Техническая область
Настоящее изобретение относится к полипептидам, содержащим Fc-область IgG, которые имеют сохраненную или сниженную активность связывания с обоими аллотипами FcγRIIa: типом H и типом R, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и которые имеют повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом из-за внесения аминокислотных замен в Fc-область IgG; к фармацевтическим композициям, содержащим полипептид; лекарственным средствам или профилактическим средствам, содержащим полипептид, при иммунологических воспалительных заболеваниях; и к способам их получения. Более того, настоящее изобретение относится к способам сохранения или снижения активности связывания с обоими аллотипами FcγRIIa: типом H и типом R, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и повышения активности связывания с FcγRIIb по сравнению с исходным полипептидом; и к способам подавления продукции антител по сравнению с исходным полипептидом при введении in vivo. Также настоящее изобретение относится к способам получения полипептида, имеющего сохраненную или сниженную активность связывания в отношении обоих аллотипов FcγRIIa: типа H и типа R, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и имеющего повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом; и к способам получения полипептида, который подавляет образование антител по сравнению с исходным полипептидом при введении in vivo.
Уровень техники
Интерес к антителам как к фармацевтическим средствам возник, поскольку они высоко стабильны в крови и вызывают мало побочных эффектов (непатентные документы - 1 и 2). Практически все фармацевтические препараты на основе антител, имеющиеся в настоящее время на рынке, представляют собой антитела подкласса IgG1 человека. Одной из известных функций антител класса IgG является антителозависимая клеточная цитотоксичность (далее обозначаемая как активность ADCC) (непатентный документ 3). Для того чтобы антитело проявляло активность ADCC, Fc-область антитела должна связываться с Fcγ-рецептором (далее обозначаемым как FcγR), который представляет собой связываемый антителом рецептор, находящийся на поверхности эффекторных клеток, таких как клетки-киллеры, натуральные клетки-киллеры и активированные макрофаги.
У человека семейство белков FcγR описано изоформами FcγRIa (CD64A), FcγRIIa (CD32A), FcγRIIb (CD32B), FcγRIIIa (CD16A) и FcγRIIIb (CD16B), а также описаны их соответствующие аллотипы (непатентный документ 7). FcγRIa, FcγRIIa и FcγRIIIa называют активирующими FcγR, поскольку они обладают иммунологически активными функциями, а FcγRIIb называют ингибиторным FcγR, поскольку он обладает иммунодепрессивными функциями (непатентный документ 8).
Было показано, что при связывании Fc-области и FcγR важны несколько аминокислотных остатков в шарнирной области и домене СН2 антитела, и цепь сахаров, связанная с Asn в положении 297 (нумерации по EU) домена СН2 (непатентные документы 4, 5 и 6). До настоящего времени исследованы различные варианты, имеющие свойства связывания FcγR, главным образом, антитела с мутациями, внесенные в эти области; и получены варианты Fc-области, имеющие более высокую активность связывания с активирующим FcγR (патентные документы 1, 2, 3 и 4).
Когда активирующий FcγR связывается с иммунным комплексом, он фосфорилирует иммунорецепторные тирозиновые активирующие мотивы (ITAM), содержащиеся во внутриклеточном домене или общей γ-цепи FcR (партнер по взаимодействию), активирует передатчик сигнала SYK, и запускает воспалительный иммунный ответ путем инициации каскада активирующего сигнала (непатентный документ 9).
FcγRIIb является единственным экспрессирующимся FcγR на B-клетках (непатентный документ 10). Описано, что взаимодействие Fc-области антитела с FcγRIIb подавляет первичный иммунный ответ B-клеток (непатентный документ 11). Более того, описано, что, когда FcγRIIb на B-клетках и B-клеточный рецептор (BCR) связываются через иммунный комплекс в крови, активация B-клеток подавляется и супрессирует продукцию антител B-клетками (непатентный документ 12). Для этой иммунодепрессивной передачи сигнала, опосредуемой BCR и FcγRIIb, необходим иммунорецепторный тирозиновый ингибирующий мотив (ITIM), содержащийся во внутриклеточном домене FcγRIIb (непатентные документы 13 и 14). Когда ITIM фосфорилируется при передаче сигнала, привлекается SH2-содержащая инозитолполифосфат-5-фосфатаза (SHIP), ингибируется передача других активирующих каскадов передачи сигнала FcγR и воспалительный иммунный ответ подавляется (непатентный документ 15). Более того, описано, что агрегация FcγRIIb отдельно временно подавляет вхождение кальция вследствие связывания BCR и пролиферации B-клеток BCR-независимым образом без индукции апоптоза продуцирующих IgM B-клеток (непатентный документ 16).
Более того, FcγRIIb также экспрессируется на дендритных клетках, макрофагах, активированных нейтрофилах, тучных клетках и базофилах. FcγRIIb ингибирует функции активирующего FcγR, такие как фагоцитоз, и высвобождает воспалительные цитокины в этих клетках и подавляет воспалительные иммунные ответы (непатентный документ 8).
Значение иммунодепрессивных функций FcγRIIb до настоящего времени устанавливали с помощью исследований с использованием мышей с нокаутом FcγRIIb. Существуют сообщения о том, что у мышей с нокаутом FcγRIIb гуморальный иммунитет не регулируется надлежащим образом (непатентный документ 17), чувствительность к индуцируемому коллагеном артриту (CIA) повышается (непатентный документ 18), присутствуют волчаноподобные симптомы, и имеются симптомы, подобные синдрому Гудпасчера (непатентный документ 19).
Более того, описано, что ненадлежащая регуляция FcγRIIb связана с аутоиммунными заболеваниями. Например, описана взаимосвязь между генетическим полиморфизмом в трансмембранной области и промоторной области FcγRIIb и частотой развития системной красной волчанки (SLE) (непатентные документы 20, 21, 22, 23 и 24), и снижением экспрессии FcγRIIb на поверхности B-клеток у пациентов с SLE (непатентный документ 25 и 26).
Исходя из моделей на мышах и собственно клинических данных, считается, что FcγRIIb играет роль в контроле аутоиммунных заболеваний и воспалительных заболеваний, главным образом, путем вовлечения B-клеток, и он является перспективной молекулой-мишенью для контроля аутоиммунных заболеваний и воспалительных заболеваний.
Известно, что IgG1, в основном используемый в качестве коммерчески доступного антительного фармацевтического средства, связывается не только с FcγRIIb, но также он сильно связывается с активирующим FcγR (непатентный документ 27). Является возможной разработка антительных фармацевтических средств, имеющих более существенные иммунодепрессивные свойства по сравнению с IgG1, путем использования Fc-области с усиленным связыванием FcγRIIb, или увеличенной селективностью связывания FcγRIIb по сравнению с активирующим FcγR. Например, было предположено, что применение антитела, имеющего вариабельную область, которая связывается с BCR, и Fc с усиленным связыванием FcγRIIb, может ингибировать активацию B-клеток (непатентный документ 28).
Описано, что связывание FcγRIIb на B-клетках и IgE, связанный с B-клеточным рецептором, подавляет дифференцировку B-клеток в плазматические клетки, что в результате вызывает подавление продуцирования IgE; и у мышей, которым трансплантировали PBMC человека, сохраняются концентрации IgG и IgM человека, в то время как концентрация IgE человека снижается (непатентный документ 29). Помимо IgE, было описано, что, когда FcγRIIb и CD79b, образующие рецепторный комплекс B-клеток, связываются антителом, пролиферация B-клеток подавляется in vitro и смягчаются симптомы в модели индуцированного коллагеном артрита (непатентный документ 30).
Помимо B-клеток, было описано, что связывание FcεRI и FcγRIIb на тучных клетках с использованием молекул, в которых Fc-часть IgG с усиленным связыванием FcγRIIb слита с Fc-частью IgE, которая связывается с IgE-рецептором FcεRI, вызывает фосфорилирование FcγRIIb посредством FcγRIIb, тем самым, подавляя зависимое от FcεRI вхождение кальция. Это указывает на то, что ингибирование дегрануляции через стимуляцию FcγRIIb возможно путем повышения связывания FcγRIIb (непатентный документ 31).
Таким образом, предполагают, что антитело, имеющее Fc с повышенной активностью связывания FcγRIIb, является перспективным в качестве лекарственного средства для лечения воспалительных заболеваний, таких как аутоиммунные заболевания.
Более того, было предположено, что мутанты с усиленным связыванием FcγRIIb являются перспективными лекарственными средствами для лечения злокачественных новообразований, а также лекарственными средствами для лечения воспалительных заболеваний, таких как аутоиммунные заболевания. На настоящий момент выявлено, что FcγRIIb играет важную роль в агонистической активности антител-агонистов против семейства рецепторов TNF. В частности, было предположено, для агонистической активности антител против CD40, DR4, DR5, CD30 и CD137, которые включены в семейство рецепторов TNF, требуется взаимодействие с FcγRIIb (непатентные документы 32, 33, 34, 35, 36 и 37). В непатентном документе 32 показано, что использование антител с усиленным связыванием FcγRIIb усиливает противоопухолевый эффект антител против CD40. Таким образом, ожидается, что антитела с усиленным связыванием FcγRIIb будут иметь эффект повышения агонистической активности антител-агонистов, включая антитела против семейства рецепторов TNF.
Описаны антитела с Fc с повышенной активностью связывания FcγRIIb (непатентный документ 28). В этом документе активность связывания FcγRIIb увеличивали путем внесения изменений, таких как S267E/L328F, G236D/S267E и S239D/S267E в Fc-область антитела. Среди них антитело, в которое внесена мутация S267E/L328F, более прочно связывается с FcγRIIb и сохраняет тот же уровень связывания с FcγRIa и FcγRIIa типа H, что и у встречающегося в природе IgG1. Однако в другом сообщении показано, что это изменение усиливает связывание с FcγRIIa типа R в несколько сот раз относительно указанного уровня связывания FcγRIIb, что означает, что селективность связывания FcγRIIb не повышается по сравнению с FcγRIIa типа R (патентный документ 5).
Даже если связывание FcγRIIb усиливается по сравнению со связыванием IgG1, считается, что только эффект усиления связывания FcγRIIa, но не усиления связывания FcγRIIb, влияет на клетки, такие как тромбоциты, которые экспрессируют FcγRIIa, но не экспрессируют FcγRIIb (непатентный документ 8). Например, известно, что группа пациентов, в которой вводили бевацизумаб, антитело против VEGF, имеет увеличенный риск тромбоэмболии (непатентный документ 38). Более того, тромбоэмболию аналогичным образом наблюдали в испытаниях по клинической разработке антител против CD40-лиганда, и клиническое испытание было прекращено (непатентный документ 39). В обоих случаях этих антител последние испытания с использованием моделей на животных и т.п. позволили предположить, что введенные антитела агрегируют тромбоциты через связывание FcγRIIa на тромбоцитах и образуют кровяные сгустки (непатентные документы 40 и 41). При системной красной волчанке, которая представляет собой аутоиммунное заболевание, тромбоциты активируются через зависимый от FcγRIIa механизм, и было описано, что активация тромбоцитов коррелирует с тяжестью симптомов (непатентный документ 42). Даже если связывание FcγRIIb усилить, введение антитела с усиленным связыванием FcγRIIa таким пациентам, которые уже имеют риск развития тромбоэмболии, увеличит риск развития тромбоэмболии и, таким образом, является чрезвычайно опасным.
Более того, описано, что антитела с усиленным связыванием FcγRIIa усиливают опосредуемый макрофагами антителозависимый фагоцитоз клеток (ADCP) (непатентный документ 43). Когда антигены антител фагоцитируются макрофагами, антитела сами по себе также фагоцитируются одновременно с ними. В этом случае пептидные фрагменты, происходящие из этих антител, также представляются в качестве антигена, и антигенность может увеличиться, тем самым, увеличивая риск продукции антител против антител (анти-антитела). Более конкретно, усиление связывания FcγRIIa увеличит риск продукции антител против антител, и это значительно снизит их ценность в качестве фармацевтических препаратов.
Более конкретно, ценность в качестве фармацевтических препаратов значительно снижается при усилении связывания FcγRIIa, что приводит к увеличенному риску образования тромбов через агрегацию тромбоцитов, более высокой антигенности и увеличенному риску продукции анти-антител.
С такой точки зрения вышеупомянутый Fc с усиленным связыванием FcγRIIb демонстрирует значительно усиленное связывание FcγRIIa типа R по сравнению со встречающимся в природе IgG1. Таким образом, его ценность в качестве фармацевтического средства для пациентов, имеющих FcγRIIa типа R, значительно снижается. Типы H и R FcγRIIa наблюдаются у европиоидов и афроамериканцев приблизительно с одной и той же частотой (непатентные документы 44 и 45). Таким образом, при использовании этого Fc для лечения аутоиммунных заболеваний количество пациентов, которые могут его безопасно использовать, одновременно получая их эффекты в качестве фармацевтического средства, будет ограничено.
Более того, описано, что в дендритных клетках с дефицитом FcγRIIb или дендритных клетках, в которых взаимодействие между FcγRIIb и Fc-частью антитела ингибируется антителом против FcγRIIb, дендритные клетки созревают самопроизвольно (непатентные документы 46 и 47). Это сообщение указывает на то, что FcγRIIb активно подавляет созревание дендритных клеток в стационарном состоянии, когда воспаление и т.п. не происходят. FcγRIIa экспрессируется на поверхности дендритных клеток в дополнение к FcγRIIb; таким образом, даже если связывание с ингибиторным FcγRIIb усиливается и если связывание с активирующим FcγR, таким как FcγRIIa, также усиливается, в результате может стимулироваться созревание дендритных клеток. Более конкретно, повышение не только активности связывания FcγRIIb, но также соотношения активности связывания FcγRIIb относительно активности связывания FcγRIIa считается важным для образования антител с иммунодепрессивным действием.
Таким образом, когда рассматривается получение фармацевтических средств, в которых используется опосредуемое связыванием FcγRIIb иммунодепрессивное действие, существует потребность в Fc, которая не только имеет повышенную активность связывания с FcγRIIb, но также имеет связывание с обоими аллотипами FcγRIIa: типом H и типом R, которое поддерживается на сходном уровне или является сниженным до более низкого уровня, чем у встречающегося в природе IgG1.
Между тем, на настоящее время описаны случаи, где аминокислотные изменения вносили в область Fc для увеличения селективности связывания FcγRIIb (непатентный документ 48). Однако все варианты, которые, как сообщалось, имеют увеличенную селективность к FcγRIIb, как описано в указанном документе, продемонстрировали сниженное связывание FcγRIIb по сравнению со связыванием встречающегося в природе IgG1. Таким образом, считается трудным для этих вариантов в действительности индуцировать опосредуемую FcγRIIb иммунодепрессивную реакцию в большей степени, чем в случае IgG1.
Более того, поскольку FcγRIIb играет важную роль в антителах-агонистах, упомянутых выше, ожидается, что повышение их активности связывания усилит агонистическую активность. Однако, когда аналогичным образом усиливается связывание FcγRIIa, проявляется непредусмотренная активность, такая как активность ADCC и активность ADCP, и это может вызвать побочные эффекты. Также, с такой точки зрения, является предпочтительным иметь возможность селективно повышать активность связывания FcγRIIb.
Исходя из этих результатов, при получении фармацевтических средств на основе антител, подлежащих применению для лечения аутоиммунных заболеваний и злокачественного новообразования с использованием FcγRIIb, важно, что по сравнению со встречающимися в природе IgG активность связывания обоих аллотипов FcγRIIa сохраняется или снижается и связывание FcγRIIb усиливается. Однако FcγRIIb обладает 93% идентичностью последовательности во внеклеточной области с FcγRIIa, который является одним из активирующих FcγR, и они являются в высокой степени структурно сходными. Существуют аллотипы FcγRIIa: тип H и тип R, в которых аминокислота в положении 131 представляет собой His (тип H) или Arg (тип R), и в то же время каждый из них реагирует по разному с антителами (непатентный документ 49). Таким образом, для получения Fc-области, которая селективно связывается с FcγRIIb, наиболее трудной проблемой может быть сообщение Fc-области антитела свойства селективного улучшения активности связывания FcγRIIb, что вовлекает различение этих гомологичных последовательностей, и снижения или не повышения активности связывания в отношении каждого аллотипа FcγRIIa, при повышении активности связывания с FcγRIIb. На настоящий момент не были получены варианты, имеющие достаточную селективность к FcγRIIb. В патентном документе 5 описаны варианты с повышенной активностью связывания FcγRIIb; однако степень усиления является низкой и существует потребность в разработке вариантов, имеющих свойства, сходные со свойствами, описанными выше.
Документы уровня техники
[Патентные документы]
[Патентный документ 1] WO 2000/42072
[Патентный документ 2] WO 2006/019447
[Патентный документ 3] WO 2004/99249
[Патентный документ 4] WO 2004/29207
[Патентный документ 5] US2009/0136485
[Непатентные документы]
[Непатентный документ 1] Nat Biotechnol, 23(9), 1073-1078, 2005
[Непатентный документ 2] Eur J Pharm Biopharm, 59(3), 389-96, 2005
[Непатентный документ 3] Chem Immunol, 65, 88-110, 1997
[Непатентный документ 4] J Biol Chem, 276(19), 16478-16483, 2001
[Непатентный документ 5] Eur J Immunol, 23(5), 1098-1104, 1993
[Непатентный документ 6] Immunology, 86(2), 319-324, 1995
[Непатентный документ 7] Immunol Lett, 82(1-2), 57-65, 2002
[Непатентный документ 8] Nat Rev Immunol, 10(5), 328-343, 2010
[Непатентный документ 9] Nat Rev Immunol, 8(1), 34-47, 2008
[Непатентный документ 10] Eur J Immunol, 19(8), 1379-1385, 1989
[Непатентный документ 11] J Exp Med, 129(6), 1183-1201, 1969
[Непатентный документ 12] Immunol Lett, 88(2), 157-161, 2003
[Непатентный документ 13] Science, 256(5065), 1808-1812, 1992
[Непатентный документ 14] Nature, 368(6466), 70-73, 1994
[Непатентный документ 15] Science, 290(5489), 84-89, 2000
[Непатентный документ 16] J Immunol, 181(8), 5350-5359 2008
[Непатентный документ 17] J Immunol, 163(2), 618-622, 1999
[Непатентный документ 18] J Exp Med, 189(1), 187-194, 1999
[Непатентный документ 19] J Exp Med, 191(5), 899-906, 2000
[Непатентный документ 20] Hum Genet, 117(2-3), 220-227, 2005
[Непатентный документ 21] J Biol Chem, 282(3), 1738-1746, 2007
[Непатентный документ 22] Arthritis Rheum, 54(12), 3908-3917, 2006
[Непатентный документ 23] Nat Med, 11(10), 1056-1058, 2005
[Непатентный документ 24] J Immunol, 176(9), 5321-5328, 2006
[Непатентный документ 25] J Exp Med, 203(9), 2157-2164, 2006
[Непатентный документ 26] J Immunol, 178(5), 3272-3280, 2007
[Непатентный документ 27] Blood, 113(16), 3716-3725, 2009
[Непатентный документ 28] Mol Immunol, 45(15), 3926-3933, 2008
[Непатентный документ 29] J Allergy Clin Immunol, 2012 Jan 16. в печати (PMID: 22257644)
[Непатентный документ 30] Arthritis Rheum, 62(7), 1933-1943, 2010
[Непатентный документ 31] Immunol Lett, 2012 Jan 25. в печати (PMID: 22305932)
[Непатентный документ 32] Science, 333(6045), 1030-1034, 2011
[Непатентный документ 33] Cancer Cell, 19(1), 101-113, 2011
[Непатентный документ 34] J Clin Invest, 2012 Feb 13. pii: 61226. doi: 10.1172/JCI61226. в печати (PMID: 22326955)
[Непатентный документ 35] J Immunol, 171(2), 562-568, 2003
[Непатентный документ 36] Blood, 108(2), 705-710, 2006
[Непатентный документ 37] J Immunol, 166(8), 4891-4898, 2001
[Непатентный документ 38] J Natl Cancer Inst, 99(16), 1232-1239, 2007
[Непатентный документ 39] Arthritis Rheum, 48(3), 719-727, 2003
[Непатентный документ 40] J Thromb Haemost, 7(1), 171-181, 2008
[Непатентный документ 41] J Immunol, 185(3), 1577-1583, 2010
[Непатентный документ 42] Sci Transl Med, 2(47), 47-63, 2010
[Непатентный документ 43] Mol Cancer Ther, 7(8), 2517-2527, 2008
[Непатентный документ 44] J Clin Invest, 97(5), 1348-1354, 1996
[Непатентный документ 45] Arthritis Rheum, 41(7), 1181-1189, 1998
[Непатентный документ 46] J Clin Invest, 115(10), 2914-2923, 2005
[Непатентный документ 47] Proc Natl Acad Sci USA, 102(8), 2910-2915, 2005
[Непатентный документ 48] Mol Immunol, 40(9), 585-593, 2003
[Непатентный документ 49] J Exp Med, 172, 19-25, 1990
Сущность изобретения
Проблемы, решаемые изобретением
Настоящее изобретение было осуществлено ввиду описанных выше обстоятельств. Задачей настоящего изобретения является предоставление полипептидов, содержащих Fc-область IgG, которые имеют сохраненную или сниженную активность связывания обоих аллотипов FcγRIIa: H-типа и R-типа, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и имеющих повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, путем внесения аминокислотных замен в Fc-область IgG; фармацевтических композиций, содержащих полипептид; терапевтических средств или профилактических средств, содержащих полипептид, против иммунологических воспалительных заболеваний; и способов их получения. Более того, задачей является предоставление способа сохранения или снижения активности связывания обоих аллотипов FcγRIIa: H-типа и R-типа, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и повышения активности связывания с FcγRIIb по сравнению с исходным полипептидом; и способа подавления продукции антител по сравнению с исходным полипептидом при введении in vivo. Кроме того, задачей является предоставление способов продуцирования полипептида, имеющего сохраненную или сниженную активность связывания обоих аллотипов FcγRIIa: типа H и типа R, в которых аминокислота в положении 131 (нумерация по EU) FcγRIIa представляет собой His (тип H) или Arg (тип R), и имеющего повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом; и способов получения полипептида, подавляющего продукцию антител по сравнению с исходным полипептидом при введении in vivo.
Средства для решения проблем
Авторы настоящего изобретения провели целенаправленное исследование в отношении полипептида, содержащего Fc-область, имеющую сниженное опосредуемое Fc связывание с FcγRIIa, и увеличенное связывание с FcγRIIb по сравнению с исходным полипептидом. В результате авторы настоящего изобретения открыли, что полипептид, содержащий Fc-область антитела, которое содержит изменение, полученное путем заменой Pro в положении 238 (нумерация по EU) на Asp или Leu в положении 328 (нумерация по EU) на Glu, повышает активность связывания с FcγRIIb и снижает опосредуемую Fc-областью активность связывания обоих аллотипов FcγRIIa: типов H и R. Более того, авторы настоящего изобретения открыли, что полипептид, содержащий Fc-область антитела, которая содержит изменение, состоящее в замене Pro в положении 238 (нумерация по EU) на Asp и несколько других изменений, повышает активность связывания с FcγRIIb и сохраняет или снижает опосредуемую Fc-областью активность связывания обоих аллотипов cγRIIa, типов H и R.
Более конкретно, настоящее изобретение относится к следующему:
[1] вариант полипептида, содержащего Fc-область антитела по меньшей мере с одним аминокислотным изменением, который имеет сохраненную или сниженную активность связывания FcγRIIa (тип R) и FcγRIIa (тип H) и повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, и где величина [величина KD варианта полипептида в отношении FcγRIIa (тип R)/[величина KD варианта полипептида в отношении FcγRIIb] составляет 1,2 или более;
[2] полипептид согласно [1], где величина [величина KD варианта полипептида в отношении FcγRIIa (тип H)]/[величина KD варианта полипептида в отношении FcγRIIb] составляет 4,2 или более;
[3] полипептид согласно [1] или [2], где величина [величина KD исходного полипептида в отношении FcγRIIb]/[величина KD варианта полипептида в отношении FcγRIIb] составляет 1,6 или более;
[4] полипептид согласно любому из [1]-[3], где величина [величина KD для более повышенной из активностей связывания варианта полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип H)]/[величина KD для более повышенной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип H)] составляет 0,7 или более;
[5] полипептид согласно любому из [1]-[4], который имеет сохраненную или сниженную активность связывания FcγRIIIa по сравнению с исходным полипептидом;
[6] полипептид согласно любому из [1]-[5], который имеет сохраненную или сниженную активность связывания FcγRIa по сравнению с исходным полипептидом;
[7] полипептид согласно любому из [1]-[6], где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu;
[8] полипептид согласно любому из [1]-[7], где аминокислотное изменение представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Pro в положении 238 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met;
[9] полипептид согласно любому из [1]-[8], где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело;
[10] полипептид согласно любому из [1]-[8], где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc;
[11] способ сохранения или снижения активности связывания в отношении FcγRIIa (тип R) и FcγRIIa (тип H) и повышения активности связывания FcγRIIb у полипептида по сравнению с исходным полипептидом, который включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела, где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu;
[12] способ подавления продукции антитела против полипептида, содержащего Fc-область антитела, по сравнению и исходным полипептидом при введении in vivo, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu;
[13] способ согласно [11] или [12], где аминокислотное изменение представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Pro в положении 238 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met;
[14] способ согласно любому из [11]-[13], где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело;
[15] способ согласно любому из [11]-[13], где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc;
[16] способ получения полипептида, имеющего сохраненную или сниженную активность связывания FcγRIIa (тип R) и FcγRIIa (тип H) и имеющего повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела, где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu;
[17] способ получения полипептида, подавляющего продукцию антител против полипептида по сравнению с исходным полипептидом при введении in vivo, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела, где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu;
[18] способ согласно [16] или [17], где аминокислотная замена представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Pro в положении 238 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met;
[19] способ согласно любому из [16]-[18], где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело;
[20] способ согласно любому из [16]-[18], где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc;
[21] полипептид, продуцируемый способом согласно любому из [16]-[20];
[22] фармацевтическая композиция, содержащая полипептид согласно любому из [1]-[10] и [21];
[23] средство для подавления активации B-клеток, тучных клеток, дендритных клеток и/или базофилов, которое содержит полипептид согласно любому из [1]-[10] и [21];
[24] средство для лечения или профилактики иммунологического воспалительного заболевания, которое содержит полипептид по любому из [1]-[10] и [21];
[25] терапевтическое средство или профилактическое средство по [24], где иммунологическое воспалительное заболевание представляет собой аутоиммунное заболевание и представляет собой заболевание, которое может быть вызвано образованием антитела против аутоантигена;
[26] средство для лечения заболевания, которое содержит полипептид согласно любому из [1]-[10] и [21], где заболевание представляет собой заболевание с дефицитом биологически необходимого белка; и
[27] противовирусное средство, содержащее полипептид согласно любому из [1]-[10] и [21].
Настоящее изобретение также относится к способам лечения или профилактики иммунологических воспалительных заболеваний, которые включают стадию введения индивидууму полипептида по настоящему изобретению или полипептида, полученного способами получения по настоящему изобретению. Более того, настоящее изобретение относится к наборам для применения в терапевтических способах или профилактических способах по настоящему изобретению, которые содержат полипептид по настоящему изобретению или полипептид, продуцируемый способами получения по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению. Настоящее изобретение также относится к применению полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению, для получения лекарственных средств или профилактических средств при иммунологических воспалительных заболеваниях. Кроме того, настоящее изобретение относится к полипептиду по настоящему изобретению или полипептиду, продуцируемому способами получения по настоящему изобретению, для применения в терапевтическом способе или профилактическом способе по настоящему изобретению. Настоящее изобретение также относится к способам подавления активации B-клеток, тучных клеток, дендритных клеток и/или базофилов, которые включают стадию введения индивидууму полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению. Настоящее изобретение относится к наборам для применения в способе ингибирования по настоящему изобретению, которые содержат полипептид по настоящему изобретению или полипептид, продуцируемый способами получения по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению. Настоящее изобретение относится к применению полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению, для получения средств, которые подавляют активацию B-клеток, тучных клеток, дендритных клеток и/или базофилов. Настоящее изобретение относится к полипептидам по настоящему изобретению или полипептидам, продуцируемым способами получения по настоящему изобретению, для применения в способах ингибирования по настоящему изобретению. Настоящее изобретение относится к способам лечения заболеваний с дефицитом биологически необходимых белков, которые включают стадию введения индивидууму полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению. Настоящее изобретение относится к наборам для применения в терапевтическом способе по настоящему изобретению, которые содержат полипептид по настоящему изобретению или полипептид, продуцируемый способами получения по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению. Настоящее изобретение относится к применению полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению, для получения лекарственных средств для лечения заболеваний с дефицитом биологически необходимых белков. Настоящее изобретение также относится к полипептиду по настоящему изобретению или полипептиду, продуцируемому способами получения по настоящему изобретению, для применения в терапевтическом способе по настоящему изобретению. Настоящее изобретение относится к способам ингибирования вирусов, которые включают стадию введения индивидууму полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению. Настоящее изобретение относится к наборам для применения в способе ингибирования по настоящему изобретению, которые содержат полипептид по настоящему изобретению или полипептид, продуцируемый способами получения по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению. Более того, настоящее изобретение относится к применению полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению, для получения противовирусного средства. Более того, настоящее изобретение относится к полипептиду по настоящему изобретению или полипептиду, продуцируемому способами получения по настоящему изобретению, для применения в способе ингибирования по настоящему изобретению.
Эффекты изобретения
Настоящее изобретение относится к полипептидам, содержащим Fc-область, имеющим сохраненную или сниженную активность связывания с обоими аллотипами FcγRIIa: типами R и H, и имеющим повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом. С использованием полипептидов с увеличенной селективностью связывания FcγRIIb относительно обоих аллотипов FcγRIIa (типы H и R), является возможным передавать ингибиторные сигналы воспалительного иммунного ответа, опосредуемого фосфорилированием ITIM в FcγRIIb, у пациентов, имеющих любой из аллотипов, тип R и тип H. Более того, путем сообщения Fc антитела свойства селективного связывания FcγRIIb, является возможным подавлять продукцию анти-антител через опосредуемое FcγRIIb иммунодепрессивное действие.
Краткое описание чертежей
На фиг.1 представлено сравнение связывания FcγRIa и связывания FcγRIIb. Связывание антитела с заменой Pro в положении 238 (нумерация по EU) на Asp и связывание антитела с заменой Leu в положении 328 (нумерация по EU) на Glu обозначено. “Мутация A” относится к изменению, полученному путем замены Pro в положении 238 (нумерация по EU) на Asp, и “мутация B” относится к изменению, полученному путем замены Leu в положении 328 (нумерация по EU) на Glu.
На фиг.2 представлено сравнение связывания FcγRIIa типа H и связывания FcγRIIb. Связывание антитела с заменой Pro в положении 238 (нумерация по EU) на Asp и связывание антитела с заменой Leu в положении 328 (нумерация по EU) на Glu обозначено. “Мутация A” относится к изменению, полученному путем замены Pro в положении 238 (нумерация по EU) на Asp, и “мутация B” относится к изменению, полученному путем замены Leu в положении 328 (нумерация по EU) на Glu.
На фиг.3 представлено сравнение связывания FcγRIIa типа R и связывания FcγRIIb. Связывание антитела с заменой Pro в положении 238 (нумерация по EU) на Asp и связывание антитела с заменой Leu в положении 328 (нумерация по EU) на Glu обозначено. “Мутация A” относится к изменению, полученному путем замены Pro в положении 238 (нумерация по EU) на Asp, и “мутация B” относится к изменению, полученному путем замены Leu в положении 328 (нумерация по EU) на Glu.
На фиг.4 представлено сравнение связывания FcγRIIa и связывания FcγRIIb. Связывание антитела с заменой Pro в положении 238 (нумерация по EU) на Asp и связывание антитела с заменой Leu в положении 328 (нумерация по EU) на Glu обозначено. “Мутация A” относится к изменению, полученному путем замены Pro в положении 238 (нумерация по EU) на Asp, и “мутация B” относится к изменению, полученному путем замены Leu в положении 328 (нумерация по EU) на Glu.
На фиг.5 представлена взаимосвязь между аминокислотными остатками, составляющими Fc-области IgG1, IgG2, IgG3 и IgG4, и нумерацией по EU (в настоящем документе также обозначаемой как индекс EU).
На фиг.6 показан график, на котором горизонтальная ось показывает относительную величину активности связывания FcγRIIb для каждого варианта PD, и на вертикальной оси показана относительная величина активности связывания FcγRIIa типа R для каждого варианта PD. Значение величины связывания каждого варианта PD с каждым FcγR делили на значение величины связывания IL6R-F652, которое представляет собой контрольное антитело, перед внесением изменения (измененная Fc с заменой Pro в положении 238 (нумерация по EU) на Asp) в каждый FcγR; а затем полученную величину умножали на 100 и использовали в качестве величины относительной активности связывания для каждого варианта PD с каждым FcγR. На графике F652 на фигуре показана величина для IL6R-F652.
На фиг.7 показан график, на котором на вертикальной оси показана относительная величина активности связывания FcγRIIb для вариантов, полученных путем внесения каждого изменения в GpH7-B3, который не имеет изменения P238D, и на горизонтальной оси показана относительная величина активности связывания FcγRIIb для вариантов, полученных путем внесения каждого изменения в IL6R-F652, который имеет изменение P238D. Значение величины связывания FcγRIIb каждого варианта делили на значение величины связывания FcγRIIb предварительно измененного антитела; а затем полученную величину умножали на 100 и использовали в качестве величины относительной активности связывания. В этом случае область A содержит изменения, которые проявляют эффект усиления связывания FcγRIIb в обоих случаях, когда изменение вносят в GpH7-B3, который не имеет P238D, и когда изменение вносят в IL6R-F652, который имеет P238D. Область B содержит изменения, которые проявляют эффект усиления связывания FcγRIIb при введении в GpH7-B3, который не имеет P238D, но не проявляют эффект усиления связывания FcγRIIb при введении в IL6R-F652, который имеет P238D.
На фиг.8 показана кристаллическая структура комплекса внеклеточной области Fc(P238D)/FcγRIIb.
На фиг.9 представлено изображение с наложением кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и модельной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, в отношении внеклеточной области FcγRIIb и домена СН2 A Fc путем аппроксимации методом наименьших квадратов на основании расстояний между парами атомов Cα.
На фиг.10 показано сравнение детальной структуры в области P238D после наложения кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и модельной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, в отношении только домена СН2 A Fc или только домена СН2 B Fc путем аппроксимации методом наименьших квадратов на основании расстояний между парами атомов Cα.
На фиг.11 показано, что водородная связь может находиться между основной цепью Gly в положении 237 (нумерация по EU) в домене CH2 A Fc и Tyr в положении 160 в FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг.12 показано, что между Asp в положении 270 (нумерация по EU) в домене CH2 B Fc и Arg в положении 131 в FcγRIIb может быть выявлено электростатическое взаимодействие в кристаллической структуре комплекса внеклеточной области Fc(P238D)/FcγRIIb.
На фиг.13 представлен график, на котором на горизонтальной оси показана относительная величина активности связывания FcγRIIb для каждого варианта 2B, и на вертикальной оси показана относительная величина активности связывания FcγRIIa типа R для каждого варианта 2B. Значение величины связывания каждого варианта 2B с каждым FcγR делили на значение величины связывания контрольного антитела перед изменением (измененный Fc с заменой Pro в положении 238 (нумерация по EU) на Asp) для каждого FcγR; а затем полученную величину умножали на 100 и использовали в качестве величины относительной активности связывания каждого варианта 2B в отношении каждого FcγR.
На фиг.14 показан Glu в положении 233 (нумерация по EU) в цепи A Fc и окружающие остатки во внеклеточной области FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг.15 показан Ala в положении 330 (нумерация по EU) в цепи A Fc и окружающие остатки во внеклеточной области FcγRIIb в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb.
На фиг.16 показаны структуры Pro в положении 271 (нумерация по EU) цепи B Fc после наложения кристаллических структур комплекса Fc(P238D)/внеклеточная область FcγRIIb и комплекса Fc(WT)/внеклеточная область FcγRIIIa путем аппроксимации методом наименьших квадратов на основе расстояний между парами атомов Cα, в отношении B-цепи Fc.
Способ осуществления изобретения
Настоящее изобретение относится к полипептидам, содержащим Fc-область IgG, которая имеет сохраненное или сниженное связывание FcγRIIa, и имеющим повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом путем внесения аминокислотой замены (замен) в Fc-область IgG.
Более конкретно, настоящее изобретение относится к полипептиду, содержащему Fc-область антитела, которая содержит замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu, и полипептиду, содержащему Fc-область антитела, которая содержит комбинацию замены Pro в положении 238 (нумерация по EU) на Asp и нескольких определенных аминокислотных замен. Более того, настоящее изобретение относится к способу сохранения или снижения активности связывания с обоими аллотипами FcγRIIa и повышения активности связывания FcγRIIb по сравнению с исходным полипептидом. Настоящее изобретение также относится к способу подавления продукции антител по сравнению с исходным полипептидом, когда полипептид вводят in vivo.
Термин “полипептиды по настоящему изобретению”, главным образом, относится к пептидам или белкам длиной приблизительно десять аминокислот или более. Более того, они, как правило, представляют собой полипептиды, происходящие из организмов, но конкретно не ограничены, и, например, они могут представлять собой полипептиды, содержащие искусственно сконструированную последовательность. Более того, они могут представлять собой любые встречающиеся в природе полипептиды, синтетические полипептиды, рекомбинантные полипептиды или сходные с ними.
Термин “Fcγ-рецепторы” (в настоящем описании обозначаемые как Fcγ-рецепторы или FcγR) относится к рецепторам, которые могут связываться с Fc-областью моноклональных антител на основе IgG1, IgG2, IgG3 и IgG4, и, конкретно, означает любого представителя семейства белков, кодируемого генами Fcγ-рецепторов. У человека это семейство включает, но не ограничивается ими, FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы H131 (тип H) и R131 (тип R)), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), включая изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2), и любые FcγR человека, изоформы или аллотипы FcγR, которые будут открыты в будущем. FcγR включает, но не ограничивается ими, происходящие из человека, мыши, крысы, кролика и обезьяны FcγR, и он может происходить из любого организма. FcγR мыши включают FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD16-2) и любые изоформы или аллотипы FcγR мыши или изоформы или аллотипы FcγR, которые будут открыты в будущем. Подходящие примеры таких Fcγ-рецепторов включают FcγRI (CD64), FcγRIIA (CD32), FcγRIIB (CD32), FcγRIIIA (CD16) и/или FcγRIIIB (CD16) человека.
Полинуклеотидная последовательность и аминокислотная последовательность FcγRI указаны в SEQ ID NO:1 (NM_000566.3) и 2 (NP_000557.1), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIA указаны в SEQ ID NO:3 (BC020823.1) и 4 (AAH20823.1), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIB указаны в SEQ ID NO:5 (BC 146678.1) и 6 (AAI46679.1), соответственно;
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIA указаны в SEQ ID NO:7 (BC033678.1) и 8 (AAH33678.1), соответственно; и
полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIB указаны в SEQ ID NO:9 (BC128562.1) и 10 (AAI28563.1), соответственно (регистрационный номер RefSeq указан внутри скобок).
В FcγRIIa существует два аллотипа: один - где аминокислота в положении 131 FcγRIIa представляет собой гистидин (тип H), и другой - где эта аминокислота замещена аргинином (тип R) (J. Exp. Med, 172:19-25, 1990).
В рамках настоящего описания “исходный полипептид” относится к полипептиду, который служит в качестве основы для получения полипептидов, содержащих Fc-область антитела, по настоящему изобретению. Более конкретно, он представляет собой полипептид, содержащий Fc-область антитела и представляет собой полипептид перед изменением по меньшей мере одной аминокислоты в Fc-области. Исходный полипептид в рамках настоящего изобретения может представлять собой, например, полипептид, содержащий Fc-область встречающегося в природе IgG, или он может представлять собой полипептид, содержащий Fc-область IgG, где во встречающийся в природе IgG внесено изменение, отличное от аминокислотных изменений по настоящему изобретению.
Термин “встречающиеся в природе IgG” относится к полипептидам, относящимся к классу антител, фактически кодируемому генами иммуноглобулинов-гамма и содержащему аминокислотную последовательность, идентичную аминокислотным последовательностям IgG, встречающихся в природе. Например, встречающийся в природе IgG человека означает встречающийся в природе IgG1 человека, встречающийся в природе IgG2 человека, встречающийся в природе IgG3 человека, встречающийся в природе IgG4 и т.п. Встречающиеся в природе IgG также включают мутантов, самопроизвольно образующихся из них.
Fc-область встречающихся в природе IgG означает Fc-область, содержащую аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, происходящей из IgG, встречающегося в природе. Fc-область встречающегося в природе IgG представлена на фиг.5 (SEQ ID NO:11-14), и, например, она относится к Fc-областям, происходящим из встречающегося в природе IgG1 человека, Fc-областям, происходящим из встречающегося в природе IgG2 человека, Fc-областям, происходящим из встречающегося в природе IgG3 человека, и Fc-областям, происходящим из встречающегося в природе IgG4 человека. Fc-области встречающихся в природе IgG также включают мутантов, самопроизвольно образующихся из них.
В рамках настоящего изобретения, определение того, повышается ли или сохраняется ли или снижается ли активность связывания каждого типа FcγR в полипептиде или Fc-области по настоящему изобретению, можно проводить, например, путем наблюдения, происходит ли снижение или увеличение величины константы диссоциации (KD), полученной из результатов анализа сенсограммы, где различные FcγR подвергают взаимодействию в качестве анализируемого соединения с антителами, иммобилизованными на сенсорных чипах или уловленными на сенсорных чипах с использованием белка A, белка L, белка A/G, белка G, антител против лямбда-цепи, антител против каппа-цепи, антигенных пептидов, антигенных белков и т.п. с использованием BIACORE, который представляет собой устройство для анализа взаимодействия, в котором используется явление поверхностного плазмонного резонанса (SPR), как показано в примерах. Альтернативно, это определение также можно проводить путем выявления того, происходит ли увеличение или снижение величины, полученной путем деления значения изменения величины в резонансных единицах (RU) на сенсограмме до и после того, как различные типы FcγR подвергают взаимодействию в качестве анализируемых соединений с антителами, иммобилизованными на сенсорные чипы или уловленными на сенсорные чипы с использованием белка A, белка L, белка A/G, белка G, антител против лямбда-цепи, антител против каппа-цепи, антигенных пептидов, антигенных белков и т.п., на значение изменения величины в резонансных единицах (RU) до и после иммобилизации или улавливания антител на сенсорных чипах. Более того, это определение можно проводить путем выявления увеличения или снижения величин константы диссоциации (KD), полученных из анализа сенсограмм, где образец, такой как антитело, подлежащее оценке, подвергают взаимодействию в качестве анализируемого соединения с использованием сенсорного чипа, на который FcγR иммобилизован прямо или через антитело против метки. Альтернативно, это определение можно проводить путем выявления того, увеличивается ли или снижается ли значение изменения величин на сенсограмме до и после того как образец, такой как антитело, подлежащее оценке, подвергают взаимодействию в качестве анализируемого соединения с сенсорным чипом, на который FcγR иммобилизован прямо или через антитело против метки.
В частности, активность связывания Fc-области с Fcγ-рецептором можно измерять с помощью скрининга с использованием гомогенного анализа усиленной за счет эффекта близости люминесценции (ALPHA), способа BIACORE, в котором используется явление поверхностного плазмонного резонанса (SPR) и т.п., в дополнение к ELISA или активированной флуоресценцией сортировке клеток (FACS) (Proc. Natl. Acad. Sci. USA (2006) 103 (11): 4005-4010).
Скрининг ALPHA проводят с помощью технологии ALPHA, в которой используется два типа гранул, донорные и акцепторные, исходя из следующих принципов. Люминесцентные сигналы выявляются, только когда молекулы, связанные с донорными гранулами, физически взаимодействуют с молекулами, связанными с акцепторными гранулами, и эти два типа гранул находятся вблизи друг друга. Возбуждаемый лазером фотосенсибилизатор в донорных гранулах преобразует кислород окружающей среды в синглетный кислород в возбужденном состоянии. Синглетный кислород распространяется среди донорных гранул, и, когда он достигает соседних акцепторных гранул, в гранулах индуцируется хемилюминесцентная реакция, и, в конечном итоге, испускается свет. Когда молекулы, связанные с донорными гранулами, не взаимодействуют с молекулами, связанными с акцепторными гранулами, хемилюминесцентная реакция не происходит, поскольку синглетный кислород, продуцируемый донорными гранулами, не достигает акцепторных гранул.
Например, биотинилированный полипептидный комплекс связывают с донорными гранулами, а Fcγ-рецептор, меченный глутатион-S-трансферазой (GST), связывают с акцепторными гранулами. В отсутствие конкурирующего полипептидного комплекса, содержащего мутантную Fc-область, полипептидный комплекс, содержащий Fc-область дикого типа, взаимодействует с Fcγ-рецептором и генерирует сигналы в области 520-620 нм. Полипептидный комплекс, содержащий немеченую мутантную Fc-область, конкурирует с полипептидным комплексом, содержащим Fc-область дикого типа, за взаимодействие с Fcγ-рецептором. Относительную активность связывания можно определять путем количественного определения снижения флуоресценции, наблюдаемого в результате конкуренции. Биотинилирование полипептидных комплексов, таких как антитела, с использованием сульфо-NHS-биотина и т.п., хорошо известно. Способ экспрессии Fcγ-рецептора и GST в клетке, содержащей слитый ген, получаемый путем слияния полинуклеотида, кодирующего Fcγ-рецептор, в рамке считывания с полинуклеотидом, кодирующим GST в экспрессирующем векторе, и проведения очистки с использованием глутатионовой колонки, соответственно, принят в качестве способа мечения Fcγ-рецептора посредством GST. Полученные сигналы предпочтительно анализируют, например, путем аппроксимации их к односторонней модели конкуренции, в которой используется нелинейный регрессионный анализ с использованием программного обеспечения, такого как GRAPHPAD PRISM (GraphPad, San Diego).
Одно из веществ (лиганд) при наблюдении взаимодействия иммобилизуют на тонкую золотую пленку на сенсорном чипе, и посредством света, светящего с обратной стороны сенсорного чипа, так чтобы общее отражение происходило на поверхности контакта между тонкой золотой пленкой и стеклом, в части отраженного света образуется часть сниженной интенсивности отражения (сигнал SPR). Когда другое из веществ (анализируемое соединение) при выявлении взаимодействия пропускают на поверхности сенсорного чипа и лиганд связывается с анализируемым соединением, масса иммобилизованной молекулы лиганда возрастает, и индекс рефракции растворителя на поверхности сенсорного чипа изменяется. Положение сигнала SPR сдвигается в результате этого изменения индекса рефракции (с другой стороны, положение сигнала возвращается, когда происходит диссоциация от связывания). Система Biacore указывает величину сдвига, упомянутую выше, или, более конкретно, временную переменную массы, путем построения графика изменения массы на поверхности сенсорного чипа по оси ординат в качестве данных измерения (сенсограмма). Из сенсограммы определяют количество анализируемого соединения, связанного с лигандом, уловленным на поверхности сенсорного чипа. Параметры кинетики, такие константы скорости ассоциации (ka) и константы скорости диссоциации (kd), определяют из кривых сенсограммы, и константы диссоциации (KD) определяют из соотношения этих констант. В способе BIACORE, предпочтительно используют способ измерения ингибирования. Пример способа измерения ингибирования описан в Proc. Natl. Acad. Sci USA (2006) 103(11): 4005-4010.
Полипептид со сниженной активностью связывания FcγR относится к полипептиду, который связывается с FcγR с по существу более низкой активностью связывания, чем у исходного полипептида, когда анализ проводят путем поддержания количества исходного полипептида и количества полипептида, содержащего по меньшей мере одно аминокислотное изменение в Fc-области исходного полипептида (также называемого полипептидным вариантом) практически на одном уровне.
Например, в величинах KD, измеренных с помощью упомянутого выше способа измерения, соотношение величин KD (величина KD варианта полипептида/величина KD исходного полипептида) предпочтительно составляет 1,25 или более, 2 или более или 3 или более, и более предпочтительно 5 или более, 10 или более, 100 или более, 1000 или более или 10000 или более.
Более того, в величинах KD, измеренных описанным выше способом измерения, величина KD предпочтительно увеличивается на 1 мкМ или более, и более предпочтительно увеличивается на 2 мкМ или более, 3 мкМ или более, 5 мкМ или более, 10 мкМ или более, 20 мкМ или более, 50 мкМ или более и 100 мкМ или более. Более того, в величинах KD, измеренных упомянутым выше способом измерения, величина KD предпочтительно составляет 0,0001 мкМ или более, и более предпочтительно 0,001 мкМ или более, 0,01 мкМ или более, 0,1 мкМ или более, 0,5 мкМ или более, 1 мкМ или более, 2 мкМ или более, 3 мкМ или более, 5 мкМ или более, 10 мкМ или более, 100 мкМ или более или 1000 мкМ или более.
Полипептид с повышенной активностью связывания FcγR относится к полипептиду, который связывается с FcγR с существенно более высокой активностью связывания, чем исходный полипептид, когда анализ проводят путем поддержания количества исходного полипептида и количества варианта полипептида практически на одном уровне.
Например, в величинах KD, измеренных с помощью описанного выше способа измерения, соотношение величин KD (величина KD исходного полипептида/величина KD варианта полипептида) предпочтительно составляет 1,25 или более, 2 или более или 3 или более, и более предпочтительно 5 или более, 10 или более, 100 или более, 1000 или более или 10000 или более.
Более того, в величинах KD, измеренных упомянутым выше способом измерения, величина KD предпочтительно снижается на 0,001 мкМ или более, и более предпочтительно снижается на 0,01 мкМ, 0,1 мкМ, 1 мкМ или более, 2 мкМ или более, 3 мкМ или более, 5 мкМ или более, 10 мкМ или более, 20 мкМ или более, 50 мкМ или более и 100 мкМ или более.
Более того, в величинах KD, измеренных упомянутым выше способом измерения, величина KD предпочтительно составляет 5 мкМ или менее, и более предпочтительно 3 мкМ или менее, 1 мкМ или менее, 0,5 мкМ или менее, 0,1 мкМ или менее, 0,01 мкМ или менее, 0,001 мкМ или менее или 0,0001 мкМ или менее.
Полипептид с неизмененной (сохраненной) активностью связывания FcγR относится к полипептиду, который связывается с FcγR с активностью связывания, практически неизмененной или эквивалентной относительно исходного полипептида, когда анализ проводят путем поддержания количества исходного полипептида и количества полипептида, содержащего по меньшей мере одно аминокислотное изменение в Fc-области исходного полипептида (также называемого вариантом полипептида) практически на одном уровне.
То, является ли полипептид полипептидом, имеющим сохраненную или сниженную активность связывания FcγRIIa и имеющим повышенную активность связывания с FcγRIIb, можно определять с использованием величины KD этого полипептида в отношении FcγRIIa и величины KD этого полипептида в отношении FcγRIIb, определенной в соответствии с упомянутыми выше примерами. Примером является случай, когда величина KD полипептида по настоящему изобретению в отношении FcγRIIb снижается по сравнению с величиной KD исходного полипептида в отношении FcγRIIb; и величина KD полипептида по настоящему изобретению в отношении FcγRIIa (тип R и тип H) увеличивается или сохраняется по сравнению с величиной KD исходного полипептида в отношении FcγRIIa (тип R и тип H). Более того, это можно определить путем комбинирования соответствующим образом величины KD полипептида в отношении FcγRIa и величины KD полипептида в отношении FcγRIIIa, которые были определены в соответствии с упомянутым выше примером.
В настоящем изобретении повышенная активность связывания FcγRIIb означает, что, например, в величинах KD, измеренных способом измерения, описанным выше, соотношение KD [величина KD исходного полипептида]/[величина KD варианта полипептида] предпочтительно составляет 1,6 или более, 2 или более или 3 или более, и более предпочтительно 5 или более, 10 или более, 20 или более, 30 или более и 50 или более.
Сохраненная или сниженная активность связывания FcγRIIa (тип R) и FcγRIIa (тип H) означает, например, что в величинах KD, измеренных способом измерения, описанным выше, соотношение KD [величина KD для более сильной из активностей связывания варианта полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип H)]/[величина KD для более сильной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип H)] предпочтительно составляет 0,7 или более, 1 или более, 2 или более или 3 или более, и более предпочтительно 5 или более, 10 или более, 20 или более, 30 или более и 50 или более.
Полипептиды по настоящему изобретению предпочтительно имеют сохраненную или сниженную активность связывания с FcγRIIa типа R и FcγIIa типа H. Более того, они предпочтительно имеют сохраненную или сниженную активность связывания с FcγIIa типа R и FcγIIa типа H, а также сохраненную или сниженную активность связывания FcγRIIIa. Кроме того, они предпочтительно имеют сохраненную или сниженную активность связывания с FcγRIa.
Сохраненная или сниженная активность связывания с FcγRIIIa или FcγRIa означает, что, например, в величинах KD, измеренных способом измерения, описанным выше, соотношение KD [величина KD варианта полипептида]/[величина KD исходного полипептида] предпочтительно составляет 1 или более, 2 или более или 3 или более, и более предпочтительно 5 или более, 10 или более, 20 или более, 30 или более и 50 или более.
Более того, определение того, является ли полипептид по настоящему изобретению полипептидом с улучшенной селективностью связывания с FcγRIIb, а не с FcγRIIa, можно определять путем сравнения соотношения величины KD в отношении FcγRIIa с величиной KD в отношении FcγRIIb у полипептида по настоящему изобретению (величина KD в отношении FcγRIIa/величина KD в отношении FcγRIIb) с соотношением величины KD в отношении FcγRIIa к величине KD в отношении FcγRIIb у исходного пептида (величина KD в отношении FcγRIIa/величина KD в отношении FcγRIIb), которую определяли в соответствии с упомянутыми выше примерами. В частности, когда величина соотношения KD полипептида по настоящему изобретению превышает эту величину у исходного полипептида, может быть определено, что полипептид по настоящему изобретению имеет повышенную селективность связывания в отношении FcγRIIb относительно FcγRIIa по сравнению с исходным полипептидом.
Селективность связывания с FcγRIIa (тип R) относительно FcγRIIb представляет собой, например, соотношение величин KD [величина KD варианта полипептида в отношении FcγRIIa (тип R)]/[величина KD варианта полипептида в отношении FcγRIIb], составляющее предпочтительно 1,2 или более, 2 или более или 3 или более для величин KD, измеренных способом измерения, описанным выше, и более предпочтительно 5 или более, 10 или более, 20 или более или 30 или более.
Селективность связывания с FcγRIIa (тип H) относительно FcγRIIb представляет собой, например, соотношение величин KD [величина KD варианта полипептида в отношении FcγRIIa (тип H)]/[величина KD варианта полипептида в отношении FcγRIIb], составляющее предпочтительно 4,2 или более, 5 или более или 10 или более для величин KD, измеренных способом измерения, описанным выше, и более предпочтительно 20 или более, 30 или более, 50 или более, 100 или более или 200 или более.
Более того, определение того, сохраняется ли, повышается ли или снижается ли активность связывания полипептидов по настоящему изобретению с различными FcγR можно проводить из увеличения или снижения величин связывания различных FcγR с полипептидами по настоящему изобретению, которые определены в соответствии с примерами, описанными выше. В рамках настоящего изобретения величина связывания различных FcγR с полипептидами относится к величинам, полученным путем определения разности величин RU на сенсограммах, которые изменялись до и после взаимодействия различных FcγR в качестве анализируемого соединения с каждым полипептидом, и деления их на разность величин RU на сенсограммах, которые изменялись до и после улавливания полипептидов на сенсорные чипы.
Определение того, являются ли полипептиды по настоящему изобретению полипептидами, имеющими сохраненную или сниженную активность связывания с FcγRIIa (тип R и тип H) и имеющими повышенную активность связывания с FcγRIIb, можно определять с использованием величины связывания полипептида с FcγRIIa и величины связывания полипептида с FcγRIIb, определенных в соответствии с примерами, описанными выше.
Примером является случай, когда величина связывания FcγRIIb у полипептида по настоящему изобретению увеличивается по сравнению с величиной связывания FcγRIIb у исходного полипептида, и величина связывания FcγRIIa (тип R и тип H) у полипептида по настоящему изобретению эквивалентна (сохраняется на уровне) или предпочтительно снижена по сравнению с величиной связывания исходного полипептида с FcγRIIa (тип R и тип H). Более того, это возможно определить путем комбинирования соответствующим образом величины связывания FcγRIa и величины связывания FcγRIIIa у полипептида, определенного в соответствии с примерами, описанными выше.
“Fc-область” относится к области, содержащей фрагмент, состоящий из шарнирной области или ее части, домена СН2 или домена CH3 в молекуле антитела. В соответствии с нумерацией по EU (в настоящем описании также называемой индексом EU) (см. фиг.5), Fc-область класса IgG относится, например, к области от цистеина в положении 226 до C-конца, или от пролина в положении 230 до C-конца, но не ограничивается ими.
Fc-область можно получать, предпочтительно путем повторного элюирования фракции, адсорбированной на колонку с белком A, после частичного расщепления моноклональных антител на основе IgG1, IgG2, IgG3, IgG4 и т.п. с использованием протеазы, такой как пепсин. Протеаза конкретно не ограничена, при условии, что она может расщеплять полноразмерное антитело, так чтобы при рестрикции образовывались Fab и F(ab')2 путем установления надлежащим образом условий ферментативной реакции, таких как pH, и ее примеры включают пепсин и папаин.
Настоящее изобретение относится к константной области антитела, содержащей Fc-область, которая содержит изменение, полученное путем замены Pro в положении 238 (нумерация по EU) на Asp или замены Leu в положении 328 (нумерация по EU) на Glu в IgG человека (IgG1, IgG2, IgG3 и IgG4). Полипептиды с сохраненной или сниженной активностью связывания с FcγRIa, FcγRIIIa и обоими аллотипами FcγRIIa: типами R и H, а также с повышенной активностью связывания FcγRIIb по сравнению с исходным полипептидом могут быть предоставлены путем внесения изменения, состоящего в замене Pro в положении 238 (нумерация по EU) на Asp или замене Leu в положении 328 (нумерация по EU) на Glu в IgG человека.
В рамках настоящего изобретения по меньшей мере одно изменение можно дополнительно вносить в Fc-область IgG человека, содержащую изменение, полученное путем замены Pro в положении 238 (нумерация по EU) на Asp или замены Leu в положении 328 (нумерация по EU) на Glu. В рамках настоящего изобретения изменение относится к любому одному из, или комбинации, замен, делеций, вставок и инсерций. Вместе с этими изменениями, кроме того, можно вносить дополнительные изменения. Дополнительное изменение может быть выбрано из любого одного из, или комбинаций, аминокислотных замен, делеций или модификаций. Например, можно вносить изменения, которые повышают активность связывания с FcγRIIb, а также сохраняют или снижают активность связывания с FcγRIIa (тип H) и FcγRIIa (тип R). Внесение таких изменений увеличивает селективность связывания с FcγRIIb, а не с FcγRIIa.
Среди них изменения, которые увеличивают селективность связывания в отношении FcγRIIb, а не FcγRIIa (тип R) являются предпочтительными, и более предпочтительными являются изменения, которые увеличивают селективность связывания в отношении FcγRIIb, а не FcγRIIa (тип H). Предпочтительные примеры изменений с заменой аминокислот включают
изменение путем замены Gly в положении 237 (нумерация по EU) на Trp,
изменение путем замены Gly в положении 237 (нумерация по EU) на Phe,
изменение путем замены Pro в положении 238 (нумерация по EU) на Phe,
изменение путем замены Asn в положении 325 (нумерация по EU) на Met,
изменение путем замены Ser в положении 267 (нумерация по EU) на Ile,
изменение путем замены Leu в положении 328 (нумерация по EU) на Asp,
изменение путем замены Ser в положении 267 (нумерация по EU) на Val,
изменение путем замены Leu в положении 328 (нумерация по EU) на Trp,
изменение путем замены Ser в положении 267 (нумерация по EU) на Gln,
изменение путем замены Ser в положении 267 (нумерация по EU) на Met,
изменение путем замены Gly в положении 236 (нумерация по EU) на Asp,
изменение путем замены Ala в положении 327 (нумерация по EU) на Asn,
изменение путем замены Asn в положении 325 (нумерация по EU) на Ser,
изменение путем замены Leu в положении 235 (нумерация по EU) на Tyr,
изменение путем замены Val в положении 266 (нумерация по EU) на Met,
изменение путем замены Leu в положении 328 (нумерация по EU) на Tyr,
изменение путем замены Leu в положении 235 (нумерация по EU) на Trp,
изменение путем замены Leu в положении 235 (нумерация по EU) на Phe,
изменение путем замены Ser в положении 239 (нумерация по EU) на Gly,
изменение путем замены Ala в положении 327 (нумерация по EU) на Glu,
изменение путем замены Ala в положении 327 (нумерация по EU) на Gly,
изменение путем замены Pro в положении 238 (нумерация по EU) на Leu,
изменение путем замены Ser в положении 239 (нумерация по EU) на Leu,
изменение путем замены Leu в положении 328 (нумерация по EU) на Thr,
изменение путем замены Leu в положении 328 (нумерация по EU) на Ser,
изменение путем замены Leu в положении 328 (нумерация по EU) на Met,
изменение путем замены Pro в положении 331 (нумерация по EU) на Trp,
изменение путем замены Pro в положении 331 (нумерация по EU) на Tyr,
изменение путем замены Pro в положении 331 (нумерация по EU) на Phe,
изменение путем замены Ala в положении 327 (нумерация по EU) на Asp,
изменение путем замены Leu в положении 328 (нумерация по EU) на Phe,
изменение путем замены Pro в положении 271 (нумерация по EU) на Leu,
изменение путем замены Ser в положении 267 (нумерация по EU) на Glu,
изменение путем замены Leu в положении 328 (нумерация по EU) на Ala,
изменение путем замены Leu в положении 328 (нумерация по EU) на Ile,
изменение путем замены Leu в положении 328 (нумерация по EU) на Gln,
изменение путем замены Leu в положении 328 (нумерация по EU) на Val,
изменение путем замены Lys в положении 326 (нумерация по EU) на Trp,
изменение путем замены Lys в положении 334 (нумерация по EU) на Arg,
изменение путем замены His в положении 268 (нумерация по EU) на Gly,
изменение путем замены His в положении 268 (нумерация по EU) на Asn,
изменение путем замены Ser в положении 324 (нумерация по EU) на Val,
изменение путем замены Val в положении 266 (нумерация по EU) на Leu,
изменение путем замены Pro в положении 271 (нумерация по EU) на Gly,
изменение путем замены Ile в положении 332 (нумерация по EU) на Phe,
изменение путем замены Ser в положении 324 (нумерация по EU) на Ile,
изменение путем замены Glu в положении 333 (нумерация по EU) на Pro,
изменение путем замены Tyr в положении 300 (нумерация по EU) на Asp,
изменение путем замены Ser в положении 337 (нумерация по EU) на Asp,
изменение путем замены Tyr в положении 300 (нумерация по EU) на Gln,
изменение путем замены Thr в положении 335 (нумерация по EU) на Asp,
изменение путем замены Ser в положении 239 (нумерация по EU) на Asn,
изменение путем замены Lys в положении 326 (нумерация по EU) на Leu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Ile,
изменение путем замены Ser в положении 239 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Phe,
изменение путем замены Lys в положении 326 (нумерация по EU) на Val,
изменение путем замены Lys в положении 326 (нумерация по EU) на Tyr,
изменение путем замены Ser в положении 267 (нумерация по EU) на Asp,
изменение путем замены Lys в положении 326 (нумерация по EU) на Pro,
изменение путем замены Lys в положении 326 (нумерация по EU) на His,
изменение путем замены Lys в положении 334 (нумерация по EU) на Ala,
изменение путем замены Lys в положении 334 (нумерация по EU) на Trp,
изменение путем замены His в положении 268 (нумерация по EU) на Gln,
изменение путем замены Lys в положении 326 (нумерация по EU) на Gln,
изменение путем замены Lys в положении 326 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Met,
изменение путем замены Val в положении 266 (нумерация по EU) на Ile,
изменение путем замены Lys в положении 334 (нумерация по EU) на Glu,
изменение путем замены Tyr в положении 300 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 334 (нумерация по EU) на Met,
изменение путем замены Lys в положении 334 (нумерация по EU) на Val,
изменение путем замены Lys в положении 334 (нумерация по EU) на Thr,
изменение путем замены Lys в положении 334 (нумерация по EU) на Ser,
изменение путем замены Lys в положении 334 (нумерация по EU) на His,
изменение путем замены Lys в положении 334 (нумерация по EU) на Phe,
изменение путем замены Lys в положении 334 (нумерация по EU) на Gln,
изменение путем замены Lys в положении 334 (нумерация по EU) на Pro,
изменение путем замены Lys в положении 334 (нумерация по EU) на Tyr,
изменение путем замены Lys в положении 334 (нумерация по EU) на Ile,
изменение путем замены Gln в положении 295 (нумерация по EU) на Leu,
изменение путем замены Lys в положении 334 (нумерация по EU) на Leu,
изменение путем замены Lys в положении 334 (нумерация по EU) на Asn,
изменение путем замены His в положении 268 (нумерация по EU) на Ala,
изменение путем замены Ser в положении 239 (нумерация по EU) на Asp,
изменение путем замены Ser в положении 267 (нумерация по EU) на Ala,
изменение путем замены Leu в положении 234 (нумерация по EU) на Trp,
изменение путем замены Leu в положении 234 (нумерация по EU) на Tyr,
изменение путем замены Gly в положении 237 (нумерация по EU) на Ala,
изменение путем замены Gly в положении 237 (нумерация по EU) на Asp,
изменение путем замены Gly в положении 237 (нумерация по EU) на Glu,
изменение путем замены Gly в положении 237 (нумерация по EU) на Leu,
изменение путем замены Gly в положении 237 (нумерация по EU) на Met,
изменение путем замены Gly в положении 237 (нумерация по EU) на Tyr,
изменение путем замены Ala в положении 330 (нумерация по EU) на Lys,
изменение путем замены Ala в положении 330 (нумерация по EU) на Arg,
изменение путем замены Glu в положении 233 (нумерация по EU) на Asp,
изменение путем замены His в положении 268 (нумерация по EU) на Asp,
изменение путем замены His в положении 268 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Asp,
изменение путем замены Lys в положении 326 (нумерация по EU) на Ser,
изменение путем замены Lys в положении 326 (нумерация по EU) на Thr,
изменение путем замены Val в положении 323 (нумерация по EU) на Ile,
изменение путем замены Val в положении 323 (нумерация по EU) на Leu,
изменение путем замены Val в положении 323 (нумерация по EU) на Met,
изменение путем замены Tyr в положении 296 (нумерация по EU) на Asp,
изменение путем замены Lys в положении 326 (нумерация по EU) на Ala,
изменение путем замены Lys в положении 326 (нумерация по EU) на Asn, и
изменение путем замены Ala в положении 330 (нумерация по EU) на Met.
Более того, примеры предпочтительных аминокислотных замен среди этих изменений включают
изменение путем замены Gly в положении 237 (нумерация по EU) на Trp,
изменение путем замены Gly в положении 237 (нумерация по EU) на Phe,
изменение путем замены Ser в положении 267 (нумерация по EU) на Val,
изменение путем замены Ser в положении 267 (нумерация по EU) на Gln,
изменение путем замены His в положении 268 (нумерация по EU) на Asn,
изменение путем замены Pro в положении 271 (нумерация по EU) на Gly,
изменение путем замены Lys в положении 326 (нумерация по EU) на Leu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Gln,
изменение путем замены Lys в положении 326 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Met,
изменение путем замены Ser в положении 239 (нумерация по EU) на Asp,
изменение путем замены Ser в положении 267 (нумерация по EU) на Ala,
изменение путем замены Leu в положении 234 (нумерация по EU) на Trp,
изменение путем замены Leu в положении 234 (нумерация по EU) на Tyr,
изменение путем замены Gly в положении 237 (нумерация по EU) на Ala,
изменение путем замены Gly в положении 237 (нумерация по EU) на Asp,
изменение путем замены Gly в положении 237 (нумерация по EU) на Glu,
изменение путем замены Gly в положении 237 (нумерация по EU) на Leu,
изменение путем замены Gly в положении 237 (нумерация по EU) на Met,
изменение путем замены Gly в положении 237 (нумерация по EU) на Tyr,
изменение путем замены Ala в положении 330 (нумерация по EU) на Lys,
изменение путем замены Ala в положении 330 (нумерация по EU) на Arg,
изменение путем замены Glu в положении 233 (нумерация по EU) на Asp,
изменение путем замены His в положении 268 (нумерация по EU) на Asp,
изменение путем замены His в положении 268 (нумерация по EU) на Glu,
изменение путем замены Lys в положении 326 (нумерация по EU) на Asp,
изменение путем замены Lys в положении 326 (нумерация по EU) на Ser,
изменение путем замены Lys в положении 326 (нумерация по EU) на Thr,
изменение путем замены Val в положении 323 (нумерация по EU) на Ile,
изменение путем замены Val в положении 323 (нумерация по EU) на Leu,
изменение путем замены Val в положении 323 (нумерация по EU) на Met,
изменение путем замены Tyr в положении 296 (нумерация по EU) на Asp,
изменение путем замены Lys в положении 326 (нумерация по EU) на Ala,
изменение путем замены Lys в положении 326 (нумерация по EU) на Asn, и
изменение путем замены Ala в положении 330 (нумерация по EU) на Met.
Изменение, упомянутое выше, может представлять собой изменение, внесенное в одно положение, и альтернативно можно комбинировать изменения в двух или более положениях. Предпочтительные примеры таких изменений включают изменения, упомянутые в таблицах 6-7 и таблицах 9-12.
Более того, например, можно вносить аминокислотные замены, которые повышают активность связывания FcRn (J. Immunol. 2006 Jan 1; 176(1): 346-56; J Biol Chem. 2006 Aug 18; 281(33): 23514-24; Int. Immunol. 2006 Dec; 18(12): 1759-69; Nat Biotechnol. 2010 Feb; 28(2): 157-9; WO 2006/019447; WO 2006/053301; и WO 2009/086320), и аминокислотные замены для повышения гетерогенности или стабильности антитела (WO 2009/041613) в константную область антитела. Альтернативно, также в изобретение включены полипептиды, полученные путем сообщения полипептидам по настоящему изобретению свойства обеспечения исчезновения антигенов, которые описаны в WO 2011/122011 или PCT/JP2011/072550, и полипептиды, полученные путем сообщения свойства многократного связывания с множеством антигенных молекул, которые описаны в WO 2009/125825 или PCT/JP2011/077619.
Предпочтительные примеры полипептидов по настоящему изобретению включают IgG-антитела. Когда в качестве антитела используют IgG-антитело, тип константной области не ограничен, и можно использовать изотипы (подклассы) IgG, такие как IgG1, IgG2, IgG3 и IgG4. IgG-антитела по настоящему изобретению предпочтительно представляют собой IgG человека, и более предпочтительно IgG1 человека и IgG4 человека. Аминокислотные последовательности константных областей тяжелой цепи IgG1 человека и IgG4 человека известны. Множество последовательностей аллотипов для константной области IgG1 человека, являющихся результатом генетических полиморфизмов, описано в Sequences of Proteins of Immunological Interest, NIH Publication № 91-3242, и в рамках настоящего изобретения можно использовать любые из этих последовательностей.
Замена
При замене аминокислотных остатков замену на отличающийся аминокислотный остаток проводят с задачей изменения аспектов, таких как (a)-(c), описанные ниже:
(a) структура полипептидного остова в области структуры слоя или структуры спирали;
(b) электрический заряд и гидрофобность в заданном участке; или
(c) размер боковой цепи.
Аминокислотные остатки классифицируют на следующие группы, исходя из их общих свойств боковой цепи:
(1) гидрофобные: норлейцин, met, ala, val, leu и ile;
(2) нейтральные гидрофильные: cys, ser, thr, asn и gln;
(3) кислотные: asp и glu;
(4) основные: his, lys и arg;
(5) остатки, которые влияют на ориентацию цепи: gly и pro; и
(6) ароматические: trp, tyr и phe.
Замену между аминокислотными остатками в каждой из этих групп аминокислот называют консервативной заменой, и замену аминокислотных остатков между различными группами называют неконсервативными заменами. Замены в рамках настоящего изобретения могут представлять собой консервативные замены или неконсервативные замены, или комбинацию консервативных замен и неконсервативных замен.
Изменения аминокислотной последовательности осуществляют различными способами, известными специалистам в данной области. Такие способы включают способ сайт-направленного мутагенеза (Hashimoto-Gotoh, T, Mizuno, T, Ogasahara, Y, and Nakagawa, M. (1995) An oligodesoxyribonucleotide-directed dual amber method for site-directed muthagenesis. Gene 152: 271-275; Zoller, M.J, and Smith, M. (1983) Oligonucleotide-directed muthagenesis of DNA fragments cloned into M13 vectors. Methods Enzymol. 100: 468-500; Kramer, W, Drutsa, V, Jansen, H.W, Kramer, B, Pflugfelder, M, and Fritz, H.J. (1984) The gapped duplex DNA approach to oligonucleotide-directed mutation construction. Nucleic Acids Res. 12: 9441-9456; Kramer W, and Fritz H.J. (1987) Oligonucleotide-directed construction of mutations via gapped duplex DNA Methods. Enzymol. 154, 350-367; and Kunkel, T.A. (1985) Rapid and efficient site-specific muthagenesis without phenotypic selection. Proc. Natl. Acad. Sci USA. 82: 488-492), способ внесения мутаций с помощью ПЦР и кассетный способ внесения мутаций, но не ограничиваются ими.
Аминокислотная модификация по настоящему изобретению включает посттрансляционную модификацию. Конкретной посттрансляционной модификацией может быть вставка или делеция цепи сахара. Например, в константной области IgG1, состоящей из аминокислотной последовательности SEQ ID NO:11, аминокислотный остаток в положении 297 (нумерация по EU) может быть модифицированным по сахарной цепи. Структура цепи сахара для модификации не ограничена. Как правило, антитела, экспрессируемые в эукариотических клетках, имеют гликозилирование в константной области. Таким образом, антитела, экспрессируемые в клетках, такие как антитела, представленные ниже, обычно модифицированы с помощью некоторого типа цепи сахара:
- антителопродуцирующие клетки млекопитающих;
- эукариотические клетки, трансформированные экспрессирующим вектором, содержащим ДНК, кодирующую антитело.
Эукариотические клетки, описанные в настоящем описании, включают клетки дрожжей и клетки животных. Например, клетки CHO и клетки HEK293H являются типичными клетками животных, используемыми для трансформации экспрессирующим вектором, содержащим ДНК, кодирующую антитело. С другой стороны, константные области без гликозилирования в этом участке также включены в константную область по настоящему изобретению. Антитела, константная область которых не гликозилирована, можно получать путем экспрессии гена, кодирующего антитело, в прокариотических клетках, таких как Escherichia coli.
В частности, например, сиаловая кислота может быть добавлена к цепи сахара Fc-области (MAbs. 2010 Sep-Oct; 2(5): 519-27).
Антитело
Более того, настоящее изобретение относится к антителам, содержащим Fc-область, в которых упомянутые выше аминокислотные последовательности изменены.
Термин “антитело/антитела” в рамках настоящего изобретения используют в наиболее широком значении, и при условии, что показана желаемая биологическая активность, он включает любое антитело, такое как моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, варианты антител, фрагменты антител, полиспецифические антитела (мультиспецифические антитела) (например, биспецифические антитела (антитела-димеры)), химерные антитела и гуманизированные антитела.
Что касается антител по настоящему изобретению, тип антигена и происхождение антитела не ограничены, и они могут представлять собой антитела любого типа. Происхождение антител конкретно не ограничено, но их примеры включают антитела человека, антитела мыши, антитела крысы и антитела кролика.
Способы получения антител хорошо известны специалистам в данной области, и, например, моноклональные антитела можно получать способом гибридом (Kohler and Milstein, Nature 256: 495 (1975)), или способом рекомбинации (патент США № 4816567). Альтернативно, их можно выделять из библиотеки фагового дисплея (Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1991)).
Гуманизированное антитело также называют реконструированным антителом человека. В частности, известны гуманизированные антитела, полученные пересадкой CDR антитела животного, не являющегося человеком, такого как антитело мыши, в антитело человека и т.п. Также известны основные способы генетической инженерии для получения гуманизированных антител. В частности, например, перекрывающаяся ПЦР с удлинением известна в качестве способа пересадки CDR антител мыши в FR человека.
Вектор для экспрессии гуманизированного антитела можно получать путем встраивания ДНК, кодирующей вариабельную область антитела, в которой три CDR и четыре FR лигированы, и ДНК, кодирующей константную область антитела человека, в экспрессирующий вектор так, чтобы эти ДНК были слиты в рамке считывания. После этого вектор для встраивания переносят хозяину для установления рекомбинантных клеток, эти клетку культивируют, и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в культуре клеток (см., публикацию патента Европы № EP 239400, и международную публикацию патента № WO 1996/002576).
При необходимости аминокислотный остаток в FR можно заменять так, чтобы CDR реконструированного антитела человека образовывала соответствующий антигенсвязывающий центр. Например, мутацию можно вносить в аминокислотную последовательность FR, применяя способ ПЦР, используемый для пересадки CDR мыши в FR человека.
Желаемое антитело человека можно получать путем иммунизации посредством ДНК с использованием трансгенного животного, имеющего полный набор генов антител человека (см. международные публикации № WO 1993/012227, WO 1992/003918, WO 1994/002602, WO 1994/025585, WO 1996/034096 и WO 1996/033735), в качестве животного для иммунизации.
Более того, известны технологии получения антител человека способом пэннинга с использованием библиотеки антител человека. Например, V-область антитела человека экспрессируют на поверхности фага в качестве одноцепочечного антитела (scFv) способом фагового дисплея. Экспрессирующий scFv фаг, который связывается с антигеном, можно отбирать. Последовательность ДНК, которая кодирует V-область связанного с антигеном антитела человека, можно определять путем анализа генов выбранного фага. После определения последовательности ДНК scFv, который связывается с антигеном, можно получать экспрессирующий вектор путем слияния последовательности V-области в рамке считывания с последовательностью C-области желаемого антитела человека, а затем встраивания их в подходящий экспрессирующий вектор. Экспрессирующий вектор вводят в подходящие экспрессирующие клетки, такие как клетки, описанные выше, и антитело человека можно получать путем экспрессии гена, кодирующего антитело человека. Эти способы уже известны (см., международные публикации № WO 1992/001047, WO 1992/020791, WO 1993/006213, WO 1993/011236, WO 1993/019172, WO 1995/001438 и WO 1995/15388).
Вариабельные области, составляющие антитела по настоящему изобретению, могут представлять собой вариабельные области, которые распознают любой антиген.
В рамках настоящего изобретения отсутствует конкретное ограничение на антиген, и он может представлять собой любой антиген. Примеры таких антигенов предпочтительно включают лиганды (цитокины, хемокины и т.п.), рецепторы, антигены злокачественной опухоли, антигены MHC, антигены дифференцировки, иммуноглобулины и иммунные комплексы, частично содержащие иммуноглобулины.
Примеры цитокинов включают интерлейкины с 1 по 18, колониестимулирующие факторы (G-CSF, M-CSF, GM-CSF и т.д.), интерфероны (IFN-α, IFN-β, IFN-γ и т.д.), факторы роста (EGF, FGF, IGF, NGF, PDGF, TGF, HGF и т.д.), факторы некроза опухоли (TNF-α и TNF-β), лимфотоксин, эритропоэтин, лептин, SCF, TPO, MCAF и BMP.
Примеры хемокинов включают CC-хемокины, такие как CCL1-CCL28, CXC-хемокины, такие как CXCL1-CXCL17, C-хемокины, такие как XCL1 и XCL2, и CX3C-хемокины, такие как CX3CL1.
Примеры рецепторов включают рецепторы, принадлежащие семействам рецепторов, таким как семейство рецепторов гемопоэтических факторов роста, семейство рецепторов цитокинов, семейство рецепторов тирозинкиназного типа, семейство рецепторов серин/треонинкиназного типа, семейство рецепторов TNF, семейство сопряженных с G-белком рецепторов, семейство рецепторов типа GPI-якоря, семейство рецепторов тирозинфосфатазного типа, семейство факторов адгезии и семейство рецепторов гормонов. Рецепторы, принадлежащие этим семействам рецепторов, и их характеристики описаны во многих документах, таких как Cooke B.A., King RJB., van der Molen H.J. ed. New Comprehesive Biochemistry V0I.I8B “Hormones and their Actions Part II” pp.1-46 (1988) Elsevier Science Publishers BV; Patthy (Cell (1990) 61(1): 13-14); Ullrich et al. (Cell (1990) 61(2): 203-212); Massague (Cell (1992) 69(6): 1067-1070); Miyajima et al. (Annu. Rev. Immunol. (1992) 10: 295-331); Taga et al. (FASEB J. (1992) 6, 3387-3396); Fantl et al. (Annu. Rev. Biochem. (1993), 62:453-481); Smith et al. (Cell (1994) 76(6): 959-962); и Flower D.R. Flower (Biochim. Biophys. Acta (1999) 1422(3): 207-234).
Примеры конкретных рецепторов, принадлежащих упомянутым выше семействам рецепторов, включают рецепторы эритропоэтина (EPO) человека или мыши (Blood (1990) 76(1): 31-35; и Cell (1989) 57(2): 277-285), рецепторы гранулоцитарного колониестимулирующего фактора (G-CSF) человека или мыши (Proc. Natl. Acad. Sci. USA. (1990) 87(22): 8702-8706, mG-CSFR; Cell (1990) 61(2): 341-350), рецепторы тромбопоэтина (TPO) человека или мыши (Proc. Natl. Acad. Sci. USA. (1992) 89(12): 5640-5644; EMBO J. (1993) 12(7): 2645-53), рецепторы инсулина человека или мыши (Nature (1985) 313(6005): 756-761), рецепторы лиганда Flt-3 человека или мыши (Proc. Natl. Acad. Sci. USA. (1994) 91(2): 459-463), рецепторы тромбоцитарного фактора роста (PDGF) (Proc. Natl. Acad. Sci. USA. (1988) 85(10): 3435-3439), рецепторы интерферона (IFN)-α и -β человека или мыши (Cell (1990) 60(2): 225-234; и Cell (1994) 77(3): 391-400), рецепторы лептина человека или мыши, рецепторы гормонов роста (GH) человека или мыши, рецепторы интерлейкина (IL)-10 человека или мыши, рецепторы инсулиноподобного фактора роста (IGF)-I человека или мыши, рецепторы фактора, ингибирующего лейкозы, (LIF) человека или мыши, и рецепторы циллиарного нейротрофического фактора (CNTF) человека или мыши.
Антигены злокачественной опухоли представляют собой антигены, которые экспрессируются, когда клетки становятся злокачественными, и их также называют опухолеспецифическими антигенами. Аномальные цепи сахаров, которые появляются на поверхностях клеток или молекулах белков, когда клетки становятся злокачественными, также являются антигенами злокачественной опухоли, и их также называют антигенами злокачественной опухоли в виде цепи сахара. Примеры антигенов злокачественной опухоли предпочтительно включают GPC3, который представляет собой рецептор, принадлежащий семейству рецепторов типа GPI-якоря, упомянутому выше, и также экспрессируется в нескольких злокачественных опухолях, включая рак печени (Int J Cancer. (2003) 103(4): 455-65), а также EpCAM, который экспрессируется в нескольких злокачественных опухолях, включая рак легкого (Proc. Natl. Acad. Sci. USA. (1989) 86(1): 27-31), CA19-9, CA15-3, и сиалил-SSEA-1 (SLX).
Антигены MHC приближенно классифицируют как антигены MHC класса I и антигены MHC класса II. Антигены MHC класса I включают HLA-A, -B, -C, -E, -F, -G и -H, и антигены MHC класса II включают HLA-DR, -DQ и -DP.
Антигены дифференцировки могут включать CD1, CD2, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15s, CD16, CD18, CD19, CD20, CD21, CD23, CD25, CD28, CD29, CD30, CD32, CD33, CD34, CD35, CD38, CD40, CD41a, CD41b, CD42a, CD42b, CD43, CD44, CD45, CD45RO, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD51, CD54, CD55, CD56, CD57, CD58, CD61, CD62E, CD62L, CD62P, CD64, CD69, CD71, CD73, CD95, CD102, CD106, CD122, CD126 и CDw130.
Иммуноглобулины включают IgA, IgM, IgD, IgG и IgE. Иммунные комплексы включают компонент по меньшей мере любого из иммуноглобулинов.
Другие антигены включают, например, представленные ниже молекулы: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, аденозиновый рецептор A1, A33, ACE, ACE-2, активин, активин A, активин AB, активин B, активин C, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM 10, ADAM 12, ADAM 15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, APE, APJ, APP, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, интегрин av/b3, Ax1, b2M, B7-1, B7-2, B7-H, B-лимфоцитарный стимулирующий фактор (BlyS), BACE, BACE-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bcl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, BMP-3 остеогенин, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, костный нейротрофический фактор, BPDE, BPDE-DNA, BTC, фактор комплемента 3 (C3), C3a, C4, C5, C5a, C10, CA125, CAD-8, кальцитонин, cAMP, карциноэмбриональный антиген (CEA), ассоциированный со злокачественной опухолью, катепсин A, катепсин B, катепсин C/DPPI, катепсин D, катепсин E, катепсин H, катепсин L, катепсин O, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок p67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (B7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, токсин Botulinum, токсин Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цитокератиновый ассоциированный с опухолью антиген, DAN, DCC, DcR3, DC-SIGN, регулирующий фактор комплемента (фактор, ускоряющий распад), дез-(1-3)-IGF-I (IGF-1 головного мозга), Dhh, дигоксин, ДНКМ-1, ДНКаза, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, фактор активации фибробластов (FAP), Fas, FcR1, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа1, GFR-альфа2, GFR-альфа3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, рилизинг-фактор гормона роста, гаптен (NP-cap или NIP-cap), HB-EGF, HCC, гликопротеин оболочки gB HCMV, гликопротеин оболочки gH HCMV, UL HCMV, гемопоэтический фактор роста (HGF), HepB gp120, гепараназа, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMW-MAA), gp120 HIV, петля V3 gp 120 IIIB HIV, HLA, HLA-DR, HM1.24, HMFG PEM, HRG, Hrk, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (HGH), HVEM, I-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, рецептор IgA, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-23, интерферон (INF)-альфа, INF-бета, INF-гамма, ингибин, iNOS, A-цепь инсулина, B-цепь инсулина, инсулиноподобный фактор роста 1, интегрин альфа-2, интегрин альфа-3, интегрин альфа-4, интегрин альфа4/бета1, интегрин альфа4/бета7, интегрин альфа5 (альфа V), интегрин альфа5/бета1, интегрин альфа5/бета3, интегрин альфа6, интегрин бета1, интегрин бета2, интерферон-гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин L1, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bpl, LBP, LDGF, LECT2, lefty, антиген Lewis-Y, ассоциированный с Lewis-Y антиген, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, белок поверхности легких, лютеинизирующий гормон, рецептор лимфотоксина-бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCAM, MCK-2, MCP, M-CSF, MDC, Mer, металлопротеазы, рецептор MGDF, MGMT, MHC (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, мюллерово ингибиторное вещество, Mug, MuSK, NAIP, NAP, NCAD, адгерин N-C, NCA90, NCAM, NCAM, неприлизин, нейротрофин-3, -4 или -6, нейротурин, фактор роста нервов (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGG1, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, плацентарная щелочная фосфатаза (PLAP), P1GF, PLP, PP14, проинсулин, прорелаксин, белок C, PS, PSA, PSCA, простат-специфический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, A-цепь релаксина, B-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, SERINE, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин-72), TARC, TCA-3, T-клеточный рецептор (например, T-клеточный рецептор альфа/бета), TdT, TECK, TEM1, TEM5, TEM7, TEM8, TERT, PLAP-подобная щелочная фосфатаза семенников, TfR, TGF, TGF-альфа, TGF-бета, панспецифический TGF-бета, TGF-бета RI (ALK-5), TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета 1, TGF-бета 2, TGF-бета 3, TGF-бета 4, TGF-бета 5, тромбин, Ck-1 тимуса, тиреотропный гормон, Tie, TIMP, TIQ, тканевой фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа-бета, TNF-бета 2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD 120a, p55-60), TNFRSF1B (TNF RII CD 120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (0X40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL Rl TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд HVEM LIGHT, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 Fas-лиганда, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TSG, TSLP, ассоциированный с опухолью антиген CA125, ассоциированный с опухолью антиген, экспрессирующий ассоциированные с Lewis-Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный антиген, VLA, VLA-1, VLA-4, интегрин VNR, фактор Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, Αβ, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин A, хромогранин B, tau, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор B, фактор D, фактор H, пропердин, склеростин, фибриноген, фибрин, протромбин, тромбин, тканевой фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Xa, фактор XI, фактор XIa, фактор XII, фактор XIIa, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и SIP; и рецепторы для гормонов и факторов роста.
В аминокислотных последовательностях, составляющих вариабельные области, допустимо одно или несколько изменений аминокислотных остатков, при условии, что активность связывания антигена сохраняется. При изменении аминокислотной последовательности вариабельной области отсутствует конкретное ограничение на область изменения и количество измененных аминокислот. Например, аминокислоты, присутствующие в CDR и/или FR, могут быть изменены соответствующим образом. При изменении аминокислот в вариабельной области активность связывания предпочтительно сохраняется без конкретного ограничения; и, например, по сравнению с тем, что было до изменения, активность связывания составляет 50% или более, предпочтительно 80% или более, и более предпочтительно 100% или более. Более того, активность связывания может быть увеличена с помощью аминокислотных изменений. Например, активность связывания может в 2, 5, 10 раз превышать активность связывания до изменения. В антителах по настоящему изобретению изменение аминокислотной последовательности может представлять собой одно из замены, вставки, делеции и модификации аминокислотного остатка.
Например, модификация N-концевого глутамина вариабельной области в пироглутаминовую кислоту путем пироглутамилирования представляет собой модификацию, хорошо известную специалистам в данной области. Таким образом, когда N-конец тяжелой цепи представляет собой глутамин, антитела по настоящему изобретению содержат вариабельные области, в которых глутамин модифицирован в пироглутаминовую кислоту.
Вариабельные области антитела по настоящему изобретению могут иметь любые последовательности, и они могут представлять собой вариабельные области антитела любого происхождения, такого как антитела мыши, антитела крысы, антитела кролика, антитела козы, антитела верблюда, гуманизированные антитела, полученные путем гуманизации этих не являющихся человеческими антител, и антитела человека. “Гуманизированные антитела”, также называемые “реконструированными антителами человека”, представляют собой антитела, в которых определяющие комплементарность области (CDR) антитела, происходящего из не являющегося человеком млекопитающего, например, антитела мыши, трансплантируют в CDR антитела человека. Способы идентификации CDR известны (Kabat et al., Sequences of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989) 342: 877). Их общие способы генетической рекомбинации также известны (см. публикацию патентной заявки Европы № EP 125023 и WO 96/02576). Более того, эти антитела могут иметь различные аминокислотные замены, внесенные в их вариабельные области, для повышения их связывания антигена, фармакокинетики, стабильности и антигенности. Вариабельные области антител по настоящему изобретению могут быть способны связывать антигены многократно благодаря их зависимости от pH при связывании антигена (WO 2009/125825).
Константные области типа κ-цепи и λ-цепи присутствуют в константных областях легкой цепи антител, однако приемлема только любая одна из константных цепей легкой цепи. Более того, константные области легкой цепи по настоящему изобретению могут представлять собой константные области легкой цепи с аминокислотными изменениями, такими как замены, делеции, вставки и/или инсерции.
Например, для константных областей тяжелой цепи антитела по настоящему изобретению можно использовать константные области тяжелой цепи IgG-антител человека и предпочтительными являются константные области тяжелой цепи IgG1-антител человека и константные области тяжелой цепи IgG4-антител человека.
Более того, полипептиды по настоящему изобретению можно преобразовывать в молекулы слитого белка Fc путем связывания с другими белками, физиологически активными пептидами и т.п.
Примеры других белков и биологически активных пептидов включают, но не ограничиваются ими, рецепторы, молекулы адгезии, лиганды и ферменты.
Предпочтительные примеры молекул слитых белков Fc по настоящему изобретению включают белки с Fc-доменом, слитым с рецепторным белком, который связывается с мишенью, и такие примеры включают слитый белок TNFR-Fc, слитый белок ILIR-Fc, слитый белок VEGFR-Fc и слитый белок CTLA4-Fc (Nat Med. 2003 Jan; 9(1): 47-52; BioDrugs. 2006; 20(3): 151-60). Более того, белок, подлежащий слиянию с полипептидом по настоящему изобретению, может представлять собой любую молекулу, при условии, что она связывается с молекулой-мишенью, и его примеры включают молекулы scFv (WO 2005/037989), однодоменные молекулы антител (WO 2004/058821; WO 2003/002609), антителоподобные молекулы (Current Opinion in Biotechnology 2006, 17: 653-658; Current Opinion in Biotechnology 2007, 18: 1-10; Current Opinion in Structural Biology 1997, 7: 463-469; и Protein Science 2006, 15: 14-27) такие как DARPins (WO 2002/020565), Affibody (WO 1995/00193 7), Avimer (WO 2004/044011; WO 2005/040229) и Adnectin (WO 2002/032925). Более того, антитела и молекулы слитого белка Fc могут представлять собой полиспецифические антитела, которые связываются с несколькими типами молекул-мишеней или эпитопов-мишеней.
Более того, антитела по настоящему изобретению включают продукты модификации антител. Такие продукты модификации антител включают, например, антитела, связанные с различными молекулами, такими как полиэтиленгликоль (PEG) и цитотоксические вещества. Такие продукты модификации антител можно получать путем химической модификации антител по настоящему изобретению. Способы модификации антител уже являются общеизвестными в данной области.
Антитела по настоящему изобретению также могут представлять собой биспецифические антитела. “Биспецифическое антитело” относится к антителу, которое имеет в единой молекуле вариабельные области, которые распознают различные эпитопы. Эпитопы могут присутствовать в одной молекуле или в разных молекулах.
Полипептиды по настоящему изобретению можно получать способами, известными специалистам в данной области. Например, антитела можно получать способами, описанными ниже, однако способы не ограничиваются ими.
Экспрессируют ДНК, кодирующую тяжелую цепь антитела, в которой один или несколько аминокислотных остатков в Fc-области заменены другими представляющими интерес аминокислотами, и ДНК, кодирующую легкую цепь антитела. ДНК, кодирующую тяжелую цепь, в которой один или несколько аминокислотных остатков в Fc-области заменены другими представляющими интерес аминокислотами, можно получать, например, путем получения ДНК, кодирующей Fc-область природной тяжелой цепи, и внесения соответствующей замены так, чтобы кодон, кодирующий конкретную аминокислоту в Fc-области, кодировал другую представляющую интерес аминокислоту.
Альтернативно, ДНК, кодирующую тяжелую цепь, в которой один или несколько аминокислотных остатков в Fc-области заменены другими представляющими интерес аминокислотами, также можно получать путем моделирования, а затем химического синтеза ДНК, кодирующей белок, в которой один или несколько аминокислотных остатков в Fc-области природной тяжелой цепи заменены другими представляющими интерес аминокислотами. Положение и тип аминокислотной замены конкретно не ограничены. Более того, изменение не ограничивается заменой, и изменение может представлять собой любое из делеции, вставки или инсерции, или их комбинации.
Альтернативно, ДНК, кодирующую тяжелую цепь, в которой один или несколько аминокислотных остатков в Fc-области заменены другими представляющими интерес аминокислотами, можно получать в качестве комбинации частичных ДНК. Такие комбинации частичных ДНК включают, например, комбинацию ДНК, кодирующей вариабельную область, и ДНК, кодирующей константную область, и комбинацию ДНК, кодирующей Fab-область, и ДНК, кодирующей Fc-область, но не ограничиваются ими. Более того, ДНК, кодирующую легкую цепь, можно аналогичным образом получать в качестве комбинации частичных ДНК.
Способы экспрессии описанных выше ДНК включают способы, описанные ниже. Например, экспрессирующий вектор тяжелой цепи конструируют путем встраивания ДНК, кодирующей вариабельную область тяжелой цепи, в экспрессирующий вектор вместе с ДНК, кодирующей константную область тяжелой цепи. Аналогично, экспрессирующий вектор легкой цепи конструируют путем встраивания ДНК, кодирующей вариабельную область легкой цепи, в экспрессирующий вектор вместе с ДНК, кодирующей константную область легкой цепи. Альтернативно, эти гены тяжелой и легкой цепей можно встраивать в один вектор.
При встраивании ДНК, кодирующей представляющее интерес антитело, в экспрессирующий вектор, ДНК встраивают так, чтобы антитело экспрессировалось под контролем регулирующей экспрессию области, такой как энхансер или промотор. Далее, клетки-хозяева трансформируют этим экспрессирующим вектором для экспрессии антитела. В таких случаях можно использовать подходящую комбинацию хозяина и экспрессирующего вектора.
Примеры векторов включают векторы M13, векторы pUC, pBR322, pBluescript и PCR-Script. Альтернативно, когда целью является субклонирование и вырезание кДНК, в дополнение к описанным выше векторам можно использовать pGEM-T, pDIRECT, pT7 и т.п.
Экспрессирующие векторы особенно пригодны при использовании векторов для получения полипептидов по настоящему изобретению. Например, когда клетка-хозяин представляет собой E. coli, такую как JM109, DH5α, HB101 и XL1-Blue, экспрессирующие векторы должны иметь промотор, который обеспечивает эффективную экспрессию в E. coli, например, промотор lacZ (Ward et al., Nature (1989) 341: 544-546; FASEB J. (1992) 6: 2422-2427; полное содержание которой включено в настоящее описание в качестве ссылки), промотор araB (Better et al., Science (1988) 240: 1041-1043; полное содержание которой включено в настоящее описание в качестве ссылки), промотор T7 и т.п. Такие векторы включают pGEX-5X-1 (Pharmacia), “QIAexpress system” (Qiagen), pEGFP или pET (в этом случае хозяином предпочтительно являются BL21, которые экспрессируют РНК-полимеразу T7) в дополнение к векторам, описанным выше.
Векторы могут содержать сигнальные последовательности для секреции полипептида. В качестве сигнальной последовательности для секреции полипептида можно использовать сигнальную последовательность pelB (Lei, S. P. et al. J. Bacteriol. (1987) 169: 4379; полное содержание которой включено в настоящее описание в качестве ссылки), когда полипептид секретируется в периплазму E. coli. Вектор можно вводить в клетки-хозяева, например, способом с липофектином, способом с фосфатом кальция и способом с DEAE-декстраном.
В дополнение к экспрессирующим векторам E. coli, векторы для продуцирования полипептидов по настоящему изобретению включают, например, экспрессирующие векторы млекопитающих (например, pcDNA3 (Invitrogen), pEGF-BOS (Nucleic Acids Res. 1990, 18(17): p5322; полное содержание которой включено в настоящее описание в качестве ссылки), pEF и pCDM8), происходящие из насекомых экспрессирующие векторы (например, “бакуловирусная экспрессирующая система Bac-to-BAC” (Gibco-BRL) и pBacPAK8), происходящие из растений экспрессирующие векторы (например, pMH1 и pMH2), происходящие из вирусов животных экспрессирующие векторы (например, pHSV, pMV и pAdexLcw), ретровирусные экспрессирующие векторы (например, pZIPneo), экспрессирующие векторы дрожжей (например, “Pichia Expression Kit” (Invitrogen), pNV11 и SP-Q01) и экспрессирующие векторы Bacillus subtilis (например, pPL608 и pKTH50).
Когда целью является экспрессия в клетках животных, таких как клетки CHO, COS и NIH3T3, векторы должны иметь промотор, необходимый для экспрессии в клетках, например, промотор SV40 (Mulligan et al., Nature (1979) 277: 108; полное содержание которой включено в настоящее описание в качестве ссылки), промотор MMTV-LTR, промотор EF1a (Mizushima et al., Nucleic Acids Res. (1990) 18: 5322; полное содержание которой включено в настоящее описание в качестве ссылки), промотор CAG (Gene. (1990) 18: 5322; полное содержание которой включено в настоящее описание в качестве ссылки) и промотор CMV, и более предпочтительно они имеют ген для селекции трансформированных клеток (например, ген устойчивости к лекарственным средствам, который позволяет оценку с использованием агента (неомицин, G418 и т.п.)). Векторы с такими характеристиками включают, например, pMAM, pDR2, pBK-RSV, pBK-CMV, pOPRSV и pOP13.
Кроме того, представленный ниже способ можно использовать для стабильной экспрессии генов и амплификации количества копий гена в клетках: в клетки CHO с дефектом в каскаде синтеза нуклеиновых кислот вводят вектор, который содержит ген DHFR, который компенсирует дефект (например, pCHOI), и вектор амплифицируют с использованием метотрексата (MTX). Альтернативно, следующий способ можно использовать для временной экспрессии гена: клетки COS с геном, экспрессирующим T-антиген SV40 на их хромосоме, трансформируют вектором с ориджином репликации SV40 (pcD и т.п.). Также можно использовать ориджины репликации, происходящие из вируса полиомы, аденовируса, вируса папилломы крупного рогатого скота (BPV) и т.п. Для амплификации количества копий генов в клетках-хозяевах, экспрессирующие векторы, кроме того, могут содержать селективные маркеры, такие как ген аминогликозидтрансферазы (APH), ген тимидинкиназы (TK), ген ксантин-гуанинфосфорибозилтрансферазы (Ecogpt) E. coli и ген дигидрофолатредуктазы (dhfr).
Антитела можно собирать, например, путем культивирования трансформированных клеток, а затем выделения антител из трансформированных клеток или из культуральной среды. Антитела можно выделять и очищать с использованием подходящей комбинации способов, таких как центрифугирование, фракционирование с сульфатом аммония, высаливание, ультрафильтрация, колонка с Ig, FcRn, белком A, белком G, аффинная хроматография, ионообменная хроматография и гель-фильтрация.
Более того, настоящее изобретение относится к способам получения полипептида, содержащего Fc-область антитела, имеющую сохраненную или сниженную активность связывания FcγRIIa и повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, которые включают внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида.
Примеры включают способы получения, включающие следующие стадии:
(a) внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептидов, содержащих Fc-область антитела;
(b) измерение активности связывания FcγRIIa и активности связывания FcγRIIb у полипептидов, измененных на стадии (a); и
(с) выбор полипептидов, имеющих сохраненную или сниженную активность связывания FcγRIIa и повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом.
Предпочтительным вариантом осуществления является способ получения полипептида, содержащего Fc-область антитела, который включает стадии:
(a) изменение нуклеиновой кислоты, кодирующей полипептид, так чтобы активность связывания FcγRIIa сохранялась или снижалась и активность связывания FcγRIIb повышалась по сравнению с исходным пептидом;
(b) введение нуклеиновой кислоты в клетки-хозяева и культивирование их для индукции экспрессии; и
(с) сбор полипептида из культуры клеток-хозяев.
Более того, антитела и молекулы слитого белка Fc, продуцируемые в этом способе получения, также охватываются настоящим изобретением.
Настоящее изобретение также относится к способам получения полипептида, где продукция антитела против полипептида подавляется по сравнению с его исходным полипептидом при введении in vivo, которые включают внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела.
Примеры включают способ получения, включающий следующие стадии:
(a) внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела; и
(b) подтверждение того, что продукция антитела подавляется, когда полипептид, измененный на стадии (a), вводят in vivo, по сравнению с исходным полипептидом.
Наличие подавления продукции антител против полипептида можно подтверждать способами введения полипептида животному и т.п. Альтернативно, подавление продукции антител можно определять путем измерения активности связывания с FcγRIIa и FcγRIIb, и выявления увеличения величины, полученной путем деления величины KD в отношении FcγRIIa на величину KD в отношении FcγRIIb. Такие полипептиды считаются полезными в качестве фармацевтических средств, поскольку они могут подавлять продукцию антител без активации активирующего FcγR.
В упомянутом выше способе продуцирования является предпочтительным повышение активности связывания FcγRIIb и сохранение или снижение активности связывания с FcγRIIa (тип R) и FcγRIIa (тип H); и, кроме того, является предпочтительным снижение активности связывания с FcγRIa и/или FcγRIIIa.
В предпочтительном варианте осуществления в упомянутом выше способе продуцирования, например, полипептид, содержащий Fc-область IgG человека, изменен так, чтобы Pro в положении 238 (нумерация по EU) был заменен на Asp или Leu в положении 328 (нумерация по EU) был заменен на Glu. Другие предпочтительные варианты осуществления включают изменение полипептида посредством по меньшей мере одной замены, выбранной из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met, в дополнение к замене Pro в положении 238 (нумерация по EU) на Asp.
Более того, настоящее изобретение относится к способам изменения полипептида для получения полипептида, имеющего сохраненную или сниженную активность связывания FcγRIIa и имеющего повышенную активность связывания с FcγRIIb по сравнению с его исходным полипептидом.
Настоящее изобретение также относится к способам изменения полипептида для получения полипептида, продукция антител против которого подавлена по сравнению с исходным полипептидом, когда его вводят in vivo.
В предпочтительном варианте осуществления, например, полипептид, содержащий Fc-область IgG человека, изменяют так, чтобы Pro в положении 238 (нумерация по EU) был заменен на Asp или Leu в положении 328 (нумерация по EU) был заменен на Glu. Другие предпочтительные варианты осуществления включают изменение полипептида путем по меньшей мере одной замены, выбранной из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met, в дополнение к замене Pro в положении 238 (нумерация по EU) на Asp.
Более того, настоящее изобретение относится к нуклеиновой кислоте, кодирующей полипептид, содержащий Fc-область антитела по меньшей мере с одним аминокислотным изменением, который имеет сохраненную или сниженную активность связывания FcγRIIa и повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом. Нуклеиновая кислота по настоящему изобретению может быть в любой форме, такой как ДНК или РНК.
Настоящее изобретение также относится к векторам, содержащим описанные выше нуклеиновые кислоты по настоящему изобретению. Тип вектора могут надлежащим образом выбрать специалисты в данной области, в зависимости от клеток-хозяев, в которые предстоит вводить вектор. Векторы включают, например, векторы, описанные выше.
Более того, настоящее изобретение относится к клеткам-хозяевам, трансформированным описанными выше векторами по настоящему изобретению. Подходящие клетки-хозяева могут быть выбраны специалистами в данной области. Клетки-хозяева включают, например, описанные выше клетки-хозяева.
Более того, настоящее изобретение относится к способам сохранения или снижения активности связывания FcγRIIa и повышения активности связывания FcγRIIb у полипептида, содержащего Fc-часть антитела, по сравнению с исходным полипептидом, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область.
Настоящее изобретение также относится к способам подавления продукции антител против полипептида по сравнению с исходным полипептидом, когда полипептид вводят in vivo, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела.
В предпочтительном варианте осуществления, например, полипептид, содержащий Fc-область IgG человека, изменен так, чтобы Pro в положении 238 (нумерация по EU) был заменен на Asp или Leu в положении 328 (нумерация по EU) был заменен на Glu. Другие предпочтительные варианты осуществления включают изменение полипептида путем по меньшей мере одной замены, выбранной из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met, в дополнение к замене Pro в положении 238 (нумерация по EU) на Asp.
В упомянутом выше способе является предпочтительным повышение активности связывания FcγRIIb и сохранение или снижение активности связывания с FcγRIIa (тип R) и FcγRIIa (тип H); и, кроме того, является предпочтительным сохранение или снижение активности связывания с FcγRIa и/или FcγRIIIa.
Полипептиды, продуцируемые любыми из упомянутых выше способов, также охватываются настоящим изобретением.
Фармацевтические композиции
Настоящее изобретение относится к фармацевтическим композициям, содержащим полипептид по настоящему изобретению.
Фармацевтические композиции по настоящему изобретению могут быть составлены, в дополнение к антителу или молекулам слитого белка Fc по настоящему изобретению, описанным выше, с фармацевтически приемлемыми носителями известными способами. Например, композиции можно использовать парентерально, когда антитела составлены в стерильном растворе или суспензии для инъекции с использованием воды или любой другой фармацевтически приемлемой жидкости. Например, композиции могут быть составлены путем комбинирования соответствующим образом антител или молекул слитых белков Fc с фармацевтически приемлемыми носителями или средами, в частности, стерильной водой или физиологический раствором, растительными маслами, эмульгаторами, суспендирующими веществами, поверхностно-активными веществами, стабилизаторами, вкусовыми добавками, эксципиентами, наполнителями, консервантами, связующими веществами и т.п., путем смешения их в единичной дозе и форме, требуемой для общепринятых фармацевтических форм. Конкретные примеры носителей включают легкую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кармеллозу кальция, кармеллозу натрия, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, поливинилацеталь диэтиламиноацетат, поливинилпирролидон, желатин, триглицериды средней цепи, отвержденное полиоксиэтиленом касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганическую соль и т.п. Содержание активного ингредиента в таком составе корректируют так, чтобы можно было получить соответствующую дозу в требуемом диапазоне.
Стерильные композиции для инъекций можно составлять с использованием носителей, таких как дистиллированная вода для инъекций, в соответствии со стандартными протоколами.
Водные растворы, используемые для инъекций, включают, например, физиологический солевой раствор и изотонические растворы, содержащие глюкозу или другие адъюванты, такие как D-сорбит, D-манноза, D-маннит и хлорид натрия. Их можно использовать вместе с подходящими солюбилизаторами, такими как спирт, в частности, этанол, полиспирты, такие как пропиленгликоль и полиэтиленгликоль, и неионные поверхностно-активные вещества, такие как Polysorbat 80TM и HCO-50.
Масла включают кунжутные масла и соевые масла, и их можно комбинировать с солюбилизаторами, такими как бензилбензоат или бензиловый спирт. Их также можно составлять с буферами, например, фосфатными буферами или буферами на основе ацетата натрия; анальгетиками, например, прокаина гидрохлоридом; стабилизаторами, например, бензиловым спиртом или фенолом; или антиоксидантами. Полученные инъекции, как правило, распределяют аликвотами в соответствующие ампулы.
Введение предпочтительно проводят парентерально, и, в частности, оно включает инъекцию, интраназальное введение, внутрилегочное введение и чрескожное введение. Например, инъекции можно проводить системно или местно путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции или подкожной инъекции.
Более того, способ введения можно соответствующим образом выбирать, исходя из возраста и симптомов пациента. Единичную дозу фармацевтической композиции, содержащей антитело или полинуклеотид, кодирующий антитело, можно выбирать, например, из диапазона от 0,0001 до 1000 мг на кг массы тела. Альтернативно, дозировка может находиться, например, в диапазоне от 0,001 до 100000 мг/пациент. Однако дозировка не ограничивается этими величинами. Дозировка и способ введения варьируют, в зависимости от массы тела пациента, возраста и симптомов, и ее могут соответствующим образом выбирать специалисты в данной области.
Упомянутые выше полипептиды по настоящему изобретению пригодны в качестве активных ингредиентов фармацевтических средств, которые подавляют активацию B-клеток, тучных клеток, дендритных клеток и/или базофилов. Полипептиды по настоящему изобретению могут подавлять активацию B-клеток, тучных клеток, дендритных клеток и/или базофилов, путем селективного воздействия на FcγRIIb без активации активирующего FcγR. Активация B-клеток включает пролиферацию, продуцирование IgE, продуцирование IgM и продуцирование IgA. Упомянутые выше полипептиды по настоящему изобретению обеспечивают связывание FcγRIIb с IgE для подавления продуцирования IgE B-клетками, с IgM для подавления продуцирования IgM B-клетками, и с IgA для подавления продуцирования IgA. Помимо указанного выше, эффекты подавления, сходные с упомянутыми выше эффектами при прямом или непрямом связывании FcγRIIb с молекулами, которые экспрессируются на B-клетках и содержат ITAM-домен внутри клетки или взаимодействуют с ITAM-доменом, такими как BCR, CD19 и CD79b. Более того, активация тучных клеток включает пролиферацию, активацию посредством IgE и т.п., и дегрануляцию. В тучных клетках упомянутые выше полипептиды по настоящему изобретению могут подавлять пролиферацию, активацию посредством IgE и т.п., и дегрануляцию путем прямого или непрямого связывания FcγRIIb с молекулами рецепторов IgE, которые экспрессируются на тучных клетках и содержат ITAM-домен или взаимодействуют с ITAM-доменом, такими как FcεRI, DAP12 и CD200R3. Активация базофилов включает пролиферацию и дегрануляцию базофилов. Также в базофилах упомянутые выше полипептиды по настоящему изобретению могут подавлять пролиферацию, активацию и дегрануляцию путем прямого или непрямого связывания FcγRIIb с молекулами на клеточной мембране, которые содержат ITAM-домен внутри клетки или взаимодействуют с ITAM-доменом. Активация дендритных клеток включает пролиферацию и дегрануляцию дендритных клеток. Также в дендритных клетках упомянутые выше полипептиды по настоящему изобретению могут подавлять активацию, дегрануляцию и пролиферацию путем прямого или непрямого связывания FcγRIIb с молекулами на клеточной мембране, которые содержат ITAM-домен внутри клетки и взаимодействуют с ITAM-доменом.
В рамках настоящего изобретения полипептиды по настоящему изобретению, упомянутые выше, пригодны в качестве активного ингредиента лекарственных средств или профилактических средств при иммунологических воспалительных заболеваниях. Как описано выше, поскольку полипептиды по настоящему изобретению могут подавлять активацию B-клеток, тучных клеток, дендритных клеток и/или базофилов, введение полипептидов по настоящему изобретению в результате может обеспечивать лечение или профилактику иммунологических воспалительных заболеваний. Не ограничиваясь этим, термин “иммунологические воспалительные заболевания” включает ревматоидный артрит, аутоиммунный гепатит, аутоиммунный тиреоидит, аутоиммунные заболевания с образованием волдырей, аутоиммунное адренокортикальное заболевание, аутоиммунную гемолитическую анемию, аутоиммунную тромбоцитопеническую пурпуру, мегалоцитарную анемию, аутоиммунный атрофический гастрит, аутоиммунную нейтропению, аутоиммунный орхит, аутоиммунный энцефаломиелит, аутоиммунное связанное с рецепторами заболевание, аутоиммунное бесплодие, хронический активный гепатит, гломерулонефрит, интерстициальный фиброз легких, рассеянный склероз, болезнь Педжета, остеопороз, множественную миелому, увеит, острый и хронический спондилит, подагрический артрит, воспалительное заболевание кишечника, респираторный дистресс-синдром взрослых (ARDS), псориаз, болезнь Крона, базедову болезнь, ювенильный диабет, болезнь Аддисона, миастению gravis, вызванный ношением линз увеит, системную красную волчанку, аллергический ринит, аллергический дерматит, язвенный колит, гиперчувствительность, мышечную дегенерацию, кахексию, системную склеродермию, локализованную склеродермию, синдром Шегрена, болезнь Бехчета, синдром Рейтера, диабет типа I и типа II, связанное с резорбцией костной ткани нарушение, реакцию "трансплантат против хозяина", повреждение при ишемии-реперфузии, атеросклероз, травму головного мозга, церебральную малярию, сепсис, септический шок, синдром токсического шока, лихорадку, миалгии вследствие физической нагрузки, апластическую анемию, гемолитическую анемию, идиопатическую тромбоцитопению, синдром Гудпасчера, синдром Гийена-Барре, тиреоидит Хашимото, пемфигус, IgA-нефропатию, поллиноз, синдром антифосфолипидных антител, полимиозит, гранулематоз Вегенера, узелковый артериит, смешанное заболевание соединительных тканей, фибромиалгию, астму, атопический дерматит, хронический атрофический гастрит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, синдром аортита, быстро прогрессирующий гломерулонефрит, мегалобластную анемию, идиопатическую тромбоцитопеническую пурпуру, первичный гипотиреоз, идиопатическую болезнь Аддисона, инсулинозависимый сахарный диабет, хроническую дискоидную красную волчанку, пемфигоид, герпес беременных, буллезный дерматоз с линейными IgA, приобретенный буллезный эпидермолиз, очаговую алопецию, витилиго обыкновенный, приобретенную центробежную лейкодерму Саттона, болезнь Харада, аутоиммунную оптическую невропатию, идиопатическую азооспермию, привычный выкидыш, гипогликемию, хроническую крапивницу, анкилозирующий спондилит, псориатический артрит, энтеропатический артрит, реактивный артрит, спондилоартропатию, энтесопатию, синдром раздраженной кишки, синдром хронической усталости, дерматомиозит, миозит с тельцами включения, синдром Шмидта, болезнь Грэйвса, пернициозную анемию, люпоидный гепатит, пресенильную деменцию, болезнь Альцгеймера, демиелинизирующее нарушение, боковой амиотрофический склероз, гипопаратиреоз, синдром Дресслера, синдром Итона-Ламберта, герпетиформный дерматит, алопецию, прогрессирующий системный склероз, синдром CREST (кальциноз, явление Рейно, нарушение моторики пищевода, склеродактилию и телеангиэктазию), саркоидоз, ревматическую атаку, полиформную эритему, синдром Кушинга, трансфузионную реакцию, болезнь Хансена, артериит Такаясу, ревматическую полимиалгию, височный артериит, гигантоклеточный артрит, экзему, лимфогранулематоз, болезнь Кавасаки, эндокардит, эндомиокардиальный фиброз, эндофтальмит, фетальный эритробластоз, эозинофильный фасциит, синдром Фелти, пурпуру Геноха-Шенлейна, отторжение трансплантата, свинку, кардиомиопатию, гнойный артрит, семейную средиземноморскую лихорадку, синдром Макла-Уэлса и синдром гипер-IgD.
Более того, при аутоиммунных заболеваниях, которые могут быть вызваны продукцией антител против аутоантигенов (аутоантител), полипептиды по настоящему изобретению, упомянутые выше, пригодны в качестве активного ингредиента фармацевтических средств для лечения или профилактики аутоиммунных заболеваний путем подавления продукции этих аутоантител. Описано, что применение молекулы, полученной путем слияния Fc-части антитела с AchR (аутоантиген миастении gravis) подавляет пролиферацию B-клеток, которые экспрессируют распознающий AchR BCR и индуцируют апоптоз (J. Neuroimmunol, 227: 35-43, 2010). Применение слитого белка, образованного между антигеном, распознаваемым аутоантителом, и Fc-областью антитела по настоящему изобретению, обеспечивает связывание FcγRIIb с BCR на B-клетке, экспрессирующей BCR для этого аутоантигена, подавление пролиферации B-клеток, экспрессирующих BCR для аутоантигена, и индукцию апоптоза. Такие аутоиммунные заболевания включают синдром Гийена-Барре, миастению gravis, хронический атрофический гастрит, аутоиммунный гепатит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, синдром аортита, синдром Гудпасчера, быстро прогрессирующий гломерулонефрит, мегалобластную анемию, аутоиммунную гемолитическую анемию, аутоиммунную нейтропению, идиопатическую тромбоцитопеническую пурпуру, базедову болезнь, тиреоидит Хашимото, первичный гипотиреоз, идиопатическую болезнь Аддисона, инсулинозависимый сахарный диабет, хроническую дискоидную красную волчанку, локализованную склеродермию, пемфигус, пемфигоид, гестационный герпес, буллезный дерматоз с линейными IgA, приобретенный буллезный эпидермолиз, очаговую алопецию, витилиго обыкновенный, приобретенную центробежную лейкодерму Саттона, болезнь Харада, аутоиммунную оптическую невропатию, идиопатическую азооспермию, привычный выкидыш, диабет типа II, гипогликемию и хроническую крапивницу; но не ограничиваются ими.
Более того упомянутые выше полипептиды по настоящему изобретению пригодны в качестве активного ингредиента в лекарственных средствах от заболеваний с дефицитом биологически необходимого белка. Для заболеваний с дефицитом биологически необходимого белка используют терапевтические способы, в которых осуществляют введение белка и дополнение белком в качестве фармацевтического средства. Однако поскольку пациент изначально лишен белка, предоставляемый извне белок распознается в качестве чужеродного агента и продукции антител против этого белка. В результате белок легко удаляется, и эффект фармацевтического средства снижается. Применение слитого белка, содержащего такой белок и Fc-область антитела, по настоящему изобретению обеспечивает связывание между FcγRIIb и BCR на B-клетках, которые распознают белок, и обеспечивает подавление продукции антител против белка. Дополнительно добавляемые белки включают фактор VIII, фактор IX, TPO, EPO, α-идуронидазу, идуронатсульфатазу, гепаран N-сульфатазу A-типа, α-N-ацетилглюкозаминидазу B-типа, ацетил-CoA C-типа, ацетилтрансферазу α-глюкозаминидазы, N-ацетилглюкозамин 6-сульфатазу D-типа, галактоза-6-сульфатазу, N-ацетилгалактозамин 4-сульфатазу, β-глюкуронидазу, α-галактозидазу, кислую α-галактозидазу и глюкоцереброзидазу. Дополнение этими белками можно осуществлять при заболеваниях, таких как гемофилия, идиопатическая тромбоцитопеническая пурпура, почечная анемия и лизосомальное заболевание (мукополисахаридоз, болезнь Фабри, болезнь Помпе и болезнь Гаухера), но не ограничиваются ими.
Более того, упомянутые выше полипептиды по настоящему изобретению пригодны в качестве активного ингредиента противовирусных средств. Антитела, которые содержат Fc-область по настоящему изобретению и представляют собой противовирусные антитела, могут подавлять антителозависимое усиление, наблюдаемое для противовирусных антител. Антителозависимое усиление представляет собой явление, где вирус использует нейтрализующие антитела против вируса, чтобы фагоцитироваться через активирующие FcγR, и инфицирует экспрессирующие FcγR клетки, так что происходит распространение инфекции. Описано, что связывание нейтрализующих антител против вируса Денге с FcγRIIb играет важную роль в подавлении антителозависимого усиления (Proc. Natl. Acad. Sci. USA, 108: 12479-12484, 2011). Связывание FcγRIIb с иммунным комплексом, содержащим вирус Денге, который образуется нейтрализующими антителами против вируса Денге, ингибирует опосредуемый FcγR фагоцитоз, что приводит к подавлению антителозависимого усиления. Примеры таких вирусов включают вирус Денге (DENV1, DENV2 и DENV4) и ВИЧ, но не ограничиваются ими.
Более того, полипептиды по настоящему изобретению, описанные выше, пригодны в качестве активного ингредиента в профилактических средствах или терапевтических средствах от артериосклероза. Антитела против окисленных ЛПНП, т.е. причины артериосклероза, которые представляют собой антитела, содержащие Fc-область по настоящему изобретению, могут предотвратить зависимую от FcγRIIa адгезию воспалительных клеток. Было описано, что, хотя антитела против окисленных ЛПНП ингибируют взаимодействие между окисленным ЛПНП и CD36, антитела против окисленного ЛПНП связываются с эндотелиальными клетками, и моноциты распознают их Fc-часть зависимым от FcγRIIa или зависимым от FcγRI образом; и это приводит к адгезии (Immunol. Lett., 108: 52-61, 2007). При использовании антител, содержащих Fc-область по настоящему изобретению, такие антитела могут ингибировать FcγRIIa-зависимое связывание и подавлять адгезию моноцитов посредством опосредуемых FcγRIIb ингибиторных сигналов.
В рамках настоящего изобретения полипептиды по настоящему изобретению, описанные выше, пригодны в качестве активного ингредиента в лекарственных средствах или профилактических средствах для лечения злокачественных новообразований. Как описано выше, показано, что усиление связывания FcγRIIb повышает агонистическую активность антитела-агониста и усиливает противоопухолевый эффект антитела. Таким образом, антитела-агонисты, в которых используется Fc-область по настоящему изобретению, пригодны для лечения или профилактики злокачественного новообразования. Fc-область по настоящему изобретению повышает агонистическую активность антител-агонистов против рецепторов из семейства рецепторов TNF, таких как Aliases, CD120a, CD120b, рецептор лимфотоксина β, CD134, CD40, FAS, TNFRSF6B, CD27, CD30, CD137, TNFRSF10A, TNFRSF10B, TNFRSF10C, TNFRSF10D, RANK, остеопротегерин, TNFRSF12A, TNFRSF13B, TNFRSF13C, TNFRSF14, рецептор фактора роста нервов, TNFRSF17, TNFRSF18, TNFRSF19, TNFRSF21, TNFRSF25 и рецептор A2 эктодисплазина. Более того, также повышается агонистическая активность антител-агонистов, отличная от агонистической активности, описанной выше. Не ограничиваясь этим, злокачественная опухоль включает рак легкого (включая мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легких и плоскоклеточную карциному легкого), рак толстого кишечника, рак прямой кишки, рак ободочной кишки, рак молочной железы, рак печени, рак желудка, рак поджелудочной железы, рак почки, рак предстательной железы, рак яичника, рак щитовидной железы, холангиокарциному, рак брюшины, мезотелиому, плоскоклеточную карциному, рак шейки матки, рак эндометрия, рак мочевого пузыря, рак пищевода, рак головы и шеи, рак носоглотки, опухоль слюнных желез, тимому, рак кожи, базально-клеточную опухоль, злокачественную меланому, рак анального канала, рак полового члена, рак яичка, опухоль Вильмса, острый миелоидный лейкоз (включая острый миелолейкоз, острый миелобластный лейкоз, острый промиелоцитарный лейкоз, острый миеломоноцитарный лейкоз и острый моноцитарный лейкоз), хронический миелогенный лейкоз, острый лимфобластный лейкоз, хронический лимфацитарный лейкоз, лимфому Ходжкина, неходжскинскую лимфому (лимфому Беркитта, хронический лимфоцитарный лейкоз, фунгоидный микоз, лимфому из клеток мантийной зоны, фолликулярную лимфому, диффузную крупноклеточную лимфому, лимфому клеток краевой зоны, пилоцитарный лейкоз, плазмацитому, лимфому периферических T-клеток и T-клеточный лейкоз/лимфому взрослых), гистиоцитоз клеток Лангерганса, множественную миелому, миелодиспластический синдром, опухоль головного мозга (включая глиому, астроглиому, глиобластому, менингиому и эпендимому), нейробластому, ретинобластому, остеосаркому, саркому Капоши, саркому Юинга, ангиосаркому и гемангиоперицитому.
Более того, настоящее изобретение относится к способам лечения или профилактики иммунологических воспалительных заболеваний, которые включают введение индивидууму (пациенту) полипептида по настоящему изобретению или полипептида, продуцированного способами получения по настоящему изобретению.
Настоящее изобретение также относится к наборам для применения в терапевтических способах или профилактических способах по настоящему изобретению, которые содержат по меньшей мере полипептид по настоящему изобретению или полипептид, продуцируемый способами получения по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению. Кроме того, в набор могут быть включены фармацевтически приемлемые носители, среды, инструкции по способу применения и т.п. Более того, настоящее изобретение относится к применению полипептида по настоящему изобретению или полипептида, продуцируемого способами получения по настоящему изобретению, для получения средств для лечения или профилактики иммунологических воспалительных заболеваний. Настоящее изобретение также относится к полипептидам по настоящему изобретению или полипептидам, продуцируемым способами получения по настоящему изобретению, для применения в терапевтических способах или профилактических способах по настоящему изобретению.
Как используют в рамках изобретения, трехбуквенные или однобуквенные коды для соответствующих аминокислот являются следующими:
Аланин: Ala (A)
Аргинин: Arg (R)
Аспарагин: Asn (N)
Аспарагиновая кислота: Asp (D)
Цистеин: Cys (C)
Глутамин: Gln (Q)
Глутаминовая кислота: Glu (E)
Глицин: Gly (G)
Гистидин: His (H)
Изолейцин: Ile (I)
Лейцин: Leu (L)
Лизин: Lys (K)
Метионин: Met (M)
Фенилаланин: Phe (F)
Пролин: Pro (P)
Серин: Ser (S)
Треонин: Thr (T)
Триптофан: Trp (W)
Тирозин: Tyr (Y)
Валин: Val (V)
Все документы уровня техники, цитированные в настоящем описании, включены в качестве ссылок в полном объеме.
Примеры
Ниже в настоящем описании настоящее изобретение дополнительно описано конкретно с помощью примеров, но его не следует считать ограничивающимся ими.
Пример 1
Детальный анализ связывания вариантов Fc с FcγR
Мутации вносили в IgG1-антитела для получения антител, которые имеют сниженное опосредуемое Fc связывание с активирующим FcγR, в частности, обоими аллотипами FcγRIIa: типы H и R, а также усиленное связывание с FcγRIIb относительно IgG1; и детально анализировали связывание с каждым FcγR.
Вариабельную область (SEQ ID NO:15) антитела против глипикана 3, содержащего CDR GpH7, которое представляет собой антитело против глипикана 3 с улучшенной кинетикой в плазме, описанное в WO 2009/041062, использовали в качестве H-цепи обычного антитела. Аналогично, в качестве L-цепи обычного антитела использовали GpL16-k0 (SEQ ID NO:16) из антитела против глипикана 3 с улучшенной кинетикой в плазме, описанного в WO 2009/041062. Более того, в качестве константной области H-цепи антитела использовали B3 (SEQ ID NO:17), где в G1d, полученный путем удаления C-концевого Gly и Lys IgG1, была внесена мутация K439E. Эта H-цепь обозначается как GpH7-B3 (SEQ ID NO:18), и L-цепь обозначается как GpL16-k0 (SEQ ID NO:16).
Что касается GpH7-B3, аминокислоты, которые считаются вовлеченными в связывание FcγR, и окружающие аминокислоты (положения 234-239, 265-271, 295, 296, 298, 300 и 324-337, в соответствии с нумерацией по EU) заменяли, соответственно, 18 типами аминокислот, исключая исходные аминокислоты и Cys. Эти Fc-варианты обозначаются как варианты B3. Варианты B3 экспрессировали и очищали с использованием способа справочного примера 1, и детально оценивали связывание с каждым FcγR (FcγRIa, FcγRIIa типа H, FcγRIIa типа R, FcγRIIb и FcγRIIIa) с использованием способа справочного примера 2.
Исходя из результатов анализа взаимодействия с каждым FcγR описанным ниже способом, строили графики. Значение величины связывания FcγR для происходящего из каждого варианта B3 антитела делили на значение величины связывания FcγR для антитела, использованного для сравнения, которое не имеет мутаций, внесенных в B3 (антитело, имеющее последовательность встречающегося в природе IgG1 человека в положениях 234-239, 265-271, 295, 296, 298, 300 и 324-337, в соответствии с нумерацией по EU). Величину, полученную путем умножения этой величины на 100, использовали в качестве индикатора относительной активности связывания FcγR для каждого варианта. На горизонтальной оси показана величина относительной активности связывания FcγRIIb для каждого варианта, и на вертикальной оси показана величина соответствующей относительной активности связывания каждого варианта в отношении активирующих FcγR: FcγRIa, FcγRIIa типа H, FcγRIIa типа R и FcγRIIIa (фиг.1, 2, 3 и 4).
Как показано с помощью обозначений на фиг.1-4, результаты показывают, что из всех изменений, только когда вносили мутации, называемые мутацией A (изменение, полученное путем замены Pro в положении 238 (нумерация по EU) на Asp) и мутацией B (изменение, полученное путем замены Leu в положении 328 (нумерация по EU) на Glu), происходило значительное усиление связывания с FcγRIIb и значительное подавление связывания с обоими типами FcγRIIa по сравнению с тем, что было до введения.
Пример 2
Анализ с использованием SPR вариантов, которые селективно связываются с FcγRIIb
Что касается изменения, идентифицированного в примере 1, где Pro в положении 238 (нумерация по EU) заменен на Asp, подробно анализировали связывание с каждым FcγR.
Вариабельную область IL6R-H (SEQ ID NO:19), которая представляет собой вариабельную область антитела против рецептора интерлейкина 6 человека, описанного в WO 2009/125825, использовали в качестве вариабельной области H-цепи антитела, и IL6R-G1d (SEQ ID NO:20), которая содержит G1d с делецией C-концевого Gly и Lys IgG1 человека, использовали в качестве константной области H-цепи человека в H-цепи IgG1. Pro в положении 238 (нумерация по EU) в IL6R-G1d заменяли на Asp с получением IL6R-G1d-v1 (SEQ ID NO:21). Далее Leu в положении 328 (нумерация по EU) в IL6R-G1d заменяли на Glu с получением IL6R-G1d-v2 (SEQ ID NO:23). Более того, для сравнения, Ser в положении 267 (нумерация по EU) заменяли на Glu и Leu в положении 328 (нумерация по EU) заменяли на Phe в IL6R-G1d с получением IL6R-G1d-v3 (SEQ ID NO:24), как описано в непатентном документе 27. IL6R-L (SEQ ID NO:22), которая представляет собой L-цепь тоцилизумаба, использовали в качестве общей L-цепи антител; и вместе с каждой H-цепью антитела экспрессировали и очищали в соответствии со способом справочного примера 1. Полученные антитела, которые содержат аминокислотную последовательность, происходящую из IL6R-G1d, IL6R-G1d-v1, IL6R-G1d-v2 или IL6R-G1d-v3 в качестве H-цепи антитела, упоминаются как IgG1, IgG1-v1, IgG1-v2 и IgG1-v3, соответственно.
Далее проводили анализ кинетики взаимодействий между этими антителами и FcγR с использованием Biacore T100 (GE Healthcare). В качестве подвижного буфера использовали HBS-EP+ (GE Healthcare), и температура измерения была установлена на 25°C. Использовали чип, полученный путем иммобилизации белка A на чип Series S Sensor Chip CM5 (GE Healthcare) способом присоединения аминов. Представляющее интерес антитело улавливали на этом чипе для взаимодействия с каждым FcγR, который был разбавлен подвижным буфером, и измеряли связывание с антителом. После измерения антитело, уловленное на чипе, смывали проведением реакции с 10 мМ глицином-HCl, pH 1,5, и чип регенерировали и использовали повторно. Сенсограммы, полученные в качестве результатов измерения, анализировали с помощью модели связывания Ленгмюра 1:1 с использованием программного обеспечения Biacore Evaluation для вычисления константы скорости связывания ka (л/моль/с) и константы скорости диссоциации kd (1/с), и из этих величин вычисляли константу диссоциации KD (моль/л).
На этот раз, поскольку связывание IgG1-v1 и IgG1-v2 с FcγRIIa типа H и с FcγRIIIa было слабым, параметры кинетики, такие как KD, нельзя был вычислять с помощью упомянутого выше аналитического способа. Что касается таких взаимодействий, величины KD вычисляли с использованием следующей модели связывания 1:1, описанной в Biacore T100 Software Handbook BR1006-48 Edition AE.
Поведение взаимодействующих молекул в соответствии с моделью связывания 1:1 в Biacore может быть описано с помощью уравнения 1, представленного ниже.
[Уравнение 1]
Req= C·Rmax/(KD+C) + RI
Req: график уровней связывания в стационарном состоянии против концентрации анализируемого соединения
C: концентрация
RI: совокупный вклад индекса рефракции в образце
Rmax: способность поверхности связывать анализируемое соединение.
После преобразования этого уравнения KD может быть выражена с помощью уравнения 2, представленного ниже.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
KD можно вычислять путем подстановки величин Rmax, RI и C в это уравнение. Из текущих условий измерения можно использовать RI=0, C=2 мкмоль/л. Более того, величину Rmax, полученную при глобальной аппроксимации сенсограммы, полученной в результате анализа взаимодействия каждого FcγR с IgG1 с использованием модели связывания Ленгмюра 1:1, делили на количество уловленного IgG1, затем умножали на величину уловленного IgG1-v1 и IgG1-v2, и полученную величину использовали в качестве Rmax. Это вычисление основано на гипотезе, что предельное количество каждого FcγR, который может связываться IgG1, остается неизмененным для всех вариантов, полученных путем внесения мутаций в IgG1, и величина Rmax в момент измерения пропорциональна количеству антитела, связанного с чипом во время измерения. Req определяли как величину связывания каждого FcγR с каждым вариантом на сенсорном чипе, наблюдаемого во время измерения.
В этих условиях измерения величина связывания (Req) IgG1-v1 и IgG1-v2 с FcγRIIa типа H составляла приблизительно 2,5 RU и 10 RU, соответственно, и величина связывания (Req) IgG1-v1 и IgG1-v2 с FcγRIIIa составляла приблизительно 2,5 RU и 5 RU, соответственно. Количество IgG1, IgG1-v1 и IgG1-v2, уловленных в анализе взаимодействия с FcγRIIa H-типа, составляло 452 RU, 469,2 RU и 444,2 RU, соответственно, и количество IgG1, IgG1-v1 и IgG1-v2, уловленных в анализе взаимодействий с FcγRIIIa, составляло 454,5 RU, 470,8 RU и 447,1 RU, соответственно. Величины Rmax, полученные путем глобальной аппроксимации сенсограмм, полученных в результате анализа взаимодействия IgG1 с FcγRIIa H-типа и FcγRIIIa с использованием модели связывания Ленгмюра 1:1, составляли 69,8 RU и 63,8 RU, соответственно. Когда использовали эти величины, вычисленные величины Rmax для IgG1-v1 и IgG1-v2 в отношении FcγRIIa типа H составляли 72,5 RU и 68,6 RU, соответственно, и вычисленные величины Rmax для IgG1-v1 и IgG1-v2 в отношении FcγRIIIa составляли 66,0 RU и 62,7 RU, соответственно. Эти величины подставляли в уравнение 2 для вычисления KD IgG1-v1 и IgG1-v2 для FcγRIIa типа H и FcγRIIIa.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
Величины KD IgG1, IgG1-v1, IgG1-v2 и IgG1-v3 для каждого FcγR (величины KD каждого антитела для каждого FcγR) представлены в таблице 1, и относительные величины KD IgG1-v1, IgG1-v2 и IgG1-v3, полученные путем деления величин KD IgG1 для каждого FcγR и на величины KD IgG1-v1, IgG1-v2 и IgG1-v3 для каждого FcγR (относительные величины KD каждого антитела для каждого FcγR), представлены в таблице 2.
В таблице 1, представленной выше, "*" означает, что величину KD вычисляли с использованием уравнения 2, поскольку не наблюдали достаточного связывания FcγR с IgG.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
Согласно таблице 2, по сравнению с IgG1, активность связывания IgG1-v1 снижалась в 0,047 раз в отношении FcγRIa, снижалась в 0,10 раз в отношении FcγRIIa типа R, снижалась в 0,014 раз в отношении FcγRIIa типа H, снижалась в 0,061 раз в отношении FcγRIIIa и увеличивалась в 4,8 раз в отношении FcγRIIb.
Более того, согласно таблице 2, по сравнению с IgG1, активность связывания IgG1-v2 снижалась в 0,74 раз в отношении FcγRIa, снижалась в 0,41 раз в отношении FcγRIIa типа R, снижалась в 0,064 раз в отношении FcγRIIa типа H, снижалась в 0,14 раз в отношении FcγRIIIa и увеличивалась в 2,3 раза в отношении FcγRIIb.
Более конкретно, эти результаты продемонстрировали, что IgG1-v1, имеющий изменение в виде замены Pro в положении 238 (нумерация по EU) на Asp, и IgG1-v2, имеющий изменение в виде замены Leu в положении 328 (нумерация по EU) на Glu, имеют одинаковые свойства ослабления связывания со всеми активирующими FcγR, включая оба аллотипа FcγRIIa, при усилении связывания с FcγRIIb, который является ингибиторным FcγR.
Далее, оценивали селективность полученного варианта в отношении FcγRIIb с использованием отношения активности связывания FcγRIIb к активности связывания с FcγRIIa типа R или типа H в качестве индикатора. В частности, I/A(R) или I/A(H), которая представляет собой величину, полученную делением величины KD в отношении FcγRIIa типа R или типа H на величину KD для FcγRIIb, использовали в качестве индикатора селективности FcγRIIb в отношении каждого FcγRIIa. Этот индикатор имеет более высокую величину, когда величина KD в отношении FcγRIIb становится меньше или когда величина KD в отношении FcγRIIa становится больше. Т.е. вариант, который демонстрирует более высокую величину, проявляет повышенную активность связывания в отношении FcγRIIb относительно FcγRIIa. Эти индикаторы обобщенно представлены в таблице 3 для каждого варианта.
Согласно результатам, представленным в таблице 3, по сравнению с IgG1, IgG1-v3, который продуцировали с использованием существующей технологии, продемонстрировал более высокую величину 1/A(H), чем IgG1, и более высокую селективность в отношении FcγRIIb, но меньшую величину 1/A(R), чем IgG1, и увеличенную селективность в отношении FcγRIIb. С другой стороны, IgG1-v1 и IgG1-v2, представленные в примерах, имеют более высокие величины I/A(R) и I/A(H), чем IgG1, и увеличенную селективность в отношении FcγRIIb относительно обоих аллотипов FcγRIIa.
До настоящего времени не были описаны изменения, имеющие такие свойства, и они в действительности являются очень редкими, как показано на фиг.1, 2, 3 и 4. Изменения, полученные путем замены Pro в положении 238 (нумерация по EU) на Asp или замены Leu в положении 328 (нумерация по EU) на Glu, очень полезны для разработки лекарственных средств при иммунологических воспалительных заболеваниях и т.п.
Более того, в таблице 2 показано, что IgG1-v3, описанный в непатентном документе 27, определенно проявляет в 408 раз усиленное связывание с FcγRIIb, в то время как связывание с FcγRIIa типа H снижено в 0,51 раза, и связывание с FcγRIIa типа R усилено в 522 раза. В соответствии этими результатами, поскольку IgG1-v1 и IgG1-v2 подавляют их связывание с обоими типами FcγRIIa: R и H и усиливают их связывание с FcγRIIb, они считаются вариантами, которые связываются с FcγRIIb с большей селективностью по сравнению с IgG1-v3. В частности, изменения, полученные путем замены Pro в положении 238 (нумерация по EU) на Asp или замены Leu в положении 328 (нумерация по EU) на Glu, в высокой степени пригодны для разработки лекарственных средств при иммунологических воспалительных заболеваниях и т.п.
Пример 3
Эффекты комбинирования изменений, обеспечивающих селективное связывание с FcγRIIb, с другими аминокислотными заменами в Fc-области
Была предпринята дальнейшая попытка усиления селективности к FcγRIIb, исходя из варианта, который имеет увеличенную селективность в отношении FcγRIIb и имеет замену Pro в положении 238 (нумерация по EU) на Asp, как показано в примерах 1 и 2.
Во-первых, в IL6R-G1d_v1 (SEQ ID NO:21), полученный путем внесения в IL6R-G1d изменения, полученного путем замены Pro в положении 238 (нумерация по EU) на Asp, вносили замену Leu в положении 328 (нумерация по EU) на Glu, как описано в примере 2, которая усиливает селективность в отношении FcγRIIb, с получением варианта IL6R-G1d-v4 (SEQ ID NO:25). Его комбинировали с IL6R-L (SEQ ID NO:22) и получали в соответствии со способом справочного примера 1. Полученное антитело, имеющее аминокислотную последовательность, происходящую из IL6R-G1d-v4 в качестве H-цепи антитела, было названо IgG1-v4. Связывающую активность IgG1, IgG1-vl, IgG1-v2, и IgG1-v4 с FcγRIIb оценивали в соответствии со способом справочного примера 2, и эти результаты представлены в таблице 4.
Из результатов, представленных в таблице 4, ожидалось, что, поскольку L328E повышает активность связывания FcγRIIb в 2,3 раза по сравнению с IgG1, то комбинирование этой замены с P238D, которая аналогично повышает активность связывания FcγRIIb в 4,8 раза по сравнению с IgG1, далее увеличит степень повышения активности связывания FcγRIIb; однако, в действительности, активность связывания FcγRIIb у варианта, содержащего комбинацию этих изменений, была снижена в 0,47 раз по сравнению с IgG1. Этот результат представляет собой эффект, который не мог быть предсказан, исходя из соответствующих изменений.
Аналогично, в IL6R-G1d-v1 (SEQ ID NO:21), полученный путем внесения в IL6R-G1d изменения путем замены Pro в положении 238 (нумерация по EU) на Asp, вносили замену Ser в положении 267 (нумерация по EU) на Glu и Leu в положении 328 (нумерация по EU) на Phe, как описано в примере 2, которые повышают активность связывания FcγRIIb, и вариант IL6R-G1d-v5 (SEQ ID NO:26) получали в соответствии со способом справочного примера 1. Полученное антитело, имеющее аминокислотную последовательность, происходящую из IL6R-G1d-v5, в качестве H-цепи антитела, было названо IgG1-v5. Активность связывания FcγRIIb у IgG1, IgG1-v1, IgG1-v3 и IgG1-v5 оценивали в соответствии со способом справочного примера 2, и эти результаты представлены в таблице 5.
Замены S267E/L328F, которые имели усиливающий эффект в отношении FcγRIIb в примере 2, вносили в вариант P238D, и оценивали его активность связывания FcγRIIb до и после внесения этого изменения. Результаты представлены в таблице 5.
Из результатов, представленных в таблице 5, ожидалось, что, поскольку S267E/L328F повышает активность связывания FcγRIIb в 408 раз по сравнению IgG1, то комбинирование этих замен с P238D, которая аналогично повышает активность связывания FcγRIIb в 4,8 раза по сравнению IgG1, далее увеличит степень повышения активности связывания FcγRIIb; однако в действительности, аналогично предшествующему примеру, активность связывания FcγRIIb у варианта, содержащего комбинацию этих изменений, повышалась только в 12 раз или около того по сравнению с IgG1. Этот результат также представляет собой эффект, который не мог быть предсказан из эффектов соответствующих изменений.
Эти результаты показали, что, хотя замена Pro в положении 238 (нумерация по EU) на Asp отдельно повышает активность связывания FcγRIIb, эффект не проявляется, когда ее комбинируют с другими изменениями, которые повышают активность связывания FcγRIIb. Причина для этого может состоять в том, что структура на поверхности взаимодействия между Fc и FcγR изменяется при внесении замены Pro в положении 238 (нумерация по EU) на Asp, и эффекты изменений, наблюдаемых во встречающемся в природе антителе, более не отражаются в результатах. Таким образом, посчитали, что крайне трудно создать Fc с превосходной селективностью в отношении FcγRIIb с использованием Fc, содержащего замены Pro в положении 238 (нумерация по EU) на Asp, в качестве матрицы, поскольку информация в отношении эффектов изменений, полученных в случае встречающихся в природе антител, не могла быть использована.
Пример 4
Детальный анализ связывания FcγRIIb для вариантов, в которые внесено изменение в шарнирной области в дополнение к изменению P238D
Как показано в примере 3, в Fc, полученной путем замены Pro в положении 238 (нумерация по EU) на Asp во встречающемся в природе IgG1 человека, ожидаемый комбинаторный эффект не мог быть достигнут даже путем комбинирования его с другим изменением, для которого предсказано, что оно далее повышает связывание FcγRIIb. Таким образом, на основе измененного Fc, полученного путем замены Pro в положении 238 (нумерация по EU) на Asp, проводили исследование путем комплексного внесения изменений в Fc, чтобы найти варианты, которые далее усиливают связывание FcγRIIb. Для H-цепей антител, IL6R-F11 (SEQ ID NO:27) получали путем внесения изменения в виде замены Met в положении 252 (нумерация по EU) на Tyr и изменения путем замены Asn в положении 434 (нумерация по EU) на Tyr в IL6R-G1d (SEQ ID NO:20), и IL6R-F652 (SEQ ID NO:28) получали путем внесения дополнительного изменения путем замены Pro в положении 238 (нумерация по EU) на Asp. Экспрессирующие плазмиды, содержащие последовательность H-цепи антитела, получали для каждой из последовательностей H-цепи антитела, полученных путем замены области вблизи остатка в положении 238 (нумерация по EU) (положения с 234 по 237, и 239 (нумерация по EU)) в IL6R-F652, в каждом случае на 18 аминокислот, за исключением исходных аминокислот и Cys. Для всех антител в качестве общей L-цепи антитела использовали IL6R-L (SEQ ID NO:22). Эти варианты экспрессировали, очищали и экспрессировали способом по справочному примеру 1. Эти варианты Fc названы вариантами PD. Взаимодействия каждого варианта PD с FcγRIIa типа R и FcγRIIb детально оценивали способом по справочному примеру 2.
Что касается результатов анализа взаимодействия с соответствующими FcγR, строили график в соответствии со следующим способом. Величину, полученную делением значения величины связывания каждого варианта PD с каждым FcγR на значение величины связывания FcγR предварительно измененного антитела, которое использовали в качестве контроля (IL6R-F652/IL6R-L, который имеет изменение в виде замены Pro в положении 238 (нумерация по EU) на Asp, а затем умножения результата на 100, использовали в качестве относительной величины активности связывания каждого варианта PD с каждым FcγR. На горизонтальной оси показаны относительные величины активности связывания FcγRIIb для каждого варианта PD, и на вертикальной оси показаны относительные величины активности связывания FcγRIIa типа R для каждого варианта PD (фиг.6).
В результате было выявлено, что одиннадцать типов изменений имеют эффекты усиления связывания FcγRIIb и сохранения или усиления связывания FcγRIIa типа R по сравнению с антителом до внесения изменений. Активность этих одиннадцати вариантов в отношении связывания FcγRIIb и FcγRIIa типа R обобщенно представлена в таблице 6. В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцениваемого варианта, и изменение относится к изменению, внесенному в IL6R-F11 (SEQ ID NO:27).

На фиг.7 показаны относительные величины для активности связывания FcγRIIb, полученной путем дальнейшего внесения этих одиннадцати изменений в вариант, имеющий изменение P238D, и относительные величины для активности связывания FcγRIIb, полученной путем внесения этих изменений в Fc, которая не содержит изменения P238D, как описано в примере 1. Эти одиннадцать изменений усиливали величину связывания FcγRIIb по сравнению с тем, что было до внесения, когда их дополнительно вносили в вариант P238D, но, напротив, наблюдали эффект снижения связывания FcγRIIb для восьми из этих изменений, за исключением G237F, G237W и S239D, когда их вносили в вариант, который не содержит P238D (GpH7-B3/GpL16-k0), использованный в примере 1. Пример 3 и эти результаты показали, что из эффектов внесения изменений во встречающийся в природе IgG1 трудно предсказать эффекты внесения тех же изменений в вариант, содержащий Fc с изменением P238D. Иными словами, невозможно выявить эти восемь изменений, идентифицированных в этот раз, без этого исследования.
Результаты измерения величин KD вариантов, указанных в таблице 6 для FcγRIa, FcγRIIaR, FcγRIIaH, FcγRIIb и FcγRIIIaV способом по справочному примеру 2, обобщенно представлены в таблице 7. В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцененного варианта, и изменение относится к изменению, внесенному в IL6R-F11 (SEQ ID NO:27). Матрица, использованная для получения IL6R-F11, IL6R-G1d/IL6R-L, обозначена звездочкой (*). Более того, KD(IIaR)/KD(IIb) и KD(IIaH)/KD(IIb) в таблице, соответственно, показывают величину, полученную путем деления величины KD каждого варианта для FcγRIIaR на величину KD каждого варианта для FcγIIb, и величины, полученной делением величины KD каждого варианта для FcγRIIaH на величину KD каждого варианта для FcγRIIb. KD(IIb) исходного полипептида/KD(IIb) измененного полипептида относится к величине, полученной путем деления величины KD исходного полипептида в отношении FcγRIIb на величину KD каждого варианта для FcγRIIb. Кроме того, в таблице 7 показаны величины KD для более сильной активности связывания FcγRIIaR и FcγRIIaH каждого варианта/величины KD для более сильной активности связывания FcγRIIaR и FcγRIIaH исходного полипептида. В этом случае исходный полипептид относится к варианту, который имеет IL6R-F11 (SEQ ID NO:27) в качестве H-цепи. Было определено, что, вследствие слабого связывания FcγR с IgG, его было невозможно точно проанализировать с помощью анализа кинетики, и, таким образом, закрашенные серым цветом ячейки в таблице 7 показывают величины, вычисленные с использованием уравнения 2 справочного примера 2.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
В таблице 7 показано, что у всех вариантов аффинность связывания с FcγRIIb повышалась по сравнению с IL6R-F11, и диапазон улучшения составлял от 1,9 раза до 5,0 раз. Соотношение величина KD каждого варианта в отношении FcγRIIaR/величина KD каждого варианта в отношении FcγRIIb и соотношение величина KD каждого варианта в отношении FcγRIIaH/величина KD каждого варианта в отношении FcγRIIb соответствуют активности связывания FcγRIIb относительно активности связывания FcγRIIaR и активности связывания FcγRIIaH, соответственно. Следовательно, эти величины показывают степень селективности связывания каждого варианта в отношении FcγRIIb, и более высокая величина указывает на более высокую селективность связывания в отношении FcγRIIb. Для исходного полипептида IL6R-F11/IL6R-L, соотношение величина KD для FcγRIIaR/величина KD для FcγRIIb и соотношение величина KD для FcγRIIaH/величина KD для FcγRIIb в обоих случаях равны 0,7, и, соответственно, все варианты, представленные в таблице 7, продемонстрировали увеличение селективности связывания в отношении FcγRIIb по сравнению с исходным полипептидом. Когда отношение величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH варианта/величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида равно 1 или более, это означает, что более сильная из активностей связывания FcγRIIaR и FcγRIIaH варианта имеет эквивалентное или сниженное связывание по сравнению со связыванием при более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида. Поскольку эта величина составляла от 0,7 до 5,0 для вариантов, полученных на этот раз, можно сказать, что связывание при более сильной из активностей связывания FcγRIIaR и FcγRIIaH вариантов, полученных на этот раз, было практически таким же или сниженным по сравнению с исходным полипептидом. Эти результаты показали, что, по сравнению с исходным полипептидом, варианты, полученные на этот раз, имеют сохраненную или сниженную активность связывания с FcγRIIa типа R и типа H, и увеличенную селективность в отношении FcγRIIb. Более того, по сравнению с IL6R-F11, все варианты имели более низкую аффинность в отношении FcγRIa и FcγRIIIaV.

Пример 5
Рентгеноструктурный анализ комплекса, образованного между Fc, содержащей P238D, и внеклеточной областью FcγRIIb
Как показано ранее в примере 3, даже несмотря на то, что в Fc, содержащую P238D, внесено изменение, которое повышает активность связывания FcγRIIb или селективность к FcγRIIb, было выявлено, что активность связывания FcγRIIb снижается, и причиной для этого может быть то, что структура на поверхности взаимодействия между Fc и FcγRIIb изменяется из-за внесения P238D. Таким образом, для установления причины этого явления определяли трехмерную структуру комплекса, образованного между Fc IgG1, содержащей мутацию P238D (далее, Fc(P238D)), и внеклеточной областью FcγRIIb, с помощью рентгеновской кристаллографии, и трехмерную структуру и способ связывания сравнивали с трехмерной структурой и способом связывания комплекса, образованного между Fc встречающегося в природе IgG1 (далее Fc(WT)) и внеклеточной областью FcγRIIb. Было сделано множество сообщений о трехмерной структуре комплекса, образованного между Fc и внеклеточной областью FcγR; и были проанализированы трехмерные структуры комплекса Fc(WT)/внеклеточная область FcγRIIIb (Nature, 2000, 400: 267-273; J. Biol. Chem. 2011, 276: 16469-16477), комплекса Fc(WT)/внеклеточная область FcγRIIIa (Proc. Natl. Acad. Sci. USA, 2011, 108: 12669-126674), и комплекса Fc(WT)/внеклеточная область FcγRIIa (J. Imunol. 2011, 187: 3208-3217). Хотя трехмерная структура комплекса Fc(WT)/внеклеточная область FcγRIIb не была проанализирована, трехмерная структура комплекса, образованного с Fc(WT), известна для FcγRIIa, и внеклеточные области FcγRIIa и FcγRIIb совпадают на 93% по аминокислотной последовательности и имеют очень высокую гомологию. Таким образом, трехмерная структура комплекса Fc(WT)/внеклеточная область FcγRIIb была предсказана путем моделирования с использованием кристаллической структуры комплекса Fc(WT)/внеклеточная область FcγRIIa.
Трехмерная структура комплекса Fc(P238D)/внеклеточная область FcγRIIb была определена с помощью рентгеновской кристаллографии при разрешении 2,6 Å. Структура, полученная в результате этого анализа, представлена на фиг.8. Внеклеточная область FcγRIIb связывается между двумя доменами СН2 Fc, и это аналогично трехмерным структурам комплексов, образованных между Fc(WT) и соответствующей внеклеточной областью FcγRIIIa, FcγRIIIb или FcγRIIa, проанализированных до настоящего времени.
Далее, для детального сравнения кристаллическую структуру комплекса Fc(P238D)/внеклеточная область FcγRIIb и структуру комплекса Fc(WT)/внеклеточная область FcγRIIb накладывали друг на друга с помощью аппроксимации методом наименьших квадратов на основе расстояний между парами атомов Cα в отношении внеклеточной области FcγRIIb и домена СН2 A Fc (фиг.9). В этом случае степень перекрывания между доменами СН2 B Fc не была удовлетворительной, и в этой части были выявлены конформационные различия. Более того, с использованием кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и модельной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb, отбирали пары атомов, которые имеют расстояние 3,7 Å или менее между внеклеточной областью FcγRIIb и доменом СН2 B Fc, и сравнивали для выявления отличий в межатомных взаимодействиях между FcγRIIb и доменом СН2 B Fc в Fc(WT) и Fc(P238D). Как показано в таблице 8, межатомные взаимодействия между доменом СН2 B Fc и FcγRIIb в Fc(P238D) и Fc(WT) не совпадают.
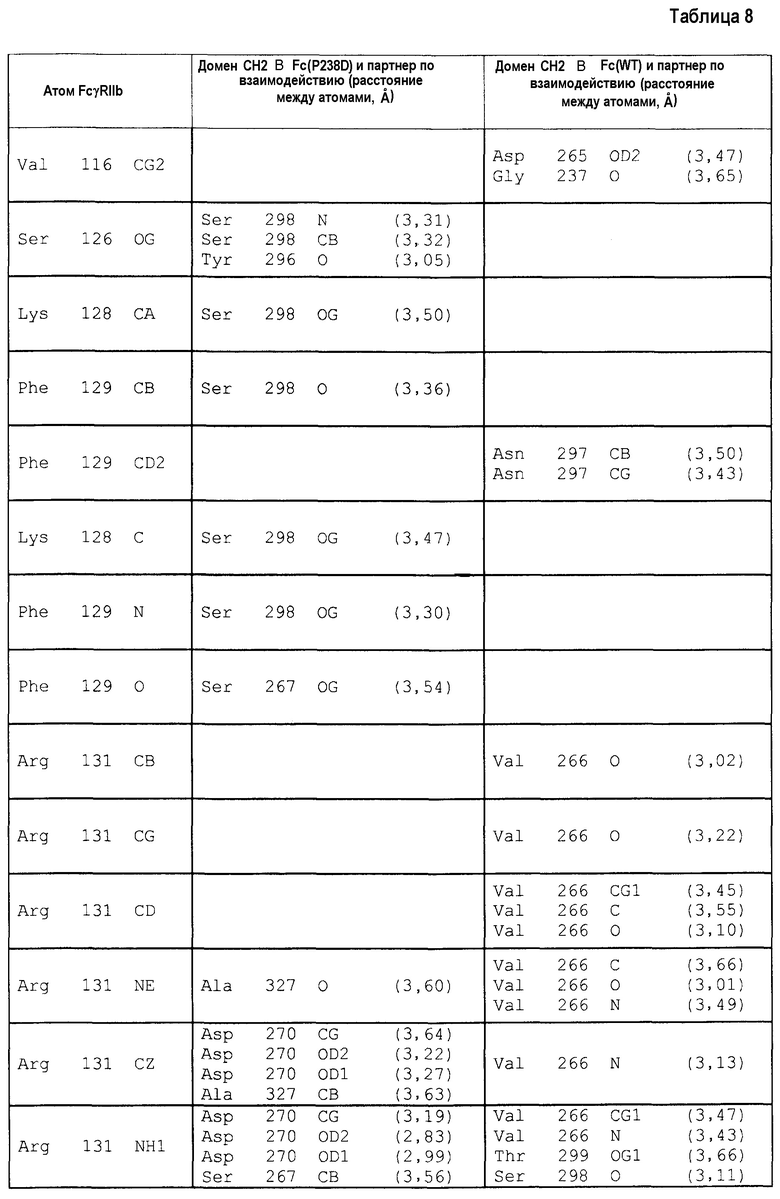

Более того, рентгеновскую кристаллическую структуру комплекса Fc(P238D)/внеклеточная область FcγRIIb и модельную структуру комплекса Fc(WT)/внеклеточная область FcγRIIb накладывали друг на друга путем аппроксимации методом наименьших квадратов на основе расстояний между парами атомов Cα в отношении только домена СН2 A Fc или только домена СН2 B Fc, и сравнивали детальные структуры вблизи P238D. Расположение аминокислотного остатка в положении 238 (нумерация по EU), которое представляет собой положение внесения мутации, изменяется между Fc(P238D) и Fc(WT), и можно видеть, что вместе с этим изменением ближайшая структура петли, исходящей из этой шарнирной области, изменяется в Fc(P238D) относительно Fc(WT) (фиг.10). Первоначально в Fc(WT) Pro в положении 238 (нумерация по EU) присутствует на внутренней стороне белка и образует гидрофобное ядро с окружающими остатками. Однако когда этот остаток изменен на заряженный или высокогидрофильный Asp, его присутствие в том же гидрофобном ядре приведет к энергетическому недостатку с точки зрения десольватации. Таким образом, в Fc(P238D) для устранения этой энергетически невыгодной ситуации аминокислотный остаток в положении 238 (нумерация по EU) изменяет его ориентацию в направлении растворителя, и это может вызвать это изменение в близлежащей структуре петли. Более того, поскольку эта петля исходит из шарнирной области с поперечными S-S связями, ее структурное изменение не ограничится локальным изменением и повлияет на относительное расположение домена СН2 A и домена B Fc. В результате межатомные взаимодействия между FcγRIIb и доменом СН2 B Fc изменяются. Таким образом, предсказанные эффекты не могли быть выявлены, когда изменения, которые повышают селективность и активность связывания FcγRIIb во встречающемся в природе IgG, комбинировали с Fc, содержащей изменение P238D.
Более того, в результате структурных изменений из-за внесения P238D в домен СН2 A Fc, была выявлена водородная связь между основной цепью Gly в соседнем положении 237 (нумерация по EU) и Tyr в положении 160 в FcγRIIb (фиг.11). Остаток в FcγRIIa, который соответствует этому Tyr160, представляет собой Phe; и, когда происходит связывание с FcγRIIa, эта водородная связь не образуется. Аминокислота в положении 160 представляет собой одно из небольшого количества отличий между FcγRIIa и FcγRIIb на границе взаимодействия с Fc, и присутствие этой водородной связи, которая является специфичной к FcγRIIb, предположительно привело к повышению активности связывания FcγRIIb и снижению активности связывания FcγRIIa в Fc(P238D) и повышению его селективности.
Более того, в домене СН2 B Fc наблюдают электростатическое взаимодействие между Asp в положении 270 (нумерация по EU) и Arg в положении 131 в FcγRIIb (фиг.12). В FcγRIIa типа H, который является одним из аллотипов FcγRIIa, соответствующий остаток представляет собой His, и, таким образом, он не может образовывать это электростатическое взаимодействие. Это может объяснить, почему активность связывания Fc(P238D) снижается в FcγRIIa типа H по сравнению с FcγRIIa типа R. Наблюдения, сделанные на основе таких результатов рентгеновской кристаллографии, показали, что изменение структуры петли рядом с P238D из-за внесения P238D и сопутствующего изменения в относительном расположении доменов вызывает формирование новых взаимодействий, не встречающихся во встречающемся в природе IgG, и это привело к селективному профилю связывания вариантов P238D в отношении FcγRIIb.
Экспрессия и очистка Fc(P238D)
Fc, содержащую изменение P238D, получали следующим образом. Сначала Cys в положении 220 (нумерация по EU) hIL6R-IgG1-v1 (SEQ ID NO:21) заменяли на Ser. Затем генетическую последовательность Fc(P238D) от Glu в положении 236 (нумерация по EU) до ее C-конца клонировали способом ПЦР. С использованием этой клонированной генетической последовательности проводили продуцирование экспрессирующих векторов и экспрессию и очистку Fc(P238D) в соответствии со способом по справочному примеру 1. Cys в положении 220 (нумерация по EU) образует дисульфидную связь с Cys L-цепи в общем IgG1. L-цепь не коэкспрессируется, когда получают Fc отдельно, и, таким образом, этот остаток заменяли на Ser, чтобы избежать образования ненужных дисульфидных связей.
Экспрессия и очистка внеклеточной области FcγRIIb
Ее получали в соответствии со способом по справочному примеру 2.
Очистка комплекса Fc(P238D)/внеклеточная область FcγRIIb
К 2 мг образца внеклеточной области FcγRIIb, полученного для кристаллизации, добавляли 0,29 мг Endo F1 (Protein Science 1996, 5: 2617-2622), экспрессированного и очищенного из Escherichia coli, в качестве слитого белка глутатион-S-трансферазы. Смеси позволяли стоять при комнатной температуре в течение трех суток в 0,1 M буфере Bis-Tris при pH 6,5, и N-связанный олигосахарид отщепляли, оставляя N-ацетилглюкозамин прямо связанным с Asn. Далее этот образец внеклеточного домена FcγRIIb, подвергнутый обработке для отщепления углевода, концентрировали путем ультрафильтрации с 5000 MWCO, и очищали гель-фильтрацией (Superdex200 10/300) с использованием колонки, уравновешенной в 20 мМ HEPS при pH 7,5, содержавшем 0,05 M NaCl. Более того, к полученной после отщепления углевода фракции внеклеточной области FcγRIIb добавляли Fc(P238D), так чтобы внеклеточная область FcγRIIb в молярном соотношении присутствовала в небольшом избытке, и после концентрирования путем ультрафильтрации с 10000 MWCO получали образец комплекса Fc(P238D)/внеклеточная область FcγRIIb путем очистки гель-фильтрацией (Superdex200 10/300) с использованием колонки, уравновешенной в 20 мМ HEPS при pH 7,5, содержащем 0,05 M NaCl.
Кристаллизация комплекса Fc(P238D)/внеклеточная область FcγRIIb
Образец комплекса Fc(P238D)/внеклеточная область FcγRIIb концентрировали до приблизительно 10 мг/мл путем ультрафильтрации с 10000 MWCO, и проводили кристаллизацию способом паровой диффузии с сидячей каплей. Для кристаллизации использовали Hydra II Plus One (MATRIX); и в качестве резервуарного раствора, содержавшего 100 мМ Bis-Tris pH 6,5, 17% PEG3350, 0,2 M ацетат аммония, и 2,7% (масс./об.) D-галактозу, кристаллизационную каплю получали путем смешения соотношения резервуарный раствор:образец для кристаллизации = 0,2 мкл:0,2 мкл, емкость закрывали, и ей позволяли стоять при 20°C, а затем успешно получали плоские кристаллы.
Измерение данных рентгенодифракции в кристалле комплекса Fc(P238D)/внеклеточная область FcγRIIb
Один из полученных монокристаллов комплекса Fc(P238D)/внеклеточная область FcγRIIb погружали в раствор 100 мМ Bis-Tris pH 6,5, 20% PEG3350, ацетат аммония, 2,7% (масс./об.) D-галактоза, 22,5% (об./об.) этиленгликоль. Кристалл вылавливали из раствора с использованием булавки с прикрепленной тонкой нейлоновой петлей и замораживали в жидком азоте; а затем измеряли данные рентгенодифракции на оборудовании с магнитотормозном излучением Photon Factory BL-1A в High Energy Accelerator Research Organization. В ходе измерения кристалл постоянно помещали в поток азота при -178°C для поддержания в замороженном состоянии, и было получено всего 225 рентгенограмм с использованием детектора Quantum 270 CCD (ADSC), подсоединенного к линии пучка, с вращением кристалла 0,8° за один раз. Определение параметров ячейки, индексирование дифракционных пятен и обработку дифракционных данных из полученных рентгенограмм проводили с использованием программы Xia2 (CCP4 Software Suite), XDS Package (Walfgang Kabsch) и Scala (CCP4 Software Suite); и, наконец, получали данные об интенсивности дифракции при разрешении вплоть до 2,46 Å. Кристалл относится к пространственной группе P21 и имеет следующие параметры ячейки: a=48,85 Å, b=76,01 Å, c=115,09 Å, a=90°, β=100,70°, γ=90°.
Рентгеновский кристаллографический анализ комплекса Fc(P238D)/внеклеточная область FcγRIIb
Кристаллическую структуру комплекса Fc(P238D)/внеклеточная область FcγRIIb определяли способом молекулярного замещения с использованием программы Phaser (CCP4 Software Suite). Из размера полученной кристаллической решетки и молекулярной массы комплекса Fc(P238D)/внеклеточная область FcγRIIb было предсказано, что количество комплексов в асимметричном элементе равно единице. Из структурных координат кода PDB: 3SGJ, который представляет собой кристаллическую структуру комплекса Fc(WT)/внеклеточная область FcγRIIIa, участки аминокислотных остатков A-цепи в положениях 239-340 и B-цепи в положениях 239-340 брали в качестве отдельных координат, и их использовали, соответственно, в качестве моделей для поиска доменов CH2 Fc. Участки аминокислотных остатков A-цепи в положениях 341-444 и B-цепи в положениях 341-443 брали в качестве единого набора координат из тех же структурных координат кода PDB: 3SGJ; и их использовали в качестве модели для поиска доменов CH3 Fc. Наконец, из структурных координат кода PDB: 2FCB, что соответствует кристаллической структуре внеклеточной области FcγRIIb, участки аминокислотных остатков A-цепи в положениях 6-178 выбирали и использовали в качестве модели для поиска внеклеточной области FcγRIIb. Ориентацию и положение каждой поисковой модели в кристаллической решетке определяли в порядке домен CH3 Fc, внеклеточная область FcγRIIb и домен CH2 Fc, исходя из функции вращения и функции трансляции, с получением первоначальной модели кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb. Когда на полученной первоначальной модели проводили уточнение жесткого тела, которое осуществляет движение двух доменов CH2 Fc, двух доменов CH3 Fc и внеклеточной области FcγRIIb, кристаллографический фактор достоверности, R-величина, становилась равной 40,4%, и свободная R-величина становилась равной 41,9% для данных интенсивности дифракции от 25 Å до 3,0 Å в этот момент времени. Более того, уточнение структуры с использованием программы Refmac5 (CCP4 Software Suite) и пересмотр модели для выявления карт электронной плотности, коэффициент которых имеет 2Fo-Fc или Fo-Fc, которые вычисляют, исходя из экспериментально определенного структурного фактора Fo, вычисленного структурного фактора Fc и вычисленной фазы с использованием модели, проводили с помощью программы Coot (Paul Emsley), и уточнение модели проводили путем повторения этих стадий. Наконец, в результате включения молекул воды в модель на основе карт электронной плотности, в которых используются 2Fo-Fc или Fo-Fc в качестве коэффициента, и последующего уточнения, кристаллографический фактор достоверности, R-величина, и свободная R-величина модели, содержащей 4846 не являющихся водородом атомами, становились равными 23,7% и 27,6% для 24291 данных интенсивности дифракции при разрешении от 25 Å до 2,6 Å, соответственно.
Получение модельной структуры комплекса Fc(WT)/внеклеточная область FcγRIIb
Исходя из структурных координат кода PDB: 3RY6, что соответствует кристаллической структуре комплекса Fc(WT)/внеклеточная область FcγRIIa, функцию Build Mutants программы Discovery Studio 3.1 (Accelrys) использовали для внесения мутаций, чтобы они соответствовали аминокислотной последовательности FcγRIIb, в FcγRIIa в этих структурных координатах. В этом случае уровень оптимизации был установлен на высокий, радиус отсечения был установлен на 4,5, получали пять моделей, и модель с наилучшей энергетической оценкой среди них использовали в качестве модельной структуры для комплекса Fc(WT)/внеклеточная область FcγRIIb.
Пример 6
Анализ связывания FcγR вариантами Fc, в которых участки изменения были определены на основе кристаллических структур
Исходя из результатов рентгеноструктурного анализа комплекса, образованного между Fc(P238D) и внеклеточной областью FcγRIIb, полученной в примере 5, вносили комплексные изменения в участки измененной Fc, имеющей замену Pro в положении 238 (нумерация по EU) на Asp, которые, как было предсказано, повлияют на взаимодействие с FcγRIIb (остатки положений 233, 240, 241, 263, 265, 266, 267, 268, 271, 273, 295, 296, 298, 300, 323, 325, 326, 327, 328, 330, 332 и 334 (нумерация по EU)), и исследовали варианты с комбинацией изменений, которые усиливают связывание FcγRIIb.
IL6R-B3 (SEQ ID NO:40) получали путем внесения в IL6R-G1d (SEQ ID NO:20), полученный согласно примеру 2, изменения, полученного путем замены Lys в положении 439 (нумерация по EU) на Glu. Далее, получали IL6R-BF648 (SEQ ID NO:41) путем внесения в IL6R-B3 замены, полученной путем замены Pro в положении 238 (нумерация по EU) на Asp. IL6R-L (SEQ ID NO:22) использовали в качестве общей L-цепи антитела для всех из антител. Эти варианты антител экспрессировали и очищали в соответствии со способом справочного примера 1, и связывание каждого из FcγR (FcγRIa, FcγRIIa типа H, FcγRIIa типа R, FcγRIIb и FcγRIIIa типа V) детально оценивали способом справочного примера 2.
В соответствии со следующим способом строили график для результатов анализа взаимодействий с соответствующими FcγR. Значение величины связывания каждого варианта с каждым FcγR делили на величину связывания предварительно измененного контрольного антитела (IL6R-BF648/IL6R-L с заменой Pro в положении 238 (нумерация по EU) на Asp) с каждым FcγR, а затем полученную величину умножали на 100 и использовали в качестве величины относительной активности связывания каждого варианта с каждым FcγR. На горизонтальной оси показана величина относительной активности связывания каждого варианта с FcγRIIb, и на вертикальной оси показана величина относительной активности связывания каждого варианта с FcγRIIa типа R (фиг.13).
Как показано на фиг.13, результаты показывают, что было выявлено, что из всех изменений 24 типа изменений обладают эффектом сохранения или усиления связывания FcγRIIb по сравнению с предварительно измененным антителом. Связывание этих вариантов с каждым из FcγR представлено в таблице 9. В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцененного варианта, и изменение относится к изменению, внесенному в IL6R-B3 (SEQ ID NO:40). Матрица, использованная для получения IL6R-B3, IL6R-G1d/IL6R-L, обозначена звездочкой (*).

Результаты измерения величин KD вариантов, представленных в таблице 9 в отношении FcγRIa, FcγRIIaR, FcγRIIaH, FcγRIIb и FcγRIIIa типа V, с помощью способа по справочному примеру 2 обобщенно представлены в таблице 10. В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцениваемого варианта, и изменение относится к изменению, внесенному в IL6R-B3 (SEQ ID NO:40). Матрица, использованная для получения IL6R-B3, IL6R-G1d/IL6R-L, обозначена звездочкой (*). Более того, KD(IIaR)/KD(IIb) и KD(IIaH)/KD(IIb) в таблице, соответственно, соответствуют величине, полученной путем деления величины KD каждого варианта для FcγRIIaR на величину KD каждого варианта для FcγRIIb, и величине, полученной делением величины KD каждого варианта для FcγRIIaH на величину KD каждого варианта для FcγRIIb. KD(IIb) исходного полипептида/KD(IIb) измененного полипептида относится к величине, полученной делением величины KD исходного полипептида в отношении FcγRIIb на величину KD каждого варианта для FcγRIIb. Кроме того, величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH каждого варианта/величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида представлена в таблице 10. Здесь исходный полипептид относится к варианту, который имеет IL6R-B3 (SEQ ID NO:40) в качестве H-цепи. Было определено, что, вследствие слабого связывания FcγR с IgG, его было невозможно точно проанализировать с помощью анализа кинетики, и, таким образом, закрашенные серым цветом ячейки в таблице 10 показывают величины, вычисленные с использованием уравнения 2 справочного примера 2.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
В таблице 10 показано, что по сравнению с IL6R-B3 все варианты показали улучшение аффинности в отношении FcγRIIb, и диапазон улучшения составлял от 2,1 раза до 9,7 раз. Соотношение величина KD каждого варианта для FcγRIIaR/величина KD каждого варианта для FcγRIIb и соотношение величина KD каждого варианта для FcγRIIaH/величина KD каждого варианта для FcγRIIb соответствуют активности связывания FcγRIIb относительно активности связывания FcγRIIaR и активности связывания FcγRIIaH, соответственно. Следовательно, эти величины показывают степень селективности связывания каждого варианта с FcγRIIb, и более высокая величина указывает на более высокую селективность связывания с FcγRIIb. Поскольку соотношение величина KD для FcγRIIaR/величина KD для FcγRIIb и соотношение величина KD для FcγRIIaH/величина KD для FcγRIIb в исходном полипептиде IL6R-B3/IL6R-L составляло 0,3 и 0,2, соответственно, все варианты в таблице 10 показали улучшение селективности связывания с FcγRIIb по сравнению с исходным полипептидом. Когда соотношение величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH варианта/величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида равно 1 или более, это означает, что более сильная из активностей связывания FcγRIIaR и FcγRIIaH варианта имеет эквивалентное или сниженное связывание по сравнению со связыванием при более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида. Поскольку эта величина составляла от 4,6 до 34,0 для вариантов, полученных на этот раз, можно сказать, что по сравнению с исходным полипептидом варианты, полученные на этот раз, имели сниженное связывание при более сильной из активностей связывания FcγRIIaR и FcγRIIaH. Эти результаты показали, что по сравнению с исходным полипептидом варианты, полученные на этот раз, имеют сохраненную или сниженную активность связывания FcγRIIa типа R и типа H, повышенную активность связывания с FcγRIIb и увеличенную селективность в отношении FcγRIIb. Более того, по сравнению с IL6R-B3 все варианты имели более низкую аффинность к FcγRIa и FcγRIIIaV.

Что касается перспективных вариантов среди полученных комбинированных вариантов, факторы, обеспечивающие их эффекты, исследовали с использованием кристаллической структуры. На фиг.14 показана кристаллическая структура комплекса Fc(P238D)/внеклеточная область FcγRIIb. На этой фигуре H-цепь, расположенная слева, представляет собой A-цепь Fc, и H-цепь, расположенная справа, представляет собой B-цепь Fc. Здесь можно видеть, что участок в положении 233 (нумерация по EU) в A-цепи Fc расположен вблизи Lys в положении 113 (нумерация по EU) FcγRIIb. Однако в этой кристаллической структуре боковая цепь E233 находится в состоянии в значительной степени высокой подвижности, и ее электронная плотность четко не выявляется. Таким образом, изменение, полученное путем замены Glu в положении 233 (нумерация по EU) на Asp, приводит к снижению степени свободы боковой цепи, поскольку боковая цепь становится на один атом углерода короче. В результате, может быть снижена потеря энтропии при формировании взаимодействия с Lys в положении 113 (нумерация по EU) FcγRIIb, и, следовательно, предположительно это приводит к повышению свободной энергии связывания.
Аналогично, на фиг.15 показано окружение вблизи участка в положении 330 (нумерация по EU) в структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb. На этой фигуре показано, что окружение участка в положении 330 (нумерация по EU) A-цепи Fc в Fc (P238D) представляет собой гидрофильное окружение, состоящее из Ser в положении 85, Glu в положении 86, Lys в положении 163 и т.п. (нумерация по EU) FcγRIIb. Таким образом, изменение, полученное путем замены Ala в положении 330 (нумерация по EU) на Lys или Arg, предположительно приводит к упрочнению взаимодействия с Ser в положении 85 (нумерация по EU) или Glu в положении 86 (нумерация по EU) в FcγRIIb.
На фиг.16 представлены структуры Pro в положении 271 (нумерация по EU) B-цепи Fc после наложения друг на друга кристаллической структуры комплекса Fc(P238D)/внеклеточная область FcγRIIb и комплекса Fc(WT)/внеклеточная область FcγRIIIa путем аппроксимации с помощью метода наименьших квадратов на основе расстояний между парами атомов Cα в отношении B-цепи Fc. Эти две структуры хорошо соответствуют, но имеют различные трехмерные структуры Pro в положении 271 (нумерация по EU). Когда также учитывают слабую электронную плотность в этой области в кристаллической структуре комплекса Fc(P238D)/внеклеточная область FcγRIIb, предполагается, что существует возможность того, что Pro в положении 271 (нумерация по EU) в Fc(P238D)/FcγRIIb вызывает большое напряжение в структуре, таким образом, нарушая равновесие структуры петли для достижения оптимальной структуры. Таким образом, изменение, полученное путем замены Pro в положении 271 (нумерация по EU) на Gly, обеспечивает подвижность этой структуры петли и предположительно приводит к усилению связывания путем снижения энергетического барьера, когда позволяют образоваться оптимальной структуре в ходе взаимодействия с FcγRIIb.
Пример 7
Исследование комбинаторного эффекта изменений, которые усиливают связывание FcγRIIb, при комбинировании с P238D
Из изменений, полученных согласно примерам 4 и 6, изменения, которые усиливали связывание FcγRIIb или сохраняли связывание FcγRIIb и продемонстрировали эффекты подавления связывания с другими FcγR, комбинировали друг с другом, и их эффекты исследовали.
Из таблиц 6 и 9 выбирали особенно удачные изменения, и их комбинировали и вносили в H-цепь антитела IL6R-BF648 аналогично способу по примеру 6. IL6R-L использовали в качестве общей L-цепи антитела для всех из антител, антитела экспрессировали и очищали в соответствии со способом справочного примера 1, и связывание с каждым из FcγR (FcγRIa, FcγRIIa H-типа, FcγRIIa R-типа, FcγRIIb и FcγRIIIa V-типа) детально оценивали способом справочного примера 2.
Относительную активность связывания вычисляли для результатов анализа взаимодействий с соответствующими FcγR в соответствии со следующим способом. Значение величины связывания каждого варианта с каждым FcγR делили на значение величины связывания предварительно измененного контрольного антитела (IL6R-BF648/IL6R-L с заменами Pro в положении 238 (нумерация по EU) на Asp с каждым FcγR, и умножали на 100; а затем эту величину использовали в качестве относительной активности связывания каждого варианта с каждым FcγR. На горизонтальной оси показана величина относительной активности связывания каждого варианта с FcγRIIb, и на вертикальной оси показана величина относительной активности связывания каждого варианта с FcγRIIa типа R (таблица 11).
В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцениваемого варианта, и изменение относится к изменению, внесенному в IL6R-B3 (SEQ ID NO:40). Матрица, использованная для получения IL6R-B3, IL6R-G1d/IL6R-L, обозначена звездочкой (*).

Результаты измерения величин KD вариантов, представленных в таблице 11, для FcγRIa, FcγRIIaR, FcγRIIaH, FcγRIIb и FcγRIIIa типа V способом справочного примера 2, обобщенно представлены в таблице 12. В таблице SEQ ID NO относится к SEQ ID NO H-цепи оцениваемого варианта и изменение относится к изменению, внесенному в IL6R-B3 (SEQ ID NO:40). Матрица, использованная для получения IL6R-B3, IL6R-G1d/IL6R-L, обозначена звездочкой (*). Более того, KD(IIaR)/KD(IIb) и KD(IIaH)/KD(IIb) в таблице, соответственно, соответствуют величине, полученной путем деления величины KD каждого варианта для FcγRIIaR на величину KD каждого варианта для FcγRIIb, и величине, полученной путем деления величины KD каждого варианта для FcγRIIaH на величину KD каждого варианта для FcγRIIb. KD(IIb) исходного полипептида/KD(IIb) измененного полипептида относится к величине, полученной путем деления величины KD исходного полипептида в отношении FcγRIIb на величину KD каждого варианта для FcγRIIb. Кроме того, в таблице 12 представлено соотношение величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH каждого варианта/величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида. Здесь, исходный полипептид относится к варианту, который имеет IL6R-B3 (SEQ ID NO:40) в качестве H-цепи. Было определено, что вследствие слабого связывания FcγR с IgG, его было невозможно точно проанализировать путем анализа кинетики, и, таким образом, закрашенные серым цветом ячейки в таблице 12 показывают величины, вычисленные с использованием уравнения 2 справочного примера 2.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
В таблице 12 показано, что по сравнению с IL6R-B3 все варианты продемонстрировали увеличение аффинности к FcγRIIb, и диапазон улучшения составлял от 3,0 раз до 99,0 раз. Соотношение величина KD каждого варианта в отношении FcγRIIaR/величина KD каждого варианта в отношении FcγRIIb и соотношение величина KD каждого варианта в отношении FcγRIIaH/величина KD каждого варианта в отношении FcγRIIb соответствуют активности связывания FcγRIIb относительно активности связывания FcγRIIaR и активности связывания FcγRIIaH, соответственно. Следовательно, эти величины показывают степень селективности связывания каждого варианта с FcγRIIb, и более высокая величина указывает на более высокую селективность связывания с FcγRIIb. Поскольку соотношение величина KD в отношении FcγRIIaR/величина KD в отношении FcγRIIb и соотношение величина KD в отношении FcγRIIaH/величина KD для в отношении FcγRIIb исходного полипептида IL6R-B3/IL6R-L составляли 0,3 и 0,2, соответственно, все варианты в таблице 12 показали увеличение селективности связывания с FcγRIIb по сравнению с исходным полипептидом. Когда соотношение величина KD для более сильной из активностей связывания FcγRIIaR и FcγRIIaH варианта/величина KD для более сильной из активностей связывания исходного полипептида равно 1 или более, это означает, что более сильная из активностей связывания FcγRIIaR и FcγRIIaH варианта имеет эквивалентное или сниженное связывание по сравнению со связыванием при более сильной из активностей связывания FcγRIIaR и FcγRIIaH исходного полипептида. Поскольку эта величина составляла от 0,7 до 29,9 для вариантов, полученных на этот раз, можно сказать, что связывание при более сильной из активностей связывания вариантов FcγRIIaR и FcγRIIaH, полученных на этот раз, практически эквивалентно или снижено по сравнению с исходным полипептидом. Эти результаты показали, что по сравнению с исходным полипептидом варианты, полученные на этот раз, имеют сохраненную или сниженную активность связывания FcγRIIa типа R и типа H, повышенную активность связывания с FcγRIIb и увеличенную селективность в отношении FcγRIIb. Более того, по сравнению с IL6R-B3 все варианты имели более низкую аффинность в отношении FcγRIa и FcγRIIIaV.


Справочный пример 1
Конструирование экспрессирующих векторов для антител; и экспрессия и очистка антител
Синтез полноразмерных генов, кодирующих нуклеотидные последовательности вариабельных областей H-цепи и L-цепи антитела, проводили способами получения, известными специалистам в данной области с использованием ПЦР со сборкой и т.п. Внесение аминокислотных замен проводили способами, известными специалистам в данной области, с использованием ПЦР и т.п. Полученный плазмидный фрагмент встраивали в экспрессирующий вектор клеток животных, и получали экспрессирующий вектор H-цепи и экспрессирующий вектор L-цепи. Нуклеотидную последовательность полученного экспрессирующего вектора определяли способами, известными специалистам в данной области. Полученные плазмиды временно вводили в клеточную линию HEK293H, происходящую из клеток рака почки эмбриона человека (Invitrogen) или в клетки FreeStyle293 (Invitrogen) для экспрессии антител. Полученный культуральный супернатант собирали, а затем пропускали через 0,22-мкм фильтр MILLEX(R)-GV (Millipore) или через 0,45-мкм фильтр MILLEX(R)-GV (Millipore) с получением культурального супернатанта. Антитела очищали из полученного культурального супернатанта способами, известными специалистам в данной области, с использованием rProtein A Sepharose Fast Flow (GE Healthcare) или Protein G Sepharose 4 Fast Flow (GE Healthcare). Для концентрирования очищенных антител измеряли их поглощение при 280 нм с использованием спектрофотометра. Коэффициент экстинкции, вычисленный на основе полученной величины способами, такими как PACE, использовали для вычисления концентрации антитела (Protein Science 1995; 4: 2411-2423).
Справочный пример 2
Способ получения FcγR и способ анализа взаимодействия между измененным антителом и FcγR
Внеклеточные домены FcγR получали следующим способом. Сначала ген внеклеточного домена FcγR синтезировали способом, хорошо известным специалистам в данной области. В то же время получали последовательность каждого FcγR, исходя из информации, зарегистрированной в NCBI. В частности, FcγRI получали на основе последовательности с номером доступа NCBI № NM_000566.3, FcγRIIa получали на основе последовательности с номером доступа NCBI № NM_001136219.1, FcγRIIb получали на основе последовательности с номером доступа NCBI № NM_004001.3, FcγRIIIa получали на основе последовательности с номером доступа NCBI № NM_001127593.1, и FcγRIIIb получали на основе последовательности с номером доступа NCBI № NM_000570.3, и к C-концу присоединяли His-метку. Более того, известен полиморфизм для FcγRIIa, FcγRIIIa и FcγRIIIb, и полиморфные участки получали согласно J. Exp. Med., 1990, 172: 19-25, для FcγRIIa; J. Clin. Invest., 1997, 100 (5): 1059-1070, для FcγRIIIa; и J. Clin. Invest., 1989, 84, 1688-1691, для FcγRIIIb.
Полученные фрагменты генов встраивали в экспрессирующий вектор для клеток животных, и продуцировали экспрессирующие векторы. Полученные экспрессирующие векторы временно вводили в происходящие из клеток рака почки эмбриона человека клетки FreeStyle293 (Invitrogen) для экспрессии представляющих интерес белков. Что касается FcγRIIb, использованного для кристаллографического анализа, представляющий интерес белок экспрессировали в присутствии Kifunensine в конечной концентрации 10 мкг/мл, так чтобы цепь сахара, добавленная к FcγRIIb, представляла собой цепь сахара с высоким содержанием маннозы. Клетки культивировали, и после сбора полученного культуральрного супернатанта их пропускали через 0,22-мкм фильтр для получения культурального супернатанта. В принципе, полученные культуральные супернатанты очищали посредством следующих четырех стадий. Проводимые стадии представляли собой катионообменную колоночную хроматографию (SP Sepharose FF) на стадии 1, аффинную колоночную хроматографию (HisTrap HP) для His-метки на стадии 2, гель-фильтрационную колоночную хроматографию (Superdex200) на стадии 3 и асептическую хроматографию на стадии 4. Однако для FcγRI в качестве стадии 1 проводили анионообменную колоночную хроматографию с использованием Q sepharose FF. Очищенные белки подвергали измерению поглощения при 280 нм с использованием спектрофотометра; и из полученных величин вычисляли концентрации очищенных белков с использованием коэффициента поглощения, вычисленного с использованием способов, таких как PACE (Protein Science 1995; 4: 2411-2423).
Анализ взаимодействия между каждым измененным антителом и Fcγ-рецептором, полученным, как указано выше, проводили с использованием Biacore T100 (GE Healthcare), Biacore T200 (GE Healthcare), Biacore A100 и Biacore 4000. В качестве подвижного буфера использовали HBS-EP+ (GE Healthcare), и температуру измерения устанавливали на 25°C. Использовали чипы, полученные путем иммобилизации антигенного пептида, белка A (Thermo Scientific), белка A/G (Thermo Scientific) и белка L (ACTIGEN или BioVision) способом присоединения аминов к сенсорному чипу CM5 Series S (GE Healthcare) или сенсорному чипу CM4 Series S (GE Healthcare), или альтернативно чипы, полученные путем обеспечения взаимодействия предварительно биотинилированных антигенных пептидов и иммобилизации на сенсорном чипе SA Series S (сертифицированный) (GE Healthcare).
После улавливания представляющих интерес антител на этих сенсорных чипах, Fcγ-рецептору, разбавленному подвижным буфером, позволяли взаимодействовать, измеряли количество, связавшееся с антителом, и антитела сравнивали. Однако, поскольку количество связанного рецептора Fcγ зависит от количества уловленных антител, количество связанного Fcγ-рецептора делили на количество каждого уловленного антитела, с получением скорректированных величин, и эти величины сравнивали. Более того, антитела, уловленные на чипы, промывали путем реакции с 10 мМ глицин-HCl, pH 1,5, и чипы регенерировали и использовали повторно.
Анализы кинетики для вычисления величин KD каждого измененного антитела для FcγR проводили согласно следующему способу. Сначала представляющие интерес антитела улавливали на упомянутых выше сенсорных чипах, и Fcγ-рецептору, разбавленному подвижным буфером, позволяли взаимодействовать. Для глобальной аппроксимации измеренных результатов к полученной сенсограмме использовали программное обеспечение Biacore Evaluation Software с применением модели связывания Ленгмюра 1:1, и вычисляли константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (1/с); и из этих результатов вычисляли константы диссоциации KD (моль/л).
Когда взаимодействие между каждым из измененных антител и FcγR было слабым, и определяли, что корректный анализ был невозможен с помощью упомянутого выше анализа кинетики, KD для таких взаимодействий вычисляли с использованием следующего уравнения модели связывания 1:1, описанного в Biacore T100 Software Handbook BR1006-48 Edition AE.
Поведение взаимодействующих молекул согласно модели связывания 1:1 в Biacore может быть описано с помощью уравнения 1, представленного ниже.
[Уравнение 1]
Req = C·Rmax/(KD+C)+RI
Req: величина из графика уровней связывания в стационарном состоянии против концентрации анализируемого соединения
C: концентрация
RI: совокупный вклад индекса рефракции в образце
Rmax: способность поверхности связывать анализируемое соединение
Путем преобразования этого уравнения KD может быть выражена с помощью уравнения 2, представленного ниже.
[Уравнение 2]
KD=C·Rmax/(Req-RI)-C
KD можно вычислять путем подстановки величин Rmax, RI и C в это уравнение. Величины RI и C могут быть определены из сенсограммы по результатам измерения и условиям измерения. Rmax вычисляли согласно следующему способу. Для сравнения, для антител, которые имели достаточно сильные взаимодействия при оценке одновременно в том же раунде измерения, величину Rmax получали путем глобальной аппроксимации с использованием модели связывания Ленгмюра 1:1, а затем ее делили на величину для сравнительного антитела, уловленного на сенсорном чипе, и умножали на количество уловленного измененного антитела, подлежащего оценке.
Промышленная применимость
Настоящее изобретение предусматривает полипептиды, содержащие Fc-область, которая имеет сохраненную или сниженную активность связывания с обоими аллотипами FcγRIIa: типы R и H, и имеющие повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом. С использованием полипептидов с усиленной селективностью связывания FcγRIIb, а не обоих аллотипов FcγRIIa (типы R и H), можно передавать ингибиторный сигнал для воспалительного иммунного ответа, опосредуемого фосфорилированием ITIM FcγRIIb у пациентов, имеющих любой из аллотипов, типы R и H. Более того, путем сообщения Fc антитела свойства селективного связывания FcγRIIb, можно подавлять продукцию анти-антител через опосредуемое FcγRIIb иммунодепрессивное действие.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ЭЛИМИНАЦИИ АНТИГЕНОВ | 2012 |
|
RU2772771C1 |
| СПОСОБ ПОЛУЧЕНИЯ ВАРИАНТОВ FcyRIIB-СПЕЦИФИЧЕСКОЙ Fc-ОБЛАСТИ | 2013 |
|
RU2820161C1 |
| ВАРИАНТ ПОЛИПЕПТИДА, КОТОРЫЙ ИМЕЕТ СОХРАНЕННУЮ ИЛИ СНИЖЕННУЮ АКТИВНОСТЬ СВЯЗЫВАНИЯ С FcγRIIa, СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2749357C2 |
| СПОСОБ ИЗМЕНЕНИЯ УДЕРЖАНИЯ В ПЛАЗМЕ И ИММУНОГЕННОСТИ АНТИГЕНСВЯЗЫВАЮЩЕЙ МОЛЕКУЛЫ | 2012 |
|
RU2799423C1 |
| ВАРИАНТЫ FcγRIIB-СПЕЦИФИЧЕСКОЙ Fc-ОБЛАСТИ | 2013 |
|
RU2729831C2 |
| ГЕТЕРОДИМЕРИЗОВАННЫЙ ПОЛИПЕПТИД | 2012 |
|
RU2719132C2 |
| ГЕТЕРОДИМЕРИЗОВАННЫЙ ПОЛИПЕПТИД | 2012 |
|
RU2641256C2 |
| ВАРИАНТ Fc-ОБЛАСТИ | 2014 |
|
RU2757124C2 |



































































































































































































































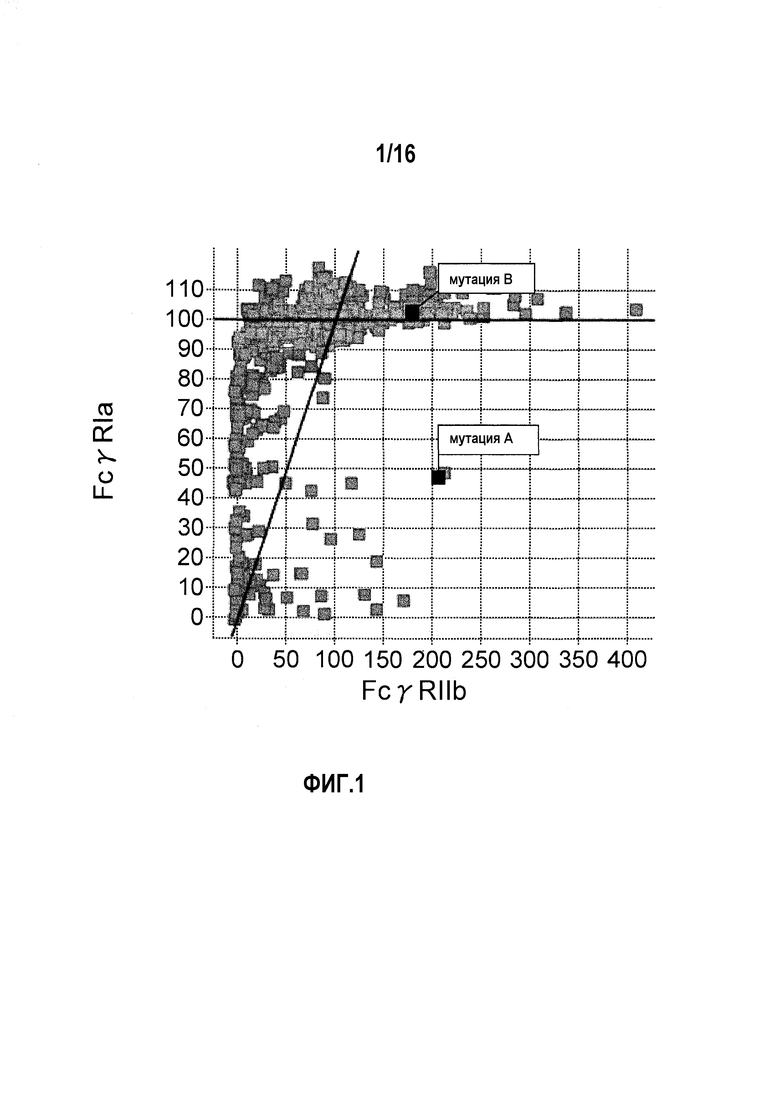

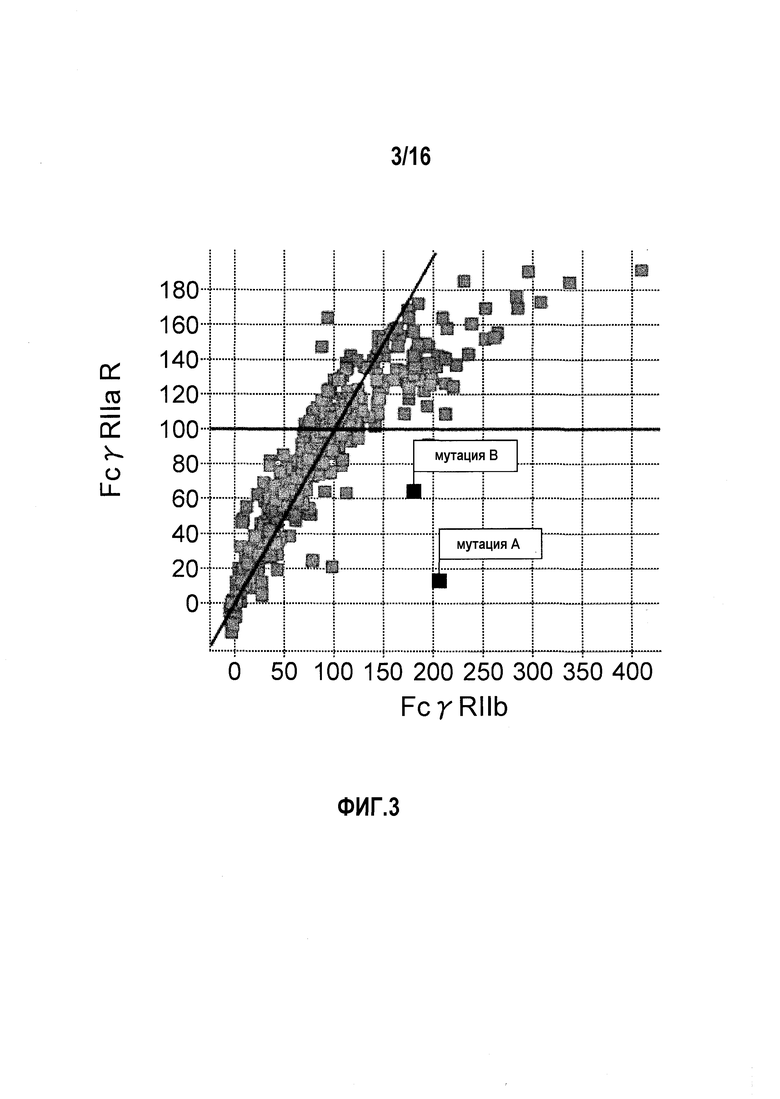










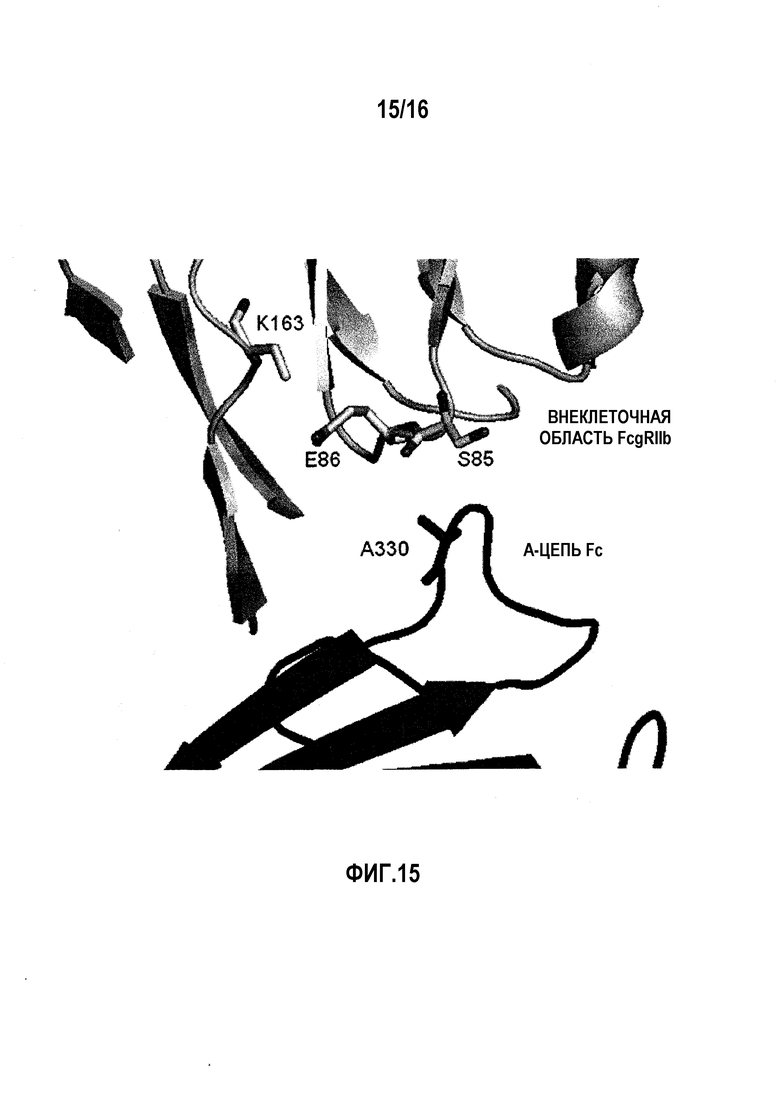

Изобретение относится к биохимии. Описан полипептид, содержащий Fc-область антитела по меньшей мере с одним аминокислотным изменением, который имеет повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, где величина [величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R)]/[величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIb] равна 1,2 или более, где величина [величина KD для более сильной из активностей связывания полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)]/[величина KD для более сильной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)] составляет 0,7 или более, и где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация EU) на Asp или замену Leu в положении 328 (нумерация EU) на Glu. Описаны способы получения такого полипептида и его применение. Изобретение расширяет арсенал терапевтических и профилактических средств. 6 н. и 14 з.п. ф-лы, 16 ил., 12 табл., 9 пр.
1. Полипептид, содержащий Fc-область антитела по меньшей мере с одним аминокислотным изменением, который имеет повышенную активность связывания с FcγRIIb по сравнению с исходным полипептидом, где величина [величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R)]/[величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIb] равна 1,2 или более,
где величина [величина KD для более сильной из активностей связывания полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)]/[величина KD для более сильной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)] составляет 0,7 или более, и
где аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация EU) на Asp или замену Leu в положении 328 (нумерация EU) на Glu.
2. Полипептид по п. 1, где величина [величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип Н)]/[величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIb] равна 4,2 или более.
3. Полипептид по п. 1, где величина [величина KD исходного полипептида в отношении FcγRIIb]/[величина KD полипептида, содержащего Fc-область антитела, в отношении FcγRIIb] равна 1,6 или более.
4. Полипептид по п. 1, который имеет сохраненную или сниженную активность связывания FcγRIIIa по сравнению с исходным полипептидом.
5. Полипептид по п. 1, который имеет сохраненную или сниженную активность связывания FcγRIa по сравнению с исходным полипептидом.
6. Полипептид по п. 1, где указанное по меньшей мере одно аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация EU) на Asp и по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met.
7. Полипептид по п. 1, где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело.
8. Полипептид по п. 1, где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc.
9. Способ повышения активности связывания с FcγRIIb у полипептида по сравнению с исходным полипептидом, который включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела, где величина [величина KD для более сильной из активностей связывания полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)]/[величина KD для более сильной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)] составляет 0,7 или более, и
где указанное по меньшей мере одно аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 328 (нумерация по EU) на Glu.
10. Способ по п. 9, где указанное по меньшей мере одно аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация EU) на Asp и по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met.
11. Способ по п. 9 или 10, где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело.
12. Способ по п. 9 или 10, где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc.
13. Способ получения полипептида, имеющего повышенную активность связывания FcγRIIb по сравнению с исходным полипептидом, где способ включает внесение по меньшей мере одного аминокислотного изменения в Fc-область полипептида, содержащего Fc-область антитела,
где величина [величина KD для более сильной из активностей связывания полипептида, содержащего Fc-область антитела, в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)]/[величина KD для более сильной из активностей связывания исходного полипептида в отношении FcγRIIa (тип R) и FcγRIIa (тип Н)] составляет 0,7 или более, и
где указанное по меньшей мере одно аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp или замену Leu в положении 32 8 (нумерация по EU) на Glu.
14. Способ по п. 17, где указанное по меньшей мере одно аминокислотное изменение представляет собой замену Pro в положении 238 (нумерация по EU) на Asp и по меньшей мере одну замену, выбранную из группы, состоящей из:
замены Gly в положении 237 (нумерация по EU) на Trp;
замены Gly в положении 237 (нумерация по EU) на Phe;
замены Ser в положении 267 (нумерация по EU) на Val;
замены Ser в положении 267 (нумерация по EU) на Gln;
замены His в положении 268 (нумерация по EU) на Asn;
замены Pro в положении 271 (нумерация по EU) на Gly;
замены Lys в положении 326 (нумерация по EU) на Leu;
замены Lys в положении 326 (нумерация по EU) на Gln;
замены Lys в положении 326 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Met;
замены Ser в положении 239 (нумерация по EU) на Asp;
замены Ser в положении 267 (нумерация по EU) на Ala;
замены Leu в положении 234 (нумерация по EU) на Trp;
замены Leu в положении 234 (нумерация по EU) на Tyr;
замены Gly в положении 237 (нумерация по EU) на Ala;
замены Gly в положении 237 (нумерация по EU) на Asp;
замены Gly в положении 237 (нумерация по EU) на Glu;
замены Gly в положении 237 (нумерация по EU) на Leu;
замены Gly в положении 237 (нумерация по EU) на Met;
замены Gly в положении 237 (нумерация по EU) на Tyr;
замены Ala в положении 330 (нумерация по EU) на Lys;
замены Ala в положении 330 (нумерация по EU) на Arg;
замены Glu в положении 233 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Asp;
замены His в положении 268 (нумерация по EU) на Glu;
замены Lys в положении 326 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ser;
замены Lys в положении 326 (нумерация по EU) на Thr;
замены Val в положении 323 (нумерация по EU) на Ile;
замены Val в положении 323 (нумерация по EU) на Leu;
замены Val в положении 323 (нумерация по EU) на Met;
замены Tyr в положении 296 (нумерация по EU) на Asp;
замены Lys в положении 326 (нумерация по EU) на Ala;
замены Lys в положении 326 (нумерация по EU) на Asn; и
замены Ala в положении 330 (нумерация по EU) на Met.
15. Способ по п. 13 или 14, где полипептид, содержащий Fc-область антитела, представляет собой IgG-антитело.
16. Способ по п. 13 или 14, где полипептид, содержащий Fc-область антитела, представляет собой молекулу слитого белка Fc.
17. Средство для подавления активации по меньшей мере одной, выбранной из группы, состоящей из В-клеток, тучных клеток, дендритных клеток и базофилов, которое содержит полипептид по любому из пп. 1-8.
18. Средство для лечения или профилактики иммунологического воспалительного заболевания, которое содержит полипептид по любому из пп. 1-8.
19. Терапевтическое средство или профилактическое средство по п. 18, где иммунологическое воспалительное заболевание представляет собой аутоиммунное заболевание и представляет собой заболевание, которое может быть вызвано образованием антитела против аутоантигена.
20. Противовирусное средство, содержащее полипептид по любому из пп. 1-8.
| US20090076251 А1, 19.03.2009 | |||
| МОДУЛЯТОРЫ FC-РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 1999 |
|
RU2236222C2 |

