Область техники
Настоящее изобретение относится к фармацевтическим композициям, содержащим в качестве активного ингредиента антигенсвязывающую молекулу, которая индуцирует иммунный ответ на антиген-мишень, или к терапевтическим способам, в которых используются эти фармацевтические композиции. Настоящее изобретение также относится к фармацевтическим композициям, содержащим в качестве активного ингредиента антигенсвязывающую молекулу, которая индуцирует упомянутый выше иммунный ответ и также обладает цитотоксичностью (цитотоксической активностью) или антипролиферативным действием (активность ингибирования пролиферации клеток) против клеток, экспрессирующих антиген-мишень, или к терапевтическим способам, в которых используются фармацевтические композиции.
Уровень техники
До настоящего времени были предприняты попытки разработать ряд терапевтических вакцин, направленных на опухолевые клетки. Это является следствием того, что считается, что существуют качественные и количественные различия между опухолевыми клетками и нормальными клетками, которые могут распознаваться иммунной системой живого организма, и иммунная система, стимулированная посредством активной и специфической сенсибилизацией вакцинами, в которых используются такие различия (неоэпитопы), может распознавать и устранять опухолевые клетки.
Для достижения такого противоопухолевого ответа должны удовлетворяться по меньшей мере два условия. Во-первых, опухолевые клетки должны экспрессировать антиген, который не встречается в нормальных клетках, или экспрессируют антиген в такой степени, чтобы нормальные клетки и опухолевые клетки могли различаться только качественно. Во-вторых, иммунная система должна активироваться вакцинами или сходными с ними для взаимодействия с представляющим интерес антигеном. Основным препятствием для иммунотерапии опухоли считается то, что иммуногенность опухолей является особенно слабой у человека.
В последние годы были открыты связанные с опухолью или опухолеспецифические антигены, включающие такие неоэпитопы, которые могут представлять собой мишени для атаки иммунной системы. Тем не менее, иммунная система не может устранить опухоли, экспрессирующие такие неоэпитопы, и это может быть следствием недостаточного иммунного ответа на эти неоэпитопы, а не следствием отсутствия неоэпитопов.
Для клеточной иммунотерапии злокачественных опухолей было разработано две общих стратегии. Одной из них является адоптивная иммунотерапия, где реактивные в отношении опухоли T-лимфоциты, увеличенные в количестве in vitro, повторно вводят пациенту, а другой является активная иммунотерапия, в которой используются опухолевые клетки для индукции системного ответа на опухоль путем запуска нового или более сильного иммунного ответа на опухолевый антиген.
Вакцины против опухоли на основе активной иммунотерапии получают различными способами. Для индукции иммунного ответа на опухолевый антиген получают облученные опухолевые клетки, смешанные со стимулирующим иммунитет адъювантом, таким как бацилла Кальмета-Герена (BCG) (непатентный документ 1), опухолевые клетки, генетически модифицированные для продуцирования, например, цитокинов (непатентный документ 2), и отчужденные аутологичные опухолевые клетки (непатентный документ 3). Однако иммуногенность опухолевых клеток является низкой, и это считается следствием количества опухолевого антигена, но не его качества.
С другой стороны, известно, что антитела индуцируют гуморальный иммунный ответ (продукция антител против антигена) и клеточный иммунный ответ (продукция CD8-положительных T-клеток против антигена) на антигены посредством перекрестного представления связанных антигенов антигенпредставляющим клеткам, и было описано, что введение антитела может индуцировать приобретенный иммунитет на антиген (непатентный документ 4). Недавно для противоопухолевого эффекта антитела против HER2 в модели на мышах in vivo было показано, что приобретенный иммунитет на антиген HER2, индуцированный введением антитела, играет более важную роль, чем прямая ADCC вводимого антитела (непатентный документ 5). В действительности, при клиническом применении герцептина, который представляет собой лекарственное средство на основе антитела подкласса IgG1 против HER2, при введении герцептина индуцировался приобретенный иммунитет, и наблюдали гуморальный иммунный ответ на HER2 (непатентный документ 6). Поскольку у пациентов, у которых было эффективным введение герцептина, в частности, был продемонстрирован увеличенный титр антител против HER2, считали, что индукция приобретенного иммунитета при введении герцептина играет важную роль в противоопухолевом эффекте.
Антитела являются высокостабильными в крови и имеют мало побочных эффектов, и, таким образом, привлекают внимание в качестве фармацевтических средств (непатентные документы 7 и 8). До настоящего времени было проведено множество исследований антителозависимой клеточной цитотоксичности (далее обозначаемой как ADCC) и комплементзависимой цитотоксичности (далее обозначаемой как CDC), которые являются эффекторными функциями антител класса IgG. Было описано, что в классе IgG человека, антитела полкласса IgG1 обладают наиболее высокой активностью ADCC и активностью CDC (непатентный документ 9). Более того, также полагают, что одной из эффекторных функций антител является антителозависимый клеточно-опосредуемый фагоцитоз (ADCP), который представляет собой фагоцитоз клеток-мишеней, опосредуемый антителами класса IgG (непатентные документы 10 и 11). Поскольку антитела подкласса IgG1 могут индуцировать эти эффекторные функции против опухолей, антитела подкласса IgG1 используют для большинства фармацевтических средств против антигенов злокачественной опухоли.
Для того, чтобы IgG-антитела опосредовали активность ADCC и ADCP, Fc-область IgG-антитела должна связываться с рецепторами антител (далее обозначаемыми как FcγR), которые присутствуют на поверхности эффекторных клеток, таких как киллерные клетки, натуральные киллерные клетки и активированные макрофаги. У человека описаны изоформы FcγRIa, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb в качестве представителей семейства FcγR, и также описаны их соответствующие аллотипы (непатентный документ 12).
Усиление цитотоксических эффекторных функций, таких как ADCC и ADCP, привлекло внимание в качестве перспективных средств для усиления противоопухолевых эффектов антител против злокачественной опухоли. Важность опосредуемых FcγR эффекторных функций, являющихся целью противоопухолевых эффектов антител, описана с использованием моделей на мышах (непатентные документы 13 и 14). Более того, было выявлено, что клинические эффекты у человека коррелировали с высокоаффинным полиморфным аллотипом (V158) и низкоаффинным полиморфным аллотипом (F158) FcγRIIIa (непатентный документ 15). В этих сообщениях указано, что антитела с Fc-областью, оптимизированной для связывания с конкретными FcγR, опосредует более мощные эффекторные функции, и, тем самым, проявляют более эффективные противоопухолевые эффекты. Баланс между аффинностью антител против активирующих рецепторов, включая FcγRIa, FcγRIIa, FcγRIIIa и FcγRIIIb, и ингибирующих рецепторов, включающих FcγRIIb, является важным фактором при оптимизации эффекторных функций антител. Повышение аффинности к активирующим рецепторам может обеспечить антитела со свойством опосредования более мощных эффекторных функций (непатентный документ 16), и, таким образом, описано в различных сообщениях до настоящего времени в качестве способа модификации с помощью инженерии антител для повышения или усиления противоопухолевой активности фармацевтических средств на основе антител против антигенов злокачественной опухоли.
Что касается связывания между Fc-областью и FcγR, было показано, что несколько остатков в шарнирной области антитела и CH2-домене антитела и цепь сахара, связанная с Asn в положении 297 (нумерация EU) в CH2-домене, являются важными (непатентные документы 9, 17 и 18). Фокусируясь на этих участках связывания, до настоящего времени проводили исследования на мутантах Fc-области, имеющих различные свойства связывания FcγR, и были получены мутанты Fc-области с более высокой аффинностью в отношении активирующего FcγR (патентные документы 1 и 2). Например, Lazar et al. успешно увеличили связывание IgG1 человека с FcγRIIIa человека (V158) приблизительно в 370 раз посредством замены Ser в положении 239, Ala в положении 330 и Ile в положении 332 (нумерация EU) IgG1 человека на Asn, Leu и Glu, соответственно (непатентный документ 19 и патентный документ 2). Соотношение связывания FcγRIIIa и FcγRIIb (соотношение A/I) для этого мутанта было приблизительно в 9 раз выше, чем для дикого типа. Более того, Shinkawa et al. успешно увеличили связывание с FcγRIIIa вплоть до приблизительно 100 раз посредством удаления фукозы из цепи сахара, связанной с Asn в положении 297 (нумерация EU) (непатентный документ 20). Эти способы могут значительно повысить активность IgG1 человека в отношении ADCC по сравнению со встречающимся в природе IgG1 человека.
Хотя, как описано выше, существует много сообщений о способах усиления ADCC с помощью модификации антител способами инженерии, до настоящего времени отсутствуют сообщения о способах модификации антител способами инженерии для усиления или улучшения индукции приобретенного иммунитета посредством введения антитела. Существует сообщение о способах индукции приобретенного иммунитета против антигена злокачественной опухоли, в которых антиген злокачественной опухоли, против которого является желательной индукция приобретенного иммунитета, подвергают слиянию с антителом, которое связывается с рецептором с высоким содержанием маннозы или DEC-205, экспрессируемым на антигенпредставляющих клетках, тем самым, ускоряя включение и представление злокачественного антигена антигенпредставляющими клетками (непатентный документ 21). Однако в этих способах мишенью для связывания антителом не является антиген злокачественной опухоли, как в случае упомянутого выше антитела против HER2. Следовательно, поскольку эти способы индуцируют приобретенный иммунитет против антигена злокачественной опухоли, слитого с самим антителом, антитело само по себе не может связываться с антигеном злокачественной опухоли, и имеет недостаток, состоящий в том, что оно не способно проявлять прямое действие на антиген злокачественной опухоли. Более того, поскольку этот способ индуцирует приобретенный иммунитет не только против антигена злокачественной опухоли, слитого с антителом, но также против самого антитела, используемого для нацеливания на антигенпредставляющие клетки, появляются антитела против лекарственного средства, и это приводит к ослаблению эффектов. Таким образом, этот способ может не быть предпочтительным для терапевтических целей.
В соответствии с вышеуказанным, хотя является желательной индукция приобретенного иммунитета против антигена-мишени посредством введения антигенсвязывающей молекулы, обладающей активностью связывания с антигеном-мишенью, отсутствуют сообщения о способах инженерии для улучшения или усиления приобретенного иммунитета такими способами.
Документы уровня техники
Патентные документы
Патентный документ 1: WO 2000/042072
Патентный документ 2: WO 2006/019447
Непатентные документы
Непатентный документ 1: Oettgen, H. F., and Old, L. J., The history of cancer immunotherapy., Biological Therapy of Cancer (1991) 87-119 DeVita et al. ed.
Непатентный документ 2: Zatloukal K, Schmidt W, Cotten M, Wagner E, Stingl G, Birnstiel ML., Somatic gene therapy for cancer: the utility of transferrinfection in generating 'tumor vaccines'., Gene (1993) 135, 199-207
Непатентный документ 3: Bronte V, Tsung K, Rao JB, Chen PW, Wang M, Rosenberg SA, Restifo NP., IL-2 enhances the function of recombinant poxvirus-based vaccines in the treatment of established pulmonary metastases., J. Immunol. (1995) 154, 5282-5292
Непатентный документ 4: Adams GP, Weiner LM., Monoclonal antibody therapy of cancer., Nat. Biotechnol. (2005) 23, 1147-1157
Непатентный документ 5: Park S, Jiang Z, Mortenson ED, Deng L, Radkevich-Brown O, Yang X, Sattar H, Wang Y, Brown NK, Greene M, Liu Y, Tang J, Wang S, Fu YX., The therapeutic effect of anti-HER2/neu antibody depends on both innate and adaptive immunity., Cancer Cell (2010) 18, 160-170 (2010)
Непатентный документ 6: Taylor C, Hershman D, Shah N, Suciu-Foca N, Petrylak DP, Taub R, Vahdat L, Cheng B, Pegram M, Knutson KL, Clynes R. Augmented HER-2 specific immunity during treatment with trastuzumab and chemotherapy., Clin. Cancer Res, (2007) 13, 5133-43
Непатентный документ 7: anice M Reichert, Clark J Rosensweig, Laura B Faden & Matthew C Dewitz, Monoclonal antibody successes in the clinic., Nat. Biotechnol. (2005) 23, 1073-1078
Непатентный документ 8: avlou AK, Belsey MJ., The therapeutic antibodies market to 2008., Eur. J. Pharm. Biopharm. (2005) 59(3), 389-396
Непатентный документ 9: Clark, M., Antibody Engineering IgG Effector Mechanisms., Chemical Immunology (1997), 65, 88-110
Непатентный документ 10: Horton HM, Bernett MJ, Pong E, Peipp M, Karki S, Chu SY, Richards JO, Vostiar I, Joyce PF, Repp R, Desjarlais JR, Zhukovsky EA., Potent in vitro and in vivo activity of an Fc-engineered anti-CD19 monoclonal antibody against lymphoma and leukemia., Cancer Res. (2008) 68, 8049-8057
Непатентный документ 11: Zalevsky J, Leung IW, Karki S, Chu SY, Zhukovsky EA, Desjarlais JR, Carmichael DF, Lawrence CE., The impact of Fc engineering on an anti-CD19 antibody: increased Fcγ receptor affinity enhances B-cell clearing in nonhuman primates., Blood (2009) 113, 3735-3743
Непатентный документ 12: Jefferis R, Lund J., Interaction sites on human IgG-Fc for FcgammaR: current models., Immunol. Lett. (2002) 82, 57-65
Непатентный документ 13: Clynes, R., Yoshizumi, T., Moroi, Y., Houghton, A.N., and Ravetch, J.V., Fc Receptors are required for passive and active immunity to melanoma., Proc. Natl. Acad. Sci. U. S. A. (1998) 95, 652-656
Непатентный документ 14: Clynes RA, Towers TL, Presta LG, Ravetch JV., Inhibitory Fc receptors modulate in vivo cytoxicity against tumor targets., Nat. Med. (2000) 6, 443-446
Непатентный документ 15: Cartron G, Dacheux L, Salles G, Solal-Celigny P, Bardos P, Colombat P, Watier H., Therapeutic activity of humanized anti-CD20 monoclonal antibody and polymorphism in IgG Fc receptor FcgammaRIIIa gene., Blood (2002) 99, 754-758
Непатентный документ 16: Nimmerjahn F, Ravetch JV., Divergent immunoglobulin g subclass activity through selective Fc receptor binding., Science (2005), 310, 1510-1512
Непатентный документ 17: Greenwood J, Clark M, Waldmann H., Structural motifs involved in human IgG antibody effector functions., Eur. J. Immunol. 23, 1098-1104 (1993)
Непатентный документ 18: Morgan A, Jones ND, Nesbitt AM, Chaplin L, Bodmer MW, Emtage JS., The N-terminal end of the CH2 domain of chimeric human IgG1 anti-HLA-DR is necessary for C1q, Fc gamma RI and Fc gamma RIII binding., Immunology (1995) 86, 319-324
Непатентный документ 19: Lazar GA, Dang W, Karki S, Vafa O, Peng JS, Hyun L, Chan C, Chung HS, Eivazi A, Yoder SC, Vielmetter J, Carmichael DF, Hayes RJ, Dahiyat BI., Engineered antibody Fc variants with enhanced effector function., Proc. Natl. Acad. Sci. U. S. A. (2006) 103, 4005-4010
Непатентный документ 20: Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, Uchida K, Anazawa H, Satoh M, Yamasaki M, Hanai N, Shitara K., The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity., J. Biol. Chem. (2003) 278, 3466-3473
Непатентный документ 21: Tsuji T, Matsuzaki J, Kelly MP, Ramakrishna V, Vitale L, He LZ, Keler T, Odunsi K, Old LJ, Ritter G, Gnjatic S., Antibody-Targeted NY-ESO-1 to Mannose Receptor or DEC-205 In Vitro Elicits Dual Human CD8+ and CD4+ T Cell Responses with Broad Antigen Specificity., J. Immunol. (2011) 186, 1218-1827
Сущность изобретения
Проблемы, решаемые с помощью изобретения
Настоящее изобретение было осуществлено с учетом описанных выше обстоятельств. Задачей настоящего изобретения является предоставление фармацевтических композиций, содержащих в качестве активного ингредиента антигенсвязывающую молекулу, которая индуцирует иммунный ответ у индивидуумов, страдающих злокачественной опухолью или инфицированных чужеродными биологическими видами при введении этим индивидуумам, или терапевтических способов, в которых используются эти фармацевтические композиции. Другой задачей является предоставление фармацевтических композиций, содержащих в качестве активного ингредиента антигенсвязывающую молекулу, которая индуцирует упомянутый выше иммунный ответ и также обладает цитотоксичностью (цитотоксическая активность) или антипролиферативным действием (активность ингибирования пролиферации клеток) против злокачественных клеток или инфицирующих чужеродных биологических видов, или терапевтических способов, в которых используются фармацевтические композиции.
Способы решения проблем
Авторы настоящего изобретения открыли, что в живых организмах, в которые вводили антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, активность связывания которого с антигеном изменяется в зависимости от условий концентрации ионов, и содержащую FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH, индуцируется иммунный ответ на антиген. Более того, авторы настоящего изобретения открыли, что в живых организмах, которым вводили антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH, индуцируется иммунный ответ на антиген, и антигенсвязывающая молекула также может обладать цитотоксичностью или антипролиферативным эффектом против злокачественных клеток, чужеродных биологических видов и т.п., которые экспрессируют антиген, с которым связывается антигенсвязывающая молекула. Исходя из этих открытий авторы настоящего изобретения установили, что антигенсвязывающие молекулы по настоящему изобретению пригодны в качестве фармацевтических композиций для индукции иммунного ответа у индивидуума, инфицированного чужеродными биологическими видами или страдающего злокачественной опухолью при введении индивидууму. Авторы настоящего изобретения также установили, что антигенсвязывающие молекулы по настоящему изобретению пригодны в качестве фармацевтических композиций, которые при введении индивидууму, инфицированному чужеродными биологическими видами или страдающему злокачественной опухолью, индуцируют иммунный ответ у индивидуума и также обладают цитотоксичностью или антипролиферативным эффектом против злокачественных клеток и чужеродных биологических видов. Также были разработаны способы получения этих фармацевтических композиций.
Более конкретно, настоящее изобретение относится к [1]-[47] ниже:
[1] фармацевтическая композиция, которая индуцирует иммунный ответ на антиген, которая содержит в качестве активного ингредиента антигенсвязывающую молекулу, где антигенсвязывающая молекула содержит антигенсвязывающий домен, активность связывания которого с антигеном изменяется в зависимости от условий концентрации ионов, и содержит FcRn-связывающий домен, обладающий активностью связывания с FcRn в диапазоне нейтральных значений pH;
[2] фармацевтическая композиция согласно [1], где концентрация ионов представляет собой концентрацию ионов кальция;
[3] фармацевтическая композиция согласно [2], где антигенсвязывающий домен представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого является более высокой в условиях высокой концентрации ионов кальция, чем в условиях низкой концентрации ионов кальция;
[4] фармацевтическая композиция согласно [1], где условия концентрации ионов представляют собой условия pH;
[5] фармацевтическая композиция согласно [4], где антигенсвязывающий домен представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого является более высокой в диапазоне нейтральных значений pH, чем в диапазоне кислых значений pH;
[6] фармацевтическая композиция согласно любому из [1]-[5], где антигенсвязывающая молекула обладает нейтрализующей активностью в отношении антигена;
[7] фармацевтическая композиция согласно любому из [1]-[6], где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген;
[8] фармацевтическая композиция согласно любому из [1]-[7], где FcRn-связывающий домен содержит Fc-область антитела;
[9] фармацевтическая композиция согласно [8], где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области;
[10] фармацевтическая композиция согласно [8] или [9], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области;
[11] фармацевтическая композиция согласно любому из [8]-[10], где активность Fc-области в отношении связывания Fcγ-рецептора является более высокой, чем у встречающейся в природе Fc-области IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[12] фармацевтическая композиция согласно [11], где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) и FcγRIIIa(F);
[13] фармацевтическая композиция согласно [11] или [12], где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области;
[14] фармацевтическая композиция согласно любому из [11]-[13], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
либо Lys, либо Tyr в положении аминокислоты 221;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 222;
любой из Phe, Trp, Glu и Lys в положении аминокислоты 223;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 224;
любой из Glu, Lys и Trp в положении аминокислоты 225;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 227;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 228;
любой из Ala, Glu, Gly и Tyr в положении аминокислоты 230;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 231;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 232;
любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 233;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 234;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 235;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 236;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 237;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 238;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 239;
любой из Ala, Ile, Met и Thr в положении аминокислоты 240;
любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении аминокислоты 241;
любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении аминокислоты 243;
His в положении аминокислоты 244;
Ala в положении аминокислоты 245;
любой из Asp, Glu, His и Tyr в положении аминокислоты 246;
любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении аминокислоты 247;
любой из Glu, His, Gln и Tyr в положении аминокислоты 249;
либо Glu, либо Gln в положении аминокислоты 250;
Phe в положении аминокислоты 251;
любой из Phe, Met и Tyr в положении аминокислоты 254;
любой из Glu, Leu и Tyr в положении аминокислоты 255;
любой из Ala, Met и Pro в положении аминокислоты 256;
любой из Asp, Glu, His, Ser и Tyr в положении аминокислоты 258;
любой из Asp, Glu, His и Tyr в положении аминокислоты 260;
любой из Ala, Glu, Phe, Ile и Thr в положении аминокислоты 262;
любой из Ala, Ile, Met и Thr в положении аминокислоты 263;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 264;
любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении аминокислоты 265;
любой из Ala, Ile, Met и Thr в положении аминокислоты 266;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 267;
любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении аминокислоты 268;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 269;
любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 270;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 271;
любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 272;
либо Phe, либо Ile в положении аминокислоты 273;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 274;
либо Leu, либо Trp в положении аминокислоты 275;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 276;
любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 278;
Ala в положении аминокислоты 279;
любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении аминокислоты 280;
любой из Asp, Lys, Pro и Tyr в положении аминокислоты 281;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 282;
любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении аминокислоты 283;
любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении аминокислоты 284;
любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении аминокислоты 285;
любой из Glu, Gly, Pro и Tyr в положении аминокислоты 286;
любой из Asn, Asp, Glu и Tyr в положении аминокислоты 288;
любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении аминокислоты 290;
любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении аминокислоты 291;
любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении аминокислоты 292;
любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 293;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 294;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 295;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении аминокислоты 296;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 297;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 298;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 299;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 300;
любой из Asp, Glu, His и Tyr в положении аминокислоты 301;
Ile в положении аминокислоты 302;
любой из Asp, Gly и Tyr в положении аминокислоты 303;
любой из Asp, His, Leu, Asn и Thr в положении аминокислоты 304;
любой из Glu, Ile, Thr и Tyr в положении аминокислоты 305;
любой из Ala, Asp, Asn, Thr, Val и Tyr в положении аминокислоты 311;
Phe в положении аминокислоты 313;
Leu в положении аминокислоты 315;
либо Glu, либо Gln в положении аминокислоты 317;
любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении аминокислоты 318;
любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 320;
любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 322;
Ile в положении аминокислоты 323;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 324;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 325;
любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 326;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 327;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 328;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 329;
любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 330;
любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 331;
любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 332;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении аминокислоты 333;
любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении аминокислоты 334;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 335;
любой из Glu, Lys и Tyr в положении аминокислоты 336;
любой из Glu, His и Asn в положении аминокислоты 337;
любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении аминокислоты 339;
либо Ala, либо Val в положении аминокислоты 376;
либо Gly, либо Lys в положении аминокислоты 377;
Asp в положении аминокислоты 378;
Asn в положении аминокислоты 379;
любой из Ala, Asn и Ser в положении аминокислоты 380;
либо Ala, либо Ile в положении аминокислоты 382;
Glu в положении аминокислоты 385;
Thr в положении аминокислоты 392;
Leu в положении аминокислоты 396;
Lys в положении аминокислоты 421;
Asn в положении аминокислоты 427;
либо Phe, либо Leu в положении аминокислоты 428;
Met в положении аминокислоты 429;
Trp в положении аминокислоты 434;
Ile в положении аминокислоты 436; и
любой из Gly, His, Ile, Leu и Tyr в положении аминокислоты 440;
в соответствии с нумерацией EU в Fc-области;
[15] фармацевтическая композиция согласно любому из [11]-[14], где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[16] фармацевтическая композиция согласно любому из [11]-[15], где Fc-область модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким;
[17] способ индукции иммунного ответа в живом организме, который включает стадию введения антигенсвязывающей молекулы согласно любому из [1]-[16] в живой организм;
[18] способ получения антигенсвязывающей молекулы, которая индуцирует иммунный ответ, который включает сообщение активности связывания FcRn в диапазоне нейтральных значений pH с FcRn-связывающим доменом, который содержится в антигенсвязывающей молекуле, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
[19] способ согласно [18], где концентрация ионов представляет собой концентрацию ионов кальция;
[20] способ согласно [19], где антигенсвязывающий домен представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого является более высокой в условиях высокой концентрации ионов кальция, чем в условиях низкой концентрации ионов кальция;
[21] способ согласно [18], где условия концентрации ионов представляют собой условия pH;
[22] способ согласно [21], где антигенсвязывающий домен представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого является более высокой в диапазоне нейтральных значений pH, чем в диапазоне кислых значений pH;
[23] способ согласно любому из [18]-[22], где антигенсвязывающая молекула обладает нейтрализующей активностью в отношении антигена;
[24] способ согласно любому из [18]-[23], где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген;
[25] способ согласно любому из [18]-[24], где FcRn-связывающий домен содержит Fc-область антитела;
[26] способ согласно [25], включающий стадию замены по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 239, 252, 257, 286, 307, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области;
[27] способ согласно [25] или [26], включающий стадию проведения по меньшей мере одной или нескольких аминокислотных замен, выбранных из группы, состоящей из:
аминокислотная замена на Ala в положении 257;
аминокислотная замена на Pro в положении 308;
аминокислотная замена на Leu в положении 428; и
аминокислотная замена на Tyr в положении 434,
в соответствии с нумерацией EU в Fc-области;
[28] способ согласно любому из [25]-[27], включающий стадию повышения активности Fc-области в отношении связывания Fcγ-рецептора по сравнению со встречающейся в природе Fc-областью IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[29] способ согласно [28], где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) и FcγRIIIa(F);
[30] способ согласно [28] или [29], включающий стадию замены по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU в Fc-области;
[31] способ согласно любому из [28]-[30], включающий стадию проведения по меньшей мере одной или нескольких замен аминокислот, выбранных из группы, состоящей из:
аминокислотная замена либо на Lys, либо на Tyr в положении 221;
аминокислотная замена на любой из Phe, Trp, Glu и Tyr в положении 222;
аминокислотная замена на любой из Phe, Trp, Glu и Lys в положении 223;
аминокислотная замена на любой из Phe, Trp, Glu и Tyr в положении 224;
аминокислотная замена на любой из Glu, Lys и Trp в положении 225;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 227;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 228;
аминокислотная замена на любой из Ala, Glu, Gly и Tyr в положении 230;
аминокислотная замена на любой из Glu, Gly, Lys, Pro и Tyr в положении 231;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 232;
аминокислотная замена на любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 233;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 234;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 235;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 236;
аминокислотная замена на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 237;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 238;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении 239;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 240;
аминокислотная замена на любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении 241;
аминокислотная замена на любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении 243;
аминокислотная замена на His в положении 244;
аминокислотная замена на Ala в положении 245;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 246;
аминокислотная замена на любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении 247;
аминокислотная замена на любой из Glu, His, Gln и Tyr в положении 249;
аминокислотная замена либо на Glu, либо на Gln в положении 250;
аминокислотная замена на Phe в положении 251;
аминокислотная замена на любой из Phe, Met и Tyr в положении 254;
аминокислотная замена на любой из Glu, Leu и Tyr в положении 255;
аминокислотная замена на любой из Ala, Met и Pro в положении 256;
аминокислотная замена на любой из Asp, Glu, His, Ser и Tyr в положении 258;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 260;
аминокислотная замена на любой из Ala, Glu, Phe, Ile и Thr в положении 262;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 263;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении 264;
аминокислотная замена на любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении 265;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 266;
аминокислотная замена на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении 267;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении 268;
аминокислотная замена на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 269;
аминокислотная замена на любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении 270;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 271;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 272;
аминокислотная замена либо на Phe, либо на Ile в положении 273;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 274;
аминокислотная замена либо на Leu, либо на Trp в положении 275;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 276;
аминокислотная замена на любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении 278;
аминокислотная замена на Ala в положении 279;
аминокислотная замена на любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении 280;
аминокислотная замена на любой из Asp, Lys, Pro и Tyr в положении 281;
аминокислотная замена на любой из Glu, Gly, Lys, Pro и Tyr в положении 282;
аминокислотная замена на любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении 283;
аминокислотная замена на любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении 284;
аминокислотная замена на любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении 285;
аминокислотная замена на любой из Glu, Gly, Pro и Tyr в положении 286;
аминокислотная замена на любой из Asn, Asp, Glu и Tyr в положении 288;
аминокислотная замена на любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении 290;
аминокислотная замена на любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении 291;
аминокислотная замена на любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении 292;
аминокислотная замена на любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 293;
аминокислотная замена на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 294;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 295;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении 296;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 297;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении 298;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении 299;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении 300;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 301;
аминокислотная замена на Ile в положении 302;
аминокислотная замена на любой из Asp, Gly и Tyr в положении 303;
аминокислотная замена на любой из Asp, His, Leu, Asn и Thr в положении 304;
аминокислотная замена на любой из Glu, Ile, Thr и Tyr в положении 305;
аминокислотная замена на любой из Ala, Asp, Asn, Thr, Val и Tyr в положении 311;
аминокислотная замена на Phe в положении 313;
аминокислотная замена на Leu в положении 315;
аминокислотная замена либо на Glu, либо на Gln в положении 317;
аминокислотная замена на любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении 318;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении 320;
аминокислотная замена на любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении 322;
аминокислотная замена на Ile в положении 323;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении 324;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 325;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении 326;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении 327;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 328;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 329;
аминокислотная замена на любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 330;
аминокислотная замена на любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении 331;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 332;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении 333;
аминокислотная замена на любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении 334;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении 335;
аминокислотная замена на любой из Glu, Lys и Tyr в положении 336;
аминокислотная замена на любой из Glu, His и Asn в положении 337;
аминокислотная замена на любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении 339;
аминокислотная замена либо на Ala, либо на Val в положении 376;
аминокислотная замена либо на Gly, либо на Lys в положении 377;
аминокислотная замена на Asp в положении 378;
аминокислотная замена на Asn в положении 379;
аминокислотная замена на любой из Ala, Asn и Ser в положении 380;
аминокислотная замена либо на Ala, либо на Ile в положении 382;
аминокислотная замена на Glu в положении 385;
аминокислотная замена на Thr в положении 392;
аминокислотная замена на Leu в положении 396;
аминокислотная замена на Lys в положении 421;
аминокислотная замена на Asn в положении 427;
аминокислотная замена либо на Phe, либо на Leu в положении 428;
аминокислотная замена на Met в положении 429;
аминокислотная замена на Trp в положении 434;
аминокислотная замена на Ile в положении 436; и
аминокислотная замена на любой из Gly, His, Ile, Leu и Tyr в положении 440,
в соответствии с нумерацией EU в Fc-области;
[32] способ согласно любому из [28]-[31], где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[33] способ согласно любому из [28]-[32], включающий стадию модификации Fc-области так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким;
[34] способ получения фармацевтической композиции, которая индуцирует иммунный ответ, который включает стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в условиях высокой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в условиях низкой концентрации ионов кальция;
(c) выбор антигенсвязывающего домена, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, выбранный согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e);
[35] способ получения фармацевтической композиции, которая индуцирует иммунный ответ, который включает стадии:
(a) определение антигенсвязывающей активности антитела в условиях высокой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антитела в условиях низкой концентрации ионов кальция;
(c) выбор антитела, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен антитела, выбранного согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e);
[36] способ получения антигенсвязывающей молекулы, который включает стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в диапазоне нейтральных значений pH;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в диапазоне кислых значений pH;
(c) выбор антигенсвязывающего домена, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, выбранный согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e);
[37] способ получения антигенсвязывающей молекулы, который включает стадии:
(a) определение антигенсвязывающей активности антитела в диапазоне нейтральных значений pH;
(b) определение антигенсвязывающей активности антитела в диапазоне кислых значений pH;
(c) выбор антитела, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен антитела, выбранного согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e);
[38] способ согласно любому из [34]-[37], где антигенсвязывающая молекула обладает нейтрализующей активностью против антигена;
[39] способ согласно любому из [34]-[38], где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген;
[40] способ согласно любому из [34]-[39], где FcRn-связывающий домен содержит Fc-область антитела;
[41] способ согласно [40], где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области;
[42] способ согласно [40] или [41], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области;
[43] способ согласно любому из [40]-[42], где активность Fc-области в отношении связывания Fcγ-рецептора является более высокой, чем у встречающейся в природе Fc-области IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[44] способ согласно [43], где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) и FcγRIIIa(F);
[45] способ согласно [43] или [44], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
либо Lys, либо Tyr в положении аминокислоты 221;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 222;
любой из Phe, Trp, Glu и Lys в положении аминокислоты 223;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 224;
любой из Glu, Lys и Trp в положении аминокислоты 225;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 227;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 228;
любой из Ala, Glu, Gly и Tyr в положении аминокислоты 230;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 231;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 232;
любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 233;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 234;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 235;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 236;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 237;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 238;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 239;
любой из Ala, Ile, Met и Thr в положении аминокислоты 240;
любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении аминокислоты 241;
любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении аминокислоты 243;
His в положении аминокислоты 244;
Ala в положении аминокислоты 245;
любой из Asp, Glu, His и Tyr в положении аминокислоты 246;
любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении аминокислоты 247;
любой из Glu, His, Gln и Tyr в положении аминокислоты 249;
либо Glu, либо Gln в положении аминокислоты 250;
Phe в положении аминокислоты 251;
любой из Phe, Met и Tyr в положении аминокислоты 254;
любой из Glu, Leu и Tyr в положении аминокислоты 255;
любой из Ala, Met и Pro в положении аминокислоты 256;
любой из Asp, Glu, His, Ser и Tyr в положении аминокислоты 258;
любой из Asp, Glu, His и Tyr в положении аминокислоты 260;
любой из Ala, Glu, Phe, Ile и Thr в положении аминокислоты 262;
любой из Ala, Ile, Met и Thr в положении аминокислоты 263;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 264;
любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении аминокислоты 265;
любой из Ala, Ile, Met и Thr в положении аминокислоты 266;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 267;
любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении аминокислоты 268;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 269;
любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 270;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 271;
любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 272;
либо Phe, либо Ile в положении аминокислоты 273;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 274;
либо Leu, либо Trp в положении аминокислоты 275;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 276;
любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 278;
Ala в положении аминокислоты 279;
любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении аминокислоты 280;
любой из Asp, Lys, Pro и Tyr в положении аминокислоты 281;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 282;
любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении аминокислоты 283;
любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении аминокислоты 284;
любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении аминокислоты 285;
любой из Glu, Gly, Pro и Tyr в положении аминокислоты 286;
любой из Asn, Asp, Glu и Tyr в положении аминокислоты 288;
любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении аминокислоты 290;
любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении аминокислоты 291;
любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении аминокислоты 292;
любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 293;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 294;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 295;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении аминокислоты 296;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 297;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 298;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 299;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 300;
любой из Asp, Glu, His и Tyr в положении аминокислоты 301;
Ile в положении аминокислоты 302;
любой из Asp, Gly и Tyr в положении аминокислоты 303;
любой из Asp, His, Leu, Asn и Thr в положении аминокислоты 304;
любой из Glu, Ile, Thr и Tyr в положении аминокислоты 305;
любой из Ala, Asp, Asn, Thr, Val и Tyr в положении аминокислоты 311;
Phe в положении аминокислоты 313;
Leu в положении аминокислоты 315;
либо Glu, либо Gln в положении аминокислоты 317;
любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении аминокислоты 318;
любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 320;
любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 322;
Ile в положении аминокислоты 323;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 324;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 325;
любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 326;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 327;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 328;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 329;
любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 330;
любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 331;
любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 332;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении аминокислоты 333;
любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении аминокислоты 334;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 335;
любой из Glu, Lys и Tyr в положении аминокислоты 336;
любой из Glu, His и Asn в положении аминокислоты 337;
любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении аминокислоты 339;
либо Ala, либо Val в положении аминокислоты 376;
либо Gly, либо Lys в положении аминокислоты 377;
Asp в положении аминокислоты 378;
Asn в положении аминокислоты 379;
любой из Ala, Asn и Ser в положении аминокислоты 380;
либо Ala, либо Ile в положении аминокислоты 382;
Glu в положении аминокислоты 385;
Thr в положении аминокислоты 392;
Leu в положении аминокислоты 396;
Lys в положении аминокислоты 421;
Asn в положении аминокислоты 427;
либо Phe, либо Leu в положении аминокислоты 428;
Met в положении аминокислоты 429;
Trp в положении аминокислоты 434;
Ile в положении аминокислоты 436; и
любой из Gly, His, Ile, Leu и Tyr в положении аминокислоты 440;
в соответствии с нумерацией EU в Fc-области;
[46] способ согласно любому из [43]-[45], где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров;
[47] способ согласно любому из [43]-[46], где Fc-область модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким.
Эффекты изобретения
Настоящее изобретение относится к фармацевтическим композициям, содержащим антигенсвязывающую молекулу, которая при введении в живой организм не только может проявлять фармакологические действия на антиген-мишень, но также индуцирует иммунный ответ на антиген-мишень, который не был возможен в случае общепринятых антител, и к способам их изготовления. Это позволяет эффективное лечение злокачественной опухоли и инфекционных заболеваний посредством индукции иммунного ответа антиген-мишень при наличии активности связывания с антигеном-мишенью и наличии цитотоксичности и антипролиферативной активности против клеток-мишеней, которые не были возможными в случае общепринятых вакцин.
Краткое описание чертежей
На фиг. 1 представлены изменения концентрации растворимого рецептора IL-6 человека в плазме мышей для группы введения антитела против CD4 мыши и группы без введения в стационарной модели для растворимого рецептора IL-6 человека. На горизонтальной оси представлено количество суток от введения антитела против CD4 мыши и на вертикальной оси представлена концентрация в плазме растворимого рецептора IL-6 человека.
На фиг. 2 представлены изменения концентрации растворимого рецептора IL-6 человека в плазме мышей для групп введения нормального антитела против рецептора IL-6 и pH-зависимого антитела против рецептора IL-6 в модели на нормальных мышах для иммунной толерантности к рецептору IL-6 человека. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6 и на вертикальной оси представлена концентрация в плазме растворимого рецептора IL-6 человека. Закрашенные круги указывают на концентрацию в плазме растворимого рецептора IL-6 человека у контрольных мышей. Незакрашенные круги указывают на концентрацию в плазме растворимого рецептора IL-6 человека у мышей, которым вводили H54/L28-IgG1, и ромбы указывают на концентрацию в плазме растворимого рецептора IL-6 человека у мышей, которым вводили Fv4-IgG1.
На фиг. 3 представлены изменения концентрации растворимого рецептора IL-6 человека у мышей для группы, в которой вводили нормальное антитело против рецептора IL-6 с усиленным связыванием FcRn при pH7,4, и группы, в которой вводили pH-зависимое антитело против рецептора IL-6 с усиленным связыванием FcRn при pH7,4, в модели на нормальных мышах для иммунной толерантности к рецептору IL-6 человека. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6, и на вертикальной оси представлена концентрация в плазме растворимого рецептора IL-6 человека. Закрашенные круги указывают на концентрацию в плазме растворимого рецептора IL-6 человека у контрольных мышей. Концентрация в плазме растворимого рецептора IL-6 человека у мышей, которым вводили H54/L28-IgG1, Fv4-IgG1, H54/L28-F157 или Fv4-F157, показана с помощью незакрашенных кругов, ромбов, незакрашенных треугольников или X и закрашенных квадратов, соответственно.
На фиг. 4 представлена неограниченная модель действия для зависимой от концентрации ионов антигенсвязывающей молекулы в отношении транспорта в лизосому растворимого антигена. В условиях концентрации ионов в плазме (в диапазоне нейральных значений pH или при высокой концентрации ионов кальция), антигенсвязывающая молекула, которая связалась с растворимым антигеном в плазме (A), захватывается в клетку посредством неспецифического эндоцитоза и т.п. (B), а затем переносится в кислую эндосому, где она связывается с FcRn, экспрессируемым в эндосоме через FcRn-связывающий домен в условиях кислых значений pH и высвобождает антиген в условиях эндосомальной концентрации ионов (в диапазоне кислых значений pH или при низкой концентрации ионов кальция) (C). Высвобожденный антиген транспортируется в лизосому, а затем деградируется (D). С другой стороны, антигенсвязывающая молекула, которая высвободила антиген, перемещается к поверхности клетки, будучи связанной с FcRn, диссоциирует от FcRn в условиях нейтральных значений pH в плазме, а затем возвращается в плазму (E).
На фиг. 5 представлена неограниченная модель действия для антигенсвязывающей молекулы, обладающей активностью связывания FcRn при pH7,4, в отношении транспорта в лизосому растворимого антигена. В условиях концентрации ионов в плазме (в диапазоне нейтральных значений pH или при высокой концентрации ионов кальция), антигенсвязывающая молекула, которая связалась с растворимым антигеном в плазме, связывается с FcRn в условиях нейтральных значений pH через FcRn-связывающий домен (A), а затем она захватывается в клетку посредством эндоцитоза (B). Антигенсвязывающая молекула, которая транспортировалась в кислую эндосому, не высвобождает антиген в условиях концентрации ионов в эндосоме (в диапазоне кислых значений pH или при низкой концентрации ионов кальция) (C), и связанная с антигеном антигенсвязывающая молекула рециклирует на клеточную поверхность, будучи связанной с FcRn (D).
На фиг. 6 представлена неограниченная модель действия для зависимой от концентрации ионов антигенсвязывающей молекулы с усиленным связыванием FcRn при pH7,4 в отношении транспорта в лизосому растворимого антигена. В условиях концентрации ионов в плазме (в диапазоне нейтральных значений pH или при высокой концентрации ионов кальция), антигенсвязывающая молекула, которая связалась с растворимым антигеном в плазме, связывается с FcRn в условиях нейтральных значений pH через FcRn-связывающий домен (A), а затем она может захватываться в клетку посредством эндоцитоза (B). Антигенсвязывающая молекула, которая транспортировалась в кислотную эндосому, высвобождает антиген в условиях концентрации ионов в эндосоме (в диапазоне кислых значений pH или при низкой концентрации ионов кальция) (C). Диссоциировавший антиген переносится в лизосому, а затем деградируется (D). С другой стороны, антигенсвязывающая молекула, которая высвободила антиген, рециклирует на клеточную поверхность, будучи связанной с FcRn (E).
На фиг. 7 представлены изменения концентрации в плазме растворимого рецептора IL-6 человека и изменения титра антитела мыши против рецептора IL-6 человека у каждой из трех мышей, которым вводили Fv4-F157 (#7, 8 и 9) в испытании 1. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6, на левой вертикальной оси представлена концентрация растворимого рецептора IL-6 человека в плазме, а на правой вертикальной оси представлена величина ECL, которая служит в качестве показателя титра антитела мыши против hsIL-6R. Сплошной линией показана концентрация растворимого рецептора IL-6 человека в плазме и пунктирной линией показаны величины ECL. Ромбами, незакрашенными квадратами и треугольниками показана концентрация в плазме растворимого рецептора IL-6 человека индивидуумов #7, 8 и 9, соответственно; и X, закрашенными квадратами и закрашенными кругами показана величина ECL у индивидуумов #7, 8 и 9, соответственно.
На фиг. 8 представлены изменения концентрации в плазме растворимого рецептора IL-6 человека и изменения титра антитела мыши против рецептора IL-6 человека у каждой из трех мышей, которым вводили Fv4-F157 (#10, 11 и 12) в испытании 2. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6, на левой вертикальной оси представлена концентрация растворимого рецептора IL-6 человека в плазме и на правой вертикальной оси представлена величина ECL, которая служит в качестве показателя титра антитела мыши против рецептора IL-6 человека. Сплошной линией показана концентрация растворимого рецептора IL-6 человека в плазме и пунктирной линией показаны величины ECL. Ромбами, незакрашенными квадратами и треугольниками показана концентрация растворимого рецептора IL-6 человека в плазме индивидуумов #10, 11 и 12, соответственно; и X, закрашенными квадратами и закрашенными кругами показаны изменения величины ECL у индивидуумов #10, 11 и 12, соответственно.
На фиг. 9 представлены изменения титра антитела мыши против hsIL-6R и антитела мыши против Fv4-F157 у каждой из трех мышей, которым вводили Fv4-F157 (#7, 8 и 9) из испытания 1. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6 и на вертикальной оси представлены величины ECL, которые служат в качестве показателей титра антител мыши против рецептора IL-6 человека и титра антитела мыши против Fv4-F157. Сплошной линией показаны изменения величины ECL, служащей показателем титра антител мыши против Fv4-F157, и пунктирными линиями показаны изменения величины ECL, служащей показателем титра антител мыши против рецептора IL-6 человека. Ромбами, незакрашенными квадратами и закрашенными треугольниками показаны изменения величины ECL, служащей показателем титра антител мыши против Fv4-F157 у индивидуумов #7, 8 и 9, соответственно; и незакрашенными квадратами, закрашенными квадратами и незакрашенными треугольниками показаны изменения величины ECL, служащей показателем титра антител мыши против рецептора IL-6 человека у индивидуумов #7, 8 и 9, соответственно.
На фиг. 10 представлены изменения титра антитела мыши против IL-6R человека и изменения титра антитела мыши против Fv4-F157 у каждой из трех мышей, которым вводили Fv4-F157 (#10, 11 и 12) из испытания 2. На горизонтальной оси представлено количество суток после введения антитела против рецептора IL-6 и на вертикальной оси представлены величины ECL, которые служат в качестве показателя титра антител мыши против рецептора IL-6 человека и показателя титра антитела мыши против Fv4-F157. Сплошными линиями показаны изменения величины ECL, служащей показателем титра антител мыши против Fv4-F157 и пунктирными линиями показаны изменения величины ECL, служащей показателем титра антител мыши против hsIL-6R. Ромбами, незакрашенными квадратами и закрашенными треугольниками показаны изменения величины ECL, служащей показателем титра антител мыши против Fv4-F157 у индивидуумов #10, 11 и 12, соответственно; и незакрашенными квадратами, закрашенными квадратами и незакрашенными треугольниками показаны изменения величины ECL, служащей показателем титра антитела мыши против рецептора IL-6 человека у индивидуумов #10, 11 и 12, соответственно.
На фиг. 11 представлена неограниченная модель механизма действия антитела, слитого с антигеном-мишенью, на злокачественные клетки и антигенпредставляющие клетки.
На фиг. 12 представлена неограниченная модель механизма действия антигенсвязывающей молекулы на злокачественные клетки и антигенпредставляющие клетки, где антигенсвязывающая молекула обладает активностью связывания FcRn в диапазоне нейтральных значений pH и обладает зависимой от концентрации ионов активностью связывания с антигеном-мишенью.
На фиг. 13 представлен способ взаимодействия между антигеном и антителом с кальций-зависимым связыванием в плазме (2 мМ Ca2+) и в эндосоме (3 мкМ Ca2+) (i) и способ взаимодействия между антигеном и антителом с pH- и кальций-зависимым связыванием в плазме (pH 7,4, 2 мМ Ca2+) и в эндосоме (pH 6,0, 3 мкМ Ca2+) (ii).
На фиг. 14 представлена ионообменная хроматограмма для антитела, содержащего последовательность k5-2 человека, и антитела, содержащего последовательность hVk5-2_L65, которая обладает модифицированной последовательностью гликозилирования в последовательности Vk5-2 человека. Сплошная линия соответствует хроматограмме для антитела, содержащего последовательность Vk5-2 человека (тяжелая цепь: CIM_H, SEQ ID NO:45; и легкая цепь: hVk5-2, SEQ ID NO:50). Прерывистая линия соответствует хроматограмме для антитела, содержащего последовательность hVk5-2_L65 (тяжелая цепь: CIM_H (SEQ ID NO:45); и легкая цепь: hVk5-2_L65 (SEQ ID NO:53)).
На фиг. 15 представлена взаимосвязь модельного распределения аминокислот (обозначенного как модель) и распределения аминокислот (указанного как библиотека) для информации о последовательности из 290 клонов, выделенных из E. coli, в которые была введена генная библиотека генов, которые связываются с антигенами Ca-зависимым образом. На горизонтальной оси показаны положения аминокислот в системе нумерации Kabat. На вертикальной оси указано % распределение аминокислот.
На фиг. 16 представлены сенсограммы для антитела против рецептора IL-6 (тоцилизумаб), антитела 6RC1IgG_010, антитела 6RC1IgG_012 и антитела 6RC1IgG_019 в условиях высокой концентрации ионов кальция (1,2 мМ).
На фиг. 17 представлены сенсограммы для антитела против рецептора IL-6 (тоцилизумаб), антитела 6RC1IgG_010, антитела 6RC1IgG_012 и антитела 6RC1IgG_019 в условиях низкой концентрации ионов кальция (3 мкМ).
На фиг. 18 представлена структура CDR3 тяжелой цепи Fab-фрагмента антитела 6RL#9, определенная посредством рентгеновской кристаллографии. Участок CDR3 тяжелой цепи в кристаллической структуре, полученный при кристаллизации в присутствии ионов кальция, представлен в (i) и участок CDR3 тяжелой цепи в кристаллической структуре, полученный при кристаллизации в отсутствие ионов кальция, представлен в (ii).
На фиг. 19 представлены изменения концентраций антител в плазме у нормальных мышей для антитела H54/L28-IgG1, антитела FH4-IgG1 и антитела 6RL#9-IgG1.
На фиг. 20 представлены изменения концентрации в плазме растворимого рецептора IL-6 человека (hsIL-6R) у нормальных мышей для антитела H54/L28-IgG1, антитела FH4-IgG1 и антитела 6RL#9-IgG1.
На фиг. 21 представлены изменения концентраций антител в плазме для антитела H54/L28-N434W, антитела FH4-N434W и антитела 6RL#9-N434W.
На фиг. 22 представлены изменения концентрации в плазме растворимого рецептора IL-6 человека (hsIL-6R) у нормальных мышей для антитела H54/L28-N434W, антитела FH4-N434W и антитела 6RL#9-N434W.
На фиг. 23 представлена взаимосвязь модельного распределения аминокислот (указанного как модель) и распределения аминокислот (указанного как библиотека) для информации о последовательности из 132 клонов, выделенных из E. coli, в которые была введена генная библиотека антител, которые связываются с антигенами pH-зависимым образом. На горизонтальной оси указаны положения аминокислот в системе нумерации Kabat. На вертикальной оси указано % распределение аминокислот.
На фиг. 24 представлены сенсограммы для антитела против рецептора IL-6 (тоцилизумаб), антитела 6RpH#01, антитела 6RpH#02 и антитела 6RpH#03 при pH 7,4. На горизонтальной оси представлено время, а на вертикальной оси представлена величина RU.
На фиг. 25 представлены сенсограммы для антитела против рецептора IL-6 (тоцилизумаб), антитела 6RpH#01, антитела 6RpH#02 и антитела 6RpH#03 при pH 6,0. На горизонтальной оси представлено время, а на вертикальной оси представлена величина RU.
На фиг. 26 представлены изменения средней концентрации в плазме hsIL-6R в группе без введения антитела и в группах введения Fv4-mIgG1, Fv4-mIgG2a, Fv4-mF3, Fv4-mFa30 и H54/L28-mF3.
На фиг. 27 представлены изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе введения Fv4-mIgG1.
На фиг. 28 представлены изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе введения Fv4-mIgG2a.
На фиг. 29 представлены изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе введения Fv4-mF3.
На фиг. 30 представлены изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе введения Fv4-mFa30.
На фиг. 31 представлены изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе введения H54/L28-mF3.
На фиг. 32A представлена схематическая диаграмма, демонстрирующая взаимосвязь между структурой геномной ДНК гена рецептора интерлейкина-6 мыши (Il6ra) (1) и вектора с нокином (Knock-in), подлежащего встраиванию (2). Вектор с нокином имеет полноразмерную кДНК рецептора интерлейкина-6 человека (hIL6R), последовательность hp7, сигнал присоединения поли-A и ген устойчивости к неомицину.
На фиг. 32B представлена схематическая диаграмма, демонстрирующая, каким образом геномная ДНК гена рецептора интерлейкина-6 мыши (a) и вектора с нокином (b) претерпевают гомологичную рекомбинацию с образованием геномной ДНК с нокином (c). Более того, на ней показан процесс завершения введенного нокином аллеля гена рецептора интерлейкина-6 человека (d) посредством обеспечения действия рекомбиназы Cre на (c) для удаления кассеты с геном устойчивости к неомицину. Стрелками на фигуре указаны положения для размещения праймеров, использованных для обнаружения подвергнутого нокину гена рецептора интерлейкина-6 человека.
На фиг. 33 представлен репрезентативный пример ПЦР, с помощью которой анализировали каждый генотип, полученный в процессе получения мышей с нокином гена рецептора интерлейкина-6 человека.
На фиг. 34 представлен профиль экспрессии гена рецептора интерлейкина-6 у мыши дикого типа и мыши с нокином гена рецептора интерлейкина-6 человека.
На фиг. 35 представлен график, на котором показаны результаты измерения концентрации в плазме растворимого рецептора интерлейкина-6 человека (hsIL-6R) у мышей дикого типа и гомозиготных и гетерозиготных мышей с нокином гена рецептора интерлейкина-6 человека. KI/KI, KI/+ и +/+ указывают на гомозиготных мышей с нокином, гетерозиготных мышей с нокином и дикий тип, соответственно.
На фиг. 36 представлен график, на котором представлена видоспецифическая реактивность в отношении интерлейкина-6 (лиганд) у мышей дикого типа и у гомозиготных мышей с нокином гена рецептора интерлейкина-6 человека.
Способ осуществления изобретения
Ниже предоставлены определения и подробное описание, чтобы облегчить понимание настоящего изобретения, проиллюстрированного в настоящем описании.
Аминокислоты
В рамках настоящего изобретения аминокислоты описаны с помощью однобуквенного или трехбуквенного кодов или обоих из них, например, Ala/A, Leu/L, Arg/R, Lys/K, Asn/N, Met/M, Asp/D, Phe/F, Cys/C, Pro/P, Gln/Q, Ser/S, Glu/E, Thr/T, Gly/G, Trp/W, His/H, Tyr/Y, Ile/I или Val/V.
Антиген
В рамках настоящего изобретения антиген конкретно не ограничен и может представлять собой любой антиген, при условии, что он представляет собой молекулу, которая может индуцировать иммунную систему организма и, тем самым, стать мишенью для иммунного ответа в этом организме. Предпочтительные примеры таких антигенов включают молекулы, которые экспрессируются специфически в опухолевых клетках, но не экспрессируются в нормальных клетках (неоэпитопы). Также являются предпочтительными молекулы, которые экспрессируются в чужеродных биологических видах, таких как бактерии и вирусы, которые инфицируют организм, но не экспрессируются в организме. Выражение "экспрессируемый специфически в опухолевых клетках, но не экспрессируемый в нормальных клетках" или "экспрессируемый в чужеродных биологических видах, которые инфицируют организм, но не экспрессируемый в организме" означает, что существует качественное или количественное отличие молекулы между "опухолевыми клетками и нормальными клетками" или "чужеродными биологическими видами, которые инфицируют организм, и организмом". Например, даже если молекула экспрессируется в нормальных клетках, когда количество молекулы, экспрессируемой в опухолевых клетках, значительно превышает количество, экспрессируемое в нормальных клетках, в рамках настоящего изобретения может быть сказано, что существует количественное отличие молекулы между опухолевыми клетками и нормальными клетками. Более того, даже если уровень экспрессии полипептида, состоящего из той же аминокислотной последовательности, сходен в опухолевых клетках и нормальных клетках, когда экспрессируемый полипептид претерпел посстрансляционную модификацию, такую как фосфорилирование в опухолевых клетках, но не в нормальных клетках, в рамках настоящего изобретения может быть сказано, что существует качественное отличие молекулы между опухолевыми клетками и нормальными клетками.
Для таких молекул предпочтительные опухолевые антигены могут включать: рецептор ALK (рецептор плейотрофина), плейотрофин; антиген карциномы поджелудочной железы KS 1/4; антиген карциномы яичника (CA125); простатический кислый фосфат; простатспецифический антиген (PSA); ассоциированный с меланомой антиген p97; антиген меланомы gp75; высокомолекулярный антиген меланомы (HMW-MAA); простатспецифический мембранный антиген; карциноэмбриональный антиген (CEA); антиген полиморфного эпиителиального муцина; антиген глобулы молочного жира человека; ассоциированные с опухолью ободочной и толстой кишки антигены, такие как CEA, TAG-72, CO17-1A, GICA 19-9, CTA-1 и LEA; антиген лимфомы Беркитта 38.13; CD19; антиген B-лимфомы человека CD20; CD33; специфические антигены меланомы, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; опухолеспецифический трансплантационный антиген клеточной поверхности (TSTA); индуцируемые вирусами опухолевые антигены, включающие T-антиген, опухолевые ДНК-вирусы и антигены оболочки опухолевых РНК-вирусов; онкофетальные антигены, такие как CEA толстого кишечника, онкофетальный трофобластный гликопротеин 5T4 и онкофетальный антиген опухоли мочевого пузыря; альфа-фетопротеин; антигены дифференцировки, такие как антигены карциномы легкого человека L6 и L20; антигены фибросаркомы; T-клеточный антиген лейкоза человека Gp37; neo гликопротеин; сфинголипиды; антигены рака молочной железы, такие как EGFR (эпидермальный фактор роста рецептор); NY-BR-16; NY-BR-16 и антиген HER2 (p185HER2); антиген полиморфного эпителиального муцина (PEM); злокачественный лимфоцитарный антиген человека APO-1; антигены дифференцировки, такие как антиген I, встречающийся в фетальных эритроцитах; антиген I первичной эндодермы, встречающийся во взрослых эритроцитах; предимплантационный эмбрион; I(Ma), встречающийся при раке желудка; M18, M39, встречающийся в эпителии молочной железы; SSEA-1, встречающийся в миелоидных клетках; VEP8; VEP9; Myl; VIM-D5; D156-22, встречающийся при раке ободочной и прямой кишки; TRA-1-85 (H групп крови); SCP-1, встречающийся при раке яичка и раке яичника; C14, встречающийся при раке толстого кишечника; F3, встречающийся при раке легкого; AH6, встречающийся при раке желудка; гаптен Y; Ley, встречающийся в клетках эмбриональной карциномы; TL5 (A групп крови); рецептор EGF, встречающийся в клетках A431; серия E1 (B групп крови), встречающийся при раке поджелудочной железы; FC10.2, встречающийся в клетках эмбриональной карциномы; антиген рака желудка; CO-514 (Lea групп крови), встречающийся при аденокарциномах; NS-10, встречающийся при аденокарциномах; CO-43 (Leb групп крови); G49, встречающийся в рецепторе EGF клеток A431; MH2 (ALeb/Ley групп крови), встречающийся при раке толстого кишечника; 19.9, встречающийся при раке толстого кишечника; муцины рака желудка; T5A7, встречающийся в миелоидных клетках; R24, встречающийся при меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и M1:22:25:8, встречающийся в клетках эмбриональной карциномы, и SSEA-3 и SSEA-4, встречающиеся в эмбрионах на стадии 4-8 клеток; подкожный антиген T-клеточной лимфомы; антиген MART-1; антиген сиалил-Tn (STn); антиген рака толстого кишечника NY-CO-45; антиген рака легкого NY-LU-12, вариант A; антиген аденокарциномы ART1; паранеопластический антиген злокачественной опухоли головного мозга-яичек (онконейрональный антиген MA2; паранеопластический нейрональный антиген); нейроонкологический вентральный антиген 2 (NOVA2); антиген гена 520 карциномы гемоцитов; ассоциированный с опухолью антиген CO-029; ассоциированные с опухолью антигены MAGE-C1 (антиген злокачественной опухоли/яичек CT7), MAGE-B1 (антиген MAGE-XP), MAGE-B2 (DAM6), MAGE-2, MAGE-4a, MAGE-4b и MAGE-X2; антиген злокачественной опухоли-яичек (NY-EOS-1); YKL-40 и фрагменты любого из упомянутых выше полипептидов или структур, получаемых посредством их модификации (например, модифицированная фосфатная группа или цепь сахаров упомянутых выше соединений).
Антигены чужеродных биологических видов включают молекулы, экспрессируемые в: Bacillus anthracis, Clostridium botulinum, Yersinia pestis, Variola major (оспа) и других поксвирусах, Francisella tularensis (туляремия) и возбудителях, которые вызывают вирусную геморрагическую лихорадку, аренавирусах, таких как LCM, вирус Хунин, вирус Мачупо, вирус Гуанарито и вирусы, которые вызывают лихорадку Ласса, буньявирусах и хантавирусах, таких как вирусы, которые вызывают лихорадку долины Рифт, кальцивирусе, вирусе гепатита A, гепатита B, гепатита C, вирусе энцефалита, таком как вирус лихорадки западного Нила, заболевания Ла-Кросс, калифорнийского энцефалита, VEE, EEE, WEE и вирус японского энцефалита, вирусе киасанурской лесной болезни, вирусе клещевой геморрагической лихорадки, вирус конго-крымской геморрагической лихорадки, вирусах клещевого энцефалита, вирусе желтой лихорадки, TB с множественной устойчивостью к лекарственным средствам, вирусе гриппа, других риккетсиях и вирусе бешенства, флавирусе, вирусе Денге, филовирусе, вирусе Эбола, Марбургских Burkholderia pseudomallei, Coxiella burnetii (лихорадка Q), видах Brucella (бруцеллез), Burkholderia mallei (сап), рициновый токсин (происходящий из Ricinus communis), эпсилон-токсин Clostridium perfringens, стафилококковый энтеротоксин B, молекулы, экспрессируемые в возбудителе сыпного тифа (Rickettsia prowazekii), переносимых с пищей и водой патогенах, бактериях, таких как вызывающие диарею E. coli, патогеннные Vibrios, виды Shigella, Salmonella, Listeria monocytogenes, Campylobacter jejuni и Yersinia enterocolitica; и простейших, таких как Cryptosporidium parvum, Cyclospora cayatanensis, Giardia lamblia, Entamoeba histolytica, Toxoplasma и Microsporidia.
Другие антигены включают, например, представленные ниже молекулы: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, аденозиновый рецептор A1, A33, ACE, ACE-2, активин, активин A, активин AB, активин B, активин C, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, APE, APJ, APP, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, интегрин av/b3, Ax1, b2M, B7-1, B7-2, B7-H, B-лимфоцитарный стимулирующий фактор (BlyS), BACE, BACE-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bсl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, BMP-3 остеогенин, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, костный нейротрофический фактор, BPDE, BPDE-DNA, BTC, фактор комплемента 3 (C3), C3a, C4, C5, C5a, C10, CA125, CAD-8, кальцитонин, cAMP, карциноэмбриональный антиген (CEA), ассоциированный со злокачественной опухолью, катепсин A, катепсин B, катепсин C/DPPI, катепсин D, катепсин E, катепсин H, катепсин L, катепсин O, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок p67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (B7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, токсин Botulinum, токсин Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цитокератиновый ассоциированный с опухолью антиген, DAN, DCC, DcR3, DC-SIGN, регулирующий фактор комплемента (фактор, ускоряющий распад), дез-(1-3)-IGF-I (IGF-1 головного мозга), Dhh, дигоксин, ДНКM-1, ДНКаза, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, фактор активации фибробластов (FAP), Fas, FcRI, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа1, GFR-альфа2, GFR-альфа3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, рилизинг-фактор гормона роста, гаптен (NP-cap или NIP-cap), HB-EGF, HCC, гликопротеин оболочки gB HCMV, гликопротеин оболочки gH HCMV, UL HCMV, гемопоэтический фактор роста (HGF), HepB gp120, гепараназа, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMW-MAA), gp120 HIV, петля V3 gp 120 IIIB HIV, HLA, HLA-DR, HM1,24, HMFG PEM, HRG, Hrk, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (HGH), HVEM, 1-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, рецептор IgA, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-23, интерферон (INF)-альфа, INF-бета, INF-гамма, ингибин, iNOS, A-цепь инсулина, B-цепь инсулина, инсулиноподобный фактор роста 1, интегрин альфа-2, интегрин альфа-3, интегрин альфа-4, интегрин альфа4/бета1, интегрин альфа4/бета7, интегрин альфа5 (альфа V), интегрин альфа5/бета1, интегрин альфа5/бета3, интегрин альфа6, интегрин бета1, интегрин бета2, интерферон-гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин LI, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bp1, LBP, LDGF, LECT2, lefty, антиген Lewis-Y, ассоциированный с Lewis-Y антиген, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, белок поверхности легких, лютеинизирующий гормон, рецептор лимфотоксина-бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCAM, MCK-2, MCP, M-CSF, MDC, Mer, металлопротеазы, рецептор MGDF, MGMT, MHC (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, мюллерово ингибиторное вещество, Mug, MuSK, NAIP, NAP, NCAD, адгерин N-C, NCA90, NCAM, NCAM, неприлизин, нейротрофин-3, -4 или -6, нейротурин, фактор роста нервов (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGGI, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, плацентарная щелочная фосфатаза (PLAP), P1GF, PLP, PP14, проинсулин, прорелаксин, белок C, PS, PSA, PSCA, простат-специфический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, A-цепь релаксина, B-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, SERINE, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин-72), TARC, TCA-3, T-клеточный рецептор (например, T-клеточный рецептор альфа/бета), TdT, TECK, TEM1, TEM5, TEM7, TEM8, TERT, PLAP-подобная щелочная фосфатаза семенников, TfR, TGF, TGF-альфа, TGF-бета, панспецифический TGF-бета, TGF-бета RI (ALK-5), TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета 1, TGF-бета 2, TGF-бета 3, TGF-бета 4, TGF-бета 5, тромбин, Ck-1 тимуса, тиреотропный гормон, Tie, TIMP, TIQ, тканевой фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа-бета, TNF-бета 2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD 120a, p55-60), TNFRSF1B (TNF RII CD 120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (0X40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL Rl TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд IVEM LIGHT I, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 Fas-лиганда, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TSG, TSLP, ассоциированный с опухолью CA125, ассоциированный с опухолью антиген, экспрессирующий ассоциированные с Lewis-Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный анотиген, VLA, VLA-1, VLA-4, интегрин VNR, фактор Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, Αβ, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин A, хромогранин B, tau, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор B, фактор D, фактор H, пропердин, склеростин, фибринoген, фибрин, протромбин, тромбин, тканевой фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Xa, фактор XI, фактор XIa, фактор XII, фактор Xlla, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA, SIP; рецептор ацетилхолина, AdipoR1, AdipoR2, ADP рибозилциклазу-1, альфа-4/бета-7 интегрин, альфа-5/бета-1 интегрин, альфа-v/бета-6 интегрин, альфа-v/бета-1 интегрин, лиганд ангиопоэтина-2, Angptl2, Anthrax, кадгерин, карбоангидразу-IX, CD105, CD155, CD158a, CD37, CD49b, CD51, CD70, CD72, клаудин 18, токсин Clostridium difficile, CS1, лиганд Delta-подобного белка 4, оксидазу DHICA, лиганд Dickkopf-1, дипептидилпептидазу IV, EPOR, F-белок RSV, фактор Ia, FasL, рецептор фолатов альфа, рецептор глюкагона, рецептор глюкагон-подобного пептида 1, глутаматкарбоксипептидаза II, GMCSFR, гликопротеин E2 вируса гепатита C, гепцидин, рецептор IL-17, рецептор IL-22, рецептор IL-23, рецептор IL-3, тирозинкиназу Kit, лейцин-блогатый альфа-2-гликопротеин 1 (LRG1), рецептор лизосфинголипидов, мембранный гликопротеин OX2, мезотелин, MET, MICA, MUC-16, ассоциированный с миелином гликопротеин, нейропилин-1, нейропилин-2, рецептор Nogo, PLXNA1, PLXNA2, PLXNA3, PLXNA4A, PLXNA4B, PLXNB1, PLXNB2, PLXNB3, PLXNC1, PLXND1, лиганд запрограммированной гибели клеток 1, конвертазу пробелков PC9, лиганд 1 гликопротеина P-селектина, RAGE, ретикулон 4, RF, RON-8, SEMA3A, SEMA3B, SEMA3C, SEMA3D, SEMA3E, SEMA3F, SEMA3G, SEMA4A, SEMA4B, SEMA4C, SEMA4D, SEMA4F, SEMA4G, SEMA5A, SEMA5B, SEMA6A, SEMA6B, SEMA6C, SEMA6D, SEMA7A, шига-подобный токсин II, рецептор 1 cфингозин-1-фосфата, ST2, стафилококковую липотейхоевую кислоту, тенасцин, TG2, тимический стромальный рецептор лимфопротеинов, рецептор 12A суперсемейства TNF, трансмембранный гликопротеин NMB, TREM-1, TREM-2, трофобластный гликопротеин, рецептор TSH, TTR, тубулин и ULBP2; и рецепторы для гормонов и факторов роста.
Эпитоп
"Эпитоп" означает антигенную детерминанту в антигене и относится к антигенному центру, с которым связывается антигенсвязывающий домен антигенсвязывающей молекулы, описанной в настоящем описании. Таким образом, например, эпитоп может быть определен в соответствии с его структурой. Альтернативно эпитоп может быть определен в соответствии с антигенсвязывающей активностью антигенсвязывающей молекулы, которая распознает эпитоп. Когда антиген представляет собой пептид или полипептид, эпитоп может быть указан с помощью аминокислотных остатков, образующих эпитоп. Альтернативно, когда эпитоп представляет собой цепь сахара, эпитоп может быть указан с помощью его определенной структуры цепи сахаров.
Линейный эпитоп представляет собой эпитоп, который содержит эпитоп, в котором распознается его первичная аминокислотная последовательность. Такой линейный эпитоп, как правило, содержит по меньшей мере три и наиболее часто по меньшей мере пять, например, приблизительно 8-10 или 6-20 аминокислот в его конкретной последовательности.
В противоположность линейному эпитопу, "конформационный эпитоп" представляет собой эпитоп, в котором первичная аминокислотная последовательность, содержащая эпитоп, не является единственной детерминантой распознаваемого эпитопа (например, первичная аминокислотная последовательность конформационного эпитопа не обязательно распознается определяющим эпитоп антителом). Конформационные эпитопы могут содержать большее количество аминокислот по сравнению с линейными эпитопами. Антитело, распознающее конформационный эпитоп, распознает трехмерную структуру пептида или белка. Например, когда молекула белка сворачивается и формирует трехмерную структуру, аминокислоты и/или основные цепи полипептида, которые образуют коноформационный эпитоп, располагаются рядом друг с другом, и эпитоп становится распознаваемым для антитела. Способы определения конформаций эпитопов включают, например, рентгеновскую кристаллографию, двумерный ядерный магнитный резонанс, сайт-специфическое спиновое мечение и электронный парамагнитный резонанс, но они не ограничиваются ими. Смотрите, например, Epitope Mapping Protocols in Methods in Molecular Biology (1996), Vol. 66, Morris (ed.).
Активность связывания
Примеры способа оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для рецептора IL-6, описаны ниже. В соответствии с примерами ниже, также можно соответствующим образом проводить способы оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, с антигеном, отличным от рецептора IL-6.
Например, то, что исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для рецептора IL-6, распознает линейный эпитоп в молекуле рецептора IL-6, можно подтвердить, например, как описано ниже. Для указанной выше цели синтезируют линейный пептид, содержащий аминокислотную последовательность, образующую внеклеточный домен рецептора IL-6. Пептид можно синтезировать химически или его можно получать способами генетической инженерии с использованием области, кодирующей аминокислотную последовательность, соответствующую внеклеточному домену в кДНК рецептора IL-6. Затем исследуемую антигенсвязывающую молекулу, содержащую антигенсвязывающий домен для рецептора IL-6, оценивают в отношении ее активности связывания с линейным пептидом, содержащим аминокислотную последовательность, образующую внеклеточный домен. Например, иммобилизованный линейный пептид можно использовать в качестве антигена в ELISA для оценки активности связывания антигенсвязывающей молекулы с пептидом. Альтернативно активность связывания с линейным пептидом можно оценивать, исходя из уровня, на котором линейный пептид ингибирует связывание антигенсвязывающей молекулы с клетками, экспрессирующими рецептор IL-6. Эти исследования могут продемонстрировать активность связывания антигенсвязывающей молекулы с линейным пептидом.
Наличие распознавания исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для рецептора IL-6, конформационного эпитопа можно оценивать следующим образом. Для указанной выше цели получают экспрессирующие рецептор IL-6 клетки. Наличие распознавания исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен рецептора IL-6, конформационного эпитопа, можно определять, когда она прочно связывается с экспрессирующими рецептор IL-6 клетками при контакте, но по существу не связывается с иммобилизованным линейным пептидом, содержащим аминокислотную последовательность, образующую внеклеточный домен рецептора IL-6. В рамках настоящего изобретения "по существу не связывает" означает, что активность связывания составляет 80% или менее, как правило, 50% или менее, предпочтительно 30% или менее, и особенно предпочтительно 15% или менее по сравнению с активностью связывания с клетками, экспрессрующими рецептор IL-6 человека.
Способы оценки активности связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для рецептора IL-6, с экспрессирующими рецептор IL-6 клетками, включают, например, способы, описанные в Antibodies: A Laboratory Manual (Ed Harlow, David Lane, Cold Spring Harbor Laboratory (1988) 359-420). В частности, оценку можно проводить на основе принципа ELISA или активированной флуоресценцией сортировки клеток (FACS) с использованием в качестве антигена экспрессирующих рецептор IL-6 клеток.
В формате ELISA активность связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для рецептора IL-6, с экспрессирующими рецептор IL-6 клетками можно оценивать количественно путем сравнения уровней сигнала, генерируемого ферментативной реакцией. В частности, исследуемый полипептидный комплекс добавляют в планшет для ELISA, на котором иммобилизованы эксперессирующие рецептор IL-6 клетки. Затем антигенсвязывающую молекулу, связанную с клетками, выявляют с использованием меченного ферментом антитела, которое распознает исследуемую антигенсвязывающую молекулу. Альтернативно, когда используют FACS, приготавливают серии разведений исследуемой антигенсвязывающей молекулы и можно определять титр связывания антитела с экспрессирующими рецептор IL-6 клетками для сравнения активности связывания исследуемой антигенсвязывающей молекулы с экспрессирующими рецептор IL-6 клетками.
Связывание исследуемой антигенсвязывающей молекулы с антигеном, экспрессируемым на поверхности клеток, суспендированных в буфере или сходных с ним, можно выявлять с использованием проточного цитометра. Известные проточные цитометры включают, например, следующие устройства:
FACSCanto™ II
FACSAria™
FACSArray™
FACSVantage™ SE
FACSCalibur™ (все являются торговыми названиями BD Biosciences)
EPICS ALTRA HyPerSort
Cytomics FC 500
EPICS XL-MCL ADC EPICS XL ADC
Cell Lab Quanta/Cell Lab Quanta SC (все являются торговыми названиями Beckman Coulter).
Предпочтительные способы оценки активности связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для рецептора IL-6, с антигеном, включают, например, следующий способ. Сначала экспрессирующие рецептор IL-6 клетки подвергают реакции с исследуемой антигенсвязывающей молекулой, а затем их окрашивают меченным FITC вторичным антителом, которое распознает антигенсвязывающую молекулу. Исследуемую антигенсвязывающую молекулу надлежащим образом разбавляют подходящим буфером с получением молекулы с желаемой концентрацией. Например, молекулу можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Затем определяют интенсивность флуоресценции и количество клеток с использованием FACSCalibur (BD). Интенсивность флуоресценции, полученную с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. среднее геометрическое значение, отражает количество антитела, связанного с клетками. Следовательно, активность связывания исследуемой антигенсвязывающей молекулы, которую отражает количество связанной исследуемой антигенсвязывающей молекулы, можно определять путем определения геометрического среднего значения.
То, обладает ли исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для рецептора IL-6, общим эпитопом с другой антигенсвязывающей молекулой, можно оценивать на основе конкуренции между двумя молекулами за один и тот же эпитоп. Конкуренцию между антигенсвязывающими молекулами можно выявлять с помощью анализа перекрестного блокирования и т.п. Например, предпочтительным анализом перекрестного блокирования является конкурентный анализ ELISA.
В частности, в анализе перекрестного блокирования белок рецептора IL-6, иммобилизованный на лунках микропланшета для титрования, предварительно инкубируют в присутствии или в отсутствие являющейся кандидатом конкурентной антигенсвязывающей молекулы, а затем к ним добавляют антигенсвязывающую молекулу. Количество исследуемой антигенсвязывающей молекулы, связавшейся с белком рецептора IL-6 в лунках, непрямо коррелирует со связывающей способностью являющейся кандидатом конкурентной антигенсвязывающей молекулы, которая конкурирует за связывание с тем же эпитопом. Следовательно, чем более высокой является аффинность конкурентной антигенсвязывающей молекулы к тому же эпитопу, тем более низкой является активность связывания исследуемой антигенсвязывающей молекулы с покрытыми белком рецептора IL-6 лунками.
Количество исследуемой антигенсвязывающей молекулы, связавшейся с лунками через рецептор IL-6, можно без труда определить путем предварительного мечения антигенсвязывающей молекулы. Например, меченную биотином антигенсвязывающую молекулу определяют с использованием конъюгата авидин/пероксидаза и соответствующего субстрата. В частности, анализ перекрестного блокирования, в котором используются ферментные метки, такие пероксидаза, называют "конкурентным анализом ELISA". Антигенсвязывающую молекулу также можно метить другими агентами для мечения, которые обеспечивают выявление или измерение. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Когда являющаяся кандидатом конкурентная антигенсвязывающая молекула может блокировать связывание исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для рецептора IL-6, по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50%, и более предпочтительно по меньшей мере на 50% по сравнению с активностью связывания в контрольном эксперименте, проведенном в отсутствие конкурентной антигенсвязывающей молекулы, то определяют, что исследуемая антигенсвязывающая молекула по существу связывается с тем же эпитопом, с которым связывается конкурентная антигенсвязывающая молекула, или конкурирует за связывание с тем же эпитопом.
Когда структура эпитопа, с которым связывается исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для рецептора IL-6, уже идентифицирована, тогда то, имеют ли исследуемые и контрольные антигенсвязывающие молекулы общий эпитоп, можно оценивать путем сравнения активности связывания двух антигенсвязывающих молекул с пептидом, полученным путем внесения аминокислотных мутаций в пептид, формирующий эпитоп.
Для измерения указанной выше активности связывания, например, активность связывания исследуемых и контрольных антигенсвязывающих молекул в отношении линейного пептида, в который внесена мутация, сравнивают в описанном выше формате ELISA. Помимо способов ELISA, активность связывания с мутантным пептидом, связанным с колонкой, можно определять путем пропускания исследуемых и контрольных антигенсвязывающих молекул через колонку, а затем количественного определения антигенсвязывающей молекулы, элюированной в элюирующем растворе. Способы адсорбции мутантного пептида на колонку, например, в форме слитого пептида с GST, известны.
Альтернативно, когда идентифицированный эпитоп представляет собой конформационный эпитоп, тогда то, обладают ли исследуемые и контрольные антигенсвязывающие молекулы общим эпитопом, можно оценивать следующим способом. Сначала получают экспрессирующие рецептор IL-6 клетки и клетки, экспрессирующие рецептор IL-6 с мутацией, внесенной в эпитоп. Исследуемые и контрольные антигенсвязывающие молекулы добавляют в суспензию клеток, полученную суспендированием этих клеток в соответствующем буфере, таком как PBS. Затем клеточные суспензии надлежащим образом промывают буфером, и к ним добавляют меченное FITC антитело, которое распознает исследуемые и контрольные антигенсвязывающие молекулы. Интенсивность флуоресценции и количество клеток, окрашенных меченым антителом, определяют с использованием FACSCalibur (BD). Исследуемые и контрольные антигенсвязывающие молекулы надлежащим образом разбавляют с использованием подходящего буфера и используют в желаемых концентрациях. Например, их можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Интенсивность флуоресценции, определенная с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. геометрическое среднее значение, отражает количество меченного антитела, связавшегося с клетками. Следовательно, активность связывания исследуемых и контрольных антигенсвязывающих молекул, которую отражает количество связавшегося меченого антитела, можно определять путем измерения геометрического среднего значения.
В описанном выше способе то, что антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный рецептор IL-6," можно оценивать, например, следующим способом. Сначала исследуемые и контрольные антигенсвязывающие молекулы, связанные с клетками, экспрессирующими мутантный рецептор IL-6, окрашивают меченым антителом. Затем определяют интенсивность флуоресценции клеток. Когда используют FACSCalibur для выявления флуоресценции проточной цитометрией, определенную интенсивность флуоресцении можно анализировать с использованием программного обеспечения CELL QUEST. Из геометрических средних значений в присутствии и в отсутствие полипептидного комплекса можно вычислять сравнительное значение (ΔGeo-Mean) по следующей формуле для определения соотношения увеличения интенсивности флуоресценции в результате связывания антигенсвязывающей молекулой.
ΔGeo-Mean=Geo-Mean (в присутствии полипептидного комплекса)/Geo-Mean (в отсутствие полипептидного комплекса)
Геометрическое среднее сравнительное значение (значение ΔGeo-Mean для мутантной молекулы рецептора IL-6), определенное с помощью описанного выше анализа, которое отражает количество исследуемой антигенсвязывающей молекулы, связавшейся с клетками, экспрессирующими мутантный рецептор IL-6, сравнивают со сравнительным значением ΔGeo-Mean, которое отражает количество исследуемой антигенсвязывающей молекулы, связавшейся с экспрессирующими рецептор IL-6 клетками. В этом случае, особенно предпочтительно, чтобы концентрации исследуемой антигенсвязывающей молекулы, используемой для определения сравнительных значений ΔGeo-Mean для экспрессирующих рецептор IL-6 клеток и клеток, экспрессирующих мутантный рецептор IL-6, были скорректированы так, чтобы они были равными или по существу равными. В качестве контрольной антигенсвязывающей молекулы используют антигенсвязывающую молекулу, для которой подтверждено, что она распознает эпитоп в рецептор IL-6.
Если сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для клеток, экспрессирующих мутантный рецептор IL-6, меньше чем сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для экспрессирующих рецептор IL-6 клеток по меньшей мере на 80%, предпочтительно на 50%, более предпочтительно на 30% и особенно предпочтительно на 15%, тогда исследуемая антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный рецептор IL-6". Формула для определения значения Geo-Mean (геометрического среднего значения) описана в руководстве пользователя программного обеспечения CELL QUEST (BD biosciences). Если сравнение показывает, что сравнительные величины по существу эквивалентны, то может быть определено, что эпитоп для исследуемой и контрольной антигенсвязывающих молекул является одним и тем же.
Антигенсвязывающий домен
"Антигенсвязывающий домен" может представлять собой любую структуру при условии, что она связывается с представляющим интерес антигеном. Такие домены предпочтительно включают, например:
вариабельные области тяжелых цепей и легких цепей антител;
модуль приблизительно из 35 аминокислот, называемый A-доменом, который содержится в мембранном белке клеток авимере in vivo (WO 2004/044011, WO 2005/040229);
аднектин, содержащий домен 10Fn3, который связывается с белковой частью фибронектина - гликопротеина, экспрессирующегося на клеточной мембране (WO 2002/032925);
аффитело, которое состоит из трехспирального пучка из 58-аминокислот на основе каркаса связывающего IgG домена белка A (WO 1995/001937);
смоделированные белки анкиринового повтора (DARPin), которые представляют собой область, экспонированную на молекулярной поверхности анкириновых повторов (AR), имеющих структуру, в которой субъединица, состоящая из поворота, содержащего 33 аминокислотных остатка, две антипараллельных спирали и петли многократно сложены в стопку (WO 2002/020565);
антикалины и т.п., которые представляют собой домены, состоящие из четырех петель, которые поддерживают одну сторону структуры бочки, состоящей из восьми расположенных по кругу антипараллельных нитей, которые являются высоко консервативными среди молекул липокалина, таких как ассоциированный с желатином липокалин нейтрофилов (NGAL) (WO 2003/029462); и
вогнутая область, образованная структурой параллельных слоев внутри структуры в форме подковы, образованной уложенными в стопку повторами модуля лейцин-богатого повтора (LRR) вариабельного рецептора лимфоцитов (VLR), который не имеет иммуноглобулиновой структуры и используется в системе приобретенного иммунитета у бесчелюстных позвоночных, таких как минога и миксина (WO 2008/016854). Предпочтительные антигенсвязывающие домены по настоящему изобретению включают, например, антигенсвязывающие домены, имеющие вариабельные области тяжелых цепей и легких цепей антител. Предпочтительные примеры антигенсвязывающих доменов включают "одноцепочечный Fv (scFv)", "одноцепочечное антитело", "Fv", "одноцепочечный Fv2 (scFv2)", "Fab" и "F(ab')2".
Антигенсвязывающие домены антигенсвязывающих молекул по настоящему изобретению могут связываться с идентичным эпитопом. Такой эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:1. Альтернативно, каждый из антигенсвязывающих доменов антигенсвязывающих молекул по настоящему изобретению может связываться с отличающимся эпитопом. В рамках настоящего изобретения, отличающийся эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:1.
Специфичность
"Специфичный" означает, что одна из молекул, которая специфически связывается, не проявляет никакого существенного связывания с молекулами, отличными от одной или нескольких молекул-партнеров по связыванию. Более того, "специфичный" также используют, когда антигенсвязывающий домен является специфичным к конкретному эпитопу среди множества эпитопов в антигене. Когда эпитоп, связываемый антигенсвязывающим доменом, содержится в нескольких различных антигенах, антигенсвязывающие молекулы, содержащие антигенсвязывающий домен, могут связываться с различными антигенами, которые имеют этот эпитоп.
Антитела
В рамках настоящего изобретения "антитело" относится к природному иммуноглобулину или иммуноглобулину, полученному частичным или полным синтезом. Антитела можно выделять из природных источников, таких как природная плазма и сыворотка, или из культуральных супернатантов продуцирующих антитела гибридом. Альтернативно антитела можно частично или полностью синтезировать с использованием способов, таких как генетическая рекомбинация. Предпочтительные антитела включают, например, антитела изотипа иммуноглобулинов или принадлежащего ему подкласса. Известные иммуноглобулины человека включают антитела следующих девяти классов (изотипов): IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE и IgM. Из этих изотипов антитела по настоящему изобретению включают IgG1, IgG2, IgG3 и IgG4.
Способы получения антитела с желаемой активностью связывания известны специалистам в данной области. Ниже приведен пример, в котором описан способ получения антитела, которое связывается с рецептором IL-6 (антитело против рецептора IL-6). Антитела, которые связываются с антигеном, отличным от рецептора IL-6, также можно получать в соответствии с примером, описанным ниже.
Антитела против рецептора IL-6 можно получать в качестве поликлональных или моноклональных антител с использованием известных способов. Продуцируемые антитела против рецептора IL-6 предпочтительно представляют собой моноклональные антитела, происходящие из млекопитающих. Такие происходящие из млекопитающих моноклональные антитела включают антитела, продуцируемые гибридомами или клетками-хозяевами, трансформированными экспрессирующим вектором, несущим ген антитела, способами генетической инженерии. Моноклональные антитела по настоящему изобретению включают "гуманизированные антитела" или "химерные антитела".
Продуцирующие моноклональные антитела гибридомы можно получать с использованием известных способов, например, как описано ниже. В частности, млекопитающих иммунизируют общепринятыми способами иммунизации с использованием белка рецептора IL-6 в качестве сенсибилизирующего антигена. Полученные иммунные клетки подвергают слиянию с известными родительскими клетками общепринятыми способами слияния клеток. Затем гибридомы, продуцирующие антитело против рецептора IL-6, можно отбирать путем скрининга продуцирующих моноклональное антитело клеток с использованием общепринятых способов скрининга.
В частности, моноклональные антитела получают так, как описано ниже. Сначала можно экспрессировать ген рецептора IL-6, нуклеотидная последовательность которого представлена в SEQ ID NO:2, для получения белка рецептора IL-6, представленного в SEQ ID NO:1, который будет использован в качестве сенсибилизирующего антигена для получения антитела. Для этого последовательность гена, кодирующего рецептор IL-6, встраивают в известный экспрессирующий вектор, и подходящие клетки-хозяева трансформируют этим вектором. Желаемый белок рецептора IL-6 человека очищают из клеток-хозяев или их культуральных супернатантов известными способами. Для получения растворимого рецептора IL-6 из культуральных супернатантов, например, белок, состоящий из аминокислот в положениях 1-357 в полипептидной последовательности рецептора IL-6 SEQ ID NO:1, такой как описан в Mullberg et al. (J. Immunol. (1994) 152 (10), 4958-4968), экспрессируют в качестве растворимого рецептора IL-6 вместо белка рецептора IL-6 SEQ ID NO:1. Также в качестве сенсибилизирующего антигена можно использовать очищенный природный белок рецептора IL-6.
Очищенный белок рецептора IL-6 можно использовать в качестве сенсибилизирующего антигена для иммунизации млекопитающих. Также в качестве сенсибилизирующего антигена можно использовать неполный пептид рецептора IL-6. В этом случае, неполный пептид можно получать путем химического синтеза на основе аминокислотной последовательности рецептора IL-6 человека, или путем встраивания неполного гена рецептора IL-6 в экспрессирующий вектор для экспрессии. Альтернативно неполный пептид можно получать путем деградации белка рецептора IL-6 протеазой. Длина и область неполного пептида рецептора IL-6 не ограничены конкретными вариантами осуществления. Предпочтительная область может быть произвольным образом выбрана из аминокислотной последовательности в положениях аминокислот 20-357 аминокислотной последовательности SEQ ID NO:1. Количество аминокислот, образующих пептид, подлежащий применению в качестве сенсибилизирующего антигена, предпочтительно составляет по меньшей мере пять или более, шесть или более, или семь или более. Более конкретно, в качестве сенсибилизирующего антигена можно использовать пептид из 8-50 остатков, более предпочтительно, из 10-30 остатков.
Альтернативно, для сенсибилизирующего антигена можно использовать слитый белок, полученный путем слияния желаемого неполного полипептида или пептида белка рецептора IL-6 с отличающимся полипептидом. Например, предпочтительно в качестве сенсибилизирующих антигенов используют Fc-фрагменты антител и пептидные метки. Векторы для экспрессии таких слитых белков можно конструировать путем слияния в рамке считывания генов, кодирующих два или более желаемых полипептидных фрагментов, и встраивания слитого гена в экспрессирующий вектор, как описано выше. Способы получения слитых белков описаны в Molecular Cloning 2nd ed. (Sambrook, J et al., Molecular Cloning 2nd ed., 9.47-9.58 (1989) Cold Spring Harbor Lab. Press). Способы получения рецептора IL-6 для применения в качестве сенсибилизирующего антигена и способы иммунизации с использованием рецептора IL-6 конкретно описаны в WO 2003/000883, WO 2004/022754, WO 2006/006693 и т.п.
Отсутствует конкретное ограничение на млекопитающих, подлежащих иммунизации сенсибилизирующим антигеном. Однако предпочтительно выбирать млекопитающих, учитывая их совместимость с родительскими клетками, используемыми для слияния клеток. Как правило, предпочтительно используют грызунов, таких как мыши, крысы и хомячки, кроликов и обезьян.
Указанных выше животных иммунизируют сенсибилизирующим антигеном известными способами. Как правило, выполняемые способы иммунизации включают, например, внутрибрюшинную или подкожную инъекцию сенсибилизирующего антигена млекопитающим. В частности, сенсибилизирующий антиген соответствующим образом разбавляют PBS (фосфатно-солевой буфер), физиологическим солевым раствором или сходными с ними. Если желательно, общепринятый адъювант, такой как полный адъювант Фрейнда, смешивают с антигеном и смесь эмульгируют. Затем сенсибилизирующий антиген вводят млекопитающем несколько раз с интервалами 4-21 суток. При иммунизации сенсибилизирующим антигеном можно использовать подходящие носители. В частности, когда в качестве сенсибилизирующего антигена используют низкомолекулярный неполный пептид, иногда желательно связывать сенсибилизирующий антигенный пептид с белком-носителем, таким как альбумин или гемоцианин лимфы улитки, для иммунизации.
Альтернативно гибридомы, продуцирующие желаемое антитело, можно получать с использованием способов иммунизации ДНК, как описано ниже. Иммунизация ДНК представляет собой способ иммунизации, который обеспечивает стимуляцию иммунной системы путем экспрессии сенсибилизирующего антигена у животного, имунизированного в результате введения векторной ДНК, сконструированной для обеспечения экспрессии гена, кодирующего антигенный белок, у животного. По сравнению с общепринятыми способами иммунизации, в которых белковый антиген вводят животным, подлежащим иммунизации, иммунизация ДНК является лучшей в том, что:
- может быть обеспечена стимуляция иммунной системы при сохранении структуры мембранного белка, такого как рецептор IL-6; и
- отсутствует необходимость в очистке антигена для иммунизации.
Для получения моноклонального антитела по настоящему изобретению с использованием иммунизации ДНК, сначала ДНК, экспрессирующую белок рецептора IL-6, вводят животному, подлежащему иммунизации. ДНК, кодирующую рецептор IL-6, можно синтезировать известными способами, такими как ПЦР. Полученную ДНК встраивают в соответствующий экспрессирующий вектор, а затем его вводят животному, подлежащему иммунизации. Предпочтительно используемые экспрессирующие векторы включают, например, коммерчески доступные экспрессирующие векторы, такие как pcDNA3.1. Векторы можно вводить в организм с использованием общепринятых способов. Например, иммунизацию ДНК проводят с использованием генной пушки для введения покрытых экспрессирующим вектором частиц золота в клетки организма животного, подлежащего иммунизации. Антитела, которые распознают рецептор IL-6. также можно получать способами, описанными в WO 2003/104453.
После иммунизации млекопитающего, как описано выше, подтверждают увеличение титра связывающего рецептор IL-6 антитела в сыворотке. Затем иммунные клетки собирают от млекопитающего, а затем подвергают слиянию клеток. В частности, предпочтительно в качестве иммунных клеток используют спленоциты.
В качестве клетки, подлежащей слиянию с упомянутыми выше иммунными клетками, используют клетки миеломы млекопитающих. Клетки миеломы предпочтительно содержат подходящий селективный маркер для скрининга. Селективный маркер придает клеткам характеристики для их выживания (или гибели) в конкретных условиях культивирования. В качестве селективных маркеров известны дефицит гипоксантин-гуанин-фосфорибозилтрансферазы (далее сокращенно обозначаемый как дефицит HGPRT) и дефицит тимидинкиназы (далее сокращенно обозначаемый как дефицит TK). Клетки с дефицитом HGPRT или TK обладают чувствительностью к гипоксантину-аминоптерину-тимидину (далее сокращенно обозначаемой чувствительностью HAT). Чувствительные к HAT клетки не могут синтезировать ДНК в селективной среде HAT, и, таким образом, погибают. Однако, когда клетки слиты с нормальными клетками, они могут продолжать синтезировать ДНК с использованием пути спасения нормальных клеток, и, таким образом, они могут расти в селективной среде HAT.
Селекцию клеток с дефицитом HGPRT и TK можно проводить в среде, содержащей 6-тиогуанин, 8-азагуанин (далее сокращенно обозначаемый как 8AG) или 5'-бромдезоксиуридин, соответственно. Нормальные клетки погибают, поскольку они включают аналоги пиримидина в их ДНК. Между тем, клетки, которые являются дефицитными по этим ферментам, могут выживать в селективной среде, поскольку они не могут включать эти аналоги пиримидина. Кроме того, селективный маркер, называемый устойчивостью к G418, обеспечиваемый геном устойчивости к неомицину, обеспечивает устойчивость к 2-дезоксистрептаминовым антибиотикам (аналоги гентамицина). Известны различные типы миеломных клеток, которые пригодны для слияния клеток.
Например, предпочтительно можно использовать миеломные клетки, включающие следующие клетки:
P3(P3x63Ag8,653) (J. Immunol. (1979) 123 (4), 1548-1550);
P3x63Ag8U.1 (Current Topics in Microbiology и Immunology (1978)81, 1-7);
NS-1 (C. Eur. J. Immunol. (1976)6 (7), 511-519);
MPC-11 (Cell (1976) 8 (3), 405-415);
SP2/0 (Nature (1978) 276 (5685), 269-270);
FO (J. Immunol. Methods (1980) 35 (1-2), 1-21);
S194/5.XX0.BU.1 (J. Exp. Med. (1978) 148 (1), 313-323);
R210 (Nature (1979) 277 (5692), 131-133), и т.д.
Слияние клеток между иммуноцитами и клетками миеломы по существу проводят с использованием известных способов, например, способа Kohler and Milstein et al. (Methods Enzymol. (1981) 73: 3-46).
Более конкретно, слияние клеток можно проводить, например, в общепринятой культуральной среде в присутствии агента, обеспечивающего слияние клеток. Агенты, обеспечивающие слияние клеток, включают, например, полиэтиленгликоль (PEG) и вирус Сендай (HVJ). Если необходимо, также можно добавлять вспомогательное вещество, такое как диметилсульфоксид, для повышения эффективности слияния.
Соотношение иммунных клеток и клеток миеломы можно определять самостоятельно, предпочтительно, например, оно составляет одну клетку миеломы на каждые от одного до десяти иммуноцитов. Культуральные среды, используемые для слияния клеток, включают, например, среды, которые пригодны для выращивания клеточных линий, такие как среда RPMI1640 и среда MEM, и другие общепринятые культуральные среды, используемые для этого типа клеточной культуры. Кроме того, предпочтительно в культуральную среду можно добавлять сывороточные добавки, такие как эмбриональная телячья сыворотка (FCS).
Для слияния клеток заданные количества указанных выше иммунных клеток и миеломных клеток смешивают в описанной выше культуральной среде. Затем добавляют раствор PEG (например, средняя молекулярная масса составляет приблизительно от 1000 до 6000), предварительно нагретый до приблизительно 37°C, в концентрации, как правило, от 30% до 60% (масс./об.). Затем эту смесь осторожно перемешивают для получения желаемых слитых клеток (гибридом). Затем к клеткам постепенно добавляют подходящую культуральную среду, упомянутую выше, и смесь многократно центрифугируют для удаления супернатанта. Таким образом, можно удалить агенты слияния клеток и т.п., которые являются неблагоприятными для роста гибридомы.
Селекцию полученных таким образом гибридом можно проводить путем культивирования с использованием общепринятой селективной среды, например, среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Клетки, отличные от желаемых гибридом (неслитые клетки) можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. Как правило, период составляет от нескольких суток до нескольких недель. Затем, гибридомы, продуцирующие желаемое антитело, подвергают скринингу и клонируют по отдельности общепринятыми способами лимитирующих разведений.
Селекцию полученных таким образом гибридом можно проводить с использованием селективной среды на основе селективного маркера, которым обладает миелома, используемая для слияния клеток. Например, селекцию клеток с дефицитом HGPRT или TK можно проводить путем культивирования с использованием среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). В частности, когда для слияния клеток используют чувствительные к HAT миеломные клетки, клетки, успешно слитые с нормальными клетками, могут селективно пролиферировать в среде HAT. Клетки, отличные от желаемых гибридом (неслитые клетки), можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. В частности, селекцию желаемых гибридом можно проводить путем культивирования, как правило, в течение от нескольких суток до нескольких недель. Затем гибридомы, продуцирующие желаемое антитело, подвергают скринингу и по отдельности клонируют общепринятыми способами лимитирующих разведений.
Желаемые антитела можно предпочтительно отбирать и по отдельности клонировать способами скрининга на основе известной реакции антиген/антитело. Например, связывающее рецептор IL-6 моноклональное антитело может связываться с рецептором IL-6, экспрессированным на поверхности клетки. Такое моноклональное антитело можно подвергать скринингу с помощью активированной флуоресценцией сортировки клеток (FACS). FACS представляет собой систему, которая оценивает связывание антитела с клеточной поверхностью путем анализа клеток, контактирующих с флуоресцентным антителом, с использованием лазерного луча и измерения флуоресценции, испускаемой индивидуальными клетками.
Для скрининга гибридом, которые продуцируют моноклональные антитело по настоящему изобретению, с помощью FACS, сначала получают экспрессирующие рецептор IL-6 клетки. Клетки, предпочтительно используемые для скрининга, представляют собой клетки млекопитающих, в которых рецептор IL-6 экспрессируется неестественным образом. В качестве контроля можно проводить селективное выявление активности антитела, которое связывается с рецептором IL-6 клеточной поверхности, с использованием в качестве клеток-хозяев нетрансформированных клеток млекопитающих. В частности, гибридомы, продуцирующие моноклональное антитело против рецептора IL-6, можно выделять путем селекции гибридом, которые продуцируют антитело, которое связывается с клетками, в которых экспрессируется рецептор IL-6 неестественным образом, но не с клетками-хозяевами.
Альтернативно активность связывания антитела с иммобилизованными экспрессирующими рецептор IL-6 клетками можно оценивать на основе принципа ELISA. Например, экспрессирующие рецептор IL-6 клетки иммобилизуют на лунках планшета для ELISA. Культуральные супернатанты гибридом контактируют с иммобилизованными клетками в лунках и выявляют антитела, которые связываются с иммобилизованными клетками. Когда моноклональные антитела происходят из мыши, антитела, связанные с клетками, можно выявлять с использованием антитела против иммуноглобулина мыши. Селекцию гибридом, продуцирующих желаемое антитело, обладающее антигенсвязывающей способностью, проводят с помощью описанного выше скрининга, и их можно клонировать способом лимитирующих разведений и т.п.
Продуцирующие моноклональное антитело гибридомы, полученные таким образом, можно пассировать в общепринятой культуральной среде и хранить в жидком азоте в течение длительного периода времени.
Описанные выше гибридомы культивируют общепринятым способом, и желаемые моноклональные антитела можно получать из культуральных супернатантов. Альтернативно гибридомы вводят совместимым млекопитающим и выращивают в них, и моноклональные антитела получают из асцитной жидкости. Первый из этих способов пригоден для получения антител с высокой чистотой.
Предпочтительно также можно использовать антитела, кодируемые генами антител, которые клонированы из антителопродуцирующих клеток, таких как описанные выше гибридомы. Клонированный ген антитела встраивают в подходящий вектор, и его вводят в клетку-хозяина для экспрессии антитела, кодируемого геном. Способы выделения генов антител, встраивания генов в векторы и трансформации клеток-хозяев, уже разработаны, например, Vandamme et al. (Eur. J. Biochem. (1990) 192(3), 767-775). Способы продуцирования рекомбинантных антител также известны, как описано ниже.
Например, кДНК, кодирующую вариабельную область (V-область) антитела против рецептора IL-6, получают из клеток гибридомы, экспрессирующих антитело против рецептора IL-6. Для этой цели сначала из гибридом экстрагируют тотальную РНК. Способы, используемые для экстракции мРНК из клеток, включают, например:
- способ ультрацентрифугирования с гуанидином (Biochemistry (1979) 18(24), 5294-5299), и
- способ AGPC (Anal. Biochem. (1987) 162(1), 156-159).
Экстрагированные мРНК можно очищать с использованием набора для очистки мРНК mRNA Purification Kit (GE Healthcare Bioscience) и т.п. Альтернативно также коммерчески доступны наборы для экстракции тотальной мРНК непосредственно из клеток, такие как набор для очистки мРНК QuickPrep mRNA Purification Kit (GE Healthcare Bioscience). мРНК можно получать из гибридом с использованием таких наборов. кДНК, кодирующие V-область антитела, можно синтезировать из полученных мРНК с использованием обратной транскриптазы. кДНК можно синтезировать с использованием набора для синтеза первой цепи кДНК с обратной транскриптазой AMV Reverse Transcriptase First-strand cDNA Synthesis Kit (Seikagaku Co.) и т.п. Более того, для синтеза и амплификации кДНК можно соответствующим образом использовать набор для амплификации кДНК SMART RACE cDNA amplification kit (Clontech) и способ 5'-RACE на основе ПЦР (Proc. Natl. Acad. Sci. USA (1988) 85(23), 8998-9002; Nucleic Acids Res. (1989) 17(8), 2919-2932). В таком способе синтеза кДНК на оба конца кДНК можно вносить подходящие участки для ферментов рестрикции, описанные ниже.
Представляющий интерес фрагмент кДНК очищают из полученного продукта ПЦР, а затем его лигируют в векторную ДНК. Таким образом, конструируют рекомбинантный вектор и вводят в E. coli и т.п. После селекции колоний из образующих колонии E. coli можно получать желаемый рекомбинантный вектор. Затем исследуют, имеет ли рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК, известным способом, таким как способ с терминацией цепи дидезоксинуклеотидами.
Способ 5'-RACE, в котором используются праймеры для амплификации гена вариабельной области, удобно использовать для выделения гена, кодирующего вариабельную область. Сначала конструируют библиотеку кДНК 5'-RACE с помощью синтеза кДНК с использованием РНК, экстрагированных из клеток гибридом в качестве матрицы. Для синтеза библиотеки кДНК 5'-RACE соответствующим образом используют коммерчески доступный набор, такой как набор для амплификации кДНК SMART RACE.
Ген антитела амплифицируют способом ПЦР с использованием полученной библиотеки кДНК 5'-RACE в качестве матрицы. Праймеры для амплификации гена антитела мыши можно конструировать на основе известных последовательностей генов антител. Нуклеотидные последовательности праймеров варьируют, в зависимости от подкласса иммуноглобулинов. Таким образом, предпочтительно, чтобы подкласс был определен заранее с использованием коммерчески доступного набора, такого как набор для изотипирования моноклональных антител мыши Iso Strip mouse monoclonal antibody isotyping kit (Roche Diagnostics).
В частности, например, праймеры, которые обеспечивают амплификацию генов, кодирующих тяжелые цепи γ1, γ2a, γ2b и γ3 и легкие цепи κ и λ, используют для выделения генов, кодирующих IgG мыши. Как правило, для амплификации гена вариабельной области gG используют праймер, который гибридизуется с областью константной области вблизи вариабельной области, в качестве 3'-праймера. Между тем, в качестве 5'-праймера используют праймер, прилагаемый к набору для конструирования библиотеки кДНК 5'-RACE.
Продукты ПЦР, амплифицированные таким образом, используют для реорганизации иммуноглобулинов, состоящих из комбинации тяжелых и легких цепей. Селекцию желаемого антитела можно проводить с использованием активности связывания рецептора IL-6 переформированного иммуноглобулина в качестве индикатора. Например, когда задачей является выделение антитела против рецептора IL-6, более предпочтительно, чтобы связывание антитела с рецептором IL-6 было специфическим. Скрининг связывающего рецептор IL-6 антитела можно проводить, например, с помощью следующих стадий:
(1) контактирование экспрессирующей рецептор IL-6 клетки с антителом, содержащим V-область, кодируемую кДНК, выделенной из гибридомы;
(2) выявление связывания антитела с эксперессирующей рецептор IL-6 клеткой; и
(3) селекция антитела, которое связывается с экспрессирующей рецептор IL-6 клеткой.
Способы выявления связывания антитела с экспрессирующими рецептор IL-6 клетками известны. В частности, связывание антитела с экспрессирующими рецептор IL-6 клетками можно выявлять с помощью описанных выше способов, таких как FACS. Для оценки активности связывания антитела соответствующим образом используют иммобилизованные образцы экспрессирующих рецептор IL-6 клеток.
Предпочтительные способы скрининга антител, в которых в качестве индикатора используется активность связывания, также включают способы пэннинга с использованием фаговых векторов. Способы скрининга с использованием фаговых векторов являются преимущественными, когда гены антител выделяют из библиотек подклассов тяжелых цепей и легких цепей из популяции клеток, экспрессирующей поликлональное антитело. Гены, кодирующие вариабельные области тяжелых цепей и легких цепей, можно связывать соответствующей линкерной последовательностью с получением одноцепочечного Fv (scFv). Фаги, представляющие scFv на их поверхности, можно получать встраиванием гена, кодирующего scFv, в фаговый вектор. Фаги контактируют с представляющим интерес антигеном. Затем ДНК, кодирующую scFv, обладающие представляющей интерес активностью связывания, можно выделять путем сбора фагов, связанных с антигеном. Этот процесс можно повторять при необходимости для увеличения содержания scFv, обладающих представляющий интерес активностью связывания.
После выделения кДНК, кодирующей V-область представляющего интерес антитела против рецептора IL-6, кДНК расщепляют ферментами рестрикции, которые распознают участки рестрикции, встроенные на обоих концах кДНК. Предпочтительные ферменты рестрикции распознают и расщепляют нуклеотидную последовательность, которая встречается в нуклеотидной последовательности гена антитела с низкой частотой. Более того, предпочтительно в представляющий интерес вектор вводят участок рестрикции для фермента, который образует липкий конец, для встраивания одной копии расщепленного фрагмента в правильной ориентации. Кодирующую кДНК V-область антитела против рецептора IL-6 расщепляют, как описано выше, и ее встраивают в соответствующий экспрессирующий вектор для конструирования экспрессирующего вектора для антитела. В этом случае, если ген, кодирующий константную область антитела (C-область) и ген, кодирующий описанную выше V-область, слить в рамке считывания, то получится химерное антитело. В рамках настоящего изобретения "химерное антитело" означает, что происхождение константной области отличается от вариабельной области. Таким образом, в дополнение к гетерохимерным антителам мыши/человека, химерные антитела по настоящему изобретению включают аллохимерные антитела человека/человека. Экспрессирующий вектор для химерного антитела можно конструировать путем встраивания гена V-области в экспрессирующий вектор, который уже имеет константную область. В частности, например, последовательность, распознаваемая первым ферментом рестрикции, который расщепляет выше гена V-области, можно соответствующим образом помещать на 5'-конец экспрессирующего вектора, содержащего ДНК, кодирующую желаемую константную область (C-область) антитела. Экспрессирующий вектор для химерного антитела конструируют путем слияния в рамке считывания двух генов, расщепленных одной и той же комбинацией ферментов рестрикции.
Для получения моноклонального антитела против рецептора IL-6, гены антител встраивают в экспрессирующий вектор так, чтобы гены экспрессировались под контролем регулирующей экспрессию области. Регулирующая экспрессию область для экспрессии антитела включает, например, энхансеры и промоторы. Более того, на N-конец можно присоединять подходящую сигнальную последовательность, так чтобы экспрессируемое антитело секретировалось наружу клеток. В примерах, описанных ниже, в качестве сигнальной последовательности используют пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO:3). Между тем, можно присоединять другие подходящие сигнальные последовательности. Экспрессированный полипептид расщепляется на С-конце описанной выше последовательности, и полученный полипептид секретируется из клеток в качестве зрелого полипептида. Затем подходящие клетки-хозяева трансформируют экспрессирующим вектором и получают рекомбинантные клетки, экспрессирущие ДНК, кодирующую антитело против рецептора IL-6.
ДНК, кодирующие тяжелую цепь (H-цепь) и легкую цепь (L-цепь) антитела по отдельности встраивают в различные экспрессирующие векторы для экспрессии гена антитела. Молекулу антитела, имеющую H- и L-цепи, можно экспрессировать путем котрансфекции одной и той же клетки-хозяина векторами, в которые соответственно встроены гены H-цепи и L-цепи. Альтернативно клетки-хозяева можно трансформировать одним экспрессирующим вектором, в который встроены ДНК, кодирующие H- и L-цепи (смотрите WO 1994/11523).
Существуют различные известные комбинации клетка-хозяин/экспрессирующий вектор для получения антител путем введения выделенных генов антител в соответствующих хозяев. Все из этих экспрессирующих систем пригодны для выделения антигенсвязывающих доменов по настоящему изобретению. Подходящие эукариотические клетки, используемые в качестве клеток-хозяев, включают клетки животных, клетки растений и клетки грибов. В частности, клетки животных включают, например, следующие клетки:
(1) клетки млекопитающих: CHO, COS, клетки миеломы, клетки почки китайского хомячка (BHK), HeLa, Vero клетки почки эмбриона человека (HEK) 293 и т.п.;
(2) клетки земноводных: ооциты Xenopus и т.п.; и
(3) клетки насекомых: sf9, sf21, Tn5 и т.п.
Кроме того, в качестве клетки растений известна экспрессирующая система для генов антител с использованием клеток, происходящих из рода Nicotiana, таких как Nicotiana tabacum. Для трансформации клеток растений можно соответствующим образом использовать культивированные клетки каллуса.
Более того, в качестве клеток грибов можно использовать следующие клетки:
- дрожжи: род Saccharomyces, как например, Saccharomyces cerevisiae, и род Pichia, как например, Pichia pastoris; и
- нитчатые грибы: род Aspergillus, как например, Aspergillus niger.
Более того, также известны экспрессирующие системы для генов антител, в которых используются прокариотические клетки. Например, при использовании бактериальных клеток, в рамках настоящего изобретения можно подходящим образом использовать клетки E. coli, клетки Bacillus subtilis и т.п. Экспрессирующие векторы, несущие представляющие интерес гены антитела, вводят в клетки путем трансфекции. Трансфицированные клетки культивируют in vitro, и из культуры трансформированных клеток можно получать желаемое антитело.
В дополнение к описанным выше клеткам-хозяевам, также для получения рекомбинантного антитела можно использовать трансгенных животных. Следовательно, антитело можно получать из животного, которому введен ген, кодирующий представляющее интерес антитело. Например, ген антитела можно конструировать в качестве слитого гена путем встраивания в рамке считывания в ген, который кодирует белок, специфически продуцируемый в молоке. В качестве белка, секретируемого в молоке, можно использовать, например, β-казеин козы и т.п. Фрагменты ДНК, содержащие слитый ген, в который встроен ген антитела, инъецируют в эмбрион козы, а затем этот эмбрион имплантируют самке козы. Желаемые антитела можно получать в качестве белка, слитого с белком молока, из молока, продуцируемого трансгенной козой, родившейся от козы, являющейся реципиентом эмбриона (или ее потомства). Кроме того, для увеличения объема молока, содержащего желаемое антитело, продуцируемого трансгенной козой, трансгенной козе при необходимости можно вводить гормоны, Ebert, K. M. et al., Bio/Technology (1994) 12 (7), 699-702).
Когда полипептидный комплекс, описанный в настоящем описании, вводят человеку, антигенсвязывающий домен, происходящий из генетически рекомбинантного антитела, которое является искусственно модифицированным для снижения гетерологической антигенности против человека и т.п., можно соответствующим образом использовать в качестве антигенсвязывающего домена комплекса. Такие генетически рекомбинантные антитела включают, например, гуманизированные антитела. Эти модифицированные антитела получают соответствующим образом известными способами.
Вариабельная область антитела, используемая для получения антигенсвязывающего домена полипептидного комплекса, описанного в настоящем описании, как правило, образована тремя определяющими комплементарность областями (CDR) которые разделены четырьмя каркасными областями (FR). CDR представляет собой область, которая по существу определяет специфичность связывания антитела. Аминокислотные последовательности CDR являются в высокой степени разнообразными. С другой стороны, образующие FR аминокислотные последовательности часто обладают высокой идентичностью, даже среди антител с отличающейся специфичностью связывания. Таким образом, как правило, специфичность связывания определенного антитела можно вносить в другое антитело путем пересадки CDR.
Гуманизированное антитело также называют переформированным антителом человека. В частности, известны гуманизированные антитела, полученные пересадкой CDR антитела не являющегося человеком животного, такого как антитело мыши, в антитело человека и т.п. Также известны стандартные способы генной инженерии для получения гуманизированных антител. В частности, например, в качестве способа пересадки CDR антитела мыши в FR человека известна перекрывающаяся ПЦР. В перекрывающейся ПЦР нуклеотидную последовательность, кодирующую CDR антитела мыши, подлежащую пересадке, добавляют к праймерам для синтеза FR антитела человека. Праймеры получают для каждой из четырех FR. Является общепризнанным, что при пересадке CDR мыши в FR человека, выбор FR человека, который имеет высокую идентичность FR мыши, является преимущественным для сохранения функции CDR. Таким образом, как правило, предпочтительно использовать FR человека, содержащую аминокислотную последовательность, которая обладает высокой идентичностью с аминокислотной последовательностью FR, соседней с CDR мыши, подлежащей пересадке.
Нуклеотидные последовательности, подлежащие лигированию, конструируют так, чтобы они были соединены друг с другом в рамке считывания. FR человека синтезируют по отдельности с использованием соответствующих праймеров. В результате получают продукты, в которых ДНК, кодирующая CDR мыши, связана с индивидуальными ДНК, кодирующими FR. Нуклеотидные последовательности, кодирующие CDR мыши, каждого продукта конструируют так, чтобы они перекрывались друг с другом. Затем проводят синтез комплементарной цепи для гибридизации перекрывающихся CDR-областей продуктов, синтезированных с использованием гена антитела человека в качестве матрицы. С помощью этой реакции FR человека лигируют через последовательности CDR мыши.
Полноразмерный ген V-области, в котором в конечном итоге лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, которые гибридизуются с его 5'- или 3'-концом, в которые добавлены подходящие последовательности, распознаваемые ферментом рестрикции. Экспрессирующий вектор для гуманизированного антитела можно получать путем встраивания ДНК, полученной, как описано выше, и ДНК, которая кодирует C-область антитела человека, в экспрессирующий вектор, так чтобы они лигировались в рамке считывания. После трансфекции рекомбинантным вектором хозяина для получения рекомбинантных клеток, рекомбинантные клетки культивируют и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в клеточной культуре (смотрите публикацию патента Европы № EP 239400 и международную публикацию патента № WO 1996/002576).
Путем качественного или количественного определения и оценки антигенсвязывающей активности гуманизированного антитела, полученного, как описано выше, можно пригодным образом отобрать FR антитела человека, которые позволяют CDR образовывать подходящий антигенсвязывающий центр при лигировании через CDR. Аминокислотные остатки в FR можно заменять при необходимости, так чтобы CDR переформированного анитетела человека образовывали соответствующий антигенсвязывающий центр. Например, мутации аминокислотной последовательности можно вносить в FR с использованием способа ПЦР, который используется для пересадки CDR мыши в FR человека. Более конкретно, в праймеры, которые гибридизуются с FR, можно вносить частичные мутации нуклеотидной последовательности. Мутации нуклеотидной последовательности вносятся в FR, синтезированные с использованием таких праймеров. Мутантные последовательности FR, имеющие желаемые характеристики, можно отбирать путем измерения и оценки активности мутанта антитела с замененной аминокислотой в отношении связывания с антигеном с помощью упомянутого выше способа (Cancer Res. (1993) 53: 851-856).
Альтернативно желаемые антитела человека можно получать путем иммунизации трансгенных животных, имеющих полный набор генов антител человека (смотрите WO 1993/012227; WO 1992/003918; WO 1994/002602; WO 1994/025585; WO 1996/034096; WO 1996/033735) путем иммунизации ДНК.
Более того, также известны способы получения антител человека путем пэннинга с использованием библиотек антител человека. Например, V-область антитела человека экспрессируют в качестве одноцепочечного антитела (scFv) на поверхности фага способом фагового дисплея. Можно отбирать фаги, экспрессирующие scFv, которые связываются с антигеном. Последовательность ДНК, кодирующую V-область антитела человека, которая связывается с антигеном, можно определять путем анализа генов выбранных фагов. Определяют последовательность ДНК scFv, которая связывается с антигеном. Получают Экспрессирующий вектор путем слияния последовательности V-области в рамке считывания с последовательностью C-области желаемого антитела человека, и встраивания ее в соответствующий экспрессирующий вектор. Экспрессирующий вектор вводят в клетки, пригодные для экспрессии, такие как клетки, описанные выше. Антитело человека можно получать путем экспрессии гена, кодирующего антитело человека, в клетках. Эти способы уже известны (смотрите WO 1992/001047; WO 1992/020791; WO 1993/006213; WO 1993/011236; WO 1993/019172; WO 1995/001438; WO 1995/015388).
В дополнение к способам, описанным выше, для выделения генов антител можно соответствующим образом использовать способы клонирования B-клеток (идентификация каждой кодирующей антитело последовательности, ее клонирование и выделение; использование при конструировании экспрессирующего вектора для получения каждого антитела (IgG1, IgG2, IgG3 или IgG4, в частности); и т.п.), как описано, например Bernasconi et al. (Science (2002) 298: 2199-2202) или в WO 2008/081008.
Система нумерации EU и система нумерации Kabat
В соответствии со способами, используемыми в рамках настоящего изобретения, положения аминокислот, приписываемые CDR и FR антитела, указаны в соответствии с нумерацией по Kabat (Sequences of Proteins of Immunological Interest (National Institute of Health, Bethesda, Md., 1987 и 1991)). В рамках настоящего изобретения, когда антигенсвязывающая молекула представляет собой антитело или антигенсвязывающий фрагмент, аминокислоты вариабельной области указаны в соответствии с нумерацией по системе Kabat, в то время как аминокислоты константной области указаны в соответствии с системой нумерации EU на основе положений аминокислот по Kabat.
Условия концентрации ионов
Условия концентрации ионов металлов
В неограничивающем варианте осуществления настоящего изобретения концентрация ионов относится к концентрации ионов металлов. "Ионы металлов" относятся к ионам элементов группы I, за исключением водорода, таких как щелочные металлы и элементы группы меди, ионам элементов группы II, таких как щелочноземельные металлы и элементы группы цинка, ионам элементов группы III, за исключением бора, ионам элементов группы IV, за исключением углерода и кремния, ионам элементов группы VIII, таких как элементы группы железа и элементы группы платины, ионам элементов, принадлежащих подгруппе A групп V, VI и VII, и элементов металлов, таких как сурьма, висмут и полоний. Атомы металлов обладают свойством высвобождения валентных электронов так, что они становятся катионами. Это называют тенденцией к ионизации. Металлы с высокой тенденцией к ионизации считаются химически активными.
В рамках настоящего изобретения предпочтительные ионы металлов включают, например, ион кальция. Ион кальция вовлечен в модулирование множества биологических явлений, включая сокращение мышц, таких как скелетные, гладкие и сердечная мышцы; активацию движения, фагоцитоза и т.п. лейкоцитов; активацию изменения формы, секреции и т.п. тромбоцитов; активацию лимфоцитов; активацию тучных клеток, включая секрецию гистамина; клеточные ответы, опосредуемые рецептором катехоламина α или рецептором ацетилхолина; экзоцитоз; высвобождение веществ-передатчиков из окончаний нейронов; и поток аксоплазмы в нейронах. Известные внутриклеточные рецепторы ионов кальция включают тропонин C, кальмодулин, парвальбумин и легкую цепь миозина, которые имеют несколько участков связывания ионов кальция и, как полагают, имеют общее происхождение с точки зрения молекулярной эволюции. Также существует множество известных кальций-связывающих мотивов. Такие хорошо известные мотивы включают, например, домены кадгерина, EF-hand кальмодулина, C2-домен протеинкиназы C, Gla-домен белка фактора свертывания крови IX, лектины C-типа рецептора ациарогликопротеинов и манноза-связывающего рецептора, A-домены рецепторов LDL, аннексин, домен тромбоспондина 3 типа и EGF-подобные домены.
В рамках настоящего изобретения, когда ион металла представляет собой ион кальция, условия концентрации ионов кальция включают низкие концентрации ионов кальция и высокие концентрации ионов кальция. "Активность связывания варьирует, в зависимости от концентраций ионов кальция" означает, что антигенсвязывающая активность антигенсвязывающей молекулы варьирует вследствие различий между условиями низкой и высокой концентрации ионов кальция. Например, антигенсвязывающая активность антигенсвязывающей молекулы может быть более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция. Альтернативно антигенсвязывающая активность антигенсвязывающей молекулы может быть более высокой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция.
В рамках настоящего изобретения высокая концентрация ионов кальция, в частности, не ограничена конкретной величиной; однако концентрация предпочтительно может быть выбрана из величин в пределах от 100 мкМ до 10 мМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 200 мкМ до 5 мМ. В альтернативном варианте осуществления концентрация может быть выбрана из величин в пределах от 500 мкМ до 2,5 мМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 200 мкМ до 2 мМ. Более того, концентрация может быть выбрана из величин в переделах от 400 мкМ до 1,5 мМ. В частности, предпочтительной является концентрация, выбранная из величин в пределах от 500 мкМ до 2,5 мМ, которая является близкой к концентрации ионов кальция в плазме (крови) in vivo.
В рамках настоящего изобретения низкая концентрация ионов кальция, в частности, не ограничена конкретной величиной; однако концентрация предпочтительно может быть выбрана из величин в пределах от 0,1 мкМ до 30 мкМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 0,2 мкМ до 20 мкМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 0,5 мкМ до 10 мкМ. В альтернативном варианте осуществления концентрация может быть выбрана из величин в пределах от 1 мкМ до 5 мкМ. Более того, концентрация может быть выбрана из величин в пределах от 2 мкМ до 4 мкМ. В частности, предпочтительной является концентрация, выбранная из величин в пределах от 1 мкМ до 5 мкМ, которая является близкой к концентрации ионизированного кальция в ранних эндосомах in vivo.
В рамках настоящего изобретения "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 0,1 мкМ до 30 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 100 мкМ до 10 мМ. Предпочтительно, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 0,5 мкМ до 10 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 200 мкМ до 5 мМ. Особенно предпочтительно, это означает, что антигенсвязывающая активность при концентрации ионов кальция в ранней эндосоме in vivo является более слабой, чем при концентрации ионов кальция в плазме in vivo; и, в частности, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 1 мкМ до 5 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 500 мкМ до 2,5 мМ.
То, изменяется ли антигенсвязывающая активность антигенсвязывающей молекулы в зависимости от концентрации ионов металлов, можно определять, например, с использованием известных способов измерения, таких как способы, описанные в разделе "Активность связывания" выше. Например, для подтверждения того, что антигенсвязывающая активность антигенсвязывающей молекулы становится более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция, сравнивают антигенсвязывающую активность антигенсвязывающей молекулы при низкой и высокой концентрациях ионов кальция.
В рамках настоящего изобретения выражение "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" также может быть выражено как "антигенсвязывающая активность антигенсвязывающей молекулы является более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция". В настоящем описании "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" иногда представлено как "антигенсвязывающая способность является более слабой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция". Также "антигенсвязывающая активность при низкой концентрации ионов кальция снижена так, чтобы она была ниже, чем антигенсвязывающая активность при высокой концентрации ионов кальция" может быть представлено как "антигенсвязывающую способность при низкой концентрации ионов кальция ослабили по сравнению с антигенсвязывающей способностью при высокой концентрации ионов кальция".
При определении антигенсвязывающей активности, условия, отличные от концентрации ионов кальция, могут быть соответствующим образом выбраны специалистами в данной области, и они конкретно не ограничены. Например, активность можно определять при 37°C в буфере HEPES. Например, для определения можно использовать Biacore (GE Healthcare) и т.п. Когда антиген представляет собой растворимый антиген, антигенсвязывающую активность антигенсвязывающей молекулы можно оценивать путем пропускания антигена в качестве анализируемого соединения над чипом, на котором иммобилизована антигенсвязывающая молекула. Когда антиген представляет собой антиген мембраны, активность связывания антигенсвязывающей молекулы с антигеном мембраны можно оценивать путем пропускания антигенсвязывающей молекулы в качестве анализируемого соединения над чипом, на котором иммобилизован антиген.
При условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению является более слабой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, соотношение антигенсвязывающей активности между низкими и высокими концентрациями ионов кальция конкретно не ограничено. Однако соотношение KD (константа диссоциации) антигенсвязывающей молекулы в отношении антигенов при низкой концентрации ионов кальция и KD при высокой концентрации ионов кальция, т.е. величина KD (3 мкМ Ca)/KD (2 мМ Ca), предпочтительно составляет 2 или более, более предпочтительно 10 или более, и еще более предпочтительно 40 или более. Верхний предел величины KD (3 мкМ Ca)/KD (2 мМ Ca) конкретно не ограничен и он может представлять собой любую величину, такую как 400, 1000 или 10000, при условии, что молекулу можно получить способами, известными специалистам в данной области. Более того, оно также может быть указано с помощью величины KD (Ca 3 мкМ)/KD (Ca 1,2 мМ). В частности, величина KD (Ca 3 мкМ)/KD (Ca 1,2 мМ) равна 2 или более, предпочтительно величина KD (Ca 3 мкМ)/KD (Ca 1,2 мМ) равна 10 или более и более предпочтительно величина KD (Ca 3 мкМ)/KD (Ca 1,2 мМ) равна 40 или более. Верхний предел величины KD (Ca 3 мкМ)/KD (Ca 1,2 мМ) конкретно не ограничен и может представлять собой любую величину, такую как 400, 1000 или 10000 при условии, что молекулу можно получить способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, для отображения антигенсвязывающей активности можно использовать KD (константа диссоциации). Между тем, когда антиген представляет собой антиген мембраны, для отображения активности можно использовать кажущуюся KD (кажущаяся константа диссоциации). KD (константа диссоциации) и кажущуюся KD (кажущаяся константа диссоциации) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или проточного цитометра.
Альтернативно, например, также в качестве показателя для отображения соотношения антигенсвязывающей активности антигенсвязывающей молекулы по настоящему изобретению при низкой и высокой концентрациях кальция предпочтительно можно использовать константу скорости диссоциации (kd). Когда константу скорости диссоциации (kd) используют вместо константы диссоциации (KD) в качестве показателя для отображения соотношение активностей связывания, соотношение констант скорости диссоциации (kd) при низкой и высокой концентрациях кальция, т.е. величина kd (низкая концентрация кальция)/kd (высокой концентрация кальция), предпочтительно равна 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более, и еще более предпочтительно 30 или более. Верхний предел величины Kd (низкая концентрация кальция)/kd (высокая концентрация кальция) конкретно не ограничен, и он может представлять собой любую величину, такую как 50, 100 или 200, при условии что можно получить молекулу способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, для отображения антигенсвязывающей активности можно использовать kd (константа скорости диссоциации). Между тем, когда антиген представляет собой антиген мембраны, для отображения антигенсвязывающей активности можно использовать кажущуюся kd (кажущуюся константу скорости диссоциации). kd (константа скорости диссоциации) и кажущуюся kd (кажущаяся константа скорости диссоциации) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare) или проточного цитометра. В рамках настоящего изобретения, когда антигенсвязывающую активность антигенсвязывающей молекулы определяют при различных концентрациях ионов кальция, предпочтительно использовать одни и те же условия, за исключением концентраций кальция.
Например, антигенсвязывающий домен или антитело, антигенсвязывающая активность которого является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антител, включающего стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена или антитела при низкой концентрации кальция;
(b) определение антигенсвязывающей активности антигенсвязывающего домена или антитела при высокой концентрации кальция; и
(c) выбор антигенсвязывающего домена или антитела, антигенсвязывающая активность которого является более низкой при низкой концентрации кальция, чем при высокой концентрации кальция.
Более того, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антител, или их библиотеки, включающего стадии:
(a) контактирование антигена с антигенсвязывающим доменом или антителом или их библиотекой при высокой концентрации кальция;
(b) инкубация при низкой концентрации кальция антигенсвязывающего домена или антитела, которые связались с антигеном на стадии (a); и
(c) выделение антигенсвязывающего домена или антитела, диссоциировавших на стадии (b).
Более того, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антител или их библиотеки, включающего стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антител при низкой концентрации кальция;
(b) выбор антигенсвязывающего домена или антитела, которые не связались с антигеном на стадии (a);
(c) позволение антигенсвязывающему домену или антителу, выбранным на стадии (b), связываться с антигеном при высокой концентрации кальция; и
(d) выделение антигенсвязывающего домена или антитела, которые связались с антигеном на стадии (c).
Кроме того, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) контактирование при высокой концентрации кальция библиотеки антигенсвязывающих доменов или антител с колонкой, на которой иммобилизован антиген;
(b) элюирование антигенсвязывающего домена или антитела, которые связались с колонкой на стадии (a), из колонки при низкой концентрации кальция; и
(c) выделение антигенсвязывающего домена или антитела, элюированных на стадии (b).
Более того, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) позволение библиотеке антигенсвязывающих доменов или антител при низкой концентрации кальция пройти через колонку, на которой иммобилизован антиген;
(b) сбор антигенсвязывающего домена или антитела, которые элюировались без связывания с колонкой на стадии (a);
(c) позволение антигенсвязывающему домену или антителу, собранным на стадии (b), связаться с антигеном при высокой концентрации кальция; и
(d) выделение антигенсвязывающего домена или антитела, которые связались с антигеном на стадии (c).
Более того, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антител при высокой концентрации кальция;
(b) получение антигенсвязывающего домена или антитела, которые связались с антигеном на стадии (a);
(c) инкубация при низкой концентрации кальция антигенсвязывающего домена или антитела, полученных на стадии (b); и
(d) выделение антигенсвязывающего домена или антитела, антигенсвязывающая активность которых на стадии (c) является более слабой, чем критерий отбора на стадии (b).
Описанные выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающим доменам или антителам, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые получают способами скрининга, которые дополнительно включают стадию повторения два или более раз стадий (a)-(c) или (a)-(d) в описанных выше способах скрининга. Количество циклов стадий (a)-(c) или (a)-(d) конкретно не ограничено, однако обычно составляет 10 или менее.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена или антитела при низкой концентрации кальция конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 0,1 мкМ до 30 мкМ, но предпочтительно она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 0,5 мкМ до 10 мкМ. Более предпочтительно, она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция в ранней эндосоме in vivo, в частности, от 1 мкМ до 5 мкМ. Между тем, антигенсвязывающая активность антигенсвязывающего домена или антитела при высокой концентрации кальция конкретно не ограничена, при условии что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 100 мкМ до 10 мМ, но предпочтительно она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 200 мкМ до 5 мМ. Более предпочтительно, она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция в плазме in vivo, в частности, от 0,5 мМ до 2,5 мМ.
Антигенсвязывающую активность антигенсвязывающего домена или антитела можно измерять способами, известными специалистам в данной области. Условия, отличные от концентрации ионизированного кальция, могут быть определены специалистами в данной области. Антигенсвязывающую активность антигенсвязывающего домена или антитела можно оценивать в качестве константы диссоциации (KD), кажущейся константы диссоциации (кажущаяся KD), константы скорости диссоциации (kd), кажущейся константы диссоциации (кажущаяся kd) и т.п. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или FACS.
В рамках настоящего изобретения стадия выбора антигенсвязывающего домена или антитела, антигенсвязывающая активность которых является более высокой при высокой концентрации кальция, чем при низкой концентрации кальция, является тождественной стадии выбора антигенсвязывающего домена или антитела, антигенсвязывающая активность которых является более низкой при низкой концентрация кальция, чем при высокой концентрации кальция.
При условии, что антигенсвязывающая активность является более высокой при высокой концентрации кальция, чем при низкой концентрации кальция, различие в антигенсвязывающей активности между высокой и низкой концентрациями концентрация конкретно не ограничено; однако антигенсвязывающая активность при высокой концентрации кальция предпочтительно в два раза или более, более предпочтительно в 10 раз или более, и еще более предпочтительно в 40 раз или более превышает низкую концентрацию кальция.
Антигенсвязывающие домены или антитела по настоящему изобретению, подлежащие скринингу способами скрининга, описанными выше, могут представлять собой любые антигенсвязывающие домены и антитела. Например, можно проводить скрининг описанных выше антигенсвязывающих доменов или антител. Например, можно проводить скрининг антигенсвязывающих доменов или антител, имеющих природные последовательности или последовательности с замещенными аминокислотами.
Библиотеки
В одном варианте осуществления антигенсвязывающий домен или антитело по настоящему изобретению можно получать из библиотеки, которая в основном состоит из множества антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены которых имеют по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентрации ионов. Концентрации ионов предпочтительно включают, например, концентрацию ионов металлов и концентрацию ионов водорода.
В рамках настоящего изобретения "библиотека" относится к множеству антигенсвязывающих молекул или множеству слитых полипептидов, содержащих антигенсвязывающие молекулы, или нуклеиновых кислот или полинуклеотидов, кодирующих их последовательности. Последовательности множества антигенсвязывающих молекул или множества слитых полипептидов, содержащих антигенсвязывающие молекулы, в библиотеке не идентичны, а отличаются друг от друга.
В рамках настоящего изобретения выражение "последовательности отличаются друг от друга" в выражении "множество антигенсвязывающих молекул, последовательности которых отличаются друг от друга," означает, что последовательности антигенсвязывающих молекул в библиотеке отличаются друг от друга. В частности, в библиотеке количество последовательностей, отличающихся друг от друга, отражает количество независимых клонов с различными последовательностями, и также может называться "размером библиотеки". Размер библиотеки для обычной библиотеки фагового дисплея находится в диапазоне от 106 до 1012. Размер библиотеки может быть увеличен вплоть до 1014 с использованием известных способов, таких как рибосомный дисплей. Однако истинное количество фаговых частиц, используемых в селекции способом пэннинга фаговой библиотеки, как правило, в 10-10000 раз превышает размер библиотеки. Эту избыточную множественность также называют "количеством эквивалентов библиотеки", и она означает, что существует от 10 до 10000 индивидуальных клонов, которые обладают одной и той же аминокислотной последовательностью. Таким образом, в рамках настоящего изобретения выражение "последовательности отличаются друг от друга" означает, что последовательности независимых антигенсвязывающих молекул в библиотеке, исключая эквиваленты библиотеки, отличаются друг от друга. Более конкретно, указанное выше означает, что существует от 106 до 1014 антигенсвязывающих молекул, последовательности которых отличаются друг от друга, предпочтительно от 107 до 1012 молекул, более предпочтительно от 108 до 1011 молекул, и особенно предпочтительно от 108 до 1012 молекул, последовательности которых отличаются друг от друга.
В рамках настоящего изобретения выражение "множество" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" в случае, например, антигенсвязывающих молекул, слитых полипептидов, полинуклеотидных молекул, векторов или вирусов по настоящему изобретению, главным образом, относится к группе из двух или более типов вещества. Например, когда два или более веществ отличаются друг от друга конкретной характеристикой, это означает, что существует два или более типов вещества. Такие примеры могут включать, например, мутантные аминокислоты, наблюдаемые в конкретных аминокислотных положениях аминокислотной последовательности. Например, когда существует две или более антигенсвязывающих молекул по настоящему изобретению, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нестрогих остатков или за исключением конкретных мутантных аминокислот в гипервариабельных положениях, экспонированных на поверхности, существует множество антигенсвязывающих молекул по настоящему изобретению. В другом примере, когда существует две или более полинуклеотидных молекулы, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нуклеотидов, кодирующих нестрогие остатки, или нуклеотидов, кодирующих мутантные аминокислоты гипервариабельных положений, экспонированных на поверхности, существует множество полинуклеотидных молекул по настоящему изобретению.
Кроме того, в рамках настоящего изобретения выражение "в основном состоит из" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" отражает количество антигенсвязывающих молекул, антигенсвязывающая активность которых варьирует, в зависимости от концентрации ионов, среди независимых клонов с отличающимися последовательностями в библиотеке. Особенно предпочтительно, чтобы в библиотеке существовало по меньшей мере 104 антигенсвязывающих молекул, имеющих такую активность связывания. Более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 105 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 106 антигенсвязывающих молекул, обладающих такой активностью связывания. Особенно предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 107 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 108 антигенсвязывающих молекул, обладающих такой активностью связывания. Альтернативно это также предпочтительно может быть выражено как соотношение количества антигенсвязывающих молекул, антигенсвязывающая активность которых варьирует, в зависимости от концентрации ионов, и количества независимых клонов, имеющих различные последовательности, в библиотеке. В частности, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в которой антигенсвязывающие молекулы, имеющие такую активность связывания, составляют от 0,1% до 80%, предпочтительно от 0,5% до 60%, более предпочтительно от 1% до 40%, еще более предпочтительно от 2% до 20%, и особенно предпочтительно от 4% до 10% независимых клонов с отличающимися последовательностями в библиотеке. В случае слитых полипептидов, полинуклеотидных молекул или векторов, могут быть возможными сходные выражения, в которых используются количества молекул или отношение к общему количеству молекул. В случае вирусов, также могут быть возможными сходные выражения, в которых используется количество вирионов или отношение к общему количеству вирионов.
Аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих доменов в зависимости от концентрации ионов кальция
Антигенсвязывающие домены или антитела по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым путем. Например, когда ион металла представляет собой ион кальция, можно использовать заранее существующие антитела, заранее существующие библиотеки (фаговая библиотека, и т.д.), антитела или библиотеки, полученные из гибридом, полученных путем иммунизации животных или из B-клеток иммунизированных животных, антитела или библиотеки, полученные внесением аминокислот, способных хелатировать кальций (например, аспарагиновая кислота и глутаминовая кислота), или неприродных аминокислотных мутаций в описанные выше антитела или библиотеки (хелатирующие кальций аминокислоты (такие как аспарагиновая кислота и глутаминовая кислота), библиотеки с увеличенным содержанием неприродных аминокислот, библиотеки полученные путем внесения хелатирующих кальций аминокислот (таких как аспарагиновая кислота и глутаминовая кислота) или неприродных аминокислотных мутаций в конкретные положения, и т.п.
Примеры аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентрации ионов кальция, как описано выше, могут представлять собой любые типы аминокислот, при условии что аминокислоты формируют кальций-связывающий мотив. Кальций-связывающие мотивы хорошо известны специалистам в данной области и подробно описаны (например, Springer et al. (Cell (2000) 102, 275-277); Kawasaki и Kretsinger (Белок Prof. (1995) 2, 305-490); Moncrief et al. (J. Mol. Evol. (1990) 30, 522-562); Chauvaux et al. (Biochem. J. (1990) 265, 261-265); Bairoch и Cox (FEBS Lett. (1990) 269, 454-456); Davis (New Biol. (1990) 2, 410-419); Schaefer et al. (Genomics (1995) 25, 638-643); Economou et al. (EMBO J. (1990) 9, 349-354); Wurzburg et al. (Structure. (2006) 14, 6, 1049-1058)). В частности, любые известные кальций-связывающие мотивы, включая лектины C-типа, такие как ASGPR, CD23, MBR и DC-SIGN, могут быть включены в антигенсвязывающие молекулы по настоящему изобретению. Предпочтительные примеры таких предпочтительных кальций-связывающих мотивов также включают, в дополнение к кальций-связывающим мотивам, описанным выше, например, кальций-связывающий мотив в антигенсвязывающем домене SEQ ID NO:4, 9 или 10.
Более того, в качестве аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция, например, предпочтительно можно использовать аминокислоты, имеющие активность хелатирования металлов. Примеры таких хелатирующих металлы аминокислот включают, например, серин (Ser(S)), треонин (Thr(T)), аспарагин (Asn(N)), глутамин (Gln(Q)), аспарагиновую кислоту (Asp(D)) и глутаминовую кислоту (Glu(E)).
Положения в антигенсвязывающих доменах, в которых содержатся описанные выше аминокислоты, конкретно не ограничены конкретными положениями, и они могут представлять собой любые положения в вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые образуют антигенсвязывающий домен, при условии, что они изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция. В частности, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены тяжелых цепей которых содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекулы в зависимости от концентраций ионов кальция. В другом неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены тяжелой цепи которых содержат упомянутые выше аминокислоты. В другом неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены тяжелых цепей которых содержат упомянутые выше аминокислоты в положениях 95, 96, 100a и/или 101, как указано в соответствии с системой нумерации Kabat.
Между тем, в неограничивающем варианте осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены легкой цепи которых содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR1-домены легкой цепи которых содержат упомянутые выше аминокислот. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR1-домены легкой цепи которых содержат упомянутые выше аминокислоты в положениях 30, 31 и/или 32, как указано в соответствии с системной нумерации Kabat.
В другом неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR2-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки. В другом варианте осуществления настоящее изобретение относится к библиотекам, в основном состоящим из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR2-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки в положении 50, как указано в соответствии с системой нумерации Kabat.
В другом неограничивающем варианте осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки. В альтернативном варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки в положении 92, как указано в соответствии с системной нумерации Kabat.
Более того, в различных вариантах осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и в которых две или три CDR, выбранных из описанных выше CDR1, CDR2 и CDR3 легкой цепи, содержат упомянутые выше аминокислотные остатки. Более того, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и легкие цепи которых содержат упомянутые выше аминокислотные остатки в любом одном или нескольких из положений 30, 31, 32, 50 и/или 92, как указано в соответствии с системной нумерации Kabat.
В особенно предпочтительном варианте осуществления каркасные последовательности вариабельной области легкой цепи и/или тяжелой цепи антигенсвязывающей молекулы предпочтительно содержат каркасные последовательности человека эмбрионального типа. Таким образом, в одном варианте осуществления настоящего изобретения, когда каркасные последовательности представляют собой полностью человеческие последовательности, ожидается, что при введении такой антигенсвязывающей молекулы по настоящему изобретению человеку (например, для лечения заболеваний), она индуцирует небольшой иммуногенный ответ или не индуцирует иммуногенного ответа. В указанном выше контексте выражение "содержащий последовательность эмбрионального типа" в рамках настоящего изобретения означает, что часть каркасных последовательностей по настоящему изобретению идентична части любых каркасных последовательностей человека эмбрионального типа. Например, когда последовательность FR2 тяжелой цепи антигенсвязывающей молекулы по настоящему изобретению представляет собой комбинацию последовательностей FR2 тяжелой цепи различных каркасных последовательностей человека эмбрионального типа, такая молекула также представляет собой антигенсвязывающую молекулу по настоящему изобретению, "содержащую последовательность эмбрионального типа".
Предпочтительные примеры каркасных областей включают, например, последовательности полностью человеческих каркасных областей человека, известные в настоящее время, которые включены в web-сайт V-Base (http://vbase.mrc-cpe.cam.ac.uk/) или другие. Эти последовательности каркасных областей можно соответствующим образом использовать в качестве последовательности эмбрионального типа, содержащейся в антигенсвязывающей молекуле по настоящему изобретению. Последовательности эмбрионального типа могут быть подразделены на категории в соответствии с их сходством (Tomlinson et al. (J. Mol. Biol. (1992) 227, 776-798); Williams и Winter (Eur. J. Immunol. (1993) 23, 1456-1461); Cox et al. (Nat. Genetics (1994) 7, 162-168)). Соответствующие последовательности эмбрионального типа могут быть выбраны из Vκ, которые подразделяются на семь подгрупп; Vλ, которые подразделяются на десять подгрупп; и VH, которые подразделяются на семь подгрупп.
Полностью человеческие последовательности VH предпочтительно включают, но не ограничиваются ими, например, последовательности VH:
подгруппы VH1 (например, VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58 и VH1-69);
подгруппы VH2 (например, VH2-5, VH2-26 и VH2-70);
подгруппы VH3 (VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73 и VH3-74);
подгруппы VH4 (VH4-4, VH4-28, VH4-31, VH4-34, VH4-39, VH4-59 и VH4-61);
подгруппы VH5 (VH5-51);
подгруппы VH6 (VH6-1); и
подгруппы VH7 (VH7-4 и VH7-81).
Они также описаны в известных документах (Matsuda et al. (J. Exp. Med. (1998) 188, 1973-1975)) и т.п., и, таким образом, специалисты в данной области могут соответствующим образом моделировать антигенсвязывающие молекулы по настоящему изобретению, исходя из информации об этих последовательностях. Также предпочтительно использовать другие полностью человеческие каркасные области или каркасные субобласти.
Полностью человеческие последовательности VK предпочтительно включают, но не ограничиваются ими, например:
A20, A30, L1, L4, L5, L8, L9, L11, L12, L14, L15, L18, L19, L22, L23, L24, O2, O4, O8, O12, O14 и O18, выделенные в подгруппу Vk1;
A1, A2, A3, A5, A7, A17, A18, A19, A23, O1 и O11, выделенные в подгруппу Vk2;
A11, A27, L2, L6, L10, L16, L20 и L25, выделенные в подгруппу Vk3;
B3, выделенная в подгруппу Vk4;
B2 (также обозначаемая как Vk5-2), выделенная в подгруппу Vk5; и
A10, A14 и A26, выделенные в подгруппу VK6.
(Kawasaki et al. (Eur. J. Immunol. (2001) 31, 1017-1028); Schable and Zachau (Biol. Chem. Hoppe Seyler (1993) 374, 1001-1022); Brensing-Kuppers et al. (Gene (1997) 191, 173-181)).
Полностью человеческие последовательности VL предпочтительно включают, но не ограничиваются ими, например:
V1-2, V1-3, V1-4, V1-5, V1-7, V1-9, V1-11, V1-13, V1-16, V1-17, V1-18, V1-19, V1-20 и V1-22, выделенные в подгруппу VL1;
V2-1, V2-6, V2-7, V2-8, V2-11, V2-13, V2-14, V2-15, V2-17 и V2-19, выделенные в подгруппу VL1;
V3-2, V3-3 и V3-4, выделенные в подгруппу VL3;
V4-1, V4-2, V4-3, V4-4 и V4-6, выделенные в подгруппу VL4; и
V5-1, V5-2, V5-4 и V5-6, выделенные в подгруппу VL5 (Kawasaki et al. (Genome Res. (1997) 7, 250-261)).
Обычно эти каркасные последовательности отличаются друг от друга одним или несколькими аминокислотными остатками. Эти каркасные последовательности можно использовать в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов" по настоящему изобретению. Другие примеры полностью человеческих каркасных областей, используемых в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов" по настоящему изобретению включают, но не ограничиваются ими, например, KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (например, Kabat et al. (1991), выше, Wu et al. (J. Exp. Med. (1970) 132, 211-250)).
Без связи с конкретной теорией полагают, что одна из причин для того, чтобы ожидать, что использование последовательностей эмбрионального типа предотвратит неблагоприятные иммунные ответы у большинства индивидуумов, состоит в следующем. В результате процесса созревания аффинности в ходе нормальных иммунных ответов, часто в вариабельных областях иммуноглобулинов происходят соматические мутации. Такие мутации чаще всего происходят в области CDR, последовательности которых являются гипервариабельными, но также затрагивают остатки каркасных областей. Такие мутации каркасной области не существуют в генах эмбрионального типа, и они также с меньшей вероятностью будут иммуногенными у пациентов. С другой стороны, в нормальной человеческой популяции присутствует большинство каркасных последовательностей, экспрессируемых с генов эмбрионального типа. В результате иммунной толерантности, ожидается, что эти каркасные области эмбрионального типа будут иметь низкую иммуногенность у пациентов или не иметь иммуногенности у пациентов. Для максимизации возможности иммунной толерантности можно выбирать гены, кодирующие вариабельную область, из группы часто встречающихся функциональных генов эмбрионального типа.
Для получения антигенсвязывающих молекул по настоящему изобретению, в которых описанные выше каркасные последовательности содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентрации ионов кальция можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР.
Например, библиотеку, которая содержит множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, где вариабельную область легкой цепи выбирают в качестве каркасной последовательности, первоначально содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. Неограничивающие примеры таких библиотек, когда концентрация ионов представляет собой концентрацию ионов кальция, включают библиотеку, в которой последовательность вариабельной области легкой цепи, принадлежащая семейству Vk5-2, соответствующая SEQ ID NO:4 (Vk5-2) комбинирована с вариабельными областями тяжелой цепи, полученными в качестве рандомизированной библиотеки последовательностей вариабельной области.
Альтернативно последовательность вариабельной области легкой цепи, выбранную в качестве каркасной области, первоначально содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы, как упоминалось выше, можно конструировать так, чтобы она содержала различные аминокислотные остатки, отличные от упомянутых выше аминокислотных остатков. В настоящем описании такие остатки называют нестрогими остатками. Количество и положение нестрогих остатков конкретно не ограничено, при условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков. Например, когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, подлежащих внесению в последовательность вариабельной области легкой цепи SEQ ID NO:4 (Vk5-2), включают аминокислотные остатки, приведенные в таблицах 1 или 2.
В настоящем описании нестрогие остатки относятся к варьированию аминокислотных остатков, имеющемуся в гипервариабельных положениях, в которых присутствует несколько различных аминокислот, в вариабельных областях легкой цепи и тяжелой цепи, когда сравнивают аминокислотные последовательности известных и/или нативных антител или антигенсвязывающих доменов. Гипервариабельные положения, как правило, расположены в областях CDR. В одном варианте осуществления для определения гипервариабельных положений в известных и/или нативных антителах пригодны данные, предоставленные Kabat, Sequences of Proteins of Immunological Interest (National Institute of Health Bethesda Md.) (1987 и 1991). Более того, в базах данных в Интернете (http://vbase.mrc-cpe.cam.ac.uk/, http://www.bioinf.org.uk/abs/index.html) предоставлены собранные последовательности многих легких цепей и тяжелых цепей человека и их положения. Информация о последовательностях и положениях пригодна для определения гипервариабельных положений в рамках настоящего изобретения. В соответствии с настоящим изобретением, когда определенное положение аминокислоты имеет предпочтительно от приблизительно 2 до приблизительно 20 возможных вариантов аминокислотных остатков, предпочтительно от приблизительно 3 до приблизительно 19, предпочтительно от приблизительно 4 до приблизительно 18, предпочтительно от 5 до 17, предпочтительно от 6 до 16, предпочтительно от 7 до 15, предпочтительно от 8 до 14, предпочтительно от 9 до 13, и предпочтительно от 10 до 12 возможных вариантов аминокислотных остатков, это положение является гипервариабельным. В некоторых вариантах осуществления определенное положение аминокислоты может иметь предпочтительно по меньшей мере приблизительно 2, предпочтительно по меньшей мере приблизительно 4, предпочтительно по меньшей мере приблизительно 6, предпочтительно по меньшей мере приблизительно 8, предпочтительно приблизительно 10 и предпочтительно приблизительно 12 вариантов аминокислотных остатков.
Альтернативно библиотеку, содержащую множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, с вариабельными областями легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов, как упоминалось выше. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают, например, библиотеки, в которых вариабельные области тяжелой цепи, полученные в качестве рандомизированной библиотеки последовательностей вариабельной области, комбинируют с последовательностями вариабельной области легкой цепи, в которых конкретный остаток(и) в последовательности эмбрионального типа, такой как SEQ ID NO:5 (Vk1), SEQ ID NO:6 (Vk2), SEQ ID NO:7 (Vk3) или SEQ ID NO:8 (Vk4) заменен по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. Неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки в CDR1 легкой цепи. Более того, неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки в CDR2 легкой цепи. Кроме того, неограничивающие примеры таких аминокислотных остатков также включают аминокислотные остатки в CDR3 легкой цепи.
Неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR1 легкой цепи, включают остатки в положениях 30, 31 и/или 32 в CDR1 вариабельной области легкой цепи, как указано согласно нумерации Kabat. Более того, неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR2 легкой цепи, включают аминокислотный остаток в положении 50 в CDR2 вариабельной области легкой цепи, как указано согласно нумерации Kabat. Более того, неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR3 легкой цепи, включают аминокислотный остаток в положении 92 в CDR3 вариабельной области легкой цепи, как указано согласно нумерации Kabat. Эти аминокислотные остатки могут содержаться отдельно или в комбинации, при условии что они образуют кальций-связывающий мотив и/или при условии что антигенсвязывающая активность антигенсвязывающей молекулы варьирует, в зависимости от концентрации ионов кальция. Между тем, поскольку известны тропонин C, кальмодулин, парвальбумин и легкая цепь миозина, которые обладают несколькими участками связывания ионов кальция и, как полагают, имеют общее происхождение с точки зрения молекулярной эволюции, можно конструировать CDR1, CDR2 и/или CDR3 легкой цепи так, чтобы они имели их связывающие мотивы. Например, можно использовать домены кадгерина, EF-hand кальмодулина, C2-домен протеинкиназы C, Gla-домен белка фактора свертывания крови IX, лектины C-типа рецептора ациарогликопротеинов и манноза-связывающего рецептора, A-домены рецепторов LDL, аннексин, домена тромбоспондина 3 типа, и EGF-подобные домены соответствующим образом для описанных выше целей.
Когда вариабельные области тяжелой цепи, полученные в качестве рандомизированной библиотеки последовательностей вариабельной области, и вариабельные области легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов, комбинируют, как описано выше, последовательности вариабельных областей легкой цепи можно конструировать так, чтобы они содержали нестрогие остатки аналогично тому, как описано выше. Количество и положение таких нестрогих остатков конкретно не ограничивается конкретными вариантами осуществления, при условии, что антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области легкой цепи, включают аминокислотные остатки, приведенные в таблицах 1 и 2.
Предпочтительные вариабельные области тяжелой цепи, подлежащие комбинированию, включают, например, рандомизированные библиотеки вариабельной области. Для получения рандомизированной библиотеки вариабельной области известные способы комбинируют соответствующим образом. В неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области предпочтительно можно использовать иммунную библиотеку, сконструированную на основе генов антител, происходящих из лимфоцитов животных, иммунизированных конкретным антигеном, пациентов с инфекциями, индивидуумов с повышенным титром антител в крови в результате вакцинации, пациентов со злокачественной опухолью или пациентов с аутоиммунным заболеванием.
В другом неограничивающем варианте осуществления настоящего изобретения, в качестве рандомизированной библиотеки вариабельной области также предпочтительно можно использовать синтетическую библиотеку, полученную путем замены последовательностей CDR V-генов в геномной ДНК или функциональных реконструированных V-генов набором синтетических олигонуклеотидов, содержащих последовательности, кодирующие наборы кодонов соответствующей длины. В этом случае, поскольку в последовательности CDR3 тяжелой цепи наблюдают разнообразие, также можно заменять только последовательность CDR3. Критерием возникновения разнообразия аминокислот в вариабельной области антигенсвязывающей молекулы является то, что разнообразие возникает у аминокислотных остатков в экспонированных на поверхности положениях антигенсвязывающей молекулы. Экспонированное на поверхности положение относится к положению, которое считается способным экспонироваться на поверхности и/или контактировать с антигеном, исходя из структуры, группы структур и/или смоделированной структуры антигенсвязывающей молекулы. Как правило, такие положения представляют собой CDR. Предпочтительно, экспонированные на поверхности положения определяют с использованием координат из трехмерной модели антигенсвязывающей молекулы с использованием компьютерной программы, такой как программа InsightII (Accelrys). Экспонированные на поверхности положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (J. Mol. Biol. (1971) 55, 379-400); Connolly (J. Appl. Cryst. (1983) 16, 548-558)). Определение экспонированных на поверхности положений можно проводить с использованием программного обеспечения, пригодного для моделирования белка, и информации о трехмерной структуре, полученной для антитела. Программное обеспечение, которое можно использовать для этих целей, предпочтительно включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Главным образом или предпочтительно, когда алгоритм требует введения пользователем параметра размера, "размер" образца, который используют в вычислении, устанавливают как радиус, равный приблизительно 1,4 Ангстрем или менее. Более того, способы определения экспонированных на поверхности областей и участков с использованием программного обеспечения для персональных компьютеров описаны Pacios (Comput. Chem. (1994) 18 (4), 377-386; J. Mol. Model. (1995) 1, 46-53).
В другом неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области также особенно предпочтительно можно использовать наивную библиотеку, которая сконструирована из генов антител, происходящих из лимфоцитов здоровых индивидуумов, и набор генов которой состоит из наивных последовательностей, которые представляют собой последовательности антител без предпочтений (Gejima et al. (Human Antibodies (2002) 11, 121-129); Cardoso et al. (Scand. J. Immunol. (2000) 51, 337-344)). В настоящем описании, аминокислотная последовательность, содержащая наивную последовательность, относится к аминокислотной последовательности, полученной из такой наивной библиотеки.
В одном варианте осуществления настоящего изобретения антигенсвязывающий домен по настоящему изобретению можно получать из библиотеки, содержащей множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, полученных путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки вариабельной области, где вариабельная область тяжелой цепи, выбрана в качестве каркасной последовательности, которая первоначально содержит "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов". Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают библиотеки, сконструированные путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки вариабельной области с последовательностью вариабельной области тяжелой цепи SEQ ID NO:9 (6RL#9-IgG1) или SEQ ID NO:10 (6KC4-1#85-IgG1). Альтернативно такую библиотеку можно конструировать путем выбора соответствующих вариабельных областей легкой цепи из вариабельных областей легкой цепи, которые имеют последовательности эмбрионального типа, вместо вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки последовательностей вариабельной области. Такие предпочтительные библиотеки включают, например, библиотеки, в которых последовательность вариабельной области тяжелой цепи SEQ ID NO:9 (6RL#9-IgG1) или SEQ ID NO:10 (6KC4-1#85-IgG1) комбинируют с вариабельными областями легкой цепи, имеющими последовательность эмбрионального типа.
Альтернативно последовательность вариабельной области тяжелой цепи, выбранную в качестве каркасной последовательности, которая первоначально содержит "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы", как упоминалось выше, можно конструировать так, чтобы она содержала нестрогие остатки. Количество и положение нестрогих остатков конкретно не ограничено, при условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области тяжелой цепи SEQ ID NO:9 (6RL#9-IgG1), включают все аминокислотные остатки CDR1 и CDR2 тяжелой цепи и аминокислотные остатки CDR3 тяжелой цепи, за исключением остатков в положениях 95, 96 и/или 100a, как указано согласно нумерации Kabat. Альтернативно неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области тяжелой цепи SEQ ID NO:10 (6KC4-1#85-IgG1), включают все аминокислотные остатки CDR1 и CDR2 тяжелой цепи и аминокислотные остатки CDR3 тяжелой цепи, за исключением остатков в положениях аминокислоты 95 и/или 101, как указано согласно нумерации Kabat.
Альтернативно библиотеку, содержащую множество антигенсвязывающих молекул, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельных областей легкой цепи, имеющих последовательности эмбрионального типа, с вариабельными областями тяжелой цепи, в которые внесен "по меньшей мере один аминокислотный остаток, ответственный за зависимое от концентрации ионов изменение антигенсвязывающей активности антигенсвязывающей молекулы". Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают библиотеки, в которых вариабельные области легкой цепи, сконструированные в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельные области легкой цепи, имеющие последовательности эмбрионального типа, комбинируют с последовательностью вариабельной области тяжелой цепи, в которой конкретный остаток(-и) заменен по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. Неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки CDR1 тяжелой цепи. Следующие неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки CDR2 тяжелой цепи. Кроме того, неограничивающие примеры таких аминокислотных остатков также включают аминокислотные остатки CDR3 тяжелой цепи. Неограничивающие примеры таких аминокислотных остатков CDR3 тяжелой цепи включают аминокислоты в положениях 95, 96, 100a и/или 101 в CDR3 вариабельной области тяжелой цепи, как указано согласно нумерации Kabat. Более того, эти аминокислотные остатки могут содержаться отдельно или в комбинации, при условии что они образуют кальций-связывающий мотив и/или антигенсвязывающая активность антигенсвязывающей молекулы варьирует в зависимости от концентрации ионов кальция.
Когда вариабельные области легкой цепи, сконструированные в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельные области легкой цепи, имеющие последовательность эмбрионального типа, комбинируют с вариабельной областью тяжелой цепи, в которую внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов, как упоминалось выше, последовательность вариабельной области тяжелой цепи также можно конструировать так, чтобы она содержала нестрогие остатки, аналогично тому, как описано выше. Количество и положение нестрогих остатков конкретно не ограничено, при условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует, в зависимости от концентраций ионов. В частности, последовательности CDR и/или FR тяжелой цепи могут содержать один или несколько нестрогих остатков. Более того, рандомизированные библиотеки вариабельной области можно предпочтительно использовать в качестве аминокислотных последовательностей CDR1, CDR2 и/или CDR3 вариабельной области тяжелой цепи, отличных от аминокислотных остатков, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы. Когда в качестве вариабельных областей легкой цепи используют последовательности эмбрионального типа, неограничивающие примеры таких последовательностей включают последовательности SEQ ID NO:5 (Vk1), SEQ ID NO:6 (Vk2), SEQ ID NO:7 (Vk3) и SEQ ID NO:8 (Vk4).
Предпочтительно можно использовать любую из описанных выше аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов кальция, при условии что они образуют кальций-связывающий мотив. В частности, такие аминокислоты включают электронодонорные аминокислоты. Предпочтительные примеры таких электронодонорных аминокислот включают серин, треонин, аспарагин, глутаминовую кислоту, аспарагиновую кислоту и глутаминовую кислоту.
Условия концентраций ионов водорода
В одном варианте осуществления настоящего изобретения условия концентраций ионов относятся к условиям концентраций ионов водорода или условиям pH. В рамках настоящего изобретения концентрация протона, т.е. ядра атома водорода, используется в качестве синонима водородного показателя (pH). Когда активность ионов водорода в водном растворе отображают как aH+, pH определяется как -log10aH+. Когда ионная сила водного раствора является низкой (например, ниже 10-3), aH+ практически равна ионной силе водорода. Например, ионное произведение воды при 25°C и 1 атмосфере составляет Kw=aH+aOH=10-14, и, таким образом, в чистой воде aH+=aOH=10-7. В этом случае, pH=7 является нейтральным; водный раствор, pH которого ниже 7, является кислым, или pH которого больше 7, является щелочным.
В рамках настоящего изобретения, когда условия pH используют в качестве условий концентрации ионов, условия pH включают высокие концентрации ионов водорода или низкие значения pH, т.е. диапазон кислых значений pH, и низкие концентрации ионов водорода или высокие значения pH, т.е. диапазон нейтральных значений pH. "Активность связывания варьирует в зависимости от условий pH" означает, что антигенсвязывающая активность антигенсвязывающей молекулы варьирует вследствие различий в условиях высокой концентрации ионов водорода или низких значений pH (диапазон кислых значений pH) и низкой концентрации ионов водорода или высоких значений pH (диапазон нейтральных значений pH). Это включает, например, случай, когда антигенсвязывающая активность антигенсвязывающей молекулы является более высокой в диапазоне нейтральных значений pH, чем в диапазоне кислых значений pH, и случай, когда антигенсвязывающая активность антигенсвязывающей молекулы является более высокой в диапазоне кислых значений pH, чем в диапазоне нейтральных значений pH.
В рамках настоящего изобретения диапазон нейтральных значений pH не ограничивается конкретной величиной и предпочтительно выбран из диапазона от 6,7 до 10,0. В другом варианте осуществления значение pH может быть выбрано из диапазона от 6,7 до 9,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от 7,0 до 9,0. В другом варианте осуществления значение pH может быть выбрано из диапазона от 7,0 до 8,0. Особенно предпочтительное значение pH включает 7,4, которое является близким к pH плазмы (крови) in vivo.
В рамках настоящего изобретения диапазон кислых значений pH не ограничивается конкретной величиной и предпочтительно выбран из диапазона от 4,0 до 6,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от 4,5 до 6,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от 5,0 до 6,5. В другом варианте осуществления pH может быть выбрано из диапазона от 5,5 до 6,5. Особенно предпочтительное значение pH включает 5,8, что является близким к концентрации ионизированного кальция в ранней эндосоме in vivo.
В рамках настоящего изобретения "антигенсвязывающая активность антигенсвязывающей молекулы при высокой концентрации ионов водорода или низком значении pH (диапазон кислых значений pH) является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH (диапазон нейтральных значений pH)" означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от 4,0 до 6,5, является более слабой, чем при значении pH, выбранном из диапазона от 6,7 до 10,0; предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от 4,5 до 6,5, является более слабой, чем при pH, выбранном из диапазона от 6,7 до 9,5; более предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от 5,0 до 6,5, является более слабой, чем при значении pH, выбранном из диапазона от 7,0 до 9,0; еще более предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от 5,5 до 6,5 является более слабой, чем при значении pH, выбранном из диапазона от 7,0 до 8,0; особенно предпочтительно это означает, что антигенсвязывающая активность при значении pH в ранней эндосоме in vivo является более слабой, чем антигенсвязывающая активность при значении pH в плазме in vivo; и, конкретно, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при pH 5,8 является более слабой, чем антигенсвязывающая активность при pH 7,4.
Определение того, изменяется ли антигенсвязывающая активность антигенсвязывающей молекулы в зависимости от условий pH можно проводить, например, с использованием известных способов измерения, таких как способы, описанные в разделе "Активность связывания" выше. В частности, активность связывания измеряют в различных условиях pH с использованием способов измерения, описанных выше. Например, антигенсвязывающую активность антигенсвязывающей молекулы сравнивают в условиях диапазона кислых значений pH и диапазона нейтральных значений pH, чтобы подтвердить, что антигенсвязывающая активность антигенсвязывающей молекулы изменяется так, что она является более высокой в условиях диапазона нейтральных значений pH, чем в условиях диапазона кислых значений pH.
Более того, в рамках настоящего изобретения выражение "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или при высоком значении pH, т.е. в диапазоне нейтральных значений pH" также может быть выражено как "антигенсвязывающая активность антигенсвязывающей молекулы при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой чем при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH". В рамках настоящего изобретения "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH" может быть описана как "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более слабой, чем антигенсвязывающая способность при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH". Альтернативно "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, снижена так, чтобы она более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH" может быть описана как "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, снижена так, чтобы она была слабее, чем антигенсвязывающая способность при низкой концентрации ионов водорода или высоких значениях pH, т.е. в диапазоне нейтральных значений pH".
Условия, отличные от концентрации ионов водорода или pH для измерения и антигенсвязывающей активности, могут быть соответствующим образом выбраны специалистами в данной области и они конкретно не ограничены. Измерение можно проводить, например, при 37°C с использованием буфера HEPES. Измерение можно проводить, например, с использованием Biacore (GE Healthcare). Когда антиген представляет собой растворимый антиген, антигенсвязывающую активность антигенсвязывающей молекулы можно определять путем оценки активности связывания с растворимым антигеном путем нанесения антигена в качестве анализируемого соединения на чип, на который иммобилизована антигенсвязывающая молекула. Когда антиген представляет собой антиген мембраны, активность связывания с антигеном мембраны можно оценивать путем нанесения антигенсвязывающей молекулы в качестве анализируемого соединения на чип, на который иммобилизован антиген.
При условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более слабой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, соотношение антигенсвязывающей активности между антигенсвязывающей активностью при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, и при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH конкретно не ограничено, и величина KD (pH 5,8)/KD (pH 7,4), которая представляет собой соотношение константы диссоциации (KD) для антигена при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, и KD при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно составляет 2 или более; более предпочтительно величина KD (pH 5,8)/KD (pH 7,4) составляет 10 или более; и еще более предпочтительно величина KD (pH 5,8)/KD (pH 7,4) составляет 40 или более. Верхний предел величины KD (pH 5,8)/KD (pH 7,4) конкретно не ограничен и он может представлять собой любую величину, такую как 400, 1000 или 10000, при условии, что молекулу можно получать способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, константу диссоциации (KD) можно использовать в качестве величины антигенсвязывающей активности. Между тем, когда антиген представляет собой мембранный антиген, можно использовать кажущуюся константу диссоциации (KD). Константу диссоциации (KD) и кажущуюся константу диссоциации (KD) можно измерять способами, известными специалистам в данной области, и можно использовать Biacore (GE healthcare), график Скэтчарда, проточный цитометр и т.п.
Альтернативно, например, константу скорости диссоциации (kd) можно соответствующим образом использовать в качестве показателя, указывающего на соотношение антигенсвязывающей активности антигенсвязывающей молекулы по настоящему изобретению между активностью при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, и при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH. Когда kd (константа скорости диссоциации) используют в качестве показателя, указывающего на соотношение активностей связывания вместо KD (константа диссоциации), величина kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH), которая представляет собой соотношение kd (константа скорости диссоциации) в отношении антигена при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений, pH и kd (константа скорости диссоциации) при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно составляет 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более и еще более предпочтительно 30 или более. Верхний предел величины kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) конкретно не ограничен, и он может представлять собой любую величину, такую как 50, 100 или 200, при условии, что можно молекулу можно получать способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, в качестве величины антигенсвязывающей активности можно использовать константу скорости диссоциации (kd), и когда антиген представляет собой антиген мембраны, можно использовать кажущуюся константу скорости диссоциации (kd). Константу скорости диссоциации (kd) и кажущуюся константу скорости диссоциации (kd) можно определять способами, известными специалистам в данной области, и можно использовать Biacore (GE healthcare), проточный цитометр и т.п. В рамках настоящего изобретения, когда антигенсвязывающую активность антигенсвязывающей молекулы измеряют при различных концентрациях ионов водорода, т.е. pH, условия, отличные от концентрации ионов водорода, т.е. pH, предпочтительно являются одинаковыми.
Например, антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются одним из вариантов осуществления, предусматриваемых настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов или антител, включающего следующие стадии (a)-(c):
(a) получение значения антигенсвязывающей активности антигенсвязывающего домена или антитела в диапазоне кислых значений pH;
(b) получение значения антигенсвязывающей активности антигенсвязывающего домена или антитела в диапазоне нейтральных значений pH; и
(c) выбор антигенсвязывающего домена или антитела, антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH.
Альтернативно антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются одним из вариантов осуществления, предусматриваемых настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов или антител или их библиотеки, включающего следующие стадии (a)-(c):
(a) контактирование антигенсвязывающего домена, или антитела, или их библиотеки, в диапазоне нейтральных значений pH с антигеном;
(b) помещение антигенсвязывающего домена или антитела, связавшихся с антигеном на стадии (a), в диапазон кислых значений pH; и
(c) выделение антигенсвязывающего домена или антитела, диссоциировавших на стадии (b).
Антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов, или антител, или их библиотеки, включающего следующие стадии (a)-(d):
(a) контактирование в диапазоне кислых значений pH антигена с библиотекой антигенсвязывающих доменов или антител;
(b) отбор антигенсвязывающего домена или антитела, которые не связываются с антигеном на стадии (a);
(c) позволение антигенсвязывающему домену или антителу, отобранным на стадии (b), связываться с антигеном в диапазоне нейтральных значений pH; и
(d) выделение антигенсвязывающего домена или антитела, связавшихся с антигеном на стадии (c).
Антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(c):
(a) контактирование в диапазоне нейтральных значений pH библиотеки антигенсвязывающих доменов или антител с колонкой, на которой иммобилизован антиген;
(b) элюирование в диапазоне кислых значений pH из колонки антигенсвязывающего домена или антитела, связавшегося с колонкой на стадии (a); и
(c) выделение антигенсвязывающего домена или антитела, элюированных на стадии (b).
Антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(d):
(a) позволение в диапазоне кислых значений pH библиотеке антигенсвязывающих доменов или антител проходить через колонку, на которую иммобилизован антиген;
(b) сбор антигенсвязывающего домена или антитела, элюированных без связывания с колонкой на стадии (a);
(c) позволение антигенсвязывающему домену или антителу, собранным на стадии (b), связываться с антигеном в диапазоне нейтральных значений pH; и
(d) выделение антигенсвязывающего домена или антитела, связавшихся с антигеном на стадии (c).
Антигенсвязывающий домен или антитело, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(d):
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антител в диапазоне нейтральных значений pH;
(b) получение антигенсвязывающего домена или антитела, связавшихся с антигеном на стадии (a);
(c) помещение антигенсвязывающего домена или антитела, полученных на стадии (b), в диапазон кислых значений pH; и
(d) выделение антигенсвязывающего домена или антитела, антигенсвязывающая активность которых на стадии (c) является более слабой, чем критерий для отбора на стадии (b).
Описанные выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающим доменам и антителам, антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH, которые получают способом скрининга, который дополнительно включает стадии повторения стадий (a)-(c) или (a)-(d) в описанных выше способах скрининга. Количество повторений стадий (a)-(c) или (a)-(d) конкретно не ограничено; однако обычно количество составляет 10 или менее.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена или антитела при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, конкретно не ограничена, при условии что она представляет собой антигенсвязывающую активность при pH от 4,0 до 6,5, и включает антигенсвязывающую активность при pH от 4,5 до 6,6 в качестве предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH от 5,0 до 6,5, и антигенсвязывающую активность при pH от 5,5 до 6,5 в качестве других предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH в ранней эндосоме in vivo в качестве более предпочтительного значения pH, и, в частности, антигенсвязывающую активность при pH 5,8. Между тем, антигенсвязывающая активность антигенсвязывающего домена или антитела при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность при pH от 6,7 до 10, и включает антигенсвязывающую активность при pH от 6,7 до 9,5 в качестве предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH от 7,0 до 9,5 и антиген связывающую активность при pH от 7,0 до 8,0 в качестве других предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH плазмы in vivo в качестве более предпочтительных значений pH, и, в частности, антигенсвязывающую активность при pH 7,4.
Антигенсвязывающую активность антигенсвязывающего домена или антитела можно измерять способами, известными специалистам в данной области. Специалисты в данной области могут соответствующим образом определить условия, отличные от концентрации ионизированного кальция. Антигенсвязывающую активность антигенсвязывающего домена или антитела можно оценивать, основываясь на константе диссоциации (KD), кажущейся константе диссоциации (KD), константе скорости диссоциации (kd), кажущейся константе скорости диссоциации (kd) и т.п. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или FACS.
В рамках настоящего изобретения стадия отбора антигенсвязывающего домена или антитела, антигенсвязывающая активность которых при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является синонимичной стадии отбора антигенсвязывающего домена или антитела, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH.
При условии, что антигенсвязывающая активность при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, отличие между антигенсвязывающей активностью при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, и антигенсвязывающей активностью при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, конкретно не ограничено; однако антигенсвязывающая активность при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно в два или более раз, более предпочтительно в 10 или более раз, и еще более предпочтительно в 40 или более раз превышает антигенсвязывающую активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH.
Антигенсвязывающий домен или антитело по настоящему изобретению, подвергаемые скринингу способами скрининга, описанными выше, могут представлять собой любой антигенсвязывающий домен или антитело, и можно проводить скрининг упомянутого выше антигенсвязывающего домена или антитела. Например, можно проводить скрининг антигенсвязывающего домена или антитела, имеющих нативную последовательность, и можно проводить скрининг антигенсвязывающего домена или антитела, в которых их аминокислотные последовательности были заменены.
Антигенсвязывающий домен или антитело по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым способом. Например, можно использовать общепринятые антитела, общепринятые библиотеки (фаговая библиотека и т.д.), антитела или библиотеки, полученные из B-клеток иммунизированных животных или из гибридом, полученных путем иммунизации животных, антитела или библиотеки (библиотеки с увеличенным содержанием аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, библиотеки, в которые внесены аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислотные мутации в конкретных положениях и т.д.), полученные путем внесения аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислотных мутаций в описанные выше антитела или библиотеки.
Способы получения антигенсвязывающего домена или антитела, антигенсвязывающая активность которых при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, из антигенсвязывающих доменов или антител, полученных из гибридом, полученных иммунизацией животных или из B-клеток иммунизированных животных, предпочтительно включают, например, антигенсвязывающую молекулу или антитело, в которых по меньшей мере одна из аминокислот антигенсвязывающего домена или антитела заменена аминокислотой с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или мутацией на неприродную аминокислоту, или антигенсвязывающий домен или антитело, в которые встроена аминокислота с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродная аминокислота, такая как описана в WO 2009/125825.
Участки внесения мутаций на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты конкретно не ограничены, и они могут представлять собой любое положение, при условии что антигенсвязывающая активность в диапазоне кислых значений pH становится более слабой, чем в диапазоне нейтральных значений pH (величина KD (в диапазоне кислых значений pH)/KD (в диапазоне нейтральных значений pH) или kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) увеличивается) по сравнению с величиной до замены или вставки. Например, когда антигенсвязывающая молекула представляет собой антитело, являются пригодными вариабельная область и CDR антитела. Специалисты в данной области могут соответствующим образом определить количество аминокислот, подлежащих замене, или количество аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, подлежащих встраиванию. Можно осуществить замену на одну аминокислоту, имеющую pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или одну неприродную аминокислоту; можно осуществить встраивание одной аминокислоты, имеющей pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или одной неприродной аминокислоты; можно осуществить замену на две или более аминокислот, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или на две или более неприродных аминокислоты; и можно осуществить встраивание двух или более аминокислот, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или двух или более неприродных аминокислот. Альтернативно другие аминокислоты можно параллельно удалять, добавлять, встраивать и/или заменять, помимо замены на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, или встраивания аминокислот, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот. Замену на или встраивание аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот можно проводить случайным образом способами, такими как сканирование гистидином, в которых аланин в способе сканирования аланином, известном специалистам в данной области, заменяют на гистидин. Антигенсвязывающие молекулы, проявляющие более высокую величину KD (в диапазоне кислых значений pH)/KD (в диапазоне нейтральных значений pH) или kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) по сравнению с величиной до внесения мутации можно выбирать из антигенсвязывающих доменов или антител, в которые внесены случайные инсерции или мутации с заменой на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты.
Предпочтительные примеры антигенсвязывающих молекул, содержащих мутацию на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, как описано выше, и антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH, включают антигенсвязывающие молекулы, антигенсвязывающая активность которых в диапазоне нейтральных значений pH после мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты сравнима с антигенсвязывающей активностью до мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты. В рамках настоящего изобретения "антигенсвязывающая молекула после мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты имеет антигенсвязывающую активность, сравнимую с антигенсвязывающей активностью до мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты" означает, что, при принятии антигенсвязывающей активности антигенсвязывающей молекулы до мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты как 100%, антигенсвязывающая активность антигенсвязывающей молекулы после мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты составляет по меньшей мере 10% или более, предпочтительно 50% или более, более предпочтительно 80% или более, и еще более предпочтительно 90% или более. Антигенсвязывающая активность после мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты при pH 7,4 может быть более высокой, чем до мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты при pH 7,4. Если антигенсвязывающая активность антигенсвязывающей молекулы снижается вследствие встраивания или замены на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, антигенсвязывающую активность можно сделать сравнимой с антигенсвязывающей активностью до встраивания или замены на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, путем внесения замены, делеции, вставки и/или инсерции одной или нескольких аминокислот антигенсвязывающей молекулы. Настоящее изобретение также включает антигенсвязывающие молекулы, активность связывания которых скорректирована так, чтобы она была сравнимой, путем замены, делеции, вставки и/или инсерции одной или нескольких аминокислот после замены на или вставки аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродной аминокислоты.
Между тем, когда антигенсвязывающая молекула представляет собой вещество, содержащее константную область антитела, предпочтительные варианты осуществления антигенсвязывающих молекул, антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH, включают способы, в которых константные области антитела, содержащиеся в антигенсвязывающих молекулах, модифицированы. Конкретные примеры модифицированных константных областей антител предпочтительно включают константные области SEQ ID NO:11, 12, 13 и 14.
Аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающего домена в зависимости от условий концентрации ионов водорода
Антигенсвязывающие домены или антитела по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым способом. Например, когда условия концентрации ионов представляют собой условия концентрации ионов водорода или условия pH, можно использовать общепринятые антитела, общепринятые библиотеки (фаговая библиотека и т.д.), антитела или библиотеки, полученные из B-клеток иммунизированных животных или из гибридом, полученных иммунизацией животных, антитела или библиотеки (библиотеки с увеличенным содержанием аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, библиотеки, в которые внесены мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты в конкретных положениях и т.д.), полученные путем внесения мутаций на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты в описанные выше антитела или библиотеки.
В одном неограничивающем варианте осуществления настоящего изобретения библиотеку, содержащую множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, также можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, с вариабельными областями легкой цепи, в которые внесен "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации ионов водорода".
Такие аминокислотные остатки включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR1 легкой цепи. Аминокислотные остатки также включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR2 легкой цепи. Аминокислотные остатки также включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR3 легкой цепи.
Описанные выше аминокислотные остатки, содержащиеся в CDR1 легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 24, 27, 28, 31, 32 и/или 34 согласно нумерации Kabat в CDR1 вариабельной области легкой цепи. Между тем, аминокислотные остатки, содержащиеся в CDR2 легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 50, 51, 52, 53, 54, 55 и/или 56 согласно нумерации Kabat в CDR2 вариабельной области легкой цепи. Более того, аминокислотные остатки в CDR3 легкой цепи включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 89, 90, 91, 92, 93, 94 и/или 95A согласно нумерации Kabat, в CDR3 вариабельной области легкой цепи. Более того, аминокислотные остатки могут содержаться отдельно или они могут содержаться в комбинации из двух или более аминокислот, при условии, что они позволяют изменять антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов водорода.
Даже когда вариабельную область тяжелой цепи, полученную в качестве рандомизированной библиотеки последовательностей вариабельной области, комбинируют с описанной выше вариабельной областью легкой цепи, в которую внесен "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации ионов водорода", ее можно конструировать так, чтобы нестрогие остатки содержались в последовательности вариабельной области легкой цепи аналогично тому, как описано выше. Количество и положение нестрогих остатков конкретно не ограничено конкретным вариантом осуществления, при условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению изменяется в зависимости от условий концентрации ионов водорода. В частности, последовательности CDR и/или FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков. Например, нестрогие остатки, подлежащие внесению в последовательности вариабельных областей легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки, приведенные в таблицах 3 и 4. Между тем, аминокислотные последовательности вариабельных областей легкой цепи, отличные от нестрогих остатков, и аминокислотные остатки, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации ионов водорода, соответственно включают, но не ограничиваются ими, последовательности эмбрионального типа, такие как Vk1 (SEQ ID NO:5), Vk2 (SEQ ID NO:6), Vk3 (SEQ ID NO:7) и Vk4 (SEQ ID NO:8).
Любой аминокислотный остаток можно соответствующим образом использовать в качестве описанных выше аминокислотных остатков, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации ионов водорода. В частности, такие аминокислотные остатки включают аминокислоты с pKa боковой цепи 4,0-8,0. Такие электроноотдающие аминокислоты предпочтительно включают, например, встречающиеся в природе аминокислоты, такие как гистидин и глутаминовая кислота, а также неприродные аминокислоты, такие как аналоги гистидина (US 20090035836), m-NO2-Tyr (pKa 7,45), 3.5-Br2-Tyr (pKa 7,21) и 3.5-I2-Tyr (pKa 7,38) (Bioorg. Med. Chem. (2003) 11 (17), 3761-2768). Особенно предпочтительные аминокислотные остатки включают, например, аминокислоты с pKa боковой цепи 6,0-7,0. Такие электроноотдающие аминокислотные остатки предпочтительно включают, например, гистидин.
Для модификации аминокислот антигенсвязывающих доменов можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которых амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
Предпочтительная вариабельная область тяжелой цепи, которую используют в комбинации, включает, например, рандомизированные библиотеки вариабельной области. Известные способы соответствующим образом комбинируют в качестве способа получения рандомизированной библиотеки вариабельной области. В неограничивающем варианте осуществления настоящего изобретения, в качестве рандомизированной библиотеки вариабельной области можно использовать иммунную библиотеку, сконструированную на основе генов антител, происходящих из животных, иммунизированных конкретными антигенами, пациентов с инфекцией или индивидуумов с повышенным титром антител в крови в результате вакцинации, пациентов со злокачественной опухолью или лимфоцитов от пациентов с иммунными заболеваниями.
В другом неограничивающем варианте осуществления настоящего изобретения, аналогично тому, как описано выше, в качестве рандомизированной библиотеки вариабельной области также пригодным образом можно использовать синтетическую библиотеку, полученную путем замены последовательностей CDR V-генов в геномной ДНК или функциональных реконструированных V-генов набором синтетических олигонуклеотидов, содержащих последовательности, кодирующие наборы кодонов соответствующей длины. В этом случае, поскольку в последовательности CDR3 тяжелой цепи наблюдают разнообразие, также можно заменять только последовательность CDR3. Основанием для получения вариантов аминокислот в вариабельной области антигенсвязывающей молекулы является получение вариантов аминокислотных остатков экспонированных на поверхности положений антигенсвязывающей молекулы. Экспонированное на поверхности положение относится к положению, которое считается способным экспонироваться на поверхности и/или контактировать с антигеном, исходя из структуры, группы структур и/или смоделированной структуры антигенсвязывающей молекулы, и, как правило, такие положения представляют собой CDR. Предпочтительно, экспонированные на поверхности положения определяют с использованием координат из трехмерной модели антигенсвязывающей молекулы с использованием компьютерной программы, такой как программа InsightII (Accelrys). Экспонированные на поверхности положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (J. Mol. Biol. (1971) 55, 379-400); Connolly (J. Appl. Cryst. (1983) 16, 548-558)). Определение экспонированных на поверхности положений можно проводить на основе информации о трехмерной структуре с использованием программного обеспечения, пригодного для моделирования белка. Программное обеспечение, которое можно использовать для этих целей, предпочтительно включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Когда алгоритм требует введения пользователем параметра размера, "размер" образца, который используют в вычислении, устанавливают как радиус, приблизительно равный 1,4 Ангстрем или менее. Более того, способы определения экспонированных на поверхности областей и участков с использованием программного обеспечения для PC описаны Pacios (Comput. Chem. (1994) 18 (4), 377-386; J. Mol. Model. (1995) 1, 46-53).
В другом неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области также особенно пригодным является использование наивной библиотеки, которая сконструирована из генов антител, происходящих из лимфоцитов здоровых индивидуумов, и набор генов которой состоит из наивных последовательностей, которые представляют собой последовательности антител без предпочтений (Gejima et al. (Human Antibodies (2002) 11, 121-129); Cardoso et al. (Scand. J. Immunol. (2000) 51, 337-344)).
FcRn
В отличие от Fcγ-рецептора, принадлежащего семейству иммуноглобулинов, FcRn, в частности, FcRn человека, структурно сходен с полипептидами главного комплекса гистосовместимости (MHC) класса I, проявляя 22%-29% идентичность последовательности с молекулами MHC класса I (Ghetie el al., Immunol. Today (1997) 18 (12): 592-598). FcRn экспрессируется в качестве гетеродимера, состоящего из растворимой β- или легкой цепи (β2-микроглобулин) в комплексе с трансмембранной α- или тяжелой цепью. Подобно MHC, α-цепь FcRn содержит три внеклеточных домена (α1, α2 и α3) и ее короткий цитоплазматический домен заякоривает белок на клеточной поверхности. α1- и α2-домены взаимодействуют с FcRn-связывающим доменом Fc-области антитела (Raghavan et al., Immunity (1994) 1: 303-315).
FcRn экспрессируется в материнской плаценте и желточном мешке млекопитающих, и он вовлечен в перенос IgG от матери к плоду. Кроме того, в тонком кишечнике новорожденных грызунов, где FcRn экспрессируется, FcRn вовлечен в перенос материнских IgG через эпителий щеточной каемки из проглоченного молозива или молока. FcRn экспрессируется во множестве других тканей и системах эндотелиальных клеток различных видов. FcRn также экспрессируется у взрослого человека в эндотелии, кровеносных сосудах мышц и синусоидных капиллярах печени. Полагают, что FcRn участвует в поддержании концентрации IgG в плазме путем опосредования рециклирования IgG в сыворотку при связывании с IgG. Как правило, связывание FcRn с молекулами IgG строго зависит от pH. Оптимальное связывание наблюдают при кислом диапазоне pH ниже 7,0.
FcRn человека, предшественником которого является полипептид, имеющий сигнальную последовательность SEQ ID NO:15 (полипептид с сигнальной последовательностью представлен в SEQ ID NO:16), образует комплекс с β2-микроглобулином человека in vivo. Как показано в справочных примерах, описанных ниже, растворимый FcRn человека в комплексе с β2-микроглобулином получают с использованием общепринятых способов рекомбинантной экспрессии. FcRn-области по настоящему изобретению можно оценивать в отношении их активности связывания с таким растворимым FcRn человека в комплексе с β2-микроглобулином. В рамках настоящего изобретения, если нет иных указаний, FcRn человека относится к форме, способной связываться с FcRn-связывающим доменом по настоящему изобретению. Примеры включают комплекс между FcRn человека и β2-микроглобулином человека.
FcRn-связывающие домены
Один вариант осуществления настоящего изобретения относится к фармацевтическим композициям, индуцирующим иммунный ответ на упомянутый выше антиген, которые включают в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH.
Антигенсвязывающие молекулы по настоящему изобретению обладают FcRn-связывающим доменом. FcRn-связывающий домен конкретно не ограничен при условии, что антигенсвязывающая молекула обладает активностью связывания FcRn в диапазоне нейтральных значений pH, и он может представлять собой домен, который обладает активностью прямого или непрямого связывания с FcRn. Предпочтительные примеры таких доменов включают Fc-области IgG-иммуноглобулинов, альбумин, домен 3 альбумина, антитела против FcRn, пептиды против FcRn, каркасные молекулы против FcRn и т.п., которые обладают активностью прямого связывания с FcRn, или молекулы, которые связываются с IgG или альбумином, которые обладают активностью прямого связывания с FcRn. Для каркаса против FcRn можно использовать домен с любой структурой упомянутых выше антигенсвязывающих доменов, характеризующийся связыванием с FcRn. В рамках настоящего изобретения, предпочтительным является домен, обладающий активностью связывания с FcRn в диапазоне кислых значений pH и в диапазоне нейтральных значений pH. Такой домен предпочтительно можно использовать как есть, если он представляет собой домен, уже обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. Когда домен не обладает или обладает слабой активностью связывания FcRn в диапазоне нейтральных значений pH, аминокислоты в антигенсвязывающей молекуле можно модифицировать для сообщения активности связывания FcRn. Альтернативно активность связывания FcRn можно усиливать посредством изменения аминокислот в домене, уже обладающем активностью связывания FcRn в диапазоне нейтральных значений pH. Желательные изменения аминокислот в FcRn-связывающем домене можно идентифицировать посредством сравнения активности связывания FcRn активность в диапазоне нейтральных значений pH до и после изменения аминокислот.
Более того, в другом варианте осуществления настоящего изобретения предпочтительно используют FcRn-связывающий домен, который обладает активностью связывания FcRn в условиях низкой концентрации ионов кальция и в условиях высокой концентрация ионов кальция. Такой домен предпочтительно можно использовать как есть, если домен уже обладает активностью связывания FcRn в условиях высокой концентрации ионов кальция. Когда домен не обладает или обладает слабой активностью связывания FcRn в условиях высокой концентрации ионов кальция, аминокислоты в антигенсвязывающей молекуле можно модифицировать для сообщения активности связывания FcRn. Альтернативно активность связывания FcRn можно увеличивать посредством изменения аминокислот в домене, уже обладающем активностью связывания FcRn в условиях высокой концентрации ионов кальция. Желательные аминокислотные изменения в FcRn-связывающем домене можно идентифицировать посредством сравнения активности связывания FcRn в условиях высокой концентрации ионов кальция до и после изменения аминокислот. FcRn-связывающие домены можно получать способами на основе способов скрининга или продуцирования антигенсвязывающих доменов, обладающих антигенсвязывающей активностью, которая изменяется в зависимости от условий концентрации ионов кальция, как упоминалось выше в разделе "Условия концентрации ионов". Примеры таких FcRn-связывающих доменов включают антитела против FcRn, пептиды против FcRn, каркасные молекулы против FcRn и т.п.
Предпочтительный FcRn-связывающий домен человека представляет собой область, которая прямо связывается с FcRn. Такие предпочтительные FcRn-связывающие домены включают, например, Fc-области антител. Между тем, области, способные связываться с полипептидом, таким как альбумин или IgG, которые обладают активностью связывания FcRn, могут непрямо связываться с FcRn через альбумин, IgG и т.п. Таким образом, для FcRn-связывающей области в рамках настоящего изобретения, предпочтительно можно использовать область, которая связывается с полипептидом, обладающим активностью связывания FcRn. Fc-область содержит аминокислотную последовательность, происходящую из константной области тяжелой цепи антитела. Fc-область представляет собой часть константной области тяжелой цепи антитела, начинающуюся с N-конца шарнирной области в участке расщепления папаином, который находится на аминокислоте приблизительно в положении 216 в соответствии с нумерацией EU, и включающую шарнирную область, домены CH2 и CH3.
Активность связывания FcRn-связывающего домена по настоящему изобретению с FcRn, в частности FcRn человека, можно измерять способами, известными специалистам в данной области, как описано в разделе "Активность связывания" выше. Специалисты в данной области могут соответствующим образом определить условия, отличные от pH. Антигенсвязывающую активность и активность связывания FcRn человека антигенсвязывающей молекулы можно оценивать, основываясь на константе диссоциации (KD), кажущейся константе диссоциации (KD), константе скорости диссоциации (kd), кажущейся константе скорости диссоциации (kd) и т.п. Их можно измерять способами, известными специалистам в данной области. Например, можно использовать Biacore (GE healthcare), график Скэтчарда или проточный цитометр.
Когда измеряют активность связывания FcRn-связывающего домена с FcRn человека, условия, отличные от pH, конкретно не ограничены, и они могут быть соответствующим образом выбраны специалистами в данной области. Измерения можно проводить, например, при 37°C с использованием буфера MES, как описано в WO 2009/125825. Альтернативно активность связывания FcRn-связывающего домена с FcRn можно измерять способами, известными специалистам в данной области, и ее можно измерять с использованием, например, Biacore (GE Healthcare) т.п. Активность связывания FcRn-связывающего домена с FcRn можно оценивать путем нанесения в качестве анализируемого образца FcRn, FcRn-связывающего домена или антигенсвязывающей молекулы по настоящему изобретению, содержащей FcRn-связывающий домен, на чип, на который иммобилизован FcRn-связывающий домен, антигенсвязывающая молекула по настоящему изобретению, содержащая FcRn-связывающий домен, или FcRn.
Диапазон кислых значений pH в качестве условий, в которых FcRn-связывающий домен в антигенсвязывающей молекуле по настоящему изобретению обладает активностью связывания FcRn, обычно относится к pH 4,0-6,5. Предпочтительно он относится к pH 5,5-6,5 и особенно предпочтительно к pH 5,8-6,0, что является близким к pH в эндосоме ранней стадии in vivo. Более того, диапазон нейтральных значений pH в качестве условий, в которых FcRn-связывающий домен в антигенсвязывающей молекуле по настоящему изобретению обладает активностью связывания FcRn, обычно относится к pH 6,7-10,0. Диапазон нейтральных значений pH предпочтительно представляет собой диапазон, определяемый любой величиной pH в пределах 7,0-8,0, и предпочтительно его выбирают из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, и особенно предпочтительно pH 7,4, что является близким к pH плазмы (крови) in vivo. Когда аффинность связывания между FcRn-связывающим доменом человека и FcRn человека трудно оценить вследствие того, что аффинность связывания при pH 7,4 является низкой, вместо pH 7,4 можно использовать pH 7,0. Что касается температуры, используемой для условий измерения, аффинность связывания между FcRn-связывающим доменом и FcRn можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, для определения аффинности связывания между FcRn-связывающим доменом и FcRn человека используют температуру от 15°C до 40°C. Более предпочтительно, для определения аффинности связывания между FcRn-связывающим доменом и FcRn сходным образом используют любую температуру от 20°C до 35°C, такую как любая температура, выбранная из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35°C. В одном варианте осуществления настоящего изобретения 25°C является неограничивающим примером такой температуры.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность встречающегося в природе IgG1 человека в отношении связывания FcRn человека составляет KD 1,7 мкМ в диапазоне кислых значений pH (pH 6,0), но практически не поддается определению в диапазоне нейтральных значений pH. Таким образом, в предпочтительном варианте осуществления можно использовать антигенсвязывающие молекулы по настоящему изобретению, обладающие активностью связывания FcRn человека в диапазоне кислых значений pH, которые включают антигенсвязывающие молекулы, активность связывания FcRn человека которых в диапазоне кислых значений pH составляет KD 20 мкМ или сильнее, и активность связывания FcRn человека которых в диапазоне нейтральных значений pH эквивалентна этой активности у встречающегося в природе IgG человека. В более предпочтительном варианте осуществления, можно использовать антигенсвязывающие молекулы по настоящему изобретению, включающие антигенсвязывающие молекулы, активность связывания FcRn человека в диапазоне кислых значений pH составляет KD 2,0 мкМ или сильнее. В еще более предпочтительном варианте осуществления можно использовать антигенсвязывающие молекулы, активность связывания FcRn человека которых активность в диапазоне кислых значений pH составляет KD 0,5 мкМ или сильнее. Описанные выше величины KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (антигенсвязывающие молекулы иммобилизуют на чип и FcRn человека позволяют протекать в качестве анализируемого соединения).
В рамках настоящего изобретения предпочтительной является Fc-область, обладающая активностью связывания FcRn в диапазоне кислых значений pH. Если такой домен представляет собой Fc-область, уже обладающую активностью связывания FcRn в диапазоне кислых значений pH, его можно использовать как есть. Если домен не обладает или обладает слабой активностью связывания FcRn в диапазоне кислых значений pH, Fc-область, обладающую желаемой активностью связывания FcRn, можно получать путем изменения аминокислот в антигенсвязывающей молекуле. Также Fc-область, обладающую желаемой или усиленной активностью связывания FcRn в диапазоне кислых значений pH, можно пригодным образом получать путем изменения аминокислот в Fc-области. Изменения аминокислот в Fc-области, которые приводят к такой желаемой активности связывания, можно определять посредством сравнения активности связывания FcRn в диапазоне кислых значений pH до и после изменения аминокислот. Специалисты в данной области могут соответствующим образом осуществлять изменение аминокислот с использованием известных способов, таких как перекрывающаяся ПЦР и сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)), аналогично упомянутым выше способам, используемым для изменения антигенсвязывающей активности.
Fc-область, обладающую активностью связывания FcRn в диапазоне кислых значений pH, которая содержится в антигенсвязывающих молекулах по настоящему изобретению, можно получать любыми способами, но, в частности, FcRn-связывающий домен, обладающий активностью связывания FcRn или усиленной активностью связывания FcRn в диапазоне кислых значений pH, можно получать путем изменения аминокислот IgG-иммуноглобулина человека, который используют в качестве исходной Fc-области. Примеры предпочтительных Fc-областей IgG-иммуноглобулинов, подлежащих изменению, включают Fc-область IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Аминокислоты в любых положениях можно заменять другими аминокислотами при условии, что Fc-область обладает активностью связывания FcRn в диапазоне кислых значений pH, или ее активность связывания FcRn человека в кислом диапазоне может быть усилена. Когда антигенсвязывающая молекула включает Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят усилению связывания FcRn в диапазоне кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Примеры аминокислот, в которые можно вносить такие изменения, предпочтительно включают, например, аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447 в соответствии с нумерацией EU, как описано в WO 2000/042072. Аналогично, примеры аминокислот, в которые можно вносить такие изменения, также предпочтительно включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, примеры аминокислот, в которые можно вносить такие изменения, также включают аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO 2004/092219. Кроме того, примеры аминокислот, в которые можно вносить такие изменения, также предпочтительно включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193. Изменение этих аминокислот усиливает связывание Fc-области IgG-иммуноглобулина с FcRn в диапазоне кислых значений pH.
В рамках настоящего изобретения предпочтительной является Fc-область, обладающая активностью связывания FcRn в диапазоне нейтральных значений pH. Если домен представляет собой Fc-область, уже обладающую активностью связывания FcRn в диапазоне нейтральных значений pH, его можно использовать как есть. Когда домен не обладает или обладает слабой активностью связывания FcRn в диапазоне нейтральных значений pH, Fc-область, обладающую желаемой активностью связывания FcRn, можно получать путем изменения аминокислот в антигенсвязывающей молекуле. Также Fc-область, обладающую желаемой или усиленной активностью связывания FcRn в диапазоне нейтральных значений pH, можно пригодным образом получать путем изменения аминокислот в Fc-области. Аминокислотные изменения в Fc-области, которые приводят к такой желаемой активности связывания, можно определять путем сравнения активности связывания FcRn в диапазоне нейтральных значений pH до и после изменения аминокислот. Специалисты в данной области могут пригодным образом осуществлять изменение аминокислот с использованием известных способов, таких как перекрывающаяся ПЦР или сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)), аналогично упомянутым выше способам, используемым для изменения антигенсвязывающей активности.
Fc-область, обладающую активностью связывания FcRn в диапазоне нейтральных значений pH, которая содержится в антигенсвязывающей молекуле по настоящему изобретению, можно получать любым способом, но, в частности, FcRn-связывающий домен, обладающий активностью связывания FcRn или усиленной активностью связывания FcRn в диапазоне нейтральных значений pH, можно получать путем модификации аминокислот IgG-иммуноглобулина человека, используемого в качестве исходной Fc-области. Примеры предпочтительных Fc-областей IgG-иммуноглобулинов, подлежащих модификации, включают Fc-область IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Аминокислоты в любых положениях можно заменять другими аминокислотами при условии, что Fc-область обладает активностью связывания FcRn в диапазоне нейтральных значений pH или ее активность связывания FcRn человека в нейтральном диапазоне может быть усилена. Когда антигенсвязывающая молекула включает Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят к усилению связывания FcRn в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Величины KD для FcRn в диапазоне нейтральных значений pH определяют, как описано выше, способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (антигенсвязывающие молекулы иммобилизуют на чип и FcRn человека позволяют протекать в качестве анализируемого соединения).
Примеры предпочтительных Fc-областей IgG-иммуноглобулинов, подлежащих изменению, включают Fc-область IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Аминокислоты в любых положениях можно заменять другими аминокислотами при условии, что Fc-область обладает активностью связывания FcRn в диапазоне нейтральных значений pH или ее активность связывания FcRn человека в нейтральном диапазоне может быть усилена. Когда антигенсвязывающая молекула включает Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят к усилению связывания FcRn в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Для получения Fc-областей, в которых проведены такие изменения, различные мутации, представленные в таблице 5, вносили в VH3-IgG1 (SEQ ID NO:17) и оценивали. Варианты (с IgG1-F1 по IgG1-F1052), каждый из которых содержал продуцированную тяжелую цепь и легкую цепь, L (WT) (SEQ ID NO:18), экспрессировали и очищали в соответствии со способами, описанными в справочном примере 1.
Связывание между антителом FcRn человека анализировали в соответствии со способом, описанным в примере 3-3. Активность связывания вариантов с FcRn человека в нейтральных условиях (pH7,0), измеренная с использованием Biacore, представлена в таблице 5 (таблицы с 5-1 по 5-33).
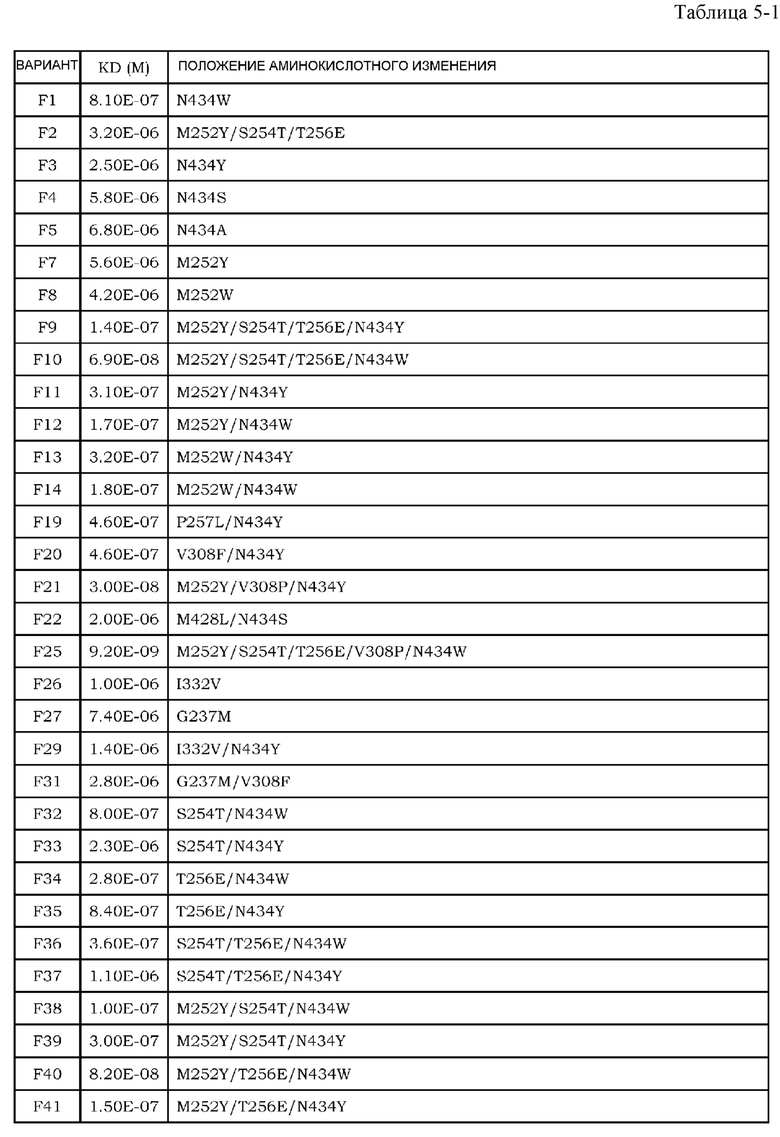
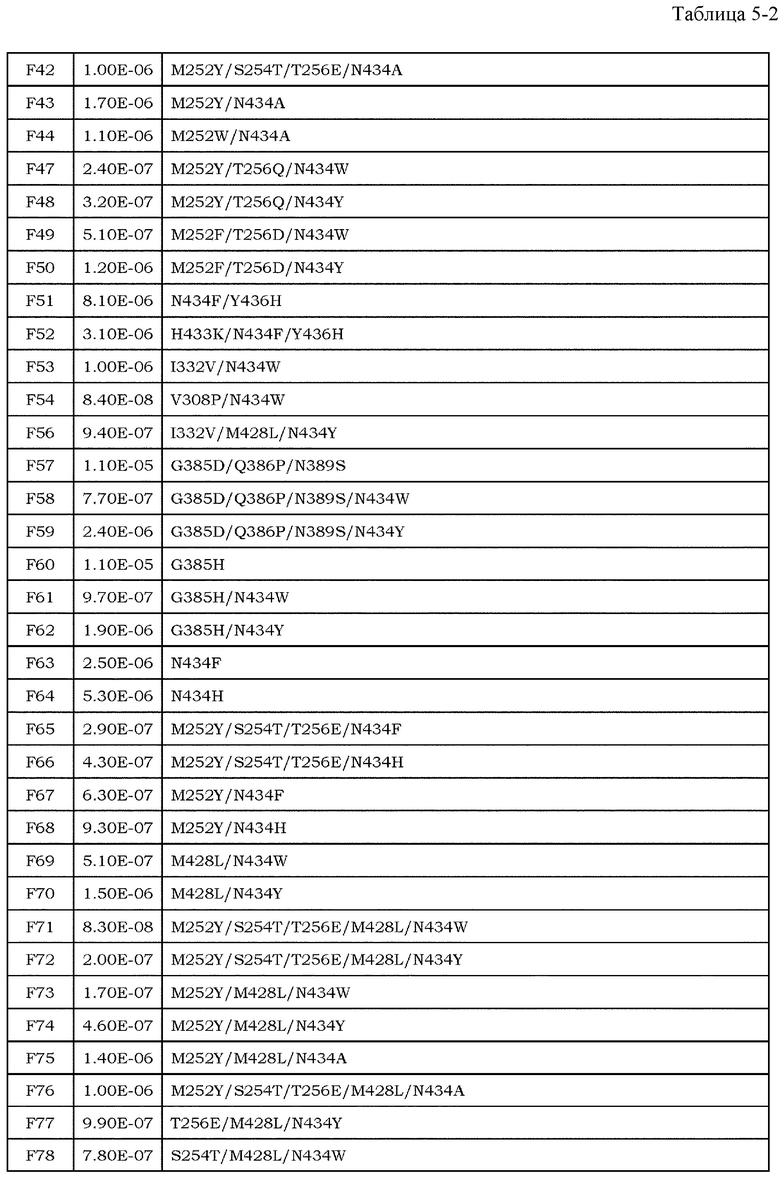
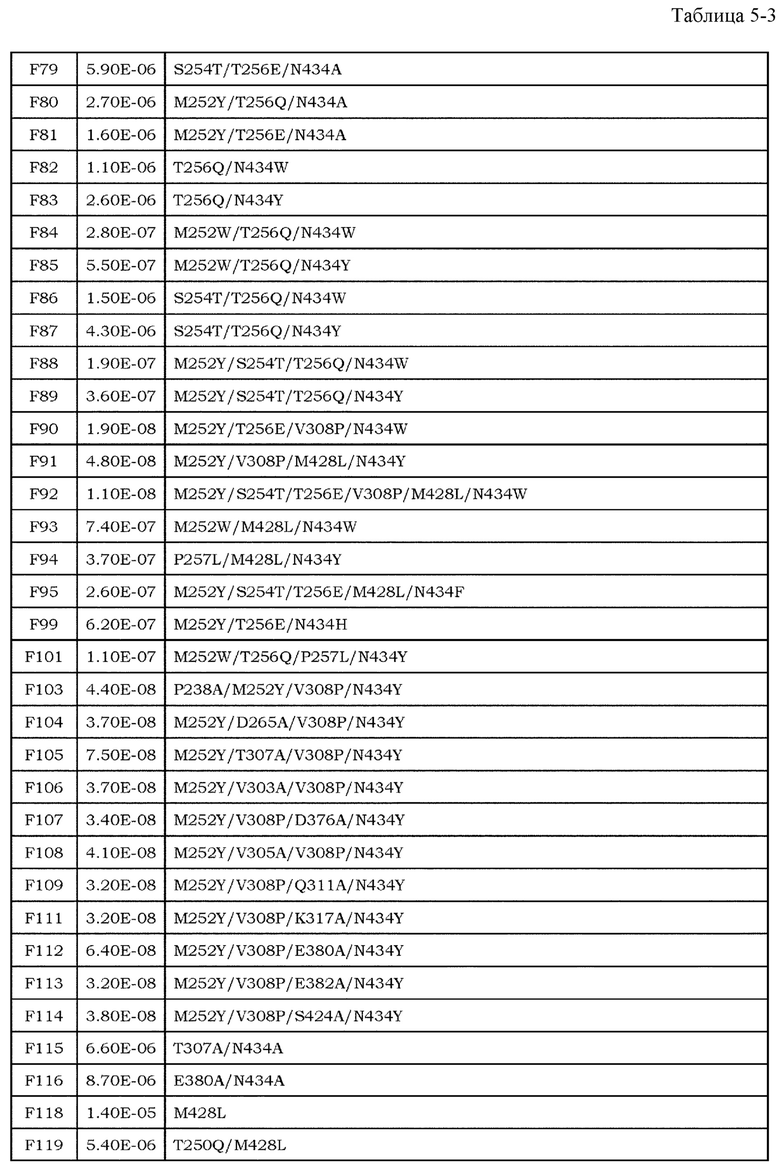
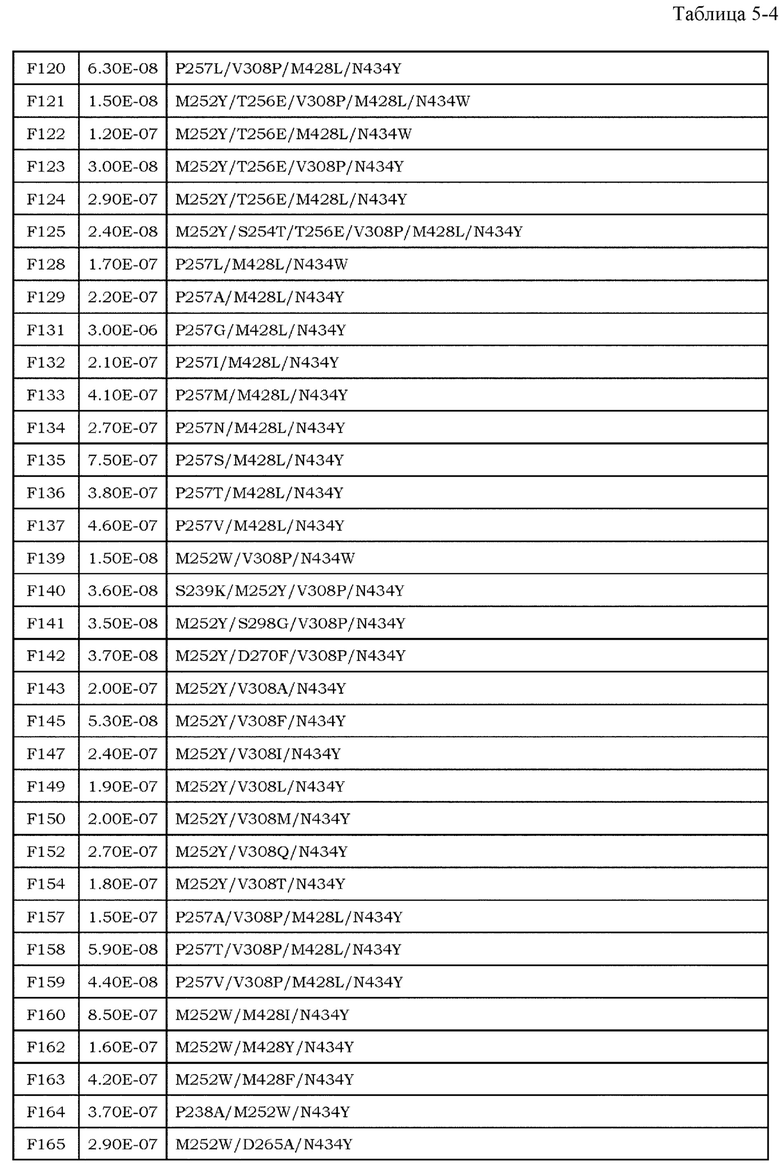

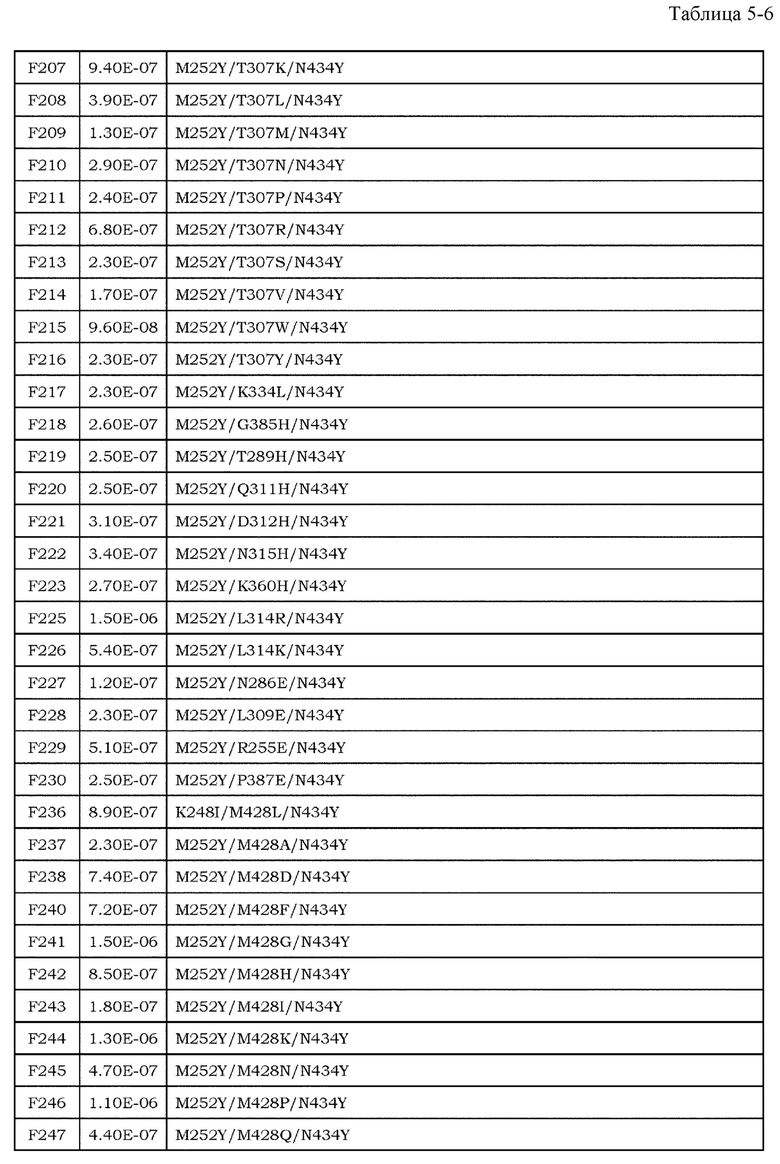




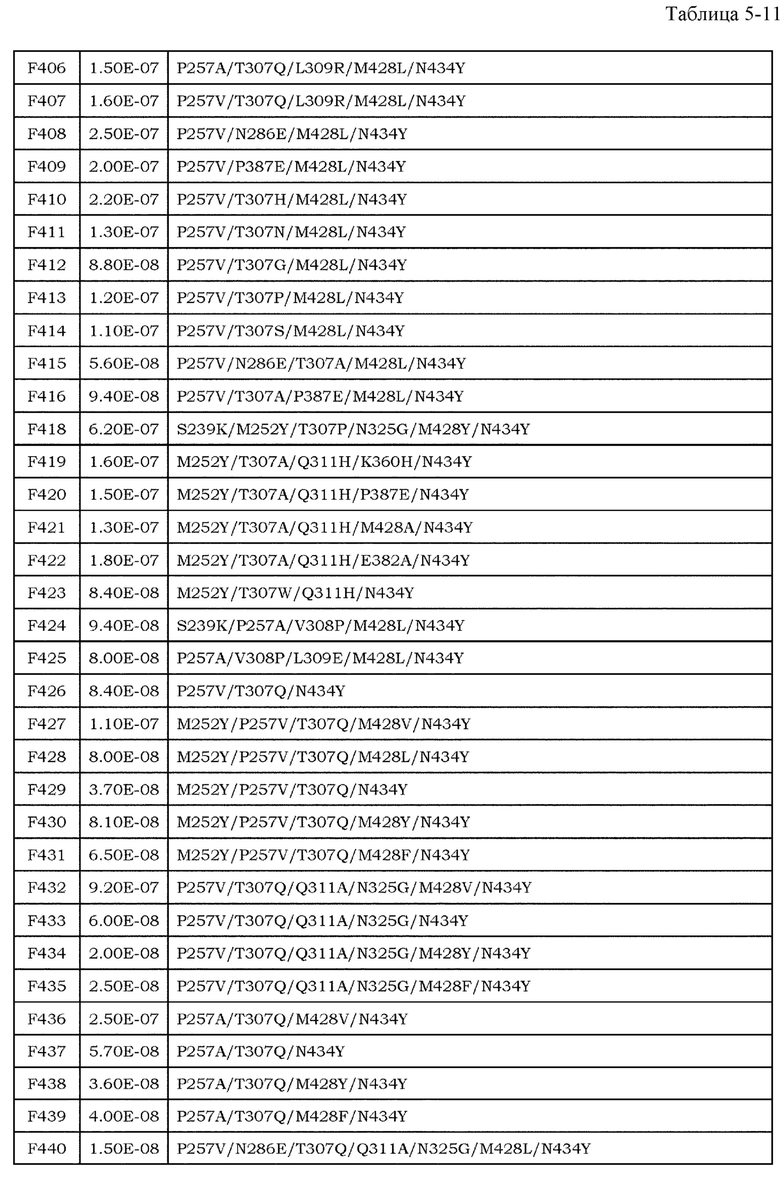
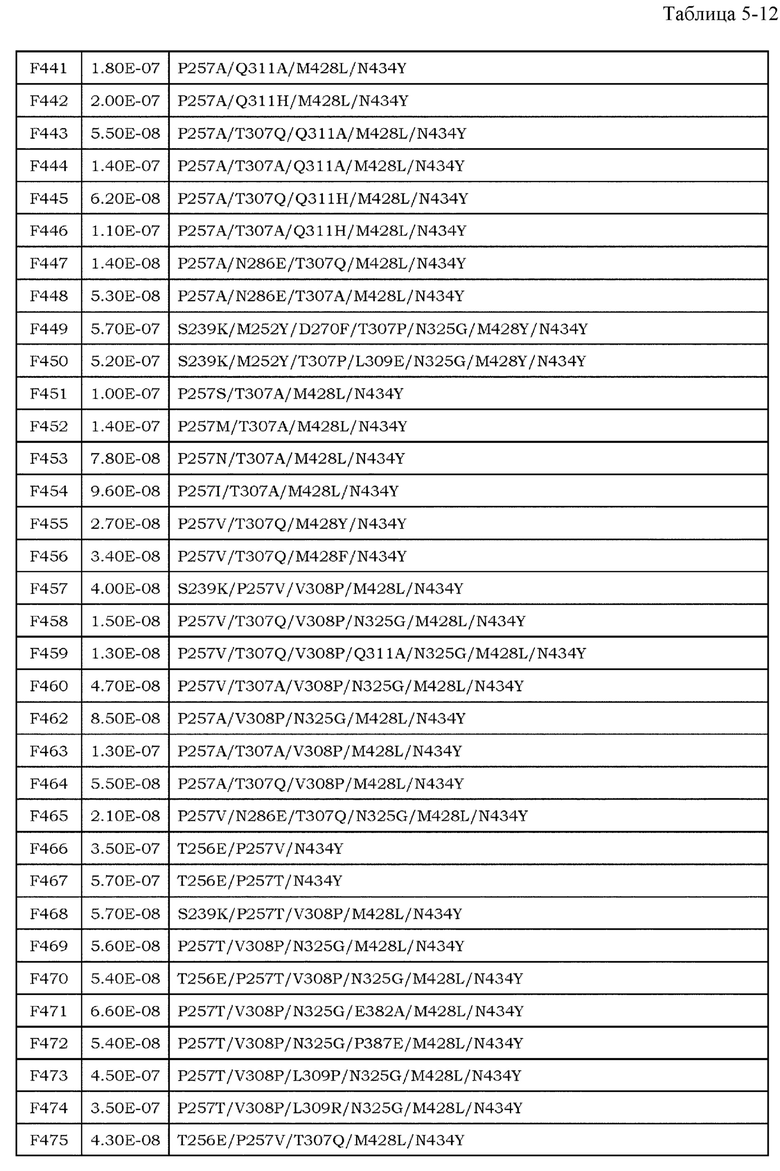






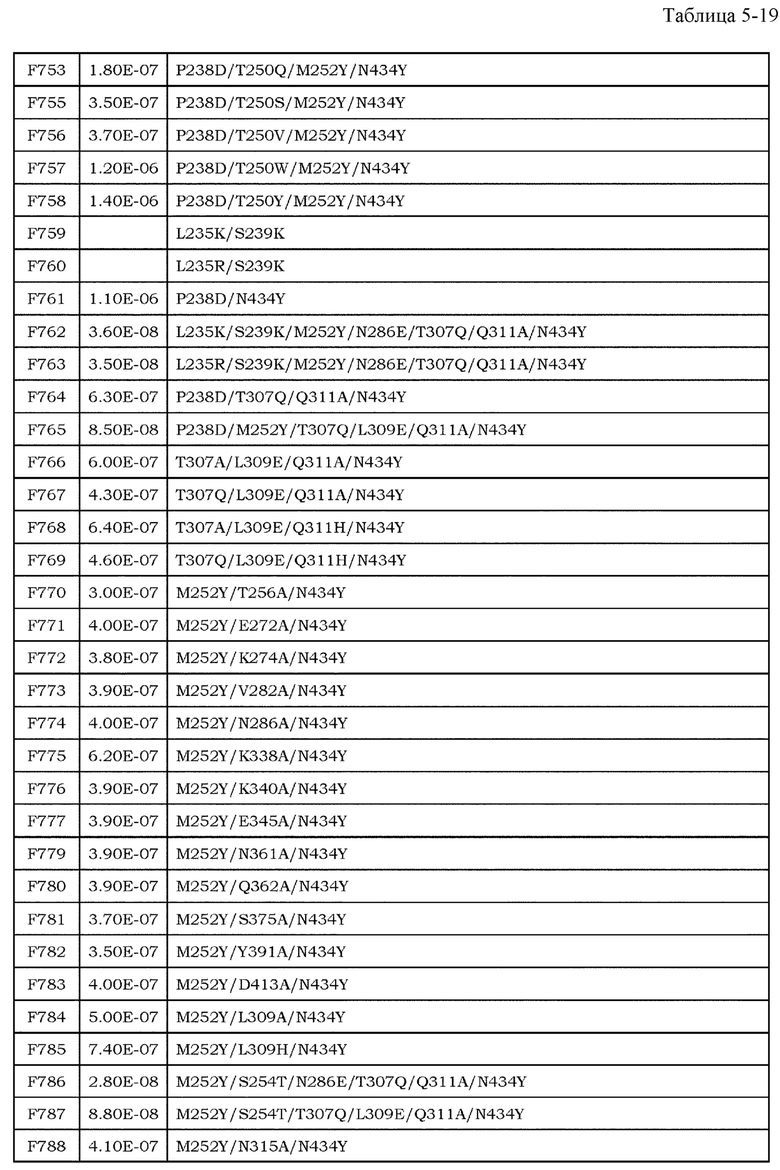
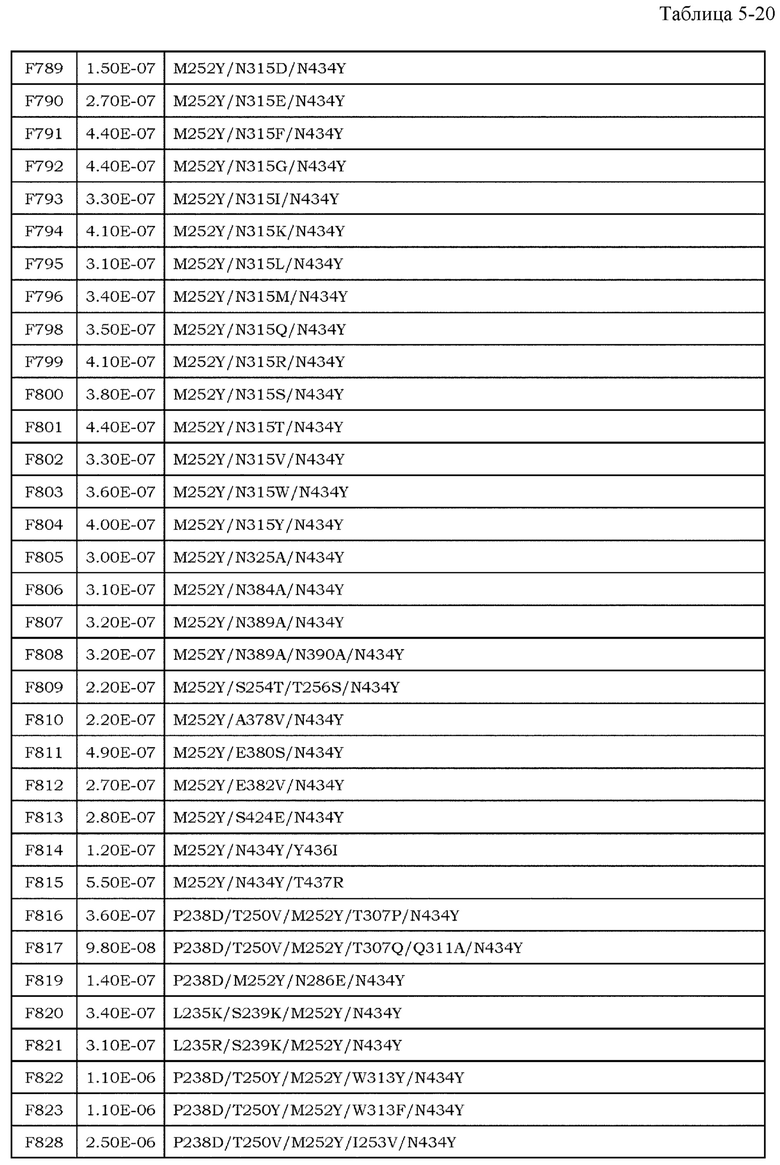
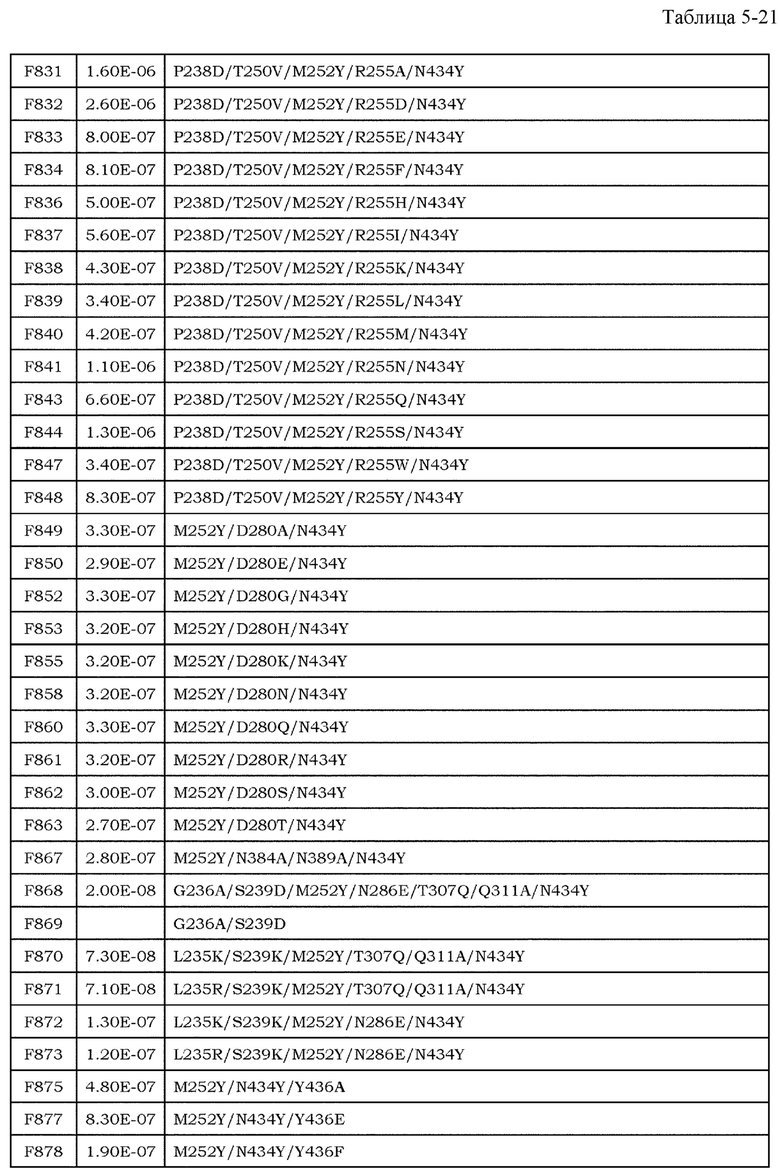
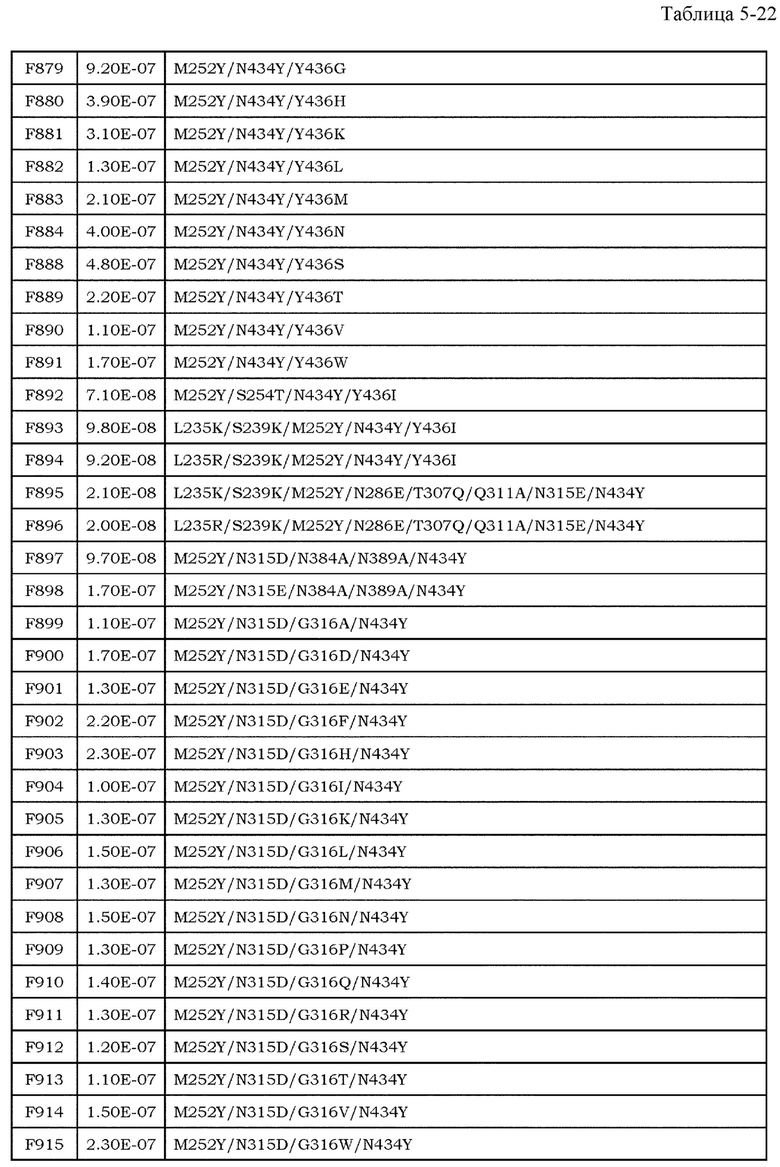
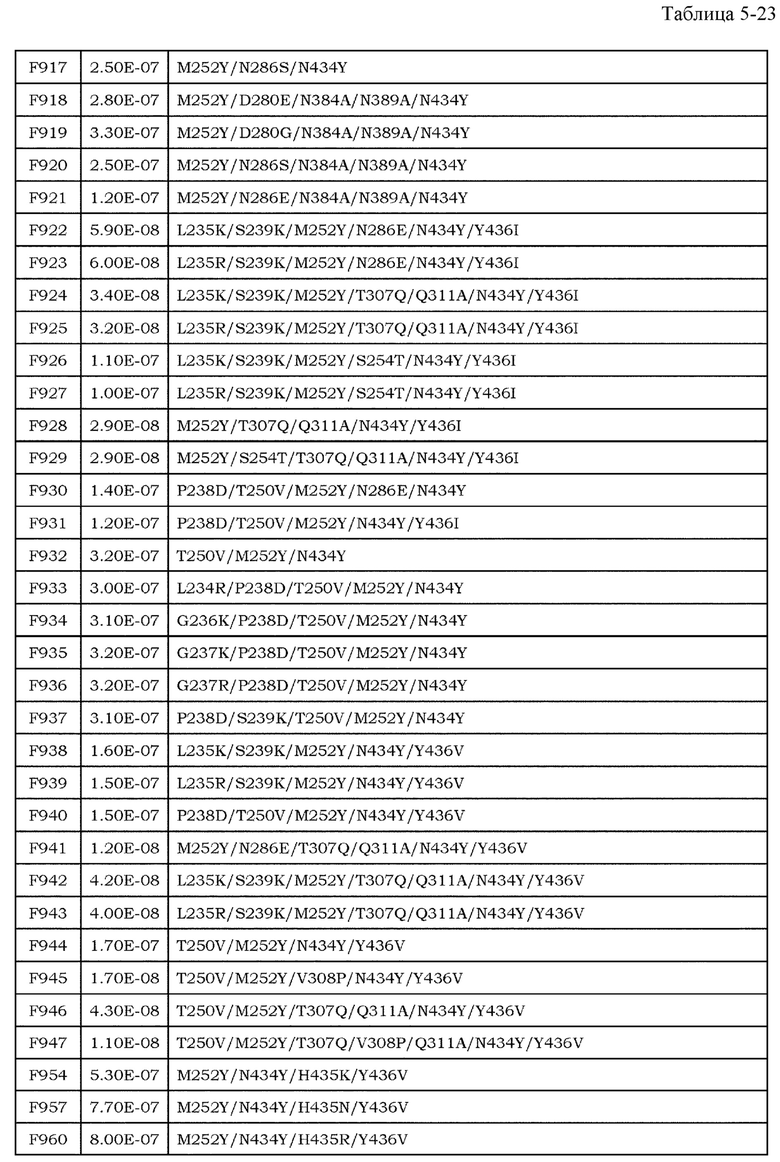
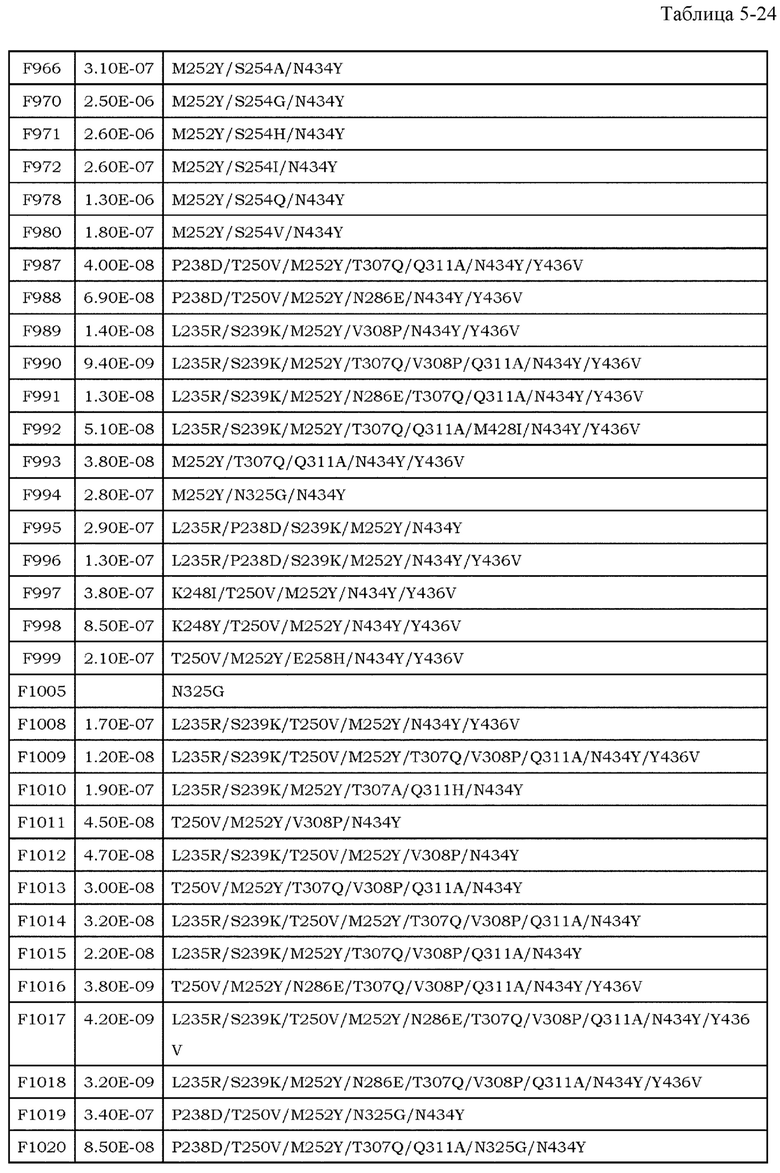
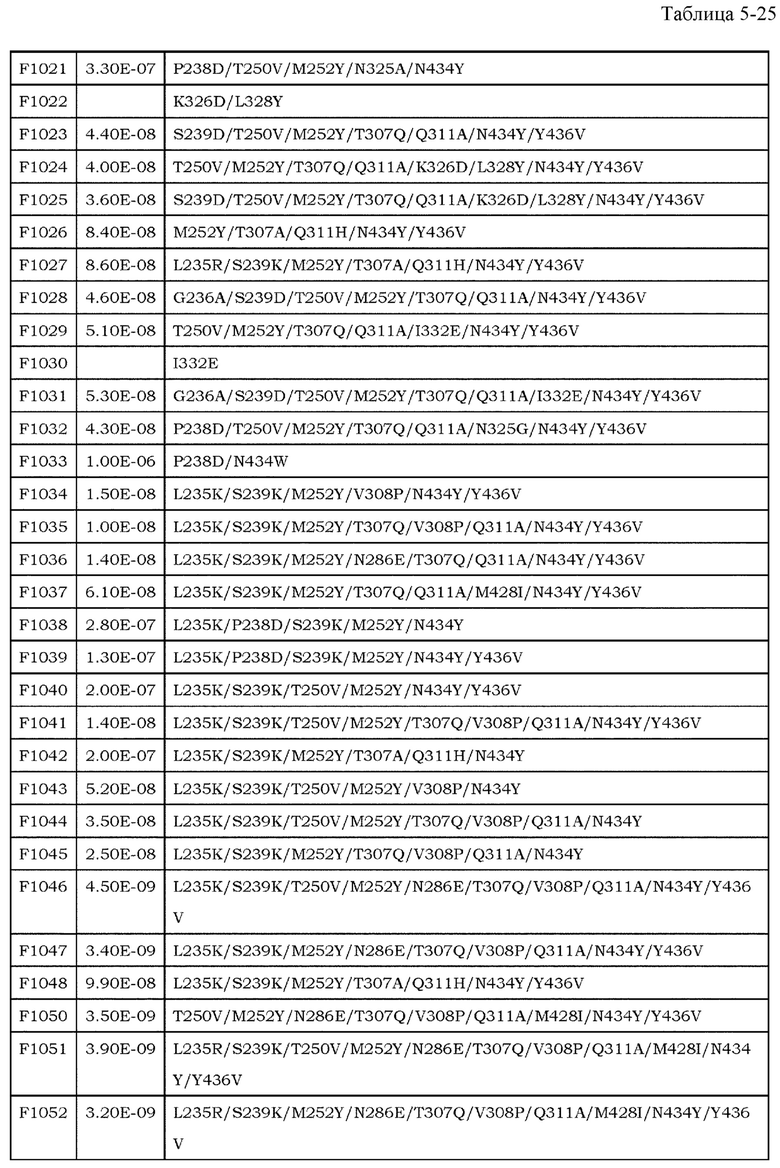

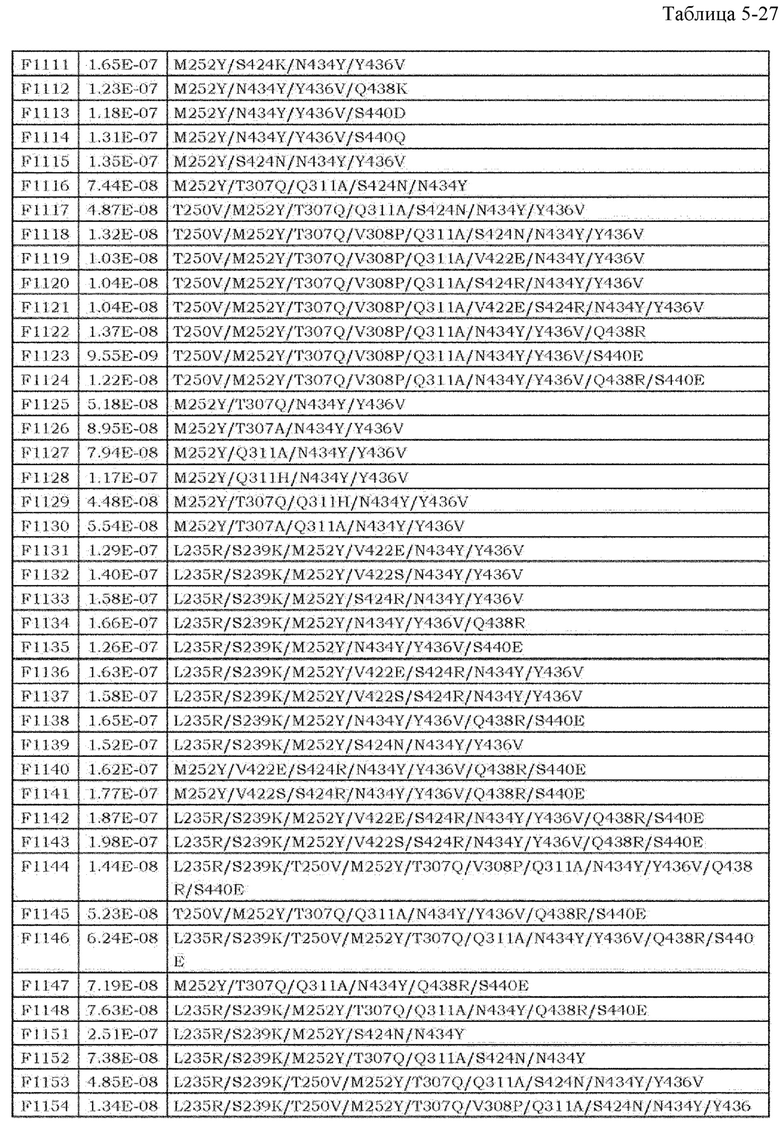

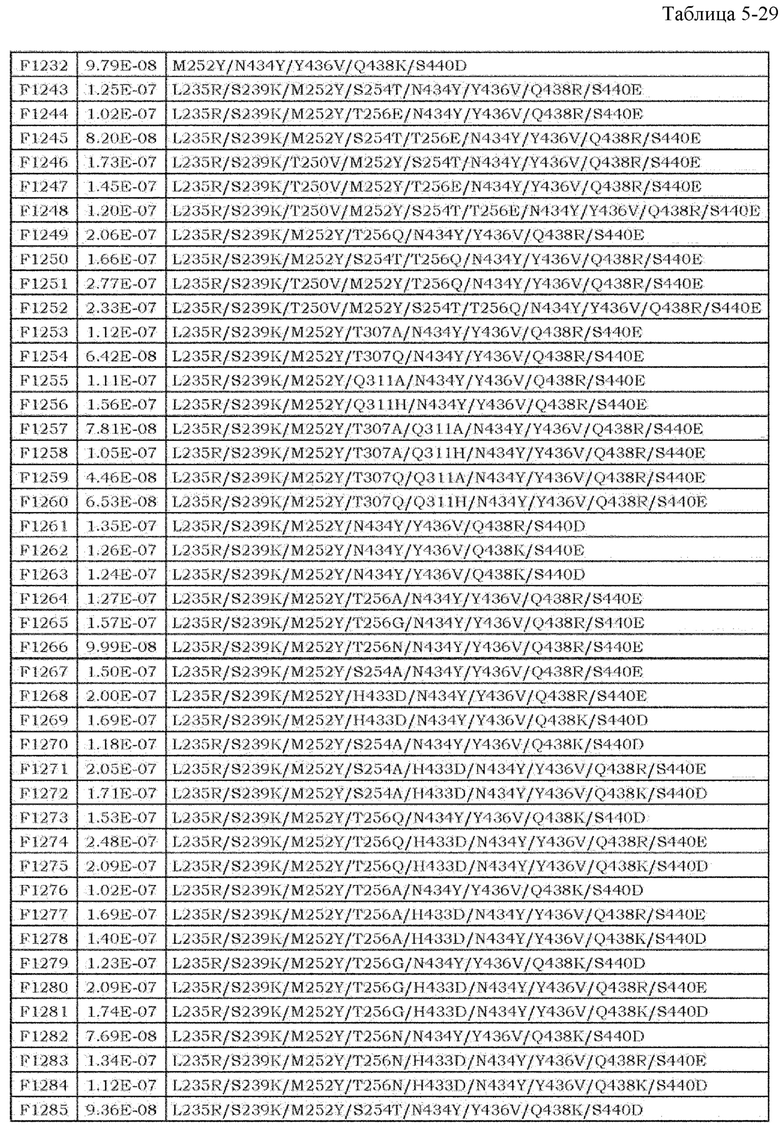

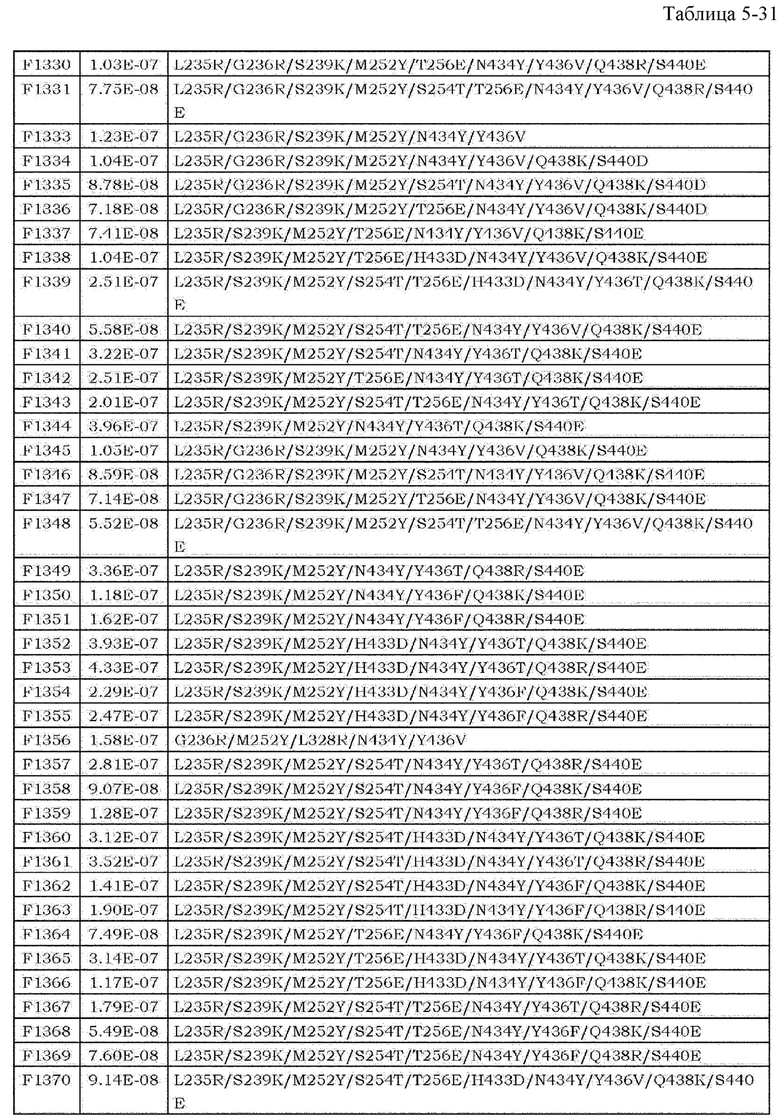
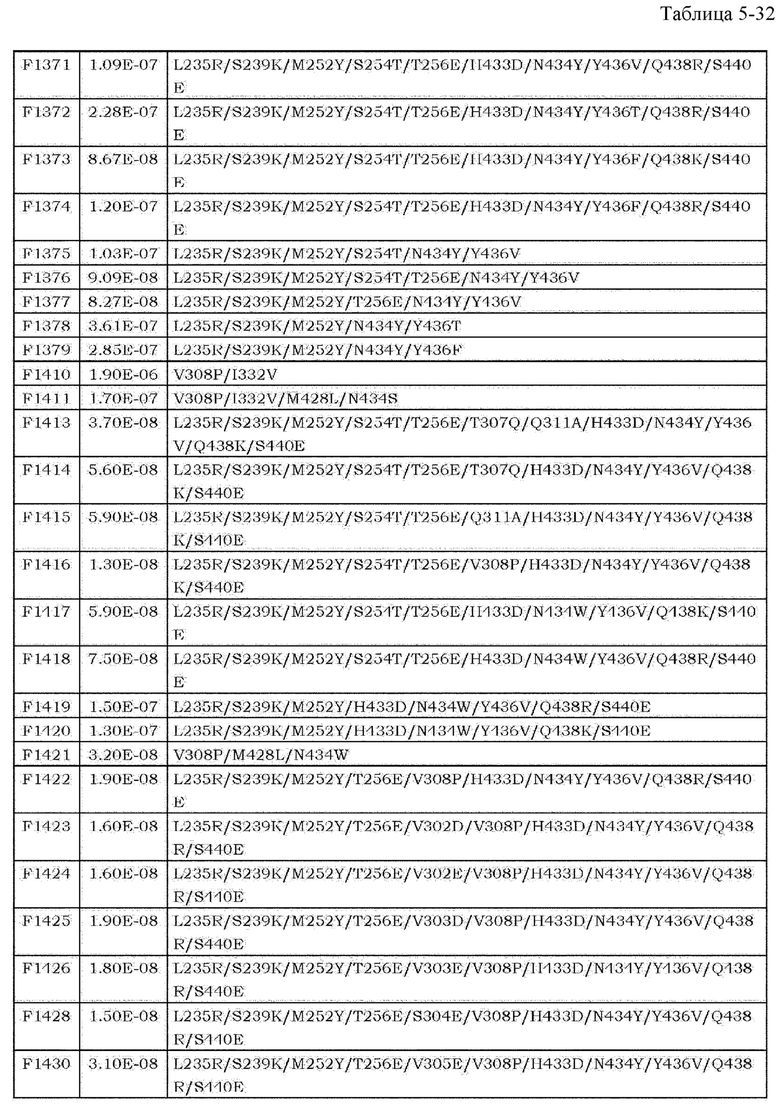

В неограничивающем варианте осуществления настоящего изобретения предпочтительно используют Fc-области, в которых по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области. Неограничивающие примеры таких Fc-областей предпочтительно включают Fc-области, содержащие по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области.
Fcγ-рецептор
Fcγ-рецептор относится к рецептору, способному связываться с Fc-областью моноклональных антител IgG1, IgG2, IgG3 или IgG4, и включает всех представителей, принадлежащих семейству белков, по существу кодируемых геном Fcγ-рецептора. У человека семейство включает FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотип H131 и R131), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16) включая изоформу FcγRIIIa (включая аллотип V158 и F158) и FcγRIIIb (включая аллотип FcγRIIIb-NA1 и FcγRIIIb-NA2); а также неидентифицированные FcγR человека, изоформы FcγR и их аллотипы. Однако Fcγ-рецептор не ограничивается этими примерами. Не ограничиваясь этим, FcγR включает FcγR человека, мыши, крысы, кролика и обезьяны. FcγR может происходить из любых организмов. FcγR мыши включает, но не ограничивается ими, FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (FcγRIV, CD16-2), а также неидентифицированные FcγR мыши, изоформы FcγR и их аллотипы. Такие предпочтительные Fcγ-рецепторы включают, например, FcγRI (CD64), FcγRIIa (CD32), FcγRIIb (CD32), FcγRIIIa (CD16) и/или FcγRIIIb (CD16) человека. Полинуклеотидная последовательность и аминокислотная последовательность FcγRI представлены в SEQ ID NO:19 (NM_00056.3) и 20 (NP_000557.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIa (аллотип H131) представлены в SEQ ID NO:21 (BC020823.1) и 27 (AAH20823.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγIIB представлены в SEQ ID NO:23 (BC146678.1) и 24 (AAI46679.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIa представлены в SEQ ID NO:22 (BC033678.1) и 26 (AAH33678.1), соответственно; и полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIb представлены в SEQ ID NO:27 (BC128562.1) и 28 (AAI28563.1), соответственно (номер доступа RefSeq представлен в каждом случае в скобках). То, обладает ли Fcγ-рецептор активностью связывания с Fc-областью моноклонального антитела IgG1, IgG2, IgG3 или IgG4 можно оценивать с помощью скрининга ALPHA (анализ усиленной за счет эффекта близости люминесценции), способа BIACORE на основе поверхностного плазмонного резонанса (SPR) и другие (Proc. Natl. Acad. Sci. USA (2006) 103(11), 4005-4010), в дополнение к описанным выше форматам FACS и ELISA.
Между тем, "Fc-лиганд" или "эффекторный лиганд" относится к молекуле и предпочтительно полипептиду, которые связываются с Fc-областью антитела, образуя комплекс Fc/Fc-лиганд. Молекула может происходить из любых организмов. Связывание Fc-лиганда с Fc предпочтительно индуцирует одну или несколько эффекторных функций. Такие Fc-лиганды включают, но не ограничиваются ими, Fc-рецепторы, FcγR, FcγR, FcγR, FcRn, C1q и C3, маннан-связывающий лектин, рецептор маннозы, белок A Staphylococcus, белок G Staphylococcus и вирусные FcγR. Fc-лиганды также включают гомологи Fc-рецептора (FcRH) (Davis et al., (2002) Immunological Reviews 190, 123-136), которые являются семейством Fc-рецепторов, гомологичных FcγR. Fc-лиганды также включают неидентифицированные молекулы, которые связываются с Fc.
В FcγRI (CD64), включая FcγRIa, FcγRIb, и FcγRIc, и FcγRIII (CD16), включающий изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2), α-цепь, которая связывается с Fc-частью IgG, ассоциирована с общей γ-цепью, имеющей ITAM, ответственный за передачу внутриклеточного активирующего сигнала. Между тем, цитоплазматический домен FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы H131 и R131) и FcγRIIc содержит ITAM. Эти рецепторы экспрессируются на многих клетках, таких как макрофаги, тучные клетки и антигенпредставляющие клетки. Активирующий сигнал, передаваемый при связывании этих рецепторов с Fc-частью IgG, приводит к усилению фагоцитарной активности макрофагов, продукции воспалительных цитокинов, дегрануляции тучных клеток и усилению функции антигенпредставляющих клеток. Fcγ-рецепторы, обладающие способность передавать активирующий сигнал, как описано выше, также называют активирующими Fcγ-рецепторами.
Между тем, внутрицитоплазматический домен FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) содержит ITIM, ответственный за передачу ингибиторных сигналов. Связывание между FcγRIIb и B-клеточным рецептором (BCR) на B-клетках подавляет активирующий сигнал из BCR, что приводит к подавлению продукции антитела через BCR. Связывание FcγRIII и FcγRIIb на макрофагах подавляет фагоцитарную активность и продукцию воспалительных цитокинов. Fcγ-рецепторы, обладающие способностью передавать ингибирующий сигнал, как описано выше, также называют ингибирующим Fcγ-рецептором.
Активность связывания с Fcγ-рецептором
Один вариант осуществления настоящего изобретения относится к фармацевтическим композициям, индуцирующим иммунный ответ, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, содержащую FcRn-связывающий домен, который содержит Fc-область, активность связывания которой с Fcγ-рецепторами человека является более высокой, чем активность связывания Fc IgG-человека Fcγ-рецепторами. То, является ли активность связывания Fc-области с любым из Fcγ-рецепторов человека: FcγRI, FcγRIIa, FcγRIb, FcγRIIIa и FcγRIIIb, чем активность связывания Fc IgG человека с этими Fcγ-рецепторами человека, можно подтверждать с помощью формата FACS или ELISA, как описано выше, и также с помощью скрининга ALPHA (анализ усиленной за счет эффекта близости люминесценции), способа BIACORE, который основан на явлении поверхностного плазмонного резонанса (SPR) и т.п. (Proc. Natl. Acad. Sci. USA (2006) 103 (11), 4005-4010).
Скрининг ALPHA проводят с помощью технологии ALPHA на основе принципа, описанного ниже, где используется два типа гранул: донорные и акцепторные. Люминесцентный сигнал выявляется только когда молекулы, связанные с донорными гранулами, биологически взаимодействуют с молекулами, связанными с акцепторными гранулами, и эти два типа гранул находятся вблизи друг друга. Возбуждаемый лазерным пучком фотосенсибилизатор в донорных гранулах преобразует кислород вокруг гранулы в возбужденный синглетный кислород. Когда синглетный кислород распространяется среди донорных гранул и достигает акцепторных гранул, расположенных близко, в акцепторных гранулах индуцируется хемилюминесцентная реакция. Это в конечном итоге приводит к испусканию света. Когда молекулы, связанные с донорными гранулами, не взаимодействуют с молекулами, связанными с акцепторными гранулами, сиглетный кислород, продуцируемый донорными гранулами, не достигает акцепторных гранул и хемилюминесцентная реакция не происходит.
Например, меченную биотином антигенсвязывающую молекулу, содержащую Fc-область, иммобилизуют на донорных гранулах, и Fcγ-рецептор, меченный глутатион-S-трансферазой (GST) иммобилизуют на акцепторных гранулах. В отсутствие конкурентной антигенсвязывающей молекулы, содержащей измененную Fc-область, Fcγ-рецептор взаимодействует с полипептидным комплексом, содержащим Fc-область дикого типа, что приводит к индукции сигнала в области 520-620 нм. Немеченая антигенсвязывающая молекула, имеющая измененную Fc-область, конкурирует с антигенсвязывающей молекулой, содержащей Fc-область дикого типа, для взаимодействия с Fcγ-рецептором. Относительную аффинность связывания можно определять путем количественного определения снижения флуоресценции в результате конкуренции. Способы биотинилирования антигенсвязывающих молекул, таких как антитела, с использованием сульфо-NHS-биотина и т.п., известны. Соответствующие способы добавления GST-метки к Fcγ-рецептору включают способы, которые вовлекают слияние полипептидов, кодирующих Fcγ и GST в рамке считывания, экспрессию слитого гена с использованием клеток, в которые введен вектор, с которым ген функционально связан, а затем очистку с использованием колонки с глутатионом. Индуцированный сигнал предпочтительно можно анализировать, например, путем аппроксимации его к односторонней модели конкуренции на основе нелинейного регрессионного анализа с использованием программного обеспечения, такого как GRAPHPAD PRISM (GraphPad, San Diego).
Одно из веществ при наблюдении их взаимодействия иммобилизуют в качестве лиганда на тонкую золотую пленку на сенсорном чипе. Когда свет падает на заднюю поверхность сенсорного чипа, так чтобы общее отражение происходило на поверхности контакта между тонкой золотой пленкой и стеклом, интенсивность отраженного света частично снижается в определенной области (сигнал SPR). Другое из веществ при выявлении взаимодействия инжектируют в качестве анализируемого соединения на поверхность сенсорного чипа. Когда анализируемое соединение связывается с лигандом, масса иммобилизованной молекулы лиганда возрастает. Изменение индекса рефракции вызывает сдвиг положения сигнала SPR (с другой стороны, диссоциация вызывает сдвиг сигнала обратно в исходное положение). В системе Biacore величину сдвига, упомянутая выше (т.е. изменение массы на поверхности сенсорного чипа) наносят на график по вертикальной оси, и, таким образом, изменение массы с течением времени показывают в качестве данных измерения (сенсограмма). Параметры кинетики (константа скорости ассоциации (ka) и константу скорости диссоциации (kd)), определяют из кривых сенсограммы, и аффинность (KD) определяют из соотношения этих двух констант. В способе BIACORE, предпочтительно используют анализ ингибирования. Примеры такого анализа ингибирования описаны в Proc. Natl. Acad. Sci USA (2006) 103 (11): 4005-4010.
Для условий pH для измерения активности связывания Fc-области и Fcγ-рецептора, содержащихся в антигенсвязывающей молекуле по настоящему изобретению, можно соответствующим образом использовать условия в диапазоне кислых значений pH или в диапазоне нейтральных значений pH. Диапазон нейтральных значений pH в качестве условий для измерения активности связывания Fc-области и Fcγ-рецептора, содержащихся в антигенсвязывающей молекуле по настоящему изобретению, как правило, обозначают как pH от 6,7 до 10,0. Предпочтительно, он представляет собой диапазон, обозначаемый с помощью произвольных величин pH от 7,0 до 8,0; и предпочтительно он выбран из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0; и особенно предпочтительно он представляет собой pH 7,4, что является близким к pH плазмы (крови) in vivo. В рамках настоящего изобретения диапазон кислых значений pH, в качестве условий наличия активности связывания Fc-области и Fcγ-рецептора, которой обладает антигенсвязывающая молекула по настоящему изобретению, как правило, обозначают как pH от 4,0 до 6,5. Предпочтительно, его обозначают как pH от 5,5 до 6,5, и особенно предпочтительно его обозначают как pH от 5,8 до 6,0, что близко к pH в ранней эндосоме in vivo. Что касается температуры, используемой в качестве условий измерения, аффинность связывания между Fc-областью и Fcγ-рецептором человека можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, для определения аффинности связывания между Fc-областью человека и Fcγ-рецептором используют температуру от 15°C до 40°C. Более предпочтительно, аналогично для определения аффинности связывания между Fc-областью и Fcγ-рецептором используют температуру от 20°C до 35°C, такую как любая температура из 20°C, 21°C, 22°C, 23°C, 24°C, 25°C, 26°C, 27°C, 28°C, 29°C, 30°C, 31°C, 32°C, 33°C, 34°C или 35°C. В одном варианте осуществления настоящего изобретения неограничивающим примером является температура, составляющая 25°C.
В настоящем описании "активность связывания Fc-области с любым из Fcγ-рецепторов человека: FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и/или FcγRIIIb является более высокой, чем активность связывания Fc-области IgG человека с этими Fcγ-рецепторами человека" означает, например, что, исходя из вышеупомянутых способов анализа, активность связывания антигенсвязывающей молекулы, содержащей исследуемую Fc-область, составляет 105% или выше, предпочтительно 110% или выше, 120% или выше, 130% или выше, 140% или выше, в особенно предпочтительно 150% или выше, 160% или выше, 170% или выше, 180% или выше, 190% или выше, 200% или выше, 250% или выше, 300% или выше, 350% или выше, 400% или выше, 450% или выше, 500% или выше, 750% или выше, является более высокой в 10 раз или выше, в 20 раз или выше, в 30 раз или выше, 40 раз или выше и в 50 раз или выше относительно активности связывания антигенсвязывающей молекулы, содержащей Fc-область IgG человека, в качестве контроля.
Fc-область с более высокой активностью связывания Fcγ-рецептора, чем активность связывания Fcγ-рецептора встречающейся в природе Fc-области, можно получать путем изменения аминокислот встречающейся в природе Fc-области. Встречающаяся в природе Fc-область, упомянутая в настоящем описании, относится к встречающейся в природе Fc-области, в которой цепь сахара в положении 297 (нумерация EU) представляет собой связанную через фукозу цепь сахаров комплексного типа. То, является ли активность связывания Fcγ-рецептора Fc-области более высокой, чем у встречающейся в природе Fc-области, можно определять соответствующим образом с использованием способов, описанных в разделе "Активность связывания", упоминаемом выше.
В рамках настоящего изобретения "изменение аминокислот" или "аминокислотные изменения" Fc-области включают изменение на аминокислотную последовательность, которая отличается от аминокислотной последовательности исходной Fc-области. В качестве исходного домена можно использовать любую Fc-область при условии, что модифицированный вариант исходной Fc-области может связываться с Fcγ-рецепторами человека в диапазоне нейтральных значений pH. Примеры исходной Fc-области предпочтительно включают Fc-область IgG-антитела человека или, более конкретно, встречающуюся в природе Fc-область, в которой цепь сахара в положении 297 (нумерация EU) представляет собой связанную через фукозу цепь сахаров комплексного типа. Более того, Fc-область, получаемую посредством дальнейшего изменения уже измененной Fc-области, используемой в качестве исходной Fc-области, также можно предпочтительно использовать в качестве Fc-области по настоящему изобретению. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, что в качестве исходной Fc-области можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или модифицированных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности с их исходной Fc-областью, меньшим чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с системой нумерации EU.
Для модификации аминокислот Fc-областей можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот аминокислотами, отличными от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно пригодным образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которой амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
Fc-область, обладающую активностью связывания Fcγ-рецептора в диапазоне нейтральных значений pH, которая содержится в антигенсвязывающих молекулах по настоящему изобретению, можно получать любым способом, но, в частности, Fc-область, обладающую активностью связывания Fcγ-рецептора в диапазоне нейтральных значений pH можно получать путем изменения аминокислот IgG-иммуноглобулин человека, используемой в качестве исходной Fc-области. Предпочтительные Fc-области IgG-иммуноглобулинов, подлежащие изменению, включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). В качестве предпочтительных примеров Fc-областей IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты), предпочтительно можно использовать Fc-области IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Структуры этих Fc-областей представлены в SEQ ID NO:11 (A добавлен на N-конец последовательности с номером доступа RefSeq AAC82527.1), SEQ ID NO:12 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59393.1), SEQ ID NO:13 (номер доступа RefSeq CAA27268.1) и SEQ ID NO:14 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59394.1). Более того, при использовании в качестве исследуемого вещества антигенсвязывающей молекулы, обладающей Fc-областью, полученную путем изменения антитела определенного изотипа, используемого в качестве исходной Fc-области, антигенсвязывающую молекулу, обладающую Fc-областью моноклонального IgG-антитела этого изотипа используют в качестве контроля для подтверждения эффектов на активность связывания с Fcγ-рецепторами антигенсвязывающей молекулы, имеющей измененную Fc-область. Антигенсвязывающие молекулы, содержащие Fc-область, для которой подтверждено, что она обладает высокой активностью связывания Fcγ-рецептора, как описано выше, отбирают соответствующим образом.
Аминокислоты в любых положениях можно изменять на другие аминокислоты при условии, что Fc-область обладает активностью связывания Fcγ-рецептора в диапазоне нейтральных значений pH или ее активность связывания Fcγ-рецептора в нейтральном диапазоне может быть усилена. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят к усилению связывания Fcγ-рецептора в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислотные изменения для усиления активности связывания Fcγ-рецептора в диапазоне нейтральных значений pH описана, например, в WO 2007/024249, WO 2007/021841, WO 2006/031370, WO 2000/042072, WO 2004/029207, WO 2004/099249, WO 2006/105338, WO 2007/041635, WO 2008/092117, WO 2005/070963, WO 2006/020114, WO 2006/116260 и WO 2006/023403.
Примеры таких аминокислот, которые могут быть изменены, включают по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU. Изменение этих аминокислот усиливает связывание Fcγ-рецептора Fc-областью IgG-иммуноглобулина в диапазоне нейтральных значений pH.
Особенно предпочтительные изменения для применения в рамках настоящего изобретения включают следующие изменения:
аминокислота в положении 221 либо на Lys, либо на Tyr;
аминокислота в положении 222 на любой из Phe, Trp, Glu и Tyr;
аминокислота в положении 223 на любой из Phe, Trp, Glu и Lys;
аминокислота в положении 224 на любой из Phe, Trp, Glu и Tyr;
аминокислота в положении 225 на любой из Glu, Lys и Trp;
аминокислота в положении 227 на любой из Glu, Gly, Lys и Tyr;
аминокислота в положении 228 на любой из Glu, Gly, Lys и Tyr;
аминокислота в положении 230 на любой из Ala, Glu, Gly и Tyr;
аминокислота в положении 231 на любой из Glu, Gly, Lys, Pro и Tyr;
аминокислота в положении 232 на любой из Glu, Gly, Lys и Tyr;
аминокислота в положении 233 на любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 234 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 235 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 236 на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 237 на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 238 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 239 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 240 на любой из Ala, Ile, Met и Thr;
аминокислота в положении 241 на любой из Asp, Glu, Leu, Arg, Trp и Tyr;
аминокислота в положении 243 на любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr;
аминокислота в положении 244 на His;
аминокислота в положении 245 на Ala;
аминокислота в положении 246 на любой из Asp, Glu, His и Tyr;
аминокислота в положении 247 на любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr;
аминокислота в положении 249 на любой из Glu, His, Gln и Tyr;
аминокислота в положении 250 либо на Glu, либо на Gln;
аминокислота в положении 251 на Phe;
аминокислота в положении 254 на любой из Phe, Met и Tyr;
аминокислота в положении 255 на любой из Glu, Leu и Tyr;
аминокислота в положении 256 на любой из Ala, Met и Pro;
аминокислота в положении 258 на любой из Asp, Glu, His, Ser и Tyr;
аминокислота в положении 260 на любой из Asp, Glu, His и Tyr;
аминокислота в положении 262 на любой из Ala, Glu, Phe, Ile и Thr;
аминокислота в положении 263 на любой из Ala, Ile, Met и Thr;
аминокислота в положении 264 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr;
аминокислота в положении 265 на любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr;
аминокислота в положении 266 на любой из Ala, Ile, Met и Thr;
аминокислота в положении 267 на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 268 на любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp;
аминокислота в положении 269 на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 270 на любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr;
аминокислота в положении 271 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 272 на любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 273 либо на Phe, либо на Ile;
аминокислота в положении 274 на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 275 либо на Leu, либо на Trp;
аминокислота в положении 276 на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 278 на любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp;
аминокислота в положении 279 на Ala;
аминокислота в положении 280 на любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr;
аминокислота в положении 281 на любой из Asp, Lys, Pro и Tyr;
аминокислота в положении 282 на любой из Glu, Gly, Lys, Pro и Tyr;
аминокислота в положении 283 на любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr;
аминокислота в положении 284 на любой из Asp, Glu, Leu, Asn, Thr и Tyr;
аминокислота в положении 285 на любой из Asp, Glu, Lys, Gln, Trp и Tyr;
аминокислота в положении 286 на любой из Glu, Gly, Pro и Tyr;
аминокислота в положении 288 на любой из Asn, Asp, Glu и Tyr;
аминокислота в положении 290 на любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr;
аминокислота в положении 291 на любой из Asp, Glu, Gly, His, Ile, Gln и Thr;
аминокислота в положении 292 на любой из Ala, Asp, Glu, Pro, Thr и Tyr;
аминокислота в положении 293 на любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 294 на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 295 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 296 на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val;
аминокислота в положении 297 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 298 на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 299 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr;
аминокислота в положении 300 на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp;
аминокислота в положении 301 на любой из Asp, Glu, His и Tyr;
аминокислота в положении 302 на Ile;
аминокислота в положении 303 на любой из Asp, Gly и Tyr;
аминокислота в положении 304 на любой из Asp, His, Leu, Asn и Thr;
аминокислота в положении 305 на любой из Glu, Ile, Thr и Tyr;
аминокислота в положении 311 на любой из Ala, Asp, Asn, Thr, Val и Tyr;
аминокислота в положении 313 на Phe;
аминокислота в положении 315 на Leu;
аминокислота в положении 317 либо на Glu, либо на Gln;
аминокислота в положении 318 на любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr;
аминокислота в положении 320 на любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 322 на любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 323 на Ile;
аминокислота в положении 324 на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 325 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 326 на любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 327 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 328 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 329 на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 330 на любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 331 на любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr;
аминокислота в положении 332 на любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr;
аминокислота в положении 333 на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr;
аминокислота в положении 334 на любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr;
аминокислота в положении 335 на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr;
аминокислота в положении 336 на любой из Glu, Lys и Tyr;
аминокислота в положении 337 на любой из Glu, His и Asn;
аминокислота в положении 339 на любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr;
аминокислота в положении 376 либо на Ala, либо на Val;
аминокислота в положении 377 либо на Gly, либо на Lys;
аминокислота в положении 378 на Asp;
аминокислота в положении 379 на Asn;
аминокислота в положении 380 на любой из Ala, Asn и Ser;
аминокислота в положении 382 либо на Ala, либо на Ile;
аминокислота в положении 385 на Glu;
аминокислота в положении 392 на Thr;
аминокислота в положении 396 на Leu;
аминокислота в положении 421 на Lys;
аминокислота в положении 427 на Asn;
аминокислота в положении 428 либо на Phe, либо на Leu;
аминокислота в положении 429 на Met;
аминокислота в положении 434 на Trp;
аминокислота в положении 436 на Ile; и
аминокислота в положении 440 на любой из Gly, His, Ile, Leu и Tyr,
в соответствии с нумерацией EU в Fc-области.
Количество аминокислот, которые изменены, конкретно не ограничено. Может быть изменена аминокислота только в одном положении или могут быть изменены аминокислоты в двух или более положениях. Примеры комбинаций изменений аминокислот в двух или более положениях включают комбинации, представленные в таблице 6 (таблицы 6-1 - 6-3).
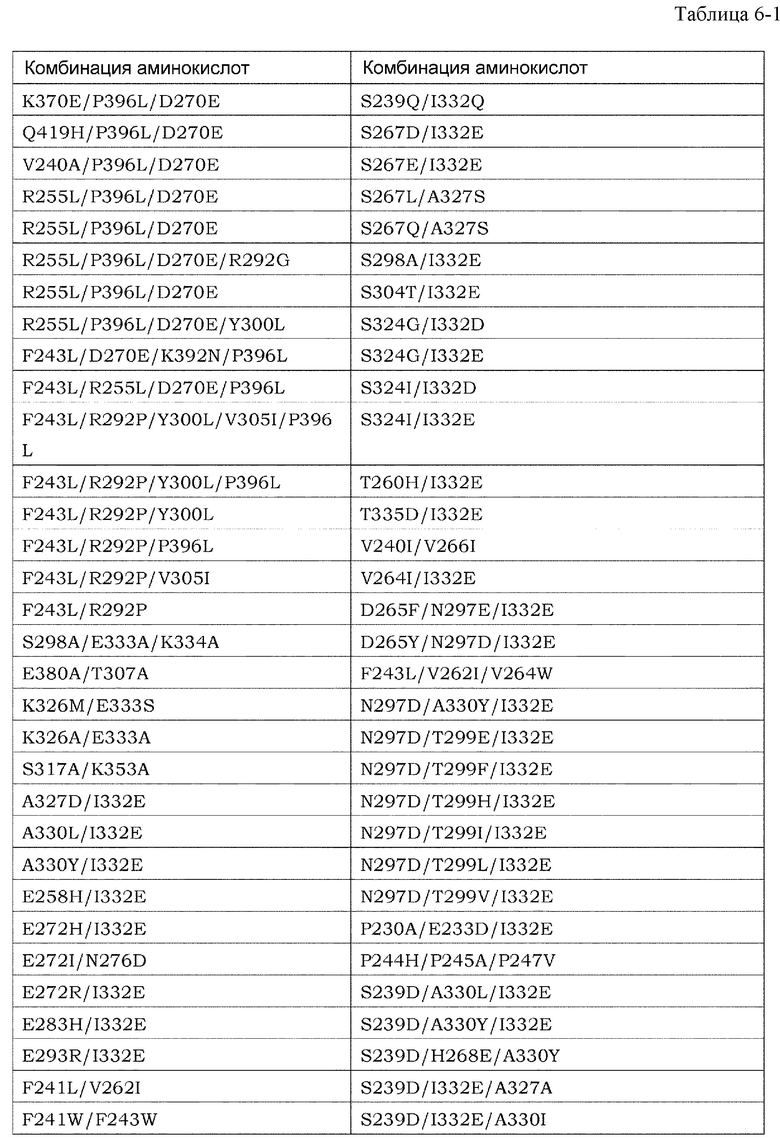
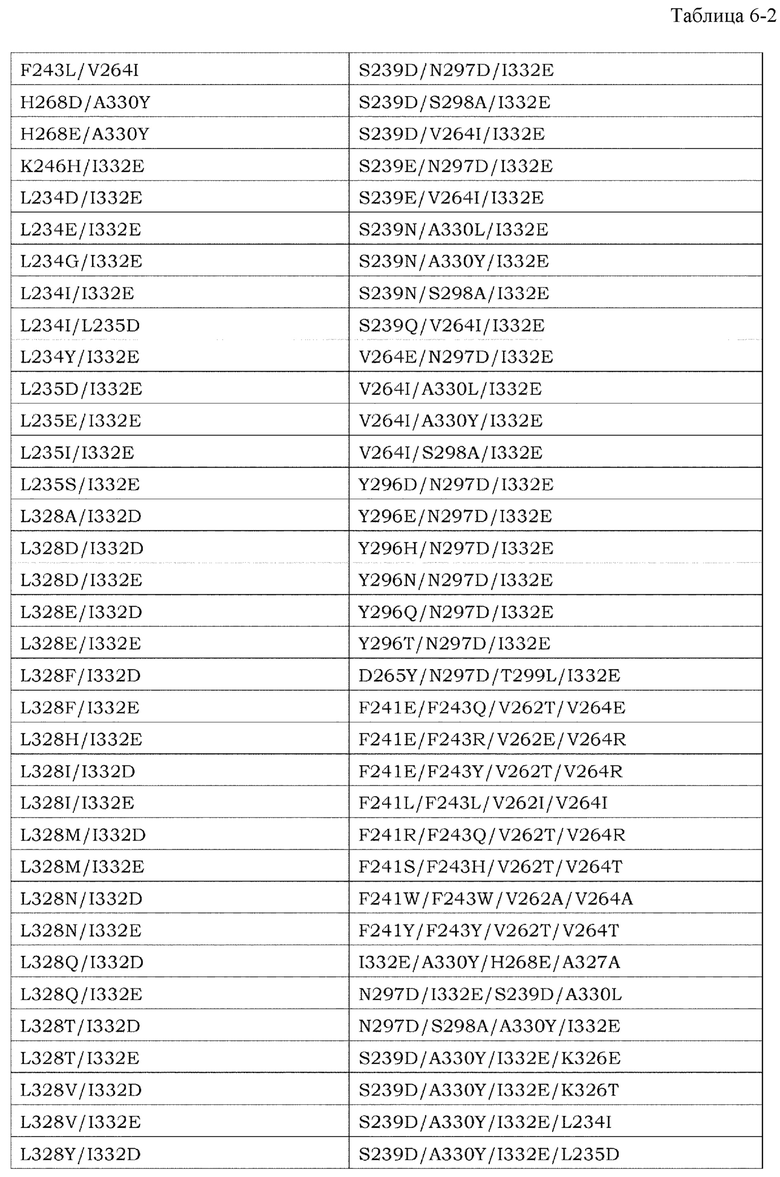
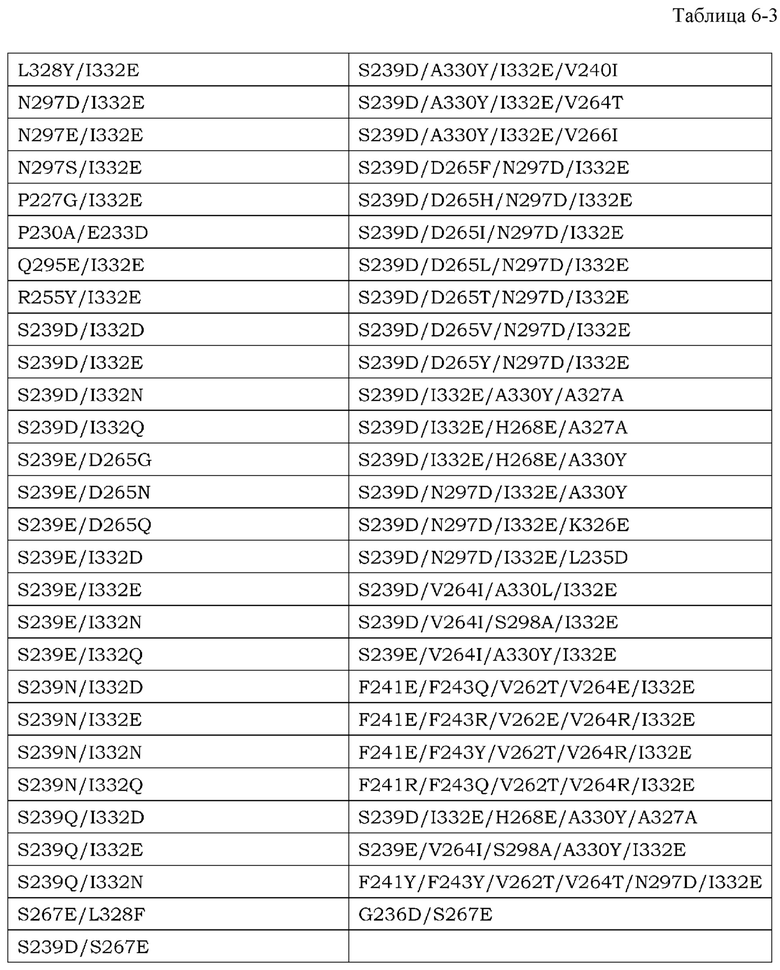
Fc-область с модифицированными цепями сахаров
Fc-области, предусматриваемые в рамках настоящего изобретения, могут включать Fc-области, которые модифицированы так, чтобы процент Fc-областей с которыми связана цепь сахаров с дефицитом фукозы, становился более высоким, или чтобы процент Fc-областей, к которым добавлен бисекторный N-ацетилглюкозамин, становился более высоким. Известно, что аффинность Fc-области антитела в отношении FcγIIIa повышается, когда остаток фукозы удаляют из N-ацетилглюкозамина на восстанавливающем конце связанной через N-гликозид цепи сахаров комплексного типа, связанной с Fc-областью антитела (непатентный документ 20). Известно, что IgG1-антитела, содержащие такие Fc-области, обладают увеличенной активностью ADCC, которая описана ниже; таким образом, антигенсвязывающие молекулы, содержащие такие Fc-области, часто пригодны в качестве антигенсвязывающих молекул, подлежащих включению в фармацевтические композиции по настоящему изобретению. Известные примеры антител, в которых из N-ацетилглюкозамина удален остаток фукозы на восстанавливающем конце связанной через N-гликозид цепи сахаров комплексного типа, связанной с Fc-областью антитела, являются следующими:
антитела с модифицированным гликозилированием (например, WO 1999/054342);
антитела, лишенные фукозы, связанной с цепями сахаров (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913);
антитела, обладающие цепью сахаров с бисекторным GlcNAc (например, WO 2002/079255) и т.п. Способы получения этих антител также можно использовать в способах получения антигенсвязывающих молекул, содержащих измененную Fc-область, модифицированную так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы, становился более высоким, или чтобы процент Fc-области, к которой добавлен бисекторный N-ацетилглюкозамин, становилтся более высоким.
Антигенсвязывающие молекулы
В рамках настоящего изобретения этот термин используют в наиболее широком значении для обозначения молекул, содержащих антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. В частности, различные типы молекул могут быть включены при условии, что они демонстрируют антигенсвязывающую активность. Молекулы, в которых антигенсвязывающий домен связан с Fc-областью, включают, например, антитела. Антитела могут включать единичные моноклональные антитела (включая антитела-агонисты и антитела-антагонисты), антитела человека, гуманизированные антитела, химерные антитела и т.п. Альтернативно при использовании в качестве фрагментов антител, они предпочтительно включают антигенсвязывающие домены и антигенсвязывающие фрагменты (например, Fab, F(ab')2, scFv и Fv). Каркасные молекулы, где трехмерные структуры, такие как уже известная стабильная белковая структура α/β-бочонок, используются в качестве каркаса (основы) и только некоторые части структур преобразуют в библиотеки для конструирования антигенсвязывающих доменов, также включены в антигенсвязывающие молекулы по настоящему изобретению.
Антигенсвязывающая молекула по настоящему изобретению может содержать по меньшей мере некоторые части Fc-области, которые опосредуют связывание с FcRn и Fcγ-рецептором. В неограничивающем варианте осуществления антигенсвязывающая молекула включает, например, антитела и слитые белки Fc. Слитый белок относится к химерному полипептиду, содержащему полипептид, имеющий первую аминокислотную последовательность, который связан с полипептидом, имеющим вторую аминокислотную последовательность, которые не связаны в природе. Например, слитый белок может содержать аминокислотную последовательность по меньшей мере части Fc-области (например, часть Fc-области, ответственная за связывание с FcRn, или часть Fc-области, ответственная за связывание с Fcγ-рецептором) и не являющийся иммуноглобулином полипептид, содержащий, например, аминокислотную последовательность лиганд-связывающего домена рецептора или рецептор-связывающего домена лиганда. Аминокислотные последовательности могут находиться в отдельных белках, которые переносят вместе в слитый белок, или, как правило, они могут находиться в одном белке, но они включены в слитый полипептид в новом порядке. Слитые белки можно получать, например, посредством химического синтеза или способами генетической рекомбинации для экспрессии полинуклеотида, кодирующего области пептидов в желаемом порядке.
Соответствующие домены по настоящему изобретению могут быь связаны вместе через линкеры или прямо через связывание полипептида.
Линкеры включают произвольные пептидные линкеры, которые можно вносить способами генетической инженерии, синтетические линкеры и линкеры, описанные, например, в Protein Engineering (1996) 9(3), 299-305. Однако пептидные линкеры являются предпочтительными в рамках настоящего изобретения. Длина пептидных линкеров конкретно не ограничена, и ее могут надлежащим образом выбирать специалисты в данной области в зависимости от назначения. Длина предпочтительно составляет пять аминокислот или более (без конкретных ограничений, верхний предел обычно составляет 30 аминокислот или менее, предпочтительно 20 аминокислот или менее), и особенно предпочтительно 15 аминокислот.
Например, такие пептидные линкеры предпочтительно включают:
Ser
Gly·Ser
Gly·Gly·Ser
Ser·Gly·Gly
Gly·Gly·Gly·Ser (SEQ ID NO:29)
Ser·Gly·Gly·Gly (SEQ ID NO:30)
Gly·Gly·Gly·Gly·Ser (SEQ ID NO:31)
Ser·Gly·Gly·Gly·Gly (SEQ ID NO:32)
Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:33)
Ser·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:34)
Gly·Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:35)
Ser·Gly·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:36)
(Gly·Gly·Gly·Gly·Ser (SEQ ID NO:31))n
(Ser·Gly·Gly·Gly·Gly (SEQ ID NO:32))n
где n представляет собой целое число, равное 1 или более. Длину или последовательности пептидных линкеров могут соответствующим образом выбирать специалисты в данной области, в зависимости от назначения.
Обычно для сшивания пептидов используют синтетические линкеры (химические сшивающие агенты), и например:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидил пропионат) (DSP),
дитиобис(сульфосукцинимидилпропионат) (DTSSP),
этиленгликоль бис(сукцинимидилсукцинат) (EGS),
этиленгликоль бис(сульфосукцинимидилсукцинат) (сульфо-EGS),
дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES), и
бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти сшивающие агенты являются коммерчески доступными.
Когда используют множество линкеров для связывания соответствующих доменов, все они могут быть одного типа или они могут быть различных типов.
В дополнение к линкерам, проиллюстрированным выше, также можно соответствующим образом использовать линкеры с пептидными метками, такими как His-метка, HA-метка, myc-метка и FLAG-метка. Более того, также можно соответствующим образом использовать образование водородных связей, образование дисульфидных связей, образование ковалентных связей, ионное взаимодействие и свойства связывания друг с другом в результате их комбинации. Например, можно использовать аффинность между CH1 и CL антитела, и также можно использовать Fc-области, происходящие из описанных выше биспецифических антител для гетероассоциации Fc-областей. Более того, также можно соответствующим образом использовать дисульфидные связи, образующиеся между доменами.
Для связывания соответствующих доменов пептидной связью, полинуклеотиды, кодирующие домены, связывают вместе в рамке считывания. Известные способы связывания полинуклеотидов в рамке считывания включают способы, такие как лигирование фрагментов рестрикции, ПЦР со слиянием и перекрывающаяся ПЦР. Такие способы можно соответствующим образом использовать отдельно или в комбинации для конструирования антигенсвязывающих молекул по настоящему изобретению. В рамках настоящего изобретения термины "связанный" и "слитый", или "связывание" и "слияние" используют взаимозаменяемо. Эти термины означают, что два или более элементов или компонентов, таких как полипептиды, слиты вместе с образованием единой структуры любыми способами, включая описанные выше способы химического связывания и способы генетической рекомбинации. Слияние в рамке считывания означает, когда два или более элементов или компонентов представляют собой полипептиды, связывание двух или более элементов рамок считывания с образованием непрерывной более длинной рамки считывания при сохранении правильных рамок считывания полипептидов. Когда две молекулы Fab используют в качестве антигенсвязывающего домена, в качестве предпочтительной антигенсвязывающей молекулы по настоящему изобретению можно использовать антитело, которое представляет собой антигенсвязывающую молекулу по настоящему изобретению, где антигенсвязывающий домен связан в рамке считывания с Fc-областью через пептидную связь без линкера.
Нейтрализующая активность
Один вариант осуществления настоящего изобретения относится к фармацевтическим композициям, которые индуцируют иммунный ответ, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, обладающую нейтрализующей активностью против антигена и содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. Главным образом, нейтрализующая активность относится к активности ингибирования биологической активности лиганда, такого как вирусы и токсины, обладающие биологической активностью в отношении клеток. Таким образом, вещества, обладающие нейтрализующей активностью, относятся к веществам, которые связываются с лигандом или рецептором, с которым связывается лиганд, и ингибируют связывание между лигандом и рецептором. Рецепторы, связывание которых с лигандом блокировано вследствие нейтрализующей активности, не способны проявлять биологическую активность через этот рецептор. Когда антигенсвязывающая молекула представляет собой антитело, такое антитело, обладающее нейтрализующей активностью, обычно называют нейтрализующим антителом. Нейтрализующую активность исследуемого вещества можно измерять путем сравнения биологической активности в присутствии лиганда между ситуациями, когда исследуемое вещество присутствует и отсутствует.
Например, основные возможные лиганды для рецептора IL-6 предпочтительно включают IL-6, как показано в SEQ ID NO:37. Рецептор IL-6, который представляет собой мембранный белок типа I, N-конец которого образует внеклеточный домен, образует гетеротетрамер с рецептором gp130, который индуцируется к димеризации посредством IL-6 (Heinrich et al. (Biochem. J. (1998) 334, 297-314)). Образование гетеротетрамера активирует Jak, которая ассоциирована с рецептором gp130. Jak претерпевает аутофосфорилирование и фосфорилирует рецептор. Участок фосфорилирования рецептора и Jak служит в качестве участка связывания для содержащих SH2 молекул, принадлежащих семейству Stat, таких как Stat3; MAP-киназа; PI3/Akt; и другие содержащие SH2 белки и адаптеры. Далее, Stat, связанный с рецептором gp130, фосфорилируется посредством Jak. Фосфорилированный Stat димеризуется и перемещается в ядро, и регулирует транскипцию генов-мишеней. Jak или Stat также могут быть вовлечены в каскады передачи сигнала через рецепторы других классов. Нарушенную регуляцию каскадов передачи сигнала IL-6 наблюдают при воспалительных и патологических состояниях аутоиммунных заболеваний и злокачественных опухолей, таких как рак предстательной железы и множественная миелома. Stat3, который может действовать в качестве онкогена, конститутивно активируется во многих злокачественных опухолях. При раке предстательной железы и множественной миеломе существует перекрестная связь между каскадом передачи сигнала через рецептор IL-6 и каскадом передачи сигнала через представителей семейства рецептора эпителиального фактора роста (EGFR) (Ishikawa et al. (J. Clin. Exp. Hematopathol. (2006) 46 (2), 55-66)).
Такие внутриклеточные каскады передачи сигнала различаются для каждого типа клеток; таким образом, соответствующие молекулы-мишени можно определять для каждой представляющей интерес клетки-мишени, и они не ограничиваются упомянутыми выше факторами. Активность нейтрализации можно оценивать путем измерения активации передачи сигнала in vivo. Более того, активацию передачи сигнала in vivo можно выявлять с использованием в качестве показателя действия индукции транскрипции гена-мишени, который располагается ниже каскада передачи сигнала in vivo. Изменение активности транскрипции гена-мишени можно выявлять по принципу репортерных анализов. В частности, репортерный ген, такой как зеленый флуоресцентный белок (GFP) или люцифераза, помещают ниже промоторной области или фактора транскрипции гена-мишени, его репортерную активность измеряют, и, тем самым, можно измерять изменение активности транскрипции в качестве активности репортера. Можно соответствующим образом использовать коммерчески доступные наборы для измерения активации передачи сигнала in vivo (например, Mercury Pathway Profiling Luciferase System (Clontech)).
Более того, для способов измерения активности нейтрализации рецепторов/лигандов семейства рецепторов EGF и т.п., которая обычно воздействует на каскады передачи сигнала, которые действуют в направлении стимуляции пролиферации клеток, активность нейтрализации нейтрализующих антител, можно оценивать путем измерения активности пролиферации клеток-мишеней. Например, когда пролиферация клеток стимулируется факторами роста семейства EGF, такими как HB-EGF, ингибиторный эффект на пролиферацию таких клеток, основанный на нейтрализующей активности антитела против HB-EGF, можно пригодным образом оценивать или измерять следующими способами: для оценки или измерения активности ингибирования пролиферации клеток in vitro используют способ измерения включения [3H]-меченного тимидина, добавленного в среду, жизнеспособными клетками в качестве показателя способности к репликации ДНК. В качестве более удобных способов используют способ исключения красителя, в котором измеряют способность клеток исключать краситель, такой как трипановый синий, из клетки под микроскопом, и способ с MTT. В последнем из этих способов используется способность живых клеток преобразовывать MTT (бромид 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия), который представляет собой соль тетразолия, в синий продукт формазан. Более конкретно, исследуемое антитело добавляют вместе с лигандом в культуральный раствор исследуемых клеток, и после определенного периода времени в культуральный раствор добавляют MTT и его оставляют стоять в течение некоторого периода времени для включения MTT в клетку. В результате, MTT, который представляет собой желтое соединение, преобразуется в синее соединение под действием сукцинатдегидрогеназы в митохондриях клеток. После растворения этого синего продукта для окрашивания, его поглощение измеряют и используют в качестве показателя количества живых клеток. В дополнение к MTT, также являются коммерчески доступными реагенты, такие как MTS, XTT, WST-1 и WST-8 (Nacalai Tesque и т.п.) и их можно пригодным образом использовать. Для измерения активности связывающее антитело, которое представляет собой антитело того же изотипа, что и антитело против HB-EGF, но не обладает активностью ингибирования пролиферации клеток, можно использовать в качестве контрольного антитела аналогично тому, как и антитело против HB-EGF, и активность можно определять, когда антитело против HB-EGF демонстрирует более высокую активность ингибирования пролиферации клеток, чем контрольное антитело.
Клетки, которые предпочтительно можно использовать для оценки активности, включают, например, клетки, пролиферация которых стимулируется посредством HB-EGF, такие как клеточная линия рака яичника RMG-1 и клетки мыши Ba/F3, трансформированные вектором для экспрессии гена, кодирующего hEGFR/mG-CSFR, который представляет собой слитый белок, в котором внеклеточный домен EGFR человека слит в рамке считывания с внутриклеточным доменом рецептора GCSF мыши. Таким образом, специалисты в данной области могут соответствующим образом выбрать клетки для применения для оценки активности и использовать их для измерения активности пролиферации клеток, как упоминалось выше.
Цитотоксическая активность
Один вариант осуществления настоящего изобретения относится к фармацевтическим композициям, которые индуцируют иммунный ответ, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, которая обладает цитотоксической активностью против клеток, экспресирующих антиген, и содержит антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. В рамках настоящего изобретения цитотоксическая активность включает, например, активность в форме антителозависимой клеточно-опосредуемой цитотоксичности (ADCC) и активность в форме комплементзависимой цитотоксичности (CDC). В рамках настоящего изобретения активность CDC относится к цитотоксической активности, опосредуемой системой комплемента. Активность ADCC относится к активности повреждения клетки-мишени, когда специфическая антигенсвязывающая молекула связывается с поверхностным антигеном экспрессирующей антиген клетки, а затем экспрессирующая Fcγ-рецептор клетка (иммунная клетка и т.п.) связывается с Fc-частью антигенсвязывающей молекулы через Fcγ-рецептор. То, обладает ли представляющая интерес антигенсвязывающая молекула активностью ADCC или активностью CDC, можно определять с использованием известных способов (например, Current Protocols in Immunology, Chapter 7. Immunologic studies in humans, Coligan et al. Ed. (1993)).
Сначала, в частности, получают эффекторные клетки, раствор комплемента и клетки-мишени.
(1) Получение эффекторных клеток
Селезенку извлекают из мыши CBA/N и т.п. и клетки селезенки выделяют в среде RPMI1640 (Invitrogen). После промывания клеток той же средой, содержащей 10% эмбриональную телячью сыворотку (FBS, HyClone), концентрацию промытых клеток селезенки можно доводить до 5×106 /мл, получая эффекторные клетки.
(2) Получение раствора комплемента
Комплемент детенышей кролика (CEDARLANE) разбавляют в 10 раз в культуральной среде (Invitrogen), содержащей 10% FBS, получая раствор комплемента.
(3) Получение клеток-мишеней
Клетки-мишени можно метить радиоактивной меткой путем культивирования клеток, экспрессирующих антиген, с 0,2 мкКи хромата натрия-51Cr (GE Healthcare Bio-Sciences) в среде DMEM, содержащей 10% FBS, в течение одного часа при 37°C. После мечения радиоактивной меткой, клетки промывают три раза средой RPMI1640, содержащей 10% FBS, и клетки-мишени можно получать путем доведения концентрации клеток до 2×105 /мл.
Активность ADCC или активность CDC можно измерять способом, описанным ниже. В случае измерения активности ADCC, 50 мкл каждых из клеток-мишеней и антигенсвязывающей молекулы добавляют в 96-луночный планшет с U-образным дном (Becton Dickinson) и позволяют им реагировать в течение 15 минут на льду. После этого в планшет добавляют 100 мкл эффекторных клеток и планшет оставляют стоять в инкубаторе с диоксидом углерода в течение четырех часов. Конечную концентрацию антитела можно доводить, например, до 0 или 10 мкг/мл. После стояния 100 мкл супернатанта собирают из каждой лунки и измеряют радиоактивность с помощью гамма-счетчика (COBRAII AUTO-GAMMA, MODEL D5005, Packard Instrument Company). Измеренную величину используют для вычисления цитотоксической активности (%) по формуле: (A-C)/(B-C)×100. A обозначает радиоактивность (cpm) в каждом образце, B обозначает радиоактивность (cpm) в образце, в который добавлен 1% NP-40 (Nacalai Tesque) и C обозначает радиоактивность (cpm) в образце, содержащем только клетки-мишени.
Между тем, в случае измерения активности CDC, 50 мкл каждых из клеток-мишеней и антигенсвязывающей молекулы добавляют в 96-луночный планшет с плоским дном (Becton Dickinson) и позволяют реагировать в течение 15 минут на льду. После этого в планшет добавляют 100 мкл раствора комплемента и планшет оставляют стоять в инкубаторе с диоксидом углерода в течение четырех часов. Конечную концентрацию антитела можно доводить, например, до 0 или 3 мкг/мл. После стояния, 100 мкл супернатанта собирают из каждой лунки и измеряют радиоактивность с помощью гамма-счетчика. Цитотоксическую активность можно вычислять аналогично измерению активности ADCC.
Иммунный ответ
Неограничивающий вариант осуществления настоящего изобретения относится к фармацевтическим композициям, индуцирующим иммунный ответ на антиген, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH.
То, индуцируется ли иммунный ответ, можно оценивать путем измерения ответа в живом организме, которому ввели фармацевтическую композицию, которая индуцирует иммунный ответ против упомянутого выше антигена и содержит антигенсвязывающую молекулу в качестве активного ингредиента. Примеры ответа в живом организме, главным образом, включают два иммунных ответа: клеточный иммунитет (индукция цитотоксических T-клеток, которые распознают пептидный фрагмент антигена, связанный с MHC класса I) и гуморальный иммунитет (индукция продуцирования антигенсвязывающих антител). Способы оценки индукции гуморального иммунитета (иммунный ответ) включают способы оценки продуцирования антител против антигена in vivo.
То, индуцируется ли гуморальный иммунитет фармацевтической композицией по настоящему изобретению, которая индуцирует иммунный ответ, можно оценивать путем введения фармацевтической композиции в живой организм и обнаружения в периферической крови, выделенной из организма, антител, индуцированных в организме против антигена, на который нацелена антигенсвязывающая молекула, содержащая фармацевтическую композицию. Титр антитела против антигена можно измерять с использованием способов количественного определения молекул, которые специфически связываются с введенной молекулой, с использованием ELISA, ECL, SPR, которые известны специалистам в данной области (J. Pharm. Biomed. Anal. 2011 Jul 15;55(5):878-88).
То, появляется ли клеточный иммунитет благодаря фармацевтической композиции по настоящему изобретению, которая индуцирует иммунный ответ, можно оценивать путем введения фармацевтической композиции в живой организм и обнаружения в периферической крови, выделенной из организма, подгруппы CD8-экспрессирующих T-клеток, которые имеют фенотип по типу памяти и являются специфичными к антигену, на который нацелена антигенсвязывающая молекула, содержащаяся в фармацевтической композиции. Популяция CD8-экспрессирующих клеток, имеющих фенотип по типу памяти, представляет собой гетерогенную клеточную популяцию. В частности, она включает центральные клетки памяти, которые быстро делятся в ответ на антиген, и эффекторные клетки памяти, которые демонстрируют память эффекторных функций, таких как цитотоксичность. Эти подгруппы не являются взаимоисключающими. Иными словами, клетки могут быстро делиться, но также могут повреждать клетки-мишени, представляющие антиген. Существует коммерчески доступный набор (Cytokine Secretion Assay - Cell Enrichment and Detection Kit (Miltenyi Biotec)) для обнаружения цитокинов, продуцируемых в результате проведения культивирования для увеличения в количестве такой подгруппы антигенспецифических CD8-экспрессирующих T-клеток, обладающих фенотипом по типу памяти. Также предоставляются протоколы выделения такой антигенспецифичевкой популяции. С использованием такого набора как антигенспецифические центральные клетки памяти, так и эффекторные T-клетки памяти можно эффективно выращивать in vitro. Антигенпредставляющие клетки, которые могут стимулировать пролиферацию подгруппы таких T-клеток, можно выделять из периферической крови, полученной из организма, в которую введена упомянутая выше индуцирующая иммунный ответ фармацевтическая композиция. В качестве антигенпредставляющих клеток можно использовать дендритные клетки, обработанные антигеном, или дендритные клетки, трансфицированные антигеном (Overes et al. (J. Immunother. (2009) 32, 539-551).
Фармацевтические композиции
В другом аспекте настоящее изобретение относится к фармацевтическим композициям, которые индуцируют иммунный ответ на антиген, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. Другой вариант осуществления настоящего изобретения относится к ингибиторам роста клеток или средствам против злокачественной опухоли, которые индуцируют иммунный ответ на упомянутый выше антиген, которые включают антигенсвязывающую молекулу в качестве активного ингредиента. Фармацевтическую композицию, ингибитор роста клеток или средство против злокачественной опухоли по настоящему изобретению предпочтительно вводят индивидууму, страдающему инфекцией чужеродными биологическими видами или злокачественной опухолью, или индивидууму, у которого может возникнуть рецидив.
В одном варианте осуществления настоящего изобретения фармацевтическая композиция, ингибитор роста клеток или средство против злокачественной опухоли, индуцирующие иммунный ответ на вышеупомянутый антиген, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH, могут символизировать применение антигенсвязывающей молекулы для получения фармацевтической композиции, ингибитора роста клеток или средства против злокачественной опухоли.
В другом варианте осуществления настоящего изобретения они также могут символизировать способ индукции иммунного ответа на антиген, который включает стадию введение фармацевтической композиции, ингибитора роста клеток или средства против злокачественной опухоли, которые содержат в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH.
В другом варианте осуществления настоящего изобретения они могут символизировать антигенсвязывающую молекулу, которая содержит антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH, для применения для индукции иммунного ответа на антиген.
В другом варианте осуществления настоящего изобретения, они могут символизировать способ получения фармацевтической композиции, ингибитора роста клеток или средства против злокачественной опухоли, индуцирующих иммунный ответ на антиген, который включает стадию использования антигенсвязывающей молекулы, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH.
В рамках настоящего изобретения выражение "содержит в качестве активного ингредиента антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH," означает содержание указанной антигенсвязывающей молекулы в качестве основного активного компонента и не ограничивает содержание антигенсвязывающей молекулы. Более того, настоящее изобретение может предусматривать фармацевтические композиции, ингибиторы роста клеток и средства против злокачественной опухоли, индуцирующие иммунный ответ на антиген, которые содержат в качестве активного ингредиента, в дополнение к антигенсвязывающей молекуле, которая не связывается с антигеном, антигенсвязывающую молекулу, которая уже связалась с антигеном. Более того, настоящее изобретение относится к способам индукции иммунного ответа на антиген, которые включают введение, в дополнение к антигенсвязывающей молекуле, которая не связана с антигеном, антигенсвязывающей молекулы, которая уже связана с антигеном.
Более того, фармацевтические композиции, ингибиторы роста клеток и средства против злокачественной опухоли по настоящему изобретению при необходимости могут включать различные антигенсвязывающие молекулы. Например, коктейль из различных антигенсвязывающих молекул по настоящему изобретению, которые связываются с одним и тем же антигеном, может усиливать действие индукции иммунного ответа, цитотоксической активности или нейтрализующей активности против клеток, экспрессирующих антиген, что приводит к увеличению терапевтических эффектов против заболеваний, вызываемых клетками, экспрессирующими антиген. Альтернативно фармацевтическую композицию, ингибитор роста клеток или средство против злокачественной опухоли по настоящему изобретению, которые содержат антигенсвязывающую молекулу по настоящему изобретению, содержащую антигенсвязывающий домен, который связывается с антигеном, экспрессируемым клетками, вызывающими заболевание, подлежащее лечению, и также содержат антигенсвязывающую молекулу по настоящему изобретению, содержащую антигенсвязывающий домен, который связывается с другим антигеном, экспрессируемым клетками, вызывающими то же заболевание, вводят для увеличения терапевтических эффектов в отношении заболевания.
Если необходимо, фармацевтическую композицию, ингибитор роста клеток или средство против злокачественной опухоли по настоящему изобретению можно инкапсулировать в микрокапсулы (микрокапсулы, изготовленные из гидроксиметилцеллюлозы, желатина, поли[метилметакрилата] и т.п.) и приготавливать в качестве коллоидных систем для доставки лекарственного средства (таких как липосомы, альбуминовые микросферы, микроэмульсия, наночастицы и нанокапсулы) (смотрите, например, "Remington's Pharmaceutical Science 16th edition", Oslo Ed. (1980)). Также известны способы получения лекарственного средства в качестве лекарственного средства с контролируемым высвобождением и такие способы можно использовать для фармацевтических композиций, ингибиторов роста клеток и средств против злокачественной опухоли по настоящему изобретению (J. Biomed. Mater. Res. (1981) 15, 267-277; Chemtech. (1982) 12: 98-105; патент США № 3773919; публикации патентов Европы EP 58481 и EP 133988; Biopolymers (1983) 22, 547-556).
Фармацевтические композиции, ингибиторы роста клеток и средства против злокачественной опухоли по настоящему изобретению можно вводить пациентам либо перорально, либо парентерально. Предпочтительным является парентеральное введение. Такие способы введения, в частности, включают введение посредством инъекции, трансназальное введение, легочное введение и трансдермальное введение. Для введения посредством инъекции, фармацевтическую композицию, ингибитор роста клеток или средство против злокачественной опухоли по настоящему изобретению можно системно или местно вводить, например, посредством внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции и подкожной инъекции. Способ введения можно соответствующим образом выбирать в зависимости от возраста и симптомов пациента. Дозу можно выбирать, например, из диапазона от 0,0001 мг до 1000 мг на килограмм массы тела на введение. Альтернативно дозу можно выбирать, например, из диапазона от 0,001 мг/организм до 100000 мг/организм на пациента. Однако фармацевтические композиции, ингибиторы роста клеток или средства против злокачественной опухоли по настоящему изобретению не ограничиваются этими дозами.
Фармацевтические композиции, ингибиторы роста клеток и средства против злокачественной опухоли по настоящему изобретению можно составлять в соответствии с общепринятыми способами (например, Remington's Pharmaceutical Science, последнее издание, Mark Publishing Company, Easton, U.S.A) и они также могут содержать фармацевтически приемлемые носители и добавки. Примеры включают поверхностно-активные вещества, эксципиенты, красители, вкусовые добавки, консерванты, стабилизаторы, буферы, суспендирующие вещества, обеспечивающие изотоничность средства, связующие вещества, дезинтегрирующие средства, смазывающие вещества, повышающие текучесть средства и корригирующие вещества. Не ограничиваясь ими, можно пригодным образом использовать другие широко используемые носители. Конкретные примеры носителей включают легкую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кармеллозу кальций, кармеллозу натрий, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, поливинилацеталь диэтиламиноацетат, поливинилпирролидон, желатин, триглицериды жирных кислот средней цепи, отвержденное полиоксиэтиленом касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганические соли и т.п.
Аминокислоты, содержащиеся в аминокислотных последовательностях в рамках настоящего изобретения, могут быть посттрансляционно модифицированными (например, модификация N-концевого глутамина в пироглутаминовую кислоту посредством пироглутамилирования хорошо известна специалистам в данной области). Естественно, такие посттрансляционно модифицированные аминокислоты включены в аминокислотные последовательности по настоящему изобретению.
Способы изготовления фармацевтических композиций
Неограничивающий вариант осуществления настоящего изобретения относится к способам получения антигенсвязывающей молекулы, которая индуцирует иммунный ответ, которые включают сообщение активности связывания FcRn в диапазоне нейтральных значений pH FcRn-связывающему домену, который содержится в антигенсвязывающей молекуле, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов.
В способах получения по настоящему изобретению, когда активность связывания FcRn FcRn-связывающего домена, который содержится в антигенсвязывающей молекуле, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, является слабой или не обнаруживается в диапазоне нейтральных значений pH, активность связывания FcRn в диапазоне нейтральных значений pH можно сообщать FcRn-связывающему домену, получая антигенсвязывающую молекулу по настоящему изобретению.
Например, когда антигенсвязывающий домен, содержащий вариабельные области тяжелой и легкой цепи антитела против FcRn используют в качестве FcRn-связывающего домена, можно получить FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH, в соответствии с упомянутым выше способом получения антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов. Когда Fc-область, активность связывания FcRn которой в диапазоне нейтральных значений pH является слабой или не обнаруживается, используют в качестве FcRn-связывающего домена, антигенсвязывающую молекулу, содержащую Fc-область с желаемой активностью связывания FcRn можно получать путем изменения аминокислот Fc-области, содержащейся в антигенсвязывающей молекуле. Аминокислотные изменения Fc-области, которые приводят к такой желаемой активности связывания, можно идентифицировать путем сравнения активности связывания FcRn в диапазоне нейтральных значений pH до и после изменения. Специалисты в данной области могут осуществить соответствующие аминокислотные изменения с использованием известных способов, таких как перекрывающаяся ПЦР или сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) аналогично упомянутым выше способам, используемым для изменения антигенсвязывающей активности.
Fc-область, обладающую активностью связывания FcRn в диапазоне нейтральных значений pH, которая содержится в антигенсвязывающей молекуле по настоящему изобретению, можно получать любыми способами, но, в частности, FcRn-связывающий домен, обладающий активностью связывания FcRn или усиленной активностью связывания FcRn в диапазоне нейтральных значений pH, можно получать путем изменения аминокислот IgG-иммуноглобулина человека, используемого в качестве исходной Fc-области. Примеры предпочтительных Fc-областей IgG-иммуноглобулинов, подлежащих изменению, включают Fc-область IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Аминокислоты в любых положениях можно заменять другими аминокислотами при условии, что Fc-область обладает активностью связывания FcRn в диапазоне нейтральных значений pH или ее активность связывания FcRn человека в нейтральном диапазоне может быть усилена. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят к усилению связывания FcRn в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Величины KD для FcRn в диапазоне нейтральных значений pH определяют, как упоминалось выше, способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (антигенсвязывающие молекулы иммобилизуют на чип и FcRn человека позволяют протекать в качестве анализируемого соединения).
Примеры предпочтительных Fc-областей IgG-иммуноглобулинов, подлежащих изменению, включают Fc-область IgG человека (IgG1, IgG2, IgG3 или IgG4 и их варианты). Аминокислоты в любых положениях можно заменять на другие аминокислоты при условии, что Fc-область обладает активностью связывания FcRn в диапазоне нейтральных значений pH или ее активностью связывания FcRn человека в нейтральном диапазоне может быть усилена. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека, предпочтительно вносить изменения, которые приводят к усилению связывания FcRn в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Примеры такой модифицированной Fc-области включают измененные Fc-области, в которых аминокислоты, такие как аминокислоты, приведенные в таблице 5 выше, изменены и которые обладают активностью связывания в диапазоне нейтральных значений pH.
Неограничивающий вариант осуществления настоящего изобретения относится к способам продуцирования фармацевтической композиции, которая индуцирует иммунный ответ, которые включают стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в условиях высокой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в условиях низкой концентрации ионов кальция;
(c) выбор антигенсвязывающего домена, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, выбранный согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клеток, в которые введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e).
Другой неограничивающий вариант осуществления настоящего изобретения также относится к способам получения фармацевтической композиции, которая индуцирует иммунный ответ, который включает стадии:
(a) определение антигенсвязывающей активности антитела в условиях высокой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антитела в условиях низкой концентрации ионов кальция;
(c) выбор антитела, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен антитела, выбранного согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клеток, в которые введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e).
Более того, неограничивающий вариант осуществления настоящего изобретения относится к способам получения антигенсвязывающей молекулы, которые включают стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в диапазоне нейтральных значений pH;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в диапазоне кислых значений pH;
(c) выбор антигенсвязывающего домена, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, выбранный согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клеток, в которые введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e).
Более того, другой неограничивающий вариант осуществления настоящего изобретения относится к способам получения антигенсвязывающей молекулы, которые включают стадии:
(a) определение антигенсвязывающей активности антитела в диапазоне нейтральных значений pH;
(b) определение антигенсвязывающей активности антитела в диапазоне кислых значений pH;
(c) выбор антитела, антигенсвязывающая активность которого, определенная согласно (a), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен антитела, выбранного согласно (c), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH;
(e) культивирование клеток, в которые введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (e).
В неограничивающем варианте осуществления настоящего изобретения антигенсвязывающий домен предпочтительно включает множество антигенсвязывающих доменов, составляющих библиотеку фрагментов антител. Более того, в неограничивающем варианте осуществления настоящего изобретения антитело предпочтительно включает панель группы антител, которые заранее преобразовали в моноклональные. Способы получения этих библиотек и антител описаны в разделе "Антитела", упоминаемом выше.
Например, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях высокой концентрации ионов водорода и низком значении pH, т.е. в диапазоне кислых значений pH, чем в условиях низкой концентрации ионов водорода или при высоком значении pH, т.е. в диапазоне нейтральных значений pH, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов в диапазоне нейтральных значений pH;
(b) помещение библиотеки антигенсвязывающих доменов, связавшихся с антигеном на стадии (a) в диапазон кислых значений pH; и
(c) выделение антигенсвязывающего домена, который диссоциировал на стадии (b).
Например, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях низкой концентрации ионов кальция, чем в условиях высокой концентрации ионов кальция, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов в условиях высокой концентрации ионов кальция;
(b) помещение библиотеки антигенсвязывающих доменов, связавшихся с антигеном на стадии (a), в условия низкой концентрации ионов кальция; и
(c) выделение антигенсвязывающего домена, который диссоциировал на стадии (b).
Более того, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях высокой концентрации ионов водорода или при низком значении pH, т.е. в диапазоне кислых значений pH, чем в условиях низкой концентрации ионов водорода или при высоком значении pH, т.е. в диапазоне нейтральных значений pH, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) контактирование в диапазоне нейтральных значений pH библиотеки антигенсвязывающих доменов с колонкой, на которой иммобилизован антиген;
(b) элюирование антигенсвязывающего домена, связавшегося с колонкой на стадии (a), из колонки в диапазоне кислых значений pH; и
(c) выделение антигенсвязывающего домена, элюированного на стадии (b).
Более того, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях низкой концентрации ионов кальция, чем в условиях высокой концентрации ионов кальция, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) контактирование в условиях высокой концентрации ионов кальция библиотеки антигенсвязывающих доменов с колонкой, на которой иммобилизован антиген;
(b) элюирование антигенсвязывающего домена, связавшегося с колонкой на стадии (a), из колонки в условиях низкой концентрации ионов кальция; и
(c) выделение антигенсвязывающего домена, элюированного на стадии (b).
Более того, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях высокой концентрации ионов водорода или при низком значении pH, т.е. в диапазоне кислых значений pH, чем в условиях низкой концентрации ионов водорода или высокого значения pH, т.е. в диапазоне нейтральных значений pH, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) пропускание в диапазоне кислых значений pH библиотеки антигенсвязывающих доменов через колонку, на которой иммобилизован антиген;
(b) сбор антигенсвязывающих доменов, элюированных без связывания с колонкой на стадии (a);
(c) обеспечение возможности антигенсвязывающим доменам, собранным на стадии (b), связаться с антигеном в диапазоне нейтральных значений pH; и
(d) выделение антигенсвязывающего домена, связавшегося с антигеном на стадии (c).
Более того, стадия получения антигенсвязывающего домена, антигенсвязывающая активность которого является более низкой в условиях низкой концентрации ионов кальция, чем в условиях высокой концентрации ионов кальция, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антигенсвязывающего домена, включающую следующие стадии:
(a) пропускание в условиях низкой концентрации ионов кальция библиотеки антигенсвязывающих доменов через колонку, на которой иммобилизован антиген;
(b) сбор антигенсвязывающих доменов, элюированных без связывания с колонкой на стадии (a);
(c) обеспечение возможности антигенсвязывающим доменам, собранным на стадии (b), связаться с антигеном в условиях высокой концентрации ионов кальция; и
(d) выделение антигенсвязывающего домена, связавшегося с антигеном на стадии (c).
Например, стадия получения антитела, антигенсвязывающая активность которого является более низкой в условиях высокой концентрации ионов водорода или при низком значении pH, т.е. в диапазоне кислых значений pH, чем в условиях низкой концентрации ионов водорода или при высоком значении pH, т.е. в диапазоне нейтральных значений pH, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антитела, включающую следующие стадии:
(a) определение антигенсвязывающей активности антитела в диапазоне кислых значений pH;
(b) определение антигенсвязывающей активности антитела в диапазоне нейтральных значений pH; и
(c) выбор антитела, антигенсвязывающая активность которого, определенная в диапазоне кислых значений pH, является более низкой, чем антигенсвязывающая активность, определенная в диапазоне нейтральных значений pH.
Например, стадия получения антитела, антигенсвязывающая активность которого является более низкой в условиях низкой концентрации ионов кальция, чем в условиях высокой концентрации ионов кальция, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антитела, включающую следующие стадии:
(a) определение антигенсвязывающей активности антитела в условиях низкой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антитела в условиях высокой концентрации ионов кальция; и
(c) выбор антитела, антигенсвязывающая активность которого, определенная в условиях низкой концентрации ионов кальция, является более низкой, чем антигенсвязывающая активность, определенная в условиях высокой концентрации ионов кальция.
Более того, стадия получения антитела, антигенсвязывающая активность которого является более низкой в условиях высокой концентрации ионов водорода или при низких значениях pH, т.е. в диапазоне кислых значений pH, чем в условиях низкой концентрации ионов водорода или при высоком значении pH, т.е. в диапазоне нейтральных значений pH, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антитела, включающую следующие стадии:
(a) контактирование антитела с антигеном в диапазоне нейтральных значений pH;
(b) получение антитела, связавшегося с антигеном на стадии (a);
(c) помещение антитела, полученного на стадии (b), в диапазон кислых значений pH; и
(d) выбор антитела, антигенсвязывающая активность которого на стадии (c) является более слабой, чем критерий отбора на стадии (b).
Более того, стадия получения антитела, антигенсвязывающая активность которого является более низкой в условиях низкой концентрации ионов кальция, чем в условиях высокой концентрации ионов кальция, которая является неограничивающим вариантом осуществления настоящего изобретения, предпочтительно включает стадию получения антитела, включающую следующие стадии:
(a) контактирование антитела с антигеном в условиях высокой концентрации ионов кальция;
(b) получение антитела, связавшегося с антигеном на стадии (a);
(c) помещение антитела, полученного на стадии (b), в условиях низкой концентрации ионов кальция; и
(d) выбор антитела, антигенсвязывающая активность которого на стадии (c) является более слабой, чем критерий отбора на стадии (b).
Упомянутые выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающим доменам или антителам, антигенсвязывающая активность которых является более низкой в диапазоне кислых значений pH, чем в диапазоне нейтральных значений pH, которые получают посредством упомянутых выше способов скрининга, которые, кроме того, включают стадию из повторяющихся стадий (a)-(c) или стадий (a)-(d) два или более раз. Количество циклов стадий (a)-(c) или (a)-(d) конкретно не ограничено, однако обычно составляет десять или менее.
В упомянутых выше стадиях антигенсвязывающая активность антигенсвязывающего домена или антитела в условиях низкой концентрации ионов водорода или при низких значениях концентрация pH, т.е. в диапазоне кислых значений pH, конкретно не ограничена при условии, что она представляет собой антигенсвязывающую активность при pH 4,0-6,5, однако антигенсвязывающая активность при pH 4,5-6,5 может быть предпочтительной. Альтернативно также может быть предпочтительной антигенсвязывающая активность при pH 5,0-6,5 или антигенсвязывающая активность при pH 5,5-6,0. Более предпочтительные значения pH включают pH в ранних эндосомах in vivo, и конкретным примером является антигенсвязывающая активность при pH 5,8. Антигенсвязывающая активность антигенсвязывающего домена или антитела в условиях высокой концентрации ионов водорода или при высоких значениях pH, т.е. в диапазоне нейтральных значений pH, конкретно не ограничена при условии, что она представляет собой антигенсвязывающую активность при pH 6,7-10, однако антигенсвязывающая активность при pH 6,7-9,5 может быть предпочтительной. Альтернативно также может быть предпочтительной антигенсвязывающая активность при pH 7,0-9,5 или антигенсвязывающая активность при pH 7,0-8,0. Более предпочтительные значения pH включают pH в плазме крови in vivo, и конкретным примером является антигенсвязывающая активность при pH 7,4.
В упомянутых выше стадиях антигенсвязывающая активность антигенсвязывающего домена или антитела в условиях низкой концентрации кальция конкретно не ограничена при условии, что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 0,1 мкМ до 30 мкМ, однако антигенсвязывающая активность при концентрации ионизированного кальция от 0,2 мкМ до 20 мкМ может быть предпочтительной. В другом варианте осуществления предпочтительной может быть антигенсвязывающая активность при от 0,5 мкМ до 10 мкМ. Более предпочтительные концентрации ионизированного кальция включают концентрацию ионизированного кальция в ранних эндосомах in vivo, и конкретные примеры включают антигенсвязывающую активность при от 1 мкМ до 5 мкМ и антигенсвязывающую активность при от 2 мкМ до 4 мкМ. Антигенсвязывающая активность антигенсвязывающего домена или антитела в условиях высокой концентрации ионов кальция конкретно не ограничена при условии, что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 100 мкМ до 10 мМ, однако предпочтительной может быть антигенсвязывающая активность при концентрации ионизированного кальция от 200 мкМ до 5 мМ. В другом варианте осуществления предпочтительной может быть антигенсвязывающая активность при от 500 мкМ до 2,5 мМ и в другом варианте осуществления предпочтительной может быть антигенсвязывающая активность при от 200 мкМ до 2 мМ. В другом варианте осуществления также может быть предпочтительной антигенсвязывающая активность при от 400 мкМ до 1,5 мМ. Более предпочтительные концентрации ионизированного кальция включают концентрацию ионизированного кальция в плазме in vivo, и конкретные примеры включают антигенсвязывающую активность при от 0,5 мМ до 2,5 мМ.
FcRn-связывающие домены, обладающие активностью связывания FcRn в диапазоне нейтральных значений pH, модифицированные Fc-области с активностью связывания Fcγ-рецептора, которая превышает активность связывания Fcγ-рецептора встречающейся в природе Fc-области, в которой фукоза связана с цепью сахаров в положении 297 в соответствии с нумерацией EU, и Fc-области, включающие такие FcRn-связывающие домены и модифицированные Fc-области, получают способами, описанными в разделах "FcRn-связывающий домены" и "FcRn-связывающий домены", упомянутых выше. Полинуклеотиды, кодирующие такие домены, можно получать известными способами генетической рекомбинации, описанными ниже в этом разделе.
Соответствующие домены по настоящему изобретению можно связывать друг с другом через линкеры или прямо через полипептидные связи. Линкеры включают произвольные пептидные линкеры, которые можно вносить способами генетической инженерии, синтетические линкеры и линкеры, описанные, например, в Protein Engineering (1996) 9(3), 299-305. Однако пептидные линкеры являются предпочтительными в рамках настоящего изобретения. Длина пептидных линкеров конкретно не ограничена, и ее могут надлежащим образом выбирать специалисты в данной области в зависимости от назначения. Длина предпочтительно составляет пять аминокислот или более (без конкретных ограничений, верхний предел обычно составляет 30 аминокислот или менее, предпочтительно 20 аминокислот или менее), и особенно предпочтительно 15 аминокислот.
Например, такие пептидные линкеры предпочтительно включают:
Ser
Gly·Ser
Gly·Gly·Ser
Ser·Gly·Gly
Gly·Gly·Gly·Ser (SEQ ID NO:29)
Ser·Gly·Gly·Gly (SEQ ID NO:30)
Gly·Gly·Gly·Gly·Ser (SEQ ID NO:31)
Ser·Gly·Gly·Gly·Gly (SEQ ID NO:32)
Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:33)
Ser·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:34)
Gly·Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:35)
Ser·Gly·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:36)
(Gly·Gly·Gly·Gly·Ser (SEQ ID NO:31))n
(Ser·Gly·Gly·Gly·Gly (SEQ ID NO:32))n
где n представляет собой целое число, равное 1 или более. Длину или последовательности пептидных линкеров могут соответствующим образом выбирать специалисты в данной области, в зависимости от назначения.
Обычно для сшивания пептидов используют синтетические линкеры (химические сшивающие агенты), и например:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидил пропионат) (DSP),
дитиобис(сульфосукцинимидилпропионат) (DTSSP),
этиленгликоль бис(сукцинимидилсукцинат) (EGS),
этиленгликоль бис(сульфосукцинимидилсукцинат) (сульфо-EGS),
дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES), и
бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти сшивающие агенты являются коммерчески доступными.
Когда используют множество линкеров для связывания соответствующих доменов, все они могут быть одного типа или они могут быть различных типов.
В дополнение к линкерам, проиллюстрированным выше, также можно соответствующим образом использовать линкеры с пептидными метками, такими как His-метка, HA-метка, myc-метка и FLAG-метка. Более того, также можно соответствующим образом использовать образование водородных связей, образование дисульфидных связей, образование ковалентных связей, ионное взаимодействие и свойства связывания друг с другом в результате их комбинации. Например, можно использовать аффинность между CH1 и CL антитела, и также можно использовать Fc-области, происходящие из описанных выше биспецифических антител для гетероассоциации Fc-областей. Более того, также можно соответствующим образом использовать дисульфидные связи, образующиеся между доменами.
Для связывания соответствующих доменов пептидной связью, полинуклеотиды, кодирующие домены, связывают вместе в рамке считывания. Известные способы связывания полинуклеотидов в рамке считывания включают способы, такие как лигирование фрагментов рестрикции, ПЦР со слиянием и перекрывающаяся ПЦР. Такие способы можно соответствующим образом использовать отдельно или в комбинации для конструирования антигенсвязывающих молекул по настоящему изобретению.
Для получения полинуклеотидов, кодирующих каждый домен, можно соответствующим образом использовать известные способы. Например, полинуклеотидную последовательность, кодирующую антигенсвязывающий домен, выделяют из фага, экспонирующего представляющий интерес антигенсвязывающий домен из библиотеки, содержащей множество антигенсвязывающих доменов, способом ПЦР с использованием праймеров, использованных для конструирования библиотеки, или праймеров, имеющих последовательность фагового вектора, использованного для конструирования библиотеки. Для получения генов, кодирующих вариабельные области антитела, удобно использовать способ 5'-RACE с использованием праймеров для амплификации генов вариабельных областей. Сначала РНК, экстрагированные из гибридомных клеток, используют в качестве матриц для синтеза кДНК, и, тем самым, получения библиотеки кДНК 5'-RACE. Для синтеза библиотеки кДНК 5'-RACE соответствующим образом используют набор для амплификации кДНК SMART RACE.
Полученную библиотеку кДНК 5'-RACE используют в качестве матрицы для амплификации генов антител с помощью ПЦР. Праймеры для амплификации генов антител мыши можно конструировать на основе известных последовательностей генов. Нуклеотидные последовательности этих праймеров варьируют. в зависимости от подкласса иммуноглобулина. Таким образом, предпочтительно определять подкласс заранее с использованием коммерчески доступного набора, такого как набор для изотипирования моноклональных антител мыши Iso Strip (Roche Diagnostics).
Более конкретно, когда намереваются получить гены, кодирующие IgG мыши, можно использовать праймеры, способные амплифицировать гены, кодирующие γ1, γ2a, γ2b и γb3 для тяжелой цепи, и гены, кодирующие цепь κ и цепь λ для легкой цепи. Для амплификации генов вариабельных областей IgG, как правило, в качестве 3'-праймера используют праймер, который подвергается отжигу с частью, соответствующей константной области вблизи вариабельной области. С другой стороны, в качестве 5'-праймера можно использовать праймер, предоставляемый в наборе для продуцирования библиотеки кДНК 5'-RACE.
После определения полинуклеотидной последовательности антигенсвязывающего домена или антитела по настоящему изобретению, выделенной, как описано выше, получают полинуклеотид, содержащий слитый ген, в котором этот полинуклеотид связан в рамке считывания с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий активностью связывания FcRn в диапазоне нейтральных значений pH. Полученный полинуклеотид, содержащий слитый ген, функционально связан с подходящим экспрессирующим вектором так, чтобы он экспрессировался в желаемых клетках.
Термины "клетка", "клеточная линия" и "клеточная культура" используют в настоящем описании синонимично и такие обозначения могут включать любого потомка клетки или клеточной линии. Таким образом, например, термины "трансформанты" и "трансформированные клетки" включают первичные рассматриваемые клетки и культуры, происходящие из них, не зависимо количества переносов. Также понятно, что все потомки могут не быть точно идентичными по содержанию ДНК вследствие преднамеренных или случайных мутаций. Также могут быть включены мутантные потомки, которые обладают по существу те и же функциями или биологической активностью, в отношении которых подвергали скрининг первоначально трансформированных клеток. Если подразумевается другое обозначение, это будет очевидно из контекста описания.
При указании на экспрессию кодирующей последовательности, термин "последовательность контроля" относится к нуклеотидным последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, пригодные, например, для прокариот, включают промотор, необязательные последовательность оператора, участок связывания рибосом и, возможно, другие последовательности, которые все еще предстоит изучить. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры для экспрессии кодирующей последовательности.
Для нуклеиновой кислоты термин "функционально связанный" означает, что нуклеиновая кислота имеет функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК препоследовательности или секреторного лидера функционально связана с ДНК полипептида, если он экспрессируется в качестве белка-предшественника, который участвует в секреции полипептида. Промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности. Участок связывания рибосом функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Обычно выражение "функционально связанный" означает, что связанные последовательности ДНК являются смежными и, в случае секреторного лидера, являются смежными и находятся в рамке считывания. Однако энхансеры не должны быть смежными. Связывание осуществляют посредством лигирования в подходящих участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой. Более того, связанные нуклеиновые кислоты можно получать с помощью упомянутой выше технологии перекрывающейся ПЦР.
"Лигирование" относится к способу формирования фосфодиэфирной связи между двумя фрагментами нуклеиновой кислоты. Для лигирования двух фрагментов концы этих фрагментов должны быть совместимыми друг с другом. В некоторых случаях эти концы обладают совместимостью сразу после расщепления эндонуклеазой. Однако может быть необходимо, чтобы сначала липкие концы, обычно образующиеся при расщеплении эндонуклеазой, были преобразованы в тупые концы, чтобы они стали совместимыми для лигирования. Для преобразования в тупые концы ДНК обрабатывают в подходящем буфере в течение по меньшей мере 15 минут при 15°C приблизительно 10 единицами фрагмента Кленова ДНК-полимеразы I или ДНК-полимеразой T4 в присутствии четырех дезоксирибонуклеотидтрифосфатов в соответствующем буферном растворе. Далее ДНК очищают экстракцией фенолом-хлороформом и преципитацией этанолом, или очисткой на диоксиде кремния. Фрагменты ДНК, подлежащие слиянию, добавляют в эквимолярных количествах к раствору. Этот раствор содержит ATP и буфер лигазы, а также приблизительно 10 единиц лигазы, такой как ДНК-лигаза T4 на 0,5 мкг ДНК. Если ДНК подлежит лигированию с вектором, вектор сначала линеаризуют посредством расщепления соответствующей эндонуклеазой(-ами) рестрикции. Затем линеаризованный фрагмент обрабатывают бактериальной щелочной фосфатазой или фосфатазой кишечника теленка, тем самым препятствуя самолигированию фрагмента на стадии лигирования.
Получение экспрессирующего вектора, содержащего полинуклеотид, кодирующий антигенсвязывающую молекулу по настоящему изобретению, введение экспрессирующего вектора в клетки, экспрессию полинуклеотида в клетках и получение экспрессированной антигенсвязывающей молекулы из культуральной жидкости клеток проводят в соответствии со способами, описанными в разделе "Антитела", упомянутом выше.
Все цитированные в настоящем описании ссылки уровня техники включены в настоящее описание в качестве ссылок.
Примеры
Ниже настоящее изобретение конкретно описано с помощью примеров, однако его не следует считать ограниченным ими.
Пример 1. Получение модельных мышей, которые являются иммунотолерантными к рецептору IL-6 человека
Поскольку рецептор IL-6 в высокой степени экспрессируется в миеломе, для которой фактором роста является лиганд IL-6, известно, что антитела против рецептора IL-6 человека проявляют противоопухолевые эффекты в модели на иммунодефицитных мышах с ксенотрансплантатом миеломы человека (Eur. J. Immunol. 22, 1989-93 (1992)). Если продукция антител мыши против рецептора IL-6 может индуцироваться путем введения мыши антител против рецептора IL-6, эти антитела против рецептора IL-6 могут индуцировать приобретенный иммунитет и могут оказаться пригодными в качестве вакцин против злокачественной опухоли.
Однако поскольку иммунные функции не работают у мышей с иммунодефицитом, которые используют в качестве противоопухолевых моделей, индукция приобретенного иммунитета невозможна. Даже если растворимые рецепторы IL-6 человека вводят нормальны мышам, антитела мыши против рецептора IL-6 человека антитела будут быстро продуцироваться против рецепторов IL-6 человека, которые являются чужеродными веществами. Таким образом, нормальных мышей нельзя использовать, поскольку они являются системами для оценки эффектов антител против рецептора IL-6 человека на продукцию антител мыши против рецептора IL-6 человека (приобретенный иммунитет к рецептору IL-6 человека).
С другой стороны, в клинических ситуациях у человека, поскольку антиген, на который нацелены молекулы антитела, представляет собой антиген человека, люди являются иммунотолерантными в отношении этого собственного антитела, на который осуществляется нацеливание. В клинических ситуациях у человека, когда индуцируется продукция аутоантител против антигена-мишени после введения антител против антигена-мишени, иммунотолерантность, по-видимому, утрачивается.
Таким образом, для оценки, предусматривающей клиническую ситуацию у человека, в отношении того, могут ли введенные мыши антитела индуцировать приобретенный иммунитет против антигена-мишени человека у мышей, необходимо, чтобы мыши находились в состоянии, близком к иммунотолерантности в отношении антигена-мишени человека (т.е. введение антигена-мишени человека мышам не приведет к продукции антител против антигена-мишени человека у мышей), и необходимо разработать аналитическую систему для оценки того, что при введении антител против антигена-мишени утрачивается практически иммунотолерантное состояние и индуцируется продукция аутоантител против антигена-мишени.
Таким образом, чтобы избежать продуцирования аутоантител против антигена-мишени человека у мышей посредством введения мышам антигена-мишени человека, конструировали аналитическую систему, в которой CD4-положительные T-клетки, необходимые для индукции продуцирования антител после представления антигена, успешно удалены с использованием антител против CD4 мыши.
Следующую тестовую модель конструировали в качестве модели для поддержания концентрации в плазме растворимого рецептора IL-6 человека, антигена-мишени человека, в стационарном состоянии (приблизительно 20 нг/мл). Инфузионный насос (MINI-OSMOTIC PUMP MODEL 2004; alzet), заполненный растворимым рецептором IL-6 человека, имплантировали под кожу спины нормальной мыши (мышь C57BL/6J, Charles River Japan) для получения модельных животных, у которых концентрация в плазме растворимого рецептора IL-6 человека поддерживалась в стационарном состоянии.
Исследование проводили в двух группах (n=4 на группу). В группе мышей, которые имитируют иммунную толерантность, вводили однократную дозу (20 мг/кг) моноклонального антитела против CD4 мыши (R&D) в хвостовую вену для ингибирования продукции антител мыши против растворимого рецептора IL-6 человека. Затем антитело аналогичным образом вводили один раз в 10 суток (далее эта группа обозначается как группа введения антитела против CD4 мыши). Другую группу использовали в качестве контрольной группы, т.е. группы без введения антитела против CD4 мыши, в которой не вводили моноклональное антитело против CD4 мыши. Затем инфузионный насос, заполненный 92,8 мкг/мл растворимого рецептора IL-6 человека, подкожно имплантировали в спину мыши. После имплантации инфузионного насоса проводили взятие образцов крови с течением времени, а затем сразу проводили центрифугирование в течение 15 минут при 4°C и 15000 об./мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до времени измерения. Концентрацию в плазме растворимого рецептора IL-6 человека (hsIL-6R) определяли способом, описанным ниже.
Концентрацию hsIL-6R в плазме мыши определяли с использованием способа электрохемилюминесценции. Образец hsIL-6R для калибровочной кривой, полученный в концентрации 2000, 1000, 500, 250, 125, 62,5 или 31,25 пг/мл, и образец плазмы мыши для измерения, разбавленный в 50 раз или выше, смешивали с моноклональным антителом против IL-6R человека (R&D), рутенированным с помощью SULFO-TAG NHS Ester (Meso Scale Discovery), биотинилированным антителом против IL-6R человека (R&D), и тоцилизумабом, с последующей реакцией в течение ночи при 37°C. Тоцилизумаб получали в конечной концентрации 333 мкг/мл. Затем реакционную жидкость распределяли в планшет со стрептавидином MA400 PR Streptavidin Plate (Meso Scale Discovery). Кроме того, после смывания реакционной жидкости, которой позволяли реагировать в течение 1 часа при комнатной температуре, распределяли буфер для считывания T (×4) (Meso Scale Discovery). Затем сразу проводили измерение в реакционной жидкости с использованием устройства для считывания SECTOR PR 400 (Meso Scale Discovery). Концентрацию hsIL-6R вычисляли из ответа калибровочной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Изменение концентрации hsIL-6R в плазме каждой нормальной мыши, которое было определено этим способом, представлено на фиг. 1.
В результате у всех мышей в группе без введения антитела против CD4 мыши, снижение концентрации hsIL-6R в плазме наблюдали через 14 дней после имплантации инфузионного насоса под кожу спины мышей. Иными словами, если антитела мыши против рецептора IL-6 человека продуцируются, концентрация hsIL-6R в плазме будет снижаться, и это показало, что антитела мыши против рецептора IL-6 человека продуцируются в этих группах. С другой стороны, снижение концентрации hsIL-6R в плазме не наблюдали у каких-либо мышей, относящихся к группе введения антител против CD4 мыши. Таким образом, это показало, что антитела мыши против рецептора IL-6 человека не продуцировались, поскольку в течение 28 суток, когда инфузионный насос был эффективным, поддерживалась практически постоянная концентрация hsIL-6R в плазме (приблизительно 20 нг/мл).
Описанные выше результаты подтвердили, что предшествующее введение антитела против CD4 мыши не приводило к индукции приобретенного иммунитета у нормальных мышей, когда рецептор IL-6 человека, который является антигеном-мишенью, вводят отдельно. В примерах, которые следуют далее, модель введения антитела против CD4 мыши использовали в качестве системы для оценки индукции приобретенного иммунитета в отношении рецептора IL-6 человека после введения антитела против рецептора IL-6 человека.
Пример 2. Сравнение эффектов нормальных антител против рецептора IL-6 и pH-зависимых антител против рецептора IL-6 в отношении индукции приобретенного иммунитета в модели на нормальных мышах с иммунной толерантностью к рецептору IL-6 человека
Для индукции приобретенного иммунитета к антигену-мишени, антиген-мишень, захваченный в антигенпредставляющие клетки, должен соответствующим образом деградироваться лизосомами в клетках и фрагментированные пептиды антигена-мишени должны претерпевать представление антигена посредством связывания с MHC класса I или MHC класса II. Считается, что чем больше количество пептидов, представляемых в качестве антигеном, тем сильнее индукция иммунитета; таким образом, в качестве способы усиления индукции приобретенного иммунитета к антигену-мишени, рассматривали способ направления большого количества антигена-мишени в антигенпредставляющие клетки.
Обычно, когда антиген включается неспецифически в антигенпредставляющую клетку, он переносится в таком виде из эндосомы в лизосому, и, таким образом, фрагментированные пептиды могут представляться в качестве антигенов. Однако, когда обычное IgG-антитело связано с антигеном, поскольку IgG-антитело рециклирует на клеточную поверхность (в плазму) из эндосомы посредством связывания FcRn, антиген, связанный с антителом, может не переноситься в лизосому и может рециклировать на поверхность (в плазму). Таким образом, введение нормальных IgG-антитела будет подавлять деградацию антигена-мишени. В результате фрагментированные пептиды антигена-мишени, которые соответствующим образом деградируются в лизосомах антигенпредставляющих клеток, не представляются как антигены, а, скорее, полагают, что индукция приобретенного иммунитета против антигена-мишени снижается. Таким образом, поскольку IgG-антитела, обладающие pH-зависимой активностью связывания, так что они связываются с антигеном-мишенью при pH 74 в плазме и высвобождают антиген в кислых условиях pH 5,0-6,5 в эндосоме (WO 2009/125825), могут диссоциировать от антигена-мишени в кислых эндосомах антигенпредставляющих клеток, было предположено, что при использовании этого IgG-антитела только антиген-мишень может транслоцироваться в лизосомы антигенпредставляющих клеток, фрагментированные пептиды антигена-мишени могут представляться в качестве антигенов, и IgG-антитела, которые высвободили антиген, могут рециклировать на поверхность клеток (в плазму) из эндосом посредством связывания с FcRn, не подвергаясь представлению антигена.
Таким образом, эффекты нормальных IgG-антител против рецептора IL-6 человека и pH-зависимых IgG-антител против рецептора IL-6 человека в отношении индукции приобретенного иммунитета оценивали с использованием модели на нормальных мышах, которые являются иммунотолерантными к рецепторам IL-6 человека, разработанной согласно примеру 1. H54/L28-IgG1, состоящее из H54-IgG1 (SEQ ID NO:38) и L28-CK (SEQ ID NO:39), которое описано в WO 2009/125825, использовали в качестве обычного IgG-антитела против рецептора IL-6 человека. Fv4-IgG1, состоящее из VH3-IgG1 (SEQ ID NO:17) и VL3-CK (SEQ ID NO:40), использовали в качестве pH-зависимого IgG-антитела против рецептора IL-6 человека. Получение антител проводили с использованием способа, описанного в справочном примере 1.
Эффекты H54/L28-IgG1 и Fv4-IgG1 в отношении индукции приобретенного иммунитета оценивали с использованием модели на мышах, иммунотолерантных к рецептору IL-6 человека, разработанной из трансгенных мышей с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 32 +/+ мышь, Jackson Laboratories (Methods Mol. Biol. (2010) 602: 93-104)). H54/L28-IgG1 и Fv4-IgG1 вводили модельным мышам, иммунотолерантным к рецептору IL-6 человека. В частности, аналогично примеру 1, через трое суток после имплантации инфузионного насоса, антитело против рецептора IL-6 человека вводили один раз в количестве 1 мг/кг в хвостовую вену. Взятие крови проводили с течением времени после введения антитела против рецептора IL-6 человека. Собранную кровь сразу центрифугировали при 15000 об./мин и 4°C в течение 15 минут с получением плазмы. Отделенную плазму хранили в морозилке, установленной при -20°C или ниже до проведения измерений. Концентрацию hsIL-6R в плазме измеряли тем же способом, который описан в примере 1.
Изменения средней концентрации hsIL-6R в плазме для каждой из контрольной группы (группа без введения антитела против рецептора IL-6 человека), группы введения H54/L28-IgG1 и группы введения Fv4-IgG1 представлены на фиг. 2. У всех животных в контрольной группе концентрация hsIL-6R в плазме поддерживалась практически на постоянном уровне, как и в примере 1, что указывает на то, что антитела мыши против рецептора IL-6 человека не продуцировались. Значительное повышение концентрации hsIL-6R в плазме наблюдали у всех животных в группе введения H54/L28-IgG1 по сравнению с контролем, и это повышенное состояние сохранялось в течение 28 суток. Полагали, что это является следствием того, что когда hsIL-6R захватывается в клетки неспецифическим образом, он прямо деградируется в лизосомах, в то время как hsIL-6R, связанный с IgG-антителом, рециклирует в плазму в качестве комплекса антитело-антиген посредством FcRn, так что выведение hsIL-6R снижается и, следовательно, концентрация hsIL-6R в плазме увеличивается. Поскольку это повышенное состояние сохранялось в течение 28 суток, это указывает на то, что аналогично контролю, продукция антител мыши против рецептора IL-6 человека не индуцировалась в группе введения H54/L28-IgG1. У всех животных в группе введения Fv4-IgG1 концентрация hsIL-6R в плазме увеличивалась по сравнению с концентрацией в контрольной группе, однако степень повышения концентрации hsIL-6R в плазме была отчетливо снижена по сравнению с группой введения H54/L28-IgG1. Это является следствием того, что антитело с pH-зависимым связыванием высвобождает антиген в эндосому, что приводит к подавлению снижения выведения hsIL-6R, что затем приводит к подавлению повышения концентрации hsIL-6R в плазме. Исходя из этих результатов, было предположено, что антитело с pH-зависимым связыванием высвобождает больше антигенов-мишеней в эндосомах антигенпредставляющих клетках по сравнению с нормальными антителами, и стимулирует перемещение антигена-мишени в лизосомы. Однако поскольку состояние повышенной концентрации hsIL-6R сохранялось в течение 28 суток аналогично H54/L28-IgG1, было показано, что Fv4-IgG1 не индуцирует продукцию антител мыши против рецептора IL-6 человека.
Эти результаты показали, что приобретенный иммунитет не может быть индуцирован против антигена-мишени с использованием обычных IgG-антител и с использованием IgG-антител с pH-зависимым связыванием, которые высвобождают антиген внутрь эндосом антигенпредставляющих клеток для переноса в лизосомы.
Пример 3. Эффекты pH-зависимого связывания рецептора IL-6 и усиления связывания FcRn при pH 7,4 на эффекты индукции приобретенного иммунитета в модели на трансгенных мышах с FcRn человека с иммунной толерантностью к рецептору IL-6 человека
(3-1) Сущность
В примере 2 было подтверждено, что обычное IgG1-антитело и IgG1-антитело с pH-зависимым связыванием не может индуцировать приобретенный иммунитет против антигена-мишени. С другой стороны, в качестве способа повышения иммуногенности пептида T-клеточного эпитопа, недавно был описан способ слияния пептида T-клеточного эпитопа с Fc, а затем усиления связывания Fc-части с FcRn при pH 7,4, так чтобы большее количество пептидов T-клеточных эпитопов переносилось в лизосомы (J. Immunol. 181, 7550-61 (2008)). Поскольку FcRn экспрессируется на антигенпредставляющих клетках, усиление связывание Fc-части с FcRn при pH 7,4 может усилить представление пептида T-клеточного эпитопа. Однако, поскольку молекула, описанная в этом способе, которая имеет антигенный пептид, прямо слитый с Fc, не может связываться с антигенами злокачественной опухоли в качестве антигенсвязывающей молекулы, эта молекула не может проявлять никакого прямого действия на злокачественные клетки. Более того, способ усиления связывания FcRn Fc-части при pH 7,4 усиливает иммуногенность пептида T-клеточного эпитоп in vitro, однако, напротив, снижает иммуногенность in vivo, и это не было эффективным in vivo. Таким образом, поскольку молекула, продуцированная посредством прямого слияния антигена-мишени с Fc, обладающей усиленным связыванием FcRn при pH 7,4, не может продемонстрировать активность связывания с антигеном-мишенью, она не может действовать прямо на антиген-мишень и, более того, усиление связывания FcRn in vivo приводило к снижению иммуногенности.
Учитывая сообщения, сделанные до настоящего времени и пример 2, антитело (H54/L28-F157), полученное путем внесения в нормальное IgG1-антитело (H54/L28-IgG1) изменений, которые усиливают связывание FcRn при pH 7,4, и антитело (Fv4-F157), полученное путем внесения в IgG1-антитело с pH-зависимым связыванием (Fv4-IgG1) изменений, которые усиливают связывание FcRn при pH 7,4, вводили для оценки того, может ли индуцироваться приобретенный иммунитет против антигена-мишени.
(3-2) Получение антител
H54/L28-F157, состоящее из H54-F157 (SEQ ID NO:41) и L28-CK (SEQ ID NO:39), использовали в качестве нормального IgG-антитела против рецептора IL-6 человека с усиленным связыванием FcRn. Fv4-F157, состоящее из VH3-F157 (SEQ ID NO:42) и VL3-CK (SEQ ID NO:40), использовали в качестве pH-зависимого IgG-антитела против рецептора IL-6 человека с усиленным связыванием FcRn. Получение антитела проводили с использованием способа согласно справочному примеру 1.
(3-3) Измерение аффинности в отношении FcRn человека
Для определения аффинности Fc-областей Fv4-IgG1 и Fv4-F157 (обозначаемых как IgG1 и F157, соответственно), полученных согласно примеру 2, к FcRn человека при pH 7,0, аффинность VH3/L(WT)-IgG1, состоящего из VH3-IgG1 (SEQ ID NO:17) и L(WT)-CK (SEQ ID NO:18), и VH3/L(WT)-F157, состоящего из VH3-F157 (SEQ ID NO:42) и L(WT)-CK (SEQ ID NO:18), в отношении FcRn человека определяли способом, представленным ниже.
Анализы кинетики между FcRn человека и антителом проводили с использованием Biacore T100 (GE Healthcare). Подходящее количество белка L (ACTIGEN) иммобилизовывали на сенсорный чип CM4 (GE healthcare) способом присоединения аминов, и представляющие интерес антитела улавливали на него. Далее, инжектировали разбавленные растворы FcRn и подвижный буфер, который представлял собой пустой образец, и FcRn человека подвергали взаимодействию с антителами, уловленными на сенсорном чипе. Использованный подвижный буфер представлял собой 50 ммоль/л фосфат натрия, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween 20, pH 7,0, и этот буфер использовали для разведений FcRn. Сенсорный чип регенерировали с использованием 10 ммоль/л глицина-HCl, pH 1,5. Все измерения проводили при 25°C. На основе константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с), которые являются параметрами кинетики, вычисленными из сенсограммы, полученной при измерении, вычисляли KD (M) (аффинность) каждого антитела в отношении FcRn человека. Каждый параметр вычисляли с использованием программного обеспечения Biacore T100 Evaluation Software (GE Healthcare).
Аффинность IgG1 и F157 в отношении FcRn человека представлена в таблице 7. Было подтверждено, что аффинность F157 в отношении FcRn человека была приблизительно в 600 раз более высокой по сравнению с IgG1.
(3-4) Изменение концентрации антигена в плазме (растворимый рецептор IL-6 человека) в тесте введения антитела
Далее, в испытании 1 использовали модель на мышах с иммунной толерантностью в отношении рецептора IL-6 человека, которая была разработана для трансгенных мышей с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 32 +/+ мышь, Jackson Laboratories, Methods Mol. Biol. (2010) 602, 93-104). H54/L28-F157 и Fv4-F157 вводили в моделях мышах, иммунотолерантных к рецептору IL-6 человека. В частности, аналогично примеру 1, через трое суток после имплантации инфузионного насоса, антитело против рецептора IL-6 человека вводили один раз в дозе 1 мг/кг в хвостовую вену (три индивидуума в каждой группе). Взятие крови проводили с течением времени после введения мышам антитела против рецептора IL-6 человека. Более того, в испытании 2 проводили взятие крови с течением времени после введения Fv4-F157 отдельно аналогичным образом. Собранную кровь сразу центрифугировали при 15000 об./мин и 4°C в течение 15 минут с получением плазмы. Отделенную плазму хранили в морозильной камере при -20°C или менее до проведения измерений. Концентрацию hsIL-6R в плазме измеряли тем же способом, который описан в примере 1.
Изменения средней концентрации hsIL-6R в плазме для каждой из групп с введением H54/L28-F157 и с введением Fv4-F157 в испытании 1 и группы введения Fv4-F157 в испытании 2 представлены на фиг. 3, вместе с изменением средней концентрации hsIL-6R в плазме для каждой из контрольной группы (группа без введения антитела), группы с введением H54/L28-IgG1 и группы с введением Fv4-IgG1, полученных согласно примеру 2. В группе с введением H54/L28-F157 было показано увеличение концентрации hsIL-6R в плазме аналогично группе введения H54/L28-IgG1, однако увеличение было временным, и через 13 суток концентрация hsIL-6R в плазме снижалась до уровня, эквивалентного уровню контроля. После этого уровень, эквивалентный уровню контроля, сохранялся в течение 28 суток; таким образом, было показано, что аналогично H54/L28-IgG1, антитела мыши против рецептора IL-6 человека не продуцировались, и было выявлено, что усиление связывания FcRn при pH 7,4 отдельно не может индуцировать приобретенный иммунитет в отношении антигена-мишени. Это не противоречит данным, что молекула, полученная слиянием пептида T-клеточного эпитопа с Fc-областью с усиленным FcRn-связыванием при pH 7,4 не может усилить индукцию приобретенного иммунитета против пептида T-клеточного эпитопа, как описано в J. Immunol. (2008), 181, 7550-7561. Напротив, в группе введения Fv4-F157 наблюдали быстрое снижение концентрации hsIL-6R в плазме после введения антитела, и через одни сутки после введения концентрация hsIL-6R снижалась до уровня ниже предела обнаружения (15,6 нг/мл или ниже).
Не ограничиваясь конкретной теорией, упомянутые выше явления могут быть описаны следующим образом. Усиление приобретенного иммунитета к антигену-мишени может зависеть от количества антигена-мишени, захватываемого антигенпредставляющими клетками, при введении антитела. Количество включенного антигена-мишени можно оценивать посредством определения того, насколько концентрации антигена-мишени в плазме снизилась. Хотя антитело с pH-зависимым связыванием (Fv4-IgG1) высвобождает антиген-мишень в кислой эндосоме после захвата в клетки, процесс включения в клетки pH-зависимого антитела, которое связалось с антигеном, зависит от неспецифического пиноцитоза, и, поскольку скорость включения является медленной, концентрация антигена-мишени в плазме не снижается до уровня, ниже чем в случае контроля (фиг. 4). Более того, антитело (H54/L28-F157), в котором только связывание FcRn при pH 7,4 усилено, активно захватывается в клетки с использованием связывания FcRn при pH 7,4; однако, поскольку большая его часть рециклирует прямо на поверхность вследствие функции, присущей FcRn, в состоянии, когда антиген-мишень все еще связан с антителом, т.е. в качестве комплекса антитела и антигена-мишени, концентрация антигена-мишени в плазме не увеличивается до уровня ниже уровня контроля (фиг. 5). Напротив, поскольку антитело с pH-зависимым связыванием с усиленным связыванием FcRn при pH 7,4 (Fv4-F157), которое активно захватывается в клетки через связывание FcRn при pH 7,4, высвобождает антиген-мишень в эндосому, антиген направляется в лизосому, в то время как антитело, которое высвободило антиген-мишень, рециклирует на клеточную поверхность вследствие функции, присущей FcRn, вновь связывается с антигеном-мишенью и может аналогичным образом также направлять антиген-мишень в лизосому (фиг. 6). Считалось, что только антитела с pH-зависимым связыванием, которые обладают усиленным связыванием FcRn при pH 7,4, способны снижать концентрацию антигена-мишени в плазме до уровня, который является значительно более низким, чем уровень в случае контроля, путем повторения этого цикла.
Концентрация hsIL-6R для каждой из шести мышей, которым вводили Fv4-F157, в испытаниях 1 и 2, представлены на фиг. 7 и 8. Среди шести мышей, которым вводили Fv4-F157, концентрация hsIL-6R была увеличена после 13 суток у трех мышей, а именно #7, #8 и #10, а затем возвращалась до уровня, эквивалентного уровню контроля, и после этого уровень, эквивалентный уровню контроля, сохранялся в течение 28 суток. С другой стороны, у остальных трех мышей, а именно #9, #11 и #12, поскольку концентрация hsIL-6R не восстанавливалась и сохранялась после этого на этом низком уровне в течение 28 суток, было сделано предположение, что у этих трех мышей продуцировались антитела против рецептора IL-6 человека, и они устраняли рецепторы IL-6 человека из плазмы. Таким образом, титр антитела против рецептора IL-6 человека определяли у шести мышей, которым вводили Fv4-F157, способом, указанным ниже.
(3-4) Изменение титра антител мыши (антитело мыши против рецептора IL-6 человека) против антигена (растворимый рецептор IL-6 человека) у мышей в испытании с введением антитела
Титр антитела мыши против рецептора IL-6 человека в плазме мыши измеряли способом хемилюминесценции. Сначала рецептор IL-6 человека распределяли в непокрытый планшет MA100 PR (Meso Scale Discovery), и, оставляя стоять в течение ночи при 4°C, получали планшет с иммобилизованным рецептором IL-6 человека. Образцы плазмы мыши для измерения, разбавленные в 50 раз, распределяли в планшет с иммобилизованным рецептором IL-6 человека, и его оставляли стоять в течение ночи при 4°C. Затем планшету позволяли реагировать с антителом против IgG мыши (целая молекула) (Sigma-Aldrich), которое было меченным рутением, с использованием SULFO-TAG NHS Ester (Meso Scale Discovery) при комнатной температуре в течение одного часа, а затем промывали. После распределения буфера для считывания T (×4) (Meso Scale Discovery) в планшет, сразу проводили измерения на устройстве для считывания SECTOR PR 400 (Meso Scale Discovery).
Изменение титра антитела мыши против рецептора IL-6 человека антитело (антитело против hsIL-6R) в группе введения Fv4-F157 представлены на фиг. 7 и 8. В результате у трех мышей, #7, #8 и #10, у которых концентрация hsIL-6R возвращалась до того же уровня, что и у контроля, повышения титра антител мыши против рецептора IL-6 человека не наблюдали; однако у трех мышей, #9, #11 и #12, у которых сохранялись низкие уровни концентрации hsIL-6R до 28 суток, наблюдали повышение титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R). Из представленных выше результатов было обнаружено, что введение Fv4-F157, которое представляет собой pH-зависимое IgG-антитело против рецептора IL-6 человека с усиленным связыванием FcRn, в модели на нормальных мышах с иммунной толерантностью к рецептору IL-6 человека способно индуцировать приобретенный иммунитет против рецептора IL-6 человека, который является антигеном-мишенью в трех из шести случаев. Таким образом, даже если нормальное IgG-антитело (H54/L28-IgG1) или IgG-антитело (Fv4-IgG1), демонстрирующее pH-зависимое связывание с антигеном-мишенью, или IgG-антитело (H54/L28-IgG1) только с усилением связывания FcRn при pH 7,4 вводят in vivo, приобретенный иммунитет против антигена-мишени не может индуцироваться, и было обнаружено, что только IgG-антитело (Fv4-F157), демонстрирующее pH-зависимое связывание антигена-мишени и усиленное связывание FcRn при pH 7,4 является способным индуцировать приобретенный иммунитет против антигена-мишени.
(3-5) Изменение титра антитела мыши (антитело мыши против Fv4-F157) против антитела (Fv4-F157), введенного мыши в испытании введения антитела
Даже если получение антител против антигена-мишени станет возможным с помощью указанного выше способа (J. Immunol. (2008) 181, 7550-7561), в котором используется в качестве фармацевтического средства молекула, полученная посредством прямого слияния антигена-мишени с Fc с усиленным связыванием FcRn при pH 7,4, поскольку антитела против антигена-мишени будут связываться с самим фармацевтическим средством, они будут действовать в качестве антител против фармацевтического средства, и это приведет к снижению действия фармацевтического средства.
Таким образом, использование молекулы, имеющей антиген-мишень, прямо слитый с фармацевтическим средством (например, соединения, описанные в J. Immunol. (2008) 181, 7550-7561 и J. Immunol. (2011), 186, 1218-1227) означает индукцию продукции антител против антигена-мишени, или, более конкретно, индукцию антител против самого фармацевтического средства, что приведет к снижению действия фармацевтического средства, и таким образом, это применение может не быть эффективным.
pH-зависимое IgG-антитело против рецептора IL-6 человека (Fv4-F157) с усиленным связыванием с FcRn представляет собой фармацевтическое средство, обладающее pH-зависимой активностью связывания с рецептором IL-6 человека, который является антигеном-мишенью, и не является молекулой с прямо слитым с ней антигеном-мишенью. Таким образом, как показано на фиг. 7 и 8, в то время как Fv4-F157 индуцировало продукцию антитела против рецептора IL-6 человека, который является антигеном-мишенью, оно не вызывало продукцию антител против самого фармацевтического средства (Fv4-F157). Таким образом, титры антитела мыши против Fv4-F157 у шести мышей, которым вводили Fv4-F157, определяли способом, описанным ниже.
Титр антитела против Fv4-F157 в плазме мыши измеряли способом электрохемилюминесценции. Сначала антитело против рецептора IL-6 человека распределяли в непокрытый планшет MA100 PR (Meso Scale Discovery), и, оставляя его стоять в течение ночи при 4°C, получали планшет с иммобилизованным на нем антителом против рецептора IL-6 человека. Образцы плазмы мыши для измерения, разбавленные в 50 раз, распределяли в планшет с иммобилизованным рецептором IL-6 человека и его оставляли стоять в течение ночи при 4°C. Затем планшету позволяли реагировать с антителом против IgG мыши (целая молекула) (Sigma-Aldrich), которое было меченным рутением, с использованием SULFO-TAG NHS Ester (Meso Scale Discovery) при комнатной температуре в течение одного часа, а затем промывали. После распределения буфера для считывания T (×4) (Meso Scale Discovery) в планшет, сразу проводили измерения на устройстве для считывания SECTOR PR 400 (Meso Scale Discovery).
Изменения титра антитела мыши против Fv4-F157 и титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) в группе введения Fv4-F157 представлены на фиг. 9 и 10. В группе введения Fv4-F157, независимо от продукции антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R), продукцию антитела мыши против антитела Fv4-F157 не наблюдали ни у одной мыши. Таким образом, поскольку pH-зависимое IgG-антитело против рецептора IL-6 человека с усиленным связыванием FcRn при pH 7,4 (Fv4-F157) индуцировало продукцию антител против антигена-мишени (рецептор IL-6 человека), но не индуцировало продукцию антител против самого фармацевтического средства (Fv4-F157), было предположено, что pH-зависимое (антитело против рецептора IL-6 человека) IgG-антитело с усиленным связыванием FcRn при pH 7,4 является в высокой степени пригодным в качестве фармацевтического средства для индукции приобретенного иммунитета в отношении антигена-мишени.
На настоящий момент, в качестве способа индукции приобретенного иммунитета на антиген злокачественной опухоли, описан способ с использованием молекулы фармацевтического средства, в котором антиген злокачественной опухоли, против которого желательна индукция приобретенного иммунитета, слит с антителом, которое связывается с рецептором, экспрессированным на антигенпредставляющих клетках (J. Immunol. (2011) 186, 1218-1227). Такая молекула фармацевтического средства не может прямо связываться с антигенами злокачественной опухоли и проявлять действие (фиг. 11). Таким образом, такая молекула фармацевтического средства не может демонстрировать прямую цитотоксическую активность в направлении клеток-мишеней или эффекты ингибирования функций антигенов злокачественной опухоли аналогично общепринятым фармацевтическим средствам на основе антител. Более того, поскольку такая молекула фармацевтического средств захватывается в антигенпредставляющие клетки целиком, а затем деградируется, фрагментированные пептиды молекулы фармацевтического средств представляются на MHC класса II и MHC класса I. Таким образом, может не только индуцироваться клеточный иммунитет и гуморальный иммунитет против злокачественных антигенов, но также может индуцироваться гуморальный иммунитет против самого фармацевтического средства, и это может быстро привести к снижению эффектов вследствие продукции антител против фармацевтического средства. Таким образом, такая молекула считается неэффективной (фиг. 11).
Напротив, антитело, выявленное в настоящих примерах, которое демонстрирует pH-зависимое связывание с антигеном-мишенью злокачественной опухоли и усиленное связывание FcRn при pH 7,4, может проявлять действие связывания прямо с антигенами злокачественной опухоли (фиг. 12). Таким образом, оно может продемонстрировать прямую цитотоксическую активность в отношении злокачественных клеток и эффекты ингибирования функций антигенов злокачественной опухоли аналогично общепринятым фармацевтическим средствам на основе антител. Более того, такое антитело высвобождает антиген-мишень злокачественной опухоли в кислую эндосому и само антитело рециклирует на клеточную поверхность, так что антиген-мишень злокачественной опухоли селективно деградируется, и поскольку пептидные фрагменты антигена-мишени представляются посредством MHC класса II и MHC класса I, подукция антител против самого антитела из фармацевтического средства на основе антител не индуцируется, и клеточный иммунитет и гуморальный может индуцироваться селективно против антигенов злокачественной опухоли (фиг. 12).
Пример 4. Эффекты pH-зависимого связывания рецептора IL-6 и усиления связывания FcRn при pH 7,4 на эффекты индукции приобретенного иммунитета против эндогенного рецептора IL-6 человека у мышей с нокином рецептора IL-6 человека
(4-1) Сущность
Как показано в примере 3, антитело (H54/L28-F157), полученное путем внесения в нормальное IgG1-антитело (H54/L28-IgG1) изменений, которые усиливают связывание FcRn при pH 7,4, не индуцировало приобретенный иммунитет против антигена-мишени у мышей, когда его вводили в модели на трансгенных мышах с FcRn человека с иммунной толерантностью в отношении рецептора IL-6 человека; однако антитело (Fv4-F157), полученное путем внесения в IgG1-антитело с pH-зависимым связыванием (Fv4-IgG1) изменений, которые усиливают связывание FcRn при pH 7,4, индуцировало приобретенный иммунитет против антигена-мишени у мышей после введения мышам.
Однако, хотя упомянутая выше модель представляет собой модель, которая индуцирует приобретенный иммунитет против антигена человека, введенного извне, при исследовании клинических применений описанной выше индукции приобретенного иммунитета предпочтительно, чтобы иммунная толерантность к эндогенному антигену, обладающему полной иммунотолерантностью, устранялась и была подтверждена индукция приобретенного иммунитета.
С использованием свойства, состоящего в том, что мыши с нокином рецептора IL-6 человека, экспрессирующие рецептор IL-6 человека, обладают иммунной толерантностью к эндогенному рецептору IL-6 человека, упомянутое выше антитело вводили мышам для оценки того, может ли индуцироваться приобретенный иммунитет против эндогенного рецептора IL-6 человека.
(4-2) Получение антитела
H54/L28-mF3, содержащее H54-mF3 (SEQ ID NO:124) и L28-mCK (SEQ ID NO:125), получали в качестве нормального антитела против рецептора IL-6 человека с усиленным связыванием с FcRn мыши. Fv4-mIgG1, содержащее VH3-mIgG1 (SEQ ID NO:126) и VL3-mCK (SEQ ID NO:127), и Fv4-mIgG2a, содержащее VH3-mIgG2a (SEQ ID NO:128) и VL3-mCK (SEQ ID NO:127), получали в качестве pH-зависимых IgG-антител против рецептора IL-6 человека. Fv4-mF3, содержащее VH3-mF3 (SEQ ID NO:129) и VL3-mCK (SEQ ID NO:127), получали в качестве pH-зависимого IgG-антитела против рецептора IL-6 человека с усиленным связыванием с FcRn мыши. Более того, Fv4-mFa30, содержащее VH3-mFa30 (SEQ ID NO:130) и VL3-mCK (SEQ ID NO:127), получали в качестве pH-зависимого IgG-антитела против рецептора IL-6 человека с усиленным связыванием с FcRn мыши, а также с усиленным связыванием с FcγR мыша. Эти антитела получали с использованием способа, описанного в справочном примере 1.
(4-3) Измерение аффинности к FcRn мыши и FcγR мыши
С использованием величин, определенных в соответствии со способом, представленным в (3-3), определяли аффинность к FcRn мыши при pH 7,0 для Fc-частей из mIgG1, mIgG2a, mF3 и mFa30 полученных Fv4-mIgG1, Fv4-mIgG2a, Fv4-mF3 и Fv4-mFa30.
Внеклеточный домен FcγR получали следующим способом. Во-первых, на основе информации о последовательности, зарегистрированной в NCBI, ген внеклеточного домена FcγR синтезировали способом, известным специалистам в данной области. В частности, получали гены, кодирующие внеклеточные домены FcγR с His-меткой, добавленной на C-конце каждого из полипептидов с номером доступа NCBI NP_034316 (версия номер NP_034316.1) для mFcγRI, номером доступа NCBI NP_034317 (версия номер NP_034317.1) для mFcγRII, номером доступа NCBI NP_034318 (версия номер NP_034318.2) для FcγRIII, и номером доступа NCBI NP_653142 (версия номер NP_653142.2) для FcγRIV.
Полученные фрагменты генов встраивали в экспрессирующий вектор клеток животных с получением экспрессирующих векторов. Экспрессирующие векторы вводили временно в клетки, происходящие из клеток рака почки эмбриона человека, FreeStyle 293 (Invitrogen), и культуральный супернатант трансдуцированных клеток, экспрессирующих представляющие интерес белки, пропускали через 0,22-мкм фильтр с получением культурального супернатанта. В качестве общего правила, внеклеточные домены каждого из FcγR очищали из полученного культурального супернатанта посредством следующих четырех стадий очистки: ионообменная колоночная хроматография на стадии 1 (стадия 1), аффинная колоночная хроматография для His-метки (HisTrap HP) (стадия 2), гель-фильтрационная колоночная хроматография (Superdex200) (стадия 3) и асептическая фильтрация (стадия 4). Колонка для ионообменной колоночной хроматографии стадии 1 представляла собой Q sepharose HP для очистки mFcγRI, SP Sepharose FF для очистки mFcγRII и mFcγRIV, и SP Sepharose HP для очистки mFcγRIII. На стадии 3 и на последующих стадиях очистки, в качестве растворителя использовали D-PBS(-), однако для очистки mFcγRIII использовали D-PBS(-), содержащий 0,1 M аргинин. Поглощение при 280 нм очищенного раствора, содержащего внеклеточный домен FcγR, измеряли с использованием спектрофотометра. Из полученных выше величин поглощения вычисляли концентрации очищенного внеклеточного домена FcγR с использованием коэффициента поглощения, вычисленного способами, такими как PACE (Protein Science (1995) 4, 2411-2423).
Анализ взаимодействия между каждым из измененных антител и внеклеточным доменом Fcγ-рецептора, полученным, как описано выше, проводили с использованием Biacore T100 (GE Healthcare), Biacore T200 (GE Healthcare), Biacore A100, и Biacore 4000. HBS-EP+ (GE Healthcare) использовали в качестве подвижного буфера и взаимодействия определяли при температуре измерения 25°C. Использовали чипы, полученные посредством иммобилизации белка L (ACTIGEN или BioVision) способом присоединения аминов на сенсорный чип CM5 Series S (GE Healthcare) или сенсорный чип CM4 Series S (GE Healthcare).
После улавливания каждого из измененных антител на эти сенсорные чипы, внеклеточному домену Fcγ-рецептора, разбавленному подвижным буфером, позволяли действовать на чипы для измерения уровня связывания каждого из доменов с каждым из антител, и сравнивали уровни связывания. Однако поскольку количество связанного внеклеточного домена Fcγ-рецептора зависит от количества антитела, иммобилизованного на сенсорный чип, количество связанного внеклеточного домена Fcγ-рецептора делили на соответствующее количество иммобилизованного антитела, получая скорректированные величины, и эти величины сравнивали. Более того, посредством реакции с 10 мМ глицин-HCl, имеющим pH 1,5, антитела, уловленные на сенсорный чип, смывали и регенерированные сенсорные чипы использовали повторно.
Величины KD для каждого из измененных антител для внеклеточного домена Fcγ-рецептора вычисляли в соответствии со следующим способом кинетического анализа. Представляющие интерес антитела иммобилизовывали на упомянутые выше сенсорные чипы, внеклеточному домену Fcγ-рецептора, разбавленному подвижным буфером, позволяли взаимодействовать, и с использованием программного обеспечения Biacore Evaluation Software для глобальной аппроксимации измеренных результатов с помощью полученной сенсограммы с использованием модели связывания Ленгмюра 1:1, вычисляли константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (1/с), и из этих величин определяли константы диссоциации KD (моль/л).
Аффинность mIgG1, mIgG2a, mF3 и mFa30 для FcRn мыши представлена в таблице 8, и их аффинность в отношении FcγR мыши представлена в таблице 9.
(4-4) Изменение концентрации в плазме антигена (растворимый рецептор IL-6 человека) в испытании с введением антитела
Далее, Fv4-mIgG1, Fv4-mIgG2a, Fv4-mF3, Fv4-mFa30 и H54/L28-mF3 вводили мышам с нокином рецептора IL-6 человека (справочный пример 25), и у этих мышей проводили взятие крови с течением времени. Собранную кровь сразу центрифугировали при 15000 об./мин и 4°C в течение 15 минут для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до проведения измерений. Концентрацию hsIL-6R в плазме измеряли те же способом, который описан в примере 1.
Изменения средней концентрации hsIL-6R в плазме для группы без введения антитела и групп введения Fv4-mIgG1, Fv4-mIgG2a, Fv4-mF3, Fv4-mFa30 и H54/L28-mF3 представлены на фиг. 26. Увеличение концентрации hsIL-6R наблюдали в плазме для групп введения Fv4-mIgG1 и Fv4-mIgG2a. С другой стороны, значимое снижение концентрации hsIL-6R по сравнению с группой без введения антитела наблюдали в плазме для групп введения Fv4-mF3, H54/L28-mF3 и Fv4-mFa30, где Fv4-mF3 и H54/L28-mF3 обладают усиленным связыванием с FcRn мыши и Fv4-mFa30 обладает усиленным связыванием с FcRn мыши и FcgR мыши.
(4-5) Изменение титра антитела мыши (антитело мыши против рецептора IL-6 человека) против антигена (растворимый рецептор IL-6 человека) у мышей в испытании с введением антитела
Титр антитела против hsIL-6R в плазме мыши измеряли посредством электролюминесценции. Образцы плазмы мыши, разбавленные в 50 раз, и идиотипическое антитело против Fv4, доведенное до 30 мкг/мл, смешивали и подвергали реакции при комнатной температуре в течение одного часа. Идиотипическое антитело получали посредством очистки сыворотки из иммунизированного Fv4-M73 (WO 2009/125825) кролика на ионообменной смоле, а затем проведения аффинной очистки на колонке, на которой было иммобилизовано Fv4-M73, и последующей адсорбции на иммобилизованную колонку с антигеном человека. К упомянутому выше смешанному раствору добавляли 50 мкг/мл раствора, содержавшего 1 мкг/мл hsIL-6R, который был биотинилирован с использованием Sulfo-NHS-биотина EZ-Link и набора для биотинилирования (Pierce), и 2 мкг/мл SULFO-антитела против IgG мыши (H+L) (BECKMAN COULTER), которое было меченным рутением с использованием SULFO-TAG NHS Ester (Meso Scale Discovery), и их перемешивали и позволяли реагировать в течение ночи при 4°C. В этих обстоятельствах для предотвращения связывания введенного образца, включенного в образец для измерения, с hsIL-6R и обнаружения ADA, направленного на введенный образец, к образцу заранее добавляли избыточное количество идиотипического антитела против Fv4. После этого упомянутый выше реакционный раствор распределяли в планшет со стрептавидином MA400 PR (Meso Scale Discovery). В каждую из лунок, которые дополнительно подвергали реакции при 25°C в течение одного часа, а затем промывали, распределяли буфер для считывания T (×4) (Meso Scale Discovery) и поглощение реакционного раствора в каждой лунке сразу измеряли с использованием устройства для считывания SECTOR PR 400 (Meso Scale Discovery).
Изменения титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) для каждого индивидуума в группе без введения антитела, и группах введения Fv4-mIgG1, Fv4-mIgG2a, Fv4-mF3, Fv4-mFa30 и H54/L28-mF3 представлены на фиг. 27-31. Увеличение титра антитела мыши против рецептора IL-6 человека (антитело против hsIL-6R) не наблюдали у индивидуумов в группе введения Fv4-mIgG1, в группе введения Fv4-mIgG2a и в группе введения H54/L28-mF3, где Fv4-mIgG1 и Fv4-mIgG2a не обладают усиленным связыванием с FcRn мыши и H54/L28-mF3 обладает усиленным связыванием с FcRn мыши, но не обладает pH-зависимым связыванием с рецептором IL-6 человека. С другой стороны, увеличение титра антитела мыши против рецептор IL-6 человека (антитело против hsIL-6R) у индивидуумов было подтверждено в группе введения Fv4-mF3 и в группе введения Fv4-mFa30, где Fv4-mF3 представляет собой антитело с pH-зависимым связыванием с усиленным связыванием с FcRn мыши, и Fv4-mFa30 представляет собой антитело с pH-зависимым связыванием с усиленным связыванием с FcRn мыши и также усиленным связыванием с FcgR мыши.
Из описанного выше, было показано, что антигенсвязывающая молекула, обладающая pH-зависимой активностью связывания антигена-мишени и усиленным связыванием FcRn при pH 7,4 способна индуцировать приобретенный иммунитет против эндогенного антигена-мишени, к которому существует иммунологическая толерантность. Таким образом, поскольку такая молекула может быть способна индуцировать приобретенный иммунитет на собственные антигены против злокачественной опухоли, она является в значительной степени перспективной в качестве лекарственного средства от злокачественной опухоли.
Справочный пример 1
Антитела экспрессировали способом, описанным ниже. Клетки клеточной линии, происходящей из рака почки эмбриона человека HEK293H (Invitrogen) суспендировали в среде DMEM (Invitrogen), дополненной 10% эмбриональной телячьей сывороткой (Invitrogen) и высевали в количестве 10 мл на чашку в чашки для прикрепляющихся клеток (диаметром 10 см; CORNING) при плотности клеток от 5-6×105 до 6×105 клеток/мл. Клетки культивировали в CO2-инкубаторе (37°C, 5% CO2) в течение всего дня и ночи, затем среду удаляли аспирированием и в чашки добавляли 6,9 мл среды CHO-S-SFM-II (Invitrogen). Полученные плазмиды вводили в клетки способом липофекции. Культуральные супернатанты собирали и центрифугировали (приблизительно 2000 g, 5 мин, комнатная температура) для удаления клеток. Культуральные супернатанты далее стерилизовали фильтрацией через 0,22-мкм фильтр MILLEX(R)-GV (Millipore) с получением культуральных супернатантов. Экспрессированные антитела очищали из полученных культуральных супернатантов способом, известным специалистам в данной области, с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Для определения концентрации очищенного антитела измеряли поглощение при 280 нм с использованием спектрофотометра, и из измеренных величин вычисляли концентрации антитела с использованием коэффициента поглощения, вычисленного способом, описанным в Protein Science (1995) 4, 2411-2423).
Справочный пример 2. Исследование последовательностей человека эмбрионального типа, которые связываются с ионами кальция
(15-1) Антитело, которое связывается с антигеном зависимым от кальция образом
Антитела, которые связываются с антигеном зависимым от Ca образом (антитела с зависимым от Ca связыванием антигена) представляют собой антитела, взаимодействия которых с антигеном изменяются, в зависимости от концентрации кальция. Полагают, что антитело с зависимым от Ca связыванием связывается с антигеном через ион кальция. Таким образом, аминокислоты, которые образуют эпитоп на одной стороне антигена представляют собой отрицательно заряженные аминокислоты, которые могут хелатировать ионы кальция, или аминокислоты, которые могут являться акцептором водородной связи. Эти свойства аминокислот, которые образуют эпитоп, позволяют нацеливание на эпитоп, отличное от связывающих молекул, которые получают внесением остатков гистидина и которые связываются с антигеном pH-зависимым образом. Боле того, как показано на фиг. 13, полагают, что применение антигенсвязывающих молекул, обладающих свойствами кальций- и pH-зависимого связывания, позволит образование антигенсвязывающих молекул, которые могут быть индивидуально нацелены на различные эпитопы, имеющие широкие свойства. Таким образом, если конструируют совокупность молекул, содержащих кальций-связывающий мотив (Ca-библиотека), из которых получают антигенсвязывающие молекулы, полагают, что будут эффективным путем получены антитела с зависимым от Ca связыванием антигена.
(2-2) Получение последовательностей человека эмбрионального типа
Примером совокупности молекул, содержащих кальций-связывающий мотив, является пример, в котором указанные молекулы представляют собой антитела. Иными словами, библиотека антител, содержащих кальций-связывающий мотив, может представлять собой Ca-библиотеку.
Антитела с зависимым от ионов кальция связыванием, содержащие последовательности человека эмбрионального типа, не были описаны. Таким образом, последовательности антител эмбрионального типа, обладающие последовательностями человека эмбрионального типа, клонировали с использованием в качестве матрицы кДНК, полученной из поли-РНК селезенки эмбриона (Clontech) для оценки того, связываются ли антитела, обладающие последовательностями человека эмбрионального типа, с ионом кальция. Клонировенные фрагменты ДНК встраивали в экспрессируюище векторы клеток животных. Нуклеотидные последовательности сконструированных экспрессирующих векторов определяли способом, известным специалистам в данной области. SEQ ID представлены в таблице 10. С помощью ПЦР полинуклеотиды, кодирующие SEQ ID NO:5 (Vk1), SEQ ID NO:6 (Vk2), SEQ ID NO:7 (Vk3), SEQ ID NO:8 (Vk4) и SEQ ID NO:43 (Vk5), связывали с полинуклеотидом, кодирующим природную константную область каппа-цепи (SEQ ID NO:44). Связанные фрагменты ДНК встраивали в экспрессирующие векторы клеток животных. Более того, полинуклеотиды, кодирующие SEQ ID NO:46 (Vk1), SEQ ID NO:47 (Vk2), SEQ ID NO:48 (Vk3), SEQ ID NO:49 (Vk4) и SEQ ID NO:45 (Vk5), связывали способом ПЦР с полинуклеотидом, кодирующим полипептид (SEQ ID NO:11), имеющий делецию двух аминокислот на C-конце IgG1. Полученные фрагменты ДНК встраивали в экспрессирующие векторы клеток животных. Последовательности сконструированных вариантов подтверждали способом, известным специалистам в данной области.
(3-3) Экспрессия и очистка антител
Сконструированные экспрессирующие векторы клеток животных, в которые встроены фрагменты ДНК, имеющие пять типов последовательностей человека эмбрионального типа, вводили в клетки животных. Экспрессию антитела проводили следующим способом. Клетки клеточной линии, происходящей из почки эмбриона человека FreeStyle 293-F (Invitrogen) суспендировали в среде FreeStyle 293 Expression Medium (Invitrogen), и высевали с плотностью клеток 1,33×106 клеток/мл (3 мл) в каждую лунку планшета с 6 ячейками. Полученные плазмиды вводили в клетки способом липофекции. Клетки культивировали в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин). Из культуральных супернатантов, полученных как описано выше, антитела очищали с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences) способом, известным специалистам в данной области. Поглощение растворов очищенного антитела при 280 измеряли с использованием спектрофотометра. Концентрации антител вычисляли из определенных величин с использованием коэффициента экстинкции, вычисленного способом PACE (Protein Science (1995) 4: 2411-2423).
(2-4) Оценка антител, имеющих последовательности человека эмбрионального типа, в отношении их активности связывания ионов кальция
Очищенные антитела оценивали в отношении их активности связывания ионов кальция. Промежуточную температуру термической денатурации (величину Tm) измеряли с помощью дифференциальной сканирующей калориметрии (DSC) в качестве показателя для исследования связывания ионов кальция с антителом (MicroCal VP-Capillary DSC, MicroCal). Промежуточная температура термической денатурации (величина Tm) является показателем стабильности. Она становится более высокой, когда белок стабилизирован путем связывания с ионами кальция, по сравнению с отсутствием связывания ионов кальция (J. Biol. Chem. (2008) 283, 37, 25140-25149). Активность связывания ионов кальция с антителом исследовали путем исследования изменений величины Tm антитела, в зависимости от изменений концентрации ионов кальция в растворе антитела. Очищенное антитело подвергали диализу (EasySEP, TOMY) с использованием внешнего раствора 20 мМ Tris-HCl, 150 мМ NaCl и 2 мМ CaCl2 (pH 7,4), или 20 мМ Tris-HCl, 150 мМ NaCl и 3 мкM CaCl2 (pH 7,4). Измерение DSC проводили при скорости нагревания 240°C/ч от 20 до 115°C с использованием раствора антитела, приготовленного в концентрации приблизительно 0,1 мг/мл с диализатом в качестве исследуемого вещества. Промежуточные температуры термической денатурации (величины Tm) Fab-доменов каждого антитела, вычисленные исходя из кривой денатурации, полученной с помощью DSC, представлены в таблице 11.
Результат показал, что величины Tm Fab-доменов антител, имеющих последовательность hVk1, hVk2, hVk3 или hVk4, не варьировали в зависимости от концентрации ионов кальция в растворах, содержащих Fab-домен. Между тем, величина Tm для Fab-домена антитела, имеющего последовательность hVk5, варьировала в зависимости от концентрации ионов кальция в растворе, содержащем Fab-домен. Это демонстрирует, что последовательность hVk5 связывается с ионами кальция.
(2-5) Оценка последовательности hVk5-2 в отношении связывания кальция
В дополнение к Vk5-2 (SEQ ID NO:50, полученное путем слияния константной области каппа-цепи SEQ ID NO:44 с SEQ ID NO:43), получали вариант 1 Vk5-2 (SEQ ID NO:51) и вариант 2 Vk5-2 (SEQ ID NO:52), классифицируемые как Vk5-2 в справочном примере 2, (2-2). Эти варианты оценивали в отношении их активности связывания кальция. Каждый из фрагментов ДНК VK5-2, варианта 1 Vk5-2 и варианта 2 Vk5-2 включали в экспрессирующие векторы для клеток животных. Нуклеотидные последовательности полученных экспрессирующих векторов определяли способом, известным специалистам в данной области. Способом, описанным в справочном примере 2, (2-3), экспрессирующие векторы клеток животных, в которые были встроены фрагменты ДНК для каждого из Vk5-2, варианта 1 Vk5-2 и варианта 2 Vk5-2, вводили в клетки животных вместе с экспресирующим вектором, содержащим вставку для экспрессии CIM_H (SEQ ID NO:45) в качестве тяжелой цепи, и антитела очищали. Очищенные антитела оценивали в отношении их активности связывания ионов кальция. Очищенные антитела подвергали диализу (EasySEP, TOMY) против внешнего раствора 20 мМ Tris-HCl, 150 мМ NaC1, 2 мМ CaCl2 (pH 7,5), или внешнего раствора 20 мМ Tris-HCl, 150 мМ NaCl (pH 7,5) (указан в качестве концентрации ионов кальция 0 мМ в таблице 12). Измерение DSC проводили при скорости повышения температуры 240°C/ч от 20°C до 115°C с использованием в качестве исследуемого вещества растворов антител, полученных в концентрации 0,1 мг/мл, с использованием того же раствора, что и раствор для диализа. На основе полученных кривых денатурации DSC, вычисляли промежуточную температуру термической денатурации (величину Tm) для Fab-домена каждого антитела, и она представлена в таблице 12.
Результат показал, что величина Tm для Fab-доменов антител, обладающих последовательностью Vk5-2, варианта 1 Vk5-2 или варианта 2 Vk5-2 варьировала в зависимости от концентрации ионов кальция в растворах, содержащих антитела, имеющие Fab-домены. Это демонстрирует, что антитела, обладающие последовательностью, классифицируемой как Vk5-2, связываются с ионами кальция.
Справочный пример 3. Оценка последовательности Vk5 человека (hVk5)
(3-1) Последовательность hVk5
Единственной последовательностью hVk5, зарегистрированной в базе данных Kabat, является последовательность hVk5-2. Далее hVk5 и hVk5-2 используются в качестве синонимов.
В WO2010/136598 описано, что относительная распространенность последовательности hVk5-2 в последовательности эмбрионального типа составляет 0,4%. Также существуют другие сообщения, согласно которым относительная распространенность последовательности hVk5-2 в последовательности эмбрионального типа составляет 0-0,06% (J. Mol. Biol. (2000) 296, 57-86; Proc. Natl. Acad. Sci. USA (2009) 106, 48, 20216-20221). Как описано выше, поскольку последовательность hVk5-2 представляет собой последовательность с низкой частотой появления в последовательности эмбрионального типа, полагали, что она является неэффективной для получения антитела с кальций-зависимым связыванием из библиотеки антител, состоящей из последовательностей человека эмбрионального типа, или B-клеток, полученных путем иммунизации мыши, экспрессирующей антитела человека. Таким образом, возможно сконструировать Ca-библиотеки, содержащие последовательность hVk5-2 человека. Между тем, описанные синтетические библиотеки антител (WO2010/105256 и WO2010/136598) не содержали последовательности hVk5. Кроме того, осуществимость этой возможности неизвестна, поскольку не было опубликовано отчета о физико-химических свойствах последовательности hVk5-2.
(3-2) Конструирование, экспрессия и очистка негликозилированной формы последовательности hVk5-2
Последовательность hVk5-2 имеет последовательность для потенциального N-гликозилирования в положении аминокислоты 20 (нумерация по Kabat). Цепи сахаров, присоединенные к белкам, проявляют гетерогенность. Таким образом, желательно избегать гликозилирования с точки зрения гомогенности вещества. В этом контексте, конструировали вариант hVk5-2_L65 (SEQ ID NO:53), в котором остаток Asn (N) в положении 20 (нумерация по Kabat) заменен на Thr (T). Аминокислотную замену проводили способом, известным специалистам в данной области, с использованием набора для сайт-направленного мутагенеза QuikChange Site-Directed Mutagenesis Kit (Stratagene). ДНК, кодирующую вариант hVk5-2_L65, встраивали в экспрессирующий вектор животных. Экспрессирующий вектор животных, в который встроена конструированная ДНК, кодирующая вариант hVk5-2_L65, в комбинации с экспрессирующим вектором животных, имеющим вставку для экспрессии CIM_H (SEQ ID NO:45) в качестве тяжелой цепи, встраивали в клетки животных способом, описанным в справочном примере 2. Антитело, содержащее hVk5-2_L65 и CIM_H, которые экспрессировались в клетках животных, в которые были введены векторы, очищали способом, описанным в справочном примере 2.
(3-3) Оценка антитела, имеющего негликозилированную последовательность hVk5-2, в отношении физико-химических свойств
Выделенное антитело, имеющее модифицированную последовательность hVk5-2_L65, анализировали ионообменной хроматографией для исследования того, является ли она менее гетерогенной, чем антитело, имеющее исходную последовательность hVk5-2, до модификации. Методика ионообменной хроматографии представлена в таблице 13. Результат анализа показал, что hVk5-2_L65, модифицированное в участке гликозилирования, было менее гетерогенным, чем исходная последовательность hVk5-2, как показано на фиг. 14.
Далее оценку того, связывается ли антитело, содержащее менее гетерогенную последовательность hVk5-2_L65, с ионом кальция, оценивали способом, описанным в справочном примере 2. Результат показал, что величина Tm для Fab-домена антитела, имеющего hVk5-2_L65 с измененным участком гликозилирования, также варьировала, в зависимости от концентрации ионов кальция в растворах антител, как показано в таблице 14. В частности, было продемонстрировано, что Fab-домен антитела, имеющего hVk5-2_L65 с измененным участком гликозилирования, связывается с ионом кальция.
Справочный пример 4. Оценка активности связывания ионов кальция молекул антител, имеющих последовательность CDR из последовательности hVk5-2
(4-1) Конструирование, экспрессия и очистка модифицированных антител, имеющих последовательность CDR из последовательности hVk5-2
Последовательность hVk5-2_L65 представляет собой последовательность с измененными аминокислотами в участке гликозилирования каркасной области последовательности Vk5-2 человека. Как описано в справочном примере 3, было продемонстрировано, что связывание ионов кальция происходило даже после изменения участка гликозилирования. Между тем, с точки зрения иммуногенности, главным образом, желательно, чтобы каркасная последовательность представляла собой последовательность эмбрионального типа. Таким образом, авторы настоящего изобретения оценивали, может ли каркасная последовательность антитела быть заменена каркасной последовательностью негликозилированной последовательности эмбрионального типа при сохранении активности антитела в отношении связывания ионов кальция.
Полинуклеотиды, кодирующие химически синтезированные последовательности, которые содержат измененную последовательность каркасной области из последовательности hVk5-2, hVk1, hVk2, hVk3 или hVk4 (CaVk1 (SEQ ID NO:54), CaVk2 (SEQ ID NO:55), CaVk3 (SEQ ID NO:56) или CaVk4 (SEQ ID NO:57), соответственно) связывали способом ПЦР с полинуклеотидом, кодирующим константную область (SEQ ID NO:44) природной каппа-цепи. Связанные фрагменты ДНК встраивали в экспрессирующие векторы клеток животных. Последовательности сконструированных вариантов подтверждали способом, известным специалистам в данной области. Каждую плазмиду, сконструированную, как описано выше, встраивали в клетки животных в комбинации с плазмидой, в которую встроен полинуклеотид, кодирующий CIM_H (SEQ ID NO:45), способом, описанным в примере 6. Экспрессированные молекулы представляющего интерес антитела очищали из культуральной среды клеток животных, в которые были введены плазмиды.
(4-2) Оценка измененных антител, имеющих последовательность CDR из последовательности hVk5-2, в отношении их активности связывания ионов кальция
Оценку того, связываются ли ионы кальция с измененными антителами, имеющими последовательность CDR из последовательности hVk5-2 и каркасные последовательности из последовательностей эмбрионального типа, отличные от hVk5-2 (hVk1, hVk2, hVk3 и hVk4), оценивали с использованием способа, описанного в справочном примере 2. Результат оценки представлен в таблице 15. Было выявлено, что величина Tm Fab-домена каждого из измененных антител варьировала, в зависимости от концентрации ионов кальция в растворах антител. Это демонстрирует, что антитела, имеющие каркасную последовательность, отличную от каркасных последовательностей из последовательности hVk5-2, также связываются с ионами кальция.
Было продемонстрировано, что температура термической денатурации (величина Tm), в качестве индикатора термической стабильности Fab-домена каждого антитела, измененного так, чтобы оно имело последовательность CDR из последовательности hVk5-2 и каркасную последовательность эмбрионального типа, отличную от последовательности hVk5-2 (hVk1, hVk2, hVk3 или hVk4) является более высокой, чем температура термической денатурации Fab-домена исходного антитела, имеющего последовательность hVk5-2. Этот результат показывает, что антитела, имеющие последовательность CDR из последовательности hVk5-2 и каркасную последовательность hVk1, hVk2, hVk3 или hVk4, не только обладают активностью связывания ионов кальция, но также являются превосходными молекулами с точки зрения термической стабильности.
Справочный пример 5. Идентификация участка связывания ионов кальция в последовательности hVk5-2 человека эмбрионального типа
(5-1) Конструирование участка мутации в последовательности CDR из последовательности hVk5-2
Как описано в справочном примере 4, также было показано, что антитела, имеющие легкую цепь, полученную в результате введения CDR-домена последовательности hVk5-2 в каркасную последовательность отличающейся последовательности эмбрионального типа, связываются с ионом кальция. Этот результат указывает на то, что в hVk5-2 участок связывания ионов кальция локализован в его CDR. Аминокислоты, которые связываются с ионом кальция, т.е. хелатируют ион кальция, включают отрицательно заряженные аминокислоты и аминокислоты, которые могут быть акцептором водородной связи. Таким образом, исследовали, связываются ли антитела, имеющие мутантную последовательность hVk5-2 с заменой остатком Ala (A) остатка Asp (D) или Glu (E) в последовательности CDR из последовательности hVk5-2, с ионом кальция.
(5-2) Конструирование последовательностей вариантов hVk5-2 с заменой на Ala, и экспрессия и очистка антител
Молекулы антител получали так, чтобы они содержали легкую цепь с заменой остатка Asp и/или Glu остатком Ala в последовательности CDR из последовательности hVk5-2. Как описано в справочном примере 3, негликозилированный вариант hVk5-2_L65 проявлял связывание ионов кальция, и было предположено, что он является эквивалентным последовательности hVk5-2 с точки зрения связывания ионов кальция. В этом справочном примере аминокислотные замены вносили в hVk5-2_L65 в качестве матричной последовательности. Сконструированные варианты представлены в таблице 16. Аминокислотные замены проводили способами, известными специалистам в данной области, такими как использование набора для сайт-направленного мутагенеза QuikChange Site-Directed Mutagenesis Kit (Stratagene), ПЦР или набора In fusion Advantage PCR Cloning Kit (TAKARA) для конструирования экспрессирующих векторов для измененных легких цепей, имеющих аминокислотную замену.
Нуклеотидные последовательности сконструированных экспрессирующих векторов подтверждали способом, известным специалистам в данной области. Экспрессирующие векторы, сконструированные для измененных легких цепей, временно вводили, в комбинации с экспрессирующим вектором для тяжелой цепи CIM_H (SEQ ID NO:45), в клетки линии, происходящей из клеток почки эмбриона человека HEK293H (Invitrogen) или FreeStyle293 (Invitrogen) для экспрессии антител. Из полученных культуральных супернатантов антитела очищали с использованием rProtein A SepharoseTM Fast Flow (GE Healthcare) способом, известным специалистам в данной области. Поглощение растворов очищенного антитела при 280 нм измеряли с использованием спектрофотометра. Концентрации антител вычисляли из определенных величин с использованием коэффициента экстинкции, вычисленного способом PACE (Protein Science (1995) 4: 2411-2423).
(5-3) Оценка антител, имеющих замену Ala в последовательности hVk5-2, в отношении активности связывания ионов кальция
Проводили исследование, связываются ли полученные очищенные антитела с ионами кальция, способом, описанным в справочном примере 2. Результат представлен в таблице 17. Было выявлено, что некоторые антитела, имеющие замену остатка Asp или Glu в последовательности CDR из последовательности hVk5-2 на остаток Ala, который не может быть вовлечен в связывание или хелатирование ионов кальция, имеют Fab-домен, Tm которого не варьировала, в зависимости от концентрации ионов кальция в растворах антител. Было показано, что участки замены, в которых замена на Ala не изменяла Tm (положения 32 и 92 (нумерация по Kabat)) являются в значительной степени важными для связывания антителом ионов кальция.
Справочный пример 6. Оценка антител, имеющих последовательность hVk1 со связывающим ионы кальция мотивом, в отношении связывания ионов кальция
(6-1) Конструирование последовательности hVk1 со связывающим ионы кальция мотивом, и экспрессия и очистка антител
Результат, описанный в справочном примере 4, в отношении активности связывания кальция в случае замены Ala, демонстрирует, что остатки Asp или Glu в последовательности CDR из последовательности hVk5-2 были важными для связывания кальция. Таким образом, авторы настоящего изобретения оценивали, может ли антитело связываться с ионом кальция, когда остатки в положениях 30, 31, 32, 50 и 92 (нумерация по Kabat) отдельно вносили в другую последовательность вариабельной области эмбрионального типа. В частности, вариант LfVk1_Ca (SEQ ID NO:66) конструировали путем замены остатками в положениях 30, 31, 32, 50 и 92 (нумерация по Kabat) в последовательности hVk5-2 остатков в положениях 30, 31, 32, 50 и 92 (нумерация по Kabat) последовательности hVk1 (последовательность человека эмбрионального типа). В частности исследовали, могут ли антитела, имеющие последовательность hVk1, в которую встроено только 5 остатков из последовательности hVk5-2, связывать кальций. Варианты получали тем же способом, который описан в справочном примере 5. Полученные варианты легкой цепи LfVk1_Ca и LfVk1, имеющие последовательность легкой цепи hVk1 (SEQ ID NO:67) совместно экспрессировали с тяжелой цепью CIM_H (SEQ ID NO:45). Антитела экспрессировали и очищали тем же способом, который описан в справочном примере 4.
(6-2) Оценка антител, имеющих последовательность hVk1 человека со связывающим ионы кальция мотивом, в отношении активности связывания ионов кальция
Оценку того, связывает ли очищенное антитело, полученное как описано выше, ионы кальция, проводили способом, описанным в справочном примере 2. Результаты представлены в таблице 18. Величина Tm для Fab-домена антитела, имеющего LfVk1 с последовательностью hVk1, не варьировала, в зависимости от концентрации кальция в растворах антител. Между тем, Tm антитела, имеющего последовательность LfVk1_Ca, сдвигалась на 1°C или более при изменении концентрации кальция в растворах антител. Таким образом, было показано, что антитело, имеющее LfVk1_Ca, связывается с кальцием. Результаты, описанные выше, демонстрирует, что полная последовательность CDR hVk5-2 не требуется, в то время как остатки, внесенные для конструирования последовательности LfVk1_Ca отдельно, являются достаточными для связывания ионов кальция.
Справочный пример 7. Конструирование совокупности молекул антител (Ca-библиотека) со связывающим ионы кальция мотивом, встроенным в вариабельную область, для эффективного получения связывающих антител, которые связываются с антигеном зависимым от концентрации Ca образом
Предпочтительные кальций-связывающие мотивы включают, например, последовательность hVk5-2 и последовательность CDR, а также остатки в положениях 30, 31, 32, 50 и 92 (нумерация Kabat). Другие кальций-связывающие мотивы включают мотив EF-hand, которым обладают кальций-связывающие белки (например, кальмодулин) и лектин C-типа (например, ASGPR).
Ca-библиотека состоит из вариабельных областей тяжелой цепи и вариабельных областей легкой цепи. Последовательности антител человека использовали в качестве вариабельных областей, и кальций-связывающий мотив вносили в вариабельные области легкой цепи. Последовательность hVk1 была выбрана в качестве матричной последовательности вариабельной области легкой цепи, которую встроен кальций-связывающий мотив. Было показано, что антитело, содержащее последовательность LfVk1_Ca, которое имеет встроенную последовательность CDR hVk5-2, одного из кальций-связывающих мотивов, в последовательность hVk1, связывается с ионом кальция, как показано в справочном примере 5. Множеству аминокислот позволяли оказаться в матричной последовательности, чтобы разнообразить антигенсвязывающие молекулы, которые составляют библиотеку. Положения, экспонированные на поверхности вариабельной области, которые, вероятно, взаимодействуют с антигеном, выбирали в качестве положений, где позволяли оказаться множеству аминокислот. В частности, в качестве нестрогих остатков были выбраны положения 30, 31, 32, 34, 50, 53, 91, 92, 93, 94 и 96 (нумерация Kabat).
Определяли тип и частоту появления аминокислотных остатков, которым впоследствии позволяли появиться. Анализировали частоту появления аминокислотных остатков в нестрогих остатках последовательностей hVk1 и hVk3, зарегистрированных в базе данных Kabat (KABAT, E.A. ET AL.: "Sequence of proteins of immunological interest", vol. 91, 1991, NIH PUBLICATION). Исходя из результатов анализа, отбирали тип аминокислот, которым позволяли появиться в Ca-библиотеке, из аминокислот с более высокой частотой появления в каждом положении. В это время, аминокислоты, для которых было определено, что частота их появления является низкой, исходя из результатов анализа, также отбирали для избежания отклонения свойств аминокислот. Частоту появления выбранных аминокислот определяли, ссылаясь на результаты анализа в базе данных Kabat.
Ca-библиотеку, содержащую кальций-связывающий мотив с акцентом на разнообразии последовательностей, чтобы она содержала множество аминокислот в каждом остатке, отличном от мотива, конструировали в качестве Ca-библиотеки с учетом аминокислот и частоты появления, как описано выше. Детальные конструкции Ca-библиотеки представлены в таблицах 1 и 2 (где положения в каждой таблице соответствуют нумерации Kabat). Когда положение 92, исходя из нумерации Kabat, представляет собой Asn (N), для частоты появления аминокислот, представленной в таблицах 1 и 2, может присутствовать Leu (L) вместо Ser (S) в положении 94.
Справочный пример 8. Получение Ca-библиотеки
Библиотеку генов вариабельной области тяжелой цепи человека амплифицировали способом ПЦР с использованием поли-A РНК, полученной из PBMC человека, или коммерчески доступной поли-A РНК человека в качестве матрицы. Что касается участков вариабельной области легкой цепи антитела, как показано в справочном примере 7, конструировали участки вариабельной области легкой цепи антитела, которые увеличивают частоту появления антител с сохранением кальций-связывающего мотива и обеспечивают связывание с антигенами зависимым от концентрации кальция образом. Кроме того, для аминокислотных остатков среди нестрогих остатков, отличных от остатков с внесенным кальций-связывающим мотивом, конструировали библиотеку вариабельных областей легкой цепи антитела с равномерно распределенными аминокислотами с высокой частотой появления в последовательностях природных антител человека, ссылаясь на информацию о частоте появления аминокислот в природных антителах человека (KABAT, E.A. ET AL.: "Seqeuences of proteins of immunological interest", vol. 91, 1991, NIH PUBLICATION). Комбинацию библиотек генов вариабельных областей тяжелой цепи и легкой цепи антител, полученных как описано выше, встраивали в фагмидный вектор для конструирования библиотеки фагового дисплея антител человека, которая представляет Fab-домены, состоящие из последовательностей антител человека (Methods Mol Biol. (2002) 178, 87-100). Последовательности участков генов антител, выделенных из E. coli, содержащих библиотеку генов антител, подтверждали. Распределение аминокислот в последовательностях полученных 290 клонов и модельное распределение аминокислот представлено на фиг. 15.
Справочный пример 9. Оценка активности молекул, включенных в Ca-библиотеку, в отношении активности связывания ионов кальция
(9-1) Активность молекул, включенных в Ca-библиотеку, в отношении связывания ионов кальция
Как показано в справочном примере 3, последовательность hVk5-2, для которой показано, что она связывается с ионами кальция, обладает низкой частотой появления в последовательностях эмбрионального типа; таким образом, попытка получить кальций-связывающие антитела из библиотек антител, состоящих из последовательностей человека эмбрионального типа или из B-клеток, полученных путем иммунизации мышей, экспрессирующих антитела человека, была сочтена неэффективной. Таким образом, конструировали Ca-библиотеку согласно справочному примеру 8. Сконструированную Ca-библиотеку оценивали в отношении присутствия клонов, демонстрирующих связывание кальция.
(9-2) Экспрессия и очистка антител антитела
Клоны Ca-библиотеки вносили в экспрессирующие плазмиды клеток животных. Антитела экспрессировали с использованием способа, описанного ниже. Клетки линии, происходящей из клеток почки эмбриона человека, FreeStyle 293-F (Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 (Invitrogen), и высевали при плотности клеток 13,3×106 клеток/мл (3 мл) в каждую лунку 6-луночного планшета. Полученные плазмиды вводили в клетки способом липофекции. Клетки культивировали в CO2-инкубаторе (37°C, 8% CO2, 90 об./мин) в течение четырех суток. Способом, известным специалистам в данной области, антитела очищали с использованием rProtein A SepharoseTM Fast Flow (Amersham Biosciences) из культуральных супернатантов, полученных, как описано выше. Поглощение растворов очищенных антител измеряли при 280 нм с использованием спектрофотометра. Концентрации антител вычисляли из измеренных величин с использованием коэффициента поглощения, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
(9-3) Оценка свойства связывания ионов кальция полученными антителами
Оценку того, связываются ли очищенные антитела, полученные, как описано выше, с ионами кальция, оценивали способом, описанным в примере 6. Результаты представлены в таблице 19. Tm Fab-доменов множества антител, включенных в Ca-библиотеку, изменялись в зависимости от концентрации ионов кальция, и это показало, что библиотека включает молекулы, связывающие ионы кальция.
Справочный пример 10. Получение антител, которые связываются с рецептором IL-6 Ca-зависимым образом
(10-1) Получение фрагментов антител, которые связываются с антигеном Ca-зависимым образом, из библиотеки с помощью пэннинга на гранулах
Первичный отбор антител, которые связываются с рецептором IL-6 Ca-зависимым образом, из сконструированной библиотеки проводили посредством обогащения фрагментами антител, которые обладают активностью связывания антигена (IL-6 рецептор).
Фаги получали с помощью E. coli, несущих фагмиды фагового дисплея. К осадку фагов, полученных с помощью E. coli, добавляли 2,5 M NaCl/10% PEG в культуральную среду для E. coli. Фракцию фагов разбавляли TBS для получения раствора с фаговой библиотекой. Затем к раствору с фаговой библиотекой добавляли BSA и CaCl2 до конечных концентраций 4% и 1,2 мМ ионов кальция, соответственно. Использованный способ пэннинга представлял собой общепринятый способ пэннинга с использованием магнитных гранул с иммобилизованным на них антигеном (J Immunol Methods. 2008 Mar 20, 332(1-2): 2-9; J Immunol Methods. 2001 Jan 1, 247(1-2): 191-203; Biotechnol Prog. 2002 Mar-Apr, 18(2): 212-20; Mol Cell Proteomics. 2003 Feb, 2(2): 61-9). Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
В частности, 250 пмоль биотинилированного антигена добавляли к полученному раствору с фаговой библиотекой. Таким образом, раствор контактировали с антигеном при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали три раза 1 мл 1,2 мМ CaCl2/TBS (TBS, содержащий 1,2 мМ CaCl2). После этого к гранулам добавляли 0,5 мл трипсина в концентрации 1 мг/мл. После диспергирования в течение 15 минут при комнатной температуре, гранулы сразу разделяли с использованием магнитного стенда для сбора суспензии фагов. Полученную суспензию фагов добавляли к 10 мл E. coli штамма ER2738 в фазе логарифмического роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагами. Инфицированные E. coli высевали в чашку (225×225 мм). Затем из культуральной среды посеянных E. coli собирали фаги для получения раствора с фаговой библиотекой.
На втором раунде пэннинга обогащение фагов проводили с использованием активности связывания антигена или активности Ca-зависимого связывания в качестве показателя.
В частности, когда проводили обогащение с использованием антигенсвязывающей способности в качестве показателя, 40 пмоль меченного биотином антигена добавляли к полученному раствору с фаговой библиотекой, чтобы обеспечить контактирование раствора с фаговой библиотекой и антигена при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и позволяли им связываться с комплексами антиген/фаг при комнатной температуре в течение 15 минут. Гранулы промывали три раза 1 мл 1,2 мМ CaCl2/TBST, а затем два раза 1,2 мМ CaCl2/TBS. Затем гранулы, к которым было добавлено 0,5 мл 1 мг/мл трипсина, суспендировали при комнатной температуре в течение 15 минут. Сразу после этого гранулы разделяли с использованием магнитного стенда для сбора раствора с фагом. Для устранения способности фагов, экспонирующих Fab, инфицировать E. coli, белок pIII (происходящий из фага-помощника белок pIII) фагов, не экспонирующих Fab, расщепляли добавлением 5 мкл 100 мг/мл трипсина к собранному раствору с фагами. Собранный раствор с фагами добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600 0,4-0,7). E. coli культивировали при осторожном перемешивании при 37°C в течение 1 часа, чтобы позволить фагам инфицировать E. coli. Инфицированные E. coli инокулировали в чашку 225×225 мм. Затем из культуральной среды E. coli после инокуляции собирали фаги, получая раствор с фаговой библиотекой.
Когда обогащение проводили с использованием способности к Ca-зависимому связыванию в качестве показателя, 40 моль меченного биотином антигена добавляли к полученному раствору с фаговой библиотекой, чтобы обеспечить контактирование раствора с фаговой библиотекой и антигена при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и позволяли им связываться с комплексами антиген/фаг при комнатной температуре в течение 15 минут. Гранулы промывали 1 мл 1,2 мМ CaCl2/TBST и 1,2 мМ CaCl2/TBS. Затем гранулы, к которым было добавлено 0,1 мл 2 мМ EDTA/TBS (TBS, содержащий 2 мМ EDTA), суспендировали при комнатной температуре. Сразу после этого гранулы разделяли с использованием магнитного стенда для сбора раствора с фагом. Для устранения способности фагов, экспонирующих Fab, инфицировать E. coli, белок pIII (происходящий из фага-помощника белок pIII) фагов, не экспонирующих Fab, расщепляли добавлением 5 мкл 100 мг/мл трипсина к собранному раствору с фагами. Собранный раствор с фагами добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600 0,4-0,7). E. coli культивировали при осторожном перемешивании при 37°C в течение 1 часа, чтобы позволить фагам инфицировать E. coli. Инфицированные E. coli инокулировали в чашку 225×225 мм. Затем из культуральной среды E. coli после инокуляции собирали фаги, получая раствор с фаговой библиотекой.
(10-2) Оценка с помощью ELISA фагов
Содержащие фаг культуральные супернатанты собирали из единичных колоний E. coli, полученных способом, описанным выше, в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145).
В содержащие фаг культуральные супернатанты добавляли BSA и CaCl2. Супернатанты подвергали ELISA по следующей методике. 96-луночные микропланшеты для титрования StreptaWell (Roche) покрывали в течение ночи с использованием 100 мкл PBS, содержащего меченный биотином антиген. Антиген удаляли из лунки путем промывания каждой лунки планшета PBST. Затем лунки блокировали 250 мкл 4% BSA-TBS в течение одного часа или более. После удаления 4% BSA-TBS в каждую лунку добавляли полученные культуральные супернатанты. Планшет инкубировали при 37°C в течение одного часа, так чтобы позволить экспонирующим антитело фагам связаться с антигеном на каждой лунке. После промывания каждой лунки 1,2 мМ CaCl2/TBST, добавляли 1,2 мМ CaCl2/TBS или 1 мМ EDTA/TBS. Планшет оставляли для инкубации при 37°C в течение 30 минут. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS при концентрации ионизированного кальция 1,2 мМ, и планшет инкубировали в течение одного часа. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли раствор только TMB (ZYMED). Хромогенную реакцию в растворе в каждой лунке останавливали добавлением серной кислоты. Затем развитие окраски оценивали путем измерения поглощения при 450 нм.
Для клонов, подвергнутых описанному выше ELISA фагов, анализировали последовательность оснований гена, амплифицированного с помощью специфических праймеров. Результаты анализа последовательности представлены в таблице 20 ниже.
(10-3) Экспрессия и очистка антител
Клоны, которые оценили как имеющие способность к Ca-зависимому связыванию с помощью ELISA фагов, вводили в экспрессирующие плазмиды клеток животных. Антитела экспрессировали с использованием следующего способа. Клетки линии, происходящей из клеток почки эмбриона FreeStyle 293-F (Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen), и аликвоты объемом 3 мл высевали в каждую ячейку планшетов с 6 ячейками с плотностью клеток 1,33×106 клеток/мл. Полученные плазмиды вводили в клетки способом липофекции. Клетки культивировали в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин) в течение четырех суток. Из полученных культуральных супернатантов антитела очищали с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences) способом, известным специалистам в данной области. Концентрации очищенных антител определяли путем измерения поглощения при 280 нм с использованием спектрофотометра. Концентрации антител вычисляли из установленных величин на основе коэффициента экстинкции, определенного способом PACE (Protein Science 1995; 4: 2411-2423).
(10-4) Оценка способности полученных антител к Ca-зависимому связыванию с рецептором IL-6 человека
Для определения того, обладают ли антитела, полученные согласно справочному примеру 9, т.е. 6RC1IgG_010 (тяжелая цепь SEQ ID NO:90, легкая цепь SEQ ID NO:91), 6RC1IgG_012 (тяжелая цепь SEQ ID NO:92, легкая цепь SEQ ID NO:93) и 6RC1IgG_019 (тяжелая цепь SEQ ID NO:94, легкая цепь SEQ ID NO:95), активностью Ca-зависимого связывания с рецептором IL-6 человека, проводили анализы взаимодействия между этими антителами и рецептором IL-6 человека с использованием Biacore T100 (GE Healthcare). Тоцилизумаб (тяжелая цепь SEQ ID NO:96, легкая цепь SEQ ID NO:97) использовали в качестве контрольного антитела, которое не обладает Ca-зависимой активностью связывания с рецептором IL-6 человека. Анализ взаимодействия проводили в растворе с концентрацией ионов кальция 1,2 мМ для условий высокой концентрации ионов кальция или 3 мкM для условий низкой концентрации ионов кальция. Подходящее количество белка A/G (Invitrogen) иммобилизовывали на сенсорном чипе CM5 (GE Healthcare) способом присоединения аминов, а затем представляющие интерес антитела улавливали на чипы. В качестве подвижного буфера использовали два типа буферов 20 мМ ACES, 150 мМ NaC1, 0,05% (масс./об.) Tween 20, 1,2 мМ CaCl2 (pH 7,4) или 20 мМ ACES, 150 мМ NaC1, 0,05% (масс./об.) Tween 20, 3 мкM CaCl2 (pH 7,4). Соответствующие буферы также использовали для разбавления рецептора IL-6 человека. Все измерения проводили при 37°C.
При проведении анализов взаимодействия на реакциях антиген-антитело с использованием антитела тоцилизумаба, которое представляет собой контрольное антитело, антитела 6RC1IgG_010, антитела 6RC1IgG_012 и антитела 6RC1IgG_019, разбавленный раствор рецептора IL-6 и подвижный буфер, который представляет собой пустой образец, инжектировали со скоростью потока 5 мкл/мин в течение трех минут, чтобы позволить рецептору IL-6 взаимодействовать с антителом тоцилизумабом, антителом 6RC1IgG_010, антителом 6RC1IgG_012 и антителом 6RC1IgG_019 уловленными на сенсорном чипе. Затем инжектировали 10 мМ глицин-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд, чтобы регенерировать сенсорный чип.
Сенсограммы при высокой концентрации ионов кальция, полученные этим способом, представлены на фиг. 16.
Сенсограммы для антитела тоцилизумаба, антитела 6RC1IgG_010, антитела 6RC1IgG_012 и антитела 6RC1IgG_019 в условиях низкой концентрации ионов кальция, также получали сходным образом. Сенсограммы, полученные при низкой концентрации ионов кальция, представлены на фиг. 17.
Из представленных выше результатов было выявлено, что способность антитела 6RC1IgG_010, антитела 6RC1IgG_012 и антитела 6RC1IgG_019 связываться с рецептором IL6 значительно снижалась при изменении концентрации ионов кальция с 1,2 мМ до 3 мкМ.
Справочный пример 11. Получение антител, которые связываются с рецептором IL-6 Ca-зависимым образом, из библиотеки антител с использованием технологии фагового дисплея
(11-1) Получение библиотеки фагового дисплея для наивных антител человека
Библиотеку фагового дисплея для антител человека, состоящую из множества фагов, представляющих Fab-домены взаимно отличающихся последовательностей антител человека, конструировали в соответствии со способом, известным специалистам в данной области, с использованием поли-A РНК, полученной из PBMC человека, и коммерческой поли-A РНК человека в качестве матрицы.
(11-2) Получение фрагментов антител, которые связываются с антигеном Ca-зависимым образом из библиотеки посредством пэннинга гранул
Сконструированную библиотеку фагового дисплея для наивных антител человека подвергали первоначальной селекции посредством концентрирования только тех фрагментов антител, которые обладают способностью связывать антиген (рецептор IL-6), или концентрирования фрагментов антител с использованием зависимой от концентрации Ca способности связывания антигена (рецептора IL-6) в качестве индикатора. Концентрирование фрагментов антител с использованием зависимой от концентрации Ca способности связывать антиген (рецептор IL-6) в качестве индикатора проводили путем элюирования фагов фаговой библиотеки, связавшихся с рецептором IL-6 в присутствии ионов Ca, с EDTA, которая хелатирует ионы Ca. В качестве антигена использовали биотинилированный рецептор IL-6.
Фаги получали из Escherichia coli, содержащих сконструированную фагмиду фагового дисплея. Раствор фаговой библиотеки получали путем разбавления с помощью TBS популяции фагов, преципитированных добавлением 2,5 M NaCl/10% PEG к культуральному раствору E. coli, в котором продуцировались фаги. Затем к раствору фаговой библиотеки добавляли BSA и CaCl2 в конечной концентрации 4% BSA и концентрации ионов кальция 1,2 мМ. Обычный способ пэниннга с использованием антигена, иммобилизованного на магнитных гранулах, был назван способом пэниннга (J. Immunol. Methods. (2008) 332 (1-2), 2-9; J. Immunol. Способы. (2001) 247 (1-2), 191-203; Biotechnol. Prog. (2002) 18(2) 212-20; Mol. Cell Proteomics (2003) 2(2), 61-9). В качестве магнитных гранул использовали покрытые нейтравидином гранулы (Sera-Mag SpeedBeads Neutravidin-coated) или покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
В частности, к полученному раствору фаговой библиотеки добавляли 250 пмоль меченного биотином антигена, чтобы обеспечить контактирование указанного раствора фаговой библиотеки с антигеном в течение 60 минут при комнатной температуре. Добавляли магнитные гранулы, блокированные с помощью BSA, чтобы они связывались с комплексами антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали один раз с помощью 1 мл 1,2 мМ CaCl2/TBS (TBS, содержащий 1,2 мМ CaCl2). Затем раствор фага собирали с помощью общего способа элюирования для концентрирования фрагмента антитела, обладающего способностью связывать рецептор IL-6, или с помощью элюирования с гранул, суспендированных в 2 мМ EDTA/TBS (TBS, содержащий 2 мМ EDTA), для концентрирования фрагмента антитела с использованием способности связывать рецептор IL-6 зависимым от концентрации Ca образом в качестве показателя. Собранный раствор фага добавляли к 10 мл штамма E. coli TG1 в логарифмической фазе роста (OD600 0,4-0,7). E. coli культивировали при осторожном перемешивании при 37°C в течение 1 часа, чтобы позволить фагам инфицировать E. coli. Инфицированные E. coli инокулировали в чашку 225×225 мм. Затем после инокуляции фаги собирали из культуральной среды E. coli для получения раствора фаговой библиотеки.
Во втором и последующем пэннинге фаги концентрировали с использованием зависимой от Ca способности связывания в качестве индикатора. В частности, 40 пмоль меченного биотином антитела добавляли к полученному раствору фаговой библиотеки, чтобы обеспечить контактирование фаговой библиотеки с антигеном в течение 60 минут при комнатной температуре. Добавляли магнитные гранулы, блокированные с помощью BSA, чтобы они связывались с комплексами антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали один раз с помощью 1 мл 1,2 мМ CaCl2/TBST и 1,2 мМ CaCl2/TBS. Затем гранулы, к которым добавляли 0,1 мл 2 мМ EDTA/TBS, суспендировали при комнатной температуре. Сразу после этого гранулы отделяли с использованием магнитного стенда для сбора раствора фага. Собранный раствор фага добавляли к 10 мл штамма E. coli TG1 в логарифмической фазе роста (OD600 0,4-0,7). E. coli культивировали при осторожном перемешивании при 37°C в течение 1 часа, чтобы позволить фагам инфицировать E. coli. Инфицированные E. coli инокулировали в чашку 225×225 мм. Затем после инокуляции фаги собирали из культуральной среды E. coli для получения раствора фаговой библиотеки. Пэннинг с использованием способности Ca-зависимого связывания в качестве показателя повторяли несколько раз.
(11-3) Исследование с помощью ELISA фагов
Фаг, содержащий культуральный супернатант, собирали общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичной колонии E. coli, полученной, как описано выше.
Культуральный супернатант, содержащий фаги, в который были добавлены BSA и CaCl2 до конечной концентрации 4% BSA и концентрации ионов кальция 1,2 мМ, подвергали ELISA, как описано ниже. На 96-луночный микропланшет для титрования StreptaWell (Roche) наносили в течение ночи 100 мкл PBS, содержащего меченный биотином антиген. Каждую лунку указанного планшета промывали PBST для удаления антигена, а затем лунки блокировали с помощью 250 мкл 4% BSA-TBS в течение 1 часа или дольше. Указанному планшету с полученным культуральным супернатантом, добавленным в каждую лунку, из которой был удален 4% BSA-TBS, позволяли стоять нетронутым при 37°C в течение 1 часа, позволяя связываться представляющему фаг антителу с антигеном, присутствующим в каждой лунке. В каждую лунку, промытую 1,2 мМ CaCl2/TBST, добавляли 1,2 мМ CaCl2/TBS или 1 мМ EDTA/TBS. Планшету позволяли стоять нетронутым в течение 30 минут при 37°C для инкубации. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли конъюгироваанное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS в конечной концентрации 4% BSA и концентрации ионизированного кальция 1,2 мМ, и планшет инкубировали в течение 1 часа. После промывания 1,2 мМ CaCl2/TBST хромогенную реакцию раствора в каждой лунке, в которую добавляли только раствор TMB (ZYMED), останавливали добавлением серной кислоты. Затем указанный цвет определяли путем измерения поглощения при 450 нм.
В результате указанного выше ELISA фагов анализировали последовательность оснований гена, амплифицированного с помощью специфических праймеров и фрагмента антитела, идентифицированного как обладающего способностью зависимого от Ca связывания антигена в качестве матрицы.
(11-4) Экспрессия и очистка антител
В результате описанного выше ELISA фагов, клон, идентифицированный как обладающий зависимой от Ca способностью связывания антигена, вводили в экспрессирующую плазмиду для клеток животных. Антитела экспрессировали, как описано ниже. Штамм FreeStyle 293-F (Invitrogen), происходящий из клеток почки эмбриона человека, суспендировали в среде FreeStyle 293 Expression Medium (Invitrogen), а затем инокулировали по 3 мл в каждую лунку планшета с 6 ячейками с плотностью клеток 1,33×106 клеток/мл. Полученную плазмиду вводили в клетки способом липофекции. Клетки культивировали в течение 4 суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин). Антитела очищали из культурального супернатанта, полученного, как описано выше, способом, известным в данной области, с использованием rProtein A Sepharose (торговый знак) Fast Flow (Amersham Biosciences). Поглощение очищенного раствора антитела измеряли при 280 нм с использованием спектрофотометра. Концентрацию антитела вычисляли из данных измерения, полученных с использованием коэффициента экстинкции, вычисленного способом PACE (Protein Science (1995) 4, 2411-2423).
Справочный пример 12. Исследование зависимой от Ca способности связывания полученных антител к рецептору IL-6 человека
Для исследования того, является ли активность связывания антител 6RL#9-IgG1 [тяжелая цепь (последовательность, в которой константная область из IgG1 связана с SEQ ID NO:98) и легкая цепь (SEQ ID NO:99)] и FH4-IgG1 [тяжелая цепь (SEQ ID NO:100) и легкая цепь (SEQ ID NO:101)], полученных согласно справочному примеру 11, с рецептором IL-6 человека зависимой от Ca, проводили кинетический анализ реакций антиген-антитело для этих антител с рецептором IL-6 человека с использованием Biacore T100 (GE Healthcare). H54/L28-IgG1 [тяжелая цепь: SEQ ID NO:103 и легкая цепь: SEQ ID NO:103], описанное в WO2009/125825, использовали в качестве контрольного антитела, которое обладает Ca-зависимой активностью связывания с рецептором IL-6 человека. Кинетический анализ реакций антиген-антитело проводили в растворах с концентрациями ионов кальция 2 мМ и 3 мкМ в качестве условий высокой и низкой концентрации ионов кальция, соответственно. Представляющее интерес антитело иммобилизовывали на сенсорном чипе CM4 (GE Healthcare), на котором было иммобилизовано подходящее количество белка A (Invitrogen) способом присоединения аминов. В качестве подвижных буферов использовали два буфера [10 мМ ACES, 150 мМ NaCl, 0,05% (масс./об.) Tween 20, и 2 мМ CaCl2 (pH 7,4) или 10 мМ ACES, 150 мМ NaCl, 0,05% (масс./об.) Tween 20, и 3 мколь/л CaCl2 (pH 7,4)]. Эти буферы использовали для разбавления рецептора IL-6 человека. Все измерения проводили при 37°C.
В кинетическом анализе реакции антиген-антитело с использованием антитела H54L28-IgG1, антителу H54L28-IgG1, уловленному на сенсорном чипе, позволяли взаимодействовать с рецептором IL-6 путем инжекции разбавителя рецептора IL-6 и подвижного буфера (пустой образец) со скоростью потока 20 мкл/мин в течение 3 минут. Затем после выявления диссоциации рецептора IL-6 с использованием подвижного буфера со скоростью потока 20 мкл/мин в течение 10 минут, сенсорный чип регенерировали путем инъекции 10 мМ глицин-HCl (pH 1,5) со скоростью потока 30 мкл/мин в течение 30 секунд. Из сенсограмм, полученных при измерении, вычисляли параметры кинетики: константу связывания (ka) (1/Мс) и константу скорости диссоциации (kd) (1/c). Эти величины использовали для вычисления константы диссоциации (KD) (M) антитела H54L28-IgG1 в отношении рецептора IL-6 человека. Каждый параметр вычисляли с использованием программного обеспечения Biacore T100 Evaluation Software (GE Healthcare).
В кинетическом анализе реакции антиген-антитело с использованием антител FH4-IgG1 и 6RL#9-IgG1, антителу FH4-IgG1 или 6RL#9-IgG1, уловленному на сенсорном чипе, позволяли взаимодействовать с рецептором IL-6 путем инжекции разбавителя рецептора IL-6 и подвижного буфера (пустой образец) со скоростью потока 5 мкл/мин в течение 15 минут. Затем сенсорный чип регенерировали путем инжекции 10 мМ глицин-HCl (pH 1,5) со скоростью потока 30 мкл/мин в течение 30 секунд. Константы диссоциации (KD) (M) вычисляли из сенсограмм при измерении с использованием стационарной модели аффинности. Каждый параметр вычисляли с использованием программного обеспечения Biacore T100 Evaluation Software (GE Healthcare). Константы диссоциации (KD) между каждым антителом и рецептором IL-6 в присутствии 2 мМ CaCl2, определенные описанным выше способом, представлены в таблице 21.
Величину KD для антитела H54/L28-IgG1 в условиях концентрации Ca 3 мкМ можно вычислять аналогично тому, как и в присутствии концентрации кальция Ca 2 мМ. В условиях концентрации Ca 3 мкМ наблюдали, что антитела FH4-IgG1 и 6RL#9-IgG1 практически не связываются с рецептором IL-6, таким образом, вычисление величин KD способом, описанным выше, является трудным. Однако величины KD для этих антител в условиях концентрации Ca 3 мкМ можно оценивать с использованием уравнения 1 (Biacore T100 Software Handbook, BR-1006-48, AE 01/2007) ниже.
Уравнение 1
Req=C×Rmax/(KD+C)+RI
Значение каждого параметра в упомянутом выше уравнении 1 является следующим:
Req (RU): уровень связывания в стационарном состоянии;
C (M): концентрация анализируемого соединения;
C: концентрация;
Rmax (RU): способность к поверхностному связыванию анализируемого соединения;
RI (RU): совокупный вклад индекса рефракции в образце;
KD (M): равновесная константа диссоциации.
Приблизительные результаты для величин константы диссоциации KD для антител и рецептора IL-6 при концентрации Ca 3 мколь/л, оцененной с использованием указанного выше уравнения 1 представлены в таблице 22. В таблице 22 величины Req, Rmax, RI и C оценены, исходя из результата анализа.
Исходя из данных, описанных выше, было предсказано, что KD между рецептором IL-6 и антителом FH4-IgG1 или антителом 6RL#9-IgG1 увеличивалась приблизительно в 60 или 120 раз (аффинность снижалась в 60 или 120 раз или более), когда концентрацию CaCl2 в буфере снижали от 2 мМ до 3 мкМ.
В таблице 23 обобщенно представлены величины KD при концентрациях CaCl2 2 мМ и 3 мкМ и зависимость от Ca для трех типов антител H54/L28-IgG1, FH4-IgG1 и 6RL#9-IgG1.
Не было выявлено отличий в связывании антитела H54/L28-IgG1 с рецептором IL-6 вследствие отличий в концентрации Ca. С другой стороны, было выявлено, что связывание антител FH4-IgG1 и 6RL#9-IgG1 с рецептором IL-6 было значительно ослаблено в условиях низкой концентрации Ca (таблица 23).
Справочный пример 13. Исследование связывания ионов кальция с полученным антителом
Затем измеряли промежуточную температуру термической денатурации (величину Tm) с помощью дифференциальной сканирующей калориметрии (DSC) в качестве показателя для исследования связывания ионов кальция с антителом (MicroCal VP-Capillary DSC, MicroCal). Промежуточная температура термической денатурации (величина Tm) является показателем стабильности. Промежуточная температура термической денатурации (величина Tm) становится более высокой, когда белок стабилизирован путем связывания с ионами кальция, по сравнению с отсутствием связывания ионов кальция (J. Biol. Chem. (2008) 283, 37, 25140-25149). Активность связывания ионов кальция с антителом исследовали путем исследования изменений величины Tm антитела, в зависимости от изменений концентрации ионов кальция в растворе антитела. Очищенное антитело подвергали диализу (EasySEP, TOMY) с использованием внешнего раствора 20 мМ Tris-HCl, 150 мМ NaCl и 2 мМ CaCl2 (pH 7,4), или 20 мМ Tris-HCl, 150 мМ NaCl и 3 мкM CaCl2 (pH 7,4). Измерение DSC проводили при скорости нагревания 240°C/ч от 20 до 115°C с использованием раствора антитела, приготовленного в концентрации приблизительно 0,1 мг/мл с диализатом в качестве исследуемого вещества. Промежуточные температуры термической денатурации (величины Tm) Fab-доменов каждого антитела, вычисленные исходя из кривой денатурации, полученной с помощью DSC, представлены в таблице 24.
Результаты, представленные в таблице 24, показывают, что величины Tm для Fab антител FH4-IgG1 и 6RL#9-IgG1, которые проявляют зависимую от кальция способность связывания, варьировали при изменении концентрации ионов кальция, в то время как величины Tm для Fab антитела H54/L28-IgG1, которое не проявляет зависимой от кальция способности связывания, не варьируют при изменении концентрации ионов кальция. Варьирование величин Tm для Fab антител FH4-IgG1 и 6RL#9-IgG1 демонстрирует, что ионы кальция, связанные с этими антителами, стабилизируют Fab-части. Описанные выше результаты показывают, что ионы кальция связывались с антителами FH4-IgG1 и 6RL#9-IgG1, в то время как ионы кальция не связывались с антителом H54/L28-IgG1.
Справочный пример 14. Идентификация участка связывания ионов кальция в антителе 6RL#9 с помощью рентгеновской кристаллографии
(14-1) Рентгеновская кристаллография
Как описано в справочном примере 13, измерение температуры термической денатурации Tm показало, что антитело 6RL#9 связывается с ионами кальция. Однако было непредсказуемо, какая часть антитела 6RL#9 связывается с ионом кальция. Следовательно, с использованием способа рентгеновской кристаллографии идентифицировали остатки антитела 6RL#9, которые взаимодействуют с ионом кальция.
(14-2) Экспрессия и очистка антитела 6RL#9
Антитело 6RL#9 экспрессировали и очищали для рентгеновской кристаллографии. В частности, плазмиды для экспрессии у животных, сконструированные так, чтобы они были способны экспрессировать тяжелую цепь (последовательность, в которой константная область, происходящая из IgG1, связана с SEQ ID NO:98) и легкую цепь (SEQ ID NO:99) антитела 6RL#9, вводили временно в клетки животных. Сконструированные плазмиды вводили способом липофекции в клетки линии, происходящей из клеток почек эмбриона человека FreeStyle 293-F (Invitrogen), суспендированные в 800 мл среды для экспрессии FreeStyle 293 Expression Medium (Invitrogen) (конечная плотность клеток: 1×106 клеток/мл). Клетки с введенной плазмидой культивировали в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин) в течение пяти суток. Из культурального супернатанта, полученного, как описано выше, антитела очищали способом, известным специалистам в данной области, с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Поглощение при 280 нм очищенных растворов антитела измеряли с использованием спектрофотометра. Из измеренных значений вычисляли концентрации антител с использованием коэффициента экстинкции, вычисленного способом PACE (Protein Science (1995) 4, 2411-2423).
(14-3) Очистка Fab-фрагмента антитела 6RL#9
Антитело 6RL#9 концентрировали до 21 мг/мл с использованием ультрафильтра с пределом молекулярной массы 10000 MWCO. Образец антитела в концентрации 5 мг/мл (2,5 мл) получали разбавлением раствора антитела с использованием буфера 4 мМ L-цистеин/5 мМ EDTA/20 мМ фосфат натрия (pH 6,5). К образцу добавляли 0,125 мг папаина (Roche Applied Science). После перемешивания образец инкубировали при 35°C в течение двух часов. После инкубации таблетку с ингибиторами протеаз Protease Inhibitor Cocktail Mini, не содержащую EDTA (Roche Applied Science) растворяли в 10 мл буфера 25 мМ MES (pH 6) и добавляли к образцу. Образец инкубировали на льду для остановки протеолитического расщепления папаином. Затем образец наносили на 1-мл катионнообменную колонку HiTrap SP HP (GE Healthcare), уравновешенную 25 мМ буфером MES (pH 6), ниже которой была последовательно подсоединена 1-мл колонка с белком A HiTrap MabSelect Sure Protein A column (GE Healthcare). Очищенную фракцию Fab-фрагмента антитела 6RL#9 получали, путем проведения элюирования с помощью линейного градиента концентрации NaCl вплоть до 300 мМ в описанном выше буфере. Затем полученную очищенную фракцию концентрировали до приблизительно 0,8 мл с использованием ультрафильтра 5000 MWCO. Концентрат наносили на колонку для гель-фильтрации Superdex 200 10/300 GL (GE Healthcare), уравновешенную 100 мМ буфером HEPES (pH 8), содержащим 50 мМ NaCl. Очищенный Fab-фрагмент антитела 6RL#9 для кристаллизации элюировали с колонки с использованием того же буфера. Все обработки колонок, описанные выше, проводили при низкой температуре от 6 до 7,5°C.
(14-4) Кристаллизация Fab-фрагмента антитела 6RL#9 в присутствии Ca
Затравочные кристаллы Fab-фрагмента 6RL#9 получали заранее согласно общим условиям. Затем очищенный Fab-фрагмент антитела 6RL#9 в 5 мМ CaCl2 концентрировали до 12 мг/мл с помощью ультрафильтра 5000 MWCO. Далее, образец, концентрированный, как описано выше, кристаллизовали способом висячей капли посредством диффузии в парах с использованием 100 мМ буфера HEPES (pH 7,5), содержавшего 20-29% PEG4000, в качестве резервуарного раствора. Описанные выше затравочные кристаллы, дробили в 100 мМ буфере HEPES (pH 7,5), содержавшем 29% PEG4000 и 5 мМ CaCl2, и проводили серийное разведение в от 100 до 10000 раз. Затем 0,2 мкл разбавленных растворов комбинировали со смесью 0,8 мкл резервуарного раствора и 0,8 мкл концентрированного образца для получения кристаллизационных капель на покровном стекле. Каплям кристаллов позволяли держаться при 20°C в течение от двух до трех суток до получения плоских кристаллов. С использованием кристаллов получали данные рентгенодиффракции.
(14-5) Кристаллизация Fab-фрагмента антитела 6RL#9 в отсутствие Ca
Очищенный Fab-фрагмент антитела 6RL#9 концентрировали до 15 мг/мл с помощью ультрафильтра 5000 MWCO. Далее, образец, концентрированный, как описано выше, кристаллизовали способом висячей капли посредством диффузии в парах с использованием 100 мМ буфера HEPES (pH 7,5), содержавшего 18-25% PEG4000, в качестве резервуарного раствора. Описанные выше затравочные кристаллы, дробили в 100 мМ буфере HEPES (pH 7,5), содержавшем 25% PEG4000, и проводили серийное разведение в от 100 до 10000 раз. Затем 0,2 мкл разбавленных растворов комбинировали со смесью 0,8 мкл резервуарного раствора и 0,8 мкл концентрированного образца для получения кристаллизационных капель на покровном стекле. Каплям кристаллов позволяли держаться при 20°C в течение от двух до трех суток до получения плоских кристаллов. С использованием кристаллов получали данные рентгенодиффракции.
(14-6) Рентгеновское кристаллографическое измерение кристалла Fab-фрагмента из антитела 6RL#9 в присутствии Ca
Кристаллы Fab-фрагмента антитела 6RL#9, полученные в присутствии Ca, смачивали 100 мМ раствором буфера HEPES (pH 7,5), содержавшим 35% PEG4000 и 5 мМ CaCl2. Удалив внешней раствор с поверхности монокристалла с помощью микробулавки в виде нейлоновой петли, монокристалл замораживали в жидком азоте. Данные рентгенодиффракции замороженного кристалла получали из линии пучка BL-17A Photon Factory в High Energy Accelerator Research Organization. Замороженный кристалл поддерживали в замороженном состоянии в ходе измерения путем постоянного помещения его в поток газообразного азота при -178°C. Всего было получено 180 дифракционных изображений с использованием CCD-детектора Quantum315r (ADSC), присоединенного к линии пучка, при вращении кристалла с интервалами 1°. Определение константы решетки, индексирование дифракционных пятен и анализ данных дифракции проводили с использованием программ Xia2 (CCP4 Software Suite), XDS Package (Walfgang Kabsch)и Scala (CCP4 Software Suite). Наконец, были получены данные интенсивности дифракции с разрешением вплоть до 2,2 Ангстрем. Кристалл принадлежит к группе симметрии P212121 с константой решетки a=45,47 Ангстрем, b=79,86 Ангстрем, c=116,25 Ангстрем, α=90°, β=90° и γ=90°.
(14-7) Рентгеновское кристаллографическое измерение кристалла Fab-фрагмента из антитела 6RL#9 в отсутствие Ca
Кристаллы Fab-фрагмента антитела 6RL#9, полученные в отсутствие Ca, смачивали 100 мМ раствором буфера HEPES (pH 7,5), содержавшим 35% PEG4000. Удалив внешней раствор с поверхности монокристалла с помощью микробулавки в виде нейлоновой петли, монокристалл замораживали в жидком азоте. Данные рентгенодиффракции замороженного кристалла получали из линии пучка BL-5A Photon Factory в High Energy Accelerator Research Organization. Замороженный кристалл поддерживали в замороженном состоянии в ходе измерения путем постоянного помещения его в поток газообразного азота при -178°C. Всего было получено 180 дифракционных изображений с использованием CCD-детектора Quantum210r (ADSC), присоединенного к линии пучка, при вращении кристалла с интервалами 1°. Определение константы решетки, индексирование дифракционных пятен и анализ данных дифракции проводили с использованием программ Xia2 (CCP4 Software Suite), XDS Package (Walfgang Kabsch)и Scala (CCP4 Software Suite). Наконец, были получены данные интенсивности дифракции с разрешением вплоть до 2,3 Ангстрем. Кристалл принадлежит к группе симметрии P212121 с константой решетки a=45,40 Ангстрем, b=79,63 Ангстрем, c=116,07 Ангстрем, α=90°, β=90° и γ=90°, и, таким образом, он структурно идентичен кристаллу, полученному в присутствии Ca.
(14-8) Измерение способом рентгеновской кристаллографии кристалла Fab-фрагмента из антитела 6RL#9 в присутствии Ca
Кристаллическую структуру Fab-фрагмента антитела 6RL#9 в присутствии Ca определяли способом молекулярного замещения с использованием программы Phaser (CCP4 Software Suite). Количество молекул в асимметричном элементе оценивали, исходя из размера кристаллической решетки и молекулярной массы Fab-фрагмента антитела 6RL#9. Исходя из гомологии первичной последовательности, часть из положений аминокислот 112-220 A-цепи и часть из положений аминокислот 116-218 B-цепи в конформационной координате PDB-кода 1ZA6, использовали в качестве модельных молекул для анализа областей CL и CH1. Затем часть из положений аминокислот 1-115 B-цепи в конформационной координате PDB-кода 1ZA6 использовали в качестве модельной молекулы для анализа VH-области. Наконец, часть из положений аминокислот 3-147 легкой цепи в конформационной координате PDB-кода 2A9M использовали в качестве модельной молекулы для анализа VL-области. Исходя из этого порядка, была получена первоначальная модель структуры Fab-фрагмента антитела 6RL#9 путем определения из функций сдвига и вращения положений и ориентаций модельных молекул для анализа кристаллической решетки. Кристаллографический фактор достоверности R для данных отражения при разрешении от 25 до 3,0 составлял 46,9% и свободный R составлял 48,6% после детализации жесткого каркаса, где каждому из VH-, VL-, CH1- и CL-доменов позволяли отклоняться от первоначальной структурной модели. Затем детализации модели достигали путем повторения структурной детализации с использованием программы Refmac5 (CCP4 Software Suite) с последующей проверкой модели, проводимой с использованием программы Coot (Paul Emsley) на основании карт электронной плотности Fo-Fc и 2Fo-F, где коэффициенты Fo-Fc и 2Fo-Fc вычисляли с использованием экспериментально определенного структурного фактора Fo, структурного фактора Fc, вычисленного на основе этой модели, и фаз. Конечную детализацию проводили с использованием программы Refmac5 (CCP4 Software Suite) на основе карт электронной плотности Fo-Fc и 2Fo-F путем добавления молекулы воды и иона Ca в модель. С помощью 21020 данных отражения при разрешении от 25 до 2,2 Ангстрем, в конечном итоге кристаллографический фактор достоверности R составил 20,0% и свободный R составил 27,9% для модели, состоящей из 3440 атомов.
(14-9) Определение данных рентгенодиффракции кристалла Fab-фрагмента антитела 6RL#9 в отсутствие of Ca
Кристаллическую структуру Fab-фрагмента антитела 6RL#9 в отсутствие Ca определяли на основе структуры кристалла, полученного в присутствии Ca. Молекулами воды и ионов Ca пренебрегали в конформационных координатах кристалла Fab-фрагмента антитела 6RL#9, полученного в присутствии Ca. Кристаллографический фактор достоверности R для данных отражения при разрешении от 25 до 3,0 Ангстрем составил 30,3% и свободный R составил 31,7% после детализации жесткого каркаса, где каждому из VH-, VL-, CH1- и CL-доменов позволяли отклоняться. Затем детализации модели достигали путем повторения структурной детализации с использованием программы Refmac5 (CCP4 Software Suite) с последующей проверкой модели, проводимой с использованием программы Coot (Paul Emsley) на основании карт электронной плотности Fo-Fc и 2Fo-F, где коэффициенты Fo-Fc и 2Fo-Fc вычисляли с использованием экспериментально определенного структурного фактора Fo, структурного фактора Fc, вычисленного на основе этой модели, и фаз. Конечную детализацию проводили с использованием программы Refmac5 (CCP4 Software Suite) на основе карт электронной плотности Fo-Fc и 2Fo-F путем добавления молекулы воды и иона Ca в модель. C помощью 18357 данных отражения при разрешении от 25 до 2,3 Ангстрем, в конечном итоге кристаллографический фактор достоверности R составил 20,9% и свободный R составил 27,7% для модели, состоящей из 3351 атомов.
(14-10) Сравнение данных рентгеновской кристаллографической дифракции Fab-фрагментов антитела 6RL#9 в присутствии и в отсутствие Ca
При сравнении кристаллографических структур Fab-фрагментов антитела 6RL#9 в присутствии и в отсутствие Ca, были выявлены значительные изменения в CDR3 тяжелой цепи. Структура CDR3 тяжелой цепи Fab-фрагмента антитела 6RL#9, определенная рентгеновской кристаллографией, представлена на фиг. 18. В частности, ион кальция располагался в центре области петли CDR3 тяжелой цепи Fab-фрагмента антитела 6RL#9, полученного в присутствии Ca. Было предположено, что ион кальция взаимодействует с положениями 95, 96 и 100a (нумерация по Kabat) CDR3 тяжелой цепи. Было предположено, что петля CDR3 тяжелой цепи, которая является важной для связывания антигена, стабилизировалась связыванием кальция в присутствии Ca, и становилась оптимальной структурой для связывания антигена. Отсутствуют сообщения, демонстрирующие, что кальций связывается с CDR3 тяжелой цепи антитела. Таким образом, связанная с кальцием структура CDR3 тяжелой цепи антитела является новой структурой.
Кальций-связывающий мотив, который, как было выявлено, существует в CDR3 тяжелой цепи, исходя из структуры Fab-фрагмента антитела 6RL#9, также может быть новым компонентом при конструировании Ca-библиотеки, такой как библиотеки, описанные в справочном примере 7. Иными словами, в то время как кальций-связывающий мотив вносили в вариабельную цепь легкой цепи в справочном примере 7, можно рассматривать, например, библиотеку, содержащую CDR3 тяжелой цепи антитела 6RL#9 и нестрогие остатки в других CDR, включая легкую цепь.
Справочный пример 15. Получение антител, которые связываются с IL-6 Ca-зависимым образом, из библиотеки антител человека с использованием способов фагового дисплея
(15-1) Конструирование библиотеки фагового дисплея наивных антител человека
Библиотеку фагового дисплея антител человека, содержавшую множество фагов, которые экспонируют различные последовательности Fab-доменов антител человека, конструировали способом, известным специалистам в данной области с использованием, в качестве матрицы, поли-A-РНК, полученной из PBMC человека, коммерчески доступной поли-A-РНК человека и т.п.
(15-2) Получение фрагментов антител, которые связываются с антигеном Ca-зависимым образом? из библиотеки с помощью пэннинга на гранулах
Первичную селекцию из сконструированных библиотек фагового дисплея наивных антител человека проводили путем обогащения фрагментами антител, которые имеют активность связывания антигена (IL-6). Использованный антиген представлял меченный биотином IL-6.
Фаги получали с помощью E. coli, несущих фагмиды фагового дисплея. К осадку фагов, полученных с помощью E. coli, добавляли 2,5 M NaCl/10% PEG в культуральную среду для E. coli. Фракцию фагов разбавляли TBS для получения раствора с фаговой библиотекой. Затем к раствору с фаговой библиотекой добавляли BSA и CaCl2 до конечных концентраций 4% и 1,2 мМ ионов кальция, соответственно. Использованный способ пэннинга представлял собой общепринятый способ пэннинга с использованием магнитных гранул с иммобилизованным на них антигеном (J Immunol Methods. 2008 Mar 20, 332(1-2): 2-9; J Immunol Methods. 2001 Jan 1, 247(1-2): 191-203; Biotechnol Prog. 2002 Mar-Apr, 18(2): 212-20; Mol Cell Proteomics. 2003 Feb, 2(2): 61-9). Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
В частности, 250 пмоль биотинилированного антигена добавляли к полученному раствору с фаговой библиотекой. Таким образом, раствор контактировали с антигеном при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали три раза 1 мл 1,2 мМ CaCl2/TBS (TBS, содержащий 1,2 мМ CaCl2). После этого к гранулам добавляли 0,5 мл трипсина в концентрации 1 мг/мл. После диспергирования в течение 15 минут при комнатной температуре, гранулы сразу разделяли с использованием магнитного стенда для сбора суспензии фагов. Полученную суспензию фагов добавляли к 10 мл E. coli штамма TG1 в фазе логарифмического роста (OD600=0,4-0,5). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагами. Инфицированные E. coli высевали в чашку (225×225 мм). Затем из культуральной среды посеянных E. coli собирали фаги для получения раствора с фаговой библиотекой.
При втором и последующих пэннингах, фаги обогащали с использованием способности к Ca-зависимому связыванию в качестве индикатора. В частности, 40 пмоль меченного биотином антигена добавляли к полученному раствору фаговой библиотеки. Таким образом, фаговую библиотеку контактировали с антигеном при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и комплексу антиген-фаг позволяли связаться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз 1 мл 1,2 мМ CaCl2/TBST (TBS содержащий 1,2 мМ CaCl2 и 0,1% Tween-20) и 1,2 мМ CaCl2/TBS. Затем к гранулам добавляли 0,1 мл 2 мМ EDTA/TBS. После диспергирования при комнатной температуре гранулы сразу отделяли с использованием магнитного стенда для сбора суспензии фагов. Белок pIII (белок pIII, происходящий из фага-помощника) отщепляли от фагов, которые не экспонировали Fab, добавлением 5 мкл трипсина в концентрации 100 мг/мл к собранной суспензии фага для устранения способности фагов, не экспонирующих Fab, инфицировать E. coli. Фаги, собранные из исходного раствора с фагом после обработки трипсином, добавляли к 10 мл клеток E. coli штамма TG1 в фазе логарифмического роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагами. Инфицированные E. coli высевали в чашку (225×225 мм). Затем из культуральной среды посеянных E. coli собирали фаги для получения жидкого исходного раствора с фаговой библиотекой. Пэннинг проводили три раза с использованием активности Ca-зависимого связывания в качестве индикатора.
(15-3) Оценка с помощью ELISA фагов
Содержащие фаг культуральные супернатанты собирали из единичных колоний E. coli, полученных способом, описанным выше, в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145). В содержащие фаг культуральные супернатанты добавляли BSA и CaCl2 до конечных концентраций 4% и 1,2 мМ ионов кальция, соответственно. Супернатанты подвергали ELISA по следующей методике. 96-луночные микропланшеты для титрования StreptaWell (Roche) покрывали в течение ночи с использованием 100 мкл PBS, содержащего меченный биотином антиген. Антиген удаляли из лунки путем промывания каждой лунки планшета PBST. Затем лунки блокировали 250 мкл 4% BSA-TBS в течение одного часа или более. После удаления 4% BSA-TBS в каждую лунку добавляли полученные культуральные супернатанты. Планшет инкубировали при 37°C в течение одного часа, так чтобы позволить экспонирующим антитело фагам связаться с антигеном на каждой лунке. После промывания каждой лунки 1,2 мМ CaCl2/TBST, добавляли 1,2 мМ CaCl2/TBS или 1 мМ EDTA/TBS. Планшет оставляли для инкубации при 37°C в течение 30 минут. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS, содержащим BSA и ионы кальция в конечных концентрациях 4% и 1,2 мМ ионов кальция, и планшет инкубировали в течение одного часа. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли раствор только TMB (ZYMED). Хромогенную реакцию в растворе в каждой лунке останавливали добавлением серной кислоты. Затем проявившийся цвет оценивали путем измерения поглощения при 450 нм.
Из 96 выделенных клонов с помощью ELISA фагов было получено антитело 6KC4-1#85, имеющее Ca-зависимую активность связывания IL-6. С использованием фрагментов антител, для которых было предсказано, что они имеют Ca-зависимую антигенсвязывающую активность, исходя из результата фагового ELISA, описанного выше, в качестве матрицы, гены амплифицировали со специфическими праймерами и их последовательности анализировали. Последовательности вариабельных областей тяжелой цепи и легкой цепи антитела 6KC4-1#85 представлены в SEQ ID NO:10 и 104, соответственно. Полинуклеотид, кодирующий вариабельную область тяжелой цепи антитела 6KC4-1#85 (SEQ ID NO:10) связывали с полинуклеотидом, кодирующим последовательность, происходящую из IgG1 способом ПЦР. Полученный фрагмент ДНК встраивали в экспрессирующий вектор клеток животных для конструирования экспрессирующего вектора для тяжелой цепи SEQ ID NO:105. Полинуклеотид, кодирующий вариабельную область легкой цепи антитела 6KC4-1#85 (SEQ ID NO:104), связывали с полинуклеотидом, кодирующим константную область природной каппа-цепи (SEQ ID NO:44) способом ПЦР. Связанный фрагмент ДНК встраивали в экспрессирующий вектор клеток животных. Последовательности сконструированных вариантов подтверждали способом, известным специалистам в данной области. Последовательности сконструированных вариантов подтверждали способом, известным специалистам в данной области.
(10-5) Экспрессия и очистка антител
Клон 6KC4-1#85, который оценили как имеющий способность к Ca-зависимому связыванию с помощью ELISA фагов, вводили в экспрессирующие плазмиды клеток животных. Антитела экспрессировали с использованием следующего способа. Клетки линии, происходящей из клеток почки эмбриона FreeStyle 293-F (Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen), и аликвоты объемом 3 мл высевали в каждую ячейку планшетов с 6 ячейками с плотностью клеток 1,33×106 клеток/мл. Полученные плазмиды вводили в клетки способом липофекции. Клетки культивировали в инкубаторе CO2 (37°C, 8% CO2, 90 об./мин) в течение четырех суток. Из полученных культуральных супернатантов антитела очищали с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences) способом, известным специалистам в данной области. Концентрации очищенных антител определяли путем измерения поглощения при 280 нм с использованием спектрофотометра. Концентрации антител вычисляли из установленных величин на основе коэффициента экстинкции, определенного способом PACE (Protein Science 1995; 4: 2411-2423).
Справочный пример 16. Оценка связывания антитела 6KC4-1#85 с ионом кальция
Антитело 6KC4-1#85 с кальций-зависимым связыванием, которое было выделено из библиотеки антител человека, оценивали в отношении связывания им кальция. Оценку того, варьирует ли измеренная величина Tm, в зависимости от концентрации ионизированного кальция, проводили способом, описанным в справочном примере 2.
Величины Tm для Fab-домена антитела 6KC4-1#85 представлены в таблице 25. Как показано в таблице 25, величина Tm для Fab-домена антитела 6KC4-1#85 варьировала, в зависимости от концентрации ионов кальция. Это демонстрирует, что антитело 6KC4-1#85 связывается с кальцием.
Справочный пример 17. Идентификация участка связывания ионов кальция в антителе 6KC4-1#85
Как показано в справочном примере 16, антитело 6KC4-1#85 связывается с ионом кальция. Однако 6KC4-1#85 не имеет кальций-связывающего мотива, такого как последовательность hVk5-2, для которой путем оценки было выявлено, что она обладает кальций-связывающим мотивом. Затем то, связывается ли ион кальция с любой или обеими из тяжелой цепи и легкой цепи антитела 6KC4-1#85, подтверждали путем оценки связывания ионов кальция измененными антителами, полученными заменой тяжелой цепи и легкой цепи в 6KC4-1#85, соответственно, на тяжелую цепь и легкую цепь антитела против глипикана 3 (последовательность тяжелой цепи GC_H (SEQ ID NO:106), последовательность легкой цепи GC_L (SEQ ID NO:107)) которое не связывает ион кальция. Величины Tm измененных антител, измеренные способом, описанным в справочном примере 2, представлены в таблице 26. Результат указывает на то, что тяжелая цепь антитела 6KC4-1#85 связывается с кальцием, поскольку величины Tm измененного антитела, имеющего тяжелую цепь антитела 6KC4-1#85, изменялись в зависимости от концентрации ионов кальция.
Таким образом, для дальнейшей идентификации остатков, ответственных за связывание ионов кальция антителом 6KC4-1#85, конструировали измененные тяжелые цепи (6_H1-11 (SEQ ID NO:108), 6_H1-12 (SEQ ID NO:109), 6_H1-13 (SEQ ID NO:110), 6_H1-14 (SEQ ID NO:111), 6_H1-15 (SEQ ID NO:112)) и измененные легкие цепи (6_L1-5 (SEQ ID NO:113) и 6_L1-6 (SEQ ID NO:114)) путем замены остатка Asp (D) в CDR антитела 6KC4-1#85 остатком Ala (A), который не участвует в связывании или хелатировании ионов кальция. Способом, описанным в справочном примере 15, измененные антитела очищали из культуральных супернатантов клеток животных, в которые введены экспрессирующие векторы, несущие гены измененных антител. Очищенные измененные антитела оценивали в отношении связывания ими кальция способом, описанным в примере 2. Результат измерения представлен в таблице 33. Как показано в таблице 27, замена остатком Ala остатка в положении 95 или 101 (нумерация Kabat) в CDR3 тяжелой цепи антитела 6KC4-1#85 приводила к утрате активности антитела 6KC4-1#85 в отношении связывания кальция. Это указывает на то, что эти остатки ответственны за связывание кальция. Кальций-связывающий мотив, расположенный в основании петли CDR3 в тяжелой цепи антитела 6KC4-1#85, который был выявлен, исходя из способности связывать кальций антитела, измененного относительно антитела 6KC4-1#85, может представлять собой новый фактор для конструирования Ca-библиотек, как описано в справочном примере 7. В справочном примере 7 кальций-связывающие мотивы вносили в вариабельную область легкой цепи. Между тем, такие библиотеки включают, например, библиотеки, содержащие CDR3 тяжелой цепи из антитело 6KC4-1#85 и нестрогие остатки в CDR, отличных от CDR3 тяжелой цепи, но включающих CDR легкой цепи.
Справочный пример 18. Исследование эффектов антитела с Ca-зависимым связыванием на удержание в плазме антигена с использованием нормальных мышей
(18-1) Исследование in vivo с использованием нормальных мышей
Нормальной мыши (мышь C57BL/6J, Charles River Japan), вводили hsIL-6R (растворимый рецептор IL-6 человека, полученный согласно справочному примеру 21) отдельно, или вводили shIL-6R и антитело против рецептора IL-6 человека одновременно для исследования кинетики shIL-6R и антитела против рецептора IL-6 человека in vivo. Однократную дозу (10 мл/кг) раствора shIL-6R (5 мкг/мл) или смеси shIL-6R и антитела против рецептора IL-6 человека вводили в хвостовую вену. Указанные выше H54/L28-IgG1, 6RL#9-IgG1 и FH4-IgG1 использовали в качестве антител против рецептора IL-6 человека.
Концентрация shIL-6R во всех смесях составляет 5 мкг/мл. Концентрации антитела против рецептора IL-6 человека варьируют для антител: 0,1 мг/мл для H54/L28-IgG1 и 10 мг/мл для 6RL#9-IgG1 и FH4-IgG1. В это время полагали, что большинство shIL-6R связываются с антителом, поскольку антитело против shIL-6R присутствует в достаточном или избыточном количестве. Взятие образцов крови проводили через 15 минут, 7 часов и 1, 2, 4, 7, 14, 21 и 28 суток после введения. Полученные образцы крови сразу центрифугировали в течение 15 минут при 4°C и 12000 об./мин для отделения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до времени измерения.
(18-2) Определение концентрации в плазме рецептора против IL-6 человека у нормальных мышей с помощью ELISA
Концентрацию антитела против рецептора IL-6 человека в плазме мыши определяли с помощью ELISA. Сначала F(ab')2-фрагмент антитела против IgG человека (специфичный к γ-цепи) (SIGMA) распределяли в планшеты Nunc-Immuno Plate, MaxiSoup (Nalge nunc International) и позволяли им стоять в течение ночи при 4°C с получением планшетов с антителом против IgG человека в твердой фазе. Образцы для калибровочной кривой, имеющие концентрации в плазме 0,64, 0,32, 0,16, 0,08, 0,04, 0,02 или 0,01 мкг/мл, и образцы плазмы мыши для измерения, разбавленные в 100 раз или более, распределяли в планшет с антителом против IgG человека на твердой фазе, а затем проводили инкубацию в течение 1 часа при 25°C. Затем планшету позволяли реагировать с биотинилированным антителом против IL-6 R человека (R&D) в течение 1 часа при 25°C, а затем проводили реакцию со стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в течение 0,5 часа при 25°C. Хромогенную реакцию проводили с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки хромогенной реакции 1 н. серной кислотой (Showa Chemical) измеряли поглощение цветного раствора при 450 нм на устройстве для считывания микропланшетов. Концентрации антител в плазме у мышей вычисляли из величин поглощения на калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения в концентрациях в плазме антител H54/L28-IgG1, 6RL#9-IgG1 и FH4-IgG1, у нормальных мышей после внутривенного введения, измеренных как описано выше, представлено на фиг. 42.
(18-3) Определение концентрации в плазме shIL-6R способом электрохемилюминесценции
Концентрацию shIL-6R в плазме мыши определяли способом электрохемилюминесценции. Образец для калибровочной кривой shIL-6R, приготовленный в концентрации 2000, 1000, 500, 250, 125, 62,5 или 31,25 пг/мл, и образец плазмы мыши для измерения, разбавленный в 50 раз или выше, смешивали с моноклональным антителом против IL-6R человека (R&D), рутенированным с помощью SULFO-TAG NHS Ester (Meso Scale Discovery), биотинилированным антителом против IL-6R человека (R&D), и тоцилизумабом (тяжелая цепь SEQ ID NO:96, легкая цепь SEQ ID NO:97), с последующей реакцией в течение ночи при 4°C. К этому времени буфер для анализа содержал 10 мМ EDTA для снижения концентрации свободного Ca в образце и диссоциации практически всех shIL-6R в образце от 6RL#9-IgG1 или FH4-IgG1 для связывания с добавленным тоцилизумабом. Затем указанную реакционную жидкость распределяли в планшет со стрептавидином MA400 PR Streptavidin Plate (Meso Scale Discovery). Кроме того, после промывания каждой лунки планшета, которому позволяли реагировать в течение 1 часа при 25°C, в каждую лунку распределяли буфер для считывания T (×4) (Meso Scale Discovery). Сразу после этого реакционную жидкость подвергали измерению с использованием устройства для считывания SECTOR PR 400 (Meso Scale Discovery). Концентрацию shIL-6R вычисляли из ответа калибровочной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Изменения концентрации в плазме shIL-6R у нормальной мыши после внутривенного введения, определенные как описано выше, представлены на фиг. 20.
В результате, исчезновение shIL-6R было очень быстрым, когда shIL-6R вводили отдельно, в то время как исчезновение shIL-6R было значительно замедленным, когда shIL-6R вводили одновременно с H54/L28-IgG1, обычным антителом, не обладающим активностью связывания Ca с shIL-6R. Напротив, исчезновение shIL-6R было значительно ускорено, когда shIL-6R вводили одновременно с 6RL#9-IgG1 или FH4-IgG1, обладающим в 100 раз или более высокой зависимой от Ca способностью связывания с shIL-6R. Концентрации в плазме shIL-6R через одни сутки после введения shIL-6R одновременно с 6RL#9-IgG1 и FH4-IgG1 были снижены в 39 раз и 2 раза, соответственно, по сравнению с одновременным введением с H54/L28-IgG1. Таким образом, было подтверждено, что антитела с зависимым от кальция связывания способны ускорять исчезновение антигена из плазмы.
Справочный пример 19. Испытания по улучшению эффекта ускорения элиминации антигена с помощью зависимого от Ca связывания антигена (получение антител)
(19-1) Связывание IgG-антитела с FcRn
IgG-антитела имеют более длительное время удержания в плазме в результате связывания FcRn. Связывание между IgG и FcRn наблюдают только в кислых условиях (pH 6,0). Напротив, связывание практически не поддается выявлению в нейтральных условиях (pH 7,4). IgG-антитело захватывается в клетки неспецифическим образом. Антитело возвращается на клеточную поверхность путем связывания с эндосомальным FcRn в кислых условиях эндосом, а затем диссоциирует от FcRn в нейтральных условиях плазмы. Когда связывание FcRn в кислых условиях утрачивается вследствие внесения мутаций в Fc-домен IgG, время удержания антитела в плазме значительно снижается, поскольку антитело более не рециклирует в плазму из эндосомы.
Описанным способом увеличения удержания в плазме IgG-антитела является усиление связывания FcRn в кислых условиях. Аминокислотные мутации вносят в Fc-домен IgG-антитела для увеличения его связывания с FcRn в кислых условиях. Это увеличивает эффективность рециклирования в плазму из эндосомы, что приводит к увеличению удержания в плазме IgG-антитела. При внесении замены считается важным не увеличить связывание с FcRn в нейтральных условиях. IgG-антитела, которые связываются с FcRn в нейральных условиях, могут возвращаться на клеточную поверхность путем связывания с FcRn в кислых условиях эндосомы, однако IgG антитела не диссоциируют от FcRn в плазме в нейтральных условиях и не рециклируют в плазму, и, таким образом, полагали, что удержание в плазме IgG-антител напротив ухудшится.
Например, как описано в Dall' Acqua et al. (J. Immunol. (2002) 169 (9), 5171-5180), удержание в плазме IgG1-антитела, которому позволяли связываться с FcRn мыши в нейтральных условиях (pH 7,4), усиливалось в результате внесения аминокислотной замены у мыши. Кроме того, как описано Yeung et al. (J. Immunol. (2009) 182 (12), 7663-7671), Datta-Mannan et al. (J. Biol. Chem. (2007) 282 (3), 1709-1717), и Dall' Acqua et al. (J. Immunol. (2002) 169 (9), 5171-5180), выявлено, что варианты IgG1-антител, связывание которых с FcRn человека в кислых условиях (pH 6,0) улучшается путем внесения аминокислотной замены, также связываются с FcRn человека в нейтральных условиях (pH 7,4). Согласно сообщениям, время удержания в плазме указанного антитела, введенного яванскому макаку, не повышалось, не демонстрируя изменений в удержании в плазме. Таким образом, в технологии инженерии антител для повышения функций антител, были предприняты попытки для повышения удержания в плазме антитела путем увеличения его связывания с FcRn человека в кислых условиях без увеличения его связывания с FcRn человека в нейтральных условиях (pH 7,4). Иными словами, не было опубликовано сообщений в отношении преимуществ IgG1-антител, активность связывания которых с FcRn человека в нейтральных условиях (pH 7,4) увеличена путем внесения аминокислотных замен в Fc-область.
Антитела, которые связываются с антигеном Ca-зависимым образом, являются чрезвычайно полезными, поскольку они обладают эффектом ускорения исчезновения растворимого антигена и повторного связывания единичной молекулы антитела с растворимым антигеном. Способ усиления связывания с FcRn в нейтральных условиях (pH 7,4) исследовали в качестве способа дальнейшего улучшения эффекта ускорения исчезновения антигена.
(19-2) Получение антител с Ca-зависимым связыванием рецептора IL-6 человека, обладающих способностью связывания FcRn в нейтральных условиях
Аминокислотную мутацию вносили в Fc-области FH4-IgG1 и 6RL#9-IgG1, обладающих способностью кальций-зависимого связывания антигена, и H54/L28-IgG1, не обладающего способностью кальций-зависимого связывания антигена (используемого в качестве контроля) с получением вариантов, обладающих способностью связываться с FcRn в нейтральных условиях (pH 7,4). Аминокислотную мутацию вносили способом, известным в данной области, с использованием ПЦР. В частности, получали FH4-N434W (тяжелая цепь SEQ ID NO:115, легкая цепь SEQ ID NO:101), 6RL#9-N434W (тяжелая цепь SEQ ID NO:116, легкая цепь SEQ ID NO:99) и H54/L28-N434W (тяжелая цепь SEQ ID NO:117, легкая цепь SEQ ID NO:39) с Asn (аминокислота в положении 434, указанном согласно нумерации EU), замененным на Trp, в константной области тяжелой цепи IgG1. Экспрессирующий вектор для клеток животных, в который был встроен полинуклеотид, кодирующий вариант с аминокислотной заменой, получали с использованием набора для сайт-направленного мутагенеза QuikChange Site-Directed Mutahenesis Kit (Stratagene) способом, описанным в прилагаемых инструкциях. Экспрессию и очистку антитела и измерение концентрации проводили в соответствии со способом, описанным в справочном примере 11.
Справочный 20. Исследование эффекта ускорения исчезновения антител с Ca-зависимым связыванием с использованием нормальных мышей
(20-1) Исследование in vivo с использованием нормальных мышей
Нормальным мышам (мышь C57BL/6J; Charles River Japan) вводили hsIL-6R (растворимый рецептор IL-6 человека, полученный, как описано в справочном примере 20) отдельно или вводили shIL-6R и антитело против IL-6 человека одновременно для исследования кинетики shIL-6R и антитела против рецептора IL-6 человека in vivo. Однократную дозу (10 мл/кг) раствора shIL-6R (5 мкг/мл) или смеси shIL-6R и антитела против IL-6 человека вводили в хвостовую вену. Описанные выше H54/L28-N434W, 6RL#9-N434W и FH4-N434W использовали в качестве антител против рецептора IL-6 человека.
Концентрация shIL-6R во всех смесях составляет 5 мкг/мл. Концентрации антитела против рецептора IL-6 человека варьируют для антител: приготавливали в концентрации 0,042 мг/мл для H54/L28-N434W, в концентрации 0,55 мг/мл для 6RL#9-N434W, и в концентрации 1 мг/мл для FH4-N434W. В это время полагали, что большая часть shIL-6R связалась с антителом, поскольку антитело против shIL-6R существует в достаточном или избыточном количестве. Взятие образцов крови проводили через 15 минут, 7 часов и 1, 2, 4, 7, 14, 21 и 28 суток после введения. Образцы крови сразу центрифугировали в течение 15 минут при 4°C и 12000 об./мин для отделения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до времени измерения.
(20-2) Определение концентрации антитела против рецептора IL-6 человека у нормальных мышей с помощью ELISA
Концентрацию в плазме антитела против рецептора IL-6 человека у мыши определяли с помощью ELISA, как описано в справочном примере 18. Изменения концентрации в плазме антител H54/L28-N434W, 6RL#9-N434W и FH4-N434W, у нормальных мышей после внутривенного введения, измеренные как описано выше, представлены на фиг. 21.
(20-3) Определение концентрации shIL-6R способом электрохемилюминесценции
Концентрацию shIL-6R в плазме мыши определяли способом электрохемилюминесценции. Образец для калибровочной кривой shIL-6R, приготовленный в концентрации 2000, 1000, 500, 250, 125, 62,5 или 31,25 пг/мл, и образец для измерения в плазме мыши, разбавленный в 50 раз или выше, смешивали с моноклональным антителом против IL-6R человека (R&D), рутенированным с помощью SULFO-TAG NHS Ester (Meso Scale Discovery) и биотинилированным антителом против IL-6R человека (R&D), с последующей реакцией в течение ночи при 4°C. К этому времени буфер для анализа содержал 10 мМ EDTA для снижения концентрации свободного Ca в образце и диссоциации практически всех shIL-6R в образце от 6RL#9-N434W или FH4-N434W, чтобы они существовали в свободном состоянии. Затем указанную реакционную жидкость распределяли в планшет со стрептавидином MA400 PR Streptavidin Plate (Meso Scale Discovery). Кроме того, после промывания каждой лунки планшета, которому позволяли реагировать в течение 1 часа при 25°C, в каждую лунку распределяли буфер для считывания T (×4) (Meso Scale Discovery). Сразу после этого реакционную жидкость подвергали измерению с использованием устройства для считывания SECTOR PR 400 (Meso Scale Discovery). Концентрацию shIL-6R вычисляли из ответа калибровочной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Изменения концентрации в плазме shIL-6R у нормальной мыши после внутривенного введения, определенные как описано выше, представлены на фиг. 22.
В результате по сравнению с введением shIL-6R отдельно, одновременное введение shIL-6R с антителом H54/L28-N434W, которое обладает активностью связывания FcRn при pH 7,4 и не обладает зависимой от Ca активностью связывания с shIL-6R, имело значительно замедленное исчезновение shIL-6R. Напротив, исчезновение shIL-6R было ускоренным, когда shIL-6R вводили одновременно с антителом 6RL#9-N434W или FH4-N434W, которое имеет в 100 раз или более превышающую зависимую от Ca способность связывания с shIL-6R и активность связывания FcRn при pH 7,4 по сравнению с введением shIL-6R отдельно. Концентрации в плазме shIL-6R через одни сутки после введения shIL-6R одновременно с антителом 6RL#9-N434W или FH4-N434W были снижены в 3 раза и 8 раз, соответственно, по сравнению с введением shIL-6R отдельно. В результате было подтверждено, что исчезновение антигена из плазмы можно далее ускорять путем сообщения активности связывания FcRn при pH 7,4 антителу, которое связывается с антигеном кальций-зависимым образом.
Было подтверждено, что антитело 6RL#9-IgG1 или FH4-IgG1, обладающее в 100 раз или более увеличенной зависимой от Ca активностью связывания с shIL-6R увеличивает исчезновение shIL-6R, по сравнению с антителом H54/L28-IgG1, не обладающим зависимой от Ca активностью связывания с shIL-6R. Было подтверждено, что антитело 6RL#9-N434W или FH4-N434W, которое обладает в 100 раз или более увеличенной Ca-зависимой активностью связывания с shIL-6R и активностью связывания FcRn при pH 7,4, в большей степени ускоряет исчезновение shIL-6R, по сравнению с сведением shIL-6R отдельно. Эти данные указывают на то, что антитело, которое связывается с антигеном зависимым от Ca образом, диссоциирует от антигена в эндосоме, аналогично антителу, которое связывается с антигеном pH-зависимым образом.
Справочный пример 21. Получение растворимого рецептора IL-6 человека (hsIL-6R)
Рекомбинантный рецептор IL-6 человека, который представляет собой антиген, получали следующим образом. Клеточную линию CHO, конститутивно экспрессирующую растворимый рецептор IL-6 человека (далее обозначаемый как hsIL-6R), имеющий аминокислотную последовательность положений 1-357 от N-конца, как описано Mullberg et al. (J. Immunol. (1994) 152, 4958-4968), получали способом, известным специалистам в данной области. Экспрессирующую линию культивировали для экспрессии hsIL-6R. hsIL-6R очищали из полученного культурального супернатанта с помощью колоночной хроматографии Blue Sepharose 6 FF и гель-фильтрационной колоночной хроматографии. Фракцию, элюированную в качестве основного пика на последней стадии, использовали в качестве конечного продукта очистки.
Справочный пример 22. Конструирование библиотеки антител с pH-зависимым связыванием
(22-1) Способ получения антител с pH-зависимым связыванием
В WO 2009/125825 описано антитело с pH-зависимым связыванием антигена, свойства которого изменяются в области нейтральных и кислых значений pH, вследствие внесения гистидина в антигенсвязывающую молекулу. Описанное антитело с pH-зависимым связыванием получают путем модификации для замены части аминокислотной последовательности представляющей интерес антигенсвязывающей молекулы гистидином. Для более эффективного получения антитела с pH-зависимым связыванием без предварительного получения представляющей интерес антигенсвязывающей молекулы, подлежащей модификации, одним из способов может быть получение антигенсвязывающей молекулы, которая связывается с желаемым антигеном из совокупности антигенсвязывающих молекул (обозначаемой как His-библиотека) с гистидином, внесенным в вариабельную область (более предпочтительно, область, потенциально вовлеченную в связывание антигена). Может быть возможным эффективное получение антигенсвязывающей молекулы, обладающей желаемыми свойствами, из библиотеки His, поскольку гистидин встречается более часто в антигенсвязывающих молекулах из His-библиотеки, чем из общепринятых библиотек антител.
(22-2) Конструирование популяции молекул антител (His-библиотеки) с остатком гистидина, внесенным в их вариабельную область, для эффективного получения связывающих антител, которые связываются с антигеном pH-зависимым образом
Сначала выбирали положения для встраивания гистидина в His-библиотеке. В WO 2009/125825 описано получение антител с pH-зависимым связыванием антигена путем замены аминокислотных остатков в последовательностях антител против рецептора IL-6, IL-6 и рецептора IL-31 на гистидин. Кроме того, получали антитела против лизоцима яичного белка (FEBS Letter 11483, 309, 1, 85-88) и против гепцидина (WO 2009/139822), обладающие способностью pH-зависимого связывания антигена, путем замены в аминокислотной последовательности антигенсвязывающей молекулы на гистидин. Положения, где вносили остатки гистидина в антитело против рецептора IL-6, антитело против IL-6, антитело против рецептора IL-31, антитело против лизоцима яичного белка и антител против гепцидина, представлены в таблице 28. Положения, представленные в таблице 28, могут быть приведены в качестве положений-кандидатов, которые контролируют связывание антиген-антитело. Кроме того, помимо положений, представленных в таблице 28, также пригодными для внесения остатков гистидина считали положения, которые вероятно контактируют с антигеном.
В His-библиотеке, состоящей из вариабельных областей тяжелой цепи и легкой цепи, последовательность антитела человека использовали в качестве вариабельной области тяжелой цепи, и в вариабельную область легкой цепи вносили остатки гистидина. Положения, приведенные выше, и положения, которые могут быть вовлечены в связывание антигена, т.е. положения 30, 32, 50, 53, 91, 92 и 93 (нумерация Kabat, Kabat EA et al. 1991. Sequences of Proteins of Immunological Interest. NIH) в легкой цепе отбирали в качестве положений для внесения остатков гистидина в His-библиотеку. Кроме того, последовательность Vk1 отбирали в качестве матричной последовательности вариабельной области легкой цепи для внесения остатков гистидина. Множеству аминокислот позволяли появляться в матричной последовательности, чтобы разнообразить антигенсвязывающие молекулы, которые составляют библиотеку. Положения, экспонированные на поверхности вариабельной области, которые вероятно будут взаимодействовать с антигеном, отбирали в качестве положений, где позволяли появиться множеству аминокислот. В частности, в качестве нестрогих остатков были выбраны положения 30, 31, 32, 34, 50, 53, 91, 92, 93, 94 и 96 легкой цепи (нумерация Kabat, Kabat EA et al. 1991. Sequences of Proteins of Immunological Interest. NIH).
Определяли тип и частоту появления аминокислотных остатков, которым впоследствии позволяли появиться. Анализировали частоту появления аминокислот в нестрогих остатках в последовательностях hVk1 и hVk3, зарегистрированных в базе данных Kabat (KABAT, E.A. ET AL.: "Sequences of proteins of immunological interest", vol. 91, 1991, NIH PUBLICATION). Исходя из результатов анализа, отбирали тип аминокислот, которым позволяли появиться в His-библиотеке, из аминокислот с более высокой частотой появления в каждом положении. В то же время, аминокислоты, для которых было определено, что частота их появления является низкой, исходя из результатов анализа, также отбирали для избежания смещения свойств аминокислот. Частоту появления выбранных аминокислот определяли, ссылаясь на результаты анализа в базе данных Kabat.
В качестве His-библиотек конструировали His-библиотеку 1, которая зафиксирована так, чтобы обязательно включать один остаток гистидина в каждую CDR, и His-библиотеку 2, которая более сфокусирована на разнообразии последовательностей, чем His-библиотека 1, учитывая аминокислоты и частоту появления, как описано выше. Детальные конструкции His-библиотек 1 и 2 представлены в таблицах 3 и 4 (причем положения в каждой таблице соответствовали нумерации Kabat). Ser (S) в положении 94 может быть исключен, если в положении 92, соответствующем нумерации Kabat, находится Asn (N), для частоты появления аминокислот, как описано в таблице 3 и 4.
Справочный пример 23. Получение библиотеки фагового дисплея для антител человека (His-библиотека 1) для получения антитела, которое связывается с антигеном pH-зависимым образом.
Библиотеку генов вариабельных областей тяжелой цепи антител амплифицировали способом ПЦР с использованием поли-A РНК, полученной из PBMC человека, и коммерческой поли-A РНК человека в качестве матрицы. Библиотеку генов вариабельных областей легкой цепи антител, сконструированную в качестве His-библиотеки 1, как описано в примере 1, амплифицировали с использованием ПЦР. Комбинацию библиотек генов вариабельных областей тяжелой цепи и легкой цепи антител, полученных как описано выше, встраивали в фагмидный вектор для конструирования библиотеки фагового дисплея антител человека, которая представляет Fab-домены, состоящие из последовательностей антител человека. Для способа конструирования использовали (Methods Mol Biol. (2002) 178, 87-100) в качестве справочных материалов. Для конструирования библиотеки использовали линкерную часть, соединяющую фагмидный Fab с белком фага pIII, и последовательности библиотеки фагового дисплея с последовательностью расщепления трипсином, встроенной между доменами N2 и CT гена белка pIII хелперного фага. Идентифицировали последовательности частей генов антител, выделенных из E. coli, в которые были введены библиотеки генов антител, и получали информацию о последовательности для 132 клонов. Намеченное распределение аминокислот и распределение аминокислот в идентифицированных последовательностях представлено на фиг. 23. Конструировали библиотеку, содержащую различные последовательности, соответствующие намеченному распределению аминокислот.
Справочный пример 24. Выделение антител, которые связываются с IL-6R pH-зависимым образом
(24-1) Выделение фрагментов антител, которые связываются с антигенами pH-зависимым образом, из библиотеки посредством пэннинга на гранулах
Первую селекцию из сконструированной His-библиотеки 1 проводили посредством обогащения только фрагментами антител, обладающих способностью к связыванию антигена (IL-6R).
Фаги получали с помощью E. coli, содержащих сконструированные фагмиды длял фагового дисплея. Для преципитации фагов в культуральную жидкость E. coli для продукции фага добавляли 2,5 M NaCl/10% PEG. Преципитированную популяцию фагов разбавляли TBS, получая раствор с фаговой библиотекой. К раствору с фаговой библиотекой добавляли BSA и CaCl2 для доведения конечной концентрации BSA до 4% и конечной концентрации ионов кальция до 1,2 мМ. Что касается способа пэннинга, авторы настоящего изобретения использовали общие способы пэннинга с использованием антигенов, иммобилизованных на магнитные гранулы (J. Immunol. Methods. (2008) 332 (1-2), 2-9; J. Immunol. Methods. (2001) 247 (1-2), 191-203; Biotechnol. Prog. (2002) 18(2) 212-20, Mol. Cell Proteomics (2003) 2 (2), 61-9). Использованные магнитные гранулы представляли собой покрытые NeutrAvidin гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) или покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
В частности, к полученному раствору фаговой библиотеки добавляли 250 пмоль меченного биотином антигена, чтобы обеспечить контактирование указанного раствора фаговой библиотеки с антигеном в течение 60 минут при комнатной температуре. Добавляли магнитные гранулы, блокированные с помощью BSA, и позволяли им связываться с комплексами антиген/фаг при комнатной температуре в течение 15 минут. Гранулы промывали три раза с помощью 1 мл 1,2 мМ CaCl2/TBST (TBS, содержащий 1,2 мМ CaCl2 и 0,1% Tween20), а затем два раза с помощью 1 мл 1,2 мМ CaCl2/TBS (pH 7,6). Затем гранулы, к которым добавили 0,5 мл трипсина в концентрации 1 мг/мл, суспендировали при комнатной температуре в течение 15 минут, а затем сразу разделяли с использованием магнитного стенда для сбора раствора с фагом. Собранный раствор с фагом добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600 0,4-0,7). E. coli инфицировали фагами посредством культивирования их при осторожном перемешивания 37°C в течение одного часа. Инфицированные E. coli высевали в чашку 225×225 мм. Затем из культуруальной жидкости посеянных E. coli собирали фаги, получая раствор с фаговой библиотекой.
Для обогащения фагами, второй и последующие раунды пэннинга проводили с использованием антигенсвязывающей способности или способности к pH-зависимому связыванию в качестве показателя. В частности, 40 пмоль меченного биотином антигена добавляли в полученный раствор с фаговой библиотекой, чтобы обеспечит контактирование раствора с фаговой библиотекой и антигеном при комнатной температуре в течение 60 минут. Добавляли блокированные BSA магнитные гранулы и позволяли им связываться с комплексами антиген/фаг комплексы при комнатной температуре в течение 15 минут. Гранулы промывали несколько раз 1 мл 1,2 мМ CaCl2/TBST и 1,2 мМ CaCl2/TBS. Затем, при обогащении фагами с использованием антигенсвязывающей способности в качестве показателя, гранулы, в которые было добавлено 0,5 мл трипсина в концентрации 1 мг/мл, суспендировали при комнатной температуре в течение 15 минут, а затем сразу разделяли с использованием магнитного стенда для сбора раствора с фагами. Альтернативно, при обогащении фагами с использованием способности к pH-зависимому связыванию антигена в качестве показателя, гранулы, в которые было добавлено 0,1 мл 50 мМ MES/1,2 мМ CaCl2/150 мМ NaCl (pH 5,5), суспендировали при комнатной температуре, а затем сразу разделяли с использованием магнитного стенда для сбора раствора с фагом. Для устранения способности фагов, не экспонирующих Fab, инфицировать E. coli, белок pIII (происходящий из фага-помощника белок pIII) фагов, не экспонирующих Fab, расщепляли добавлением 5 мкл трипсина в концентрации 100 мг/мл к собранному раствору с фагом. Собранные фаги добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600 0,4-0,7). E. coli инфицировали фагами посредством культивирования их при осторожном перемешивании при 37°C в течение одного часа. Инфицированные E. coli высевали в планшет 225×225 мм. Затем фаги собирали из культуральной среды посеянных E. coli для сбора раствора с библиотекой фагов. Пэннинг с использованием антигенсвязывающей способности или способности к pH-зависимому связыванию в качестве показателя повторяли два раза.
(24-2) Оценка с помощью ELISA фагов
Содержание фаг культуральные супернатанты собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из отдельных колоний E. coli, полученных способом, описанным выше.
К содержащим фаг культуральным супернатантам добавляли BSA и CaCl2 в конечной концентрации 4% BSA и в конечной концентрации ионов кальция 1,2 мМ. Эти содержащие фаг культуральные супернатанты подвергали ELISA по следующей методике. Микропланшет для титрования StreptaWell 96 (Roche) покрывали в течение ночи 100 мкл PBS, содержавшего меченный биотином антиген. После промывания каждой лунки планшета посредством PBST (PBS, содержащий 0,1% Tween 20) для удаления антигена, лунки блокировали 250 мкл 4% BSA/TBS в течение одного часа или более. После удаления 4% BSA/TBS полученные супернатанты добавляли в каждую лунку. Антителам, находящимся на фагах, позволяли связаться с антигенами на каждой лунке посредством инкубации планшета при 37°C в течение одного часа. После промывания 1,2 мМ CaCl2/TBST, 12, в каждую лунку добавляли мМ CaCl2/TBS (pH 7,6) или 1,2 мМ CaCl2/TBS (pH 5,5). Планшет инкубировали при 37°C в течение 30 минут. После промывания 1,2 мМ CaCl2/TBST (pH 7,6), в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS, содержащее 4% BSA и 1,2 мМ ионизированный кальций. Планшет инкубировали в течение одного часа. После промывания 1,2 мМ CaCl2/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты, а затем измеряли поглощение при 450 нм для оценки развития окраски.
Когда обогащение проводили с использованием антигенсвязывающей способности в качестве показателя, ELISA фагов после двух раундов пэннинга показал, что 17 из 96 клонов были положительными в ELISA антигенспецифическим образом. Таким образом, клоны анализировали после трех раундов пэннинга. Между тем, когда обогащение проводили с использованием способности к pH-зависимому связыванию антигена в качестве показателя, ELISA фагов после двух раундов пэннинга показал, что 70 из 94 клонов были положительными в ELISA. Таким образом, клоны анализировали после двух раундов пэннинга.
Для клонов, подвергнутых описанному выше ELISA фагов, анализировали последовательность оснований гена, амплифицированного с помощью специфических праймеров. Результаты ELISA фагов и анализа последовательности представлены в таблице 29 ниже.
Тем же способом антитела со способностью к pH-зависимому связыванию антигена выделяли из библиотеки фагового дисплея нативных антител человека. Когда обогащение проводили с использованием антигенсвязывающей способности в качестве показателя, 13 типов антител с pH-зависимым связывванием было выделено из 88 исследованных клонов. Между тем, когда обогащение проводили с использованием способности к pH-зависимому связыванию антигена в качестве показателя, 27 типов антител с pH-зависимым связыванием было выделено из 83 исследованных клонов.
Описанный выше результат продемонстрировал, что варьирование клонов со способностью к pH-зависимому связыванию антигена из His-библиотеки 1 было более высоким по сравнению с библиотекой фагового дисплея наивных антител человека.
(24-3) Экспрессия и очистка антител
Клоны, которые оценили как имеющие способность к pH-зависимому связыванию, исходя из результатов ELISA фагов, вводили в экспрессирующие плазмиды клеток животных. Антитела экспрессировали с использованием описанного ниже способа. Клетки линии, происходящей из клеток почки эмбриона FreeStyle 293-F (Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen), и высевали при плотности клеток 1,33×106 клеток/мл (3 мл) в каждую лунку планшета с 6-ячейками. Полученные плазмиды вводили в клетки способом липофекции. Клетки культивировали в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин) в течение четырех суток. Из культуральных супернатантов, полученных как описано выше, антитела очищали с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences) способом, известным специалистам в данной области. Поглощение растворов очищенных антител измеряли при 280 нм с использованием спектрофотометра. Концентрации антител вычисляли из измеренных величин с использованием коэффициента поглощения, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
(24-4) Оценка выделенных антител в отношении их способности к pH-зависимому связыванию с рецептором IL-6 человека
Антитела 6RpH#01 (тяжелая цепь SEQ ID NO:118; легкая цепь SEQ ID NO:119), 6RpH#02 (тяжелая цепь SEQ ID NO:120; легкая цепь SEQ ID NO:121) и 6RpH#03 (тяжелая цепь SEQ ID NO:122; легкая цепь SEQ ID NO:123), выделенные, как описано в (24-3), оценивали в отношении зависимости от pH их активности связывания рецептора IL-6 человека посредством анализа взаимодействия между антителами и рецептором IL-6 человека с использованием Biacore T100 (GE Healthcare). Тоцилизумаб (тяжелая цепь SEQ ID NO:60; легкая цепь SEQ ID NO:61) использовали в качестве контрольного антитела, которое не обладает pH-зависимой активностью связывания с рецептором IL-6 человека. Взаимодействие для реакции антиген-антитело анализировали в растворах при pH 7,4 и pH 6,0, соответствующих условиям нейтральных pH и кислых pH, соответственно. Соответствующее количество белка A/G (Invitrogen) иммобилизовывали на сенсорный чип CM5 (GE Healthcare) способом присоединения аминов и приблизительно 300 RU каждого из представляющих интерес антител улавливали на чипе. Два типа используемых подвижных буфера представляли собой: 20 мМ ACES/150 мМ NaCl/0,05% (масс./об.) Tween20/1,2 мМ CaCl2 (pH 7,4); и 20 мМ ACES/150 мМ NaCl/0,05% (масс./об.) Tween20/1,2 мМ CaCl2 (pH 6,0). Каждый из этих буферов использовали для разбавления рецептора IL-6 человека. Все измерения проводили при 37°C.
В анализе взаимодействия в реакции антиген-антитело с использованием тоцилизумаба в качестве контрольного антитела и антител 6RpH#01, 6RpH#02 и 6RpH#03, разбавленный раствор рецептора IL-6 и подвижный буфер в качестве пустого образца инжектировали при скорости потока 5 мкл/мин в течение трех минут, чтобы позволить рецептору IL-6 взаимодействовать с антителами тоцилизумабом, 6RpH#01, 6RpH#02 и 6RpH#03, уловленными на сенсорном чипе. Затем инъецировали 10 мМ глицин-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд для регенерации сенсорного чипа.
Сенсограммы при pH 7,4, полученные при измерении с использованием способа, описанного выше, представлены на фиг. 24. Сенсограммы в условиях pH 6,0, полученные тем же способом, представлены на фиг. 25.
Описанный выше результат показывает, что способность к связыванию рецептора IL-6 антител 6RpH#01, 6RpH#02 и 6RpH#03 была значительно снижена, когда pH буфера изменяли от pH 7,4 до pH 6,0.
Справочный пример 25. Получение мышей с нокином рецептора IL-6 человека
(25-1) Конструирование вектора для нокина
Использовали клон бактериальной искусственной хромосомы (BAC), в которую была клонирована геномная область гена интерлейкина-6 мыши (Il6ra). Фрагмент ДНК, в котором кодирующая последовательность гена рецептора интерлейкина-6 человека (GenBank# NM_000565_), последовательность hp7, сигнал присоединения поли-A, последовательность loxP, кассета гена устойчивости к неомицину (neo) и loxP были последовательно связаны, встраивали в заданную область гена Il6ra мыши на этой BAC посредством гомологичной рекомбинации с использованием системы Red/ET (GeneBridges). В этом случае, встраивание проводили посредством совмещения участка инициации трансляции, расположенного в экзоне 1 гена Il6ra мыши на BAC, с участком инициации трансляциии гена IL6R человека, и удаляли только 40 пар оснований нуклеотидной последовательности после участка инициации трансляции в экзоне 1 гена Il6ra мыши. К neo, который является геном устойчивости к лекарственному средству, был добавлен промотор гена pkg, так чтобы ген neo экспрессировался в клетках ES. однако, согласно прогнозам, ген neo, возможно, будет подавлять экспрессию гена hIL6R, встроенную в вышележащую область. Таким образом, для обеспечения удаления гена neo на обеих сторонах гена neo помещали последовательность loxP (ATAACTTCGTATAGCATACATTATACGAAGTTAT (SEQ ID NO:131)). Это обеспечивало систему, где ген neo, расположенный между последовательностями loxP, будет удаляться посредством рекомбинации при воздействии на него Cre. Далее, для обеспечения линеаризации вектора для нокина, в 5'-вышележащую область гена Il6ra мыши на BAC встраивали последовательность распознавания ферментом рестрикции NotI (GCGGCCGC) вместе с геном устойчивости к ампициллину.
(25-2) Введение в клетки ES
Упомянутый выше вектор для нокина hIL6R вводили электропорицией в клетки ES (происходящие из мыши клетки 129SvEv), и после селективного культивирования с G418 получали клоны, устойчивые к лекарственному средству. Среди этих клонов проводили скрининг гомологичных рекомбинантов способом ПЦР. 60 мкг вектора для нокина линеаризовывали с помощью NotI, экстрагировали фенолом/хлороформом, преципитировали этанолом, а затем растворяли в PBS.
Для получения образцов для ПЦР клетки ES, подлежащие применению в скрининге, культивировали на 96-луночном планшете, промывали два раза с использованием 200 мкл PBS на лунку, а затем к ним добавляли буфер для лизиса клеток, имеющий следующий состав (5 мкл 10× буфера LA II (TAKARA LA для Taq), 5 мкл 5% NP-40, 4 мкл протеазы K (TAKARA) (20 мг/мл) и 36 мкл дистиллированной воды) для обработки при 55°C в течение двух часов, с последующей обработкой при 95°C в течение 15 минут для инактивации протеазы K.
Общий объем реакционной смеси для ПЦР, равный 25 мкл, получали посредством смешения 1 мкл образца, 2,5 мкл 10× буфера LA II, 2,5 мкл 25 мМ MgCl2, 4 мкл dNTP (включающие 2,5 мМ каждого из dATP, dCTP, dGTP и dTTP), 0,2 мкл каждого из праймеров (по 50 мкМ каждого), 0,25 мкл LA Taq (TAKARA) и 14,35 мкл дистиллированной воды. Условия ПЦР представляли собой: предварительное нагревание при 94°C в течение пяти минут, 35 циклов амплификации, состоящих из 98°C в течение десяти секунд и 68°C в течение 3 минут 30 секунд, а также нагревание при 68°C в течение семи минут.
В качестве праймеров использовали P6Ra1 (прямой) 5'-ACAGGGCCTTAGACTCACAGC-3' (SEQ ID NO:132) и hRLI6 11638R (обратный) 5'-AACTTGCTCCCGACACTACTGG-3' (SEQ ID NO:133). Праймер P6Ra1 располагался в области генома Il6ra мыши с 5'-стороны, выше гомологичного элемента вектора для нокина, и hRLI6 11638R располагался внутри кДНК hIL6R (смотрите фиг. 32). В образцах клеток ES, которые претерпели гомологичную рекомбинацию, амплифицируется фрагмент, соответствующий полосе на уровне приблизительно 2,2 т.п.н.
(3) Получение мышей с нокином
Гомологичные рекомбинантные клоны ES суспендировали посредством обработки трипсином и промывали клеточной средой для ES. Пять МЕ хорионического гонадотропина лошадей (eCG) и хорионического гонадотропина человека (hCG) вводили внутрибрюшинно с интервалами 48 часов самкам мышей C57BL/6J(B6) для достижения гиперовуляции, и этих самок мышей скрещивали с самцами мышей той же линии. Сутки, когда у самок мышей подтверждали пробку, считали сутками 0,5. На сутки гестации 2,5 матку и яйцеводы подвергали перфузии и эмбрионы на стадии от 8-клеток до морулы собирали. Собранные эмбрионы инкубировали в течение ночи при 37°C, эмбрионы, которые развивались в бластоцисты, использовали в качестве эмбрионов-хозяев и в них инъецировали 10-15 клеток ES. Эмбрионы после инъекции трансплантировали в матку псевдобеременных рецепиентных самок типа ICR на сутки гестации 2,5, и их потомков наблюдали через 17 суток. На основе различий масти потомков, полученных посредством инъекции клеток ES в бластоцисты, получали химерных мышей со смешенным присутствием рекомбинантных клеток ES (цвет дикого типа) и клеток, происходящих из бластоцисты хозина (черный). После созревания самцов мышей скрещивали с самками мышей B6, и передачу введенного нокином аллеля мышам следующего поколения подтверждали способом ПЦР с использованием геномной ДНК, извлеченной из хвоста мышей следующего поколения в качестве матрицы. ПЦР проводили способом, использованным при скрининге упомянутых выше гомологичных рекомбинантных клеток ES. В результате получали индивидуумов, у которых был обнаружен сигнал на уровне 2,2 т.п.н. и было подтверждено, что введенный нокином аллель передался этим индивидуумам.
(4) Удаление гена neo
Кассету гена neo удаляли посредством микроинъекциии вектора, экспрессирующего рекомбиназу Cre, в пронуклеус оплодотворенной яйцеклетки, полученного путем размножения индивидуумов, у которых был подтвержден введенный нокином аллель. Следовательно, посредством временной экспрессии Cre индуцировали рекомбинацию между двумя участками loxP, помещенными в введенный нокином аллель, и удаляли кассету гена neo. Оплодотворенная яйцеклетка, в которую был введен микроинъекцией экспрессирующий вектор для Cre, переносили в яйцеводы псевдобеременных реципиентных самок ICR на сутки гестации 0,5, и через 19 суток рождались потомки. Удаление кассеты гена neo подтверждали способом ПЦР с использованием геномной ДНК, экстрагированной из хвостов, собранных после отлучения потомков.
Раствор для реакции ПЦР состоял из 1 мкл образца, 12, мкл 2× GC буфера I, 4 мкл dNTP (включающих 2,5 мМ каждого из dATP, dCTP, dGTP и dTTP), 0,25 мкл каждого из праймеров (по 50 мкМ каждого), 0,25 мкL LA Taq (TAKARA) и 6,75 мкл дистиллированной воды, и после их смешения общее количество составляло 25 мкл. Условия реакции ПЦР представляли собой: предварительное нагревание при 94°C в течение четырех минут, 35 циклов амплификации, состоящих из 94°C в течение 30 секунд, 62°C в течение 30 секунд, и 72°C в течение трех минут, а также нагревание при 72°C в течение семи минут.
Положения, где располагаются праймеры, представлены на фиг. 32B. В качестве праймеров использовали mRLI6 10355 (5'-TCTGCAGTAGCCTTCAAAGAGC-3' (SEQ ID NO:134)) и mRLI6 11166R (5'-AACCAGACAGTGTCACATTCC-3' (SEQ ID NO:135)). В образцах от индивидуумов до удаления кассеты neo, способом ПЦР обнаруживали сигнал, соответствующий 4,2 т.п.н., в качестве продукта амплификации, происходящего из введенного нокином аллеля, и сигнал, соответствующий 0,8 т.п.н., происходящий из аллеля дикого типа, в то время как в образцах, в которых кассета neo была удалена, обнаруживали сигнал, соответствующий приблизительно 2,7 т.п.н., и сигнал, соответствующий 0,8 т.п.н. происходящий из аллеля дикого типа (фиг. 33).
(6) Подтверждение экспрессии IL6R человека и экспрессии IL6ra мыши
(6-1) Подтверждение способом ОТ-ПЦР с использованием тканевой РНК
Анализы экспрессии IL6R человека и IL6ra мыши проводили способом ОТ-ПЦР с использованием тканевой РНК гомозиготных мышей с нокином и мышей дикого типа. Тканевую РНК получали из печени, селезенки, тимуса, почки, сердца и легкого. С использованием 1 мкг каждой из тканевых РНК в качестве матриц, синтезировали кДНК посредством проведения реакции обратной транскрипции с использованием набора для синтеза первой цепи кДНК SuperScript II First Strand cDNA Synthesis Kit (Invitrogen) и олиго-dT (20) праймера. IL6R человека и IL6ra мыши выявляли посредством проведения ПЦР с использованием синтезированных кДНК в качестве матриц. IL6R человека выявляли с использованием комбинации прямого праймера 6RIK-s1 (5'-CCCGGCTGCGGAGCCGCTCTGC-3' (SEQ ID NO:136)), располагающегося в 5'-нетранслируемой области выше участка инициации трансляции, который представляет собой положение вставки гена hIL6R в введенном нокином аллеле, и специфичного к IL6R обратного праймера RLI6-a1 (5'-ACAGTGATGCTGGAGGTCCTT-3' (SEQ ID NO:137)). С другой стороны, IL6ra мыши выявляли с использованием комбинации упомянутого выше прямого праймера 6RIK-s1 и обратного праймера 6RLIcA2 (5'-AGCAACACCGTGAACTCCTTTG-3' (SEQ ID NO:138)), который является специфичным к IL6ra мыши. Раствор для реакции ПЦР состоял из 12,5 мкл образца, 12,5 мкл 2× буфера GC I, 4 мкл dNTP (включающих 2,5 мМ каждого из dATP, dCTP, dGTP, и dTTP), 0,25 мкл каждого из праймеров (по 50 мкМ каждого), 0,25 мкл LA Taq (TAKARA) и 6,75 мкл дистиллированной воды, и при их смешении общее количество составляло 25 мкл. Условия ПЦР представляли собой: предварительное нагревание при 94°C в течение двух минут, 30 циклов амплификации, состоящих из 94°C в течение 30 секунд, 62°C в течение 30 секунд и 72°C в течение одной минуты, а также нагревание при 72°C в течение пяти минут. Хотя продукт амплификации IL6R человека обнаруживается на уровне 880 п.н. и продукт амплификации IL6ra мыши обнаруживается на уровне 846 п.н., только IL6R человека обнаруживался в каждой из тканей гомозиготных мышей с нокином hIL6R, и IL6ra мыши не обнаруживался. Между тем, IL6R человека не обнаруживался в каждой из тканей мышей дикого типа и обнаруживался только IL6ra мыши (фиг. 34). Эти результаты подтвердили, что вектор с нокином претерпел гомологичную рекомбинацию, как и было намечено, обеспечив мышей, которые экспрессируют IL6R человека вместо IL6ra мыши.
(6-2) измерение концентрации IL-6R человека в плазме
Лапаротомию проводили под ингаляционной анестезией изофлураном и концентрацию растворимого IL-6R человека в плазме, выделенной из крови, собранной из большой абдоминальной вены измеряли с использованием набора Quantikine Human IL-6sR Immunoassay Kit (R&D Systems). В результате концентрация растворимого hIL-6R в плазме составляла 22,1±5,0 нг/мл для гомозиготных мышей с нокином, и 11,5±4,1 нг/мл для гетерозиготных мышей с нокином. Растворимый hIL-6R не обнаруживался в плазме мышей дикого типа (фиг. 35). Концентрация у гомозиготных мышей с нокином была эквивалентной концентрации в крови, сообщенной для человека (Blood (2008) 112, 3959-3969).
(6-3) Подтверждение видоспецифической реактивности лиганда
IL-6 мыши или IL-6 человека вводили внутрибрюшинно гомозиготным мышам с нокином и диким мышам в количестве 4 мкг на кг массы тела, через шесть часов проводили взятие крови и концентрацию сывороточного амилоида A (SAA) в крови количественно определяли с использованием набора ELISA для SAA (Invitrogen). Раствор, полученный добавлением в фосфатно-солевой буфер (PBS) плазмы мыши, так чтобы ее концентрация становилась равной 0,5%, использовали в качестве растворителя IL-6 для введения. Получали контрольную группу, в которой вводили только растворитель. В результате гомозиготная мышь с нокином отвечала только на IL-6 человека и уровень SAA в плазме возрастал, однако она не продемонстрировала никакого ответа на IL-6 мыши (фиг. 36). С другой стороны, мыши дикого типа отвечали на IL-6 человека и IL-6 мыши, и продемонстрировали увеличение уровня SAA в плазме (фиг. 36). Известно, что в то время как IL-6ra мыши связывается с IL-6 мыши, а также с IL-6 человека, IL-6R человека связывается с IL-6 человека, но не связывается с IL-6 мыши, и результаты этого эксперименты согласовывались с этой информацией. Таким образом, было выявлено, что у гомозиготных мышей с нокином IL6ra мыши не экспрессировался, а вместо этого экспрессировался IL6R, и он функционирует намеченным образом.
Поскольку мРНК гена hIL6R, транскрибированного с введенного нокином аллеля по настоящему изобретению, имеет структуру, которая не вырезается посредством сплайсинга, она не деградируется механизмом NMD, но с другой стороны известно, что уровень экспрессии генов, которые не вырезаются посредством сплайсинга, становится ниже. Однако у мышей с нокином hIL6R по настоящему изобретению концентрация растворимого hIL-6R в крови является такой же, как и у здоровых индивидуумов, и, более того, мыши в достаточной степени отвечают на введенный IL-6 человека, и продукция SAA была подтверждена. Это демонстрирует, что hp7, встроенный вместе с сигналом присоединения поли-A, участвовал в стабилизации уровня экспрессии hIL6R, который обычно снижался вследствие того, что его структура не вырезалась посредством сплайсинга.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА
АНТИГЕН-МИШЕНЬ
<130> C1-A1110P
<150> JP 2011-216958
<151> 2011-09-30
<160> 138
<170> PatentIn version 3.5
<210> 1
<211> 468
<212> Белок
<213> Homo sapiens
<400> 1
Met Leu Ala Val Gly Cys Ala Leu Leu Ala Ala Leu Leu Ala Ala Pro
1 5 10 15
Gly Ala Ala Leu Ala Pro Arg Arg Cys Pro Ala Gln Glu Val Ala Arg
20 25 30
Gly Val Leu Thr Ser Leu Pro Gly Asp Ser Val Thr Leu Thr Cys Pro
35 40 45
Gly Val Glu Pro Glu Asp Asn Ala Thr Val His Trp Val Leu Arg Lys
50 55 60
Pro Ala Ala Gly Ser His Pro Ser Arg Trp Ala Gly Met Gly Arg Arg
65 70 75 80
Leu Leu Leu Arg Ser Val Gln Leu His Asp Ser Gly Asn Tyr Ser Cys
85 90 95
Tyr Arg Ala Gly Arg Pro Ala Gly Thr Val His Leu Leu Val Asp Val
100 105 110
Pro Pro Glu Glu Pro Gln Leu Ser Cys Phe Arg Lys Ser Pro Leu Ser
115 120 125
Asn Val Val Cys Glu Trp Gly Pro Arg Ser Thr Pro Ser Leu Thr Thr
130 135 140
Lys Ala Val Leu Leu Val Arg Lys Phe Gln Asn Ser Pro Ala Glu Asp
145 150 155 160
Phe Gln Glu Pro Cys Gln Tyr Ser Gln Glu Ser Gln Lys Phe Ser Cys
165 170 175
Gln Leu Ala Val Pro Glu Gly Asp Ser Ser Phe Tyr Ile Val Ser Met
180 185 190
Cys Val Ala Ser Ser Val Gly Ser Lys Phe Ser Lys Thr Gln Thr Phe
195 200 205
Gln Gly Cys Gly Ile Leu Gln Pro Asp Pro Pro Ala Asn Ile Thr Val
210 215 220
Thr Ala Val Ala Arg Asn Pro Arg Trp Leu Ser Val Thr Trp Gln Asp
225 230 235 240
Pro His Ser Trp Asn Ser Ser Phe Tyr Arg Leu Arg Phe Glu Leu Arg
245 250 255
Tyr Arg Ala Glu Arg Ser Lys Thr Phe Thr Thr Trp Met Val Lys Asp
260 265 270
Leu Gln His His Cys Val Ile His Asp Ala Trp Ser Gly Leu Arg His
275 280 285
Val Val Gln Leu Arg Ala Gln Glu Glu Phe Gly Gln Gly Glu Trp Ser
290 295 300
Glu Trp Ser Pro Glu Ala Met Gly Thr Pro Trp Thr Glu Ser Arg Ser
305 310 315 320
Pro Pro Ala Glu Asn Glu Val Ser Thr Pro Met Gln Ala Leu Thr Thr
325 330 335
Asn Lys Asp Asp Asp Asn Ile Leu Phe Arg Asp Ser Ala Asn Ala Thr
340 345 350
Ser Leu Pro Val Gln Asp Ser Ser Ser Val Pro Leu Pro Thr Phe Leu
355 360 365
Val Ala Gly Gly Ser Leu Ala Phe Gly Thr Leu Leu Cys Ile Ala Ile
370 375 380
Val Leu Arg Phe Lys Lys Thr Trp Lys Leu Arg Ala Leu Lys Glu Gly
385 390 395 400
Lys Thr Ser Met His Pro Pro Tyr Ser Leu Gly Gln Leu Val Pro Glu
405 410 415
Arg Pro Arg Pro Thr Pro Val Leu Val Pro Leu Ile Ser Pro Pro Val
420 425 430
Ser Pro Ser Ser Leu Gly Ser Asp Asn Thr Ser Ser His Asn Arg Pro
435 440 445
Asp Ala Arg Asp Pro Arg Ser Pro Tyr Asp Ile Ser Asn Thr Asp Tyr
450 455 460
Phe Phe Pro Arg
465
<210> 2
<211> 1407
<212> ДНК
<213> Homo sapiens
<400> 2
atgctggccg tcggctgcgc gctgctggct gccctgctgg ccgcgccggg agcggcgctg 60
gccccaaggc gctgccctgc gcaggaggtg gcgagaggcg tgctgaccag tctgccagga 120
gacagcgtga ctctgacctg cccgggggta gagccggaag acaatgccac tgttcactgg 180
gtgctcagga agccggctgc aggctcccac cccagcagat gggctggcat gggaaggagg 240
ctgctgctga ggtcggtgca gctccacgac tctggaaact attcatgcta ccgggccggc 300
cgcccagctg ggactgtgca cttgctggtg gatgttcccc ccgaggagcc ccagctctcc 360
tgcttccgga agagccccct cagcaatgtt gtttgtgagt ggggtcctcg gagcacccca 420
tccctgacga caaaggctgt gctcttggtg aggaagtttc agaacagtcc ggccgaagac 480
ttccaggagc cgtgccagta ttcccaggag tcccagaagt tctcctgcca gttagcagtc 540
ccggagggag acagctcttt ctacatagtg tccatgtgcg tcgccagtag tgtcgggagc 600
aagttcagca aaactcaaac ctttcagggt tgtggaatct tgcagcctga tccgcctgcc 660
aacatcacag tcactgccgt ggccagaaac ccccgctggc tcagtgtcac ctggcaagac 720
ccccactcct ggaactcatc tttctacaga ctacggtttg agctcagata tcgggctgaa 780
cggtcaaaga cattcacaac atggatggtc aaggacctcc agcatcactg tgtcatccac 840
gacgcctgga gcggcctgag gcacgtggtg cagcttcgtg cccaggagga gttcgggcaa 900
ggcgagtgga gcgagtggag cccggaggcc atgggcacgc cttggacaga atccaggagt 960
cctccagctg agaacgaggt gtccaccccc atgcaggcac ttactactaa taaagacgat 1020
gataatattc tcttcagaga ttctgcaaat gcgacaagcc tcccagtgca agattcttct 1080
tcagtaccac tgcccacatt cctggttgct ggagggagcc tggccttcgg aacgctcctc 1140
tgcattgcca ttgttctgag gttcaagaag acgtggaagc tgcgggctct gaaggaaggc 1200
aagacaagca tgcatccgcc gtactctttg gggcagctgg tcccggagag gcctcgaccc 1260
accccagtgc ttgttcctct catctcccca ccggtgtccc ccagcagcct ggggtctgac 1320
aatacctcga gccacaaccg accagatgcc agggacccac ggagccctta tgacatcagc 1380
aatacagact acttcttccc cagatag 1407
<210> 3
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 3
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 4
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 4
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 5
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 6
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 6
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Asp Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Val
85 90 95
Leu Arg Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Gln
100 105 110
<210> 7
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 7
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 8
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 8
Asp Ile Val Met Thr Gln Ser Pro Glu Ser Leu Val Leu Ser Leu Gly
1 5 10 15
Gly Thr Ala Thr Ile Asn Cys Arg Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Thr Leu Leu Phe Ser Trp Ala Ser Ile Arg Asp Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Ala Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
65 70 75 80
Ile Ser Asp Leu Gln Ala Glu Asp Ala Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Arg Ala Pro Ser Phe Gly Gln Gly Thr Lys Leu Gln Ile Lys
100 105 110
<210> 9
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 9
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asp Pro Gly Gly Gly Glu Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 10
<211> 126
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 11
<211> 330
<212> Белок
<213> Homo sapiens
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 12
<211> 326
<212> Белок
<213> Homo sapiens
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 13
<211> 377
<212> Белок
<213> Homo sapiens
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 14
<211> 327
<212> Белок
<213> Homo sapiens
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 15
<211> 365
<212> Белок
<213> Homo sapiens
<400> 15
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 16
<211> 119
<212> Белок
<213> Homo sapiens
<400> 16
Met Ser Arg Ser Val Ala Leu Ala Val Leu Ala Leu Leu Ser Leu Ser
1 5 10 15
Gly Leu Glu Ala Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Ala Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser
35 40 45
Gly Phe His Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu
50 55 60
Arg Ile Glu Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp
85 90 95
Glu Tyr Ala Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile
100 105 110
Val Lys Trp Asp Arg Asp Met
115
<210> 17
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 17
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 18
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 19
<211> 1125
<212> ДНК
<213> Homo sapiens
<400> 19
atgtggttct tgacaactct gctcctttgg gttccagttg atgggcaagt ggacaccaca 60
aaggcagtga tcactttgca gcctccatgg gtcagcgtgt tccaagagga aaccgtaacc 120
ttgcactgtg aggtgctcca tctgcctggg agcagctcta cacagtggtt tctcaatggc 180
acagccactc agacctcgac ccccagctac agaatcacct ctgccagtgt caatgacagt 240
ggtgaataca ggtgccagag aggtctctca gggcgaagtg accccataca gctggaaatc 300
cacagaggct ggctactact gcaggtctcc agcagagtct tcacggaagg agaacctctg 360
gccttgaggt gtcatgcgtg gaaggataag ctggtgtaca atgtgcttta ctatcgaaat 420
ggcaaagcct ttaagttttt ccactggaat tctaacctca ccattctgaa aaccaacata 480
agtcacaatg gcacctacca ttgctcaggc atgggaaagc atcgctacac atcagcagga 540
atatctgtca ctgtgaaaga gctatttcca gctccagtgc tgaatgcatc tgtgacatcc 600
ccactcctgg aggggaatct ggtcaccctg agctgtgaaa caaagttgct cttgcagagg 660
cctggtttgc agctttactt ctccttctac atgggcagca agaccctgcg aggcaggaac 720
acatcctctg aataccaaat actaactgct agaagagaag actctgggtt atactggtgc 780
gaggctgcca cagaggatgg aaatgtcctt aagcgcagcc ctgagttgga gcttcaagtg 840
cttggcctcc agttaccaac tcctgtctgg tttcatgtcc ttttctatct ggcagtggga 900
ataatgtttt tagtgaacac tgttctctgg gtgacaatac gtaaagaact gaaaagaaag 960
aaaaagtggg atttagaaat ctctttggat tctggtcatg agaagaaggt aatttccagc 1020
cttcaagaag acagacattt agaagaagag ctgaaatgtc aggaacaaaa agaagaacag 1080
ctgcaggaag gggtgcaccg gaaggagccc cagggggcca cgtag 1125
<210> 20
<211> 374
<212> Белок
<213> Homo sapiens
<400> 20
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
Glu Pro Gln Gly Ala Thr
370
<210> 21
<211> 951
<212> ДНК
<213> Homo sapiens
<400> 21
atgactatgg agacccaaat gtctcagaat gtatgtccca gaaacctgtg gctgcttcaa 60
ccattgacag ttttgctgct gctggcttct gcagacagtc aagctgctcc cccaaaggct 120
gtgctgaaac ttgagccccc gtggatcaac gtgctccagg aggactctgt gactctgaca 180
tgccaggggg ctcgcagccc tgagagcgac tccattcagt ggttccacaa tgggaatctc 240
attcccaccc acacgcagcc cagctacagg ttcaaggcca acaacaatga cagcggggag 300
tacacgtgcc agactggcca gaccagcctc agcgaccctg tgcatctgac tgtgctttcc 360
gaatggctgg tgctccagac ccctcacctg gagttccagg agggagaaac catcatgctg 420
aggtgccaca gctggaagga caagcctctg gtcaaggtca cattcttcca gaatggaaaa 480
tcccagaaat tctcccattt ggatcccacc ttctccatcc cacaagcaaa ccacagtcac 540
agtggtgatt accactgcac aggaaacata ggctacacgc tgttctcatc caagcctgtg 600
accatcactg tccaagtgcc cagcatgggc agctcttcac caatgggggt cattgtggct 660
gtggtcattg cgactgctgt agcagccatt gttgctgctg tagtggcctt gatctactgc 720
aggaaaaagc ggatttcagc caattccact gatcctgtga aggctgccca atttgagcca 780
cctggacgtc aaatgattgc catcagaaag agacaacttg aagaaaccaa caatgactat 840
gaaacagctg acggcggcta catgactctg aaccccaggg cacctactga cgatgataaa 900
aacatctacc tgactcttcc tcccaacgac catgtcaaca gtaataacta a 951
<210> 22
<211> 316
<212> Белок
<213> Homo sapiens
<400> 22
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 23
<211> 876
<212> ДНК
<213> Homo sapiens
<400> 23
atgggaatcc tgtcattctt acctgtcctt gccactgaga gtgactgggc tgactgcaag 60
tccccccagc cttggggtca tatgcttctg tggacagctg tgctattcct ggctcctgtt 120
gctgggacac ctgcagctcc cccaaaggct gtgctgaaac tcgagcccca gtggatcaac 180
gtgctccagg aggactctgt gactctgaca tgccggggga ctcacagccc tgagagcgac 240
tccattcagt ggttccacaa tgggaatctc attcccaccc acacgcagcc cagctacagg 300
ttcaaggcca acaacaatga cagcggggag tacacgtgcc agactggcca gaccagcctc 360
agcgaccctg tgcatctgac tgtgctttct gagtggctgg tgctccagac ccctcacctg 420
gagttccagg agggagaaac catcgtgctg aggtgccaca gctggaagga caagcctctg 480
gtcaaggtca cattcttcca gaatggaaaa tccaagaaat tttcccgttc ggatcccaac 540
ttctccatcc cacaagcaaa ccacagtcac agtggtgatt accactgcac aggaaacata 600
ggctacacgc tgtactcatc caagcctgtg accatcactg tccaagctcc cagctcttca 660
ccgatgggga tcattgtggc tgtggtcact gggattgctg tagcggccat tgttgctgct 720
gtagtggcct tgatctactg caggaaaaag cggatttcag ccaatcccac taatcctgat 780
gaggctgaca aagttggggc tgagaacaca atcacctatt cacttctcat gcacccggat 840
gctctggaag agcctgatga ccagaaccgt atttag 876
<210> 24
<211> 291
<212> Белок
<213> Homo sapiens
<400> 24
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
Asn Arg Ile
290
<210> 25
<211> 765
<212> ДНК
<213> Homo sapiens
<400> 25
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagggt gctcgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gttgacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaagg caggaagtat tttcatcata attctgactt ctacattcca 480
aaagccacac tcaaagacag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tgtcagtgtc aaccatctca 600
tcattctttc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattc gaagctcaac aagagactgg 720
aaggaccata aatttaaatg gagaaaggac cctcaagaca aatga 765
<210> 26
<211> 254
<212> Белок
<213> Homo sapiens
<400> 26
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 27
<211> 702
<212> ДНК
<213> Homo sapiens
<400> 27
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagcgt gcttgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gtcaacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaaga caggaagtat tttcatcata attctgactt ccacattcca 480
aaagccacac tcaaagatag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tggcagtgtc aaccatctca 600
tcattctctc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattt ga 702
<210> 28
<211> 233
<212> Белок
<213> Homo sapiens
<400> 28
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 29
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 29
Gly Gly Gly Ser
1
<210> 30
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 30
Ser Gly Gly Gly
1
<210> 31
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 31
Gly Gly Gly Gly Ser
1 5
<210> 32
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 32
Ser Gly Gly Gly Gly
1 5
<210> 33
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 33
Gly Gly Gly Gly Gly Ser
1 5
<210> 34
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 34
Ser Gly Gly Gly Gly Gly
1 5
<210> 35
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 35
Gly Gly Gly Gly Gly Gly Ser
1 5
<210> 36
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 36
Ser Gly Gly Gly Gly Gly Gly
1 5
<210> 37
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 37
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 38
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 38
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 39
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 39
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 40
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 41
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 41
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Ala Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Pro Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Tyr His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 42
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 42
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Ala Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Pro Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Tyr His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 43
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 43
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 44
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 44
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 45
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Ile Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Asp Tyr Asn Pro Gln Phe
50 55 60
Gln Asp Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Asp Asp Gly Pro Tyr Thr Leu Glu Thr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys
210 215 220
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
290 295 300
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 46
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 46
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Thr Leu Tyr Asp Phe Trp Ser Gly Tyr Tyr Ser Tyr
100 105 110
Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 47
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 47
Gln Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Asp Thr Gly Pro Tyr Tyr Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 48
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 48
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Pro Val Pro Gly Val Tyr Tyr Tyr Tyr Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 49
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg His Arg Ala Gly Asp Leu Gly Gly Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 50
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 50
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 51
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 51
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 52
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 52
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Glu Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 53
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 53
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 54
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Thr Thr Leu Val Pro Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 55
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Glu Ala Thr Thr Leu Val Pro Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
65 70 75 80
Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 56
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 56
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Glu Ala Thr Thr Leu Val Pro Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 57
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 57
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Thr Thr Leu Val Pro Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Val Tyr Tyr Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 58
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 58
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Ala Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 59
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 59
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Ala Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 60
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 60
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Ala
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 61
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 61
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Ala Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 62
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 62
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Ala Asp Ala
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 63
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 63
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Ala Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Ala Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 64
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 64
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Ala Asp Ala
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Ala Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 65
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 65
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Thr Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Ala Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 66
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 66
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asp Asp Asp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asp Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 67
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 68
<211> 456
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 68
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Val Pro Pro Tyr Ser Ser Ser Ser Tyr Tyr Tyr Tyr Tyr
100 105 110
Tyr Tyr Met Asp Val Trp Gly Lys Gly Thr Thr Val Thr Val Ser Ser
115 120 125
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
130 135 140
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
145 150 155 160
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
165 170 175
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
180 185 190
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
195 200 205
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
210 215 220
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro
450 455
<210> 69
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ala Pro Gly Ile Gln Leu Trp Leu Arg Pro Ser Tyr Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro
450
<210> 70
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 70
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Trp Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ala Gly Asp Ser Ile Lys Tyr Ser
20 25 30
Ser Asp Tyr Trp Gly Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ser Tyr Leu Ser Gly Thr Thr Gln Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Met Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Arg Gly Pro Thr Gly Val Asp Gln Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 71
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 71
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Thr Leu Ser Cys Val Gly Tyr Gly Phe Thr Phe His Glu Asn
20 25 30
Asp Met His Trp Leu Arg Gln Pro Leu Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser His Ile Gly Trp Asn Asn Asn Arg Val Ala Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Val Ser Arg Asp Asn Ala Lys Asn Ser Leu Phe
65 70 75 80
Leu His Met Asn Ser Leu Arg Pro Asp Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Asp Leu Gly Asn Pro Ile Tyr Asp Val Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 72
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 72
Gln Pro Ala Leu Ala Gln Met Gln Leu Val Glu Ser Gly Gly Gly Leu
1 5 10 15
Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
20 25 30
Thr Phe Asp Asp Tyr Ala Met His Trp Val Arg Gln Ala Pro Gly Lys
35 40 45
Gly Leu Glu Trp Val Ser Gly Ile Ser Trp Asn Ser Gly Ser Ile Gly
50 55 60
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
65 70 75 80
Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
85 90 95
Ala Leu Tyr Tyr Cys Ala Arg Glu Gly Val Leu Gly Asp Ala Phe Asp
100 105 110
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro
450
<210> 73
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 73
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
Ser Ala Ala Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Ser Lys Trp Tyr Asn Asp Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Asn
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Arg Val Arg Ser Gly Ser Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 74
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 74
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
Ser Ala Ala Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Ser Lys Trp Tyr Asn Asp Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Ile Pro Asp Thr Ser Lys Asn
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Lys Asp Pro Arg Val Trp Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 75
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 75
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Val Leu Ala Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr His Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Gly Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Gly Tyr Ser Ala Gly Tyr Gly Gly Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 76
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 76
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Lys Thr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Arg Ala Asp Gly Gly Gln Met Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asp Ser Leu Tyr
65 70 75 80
Leu Gln Met Ile Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Phe Ala Ser Gly Gly Leu Asp Gln Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 77
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 77
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Glu Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Ser His Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Val Arg Tyr Phe Asp Ser Tyr Gly Met Asp Val Trp
100 105 110
Gly His Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 78
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 78
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Phe
20 25 30
Trp Ile Asn Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asp Pro Ser Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly His Val Ala Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu His Trp Asn Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg His Arg Tyr Leu Gly Gln Leu Ala Pro Phe Asp Pro Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 79
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Asp Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 80
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 80
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asp Ser Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 81
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Ser Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 82
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 82
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asp Gly Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 83
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 83
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Ser Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 84
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 85
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Ser His Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Asn Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 86
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Asn Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 87
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Asp Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 88
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 88
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Asp Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 89
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 89
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asp Asn Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 90
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 90
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Ala Phe Leu Glu Trp Pro Ile Trp Gly Ser Glu Tyr Phe
100 105 110
Gln His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 91
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 91
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Asp Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 92
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 92
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Thr Tyr Tyr Tyr Asp Ser Ser Ala Pro Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 93
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asp Asp Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 94
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 94
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Gly Ser Gly Phe Asn Trp Gly Asn Tyr Ala Phe Asp
100 105 110
Ile Trp Gly Gln Gly Phe Ser Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 95
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 95
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Arg Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 96
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 96
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 97
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 97
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 98
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 98
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asp Pro Gly Gly Gly Glu Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 99
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 99
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Arg Ser Asn Met Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Leu Leu Pro Gly Ala Ala Pro Lys Leu
35 40 45
Leu Ile Ser His Asn Thr His Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Ala Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser His Asp Ser Ser
85 90 95
Leu Ser Ala Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ser
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 100
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 100
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Asp Tyr Tyr Asp Ser Ser Gly Tyr Tyr Asp Ala Phe
100 105 110
Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 101
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 101
Glu Thr Thr Val Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ile Thr Thr Thr Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Phe Gln Gln Glu Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Glu Gly Asn Ile Leu Arg Pro Gly Val Pro Ser Arg Phe Ser Ser
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Lys Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Ser Asp Asn Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 102
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 102
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 103
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 103
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 104
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 104
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Phe Asp Ala Ser Asn Arg Ala Ala Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Lys Trp Val Thr
85 90 95
Phe Gly Gly Gly Thr Thr Val Glu Ile Arg Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 105
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 105
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 106
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 106
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 107
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 107
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 108
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Ala Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 109
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 110
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 110
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Ala Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 111
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Ala Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 112
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 112
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Ala Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro
450
<210> 113
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 113
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Phe Ala Ala Ser Asn Arg Ala Ala Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Lys Trp Val Thr
85 90 95
Phe Gly Gly Gly Thr Thr Val Glu Ile Arg Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 114
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 114
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Phe Asp Ala Ser Asn Arg Ala Ala Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Ala Lys Trp Val Thr
85 90 95
Phe Gly Gly Gly Thr Thr Val Glu Ile Arg Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 115
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 115
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Asp Tyr Tyr Asp Ser Ser Gly Tyr Tyr Asp Ala Phe
100 105 110
Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Trp His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro
450
<210> 116
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 116
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asp Pro Gly Gly Gly Glu Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 117
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 117
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 118
<211> 458
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 118
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Ser Ile Tyr Cys Ser Ser Thr Ser Cys Tyr Glu Pro Pro
100 105 110
Tyr Tyr Tyr Tyr Met Asp Val Trp Gly Lys Gly Thr Met Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
130 135 140
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
225 230 235 240
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
245 250 255
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
260 265 270
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
275 280 285
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
290 295 300
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
305 310 315 320
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
325 330 335
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
340 345 350
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
355 360 365
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
370 375 380
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
385 390 395 400
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
405 410 415
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
420 425 430
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
435 440 445
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455
<210> 119
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser His
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser His Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 120
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 120
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Ile Asn Gly Val Trp Glu Gly Gly Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 121
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 121
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile His Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Ser Lys Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Gly Asn Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 122
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 122
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Val Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asp Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Ala Asp Val Pro Ala Ser Asn Pro Tyr Gly Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 123
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 123
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser His Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asn Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 124
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 124
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Tyr Ile Thr Leu Glu Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu Lys Phe His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 125
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 125
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 126
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 126
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 127
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 127
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 128
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 128
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Val Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser
180 185 190
Thr Trp Pro Ser Gln Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys
210 215 220
Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser
245 250 255
Leu Ser Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp
260 265 270
Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr
275 280 285
Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val
290 295 300
Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu
305 310 315 320
Phe Lys Cys Lys Val Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr
355 360 365
Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr
370 375 380
Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys
405 410 415
Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu
420 425 430
Gly Leu His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly
435 440 445
Lys
<210> 129
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 129
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Tyr Ile Thr Leu Glu Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu Lys Phe His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 130
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 130
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Val Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser
180 185 190
Thr Trp Pro Ser Gln Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys
210 215 220
Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro
225 230 235 240
Asp Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Tyr Ile Ser
245 250 255
Leu Glu Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp
260 265 270
Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr
275 280 285
Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val
290 295 300
Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu
305 310 315 320
Phe Lys Cys Lys Val Asn Asn Asp Asp Tyr Pro Ala Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr
355 360 365
Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr
370 375 380
Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys
405 410 415
Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu
420 425 430
Gly Leu Lys Phe His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly
435 440 445
Lys
<210> 131
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 131
ataacttcgt atagcataca ttatacgaag ttat 34
<210> 132
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 132
acagggcctt agactcacag c 21
<210> 133
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 133
aacttgctcc cgacactact gg 22
<210> 134
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 134
tctgcagtag ccttcaaaga gc 22
<210> 135
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 135
aaccagacag tgtcacattc c 21
<210> 136
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 136
cccggctgcg gagccgctct gc 22
<210> 137
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 137
acagtgatgc tggaggtcct t 21
<210> 138
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 138
agcaacaccg tgaactcctt tg 22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ МНОГОКРАТНО СВЯЗЫВАТЬСЯ С МНОЖЕСТВОМ АНТИГЕННЫХ МОЛЕКУЛ | 2018 |
|
RU2757786C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ К МНОГОКРАТНОМУ СВЯЗЫВАНИЮ ДВУХ ИЛИ БОЛЕЕ МОЛЕКУЛ АНТИГЕНА | 2014 |
|
RU2756029C2 |
| АНТИТЕЛА С МОДИФИЦИРОВАННОЙ АФФИННОСТЬЮ К FcRn, КОТОРЫЕ УВЕЛИЧИВАЮТ КЛИРЕНС АНТИГЕНОВ | 2011 |
|
RU2810471C1 |
| СОДЕРЖАЩИЙ ЛЕКАРСТВЕННОЕ СРЕДСТВО ПЕРЕНОСЧИК В КЛЕТКУ ДЛЯ ФОРМИРОВАНИЯ ИММУННОГО КОМПЛЕКСА | 2012 |
|
RU2739792C1 |
| СПЕЦИФИЧНАЯ К ТКАНИ-МИШЕНИ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2013 |
|
RU2743463C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ К МНОГОКРАТНОМУ СВЯЗЫВАНИЮ ДВУХ ИЛИ БОЛЕЕ МОЛЕКУЛ АНТИГЕНА | 2021 |
|
RU2841525C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| БИБЛИОТЕКА ЗАВИСИМЫХ ОТ КОНЦЕНТРАЦИИ ИОНОВ СВЯЗЫВАЮЩИХ МОЛЕКУЛ | 2012 |
|
RU2732151C2 |
| АНТИТЕЛА С МОДИФИЦИРОВАННОЙ АФФИННОСТЬЮ К FcRn, КОТОРЫЕ УВЕЛИЧИВАЮТ КЛИРЕНС АНТИГЕНОВ | 2011 |
|
RU2745989C2 |
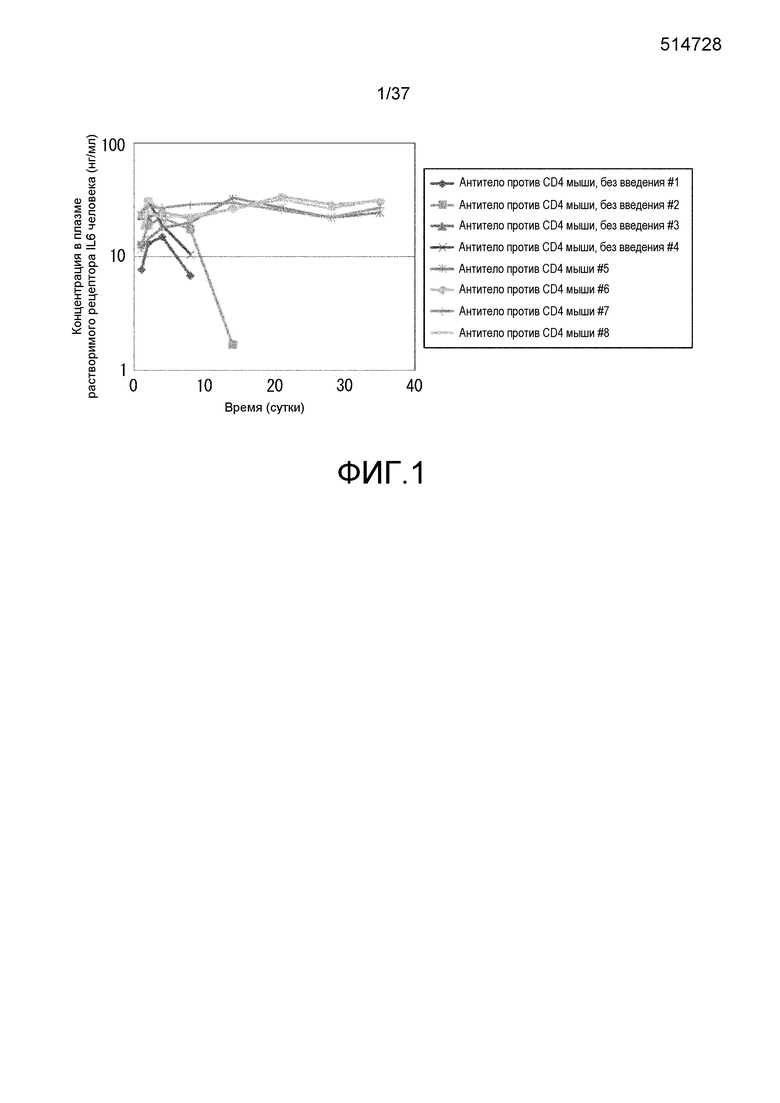
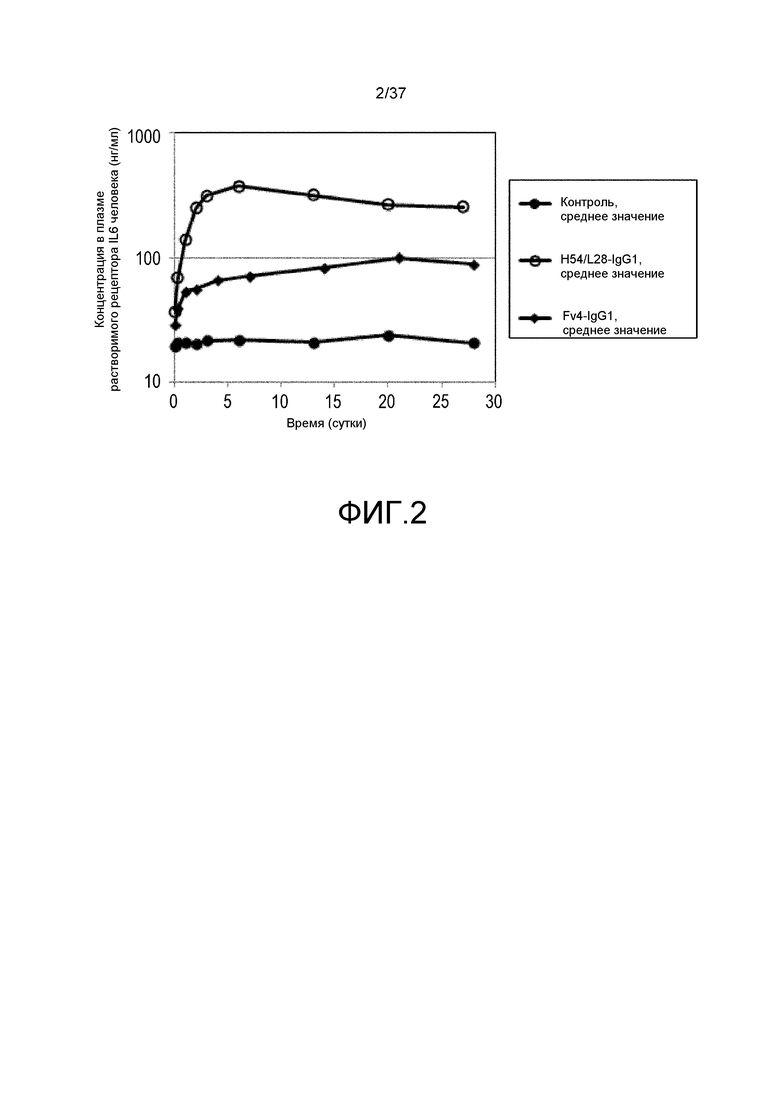
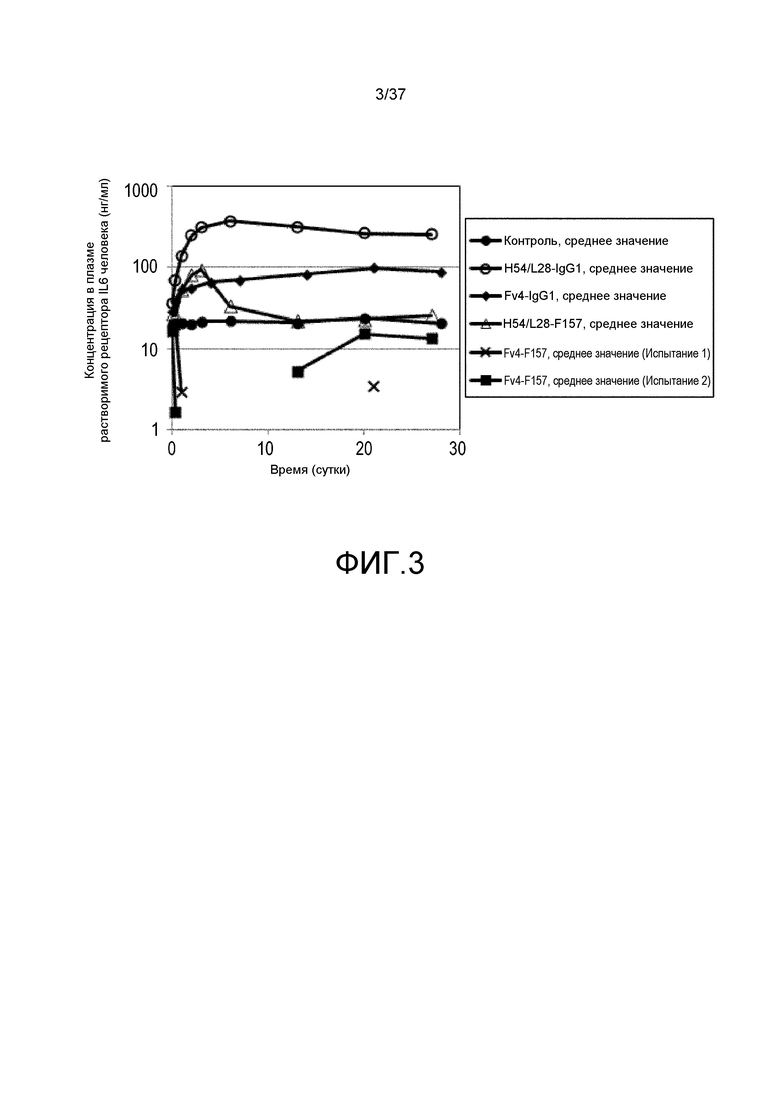
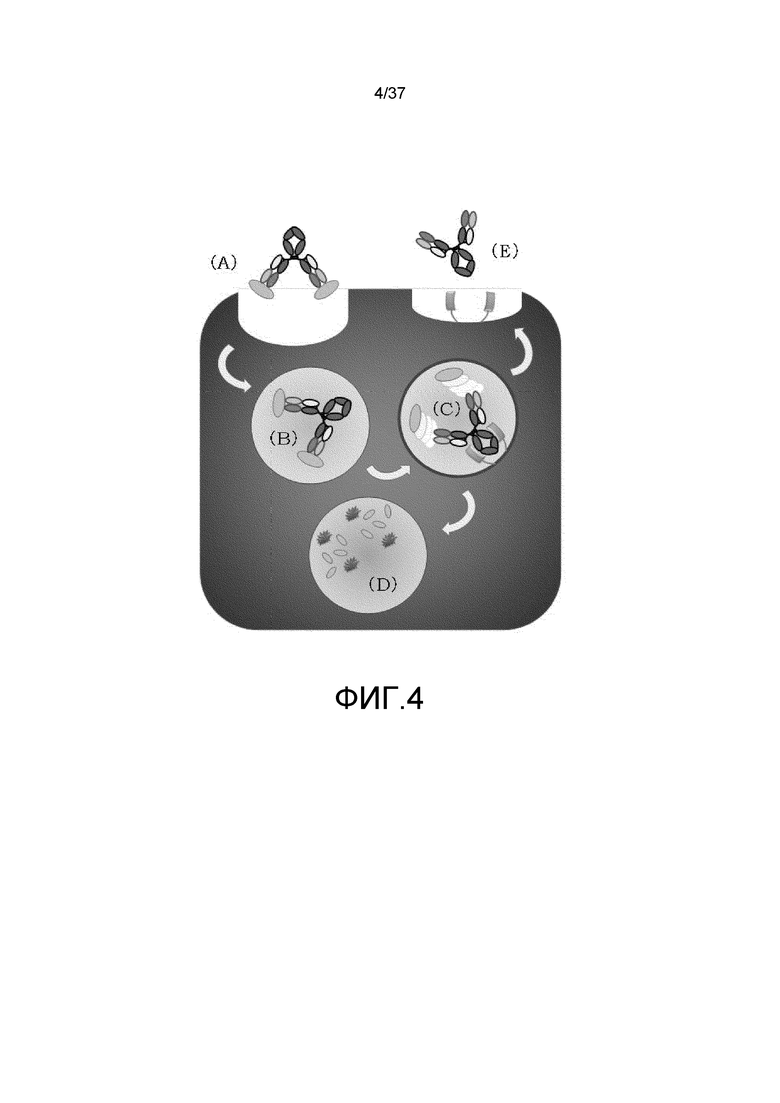
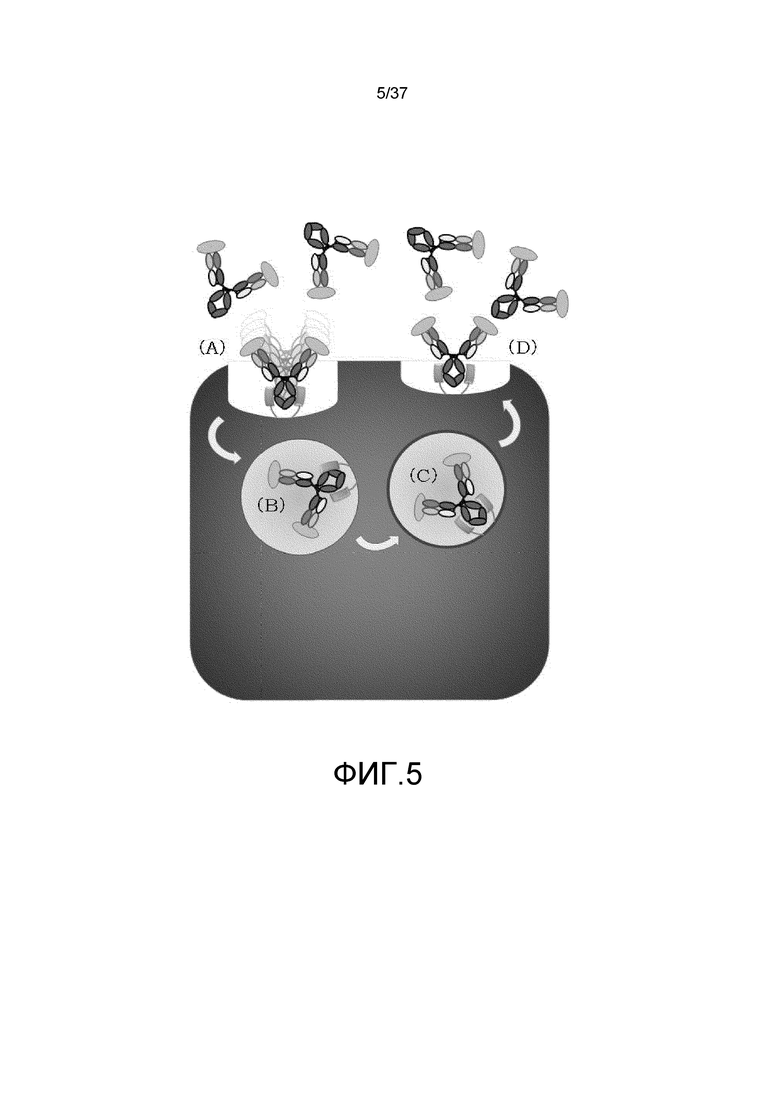
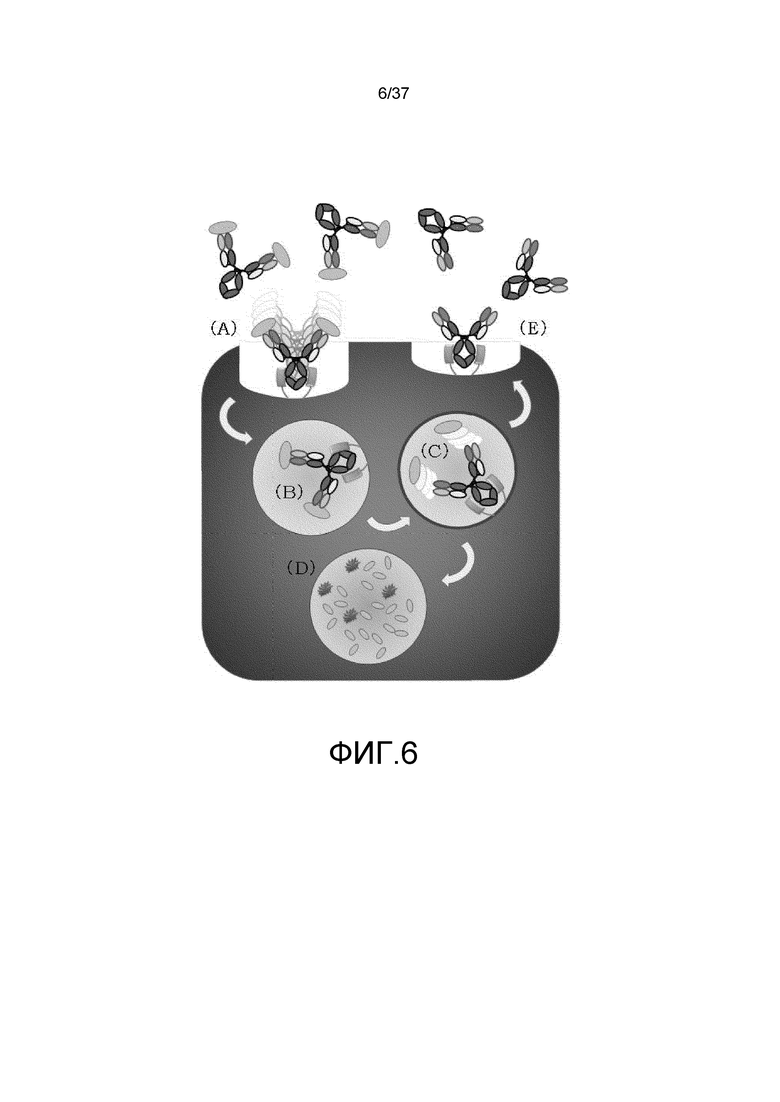
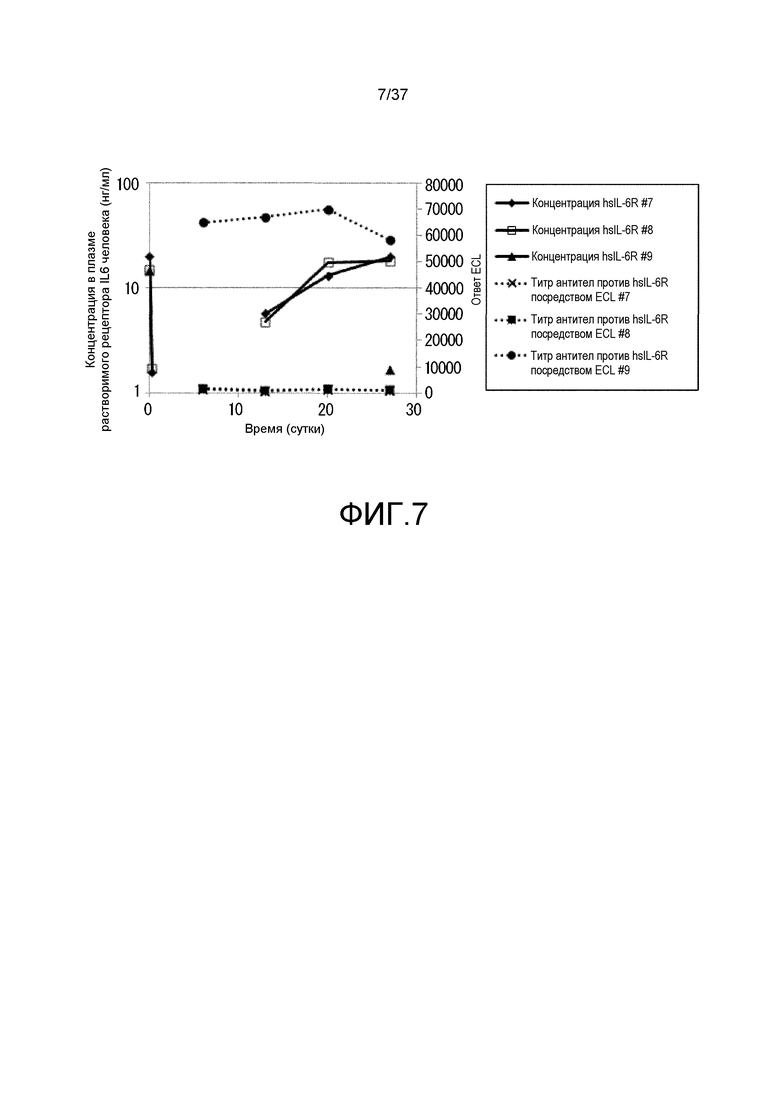
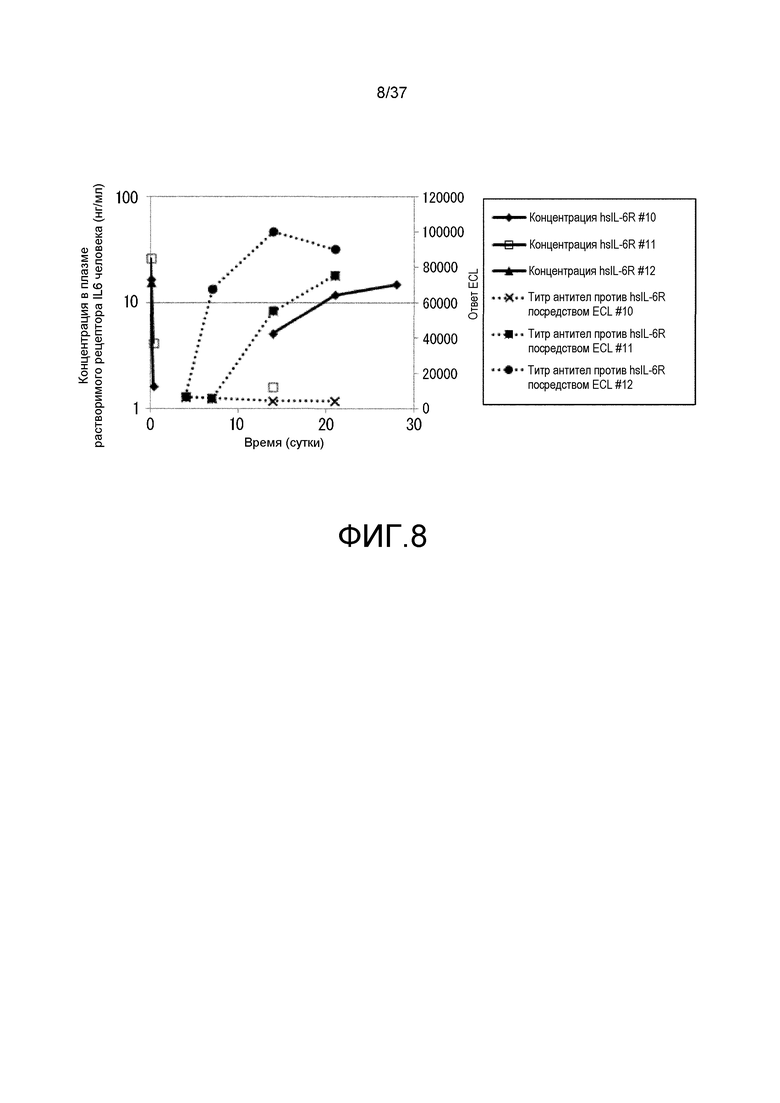
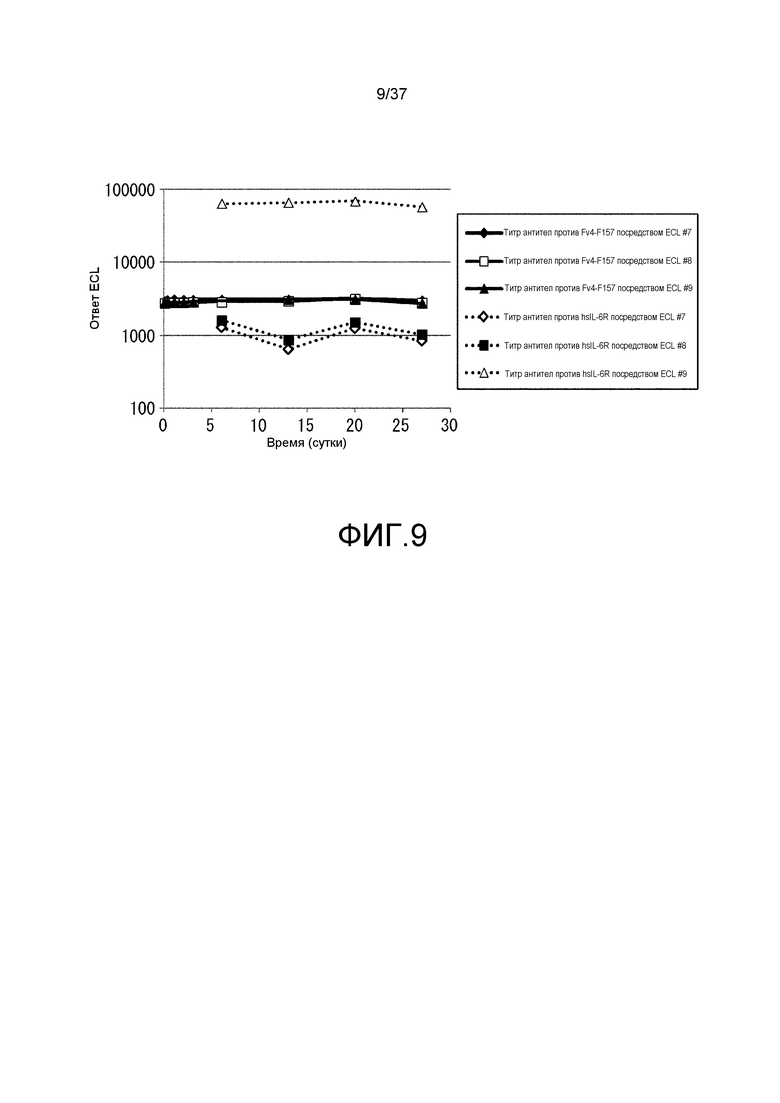
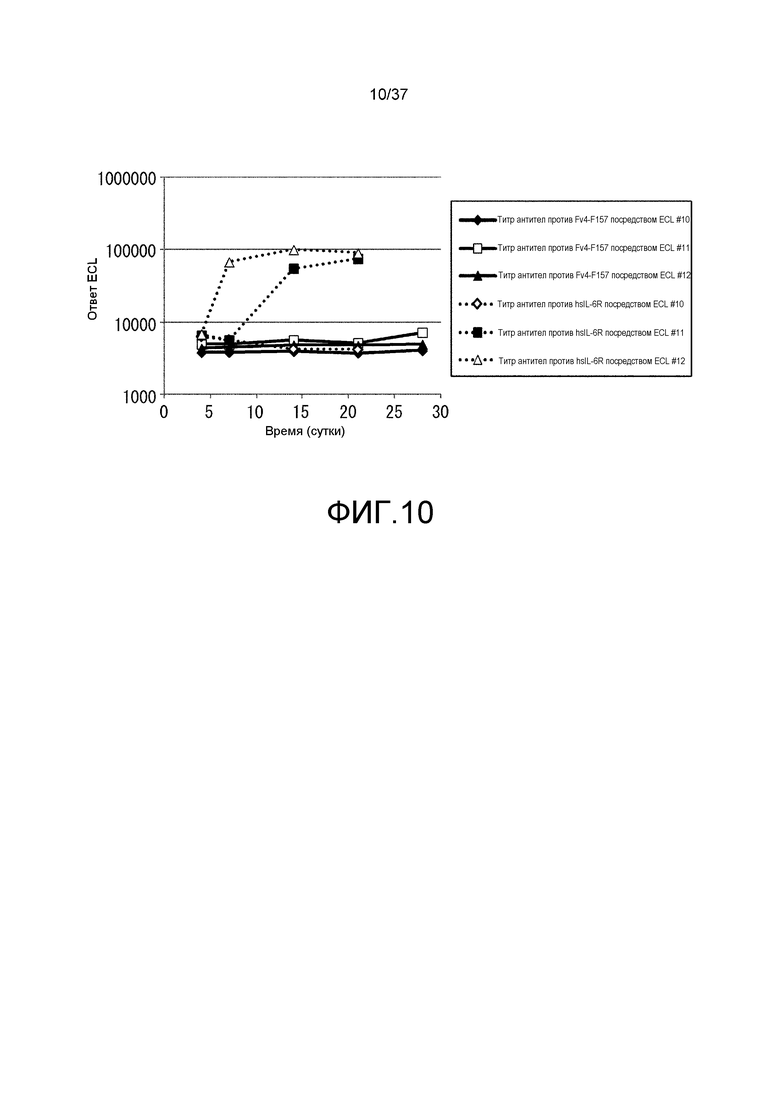
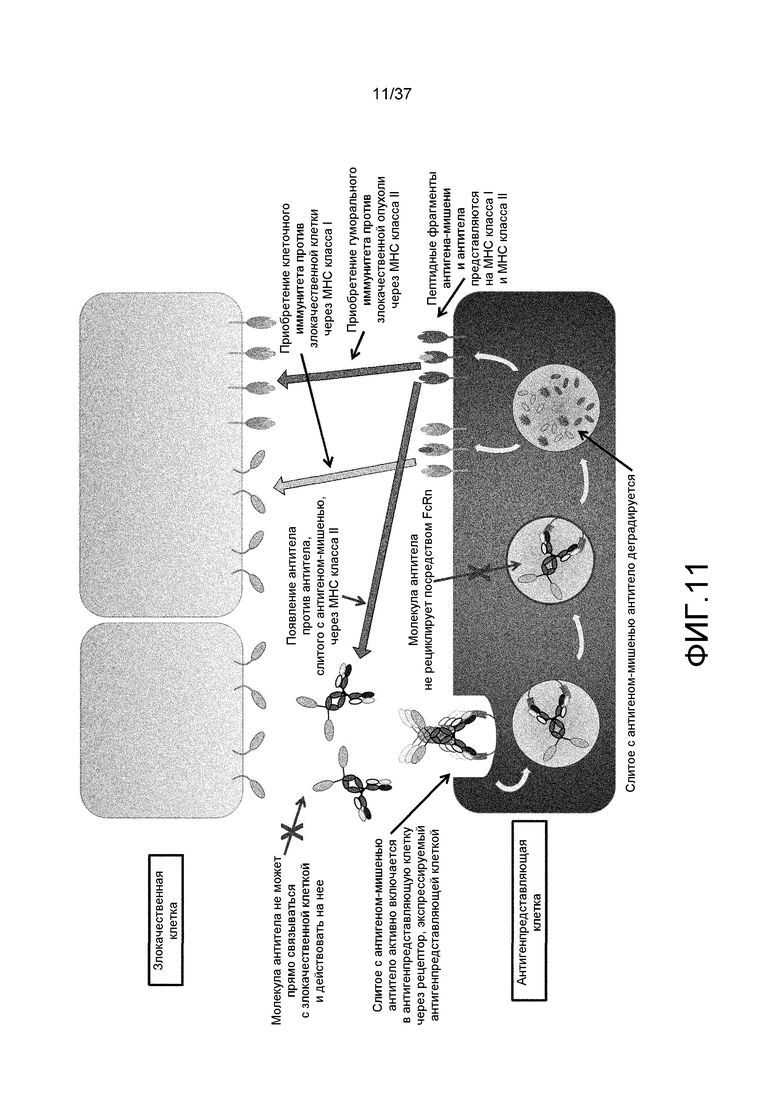
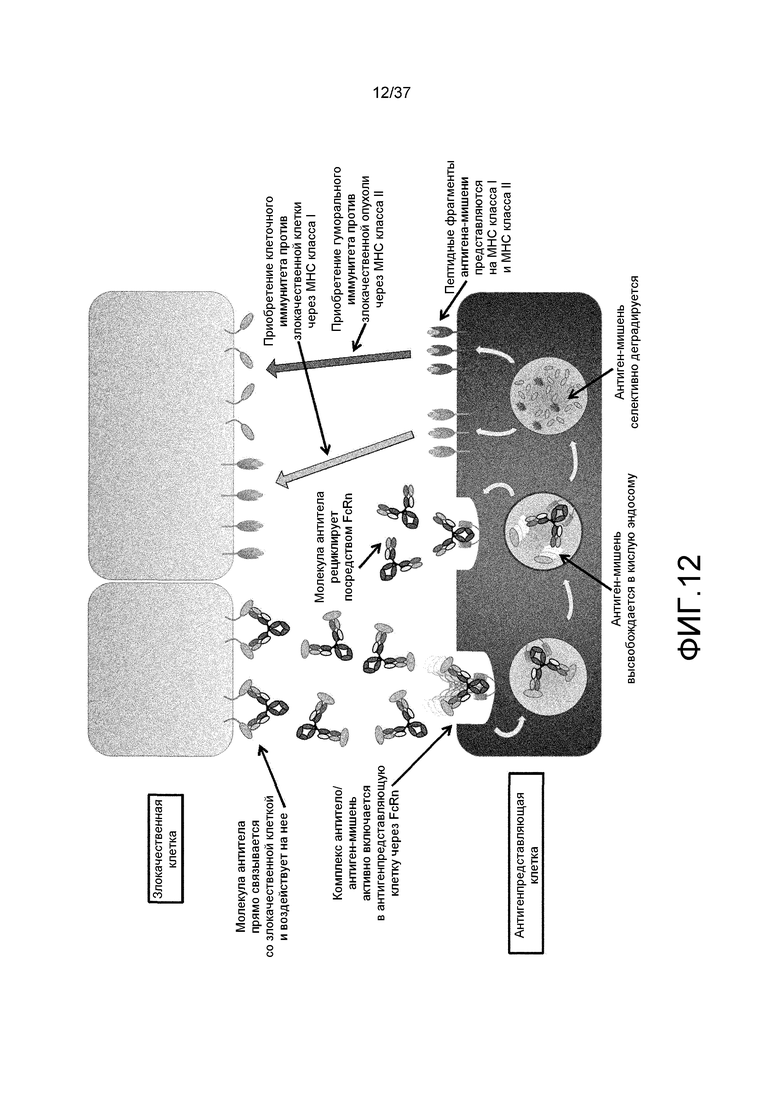
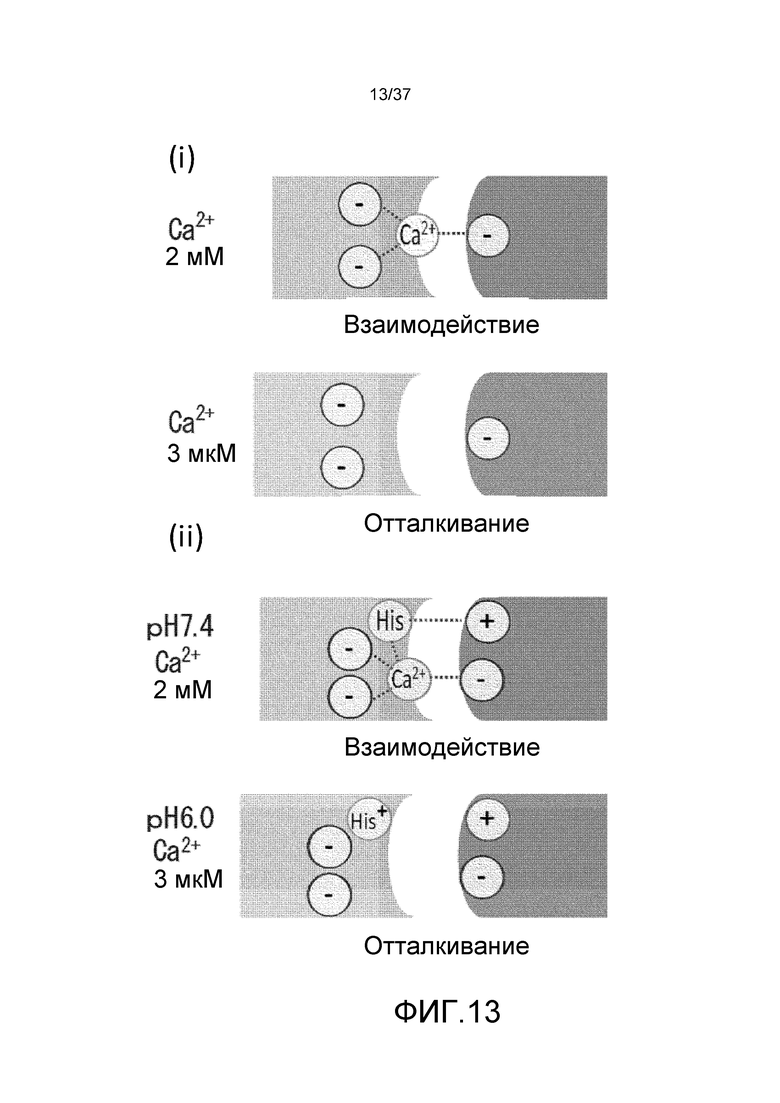
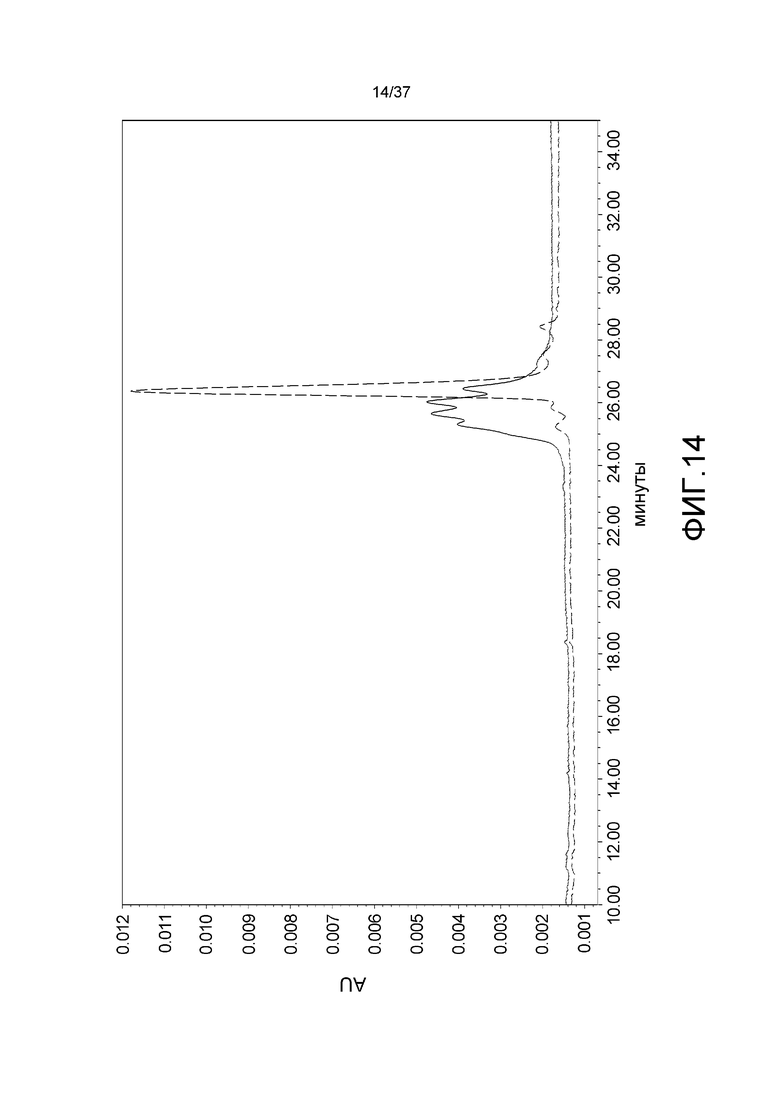
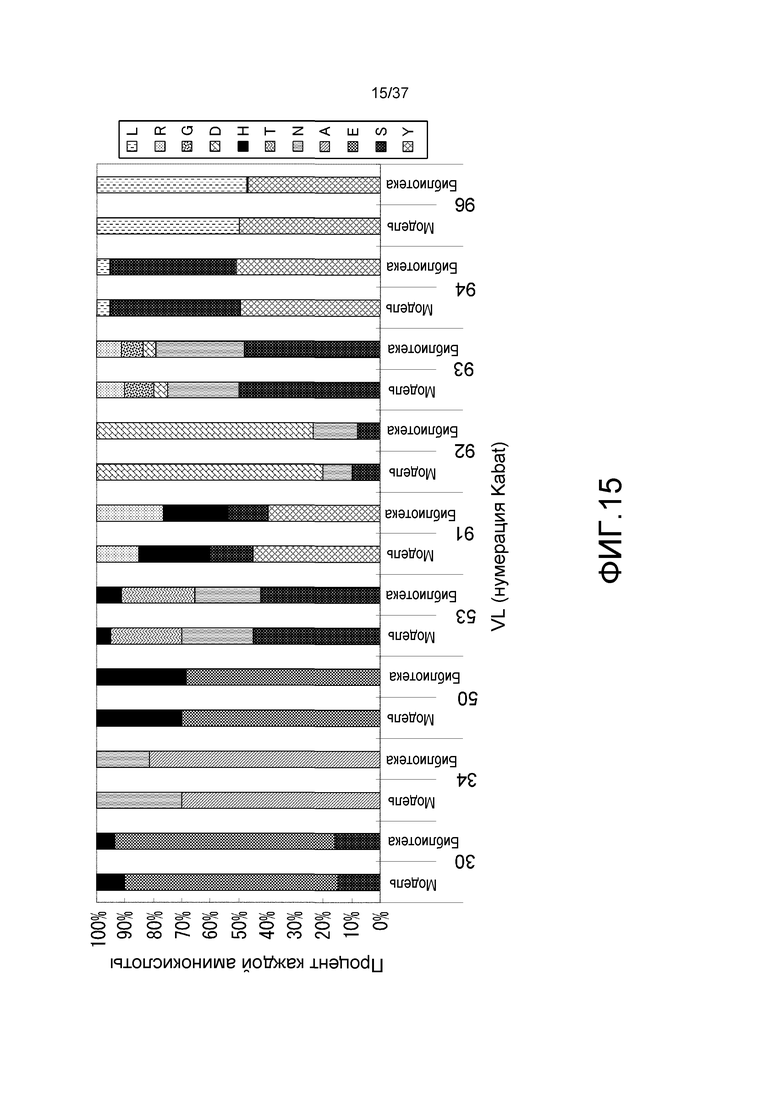
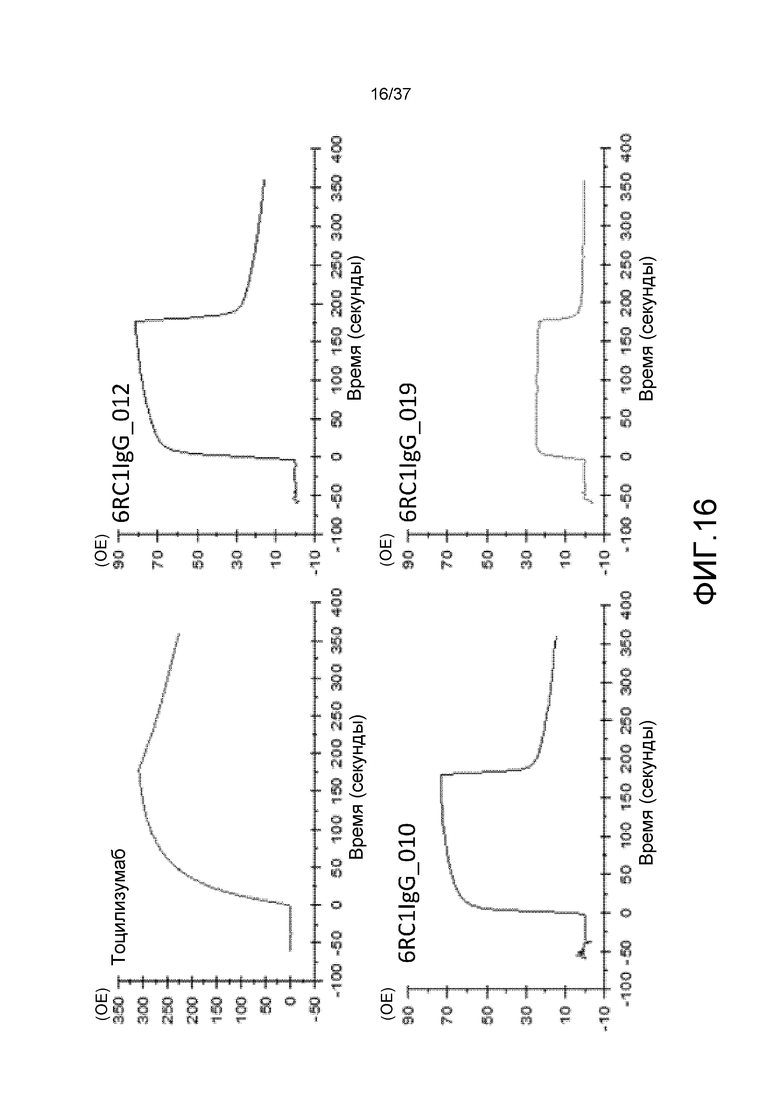
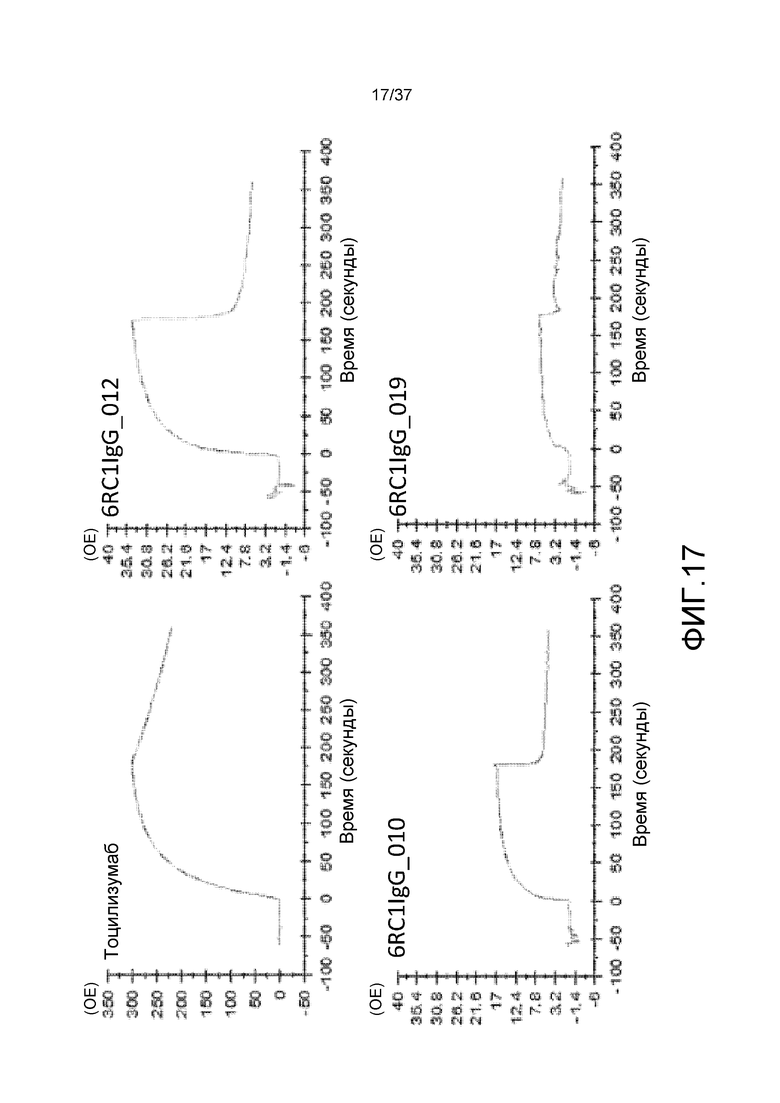
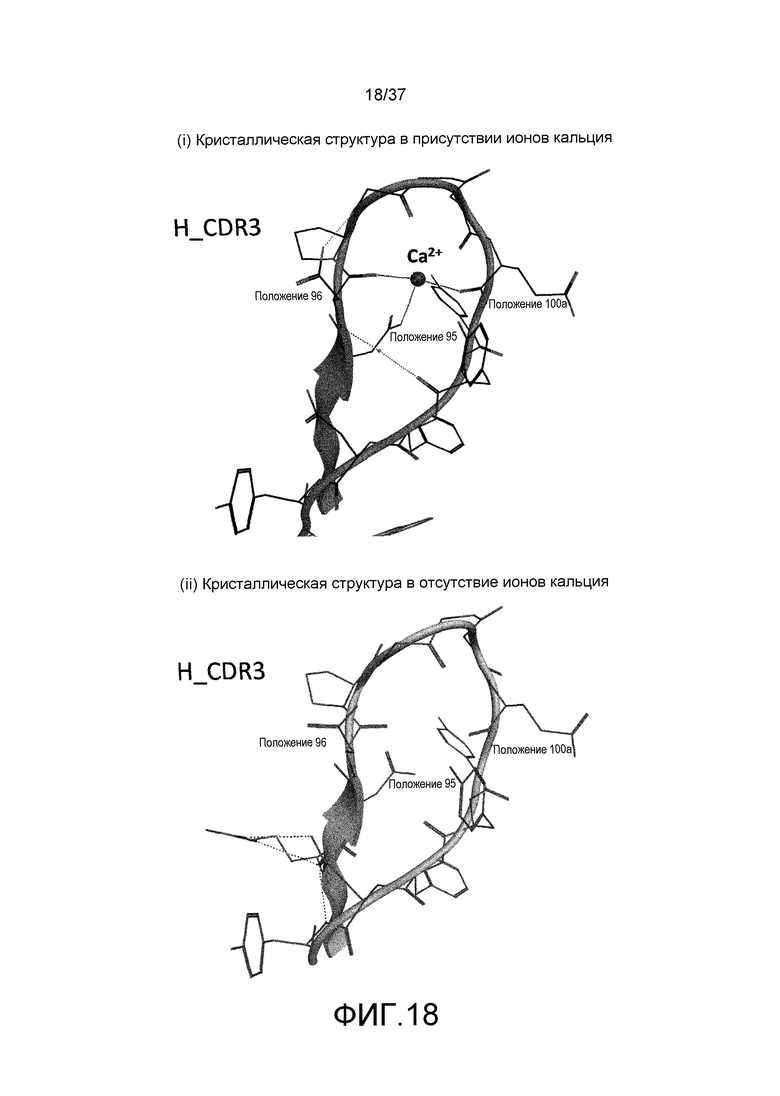
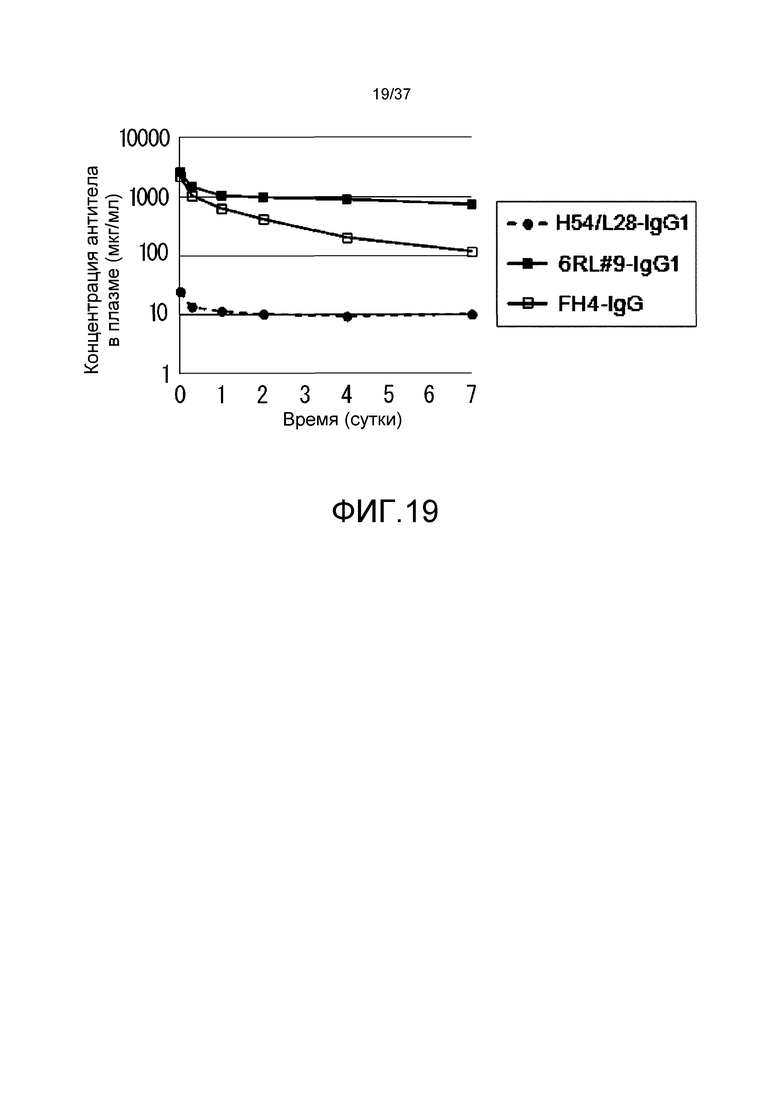
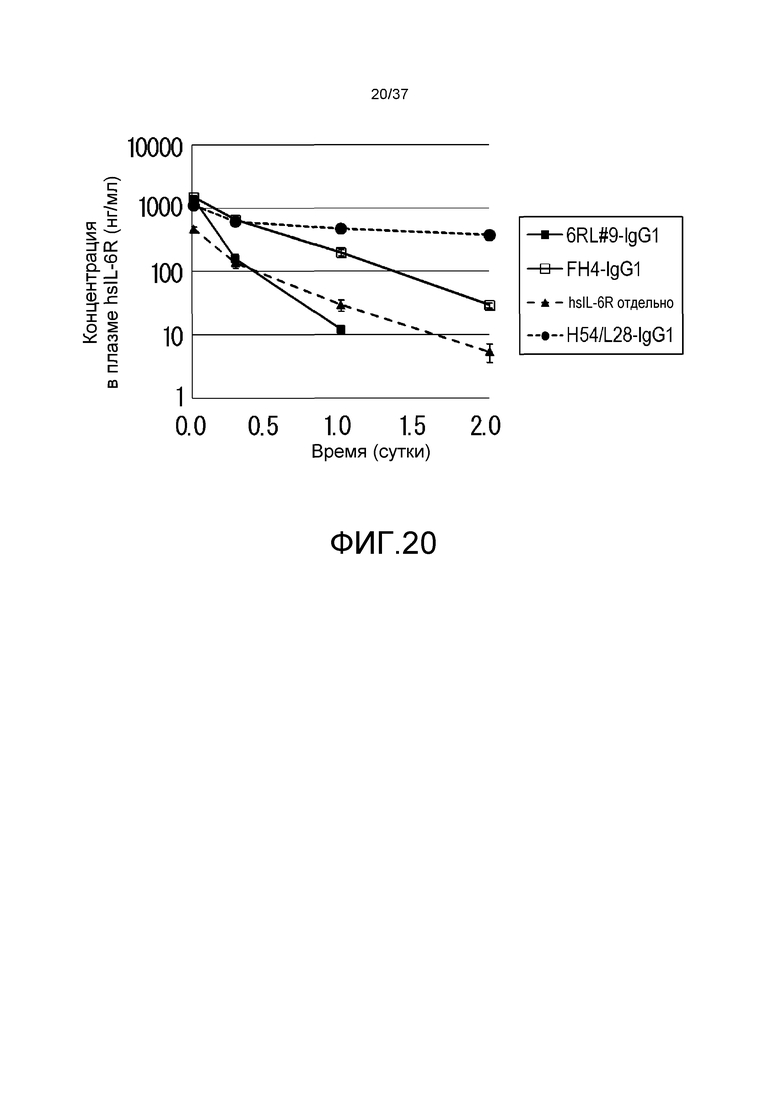
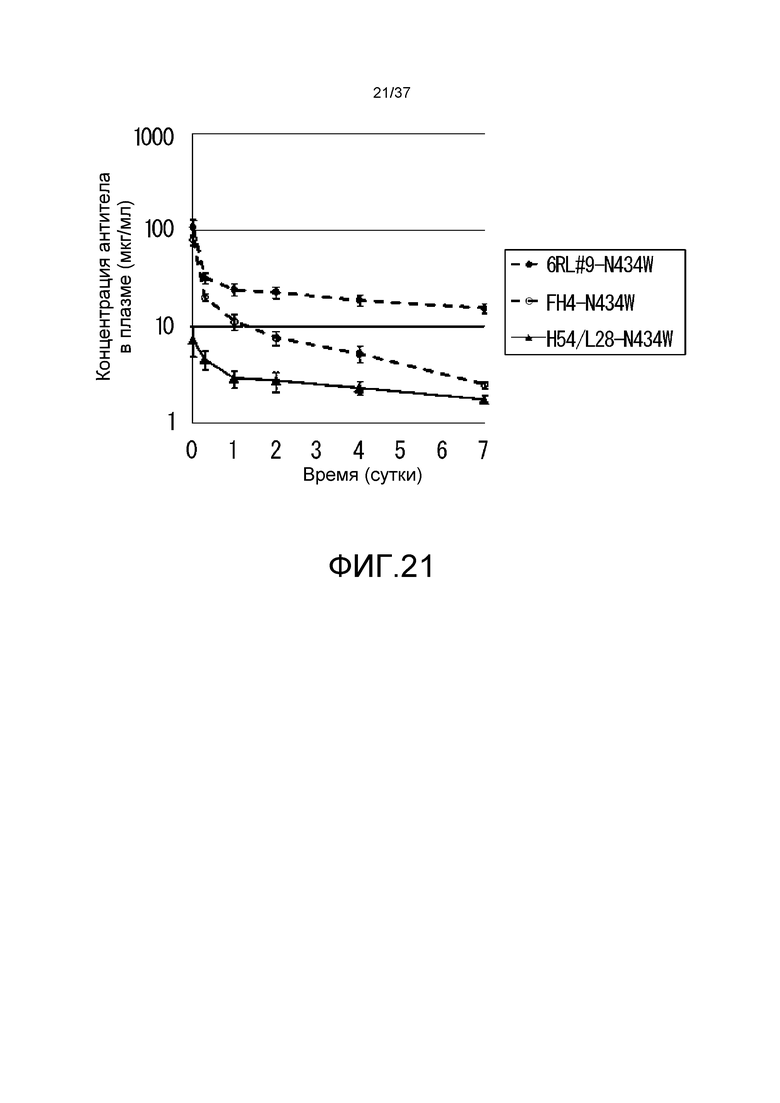
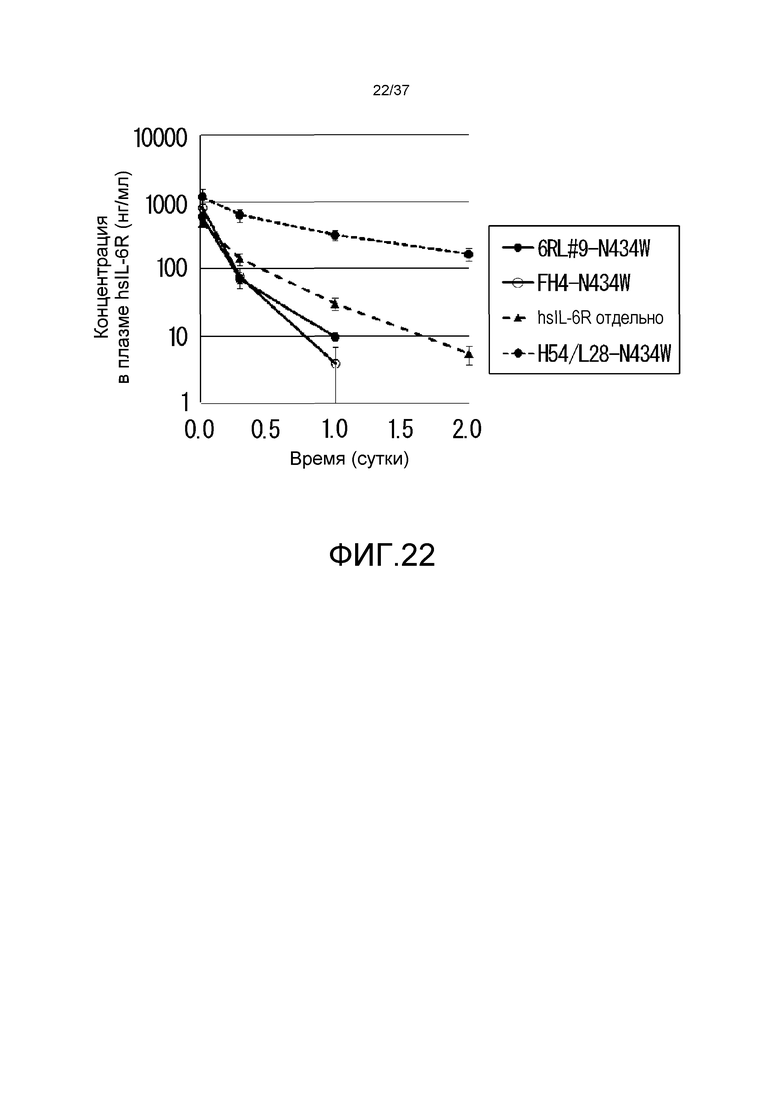
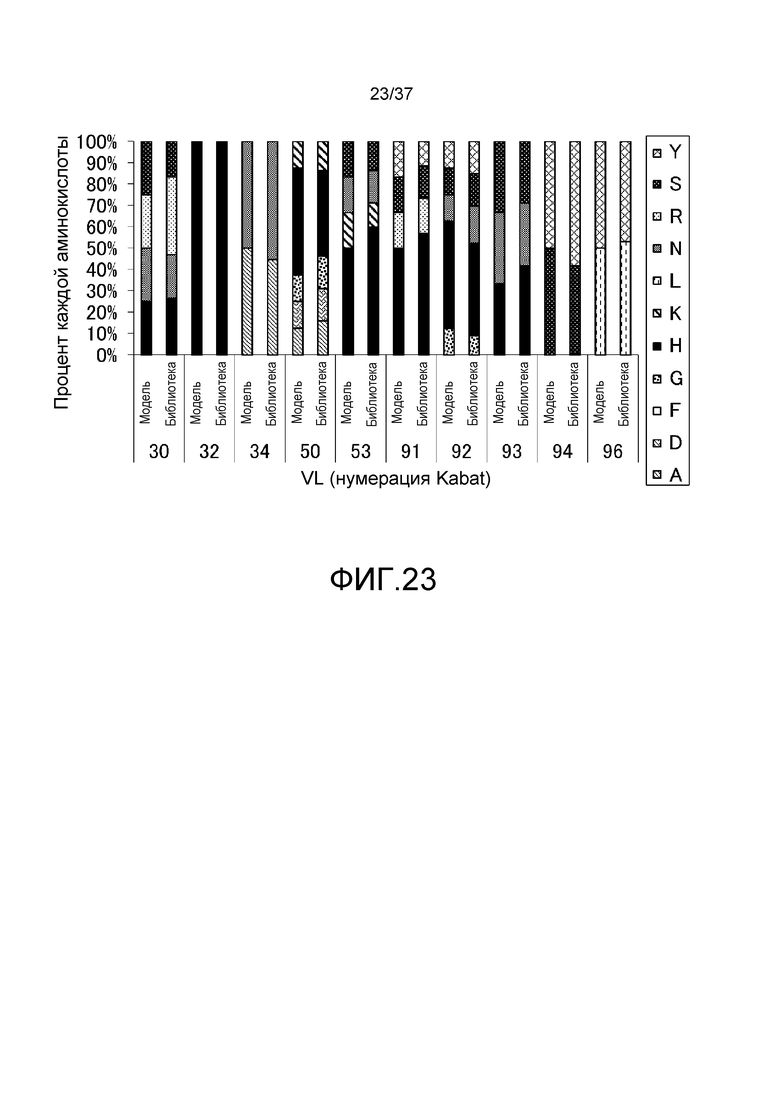
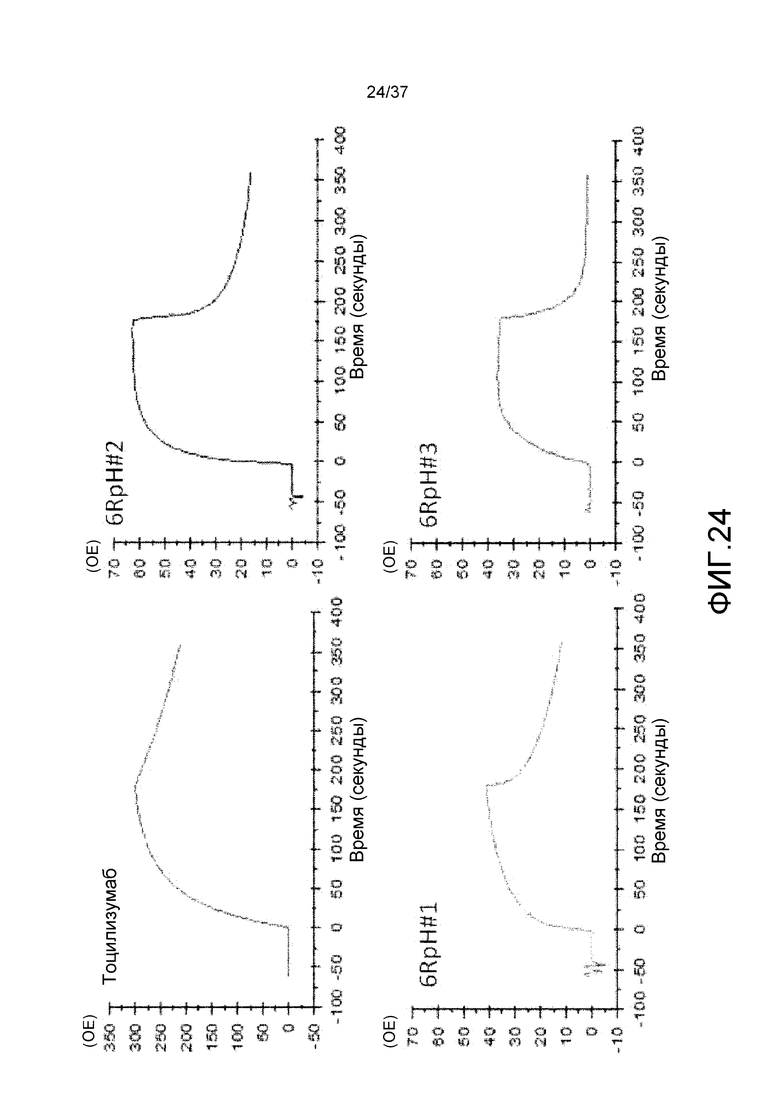
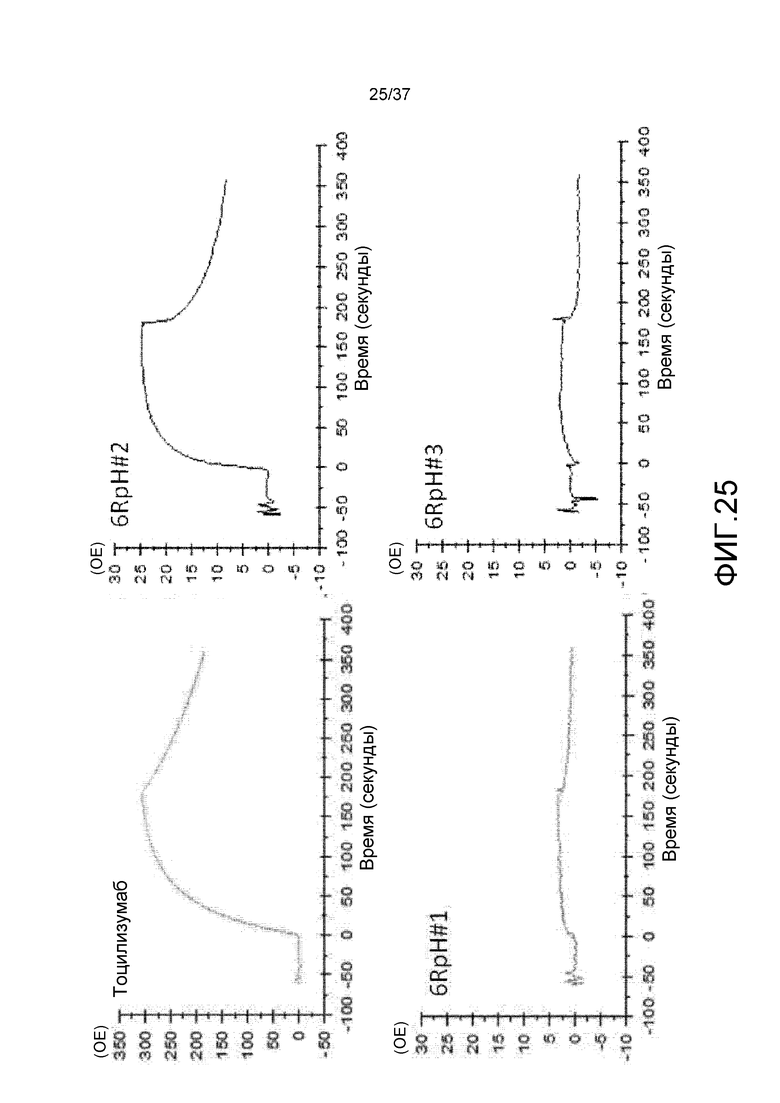
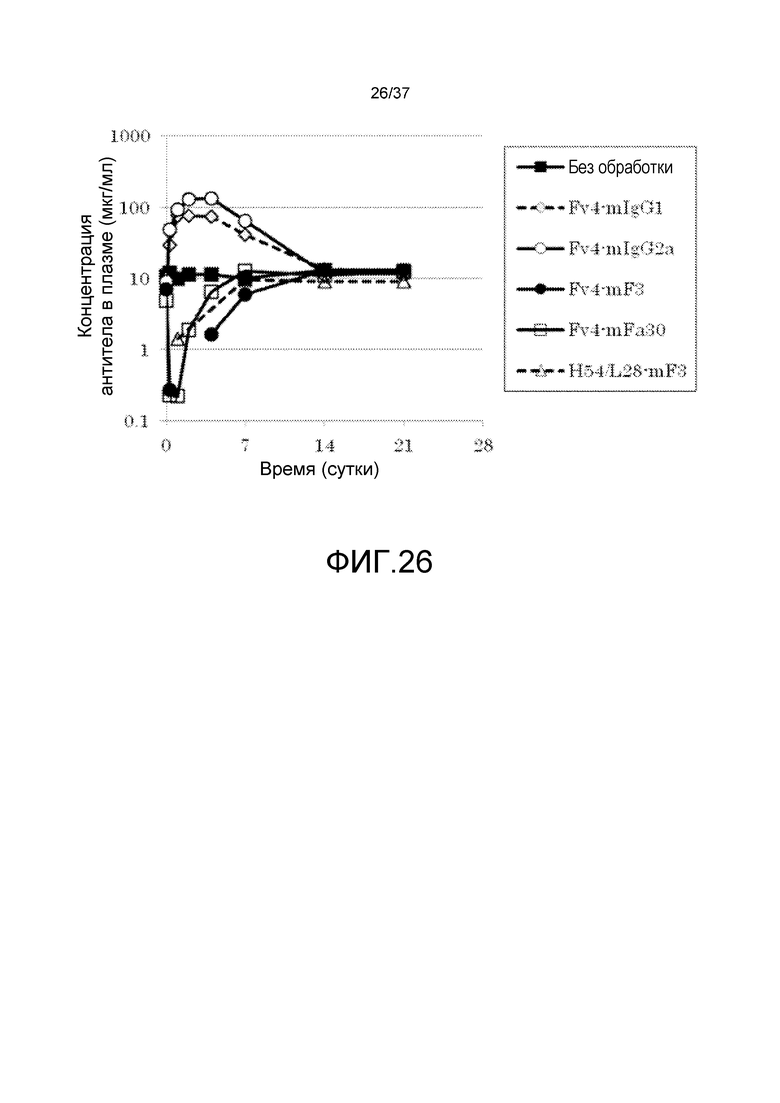
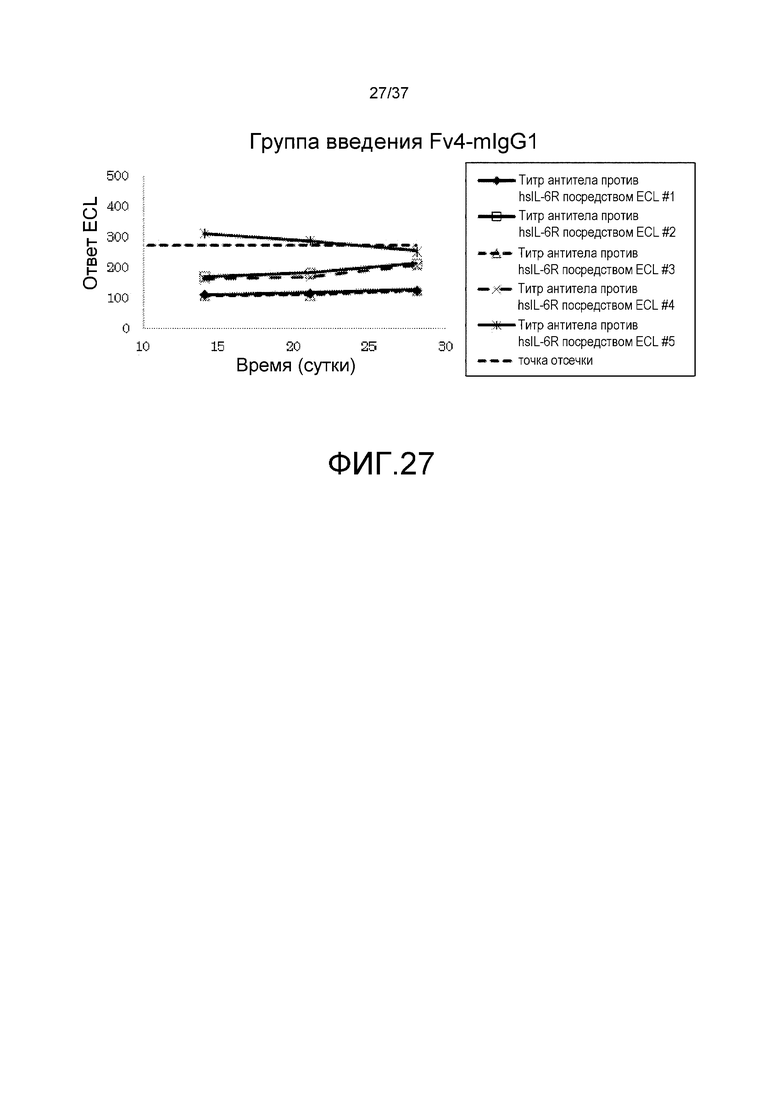
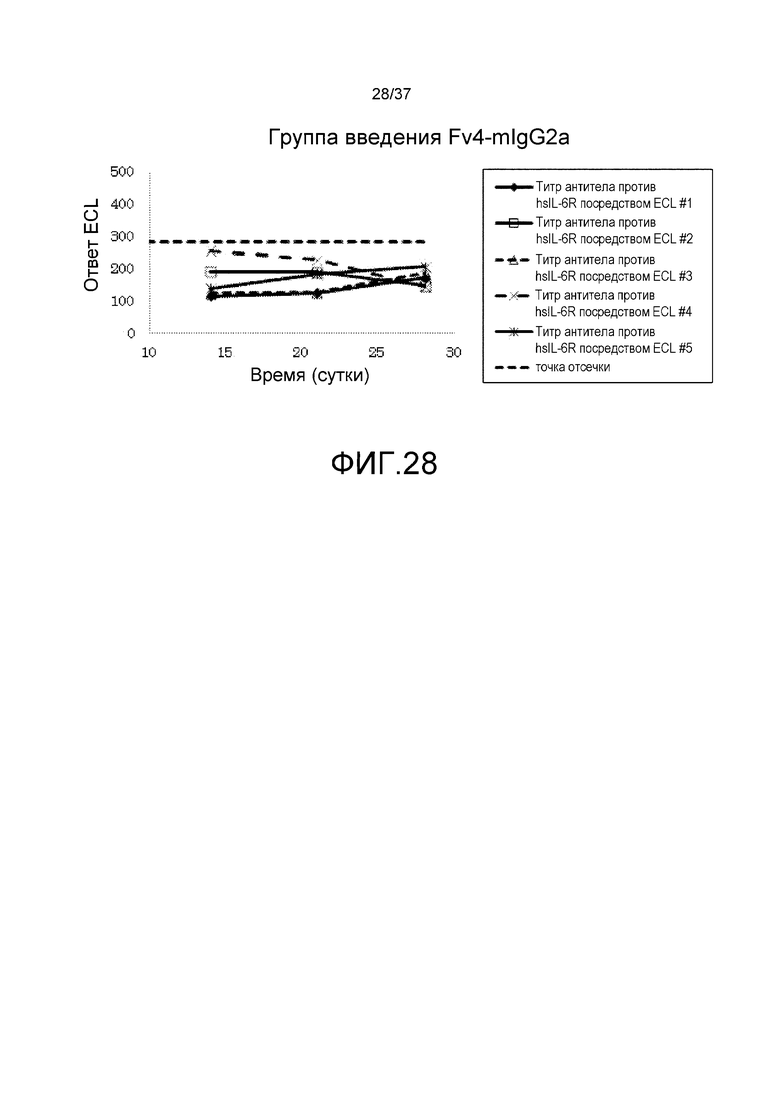
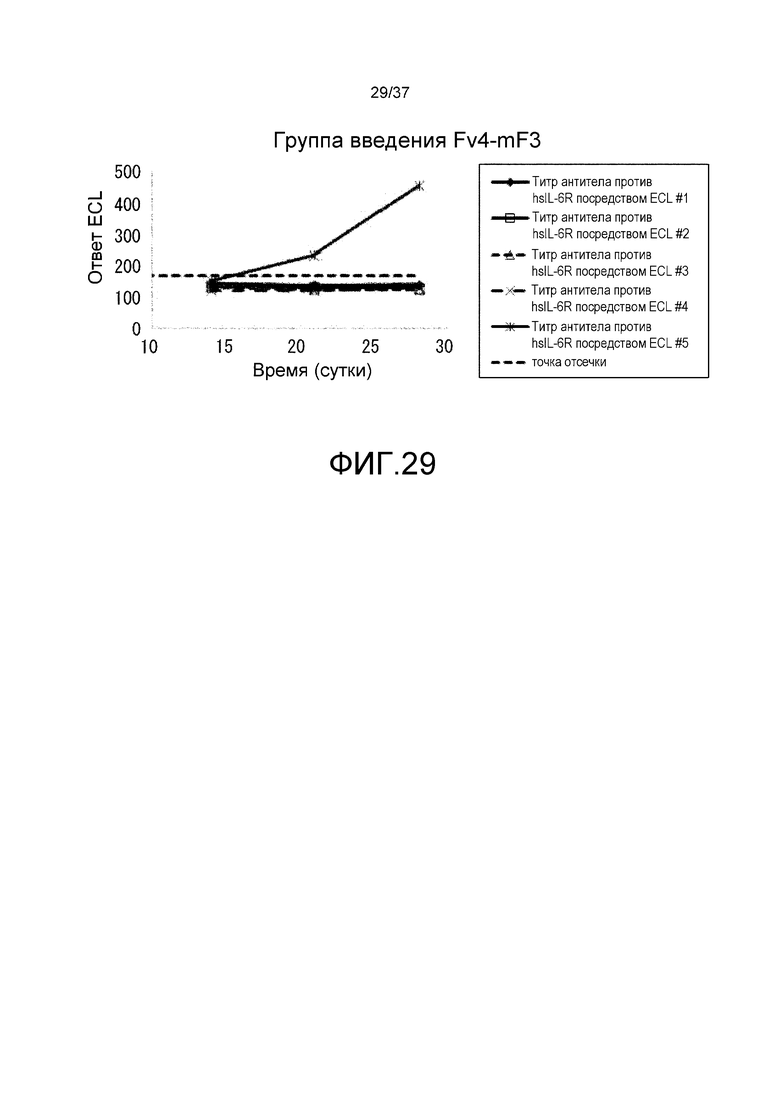
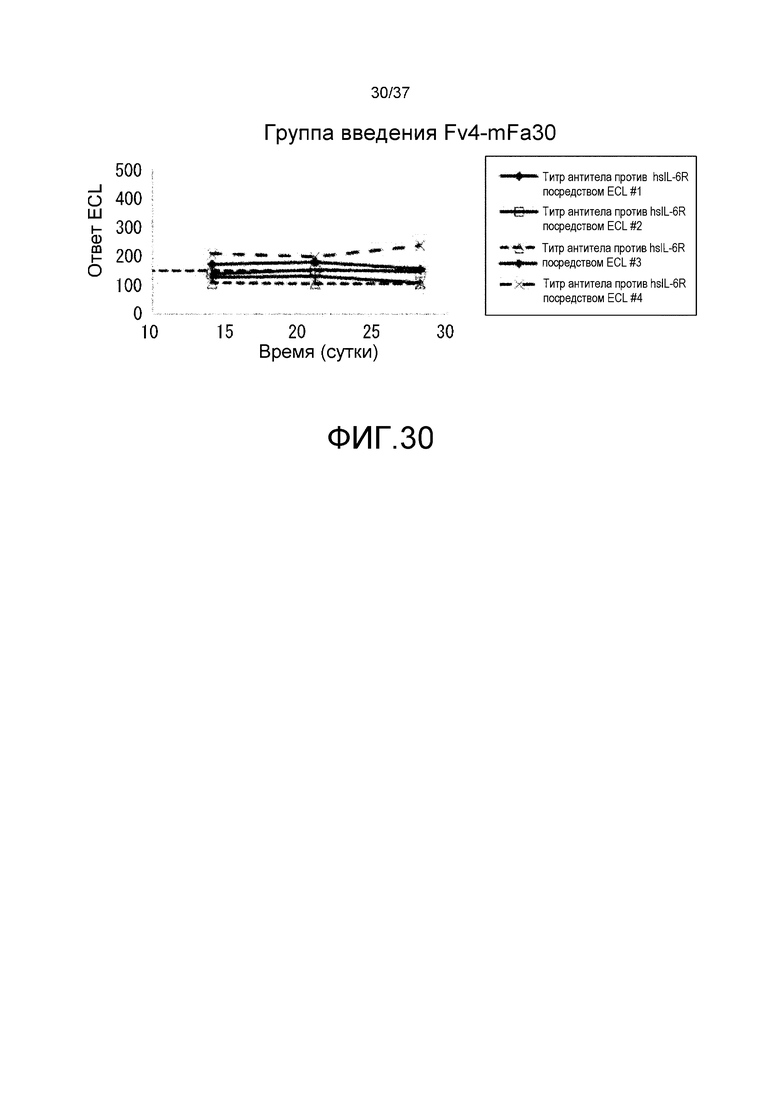
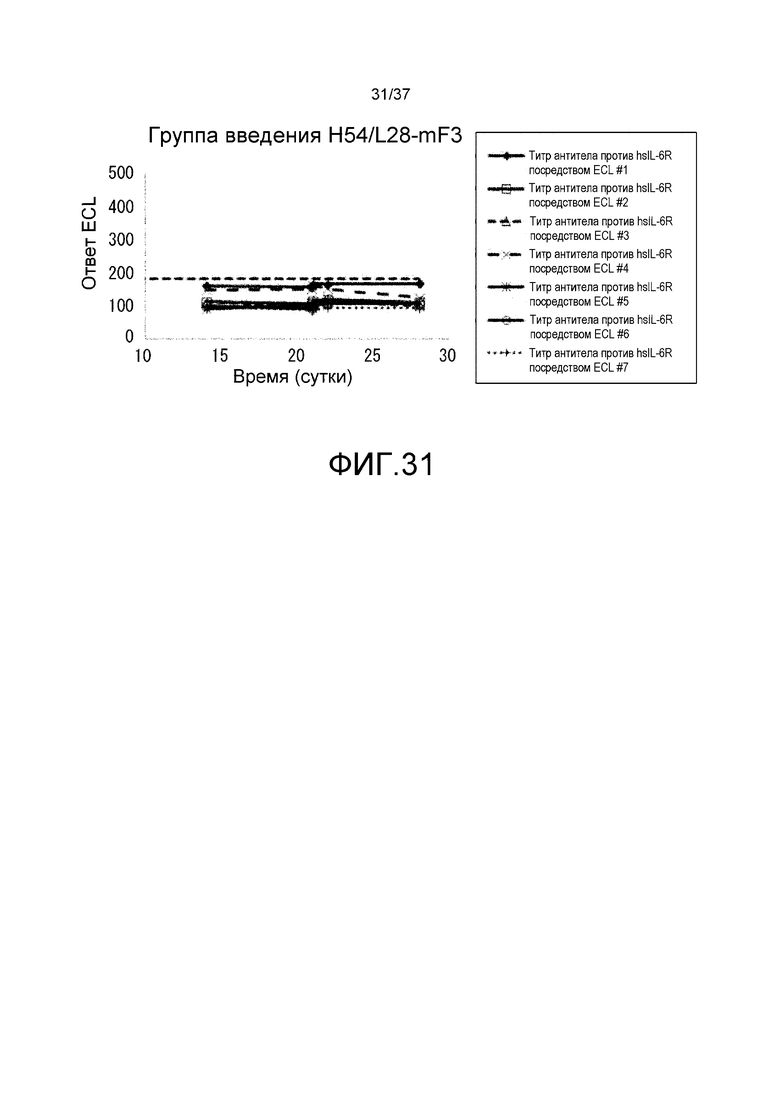
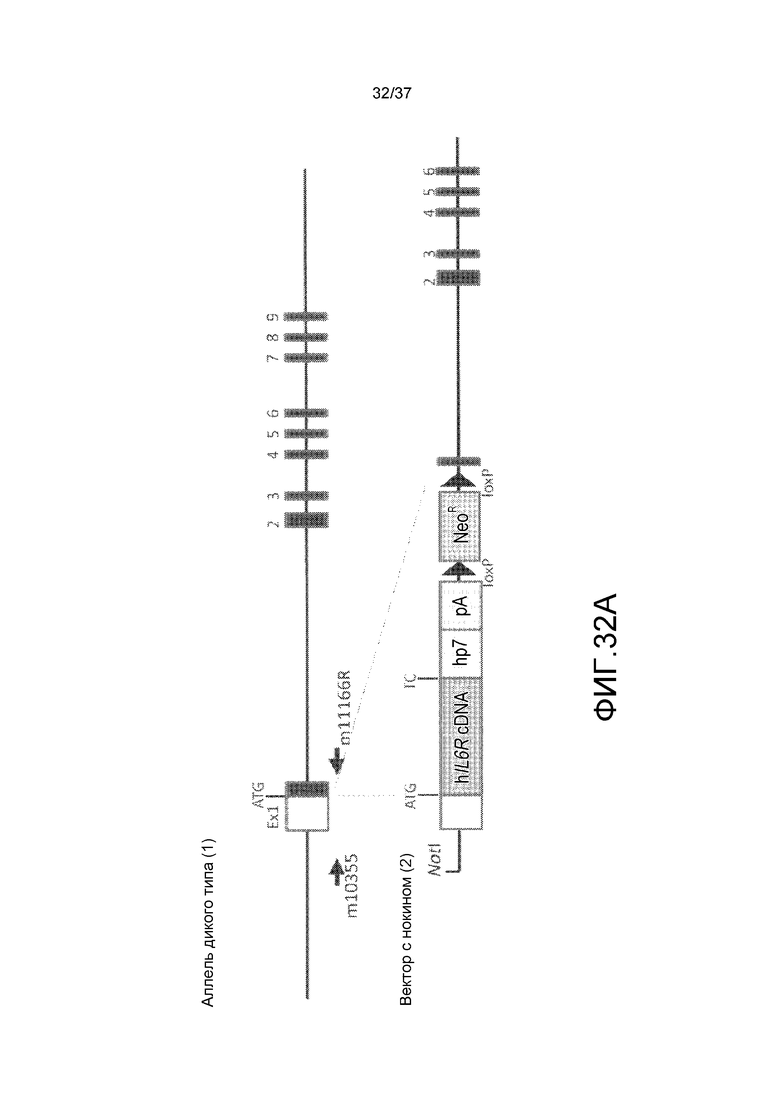
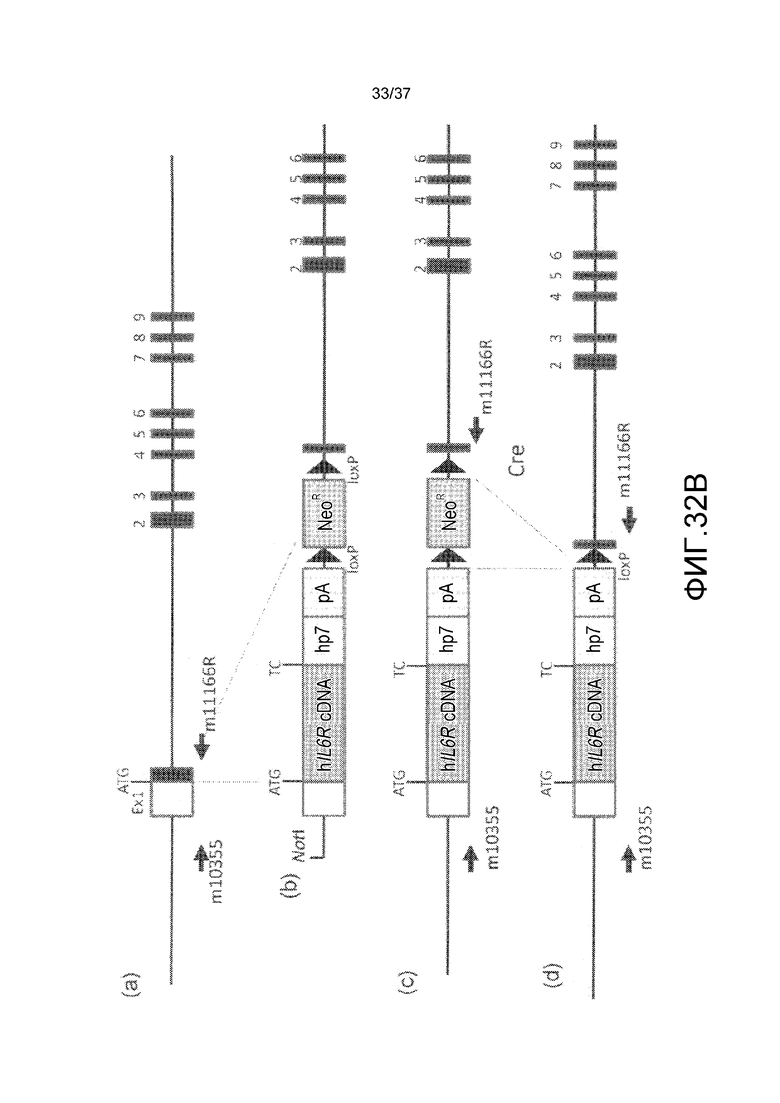
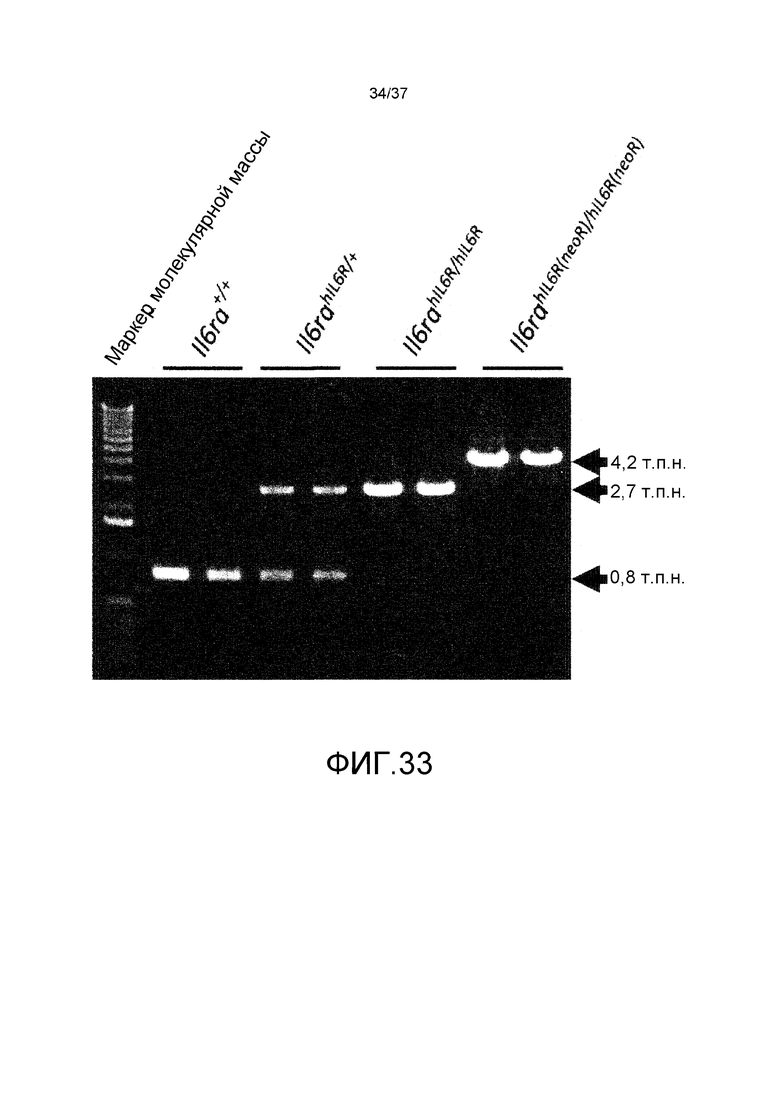
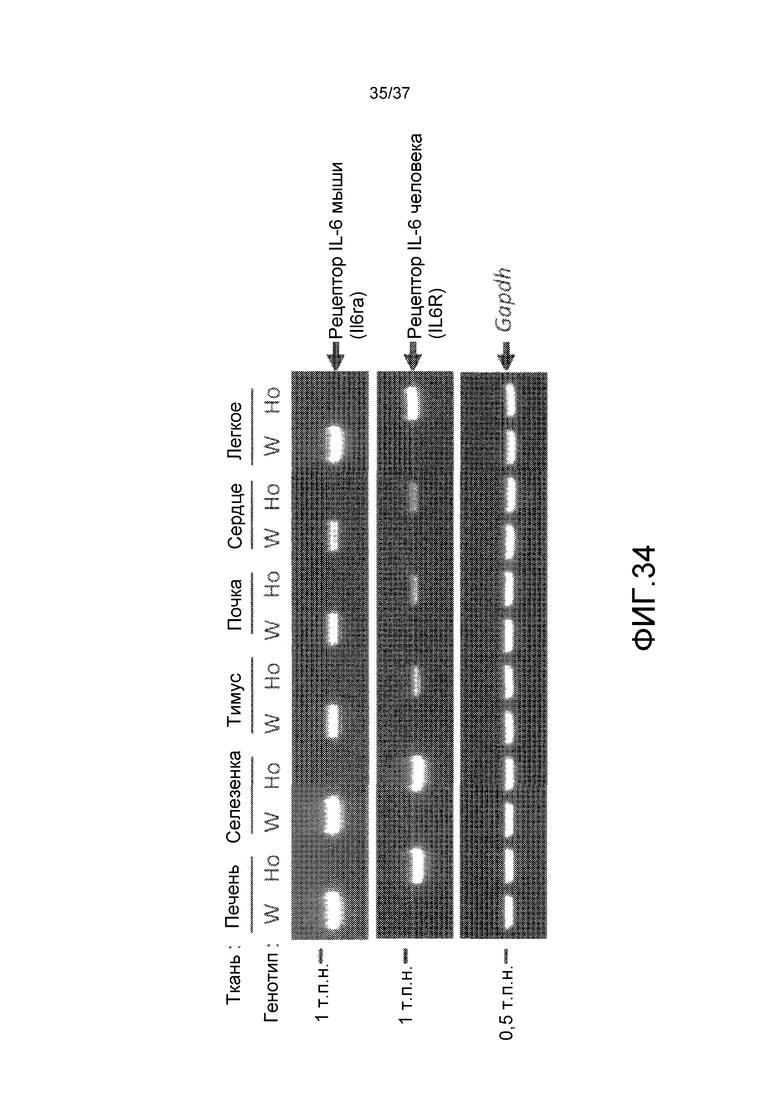
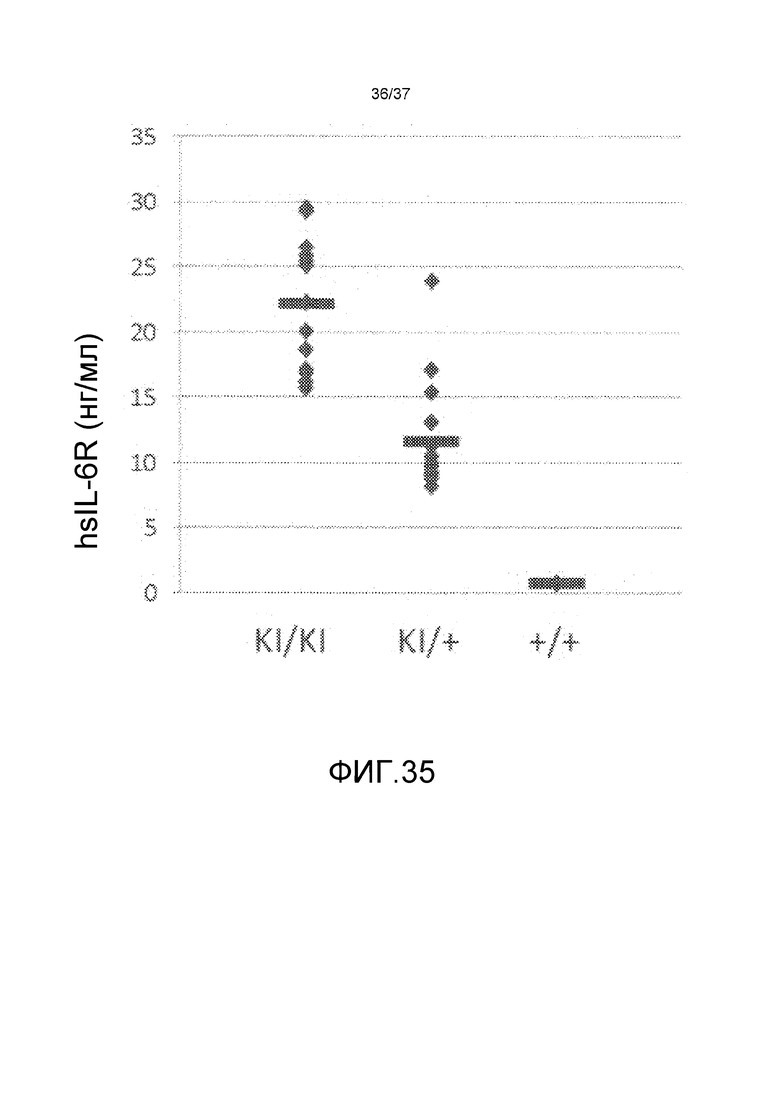
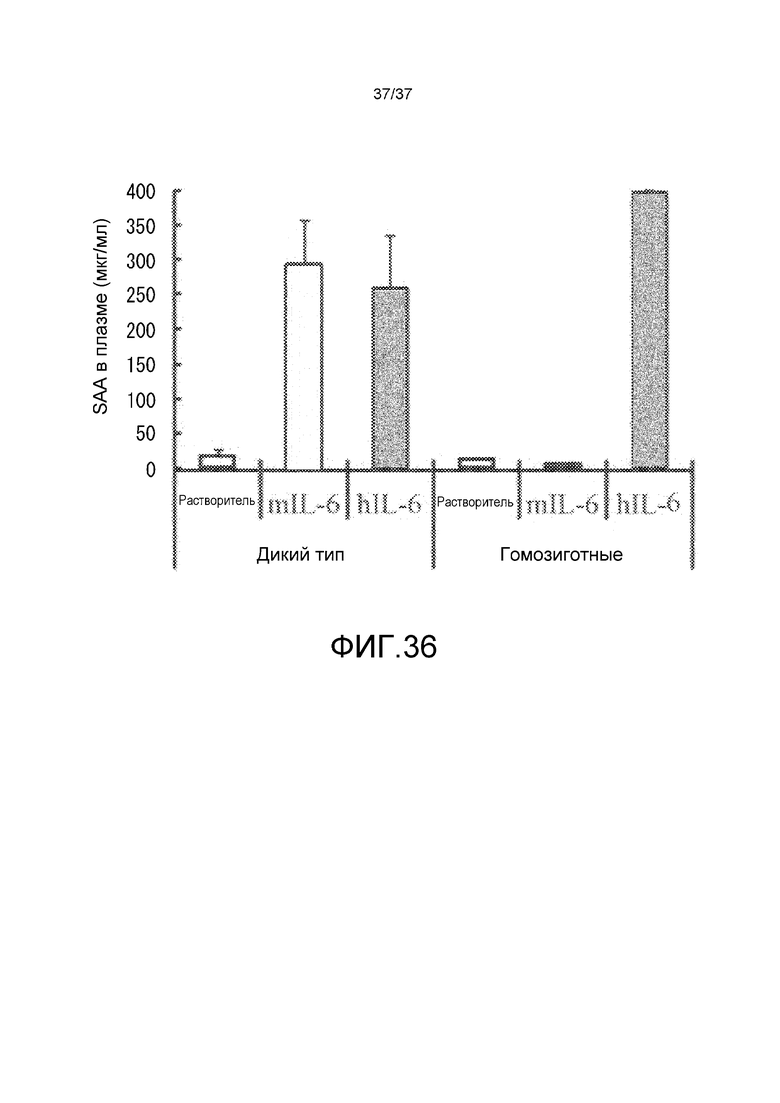
Изобретение относится к области биохимии, в частности к композиции, которая индуцирует приобретенный иммунный ответ на антиген, которая содержит в качестве активного ингредиента эффективное количество антигенсвязывающей молекулы, а также к способу ее получения. Также раскрыт способ индукции приобретенного иммунного ответа к антигену в живом организме, который включает стадию введения вышеуказанной композиции в живой организм. Изобретение позволяет эффективно индуцировать приобретенный иммунный ответ на антиген в живом организме. 5 н. и 34 з.п. ф-лы, 36 ил., 29 табл., 18 пр.
1. Композиция, которая индуцирует приобретенный иммунный ответ на антиген, которая содержит в качестве активного ингредиента эффективное количество антигенсвязывающей молекулы, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен, обладающий FcRn-связывающей активностью в диапазоне нейтральных значений рН, и где антигенсвязывающий домен выбран из (1) или (2), представленных ниже:
1) антигенсвязывающий домен, антигенсвязывающая активность которого против указанного антигена выше в условиях высокой концентрации ионов кальция, чем в условиях низкой концентрации ионов кальция, причем антигенсвязывающий домен содержит вариабельную область антитела, в которой по меньшей мере одна или более аминокислот обладают металл-хелатирующей активностью;
2) антигенсвязывающий домен, антигенсвязывающая активность которого против указанного антигена выше в диапазоне нейтральных значений рН, чем в диапазоне кислых значений рН, причем антигенсвязывающий домен содержит вариабельную область антитела, в которой по меньшей мере одна или более аминокислот являются His;
где указанная высокая концентрация ионов кальция представляет собой концентрацию между 100 мкМ и 10 мМ; где указанная низкая концентрация ионов кальция представляет собой концентрацию между 0,1 мкМ и 30 мкМ; где указанный диапазон нейтральных значений рН находится между рН 7,0 и рН 8,0 и где указанный диапазон кислых значений рН находится между рН 5,5 и рН 6,5,
где FcRn-связывающий домен содержит Fc-область антитела, и где Fc-область получена из Fc-области IgG и имеет FcRn-связывающую активность в диапазоне нейтральных значений рН больше, чем FcRn-связывающая активность интактного человеческого IgG, и где Fc-область является Fc-областью, в которой по меньшей мере одна или более аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области IgG человека.
2. Композиция по п. 1, где антигенсвязывающая молекула обладает нейтрализующей активностью в отношении антигена.
3. Композиция по любому из пп. 1, 2, где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген.
4. Композиция по п. 1, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области.
5. Композиция по п. 1, где активность Fc-области в отношении связывания Fcγ-рецептора является более высокой, чем у встречающейся в природе Fc-области IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
6. Композиция по п. 5, где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) или FcγRIIIa(F).
7. Композиция по п. 5 или 6, где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области.
8. Композиция по п. 5 или 6, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
либо Lys, либо Tyr в положении аминокислоты 221;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 222;
любой из Phe, Trp, Glu и Lys в положении аминокислоты 223;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 224;
любой из Glu, Lys и Trp в положении аминокислоты 225;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 227;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 228;
любой из Ala, Glu, Gly и Tyr в положении аминокислоты 230;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 231;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 232;
любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 233;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 234;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 235;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 236;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 237;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 238;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 239;
любой из Ala, Ile, Met и Thr в положении аминокислоты 240;
любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении аминокислоты 241;
любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении аминокислоты 243;
His в положении аминокислоты 244;
Ala в положении аминокислоты 245;
любой из Asp, Glu, His и Tyr в положении аминокислоты 246;
любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении аминокислоты 247;
любой из Glu, His, Gln и Tyr в положении аминокислоты 249;
либо Glu, либо Gln в положении аминокислоты 250;
Phe в положении аминокислоты 251;
любой из Phe, Met и Tyr в положении аминокислоты 254;
любой из Glu, Leu и Tyr в положении аминокислоты 255;
любой из Ala, Met и Pro в положении аминокислоты 256;
любой из Asp, Glu, His, Ser и Tyr в положении аминокислоты 258;
любой из Asp, Glu, His и Tyr в положении аминокислоты 260;
любой из Ala, Glu, Phe, Ile и Thr в положении аминокислоты 262;
любой из Ala, Ile, Met и Thr в положении аминокислоты 263;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 264;
любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении аминокислоты 265;
любой из Ala, Ile, Met и Thr в положении аминокислоты 266;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 267;
любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении аминокислоты 268;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 269;
любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 270;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 271;
любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 272;
либо Phe, либо Ile в положении аминокислоты 273;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 274;
либо Leu, либо Trp в положении аминокислоты 275;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 276;
любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 278;
Ala в положении аминокислоты 279;
любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении аминокислоты 280;
любой из Asp, Lys, Pro и Tyr в положении аминокислоты 281;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 282;
любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении аминокислоты 283;
любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении аминокислоты 284;
любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении аминокислоты 285;
любой из Glu, Gly, Pro и Tyr в положении аминокислоты 286;
любой из Asn, Asp, Glu и Tyr в положении аминокислоты 288;
любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении аминокислоты 290;
любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении аминокислоты 291;
любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении аминокислоты 292;
любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 293;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 294;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 295;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении аминокислоты 296;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 297;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 298;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 299;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 300;
любой из Asp, Glu, His и Tyr в положении аминокислоты 301;
Ile в положении аминокислоты 302;
любой из Asp, Gly и Tyr в положении аминокислоты 303;
любой из Asp, His, Leu, Asn и Thr в положении аминокислоты 304;
любой из Glu, Ile, Thr и Tyr в положении аминокислоты 305;
любой из Ala, Asp, Asn, Thr, Val и Tyr в положении аминокислоты 311;
Phe в положении аминокислоты 313;
Leu в положении аминокислоты 315;
либо Glu, либо Gln в положении аминокислоты 317;
любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении аминокислоты 318;
любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 320;
любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 322;
Ile в положении аминокислоты 323;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 324;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 325;
любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 326;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 327;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 328;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 329;
любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 330;
любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 331;
любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 332;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении аминокислоты 333;
любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении аминокислоты 334;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 335;
любой из Glu, Lys и Tyr в положении аминокислоты 336;
любой из Glu, His и Asn в положении аминокислоты 337;
любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении аминокислоты 339;
либо Ala, либо Val в положении аминокислоты 376;
либо Gly, либо Lys в положении аминокислоты 377;
Asp в положении аминокислоты 378;
Asn в положении аминокислоты 379;
любой из Ala, Asn и Ser в положении аминокислоты 380;
либо Ala, либо Ile в положении аминокислоты 382;
Glu в положении аминокислоты 385;
Thr в положении аминокислоты 392;
Leu в положении аминокислоты 396;
Lys в положении аминокислоты 421;
Asn в положении аминокислоты 427;
либо Phe, либо Leu в положении аминокислоты 428;
Met в положении аминокислоты 429;
Trp в положении аминокислоты 434;
Ile в положении аминокислоты 436; и
любой из Gly, His, Ile, Leu и Tyr в положении аминокислоты 440; в соответствии с нумерацией EU в Fc-области.
9. Композиция по п. 5 или 6, где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой содержащую фукозу цепь сахаров.
10. Композиция по п. 5 или 6, где Fc-область модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким.
11. Способ индукции приобретенного иммунного ответа к антигену в живом организме, который включает стадию введения композиции по любому из пп. 1-10 в живой организм.
12. Способ получения композиции, которая индуцирует приобретенный иммунный ответ к антигену и содержит в качестве активного ингридиента в эффективном количестве антигенсвязывающую молекулу,
где антигенсвязывающая молекула включает антигенсвязывающий домен, антигенсвязывающая активность которого зависит от условия - концентрации ионов, и FcRn-связывающий домен, содержащий Fc-область антитела;
в котором способ включает сообщение FcRn-связывающей активности в диапазоне нейтральных значений рН Fc-области антитела, содержащейся в FcRn-связывающем домене, который содержится в антигенсвязывающей молекуле, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, с помощью замены по меньшей мере одной или более аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области,
где антигенсвязывающий домен выбран из (1) или (2), представленных ниже:
1) антигенсвязывающий домен, антигенсвязывающая активность которого к указанному антигену выше в условиях высокой концентрации ионов кальция, чем в условиях низкой концентрации ионов кальция и антигенсвязывающий домен содержит тяжелую цепь вариабельной области антитела и легкую цепь вариабельной области антитела, в которых по меньшей мере одна или более аминокислот обладают металл-хелатирующей активностью;
2) антигенсвязывающий домен, антигенсвязывающая активность которого к указанному антигену выше в диапазоне нейтральных значений рН, чем в диапазоне кислых значений рН и антигенсвязывающий домен содержит тяжелую цепь вариабельной области антитела и легкую цепь вариабельной области антитела, в которых по меньшей мере одна или более аминокислот являются His;
где указанная высокая концентрация ионов кальция представляет собой концентрацию между 100 мкМ и 10 мМ; где указанная низкая концентрация ионов кальция представляет собой концентрацию между 0,1 мкМ и 30 мкМ; где указанный диапазон нейтральных значений рН находится между рН 7,0 и рН 8,0 и где указанный диапазон кислых значений рН находится между рН 5,5 и рН 6,5.
13. Способ по п. 12, где антигенсвязывающая молекула обладает нейтрализующей активностью в отношении антигена.
14. Способ по п. 12, где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген.
15. Способ по любому из пп. 12-14, где заменой(ми) аминокислот(ы) в Fc-области являются по меньшей мере одна или несколько аминокислотных замен, выбранных из группы, состоящей из:
аминокислотная замена на Ala в положении 257;
аминокислотная замена на Pro в положении 308;
аминокислотная замена на Leu в положении 428; и
аминокислотная замена на Tyr в положении 434,
в соответствии с нумерацией EU в Fc-области.
16. Способ по п. 12, включающий стадию повышения активности Fc-области в отношении связывания Fcγ-рецептора по сравнению со встречающейся в природе Fc-областью IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
17. Способ по п. 16, где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) или FcγRIIIa(F).
18. Способ по п. 16 или 17, включающий стадию замены по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU в Fc-области.
19. Способ по п. 16 или 17, включающий стадию проведения по меньшей мере одной или нескольких замен аминокислот, выбранных из группы, состоящей из:
аминокислотная замена либо на Lys, либо на Tyr в положении 221;
аминокислотная замена на любой из Phe, Trp, Glu и Tyr в положении 222;
аминокислотная замена на любой из Phe, Trp, Glu и Lys в положении 223;
аминокислотная замена на любой из Phe, Trp, Glu и Tyr в положении 224;
аминокислотная замена на любой из Glu, Lys и Trp в положении 225;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 227;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 228;
аминокислотная замена на любой из Ala, Glu, Gly и Tyr в положении 230;
аминокислотная замена на любой из Glu, Gly, Lys, Pro и Tyr в положении 231;
аминокислотная замена на любой из Glu, Gly, Lys и Tyr в положении 232;
аминокислотная замена на любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 233;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 234;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 235;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 236;
аминокислотная замена на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 237;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 238;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении 239;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 240;
аминокислотная замена на любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении 241;
аминокислотная замена на любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении 243;
аминокислотная замена на His в положении 244;
аминокислотная замена на Ala в положении 245;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 246;
аминокислотная замена на любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении 247;
аминокислотная замена на любой из Glu, His, Gln и Tyr в положении 249;
аминокислотная замена либо на Glu, либо на Gln в положении 250;
аминокислотная замена на Phe в положении 251;
аминокислотная замена на любой из Phe, Met и Tyr в положении 254;
аминокислотная замена на любой из Glu, Leu и Tyr в положении 255;
аминокислотная замена на любой из Ala, Met и Pro в положении 256;
аминокислотная замена на любой из Asp, Glu, His, Ser и Tyr в положении 258;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 260;
аминокислотная замена на любой из Ala, Glu, Phe, Ile и Thr в положении 262;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 263;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении 264;
аминокислотная замена на любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении 265;
аминокислотная замена на любой из Ala, Ile, Met и Thr в положении 266;
аминокислотная замена на любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении 267;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении 268;
аминокислотная замена на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 269;
аминокислотная замена на любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении 270;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 271;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 272;
аминокислотная замена либо на Phe, либо на Ile в положении 273;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 274;
аминокислотная замена либо на Leu, либо на Trp в положении 275;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 276;
аминокислотная замена на любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении 278;
аминокислотная замена на Ala в положении 279;
аминокислотная замена на любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении 280;
аминокислотная замена на любой из Asp, Lys, Pro и Tyr в положении 281;
аминокислотная замена на любой из Glu, Gly, Lys, Pro и Tyr в положении 282;
аминокислотная замена на любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении 283;
аминокислотная замена на любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении 284;
аминокислотная замена на любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении 285;
аминокислотная замена на любой из Glu, Gly, Pro и Tyr в положении 286;
аминокислотная замена на любой из Asn, Asp, Glu и Tyr в положении 288;
аминокислотная замена на любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении 290;
аминокислотная замена на любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении 291;
аминокислотная замена на любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении 292;
аминокислотная замена на любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 293;
аминокислотная замена на любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 294;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 295;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении 296;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 297;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении 298;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении 299;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении 300;
аминокислотная замена на любой из Asp, Glu, His и Tyr в положении 301;
аминокислотная замена на Ile в положении 302;
аминокислотная замена на любой из Asp, Gly и Tyr в положении 303;
аминокислотная замена на любой из Asp, His, Leu, Asn и Thr в положении 304;
аминокислотная замена на любой из Glu, Ile, Thr и Tyr в положении 305;
аминокислотная замена на любой из Ala, Asp, Asn, Thr, Val и Tyr в положении 311;
аминокислотная замена на Phe в положении 313;
аминокислотная замена на Leu в положении 315;
аминокислотная замена либо на Glu, либо на Gln в положении 317;
аминокислотная замена на любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении 318;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении 320;
аминокислотная замена на любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении 322;
аминокислотная замена на Ile в положении 323;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении 324;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 325;
аминокислотная замена на любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении 326;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении 327;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 328;
аминокислотная замена на любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 329;
аминокислотная замена на любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении 330;
аминокислотная замена на любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении 331;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении 332;
аминокислотная замена на любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении 333;
аминокислотная замена на любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении 334;
аминокислотная замена на любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении 335;
аминокислотная замена на любой из Glu, Lys и Tyr в положении 336;
аминокислотная замена на любой из Glu, His и Asn в положении 337;
аминокислотная замена на любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении 339;
аминокислотная замена либо на Ala, либо на Val в положении 376;
аминокислотная замена либо на Gly, либо на Lys в положении 377;
аминокислотная замена на Asp в положении 378;
аминокислотная замена на Asn в положении 379;
аминокислотная замена на любой из Ala, Asn и Ser в положении 380;
аминокислотная замена либо на Ala, либо на Ile в положении 382;
аминокислотная замена на Glu в положении 385;
аминокислотная замена на Thr в положении 392;
аминокислотная замена на Leu в положении 396;
аминокислотная замена на Lys в положении 421;
аминокислотная замена на Asn в положении 427;
аминокислотная замена либо на Phe, либо на Leu в положении 428;
аминокислотная замена на Met в положении 429;
аминокислотная замена на Trp в положении 434;
аминокислотная замена на Ile в положении 436; и
аминокислотная замена на любой из Gly, His, Ile, Leu и Tyr в положении 440, в соответствии с нумерацией EU в Fc-области.
20. Способ по п. 16 или 17, где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
21. Способ по п. 16 или 17, включающий стадию модификации Fc-области так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким.
22. Способ получения композиции, которая индуцирует приобретенный иммунный ответ к антигену и включает в качестве активного ингредиента в эффективном количестве антигенсвязывающую молекулу, где способ включает стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в условиях высокой концентрации ионов кальция;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в условиях низкой концентрации ионов кальция;
(c) выбор антигенсвязывающего домена, антигенсвязывающая активность которого, определенная согласно (а), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, выбранный согласно (с), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий FcRn-связывающей активностью в диапазоне нейтральных значений рН;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (е),
где указанная высокая концентрация ионов кальция представляет собой концентрацию между 100 мкМ и 10 мМ; где указанная низкая концентрация ионов кальция представляет собой концентрацию между 0,1 мкМ и 30 мкМ; где указанный диапазон нейтральных значений рН находится между рН 7,0 и рН 8,0;
где FcRn-связывающий домен содержит Fc-область антитела, и где Fc-область получена из Fc-области IgG и имеет FcRn-связывающую активность в диапазоне нейтральных значений рН больше, чем FcRn-связывающая активность интактного человеческого IgG, и где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области IgG.
23. Способ по п. 22, где антигенсвязывающая молекула обладает нейтрализующей активностью против антигена.
24. Способ по п. 22, где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген.
25. Способ по любому из п. 22-24, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области.
26. Способ по п. 22, где активность Fc-области в отношении связывания Fcγ-рецептора является более высокой, чем у встречающейся в природе Fc-области IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
27. Способ по п. 26, где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) или FcγRIIIa(F).
28. Способ по п. 26 или 27, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
либо Lys, либо Tyr в положении аминокислоты 221;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 222;
любой из Phe, Trp, Glu и Lys в положении аминокислоты 223;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 224;
любой из Glu, Lys и Trp в положении аминокислоты 225;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 227;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 228;
любой из Ala, Glu, Gly и Tyr в положении аминокислоты 230;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 231;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 232;
любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 233;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 234;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 235;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 236;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 237;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 238;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 239;
любой из Ala, Ile, Met и Thr в положении аминокислоты 240;
любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении аминокислоты 241;
любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении аминокислоты 243;
His в положении аминокислоты 244;
Ala в положении аминокислоты 245;
любой из Asp, Glu, His и Tyr в положении аминокислоты 246;
любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении аминокислоты 247;
любой из Glu, His, Gln и Tyr в положении аминокислоты 249;
либо Glu, либо Gln в положении аминокислоты 250;
Phe в положении аминокислоты 251;
любой из Phe, Met и Tyr в положении аминокислоты 254;
любой из Glu, Leu и Tyr в положении аминокислоты 255;
любой из Ala, Met и Pro в положении аминокислоты 256;
любой из Asp, Glu, His, Ser и Tyr в положении аминокислоты 258;
любой из Asp, Glu, His и Tyr в положении аминокислоты 260;
любой из Ala, Glu, Phe, Ile и Thr в положении аминокислоты 262;
любой из Ala, Ile, Met и Thr в положении аминокислоты 263;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 264;
любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении аминокислоты 265;
любой из Ala, Ile, Met и Thr в положении аминокислоты 266;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 267;
любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении аминокислоты 268;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 269;
любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 270;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 271;
любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 272;
либо Phe, либо Ile в положении аминокислоты 273;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 274;
либо Leu, либо Trp в положении аминокислоты 275;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 276;
любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 278;
Ala в положении аминокислоты 279;
любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении аминокислоты 280;
любой из Asp, Lys, Pro и Tyr в положении аминокислоты 281;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 282;
любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении аминокислоты 283;
любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении аминокислоты 284;
любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении аминокислоты 285;
любой из Glu, Gly, Pro и Tyr в положении аминокислоты 286;
любой из Asn, Asp, Glu и Tyr в положении аминокислоты 288;
любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении аминокислоты 290;
любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении аминокислоты 291;
любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении аминокислоты 292;
любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 293;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 294;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 295;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении аминокислоты 296;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 297;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 298;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 299;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 300;
любой из Asp, Glu, His и Tyr в положении аминокислоты 301;
Ile в положении аминокислоты 302;
любой из Asp, Gly и Tyr в положении аминокислоты 303;
любой из Asp, His, Leu, Asn и Thr в положении аминокислоты 304;
любой из Glu, Ile, Thr и Tyr в положении аминокислоты 305;
любой из Ala, Asp, Asn, Thr, Val и Tyr в положении аминокислоты 311;
Phe в положении аминокислоты 313;
Leu в положении аминокислоты 315;
либо Glu, либо Gln в положении аминокислоты 317;
любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении аминокислоты 318;
любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 320;
любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 322;
Ile в положении аминокислоты 323;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 324;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 325;
любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 326;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 327;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 328;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 329;
любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 330;
любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 331;
любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 332;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении аминокислоты 333;
любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении аминокислоты 334;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 335;
любой из Glu, Lys и Tyr в положении аминокислоты 336;
любой из Glu, His и Asn в положении аминокислоты 337;
любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении аминокислоты 339;
либо Ala, либо Val в положении аминокислоты 376;
либо Gly, либо Lys в положении аминокислоты 377;
Asp в положении аминокислоты 378;
Asn в положении аминокислоты 379;
любой из Ala, Asn и Ser в положении аминокислоты 380;
либо Ala, либо Ile в положении аминокислоты 382;
Glu в положении аминокислоты 385;
Thr в положении аминокислоты 392;
Leu в положении аминокислоты 396;
Lys в положении аминокислоты 421;
Asn в положении аминокислоты 427;
либо Phe, либо Leu в положении аминокислоты 428;
Met в положении аминокислоты 429;
Trp в положении аминокислоты 434;
Ile в положении аминокислоты 436; и
любой из Gly, His, Ile, Leu и Tyr в положении аминокислоты 440; в соответствии с нумерацией EU в Fc-области.
29. Способ по п. 26 или 27, где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
30. Способ по п. 26 или 27, где Fc-область модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким.
31. Способ получения композиции, которая вызывает приобретенный иммунный ответ к антигену и содержит в качестве активного ингридиента в эффективном количестве антигенсвязывающую молекулу, где способ включает стадии:
(a) определение антигенсвязывающей активности антитела в диапазоне нейтральных значений рН;
(b) определение антигенсвязывающей активности антитела в диапазоне кислых значений рН;
(c) выбор антитела, антигенсвязывающая активность которого, определенная согласно (а), является более высокой, чем антигенсвязывающая активность, определенная согласно (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен антитела, выбранного согласно (с), с полинуклеотидом, кодирующим FcRn-связывающий домен, обладающий FcRn-связывающей активностью в диапазоне нейтральных значений рН;
(e) культивирование клетки, в которую введен вектор, с которым функционально связан полинуклеотид, полученный согласно (d); и
(f) сбор антигенсвязывающей молекулы из культуральной жидкости клеток, культивированных согласно (е),
где указанный диапазон нейтральных значений рН находится между рН 7,0 и рН 8,0 и где указанный диапазон кислых значений рН находится между рН 5,5 и рН 6,5,
где FcRn-связывающий домен содержит Fc-область антитела, и где Fc-область получена из Fc-области IgG и имеет FcRn-связывающую активность в диапазоне нейтральных значений рН больше, чем FcRn-связывающая активность интактного человеческого IgG, и где Fc-область представляет собой Fc-область, в которой по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 257, 308, 428 и 434 в соответствии с нумерацией EU в Fc-области, отличаются от аминокислот в соответствующих положениях во встречающейся в природе Fc-области IgG.
32. Способ по п. 31, где антигенсвязывающая молекула обладает нейтрализующей активностью против антигена.
33. Способ по п. 31, где антигенсвязывающая молекула обладает цитотоксической активностью в отношении клетки, экспрессирующей антиген.
34. Способ по любому из п. 31-33, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala в положении аминокислоты 257;
Pro в положении аминокислоты 308;
Leu в положении аминокислоты 428; и
Tyr в положении аминокислоты 434,
в соответствии с нумерацией EU в Fc-области.
35. Способ по п. 31, где активность связывания Fcγ-рецептора Fc-областью выше по сравнению со встречающейся в природе Fc-областью IgG человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
36. Способ по п. 35, где Fcγ-рецептор представляет собой FcγRIa, FcγRIIa(R), FcγRIIa(H), FcγRIIb, FcγRIIIa(V) или FcγRIIIa(F).
37. Способ по п. 35 или 36, где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
либо Lys, либо Tyr в положении аминокислоты 221;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 222;
любой из Phe, Trp, Glu и Lys в положении аминокислоты 223;
любой из Phe, Trp, Glu и Tyr в положении аминокислоты 224;
любой из Glu, Lys и Trp в положении аминокислоты 225;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 227;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 228;
любой из Ala, Glu, Gly и Tyr в положении аминокислоты 230;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 231;
любой из Glu, Gly, Lys и Tyr в положении аминокислоты 232;
любой из Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 233;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 234;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 235;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 236;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 237;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 238;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 239;
любой из Ala, Ile, Met и Thr в положении аминокислоты 240;
любой из Asp, Glu, Leu, Arg, Trp и Tyr в положении аминокислоты 241;
любой из Leu, Glu, Leu, Gln, Arg, Trp и Tyr в положении аминокислоты 243;
His в положении аминокислоты 244;
Ala в положении аминокислоты 245;
любой из Asp, Glu, His и Tyr в положении аминокислоты 246;
любой из Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val и Tyr в положении аминокислоты 247;
любой из Glu, His, Gln и Tyr в положении аминокислоты 249;
либо Glu, либо Gln в положении аминокислоты 250;
Phe в положении аминокислоты 251;
любой из Phe, Met и Tyr в положении аминокислоты 254;
любой из Glu, Leu и Tyr в положении аминокислоты 255;
любой из Ala, Met и Pro в положении аминокислоты 256;
любой из Asp, Glu, His, Ser и Tyr в положении аминокислоты 258;
любой из Asp, Glu, His и Tyr в положении аминокислоты 260;
любой из Ala, Glu, Phe, Ile и Thr в положении аминокислоты 262;
любой из Ala, Ile, Met и Thr в положении аминокислоты 263;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 264;
любой из Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Val, Trp и Tyr в положении аминокислоты 265;
любой из Ala, Ile, Met и Thr в положении аминокислоты 266;
любой из Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 267;
любой из Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val и Trp в положении аминокислоты 268;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 269;
любой из Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp и Tyr в положении аминокислоты 270;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 271;
любой из Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 272;
либо Phe, либо Ile в положении аминокислоты 273;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 274;
либо Leu, либо Trp в положении аминокислоты 275;
любой из Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 276;
любой из Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 278;
Ala в положении аминокислоты 279;
любой из Ala, Gly, His, Lys, Leu, Pro, Gln, Trp и Tyr в положении аминокислоты 280;
любой из Asp, Lys, Pro и Tyr в положении аминокислоты 281;
любой из Glu, Gly, Lys, Pro и Tyr в положении аминокислоты 282;
любой из Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg и Tyr в положении аминокислоты 283;
любой из Asp, Glu, Leu, Asn, Thr и Tyr в положении аминокислоты 284;
любой из Asp, Glu, Lys, Gln, Trp и Tyr в положении аминокислоты 285;
любой из Glu, Gly, Pro и Tyr в положении аминокислоты 286;
любой из Asn, Asp, Glu и Tyr в положении аминокислоты 288;
любой из Asp, Gly, His, Leu, Asn, Ser, Thr, Trp и Tyr в положении аминокислоты 290;
любой из Asp, Glu, Gly, His, Ile, Gln и Thr в положении аминокислоты 291;
любой из Ala, Asp, Glu, Pro, Thr и Tyr в положении аминокислоты 292;
любой из Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 293;
любой из Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 294;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 295;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr и Val в положении аминокислоты 296;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 297;
любой из Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 298;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 299;
любой из Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val и Trp в положении аминокислоты 300;
любой из Asp, Glu, His и Tyr в положении аминокислоты 301;
Ile в положении аминокислоты 302;
любой из Asp, Gly и Tyr в положении аминокислоты 303;
любой из Asp, His, Leu, Asn и Thr в положении аминокислоты 304;
любой из Glu, Ile, Thr и Tyr в положении аминокислоты 305;
любой из Ala, Asp, Asn, Thr, Val и Tyr в положении аминокислоты 311;
Phe в положении аминокислоты 313;
Leu в положении аминокислоты 315;
либо Glu, либо Gln в положении аминокислоты 317;
любой из His, Leu, Asn, Pro, Gln, Arg, Thr, Val и Tyr в положении аминокислоты 318;
любой из Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 320;
любой из Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 322;
Ile в положении аминокислоты 323;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 324;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 325;
любой из Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 326;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 327;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 328;
любой из Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 329;
любой из Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 330;
любой из Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp и Tyr в положении аминокислоты 331;
любой из Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp и Tyr в положении аминокислоты 332;
любой из Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val и Tyr в положении аминокислоты 333;
любой из Ala, Glu, Phe, Ile, Leu, Pro и Thr в положении аминокислоты 334;
любой из Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp и Tyr в положении аминокислоты 335;
любой из Glu, Lys и Tyr в положении аминокислоты 336;
любой из Glu, His и Asn в положении аминокислоты 337;
любой из Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser и Thr в положении аминокислоты 339;
либо Ala, либо Val в положении аминокислоты 376;
либо Gly, либо Lys в положении аминокислоты 377;
Asp в положении аминокислоты 378;
Asn в положении аминокислоты 379;
любой из Ala, Asn и Ser в положении аминокислоты 380;
либо Ala, либо Ile в положении аминокислоты 382;
Glu в положении аминокислоты 385;
Thr в положении аминокислоты 392;
Leu в положении аминокислоты 396;
Lys в положении аминокислоты 421;
Asn в положении аминокислоты 427;
либо Phe, либо Leu в положении аминокислоты 428;
Met в положении аминокислоты 429;
Trp в положении аминокислоты 434;
Ile в положении аминокислоты 436; и
любой из Gly, His, Ile, Leu и Tyr в положении аминокислоты 440;
в соответствии с нумерацией EU в Fc-области.
38. Способ по п. 35 или 36, где встречающаяся в природе Fc-область представляет собой Fc-область любого из IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека, в которой цепь сахаров, присоединенная в положении 297 в соответствии с нумерацией EU, представляет собой цепь сахаров, содержащую фукозу.
39. Способ по п. 35 или 36, где Fc-область модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или к которым добавлен бисекторный N-ацетилглюкозамин в положении 297 в соответствии с нумерацией EU в Fc-области, стал более высоким.
| WO2010088444 A1, 05.08.2010 | |||
| EP22275443 A1, 19.01.2011 | |||
| MI WENTAO et al., Targeting the Neonatal Fc Receptor for Antigen DeliveryUsing Engineered Fc Fragments, J Immunol, 2008, 181, pp.7550-7561 | |||
| АНТИТЕЛА ПРОТИВ IL-6, КОМПОЗИЦИИ, СПОСОБЫ И ПРИМЕНЕНИЕ | 2002 |
|
RU2318829C2 |

