могут быть приготовлены путем соответствующего объединения молекул антител или слитого белка Fc с фармацевтически приемлемыми носителями или средами, а в частности, со стерильной водой или физиологическим раствором, растительными маслами, эмульгаторами, суспендирующими агентами, поверхностно-активными веществами, стабилизаторами, ароматизаторами, наполнителями, носителями, консервантами, связывающими агентами и т.п., путем их смешивания с получением унифицированной дозы в форме, отвечающей общеизвестным фармацевтическим требованиям. Конкретными примерами таких носителей являются легкая безводная кремневая кислота, лактоза, кристаллическая целлюлоза, маннит, крахмал, кальций-содержащая кармеллоза, натрий-содержащая кармеллоза, гидроксипропилцеллюлоза, гидроксипропилметилцеллюлоза, диэтиламиноацетат поливинилацетала, поливинилпирролидон, желатин, триглицерид со средней длиной цепи, отвержденное полиоксиэтилен-содержащее касторовое масло 60, сахароза, карбоксиметилцеллюлоза, кукурузный крахмал, неорганическая соль и т.п. Содержание активного ингредиента в такой композиции корректируют так, чтобы получить его соответствующую дозу в требуемом интервале.
Стерильные композиции для инъекции могут быть приготовлены с использованием носителей, таких как дистиллированная вода для инъекций, в соответствии со стандартными протоколами.
Водными растворами для инъекций являются, например, физиологический раствор и изотонические растворы, содержащие глюкозу или другие адъюванты, такие как D-сорбит, D-манноза, D-маннит и хлорид натрия. Эти растворы могут быть использованы в комбинации с подходящими солюбилизаторами, такими как спирт, а в частности, этанол, многоатомные спирты, такие как пропиленгликоль и полиэтиленгликоль, и неионные поверхностно-активные вещества, такие как полисорбат 80(TM) и HCO-50.
Маслами являются кунжутное масло и соевое масло, и эти масла могут быть объединены с солюбилизаторами, такими как бензилбензоат или бензиловый спирт. Эти композиции могут быть также приготовлены с использованием буферов, например, фосфатных буферов и натрий-ацетатных буферов; аналгетиков, например, гидрохлорида прокаина; стабилизаторов, например, бензилового спирта или фенола; или антиоксидантов. Приготовленные инъекции обычно разделяют на аликвоты и разливают по соответствующим ампулам.
Предпочтительным способом введения является парентеральное введение, а в частности, введение путем инъекции, интраназальное введение, внутрилегочное введение и чрескожное введение. Так, например, инъекции могут быть введены системно или местно путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции или подкожной инъекции.
Кроме того, способ введения может быть соответствующим образом выбран в зависимости от возраста и симптомов пациента. Разовая доза фармацевтической композиции, содержащая антитело или полинуклеотид, кодирующий антитело, может быть выбрана, например, так, чтобы она составляла, например, в пределах от 0,0001 до 1000 мг/кг массы тела. Альтернативно, такая доза может, например, составлять в пределах от 0,001 до 100000 мг/пациента. Однако эта доза не ограничивается указанными величинами. Доза и способ введения варьируются в зависимости от массы тела и возраста пациента, а также от симптомов его заболевания, и могут быть соответствующим образом выбраны самим специалистом.
Вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активных ингредиентов фармацевтических средств, которые подавляют активацию В-клеток, тучных клеток, дендритных клеток и/или базофилов. Полипептиды, содержащие вариант Fc-области согласно изобретению, могут подавлять активацию В-клеток, тучных клеток, дендритных клеток и/или базофилов, посредством селективного воздействия на FcγRIIb без активации активирующего FcγR. Активация В-клеток включает пролиферацию, продуцирование IgЕ, продуцирование IgM и продуцирование IgA. Вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, перекрестно связываются с FcγRIIb и IgЕ, что приводит к подавлению продуцирования IgЕ В-клетками; перекрестно связываются с FcγRIIb и IgМ, что приводит к подавлению продуцирования IgМ В-клетками; и перекрестно связываются с FcγRIIb и IgА, что приводит к подавлению продуцирования IgА В-клетками. Помимо этого, ингибирующее действие, аналогичное вышеупомянутому действию, осуществляется посредством прямого или опосредованного перекрестного связывания FcγRIIb с молекулами, которые экспрессируются на В-клетках и содержат внутриклеточный домен ITAM или взаимодействуют с доменом ITAM, таким как BCR, CD19 и CD79b. Кроме того, активация тучных клеток включает пролиферацию, активацию посредством IgE и т.п. и дегрануляцию. В тучных клетках вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут подавлять пролиферацию, активацию посредством IgE и т.п. и дегрануляцию посредством прямого или опосредованного перекрестного связывания FcγRIIb с молекулами рецептора IgЕ, которые экспрессируются на тучных клетках и содержат домен ITAM или взаимодействуют с доменом ITAM, таким как FcεRI, DAP12 и CD200R3. Активация базофилов включает их пролиферацию и дегрануляцию. В базофилах вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут подавлять пролиферацию, активацию и дегрануляцию посредством прямого или опосредованного перекрестного связывания FcγRIIb с молекулами на клеточной мембране, которые содержат внутриклеточный домен ITAM или взаимодействуют с доменом ITAM. Активация дендритных клеток включает их пролиферацию и дегрануляцию. В дендритных клетках вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут также подавлять активацию, дегрануляцию и пролиферацию посредством прямого или опосредованного перекрестного связывания FcγRIIb с молекулами на клеточной мембране, которые содержат внутриклеточный домен ITAM или взаимодействуют с доменом ITAM.
В настоящем изобретении вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента в терапевтических или профилактических средствах для лечения иммунных воспалительных заболеваний. Поскольку, как описано выше, полипептиды, содержащие вариант Fc-области согласно изобретению, могут подавлять активацию В-клеток, тучных клеток, дендритных клеток и/или базофилов, то введение полипептидов, содержащих вариант Fc-области согласно изобретению, может быть осуществлено для лечения или предупреждения иммунных воспалительных заболеваний. Термин «иммунные воспалительные заболевания» включает, но не ограничивается ими, ревматоидный артрит, аутоиммунный гепатит, аутоиммунный тиреоидит, буллезные аутоиммунные заболевания, аутоиммунное заболевание коры надпочечника, аутоиммунную гемолитическую анемию, аутоиммунную тромбоцитопеническую пурпуру, мегалоцитарную анемию, аутоиммунный атрофический гастрит, аутоиммунную нейтропению, аутоиммунный орхит, аутоиммунный энцефаломиелит, аутоиммунное заболевание, ассоциированное с рецепторными функциями, аутоиммунное бесплодие, хронический активный гепатит, гломерулонефрит, интерстициальный фиброз легких, рассеянный склероз, болезнь Пэджета, остеопороз, множественную миелому, увеит, острый и хронический спондилит, подагрический артрит, воспалительное заболевание кишечника, респираторный дистресс-синдром взрослых (РДСВ), псориаз, болезнь Крона, базедову болезнь, ювенильный диабет, болезнь Аддисона, тяжелую миастению, увеит, вызываемый поражением хрусталика, системную красную волчанку, аллергический ринит, аллергический дерматит, язвенный колит, гиперчувствительность, мышечную дегенерацию, кахексию, системную склеродермию, локализованную склеродермию, синдром Сьегрена, болезнь Бехчета, синдром Рейтера, диабет типа I и типа II, расстройства, ассоциированные с резорбцией кости, реакцию «трансплантат против хозяина», ишемическое реперфузионное повреждение, атеросклероз, травму головного мозга, церебральную малярию, сепсис, септический шок, синдром токсического шока, лихорадку, миалгии, ассоциированные с окрашиванием, апластическую анемию, гемолитическую анемию, идиопатическую тромбоцитопению, синдром Гудпасчера, синдром Гийена-Барре, тиреоидит Хашимото, пузырчатку, IgA-нефропатию, поллиноз, антифосфолипидный синдром, полимиозит, гранулематоз Вегенера, нодозный артерит, смешанное заболевание соединительных тканей, фибромиалгию, астму, атопический дерматит, хронический атрофический гастрит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, аортит, быстро прогрессирующий гломерулонефрит, мегалобластную анемию, идиопатическую тромбоцитопеническую пурпуру, первичный гипотиреоидит, идиопатическую болезнь Аддисона, инсулинозависимый сахарный диабет, хроническую дисковидную красную волчанку, пемфигоид, герпес беременных, буллезный дерматоз, вызываемый линейными цепями IgA, приобретенный буллезный эпидермолиз, гнездную алопецию, вульгарное витилиго, центробежную приобретенную лейкодермию Саттона, болезнь Харады, аутоиммунную невропатию зрительного нерва, идиопатическую азооспермию, привычный выкидыш, гипогликемию, хроническую крапивницу, анкилозирующий спондилит, псориатический артрит, энтеропатический артрит, реактивный артрит, спондилоартропатию, энтезопатию, синдром раздраженного кишечника, синдром хронической усталости, дерматомиозит, миозит, вызываемый тельцами включения, синдром Шмидта, болезнь Грэйвса, пернициозную анемию, волчаночный гепатит, пресенильную деменцию, болезнь Альцгеймера, демиелинизирующее расстройство, амиотрофический боковой склероз, гипопаратиреоидит, синдром Дресслера, синдром Итона-Ламберта, герпетиформный дерматит, алопецию, прогрессирующий системный склероз, синдром CREST (кальциноз, феномен Рейно, нарушение перистальтики пищевода, склеродактилия и телангиэктазия), саркоидоз, ревматическую лихорадку, мультиформную эритему, синдром Кушинга, заболевание, вызываемое переливанием крови, болезнь Хансена, артерит Такаясу, ревматическую полимиалгию, височный артерит, гигантоклеточный артрит, экзему, лимфоматозный гранулематоз, болезнь Кавазаки, эндокардит, фиброз эндомиокарда, эндофтальмит, эритробластоз плода, эозинофильный фасцит, синдром Фэлти, пурпуру Геноха-Шенлейна, отторжение трансплантата, паротит, кардиомиопатию, гнойный артрит, наследственную средиземноморскую лихорадку, синдром Макла-Уэлса и гипер-IgD-синдром.
Кроме того, при аутоиммунных заболеваниях, которые могут быть вызваны продуцированием антител против аутоантигенов (аутоантител), полипептиды, содержащие вышеупомянутый вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента в фармацевтических средствах для лечения или предупреждения аутоиммунных заболеваний посредством подавления продуцирования этих аутоантител. Сообщалось, что использование молекулы, продуцируемой путем присоединения Fc-части антитела к AchR (аутоантигену тяжелой миастении), позволяет ингибировать пролиферацию В-клеток, которые экспрессируют AchR-распознающий BCR и индуцируют апоптоз (J. Neuroimmunol, 227, 35-43, 2010). С использованием слитого белка, образованного антигеном, распознаваемым аутоантителом и Fc-областью антитела согласно изобретению, можно осуществлять перекрестное связывание FcγRIIb с BCR на B-клетках, экспрессирующих BCR для этого аутоантигена, подавлять пролиферацию В-клеток, экспрессирующих BCR для этого аутоантигена, и индуцировать их апоптоз. Такими аутоиммунными заболеваниями являются, но не ограничиваются ими, синдром Гийена-Барре, тяжелая миастения, хронический атрофический гастрит, аутоиммунный гепатит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, аортит, синдром Гудпасчера, быстро прогрессирующий гломерулонефрит, мегалобластная анемия, аутоиммунная гемолитическая анемия, аутоиммунная нейтропения, идиопатическая тромбоцитопеническая пурпура, базедова болезнь, тиреоидит Хашимото, первичный гипотиреоидит, идиопатическая болезнь Аддисона, инсулинозависимый сахарный диабет, хроническая дисковидная красная волчанка, локализованная склеродермия, пузырчатка, пемфигоид, герпес беременных, буллезный дерматоз, вызываемый линейными цепями IgA, приобретенный буллезный эпидермолиз, гнездная алопеция, вульгарное витилиго, центробежная приобретенная лейкодермия Саттона, болезнь Харады, аутоиммунная невропатия зрительного нерва, идиопатическая азооспермия, привычный выкидыш, диабет типа II, гипогликемия и хроническая крапивница.
Кроме того, вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента в терапевтических средствах для лечения заболеваний, ассоциированных с дефицитом жизненно необходимых белков. Для лечения заболеваний, ассоциированных с дефицитом жизненно необходимых белков, могут быть применены методы терапии, включающие введение белка в качестве фармацевтического средства и в качестве добавки. Однако, поскольку у пациента изначально отсутствует такой белок, то экзогенно вводимый белок будет распознаваться как чужеродное вещество, в результате чего будут продуцироваться антитела против этого белка. Поэтому такой белок становится легко удаляемым, что будет приводить к снижению эффекта фармацевтического средства. С использованием слитого белка, содержащего указанный белок и Fc-область антитела согласно изобретению, можно осуществлять перекрестное связывание FcγRIIb с BCR на B-клетках, распознающих этот белок, и подавлять продуцирование антитела против указанного белка. Белками, вводимыми в качестве добавки, являются фактор VIII, фактор IX, TPO, EPO, α-идуронидаза, идуронат-сульфатаза, гепаран-N-сульфатаза типа A, α-N-ацетилглюкозаминидаза типа В, ацетил-СоА-α-глюкозаминидаза-ацетилтрансфераза типа С, N-ацетилглюкозамин-6-сульфатаза типа D, галактоза-6-сульфатаза, N-ацетилгалактозамин-4-сульфатаза, β-глюкуронидаза, α-галактозидаза, кислотная α-галактозидаза и глюкоцереброзидаза. Эти белки могут быть введены в качестве добавок для лечения заболеваний, таких как, но не ограничивающихся ими, гемофилия, идиопатическая тромбоцитопеническая пурпура, почечная анемия и заболевание, ассоциированное с лизосомами (мукополисахаридоз, болезнь Фабри, болезнь Помпе и болезнь Гуше).
Кроме того, вышеупомянутые полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента противовирусных средств. Антитела, содержащие Fc-область согласно изобретению, и антивирусные антитела могут подавлять антитело-зависимое иммунологическое усиление, наблюдаемое при использовании антивирусных антител. Антитело-зависимое иммунологическое усиление представляет собой явление, при котором вирус вырабатывает нейтрализующие антитела против этого вируса и подвергается фагоцитозу посредством активирующих FcγR, что приводит к инфицированию FcγR-экспрессирующих клеток и к дальнейшему распространению этой инфекции. Сообщалось, что связывание нейтрализующих антител против вируса денге с FcγRIIb играет важную роль в ингибировании антитело-зависимого иммунологического усиления (Proc. Natl. Acad. Sci. USA, 108, 12479-12484, 2011). Перекрестное связывание FcγRIIb с иммунным комплексом вируса денге, образованным нейтрализующими антителами против вируса денге, ингибирует FcγR-опосредуемый фагоцитоз и подавляет антитело-зависимое иммунологическое усиление. Примерами таких вирусов являются, но не ограничиваются ими, вирус денге (DENV1, DENV2 и DENV4) и ВИЧ.
Кроме того, вышеописанные полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента в профилактических или терапевтических средствах для лечения артериосклероза. Антитела против окисленного ЛНП, вызывающего артериосклероз, то есть антитела, содержащие Fc-область согласно изобретению, могут способствовать предупреждению FcγRIIа-зависимой адгезии воспалительных клеток. Сообщалось, что антитела против окисленного ЛНП ингибируют взаимодействие окисленного ЛНП с CD36 и связываются с эндотелиальными клетками, а моноциты распознают Fc-часть этих антител по FcγRIIа-зависимому или FcγRI-зависимому механизму, что приводит к адгезии моноцитов (Immunol. Lett., 108, 52-61, 2007). С использованием антител, содержащих Fc-область согласно изобретению, можно ингибировать FcγRIIа-зависимое связывание и подавлять адгезию моноцитов под действием FcγRIIb-опосредуемых ингибирующих сигналов.
В настоящем изобретении вышеописанные полипептиды, содержащие вариант Fc-области согласно изобретению, могут быть использованы в качестве активного ингредиента в терапевтических или профилактических средствах для лечения рака. Как описано выше, известно, что повышение активности связывания с FcγRIIb усиливает агонистическую активность антитела-агониста и повышает противоопухолевый эффект антитела. Поэтому антитела-агонисты, содержащие вариант Fc-области согласно изобретению, могут быть использованы для лечения или предупреждения рака. В частности, вариант Fc-области согласно изобретению усиливает агонистическую активность антител-агонистов, направленных, например, против рецепторов, принадлежащих к семейству рецепторов TNF, таких как A-лиазы, CD120a, CD120b, рецептор лимфотоксина β, CD134, CD40, FAS, TNFRSF6B, CD27, CD30, CD137, TNFRSF10A, TNFRSF10B, TNFRSF10C, TNFRSF10D, RANK, остеопротегерин, TNFRSF12A, TNFRSF13B, TNFRSF13C, TNFRSF14, рецептор фактора роста нервных клеток, TNFRSF17, TNFRSF18, TNFRSF19, TNFRSF21, TNFRSF25 и рецептор эктодисплазина A2, и может быть использован для лечения или предупреждения рака. Кроме того, в дополнение к вышеописанному можно сказать, что наблюдается также усиление агонистической активности антител-агонистов против молекул, которым, для их агонистической активности, необходимо взаимодействие с FcγRIIb. Кроме того, при введении варианта Fc-области согласно изобретению в полипептид, обладающий активностью связывания с молекулой, такой как Kit, то есть тирозинкиназный рецептор (RTK), который подавляет пролиферацию клеток после перекрестного связывания с FcγRIIb, ингибирующий эффект, направленный против клеток, экспрессирующих такую молекулу, может усиливаться. Раковыми заболеваниями являются, но не ограничиваются ими, рак легких (включая мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и плоскоклеточную карциному легких), рак толстого кишечника, рак прямой кишки, рак толстой кишки, рак молочной железы, рак печени, рак желудка, рак поджелудочной железы, рак почек, рак предстательной железы, рак яичника, рак щитовидной железы, холангиокарцинома, рак брюшной полости, мезотелиома, плоскоклеточная карцинома, рак шейки матки, рак эндометрия, рак мочевого пузыря, рак пищевода, рак головы и шеи, рак носоглотки, опухоль слюнных желез, тимома, рак кожи, базальноклеточная опухоль, злокачественная меланома, рак заднего прохода, рак пениса, рак яичек, опухоль Вильмса, острый миелоидный лейкоз (включая острый миелолейкоз, острый миелобластный лейкоз, острый промиелоцитарный лейкоз, острый миеломоноцитарный лейкоз и острый моноцитарный лейкоз), хронический миелогенный лейкоз, острый лимфобластный лейкоз, хронический лимфолейкоз, ходжкинская лимфома, неходжкинская лимфома (лимфома Беркитта, хронический лимфоцитарный лейкоз, грибовидный микоз, лимфома клеток коры головного мозга, фолликулярная лимфома, диффузная крупноклеточная лимфома, лимфома маргинальной зоны, пилоцитарная лейкоплазмацитома, лимфома периферических Т-клеток и Т-клеточный лейкоз/Т-клеточная лимфома взрослых), гистиоцитоз клеток Лангерганса, множественная миелома, миелодиспластический синдром, опухоль головного мозга (включая глиому, астроглиому, глиобластому, менингиому и эпендимому), нейробластома, ретинобластома, остеосаркома, саркома Капоши, саркома Юинга, ангиосаркома и гемангиоперицитома.
Кроме того, настоящее изобретение относится к способам лечения или предупреждения иммунных воспалительных заболеваний, где указанные способы включают стадию введения индивидууму (пациенту) полипептида, содержащего вариант Fc-области согласно изобретению, или полипептида, содержащего вариант Fc-области и полученного способами продуцирования согласно изобретению.
Настоящее изобретение также относится к наборам, используемым в способах терапии или профилактики согласно изобретению, где указанные наборы содержат по меньшей мере полипептид, содержащий вариант Fc-области согласно изобретению, или полипептид, содержащий вариант Fc-области и полученный способами продуцирования согласно изобретению, или фармацевтическую композицию согласно изобретению. Кроме того, в набор могут быть включены фармацевтически приемлемые носители и среды, а также к нему могут прилагаться инструкции по применению такого набора. Кроме того, настоящее изобретение относится к применению полипептида, содержащего вариант Fc-области согласно изобретению, или полипептида, содержащего вариант Fc-области и полученного способами продуцирования согласно изобретению, в целях приготовления средств для лечения или предупреждения иммунных воспалительных заболеваний. Настоящее изобретение также относится к полипептиду, содержащему вариант Fc-области согласно изобретению, или полипептиду, содержащему вариант Fc-области и полученному способами продуцирования согласно изобретению, где указанные полипептиды могут быть применены в способах терапии или профилактики согласно изобретению.
Используемые здесь аминокислоты имеют нижеследующие трехбуквенные и однобуквенные обозначения:
Аланин: Ala (A)
Аргинин: Arg (R)
Аспарагин: Asn (N)
Аспарагиновая кислота: Asp (D)
Цистеин: Cys (C)
Глутамин: Gln (Q)
Глутаминовая кислота: Glu (E)
Глицин: Gly (G)
Гистидин: His (H)
Изолейцин: Ile (I)
Лейцин: Leu (L)
Лизин: Lys (K)
Метионин: Met (M)
Фенилаланин: Phe (F)
Пролин: Pro (P)
Серин: Ser (S)
Треонин: Thr (T)
Триптофан: Trp (W)
Тирозин: Tyr (Y)
Валин: Val (V)
Все цитируемые здесь предшествующие документы во всей своей полноте вводятся в настоящее описание посредством ссылки.
Примеры
Ниже приводится конкретные примеры осуществления настоящего изобретения, которые не должны рассматриваться как ограничение объема настоящего изобретения.
[Пример 1]. Оценка способности антител, содержащих Fc-вариант, обладающий повышенной активностью связывания с FcγRIIb, к агрегации тромбоцитов
Как показано в таблице 16 сравнительного примера 4, существующий метод повышения активности связывания с FcγRIIb, в котором осуществляют модификации, а именно, замену Ser на Glu в положении 267 и замену Leu на Phe в положении 328 (в соответствии с Европейской нумерацией) в нативном человеческом IgG1 (непатентный документ 28), позволяет в 408 раз повышать активность связывания с FcγRIIb и в 0,51 раз снижать активность связывания с FcγRIIаН, а также в 522 раза повышать активность связывания с FcγRIIаR по сравнению с активностью IgG1. Как описано в разделе «Предшествующий уровень техники», даже при повышении активности связывания с FcγRIIb, которое происходит в клетках, таких как тромбоциты, экспрессирующие только FcγRIIа, такие эффекты усиления могут наблюдаться только для FcγRIIа. То есть, существующие методы повышения активности связывания с FcγRIIаR повышают риск агрегации тромбоцитов и риск развития тромбоза. Для подтверждения этого факта необходимо провести анализ для того, чтобы определить, может ли повышаться активность в агрегации тромбоцитов при повышении активности связывания антитела с FcγRIIа.
С применением метода, описанного в сравнительном примере 1, омализумаб_VH-G1d (SEQ ID NO:25) был получен в виде тяжелой цепи, а омализумаб_VL-CK (SEQ ID NO:26) был получен в виде легкой цепи человеческого антитела IgG1, которое связывается с IgЕ. Кроме того, для повышения активности связывания омализумаб_VH-G1d с человеческим FcγRIIb был получен омализумаб_VH-G1d-v3 путем замены Ser на Glu в положении 267 и замены Leu на Phe в положении 328 омализумаб_VH-G1d в соответствии с Европейской нумерацией. С применением метода, описанного в сравнительном примере 1, были получены омализумаб_VH-G1d-v3 в виде тяжелой цепи и омализумаб_VL-CK в виде легкой цепи. С использованием этого антитела была оценена способность к агрегации тромбоцитов.
Агрегацию тромбоцитов оценивали на агрегометре тромбоцитов HEMATRACER 712 (LMS Co.). Сначала, приблизительно 50 мл цельной крови собирали в фиксированном количестве в 4,5-миллилитровые пробирки для забора крови с откаченным воздухом, содержащие 0,5 мл 3,8% цитрата натрия, а затем эти пробирки центрифугировали при 200×g в течение 15 минут. Полученный супернатант собирали и использовали в качестве плазмы, богатой тромбоцитами (PRP). После этого, PRP промывали буфером 1 (137 мМ NaCl, 2,7 мМ KCl, 12 мМ NaHCO3, 0,42 мМ NaH2PO4, 2 мМ MgCl2, 5 мМ HEPES, 5,55 мМ декстрозы, 1,5 ед/мл апиразы, 0,35% BSA), а затем этот буфер заменяли на буфер 2 (137 мМ NaCl, 2,7 мМ KCl, 12 мМ NaHCO3, 0,42 мМ NaH2PO4, 2 мМ MgCl2, 5 мМ HEPES, 5,55 мМ декстрозы, 2 мМ CaCl2, 0,35% BSA) и получали приблизительно 300000/мкл промытых тромбоцитов. 156 мкл промытых тромбоцитов разделяли на аликвоты и помещали в аналитические кюветы агрегометра для тромбоцитов, содержащие стержневую мешалку. Тромбоциты перемешивали стержневой мешалкой со скоростью 1000 об/мин в кюветах агрегометра для тромбоцитов при 37,0°C. Затем в кюветы добавляли 44 мкл иммунного комплекса омализумаб-G1d-v3 и IgЕ в молярном соотношении 1:1 (в конечных концентрациях 600 мкг/мл и 686 мкг/мл, соответственно). Тромбоциты подвергали реакции с иммунным комплексом в течение пяти минут. После этого к реакционной смеси добавляли аденозиндифосфат (ADP, SIGMA) в концентрации, при которой не наблюдалась вторичная агрегация тромбоцитов, для того чтобы определить, происходит ли повышение уровня такой агрегации.
Результаты для каждого донора с полиморфной формой FcγRIIа (H/H или R/H), полученные в вышеописанном анализе, представлены на фиг. 1 и 2. Результат, представленный на фиг. 1, показал, что уровень агрегации тромбоцитов повышается в случае полиморфной формы FcγRIIа (R/H) при добавлении иммунного комплекса. С другой стороны, как показано на фиг. 2, полиморфизм FcγRIIа (H/H) не приводил к повышению уровня агрегации тромбоцитов.
Затем оценивали активацию тромбоцитов с использованием маркеров активации. Активация тромбоцитов может быть оценена по повышению уровней экспрессии маркера активации, такого как CD62p (р-селектин) или активный интегрин на мембранной поверхности тромбоцитов. К 7,7 мкл промытых тромбоцитов, полученных описанным выше методом, добавляли 2,3 мкл иммунного комплекса. После прохождения реакции в течение пяти минут при комнатной температуре, активацию индуцировали путем добавления ADP в конечной концентрации 30 мкМ и определяли, может ли иммунный комплекс усиливать ADP-зависимую активацию. Образец, в который был добавлен фосфатный буфер (pH7,4, Gibco) вместо иммунного комплекса, использовали в качестве негативного контроля. Окрашивание осуществляли путем добавления в образец, полученный после реакции, ФЭ-меченного анти-CD62 антитела (BECTON DICKINSON), PerCP-меченного анти-CD61 антитела и ФИТЦ-меченного антитела PAC-1 (BD bioscience). Все величины интенсивности флуоресценции измеряли на проточном цитометре (FACS CantoII, BD bioscience).
Данные по экспрессии CD62p, полученные вышеописанным аналитическим методом, представлены на фиг. 3. Данные по экспрессии активированного интегрина представлены на фиг. 4. Используемые промытые тромбоциты были взяты у здорового индивидуума с полиморфной формой FcγRIIа R/H. Экспрессия CD62p и активированного интегрина на мембранной поверхности тромбоцитов, индуцированная посредством ADP-стимуляции, повышалась в присутствии иммунного комплекса.
Эти результаты показали, что в имеющихся антителах, содержащих Fc-вариант, обладающий повышенной активностью связывания с человеческим FcγRIIb и полученный путем замены Ser на Glu в положении 267 и замены Leu на Phe в положении 328 (в соответствии с Европейской нумерацией) в Fc IgG1, генетические полиморфы FcγRIIа, в которых аминокислотой в положении 131 является R, обладали повышенной активностью в агрегации тромбоцитов, чем полиморфы, в которых аминокислотой в положении 131 являеся Н. То есть, можно предположить, что имеющиеся антитела, содержащие Fc-вариант, обладающий повышенной активностью связывания с человеческим FcγRIIb, представляют повышенный риск развития тромбоза, что обусловлено агрегацией тромбоцитов у человека, несущего FcγRIIа R-типа, а поэтому очевидно, что такая проблема может быть решена с использованием Fc-варианта, обладающего повышенной селективностью связывания с FcγRIIb.
[Пример 2]. Продуцирование вариантов, обладающих повышенной активностью связывания с FcγRIIb
Как показано в примере 1, если требуется повышение активности связывания с FcγRIIb, то при этом также необходимо ингибировать связывание с другими активирующими FcγR, если это возможно. Поэтому продуцирование вариантов, обладающих повышенной активностью или селективностью связывания с FcγRIIb, было оценено путем объединения замен, которые способствуют повышению активности связывания или селективности связывания с FcγRIIb. В частности, взятая за основу модификация P238D, которая дает превосходный эффект в отношении повышения активности и селективности связывания с FcγRIIb, может быть также объединена с модификациями, которые, как было обнаружено, являются эффективными для такого повышения активности и селективности связывания, как описано в сравнительных примерах 6, 8 и 9.
Вариабельная область IL6R-H (SEQ ID NO:18), которая описана в WO 2009/125825 и которая представляет собой вариабельную область антитела против рецептора человеческого интерлейкина 6, была получена как вариабельная область Н-цепи антитела, а IL6R-G1d (SEQ ID NO:19), которая имеет G1d, продуцированную путем удаления С-концевого Gly и Lys человеческого IgG1, была получена как константная область Н-цепи антитела. Кроме того, IL6R-B3 (SEQ ID NO:23) получали путем введения K439E в IL6R-G1d. Затем были получены варианты IL6R-B3 путем объединения E233D, L234Y, G237D, S267Q, H268D, P271G, Y296D, K326D, K326A, A330R, A330K, которые, как было обнаружено, являются эффективными при их использовании в комбинации с P238D, как описано в сравнительных примерах 6, 8 и 9. IL6R-L (SEQ ID NO:21) обычно используют как L-цепь антитела. Эти варианты были использованы для экспрессии и очистки антитела методом, описанным в сравнительном примере 1, а связывание с каждым из FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 1. В данной таблице «модификации» означают модификации, вводимые в IL6R-B3 (SEQ ID NО:23). При этом IL6R-B3/IL6R-L, которые используются в качестве матрицы для продуцирования каждого из вариантов, отмечены звездочкой (*). В этой таблице «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность к FcγRIIb. «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIаR. В таблице 1 закрашенные серым ячейки относятся к величинам, вычисленным по следующему уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2, где, как было установлено, связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа.

Если принять активность связывания IL6R-B3/IL6R-L, продуцируемого путем введения K439E в IL6R-G1d/IL6R-L, с соответствующими FcγR за 1, то активность связывания IL6R-G1d/IL6R-L, который несет нативную последовательность человеческого IgG1, будет в 1,3 раза выше для FcγRIа, в 1,1 раза выше для FcγRIIаR, 1,1 раза выше для FcγRIIаН, в 1,2 раза выше для FcγRIIb и в 0,9 раза выше для FcγRIIIаV, а все активности связывания с FcγR будут эквивалентны активности связывания с IL6R-G1d/IL6R-L. Поэтому сравнение связывания каждого варианта с IL6R-B3/IL6R-L до модификации может рассматриваться как эквивалентное сравнению активности связывания каждого варианта с антителом IL6R-G1d/IL6R-L, имеющим нативную последовательность человеческого IgG1. В соответствии с этим, как описано ниже в примерах, активность связывания каждого варианта сравнивают с активностью связывания IL6R-B3/IL6R-L до модификации.
В таблице 1 показано, что все варианты обладают повышенной аффинностью связывания с FcγRIIb по сравнению с аффинностью связывания с IL6R-B3 до модификации, причем IL6R-BF648/IL6R-L обладает в 2,6 раза большей аффинностью связывания, и эта величина является наименьшей, а IL6R-BP230/IL6R-L обладает в 147,6 раз большей аффинностью связывания, и эта величина является наибольшей. Кроме того, величина KD(IIaR)/KD(IIb), которая определяет степень селективности, составляет 10,0 для IL6R-BP234/IL6R-L, где такая величина является наименьшей, и 32,2 для IL6R-BP231/IL6R-L, где такая величина является наибольшей, причем все варианты обладают повышенной селективностью связывания по сравнению с IL6R-B3/IL6R-L до модификации, и такая величина составляет 0,3. Все варианты обладают более низкой активностью связывания с FcγRIа, FcγRIIаН и FcγRIIIаV по сравнению с активностью IL6R-B3/IL6R-L до модификации.
[Пример 3] Рентгеноструктурный анализ комплекса, образованного между Fc с повышенной активностью связывания с FcγRIIb и внеклеточной областью FcγRIIb, и комплекса, образованного между этим Fc и внеклеточной областью FcγRIIаR
Вариант IL6R-BP230/IL6R-L, имеющий наибольшую активность связывания с FcγRIIb, как описано в примере 2, обладал приблизительно в 150 раз большей активностью связывания с FcγRIIb, а активность связывания с FcγRIIа R-типа ингибировалась до уровня активности, который был приблизительно в 1,9 раз выше активности IL6R-B3/IL6R-L до модификации. Поэтому IL6R-BP230/IL6R-L представляет собой превосходный вариант с точки зрения активности и селективности связывания с FcγRIIb, однако, для получения еще лучших вариантов, предпочтительно, чтобы активность связывания с FcγRIIb была еще выше, а активность связывания с FcγRIIаR была как можно ниже.
Как показано на фиг. 28 сравнительного примера 7, в Fc с модификацией P238D, сильное электростатическое взаимодействие наблюдалось между Asp в положении 270 (в соответствии с Европейской нумерацией) CH2-домена B и Arg в положении 131 FcγRIIb. В FcγRIIIа и FcγRIIа Н-типа, таким остатком в положении 131 является His, а в FcγRIIа R-типа, как и в случае FcγRIIb, таким остатком является Arg. В результате было обнаружено, что какого-либо отличия во взаимодействиях в этой части не наблюдалось, и по этой причине трудно определить селективность связывания с FcγRIIа R-типа.
С другой стороны, внеклеточные области FcγRIIа и FcγRIIb имеют аминокислотные последовательности с гомологией, составляющей 93%, то есть с очень высокой гомологией. Анализ кристаллической структуры комплекса, образованного между нативным Fc IgG1 (далее обозначаемым «Fc(WT)») и внеклеточной областью FcγRIIа R-типа (J. Imunol. 2011, 187, 3208-3217), показал, что на границе взаимодействия между FcγRIIа R-типа и FcγRIIb, аминокислоты отличались лишь в трех положениях (Gln127, Leu132 и Phe160), а поэтому, как и ожидалось, очень трудно определить повышение селективности связывания по отношению к FcγRIIа R-типа.
Поэтому авторы настоящего изобретения считают, что для дополнительного повышения селективности и активности связывания с FcγRIIb, вводимые аминокислотные мутации должны быть детально оценены с помощью конформационного анализа тонких различий во взаимодействиях, вызываемых различиями в рецепторах путем получения не только трехмерной структуры комплекса, образованного между Fc, обладающим повышенной активностью связывания с FcγRIIb, и внеклеточной областью FcγRIIb, но также одновременно получения трехмерной структуры комплекса, образованного внеклеточной областью FcγRIIа R-типа, для которой очень трудно определить повышение селективности. Затем были проведены рентгеноструктурные анализы комплекса, образованного между внеклеточной областью FcγRIIb и Fc(Р208), где модификация K439E была удалена из Fc комплекса IL6R-BP208/IL6R-L (полученного как описано в сравнительном примере 9), который представляет собой вариант, взятый за основу для продуцирования IL6R-BP230/IL6R-L, и комплекса, образованного между внеклеточной областью FcγRIIа R-типа и Fc(P208).
3-1. Рентгеноструктурный анализ комплекса, образованного между Fc(P208) и внеклеточной областью FcγRIIb
Трехмерную структуру комплекса внеклеточной области Fc(P208)/FcγRIIb определяли с помощью рентгеноструктурного анализа с разрешением 2,81 Å. Структура, полученная в результате такого анализа, представлена на фиг. 5. Внеклеточная область FcγRIIb расположена между двумя СН2-доменами Fc, структура которых аналогична трехмерным структурам комплексов, образованных между областью Fc(WT), представляющей собой Fc нативного IgG, и соответствующей внеклеточной областью FcγRIIIа (Proc. Natl. Acad. Sci. USA, 2011, 108, 12669-126674), FcγRIIIb (Nature, 2000, 400, 267-273; J. Biol. Chem 2011, 276, 16469-16477) или FcγRIIа, проанализированных ранее.
Однако детальный анализ показал, что в комплексе Fc(P208)/внеклеточная область FcγRIIb петлевая структура в положениях 233-239 (в соответствии с Европейской нумерацией), происходящая от шарнирной области CH2-домена А Fc, отличалась от петлевой структуры комплекса Fc(WT)/внеклеточная область FcγRIIа R-типа, что было обусловлено введением замен G237D и P238D (фиг. 6). В результате, между главной цепью Asp в положении 237 (в соответствии с Европейской нумерацией) Fc(P208) и боковой цепью Tyr в положении 160 FcγRIIb (фиг. 7) образовывалась сильная водородная связь. В случае, когда в FcγRIIа H-типа и R-типа, Tyr160 заменяли на Phe, то водородная связь не образовывалась. Поэтому можно сказать, что такая водородная связь играет важную роль в сообщении селективности, которая обеспечивает повышение активности связывания с FcγRIIb и снижение активности связывания с FcγRIIа.
С другой стороны, боковая цепь Asp в положении 237 (в соответствии с Европейской нумерацией) Fc(P208) не обнаруживала каких-либо явных взаимодействий с FcγRIIb, при этом также не наблюдалось взаимодействия с остатками в Fc. В Fc Ile в положении 332 (в соответствии с Европейской нумерацией), Glu в положении 333 (в соответствии с Европейской нумерацией) и Lys в положении 334 (в соответствии с Европейской нумерацией) расположены в непосредственной близости от остатка Asp в положении 237 (в соответствии с Европейской нумерацией) (фиг. 8). Если петлевая структура была стабилизирована путем замены остатков в этих положениях гидрофильными остатками, что обеспечивало взаимодействие с боковой цепью Asp в положении 237 (в соответствии с Европейской нумерацией), то потеря энергии в результате ее энтропии после образования водородной связи с Tyr в положении 160 FcγRIIb может быть уменьшена, что может приводить к повышению свободной энергии связывания и, тем самым, к усилению активности связывания.
Сравнение кристаллической структуры комплекса, образованного между Fc(P238D), имеющего модификацию P238D, и внеклеточной областью FcγRIIb, которая описана в сравнительном примере 7, и кристаллической структуры комплекса, образованного между Fc(P208) и внеклеточной областью FcγRIIb, где указанное сравнение проводили с помощью рентгеноструктурного анализа, показало, что Fc(P208) по сравнению с Fc(P238D) содержит пять новых мутаций, и большинство из них представляет собой замены только в боковой цепи. Однако в результате замены Pro в положении 271 (в соответствии с Европейской нумерацией) на Gly в СН2-домене В Fc происходит изменение локализации на уровне главной цепи, и такие структурные изменения наблюдаются в области выше петли, образованной остатками в положениях 266-270 (в соответствии с Европейской нумерацией)(фиг. 9). Как указывается в сравнительном примере 8 для Fc(P238D), было высказано предположение, что Pro-часть в положении 271 (в соответствии с Европейской нумерацией) может иметь стереохимические затруднения, если Asp в положении 270 (в соответствии с Европейской нумерацией) вступает в сильное электростатическое взаимодействие с Arg в положении 131 FcγRIIb. Структурные изменения, наблюдаемые после введения Gly в положении 271 (в соответствии с Европейской нумерацией), могут быть обусловлены снятием структурного напряжения, аккумулированного в Pro-части до модификации, и при этом считается, что количество такого снятия напряжения приводит к повышению свободной энергии связывания с FcγRIIb, то есть к повышению активности связывания.
Кроме того, было подтверждено, что Arg в положении 292 (в соответствии с Европейской нумерацией) подвергается структурным изменениям, принимая две формы в результате структурных изменений петли в положениях 266-271 (в соответствии с Европейской нумерацией). В данном случае Arg в положении 292 (в соответствии с Европейской нумерацией) вступает в электростатическое взаимодействие с аминокислотой Asp в положении 268 (в соответствии с Европейской нумерацией), которая представляет собой один из других модифицированных остатков в Fc(P208) (фиг. 9), что может способствовать стабилизации этой петлевой структуры. Электростатическое взаимодействие между Asp в положении 270 (в соответствии с Европейской нумерацией) этой петли и Arg в положении 131 FcγRIIb оказывает значительное влияние на активность связывания с FcγRIIb, а поэтому введение модификации H268D может приводить к снижению потери энергии в результате ее энтропии после связывания и к повышению свободной энергии связывания, то есть к усилению активности связывания благодаря стабилизации конформации этой петлевой структуры в FcγRIIb-связанной форме.
После проведения дополнительных исследований на возможность повышения активности посредством введения модификаций, осуществляемых на основе имеющихся результатов структурного анализа, было обнаружено, что Ser в положении 239 (в соответствии с Европейской нумерацией) является одним из положений-кандидатов на введение модификаций. Как показано на фиг. 10, этот остаток Ser в положении 239 (в соответствии с Европейской нумерацией) CH2-домена B расположен в области, в которой остаток Lys в положении 117 FcγRIIb простирается в направлении области, имеющей структуру, наиболее близкую к природной. Однако, поскольку электронная плотность Lys в положении 117 FcγRIIb не была детектирована в данном анализе, то этот остаток Lys не образует какую-либо стабильную структуру. Таким образом, в настоящее время считается, что участие этого остатка Lys во взаимодействии с Fc(P208) имеет определенные ограничения, однако, если этот остаток Ser в положении 239 (в соответствии с Европейской нумерацией) CH2-домена B заменить на отрицательно заряженный остаток Asp или Glu, то можно ожидать возникновение электростатического взаимодействия с положительно заряженным остатком Lys в положении 117 FcγRIIb, которое, как предполагается, будет способствовать усилению активности связывания с FcγRIIb.
С другой стороны, анализ структуры Ser в положении 239 (в соответствии с Европейской нумерацией) в CH2-домене А показал, что боковая цепь этой аминокислоты образует водородные связи с главной цепью Gly в положении 236 (в соответствии с Европейской нумерацией) и стабилизирует петлевую структуру в положениях 233-239, которые являются продолжением шарнирной области и включают остаток Asp в положении 237 (в соответствии с Европейской нумерацией), образующий водородную связь с боковой цепью Tyr160 FcγRIIb (фиг. 7). Стабилизация этой петлевой структуры с образованием нужной конформации во время связывания может приводить к снижению потери энергии в результате ее энтропии после связывания и, тем самым, к повышению свободной энергии связывания, то есть, к усилению активности связывания. С другой стороны, если этот остаток Ser в положении 239 (в соответствии с Европейской нумерацией) в CH2-домене А заменить на Asp или Glu, то водородная связь с главной цепью Gly в положении 239 (в соответствии с Европейской нумерацией) может быть потеряна, что, в свою очередь, может приводить к электростатическому отталкиванию между остатком Glu и остатком Asp в положении 265 (в соответствии с Европейской нумерацией), который присутствует в непосредственной близости от указанного остатка Gly, и, тем самым, будет способствовать значительной дестабилизации петлевой структуры. Такая энергия дестабилизации будет снижать свободную энергию связывания с FcγRIIb, что может приводить к снижению активности связывания.
[Экспрессия и очистка Fc(P208)]
Fc(P208) получали следующим образом. Сначала получали IL6R-BP208 путем замены Glu в положении 439 (в соответствии с Европейской нумерацией) IL6R-BP208 (SEQ ID NO:24) на Lys, где указанная последовательность представляет собой последовательность нативного человеческого IgG1. Затем Cys в положении 220 (в соответствии с Европейской нумерацией) IL6R-P208 заменяли на Ser. После этого генную последовательность Fc(P208), в которой у С-конца в положении 216 присутствует Glu (в соответствии с Европейской нумерацией), клонировали с помощью ПЦР. С использованием этой клонированной генной последовательности получали экспрессионные векторы, а также осуществляли экспрессию и очистку Fc(P208) методом, описанным в сравнительном примере 1. Cys в положении 220 (в соответствии с Европейской нумерацией) образует дисульфидную связь с Cys L-цепи в природном IgG1. При получении одного Fc, L-цепь не экспрессировалась, а поэтому, для предотвращения образования нежелательных дисульфидных связей, этот остаток заменяли на Ser.
[Экспрессия и очистка внеклеточной области FcγRIIb]
Экспрессия и очистка были проведены методом, описанным в сравнительном примере 2.
[Очистка комплекса Fc(P208)/внеклеточная область FcγRIIb]
К 1,5 мг образца внеклеточной области FcγRIIb, полученного в целях кристаллизации, добавляли 0,15 мг Endo F1 (Protein Science 1996, 5, 2617-2622), экспрессированного и очищенного из Escherichia coli в виде слитого белка глутатион-S-трансферазы. Этот белок выдерживали при комнатной температуре в течение трех дней в 0,1 M бис-трис-буфере при pH 6,5, и N-связанный олигосахарид отделяли, в результате чего оставался N-ацетилглюкозамин, непосредственно связанный с Asn. Затем этот образец внеклеточной области FcγRIIb, подвергнутый реакции отщепления углевода, концентрировали путем ультрафильтрации с отсечкой молекулярной массы 5000, после чего его очищали с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 20 мM HEPES при pH 7,5, содержащем 0,1M NaCl. Кроме того, к полученной фракции внеклеточной области FcγRIIb, от которой был отщеплен углевод, добавляли Fc(P208) так, чтобы молярное отношение внеклеточной области FcγRIIb было слегка повышенным, и после концентрирования этого образца путем ультрафильтрации с отсечкой молекулярной массы 10000 получали образец комплекса Fc(P208)/внеклеточная область FcγRIIb с последующей очисткой с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 25 мM HEPES при pH 7,5, содержащего 0,1M NaCl.
[Кристаллизация комплекса Fc(P208)/внеклеточная область FcγRIIb]
Образец комплекса Fc(P208)/внеклеточная область FcγRIIb концентрировали с получением концентрации приблизительно 10 мг/мл с использованием фильтра для ультрафильтрации с отсечкой молекулярной массы 10000, и кристаллизовали методом паровой диффузии с висячей каплей в комбинации с методом затравки кристалла. Для кристаллизации использовали пластину VDXm (Hampton Research). С использованием резервуарного раствора, содержащего 0,1M бис-трис (pH 6,5), 19% (масс/об) PEG3350 и 0,2M двухосновного фосфата калия, капли для инициации кристаллизации были получены путем смешивания резервуарного раствора и образца кристаллизации в отношении 0,85 мкл:0,85 мкл. Кристаллы комплекса, полученного в тех же условиях, измельчали с использованием сфер Seed Bead (Hampton Research) для получения раствора затравочного кристалла. 0,15 мкл разведенного раствора, полученного из раствора затравочного кристалла, добавляли к каплям для инициации кристаллизации, которые были герметично закрыты в лунках, находящихся в резервуарах, и оставляли при 20°C. В результате этого были успешно получены пластинчатые кристаллы.
[Получение рентгенографических данных для кристалла комплекса Fc(P208)/внеклеточная область FcγRIIb]
Один из полученных монокристаллов комплекса Fc(P208)/ внеклеточной области FcγRIIb заливали в раствор 0,1M бис-трис (pH 6,5), 24% (масс/об) PEG3350, 0,2M двухосновного фосфата калия, 20% (об/об) этиленгликоля. Кристалл промывали раствором с помощью иглы, к которой была подсоединена тонкая нейлоновая петля, и замораживали в жидком азоте, а затем проводили рентгенографический анализ с использованием BL32XU на приборе Spring-8. Во время измерений кристалл постоянно выдерживали в потоке азота при -178°C для поддержания этого кристалла в замороженном состоянии, и все 300 дифракционных изображений получали на ПЗС-детекторе MX-225HE (RAYONIX), подсоединенном к источнику луча света, с одновременным вращением кристалла под углом 0,6°. Определение параметров ячейки, идентификацию пятен дифракции и обработку рентгенографических данных после получения дифракционных изображений проводили с использованием программы Xia2 (J. Appl. Cryst. 2010, 43, 186-190), пакета программ XDS (Acta Cryst. 2010, D66: 125-132) и программы Scala (Acta Cryst. 2006, D62:72-82), и наконец, получали данные интенсивности дифракции с разрешением 2,81 Å. Этот кристалл принадлежал к пространственной группе C2221 и имел следующие параметры ячейки: а=156,69 Å, b=260,17 Å, c=56,85 Å, α=90°, β=90°, γ=90°.
[Рентгеноструктурный анализ комплекса Fc(P208)/внеклеточная область FcγRIIb]
Кристаллическую структуру комплекса Fc(P208)/внеклеточная область FcγRIIb определяли методом молекулярной замены с использованием программы Phaser (J. Appl. Cryst. 2007, 40, 658-674). Исходя из размера полученной кристаллической решетки и молекулярной массы комплекса Fc(P208)/внеклеточная область FcγRIIb было предположено, что число комплексов в асимметрической ячейке равно 1. Исходя из структурных координат кода PDB: 3SGJ, который представляет собой кристаллическую структуру комплекса Fc(WT)/внеклеточная область FcγRIIIа, были определены отдельные координаты аминокислотных остатков части А-цепи в положениях 239-340 и В-цепи в положениях 239-340, а затем эти координаты были использованы, соответственно, в качестве моделей для поиска CH2-доменов Fc. Исходя из тех же самых структурных координат кода PDB: 3SGJ, были определены отдельные координаты аминокислотных остатков части А-цепи в положениях 341-444 и В-цепи в положениях 341-443, а затем эти координаты были использованы, соответственно, в качестве моделей для поиска CH3-доменов Fc. И наконец, исходя из структурных координат кода PDB: 2FCB, который представляет собой кристаллическую структуру внеклеточной области FcγRIIb, были определены отдельные координаты аминокислотных остатков части А-цепи в положениях 6-178, а затем эти координаты были использованы в качестве моделей для поиска Fc(P208). Авторами настоящего изобретения были предприняты попытки определить ориентации и положения в качестве модели для поиска каждого из СН3-доменов Fc-области, внеклеточной области FcγRIIb и CH2-домена Fc-области в кристаллических решетках с использованием функций вращения и функций трансляции, однако, попытка определить положение одного из CH2-доменов оказалась безуспешной. Затем, исходя из кристаллической структуры комплекса Fc(WT)/внеклеточная область FcγRIIIа, было определено положение последнего CH2-домена по карте электронной плотности, которую вычисляли исходя из фазы, определенной из трех остальных частей. Таким образом, авторами настоящего изобретения была получена первоначальная модель кристаллической структуры комплекса «Fc(P208)/внеклеточная область FcγRIIb». Если уточнение параметров твердого тела, относящегося к двум CH2-доменам Fc, двум CH3-доменам Fc и внеклеточной области FcγRIIb, осуществляли исходя из полученной первоначальной модели, то фактор кристаллографической достоверности, то есть R-фактор, составлял 42,6%, а свободный R-фактор (R-free) составлял 43,7%, как было определено по данным интенсивности дифракции с разрешением от 25 Å до 3,0 Å. Кроме того, уточнение структуры проводили с использованием программы REFMAC5 (Acta Cryst. 2011, D67, 355-367), коррекцию модели для наблюдения карт электронной плотности с коэффициентами 2Fo-Fc или Fo-Fc, которые вычисляли исходя из экспериментально определенного структурного фактора Fo, определенного фактора Fc и фазы, определенной с использованием такой модели, осуществляли с помощью программы Сооt (Acta Cryst. 2010, D66: 486-501), а уточнение модели осуществляли путем повторения этих стадий. И наконец, в результате введения молекул воды в модель, полученную исходя из карты электронной плотности, где 2Fo-Fc и Fo-Fc использовали в качестве коэффициента, и после уточнения был получен фактор кристаллографической достоверности, то есть R-фактор и R-free модели, содержащей 4786 не-водородных атомов, которые составляли 24,5% и 28,2% соответственно, как было определено по 27259 данным интенсивности дифракции с разрешением от 25 Å до 3,0 Å, соответственно.
3-2. Рентгеноструктурный анализ комплекса, образованного между Fc(P208) и внеклеточной областью FcγRIIа R-типа
В результате структурного анализа была определена кристаллическая структура комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа с разрешением 2,87 Å. Кристаллическую структуру комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа сравнивали с кристаллической структурой комплекса Fc(P208)/внеклеточная область FcγRIIb, описанного в примере 3-1, в результате чего было обнаружено, что эти два рецептора имеют очень высокую гомологию аминокислот, причем во всех этих структурах не наблюдалось почти никаких различий (фиг. 11). Однако при детальном анализе этой структуры на уровне электронной плотности были обнаружены различия, которые могут быть использованы для повышения селективности. В FcγRIIа R-типа, остатком в положении 160 является Phe вместо Tyr, и как показано на фиг. 12, этот Phe не может образовывать водородную связь с главной цепью аминокислотного остатка в положении 237 (в соответствии с Европейской нумерацией) СН2-домена А Fc, который участвует в связывании между FcγRIIb и Fc, содержащим модификацию P238D. Хотя введение модификации P238D является главным фактором в повышении селективности связывания с FcγRIIа R-типа, однако, дополнительное сравнение на уровне электронной плотности показало, что в комплексе, образованном FcγRIIb, электронная плотность боковых цепей Leu в положении 235 (в соответствии с Европейской нумерацией) или Leu в положении 234 (в соответствии с Европейской нумерацией) в СН2-домене А Fc может быть зарегистрирована, тогда как электронные плотности этих боковых цепей в комплексе, образованном FcγRIIа R-типа, не были детектированы, а петля, расположенная рядом с положением 237 (в соответствии с Европейской нумерацией), может подвергаться флуктуации, что обусловлено снижением степени взаимодействия с FcγRIIа R-типа вокруг этой области. С другой стороны, при сравнении структур той же самой области со структурой СН2-домена В (фиг. 13) было обнаружено, что электронная плотность Asp в положении 237 (в соответствии с Европейской нумерацией) может быть подтверждена в структуре комплекса, образованного FcγRIIb, тогда как электронная плотность приблизительно трех остатков, расположенных выше Asp в положении 237 (в соответствии с Европейской нумерацией), может быть подтверждена для комплекса, образованного FcγRIIа R-типа, и по сравнению с уровнем связывания с FcγRIIb, эта область, вероятно, является более широкой для взаимодействия. Полученные данные позволяют предположить, что в области, простирающейся от положения 234 до положения 238 (в соответствии с Европейской нумерацией) в Fc(Р208), боковая цепь СН2-домена А может вносить значительный вклад в связывание с FcγRIIb, а боковая цепь СН2-домена В может вносить значительный вклад в связывание с FcγRIIaR.
[Экспрессия и очистка внеклеточной области FcγRIIа R-типа]
Экспрессия и очистка были проведены методом, описанным в сравнительном примере 2.
[Очистка комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа]
К 1,5 мг очищенного образца внеклеточной области FcγRIIа R-типа добавляли 0,15 мг Endo F1 (Protein Science 1996, 5, 2617-2622), экспрессированного и очищенного из Escherichia coli в виде слитого белка глутатион-S-трансферазы, 20 мкл 5 ед/мл Endo F2 (QA-bio) и 20 мкл 5 ед/мл Endo F3 (QA-bio). Этот белок выдерживали при комнатной температуре в течение девяти дней в 0,1 M натрий-ацетатном буфере (pH 4,5), а затем добавляли 0,07 мг Endo F1 (Protein Science 1996, 5, 2617-2622), экспрессированного и очищенного из Escherichia coli в виде слитого белка глутатион-S-трансферазы, 7,5 мкл 5 ед/мл Endo F2 (QA-bio) и 7,5 мкл 5 ед/мл Endo F3 (QA-bio). Этот белок выдерживали при комнатной температуре в течение еще трех дней, и N-связанный олигосахарид отделяли, в результате чего оставался N-ацетилглюкозамин, непосредственно связанный с Asn. Затем этот образец внеклеточного домена FcγRIIа R-типа, подвергнутый реакции отщепления сахарной цепи, концентрировали путем ультрафильтрации с отсечкой молекулярной массы 10000, после чего его очищали с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 25 мM HEPES (pH 7), содержащем 0,1M NaCl. Кроме того, к полученной фракции внеклеточной области FcγRIIа R-типа, от которой была отщеплена сахарная цепь, добавляли Fc(P208) так, чтобы молярное соотношение внеклеточной области FcγRIIа R-типа было слегка повышенным, и после концентрирования этого образца путем ультрафильтрации с отсечкой молекулярной массы 10000 получали образец комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа с последующей очисткой с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 25 мM HEPES (pH 7), содержащем 0,1M NaCl.
[Кристаллизация комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа]
Образец комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа, концентрированный приблизительно до 10 мг/мл с помощью фильтра для ультрафильтрации с отсечкой молекулярной массы 10000, кристаллизовали методом паровой диффузии с сидячей каплей. С использованием резервуарного раствора, содержащего 0,1M бис-трис (pH 7,5), 26% (масс/об) PEG3350 и 0,2M сульфата аммония, капли для инициации кристаллизации были получены путем смешивания резервуарного раствора и образца кристаллизации в соотношении 0,8 мкл:1,0 мкл. Затем капли герметично закрывали и оставляли при 20°C. В результате этого были успешно получены пластинчатые кристаллы.
[Данные рентгенографических анализов для кристалла комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа]
Полученный монокристалл комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа заливали в раствор 0,1M бис-трис (pH 7,5), 27,5% (масс/об) PEG3350, 0,2M сульфата аммония, 20% (об/об) глицерина. Затем кристалл промывали раствором с помощью иглы, к которой была подсоединена тонкая нейлоновая петля, и замораживали в жидком азоте. Рентгенографический анализ монокристалла проводили на синхротронном излучателе Photon Factory BL-17A в научно-исследовательском институте высоких энергий. Во время измерений кристалл постоянно выдерживали в потоке азота при -178°C для его поддержания в замороженном состоянии. Все 225 дифракционных изображений этого монокристалла получали на ПЗС-детекторе Quantum 315r (ADSC), подсоединенном к источнику луча света, с одновременным вращением кристалла под углом 0,6°. Исходя из полученных дифракционных изображений, определение констант ячейки, идентификацию пятен дифракции и обработку рентгенографических данных проводили с использованием программы Xia2 (J. Appl. Cryst. (2010), 43, 186-190), пакета программ XDS (Acta Cryst. (2010), D66: 125-132) и программы Scala (Acta Cryst. (2006), D62:72-82). И наконец, получали данные интенсивности дифракции с разрешением 2,87 Å. Этот кристалл принадлежал к пространственной группе C2221 и имел следующие параметры ячейки: а=154,31 Å, b=257,61 Å, c=56,19 Å, α=90°, β=90°, γ=90°.
[Рентгеноструктурный анализ комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа]
Структуру комплекса определяли методом молекулярной замены с использованием программы Phaser (J. Appl. Cryst. (2007), 40, 658-674). Исходя из размера полученной кристаллической решетки и молекулярной массы комплекса Fc(P208)/внеклеточная область FcγRIIа R-типа было предположено, что число комплексов в асимметрической ячейке равно 1. С использованием кристаллографической структуры комплекса Fc(P208)/внеклеточная область FcγRIIb в качестве поисковой модели, полученной как описано в примере 3-1, были определены ориентация и положение в в кристаллических решетках с использованием функций вращения и функций трансляции. После уточнения параметров твердого тела, относящегося к двум CH2-доменам Fc, двум CH3-доменам Fc и внеклеточной области FcγRIIа R-типа, исходя из полученной первоначальной модели, определяли фактор кристаллографической достоверности, то есть R-фактор, который составлял 38,4%, а свободный R-фактор (R-free) составлял 38,0%, как было определено по данным интенсивности дифракции с разрешением от 25 Å до 3,0 Å. Затем уточнение структурной модели проводили путем повторения уточнений структуры с использованием программы REFMAC5 (Acta Cryst. (2011) D67, 355-367), с последующей коррекцией модели, проводимой с использованием программы Сооt (Acta Cryst. 2010, D66: 486-501) по картам электронной плотности с коэффициентами 2Fo-Fc и Fo-Fc, которые вычисляли исходя из экспериментально определенного структурного фактора Fo, определенного фактора Fc и фазы, определенной с использованием такой модели. И наконец, в результате введения молекул воды в модель, полученную исходя из карты электронной плотности, где 2Fo-Fc или Fo-Fc использовали в качестве коэффициента, и последующего уточнения получали фактор кристаллографической достоверности, то есть R-фактор и R-free модели, содержащей 4758 не-водородных атомов, которые составляли 26,3% и 29,8% соответственно, как было определено по 24838 данным интенсивности дифракции с разрешением от 25 Å до 2,87 Å, соответственно.
[Пример 4] Fc-варианты, в которых сайты с модификациями были определены исходя из кристаллических структур
Как указывалось в примере 3, предполагается, что электростатическое взаимодействие между Asp в положении 268 (в соответствии с Европейской нумерацией) и Arg в положении 292 (Европейская нумерация) является результатом структурных изменений, которые, вероятно, обусловлены введением модификации P271G в CH2-домен B варианта Fc(Р208) с повышенной активностью связывания с FcγRIIb (фиг. 9). Такое взаимодействие способствует стабилизации петлевой структуры в положениях 266-271 (в соответствии с Европейской нумерацией) и может вносить свой вклад в повышение активности связывания с FcγRIIb. В соответствии с этим, был проведен анализ для того, чтобы определить, может ли такое усиление электростатического взаимодействия с Arg в положении 292 (Европейская нумерация) стабилизировать эту петлевую структуру посредством замены Asp в положении 268 (в соответствии с Европейской нумерацией) на Glu и повышать активность взаимодействия с FcγRIIb. Кроме того, как показано на фиг. 8, Tyr в положении 160 (в соответствии с Европейской нумерацией) FcγRIIb образует водородные связи с главной цепью Asp в положении 237 (в соответствии с Европейской нумерацией) CH2-домена А Fc(P208) и играет важную роль в связывании с FcγRIIb. Хотя часть боковой цепи Asp в положении 237 (в соответствии с Европейской нумерацией) не вступает в какое-либо конкретное взаимодействие, однако, Ile в положении 332 (в соответствии с Европейской нумерацией), Glu в положении 333 (в соответствии с Европейской нумерацией) и Lys в положении 334 (в соответствии с Европейской нумерацией) расположены в непосредственной близости в этой молекуле. Был проведен анализ для того, чтобы определить, может ли замена остатков в этих сайтах гидрофильными остатками усиливать взаимодействие с Asp в положении 237 (в соответствии с Европейской нумерацией) и стабилизировать петлевую структуру, находящуюся поблизости от этого остатка, а также усиливать взаимодействие с Tyr в положении 160 FcγRIIb.
Варианты были получены путем введения замен H268E, I332T, I332S, I332E, I332K, E333K, E333R, E333S, E333T, K334S, K334T и K334E в антитело IL6R-BP230/IL6R-L (SEQ ID NO:27/SEQ ID NO:21), которое было продуцировано, как описано в примере 2. IL6R-L (SEQ ID NО: 21) обычно используют как L-цепь антитела. Эти варианты были использованы для экспрессии и очистки антител методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 2. В данной таблице «модификации» означают модификации, вводимые в IL6R-BР3 (SEQ ID NО:23). IL6R-B3/IL6R-L, которые используются в качестве матрицы для продуцирования IL6R-BР230, отмечены звездочкой (*). В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 2 закрашенные серым ячейки относятся к величинам, указывающим на то, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа, а поэтому они были вычислены по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2.
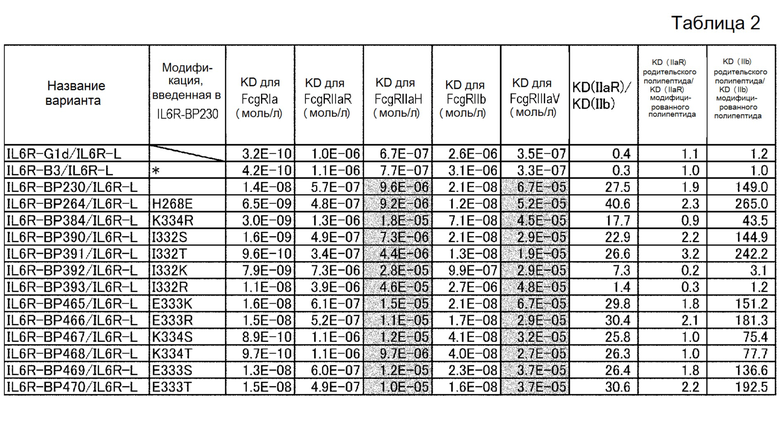
Активность связывания IL6R-BP264/IL6R-L, IL6R-BP465/IL6R-L, IL6R-BP466/IL6R-L и IL6R-BP470 с FcγRIIb и селективность связывания этих антител с FcγRIIb после введения модификаций H268E, E333K, E333R и E333T, соответственно, в IL6R-BP230/IL6R-L повышались по сравнению с активностью и селективностью IL6R-BP230/IL6R-L. Селективность связывания IL6R-BP391/IL6R-L с FcγRIIb после введения модификации I332T снижалась, а активность связывания этого антитела с FcγRIIb повышалась по сравнению с активностью и селективностью связывания IL6R-BP230/IL6R-L.
[Пример 5] Введение всех возможных модификаций в область, находящуюся возле положения 271 (в соответствии с Европейской нумерацией)
При сравнении рентгенографической кристаллической структуры комплекса, образованного между Fc(P238D), несущего модификацию P238D, и внеклеточной областью FcγRIIb, и рентгенографической кристаллической структуры комплекса, образованного между Fc(Р208) и внеклеточной областью FcγRIIb, было обнаружено, что структура, находящаяся возле положения 271 (в соответствии с Европейской нумерацией), подвергается наибольшему изменению (фиг. 9). Как указывалось в сравнительном примере 8, предполагается, что в Fc(P238D), если Asp в положении 270 (в соответствии с Европейской нумерацией) вступает в сильное электростатическое взаимодействие с Arg в положении 131 FcγRIIb, то Pro-часть в положении 271 (в соответствии с Европейской нумерацией) может иметь стереохимические затруднения. В структуре Fc(P208)/FcγRIIb введение модификации P271G приводит к позитивным изменениям на уровне главной цепи и способствует устранению такого структурного затруднения. В результате, структура, находящаяся поблизости от положения 271 (в соответствии с Европейской нумерацией), может подвергаться значительным изменениям. При эффективном введении модификаций, которые стабилизируют такую измененную структуру, потеря энергии в результате ее энтропии после электростатического взаимодействия с Arg в положении 131 FcγRIIb может быть уменьшена, что будет способствовать повышению активности связывания. В соответствии с этим, в область, расположенную возле положения 271 (в соответствии с Европейской нумерацией), были введены все возможные модификации для проведения скрининга на модификации, которые будут способствовать повышению активности связывания с FcγRIIb или селективности связывания с FcγRIIb.
Антитело IL6R-BP267 (SEQ ID NO:29), полученное путем введения E233D, G237D, P238D, H268E и P271G в IL6R-B3 (SEQ ID NO:23), использовали в качестве матрицы для введения всех возможных модификаций. Аминокислоты в положениях 264, 265, 266, 267, 269 и 272 (в соответствии с Европейской нумерацией) в IL6R-BP267 были по отдельности заменены любой из 18 аминокислот, не являющихся исходными аминокислотами и Cys. IL6R-L (SEQ ID NО: 21) обычно используют как L-цепь антитела. Эти варианты были использованы для экспрессии и очистки антител методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2. Все полученные варианты, обладающие повышенной активностью или селективностью связывания с FcγRIIb, по сравнению с вариантом IL6R-BP267/IL6R-L, в который не была введена модификация, систематизированы в таблице 3.

Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 3. В данной таблице «модификация, введенная в IL6R-BP267» означает модификацию, введенную в антитело IL6R-BР267 (SEQ ID NО:29), которое используется в качестве матрицы. IL6R-B3/IL6R-L, которое используется в качестве родительского антитела для продуцирования IL6R-B3, отмечено звездочкой (*). В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 3 закрашенные серым ячейки относятся к величинам, указывающим на то, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа, а поэтому они были вычислены по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2.
Все активности связывания вариантов, представленных в таблице 3, с FcγRIа, FcγRIIаН и FcγRIIIаV, были аналогичны активностям IL6R-B3/IL6R-L или ниже этих активностей. Кроме того, FcγRIIb-связывающая активность вариантов, полученных путем введения модификаций S267A, V264I, E269D, S267E, V266F, S267G и V266M, соответственно, в IL6R-BP267/IL6R-L, превышала активность IL6R-BP267/IL6R-L до введения модификации. Кроме того, величины KD(IIaR)/KD(IIb) для вариантов, полученных путем введения S267A, S267G, E272M, E272Q, D265E, E272D, E272N, V266L, E272I и E272F, соответственно, в IL6R-BP267/IL6R-L, превышали величины для варианта IL6R-BP267/IL6R-L до введения модификации, что указывало на повышение селективности связывания с FcγRIIb.
[Пример 6] Повышение активности связывания с FcγRIIb путем введения модификаций в СН3-область
Сообщалось, что замена Pro на Leu в положении 396 (в соответствии с Европейской нумерацией) приводит к повышению активности связывания с FcγRIIb (Cancer Res., (2007) 67, 8882-8890). Аминокислота в положении 396 (в соответствии с Европейской нумерацией) не принимает непосредственного участия во взаимодействии с FcγR. Однако было высказано предположение, что такая аминокислота может влиять на взаимодействие с FcγR после изменения структуры антитела. Таким образом, авторами настоящего изобретения был проведен анализ для того, чтобы определить, может ли повышение активности связывания с FcγRIIb или повышение селективности связывания с FcγRIIb стимулироваться введением всех возможных аминокислотных модификаций в положение 396 (в соответствии с Европейской нумерацией).
Антитело IL6R-BP423 (SEQ ID NO:33), полученное путем введения модификаций E233D, G237D, P238D, S267A, H268E, P271G и A330R в IL6R-B3 (SEQ ID NO:23), использовали в качестве матрицы. Варианты были получены путем замены аминокислоты в положении 396 (в соответствии с Европейской нумерацией) в IL6R-BP423 любой из 18 аминокислот, не являющихся исходной аминокислотой и цистеином. IL6R-L (SEQ ID NО: 21) обычно используют как L-цепь антитела. Эти варианты были использованы для экспрессии и очистки антител методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2. Связывание полученных вариантов с каждым FcγR систематизировано в таблице 4.
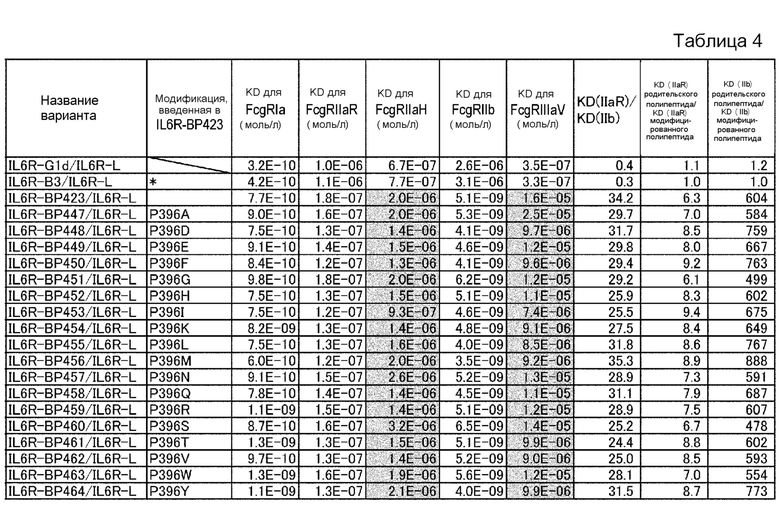
В этой таблице термин «модификация, введенная в IL6R-BP423» означает модификацию, введенную в антитело IL6R-BР423. Антитело IL6R-B3/IL6R-L, которое используется в качестве матрицы для продуцирования IL6R-BР423, отмечено звездочкой (*). В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 4 закрашенные серым ячейки относятся к величинам, указывающим на то, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа, а поэтому они были вычислены по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2.
Результаты, представленные в таблице 4, продемонстрировали, что FcγRIIb-связывающая активность антитела IL6R-BP456/IL6R-L, полученного путем введения P396M в IL6R-BP423/IL6R-L; антитела IL6R-BP455/IL6R-L, полученного путем введения P396L в IL6R-BP423/IL6R-L; антитела IL6R-BP464/IL6R-L, полученного путем введения P396Y в IL6R-BP423/IL6R-L; антитела IL6R-BP450/IL6R-L, полученного путем введения P396F в IL6R-BP423/IL6R-L; антитела IL6R-BP448/IL6R-L, полученного путем введения P396D в IL6R-BP423/IL6R-L; антитела IL6R-BP458/IL6R-L, полученного путем введения P396Q в IL6R-BP423/IL6R-L; антитела IL6R-BP453/IL6R-L, полученного путем введения P396I в IL6R-BP423/IL6R-L; антитела IL6R-BP449/IL6R-L, полученного путем введения P396E в IL6R-BP423/IL6R-L; антитела IL6R-BP454/IL6R-L, полученного путем введения P396K в IL6R-BP423/IL6R-L, и антитела IL6R-BP459/IL6R-L, полученного путем введения P396R в IL6R-BP423/IL6R-L, превышала активность IL6R-BP423/IL6R до введения модификаций. Кроме того, величина KD(IIaR)/KD(IIb) для IL6R-BP456/IL6R-L, полученного путем введения P396M в IL6R-BP423/IL6R-L, превышала величину для IL6R-BP423/IL6R-L до введения модификации, что указывало на повышение селективности связывания с FcγRIIb. Как видно из таблицы 4, аффинность связывания полученных вариантов с FcγRIа, FcγRIIаН и FcγRIIIаV была ниже, чем аффинность IL6R-B3/IL6R-L, который представляет собой родительский полипептид.
[Пример 7] Получение вариантов с повышенной активностью связывания с FcγRIIb с использованием последовательностей различных подклассов
Человеческие IgG имеют различные подклассы, и профили их связывания с FcγR варьируются. Авторами настоящего изобретения был проведен анализ для того, чтобы определить, может ли различие в аффинностях связывания IgG1 и IgG4 с каждым FcγR приводить к повышению активности и/или селективности связывания с FcγRIIb.
Сначала, IgG1 и IgG4 были проанализированы на их аффинность связывания с каждым из FcγR. Антитело IL6R-G4d (SEQ ID NO:30), содержащее G4d, было сконструировано в виде Н-цепи. В G4d отсутствуют С-концевые Gly и Lys, а в человеческом IgG4 Ser заменен на Pro в положении 228 (в соответствии с Европейской нумерацией). IL6R-L (SEQ ID NО: 21) обычно используют как L-цепь антитела. IL6R-G1d/IL6R-L и IL6R-G4d/IL6R-L были использованы для экспрессии и очистки методом, описанным в сравнительном примере 1. Эти варианты были оценены на связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb или FcγRIIIа V-типа) методом, описанным в сравнительном примере 2. Связывание полученных вариантов с каждым FcγR систематизировано в таблице 5.

При сравнении IL6R-G4d/IL6R-L с IL6R-G4d/IL6R-L было обнаружено, что IL6R-G4d/IL6R-L обладает в 1,5 раза большей активностью связывания с FcγRIIb и в 2,2 раза меньшей активностью связывания с FcγRIIаR. Кроме того, IL6R-G4d/IL6R-L, по сравнению с IL6R-G1d/IL6R-L, обладал меньшей аффинностью связывания с FcγRIа, FcγRIIаН и FcγRIIIаV. Вышеупомянутые результаты показали, что IL6R-G4d, по сравнению с IL6R-G1d, обладает превосходной селективностью и активностью связывания с FcγRIIb.
На фигуре 14 проиллюстрировано сравнение последовательностей G1d и G4d, простирающихся от СН1-домена и до С-конца (в положениях 118-445 (в соответствии с Европейской нумерацией)). Представленные в рамках аминокислоты на фиг. 14 означают остатки, которые отличаются в G1d и G4d. Несколько сайтов, которые, как было предсказано, участвуют во взаимодействии с FcγR, были выбраны из сайтов, имеющих различные аминокислоты, и был проведен анализ для того, чтобы определить, можно ли достичь еще большего повышения селективности и активности связывания путем переноса последовательности G4d, обладающей превосходной селективностью и активностью связывания с FcγRIIb, в варианты с повышенной активностью связывания с FcγRIIb.
В частности, антитело IL6R-BP473 было получено путем введения A327G в IL6R-BP230; антитело IL6R-BP472 было получено путем введения A330S в IL6R-BP230; антитело IL6R-BP471 было получено путем введения P331S в IL6R-BP230; антитело IL6R-BP474 было получено путем введения A330S и P331S в IL6R-BP230; антитело IL6R-BP475 было получено путем введения A327G и A330S в IL6R-BP230; антитело IL6R-BP476 было получено путем введения A327G, A330S и P331S в IL6R-BP230; а антитело IL6R-BP477 было получено путем введения A327G и P331S в IL6R-BP230. Кроме того, IL6R-BP478 (SEQ ID NO:31) получали путем замены последовательности, простирающейся от Ala в положении 118 и до Thr в положении 225 (в соответствии с Европейской нумерацией), в IL6R-BP230, последовательностью G4d (с Ala в положении 118 до Pro в положении 222 (в соответствии с Европейской нумерацией)). IL6R-L (SEQ ID NО: 21) обычно используют как L-цепь антитела. Эти варианты были использованы для экспрессии и очистки методом, описанным в сравнительном примере 1, а связывание этих вариантов с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb или FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым из FcγR представлены в таблице 6. В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Термин «модификация(и), введенная(ые) в IL6R-BP230» означает модификацию, введенную в антитело IL6R-BР230. IL6R-B3/IL6R-L, которое используется в качестве матрицы для продуцирования IL6R-BР230, отмечено *1. Кроме того, антитело IL6R-B478 (SEQ ID NО: 31), в котором сегмент, простирающийся от Ala в положении 118 до Thr в положении 225 (в соответствии с Европейской нумерацией) в IL6R-BP230, заменен последовательностью G4d (от Ala в положении 118 до Pro в положении 222 (в соответствии с Европейской нумерацией)), обозначено *2. Термин «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 6 закрашенные серым ячейки относятся к величинам, указывающим на то, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа, а поэтому они были вычислены по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2.
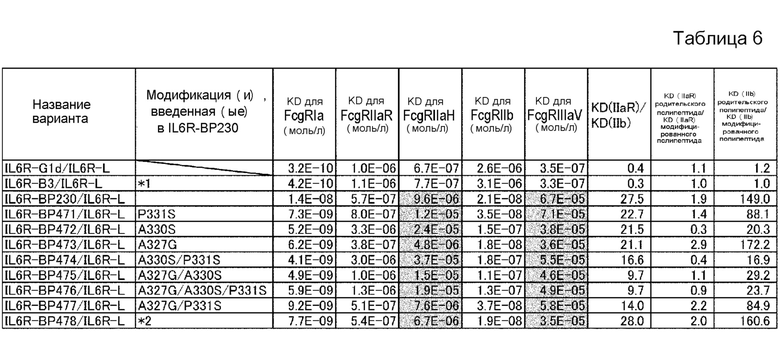
Из вариантов, описанных в таблице 6, антитело IL6R-BP473/IL6R-L, в которое была введена замена A327G, обнаруживало активность связывания с FcγRIIb, которая в 1,2 раза превышала активность IL6R-BP230/IL6R-L. По сравнению с IL6R-BP230/IL6R-L, антитело IL6R-BP478/IL6R-L, полученное путем замены последовательности, простирающейся от Ala в положении 118 и до Thr в положении 225 (в соответствии с Европейской нумерацией) в IL6R-BP230, последовательностью G4d (от Ala в положении 118 до Pro в положении 222 (в соответствии с Европейской нумерацией)), обнаруживало 1,1-кратное увеличение активности связывания с FcγRIIb и с FcγRIIаR. По сравнению с антителом IL6R-B3/IL6R-L, которое представляет собой родительский пептид, все варианты обладали более низкой аффинностью связывания с FcγRIа, FcγRIIаН и FcγRIIIаV.
Кроме того, как показано на фиг. 14, другие сайты, в которых аминокислоты отличаются в последовательности G1d и G4d, включают положения 268, 274, 296, 355, 356, 358, 409, 419 и 445 (в соответствии с Европейской нумерацией). Поэтому, в результате замены этих сайтов аминокислотами, происходящими от IgG4, может быть повышена селективность и активность связывания с FcγRIIb.
Проведенные ранее исследования показали, что введение замены A327G, которая имеется в последовательности человеческого IgG4, в вариант IL6R-BP230/IL6R-L приводило к повышению активности связывания с FcγRIIb. Части, которые не соответствовали последовательностям IgG4 и IgG1, были подвергнуты дополнительному анализу.
В частности, были получены варианты путем введения в антитело IL6R-BP230, которое представляет собой Н-цепь антитела, следующих модификаций: K274Q, введенной для продуцирования IL6R-BP541; Y296F, введенной для продуцирования IL6R-BP542; H268Q, введенной для продуцирования IL6R-BP543; R355Q, введенной для продуцирования IL6R-BP544; D356E, введенной для продуцирования IL6R-BP545; L358M, введенной для продуцирования IL6R-BP546; K409R, введенной для продуцирования IL6R-BP547; и Q419E, введенной для продуцирования IL6R-BP548. Кроме того, IL6R-L обычно используют как L-цепь антитела. Антитела, содержащие вышеуказанный вариант тяжелой цепи и IL6R-L легкой цепи, очищали методами, описанными в сравнительном примере 1. Очищенные антитела оценивали на их связывание с каждым FcγR (FcγRIа, FcγRIIаН, FcγRIIаR, FcγRIIb или FcγRIIIаV) методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 7. В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. В этой таблице «модификации, введенные в IL6R-BP230» означают модификации, введенные в антитело IL6R-BР230. Кроме того, IL6R-B3/IL6R-L, которое используется в качестве матрицы для продуцирования IL6R-BР230, отмечено *1. «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 7 закрашенные серым ячейки относятся к величинам, вычисленным по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2, где было показано, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа.
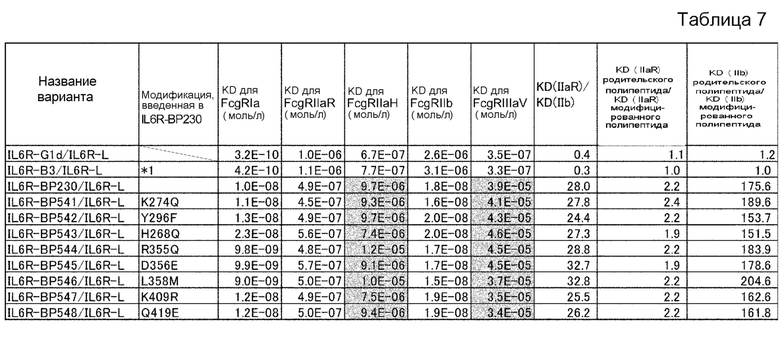
Как показано в таблице 7, по сравнению с вариантом IL6R-BP230/IL6R-L до модификации, вариант IL6R-BP541/IL6R-L, полученный путем введения K274Q в IL6R-BP230/IL6R-L; вариант IL6R-BP544/IL6R-L, полученный путем введения R355Q в IL6R-BP230/IL6R-L; вариант IL6R-BP545/IL6R-L, полученный путем введения D356E в IL6R-BP230/IL6R-L; и вариант IL6R-BP546/IL6R-L, полученный путем введения L358M в IL6R-BP230/IL6R-L, обладали повышенной активностью связывания с FcγRIIb. Из этих вариантов вариант IL6R-BP544/IL6R-L, полученный путем введения R355Q в IL6R-BP230/IL6R-L; вариант IL6R-BP545/IL6R-L, полученный путем введения D356E в IL6R-BP230/IL6R-L; и вариант IL6R-BP546/IL6R-L, полученный путем введения L358M в IL6R-BP230/IL6R-L, имели более высокие величины KD(IIaR)/KD(IIb) по сравнению с величинами для IL6R-BP230/IL6R-L до модификации, что указывает на то, что такие модификации также повышают селективность связывания с FcγRIIb.
[Пример 8] Оценка комбинаций модификаций, которые приводят к повышению селективности и активности связывания с FcγRIIb
Комбинации модификаций, которые, как было обнаружено в предыдущих исследованиях, повышают селективность или активность связывания с FcγRIIb, оценивали для дополнительной оптимизации.
Комбинацию модификаций, которая повышает активность и/или селективность связывания с FcγRIIb, как было обнаружено в предыдущих исследованиях, вводили в IL6R-B3. Для сравнения с контролем был продуцирован вариант IL6R-BP253 путем введения модификаций S267E и L328F, которые, как известно, повышают активность связывания с FcγRIIb (Seung et al., Mol. Immunol. (2008) 45, 3926-3933), в IL6R-B3. IL6R-L использовали в качестве L-цепи антитела. Антитело, содержащее вышеупомянутый вариант тяжелой цепи и легкой цепи IL6R-L, который был экспрессирован методом, описанным в сравнительном примере 1, очищали. Связывание очищенного антитела с каждым FcγR (FcγRIа, FcγRIIаН, FcγRIIаR, FcγRIIb и FcγRIIIаV) оценивали методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 8. В этой таблице «модификация» означает модификации, введенные в IL6R-B3. Кроме того, антитело IL6R-B3/IL6R-L, которое используется в качестве матрицы для продуцирования каждого из вариантов, отмечено звездочкой (*). «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания соответствующего варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого соответствующего варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb по сравнению с FcγRIIаR. «KD(IIaН)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаН на величину KD для связывания этого соответствующего варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb по сравнению с FcγRIIаН. В таблице 8 числа в закрашенных серым ячейках относятся к величинам, вычисленным по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2, где было показано, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа.

Из вариантов, описанных в таблице 8, антитело IL6R-BP253/IL6R-L, полученное путем введения известных модификаций, повышающих активность связывания с FcγRIIb, обладало активностью связывания с FcγRIIb и FcγRIIаR, которая, соответственно, в 277 раз и 529 раз превышала активность связывания IL6R-B3/IL6R-L до модификации. Кроме того, FcγRIа-связывающая активность IL6R-BP253/IL6R-L также превышала активность IL6R-B3/IL6R-L. С другой стороны, активность связывания IL6R-BP253/IL6R-L с FcγRIIаН и FcγRIIIаV была ниже активности IL6R-B3/IL6R-L. Что касается других вариантов, то активность связывания IL6R-BP436/IL6R-L, IL6R-BP438/IL6R-L, IL6R-BP567/IL6R-L и IL6R-BP568/IL6R-L с FcγRIа слегка превышала активность связывания IL6R-B3/IL6R-L до модификации, однако, все другие варианты имели пониженную активность связывания с FcγRIа. Кроме того, активность связывания всех вариантов с FcγRIIаН и FcγRIIIаV была ниже, чем активность связывания IL6R-B3/IL6R-L.
Проводимое в этом исследовании сравнение полученных вариантов с существующим вариантом IL6R-BP253/IL6R-L, обладающим повышенной активностью связывания с FcγRIIb, показало, что величина KD(IIaH)/KD(IIb) для IL6R-BР480/IL6R-L составляла 107,7 и является наименьшей величиной, а для IL6R-BР426/IL6R-L она составляла 8362 и является наибольшей величиной, причем величины для всех вариантов превышали величину 107,1 для IL6R-BP253/IL6R-L. Кроме того, величина KD(IIaR)/KD(IIb) для IL6R-BР479/IL6R-L составляла 16,1 и является наименьшей величиной, а для IL6R-BР567/IL6R-L она составляла 64,4 и является наибольшей величиной, причем величины для всех вариантов превышали величину 0,2 для IL6R-BP253/IL6R-L. Исходя из этих результатов было установлено, что все варианты, представленные в таблице 8, обладают повышенной селективностью связывания с FcγRIIb по сравнению с существующим вариантом, в который была(и) введена(ы) модификация(и) для повышения активности связывания с FcγRIIb. В частности, все варианты IL6R-BP559/IL6R-L, IL6R-BP493/IL6R-L, IL6R-BP557/IL6R-L, IL6R-BP492/IL6R-L, IL6R-BP500/IL6R-L и IL6R-BP567/IL6R-L обладали активностью связывания с FcγRIIаR, которая не более чем в 1,5 раза превышала активность IL6R-B3/IL6R-L, и в то же время активность связывания с FcγRIIb была выше в 100 раз или более, а поэтому эти варианты, как и предполагалось, обладали повышенной активностью связывания с FcγRIIb, но при этом не давали побочных эффектов, вызываемых повышением активности связывания с FcγRIIаR.
Кроме того, что касается вариантов IL6R-BP489/IL6R-L, IL6R-BP487/IL6R-L, IL6R-BP499/IL6R-L, IL6R-BP498/IL6R-L, IL6R-BP503/IL6R-L, IL6R-BP488/IL6R-L, IL6R-BP490/IL6R-L, IL6R-BP445/IL6R-L, IL6R-BP552/IL6R-L, IL6R-BP507/IL6R-L, IL6R-BP536/IL6R-L, IL6R-BP534/IL6R-L, IL6R-BP491/IL6R-L, IL6R-BP553/IL6R-L, IL6R-BP532/IL6R-L, IL6R-BP506/IL6R-L, IL6R-BP511/IL6R-L, IL6R-BP502/IL6R-L, IL6R-BP531/IL6R-L, IL6R-BP510/IL6R-L, IL6R-BP535/IL6R-L, IL6R-BP497/IL6R-L, IL6R-BP533/IL6R-L, IL6R-BP555/IL6R-L, IL6R-BP554/IL6R-L, IL6R-BP436/IL6R-L, IL6R-BP423/IL6R-L, IL6R-BP440/IL6R-L, IL6R-BP538/IL6R-L, IL6R-BP429/IL6R-L, IL6R-BP438/IL6R-L, IL6R-BP565/IL6R-L, IL6R-BP540/IL6R-L, IL6R-BP426/IL6R-L, IL6R-BP437/IL6R-L, IL6R-BP439/IL6R-L, IL6R-BP551/IL6R-L, IL6R-BP494/IL6R-L, IL6R-BP537/IL6R-L, IL6R-BP550/IL6R-L, IL6R-BP556/IL6R-L, IL6R-BP539/IL6R-L, IL6R-BP558/IL6R-L, IL6R-BP425/IL6R-L и IL6R-BP495/IL6R-L, то их активность связывания с FcγRIIb превышает активность связывания существующего варианта IL6R-BP253/IL6R-L, в который была введена модификация для повышения активности связывания с FcγRIIb. Кроме того, активность связывания с FcγRIIb для вариантов от IL6R-BP489/IL6R-L до IL6R-BP495/IL6R-L была выше, чем активность связывания IL6R-B3/IL6R-L (которая принималась за 1), и в 321 раз (наименьшая) - 3100 раз (наибольшая) превышала активность связывания IL6R-B3/IL6R-L, соответственно. Поэтому можно сказать, что, с точки зрения селективности и активности связывания с FcγRIIb, эти варианты являются наилучшими из всех вариантов, полученных по существующей технологии.
В настоящем изобретении варианты, родственные варианту IL6R-BP567/IL6R-L, который рассматривается как наилучший с точки зрения селективности и активности связывания с FcγRIIb, были исследованы на их перспективность в отношении иммуногенности. Модификация Y296D была введена в вариант IL6R-BP567/IL6R-L, который обнаруживал наивысшую селективность связывания, и в вариант IL6R-BP493/IL6R-L, который обнаруживал активность связывания с FcγRIIаR, полностью эквивалентную активности нативной формы и в 147 раз превышающую активность связывания с FcγRIIb. Сообщалось, что в последовательность Tregitope была введена модификация Y296 (De Groot et al. Blood (2008) 112, 3303-3311), и введение модификаций в этот сайт может приводить к потере иммуносупрессорных функций, которыми обычно обладает нативный IgG1. Поэтому варианты, в которые не была введена модификация Y296D, являются более предпочтительными с точки зрения возможной иммуногенности. IL6R-BP568/IL6R-L и IL6R-BP492/IL6R-L были получены путем удаления модификации Y296D из IL6R-BP567/IL6R-L и IL6R-BP493/IL6R-L, соответственно. Принимая во внимание селективность и активность связывания с FcγRIIb, следует отметить, что удаление модификации Y296D из IL6R-BP492/IL6R-L и IL6R-BP568/IL6R-L будет приводить к снижению селективности и активности связывания по сравнению с селективностью и активностью связывания варианта, имеющего Y296D. Однако, по сравнению с нативной формой, активность связывания IL6R-BP568/IL6R-L с FcγRIIаR была в 1,6 раза выше, а с FcγRIIb - в 211 раз выше; а активность связывания IL6R-BP492/IL6R-L с FcγRIIаR была в 1,2 раза выше, а с FcγRIIb - в 131 раз выше; и отсюда следует, что высокая активность и селективность связывания этих вариантов все еще сохранялась. Полученные результаты позволяют сделать вывод, что IL6R-BP568/IL6R-L и IL6R-BP492/IL6R-L являются превосходными вариантами не только с точки зрения селективности и активности связывания с FcγRIIb, но также и с точки зрения иммуногенности.
[Пример 9] Повышение активности связывания гетеродимеризованного антитела с FcγRIIb
9-1. Оценка результатов введения P238D только в одну из цепей
Как показано на фиг. 26 в сравнительном примере 7, причина того, что Fc(P238D) приобретает высокую активность связывания с FcγRIIb после введения модификации P238D, заключается в том, что область, которая имеет гидрофобную сердцевину с окружающими ее остатками в случае Pro, не может существовать в гидрофобной сердцевине после замены этого остатка на Asp и расширяется в сторону растворителя, что приводит к значительному изменению петлевой структуры домена А. Однако, следует проверить, необходимо ли введение модификации P238D в обе цепи и допустимо ли введение этой модификации в одну из цепей, и других модификаций - в другую цепь. В соответствии с этим, для такого подтверждения были использованы гетеродимеризованные антитела с различными модификациями, вводимыми в каждую Н-цепь антитела.
Вариабельная область (SEQ ID NO:15) антитела против глипикана 3, содержащая CDR антитела GpH7, которое представляет собой антитело против глипикана 3, обладающее повышенной кинетикой в плазме, как описано в WO 2009/041062, была использована в качестве Н-цепи антитела. Было использовано антитело GpH7-A5 (SEQ ID NО: 35), полученное путем введения модификаций D356K и H435R в GpH7-G1d (SEQ ID NO:34), где Gly и Lys были удалены из C-конца IgG1, несущего GpH7 в качестве вариабельной области, и антитело GpH7-B3 (SEQ ID NО:17), полученное путем введения модификации K439E в GpH7-G1d. Модификации D356K и K439E были введены в соответствующие Н-цепи для более эффективного образования гетеродимеров в каждой Н-цепи при продуцировании гетеродимеризованных антител, содержащих две Н-цепи (WO 2006/106905). H435R представляет собой модификацию, которая ингибирует связывание с белком A, и которая была введена для эффективного отделения димерного гетеромера, содержащего две Н-цепи, в каждую из которых были введены различные модификации, от димерного гомомера, содержащего две Н-цепи, в каждую из которых были введены одинаковые модификации. Варианты, в которых аминокислоты в положениях 236, 237 и 238 (Европейская нумерация) антитела GpH7-B3 (SEQ ID NO:17), полученного по сравнительному примеру 3, были заменены любой из 18 аминокислот, отличающейся от исходной аминокислоты и Cys, использовали в качестве одной из Н-цепей. Антитело GpH7-AP001, полученное путем введения P238D в GpH7-A5 (SEQ ID NO:35), использовали в качестве другой цепи. Вариант GpL16-k0 (SEQ ID NO:16) антитела против глипикана 3, обладающего повышенной кинетикой в плазме, как описано в WO 2009/041062, был использован в качестве L-цепи антитела. Эти варианты были экспрессированы и очищены методом, описанным в сравнительном примере 1, а активность связывания с FcγRIIа R-типа и с FcγRIIb оценивали методом, описанным в сравнительном примере 2. Активность связывания каждого варианта с FcγR представлена на фиг. 15.
Модификации G237W, G237F, G236N, P238G, P238N, P238E и P238D, представленные на фиг. 15, означают модификации, вводимые в GpH7-B3. A5/B3 означает вариант GpH7-A5/GpH7-B3/GpL16-k0, в который не были введены модификации ни в одну из цепей, а вариант, содержащий P238D только в одной из этих цепей, обозначен GpH7-A5/GpH7-BF648/GpL16-k0. Полученные результаты представлены в таблице 9.
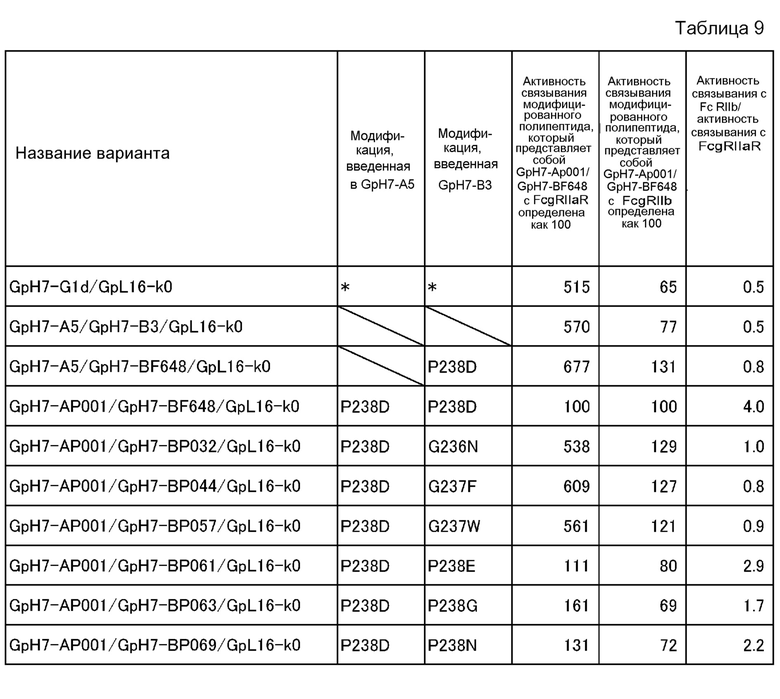
В таблице 9 «активность связывания с FcγRIIb/активность связывания с FcγRIIаR» означает величины, полученные путем деления активности связывания каждого варианта с FcγRIIb на активность связывания каждого варианта с FcγRIIаR, и, как было показано, чем выше эта величина, тем выше селективность связывания с FcγRIIb. Кроме того, «модификация, введенная в GpH7-A5» и «модификация, введенная в GpH7-В3» означает модификации, введенные в GpH7-A5 и GpH7-B3, соответственно, а вариант GpH7-G1d, который был использован в качестве матрицы при продуцировании GpH7-A5 и GpH7-B3, показан звездочкой (*). Результаты, представленные в таблице 9, показали, что вариант GpH7-AP001/GpH7-BF648/GpL16-k0, имеющий модификацию P238D, введенную в обе цепи, имеет наивысшую селективность связывания с FcγRIIb. Варианты GpH7-AP001/GpH7-BP061/GpL16-k0, GpH7-AP001/GpH7-BP069/GpL16-k0 и GpH7-AP001/GpH7-BP063/GpL16-k0, которые содержат модификации P238E, P238N и P238G в другой цепи, имеют величины «активности связывания с FcγRIIb/активности связывания с FcγRIIаR», составляющие 2,9, 2,2 и 1,7, соответственно, и эти варианты сохраняют высокую селективность связывания с FcγRIIb даже по сравнению с селективностью связывания вариантов GpH7-AP001/GpH7-BF648/GpL16-k0, имеющих модификацию P238D в обеих цепях. Кроме того, поскольку вариант GpH7-AP001/GpH7-BF648/GpL16-k0, имеющий модификацию P238D в обеих цепях, также сохраняет 69% аффинности связывания с FcγRIIb или более, то можно сказать, что, если модификация P238D присутствует в одной из этих цепей, то другая цепь может иметь модификацию P238Е, P238N или P238G. Кроме того, что касается активности связывания с FcγRIIb, то по сравнению с вариантом GpH7-AP001/GpH7-BF648/GpL16-k0, имеющим модификацию P238D в обеих цепях, вариант GpH7-A5/GpH7-BF648/GpL16-k0, который имеет модификацию P238D только в одной из цепей и не имеет модификации в другой цепи, обладает более высокой активностью с FcγRIIb, а варианты GpH7-AP001/GpH7-BP032/GpL16-k0, GpH7-AP001/GpH7-BP044/GpL16-k0 и GpH7-AP001/GpH7-BP057/GpL16-k0, которые имеют модификацию P238D в одной из цепей и модификации G236N, G237F и G237W, соответственно, в другой цепи, обладают еще большей активностью связывания с FcγRIIb.
9-2. Подтверждение модификаций на основе информации о структуре Fc(P208)/FcγRIIb
Как показано на фиг. 10, в кристаллической структуре Fc(P208)/FcγRIIb электронная плотность Lys в положении 117 FcγRIIb не наблюдалась, и этот остаток не рассматривался как остаток, играющий важную роль в связывании с Fc(P208), однако, после замены Ser на Asp или Glu в положении 239 (в соответствии с Европейской нумерацией) СН2-домена В, который расположен поблизости, этот остаток Lys может вступать в электростатическое взаимодействие в положении 117 FcγRIIb. С другой стороны, как показано на фиг. 7, в СН2-домене А остаток Ser в положении 239 (в соответствии с Европейской нумерацией) образует водородную связь с Gly в положении 236 (в соответствии с Европейской нумерацией), а стабилизация петлевой структуры в положениях 233-239 (в соответствии с Европейской нумерацией) может вносить свой вклад в повышение активности связывания с Tyr в положении 160 (в соответствии с Европейской нумерацией), и введение замен в эту часть, как было предсказано, будет способствовать снижению активности связывания и будет приводить к дестабилизации петлевой структуры в СН2-домене А, где такие эффекты, как было предсказано, будут подавлять друг друга в случае гомологичных модификаций. В соответствии с этим, в данном исследовании модификацию S239D или модификацию S239E вводили только в одну из цепей посредством гетеродимеризации, а затем оценивали эффекты повышения активности связывания с FcγRIIb.
Антитела IL6R-BP256 и IL6R-BP257, используемые в качестве одной из Н-цепей, продуцировали путем введения модификаций S239D и S239E, соответственно, в IL6R-BP208 (SEQ ID NO:24). Аналогичным образом, модификацию S239D вводили в IL6R-BP230 (SEQ ID NO:27) с получением IL6R-BP259, а модификацию S239E вводили в IL6R-BP230 с получением IL6R-BP260. IL6R-AP002 получали путем введения тех же самых модификаций, которые были введены в СН2 IL6R-BP208, то есть E233D, G237D, P238D, H268D, P271G и A330R в IL6R-A5 (SEQ ID NO:69), а IL6R-AP009 получали путем введения тех же самых модификаций, которые были введены в СН2 IL6R-BP230, то есть E233D, G237D, P238D, H268D, P271G, Y296D и A330R в IL6R-A5, и эти варианты были использованы в качестве другой Н-цепи антитела. В противоположность этому, IL6R-BP253 (SEQ ID NO:32) получали путем введения S267E и L328F в IL6R-B3, и такое введение является известным методом повышения активности связывания с FcγRIIb (непатентный документ 28). IL6R-L (SEQ ID NO:21) обычно используется как L-цепь антитела. Антитела экспрессировали с использованием этих вариантов, а затем очищали методом, описанным в сравнительном примере 1, и их связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb или FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2.
Величины KD для связывания каждого варианта с каждым FcγR представлены в таблице 10. «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 10 закрашенные серым ячейки относятся к величинам, вычисленным по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2, где было показано, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа.
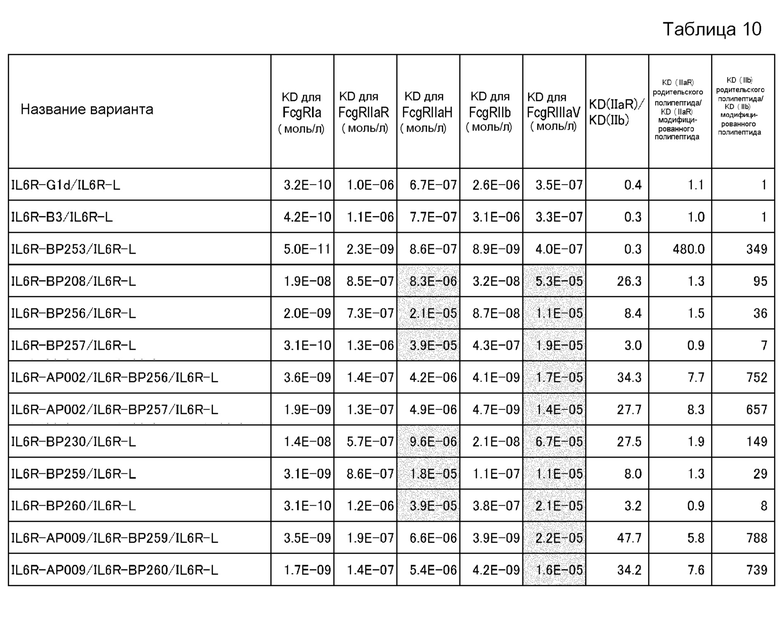
Как показано в таблице 10, по сравнению с вариантом IL6R-BP208/IL6R-L, оба варианта IL6R-AP002/IL6R-BP256/IL6R-L и IL6R-AP002/IL6R-BP257/IL6R-L, продуцированные путем введения модификаций S239D и S239E, соответственно, в одну из цепей IL6R-BP208/IL6R-L, обладали повышенной активностью связывания с FcγRIIb. Кроме того, величина KD (IIaR)/KD(IIb) этого варианта превышала величину для IL6R-BP256/IL6R-L, и его селективность связывания с FcγRIIb также была более высокой. С другой стороны, по сравнению с IL6R-BP208/IL6R-L, вариант IL6R-BP256/IL6R-L, продуцированный путем введения S239D в обе цепи IL6R-BP208/IL6R-L, и вариант IL6R-BP257/IL6R-L, продуцированный путем введения S239E в обе цепи IL6R-BP208/IL6R-L, обладали значительно более низкой селективностью и активностью связывания с FcγRIIb. Таким образом, при введении модификаций S239D или S239E только в одну из цепей наблюдалось повышение уровня связывания с FcγRIIb, тогда как при введении модификаций S239D или S239E в обе цепи, активность связывания с FcγRIIb значительно снижалась. Главной причиной такого эффекта может быть дестабилизация петлевой структуры в СН2-домене А, описанная выше. Аналогичные результаты были получены при введении модификаций S239D или S239E в IL6R-BP230/IL6R-L, используемый в качестве матрицы. Варианты IL6R-AP009/IL6R-BP259/IL6R-L и AP009/IL6R-BP260/IL6R-L, продуцированные путем введения модификаций S239D или S239E, соответственно, в одну из цепей IL6R-BP230/IL6R-L, обладали более высокой селективностью и активностью связывания с FcγRIIb, чем IL6R-BP230/IL6R-L. С другой стороны, варианты IL6R-BP259/IL6R-L и IL6R-BP260/IL6R-L, продуцированные путем введения модификаций S239D или S239E, соответственно, в обе цепи, обладали значительно меньшей селективностью и активностью связывания с FcγRIIb по сравнению с IL6R-BP230/IL6R-L. Кроме того, все варианты, продуцированные путем введения модификаций S239D или S239E, соответственно, в одну из цепей IL6R-BP208/IL6R-L и IL6R-BP230/IL6R-L, обладали повышенной селективностью и активностью связывания с FcγRIIb по сравнению с IL6R-BP253/IL6R-L, как было определено с применением известного метода повышения активности связывания с FcγRIIb.
9-3. Подтверждение модификаций на основе информации о структуре Fc(P208)/FcγRIIaR
Сравнение кристаллических структур Fc(P208) с FcγRIIb и FcγRIIаR, как описано в примере 3, указывало на различие электронной плотности возле положения 237 (в соответствии с Европейской нумерацией), где водородная связь образовывалась с остатком Tyr в положении 160 FcγRIIb, и было высказано предположение, что боковая цепь СН2-домена А вносит значительный вклад в связывание с FcγRIIb, а боковая цепь СН2-домена В вносит значительный вклад в связывание с FcγRIIаR (фиг. 12 и 13). Так, например, исходя из оценки электронной плотности при связывании с FcγRIIа R-типа, было высказано предположение, что Leu в положении 234 и Leu в положении 235 (в соответствии с Европейской нумерацией) в СН2-домене В участвуют в связывании с рецептором, тогда как эти остатки могут оказывать лишь незначительное влияние на связывание с FcγRIIb. Поэтому, в результате замены этих двух остатков другими остатками, не являющимися гидрофобными остатками, взаимодействие с FcγRIIа R-типа может быть значительно снижено. Однако считается, что в боковой цепи СН2-домена А остатки Leu в положении 234 и Leu в положении 235 (в соответствии с Европейской нумерацией) вносят определенный вклад в стабилизацию петлевой структуры возле положения 237 (в соответствии с Европейской нумерацией), а в частности, существует большая вероятность того, что они будут принимать большее участие в связывании с FcγRIIb. Поэтому замена этих остатков другими остатками, не являющимися гидрофобными остатками, будет приводить к снижению взаимодействия СН2-домена А с FcγRIIb. В частности, остаток Leu в положении 235 (в соответствии с Европейской нумерацией) способствует гидрофобному взаимодействию в СН2-домене А комплексной структуры, образованной FcγRIIb, и поскольку такой остаток вносит значительный вклад в стабилизацию петлевой структуры возле положения 237 (в соответствии с Европейской нумерацией), этот остаток был заменен негидрофобным остатком только в одной из цепей. В частности, если гидрофобное взаимодействие в СН2-домене А может еще больше стабилизировать петлевую структуру возле положения 237 (в соответствии с Европейской нумерацией) после замены Leu в положении 235 (в соответствии с Европейской нумерацией) гидрофобными аминокислотами, не являющимися Leu в обеих цепях, то это может также приводить к снижению потери энергии в результате ее энтропии с последующим образованием водородной связи с Tyr в положении 160 FcγRIIb и способствовать повышению селективности и активности связывания с FcγRIIb, как было определено в данном анализе.
Антитело IL6R-BP264 (SEQ ID NO:28), используемое как Н-цепь, было продуцировано путем введения модификаций E233D, G237D, P238D, H268E, P271G, Y296D и A330R в IL6R-B3 (SEQ ID NO:23) и служило в качестве матрицы. Были продуцированы варианты, в которых остаток Leu в положении 234 (в соответствии с Европейской нумерацией) IL6R-BP264 был заменен остатками Asn, Ser, Asp, Gln, Glu, Thr, Arg, His, Gly, Lys и Tyr. Были также продуцированы варианты, в которых аминокислота в положении 235 (в соответствии с Европейской нумерацией) в IL6R-BP264 была заменена любой из 18 аминокислот, не являющихся исходной аминокислотой и Cys. Вариант IL6R-AP029 (SEQ ID NO:42), который используется в качестве другой Н-цепи антитела, был продуцирован путем введения модификаций E233D, G237D, P238D, H268E, P271G, Y296D и A330R в IL6R-A5 (SEQ ID NO:69). IL6R-L (SEQ ID NO:21) обычно использовали как L-цепь антитела. Варианты, продуцированные путем введения модификаций L234N, L234S, L234D, L234Q, L234E, L234T, L234R, L234H, L234G, L234K и L234Y в IL6R-BP264, и варианты, продуцированные путем введения модификаций L235W, L235M, L235P, L235F, L235A, L235V и L235I, соответственно, в IL6R-BP264, получали в виде гомологичных антител, содержащих одинаковые модификации в обеих цепях, а варианты, продуцированные путем введения комбинации модификаций L235N, L235S, L235D, L235Q, L235E, L235T, L235R, L235H, L235G, L235K и L235Y, соответственно, в IL6R-AP029, получали в виде гетеродимерных антител, а затем эти антитела анализировали.
Эти варианты использовали для экспрессии и очистки антител методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) оценивали методом, описанным в сравнительном примере 2. На фиг. 16 представлен график, в котором на горизонтальной оси отложены величины KD для связывания каждого варианта с FcγRIIb, а на вертикальной оси отложены величины KD для связывания каждого варианта с FcγRIIаR.
Как показано на фиг. 16, вариант IL6R-BP404/IL6R-L, продуцированный путем введения модификации L234Y в обе цепи IL6R-BP264/IL6R-L, обладал несколько более высокой активностью связывания с FcγRIIb по сравнению с активностью IL6R-BP264/IL6R-L до модификации.
Из этих вариантов, систематизированных в таблице 11, вариант IL6R-BP404/IL6R-L обладал повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb. В этой таблице «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. «KD(IIaR) родительского полипептида/KD(IIaR) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания IL6R-B3/IL6R-L с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIаR. «KD(IIaR)/KD(IIb)» означает величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания этого варианта с FcγRIIb. Чем выше эта величина, тем выше селективность связывания с FcγRIIb. В таблице 11 закрашенные серым ячейки относятся к величинам, вычисленным по уравнению:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2],
описанному в сравнительном примере 2, где было показано, что связывание FcγR с IgG является слишком слабым для его точной оценки с помощью кинетического анализа.
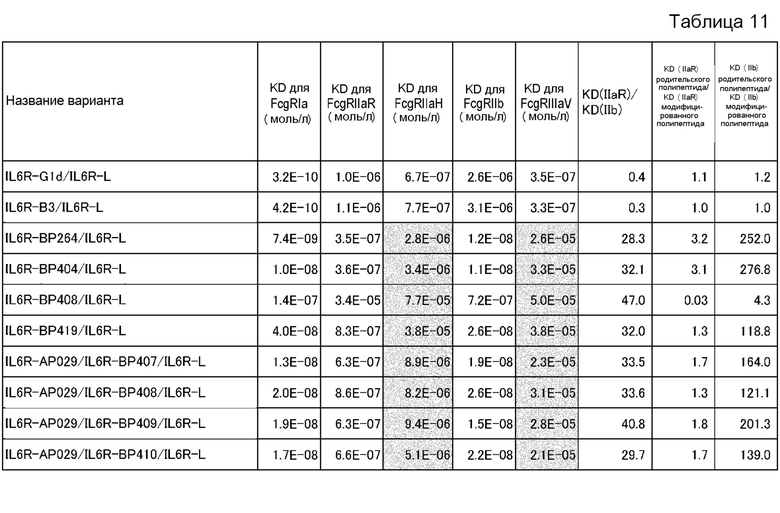
Как показано в таблице 11, вариант IL6R-BP404/IL6R-L, продуцированный путем введения модификации L234Y в обе цепи IL6R-BP264/IL6R-L, обнаруживал 1,1-кратное увеличение активности связывания с FcγRIIb по сравнению с активностью IL6R-BP264/IL6R-L до введения модификаций. Вариант IL6R-BP408/IL6R-L, продуцированный путем введения модификации L235Q в обе цепи IL6R-BP264/IL6R-L; вариант IL6R-BP419/IL6R-L, продуцированный путем введения модификации L235F в обе цепи IL6R-BP264/IL6R-L; вариант IL6R-AP029/IL6R-BP407/IL6R-L, продуцированный путем введения модификации L235D в одну из цепей IL6R-BP264/IL6R-L; вариант AP029/IL6R-BP408/IL6R-L, продуцированный путем введения модификации L235Q в одну из цепей IL6R-BP264/IL6R-L; вариант IL6R-AP029/IL6R-BP409/IL6R-L, продуцированный путем введения модификации L235Е в одну из цепей IL6R-BP264/IL6R-L; и вариант IL6R-AP029/IL6R-BP410/IL6R-L, продуцированный путем введения модификации L235Т в одну из цепей IL6R-BP264/IL6R-L, имели величины KD(IIaR)/KD(IIb), которые были выше, чем величины для IL6R-BP264/IL6R-L до введения модификации, и эти варианты обладали повышенной селективностью связывания с FcγRIIb.
[Пример 10] Оценка иммуногенности Fc-вариантов, обладающих повышенной активностью связывания с FcγRIIb, с использованием программы по предсказанию иммуногенности in silico
При использовании Fc-вариантов, описанных в этом примере, в качестве терапевтических антител предпочтительно, чтобы не происходило продуцирование антитела против лекарственных средств, которое ослабляет фармакологический эффект лекарственного средства. Поскольку антитела с высокой иммуногенностью имеют тенденцию к образованию антител против лекарственного средства, то иммуногенность терапевтических антител предпочтительно должна быть как можно более низкой. В целях предотвращения, насколько это возможно, повышения иммуногенности этих вариантов, могут быть использованы программы для предсказания иммуногенности in silico, например, программы для предсказания Т-клеточных эпитопов, такие как Epibase и EpiMatrix. Epibase Light (Lonza) представляет собой программу для предсказания иммуногенности in silico, позволяющую определить способность к связыванию 9-мерного пептида с MHC класса II, содержащего главные аллели DRB1, с использованием алгоритма FASTER (Expert Opin Biol Ther., 2007, Mar; 7 (3): 405-18). Эта программа позволяет идентифицировать Т-клеточные эпитопы с высоким уровнем связывания (сильные эпитопы) и средним уровнем связывания (слабые эпитопы) с МНС класса II.
Для вычисления использовали частоту встречаемости аллотипа DRB1 у группы индивидуумов, то есть частоту встречаемости этого аллотипа у группы индивидуумов индоевропейской расы, как показано ниже в таблице 12.
Эта программа была использована для сравнения общего числа Т-клеточных эпитопов с высокой и низкой связывающей способностью, которые были включены в последовательности (последовательность, простирающуюся от положения 118 до С-конца (в соответствии с Европейской нумерацией)) различных Fc-вариантов, описанных в литературе, и Fc-вариантов, обладающих повышенной селективностью связывания с FcγRIIb, как описано в примере. В частности, в качестве сравниваемого контроля, используемого для проведения анализа уже известными методами, были продуцированы следующие антитела: антитело Fc(DLE) (SEQ ID NО: 78), представляющее собой Fc-область антитела с введенными в нее модификациями S239D, A330L и I332E, которые, как уже сообщалось, повышают активность связывания с FcγRIIIа (Proc. Natl. Acad. Sci. USA. 2006, 103: 4005-10); антитело Fc(YTE) (SEQ ID NO:79), представляющее собой Fc-область антитела с введенными в нее модификациями M252Y, S254T и T256E, которые, как сообщалось, повышают активность связывания с FcRn (J. Biol. Chem. 2006, 281: 23514-24); антитело Fc(EF) (SEQ ID NO:80), представляющее собой Fc-область антитела с введенными в нее модификациями S267E и L328F, которые, как сообщалось, повышают активность связывания с FcγRIIb (Mol. Immunol. 2008, 45: 3926-33); и антитело Fc(P208) (SEQ ID NO:81), представляющее собой Fc-область антитела с введенными в нее модификациями E233D, G237D, P238D, H268D, P271G и A330R, которые, как сообщалось, повышают активность связывания с FcγRIIb и описаны в WO2012/115241. Кроме того, было получено антитело Fc(P587) (SEQ ID NO:70), которое представляет собой Fc-область антитела с введенными в нее модификациями Е233D, P238D, S264I, S267A, H268E и P271G, аналогичными модификациям варианта ВР568, способствующим повышению активности связывания с FcγRIIb и описанным в примерах, и антитело Fc(P588) (SEQ ID NO:71), которое представляет собой Fc-область антитела с введенными в нее модификациями P238D, S264I, S267A, H268E и P271G, аналогичными модификациям варианта BP492. Общее число эпитопов с сильной и слабой активностью связывания в этих Fc-вариантах сравнивали с использованием программы Epibase. Результаты представлены в таблице 13.
Эти результаты показали, что из уже существующих Fc-вариантов, варианты Fc(P587) и Fc(P588), которые представляют собой Fc-области вариантов, описанных в примерах, имеют небольшое число Т-клеточных эпитопов и низкий риск развития иммуногенности. Это свойство указывает на то, что, при использовании этих вариантов в качестве фармацевтических средств, вероятность продуцирования антител против лекарственных средств будет мала, и что эти варианты обладают превосходными функциями.
[Пример 11] Оценка кинетики Fc-вариантов с повышенной активностью связывания с человеческим FcγRIIb в кровотоке, проводимая с использованием трансгенных мышей, несущих человеческий FcγRIIb
(11-1) Описание метода оценки
Как указывается в WO 2013/047752, по сравнению с нативным человеческим IgG, концентрация растворимого антигена-мишени в плазме живого организма может быть значительно снижена путем введения антигенсвязывающей молекулы, которая обладает активностью связывания с человеческим FcRn при кислотных значениях рН и содержит антигенсвязывающий домен, где антигенсвязывающая активность такой антигенсвязывающей молекулы изменяется в зависимости от концентрации ионов, и которая также содержит FcγR-связывающий домен с более высокой FcγR-связывающей активностью, чем FcγR-связывающий домен Fc-области нативного человеческого IgG, где сахарная цепь, присоединенная в положении 297 (в соответствии с Европейской нумерацией), представляет собой сахарную цепь, содержащую фукозу. Также сообщалось, что при введении in vivo антигенсвязывающей молекулы, которая обладает повышенной активностью связывания с FcγRIIb по сравнению с другими FcγR, скорость элиминации растворимых антигенов из плазмы увеличивается, а концентрация растворимых антигенов в плазме может значительно снижаться. В этом примере Fc-вариант с повышенной активностью связывания с человеческим FcγRIIb вводили генетически модифицированным трансгенным мышам, несущим человеческий FcγRIIb, для того чтобы определить, может ли скорость элиминации растворимых антигенов-мишеней увеличиваться под действием Fc-варианта, который фактически обладает повышенной активностью связывания с описанным здесь человеческим FcγRIIb.
(11-2) Получение антител с повышенной активностью связывания с FcγRIIb
Нижеследующие антитела были использованы в качестве Fc-вариантов с повышенной активностью связывания с человеческим FcγRIIb.
Вариант IL6R-P587 был получен путем введения модификаций E233D, P238D, S264I, S267A, H268E и P271G, аналогичных модификациям, вводимым в BP568 варианта IL6R-G1d (SEQ ID NO:19), состоящего из константной области антитела G1d, которое имеет С-концевые Gly и Lys, удаленные из человеческого IgG1, и вариабельные области антитела против рецептора человеческого интерлейкина-6 (человеческого IL-6R), описанного в WO 2009/125825. Вариант Fv4-P587, содержащий IL-6R-P587 в качестве Н-цепи антитела, и вариант IL6R-L2 (SEQ ID NO:74), который представляет собой L-цепь антитела против человеческого IL-6R, описанного в WO 2009/125825 и используемого в качестве L-цепи антитела, получали методом, описанным в сравнительном примере 1. В качестве контроля для сравнения, вариант Fv4-IgG1, содержащий антитела IL6R-G1d (SEQ ID NO:19) и IL6R-L2 (SEQ ID NO:74), используемые в качестве Н-цепи и L-цепи антитела, соответственно, получали методом, аналогичным методу, описанному в сравнительном примере 1. Как описано в WO 2009/125825, варианты Fv4-G1d и Fv4-P587, полученные в настоящем изобретении, содержат антигенсвязывающий домен антигенсвязывающей молекулы, антигенсвязывающая активность которого изменяется в зависимости от концентрации протонов, то есть антигенсвязывающая активность этого антигенсвязывающего домена, связывающегося с человеческим IL-6R (антигеном) при кислотном рН, ниже, чем соответствующая активность при нейтральном рН.
(11-3) Получение трансгенных мышей, имеющих человеческий FcγRIIb
Трансгенные мыши, имеющие человеческий FcγRIIb, получали нижеследующим методом.
Трансгенные мыши были получены путем введения мышам C57BL/6(B6) человеческого гена FcγRIIb. Получение трансгенных мышей осуществляли в соответствии с процедурой, описанной в публикациях «Nagy et al. (Manipulating the mouse embryo, CSHL press. (2003) 399-506)» и «Ueda, et al., (Latest technology of gene targeting, Yodosha. (2000) 190-207)». Более конкретно, трансгенных мышей получали путем микроинъекции в пронуклеарные оплодотворенные яйца мышей B6 бактериальной искусственной хромосомы, в которую была клонирована геномная область человеческого гена FcγRIIb (GeneBank # NW_004077999:18307411-18381603). Мышей, которым был перенесен человеческий ген FcγRIIb, отбирали путем Саузерн-блот-анализа с использованием зонда, который специфически гибридизуется с человеческим геном FcγRIIb, и с помощью ПЦР. У трансгенных мышей с человеческим FcγRIIb брали кровь и печень, и экспрессию гена человеческого FcγRIIb подтверждали с помощью полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР) с использованием праймеров, которые специфически амплифицируют человеческий ген FcγRIIb. В результате была детектирована экспрессия человеческого гена FcγRIIb. Кроме того, мышиные мононуклеарные клетки периферической крови (МКПК) выделяли из крови трансгенных мышей с человеческим FcγRIIb, и экспрессию человеческого FcγRIIb в МКПК подтверждали с помощью анализов методом клеточного сортинга с активацией флуоресценции (FACS). В результате была детектирована экспрессия человеческого FcγRIIb. Вышеописанный анализ подтвердил получение трансгенных мышей с человеческим FcγRIIb, экспрессирующих человеческий FcγRIIb.
(11-4) In vivo тест на одновременное введение антигенов и антител трансгенным мышам с человеческим FcγRIIb
Трансгенным мышам с человеческим FcγRIIb, полученным как описано в (11-3), одновременно вводили растворимый человеческий IL-6R в качестве антигена и антитело против человеческого IL-6R, полученное как описано в (11-2), и после их введения оценивали концентрации растворимого человеческого IL-6R и антитела против человеческого IL-6R в плазме.
В хвостовую вену вводили смешанный раствор растворимого человеческого IL-6R и антитела против человеческого IL-6R (5 мкг/мл и 0,1 мг/мл, соответственно) в одной дозе, составляющей 10 мл/кг. Поскольку в этом случае антитело против человеческого IL-6R присутствует в значительном избытке по сравнению с растворимым человеческим IL-6R, то считается, что почти весь растворимый человеческий IL-6R связывался с антителом. Кровь брали через 5 минут, 1 час, 4 часа, 7 часов, 1 день, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Взятую кровь сразу центрифугировали при 4°C и при 15000 об/мин в течение 15 минут с получением плазмы. Выделенную плазму хранили в холодильнике при температуре -20°C или менее до начала проведения анализа. Вышеописанные Fv4-P587 и Fv4-IgG1 использовали в качестве антитела против человеческого IL-6R.
(11-5) Измерение концентрации антитела против человеческого IL-6R в плазме с помощью ELISA
Концентрацию антитела против человеческого IL-6R в плазме мышей измеряли с помощью ELISA. Сначала F(ab')2-фрагмент антитела против человеческого IgG (специфичного к γ-цепи) (Sigma) разделяли на аликвоты и помещали в планшет Nunc-ImmunoPlate, MaxiSorp (Nalge Nunc International), а затем оставляли на ночь при 4°C для приготовления планшета, на котором было иммобилизовано антитело против человеческого IgG. Затем строили калибровочные кривые для образцов плазмы при концентрациях 0,8, 0,4, 0,2, 0,1, 0,05, 0,025 и 0,0125 мкг/мл, и образцы мышиной плазмы для анализа разводили 100-кратно или более. Затем смеси, полученные путем добавления 200 мкл 20 нг/мл растворимого человеческого IL-6R к 100 мкл образцов для построения калибровочных кривых или образцов для анализа плазмы, перемешивали в течение 1 часа при комнатной температуре. Затем, планшет с иммобилизованным антителом против человеческого IgG, в который были добавлены смеси, снова перемешивали в течение одного часа при комнатной температуре. После этого биотинилированное антитело против человеческого IL-6R (R&D) подвергали реакции взаимодействия с образцами при комнатной температуре в течение одного часа, и стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) также подвергали реакции взаимодействия с образцами при комнатной температуре в течение одного часа. Хромогенную реакцию осуществляли с использованием микролуночного субстрата, конъюгированного с ПХ и содержащего один компонент TMB (BioFX Laboratories). После этого реакцию завершали путем добавления 1н серной кислоты (Showa Chemical), и измеряли оптическую плотность на 450 нм на микропланшете-ридере. Концентрации антитела в мышиной плазме вычисляли исходя из величин оптической плотности по калибровочной кривой с использованием программы для анализа SOFTmax PRO (Molecular Devices). После внутривенного введения антитела трансгенным мышам с человеческим FcγRIIb измеряли концентрацию антитела в плазме мышей в зависимости от времени в соответствии с методом, описанным на фиг. 33.
(11-5) Измерение концентрации человеческого IL-6R в плазме с помощью электрохемилюминесценции
Концентрацию человеческого IL-6R в мышиной плазме измеряли методом электрохемилюминесценции. Затем строили калибровочные кривые для образцов человеческого IL-6R в плазме при концентрациях 12,5, 6,25, 3,13, 1,56, 0,781, 0,391 и 0,195 нг/мл, и приготавливали образцы мышиной плазмы для анализа путем их 50-кратного разведения или более. Моноклональное антитело против человеческого IL-6R (R&D) было помечено рутением с использованием сложного эфира SULFO-TAG NHS (Meso Scale Discovery), а затем биотинилированное антитело против человеческого IL-6R (R&D) и раствор тоцилизумаба перемешивали и оставляли на ночь при 37°C для прохождения реакции. Затем смешанный раствор разделяли на аликвоты и помещали в покрытый стрептавидином планшет Streptavidin Gold Multi-ARRAY (Meso Scale Discovery), после чего блокировали с использованием раствора PBS-Tween, содержащего 0,5% BSA (масс/об), в течение ночи при 5°C. После реакции в течение более двух часов при комнатной температуре, планшет промывали. Сразу после промывки, в планшет добавляли аликвоты буфера Read Buffer T(х2)(Meso Scale Discovery), и проводили измерения на устройстве SECTOR Imager 2400 (Meso Scale Discovery). Концентрацию hSIL-6R вычисляли по калибровочной кривой с использованием аналитической компьютерной программы SOFTmax PRO (Molecular Devices). Концентрацию растворимого человеческого IL-6R в плазме в зависимости от времени у трансгенных мышей с человеческим FcγRIIb после внутривенного введения антитела измеряли методом, описанным на фиг. 34.
(11-6) Эффекты повышения уровня связывания с человеческим FcγRIIb
Проводили сравнение результатов in vivo теста, полученных для Fv4-P587, который обладал повышенной активностью связывания с человеческим FcγRIIb, и для Fv4-IgG1. Как показано на фиг. 33, время нахождения обоих антител в плазме было почти одинаковым, однако, как показано на фиг. 34, было подтверждено, что элиминация человеческого IL-6R была выше в случае, когда человеческий IL-6R вводили одновременно с Fv4-P587, обладающим повышенной активностью связывания с человеческим FcγRIIb, чем элиминация при одновременном введении человеческого IL-6R с Fv4-IgG1. Более конкретно, было обнаружено, что антитело, которое связывается с человеческим IL-6R в зависимости от рН, способно снижать концентрацию растворимого человеческого IL-6R благодаря повышению способности связываться с человеческим FcγRIIb.
Не ограничиваясь какой-либо конкретной теорией, авторы лишь отмечают, что, как показали результаты, представленные на фиг. 35, растворимые антигены плазмы, которые связываются с этим антителом, элиминируются в результате их включения в FcγRIIb-экспрессирующие клетки посредством человеческого FcγRIIb.
Растворимый человеческий IL-6R, связанный с антителом, которое связывается с растворимым человеческим IL-6R, подвергается рециклингу в плазме посредством FcRn вместе с антителом. В противоположность этому, Fv4-IgG1, которое представляет собой антитело, связывающееся с растворимым человеческим IL-6R в зависимости от рН, подвергается реакции диссоциации с растворимым человеческим IL-6R, связанным с антителом, в эндосоме в кислотных условиях. Поскольку диссоциированный растворимый человеческий IL-6R разлагается под действием лизосомы, то элиминация растворимого человеческого IL-6R может значительно повышаться, а Fv4-IgG1, которое представляет собой антитело, связывающееся с растворимым человеческим IL-6R в зависимости от рН, связывается с FcRn в эндосоме, а затем подвергается рециклингу в плазме. Это подвергнутое рециклингу антитело может снова связываться с растворимым человеческим IL-6R, а поэтому связывание с антигеном (с растворимым человеческим IL-6R) и рециклинг в плазме под действием FcRn могут повторяться. В результате можно сказать, что одна молекула антитела может связываться с растворимым человеческим IL-6R несколько раз. Кроме того, считается, что благодаря повышению активности связывания Fv4-IgG1 с FcγRIIb, то есть связывания с антигеном в зависимости от рН, комплекс, образованный антителом, которое связывается с растворимым человеческим IL-6R, и с растворимым человеческим IL-6R, быстро включается в клетки посредством FcγRIIb, что приводит к более быстрому снижению концентрации растворимого человеческого IL-6R (фиг. 35).
[Пример 12] Оценка кинетики Fc-вариантов с повышенной активностью связывания с человеческим FcγRIIb в кровотоке, проводимая с использованием трансгенных мышей, несущих человеческий FcγRIIb и человеческий FcRn
(12-1) Описание метода оценки
Как указывается в WO 2013/047752, использование антигенсвязывающей молекулы, обладающей более высокой FcγR-связывающей активностью по сравнению с Fc-областью нативного IgG и более высокой активностью связывания с человеческим FcRn при кислотных значениях рН, подтвердило, что такая молекула имеет повышенную способность сохраняться в плазме по сравнению с антигенсвязывающими молекулами, которые не обладают повышенной активностью связывания с человеческим FcRn при кислотных значениях рН. С другой стороны, сообщалось, что антигенсвязывающая молекула, обладающая более высокой FcγR-связывающей активностью по сравнению с Fc-областью нативного человеческого IgG и более высокой активностью связывания с человеческим FcRn при кислотных значениях рН, способствовала снижению концентрации антигена-мишени в плазме по сравнению с антигенсвязывающей молекулой, которая обладает более высокой активностью связывания с FcγR, чем Fc-область нативного человеческого IgG и которая не обладает повышенной активностью связывания с человеческим FcRn при кислотных значениях рН. В соответствии с этим, было проведено исследование для того, чтобы определить, имеют ли антигенсвязывающие молекулы, несущие варианты Fc-области с повышенной активностью связывания с человеческим FcγRIIb, как описано в примерах, одинаковые свойства.
(12-2) Получение антигенсвязывающей молекулы, обладающей более высокой FcγR-связывающей активностью по сравнению с Fc-областью нативного человеческого IgG и более высокой активностью связывания с человеческим FcRn при кислотных значениях рН
Помимо Fv4-IgG1 и Fv4-P587, описанных в примере 11-2, был получен вариант Fv4-P587-LS методом, описанным в сравнительном примере 1, где Fv4-P587-LS содержит IL6R-L2 в качестве L-цепи антитела и IL6R-P587-LS (SEQ ID NО:73) в качестве Н-цепи антитела. IL6R-P587-LS получали путем введения в IL6R-P587, то есть в Н-цепь Fv4-P587, модификаций, состоящих из замены Met в положении 428 на Leu и замены Asn в положении 434 на Ser в соответствии с Европейской нумерацией, где указанные замены, как уже сообщалось, способствуют повышению кинетики антител в кровотоке (Nat. Biotechnol. 2010, 28: 157-159).
(12-3) Анализ взаимодействия с человеческим FcRn
Анализ взаимодействия полученного антитела и человеческого FcRn осуществляли с использованием Biacore T200. Соответствующее количество белка L (BioVision) иммобилизовали на сенсорном чипе CM4 (GE Healthcare) методом связывания с амином, а затем на этом чипе иммобилизовали представляющие интерес антитела. После этого впрыскивали разведенный FcRn и рабочий буфер (используемый в качестве контрольного раствора) в целях осуществления взаимодействия антител, иммобилизованных на этом сенсорном чипе, с человеческим FcRn. В качестве рабочего буфера использовали 50 ммоль/л фосфата натрия, 150 ммоль/л NaCl и 0,05% (масс/об) твина 20 (рН 6,0), и этот рабочий буфер также использовали для разведения FcRn. Для регенерации чипа использовали 10 ммоль/л глицина-HCl (pH 1,5). Все измерения проводили при 25°C. Кинетические параметры, такие как константы скорости ассоциации ka (1/Mс) и константы скорости диссоциации kd (1/с), определяли по сенсограмме, построенной по данным этих измерений, а KD (M) для связывания каждого антитела с человеческим FcRn вычисляли по величинам этих констант. Вычисление каждого параметра проводили с помощью компьютерной программы Biacore T200 Evaluation Software (GE Healthcare). Величины KD для связывания полученных антител с человеческим FcRn вычисляли методом, описанным в таблице 14. Как показано в таблице 14, было подтверждено, что Fv4-P587-LS, по сравнению с Fv4-P587, обладал повышенной активностью связывания с FcRn в кислотных условиях.
(12-4) Получение трансгенных мышей с человеческим FcγRIIb и человеческим FcRn
Трансгенных мышей с человеческим FcγRIIb и человеческим FcRn и мышей, дефицитных по мышиному FcRn (нокаут-мышей), получали нижеследующим методом.
Сначала получали мышей, дефицитных по мышиному FcRn. Получение нокаут-мышей осуществляли методом, описанным в публикации «Nagy et al. (Manipulating the mouse embryo, CSHL press. (2003) 399-506)». Более конкретно, был получен нацеливающий вектор для разрушения мышиного гена FcRn, а затем этот вектор вводили в клетки ES (взятые у мышей C57BL/6) для разрушения мышиного гена FcRn посредством гомологичной рекомбинации. РНК экстрагировали из печени выведенных мышей с дефицитом мышиного FcRn, и на основе кДНК, синтезированной из этой РНК и используемой в качестве матрицы, осуществляли ОТ-ПЦР с помощью праймеров, которые специфически амплифицируют мышиный FcRn. В результате, у мышей с дефицитом мышиного FcRn, этот мышиный ген FcRn не детектировался. Затем получали трансгенных мышей путем введения человеческих генов FcγRIIb и FcRn мышам, дефицитным по мышиному FcRn. Трансгенных мышей получали в соответствии с процедурами, описанными в публикациях «Nagy et al. (Manipulating the mouse embryo, CSHL press. (2003) 399-506)» и «Ueda, et al., (Latest technology of gene targeting, Yodosha. (2000) 190-207)». Более конкретно, мышей получали путем микроинъекции в пронуклеарные оплодотворенные яйца мышей, дефицитных по мышиному FcRn, бактериальной искусственной хромосомы, в которую были клонированы геномные области человеческого гена FcRn (GeneBank # NC_000019.9:50000108-50039865) и человеческого гена FcγRIIb (GeneBank # NW_004077999:18307411-18381603). Мышей, которым были перенесены человеческий ген FcRn и человеческий ген FcγRIIb и которые были гомозиготными по аллелю, дефицитному по мышиному FcRn, отбирали с помощью Саузерн-блот-анализа с использованием зонда, который специфически гибридизуется с каждым из этих генов, и с помощью ПЦР. У трансгенных мышей с человеческим FcγRIIb и человеческим FcRn и мышей, дефицитных по мышиному FcRn, брали кровь, и экспрессию человеческого гена FcRn и человеческого гена FcγRIIb подтверждали с помощью ОТ-ПЦР с использованием праймеров, которые специфически амплифицируют человеческий ген FcRn и человеческий ген FcγRIIb. В результате была детектирована экспрессия человеческого гена FcRn и человеческого гена FcγRIIb. Полученные результаты подтвердили получение трансгенных мышей с человеческим FcγRIIb и человеческим FcRn и мышей с дефицитом мышиного FcRn, у которых экспрессировались человеческий FcRn и человеческий FcγRIIb, но не экспрессировался мышиный FcRn.
(12-5) Улучшение фармакокинетики путем повышения активности связывания с человеческим FcRn при кислотных значениях рН
Анализ in vivo осуществляли методом, аналогичным методу, описанному в примере 11, путем введения Fv4-IgG1, Fv4-P587 и Fv4-P587-LS трансгенным мышам с человеческим FcγRIIb и человеческим FcRn, и у мышей этих групп измеряли концентрацию растворимого IL-6R и антитела против человеческого IL-6R в плазме. Результаты измерения концентраций антитела против человеческого IL-6R и растворимого IL-6R в плазме представлены на фиг. 36 и фиг. 37, соответственно.
Было обнаружено, что у мышей, которым вводили Fv4-P587-LS и у которых наблюдалась повышенная активность связывания Fv4-P587 с человеческим FcRn при кислотных значениях рН, время пребывания антител в плазме превышало время пребывания антител в плазме у группы мышей, которым вводили Fv4-P587. Кроме того, Fv4-P587-LS имело более длительное время пребывания в плазме, чем Fv4-IgG1. С другой стороны, концентрация растворимого IL-6R в плазме у группы мышей, которым вводили Fv4-P587-LS, была эквивалентна концентрации растворимого IL-6R у группы мышей, которым вводили Fv4-P587. У группы мышей, которым вводили Fv4-P587-LS или Fv4-P587, концентрация растворимого IL-6R в плазме была ниже, чем у группы мышей, которым вводили Fv4-IgG1.
В соответствии с этим, введение антитела, то есть антигенсвязывающей молекулы с повышенной активностью связывания с человеческим FcRn при кислотных значениях рН, где активность связывания антигенсвязывающей молекулы с человеческим FcγRIIb была выше, чем активность связывания Fc-области нативного человеческого IgG, показало, что время удерживания вводимой молекулы антитела в плазме в живом организме может увеличиваться после такого введения. Кроме того, было обнаружено, что, даже при увеличении времени пребывания антигенсвязывающей молекулы в плазме живого организма, антиген-элиминирующий эффект у этого живого организма не снижался, а сохранялся на обычном уровне.
Модификации, используемые для повышения активности связывания с человеческим FcRn при кислотных значениях рН, не имеют конкретных ограничений, и такие модификации могут быть введены в IgG путем замены Met на Leu в положении 428 и замены Asn на Ser в положении 434 (в соответствии с Европейской нумерацией) (Nat. Biotechnol. (2010) 28: 157-159); замены Asn на Ala в положении 434 (Drug. Metab. Dispos. (2010) Apr.; 38(4): 600-605); замены Met на Thr в положении 252, замены Ser на Thr в положении 254 и замены Thr на Glu в положении 256 (J. Biol. Chem. (2006) 281: 23514-23524); замены Thr на Gln в положении 250 и замены Met на Leu в положении 428 (J. Immunol. (2006), 176(1): 346-356); замены Asn на His в положении 434 (Clin. Pharm. & Ther. (2011) 89(2): 283-290) или замен, описанных в WO 2010/106180, WO 2010/045193, WO 2009/058492, WO 2008/022152, WO 2006/050166, WO 2006/053301, WO 2006/031370, WO 2005/123780, WO 2005/047327, WO 2005/037867, WO 2004/035752, WO 2002/060919 и т.п.
[Сравнительный пример 1] Конструирование антитело-экспрессирующих векторов; и экспрессия и очистка антител
Синтез полноразмерных генов, содержащих нуклеотидные последовательности Н-цепи и L-цепи вариабельных областей антитела, осуществляли методами продуцирования, известными специалистам, посредством ПЦР-сборки и т.п. Введение аминокислотных замен осуществляли методами, известными специалистам, с помощью ПЦР или т.п. Полученный плазмидный фрагмент встраивали в экспрессионный вектор клеток животных и получали экспрессионный вектор для Н-цепи и экспрессионный вектор для L-цепи. Нуклеотидную последовательность полученного экспрессионного вектора определяли методами, известными специалистам. Полученные плазмиды транзиентно вводили в клеточную линию HEK293H, происходящую от раковых клеток почек человеческого эмбриона (Invitrogen), или в клетки FreeStyle293 (Invitrogen) для экспрессии антитела. Полученный супернатант культуры собирали, а затем пропускали через 0,22-мкм фильтр MILLEX(R)-GV (Millipore) или через 0,45-мкм фильтр MILLEX (R)-GV (Millipore). Антитела очищали из полученного супернатанта культуры методами, известными специалистам, с использованием рекомбинантного белка А-сефарозы Fast Flow (GE Healthcare) или G-белка-сефарозы 4 Fast Flow (GE Healthcare). Концентрацию очищенных антител определяли по оптической плотности на 280 нм, измеренной на спектрофотометре. По полученному значению вычисляли коэффициент возбуждения методами, такими как PACE, а затем этот коэффициент использовали для оценки концентрации антитела (Protein Science 1995; 4: 2411-2423).
[Сравнительный пример 2] Метод получения FcγR и метод анализа взаимодействия между модифицированным антителом и FcγR
Внеклеточные домены FcγR получали следующим методом. Сначала, ген внеклеточного домена FcγR синтезировали методом, хорошо известным специалистам. Затем получали последовательность каждого FcγR на основе информации, имеющейся в NCBI. В частности, FcγRI продуцировали на основе последовательности NCBI рег. № NM_000566.3, FcγRIIа продуцировали на основе последовательности NCBI рег. № NM_001136219.1, FcγRIIb продуцировали на основе последовательности NCBI рег. № NM_004001.3, FcγRIIIа продуцировали на основе последовательности NCBI рег. № NM_001127593.1, а FcγRIIIb продуцировали на основе последовательности NCBI рег. № NM_000570.3, и к С-концу присоединяли His-метку. Кроме того, известно, что существует полиморфизм между FcγRIIа, FcγRIIIа и FcγRIIIb, и эти полиморфные сайты были получены, как описано в публикациях J. Exp. Med, 1990, 172: 19-25, для FcγRIIа; J. Clin Invest 1997, 100(5): 1059-1070, для FcγRIIIа; и J. Clin Invest, 1989, 84, 1688-1691, для FcγRIIIb.
Полученные генные фрагменты встраивали в экспрессионный вектор для клеток животных и получали нужные экспрессионные векторы. Полученные экспрессионные векторы транзиентно вводили в клетки FreeStyle293, происходящие от раковых клеток почек человеческого эмбриона (Invitrogen), для экспрессии представляющих интерес белков. В случае FcγRIIb, используемого для кристаллографического анализа, представляющий интерес белок экспрессировали в присутствии кифуненсина в конечной концентрации 10 мкг/мл, так, чтобы сахарная цепь, присоединенная к FcγRIIb, имела высокое содержание маннозы. Клетки культивировали, и после сбора полученного супернатанта культуры его пропускали через 0,22-мкм фильтр. В принципе, полученные супернатанты культуры были очищены за нижеследующие четыре стадии. Эти стадии проводили следующим образом, а именно, в первой стадии осуществляли катионообменную колоночную хроматографию (SP Sepharose FF), во второй стадии проводили аффинную колоночную хроматографию (HisTrap HP) для присоединения His-метки, в третьей стадии проводили колоночную гель-фильтрацию (Superdex200), а в четвертой стадии проводили хроматографию в асептических условиях. Однако, для FcγRI, анионообменную колоночную хроматографию, проводимую с использованием сефарозы (Q sepharose FF), осуществляли, как описано в стадии 1. Очищенные белки оценивали путем измерения оптической плотности на 280 нм на спектрофотометре, и по полученным величинам вычисляли концентрации очищенных белков с использованием коэффициента поглощения, определенного таким методом, как PACE (Protein Science 1995; 4: 2411-2423).
Анализ взаимодействия между модифицированным антителом и Fcγ-рецептором, полученными как описано выше, осуществляли с использованием Biacore T100 (GE Healthcare), Biacore T200 (GE Healthcare), Biacore A100 и Biacore 4000. В качестве рабочего буфера использовали HBS-EP+ (GE Healthcare), а контрольную температуру устанавливали на 25°C. Были использованы чипы, полученные путем иммобилизации антигенного пептида, белка А (Thermo Scientific), белка А/G (Thermo Scientific) и белка L (ACTIGEN или BioVision), методом присоединения амина к сенсорным чипам серии S CM5 (GE Healthcare) или серии S CM4 (GE Healthcare), или, альтернативно, были использованы чипы, полученные путем предварительного взаимодействия биотинилированных антигенных пептидов с сенсорным чипом серии S SA (сертифицированным) (GE Healthcare) и их иммобилизации на этом чипе.
После захвата представляющих интерес антител на этих сенсорных чипах, Fcγ-рецептор, разведенный рабочим буфером, оставляли для взаимодействия с антителом, после чего определяли количество рецептора, связанного с антителами, и антитела сравнивали. Однако, поскольку количество связанного Fcγ-рецептора зависит от количества захваченных антител, то количество связанного Fcγ-рецептора делили на количество каждого захваченного антитела и получали скорректированные величины, которые затем сравнивали. Кроме того, антитела, иммобилизованные на чипах, смывали путем реакции с 10 мМ глицина-HCl, pH 1,5, и эти чипы регенерировали и использовали повторно.
Кинетические анализы для вычисления величин KD для связывания каждого модифицированного антитела с FcγR осуществляли описанным ниже методом. Сначала представляющие интерес антитела иммобилизовали на вышеупомянутых сенсорных чипах, а затем Fcγ-рецептор, разведенный рабочим буфером, оставляли для взаимодействия. Для построения общей кривой зависимости использовали программу Biacore Evaluation Software, и по полученным результатам строили сенсограмму с использованием лангмюровской модели связывания 1:1, а затем вычисляли константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (1/с), и по этим величинам вычисляли константы диссоциации KD (моль/л).
Если взаимодействие каждого из модифицированных антител и FcγR было слабым, и если было установлено, что коррекция не может быть проведена с помощью вышеупомянутого кинетического анализа, то KD для таких взаимодействий вычисляли по нижеследующему уравнению модели связывания 1:1, как описано в руководстве Biacore T100 Software Handbook BR1006-48 Edition AE.
Характер взаимодействия молекул в соответствии с моделью связывания 1:1 на Biacore может быть представлен нижеследующим уравнением 1.
[Уравнение 1]
Req=C ⋅ Rmax/(KD + C) + RI, где
Req: график зависимости устойчивых уровней связывания от концентрации аналита
С: концентрация
RI: вклад объемного показателя преломления в образце
Rmax: способность аналита связываться с поверхностью чипа
При преобразовании этого уравнения, KD может быть вычислена по уравнению 2, представленному ниже.
[Уравнение 2]
KD=C⋅Rmax/(Req-RI)-C
KD может быть вычислена путем замены величин Rmax, RI и C в этом уравнении. Величины RI и C могут быть определены из сенсограммы, полученной по результатам и условиям измерений. Rmax вычисляли следующим методом. В качестве объекта для сравнения были выбраны антитела, которые обнаруживали достаточно сильное взаимодействие, как было оценено одновременно в одном и том же раунде измерений, и величину Rmax определяли путем построения общей кривой с использованием лангмюровской модели связывания 1:1, а затем полученную величину делили на количество сравниваемого антитела, иммобилизованного на сенсорном чипе, и результат умножали на оцениваемое количество иммобилизованного модифицированного антитела.
[Сравнительный пример 3] Детальный анализ связывания Fc-вариантов с FcγR
В антитела IgG1 были введены мутации для получения антител, которые обладают пониженной Fc-опосредуемой активностью связывания с активирующим FcγR, а в частности, с обоими аллотипами FcγRIIа Н-типа и R-типа, и повышенной активностью связывания с FcγRIIb по сравнению с антителом IgG1, а затем был проведен детальный анализ связывания с каждым FcγR.
Вариабельная область (SEQ ID NO:15) антитела против глипикана 3, содержащая CDR антитела GpH7, которое представляет собой антитело против глипикана 3, обладающее повышенной кинетикой в плазме, как описано в WO2009/041062, была использована в качестве Н-цепи природного антитела. Аналогичным образом, в качестве L-цепи природного антитела использовали GpL16-k0 (SEQ ID NO:16) антитела против глипикана 3, обладающего повышенной кинетикой в плазме, как описано в WO 2009/041062. Кроме того, антитело В3, содержащее область G1d, в которую была введена мутация K439E и которая была продуцирована путем удаления С-концевого Gly и Lys IgG1, использовали в качестве константной области Н-цепи антитела. Эта Н-цепь обозначается GpH7-B3 (SEQ ID NО:17), а L-цепь обозначается GpL16-k0 (SEQ ID NO:16).
В GpH7-B3 аминокислоты, которые, как считается, участвуют в связывании с FcγR, и окружающие их аминокислоты (в положениях 234-239, 265-271, 295, 296, 298, 300 и 324-337 в соответствии с Европейской нумерацией) заменяли, соответственно, аминокислотами 18 типов, за исключением исходных аминокислот и Cys. Эти варианты называются вариантами В3. Варианты В3 были экспрессированы и очищены методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа) детально анализировали методом, описанным в сравнительном примере 2.
Результаты анализа на взаимодействие с каждым FcγR, проводимого методом, описанным ниже, были представлены на
графиках. Величину количества FcγR, связывающегося с каждым антителом, происходящим от варианта В3, делили на величину количества FcγR, связывающегося с антителом, которое было использовано для сравнения и которое не имело мутаций, введенных в В3 (антитело, имеющее последовательность природного человеческого IgG1 в положениях 234-239, 265-271, 295, 296, 298, 300 и 324-337 в соответствии с Европейской нумерацией). Величину, полученную путем умножения данного результата на 100, использовали в качестве показателя относительной активности связывания FcγR с каждым вариантом. На горизонтальной оси отложены величины относительной активности связывания FcγRIIb с каждым вариантом, а на вертикальной оси отложены величины относительной активности связывания каждого варианта с активирующими FcγR: FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа и FcγRIIIа (фиг. 17, 18, 19 и 20).
Как отмечено на фиг. 17-20, результаты показали, что все модификации, а именно, только введенные мутации, называемые мутацией А (модификацией, полученной путем замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp) и мутацией В (модификацией, полученной путем замены Leu в положении 328 (в соответствии с Европейской нумерацией) на Glu), приводили к заметному повышению активности связывания с FcγRIIb и к заметному ингибированию связывания с FcγRIIа обоих типов по сравнению с активностью связывания до введения модификаций.
[Сравнительный пример 4] ППР-анализ вариантов, селективно связывающихся в FcγRIIb
После введения модификации, описанной в примере 1, а именно, замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, проводили детальный анализ активности связывания с каждым FcγR.
Вариабельную область IL6R-H (SEQ ID NO:18), которая представляет собой вариабельную область антитела против рецептора человеческого интерлейкина-6, описанного в WO 2009/125825, использовали в качестве вариабельной области Н-цепи антитела, а IL6R-G1d (SEQ ID NO:19), которая содержит G1d с делецией С-концевого Gly и Lys человеческого IgG1, использовали в качестве константной области Н-цепи антитела в Н-цепи IgG1. В IL6R-G1d Pro в положении 238 (в соответствии с Европейской нумерацией) заменяли на Asp с получением IL6R-G1d-v1 (SEQ ID NO:20). Затем аминокислоту Leu в положении 328 (в соответствии с Европейской нумерацией) в IL6R-G1d заменяли аминокислотой Glu с получением IL6R-G1d-v2. Кроме того, для сравнения, аминокислоту Ser в положении 267 (в соответствии с Европейской нумерацией) заменяли на Glu, а аминокислоту Leu в положении 328 (в соответствии с Европейской нумерацией) заменяли на Phe в IL6R-G1d с получением IL6R-G1d-v3, как описано в непатентном документе 28. Антитело IL6R-L (SEQ ID NO:21), которое представляет собой L-цепь тоцилизумаба, использовали как L-цепь общего антитела, и эти антитела, вместе с каждой Н-цепью, экспрессировали и очищали методом, описанным в сравнительном примере 1. Полученные антитела, которые содержат в качестве Н-цепи аминокислотную последовательность, происходящую от IL6R-G1d, IL6R-G1d-v1, IL6R-G1d-v2 или IL6R-G1d-v3, обозначаются IgG1, IgG1-v1, IgG1-v2 и IgG1-v3, соответственно.
Затем проводили кинетический анализ взаимодействия между этими антителами и FcγR с использованием Biacore T100 (GE Healthcare). В качестве рабочего буфера использовали HBS-EP+ (GE Healthcare), а контрольную температуру устанавливали на 25°C. Был использован чип, полученный путем иммобилизации белка А на сенсорном чипе серии S CM5 (GE Healthcare) методом присоединения амина. Представляющее интерес антитело иммобилизовали на этом чипе для взаимодействия с каждым FcγR, который был разведен рабочим буфером, и оценивали активность связывания с антителом. После оценки активности связывания, антитело, иммобилизованное на чипе, смывали путем реакции с 10 мМ глицин-HCl, pH 1,5, и этот чип регенерировали и использовали повторно. Сенсограммы, построенные по результатам измерений, анализировали с использованием лангмюровской модели связывания 1:1 с помощью компьютерной программы Biacore Evaluation Software, и вычисляли константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (1/с), а затем по этим величинам вычисляли константу диссоциации KD (моль/л).
Если связывание IgG1-v1 и IgG1-v2 с FcγRIIа Н-типа и FcγRIIIа было слабым, то кинетические параметры, такие как KD, не могли быть вычислены с применением вышеупомянутого аналитического метода. Величины KD для таких взаимодействий вычисляли с использованием нижеследующей модели связывания 1:1, как описано в руководстве Biacore T100 Software Handbook BR1006-48 Edition AE.
Характер взаимодействия молекул в соответствии с моделью связывания 1:1 на Biacore может быть представлен нижеследующим уравнением 1.
[Уравнение 1]
Req=C⋅Rmax/(KD+C)+RI, где
Req: график зависимости устойчивых уровней связывания от концентрации аналита
С: концентрация
RI: вклад объемного показателя преломления в образце
Rmax: способность аналита связываться с поверхностью чипа
При преобразовании этого уравнения, KD может быть вычислена по уравнению 2, представленному ниже.
[Уравнение 2]
KD=C⋅Rmax/(Req-RI)-C
KD может быть вычислена путем замены величин Rmax, RI и C в этом уравнении. Исходя из условий данного измерения могут быть использованы величины RI=0, C=2 мкмоль/л. Кроме того, величину Rmax, полученную путем построения общей кривой сенсограммы по результатам анализа взаимодействия каждого FcγR с IgG1 с использованием лангмюровской модели связывания 1:1, делили на количество иммобилизованного IgG1, и результат умножали на количество иммобилизованного IgG1-v1 и IgG1-v2, а затем полученную величину использовали как Rmax. Это вычисление проводили исходя из гипотезы, что ограниченное количество каждого FcγR, который может быть связан с IgG1, остается неизменным для всех вариантов, полученных путем введения мутаций в IgG1, а величина Rmax пропорциональна количеству антитела, связанного с чипом на время проведения измерений. Величина Req определяется как количество каждого FcγR, связывающегося с каждым вариантом на сенсорном чипе на время проведения измерений.
В этих условиях измерения уровень связывания (Req) IgG1-v1 и IgG1-v2 с FcγRIIа Н-типа составляет приблизительно 2,5 RU и 10 RU (RU=относительная единица), соответственно, а уровень связывания (Req) IgG1-v1 и IgG1-v2 с FcγRIIIа составляет приблизительно 2,5 RU и 5 RU, соответственно. Уровень связывания иммобилизованных IgG1, IgG1-v1 и IgG1-v2 в анализе на взаимодействие с FcγRIIа Н-типа составляет 452 RU, 469,2 RU и 444,2 RU, соответственно, а уровень связывания иммобилизованных IgG1, IgG1-v1 и IgG1-v, в анализе взаимодействия с FcγRIIIа составляет 454,5 RU, 470,8 RU и 447,1 RU, соответственно. Величины Rmax, полученные путем построения общей кривой сенсограммы по результатам анализа на взаимодействие IgG1 с FcγRIIа Н-типа и FcγRIIIа с использованием лангмюровской модели связывания 1:1, составляли 69,8 RU и 63,8 RU, соответственно. Вычисленные величины Rmax для связывания IgG1-v1 и IgG1-v2 с FcγRIIа Н-типа составляли 72,5 RU и 68,6 RU, соответственно, а вычисленные величины Rmax для связывания IgG1-v1 и IgG1-v2 с FcγRIIIа составляли 66,0 RU и 62,7 RU, соответственно. Эти величины были заменены в уравнении 2 для вычисления KD для связывания IgG1-v1 и IgG1-v2 с FcγRIIа Н-типа и FcγRIIIа.
[Уравнение 2]
http://patentimages.storage.googleapis.com/WO2014030728A1/JPOXMLDOC01-appb-I000003.png
KD=C⋅Rmax/(Req-RI)-C
Величины KD для связывания IgG1, IgG1-v1, IgG1-v2 и IgG1-v3 с каждым FcγR (величины KD для связывания каждого антитела с каждым FcγR) представлены в таблице 15, а относительные величины KD для IgG1-v1, IgG1-v2 и IgG1-v3, полученные путем деления величин KD для связывания IgG1 с каждым FcγR на величины KD для связывания IgG1-v1, IgG1-v2 и IgG1-v3 с каждым FcγR (относительные величины KD для связывания каждого антитела с каждым FcγR), представлены в таблице 16.
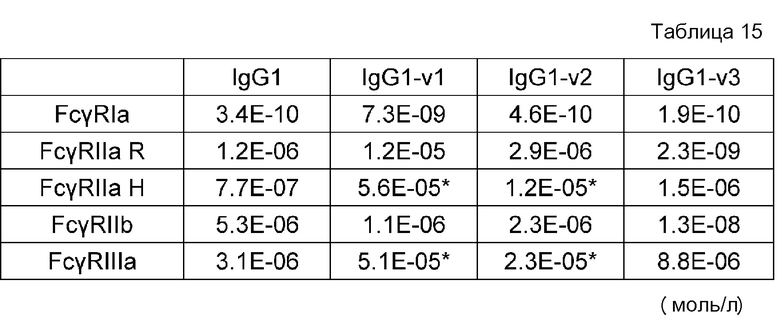
В этой таблице 15 цифры, отмеченные «*», означают, что величина KD была вычислена по уравнению 2, поскольку какого-либо значимого уровня связывания FcγR с IgG не наблюдалось.
[Уравнение 2]
http://patentimages.storage.googleapis.com/WO2014030728A1/JPOXMLDOC01-appb-I000003.png
KD=C⋅Rmax/(Req-RI)-C
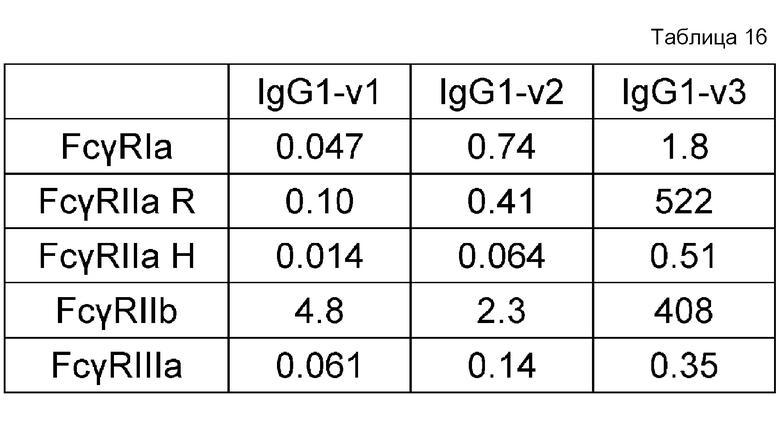
(Величина, полученная путем деления величины KD для связывания IgG1 с каждым FcγR на величину KD для связывания каждого антитела IgG1 с каждым FcγR).
Как показано в таблице 16, по сравнению с IgG1, активность связывания IgG1-v1 снижалась в 0,047 раза для FcγRIа; снижалась в 0,10 раза для FcγRIIа R-типа; снижалась в 0,014 раза для FcγRIIа Н-типа; снижалась в 0,061 раза для FcγRIIIа и увеличивалась в 4,8 раза для FcγRIIb.
Кроме того, как показано в таблице 16, по сравнению с IgG1, активность связывания IgG1-v2 снижалась в 0,74 раза для FcγRIа; снижалась в 0,41 раза для FcγRIIа R-типа; снижалась в 0,064 раза для FcγRIIа Н-типа; снижалась в 0,14 раза для FcγRIIIа и увеличивалась в 2,3 раза для FcγRIIb.
Более конкретно, полученные результаты продемонстрировали, что IgG1-v1, в котором остаток Pro в положении 238 (в соответствии с Европейской нумерацией) заменен Asp, и что IgG1-v2, в котором остаток Leu в положении 328 (в соответствии с Европейской нумерацией) заменен Glu, обладают более слабой активностью связывания со всеми активирующими FcγR, включая FcγRIIа обоих аллотипов, и повышенной активностью связывания с FcγRIIb, который представляет собой ингибирующий FcγR.
Затем оценивали селективность связывания полученного варианта с FcγRIIb с использованием отношения FcγRIIb-связывающей активности к активности связывания с FcγRIIа R-типа или Н-типа, где указанное соотношение служило в качестве индикатора. В частности, величина I/A(R) или I/A(H), которая представляет собой величину, полученную путем деления величины KD для FcγRIIа R-типа или Н-типа на величину KD для FcγRIIb, использовали в качестве показателя селективности связывания FcγRIIb с каждым FcγRIIа. Этот показатель увеличивается, если величина KD для FcγRIIb снижается, или если KD для FcγRIIа увеличивается. То есть, вариант, имеющий большую величину, обладает повышенной активностью связывания с FcγRIIb по сравнению с FcγRIIа. Эти показатели систематизированы в таблице 17 для каждого варианта.
В соответствии с результатами, представленными в таблице 17, по сравнению с IgG1, вариант IgG1-v3, который был получен с применением известной технологии, имел более высокую величину I/A(Н), чем IgG1, и более высокую селективность связывания с FcγRIIb, но меньшую величину I/A(R), чем IgG1, и более высокую селективность связывания с FcγRIIb. С другой стороны, как описано в примерах, IgG1-v1 и IgG1-v2, как было обнаружено, имеет большие величины I/A(R) и I/A(Н), чем IgG1, и более высокую селективность связывания с FcγRIIb по сравнению с FcγRIIа обоих аллотипов.
До сих пор не было получено каких-либо данных о модификациях, обладающих такими свойствами, и действительно, такие модификации являются очень редкими, как показано на фиг. 17, 18, 19, и 20. Модификации, вводимые путем замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp или замены Leu в положении 328 (в соответствии с Европейской нумерацией) на Glu, являются наиболее подходящими при получении терапевтических средств для лечения иммунных воспалительных заболеваний и т.п.
Кроме того, в таблице 16 было убедительно продемонстрировано, что IgG1-v3, описанный в непатентном документе 28, обладает в 408 раз большей активностью связывания с FcγRIIb, в 0,51 раза более низкой активностью связывания с FcγRIIа Н-типа и в 522 раза большей активностью связывания с FcγRIIа R-типа. Эти результаты показали, что поскольку IgG1-v1 и IgG1-v2 обладают пониженной активностью связывания с FcγRIIа R-типа и Н-типа и повышенной активностью связывания с FcγRIIb, то считается, что эти варианты связываются с FcγRIIb с большей селективностью, чем IgG1-v3. В частности, модификации, вводимые путем замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp или замены Leu в положении 328 (в соответствии с Европейской нумерацией) на Glu, являются наиболее подходящими при получении терапевтических средств для лечения иммунных воспалительных заболеваний и т.п.
[Сравнительный пример 5] Эффекты объединения модификаций и аминокислотных замен в другой Fc-области, влияющих на селективность связывания с FcγRIIb,
Были предприняты попытки достичь еще большего повышения селективности связывания с FcγRIIb с использованием варианта, который обладал повышенной селективностью связывания с FcγRIIb и имел замену Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, как описано в сравнительных примерах 3 и 4.
Сначала в вариант IL6R-G1d_v1 (SEQ ID NO:20) вводили в IL6R-G1d модификации, а именно, замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp и замены Leu в положении 328 (в соответствии с Европейской нумерацией) на Glu, как описано в сравнительном примере 4, которые повышают селективность связывания с FcγRIIb, в результате чего получали вариант IL6R-G1d-v4. Полученный вариант объединяли с IL6R-L (SEQ ID NO:21) методом, описанным в сравнительном примере 1. Полученное антитело, имеющее аминокислотную последовательность, происходящую от IL6R-G1d-v4, и служащее в качестве Н-цепи антитела, было обозначено IgG1-v4. Активности связывания IgG1, IgG1-v1, IgG1-v2 и IgG1-v4 с FcγRIIb оценивали методом, описанным в сравнительном примере 2, и полученные результаты представлены в таблице 18.
Исходя из результатов таблицы 18, следует отметить, что поскольку модификация L328E приводит к 2,3-кратному повышению активности связывания с FcγRIIb по сравнению с активностью IgG1, то можно предположить, что комбинация этой модификации с заменой P238D, которая также приводит к 4,8-кратному повышению активности связывания с FcγRIIb по сравнению с активностью IgG1, будет еще больше повышать активность связывания с FcγRIIb, однако, фактически, FcγRIIb-связывающая активность варианта, содержащего комбинацию этих модификаций, была в 0,47 раза ниже активности IgG1. Этот результат указывает на то, что эффект от введения этих конкретных модификаций не может быть предсказан.
Аналогичным образом, IL6R-G1d-v1 (SEQ ID NO:20) получали путем введения в IL6R-G1d модификации, а именно, замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, замены Ser в положении 267 (в соответствии с Европейской нумерацией) на Glu и замены Leu в положении 328 (в соответствии с Европейской нумерацией) на Phe, как описано в сравнительном примере 4, которые повышают активность связывания с FcγRIIb, в результате чего был получен вариант IL6R-G1d-v5 методом, описанным в сравнительном примере 1. Полученное антитело, имеющее аминокислотную последовательность, происходящую от IL6R-G1d-v5, и служащее в качестве Н-цепи антитела, было обозначено IgG1-v5. Активности связывания IgG1, IgG1-v1, IgG1-v3 и IgG1-v5 с FcγRIIb оценивали методом, описанным в сравнительном примере 2, и полученные результаты представлены в таблице 19.
Модификацию S267E/L328F, повышающую активность связывания с FcγRIIb, как описано в сравнительном примере 4, вводили в P238D-вариант, а затем оценивали активность его связывания с FcγRIIb до и после введения такой модификации. Результаты представлены в таблице 19.
Исходя из результатов таблицы 19, следует отметить, что поскольку модификация S267E/L328F приводит к 408-кратному повышению активности связывания с FcγRIIb по сравнению с активностью IgG1, то можно предположить, что комбинация этой модификации с модификацией P238D, которая также приводит к 4,8-кратному повышению активности связывания с FcγRIIb по сравнению с активностью IgG1, будет еще больше повышать активность связывания с FcγRIIb, однако, фактически, как описано в предыдущем примере, FcγRIIb-связывающая активность варианта, содержащего комбинацию этих модификаций, была только в 12 раз выше или т.п. по сравнению с активностью IgG1. Этот результат также указывает на то, что эффект от введения комбинации модификаций не может быть предсказан исходя из эффектов конкретных модификаций.
Эти результаты показали, что хотя одна замена Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp приводит к повышению активности связывания с FcγRIIb, однако, этот эффект не наблюдается при объединении с другими модификациями, которые повышают активность связывания с FcγRIIb. Причиной этого может быть то, что структура в пограничной области взаимодействия между Fc и FcγR после замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp изменяется, и эффекты таких модификаций, наблюдаемые в природном антителе, уже не дают ожидаемых результатов. В соответствии с этим, очевидно, что, с использованием в качестве матрицы Fc-области, содержащей замену Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, очень трудно получить Fc-область, которая обладала бы превосходной селективностью связывания с FcγRIIb, поскольку информация об эффектах модификаций, полученная для природных антител, не может быть применена.
[Сравнительный пример 6] Детальный анализ FcγRIIb-связывающей активности вариантов, в которые, помимо модификации P238D, была введена модификация в шарнирную область
Как было показано в примере 5, в Fc-области, полученной путем замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp в природном человеческом IgG1, ожидаемый комбинаторный эффект не мог быть получен даже при объединении этой модификации с другой модификацией, которая, как предполагалось, повышает активность связывания с FcγRIIb. Поэтому, исходя из Fc-варианта, полученного путем замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, был проведен детальный анализ путем введения модификаций в Fc для поиска вариантов, которые также обладали бы повышенной активностью связывания с FcγRIIb. Для Н-цепей антитела получали вариант IL6R-F11 (SEQ ID NО: 22) путем введения модификации, а именно, замены Met в положении 252 (в соответствии с Европейской нумерацией) на Tyr и замены Asn в положении 434 (в соответствии с Европейской нумерацией) на Tyr в IL6R-G1d (SEQ ID NO:19), и вариант IL6R-F652 путем введения дополнительной замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp. Экспрессионные плазмиды, содержащие последовательность Н-цепи антитела, были получены для каждой последовательности Н-цепи антитела путем замены области возле остатка в положении 238 (в соответствии с Европейской нумерацией)(в положениях 234-237 и 239 (в соответствии с Европейской нумерацией)) в IL6R-F652 каждой из 18 аминокислот, за исключением исходных аминокислот и цистеина. IL6R-L (SEQ ID NO:21) использовали в качестве общей L-цепи для всех антител. Эти варианты были экспрессированы и очищены методом, описанным в сравнительном примере 1. Эти Fc-варианты были названы PD-вариантами. Взаимодействие каждого PD-варианта с FcγRIIа R-типа и с FcγRIIb детально оценивали методом, описанным в сравнительном примере 2.
Результаты анализа взаимодействия с соответствующими FcγR, представленные на графике, были получены следующим методом. Величину, полученную путем деления величины активности связывания каждого PD-варианта с каждым FcγR на количество FcγR, связывающегося с предварительно модифицированным антителом, которое использовали в качестве контроля (антитело IL6R-F652/IL6R-L, которое имеет замену Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp), и умножения полученного результата на 100, использовали как относительную величину активности связывания каждого PD-варианта с каждым FcγR. На горизонтальной оси отложены относительные величины активности связывания FcγRIIb с каждым PD-вариантом, а на вертикальной оси отложены относительные величины активности связывания FcγRIIа R-типа с каждым PD-вариантом (фиг. 22).
В результате было обнаружено, что модификации одиннадцати типов обладали действием, усиливающим активность связывания с FcγRIIb и сохраняющим или усиливающим активность связывания с FcγRIIа R-типа по сравнению с активностью антитела до введения в него модификаций. Активности связывания этих одиннадцати вариантов с FcγRIIb и FcγRIIаR систематизированы в таблице 20. В этой таблице «модификация» означает модификацию, введенную в IL6R-F11 (SEQ ID NO:22).
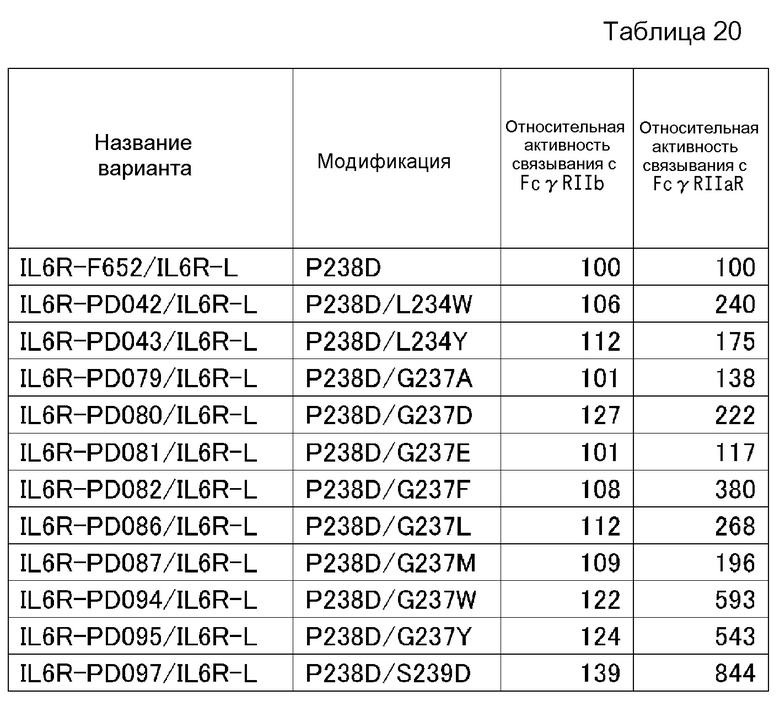
На фиг. 23 представлены относительные величины активности связывания с FcγRIIb, полученные путем дополнительного введения этих одиннадцати модификаций в вариант, имеющий модификацию P238D, и относительные величины активности связывания с FcγRIIb, полученные путем введения этих модификаций в Fc-область, которая не содержит модификации P238D, как описано в сравнительном примере 3. Эти одиннадцать модификаций повышали активность связывания с FcγRIIb, если они были дополнительно введены в вариант с P238D, по сравнению с активностью до введения, однако, с другой стороны, в случае введения этих восьми модификаций, за исключением G237F, G237W и S239D, эффект снижения активности связывания с FcγRIIb наблюдался, только когда эти модификации были введены в вариант, не содержащий P238D (GpH7-B3/GpL16-k0), как описано в сравнительном примере 3.
Результаты, описанные в сравнительном примере 5, показали, что исходя из эффектов введения модификаций в природный IgG1 трудно предсказать эффекты введения тех же самых модификаций в вариант, содержащий Fc с модификацией P238D. Другими словами, эффект от введения этих восьми идентифицированных здесь модификаций невозможно предсказать без проведения описанного исследования.
Результаты измерения величин KD для связывания вариантов, указанных в таблице 20, с FcγRIа, FcγRIIаR, FcγRIIаН, FcγRIIb и FcγRIIIаV методом, описанным в сравнительном примере 2, систематизированы в таблице 21. В этой таблице «модификация» означает модификацию, введенную в IL6R-F11 (SEQ ID NО:22). Матрица, используемая для продуцирования IL6R-F11, IL6R-G1d/IL6R-L, отмечена звездочкой (*). Кроме того, в таблице «KD(IIaR)/KD(IIb)» и «KD(IIaН)/KD(IIb)», соответственно, означают величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIb, и величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаН на величину KD для связывания каждого варианта с FcγRIIb. «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания родительского полипептида с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, в таблице 21 указаны величины KD для более высокой активности связывания каждого варианта с FcγRIIаR и FcγRIIаН/величины KD для более высокой активности связывания родительского полипептида с FcγRIIаR и FcγRIIаН. В данном случае «родительский полипептид» означает вариант, который содержит IL6R-F11 (SEQ ID NО:22) в качестве Н-цепи. Было определено, что из-за слабого связывания FcγR с IgG невозможно провести точный кинетический анализ, а поэтому числа в закрашенных серым ячейках в таблице 21 относятся к величинам, вычисленным по уравнению 2 в сравнительном примере 2:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2].
В таблице 21 показано, что все варианты обладали более высокой аффинностью связывания с FcγRIIb по сравнению с аффинностью IL6R-F11, и эта аффинность была выше в 1,9-5,0 раза. Отношение величины KD для связывания каждого варианта с FcγRIIаR/величины KD для связывания каждого варианта с FcγRIIb, и отношение величины KD для связывания каждого варианта с FcγRIIаН/величины KD для связывания каждого варианта с FcγRIIb означает отношение активности связывания с FcγRIIb к активности связывания с FcγRIIаR и активности связывания с FcγRIIаН, соответственно. То есть, эти величины указывают на степень селективности связывания каждого варианта с FcγRIIb, и чем выше эта величина, тем выше селективность связывания с FcγRIIb. Для родительского полипептида IL6R-F11/IL6R-L, отношение величины KD для FcγRIIаR/величины KD для FcγRIIb и отношение величины KD для FcγRIIаН/величины KD для FcγRIIb составляют 0,7, и, соответственно, все варианты, представленные в таблице 21, обладают повышенной селективностью связывания с FcγRIIb по сравнению с родительским полипептидом. Если отношение «величина KD для более высокой активности связывания варианта с FcγRIIаR и с FcγRIIаН/величина KD для более высокой активности связывания родительского полипептида с FcγRIIаR и с FcγRIIаН» составляет 1 или более, то это означает, что более сильные FcγRIIаR-связывающие и FcγRIIаН-связывающие активности варианта являются эквивалентными FcγRIIаR-связывающей и FcγRIIаН-связывающей активности родительского полипептида или более низкими, чем активность родительского полипептида. Поскольку эта величина составляет от 0,7 до 5,0 для полученных здесь вариантов, то можно сказать, что связывание этих вариантов, имеющих более высокие FcγRIIаR-связывающие и FcγRIIаН-связывающие активности, являются почти аналогичными соответствующей активности родительского полипептида или более низкими, чем активность родительского полипептида. Эти результаты показали, что полученные варианты, по сравнению с родительским полипептидом, обладают повышенной активностью связывания с FcγRIIb, но обладают аналогичной или пониженной активностью связывания с FcγRIIа R-типа и Н-типа, а поэтому эти варианты обладают повышенной селективностью связывания с FcγRIIb. Кроме того, все эти варианты, по сравнению с IL6R-F11, обладают пониженной аффинностью связывания с FcγRIа и FcγRIIIаV.
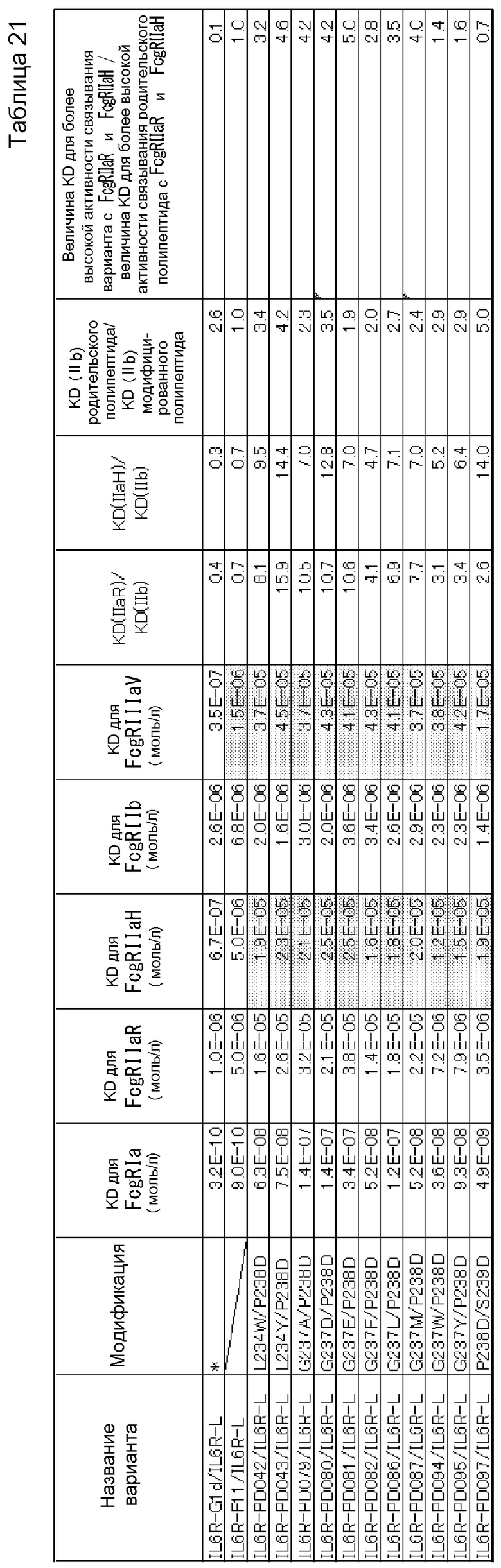
[Сравнительный пример 7] Рентгеноструктурный анализ комплекса, образованного Fc-областью, содержащей P238D, и внеклеточной областью FcγRIIb
Как показано в сравнительном примере 5, даже если было обнаружено, что при введении в P238D-содержащую Fc-область модификации, которая повышает активность или селективность связывания с FcγRIIb, наблюдается снижение этой активности, то это может быть обусловлено тем, что структура пограничной области взаимодействия Fc и FcγRIIb изменяется в результате введения модификации P238D. Поэтому, чтобы понять причину такого явления, трехмерную структуру комплекса, образованного Fc-областью IgG1, содержащей мутацию P238D (далее обозначаемую Fc(P238D)), и внеклеточной областью FcγRIIb, оценивали с помощью рентгеноструктурного анализа кристаллической структуры, а затем эту трехмерную структуру и характер связывания сравнивали с трехмерной структурой и характером связывания комплекса, образованного Fc-областью природного IgG1 (далее обозначаемой Fc(WT)) и внеклеточной областью FcγRIIb. Имеется множество данных о трехмерной структуре комплекса, образованного Fc и внеклеточной областью FcγR, и был проведен анализ трехмерных структур комплекса Fc(WT)/внеклеточная область FcγRIIIb (Nature, 2000, 400, 267-273; J.Biol. Chem. 2011, 276, 16469-16477), комплекса Fc(WT)/внеклеточная область FcγRIIIа (Proc. Natl. Acad. Sci. USA, 2011, 108, 12669-126674) и комплекса Fc(WT)/внеклеточная область FcγRIIа (J. Imunol. 2011, 187, 3208-3217). Хотя трехмерная структура комплекса Fc(WT)/внеклеточная область FcγRIIb не была проанализирована, однако, трехмерная структура комплекса, образованного Fc(WT) и FcγRIIа, является известной, и также известно, что внеклеточые области FcγRIIа и FcγRIIb имеют аминокислотные последовательности, идентичные на 93%, то есть они имеют очень высокую гомологию. Таким образом, трехмерная структура комплекса Fc(WT)/внеклеточная область FcγRIIb была предсказана путем моделирования с использованием кристаллической структуры комплекса Fc(WT)/внеклеточная область FcγRIIа.
Трехмерная структура комплекса Fc(P238D)/внеклеточная область FcγRIIb была определена с помощью рентгеноструктурного анализа кристаллической структуры с разрешением 2,6 Å. Структура, полученная в результате такого анализа, представлена на фиг. 24. В настоящее время уже была проанализирована внеклеточная область FcγRIIb, образованная двумя СН2-доменами Fc, и было обнаружено, что структура этой области аналогична трехмерной структуре комплексов, образованных Fc(WT) и соответствующими внеклеточыми областями FcγRIIIа, FcγRIIIb или FcγRIIа.
Затем, для детального сравнения, осуществляли суперпозицию кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и смоделированной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, где такое сравнение проводили методом наименьших квадратов, исходя из расстояния между парами атомов Cα для внеклеточной области FcγRIIb и CH2-домена А Fc (фиг. 25). В этом случае степень перекрывания между CH2-доменами В Fc была неудовлетворительной, и в этой части наблюдались конформационные различия. Кроме того, с использованием кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и смоделированной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, пары атомов, которые имеют расстояния 3,7 Å или менее между внеклеточной областью FcγRIIb и CH2-доменом В Fc, исключали, и проводили сравнение для выявления различий в межатомных взаимодействиях между FcγRIIb и CH2-доменом В Fc в Fc(WT) и Fc(P238D). Как показано в таблице 22, межатомные взаимодействия между CH2-доменом В Fc и FcγRIIb в Fc(P238D) и в Fc(WT) не соответствовали друг другу.
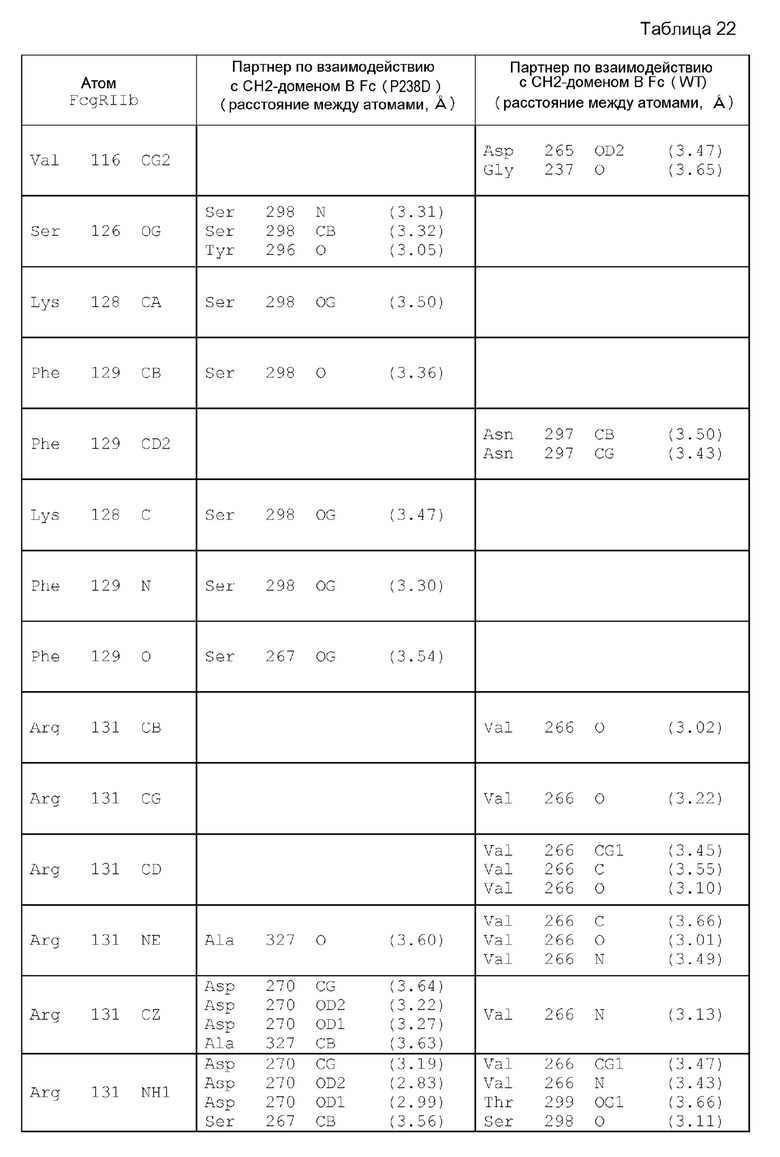
Кроме того, осуществляли суперпозицию кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и смоделированной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, определенной с помощью рентгеноструктурного анализа, и проводили сравнение методом наименьших квадратов, исходя из расстояния между парами атомов Cα только для CH2-домена А Fc или только для CH2-домена В Fc, а затем проводили детальное сравнение структур, находящихся возле P238D. Локализации аминокислотного остатка в положении 238 (в соответствии с Европейской нумерацией), которое представляет собой положение вводимой мутации, отличались для Fc(P238D) и в Fc(WT), то есть можно видеть, что наряду с таким изменением находящаяся рядом петлевая структура, являющаяся продолжением шарнирной области, отличалась для Fc(P238D) и Fc(WT) (фиг. 26). В природном Fc(WT) Pro в положении 238 (в соответствии с Европейской нумерацией) присутствует на внутренней стороне белка и образует гидрофобную сердцевину с окружающими ее остатками. Однако, при замене этого остатка заряженным и в высокой степени гидрофильным Asp, присутствие такого остатка в той же самой гидрофобной сердцевине может вызывать энергетически неблагоприятную ситуацию, называемую десольватацией. Поэтому, в Fc(P238D), для устранения такой неблагоприятной энергетической ситуации, аминокислотный остаток в положении 238 (в соответствии с Европейской нумерацией) заменяют так, чтобы он был ориентирован в сторону растворителя, и это может вызывать изменение расположенной рядом петлевой структуры. Кроме того, поскольку такая петля является продолжением шарнирной области, перекрестно связанной связью S-S, то такое структурное изменение не будет ограничиваться изменением в какой-либо конкретной области, а будет влиять на относительное расположение CH2-домена А Fc и CH2-домена В Fc. В результате происходит изменение межатомных взаимодействий между FcγRIIb и CH2-доменом В Fc. Поэтому предсказанные эффекты не наблюдались при введении модификаций, которые повышают селективность и активность связывания с FcγRIIb в природном IgG, в комбинации с модификациями в Fc-области, содержащей модификацию P238D.
Кроме того, в результате структурных изменений, обусловленных введением модификации P238D в CH2-домен А Fc, образовывалась водородная связь между главной цепью Gly в соседнем положении 237 (в соответствии с Европейской нумерацией) и Tyr в положении 160 FcγRIIb (фиг. 27). Остатком в FcγRIIа, который соответствует этому остатку Tyr 160, является Phe, а при связывании этого остатка с FcγRIIа, такая водородная связь не образуется. Аминокислота в положении 160 представляет собой одну из нескольких аминокислот, отличающихся в FcγRIIа и в FcγRIIb в пограничной области взаимодействия с Fc, и было высказано предположение, что такая водородная связь, которая является специфичной для FcγRIIb, будет способствовать повышению FcγRIIb-связывающей активности и снижению FcγRIIа-связывающей активности Fc(P238D), а также к повышению селективности такого связывания. Кроме того, в CH2-домене В Fc-области в FcγRIIb наблюдалось электростатическое взаимодействие между остатком Asp в положении 270 (в соответствии с Европейской нумерацией) и остатком Arg в положении 131 (фиг. 28). В FcγRIIа Н-типа, который представляет собой один из аллотипов FcγRIIа, соответствующим остатком является His, а поэтому этот остаток не может вступать в такое электростатическое взаимодействие. Этим объясняется снижение активности связывания Fc(P238D) с FcγRIIа Н-типа, а не с FcγRIIа R-типа. Оценка таких результатов рентгеноструктурного анализа кристаллической структуры показала, что изменение петлевой структуры после введения модификации P238D с последующим изменением относительной локализации домена будет приводить к новым взаимодействиям, которые не наблюдаются в природном IgG, и к селективному характеру связывания P238D-вариантов с FcγRIIb.
[Экспрессия и очистка Fc(P238D)]
Fc-область, содержащую модификацию P238D, получали следующим образом. Сначала получали hIL6R-IgG1-v1 (SEQ ID NO:20) путем замены Cys в положении 220 (в соответствии с Европейской нумерацией) на Ser. Затем генную последовательность Fc(P238D), в которой у С-конца в положении 216 присутствует Glu (в соответствии с Европейской нумерацией), клонироваали с помощью ПЦР. С использованием этой клонированной генной последовательности осуществляли конструирование экспрессионных векторов, и проводили экспрессию и очистку Fc(P238D) методом, описанным в сравнительном примере 1. Cys в положении 220 (в соответствии с Европейской нумерацией) образует дисульфидную связь с Cys L-цепи в природном IgG1. При получении одного Fc, L-цепь не экспрессируется, а поэтому, для предотвращения образования нежелательных дисульфидных связей, этот остаток заменяют на Ser.
[Экспрессия и очистка внеклеточной области FcγRIIb]
Экспрессия и очистка были проведены методом, описанным в сравнительном примере 2.
[Очистка комплекса Fc(P238D))/внеклеточная область FcγRIIb]
К 2 мг образца внеклеточной области FcγRIIb, полученного в целях кристаллизации, добавляли 0,29 мг Endo F1 (Protein Science 1996, 5, 2617-2622), экспрессированного и очищенного из Escherichia coli в виде слитого белка глутатион-S-трансферазы. Этот белок выдерживали при комнатной температуре в течение трех дней в 0,1 M бис-трис-буфера при pH 6,5, и N-связанный олигосахарид отделяли, в результате чего оставался N-ацетилглюкозамин, непосредственно связанный с Asn. Затем этот образец внеклеточного домена FcγRIIb, подвергнутый реакции отщепления углевода, концентрировали путем ультрафильтрации с отсечкой молекулярной массы 5000, после чего его очищали с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 20 мM HEPES при pH 7,5, содержащем 0,05M NaCl. Кроме того, к полученной фракции внеклеточной области FcγRIIb, от которой был отщеплен углевод, добавляли Fc(P238D) так, чтобы молярное отношение внеклеточной области FcγRIIb было слегка повышенным, и после концентрирования этого образца путем ультрафильтрации с отсечкой молекулярной массы 10000 получали образец комплекса Fc(P238D)/внеклеточная область FcγRIIb с последующей очисткой с помощью гель-фильтрации (Superdex 200 10/300) на колонке, уравновешенной в 20 мM HEPES при pH 7,5, содержащем 0,05M NaCl.
[Кристаллизация комплекса Fc(P238D)/внеклеточная область FcγRIIb]
Образец комплекса Fc(P238D)/внеклеточная область FcγRIIb концентрировали до получения концентрации приблизительно 10 мг/мл с использованием фильтра для ультрафильтрации с отсечкой молекулярной массы 10000, и кристаллизовали методом паровой диффузии с сидячей каплей. Для кристаллизации использовали Hydra II Plus One (MATRIX), а для резервуарного раствора, содержащего 100 мM бис-трис, pH 6,5, 17% PEG3350, 0,2М ацетата аммония и 2,7% (масс/об) D-галактозы, каплю для инициации кристаллизации получали путем смешивания резервуарного раствора и образца кристаллизации в соотношении 0,2 мкл:0,2 мкл, а затем этот раствор герметично закрывали и оставляли при температуре 20°C, в результате чего были успешно получены тонкие пластинчатые кристаллы.
[Получение рентгенографических данных для кристалла комплекса Fc(P238D)/внеклеточная область FcγRIIb]
Один из полученных монокристаллов комплекса Fc(P238D)/внеклеточная область FcγRIIb заливали в раствор 100 мM бис-трис (pH 6,5), 20% PEG3350, ацетата аммония, 2,7% (масс/об) D-галактозы и 22,5% (об/об) этиленгликоля. Кристалл промывали раствором с помощью иглы, к которой была подсоединена тонкая нейлоновая петля, и замораживали в жидком азоте, а затем проводили рентгенографический анализ на синхротронном излучателе Photon Factory BL-1A в научно-исследовательском институте высоких энергий. Во время измерений кристалл постоянно выдерживали в потоке азота при -178°C для его поддержания в замороженном состоянии, и все 225 дифракционных изображений этого монокристалла получали на ПЗС-детекторе Quantum 270 (ADSC), подсоединенном к источнику луча света, с одновременным вращением кристалла под углом 0,8°. Исходя из полученных дифракционных изображений, определение констант ячейки, идентификацию пятен дифракции и обработку рентгенографических данных проводили с использованием программы Xia2 (пакета программ CCP4), программы XDS (Walfgang Kabsch) и Scala (пакета программ CCP4), и наконец, получали данные интенсивности дифракции с разрешением 2,46 Å. Этот кристалл принадлежал к пространственной группе Р21 и имел следующие параметры ячейки: а=48,85 Å, b=76,01 Å, c=115,09 Å, α=90°, β=100,70°, γ=90°.
[Рентгеноструктурный анализ комплекса Fc(P238D)/внеклеточная область FcγRIIb]
Кристаллическую структуру комплекса Fc(P238D)/внеклеточная область FcγRIIb определяли методом молекулярной замены с использованием программы Phaser (пакета программ CCP4). Исходя из размера полученной кристаллической решетки и молекулярной массы комплекса Fc(P238D)/внеклеточная область FcγRIIb было предсказано, что число комплексов в асимметрической ячейке равно 1. Исходя из структурных координат кода PDB: 3SGJ, который представляет собой кристаллическую структуру комплекса Fc(WT)/внеклеточная область FcγRIIIа, были определены отдельные координаты аминокислотных остатков части А-цепи в положениях 239-340 и В-цепи в положениях 239-340, а затем эти координаты были использованы, соответственно, в качестве моделей для поиска CH2-доменов Fc. Исходя из тех же самых структурных координат кода PDB: 3SGJ, был определен отдельный ряд координат аминокислотных остатков части А-цепи в положениях 341-444 и В-цепи в положениях 341-444, а затем эти координаты были использованы, соответственно, в качестве моделей для поиска CH3-доменов Fc. И наконец, исходя из структурных координат кода PDB: 2FCB, который представляет собой кристаллическую структуру внеклеточной области FcγRIIb, были определены отдельные координаты аминокислотных остатков части А-цепи в положениях 6-178, а затем эти координаты были использованы в качестве моделей для поиска внеклеточной области FcγRIIb. Затем определяли ориентацию и положение CH3-доменов Fc, внеклеточной области FcγRIIb и CH2-домена Fc для каждой модели поиска в кристаллических решетках с использованием функций вращения и функций трансляции, в результате чего была получена первоначальная модель кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb. Если уточнение параметров твердого тела, относящегося к двум CH2-доменам Fc, двум CH3-доменам Fc и внеклеточной области FcγRIIb, осуществляли исходя из полученной первоначальной модели, то фактор кристаллографической достоверности, то есть R-фактор, составлял 40,4%, а свободный R-фактор (R-free) составлял 41,9%, как было определено по данным интенсивности дифракции с разрешением от 25 Å до 3,0 Å. Кроме того, уточнение структуры проводили с использованием программы REFMAC5 (пакета программ CCP4), а коррекцию модели с наблюдением карт электронной плотности с коэффициентами 2Fo-Fc или Fo-Fc, которые вычисляли исходя из экспериментально определенного структурного фактора Fo, вычисленного структурного фактора Fc и фазы, вычисленной с использованием такой модели, осуществляли с помощью программы Сооt (Paul Emsley), а затем проводили уточнение модели путем повторения этих стадий. И наконец, в результате введения молекул воды в модель, полученную исходя из карты электронной плотности, где 2Fo-Fc и Fo-Fc использовали в качестве коэффициента, и после уточнения был получен фактор кристаллографической достоверности, то есть R-фактор и R-free модели, содержащей 4846 не-водородных атомов, которые составляли 23,7% и 27,6%, соответственно, как было определено по 24291 данным интенсивности дифракции с разрешением от 25 Å до 2,6 Å, соответственно.
[Создание модели структуры комплекса Fc(WT)/внеклеточная область FcγRIIb]
Исходя из структурных координат кода PDB: 3RY6, который представляет собой кристаллическую структуру комплекса Fc(WT)/внеклеточная область FcγRIIа, функцию сконструированных мутантов, заложенную в программу Disсovery Studio 3.1 (Accelrys), использовали для введения мутаций в FcγRIIа в целях поиска соответствий аминокислотной последовательности FcγRIIb и FcγRIIа в этих структурных координатах. В данном случае уровень оптимизации ставили на «высокий», а радиус отсечки устанавливали на 4,5, при этом получали пять моделей, и одну из этих моделей с наилучшей энергетической оценкой использовали в качестве смоделированной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb.
[Сравнительный пример 8] Анализ на FcγR-связывание Fc-вариантов, в которых сайты модификаций были определены исходя из кристаллических структур
На основе результатов рентгеноструктурного анализа комплекса, образованного Fc(P238D) и внеклеточной областью FcγRIIb, полученного как описано в сравнительном примере 7, было предсказано, что всевозможные модификации, которые были введены в сайты Fc-варианта, имеющего замену Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp, влияют на взаимодействие с FcγRIIb (в положениях остатков 233, 240, 241, 263, 265, 266, 267, 268, 271, 273, 295, 296, 298, 300, 323, 325, 326, 327, 328, 330, 332 и 334 (в соответствии с Европейской нумерацией)), и были оценены варианты, имеющие комбинацию модификаций, повышающих активность связывания с FcγRIIb.
IL6R-B3 (SEQ ID NO:23) получали путем введения в вариант IL6R-G1d (SEQ ID NO:19), полученный как описано в сравнительном примере 4, модификации, а именно замены Lys в положении 439 (в соответствии с Европейской нумерацией) на Glu. Затем получали IL6R-BF648 путем введения в вариант IL6R-B3 модификации, а именно, замены Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp. IL6R-L (SEQ ID NO:21) использовали в качестве L-цепи антитела, которая является общей для всех антител. Эти варианты антител были экспрессированы и очищены методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) детально анализировали методом, описанным в сравнительном примере 2.
Величины активности связывания вычисляли по результатам анализа на взаимодействие с соответствующим FcγR методом, описанным ниже. Величину активности связывания каждого варианта с каждым FcγR делили на величину активности связывания предварительно модифицированного контрольного антитела (IL6R-BF648/IL6R-L с заменой Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp) с каждым FcγR, и полученный результат умножали на 100, в результате чего получали относительную активность связывания каждого варианта с каждым FcγR. На горизонтальной оси отложены относительные величины активности связывания каждого варианта с FcγRIIb, а на вертикальной оси отложены относительные величины активности связывания каждого варианта с FcγRIIа R-типа (фиг. 29).
Результаты, представленные на фиг. 29, показали, что из всех модификаций модификации 24 типов способны сохранять или повышать активность связывания с FcγRIIb по сравнению с предварительно модифицированным антителом. Связывание этих вариантов с каждым FcγR проиллюстрировано в таблице 23. В этой таблице «модификация» означает модификацию, введенную в IL6R-B3 (SEQ ID NО: 23; IL6R-2B999 в таблице 23). Матрица, используемая для продуцирования IL6R-B3, IL6R-G1d/IL6R-L, показана звездочкой (*).
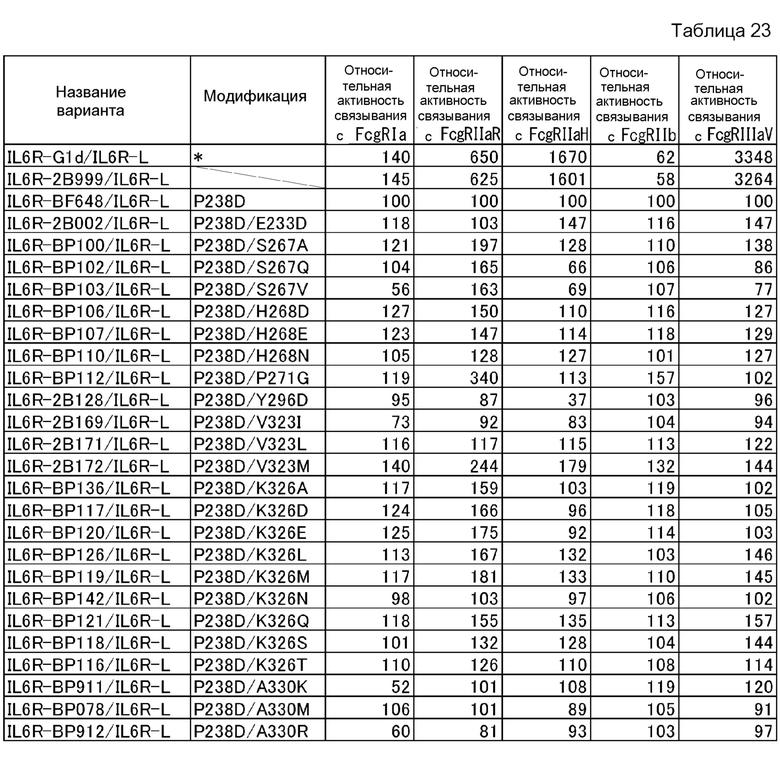
Результаты измерения величин KD для связывания вариантов, указанных в таблице 23, с FcγRIа, FcγRIIаR, FcγRIIаН, FcγRIIb и FcγRIIIа V-типа, проводимого методом, описанным в сравнительном примере 2, систематизированы в таблице 24. В этой таблице «модификация» означает модификацию, введенную в IL6R-В3 (SEQ ID NО:23). Матрица, используемая для продуцирования IL6R-В3, IL6R-G1d/IL6R-L, отмечена звездочкой (*). Кроме того, в этой таблице «KD(IIaR)/KD(IIb)» и «KD(IIaН)/KD(IIb)», соответственно, означают величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIb, и величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаН на величину KD для связывания каждого варианта с FcγRIIb. «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания родительского полипептида с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, в таблице 24 указаны соотношения «величины KD для более высокой активности связывания каждого варианта с FcγRIIаR и FcγRIIаН/величины KD для более высокой активности связывания родительского полипептида с FcγRIIаR и FcγRIIаН». В данном случае родительский полипептид означает вариант, который содержит IL6R-В3 (SEQ ID NО:23) в качестве Н-цепи. Было определено, что из-за слабого связывания FcγR с IgG невозможно провести точный кинетический анализ, а поэтому числа в закрашенных серым ячейках, как показано в таблице 24, относятся к величинам, вычисленным по уравнению 2 в сравнительном примере 2:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2].
В таблице 24 показано, что все варианты обладали более высокой аффинностью связывания с FcγRIIb по сравнению с аффинностью IL6R-В3 (IL6R-2B999 в таблице 24), и эта аффинность была выше в 2,1-9,7 раза. Отношение величины KD для связывания каждого варианта с FcγRIIаR/величины KD для связывания каждого варианта с FcγRIIb и отношение величины KD для связывания каждого варианта с FcγRIIаН/величины KD для связывания каждого варианта с FcγRIIb означает отношение активности связывания с FcγRIIb к активности связывания с FcγRIIаR и к активности связывания с FcγRIIаН, соответственно. То есть, эти величины указывают на степень селективности связывания каждого варианта с FcγRIIb, и чем выше эта величина, тем выше селективность связывания с FcγRIIb. Поскольку для родительского полипептида IL6R-B3/IL6R-L отношение величины KD для FcγRIIаR/величины KD для FcγRIIb и отношение величины KD для FcγRIIаН/величины KD для FcγRIIb составляют 0,3 и 0,2, соответственно, то все варианты, представленные в таблице 24, обладают повышенной селективностью связывания с FcγRIIb по сравнению с родительским полипептидом. Если отношение «величина KD для более высокой активности связывания варианта с FcγRIIаR и с FcγRIIаН/величина KD для более высокой активности связывания родительского полипептида с FcγRIIаR и с FcγRIIаН» составляет 1 или более, то это означает, что более сильные FcγRIIаR-связывающие и FcγRIIаН-связывающие активности варианта эквивалентны или ниже по сравнению с более высокими FcγRIIаR-связывающей и FcγRIIаН-связывающей активностями родительского полипептида. Поскольку эта величина составляет от 4,6 до 34,0 для полученных здесь вариантов, то можно сказать, что, по сравнению с родительским полипептидом, полученные здесь варианты имели более низкую активность связывания с FcγRIIаR и с FcγRIIаН. Эти результаты показали, что полученные варианты, по сравнению с родительским полипептидом, обладают одинаковой или пониженной активностью связывания с FcγRIIа R-типа и Н-типа, повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb. Кроме того, все эти варианты, по сравнению с IL6R-В3, обладают пониженной аффинностью связывания с FcγRIа и FcγRIIIаV.
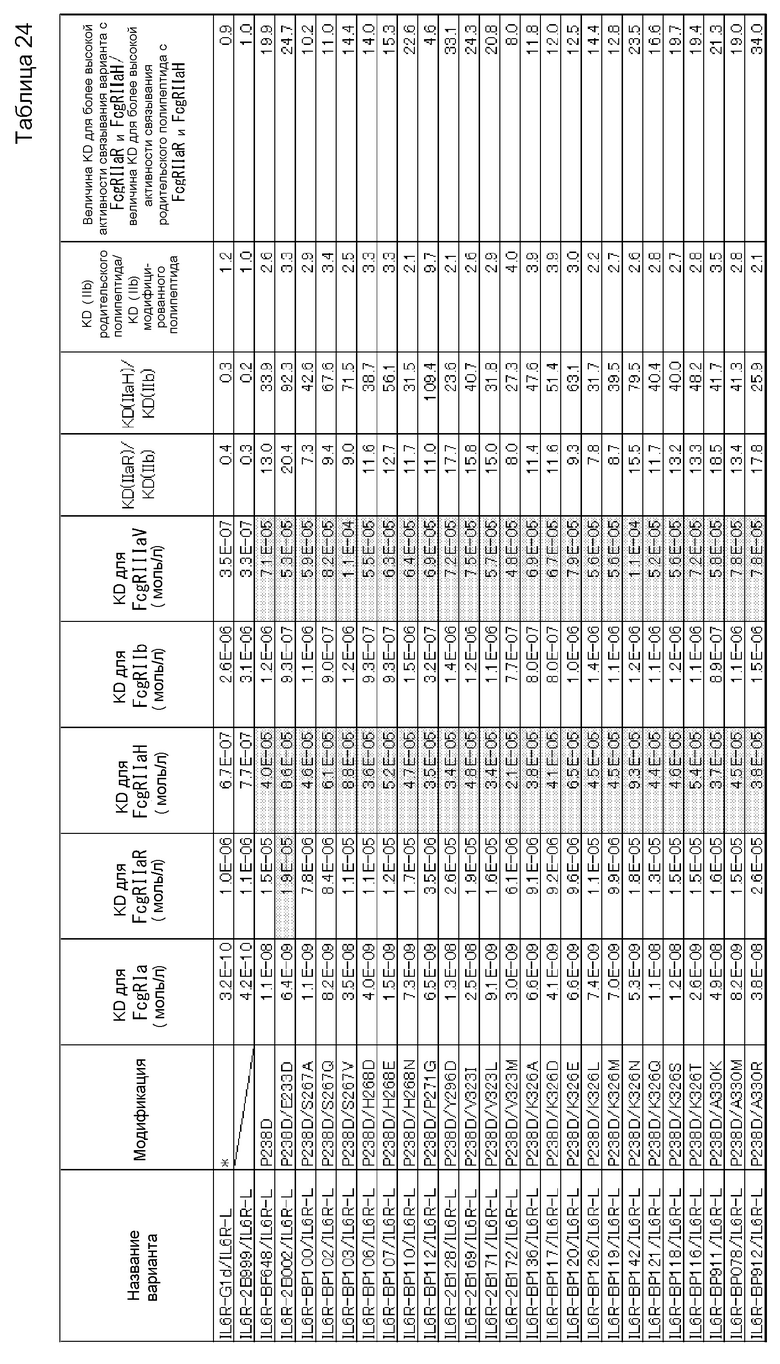
При рассмотрении наиболее перспективных вариантов из числа полученных вариантов, имеющих комбинации модификаций, факторы, обеспечивающие действие этих вариантов, были исследованы с использованием данных кристаллической структуры. На фиг. 30 показана кристаллическая структура комплекса Fc(P238D)/внеклеточная область FcγRIIb. На этой фигуре, Н-цепь, расположенная слева, представляет собой цепь А Fc, а Н-цепь, расположенная справа, представляет собой цепь В Fc. Отсюда можно видеть, что сайт в положении 233 (в соответствии с Европейской нумерацией) в цепи А Fc расположен рядом с Lys в положении 113 (в соответствии с Европейской нумерацией) FcγRIIb. Однако, в этой кристаллической структуре, боковая цепь E233 находится в состоянии довольно высокой подвижности, а электронная плотность этой цепи не была точно определена. Таким образом, модификация, вводимая путем замены Glu в положении 233 (в соответствии с Европейской нумерацией) на Asp, приводила к уменьшению степени свободы боковой цепи, поскольку боковая цепь становилась короче на один атом углерода. В результате этого, потеря энтропии при взаимодействии с Lys в положении 113 (в соответствии с Европейской нумерацией) в FcγRIIb может снижаться, что дает основание предположить, что указанный фактор может вносить свой вклад в увеличение свободной энергии связывания.
Аналогичным образом, на фиг. 31 показано окружение сайта в положении 330 (в соответствии с Европейской нумерацией) в структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb. На этой фигуре показано, что окружение сайта в положении 330 (в соответствии с Европейской нумерацией) в цепи А Fc(P238D) представляет собой гидрофильное окружение, состоящее из Ser в положении 85, Glu в положении 86, Lys в положении 163 и т.п. (в соответствии с Европейской нумерацией) в FcγRIIb. Таким образом, было высказано предположение, что модификация, полученная путем замены Ala в положении 330 (в соответствии с Европейской нумерацией) на Lys или Arg, будет вносить свой вклад в усиление взаимодействия с Ser в положении 85 (в соответствии с Европейской нумерацией) или Glu в положении 86 (в соответствии с Европейской нумерацией) в FcγRIIb.
На фиг. 32 показаны структуры с Pro в положении 271 (в соответствии с Европейской нумерацией) в цепи В Fc после суперпозиции кристаллических структур комплекса Fc(P238D)/внеклеточная область FcγRIIb и комплекса Fc(WT)/внеклеточная область FcγRIIIa со структурой цепи В Fc, где указанную суперпозицию анализировали методом наименьших квадратов исходя из расстояния между парами атомов Cα. Эти две структуры имеют хорошее соответствие, но они имеют различия в трехмерной структуре у Pro в положении 271 (в соответствии с Европейской нумерацией). Если принять во внимание слабую электронную плотность вокруг этой области в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb, то можно предположить, что существует вероятность того, что остаток Pro в положении 271 (в соответствии с Европейской нумерацией) в Fc(P238D)/FcγRIIb будет создавать значительные стерические затруднения в этой структуре, а значит он будет препятствовать оптимизации петлевой структуры. Следовательно, можно сказать, что модификация, осуществляемая путем замены Pro в положении 271 (в соответствии с Европейской нумерацией) на Gly, будет придавать этой петлевой структуре большую гибкость, а поэтому будет вносить свой вклад в повышение активности связывания благодаря снижению энергетического барьера, что даст возможность получить оптимальную структуру после взаимодействия с FcγRIIb.
[Сравнительный пример 9] Оценка комбинаторного эффекта модификаций, которые, при их объединении с P238D, будут повышать активность связывания с FcγRIIb
Из всех модификаций, полученных, как описано в сравнительных примерах 6 и 8, те модификации, которые повышали или сохраняли активность связывания с FcγRIIb и подавляли активность связывания с другими FcγR, объединяли друг с другом, а затем оценивали их эффекты.
В частности, хорошие модификации были выбраны из таблиц 19 и 22, и эти модификации были объединены и введены в Н-цепь антитела IL6R-BF648 методом, аналогичным методу, описанному в сравнительном примере 8. IL6R-L использовали в качестве L-цепи антитела, которая является общей для всех антител, и эти антитела были экспрессированы и очищены методом, описанным в сравнительном примере 1, а связывание с каждым FcγR (FcγRIа, FcγRIIа Н-типа, FcγRIIа R-типа, FcγRIIb и FcγRIIIа V-типа) детально анализировали методом, описанным в сравнительном примере 2.
Относительные активности связывания вычисляли по результатам анализа на взаимодействие с соответствующими FcγR методом, описанным ниже. Величину активности связывания каждого варианта с каждым FcγR делили на величину активности связывания контрольного предварительно модифицированного антитела (IL6R-BF648/IL6R-L с заменой Pro в положении 238 (в соответствии с Европейской нумерацией) на Asp) с каждым FcγR, и полученный результат умножали на 100, в результате чего получали относительную активность связывания каждого варианта с каждым FcγR. На горизонтальной оси отложены относительные величины активности связывания каждого варианта с FcγRIIb, а на вертикальной оси отложены относительные величины активности связывания каждого варианта с FcγRIIа R-типа (таблица 25).
В этой таблице «модификация» означает модификацию, введенную в IL6R-B3 (SEQ ID NО:23). Матрица, используемая для продуцирования IL6R-B3, IL6R-G1d/IL6R-L, показана звездочкой (*).
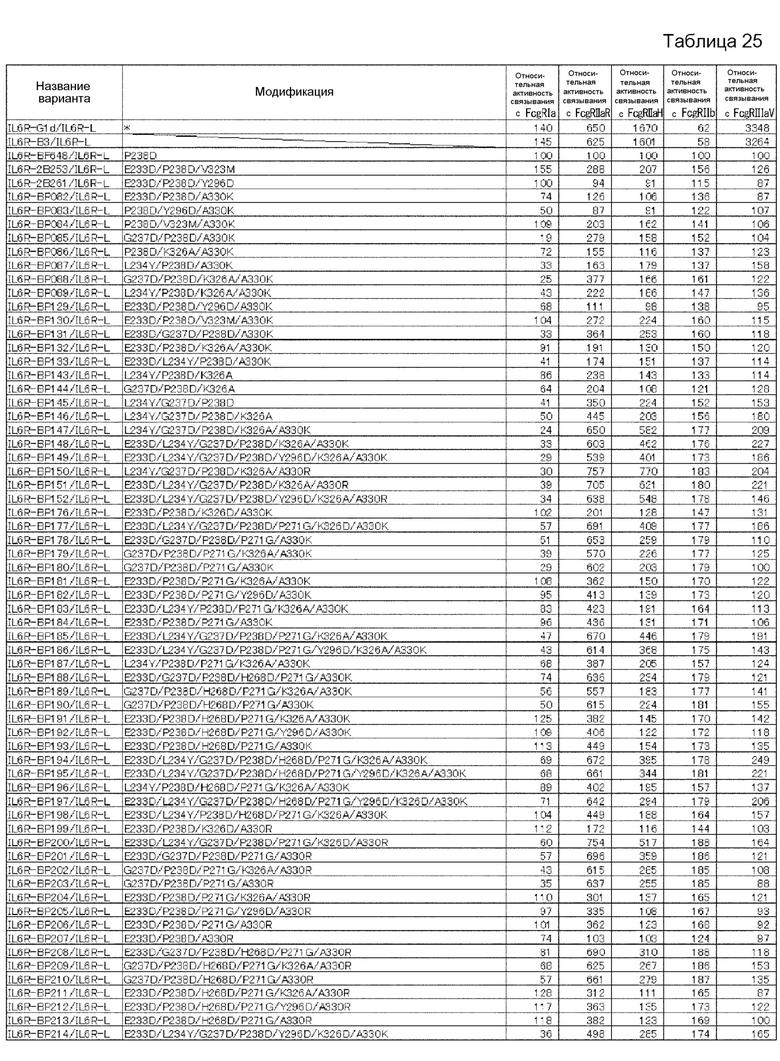
Результаты измерения величин KD для связывания вариантов, указанных в таблице 25, с FcγRIа, FcγRIIаR, FcγRIIаН, FcγRIIb и FcγRIIIа V-типа методом, описанным в сравнительном примере 2, систематизированы в таблице 26. В этой таблице «модификация» означает модификацию, введенную в IL6R-В3 (SEQ ID NО:23). Матрица, используемая для продуцирования IL6R-В3, IL6R-G1d/IL6R-L, отмечена звездочкой (*). Кроме того, в таблице «KD(IIaR)/KD(IIb)» и «KD(IIaН)/KD(IIb)», соответственно, означают величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаR на величину KD для связывания каждого варианта с FcγRIIb, и величину, полученную путем деления величины KD для связывания каждого варианта с FcγRIIаН на величину KD для связывания каждого варианта с FcγRIIb. «KD(IIb) родительского полипептида/KD(IIb) модифицированного полипептида» означает величину, полученную путем деления величины KD для связывания родительского полипептида с FcγRIIb на величину KD для связывания каждого варианта с FcγRIIb. Кроме того, в таблице 24 указаны отношения «величины KD для более высокой активности связывания каждого варианта с FcγRIIаR и FcγRIIаН/величины KD для более высокой активности связывания родительского полипептида с FcγRIIаR и FcγRIIаН». В данном случае родительский полипептид означает вариант, который содержит IL6R-В3 (SEQ ID NО:23) в качестве Н-цепи. Было определено, что из-за слабого связывания FcγR с IgG невозможно провести точный кинетический анализ, а поэтому числа в закрашенных серым ячейках, как показано в таблице 26, относятся к величинам, вычисленным по уравнению 2 в сравнительном примере 2:
KD=C⋅Rmax/(Req-RI)-C [Уравнение 2].
В таблице 26 показано, что все варианты обладали более высокой аффинностью связывания с FcγRIIb по сравнению с аффинностью IL6R-В3, и эта аффинность была выше в 3,0-99,0 раз. Отношение величины KD для связывания каждого варианта с FcγRIIаR/величины KD для связывания каждого варианта с FcγRIIb и отношение величины KD для связывания каждого варианта с FcγRIIаН/величины KD для связывания каждого варианта с FcγRIIb означает отношение активности связывания с FcγRIIb к активности связывания с FcγRIIаR и активности связывания с FcγRIIаН, соответственно. То есть, эти величины указывают на степень селективности связывания каждого варианта с FcγRIIb, и чем выше эта величина, тем выше селективность связывания с FcγRIIb. Поскольку для родительского полипептида IL6R-B3/IL6R-L отношение величины KD для FcγRIIаR/величины KD для FcγRIIb и отношение величины KD для FcγRIIаН/величины KD для FcγRIIb составляют 0,3 и 0,2, соответственно, то все варианты, представленные в таблице 26, обладают повышенной селективностью связывания с FcγRIIb по сравнению с родительским полипептидом. Если отношение «величина KD для более высокой активности связывания варианта с FcγRIIаR и с FcγRIIаН/величина KD для более высокой активности связывания родительского полипептида с FcγRIIаR и с FcγRIIаН» составляет 1 или более, то это означает, что более сильные FcγRIIаR-связывающие и FcγRIIаН-связывающие активности варианта эквивалентны или ниже FcγRIIаR-связывающей и FcγRIIаН-связывающей активности связывания родительского полипептида. Поскольку эта величина составляет от 0,7 до 29,9 для полученных здесь вариантов, то можно сказать, что полученные здесь варианты имели почти эквивалентную или более низкую активность связывания по сравнению с более сильной FcγRIIаR-связывающей и FcγRIIаН-связывающей активностью родительского полипептида. Эти результаты показали, что полученные варианты, по сравнению с родительским полипептидом, обладают одинаковой или пониженной активностью связывания с FcγRIIа R-типа и Н-типа, повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb. Кроме того, все эти варианты, по сравнению с IL6R-В3, обладают пониженной аффинностью связывания с FcγRIа и FcγRIIIаV.
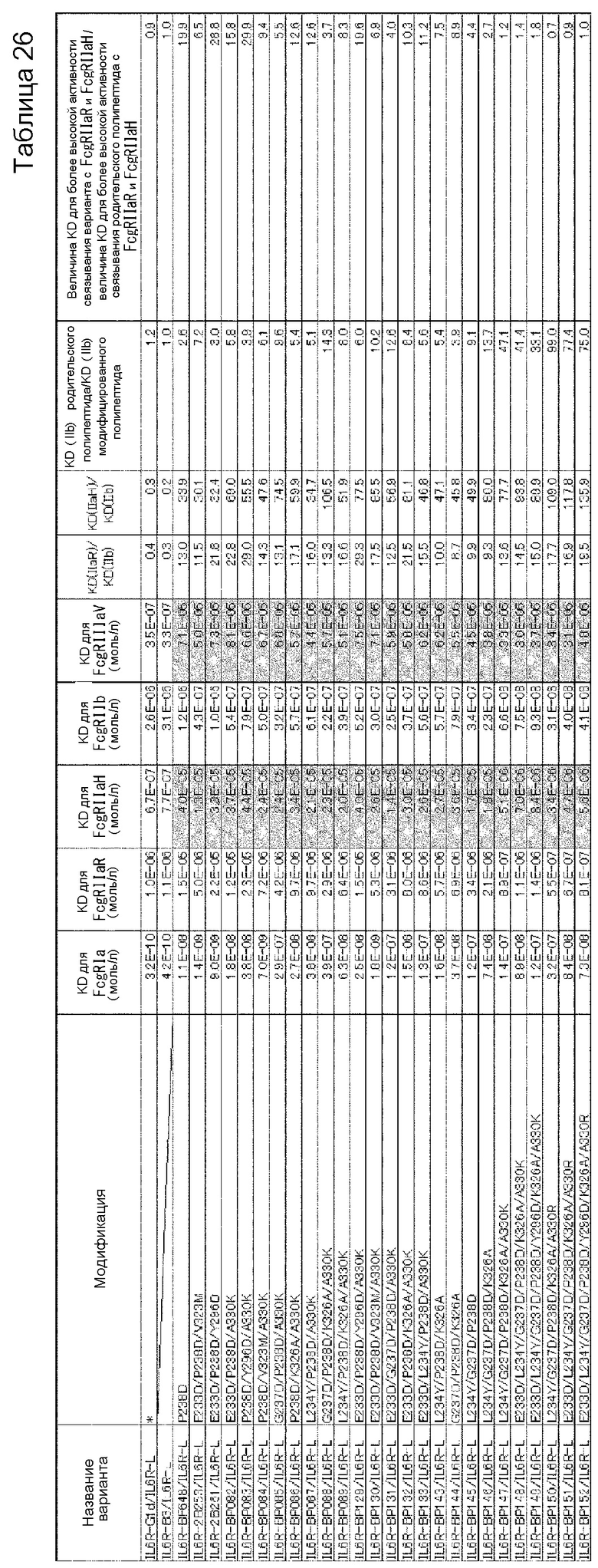
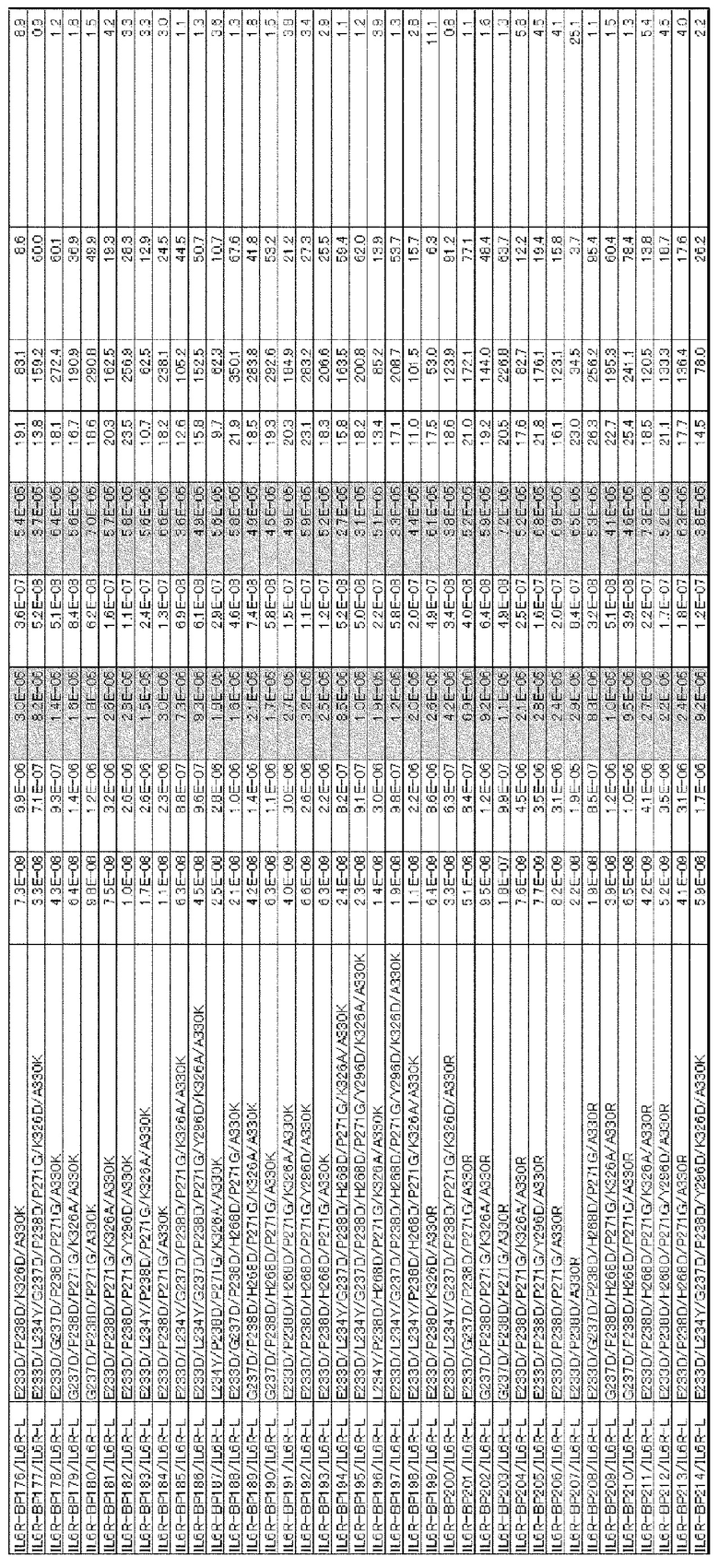
Вариабельная область и константная область в соответствующих последовательностях SEQ ID NO систематизированы в нижеследующей таблице. В этой таблице «В3» означает «2B999(B30)», «omlizH» означает «омализумаб_VH», а «omlizL» означает «омализумаб_VL».
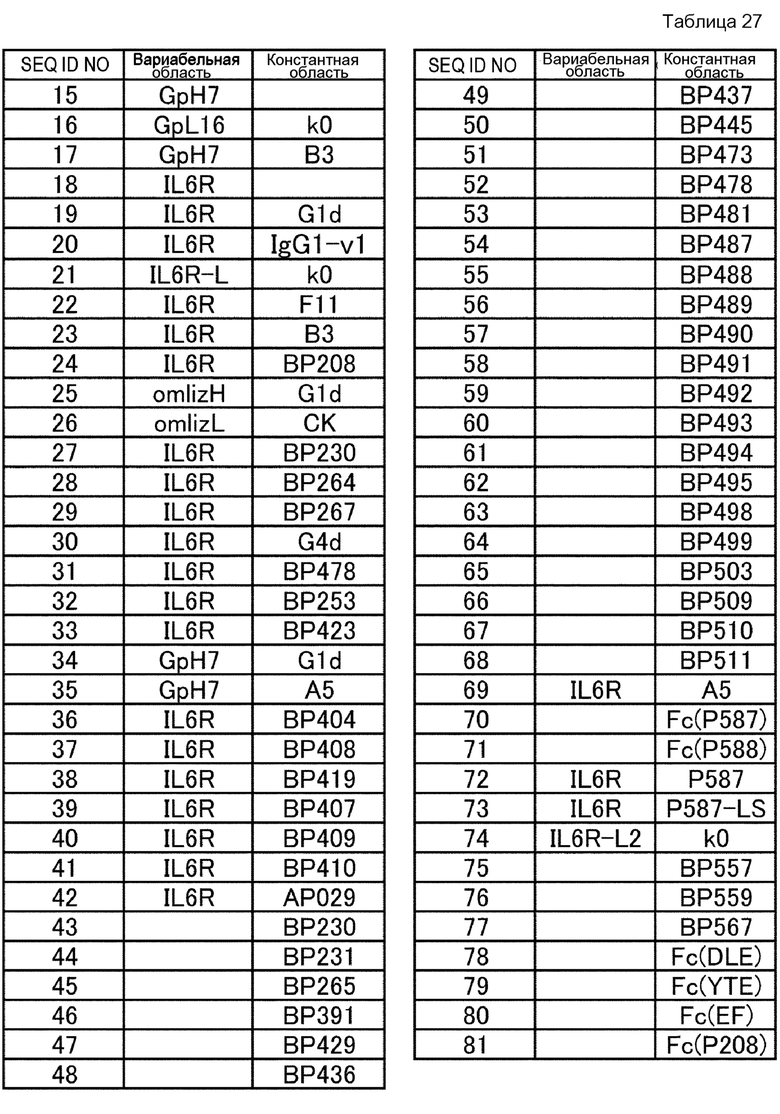
Промышленное применение
Настоящее изобретение относится к варианту Fc-области с повышенной активностью связывания с FcγRIIb и повышенной селективностью связывания с FcγRIIb по сравнению с активностью и селективностью с FcγRIIа (типа R), в отличие от полипептида, содержащего Fc-область, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и); и к полипептиду, содержащему вариант Fc-области. Применение указанного полипептида позволяет осуществлять передачу сигнала, ингибирующего воспалительный иммунный ответ, опосредуемый фосфорилированием ITIM FcγRIIb. Кроме того, в результате сообщения Fc-части антитела способности селективно связываться с FcγRIIb, можно ингибировать продуцирование антитела против лекарственного средства благодаря FcγRIIb-опосредуемому иммуносупрессорному действию.
--->
Список последовательностей
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> Варианты FcγRIIB-специфической Fc-области
<130> C1-A1212P
<150> JP 2012-185868
<151> 2012-08-24
<160> 81
<170> PatentIn version 3.5
<210> 1
<211> 1125
<212> ДНК
<213> Homo sapiens
<400> 1
atgtggttct tgacaactct gctcctttgg gttccagttg atgggcaagt ggacaccaca 60
aaggcagtga tcactttgca gcctccatgg gtcagcgtgt tccaagagga aaccgtaacc 120
ttgcactgtg aggtgctcca tctgcctggg agcagctcta cacagtggtt tctcaatggc 180
acagccactc agacctcgac ccccagctac agaatcacct ctgccagtgt caatgacagt 240
ggtgaataca ggtgccagag aggtctctca gggcgaagtg accccataca gctggaaatc 300
cacagaggct ggctactact gcaggtctcc agcagagtct tcacggaagg agaacctctg 360
gccttgaggt gtcatgcgtg gaaggataag ctggtgtaca atgtgcttta ctatcgaaat 420
ggcaaagcct ttaagttttt ccactggaat tctaacctca ccattctgaa aaccaacata 480
agtcacaatg gcacctacca ttgctcaggc atgggaaagc atcgctacac atcagcagga 540
atatctgtca ctgtgaaaga gctatttcca gctccagtgc tgaatgcatc tgtgacatcc 600
ccactcctgg aggggaatct ggtcaccctg agctgtgaaa caaagttgct cttgcagagg 660
cctggtttgc agctttactt ctccttctac atgggcagca agaccctgcg aggcaggaac 720
acatcctctg aataccaaat actaactgct agaagagaag actctgggtt atactggtgc 780
gaggctgcca cagaggatgg aaatgtcctt aagcgcagcc ctgagttgga gcttcaagtg 840
cttggcctcc agttaccaac tcctgtctgg tttcatgtcc ttttctatct ggcagtggga 900
ataatgtttt tagtgaacac tgttctctgg gtgacaatac gtaaagaact gaaaagaaag 960
aaaaagtggg atttagaaat ctctttggat tctggtcatg agaagaaggt aatttccagc 1020
cttcaagaag acagacattt agaagaagag ctgaaatgtc aggaacaaaa agaagaacag 1080
ctgcaggaag gggtgcaccg gaaggagccc cagggggcca cgtag 1125
<210> 2
<211> 374
<212> PRT
<213> Homo sapiens
<400> 2
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
Glu Pro Gln Gly Ala Thr
370
<210> 3
<211> 951
<212> ДНК
<213> Homo sapiens
<400> 3
atgactatgg agacccaaat gtctcagaat gtatgtccca gaaacctgtg gctgcttcaa 60
ccattgacag ttttgctgct gctggcttct gcagacagtc aagctgctcc cccaaaggct 120
gtgctgaaac ttgagccccc gtggatcaac gtgctccagg aggactctgt gactctgaca 180
tgccaggggg ctcgcagccc tgagagcgac tccattcagt ggttccacaa tgggaatctc 240
attcccaccc acacgcagcc cagctacagg ttcaaggcca acaacaatga cagcggggag 300
tacacgtgcc agactggcca gaccagcctc agcgaccctg tgcatctgac tgtgctttcc 360
gaatggctgg tgctccagac ccctcacctg gagttccagg agggagaaac catcatgctg 420
aggtgccaca gctggaagga caagcctctg gtcaaggtca cattcttcca gaatggaaaa 480
tcccagaaat tctcccattt ggatcccacc ttctccatcc cacaagcaaa ccacagtcac 540
agtggtgatt accactgcac aggaaacata ggctacacgc tgttctcatc caagcctgtg 600
accatcactg tccaagtgcc cagcatgggc agctcttcac caatgggggt cattgtggct 660
gtggtcattg cgactgctgt agcagccatt gttgctgctg tagtggcctt gatctactgc 720
aggaaaaagc ggatttcagc caattccact gatcctgtga aggctgccca atttgagcca 780
cctggacgtc aaatgattgc catcagaaag agacaacttg aagaaaccaa caatgactat 840
gaaacagctg acggcggcta catgactctg aaccccaggg cacctactga cgatgataaa 900
aacatctacc tgactcttcc tcccaacgac catgtcaaca gtaataacta a 951
<210> 4
<211> 316
<212> PRT
<213> Homo sapiens
<400> 4
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 5
<211> 876
<212> ДНК
<213> Homo sapiens
<400> 5
atgggaatcc tgtcattctt acctgtcctt gccactgaga gtgactgggc tgactgcaag 60
tccccccagc cttggggtca tatgcttctg tggacagctg tgctattcct ggctcctgtt 120
gctgggacac ctgcagctcc cccaaaggct gtgctgaaac tcgagcccca gtggatcaac 180
gtgctccagg aggactctgt gactctgaca tgccggggga ctcacagccc tgagagcgac 240
tccattcagt ggttccacaa tgggaatctc attcccaccc acacgcagcc cagctacagg 300
ttcaaggcca acaacaatga cagcggggag tacacgtgcc agactggcca gaccagcctc 360
agcgaccctg tgcatctgac tgtgctttct gagtggctgg tgctccagac ccctcacctg 420
gagttccagg agggagaaac catcgtgctg aggtgccaca gctggaagga caagcctctg 480
gtcaaggtca cattcttcca gaatggaaaa tccaagaaat tttcccgttc ggatcccaac 540
ttctccatcc cacaagcaaa ccacagtcac agtggtgatt accactgcac aggaaacata 600
ggctacacgc tgtactcatc caagcctgtg accatcactg tccaagctcc cagctcttca 660
ccgatgggga tcattgtggc tgtggtcact gggattgctg tagcggccat tgttgctgct 720
gtagtggcct tgatctactg caggaaaaag cggatttcag ccaatcccac taatcctgat 780
gaggctgaca aagttggggc tgagaacaca atcacctatt cacttctcat gcacccggat 840
gctctggaag agcctgatga ccagaaccgt atttag 876
<210> 6
<211> 291
<212> PRT
<213> Homo sapiens
<400> 6
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
Asn Arg Ile
290
<210> 7
<211> 765
<212> ДНК
<213> Homo sapiens
<400> 7
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagggt gctcgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gttgacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaagg caggaagtat tttcatcata attctgactt ctacattcca 480
aaagccacac tcaaagacag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tgtcagtgtc aaccatctca 600
tcattctttc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattc gaagctcaac aagagactgg 720
aaggaccata aatttaaatg gagaaaggac cctcaagaca aatga 765
<210> 8
<211> 254
<212> PRT
<213> Homo sapiens
<400> 8
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 9
<211> 702
<212> ДНК
<213> Homo sapiens
<400> 9
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagcgt gcttgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gtcaacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaaga caggaagtat tttcatcata attctgactt ccacattcca 480
aaagccacac tcaaagatag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tggcagtgtc aaccatctca 600
tcattctctc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattt ga 702
<210> 10
<211> 233
<212> PRT
<213> Homo sapiens
<400> 10
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 11
<211> 330
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 12
<211> 326
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 13
<211> 377
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 14
<211> 327
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 15
<211> 115
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Thr Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Glu Ser Phe
50 55 60
Gln Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 16
<211> 219
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 16
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Gln Ala Ser Glu Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 17
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Thr Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Glu Ser Phe
50 55 60
Gln Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440
<210> 18
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 18
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 19
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 19
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 20
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 20
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 21
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 21
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 22
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 22
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Tyr His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 23
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 23
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 24
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 24
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Asp Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 25
<211> 449
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 26
<211> 218
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 26
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 27
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 27
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Asp Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 28
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 28
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 29
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 29
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 30
<211> 444
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 30
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440
<210> 31
<211> 444
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 31
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440
<210> 32
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 32
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 33
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 33
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ala Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 34
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Thr Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Glu Ser Phe
50 55 60
Gln Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 35
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 35
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Thr Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Glu Ser Phe
50 55 60
Gln Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Lys Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 36
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 36
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Tyr Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 37
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 37
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Gln Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 38
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 38
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Phe Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 39
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 39
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Asp Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 40
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 40
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Glu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 41
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 41
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Thr Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440 445
<210> 42
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 42
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Asp Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Arg Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Lys Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 43
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 43
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 44
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 45
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 45
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 46
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 46
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Thr Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 47
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 47
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Gly Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 48
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 48
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Thr Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 49
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 49
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Thr Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 50
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 50
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 51
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 51
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Gly Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 52
<211> 325
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 52
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Asp
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
195 200 205
Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 53
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 53
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 54
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 54
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 55
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 55
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 56
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 56
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 57
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 57
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 58
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 58
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 59
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 59
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 60
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 60
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 61
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 61
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 62
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 62
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Gly Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 63
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 63
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Leu Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 64
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 64
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Leu Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 65
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 65
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Gly Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 66
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 66
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Asp Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 67
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 67
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Pro Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 68
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 68
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Pro Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 69
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 69
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Lys Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 70
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 70
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 71
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 71
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 72
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 72
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Gly Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Ile Asp Val Ala Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 73
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 73
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Asp Leu Leu Gly Gly Asp
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Ile Asp Val Ala Glu Glu Asp
260 265 270
Gly Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 74
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 74
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 75
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 75
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Gly Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 76
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 76
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Asp Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 77
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 77
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Asp Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 78
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 78
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Asp Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Leu Pro Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 79
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 79
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 80
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 80
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 81
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> Искусственно полученная последовательность
<400> 81
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Arg Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<---
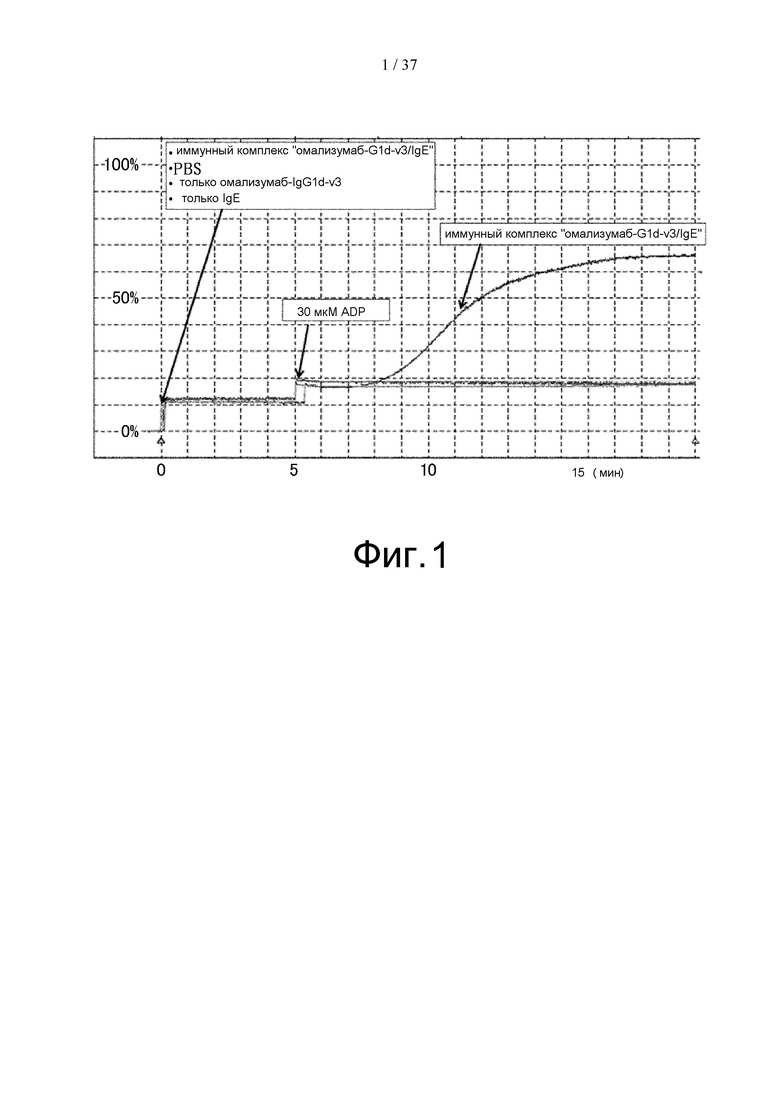
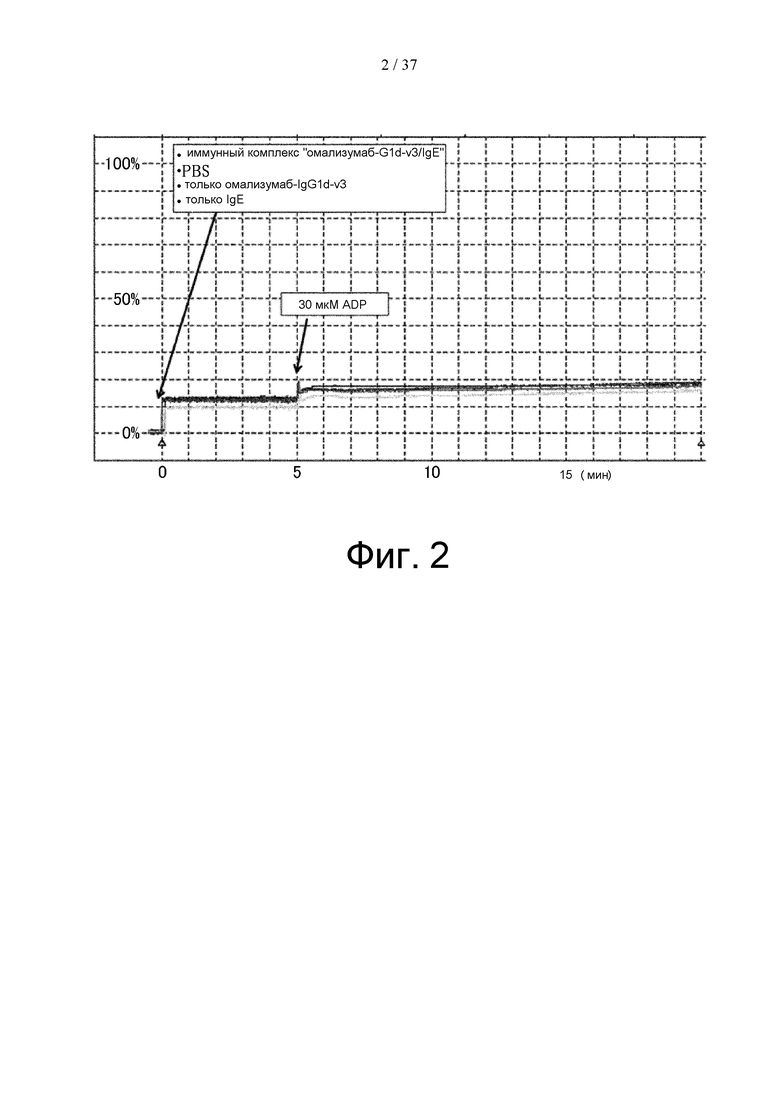
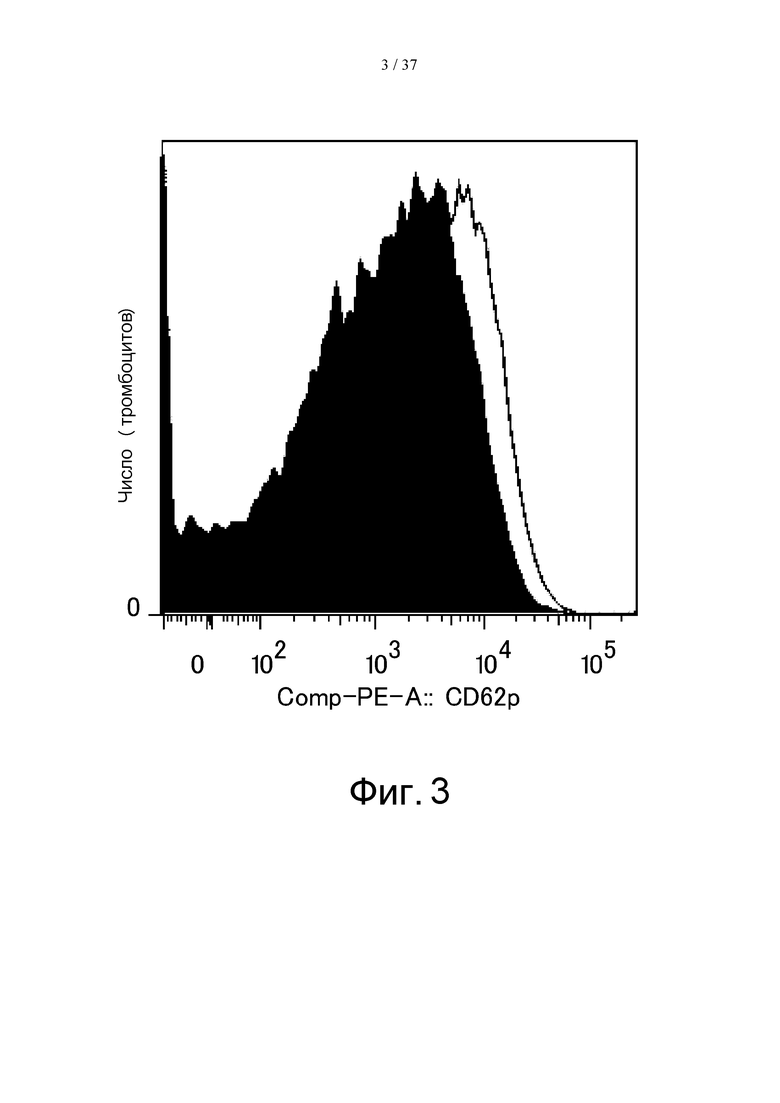
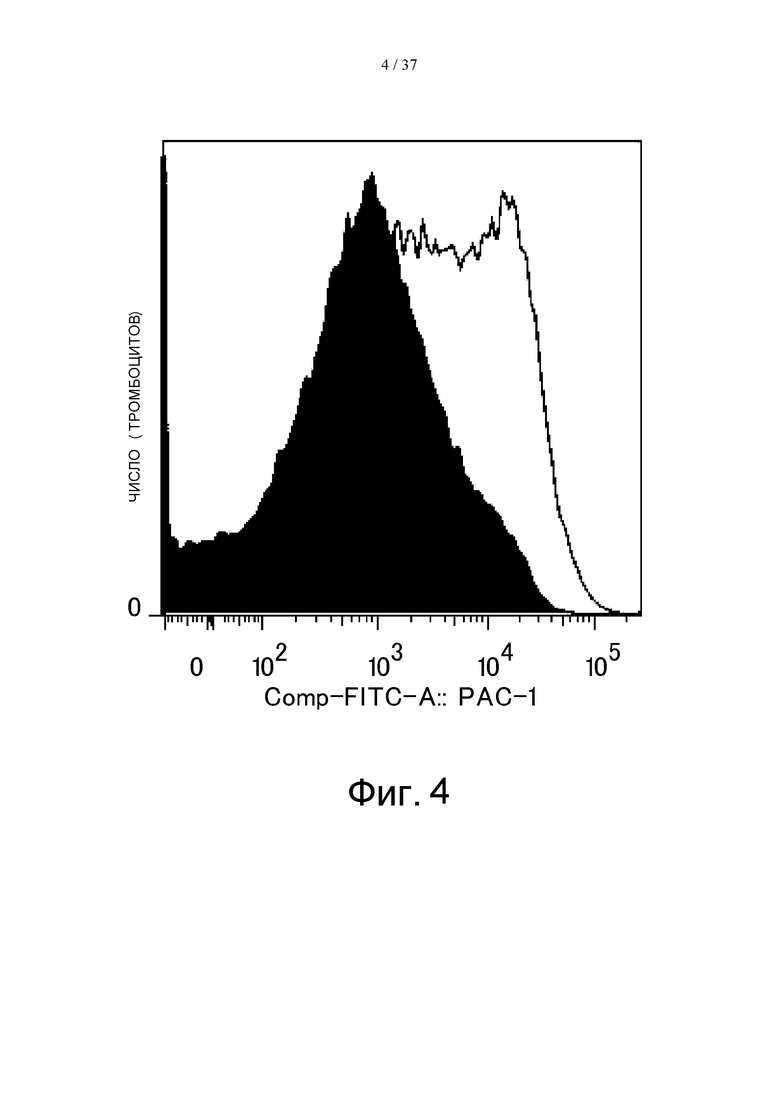
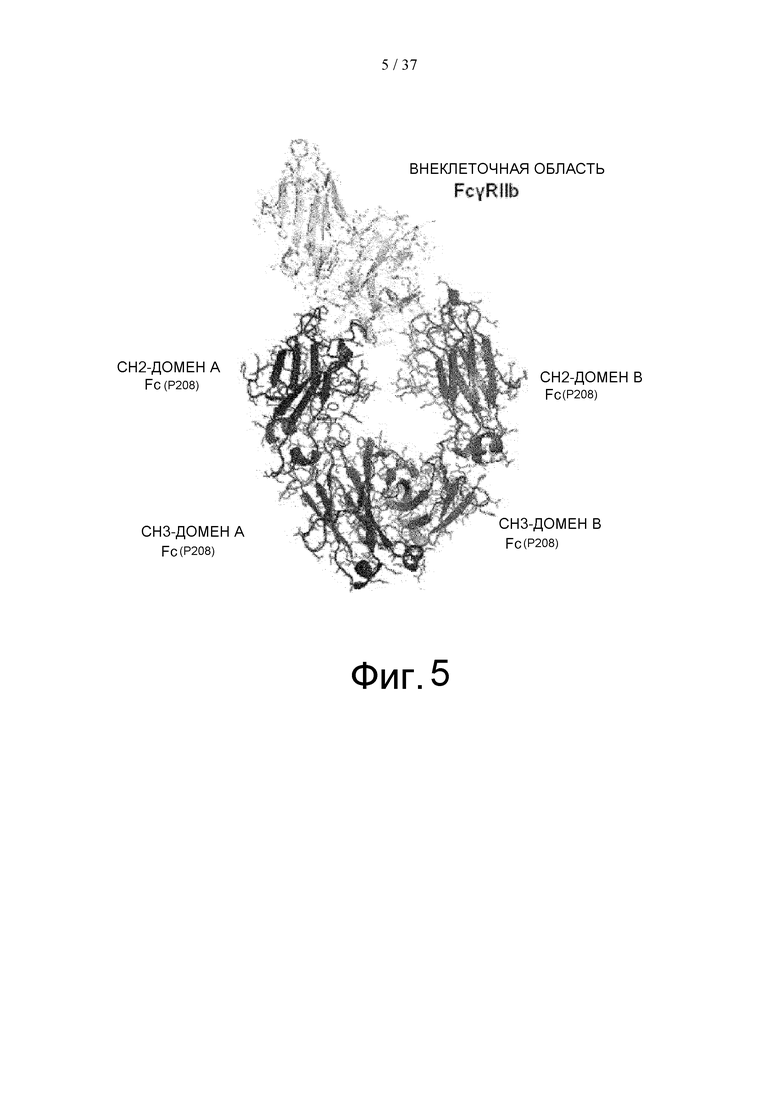
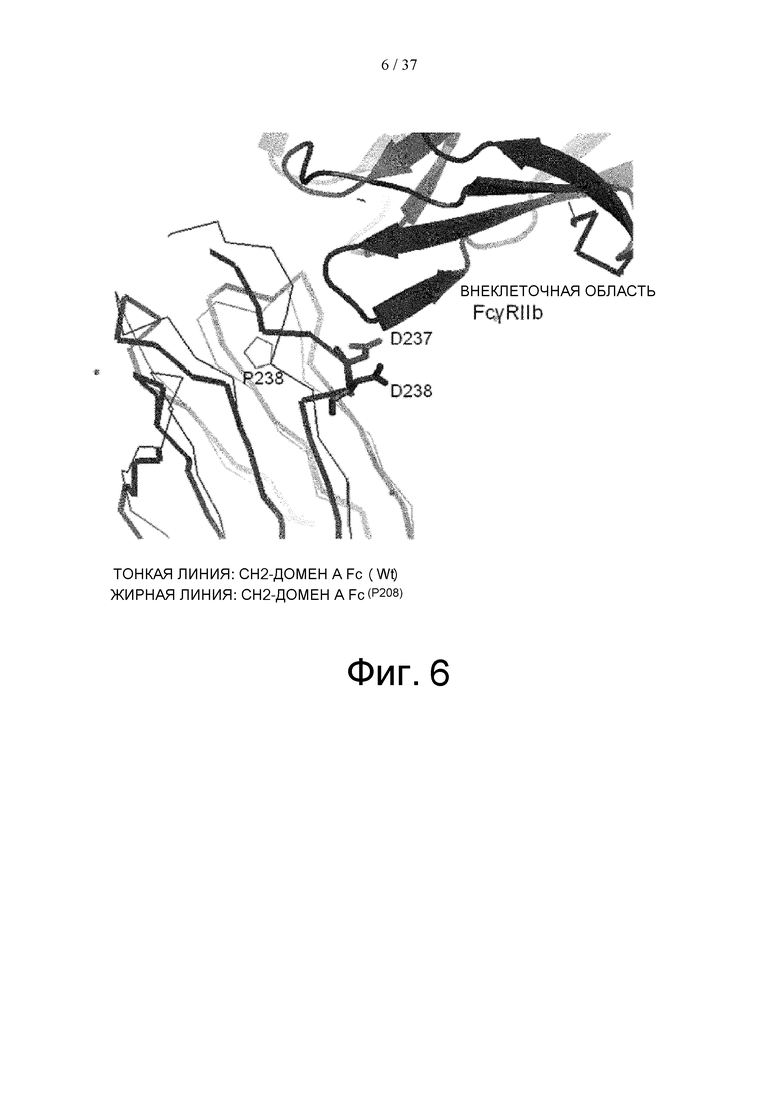
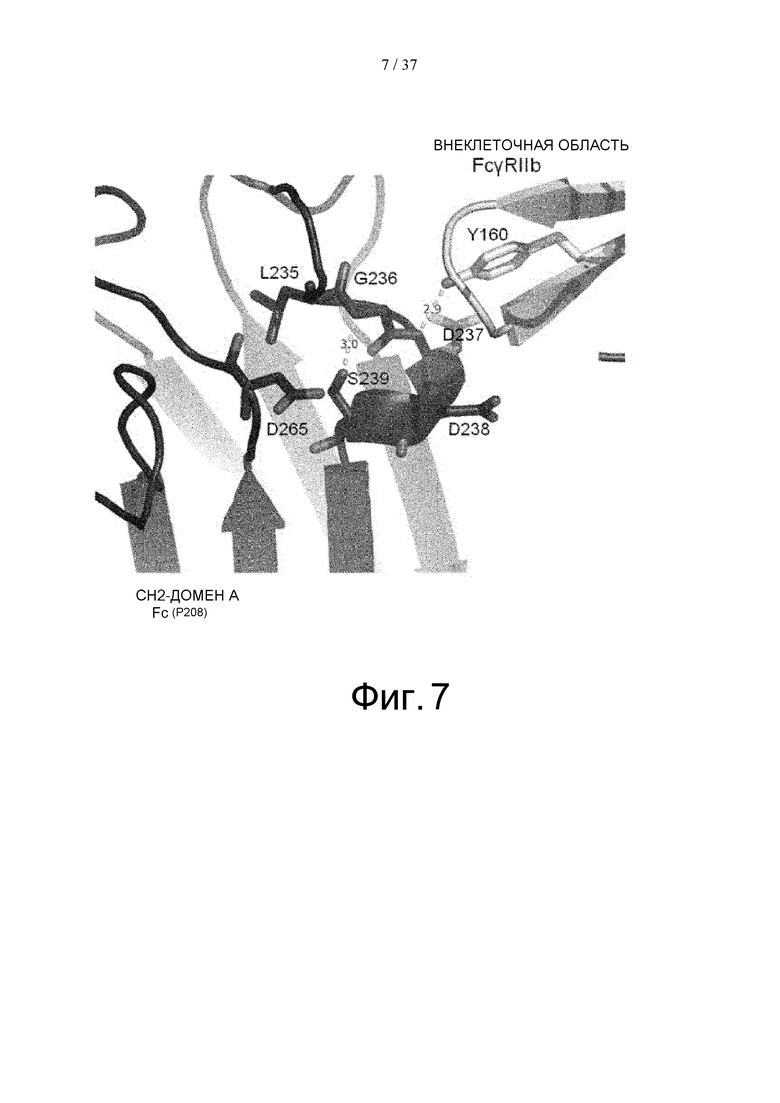
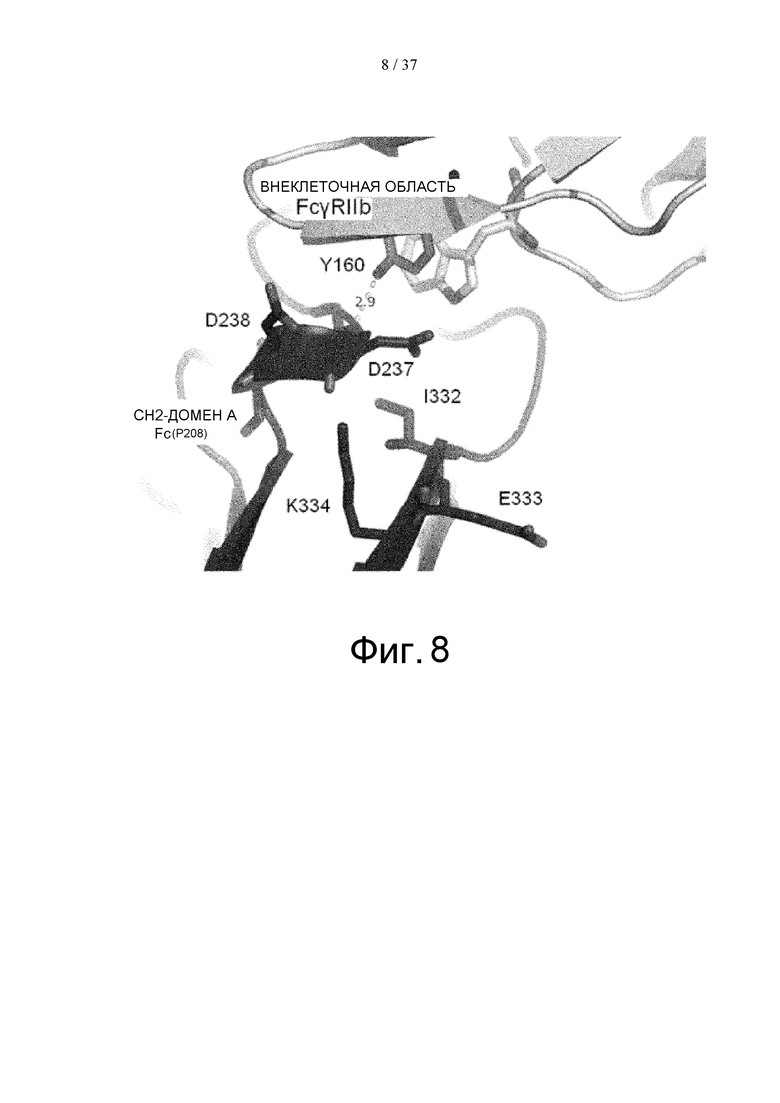
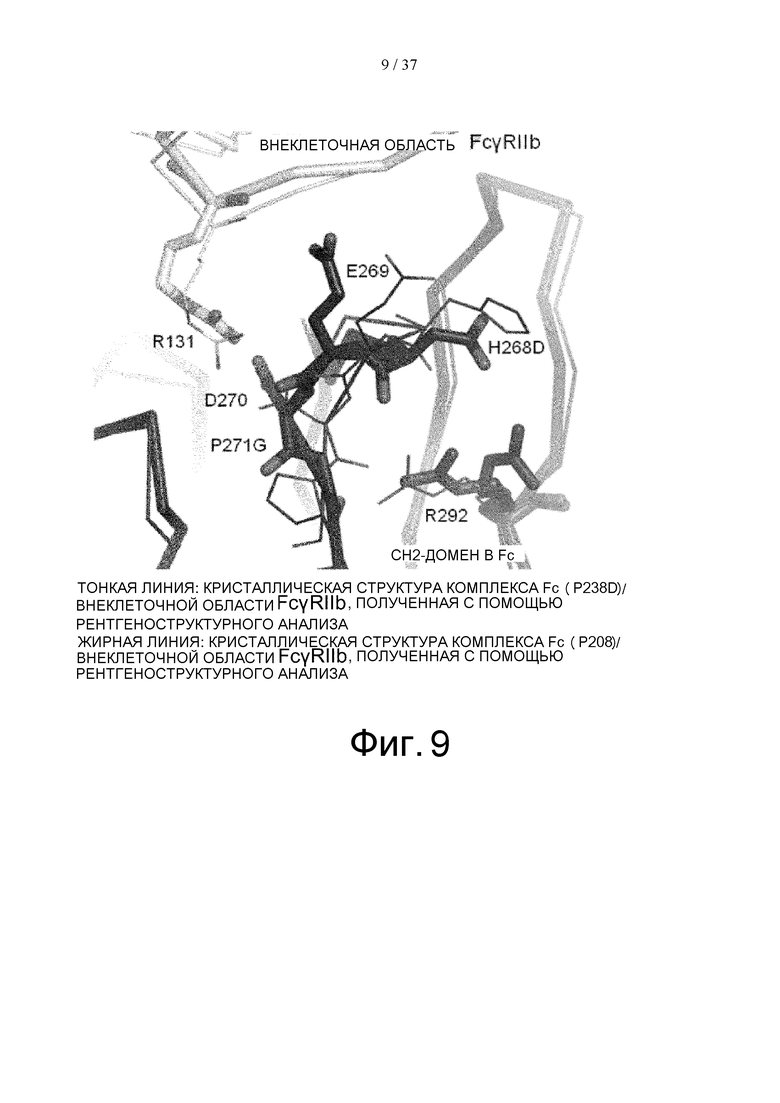
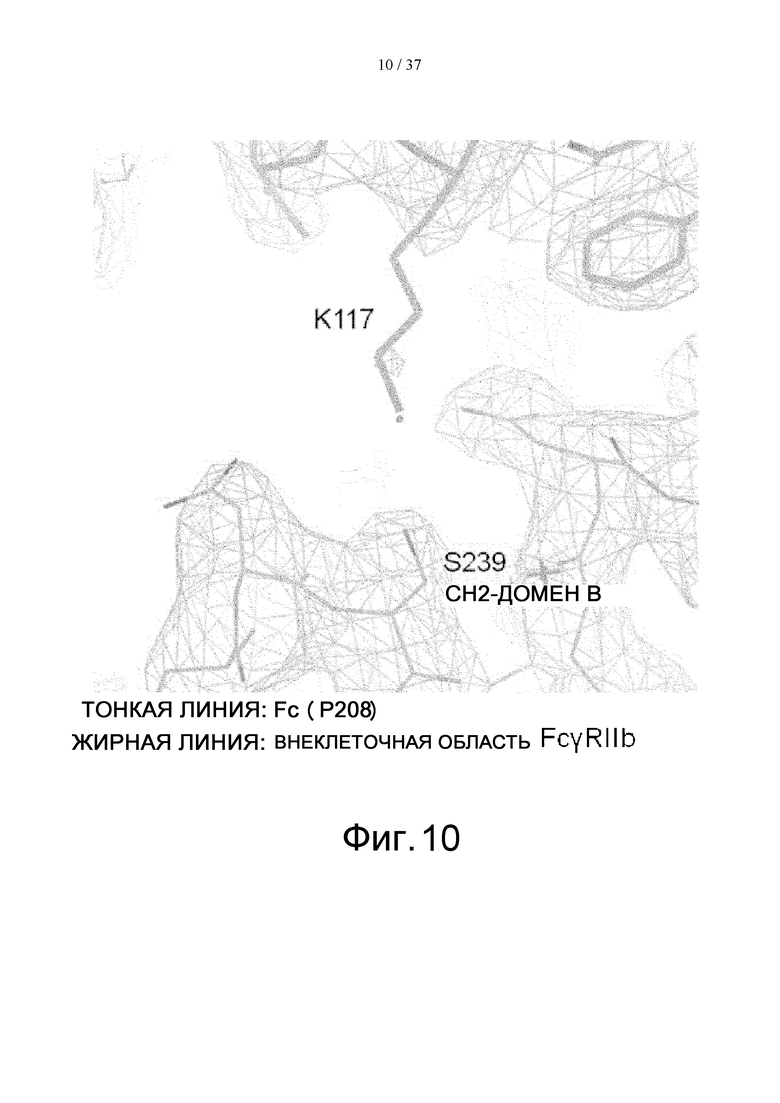
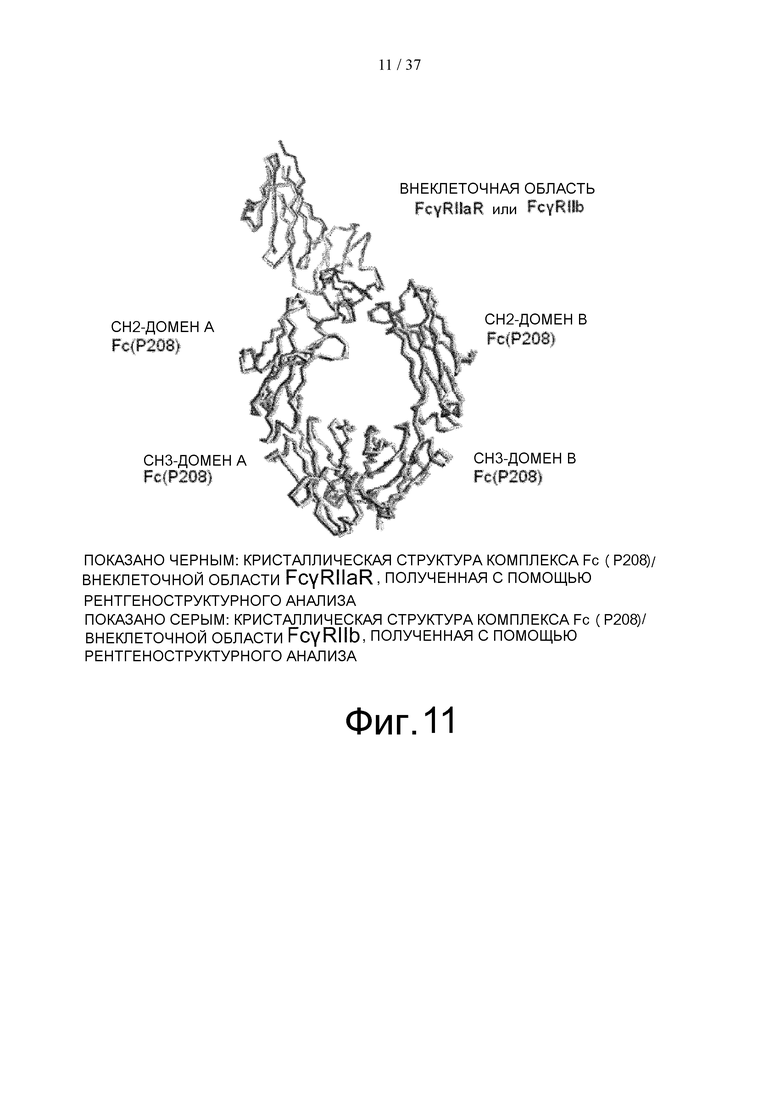
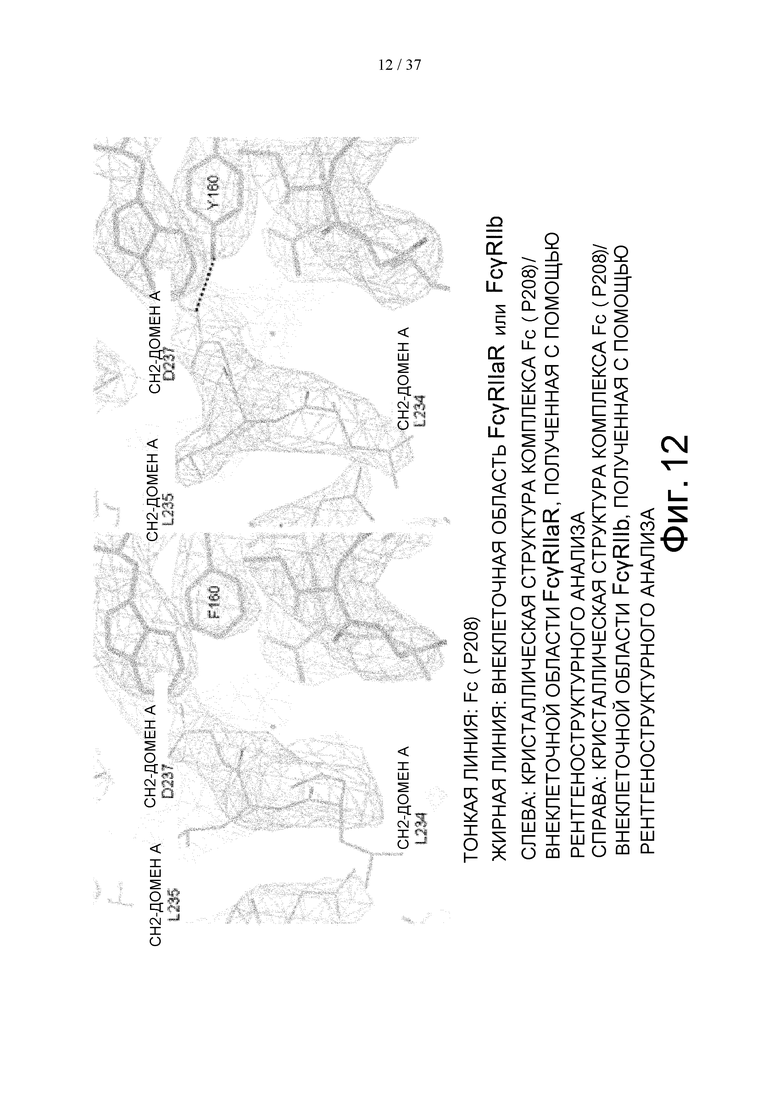

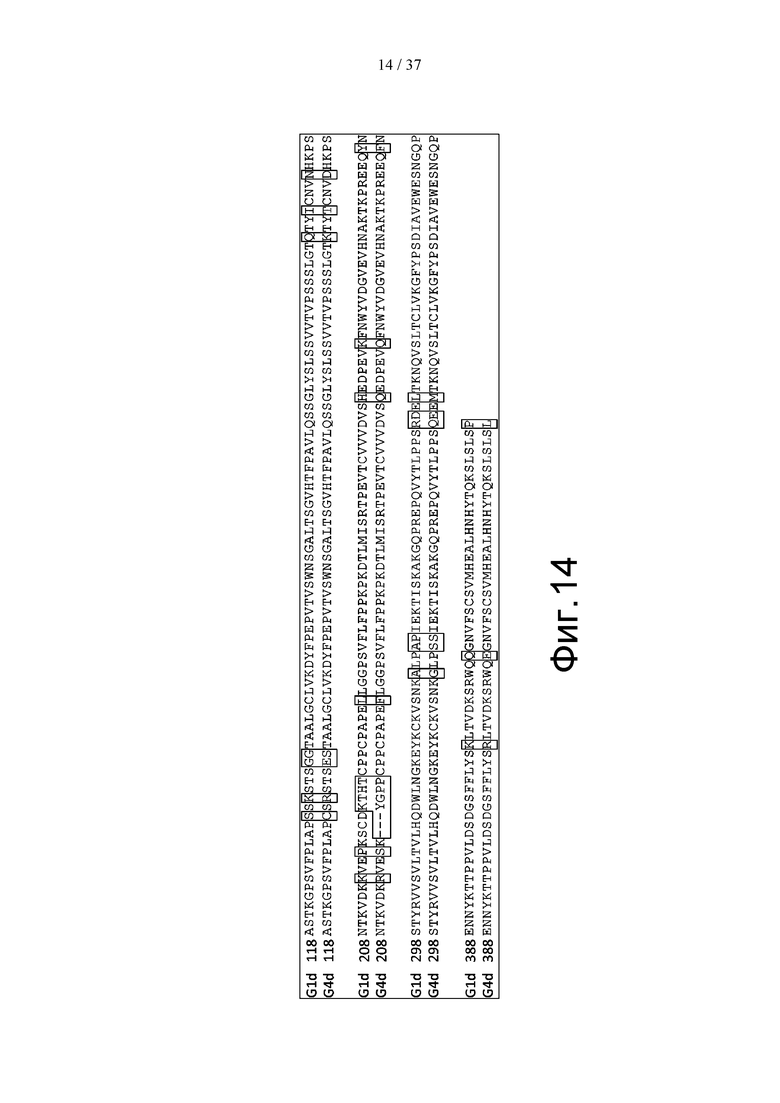
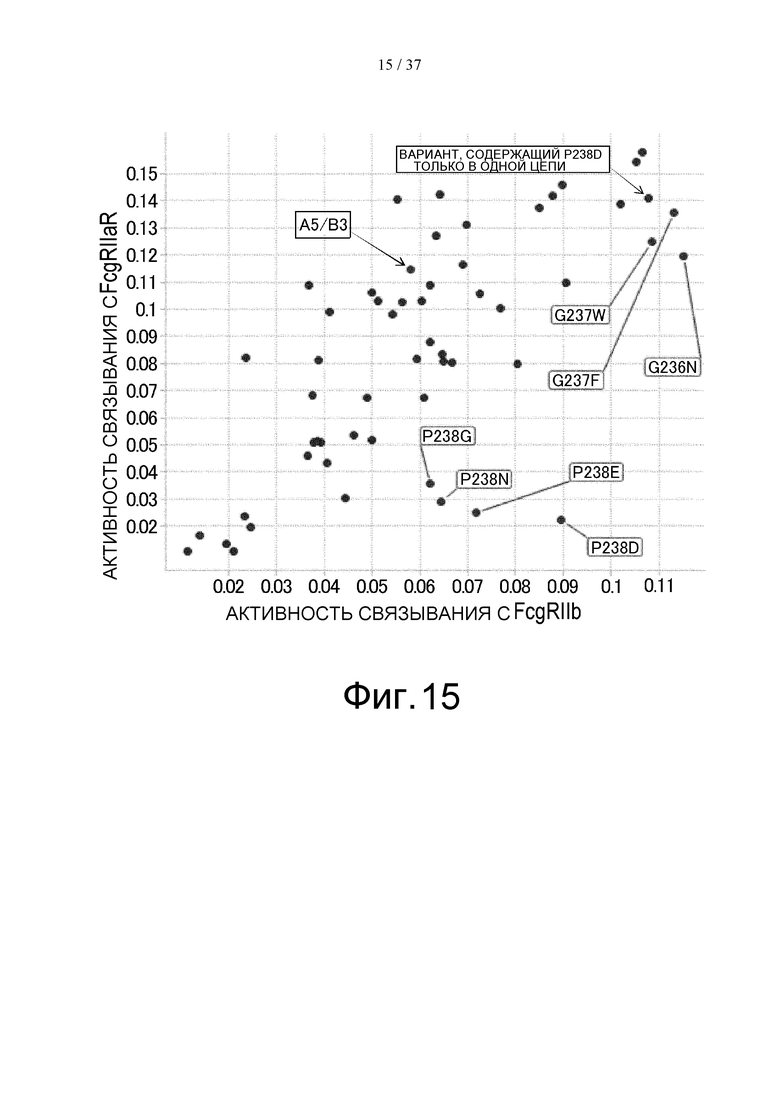
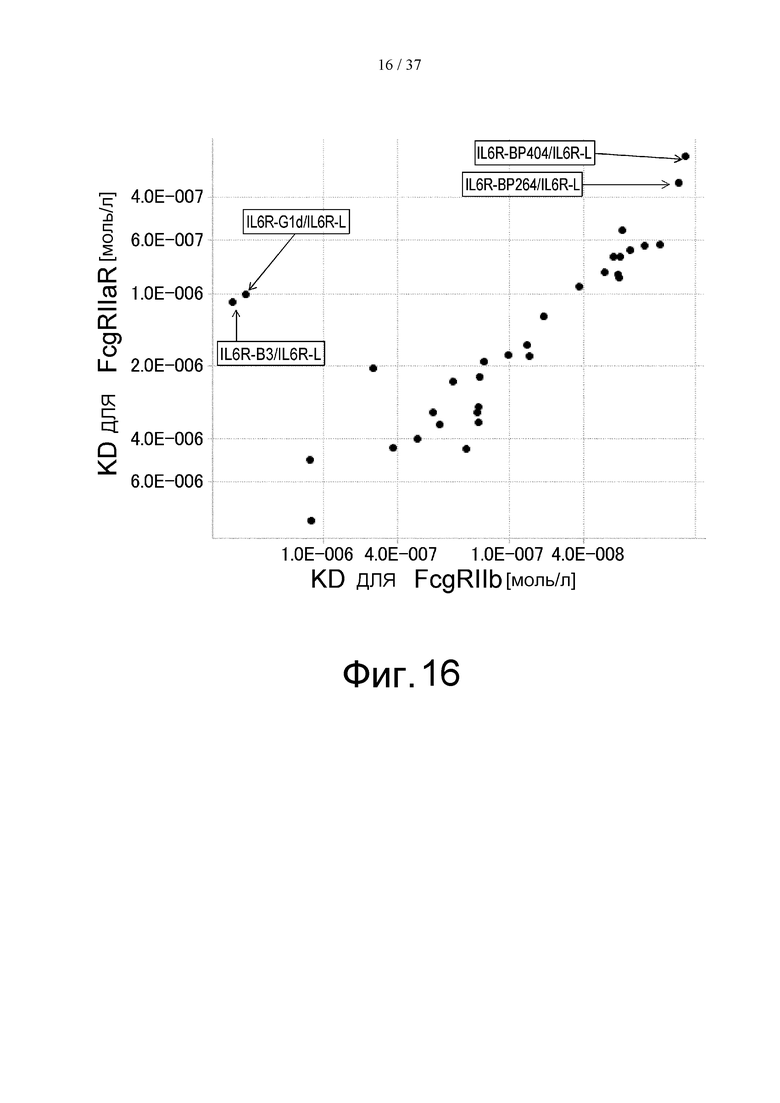
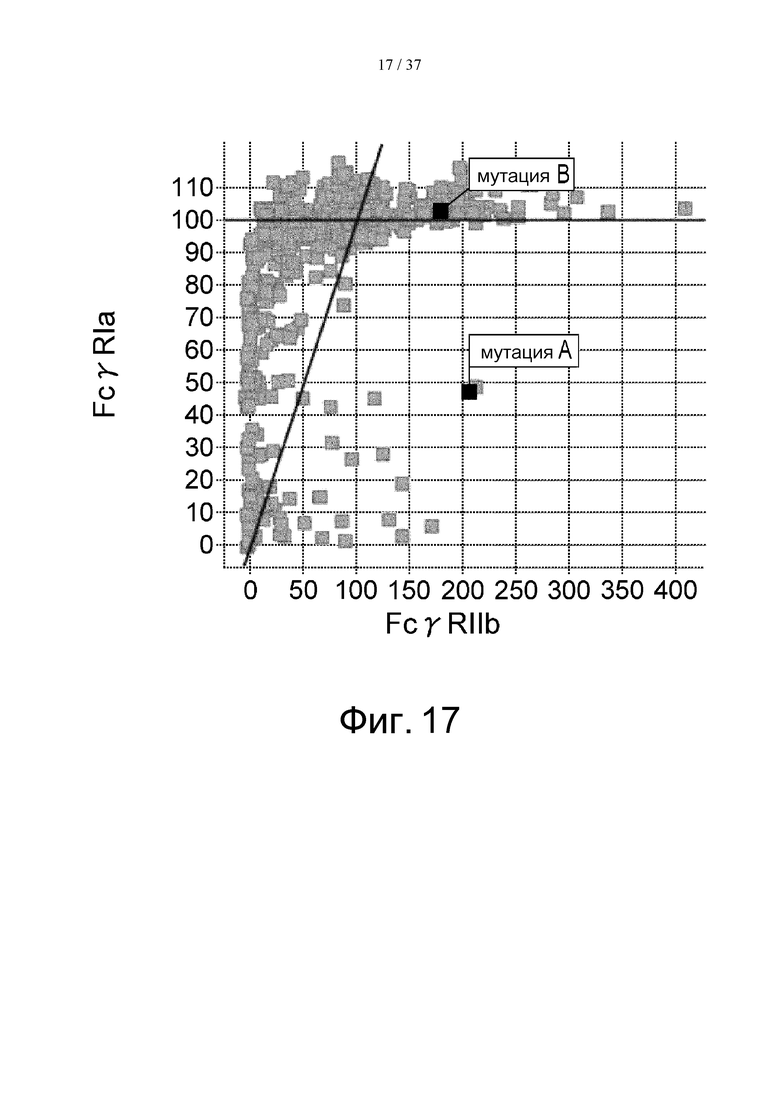
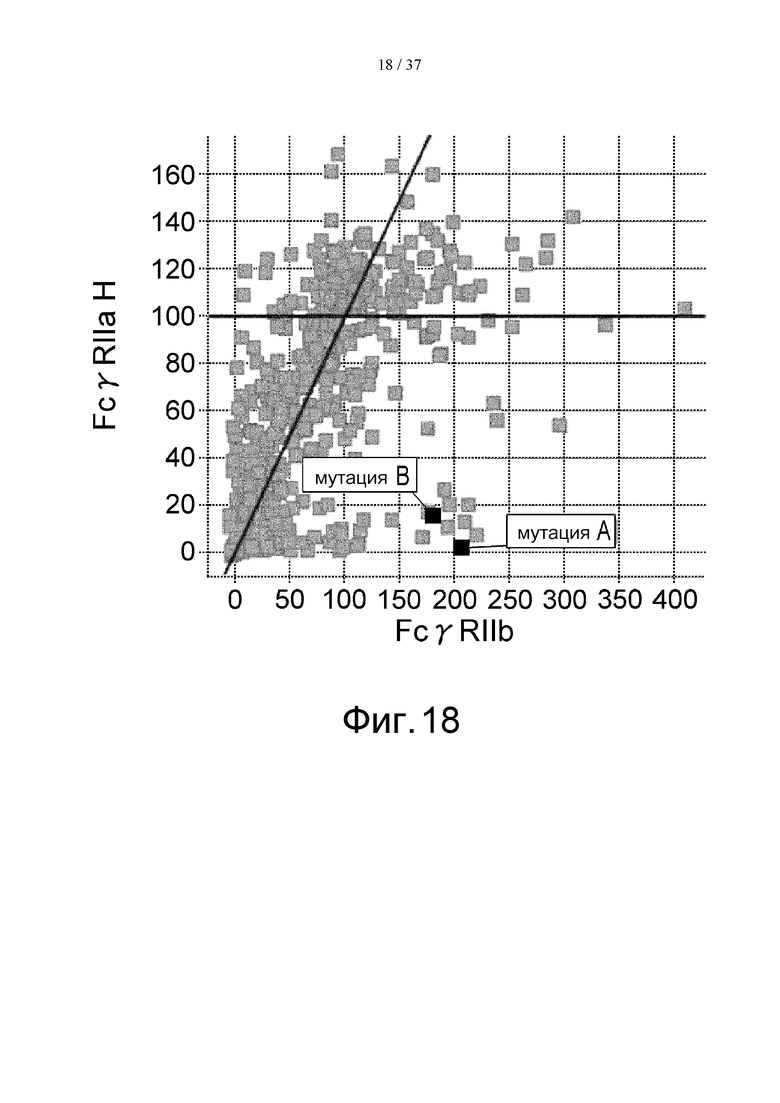
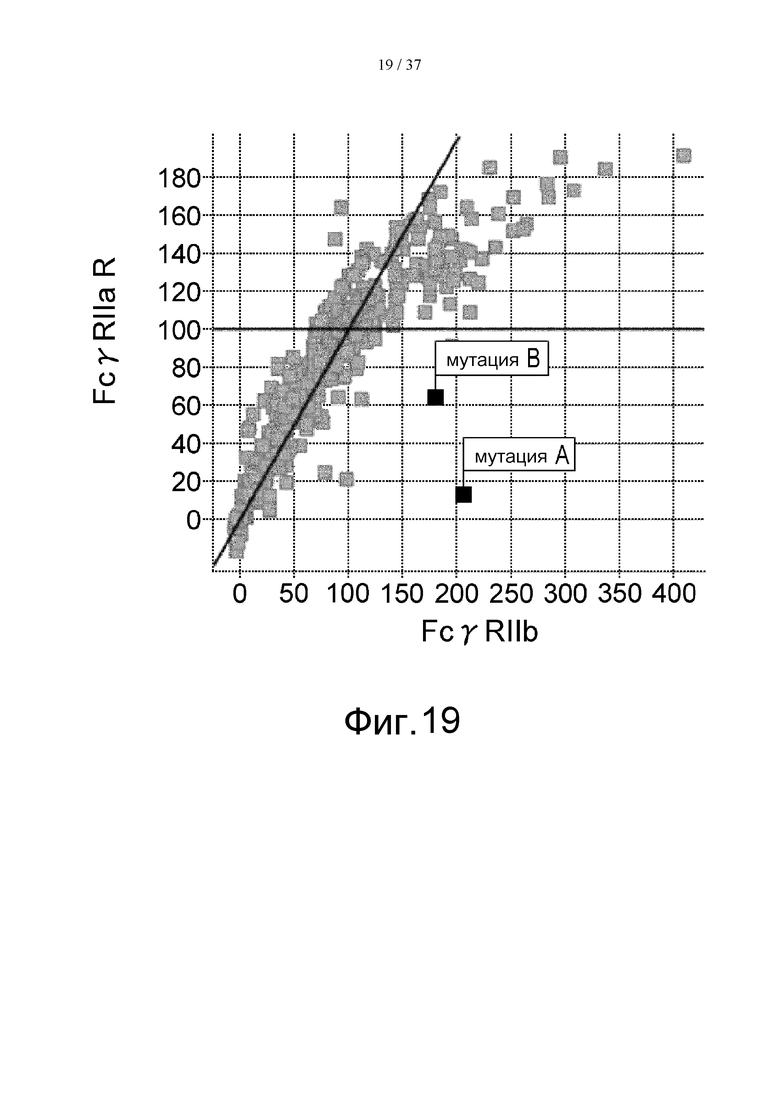
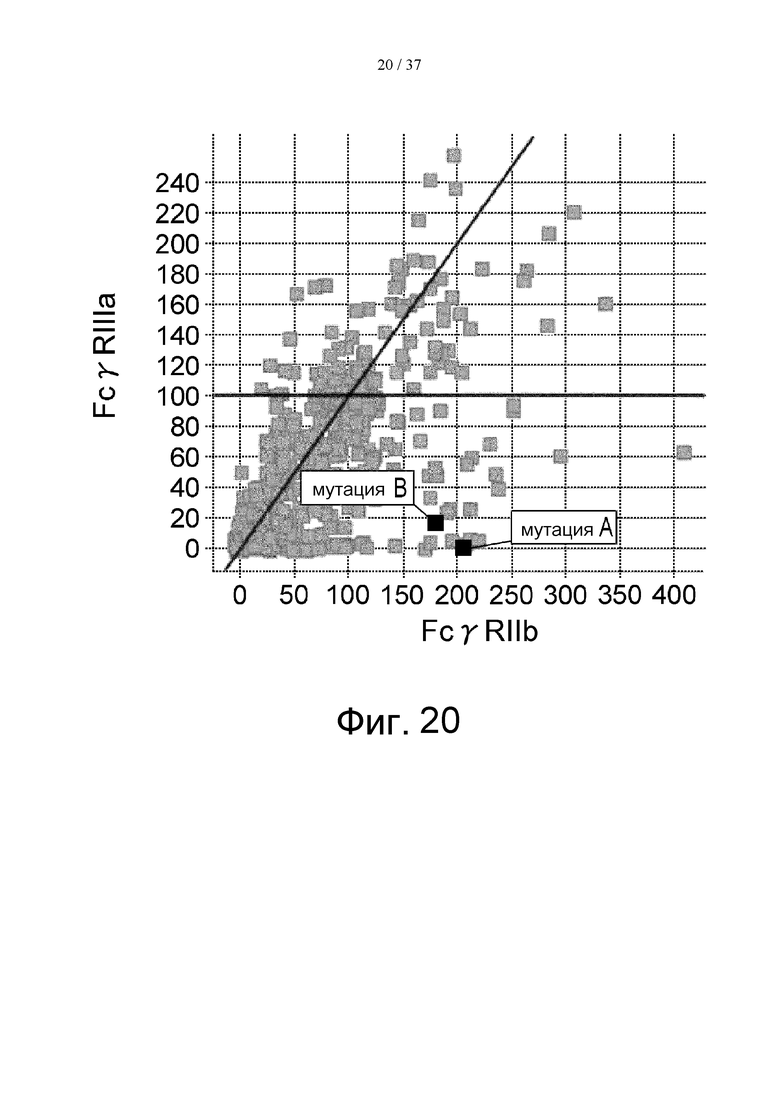
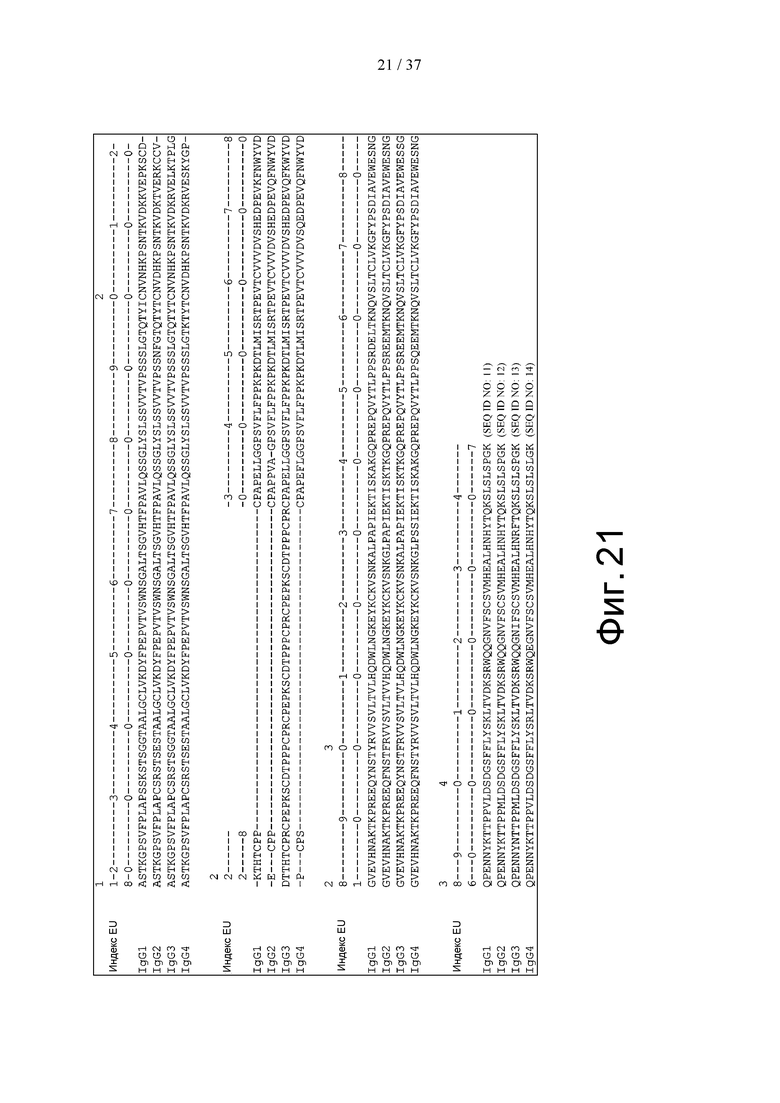
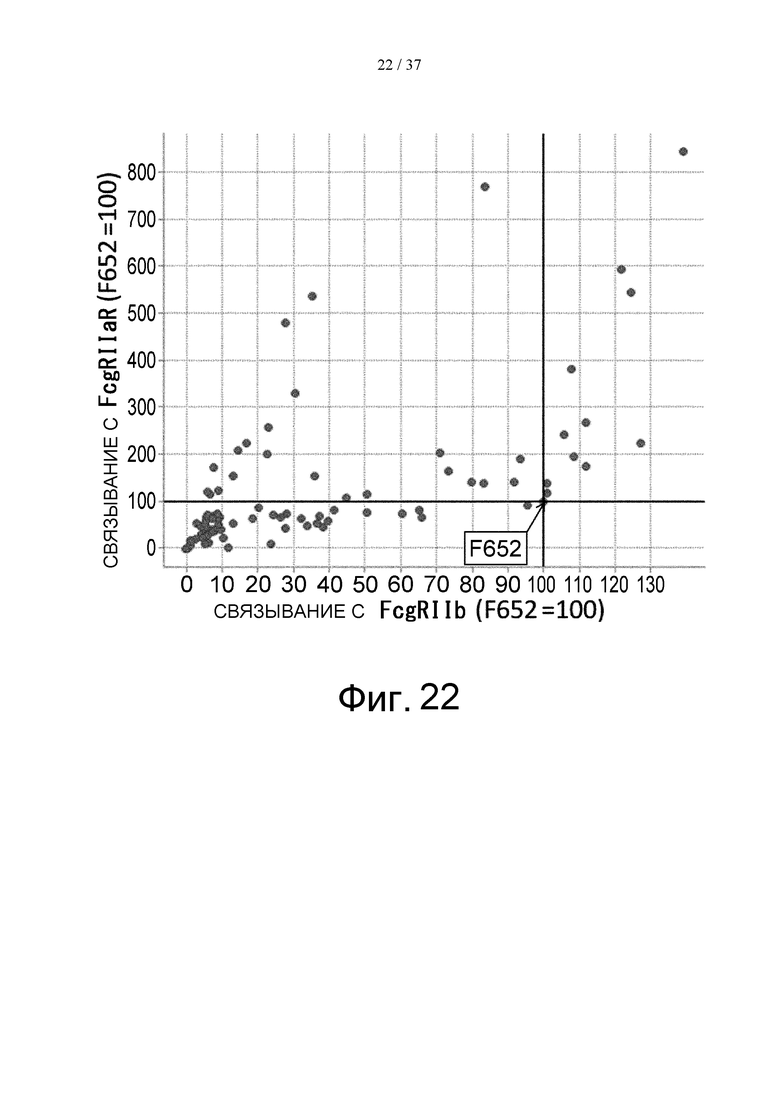
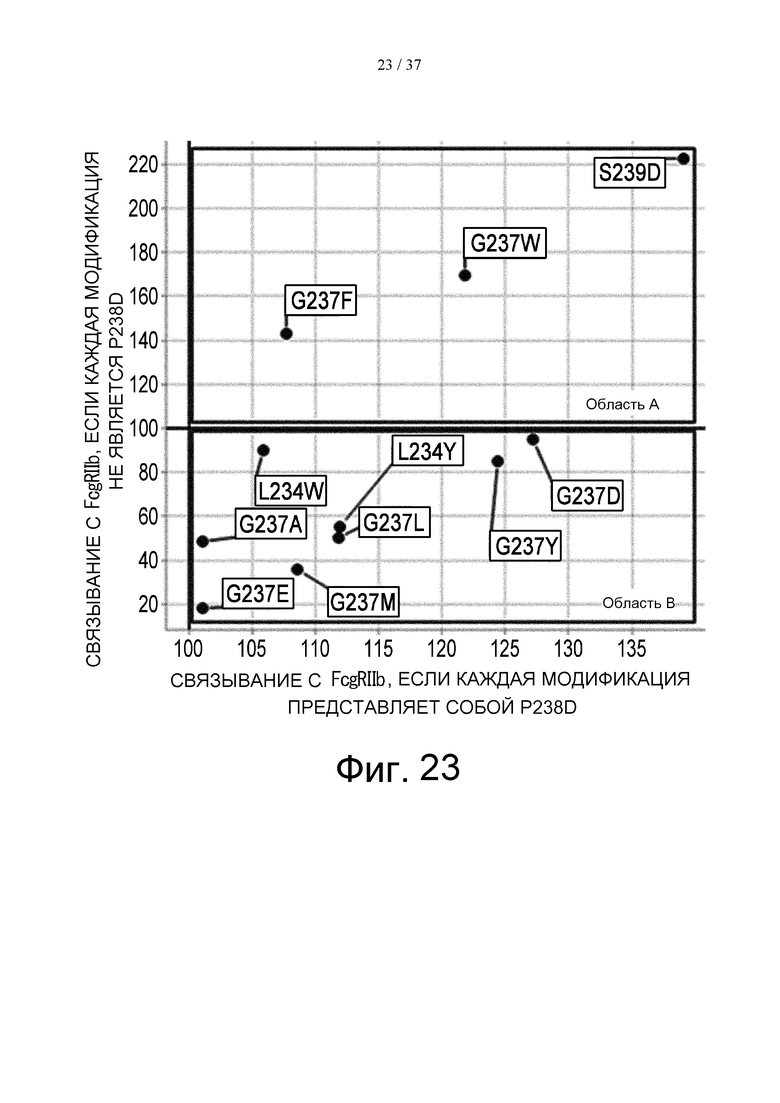
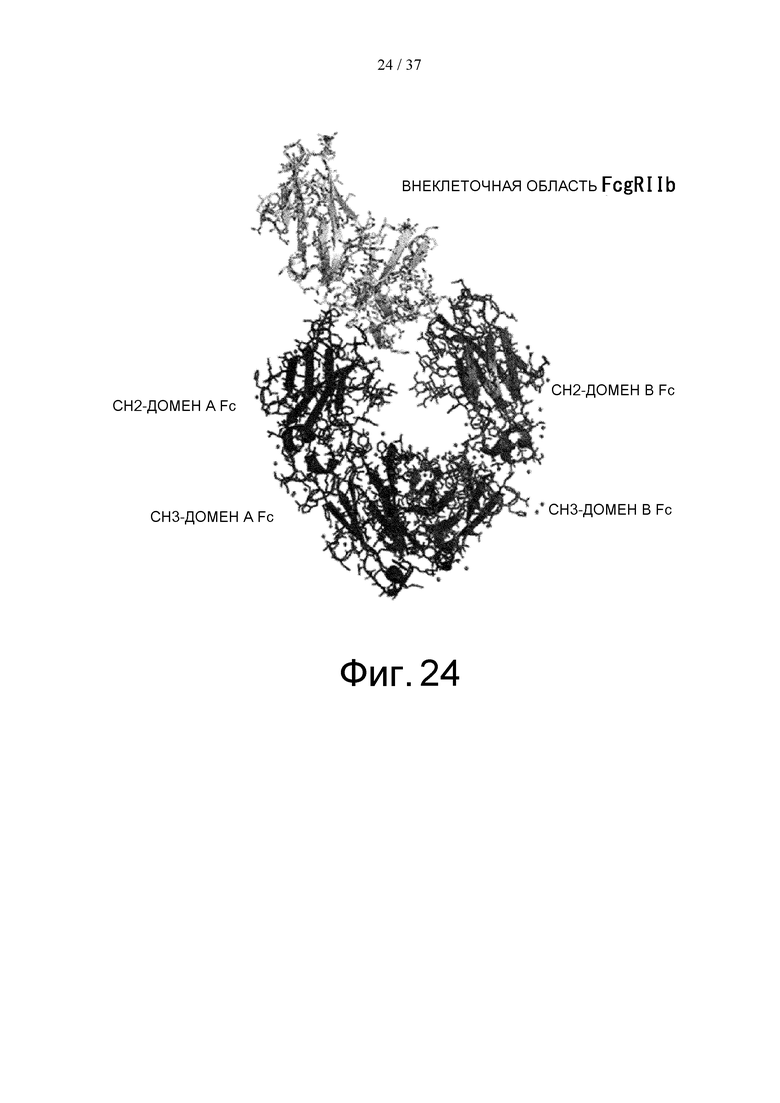


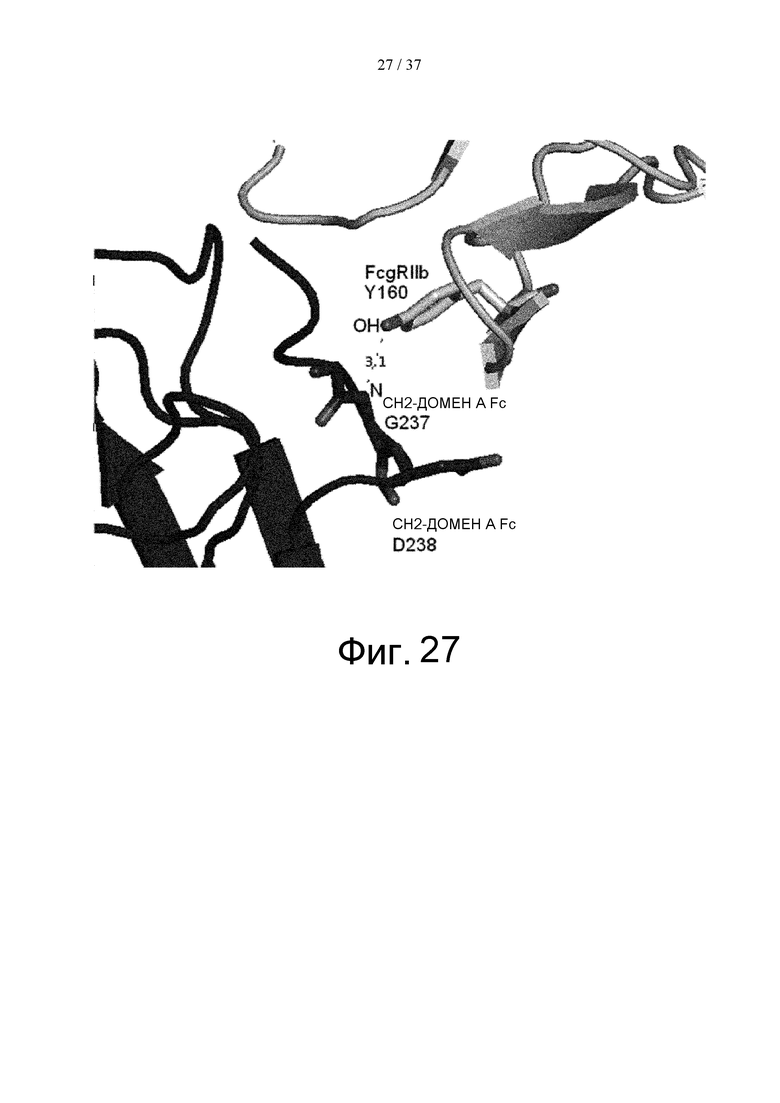

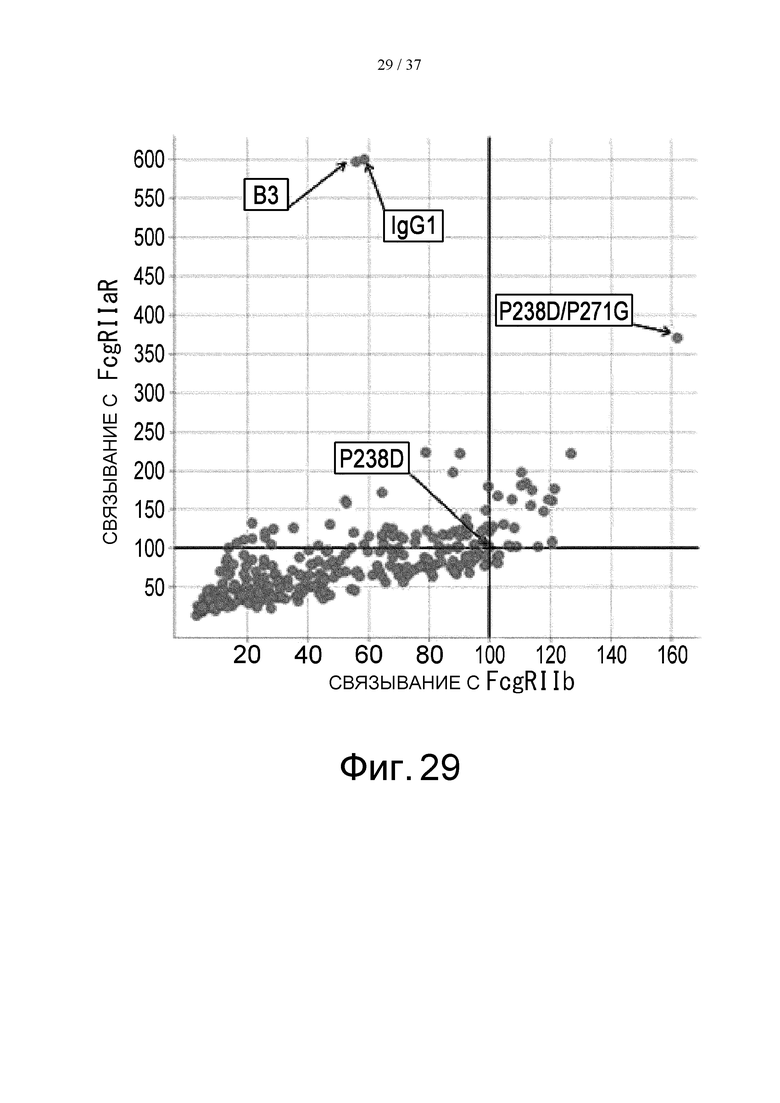
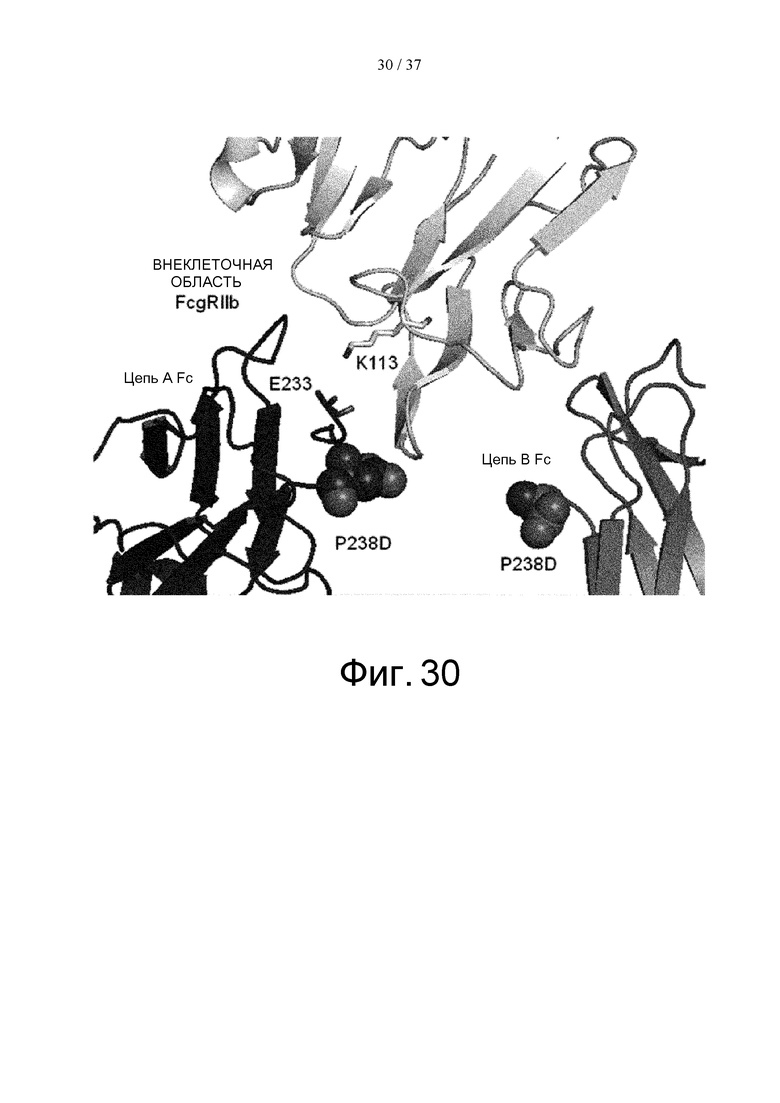
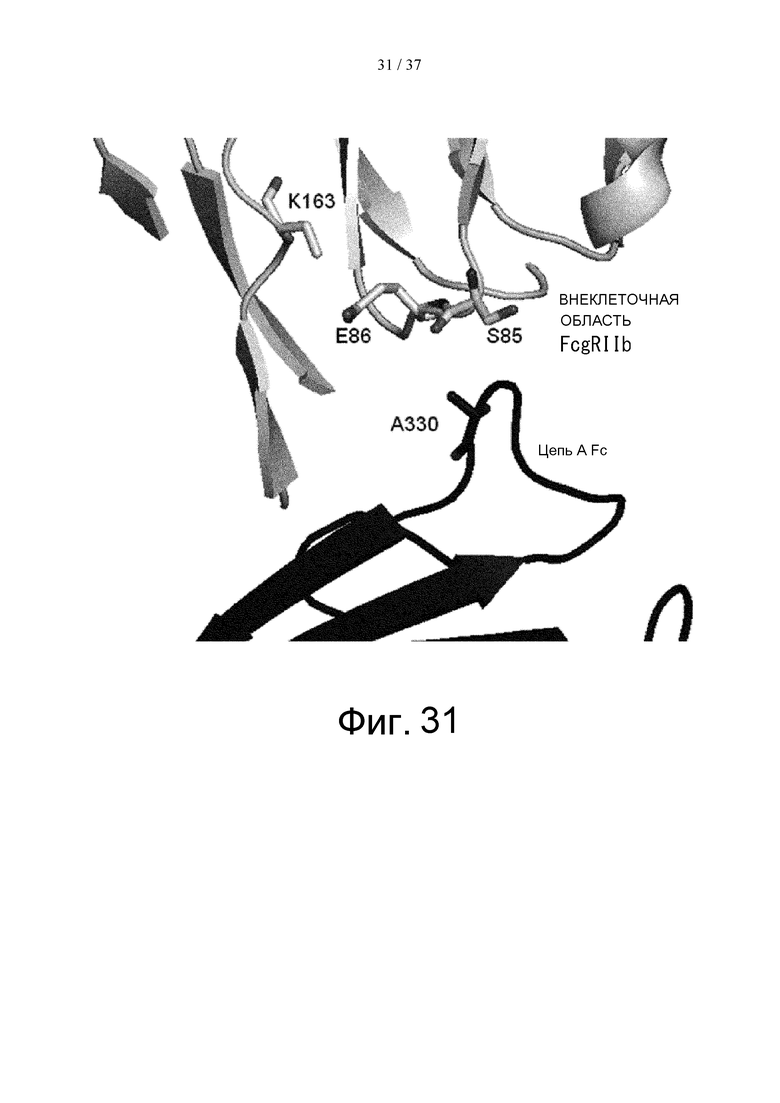
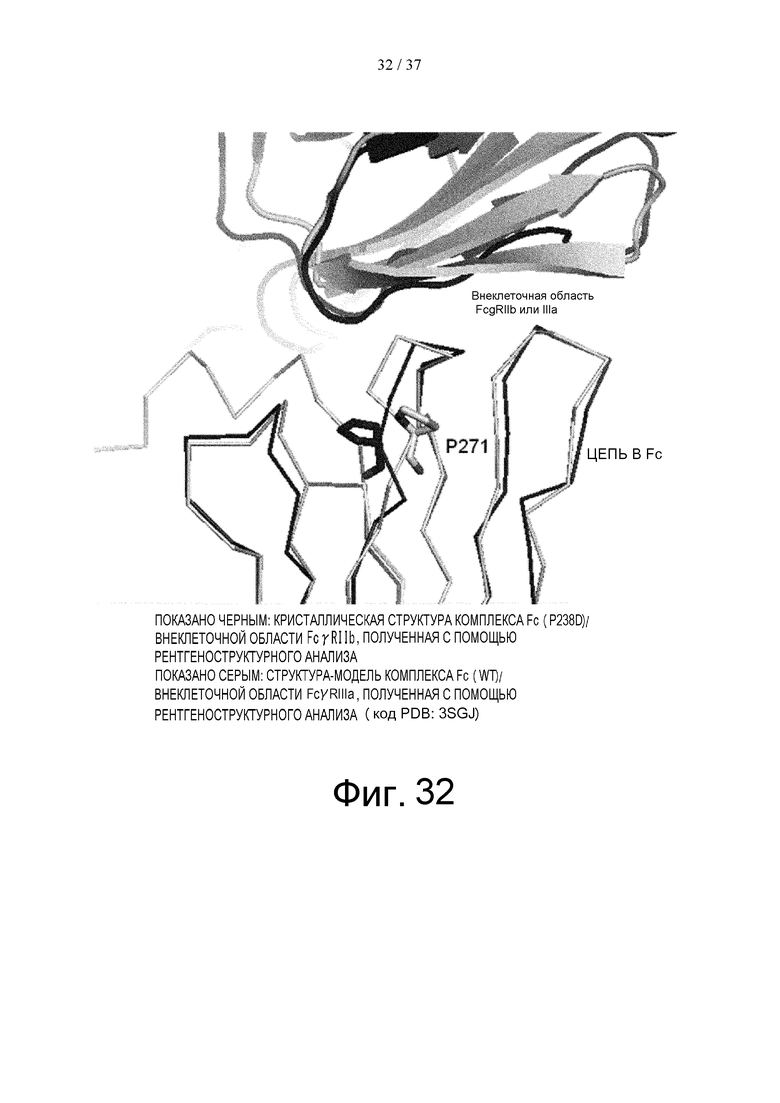
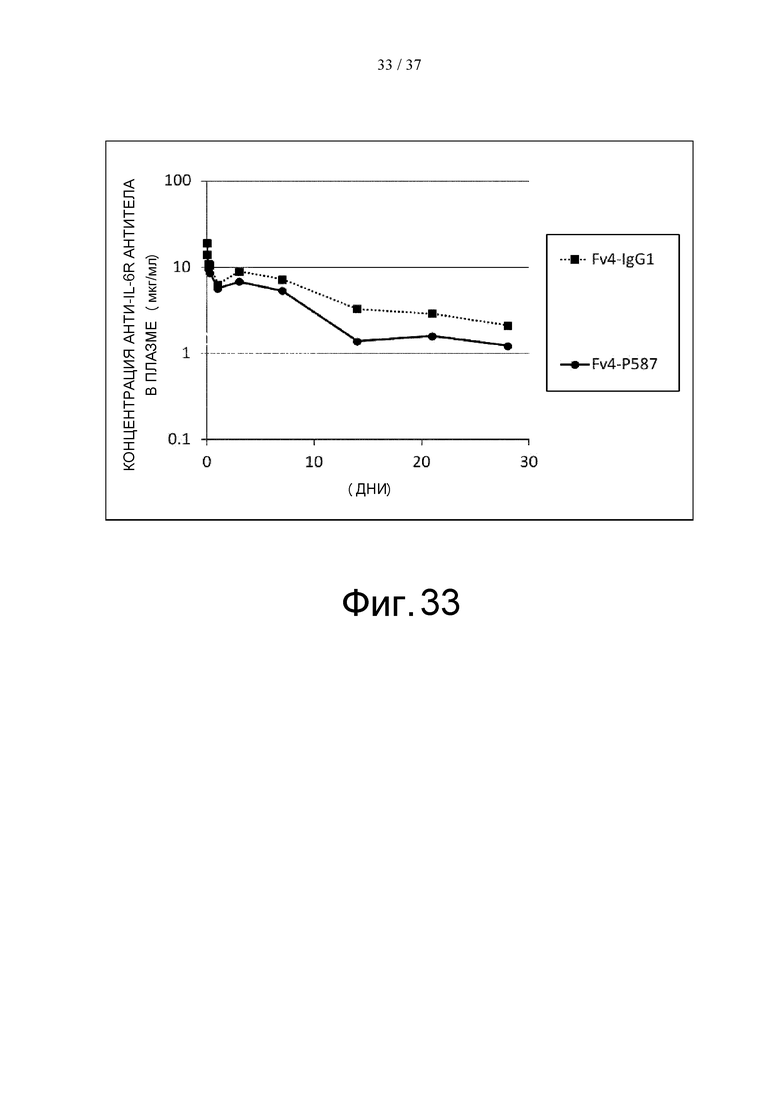
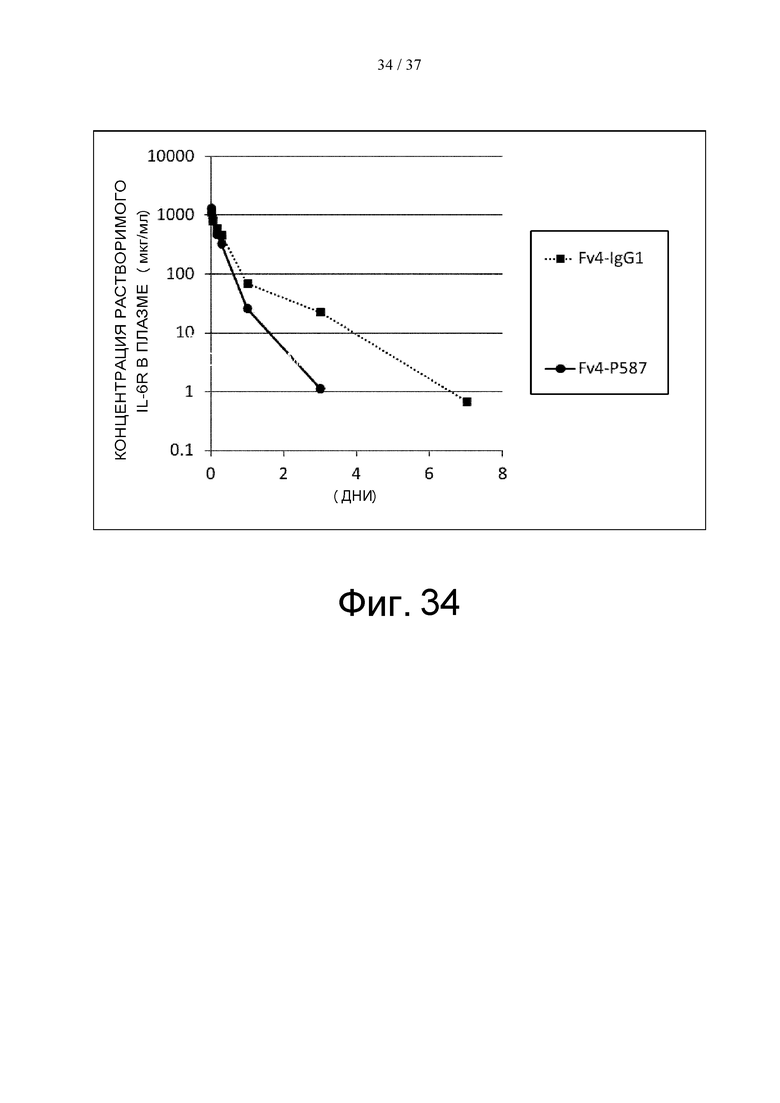
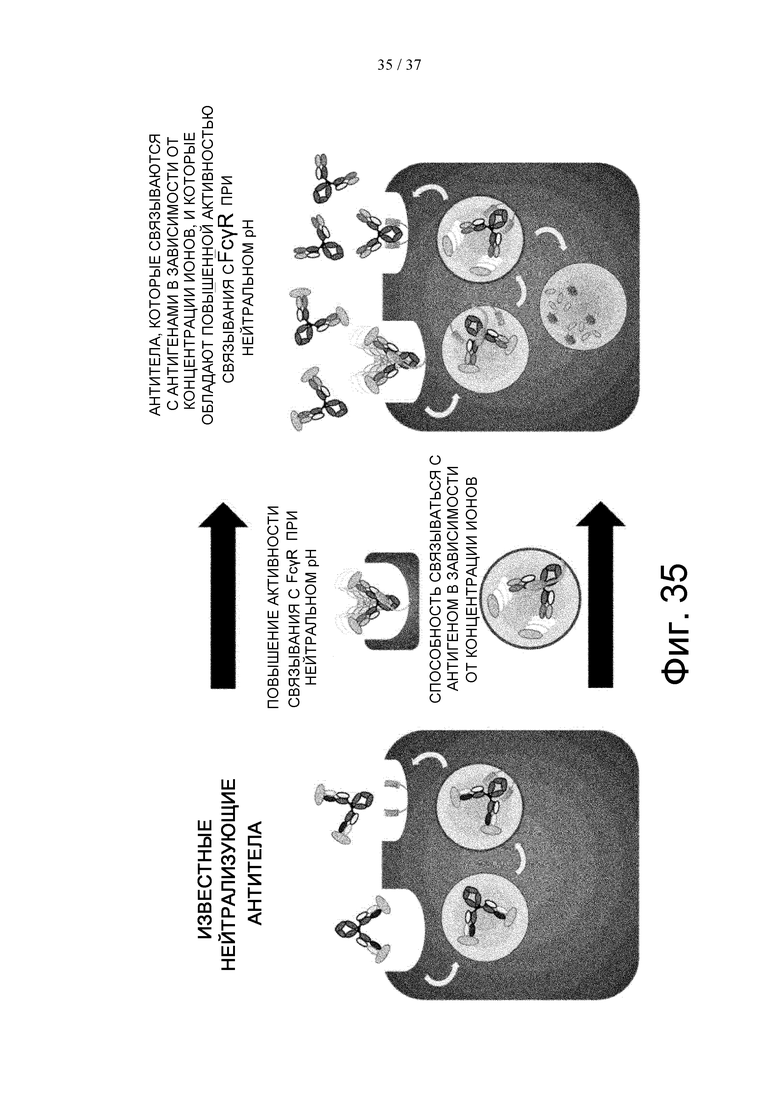
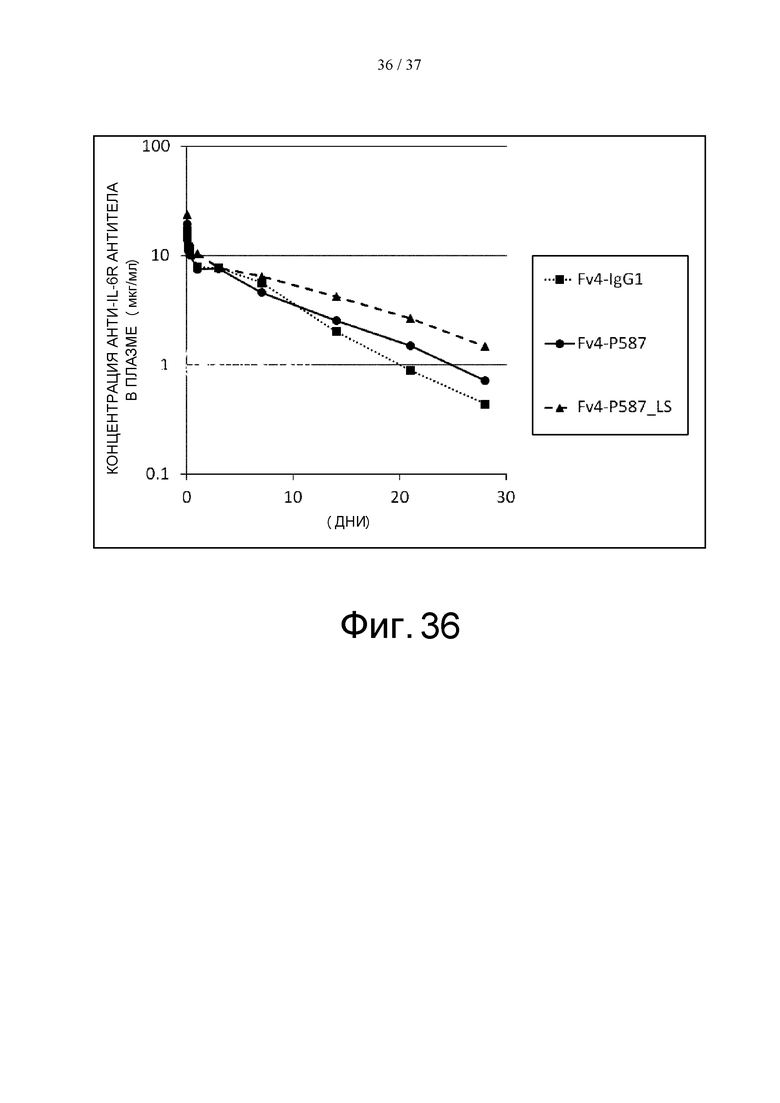
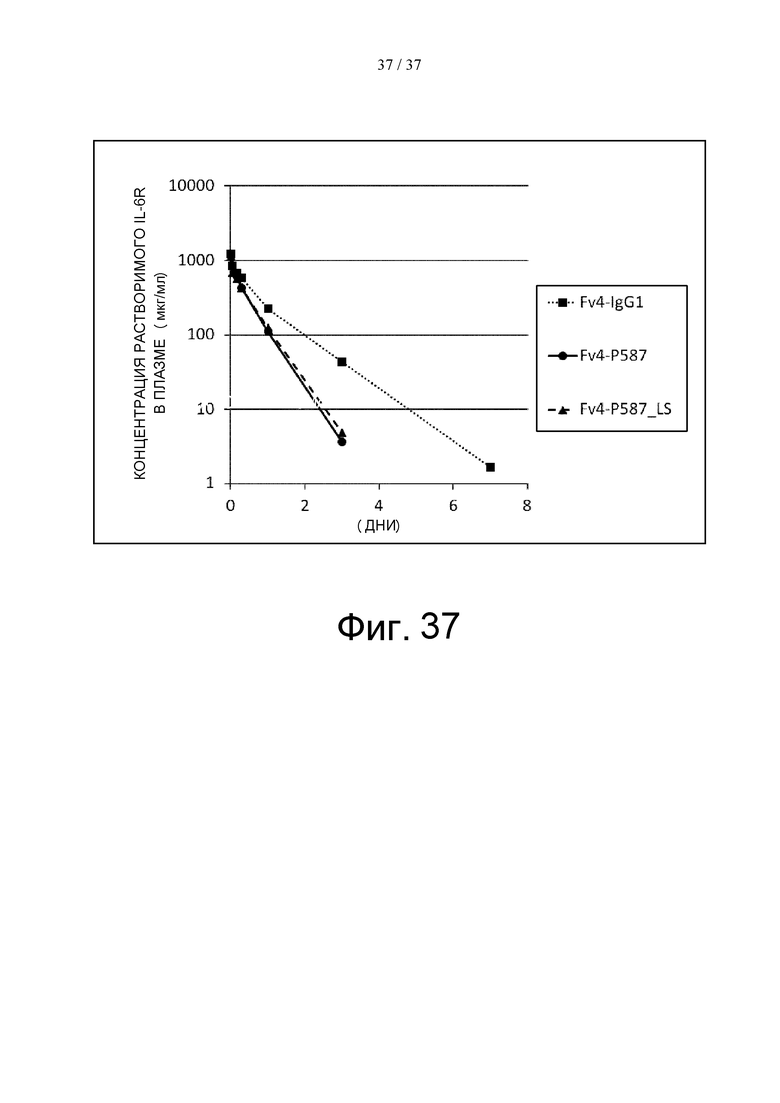
Изобретение относится к биотехнологии. Описаны варианты Fc-области, обладающие повышенной активностью и/или селективностью связывания с FcγRIIb, по сравнению с FcγRIIа (R-типа), по сравнению с активностью и/или селективностью связывания полипептида, содержащего Fc-область, в которую не была(и) введена(ы) аминокислотная(ые) модификация(и); полипептид, включающий указанный вариант Fc-области. Также представлена соответствующая фармацевтическая композиция для лечения или предупреждения воспалительных заболеваний или онкологических заболеваний. 4 н. и 11 з.п. ф-лы, 37 ил., 27 табл., 9 пр.
1. Вариант Fc-области для получения антигенсвязывающей молекулы с улучшенной связывающей активностью к FcγRIIb, где вариант Fc-области обладает улучшенной связывающей активностью к FcγRIIb и включает Asp в положении 238 в соответствии с нумерацией EU и по меньшей мере две аминокислоты, выбранные из группы аминокислот, состоящей из аминокислоты Tyr в положении 234, аминокислоты Phe в положении 235, аминокислоты Ile в положении 264, аминокислоты Glu в положении 265, аминокислот Phe, Leu или Met в положении 266, аминокислот Ala, Glu, Gly или Gln в положении 267, аминокислот Gln или Glu в положении 268, аминокислоты Asp в положении 269, аминокислот Asp, Phe, Ile, Met, Asn, Pro или Gln в положении 272, аминокислоты Gln в положении 274, аминокислоты Phe в положении 296, аминокислоты Asp в положении 326, аминокислоты Gly в положении 327, аминокислот Lys, Arg или Ser в положении 330, аминокислоты Ser в положении 331, аминокислот Lys, Arg, Ser или Thr в положении 332, аминокислот Lys, Arg, Ser или Thr в положении 333, аминокислот Arg, Ser или Thr в положении 334, аминокислот Ala или Gln в положении 355, аминокислоты Glu в положении 356, аминокислоты Met в положении 358, аминокислот Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в положении 396, аминокислоты Arg в положении 409 и аминокислоты Glu в положении 419, где величина отношения [KD для FcγRIIa (тип R) полипептида, содержащего вариант Fc-области]/[KD для FcγRIIb полипептида, содержащего вариант Fc-области] равна 20 или более.
2. Вариант Fc-области по п. 1, где активность связывания варианта с Fcy-рецепторами [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 15,0 или более.
3. Вариант Fc-области по п. 1 или 2, в котором указанная величина [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 50,0 или более.
4. Вариант Fc-области по любому из пп. 1-3, в котором указанная величина [величина KD для FcγRIIb полипептида, содержащего Fc-область, в которую не была (были) введена(ы) аминокислотная(ые) модификация(и)]/[величина KD для FcγRIIb полипептида, содержащего вариант Fc-области] имеет значение, равное 100,0 или более.
5. Вариант Fc-области по любому из пп. 1-4, в котором указанный вариант Fc-области содержит любую из нижеследующих групп аминокислотных модификаций (а)-(х), таких как:
(a) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(b) аминокислотные модификации в положениях 238, 237, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(c) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(d) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(e) аминокислотные модификации в положениях 238, 233, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(f) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296, 330 и 332 (в соответствии с Европейской нумерацией) Fc-области;
(g) аминокислотные модификации в положениях 238, 233, 237, 268, 271, 296, 327 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(h) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(i) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(j) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(k) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(l) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(m) аминокислотные модификации в положениях 238, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(n) аминокислотные модификации в положениях 238, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(о) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(р) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(q) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 296, 327, 330 и 396 (в соответствии с Европейской нумерацией) Fc-области;
(r) аминокислотные модификации в положениях 238, 233, 237, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области;
(s) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(t) аминокислотные модификации в положениях 238, 237, 264, 267, 268, 271, 272, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(u) аминокислотные модификации в положениях 238, 233, 264, 267, 268 и 271 (в соответствии с Европейской нумерацией) Fc-области;
(v) аминокислотные модификации в положениях 238, 237, 267, 268, 271, 296 и 330 (в соответствии с Европейской нумерацией) Fc-области;
(w) аминокислотные модификации в положениях 238, 264, 267, 268, 271, 272 и 296 (в соответствии с Европейской нумерацией) Fc-области; и
(х) аминокислотные модификации в положениях 238, 233, 264, 267, 268, 271 и 296 (в соответствии с Европейской нумерацией) Fc-области.
6. Вариант Fc-области по любому из пп. 1-5, в котором указанный вариант Fc-области содержит любую из нижеследующих аминокислотных последовательностей (b)-(х):
(b) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp или Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(c) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(d) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Gly или Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(e) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(f) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 332 является Thr в соответствии с Европейской нумерацией;
(g) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(h) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(i) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(j) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(k) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(l) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(m) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(n) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(о) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Ala или Gly, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(р) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met или Leu в соответствии с Европейской нумерацией;
(q) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, аминокислотой в положении 327 является Gly, аминокислотой в положении 330 является Arg, а аминокислотой в положении 396 является Met в соответствии с Европейской нумерацией;
(r) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией;
(s) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(t) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Pro, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(u) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, а аминокислотой в положении 271 является Gly в соответствии с Европейской нумерацией;
(v) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 237 является Asp, аминокислотой в положении 267 является Gly, аминокислотой в положении 268 является Asp, аминокислотой в положении 271 является Gly, аминокислотой в положении 296 является Asp, а аминокислотой в положении 330 является Arg в соответствии с Европейской нумерацией;
(w) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, аминокислотой в положении 272 является Asp, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией; и
(х) аминокислотная последовательность Fc-области, в которой аминокислотой в положении 238 является Asp, аминокислотой в положении 233 является Asp, аминокислотой в положении 264 является Ile, аминокислотой в положении 267 является Ala, аминокислотой в положении 268 является Glu, аминокислотой в положении 271 является Gly, а аминокислотой в положении 296 является Asp в соответствии с Европейской нумерацией.
7. Вариант Fc-области для получения антигенсвязывающей молекулы с улучшенной связывающей активностью к FcγRIIb, где вариант Fc-области обладает улучшенной связывающей активностью к FcγRIIb и состоит из любой аминокислотной последовательности, выбранной из SEQ ID NO: 43-68, SEQ ID NO: 70, SEQ ID NO: 71 и SEQ ID NO: 75-77.
8. Антигенсвязывающая молекула с улучшенной связывающей активностью к FcγRIIb, где антигенсвязывающая молекула содержит по меньшей мере два варианта Fc-области по любому из пп. 1-7, где указанные два варианта Fc-области связаны.
9. Антигенсвязывающая молекула по п. 8, в которой аминокислотные последовательности двух указанных связанных вариантов Fc-области в антигенсвязывающей молекуле являются одинаковыми.
10. Антигенсвязывающая молекула по п. 8, в которой аминокислотные последовательности двух указанных связанных вариантов Fc-области в антигенсвязывающей молекуле являются различными.
11. Антигенсвязывающая молекула по п. 10, в которой аминокислотные последовательности двух указанных связанных вариантов Fc-области имеют отличающуюся(иеся) аминокислоту(ы) по меньшей мере в одном положении, выбранном из положений 235, 236, 237, 238 и 239 в соответствии с Европейской нумерацией в указанном варианте Fc-области.
12. Антигенсвязывающая молекула по п. 11, в которой одной из аминокислотных последовательностей двух связанных вариантов Fc-области является аминокислотная последовательность, содержащая по меньшей мере одну аминокислоту, выбранную из аминокислот Asp, Gln, Glu или Thr в положении 235, аминокислоты Asn в положении 236, аминокислот Phe или Trp в положении 237, аминокислот Glu, Gly или Asn в положении 238 и аминокислот Asp или Glu в положении 239 в соответствии с Европейской нумерацией.
13. Антигенсвязывающая молекула по любому из пп. 8-12, в которой антигенсвязывающая молекула, содержащая вариант Fc-области, является антителом IgG.
14. Антигенсвязывающая молекула по любому из пп. 8-12, в которой антигенсвязывающая молекула, содержащая вариант Fc-области, является молекулой слитого Fc-белка.
15. Фармацевтическая композиция для лечения воспалительных или онкологических заболеваний, содержащая терапевтически эффективное количество антигенсвязывающей молекулы по любому из пп. 8-14 и фармацевтически приемлемый носитель.
| WO2008091954 А2, 23.01.2008 | |||
| ОПТИМИЗИРОВАННЫЕ Fc-ВАРИАНТЫ, ИМЕЮЩИЕ ИЗМЕНЕННОЕ СВЯЗЫВАНИЕ С FcγR, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2337107C2 |
| WO2008002933 А2, 03.01.2008 | |||
| US20100184959 А1, 22.07.2010. | |||

