Ссылка на связанные заявки
[0001] Согласно настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой США с серийным номером 62/438516 (поданной 23 декабря 2016 года; в стадии рассмотрения), указанная заявка включена в данный документ посредством ссылки в полном объеме.
Ссылка на перечень последовательностей
[0002] Настоящая заявка включает в себя одну или несколько перечней последовательностей, соответственно п. 1.821 и далее раздела 37 Свода федеральных нормативных актов США Правил, которые раскрыты в машиночитаемых носителях (название файла: 1301.0147PCT_Sequence_Listing_ST25.txt, созданного 29 ноября 2017 года и имеющего размер 175962 байтов), указанный файл включен в данный документ посредством ссылки в полном объеме.
Область техники, к которой относится настоящее изобретение
[0003] Настоящее изобретение направлено на молекулы, такие как моноспецифические антитела и биспецифические, триспецифические или мультиспецифические молекулы, в том числе диатела, BiTE и антитела, которые способны специфически связываться с «белком 9, содержащим домен дезинтегрина и металлопротеиназы» («ADAM9»). В настоящем изобретении, в частности, рассматриваются такие связывающие молекулы, которые способны проявлять высокую аффинность связывания с ADAM9 человека и не относящимся к человеку ADAM9. Настоящее изобретении дополнительно, в частности, связано с такими молекулами, которые, таким образом, являются перекрестно-реактивными в отношении ADAM9 человека и ADAM9 не относящегося к человеку примата (например, яванского макака). Настоящее изобретение, помимо этого, относится ко всем таким молекулам, связывающим ADAM9, которые содержат вариабельный домен легкой цепи (VL) и/или вариабельный домен тяжелой цепи (VH), которые были гуманизированы и/или деиммунизированы таким образом, чтобы проявлять ослабленную иммуногенность при введении такой молекулы, связывающей ADAM9, субъекту-реципиенту. Настоящее изобретение также направлено на фармацевтические композиции, которые содержат любую из таких молекул, связывающих ADAM9, а также способы, предусматривающие применение любой из таких молекул, связывающих ADAM9, в лечении рака и других заболеваний и патологических состояний.
Предшествующий уровень техники настоящего изобретения
[0004] ADAM представляет собой семейство белков, участвующих в различных физиологических и патологических процессах (Amendola, R.S. et al. (2015) “ADAM9 Disintegrin Domain Activates Human Neutrophils Through An Autocrine Circuit Involving Integrins And CXCR2,” J. Leukocyte Biol. 97(5):951-962; Edwars, D.R. et al. (2008) “The ADAM Metalloproteases,” Molec. Aspects Med. 29:258-289). По меньшей мере 40 генов-членов семейства были идентифицированы и считается, что по меньшей мере 21 из таких членов является функциональным у человека (Li, J. et al. (2016) “Overexpression of ADAM9 Promotes Colon Cancer Cells Invasion,” J. Invest. Surg. 26(3):127-133; Duffy, M.J. et al. (2011) “The ADAMs Family Of Proteases: New Biomarkers And Therapeutic Targets For Cancer?,” Clin. Proteomics 8:9:1-13; см. также публикацию патента США № 2013/0045244).
[0005] Члены семейства ADAM имеют высококонсервативную структуру с 8 доменами, среди которых присутствуют домен металлопротеиназы и домен, связывающий интегрин (дезинтегрин) (Duffy, M.J. et al. (2009) “The Role Of ADAMs In Disease Pathophysiology,” Clin. Chim. Acta 403:31-36). Домен металлопротеиназы ADAM выступает в качестве шеддазы и, как было описано, модулирует ряд биологических процессов в результате расщепления трансмембранных белков, которые затем выступают в качестве растворимых лигандов и регулируют передачу сигналов в клетке (Amendola, R.S. et al. (2015) “ADAM9 Disintegrin Domain Activates Human Neutrophils Through An Autocrine Circuit Involving Integrins And CXCR2,” J. Leukocyte Biol. 97(5):951-962; Ito, N. et al. (2004) “ADAMs, A Disintegrin And Metalloproteinases, Mediate Shedding Of Oxytocinase,” Biochem. Biophys. Res. Commun. 314 (2004) 1008-1013).
[0006] ADAM9 представляет собой члена семейство молекул ADAM. Он синтезируется в виде неактивной формы, которая протеолитически расщепляется с образованием активного фермента. Процессинг в вышерасположенной части является особенно важным для активации профермента. ADAM9 экспрессируется в фибробластах (Zigrino, P. et al. (2011) “The Disintegrin-Like And Cysteine-Rich Domains Of ADAM-9 Mediate Interactions Between Melanoma Cells And Fibroblasts,” J. Biol. Chem. 286:6801-6807), активированных клетках гладкой мускулатуры сосудов (Sun, C. et al. (2010) “ADAM15 Regulates Endothelial Permeability And Neutrophil Migration Via Src/ERK1/2 Signalling,” Cardiovasc. Res. 87:348-355), моноцитах (Namba, K. et al. (2001) “Involvement Of ADAM9 In Multinucleated Giant Cell Formation Of Blood Monocytes,” Cell. Immunol. 213:104-113), активированных макрофагах (Oksala, N. et al. (2009) “ADAM-9, ADAM-15, And ADAM-17 Are Upregulated In Macrophages In Advanced Human Atherosclerotic Plaques In Aorta And Carotid And Femoral Arteries - Tampere Vascular Study,” Ann. Med. 41:279-290).
[0007] Активность металлопротеиназы ADAM9 проявляется в разрушении компонентов матрикса, тем самым, способствуя, миграции опухолевых клеток (Amendola, R.S. et al. (2015) “ADAM9 Disintegrin Domain Activates Human Neutrophils Through An Autocrine Circuit Involving Integrins And CXCR2,” J. Leukocyte Biol. 97(5):951-962). Его домен дезинтегрина, который является высокогомологичным по отношению ко многим дезинтегринам змеиного яда, способствует взаимодействию между ADAM9 и интегринами, а также позволяет ADAM9 модулировать, положительно или отрицательно, явления, связанные с клеточной адгезией (Zigrino, P. et al. (2011) “The Disintegrin-Like And Cysteine-Rich Domains Of ADAM-9 Mediate Interactions Between Melanoma Cells And Fibroblasts,” J. Biol. Chem. 286:6801-6807; Karadag, A. et al. (2006) “ADAM-9 (MDC-9/Meltringamma), A Member Of The A Disintegrin And Metalloproteinase Family, Regulates Myeloma-Cell-Induced Interleukin-6 Production In Osteoblasts By Direct Interaction With The Alpha(v)Beta5 Integrin,” Blood 107:3271-3278; Cominetti, M.R. et al. (2009) “Inhibition Of Platelets And Tumor Cell Adhesion By The Disintegrin Domain Of Human ADAM9 To Collagen I Under Dynamic Flow Conditions,” Biochimie 91:1045-1052). Было показано, что домен дезинтегрина ADAM9 взаимодействует с интегринами α6β1, α6β4, αvβ5 и α9β1.
[0008] Было обнаружено, что экспрессия ADAM9 связано с заболеваниями, особенно раком. Было обнаружено, что ADAM9 расщепляет и способствует высвобождению ряда молекул с важными функциями в онкогенезе и ангиогенезе, таких как TEK, KDR, EPHB4, CD40, VCAM1 и CDH5. ADAM9 экспрессируется многими типами опухолевых клеток, в том числе опухолевых клеток видов рака молочной железы, видов рака толстой кишки, видов рака желудка, глиом, видов рака печени, видов немелкоклеточного рака легких, меланом, миелом, видов рака поджелудочной железы и видов рака предстательной железы (Yoshimasu, T. et al. (2004) “Overexpression Of ADAM9 In Non-Small Cell Lung Cancer Correlates With Brain Metastasis,” Cancer Res. 64:4190-4196; Peduto, L. et al. (2005) “Critical Function For ADAM9 In Mouse Prostate Cancer,” Cancer Res. 65:9312-9319; Zigrino, P. et al. (2005) “ADAM-9 Expression And Regulation In Human Skin Melanoma And Melanoma Cell Lines,” Int. J. Cancer 116:853-859; Fritzsche, F.R. et al. (2008) “ADAM9 Is Highly Expressed In Renal Cell Cancer And Is Associated With Tumour Progression,” BMC Cancer 8:179:1-9; Fry, J.L. et al. (2010) “Secreted And Membrane-Bound Isoforms Of Protease ADAM9 Have Opposing Effects On Breast Cancer Cell Migration,” Cancer Res. 70, 8187-8198; Chang, L. et al. (2016) “Combined Rnai Targeting Human Stat3 And ADAM9 As Gene Therapy For Non-Small Cell Lung Cancer,” Oncology Letters 11:1242-1250; Fan, X. et al. (2016) “ADAM9 Expression Is Associate with Glioma Tumor Grade and Histological Type, and Acts as a Prognostic Factor in Lower-Grade Gliomas,” Int. J. Mol. Sci. 17:1276:1-11).
[0009] Было обнаружено, что значительно повышенная экспрессия ADAM9 коррелирует со злокачественным характером опухолей и метастатическим потенциалом (Amendola, R.S. et al. (2015) “ADAM9 Disintegrin Domain Activates Human Neutrophils Through An Autocrine Circuit Involving Integrins And CXCR2,” J. Leukocyte Biol. 97(5):951-962; Fan, X. et al. (2016) “ADAM9 Expression Is Associate with Glioma Tumor Grade and Histological Type, and Acts as a Prognostic Factor in Lower-Grade Gliomas,” Int. J. Mol. Sci. 17:1276:1-11; Li, J. et al. (2016) “Overexpression of ADAM9 Promotes Colon Cancer Cells Invasion,” J. Invest. Surg. 26(3):127-133). Помимо этого, по-видимому, ADAM9 и его секретируемая растворимая изоформа играют важнейшую роль в диссеминировании раковых клеток (Amendola, R.S. et al. (2015) “ADAM9 Disintegrin Domain Activates Human Neutrophils Through An Autocrine Circuit Involving Integrins And CXCR2,” J. Leukocyte Biol. 97(5):951-962; Fry, J.L. et al. (2010) “Secreted And Membrane-Bound Isoforms Of Protease ADAM9 Have Opposing Effects On Breast Cancer Cell Migration,” Cancer Res. 70, 8187-8198; Mazzocca, A. (2005) “A Secreted Form Of ADAM9 Promotes Carcinoma Invasion Through Tumor-Stromal Interactions,” Cancer Res. 65:4728-4738; см. также патенты США №№ 9150656; 7585634; 7829277; 8101361; и 8445198, а также публикацию патента США № 2009/0023149).
[0010] Таким образом, в ряде исследований ADAM9 идентифицировали в качестве потенциальной мишени противораковой терапии (Peduto, L. (2009) “ADAM9 As A Potential Target Molecule In Cancer,” Curr. Pharm. Des. 15:2282-2287; Duffy, M.J. et al. (2009) “Role Of ADAMs In Cancer Formation And Progression,” Clin. Cancer Res. 15:1140-1144; Duffy, M.J. et al. (2011) “The ADAMs Family Of Proteases: New Biomarkers And Therapeutic Targets For Cancer?” Clin. Proteomics 8:9:1-13; Josson, S. et al. (2011) “Inhibition of ADAM9 Expression Induces Epithelial Phenotypic Alterations and Sensitizes Human Prostate Cancer Cells to Radiation and Chemotherapy,” Prostate 71(3):232-240; см. также публикации патента США №№ 2016/0138113, 2016/0068909, 2016/0024582, 2015/0368352, 2015/0337356, 2015/0337048, 2015/0010575, 2014/0342946, 2012/0077694, 2011/0151536, 2011/0129450, 2010/0291063, 2010/0233079, 2010/0112713, 2009/0285840, 2009/0203051, 2004/0092466, 2003/0091568 и 2002/0068062, а также публикации согласно PCT №№ WO 2016/077505, WO 2014/205293, WO 2014/186364, WO 2014/124326, WO 2014/108480, WO 2013/119960, WO 2013/098797, WO 2013/049704 и WO 2011/100362). Помимо этого, также было обнаружено, что экспрессия ADAM9 связана с заболеванием и воспалением легких (см., например, публикации патентов США №№ 2016/0068909; 2012/0149595; 2009/0233300; 2006/0270618; и 2009/0142301). Антитела, которые связываются с ADAM9, являются коммерчески доступными от Abcam, Thermofisher, Sigma-Aldrich и других компаний.
[0011] В то же время, несмотря на все предыдущие успехи, сохраняется потребность в высокоаффинных молекулах, связывающих ADAM9, которые характеризуются минимальным связыванием с нормальными тканями и способны связываться с ADAM9 человека и не относящимся к человеку ADAM9 с аналогичной высокой аффинностью. Настоящее изобретение относится к этой потребности и потребности в улучшенных терапевтических препаратах для лечения рака.
Краткое раскрытие настоящего изобретения
[0012] Настоящее изобретение направлено на молекулы, такие как моноспецифические антитела и биспецифические, триспецифические или мультиспецифические молекулы, в том числе диатела, BiTE и антитела, которые способны специфически связываться с «белком 9, содержащим домен дезинтегрина и металлопротеиназы» («ADAM9»). В настоящем изобретении, в частности, рассматриваются такие связывающие молекулы, которые способны проявлять высокую аффинность связывания с ADAM9 человека и не относящимся к человеку ADAM9. Настоящее изобретении дополнительно, в частности, связано с такими молекулами, которые, таким образом, являются перекрестно-реактивными в отношении ADAM9 человека и ADAM9 не относящегося к человеку примата (например, яванского макака). Настоящее изобретение, кроме того, относится ко всем таким молекулам, связывающим ADAM9, которые содержат вариабельный домен легкой цепи (VL) и/или вариабельный домен тяжелой цепи (VH), которые были гуманизированы и/или деиммунизированы таким образом, чтобы проявлять ослабленную иммуногенность при введении такой молекулы, связывающей ADAM9, субъекту-реципиенту. Настоящее изобретение также направлено на фармацевтические композиции, которые содержат любую из таких молекул, связывающих ADAM9, а также способы, предусматривающие применение любой из таких молекул, связывающих ADAM9, в лечении рака и других заболеваний и патологических состояний.
[0013] Более конкретно, в настоящем изобретении предусмотрена молекула, связывающая ADAM9, которая содержит домен, связывающий ADAM9, где такой домен, связывающий ADAM9, содержит вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH), при этом такой вариабельный домен тяжелой цепи содержит домен CDRH1, домен CDRH2 и домен CDRH3, и такой вариабельный домен легкой цепи содержит домен CDRL1, домен CDRL2 и домен CDRL3, где:
(A) такой домен CDRH1, домен CDRH2 и домен CDRH3 имеют аминокислотную последовательность домена CDRH1, домена CDRH2 и домена CDRH3 вариабельного домена тяжелой цепи (VH) оптимизированного варианта MAB-A; и такой домен CDRL1, домен CDRL2 и домен CDRL3 имеют аминокислотную последовательность домена CDRL1, домена CDRL2 и домена CDRL3 вариабельного домена легкой цепи (VL) MAB-A; или
(B) такой домен CDRH1, домен CDRH2 и домен CDRH3 имеют аминокислотную последовательность домена CDRH1, домена CDRH2 и домена CDRH3 вариабельного домена тяжелой цепи (VH) MAB-A; и такой домен CDRL1, домен CDRL2 и домен CDRL3 имеют аминокислотную последовательность домена CDRL1, домена CDRL2 и домена CDRL3 вариабельного домена легкой цепи (VL) оптимизированного варианта MAB-A; или
(С) такой домен CDRH1, домен CDRH2 и домен CDRH3 имеют аминокислотную последовательность домена CDRH1, домена CDRH2 и домена CDRH3 вариабельного домена тяжелой цепи (VH) оптимизированного варианта MAB-A; и такой домен CDRL1, домен CDRL2 и домен CDRL3 имеют аминокислотную последовательность домена CDRL1, домена CDRL2 и домена CDRL3 вариабельного домена легкой цепи (VL) оптимизированного варианта MAB-A.
[0014] В настоящем изобретении, в частности, рассматривает вариант осуществления таких молекул, связывающих ADAM9, при этом такие домены, связывающие ADAM9, имеют:
(A) (1) домен CDRH1, домен CDRH2 и домен CDRH3 вариабельного домена тяжелой цепи (VH) MAB-A; и
(2) FR1, FR2, FR3 и FR4 домена VH гуманизированного варианта MAB-A; или
(B) (1) домен CDRL1, домен CDRL2 и домен CDRL3 вариабельного домена легкой цепи (VL) MAB-A; и
(2) FR1, FR2, FR3 и FR4 домена VL гуманизированного варианта MAB-A; или
(С) (1) домен CDRH1, домен CDRH2 и домен CDRH3 вариабельного домена тяжелой цепи (VH) оптимизированного варианта MAB-A; и
(2) FR1, FR2, FR3 и FR4 домена VH гуманизированного варианта MAB-A; или
(D) (1) домен CDRL1, домен CDRL2 и домен CDRL3 вариабельного домена легкой цепи (VL) оптимизированного варианта MAB-A; и
(2) FR1, FR2, FR3 и FR4 домена VL гуманизированного варианта MAB-A; или
(E) (1) вариабельный домен тяжелой цепи (VH) гуманизированного/оптимизированного MAB-A; и
(2) вариабельный домен легкой цепи (VL) гуманизированного/оптимизированного варианта MAB-A.
[0015] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такой вариабельный домен тяжелой цепи (VH) такого оптимизированного варианта MAB-A содержит аминокислотную последовательность SEQ ID NO:15:
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWX1HWVRQA
PGKGLEWVGE IIPIX2GHTNY NEX3FX4X5RFTI SLDNSKNTLY
LQMGSLRAED TAVYYCARGG YYYYX6X7X8X9X10X11
DYWGQGTTVT VSS,
где: X1, X2, X3, X4, X5 и X6 выбирают независимо,
где: X1 представляет собой M или I; X2 представляет собой N или F;
X3 представляет собой K или R; X4 представляет собой K или Q;
X5 представляет собой S или G, и X6 представляет собой P, F, Y, W, I, L, V, T, G или D;
где: X7, X8, X9, X10 и X11 выбирают независимо, таким образом, что:
если X6 представляет собой P; то X7 представляет собой K или R; X8 представляет собой F или M; X9 представляет собой G; X10 представляет собой W или F; и X11 представляет собой M, L или K;
если X6 представляет собой F, Y или W; то X7 представляет собой N или H; X8 представляет собой S или K; X9 представляет собой G или A; X10 представляет собой T или V; и X11 представляет собой M, L или K;
если X6 представляет собой I, L или V; X7 представляет собой G; X8 представляет собой K; X9 представляет собой G или A; X10 представляет собой V; и X11 представляет собой M, L или K;
если X6 представляет собой T; то X7 представляет собой G; X8 представляет собой K, M или N; X9 представляет собой G; X10 представляет собой V или T; и X11 представляет собой L или M;
если X6 представляет собой G; то X7 представляет собой G; X8 представляет собой S; X9 представляет собой G; X10 представляет собой V; и X11 представляет собой L;
если X6 представляет собой D; то X7 представляет собой S; X8 представляет собой N; X9 представляет собой A; X10 представляет собой V; и X11 представляет собой L.
[0016] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такой домен CDRH1, домен CDRH2 и домен CDRH3 такого вариабельного домена тяжелой цепи (VH) такого оптимизированного варианта MAB-A соответственно имеют аминокислотные последовательности:
(1) SEQ ID NO:47 (SYWX1H),
где: X1 представляет собой M или I;
(2) SEQ ID NO:48 (EIIPIX2GHTNYNEX3FX4X5),
где: X2, X3, X4 и X5 выбирают независимо, и
где: X2 представляет собой N или F; X3 представляет собой K или R; X4 представляет собой K или Q; и X5 представляет собой S или G; и
(3) SEQ ID NO:49 (GGYYYYX6X7X8X9X10X11DY),
где: X6 представляет собой P, F, Y, W, I, L, V, T, G или D, и X7, X8, X9, X10 и X11 выбирают таким образом, что:
(A) если X6 представляет собой P, то:
X7 представляет собой K или R; X8 представляет собой F или M; X9 представляет собой G;
X10 представляет собой W или F; и X11 представляет собой M, L или K;
(B) если X6 представляет собой F, Y или W, то:
X7 представляет собой N или H; X8 представляет собой S или K; X9 представляет собой G или A;
X10 представляет собой T или V; и X11 представляет собой M, L или K;
(C) если X6 представляет собой I, L или V, то:
X7 представляет собой G; X8 представляет собой K; X9 представляет собой G или A;
X10 представляет собой V; и X11 представляет собой M, L или K;
(D) если X6 представляет собой T, то:
X7 представляет собой G; X8 представляет собой K, M или N; X9 представляет собой G;
X10 представляет собой V или T; и X11 представляет собой L или M;
(E) если X6 представляет собой G, то:
X7 представляет собой G; X8 представляет собой S; X9 представляет собой G;
X10 представляет собой V; и X11 представляет собой L; и
(F) если X6 представляет собой D, то:
X7 представляет собой S; X8 представляет собой N; X9 представляет собой A;
X10 представляет собой V; и X11 представляет собой L.
[0017] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такой вариабельный домен тяжелой цепи (VH) такого оптимизированного варианта MAB-A выбирают из группы, состоящей из:
(1) hMAB-A VH(1) (SEQ ID NO:16);
(2) hMAB-A VH(2) (SEQ ID NO:17);
(3) hMAB-A VH(3) (SEQ ID NO:18);
(4) hMAB-A VH(4) (SEQ ID NO:19);
(5) hMAB-A VH(2A) (SEQ ID NO:20);
(6) hMAB-A VH(2B) (SEQ ID NO:21);
(7) hMAB-A VH(2C) (SEQ ID NO:22);
(8) hMAB-A VH(2D) (SEQ ID NO:23);
(9) hMAB-A VH(2E) (SEQ ID NO:24);
(10) hMAB-A VH(2F) (SEQ ID NO:25);
(11) hMAB-A VH(2G) (SEQ ID NO:26);
(12) hMAB-A VH(2H) (SEQ ID NO:27);
(13) hMAB-A VH(2I) (SEQ ID NO:28); и
(14) hMAB-A VH(2J) (SEQ ID NO:29).
[0018] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такой вариабельный домен легкой цепи (VL) содержит аминокислотную последовательность SEQ ID NO:53:
DIVMTQSPDS LAVSLGERAT ISCX12ASQSVD
YX13GDSYX14NWY QQKPGQPPKL LIYAASDLES
GIPARFSGSG SGTDFTLTIS SLEPEDFATY
YCQQSX15X16X17PF TFGQGTKLEI K,
где: X12, X13, X14, X15, X16 и X17 выбирают независимо, и
где: X12 представляет собой K или R; X13 представляет собой D или S;
X14 представляет собой M или L; X15 представляет собой H или Y;
X16 представляет собой E или S; и X17 представляет собой D или T.
[0019] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такой домен CDRL1, домен CDRL2 и домен CDRL3 такого вариабельного домена легкой цепи (VH) такого оптимизированного варианта MAB-A соответственно имеют аминокислотные последовательности:
(1) SEQ ID NO:66 (X12ASQSVDYX13GDSYX14N),
где: X12, X13, X14 выбирают независимо, и
где: X12 представляет собой K или R; X13 представляет собой D или S; и X14 представляет собой M или L;
(2) SEQ ID NO:13 (AASDLES); и
(3) SEQ ID NO:67 (QQSX15X16X17PFT),
где: X15, X16 и X17 выбирают независимо, и
где: X15 представляет собой H или Y; X16 представляет собой E или S; и X17 представляет собой D или T.
[0020] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой вариабельный домен легкой цепи (VL) такого оптимизированного варианта MAB-A выбирают из группы, состоящей из:
(1) hMAB-A VL(1) (SEQ ID NO:54);
(2) hMAB-A VL(2) (SEQ ID NO:55);
(3) hMAB-A VL(3) (SEQ ID NO:56);
(4) hMAB-A VL(4) (SEQ ID NO:57);
(5) hMAB-A VL(2A) (SEQ ID NO:20).
[0021] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом домен, связывающий ADAM9, содержит:
(A) (1) домен CDRH1, который содержит аминокислотную последовательность SYWMH (SEQ ID NO:8);
(2) домен CDRH2, который содержит аминокислотную последовательность EIIPIFGHTNYNEKFKS (SEQ ID NO:35); или
(3) домен CDRH3, который содержит аминокислотную последовательность GGYYYYPRQGFLDY (SEQ ID NO:45);
или
(B) (1) домен CDRL1, который содержит аминокислотную последовательность KASQSVDYSGDSYMN (SEQ ID NO:62);
(2) домен CDRL2, который содержит аминокислотную последовательность AASDLES (SEQ ID NO:13); или
(3) домен CDRL3, который содержит аминокислотную последовательность QQSHEDPFT (SEQ ID NO:14).
[0022] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом домен, связывающий ADAM9, содержит домен CDRH1, который содержит аминокислотную последовательность SYWMH (SEQ ID NO:8), домен CDRH2, который содержит аминокислотную последовательность EIIPIFGHTNYNEKFKS (SEQ ID NO:35), и домен CDRH3, который содержит аминокислотную последовательность GGYYYYPRQGFLDY (SEQ ID NO:45).
[0023] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом домен, связывающий ADAM9, содержит домен CDRH1, который содержит аминокислотную последовательность KASQSVDYSGDSYMN (SEQ ID NO:62), домен CDRL2, который содержит аминокислотную последовательность AASDLES (SEQ ID NO:13), и домен CDRL3, который содержит аминокислотную последовательность QQSHEDPFT (SEQ ID NO:14).
[0024] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит:
(A) вариабельный домен тяжелой цепи (VH) hMAB-A (2I.2) (SEQ ID NO:28); или
(В) вариабельный домен легкой цепи (VL) hMAB-A (2I.2) (SEQ ID NO:55); или
(C) вариабельный домен тяжелой цепи (VH) hMAB-A (2I.2) (SEQ ID NO:28) вариабельный домен легкой цепи (VL) hMAB-A (2I.2) (SEQ ID NO:55).
[0025] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит домен CDRH1, домен CDRH2 и домен CDRH3, и при этом домен CDRL1, домен CDRL2 и домен CDRL3 имеют последовательности, выбранные из группы, состоящей из:
(a) SEQ ID NO:8, 35 и 10 и SEQ ID NO:62, 13 и 14 соответственно;
(b) SEQ ID NO:8, 35 и 10 и SEQ ID NO:63, 13 и 14 соответственно;
(c) SEQ ID NO:8, 36 и 10 и SEQ ID NO:63, 13 и 14 соответственно; и
(d) SEQ ID NO:34, 36 и 10 и SEQ ID NO:64, 13 и 65 соответственно.
[0026] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, которые на по меньшей мере 90%, на по меньшей мере 95% или на по меньшей мере 99% идентичны последовательности выбранный из группы, состоящей из:
(a) SEQ ID NO:17 и SEQ ID NO:55 соответственно;
(b) SEQ ID NO:17 и SEQ ID NO:56 соответственно;
(с) SEQ ID NO:18 и SEQ ID NO:56 соответственно; и
(d) SEQ ID NO:19 и SEQ ID NO:57 соответственно.
[0027] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, выбранные из группы, состоящей из:
(a) SEQ ID NO:17 и SEQ ID NO:55 соответственно;
(b) SEQ ID NO:17 и SEQ ID NO:56 соответственно;
(с) SEQ ID NO:18 и SEQ ID NO:56 соответственно; и
(d) SEQ ID NO:19 и SEQ ID NO:57 соответственно.
[0028] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, характеризуется по меньшей мере 150-кратным усилением аффинности связывания с ADAM9 яванского макака и сохраняет высокую аффинность связывания с ADAM9 человека по сравнению с MAB-A.
[0029] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит домен CDRH1, домен CDRH2 и домен CDRH3, и при этом домен CDRL1, домен CDRL2 и домен CDRL3 имеют последовательности, выбранные из группы, состоящей из:
(a) SEQ ID NO:8, 35 и 37 и SEQ ID NO:62, 13 и 14 соответственно;
(b) SEQ ID NO:8, 35 и 38 и SEQ ID NO:62, 13 и 14 соответственно;
(c) SEQ ID NO:8, 35 и 39 и SEQ ID NO:62, 13 и 14 соответственно;
(d) SEQ ID NO:8, 35 и 40 и SEQ ID NO:62, 13 и 14 соответственно;
(e) SEQ ID NO:8, 35 и 41 и SEQ ID NO:62, 13 и 14 соответственно;
(f) SEQ ID NO:8, 35 и 42 и SEQ ID NO:62, 13 и 14 соответственно;
(g) SEQ ID NO:8, 35 и 43 и SEQ ID NO:62, 13 и 14 соответственно;
(h) SEQ ID NO:8, 35 и 44 и SEQ ID NO:62, 13 и 14 соответственно;
(i) SEQ ID NO:8, 35 и 45 и SEQ ID NO:62, 13 и 14 соответственно; и
(j) SEQ ID NO:8, 35 и 46 и SEQ ID NO:62, 13 и 14 соответственно.
[0030] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, которые на по меньшей мере 90%, на по меньшей мере 95% или на по меньшей мере 99% идентичны последовательности выбранный из группы, состоящей из:
(a) SEQ ID NO:20 и SEQ ID NO:55 соответственно;
(b) SEQ ID NO:21 и SEQ ID NO:55 соответственно;
(c) SEQ ID NO:22 и SEQ ID NO:55 соответственно;
(d) SEQ ID NO:23 и SEQ ID NO:55 соответственно;
(e) SEQ ID NO:24 и SEQ ID NO:55 соответственно;
(f) SEQ ID NO:25 и SEQ ID NO:55 соответственно;
(g) SEQ ID NO:26 и SEQ ID NO:55 соответственно;
(h) SEQ ID NO:27 и SEQ ID NO:55 соответственно;
(i) SEQ ID NO:28 и SEQ ID NO:55 соответственно; и
(j) SEQ ID NO:29 и SEQ ID NO:55 соответственно.
[0031] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой домен, связывающий ADAM9, содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, выбранные из группы, состоящей из:
(a) SEQ ID NO:20 и SEQ ID NO:55 соответственно;
(b) SEQ ID NO:21 и SEQ ID NO:55 соответственно;
(c) SEQ ID NO:22 и SEQ ID NO:55 соответственно;
(d) SEQ ID NO:23 и SEQ ID NO:55 соответственно;
(e) SEQ ID NO:24 и SEQ ID NO:55 соответственно;
(f) SEQ ID NO:25 и SEQ ID NO:55 соответственно;
(g) SEQ ID NO:26 и SEQ ID NO:55 соответственно;
(h) SEQ ID NO:27 и SEQ ID NO:55 соответственно;
(i) SEQ ID NO:28 и SEQ ID NO:55 соответственно; и
(j) SEQ ID NO:29 и SEQ ID NO:55 соответственно.
[0032] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такая молекула представляет собой моноспецифическое антитело, связывающее ADAM9 или его фрагмент, связывающий ADAM9, или при этом такая молекула представляет собой биспецифическое антитело.
[0033] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такая молекула представляет собой диатело, при этом такое диатело представляет собой ковалентно связанный комплекс, который содержит две, три, четыре или пять полипептидных цепей.
[0034] В настоящем изобретении дополнительно рассматривается вариант осуществления всех таких молекул, связывающих ADAM9, при этом такая молекула представляет собой трехвалентную связывающую молекулу, при этом такая трехвалентная связывающая молекула представляет собой ковалентно связанный комплекс, который содержит три, четыре, пять или более полипептидных цепей.
[0035] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула содержит домен, связывающий альбумин (ABD).
[0036] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула содержит Fc-область, и, в частности, вариант осуществления, где такая Fc-область представляет собой вариантную Fc-область, которая содержит:
(a) одну или несколько аминокислотных модификаций, которые снижают аффинность вариантной Fc-области в отношении FcγR; и/или
(b) одну или несколько аминокислотных модификаций, которые усиливают период полужизни в сыворотке крови такой молекулы, связывающей ADAM9.
[0037] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая одна или несколько аминокислотных модификаций, которые снижают аффинность вариантной Fc-области по отношению к FcγR, содержат:
(A) L234A;
(B) L235A; или
(C) L234A и L235A;
где такая нумерация представляет собой нумерацию EU-индекса в соответствии с Кабат.
[0038] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая одна или несколько аминокислотных модификаций, которые повышают период полужизни в сыворотке крови такой молекулы, связывающей ADAM9, содержат:
(A) M252Y;
(B) M252Y и S254T;
(C) M252Y и T256E;
(D) M252Y, S254T и T256E; или
(E) K288D и H435K;
где такая нумерация представляет собой нумерацию EU-индекса в соответствии с Кабат.
[0039] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула является биспецифической и содержит эпитопсвязывающий сайт, способный к иммуноспецифическому связыванию эпитопа ADAM9, и эпитопсвязывающий сайт, способный к иммуноспецифическому связыванию эпитопа молекулы, присутствующей на поверхности эффекторной клетки.
[0040] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула содержит два эпитопсвязывающих сайта, способных к иммуноспецифическому связыванию эпитопа(эпитопов) ADAM9, и два эпитопсвязывающих сайта, способных к иммуноспецифическому связыванию эпитопа(эпитопов) молекулы, присутствующей на поверхности эффекторной клетки.
[0041] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такой молекула является триспецифической и содержит:
(a) один эпитопсвязывающий сайт, способный к иммуноспецифическому связыванию с эпитопом ADAM9;
(b) один эпитопсвязывающий сайт, способный к иммуноспецифическому связыванию с эпитопом первой молекулы, присутствующей на поверхности эффекторной клетки; и
(c) один эпитопсвязывающий сайт, способный к иммуноспецифическому связыванию с эпитопом второй молекулы, присутствующей на поверхности эффекторной клетки.
[0042] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула способна одновременно связываться с ADAM9 и такой молекулой, присутствующей на поверхности эффекторной клетки.
[0043] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула, присутствующая на поверхности эффекторной клетки представляет собой CD2, CD3, CD8, TCR или NKG2D.
[0044] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая эффекторная клетка представляет собой цитотоксическую Т-клетку или естественный киллер (NK).
[0045] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая первая молекула, присутствующая на поверхности эффекторной клетки, представляет собой CD3, и такая вторая молекула, присутствующая на поверхности эффекторной клетки, представляет собой CD8.
[0046] В настоящем изобретении дополнительно рассматривается вариант осуществления таких молекул, связывающих ADAM9, при этом такая молекула, связывающая ADAM9, опосредует координированное связывание клетки, экспрессирующей ADAM9, и цитотоксической T-клетки.
[0047] В настоящем изобретении дополнительно рассматривается фармацевтическая композиция, которая содержит эффективное количество любой из вышеописанных молекул, связывающих ADAM9, и фармацевтически приемлемого носителя, наполнителя или разбавителя.
[0048] В настоящем изобретении дополнительно рассматривается применение любой из вышеописанных молекул, связывающих ADAM9, или применение вышеописанной фармацевтической композиции в лечении заболевания или патологического состояния, ассоциированного с экспрессией ADAM9 или характеризующегося ей.
[0099] В настоящем изобретении, в частности, рассматривается такое применение, при этом такое заболевание или патологического состояние, ассоциированной с экспрессией ADAM9, или характеризующееся ей, представляет собой рак, и, особенно при этом такой рак выбирают из группы, состоящей из рака мочевого пузыря, рака молочной железы, рака шейки матки, колоректального рака (особенно аденокарциномы, желудочно-кишечных карциноидных опухолей, желудочно-кишечных стромальных опухолей, первичной колоректальной лимфомы, лейомиосаркомы, меланомы или плоскоклеточной карциномы), рака пищевода, рака желудка, рака головы и шеи, рака печени, немелкоклеточного рака легких (особенно плоскоклеточной карциномы, аденокарциномы или крупноклеточной недифференцированной карциномы), рака миелоидного происхождения, рака яичников, рака поджелудочной железы, рака предстательной железы, почечно-клеточной карциномы, рака щитовидной железы, рака яичка и рака матки.
[0050] В настоящем изобретении дополнительно рассматривается способ лечения заболевания или патологического состояния, ассоциированного с экспрессией ADAM9 или характеризующегося ей, у субъекта, предусматривающий введение такому субъекту эффективного количества любой из вышеописанных молекул, связывающих ADAM9, или любых из вышеуказанных фармацевтических композиций.
[0051] В настоящем изобретении, в частности, рассматривается такой способ, при этом такое заболевание или патологического состояние, ассоциированной с экспрессией ADAM9, или характеризующееся ей, представляет собой рак, и, особенно при этом такой рак выбирают из группы, состоящей из рака мочевого пузыря, рака молочной железы, рака шейки матки, колоректального рака (особенно аденокарциномы, желудочно-кишечных карциноидных опухолей, желудочно-кишечных стромальных опухолей, первичной колоректальной лимфомы, лейомиосаркомы, меланомы или плоскоклеточной карциномы), рака пищевода, рака желудка, рака головы и шеи, рака печени, немелкоклеточного рака легких (особенно плоскоклеточной карциномы, аденокарциномы или крупноклеточной недифференцированной карциномы), рака миелоидного происхождения, рака яичников, рака поджелудочной железы, рака предстательной железы, почечно-клеточной карциномы, рака щитовидной железы, рака яичка и рака матки.
Краткое описание чертежей
[0052] На Фиг. 1 представлена схема иллюстративного ковалентно связанного диатела, имеющего два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, при этом каждая из них имеет E-петлевой или K-петлевой активирующий сборку гетеродимера домен (альтернативные домены, активирующие сборку гетеродимера, представлены ниже). Цистеиновый остаток может присутствовать в линкере и/или в активирующем сборку гетеродимера домене, как показано на Фиг. 3B. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки.
[0053] На Фиг. 2 представлена схема иллюстративной ковалентно связанной молекулы диатела, имеющей два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, при этом каждая из них содержит домен CH2 и CH3, таким образом, что ассоциированные цепи образуют всю или часть Fc-области. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки.
[0054] На Фиг. 3A-3C представлены схемы, на которых показаны иллюстративные ковалентно связанные тетравалентные диатела, имеющие четыре эпитопсвязывающих сайта, состоящие из двух пар полипептидных цепей (т.е., в общей сложности четырех полипептидных цепей). Один полипептид каждой пары имеет домен CH2 и CH3, таким образом, что ассоциированные цепи образуют всю или часть Fc-области. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки. Две пары полипептидных цепей могут быть одинаковыми. В таких вариантах осуществления, где две пары полипептидных цепей являются одинаковыми, а домены VL и VH распознают различные эпитопы (как показано на Фиг. 3A-3B), образующаяся молекула имеет четыре эпитопсвязывающих сайта и является биспецифической и трехвалентной по отношению к каждому связанному эпитопу. В таких вариантах осуществления, где домены VL и VH распознают один и тот же эпитоп (например, CDR одного и того же домена VL или CDR одного и того же домена VH используются в обоих цепях), образующаяся молекула имеет четыре эпитопсвязывающих сайта и является моноспецифической и четырехвалентной по отношению к одному эпитопу. В альтернативном случае две пары полипептидов могут быть различными. В таких вариантах осуществления, где две пары полипептидных цепей являются различными, а домены VL и VH каждой пары полипептидов распознают различные эпитопы (как показано с помощью различных вариантов штриховки и заливки на Фиг. 3C), образующаяся молекула имеет четыре эпитопсвязывающих сайта и является тетраспецифической и моновалентной по отношению к каждому связанному эпитопу. На Фиг. 3A показано диатело, содержащее Fc-область, которая содержит пептидный активирующий сборку гетеродимера домен, содержащий цистеиновый остаток. На Фиг. 3B показано диатело, содержащее Fc-область, которая содержит E-петлевые и K-петлевые активирующие сборку гетеродимера домены, содержащего цистеиновый остаток и линкер (с необязательным цистеиновым остатком). На Фиг. 3C показано диатело, содержащее Fc-область, которое содержит домены CH1 и CL.
[0055] На Фиг. 4A-4B представлена схема иллюстративной ковалентно связанной молекулы диатела, имеющей два эпитопсвязывающих сайта, состоящих из трех полипептидных цепей. Две из полипептидных цепей имеют домен CH2 и CH3, таким образом, что ассоциированные цепи образуют всю или часть Fc-области. Полипептидные цепи, содержащие домен VL и VH, дополнительно содержат активирующий сборку гетеродимера домен. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки.
[0056] На Фиг. 5 представлена схема иллюстративной ковалентно связанной молекулы диатела, имеющей четыре эпитопсвязывающих сайта, состоящих из пяти полипептидных цепей. Две из полипептидных цепей имеют домен CH2 и CH3, таким образом, что ассоциированные цепи образуют Fc-область, которая представляет собой всю или часть Fc-области. Полипептидные цепи, содержащие связанные домены VL и VH, дополнительно содержат активирующий сборку гетеродимера домен. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки.
[0057] На Фиг. 6A-6F представлены схемы иллюстративных трехвалентных связывающих молекул, содержащих Fc-область, имеющих три эпитопсвязывающих сайта. На Фиг. 6A и 6B соответственно схематически иллюстрируются домены трехвалентных связывающих молекул, содержащих два связывающих домена по типу диатела и связывающий домен Fab-типа, имеющих различные ориентации доменов, в которых связывающие домены по типу диатела являются N-концевыми или C-концевыми по отношению к Fc-области. Молекулы на Фиг. 6A и 6B содержат четыре цепи. На Фиг. 6C и 6D соответственно схематически иллюстрируются домены трехвалентных связывающих молекул, содержащих два связывающих домена по типу диатела, N-концевых по отношению к Fc-области, и связывающий домен Fab-типа, в котором легкая и тяжелая цепь связаны посредством полипептидного спейсера, или связывающий домен scFv-типа. Трехвалентные связывающие молекулы на Фиг. 6E и 6F соответственно схематически иллюстрируют домены трехвалентных связывающих молекул, содержащих два связывающих домена по типу диатела, С-концевых по отношению к Fc-области, и связывающий домен Fab-типа, в котором легкая и тяжелая цепь связаны посредством полипептидного спейсера, или связывающий домен scFv-типа. Трехвалентные связывающие молекулы на Фиг. 6C-6F содержат три цепи. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с помощью одного и того же варианта штриховки или заливки.
[0058] На Фиг. 7A-7C представлены результаты иммуногистохимических (IHC) исследований и показана способность MAB-A специфически метить клетки ряда типов немелкоклеточного рака легких (Фиг. 7A, панели 1-8), клетки рака молочной железы, клетка рака предстательной железы, клетки рака желудка (Фиг. 7B, панели 1-6) и клетки рака толстой кишки (Фиг. 7C, панели 1-8), в то время как изотипический контроль не мог специфически метить никакие из этих типов раковых клеток (Фиг. 7A-7C).
[0059] На Фиг. 8A-8B представлены результаты исследований по окрашиванию клеток и показано, что MAB-A связывается с ADAM9 человека, и в меньшей степени, ADAM9 яванского макака, временно экспрессируемым на поверхности клеток 293-FT и CHO-K (Фиг. 8A и Фиг. 8B соответственно).
[0060] На Фиг. 9A-9B представлены аминокислотные последовательности мышиного анти-ADAM9-VH-домена, выравненного по отношению к гуманизированным/оптимизированным вариантам MAB-A (Фиг. 9A, SEQ ID NO:7, 16, 17, 18, 19, 21, 22, 23 и 28) и мышиного анти-ADAM9-VL-домена, выравненные по отношению к гуманизированным/оптимизированным вариантам MAB-A (Фиг. 9B, SEQ ID NO:11, 51, 52, 53 и 54). Положения, замещенные в CDR во время исходной оптимизации, подчеркнуты следующим образом: потенциальные сайты дезамидирования и изомерации обозначены одним подчеркиванием, лизиновые остатки обозначены двойным подчеркиванием, дополнительные лабильные остатки обозначены двойным пунктирным подчеркиванием.
[0061] На Фиг. 10A-10B представлены кривые связывания ELISA десяти выбранных оптимизированных клонов hMAB-A, содержащих варианты CDRH3, исходное hMAB-A (2.2) и антитело изотипического контроля. На Фиг. 10A представлены кривые связывания для cynoADAM9 и на Фиг. 10B представлены кривые связывания для huADAM9.
Подробно раскрытие настоящего изобретения
[0062] Настоящее изобретение направлено на молекулы, такие как моноспецифические антитела и биспецифические, триспецифические или мультиспецифические молекулы, в том числе диатела, BiTE и антитела, которые способны специфически связываться с «белком 9, содержащим домен дезинтегрина и металлопротеиназы» («ADAM9»). В настоящем изобретении, в частности, рассматриваются такие связывающие молекулы, которые способны проявлять высокую аффинность связывания с ADAM9 человека и не относящимся к человеку ADAM9. Настоящее изобретение дополнительно, в частности, связано с такими молекулами, которые, таким образом, являются перекрестно-реактивными в отношении ADAM9 человека и ADAM9 не относящегося к человеку примата (например, яванского макака). Настоящее изобретение, кроме того, относится ко всем таким молекулам, связывающим ADAM9, которые содержат вариабельный домен легкой цепи (VL) и/или вариабельный домен тяжелой цепи (VH), которые были гуманизированы и/или деиммунизированы таким образом, чтобы проявлять ослабленную иммуногенность при введении такой молекулы, связывающей ADAM9, субъекту-реципиенту. Настоящее изобретение также направлено на фармацевтические композиции, которые содержат любую из таких молекул, связывающих ADAM9, а также способы, предусматривающие применение любой из таких молекул, связывающих ADAM9, в лечении рака и других заболеваний и патологических состояний.
I. Антитела и их связывающие домены
[0063] Антитела по настоящему изобретению представляют собой иммуноглобулиновые молекулы, способные специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., посредством по меньшей мере одного сайта распознавания антигена, расположенного в вариабельном домене иммуноглобулиновой молекулы. Используемые в данном документе термины «антитело» и «антитела» относятся к моноклональным антителам, мультиспецифическим антителам, человеческим антителам, гуманизированным антителам, синтетическим антителам, химерным антителам, поликлональным антителам, верблюжьим антителам, одноцепочечным Fv (scFv), одноцепочечным антителам, Fab-фрагментам, F(ab’)-фрагментам, связанным дисульфидными связями биспецифическим Fv (sdFv), интрателам и эпитопсвязывающим фрагментам любого из вышеуказанного. В частности, термин «антитело» включает в себя иммуноглобулиновые молекулы и иммунологически активные фрагменты иммуноглобулиновых молекул, т.е., молекулы, которые содержат один эпитопсвязывающий сайт. Иммуноглобулиновые молекулы могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса. В последние несколько десятилетий наблюдается оживление интереса к терапевтическому потенциалу антител и антитела стали одним из ведущих классов лекарственных препаратов, получаемых биотехнологическим путем (Chan, C.E. et al. (2009) “The Use Of Antibodies In The Treatment Of Infectious Diseases,” Singapore Med. J. 50(7):663-666). В дополнение к их применению в диагностике было показано, что антитела являются пригодными в качестве терапевтических средств. Свыше 200 лекарственных препаратов на основе антител были одобрены для применения или находятся в разработке.
[0064] Антитела способны «иммуноспецифически связываться» с полипептидом или белком или небелковой молекулой вследствие наличия на такой молекуле определенного домена или фрагмента или конформации («эпитопа»). Молекула, содержащая эпитоп, может характеризоваться иммуногенной активностью, таким образом, что она вызывает ответ в виде продуцирования антитела; такие молекулы называется «антигены». Используемые в данном документе антитело, диатело или другая эпитопсвязывающая молекула «иммуноспецифически» связывается с участком другой молекулы (т.е., эпитопом), если она реагирует или ассоциируется более часто, более быстро, с большей продолжительностью и/или с большей аффинностью с данным эпитопом по сравнению с альтернативными эпитопами. Например, антитело, которое иммуноспецифически связывается с вирусным эпитопом, представляет собой антитело, которое связывает этот вирусный эпитоп с более высокой аффинностью, авидностью, более легко и/или с большей продолжительностью, чем оно иммуноспецифически связывается с другими вирусными эпитопами или невирусными эпитопами. Также подразумевается при трактовке данного определения, что, например, антитело (или фрагмент или эпитоп), которое иммуноспецифически связывается с первой мишенью, может или не может специфически или предпочтительно связываться со второй мишенью. Соответственно, «иммуноспецифическое связывание» с определенным эпитопом не обязательно требует (хотя оно может включать) исключительное связывание с этим эпитопом. Как правило, но необязательно, отсылка на связывания означает «иммуноспецифическое» связывание. Считается, что две молекулы способны связываться друг с другом «физиоспецифическим» образом, если такое связывание характеризуется специфичностью, с которой рецепторы связываются со своими соответствующими лигандами.
[0065] Термин «моноклональное антитело» относится к популяции гомогенных антител, при этом моноклональное антитело состоит из аминокислот (встречающихся в природе или не встречающихся в природе), которые участвуют в селективном связывании антигена. Моноклональные антитела являются высокоспецифическими, при этом они направлены на один эпитоп (или антигенный сайт). Термин «моноклональное антитело» охватывает не только интактные моноклональные антитела и полноразмерные моноклональные антитела, но также и их фрагменты (такие как Fab, Fab', F(ab')2 Fv), одноцепочечные scFv, их мутантные формы, слитые белки, содержащие участок антитела, гуманизированные моноклональные антитела, химерные моноклональные антитела и любую другую модифицированную конфигурацию иммуноглобулиновой молекулы, которая содержит сайт распознавания антигена с необходимой специфичностью и способностью связываться с антигеном. Термин не предполагает ограничение по отношению к источнику антитела или способу, которым оно получено (например, с помощью гибридомы, селекции с помощью фагового дисплея, рекомбинантной экспрессии, трансгенных животных и т.д.). Термин включает в себя целые иммуноглобулины, а также фрагменты, например, описанные выше под определением «антитело». Способы получения моноклональных антител известны в данной области техники. Один из способов, который может быть использован, представляет собой способ Kohler, G. et al. (1975) “Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity,” Nature 256:495-497, или его модификация. В типичном случае моноклональные антитела развиваются в мышах, крысах или кроликах. Антитела продуцируются в результате иммунизации животного иммуногенным количеством клеток, клеточных экстрактов или белковых препаратов, которые содержат необходимый эпитоп. Иммуноген может представлять собой без ограничения первичные клетки, культивированные клеточные линии, раковые клетки, белки, пептиды, нуклеиновые кислоты или ткани. Клетки, используемые для иммунизации, можно культивировать в течение периода времени (например, по меньшей мере 24 часов) до их применения в качестве иммуногена. Клетки могут быть использованы в качестве иммуногена сами по себе или в комбинации с неденатурирующим адъювантом, таким как Ribi (см., например, Jennings, V.M. (1995) “Review of Selected Adjuvants Used in Antibody Production,” ILAR J. 37(3):119-125). Как правило, клетки должны содержаться интактными и предпочтительно жизнеспособными при использовании в качестве иммуногенов. Интактные клетки могут способствовать более эффективному выявлению антигенов, чем разрушенные иммунизированным животным клетки. Применение денатурирующих или агрессивных адъювантов, таких как адъювант Фрейнда, может разрушать клетки и, таким образом, является нежелательным. Иммуноген можно вводить много раз через периодические интервалы, например, два раза в неделю или еженедельно, или можно вводить таким образом, чтобы поддерживать жизнеспособность животного (например, рекомбинантным в отношении ткани образом). В альтернативном случае существующие моноклональные антитела и любые другие эквивалентные антитела, которые являются иммуноспецифическими по отношению к необходимому патогенному эпитопу, могут быть секвенированы и получены рекомбинантно с помощью любых средств, известных в данной области техники. В соответствии с одним вариантом осуществления такое антитело секвенируют и полинуклеотидную последовательность затем клонируют в вектор для экспрессии или размножения. Последовательность, кодирующая антитело, представляющее интерес, может поддерживаться в векторе в клетке-хозяине и клетку-хозяина затем можно размножать и замораживать для будущего применения. Полинуклеотидная последовательность таких антител может быть использована для генетической манипуляции с получением моноспецифических или мультиспецифических (например, биспецифических, триспецифических и тетраспецифических) молекул по настоящему изобретению, а также оптимизированного по аффинности, химерного антитела, гуманизированного антитела и/или канинизированного антитела, с целью улучшения аффинности или других характеристик антитела. Общий принцип гуманизации антитела включает в себя сохранение основной последовательности антигенсвязывающего участка антитела, при этом в то же время происходит обмен не относящегося к человеческому остатку антитела последовательностями человеческих антител.
[0066] Природные антитела (такие как природные антитела IgG) состоят из двух «легких цепей», образующих комплекс с двумя «тяжелыми цепями». Каждая легкая цепь содержит вариабельный домен («VL») и константный домен («CL»). Каждая тяжелая цепь содержит вариабельный домен («VH»), три константных домена («CH1», «CH2» и «CH3») и «шарнирную» область («H»), расположенную между доменами CH1 и CH2. Основной структурной единицей встречающихся в природе иммуноглобулинов (например, IgG) является, таким образом, тетрамер, имеющий две легкие цепи и две тяжелые цепи, обычно экспрессируемые в виде гликопротеина, составляющего приблизительно 150000 Да. Аминоконцевой («N-концевой») участок каждой цепи включает в себя вариабельный домен из приблизительно 100-110 или более аминокислот, главным образом отвечающих за распознавание антигена. Карбоксиконцевой («C-концевой») участок каждой цепи определяет константную область, при этом легкие цепи имеют один константный домен и тяжелые цепи обычно имеют три константных домена и шарнирную область. Таким образом, структура легких цепей молекулы IgG представляет собой n-VL-CL-c, а структура тяжелой цепи IgG представляет собой n-VH-CH1-H-CH2-CH3-c (где n и c представляют собой соответственно N-конец и C-конец полипептида). Вариабельные домены молекулы IgG состоят из 1, 2 и наиболее часто 3 областей, определяющих комплементарность («CDR», т.е., CDR1, CDR2 и CDR3 соответственно), которые содержат остатки в контакте с эпитопом, и не относящиеся к CDR сегменты, обозначаемые как каркасные области («FR»), которые, как правило, поддерживают структуру и определяют расположение областей CDR таким образом, чтобы обеспечивать такое контактирование (хотя определенные каркасные остатки могут также контактировать с эпитопом). Таким образом, домены VL и VH в типичном случае имеют структуру: n-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4-c (где «n» обозначает N-конец и «c» обозначает C-конец). Полипептиды, которые представляют собой первую, вторую, третью и четвертую FR легкой цепи антитела (или могут выступать в качестве их) в данном документе соответственно обозначаются как: домен FRL1, домен FRL2, домен FRL3 и домен FRL4. Аналогичным образом, полипептиды, которые представляют собой первую, вторую, третью и четвертую FR тяжелой цепи антитела (или могут выступать в качестве их) в данном документе соответственно обозначаются как: домен FRH1, домен FRH2, домен FRH3 и домен FRH4. Полипептиды, которые представляют собой первую, вторую и третью CDR легкой цепи антитела (или могут выступать в качестве их) в данном документе соответственно обозначаются как: домен CDRL1, домен CDRL2 и домен CDRL3. Аналогичным образом, полипептиды, которые представляют собой первую, вторую и третью CDR тяжелой цепи антитела (или могут выступать в качестве их) в данном документе соответственно обозначаются как: домен CDRH1, домен CDRH2 и домен CDRH3. Таким образом, термины домен CDRL1, домен CDRL2, домен CDRL3, домен CDRH1, домен CDRH2 и домен CDRH3 направлены на полипептиды, которые при введении в белок приводят к тому, что этот белок способен связываться со специфическим эпитопом, вне зависимости от того, представляет ли собой такой белок антитело, имеющее легкие и тяжелые цепи, или диатело или одноцепочечную связывающую молекулу (например, scFv, BiTe и т.д.), или другой тип белка.
[0067] Соответственно, используемый в данном документе термин «эпитопсвязывающий фрагмент» означает фрагмент антитела, способного иммуноспецифически связываться с эпитопом, а термин «эпитопсвязывающий сайт» относится к участку молекулы, содержащей эпитопсвязывающий фрагмент. Эпитопсвязывающий фрагмент может содержать 1, 2, 3, 4 или 5 доменов CDR антитела, или может содержать все 6 доменов CDR антитела, и несмотря на то, что при этом он способен иммуноспецифически связываться с таким эпитопом, он может характеризоваться иммуноспецифичностью, аффинностью или селективностью, которые отличаются от таковых такого антитела. Однако предпочтительно, чтобы эпитопсвязывающий фрагмент содержал все 6 доменов CDR такого антитела. Эпитопсвязывающий фрагмент антитела может представлять собой одну полипептидную цепь (например, scFv) или может содержать две или более полипептидные цепи, при этом каждая из них имеет аминоконец и карбоксиконец (например, диатело, Fab-фрагмент, Fab2-фрагмент и др.). Если особо не указано, порядок доменов белковых молекул, описанных в данном документе, следует в направлении «N-конец-C-конец».
[0068] Настоящее изобретение, в частности, охватывает одноцепочечные фрагменты вариабельных доменов (“scFv”), содержащие анти-ADAM9-VL- и/или VH-домен по настоящему изобретению, а также мультиспецифические связывающие молекулы, содержащие такие анти-ADAM9-VL- и/или VH-домены. Одноцепочечные фрагменты вариабельных доменов содержат домены VL и VH, которые связаны совместно с помощью короткого «линкерного» пептида. Такие линкеры можно модифицировать с получением дополнительных функций, например, обеспечения прикрепления лекарственного средства или обеспечения прикрепления к твердой подложке. Одноцепочечные варианты можно получать либо рекомбинантно, либо синтетически. Для синтетического получения scFv может быть использован синтезатор. Для рекомбинантного получения scFv подходящая плазмида, содержащая полинуклеотид, который кодирует scFv, может быть введена в подходящую клетку-хозяина, либо эукариотическую, такую как клетки дрожжевых грибов, растений, насекомых или млекопитающих, либо прокариотическую, такие как E. coli. Полинуклеотиды, кодирующие scFv, представляющие интерес, могут быть получены с помощью стандартных манипуляций, таких как лигирование полинуклеотидов. Образующийся в результате scFv может быть выделен с помощью стандартных методик очистки белка, известных в данной области техники.
[0069] Настоящее изобретение, также, в частности, охватывает домены CDRH1, CDRH2, CDRH3, CDRL1, CDRL2 и CDRL3 гуманизированных вариантов антител к ADAM9 по настоящему изобретению, а также домены VL, которые которые содержат любые 1, 2 или 3 из таких CDRL, и домены VH, которые содержат любые 1, 2 или 3 из таких CDRH, а также мультиспецифические связывающие молекулы, содержащие их. Термин «гуманизированное» антитело относится к химерной молекуле, имеющей эпитопсвязывающие сайты иммуноглобулина от не относящегося к человеку вида и оставшуюся иммуноглобулиновую структуру, которая основана на структуре и/или последовательности человеческого иммуноглобулина. Гуманизированные антитела, как правило, получают с помощью рекомбинантных методик. Антитела к ADAM9 по настоящему изобретению включают в себя гуманизированные, химерные или канинизированные варианты антитела, которые обозначаются в данном документе как «MAB-A». Полинуклеотидные последовательности, которые кодируют вариабельные домены MAB-A, могут быть использованы для генетической манипуляции с целью получения производных MAB-A, обладающих улучшенными или измененными характеристиками (например, аффинностью, перекрестной реактивностью, специфичностью и т.д.). Общий принцип гуманизации антитела включает в себя сохранение основной последовательности эпитопсвязывающего участка антитела, при этом в то же время происходит обмен не относящегося к человеческому остатку антитела последовательностями человеческих антител. Существуют четыре основные стадии гуманизации моноклонального антитела. Они представляют собой: (1) определение нуклеотидной и предполагаемой аминокислотной последовательности вариабельных доменов легкой и тяжелой цепи исходного антитела; (2) конструирование гуманизированного или канинизированного антитела, т.е., решение вопроса о том, какую каркасную область антитела использовать во время процесса гуманизации или канинизации; (3) использование фактических методологий/методик гуманизации или канинизации; и (4) трансфекцию и экспрессию гуманизированного антитела. См., например, патенты США №№ 4816567; 5807715; 5866692; и 6331415. Термин «оптимизированное» антитело относится к антителу, имеющему по меньшей мере одну аминокислоту, которая отличается от исходного антитела, на по меньшей мере одну область, определяющую комплементарность (CDR), в вариабельной области легкой или тяжелой цепи, что придает более высокую аффинность (например, в 2 раза больше или более раз), более высокую аффинность связывания по отношению к ADAM9 человека и/или ADAM9 яванского макака по сравнению с исходным антителом. Будет понятно из описания, предусмотренного в данном документе, что антитела по настоящему изобретению можно гуманизировать, оптимизировать или либо гуманизировать, либо оптимизировать.
[0070] Эпитопсвязывающие сайты могут содержать либо полный вариабельный домен, слитый с одним или несколькими константными доменами, либо только CDR такого вариабельного домена, привитого на подходящие каркасные области. Эпитопсвязывающие сайты могут быть дикого типа или могут быть модифицированы с помощью одной или нескольких аминокислотных замен, вставок или делеций. Такое действие частично или полностью устраняет способность константной области выступать в качестве иммуногена у реципиентов (например, индивидов-людей), однако, возможность иммунного ответа на чужеродный вариабельный домен сохраняется (LoBuglio, A.F. et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход сосредоточен не только на получении константных областей человеческого происхождения, но и на модифицировании вариабельных доменов, а также на преобразовании их как можно ближе к форме, встречающейся в человеческих иммуноглобулинах. Известно, что вариабельные домены как тяжелых, так и легких цепей антител содержат три CDR, которые варьируются в ответ на рассматриваемые антигены и определяют связывающую способность, фланкируются четырьмя каркасными областями, которые являются относительно консервативными у определенного вида и которые предположительно обеспечивают каркас для CDR. Если не относящиеся к человеку антитела получают в отношении определенного антигена, то вариабельные домены могут быть «преобразованы» или «гуманизированы» с помощью привития CDR, происходящих из не относящихся к человеку антитела, на FR, присутствующие в человеческом антителе, подлежащем модификации. Применение данного подхода к различным антителам, было описано Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154. В соответствии с некоторыми вариантами осуществления гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное мышиное антитело, которое содержит все шесть из CDR, присутствующих в мышином антителе). В соответствии с другими вариантами осуществления гуманизированные антитела имеют одну или несколько CDR (один, два, три, четыре, пять или шесть), которые отличаются последовательностью по отношению к CDR исходного антитела.
[0071] Были описаны несколько гуманизированных молекул антител, содержащих эпитопсвязывающие сайты, происходящие из не относящегося к человеку иммуноглобулина, в том числе химерные антитела, имеющие вариабельный домен грызуна или модифицированный домен грызуна и их соответствующие области, определяющие комплементарность (CDR), слитые с человеческими константными доменами (см., например, Winter et al. (1991) “Man-made Antibodies,” Nature 349:293-299; Lobuglio et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224; Shaw et al. (1987) “Characterization Of A Mouse/Human Chimeric Monoclonal Antibody (17-1A) To A Colon Cancer Tumor-Associated Antigen,” J. Immunol. 138:4534-4538; и Brown et al. (1987) “Tumor-Specific Genetically Engineered Murine/Human Chimeric Monoclonal Antibody,” Cancer Res. 47:3577-3583). В других литературных источниках описаны CDR грызунов, привитые на человеческие поддерживающие каркасные области (FR) до слияния с соответствующим константным доменом человеческого антитела (см., например, Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; и Jones et al. (1986) “Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse,” Nature 321:522-525). В другом литературном источнике описаны CDR грызунов, поддерживаемые рекомбинантно венированными каркасными областями грызунов (см., например, публикацию Европейского патента № 519596). Эти «гуманизированные» молекулы конструируют с целью сведения к минимуму нежелательного иммунологического ответа по отношению к молекулам антител грызунов к человеческим антителам, что ограничивает продолжительность и эффективность терапевтических областей применения таких фрагментов у реципиентов-людей. Другие способы гуманизации антител, которые также могут быть использованы, раскрыты Daugherty et al. (1991) “Polymerase Chain Reaction Facilitates The Cloning, CDR-Grafting, And Rapid Expression Of A Murine Monoclonal Antibody Directed Against The CD18 Component Of Leukocyte Integrins,” Nucl. Acids Res. 19:2471-2476 и в патентах США №№ 6180377; 6054297; 5997867; и 5866692.
II. Fcγ-рецепторы (FcγR)
[0072] Домены CH2 и CH3 двух тяжелых цепей антитела взаимодействуют с образованием «Fc-области», которая представляет собой домен, который распознается клеточными «Fc-рецепторами», в том числе без ограничения Fc-гамма-рецепторами («FcγR»). Используемый в данном документе термин «Fc-область» используется для определения C-концевой области тяжелой цепи IgG, которая содержит домены CH2 и CH3 этой цепи. Считается, что Fc-область имеет определенный изотип, класс или подкласс IgG, если ее аминокислотная последовательность является наиболее гомологичной в отношении этого изотипа по сравнению с другими изотипами IgG.
[0073] Аминокислотная последовательность домена CH2-CH3 иллюстративного IgG1 человека представляет собой (SEQ ID NO:1):

пронумерованную с помощью EU-индекса в соответствии с Кабат, где X представляет собой (K) или отсутствует.
[0074] Аминокислотная последовательность домена CH2-CH3 иллюстративного IgG2 человека представляет собой (SEQ ID NO:2):

пронумерованную с помощью EU-индекса в соответствии с Кабат, где X представляет собой (K) или отсутствует.
[0075] Аминокислотная последовательность домена CH2-CH3 иллюстративного IgG3 человека представляет собой (SEQ ID NO:3):

пронумерованную с помощью EU-индекса в соответствии с Кабат, где X представляет собой (K) или отсутствует.
[0076] Аминокислотная последовательность домена CH2-CH3 иллюстративного IgG4 человека представляет собой (SEQ ID NO:4):

пронумерованную с помощью EU-индекса в соответствии с Кабат, где X представляет собой (K) или отсутствует.
[0077] Во всем описании настоящего изобретения нумерация остатков в константной области тяжелой цепи IgG представлена с помощью EU-индекса, как в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, NH1, MD (1991) (“Kabat”), явно включенного в данный документ посредством ссылки. Термин «EU-индекс в соответствии с Кабат» относится к нумерации EU константных доменов антитела IgG1 человека, представленного в Kabat. Аминокислоты из вариабельных доменов зрелых тяжелых и легких цепей иммуноглобулинов обозначены с помощью положения аминокислот в цепи. Kabat описал многочисленные аминокислотные последовательности антител, идентифицировал аминокислотную консенсусную последовательность для каждой подгруппы и присвоил номер остатка каждой аминокислоте, и CDR идентифицируют, как описано в Kabat (будет понятно, что CDRH1, обозначаемая в соответствии с Chothia, C. & Lesk, A. M. ((1987) “Canonical structures for the hypervariable regions of immunoglobulins,” J. Mol. Biol. 196:901-917), начинается на пять остатков ранее). Схема нумерации в соответствии с Кабат, распространяется на антитела, не включенные в его компендиум, в результате выравнивания рассматриваемого антитела по отношению к одной из консенсусных последовательностей в нумерации Кабат с помощью отсылки на консервативные аминокислоты. Данный способ присвоения номеров остатков стал стандартом в данной области техники и с его помощью легко идентифицируют аминокислоты в эквивалентных положениях в других антителах, в том числе химерных или гуманизированных вариантах. Например, аминокислота в положении 50 легкой цепи человеческого антитела занимает эквивалентное положение по отношению к положению 50 легкой цепи мышиного антитела.
[0078] Наблюдали полиморфизмы по ряду различных положений в константных областях аминокислот (например, Fc-положений, в том без ограничения положений 270, 272, 312, 315, 356 и 358, пронумерованных с помощью EU-индекса в соответствии с Кабат), и, следовательно, могут существовать незначительные различия между представленной последовательностью и последовательностями из уровня техники. Полиморфные формы человеческих иммуноглобулинов были хорошо охарактеризованы. В настоящее время известны 18 аллотипов Gm: G1m (1, 2, 3, 17) или G1m (a, ×, f, z), G2m (23) или G2m (n), G3m (5, 6, 10, 11, 13, 14, 15, 16, 21, 24, 26, 27, 28) или G3m (b1, c3, b3, b0, b3, b4, s, t, g1, c5, u, v, g5) (Lefranc, et al., “The Human IgG Subclasses: Molecular Analysis of Structure, Function And Regulation.” Pergamon, Oxford, pp. 43-78 (1990); Lefranc, G. et al., 1979, Hum. Genet.: 50, 199-211). В частности, предполагается, что антитела по настоящему изобретению могут включать в себя любой аллотип, изоаллотип или гаплотип любого гена иммуноглобулина, и не ограничены аллотипом, изоаллотипом или гаплотипом последовательностей, предусмотренных в данном документе. Кроме того, в некоторых системах экспрессии C-концевой аминокислотный остаток (выделен жирным выше) домена CH3 может быть удален посттрансляционно. Соответственно, C-концевой остаток домена CH3 представляет собой необязательный аминокислотный остаток в молекулах, связывающих ADAM9, по настоящему изобретению. В частности, настоящим изобретением охвачены молекулы, связывающие ADAM9, не имеющие C-концевого остатка домена CH3. Также, в частности, настоящим изобретением охвачены такие конструкции, содержащие C-концевой лизиновый остаток домена CH3.
[0079] Как указано выше, Fc-область природных антител IgG способна связываться с клеточными Fc-гамма-рецепторами (FcγR). Такое связывание приводит к передаче активирующих и ингибирующих сигналов в иммунной системе. Способность такого связывания приводить к диаметрально противоположным функциям отражает структурные различия среди различных FcγR, и, в частности, отражает то, обладает ли связанный FcγR иммунорецепторным тирозиновым активирующим мотивом («ITAM») или иммунорецепторным тирозиновым ингибирующим мотивом («ITIM»). Рекрутинг различных цитоплазматических ферментов в этим структурам обусловливает результат FcγR-опосредованных клеточных ответов. ITAM-содержащие FcγR включают в себя FcγRI, FcγRIIA, FcγRIIIA и активируют иммунную систему при связывании с Fc-областями (например, агрегированными Fc-областями, присутствующими в иммунном комплексе). FcγRIIB представляет собой единственный известный в настоящее время ITIM-содержащий FcγR; он функционирует с целью ослабления или ингибирования иммунной системы при связывании с агрегированными Fc-областями. Нейтрофилы человека экспрессируют ген FcγRIIA. Кластеризация FcγRIIA посредством иммунных комплексов или специфическое перекрестное связывание антител необходимо для агрегации ITAM с ассоциированными с рецепторами киназами, которые облегчает фосфорилирование ITAM. Фосфорилирование ITAM выступает в качестве сайта докинга для Syk-киназы, активация которой приводит к активации расположенных далее субстратов (например, PI3K). Клеточная активация приводит к высвобождению провоспалительных медиаторов. Ген FcγRIIB экспрессируется на В-лимфоцитах; его внеклеточный домен является на 96% идентичным по отношению к FcγRIIA и связывается с комплексами IgG неразличимым образом. Наличие ITIM в цитоплазматическом домене FcγRIIB определяет этот ингибирующий класс FcγR. В последнее время была установлена молекулярная основа данного ингибирования. При колигировании совместно с активирующим FcγR ITIM в FcγRIIB становится фосфорилированным и притягивает домен SH2 инозитолполифосфат-5'-фосфатазы (SHIP), которая гидролизует фосфоинозитоловые посредники, высвобождаемые вследствие активации ITAM-содержащей FcγR-опосредованной тирозинкиназы, впоследствии предупреждая приток внутриклеточного Ca++. Таким образом, перекрестное связывание FcγRIIB ослабляет активирующий ответ в отношении лигирования FcγR и ингибирует восприимчивость клетки. Таким образом, прекращается B-клеточная активация, B-клеточная пролиферация и секреция антител.
III. Биспецифические антитела, мультиспецифические диатела и диатела DART®
[0080] Способность антитела связывать эпитоп антигена зависит от наличия и аминокислотной последовательности доменов VL и VH антитела. Взаимодействия легкой цепи и тяжелой цепи антитела, в частности, взаимодействие доменов VL и VH, приводит к образованию одного из двух эпитопсвязывающих сайтов природного антитела, такого как IgG. Природные антитела способны связываться только с одной молекулой эпитопа (т.е., они являются моноспецифическими), хотя они могут связывать многочисленные копии молекул этого эпитопа (т.е., характеризоваться бивалентностью или мультивалентностью).
[0081] Функциональность антител может быть повышена в результате создания мультиспецифических молекул на основе антител, которые могут одновременно связывать два отдельных и различных антигена (или различные эпитопы одного и того же антигена) и/или в результате создания молекул на основе антител, имеющих более высокую валентность (т.е., более чем два связывающих сайта) по отношению к одному и то же эпитопу и/или антигену.
[0082] С целью получения молекул, имеющих более высокую способность, чем у природных антител, было разработано большое разнообразие форматов биспецифических антител (см., например, публикации в соответствии с PCT №№ WO 2008/003116, WO 2009/132876, WO 2008/003103, WO 2007/146968, WO 2009/018386, WO 2012/009544, WO 2013/070565), в большинстве из которых используются линкерные пептиды для слияния дополнительного эпитопсвязывающего фрагмента (например, scFv, VL, VH и т.д.) с кором антитела или в его пределах (IgA, IgD, IgE, IgG or IgM), или для слияния множественных эпитопсвязывающих фрагментов (например, двух Fab-фрагментов или scFv). В альтернативных форматах используются линкеры для слияния эпитопсвязывающего фрагмента (например, scFv, VL, VH и др.) с доменом димеризации, такими как домен CH2-CH3, или альтернативные пептиды (см., например, публикации согласно PCT №№ WO 2005/070966, WO 2006/107786A WO 2006/107617A, WO 2007/046893). В публикациях согласно PCT №№ WO 2013/174873, WO 2011/133886 и WO 2010/136172 раскрыто триспецифическое антитело, в котором домены CL и CH1 переключены из своих соответствующих природных положений, а домены VL и VH были диверсифицированы (см., например, публикации согласно PCT №№ WO 2008/027236; WO 2010/108127) с целью обеспечения их связывания с более чем одним антигеном. В публикациях согласно PCT №№ WO 2013/163427 и WO 2013/119903 раскрыто модифицирование домена CH2 с целью содержания аддукта слитого белка, содержащего связывающий домен. В публикациях согласно PCT №№ WO 2010/028797, WO 2010/028796 и WO 2010/028795 раскрыты рекомбинантные антитела, Fc-области которых были замещены дополнительными доменами VL и VH, таким образом, чтобы образовать трехвалентные связывающие молекулы. В публикациях согласно PCT №№ WO 2003/025018 и WO 2003/012069 раскрыты рекомбинантные диатела, отдельные цепи которых содержат домены scFv. В публикации согласно PCT № WO 2013/006544 раскрыты многовалентные Fab-молекулы, которые синтезируются в виде одной полипептидной цепи и затем подвергаются протеолизу с образованием гетеродимерных структур. В публикациях согласно PCT №№ WO 2014/022540, WO 2013/003652, WO 2012/162583, WO 2012/156430, WO 2011/086091, WO 2008/024188, WO 2007/024715, WO 2007/075270, WO 1998/002463, WO 1992/022583 и WO 1991/003493 раскрыто добавление дополнительных связывающих доменов или функциональных групп с антителом или участком антитела (например, добавление диатела к легкой цепи антитела, или добавление дополнительных доменов VL и VH к легким и тяжелым цепям антитела, или добавление гетерологичного слитого белка, или соединение нескольких Fab-доменов друг с другом в виде цепи).
[0083] Конструирование диатела основано на производном антитела, известном как одноцепочечный фрагмент вариабельного домена (scFv). Такие молекулы образуются в результате связывания вариабельных доменов легких и/или тяжелых цепей с помощью короткого связывающего пептида. Bird, R.E. et al. (1988) (“Single-Chain Antigen-Binding Proteins,” Science 242:423-426) описывает примеры связывающих пептидов, которые выступают в качестве мостиков, составляющих примерно 3,5 нм, между карбоксиконцом одного вариабельного домена и аминоконцом другого вариабельного домена. Были сконструированы и использованы линкеры других последовательностей (Bird et al. (1988) “Single-Chain Antigen-Binding Proteins,” Science 242:423-426). Линкеры, в свою очередь, можно модифицировать с целью дополнительных функций, таких как прикрепление лекарственных средств или прикрепление к твердым подложкам. Одноцепочечные варианты можно получать либо рекомбинантно, либо синтетически. Для синтетического получения scFv может быть использован синтезатор. Для рекомбинантного получения scFv подходящая плазмида, содержащая полинуклеотид, который кодирует scFv, может быть введена в подходящую клетку-хозяина, либо эукариотическую, такую как клетки дрожжевых грибов, растений, насекомых или млекопитающих, либо прокариотическую, такие как E. coli. Полинуклеотиды, кодирующие scFv, представляющие интерес, могут быть получены с помощью стандартных манипуляций, таких как лигирование полинуклеотидов. Образующийся в результате scFv может быть выделен с помощью стандартных методик очистки белка, известных в данной области техники.
[0084] В данной области техники была отмечена возможность получать диатела, которые отличаются от таких природных антител способностью связывать две или более различных молекул эпитопов (т.е., характеризующихся биспецифичностью или мультиспецифичностью, помимо бивалентности или мультивалентности) (см., например, Holliger, P. et al. (1993) “’Diabodies’: Small Bivalent And Bispecific Antibody Fragments,” Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; US 2004/0058400 (Hollinger et al.); US 2004/0220388 / WO 02/02781 (Mertens et al.); Alt et al. (1999) FEBS Lett. 454(1-2):90-94; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672; WO 02/02781 (Mertens et al.); Olafsen, T. et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Protein Eng. Des. Sel. 17(1):21-27; Wu, A. et al. (2001) “Multimerization Of A Chimeric Anti-CD20 Single Chain Fv-Fv Fusion Protein Is Mediated Through Variable Domain Exchange,” Protein Engineering 14(2):1025-1033; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Baeuerle, P.A. et al. (2009) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944).
[0085] Получение биспецифических связывающих молекул (например, не являющихся моноспецифическими диател) обеспечивает значительное преимущество над антителами, в том числе без ограничения способность связываться в «транс»-положении, достаточную для колигирования и/или колокализации различных клеток, которые экспрессируют различные эпитопы, и/или способность связываться в «цис»-положении, достаточную для колигирования и/или колокализации различных молекул, экспрессируемых одной и той же клеткой. Биспецифические связывающие молекулы (например, не являющиеся моноспецифическими диатела), таким образом, имеют области применения в широком диапазоне, включая терапию и иммунодиагностику. Биспецифичность способствует более высокой гибкости в конструировании и разработке диатела в различных областях применения, обеспечивая повышенную авидность в отношении мультимерных антигенов, перекрестное связывание различных антигенов и направленное целенаправленное воздействие в отношении специфических типов клеток на основании наличия обоих целевых антигенов. В связи со своей повышенной валентностью, низкими скоростями диссоциации и быстрым клиренсом из кровотока (в случае диател небольшого размера при ~50 кДа или менее) молекулы диател, известные в данной области техники, также показали определенное применение в области визуализации опухолей (Fitzgerald et al. (1997) “Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris,” Protein Eng. 10:1221-1225).
[0086] Возможность получать биспецифические диатела привела к их применению (в «транс»-положении) с целью колигирования двух клеток вместе, например, с помощью колигирующих рецепторов, которые присутствуют на поверхности различных клеток (например, перекрестного связывания цитотоксических T-клеток с опухолевыми клетками) (Staerz et al. (1985) “Hybrid Antibodies Can Target Sites For Attack By T Cells,” Nature 314:628-631; Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305; и Marvin et al. (2005) “Recombinant Approaches To IgG-Like Bispecific Antibodies,” Acta Pharmacol. Sin. 26:649-658). В альтернативном случае (или дополнительно) биспецифические (или три- или мультиспецифические) диатела могут быть использованы (в «цис»-положении) с целью колигирования молекул, таких как рецепторы и т.д., которые присутствуют на поверхности одной и той же клетки. Колигирование различных клеток и/или рецепторов является целесообразным для модулирования эффекторных функций и/или передачи сигналов с участием иммунных клеток. Мультиспецифические молекулы (например, биспецифические диатела), содержащие эпитопсвязывающие сайты, могут быть направлены на поверхностную детерминанту любой иммунной клетки, такую как CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д., которые экспрессируются на T-лимфоцитах, естественных киллерах (NK), антигенпрезентирующих клетках или других мононуклеарных клетках. В частности, эпитопсвязывающие сайты, направленные на рецептор клеточной поверхности, который присутствует на иммунных эффекторных клетках, являются пригодными при получении мультиспецифических связывающих молекул, способных опосредовать перенаправленное уничтожение клеток.
[0087] В то же время, вышеуказанные преимущества сопряжены с существенной стоимостью. Образование таких не являющихся моноспецифическими диател требует успешной сборки двух или более отдельных и различных полипептидов (т.е., такое образование требует, чтобы диатела образовывались посредством гетеродимеризации различных молекул полипептиных цепей). Данный факт отличается от моноспецифических диател, которые образуются посредством гомодимеризации идентичных полипептидных цепей. Поскольку по меньшей мере два различных полипептида (т.е., две полипептидные молекулы) должны быть предусмотрены для образования не являющегося моноспецифическим диатела и поскольку гомодимеризация таких полипептидов приводит к образованию неактивных молекул (Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588), получение таких полипептидов должно осуществляться таким образом, чтобы предупреждать ковалентное связывание между полипептидами одной и той же молекулы (т.е., чтобы предупреждать гомодимеризацию) (Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588). В данной области техники, таким образом, описана нековалентная ассоциация таких полипептидов (см., например, Olafsen et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Prot. Engr. Des. Sel. 17:21-27; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; и Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
[0088] В то же время, в данной области техники признано, что биспецифические диатела, состоящие из нековалентно ассоциированных полипептидов, являются нестабильными и легко диссоциируют на нефункциональные мономеры (см., например, Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
[0089] Учитывая эту проблему, в данной области техники преуспели в разработке стабильных, ковалентно связанных гетеродимерных не являющихся специфическими диател, называемых диатела DART®; см., например, патенты США №№ 9296816 и 9284375 и публикации патента США №№ 2015/0175697; 2014/0255407; 2014/0099318; 2013/0295121; WO 2012/018687; WO 2012/162068; 2010/0174053; WO 2010/080538; 2009/0060910; 2007-0004909; публикации Европейских патентов №№ EP 2714079; EP 2601216; EP 2376109; EP 2158221; EP 1868650; и публикации согласно PCT WO 2012/162068; WO 2012/018687; WO 2010/080538; WO 2006/113665; и Sloan, D.D. et al. (2015) “Targeting HIV Reservoir in Infected CD4 T Cells by Dual-Affinity Re-targeting Molecules (DARTs) that Bind HIV Envelope and Recruit Cytotoxic T Cells,” PLoS Pathog. 11(11):e1005233. doi: 10.1371/journal.ppat.1005233; Al Hussaini, M. et al. (2015) “Targeting CD123 In AML Using A T-Cell Directed Dual-Affinity Re-Targeting (DART®) Platform,” Blood pii: blood-2014-05-575704; Chichili, G.R. et al. (2015) “A CD3xCD123 Bispecific DART For Redirecting Host T Cells To Myelogenous Leukemia: Preclinical Activity And Safety In Nonhuman Primates,” Sci. Transl. Med. 7(289):289ra82; Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T-Cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Veri, M.C. et al. (2010) “Therapeutic Control Of B Cell Activation Via Recruitment Of Fcgamma Receptor IIb (CD32B) Inhibitory Function With A Novel Bispecific Antibody Scaffold,” Arthritis Rheum. 62(7):1933-1943; и Johnson, S. et al. (2010) “Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And in vivo B-Cell Depletion,” J. Mol. Biol. 399(3):436-449). Такие диатела содержат два или более ковалентно связанных в виде комплекса полипептида и предусматривают разработку одного или нескольких цистеиновых остатка в каждой из используемых полипептидных молекул, которые обеспечивают образование дисульфидных связей и, тем самым, ковалентное связывание одной или нескольких пар таких полипептидных цепей друг с другом. Например, было показано, что добавление цистеинового остатка к C-концу таких конструкций способствует дисульфидному связыванию между участвующими полипептидными связями, стабилизируя образующееся в результате диатело без воздействия на характеристики связывания диатела. Такие молекулы могут быть образованы таким образом, чтобы они были биспецифическими (или мультиспецифическими) и, таким образом, могут быть образованы таким образом ,чтобы колигировать две или более молекул. Такое колигирование способствует обеспечению усиленной иммунотерапии. Помимо этого, поскольку отдельные полипептидные цепи таких молекул образуют ковалентно связанный комплекс, молекулы характеризуются намного большей стабильностью, чем диатела, включающие не связанные ковалентно полипептидные цепи.
[0090] В последнее время были описаны трехвалентные и многовалентные молекулы, включающие два связывающих домена по типу диатела и один домен не по типу диатела, и Fc-область (см., например, публикации согласно PCT №№ WO 2015/184207 и WO 2015/184203). Такие связывающие молекулы могут быть использованы для получения моноспецифических, биспецифических или триспецифических молекул. Способность связывать три различные эпитопы обеспечивает усиленные способности.
[0091] Альтернативные конструкции известны в данной области техники для областей применения, где четырехвалентная молекула является предпочтительной, однако Fc не требуется, в том числе без ограничения четырехвалентные тандемные антитела, также обозначаемые как «TandAb» (см., например, публикации патентов США №№ 2005-0079170, 2007-0031436, 2010-0099853, 2011-020667 2013-0189263; публикации Европейских патентов №№ EP 1078004, EP 2371866, EP 2361936 и EP 1293514; публикации согласно PCT №№ WO 1999/057150, WO 2003/025018 и WO 2013/013700), которые образуются в результате гомодимеризации двух идентичных полипептидных цепей, каждая из которых имеет домен VH1, VL2, VH2 и VL2.
IV. ADAM9
[0092] Иллюстративный полипептид ADAM9 человека (последовательность в NCBI NP_003807, включая сигнальную последовательность из 28 аминокислотных остатков, показанную подчеркнутой) имеет аминокислотную последовательность (SEQ ID NO:5):
MGSGARFPSG TLRVRWLLLL GLVGPVLGAA RPGFQQTSHL SSYEIITPWR LTRERREAPR PYSKQVSYVI QAEGKEHIIH LERNKDLLPE DFVVYTYNKE GTLITDHPNI QNHCHYRGYV EGVHNSSIAL SDCFGLRGLL HLENASYGIE PLQNSSHFEH IIYRMDDVYK EPLKCGVSNK DIEKETAKDE EEEPPSMTQL LRRRRAVLPQ TRYVELFIVV DKERYDMMGR NQTAVREEMI LLANYLDSMY IMLNIRIVLV GLEIWTNGNL INIVGGAGDV LGNFVQWREK FLITRRRHDS AQLVLKKGFG GTAGMAFVGT VCSRSHAGGI NVFGQITVET FASIVAHELG HNLGMNHDDG RDCSCGAKSC IMNSGASGSR NFSSCSAEDF EKLTLNKGGN CLLNIPKPDE AYSAPSCGNK LVDAGEECDC GTPKECELDP CCEGSTCKLK SFAECAYGDC CKDCRFLPGG TLCRGKTSEC DVPEYCNGSS QFCQPDVFIQ NGYPCQNNKA YCYNGMCQYY DAQCQVIFGS KAKAAPKDCF IEVNSKGDRF GNCGFSGNEY KKCATGNALC GKLQCENVQE IPVFGIVPAI IQTPSRGTKC WGVDFQLGSD VPDPGMVNEG TKCGAGKICR NFQCVDASVL NYDCDVQKKC HGHGVCNSNK NCHCENGWAP PNCETKGYGG SVDSGPTYNE MNTALRDGLL VFFFLIVPLI VCAIFIFIKR DQLWRSYFRK KRSQTYESDG KNQANPSRQP GSVPRHVSPV TPPREVPIYA NRFAVPTYAA KQPQQFPSRP PPPQPKVSSQ GNLIPARPAP APPLYSSLT
Из 819 аминокислотных остатков ADAM9 (SEQ ID NO:5) остатки 1-28 представят собой сигнальную последовательность, остатки 29-697 представляют собой внеклеточный домен, остатки 698-718 представляют собой трансмембранный домен и остатки 719-819 представляют собой внутриклеточный домен. Три структурные домена расположены во внеклеточном домене: домен репролизина (M12B) семейства цинковых металлопротеиназ (примерно в остатках 212-406); домен дезинтегрина (примерно в остатках 423-497); и EGF-подобный домен (примерно в остатках 644-697). Ряд посттрансляционных модификаций и изоформ были идентифицированы, а белок протеолитически расщепляется в сети транс-Гольджи перед тем, как он достигает плазматической мембраны с образованием зрелого белка. Удаление продомена происходит в результате расщепления в двух различных сайтах. Процессинг вероятнее всего происходит с помощью пробелковой конвертазы, такой как фурин, на границе между продоменом и каталитическим доменом (Arg-205/Ala-206). Дополнительный вышерасположенный сайт расщепления пробелковой конвертазы (Arg-56/Glu-57) играет важную роль в активации ADAM9.
[0093] Иллюстративный полипептид ADAM9 яванского макака (последовательность в NCBI XM_005563126.2, включая возможную сигнальную последовательность из 28 аминокислотных остатков, показанную подчеркнутой) имеет аминокислотную последовательность (SEQ ID NO:6):
MGSGVGSPSG TLRVRWLLLL CLVGPVLGAA RPGFQQTSHL SSYEIITPWR LTRERREAPR PYSKQVSYLI QAEGKEHIIH LERNKDLLPE DFVVYTYNKE GTVITDHPNI QNHCHFRGYV EGVYNSSVAL SNCFGLRGLL HLENASYGIE PLQNSSHFEH IIYRMDDVHK EPLKCGVSNK DIEKETTKDE EEEPPSMTQL LRRRRAVLPQ TRYVELFIVV DKERYDMMGR NQTAVREEMI LLANYLDSMY IMLNIRIVLV GLEIWTNGNL INIAGGAGDV LGNFVQWREK FLITRRRHDS AQLVLKKGFG GTAGMAFVGT VCSRSHAGGI NVFGHITVET FASIVAHELG HNLGMNHDDG RDCSCGAKSC IMNSGASGSR NFSSCSAEDF EKLTLNKGGN CLLNIPKPDE AYSAPSCGNK LVDAGEECDC GTPKECELDP CCEGSTCKLK SFAECAYGDC CKDCRFLPGG TLCRGKTSEC DVPEYCNGSS QFCQPDVFIQ NGYPCQNNKA YCYNGMCQYY DAQCQVIFGS KAKAAPKDCF IEVNSKGDRF GNCGFSGNEY KKCATGNALC GKLQCENVQE IPVFGIVPAI IQTPSRGTKC WGVDFQLGSD VPDPGMVNEG TKCGADKICR NFQCVDASVL NYDCDIQKKC HGHGVCNSNK NCHCENGWAP PNCETKGYGG SVDSGPTYNE MNTALRDGLL VFFFLIVPLI VCAIFIFIKR DQLWRRYFRK KRSQTYESDG KNQANPSRQP VSVPRHVSPV TPPREVPIYA NRFPVPTYAA KQPQQFPSRP PPPQPKVSSQ GNLIPARPAP APPLYSSLT
Домен репролизина (M12B) семейства цинковых металлопротеиназ белка находится примерно в остатках 212-406; домен дезинтегрина белка находится примерно в остатках 423-497.
[0094] В соответствии с определенными вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению (например, scFv, антитела, биспецифические диатела и др.) характеризуются одним, двумя, тремя, четырьмя, пятью, шестью, семью или восемью из следующих критериев:
(1) способность иммуноспецифически связывать ADAM9 человека в виде эндогенно экспрессируемого на поверхности раковой клетки;
(2) специфически связывает ADAM9 человека и ADAM9 не относящегося к человеку примата (например, ADAM9 яванского макака) с аналогичной аффинностью связывания;
(3) специфически связывает ADAM9 человека с равновесной константой связывания (KD) 4 нМ или менее;
(4) специфически связывает ADAM9 не относящегося к человеку примата с равновесной константой связывания (KD) 4 нМ или менее;
(5) специфически связывает ADAM9 человека со скоростью ассоциации (ka) 5 x 105 M-1мин-1 или более;
(6) специфически связывает ADAM9 не относящегося к человеку примата со скоростью ассоциации (ka) 1 x 106 M-1мин-1 или более;
(7) специфически связывает ADAM9 человека со скоростью диссоциации (kd) 1 x 10-3 мин-1 или менее;
(8) специфически связывает ADAM9 не относящегося к человеку примата со скоростью диссоциации (kd) 9 x 10-4 мин-1 или менее;
(9) оптимизирована с целью наличия по меньшей мере 100-кратного усиления (например, по меньшей мере 100-кратного, меньшей мере 150-кратного, меньшей мере 200-кратного, меньшей мере 250-кратного, меньшей мере 300-кратного, меньшей мере 350-кратного, меньшей мере 400-кратного, меньшей мере 450-кратного, меньшей мере 500-кратного, меньшей мере 550-кратного или меньшей мере 600-кратного усиления) аффинности связывания (например, измеряемой с помощью анализа BIACORE®) с ADAM9 яванского макака и сохраняет высокую аффинность связывания с ADAM9 человека (например, измеряемую с помощью анализа BIACORE®) по сравнению с химерным или мышиным исходным антителом.
[0095] Описанные в данном документе константы связывания молекулы, связывающей ADAM9, могут быть определены с помощью поверхностного плазмонного резонанса, например, с помощью анализа BIACORE®. Данные, полученные в результате поверхностного плазмонного резонанса, могут быть подогнаны в соответствии с моделью связывания Ленгмюра 1:1 (одновременно ka и kd) и равновесной константой связывания KD, рассчитанной из соотношения констант скоростей kd/ka. Такие константы связывания могут быть определены для моновалентной молекулы, связывающей ADAM9 (т.е., молекулы, содержащей один эпитопсвязывающий сайт ADAM9), бивалентной молекулы, связывающей ADAM9 (т.е., молекулы, содержащей два эпитопсвязывающих сайта ADAM9 ), или молекулы, связывающей ADAM9, имеющий более высокую валентность (например, молекулы, содержащей три, четыре или пять эпитопсвязывающих сайта ADAM9).
[0096] Настоящее изобретение, в частности, охватывает молекулы, связывающие ADAM9 (например, антитела, диатела, трехвалентные связывающие молекулы и т.д.), содержащие анти-ADAM9 вариабельный(вариабельные) домен(домены) легкой цепи (VL) и анти-ADAM9 вариабельный(вариабельные) домен(домены) тяжелой цепи (VH), которые иммуноспецифически связываются с эпитопом полипептида ADAM9 человека. Если не указано иное, то все такие молекулы, связывающие ADAM9, способны иммуноспецифически связываться с ADAM9 человека. Как используется в данном документе, такие вариабельные домены ADAM9 обозначаются как «анти-ADAM9-VL» и «анти-ADAM9-VH» соответственно.
V. Мышиные антитела к ADAM9 человека
[0097] Было идентифицировано мышиное антитело к ADAM9 человека, которое блокирует активность целевого белка, связанную с процессингом, ADAM9, интернализируется и характеризуется противоопухолевой активностью (см., например, патент США № 8361475). Это антитело, обозначенное в патентах США №№ 7674619 и 8361475 как «антитело к KID24», продуцируемое клоном гибридомы ATCC PTA-5174, обозначается в данном документе как «MAB-A». MAB-A характеризуется выраженным предпочтительным связыванием с опухолями по сравнению с нормальными тканями (см., Фиг. 7A-7C). MAB-A характеризуется незначительным окрашиванием или его отсутствием в значительной панели нормальных типов клеток (Табл. 1).
Клетки (предпочтительно главные клетки) 2+ (m,c) 5% 1+ (m,c) 10%, главным образом апикальные
Мезенхимальные клетки хориальной пластинки 1-2+ (gr c), 5%
Интерстициальные клетки (возможно клетки Лейдига) 2-3+ (gr c) <5% и
1+ (gr c) 10%
Эндотелиальные клетки 1+ (c) <5%
[0098] Как показано на Фиг. 8A-8B, MAB-A связывает ADAM9 человека с высокой аффинностью, однако связывает ADAM9 не относящегося к человеку примата (например, яванского макака) в меньшей степени.
[0099] Аминокислотные последовательности доменов VL и VH MAB-A представлены ниже. Домены VH и VL MAB-A гуманизировали, а CDR оптимизировали с целью повышения аффинности и/или устранения потенциальных аминокислотных отклонений. CDRH3 дополнительно оптимизировали с целью усиления связывания с ADAM9 не относящегося к человеку примата, при этом сохраняя его высокую аффинность в отношении ADAM9 человека.
[00100] Предпочтительные молекулы, связывающие ADAM9 человека, по настоящему изобретению, имеют 1, 2 или все 3 из CDRH домена VH и/или 1, 2 или все 3 из CDRL домена VL оптимизированного варианта MAB-A, и предпочтительно дополнительно имеют гуманизированные каркасные области («FR») доменов VH и/или VL гуманизированного MAB-A. Другие предпочтительные молекулы, связывающие ADAM9, по настоящему изобретению, имеют целые домены VH и/или VL гуманизированного/оптимизированного варианта MAB-A. Такие предпочтительные молекулы, связывающие ADAM9, включают в себя антитела, биспецифические (или мультиспецифические) антитела, химерные или гуманизированные антитела, BiTe, диатела и др., а также такие связывающие молекулы, которые дополнительно содержат встречающуюся в природе или вариантную Fc-область.
[00101] Настоящее изобретение, в частности, относится к молекулам, связывающим ADAM9, содержащим домен, связывающий ADAM9, который имеет:
(A) (1) три CDRH домена VH MAB-A; и
(2) четыре FR домена VH гуманизированного варианта MAB-A; или
(B) (1) три CDRL домена VL MAB-A; и
(2) четыре FR домена VL гуманизированного варианта MAB-A; или
(C) три CDRH домена VH оптимизированного варианта MAB-A; и три CDRL домена VL MAB-A; или
(D) три CDRH домена VH MAB-A; и три CDRL домена VL оптимизированного варианта MAB-A; или
(Е) три CDRH домена VH оптимизированного варианта MAB-A; и три CDRL домена VL оптимизированного варианта MAB-A; или
(F) (1) три CDRH домена VH оптимизированного варианта MAB-A; и
(2) четыре FR домена VH гуманизированного варианта MAB-A; или
(G) (1) три CDRL домена VL оптимизированного варианта MAB-A; и
(2) четыре FR домена VL гуманизированного варианта MAB-A; или
(H) (1) домен VH гуманизированного/оптимизированного варианта MAB-A; и
(2) домен VL гуманизированного/оптимизированного варианта MAB-A. «MAB-A» означает мышиное антитело.
[00102] Аминокислотная последовательность домена VH мышиного антитела к ADAM9 MAB-A представляет собой SEQ ID NO:7 (остатки CDRH показаны подчеркнутыми):
QVQLQQPGAE LVKPGASVKL SCKASGYTFT SYWMHWVKQR PGQGLEWIGE IIPINGHTNY NEKFKSKATL TLDKSSSTAY MQLSSLASED SAVYYCARGG YYYYGSRDYF DYWGQGTTLT VSS
[00103] Аминокислотная последовательность домена СDRH1 MAB-A представляет собой (SEQ ID NO:8): SYWMH.
[00104] Аминокислотная последовательность домена СDRH2 MAB-A представляет собой (SEQ ID NO:9): EIIPINGHTNYNEKFKS.
[00105] Аминокислотная последовательность домена СDRH3 MAB-A представляет собой (SEQ ID NO:10): GGYYYYGSRDYFDY.
[00106] Аминокислотная последовательность домена VL мышиного антитела к ADAM9 MAB-A представляет собой SEQ ID NO:11 (остатки CDRL показаны подчеркнутыми):
DIVLTQSPAS LAVSLGQRAT ISCKASQSVD YDGDSYMNWY QQIPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLNIH PVEEEDAATY YCQQSHEDPF TFGGGTKLEI K
[00107] Аминокислотная последовательность домена СDRL1 MAB-A представляет собой (SEQ ID NO:12): KASQSVDYDGDSYMN.
[00108] Аминокислотная последовательность домена СDRL2 MAB-A представляет собой (SEQ ID NO:13): AASDLES.
[00109] Аминокислотная последовательность домена СDRL3 MAB-A представляет собой (SEQ ID NO:14): QQSHEDPFT.
VI. Иллюстративные гуманизированные/оптимизированные анти-ADAM9-VH-домены и домены VL
1. Вариантные домены VH MAB-A
[00110] Аминокислотные последовательности определенных предпочтительных гуманизированных/оптимизированных анти-ADAM9-VH-доменов MAB-A представляют собой варианты ADAM9-VH-домена MAB-A (SEQ ID NO:7) и представлены SEQ ID NO:15 (остатки CDRH показаны подчеркнутыми):
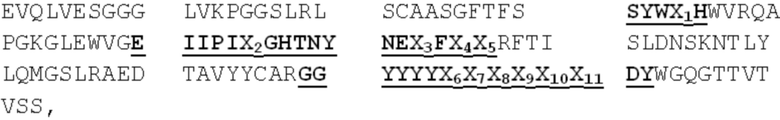
где: X1, X2, X3, X4, X5 и X6 выбирают независимо,
где: X1 представляет собой M или I; X2 представляет собой N или F;
X3 представляет собой K или R; X4 представляет собой K или Q;
X5 представляет собой S или G, и X6 представляет собой P, F, Y, W, I, L, V, T, G или D;
где: X7, X8, X9, X10 и X11 выбирают независимо, таким образом, что:
X7 представляет собой K или R;
X8 представляет собой F или M;
X9 представляет собой G;
X10 представляет собой W или F; и
X11 представляет собой M, L или K;
X7 представляет собой N или H;
X8 представляет собой S или K;
X9 представляет собой G или A;
X10 представляет собой T или V; и
X11 представляет собой M, L или K;
X7 представляет собой G;
X8 представляет собой K;
X9 представляет собой G или A;
X10 представляет собой V; и
X11 представляет собой M, L или K;
X7 представляет собой G;
X8 представляет собой K, M или N;
X9 представляет собой G;
X10 представляет собой V или T; и
X11 представляет собой L или M;
X7 представляет собой G;
X8 представляет собой S;
X9 представляет собой G;
X10 представляет собой V; и
X11 представляет собой L;
X7 представляет собой S;
X8 представляет собой N;
X9 представляет собой A;
X10 представляет собой V; и
X11 представляет собой L.
[00111] Аминокислотные последовательности предпочтительного гуманизированного анти-ADAM9 VH-домена MAB-A: hMAB-A VH(1) (SEQ ID NO:16) и определенных предпочтительных гуманизированных/оптимизированных анти-ADAM9-VH-доменов MAB-A:
hMAB-A VH(2) (SEQ ID NO:17) hMAB-A VH(2D) (SEQ ID NO:23)
hMAB-A VH(3) (SEQ ID NO:18) hMAB-A VH(2E) (SEQ ID NO:24)
hMAB-A VH(4) (SEQ ID NO:19) hMAB-A VH(2F) (SEQ ID NO:25)
hMAB-A VH(2A) (SEQ ID NO:20) hMAB-A VH(2G) (SEQ ID NO:26)
hMAB-A VH(2B) (SEQ ID NO:21) hMAB-A VH(2H) (SEQ ID NO:27)
hMAB-A VH(2C) (SEQ ID NO:22) hMAB-A VH(2I) (SEQ ID NO:28)
и hMAB-A VH(2J) (SEQ ID NO:29)
представлены ниже (остатки CDRH показаны одним подчеркиванием; различия по отношению к hMAB-A VH(1) (SEQ ID NO:7) показаны двойным подчеркиванием).
hMAB-A VH(1) (SEQ ID NO:16):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPINGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGSRDYF DYWGQGTTVT VSS
hMAB-A VH(2) (SEQ ID NO:17):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGSRDYF DYWGQGTTVT VSS
hMAB-A VH(3) (SEQ ID NO:18):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NERFQGRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGSRDYF DYWGQGTTVT VSS
hMAB-A VH(4) (SEQ ID NO:19):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWIHWVRQA PGKGLEWVGE IIPIFGHTNY NERFQGRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGSRDYF DYWGQGTTVT VSS
hMAB-A VH(2A) (SEQ ID NO:20):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYFNSGTL DYWGQGTTVT VSS
hMAB-A VH(2B) (SEQ ID NO:21):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYIGKGVL DYWGQGTTVT VSS
hMAB-A VH(2C) (SEQ ID NO:22):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYPRFGWL DYWGQGTTVT VSS
hMAB-A VH(2D) (SEQ ID NO:23):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYTGKGVL DYWGQGTTVT VSS
hMAB-A VH(2E) (SEQ ID NO:24):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYDSNAVL DYWGQGTTVT VSS
hMAB-A VH(2F) (SEQ ID NO:25):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYFHSGTL DYWGQGTTVT VSS
hMAB-A VH(2G) (SEQ ID NO:26):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYFNKAVL DYWGQGTTVT VSS
hMAB-A VH(2H) (SEQ ID NO:27):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGGSGVL DYWGQGTTVT VSS
hMAB-A VH(2I) (SEQ ID NO:28):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYPRQGFL DYWGQGTTVT VSS
hMAB-A VH(2J) (SEQ ID NO:29):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYYNSGTL DYWGQGTTVT VSS
[00112] Подходящие человеческие аминокислотные последовательности для FR гуманизированного и/или оптимизированного анти-ADAM9-VH-домена MAB-A представляют собой:
Домен FRH1 (SEQ ID NO:30): EVQLVESGGGLVKPGGSLRLSCAASGFTFS
Домен FRH2 (SEQ ID NO:31): WVRQAPGKGLEWVG
Домен FRH3 (SEQ ID NO:32): RFTISLDNSKNTLYLQMGSLRAEDTAVYYCAR
Домен FRH4 (SEQ ID NO:33): WGQGTTVTVSS
[00113] Подходящие альтернативные аминокислотные последовательности для домена CDRH1 анти-ADAM9-VH-домена MAB-A включают в себя:
SEQ ID NO:8: SYWMH
SEQ ID NO:34: SYWIH
[00114] Подходящие альтернативные аминокислотные последовательности для домена CDRH2 анти-ADAM9-VH-домена MAB-A включают в себя:
SEQ ID NO:9: EIIPINGHTNYNEKFKS
SEQ ID NO:35: EIIPIFGHTNYNEKFKS
SEQ ID NO:36: EIIPIFGHTNYNERFQG
Подходящие альтернативные аминокислотные последовательности для домена CDRH3 анти-ADAM9-VH-домена MAB-A включают в себя:
SEQ ID NO:10: GGYYYYGSRDYFDY
SEQ ID NO:37: GGYYYYFNSGTLDY
SEQ ID NO:38: GGYYYYIGKGVLDY
SEQ ID NO:39: GGYYYYPRFGWLDY
SEQ ID NO:40: GGYYYYTGKGVLDY
SEQ ID NO:41: GGYYYYDSNAVLDY
SEQ ID NO:42: GGYYYYFHSGTLDY
SEQ ID NO:43: GGYYYYFNKAVLDY
SEQ ID NO:44: GGYYYYGGSGVLDY
SEQ ID NO:45: GGYYYYPRQGFLDY
SEQ ID NO:46: GGYYYYYNSGTLDY
[00116] Соответственно, настоящее изобретение охватывает молекулы, связывающие ADAM9, имеющие VH-домен, содержащий:
(1) домен CDRH1, имеющий аминокислотную последовательность:
SEQ ID NO:47: SYWX1H,
где: X1 представляет собой M или I;
(2) домен CDRH2, имеющий аминокислотную последовательность:
SEQ ID NO:48: EIIPIX2GHTNYNEX3FX4X5,
где: X2, X3, X4 и X5 выбирают независимо, и
где: X2 представляет собой N или F; X3 представляет собой K или R;
X4 представляет собой K или Q; и X5 представляет собой S или G.
и
(3) домен CDRH3, имеющий аминокислотную последовательность:
SEQ ID NO:49: GGYYYYX6X7X8X9X10X11DY,
где: X6 представляет собой P, F, Y, W, I, L, V, T, G или D, и X7, X8, X9, X10 и X11 выбирают таким образом, что:
X7 представляет собой K или R;
X8 представляет собой F или M;
X9 представляет собой G;
X10 представляет собой W или F; и
X11 представляет собой M, L или K;
X7 представляет собой N или H;
X8 представляет собой S или K;
X9 представляет собой G или A;
X10 представляет собой T или V; и
X11 представляет собой M, L или K;
X7 представляет собой G;
X8 представляет собой K;
X9 представляет собой G или A;
X10 представляет собой V; и
X11 представляет собой M, L или K;
X7 представляет собой G;
X8 представляет собой K, M или N;
X9 представляет собой G;
X10 представляет собой V или T; и
X11 представляет собой L или M;
X7 представляет собой G;
X8 представляет собой S;
X9 представляет собой G;
X10 представляет собой V; и
X11 представляет собой L;
X7 представляет собой S;
X8 представляет собой N;
X9 представляет собой A;
X10 представляет собой V; и
X11 представляет собой L.
[001117] Тяжелая цепь первого иллюстративного гуманизированного/оптимизированного IgG1 производного/варианта MAB-A содержит домен hMAB-A VH (2) (SEQ ID NO:17) и имеет аминокислотную последовательность (SEQ ID NO:50):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYGSRDYF DYWGQGTTVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEL LGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LTCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGX,
где X представляет собой лизин (K) или отсутствует.
[00118] Тяжелая цепь второго иллюстративного гуманизированного/оптимизированного IgG1 производного/варианта MAB-A содержит домен hMAB-A VH (2С) (SEQ ID NO:22) и имеет аминокислотную последовательность (SEQ ID NO:51):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYPRFGWL DYWGQGTTVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEL LGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LTCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGX,
где X представляет собой лизин (K) или отсутствует.
[00119] Тяжелая цепь третьего иллюстративного гуманизированного/оптимизированного я IgG1 производного/варианта MAB-A содержит домен hMAB-A VH (2I) (SEQ ID NO:28) и имеет аминокислотную последовательность (SEQ ID NO:52):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYPRQGFL DYWGQGTTVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEL LGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LTCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGX,
где X представляет собой лизин (K) или отсутствует.
[00120] Предусмотренные в данном документе домены CH2-CH3 Fc-области могут быть сконструированы, например, с целью ослабления эффекторной функции. В соответствии с определенными вариантами осуществления домены CH2-CH3 иллюстративных гуманизированных/оптимизированных тяжелых цепей IgG1 по настоящему изобретению содержат один или несколько замен, выбранных из L234A и L235A.
[00121] Таким образом, тяжелая цепь четвертого иллюстративного гуманизированного/оптимизированного IgG1 производного/варианта MAB-A содержит домен hMAB-A VH (2I) (SEQ ID NO:28) и дополнительно содержит замены L234A и L235A в доменах CH2-CH3 Fc-области (SEQ ID NO:106), подчеркнутые ниже) и имеет аминокислотную последовательность (SEQ ID NO:202):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS SYWMHWVRQA PGKGLEWVGE IIPIFGHTNY NEKFKSRFTI SLDNSKNTLY LQMGSLRAED TAVYYCARGG YYYYPRQGFL DYWGQGTTVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LTCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGX,
где X представляет собой лизин (K) или отсутствует.
2. Вариантные домены VL MAB-A
[00122] Аминокислотные последовательности предпочтительных гуманизированных/оптимизированных анти-ADAM9-VL-доменов MAB-A представляют собой варианты ADAM9-VL-домена MAB-A (SEQ ID NO:11) и представлены SEQ ID NO:53 (остатки CDRL показаны подчеркнутыми):
DIVMTQSPDS LAVSLGERAT ISCX12ASQSVD YX13GDSYX14NWY
QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS
SLEPEDFATY YCQQSX15X16X17PF TFGQGTKLEI K,
где: X12, X13, X14, X15, X16 и X17 выбирают независимо, и
где: X12 представляет собой K или R; X13 представляет собой D или S;
X14 представляет собой M или L; X15 представляет собой H или Y;
X16 представляет собой E или S; и X17 представляет собой D или T.
[00123] Аминокислотные последовательности предпочтительного гуманизированного анти-ADAM9-VL-домена MAB-A: hMAB-A VL(1) (SEQ ID NO:54), и определенных предпочтительных гуманизированных/оптимизированных анти-ADAM9-VL-доменов MAB-A: hMAB-A VL(2) (SEQ ID NO:55), hMAB-A VL(3) (SEQ ID NO:56) и hMAB-A VL(4) (SEQ ID NO:57) представлены ниже (остатки CDRL показаны одним подчеркиванием; различия по отношению к hMAB-A VL(1) (SEQ ID NO:54) показаны двойным подчеркиванием).
hMAB-A VL(1) (SEQ ID NO:54):
DIVMTQSPDS LAVSLGERAT ISCKASQSVD YDGDSYMNWY QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS SLEPEDFATY YCQQSHEDPF TFGQGTKLEI K
hMAB-A VL(2) (SEQ ID NO:55):
DIVMTQSPDS LAVSLGERAT ISCKASQSVD YSGDSYMNWY QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS SLEPEDFATY YCQQSHEDPF TFGQGTKLEI K
hMAB-A VL(3) (SEQ ID NO:56):
DIVMTQSPDS LAVSLGERAT ISCRASQSVD YSGDSYMNWY QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS SLEPEDFATY YCQQSHEDPF TFGQGTKLEI K
hMAB-A VL(4) (SEQ ID NO:57):
DIVMTQSPDS LAVSLGERAT ISCRASQSVD YSGDSYLNWY QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS SLEPEDFATY YCQQSYSTPF TFGQGTKLEI K
[00124] Соответственно, подходящие человеческие аминокислотные последовательности для FR гуманизированного и/или оптимизированного анти-ADAM9-VL-домена MAB-A представляют собой:
Домен FRL1 (SEQ ID NO:58): DIVMTQSPDSLAVSLGERATISC
Домен FRL2 (SEQ ID NO:59): WYQQKPGQPPKLLIY
Домен FRL3 (SEQ ID NO:60): GIPARFSGSGSGTDFTLTISSLEPEDFATYYC
Домен FRL4 (SEQ ID NO:61): FGQGTKLEIK
[00125] Подходящие альтернативные аминокислотные последовательности для домена CDRL1 анти-ADAM9-VL-домена включают в себя:
SEQ ID NO:12: KASQSVDYDGDSYMN
SEQ ID NO:62: KASQSVDYSGDSYMN
SEQ ID NO:63: RASQSVDYSGDSYMN
SEQ ID NO:64: RASQSVDYSGDSYLN
[00126] Подходящие альтернативные аминокислотные последовательности для домена CDRL3 анти-ADAM9-VL-домена включают в себя:
SEQ ID NO:14: QQSHEDPFT
SEQ ID NO:65: QQSYSTPFT
[00127] Соответственно, настоящее изобретение охватывает домен VL антитела к ADAM9, содержащий:
(1) домен CDRL1, имеющий аминокислотную последовательность:
SEQ ID NO:66: X12ASQSVDYX13GDSYX14N,
где: X12, X13, X14 выбирают независимо, и
где: X12 представляет собой K или R; X13 представляет собой D или S; и X14 представляет собой M или L;
(2) домен CDRL2, имеющий аминокислотную последовательность:
SEQ ID NO:13: AASDLES
и
(3) домен CDRL3, имеющий аминокислотную последовательность:
SEQ ID NO:67: QQSX15X16X17PFT,
где: X15, X16 и X17 выбирают независимо, и
где: X15 представляет собой H или Y; X16 представляет собой E или S; и X17 представляет собой D или T.
[00128] Легкая цепь иллюстративного гуманизированного/оптимизированного IgG1 производного/варианта MAB-A содержит домен hMAB-A VL (2) (SEQ ID NO:55) и имеет аминокислотную последовательность (SEQ ID NO:68):
DIVMTQSPDS LAVSLGERAT ISCKASQSVD YSGDSYMNWY QQKPGQPPKL LIYAASDLES GIPARFSGSG SGTDFTLTIS SLEPEDFATY YCQQSHEDPF TFGQGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
[00129] Таким образом, в настоящем изобретении явно предусмотрены молекулы, связывающие ADAM9 (например, антитела, диатела, трехвалентные связывающие молекулы и т.д.), которые иммуноспецифически связываются с эпитопом полипептида ADAM9 человека и которые содержат любую из предусмотренных выше CDRH1, CDRH2, CDRH3, CDRL1, CDRL2 или CDRL3 MAB-A и, в частности, предусмотрены такие молекулы, связывающие ADAM9, которые содержит одну из предусмотренных выше CDRH1 MAB-A, одну из предусмотренных выше CDRH2 MAB-A, одну из предусмотренных выше CDRH3 MAB-A, одну из предусмотренных выше CDRL1 MAB-A, одну из предусмотренных выше CDRL2 MAB-A и одну из предусмотренных выше CDRL3 MAB-A.
[00130] В настоящем изобретении дополнительно предусмотрены такие молекулы, связывающие ADAM9, которые дополнительно содержат любую из предусмотренных выше гуманизированных FRH1, FRH2, FRH3 или FRH4, FRL1, FRL2, FRL3 или FRL4 MAB-A и, в частности, предусмотрены такие молекулы, связывающие ADAM9, которые содержат FRH1, FRH2, FRH3 и FRH4, и/или которые содержат FRL1, FRL2, FRL3, FRL4 и FRH1.
[00131] В соответствии с некоторыми вариантами осуществления молекулы, связывающие ADAM9, содержат домен CDRH1, домен CDRH2 и домен CDRH3 и домен CDRL1, домен CDRL2 и домен CDRL3, имеющие последовательности, выбранные из группы, состоящей из:
(a) SEQ ID NO:8, 35 и 10 и SEQ ID NO:62, 13 и 14 соответственно;
(b) SEQ ID NO:8, 35 и 10 и SEQ ID NO:63, 13 и 14 соответственно;
(c) SEQ ID NO:8, 36 и 10 и SEQ ID NO:63, 13 и 14 соответственно;
(d) SEQ ID NO:34, 36 и 10 и SEQ ID NO:64, 13 и 65 соответственно;
(e) SEQ ID NO:8, 35 и 37 и SEQ ID NO:62, 13 и 14 соответственно;
(f) SEQ ID NO:8, 35 и 38 и SEQ ID NO:62, 13 и 14 соответственно;
(g) SEQ ID NO:8, 35 и 39 и SEQ ID NO:62, 13 и 14 соответственно;
(h) SEQ ID NO:8, 35 и 40 и SEQ ID NO:62, 13 и 14 соответственно;
(i) SEQ ID NO:8, 35 и 41 и SEQ ID NO:62, 13 и 14 соответственно;
(j) SEQ ID NO:8, 35 и 42 и SEQ ID NO:62, 13 и 14 соответственно;
(k) SEQ ID NO:8, 35 и 43 и SEQ ID NO:62, 13 и 14 соответственно;
(l) SEQ ID NO:8, 35 и 44 и SEQ ID NO:62, 13 и 14 соответственно;
(m) SEQ ID NO:8, 35 и 45 и SEQ ID NO:62, 13 и 14 соответственно; и
(n) SEQ ID NO:8, 35 и 46 и SEQ ID NO:62, 13 и 14 соответственно.
[00132] В соответствии с определенными вариантами осуществления молекулы, связывающие ADAM9, содержат домен CDRH1, домен CDRH2 и домен CDRH3 и домен CDRL1, домен CDRL2 и домен CDRL3, имеющие последовательности SEQ ID NO:8, 35 и 45 и SEQ ID NO:62, 13 и 14 соответственно.
[00133] В соответствии с некоторыми вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, которые на по меньшей мере 90%, на по меньшей мере 95%, на по меньшей мере 99% или на 100% идентичны следующим последовательностям:
SEQ ID NO:17 и SEQ ID NO:55 соответственно;
SEQ ID NO:17 и SEQ ID NO:56 соответственно;
SEQ ID NO:18 и SEQ ID NO:56 соответственно;
SEQ ID NO:19 и SEQ ID NO:57 соответственно;
SEQ ID NO:20 и SEQ ID NO:55 соответственно;
SEQ ID NO:21 и SEQ ID NO:55 соответственно;
SEQ ID NO:22 и SEQ ID NO:55 соответственно;
SEQ ID NO:23 и SEQ ID NO:55 соответственно;
SEQ ID NO:24 и SEQ ID NO:55 соответственно;
SEQ ID NO:25 и SEQ ID NO:55 соответственно;
SEQ ID NO:26 и SEQ ID NO:55 соответственно;
SEQ ID NO:27 и SEQ ID NO:55 соответственно;
SEQ ID NO:28 и SEQ ID NO:55 соответственно; и
SEQ ID NO:29 и SEQ ID NO:55 соответственно.
[00134] Под «по сути идентичный» или «идентичный» подразумевается полипептид, на по меньшей мере 50% идентичный эталонной аминокислотной последовательности (например, любой из аминокислотных последовательностей, описанных в данном документе). Предпочтительно, такая последовательность на по меньшей мере 60%, более предпочтительно на по меньшей мере 80% или на по меньшей мере 85%, и более предпочтительно на по меньшей мере 90%, на по меньшей мере 95%, на по меньшей мере 99% или даже 100% идентична по аминокислотному уровню полипептидной последовательности, используемой для сравнения.
[00135] Идентичность последовательностей в типичном случае измеряется с помощью компьютерной программы анализа последовательностей (например, пакета компьютерных программ для анализа последовательностей Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, Wis. 53705, программы BLAST, BESTFIT, GAP или PILEUP/PRETTYBOX). Такая компьютерная программа сопоставляет идентичные или аналогичные последовательности с помощью присваивания степеней гомологии различным заменам, делециям и/или другим модификациям. Консервативные замены в типичном случае включают в себя замены в пределах следующих групп: глицин, аланин; валин, изолейцин, лейцин; аспарагиновая кислота, глутаминовая кислота, аспарагин, глутамин; серин, треонин; лизин, аргинин; и фенилаланин, тирозин. В иллюстративном подходе для определения степени идентичности может быть использована программа BLAST, с индексом вероятности от e-3 до e-100, указывающим на близкородственную последовательность.
[00136] В соответствии с определенными вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющие последовательности, которые на по меньшей мере 90%, на по меньшей мере 95%, на по меньшей мере 99% или на 100% идентичны последовательностям SEQ ID NO:28 и SEQ ID NO:55 соответственно.
[00137] В соответствии с определенными последовательностями молекулы, связывающие ADAM9, по настоящему изобретению, содержат следующую последовательность тяжелой цепи и легкой цепи:
SEQ ID NO:50 и SEQ ID NO:68 соответственно;
SEQ ID NO:51 и SEQ ID NO:68 соответственно;
SEQ ID NO:52 и SEQ ID NO:68 соответственно; и
SEQ ID NO:202 и SEQ ID NO:68 соответственно.
[00138] В настоящем изобретении также явно предусмотрены молекулы, связывающие ADAM9 (например, антитела, диатела, трехвалентные связывающие молекулы и т.д.), которые иммуноспецифически связываются с эпитопом полипептида ADAM9 человека и которые содержат любые из предусмотренных выше гуманизированных/оптимизированных ADAM9 VL- или VH-доменов MAB-A. В настоящем изобретении, в частности, предусмотрены такие молекулы, связывающие ADAM9, которые содержат любую из следующих комбинаций гуманизированных анти-ADAM9 VL- или VH-доменов:
[00139] Настоящее изобретение особо охватывает молекулы, связывающие ADAM9, содержащие (i) гуманизированный/оптимизированный анти-ADAM9-VL- и/или VH-домен, предусмотренный выше, и (ii) Fc-область. В соответствии с конкретными вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, представляют собой моноклональные антитела, содержащие (i) гуманизированный/оптимизированный анти-ADAM9-VL- и/или VH-домен, предусмотренный выше, и (ii) Fc-область. В соответствии с другими вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, выбирают из группы, состоящей из моноклональных антител, мультиспецифических антител, синтетических антител, химерных антител, одноцепочечных Fv (scFv), одноцепочечных антител, Fab-фрагментов, F(ab’)-фрагментов, связанных дисульфидными связями биспецифических Fv (sdFv), BiTE, диател и трехвалентных связывающих молекул.
[00140] Несмотря на то, что определенные модификации анти-ADAM9 VH- и VL-доменов представлены выше и сравниваются на Фиг. 9A-9B, необязательно модифицировать все или большую часть этих остатков при разработке гуманизированного и/или оптимизированного анти-ADAM9-VH- или VL-домена по настоящему изобретению. Настоящее изобретение также охватывает незначительные вариации этих последовательностей VH и VL, в том числе, например, аминокислотные замены C-коневых и/или N-концевых аминокислотных остатков, которые могут быть включены для облегчения субклонирования.
VII. Химерные антигенные рецепторы
[00141] Молекулы, связывающие ADAM9, по настоящему изобретению, могут представлять собой моноспецифические одноцепочечные молекулы, такие как анти-ADAM9 одноцепочечные вариабельные фрагменты («анти-ADAM9-scFv») или анти-ADAM9 химерные антигенные рецепторы («анти-ADAM9-CAR»). Как описано выше, scFv образуются в результате связывания вариабельных доменов легких и тяжелых цепей совместно посредством короткого связывающего пептида. Химерные антигенные рецепторы первого поколения («CAR») в типичном случае содержат внутриклеточный домен из ζ-цепи CD3, который представляет собой первичный переносчик сигналов от эндогенных T-клеточных рецепторов («TCR»). CAR второго поколения имеют дополнительные внутриклеточные сигнальные домены от костимулирующих белковых рецепторов (например, CD28, 41BB, ICOS и т.д.), слитых с цитоплазматическим хвостом CAR с целью получения дополнительных сигналов по отношению T-клетке. CAR третьего поколения сочетают многочисленные сигнальные домены, такие как CD3ζ-CD28-41BB или CD3ζ-CD28-OX40, с целью усиления их активности (Tettamanti, S. et al. (2013) “Targeting Of Acute Myeloid Leukaemia By Cytokine-Induced Killer Cells Redirected With A Novel CD123-Specific Chimeric Antigen Receptor,” Br. J. Haematol. 161:389-401; Gill, S. et al. (2014) “Efficacy Against Human Acute Myeloid Leukemia And Myeloablation Of Normal Hematopoiesis In A Mouse Model Using Chimeric Antigen Receptor-Modified T Cells,” Blood 123(15): 2343-2354; Mardiros, A. et al. (2013) “T Cells Expressing CD123-Specific Chimeric Antigen Receptors Exhibit Specific Cytolytic Effector Functions And Antitumor Effects Against Human Acute Myeloid Leukemia,” Blood 122:3138-3148; Pizzitola, I. et al. (2014) “Chimeric Antigen Receptors Against CD33/CD123 Antigens Efficiently Target Primary Acute Myeloid Leukemia Cells in vivo,” Leukemia doi:10.1038/leu.2014.62). Химерные антигенные рецепторы описаны в патентах США №№ 9447194; 9266960; 9212229; 9074000; 8822196; и 8,465,743, и в публикациях патентов №№ 2016/0272718, 2016/0144026, 2016/0130357, 2016/0081314, 2016/0075784, 2016/0058857, 2016/0046729, 2016/0046700, 2016/0045551, 2016/0015750, 2015/0320799, 2015/0307623, 2015/0307564, 2015/0038684, 2014/0134142 и 2013/0280285.
[00142] Анти-ADAM9-CAR по настоящему изобретению содержат анти-ADAM9-scFv, слитые с внутриклеточным доменом рецептора. Вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH) анти-ADAM9-scFv выбирают из любого из гуманизированных анти-ADAM9-VL- и анти-ADAM9-VH-доменов, раскрытых в данном документе. Предпочтительно домен VH выбирают из группы, состоящей из: hMAB-A VH(1) (SEQ ID NO:16), hMAB-A VH(2) (SEQ ID NO:17), hMAB-A VH(3) (SEQ ID NO:18), hMAB-A VH(4) (SEQ ID NO:19), hMAB-A VH(2A) (SEQ ID NO:20), hMAB-A VH(2B) (SEQ ID NO:21), hMAB-A VH(2C) (SEQ ID NO:22), hMAB-A VH(2D) (SEQ ID NO:23), hMAB-A VH(2E) (SEQ ID NO:24), hMAB-A VH(2F) (SEQ ID NO:25), hMAB-A VH(2G) (SEQ ID NO:26), hMAB-A VH(2H) (SEQ ID NO:27), hMAB-A VH(2I) (SEQ ID NO:28) и hMAB-A VH(2J) (SEQ ID NO:29), а домен VL выбирают из группы, состоящей из: hMAB-A VL(1) (SEQ ID NO:54), hMAB-A VL(2) (SEQ ID NO:55), hMAB-A VL(3) (SEQ ID NO:56) и hMAB-A VL(4) (SEQ ID NO:57). Комбинации гуманизированных/оптимизированных анти-ADAM9-VL- и анти-ADAM9-VH-доменов и комбинации CDRH и CDRL, которые могут быть использованы для образования таких химерных антигенных рецепторов представлены выше.
[00143] Внутриклеточный домен анти-ADAM9-CAR по настоящему изобретению предпочтительно выбирают из внутриклеточного домена любого из: 41BB-CD3ζ, b2c-CD3ζ, CD28, CD28-4-1BB-CD3ζ, CD28-CD3ζ, CD28-FcεRIγ, CD28mut-CD3ζ, CD28-OX40-CD3ζ, CD28-OX40-CD3ζ, CD3ζ, CD4-CD3ζ, CD4-FcεRIγ, CD8-CD3ζ, FcεRIγ, FcεRIγCAIX, герегулин-CD3ζ, IL-13-CD3ζ или Ly49H-CD3ζ (Tettamanti, S. et al. (2013) “Targeting Of Acute Myeloid Leukaemia By Cytokine-Induced Killer Cells Redirected With A Novel CD123-Specific Chimeric Antigen Receptor,” Br. J. Haematol. 161:389-401; Gill, S. et al. (2014) “Efficacy Against Human Acute Myeloid Leukemia And Myeloablation Of Normal Hematopoiesis In A Mouse Model Using Chimeric Antigen Receptor-Modified T Cells,” Blood 123(15): 2343-2354; Mardiros, A. et al. (2013) “T Cells Expressing CD123-Specific Chimeric Antigen Receptors Exhibit Specific Cytolytic Effector Functions And Antitumor Effects Against Human Acute Myeloid Leukemia,” Blood 122:3138-3148; Pizzitola, I. et al. (2014) “Chimeric Antigen Receptors Against CD33/CD123 Antigens Efficiently Target Primary Acute Myeloid Leukemia Cells in vivo,” Leukemia doi:10.1038/leu.2014.62).
VIII. Мультиспецифические молекулы, связывающие ADAM9
[00144] Настоящее изобретение также направлено на мультиспецифические (например, биспецифические, триспецифические и т.д.) молекулы, связывающие ADAM9, содержащие эпитопсвязывающие сайты (предпочтительно содержащие 1, 2 или все 3 из CDRH анти-ADAM9-VH-домена по настоящему изобретению и/или 1, 2 или все 3 из CDRL анти-ADAM9-VL-домена по настоящему изобретению, или такой анти-ADAM9-VH-домен и/или такой анти-ADAM9-VL-домен), и дополнительно содержащие второй эпитопсвязывающий сайт, который иммуноспецифически связывается со вторым эпитопом, при этом такой второй эпитоп представляет собой (i) другой эпитоп ADAM9, или (ii) эпитоп молекулы, которая не представляет собой ADAM9. Такие мультиспецифические молекулы, связывающие ADAM9, предпочтительно содержат комбинацию эпитопсвязывающих сайтов, которые распознают совокупность антигенов, уникальных по отношению к целевым клеткам или типу тканей. В частности, настоящее изобретение относится к мультиспецифическим молекулам, связывающим ADAM9, которые способны связываться с эпитопом ADAM9 и эпитопом молекулы, присутствующей на поверхности эффекторной клетки, особенно T-лимфоцита, естественного киллера (NK) или другой мононуклеарной клетки. Например, такие молекулы, связывающие ADAM9, по настоящему изобретению, могут быть сконструированы таким образом, чтобы содержать эпитопсвязывающий сайт, который иммуноспецифически связывает CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR) или NKG2D.
[00145] Один вариант настоящего изобретения относится к биспецифическим молекулам, связывающим ADAM9, которые способны связываться с «первым эпитопом» и «вторым эпитопом», при этом такие эпитопы не являются идентичными друг другу. Такие биспецифические молекулы содержат домены «VL1»/«VH1», которые способны связываться с первым эпитопом, и домены «VL2»/«VH2», которые способны связываться со вторым эпитопом. Обозначение «VL1» и «VH1» означает соответственно вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связываются с «первым» эпитопом таких биспецифических молекул. Аналогичным образом, обозначение «VL2» и «VH2» означает соответственно вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связываются со «вторым» эпитопом таких биспецифических молекул. Не имеет значения, обозначен ли определенный эпитоп как первый или второй эпитоп; такое обозначение имеет значение только в связи с наличием или ориентацией доменов полипептидных цепей связывающих молекул по настоящему изобретению. В соответствии с одним вариантом осуществления один из таких эпитопов представляет собой эпитоп ADAM9 человека, а другой представляет собой другой эпитоп ADAM9, или представляет собой эпитоп молекулы, которая не представляет собой ADAM9. В соответствии с определенными вариантами осуществления один из таких эпитопов представляет собой эпитоп ADAM9 человека, а другой представляет собой эпитоп молекулы (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующей на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. В соответствии с определенными вариантами осуществления биспецифическая молекула содержит более двух эпитопсвязывающих сайтов. Такие биспецифические молекулы будут связывать по меньшей мере один эпитоп ADAM9 и по меньшей мере один эпитоп молекулы, которая не представляет собой ADAM9, и может дополнительно связывать дополнительные эпитопы ADAM9 и/или дополнительные эпитопы молекулы, которая не представляет собой ADAM9.
[00146] Настоящее изобретение, в частности, относится к биспецифическим, триспецифическим и мультиспецифическим молекулам, связывающим ADAM9 (например, биспецифическим антителам, биспецифическим диателам, трехвалентных связывающим молекулам и т.д.), которые имеют эпитопсвязывающие фрагменты антител (например, домены VL и VH), которые способствуют их координированному связыванию по меньшей мере с одним эпитопом ADAM9 и по меньшей мере с одним эпитопом второй молекулы, которая не представляет собой ADAM9. Выбор доменов VL и VH полипептидов таких молекул координируется таким образом, что полипептидные цепи, которые образуют такие мультиспецифические молекулы, связывающие ADAM9, собираются с образованием по меньшей мере одного функционального эпитопсвязывающего сайта, который является специфическим по меньшей мере для одного эпитопа ADAM9, и по меньшей мере одного функционального эпитопсвязывающего сайта который является специфическим по меньшей мере для одного эпитопа молекулы, которая не представляет собой ADAM9. Предпочтительно мультиспецифические молекулы, связывающие ADAM9, содержат 1, 2 или все 3 из CDRH анти-ADAM9-VH-домена по настоящему изобретению и/или 1, 2 или все 3 из CDRL анти-ADAM9-VL-домена по настоящему изобретению, или такой анти-ADAM9-VH-домен и/или такой анти-ADAM9-VL-домен, предусмотренные в данном документе.
Биспецифические антитела
[00147] Настоящее изобретение охватывает биспецифические антитела, способные одновременно связываться с эпитопом ADAM9 и эпитопом молекулы, которая не представляет собой ADAM9. В соответствии с некоторыми вариантами осуществления биспецифическое антитело, способное одновременно связываться с ADAM9 и второй молекулой, которая не представляет собой ADAM9, получают с помощью способов, описанных в публикациях согласно PCT №№ WO 1998/002463, WO 2005/070966, WO 2006/107786 WO 2007/024715, WO 2007/075270, WO 2006/107617, WO 2007/046893, WO 2007/146968, WO 2008/003103, WO 2008/003116, WO 2008/027236, WO 2008/024188, WO 2009/132876, WO 2009/018386, WO 2010/028797, WO2010028796, WO 2010/028795, WO 2010/108127, WO 2010/136172, WO 2011/086091, WO 2011/133886, WO 2012/009544, WO 2013/003652, WO 2013/070565, WO 2012/162583, WO 2012/156430, WO 2013/174873 и WO 2014/022540, каждая из которых тем самым включена в данный документ посредством ссылки в полном объеме.
B. Биспецифические диатела, не имеющие Fc-областей
[00148] Один вариант осуществления настоящего изобретения относится к биспецифическим диателам, которые способны связываться с первым эпитопом и вторым эпитопом, при этом первый эпитоп представляет собой эпитоп ADAM9 человека, а второй представляет собой эпитоп молекулы, которая не представляет собой ADAM9, предпочтительно молекулы (например, CD2, CD3, CD8, CD16, T-клеточного рецептора (TCR), NKG2D и т.д.), присутствующей на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. Такие диатела содержат первую полипептидную цепь и вторую полипептидную цепь, и предпочтительно состоят из них, последовательности которых обеспечивают ковалентное связывание друг с другом с образованием ковалентно ассоциированного диатела, которое способно одновременно связываться с эпитопом ADAM9 и вторым эпитопом.
[00149] Первая полипептидная цепь такого варианта осуществления биспецифических диател содержит в направлении от N-конца до C-конца: N-конец, домен VL моноклонального антитела, способный связываться либо с первым, либо со вторым эпитопом (например, либо VLanti-ADAM9-VL либо VLEpitope 2), первый промежуточный спейсерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо со вторым эпитопом (если такая первая полипептидная цепь содержит VLanti-ADAM9-VL), либо с ADAM9 (если такая первая полипептидная цепь содержит VLEpitope 2), второй промежуточный спейсерный пептид (линкер 2), необязательно содержащий цистеиновый остаток, активирующий сборку гетеродимера домен и C-конец (Фиг. 1).
[00150] Вторая полипептидная цепь данного варианта осуществления биспецифических диател содержит в направлении от N-конца до C-конца: N-конец, домен VL моноклонального антитела, способный связываться либо с первым, либо со вторым эпитопом (например, либо VLanti-ADAM9-VL либо VLEpitope 2, и при этом домен VL не выбирают для вставки в первую полипептидную цепь диатела), первый промежуточный спейсерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо со вторым эпитопом (если такая вторая полипептидная цепь содержит VLanti-ADAM9-VL), либо с ADAM9 (если такая вторая полипептидная цепь содержит VLEpitope 2), второй промежуточный спейсерный пептид (линкер 2), необязательно содержащий цистеиновый остаток, активирующий сборку гетеродимера домен и C-конец (Фиг. 1).
[00151] Домен VL первой полипептидной цепи взаимодействует с доменом VH второй полипептидной цепи с образованием первого функционального эпитопсвязывающего сайта, который является специфическим для первого антигена (т.е., либо ADAM9, либо молекулы, которая содержит второй эпитоп). Аналогичным образом, домен VL второй полипептидной цепи взаимодействует с доменом VH первой полипептидной цепи с образованием второго функционального эпитопсвязывающего сайта, который является специфическим для второго антигена (например, либо молекулы, которая содержит второй эпитоп, либо ADAM9). Таким образом, выбор доменов VL и VH первой и второй полипептидных цепей координируется таким образом, что две полипептидные цепи диатела вместе содержат домены VL и VH, способные связывать как эпитоп ADAM9, таки второй эпитоп (т.е., они вместе содержат VLanti-ADAM9-VL/VHanti-ADAM9-VH и VLEpitope 2/VHEpitope 2).
[00152] Более предпочтительно длину промежуточного спейсерного пептида (т.е., «линкера 1» который разделяет такие домены VL и VH) выбирают таким образом, что значительно или полностью предупреждать связывание доменов VL и VH полипептида друг с другом (например, состоит из 0, 1, 2, 3, 4, 5, 6, 7, 8 или 9 аминокислотных остатков промежуточных линкеров). Таким образом, домены VL и VH первой полипептидной цепи по сути или полностью неспособны связываться друг с другом. Аналогичным образом, домены VL и VH второй полипептидной цепи по сути или полностью неспособны связываться друг с другом. Предпочтительный промежуточный спейсерный пептид (линкер 1) имеет последовательность (SEQ ID NO:69): GGGSGGGG.
[00153] Длину и состав второго промежуточного спейсерного пептида («линкер 2») выбирают на основе выбора одного или нескольких полипептидных доменов, которые активируют такую димеризацию (т.е., «активирующего сборку гетеродимера домена»). В типичном случае второй промежуточный спейсерный пептид (линкер 2) будет содержать 3-20 аминокислотные остатков. В частности, в случае, если используемый(используемые) активирующий(активирующие) сборку гетеродимера домен(домены), не содержит(не содержат) цистеиновый остаток, используется содержащий цистеин второй промежуточный спейсерный пептид (линкер 2). Содержащий цистеин второй промежуточный спейсерный пептид (линкер 2) будет содержать 1, 2, 3 или более цистеинов. Предпочтительный содержащий цистеин спейсерный пептид (линкер 2) имеет последовательность GGCGGG (SEQ ID NO:70). В альтернативном случае линкер 2 не содержит цистеин (например, GGG, GGGS (SEQ ID NO:71), LGGGSG (SEQ ID NO:72), GGGSGGGSGGG (SEQ ID NO:73), ASTKG (SEQ ID NO:74), LEPKSS (SEQ ID NO:75), APSSS (SEQ ID NO:76) и т.д.) и используется содержащий цистеин активирующий сборку гетеродимера домен, описанный ниже. Необязательно используют как содержащий цистеин линкер 2, так и содержащий цистеин активирующий сборку гетеродимера домен.
[00154] Активирующие сборку гетеродимера домены могут представлять собой GVEPKSC (SEQ ID NO:77) или VEPKSC (SEQ ID NO:78) или AEPKSC (SEQ ID NO:79) в одной полипептидной цепи и GFNRGEC (SEQ ID NO:80) или FNRGEC (SEQ ID NO:81) в другой полипептидной цепи (см., патент США № 9296816).
[00155] В соответствии с предпочтительным вариантом осуществления активирующие сборку гетеродимера домены будут содержать тандемно повторяющиеся петлевые домены противоположного заряда, например, «E-петлевые» спиральные домены (SEQ ID NO:82): EVAALEK-EVAALEK-EVAALEK-EVAALEK), глутаматные остатки которых будут образовывать отрицательный заряд при pH 7, и «K-петлевые» домены (SEQ ID NO:83): KVAALKE-KVAALKE-KVAALKE-KVAALKE), лизиновые остатки которых будут образовывать положительный заряд при pH 7. Наличие таких заряженных доменов активирует ассоциацию между первым и вторым полипептидами, и, таким образом, содействует образованию гетеродимеров. Могут быть использованы активирующие сборку гетеродимера домены, которые содержат модификации вышеописанных E-петлевых и K-петлевых последовательностей с целью включения одного или нескольких цистеиновых остатков. Наличие таких цистеиновых остатков способствует тому, что петля, присутствующая в полипептидной цепи, становится ковалентно связанной с комплементарной петлей, присутствующей в другой полипептидной цепи, тем самым, ковалентно связывая друг с другом полипептидные цепи и повышая стабильность диатела. Примеры таких особо предпочтительных активирующих сборку гетеродимера доменов включают в себя модифицированную E-петлю, имеющую аминокислотную последовательность EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO:84), и модифицированную K-петлю, имеющую аминокислотную последовательность KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO:85).
[00156] Как раскрыто в публикации согласно № WO 2012/018687, с целью улучшения фармакокинетических свойств диател in vivo, диатело можно модифицировать таким образом, чтобы оно содержало полипептидный участок сывороточного связывающегося белка на одном или более концах диатела. Наиболее предпочтительно, чтобы такой полипептидный участок сывороточного связывающегося белка был встроен на C-конце полипептидной цепи диатела. Альбумин представляет собой наиболее распространенный белок в плазме крови и имеет период полужизни, составляющий 19 дней, у человека. Альбумин имеет несколько низкомолекулярных связывающих сайтов, которые обеспечивают его нековалентное связывание с другими белками и, тем самым, продлевают их периоды полужизни в сыворотке крови. Домен 3, связывающий альбумин (ABD3), белка G штамма Streptococcus G148 состоит из 46 аминокислотных остатков, образующих стабильный трехспиральный пучок и характеризуется широкой специфичностью связывания с альбумином (Johansson, M.U. et al. (2002) “Structure, Specificity, And Mode Of Interaction For Bacterial Albumin-Binding Modules,” J. Biol. Chem. 277(10):8114-8120). Таким образом, особо предпочтительным полипептидным участком сывороточного связывающего белка для улучшения фармакокинетических свойств диатела in vivo является домен, связывающий альбумин (ABD), из белка стрептококков G, и более предпочтительно домен 3, связывающий альбумин (ABD3), белка G штамма Streptococcus G148 (SEQ ID NO:86): LAEAKVLANR ELDKYGVSDY YKNLIDNAKS AEGVKALIDE ILAALP.
[00157] Как раскрыто в публикации согласно PCT № WO 2012/162068 (включенной в данный документ посредством ссылки), «деиммунизированные» варианты SEQ ID NO:86 имеют способность ослаблять или устранять связывание с MHC класса II. На основании комбинационных результатов мутаций, следующие комбинации замен считаются предпочтительными заменами для образования такого деиммунизированного ABD: 66D/70S +71A; 66S/70S +71A; 66S/70S +79A; 64A/65A/71A; 64A/65A/71A+66S; 64A/65A/71A+66D; 64A/65A/71A+66E; 64A/65A/79A+66S; 64A/65A/79A+66D; 64A/65A/79A+66E. Вариантные ABD имеют модификации L64A, I65A и D79A или модификации N66S, T70S и D79A. Вариантные деиммунизированные ABD, имеющие аминокислотную последовательность:
LAEAKVLANR ELDKYGVSDY YKNLID66NAKS70 A71EGVKALIDE ILAALP (SEQ ID NO:87),
или аминокислотную последовательность:
LAEAKVLANR ELDKYGVSDY YKNA64A65NNAKT VEGVKALIA79E ILAALP (SEQ ID NO:88),
или аминокислотную последовательность:
LAEAKVLANR ELDKYGVSDY YKNLIS66NAKS70 VEGVKALIA79E ILAALP (SEQ ID NO:89)
являются особо предпочтительными, поскольку такие деиммунизированные ABD характеризуются по сути связыванием дикого типа, при этом обеспечивая ослабленное связывание с MHC класса II. Таким образом, первая полипептидная цепь такого диатела, имеющего ABD, содержит третий линкер (линкер 3), предпочтительно расположенный в C-концевом направлении по отношению к E-петлевому (или K-петлевому) домену такой полипептидной цепи, таким образом, чтобы располагаться между E-петлевым (или K-петлевым) доменом и ABD (который предпочтительно представляет собой деиммунизированный ABD). Предпочтительная последовательность в случае такого линкера 3 представляет собой GGGS (SEQ ID NO:71).
C. Мультиспецифические диатела, содержащие Fc-области
[00158] Один вариант осуществления настоящего изобретения относится к мультиспецифическим диателам, способным одновременно связываться с эпитопом ADAM9 и вторым эпитопом (т.е., другим эпитопом ADAM9 или эпитопом молекулы, которая не представляет собой ADAM9), которые содержат Fc-область. Добавление домена CH2-CH3 IgG к одному или обоим полипептидным цепям диатела, в результате чего связывание в комплекс цепей диатела приводит к образованию Fc-области, повышает биологический период полужизни и/или изменит валентность диатела. Такие диатела содержат две или более полипептидных цепей, последовательности которых обеспечивают ковалентное связывание друг с другом с образованием ковалентно ассоциированного диатела, которое способно одновременно связываться с эпитопом ADAM9 и вторым эпитопом. Введение доменов CH2-CH3 IgG в оба полипептида диатела будут обеспечивать образование двухцепочечного биспецифического диатела, содержащего Fc-область (Фиг. 2).
[00159] В альтернативном случае введение доменов CH2-CH3 IgG только в один из полипептидов диатела будут обеспечивать образование более сложного четырехцепочечного биспецифического диатела, содержащего Fc-область (Фиг. 3А-3С). На Фиг. 3C показано иллюстративное четырехцепочечное диатело, имеющее константный домен легкой цепи (CL) и константный домен тяжелой цепи CH1, однако фрагменты таких доменов, а также другие полипептиды могут быть использованы альтернативно (см., например, Фиг. 3A и 3B, публикации патента США №№ 2013-0295121; 2010-0174053 и 2009-0060910; публикации Европейских патентов №№ EP 2714079; EP 2601216; EP 2376109; EP 2158221, и публикации согласно PCT №№ WO 2012/162068; WO 2012/018687; WO 2010/080538). Таким образом, например, вместо домена CH1 можно использовать пептид, имеющий аминокислотную последовательность GVEPKSC (SEQ ID NO:77), VEPKSC (SEQ ID NO:78) или AEPKSC (SEQ ID NO:79), происходящую из шарнирной области IgG человека, а вместо домена CL можно использовать C-концевые 6 аминокислоты легкой каппа-цепи человека, GFNRGEC (SEQ ID NO:80) или FNRGEC (SEQ ID NO:81). Иллюстративный пептид, содержащий четырехцепочечное диатело, показан на Фиг. 3A. В альтернативном случае или дополнительно можно использовать пептид, содержащий тандемные петлевые домены противоположного заряда, такие как «E-петлевые» спиральные домены (SEQ ID NO:82): EVAALEK-EVAALEK-EVAALEK-EVAALEK или SEQ ID NO:84): EVAACEK-EVAALEK-EVAALEK-EVAALEK); и «K-петлевые» домены (SEQ ID NO:83): KVAALKE-KVAALKE-KVAALKE-KVAALKE или SEQ ID NO:85): KVAACKE-KVAALKE-KVAALKE-KVAALKE). Иллюстративный петлевой домен, содержащий четырехцепочечное диатело, показан на Фиг. 3В.
[00160] Молекулы, содержащие Fc-область, по настоящему изобретению, могут содержать дополнительные промежуточные спейсерные пептиды (линкеры), как правило, такие линкеры будут встроены между активирующим сборку гетеродимера доменом (например, E-петлей или K-петлей) и доменом CH2-CH3 и/или между доменом CH2-CH3 и вариабельным доменом (т.е., VH или VL). В типичном случае дополнительные линкеры будут содержать 3-20 аминокислотных остатка и могут необязательно содержать всю или часть шарнирной области IgG (предпочтительно содержащую цистеин часть шарнирной области IgG). Линкеры, которые могут быть использованы в биспецифических молекулах диател, содержащих Fc-область, по настоящему изобретению, включат в себя: GGGS (SEQ ID NO:71), LGGGSG (SEQ ID NO:72), GGGSGGGSGGG (SEQ ID NO:73), ASTKG (SEQ ID NO:74), LEPKSS (SEQ ID NO:75), APSSS (SEQ ID NO:76), APSSSPME (SEQ ID NO:90), VEPKSADKTHTCPPCP (SEQ ID NO:91), LEPKSADKTHTCPPC (SEQ ID NO:92), DKTHTCPPCP (SEQ ID NO:93), GGC и GGG. LEPKSS (SEQ ID NO:75) могут быть использованы вместо GGG или GGC для облегчения клонирования. Помимо этого, после аминокислот GGG или LEPKSS (SEQ ID NO:75) могут следовать непосредственно DKTHTCPPCP (SEQ ID NO:93) с образованием альтернативных линкеров: GGGDKTHTCPPCP (SEQ ID NO:94); и LEPKSSDKTHTCPPCP (SEQ ID NO:95). Биспецифические молекулы, содержащие Fc-область, по настоящему изобретению, могут включать шарнирную область IgG в дополнение к линкеру или вместо него. Иллюстративные шарнирные области включают в себя: EPKSCDKTHTCPPCP (SEQ ID NO:96) из IgG1, ERKCCVECPPCP (SEQ ID NO:97) из IgG2, ELKTPLGDTT HTCPRCPEPK SCDTPPPCPR CPEPKSCDTP PPCPRCPEPK SCDTPPPCPR CP (SEQ ID NO:206) из IgG3, ESKYGPPCPSCP (SEQ ID NO:98) из IgG4 и ESKYGPPCPPCP (SEQ ID NO:99), который представляет собой вариант шарнира IgG4, содержащего стабилизирующую замену S228P (подчеркнуто) (пронумерованную с помощью EU-индекса в соответствии с нумерацией Кабат) с целью снижения обмена нитями.
[00161] Как представлено на Фиг. 3A-3C, диатела, содержащие Fc-область, по настоящему изобретению, могут содержать четыре цепи. Первая и третья полипептидные цепи такого диатела содержат три домена: (i) домен, содержащий VL1, (ii) домен, содержащий VH2, (iii) активирующий сборку гетеродимера домен, и (iv) домен, содержащий последовательность CH2-CH3. Вторая и четвертая полипептидные цепи содержат: (i) домен, содержащий VL2, (ii) домен, содержащий VH1, и (iii) активирующий сборку гетеродимера домен, при этом активирующие сборку гетеродимера домены активируют димеризацию первой/третьей полипептидных цепей со второй/четвертой полипептидными цепями. Домены VL и/или VH третьей и четвертой полипептидных цепей и домены VL и/или VH первой и второй полипептидных цепей могут быть одинаковыми или разными, с тем, чтобы обеспечивать четырехвалентное связывание, которое является моноспецифическим, биспецифическим или тетраспецифическим. Обозначение «VL3» и «VH3» означает соответственно вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связываются с «третьим» эпитопом такого диатела. Аналогичным образом, обозначение «VL4» и «VH4» означает соответственно вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связываются с «четвертым» эпитопом такого диатела. Общая структура полипептидных цепей иллюстративных четырехцепочечных биспецифических диател, содержащих Fc-область, по настоящему изобретению, представлена в Табл. 2.
HPD = активирующий сборку гетеродимера домен
[00162] В соответствии с конкретным вариантом осуществления диатела по настоящему изобретению представляют собой биспецифические тетравалентные (т.е., имеют четыре эпитопсвязывающих сайта) диатела, содержащие Fc-область, которые состоят в общей сложности из четырех полипептидных цепей (Фиг. 3A-3C). Биспецифические тетравалентные диатела, содержащие Fc-область, по настоящему изобретению, содержат два эпитопсвязывающих сайта, иммуноспецифических в отношении ADAM9 (которые могут быть способны связываться с одним и тем же эпитопом ADAM9 или с разными эпитопами ADAM9), и два эпитопсвязывающих сайта, иммуноспецифических в отношении второй молекулы (которые могут быть способны связываться с одним и тем же эпитопом второй молекулы или с различными эпитопами второй молекулы). Предпочтительно вторая молекула представляет собой молекулу (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующую на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка.
[00163] В соответствии с дополнительным вариантом осуществления диатела, содержащие Fc-область, по настоящему изобретению, могут содержать три полипептидные цепи. Первый полипептид такого диатела содержит три домена: (i) домен, содержащий VL1, (ii) домен, содержащий VH2 и (iii) домен, содержащий последовательность CH2-CH3. Второй полипептид такого диатела содержит: (i) домен, содержащий VL2, (ii) домен, содержащий VH1, и (iii) домен, который активирует гетеродимеризацию и ковалентное связывание с первой полипептидной цепью диатела. Третий полипептид такого диатела содержит последовательность CH2-CH3. Таким образом, первая и вторая полипептидные цепи такого диатела ассоциируются вместе с образованием эпитопсвязывающего сайта VL1/VH1, который способен связываться с первым антигеном (т.е., либо ADAM9, либо молекулой, которая содержит второй эпитоп), а также эпитопсвязывающего сайта VL2/VH2, который способен связываться со вторым антигеном (т.е., либо молекулой, которая содержит второй эпитоп, либо ADAM9). Первый и второй полипептиды связываются друг с другом посредством дисульфидной связи, включающей цистеиновые остатки в их соответствующих третьих доменах. Примечательно, что первая и третья полипептидные цепи образуют комплекс друг с другом с образованием Fc-области, которая стабилизируется посредством дисульфидной связи. Такие биспецифические диатела имеют повышенную активность. На Фиг. 4A и 4B изображены структуры таких диател. Такие диатела, содержащие Fc-область, могут иметь любую из двух ориентаций (Табл. 3):
ориентация
ориентация
HPD = активирующий сборку гетеродимера домен
[00164] В соответствии с конкретным вариантом осуществления диатела по настоящему изобретению представляют собой биспецифические бивалентные (т.е., имеют два эпитопсвязывающих сайта) диатела, содержащие Fc-область, которые состоят в общей сложности из трех полипептидных цепей (Фиг. 4A-4B). Биспецифические бивалентные диатела, содержащие Fc-область, по настоящему изобретению, содержат один эпитопсвязывающий сайт, иммуноспецифический в отношении ADAM9, и один эпитопсвязывающий сайт, иммуноспецифический в отношении второй молекулы. Предпочтительно вторая молекула представляет собой молекулу (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующую на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка.
[00165] В соответствии с дополнительным вариантом осуществления диатела, содержащие Fc-область, могут содержать в общей сложности пять полипептидных цепей. В соответствии с конкретным вариантом осуществления две из указанных пяти полипептидных цепей имеют одинаковую аминокислотную последовательность. Первая полипептидная цепь такого диатела содержит: (i) домен, содержащий VH1, (ii) домен, содержащий CH1 и (iii) домен, содержащий последовательность CH2-CH3. Первая полипептидная цепь может представлять собой тяжелую цепь антитела, которая содержит VH1 и константную область тяжелой цепи. Вторая и пятая полипептидные цепи такого диатела содержат: (i) домен, содержащий VL1, и (ii) домен, содержащий CL. Вторая и/или пятая полипептидные цепи такого диатела могут представлять собой легкие цепи, которые содержат VL1, комплементарный к VH1 первой/третьей полипептидной цепи. Первая, вторая и/или пятая полипептидные цепи могут быть выделены из встречающегося в природе антитела. В альтернативном случае они могут быть сконструированы рекомбинантно. Третья полипептидная цепь такого диатела содержит: (i) домен, содержащий VH1, (ii) домен, содержащий CH1, (iii) домен, содержащий последовательность CH2-CH3, (iv) домен, содержащий VL2, (v) домен, содержащий VH3, и (vi) активирующий сборку гетеродимера домен, при этом активирующие сборку гетеродимера домены активируют димеризацию третьей цепи с четвертой цепью. Четвертый полипептид таких диател содержит: (i) домен, содержащий VL3, (ii) домен, содержащий VH2, и (iii) домен, который активирует гетеродимеризацию и ковалентное связывание с третьей полипептидной цепью диатела.
[00166] Таким образом, первая и вторая, а также третья и четвертая полипептидные цепи таких диател ассоциируются вместе с образованием эпитопсвязывающих сайтов VL1/VH1, способных связываться с первым эпитопом. Третья и четвертая полипептидные цепи таких диател ассоциируются вместе с образованием эпитопсвязывающего сайта VL2/VH2, который способен связываться со вторым эпитопом, а также связывающего сайта VL3/VH3, который способен связываться с третьим эпитопом. Первый и третий полипептиды связываются друг с другом посредством дисульфидной связи, включающей цистеиновые остатки в их соответствующих константных областях. Примечательно, что первая и третья полипептидные цепи образуют комплекс друг с другом с образованием Fc-области. Такие мультиспецифические диатела имеют повышенную активность. На Фиг. 5 изображена структура таких диател. Будет понятно, что домены VL1/VH1, VL2/VH2 и VL3/VH3 могут быть одинаковыми или разными, с тем, чтобы обеспечивать связывание, которое является моноспецифическим, биспецифическим или триспецифическим. Как предусмотрено в данном документе, эти домены предпочтительно выбирают таким образом, чтобы связывать эпитоп ADAM9, эпитоп второй молекулы и необязательно эпитоп третьей молекулы.
[00167] Домены VL и VH полипептидных цепей выбирают таким образом ,чтобы образовались связывающие сайты VL/VH, специфические в отношении предпочтительно эпитопа. Связывающие сайты VL/VH, образованные с помощью ассоциации полипептидных цепей, могут быть одинаковыми или разными, с тем, чтобы обеспечивать тетравалентное связывание, которое может быть моноспецифическим, биспецифическим или тетраспецифическим. В частности, домены VL и VH могут быть выбраны таким образом, что мультивалентное диатело может содержать два связывающих сайта в отношении первого эпитопа и два связывающих сайта в отношении второго эпитопа, или три связывающих сайта в отношении первого эпитопа и один связывающий сайт в отношении второго эпитопа, или два связывающих сайта в отношении первого эпитопа, один связывающий сайт в отношении второго эпитопа и один связывающий сайт в отношении третьего эпитопа (как показано на Фиг. 5). Общая структура полипептидных цепей иллюстративных пятицепочечных диател, содержащих Fc-область, по настоящему изобретению, представлена в Табл. 4.
HPD = активирующий сборку гетеродимера домен
[00168] В соответствии с конкретным вариантом осуществления диатела по настоящему изобретению представляют собой биспецифические тетравалентные (т.е., имеют четыре эпитопсодержащих сайта) диатела, содержащие Fc-область, которые состоят в общей сложности из пяти полипептидных цепей, имеющих два эпитопсвязывающих сайта, иммуноспецифических в отношении ADAM9 (которые могут быть способны связываться с одним и тем же эпитопом ADAM9 или с разными эпитопами ADAM9), и два эпитопсвязывающих сайта, специфических в отношении второй молекулы (которые могут быть способны связываться с одним и тем же эпитопом второй молекулы или с различными эпитопами второй молекулы). В соответствии с другим вариантом осуществления биспецифические тетравалентные диатела, содержащие Fc-область, по настоящему изобретению, содержат три эпитопсвязывающих сайта, иммуноспецифических в отношении ADAM9 (которые могут быть способны связываться с одним и тем же эпитопом ADAM9 или двумя или тремя разными эпитопами ADAM9), и один эпитопсвязывающий сайт, специфический в отношении второй молекулы. В соответствии с другим вариантом осуществления биспецифические тетравалентные диатела, содержащие Fc-область, по настоящему изобретению, содержат один эпитопсвязывающий сайт, иммуноспецифический в отношении ADAM9, и три эпитопсвязывающих сайта, специфических в отношении второй молекулы (которые могут быть способны связываться с одним и тем же эпитопом второй молекулы или с двумя или тремя разными эпитопами второй молекулы). Как представлено выше, домены VL и VH могут быть выбраны для обеспечения триспецифического связывания. Соответственно, настоящее изобретение также охватывает триспецифические тетравалентные диатела, содержащие Fc-область. Триспецифические тетравалентные диатела, содержащие Fc-область, по настоящему изобретению, содержат два эпитопсвязывающих сайта, иммуноспецифические в отношении ADAM9, один эпитопсвязывающий сайт, иммуноспецифический в отношении второй молекулы, и один эпитопсвязывающий сайт, иммуноспецифический в отношении третьей молекулы. В соответствии с определенными вариантами осуществления вторая молекула представляет собой молекулу (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующую на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. В соответствии с определенными вариантами осуществления вторая молекула представляет собой CD3 и третья молекула представляет собой CD8.
D. Трехвалентные связывающие молекулы, содержащие Fc-области
[00169] Дополнительный вариант осуществления настоящего изобретения относится к трехвалентным связывающим молекулам, содержащим Fc-область, способным связывать одновременно первый эпитоп, второй эпитоп и третий эпитоп, при этом по меньшей мере один из таких эпитопов не является идентичным другому. Такие трехвалентные связывающие молекулы содержат три эпитопсвязывающих сайта, два из которых представляют собой связывающие домены по типу диатела, которые обеспечивают связывание сайта A и связывание сайта B, и один из которых представляет собой связывающий домен Fab-типа или связывающий домен scFv-типа, который обеспечивает связывание сайта C (см., например, Фиг. 6A-6F, и публикации согласно PCT №№: WO 2015/184207; и WO 2015/184203). Таким образом, такие трехвалентные связывающие молекулы содержат домены «VL1»/«VH1», которые способны связываться с первым эпитопом, домены «VL2»/«VH2», которые способны связываться со вторым эпитопом, и домены «VL3» и «VH3», которые способны связываться с «третьим» эпитопом такой трехвалентной связывающей молекулы. «Связывающий домен типа диатела» представляет собой тип эпитопсвязывающего сайта, присутствующего в диателе, и, в частности, диателе DART®, описанном выше. Каждый из «связывающих доменов Fab-типа» и «связывающих доменов scFv-типа» представляют собой эпитопсвязывающие сайты, которые образуются в результате взаимодействия домена VL легкой цепи иммуноглобулина и комплементирующего домена VH тяжелой цепи иммуноглобулина. Связывающие домены Fab-типа отличаются от связывающих доменов по типу диатела тем, что две полипептидные цепи, которые образуют связывающий домен Fab-типа, содержат только один эпитопсвязывающий сайт, в то время как две полипептидные цепи, которые образуют связывающий домен по типу диатела, образуют по меньшей мере два эпитопсвязывающих сайта. Аналогичным образом, связывающие домены scFv-типа также отличаются от связывающих доменов по типу диатела тем, что они содержат только один эпитопсвязывающий сайт. Таким образом, используемые в данном документе связывающие домены Fab-типа и scFv-типа отличаются от связывающих доменов по типу диатела.
[00170] В типичном случае трехвалентные связывающие молекулы по настоящему изобретению содержат четыре различные полипептидные цепи (см. Фиг. 6A-6B), однако данные молекулы могут содержать меньшее или большее число полипептидных цепей, например, в результате слияния таких полипептидных цепей друг с другом (например, посредством пептидной связи) или в результате разделения таких полипептидных цепей с образованием дополнительных полипептидных цепей, или в результате ассоциации меньшего количества или дополнительных полипептидных цепей посредством дисульфидных связей. На Фиг. 6C-6F продемонстрирован данный аспект настоящего изобретения с помощью схематического изображения таких молекул, имеющих три полипептидные цепи. Как представлено на Фиг. 6A-6F, трехвалентные связывающие молекулы по настоящему изобретению могут иметь альтернативные ориентации, в которых связывающие домены по типу диатела расположены в N-концевом (Фиг. 6A, 6C и 6D) или C-концевом (Фиг. 6B, 6E и 6F) направлении по отношению к Fc-области.
[00171] В соответствии с определенными вариантами осуществления первая полипептидная цепь таких трехвалентных связывающих молекул по настоящему изобретению содержит: (i) домен, содержащий VL1, (ii) домен, содержащий VH2, (iii) активирующий сборку гетеродимера домен, и (iv) домен, содержащий последовательность CH2-CH3. Домены VL1 и VL2 расположены в N-концевом или C-концевом направлении по отношению к домену, содержащему CH2-CH3, как представлено в Табл. 4 (также см. Фиг. 6A и 6B). Вторая полипептидная цепь таких вариантов осуществления содержит: (i) домен, содержащий VL2, (ii) домен, содержащий VH1 и (iii) активирующий сборку гетеродимера домен. Третья полипептидная цепь таких вариантов осуществления содержит: (i) домен, содержащий VH3, (ii) домен, содержащий CH1 и (iii) домен, содержащий последовательность CH2-CH3. Третья полипептидная цепь может представлять собой тяжелую цепь антитела, которая содержит VH3 и константную область тяжелой цепи, или полипептид, который содержит такие домены. Четвертый полипептид таких вариантов осуществления содержит: (i) домен, содержащий VL3, и (ii) домен, содержащий CL. Четвертая полипептидная цепь может представлять собой легкую цепь антитела, которая содержит VL3, комплементарный третьей полипептидной цепи, или полипептид, который содержит такие домены. Третья или четвертая полипептидные цепи могут быть выделены из встречающихся в природе антител. В альтернативном случае они могут быть сконструированы рекомбинантно, синтетически или с помощью других средств.
[00172] Вариабельные домены легкой цепи первых и вторых полипептидных цепей отделяются от вариабельных доменов тяжелой цепи с помощью промежуточного спейсерного пептида, имеющего длину, которая является слишком короткой для того, чтобы обеспечивать ассоциацию доменов VL1/VH2 (или VL2/VH1) вместе с образованием эпитопсвязывающего сайта, способного связываться либо с первым, либо со вторым эпитопом. Предпочтительный промежуточный спейсерный пептид (линкер 1) для этой цели имеет последовательность (SEQ ID NO:69): GGGSGGGG. Другие домены трехвалентных связывающих молекул могут быть отделены одним или несколькими промежуточными спейсерными пептидами (линкерами), необязательно содержащими цистеиновый остаток. В частности, как предусмотрено выше, такие линкеры в типичном случае будут встроены между вариабельными доменами (т.е., VH или VL) и пептидными активирующими сборку гетеродимера доменами (например, E-петлей или K-петлей) и между такими пептидными активирующими сборку гетеродимера доменами (например, E-петлей или K-петлей) и доменами CH2-CH3. Иллюстративные линкеры, пригодные для получения трехвалентных связывающих молекул, представлены выше и также предусмотрены в публикациях согласно PCT №№ PCT/US15/33081; и PCT/US15/33076. Таким образом, первая и вторая полипептидные цепи таких трехвалентных связывающих молекул ассоциируются вместе с образованием связывающего сайта VL1/VH1, который способен связываться с первым эпитопом, а также связывающего сайта VL2/VH2, который способен связываться со вторым эпитопом. Третья и четвертая полипептидные цепи таких трехвалентных связывающих молекул ассоциируются вместе с образованием связывающего сайта VL3/VH3, который способен связываться с третьим эпитопом.
[00173] Как описано выше, трехвалентные связывающие молекулы по настоящему изобретению могут содержать три полипептида. Трехвалентные связывающие молекулы, содержащие три полипептидные цепи, могут быть получены в результате связывания доменов четвертого полипептида в N-концевом направлении с доменом, содержащим VH3, третьего полипептида (т.е., с помощью промежуточного спейсерного пептида (линкер 4)). В альтернативном случае используют третью полипептидная цепь трехвалентной связывающей молекулы по настоящему изобретению, содержащую следующие домены: (i) домен, содержащий VL3, (ii) домен, содержащий VH3, и (iii) домен, содержащий последовательность CH2-CH3, при этом VL3 и VH3 разделены друг от друга с помощью промежуточного спейсерного пептида, который является достаточно длинным (по меньшей мере 9 или более аминокислотных остатков) с тем, чтобы обеспечивать ассоциацию этих доменов с образованием эпитопсвязывающего сайта. Один предпочтительный промежуточный спейсерный пептид для этой цели имеет последовательность: GGGGSGGGGSGGGGS (SEQ ID NO:100).
[00174] Будет понятно, что домены VL1/VH1, VL2/VH2 и VL3/VH3 таких трехвалентных связывающих молекул могут быть разными, с тем, чтобы обеспечивать связывание, которое является моноспецифическим, биспецифическим или триспецифическим. В частности, домены VL и VH могут быть выбраны таким образом, что трехвалентная связывающая молекула содержит два связывающих сайта в отношении первого эпитопа и один связывающий сайт в отношении второго эпитопа, или один связывающий сайт в отношении первого эпитопа и два связывающих сайта в отношении второго эпитопа, или один связывающий сайт в отношении первого эпитопа, один связывающий сайт в отношении второго эпитопа и один связывающий сайт в отношении третьего эпитопа.
[00175] В то же время, как предусмотрено в данном документе, эти домены предпочтительно выбирают таким образом, чтобы связывать эпитоп ADAM9, эпитоп второй молекулы и эпитоп третьей молекулы. В соответствии с определенными вариантами осуществления вторая молекула представляет собой молекулу (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующую на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. В соответствии с определенными вариантами осуществления третья молекула представляет собой CD8.
[00176] Общая структура полипептидных цепей иллюстративных трехвалентных связывающих молекул по настоящему изобретению представлена на Фиг. 6A-6F и в Табл. 5.
1-я ориентация
2-я ориентация
1-я ориентация
2-я ориентация
HPD = активирующий сборку гетеродимера домен
[00177] Один вариант осуществления настоящего изобретения относится к трехвалентным связывающим молекулам, которые содержат два эпитопсвязывающих сайта в отношении ADAM9 и один эпитопсвязывающий сайт в отношении второй молекулы. Два эпитопсвязывающих сайта в отношении ADAM9 могут связывать один и тот же эпитоп или разные эпитопы. Другой вариант осуществления настоящего изобретения относится к трехвалентным связывающим молекулам, которые содержат один эпитопсвязывающий сайт в отношении ADAM9 и два эпитопсвязывающих сайта в отношении второй молекулы. Два эпитопсвязывающих сайта в отношении второй молекулы могут связывать один и тот же эпитоп или разные эпитопы второй молекулы. Дополнительный вариант осуществления настоящего изобретения относится к триспецифическим трехвалентным связывающим молекулам, которые содержат один эпитопсвязывающий сайт в отношении ADAM9, один эпитопсвязывающий сайт в отношении второй молекулы и один эпитопсвязывающий сайт в отношении третьей молекулы. В соответствии с определенными вариантами осуществления вторая молекула представляет собой молекулу (например, CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR), NKG2D и т.д.), присутствующую на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. В соответствии с определенными вариантами осуществления вторая молекула представляет собой CD3 и третья молекула представляет собой CD8. Как предусмотрено в данном документе, такие трехвалентные связывающие молекулы могут содержать три, четыре, пять или более полипептидных цепей.
Константные домены и вариантные Fc-области
[00178] В данном документе предусмотрены «константные домены» антител, пригодные в получении молекул, связывающих ADAM9 (например, антител, диател, трехвалентных связывающих молекул и т.д.), по настоящему изобретению.
[00179] Предпочтительный домен CL представляет собой домен CL каппа-цепи IgG человека. Аминокислотная последовательность иллюстративного домена CL каппа-цепи человека представляет собой (SEQ ID NO:101):
RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT HQGLSSPVTK SFNRGEC
[00180] В альтернативном случае иллюстративный домен CL представляет собой домен CL лямбда-цепи IgG человека. Аминокислотная последовательность иллюстративного домена CL лямбда-цепи человека представляет собой (SEQ ID NO:102):
QPKAAPSVTL FPPSSEELQA NKATLVCLIS DFYPGAVTVA WKADSSPVKA GVETTPSKQS NNKYAASSYL SLTPEQWKSH RSYSCQVTHE GSTVEKTVAP
TECS
[00181] Предусмотренные в данном документе молекулы, связывающие ADAM9, по настоящему изобретению, могут содержать Fc-область. Fc-область таких молекул по настоящему изобретению может быть любого типа (например, IgG1, IgG2, IgG3 или IgG4). Молекулы, связывающие ADAM9, по настоящему изобретению, могут дополнительно содержать домен CH1 и/или шарнирную область. В случае наличия, домен CH1 и/или шарнирная область могут быть любого изотипа (например, IgG1, IgG2, IgG3 или IgG4), и предпочтительно одного и того же изотипа, что и предпочтительная Fc-область.
[00182] Иллюстративный домен CH1 представляет собой домен CH1 IgG1. Аминокислотная последовательность иллюстративного домена CH1 IgG1 человека представляет собой (SEQ ID NO:103):
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKRV
[00183] Иллюстративный домен CH1 представляет собой домен CH1 IgG2. Аминокислотная последовательность иллюстративного домена CH1 IgG2 человека представляет собой (SEQ ID NO:104):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSNFGTQT YTCNVDHKPS NTKVDKTV
[00184] Иллюстративный домен CH1 представляет собой домен CH1 IgG3. Аминокислотная последовательность иллюстративного домена CH1 IgG3 человека представляет собой (SEQ ID NO:207):
ASTKGPSVFP LAPCSRSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YTCNVNHKPS NTKVDKRV
[00185] Иллюстративный домен CH1 представляет собой домен CH1 IgG4. Аминокислотная последовательность иллюстративного домена CH1 IgG4 человека представляет собой (SEQ ID NO:105):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTKT YTCNVDHKPS NTKVDKRV
[00186] Одна иллюстративная шарнирная область представляет собой шарнирную область IgG1 человека. Аминокислотная последовательность иллюстративной шарнирной области IgG1 человека представляет собой (SEQ ID NO:96): EPKSCDKTHTCPPCP.
[00187] Другая иллюстративная шарнирная область представляет собой шарнирную область IgG2 человека. Аминокислотная последовательность иллюстративной шарнирной области IgG2 человека представляет собой (SEQ ID NO:97): ERKCCVECPPCP.
[00188] Другая иллюстративная шарнирная область представляет собой шарнирную область IgG4 человека. Аминокислотная последовательность иллюстративной шарнирной области IgG4 человека представляет собой (SEQ ID NO:98): ESKYGPPCPSCP. Как описано выше, шарнирная область IgG4 может содержать стабилизующую мутацию, такую как замена S228P. Аминокислотная последовательность иллюстративной шарнирной области стабилизированного IgG4 представляет собой (SEQ ID NO:99): ESKYGPPCPPCP.
[00189] Fc-область молекул, содержащих Fc-область (например, антител, диател, трехвалентных связывающих молекул и т.д.), по настоящему изобретению может представлять собой либо полную Fc-область (например, полную Fc-область IgG), либо только фрагмент Fc-области. Необязательно Fc-область молекул, содержащих Fc-область, по настоящему изобретению, не имеет C-концевого лизинового аминокислотного остатка.
[00190] При традиционной иммунной функции взаимодействие комплексов антитело-антиген с клетками иммунной системы приводит к широкому разнообразию ответов, находящихся в диапазоне от эффекторных функций, таких как антителозависимая цитотоксичность, дегрануляция тучных клеток и фагоцитоз, до иммуномодулирующих сигналов, таких как регуляция пролиферации лимфоцитов и секреция антител. Все из этих взаимодействий инициируются посредством связывания Fc-области антител или иммунных комплексов со специализированными рецепторами клеточной поверхности на гематопоэтических клетках. Разнообразие клеточных ответов, вызываемых антителами и иммунными комплексами, является результатом структурой гетерогенности трех Fc-рецепторов: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). FcγRI (CD64), FcγRIIA (CD32A) и FcγRIII (CD16) представляют собой активирующие (т.е., усиливающие иммунную систему) рецепторы; FcγRIIB (CD32B) представляет собой ингибирующий (т.е., подавляющий иммунную систему) рецептор. Помимо этого, взаимодействие с неонатальным Fc-рецептором (FcRn) опосредует рециркулирование молекул IgG из эндосомы к клеточной поверхности и высвобождению в кровь. Аминокислотная последовательность иллюстративного IgG1 дикого типа (SEQ ID NO:1), IgG2 (SEQ ID NO:2), IgG3 (SEQ ID NO:3) и IgG4 (SEQ ID NO:4) представлена выше.
[00191] Модификация Fc-области может приводить к измененному фенотипу, например, измененному периоду полужизни в сыворотке крови, измененной стабильности, измененной восприимчивости к клеточным ферментам или измененной эффекторной функции. Таким образом, это может быть желательным для модификации содержащей Fc-область молекулы, связывающей ADAM9, по настоящему изобретению, в отношении эффекторной функции, например, с тем, чтобы усиливать эффективность такой молекулы при лечении рака. В определенных случаях ослабление или устранение эффекторной функции является желательной, например, в случае антител, механизм действия которых включает в себя блокирование или антагонизм, но не уничтожение клеток, несущих целевой антиген. Повышенная эффекторная функция, как правило, является желательной в случае направленности на патогенные клетки, такие как опухолевые и чужеродные клетки, когда FcγR экспрессируются при низких уровнях, например, опухольспецифические B-клетки с низкими уровнями FcγRIIB (например неходжкинская лимфома, CLL и лимфома Беркитта). Молекулы по настоящему изобретению, характеризующиеся такой придаваемой или измененной эффекторной функциональной активностью, являются пригодными для лечения и/или предупреждения заболевания, нарушения или инфекции, при которых усиленная эффективность эффекторной функциональной активности является желательной.
[00192] Соответственно, в соответствии с определенными вариантами осуществления Fc-область молекул, содержащих Fc-область, по настоящему изобретению, могут представлять собой разработанную вариантную Fc-область. Несмотря на то, что Fc-область биспецифических молекул, содержащих Fc-область, по настоящему изобретению, может характеризоваться способностью связываться с одним или несколькими Fc-рецепторами (например, FcγR), более предпочтительно такая измененная Fc-область характеризуется измененным связыванием с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по отношению к связыванию, проявляемому Fc-областью дикого типа), например, будет характеризоваться повышенным связыванием с активирующим рецептором и/или будет характеризоваться значительно сниженной или отсутствующей способностью связываться с ингибирующим(ингибирующими) рецептором(рецепторами). Таким образом, Fc-область молекул, содержащих Fc-область, по настоящему изобретению, может содержать весь домен CH2 и/или некоторую часть или весь домен CH3 полной Fc-области, или может содержать последовательность вариантного CH2 и/или вариантного CH3 (которые могут содержать, например, одну или несколько вставок и/или одну или несколько делеций по отношению к доменам CH2 или CH3 полной Fc-области). Такие Fc-области могут содержать не относящиеся к Fc-области полипептидные участки, или могут содержать не встречающиеся в природе полные Fc-области, или могут содержать не встречающиеся в природе ориентации доменов CH2 и/или CH3 (такие как, например, два домена CH2 или два домена CH3, или в направлении от N-конца к C-концу домен CH3, связанный с доменом CH2 и т.д.).
[00193] Модификации Fc-области, идентифицированные в виде измененной эффекторной функции, известны в данной области техники, в том числе модификации, которые повышают связывание с активирующими рецепторами (например, FcγRIIA (CD16A) и ослабляют связывание с ингибирующими рецепторами (например, FcγRIIB (CD32B) (см., например, Stavenhagen, J.B. et al. (2007) “Fc Optimization Of Therapeutic Antibodies Enhances Their Ability To Kill Tumor Cells In Vitro And Controls Tumor Expansion In Vivo Via Low-Affinity Activating Fcgamma Receptors,” Cancer Res. 57(18):8882-8890). В Табл. 6 приведены иллюстративные одиночные, двойные, тройные, четверные и пятерные замены (нумерация представлена с помощью EU-индекса в соответствии с Кабат, и замены представлены по отношению к аминокислотной последовательности SEQ ID NO:1) иллюстративной модификации, которая повышает связывание с активирующими рецепторами и/или ослабляет связывание с ингибирующими рецепторами.
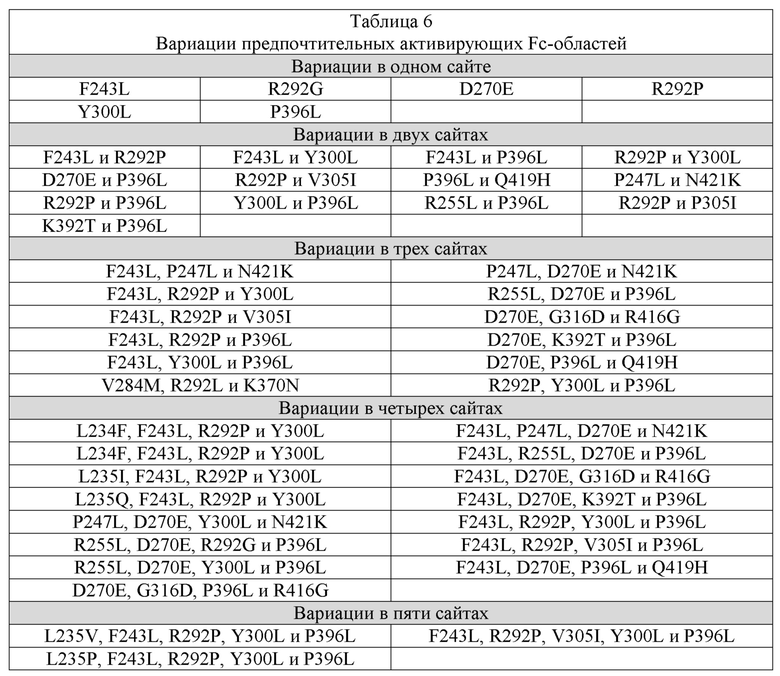
[00194] Иллюстративные варианты Fc-областей IgG1 человека с ослабленным связыванием с CD32B и/или повышенным связыванием с CD16A, содержат замены F243L, R292P, Y300L, V305I или P396L, при этом нумерация представлена с помощью EU-индекса в соответствии с Кабат. Эти аминокислотные замены могут присутствовать в Fc-области IgG1 человека в любой комбинации. В соответствии с одним вариантом осуществления вариантная Fc-область IgG1 человека содержит замену F243L, R292P и Y300L. В соответствии с другим вариантом осуществления вариантная Fc-область IgG1 человека содержит замену F243L, R292P, Y300L, V305I и P396L.
[00195] В соответствии с определенными вариантами осуществления предпочтительно, чтобы Fc-области молекул, связывающих ADAM9, по настоящему изобретению, характеризовались сниженным (или по сути отсутствующим) связыванием с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по отношению к связыванию, проявляемому Fc-областью IgG1 дикого типа (SEQ ID NO:1). В соответствии с конкретным вариантом осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат Fc-область IgG, которая характеризуется ослабленной эффекторной функцией ADCC. В соответствии с предпочтительным вариантом осуществления домены CH2-CH3 таких молекул, связывающих ADAM9, содержат любые 1, 2, 3 или 4 из замен: L234A, L235A, D265A, N297Q и N297G, при этом нумерация представлена с помощью EU-индекса в соответствии с Кабат. В соответствии с другим вариантом осуществления домены CH2-CH3 содержат замену N297Q, замену N297G, замены L234A и L235A или замену D265A, поскольку эти мутацию устраняют связывание FcR. В альтернативном случае используют домен CH2-CH3 встречающей в природе Fc-области, которая изначально характеризуется сниженным (или по сути отсутствующим) связыванием с FcγRIIIA (CD16a) и/или ослабленной эффекторной функцией (по отношению к связыванию и эффекторной функции, проявляемыми Fc-областью IgG1 дикого типа (SEQ ID NO:1)). В соответствии с конкретным вариантом осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат Fc-область IgG2 (SEQ ID NO:2) или Fc-область IgG4 (SEQ ID:NO:4). При использовании Fc-области IgG4 настоящее изобретение также охватывает включение стабилизирующей мутации, такой как замена S228P шарнирной области выше (см., например, SEQ ID NO:99). Поскольку замены N297G, N297Q, L234A, L235A и D265A устраняют эффекторную функцию, в случаях, когда эффекторная функция является желательной, эти замены предпочтительно не используют.
[00196] Предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, имеющих ослабленную или устраненную эффекторную функцию, будет содержать замены L234A/L235A (показанные подчеркнутыми) (SEQ ID NO:106):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00197] Вторая предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, содержит замену S442C (показанную подчеркнутой), с тем, чтобы обеспечивать ковалентное связывание доменов CH3 друг с другом посредством дисульфидной связи или чтобы обеспечивать конъюгацию лекарственного фрагмента. Аминокислотная последовательность такой молекулы представляет собой (SEQ ID NO:107):
APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LCLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00198] Третья предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, содержит замены L234A/L235A (показаны подчеркнутыми), которые ослабляют или устраняют эффекторную функцию, и замену S442C (показана подчеркнутой), которая обеспечивает ковалентное связывание двух доменов CH3 друг с другом посредством дисульфидной связи или конъюгацию лекарственного фрагмента. Аминокислотная последовательность такой молекулы представляет собой (SEQ ID NO:108):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LCLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00199] Период полужизни в сыворотке крови белков, содержащих Fc-области, может быть увеличено в результате повышения аффинности связывания Fc-области в отношении FcRn. Термин «период полужизни», используемый в данном документе, означает фармакокинетическое свойство молекулы, которое представляет собой величину среднего времени жизни молекул после их введения. Период полужизни можно выразить в виде времени, требующегося для выведения пятидесяти процентов (50%) известного количества молекулы из организма субъекта (например, пациента-человека или другого млекопитающего) или его определенного компартмента, например, измеряемого в сыворотки крови, т.е., период полужизни в кровотоке или в других тканях. Как правило, повышение периода полужизни приводит к повышению среднего времени удержания (MRT) в кровотоке в случае вводимой молекулы.
[00200] В соответствии с некоторыми вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат вариантную Fc-область, которая содержит по меньшей мере одну аминокислотную модификацию по отношению к Fc-области дикого типа, таким образом, что указанная молекула характеризуется повышенным периодом полужизни (по отношению к молекуле, содержащей Fc-область дикого типа). В соответствии с некоторыми вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, содержат вариантную Fc-область IgG, при этом указанная вариантная Fc-область содержит продлевающую период полужизни аминокислотную замену, в одном или нескольких положениях, выбранных из группы, состоящей из 238, 250, 252, 254, 256, 257, 256, 265, 272, 286, 288, 303, 305, 307, 308, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424, 428, 433, 434, 435 и 436, при этом нумерация представлена с помощью EU-индекса в соответствии с Кабат. Многочисленные мутации, способные повышать период полужизни молекулы, содержащей Fc-область, известны в данной области техники и включают в себя, например, M252Y, S254T, T256E и их комбинации. Например, см. мутации, описанные в патентах США №№ 6277375, 7083784; 7217797, 8088376; публикациях патентов США №№ 2002/0147311; 2007/0148164; и публикациях согласно PCT №№ WO 98/23289; WO 2009/058492; и WO 2010/033279, которые включены в данный документ посредством ссылки в полном объеме. Молекулы, связывающие ADAM9, с повышенным периодом полужизни, также включают таковые, имеющие вариантные Fc-области, содержащие замены в двух или более из остатков Fc-областей 250, 252, 254, 256, 257, 288, 307, 308, 309, 311, 378, 428, 433, 434, 435 и 436. В частности, две или более замены выбирают из: T250Q, M252Y, S254T, T256E, K288D, T307Q, V308P, A378V, M428L, N434A, H435K и Y436I, при этом нумерация представлена с помощью EU-индекса в соответствии с Кабат.
[00201] В соответствии с конкретным вариантом осуществления молекула, связывающая ADAM9, по настоящему изобретению, имеет вариантную Fc-область IgG, содержащую замены:
(A) M252Y, S254T и T256E;
(B) M252Y и S254T;
(C) M252Y и T256E;
(D) T250Q и M428L;
(E) T307Q и N434A;
(F) A378V и N434A;
(G) N434A и Y436I;
(H) V308P и N434A; или
(I) K288D и H435K.
[00202] В соответствии с предпочтительным вариантом осуществления молекула, связывающая ADAM9, по настоящему изобретению, имеет вариантную Fc-область IgG, содержащую любую 1, 2 или 3 из замен: M252Y, S254T и T256E. Настоящее изобретение дополнительно охватывает молекулы, связывающие ADAM9, имеющие вариантные Fc-области, содержащие:
(A) одну или несколько мутаций, которые изменяют эффекторную функцию и/или FcγR; и
(B) одну или несколько мутаций, которые продлевают период полужизни в сыворотке крови.
[00203] Четвертая предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, содержит замены M252Y, S254T и T256E (показаны подчеркнутыми), с тем, чтобы продлевать период полужизни в сыворотке крови. Аминокислотная последовательность такой молекулы представляет собой (SEQ ID NO:200):
APELLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00204] Пятая предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, содержит замены L234A/L235A (показаны подчеркнутыми), которые ослабляют или устраняют эффекторную функцию, и замены M252Y, S254T и T256E (показаны подчеркнутыми), с тем, чтобы продлевать период полужизни в сыворотке крови. Аминокислотная последовательность такой молекулы представляет собой (SEQ ID NO:201):
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00205] Шестая предпочтительная последовательность IgG1 в случае доменов CH2 и CH3 молекул, содержащих Fc-область, по настоящему изобретению, содержит замены L234A/L235A (показаны подчеркнутыми), которые ослабляют или устраняют эффекторную функцию, и замены M252Y, S254T и T256E (показаны подчеркнутыми), с тем, чтобы продлевать период полужизни в сыворотке крови, а также замену S442C (показана подчеркнутой), с тем, чтобы обеспечивать ковалентное связывание двух доменов CH3 друг с другом посредством дисульфидной связи или чтобы обеспечивать конъюгацию лекарственного фрагмента. Аминокислотная последовательность такой молекулы представляет собой (SEQ ID NO:203):
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LCLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00206] В случае определенных антител, диател и трехвалентных связывающих молекул, содержащие Fc-область первые и третьи полипептидные цепи которых не являются идентичными, желательно ослаблять или предупреждать возникновение гомодимеризации между доменами CH2-CH3 двух первых полипептидных цепей или между доменами CH2-CH3 двух третьих полипептидных цепей. Домены CH2 и/или CH3 таких полипептидных цепей не должны быть идентичными в последовательности и предпочтительно их модифицируют, чтобы способствовать образованию комплекса между двумя полипептидными цепями. Например, аминокислотная замена (предпочтительно замена аминокислотной, содержащей объемную боковую группу, образующую «выступ», например, триптофаном) может быть введена в домен CH2 или CH3 таким образом, что стерическое влияние будет препятствовать взаимодействию с аналогичным мутантным доменом и будет способствовать тому, чтобы мутантный домен связывался с доменом, в котором комплементарная или аккомодирующая мутация была разработана, т.е., «впадиной» (например, заменой глицином). Такие совокупности мутаций могут быть разработаны в любой паре полипептидов, содержащих домены CH2-CH3, которые образуют Fc-область, с целью содействия гетеродимерацизии. Способы разработки белков, которые содействуют преобладанию гетеродимеризации над гомодимеразацией, хорошо известны в данной области техники, в частности, в связи с разработкой иммуноглобулиноподобных молекул, и охвачены в данном документе (см., например, Ridgway et al. (1996) “‘Knobs-Into-Holes’ Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization,” Protein Engr. 9:617-621; Atwell et al. (1997) “Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library,” J. Mol. Biol. 270: 26-35; и Xie et al. (2005) “A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis,” J. Immunol. Methods 296:95-101; каждая из которых тем самым включена в данный документ посредством ссылки в полном объеме).
[00207] Предпочтительный выступ создают с помощью модификации Fc-области IgG с содержанием модификации T366W. Предпочтительную впадину создают с помощью модификации Fc-области IgG с содержанием модификации T366S, L368A и Y407V. С целью облегчения очистки гомодимера несущей впадину третьей полипептидной цепи от конечной биспецифической гетеродимерной содержащей Fc-область молекулы, связывающий сайт белка A несущих впадину доменов CH2 и CH3 третьей полипептидной цепи предпочтительно подвергают мутированию посредством аминокислотной замены в положении 435 (H435R). Таким образом, гомодимер несущей впадину третьей полипептидной цепи не будет связываться с белком A, в то время как биспецифический димер будет сохранять свою способность связывать белок A посредством связывающего сайта белка A в первой полипептидной цепи. В соответствии с альтернативным вариантом осуществления несущая впадину третья полипептидная цепь может включать аминокислотные замены в положениях 434 и 435 (N434A/N435K).
[00208] Предпочтительная аминокислотная последовательность IgG в случае доменов CH2 и CH3 первой полипептидной цепи молекулы, содержащей Fc-область, по настоящему изобретению, будет иметь «несущую выступ» последовательность (SEQ ID NO:109):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00209] Предпочтительная аминокислотная последовательность IgG в случае доменов CH2 и CH3 второй полипептидной цепи молекулы, содержащий Fc-область, по настоящему изобретению, имеющий две полипептидные цепи (или третей полипептидной цепи молекулы, содержащей Fc-область, имеющую три, четыре или пять полипептидных цепей), будет иметь «несущую впадину» последовательность (SEQ ID NO:110):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00210] Как будет отмечено, домены CH2-CH3 SEQ ID NO:109 и SEQ ID NO:110 включают в себя замену в положении 234 аланином и 235 аланином, и, таким образом, образуют Fc-область, которая проявляет снижение (или по сути отсутствие) связывания с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по отношению к связыванию, проявляемому Fc-областью дикого типа (SEQ ID NO:1). Настоящее изобретение также охватывает такие домены CH2-CH3, которые содержат аланиновые остатки дикого типа, альтернативные и/или дополнительные замены, которые модифицируют эффекторную функцию и/или активность связывания FγR Fc-области.
[00211] Настоящее изобретение также охватывает такие домены CH2-CH3, которые дополнительно содержат одну или несколько аминокислотных замен, продлевающих период полужизни. В частности, настоящее изобретение охватывает такие несущие впадину и такие несущие выступ домены CH2-CH3, которые дополнительно содержат замены M252Y/S254T/T256E.
[00212] Иллюстративные несущие выступ домены CH2 и CH3, содержащие замены L234A и L235A, и дополнительно содержащие замены M252Y, S254T и T256E, представлены ниже (SEQ ID NO:204):
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00213] Иллюстративные несущие впадину домены CH2 и CH3, содержащие замены L234A и L235A, и дополнительно содержащие замены M252Y, S254T и T256E, представлены ниже (SEQ ID NO:205):
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00214] Предпочтительно, чтобы первая полипептидная цепь имела «несущую выступ» последовательность CH2-CH3, такую как таковая SEQ ID NO:109. В то же время, как будет признано, «несущий впадину» домен CH2-CH3 (например, SEQ ID NO:110 мог быть использован в первой полипептидной цепи, в случае чего «несущий выступ» домен CH2-CH3 (например, SEQ ID NO:109) был бы использован во второй полипептидной цепи молекулы, содержащей Fc-область, по настоящему изобретению, имеющей две полипепитдные цепи (или в третьей полипептидной цепи молекулы, содержащей Fc-область, имеющей три, четыре или пять полипептидных цепей).
[00215] В соответствии с другими вариантами осуществления настоящее изобретение охватывает молекулы, связывающие ADAM9, содержащие домены CH2 и/или CH3, которые были разработаны для преобладания гетеродимеризации над гомодимеризаций, с использованием мутаций, известных в данной области техники, таких как таковые, раскрытые в публикациях согласно PCT №№ WO 2007/110205; WO 2011/143545; WO 2012/058768; и WO 2013/06867, все из которых включены в данный документ посредством ссылки в полном объеме.
X. Связывающие свойства эффекторных клеток
[00216] Предусмотренные в данном документе молекулы, связывающие ADAM9, по настоящему изобретению, могут быть разработаны таким образом, чтобы содержать комбинацию эпитопсвязывающих сайтов, которые распознают совокупность антигенов, уникальных по отношению к целевой клетке или типу тканей. В частности, настоящее изобретение относится к мультиспецифическим молекулам, связывающим ADAM9, которые способны связываться с эпитопом ADAM9 и эпитопом молекулы, присутствующей на поверхности эффекторной клетки, такой как T-лимфоцит, естественный киллер (NK) или другая мононуклеарная клетка. Например, молекулы, связывающие ADAM9, по настоящему изобретению, могут быть сконструированы таким образом, чтобы содержать эпитопсвязывающий сайт, который иммуноспецифически связывает CD2, CD3, CD8, CD16, T-клеточный рецептор (TCR) или NKG2D. Настоящее изобретение также относится к триспецифическим молекулам, связывающим ADAM9, которые способны связываться с эпитопом CD3 и эпитопом CD8 (см., например, публикацию согласно PCT № WO 2015/026894).
A. Связывающие способности CD2
[00217] В соответствии с одним вариантом осуществления биспецифические, триспецифические или мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп CD2. CD2 представляет собой молекулу клеточной адгезии, встречающуюся на поверхности T-клеток и естественных киллеров (NK). CD2 усиливает цитотоксичность NK-клеток, возможно, в качестве промотора образования нанотрубочек NK-клеток (Mace, E.M. et al. (2014) “Cell Biological Steps and Checkpoints in Accessing NK Cell Cytotoxicity,” Immunol. Cell. Biol. 92(3):245-255; Comerci, C.J. et al. (2012) “CD2 Promotes Human Natural Killer Cell Membrane Nanotube Formation,” PLoS One 7(10):e47664:1-12). Молекулы, которые специфически связывают CD2, включают антитело к CD2 «Lo-CD2a».
[00218] Аминокислотная последовательность домена VH Lo-CD2a (№ доступа в ATCC: 11423; SEQ ID NO:111) показана ниже (остатки CDRH показаны подчеркнутыми):
EVQLQQSGPE LQRPGASVKL SCKASGYIFT EYYMYWVKQR PKQGLELVGR IDPEDGSIDY VEKFKKKATL TADTSSNTAY MQLSSLTSED TATYFCARGK FNYRFAYWGQ GTLVTVSS
[00219] Аминокислотная последовательность домена VL Lo-CD2a (№ доступа в ATCC: 11423; SEQ ID NO:112) показана ниже (остатки CDRL показаны подчеркнутыми):
DVVLTQTPPT LLATIGQSVS ISCRSSQSLL HSSGNTYLNW LLQRTGQSPQ PLIYLVSKLE SGVPNRFSGS GSGTDFTLKI SGVEAEDLGV YYCMQFTHYP YTFGAGTKLE LK
B. Связывающие способности CD3
[00220] В соответствии с одним вариантом осуществления биспецифические, триспецифические или мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп CD3. CD3 представляет собой T-клеточный корецептор, состоящий из четырех различных цепей (Wucherpfennig, K.W. et al. (2010) “Structural Biology Of The T-Cell Receptor: Insights Into Receptor Assembly, Ligand Recognition, And Initiation Of Signaling,” Cold Spring Harb. Perspect. Biol. 2(4):a005140; pages 1-14). У млекопитающих данный комплекс содержит CD3γ-цепь, CD3δ-цепь и две CD3ε-цепи. Эти цепи ассоциируют с молекулой, известной в качестве T-клеточного рецептора (TCR) с целью генерации активирующего сигнала в T-лимфоцитах. В отсутствии CD3 TCR не собираются должным образом и распадаются (Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177). CD3 встречается связанным с мембранами всех взрослых T-клеток и фактически не встречается ни у какого другого типа клеток (см., Janeway, C.A. et al. (2005) In: Immunobiology: The Immune System In Health And Disease,” 6th ed. Garland Science Publishing, NY, pp. 214- 216; Sun, Z. J. et al. (2001) “Mechanisms Contributing To T Cell Receptor Signaling And Assembly Revealed By The Solution Structure Of An Ectodomain Fragment Of The CD3ε:γ Heterodimer,” Cell 105(7):913-923; Kuhns, M.S. et al. (2006) “Deconstructing The Form And Function Of The TCR/CD3 Complex,” Immunity. 2006 Feb;24(2):133-139). Молекулы, которые специфически связывают CD3, включают в себя антитела к CD3 «CD3 mAb-1» и «OKT3». Антитело к CD3 CD3 mAb-1 способно к связыванию у не относящихся к человеку приматов (например, яванского макака).
[00221] Аминокислотная последовательность домена VH CD3 mAb-1 VH(1) (SEQ ID NO:113) показана ниже (остатки CDRH показаны подчеркнутыми):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKDRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSS
[00222] Аминокислотная последовательность альтернативного домена VH CD3 mAb-1 VH(2) (SEQ ID NO:114) показана ниже (остатки CDRH показаны одним подчеркиванием; различия по отношению к домену VH CD3 mAb-1 VH(1) (SEQ ID NO:92) показаны двойным подчеркиванием).
EVQLVESGGG LVQPGGSLRL SCAASGFTFN TYAMNWVRQA PGKGLEWVAR IRSKYNNYAT YYADSVKDRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSS
[00223] Аминокислотная последовательность домена VL CD3 mAb-1 (SEQ ID NO:115) показана ниже (остатки CDRL показаны подчеркнутыми):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG
[00224] Домен VH CD3 mAb-1 VH(1) (SEQ ID NO:) может быть использован вместе с доменом VL CD3 mAb-1 (SEQ ID NO:) с образованием функциональной CD3-связывающей молекулы в соответствии с настоящим изобретением. Аналогичным образом, домен VH CD3 mAb-1 VH(2) (SEQ ID NO:) может быть использован вместе с доменом VL CD3 mAb-1 (SEQ ID NO:) с образованием функциональной CD3-связывающей молекулы в соответствии с настоящим изобретением.
[00225] Как описано ниже, с целью иллюстрации настоящего изобретения получают биспецифические ADAM9 × CD3-связывающие молекулы. В некоторых из конструкций ADAM9 × CD3 был использован вариант CD3 mAb-1. Вариант «CD3 mAb-1 (D65G)» содержит домен VL CD3 mAb-1 (SEQ ID NO:115) и домен VH CD3 mAb-1, имеющий замену D65G (положение 65 в соответствии с нумерацией Кабат, соответствующее остатку 68 SEQ ID NO:113).
[00226] Аминокислотная последовательность домена VH CD3 mAb-1 (D65G) (SEQ ID NO:116) показана ниже (остатки CDRH показаны подчеркнутыми, замещенное положение (D65G) показано двойным подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSS
[00227] В альтернативном случае может быть встроен аффинный вариант CD3 mAb-1. Варианты включают в себя низкоаффинный вариант, обозначаемый «CD3 mAb-1 Low», и вариант, имеющий более быструю скорость диссоциации, обозначаемый «CD3 mAb-1 Fast». Домен VL CD mAb1 (SEQ ID NO:115) является распространенным в отношении к CD3 mAb-1 Low и CD3 mAb1 Fast и представлен выше. Аминокислотные последовательности доменов VH каждого из CD3 mAb-1 Low и CD3 mAb-1 Fast представлены ниже.
[00228] Аминокислотная последовательность домена VH CD3 mAb-1 Low человека (SEQ ID NO:117) показана ниже (остатки CDRH показаны подчеркиванием; различия по отношению к домену VH CD3 mAb-1 VH(1) (SEQ ID NO:113) показаны двойным подчеркиванием).
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVT WFAYWGQGTL VTVSS
[00229] Аминокислотная последовательность домена VH CD3 mAb-1 Fast человека (SEQ ID NO:118) показана ниже (остатки CDRH показаны подчеркиванием; различия по отношению к домену VH CD3 mAb-1 VH(1) (SEQ ID NO:113) показаны двойным подчеркиванием).
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HKNFGNSYVT WFAYWGQGTL VTVSS
[00230] Другим антителом к CD3, которое может быть использовано, является антитело муромонаб-CD3 «OKT3» (Xu et al. (2000) “In Vitro Characterization Of Five Humanized OKT3 Effector Function Variant Antibodies,” Cell. Immunol. 200:16-26; Norman, D.J. (1995) “Mechanisms Of Action And Overview Of OKT3,” Ther. Drug Monit. 17(6):615-620; Canafax, D.M. et al. (1987) “Monoclonal Antilymphocyte Antibody (OKT3) Treatment Of Acute Renal Allograft Rejection,” Pharmacotherapy 7(4):121-124; Swinnen, L.J. et al. (1993) “OKT3 Monoclonal Antibodies Induce Interleukin-6 And Interleukin-10: A Possible Cause Of Lymphoproliferative Disorders Associated With Transplantation,” Curr. Opin. Nephrol. Hypertens. 2(4):670-678).
[00231] Аминокислотная последовательность домена VH OKT3 (SEQ ID NO:119) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLQQSGAE LARPGASVKM SCKASGYTFT RYTMHWVKQR PGQGLEWIGY INPSRGYTNY NQKFKDKATL TTDKSSSTAY MQLSSLTSED SAVYYCARYY DDHYCLDYWG QGTTLTVSS
[00232] Аминокислотная последовательность домена VL OKT3 (SEQ ID NO:120) показана ниже (остатки CDRL показаны подчеркнутыми):
QIVLTQSPAI MSASPGEKVT MTCSASSSVS YMNWYQQKSG TSPKRWIYDT SKLASGVPAH FRGSGSGTSY SLTISGMEAE DAATYYCQQW SSNPFTFGSG TKLEINR
[00233] Дополнительные антитела к CD3, которые могут быть использованы, включают в себя без ограничения таковые, описанные в публикациях согласно PCT №№ WO 2008/119566; и WO 2005/118635.
C. Связывающие способности CD8
[00234] В соответствии с одним вариантом осуществления биспецифические, триспецифические или мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп CD8. CD8 представляет собой T-клеточный корецептор, состоящий из двух различных цепей (Leahy, D.J., (1995) “A Structural View of CD4 and CD8,” FASEB J., 9:17-25), который экспрессируется на цитотоксических T-клетках. Было обнаружено, что активация CD8+ T-клеток опосредуется посредством костимулирующих взаимодействий комплекса антиген:молекула главного комплекса гистосовместимости класса I (MHC I), который располагается на поверхности целевой клетки, и комплекса CD8 и T-клеточного рецептора, которые располагаются на поверхности CD8+ T-клетки (Gao, G., and Jakobsen, B., (2000). "Molecular interactions of coreceptor CD8 and MHC class I: the molecular basis for functional coordination with the T-Cell Receptor". Immunol Today 21: 630-636). В отличие от молекул MHC II, которые экспрессируются только определенными клетками иммунной системы, молекулы MHC I являются широко распространенными. Таким образом, цитотоксические T-клетки способны связываться с большим разнообразием типов клеток. Активированные цитотоксические T-клетки опосредуют уничтожение клеток посредством высвобождения ими цитотоксинов перфорина, гранзимов и гранулизина. Антитела, которые специфически связывают CD8, включают в себя антитела к CD8 «OKT8» и «TRX2».
[00235] Аминокислотная последовательность домена VH OKT8 (SEQ ID NO:121) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLLESGPE LLKPGASVKM SCKASGYTFT DYNMHWVKQS HGKSLEWIGY IYPYTGGTGY NQKFKNKATL TVDSSSSTAY MELRSLTSED SAVYYCARNF RYTYWYFDVW GQGTTVTVSS
[00236] Аминокислотная последовательность домена VL OKT8 (SEQ ID NO:122) показана ниже (остатки CDRL показаны подчеркнутыми):
DIVMTQSPAS LAVSLGQRAT ISCRASESVD SYDNSLMHWY QQKPGQPPKV LIYLASNLES GVPARFSGSG SRTDFTLTID PVEADDAATY YCQQNNEDPY TFGGGTKLEI KR
[00237] Аминокислотная последовательность домена VH TRX2 (SEQ ID NO:123) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLVESGGG VVQPGRSLRL SCAASGFTFS DFGMNWVRQA PGKGLEWVAL IYYDGSNKFY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKPH YDGYYHFFDS WGQGTLVTVS S
[00238] Аминокислотная последовательность домена VL TRX2 (SEQ ID NO:124) показана ниже (остатки CDRL показаны подчеркнутыми):
DIQMTQSPSS LSASVGDRVT ITCKGSQDIN NYLAWYQQKP GKAPKLLIYN TDILHTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCYQ YNNGYTFGQG TKVEIK
D. Связывающие способности CD16
[00239] В соответствии с одним вариантом осуществления мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп CD16. CD16 представляет собой рецептор FcγRIIIA. CD16 экспресссируется нейтрофилами, эозинофилами, естественными киллерами (NK) и тканевыми макрофагами, которые связывают агрегированный, но не мономерный IgG человека (Peltz, G.A. et al. (1989) “Human Fc Gamma RIII: Cloning, Expression, And Identification Of The Chromosomal Locus Of Two Fc Receptors For IgG,” Proc. Natl. Acad. Sci. (U.S.A.) 86(3):1013-1017; Bachanova, V. et al. (2014) “NK Cells In Therapy Of Cancer,” Crit. Rev. Oncog. 19(1-2):133-141; Miller, J.S. (2013) “Therapeutic Applications: Natural Killer Cells In The Clinic,” Hematology Am. Soc. Hematol. Educ. Program. 2013:247-253; Youinou, P. et al. (2002) “Pathogenic Effects Of Anti-Fc Gamma Receptor IIIB (CD16) On Polymorphonuclear Neutrophils In Non-Organ-Specific Autoimmune Diseases,” Autoimmun Rev. 1(1-2):13-19; Peipp, M. et al. (2002) “Bispecific Antibodies Targeting Cancer Cells,” Biochem. Soc. Trans. 30(4):507-511). Молекулы, которые специфически связывают CD16, включают в себя антитела к CD16 «3G8» и «A9». Гуманизированные антитела A9 описаны в публикации согласно PCT № WO 03/101485.
[00240] Аминокислотная последовательность домена VH 3G8 (SEQ ID NO:125) показана ниже (остатки CDRH показаны подчеркнутыми):
QVTLKESGPG ILQPSQTLSL TCSFSGFSLR TSGMGVGWIR QPSGKGLEWL AHIWWDDDKR YNPALKSRLT ISKDTSSNQV FLKIASVDTA DTATYYCAQI NPAWFAYWGQ GTLVTVSA
[00241] Аминокислотная последовательность домена VL 3G8 (SEQ ID NO:126) показана ниже (остатки CDRL показаны подчеркнутыми):
DTVLTQSPAS LAVSLGQRAT ISCKASQSVD FDGDSFMNWY QQKPGQPPKL LIYTTSNLES GIPARFSASG SGTDFTLNIH PVEEEDTATY YCQQSNEDPY TFGGGTKLEI K
[00242] Аминокислотная последовательность домена VH A9 (SEQ ID NO:127) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLQQSGAE LVRPGTSVKI SCKASGYTFT NYWLGWVKQR PGHGLEWIGD IYPGGGYTNY NEKFKGKATV TADTSSRTAY VQVRSLTSED SAVYFCARSA SWYFDVWGAR TTVTVSS
[00243] Аминокислотная последовательность домена VL A9 (SEQ ID NO:128) показана ниже (остатки CDRL показаны подчеркнутыми):
DIQAVVTQES ALTTSPGETV TLTCRSNTGT VTTSNYANWV QEKPDHLFTG LIGHTNNRAP GVPARFSGSL IGDKAALTIT GAQTEDEAIY FCALWYNNHW VFGGGTKLTVL
[00244] Дополнительные антитела к CD16, которые могут быть использованы, включают в себя без ограничения таковые, описанные в публикациях согласно PCT №№ WO 03/101485; и WO 2006/125668.
E. Связывающие способности TCR
[00245] В соответствии с одним вариантом осуществления биспецифические, триспецифические или мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп T-клеточного рецептора (TCR). T-клеточный рецептор естественным образом экспрессируется CD4+ или CD8+ T-клетками и способствует распознаванию такими клетками антигенных пептидов, которые связываются и презентируются белками MHC I класса или II класса антигенпрезентирующих клеток. Распознавание комплекса pMHC (пептид-MHC) TCR инициирует передачу клеточного иммунного ответа, который приводит к продуцированию цитокинов и лизису антигенпрезентирующей клетки (см., например, Armstrong, K.M. et al. (2008) “Conformational Changes And Flexibility In T-Cell Receptor Recognition Of Peptide-MHC Complexes,” Biochem. J. 415(Pt 2):183-196; Willemsen, R. (2008) “Selection Of Human Antibody Fragments Directed Against Tumor T-Cell Epitopes For Adoptive T-Cell Therapy,” Cytometry A. 73(11):1093-1099; Beier, K.C. et al. (2007) “Master Switches Of T-Cell Activation And Differentiation,” Eur. Respir. J. 29:804-812; Mallone, R. et al. (2005) “Targeting T Lymphocytes For Immune Monitoring And Intervention In Autoimmune Diabetes,” Am. J. Ther. 12(6):534-550). CD3 представляет собой рецептор, который связывается с TCR (Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177; Guy, C.S. et al. (2009) “Organization Of Proximal Signal Initiation At The TCR:CD3 Complex,” Immunol. Rev. 232(1):7-21; St. Clair, E.W. (Epub 2009 Oct 12) “Novel Targeted Therapies For Autoimmunity,” Curr. Opin. Immunol. 21(6):648-657; Baeuerle, P.A. et al. (Epub 2009 Jun 9) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944; Smith-Garvin, J.E. et al. (2009) “T Cell Activation,” Annu. Rev. Immunol. 27:591-619; Renders, L. et al. (2003) “Engineered CD3 Antibodies For Immunosuppression,” Clin. Exp. Immunol. 133(3):307-309).
[00246] Молекулы, которые специфически связываются с T-клеточным рецептором, включают в себя антитело к TCR «BMA 031» (EP 0403156; Kurrle, R. et al. (1989) “BMA 031 - A TCR-Specific Monoclonal Antibody For Clinical Application,” Transplant Proc. 21(1 Pt 1):1017-1019; Nashan, B. et al. (1987) “Fine Specificity Of A Panel Of Antibodies Against The TCR/CD3 Complex,” Transplant Proc. 19(5):4270-4272; Shearman, C.W. et al. (1991) “Construction, Expression, And Biologic Activity Of Murine/Human Chimeric Antibodies With Specificity For The Human α/β T Cell,” J. Immunol. 146(3):928-935; Shearman, C.W. et al. (1991) “Construction, Expression And Characterization of Humanized Antibodies Directed Against The Human α/β T Cell Receptor,” J. Immunol. 147(12):4366-4373; и публикация согласно PCT № WO 2010/027797).
[00247] Аминокислотная последовательность домена VH BMA 031 (SEQ ID NO:129) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLVQSGAE VKKPGASVKV SCKASGYKFT SYVMHWVRQA PGQGLEWIGY INPYNDVTKY NEKFKGRVTI TADKSTSTAY LQMNSLRSED TAVHYCARGS YYDYDGFVYW GQGTLVTVSS
[00248] Аминокислотная последовательность домена VL BMA 031 (SEQ ID NO:130) показана ниже (остатки CDRL показаны подчеркнутыми):
EIVLTQSPAT LSLSPGERAT LSCSATSSVS YMHWYQQKPG KAPKRWIYDT SKLASGVPSR FSGSGSGTEF TLTISSLQPE DFATYYCQQW SSNPLTFGQG TKLEIK
F. Связывающие способности NKG2D
[00249] В соответствии с одним вариантом осуществления мультиспецифические молекулы, связывающие ADAM9, по настоящему изобретению, способны связывать эпитоп ADAM9 и эпитоп рецептора NKG2D. Рецептор NKG2D экспрессируется на всех естественных киллерах человека (и других млекопитающих) (Bauer, S. et al. (1999) “Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA,” Science 285(5428):727-729; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29), а также на всех CD8+ T-клетках (Groh, V. et al. (2001) “Costimulation Of CD8αβ T Cells By NKG2D Via Engagement By MIC Induced On Virus-Infected Cells,” Nat. Immunol. 2(3):255-260; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29). Такие связывающие лиганды, и, в частности, таковые, которые не экспрессируются на нормальных клетках, включают в себя молекулу гистосовместимости 60 (H60), продукт раннего гена 1, индуцируемый ретиноевой кислотой (RAE-1), и мышиный UL16-связывающий протеинподобный транскрипт 1 (MULT1) (Raulet D.H. (2003) “Roles Of The NKG2D Immunoreceptor And Its Ligands,” Nature Rev. Immunol. 3:781-790; Coudert, J.D. et al. (2005) “Altered NKG2D Function In NK Cells Induced By Chronic Exposure To Altered NKG2D Ligand-Expressing Tumor Cells,” Blood 106:1711-1717). Молекулы, которые специфически связываются с рецептором NKG2D, включают в себя антитела к NKG2D «KYK-1.0» и «KYK-2.0» (Kwong, KY et al. (2008) “Generation, Affinity Maturation, And Characterization Of A Human Anti-Human NKG2D Monoclonal Antibody With Dual Antagonistic And Agonistic Activity,” J. Mol. Biol. 384:1143-1156).
[00250] Аминокислотная последовательность домена VH KYK-1.0 (SEQ ID NO:131) показана ниже (остатки CDRH показаны подчеркнутыми):
EVQLVESGGG VVQPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF IRYDGSNKYY ADSVKGRFTI SRDNSKNTKY LQMNSLRAED TAVYYCAKDR FGYYLDYWGQ GTLVTVSS
[00251] Аминокислотная последовательность домена VL KYK-1.0 (SEQ ID NO:132) показана ниже (остатки CDRL показаны подчеркнутыми):
QPVLTQPSSV SVAPGETARI PCGGDDIETK SVHWYQQKPG QAPVLVIYDD DDRPSGIPER FFGSNSGNTA TLSISRVEAG DEADYYCQVW DDNNDEWVFG GGTQLTVL
[00252] Аминокислотная последовательность домена VH KYK-2.0 (SEQ ID NO:133) показана ниже (остатки CDRH показаны подчеркнутыми):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF IRYDGSNKYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKDR GLGDGTYFDY WGQGTTVTVS S
[00253] Аминокислотная последовательность домена VL KYK-2.0 (SEQ ID NO:134) показана ниже (остатки CDRL показаны подчеркнутыми):
QSALTQPASV SGSPGQSITI SCSGSSSNIG NNAVNWYQQL PGKAPKLLIY YDDLLPSGVS DRFSGSKSGT SAFLAISGLQ SEDEADYYCA AWDDSLNGPV FGGGTKLTVL
XI. Мультиспецифические молекулы, связывающие ADAM9
A. Биспецифические двухцепочечные диатела ADAM9 × CD3
[00254] Домены VL и VH вышеописанного оптимизированного гуманизированного антитела к ADAM9 MAB-A используются для конструирования биспецифических диател ADAM9 × CD3, состоящих из двух ковалентно связанных полипептидных цепей и содержащих вышеописанные оптимизированные домены VL и VH MAB-A. Общая структура аминокислотных последовательностей таких биспецифических диател ADAM9 × CD3 представлена ниже.
[00255] Первая полипептидная цепь одного иллюстративного биспецифического двухцепочечного диатела ADAM9 × CD3 содержит в направлении от N-конца к C-концу: N-конец; домен VL антитела к ADAM9 (например, hMAB-A VL (2) (SEQ ID NO:55); промежуточный спейсерный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH антитела к CD3 (например, CD3 mAb 1 (D65G) (SEQ ID NO:116)); содержащий цистеин промежуточный спейсерный пептид (линкер 2: GGCGGG (SEQ ID NO:70)); активирующий сборку гетеродимера домен (например, E-петля) (EVAALEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO:82)); и C-конец.
[00256] Вторая полипептидная цепь такого иллюстративного биспецифического двухцепочечного диатела ADAM9 × CD3 содержит в направлении от N-конца к C-концу: N-конец; домен VL соответствующего антитела к CD3 (например, домен VL, который в ассоциации с доменом VH первой полипептидной цепи образует CD3-связывающий сайт, например, домен VL CD3 mAb-1 (SEQ ID NO:115); промежуточный спейсерный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH соответствующего антитела к ADAM9 (например, домен VH, который в ассоциации с доменом VL первой полипептидной цепи образует ADAM9-связывающий сайт, например, hMAB-A VH (2) (SEQ ID NO:17); содержащий цистеин промежуточный спейсерный пептид (линкер 2: GGCGGG (SEQ ID NO:70)); активирующий сборку гетеродимера домен (например, К-петля) (KVAALKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO:83)); и C-конец.
[00257] Предусмотренные в данном документе альтернативные линкеры и/или альтернативные активирующие сборку гетеродимера домены могут быть использованы в получении таких диател. Например, первая полипептидная цепь альтернативного иллюстративного биспецифического двухцепочечного диатела ADAM9 × CD3 может содержать в направлении от N-конца к C-концу: N-конец; домен VL антитела к ADAM9; промежуточный спейсерный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH антитела к CD3 или соответствующего антитела к CD3; промежуточный спейсерный пептид (линкер 2: ASTKG (SEQ ID NO:74)); содержащий цистеин активирующий сборку гетеродимера домен (например, К-петля) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO:85)); и C-конец. Вторая полипептидная цепь такого альтернативного иллюстративного диатела может содержать в направлении от N-конца к C-концу: N-конец; домен VL соответствующего антитела к CD3; промежуточный спейсерный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH соответствующего антитела к ADAM9; промежуточный спейсерный пептид (линкер 2: ASTKG (SEQ ID NO:74)); содержащий цистеин активирующий сборку гетеродимера домен (например, Е-петля) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO:84)); и C-конец.
[00258] Сконструировано иллюстративное биспецифическое двухцепочечное диатело ADAM9 × CD3 («DART-1»), содержащее домены VH и VL hMAB-A (2.2) и домены VH и VL CD3 mAb-1.
[00259] Аминокислотная последовательность первой полипептидной цепи DART1 (SEQ ID NO:135) показана ниже (последовательность домена hMAb-A VL(2) (SEQ ID NO:55) подчеркнута; последовательность CD3 mAb-1 (D65G) VH-домена (SEQ ID NO:116) представлена курсивом):

[00260] Аминокислотная последовательность второй полипептидной цепи DART1 (SEQ ID NO:136) показана ниже (последовательность домена hMAB-A VH (2) (SEQ ID NO:17) подчеркнута; последовательность CD3 mAb-1 VL -домена (SEQ ID NO:115) представлена курсивом):

B. Биспецифические трехцепочечные диатела ADAM9 × CD3
[00261] Получено диатело ADAM9 × CD3, имеющее три цепи и имеющее Fc-область, имеющее один связывающий сайт, специфический в отношении ADAM9 (содержащий гуманизированные/оптимизированные домены VH и VL hMAB-A) и один связывающий сайт, специфический в отношении CD3 (содержащий домены VL и VH CD3 mAb 1 (D65G)). Диатело называется «DART-2».
[00262] Первая полипептидная цепь иллюстративного биспецифического трехцепочечного диатела ADAM9 × CD3 содержит в направлении от N-конца к C-концу: N-конец; домен VL антитела к ADAM9 (например, hMAB-A VL (2) (SEQ ID NO:55); промежуточный спейсерный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH CD3 mAb 1 (D65G) (SEQ ID NO:116); промежуточный спейсерный пептид (линкер 2: ASTKG (SEQ ID NO:74)); содержащий цистеин активирующий сборку гетеродимера домен (Е-петля) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO:82)); промежуточный спейсерный пептид (линкер 3: GGGDKTHTCPPCP (SEQ ID NO:94)); несущий выступ домен CH2-CH3 IgG1 (SEQ ID NO:109); и C-конец. Полинуклеотиды, кодирующие эту полипептидную цепь, могут кодировать C-концевой лизиновый остаток SEQ ID NO:109 (т.е., X SEQ ID NO:109), однако, как описано выше, этот лизиновый остаток может быть пострансляционно удален в некоторых системах экспрессии. Соответственно, настоящее изобретение охватывает такую первую полипептидную цепь, которая содержит такой лизиновый остаток (т.е., SEQ ID NO:109, где X представляет собой лизин), а также первую полипептидную целью, которая не имеет такого лизинового остатка (т.е., SEQ ID NO:109, где X отсутствует). Аминокислотная последовательность такой первой полипептидной цепи DART-2 (SEQ ID NO:137) представлена ниже (последовательность домена hMAB-A VL(2) (SEQ ID NO:55) подчеркнута; последовательность CD3 mAb-1 (D65G) VH-домена (SEQ ID NO:116) представлена курсивом):

где X представляет собой лизин (K) или отсутствует.
[00263] Вторая полипептидная цепь иллюстративного биспецифического трехцепочечного диатела ADAM9 × CD3 содержит в направлении от N-конца к C-концу: N-конец; домен VL антитела к CD3 mAb 1 (SEQ ID NO:115); промежуточный спейсперный пептид (линкер 1: GGGSGGGG (SEQ ID NO:69)); домен VH антитела к ADAM9 (например, hMAB-A VH (2) (SEQ ID NO:17)); промежуточный спейсерный пептид (линкер 2: ASTKG (SEQ ID NO:74)); содержащий цистеин активирующий сборку гетеродимера домен (К-петля) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO:85)); и C-конец. Аминокислотная последовательность такой второй полипептидной цепи DART-2 (SEQ ID NO:138) представлена ниже (последовательность домена hMAB-A VH (2) (SEQ ID NO:17) подчеркнута; последовательность CD3 mAb-1 VL -домена (SEQ ID NO:115) представлена курсивом):

[00264] Третья полипептидная цепь иллюстративного биспецифического трехцепочечного диатела ADAM9 × CD3 содержит в направлении от N-конца к C-концу: N-конец; спейсерный пептид (DKTHTCPPCP (SEQ ID NO:93)); несущий выступ домен CH2-CH3 IgG1 (SEQ ID NO:110); и C-конец. Полинуклеотиды, кодирующие эту полипептидную цепь, могут кодировать C-концевой лизиновый остаток SEQ ID NO:110 (т.е., X SEQ ID NO:110), однако, как описано выше, этот лизиновый остаток может быть пострансляционно удален в некоторых системах экспрессии. Соответственно, настоящее изобретение охватывает такую третью полипептидную цепь, которая содержит такой лизиновый остаток (т.е., SEQ ID NO:110, где X представляет собой лизин), а также третью полипептидную цепь, которая не имеет такого лизинового остатка (т.е., SEQ ID NO:110, где X отсутствует). Аминокислотная последовательность такой третьей полипептидной цепи (SEQ ID NO:139) представлена ниже:
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGX,
где X представляет собой лизин (K) или отсутствует.
[00265] Принимая во внимание идеи, предусмотренные в данном документе, будет понятно, что различные ориентации доменов, доменов VH, доменов VL, линкеров и/или активирующих сборку гетеродимера доменов, могут быть использованы для получения альтернативных биспецифических трехцепочечных диател ADAM9 × CD3. В частности, могут быть использованы домен VH и домен VL различных вариантов hMAB-A.
C. Трехвалентные связывающие молекулы ADAM9 × CD3 × CD8
[00266] Предусмотрены иллюстративные трехвалентные связывающие молекулы «ADAM9 × CD3 × CD8», имеющие один связывающий сайт, специфический в отношении ADAM9 (содержащий исходный и/или гуманизированный анти-ADAM9-VL-домен и соответствующий анти-ADAM9-VH-домен, описанные выше), один связывающий сайт, специфический в отношении CD3 (содержащий, например, домен VL CD3 mAb-1 (SEQ ID NO:115) и домен VH антитела к CD3 (например, CD3 mAb 1 (D65G) (SEQ ID NO:116)), и один связывающий сайт, специфический в отношении CD8 (содержащий, например, домены VH и VL TRX2 (SEQ ID NO:123 и 124 соответственно). Такие трехвалентные связывающие молекулы могут иметь две полипептидные цепи (см., например, Фиг. 6E и Фиг. 6F), три полипептидные цепи (см., например, Фиг. 6C и Фиг. 6D), четыре полипептидные цепи (см., например, Фиг. 6A и Фиг. 6B) или пять полипептидных цепей (см., например, Фиг. 5).
XII. Способы получения
[00267] Молекулы, связывающие ADAM9, по настоящему изобретению, наиболее предпочтительно получают посредством рекомбинантной экспрессии молекул нуклеиновой кислоты, которые кодируют такие полипептиды, как хорошо известно в данной области техники.
[00268] Полипептиды по настоящему изобретению могут быть стандартно получены с помощью твердофазного пептидного синтеза (Merrifield, B. (1986) “Solid Phase Synthesis,” Science 232(4748):341-347; Houghten, R.A. (1985) “General Method For The Rapid Solid-Phase Synthesis Of Large Numbers Of Peptides: Specificity Of Antigen-Antibody Interaction At The Level Of Individual Amino Acids,” Proc. Natl. Acad. Sci. (U.S.A.) 82(15):5131-5135; Ganesan, A. (2006) “Solid-Phase Synthesis In The Twenty-First Century,” Mini Rev. Med. Chem. 6(1):3-10).
[00269] В альтернативном случае антитела могут быть получены рекомбинантно и экспрессироваться с помощью любого способа, известного в данной области техники. Антитела могут быть получены рекомбинантно, вначале в результате выделения антител, полученных из других животных-хозяев, получения последовательности генов и использования последовательности генов для рекомбинантной экспрессии антитела в клетках-хозяевах (например, клетках CHO). Другой способ, который может быть использован, представляет собой экспрессию последовательности антитела в растениях (например, табаке) или трансгенном молоке. Были раскрыты подходящие способы рекомбинантной экспрессии антител в растениях или молоке (см., например, Peeters et al. (2001) “Production Of Antibodies And Antibody Fragments In Plants,” Vaccine 19:2756; Lonberg, N. et al. (1995) “Human Antibodies From Transgenic Mice,” Int. Rev. Immunol 13:65-93; и Pollock et al. (1999) “Transgenic Milk As A Method For The Production Of Recombinant Antibodies,” J. Immunol Methods 231:147-157). Подходящие способы получения производных антител, например, гуманизированных, одноцепочечных и т.д., известны в данной области техники и были описаны выше. В альтернативном случае антитела могут быть получены рекомбинантно с помощью технологии фагового дисплея (см., например, патенты США №№ 5565332; 5580717; 5733743; 6265150; и Winter, G. et al. (1994) “Making Antibodies By Phage Display Technology,” Annu. Rev. Immunol. 12,433-455).
[00270] Векторы, содержащие полинуклеотиды, представляющие интерес (например, полинуклеотиды, кодирующие полипептидные цепи молекул, связывающих ADAM9, по настоящему изобретению), могут быть введены в клетку-хозяина с помощью любого из ряда подходящих средств, в том числе элетропорации, трансфекции с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировки микрочастицами; липофекции; и инфицирования (например, если вектор представляет собой инфекционный агент, такой как вирус осповакцины). Выбор вводимых векторов или полинуклеотидов будет часто зависеть от свойств клетки-хозяина.
[00271] Любая клетка-хозяин, способная сверхэкспрессировать гетерологичные ДНК, может быть использована для целей экспрессии полипептида или белка, представляющих интерес. Неограничивающие примеры подходящих клеток-хозяев млекопитающих включают в себя без ограничения клетки COS, HeLa и CHO.
[00272] В настоящем изобретении также предусмотрены полипептиды, содержащие аминокислотную последовательность молекулы, связывающей ADAM9, по настоящему изобретению. Полипептиды по настоящему изобретению, могут быть получены с помощью процедур, известных в данной области техники. Полипептиды могут быть получены с помощью протеолитического или другого расщепления антител, с помощью рекомбинантных способов (т.е., одиночные или слитые пептиды), как описано выше, или с помощью химического синтеза. Полипептиды антител, особенно более короткие полипептиды, размером до приблизительно 50 аминокислот, легко получают с помощью химического синтеза. Способы химического синтеза известны в данной области техники и являются коммерчески доступными.
[00273] В настоящем изобретении также предусмотрены варианты молекул, связывающих ADAM9, в том числе функционально эквивалентных полипептидов, которые существенно не влияют на свойства таких молекул, а также варианты, которые характеризуются повышенной или сниженной активностью. Модификация полипептидов представляет собой стандартную практику в данной области техники и не нуждается в подробном описании в данном документе. Примеры модифицированных полипептидов включают в себя полипептиды с консервативными заменами аминокислотных остатков, одной или несколькими делециями или добавлениями аминокислот, которые существенно или отрицательным образом не изменяют функциональную активность, или применение химических аналогов. Аминокислотные остатки, которые могут быть консервативно замещены друг на друга, включают в себя без ограничения: глицин/аланин; серин/треонин; валин/изолейцин/лейцин; аспарагин/глутамин; аспарагиновую кислоту/глутаминовую кислоту; лизин/аргинин; и фенилаланин/тирозин. Эти полипептиды также включают в себя гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование с различными сахарами, ацетилирование и фосфорилирование. Предпочтительно, чтобы аминокислотные замены были консервативными, т.е., замещенная аминокислот имела бы аналогичные химические свойства, что и исходная аминокислота. Такие консервативные замены известны в данной области техники, а примеры были представлены выше. Аминокислотные модификации могут находиться в диапазоне от изменения или модификации одной или нескольких аминокислот до полного изменения конструкции области, такой как вариабельный домен. Изменения в вариабельном домене могут изменять аффинность и/или специфичность связывания. Другие способы модификации включают в себя применение методик связывания, известных в данной области техники, в том числе без ограничения ферментативных средств, окислительной замены и хелатирования. Модификации могут быть использованы, например, для присоединения меток для иммунологического анализа, например, присоединения радиоактивных фрагментов для радиоиммунологического анализа. Модифицированные полипептиды получают с помощью разработанных процедур в данной области техники и могут быть подвергнуты скринингу с помощью стандартных анализов, известных в данной области техники.
[00274] Настоящее изобретение охватывает слитые белки, содержащие один или несколько анти-ADAM9-VL- и/или VH-доменов по данному изобретению. В соответствии с одним вариантом осуществления предусмотрен слитый полипептид, который содержит легкую цепь, тяжелую цепь или как легкую цепь, так и тяжелую цепь. В соответствии с другим вариантом осуществления слитый полипептид содержит константную область гетерологичного иммуноглобулина. В соответствии с другим вариантом осуществления слитый полипептид содержит вариабельный домен легкой цепи и вариабельный домен тяжелой цепи антитела, продуцируемого из депонируемой в открытом доступе гибридомы. Для целей настоящего изобретения слитый белок на основе антитела содержит один или несколько полипептидных доменов, которые специфически связываются ADAM9, и другую аминокислотную последовательность, которая не присоединена в нативной молекуле, например, гетерологичную последовательность или гомологичную последовательность из другой области.
[00275] Настоящее изобретение, в частности, охватывает молекулы, связывающие ADAM9 (например, антитела, диатела, трехвалентные связывающие молекулы и т.д.), конъюгированные с диагностическим или терапевтическим фрагментом. Для диагностических целей молекулы, связывающие ADAM9, по настоящему изобретению, могут быть связаны с детектируемым веществом. Такие молекулы, связывающие ADAM9, являются пригодными для мониторинга и/или прогнозирования развития или прогрессирования заболевания в виде части процедуры клинического исследования, такой как определение эффективности определенной терапии. Примеры детектируемых веществ включают в себя различные ферменты (например, пероксидазу хрена, бета-галактозидазу и т.д.), простетические группы (например, авидин/биотин), флуоресцентные вещества (например, умбеллиферон, флуоресцеин или фикоэритрин), люминесцентные вещества (например, люминол), биолюминесцентные вещества (например, люциферазу или экворин), радиоактивные вещества (например, углерод-14, марганец-54, стронций-85 или цинк-65), позитрон-эмиттирующие металлы и ионы нерадиоактивных парамагнитных металлов. Детектируемое вещество можно связать или конъюгировать либо непосредственно с молекулой, связывающей ADAM9, либо косвенно, с помощью посредника (например, линкера) с использованием методик, известных в данной области техники.
[00276] Для терапевтических целей молекулы, связывающие ADAM9, по настоящему изобретению, можно конъюгировать с терапевтическим фрагментом, таким как цитотоксин (например, цитостатическое или цитотоксическое средство), терапевтическое средство или ион радиоактивного металла, например, альфа-излучатели. Цитотоксин или цитотоксическое средство включает в себя любое средство, которое является вредным для клеток, такое как, например, экзотоксин синегнойной палочки, дифтерийный токсин, ботулинический токсин от типа А до типа F, рицин, абрин, сапорин и цитотоксические фрагменты таких средств. Терапевтическое средство включает в себя любое средство, оказывающее терапевтический эффект для профилактического или терапевтического лечения нарушения. Такие терапевтические средства могут представлять собой химические терапевтические средства, белковые или полипептидные терапевтические средства, и включают в себя терапевтические средства, которые характеризуются необходимой биологической активностью и/или модифицируют определенный биологический ответ. Примеры терапевтических средств включают в себя алкилирующие средства, ингибиторы ангиогенеза, антимитотические средства, средства гормональной терапии и антитела, пригодные для лечения нарушений клеточной пролиферации. Терапевтический фрагмент можно связать или конъюгировать либо непосредственно с молекулой, связывающей ADAM9, либо косвенно, с помощью посредника (например, линкера) с использованием методик, известных в данной области техники.
XIII. Применение молекул, связывающих ADAM9, по настоящему изобретению
[00277] Настоящее изобретение охватывает композиции, в том числе фармацевтические композиции, содержащие молекулы, связывающие ADAM9, по настоящему изобретению (например, антитела, биспецифические диатела, трехвалентные связывающие молекулы и т.д.), полипептиды, происходящие из таких молекул, полинуклеотиды, содержащие последовательности, кодирующие такие молекулы или полипептиды, а также другие средства, описанные в данном документе.
[00278] Как предусмотрено в данном документе, молекулы, связывающие ADAM9, по настоящему изобретению, содержащие анти-ADAM9-VL- и/или VH-домены, предусмотренные в данном документе, характеризуются способностью связывать ADAM9, присутствующий и на поверхности клетки, и индуцировать антителозависимую клеточноопосредованную цитотоксичность (ADCC) и/или комплементзависимую цитотоксичность (CDC) и/или опосредовать перенаправленное уничтожение клеток (например, перенаправленную T-клеточную цитотоксичность).
[00279] Таким образом, молекулы, связывающие ADAM9, по настоящему изобретению, содержащие анти-ADAM9-VL- и/или VH-домены, предусмотренные в данном документе, характеризуются способностью лечить любое заболевание или патологическое состояние, ассоциированное с экспрессией ADAM9 или характеризующееся ей. Как описано выше, ADAM9 представляет собой онкоэмбриональный антиген, экпрессируемый в многочисленных злокачественных новообразованиях крови и солидных злокачественных новообразованиях, который ассоциируется с опухолями высокой степени злокачественности, характеризующимися менее дифференцированной морфологией, и коррелирующий с неблагоприятными клиническими исходами. Таким образом, без ограничения молекулы, связывающие ADAM9, по настоящему изобретению, могут быть использованы в диагностике или лечении рака, в частности, рака, характеризующегося экспрессией ADAM9.
[00280] В частности, молекулы, связывающие ADAM9, по настоящему изобретению, могут быть использованы в лечении рака мочевого пузыря, рака молочной железы, рака шейки матки, колоректального рака, рака пищевода, рака желудка, рака головы и шеи, рака печени, рака лимфоидного происхождения, немелкоклеточного рака легких, рака миелоидного происхождения, рака яичников, рака поджелудочной железы, рака предстательной железы, почечно-клеточной карциномы, рака щитовидной железы, рака яичка и рака матки.
[00281] В соответствии с дополнительными вариантами осуществления молекулы, связывающие ADAM9, по настоящему изобретению, могут быть пригодны в лечении немелкоклеточного рака легких (плоскоклеточной карциномы, аденокарциномы или крупноклеточной недифференцированной карциномы) и колоректального рака (аденокарциномы, желудочно-кишечных карциноидных опухолей, желудочно-кишечных стромальных опухолей, первичной колоректальной лимфомы, лейомиосаркомы, меланомы или плоскоклеточной карциномы).
[00282] Биспецифические молекулы, связывающие ADAM9, по настоящему изобретению, усиливают терапию рака, обусловленную ADAM9 в результате перенаправленного уничтожения опухолевых клеток, которые экспрессируют вторую специфичность таких молекул (например, CD2, CD3, CD8, CD16, T-клеточного рецептора (TCR), NKG2D и т.д.). Такие молекулы, связывающие ADAM9, являются особо пригодными для лечения рака.
[00283] Помимо своего применения в терапии, молекулы, связывающие ADAM9, по настоящему изобретению, могут быть мечены детектируемым образом и использованы в диагностике рака или в визуализации опухолей и опухолевых клеток.
XIV. Фармацевтические композиции
[00284] Композиции по настоящему изобретению могут включать в себя композиции из нерасфасованной лекарственной формы, пригодные при производстве фармацевтических композиций (например, неочищенных или нестерильных композиций), и фармацевтические композиции (т.e., композиции, которые являются подходящими для введения пациенту или субъекту), которые можно применять при получении дозированных лекарственных форм. Такие композиции содержат профилактически или терапевтически эффективное количество молекул, связывающих ADAM9, по настоящему изобретению, или комбинации таких средств и фармацевтически приемлемый носитель. Предпочтительно композиции по настоящему изобретению содержат профилактически или терапевтически эффективное количество молекул, связывающих ADAM9, по настоящему изобретению, и фармацевтически приемлемый носитель. Настоящее изобретение также охватывает такие фармацевтические композиции, которые дополнительно содержат второе терапевтическое антитело (например, опухольспецифическое моноклональное антитело), которое является специфическим в отношении определенного ракового антигена, и фармацевтически приемлемый носитель.
[00285] В соответствии с конкретным вариантом осуществления термин «фармацевтически приемлемый» означает разрешенный органом регулирования федерального правительства или правительства штата или указанный в Фармакопее США или другой общепризнанной фармакопее для применения у животных, и, в частности, у человека. Термин «носитель» означает разбавитель, адъювант (например, адъювант Фрейнда (полный и неполный), вспомогательное вещество или носитель, с которым терапевтическое средство вводится. Как правило, ингредиенты композиций по настоящему изобретению могут поставляться либо раздельно, либо смешиваться совместно в дозированной лекарственной форме, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметически запечатанном контейнере, таком как ампула или саше с указанием количества действующего вещества. Если композицию необходимо вводить в виде инфузии, ее можно диспергировать в инфузионном флаконе, содержащем стерильную воду фармацевтической степени чистоты или физиологический раствор. Если композицию вводят путем инъекции, может быть предусмотрена ампула стерильной воды для инъекции или физиологического раствора для того, чтобы перед введением ингредиенты можно было смешать.
[00286] В настоящем изобретении также предусмотрены фармацевтическая упаковка или набор, содержащие один или несколько контейнеров, заполненных молекулой, связывающей ADAM9, по настоящему изобретению, в отдельности или вместе с таким фармацевтически приемлемым носителем. Помимо этого, одно или несколько других профилактических или терапевтических средств, пригодных для лечения заболевания, также могут быть включены в фармацевтическую упаковку или или набор. В настоящем изобретении также предусмотрены фармацевтическая упаковка или набор, заполненные одним или несколькими из ингредиентов фармацевтических композиций по настоящему изобретению. Необязательно вместе с таким контейнером(контейнерами) может находиться уведомление в форме, предписанной государственным органом, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, при этом уведомление указывает на разрешение органом производства, применения или продажи для введения человеком.
[00287] В настоящем изобретении предусмотрены наборы, которые могут быть использованы в вышеописанных способах. Набор может содержать любые из молекул, связывающих ADAM9, по настоящему изобретению. Набор может дополнительно содержать одно или несколько других профилактических и/или терапевтических средств, пригодных для лечения рака, в одном или нескольких контейнерах.
XV. Способы введения
[00288] Композиции по настоящему изобретению могут быть предусмотрены для лечения, профилактики и нормализации одного или нескольких симптомов, ассоциированных с заболеванием, нарушением или инфекцией, путем введения субъекту эффективного количества молекулы, связывающей ADAM9 (например, антитела, биспецифического диатела, трехвалентной молекулы, слитого белка и т.д.), или конъюгированной молекулы, связывающей ADAM9, по настоящему изобретению, или фармацевтической композиции, содержащей молекулу, связывающую ADAM9, или конъюгированной молекулы, связывающей ADAM9, по настоящему изобретению. В соответствии с предпочтительным аспектом такие композиции являются по сути очищенными (т.е., по сути не содержащими веществ, которые ограничивают его влияние или приводят к нежелательным побочным эффектам). В соответствии с конкретным вариантом осуществления субъект представляет собой животное, предпочтительно млекопитающее, такое как не относящееся к примату (например, корова, лошадь, кошка, собака, грызун и т.д.) или примат (например, обезьяна, такая как яванский макак, человек и т.д.). В соответствии с предпочтительным вариантом осуществления субъектом является человек.
[00289] Различные системы доставки известны и могут быть использованы для введения композиций по настоящему изобретению, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать антитело или слитый белок, опосредованный рецепторами эндоцитоз (см., например, Wu et al. (1987) “Receptor-Mediated In Vitro Gene Transformation By A Soluble DNA Carrier System,” J. Biol. Chem. 262:4429-4432), конструирование нуклеиновой кислоты в виде части ретровирусного или другого вектора и т.д.
[00290] Способы введения молекулы, связывающей ADAM9, по настоящему изобретению, включают в себя без ограничения парентеральное введение (например, внутрикожное, внутримышечное, интраперитонеальное, внутривенное и подкожное), эпидуральное и слизистое (например, интраназальные и пероральный пути). В соответствии с конкретным вариантом осуществления молекулы, связывающие ADAM9, по настоящему изобретению, вводят внутримышечно, внутривенно или подкожно. Композиции могут быть введены любым удобным путем, например, с помощью инфузии или болюсной инъекции, с помощью всасывания через эпителиальные или кожно-слизистые покровы (например, слизистую ротовой полости, слизистую прямой кишки и кишечника и т.д.) и могут быть введены совместно с другими биологически активными веществами. Введение может быть системным или локальным. Помимо этого, также может быть использовано легочное введение, например, с применением ингалятора или небулайзера, и состава с аэрозольным средством. См., например, патенты США №№ 6019968; 5985320; 5985309; 5934272; 5874064; 5855913; 5290540 и 4880078; и публикации согласно PCT №№ WO 92/19244; WO 97/32572; WO 97/44013; WO 98/31346 и WO 99/66903, каждое из которых включено в данный документ посредством ссылки в полном объеме.
[00291] В настоящем изобретении также предусмотрено, что препараты молекул, связывающих ADAM9, по настоящему изобретению, упакованы в герметически запечатанный контейнер, такой как ампула или саше, с указанием количества молекулы. В соответствии с одним вариантом осуществления такие молекулы поставляют в виде сухого стерилизованного лиофилизованного порошка или безводного концентрата в герметически запечатанном контейнере и могут быть разведены, например, водой или солевым раствором до соответствующей концентрации для введения субъекту. Предпочтительно молекулы, связывающие ADAM9, по настоящему изобретению, поставляют в виде сухого стерильного лиофилизированного порошка в герметически запечатанном контейнере.
[00292] Лиофилизированные препараты молекул, связывающих ADAM9, по настоящему изобретению, необходимо хранить при от 2°C до 8°C в их оригинальном контейнере и молекулы необходимо вводить в течение 12 часов, предпочтительно, в течение 6 часов, в течение 5 часов, в течение 3 часов или в течение 1 часа после разведения. В соответствии с альтернативным вариантом осуществления такие молекулы поставляют в жидкой форме в герметически запечатанном контейнере с указанием количества и концентрации молекулы, слитого белка или конъюгированной молекулы. Предпочтительно такие молекулы, связывающие ADAM9, при введении в жидкой форме, поставляют в герметически запечатанном контейнере.
[00293] Используемый в данном документе термин «эффективное количество» фармацевтической композиции представляет собой количество, достаточное для полезных или желательных результатов, в том числе без ограничения клинических результатов, таких как уменьшение симптомов, возникающих в результате заболевания, ослабления симптома инфекции (т.е., вирусной нагрузки, лихорадки, боли, сепсиса и т.д.) или симптома рака (например, пролиферации раковых клеток, наличия опухоли, метастазирования опухоли и т.д.), тем самым повышая качество жизни страдающих от этого заболевания, снижая дозу других препаратов, требуемых для лечения заболевания, усиления эффекта другого препарата, например, посредством целевого воздействия и/или интернализации, задержки прогрессирования заболевания и/или продления выживаемости индивидов.
[00294] Эффективное количество может быть введено в одном или нескольких введениях. Для целей настоящего изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для: устранения и/или ослабления пролиферации раковых клеток, и/или для устранения, ослабления и/или задержки развития метастазирования из первичного очага рака. В соответствии с некоторыми вариантами осуществления эффективное количество лекарственного средства, соединения или фармацевтической композиции может быть или не может быть получено в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективное количество» может рассматриваться в контексте введения одного или нескольких терапевтических средств, и можно считать, что одно средство вводят в эффективном количестве, если, в сочетании с одним или несколькими другими средствами может быть достигнут или является достигнутым.
[00295] В случае молекул, связывающих ADAM9, охваченных настоящим изобретением, дозу, вводимую пациенту, предпочтительно определяют на основе массы тела (кг) субъекта-реципиента.
[00296] Доза и частота введения молекул, связывающих ADAM9, по настоящему изобретению, могут быть снижены или изменены в результате повышения захвата и проникновения в ткани молекулы в результате модификаций, таких как, например, липидация.
[00297] Доза молекулы, связывающей ADAM9, по настоящему изобретению, вводимая пациенту, может быть рассчитана для применения в качестве монотерапии. В альтернативном случае молекула может быть использована в комбинации с другими терапевтическими композициями и доза, вводимая пациенту, является более низкой, чем тогда, когда указанные молекулы используются в качестве монотерапии.
[00298] Фармацевтические композиции по настоящему изобретению могут быть введены локально в необходимую для лечения область; этого можно достигнуть, например, и без ограничения с помощью локальной инфузии, с помощью инъекции или с помощью импланта, при этом указанный имплант представляет собой пористое непористое или гелеобразное вещество, в том числе мембраны, такие как мембраны SILASTIC®, или волокна. Предпочтительно при введении молекулы по настоящему изобретению необходимо с осторожностью использовать вещества, к которым молекула не абсорбируется.
[00299] Композиции по настоящему изобретению могут быть доставлены в везикуле, в частности, в липосоме (см., Langer (1990) “New Methods Of Drug Delivery,” Science 249:1527-1533); Treat et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.), Liss, New York, pp. 353- 365 (1989); Lopez-Berestein, ibid., pp. 3 17-327).
[00300] Если композиция по настоящему изобретению представляет собой нуклеиновую кислоту, кодирующую молекулу, связывающую ADAM9, по настоящему изобретению, то нуклеиновую кислоту можно вводить in vivo для активации экспрессии кодируемой ею молекулы, связывающей ADAM9, с помощью конструирования ее в виде части соответствующего экспрессионного вектора нуклеиновой кислоты и введения его таким образом, что он становится внутриклеточным, например, с помощью ретровирусного вектора (см. патент США № 4980286), или с помощью прямой инъекции или с помощью бомбардировки микрочастицами (например, генной пушки; Biolistic, Dupont), или покрытия липидами или рецепторами клеточной поверхности или трансфицирующими средствами, или путем введения в соединении с гомеобокс-пептидом, который, как известно, проникает в ядро (см., например, Joliot et al. (1991) “Antennapedia Homeobox Peptide Regulates Neural Morphogenesis,” Proc. Natl. Acad. Sci. (U.S.A.) USA 88:1864-1868) и т.д. В альтернативном случае нуклеиновую кислоту можно вводить внутриклеточно и включать для экспрессии в ДНК клетки-хозяина с помощью гомологической рекомбинации.
Примеры
[00301] После описания в данном документе изобретения в целом, то же самое будет более легко понять посредством отсылки на следующие примеры. Следующие примеры иллюстрируют различные способы в случае композиций в диагностических способах или способах лечения по настоящему изобретению. Примеры предполагают иллюстрировать, но никоим образом не ограничивать объем настоящего изобретения.
Пример 1
Специфичность антитела к ADAM9 MAB-A в отношении опухолевых клеток
[00302] Идентифицировали мышиное антитело к ADAM9 (обозначенное в данном документе как MAB-A), которое: (1) блокирует активность целевого белка, связанную с процессингом, ADAM9; (2) интернализируется; и (3) имеет противоопухолевую активность (см., например, патент США № 8361475). Специфичность MAB-A в отношении опухолевых клеток изучали с помощью IHC. Опухолевую ткань приводили в контакт с MAB-A (0,4 мкг/мл) или изотипическим контролем (0,4 мкг/мл) и степень окрашивания визуализировали. Было обнаружено, что MAB-A в значительной степени метит типы клеток немелкоклеточного рака легких, относящиеся к крупноклеточной карциноме, плоскоклеточной карциноме и аденокарциноме (Фиг. 7A, панели 1-8), клетки рака молочной железы, клетки рака предстательной железы, клетки рака желудка (Фиг. 7B, панели 1-6), а также образцы рака толстой кишки (Фиг. 7C, панели 1-8). Нормальную ткань приводили в контакт с MAB-A (1,25 мкг/мл) и степень окрашивания визуализировали. Как представлено в Табл. 1 выше, MAB-A характеризовались незначительным или отсутствием окрашивания в отношении большого разнообразия нормальных клеток. Следует отметить, что концентрация MAB-A, используемая в данных исследованиях, было примерно в 3 раза большей по сравнению с используемой для окрашивания опухолевых клеток. Результаты этих исследований на основе IHC указывают на то, что MAB-A характеризуется значительным предпочтительным связыванием с опухолевыми клетками по сравнению с нормальными клетками.
Пример 2
Межвидовая перекрестная реактивность
[00303] Исследовали связывание MAB-A с ADAM9 человека (huADAM9) и ADAM9 яванского макака (cynoADAM9). Вкратце, клетки 293-FT и CHO-K, временно экспрессирующие huADAM9, cynoADAM9 и не родственный антиген, или нетрансфицированные исходные клетки инкубировали с MAB-A, после чего с козьим меченым PE вторичным антителом к мышиному антителу и анализировали с помощью FACS. Как показано на Фиг. 8A-8B, MAB-A характеризовалось значительным связыванием с huADAM9, временно экспрессируемым на обоих типах клеток. MAB-A характеризовалось слабым связыванием с cynoADAM9. MAB-A не связывалось с исходными клетками или клетками, экспрессирующими нерелевантный антиген. Аналогичное слабое связывание с cynoADAM наблюдали в анализах ELISA.
Пример 3
Гуманизация и исходная оптимизация
[00304] Гуманизация MAB-A приводила к образованию гуманизированного домена VH, обозначаемого в данном документе как «hMAB-A VH(1)» и гуманизированного домена VL, обозначаемого в данном документе как «hMAB-A VL(1)». Гуманизированные вариабельные домены затем оптимизировали с целью усиления связывающей активности и/или устранения потенциально лабильных аминокислотных остатков, как описано более подробно ниже. Первый раунд оптимизации приводил к образованию трех дополнительных гуманизированных доменов VH, обозначаемых в данном документе как «hMAB-A VH(2)», «hMAB-A VH(3)» и «hMAB-A VH(4)», и трех дополнительных гуманизированных доменов VL, обозначаемых в данном документе как «hMAB-A VL(2)», «hMAB-A VL(3)» и «hMAB-A VL(4)». Помимо этого, получали химерную версию MAB-A («chMAB-A»), имеющего мышиные домены VH и VL и человеческие константные области. Аминокислотные последовательности мышиных и гуманизированных/оптимизированных доменов VH и VL представлены выше, выравнивание представлено на Фиг. 9A и 9B. Консенсусная последовательность этих гуманизированных/оптимизированных доменов VH и VL представлена выше. При получении гуманизированных вариабельных доменов гуманизированные вариабельные домены тяжелой и легкой цепи определенного антитела к ADAM9 (например, MAB-A) могли использовать в любой комбинации и определенные комбинации гуманизированных цепей обозначаются посредством отсылки на конкретные домены VH/VL, например, молекула (например, антитело или диатело), содержащая hMAB-A VH(1) и hMAB-A VL(2), специфически обозначается как «hMAB-A (1.2).»
[00305] hMAB-A VH(1) получали с наличием каркасных областей, происходящих из зародышевых линий человека VH3-21 и VH3-64, а hMAB-A VL(1) получали с наличием каркасных областей, происходящих из зародышевых линий человека B3 и L6. Мышиные CDR в этих гуманизированных вариабельных доменах сохраняли.
[00306] Потенциальный сайт дезамидирования идентифицировали в CDRH2 (показано одним подчеркиванием на Фиг. 9A), а потенциальный сайт изомеризации аспарагиновой кислоты идентифицировали в CDRL1 (показано одним подчеркиванием на Фиг. 9B). Аминокислотные замены в этих положениях исследовали для идентификации замен с целью удаления этих сайтов, при этом сохраняя аффинность связывания. Выбирали замену фенилаланина в положении 54 (N54F) CDRH2 (присутствующего в hMAB-A VH(2)) и серина в положении 28 (D28S) CDRL1 (присутствующего в hMAB-A VL(2)), при этом нумерация соответствовала Кабат. Идентифицированные замены могли использовать отдельно или в комбинации. Неожиданным образом было обнаружено, что антитела, содержащие замену N54F, характеризуются приблизительно 2-кратным повышением аффинности в отношении ADAM9 человека, а также характеризуются слегка повышенным связыванием в отношении ADAM9 яванского макака.
[00307] Дополнительные оптимизированные варианты получали с целью сведения к минимуму числа лизиновых остатков, присутствующих в CDR. Лизиновые остатки присутствовали в CDRH2 (указано двойным подчеркиванием на Фиг. 9A), а один лизин присутствовал в CDRL1 (указано двойным подчеркиванием на Фиг. 9B). Аминокислотные замены в этих положениях исследовали для идентификации замен, которые сохраняли аффинность связывания. Замены аргинина в положении 62 (K62R), глутамина в положении 64 (K64Q) и серина в положении 65 (S65G) выбирали в случае CDRH2 (присутствующие в hMAB-A VH(3)), при этом нумерации соответствовала Кабат. Замену аргинина в положении 24 (K24R) выбирали в случае CDRL1 (присутствующего в hMAB-A VL(3)). Идентифицированные замены могли использовать отдельно или в комбинации.
[00308] Идентифицировали другие потенциально лабильные остатки в CDR (указанные пунктирным подчеркиванием на Фиг. 9A-9B), один метиониновый остаток в CDRH1 в положении 34 (M34), один метиониновый остаток в CDRL1 в положении 33 (M33) и остатки гистидина, глутаминовой кислоты и аспарагиновой кислоты в положениях 92 (H93), 93 (E93) и 94 (D94) в CDRL3, при этом нумерация соответствовала Кабат. Аминокислотные замены в этих положениях исследовали для идентификации замен, которые сохраняли аффинность связывания. Замену изолейцина в положении 34 (M34I) выбирали в случае CDRH1 и замены лейцина, серина и треонина выбирали в случае положений 33 (M33L), 92 (H93Y), 93 (E93S) и 94 (D94T) CDRL3, при этом нумерация соответствовала Кабат. Каждое из этих положений можно было легко заменить в комбинации со всеми из замен, подробно описанных выше, с целью получения hMAB-A VH(4) и hMAB-A VL(4), которые затем связывали вместе с получением антитела, которое сохраняло незначительное повышение аффинности по сравнению исходным мышиным антителом, и которое характеризовалось значительно сниженным потенциалом дезамидирования или окисления и отсутствием лизиновых остатков в CDR.
[00309] Относительную аффинность связывания гуманизированных/оптимизированных антител hMAB-A (1.1), hMAB-A (2.2), hMAB-A (2.3), hMAB-A (3.3), hMAB-A (4.4) и химерного антитела chMAB-A (имеющего мышиные домены VH/VL) с huADAM изучали с помощью анализа BIACORE®, в котором меченый His растворимый ADAM9 человека («shADAM9-His», содержащий внеклеточный участок ADAM9 человека, слитый с содержащим гистидин пептидом) пропускали через поверхность, покрытую иммобилизированным антителом. Вкратце, каждое антитело захватывали на поверхности, покрытой козьим Fab2-антителом к человеческому Fc-антителу и затем инкубировали в присутствии различных концентраций (6,25-100 нМ) пептида shADAM9-His. Кинетику связывания определяли с помощью анализа связывания BIACORE® (нормализованного в соответствии с моделью связывания Ленгмюра 1:1). Рассчитанные значения ka, kd и KD на основе этих исследований представлены в Табл. 7. Связывание с cynoADAM9 исследовали с помощью FACS, как описано выше, и с помощью ELISA.
[00310] Результаты этих исследований показывают, что гуманизированные/оптимизированные антитела характеризуются такой же самой или более высокой аффинностью связывания с ADAM9 человека, чем исходное мышиное антитело. В частности, было отмечено, что введение мутации N54F в гуманизированные антитела приводило к повышенному связыванию с huADAM9 (т.е., hMAB-A (2.2), hMAB-A (2.3) и hMAB-A (3.3)). Данная мутация также предусматривала незначительное повышение связывания с cynoADAM9, определяемое с помощью FACS и ELISA, однако эти антитела продолжали характеризоваться слабым связыванием с cynoADAM9. В этих исследованиях также идентифицировали дополнительные замены, которые могли вводить для удаления лизиновых остатков из CDR без снижения аффинности. Дополнительные замены идентифицировали с целью удаления других потенциально лабильных остатков при минимального влиянии на аффинность.
Пример 4
Оптимизация связывания с ADAM9 не относящегося к человека примата
[00311] Случайный мутагенез использовали для введения замен в домены CDRH2 (положения в соответствии с Кабат 53-58) и CDRH3 (положения в соответствии с Кабат 95-100 и 100a-100f) тяжелой цепи hMAB-A (2.2). Мутанты подвергали скринингу с целью идентификации клонов, характеризующихся повышенным связыванием с ADAM9 не относящегося к примату человека (например, cynoADAM9), и которые сохраняли высокую аффинность связывания с huADAM9. 48 клонов отбирали в результате двух независимых скринингов мутаций в CDRH3 (положения в соответствии с Кабат 100a-100f). В Табл. 8 представлено выравнивание аминокислотной последовательности остаток 100a-f CDRH3 в соответствии с Кабат из клонов hMAB-A (2.2), выбранных в связи с повышенным связыванием с cynoADAM9 в результате двух независимых скринингов. Дополнительные выравнивания в клонах представлены в Табл. 9. Как указано в таких таблицах, аналогичные клоны образовывались в каждом эксперименте, которые подпадали в отдельные паттерны замен.
Замены в субдомене CDRH3 тяжелой цепи MAB-A
(положения в соответствии с Кабат 100a-100f)
Замены в субдомене CDRH3 тяжелой цепи MAB-A
(положения в соответствии с Кабат 100a-100f)
[00312] В случае всех исследуемых клонов Gly и Ala представляли собой предпочтительные аминокислотные остатки в положении 4 (P4) и Leu, Met и Phe представляли собой предпочтительные аминокислотные остатки в положении 6 (P6). Предпочтительные аминокислотные остатки в других положениях (например, положении 2 (P2), положении 3 (P3) и положении 5 (P5)) зависели от аминокислотного остатка, встречающегося в P1. В случае клонов, имеющих остаток Pro в положении 1 (P1), Lys и Arg были предпочтительными в P2, Phe и Met -в P3, Gly -в P4 и Trp или Phe - в P5. В случае клонов, имеющих Phe, Tyr или Trp в P1, Asn и His были предпочтительными в P2, Ser и His - в P3 и Leu - в P6. В случае клонов, имеющих Ile, Leu или Val в P1, Gly был предпочтительным в P2, Lys - в P3, Val - в P5 и гидрофобный остаток - в P6. Помимо этого, как можно заметить в Табл. 8, в случае клонов, имеющих остаток Thr в P1, Gly был предпочтительным в P2, Lys, Met и Asn были предпочтительными в P3, Gly был предпочтительным в P4, Val или Thr были предпочтительными в P5 и Leu и Met - в P6. Дополнительные клоны, имеющие остаток Asp, Gly, Arg, His или Ser в P1 также идентифицировали при более низких частотах (см. Табл. 8 и Табл. 9).
[00313] Домен VH десяти клонов, показанных в Табл. 9, использовали с целью получения дополнительных оптимизированных вариантов hMAB-A (2.2), обозначенных hMAB-A (2A.2). Связывание выбранных клонов исследовали с помощью анализа ELISA. Вкратце, антитела, которые связывались с содержащими гистидин пептидами, и которые наносили на микротитровальные планшеты, использовали для захвата меченого His пептидом растворенного cynoADAM9 («cynoADAM9-His») (1 мкг/мл) или меченого His пептидом растворенного huADAM9 (1 мкг/мл), и исследовали связывание последовательных разведений исходного hMAB-A (2.2) и десяти вариантов CDRH3 hMAB-A (2A.2). Кривые связывания в случае cynoADAM9 и huADAM9 представлены на Фиг. 10A и Фиг. 10B соответственно. Варианты hMAB-A (2A.2), содержащие каждый из выбранных доменов VH, характеризовались повышенным связыванием с cynoADAM9 в случае MAB-A VH(2B), MAB-A VH(2C), MAB-A VH(2D) и MAB-A VH(2I), показывая наибольшее повышение связывания с cynoADAM9, при этом сохраняя аналогичное связывание с huADAM9 в качестве исходного антитела hMAB-A (2.2).
[00314] Относительную аффинность связывания гуманизированных/дополнительно оптимизированных антител MAB-A VH(2B.2), MAB-A VH(2C.2), MAB-A VH(2D.2) и MAB-A VH(2I.2), а также исходного hMAB-A (2.2) с huADAM9-His и cynoADAM9-His исследовали с помощью анализа BIACORE® по сути, как описано выше. Рассчитанные значения ka, kd и KD на основе этих исследований представлены в Табл. 10.
(M-1с-1)
(с-1)
(нM)
(M-1с-1)
(с-1)
(нM)
[00315] Исследования связывания продемонстрировали, что четыре расположенных в верхних позициях клона характеризовались 150-550-кратным усилением аффинности связывания в отношении cynoADAM9, при этом сохраняя ту же самую аффинность связывания в отношении huADAM9, что и исходное антитело. hMAB-A (2C.2) и hMAB-A (2I.2) выбирали для дальнейших исследований.
Пример 5
Иммуногистохимическое исследование антитела hMAB-A (2I.2)
[00316] Специфичность hMAB-A (2I.2) в отношении клеток изучали с помощью IHC. Клетки положительного и отрицательного контроля, а также нормальные ткани человека и яванского макака приводили в контакт с hMAB-A (2I.2) (2,5 мкг/мл) или изотипическим контролем (2,5 мкг/мл) и степень окрашивания визуализировали. Результаты исследования представлены в Табл. 11.
[00317] Исследования на основе IHC также проводили для оценки связывания гуманизированного/оптимизированного hMAB-A (2I.2) при концентрации 12,5 мкг/мл (5x оптимальная концентрация при окрашивании). В данном исследовании использовали клетки положительного и отрицательного контроля, нормальные ткани человека и ткани яванского макака использовали. Результаты исследования представлены в Табл. 12.
[00318] Сравнительное исследование на основе IHC проводили с целью оценки различий в связывании hMAB-A (2.2), hMAB-A (2.3), hMAB-A (2C.2) и hMAB-A (2I.2) при 2,5 мкг/мл или 5 мкг/мл. В данном исследовании использовали клетки положительного и отрицательного контроля, нормальные ткани человека и ткани яванского макака использовали. Результаты исследования представлены в Табл. 13.
5 мкг/мл
2,5 мкг/мл
2,5 мкг/мл
2,5 мкг/мл
5 мкг/мл
[00319] Дополнительное сравнительное исследование на основе IHC проводили с целью оценки различий в связывании hMAB-A (2.2), hMAB-A (2.3), hMAB-A (2C.2) и hMAB-A (2I.2), а также мышиного MAB-A при 2,5 мкг/мл или 12,5 мкг/мл. В данном исследовании использовали клетки положительного и отрицательного контроля, нормальные ткани человека и ткани яванского макака использовали. Результаты исследования представлены в Табл. 14.
5 мкг/мл
2,5 мкг/мл
2,5 мкг/мл
12,5 мкг/мл
5 мкг/мл
MG06-CHTN-96 B
MG06-CHtN-162B1 A
ILS11103 B
ILS10266
ILS10241 B
ILSD8011 J
яванского макака
[00320] Таким образом, результаты продемонстрировали, что hMAB-A (2.2) характеризовалось в целом окрашиванием низкого уровня в отношении гепатоцитов и почечных канальцев человека при оптимальной концентрации, при более низкой интенсивности окрашивания/частоте реактивности в гепатоцитах и почечных канальцах, наблюдаемых в отрицательном контроле. hMAB-A (2.2) характеризовалось аналогичным окрашиванием низкого уровня в отношении гепатоцитов и почечных канальцев яванского макака при оптимальной концентрации, при более низкой интенсивности окрашивания/частоте реактивности в почечных канальцах, наблюдаемых в отрицательном контроле.
[00321] Результаты также продемонстрировали, что hMAB-A (2С.2) характеризовалось в целом окрашиванием низкого уровня в отношении гепатоцитов и почечных канальцев человека при оптимальной концентрации, при более низкой интенсивности окрашивания/частоте реактивности в гепатоцитах и почечных канальцах, наблюдаемых в отрицательном контроле. hMAB-A (2C.2) характеризовалось аналогичным окрашиванием низкого уровня в гепатоцитах и почечных канальцах яванского макака при оптимальном концентрации. Дополнительные минимальные результаты в легочном эпителии, островках/эпителии поджелудочной железы и эпителии мочевого пузыря яванского макака в случае hMAB-A (2C.2) не наблюдали в соответствующей ткани человека; более низкую интенсивность окрашивания/частоту реактивности наблюдали в легочном эпителии, почечных канальцах, эпителии мочевого пузыря в отрицательном контроле.
[00322] Результаты также продемонстрировали, что hMAB-A (2I.2) характеризовалось отсутствием окрашивания тканей человека или яванского макака при оптимальной концентрации, при редком +/- окрашивании клеток переходного эпителия мочевого пузыря. hMAB-A (2I.2) также характеризовалось в целом окрашиванием низкого уровня и частотой альвеолярных клеток легких, протокового эпителия поджелудочной железы, почечных канальцев, клеток переходного эпителия мочевого пузыря человека при 5x оптимальной концентрации, и в целом окрашиванием низкого уровня эпителия бронхов и клеток переходного эпителия мочевого пузыря яванского макака при 5x оптимальной концентрации. hMAB-A (2I.2) характеризовалось в целом благоприятным профилем IHC в нормальных исследуемых тканях человека и аналогичным профилем в соответствующих тканях яванского макака.
[00323] Все публикации и патенты, упомянутые в данном описании, включены в данный документ посредством ссылки в той же степени, в которой каждая отельная публикация или патентная заявка была бы конкретно и отдельно указана для включения посредством ссылки в полном объеме. В то время как настоящее изобретение было описано в связи с конкретными вариантами его осуществления, будет понятно, что возможны дополнительные модификации, и данная заявка предполагает включение любых вариаций, областей применения или адаптаций настоящего изобретения, в целом в соответствии с принципами настоящего изобретения и включая такие отступления от настоящего раскрытия, которые подпадают под известную или обычную практику в пределах области техники, к которой настоящее изобретение принадлежит, и которые могут применяться по отношению к обязательным особенностям, изложенным ранее в данном документе.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> МакроДженикс, Инк.
Дерик Лу
Джунипер Шрибнер
Бхасвати Барат
Гундо Дидрих
Лесли Джонсон
Эзио Бонвини
<120> Молекулы, связывающие ADAM9, и способы их применения
<130> 1301.0147PCT
<150> US 62/438516
<151> 23 декабря 2016 г.
<160> 207
<170> Версия PatentIn 3.5
<210> 1
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(200)
<223> Домен CH2-CH3 IgG1 человека
<220>
<221> MISC_FEATURE
<222> (200)..(200)
<223> Xaa представляет собой лизин (K) или отсутствует
<220>
<221> misc_feature
<222> (217)..(217)
<223> Xaa может представлять собой любую встречающуюся в природе
аминокислоту
<400> 1
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 2
<211> 216
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(216)
<223> Домен CH2-CH3 IgG2 человека
<220>
<221> MISC_FEATURE
<222> (216)..(216)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 2
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
1 5 10 15
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
20 25 30
Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
35 40 45
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
50 55 60
Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln
65 70 75 80
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
85 90 95
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro
100 105 110
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
115 120 125
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
130 135 140
Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
145 150 155 160
Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
165 170 175
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
180 185 190
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
195 200 205
Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 3
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(217)
<223> Домен CH2-CH3 IgG3 человека
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 3
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 4
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(217)
<223> Домен CH2-CH3 IgG4 человека
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 4
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Xaa
210 215
<210> 5
<211> 819
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(819)
<223> Иллюстративный полипептид ADAM9 человека (№ последовательности в
NCBI NP_003807)
<220>
<221> SIGNAL
<222> (1)..(28)
<223> Сигнальная последовательность
<400> 5
Met Gly Ser Gly Ala Arg Phe Pro Ser Gly Thr Leu Arg Val Arg Trp
1 5 10 15
Leu Leu Leu Leu Gly Leu Val Gly Pro Val Leu Gly Ala Ala Arg Pro
20 25 30
Gly Phe Gln Gln Thr Ser His Leu Ser Ser Tyr Glu Ile Ile Thr Pro
35 40 45
Trp Arg Leu Thr Arg Glu Arg Arg Glu Ala Pro Arg Pro Tyr Ser Lys
50 55 60
Gln Val Ser Tyr Val Ile Gln Ala Glu Gly Lys Glu His Ile Ile His
65 70 75 80
Leu Glu Arg Asn Lys Asp Leu Leu Pro Glu Asp Phe Val Val Tyr Thr
85 90 95
Tyr Asn Lys Glu Gly Thr Leu Ile Thr Asp His Pro Asn Ile Gln Asn
100 105 110
His Cys His Tyr Arg Gly Tyr Val Glu Gly Val His Asn Ser Ser Ile
115 120 125
Ala Leu Ser Asp Cys Phe Gly Leu Arg Gly Leu Leu His Leu Glu Asn
130 135 140
Ala Ser Tyr Gly Ile Glu Pro Leu Gln Asn Ser Ser His Phe Glu His
145 150 155 160
Ile Ile Tyr Arg Met Asp Asp Val Tyr Lys Glu Pro Leu Lys Cys Gly
165 170 175
Val Ser Asn Lys Asp Ile Glu Lys Glu Thr Ala Lys Asp Glu Glu Glu
180 185 190
Glu Pro Pro Ser Met Thr Gln Leu Leu Arg Arg Arg Arg Ala Val Leu
195 200 205
Pro Gln Thr Arg Tyr Val Glu Leu Phe Ile Val Val Asp Lys Glu Arg
210 215 220
Tyr Asp Met Met Gly Arg Asn Gln Thr Ala Val Arg Glu Glu Met Ile
225 230 235 240
Leu Leu Ala Asn Tyr Leu Asp Ser Met Tyr Ile Met Leu Asn Ile Arg
245 250 255
Ile Val Leu Val Gly Leu Glu Ile Trp Thr Asn Gly Asn Leu Ile Asn
260 265 270
Ile Val Gly Gly Ala Gly Asp Val Leu Gly Asn Phe Val Gln Trp Arg
275 280 285
Glu Lys Phe Leu Ile Thr Arg Arg Arg His Asp Ser Ala Gln Leu Val
290 295 300
Leu Lys Lys Gly Phe Gly Gly Thr Ala Gly Met Ala Phe Val Gly Thr
305 310 315 320
Val Cys Ser Arg Ser His Ala Gly Gly Ile Asn Val Phe Gly Gln Ile
325 330 335
Thr Val Glu Thr Phe Ala Ser Ile Val Ala His Glu Leu Gly His Asn
340 345 350
Leu Gly Met Asn His Asp Asp Gly Arg Asp Cys Ser Cys Gly Ala Lys
355 360 365
Ser Cys Ile Met Asn Ser Gly Ala Ser Gly Ser Arg Asn Phe Ser Ser
370 375 380
Cys Ser Ala Glu Asp Phe Glu Lys Leu Thr Leu Asn Lys Gly Gly Asn
385 390 395 400
Cys Leu Leu Asn Ile Pro Lys Pro Asp Glu Ala Tyr Ser Ala Pro Ser
405 410 415
Cys Gly Asn Lys Leu Val Asp Ala Gly Glu Glu Cys Asp Cys Gly Thr
420 425 430
Pro Lys Glu Cys Glu Leu Asp Pro Cys Cys Glu Gly Ser Thr Cys Lys
435 440 445
Leu Lys Ser Phe Ala Glu Cys Ala Tyr Gly Asp Cys Cys Lys Asp Cys
450 455 460
Arg Phe Leu Pro Gly Gly Thr Leu Cys Arg Gly Lys Thr Ser Glu Cys
465 470 475 480
Asp Val Pro Glu Tyr Cys Asn Gly Ser Ser Gln Phe Cys Gln Pro Asp
485 490 495
Val Phe Ile Gln Asn Gly Tyr Pro Cys Gln Asn Asn Lys Ala Tyr Cys
500 505 510
Tyr Asn Gly Met Cys Gln Tyr Tyr Asp Ala Gln Cys Gln Val Ile Phe
515 520 525
Gly Ser Lys Ala Lys Ala Ala Pro Lys Asp Cys Phe Ile Glu Val Asn
530 535 540
Ser Lys Gly Asp Arg Phe Gly Asn Cys Gly Phe Ser Gly Asn Glu Tyr
545 550 555 560
Lys Lys Cys Ala Thr Gly Asn Ala Leu Cys Gly Lys Leu Gln Cys Glu
565 570 575
Asn Val Gln Glu Ile Pro Val Phe Gly Ile Val Pro Ala Ile Ile Gln
580 585 590
Thr Pro Ser Arg Gly Thr Lys Cys Trp Gly Val Asp Phe Gln Leu Gly
595 600 605
Ser Asp Val Pro Asp Pro Gly Met Val Asn Glu Gly Thr Lys Cys Gly
610 615 620
Ala Gly Lys Ile Cys Arg Asn Phe Gln Cys Val Asp Ala Ser Val Leu
625 630 635 640
Asn Tyr Asp Cys Asp Val Gln Lys Lys Cys His Gly His Gly Val Cys
645 650 655
Asn Ser Asn Lys Asn Cys His Cys Glu Asn Gly Trp Ala Pro Pro Asn
660 665 670
Cys Glu Thr Lys Gly Tyr Gly Gly Ser Val Asp Ser Gly Pro Thr Tyr
675 680 685
Asn Glu Met Asn Thr Ala Leu Arg Asp Gly Leu Leu Val Phe Phe Phe
690 695 700
Leu Ile Val Pro Leu Ile Val Cys Ala Ile Phe Ile Phe Ile Lys Arg
705 710 715 720
Asp Gln Leu Trp Arg Ser Tyr Phe Arg Lys Lys Arg Ser Gln Thr Tyr
725 730 735
Glu Ser Asp Gly Lys Asn Gln Ala Asn Pro Ser Arg Gln Pro Gly Ser
740 745 750
Val Pro Arg His Val Ser Pro Val Thr Pro Pro Arg Glu Val Pro Ile
755 760 765
Tyr Ala Asn Arg Phe Ala Val Pro Thr Tyr Ala Ala Lys Gln Pro Gln
770 775 780
Gln Phe Pro Ser Arg Pro Pro Pro Pro Gln Pro Lys Val Ser Ser Gln
785 790 795 800
Gly Asn Leu Ile Pro Ala Arg Pro Ala Pro Ala Pro Pro Leu Tyr Ser
805 810 815
Ser Leu Thr
<210> 6
<211> 819
<212> PRT
<213> Macaca fascicularis
<220>
<221> MISC_FEATURE
<222> (1)..(819)
<223> Иллюстративный полипептид ADAM9 яванского макака
(№ последовательности в NCBI XM_005563126.2)
<220>
<221> SIGNAL
<222> (1)..(28)
<223> Возможная сигнальная последовательность
<400> 6
Met Gly Ser Gly Val Gly Ser Pro Ser Gly Thr Leu Arg Val Arg Trp
1 5 10 15
Leu Leu Leu Leu Cys Leu Val Gly Pro Val Leu Gly Ala Ala Arg Pro
20 25 30
Gly Phe Gln Gln Thr Ser His Leu Ser Ser Tyr Glu Ile Ile Thr Pro
35 40 45
Trp Arg Leu Thr Arg Glu Arg Arg Glu Ala Pro Arg Pro Tyr Ser Lys
50 55 60
Gln Val Ser Tyr Leu Ile Gln Ala Glu Gly Lys Glu His Ile Ile His
65 70 75 80
Leu Glu Arg Asn Lys Asp Leu Leu Pro Glu Asp Phe Val Val Tyr Thr
85 90 95
Tyr Asn Lys Glu Gly Thr Val Ile Thr Asp His Pro Asn Ile Gln Asn
100 105 110
His Cys His Phe Arg Gly Tyr Val Glu Gly Val Tyr Asn Ser Ser Val
115 120 125
Ala Leu Ser Asn Cys Phe Gly Leu Arg Gly Leu Leu His Leu Glu Asn
130 135 140
Ala Ser Tyr Gly Ile Glu Pro Leu Gln Asn Ser Ser His Phe Glu His
145 150 155 160
Ile Ile Tyr Arg Met Asp Asp Val His Lys Glu Pro Leu Lys Cys Gly
165 170 175
Val Ser Asn Lys Asp Ile Glu Lys Glu Thr Thr Lys Asp Glu Glu Glu
180 185 190
Glu Pro Pro Ser Met Thr Gln Leu Leu Arg Arg Arg Arg Ala Val Leu
195 200 205
Pro Gln Thr Arg Tyr Val Glu Leu Phe Ile Val Val Asp Lys Glu Arg
210 215 220
Tyr Asp Met Met Gly Arg Asn Gln Thr Ala Val Arg Glu Glu Met Ile
225 230 235 240
Leu Leu Ala Asn Tyr Leu Asp Ser Met Tyr Ile Met Leu Asn Ile Arg
245 250 255
Ile Val Leu Val Gly Leu Glu Ile Trp Thr Asn Gly Asn Leu Ile Asn
260 265 270
Ile Ala Gly Gly Ala Gly Asp Val Leu Gly Asn Phe Val Gln Trp Arg
275 280 285
Glu Lys Phe Leu Ile Thr Arg Arg Arg His Asp Ser Ala Gln Leu Val
290 295 300
Leu Lys Lys Gly Phe Gly Gly Thr Ala Gly Met Ala Phe Val Gly Thr
305 310 315 320
Val Cys Ser Arg Ser His Ala Gly Gly Ile Asn Val Phe Gly His Ile
325 330 335
Thr Val Glu Thr Phe Ala Ser Ile Val Ala His Glu Leu Gly His Asn
340 345 350
Leu Gly Met Asn His Asp Asp Gly Arg Asp Cys Ser Cys Gly Ala Lys
355 360 365
Ser Cys Ile Met Asn Ser Gly Ala Ser Gly Ser Arg Asn Phe Ser Ser
370 375 380
Cys Ser Ala Glu Asp Phe Glu Lys Leu Thr Leu Asn Lys Gly Gly Asn
385 390 395 400
Cys Leu Leu Asn Ile Pro Lys Pro Asp Glu Ala Tyr Ser Ala Pro Ser
405 410 415
Cys Gly Asn Lys Leu Val Asp Ala Gly Glu Glu Cys Asp Cys Gly Thr
420 425 430
Pro Lys Glu Cys Glu Leu Asp Pro Cys Cys Glu Gly Ser Thr Cys Lys
435 440 445
Leu Lys Ser Phe Ala Glu Cys Ala Tyr Gly Asp Cys Cys Lys Asp Cys
450 455 460
Arg Phe Leu Pro Gly Gly Thr Leu Cys Arg Gly Lys Thr Ser Glu Cys
465 470 475 480
Asp Val Pro Glu Tyr Cys Asn Gly Ser Ser Gln Phe Cys Gln Pro Asp
485 490 495
Val Phe Ile Gln Asn Gly Tyr Pro Cys Gln Asn Asn Lys Ala Tyr Cys
500 505 510
Tyr Asn Gly Met Cys Gln Tyr Tyr Asp Ala Gln Cys Gln Val Ile Phe
515 520 525
Gly Ser Lys Ala Lys Ala Ala Pro Lys Asp Cys Phe Ile Glu Val Asn
530 535 540
Ser Lys Gly Asp Arg Phe Gly Asn Cys Gly Phe Ser Gly Asn Glu Tyr
545 550 555 560
Lys Lys Cys Ala Thr Gly Asn Ala Leu Cys Gly Lys Leu Gln Cys Glu
565 570 575
Asn Val Gln Glu Ile Pro Val Phe Gly Ile Val Pro Ala Ile Ile Gln
580 585 590
Thr Pro Ser Arg Gly Thr Lys Cys Trp Gly Val Asp Phe Gln Leu Gly
595 600 605
Ser Asp Val Pro Asp Pro Gly Met Val Asn Glu Gly Thr Lys Cys Gly
610 615 620
Ala Asp Lys Ile Cys Arg Asn Phe Gln Cys Val Asp Ala Ser Val Leu
625 630 635 640
Asn Tyr Asp Cys Asp Ile Gln Lys Lys Cys His Gly His Gly Val Cys
645 650 655
Asn Ser Asn Lys Asn Cys His Cys Glu Asn Gly Trp Ala Pro Pro Asn
660 665 670
Cys Glu Thr Lys Gly Tyr Gly Gly Ser Val Asp Ser Gly Pro Thr Tyr
675 680 685
Asn Glu Met Asn Thr Ala Leu Arg Asp Gly Leu Leu Val Phe Phe Phe
690 695 700
Leu Ile Val Pro Leu Ile Val Cys Ala Ile Phe Ile Phe Ile Lys Arg
705 710 715 720
Asp Gln Leu Trp Arg Arg Tyr Phe Arg Lys Lys Arg Ser Gln Thr Tyr
725 730 735
Glu Ser Asp Gly Lys Asn Gln Ala Asn Pro Ser Arg Gln Pro Val Ser
740 745 750
Val Pro Arg His Val Ser Pro Val Thr Pro Pro Arg Glu Val Pro Ile
755 760 765
Tyr Ala Asn Arg Phe Pro Val Pro Thr Tyr Ala Ala Lys Gln Pro Gln
770 775 780
Gln Phe Pro Ser Arg Pro Pro Pro Pro Gln Pro Lys Val Ser Ser Gln
785 790 795 800
Gly Asn Leu Ile Pro Ala Arg Pro Ala Pro Ala Pro Pro Leu Tyr Ser
805 810 815
Ser Leu Thr
<210> 7
<211> 123
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(123)
<223> Домен VH мышиного антитела к ADAM9 MAB-A
<400> 7
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Ile Pro Ile Asn Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Lys Ala Thr Leu Thr Leu Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
115 120
<210> 8
<211> 5
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(5)
<223> Домен CDRH1 антитела к ADAM9 MAB-A
<400> 8
Ser Tyr Trp Met His
1 5
<210> 9
<211> 17
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(17)
<223> Домен CDRH2 антитела к ADAM9 MAB-A
<400> 9
Glu Ile Ile Pro Ile Asn Gly His Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Ser
<210> 10
<211> 14
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(14)
<223> Домен CDRH3 антитела к ADAM9 MAB-A
<400> 10
Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
1 5 10
<210> 11
<211> 111
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(111)
<223> Домен VL мышиного антитела к ADAM9 MAB-A
<400> 11
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Ile Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 12
<211> 15
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(15)
<223> Домен CDRL1 антитела к ADAM9 MAB-A
<400> 12
Lys Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 13
<211> 7
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(7)
<223> Домен CDRL2 антитела к ADAM9 MAB-A
<400> 13
Ala Ala Ser Asp Leu Glu Ser
1 5
<210> 14
<211> 9
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(9)
<223> Домен CDRL3 антитела к ADAM9 MAB-A
<400> 14
Gln Gln Ser His Glu Asp Pro Phe Thr
1 5
<210> 15
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH гуманизированного/оптимизированного мышиного антитела
к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (34)..(34)
<223> XAA представляет собой метионин (M) или изолейцин (I)
<220>
<221> MISC_FEATURE
<222> (55)..(55)
<223> XAA представляет собой аспарагин (N) или фенилаланин (F)
<220>
<221> MISC_FEATURE
<222> (63)..(63)
<223> XAA представляет собой лизин (K) или аргинин (R)
<220>
<221> MISC_FEATURE
<222> (65)..(65)
<223> XAA представляет собой лизин (K) или глутамин (Q)
<220>
<221> MISC_FEATURE
<222> (66)..(66)
<223> XAA представляет собой серин (S) или глицин (E)
<220>
<221> MISC_FEATURE
<222> (105)..(105)
<223> XAA представляет собой пролин (P), фенилаланин (F), тирозин (Y),
триптофан (W), изолейцин (I), лейцин (L), валин (V), треонин (T),
глицин (G) или аспартат (D)
<220>
<221> MISC_FEATURE
<222> (106)..(106)
<223> XAA представляет собой лизин (K), аргинин (R), глицин (G),
аспарагин (N), гистидин (H) или серин (S)
<220>
<221> MISC_FEATURE
<222> (107)..(107)
<223> XAA представляет собой фенилаланин (F), метионин (M), серин (S),
лизин (K) или аспарагин (N)
<220>
<221> MISC_FEATURE
<222> (108)..(108)
<223> XAA представляет собой глицин (G) или аланин (A)
<220>
<221> MISC_FEATURE
<222> (109)..(109)
<223> XAA представляет собой валин (V), треонин (T), триптофан (W)
или фенилаланин (F)
<220>
<221> MISC_FEATURE
<222> (109)..(109)
<223> XAA представляет собой метионин (M), лейцин (L) или лизин (K)
<220>
<221> misc_feature
<222> (110)..(110)
<223> Xaa может представлять собой любую встречающуюся в природе
аминокислоту
<400> 15
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Xaa His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Xaa Gly His Thr Asn Tyr Asn Glu Xaa Phe
50 55 60
Xaa Xaa Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Xaa Xaa Xaa Xaa Xaa Xaa Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 16
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(1) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(1))
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Asn Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 17
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2))
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 18
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(3) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(3))
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Arg Phe
50 55 60
Gln Gly Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 19
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(4) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(4))
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Arg Phe
50 55 60
Gln Gly Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 20
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2A) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2A))
<400> 20
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Phe Asn Ser Gly Thr Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 21
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2B) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2B))
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Ile Gly Lys Gly Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 22
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2C) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2C))
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Pro Arg Phe Gly Trp Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 23
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2D) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2D))
<400> 23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Thr Gly Lys Gly Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 24
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2E) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2E))
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Asp Ser Asn Ala Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 25
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2F) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2F))
<400> 25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Phe His Ser Gly Thr Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 26
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2G) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2G))
<400> 26
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Phe Asn Lys Ala Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 27
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2H) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2H))
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Gly Ser Gly Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 28
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2I) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2I))
<400> 28
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Pro Arg Gln Gly Phe Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 29
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2J) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VH(2J))
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Tyr Asn Ser Gly Thr Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 30
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRH1 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 30
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 31
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRH2 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 31
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly
1 5 10
<210> 32
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRH3 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 32
Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 33
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRH4 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 33
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 34
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH1 антитела к ADAM9 MAB-A
<400> 34
Ser Tyr Trp Ile His
1 5
<210> 35
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH2 антитела к ADAM9 MAB-A
<400> 35
Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Ser
<210> 36
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH2 антитела к ADAM9 MAB-A
<400> 36
Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Arg Phe Gln
1 5 10 15
Gly
<210> 37
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 37
Gly Gly Tyr Tyr Tyr Tyr Phe Asn Ser Gly Thr Leu Asp Tyr
1 5 10
<210> 38
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 38
Gly Gly Tyr Tyr Tyr Tyr Ile Gly Lys Gly Val Leu Asp Tyr
1 5 10
<210> 39
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 39
Gly Gly Tyr Tyr Tyr Tyr Pro Arg Phe Gly Trp Leu Asp Tyr
1 5 10
<210> 40
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 40
Gly Gly Tyr Tyr Tyr Tyr Thr Gly Lys Gly Val Leu Asp Tyr
1 5 10
<210> 41
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 41
Gly Gly Tyr Tyr Tyr Tyr Asp Ser Asn Ala Val Leu Asp Tyr
1 5 10
<210> 42
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 42
Gly Gly Tyr Tyr Tyr Tyr Phe His Ser Gly Thr Leu Asp Tyr
1 5 10
<210> 43
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 43
Gly Gly Tyr Tyr Tyr Tyr Phe Asn Lys Ala Val Leu Asp Tyr
1 5 10
<210> 44
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 44
Gly Gly Tyr Tyr Tyr Tyr Gly Gly Ser Gly Val Leu Asp Tyr
1 5 10
<210> 45
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 45
Gly Gly Tyr Tyr Tyr Tyr Pro Arg Gln Gly Phe Leu Asp Tyr
1 5 10
<210> 46
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRH3 антитела к ADAM9 MAB-A
<400> 46
Gly Gly Tyr Tyr Tyr Tyr Tyr Asn Ser Gly Thr Leu Asp Tyr
1 5 10
<210> 47
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Консенсусный домен CDRH1 антитела к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa представляет собой метионин (M) или серин (S)
<400> 47
Ser Tyr Trp Xaa His
1 5
<210> 48
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Консенсусный домен CDRH2 антитела к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa представляет собой аспарагин (N) или фенилаланин (F)
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa представляет собой лизин (K) или аргинин (R)
<220>
<221> MISC_FEATURE
<222> (16)..(16)
<223> Xaa представляет собой лизин (K) или Gluatamine (Q)
<220>
<221> MISC_FEATURE
<222> (17)..(17)
<223> Xaa представляет собой серин (S) или глицин (G)
<400> 48
Glu Ile Ile Pro Ile Xaa Gly His Thr Asn Tyr Asn Glu Xaa Phe Xaa
1 5 10 15
Xaa
<210> 49
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Консенсусный домен CDRH3 антитела к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa представляет собой пролин (P), фенилаланин (F), тирозин (Y),
триптофан (W), изолейцин (I), лейцин (L), валин (V), треонин (T),
глицин (G) или аспартат (D)
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> Xaa представляет собой лизин (K), аргинин (R), глицин (G),
аспарагин (N), гистидин (H), или серин (S)
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa представляет собой фенилаланин (F), метионин (M), серин (S),
лизин (K) или аспарагин (N)
<220>
<221> MISC_FEATURE
<222> (10)..(10)
<223> Xaa представляет собой глицин (G) или аланин (A)
<220>
<221> MISC_FEATURE
<222> (11)..(11)
<223> Xaa представляет собой валин (V), треонин (T), триптофан (W)
или фенилаланин (F)
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa представляет собой метионин (M), лейцин (L) или лизин (K)
<400> 49
Gly Gly Tyr Tyr Tyr Tyr Xaa Xaa Xaa Xaa Xaa Xaa Asp Tyr
1 5 10
<210> 50
<211> 453
<212> PRT
<213> Искусственная последовательность
<220>
<223> Иллюстративная гуманизированная/оптимизированная тяжелая цепь
IgG1 производного/варианта MAB-A, содержащая домен hMAB-A VH (2)
<220>
<221> MISC_FEATURE
<222> (453)..(453)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser Arg Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Xaa
450
<210> 51
<211> 453
<212> PRT
<213> Искусственная последовательность
<220>
<223> <223> Иллюстративная гуманизированная/оптимизированная тяжелая
цепь IgG1 производного/варианта MAB-A, содержащая домен hMAB-A VH (2С)
<220>
<221> MISC_FEATURE
<222> (453)..(453)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Pro Arg Phe Gly Trp Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Xaa
450
<210> 52
<211> 453
<212> PRT
<213> Искусственная последовательность
<220>
<223> <223> Иллюстративная гуманизированная/оптимизированная тяжелая
цепь IgG1 производного/варианта MAB-A, содержащего домен hMAB-A VH (2I)
<220>
<221> MISC_FEATURE
<222> (453)..(453)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Pro Arg Gln Gly Phe Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Xaa
450
<210> 53
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гуманизированный/оптимизированный домен VL мышиного антитела
к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (24)..(24)
<223> Xaa представляет собой лизин (K) или аргинин (R)
<220>
<221> MISC_FEATURE
<222> (32)..(32)
<223> Xaa представляет собой аспартат (D) или серин (S))
<220>
<221> MISC_FEATURE
<222> (37)..(37)
<223> Xaa представляет собой метионин (M) или лейцин (L)
<220>
<221> MISC_FEATURE
<222> (96)..(96)
<223> Xaa представляет собой гистидин (H) или тирозин (Y)
<220>
<221> MISC_FEATURE
<222> (97)..(97)
<223> Xaa представляет собой глутамат (E) или серин (S)
<220>
<221> MISC_FEATURE
<222> (98)..(98)
<223> Xaa представляет собой аспартат (D) или треонин (T)
<400> 53
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Xaa Ala Ser Gln Ser Val Asp Tyr Xaa
20 25 30
Gly Asp Ser Tyr Xaa Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Xaa
85 90 95
Xaa Xaa Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 54
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL(1) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VL(1))
<400> 54
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 55
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL(2) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VL(2))
<400> 55
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 56
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL(3) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VL(3))
<400> 56
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 57
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL(4) гуманизированного/оптимизированного антитела
к ADAM9 hMAB-A (hMAB-A VL(4))
<400> 57
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr
85 90 95
Ser Thr Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 58
<211> 23
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRL1 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 58
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys
20
<210> 59
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRL2 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 59
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 60
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRL3 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 60
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 61
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен FRL4 гуманизированного/оптимизированного мышиного
антитела к ADAM9 MAB-A
<400> 61
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 62
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRL1 антитела к ADAM9 MAB-A
<400> 62
Lys Ala Ser Gln Ser Val Asp Tyr Ser Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 63
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRL1 антитела к ADAM9 MAB-A
<400> 63
Arg Ala Ser Gln Ser Val Asp Tyr Ser Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 64
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRL1 антитела к ADAM9 MAB-A
<400> 64
Arg Ala Ser Gln Ser Val Asp Tyr Ser Gly Asp Ser Tyr Leu Asn
1 5 10 15
<210> 65
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CDRL2 антитела к ADAM9 MAB-A
<400> 65
Gln Gln Ser Tyr Ser Thr Pro Phe Thr
1 5
<210> 66
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Консенсусный домен CDRL1 антитела к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa представляет собой лизин (K) или аргинин (R)
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa представляет собой аспартат (D) или серин (S)
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa представляет собой метионин (M) или лейцин (L)
<400> 66
Xaa Ala Ser Gln Ser Val Asp Tyr Xaa Gly Asp Ser Tyr Xaa Asn
1 5 10 15
<210> 67
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Консенсусный домен CDRL3 антитела к ADAM9 MAB-A
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa представляет собой гистидин (H) или тирозин (Y)
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa представляет собой глутамат (E) или серин (S)
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa представляет собой аспартат (D) или треонин (T)
<400> 67
Gln Gln Ser Xaa Xaa Xaa Pro Phe Thr
1 5
<210> 68
<211> 218
<212> PRT
<213> Искусственная последовательность
<220>
<223> Иллюстративная гуманизированная/оптимизированная легкая цепь
IgG1 производного/варианта MAB-A, содержащего домен hMAB-A VL (2)
<400> 68
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 69
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Предпочтительный промежуточный спейсерный пептид (линкер 1)
<400> 69
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 70
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Предпочтительный содержащий цистеин спейсерный пептид (линкер 2)
<400> 70
Gly Gly Cys Gly Gly Gly
1 5
<210> 71
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 71
Gly Gly Gly Ser
1
<210> 72
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 72
Leu Gly Gly Gly Ser Gly
1 5
<210> 73
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 73
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly
1 5 10
<210> 74
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 74
Ala Ser Thr Lys Gly
1 5
<210> 75
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 75
Leu Glu Pro Lys Ser Ser
1 5
<210> 76
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альтернативный спейсерный пептид (линкер 2)
<400> 76
Ala Pro Ser Ser Ser
1 5
<210> 77
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Активирующий сборку гетеродимера домен
<400> 77
Gly Val Glu Pro Lys Ser Cys
1 5
<210> 78
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Активирующий сборку гетеродимера домен
<400> 78
Val Glu Pro Lys Ser Cys
1 5
<210> 79
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Активирующий сборку гетеродимера домен
<400> 79
Ala Glu Pro Lys Ser Cys
1 5
<210> 80
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Активирующий сборку гетеродимера домен
<400> 80
Gly Phe Asn Arg Gly Glu Cys
1 5
<210> 81
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Активирующий сборку гетеродимера домен
<400> 81
Phe Asn Arg Gly Glu Cys
1 5
<210> 82
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> E-петлевой активирующий сборку гетеродимера домен
<400> 82
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
1 5 10 15
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
20 25
<210> 83
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> K-петлевой активирующий сборку гетеродимера домен
<400> 83
Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val
1 5 10 15
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
20 25
<210> 84
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> Содержащий цистеин E-петлевой активирующий сборку гетеродимера
домен
<400> 84
Glu Val Ala Ala Cys Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
1 5 10 15
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
20 25
<210> 85
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> Содержащий цистеин К-петлевой активирующий сборку гетеродимера
домен
<400> 85
Lys Val Ala Ala Cys Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val
1 5 10 15
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
20 25
<210> 86
<211> 46
<212> PRT
<213> Streptococcus agalactiae
<220>
<221> MISC_FEATURE
<222> (1)..(46)
<223> Домен 3, связывающий альбумин (ABD3) белка G штамма
Streptococcus G148
<400> 86
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
1 5 10 15
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asp Asn Ala Lys Ser Ala Glu
20 25 30
Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 87
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Деиммунизированный домен 3, связывающий альбумин (ABD3)
белка G штамма Streptococcus G148
<400> 87
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
1 5 10 15
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asp Asn Ala Lys Ser Ala Glu
20 25 30
Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 88
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Деиммунизированный домен 3, связывающий альбумин (ABD3)
белка G штамма Streptococcus G148
<400> 88
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
1 5 10 15
Val Ser Asp Tyr Tyr Lys Asn Ala Ala Asn Asn Ala Lys Thr Val Glu
20 25 30
Gly Val Lys Ala Leu Ile Ala Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 89
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Деиммунизированный домен 3, связывающий альбумин (ABD3)
белка G штамма Streptococcus G148
<400> 89
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
1 5 10 15
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Ser Asn Ala Lys Ser Val Glu
20 25 30
Gly Val Lys Ala Leu Ile Ala Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 90
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 90
Ala Pro Ser Ser Ser Pro Met Glu
1 5
<210> 91
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 91
Val Glu Pro Lys Ser Ala Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 92
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 92
Leu Glu Pro Lys Ser Ala Asp Lys Thr His Thr Cys Pro Pro Cys
1 5 10 15
<210> 93
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 93
Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10
<210> 94
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 94
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10
<210> 95
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 95
Leu Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 96
<211> 15
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(15)
<223> Шарнирный домен IgG1
<400> 96
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 97
<211> 12
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(12)
<223> Шарнирный домен IgG2
<400> 97
Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 98
<211> 12
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(12)
<223> Шарнирный домен IgG4
<400> 98
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 99
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариантный шарнирный домен IgG4
<400> 99
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 100
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Промежуточный спейсерный пептид
<400> 100
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 101
<211> 107
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(107)
<223> Домен CL каппа-цепи IgG человека
<400> 101
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 102
<211> 104
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(104)
<223> Домен CL лямбда-цепи IgG человека
<400> 102
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
50 55 60
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
65 70 75 80
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
85 90 95
Thr Val Ala Pro Thr Glu Cys Ser
100
<210> 103
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Домен CH1 IgG1
<400> 103
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 104
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Домен CH1 IgG2
<400> 104
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 105
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Домен CH1 IgG4 человека
<400> 105
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 106
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1 человека, содержащий замены L234A и L235A
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 106
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 107
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1, содержащий замену S442C
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 107
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Cys Leu Ser Pro Gly Xaa
210 215
<210> 108
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1, содержащий замены L234A, L235A и S442C
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 108
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Cys Leu Ser Pro Gly Xaa
210 215
<210> 109
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> «Несущий выступ» вариантный домен CH2-CH3 IgG1 человека,
содержащий замены L234A и L235A
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 109
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 110
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> «Несущий впадину» вариантный домен CH2-CH3 IgG1 человека,
содержащий замены L234A и L235A
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 110
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 111
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD2 человека Lo-CD2a (№ доступа в ATCC
11423)
<400> 111
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Gln Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Glu Tyr
20 25 30
Tyr Met Tyr Trp Val Lys Gln Arg Pro Lys Gln Gly Leu Glu Leu Val
35 40 45
Gly Arg Ile Asp Pro Glu Asp Gly Ser Ile Asp Tyr Val Glu Lys Phe
50 55 60
Lys Lys Lys Ala Thr Leu Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Lys Phe Asn Tyr Arg Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 112
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD2 человека Lo-CD2a (№ доступа в ATCC
11423)
<400> 112
Asp Val Val Leu Thr Gln Thr Pro Pro Thr Leu Leu Ala Thr Ile Gly
1 5 10 15
Gln Ser Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Ser Gly Asn Thr Tyr Leu Asn Trp Leu Leu Gln Arg Thr Gly Gln Ser
35 40 45
Pro Gln Pro Leu Ile Tyr Leu Val Ser Lys Leu Glu Ser Gly Val Pro
50 55 60
Asn Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Gly Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Met Gln Phe
85 90 95
Thr His Tyr Pro Tyr Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 113
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(1) антитела к CD3 человека CD3 mAb-1
(CD3 mAb-1VH(1))
<400> 113
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 114
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH(2) антитела к CD3 человека CD3 mAb-1
(CD3 mAb-1VH(2))
<400> 114
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 115
<211> 110
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD3 человека CD3 mAb-1
<400> 115
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 116
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD3 человека CD3 mAb-1(D65G)
<400> 116
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 117
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD3 человека mAb-1 Low
<400> 117
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Thr Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 118
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD3 человека mAb-1 Fast
<400> 118
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Lys Asn Phe Gly Asn Ser Tyr Val Thr Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 119
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD3 человека mAb-1 OKT3
<400> 119
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 120
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD3 человека mAb-1 OKT3
<400> 120
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
100 105
<210> 121
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD8 человека mAb-1 OKT8
<400> 121
Gln Val Gln Leu Leu Glu Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Tyr Thr Gly Gly Thr Gly Tyr Asn Gln Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Ser Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Phe Arg Tyr Thr Tyr Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 122
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD8 человека mAb-1 OKT8
<400> 122
Asp Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Asp Asn Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Val Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
<210> 123
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD8 человека mAb-1 TRX2
<400> 123
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Tyr Tyr Asp Gly Ser Asn Lys Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Pro His Tyr Asp Gly Tyr Tyr His Phe Phe Asp Ser Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD8 человека mAb-1 TRX2
<400> 124
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Gly Ser Gln Asp Ile Asn Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Thr Asp Ile Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Tyr Gln Tyr Asn Asn Gly Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 125
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD16 человека 3G8
<400> 125
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Arg Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Ala Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala
115
<210> 126
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD16 человека 3G8
<400> 126
Asp Thr Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 127
<211> 117
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к CD16 человека A9
<400> 127
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Leu Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Gly Gly Gly Tyr Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Val Thr Ala Asp Thr Ser Ser Arg Thr Ala Tyr
65 70 75 80
Val Gln Val Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser Ala Ser Trp Tyr Phe Asp Val Trp Gly Ala Arg Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 128
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к CD16 человека A9
<400> 128
Asp Ile Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro
1 5 10 15
Gly Glu Thr Val Thr Leu Thr Cys Arg Ser Asn Thr Gly Thr Val Thr
20 25 30
Thr Ser Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe
35 40 45
Thr Gly Leu Ile Gly His Thr Asn Asn Arg Ala Pro Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr
65 70 75 80
Gly Ala Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr
85 90 95
Asn Asn His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 129
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к TCR человека BMA 031
<400> 129
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Lys Phe Thr Ser Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Val Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Val His Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Tyr Asp Tyr Asp Gly Phe Val Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 130
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к TCR человека BMA 031
<400> 130
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Thr Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 131
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к NKG2D человека KYK-1.0
<400> 131
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Lys Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Tyr Tyr Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 132
<211> 108
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к NKG2D человека KYK-1.0
<400> 132
Gln Pro Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Glu
1 5 10 15
Thr Ala Arg Ile Pro Cys Gly Gly Asp Asp Ile Glu Thr Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Phe Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Ser Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asp Asn Asn Asp Glu
85 90 95
Trp Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu
100 105
<210> 133
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VH антитела к NKG2D человека KYK-2.0
<400> 133
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Gly Leu Gly Asp Gly Thr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 134
<211> 110
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен VL антитела к NKG2D человека KYK-2.0
<400> 134
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Asp Leu Leu Pro Ser Gly Val Ser Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Phe Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 135
<211> 278
<212> PRT
<213> Искусственная последовательность
<220>
<223> Первая полипептидная цепь биспецифического двухцепочечного
диатела ADAM9 x CD3 («DART-1»)
<400> 135
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Thr Tyr Ala Met Asn Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Gly Arg Ile Arg Ser Lys Tyr Asn Asn
165 170 175
Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asp Ser Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys
195 200 205
Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly
210 215 220
Asn Ser Tyr Val Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu
245 250 255
Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu
260 265 270
Val Ala Ala Leu Glu Lys
275
<210> 136
<211> 275
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вторая полипептидная цепь биспецифического двухцепочечного
диатела ADAM9 x CD3 («DART-1»)
<400> 136
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Gly Glu Ile Ile Pro Ile Phe Gly His Thr
165 170 175
Asn Tyr Asn Glu Lys Phe Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn
180 185 190
Ser Lys Asn Thr Leu Tyr Leu Gln Met Gly Ser Leu Arg Ala Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser
210 215 220
Arg Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
225 230 235 240
Ser Gly Gly Cys Gly Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val
245 250 255
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala
260 265 270
Leu Lys Glu
275
<210> 137
<211> 507
<212> PRT
<213> Искусственная последовательность
<220>
<223> Первая полипептидная цепь биспецифического трехцепочечного
диатела ADAM9 x CD3 («DART-2»)
<220>
<221> MISC_FEATURE
<222> (507)..(507)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 137
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Ser
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asp Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Thr Tyr Ala Met Asn Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Gly Arg Ile Arg Ser Lys Tyr Asn Asn
165 170 175
Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asp Ser Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys
195 200 205
Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly
210 215 220
Asn Ser Tyr Val Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val
225 230 235 240
Thr Val Ser Ser Ala Ser Thr Lys Gly Glu Val Ala Ala Cys Glu Lys
245 250 255
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
260 265 270
Ala Ala Leu Glu Lys Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro
275 280 285
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
290 295 300
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
305 310 315 320
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
325 330 335
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
340 345 350
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
355 360 365
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
370 375 380
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
385 390 395 400
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
405 410 415
Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
420 425 430
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
435 440 445
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
450 455 460
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
465 470 475 480
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
485 490 495
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Xaa
500 505
<210> 138
<211> 274
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вторая полипептидная цепь биспецифического трехцепочечного
диатела ADAM9 x CD3 («DART-2»)
<400> 138
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Gly Glu Ile Ile Pro Ile Phe Gly His Thr
165 170 175
Asn Tyr Asn Glu Lys Phe Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn
180 185 190
Ser Lys Asn Thr Leu Tyr Leu Gln Met Gly Ser Leu Arg Ala Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Ala Arg Gly Gly Tyr Tyr Tyr Tyr Gly Ser
210 215 220
Arg Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
225 230 235 240
Ser Ala Ser Thr Lys Gly Lys Val Ala Ala Cys Lys Glu Lys Val Ala
245 250 255
Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu
260 265 270
Lys Glu
<210> 139
<211> 227
<212> PRT
<213> Искусственная последовательность
<220>
<223> Третья полипептидная цепь биспецифического трехцепочечного
диатела ADAM9 x CD3 («DART-2»)
<220>
<221> MISC_FEATURE
<222> (227)..(227)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 139
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Xaa
225
<210> 140
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Субдоменный участок домена CDRH3 антитела к ADAM9 человека
MAB-A (2.2)
<400> 140
Gly Ser Arg Asp Tyr Phe
1 5
<210> 141
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 141
Asp Gly Glu Gly Val Met
1 5
<210> 142
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 142
Phe His Ser Gly Leu Leu
1 5
<210> 143
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 143
Phe Asn Ser Ala Thr Leu
1 5
<210> 144
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 144
Phe Asn Ser Gly Thr Leu
1 5
<210> 145
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 145
Phe Asn Ser Ser Thr Leu
1 5
<210> 146
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 146
Gly Lys Ser Lys Trp Leu
1 5
<210> 147
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 147
Gly Met Gly Gly Thr Leu
1 5
<210> 148
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 148
His Ala Lys Gly Gly Met
1 5
<210> 149
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 149
Ile Gly Glu Ala Val Leu
1 5
<210> 150
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 150
Ile Gly Lys Gly Val Phe
1 5
<210> 151
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 151
Ile Gly Lys Gly Val Leu
1 5
<210> 152
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 152
Lys His Asp Ser Val Leu
1 5
<210> 153
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 153
Leu Asn Thr Ala Val Met
1 5
<210> 154
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 154
Asn Gly Glu Gly Thr Leu
1 5
<210> 155
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 155
Asn Gly Lys Asn Thr Leu
1 5
<210> 156
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 156
Asn Ser Ala Gly Ile Leu
1 5
<210> 157
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 157
Pro Lys Glu Gly Trp Met
1 5
<210> 158
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 158
Pro Lys Phe Gly Trp Lys
1 5
<210> 159
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 159
Pro Lys Met Gly Trp Val
1 5
<210> 160
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 160
Pro Arg Leu Gly His Leu
1 5
<210> 161
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 161
Pro Ser Phe Gly Trp Ala
1 5
<210> 162
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 162
Gln Ala Lys Gly Thr Met
1 5
<210> 163
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 163
Arg Gly Met Gly Val Met
1 5
<210> 164
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 164
Arg Lys Glu Gly Trp Met
1 5
<210> 165
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 165
Thr Gly Lys Gly Val Leu
1 5
<210> 166
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 166
Thr Gly Met Gly Thr Leu
1 5
<210> 167
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 167
Thr Gly Asn Gly Val Met
1 5
<210> 168
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 168
Trp Asn Ala Gly Thr Phe
1 5
<210> 169
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 169
Tyr His His Thr Pro Leu
1 5
<210> 170
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 170
Tyr Gln Ser Ala Thr Leu
1 5
<210> 171
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 171
Asp Gly Lys Ala Val Leu
1 5
<210> 172
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 172
Phe Asn Lys Ala Val Leu
1 5
<210> 173
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 173
Phe Asn Ser Gly Thr Trp
1 5
<210> 174
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 174
Phe Asn Thr Gly Val Phe
1 5
<210> 175
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 175
Gly Lys Ser Arg Phe His
1 5
<210> 176
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 176
Ile Gly Lys Asn Val Tyr
1 5
<210> 177
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 177
Met Gly Lys Gly Val Met
1 5
<210> 178
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 178
Asn Gly Glu Ser Val Phe
1 5
<210> 179
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 179
Pro Asp Phe Gly Trp Met
1 5
<210> 180
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 180
Pro Gly Ser Gly Val Met
1 5
<210> 181
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека MAB-A
<400> 181
Pro Lys Asp Ala Trp Leu
1 5
<210> 182
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека MAB-A
<400> 182
Pro Lys Phe Gly Trp Leu
1 5
<210> 183
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека MAB-A
<400> 183
Pro Lys Ile Gly Trp His
1 5
<210> 184
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 184
Pro Lys Met Gly Trp Ala
1 5
<210> 185
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 185
Pro Lys Met Gly Trp Met
1 5
<210> 186
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 186
Pro Gln Met Gly Trp Leu
1 5
<210> 187
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 187
Pro Arg Phe Gly Trp Leu
1 5
<210> 188
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 188
Pro Arg Met Gly Phe Leu
1 5
<210> 189
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 189
Pro Arg Met Gly Phe Met
1 5
<210> 190
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 190
Pro Ser Phe Gly Trp Met
1 5
<210> 191
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 191
Arg Arg Glu Gly Trp Met
1 5
<210> 192
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 192
Ser Gly Glu Gly Val Leu
1 5
<210> 193
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 193
Ser Gly Asn Gly Val Met
1 5
<210> 194
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A (2.2)
<400> 194
Val Gly Lys Ala Val Leu
1 5
<210> 195
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A VH (2E)
<400> 195
Asp Ser Asn Ala Val Leu
1 5
<210> 196
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A VH (2F)
<400> 196
Phe His Ser Gly Thr Leu
1 5
<210> 197
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A VH (2H)
<400> 197
Gly Gly Ser Gly Val Leu
1 5
<210> 198
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A VH (2I)
<400> 198
Pro Arg Gln Gly Phe Leu
1 5
<210> 199
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариант субдоменного участка домена CDRH3 антитела к ADAM9
человека hMAB-A VH (2J)
<400> 199
Tyr Asn Ser Gly Thr Leu
1 5
<210> 200
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1 человека, содержащий замены M252Y/S254T/T256E
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 200
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 201
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1 человека, содержащий замены
L234A/L235A/M252Y/S254T/T256E
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 201
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 202
<211> 453
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гуманизированная/оптимизированная тяжелая цепь IgG1 hMAB-A VH (2I),
содержащего замены L234A/L235A
<220>
<221> MISC_FEATURE
<222> (453)..(453)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 202
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Glu Ile Ile Pro Ile Phe Gly His Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Phe Thr Ile Ser Leu Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Tyr Tyr Pro Arg Gln Gly Phe Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Xaa
450
<210> 203
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен CH2-CH3 IgG1, содержащий замены L234A, L235A, M252Y,
S254T, T256E и S442C
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 203
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Cys Leu Ser Pro Gly Xaa
210 215
<210> 204
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> «Несущий выступ» вариантный домен CH2-CH3 IgG1 человека,
содержащий замены L234A, L235A, M252Y, S254T и T256E
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 204
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 205
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> «Несущий впадину» вариантный домен CH2-CH3 IgG1 человека,
содержащий замены L234A, L235A, M252Y, S254T и T256E
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> Xaa представляет собой лизин (K) или отсутствует
<400> 205
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 206
<211> 62
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(52)
<223> Шарнирный домен IgG3
<400> 206
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro Arg Cys
1 5 10 15
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
20 25 30
Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro Glu
35 40 45
Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
50 55 60
<210> 207
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Домен IgG3 человека
<400> 207
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| АНТИ-ROR1/АНТИ-CD3 БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2831608C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| ИММУНОКОНЪЮГАТЫ IL2 И МУТАНТНОГО TNF | 2017 |
|
RU2758139C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2019 |
|
RU2810748C2 |
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
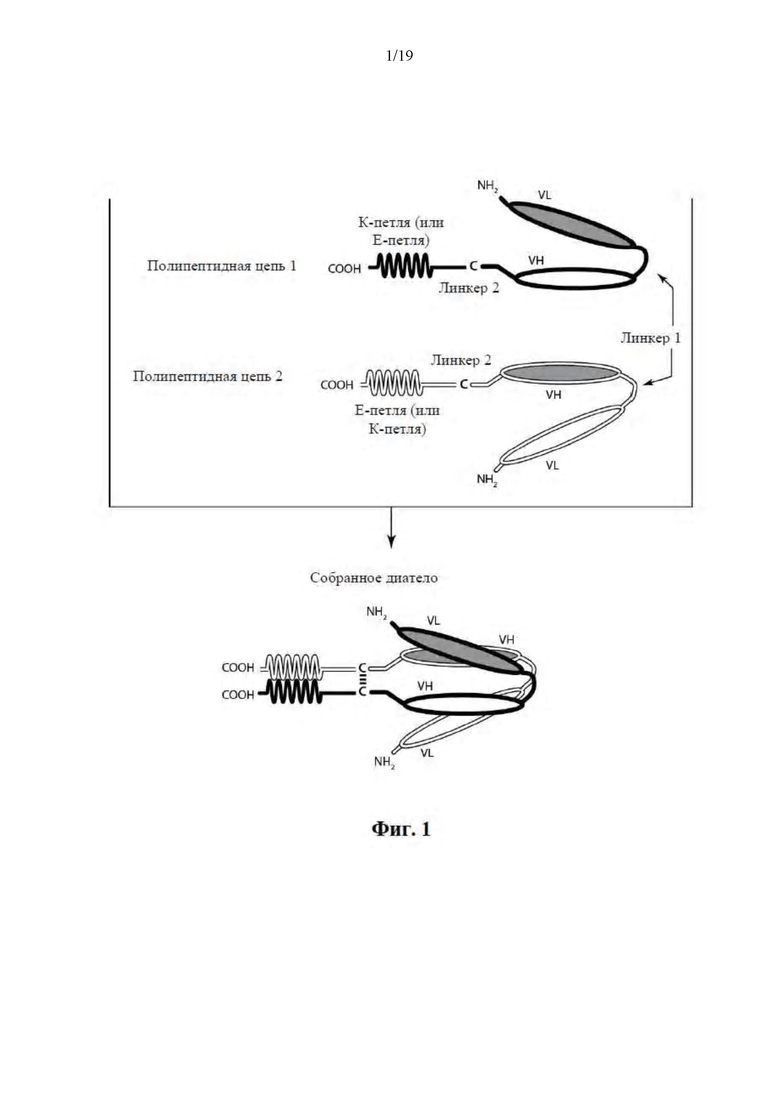
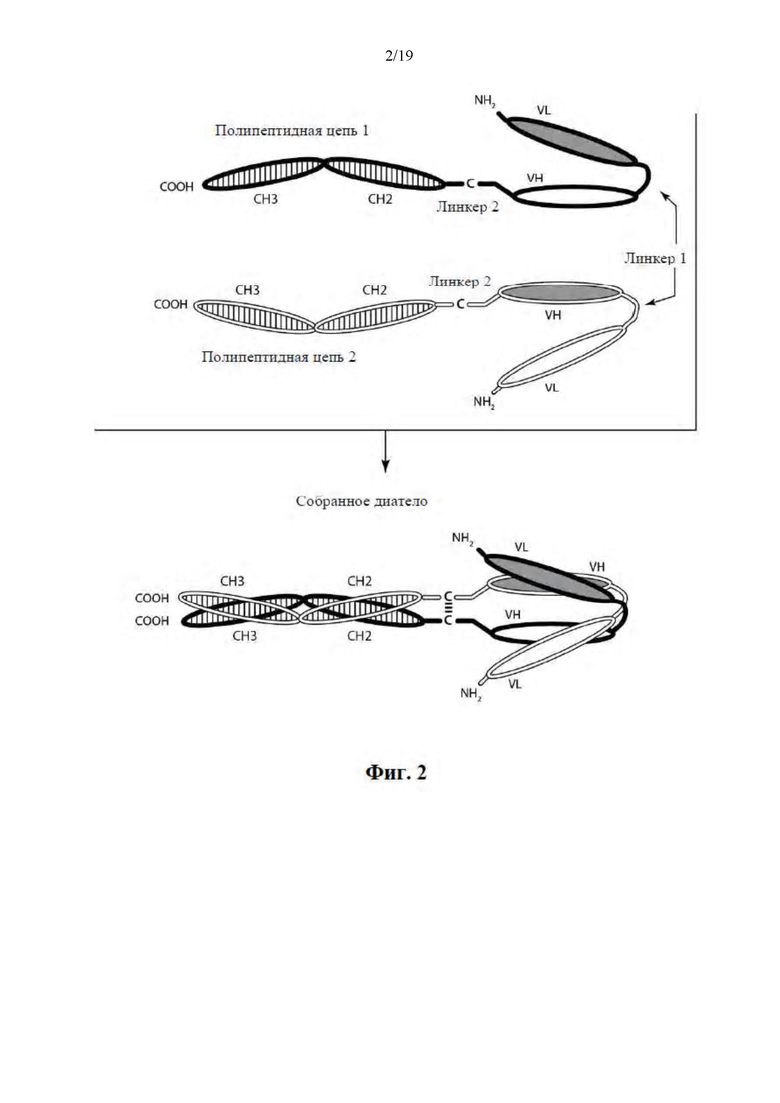
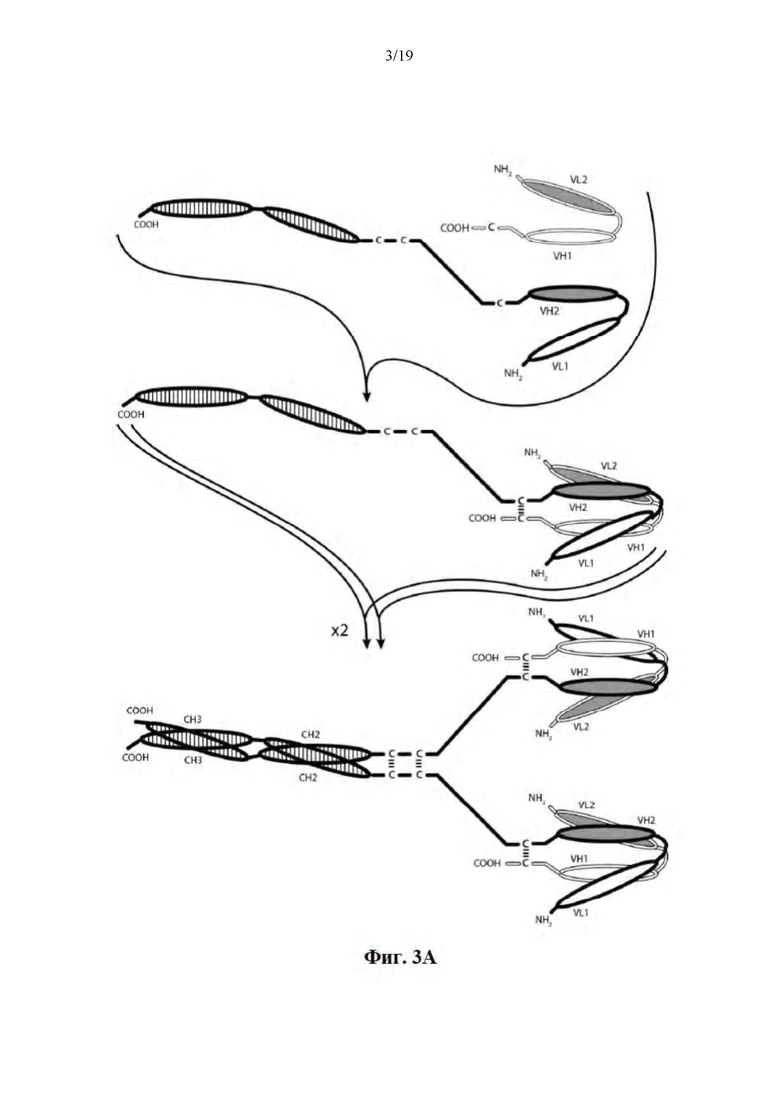
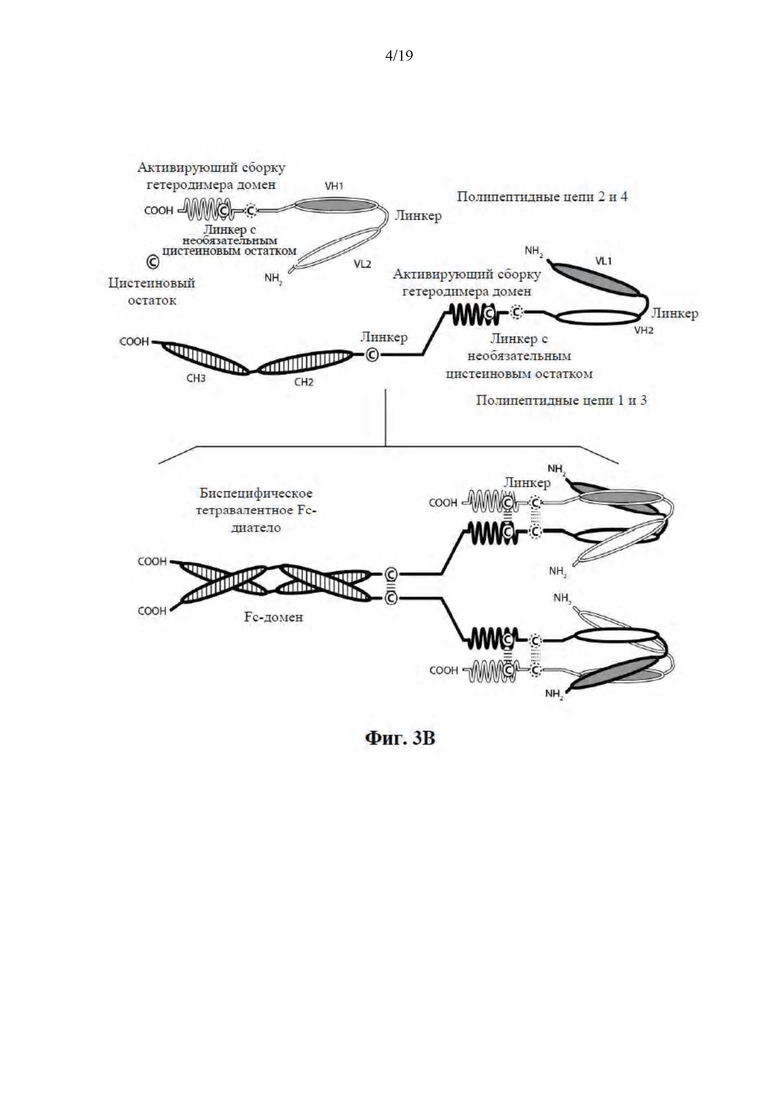
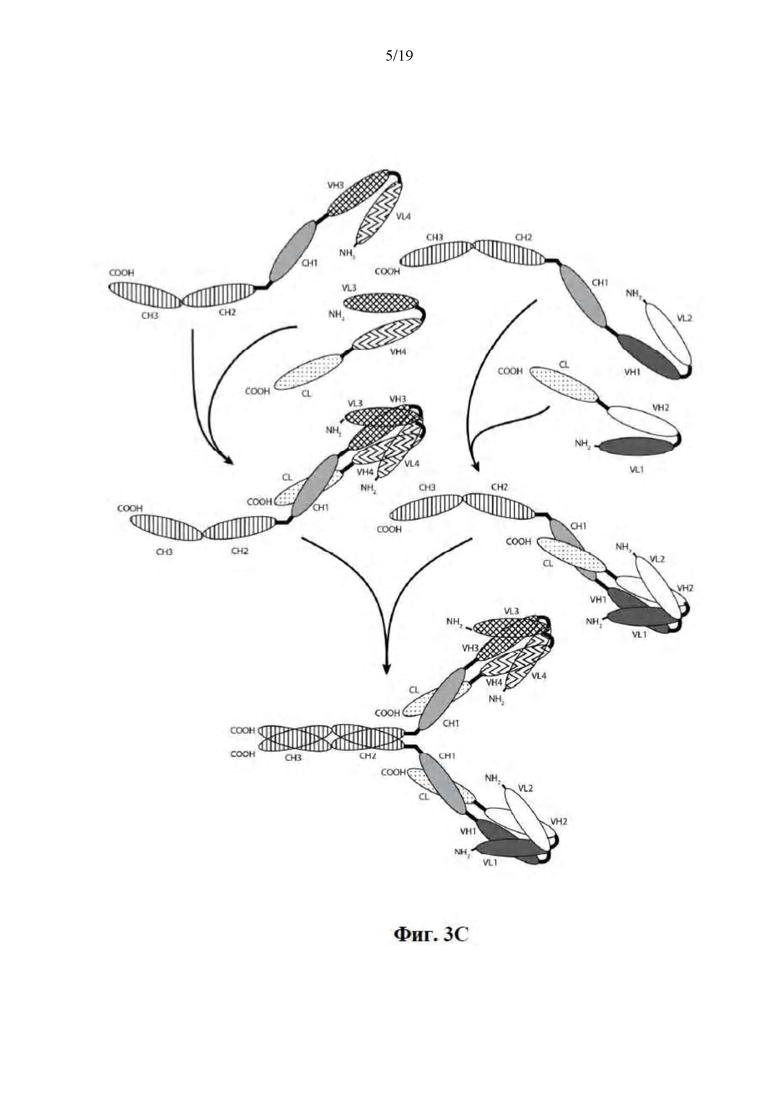
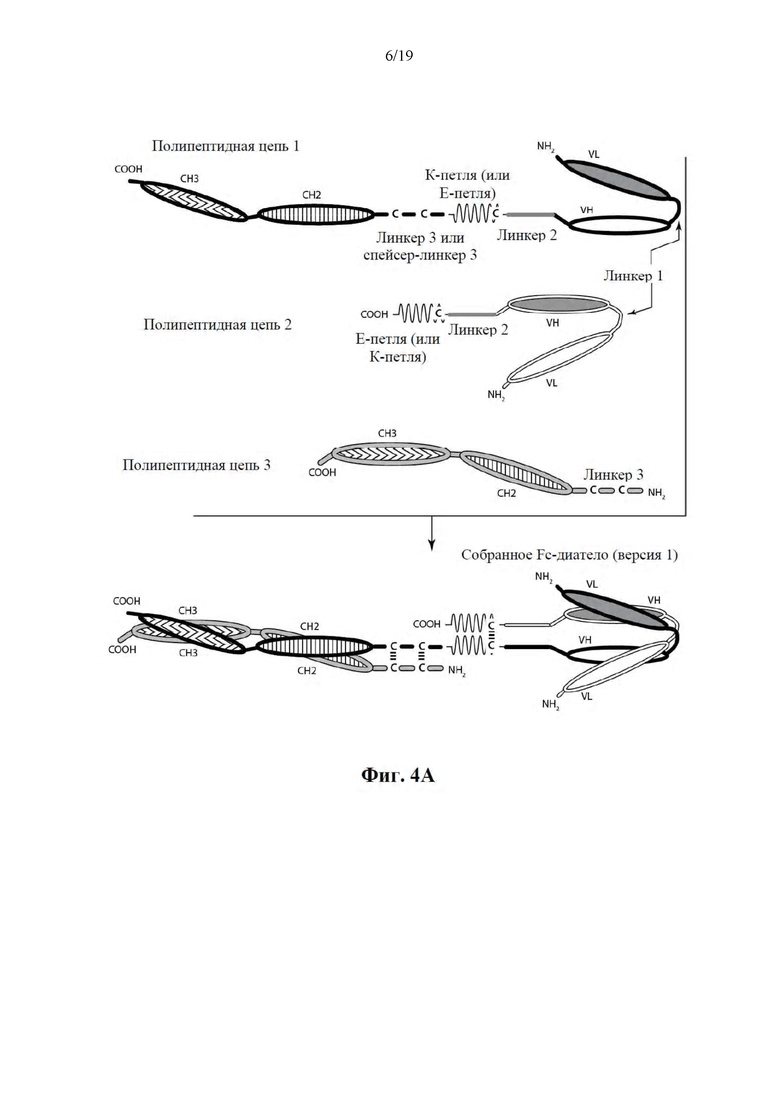
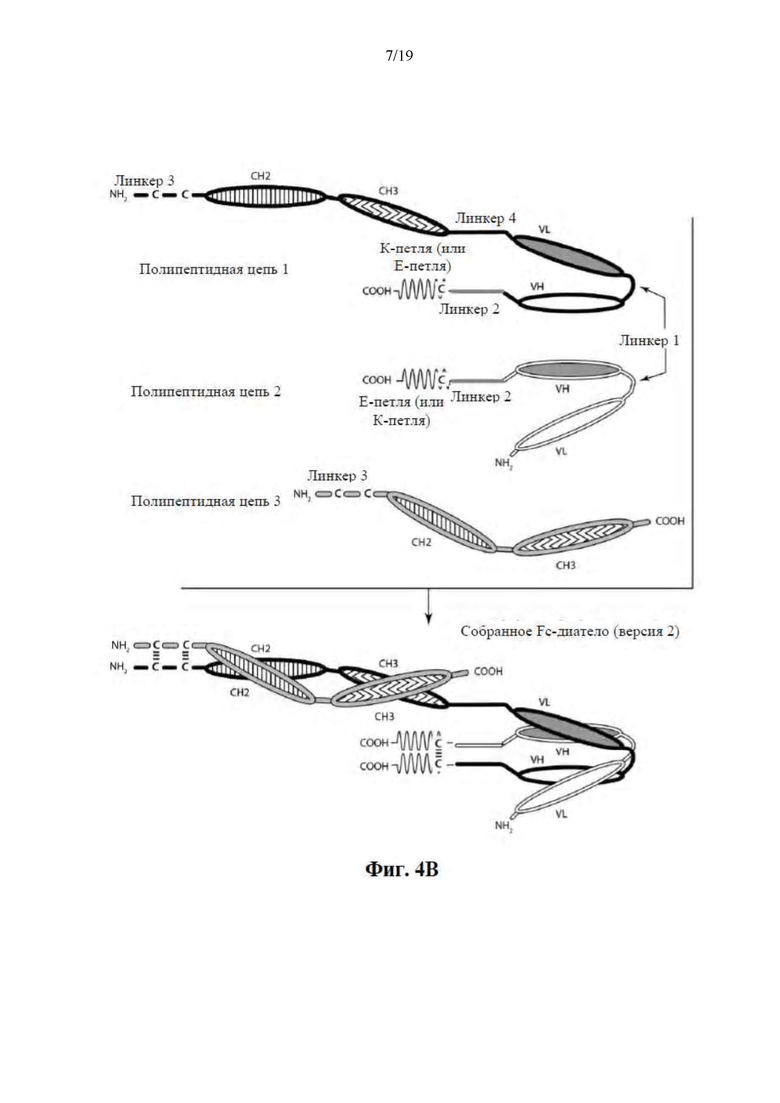
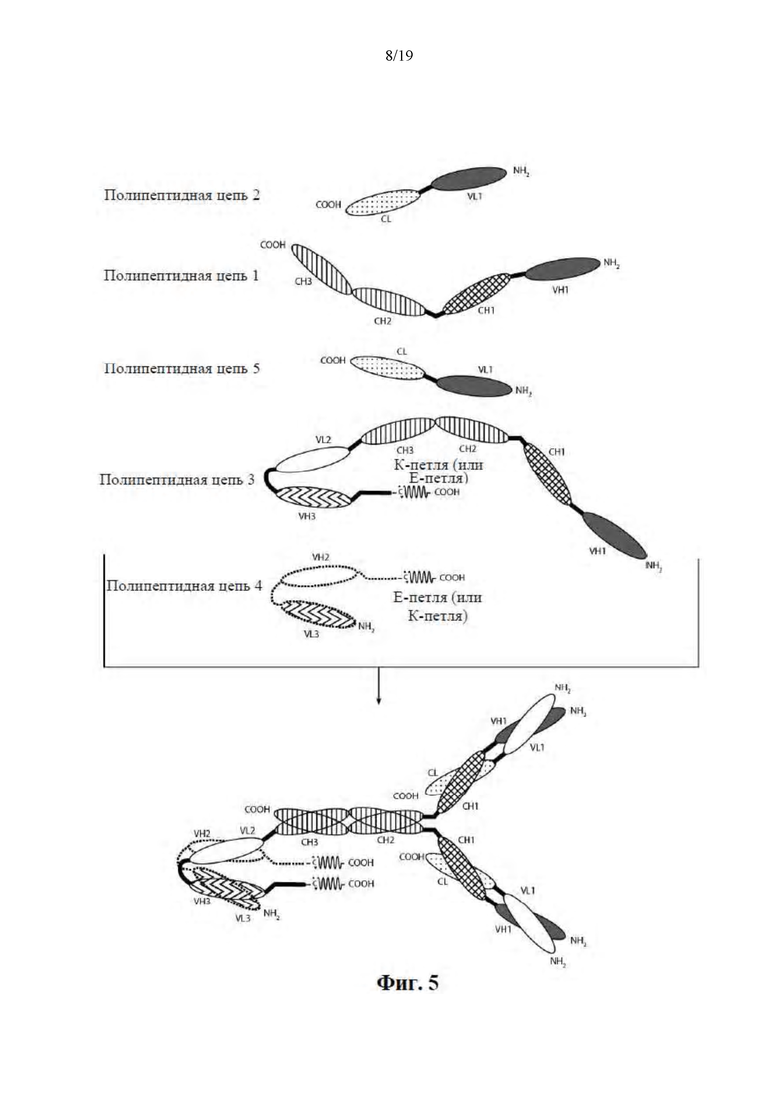
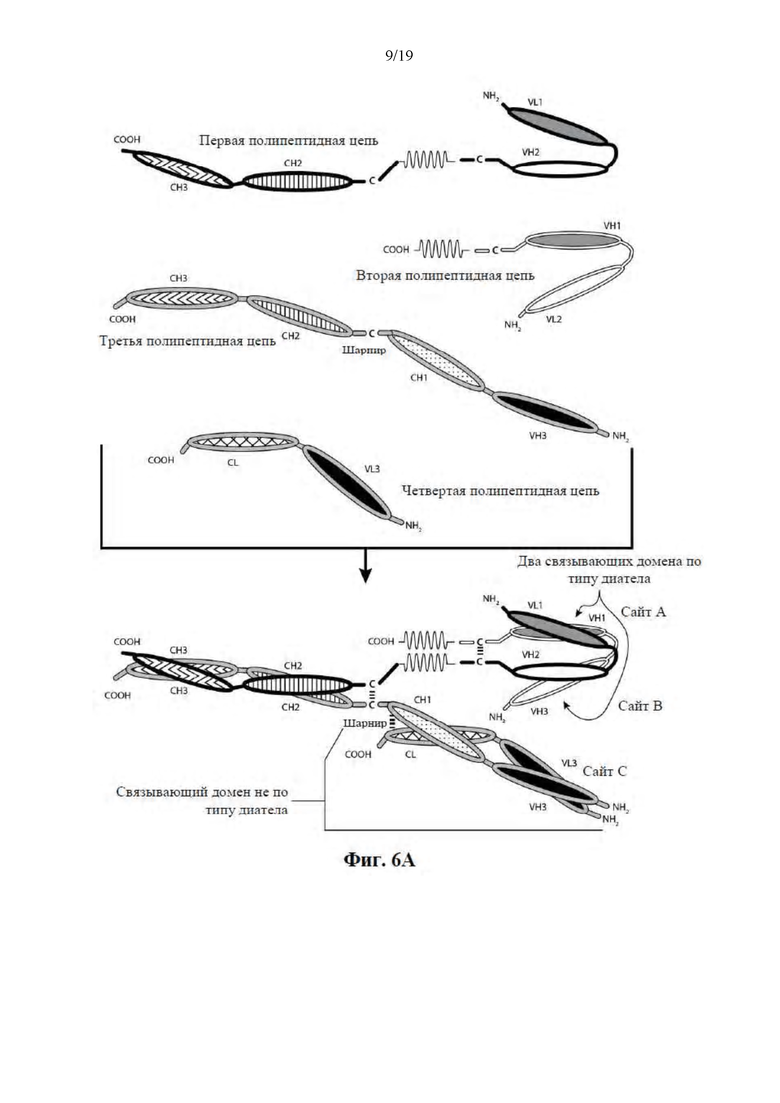
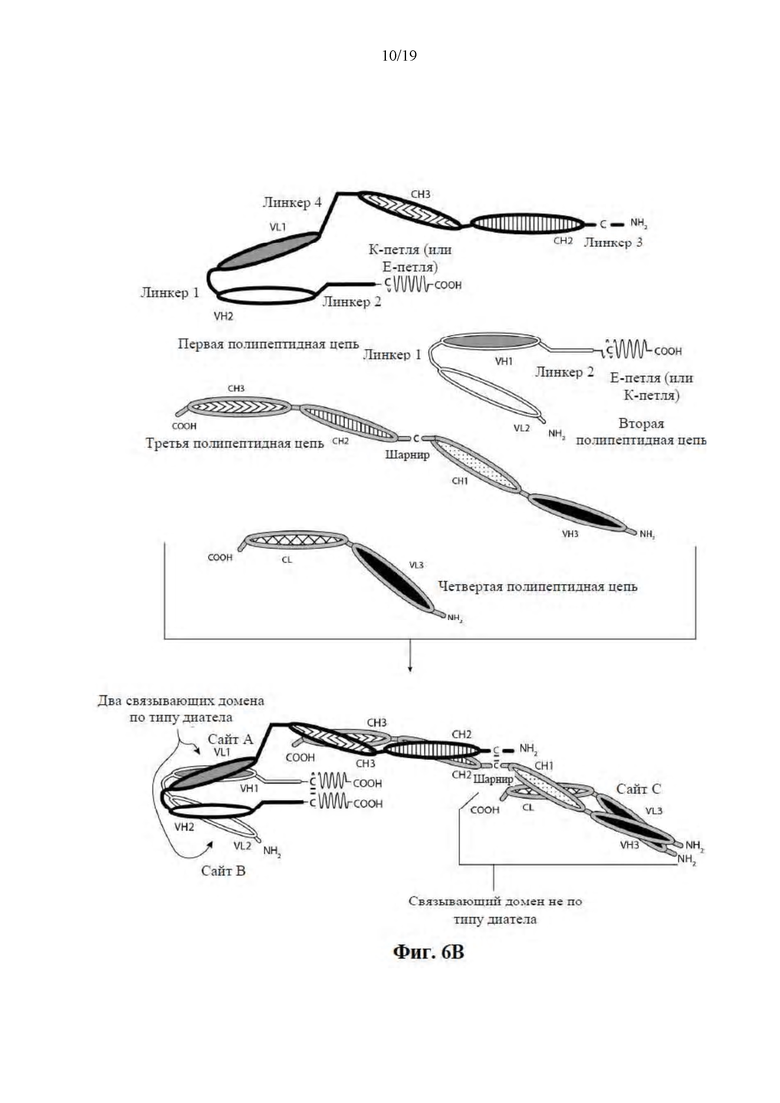
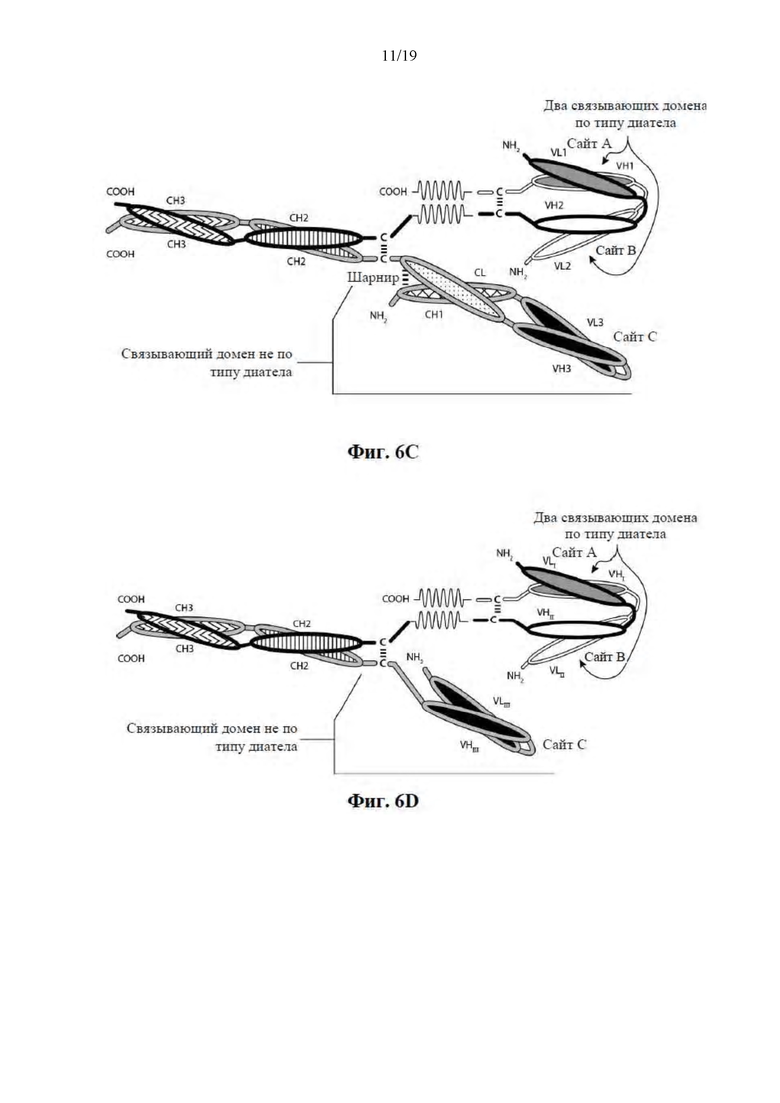
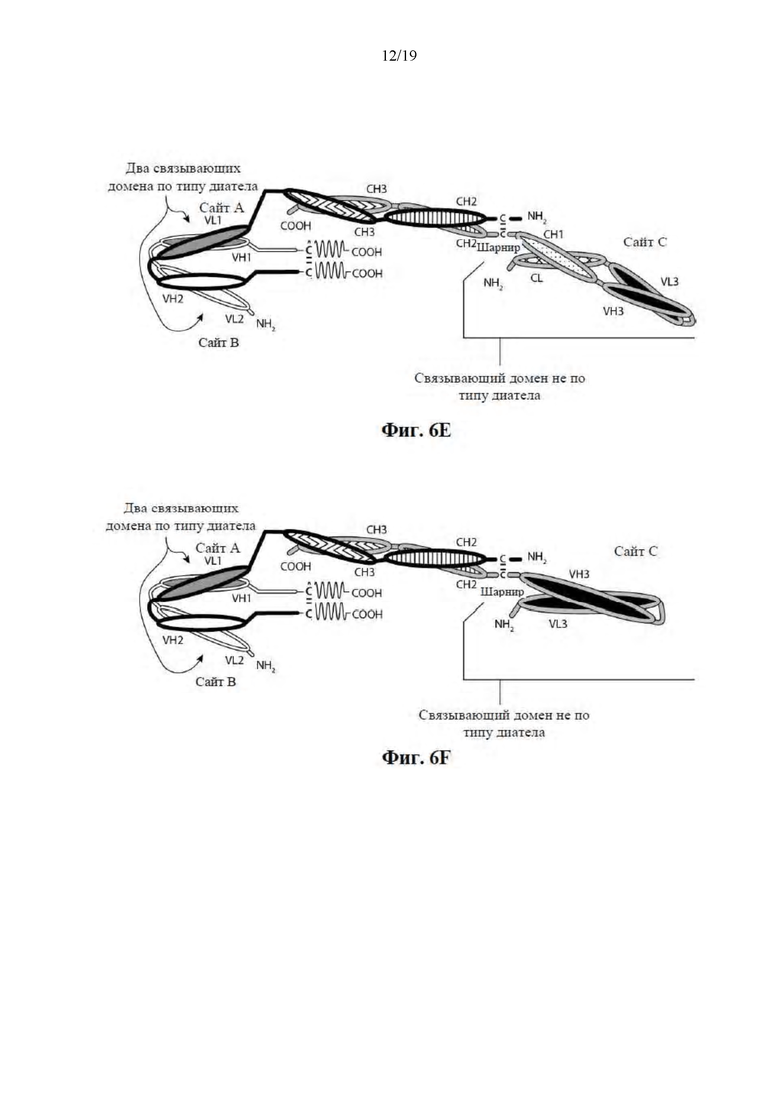
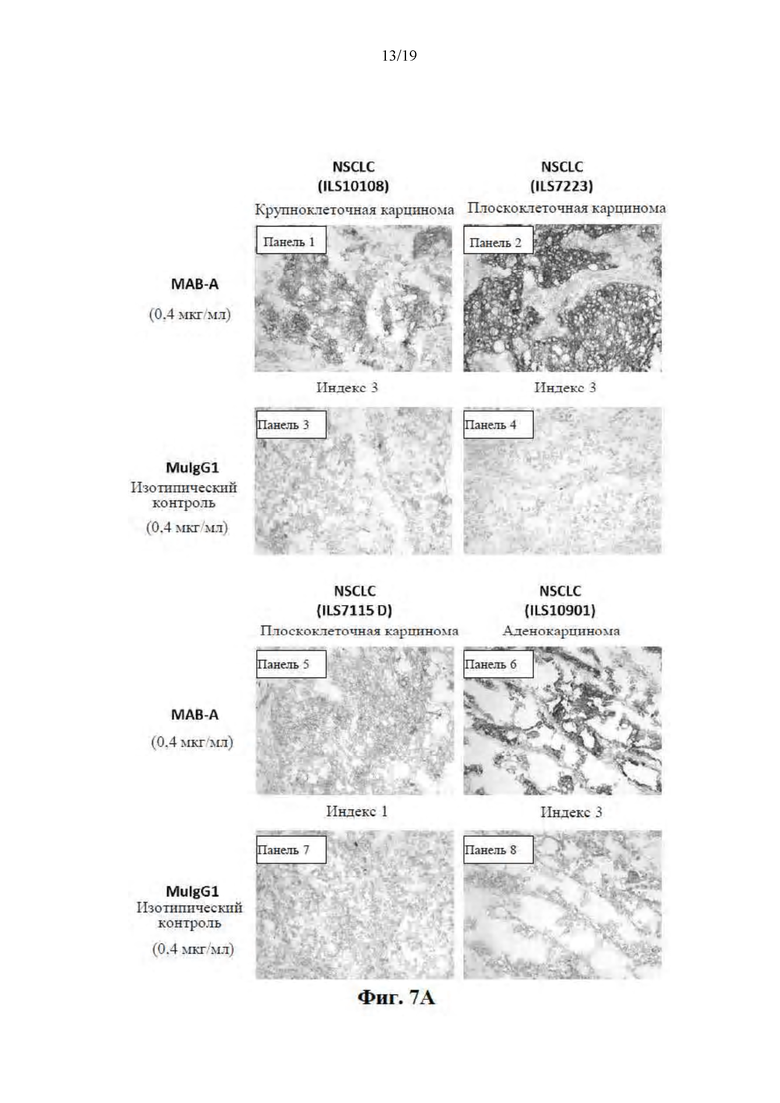

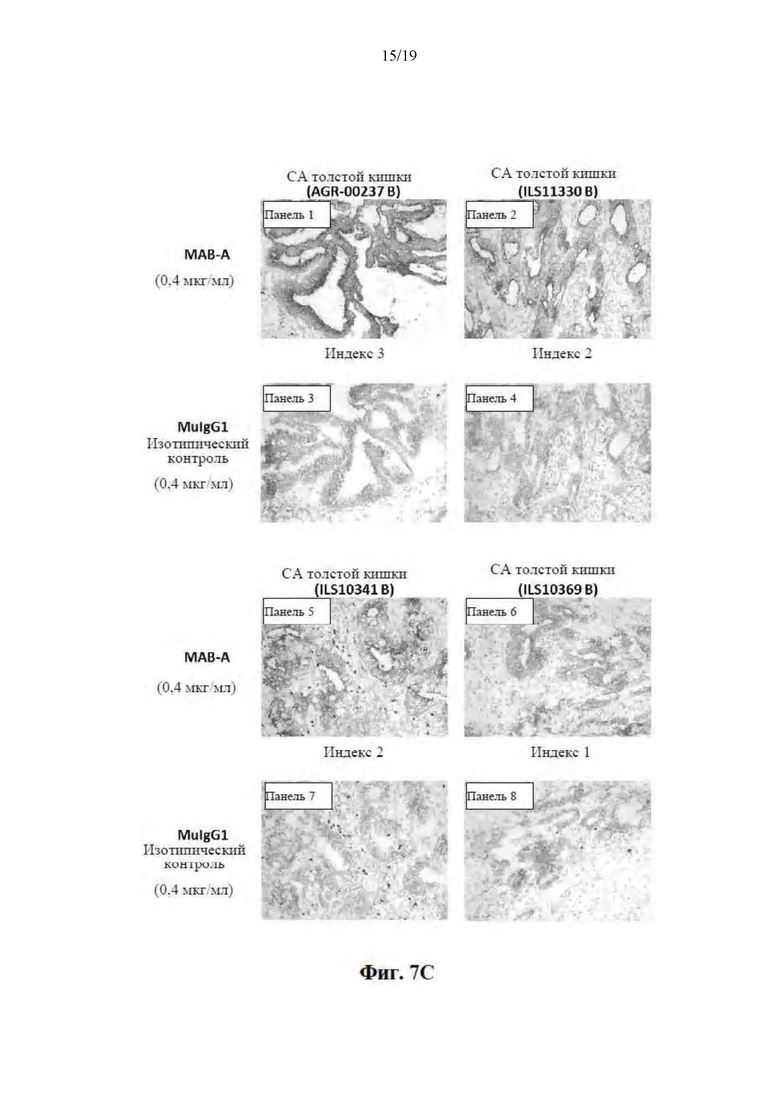
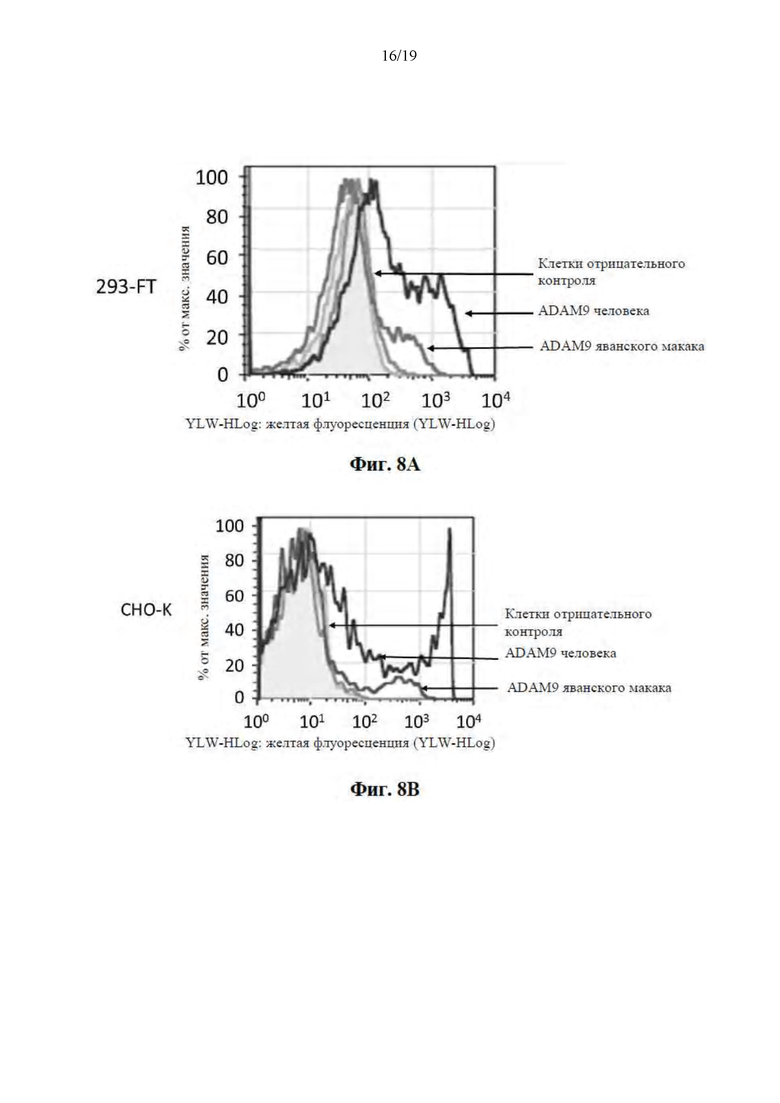


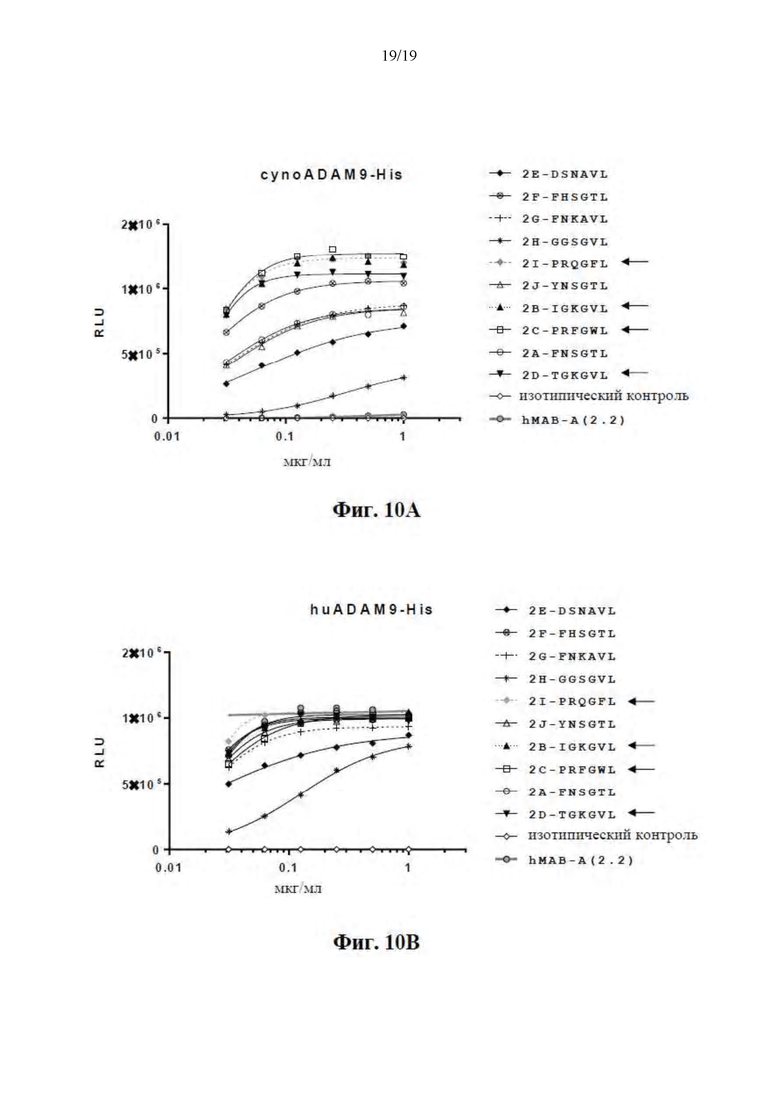
Изобретение относится к области биотехнологии, в частности к молекуле, связывающей ADAM9, а также содержащей ее композиции. Изобретение эффективно для лечения заболевания или патологического состояния, ассоциированного с экспрессией ADAM9 или характеризующегося ей. 5 н. и 14 з.п. ф-лы, 10 ил., 14 табл., 5 пр.
1. Молекула, связывающая ADAM9, содержащая эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9 и содержит вариабельный домен тяжелой цепи (VH), который содержит домен CDRH1, домен CDRH2 и домен CDRH3, и вариабельный домен легкой цепи (VL), который содержит домен CDRL1, домен CDRL2 и домен CDRL3, где:
(A) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 45 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(B) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 37 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(C) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 38 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(D) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 39 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(E) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 40 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(F) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 41 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(G) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 42 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(H) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 43 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(I) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 44 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(J) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 46 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(K) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(L) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(М) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(N) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 34, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 64, 13 и 65 соответственно.
2. Молекула, связывающая ADAM9, где указанная молекула представляет собой диатело, при этом указанное диатело представляет собой ковалентно связанный комплекс, который содержит два, три, четыре или пять полипептидных цепей, причем указанное диатело содержит:
I) эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9 и содержит вариабельный домен тяжелой цепи (VH), который содержит домен CDRH1, домен CDRH2 и домен CDRH3, и вариабельный домен легкой цепи (VL), который содержит домен CDRL1, домен CDRL2 и домен CDRL3, где:
(A) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 45 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(B) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 37 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(C) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 38 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(D) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 39 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(E) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 40 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(F) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 41 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(G) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 42 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(H) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 43 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(I) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 44 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(J) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 46 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(K) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(L) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(М) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(N) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 34, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 64, 13 и 65 соответственно, и
II) эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп молекулы, присутствующей на поверхности эффекторной клетки, где указанная молекула, присутствующая на поверхности эффекторной клетки, представляет собой CD2, CD3, CD8, TCR или NKG2D.
3. Молекула, связывающая ADAM9, по п. 2, где указанная молекула содержит два эпитопсвязывающих сайта, способных к иммуноспецифическому связыванию эпитопа (эпитопов) ADAM9, и два эпитопсвязывающих сайта, способных к иммуноспецифическому связыванию эпитопа (эпитопов) молекулы, присутствующей на поверхности эффекторной клетки.
4. Молекула, связывающая ADAM9, где указанная молекула представляет собой тривалентную связывающую молекулу, при этом указанная тривалентная связывающая молекула представляет собой ковалентно связанный комплекс, который содержит три, четыре, пять или более полипептидных цепей, причем указанная тривалентная связывающая молекула содержит:
I) эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9 и содержит вариабельный домен тяжелой цепи (VH), который содержит домен CDRH1, домен CDRH2 и домен CDRH3, и вариабельный домен легкой цепи (VL), который содержит домен CDRL1, домен CDRL2 и домен CDRL3, где:
(A) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 45 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(B) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 37 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(C) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 38 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(D) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 39 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(E) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 40 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(F) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 41 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(G) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 42 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(H) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 43 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(I) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 44 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(J) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 46 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(K) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно; или
(L) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(М) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 8, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 63, 13 и 14 соответственно; или
(N) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3 содержат аминокислотные последовательности SEQ ID NOs: 34, 36 и 10 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3 содержат аминокислотные последовательности SEQ ID NOs: 64, 13 и 65 соответственно, и
II) эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп первой молекулы, присутствующей на поверхности эффекторной клетки, где указанная первая молекула, присутствующая на поверхности эффекторной клетки, представляет собой CD2, CD3, CD8, TCR или NKG2D, и
III) эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп второй молекулы, присутствующей на поверхности эффекторной клетки, где указанная вторая молекула, присутствующая на поверхности эффекторной клетки, представляет собой CD2, CD3, CD8, TCR или NKG2D.
5. Молекула, связывающая ADAM9, по любому из пи. 1-4, где указанный эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9, содержит:
(A) (1) указанный домен CDRH1, который содержит аминокислотную последовательность SEQ ID NOs: 8;
(2) указанный домен CDRH2, который содержит аминокислотную последовательность SEQ ID NOs: 35; и
(3) указанный домен CDRH3, который содержит аминокислотную последовательность SEQ ID NOs: 37, 38, 39, 40, 41, 42, 43, 44, 45 или 46; и
(B) (1) указанный домен CDRL1, который содержит аминокислотную последовательность SEQ ID NOs: 62;
(2) указанный домен CDRL2, который содержит аминокислотную последовательность SEQ ID NOs: 13; и
(3) указанный домен CDRL3, который содержит аминокислотную последовательность SEQ ID NOs: 14.
6. Молекула, связывающая ADAM9, по п. 5, где указанный эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9, содержит:
(A) (1) указанный домен CDRH1, который содержит аминокислотную последовательность SYWMH (SEQ ID NO: 8);
(2) указанный домен CDRH2, который содержит аминокислотную последовательность EIIPIFGHTNYNEKFKS (SEQ ID NO: 35); и
(3) указанный домен CDRH3, который содержит аминокислотную последовательность GGYYYYPRQGFLDY (SEQ ID NO: 45); и
(B) (1) указанный домен CDRL1, который содержит аминокислотную последовательность KASQSVDYSGDSYMN (SEQ ID NO: 62);
(2) указанный домен CDRL2, который содержит аминокислотную последовательность AASDLES (SEQ ID NO: 13); и
(3) указанный домен CDRL3, который содержит аминокислотную последовательность QQSHEDPFT (SEQ ID NO: 14).
7. Молекула, связывающая ADAM9, по любому из пи. 1-6, где указанный эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9, содержит:
(A) указанный домен VH, который содержит аминокислотную последовательность SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 или SEQ ID NO: 29; и
(B) указанный домен VL, который содержит аминокислотную последовательность SEQ ID NO: 55.
8. Молекула, связывающая ADAM9, по любому из пи. 1-6, где указанный эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9, содержит:
(A) указанный домен VH, который содержит аминокислотную последовательность SEQ ID NO: 17, и указанный домен VL, который содержит аминокислотную последовательность SEQ ID NO: 55; или
(B) указанный домен VH, который содержит аминокислотную последовательность SEQ ID NO: 17, и указанный домен VL, который содержит аминокислотную последовательность SEQ ID NO: 56; или
(C) указанный домен VH, который содержит аминокислотную последовательность SEQ ID NO: 18, и указанный домен VL, который содержит аминокислотную последовательность SEQ ID NO: 56; или
(D) указанный домен VH, который содержит аминокислотную последовательность SEQ ID NO: 19, и указанный домен VL, который содержит аминокислотную последовательность SEQ ID NO: 57.
9. Молекула, связывающая ADAM9, по п. 7, где указанный эпитопсвязывающий сайт, который иммуноспецифически связывает эпитоп ADAM9, содержит домен VH SEQ ID NO: 28.
10. Молекула, связывающая ADAM9, по п. 1, где указанная молекула представляет собой моноспецифическое антитело, связывающее ADAM9 или его фрагмент, связывающий ADAM9.
11. Молекула, связывающая ADAM9, по любому из пп. 1-10, где указанная молекула содержит Fc-область, выбранную из группы, состоящей из IgG1, IgG2, IgG3 и IgG4.
12. Молекула, связывающая ADAM9, по п. 11, где указанная Fc-область представляет собой вариантную Fc-область, которая содержит, по отношению к Fc-области дикого типа, одну или несколько аминокислотных модификаций, которые:
(a) снижают аффинность вариантной Fc-области в отношении FcγR по отношению к аффинности, проявляемой Fc-областью дикого типа; и/или
(b) усиливают период полужизни в сыворотке крови указанной молекулы, связывающей ADAM9 по отношению к сходной молекуле, содержащей Fc-область дикого типа.
13. Молекула, связывающая ADAM9, по п. 12, где указанная одна или несколько аминокислотных модификаций, которые снижают аффинность вариантной Fc-области в отношении FcγR, содержат:
(A) L234A;
(B) L235A; или
(C) L234A и L235A;
где указанная нумерация представляет собой нумерацию EU-индекса в соответствии с Кабат.
14. Молекула, связывающая ADAM9, по любому из пп. 12, 13, где указанная одна или несколько аминокислотных модификаций, которые усиливают период полужизни в сыворотке крови указанной молекулы, связывающей ADAM9, содержат:
(A) M252Y;
(B) M252Y и S254T;
(C) M252Y и Т256Е;
(D) M252Y, S254T и Т256Е; или
(E) K288D и H435K;
где указанная нумерация представляет собой нумерацию EU-индекса в соответствии с Кабат.
15. Молекула, связывающая ADAM9, по любому из пп. 1-14, где указанная молекула содержит константный домен каппа CL IgG человека, имеющий аминокислотную последовательность SEQ ID NO: 101.
16. Молекула, связывающая ADAM9, по п. 15, где указанная молекула содержит:
(a) указанный домен CDRH1, указанный домен CDRH2 и указанный домен CDRH3, которые содержат аминокислотные последовательности SEQ ID NOs: 8, 35 и 45 соответственно; и указанный домен CDRL1, указанный домен CDRL2 и указанный домен CDRL3, которые содержат аминокислотные последовательности SEQ ID NOs: 62, 13 и 14 соответственно;
(b) Fc область IgG1, где указанная Fc область представляет собой вариантную Fc область, которая содержит модификации аминокислот M252Y, S254T и Т256Е,
где указанная нумерация представляет собой нумерацию EU-индекса в соответствии с Кабат.
17. Фармацевтическая композиция для лечения заболевания или патологического состояния, ассоциированного с экспрессией ADAM9 или характеризующегося ей, указанная фармацевтическая композиция содержит эффективное количество молекулы, связывающей ADAM9, по любому из пп. 1-16 и фармацевтически приемлемый носитель, наполнитель или разбавитель.
18. Применение молекулы, связывающей ADAM9, по любому из пп. 1-16 или фармацевтической композиции по п. 17 в лечении заболевания или патологического состояния, ассоциированного с экспрессией ADAM9 или характеризующегося ей.
19. Применение по п. 18, где указанное заболевание или патологическое состояние, ассоциированное с экспрессией ADAM9 или характеризующееся ей, представляет собой рак, выбранный из группы, состоящей из рака мочевого пузыря, рака молочной железы, рака шейки матки, колорекательного рака, рака пищевода, рака желудка, рака головы и шеи, рака печени, немелкоклеточного рака легких, рака миелоидного происхождения, рака яичников, рака поджелудочной железы, рака предстательной железы, почечно-клеточной карциномы, рака щитовидной железы, рака яичка и рака матки.
| DE 10337368 A1, 03.03.2005 | |||
| WO 2006084075 A2, 10.08.2006 | |||
| LUCIE PEDUTO, ADAM9 as a Potential Target Molecule in Cancer, Current Pharmaceutical Design, 2009, 15, 2282-2287 | |||
| KOHLER G | |||
| et al., Continuous cultures of fused cells secreting antibody of predefined specificity, Nature, 1975, Vol.256, N.5517, pp.495-497 | |||
| СТАБИЛЬНЫЕ И РАСТВОРИМЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ VEGF | 2009 |
|
RU2588467C2 |

