ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области анализа несбалансированных хромосомных нарушений с использованием конкурентной геномной гибридизации (CGH).
УРОВЕНЬ ТЕХНИКИ
В области сравнительной геномной гибридизации (CGH) появление микроматриц обеспечило явное преимущество по сравнению с традиционным цитогенетическим анализом, поскольку микроматрицы позволяют детектировать большую часть микроскопических и субмикроскопических хромосомных нарушений во всем исследуемом геноме.1 Хромосомный анализ может быть проведен в рамках одного эксперимента на клетках, полученных из множества источников, включая лимфоциты периферической крови или кровь плода, ооциты или полученные полярное тельце и сперматозоиды, бластомеры эмбрионов и любой связанный биоптат (биопсийный материал), ворсинки хориона, клетки амниотической жидкости, кожу, костный мозг, ткани плода (например, легкое, печень), солидные опухоли и асциты. Существующие способы, связанные с данной методикой, подразумевают сначала выделение генетического материала из этих клеток.2,3
Обладающие высокой пропускной способностью методики, в которых применяются микроматрицы, можно также потенциально применять в ветеринарии для развития диагностических и прогностических тестов и для идентификации новых терапевтических мишеней.4
Многочисленные исследования показали, что наследственная хромосомная аберрация может влиять на эффективность репродукции, ставя под угрозу успех, достигнутый за многие годы в области селекции животных. За последние 40 лет были опубликованы сотни научных публикаций, сообщающие о новых хромосомных нарушениях, главным образом связанных с клиническими нарушениями (по большей части снижением фертильности). Каждый год по всему миру проводится приблизительно 8000 и 10000 хромосомных анализов в основном у крупного рогатого скота5,6, свиней7 и лошадей. Например, цитогенетические анализы у лошадей оказались выгодными для коневодства благодаря идентификации хромосомных аберраций, вызывающих врожденные аномалии, потери эмбриона и бесплодие, что является подверждением современной концепции о роли хромосом также и в репродукции животных.8,9 Кроме того, многие заболевания животных и человека имеют общую патогенетическую основу, и таким образом, представляют собой хорошие сравнительные модели для заболеваний человека, давая начало концепции «единой медицины», где существует особая взаимосвязанная область исследования, причем одной из самых близких является онкология.10,11
Отклонения по числу хромосом (анеуплоидии) особенно часто встречаются у культивируемых стаминальных, раковых клеток и, в частности, у ооцитов и эмбрионов человека и связаны с различными негативными последствиями как для природных циклов, так и для циклов, в которых используют технологии искусственного оплодотворения. Эмбрионы, содержащие неправильное число хромосом (анеуплоидия), могут быть не способны имплантироваться в матку, могут быть не выношены или могут приводить к появлению новорожденных с серьезными медицинскими проблемами. Поэтому представляется разумным задуматься о надежном способе, позволяющем отличать нормальные эмбрионы от эмбрионов с анеуплоидией, в качестве полезного инструмента для врачей и эмбриологов, помогающего им сделать правильный выбор. Эта концепция привела к разработке целого ряда различных способов детекции хромосомных нарушений в различных образцах, связанных с областью оплодотворения in vitro (IVF), чаще всего называемых предимплантационным генетическим скринингом (ПГС).12,13
ПГС представляет собой способ, который предназначен для улучшения конечных результатов применения вспомогательных репродуктивных технологий, таких как IVF, за счет обеспечения того, что эмбрионы, выбранные для переноса в матку, являются нормальными в отношении хромосом. Объединение достижений в генетике и эмбриологии, по всей видимости, готово дать толчок к вступлению в новую эру лечения бесплодия. В частности, тест на анеуплоидию представляет собой термин, применяемый для описания процесса подсчета числа хромосом, присутствующих в клетке, и CGH представляет собой методику, которую используют для обнаружения их наличия и идентификации локализации в геноме амплифицированных или удаленных ДНК-последовательностей. Если клетка не содержит ровно 46 хромосом, ее определяют как «анеуплоидную».14,15,16
Кроме наличия дополнительных хромосом, в эмбрионе могут отсутствовать хромосомы (приобретение (GAIN) или потеря (LOSS) являются названиями для описания каждого явления, соответственно). В большинстве случаев, потеря или приобретение хромосом будет приводить к появлению эмбриона, который не будет имплантироваться или нормально расти. Тестирование эмбрионов с использованием CGH позволяет определять число хромосом в каждом эмбрионе перед тем, как их перенесут в матку матери или заморозят для применения в будущем.
Можно протестировать каждый отдельный эмбрион, созданный путем IVF, и подсчитать число присутствующих хромосом. Это осуществляют путем отбора клеток из эмбриона на 3 или 5 день развития и тестированием этих клеток для анализа их генетического статуса. Эмбрион может легко компенсировать удаление клеток за счет нескольких клеточных делений. При компетентном подходе биопсия эмбриона незначительно влияет на его рост, хотя может немного замедлить его.17,18
Помимо ПГС, целью предимплантационной генетической диагностики (ПГД) является идентификация эмбрионов, не содержащих наследственные генетические заболевания, связанные либо с мутациями, влияющими на функцию генов, либо с нарушениями, влияющими на число копий целых хромосом или хромосомных областей. Выбор и предпочтительный перенос генетически нормальных эмбрионов обеспечивает высокую вероятность того, что любая развившаяся беременность будет здоровой, снижая вероятность того, что может понадобиться рассмотрение необходимости прерывания беременности, и в определенных случаях способствуя устранению наследственного заболевания в семье.19 Обе методики, ПГС и ПГД, включают получение эмбрионов с использованием методик IVF с последующей биопсией и тестированием эмбрионального материала.
Поскольку маловероятно, что перенос эмбриона с хромосомными нарушениями приведет к рождению здорового живого ребенка, было предложено направить усилия на идентификацию и предпочтительный перенос эуплоидных эмбрионов. В теории, такая практика должна увеличить частоту имплантаций и беременностей, достигаемых после IVF.
За последние 10 лет увеличивающиеся количество клиник репродуктивного здоровья внедрили в практику стратегии хромосомного скрининга, чтобы облегчить идентификацию и перенос жизнеспособных эмбрионов. Этот подход чаще всего применяют у пациентов, у которых считается, что присутствует повышенный риск формирования анеуплоидных ооцитов и/или эмбрионов, особенно у пар, у которых были повторяющиеся неудачные попытки имплантации (RIF) или повторяющееся невынашивание беременности, или если партнер-женщина находится в позднем репродуктивном возрасте (ARA) и где, соответственно, присутствует мужской фактор.20,21,22
Несмотря на то, что ПГС тесно связан с ПГД, его клиническое применение оказалось намного более спорным. В течение последних нескольких лет возникали споры о диагностической значимости ПГС с использованием флуоресцентной гибридизации in situ, влиянии однократной или двойной биопсии бластомера на жизнеспособность эмбриона и влиянии мозаицизма на стадии дробления на точную диагностику эуплоидии/анеуплоидии.23,24
Флуоресцентная гибридизация in situ (FISH) является предпочтительным способом исследования хромосом в отобранном биоптате (биопсийном материале), при этом описаны различные протоколы, позволяющие оценить от 5 до 12 хромосом на ооцит или эмбрион. Данный способ не позволяет осуществлять полное кариотипирование всех 23 хромосомных пар.25,26
Методика анализа одиночного нуклеотидного полиморфизма (SNP) на матрицах отличается от анализа CGH на матрицах и также может быть применен для определения нарушений числа копий в клетках в образцах ДНК. Применяемый механизм отличается, при этом не требуется конкурентного сравнения, способ основывается на количественном определении отдельных аллелей и последующем радиометрическом расчете. Недостатки включают повышенный уровень шума, более длительный протокол проведения анализа, сложность интерпретации данных и ограниченную применимость в отношении диплоидных образцов.27
Недавнее введение более перспективных способов, таких как CGH, спектральное кариотипирование, мечение праймерами in situ (PRINS) и способы анализа пептидно-нуклеиновой кислоты (ПНК), сделало возможным кариотипирование всех 23 хромосомных пар. Эти новые подходы оказались довольно затратными по времени, техническим средствам и стоимости.28
Кроме этого, применение ДНК-микроматрицы обеспечивает высококачественный твердый носитель для этого анализа и представляет собой незаменимый способ для крупномасштабного анализа. В двухканальном анализе на ДНК-микроматрице ДНК, выделенную из тестируемых клеток и нормальных эталонных клеток, метят соответственно двумя разными флуоресцентными красителями перед совместной гибридизацией с иммобилизованными ДНК-зондами на пластине микроматрицы. В ходе гибридизации две флуоресцентно меченые ДНК (тестируемая и эталонная) конкурируют за гибридизацию с цепями иммобилизованных ДНК-зондов, и повторяющиеся последовательности удаляют или снижают их количество тем или иным способом. Реакции гибридизации между комплементарными цепями происходят только между меченой антисмысловой цепью и иммобилизованной смысловой цепью.
Отношение интенсивностей двух флуоресцентно меченых ДНК используют для количественного определения относительных уровней геномных областей в образцах. Этот способ хорошо подходит для попарного сравнения уровней вариаций числа копий (CNV) в образцах.29 Учитывая, что более тысячи различных ДНК-зондов иммобилизованы на микроматрице, можно получить относительный уровень CNV всех хромосом в образце за один анализ.30,31,32
ДНК-микроматрицы нашли применение в области открытия новых генов, диагностики заболеваний, рака, фармакогеномики и исследованиях токсикологии. Их все чаще используют для анализа серий родственных образцов, для которых желательно осуществление сравнения по всем образцам.33,34
Крайне распространено в области экспрессии генов сравнение образцов после обработки с контрольными образцами, где самым естественным эталоном является дикий тип или биологические контрольные образцы, которые, как правило, являются наиболее распространенными. Однако если целью исследования является сравнение каждого образца с другими, то естественного контроля не существует. Надежной альтернативой является объединенная в пул эталонная ДНК, которая снижает количество больших ошибок. Некоторые исследователи развивают данный подход далее и создают «универсальный эталон», полученный из нескольких стандартных клеточных линий, которые чаще всего используются в опытах указанными исследователями. Применение универсального эталона позволяет указанным исследователям сравнивать результаты для всех их экспериментов.35
Для сравнения ряда образцов одинакового высокого качества и представляющих одинаковый интерес каждый образец гибридизируют с каждым из двух различных образцов в двух различных ориентациях красителя. Данная схема приводит к уменьшению в два раза дисперсии на одно определение, потому что каждый образец участвует дважды, а не один раз, за счет только еще одной матрицы. Недостаток заключается в том, что если одна матрица не будет работать или будет низкого качества, тогда дисперсия ошибки для всех определений удваивается. С другой стороны, тот факт, что только два состояния могут быть применены к каждой матрице, усложняет указанную схему, поскольку, как правило, существует больше чем два состояния, или потому что не рекомендуется напрямую гибридизировать два образца на одной матрице, так как это приводит к образованию искусственных пар ДНК.
Самый распространенная схема для двухцветных (конкурентно гибридизируемых точечных) матриц представляет собой «схему, включающую эталон». В простейшем варианте каждый экспериментальный образец гибридизируют против стандартного эталонного образца.
Тогда как применение неамплифицированной геномной ДНК в качестве эталона является крайне распространенным в CGH на матрицах, соответствующее применение в ПГМ/ПГД приводит к высокому уровню шума по причине отсутствия баланса между двумя препаратами ДНК, амплифицированным образцом и неамплифицированным эталоном. Соответственно, этот дисбаланс увеличивается, если в этом контексте используют в качестве эталона пул ДНК.
Эталонный материал, часто применяемый в этом контексте, представляет собой образец нормальной объединенной в пул ДНК, разведенный до количества, приблизительно сравнимого с количеством ДНК малого количества одиночных клеток. Разведенную до соответствующего уровня эталонную ДНК затем амплифицируют параллельно, рядом с образцом ДНК. Несмотря на применение этих подходов молекулярной биологии, приведение в соответствие свойств тестируемой и эталонной ДНК не всегда является эффективным и часто ясность результатов может варьировать неопределенным образом. Наиболее распространенный подход для демонстрации успешного осуществления эксперимента при диагностическом применении представляет собой применение внутренних контролей и применение в отдельном анализе эталонного образца с известными вариациями числа копий.36
Чаще всего эталонный образец не совпадает с половыми хромосомами, и, следовательно, можно гарантировать измерение динамического диапазона. Несмотря на то, что этот подход во многих случаях доступен, к сожалению, он не применим в ПГС из-за того, что пол, к которому принадлежит образец неизвестен, особенно при анализе биопсии бластомера или бластоцисты, которые могут быть любого пола.
Дополнительно, Svetlana А. с соавторами в своей работе продемонстрировали, что CGH на основе микроматриц с использованием совпадающей по полу эталонной ДНК обеспечивает большую чувствительность для детекции несбалансированных нарушений половых хромосом, чем анализ CGH на матрицах с не совпадающей по полу эталонной ДНК. Применение не совпадающих по полу эталонных ДНК при CGH на матрицах может давать ложноположительные, ложноотрицательные и неопределенные результаты для зондов, специфичных к половым хромосомам, таким образом, маскируя потенциальные патогенетические геномные нарушения. Соответственно, не рекомендуется применение эталонной ДНК, несущей известные изменения в количестве копий, поскольку те же самые изменения в эмбрионе/ооците являются нестабильными и, следовательно, высоко вариабельными.37
В качестве альтернативы существует возможность применения контрольных последовательностей, не принадлежащих человеку, что в теории представляет собой идеальный подход, но в практическом плане выбор геномных последовательностей, способных имитировать характеристики последовательностей человека, должен быть точным, и дополнительно указанные последовательности могут испытывать те же указанные ошибки амплификации.
Остающееся решение состоит в проведении анализа тестируемого образца посредством двух традиционных CGH экспериментов, первый раз против мужской эталонной ДНК и второй раз против женской эталонной ДНК, подход, который характеризуется высокой стоимостью применения.
Buffart с соавторами38 предложили в качестве улучшения данного способа модифицированную CGH на матрицах, названную «перекрестная гибридизация на матрицах», при которой сравнивают тестируемую ДНК и образец ДНК с другой матрицы. CGH на матрицах осуществляли с использованием эталонного пула, анализируемого на другом канале матрицы. В то время как второй канал на контрольной матрице больше не используют для гибридизации нормальной эталонной ДНК, этот канал можно использовать для гибридизации второго образца с той же матрицей. Хотя указанный способ обеспечивает преимущество в части стоимости и потенциально качества данных, потому что он устраняет ошибки, связанные с красителем, указанный способ не рекомендуется для прямой гибридизации двух образцов на одной матрице из-за образования искусственных пар ДНК, как упоминалось выше.
Для того чтобы применять анализ CGH в этом контексте, необходима амплификация целого генома для увеличения малого количества ДНК, которое можно получить из клетки (5-10 пг), до уровня, подходящего для анализа. Широко применяемые способы амплификации включают ПЦР с вырожденными олигонуклеотидными праймерами (DOP-PCR) (стратегия применения различных случайных праймеров) или более современных наборов для получения библиотек путем плексиса, таких как Genomeplex (Rubicon Genomics) или ПЦР с разветвленными зондами с использованием набора Phy polymerase kit (Repli-G, Qiagen).
Для получения оптимальных результатов CGH на матрицах требуется, чтобы обе ДНК, эталонная ДНК и ДНК образца, были уравновешены в отношении качества и концентрации. Стоит отметить, что в этом контексте генетический материал является ключевым моментом для любого анализа, потому что он создает представление о каждой анализируемой клетке. В настоящее время, можно выделить ДНК из полярного тельца, бластомера (стадия дробления эмбриона) или образца биопсии трофэктодермы (5-6 день стадии развития эмбриона in vitro).
Полярные тельца представляют собой дополнительные клетки ооцита, отделяющиеся в два различных момента времени жизненного цикла яйцеклетки. Для полярных телец не известна другая функция, кроме как содействие делению клеток. Они представляют собой «побочные продукты» деления яйцеклетки. После того, как происходит имплантация, полярные тельца распадаются и больше не являются частью развивающегося зародыша.39
Биопсия эмбриона представляет собой способ, который осуществляют в ходе методик IVF, когда эмбрион достигает стадии шести-восьми клеток (приблизительно через 72 часа или на 3 день культивирования эмбриона). Одну или две клетки, или бластомеры, отделяют от остального зародыша и удаляют из блестящей оболочки, которая представляет собой оболочку, окружающую развивающийся эмбрион. После удаления клетки (клеток) развивающийся эмбрион помещают обратно в культуральную среду и возвращают в инкубатор, где он может продолжить свой нормальный рост и развитие.40
Биопсия трофобласта представляет собой новейший способ для получения клеток от развивающего эмбриона для генетического тестирования. К 5 дню развития эмбрион разделен на приблизительно 100 клеток внутренней клеточной массы и клеток трофобласа. Преимущество биопсии трофобласта состоит в том, что несколько клеток можно получить, не оказывая критическое влияние на развитие эмбриона. При получении нескольких клеток становится доступным большее количество генетической информации. Это будет повышать точность результатов.41
Однако применение ПГС сопряжено с уникальными техническими трудностями, в некоторых вариантах реализации оценка содержания хромосом в клетке может быть осуществлена путем извлечения клеток после оплодотворения или косвенного измерения содержания генетического материала, проводя анализ полярных телец и, таким образом, ооцитов, составляющих эмбрион. В других вариантах реализации указанное применение имеет единственную цель - изучить ооцит, при этом не обязательно получают эмбрион. Эта процедура обычна в случае, если образцы собирают в криобанки.
Доступные способы CGH на матрицах требуют большого числа гибридизационных тестов в случаях, когда целью является детекция несбалансированных хромосомных нарушений, особенно, когда пол образца неизвестен заранее (в таких случаях, как например, когда эмбрионы подвергаются ПГС): это зависит от того факта, что тестируемая ДНК должна быть гибридизирована отдельно как с мужскими, так и с женскими эталонными ДНК для получения надежной информации о возможных несбалансированных хромосомных нарушениях; это дублирование числа проводимых тестов дополнительно умножается на число образов анализируемой ДНК. Таким образом, существует острая потребность в новых способах CGH на матрицах, способных снижать число необходимых гибридизационных тестов, в идеале применяя только одну матрицу для каждой тестируемой ДНК.
Идентификация вариации числа копий находит широкое применение в детекции аберрации любого размера по всему геному, однако, к настоящему времени были описаны различные отклонения результата определения в отношении параллельных образцов даже при измерении на одной и той же платформе.42 Кроме того, различные алгоритмы могут давать различный результат определения на одном и том же образце. Таким образом, существует дополнительная потребность в новых цепочках процессов (пайплайнах) анализа данных CGH на матрицах, обеспечивающих набор реализуемых алгоритмов, которые могут устранять специфические для матриц ошибки и автоматически определять области приобретения (gain) или потерь (loss) на протяжении всего генома, снижая риск ошибки (систематической ошибки) способа в части ложноположительных (ЛП) и/или ложноотрицательных (ЛО) результатов43,44
Дополнительная вариабельность в тестировании путем CGH обусловлена вариабельностью качества эталонной ДНК, применяемой для конкурентной гибридизации с тестируемой ДНК. Таким образом, существует дополнительная потребность в новых способах CGH, включающих внутренний контроль качества для применяемой эталонной ДНК.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали новый способ определения несбалансированных хромосомных нарушений в образце тестируемой ДНК, особенно в случае, когда пол неизвестен (то есть когда пол донора указанной тестируемой ДНК неизвестен), что обычно происходит в случае эмбрионов, проходящих генетический скрининг перед имплантацией. Следует отметить, что указанный способ можно использовать для определения несбалансированных хромосомных нарушений в обоих типах хромосом, половых хромосомах (гоносомах) и неполовых хромосомах (аутосомах), хотя он имеет особое преимущество при применении для определения несбалансированных нарушений ДНК в гоносомах. Основной признак настоящего способа, основанного на технологии CGH на матрицах, состоит в устранении необходимости применения двойной гибридизации тестируемой ДНК с эталонными ДНК, полученной от обоих полов (то есть эталонной ДНК от донора-мужчины + эталонной ДНК от донора-женщины), требуемой до настоящего времени. Таким образом, настоящий способ осуществляют с использованием одной матрицы для гибридизации (в настоящем документе называемого тестовой матрицей для гибридизации), на котором тестируемую ДНК конкурентно гибридизируют против только одной эталонной ДНК (без разницы от донора-мужчины или от донора-женщины); результат, получаемый от указанной тестовой матрицы для гибридизации, затем объединяют с результатом, полученным от эталонной матрицы для гибридизации, на котором конкурентно гибридизируют эталонные ДНК, полученные от доноров двух противоположных полов. Объединение результатов от тестовой и эталонной матриц для гибридизации обеспечивает конечное число несбалансированных хромосомных нарушений (вариаций числа копий, в настоящем документе обозначаемых как «CNV») в тестируемой ДНК. Следует отметить, что эталонную гибридизацию не повторяют n раз, в случае, когда и образцов тестируемой ДНК подвергают анализу согласно настоящему способу: вместо этого, она может быть осуществлена только один раз, чтобы получить соответствующий результат; такой результат, сохраненный подходящим образом, может быть дополнительно объединен с каждым из результатов, полученным для и образцов тестируемой ДНК. Таким образом, для каждой отдельной тестируемой ДНК, которую анализируют, применяют только одну матрицу для гибридизации.
Таким образом, целью настоящего изобретения является способ CGH на одной матрице для оценки наличия и количества вариаций числа копий [в настоящем документе обозначаемых как «CNV»] во всей гоносомальной и аутосомальной геномной ДНК из тестируемого образца [в настоящем документе обозначаемой как «тест-ДНК»] с использованием способа CGH путем сравнения указанной тестируемой ДНК с эталонной геномной ДНК от донора-мужчины (или донора-женщины) [в настоящем документе обозначаемой как «эталонная ДНК»], указанный способ включает следующие этапы:
а) Мечение первым маркером указанной тестируемой ДНК, таким образом, получая меченную тестируемую ДНК;
b) Мечение вторым маркером, отличным от первого, мужской (или женской) эталонной ДНК, таким образом, получая меченую мужскую (или женскую) эталонную ДНК;
c) Гибридизация ДНК, полученных на этапах а) и b) с тестовой матрицей для гибридизации, таким образом, получая объединенную картину распределения интенсивностей сигналов от обоих маркеров, полученных от соответствующих гибридизованных молекул ДНК;
d) Из объединенной картины распределения, полученной на этапе с) определяют отдельные интенсивности сигналов, создаваемых указанными первым и вторым маркерами;
e) Проводят первый расчет CNV в указанной тестируемой ДНК путем сравнения друг с другом двух интенсивностей сигнала, полученных на этапе d);
f) Мечение указанным первым маркером эталонной ДНК, противоположной по полу той, что применяли на этапе b), таким образом, получая эталонную ДНК, меченную указанным первым маркером;
g) Мечение указанным вторым маркером указанной эталонной ДНК, противоположной по полу той, что применяли на этапе b), таким образом, получая эталонную ДНК, меченную указанным вторым маркером;
h) Гибридизация обоих ДНК, полученных на этапах f) и g) с эталонной матрицей для гибридизации, таким образом, получая объединенную картину распределения сигналов от обоих маркеров, полученных от соответствующих гибридизованных молекул ДНК;
i) Из объединенной картины распределения, полученной на этапе h) определяют отдельные интенсивности сигналов, создаваемых указанными первым и вторым маркерами;
j) Проводят второй расчет CNV в указанной тестируемой ДНК путем сравнения друг с другом:
- интенсивности сигнала указанной тестируемой ДНК, меченной указанным первым маркером, как определено на этапе d),
- с интенсивностью сигнала указанной эталонной ДНК, меченной указанным вторым маркером, как определено на этапе i);
k) Объединение значений CNV, полученных на этапах е) и j) с получением конечного результата определения CNV, относящегося к гоносомальной и аутосомальной геномной ДНК, таким образом, получая конечное значение CNV, относящееся к цельной геномной ДНК, подвергнутой анализу.
Настоящий способ позволяет вдвое снизить количество матриц, необходимых для обеспечения CGH-тестирования n образцов ДНК и, следовательно, времени, необходимого для такого анализа. Таким образом, настоящее изобретение распространяется на новые, более компактные носители для CGH-анализа, где обеспечивают n тестовых матриц для гибридизации для тестирования n образцов ДНК согласно этапам а)-е) настоящего способа и одну эталонную матрицу для гибридизации для осуществления этапа h), таким образом, избегая дублирования матриц, необходимого в известном уровне техники. Настоящее изобретение также распространяется на набор для осуществления такого способа. Настоящее изобретение дополнительно включает возможность снижения ошибки в интенсивностях сигналов, создаваемых гибридизацией меченых молекул ДНК, с помощью подходящей обработки данных. Настоящее изобретение дополнительно включает внутренний контроль качества эталонной ДНК, применяемой для гибридизации.
КРАТКОЕ ОПИСАНИЕ ФИГУР
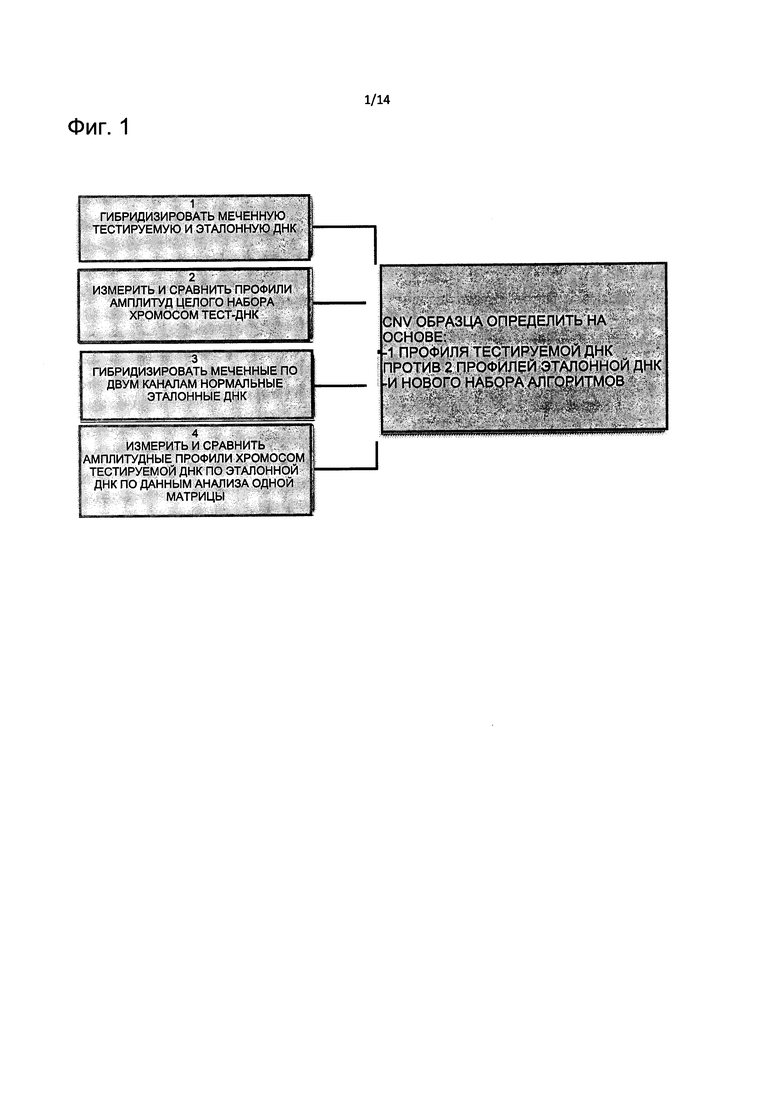
На Фиг. 1 представлена блок-схема, описывающая новый способ CGH на микроматрицах для определения CNV согласно настоящему изобретению.
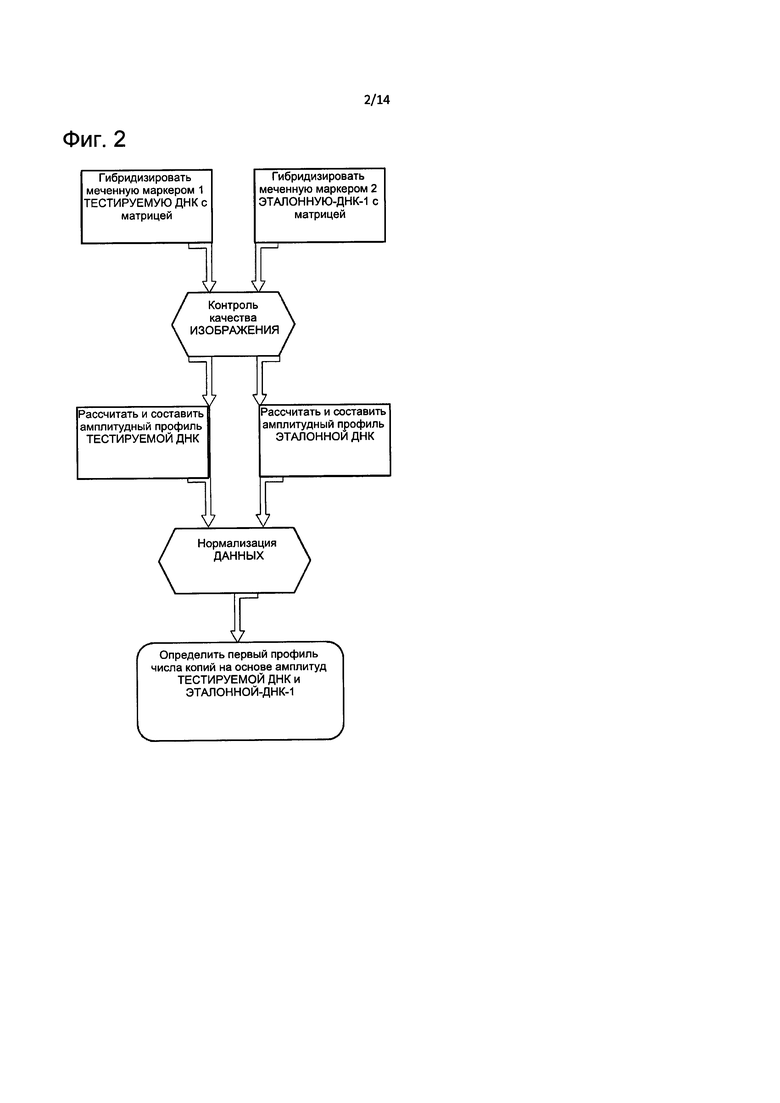
На Фиг. 2 представлена блок-схема, описывающая способ определения CNV, основанный на первом профиле амплитуд ДНК, согласно настоящему изобретению.
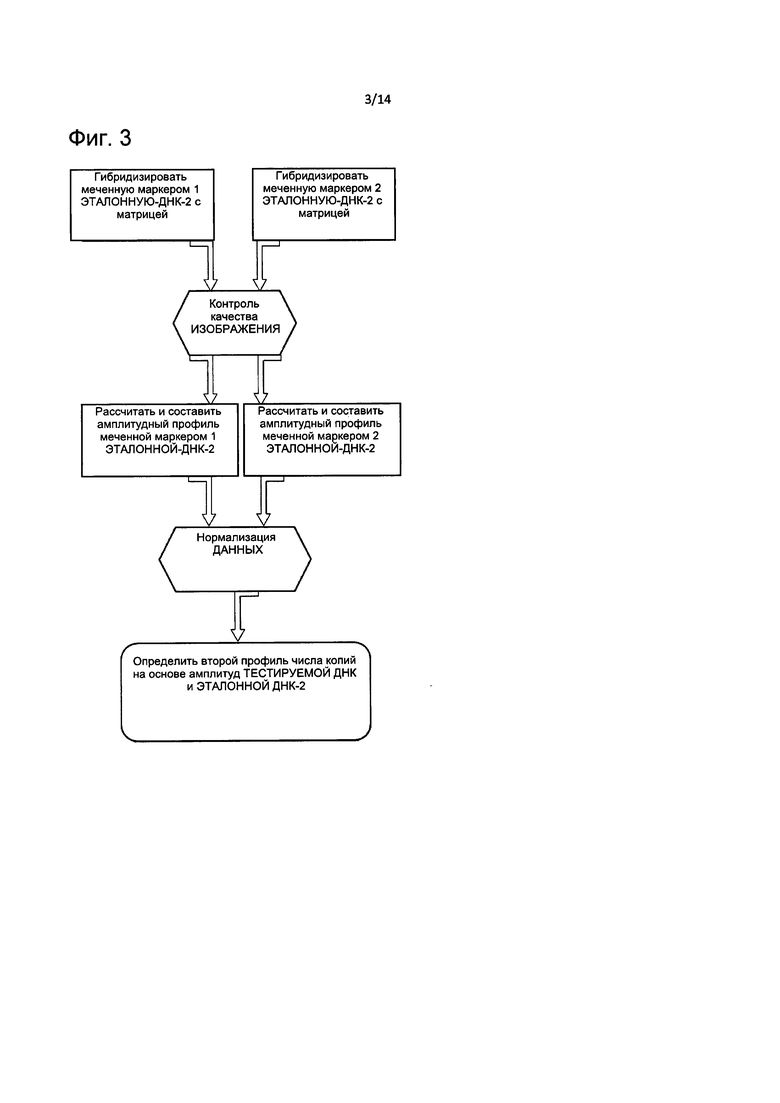
На Фиг. 3 представлена блок-схема, описывающая способ определения CNV, основанный на втором профиле амплитуд ДНК, согласно настоящему изобретению.
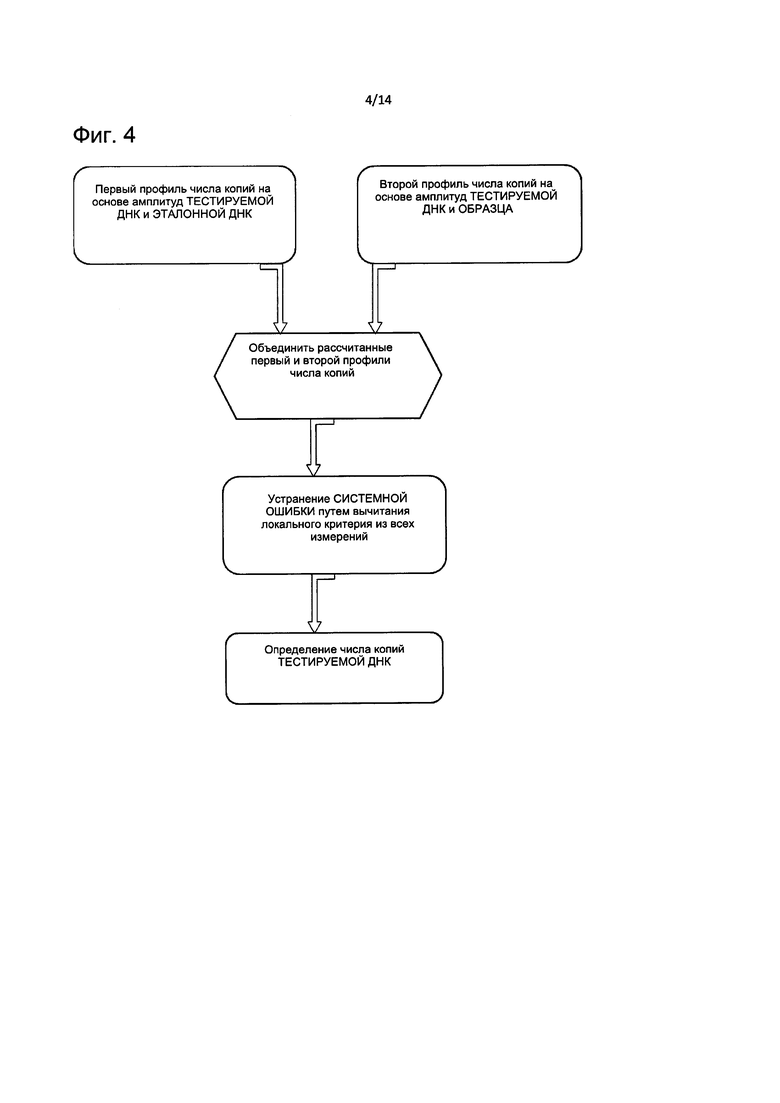
На Фиг. 4 представлена блок-схема, описывающая способ определения конечного значения CNV на основе первого и второго профиля амплитуд ДНК согласно настоящему изобретению.
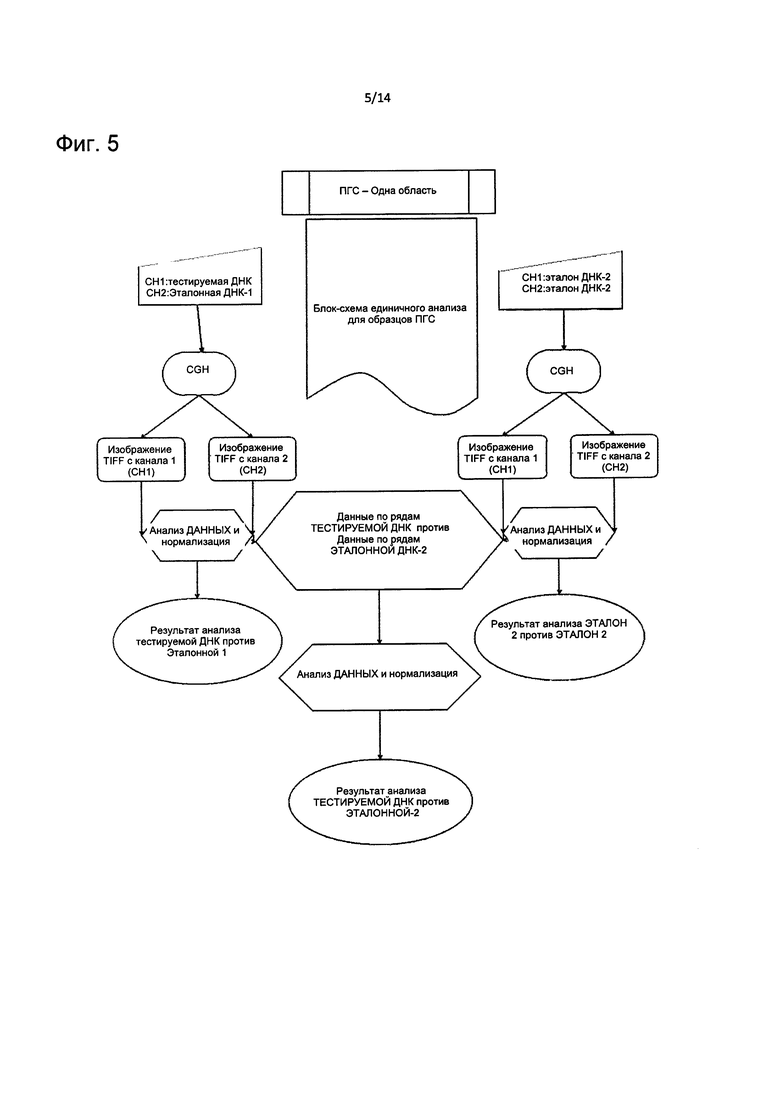
На Фиг. 5 представлено краткое описание способа определения CNV на основе первого и второго профиля амплитуд ДНК с использованием отдельной области для каждого теста, как описано в настоящем изобретении.
На Фиг. 6 представлена блок-схема, описывающая способ контроля качества изображения.
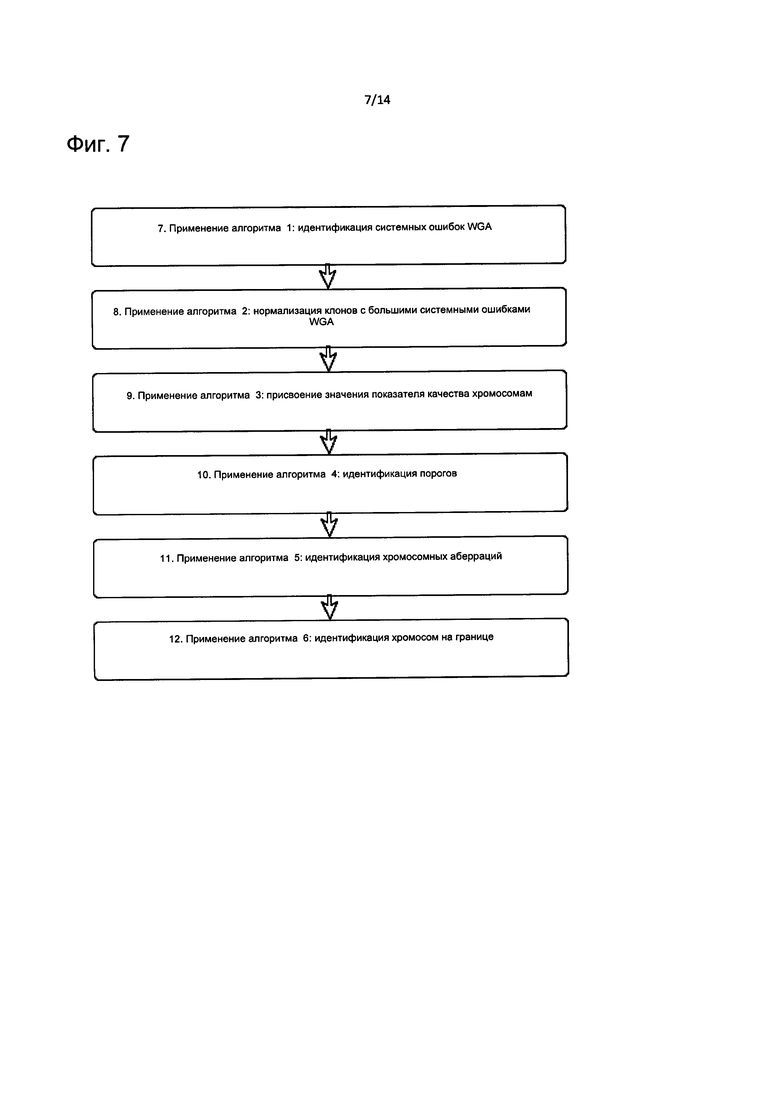
На Фиг. 7 представлена блок-схема, описывающая способ вычитания шума изображения и применения серии алгоритмов для первого расчета CNV.
На Фиг. 8 представлена блок-схема, описывающая способ расчета второго и конечного определения CNV.
На Фиг. 8а представлена таблица, сравнивающая результаты, полученные из серии анализа CNV, в отношении приобретения (GAIN) и потери (LOSS) хромосом, между тестируемой ДНК и мужской или женской эталонной ДНК; женская эталонная ДНК была представлена как от контроля качества, так и от эталона, напрямую измеряемого с помощью микроматрицы. Согласно результатам приобретения (GAIN) и потери (LOSS) аутосом и гоносом, рассчитывали кариотип ПГС и сравнивали с контрольным кариотипом. Конкордантность женских эталонов (от REF (а) до (е)) сравнивали с контролем качества и соответствующие значения конкордантности также были представлены.
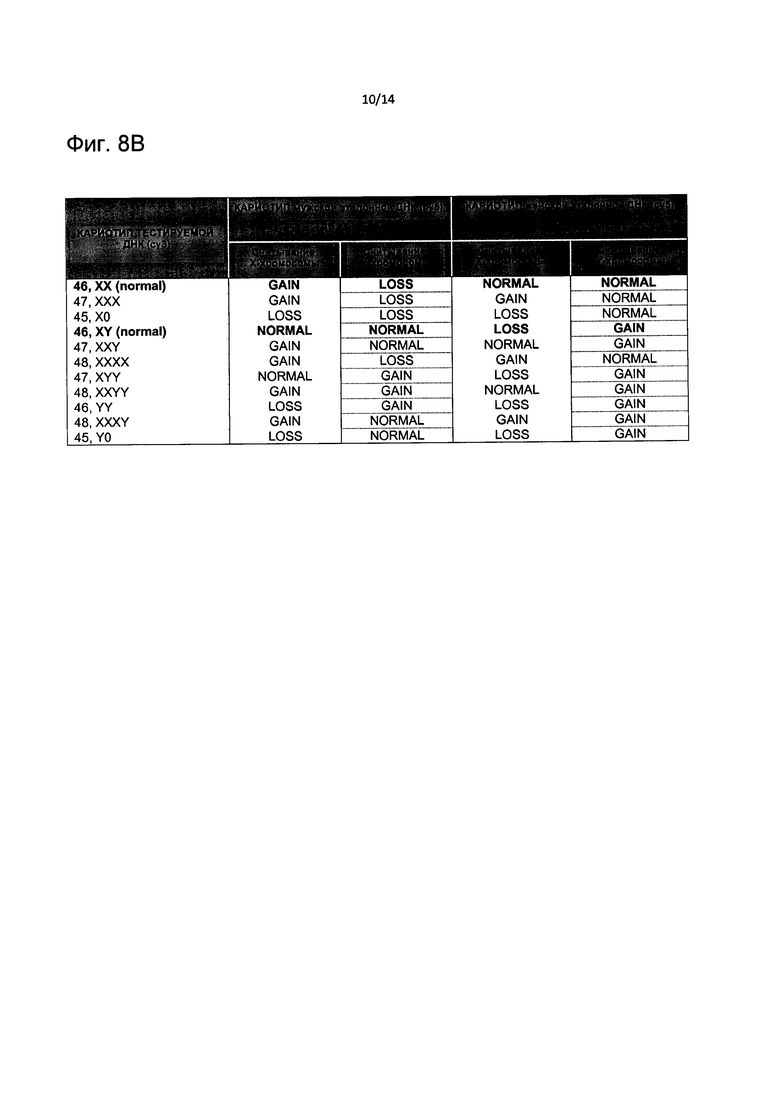
На Фиг. 8В представлены результаты считывания CNV у различных нормальных и ненормальных потенциальных генотипов в сравнении с нормальным мужским или женским эталоном, относящимся к гоносомам.
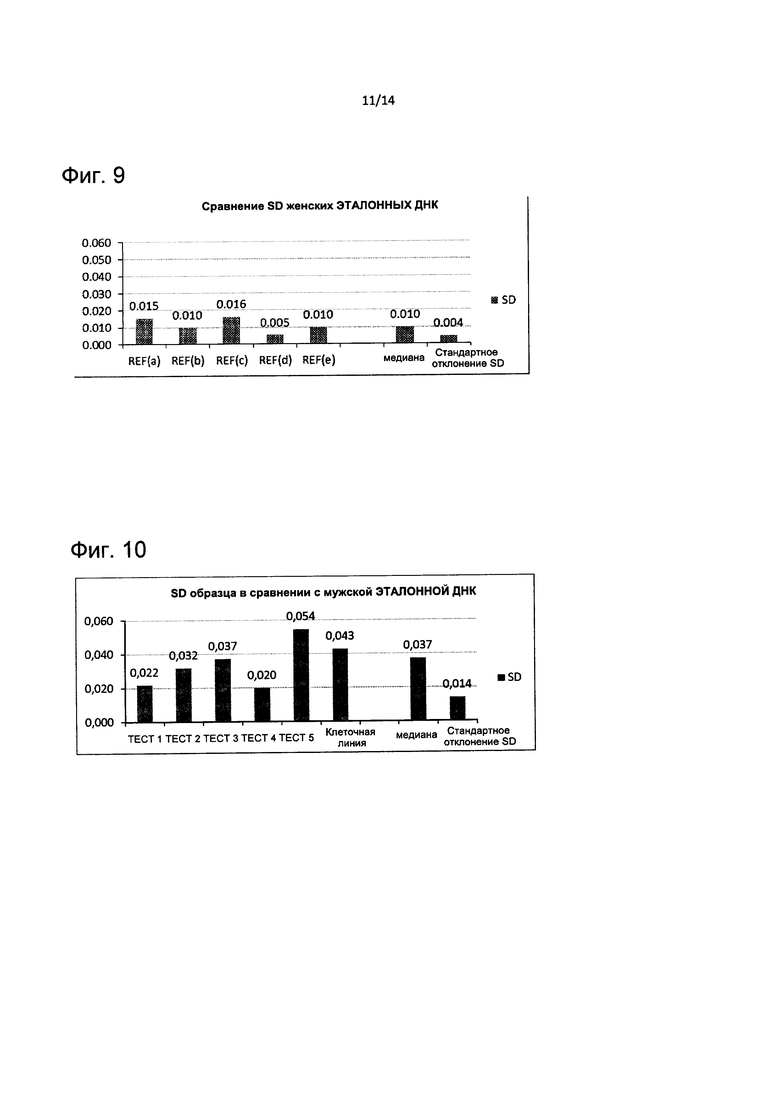
На Фиг. 9 представлено сравнение стандартного отклонения женского эталона, измеренного на различных микроматрицах. Оценка медианы и стандартного отклонения этих значений также представлена.
На Фиг. 10 представлено стандартное отклонение анализа тестируемой ДНК. Оценка медианы и стандартного отклонения этих значений также представлена.
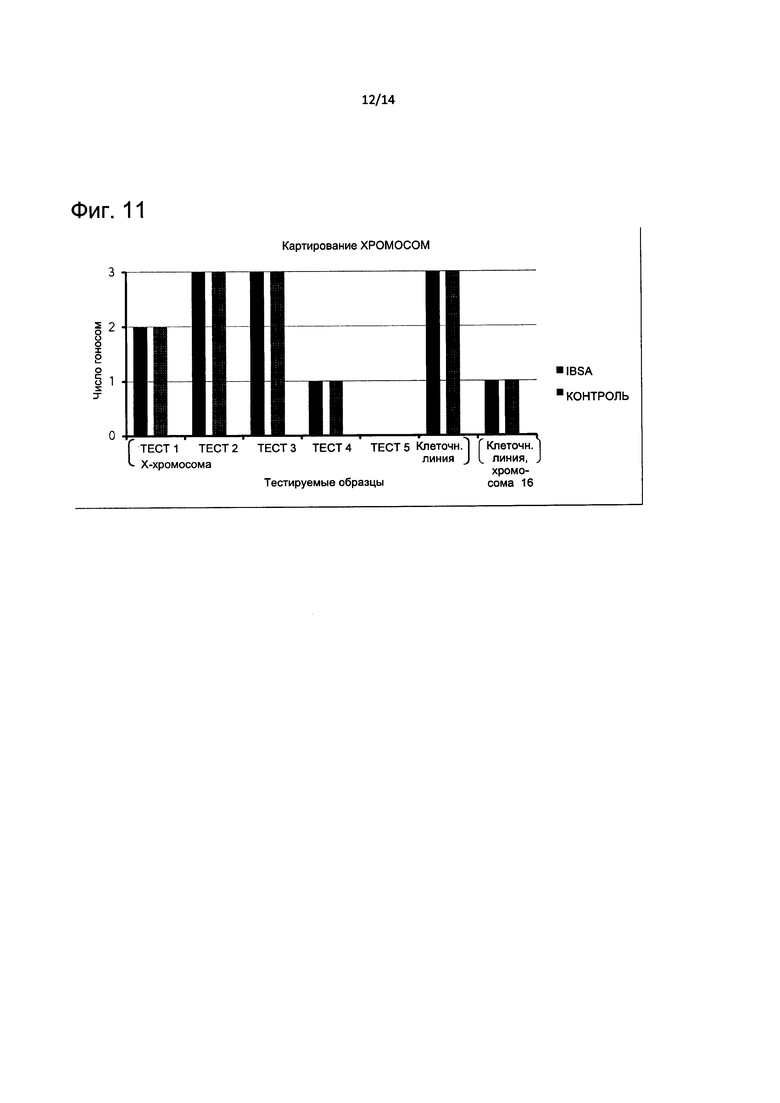
На Фиг. 11 представлено хромосомное картирование тестируемой ДНК против контроля.
На Фиг. 12 представлен тренд женского эталона при измерении тестируемых образцов против контроля качества IBSA.
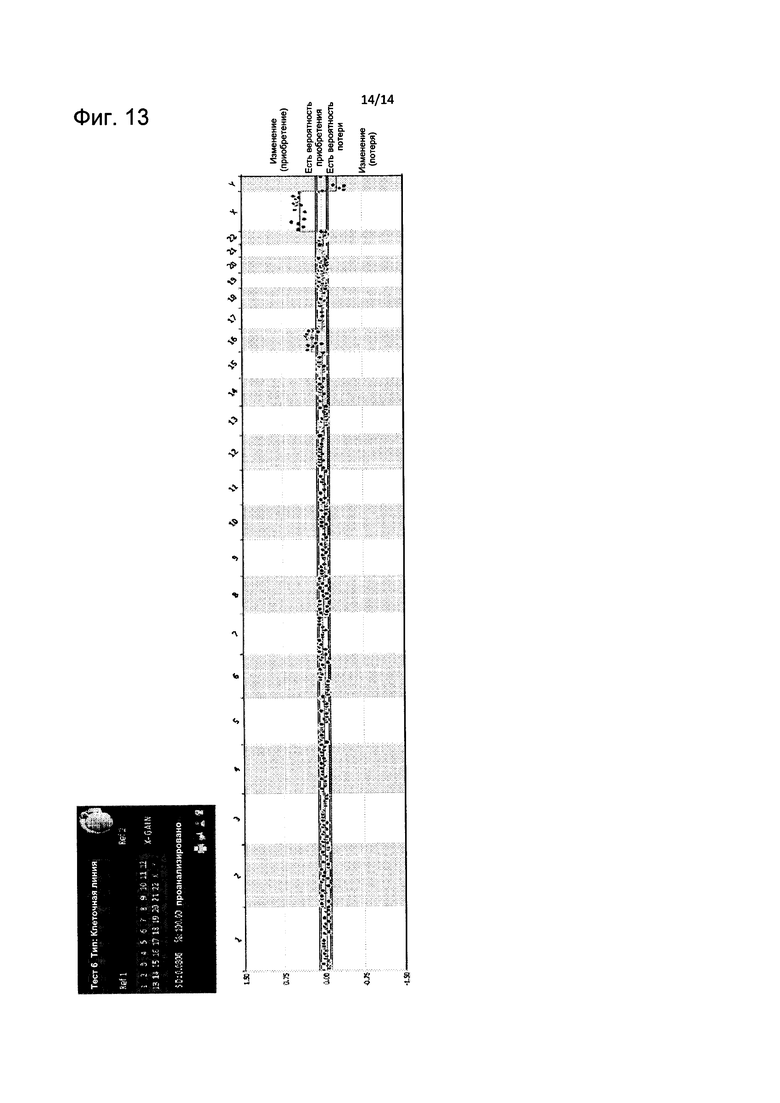
На Фиг. 13 представлен пример детекции единичной области CNV согласно настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем описании и формуле изобретении, если не указано иное, применяемая терминология должна иметь такое же значение, как обычно ей приписывается специалистом в данной области техники. Кроме того, по всему тексту описания и формулы изобретения:
«CGH» обозначает сравнительную геномную гибридизацию.
«Тестируемая ДНК» («тест-ДНК») обозначает анализируемую ДНК, в которой, в частности, необходимо определить количество вариаций числа копий.
«Вариации числа копий» или «CNV» обозначает форму структурной вариации, то есть изменений ДНК генома, которое приводит к тому, что клетка имеет ненормальное количество копий одного или более участка ДНК.
«GAIN/LOSS» означает положительные (gain-приобретение) или отрицательные (loss-потеря) значения.
«Эталонная ДНК» («референс-ДНК») обозначает ДНК, которую конкурентно гибридизируют с тестируемой ДНК на микроматрицах, применяемых в настоящем способе.
«Стандартная ДНК» обозначает ДНК, иммобилизованную на матрицах, с которой конкурентно гибридизируются как тестируемая ДНК, так и эталонная ДНК.
«Тестовая матрица для гибридизации» обозначает матрицу(ы), на которой образец (образцы) тестируемой ДНК гибридизируют на этапе с) согласно настоящему способу.
«Эталонная матрица для гибридизации» обозначает матрицу, на которой гибридизируют эталонные ДНК на этапе h) согласно настоящему способу.
«Матрицы» (или «микроматрицы», названия используются в настоящем документе взаимозаменяемо) обозначают материал-носитель (такой как стекло или пластиковая пластина) с покрытой поверхностью, к которому прикреплены многочисленные молекулы или фрагменты известных последовательностей обычно одноцепочечной ДНК или белка в виде определенной картины распределения для применения в биохимическом или генетическом анализе. В настоящем способе применяемые матрицы имеют одинаковую структуру и композицию, в независимости от того, предназначены ли они для гибридизации тестируемой ДНК или эталонных ДНК.
Тестируемая ДНК, применяемая в настоящем способе, представляет собой ДНК, полученную от любого организма, в частности, донора-мужчины или донора-женщины, в которой необходимо определить CNV. При осуществлении способа согласно настоящему изобретению пол донора тестируемой ДНК может быть известен или неизвестен. Тестируемая ДНК может нести увеличенное или сниженное число геномных фрагментов. Организм, из которого получена тестируемая ДНК, может представлять собой, например, по меньшей мере одно полярное тельце 1 и 2 из ооцитов, бластомеры из эмбриона, сперматозоиды, клетки, полученные от одного или более из следующих источников: лимфоцитов периферической крови, фетальной крови, ворсинок хориона, клеток амниотической жидкости, кожи, костного мозга, тканей плода, легкого, печени, солидных опухолей или асцитов и т.д.
Способ описан в настоящем документе со ссылкой на «тестируемую ДНК». Однако способ согласно настоящему изобретению не ограничен тестированием одиночных ДНК друг за другом: он может быть осуществлен одновременно на n-ом количестве тестируемых ДНК (от одного или разных доноров) в тех рамках, пока обеспечен подходящий носитель для CGH: такой носитель для CGH должен включать n тестовых матриц для гибридизации, на которых каждую из n тестируемых ДНК анализируют согласно настоящему изобретению. Этот вариант реализации является неотъемлемой частью способа согласно настоящему изобретению. Наконец, несмотря на то, что настоящий способ был разработан для тестирования образцов ДНК, он может быть функциональным для всех случаев, в которых может быть применен анализ на двухканальных микроматрицах.
Эталонные ДНК, применяемые в настоящем способе, можно получить от отдельных доноров или, в качестве альтернативы, от множества доноров-мужчин или от множества доноров-женщин; в случае множества доноров соответствующие генотипы могут быть также представлены в различных концентрациях. Применение эталонной ДНК с этими вариациями имеет преимущество в снижении погрешности, связанной с генотипом. Эталонную ДНК, применяемую в настоящем способе, можно получать экстемпорально или применять как коммерчески доступный вариант. Соответственно, в соответствии с настоящим изобретением термин «мужская эталонная ДНК» обозначает эталонную ДНК, полученную от одного или более доноров-мужчин; термин «женская эталонная ДНК» означает эталонную ДНК, полученную от одного или более доноров-женщин. Выражение «эталонная ДНК, противоположная по полу» (где «противоположная» используется в качестве противоположности дополнительной эталонной ДНК) обозначает эталонную ДНК, полученную от донора, чей пол является противоположным полу донора дополнительной эталонной ДНК.
В диагностической практике тестируемая ДНК, которую анализируют, часто представляет собой одну молекулу ДНК (например, полученную от одной клетки). Такая одна молекула ДНК может быть не способна создать достаточно интенсивные сигналы для CGH-анализа. В этих случаях, молекулу тестируемой ДНК можно амплифицировать подходящим способом с использованием традиционных способов, хорошо известных в данной области техники. Не ограничивающими примерами методик амплификации являются DOP-PCR (стратегия применения различных случайных праймеров), набор для создания библиотек путем плексиса, такой как Genomeplex (Rubicon Genomics), ПЦР с разветвленными зондами с использованием набора для полимеризации Phy (Repli-G, Qiagen) и т.д. Дополнительные примеры методик амплификации представлены в экспериментальном разделе настоящего описания. Степень амплификации мишени должна быть такой, чтобы быть способной обеспечить уровни сигнала, которые являются равными или по меньшей мере схожими с уровнями сигнала эталонных ДНК.
Потребность в амплифицикации эталонной ДНК является не настолько частой: фактически, эталонные ДНК, как правило, представляют собой образцы из множества молекул со степенью амплификации достаточно высокой для проведения CGH; однако настоящее изобретение не исключает возможность амплификации эталонных ДНК при необходимости (всегда с учетом получения сопоставимых сигналов с тестируемой ДНК). Амплификация ДНК (тестируемых и/или эталонных) можно осуществить за один этап или в ходе отдельных последовательных этапов, например, двух последовательных этапов амплификации, при которых получают вполне достаточное количество ДНК для мечения в ходе второго этапа амплификации.
Различные этапы от а) до k) согласно настоящему способу можно описать более подробно следующим образом:
Этап а)
Маркер, применяемый для мечения тестируемой ДНК, как правило, представляет собой флуоресцентный маркер. Не ограничивающими примерами подходящих маркеров, хорошо известных в данной области техники, являются су3- или су5-модифицированные дидеоксинуклеотиды (Су3- и су5-дЦТФ, GE Healthcare или Perkin Helmer). Также можно применять альтернативные маркеры как, например, окрашенные нефлуоресцентные маркеры, радиомаркеры и т.д. Мечения можно добиться с помощью известной методики, например, мечения со случайными праймерами с использованием фрагмента Кленова ДНК-полимеразы I E. Coli, или эквивалентными способами, известными в данной области техники.
Этап b)
Эталонная ДНК, применяемая на этом этапе, может иметь либо мужское, либо женское происхождение, это не имеет значения, так как не влияет на конечные результаты настоящего способа. Однако тип эталонной ДНК, применяемой на этом этапе определяет тип эталонных ДНК, применяемых позже на этапах f) и g). Мужская эталонная ДНК, применяемая на этапе b), потребует применения женских эталонных ДНК (таким образом, противоположных по полу) на этапах f) и g); наоборот, женская эталонная ДНК, применяемая на этапе b), потребует применения мужских эталонных ДНК на этапах f) и g). Маркер, применяемый на этом этапе, можно выбрать среди описанных на этапе а) при условии, что два маркера отличаются друг от друга, как требуется для CGH-анализа. Как правило, применяют маркеры с различными флуорофорами, различающимися по своему цвету/волне испускания. Предпочтительно два маркера относятся к одной и той же семье маркеров, например, «флуоресцентных» или «радио», чтобы было возможно провести разделение соответствующих меченых ДНК с помощью одного и того же прибора для детекции. Два маркера применяют подходящим образом в равных пропорциях.
Этап b) в настоящем документе обозначен в широком смысле, также включая возможность применения предварительно меченой коммерческой эталонной ДНК; в этом случае, процесс физического мечения эталонной ДНК заменен на получение предварительной меченого маркера из коммерческого источника: этот вариант реализации предполагается включенным в этап b) настоящего способа.
Этап с)
Этот этап представляет собой единственную реакцию гибридизации, которой подвергается тестируемая ДНК в ходе всего способа: по этой причине настоящий способ характеризуется как «одноматричный» (осуществляемый на одной матрице). На этой стадии меченую ДНК, полученную на этапах а) и b) (тестируемую и эталонную) конкурентно гибридизируют на одной и той же тестовой матрице для гибридизации. Гибридизацию осуществляют способами, известными в данной области техники: как правило, меченые ДНК растворяют в подходящем растворе для гибридизации; денатурируют ДНК, чтобы обеспечить доступность одиночных цепей для гибридизации с ДНК-зондами матрицы; конечный раствор затем распределяют на матрице, таким образом, запуская реакцию гибридизации. Как правило, в CGH, тестируемая и эталонная ДНК конкурируют за гибридизацию в отношении каждого из различных ДНК-клонов (бактериальная искусственная хромосома (ВАС)/зондов), присутствующих на матрице: любая возможная несбалансированная гибридизация тестируемой/эталонной ДНК на отдельных ВАС сигнализирует о возможном нарушении тестируемой ДНК в соответствующей ее части. После подходящего времени инкубации матрицу промывают для удаления несвязавшейся ДНК и высушивают.
Этап d)
На этом этапе уровни гибридизации тестируемой и эталонной ДНК измеряют путем оценки в каждой точке матрицы интенсивности сигналов меченых ДНК, связанных с ним. Подходящий прибор для считывания, оснащенный программным обеспечением, (например, лазерный сканер) различает и количественно определяет для каждой точки матрицы вклад двух маркеров в измеряемую интенсивность сигнала. Конечную интенсивность сигнала записывают и сохраняют для дальнейшего применения в способе.
Этап е)
CNV в тестируемой ДНК определяют путем детекции нарушений в уровнях гибридизации в соответствии со специфическими точками на матрице; как правило, при CGH несбалансированные нарушения можно детектировать как потерю/приобретение (loss/gain) сигналов эмиссии в специфических точках матрицы. В частности, когда логарифм (log2) отношения между интенсивностями двух сигналов (тестируемой ДНК и эталонной ДНК) отражается как положительное (gain) или отрицательное (loss) значение, тогда можно определить CNV. Расчет CNV осуществляют путем обработки сигналов с помощью программного обеспечения согласно способам, хорошо известным в данной области техники: это включает, например, количественное определение интенсивности сигнала, нормализацию данных, статистический анализ, снижение ошибки, расчет связанных с хромосомами интенсивностей, формирование связанных с хромосомами ДНК-сегментов и т.д. Примером подходящего программного обеспечения для этой цели является программное обеспечение, описанное в заявке на патент WO 2013/171565 А2. Дополнительные подробности о программном обеспечении и алгоритмах для этого расчета представлены в экспериментальном разделе данного описания. Значения считываний CNV хромосом на этом этапе являются финальными в отношении неполовых хромосом (аутосом), в то же время являются предварительными в отношении половых хромосом (гоносом). Причина этого в том, что когда неизвестно к какому полу принадлежит анализируемая тестируемая ДНК (то есть пол донора указанной тестируемой ДНК неизвестен), ее гибридизации с одной (женской или мужской) эталонной ДНК будет недостаточно, чтобы окончательно провести различие между CNV, представленными в нормальной и в конечном счете ненормальной половой хромосоме.
Этап f)
Эталонная ДНК, применяемая на этом этапе, имеет противоположный пол с той эталонной ДНК, примененной на этапе b): таким образом, она будет женской/мужской эталонной ДНК, если мужскую/женскую эталонную ДНК применяли на этапе b). Первую аликвоту такой ДНК метят первым маркером. Предпочтительно маркер будет одним из двух, уже примененных на этапах а) и b).
Этап g)
На этом этапе вторую аликвоту эталонной ДНК, примененной для мечения на этапе f), метят вторым маркером, отличным от первого. Применяемый в этом случае маркер будет предпочтительно тем, который был применен на этапах а) или b), который при этом не был выбран для этапа f).
Этап h)
Гибридизацию осуществляют в нормальных условиях для CGH, как описано для этапа с).
Этап i)
Уровень гибридизации двух эталонных ДНК на каждом ВАС матрицы можно измерить способами, описанными на этапе d). Относительные интенсивности сигналов записывают и сохраняют, чтобы использовать в последующем в настоящем способе. Если эти интенсивности уже были получены и доступны и сохранены на подходящих средствах хранения данных (например, в результате предыдущего выполнения способа или из коллекции базы данных), их можно использовать напрямую из этого источника как «архивные данные о гибридизации ДНК», исключая повторное выполнение подпоследовательности этапов от f) до i).
Этап j)
В отличие от стандартной CGH за гибридизацией на этапе i) не следует определение CNV напрямую на той же матрицы для гибридизации; это логично, так как на этапе i) та же самая эталонная ДНК конкурирует сама с собой, то не должно ожидаться никаких значимых CNV на этом этапе. На настоящем этапе j) вместо этого сравнивают интенсивности сигналов, полученных на этапе i) с интенсивностями сигнала, полученными на этапе d), и получают их них вторую оценку CNV в тестируемой ДНК. Интенсивности сигналов, сравниваемые на этапе j), представляют собой интенсивности сигналов, полученные от обоих типов хромосом, в частности, интенсивности сигналов, полученные от половых хромосом; таким образом, любое приобретение (gain) X-хромосомы и потеря (loss) Y-хромосомы (или наоборот), присутствующие в сравниваемых интенсивностях сигнала, будут представлять особый интерес. На этапе j) сравнивают сигналы гибридизации тестируемой ДНК с сигналами от обеих мужской и женской эталонных ДНК, таким образом, считывание CNV на этом этапе эффективно дополняет предварительную информацию о CNV, полученную на этапе е) с использованием эталонной ДНК одного пола.
Этап k)
На этом этапе с помощью программного обеспечения CNV, полученные на этапах е) и j), объединяют с получением полного и конечного профиля CNV целой тестируемой ДНК, подвергнутой анализу.
Как очевидно из вышеизложенного, подпоследовательность этапов а)-е) и подпоследовательность f)-i) относятся к двум различным и независимым анализам гибридизации. Порядок осуществления этих двух подпоследовательностей, как показано выше, является чисто иллюстративным и не ограничивает настоящий способ: последний может быть осуществлен без разницы путем инвертирования их порядка, то есть осуществляя сначала этапы f)-i), затем этапы а)-е), а затем завершая способ этапами j) и k); этот вариант реализации является неотъемлемой частью способа согласно настоящему изобретению.
Способ согласно настоящему изобретению дополнительно включает некоторые предпочтительные варианты реализации, которые способны дополнительно увеличить точность оценки CNV.
Согласно первому предпочтительному варианту реализации, методику гибридизации эталонных ДНК (этапы f-g-h-I способа) повторяют большее количество раз с использованием соответствующего числа образцов тех же эталонных ДНК; такие повторяющиеся гибридизации осуществляют на микроматрицах из различных производственных серий; затем интенсивности сигналов, полученные в результате каждой гибридизации, обрабатывают с помощью программного обеспечения с целью устранить/снизить возможную ошибку и неспецифические сигналы. Описание с примерами такой обработки более подробно представлено в экспериментальном разделе настоящего описания. Обработанные таким образом интенсивности сигналов затем обозначают как «результат этапа i)» и далее обрабатывают в соответствии со способом согласно настоящему изобретению, описанным выше.
Согласно второму предпочтительному варианту реализации интенсивности сигналов, полученные из первого варианта реализации, хранят и используют в качестве «контрольного стандарта качества» для любых дополнительных эталонных ДНК, применяемых в настоящем способе. Согласно этому варианту реализации интенсивности сигналов, полученные на этапе i) от применяемой эталонной ДНК, сравнивают с интенсивностями сигналов, соответствующих «контрольному стандарту качества» с целью определить/проконтролировать уровень качества применяемой эталонной ДНК: чем ближе ее интенсивности сигналов к интенсивностям сигналов «контрольного стандарта качества», тем выше ее качество.
Избегая повторных гибридизаций тестируемой ДНК с эталонными ДНК обоих полов, настоящий способ вводит важное упрощение в определение CNV образцов тестируемой ДНК, чей пол неизвестен заранее (то есть, когда пол донора указанного образца тестируемой ДНК неизвестен), например, это является типичным случаем в предимплантационной диагностике в протоколах оплодотворения in vitro (IVF). Это упрощение означает значительное снижение времени анализа (в особенности, когда необходимо проанализировать множество тестируемых ДНК), а также в снижении числа требуемых матриц для CGH. Определение CNV позволяет выбирать оптимальный (лучший) ооцит, сперматозоид или правильный эмбрион по правильному хромосомному статусу и т.д.
Снижение количества матриц, лежащее в основе настоящего способа, позволяет обеспечивать новые, более компактные носители для CGH, характеризующиеся тем, что содержат, нанесенные слоем на подходящий носитель: по меньшей мере одну матрицу для гибридизации для осуществления указанных этапов а)-е) на одной или более тестируемой ДНК, и одну матрицу для осуществления указанного этапа h): последний можно идентифицировать на носителе по подходящему печатному указанию на самом носителе или помещая его в конкретную область носителя, тем самым отличая его от области, содержащей матрицу(ы), необходимые для гибридизации тестируемой (тестируемых) ДНК. Эти новые, более компактные носители образуют сами по себе дополнительный вариант реализации настоящего изобретения.
Дополнительная цель настоящего изобретения представляет собой набор для осуществления способа, описанного выше, содержащий: а) по меньшей мере одну матрицу для гибридизации для осуществления указанных этапов а)-е) на одной или более тестируемой ДНК; (b) эталонные интенсивности сигналов этапа i), записанные на подходящем средстве хранения информации или, в качестве альтернативы, эталонную матрицу для осуществления указанного этапа h) и необходимые меченые мужские и женские эталонные ДНК для его осуществления, необязательно связанные с контролем качества указанных эталонных ДНК.
Настоящее изобретение теперь дополнительно будет описано со ссылкой на следующие не ограничивающие примеры.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Следующая методика поясняет этапы гибридизации дифференциально меченых образцов ДНК-зондов человека с матрицей, содержащей ВАС с фрагментами ДНК человека, прикрепленные к стеклянной пластине с покрытием. Надежность методики была доказана в лаборатории авторов изобретения и в клинических центрах.
Матрицы на основе бактериальных искусственных хромосом (ВАС) состоят из тысячи точек, каждая из которых содержит фрагменты ДНК, покрывающие относительно большие хромосомные области (размером приблизительно 150-200 тыс. оснований) и являются представителями одного ВАС-клона. Каждый ДНК-клон ВАС является специфичным к различной хромосомной области и полный набор клонов был картирован на соответствующие области хромосом. Каждый клон представлен по меньшей мере двумя повторами в каждом ряде пластины микроматрицы, чтобы избежать ошибок эксперимента.
В данном случае матрицы на основе ВАС содержат специфический дизайн генома человека со специальным числом ВАС-клонов, каждый из которых представлен в виде отдельных зондов, при этом была доказана его пригодность для основной цели скрининга хромосом клетки: детекции целых хромосом, которые были приобретены (gain) и/или потеряны (loss).
Потерю или приобретение хромосомы выявляют по маркеру, принятому каждой точкой после гибридизации двух различных ДНК, такому как отношение интенсивности флуоресценции для двух контрастных маркеров.
CGH на микроматрицах успешно применяют для детекции анеуплоидов в отдельных клетках после амплификации целого генома (WGA) с использованием одной матрицы. Например, весь подход позволяет проводить исчерпывающий хромосомный анализ за срок, необходимый для скрининга ооцита или эмбриона на стадии дробления и, кроме того, был успешно достигнут генетический анализ клеточной линии.
Микроматрица была разработана для применения с амплифицированной ДНК, тогда как амплификация целого генома (WGA) необходима для образцов с низкими количествами геномных ДНК и методика амплификации (WGA) необходима для получения достаточного количества материала для генетического анализа с помощью ДНК-микроматриц.
Схема исследования - Целью этого исследования было верифицировать и валидировать способ детекции CNV, описанный в настоящем документе, с использованием по меньшей мере двух различных матриц эуплоидных или анеуплоидных образцов, собранных в ходе методик IFV или от культивированных клеточных линий. Для первой матрицы осуществляли ретроспективное исследование, собирая набор образцов из ежедневной практики клинических центров по оплодотворению in vitro (IFV). Эуплоидный или анеуплоидный статус указанных образцов уже был предсказан по оценке хромосом, выполненной на полярных тельцах 1 или 2 или бластомерах с использованием альтернативных способов. Для второй матрицы лимфобластные клеточные линии человека, несущие по меньшей мере одну известную хромосомную аберрацию, выбирали из открытого хранилища для целей данного исследования.
Тестируемые ДНК сначала сравнивали с мужской эталонной ДНК для расчета первого порядка анеуплоидного статуса клетки. Для этого применяли специфический набор алгоритмов выбора (алгоритмы от 1 до 6) и рассчитывали первую CNV хромосом. Интенсивность точек в электронном файле тестируемой ДНК сохраняли для последующего сравнения с данными об изображении обоих эталонных ДНК, эталонной женской ДНК, измеренной на пяти микроматрицых, использованных в этом исследовании, и той же эталонной женской ДНК из контроля качества.
Второе измерение CNV хромосом получали с использованием такого же алгоритма выбора. Затем два измерения CNV хромосом сравнивали со специфическим алгоритмом (алгоритм 7), чтобы рассчитать и иметь общую оценку числа копий тестируемой ДНК.
Каждую из пяти эталонных женских ДНК (от REF1 до REF5), меченных двумя контрастными флуорофорами, гибридизировали на 5 различных микроматрицах. Соответствующий электронный файл сохраняли и сравнивали с сохраненным изображением набора данных тестируемой ДНК.
Контроль качества эталонной женской ДНК представляет собой сохраненные данные об изображении, полученные из CGH-анализа, выполненного на каждой производственной серии микроматриц, и представляющего собой анализ самого высокого качества женской или мужской эталонной ДНК.
Для оценки результатов тестов использовали два параметра, плоидность клетки и конкордантность хромосом. Первое определяет статус эуплоидности или анеуплоидности клеток, тогда как последнее определяет соответствие исследованных хромосом между тестом и контролем с известными CNV хромосом.
При применении эталонной ДНК предполагается, что большинство позиций клонов является эуплоидными и большинство log2 отношений должно колебаться около нуля. ДНК при применении в исследованиях на микроматрицах может вводить систематическую специфичную для матрицы ошибку в измеренные log2 отношения, из-за чего нормальное измерение CNV в ходе теста CGH на матрицах может привести к неправильной интерпретации данных.
Для того, чтобы снизить это явление авторы настоящего изобретения предлагают «общую нормализацию» для корректировки матриц с эталонной ДНК и три этапа контроля качества данных изображения: показатели контроля качества морфологии точек и интенсивности сигналов, контроль качества повторов клона, контроль качества клонов.
Вкратце, в ходе контроля качества эталонную ДНК анализируют на матрице каждой серии в условиях совпадения по полу и несовпадения по полу и оценивают по специфическим параметрам: % включенных клонов, экспериментальное стандартное отклонение, отношение сигнал/шум и способность к CNV для Х- и Y-хромосом. Когда эталонную ДНК анализируют в ходе анализа тестируемого образца, ее сначала сравнивают с соответствующим контролем качества эталона, чтобы оценить ее экспериментальную применимость перед последующими измерениями.
В этом исследовании каждую эталонную ДНК измеряют отдельно и сравнивают с контролем качества эталона и учитывают экспериментальное стандартное отклонение (SD) в качестве параметра оценки.
Получение тестируемой ДНК и эталонной ДНК - Для матрицы проводили амплификацию ДНК цельного генома (WGA) с использованием набора PicoPlex Single Cell WGA Kit (Rubicon Genomics Inc.), поскольку все внутренние валидационные исследования были осуществлены с использованием этого набора. Однако для амплификации ДНК можно использовать любую подходящую систему амплификации, потому что природа амплифицированного продукта не имеет такого критичного значения, как природа неамплифицированного продукта. Тестируемую ДНК и эталонную ДНК (мужскую и женскую) амплифицировали в ходе WGA и в конце разрезали на фрагменты. Оценивали качество амплифицированного продукта и продукт одной из всех десяти реакций амплификаций метили и делали пригодным для CGH на мавтрице.
Мечение амплифицированных тестируемой и эталонной ДНК - Применяли методику мечения со случайными праймерами, которая включает фрагмент Кленова ДНК-полимеразы I E. Coli, чтобы независимо пометить двумя различными маркерами тестируемую и эталонную ДНК с дополнительным конечным разрезанием на фрагменты. Две указанные ДНК можно метить, например, с использованием су3- или су5-модифицированных дидеоксинуклеотидов и смешивать в равных пропорциях для осуществления CGH-теста. Это позволяет достигать в ходе сканирования матрицы двух или одного составного изображения, соответственно связанного с образцом ДНК и контролем, и измерять интенсивность сигнала, связанную с конкурентной гибридизацией двух ДНК, для каждой точки, расположенной на пластине.
Преципитация меченой тестируемой и эталонной ДНК - Указанные меченые ДНК совместно осаждали вместе с блокирующим раствором, чтобы избежать любых ошибок, связанных с перекрестной гибридизацией высокоповторяющихся последовательностей, которые могут помешать расчету CNV. Меченые ДНК впоследствии высушивали, чтобы меченые зонды стали доступными для специфического гибридизационного буфера для переноса.
Гибридизация меченых тестируемой и эталонной ДНК - Высушенный осадок растворяли в подходящем объеме раствора для гибридизации, входящего в набор. ДНК денатурировали, чтобы одиночные цепи стали доступными для процесса гибридизации, и наносили на микроматрицу, обеспечивая возможность контакта с зондами матрицы и возникновения гибридизации. Контролировали температуру системы, гибридизацию в объеме осуществляли в соответствующих кассетах с использованием водяной бани. После гибридизации стеклянные пластины очень тщательно промывали, чтобы удалить любую несвязавшуюся меченую ДНК или любые неспецифические нуклеиновые кислоты или комплексы нуклеиновых кислот более высокого порядка, которые могут помешать измерению амплитуды сигнала.
Выделение интенсивности сигнала - После гибридизации и промывки можно считать информацию с пластинок микроматриц, информацию с микроматриц фиксируют и хранят в виде изображений с использованием двойного лазерного сканера (Innoscan 710A и последующие версии, Innopsys). Считывание микроматрицы имеет специально определенную конфигурацию, что улучшает качество данных. Лазер возбуждает флуоресцентную метку, и фотоумножитель увеличивает интенсивность сигнала на специально определенную величину, получая специфическое значение отношения. Интенсивность исходящего сигнала пропорциональна числу молекул-мишеней, которые связаны в каждой точке на матрице. Сканер имел конфигурацию для создания двух цифровых изображений для каждого канала матрицы, и информацию, полученную от каждой точки для каждого канала, хранили и дополнительно анализировали с применением изображений, полученных при применении программного обеспечения, для выделения интенсивности и расчета для каждой точки.
Анализ данных - Получают первичные данные анализа на микроматрице для гибридизации набора из шести тестируемых ДНК против мужской эталонной ДНК, состоящие из значений интенсивности перечисленных точек на канал, и можно рассчитать отношение интенсивностей между каналами для каждого признака. Отношение интенсивности обоих каналов соответствует относительной распространенности молекулы-мишени на каждой тестируемой ДНК относительно эталонной ДНК. Интерпретация данных подразумевает разработку специально определенных статистических способов, адаптированных к анализу на микроматрицах, и специализированного программного обеспечения для визуализации этих результатов анализа на микроматрице. Логическая последовательность основных этапов описана (Фиг. 6, 7 и 8).
Подробности методики следующие:
Измеряют амплитуды сигнала гибридизации тестируемой ДНК и эталонной ДНК на каждой позиции клона микроматрицы, предварительно обрабатывают и объединяют как log2 отношения. Предполагают, что этот сигнал является пропорциональным log2 отношения изменений числа копий тестируемой ДНК и эталонной ДНК в соответствующей области генома. Если эталонные ДНК выбраны эуплоидными, можно получить информацию об изменении числа копий в тестируемом образце. Первичные данные, полученные от одного CGH-эксперимента на матрице, состоят из нескольких тысяч клон-специфичных log2 отношений.
Реализация алгоритмов согласно настоящему изобретению может выполнять анализ одного и множества матриц и оказывать поддержку связанным статистическим исследованиям. Фиг. 5 дает схематическое представление об аналитических этапах в целом и каждый отдельный этап подробно представлен на Фиг. 2, 3 и 4.
С целью дополнительно повысить чувствительность способа, можно применять алгоритмы контроля качества изображения для расчета и снижения фонового шума. В этом случае после выделения интенсивности изображения осуществляют контроль качества морфологии и интенсивности сигнала точки в ходе трех различных этапов (Фиг. 6), и на Фиг. 7 применяют два разных алгоритма (алгоритм 1 и 2) для расчета и снижения более высокого шума.
Авторы настоящего изобретения также реализовали набор специфических алгоритмов, направленных на определение сегментов соседних клонов с таким же числом копий и для присвоения значения параметра качества каждой хромосоме (алгоритм 3). Полученные сегменты классифицируют на сегменты с приобретениями (gain), потерями (loss) и сегменты с нормальным числом копий (normal). Применяют следующий алгоритм (алгоритм 4) для идентификации пределов и дифференциации внутри категории потери или приобретения сегментов. В этой методике одинаково установлен фиксированный предел по шкале log2 отношения для всех матриц, потому что применение алгоритмов 1, 2 и 3 обеспечивает робастную оценку шума.
Если уровень сегмента выше предела, то это классифицируют как приобретение (gain), сегмент, который ниже отрицательного значения предела, считают потерей (loss), и сегменты на границе предела классифицируют как предупреждающие случаи. На этом этапе алгоритмы 5 и 6 могут идентифицировать соответственно истинные хромосомные аберрации и, в конечном итоге, хромосомы на границе (Фиг. 7).
Анализ второй эталонной ДНК проходит ту же самую обработку изображения: три этапа контроля качества изображения с последующим применением алгоритмов от 1 до 6. Результаты указанных контролен качества выделены на Фиг. 9, где представлено сравнение между SD каждого эксперимента с женской эталонной ДНК с соответствующими подписями данных. Показано, что общее значение SD является гомогенным, и вариация среди стандартных отклонений (SD приблизительно 0,004) является очень низкой, эти данные подтверждают точность приготовления эталонной ДНК и делают эту методику достаточной робастной для интерпретации CGH на матрицах тестируемых ДНК с соответствующей пропорцией аберрантных клонов.
Результаты теста показаны в таблице на Фиг. 8А, где представлены соответствующие приобретения и потери хромосом (gain/loss), проанализированные в сравнении как с мужскими, так и с женскими эталонными ДНК. Профиль амплитуд сигналов тестируемых ДНК сравнивали с контролем качества и с пятью различными гибридизациями на матрицах женской эталонной ДНК. Соответствующие значения SD описаны на Фиг. 10, дополненные их медианной и общими значениями SD (0,037 и 0,014 соответственно). Хотя анализ тестируемой ДНК привел к противоречивым значениям между SD, общая нормализация всех данных делает считывание данных менее подверженным ошибкам и менее затратным по времени, что приводит к успеху общей методики детектирования CNV. Фактически, только в 1 из 5 случаев анализа клеточной линии не смогли осуществить детекцию, несмотря на другие тесты.
Детекцию CNV тестируемой ДНК против мужской эталонной ДНК (ДНК-REF1) (представленной как приобретение/потеря (gain/loss) хромосом) затем сравнивают с детекцией CNV тестируемой ДНК против «контрольного стандарта качества» женской эталонной ДНК или стандартной женской эталонной ДНК (ДНК-REF 2), как представлено в примере на Фиг. 8А (в этом эксперименте для целей стандартизации гибридизация с участием ДНК-REF 2 была повторена на пяти разных матрицах для гибридизации и полученные соответствующие CNV, представленные как приобретение/потеря (gain/loss) хромосом, указаны под заголовками REF(a)-(e); определение осуществляли одновременно на шести тестируемых ДНК различного происхождения, указанных на Фиг. 8А как ТЕСТ 1-ТЕСТ 6). Этот результат получают с использованием той же методики получения изображения, применявшейся на предыдущих этапах, с целью получить два набора данных, один, связанный с тестируемой ДНК, и второй, связанный с ДНК-REF 2, где специфический алгоритм (алгоритм 7) сравнивает и нормализует два набора данных (Фиг. 8). Измерения устойчивых хромосомных аберраций получают посредством объединения предварительных результатов от первого эксперимента (тестируемая ДНК против эталонной ДНК 1) и результатов контроля качества женской эталонной ДНК и каждого из второго CGH-анализа на матрице (тестируемая ДНК против эталонной ДНК 2). На Фиг. 13 показан графический пример, описывающий результат анализа одиночной области, где тестируемый образец сравнивают с мужской эталонной ДНК и в тех же окнах также представлены результаты тестируемого образца в сравнении женской эталонной ДНК.
Соответствующий кариотип, полученный из контрольного анализа, сравнивали с кариотипом ПГС в соответствии с результатами считывания CNV на Фиг. 8В, карта хромосом, разработанная на Фиг. 11, подчеркивает полное соответствие для всех изученных хромосом.
Конкордантность детекции CNV между каждым набором женских эталонных ДНК и контролем качества также рассчитывали и тренд CNV, измеренный в отношении значений «gain», «loss», «normal» и пограничных значений показан на Фиг. 12. Результаты четко показывают 100% конкордантность от тестируемой ДНК 1 до тестируемой ДНК 5, тогда как женский REF1 не смог детектировать правильную Х-хромосому только в одном случае (клеточная линия), a REF3 показал в том же случае определение Х-хромосомы на пограничном уровне. Несмотря на то, что это явление обычно бывает вызван природой лимфобластных клеточных линий, общая конкордантность составила приблизительно 60%.
В заключении, конкордантность общего экспериментального набора из тридцати экспериментов составила 93% и конкордантность в 100% была в отношении образцов IVF.
ССЫЛОЧНЫЕ МАТЕРИАЛЫ
(1) Joo Wook Ahn et al. Array CGH as a first line diagnostic test in place of karyotyping for postnatal referrals - results from four years' clinical application for over 8,700 patients. Molecular Cytogenetics 2013, 6:16
(2) Hassold T. et al. The origin of human aneuploidy: where we have been, where we are going Human Mol Genetics. 2007 Aug; 16(2):R203-R20
(3) Fragouli E. et al. Aneuploidy screening for embryo selection. Semin Reprod Med. 2012 Aug; 30(4):289-301
(4) Yimer, N. and Rosnina. Chromosomal Anomalies and Infertility in Farm Animals: A Review Y. (2014) Pertanika J. Trop. Agric. Sci. 37 (1): 1-18
(5) Blazak, W.F., & Eldridge, F.E. (1977). A Robertsonian translocation and its effect upon fertility in Brown Swiss cattle. Journal of Dairy Science, 60(7), 1133-1142.
(6) S.M. Schmutz et al. Chromosomal aneuploidy associated with spontaneous abortions and neonatal losses in cattle. J Vet Diagn Invest 8:91-95 (1996)
(7) Popescu, C.P., Bonneau, M., Tixier, M., Bahri, I., & Boscher, J. (1984). Reciprocal translocations in pigs. The Journal of Heredity, 75, 448-452.
(8) Lear TL, Bailey E. Equine clinical cytogenetics: the past and future. Cytogenet Genome Res. 2008; 120(1-2):42-9
9 Hainan CR. Equine cytogenetics: role in equine veterinary practice. Equine Vet J. 1985 May; 17(3): 173-7
(10) Matthew Breen, PhD C. Biol M.I. Biol. Update on Genomics in Veterinary Oncology. Top Companion Anim Med. 2009 August; 24(3): 113-121. doi:10.1053/j.tcam.2009.03.002.
(11) Matthew Breen, PhD C. Biol M.I. Biol, Update on Genomics in Veterinary Oncology. Top Companion Anim Med. 2009 August; 24(3): 113-121.
(12) Brezina PR. et al. Pre implantation Genetic Screening: A Practical Guide. Clin Med Insights Reprod Health. 2013 Feb 27; 7:37-42.
(13) Robert G. EDWARDS. An astonishing journey into reproductive genetics since the 1950's. Reprod. Nutr. Dev. 45 (2005) 299-306
(14) Norbert Gleicher et al. Pre implantation genetic screening (PGS) still in search of a clinical application: a systematic review. Reproductive Biology and Endocrinology 2014, 12:22
(15) Mastenbroek S, Twisk M, van Echten-Arends J, Sikkema-Raddatz B, Korevaar JC, Verhoeve HR, Vogel NE, Arts EG, de Vries JW, Bossuyt PM, Buys CH, Heineman MJ, Repping S, van der Veen F: In vitro fertilization with pre implantation genetic screening. N Engl J Med 2007, 357:9-17.
(16) Anderson RA, Pickering S: The current status of preimplantation genetic screening: British fertility society policy and practice guidelines. Hum Fertil (Camb) 2008, 11:71-75.
(17) Geraedts J et al. Polar body array CGH for prediction of the status of the corresponding oocyte. Part I: clinical results.
(18) Magli MC. et al. Polar body array CGH for prediction of the status of the corresponding oocyte. Part II: Technical aspects. Hum Reprod. 2011 Aug; 16(11):3181-85
(19) Kuliev A, Verlinsky Y: Place of preimplantation genetic diagnosis in genetic practice. Am J Med Genet A 2005, 134A: 105-110.
(20) Hardarson T, Hanson C, Ludin K, Hillensio T, Nilsson L, Stevic J, Reismer E, Borg K, Wikland M, Bergh C: Preimplantation genetic screening in women of advanced maternal age causes a decrease in clinical pregnancy rate: a randomized controlled trial. Hum Reprod 2008, 23:2806-2812.
(21) Mausumi Das, M.D., and Hananel E.G. Holzer, M.D. Recurrent implantation failure: gamete and embryo factors. Fertility and Sterility Vol. 97, No. 5, May 2012.
22 Raziel A, Friedler S, Schachter M, Kasterstein E, Strassburger D, Ron-El R. Increased frequency of female partner chromosomal abnormalities in patients with high-order implantation failure after in vitro fertilization. Fertil Steril 2002; 78:515-9.
(23) Ginsburg ES, Baker VL, Racowsky C, Wantman E, Goldfarb J, Stern JE: Use of preimplantation genetic diagnosis and preimplantation genetic screening in the United States: a Society for Assisted Reproductive Technology Writing Group paper. Fertil Steril 2011, 96:865-868.
(24) Bassem A. Bejjani and Lisa G. Shaffer. Application of Array-Based Comparative Genomic Hybridization to Clinical Diagnostics. Journal of Molecular Diagnostics, Vol. 8, No. 5, November 2006
(25) Stephanie N. Dorman et al. Expanding probe repertoire and improving reproducibility in human genomic hybridization. Nucleic Acids Research, 2013, Vol. 41, No. 7 e81 doi:10.1093/nar/gkt048
(26) Pinkel, D., Landegent, J., Collins, C., Fuscoe, J., Segraves, R., Lucas, J. and Gray, J. (1988) Fluorescence in situ hybridization with human chromosome specific libraries: detection of trisomy 21 and translocations of chromosome 4. Proc. Natl Acad. Sci. USA, 85, 9138-9142.
(27) Andy W Pang. Towards a comprehensive structural variation map of an individual human genome. Genome Biology 2010, 11:R52
(28) Malcolm A. Ferguson-Smith, MBChB, FRCPath, FRS. Cytogenetics and the evolution of medical genetics. Genet Med 2008:10(8):553-559.
(29) Conrad, D.F., Pinto, D., Redon, R., Feuk, L., Gokcumen, O., Zhang,Y., Aerts,J., Andrews, T.D., Bames, C., Campbell, P. et al. (2010) Origins and functional impact of copy number variation in the human genome. Nature, 464, 704-712.
(30) Wells D. et al. Use of comprehensive chromosomal screening for embryo assessment: microarrays and CGH. Mol Hum Reprod. 2008 Dec; 14(12):703-10
(31) Vanneste E. et al. New array approaches to explore single cells genomes. Front Genet. 2012 Mar 27; 3:44.
(32) Pinkel, D., Segraves, R., Sudar, D., Clark, S., Poole, I., Kowbel, D., Collins, C., Kuo, W.L., Chen, C., Zhai, Y. et al. (1998) High resolution analysis of DNA copy number variation using comparative genomic hybridization to microarrays. Nat. Genet., 20, 207-211.
(33) Gray, J.W. and Pinkel, D. (1992) Molecular cytogenetics in human cancer diagnosis. Cancer, 69, 1536-1542
(34) Shinawi, M. and Cheung, S.W. (2008) The array CGH and its clinical applications. Drug Discov. Today, 13, 760-770
(35) Vikas Bansal. A statistical method for the detection of variants from next-generation resequencing of DNA pools. Bioinformatics. 2010 Jun 15; 26(12): i318-i324.
(36) Janus S Jakobsen. Amplification of pico-scale DNA mediated by bacterial carrier DNA for small-cell-number transcription factor ChIP-seq. BMC Genomics (2015) 16:46.
(37) Svetlana A, et al. Microarray-Based Comparative Genomic Hybridization Using Sex-Matched Reference DNA Provides Greater Sensitivity for Detection of Sex Chromosome Imbalances than Array-Comparative Genomic Hybridization with Sex-Mismatched Reference DNA. Volume 11, Issue 3, May 2009, Pages 226-237
(38) Tineke E. Buffart, Danielle Israeli, Marianne Tijssen, Sjoerd J. Vosse, Alan Mrsic, Gerrit A. Meijer and Bauke Ylstra. Across array comparative genomic hybridization: A strategy to reduce reference channel hybridizations. Volume 47, Issue 11, pages 994-1004, November 2008.
(39) David Greenstein. Control of oocyte meiotic maturation and fertilization. (December 28, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.53.1
(40) G.L. Harton et al. ESHRE PGD Consortium/Embryology Special Interest Group-best practice guidelines for polar body and embryo biopsy for preimplantation genetic diagnosis/screening (PGD/PGS). Human Reproduction, Vol. 0, No. 0 pp. 1-6, 2010
(41) Priscila Ramos-Ibeas. An Efficient System to Establish Biopsy-Derived Trophoblastic Cell Lines from Bovine Embryos. BIOLOGY OF REPRODUCTION (2014) 91(1):15, 1-10.
(42) Pinto, D., Darvishi, K., Shi, X., Rajan, D., Rigler. D., Fitzgerald, T., Lionel, A.C., Thiruvahindrapuram, B., Macdonald, J.R., Mills, R. et al. (2011) Comprehensive assessment of array-based platforms and calling algorithms for detection of copy number variants. Nat. Biotechnol., 29, 512-520.
(43) Dellinger, A.E., Saw, S.-M., Goh, L.K., Seielstad, M., Young, T.L. and Li, Y.-J. (2010) Comparative analyses of seven algorithms for copy number variant identification from single nucleotide polymorphism arrays. Nucleic Acids Res., 38, e105.
(44) Lai, W.R., Johnson, M.D., Kucherlapati, R. and Park, P.J. (2005) Comparative analysis of algorithms for identifying amplifications and deletions in array CGH data. Bioinformatics, 21, 3763-3770
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОВЕДЕНИЯ ПРЕНАТАЛЬНОЙ И ПОСТНАТАЛЬНОЙ ГЕНЕТИЧЕСКОЙ ДИАГНОСТИКИ ЧИСЛОВЫХ ХРОМОСОМНЫХ АНОМАЛИЙ У ЭМБРИОНОВ ЧЕЛОВЕКА И ДНК-МИКРОЧИП НА ОСНОВЕ ПОЛНОХРОМНЫХ ДНК-ЗОНДОВ | 2019 |
|
RU2725488C1 |
| ОПРЕДЕЛЕНИЕ ХРОМОСОМНЫХ АНОМАЛИЙ В КЛЕТКАХ, ПОЛУЧЕННЫХ ИЗ ФОЛЛИКУЛЯРНОЙ ЖИДКОСТИ | 2005 |
|
RU2395584C2 |
| СПОСОБЫ НЕИНВАЗИВНОГО ПРЕНАТАЛЬНОГО УСТАНОВЛЕНИЯ ПЛОИДНОСТИ | 2011 |
|
RU2671980C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ВЫСОКОМУЛЬТИПЛЕКСНОЙ ПЦР | 2012 |
|
RU2650790C2 |
| ВИРТУАЛЬНЫЕ НАБОРЫ ФРАГМЕНТОВ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2004 |
|
RU2390561C2 |
| СПОСОБ МАРКИРОВАНИЯ 7-Й ХРОМОСОМЫ ЧЕЛОВЕКА В МЕТАФАЗНЫХ И ИНТЕРФАЗНЫХ КЛЕТКАХ | 2009 |
|
RU2425890C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК рYA11-39, ПРЕДНАЗНАЧЕННАЯ ДЛЯ МАРКИРОВАНИЯ 4-Й И 9-Й ХРОМОСОМ ЧЕЛОВЕКА В МЕТАФАЗНЫХ И ИНТЕРФАЗНЫХ КЛЕТКАХ | 2006 |
|
RU2325441C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК АЛЬФА R-12, ПРЕДНАЗНАЧЕННАЯ ДЛЯ МАРКИРОВАНИЯ 6-Й ХРОМОСОМЫ ЧЕЛОВЕКА В МЕТАФАЗНЫХ И ИНТЕРФАЗНЫХ КЛЕТКАХ | 2004 |
|
RU2265060C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PYAI 960, ПРЕДНАЗНАЧЕННАЯ ДЛЯ МАРКИРОВАНИЯ 18-Й ХРОМОСОМЫ ЧЕЛОВЕКА | 2000 |
|
RU2161199C1 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ГАМЕТ У ПТИЦ | 2001 |
|
RU2294099C2 |




Изобретение относится к области биотехнологии. Предложен способ сравнительной геномной гибридизации (CGH) на одной матрице для оценки наличия и количества вариаций числа копий (CNV) в тестируемой ДНК, причём пол донора указанной тестируемой ДНК не известен, а также соответствующее применение и набор. Изобретение может быть использовано для улучшения вспомогательных репродуктивных технологий. 3 н. и 13 з.п. ф-лы, 13 ил.
1. Способ сравнительной геномной гибридизации (CGH) на одной матрице для оценки наличия и количества вариаций числа копий (CNV) в тестируемой ДНК, причём пол донора указанной тестируемой ДНК не известен, при этом указанный способ включает следующие этапы:
a) мечение первым маркером указанной тестируемой ДНК, что приводит к получению меченой тестируемой ДНК;
b) мечение вторым маркером, отличным от первого, мужской (или женской) эталонной ДНК, что приводит к получению меченой мужской (или женской) эталонной ДНК;
c) гибридизация ДНК, полученных на этапах a) и b) с тестовой матрицей для гибридизации, что приводит к получению объединенной картины распределения интенсивностей сигналов от обоих маркеров, происходящих от соответствующих гибридизованных молекул ДНК;
d) на основе объединенной картины распределения, полученной на этапе c), определение интенсивности отдельных сигналов, создаваемых указанными первым и вторым маркерами;
e) обеспечение первой оценки CNV в указанной тестируемой ДНК путем сравнения друг с другом интенсивностей двух сигналов, полученных на этапе d);
f) мечение указанным первым маркером эталонной ДНК, противоположной по полу той, что применяли на этапе b), что приводит к получению эталонной ДНК, меченной указанным первым маркером;
g) мечение указанным вторым маркером, отличным от первого, указанной эталонной ДНК, противоположной по полу той, что применяли на этапе b), что приводит к получению эталонной ДНК, меченной указанным вторым маркером;
h) гибридизация обеих ДНК, полученных на этапах f) и g), с эталонной матрицей для гибридизации, что приводит к получению объединенной картины распределения интенсивностей сигналов от обоих маркеров, происходящих от соответствующих гибридизованных молекул ДНК;
i) на основе объединенной картины распределения, полученной на этапе h), определение интенсивности отдельных сигналов, создаваемых указанными первым и вторым маркерами;
j) обеспечение второй оценки CNV в указанной тестируемой ДНК путем сравнения друг с другом:
- интенсивности сигнала указанной тестируемой ДНК, меченной указанным первым маркером, как определено на этапе d),
- с интенсивностью сигнала указанной эталонной ДНК, меченной указанным вторым маркером, как определено на этапе i);
k) объединение значений CNV, полученных на этапах e) и j), с получением конечного результата определения CNV, получая значение CNV, относящееся к целой тестируемой ДНК, подвергнутой анализу,
при этом в качестве альтернативы при осуществлении указанных этапов f)-i) интенсивности сигналов, упоминаемые на этапе i), можно получить с носителя для хранения данных, на который указанные значения были ранее записаны,
и при этом указанную мужскую эталонную ДНК получена от по меньшей мере одного мужчины-донора и/или указанную женскую эталонную ДНК получена от по меньшей мере одной женщины-донора.
2. Способ по п.1, отличающийся тем, что указанная тестируемая ДНК и/или эталонная ДНК состоит из одной молекулы ДНК.
3. Способ по п.1, отличающийся тем, что указанная тестируемая ДНК и/или эталонная ДНК состоит из более чем одной молекулы ДНК.
4. Способ по пп. 1-3, отличающийся тем, что указанная тестируемая ДНК получена из одной клетки или смеси клеток одного и того же индивидуума.
5. Способ по пп. 1-4, отличающийся тем, что указанные эталонные ДНК представляют собой смесь ДНК с по меньшей мере одним изменением числа копий в по меньшей мере одной предварительно определенной области генома с последовательным разведением.
6. Способ по пп. 1-5, отличающийся тем, что на этапе j) указанная интенсивность сигнала указанной тестируемой ДНК, меченной указанным первым маркером, определенным на этапе d), представляет собой интенсивность, получаемую от генотипов, демонстрирующих приобретение Х-хромосомы или потерю Y-хромосомы.
7. Способ по пп. 1-6, отличающийся тем, что на этапах e) и j) оценку CNV осуществляют с помощью способов с применением программного обеспечения, включающих количественное определение интенсивности сигнала, нормализацию данных, статистический анализ, сокращение систематической ошибки, расчет связанных с хромосомами интенсивностей сигнала и создание связанных с хромосомами ДНК-сегментов.
8. Способ по пп. 1-7, отличающийся тем, что указанные амплифицированные эталонные ДНК подвергают второй амплификации с получением соответствующих меченых ДНК и указанную амплифицированную тестируемую ДНК подвергают такой же второй амплификации с получением соответствующей меченой ДНК.
9. Способ по пп. 1-8, отличающийся тем, что указанные эталонные ДНК представляют собой смесь ДНК различных генотипов в различных концентрациях.
10. Способ по пп. 1-9, отличающийся тем, что указанную тестируемую ДНК получают из одной клетки или биоптата человека или животного.
11. Способ по пп. 1-10, отличающийся тем, что оценка указанных вариаций числа копий позволяет выбрать оптимальный ооцит, сперматозоид или эмбрион, подходящий по правильному хромосомному статусу.
12. Способ по пп. 1-11, дополнительно включающий оплодотворение ооцитов и перенос эмбриона в рамках программ по оплодотворению in vitro (IVF).
13. Способ по пп. 1-12, отличающийся тем, что интенсивности сигналов, упоминаемые на этапе i), не содержат систематических ошибок и неспецифических сигналов, при этом указанные интенсивности получены посредством следующей методики:
(I) указанные этапы f), g), h), i) проводят более одного раза, гибридизируя соответствующие эталонные ДНК на микроматрицах различных производственных серий, и
(II) полученные результаты обрабатывают с помощью программного обеспечения с целью получения указанных интенсивностей сигналов, не содержащих системную ошибку и неспецифические сигналы.
14. Способ по пп. 1-13, включающий осуществление «контроля качества» интенсивностей сигналов, упоминаемых на этапе i), путем сравнения их с соответствующими интенсивностями сигналов, не содержащими ошибку и неспецифические сигналы, полученными на этапах (I) и (II) согласно п. 13.
15. Продукт для осуществления способа по пп. 1-14 для оценки наличия и количества вариаций числа копий в тестируемой ДНК, причём пол донора указанной тестируемой ДНК не известен, указанный продукт представляет собой:
- по меньшей мере одну матрицу для гибридизации для осуществления указанных этапов a)-e) с по меньшей мере одной тестируемой ДНК, и
- одну матрицу для осуществления указанного этапа h),
причём указанные матрицы нанесены слоем на подходящий носитель.
16. Набор для осуществления способа по пп. 1-14 для оценки наличия и количества вариаций числа копий в тестируемой ДНК, причём пол донора указанной тестируемой ДНК не известен, содержащий:
- по меньшей мере одну матрицу для гибридизации для осуществления указанных этапов a)-e) с использованием по меньшей мере одной тестируемой ДНК,
- эталонные интенсивности сигналов согласно этапу i), записанные на подходящем носителе для хранения информации, или в качестве альтернативы эталонную матрицу для осуществления указанного этапа h) и необходимые эталонные меченые мужские и женские ДНК для осуществления указанного этапа, необязательно связанные с контролем качества указанных эталонных ДНК.
| A | |||
| YATSENKO et al | |||
| Microarray based comparative genomic hybridization using sex-matched reference DNA provides greater sensitivity for detection of sex chromosome imbalances than array-comparative genomic hybridization with sex-mismatched reference DNA | |||
| J Mol Diagn | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| WO | |||

