Область техники
Настоящее изобретение относится к применению антигенсвязывающих молекул для элиминации антигенов из плазмы; способам элиминации антигенов из плазмы, которые включают введение антигенсвязывающих молекул; фармацевтическим композициям, содержащим антигенсвязывающие молекулы, которые способны элиминировать антигены из плазмы; способам скрининга антигенсвязывающих молекул для элиминации антигенов из плазмы и способам получения антигенсвязывающих молекул для элиминации антигенов из плазмы.
Уровень техники
Антитела привлекают внимание в качестве лекарственных средств, так как они обладают высокой стабильностью в плазме и имеют малое количество побочных эффектов. В настоящее время ряд подобных фармацевтических средств IgG-типа являются коммерчески доступными и на сегодняшний день разрабатываются многие лекарственные средства на основе антител (непатентные документы 1 и 2). Между тем, описаны различные способы, применимые для второго поколения лекарственных средств на основе антител, включающие способы, которые усиливают эффекторную функцию, антигенсвязывающую способность, фармакокинетику и стабильность, и способы, которые снижают риск развития иммуногенности (непатентный документ 3). Как правило, требуемая доза лекарственного средства на основе антитела является очень высокой. Это, в свою очередь, привело к возникновению проблем, таких как высокая стоимость производства, а также к трудностям при получении подкожных составов. В теории, доза фармацевтического средства на основе антитела может быть снижена путем улучшения фармакокинетики антитела или повышения аффинности между антителами и антигенами.
В литературе описаны способы улучшения фармакокинетики антител с использованием искусственных замен аминокислот в константных областях (непатентные документы 4 и 5). Аналогично описано созревание аффинности как способ улучшения антигенсвязывающей способности или активности нейтрализации антигена (непатентный документ 6). Этот способ дает возможность увеличения антигенсвязывающей способности посредством внесения аминокислотной мутации в область CDR вариабельной области или сходные с ней. Повышение антигенсвязывающей способности обеспечивает улучшение биологической активности in vitro или обеспечивает возможность снижения дозы и дополнительно обеспечивает повышенную эффективность in vivo (в организме) (непатентный документ 7).
Между тем, способность одной молекулы антитела нейтрализовать антиген зависит от ее аффинности. Путем увеличения аффинности антиген можно нейтрализовать меньшим количеством антитела. Для увеличения аффинности антитела можно использовать различные способы (непатентный документ 6). Кроме того, если бы аффинность можно было сделать бесконечной посредством ковалентного связывания антитела с антигеном, одна молекула антитела могла бы нейтрализовать одну молекулу антигена (двухвалентное антитело могло бы нейтрализовать две молекулы антигена). Однако общепринятые способы имеют ограничение, состоящее в том, что одна молекула антитела связывается с одной молекулой антигена (двумя антигенами, когда она является двухвалентной). Однако недавно было описано, что одна антигенсвязывающая молекула может связываться с несколькими молекулами антигена при использовании антигенсвязывающей молекулы, которая связывается с антигеном pH-зависимым образом (патентный документ 1 и непатентный документ 8). Молекула с pH-зависимым связыванием антигена связывается прочно с антигеном в нейтральных условиях в плазме и высвобождает антиген в кислых условиях в эндосоме. Более того, после высвобождения антигена антигенсвязывающая молекула высвобождается в плазму через FcRn и вновь связывается с антигеном; таким образом, одна молекула с pH-зависимым связыванием антигена может неоднократно связываться с множеством антигенов.
Более того, поскольку молекулы с pH-зависимым связыванием антигена, которые были модифицированы так, чтобы усилить связывание FcRn в нейтральных условиях (pH 7,4), имеют преимущество, состоящее в том, что они способны неоднократно связываться с антигенами и обеспечивать элиминацию антигенов из плазмы, сообщалось, что введение таких антигенсвязывающих молекул обеспечивает элиминацию антигена из плазмы (патентный документ 2). Традиционные молекулы с pH-зависимым связыванием антигена, содержащие Fc-область IgG-антитела, практически не демонстрируют связывания FcRn в нейтральных условиях. Таким образом, захват комплексов, образовавшихся между антигенсвязывающей молекулой и антигеном, в клетки может осуществляться в основном через неспецифический захват. В соответствии с этим сообщением, молекулы с pH-зависимым связыванием антигена, которые модифицированы так, чтобы усиливалось связывание FcRn в нейтральных условиях (pH 7,4), могут ускорять элиминацию антигена в большей степени, чем традиционные молекулы с pH-зависимым связыванием антигена, содержащие Fc-область IgG-антитела (патентный документ 2).
Поскольку антигены имеют очень короткое время удержания в плазме по сравнению с антителами, имеющими механизм опосредуемого FcRn рециклирования, связывание антигена в плазме с антителом, имеющим механизм рециклирования (где связывание не является pH-зависимым) увеличивает нормальное время удержания в плазме и повышает концентрацию антигена в плазме. Например, когда антиген в плазме имеет несколько типов физиологических функций, даже если один тип физиологической активности блокируется при связывании антитела, концентрация антигена в плазме может обострить симптомы, вызываемые другими физиологическими функциями, вследствие связывания с антителом. С такой точки зрения, элиминация антигенов из плазмы иногда является полезной, и были описаны способы, сходные со способами, описанными выше, для внесения модификаций в Fc-область для усиления связывания FcRn с целью ускорения элиминации антигена, однако до настоящего времени не были описаны другие способы ускорения элиминации антигена.
Документы уровня техники настоящего изобретения представлены ниже.
[Документы уровня техники]
[Патентные документы]
[Патентный документ 1] Международная публикация WO2009/125825.
[Патентный документ 2] Международная публикация WO2011/122011.
[Непатентные документы]
[Непатентный документ 1] Monoclonal antibody successes in the clinic, Janice M Reichert, Clark J Rosensweig, Laura B Faden & Matthew C Dewitz, Nat. Biotechnol. (2005) 23, 1073-1078.
[Непатентный документ 2] Pavlou AK, Belsey MJ., The therapeutic antibodies market to 2008, Eur. J. Pharm. Biopharm. (2005) 59 (3), 389-396.
[Непатентный документ 3] Kim SJ, Park Y, Hong HJ., Antibody engineering for the development of therapeutic antibodies, Mol. Cells. (2005) 20 (1), 17-29.
[Непатентный документ 4] Hinton PR, Xiong JM, Johlfs MG, Tang MT, Keller S, Tsurushita N, J. Immunol. (2006) 176 (1), 346-356.
[Непатентный документ 5] Ghetie V, Popov S, Borvak J, Radu C, Matesoi D, Medesan C, Ober RJ, Ward ES., Nat. Biotechnol. (1997) 15 (7), 637-640.
[Непатентный документ 6] Rajpal A, Beyaz N, Haber L, Cappuccilli G, Yee H, Bhatt RR, Takeuchi T, Lerner RA, Crea R., Proc. Natl. Acad. Sci. USA. (2005) 102 (24), 8466-8471.
[Непатентный документ 7] Wu H, Pfarr DS, Johnson S, Brewah YA, Woods RM, Patel NK, White WI, Young JF, Kiener PA., J. Mol. Biol. (2007) 368, 652-665.
[Непатентный документ 8] Igawa T, et al., Nat. Biotechnol. (2010) 28, 1203-1207.
Сущность изобретения
[Проблемы, решаемые с помощью изобретения]
Настоящее изобретение было осуществлено ввиду указанных выше обстоятельств. Задачей настоящего изобретения является обеспечение применений антигенсвязывающих молекул для элиминации антигенов из плазмы; способов элиминации антигенов из плазмы, которые включают введение антигенсвязывающих молекул; фармацевтических композиций, содержащих антигенсвязывающие молекулы, которые способны элиминировать антигены из плазмы; способов скрининга антигенсвязывающих молекул для элиминации антигенов из плазмы; и способов получения антигенсвязывающих молекул для элиминации антигенов из плазмы.
[Способы решения проблем]
Авторы настоящего изобретения провели тщательные исследования для достижения описанных выше задач. В результате авторы настоящего изобретения получили антигенсвязывающие молекулы, содержащие (i) Fc-область и (ii) два или более антигенсвязывающих доменов, где по меньшей мере один из доменов представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от концентрации ионов, которые представляют собой антигенсвязывающие молекулы, которые могут образовывать иммунный комплекс, содержащий (a) две или более антигенсвязывающих молекул и (b) две или более молекул антигена, содержащих два или более антигенных элементов связывания. Более того, авторы настоящего изобретения обнаружили, что антигенсвязывающие молекулы можно использовать для элиминации антигенов из плазмы. Более того, авторы настоящего изобретения обнаружили, что антигенсвязывающие молекулы являются пригодными в качестве фармацевтических композиций, и они разработали способы элиминации антигенов из плазмы, которые включают введение антигенсвязывающих молекул. Более того, авторы настоящего изобретения разработали способы скрининга антигенсвязывающих молекул, имеющих упомянутые выше свойства, и разработали способы получения этих молекул, тем самым осуществив изобретение.
Более конкретно, настоящее изобретение относится к следующему:
[1] применение антигенсвязывающей молекулы, содержащей
(i) Fc-область и
(ii) два или более антигенсвязывающих доменов,
где по меньшей мере один из доменов представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и
где антигенсвязывающая молекула может образовывать иммунный комплекс, который содержит
(a) две или более антигенсвязывающих молекул и
(b) два или более антигенов, где антигены содержат два или более антигенных элементов связывания,
для элиминации антигенов из плазмы;
[2] применение согласно [1], где условия концентрации ионов представляют собой условия концентрации ионов кальция;
[3] применение согласно [2], где антигенсвязывающий домен обладает антигенсвязывающей активностью в условиях низкой концентрации ионов кальция, которая является более низкой, чем антигенсвязывающая активность в условиях высокой концентрации ионов кальция;
[4] применение согласно любому из [1]-[3], где условия концентрации ионов представляют собой условия pH;
[5] применение согласно [4], где антигенсвязывающий домен обладает антигенсвязывающей активностью в диапазоне кислых значений pH, которая является более низкой, чем антигенсвязывающая активность в условиях диапазона нейтральных значений pH;
[6] применение согласно любому из [1]-[5], где антигены, содержащие два или более антигенных элементов связывания, являются мультимерами;
[7] применение согласно [6], где антиген представляет собой любой из GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), TNF, TNF-альфа, TNF-альфа-бета, TNF-бета-2, TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 лиганда Fas, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), VEGF, IgE, IgA, IgG, IgM, RANKL, TGF-альфа, TGF-бета, панспецифического TGF-бета и IL-8;
[8] применение согласно любому из [1]-[5], где антигены, содержащие два или более антигенных элементов связывания, являются мономерами;
[9] применение согласно любому из [1]-[8], где антигенсвязывающие молекулы представляют собой мультиспецифическую или мультипаратопную антигенсвязывающую молекулу, или смесь антигенсвязывающих молекул;
[10] применение согласно любому из [1]-[9], где Fc-область соответствует любой из SEQ ID NO:13, 14, 15 и 16;
[11] применение согласно любому из [1]-[9], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[12] применение согласно [11], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442 и 447 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[13] применение согласно [12], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442; Ile, Pro или Thr в качестве аминокислоты в положении 308;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[14] применение согласно любому из [1]-[9], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[15] применение согласно [14], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из положений 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[16] применение согласно [15], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Met в качестве аминокислоты в положении 237;
Ile в качестве аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 250;
Phe, Trp или Tyr в качестве аминокислоты в положении 252;
Thr в качестве аминокислоты в положении 254;
Glu в качестве аминокислоты в положении 255;
Asp, Asn, Glu или Gln в качестве аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в качестве аминокислоты в положении 257;
His в качестве аминокислоты в положении 258;
Ala в качестве аминокислоты в положении 265;
Ala или Glu в качестве аминокислоты в положении 286;
His в качестве аминокислоты в положении 289;
Ala в качестве аминокислоты в положении 297;
Ala в качестве аминокислоты в положении 303;
Ala в качестве аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в качестве аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg в качестве аминокислоты в положении 309;
Ala, His или Ile в качестве аминокислоты в положении 311;
Ala или His в качестве аминокислоты в положении 312;
Lys или Arg в качестве аминокислоты в положении 314;
Ala, Asp или His в качестве аминокислоты в положении 315;
Ala в качестве аминокислоты в положении 317;
Val в качестве аминокислоты в положении 332;
Leu в качестве аминокислоты в положении 334;
His в качестве аминокислоты в положении 360;
Ala в качестве аминокислоты в положении 376;
Ala в качестве аминокислоты в положении 380;
Ala в качестве аминокислоты в положении 382;
Ala в качестве аминокислоты в положении 384;
Asp или His в качестве аминокислоты в положении 385;
Pro в качестве аминокислоты в положении 386;
Glu в качестве аминокислоты в положении 387;
Ala или Ser в качестве аминокислоты в положении 389;
Ala в качестве аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 428;
Lys в качестве аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr в качестве аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val в качестве аминокислоты в положении 436;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[17] применение согласно любому из [1]-[13], где Fc-область включает Fc-область, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека;
[18] применение согласно [17], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, которые отличаются от аминокислот нативной Fc-области IgG человека, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU);
[19] применение согласно [18], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU;
[20] применение согласно любому из [1]-[16], где Fc-область обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором;
[21] применение согласно [20], где ингибиторный Fcγ-рецептор представляет собой FcγRIIb человека;
[22] применение согласно [20] или [21], где активирующий Fcγ-рецептор представляет собой FcγRIa человека, FcγRIIa (R)человека, FcγRIIa (H) человека, FcγRIIIa (V) человека или FcγRIIIa (F) человека;
[23] применение согласно любому из [20]-[22], где аминокислота в положении 238 или 328 (нумерация EU) в Fc-области отличается от аминокислоты в нативной Fc-области IgG человека;
[24] применение согласно [23], где аминокислота в положении 238 Fc-области представляет собой Asp или аминокислота в положении 328 Fc-области представляет собой Glu, как указано в соответствии с нумерацией EU;
[25] применение согласно [23] или [24], где аминокислотная последовательность Fc-области содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Asp в качестве аминокислоты в положении 233;
Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Leu, Met, Phe, Trp или Tyr в качестве аминокислоты в положении 237;
Asp в качестве аминокислоты в положении 239;
Ala, Gln или Val в качестве аминокислоты в положении 267;
Asn, Asp или Glu в качестве аминокислоты в положении 268;
Gly в качестве аминокислоты в положении 271;
Ala, Asn, Asp, Gln, Glu, Leu, Met, Ser или Thr в качестве аминокислоты в положении 326;
Arg, Lys или Met в качестве аминокислоты в положении 330;
Ile, Leu или Met в качестве аминокислоты в положении 323; и
Asp в качестве аминокислоты в положении 296;
как указано в соответствии с нумерацией EU;
[26] способ скрининга антигенсвязывающей молекулы, обладающей функцией элиминации антигена из плазмы, где способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше;
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше;
(f) контактирование антигенсвязывающей молекулы, полученной согласно (e) выше, с антигеном; и
(g) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген;
[27] способ получения антигенсвязывающей молекулы, обладающей функцией элиминации антигена из плазмы, где способ включает:
(a) контактирование антигена с антигенсвязывающей молекулой, содержащей Fc-область и два или более антигенсвязывающих доменов, где по меньшей мере один из антигенсвязывающих доменов обладает антигенсвязывающей активностью, которая изменяется в зависимости от условий концентрации ионов;
(b) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген;
(c) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антигенсвязывающую молекулу, для которой подтверждено, что она образует иммунный комплекс, согласно (b) выше; и
(d) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (c) выше;
[28] способ получения антигенсвязывающей молекулы, обладающей функцией элиминации антигена из плазмы, где способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше;
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше;
(f) контактирование антигенсвязывающей молекулы, полученной согласно (e) выше, с антигеном;
(g) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген;
(h) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антигенсвязывающую молекулу, для которой подтверждено, что она образует иммунный комплекс, согласно (g) выше; и
(i) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (h) выше; и
[29] способ получения антигенсвязывающей молекулы, обладающей функцией элиминации антигена из плазмы, где способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше; и
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше; и
где способ дополнительно включает контактирование антигенсвязывающей молекулы, полученной способом получения, с антигеном, и оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген.
Упомянутые выше [1]-[25] могут быть переформулированы следующим образом:
[1’] фармацевтическая композиция для элиминации антигена из плазмы, где фармацевтическая композиция содержит антигенсвязывающую молекулу, содержащую
(i) Fc-область и
(ii) два или более антигенсвязывающих доменов,
где по меньшей мере один из доменов представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и
где антигенсвязывающая молекула может образовывать иммунный комплекс, который содержит
(a) две или более антигенсвязывающих молекул и
(b) два или более антигенов, где антигены содержат два или более антигенных элементов связывания;
[2’] фармацевтическая композиция согласно [1’], где условия концентрации ионов представляют собой условия концентрации ионов кальция;
[3’] фармацевтическая композиция согласно [2’], где антигенсвязывающий домен обладает антигенсвязывающей активностью в условиях низкой концентрации ионов кальция, которая является более низкой, чем антигенсвязывающая активность в условиях высокой концентрации ионов кальция;
[4’] фармацевтическая композиция согласно любому из [1’]-[3’], где условия концентрации ионов представляют собой условия pH;
[5’] фармацевтическая композиция согласно [4’], где антигенсвязывающий домен обладает антигенсвязывающей активностью в диапазоне кислых значений pH, которая является более низкой, чем антигенсвязывающая активность в условиях диапазона нейтральных значений pH;
[6’] фармацевтическая композиция согласно любому из [1’]-[5’], где антигены, содержащие два или более антигенных элементов связывания, являются мультимерами;
[7’] фармацевтическая композиция согласно [6’], где антиген представляет собой любой из GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), TNF, TNF-альфа, TNF-альфа-бета, TNF-бета-2, TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 лиганда Fas, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), VEGF, IgE, IgA, IgG, IgM, RANKL, TGF-альфа, TGF-бета, панспецифического TGF-бета и IL-8;
[8’] фармацевтическая композиция согласно любому из [1’]-[5’], где антигены, содержащие два или более антигенных элементов связывания, являются мономерами;
[9’] фармацевтическая композиция согласно любому из [1’]-[8’], где антигенсвязывающие молекулы представляют собой мультиспецифическую или мультипаратопную антигенсвязывающую молекулу, или смесь антигенсвязывающих молекул;
[10’] фармацевтическая композиция согласно любому из [1’]-[9’], где Fc-область соответствует любой из SEQ ID NO:13, 14, 15 и 16;
[11’] фармацевтическая композиция согласно любому из [1’]-[9’], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[12’] фармацевтическая композиция согласно [11’], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442 и 447 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[13’] фармацевтическая композиция согласно [12’], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442; Ile, Pro или Thr в качестве аминокислоты в положении 308;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[14’] фармацевтическая композиция согласно любому из [1’]-[9’], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[15’] фармацевтическая композиция согласно [14’], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из положений 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[16’] фармацевтическая композиция согласно [15’], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Met в качестве аминокислоты в положении 237;
Ile в качестве аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 250;
Phe, Trp или Tyr в качестве аминокислоты в положении 252;
Thr в качестве аминокислоты в положении 254;
Glu в качестве аминокислоты в положении 255;
Asp, Asn, Glu или Gln в качестве аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в качестве аминокислоты в положении 257;
His в качестве аминокислоты в положении 258;
Ala в качестве аминокислоты в положении 265;
Ala или Glu в качестве аминокислоты в положении 286;
His в качестве аминокислоты в положении 289;
Ala в качестве аминокислоты в положении 297;
Ala в качестве аминокислоты в положении 303;
Ala в качестве аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в качестве аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg в качестве аминокислоты в положении 309;
Ala, His или Ile в качестве аминокислоты в положении 311;
Ala или His в качестве аминокислоты в положении 312;
Lys или Arg в качестве аминокислоты в положении 314;
Ala, Asp или His в качестве аминокислоты в положении 315;
Ala в качестве аминокислоты в положении 317;
Val в качестве аминокислоты в положении 332;
Leu в качестве аминокислоты в положении 334;
His в качестве аминокислоты в положении 360;
Ala в качестве аминокислоты в положении 376;
Ala в качестве аминокислоты в положении 380;
Ala в качестве аминокислоты в положении 382;
Ala в качестве аминокислоты в положении 384;
Asp или His в качестве аминокислоты в положении 385;
Pro в качестве аминокислоты в положении 386;
Glu в качестве аминокислоты в положении 387;
Ala или Ser в качестве аминокислоты в положении 389;
Ala в качестве аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 428;
Lys в качестве аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr в качестве аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val в качестве аминокислоты в положении 436;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[17’] фармацевтическая композиция согласно любому из [1’]-[13’], где Fc-область включает Fc-область, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека;
[18’] фармацевтическая композиция согласно [17’], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, которые отличаются от аминокислот нативной Fc-области IgG человека, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU);
[19’] фармацевтическая композиция согласно [18’], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU;
[20’] фармацевтическая композиция согласно любому из [1’]-[16’], где Fc-область обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором;
[21’] фармацевтическая композиция согласно [20’], где ингибиторный Fcγ-рецептор представляет собой FcγRIIb человека;
[22’] фармацевтическая композиция согласно [20’] или [21’], где активирующий Fcγ-рецептор представляет собой FcγRIa человека, FcγRIIa (R)человека, FcγRIIa (H) человека, FcγRIIIa (V) человека или FcγRIIIa (F) человека;
[23’] фармацевтическая композиция согласно любому из [20’]-[22’], где аминокислота в положении 238 или 328 (нумерация EU) в Fc-области отличается от аминокислоты в нативной Fc-области IgG человека;
[24’] фармацевтическая композиция согласно [23’], где аминокислота в положении 238 Fc-области представляет собой Asp или аминокислота в положении 328 Fc-области представляет собой Glu, как указано в соответствии с нумерацией EU; и
[25’] фармацевтическая композиция согласно [23’] или [24’], где аминокислотная последовательность Fc-области содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Asp в качестве аминокислоты в положении 233;
Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Leu, Met, Phe, Trp или Tyr в качестве аминокислоты в положении 237;
Asp в качестве аминокислоты в положении 239;
Ala, Gln или Val в качестве аминокислоты в положении 267;
Asn, Asp или Glu в качестве аминокислоты в положении 268;
Gly в качестве аминокислоты в положении 271;
Ala, Asn, Asp, Gln, Glu, Leu, Met, Ser или Thr в качестве аминокислоты в положении 326;
Arg, Lys или Met в качестве аминокислоты в положении 330;
Ile, Leu или Met в качестве аминокислоты в положении 323; и
Asp в качестве аминокислоты в положении 296;
как указано в соответствии с нумерацией EU.
Более того, упомянутые выше [1]-[25] также могут быть переформулированы следующим образом:
[1"] способ элиминации антигена из плазмы индивидуума, где способ включает введение индивидууму антигенсвязывающей молекулы, содержащей
(i) Fc-область и
(ii) два или более антигенсвязывающих доменов,
где по меньшей мере один из доменов представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и
где антигенсвязывающая молекула может образовывать иммунный комплекс, который содержит
(a) две или более антигенсвязывающих молекул и
(b) два или более антигенов, где антигены содержат два или более антигенных элементов связывания;
[2"] способ согласно [1"], где условия концентрации ионов представляют собой условия концентрации ионов кальция;
[3"] способ согласно [2"], где антигенсвязывающий домен обладает антигенсвязывающей активностью в условиях низкой концентрации ионов кальция, которая является более низкой, чем антигенсвязывающая активность в условиях высокой концентрации ионов кальция;
[4"] способ согласно любому из [1"]-[3"], где условия концентрации ионов представляют собой условия pH;
[5"] способ согласно [4"], где антигенсвязывающий домен обладает антигенсвязывающей активностью в диапазоне кислых значений pH, которая является более низкой, чем антигенсвязывающая активность в условиях диапазона нейтральных значений pH;
[6"] способ согласно любому из [1"]-[5"], где антигены, содержащие два или более антигенных элементов связывания, являются мультимерами;
[7"] способ согласно [6"], где антиген представляет собой любой из GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), TNF, TNF-альфа, TNF-альфа-бета, TNF-бета-2, TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 лиганда Fas, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), VEGF, IgE, IgA, IgG, IgM, RANKL, TGF-альфа, TGF-бета, панспецифического TGF-бета и IL-8;
[8"] способ согласно любому из [1"]-[5"], где антигены, содержащие два или более антигенных элементов связывания, являются мономерами;
[9"] способ согласно любому из [1"]-[9"], где антигенсвязывающие молекулы представляют собой мультиспецифическую или мультипаратопную антигенсвязывающую молекулу, или смесь антигенсвязывающих молекул;
[10"] способ согласно любому из [1"]-[9"], где Fc-область соответствует любой из SEQ ID NO:13, 14, 15 и 16;
[11"] способ согласно любому из [1"]-[9"], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[12"] способ согласно [11"], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442 и 447 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[13"] способ согласно [12"], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442; Ile, Pro или Thr в качестве аминокислоты в положении 308;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[14"] способ согласно любому из [1"]-[9"], где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[15"] способ согласно [14"], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из положений 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[16"] способ согласно [15"], где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Met в качестве аминокислоты в положении 237;
Ile в качестве аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 250;
Phe, Trp или Tyr в качестве аминокислоты в положении 252;
Thr в качестве аминокислоты в положении 254;
Glu в качестве аминокислоты в положении 255;
Asp, Asn, Glu или Gln в качестве аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в качестве аминокислоты в положении 257;
His в качестве аминокислоты в положении 258;
Ala в качестве аминокислоты в положении 265;
Ala или Glu в качестве аминокислоты в положении 286;
His в качестве аминокислоты в положении 289;
Ala в качестве аминокислоты в положении 297;
Ala в качестве аминокислоты в положении 303;
Ala в качестве аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в качестве аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg в качестве аминокислоты в положении 309;
Ala, His или Ile в качестве аминокислоты в положении 311;
Ala или His в качестве аминокислоты в положении 312;
Lys или Arg в качестве аминокислоты в положении 314;
Ala, Asp или His в качестве аминокислоты в положении 315;
Ala в качестве аминокислоты в положении 317;
Val в качестве аминокислоты в положении 332;
Leu в качестве аминокислоты в положении 334;
His в качестве аминокислоты в положении 360;
Ala в качестве аминокислоты в положении 376;
Ala в качестве аминокислоты в положении 380;
Ala в качестве аминокислоты в положении 382;
Ala в качестве аминокислоты в положении 384;
Asp или His в качестве аминокислоты в положении 385;
Pro в качестве аминокислоты в положении 386;
Glu в качестве аминокислоты в положении 387;
Ala или Ser в качестве аминокислоты в положении 389;
Ala в качестве аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 428;
Lys в качестве аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr в качестве аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val в качестве аминокислоты в положении 436;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16;
[17"] способ согласно любому из [1"]-[13"], где Fc-область включает Fc-область, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека;
[18"] способ согласно [17"], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, которые отличаются от аминокислот нативной Fc-области IgG человека, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU);
[19"] способ согласно [18"], где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU;
[20"] способ согласно любому из [1"]-[16"], где Fc-область обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором;
[21"] способ согласно [20"], где ингибиторный Fcγ-рецептор представляет собой FcγRIIb человека;
[22"] способ согласно [20"] или [21"], где активирующий Fcγ-рецептор представляет собой FcγRIa человека, FcγRIIa (R)человека, FcγRIIa (H) человека, FcγRIIIa (V) человека или FcγRIIIa (F) человека;
[23"] способ согласно любому из [20"]-[22"], где аминокислота в положении 238 или 328 (нумерация EU) в Fc-области отличается от аминокислоты в нативной Fc-области IgG человека;
[24"] способ согласно [23"], где аминокислота в положении 238 Fc-области представляет собой Asp или аминокислота в положении 328 Fc-области представляет собой Glu, как указано в соответствии с нумерацией EU; и
[25"] способ согласно [23"] или [24"], где аминокислотная последовательность Fc-области содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Asp в качестве аминокислоты в положении 233;
Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Leu, Met, Phe, Trp или Tyr в качестве аминокислоты в положении 237;
Asp в качестве аминокислоты в положении 239;
Ala, Gln или Val в качестве аминокислоты в положении 267;
Asn, Asp или Glu в качестве аминокислоты в положении 268;
Gly в качестве аминокислоты в положении 271;
Ala, Asn, Asp, Gln, Glu, Leu, Met, Ser или Thr в качестве аминокислоты в положении 326;
Arg, Lys или Met в качестве аминокислоты в положении 330;
Ile, Leu или Met в качестве аминокислоты в положении 323; и
Asp в качестве аминокислоты в положении 296;
как указано в соответствии с нумерацией EU.
Краткое описание чертежей
На фиг. 1 представлена диаграмма, на которой показано, что антитело с pH-зависимым связыванием многократно связывается с растворимыми антигенами. (i) антитело связывается с растворимыми антигенами; (ii) антитело неспецифически интернализуется в клетку путем пиноцитоза; (iii) антитело связывается с FcRn в эндосомах, а затем растворимые антигены диссоциируют от антитела; (iv) растворимые антигены переносятся в лизосомы и деградируются; (v) после диссоциации от растворимых антигенов антитело рециклирует в плазму через FcRn; (vi) рециклированное антитело может вновь связываться с растворимыми антигенами.
На фиг. 2 представлена диаграмма, на которой показано, что усиление связывания FcRn в нейтральных условиях приводит к улучшению эффекта антитела с pH-зависимым связыванием в отношении многократного связывания с антигенами: (i) антитело связывается с растворимыми антигенами; (ii) антитело интернализуется в клетку посредством пиноцитоза через FcRn; (iii) растворимые антигены диссоциируют от антитела в эндосоме; (iv) растворимые антигены переносятся в лизосому и деградируются; (v) после диссоциации растворимых антигенов антитело рециклирует в плазму через FcRn; и (vi) рециклировавшее антитело вновь может связываться с растворимыми антигенами.
На фиг. 3 представлены сенсограммы Biacore, демонстрирующие взаимодействие антител против IgA человека с IgA человека в условиях 1,2 мМ Ca2+ и 3 мкМ Ca2+.
На фиг. 4 представлен график, демонстрирующий изменения с течением времени концентрации антигена в плазме у нормальных мышей для группы введения IgA человека + антитело GA1-IgG1, группы введения IgA человека + антитело GA2-IgG1, для групп введения IgA человека + антитело GA2-FcγR(-) или GA2-N434W.
На фиг. 5 представлен график, демонстрирующий изменения с течением времени концентрации в плазме IgA человека у нормальных мышей для группы введения IgA человека отдельно, группы введения IgA человека + антитело GA1-IgG1, группы введения IgA человека + антитело GA2-IgG1, для группы введения IgA человека + антитело GA2-FcγR(-) и для группы введения IgA человека + антитело GA2-N434W.
На фиг. 6 представлен график, демонстрирующий изменения с течением времени концентрации в плазме несвязанного IgA человека у нормальных мышей для группы введения IgA человека + антитело GA1-IgG1, для группы введения IgA человека + антитело GA2-IgG1, для группы введения IgA человека + антитело GA2-FcγR(-) и для группы введения IgA человека + антитело GA2-N434W.
На фиг. 7 представлена иллюстративная диаграмма, на которой показана эффективность элиминации антигена на молекулу антитела для pH/Ca-зависимого антитела, имеющего константную область природного IgG1, которое формирует крупный иммунный комплекс с мультимерным антигеном.
На фиг. 8 представлена иллюстративная диаграмма, на которой показана эффективность элиминации антигена на молекулу антитела для pH/Ca-зависимого полиспецифического антитела, которое распознает два или более эпитопов в мономерном антигене и пригодно для образования крупного иммунного комплекса.
На фиг. 9 представлены хроматографических анализов методом гель-фильтрации, которые подтвердили, что IgE человека и клон 278, который представляет собой pH-зависимое антитело против IgE, образуют крупные иммунные комплексы pH-зависимым образом.
На фиг. 10 представлен график, демонстрирующий изменения с течением времени концентрации антитела в плазме у нормальных мышей для группы введения IgE человека + клон 278 и группы введения IgE человека + антитело Xolair.
На фиг. 11 представлен график, демонстрирующий изменения с течением времени концентрации в плазме IgE человека у нормальных мышей для группы введения IgE человека отдельно, для группы введения IgE человека + антитело клона 278 и для группы введения IgE человека + антитело клона 278.
На фиг. 12 представлен график, демонстрирующий изменения с течением времени концентраций в плазме антител GA2-IgG1 и GA2-F1087 у нормальных мышей.
На фиг. 13 представлен график, на котором показаны изменения с течением времени концентрации hIgA в плазме у нормальных мышей, которым вводили GA2-IgG1 или GA2-F1087.
На фиг. 14 представлен график, демонстрирующий изменения с течением времени концентраций в плазме антител 278-IgG1 и 278-F1087 у мышей C57BL/6.
На фиг. 15 представлен график, демонстрирующий изменения с течением времени концентрации в плазме hIgE(Asp6) у мышей C57BL/6J, которым вводили 278-IgG1 или 278-F1087.
На фиг. 16 представлен график, на котором показаны изменения с течением времени концентрации в плазме GA2-F760 или GA2-F1331 у мышей с трансгенным FcRn человека, которым вводили GA2-F760 или GA2-F1331.
На фиг. 17 представлен график, демонстрирующий изменения с течением времени концентрации в плазме IgA человека у мышей с трансгенным FcRn человека, которым вводили GA2-F760 или GA2-F1331.
На фиг. 18 представлен график, на котором показаны изменения с течением времени концентрации в плазме 278-F760 или 278-F1331 у мышей с трансгенным FcRn человека, которым вводили 278-F760 или 278-F1331.
На фиг. 19 представлен график, демонстрирующий изменения с течением времени концентрации в плазме IgE человека у мышей с трансгенным FcRn человека, которым вводили 278-F760 или 278-F1331.
На фиг. 20 представлены сенсограммы, демонстрирующие взаимодействие PHX-IgG1 с hsIL-6R при pH7,4 и pH6,0.
На фиг. 21 представлен результат оценки одновременного связывания Fv4-IgG1 и PHX-F29 с IL6R способом электрохемилюминесценции (ECL).
На фиг. 22 представлен график, демонстрирующий изменения с течением времени концентрации антитела против IL6R в плазме нормальных мышей для группы введения hsIL6R + Fv4-IgG1 и для групп введения hsIL6R + Fv4-IgG1 + PHX-IgG1.
На фиг. 23 представлен график, демонстрирующий изменения с течением времени концентрации IL6R человека в плазме нормальных мышей для группы введения hsIL6R + Fv4-IgG1 и для групп введения hsIL6R + Fv4-IgG1 + PHX-IgG1.
На фиг. 24 представлены сенсограммы Biacore, полученные, когда раствор антитела отдельно или смешанный раствор антитело-антиген наносили на FcγR. Пунктирной линией показана сенсограмма, полученная, когда наносили раствор антитела отдельно, и сплошной линией показана сенсограмма, полученная, когда наносили смешанный раствор антитело-антиген.
На фиг. 25 представлены сенсограммы Biacore, полученные, когда раствор антитела отдельно или смешанный раствор антитело-антиген наносили на FcRn человека. Сплошная линия демонстрирует сенсограмму, полученную, когда наносили раствор антитела отдельно, и пунктирной линией показана сенсограмма, полученная, когда наносили смешанный раствор антитело-антиген.
На фиг. 26 представлены сенсограммы Biacore, полученные, когда раствор антитела отдельно или смешанный раствор антитело-антиген наносили на FcRn мыши. Пунктирной линией показана сенсограмма, полученная, когда наносили раствор антитела отдельно, и сплошной линией показана сенсограмма, полученная, когда наносили смешанный раствор антитело-антиген.
На фиг. 27 представлены сенсограммы Biacore, полученные, когда раствор антитела отдельно или смешанный раствор антитело-антиген наносили на FcRn мыши. Пунктирной линией показана сенсограмма, полученная, когда наносили раствор антитела отдельно, и сплошной линией показана сенсограмма, полученная, когда наносили смешанный раствор антитело-антиген.
Способ осуществления изобретения
Определения и подробное описание ниже предоставлены для того, чтобы помочь понять настоящее изобретение, проиллюстрированное в настоящем описании.
Аминокислоты
В рамках настоящего изобретения аминокислоты описаны с помощью однобуквенного или трехбуквенного кодов или обоих из них, например, Ala/A, Leu/L, Arg/R, Lys/K, Asn/N, Met/M, Asp/D, Phe/F, Cys/C, Pro/P, Gln/Q, Ser/S, Glu/E, Thr/T, Gly/G, Trp/W, His/H, Tyr/Y, Ile/I или Val/V.
Модификация аминокислот
Для модификации аминокислот в аминокислотной последовательности антигенсвязывающей молекулы можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР. Более того, различные известные способы также можно использовать в качестве способа изменения аминокислот для замены на неприродные аминокислоты (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, которая имеет неприродную аминокислоту, связанную с комплементарной амбер-супрессорной тРНК кодона UAG (амбер-кодон), который является одним из стоп-кодонов.
В настоящем описании значение термина "и/или" при описании участка изменения аминокислот включает каждую комбинацию, где "и" и "или" соответствующим образом скомбинированы. В частности, например, "аминокислоты в положениях 33, 55 и/или 96 являются замещенными" включает следующие варианты аминокислотных модификаций:
аминокислота(ы) в (a) положении 33, (b) положении 55, (c) положении 96, (d) положениях 33 и 55, (e) положениях 33 и 96, (f) положениях 55 и 96, и (g) положениях 33, 55 и 96.
Антигенные элементы связывания
В настоящем описании "антигенные элементы связывания" относятся к "количеству эпитопов, присутствующих в одной молекуле антигена в форме, обычно присутствующей в плазме в отсутствие антигенсвязывающей молекулы, где молекула антигена содержит эпитоп, который связывается с одновалентным элементом связывания антигенсвязывающего домена, содержащегося в антигенсвязывающей молекуле по настоящему изобретению". Примеры антигена, которые имеют два антигенных элемента связывания, представляют собой антигены, включающие мультимеры, обычно присутствующие в плазме в форме гомодимеров, такие как GDF, PDGF или VEGF. Например, в гомодимеризованных молекулах GDF существует два элемента эпитопов, каждый из которых связывается одновалентным элементом связывания вариабельной области, содержащимся в молекуле антитела против GDF, которая содержит две вариабельных области, имеющих одинаковую последовательность (т.е. не биспецифическое антитело, описанное ниже). Также примеры антигенов с двумя антигенными элементами связывания включают молекулы иммуноглобулинов, такие как IgE. IgE обычно существует в плазме в качестве тетрамера, содержащего димер тяжелой цепи и димер легкой цепи; и в тетрамере присутствует два элемента эпитопов, каждый из которых связывается посредством одной валентности вариабельной области, содержащейся в молекуле антитела против IgE, которая содержит две вариабельных области, имеющих одну и ту же последовательность (т.е. не описанное ниже биспецифическое антитело). IgA обычно существует в плазме в двух формах: тетрамер, содержащий димер тяжелой цепи и димер легкой цепи, и октамер, образующийся, когда тетрамеры далее образуют комплекс через J-цепь; и тетрамер и октамер, соответственно, имеют два элемента и четыре элемента эпитопов, каждый из которых связывается посредством одной валентности вариабельной области, содержащейся в молекуле антитела против IgA, которая содержит две вариабельные области, имеющие ту же последовательность (т.е. не описанное ниже биспецифическое антитело). Примеры антигенов с тремя антигенными элементами связывания представляют собой антигены, включающие мультимеры, обычно присутствующие в плазме в форме гомотримеров, такие как TNFα, RANKL или CD154 суперсемейства TNF.
Примерами молекул антигенов с одним антигенным элементом связывания являются молекулы, обычно присутствующие в плазме в форме мономеров, такие как растворимый рецептор IL-6 (далее также обозначаемый как sIL-6R), IL-6, HMGB-1 и CTGF. Гетеродимеры, такие как IL-12, содержащий IL-12p40 и IL-12p35, IL-23, содержащий IL-12p40 и IL-23p19 (также обозначаемый как IL-30B), IL-23, содержащий EBI3 и IL27p28, и IL-35, содержащий IL-12p35 и EBI3, содержат две структурно различающихся молекулы субъединиц. В молекуле антитела против субъединицы, содержащей две вариабельных области, которые связываются с одной из этих субъединиц (т.е. не описанное ниже биспецифическое антитело), эпитоп, связываемый посредством одной валентности вариабельных областей, представляет собой один элемент; таким образом, имеется один антигенный элемент связывания этих гетеродимеров. Антигенный элемент связывания гетеротримеров, таких как мультисубъединичный комплекс TNFα-TNFβ-hCG аналогично представляет собой один элемент.
Поскольку "антигенный элемент связывания" означает количество эпитопов, связываемых одновалентным элементом связывания антигенсвязывающего домена, когда в антигенсвязывающей молекуле присутствует один тип паратопа или более типов паратопов, антигенный элемент связывания антигена будет отличаться, даже если антиген, связываемый этими антигенсвязывающими молекулами, представляет собой один и тот же антиген. Например, в случае упомянутых выше гетеродимеров, когда одновалентный элемент связывания в антигенсвязывающем домене, содержащийся в антигенсвязывающей молекуле, связывается с одним типом субъединицы в гетеродимере, имеется один антигенный элемент связывания этого антигена, в то время как когда антигенсвязывающая молекула является бипаратопной или биспецифической антигенсвязывающей молекулой, содержащей два моновалентных элемента связывания, которые связываются с каждой из субъединиц, образующих гетеродимер, имеется два антигенных элемента связывания этого антигена.
Мультимеры
В настоящем описании, когда просто приведено выражение "два или более мультимеров", значение термина "мультимер" включает как гомомультимер, так и гетеромультимер. Типами связывания между субъединицами, содержащимися в мультимере, являются ковалентные связи, такие как пептидные связи или дисульфидные связи, и стабильные нековалентные связи, такие как ионные связи, ван-дер-ваальсовы связи и водородные связи, но они не ограничиваются ими. Гомомультимеры включают множество идентичных субъединиц, в то время как гетеромультимеры содержат множество различающихся субъединиц. Например, значение термина "димер" включает как гомодимер, так и гетеродимер; и в то время как гомодимер содержит две идентичных субъединицы, гетеродимер содержит две различных субъединицы. Более того, в случае полипептида, термин "мономер", который выражает каждую из единиц, образующих мультимер, относится к каждой из непрерывных структурных единиц, связанных пептидными связями.
Антигены
В рамках настоящего изобретения "антигены" конкретно не ограничены по их структуре, при условии, что они содержат эпитопы, с которыми связываются антигенсвязывающие домены. Другие антигены включают, например, представленные ниже молекулы: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, аденозиновый рецептор A1, A33, ACE, ACE-2, активин, активин A, активин AB, активин B, активин C, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, APE, APJ, APP, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, интегрин av/b3, Ax1, b2M, B7-1, B7-2, B7-H, B-лимфоцитарный стимулирующий фактор (BlyS), BACE, BACE-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bсl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, BMP-3 остеогенин, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, костный нейротрофический фактор, BPDE, BPDE-DNA, BTC, фактор комплемента 3 (C3), C3a, C4, C5, C5a, C10, CA125, CAD-8, кальцитонин, cAMP, карциноэмбриональный антиген (CEA), ассоциированный со злокачественной опухолью, катепсин A, катепсин B, катепсин C/DPPI, катепсин D, катепсин E, катепсин H, катепсин L, катепсин O, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок p67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (B7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, токсин Botulinum, токсин Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цитокератиновый ассоциированный с опухолью антиген, DAN, DCC, DcR3, DC-SIGN, регулирующий фактор комплемента (фактор, ускоряющий распад), дез-(1-3)-IGF-I (IGF-1 головного мозга), Dhh, дигоксин, ДНКM-1, ДНКаза, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, фактор активации фибробластов (FAP), Fas, FcRI, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP- 13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа1, GFR-альфа2, GFR-альфа3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp 130, gp72, GRO, рилизинг-фактор гормона роста, гаптен (NP-cap или NIP-cap), HB-EGF, HCC, гликопротеин оболочки gB HCMV, гликопротеин оболочки gH HCMV, UL HCMV, гемопоэтический фактор роста (HGF), HepB gp120, гепараназа, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMW-MAA), gp120 HIV, петля V3 gp 120 IIIB HIV, HLA, HLA-DR, HM1,24, HMFG PEM, HRG, Hrk, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (HGH), HVEM, 1-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, рецептор IgA, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-23, интерферон (INF)-альфа, INF-бета, INF-гамма, ингибин, iNOS, A-цепь инсулина, B-цепь инсулина, инсулиноподобный фактор роста 1, интегрин альфа-2, интегрин альфа-3, интегрин альфа-4, интегрин альфа4/бета1, интегрин альфа4/бета7, интегрин альфа5 (альфа V), интегрин альфа5/бета1, интегрин альфа5/бета3, интегрин альфа6, интегрин бета1, интегрин бета2, интерферон-гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин LI, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bpl, LBP, LDGF, LECT2, lefty, антиген Lewis-Y, ассоциированный с Lewis-Y антиген, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, белок поверхности легких, лютеинизирующий гормон, рецептор лимфотоксина-бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCAM, MCK-2, MCP, M-CSF, MDC, Mer, металлопротеазы, рецептор MGDF, MGMT, MHC (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, мюллерово ингибиторное вещество, Mug, MuSK, NAIP, NAP, NCAD, адгерин N-C, NCA90, NCAM, NCAM, неприлизин, нейротрофин-3, -4 или -6, нейротурин, фактор роста нервов (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGGI, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, плацентарная щелочная фосфатаза (PLAP), P1GF, PLP, PP14, проинсулин, прорелаксин, белок C, PS, PSA, PSCA, простат-специфический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, A-цепь релаксина, B-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, SERINE, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин-72), TARC, TCA-3, T-клеточный рецептор (например, T-клеточный рецептор альфа/бета), TdT, TECK, TEM1, TEM5, TEM7, TEM8, TERT, PLAP-подобная щелочная фосфатаза семенников, TfR, TGF, TGF-альфа, TGF-бета, панспецифического TGF-бета, TGF-бета RI (ALK-5), TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета 1, TGF-бета 2, TGF-бета 3, TGF-бета 4, TGF-бета 5, тромбин, Ck-1 тимуса, тиреотропный гормон, Tie, TIMP, TIQ, тканевой фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа-бета, TNF-бета 2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD 120a, p55-60), TNFRSF1B (TNF RII CD 120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (0X40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL Rl TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд IVEM LIGHT I, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 Fas-лиганда, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TSG, TSLP, ассоциированный с опухолью CA125, ассоциированный с опухолью антиген, экспрессирующий ассоциированные с Lewis-Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный анотиген, VLA, VLA-1, VLA-4, интегрин VNR, фактор Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, Αβ, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин A, хромогранин B, tau, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор B, фактор D, фактор H, пропердин, склеростин, фибринoген, фибрин, протромбин, тромбин, тканевой фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Xa, фактор XI, фактор XIa, фактор XII, фактор Xlla, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и SIP; и рецепторы для гормонов и факторов роста.
Как описано ниже, когда антигенсвязывающая молекула связывается с несколькими эпитопами в молекуле антигена, как в случае биспецифического антитела и т.п., антиген, который может образовывать комплекс с антигенсвязывающей молекулой, может представлять собой любой из антигенов, проиллюстрированных выше, или их комбинацию, т.е. мономеры или гетеромультимеры. Неограничивающие примеры гетеромультимеров включают гетеродимеры, такие как IL-12, содержащий IL-12p40 и IL-12p35, IL-23, содержащий IL-12p40 и IL-23p19 (также обозначаемый как IL-30B), IL-23, содержащий EBI-3 и IL27p28, и IL-35, содержащий IL-12p35 и EBI-3.
Хотя рецепторы приведены в качестве примеров упомянутых выше антигенов, когда рецепторы существуют в растворимых формах в биологических жидкостях, таких как плазма, они могут образовывать комплексы с антигенсвязывающими молекулами по настоящему изобретению. Таким образом, при условии, что упомянутые выше рецепторы существуют в их растворимых формах в биологических жидкостях, таких как плазма, их можно использовать в качестве антигенов, которые могут образовывать комплексы по настоящему изобретению, путем связывания с антигенсвязывающей молекулой по настоящему изобретению. Примером неограничивающего варианта осуществления такого растворимого рецептора является растворимый IL-6R, который представляет собой белок, состоящий из аминокислот в положениях 1-357 в полипептидной последовательности IL-6R SEQ ID NO:1, как описано в Mullberg et al. (J. Immunol. (1994) 152 (10), 4958-4968).
В качестве примеров упомянутых выше антигенов приводятся растворимые антигены, и растворы, в которых существуют антигены, не ограничены. Растворимые антигены могут существовать в биологических жидкостях или, более конкретно, во всех жидкостях, заполняющих пространство между тканями и клетками или сосудами в организмах. В неограничивающем варианте осуществления антигены, с которыми антигенсвязывающие молекулы по настоящему изобретению связываются, могут присутствовать во внеклеточных жидкостях. У позвоночных внеклеточная жидкость является общим термином для плазмы, интерстициальной жидкости, лимфы, плотной соединительной ткани, цереброспинальной жидкости, спинномозговой жидкости, полученной пункцией жидкости, синовиальной жидкости и сходных компонентов кости и хряще, альвеолярной жидкости (жидкости бронхоальвеолярного лаважа), перитонеальной жидкости, плевральной жидкости, перикардиальной жидкости, жидкости кисты, внутриглазной жидкости (водянистой влаги) или сходных трансцеллюлярных жидкостей (различные жидкости в полостях желез и жидкости в полости пищеварительного тракта и жидкости других полостей организма, продуцируемые в результате активного транспорта/секреторной активности клеток).
Когда эпитоп, с которым связывается антигенсвязывающий домен, содержащийся в антигенсвязывающей молекуле, представляет собой единичный эпитоп, неограничивающий вариант осуществления антигена, который может образовывать комплекс по настоящему изобретению при связывании с антигенсвязывающей молекулой, включает молекулы, проиллюстрированные ниже, которые включают гомодимер, гомотример и т.п., в качестве антигенного элемента связывания: GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), TNF, TNF-альфа, TNF-альфа-бета, TNF-бета 2, TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 лиганда Fas, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), VEGF, IgE, IgA, IgG, IgM, RANKL, TGF-альфа, TGF-бета, панспецифический TGF-бета и IL-8.
Эпитопы
"Эпитоп" означает антигенную детерминанту в антигене и относится к антигенному центру, с которым связывается антигенсвязывающий домен антигенсвязывающей молекулы, описанной в настоящем описании. Таким образом, например, эпитоп может быть определен в соответствии с его структурой. Альтернативно эпитоп может быть определен в соответствии с антигенсвязывающей активностью антигенсвязывающей молекулы, которая распознает эпитоп. Когда антиген представляет собой пептид или полипептид, эпитоп может быть указан с помощью аминокислотных остатков, образующих эпитоп. Альтернативно, когда эпитоп представляет собой цепь сахара, эпитоп может быть указан с помощью его определенной структуры цепи сахаров.
Линейный эпитоп представляет собой эпитоп, который содержит эпитоп, в котором распознается его первичная аминокислотная последовательность. Такой линейный эпитоп, как правило, содержит по меньшей мере три и наиболее часто по меньшей мере пять, например, приблизительно 8-10 или 6-20 аминокислот в его конкретной последовательности.
В противоположность линейному эпитопу, "конформационный эпитоп" представляет собой эпитоп, в котором первичная аминокислотная последовательность, содержащая эпитоп, не является единственной детерминантой распознаваемого эпитопа (например, первичная аминокислотная последовательность конформационного эпитопа не обязательно распознается определяющим эпитоп антителом). Конформационные эпитопы могут содержать большее количество аминокислот по сравнению с линейными эпитопами. Антитело, распознающее конформационный эпитоп, распознает трехмерную структуру пептида или белка. Например, когда молекула белка сворачивается и формирует трехмерную структуру, аминокислоты и/или основные цепи полипептида, которые образуют коноформационный эпитоп, располагаются рядом друг с другом, и эпитоп становится распознаваемым для антитела. Способы определения конформаций эпитопов включают, например, рентгеновскую кристаллографию, двумерный ядерный магнитный резонанс, сайт-специфическое спиновое мечение и электронный парамагнитный резонанс, он они не ограничиваются ими. См., например, Epitope Mapping Protocols in Methods in Molecular Biology (1996), Vol. 66, Morris (ed.).
Структура антигенсвязывающего домена, которая связывается с эпитопом, называется паратопом. Эпитоп и паратоп стабильно связываются под действием водородных связей, электростатической силы, ван-дер-ваальсовых силы, гидрофобных связей и т.п. между эпитопом и паратопом. Эта прочность связывания между эпитопом и паратопом называется аффинностью. Общая сумма прочностей связывания, когда связывается несколько эпитопов и несколько паратопов, называется авидностью. Когда антитело, содержащее несколько паратопов (т.е. поливалентное антитело) и т.п. связывается с несколькими эпитопами, аффинность действует аддитивно или синергично, и, таким образом, авидность превышает аффинность.
Активность связывания
Примеры способа оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, направленный на IgA, описаны ниже. В соответствии с примерами ниже, также можно соответствующим образом выполнять способы оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, с антигеном, отличным от IgA.
Например, то, что исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен против IgA, распознает линейный эпитоп в молекуле IgA, можно подтвердить, например, как описано ниже. Например, для указанной выше цели синтезируют линейный пептид, содержащий аминокислотную последовательность, образующую константную область IgA. Пептид можно синтезировать химически или его можно получать способами генетической инженерии с использованием области, кодирующей аминокислотную последовательность, соответствующую константной области в кДНК IgA. Затем исследуемую антигенсвязывающую молекулу, содержащую антигенсвязывающий домен, направленный на IgA, оценивают в отношении ее активности связывания с линейным пептидом, содержащим аминокислотную последовательность, образующую константную область. Например, можно проводить ELISA с иммобилизованным линейным пептидом в качестве антигена для оценки активности связывания антигенсвязывающей молекулы с пептидом. Альтернативно активность связывания с линейным пептидом можно оценивать, исходя из уровня ингибирования линейным пептидом связывания антигенсвязывающей молекулы с клетками с IgA. Эти исследования могут продемонстрировать активность связывания антигенсвязывающей молекулы с линейным пептидом.
Распознавание конформационного эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, нацеленный на белок IgA, можно подтверждать, как указано ниже. Для упомянутой выше задачи, как описано в настоящем описании, используют общий способ генетической рекомбинации для переноса кодирующего IgA рекомбинантного гена в клетки-хозяева (например, клетки животных, клетки насекомых или клетки дрожжей), которые обеспечивают образование нативного конформационного эпитопа в белке IgA. IgA, содержащий конформационный эпитоп, получают из культуры рекомбинантных клеток, полученных таким путем. Распознавание конформационного эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, нацеленный на IgA, происходит, например, когда исследуемая антигенсвязывающая молекула связывается прочно с молекулой IgA, когда она контактирует с иммобилизованным IgA, содержащим конформационный эпитоп, в то время как антигенсвязывающая молекула по существу не связывается с линейным пептидом, содержащим аминокислотную последовательность, составляющую аминокислотную последовательность иммобилизованного IgA. Альтернативно также можно использовать, вместо упомянутого выше линейного пептида, исследуемую нацеленную на IgA антигенсвязывающую молекулу, которая денатурирована восстановителем, который расщепляет дисульфидные связи, таким как дитиотреитол, дитиоэритрит, β-меркаптоэтанол, фосфины и боргидрид натрия, и/или хаотропными агентами, такими как поверхностно-активные вещества, включающие гидрохлорид гуанидина, мочевину и лаурилсульфат натрия. В данном случае "по существу не связывает" относится к активности связывания, не превышающей 80%, обычно не превышающей 50%, предпочтительно не превышающей 30% или особенно предпочтительно не превышающей 15% от активности связывания IgA человека.
Способы оценки активности связывания в отношении IgA исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен против IgA, включают, например, способы, описанные в Antibodies: A Laboratory Manual (Ed Harlow, David Lane, Cold Spring Harbor Laboratory (1988) 359-420). В частности, оценку можно проводить на основе принципа ELISA или EIA с использованием IgA в качестве антигена.
В формате ELISA активность связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для IgA, с IgA можно оценивать количественно путем сравнения уровней сигнала, генерируемого ферментативной реакцией. В частности, исследуемую антигенсвязывающую молекулу добавляют в планшет для ELISA, на котором иммобилизован IgA. Затем исследуемую антигенсвязывающую молекулу, которая связалась с IgA, иммобилизованном на планшете, выявляют с использованием меченного ферментом антитела, которое распознает исследуемую антигенсвязывающую молекулу. В ELISA можно получать серийные разведения исследуемой антигенсвязывающей молекулы и можно определять титр связывания антитела с IgA для сравнения активности связывания исследуемой антигенсвязывающей молекулы с IgA.
Связывание исследуемой антигенсвязывающей молекулы с антигеном, экспрессируемым на поверхности клеток, суспендированных в буфере или сходных с ним, можно выявлять с использованием проточного цитометра. Известные проточные цитометры включают, например, следующие устройства:
FACSCantoTM II,
FACSAriaTM,
FACSArrayTM,
FACSVantageTM SE,
FACSCaliburTM (все являются торговыми названиями BD Biosciences),
EPICS ALTRA HyPerSort,
Cytomics FC 500,
EPICS XL-MCL ADC EPICS XL ADC,
Cell Lab Quanta/Cell Lab Quanta SC (все являются торговыми названиями Beckman Coulter).
Предпочтительные способы оценки активности связывания в отношении антигена исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен против IgA, с антигеном, включают, например, следующий способ. Сначала экспрессирующие IgA клетки подвергают реакции с исследуемой антигенсвязывающей молекулой, а затем их окрашивают меченным FITC вторичным антителом, которое распознает антигенсвязывающую молекулу. Исследуемую антигенсвязывающую молекулу надлежащим образом разбавляют подходящим буфером с получением молекулы с желаемой концентрацией. Например, молекулу можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Затем определяют интенсивность флуоресценции и количество клеток с использованием FACSCalibur (BD). Интенсивность флуоресценции, полученную с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. среднее геометрическое значение, отражает количество антитела, связанного с клетками. Следовательно, активность связывания исследуемой антигенсвязывающей молекулы, которую отражает количество связанной исследуемой антигенсвязывающей молекулы, можно определять путем определения геометрического среднего значения.
То, обладает ли исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для IgA, общим эпитопом с другой антигенсвязывающей молекулой, можно оценивать на основе конкуренции между двумя молекулами за один и тот же эпитоп. Конкуренцию между антигенсвязывающими молекулами можно выявлять с помощью анализа перекрестного блокирования или сходных с ним. Например, предпочтительным анализом перекрестного блокирования является конкурентный анализ ELISA.
В частности, в анализе перекрестного блокирования белок IgA, иммобилизованный на лунках микропланшета для титрования, предварительно инкубируют в присутствии или в отсутствие являющейся кандидатом конкурентной антигенсвязывающей молекулы, а затем к ним добавляют антигенсвязывающую молекулу. Количество исследуемой антигенсвязывающей молекулы, связавшейся с белком IgA в лунках, непрямо коррелирует со связывающей способностью являющейся кандидатом конкурентной антигенсвязывающей молекулы, которая конкурирует за связывание с тем же эпитопом. Следовательно, чем более высокой является аффинность конкурентной антигенсвязывающей молекулы к тому же эпитопу, тем более низкой является активность связывания исследуемой антигенсвязывающей молекулы с покрытыми белком IgA лунками.
Количество исследуемой антигенсвязывающей молекулы, связанной с лунками через IgA, можно легко определить путем предварительного мечения антигенсвязывающей молекулы. Например, меченную биотином антигенсвязывающую молекулу определяют с использованием конъюгата авидин/пероксидаза и соответствующего субстрата. В частности, анализ перекрестного блокирования, в котором используются ферментные метки, такие пероксидаза, называют "конкурентным анализом ELISA". Антигенсвязывающую молекулу также можно метить другими агентами для мечения, которые обеспечивают выявление или измерение. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Когда являющаяся кандидатом конкурентная антигенсвязывающая молекула может блокировать связывание исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для IgA, по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50%, и более предпочтительно по меньшей мере на 50% по сравнению с активностью связывания в контрольном эксперименте, исследованном в отсутствие конкурентной антигенсвязывающей молекулы, то определяют, что исследуемая антигенсвязывающая молекула по существу связывается с тем же эпитопом, с которым связывается конкурентная антигенсвязывающая молекула, или конкурирует за связывание с тем же эпитопом.
Когда структура эпитопа, с которым связывается исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для IgA, уже идентифицирована, тогда то, имеют ли исследуемые и контрольные антигенсвязывающие молекулы общий эпитоп, можно оценивать путем сравнения активности связывания двух антигенсвязывающих молекул с пептидом, полученным путем внесения аминокислотных мутаций в пептид, формирующий эпитоп.
Для измерения указанной выше активности связывания, например, активность связывания исследуемых и контрольных антигенсвязывающих молекул в отношении линейного пептида, в который внесена мутация, сравнивают в описанном выше формате ELISA. Помимо способов ELISA, активность связывания с мутантным пептидом, связанным с колонкой, можно определять путем пропускания исследуемых и контрольных антигенсвязывающих молекул через колонку, а затем количественного определения антигенсвязывающей молекулы, элюированной в элюирующем растворе. Способы адсорбции мутантного пептида на колонку, например, в форме слитого пептида с GST, известны.
Альтернативно, когда идентифицированный эпитоп в антигене, экспрессируемом на клетке, представляет собой конформационный эпитоп, тогда то, обладают ли исследуемые и контрольные антигенсвязывающие молекулы общим эпитопом, можно оценивать, например, следующим способом. Ниже предоставлено пояснение с использованием случая с IgE в качестве примера. Сначала получают экспрессирующие IL-6R клетки и клетки, экспрессирующие IL-6R с мутацией, внесенной в эпитоп. Исследуемые и контрольные антигенсвязывающие молекулы добавляют в суспензию клеток, полученную суспендированием этих клеток в соответствующем буфере, таком как PBS. Затем клеточные суспензии надлежащим образом промывают буфером, и к ним добавляют меченное FITC антитело, которое распознает исследуемые и контрольные антигенсвязывающие молекулы. Интенсивность флуоресценции и количество клеток, окрашенных меченым антителом, определяют с использованием FACSCalibur (BD). Исследуемые и контрольные антигенсвязывающие молекулы надлежащим образом разбавляют с использованием подходящего буфера и используют в желаемых концентрациях. Например, их можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Интенсивность флуоресценции, определенная с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. геометрическое среднее значение, отражает количество меченного антитела, связанного с клетками. Следовательно, активность связывания исследуемых и контрольных антигенсвязывающих молекул, которую отражает количество связавшегося меченого антитела, можно определять путем измерения геометрического среднего значения.
В описанном выше способе то, что антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный IgE," можно оценивать, например, следующим способом. Сначала исследуемые и контрольные антигенсвязывающие молекулы, связанные с клетками, экспрессирующими мутантный IgE, окрашивают меченым антителом. Затем определяют интенсивность флуоресценции клеток. Когда используют FACSCalibur для выявления флуоресценции проточной цитометрией, определенную интенсивность флуоресцении можно анализировать с использованием программного обеспечения CELL QUEST. Из геометрических средних значений в присутствии и в отсутствие полипептидного комплекса, можно вычислять сравнительное значение (ΔGeo-Mean) по следующей формуле для определения соотношения увеличения интенсивности флуоресценции в результате связывания антигенсвязывающей молекулой.
ΔGeo-Mean = Geo-Mean (в присутствии полипептидного комплекса)/Geo-Mean (в отсутствие полипептидного комплекса).
Геометрическое среднее сравнительное значение (значение ΔGeo-Mean для мутантной молекулы IgE), определенное с помощью описанного выше анализа, которое отражает количество исследуемой антигенсвязывающей молекулы, связанной с клетками, экспрессирующими мутантный IgE, сравнивают со сравнительным значением ΔGeo-Mean, которое отражает количество исследуемой антигенсвязывающей молекулы, связавшейся с экспрессирующими IgE клетками. В этом случае, особенно предпочтительно, чтобы концентрации исследуемой антигенсвязывающей молекулы, используемой для определения сравнительных значений ΔGeo-Mean для экспрессирующих IgE клеток и клеток, экспрессирующих мутантный IgE, были скорректированы так, чтобы они были равными или по существу равными. В качестве контрольной антигенсвязывающей молекулы используют антигенсвязывающую молекулу, для которой подтверждено, что она распознает эпитоп в IgE.
Если сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для клеток, экспрессирующих мутантный IgE, меньше, чем сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для экспрессирующих IgE клеток, по меньшей мере на 80%, предпочтительно на 50%, более предпочтительно на 30%, и особенно предпочтительно на 15%, тогда исследуемая антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный IgE". Формула для определения значения Geo-Mean (геометрического среднего значения) описана в руководстве пользователя программного обеспечения CELL QUEST (BD biosciences). Если сравнение показывает, что сравнительные величины по существу эквивалентны, то может быть определено, что эпитоп для исследуемой и контрольной антигенсвязывающих молекул является одним и тем же.
Антигенсвязывающий домен
В рамках настоящего изобретения "антигенсвязывающий домен" может представлять собой любую структуру, при условии, что она связывается с представляющим интерес антигеном. Такие домены предпочтительно включают, например:
вариабельные области тяжелых цепей и легких цепей антител;
модуль приблизительно из 35 аминокислот, называемый A-доменом, который содержится в мембранном белке клеток авимере in vivo (международная публикация № WO 2004/044011, международная публикация № WO 2005/040229);
аднектин, содержащий домен 10Fn3, который связывается с белковой частью фибронектина - гликопротеина, экспрессирующегося на клеточной мембране (международная публикация № WO 2002/032925);
аффитело, которое состоит из трехспирального пучка из 58-аминокислот на основе каркаса связывающего IgG домена белка A (международная публикация № WO 1995/001937);
Смоделированные белки анкиринового повтора (DARPin), которые представляют собой область, экспонированную на молекулярной поверхности анкириновых повторов (AR), имеющих структуру, в которой субъединица, состоящая из поворота, содержащего 33 аминокислотных остатка, две антипараллельных спирали и петлю многократно сложены в стопку (международная публикация № WO 2002/020565);
антикалины и т.п., которые представляют собой домены, состоящие из четырех петель, которые поддерживают одну сторону структуры бочки, состоящей из восьми расположенных по кругу антипараллельных нитей, которые являются высоко консервативными среди молекул липокалина, таких как ассоциированный с желатиной липокалин нейтрофилов (NGAL) (международная публикация № WO 2003/029462); и
вогнутая область, образованная структурой параллельных слоев внутри структуры в форме подковы, образованной уложенными в стопку повторами модуля лейцин-богатого повтора (LRR) вариабельного рецептора лимфоцитов (VLR), который не имеет иммуноглобулиновой структуры и используется в системе приобретенного иммунитета у бесчелюстных позвоночных, таких как минога и миксина (WO 2008/016854). Предпочтительные антигенсвязывающие домены по настоящему изобретению включают, например, антигенсвязывающие домены, имеющие вариабельные области тяжелых цепей и легких цепей антител. Предпочтительные примеры антигенсвязывающих доменов включают "одноцепочечный Fv (scFv)", "одноцепочечное антитело", "Fv", "одноцепочечный Fv2 (scFv2)", "Fab" и "F(ab’)2".
Антигенсвязывающие домены антигенсвязывающих молекул по настоящему изобретению могут связываться с идентичным эпитопом. Такой идентичный эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:2. Альтернативно, каждый из антигенсвязывающих доменов антигенсвязывающих молекул по настоящему изобретению может связываться с отличающимся эпитопом. В рамках настоящего изобретения, отличающийся эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:2.
Иммунный комплекс
Иммунный комплекс относится к относительно стабильной структуре, образующейся, когда по меньшей мере один антиген и по меньшей мере одна антигенсвязывающая молекула связываются друг с другом с образованием комплекса с большей молекулярной массой. Неограничивающим вариантом осуществления иммунного комплекса является, например, агрегат антиген-антитело. Способ оценки образования иммунных комплексов, содержащих две или более антигенсвязывающих молекул и два или более антигенных элементов связывания, описан ниже в настоящем описании.
Специфичность
"Специфичный" означает, что одна из молекул, которая специфически связывается, не проявляет никакого существенного связывания с молекулами, отличными от одной или нескольких молекул-партнеров, с которыми она связывается. Более того, "специфичный" также используют, когда антигенсвязывающий домен является специфичным к конкретному эпитопу среди множества эпитопов в антигене. Когда эпитоп, связываемый антигенсвязывающим доменом, содержится в нескольких различных антигенах, антигенсвязывающие молекулы, содержащие антигенсвязывающий домен, могут связываться с различными антигенами, которые имеют этот эпитоп. В данном случае, "не проявляет никакого существенного связывания" означает демонстрацию не более чем 50%, как правило, не более чем 30%, предпочтительно не более чем 15%, особенно предпочтительно не более чем 10%, и еще более предпочтительно не более чем 5% активности связывания с молекулой-партнером относительно молекул, отличных от молекулы-партнера.
Антитело
В рамках настоящего изобретения "антитело" относится к природному иммуноглобулину или иммуноглобулину, полученному частичным или полным синтезом. Антитела можно выделять из природных источников, таких как природная плазма и сыворотка, или из культуральных супернатантов продуцирующих антитела гибридом. Альтернативно антитела можно частично или полностью синтезировать с использованием способов, таких как генетическая рекомбинация. Предпочтительные антитела включают, например, антитела изотипа иммуноглобулинов или принадлежащего ему подкласса. Известные иммуноглобулины человека включают антитела следующих девяти классов (изотипов): IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE и IgM. Из этих изотипов антитела по настоящему изобретению включают IgG1, IgG2, IgG3 и IgG4. Ряд аллотипических последовательностей константных областей IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека вследствие полиморфизмов генов описан в "Sequences of proteins of immunological interest", NIH Publication No. 91-3242. В рамках настоящего изобретения можно использовать любую из таких последовательностей. В частности, для последовательности IgG1 человека, аминокислотная последовательность в положениях 356-358, как указано в соответствии с нумерацией EU, может представлять собой DEL или EEM. Несколько аллотипических последовательностей вследствие генетических полиморфизмов описано в "Sequences of proteins of immunological interest", NIH Publication No. 91-3242. Для константной области Igκ (каппа) человека и константной области Igλ (лямбда) человека в рамках настоящего изобретения можно использовать любую из последовательностей.
Способы получения антитела с желаемой активностью связывания известны специалистам в данной области. Ниже приведен пример, в котором описан способ получения антитела, которое связывается с IgA (антитело против IgA). Антитела, которые связываются с антигеном, отличным от IgA, также можно получать в соответствии с примером, описанным ниже.
Антитела против IgA можно получать в качестве поликлональных или моноклональных антител с использованием известных способов. Продуцируемые антитела против IgA предпочтительно представляют собой моноклональные антитела, происходящие из млекопитающих. Такие происходящие из млекопитающих моноклональные антитела включают антитела, продуцируемые гибридомами или клетками-хозяевами, трансформированными экспрессирующим вектором, несущим ген антитела, способами генетической инженерии. Между тем, моноклональные антитела по настоящему изобретению включают "гуманизированные антитела" или "химерные антитела".
Продуцирующие моноклональные антитела гибридомы можно получать с использованием известных способов, например, как описано ниже. В частности, млекопитающих иммунизируют общепринятыми способами иммунизации с использованием белка IgA в качестве сенсибилизирующего антигена. Полученные иммунные клетки подвергают слиянию с известными родительскими клетками общепринятыми способами слияния клеток. Затем гибридомы, продуцирующие антитело против IgA, можно отбирать путем скрининга продуцирующих моноклональное антитело клеток с использованием общепринятых способов скрининга.
В частности, получение моноклонального антитела проводят, например, как представлено ниже. Например, очищенный нативный белок IgA можно использовать в качестве сенсибилизирующего антигена. Более того, рекомбинантный белок, содержащий последовательность полипептида IgA SEQ ID NO:2, которая зарегистрирована в качестве L00022|IGHE*02 в IMGT/GENE-DB, очищают с получением растворимого IgA из культурального супернатанта. Для экспрессии рекомбинантного белка сначала ген константной области тяжелой цепи IgA, нуклеотидная последовательность которого представлена в SEQ ID NO:3, можно экспрессировать с получением белка IgA, представленного в SEQ ID NO:2, который используют в качестве сенсибилизирующего антигена для получения антител. Таким образом, подходящие клетки-хозяева трансформируют путем вставки кодирующей последовательности гена IgA в известный экспрессирующий вектор. Желаемый белок IgA очищают известными способами из клеток-хозяев или из их культурального супернатанта. Для экспрессии гена константной области тяжелой цепи IgA последовательность полинуклеотида SEQ ID NO:3 можно функционально связывать с 3'-концом сигнальной последовательности. В другом неограничивающем варианте осуществления полинуклеотидную последовательность SEQ ID NO:3 можно функционально связывать с 3'-концом полинуклеотидной последовательности, кодирующей вариабельную область тяжелой цепи, содержащую сигнальную последовательность на 5'-конце. Более того, для очистки рекомбинантного белка к N-концу или С-концу SEQ ID NO:2 можно соответствующим образом добавлять пептид-метку для очистки. Примером его неограничивающего варианта осуществления является GLNDIFEAQKIEWHE (SEQ ID NO:4), который представляет собой Avi-метку, слитую через линкер GGGGS (SEQ ID NO:31). Неограничивающим примером получения рекомбинантного белка, сходного с упомянутым выше, является такой способ, как способ, описанный в примере 1, где рекомбинантный белок IgA, экспрессированный в комбинации с легкой цепью, собирают из культуры рекомбинантных клеток-хозяев, а затем очищают.
Очищенный белок IgA можно использовать в качестве сенсибилизирующего антигена для иммунизации млекопитающих. Также в качестве сенсибилизирующего антигена можно использовать неполный пептид IgA. В этом случае, неполный пептид можно получать путем химического синтеза на основе аминокислотной последовательности IgA человека, или путем встраивания неполного гена IgA в экспрессирующий вектор для экспрессии. Альтернативно неполный пептид можно получать путем деградации белка IgA протеазой. Длина и область неполного пептида IgA не ограничены конкретными вариантами осуществления. Предпочтительная область может быть произвольным образом выбрана из аминокислотной последовательности SEQ ID NO:2. Количество аминокислот, образующих пептид, подлежащий применению в качестве сенсибилизирующего антигена, предпочтительно составляет по меньшей мере пять или более, шесть или более, или семь или более. Более конкретно, в качестве сенсибилизирующего антигена можно использовать пептид из 8-50 остатков, более предпочтительно, из 10-30 остатков.
Альтернативно, для сенсибилизирующего антигена можно использовать слитый белок, полученный путем слияния желаемого неполного полипептида или пептида белка IgA с отличающимся полипептидом. Например, предпочтительно в качестве сенсибилизирующих антигенов используют Fc-фрагменты антител и пептидные метки. Векторы для экспрессии таких слитых белков можно конструировать путем слияния в рамке считывания генов, кодирующих два или более желаемых полипептидных фрагментов, и встраивания слитого гена в экспрессирующий вектор, как описано выше. Способы получения слитых белков описаны в Molecular Cloning 2nd ed. (Sambrook, J et al., Molecular Cloning 2nd ed., 9.47-9.58 (1989) Cold Spring Harbor Lab. Press). Способы получения IgA для применения в качестве сенсибилизирующего антигена и способы иммунизации с использованием IgA конкретно описаны в PCT/JP2011/077619 и т.п.
Отсутствуют конкретное ограничение на млекопитающих, подлежащих иммунизации сенсибилизирующим антигеном. Однако предпочтительно выбирать млекопитающих, учитывая их совместимость с родительскими клетками, используемыми для слияния клеток. Как правило, предпочтительно используют грызунов, таких как мыши, крысы и хомячки, кроликов и обезьян.
Указанных выше животных иммунизируют сенсибилизирующим антигеном известными способами. Как правило, выполняемые способы иммунизации включают, например, внутрибрюшинную или подкожную инъекцию сенсибилизирующего антигена млекопитающим. В частности, сенсибилизирующий антиген соответствующим образом разбавляют PBS (фосфатно-солевой буфер), физиологическим солевым раствором или сходными с ними. Если желательно, общепринятый адъювант, такой как полный адъювант Фрейнда, смешивают с антигеном и смесь эмульгируют. Затем сенсибилизирующий антиген вводят млекопитающему несколько раз с интервалами 4-21 суток. При иммунизации сенсибилизирующим антигеном можно использовать подходящие носители. В частности, когда в качестве сенсибилизирующего антигена используют низкомолекулярный неполный пептид, иногда желательно связывать сенсибилизирующий антигенный пептид с белком-носителем, таким как альбумин или гемоцианин лимфы улитки, для иммунизации.
Альтернативно гибридомы, продуцирующие желаемое антитело, можно получать с использованием способов иммунизации ДНК, как описано ниже. Иммунизация ДНК представляет собой способ иммунизации, который обеспечивает стимуляцию иммунной системы путем экспрессии сенсибилизирующего антигена у животного, иммунизированного в результате введения векторной ДНК, сконструированной для обеспечения экспрессии гена, кодирующего антигенный белок, у животного. По сравнению с общепринятыми способами иммунизации, в которых белковый антиген вводят животным, подлежащим иммунизации, иммунизация ДНК является лучшей в том, что:
- в случае, когда антиген представляет собой мембранный белок, может быть обеспечена стимуляция иммунной системы при сохранении структуры мембранного белка; и
- отсутствует необходимость в очистке антигена для иммунизации.
Для получения моноклонального антитела по настоящему изобретению с использованием иммунизации ДНК, сначала ДНК, экспрессирующую белок IgA, вводят животному, подлежащему иммунизации. ДНК, кодирующую IgA, можно синтезировать известными способами, такими как ПЦР. Полученную ДНК встраивают в соответствующий экспрессирующий вектор, а затем его вводят животному, подлежащему иммунизации. Предпочтительно используемые экспрессирующие векторы включают, например, коммерчески доступные экспрессирующие векторы, такие как pcDNA3.1. Векторы можно вводить в организм с использованием общепринятых способов. Например, иммунизацию ДНК проводят с использованием генной пушки для введения покрытых экспрессирующим вектором частиц золота в клетки организма животного, подлежащего иммунизации. Антитела, которые распознают IgA, также можно получать способами, описанными в PCT/JP2011/077619.
После иммунизации млекопитающего, как описано выше, увеличение титра связывающего IgA антитела подтверждают в сыворотке. Затем иммунные клетки собирают от млекопитающего, а затем подвергают слиянию клеток. В частности, предпочтительно в качестве иммунных клеток используют спленоциты.
В качестве клетки, подлежащей слиянию с упомянутыми выше иммунными клетками, используют клетки миеломы млекопитающих. Клетки миеломы предпочтительно содержат подходящий селективный маркер для скрининга. Селективный маркер придает клеткам характеристики для их выживания (или гибели) в конкретных условиях культивирования. В качестве селективных маркеров известны дефицит гипоксантин-гуанин-фосфорибозилтрансферазы (далее сокращенно обозначаемый как дефицит HGPRT) и дефицит тимидинкиназы (далее сокращенно обозначаемый как дефицит TK). Клетки с дефицитом HGPRT или TK обладают чувствительностью к гипоксантину-аминоптерину-тимидину (далее сокращенно обозначаемой чувствительностью HAT). Чувствительные к HAT клетки не могут синтезировать ДНК в селективной среде HAT, и, таким образом, погибают. Однако когда клетки слиты с нормальными клетками, они могут продолжать синтезировать ДНК с использованием пути спасения нормальных клеток, и, таким образом, они могут расти в селективной среде HAT.
Селекцию клеток с дефицитом HGPRT и TK можно проводить в среде, содержащей 6-тиогуанин, 8-азагуанин (далее сокращенно обозначаемый как 8AG) или 5’-бромдезоксиуридин, соответственно. Нормальные клетки погибают, поскольку они включают аналоги пиримидина в их ДНК. Между тем, клетки, которые являются дефицитными по этим ферментам, могут выживать в селективной среде, поскольку они не могут включать эти аналоги пиримидина. Кроме того, селективный маркер, называемый устойчивостью к G418, обеспечиваемый геном устойчивости к неомицину, обеспечивает устойчивость к 2-дезоксистрептаминовым антибиотикам (аналоги гентамицина). Известны различные типы миеломных клеток, которые пригодны для слияния клеток.
Например, предпочтительно можно использовать миеломные клетки, включающие следующие клетки:
P3(P3x63Ag8,653) (J. Immunol. (1979) 123 (4), 1548-1550);
P3x63Ag8U.1 (Current Topics in Microbiology и Immunology (1978)81, 1-7);
NS-1 (C. Eur. J. Immunol. (1976)6 (7), 511-519);
MPC-11 (Cell (1976) 8 (3), 405-415);
SP2/0 (Nature (1978) 276 (5685), 269-270);
FO (J. Immunol. Methods (1980) 35 (1-2), 1-21);
S194/5.XX0.BU.1 (J. Exp. Med. (1978) 148 (1), 313-323);
R210 (Nature (1979) 277 (5692), 131-133), и т.д.
Слияние клеток между иммуноцитами и клетками миеломы по существу проводят с использованием известных способов, например, способа Kohler and Milstein et al. (Methods Enzymol. (1981) 73: 3-46).
Более конкретно, слияние клеток можно проводить, например, в общепринятой культуральной среде в присутствии агента, обеспечивающего слияние клеток. Агенты, обеспечивающие слияние клеток, включают, например, полиэтиленгликоль (PEG) и вирус Сендай (HVJ). Если необходимо, также можно добавлять вспомогательное вещество, такое как диметилсульфоксид, для повышения эффективности слияния.
Соотношение иммунных клеток к клеткам миеломы можно определять самостоятельно, предпочтительно, например, оно составляет одну клетку миеломы на каждые от одного до десяти иммуноцитов. Культуральные среды, используемые для слияния клеток, включают, например, среды, которые пригодны для выращивания клеточных линий, такие как среда RPMI1640 и среда MEM, и другие общепринятые культуральные среды, используемые для этого типа клеточной культуры. Кроме того, предпочтительно в культуральную среду можно добавлять сывороточные добавки, такие как эмбриональная телячья сыворотка (FCS).
Для слияния клеток заданные количества указанных выше иммунных клеток и миеломных клеток смешивают в описанной выше культуральной среде. Затем добавляют раствор PEG (например, средняя молекулярная масса составляет приблизительно от 1000 до 6000), предварительно нагретый до приблизительно 37°C, в концентрации, как правило, от 30% до 60% (масс./об.). Затем эту смесь осторожно перемешивают для получения желаемых слитых клеток (гибридом). Затем к клеткам постепенно добавляют подходящую культуральную среду, упомянутую выше, и смесь многократно центрифугируют для удаления супернатанта. Таким образом, можно удалить агенты слияния клеток и т.п., которые являются неблагоприятными для роста гибридомы.
Селекцию полученных таким образом гибридом можно проводить путем культивирования с использованием общепринятой селективной среды, например, среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Клетки, отличные от желаемых гибридом (неслитые клетки) можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. Как правило, период составляет от нескольких суток до нескольких недель. Затем, гибридомы, продуцирующие желаемое антитело, подвергают скринингу и клонируют по отдельности общепринятыми способами лимитирующих разведений.
Селекцию полученных таким образом гибридом можно проводить с использованием селективной среды на основе селективного маркера, которым обладает миелома, используемая для слияния клеток. Например, селекцию клеток с дефицитом HGPRT или TK можно проводить путем культивирования с использованием среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). В частности, когда для слияния клеток используют чувствительные к HAT миеломные клетки, клетки, успешно слитые с нормальными клетками, могут селективно пролиферировать в среде HAT. Клетки, отличные от желаемых гибридом (неслитые клетки), можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. В частности, селекцию желаемых гибридом можно проводить путем культивирования, как правило, в течение от нескольких суток до нескольких недель. Затем гибридомы, продуцирующие желаемое антитело, подвергают скринингу и по отдельности клонируют общепринятыми способами лимитирующих разведений.
Желаемые антитела можно предпочтительно отбирать и по отдельности клонировать способами скрининга на основе известной реакции антиген/антитело. Например, активность связывания антитела с иммобилизованным IgA можно оценивать на основе принципа ELISA. Например, IgA иммобилизуют на лунках планшета для ELISA. Культуральные супернатанты гибридом контактируют с IgA в лунках, и выявляют антитела, которые связываются с IgA. Когда моноклональные антитела происходят из мыши, антитела, связанные с клетками, можно выявлять с использованием антитела против иммуноглобулина мыши. Селекцию гибридом, продуцирующих желаемое антитело, обладающее антигенсвязывающей способностью, проводят с помощью описанного выше скрининга, и их можно клонировать способом лимитирующих разведений и т.п.
Продуцирующие моноклональное антитело гибридомы, полученные таким образом, можно пассировать в общепринятой культуральной среде и хранить в жидком азоте в течение длительного периода времени.
Описанные выше гибридомы культивируют общепринятым способом, и желаемые моноклональные антитела можно получать из культуральных супернатантов. Альтернативно гибридомы вводят совместимым млекопитающим и выращивают в них, и моноклональные антитела получают из асцитной жидкости. Первый из этих способов пригоден для получения антител с высокой чистотой.
Предпочтительно также можно использовать антитела, кодируемые генами антител, которые клонированы из антителопродуцирующих клеток, таких как описанные выше гибридомы. Клонированный ген антитела встраивают в подходящий вектор, и его вводят в клетку-хозяина для экспрессии антитела, кодируемого геном. Способы выделения генов антител, встраивания генов в векторы и трансформации клеток-хозяев, уже разработаны, например, Vandamme et al. (Eur. J. Biochem. (1990) 192(3), 767-775). Способы продуцирования рекомбинантных антител также известны, как описано ниже.
Например, кДНК, кодирующую вариабельную область (V-область) антитела против IgA, получают из клеток гибридомы, продуцирующих антитело против IgA. Для этой цели сначала из гибридом экстрагируют тотальную РНК. Способы, используемые для экстракции мРНК из клеток, включают, например:
- способ ультрацентрифугирования с гуанидином (Biochemistry (1979) 18(24), 5294-5299), и
- способ AGPC (Anal. Biochem. (1987) 162(1), 156-159)
Экстрагированные мРНК можно очищать с использованием набора для очистки мРНК mRNA Purification Kit(GE Healthcare Bioscience) и т.п. Альтернативно также коммерчески доступны наборы для экстракции тотальной мРНК непосредственно из клеток, такие как набор для очистки мРНК QuickPrep mRNA Purification Kit (GE Healthcare Bioscience). мРНК можно получать из гибридом с использованием таких наборов. кДНК, кодирующие V-область антитела, можно синтезировать из полученных мРНК с использованием обратной транскриптазы. кДНК можно синтезировать с использованием набора для синтеза первой цепи кДНК с обратной транскриптазой AMV Reverse Transcriptase First-strand cDNA Synthesis Kit (Seikagaku Co.) и т.п. Более того, для синтеза и амплификации кДНК можно соответствующим образом использовать набор для амплификации кДНК SMART RACE cDNA amplification kit (Clontech) и способ 5’-RACE на основе ПЦР (Proc. Natl. Acad. Sci. USA (1988) 85(23), 8998-9002; Nucleic Acids Res. (1989) 17(8), 2919-2932). В таком способе синтеза кДНК на оба конца кДНК можно вносить подходящие участки для ферментов рестрикции, описанные ниже.
Представляющий интерес фрагмент кДНК очищают из полученного продукта ПЦР, а затем его лигируют в векторную ДНК. Таким образом, конструируют рекомбинантный вектор и вводят в E. coli и т.п. После селекции колоний из образующих колонии E. coli можно получать желаемый рекомбинантный вектор. Затем исследуют, имеет ли рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК, известным способом, таким как способ с терминацией цепи дидезоксинуклеотидами.
Способ 5’-RACE, в котором используются праймеры для амплификации гена вариабельной области, удобно использовать для выделения гена, кодирующего вариабельную область. Сначала конструируют библиотеку кДНК 5’-RACE с помощью синтеза кДНК с использованием РНК, экстрагированных из клеток гибридом в качестве матрицы. Для синтеза библиотеки кДНК 5’-RACE соответствующим образом используют коммерчески доступный набор, такой как набор для амплификации кДНК SMART RACE.
Ген антитела амплифицируют способом ПЦР с использованием полученной библиотеки кДНК 5’-RACE в качестве матрицы. Праймеры для амплификации гена антитела мыши можно конструировать на основе известных последовательностей генов антител. Нуклеотидные последовательности праймеров варьируют, в зависимости от подкласса иммуноглобулинов. Таким образом, предпочтительно, чтобы подкласс был определен заранее с использованием коммерчески доступного набора, такого как набор для изотипирования моноклональных антител мыши Iso Strip mouse monoclonal antibody isotyping kit (Roche Diagnostics).
В частности, например, праймеры, которые обеспечивают амплификацию генов, кодирующих тяжелые цепи γ1, γ2a, γ2b и γ3 и легкие цепи κ и λ, используют для выделения генов, кодирующих IgG мыши. Как правило, для амплификации гена вариабельной области gG используют праймер, который гибридизуется с областью константной области вблизи вариабельной области, в качестве 3’-праймера. Между тем, в качестве 5’-праймера используют праймер, прилагаемый к набору для конструирования библиотеки кДНК 5’-RACE.
Продукты ПЦР, амплифицированные таким образом, используют для переформировывания иммуноглобулинов, состоящих из комбинации тяжелых и легких цепей. Селекцию желаемого антитела можно проводить с использованием активности связывания IgA переформированного иммуноглобулина в качестве индикатора. Например, когда задачей является выделение антитела против IgA, более предпочтительно, чтобы связывание антитела с IgA было специфическим. Скрининг связывающего IgA антитела можно проводить, например, с помощью следующих стадий:
(1) контактирование IgA с антителом, содержащим V-область, кодируемую кДНК, выделенной из гибридомы;
(2) выявление связывания антитела с IgA; и
(3) селекция антитела, которое связывается с IgA.
Способы выявления связывания антитела с IgA известны. В частности, связывание антитела с IgA можно выявлять с помощью описанных выше способов, таких как ELISA.
Предпочтительные способы скрининга антител, в которых в качестве индикатора используется активность связывания, также включают способы пэннинга с использованием фаговых векторов. Способы скрининга с использованием фаговых векторов являются преимущественными, когда гены антител выделяют из библиотек подклассов тяжелых цепей и легких цепей из популяции клеток, экспрессирующей поликлональное антитело. Гены, кодирующие вариабельные области тяжелых цепей и легких цепей, можно связывать соответствующей линкерной последовательностью с получением одноцепочечного Fv (scFv). Фаги, представляющие scFv на их поверхности, можно получать встраиванием гена, кодирующего scFv, в фаговый вектор. Фаги контактируют с представляющим интерес антигеном. Затем ДНК, кодирующую scFv, обладающие представляющей интерес активностью связывания, можно выделять путем сбора фагов, связанных с антигеном. Этот процесс можно повторять при необходимости для увеличения содержания scFv, обладающих представляющий интерес активностью связывания.
После выделения кДНК, кодирующей V-область представляющего интерес антитела против IgA, кДНК расщепляют ферментами рестрикции, которые распознают участки рестрикции, встроенные на обоих концах кДНК. Предпочтительные ферменты рестрикции распознают и расщепляют нуклеотидную последовательность, которая встречается в нуклеотидной последовательности гена антитела с низкой частотой. Более того, предпочтительно в представляющий интерес вектор вводят участок рестрикции для фермента, который образует липкий конец, для встраивания одной копии расщепленного фрагмента в правильной ориентации. Кодирующую кДНК V-область антитела против IgA расщепляют, как описано выше, и ее встраивают в соответствующий экспрессирующий вектор для конструирования экспрессирующего вектора для антитела. В этом случае, если ген, кодирующий константную область антитела (C-область) и ген, кодирующий описанную выше V-область, слить в рамке считывания, то получат химерное антитело. В рамках настоящего изобретения "химерное антитело" означает, что происхождение константной области отличается от вариабельной области. Таким образом, в дополнение к гетерохимерным антителам мыши/человека, химерные антитела по настоящему изобретению включают аллохимерные антитела человека/человека. Экспрессирующий вектор для химерного антитела можно конструировать путем встраивания гена V-области в экспрессирующий вектор, который уже имеет константную область. В частности, например, последовательность, распознаваемая первым ферментом рестрикции, который расщепляет выше гена V-области, можно соответствующим образом помещать на 5’-конец экспрессирующего вектора, содержащего ДНК, кодирующую желаемую константную область (C-область) антитела. Экспрессирующий вектор для химерного антитела конструируют путем слияния в рамке считывания двух генов, расщепленных одной и той же комбинацией ферментов рестрикции.
Для получения моноклонального антитела против IgA, гены антител встраивают в экспрессирующий вектор так, чтобы гены экспрессировались под контролем регулирующей экспрессию области. Регулирующая экспрессию область для экспрессии антитела включает, например, энхансеры и промоторы. Более того, на N-конец можно присоединять подходящую сигнальную последовательность, так чтобы экспрессируемое антитело секретировалось наружу клеток. В примерах, описанных ниже, в качестве сигнальной последовательности используют пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO:5). Между тем, можно присоединять другие подходящие сигнальные последовательности. Экспрессированный полипептид расщепляется на С-конце описанной выше последовательности, и полученный полипептид секретируется из клеток в качестве зрелого полипептида. Затем подходящие клетки-хозяева трансформируют экспрессирующим вектором и получают рекомбинантные клетки, экспрессирущие ДНК, кодирующую антитело IgA.
ДНК, кодирующие тяжелую цепь (H-цепь) и легкую цепь (L-цепь) антитела по отдельности встраивают в различные экспрессирующие векторы для экспрессии гена антитела. Молекулу антитела, имеющую H- и L-цепи, можно экспрессировать путем котрансфекции одной и той же клетки-хозяина векторами, в которые соответственно встроены гены H-цепи и L-цепи. Альтернативно клетки-хозяева можно трансформировать одним экспрессирующим вектором, в который встроены ДНК, кодирующие H- и L-цепи (см. международную публикацию № WO 1994/011523).
Существуют различные известные комбинации клетка-хозяин/экспрессирующий вектор для получения антител путем введения выделенных генов антител в соответствующих хозяев. Все из этих экспрессирующих систем пригодны для выделения антигенсвязывающих молекул по настоящему изобретению. Подходящие эукариотические клетки, используемые в качестве клеток-хозяев, включают клетки животных, клетки растений и клетки грибов. В частности, клетки животных включают, например, следующие клетки.
(1) клетки млекопитающих: CHO (клетки яичника китайского хомячка), COS (линия клеток почки обезьяны), BHK (линия клеток детенышей хомячка), HEK293 (линия клеток почки эмбриона человека с расщепленной аденовирусной (Ad)5 ДНК), клетки PER.C6 (линия клеток сетчатки эмбриона человека, трансформированных генами E1A и E1B аденовируса типа 5 (Ad5)), HeLa, Vero, HEK293 (линия клеток почки эмбриона человека с расщепленной аденовирусной (Ad)5 ДНК) и т.п. (Current Protocols in Protein Science (май 2001 года, раздел 5.9, таблица 5.9.1));
(2) клетки земноводных: ооциты Xenopus и т.п.; и
(3) клетки насекомых: sf9, sf21, Tn5 и т.п.
Кроме того, в качестве клетки растений известна экспрессирующая система для генов антител с использованием клеток, происходящих из рода Nicotiana, таких как Nicotiana tabacum. Для трансформации клеток растений можно соответствующим образом использовать культивированные клетки каллуса.
Более того, в качестве клеток грибов можно использовать следующие клетки:
- дрожжи: род Saccharomyces, как например, Saccharomyces serevisiae, и род Pichia, как например, Pichia pastoris; и
- нитчатые грибы: род Aspergillus, как например, Aspergillus niger.
Более того, также известны экспрессирующие системы для генов антител, в которых используются прокариотические клетки. Например, при использовании бактериальных клеток, в рамках настоящего изобретения можно подходящим образом использовать клетки E. coli, клетки Bacillus subtilis и т.п. Экспрессирующие векторы, несущие представляющие интерес гены антитела, вводят в клетки путем трансфекции. Трансфицированные клетки культивируют in vitro, и из культуры трансформированных клеток можно получать желаемое антитело.
В дополнение к описанным выше клеткам-хозяевам, также для получения рекомбинантного антитела можно использовать трансгенных животных. Следовательно, антитело можно получать из животного, которому введен ген, кодирующий представляющее интерес антитело. Например, ген антитела можно конструировать в качестве слитого гена путем встраивания в рамке считывания в ген, который кодирует белок, специфически продуцируемый в молоке. В качестве белка, секретируемого в молоке, можно использовать, например β-казеин козы и т.п. Фрагменты ДНК, содержащие слитый ген, в который встроен ген антитела, инъецируют в эмбрион козы, а затем этот эмбрион вводят в самку козы. Желаемые антитела можно получать в качестве белка, слитого с белком молока, из молока, продуцируемого трансгенной козой, родившейся от козы, являющейся реципиентом эмбриона (или ее потомства). Кроме того, для увеличения объема молока, содержащего желаемое антитело, продуцируемого трансгенной козой, трансгенной козе при необходимости можно вводить гормоны, Ebert, K. M. et al., Bio/Technology (1994) 12 (7), 699-702).
Когда антигенсвязывающую молекулу, описанную в настоящем описании, вводят человеку, антигенсвязывающий домен, происходящий из генетически рекомбинантного антитела, которое является искусственно модифицированным для снижения гетерологической антигенности против человека и т.п., можно соответствующим образом использовать в качестве антигенсвязывающего домена антигенсвязывающей молекулы. Такие генетически рекомбинантные антитела включают, например, гуманизированные антитела. Эти модифицированные антитела получают соответствующим образом известными способами.
Вариабельная область антитела, используемая для получения антигенсвязывающего домена антигенсвязывающей молекулы, описанной в настоящем описании, как правило, образована тремя определяющими комплементарность областями (CDR) которые разделены четырьмя каркасными областями (FR). CDR представляет собой область, которая по существу определяет специфичность связывания антитела. Аминокислотные последовательности CDR являются в высокой степени разнообразными. С другой стороны, образующие FR аминокислотные последовательности часто обладают высокой идентичностью, даже среди антител с отличающейся специфичностью связывания. Таким образом, как правило, специфичность связывания определенного антитела можно вносить в другое антитело путем пересадки CDR.
Гуманизированное антитело также называют переформированным антителом человека. В частности, известны гуманизированные антитела, полученные с использованием технологии пересадки CDR, посредством которой пересаживают CDR антитела не являющегося человеком животного, такого как антитело мыши, в антитело человека и т.п. Также известны стандартные способы генной инженерии для получения гуманизированных антител. В частности, например, в качестве способа пересадки CDR антитела мыши в FR человека известна перекрывающаяся ПЦР с удлинением. В перекрывающейся ПЦР с удлинением, нуклеотидную последовательность, кодирующую CDR антитела мыши, подлежащую пересадке, добавляют к праймерам для синтеза FR антитела человека. Праймеры получают для каждой из четырех FR. Является общепризнанным, что при пересадке CDR мыши в FR человека, выбор FR человека, который имеет высокую идентичность FR мыши, является преимущественным для сохранения функции CDR. Таким образом, как правило, предпочтительно использовать FR человека, содержащую аминокислотную последовательность, которая обладает высокой идентичностью с аминокислотной последовательностью FR, соседней с CDR мыши, подлежащей пересадке.
Нуклеотидные последовательности, подлежащие лигированию, конструируют так, чтобы они были соединены друг с другом в рамке считывания. FR человека синтезируют по отдельности с использованием соответствующих праймеров. В результате получают продукты, в которых ДНК, кодирующая CDR мыши, связана с индивидуальными ДНК, кодирующими FR. Нуклеотидные последовательности, кодирующие CDR мыши, каждого продукта конструируют так, чтобы они перекрывались друг с другом. Затем проводят синтез комплементарной цепи для гибридизации перекрывающихся CDR-областей продуктов, синтезированных с использованием гена антитела человека в качестве матрицы. С помощью этой реакции FR человека лигируют через последовательности CDR мыши.
Полноразмерный ген V-области, в котором в конечном итоге лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, которые гибридизуются с его 5’- или 3’-концом, в которые добавлены подходящие последовательности, распознаваемые ферментом рестрикции. Экспрессирующий вектор для гуманизированного антитела можно получать путем встраивания ДНК, полученной, как описано выше, и ДНК, которая кодирует C-область антитела человека, в экспрессирующий вектор, так чтобы они лигировались в рамке считывания. После трансфекции рекомбинантным вектором хозяина для получения рекомбинантных клеток, рекомбинантные клетки культивируют и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в клеточной культуре (см., публикацию патента Европы № EP 239400 и международную публикацию патента № WO 1996/002576).
Путем качественного или количественного определения и оценки антигенсвязывающей активности гуманизированного антитела, полученного, как описано выше, можно пригодным образом отобрать FR антитела человека, которые позволяют CDR образовывать подходящий антигенсвязывающий центр при лигировании через CDR. Аминокислотные остатки в FR можно заменять при необходимости, так чтобы CDR переформированного анитетела человека образовывали соответствующий антигенсвязывающий центр. Например, мутации аминокислотной последовательности можно вносить в FR с использованием способа ПЦР, который используется для пересадки CDR мыши в FR человека. Более конкретно, в праймеры, которые гибридизуются с FR, можно вносить частичные мутации нуклеотидной последовательности. Мутации нуклеотидной последовательности вносятся в FR, синтезированные с использованием таких праймеров. Мутантные последовательности FR, имеющие желаемые характеристики, можно отбирать путем измерения и оценки активности мутанта антитела с замененной аминокислотой в отношении связывания с антигеном с помощью упомянутого выше способа (Cancer Res. (1993) 53: 851-856).
Альтернативно желаемые антитела человека можно получать путем иммунизации трансгенных животных, имеющих полный набор генов антител человека (см. международные публикации № WO 1993/012227; WO 1992/003918; WO 1994/002602; WO 1994/025585; WO 1996/034096; WO 1996/033735) путем иммунизации ДНК.
Более того, также известны способы получения антител человека путем пэннинга с использованием библиотек антител человека. Например, V-область антитела человека экспрессируют в качестве одноцепочечного антитела (scFv) на поверхности фага способом фагового дисплея. Можно отбирать фаги, экспрессирующие scFv, которые связываются с антигеном. Последовательность ДНК, кодирующую V-область антитела человека, которая связывается с антигеном, можно определять путем анализа генов выбранных фагов. Определяют последовательность ДНК scFv, которая связывается с антигеном. Получают экспрессирующий вектор путем слияния последовательности V-области в рамке считывания с последовательностью C-области желаемого антитела человека, и встраивания ее в соответствующий экспрессирующий вектор. Экспрессирующий вектор вводят в клетки, пригодные для экспрессии, такие как клетки, описанные выше. Антитело человека можно получать путем экспрессии гена, кодирующего антитело человека, в клетках. Эти способы уже известны (см. международные публикации WO 1992/001047; WO 1992/020791; WO 1993/006213; WO 1993/011236; WO 1993/019172; WO 1995/001438; WO 1995/015388).
В дополнение к способам, описанным выше, для выделения генов антител можно соответствующим образом использовать способы клонирования B-клеток (идентификация каждой кодирующей антитело последовательности, ее клонирование и выделение; использование при конструировании экспрессирующего вектора для получения каждого антитела (IgG1, IgG2, IgG3 или IgG4, в частности); и т.п.), как описано, например Bernasconi et al. (Science (2002) 298: 2199-2202) или в международной публикации № WO 2008/081008.
Система нумерации EU и система нумерации Kabat
В соответствии со способами, используемыми в рамках настоящего изобретения, положения аминокислот, приписываемые CDR и FR антитела, указаны в соответствии с нумерацией по Kabat (Sequences of Proteins of Immunological Interest (National Institute of Health, Bethesda, Md., 1987 и 1991)). В рамках настоящего изобретения, когда антигенсвязывающая молекула представляет собой антитело или антигенсвязывающий фрагмент, аминокислоты вариабельной области указаны в соответствии с нумерацией по системе Kabat (нумерация Kabat), в то время как аминокислоты константной области указаны в соответствии с системой нумерации EU на основе положений аминокислот по Kabat.
Условия концентрации ионов
Условия концентрации ионов металлов
В одном варианте осуществления настоящего изобретения концентрация ионов относится к концентрации ионов металлов. "Ионы металлов" относятся к ионам элементов группы I, за исключением водорода, таких как щелочные металлы и элементы группы меди, ионам элементов группы II, таких как щелочноземельные металлы и элементы группы цинка, ионам элементов группы III, за исключением бора, ионам элементов группы IV, за исключением углерода и кремния, ионам элементов группы VIII, таких как элементы группы железа и элементы группы платины, ионам элементов, принадлежащих подгруппе A групп V, VI и VII, и элементов металлов, таких как сурьма, висмут и полоний. Атомы металлов обладают свойством высвобождения валентных электронов так, что они становятся катионами. Это называют тенденцией к ионизации. Металлы с высокой тенденцией к ионизации считаются химически активными.
В рамках настоящего изобретения предпочтительные ионы металлов включают, например, ион кальция. Ион кальция вовлечен в модулирование множества биологических явлений, включая сокращение мышц, таких как скелетные, гладкие и сердечная мышцы; активацию движения, фагоцитоза и т.п. лейкоцитов; активацию изменения формы, секреции и т.п. тромбоцитов; активацию лимфоцитов; активацию тучных клеток, включая секрецию гистамина; клеточные ответы, опосредуемые рецептором катехоламина α или рецептором ацетилхолина; экзоцитоз; высвобождение веществ-передатчиков из окончаний нейронов; и поток аксоплазмы в нейронах. Известные внутриклеточные рецепторы ионов кальция включают тропонин C, кальмодулин, парвальбумин и легкую цепь миозина, которые имеют несколько участков связывания ионов кальция и, как полагают, имеют общее происхождение с точки зрения молекулярной эволюции. Также существует множество известных кальций-связывающих мотивов. Такие хорошо известные мотивы включают, например, домены кадгерина, EF-hand кальмодулина, C2-домен протеинкиназы C, Gla-домен белка фактора свертывания крови IX, лектины C-типа рецептора ациарогликопротеинов и манноза-связывающего рецептора, A-домены рецепторов LDL, аннексин, домен тромбоспондина 3 типа и EGF-подобные домены.
В рамках настоящего изобретения, когда ион металла представляет собой ион кальция, условия концентрации ионов кальция включают низкие концентрации ионов кальция и высокие концентрации ионов кальция. "Активность связывания варьирует, в зависимости от концентраций ионов кальция" означает, что антигенсвязывающая активность антигенсвязывающей молекулы варьирует вследствие различий между условиями низкой и высокой концентрации ионов кальция. Например, антигенсвязывающая активность антигенсвязывающей молекулы может быть более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция. Альтернативно антигенсвязывающая активность антигенсвязывающей молекулы может быть более высокой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция.
В рамках настоящего изобретения высокая концентрация ионов кальция, в частности, не ограничена конкретной величиной; однако концентрация предпочтительно может быть выбрана из величин в пределах от 100 мкМ до 10 мМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 200 мкМ до 5 мМ. В альтернативном варианте осуществления концентрация может быть выбрана из величин в пределах от 400 мкМ до 3 мМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 200 мкМ до 2 мМ. Более того, концентрация может быть выбрана из величин в переделах от 400 мкМ до 1 мМ. В частности, предпочтительной является концентрация, выбранная из величин в пределах от 500 мкМ до 2,5 мМ, которая является близкой к концентрации ионов кальция в плазме (крови) in vivo.
В рамках настоящего изобретения низкая концентрация ионов кальция, в частности, не ограничена конкретной величиной; однако концентрация предпочтительно может быть выбрана из величин в пределах от 0,1 мкМ до 30 мкМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 0,2 мкМ до 20 мкМ. В другом варианте осуществления концентрация может быть выбрана из величин в пределах от 0,5 мкМ до 10 мкМ. В альтернативном варианте осуществления концентрация может быть выбрана из величин в пределах от 1 мкМ до 5 мкМ. Более того, концентрация может быть выбрана из величин в пределах от 2 мкМ до 4 мкМ. В частности, предпочтительной является концентрация, выбранная из величин в пределах от 1 мкМ до 5 мкМ, которая является близкой к концентрации ионизированного кальция в ранних эндосомах in vivo.
В рамках настоящего изобретения "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 0,1 мкМ до 30 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 100 мкМ до 10 мМ. Предпочтительно, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 0,5 мкМ до 10 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 200 мкМ до 5 мМ. Особенно предпочтительно, это означает, что антигенсвязывающая активность при концентрации ионов кальция в ранней эндосоме in vivo является более слабой, чем при концентрации ионов кальция в плазме in vivo; и, в частности, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы является более слабой при концентрации ионов кальция, выбранной из величин в пределах от 1 мкМ до 5 мкМ, чем при концентрации ионов кальция, выбранной из величин в пределах от 500 мкМ до 2,5 мМ.
То, изменяется ли антигенсвязывающая активность антигенсвязывающей молекулы в зависимости от концентрации ионов металлов, можно определять, например, с использованием известных способов измерения, таких как способы, описанные в разделе "Активность связывания" выше. Например, для подтверждения того, что антигенсвязывающая активность антигенсвязывающей молекулы становится более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция, сравнивают антигенсвязывающую активность антигенсвязывающей молекулы при низкой и высокой концентрациях ионов кальция.
В рамках настоящего изобретения выражение "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" также может быть выражено как "антигенсвязывающая активность антигенсвязывающей молекулы является более высокой при высокой концентрации ионов кальция, чем при низкой концентрации ионов кальция". В настоящем описании "антигенсвязывающая активность является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция" иногда представлено как "антигенсвязывающая способность является более слабой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция". Также "антигенсвязывающая активность при низкой концентрации ионов кальция снижена так, чтобы она была ниже, чем антигенсвязывающая активность при высокой концентрации ионов кальция" может быть представлено как "антигенсвязывающую способность при низкой концентрации ионов кальция ослабили по сравнению с антигенсвязывающей способностью при высокой концентрации ионов кальция".
При определении антигенсвязывающей активности, условия, отличные от концентрации ионов кальция, могут быть соответствующим образом выбраны специалистами в данной области, и они конкретно не ограничены. Например, активность можно определять при 37°C в буфере HEPES. Например, для определения можно использовать Biacore (GE Healthcare) и т.п. Когда антиген представляет собой растворимый антиген, антигенсвязывающую активность антигенсвязывающей молекулы можно оценивать путем пропускания антигена в качестве анализируемого соединения над чипом, на котором иммобилизована антигенсвязывающая молекула. Когда антиген представляет собой антиген мембраны, активность связывания антигенсвязывающей молекулы с антигеном мембраны можно оценивать путем пропускания антигенсвязывающей молекулы в качестве анализируемого соединения над чипом, на котором иммобилизован антиген.
При условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению является более слабой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, соотношение антигенсвязывающей активности между низкими и высокими концентрациями ионов кальция конкретно не ограничено. Однако соотношение KD (константа диссоциации) антигенсвязывающей молекулы в отношении антигенов при низкой концентрации ионов кальция и KD при высокой концентрации ионов кальция, т.е. величина KD (3 мкМ Ca)/KD (2 мМ Ca), предпочтительно составляет 2 или более, более предпочтительно 10 или более, и еще более предпочтительно 40 или более. Верхний предел величины KD (3 мкМ Ca)/KD (2 мМ Ca) конкретно не ограничен и он может представлять собой любую величину, такую как 400, 1000 или 10000, при условии, что молекулу можно получить способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, для отображения антигенсвязывающей активности можно использовать KD (константа диссоциации). Между тем, когда антиген представляет собой антиген мембраны, для отображения активности можно использовать кажущуюся KD (кажущаяся константа диссоциации). KD (константа диссоциации) и кажущуюся KD (кажущаяся константа диссоциации) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или проточного цитометра.
Альтернативно, например, также в качестве показателя для отображения соотношения антигенсвязывающей активности антигенсвязывающей молекулы по настоящему изобретению при низкой и высокой концентрациях кальция предпочтительно можно использовать константу скорости диссоциации (kd). Когда константу скорости диссоциации (kd) используют вместо константы диссоциации (KD) в качестве показателя для отображения соотношение активностей связывания, соотношение констант скорости диссоциации (kd) при низкой и высокой концентрациях кальция, т.е. величина kd (низкая концентрация кальция)/kd (высокой концентрация кальция), предпочтительно равна 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более, и еще более предпочтительно 30 или более. Верхний предел величины Kd (низкая концентрация кальция)/kd (высокая концентрация кальция) конкретно не ограничен, и он может представлять собой любую величину, такую как 50, 100 или 200, при условии что можно получить молекулу способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, для отображения антигенсвязывающей активности можно использовать kd (константа скорости диссоциации). Между тем, когда антиген представляет собой антиген мембраны, для отображения антигенсвязывающей активности можно использовать кажущуюся kd (кажущуюся константу скорости диссоциации). kd (константа скорости диссоциации) и кажущуюся kd (кажущаяся константа скорости диссоциации) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare) или проточного цитометра. В рамках настоящего изобретения, когда антигенсвязывающую активность антигенсвязывающей молекулы определяют при различных концентрациях ионов кальция, предпочтительно использовать одни и те же условия, за исключением концентраций кальция.
Например, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антител, включающего стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена или антигенсвязывающей молекулы при низкой концентрации кальция;
(b) определение антигенсвязывающей активности антигенсвязывающего домена или антигенсвязывающей молекулы при высокой концентрации кальция; и
(c) выбор антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которого является более низкой при низкой концентрации кальция, чем при высокой концентрации кальция.
Более того, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антигенсвязывающих молекул, или их библиотеки, включающего стадии:
(a) контактирование антигена с антигенсвязывающим доменом или антигенсвязывающей молекулой или их библиотекой при высокой концентрации кальция;
(b) инкубация при низкой концентрации кальция антигенсвязывающего домена или антигенсвязывающей молекулы, которые связались с антигеном на стадии (a); и
(c) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциировавших на стадии (b).
Более того, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга антигенсвязывающих доменов или антигенсвязывающих молекул или их библиотеки, включающего стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул при низкой концентрации кальция;
(b) выбор антигенсвязывающего домена или антигенсвязывающей молекулы, которые не связались с антигеном на стадии (a);
(c) позволение антигенсвязывающему домену или антигенсвязывающей молекуле, выбранным на стадии (c), связываться с антигеном при высокой концентрации кальция; и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, которые связались с антигеном на стадии (c).
Кроме того, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) контактирование при высокой концентрации кальция библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с колонкой, на которой иммобилизован антиген;
(b) элюирование антигенсвязывающего домена или антигенсвязывающей молекулы, которые связались с колонкой на стадии (a), из колонки при низкой концентрации кальция; и
(c) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, элюированных на стадии (b).
Более того, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) позволение библиотеке антигенсвязывающих доменов или антигенсвязывающей молекуле при низкой концентрации кальция пройти через колонку, на которой иммобилизован антиген;
(b) сбор антигенсвязывающего домена или антигенсвязывающей молекулы, которые элюировались без связывания с колонкой на стадии (a);
(c) позволение антигенсвязывающему домену или антигенсвязывающей молекуле, собранным на стадии (b), связаться с антигеном при высокой концентрации кальция; и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, которые связались с антигеном на стадии (c).
Более того, антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые являются одним из вариантов осуществления настоящего изобретения, можно получать путем скрининга, включающего стадии:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул при высокой концентрации кальция;
(b) получение антигенсвязывающего домена или антигенсвязывающей молекулы, которые связались с антигеном на стадии (a);
(c) инкубация при низкой концентрации кальция антигенсвязывающего домена или антигенсвязывающей молекулы, полученных на стадии (b); и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых на стадии (c) является более слабой, чем критерий для выбора на стадии (b).
Описанные выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающим доменам или антигенсвязывающим молекулам, антигенсвязывающая активность которых является более низкой при низкой концентрации ионов кальция, чем при высокой концентрации ионов кальция, которые получают способами скрининга, которые дополнительно включают стадию повторения два или более раз стадий (a)-(c) или (a)-(d) в описанных выше способах скрининга. Количество циклов стадий (a)-(c) или (a)-(d) конкретно не ограничено, однако обычно составляет 10 или менее.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена или антигенсвязывающей молекулы при низкой концентрации кальция конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 0,1 мкМ до 30 мкМ, но предпочтительно она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 0,5 мкМ до 10 мкМ. Более предпочтительно, она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция в ранней эндосоме in vivo, в частности, от 1 мкМ до 5 мкМ. Между тем, антигенсвязывающая активность антигенсвязывающего домена или антигенсвязывающей молекулы при высокой концентрации кальция конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 100 мкМ до 10 мМ, но предпочтительно она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция от 200 мкМ до 5 мМ. Более предпочтительно, она представляет собой антигенсвязывающую активность при концентрации ионизированного кальция в плазме in vivo, в частности, от 0,5 мМ до 2,5 мМ.
Антигенсвязывающую активность антигенсвязывающего домена или антигенсвязывающей молекулы можно измерять способами, известными специалистам в данной области. Условия, отличные от концентрации ионизированного кальция, могут быть определены специалистами в данной области. Антигенсвязывающую активность антигенсвязывающего домена или антигенсвязывающей молекулы можно оценивать в качестве константы диссоциации (KD), кажущейся константы диссоциации (кажущаяся KD), константы скорости диссоциации (kd), кажущейся константы диссоциации (кажущаяся kd) и т.п. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или FACS.
В рамках настоящего изобретения стадия выбора антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых является более высокой при высокой концентрации кальция, чем при низкой концентрации кальция, является тождественной стадии выбора антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых является более низкой при низкой концентрация кальция, чем при высокой концентрации кальция.
При условии, что антигенсвязывающая активность является более высокой при высокой концентрации кальция, чем при низкой концентрации кальция, различие в антигенсвязывающей активности между высокой и низкой концентрациями концентрация конкретно не ограничено; однако антигенсвязывающая активность при высокой концентрации кальция предпочтительно в два раза или более, более предпочтительно в 10 раз или более, и еще более предпочтительно в 40 раз или более превышает низкую концентрацию кальция.
Антигенсвязывающие домены или антигенсвязывающие молекулы по настоящему изобретению, подлежащие скринингу способами скрининга, описанными выше, могут представлять собой любые антигенсвязывающие домены и антигенсвязывающие молекулы. Например, можно проводить скрининг описанных выше антигенсвязывающих доменов или антигенсвязывающих молекул. Например, можно проводить скрининг антигенсвязывающих доменов или антигенсвязывающих молекул, имеющих природные последовательности или последовательности с замещенными аминокислотами.
Библиотеки
В одном варианте осуществления антигенсвязывающий домен или антигенсвязывающую молекулу по настоящему изобретению можно получать из библиотеки, которая в основном состоит из множества антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены которых имеют по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих молекул, в зависимости от концентрации ионов. Концентрации ионов предпочтительно включают, например, концентрацию ионов металлов и концентрацию протонов.
В рамках настоящего изобретения "библиотека" относится к множеству антигенсвязывающих молекул или множеству слитых полипептидов, содержащих антигенсвязывающие молекулы, или нуклеиновых кислот или полинуклеотидов, кодирующих их последовательности. Последовательности множества антигенсвязывающих молекул или множества слитых полипептидов, содержащих антигенсвязывающие молекулы, в библиотеке не идентичны, а отличаются друг от друга.
В рамках настоящего изобретения выражение "последовательности отличаются друг от друга" в выражении "множество антигенсвязывающих молекул, последовательности которых отличаются друг от друга", означает, что последовательности антигенсвязывающих молекул в библиотеке отличаются друг от друга. В частности, в библиотеке количество последовательностей, отличающихся друг от друга, отражает количество независимых клонов с различными последовательностями, и также может называться "размером библиотеки". Размер библиотеки для обычной библиотеки фагового дисплея находится в диапазоне от 106 до 1012. Размер библиотеки может быть увеличен вплоть до 1014 с использованием известных способов, таких как рибосомный дисплей. Однако истинное количество фаговых частиц, используемых в селекции способом пэннинга фаговой библиотеки, как правило, в 10-10000 раз превышает размер библиотеки. Эту избыточную множественность также называют "количеством эквивалентов библиотеки", и она означает, что существует от 10 до 10000 индивидуальных клонов, которые обладают одной и той же аминокислотной последовательностью. Таким образом, в рамках настоящего изобретения выражение "последовательности отличаются друг от друга" означает, что последовательности независимых антигенсвязывающих молекул в библиотеке, исключая эквиваленты библиотеки, отличаются друг от друга. Более конкретно, указанное выше означает, что существует от 106 до 1014 антигенсвязывающих молекул, последовательности которых отличаются друг от друга, предпочтительно от 107 до 1012 молекул, более предпочтительно от 108 до 1011 молекул, и особенно предпочтительно от 108 до 1010 молекул, последовательности которых отличаются друг от друга.
В рамках настоящего изобретения выражение "множество" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" в случае, например, антигенсвязывающих молекул, слитых полипептидов, полинуклеотидных молекул, векторов или вирусов по настоящему изобретению, главным образом, относится к группе из двух или более типов вещества. Например, когда два или более вещества отличаются друг от друга конкретной характеристикой, это означает, что существует два или более типов вещества. Такие примеры могут включать, например, мутантные аминокислоты, наблюдаемые в конкретных аминокислотных положениях аминокислотной последовательности. Например, когда существует две или более антигенсвязывающих молекул по настоящему изобретению, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нестрогих остатков или за исключением конкретных мутантных аминокислот в гипервариабельных положениях, экспонированных на поверхности, существует множество антигенсвязывающих молекул по настоящему изобретению. В другом примере, когда существует две или более полинуклеотидных молекулы, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нуклеотидов, кодирующих нестрогие остатки, или нуклеотидов, кодирующих мутантные аминокислоты гипервариабельных положений, экспонированных на поверхности, существует множество полинуклеотидных молекул по настоящему изобретению.
Кроме того, в рамках настоящего изобретения выражение "в основном состоит из" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" отражает количество антигенсвязывающих молекул, антигенсвязывающая активность которых варьирует, в зависимости от концентрации ионов, среди независимых клонов с отличающимися последовательностями в библиотеке. Особенно предпочтительно, чтобы в библиотеке существовало по меньшей мере 104 антигенсвязывающих молекул, имеющих такую активность связывания. Более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 105 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 106 антигенсвязывающих молекул, обладающих такой активностью связывания. Особенно предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 107 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 108 антигенсвязывающих молекул, обладающих такой активностью связывания. Альтернативно это также предпочтительно может быть выражено как соотношение количества антигенсвязывающих молекул, антигенсвязывающая активность которых варьирует, в зависимости от концентрации ионов, и количества независимых клонов, имеющих различные последовательности, в библиотеке. В частности, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в которой антигенсвязывающие молекулы, имеющие такую активность связывания, составляют от 0,1% до 80%, предпочтительно от 0,5% до 60%, более предпочтительно от 1% до 40%, еще более предпочтительно от 2% до 20%, и особенно предпочтительно от 4% до 10% независимых клонов с отличающимися последовательностями в библиотеке. В случае слитых полипептидов, полинуклеотидных молекул или векторов, могут быть возможными сходные выражения, в которых используются количества молекул или отношение к общему количеству молекул. В случае вирусов, также могут быть возможными сходные выражения, в которых используется количество вирионов или отношение к общему количеству вирионов.
Аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих доменов в зависимости от концентрации ионов кальция
Антигенсвязывающие домены или антигенсвязывающие молекулы по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым путем. Например, когда ион металла представляет собой ион кальция, можно использовать ранее существующие антигенсвязывающие молекулы, ранее существующие библиотеки (фаговая библиотека, и т.д.), антитела или библиотеки, полученные из гибридом, полученных путем иммунизации животных или из B-клеток иммунизированных животных, антитела или библиотеки, полученные внесением аминокислот, способных хелатировать кальций (например, аспарагиновая кислота и глутаминовая кислота), или неприродных аминокислотных мутаций в описанные выше антитела или библиотеки (хелатирующие кальций аминокислоты (такие как аспарагиновая кислота и глутаминовая кислота), библиотеки с увеличенным содержанием неприродных аминокислот, библиотеки полученные путем внесения хелатирующих кальций аминокислот (таких как аспарагиновая кислота и глутаминовая кислота) или неприродных аминокислотных мутаций в конкретные положения, и т.п.
Примеры аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентрации ионов кальция, как описано выше, могут представлять собой любые типы аминокислот, при условии что аминокислоты формируют кальций-связывающий мотив. Кальций-связывающие мотивы хорошо известны специалистам в данной области и подробно описаны (например, Springer et al. (Cell (2000) 102, 275-277); Kawasaki и Kretsinger (Protein Prof. (1995) 2, 305-490); Moncrief et al. (J. Mol. Evol. (1990) 30, 522-562); Chauvaux et al. (Biochem. J. (1990) 265, 261-265); Bairoch и Cox (FEBS Lett. (1990) 269, 454-456); Davis (New Biol. (1990) 2, 410-419); Schaefer et al. (Genomics (1995) 25, 638-643); Economou et al. (EMBO J. (1990) 9, 349-354); Wurzburg et al. (Structure. (2006) 14, 6, 1049-1058)). В частности, любые известные кальций-связывающие мотивы, включая лектины C-типа, такие как ASGPR, CD23, MBR и DC-SIGN, могут быть включены в антигенсвязывающие молекулы по настоящему изобретению. Предпочтительные примеры таких предпочтительных кальций-связывающих мотивов также включают, в дополнение к кальций-связывающим мотивам, описанным выше, например, кальций-связывающий мотив в антигенсвязывающем домене SEQ ID NO:6.
Более того, в качестве аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция, например, предпочтительно можно использовать аминокислоты, имеющие активность хелатирования металлов. Примеры таких хелатирующих металлы аминокислот включают, например, серин (Ser(S)), треонин (Thr(T)), аспарагин (Asn(N)), глутамин (Gln(Q)), аспарагиновую кислоту (Asp(D)) и глутаминовую кислоту (Glu(E)).
Положения в антигенсвязывающих доменах, в которых содержатся описанные выше аминокислоты, конкретно не ограничены конкретными положениями, и они могут представлять собой любые положения в вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые образуют антигенсвязывающий домен, при условии, что они изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция. В частности, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены тяжелых цепей которых содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекулы в зависимости от концентраций ионов кальция. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены тяжелой цепи которых содержат упомянутые выше аминокислоты. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены тяжелых цепей которых содержат упомянутые выше аминокислоты в положениях 95, 96, 100a и/или 101, как указано в соответствии с системой нумерации Kabat.
Между тем, в одном варианте осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и антигенсвязывающие домены легкой цепи которых содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов кальция. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR1-домены легкой цепи которых содержат упомянутые выше аминокислот. В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR1-домены легкой цепи которых содержат упомянутые выше аминокислоты в положениях 30, 31 и/или 32, как указано в соответствии с системной нумерации Kabat.
В другом варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR2-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки. В другом варианте осуществления настоящее изобретение относится к библиотекам, в основном состоящим из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR2-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки в положении 50, как указано в соответствии с системой нумерации Kabat.
В другом варианте осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки. В альтернативном варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и CDR3-домены легкой цепи которых содержат упомянутые выше аминокислотные остатки в положении 92, как указано в соответствии с системной нумерации Kabat.
Более того, в различных вариантах осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и в которых две или три CDR, выбранных из описанных выше CDR1, CDR2 и CDR3 легкой цепи, содержат упомянутые выше аминокислотные остатки. Более того, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном состоящей из антигенсвязывающих молекул, последовательности которых отличаются друг от друга и легкие цепи которых содержат упомянутые выше аминокислотные остатки в любом одном или нескольких из положений 30, 31, 32, 50 и/или 92, как указано в соответствии с системной нумерации Kabat.
В особенно предпочтительном варианте осуществления каркасные последовательности вариабельной области легкой цепи и/или тяжелой цепи антигенсвязывающей молекулы предпочтительно содержат каркасные последовательности человека эмбрионального типа. Таким образом, в одном варианте осуществления настоящего изобретения, когда каркасные последовательности представляют собой полностью человеческие последовательности, ожидается, что при введении такой антигенсвязывающей молекулы по настоящему изобретению человеку (например, для лечения заболеваний), она индуцирует небольшой иммуногенный ответ или не индуцирует иммуногенного ответа. В указанном выше контексте выражение "содержащий последовательность эмбрионального типа" в рамках настоящего изобретения означает, что часть каркасных последовательностей по настоящему изобретению идентична части любых каркасных последовательностей человека эмбрионального типа. Например, когда последовательность FR2 тяжелой цепи антигенсвязывающей молекулы по настоящему изобретению представляет собой комбинацию последовательностей FR2 тяжелой цепи различных каркасных последовательностей человека эмбрионального типа, такая молекула также представляет собой антигенсвязывающую молекулу по настоящему изобретению, "содержащую последовательность эмбрионального типа".
Предпочтительные примеры каркасных областей включают, например, последовательности полностью человеческих каркасных областей человека, известные в настоящее время, которые включены в web-сайт V-Base (http://vbase.mrc-cpe.cam.ac.uk/) или другие. Эти последовательности каркасных областей можно соответствующим образом использовать в качестве последовательности эмбрионального типа, содержащейся в антигенсвязывающей молекуле по настоящему изобретению. Последовательности эмбрионального типа могут быть подразделены на категории в соответствии с их сходством (Tomlinson et al. (J. Mol. Biol. (1992) 227, 776-798); Williams и Winter (Eur. J. Immunol. (1993) 23, 1456-1461); Cox et al. (Nat. Genetics (1994) 7, 162-168)). Соответствующие последовательности эмбрионального типа могут быть выбраны из Vκ, которые подразделяются на семь подгрупп; Vλ, которые подразделяются на десять подгрупп; и VH, которые подразделяются на семь подгрупп.
Полностью человеческие последовательности VH предпочтительно включают, но не ограничиваются ими, например, последовательности VH:
подгруппы VH1 (например, VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58 и VH1-69);
подгруппы VH2 (например, VH2-5, VH2-26 и VH2-70);
подгруппы VH3 (VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73 и VH3-74);
подгруппы VH4 (VH4-4, VH4-28, VH4-31, VH4-34, VH4-39, VH4-59 и VH4-61);
подгруппы VH5 (VH5-51);
подгруппы VH6 (VH6-1); и
подгруппы VH7 (VH7-4 и VH7-81).
Они также описаны в известных документах (Matsuda et al. (J. Exp. Med. (1998) 188, 1973-1975)) и т.п., и, таким образом, специалисты в данной области могут соответствующим образом моделировать антигенсвязывающие молекулы по настоящему изобретению, исходя из информации об этих последовательностях. Также предпочтительно использовать другие полностью человеческие каркасные области или каркасные субобласти.
Полностью человеческие последовательности Vk предпочтительно включают, но не ограничиваются ими, например:
A20, A30, L1, L4, L5, L8, L9, L11, L12, L14, L15, L18, L19, L22, L23, L24, O2, O4, O8, O12, O14 и O18, выделенные в подгруппу Vk1;
A1, A2, A3, A5, A7, A17, A18, A19, A23, O1 и O11, выделенные в подгруппу Vk2;
A11, A27, L2, L6, L10, L16, L20 и L25, выделенные в подгруппу Vk3;
B3, выделенная в подгруппу Vk4;
B2 (также обозначаемая как Vk5-2), выделенная в подгруппу Vk5; и
A10, A14 и A26, выделенные в подгруппу Vk6.
(Kawasaki et al. (Eur. J. Immunol. (2001) 31, 1017-1028); Schable and Zachau (Biol. Chem. Hoppe Seyler (1993) 374, 1001-1022); Brensing-Kuppers et al. (Gene (1997) 191, 173-181)).
Полностью человеческие последовательности VL предпочтительно включают, но не ограничиваются ими, например:
V1-2, V1-3, V1-4, V1-5, V1-7, V1-9, V1-11, V1-13, V1-16, V1-17, V1-18, V1-19, V1-20 и V1-22, выделенные в подгруппу VL1;
V2-1, V2-6, V2-7, V2-8, V2-11, V2-13, V2-14, V2-15, V2-17 и V2-19, выделенные в подгруппу VL1;
V3-2, V3-3 и V3-4, выделенные в подгруппу VL3;
V4-1, V4-2, V4-3, V4-4 и V4-6, выделенные в подгруппу VL4; и
V5-1, V5-2, V5-4 и V5-6, выделенные в подгруппу VL5 (Kawasaki et al. (Genome Res. (1997) 7, 250-261)).
Обычно эти каркасные последовательности отличаются друг от друга одним или несколькими аминокислотными остатками. Эти каркасные последовательности можно использовать в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов" по настоящему изобретению. Другие примеры полностью человеческих каркасных областей, используемых в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов" по настоящему изобретению включают, но не ограничиваются ими, например, KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (например, Kabat et al. (1991) выше, Wu et al. (J. Exp. Med. (1970) 132, 211-250)).
Без связи с конкретной теорией полагают, что одна из причин для того, чтобы ожидать, что использование последовательностей эмбрионального типа предотвратит неблагоприятные иммунные ответы у большинства индивидуумов, состоит в следующем. В результате процесса созревания аффинности в ходе нормальных иммунных ответов, часто в вариабельных областях иммуноглобулинов происходят соматические мутации. Такие мутации чаще всего происходят в области CDR, последовательности которых являются гипервариабельными, но также затрагивают остатки каркасных областей. Такие мутации каркасной области не существуют в генах эмбрионального типа, и они также с меньшей вероятностью будут иммуногенными у пациентов. С другой стороны, в нормальной человеческой популяции присутствует большинство каркасных последовательностей, экспрессируемых с генов эмбрионального типа. В результате иммунной толерантности, ожидается, что эти каркасные области эмбрионального типа будут иметь низкую иммуногенность у пациентов или не иметь иммуногенности у пациентов. Для максимизации возможности иммунной толерантности можно выбирать гены, кодирующие вариабельную область, из группы часто встречающихся функциональных генов эмбрионального типа.
Для получения антигенсвязывающих молекул по настоящему изобретению, в которых описанные выше последовательности вариабельной области, последовательности вариабельной области тяжелой или легкой цепи, последовательности CDR или каркасные последовательности содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентрации ионов кальция можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением.
Например, библиотеку, которая содержит множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, где вариабельную область легкой цепи выбирают в качестве каркасной последовательности, первоначально содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. В качестве неограничивающего примера, когда концентрация ионов представляет собой концентрацию ионов кальция, такие предпочтительные библиотеки включают, например, библиотеки, сконструированные путем комбинирования последовательности вариабельной области легкой цепи SEQ ID NO:6 (Vk5-2) и вариабельной области тяжелой цепи, полученной в качестве рандомизированной библиотеки последовательностей вариабельной области.
Альтернативно последовательность вариабельной области легкой цепи, выбранную в качестве каркасной области, первоначально содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы, как упоминалось выше, можно конструировать так, чтобы она содержала различные аминокислотные остатки, отличные от упомянутых выше аминокислотных остатков. В настоящем описании такие остатки называют нестрогими остатками. Количество и положение нестрогих остатков конкретно не ограничено, при условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или более нестрогих остатков. Например, когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, подлежащих внесению в последовательность вариабельной области легкой цепи SEQ ID NO:6 (Vk5-2), включают аминокислотные остатки, приведенные в таблицах 1 или 2.
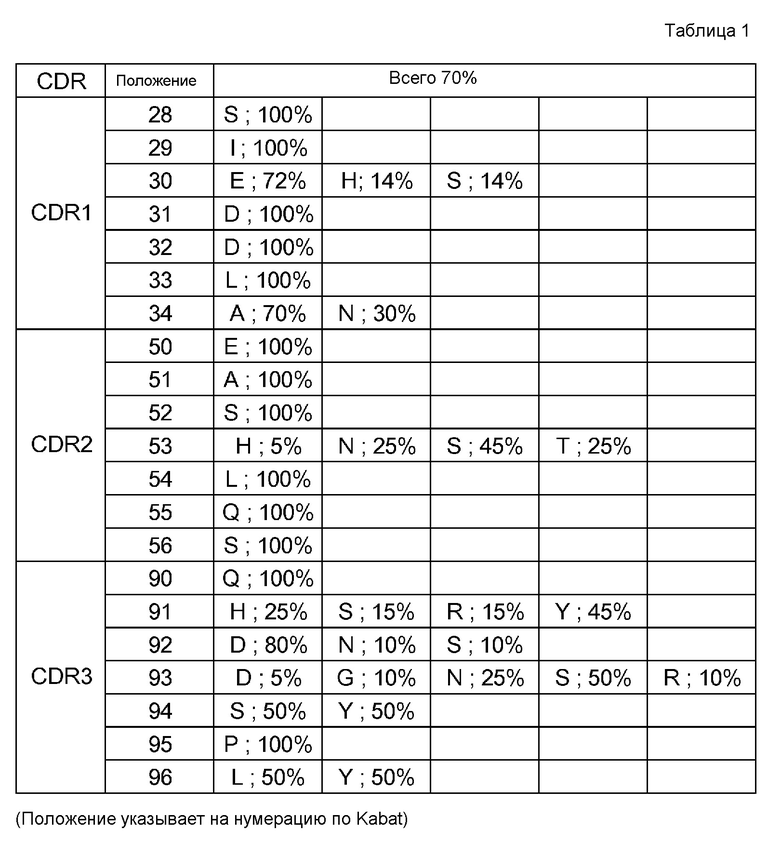
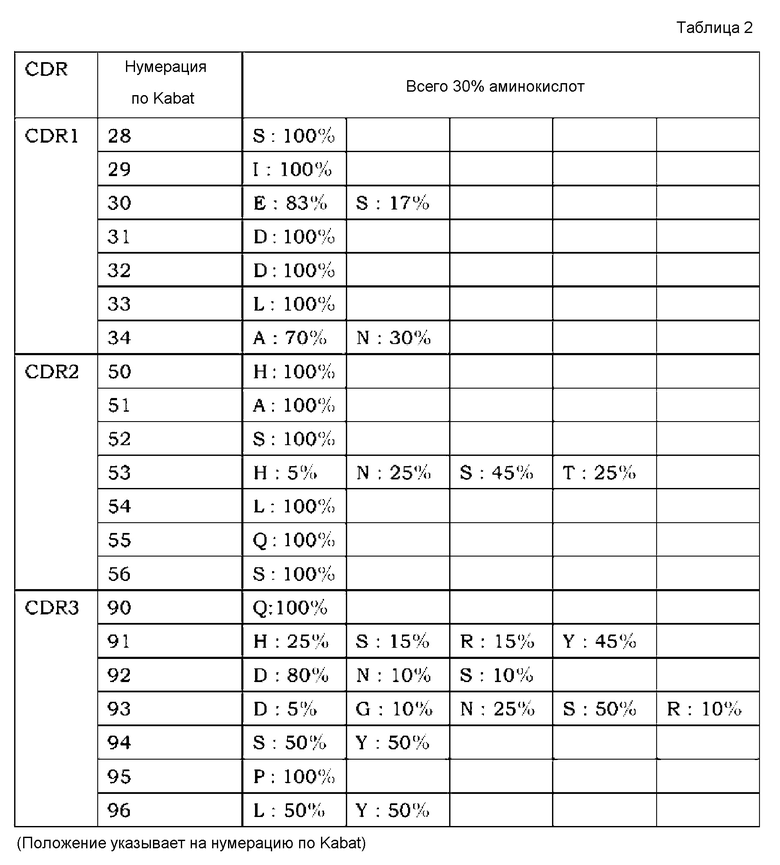
В настоящем описании нестрогие остатки относятся к варьированию аминокислотных остатков, имеющемуся в гипервариабельных положениях, в которых присутствует несколько различных аминокислот, в вариабельных областях легкой цепи и тяжелой цепи, когда сравнивают аминокислотные последовательности известных и/или нативных антител или антигенсвязывающих доменов. Гипервариабельные положения, как правило, расположены в областях CDR. В одном варианте осуществления для определения гипервариабельных положений в известных и/или нативных антителах пригодны данные, предоставленные Kabat, Sequences of Proteins of Immunological Interest (National Institute of Health Bethesda Md.) (1987 и 1991). Более того, в базах данных в Интернете (http://vbase.mrc-cpe.cam.ac.uk/, http://www.bioinf.org.uk/abs/ index.html) предоставлены собранные последовательности многих легких цепей и тяжелых цепей человека и их положения. Информация о последовательностях и положениях пригодна для определения гипервариабельных положений в рамках настоящего изобретения. В соответствии с настоящим изобретением, когда определенное положение аминокислоты имеет предпочтительно от приблизительно 2 до приблизительно 20 возможных вариантов аминокислотных остатков, предпочтительно от приблизительно 3 до приблизительно 19, предпочтительно от приблизительно 4 до приблизительно 18, предпочтительно от 5 до 17, предпочтительно от 6 до 16, предпочтительно от 7 до 15, предпочтительно от 8 до 14, предпочтительно от 9 до 13, и предпочтительно от 10 до 12 возможных вариантов аминокислотных остатков, это положение является гипервариабельным. В некоторых вариантах осуществления определенное положение аминокислоты может иметь предпочтительно по меньшей мере приблизительно 2, предпочтительно по меньшей мере приблизительно 4, предпочтительно по меньшей мере приблизительно 6, предпочтительно по меньшей мере приблизительно 8, предпочтительно приблизительно 10 и предпочтительно приблизительно 12 вариантов аминокислотных остатков.
Альтернативно библиотеку, содержащую множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, с вариабельными областями легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих молекул в зависимости от концентраций ионов, как упоминалось выше. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают, например, библиотеки, в которых вариабельные области тяжелой цепи, полученные в качестве рандомизированной библиотеки последовательностей вариабельной области, комбинируют с последовательностями вариабельной области легкой цепи, в которых конкретный остаток(и) в последовательности эмбрионального типа, такой как SEQ ID NO:7 (Vk1), SEQ ID NO:8 (Vk2), SEQ ID NO:9 (Vk3) или SEQ ID NO:10 (Vk4) заменен по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. Неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки в CDR1 легкой цепи. Более того, неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки в CDR2 легкой цепи. Кроме того, неограничивающие примеры таких аминокислотных остатков также включают аминокислотные остатки в CDR3 легкой цепи.
Неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR1 легкой цепи, включают остатки в положениях 30, 31 и/или 32 в CDR1 вариабельной области легкой цепи, как указано согласно нумерации EU. Более того, неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR2 легкой цепи, включают аминокислотный остаток в положении 50 в CDR2 вариабельной области легкой цепи, как указано согласно нумерации Kabat. Более того, неограничивающие примеры таких аминокислотных остатков, содержащихся в CDR3 легкой цепи, включают аминокислотный остаток в положении 92 в CDR3 вариабельной области легкой цепи, как указано согласно нумерации Kabat. Эти аминокислотные остатки могут содержаться отдельно или в комбинации, при условии что они образуют кальций-связывающий мотив и/или при условии что антигенсвязывающая активность антигенсвязывающей молекулы варьирует, в зависимости от концентрации ионов кальция. Между тем, поскольку известны тропонин C, кальмодулин, парвальбумин и легкая цепь миозина, которые обладают несколькими участками связывания ионов кальция и, как полагают, имеют общее происхождение с точки зрения молекулярной эволюции, можно конструировать CDR1, CDR2 и/или CDR3 легкой цепи так, чтобы они имели их связывающие мотивы. Например, можно использовать домены кадгерина, EF-hand кальмодулина, C2-домен протеинкиназы C, Gla-домен белка фактора свертывания крови IX, лектины C-типа рецептора ациарогликопротеинов и манноза-связывающего рецептора, A-домены рецепторов LDL, аннексин, домена тромбоспондина 3 типа, и EGF-подобные домены соответствующим образом для описанных выше целей.
Когда вариабельные области тяжелой цепи, полученные в качестве рандомизированной библиотеки последовательностей вариабельной области, и вариабельные области легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов, комбинируют, как описано выше, последовательности вариабельных областей легкой цепи можно конструировать так, чтобы они содержали нестрогие остатки аналогично тому, как описано выше. Количество и положение таких нестрогих остатков конкретно не ограничивается конкретными вариантами осуществления, при условии что антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или более нестрогих остатков. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области легкой цепи, включают аминокислотные остатки, приведенные в таблицах 1 и 2.
Предпочтительные вариабельные области тяжелой цепи, подлежащие комбинированию, включают, например, рандомизированные библиотеки вариабельной области. Для получения рандомизированной библиотеки вариабельной области известные способы комбинируют соответствующим образом. В неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области предпочтительно можно использовать иммунную библиотеку, сконструированную на основе генов антител, происходящих из лимфоцитов животных, иммунизированных конкретным антигеном, пациентов с инфекциями, индивидуумов с повышенным титром антител в крови в результате вакцинации, пациентов со злокачественной опухолью или пациентов с аутоиммунным заболеванием.
В другом неограничивающем варианте осуществления настоящего изобретения, в качестве рандомизированной библиотеки вариабельной области также предпочтительно можно использовать синтетическую библиотеку, полученную путем замены последовательностей CDR V-генов в геномной ДНК или функциональных реконструированных V-генов набором синтетических олигонуклеотидов, содержащих последовательности, кодирующие наборы кодонов соответствующей длины. В этом случае, поскольку в последовательности CDR3 тяжелой цепи наблюдают разнообразие, также можно заменять только последовательность CDR3. Критерием возникновения разнообразия аминокислот в вариабельной области антигенсвязывающей молекулы является то, что разнообразие возникает у аминокислотных остатков в экспонированных на поверхности положениях антигенсвязывающей молекулы. Экспонированное на поверхности положение относится к положению, которое считается способным экспонироваться на поверхности и/или контактировать с антигеном, исходя из структуры, группы структур и/или смоделированной структуры антигенсвязывающей молекулы. Как правило, такие положения представляют собой CDR. Предпочтительно, экспонированные на поверхности положения определяют с использованием координат из трехмерной модели антигенсвязывающей молекулы с использованием компьютерной программы, такой как программа InsightII (Accelrys). Экспонированные на поверхности положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (J. Mol. Biol. (1971) 55, 379-400); Connolly (J. Appl. Cryst. (1983) 16, 548-558)). Определение экспонированных на поверхности положений можно проводить с использованием программного обеспечения, пригодного для моделирования белка, и информации о трехмерной структуре, полученной для антитела. Программное обеспечение, которое можно использовать для этих целей, предпочтительно включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Главным образом или предпочтительно, когда алгоритм требует введения пользователем параметра размера, "размер" образца, который используют в вычислении, устанавливают как радиус, равный приблизительно 1,4 Ангстрем или менее. Более того, способы определения экспонированных на поверхности областей и участков с использованием программного обеспечения для персональных компьютеров описаны Pacios (Comput. Chem. (1994) 18 (4), 377-386; J. Mol. Model. (1995) 1, 46-53).
В другом неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области также особенно предпочтительно можно использовать наивную библиотеку, которая сконструирована из генов антител, происходящих из лимфоцитов здоровых индивидуумов, и набор генов которой состоит из наивных последовательностей, которые представляют собой последовательности антител без предпочтений (Gejima et al. (Human Antibodies (2002) 11, 121-129); Cardoso et al. (Scand. J. Immunol. (2000) 51, 337-344)). В настоящем описании, аминокислотная последовательность, содержащая наивную последовательность, относится к аминокислотной последовательности, полученной из такой наивной библиотеки.
В одном варианте осуществления настоящего изобретения антигенсвязывающий домен по настоящему изобретению можно получать из библиотеки, содержащей множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, полученных путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки вариабельной области, где вариабельная область тяжелой цепи выбрана в качестве каркасной последовательности, которая первоначально содержит "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов". Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают библиотеки, сконструированные путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки вариабельной области с последовательностью вариабельной области тяжелой цепи SEQ ID NO:11 (6RL#9-IgG1) или SEQ ID NO:12 (6KC4-1#85-IgG1). Альтернативно такую библиотеку можно конструировать путем выбора соответствующих вариабельных областей легкой цепи из вариабельных областей легкой цепи, которые имеют последовательности эмбрионального типа, вместо вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки последовательностей вариабельной области. Такие предпочтительные библиотеки включают, например, библиотеки, в которых последовательность вариабельной области тяжелой цепи SEQ ID NO:11 (6RL#9-IgG1) или SEQ ID NO:12 (6KC4-1#85-IgG1) комбинируют с вариабельными областями легкой цепи, имеющими последовательность эмбрионального типа.
Альтернативно последовательность вариабельной области тяжелой цепи, выбранную в качестве каркасной последовательности, которая первоначально содержит "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы", как упоминалось выше, можно конструировать так, чтобы она содержала нестрогие остатки. Количество и положение нестрогих остатков конкретно не ограничено, при условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует в зависимости от концентраций ионов. В частности, последовательности CDR и/или FR тяжелой цепи и/или легкой цепи могут содержать один или более нестрогих остатков. Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области тяжелой цепи SEQ ID NO:11 (6RL#9-IgG1), включают все аминокислотные остатки CDR1 и CDR2 тяжелой цепи и аминокислотные остатки CDR3 тяжелой цепи, за исключением остатков в положениях 95, 96 и/или 100a. Альтернативно неограничивающие примеры нестрогих остатков, вносимых в последовательность вариабельной области тяжелой цепи SEQ ID NO:12 (6KC4-1#85-IgG1), включают все аминокислотные остатки CDR1 и CDR2 тяжелой цепи и аминокислотные остатки CDR3 тяжелой цепи, за исключением остатков в положениях аминокислоты 95 и/или 101.
Альтернативно библиотеку, содержащую множество антигенсвязывающих молекул, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей легкой цепи, сконструированных в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельных областей легкой цепи, имеющих последовательности эмбрионального типа, с вариабельными областями тяжелой цепи, в которые внесен "по меньшей мере один аминокислотный остаток, ответственный за зависимое от концентрации ионов изменение антигенсвязывающей активности антигенсвязывающей молекулы". Когда концентрация ионов представляет собой концентрацию ионов кальция, неограничивающие примеры таких библиотек предпочтительно включают библиотеки, в которых вариабельные области легкой цепи, сконструированные в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельные области легкой цепи, имеющие последовательности эмбрионального типа, комбинируют с последовательностью вариабельной области тяжелой цепи, в которой конкретный остаток(остатки) заменен по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов кальция. Неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки CDR1 тяжелой цепи. Следующие неограничивающие примеры таких аминокислотных остатков включают аминокислотные остатки CDR2 тяжелой цепи. Кроме того, неограничивающие примеры таких аминокислотных остатков также включают аминокислотные остатки CDR3 тяжелой цепи. Неограничивающие примеры таких аминокислотных остатков CDR3 тяжелой цепи включают аминокислоты в положениях 95, 96, 100a и/или 101 в CDR3 вариабельной области тяжелой цепи, как указано согласно нумерации Kabat. Более того, эти аминокислотные остатки могут содержаться отдельно или в комбинации, при условии что они образуют кальций-связывающий мотив и/или антигенсвязывающая активность антигенсвязывающей молекулы варьирует в зависимости от концентрации ионов кальция.
Когда вариабельные области легкой цепи, сконструированные в качестве рандомизированной библиотеки последовательностей вариабельной области, или вариабельные области легкой цепи, имеющие последовательность эмбрионального типа, комбинируют с вариабельной областью тяжелой цепи, в которую внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации ионов, как упоминалось выше, последовательность вариабельной области тяжелой цепи также можно конструировать так, чтобы она содержала нестрогие остатки, аналогично тому, как описано выше. Количество и положение нестрогих остатков конкретно не ограничено, при условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует, в зависимости от концентраций ионов. В частности, последовательности CDR и/или FR тяжелой цепи могут содержать один или более нестрогих остатков. Более того, рандомизированные библиотеки вариабельной области можно предпочтительно использовать в качестве аминокислотных последовательностей CDR1, CDR2 и/или CDR3 вариабельной области тяжелой цепи, отличных от аминокислотных остатков, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы. Когда в качестве вариабельных областей легкой цепи используют последовательности эмбрионального типа, неограничивающие примеры таких последовательностей включают последовательности SEQ ID NO:7 (Vk1), SEQ ID NO:8 (Vk2), SEQ ID NO:9 (Vk3) и SEQ ID NO:10 (Vk4).
Предпочтительно можно использовать любую из описанных выше аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентраций ионов кальция, при условии что они образуют кальций-связывающий мотив. В частности, такие аминокислоты включают электронодонорные аминокислоты. Предпочтительные примеры таких электронодонорных аминокислот включают серин, треонин, аспарагин, глутаминовую кислоту, аспарагиновую кислоту и глутаминовую кислоту.
Условия концентраций протонов
В одном варианте осуществления настоящего изобретения условия концентраций ионов относятся к условиям концентраций протонов или условиям pH. В рамках настоящего изобретения концентрация протона, т.е. ядра атома водорода, используется в качестве синонима водородного показателя (pH). Когда активность протонов в водном растворе отображают как aH+, pH определяется как -log10aH+. Когда ионная сила водного раствора является низкой (например, ниже 10-3), aH+ практически равна ионной силе протонов. Например, ионное произведение воды при 25°C и 1 атмосфере составляет Kw=aH+aOH=10-14, и, таким образом, в чистой воде aH+=aOH=10-7. В этом случае, pH=7 является нейтральным; водный раствор, pH которого ниже 7, является кислым, или pH которого больше 7, является щелочным.
В рамках настоящего изобретения, когда условия pH используют в качестве условий концентрации ионов, условия pH включают высокие концентрации протонов или низкие значения pH, т.е. диапазон кислых значений pH, и низкие концентрации протонов или высокие значения pH, т.е. диапазон нейтральных значений pH. "Активность связывания варьирует в зависимости от условий pH" означает, что антигенсвязывающая активность антигенсвязывающей молекулы варьирует вследствие различий в условиях высокой концентрации протонов или низких значений pH (диапазон кислых значений pH) и низкой концентрации протонов или высоких значений pH (диапазон нейтральных значений pH). Это включает, например, случай, когда антигенсвязывающая активность антигенсвязывающей молекулы является более высокой в диапазоне нейтральных значений pH, чем в диапазоне кислых значений pH, и случай, когда антигенсвязывающая активность антигенсвязывающей молекулы является более высокой в диапазоне кислых значений pH, чем в диапазоне нейтральных значений pH.
В настоящем описании диапазон нейтральных значений pH не ограничивается конкретной величиной и предпочтительно выбран из диапазона от pH 6,7 до pH 10,0. В другом варианте осуществления значение pH может быть выбрано из диапазона от pH 6,7 до pH 9,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от pH 7,0 до pH 9,0. В другом варианте осуществления значение pH может быть выбрано из диапазона от pH 7,0 до pH 8,0. Особенно предпочтительное значение pH включает pH 7,4, которое является близким к pH плазмы (крови) in vivo.
В настоящем описании диапазон кислых значений pH не ограничивается конкретной величиной и предпочтительно выбран из диапазона от pH 4,0 до pH 6,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от pH 4,5 до pH 6,5. В другом варианте осуществления значение pH может быть выбрано из диапазона от pH 5,0 до pH 6,5. В другом варианте осуществления pH может быть выбрано из диапазона от pH 5,5 до pH 6,5. Особенно предпочтительное значение pH включает pH 5,8, что является близким к концентрации протонов в ранней эндосоме in vivo.
В рамках настоящего изобретения "антигенсвязывающая активность антигенсвязывающей молекулы при высокой концентрации протонов или низком значении pH (диапазон кислых значений pH) является более низкой, чем при низкой концентрации протонов или высоком значении pH (диапазон нейтральных значений pH)" означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от pH 4,0 до pH 6,5, является более слабой, чем при значении pH, выбранном из диапазона от pH 6,7 до pH 10,0; предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от pH 4,5 до pH 6,5, является более слабой, чем при pH, выбранном из диапазона от pH 6,7 до pH 9,5; более предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от pH 5,0 до pH 6,5, является более слабой, чем при значении pH, выбранном из диапазона от pH 7,0 до pH 9,0; еще более предпочтительно это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при значении pH, выбранном из диапазона от pH 5,5 до pH 6,5 является более слабой, чем при значении pH, выбранном из диапазона от pH 7,0 до pH 8,0; особенно предпочтительно это означает, что антигенсвязывающая активность при значении pH в ранней эндосоме in vivo является более слабой, чем антигенсвязывающая активность при значении pH в плазме in vivo; и, конкретно, это означает, что антигенсвязывающая активность антигенсвязывающей молекулы при pH 5,8 является более слабой, чем антигенсвязывающая активность при pH 7,4.
Определение того, изменяется ли антигенсвязывающая активность антигенсвязывающей молекулы в зависимости от условий pH можно проводить, например, с использованием известных способов измерения, таких как способы, описанные в разделе "Активность связывания" выше. В частности, активность связывания измеряют в различных условиях pH с использованием способов измерения, описанных выше. Например, антигенсвязывающую активность антигенсвязывающей молекулы сравнивают в условиях диапазона кислых значений pH и диапазона нейтральных значений pH, чтобы подтвердить, что антигенсвязывающая активность антигенсвязывающей молекулы изменяется так, что она является более высокой в условиях диапазона нейтральных значений pH, чем в условиях диапазона кислых значений pH.
Более того, в рамках настоящего изобретения выражение "антигенсвязывающая активность при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или при высоком значении pH, т.е. в диапазоне нейтральных значений pH" также может быть выражено как "антигенсвязывающая активность антигенсвязывающей молекулы при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой чем при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH". В рамках настоящего изобретения "антигенсвязывающая активность при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH" может быть описана как "антигенсвязывающая активность при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более слабой, чем антигенсвязывающая способность при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH". Альтернативно "антигенсвязывающая активность при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, снижена так, чтобы она более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH" может быть описана как "антигенсвязывающая активность при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, снижена так, чтобы она была слабее, чем антигенсвязывающая способность при низкой концентрации протонов или высоких значениях pH, т.е. в диапазоне нейтральных значений pH".
Условия, отличные от концентрации протонов или pH для измерения и антигенсвязывающей активности, могут быть соответствующим образом выбраны специалистами в данной области и они конкретно не ограничены. Измерение можно проводить, например, при 37°C с использованием буфера HEPES. Измерение можно проводить, например, с использованием Biacore (GE Healthcare). Когда антиген представляет собой растворимый антиген, антигенсвязывающую активность антигенсвязывающей молекулы можно определять путем оценки активности связывания с растворимым антигеном путем нанесения антигена в качестве анализируемого соединения на чип, на который иммобилизована антигенсвязывающая молекула. Когда антиген представляет собой антиген мембраны, активность связывания с антигеном мембраны можно оценивать путем нанесения антигенсвязывающей молекулы в качестве анализируемого соединения на чип, на который иммобилизован антиген.
При условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более слабой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, соотношение антигенсвязывающей активности между антигенсвязывающей активностью при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, и при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH конкретно не ограничено, и величина KD (pH 5,8)/KD (pH 7,4), которая представляет собой соотношение константы диссоциации (KD) для антигена при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, и KD при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно составляет 2 или более; более предпочтительно величина KD (pH 5,8)/KD (pH 7,4) составляет 10 или более; и еще более предпочтительно величина KD (pH 5,8)/KD (pH 7,4) составляет 40 или более. Верхний предел величины KD (pH 5,8)/KD (pH 7,4) конкретно не ограничен и он может представлять собой любую величину, такую как 400, 1000 или 10000, при условии что молекулу можно получать способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, константу диссоциации (KD) можно использовать в качестве величины антигенсвязывающей активности. Между тем, когда антиген представляет собой мембранный антиген, можно использовать кажущуюся константу диссоциации (KD). Константу диссоциации (KD) и кажущуюся константу диссоциации (KD) можно измерять способами, известными специалистам в данной области, и можно использовать Biacore (GE healthcare), график Скэтчарда, проточный цитометр и т.п.
Альтернативно, например, константу скорости диссоциации (kd) можно соответствующим образом использовать в качестве показателя, указывающего на соотношение антигенсвязывающей активности антигенсвязывающей молекулы по настоящему изобретению между активностью при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, и при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH. Когда kd (константа скорости диссоциации) используют в качестве показателя, указывающего на соотношение активностей связывания вместо KD (константа диссоциации), величина kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH), которая представляет собой соотношение kd (константа скорости диссоциации) в отношении антигена при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений, pH и kd (константа скорости диссоциации) при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно составляет 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более и еще более предпочтительно 30 или более. Верхний предел величины kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) конкретно не ограничен, и он может представлять собой любую величину, такую как 50, 100 или 200, при условии, что можно молекулу можно получать способами, известными специалистам в данной области.
Когда антиген представляет собой растворимый антиген, в качестве величины антигенсвязывающей активности можно использовать константу скорости диссоциации (kd), и когда антиген представляет собой антиген мембраны, можно использовать кажущуюся константу скорости диссоциации (kd). Константу скорости диссоциации (kd) и кажущуюся константу скорости диссоциации (kd) можно определять способами, известными специалистам в данной области, и можно использовать Biacore (GE healthcare), проточный цитометр и т.п. В рамках настоящего изобретения, когда антигенсвязывающую активность антигенсвязывающей молекулы измеряют при различных концентрациях протонов, т.е. pH, условия, отличные от концентрации протонов, т.е. pH, предпочтительно являются одинаковыми.
Например, антигенсвязывающий домен или антигенсвязывающая молекула, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются одним из вариантов осуществления, предусматриваемых настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов или антигенсвязывающих молекул, включающего следующие стадии (a)-(c):
(a) получение значения антигенсвязывающей активности антигенсвязывающего домена или антигенсвязывающей молекулы в диапазоне кислых значений pH;
(b) получение значения антигенсвязывающей активности антигенсвязывающего домена или антигенсвязывающей молекулы в диапазоне нейтральных значений pH; и
(c) выбор антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH.
Альтернативно антигенсвязывающий домен или антигенсвязывающая молекула, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются одним из вариантов осуществления, предусматриваемых настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов или антигенсвязывающих молекул или их библиотеки, включающего следующие стадии (a)-(c):
(a) контактирование антигенсвязывающего домена, или антигенсвязывающей молекулы, или их библиотеки, в диапазоне нейтральных значений pH с антигеном;
(b) помещение антигенсвязывающего домена или антигенсвязывающей молекулы, связавшихся с антигеном на стадии (a), в диапазон кислых значений pH; и
(c) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциировавших на стадии (b).
Антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать путем скрининга антигенсвязывающих доменов, или антигенсвязывающих молекул, или их библиотеки, включающего следующие стадии (a)-(d):
(a) контактирование в диапазоне кислых значений pH антигена с библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул;
(b) отбор антигенсвязывающего домена или антигенсвязывающей молекулы, которые не связываются с антигеном на стадии (a);
(c) позволение антигенсвязывающему домену или антигенсвязывающей молекуле, отобранным на стадии (b), связываться с антигеном в диапазоне нейтральных значений pH; и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, связавшихся с антигеном на стадии (c).
Антигенсвязывающий домен или антигенсвязывающая молекула, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(c):
(a) контактирование в диапазоне нейтральных значений pH библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с колонкой, на которой иммобилизован антиген;
(b) элюирование в диапазоне кислых значений pH из колонки антигенсвязывающего домена или антигенсвязывающей молекулы, связавшихся с колонкой на стадии (a); и
(c) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, элюированных на стадии (b).
Антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых при высокой концентрации ионов водорода или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(d):
(a) позволение в диапазоне кислых значений pH библиотеке антигенсвязывающих доменов или антигенсвязывающих молекул проходить через колонку, на которую иммобилизован антиген;
(b) сбор антигенсвязывающего домена или антигенсвязывающей молекулы, элюированных без связывания с колонкой на стадии (a);
(c) позволение антигенсвязывающему домену или антигенсвязывающей молекуле, собранным на стадии (b), связываться с антигеном в диапазоне нейтральных значений pH; и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, связавшихся с антигеном на стадии (c).
Антигенсвязывающий домен или антигенсвязывающую молекулу, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, которые являются другим вариантом осуществления, предусматриваемым настоящим изобретением, можно получать способом скрининга, включающим следующие стадии (a)-(d):
(a) контактирование антигена с библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул в диапазоне нейтральных значений pH;
(b) получение антигенсвязывающего домена или антигенсвязывающей молекулы, связавшихся с антигеном на стадии (a);
(c) помещение антигенсвязывающего домена или антигенсвязывающей молекулы, полученных на стадии (b), в диапазон кислых значений pH; и
(d) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых на стадии (c) является более слабой, чем у стандарта, выбранного на стадии (b).
Описанные выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающим доменам и антигенсвязывающим молекулам, антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH, которые получают способом скрининга, который дополнительно включает стадии повторения стадий (a)-(c) или (a)-(d) в описанных выше способах скрининга. Количество повторений стадий (a)-(c) или (a)-(d) конкретно не ограничено; однако обычно количество составляет 10 или менее.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена или антигенсвязывающей молекулы при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, конкретно не ограничена, при условии что она представляет собой антигенсвязывающую активность при pH от 4,0 до 6,5, и включает антигенсвязывающую активность при pH от 4,5 до 6,6 в качестве предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH от 5,0 до 6,5, и антигенсвязывающую активность при pH от 5,5 до 6,5 в качестве других предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH в ранней эндосоме in vivo в качестве более предпочтительного значения pH, и, в частности, антигенсвязывающую активность при pH 5,8. Между тем, антигенсвязывающая активность антигенсвязывающего домена или антигенсвязывающей молекулы при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность при pH от 6,7 до 10, и включает антигенсвязывающую активность при pH от 6,7 до 9,5 в качестве предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH от 7,0 до 9,5 и антиген связывающую активность при pH от 7,0 до 8,0 в качестве других предпочтительных значений pH. Антигенсвязывающая активность также включает антигенсвязывающую активность при pH плазмы in vivo в качестве более предпочтительных значений pH, и, в частности, антигенсвязывающую активность при pH 7,4.
Антигенсвязывающую активность антигенсвязывающего домена или антигенсвязывающей молекулы можно измерять способами, известными специалистам в данной области. Специалисты в данной области могут соответствующим образом определить условия, отличные от концентрации ионизированного кальция. Антигенсвязывающую активность антигенсвязывающего домена или антигенсвязывающей молекулы можно оценивать, основываясь на константе диссоциации (KD), кажущейся константе диссоциации (KD), константе скорости диссоциации (kd), кажущейся константе скорости диссоциации (kd) и т.п. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE healthcare), графика Скэтчарда или FACS.
В рамках настоящего изобретения стадия отбора антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является синонимичной стадии отбора антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, является более низкой, чем при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH.
При условии, что антигенсвязывающая активность при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, отличие между антигенсвязывающей активностью при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, и антигенсвязывающей активностью при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, конкретно не ограничено; однако антигенсвязывающая активность при низкой концентрации ионов водорода или высоком значении pH, т.е. в диапазоне нейтральных значений pH, предпочтительно в два или более раз, более предпочтительно в 10 или более раз, и еще более предпочтительно в 40 или более раз превышает антигенсвязывающую активность при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH.
Антигенсвязывающий домен или антигенсвязывающая молекула по настоящему изобретению, подвергаемые скринингу способами скрининга, описанными выше, могут представлять собой любой антигенсвязывающий домен или антигенсвязывающую молекулу, и, например, можно проводить скрининг упомянутого выше антигенсвязывающего домена или антигенсвязывающей молекулы. Например, можно проводить скрининг антигенсвязывающего доменов или антигенсвязывающих молекул, имеющих нативную последовательность, и также можно проводить скрининг антигенсвязывающего домена или антигенсвязывающей молекулы, в которых их аминокислотные последовательности были заменены.
Антигенсвязывающий домен или антигенсвязывающую молекулу по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым способом. Например, можно использовать существующие антигенсвязывающие молекулы, существующие библиотеки (фаговая библиотека и т.д.), антитела или библиотеки, полученные из B-клеток иммунизированных животных или из гибридом, полученных путем иммунизации животных, антитела или библиотеки (библиотеки с увеличенным содержанием аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, библиотеки, в которые внесены аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислотные мутации в конкретных положениях и т.д.), полученные путем внесения аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислотных мутаций в описанные выше антитела или библиотеки.
Способы получения антигенсвязывающего домена или антигенсвязывающей молекулы, антигенсвязывающая активность которых при низкой концентрации протонов или высоком значении pH, т.е. в диапазоне нейтральных значений pH, является более высокой, чем при высокой концентрации протонов или низком значении pH, т.е. в диапазоне кислых значений pH, из антигенсвязывающих доменов или антигенсвязывающих молекул, полученных из гибридом, полученных иммунизацией животных или из B-клеток иммунизированных животных, предпочтительно включают, например, антигенсвязывающий домен или антигенсвязывающую молекулу, в которых по меньшей мере одна из аминокислот антигенсвязывающего домена или антигенсвязывающей молекулы заменена аминокислотой с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или мутацией на неприродную аминокислоту, или антигенсвязывающий домен или антигенсвязывающую молекулу, в которые встроена аминокислота с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродная аминокислота, такая как описана в международной публикации № WO 2009/125825.
Участки внесения мутаций на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты конкретно не ограничены, и они могут представлять собой любое положение, при условии что антигенсвязывающая активность в диапазоне кислых значений pH становится более слабой, чем в диапазоне нейтральных значений pH (величина KD (в диапазоне кислых значений pH)/KD (в диапазоне нейтральных значений pH) или kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) увеличивается) по сравнению с величиной до замены или вставки. Например, когда антигенсвязывающая молекула представляет собой антитело, являются пригодными вариабельная область и CDR антитела. Специалисты в данной области могут соответствующим образом определить количество аминокислот, подлежащих замене, или количество аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, подлежащих встраиванию. Можно осуществить замену на одну аминокислоту, имеющую pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или одну неприродную аминокислоту; можно осуществить встраивание одной аминокислоты, имеющей pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или одной неприродной аминокислоты; можно осуществить замену на две или более аминокислоты, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или на две или более неприродных аминокислоты; и можно осуществить встраивание двух или более аминокислот, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или двух или более неприродных аминокислот. Альтернативно другие аминокислоты можно параллельно удалять, добавлять, встраивать и/или заменять, помимо замены на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, или встраивания аминокислот, имеющих pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот. Замену на или встраивание аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот можно проводить случайным образом способами, такими как сканирование гистидином, в которых аланин в способе сканирования аланином, известном специалистам в данной области, заменяют на гистидин. Антигенсвязывающие молекулы, проявляющие более высокую величину KD (в диапазоне кислых значений pH)/KD (в диапазоне нейтральных значений pH) или kd (в диапазоне кислых значений pH)/kd (в диапазоне нейтральных значений pH) по сравнению с величиной до внесения мутации можно выбирать из антигенсвязывающих доменов или антител, в которые внесены случайные инсерции или мутации с заменой на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты.
Предпочтительные примеры антигенсвязывающих молекул, содержащих мутацию на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, как описано выше, и антигенсвязывающая активность которых в диапазоне кислых значений pH является более низкой, чем в диапазоне нейтральных значений pH, включают антигенсвязывающие молекулы, антигенсвязывающая активность которых в диапазоне нейтральных значений pH после мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты сравнима с антигенсвязывающей активностью до мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты. В рамках настоящего изобретения "антигенсвязывающая молекула после мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты имеет антигенсвязывающую активность, сравнимую с антигенсвязывающей активностью до мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты" означает, что, при принятии антигенсвязывающей активности антигенсвязывающей молекулы до мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты как 100%, антигенсвязывающая активность антигенсвязывающей молекулы после мутации на аминокислоты, имеющие pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота), или неприродные аминокислоты составляет по меньшей мере 10% или более, предпочтительно 50% или более, более предпочтительно 80% или более, и еще более предпочтительно 90% или более. Антигенсвязывающая активность после мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты при pH 7,4 может быть более высокой, чем до мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты при pH 7,4. Если антигенсвязывающая активность антигенсвязывающей молекулы снижается вследствие встраивания или замены на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, антигенсвязывающую активность можно сделать сравнимой с антигенсвязывающей активностью до встраивания или замены на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты, путем внесения замены, делеции, вставки и/или инсерции одной или нескольких аминокислот антигенсвязывающей молекулы. Настоящее изобретение также включает антигенсвязывающие молекулы, активность связывания которых скорректирована так, чтобы она была сравнимой, путем замены, делеции, вставки и/или инсерции одной или нескольких аминокислот после замены на или вставки аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродной аминокислоты.
Аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающего домена в зависимости от условий концентрации протонов
Антигенсвязывающие домены или антигенсвязывающие молекулы по настоящему изобретению, подлежащие скринингу описанными выше способами скрининга, можно получать любым способом. Например, когда условия концентрации ионов представляют собой условия концентрации протонов или условия pH, можно использовать существующие антигенсвязывающие молекулы, существующие библиотеки (фаговая библиотека и т.д.), антитела или библиотеки, полученные из B-клеток иммунизированных животных или из гибридом, полученных иммунизацией животных, антитела или библиотеки (библиотеки с увеличенным содержанием аминокислот с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродных аминокислот, библиотеки, в которые внесены мутации на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты в конкретных положениях и т.д.), полученные путем внесения мутаций на аминокислоты с pKa боковой цепи 4,0-8,0 (например, гистидин и глутаминовая кислота) или неприродные аминокислоты в описанные выше антитела или библиотеки.
В одном варианте осуществления настоящего изобретения библиотеку, содержащую множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, также можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области, с вариабельными областями легкой цепи, в которые внесен "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации протонов".
Такие аминокислотные остатки включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR1 легкой цепи. Аминокислотные остатки также включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR2 легкой цепи. Аминокислотные остатки также включают, но не ограничиваются ими, например, аминокислотные остатки, содержащиеся в CDR3 легкой цепи.
Описанные выше аминокислотные остатки, содержащиеся в CDR1 легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 24, 27, 28, 31, 32 и/или 34 согласно нумерации Kabat в CDR1 вариабельной области легкой цепи. Между тем, аминокислотные остатки, содержащиеся в CDR2 легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 50, 51, 52, 53, 54, 55 и/или 56 согласно нумерации Kabat в CDR2 вариабельной области легкой цепи. Более того, аминокислотные остатки в CDR3 легкой цепи включают, но не ограничиваются ими, например, аминокислотные остатки в положениях 89, 90, 91, 92, 93, 94 и/или 95A согласно нумерации Kabat, в CDR3 вариабельной области легкой цепи. Более того, аминокислотные остатки могут содержаться отдельно или они могут содержаться в комбинации из двух или более аминокислот, при условии, что они позволяют изменять антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации протонов.
Даже когда вариабельную область тяжелой цепи, полученную в качестве рандомизированной библиотеки последовательностей вариабельной области, комбинируют с описанной выше вариабельной областью легкой цепи, в которую внесен "по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации протонов", ее можно конструировать так, чтобы нестрогие остатки содержались в последовательности вариабельной области легкой цепи аналогично тому, как описано выше. Количество и положение нестрогих остатков конкретно не ограничено конкретным вариантом осуществления, при условии что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению изменяется в зависимости от условий концентрации протонов. В частности, последовательности CDR и/или FR тяжелой цепи и/или легкой цепи могут содержать один или более нестрогих остатков. Например, нестрогие остатки, подлежащие внесению в последовательности вариабельных областей легкой цепи, включают, но не ограничиваются ими, например, аминокислотные остатки, приведенные в таблице 3 и 4. Между тем аминокислотные последовательности вариабельных областей легкой цепи, отличные от нестрогих остатков, и аминокислотные остатки, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации протонов, соответственно включают, но не ограничиваются ими, последовательности эмбрионального типа, такие как Vk1 (SEQ ID NO:7), Vk2 (SEQ ID NO:8), Vk3 (SEQ ID NO:9) и Vk4 (SEQ ID NO:10).
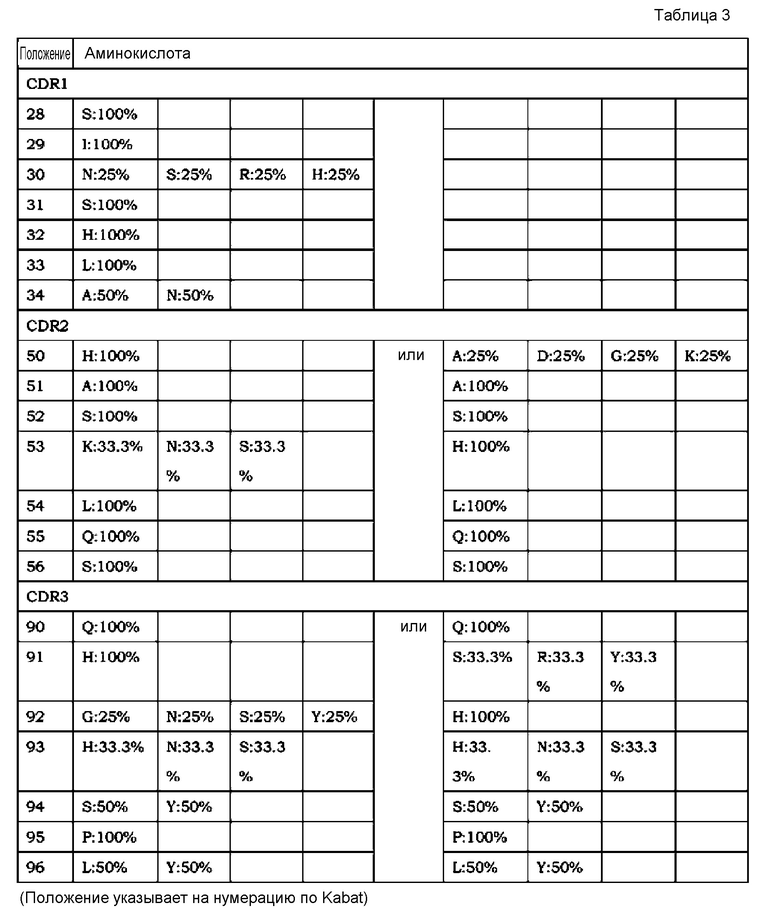
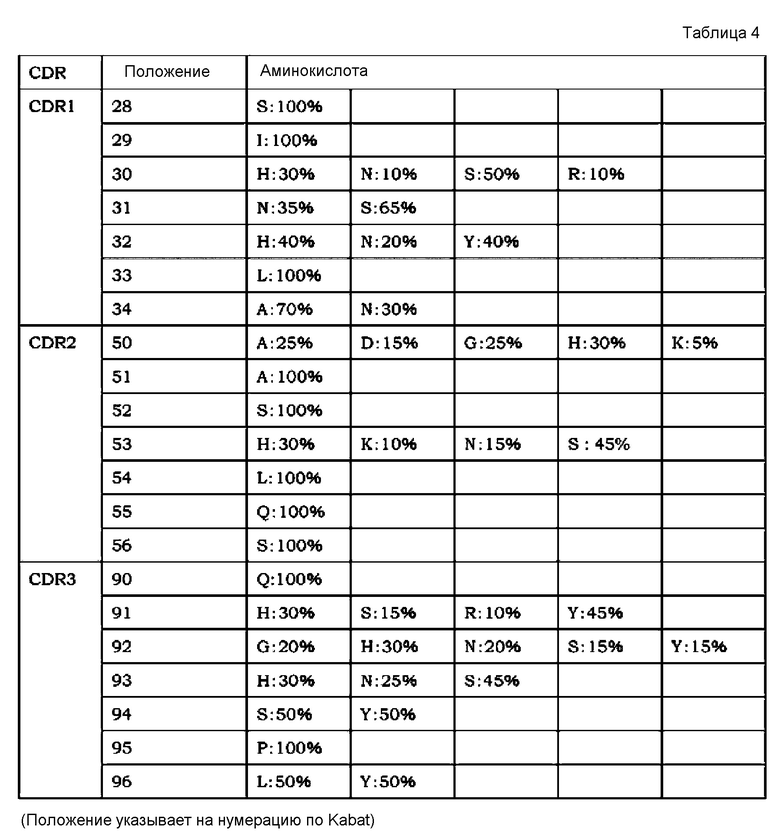
Любой аминокислотный остаток можно соответствующим образом использовать в качестве описанных выше аминокислотных остатков, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от условий концентрации протонов. В частности, такие аминокислотные остатки включают аминокислоты с pKa боковой цепи 4,0-8,0. Такие электроноотдающие аминокислоты предпочтительно включают, например, встречающиеся в природе аминокислоты, такие как гистидин и глутаминовая кислота, а также неприродные аминокислоты, такие как аналоги гистидина (US2009/0035836), m-NO2-Tyr (pKa 7,45), 3.5-Br2-Tyr (pKa 7,21) и 3.5-I2-Tyr (pKa 7,38) (Bioorg. Med. Chem. (2003) 11 (17), 3761-3768). Особенно предпочтительные аминокислотные остатки включают, например, аминокислоты с pKa боковой цепи 6,0-7,0. Такие электроноотдающие аминокислотные остатки предпочтительно включают, например, гистидин.
Для модификации аминокислот антигенсвязывающих доменов можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которых амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
Предпочтительная вариабельная область тяжелой цепи, которую используют в комбинации, включает, например, рандомизированные библиотеки вариабельной области. Известные способы соответствующим образом комбинируют в качестве способа получения рандомизированной библиотеки вариабельной области. В неограничивающем варианте осуществления настоящего изобретения, в качестве рандомизированной библиотеки вариабельной области можно использовать иммунную библиотеку, сконструированную на основе генов антител, происходящих из животных, иммунизированных конкретными антигенами, пациентов с инфекцией или индивидуумов с повышенным титром антител в крови в результате вакцинации, пациентов со злокачественной опухолью или лимфоцитов от пациентов с иммунными заболеваниями.
В другом неограничивающем варианте осуществления настоящего изобретения, аналогично тому, как описано выше, в качестве рандомизированной библиотеки вариабельной области также пригодным образом можно использовать синтетическую библиотеку, полученную путем замены последовательностей CDR V-генов в геномной ДНК или функциональных реконструированных V-генов набором синтетических олигонуклеотидов, содержащих последовательности, кодирующие наборы кодонов соответствующей длины. В этом случае, поскольку в последовательности CDR3 тяжелой цепи наблюдают разнообразие, также можно заменять только последовательность CDR3. Основанием для получения вариантов аминокислот в вариабельной области антигенсвязывающей молекулы является получение вариантов аминокислотных остатков экспонированных на поверхности положений антигенсвязывающей молекулы. Экспонированное на поверхности положение относится к положению, которое считается способным экспонироваться на поверхности и/или контактировать с антигеном, исходя из структуры, группы структур и/или смоделированной структуры антигенсвязывающей молекулы, и, как правило, такие положения представляют собой CDR. Предпочтительно, экспонированные на поверхности положения определяют с использованием координат из трехмерной модели антигенсвязывающей молекулы с использованием компьютерной программы, такой как программа InsightII (Accelrys). Экспонированные на поверхности положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (J. Mol. Biol. (1971) 55, 379-400); Connolly (J. Appl. Cryst. (1983) 16, 548-558)). Определение экспонированных на поверхности положений можно проводить на основе информации о трехмерной структуре с использованием программного обеспечения, пригодного для моделирования белка. Программное обеспечение, которое можно использовать для этих целей, предпочтительно включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Когда алгоритм требует введения пользователем параметра размера, "размер" образца, который используют в вычислении, устанавливают как радиус, приблизительно равный 1,4 Ангстрем или менее. Более того, способы определения экспонированных на поверхности областей и участков с использованием программного обеспечения для PC описаны Pacios (Comput. Chem. (1994) 18 (4), 377-386; J. Mol. Model. (1995) 1, 46-53).
В другом неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области также особенно пригодным является использование наивной библиотеки, которая сконструирована из генов антител, происходящих из лимфоцитов здоровых индивидуумов, и набор генов которой состоит из наивных последовательностей, которые представляют собой последовательности антител без предпочтений (Gejima et al. (Human Antibodies (2002) 11, 121-129); Cardoso et al. (Scand. J. Immunol. (2000) 51, 337-344)).
Fc-область
Fc-область содержит аминокислотную последовательность, происходящую из константной области тяжелой цепи антитела. Fc-область представляет собой часть константной области тяжелой цепи антитела, начинающуюся с N-конца шарнирной области, которая соответствует участку расщепления папаином у аминокислоты вблизи положения 216 согласно системе нумерации EU, и содержит шарнирную область, CH2- и CH3-домены. Хотя Fc-область можно получать из IgG1 человека, она не ограничивается конкретным подклассом IgG. Как описано ниже, подходящим примером Fc-области является Fc-область, которая обладает активностью связывания FcRn в диапазоне кислых значений pH. Более того, подходящим примером Fc-области является Fc-область, которая обладает активностью связывания Fcγ-рецептора, как описано ниже. Неограничивающим вариантом осуществления такой Fc-области является, например, Fc-область IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека.
FcRn
В отличие от Fcγ-рецептора, принадлежащего семейству иммуноглобулинов, FcRn, в частности, FcRn человека, структурно сходен с полипептидами главного комплекса гистосовместимости (MHC) класса I, проявляя 22%-29% идентичность последовательности с молекулами MHC класса I (Ghetie el al., Immunol. Today (1997) 18 (12): 592-598). FcRn экспрессируется в качестве гетеродимера, состоящего из растворимой β- или легкой цепи (β2-микроглобулин) в комплексе с трансмембранной α- или тяжелой цепью. Подобно MHC, α-цепь FcRn содержит три внеклеточных домена (α1, α2 и α3) и ее короткий цитоплазматический домен заякоривает белок на клеточной поверхности. α1- и α2-домены взаимодействуют с FcRn-связывающим доменом Fc-области антитела (Raghavan et al., Immunity (1994) 1: 303-315).
FcRn экспрессируется в материнской плаценте и желточном мешке млекопитающих, и он вовлечен в перенос IgG от матери к плоду. Кроме того, в тонком кишечнике новорожденных грызунов, где FcRn экспрессируется, FcRn вовлечен в перенос материнских IgG через эпителий щеточной каемки из проглоченного молозива или молока. FcRn экспрессируется во множестве других тканей и системах эндотелиальных клеток различных видов. FcRn также экспрессируется у взрослого человека в эндотелии, кровеносных сосудах мышц и синусоидных капиллярах печени. Полагают, что FcRn участвует в поддержании концентрации IgG в плазме путем опосредования рециклирования IgG в сыворотку при связывании с IgG. Как правило, связывание FcRn с молекулами IgG строго зависит от pH. Оптимальное связывание наблюдают при кислом диапазоне pH ниже 7,0.
FcRn человека, предшественником которого является полипептид, имеющий сигнальную последовательность SEQ ID NO:17 (полипептид с сигнальной последовательностью представлен в SEQ ID NO:18), образует комплекс с β2-микроглобулином человека in vivo. Растворимый FcRn человека в комплексе с β2-микроглобулином получают с использованием общепринятых способов рекомбинантной экспрессии. FcRn-области по настоящему изобретению можно оценивать в отношении их активности связывания с таким растворимым FcRn человека в комплексе с β2-микроглобулином. В рамках настоящего изобретения, если нет иных указаний, FcRn человека относится к форме, способной связываться с FcRn-связывающим доменом по настоящему изобретению. Примеры включают комплекс между FcRn человека и β2-микроглобулином человека. Неограничивающий вариант осуществления иллюстрируется комплексом, образованным между FcRn мыши (SEQ ID NO:73) и β2-микроглобулином мыши (SEQ ID NO:74).
Активность связывания Fc-области с FcRn, в частности, FcRn человека
Активность связывания Fc-области по настоящему изобретению с FcRn, в частности FcRn человека, можно измерять способами, известными специалистам в данной области, как описано в разделе "Активность связывания" выше. Специалисты в данной области могут соответствующим образом определить условия, отличные от pH. Антигенсвязывающую активность и активность связывания FcRn человека антигенсвязывающей молекулы можно оценивать, основываясь на константе диссоциации (KD), кажущейся константе диссоциации (KD), константе скорости диссоциации (kd), кажущейся константе скорости диссоциации (kd) и т.п. Их можно измерять способами, известными специалистам в данной области. Например, можно использовать Biacore (GE healthcare), график Скэтчарда или проточный цитометр.
Когда измеряют активность связывания FcRn человека Fc-области по настоящему изобретению, условия, отличные от pH, конкретно не ограничены, и их могут соответствующим образом выбирать специалисты в данной области. Измерения можно проводить, например, при 37°C с использованием буфера MES, как описано в международной публикации № WO 2009125825. Альтернативно активность связывания FcRn человека Fc-области по настоящему изобретению можно измерять способами, известными специалистам в данной области, и ее можно измерять с использованием, например, Biacore (GE Healthcare) и т.п. Активность Fc-области в отношении связывания по настоящему изобретению с FcRn человека можно оценивать путем нанесения в качестве анализируемого соединения FcRn человека, Fc-области или антигенсвязывающей молекулы по настоящему изобретению, содержащей Fc-область, на чип, на который иммобилизована Fc-область, антигенсвязывающая молекула по настоящему изобретению, содержащая Fc-область, или FcRn человека.
Диапазон нейтральных значений pH в качестве условий, где Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, имеет активность связывания FcRn, как правило, означает от pH 6,7 до pH 10,0. Предпочтительно, диапазон нейтральных значений pH представляет собой диапазон, указанный с помощью произвольных величин pH от pH 7,0 до pH 8,0, и предпочтительно он выбран из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, и особенно предпочтительно pH 7,4, что близко к значению pH плазмы (крови) in vivo. Когда аффинность связывания между FcRn-связывающим доменом человека и FcRn человека при pH 7,4 является слишком низкой, чтобы ее оценить, вместо pH 7,4 можно использовать pH 7,0. В рамках изобретения диапазон кислых значений pH в качестве условий, где Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, имеет активность связывания FcRn, как правило, означает от pH 4,0 до pH 6,5. Предпочтительно, диапазон кислых значений pH означает от pH 5,5 до pH 6,5, особенно предпочтительно от pH 5,8 до pH 6,0, что близко к pH в ранней эндосоме in vivo. что касается температуры, используемой в качестве условий измерения, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека можно определять при от 15°C до 40°C. Более предпочтительно, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека, можно определять аналогичным образом при произвольной температуре от 20°C до 35°C, такой как любая температура из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35°C. В одном варианте осуществления настоящего изобретения температура включает, но не ограничивается ими, например, 25°C.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность связывания нативного IgG1 человека с FcRn человека в диапазоне кислых значений pH (pH 6,0) составляет KD 1,7 мкМ, и эта активность практически не поддается выявлению в диапазоне нейтральных значений pH. Таким образом, в предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 20 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 2,0 мкМ (KD) или сильнее. В еще более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 0,5 мкМ (KD) или сильнее. Упомянутые выше величины KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизации антигенсвязывающей молекулы на чип и нанесения FcRn человека в качестве анализируемого соединения).
В рамках настоящего изобретения предпочтительные Fc-области имеют активность связывания FcRn в условиях диапазона кислых значений pH. Когда Fc-область изначально обладает активностью связывания FcRn в условиях диапазона кислых значений pH, этот домен можно использовать как есть. Когда домен обладает слабой активностью связывания FcRn в условиях диапазона кислых значений pH или не обладает ей, Fc-область, обладающую желаемой активностью связывания FcRn, можно получать путем модификации аминокислот антигенсвязывающей молекулы. Fc-области, имеющие желаемую или усиленную активность связывания FcRn в условиях диапазона кислых значений pH, также можно пригодным образом получать путем модификации аминокислот Fc-области. Аминокислотные модификации Fc-области, которые приводят к такой желаемой активности связывания, можно выявлять путем сравнения активности связывания FcRn в условиях диапазона кислых значений pH до и после модификации аминокислот. Специалисты в данной области могут соответствующим образом модифицировать аминокислоты с использованием известных способов, сходных с упомянутыми выше способами, используемыми для модификации активности связывания Fcγ-рецептора.
Fc-области, содержащиеся в антигенсвязывающих молекулах по настоящему изобретению, которые обладают активностью связывания FcRn в условиях диапазона кислых значений pH, можно получать любым способом. В частности, FcRn-связывающие домены, обладающие активностью связывания FcRn или усиленной активностью связывания FcRn в условиях диапазона кислых значений pH, можно получать путем модификации аминокислот иммуноглобулина человека IgG-типа, используемого в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулина IgG-типа для модификации включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3 и IgG4, и их варианты). При условии, что Fc-область обладает активностью связывания FcRn в условиях диапазона кислых значений pH или может увеличивать активность связывания FcRn человека в условиях диапазона кислых значений pH, аминокислоты в любом положении можно модифицировать на другие аминокислоты. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области, предпочтительно, чтобы полученная Fc-область содержала модификацию, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые позволяют такую модификацию, включают, например, аминокислоты в положениях 252, 254, 256, 309, 311, 315, 433 и/или 434 в соответствии с нумерацией EU и связанные с ними аминокислоты в положениях 253, 310, 435 и/или 426, как описано в WO 1997/034631. Подходящие примеры включают аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447, как указано в соответствии с нумерацией EU, как описано в WO 2000/042072. Аналогично, подходящие примеры аминокислот, которые позволяют такую модификацию, включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, аминокислоты, которые позволяют такую модификацию, включают, например, аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO2004/092219. Кроме того, подходящие примеры аминокислот, которые позволяют такую модификацию, включают аминокислоты в положениях 238, 244, 245, 249, 252, 256, 257, 258, 260, 262, 270, 272, 279, 283, 285, 286, 288, 293, 307, 311, 312, 316, 317, 318, 332, 339, 341, 343, 375, 376, 377, 378, 380, 382, 423, 427, 430, 431, 434, 436, 438, 440 и/или 442, как описано в WO 2006/020114. Более того, подходящие примеры аминокислот, которые позволяют такую модификацию, включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193. Модификация этих аминокислот усиливает связывание Fc-области иммуноглобулин IgG-типа с FcRn в условиях диапазона кислых значений pH.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает по меньшей мере одну или более аминокислотных модификаций, выбранных из группы, состоящей из:
Arg или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu или Thr в качестве аминокислоты в положении 256;
Ile или Thr в качестве аминокислоты в положении 308;
Pro в качестве аминокислоты в положении 309;
Glu, Leu или Ser в качестве аминокислоты в положении 311;
Ala или Asp в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
His, Phe или Tyr в качестве аминокислоты в положении 434; и
Arg, Asn, His, Lys, Met или Thr в качестве аминокислоты в положении 436, как указано в соответствии с нумерацией EU. Между тем, количество аминокислот, подлежащих модификации, конкретно не ограничено; и аминокислоту можно модифицировать только в одном участке или можно модифицировать аминокислоты в двух или более участков.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающим вариантом осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, могут быть модификации, включающие Ile в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309 и/или Glu в качестве аминокислоты в положении 311 в соответствии с нумерацией EU. Другой неограничиващий вариант осуществления этой модификации может включать Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Leu в качестве аминокислоты в положении 311, Ala в качестве аминокислоты в положении 312, и/или Ala в качестве аминокислоты в положении 314. Более того, другой неограничивающий вариант осуществления этой модификации может включать Ile или Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Glu, Leu или Ser в качестве аминокислоты в положении 311, Ala в качестве аминокислоты в положении 312, и/или Ala или Leu в качестве аминокислоты в положении 314. Другой неограничивающий вариант осуществления этой модификации может включать Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Ser в качестве аминокислоты в положении 311, Asp в качестве аминокислоты в положении 312, и/или Leu в качестве аминокислоты в положении 314.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Leu в качестве аминокислоты в положении 251, Tyr в качестве аминокислоты в положении 252, Ser или Thr в качестве аминокислоты в положении 254, Arg в качестве аминокислоты в положении 255, и/или Glu в качестве аминокислоты в положении 256 в соответствии с нумерацией EU.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428, Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433, His, Phe или Tyr в качестве аминокислоты в положении 434, и/или Arg, Asn, His, Lys, Met или Thr в качестве аминокислоты в положении 436 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать His или Met в качестве аминокислоты в положении 428, и/или His или Met в качестве аминокислоты в положении 434.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Arg в качестве аминокислоты в положении 385, Thr в качестве аминокислоты в положении 386, Arg в качестве аминокислоты в положении 387, и/или Pro в качестве аминокислоты в положении 389 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать Asp в качестве аминокислоты в положении 385, Pro в качестве аминокислоты в положении 386, и/или Ser в качестве аминокислоты в положении 389.
Более того, когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает по меньшей мере одну или более аминокислотных модификаций, выбранных из группы, состоящей из: Gln или Glu в качестве аминокислоты в положении 250; и Leu или Phe в качестве аминокислоты в положении 428 в соответствии с нумерацией EU. Количество аминокислоты, подлежащих модификации, конкретно не ограничено; и аминокислоту можно модифицировать только в одном участке или аминокислоты можно модифицировать в двух участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Gln в качестве аминокислоты в положении 250, и/или Leu или Phe в качестве аминокислоты в положении 428 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать Glu в качестве аминокислоты в положении 250, и/или Leu или Phe в качестве аминокислоты в положении 428.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека включает по меньшей мере две или более модификаций аминокислот, выбранных из группы, состоящей из:
Asp или Glu в качестве аминокислоты в положении 251;
Tyr в качестве аминокислоты в положении 252;
Gln в качестве аминокислоты в положении 307;
Pro в качестве аминокислоты в положении 308;
Val в качестве аминокислоты в положении 378;
Ala в качестве аминокислоты в положении 380;
Leu в качестве аминокислоты в положении 428;
Ala или Lys в качестве аминокислоты в положении 430;
Ala, His, Ser или Tyr в качестве аминокислоты в положении 434; и
Ile в качестве аминокислоты в положении 436, как указано в соответствии с нумерацией EU. Количество аминокислоты, подлежащих модификации, конкретно не ограничено; и аминокислоту можно модифицировать только в двух участках или аминокислоты можно модифицировать в трех или более участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Gln в качестве аминокислоты в положении 307, и Ala или Ser в качестве аминокислоты в положении 434 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать Pro в качестве аминокислоты в положении 308, и Ala в качестве аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этой модификации может включать Tyr в качестве аминокислоты в положении 252, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этой модификации может включать Val в качестве аминокислоты в положении 378, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этой модификации может включать Leu в качестве аминокислоты в положении 428, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этой модификации может включать Ala в качестве аминокислоты в положении 434, и Ile в качестве аминокислоты в положении 436. Более того, другой неограничивающий вариант осуществления этой модификации может включать Pro в качестве аминокислоты в положении 308, и Tyr в качестве аминокислоты в положении 434. Кроме того, другой неограничивающий вариант осуществления этой модификации может включать Gln в качестве аминокислоты в положении 307, и Ile в качестве аминокислоты в положении 436.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие любую из Gln в качестве аминокислоты в положении 307, Ala в качестве аминокислоты в положении 380, и Ser в качестве аминокислоты в положении 434 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать Gln в качестве аминокислоты в положении 307, Ala в качестве аминокислоты в положении 380, и Ala в качестве аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этой модификации может включать Tyr в качестве аминокислоты в положении 252, Pro в качестве аминокислоты в положении 308, и Tyr в качестве аминокислоты в положении 434. Отличающийся неограничивающий вариант осуществления этой модификации может включать Asp в качестве аминокислоты в положении 251, Gln в качестве аминокислоты в положении 307, и His в качестве аминокислоты в положении 434.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает модификацию по меньшей мере двух или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Tyr в качестве аминокислоты в положении 252;
Pro в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu или Met в качестве аминокислоты в положении 307;
Ala, Ile, Lys, Leu, Met, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 312;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp или Asn в качестве аминокислоты в положении 378;
Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Phe, Gly, His, Trp или Tyr в качестве аминокислоты в положении 434;
Ile, Leu или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442 в соответствии с нумерацией EU. Количество аминокислоты, подлежащих модификации, конкретно не ограничено и могут быть модифицированы аминокислоты только в двух участках и могут быть модифицированы аминокислоты в трех или более участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой модификации, включающие Ile в качестве аминокислоты в положении 257, и Ile в качестве аминокислоты в положении 311 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этой модификации может включать Ile в качестве аминокислоты в положении 257, и His в качестве аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой модификации может включать Val в качестве аминокислоты в положении 376, и His в качестве аминокислоты в положении 434.
Более того, в другом неограничивающем варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область с характеристикой наличия активности связывания FcRn человека в диапазоне нейтральных значений pH вместо упомянутой выше характеристики наличия активности связывания FcRn человека в диапазоне кислых значений pH. В предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 40 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH равна 15 мкМ (KD) или сильнее.
Более того, в другом неограничивающем варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область с характеристикой наличия активности связывания FcRn человека в диапазоне нейтральных значений pH вместо упомянутой выше характеристики наличия активности связывания FcRn человека в диапазоне кислых значений pH. В предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 40 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH равна 15 мкМ (KD) или сильнее.
В рамках настоящего изобретения предпочтительные Fc-области имеют активность связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH. Когда Fc-область первоначально обладает активностью связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, ее можно использовать как есть. Когда Fc-область имеет слабую активность связывания FcRn человека или не имеет активности связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, антигенсвязывающие молекулы, содержащие Fc-область, имеющую желаемую активность связывания FcRn человека, можно получать путем модификации аминокислот Fc-области, содержащейся в антигенсвязывающих молекулах. Fc-области, имеющие желаемую активность связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, также можно пригодным образом получать путем модификации аминокислот Fc-области человека. Альтернативно антигенсвязывающие молекулы, содержащие Fc-область, имеющую желаемую активность связывания FcRn человека, можно получать путем модификации аминокислот Fc-области, которые первоначально обладают активностью связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH. Аминокислотные модификации Fc-области человека, которые приводят к такой желаемой активности связывания, могут быть выявлены путем сравнения активности связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH до и после модификации аминокислот. Специалисты в данной области могут соответствующим образом модифицировать аминокислоты с использованием известных способов.
Более того, также в качестве измененной Fc-области по настоящему изобретению предпочтительно можно использовать Fc-область, модифицированную из исходной Fc-области, которая уже модифицирована. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, Fc-область, соответствующую IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или модифицированных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; международных публикациях № WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности с их исходной Fc-областью, меньшим чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с системой нумерации EU.
Для модификации аминокислот антигенсвязывающих доменов можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которых амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
Fc-области, содержащиеся в антигенсвязывающих молекулах по настоящему изобретению, которые обладают активностью связывания FcRn человека в диапазоне кислых значений pH, можно получать любым способом. В частности, можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 20 мкМ (KD) или сильнее; в более предпочтительном варианте осуществления Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 2,0 мкМ (KD) или сильнее; и в еще более предпочтительном варианте осуществления Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 0,5 мкМ (KD) или сильнее в результате модификации аминокислот иммуноглобулина IgG-типа человека, используемого в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типов для модификации включают, например, Fc-области IgG человека, таких как IgG1, IgG2, IgG3 и IgG4, представленные в SEQ ID NO:13, 14, 15 и 15, соответственно, и их варианты.
Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области, пригодные примеры аминокислот, которые можно модифицировать для достижения упомянутых выше желаемых эффектов в отношении связывания FcRn в условиях диапазона кислых значений pH путем модификации аминокислот иммуноглобулина человека IgG-типа в качестве исходной Fc-области, включают аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447 в соответствии с нумерацией EU, как описано в WO 2000/042072. Аналогично, подходящие примеры аминокислот, которые позволяют такую модификацию, включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, аминокислоты, которые позволяют такую модификацию, включают, например, аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO2004/092219. Более того, подходящие примеры аминокислот, которые позволяют такую модификацию, включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193. Модификация этих аминокислот усиливает связывание Fc-области иммуноглобулина человека IgG-типа с FcRn в условиях диапазона кислых значений pH.
Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, также можно получать путем модификации аминокислот иммуноглобулина человека IgG-типа, используемого в качестве исходной Fc-области. Fc-области иммуноглобулинов IgG-типа, подходящие для модификации, включают, например, Fc-области IgG человека (такого как IgG1, IgG2, IgG3 и IgG4, соответственно, соответствующие SEQ ID NO:13, 14, 15 и 16, их модифицированные формы). Аминокислоты в любых положениях можно изменять на другие аминокислоты, при условии, что полученные области обладают активностью связывания FcRn человека в диапазоне нейтральных значений pH или увеличенной активностью связывания FcRn человека в нейтральном диапазоне. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, предпочтительно, чтобы полученная область содержала изменение, которое приводит к эффекту усиления связывания FcRn человека в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые допускают такие изменения, включают, например, одну или более аминокислот, выбранных из группы положений 221-225, 227, 228, 230, 232, 233-241, 243-252, 254-260, 262-272, 274, 276, 278-289, 291-312, 315-320, 324, 325, 327-339, 341, 343, 345, 360, 362, 370, 375-378, 380, 382, 385-387, 389, 396, 414, 416, 423, 424, 426-438, 440 и 442 (указанные согласно нумерации EU). Изменение этих аминокислот усиливает связывание с FcRn человека Fc-области иммуноглобулина IgG-типа в диапазоне нейтральных значений pH.
Среди описанных выше, для применения в рамках настоящего изобретения выбирают подходящие изменения, которые усиливают связывание FcRn человека в диапазоне нейтральных значений pH. Особенно предпочтительные аминокислоты для таких вариантов Fc-области включают, например, аминокислоты в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (указанные согласно нумерации EU). Активность связывания FcRn человека Fc-области, включенной в антигенсвязывающую молекулу, можно увеличивать в диапазоне нейтральных значений pH путем замены по меньшей мере одной аминокислоты другой аминокислотой.
Особенно предпочтительные изменения в Fc-области включают, например, замены:
Met вместо аминокислоты в положении 237;
Ile вместо аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr вместо аминокислоты в положении 250;
Phe, Trp или Tyr вместо аминокислоты в положении 252;
Thr вместо аминокислоты в положении 254;
Glu вместо аминокислоты в положении 255;
Asp, Asn, Glu или Gln вместо аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val вместо аминокислоты в положении 257;
His вместо аминокислоты в положении 258;
Ala вместо аминокислоты в положении 265;
Ala или Glu вместо аминокислоты в положении 286;
His вместо аминокислоты в положении 289;
Ala вместо аминокислоты в положении 297;
Ala вместо аминокислоты в положении 303;
Ala вместо аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr вместо аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg вместо аминокислоты в положении 309;
Ala, His или Ile вместо аминокислоты в положении 311;
Ala или His вместо аминокислоты в положении 312;
Lys или Arg вместо аминокислоты в положении 314;
Ala, Asp или His вместо аминокислоты в положении 315;
Ala вместо аминокислоты в положении 317;
Val вместо аминокислоты в положении 332;
Leu вместо аминокислоты в положении 334;
His вместо аминокислоты в положении 360;
Ala вместо аминокислоты в положении 376;
Ala вместо аминокислоты в положении 380;
Ala вместо аминокислоты в положении 382;
Ala вместо аминокислоты в положении 384;
Asp или His вместо аминокислоты в положении 385;
Pro вместо аминокислоты а в положении 386;
Glu for вместо аминокислоты в положении 387;
Ala или Ser вместо аминокислоты в положении 389;
Ala вместо аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 428;
Lys вместо аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr вместо аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val вместо аминокислоты в положении 436 в системе нумерации EU. Между тем, количество измененных аминокислот конкретно не ограничено; такие аминокислотные изменения включают единичное изменение аминокислоты и изменение аминокислот в двух или более участков. Комбинации этих модификаций аминокислот включают, например, модификации аминокислот, представленные в таблицах с 5-1 по 5-33.

Таблица 5-2 является продолжением таблицы 5-1.

Таблица 5-3 является продолжением таблицы 5-2.

Таблица 5-4 является продолжением таблицы 5-3.

Таблица 5-5 является продолжением таблицы 5-4.
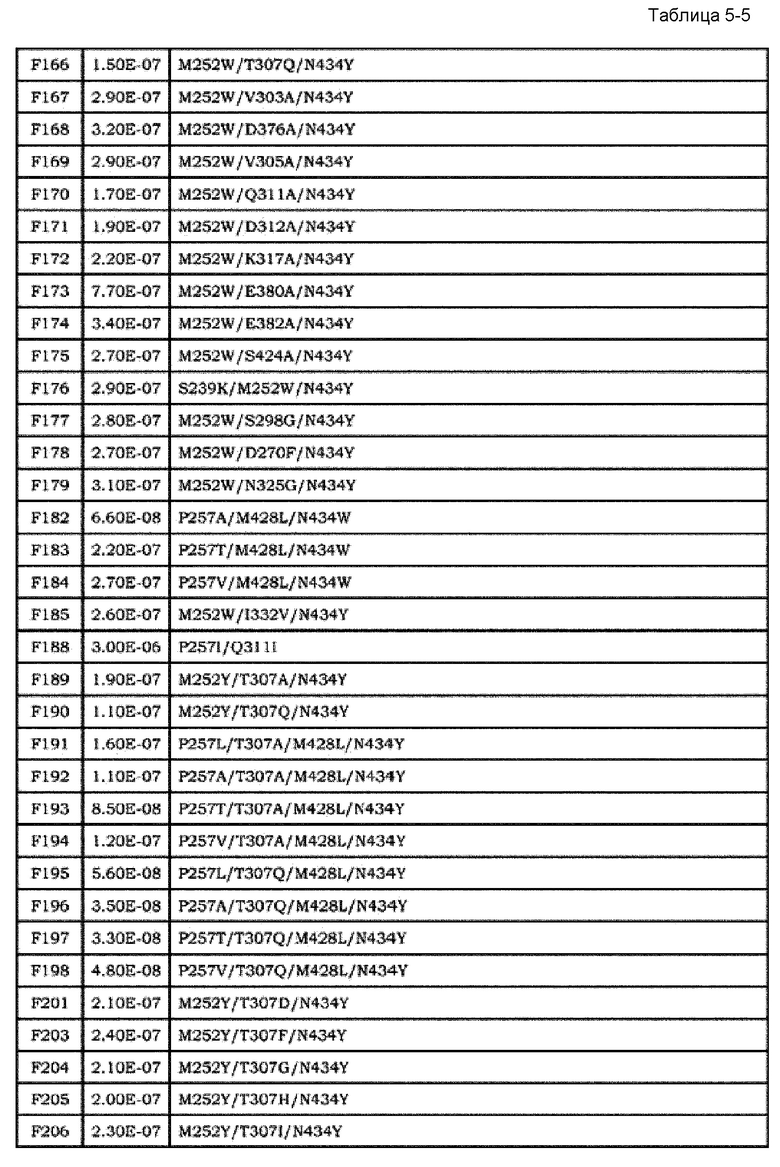
Таблица 5-6 является продолжением таблицы 5-5.
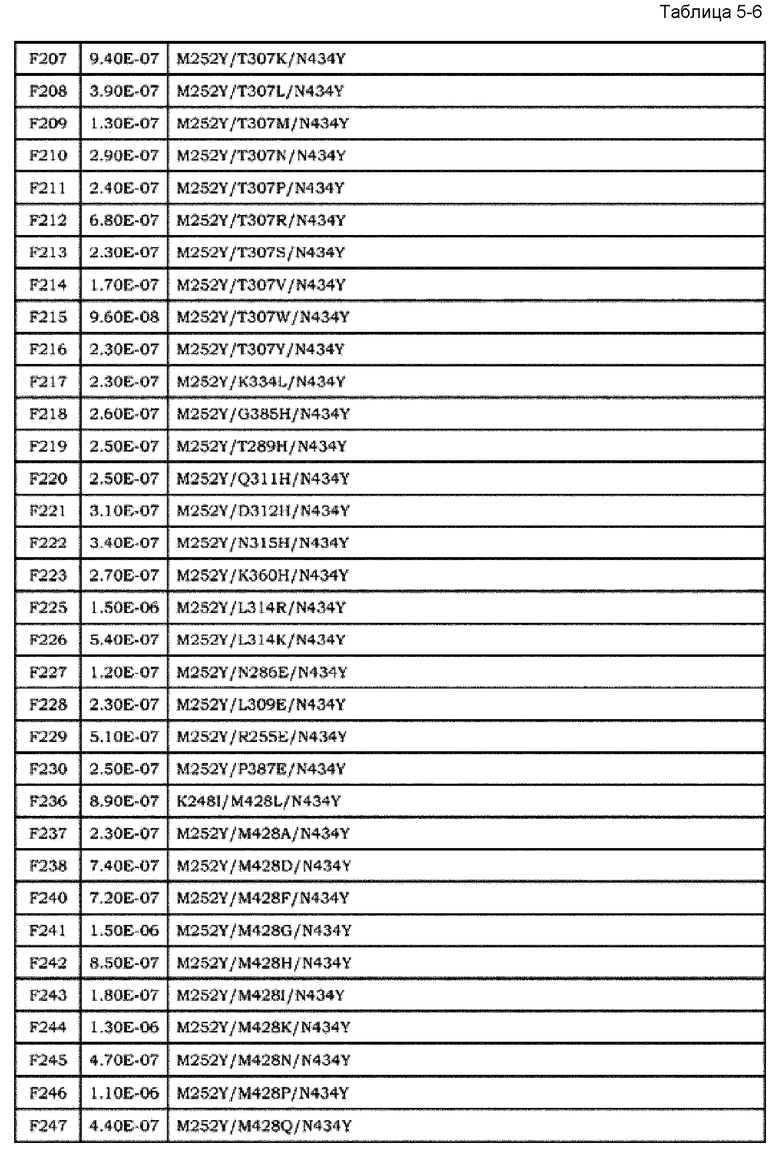
Таблица 5-7 является продолжением таблицы 5-6.

Таблица 5-8 является продолжением таблицы 5-7.

Таблица 5-9 является продолжением таблицы 5-8.

Таблица 5-10 является продолжением таблицы 5-9.

Таблица 5-11 является продолжением таблицы 5-10.

Таблица 5-12 является продолжением таблицы 5-11.

Таблица 5-13 является продолжением таблицы 5-12.

Таблица 5-14 является продолжением таблицы 5-13.
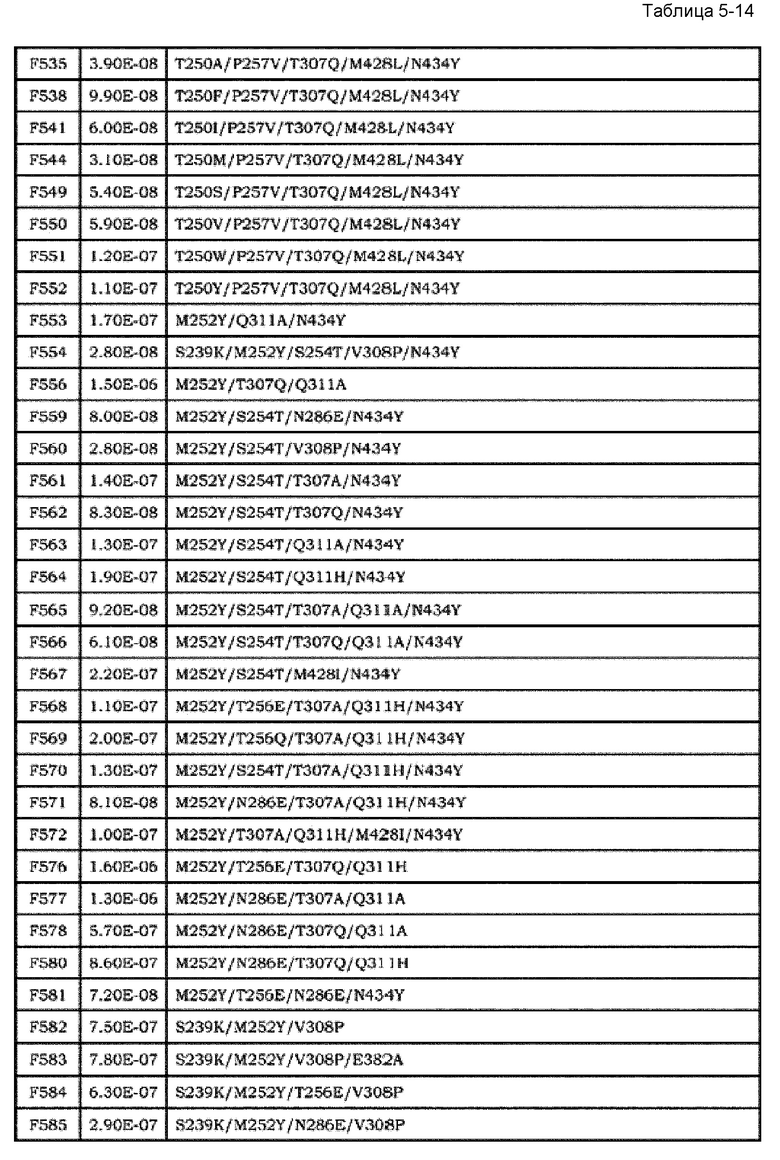
Таблица 5-15 является продолжением таблицы 5-14.

Таблица 5-16 является продолжением таблицы 5-15.

Таблица 5-17 является продолжением таблицы 5-16.

Таблица 5-18 является продолжением таблицы 5-17.
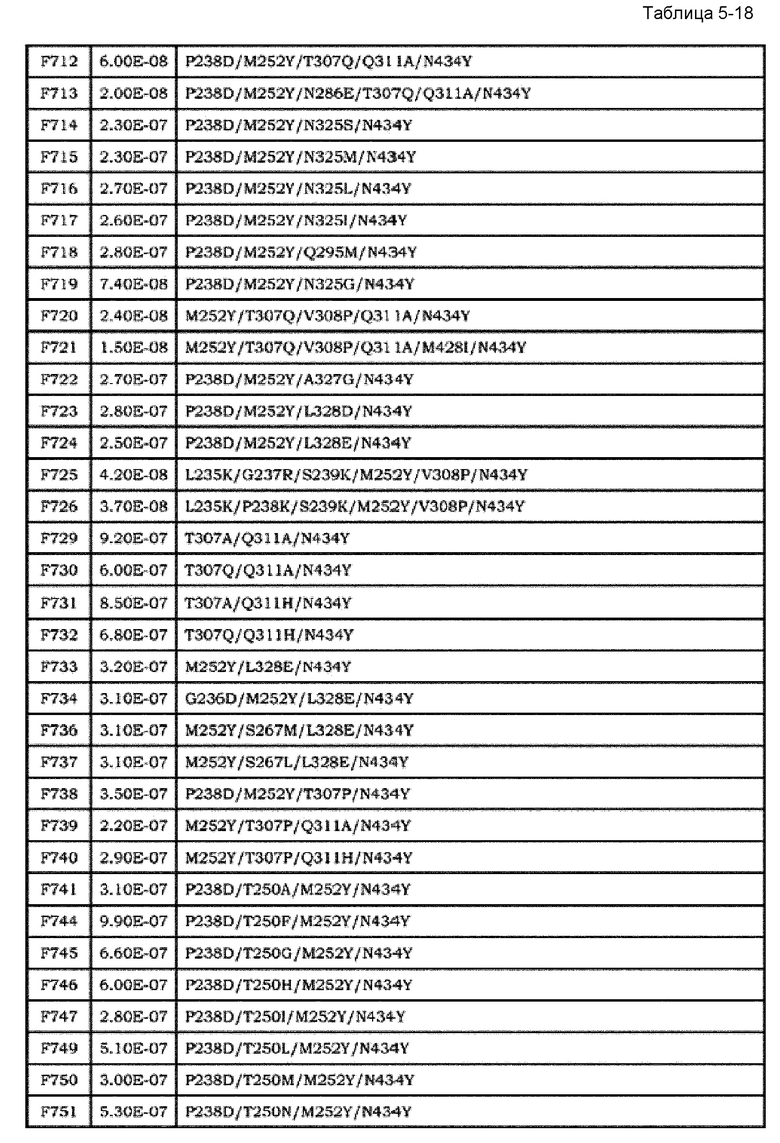
Таблица 5-19 является продолжением таблицы 5-18.
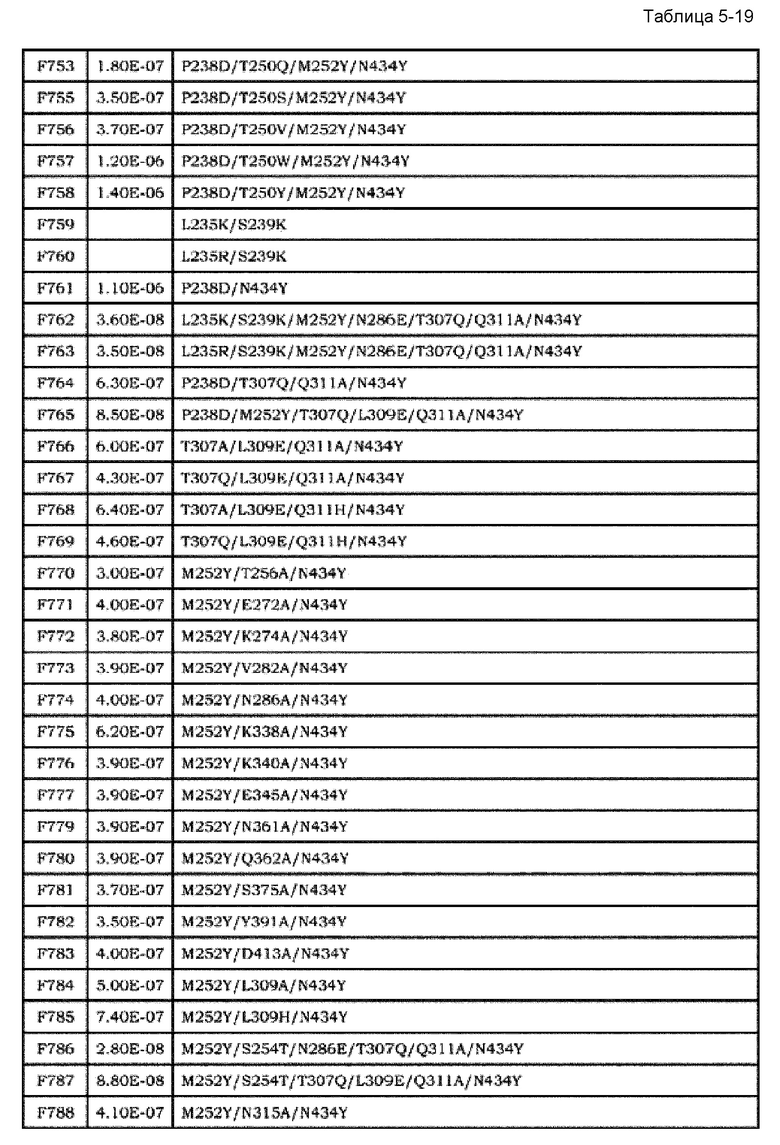
Таблица 5-20 является продолжением таблицы 5-19.
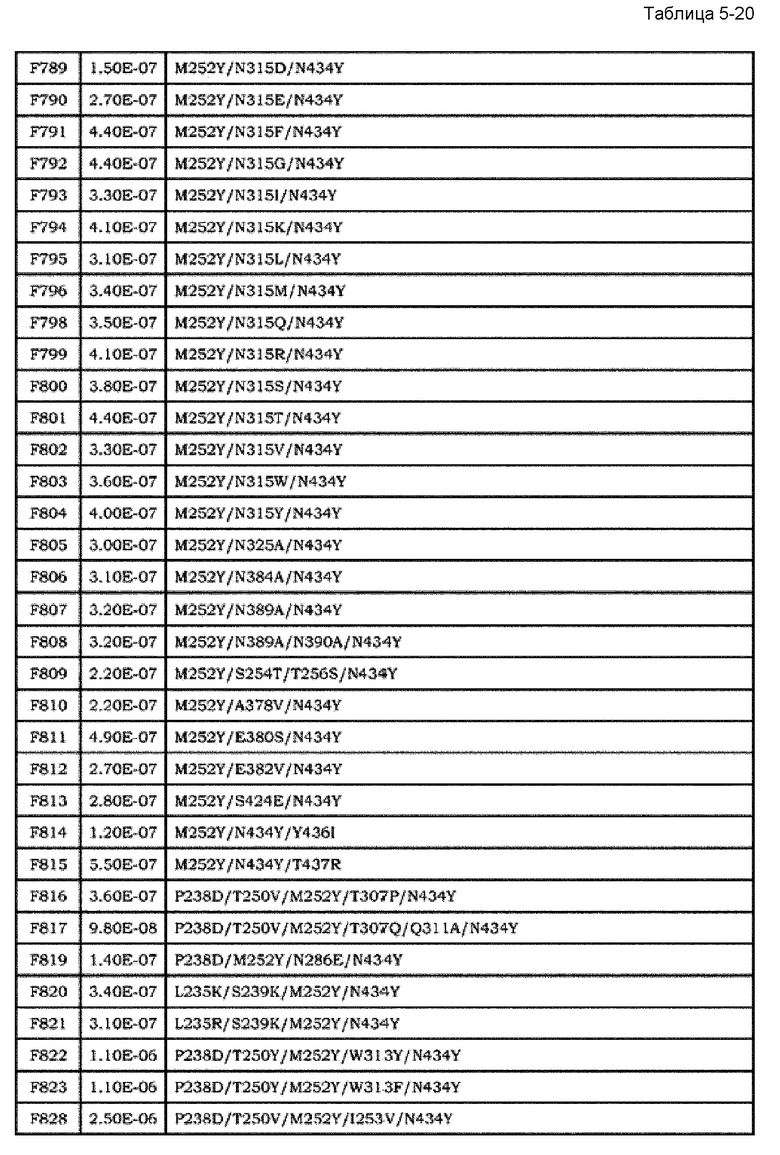
Таблица 5-21 является продолжением таблицы 5-20.
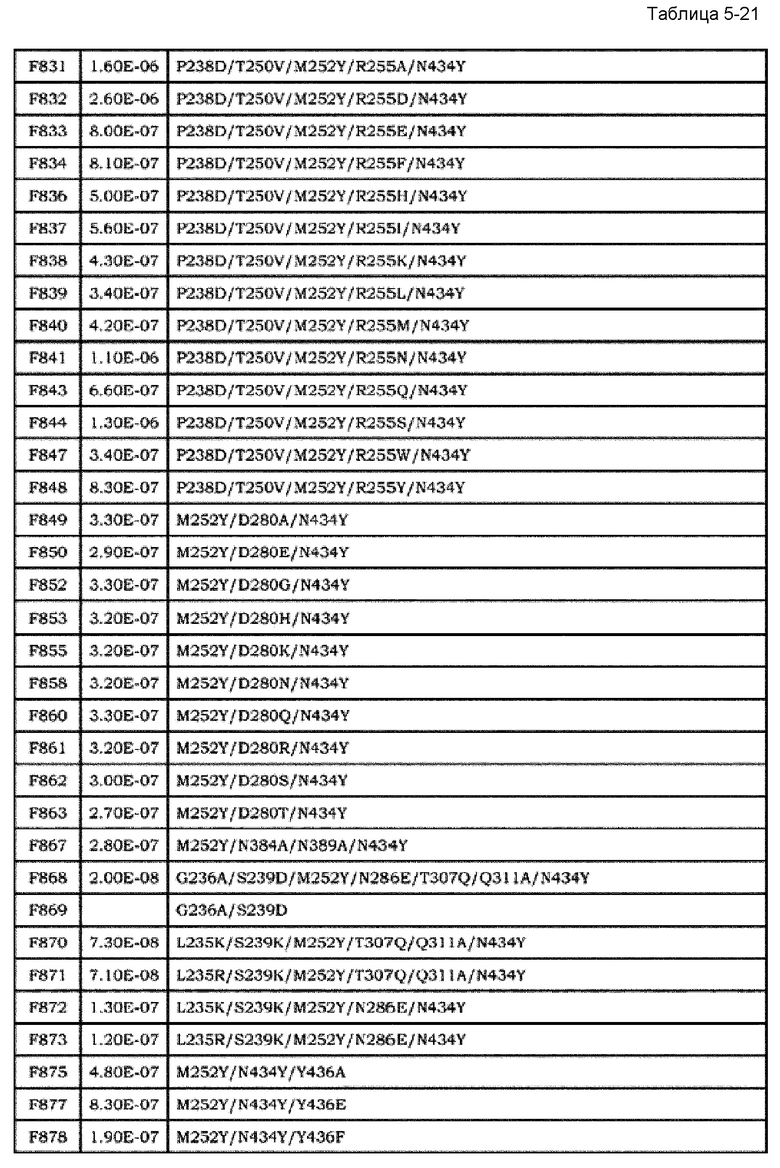
Таблица 5-22 является продолжением таблицы 5-21.
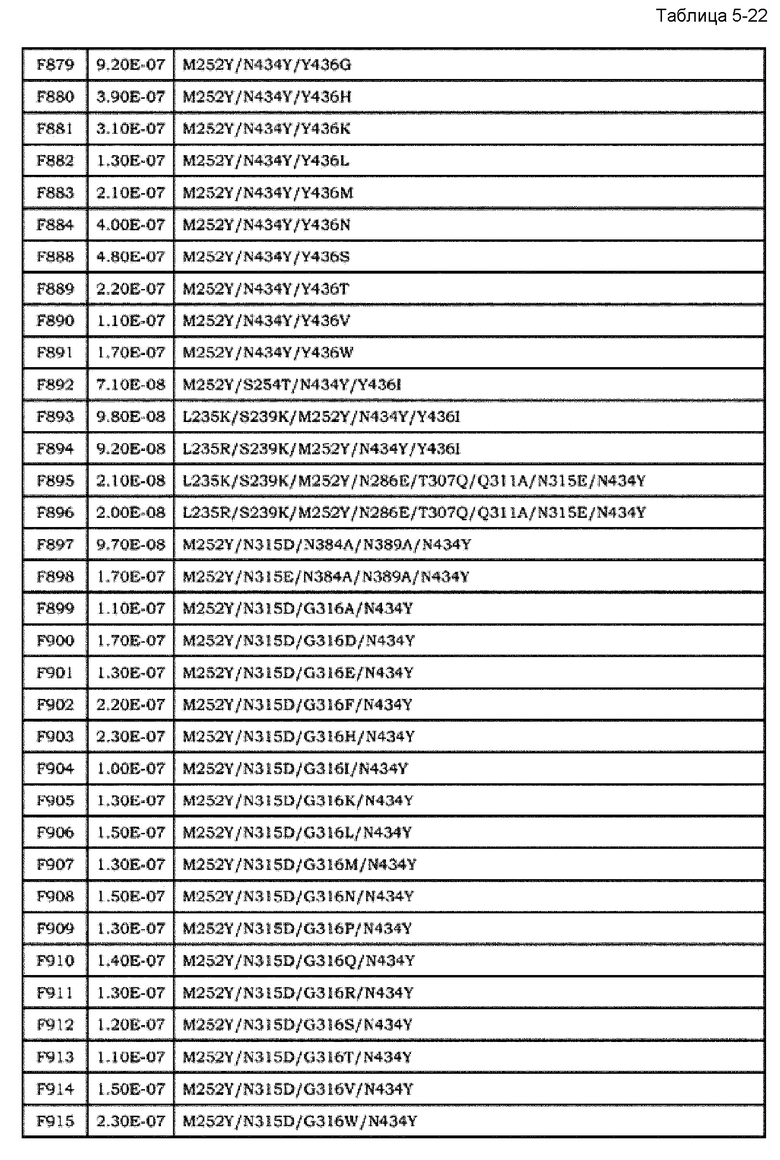
Таблица 5-23 является продолжением таблицы 5-22.

Таблица 5-24 является продолжением таблицы 5-23.

Таблица 5-25 является продолжением таблицы 5-24.

Таблица 5-26 является продолжением таблицы 5-25.
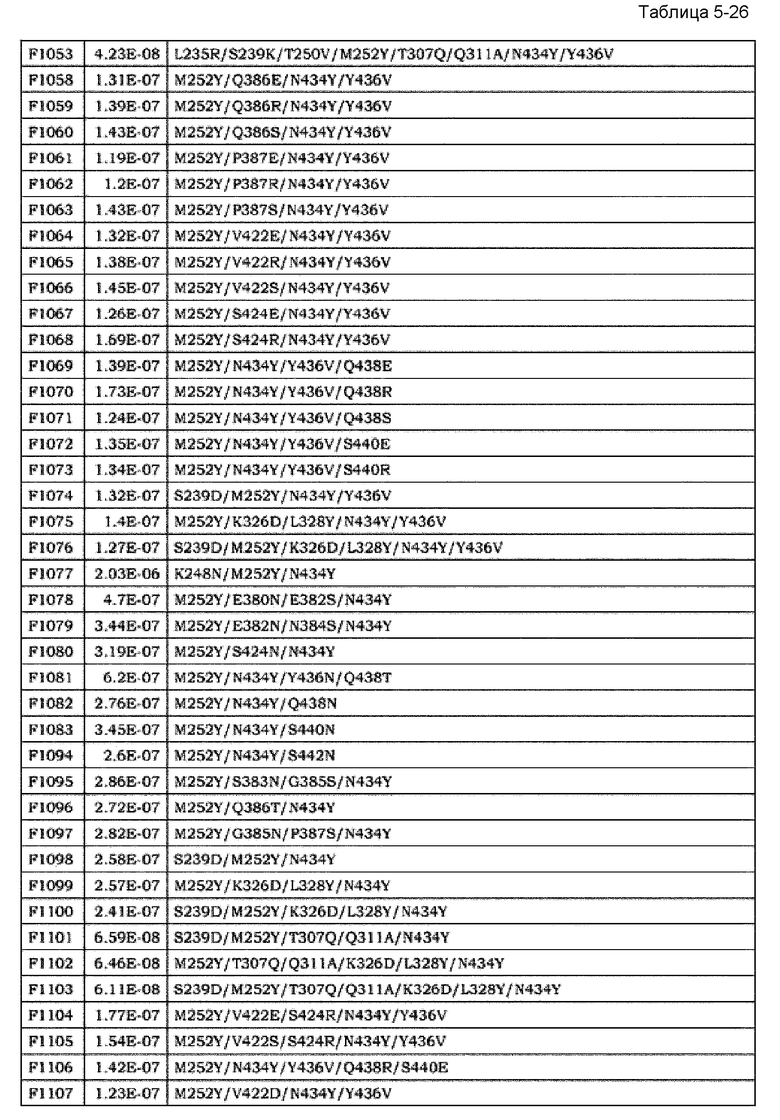
Таблица 5-27 является продолжением таблицы 5-26.

Таблица 5-28 является продолжением таблицы 5-27.
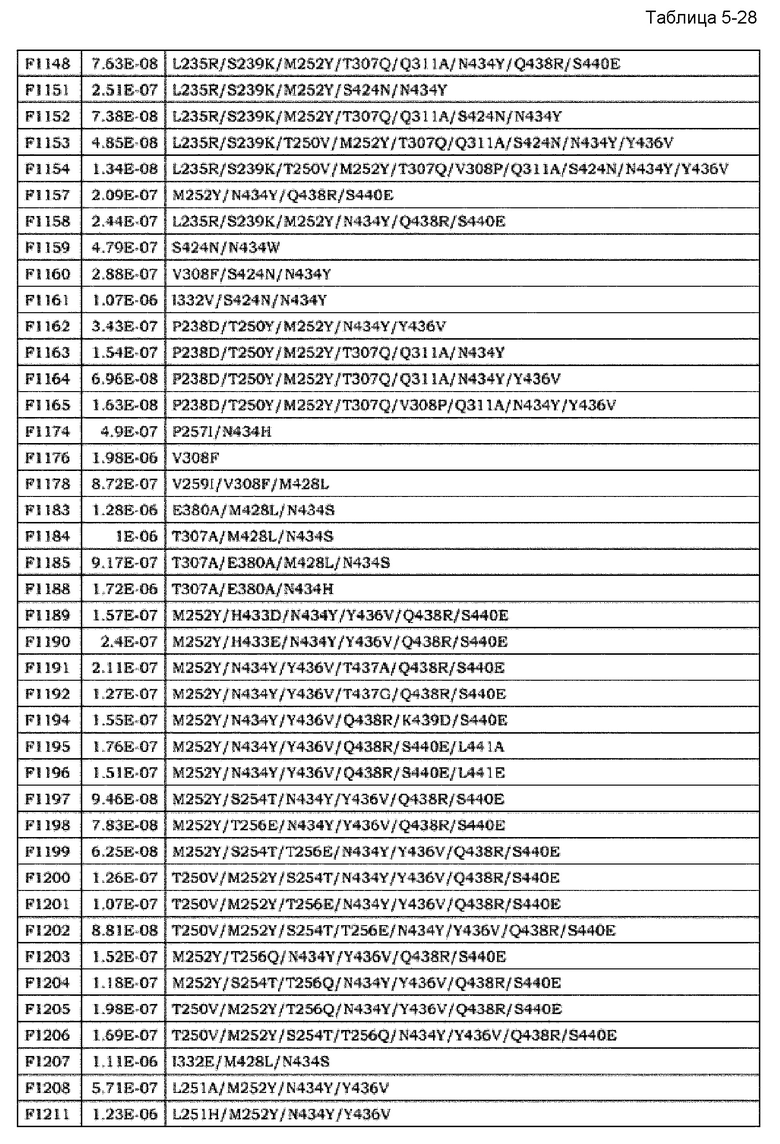
Таблица 5-29 является продолжением таблицы 5-28.

Таблица 5-30 является продолжением таблицы 5-29.
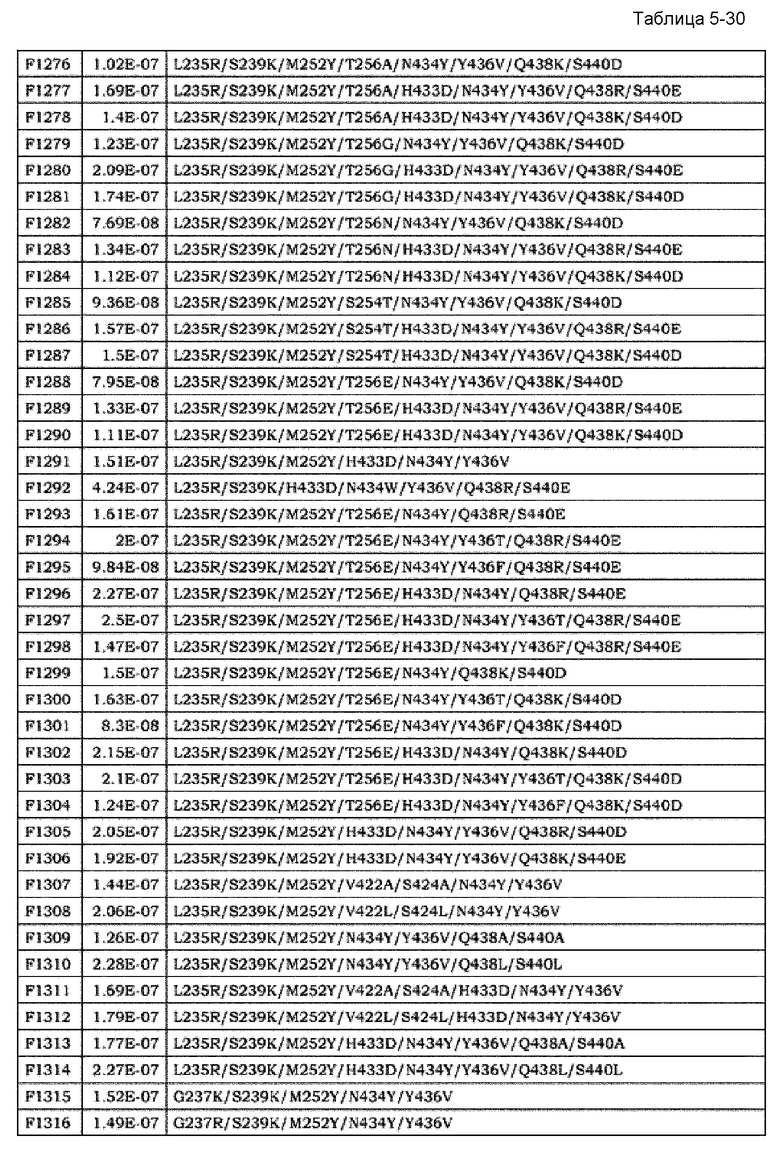
Таблица 5-31 является продолжением таблицы 5-30.

Таблица 5-32 является продолжением таблицы 5-31.
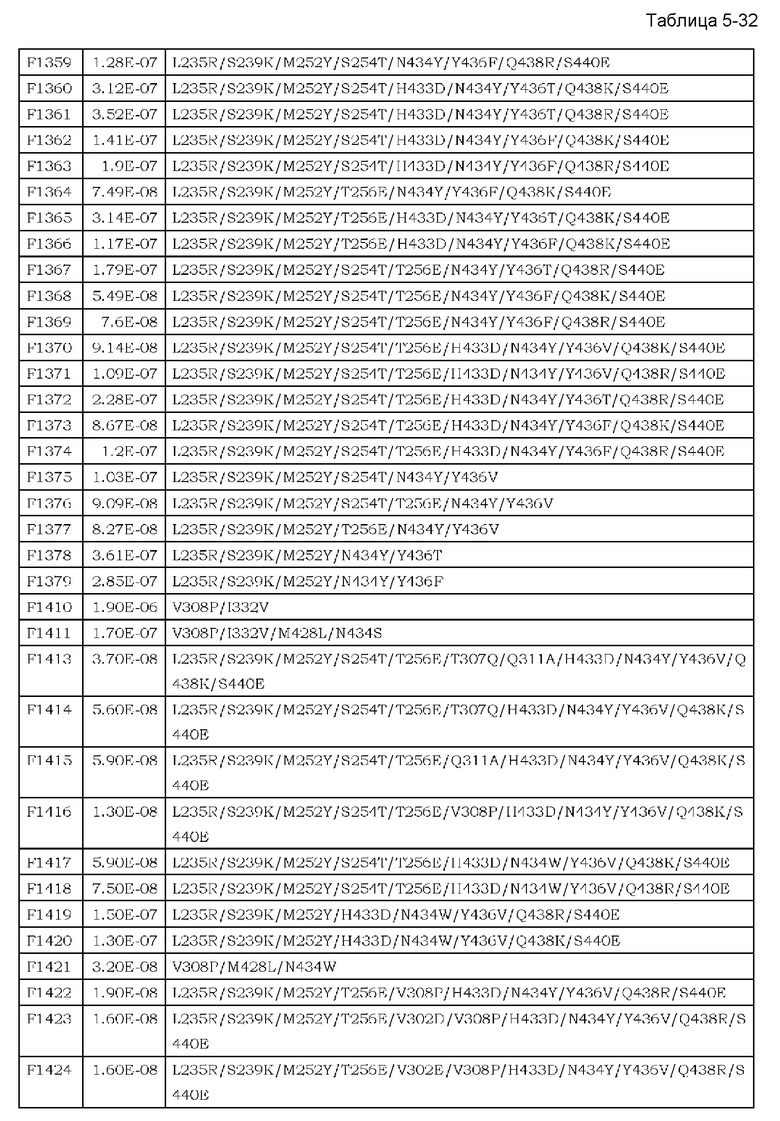
Таблица 5-33 является продолжением таблицы 5-32.

Fcγ-рецептор
Fcγ-рецептор (также описываемый как FcγR) относится к рецептору, способному связываться с Fc-областью моноклональных антител IgG1, IgG2, IgG3 или IgG4, и включает всех представителей, принадлежащих семейству белков, по существу кодируемых геном Fcγ-рецептора. У человека семейство включает FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы H131 и R131, т.е. FcγRIIa (H) и FcγRIIa (R)), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2), и FcγRIIc; и FcγRIII (CD16) включая изоформы FcγRIIIa (включая аллотипы V158 и F158, т.е. FcγRIIIa (V) и FcγRIIIa (F)) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2); а также неидентифицированные FcγR человека, изоформы FcγR и их аллотипы. Однако Fcγ-рецептор не ограничивается этими примерами. Не ограничиваясь этим, FcγR включает FcγR человека, мыши, крысы, кролика и обезьяны. FcγR может происходить из любых организмов. FcγR мыши включает, но не ограничивается ими, FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (FcγRIV, CD16-2), а также неидентифицированные FcγR мыши, изоформы FcγR и их аллотипы. Такие предпочтительные Fcγ-рецепторы включают, например, FcγRI (CD64), FcγRIIa (CD32), FcγRIIb (CD32), FcγRIIIa (CD16) и/или FcγRIIIb (CD16) человека. Полинуклеотидная последовательность и аминокислотная последовательность FcγRI представлены в SEQ ID NO:19 (NM_000566.3) и 20 (NP_000557.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIa (аллотип H131) представлены в SEQ ID NO:21 (BC020823.1) и 22 (AAH20823.1) (аллотип R131 представляет собой последовательность, в которой аминокислота в положении 166 SEQ ID NO:22 заменена на Arg), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγIIB представлены в SEQ ID NO:23 (BC146678.1) и 24 (AAI46679.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIa представлены в SEQ ID NO:25 (BC033678.1) и 26 (AAH33678.1), соответственно; и полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIb представлены в SEQ ID NO:27 (BC128562.1) и 28 (AAI28563.1), соответственно (номер доступа RefSeq представлен в каждом случае в скобках). Например, как описано для FcγRIIIaV, когда используют аллотип V158, если не указано иное, используют и аллотип F158; однако аллотип изоформы FcγRIIIa, описанной в настоящем описании, не следует интерпретировать как конкретно ограниченный.
В FcγRI (CD64), включая FcγRIa, FcγRIb, и FcγRIc, и FcγRIII (CD16), включающий изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2), α-цепь, которая связывается с Fc-частью IgG, ассоциирована с общей γ-цепью, имеющей ITAM, ответственный за передачу внутриклеточного активирующего сигнала. Между тем цитоплазматический домен FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы H131 и R131) и FcγRIIc содержит ITAM. Эти рецепторы экспрессируются на многих клетках, таких как макрофаги, тучные клетки и антигенпредставляющие клетки. Активирующий сигнал, передаваемый при связывании этих рецепторов с Fc-частью IgG, приводит к усилению фагоцитарной активности макрофагов, продукции воспалительных цитокинов, дегрануляции тучных клеток и усилению функции антигенпредставляющих клеток. Fcγ-рецепторы, обладающие способностью передавать активирующий сигнал, как описано выше, также называют активирующими Fcγ-рецепторами.
Между тем внутрицитоплазматический домен FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) содержит ITIM, ответственный за передачу ингибиторных сигналов. Связывание между FcγRIIb и B-клеточным рецептором (BCR) на B-клетках подавляет активирующий сигнал из BCR, что приводит к подавлению продукции антитела через BCR. Связывание FcγRIII и FcγRIIb на макрофагах подавляет фагоцитарную активность и продукцию воспалительных цитокинов. Fcγ-рецепторы, обладающие способностью передавать ингибирующий сигнал, как описано выше, также называют ингибирующим Fcγ-рецептором.
Активность связывания Fc-области с FcγR
Как упоминалось выше, Fc-области, обладающие активностью связывания Fcγ-рецептора, являются примерами Fc-областей, содержащихся в антигенсвязывающих молекулах по настоящему изобретению. Неограничивающий вариант осуществления такой Fc-области включает Fc-область IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека. То, обладает ли Fcγ-рецептор активностью связывания с Fc-областью моноклонального антитела IgG1, IgG2, IgG3 или IgG4, можно оценивать с помощью скрининга ALPHA (анализ усиленной за счет эффекта близости люминесценции), способа BIACORE на основе поверхностного плазмонного резонанса (SPR) и другие (Proc. Natl. Acad. Sci. USA (2006) 103(11), 4005-4010), в дополнение к описанным выше форматам FACS и ELISA.
Скрининг ALPHA проводят с помощью технологии ALPHA на основе принципа, описанного ниже, где используется два типа гранул: донорные и акцепторные. Люминесцентный сигнал выявляется только когда молекулы, связанные с донорными гранулами, биологически взаимодействуют с молекулами, связанными с акцепторными гранулами, и эти два типа гранул находятся вблизи друг друга. Возбуждаемый лазерным пучком фотосенсибилизатор в донорных гранулах преобразует кислород вокруг гранулы в возбужденный синглетный кислород. Когда синглетный кислород распространяется среди донорных гранул и достигает акцепторных гранул, расположенных близко, в акцепторных гранулах индуцируется хемилюминесцентная реакция. Это в конечном итоге приводит к испусканию света. Когда молекулы, связанные с донорными гранулами, не взаимодействуют с молекулами, связанными с акцепторными гранулами, сиглетный кислород, продуцируемый донорными гранулами, не достигает акцепторных гранул и хемилюминесцентная реакция не происходит.
Например, меченную биотином антигенсвязывающую молекулу, содержащую Fc-область, иммобилизуют на донорных гранулах, и Fcγ-рецептор, меченный глутатион-S-трансферазой (GST) иммобилизуют на акцепторных гранулах. В отсутствие антигенсвязывающей молекулы, содержащей конкурирующий полипептидный вариант Fc-области, Fcγ-рецептор взаимодействует с полипептидным комплексом, содержащим Fc-область дикого типа, что приводит к индукции сигнала в области 520-620 нм. Антигенсвязывающая молекула, имеющая немеченый вариант Fc-области, конкурирует с антигенсвязывающей молекулой, содержащей нативную Fc-область, для взаимодействия с Fcγ-рецептором. Относительную аффинность связывания можно определять путем количественного определения снижения флуоресценции в результате конкуренции. Способы биотинилирования антигенсвязывающих молекул, таких как антитела, с использованием сульфо-NHS-биотина и т.п., известны. Соответствующие способы добавления GST-метки к Fcγ-рецептору включают способы, которые вовлекают слияние полипептидов, кодирующих Fcγ и GST в рамке считывания, экспрессию слитого гена с использованием клеток, в которые введен вектор, с которым ген функционально связан, а затем очистку с использованием колонки с глутатионом. Индуцированный сигнал предпочтительно можно анализировать, например, путем аппроксимации его к односторонней модели конкуренции на основе нелинейного регрессионного анализа с использованием программного обеспечения, такого как GRAPHPAD PRISM (GraphPad, San Diego).
Одно из веществ при наблюдении их взаимодействия иммобилизуют в качестве лиганда на тонкую золотую пленку на сенсорном чипе. Когда свет падает на заднюю поверхность сенсорного чипа, так чтобы общее отражение происходило на поверхности контакта между тонкой золотой пленкой и стеклом, интенсивность отраженного света частично снижается в определенной области (сигнал SPR). Другое из веществ при выявлении взаимодействия инжектируют в качестве анализируемого соединения на поверхность сенсорного чипа. Когда анализируемое соединение связывается с лигандом, масса иммобилизованной молекулы лиганда возрастает. Изменение индекса рефракции вызывает сдвиг положения сигнала SPR (с другой стороны, диссоциация вызывает сдвиг сигнала обратно в исходное положение). В системе Biacore величину сдвига, упомянутую выше (т.е. изменение массы на поверхности сенсорного чипа) наносят на график по вертикальной оси, и, таким образом, изменение массы с течением времени показывают в качестве данных измерения (сенсограмма). Параметры кинетики (константа скорости ассоциации (ka) и константу скорости диссоциации (kd)), определяют из кривых сенсограммы, и аффинность (KD) определяют из соотношения этих двух констант. В способе BIACORE, предпочтительно используют анализ ингибирования. Примеры такого анализа ингибирования описаны в Proc. Natl. Acad. Sci USA (2006) 103 (11): 4005-4010.
В дополнение к Fc-области IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека, Fc-область с модифицированным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, можно соответствующим образом использовать в качестве Fc-области, включенной в рамках настоящего изобретения. В рамках настоящего изобретения, "Fc-область нативного IgG человека" относится к Fc-области, в которой цепь сахаров, связанная с положением 297 (нумерация EU) Fc-области IgG1, IgG2, IgG3 или IgG4 человека, представленной в SEQ ID NO:13, 14, 15 или 16, представляет собой содержащую фукозу цепь сахаров. Такие Fc-области с модифицированным связыванием FcγR можно получать путем модификации аминокислот Fc-области нативного IgG человека. То, является ли активность связывания Fc-области с модифицированным связыванием FcγR с FcγR более высокой, чем у Fc-области нативного IgG человека, можно определять соответствующим образом с использованием способов, описанных в упомянутом выше разделе, касающемся активности связывания.
В рамках настоящего изобретения "модификация аминокислот" или "аминокислотная модификация" Fc-области включает модификацию до аминокислотной последовательности, которая отличается от исходной Fc-области. Исходная Fc-область может представлять собой любую Fc-область, при условии, что варианты, полученные путем модификации исходной Fc-области, могут связываться с FcRn человека в диапазоне нейтральных значений pH. Более того, также в качестве измененной Fc-области по настоящему изобретению предпочтительно можно использовать Fc-область, модифицированную из исходной Fc-области, которая уже модифицирована. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, что в качестве исходной Fc-области можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или модифицированных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; международные публикации № WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности с их исходной Fc-областью, меньшим чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с системой нумерации EU.
Для изменения аминокислот Fc-областей можно использовать подходящие известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением. Более того, также можно использовать различные известные способы в качестве способа изменения аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, также предпочтительно использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которой неприродная аминокислота связана с амбер-супрессорной тРНК, которая комплементарна стоп-кодону UAG (амбер-кодон).
Включенную в антигенсвязывающие молекулы по настоящему изобретению Fc-область с модифицированным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека (Fc-область с модифицированным связыванием FcγR), можно получать любым способом. В частности, Fc-область с модифицированным связыванием FcγR можно получать путем модификации аминокислот иммуноглобулина человека IgG-типа, используемого в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типа для модификации включают, например, Fc-области IgG человека, представленные в SEQ ID NO:13, 14, 15 или 16 (IgG1, IgG2, IgG3 или IgG4, соответственно, и их варианты).
Аминокислоты в любых положениях можно модифицировать на другие аминокислоты при условии, что активность связывания с Fcγ-рецептором является более высокой, чем у Fc-области нативного IgG человека. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, она предпочтительно содержит модификацию, которая обеспечивает эффект более высокой активности связывания Fcγ-рецептора, чем у Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров. Такие аминокислотные модификации описаны, например, в международных публикациях, таких как WO2007/024249, WO2007/021841, WO2006/031370, WO2000/042072, WO2004/029207, WO2004/099249, WO2006/105338, WO2007/041635, WO2008/092117, WO2005/070963, WO2006/020114, WO2006/116260 и WO2006/023403.
Примеры таких аминокислот, которые можно модифицировать, включают по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU). Fc-область (Fc-область с модифицированным связыванием FcγR), обладающую более высокой активностью связывания Fcγ-рецептора, чем у Fc-области нативного IgG человека, можно получать путем модификации этих аминокислот.
Примеры особенно предпочтительных модификаций для применения в рамках настоящего изобретения включают по меньшей мере одну или более аминокислотных модификаций, выбранных из группы, состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU. Количество аминокислот, подлежащих модификации, конкретно не ограничено; и аминокислоту можно модифицировать только в одном участке или аминокислоты можно модифицировать в двух или более участков. Примеры комбинаций аминокислотных модификаций в двух или более участков включают комбинации аминокислотных модификаций, описанные в таблице 6 (таблицы с 6-1 по 6-3).
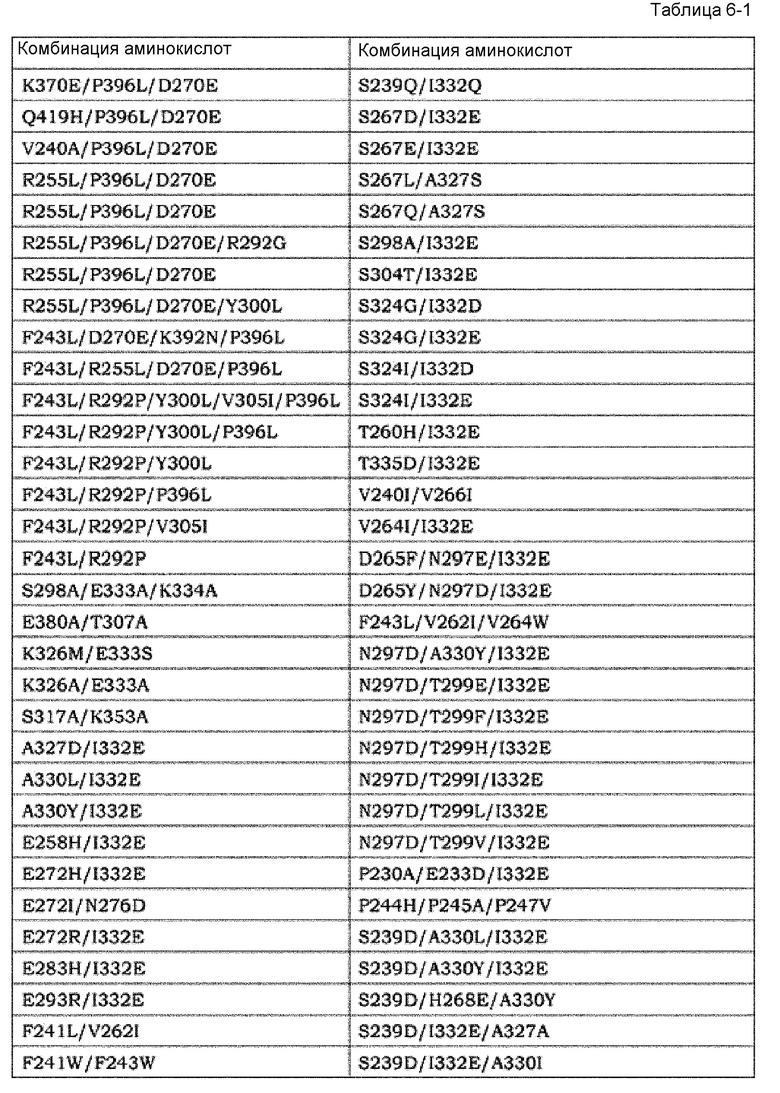
Таблица 6-2 является продолжением таблицы 6-1.
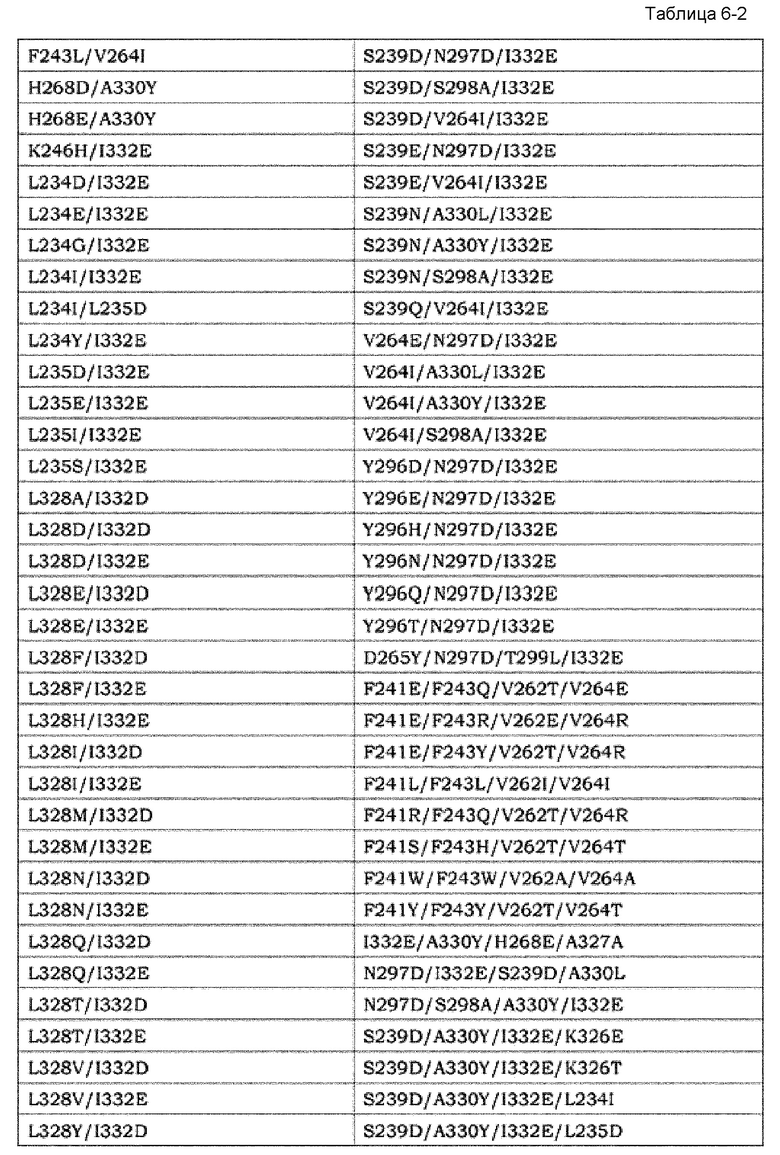
Таблица 6-3 является продолжением таблицы 6-2.
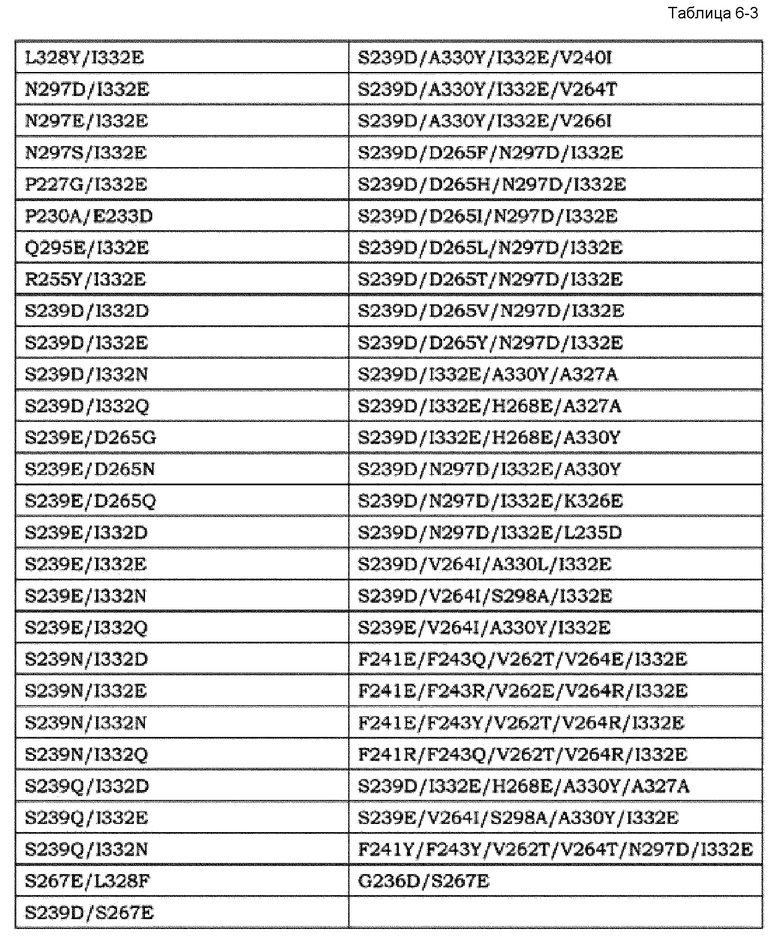
Для условий pH для измерения активности связывания Fc-области и Fcγ-рецептора, содержащихся в антигенсвязывающей молекуле по настоящему изобретению, можно соответствующим образом использовать условия в диапазоне кислых значений pH или в диапазоне нейтральных значений pH. Диапазон нейтральных значений pH в качестве условий для измерения активности связывания Fc-области и Fcγ-рецептора, содержащихся в антигенсвязывающей молекуле по настоящему изобретению, как правило, обозначают как от pH 6,7 до pH 10,0. Предпочтительно, он представляет собой диапазон, обозначаемый с помощью произвольных величин pH от pH 7,0 до pH 8,0; и предпочтительно, он выбран из pH 7,0, pH 7,1, pH 7,2, pH 7,3, pH 7,4, pH 7,5, pH 7,6, pH 7,7, pH 7,8, pH 7,9 и pH 8,0; и особенно предпочтительно он представляет собой pH 7,4, что является близким к pH плазмы (крови) in vivo. В рамках настоящего изобретения диапазон кислых значений pH, в качестве условий наличия активности связывания Fc-области и Fcγ-рецептора, которой обладает антигенсвязывающая молекула по настоящему изобретению, как правило, обозначают как от pH 4,0 до pH 6,5. Предпочтительно, его обозначают как от pH 5,5 до pH 6,5, и особенно предпочтительно его обозначают как от pH 5,8 до pH 6,0, что близко к pH в ранней эндосоме in vivo. Что касается температуры, используемой в качестве условий измерения, аффинность связывания между Fc-областью и Fcγ-рецептором можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, для определения аффинности связывания между Fc-областью и Fcγ-рецептором используют температуру от 15°C до 40°C. Более предпочтительно, аналогично для определения аффинности связывания между Fc-областью и Fcγ-рецептором используют температуру от 20°C до 35°C, такую как любая температура из 20°C, 21°C, 22°C, 23°C, 24°C, 25°C, 26°C, 27°C, 28°C, 29°C, 30°C, 31°C, 32°C, 33°C, 34°C и 35°C. В одном варианте осуществления настоящего изобретения неограничивающим примером является температура, составляющая 25°C.
В настоящем описании "Fc-область с модифицированным связыванием FcγR обладает более высокой активностью связывания Fcγ-рецептора, чем нативная Fc-область" означает, что активность связывания с Fcγ-рецептором Fc-области с модифицированным связыванием FcγR в отношении любого из Fcγ-рецепторов человека из FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и/или FcγRIIIb является более высокой, чем активность связывания нативной Fc-области в отношении этих Fcγ-рецепторов человека. Например, это означает, что на основе описанного выше аналитического способа, по сравнению с активностью связывания антигенсвязывающей молекулы, содержащей нативную Fc-область IgG человека в качестве контроля, активность связывания антигенсвязывающей молекулы, содержащей Fc-область с модифицированным связыванием FcγR, составляет 105% или более, предпочтительно 110% или более, 115% или более, 120% или более, 125% или более, особенно предпочтительно 130% или более, 135% или более, 140% или более, 145% или более, 150% или более, 155% или более, 160% или более, 165% или более, 170% или более, 175% или более, 180% или более, 185% или более, 190% или более, 195% или более, является более высокой в 2 раза или более, в 2,5 раза или более, в 3 раза или более, в 3,5 раза или более, в 4 раза или более, в 4,5 раза или более, в 5 раз или более, в 7,5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, в 90 раз или более или в 100 раз или более. Исходную Fc-область можно использовать в качестве нативной Fc-области, и также можно использовать нативные Fc-области антител того же подкласса.
В рамках настоящего изобретения, в частности, Fc-область нативного IgG человека, в которой цепь сахаров, связанная с аминокислотой в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров, пригодным образом используют в качестве Fc-области нативного IgG человека, подлежащей применению в качестве контроля. То, является ли цепь сахаров, связанная с аминокислотой в положении 297 (нумерация EU) содержащей фукозу цепью сахаров, можно определять с использованием способа, описанного в непатентном документе 6. Например, с помощью такого способа, как указано ниже, возможно определить то, является ли цепь сахаров, связанная с нативной Fc-областью IgG человека, содержащей фукозу цепью сахаров. Посредством реакции N-гликозидазы F (Roche diagnostics) с нативным IgG человека, подлежащим исследованию, цепь сахаров диссоциирует от нативного IgG человека (Weitzhandler et al. (J. Pharma. Sciences (1994) 83, 12, 1670-1675)). Далее высушенный концентрат реакционного раствора, из которого были удалены белки посредством реакции с этанолом (Schenk et al. (J. Clin. Investigation (2001) 108 (11) 1687-1695)), метят флуоресцентной меткой 2-аминопиридином или 2-аминобензамидом (Bigge et al. (Anal. Biochem. (1995) 230 (2) 229-238)). Реагенты удаляют с помощью твердофазной экстракции с использованием кассеты с целлюлозой, и флуоресцентно меченную модифицированную посредством 2-AP или 2-AB цепь сахаров анализируют с помощью нормально-фазовой хроматографии. Путем исследования выявленных пиков на хроматограмме можно определить, является ли цепь сахаров, связанная с нативной Fc-областью IgG человека, содержащей фукозу цепью сахаров.
В качестве антигенсвязывающей молекулы, содержащей Fc-область нативного антитела того же подкласса, которое используют в качестве контроля, можно пригодным образом использовать антигенсвязывающую молекулу, имеющую Fc-область моноклонального IgG-антитела. Структуры таких Fc-областей представлены в SEQ ID NO:13 (A добавлен на N-конец последовательности с номером доступа RefSeq AAC82527.1), SEQ ID NO:14 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59393.1), SEQ ID NO:15 (номер доступа RefSeq CAA27268.1) и SEQ ID NO:16 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59394.1). Более того, когда антигенсвязывающую молекулу, содержащую Fc-область конкретного изотипа антитела, используют в качестве исследуемого вещества, эффект антигенсвязывающей молекулы, содержащей исследуемую Fc-область, в отношении активности связывания Fcγ-рецептора, исследуют с использованием в качестве контроля антигенсвязывающей молекулы, имеющей Fc-область моноклонального IgG-антитела этого конкретного изотипа. Таким путем пригодным образом выбирают антигенсвязывающие молекулы, содержащие Fc-область, для которой продемонстрировано, что ее активность связывания Fcγ-рецептора является высокой.
Fc-области, обладающие активностью селективного связывания с Fcγ-рецептором
Примеры Fc-областей, пригодных для применения в рамках настоящего изобретения, включают Fc-области, обладающие более высокой активностью связывания с конкретным Fcγ-рецептором, чем с другими Fcγ-рецепторами (Fc-области, обладающие активностью селективного связывания с Fcγ-рецептором). Когда антитело используют в качестве антигенсвязывающей молекулы, одна молекула антитела может связывать только одну молекулу Fcγ-рецептора. Таким образом, одна антигенсвязывающая молекула не может связываться с другими активирующими FcγR в связанном с ингибиторным Fcγ-рецептором состоянии и не может связываться с другими активирующими Fcγ-рецепторами или ингибиторными Fcγ-рецепторами в связанном с активирующим Fcγ-рецептором состоянии.
Как описано выше, подходящие примеры активирующих Fcγ-рецепторов включают FcγRI (CD64), который включает FcγRIa, FcγRIb, и FcγRIc; FcγRIII (CD16), который включает изоформы FcγRIIIa (включая аллотипы V158 или F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2); и FcγRIIa (включая аллотипы H131 или R131). Между тем, подходящие примеры ингибиторных Fcγ-рецепторов включают FcγRIIb (включая FcγRIIb-1 или FcγRIIb-2).
Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту Fc-область, могут иметь сохраненную или сниженную активность связывания с активирующим FcγR (FcγRIa, FcγRIb, FcγRIc, FcγRIIIa, включая аллотип V158, FcγRIIIa, включая аллотип F158, FcγRIIIb, включая аллотип FcγRIIIb-NA1, FcγRIIIb, включая аллотип FcγRIIIb-NA2, FcγRIIa, включая аллотип H131, FcγRIIa, включая аллотип R131 и/или FcγRIIc), по сравнению с Fc-областью IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека (далее обозначаемой как Fc-область дикого типа) и антигенсвязывающей молекулой, содержащей эту Fc-область дикого типа.
По сравнению с Fc-областью и антигенсвязывающей молекулой, содержащей Fc-область дикого типа, Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту область Fc-область, имеют сниженную активность связывания с упомянутым выше активирующим FcγR на уровне, например, 99% или менее, 98% или менее, 97% или менее, 96% или менее, 95% или менее, 94% или менее, 93% или менее, 92% или менее, 91% или менее, 90% или менее, 88% или менее, 86% или менее, 84% или менее, 82% или менее, 80% или менее, 78% или менее, 76% или менее, 74% или менее, 72% или менее, 70% или менее, 68% или менее, 66% или менее, 64% или менее, 62% или менее, 60% или менее, 58% или менее, 56% или менее, 54% или менее, 52% или менее, 50% или менее, 45% или менее, 40% или менее, 35% или менее, 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее, 5% или менее, 4% или менее, 3% или менее, 2% или менее, 1% или менее, 0,5% или менее, 0,4% или менее, 0,3% или менее, 0,2% или менее, 0,1% или менее, 0,05% или менее, 0,01% или менее или 0,005% или менее.
Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту Fc-область, могут иметь усиленную активность связывания с ингибиторным FcγR (FcγRIIb-1 и/или FcγRIIb-2) по сравнению с Fc-областью IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека (далее обозначаемой как Fc-область дикого типа) и антигенсвязывающей молекулой, содержащей Fc-область дикого типа.
По сравнению с Fc-областью дикого типа и антигенсвязывающей молекулой, содержащей Fc-область дикого типа, Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту Fc-область, имеют усиленную активность связывания с упомянутым выше ингибиторным FcγR на уровне, например, 101% или более, 102% или более, 103% или более, 104% или более, 105% или более, 106% или более, 107% или более, 108% или более, 109% или более, 110% или более, 112% или более, 114% или более, 116% или более, 118% или более, 120% или более, 122% или более, 124% или более, 126% или более, 128% или более, 130% или более, 132% или более, 134% или более, 136% или более, 138% или более, 140% или более, 142% или более, 144% или более, 146% или более, 148% или более, 150% или более, 155% или более, 160% или более, 165% или более, 170% или более, 175% или более, 180% или более, 185% или более, 190% или более, 195% или более, или она является более высокой в 2 раза или более, в 3 раза или более, в 4 раза или более, в 5 раз или более, в 6 раз или более, в 7 раз или более, в 8 раз или более, в 9 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, в 90 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 400 раз или более, в 500 раз или более, в 600 раз или более, в 700 раз или более, в 800 раз или более, в 900 раз или более, в 1000 раз или более, в 10000 раз или более или в 100000 раз или более.
Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту Fc-область, могут иметь сохраненную или сниженную активность связывания с активирующим FcγR (FcγRIa, FcγRIb, FcγRIc, FcγRIIIa, включая аллотип V158, FcγRIIIa, включая аллотип F158, FcγRIIIb, включая аллотип FcγRIIIb-NA1, FcγRIIIb, включая аллотип FcγRIIIb-NA2, FcγRIIa, включая аллотип H131, FcγRIIa, включая аллотип R131, и/или FcγRIIc), по сравнению с Fc-областью IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека (далее обозначаемой как Fc-область дикого типа), и антигенсвязывающей молекулой, содержащей Fc-область дикого типа; и усиленную активность связывания с ингибиторным FcγR (FcγRIIb-1 и/или FcγRIIb-2) по сравнению с Fc-областью IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека (далее обозначаемой как Fc-область дикого типа) и антигенсвязывающей молекулой, содержащей Fc-область дикого типа.
По сравнению с Fc-областью IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека (далее обозначаемой как Fc-область дикого типа) и антигенсвязывающей молекулой, содержащей Fc-область дикого типа, Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, которая содержит селективный FcγR-связывающий домен, и антигенсвязывающая молекула, содержащая эту Fc-область, имеют более высокий уровень усиления их активности связывания с ингибиторным FcγR (FcγRIIb-1 и/или FcγRIIb-2), чем уровень усиления их активности связывания с активирующим Fcγ-рецептором (FcγRIa, FcγRIb, FcγRIc, FcγRIIIa, включая аллотип V158, FcγRIIIa, включая аллотип F158, FcγRIIIb, включая аллотип FcγRIIIb-NA1, FcγRIIIb, включая аллотип FcγRIIIb-NA2, FcγRIIa, включая аллотип H131, и FcγRIIa, включая аллотип R131).
В рамках настоящего изобретения можно добавить по меньшей мере одну другую модификацию в Fc-область, у которой аминокислота в положении 238 (нумерация EU) представляет собой Asp, или в Fc-область, у которой аминокислота в положении 328 (нумерация EU) представляет собой Glu в соответствии с вариантами осуществления и т.п., описанными в упомянутом разделе, касающемся аминокислотных модификаций. В дополнение к этим модификациям, также могут быть включены дополнительные модификации. Дополнительные модификации могут быть выбраны, например, из любых из аминокислотных замен, делеций и модификаций, и их комбинаций. Например, можно добавлять модификации, которые усиливают активность связывания FcγRIIb при сохранении или снижении активности связывания с FcγRIIa (H-тип) и FcγRIIa (R-тип). Добавление таких модификаций повышает селективность связывания с FcγRIIb относительно FcγRIIa.
Более того, определение того, сохраняется, усиливается или снижается ли активность связывания полипептидов по настоящему изобретению с различными FcγR, можно проводить, исходя из увеличения или уменьшения уровня связывания различных FcγR с полипептидами по настоящему изобретению, определенного в соответствии с примерами, описанными выше. В данном случае, уровень связывания различных FcγR с полипептидами относится к величинам, полученным путем определения разностей величин RU на сенсограммах до и после взаимодействия после различных FcγR в качестве анализируемого соединения с каждым полипептидом, и деления их на разности величин RU на сенсограммах до и после улавливания полипептидов на сенсорных чипах.
Является возможным определение того, сохраняется или снижается ли активность связывания FcγRIIa (R-тип и H-тип) у полипептидов по настоящему изобретению, и определение того, являются ли пептиды пептидами с усиленной активностью связывания FcγRIIb можно проводить, исходя из уровня связывания полипептидов с FcγRIIa и уровня связывания полипептидов с FcγRIIb в соответствии с упомянутыми выше примерами.
Иллюстративным случаем является то, когда количество полипептида по настоящему изобретению, связавшегося с FcγRIIb, является увеличенным по сравнению с уровнем связывания с FcγRIIb исходного полипептида, и количество полипептида по настоящему изобретению, связавшегося с FcγRIIa (R-тип и H-тип), является эквивалентным (сохраненным) или предпочтительно сниженным относительно уровня связывания исходного полипептида с FcγRIIa (R-тип и H-тип). Это также можно определить путем комбинирования соответствующим образом уровня связывания FcγRIa и уровня связывания FcγRIIIa у полипептида, определенного с помощью следующих упомянутых выше примеров.
В неограничивающем варианте осуществления настоящего изобретения примером Fc-области, имеющей более высокую активность связывания ингибиторного Fcγ-рецептора, чем активность связывания активирующего Fcγ-рецептора (обладающего активностью селективного связывания в отношении ингибиторного Fcγ-рецептора), является Fc-область, представленная в US2009/0136485 или в WO 2012/115241; и другим подходящим примером является Fc-область, в которой аминокислота в положении 238 или 328, как указано в соответствии с нумерацией EU, в упомянутой выше Fc-области, модифицирована на аминокислоту, отличающуюся от аминокислоты нативной Fc-области.
В неограничивающем варианте осуществления настоящего изобретения подходящим примером Fc-области, которая обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором (т.е. обладающая активностью селективного связывания с ингибиторным Fcγ-рецептором) является Fc-область с одной или несколькими из следующих модификаций аминокислот (указанных в соответствии с нумерацией EU) упомянутой выше Fc-области: аминокислота в положении 238 модифицирована на Asp и аминокислота в положении 328 модифицирована на Glu. Fc-области и модификации, описанные в US2009/0136485 или WO 2012/115241, можно выбирать соответствующим образом в качестве Fc-области, обладающей активностью селективного связывания с ингибиторным Fcγ-рецептором.
В неограничивающем варианте осуществления настоящего изобретения подходящим примером является Fc-область, в которой одна или более аминокислот, указанных в соответствии с нумерацией EU в положениях 238 и 328 в соответствии с нумерацией EU, соответственно модифицированы на Asp или Glu в упомянутой выше Fc-области.
Более того, в неограничивающем варианте осуществления настоящего изобретения подходящими примерами Fc-областей являются Fc-области с заменой Pro на Asp в положении 238 (нумерация EU), и одной или несколькими модификациями, выбранными из Trp в качестве аминокислоты в положении 237, Phe в качестве аминокислоты в положении 237, Val в качестве аминокислоты в положении 267, Gln в качестве аминокислоты в положении 267, Asn в качестве аминокислоты в положении 268, Gly в качестве аминокислоты в положении 271, Leu в качестве аминокислоты в положении 326, Gln в качестве аминокислоты в положении 326, Glu в качестве аминокислоты в положении 326, Met в качестве аминокислоты в положении 326, Asp в качестве аминокислоты в положении 239, Ala в качестве аминокислоты в положении 267, Trp в качестве аминокислоты в положении 234, Tyr в качестве аминокислоты в положении 234, Ala в качестве аминокислоты в положении 237, Asp в качестве аминокислоты в положении 237, Glu в качестве аминокислоты в положении 237, Leu в качестве аминокислоты в положении 237, Met в качестве аминокислоты в положении 237, Tyr в качестве аминокислоты в положении 237, Lys в качестве аминокислоты в положении 330, Arg в качестве аминокислоты в положении 330, Asp в качестве аминокислоты в положении 233, Asp в качестве аминокислоты в положении 268, Glu в качестве аминокислоты в положении 268, Asp в качестве аминокислоты в положении 326, Ser в качестве аминокислоты в положении 326, Thr в качестве аминокислоты в положении 326, Ile в качестве аминокислоты в положении 323, Leu в качестве аминокислоты в положении 323, Met в качестве аминокислоты в положении 323, Asp в качестве аминокислоты в положении 296, Ala в качестве аминокислоты в положении 326, Asn в качестве аминокислоты в положении 326, и Met в качестве аминокислоты в положении 330, в соответствии с нумерацией EU.
Антигенсвязывающая молекула
В рамках настоящего изобретения, термин "антигенсвязывающая молекула" используют в наиболее широком значении для обозначения молекулы, содержащей антигенсвязывающий домен и Fc-область. В частности, антигенсвязывающие молекулы включают различные типы молекул, при условии, что они проявляют антигенсвязывающую активность. Молекулы, в которых антигенсвязывающий домен связан с Fc-областью, включают, например, антитела. Антитела могут включать единичные моноклональные антитела (включая антитела-агонисты и антитела-антагонисты), антитела человека, гуманизированные антитела, химерные антитела и т.п. Альтернативно при использовании в качестве фрагментов антител, они предпочтительно включают антигенсвязывающие домены и антигенсвязывающие фрагменты (например, Fab, F(ab')2, scFv и Fv). Каркасные молекулы, где трехмерные структуры, такие как уже известная стабильная белковая структура α/β-бочонок, используются в качестве каркаса (основы) и только некоторые части структур преобразуют в библиотеки для конструирования антигенсвязывающих доменов, также включены в антигенсвязывающие молекулы по настоящему изобретению.
Антигенсвязывающая молекула по настоящему изобретению может содержать по меньшей мере некоторые части Fc-области, которые опосредуют связывание с FcRn и Fcγ-рецептором. В неограничивающем варианте осуществления антигенсвязывающая молекула включает, например, антитела и слитые белки Fc. Слитый белок относится к химерному полипептиду, содержащему полипептид, имеющий первую аминокислотную последовательность, который связан с полипептидом, имеющим вторую аминокислотную последовательность, которые не связаны в природе. Например, слитый белок может содержать аминокислотную последовательность по меньшей мере части Fc-области (например, часть Fc-области, ответственная за связывание с FcRn, или часть Fc-области, ответственная за связывание с Fcγ-рецептором) и не являющийся иммуноглобулином полипептид, содержащий, например, аминокислотную последовательность лиганд-связывающего домена рецептора или рецептор-связывающего домена лиганда. Аминокислотные последовательности могут находиться в отдельных белках, которые переносят вместе в слитый белок, или, как правило, они могут находиться в одном белке, но они включены в слитый полипептид в новом порядке. Слитые белки можно получать, например, посредством химического синтеза или способами генетической рекомбинации для экспрессии полинуклеотида, кодирующего области пептидов в желаемом порядке.
Любые из доменов антигенсвязывающего домена, Rc-области и т.п. по настоящему изобретению могут быть связаны через линкеры или прямо через полипептидную связь. Линкеры включают произвольные пептидные линкеры, которые можно вносить способами генетической инженерии, синтетические линкеры и линкеры, описанные, например, в Protein Engineering (1996) 9(3), 299-305. Однако пептидные линкеры являются предпочтительными в рамках настоящего изобретения. Длина пептидных линкеров конкретно не ограничена, и ее могут надлежащим образом выбирать специалисты в данной области в зависимости от назначения. Длина предпочтительно составляет пять аминокислот или более (без конкретных ограничений, верхний предел обычно составляет 30 аминокислоты или менее, предпочтительно 20 аминокислоты или менее), и особенно предпочтительно 15 аминокислот.
Например, такие пептидные линкеры предпочтительно включают:

где n представляет собой целое число, равное 1 или более. Длину или последовательности пептидных линкеров могут соответствующим образом выбирать специалисты в данной области, в зависимости от назначения.
Обычно для сшивания пептидов используют синтетические линкеры (химические сшивающие агенты), и например:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидил пропионат) (DSP),
дитиобис(сульфосукцинимидилпропионат) (DTSSP),
этиленгликоль бис(сукцинимидилсукцинат) (EGS),
этиленгликоль бис(сульфосукцинимидилсукцинат) (сульфо-EGS),
дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES),
и бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти сшивающие агенты являются коммерчески доступными.
Когда используют множество линкеров для связывания соответствующих доменов, все они могут быть одного типа или они могут быть различных типов.
В дополнение к линкерам, проиллюстрированным выше, также можно соответствующим образом использовать линкеры с пептидными метками, такими как His-метка, HA-метка, myc-метка и FLAG-метка. Более того, также можно соответствующим образом использовать образование водородных связей, образование дисульфидных связей, образование ковалентных связей, ионное взаимодействие и свойства связывания друг с другом в результате их комбинации. Например, можно использовать аффинность между CH1 и CL антитела, и также можно использовать Fc-области, происходящие из описанных выше биспецифических антител для гетероассоциации Fc-областей. Более того, также можно соответствующим образом использовать дисульфидные связи, образующиеся между доменами.
Для связывания соответствующих доменов пептидной связью, полинуклеотиды, кодирующие домены, связывают вместе в рамке считывания. Известные способы связывания полинуклеотидов в рамке считывания включают способы, такие как лигирование фрагментов рестрикции, ПЦР со слиянием и перекрывающаяся ПЦР. Такие способы можно соответствующим образом использовать отдельно или в комбинации для получения антигенсвязывающих молекул по настоящему изобретению. В рамках настоящего изобретения термины "связанный" и "слитый", или "связывание" и "слияние" используют взаимозаменяемо. Эти термины означают, что два или более элементов или компонентов, таких как полипептиды, слиты вместе с образованием единой структуры любыми способами, включая описанные выше способы химического связывания и способы генетической рекомбинации. Когда два или более доменов, элементов или компонентов представляют собой полипептиды, связывание в рамке считывания означает связывание двух или более элементов рамок считывания с образованием более длинной непрерывной рамки считывания при сохранении правильных рамок считывания полипептидов. Когда две молекулы Fab используют в качестве антигенсвязывающего домена, в качестве подходящей антигенсвязывающей молекулы настоящей заявки можно использовать антитело, которое представляет собой антигенсвязывающую молекулу по настоящему изобретению, в которой антигенсвязывающий домен связан в рамке считывания с Fc-областью через пептидную связь, но не через линкер.
Комплекс, содержащий две или более антигенсвязывающих молекул и два или более антигенных элементов связывания
Как описано в примере 1 (как показано в WO 2011/122011), Fv4-IgG1, у которого активность связывания sIL-6R изменяется в зависимости от условий pH в большей степени, чем у H54/L28-IgG1, ускоряет элиминацию sIL-6R, но не ускоряет элиминацию в большей степени, чем когда sIL-6R используют отдельно. Для ускорения элиминации в большей степени, чем когда sIL-6R используют отдельно, необходимо использовать антигенсвязывающие молекулы, содержащие Fc-область с усиленным связыванием FcRn в диапазоне нейтральных значений pH (например, Fv4-IgG1-v2 и т.п. примера 1).
Между тем, неожиданно было обнаружено, что GA2-IgG1, у которого активность связывания IgA человека варьирует в зависимости от концентрации ионов Ca, ускоряет элиминацию IgA человека в большей степени, чем когда IgA человека используют отдельно, даже несмотря на то, что оно содержит Fc-область, происходящую из нативного IgG1, у которого связывание FcRn в диапазоне нейтральных значений pH не усилено. Аналогично, было выявлено, что клон 278, у которого активность связывания IgE человека варьирует в зависимости от условий pH, ускоряет элиминацию IgE человека в большей степени, чем когда IgE человека используют отдельно, даже несмотря на то, что он содержит Fc-область, происходящую из нативного IgG1, который не обладает усиленным связыванием FcRn в диапазоне нейтральных значений pH. Без ограничения конкретной теорией, полагают, что следующий иллюстративный механизм объясняет то, что происходит в случае GA2-IgG1 и клона 278.
Когда антигенный элемент связывания представляет собой один элемент (т.е. гомомономер), такой как в sIL-6R, две молекулы (т.е. два антигенных элемента связывания) антигенов связываются с одной молекулой антитела, содержащей двухвалентный антигенсвязывающий домен, и образуется комплекс одной молекулы антитела против sIL-6R и двух молекул антигена, содержащих два антигенных элемента связывания. Таким образом, этот тип комплекса антиген-антитело имеет только одну Fc-область (нативную Fc-область IgG1), как показано на фиг. 1. Поскольку этот комплекс связывается с двумя молекулами FcRn или одной молекулой FcγR через одну Fc-область, аффинность в отношении этих рецепторов является такой же, как и аффинность обычного IgG-антитела, и полагают, что захват в клетки происходит по большей части неспецифически.
С другой стороны, когда антигенный элемент связывания представляет собой два элемента, как в IgA человека, который является димером гетерокомплекса тяжелых цепей и легких цепей, существует два элемента эпитопов, с которыми антигенсвязывающие домены будут связываться в антигенном элементе связывания. Однако, когда двухвалентное антитело против IgA (т.е. антигенсвязывающие домены, содержащиеся в одной молекуле антитела против IgA связываются с тем же эпитопом) связывается с его антигеном IgA, полагают, что связывание каждого из двухвалентных антигенсвязывающих доменов, содержащихся в одной молекуле антитела против IgA с каждым из двух элементов эпитопов, присутствующих на одной молекуле IgA, является затрудненным ввиду расположения эпитопов. В результате полагают, что отдельные молекулы антитела против IgA связываются с двумя антигенными элементами связывания, присутствующими в двух молекулах IgA, которые связываются с двухвалентными антигенсвязывающими доменами, присутствующими в одной молекуле антитела против IgA, и, следовательно, образуются комплексы антиген-антитело (иммунные комплексы), содержащие по меньшей мере четыре молекулы (т.е. две молекулы IgA, которые представляют собой молекулу антигена, и две молекулы антитела против IgA, которые являются антигенсвязывающими молекулами).
Когда антигенсвязывающая молекула, такая как антитело, связывающееся с молекулой антигена, который содержит два или более антигенных элементов связывания, образует крупный иммунный комплекс, который является по меньшей мере тетрамером, иммунный комплекс может связываться прочно с авидностью через по меньшей мере две или более поливалентных Fc-области с FcγR, FcRn, рецепторами комплемента и т.п. Таким образом, как показано на фиг. 7, комплекс захватывается в клетки, экспрессирующие эти рецепторы, с более высокой эффективностью, чем нативный IgG1. С другой стороны, поскольку опосредуемая Fc-областью аффинность в отношении этих рецепторов у иммунных комплексов, образованных из молекул антигена и антигенсвязывающих молекул, которые, например, связываются с (мономерными) молекулами антигена, содержащими один антигенный элемент связывания, является недостаточной, как упоминалось выше, иммунные комплексы захватываются по большей неспецифически (менее эффективно по сравнению с захватом, опосредуемым связыванием с авидностью) в клетки, экспрессирующие эти рецепторы, как показано на фиг. 1. Таким образом, захват является в большей степени неэффективным, чем захват, опосредуемый связыванием с авидностью.
Когда антигенсвязывающая молекула, такая как антитело, которое связывается с молекулой антигена, содержащей два или более антигенных элементов связывания, представляет собой антитело, которое содержит антигенсвязывающие домены, у которых активность связывания с антигеном варьирует в зависимости от условий концентрации ионов, как например, в случае pH- или Ca-зависимого связывания, и которые образуют комплекс антиген-антитело (иммунный комплекс), содержащий по меньшей мере четыре молекулы (две молекулы антигена и две молекулы антитела) в плазме, после захвата иммунного комплекса в клетки антигены диссоциируют от антител в эндосомах, где условия концентрации ионов отличаются от условий концентрации ионов в плазме. Таким образом, образованный иммунный комплекс распадается в эндосомах клеток, которые захватили иммунные комплексы. Поскольку диссоциированные антигены не могут связываться с FcRn в эндосомах, они деградируют после перемещения в лизосомы. С другой стороны, полагают, что антитела после диссоциации антигена рециклируют в плазму после связывания с FcRn в эндосомах (фиг. 7).
Как описано выше, если антитело, которое содержит нативную константную область IgG1-типа, против мультимерного антигена, содержащего два или более антигенных элементов связывания, и демонстрирует pH- или Ca-зависимое связывание, может образовывать крупный иммунный комплекс и связывается с FcγR, FcRn, рецепторами комплемента и т.п. с авидностью, полагают, что элиминация антигена отдельно может селективно и в значительной степени ускоряться. Полагают, что, когда вводят GA2-IgG1, которое связывается с IgA человека, образуются такие крупные иммунные комплексы. Действительно, как показано в примере 3, GA2-IgG1-FcγR(-), полученное путем внесения в GA2-IgG1 модификаций, которые ухудшают связывание с FcγR мыши, по существу не могут ускорять элиминацию IgA человека подобно GA2-IgG1 по сравнению с IgA человека отдельно, и они продемонстрировали эквивалентный уровень элиминации с IgA человека отдельно. Таким образом, причиной, по которой GA2-IgG1 могло ускорять элиминацию IgA человека, является то, что иммунный комплекс, содержащий GA2-IgG1 и IgA человека, который является мультимерным антигеном, содержащим два или более антигенных элементов связывания, связывается с авидностью с FcγR и быстро захватывается в клетки, экспрессирующие FcγR. IgA, который диссоциирует от иммунного комплекса в эндосомах клеток, которые захватили иммунный комплекс, деградируется в лизосомах. В то же время диссоциировавшее от IgA антитело, которое связалось с FcRn в эндосомах, впоследствии рециклирует в плазму и может вновь связываться с IgA в плазме. Полагают, что элиминация IgA в плазме значительно ускоряется таким образом. Способ использования Fc-области с вариантом аминокислоты, которая связывается с FcRn в диапазоне нейтральных значений pH, описан в WO2011/122011 в качестве способа ускорения элиминации антигенов из плазмы. Настоящее изобретение является пригодным в качестве способа ускорения элиминации из плазмы мультимерных антигенов, содержащих два или более антигенных элементов связывания без использования упомянутых выше вариантов, и как показано с использованием GA2-N434W, может далее ускорять элиминацию мультимерных антигенов, содержащих два или более антигенных элементов связывания из плазмы посредством комбинирования с упомянутыми выше вариантами.
Способ оценки образования иммунных комплексов, содержащих две или более антигенсвязывающих молекул и два или более антигенных элементов связывания
Для достижения эффекта дальнейшего ускорения элиминации мультимерных антигенов, содержащих два или более антигенных элементов связывания, из плазмы, как описано выше, считается предпочтительным, чтобы образовывался крупный иммунный комплекс антигенсвязывающих молекул и антигенов, и Fc-области, содержащиеся в антигенсвязывающих молекулах, прочно связывались с FcγR и/или FcRn с авидностью. Когда антиген содержит два или более антигенных элементов связывания, может образовываться крупный иммунный комплекс, содержащий две или более антигенсвязывающих молекул и два или более антигенных элементов связывания. Таким образом, посредством скрининга антигенсвязывающих молекул, у которых активность связывания с мультимерными антигенами, содержащими два или более антигенных элементов связывания, изменяется в зависимости от концентрации ионов и которые связываются с авидностью с упомянутыми выше рецепторами, можно получать антигенсвязывающие молекулы, которые далее ускоряют элиминацию мультимерных антигенов, содержащих два или более антигенных элементов связывания, из плазмы.
Способ оценки образования комплекса
Примеры способов оценки образования иммунных комплексов, содержащих антигенсвязывающие молекулы и антигены, включают способы аналитической химии, включая способы, в которых используется свойство, состоящее в том, что иммунные комплексы становятся более крупными молекулами, чем антигенсвязывающая молекула отдельно или молекула антигена отдельно, такие как эксклюзионная хроматография (гель-фильтрация), способ анализа с использованием ультрацентрифугирования, способ с использованием рассеяния света, электронная микроскопия и масс-спектрометрия (Molecular Immunology (2002), 39, 77-84; Molecular Immunology (2009), 47, 357-364). Например, когда используют эксклюзионную хроматографию (гель-фильтрацию), как показано на фиг. 9, оценку того, образуется ли иммунный комплекс, проводят путем наблюдения того, существуют ли молекулярные структуры, более крупные, чем молекулярные структуры в анализе молекулы антигена отдельно или антигенсвязывающей молекулы отдельно.
Более того, когда антигенсвязывающая молекула или антиген имеют константную область иммуноглобулина, примеры включают иммунохимические способы, включающие способы, в которых используется свойство иммунного комплекса, состоящее в более прочном связывании с Fc-рецептором или компонентом комплемента, чем у антигенсвязывающей молекулы отдельно или антигена отдельно, такие как способы ELISA, FACS или SPR (например, способы с использованием Biacore) (The Journal of Biological Chemistry (2001) 276 (9), 6591-6604; Journal of Immunological Methods (1982) 50, 109-114; Journal of Immunology (2010) 184 (4), 1968-1976; mAbs (2009) 1 (5) 491-504). Например, когда ELISA проводят путем иммобилизации Fc-рецептора, образование иммунного комплекса оценивают путем наблюдения того, увеличен ли обнаруженный сигнал по сравнению с антигеном отдельно или с антигенсвязывающей молекулой отдельно.
Способ ускорения элиминации мономерных антигенов из плазмы с использованием смеси антигенсвязывающих молекул
Как описано выше, когда антиген представляет собой мультимерный антиген (неограничивающим примером является иммуноглобулин, такой как IgA или IgE, или представитель суперсемейства TNF, такой как TNF или CD154), может образовываться крупный иммунный комплекс, содержащий две или более антигенсвязывающих молекул и два или более антигенных элементов связывания. С другой стороны, даже когда антиген является мономерным, смесь двух или более соответствующих антигенсвязывающих молекул, каждая из которых связывается с отдельным эпитопом, присутствующим на мономерном антигене, где связывание с эпитопами варьирует, в зависимости от условий концентрации ионов (таких как pH или концентрация Ca) также может образовывать крупный иммунный комплекс, содержащий две или более антигенсвязывающих молекул и два или более антигенных элементов связывания (мономерные антигены). В рамках настоящего изобретения, смесь двух или более соответствующих антигенсвязывающих молекул, каждая из которых связывается с отдельным эпитопом, присутствующим на мономерном антигене, где связывание с эпитопами варьирует в зависимости от условий концентрации ионов (таких как pH или концентрация Ca) называют смесью антигенсвязывающих молекул. Среди этих антигенсвязывающих молекул по меньшей мере одна из (с содержащимися в ней антигенсвязывающими доменами) антигенсвязывающих молекул, образующих иммунный комплекс, должна иметь антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов.
Способ ускорения элиминации мономерных антигенов из плазмы с использованием мультиспецифических или мультипаратопных антигенсвязывающих молекул
Даже в случае мономерного антигена, антигенсвязывающая молекула, содержащая антигенсвязывающие домены, каждый из которых связывается с отдельным эпитопом, присутствующим на мономерном антигене, где связывание антигенсвязывающих доменов с соответствующими эпитопами варьирует в зависимости от условий концентрации ионов (таких как pH или концентрация Ca) также могли образовывать крупный иммунный комплекс, содержащий две или более антигенсвязывающих молекул и два или более антигенных элементов связывания (мономерные антигены). Примерами неограничивающего варианта осуществления такой антигенсвязывающей молекулы являются мультипаратопные антитела или мультиспецифические антитела, содержащие соответствующие вариабельные области, которые связываются с эпитопами, присутствующими на мономерном антигене, которые отличаются друг от друга. В качестве неограничивающего варианта осуществления таких мультиспецифических антител или мультпаратопных антител, антитела, у которых вариабельные области демонстрируют pH- или Ca-зависимое связывание (биспецифические антитела или бипаратопные антитела, содержащие вариабельную область правого плеча, которая распознает эпитоп A, и вариабельную область левого плеча, которая распознает эпитоп B, как показано на фиг. 8), также могут образовывать крупный иммунный комплекс, содержащий два или более антитела и два или более антигенных элементов связывания (мономерные антигены).
Антигенсвязывающие молекулы, которые далее ускоряют элиминацию мономерных антигенов из плазмы, можно получать путем скрининга комбинаций антигенсвязывающих доменов, которые нацелены на различные эпитопы мономерного антигена, где активность связывания с каждым из эпитопов варьирует в зависимости от условий концентрации ионов, и которые могут связываться с авидностью с упомянутыми выше рецепторами. Условия концентрации ионов, которые изменяют активность связывания мультиспецифического или мультипаратопного антигенсвязывающего домена в отношении каждого из эпитопов, могут представлять собой те же самые условия концентрации ионов или отличающиеся условия концентрации ионов. Антигенсвязывающая молекула, содержащая биспецифические или бипаратопные антигенсвязывающие домены, где активность связывания эпитопа одного из биспецифических или бипаратопных антигенсвязывающих доменов варьирует в зависимости от условий pH или условий концентрации ионов металла, таких как концентрация ионов Ca, может быть предоставлена в качестве примера неограничивающего варианта осуществления антигенсвязывающих молекул по настоящему изобретению. Более того, антигенсвязывающая молекула, содержащая биспецифические или бипаратопные антигенсвязывающие домены, где активность связывания эпитопа одного из биспецифических или бипаратопных антигенсвязывающих доменов варьирует в зависимости от условий pH и активность связывания эпитопа другого антигенсвязывающего домена варьирует в зависимости от условий концентрации ионов металла, таких как концентрация ионов Ca, также могут быть представлены в качестве примера неограничивающего варианта осуществления антигенсвязывающих молекул по настоящему изобретению. Антигенсвязывающая молекула, содержащая биспецифические или бипаратопные антигенсвязывающие домены, где активность связывания эпитопа одного из биспецифических или бипаратопных антигенсвязывающих доменов варьирует в зависимости от условий pH, и активность связывания эпитопа другого антигенсвязывающего домена также варьирует в зависимости от условий pH, также может быть предоставлена в качестве примера неограничивающего варианта осуществления антигенсвязывающих молекул по настоящему изобретению. Антигенсвязывающая молекула, содержащая биспецифические или бипаратопные антигенсвязывающие домены, где активность связывания эпитопа одного из биспецифических или бипаратопных антигенсвязывающих доменов варьирует в зависимости от условий концентрации ионов металлов, таких как концентрация ионов Ca, и активность связывания эпитопа другого антигенсвязывающего домена также варьирует в зависимости от условий концентрации ионов металлов, таких как концентрация ионов Ca, также может быть предоставлена в качестве примера неограничивающего варианта осуществления антигенсвязывающих молекул по настоящему изобретению.
Мультиспецифические антигенсвязывающие молекулы и мультипаратопные антигенсвязывающие молекулы
Антигенсвязывающая молекула, содержащая по меньшей мере два антигенсвязывающих домена, где по меньшей мере один из антигенсвязывающих доменов связывается с первым эпитопом в молекуле антигена и по меньшей мере один другой из антигенсвязывающих доменов связывается со вторым эпитопом в молекуле антигена, называют мультиспецифической антигенсвязывающей молекулой с точки зрения ее реакционной специфичности. Когда одна антигенсвязывающая молекула связывается с двумя различными эпитопами через два типа антигенсвязывающих доменов, содержащихся в антигенсвязывающей молекуле, эту антигенсвязывающую молекулу называют биспецифической антигенсвязывающей молекулой. Когда одна антигенсвязывающая молекула связывается с тремя различными эпитопами через три типа антигенсвязывающих домена, содержащихся в антигенсвязывающей молекуле, эту антигенсвязывающую молекулу называют триспецифической антигенсвязывающей молекулой.
Паратоп в антигенсвязывающем домене, который связывается с первым эпитопом в молекуле антигена, имеет структуру, отличающуюся от структуры паратопа в антигенсвязывающем домене, который связывается со вторым эпитопом, который структурно отличается от первого эпитопа. Таким образом, антигенсвязывающая молекула, содержащая по меньшей мере два антигенсвязывающих домена, где по меньшей мере один из антигенсвязывающих доменов связывается с первым эпитопом в молекуле антигена и по меньшей мере один другой из антигенсвязывающих доменов связывается со вторым эпитопом в молекуле антигена, называют мультипаратопной антигенсвязывающей молекулой с точки зрения ее структуры и специфичности. Когда одна антигенсвязывающая молекула связывается с двумя различными эпитопами через два типа антигенсвязывающих домена, содержащихся в антигенсвязывающей молекуле, эту антигенсвязывающую молекулу называют бипаратопной антигенсвязывающей молекулой. Когда антигенсвязывающая молекула связывается с тремя различными эпитопами через три типа антигенсвязывающих домена, содержащихся в антигенсвязывающей молекуле, эту антигенсвязывающую молекулу называют трипаратопной антигенсвязывающей молекулой.
Поливалентные мультиспецифические или мультипаратопные антигенсвязывающие молекулы, содержащие один или более антигенсвязывающих доменов, и способы их получения описаны в непатентных документах, таких как Conrath et al. (J.Biol.Chem. (2001) 276 (10) 7346-7350), Muyldermans (Rev. Mol. Biotech. (2001) 74, 277-302), и Kontermann R.E. (2011) Bispecific Antibodies (Springer-Verlag) и в патентных документах, таких как WO 1996/034103 и WO 1999/023221. Антигенсвязывающие молекулы по настоящему изобретению можно получать с использованием мультиспецифических или мультипаратопных антигенсвязывающих молекул и способов их получения, описанных в этих документах.
Биспецифические антитела и способы их получения
Биспецифические антитела и способы их получения представлены ниже в качестве примеров варианта осуществления упомянутых выше мультиспецифических или мультипаратопных антигенсвязывающих молекул и способов их получения. Биспецифические антитела представляют собой антитела, содержащие два типа вариабельных областей, которые специфически связываются с различными эпитопами. Биспецифические антитела IgG-типа можно секретировать из гибридной гибридомы (квадромы), полученной путем слияния двух типов гибридом, которые продуцируют IgG-антитела (Milstein et al., Nature (1983) 305, 537-540).
Когда биспецифическое антитело получают с использованием рекомбинантных способов, таких как способы, описанные в упомянутом выше разделе, касающемся антител, можно использовать способ, который вводит гены, кодирующие тяжелые цепи, содержащие два типа представляющих интерес вариабельных областей, в клетки для их коэкспрессии. Однако при рассмотрении только комбинации тяжелых цепей такой способ будет обеспечивать смесь (i) комбинации пары тяжелых цепей, в которых одна из тяжелых цепей содержит вариабельную область, которая связывается с первым эпитопом, а другая тяжелая цепь содержит вариабельную область, которая связывается со вторым эпитопом, (ii) комбинации пары тяжелых цепей, которые включают только тяжелые цепи, содержащие вариабельную область, которая связывается с первым эпитопом, и (iii) комбинации пары тяжелых цепей, которые включают только тяжелые цепи, содержащие вариабельную область, которая связывается со вторым эпитопом, которые присутствуют в молекулярном соотношении 2:1:1. Трудно очистить антигенсвязывающие молекулы, содержащие желаемую комбинацию тяжелых цепей, из смеси трех типов комбинаций тяжелых цепей.
При получении биспецифических антител с использованием таких рекомбинантных способов, биспецифические антитела, содержащие гетеромерную комбинацию Fc-областей, можно предпочтительно секретировать путем добавления соответствующих аминокислотных замен в CH3-домены, составляющие тяжелые цепи. В частности, этот способ проводят путем замены аминокислотой, имеющей более крупную боковую цепь (выступ (что означает "выпуклая часть")) аминокислоты в CH3-домене одной из тяжелых цепей, и замены аминокислотой, имеющей меньшую боковую цепь (углубление (что означает "полость")) аминокислоты в CH3-домене другой тяжелой цепи, так чтобы выступ размещался в полости. Это обеспечивает образование гетеромерной тяжелой цепи и одновременно ингибирует образование гомомерной тяжелой цепи (международная публикация № WO 1996027011; Ridgway et al., Protein Engineering (1996) 9, 617-621; Merchant et al., Nature Biotechnology (1998) 16, 677-681).
Более того, также существуют известные способы получения биспецифического антитела с использованием способов контроля ассоциации полипептидов или ассоциации образованных полипептидом гетеромерных мультимеров для ассоциации между тяжелыми цепями. В частности, способы контроля образования тяжелых цепей можно использовать для получения биспецифического антитела (международная публикация № WO 2006/106905), в котором аминокислотные остатки, образующие поверхность контакта между тяжелыми цепями, которые образуют Fc-область, изменяют для ингибирования ассоциации между тяжелыми цепями, имеющими ту же последовательность, и для обеспечения образования тяжелых цепей с различными последовательностями. Такие способы можно использовать для получения биспецифических антител.
В неограничивающем варианте осуществления настоящего изобретения два полипептида, образующих Fc-область, происходящие из биспецифического антитела, описанного выше, можно соответствующим образом использовать в качестве Fc-области, содержащейся в антигенсвязывающей молекуле. Более конкретно, можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых, в аминокислотной последовательности одного из полипептидов, аминокислота в положении 349, как указано согласно нумерации EU, представляет собой Cys и аминокислота в положении 366 представляет собой Trp, и в аминокислотной последовательности других полипептидов аминокислота в положении 356, как указано согласно нумерации EU, представляет собой Cys, аминокислота в положении 366 представляет собой Ser, аминокислота в положении 368 представляет собой Ala и аминокислота в положении 407 представляет собой Val.
В другом неограничивающем варианте осуществления настоящего изобретения, в качестве Fc-области можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых, в аминокислотной последовательности одного из полипептидов, аминокислота в положении 409 согласно нумерации EU представляет собой Asp, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 399 согласно нумерации EU представляет собой Lys. В описанном выше варианте осуществления аминокислота в положении 409 может представлять собой Glu вместо Asp, и аминокислота в положении 399 может представлять собой Arg вместо Lys. Более того, в дополнение к аминокислоте Lys в положении 399, Asp можно соответствующим образом добавлять в качестве аминокислоты в положении 360 или Asp можно соответствующим образом добавлять в качестве аминокислоты в положении 392.
В другом неограничивающем варианте осуществления настоящего изобретения в качестве Fc-области можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 370 согласно нумерации EU представляет собой Glu, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 357 согласно нумерации EU представляет собой Lys.
В другом неограничивающем варианте осуществления настоящего изобретения в качестве Fc-области можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 439 согласно нумерации EU представляет собой Glu, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 356 согласно нумерации EU представляет собой Lys.
В другом неограничивающем варианте осуществления настоящего изобретения в качестве Fc-области можно соответствующим образом использовать любой из вариантов осуществления, указанных ниже, в которых вышеуказанное скомбинировано:
(i) два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 409 согласно нумерации EU представляет собой Asp и аминокислота в положении 370 представляет собой Glu, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 399 согласно нумерации EU представляет собой Lys и аминокислота в положении 357 представляет собой Lys (в этом варианте осуществления аминокислота в положении 370 согласно нумерации EU может представлять собой Asp вместо Glu, и аминокислоту Asp в положении 392 согласно нумерации EU можно использовать вместо аминокислоты Glu в положении 370 согласно нумерации EU);
(ii) два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 409 согласно нумерации EU представляет собой Asp и аминокислота в положении 439 представляет собой Glu, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 399 согласно нумерации EU представляет собой Lys и аминокислота в положении 356 представляет собой Lys (в этом варианте осуществления аминокислоту Asp в положении 360 согласно нумерации EU, аминокислоту Asp в положении 392 согласно нумерации EU или аминокислоту Asp в положении 439 согласно нумерации EU можно использовать вместо аминокислоты Glu в положении 439 согласно нумерации EU);
(iii) два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 370 согласно нумерации EU представляет собой Glu и аминокислота в положении 439 представляет собой Glu, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 357 согласно нумерации EU представляет собой Lys и аминокислота в положении 356 представляет собой Lys; и
два полипептида, образуюзщих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 409 согласно нумерации EU представляет собой Asp, аминокислота в положении 370 представляет собой Glu, и аминокислота в положении 439 представляет собой Glu, и в аминокислотной последовательности другого из полипептидов, аминокислота в положении 399 согласно нумерации EU представляет собой Lys, аминокислота в положении 357 представляет собой Lys, и аминокислота в положении 356 представляет собой Lys (в этом варианте осуществления аминокислота в положении 370 согласно нумерации EU может быть не заменена на Glu, и, более того, когда аминокислота в положении 370 не заменена на Glu, аминокислота в положении 439 может представлять собой Asp вместо Glu, или аминокислоту Asp в положении 392 можно использовать вместо аминокислоты Glu в положении 439).
Кроме того, в другом неограничивающем варианте осуществления настоящего изобретения также можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов, аминокислота в положении 356 согласно нумерации EU представляет собой Lys, и в аминокислотной последовательности другого из полипептидов аминокислота в положении 435 согласно нумерации EU представляет собой Arg и аминокислота в положении 439 представляет собой Glu.
В другом неограничивающем варианте осуществления настоящего изобретения можно соответствующим образом использовать два полипептида, образующих Fc-область, в которых в аминокислотной последовательности одного из полипептидов аминокислота в положении 356 согласно нумерации EU представляет собой Lys и аминокислота в положении 357 представляет собой Lys, в аминокислотной последовательности другого из полипептидов аминокислота в положении 370 согласно нумерации EU представляет собой Glu, аминокислота в положении 435 представляет собой Arg, и аминокислота в положении 439 представляет собой Glu.
Более того, в дополнение к упомянутому выше способу связывания гетерологичных тяжелых цепей, также для получения мультиспецифических или мультипаратопных антигенсвязывающих молекул, предусматриваемых в рамках настоящего изобретения, можно использовать технологию CrossMab, которая известна в качестве технологии связывания гетерологичных легких цепей, в которой легкую цепь, образующую вариабельную область, которая связывается с первым эпитопом, и легкую цепь, образующую вариабельную область, которая связывается со вторым эпитопом, соответственно связывают с тяжелой цепью, образующей вариабельную область, которая связывается с первым эпитопом, и тяжелой цепью, образующей вариабельную область, которая связывается с вторым эпитопом (Scaefer et al. (Proc. Natl. Acad. Sci. U.S.A. (2011) 108, 11187-11192)).
Применение антигенсвязывающей молекулы для элиминации антигенов, содержащих два или более антигенных элементов связывания, из плазмы
Настоящее изобретение относится к применению антигенсвязывающей молекулы, содержащей (i) Fc-область и (ii) антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов; где антигенсвязывающая молекула может образовывать иммунный комплекс, содержащий (a) две или более антигенсвязывающих молекул и (b) две или более молекул антигена при условии, что антигены содержат два или более антигенных элементов связывания, для элиминации антигенов из плазмы.
В рамках настоящего изобретения варианты осуществлении применения антигенсвязывающей молекулы конкретно не ограничены при условии, что антигены могут элиминироваться из плазмы. Примерами неограничивающего варианта осуществления такого применения являются фармацевтические композиции, содержащие антигенсвязывающую молекулу, предусматриваемую настоящим изобретением, и способы, включающие введение индивидууму антигенсвязывающей молекулы, предусматриваемой настоящим изобретением. Примером другого неограничивающего варианта осуществления является применение антигенсвязывающей молекулы в способе элиминации антигенов из плазмы ex vivo, который включает контактирование иммунного комплекса, образовавшегося при контакте антигенсвязывающей молекулы по настоящему изобретению, с плазмой, взятой от индивидуума, и содержащего две или более антигенсвязывающих молекул и два или более антигенов (при условии, что антигены включают два или более антигенных элементов связывания) с клетками, экспрессирующими FcRn или Fcγ-рецептор.
Улучшение фармакокинетики
В рамках настоящего изобретения "способность элиминировать антигены из плазмы" относится к способности элиминировать из плазмы антигены, которые присутствуют в плазме, когда антигенсвязывающие молекулы вводят in vivo или когда антигенсвязывающие молекулы секретируются in vivo. Таким образом, в рамках настоящего изобретения выражение "способность антигенсвязывающей молекулы элиминировать антиген из плазмы увеличивается" можно использовать, когда скорость элиминации антигена из плазмы ускоряется при введении антигенсвязывающей молекулы, по сравнению с введением антигенсвязывающих молекул, которые не могут образовывать иммунные комплексы, описанные в рамках настоящего изобретения, антигенсвязывающих молекул, содержащих антигенсвязывающие домены, у которых антигенсвязывающая активность не зависит от концентрации ионов, или антигенсвязывающих молекул, содержащих Fc-области с нарушенной активностью связывания с FcγR или FcRn. То, увеличивается ли способность антигенсвязывающей молекулы элиминировать антиген из плазмы, можно оценивать, например, путем введения растворимого антигена и антигенсвязывающей молекулы in vivo и измерения концентрации в плазме растворимого антигена после введения. Способность антигенсвязывающей молекулы элиминировать антиген из плазмы считают увеличенной, если концентрация растворимого антигена в плазме снижается после введения растворимого антигена и антигенсвязывающей молекулы, которая представляет собой антигенсвязывающую молекулу, способную образовывать иммунный комплекс, содержащий две или более антигенсвязывающих молекул и антигены с двумя или более антигенных элементов связывания, и антигенсвязывающая активность которых изменяется в зависимости от концентрации ионов, как например, снижение антигенсвязывающей активности в диапазоне кислых значений pH по сравнению с диапазоном нейтральных значений pH (или снижение антигенсвязывающей активности при низкой концентрации ионов кальция по сравнению с высокой концентрации ионов кальция). Растворимый антиген может представлять собой антиген, который связан с антигенсвязывающей молекулой, или антиген, который не связан с антигенсвязывающей молекулой в плазме, и его концентрацию можно определить как "концентрацию в плазме антигена, связанного с антигенсвязывающей молекулой" или как "концентрацию в плазме антигена, не связанного с антигенсвязывающей молекулой", соответственно (последнее является синонимом "концентрации свободного антигена в плазме"). "Общая концентрация антигена в плазме" означает сумму концентраций связанного антигенсвязывающей молекулой антигена и антигена, не связанного антигенсвязывающей молекулой или "концентрации свободного антигена в плазме", которая представляет собой концентрацию антигена, не связанного антигенсвязывающей молекулой. Таким образом, концентрацию растворимого антигена можно определить как "общую концентрацию антигена в плазме". Различные способы измерения "общей концентрации антигена в плазме" или "концентрации свободного антигена в плазме" хорошо известны в данной области, как описано в настоящем описании ниже.
В настоящем описании "усиление фармакокинетики", "улучшение фармакокинетики" и "превосходящая фармакокинетика" можно переформулировать как "усиление удержание в плазме (крови)", "улучшение удержания в плазме (крови)", "превосходящее удержание в плазме (крови)", и "пролонгированное удержание в плазме (крови)". Эти термины являются синонимами.
В настоящем описании "улучшение фармакокинетики" означает не только пролонгирование периода времени до элиминации из плазмы (например, пока антигенсвязывающая молекула не деградирует внутреклеточно и т.п. и может вернуться в плазму) после введения антигенсвязывающей молекулы человеку или не являющимся человеком животным, таким как мыши, крысы, обезьяны, кролики и собаки, но также пролонгирование удержания в плазме антигенсвязывающих молекул в форме, которая обеспечивает связывание антигена (например, в свободной от антигена форме антигенсвязывающей молекулы) в ходе периода введения до элиминации вследствие деградации. IgG человека, имеющий Fc-область дикого типа, может связываться с FcRn из не являющихся человеком животных. Например, мышь можно предпочтительно использовать для введения в целях подтверждения свойства антигенсвязывающей молекулы по изобретению, поскольку IgG человека, имеющий Fc дикого типа, может связываться с FcRn мыши прочнее, чем с FcRn человека (Int Immunol. (2001) 13(12): 1551-1559). В качестве другого примера, также для введения в целях подтверждения свойства антигенсвязывающей молекулы по изобретению, описанного далее, предпочтительно можно использовать мышь, у которой ее нативные гены FcRn разрушены и содержится трансген для гена FcRn человека (Methods Mol Biol. 2010; 602: 93-104). В частности, "улучшение фармакокинетики" также включает пролонгирование периода времени до элиминации вследствие деградации антигенсвязывающей молекулы, не связанной с антигенами (свободная от антигена форма антигенсвязывающей молекулы). Антигенсвязывающая молекула в плазме не может связываться с новым антигеном, если антигенсвязывающая молекула уже связана с антигеном. Таким образом, чем более длительным является период времени, когда антигенсвязывающая молекула не связана с антигеном, тем более длительным является период времени, когда она может связываться с новым антигеном (более высокая вероятность связывания с другим антигеном). Это обеспечивает уменьшение периода времени, когда антиген свободен от антигенсвязывающей молекулы in vivo, и продление периода времени, когда антиген связан с антигенсвязывающей молекулой. Концентрация в плазме свободной от антигена формы антигенсвязывающей молекулы может увеличиваться и период времени, когда антиген связан с антигенсвязывающей молекулой, может продлеваться путем ускорения элиминации антигена из плазмы посредством введения антигенсвязывающей молекулы. В частности, в настоящем описании "улучшение фармакокинетики антигенсвязывающей молекулы" включает улучшение фармакокинетического параметра свободной от антигена формы антигенсвязывающей молекулы (любое из продления времени полужизни в плазме, продления среднего времени удержания в плазме и снижения клиренса из плазмы), продление периода времени, когда антиген связан с антигенсвязывающей молекулой, после введения антигенсвязывающей молекулы, и ускорение опосредуемой антигенсвязывающей молекулой элиминации из плазмы. Улучшение фармакокинетики антигенсвязывающей молекулы можно оценивать путем определения любого из параметров: время полужизни в плазме, среднее время удержания в плазме и клиренс из плазмы, для антигенсвязывающей молекулы или ее свободной от антигена формы ("Pharmacokinetics: Enshu-niyoru Rikai (Understanding through practice)" Nanzando). Например, концентрацию в плазме антигенсвязывающей молекулы или ее свободной от антигена формы определяют после введения антигенсвязывающей молекулы мышам, крысам, обезьянам, кроликам, собакам или человеку. Затем определяют каждый параметр. Когда время полужизни в плазме или среднее время удержания в плазме пролонгируются, можно считать, что фармакокинктика антигенсвязывающей молекулы в плазме повышается. Параметры можно определять способами, известными специалистам в данной области. Параметры можно оценивать соответствующим образом, например, с помощью некомпартментного анализа с использованием программного обеспечения для фармакокинетического анализа WinNonlin (Pharsight) в соответствии с инструкцией по применению. Концентрацию в плазме свободной от антигена антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области, например, с использованием способа анализа, описанного в Clin Pharmacol. 2008 Apr; 48(4): 406-417.
В рамках настоящего изобретения "улучшение фармакокинетики" также включает пролонгирование периода, когда антиген связан с антигенсвязывающей молекулой, после введения антигенсвязывающей молекулы. Оценку того, пролонгируется ли период, когда антиген связан с антигенсвязывающей молекулой после введения антигенсвязывающей молекулы, можно проводить путем определения концентрации в плазме свободного антигена. Заключение о пролонгировании можно делать, исходя из определенной концентрации в плазме свободного антигена или периода времени, требуемого для увеличения соотношения концентрации свободного антигена и концентрации общего антигена.
Концентрацию свободного антигена, не связанного с антигенсвязывающей молекулой, или соотношение концентрации свободного антигена и концентрации общего антигена можно определять способами, известными специалистам в данной области, например, способом, используемым в Pharm Res. 2006 Jan; 23 (1): 95-103. Альтернативно, когда антиген проявляет конкретную функцию in vivo, оценку того, связан ли антиген с антигенсвязывающей молекулой, которая нейтрализует функцию антигена (антагонистическая молекула), можно проводить путем исследования того, нейтрализуется ли функция антигена. Оценку того, нейтрализуется ли функция антигена, можно проводить путем анализа маркера in vivo, который отражает функцию антигена. Оценку того, связан ли антиген с антигенсвязывающей молекулой, которая активирует функцию антигена (молекула-агонист), можно проводить путем анализа маркера in vivo, который отражает функцию антигена.
Определение концентрации в плазме свободного антигена и соотношения количества свободного антигена в плазме и количества общего антигена в плазме, анализ маркеров in vivo и сходные способы определения конкретно не ограничены; однако анализы предпочтительно проводят после того, как проходит определенный период времени после введения антигенсвязывающей молекулы. В рамках настоящего изобретения период после введения антигенсвязывающей молекулы конкретно не ограничен; специалисты в данной области могут определить соответствующий период, в зависимости от свойств и т.п. вводимой антигенсвязывающей молекулы. Такие периоды включают, например, одни сутки после введения антигенсвязывающей молекулы, трое суток после введения антигенсвязывающей молекулы, семь суток после введения антигенсвязывающей молекулы, 14 суток после введения антигенсвязывающей молекулы и 28 суток после введения антигенсвязывающей молекулы. В настоящем описании понятие "концентрация антигена в плазме" включает как "общую концентрацию антигена в плазме", которая представляет собой сумму концентрации антигена, связанного и не связанного антигенсвязывающей молекулой, так и "концентрацию свободного антигена в плазме", которая представляет собой концентрацию антигена, не связанного антигенсвязывающей молекулой.
Путем введения, в качестве антигенсвязывающей молекулы, антигенсвязывающей молекулы по настоящему изобретению общую концентрацию антигена в плазме можно снижать в 2 раза, в 5 раз, в 10 раз, в 20 раз, в 50 раз, в 100 раз, в 200 раз, в 500 раз, в 1000 раз или даже больше по сравнению с введением антигенсвязывающей молекулы, содержащей антигенсвязывающий домен, антигенсвязывающая активность которого не зависит от концентрации ионов, или антигенсвязывающей молекулы, содержащей Fc-область со сниженнйо активностью FcγR, или по сравнению с тем, когда молекулу с антигенсвязывающим доменом по настоящему изобретению не вводят.
Молярное соотношение антиген/антигенсвязывающая молекула можно вычислять, как показано ниже;
величина A: Молярная концентрация антигена в каждый момент времени;
величина B: Молярная концентрация антигенсвязывающей молекулы в каждый момент времени;
величина C: Молярная концентрация антигена на молярную концентрацию антигенсвязывающей молекулы (молярное соотношение антиген/антигенсвязывающая молекула) в каждый момент времени
C=A/B.
Меньшая величина C указывает на более высокую эффективность элиминации антигена на антигенсвязывающую молекулу, в то время как более высокая величина C указывает на более низкую эффективность элиминации антигена на антигенсвязывающую молекулу.
Молярное соотношение антиген/антигенсвязывающая молекула можно вычислять, как описано выше.
Введение антигенсвязывающей молекулы по настоящему изобретению может снизить молярное соотношение антиген/антигенсвязывающая молекула в 2 раза, в 5 раз, в 10 раз, в 20 раз, в 50 раз, в 100 раз, в 200 раз, в 500 раз, в 1000 раз или более по сравнению с введением антигенсвязывающих молекул, которые не могут образовывать иммунные комплексы, описанные в рамках настоящего изобретения, антигенсвязывающих молекул, содержащих антигенсвязывающие домены, антигенсвязывающая активность которых зависит от концентраций ионов, или антигенсвязывающих молекул, содержащих Fc-области с нарушенной активностью связывания с FcγR или FcRn.
В рамках настоящего изобретения, эталоном для сравнения с антигенсвязывающими молекулами по настоящему изобретению являются антигенсвязывающие молекулы, которые не могут образовывать иммунные комплексы, описанные в рамках настоящего изобретения, антигенсвязывающие молекулы, содержащие антигенсвязывающие домены, антигенсвязывающая активность которых не зависит от концентрации ионов, или антигенсвязывающие молекулы, содержащие Fc-области с нарушенной активностью связывания с FcγR или FcRn.
Когда для включения антигенсвязывающих молекул по настоящему изобретению в клетки используют опосредуемый FcRn каскад, снижение общей концентрации в плазме или молярного соотношения антиген/антитело также можно оценивать с использованием трансгенной линии мышей с FcRn человека 32 или 276 (Jackson Laboratories, Methods Mol Biol. 2010; 602: 93-104), либо с помощью модели совместной инъекции антиген-антитело, либо стационарной инфузионной модели, когда антигенсвязывающая молекула не реагирует перекрестно с антигеном-аналогом мыши. Когда антигенсвязывающая молекула перекрестно реагирует с аналогом мыши, их также можно оценивать путем простой инъекции антигенсвязывающей молекулы мышам линии трансгенных мышей с FcRn человека 32 или 276 (Jackson Laboratories). В модели совместной инъекции мышам вводят смесь антигенсвязывающей молекулы и антигена. В стационарной инфузионной модели, мышам имплантируют инфузионный насос, содержащий раствор антигена, для достижения постоянной концентрации антигена в плазме, а затем мышам инъецируют антигенсвязывающую молекулу. Исследуемые антигенсвязывающие молекулы вводят в той же дозировке. Общую концентрацию антигена в плазме, концентрацию свободного антигена в плазме и концентрацию антигенсвязывающей молекулы в плазме измеряют в соответствующие моменты времени с использованием способа, известного специалистам в данной области.
Когда для включения антигенсвязывающих молекул по настоящему изобретению из плазмы в клетки используют опосредуемый FcγR каскад, снижение общей концентрации антигена в плазме или молярного соотношения антиген/антитело можно оценивать либо в модели совместной инъекции антиген-антитело, либо в модели инфузии антигена в стационарном состоянии с использованием обычно используемых мышей C57BL/6J (Charles River Japan), когда антигенсвязывающая молекула не реагирует перекрестно с антигеном-аналогом мыши. Если антигенсвязывающая молекула перекрестно реагирует с аналогом мыши, оценку можно проводить просто путем инъекции антигенсвязывающей молекулы обычно используемым мышам C57BL/6J (Charles River Japan).
В модели совместной инъекции смесь антигенсвязывающей молекулы и антигена вводят мышам. В модели инфузии антигена в стационарном состоянии, инфузионный насос, заполненный раствором антигена, вводят мыши для достижения постоянной концентрации антигена в плазме, а затем антигенсвязывающую молекулу инъецируют мышам. Исследуемые антигенсвязывающие молекулы вводят в одинаковой дозе. Общую концентрацию антигена в плазме, концентрацию свободного антигена в плазме и концентрацию антигенсвязывающей молекулы концентрация в плазме измеряют в соответствующие моменты времени с использованием способов, известных специалистам в данной области.
Концентрацию общего или свободного антигена в плазме и молярное соотношение антиген/антигенсвязывающая молекула можно измерять через 2, 4, 7, 14, 28, 56 или 84 суток после введения для оценки долговременного эффекта настоящего изобретения. Иными словами, долговременную концентрацию антигена в плазме определяют путем измерения концентрации общего или свободного антигена в плазме и молярного соотношения антиген/антигенсвязывающая молекула через 2, 4, 7, 14, 28, 56 или 84 суток после введения антигенсвязывающей молекулы для оценки свойства антигенсвязывающей молекулы по настоящему изобретению. То, достигается ли с помощью антигенсвязывающей молекулы, описанной в рамках настоящего изобретения, снижение концентрации антигена в плазме или молярного соотношения антиген/антигенсвязывающая молекула, можно определять путем оценки снижения в любой один или более из моментов времени, описанных выше.
Концентрацию общего или свободного антигена в плазме и молярное соотношение антиген/антигенсвязывающая молекула можно измерять через 15 мин, 1, 2, 4, 8, 12 или 24 часов после введения для оценки кратковременного эффекта настоящего изобретения. Иными словами, кратковременную концентрацию антигена в плазме определяют путем измерения концентрации общего или свободного антигена в плазме и молярного соотношения антиген/антигенсвязывающая молекула через 15 мин, 1, 2, 4, 8, 12 или 24 часов после введения антигенсвязывающей молекулы для оценки свойства антигенсвязывающей молекулы по настоящему изобретению.
Пути введения антигенсвязывающей молекулы по настоящему изобретению можно выбирать из внутрикожной, внутривенной, проводимой в стекловидное тело, подкожной, внутрибрюшинной, парентеральной и внутримышечной инъекции.
В рамках настоящего изобретение предпочтительно, чтобы фармакокинетика антигенсвязывающей молекулы у человека была улучшенной. Когда удержание в плазме у человека трудно определить, его можно предсказать, исходя из удержания в плазме у мышей (например, здоровых мышей, трансгенных мышей, экспрессирующих антиген человека, трансгенных мышей, экспрессирующих FcRn человека) или обезьян (например, яванские макаки).
В настоящем описании "улучшение фармакокинетики и пролонгированное удержание в плазме антигенсвязывающих молекул" означает улучшение любого параметра фармакокинетики (любой из пролонгирования времени полужизни в плазме, пролонгирования среднего удержания в плазме, снижения клиренса из плазмы и биодоступности) после введения in vivo антигенсвязывающей молекулы, или увеличение концентрации антигенсвязывающей молекулы в плазме за соответствующий период времени после введения. Его можно определять путем измерения любого параметра, такого как время полужизни в плазме, среднее время удержания в плазме, клиренс из плазмы и биодоступность антигенсвязывающей молекулы (Pharmacokinetics: Enshu-niyoru Rikai (Understanding through practice), (Nanzando)). Например, когда антигенсвязывающую молекулу вводят мышам (нормальным мышам и трансгенным мышам с FcRn человека), крысам, обезьянам, кроликам, собакам, человеку и т.д., и определяют концентрацию антигенсвязывающей молекулы в плазме и каждый из параметров вычисляют, можно считать, что фармакокинетика антигенсвязывающей молекулы улучшается, когда время полужизни в плазме и среднее время удержания в плазме пролонгируются. Эти параметры можно определять способами, известными специалистам в данной области. Например, параметры можно соответствующим образом оценивать с помощью некомпартментного анализа с использованием программного обеспечения для анализа фармакокинетики WinNonlin (Pharsight) в соответствии с прилагаемыми инструкциями.
Без ограничения конкретной теорией, следующий механизм является примером механизма, который позволяет элиминацию антигенных элементов связывания из плазмы с помощью антигенсвязывающей молекулы по настоящему изобретению, которая содержит (i) Fc-область и (ii) антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, где антигенсвязывающая молекула может образовывать иммунный комплекс, содержащий две или более антигенсвязывающих молекул и антигены, содержащие два или более антигенных элементов связывания. Когда антигенный элемент связывания представляет собой один элемент (т.е. мономер), такой как в sIL-6R, две молекулы ((т.е. два антигенных элемента связывания) антигена связываются с одной молекулой антитела, содержащей двухвалентный антигенсвязывающий домен, и образуется комплекс одной молекулы антитела против sIL-6R и двух молекул антигена, содержащих два антигенных элемента связывания. Таким образом, этот тип комплекса антиген-антитело имеет только одну Fc-область (нативную Fc-область IgG1), как показано на фиг. 1. Поскольку этот комплекс связывается с двумя молекулами FcRn или одной молекулой FcγR через одну Fc-область, аффинность в отношении этих рецепторов является такой же, как и аффинность обычного IgG-антитела, и полагают, что захват в клетки происходит по большей части неспецифически.
С другой стороны, когда антигенный элемент связывания представляет собой два элемента, как в IgA человека, который является димером гетерокомплекса тяжелых цепей и легких цепей, существует два элемента эпитопов, с которыми антигенсвязывающие домены будут связываться в антигенном элементе связывания. Однако, когда двухвалентное антитело против IgA (т.е. антигенсвязывающие домены, содержащиеся в одной молекуле антитела против IgA связываются с тем же эпитопом) связывается с его антигеном IgA, полагают, что связывание каждого из двухвалентных антигенсвязывающих доменов, содержащихся в одной молекуле антитела против IgA с каждым из двух элементов эпитопов, присутствующих на одной молекуле IgA, является затрудненным ввиду расположения эпитопов. В результате, полагают, что отдельные молекулы антитела против IgA связываются с двумя антигенными элементами связывания, присутствующими в двух молекулах IgA, которые связываются с двухвалентными антигенсвязывающими доменами, присутствующими в одной молекуле антитела против IgA, и, следовательно, образуются комплексы антиген-антитело (иммунные комплексы), содержащие по меньшей мере четыре молекулы (т.е. две молекулы IgA, которые представляют собой молекулу антигена, и две молекулы антитела против IgA, которые являются антигенсвязывающими молекулами).
Когда антигенсвязывающая молекула, такая как антитело, связывающееся с молекулой антигена, который содержит два или более антигенных элементов связывания, образует крупный иммунный комплекс, который является по меньшей мере тетрамером, иммунный комплекс может связываться прочно с авидностью через по меньшей мере две или более поливалентных Fc-области с FcγR, FcRn, рецепторами комплемента и т.п. Таким образом, как показано на фиг. 7, комплекс захватывается в клетки, экспрессирующие эти рецепторы, с более высокой эффективностью, чем нативный IgG1. С другой стороны, поскольку опосредуемая Fc-областью аффинность в отношении этих рецепторов у иммунных комплексов, образованных из молекул антигена и антигенсвязывающих молекул, которые, например, связываются с (мономерными) молекулами антигена, содержащими один антигенный элемент связывания, является недостаточной, как упоминалось выше, иммунные комплексы захватываются по большей неспецифически (менее эффективно по сравнению с захватом, опосредуемым связыванием с авидностью) в клетки, экспрессирующие эти рецепторы, как показано на фиг. 1. Таким образом, захват является в большей степени неэффективным, чем захват, опосредуемый связыванием с авидностью.
Когда антигенсвязывающая молекула, такая как антитело, которое связывается с молекулой антигена, содержащей два или более антигенных элементов связывания, представляет собой антитело, которое содержит антигенсвязывающие домены, у которых активность связывания с антигеном варьирует в зависимости от условий концентрации ионов, как например, в случае pH- или Ca-зависимого связывания, и которые образуют комплекс антиген-антитело (иммунный комплекс), содержащий по меньшей мере четыре молекулы (две молекулы антигена и две молекулы антитела) в плазме, после захвата иммунного комплекса в клетки антигены диссоциируют от антител в эндосомах, где условия концентрации ионов отличаются от условий концентрации ионов в плазме. Таким образом, образованный иммунный комплекс распадается в эндосомах клеток, которые захватили иммунные комплексы. Поскольку диссоциированные антигены не могут связываться с FcRn в эндосомах, они деградируют после перемещения в лизосомы. С другой стороны, полагают, что антитела после диссоциации антигена рециклируют в плазму после связывания с FcRn в эндосомах (фиг. 7).
Как описано выше, если антитело, которое содержит нативную константную область IgG1-типа, против мультимерного антигена, содержащего два или более антигенных элементов связывания, и демонстрирует pH- или Ca-зависимое связывание, может образовывать крупный иммунный комплекс и связывается с FcγR, FcRn, рецепторами комплемента и т.п. с авидностью, полагают, что элиминация антигена отдельно может селективно и в значительной степени ускоряться. Полагают, что, когда вводят GA2-IgG1, которое связывается с IgA человека, образуются такие крупные иммунные комплексы. Действительно, как показано в примере 3, GA2-IgG1-FcγR(-), полученное путем внесения в GA2-IgG1 модификаций, которые ухудшают связывание с FcγR мыши, по существу не могут ускорять элиминацию IgA человека подобно GA2-IgG1 по сравнению с IgA человека отдельно, и они продемонстрировали эквивалентный уровень элиминации с IgA человека отдельно. Таким образом, причиной, по которой GA2-IgG1 могло ускорять элиминацию IgA человека, является то, что иммунный комплекс, содержащий GA2-IgG1 и IgA человека, который является мультимерным антигеном, содержащим два или более антигенных элементов связывания, связывается с авидностью с FcγR и быстро захватывается в клетки, экспрессирующие FcγR. IgA, который диссоциирует от иммунного комплекса в эндосомах клеток, которые захватили иммунный комплекс, деградируется в лизосомах. В то же время, диссоциировавшее от IgA антитело, которое связалось с FcRn в эндосомах, впоследствии рециклирует в плазму и может вновь связываться с IgA в плазме. Полагают, что элиминация IgA в плазме значительно ускоряется таким образом. Способ использования Fc-области с вариантом аминокислоты, которая связывается с FcRn в диапазоне нейтральных значений pH, описан в WO2011/122011 в качестве способа ускорения элиминации антигенов из плазмы. Настоящее изобретение является пригодным в качестве способа ускорения элиминации из плазмы мультимерных антигенов, содержащих два или более антигенных элементов связывания без использования упомянутых выше вариантов, и может далее ускорять элиминацию мультимерных антигенов, содержащих два или более антигенных элементов связывания из плазмы посредством комбинирования с упомянутыми выше вариантами.
Способ еx vivo для элиминации антигенов из плазмы
Пример неограничивающего варианта осуществления применения антигенсвязывающей молекулы в способе, предусматриваемом настоящим изобретением, для элиминации антигенов из плазмы включает применение антигенсвязывающей молекулы в так называемом способе элиминации антигенов ex vivo из плазмы, который включает формирование иммунного комплекса, содержащего две или более антигенсвязывающих молекул и две или более молекул антигена (при условии, что антигены содержат два или более антигенных элементов связывания) путем позволения антигенсвязывающим молекулам по настоящему изобретению контактировать с плазмой, взятой от индивидуума, и позволения иммунному комплексу контактировать с клетками, экспрессирующими FcRn и/или Fcγ-рецепторы. Скорость элиминации антигена из плазмы также можно увеличивать путем замены или комбинирования способа введения антигенсвязывающих молекул in vivo с так называемым способом ex vivo, в котором плазму, содержащую антигенсвязывающие молекулы и антигены, которые связываются с антигенсвязывающими молекулами, временно извлекают из организма, а затем контактируют с клетками, экспрессирующими FcRn и/или Fcγ-рецепторы, в течение определенного периода времени, и плазму, содержащую внеклеточно рециклированные или повторно секретированные или рециркулированные) антигенсвязывающие молекулы, которые не связались с антигеном, возвращают в организм.
Более того, пример неограничивающего варианта осуществления применения антигенсвязывающей молекулы в способе, предусматриваемом настоящим изобретением, для элиминации антигенов из плазмы включает применение антигенсвязывающей молекулы в так называемом способе ex vivo для элиминации антигенов из плазмы, который включает контактирование иммунного комплекса, содержащего две или более антигенсвязывающих молекул и две или более молекул антигена (при условии, что антигены содержат два или более антигенных элементов связывания), присутствующих в плазме, выделенной от индивидуума, которому антигенсвязывающие молекулы по настоящему изобретению вводят с клетками, экспрессирующими FcRn и/или Fcγ-рецепторы.
Подтверждение элиминации антигена из плазмы можно проводить, например, путем оценки того, увеличивается ли скорость элиминации антигена из упомянутой выше плазмы с использованием в качестве контроля антигенсвязывающих молекул, которые не могут образовывать иммунные комплексы, описанные в рамках настоящего изобретения, антигенсвязывающих молекул, содержащих антигенсвязывающие домены с независимой от концентрации ионов антигенсвязывающей активностью, или антигенсвязывающих молекул, содержащих Fc-область с нарушенной активностью связывания с FcγR или FcRn, вместо антигенсвязывающих молекул по настоящему изобретению.
Способ скрининга антигенсвязывающих молекул, содержащих Fc-область и антигенсвязывающий домен, антигенсвязывающая активность которого зависит от концентрации ионов
Настоящее изобретение относится к способу скрининга антигенсвязывающей молекулы, обладающей функцией элиминации антигенов из плазмы, причем способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше;
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше;
(f) контактирование антигенсвязывающей молекулы, полученной согласно (e) выше, с антигеном; и
(g) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген.
В неограничивающем варианте осуществления настоящего изобретения после выделения полинуклеотида, кодирующего антигенсвязывающий домен, активность связывания которого изменяется в зависимости от условий, выбранных, как описано выше, полинуклеотид встраивают в соответствующий экспрессирующий вектор. Например, когда антигенсвязывающий домен представляет собой вариабельную область антитела, после получения кДНК, кодирующей вариабельную область, кДНК расщепляют ферментами рестрикции, которые распознают участки рестрикции, встроенные на обоих концах кДНК. Предпочтительно, ферменты рестрикции распознают и расщепляют нуклеотидную последовательность, которая встречается с низкой частотой в нуклеотидной последовательности, составляющей ген антигенсвязывающей молекулы. Более того, предпочтительно встраивают ферменты рестрикции, которые обеспечивают липкие концы, чтобы встроить единичную копию расщепленного фрагмента в вектор в правильной ориентации. кДНК, кодирующую вариабельную область антигенсвязывающей молекулы, расщепленную как описано выше, встраивают в соответствующий экспрессирующий вектор с получением экспрессирующего вектора для антигенсвязывающей молекулы по настоящему изобретению. В это время ген, кодирующий константную область (C-область) антитела, можно подвергать слиянию в рамке считывания с геном, кодирующим вариабельную область.
Для получения представляющей интерес антигенсвязывающей молекулы, полинуклеотид, кодирующий антигенсвязывающую молекулу, встраивают так, чтобы он был функционально связан с регуляторной последовательностью, в экспрессирующий вектор. Регуляторные последовательности включают, например, энхансеры и промоторы. Более того, с N-концом может быть связана соответствующая сигнальная последовательность, чтобы экспрессированная антигенсвязывающая молекула секретировалось во внешнюю среду клеток. В качестве сигнальной последовательности используют, например, пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO:3); однако также можно присоединять другие соответствующие сигнальные последовательности. Экспрессированный полипептид расщепляется на С-конце описанной выше последовательности, и расщепленный полипептид секретируется в качестве зрелого полипептида во внешнюю среду клеток. Затем соответствующие клетки-хозяева трансформируют этим экспрессирующим вектором так, чтобы можно было получить рекомбинантные клетки, экспрессирующие полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу. Антигенсвязывающие молекулы по настоящему изобретению можно получать из рекомбинантных клеток посредством способов, описанных выше в разделе об антителах.
Для нуклеиновой кислоты "функционально связанный" означает, что нуклеиновая кислота имеет функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК, кодирующая препоследовательность или секреторную лидерную последовательность, функционально связана с ДНК, кодирующей определенный полипептид, если она подлежит экспрессии в качестве белка-предшественника, вовлеченного в секрецию полипептида. Промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию кодирующей последовательности. Участок связывания рибосомы функционально связан с кодирующей последовательностью, если он находится в положении, которое облегчает трансляцию. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются соседними и в случае секреторной лидерной последовательности, это означает, что связанные последовательности ДНК являются соседними и находятся в рамке считывания. Однако энхансеры не должны быть соседними. Связывание проводят путем лигирования в подходящих участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидые адаптеры или линкеры в соответствии с общепринятой практикой. Более того, связанные нуклеиновые кислоты можно получать упомянутым выше способом перекрывающейся ПЦР.
В неограничивающем варианте осуществления настоящего изобретения после выделения полинуклеотида, кодирующего описанную выше антигенсвязывающую молекулу, активность связывания которой варьирует, в зависимости от выбранных условий, вариант полинуклеотида встраивают в соответствующий экспрессирующий вектор. Такие варианты предпочтительно включают варианты, полученные путем гуманизации, на основе полинуклеотидной последовательности, кодирующей антигенсвязывающую молекулу по настоящему изобретению, полученную путем скрининга в качестве рандомизированной библиотеки вариабельной области сконструированной синтетической библиотеки или иммунной библиотеки, происходящей из не являющихся человеком животных. Те же способы, которые описаны выше для получения описанных выше гуманизированных антител, можно использовать в качестве способа получения гуманизированных вариантов антигенсвязывающей молекулы.
В другом варианте осуществления такие варианты предпочтительно включают варианты, полученные путем внесения изменения, которое увеличивает аффинность к антигену (созревание аффинности) антигенсвязывающей молекулы по настоящему изобретению, в выделенную полинуклеотидную последовательность для молекулы, полученной путем скрининга с использованием синтетической библиотеки или наивной библиотеки в качестве рандомизированной библиотеки вариабельной области. Такие варианты можно получать различными известными способами созревания аффинности, включая мутагенез CDR (Yang et al. (J. Mol. Biol. (1995) 254, 392-403)), шаффлинг цепей (Marks et al. (Bio/Technology (1992) 10, 779-783)), использование мутантных штаммов E. coli (Low et al. (J. Mol. Biol. (1996) 250, 359-368)), шаффлинг ДНК (Patten et al. (Curr. Opin. Biotechnol. (1997) 8, 724-733)), фаговый дисплей (Thompson et al. (J. Mol. Biol. (1996) 256, 77-88)) и половую ПЦР (Clameri et al. (Nature (1998) 391, 288-291)).
Как описано выше, антигенсвязывающие молекулы, которые получают способами получения по настоящему изобретению, включают антигенсвязывающие молекулы, имеющие Fc-область. В качестве Fc-областей можно использовать различные варианты. В одном варианте осуществления варианты по настоящему изобретению предпочтительно включают полинуклеотиды, кодирующие антигенсвязывающие молекулы, имеющие тяжелую цепь, в которой полинуклеотид, кодирующий вариант Fc-области, как описано выше, связан в рамке считывания с полинуклеотидом, кодирующим описанную выше антигенсвязывающую молекулу, активность связывания которой варьирует в зависимости от выбранных условий.
В неограничивающем варианте осуществления настоящего изобретения Fc-области предпочтительно включают, например, константные области Fc антител, таких как IgG1 SEQ ID NO:13 (Ala добавлен на N-конец AAC82527.1), IgG2 SEQ ID NO:14 (Ala добавлен на N-конец AAB59393.1), IgG3 SEQ ID NO:15 (CAA27268.1) и IgG4 SEQ ID NO:16 (Ala добавлен на N-конец AAB59394.1). Время удержания молекул IgG в плазме является относительно длительным (элиминация из плазмы является медленной), поскольку FcRn, в частности FcRn человека, функционирует в качестве рецептора спасения для молекул IgG. Молекулы IgG, включенные в эндосомы путем пиноцитоза, связываются в кислых условиях эндосом с FcRn, в частности с FcRn человека, экспрессируемым в эндосомах. Молекулы IgG, которые не могут связываться с FcRn, в частности FcRn человека, переносятся в лизосомы и деградируются в них. Между тем, молекулы IgG, связанные в FcRn, в частности FcRn человека, переносятся на клеточную поверхность, а затем возвращаются в плазму в результате диссоциации от FcRn, в частности FcRn человека, в нейтральных условиях в плазме.
Поскольку антитела, содержащие типичную Fc-область, не обладают активностью связывания с FcRn, в частности с FcRn человека, в условиях диапазона нейтральных значений pH, типичные антитела и комплексы антитело-антиген включаются в клетки посредством неспецифического эндоцитоза и переносятся на клеточную поверхность путем связывания с FcRn, в частности FcRn человека, в условиях диапазона эндосомальных кислых значений pH. FcRn, в частности FcRn человека, транспортирует антитела из эндосомы на клеточную поверхность. Таким образом, полагают, что некоторые из FcRn, в частности FcRn человека, также присутствуют на клеточной поверхности. Однако антитела рециклируют в плазму, поскольку они диссоциируют от FcRn, в частности FcRn человека, в условиях диапазона нейтральных значений pH на поверхности клетки.
Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, которые включены в антигенсвязывающие молекулы по настоящему изобретению, можно получать любым способом. В частности, Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, можно получать путем изменения аминокислот иммуноглобулина человека IgG-типа в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулина человека IgG-типа включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3 и IgG4, и их вариантов). Аминокислоты в любых положениях можно изменять на другие аминокислоты, при условии, что полученные области обладают активностью связывания FcRn человека в диапазоне нейтральных значений pH или увеличенной активностью связывания FcRn человека в нейтральном диапазоне. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, предпочтительно, чтобы полученная область содержала изменение, которое приводит к эффекту усиления связывания FcRn человека в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые допускают такие изменения, включают, например, аминокислоты в положениях 221-225, 227, 228, 230, 232, 233-241, 243-252, 254-260, 262-272, 274, 276, 278-289, 291-312, 315-320, 324, 325, 327-339, 341, 343, 345, 360, 362, 370, 375-378, 380, 382, 385-387, 389, 396, 414, 416, 423, 424, 426-438, 440 и 442 (указанные согласно нумерации EU). Более конкретно, такие изменения аминокислот включают изменения, приведенные в таблице 5. Изменение этих аминокислот усиливает связывание с FcRn человека Fc-области иммуноглобулина IgG-типа в диапазоне нейтральных значений pH.
Среди описанных выше, для применения в рамках настоящего изобретения выбирают подходящие изменения, которые усиливают связывание FcRn человека в диапазоне нейтральных значений pH. Особенно предпочтительные аминокислоты для таких вариантов Fc-области включают, например, аминокислоты в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (указанные согласно нумерации EU). Активность связывания FcRn человека Fc-области, включенной в антигенсвязывающую молекулу, можно увеличивать в диапазоне нейтральных значений pH путем замены по меньшей мере одной аминокислоты другой аминокислотой.
Особенно предпочтительные изменения в Fc-области включают, например, замены:
Met вместо аминокислоты в положении 237;
Ile вместо аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr вместо аминокислоты в положении 250;
Phe, Trp или Tyr вместо аминокислоты в положении 252;
Thr вместо аминокислоты в положении 254;
Glu вместо аминокислоты в положении 255;
Asp, Asn, Glu или Gln вместо аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val вместо аминокислоты в положении 257;
His вместо аминокислоты в положении 258;
Ala вместо аминокислоты в положении 265;
Ala или Glu вместо аминокислоты в положении 286;
His вместо аминокислоты в положении 289;
Ala вместо аминокислоты в положении 297;
Ala вместо аминокислоты в положении 303;
Ala вместо аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr вместо аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg вместо аминокислоты в положении 309;
Ala, His или Ile вместо аминокислоты в положении 311;
Ala или His вместо аминокислоты в положении 312;
Lys или Arg вместо аминокислоты в положении 314;
Ala, Asp или His вместо аминокислоты в положении 315;
Ala вместо аминокислоты в положении 317;
Val вместо аминокислоты в положении 332;
Leu вместо аминокислоты в положении 334;
His вместо аминокислоты в положении 360;
Ala вместо аминокислоты в положении 376;
Ala вместо аминокислоты в положении 380;
Ala вместо аминокислоты в положении 382;
Ala вместо аминокислоты в положении 384;
Asp или His вместо аминокислоты в положении 385;
Pro вместо аминокислоты а в положении 386;
Glu for вместо аминокислоты в положении 387;
Ala или Ser вместо аминокислоты в положении 389;
Ala вместо аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 428;
Lys вместо аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr вместо аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val вместо аминокислоты в положении 436 в системе нумерации EU. Между тем, количество измененных аминокислот конкретно не ограничено; такие аминокислотные изменения включают единичное изменение аминокислоты и изменение аминокислот в двух или более участков. Комбинации изменений аминокислот в двух или более участков включают, например, комбинации изменений, описанные в таблицах с 5-1 по 5-32.
В дополнение к Fc-области IgG1(SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека, в качестве Fc-областей, включенных в рамках настоящего изобретения, можно пригодным образом использовать Fc-области с модифицированным связыванием FcγR, которые обладают более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, в которой цепь сахаров, связанная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров. Такие Fc-области с модифицированным связыванием FcγR можно получать путем модификации аминокислот в Fc-области нативного IgG человека. То, является ли активность Fc-области в отношении связывания FcγR более высокой, чем у Fc-области нативного IgG человека, в котором цепь сахаров, связанная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров, можно подходящим образом определять с использованием способов, таких как способы, описанные выше.
В рамках настоящего изобретения "модификация аминокислот" или "аминокислотная модификация" Fc-области включает модификацию до аминокислотной последовательности, которая отличается от исходной Fc-области. Исходная Fc-область может представлять собой любую Fc-область, при условии, что варианты, полученные путем модификации исходной Fc-области, могут связываться с FcRn человека в диапазоне нейтральных значений pH. Более того, также в качестве измененной Fc-области по настоящему изобретению предпочтительно можно использовать Fc-область, модифицированную из исходной Fc-области, которая уже модифицирована. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, что в качестве исходной Fc-области можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или модифицированных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности с их исходной Fc-областью, меньшим чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с системой нумерации EU.
Fc-область с модифицированным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, в которой цепь сахаров, связанная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров, содержащуюся в антигенсвязывающих молекулах по настоящему изобретению, можно получать любым способом. В частности, Fc-область с модифицированным связыванием FcγR можно получать путем модификации аминокислот в иммуноглобулине человека IgG-типа, и использовать в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типа для модификации включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3, IgG4 и их варианты).
Аминокислоты в любых положениях можно модифицировать на другие аминокислоты при условии, что активность связывания с рецептором Fcγ является более высокой, чем у Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека с качестве Fc-области человека, она предпочтительно содержит модификацию, которая обеспечивает эффект более высокой активности связывания Fcγ-рецептора, чем у Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров. Такие модификации аминокислот описаны, например, в международных публикациях, таких как WO2007/024249, WO2007/021841, WO2006/031370, WO2000/042072, WO2004/029207, WO2004/099249, WO2006/105338, WO2007/041635, WO2008/092117, WO2005/070963, WO2006/020114, WO2006/116260 и WO2006/023403.
Примеры таких аминокислот, которые можно модифицировать, включают по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU). Модификация этих аминокислот может обеспечить Fc-области (Fc-области с модифицированным связыванием FcγR), обладающие более высокой активностью связывания Fcγ-рецептора, чем активность связывания Fcγ-рецептора Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров.
Примеры особенно предпочтительных модификаций для применения в рамках настоящего изобретения включают по меньшей мере одну или более аминокислотных модификаций в Fc-области, выбранных из группы:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU. Между тем, количество аминокислот, подлежащих модификации, конкретно не ограничено, и можно модифицировать аминокислоту только в одном участке или можно модифицировать аминокислоты в двух или более участков. Примеры комбинаций для модификаций аминокислот в двух или более участков включают комбинации, описанные в таблице 6 (таблицы с 6-1 по 6-3).
Среди Fc-областей, пригодных для применения в рамках настоящего изобретения, пригодным примером Fc-области, которая обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором (т.е. имеющей активность селективного связывания с ингибиторным Fcγ-рецептором), которую используют в качестве неограничивающего варианта осуществления Fc-области со свойством наличия более высокой активности связывания со специфическим Fcγ-рецептором, чем с другими Fcγ-рецепторами (т.е. Fc-область, имеющая селективную активность связывания с Fcγ-рецептором), представляет собой Fc-область с одной или несколькими из следующих модификаций аминокислот (указанных посредством нумерации EU) упомянутой выше Fc-области: аминокислота в положении 238 модифицирована на Asp и аминокислота в положении 328 модифицирована на Glu. Fc-области и модификации, описанные в US2009/0136485, можно соответствующим образом выбирать в качестве Fc-области, имеющей активность селективного связывания с ингибиторным Fcγ-рецептором.
В неограничивающем варианте осуществления настоящего изобретения подходящим примером является Fc-область, в которой одна или более аминокислот, указанных посредством нумерации EU в положениях 238 и 328 в соответствии с нумерацией EU, соответственно модифицированы на Asp или Glu в упомянутой выше Fc-области.
Более того, в неограничивающем варианте осуществления настоящего изобретения, подходящими примерами Fc-областей являются Fc-области с заменой Pro на Asp в положении 238 (нумерация EU), и одной или несколькими модификациями, выбранными из Trp в качестве аминокислоты в положении 237, Phe в качестве аминокислоты в положении 237, Val в качестве аминокислоты в положении 267, Gln в качестве аминокислоты в положении 267, Asn в качестве аминокислоты в положении 268, Gly в качестве аминокислоты в положении 271, Leu в качестве аминокислоты в положении 326, Gln в качестве аминокислоты в положении 326, Glu в качестве аминокислоты в положении 326, Met в качестве аминокислоты в положении 326, Asp в качестве аминокислоты в положении 239, Ala в качестве аминокислоты в положении 267, Trp в качестве аминокислоты в положении 234, Tyr в качестве аминокислоты в положении 234, Ala в качестве аминокислоты в положении 237, Asp в качестве аминокислоты в положении 237, Glu в качестве аминокислоты в положении 237, Leu в качестве аминокислоты в положении 237, Met в качестве аминокислоты в положении 237, Tyr в качестве аминокислоты в положении 237, Lys в качестве аминокислоты в положении 330, Arg в качестве аминокислоты в положении 330, Asp в качестве аминокислоты в положении 233, Asp в качестве аминокислоты в положении 268, Glu в качестве аминокислоты в положении 268, Asp в качестве аминокислоты в положении 326, Ser в качестве аминокислоты в положении 326, Thr в качестве аминокислоты в положении 326, Ile в качестве аминокислоты в положении 323, Leu в качестве аминокислоты в положении 323, Met в качестве аминокислоты в положении 323, Asp в качестве аминокислоты в положении 296, Ala в качестве аминокислоты в положении 326, Asn в качестве аминокислоты в положении 326, и Met в качестве аминокислоты в положении 330, в соответствии с нумерацией EU.
Для модификации аминокислот антигенсвязывающих доменов можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которых амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
Как описано выше, антигенсвязывающие молекулы, выделенные из культурального раствора клеток-хозяев, содержащих функционально связанные гены, кодирующие антигенсвязывающие молекулы, контактируют с антигенами в способе скрининга по настоящему изобретению. В неограничивающем варианте осуществления стадии контактирования можно соответствующим образом использовать условия, описанные, например, в упомянутом выше разделе, касающемся связывания антигенов.
Далее, проводят оценку того, образуются ли иммунные комплексы, содержащие исследуемую антигенсвязывающую молекулу и исследуемый антиген. Примеры способов оценки образования иммунных комплексов, содержащих антигенсвязывающие молекулы и антигены, включают способы, в которых используется свойство, состоящее в том, что иммунные комплексы становятся более крупными молекулами, чем антигенсвязывающая молекула отдельно или молекула антигена отдельно, такие как эксклюзионная хроматография (гель-фильтрация), способ анализа с использованием ультрацентрифугирования, способ с использованием рассеяния света, электронная микроскопия и масс-спектрометрия (Molecular Immunology (2002), 39, 77-84; Molecular Immunology (2009), 47, 357-364). Например, когда используют эксклюзионную хроматографию (гель-фильтрацию), как показано на фиг. 9, оценку того, образуется ли иммунный комплекс, проводят путем наблюдения того, существуют ли молекулярные структуры, более крупные, чем молекулярные структуры в анализе молекулы антигена отдельно или антигенсвязывающей молекулы отдельно.
Более того, когда антигенсвязывающая молекула или антиген имеют константную область иммуноглобулина, примеры также включают включают ELISA и FACS, в которых используется свойство иммунного комплекса, состоящее в более прочном связывании с Fc-рецептором или компонентом комплемента, чем у антигенсвязывающей молекулы отдельно или антигена отдельно (The Journal of Biological Chemistry (2001) 276 (9), 6591-6604; Journal of Immunological Methods (1982) 50, 109-114). Например, когда ELISA проводят путем иммобилизации Fc-рецептора, образование иммунного комплекса оценивают путем наблюдения того, увеличен ли обнаруженный сигнал по сравнению с антигеном отдельно или с антигенсвязывающей молекулой отдельно.
Способы получения антигенсвязывающей молекулы, содержащей Fc-область и антигенсвязывающий домен, демонстрирующий независимую от концентрации ионов антигенсвязывающую активность
Настоящее изобретение относится к способу получения антигенсвязывающих молекул, обладающих функцией элиминации антигенов из плазмы, причем способ включает:
(a) контактирование антигена с антигенсвязывающей молекулой, содержащей Fc-область и два или более антигенсвязывающих доменов, где по меньшей мере один из антигенсвязывающих доменов обладает антигенсвязывающей активностью, которая изменяется в зависимости от условий концентрации ионов;
(b) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген;
(c) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антигенсвязывающий домен, для которого подтверждено, что он образует иммунный комплекс согласно (b) выше; и
(d) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (c) выше.
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, обладающей функцией элиминации антигенов из плазмы, причем способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше;
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше;
(f) контактирование антигенсвязывающей молекулы, полученной согласно (e) выше, с антигеном;
(g) оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген;
(h) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антигенсвязывающий домен, для которого подтверждено, что он образует иммунный комплекс согласно (g) выше; и
(i) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (h) выше.
Кроме того, настоящее изобретение относится к способу получения антигенсвязывающей молекулы, обладающей функцией элиминации антигенов из плазмы, причем способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше; и
(e) выделение антигенсвязывающей молекулы из культурального раствора, полученного согласно (d) выше; и
где способ дополнительно включает контактирование антигенсвязывающей молекулы, полученной способом получения, с антигеном, и оценку образования иммунного комплекса, содержащего антигенсвязывающую молекулу и антиген.
В неограничивающем варианте осуществления настоящего изобретения после выделения полинуклеотида, кодирующего антигенсвязывающий домен, активность связывания которого изменяется в зависимости от условий, выбранных, как описано выше, полинуклеотид встраивают в соответствующий экспрессирующий вектор. Например, когда антигенсвязывающий домен представляет собой вариабельную область антитела, после получения кДНК, кодирующей вариабельную область, кДНК расщепляют ферментами рестрикции, которые распознают участки рестрикции, встроенные на обоих концах кДНК. Предпочтительно, ферменты рестрикции распознают и расщепляют нуклеотидную последовательность, которая встречается с низкой частотой в нуклеотидной последовательности, составляющей ген антигенсвязывающей молекулы. Более того, предпочтительно встраивают ферменты рестрикции, которые обеспечивают липкие концы, чтобы встроить единичную копию расщепленного фрагмента в вектор в правильной ориентации. кДНК, кодирующую вариабельную область антигенсвязывающей молекулы, расщепленную как описано выше, встраивают в соответствующий экспрессирующий вектор с получением экспрессирующего вектора для антигенсвязывающей молекулы по настоящему изобретению. В это время ген, кодирующий константную область (C-область) антитела, можно подвергать слиянию в рамке считывания с геном, кодирующим вариабельную область.
Для получения представляющей интерес антигенсвязывающей молекулы, полинуклеотид, кодирующий антигенсвязывающую молекулу, встраивают так, чтобы он был функционально связан с регуляторной последовательностью, в экспрессирующий вектор. Регуляторные последовательности включают, например, энхансеры и промоторы. Более того, с N-концом может быть связана соответствующая сигнальная последовательность, чтобы экспрессированная антигенсвязывающая молекула секретировалось во внешнюю среду клеток. В качестве сигнальной последовательности используют, например, пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO:3); однако также можно присоединять другие соответствующие сигнальные последовательности. Экспрессированный полипептид расщепляется на С-конце описанной выше последовательности, и расщепленный полипептид секретируется в качестве зрелого полипептида во внешнюю среду клеток. Затем соответствующие клетки-хозяева трансформируются этим экспрессирующим вектором так, чтобы можно было получить рекомбинантные клетки, экспрессирующие полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу. Антигенсвязывающие молекулы по настоящему изобретению можно получать из рекомбинантных клеток посредством способов, описанных выше в разделе об антителах.
Для нуклеиновой кислоты "функционально связанный" означает, что нуклеиновая кислота имеет функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК, кодирующая препоследовательность или секреторную лидерную последовательность, функционально связана с ДНК, кодирующей определенный полипептид, если она подлежит экспрессии в качестве белка-предшественника, вовлеченного в секрецию полипептида. Промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию кодирующей последовательности. Участок связывания рибосомы функционально связан с кодирующей последовательностью, если он находится в положении, которое облегчает трансляцию. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются соседними и в случае секреторной лидерной последовательности, это означает, что связанные последовательности ДНК являются соседними и находятся в рамке считывания. Однако энхансеры не должны быть соседними. Связывание проводят путем лигирования в подходящих участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидые адаптеры или линкеры в соответствии с общепринятой практикой. Более того, связанные нуклеиновые кислоты можно получать упомянутым выше способом перекрывающейся ПЦР.
В неограничивающем варианте осуществления настоящего изобретения после выделения полинуклеотида, кодирующего описанную выше антигенсвязывающую молекулу, активность связывания которой варьирует, в зависимости от выбранных условий, вариант полинуклеотида встраивают в соответствующий экспрессирующий вектор. Такие варианты предпочтительно включают варианты, полученные путем гуманизации, на основе полинуклеотидной последовательности, кодирующей антигенсвязывающую молекулу по настоящему изобретению, полученную путем скрининга в качестве рандомизированной библиотеки вариабельной области сконструированной синтетической библиотеки или иммунной библиотеки, происходящей из не являющихся человеком животных. Те же способы, которые описаны выше для получения описанных выше гуманизированных антител, можно использовать в качестве способа получения гуманизированных вариантов антигенсвязывающей молекулы.
В другом варианте осуществления такие варианты предпочтительно включают варианты, полученные путем внесения изменения, которое увеличивает аффинность к антигену (созревание аффинности) антигенсвязывающей молекулы по настоящему изобретению, в выделенную полинуклеотидную последовательность для молекулы, полученной путем скрининга с использованием синтетической библиотеки или наивной библиотеки в качестве рандомизированной библиотеки вариабельной области. Такие варианты можно получать различными известными способами созревания аффинности, включая мутагенез CDR (Yang et al. (J. Mol. Biol. (1995) 254, 392-403)), шаффлинг цепей (Marks et al. (Bio/Technology (1992) 10, 779-783)), использование мутантных штаммов E. coli (Low et al. (J. Mol. Biol. (1996) 250, 359-368)), шаффлинг ДНК (Patten et al. (Curr. Opin. Biotechnol. (1997) 8, 724-733)), фаговый дисплей (Thompson et al. (J. Mol. Biol. (1996) 256, 77-88)) и половую ПЦР (Clameri et al. (Nature (1998) 391, 288-291)).
Как описано выше, антигенсвязывающие молекулы, которые получают способами получения по настоящему изобретению, включают антигенсвязывающие молекулы, имеющие Fc-область. В качестве Fc-областей можно использовать различные варианты. В одном варианте осуществления варианты по настоящему изобретению предпочтительно включают полинуклеотиды, кодирующие антигенсвязывающие молекулы, имеющие тяжелую цепь, в которой полинуклеотид, кодирующий вариант Fc-области, как описано выше, связан в рамке считывания с полинуклеотидом, кодирующим описанную выше антигенсвязывающую молекулу, активность связывания которой варьирует в зависимости от выбранных условий.
В неограничивающем варианте осуществления настоящего изобретения Fc-области предпочтительно включают, например, константные области Fc антител, таких как IgG1 SEQ ID NO:11 (Ala добавлен на N-конец AAC82527.1), IgG2 SEQ ID NO:12 (Ala добавлен на N-конец AAB59393.1), IgG3 SEQ ID NO:13 (CAA27268.1) и IgG4 SEQ ID NO:14 (Ala добавлен на N-конец AAB59394.1). Время удержания молекул IgG в плазме является относительно длительным (элиминация из плазмы является медленной), поскольку FcRn, в частности FcRn человека, функционирует в качестве рецептора спасения для молекул IgG. Молекулы IgG, включенные в эндосомы путем пиноцитоза, связываются в кислых условиях эндосом с FcRn, в частности с FcRn человека, экспрессируемым в эндосомах. Молекулы IgG, которые не могут связываться с FcRn, в частности FcRn человека, переносятся в лизосомы и деградируются в них. Между тем, молекулы IgG, связанные в FcRn, в частности FcRn человека, переносятся на клеточную поверхность, а затем возвращаются в плазму в результате диссоциации от FcRn, в частности FcRn человека, в нейтральных условиях в плазме.
Поскольку антитела, содержащие типичную Fc-область, не обладают активностью связывания с FcRn, в частности с FcRn человека, в условиях диапазона нейтральных значений pH, типичные антитела и комплексы антитело-антиген включаются в клетки посредством неспецифического эндоцитоза и переносятся на клеточную поверхность путем связывания с FcRn, в частности FcRn человека, в условиях диапазона эндосомальных кислых значений pH. FcRn, в частности FcRn человека, транспортирует антитела из эндосомы на клеточную поверхность. Таким образом, полагают, что некоторые из FcRn, в частности FcRn человека, также присутствуют на клеточной поверхности. Однако антитела рециклируют в плазму, поскольку они диссоциируют от FcRn, в частности FcRn человека, в условиях диапазона нейтральных значений pH на поверхности клетки.
Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, которые включены в антигенсвязывающие молекулы по настоящему изобретению, можно получать любым способом. В частности, Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, можно получать путем изменения аминокислот иммуноглобулина человека IgG-типа в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулина человека IgG-типа включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3 и IgG4, и их вариантов). Аминокислоты в любых положениях можно изменять на другие аминокислоты, при условии, что полученные области обладают активностью связывания FcRn человека в диапазоне нейтральных значений pH или увеличенной активностью связывания FcRn человека в нейтральном диапазоне. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, предпочтительно, чтобы полученная область содержала изменение, которое приводит к эффекту усиления связывания FcRn человека в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые допускают такие изменения, включают, например, аминокислоты в положениях 221-225, 227, 228, 230, 232, 233-241, 243-252, 254-260, 262-272, 274, 276, 278-289, 291-312, 315-320, 324, 325, 327-339, 341, 343, 345, 360, 362, 370, 375-378, 380, 382, 385-387, 389, 396, 414, 416, 423, 424, 426-438, 440 и 442 (указанные согласно нумерации EU). Более конкретно, такие изменения аминокислот включают изменения, приведенные в таблице 5. Изменение этих аминокислот усиливает связывание с FcRn человека Fc-области иммуноглобулина IgG-типа в диапазоне нейтральных значений pH.
Среди описанных выше, для применения в рамках настоящего изобретения выбирают подходящие изменения, которые усиливают связывание FcRn человека в диапазоне нейтральных значений pH. Особенно предпочтительные аминокислоты для таких вариантов Fc-области включают, например, аминокислоты в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (указанные согласно нумерации EU). Активность связывания FcRn человека Fc-области, включенной в антигенсвязывающую молекулу, можно увеличивать в диапазоне нейтральных значений pH путем замены по меньшей мере одной аминокислоты другой аминокислотой.
Особенно предпочтительные изменения в Fc-области включают, например, замены:
Met вместо аминокислоты в положении 237;
Ile вместо аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr вместо аминокислоты в положении 250;
Phe, Trp или Tyr вместо аминокислоты в положении 252;
Thr вместо аминокислоты в положении 254;
Glu вместо аминокислоты в положении 255;
Asp, Asn, Glu или Gln вместо аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val вместо аминокислоты в положении 257;
His вместо аминокислоты в положении 258;
Ala вместо аминокислоты в положении 265;
Ala или Glu вместо аминокислоты в положении 286;
His вместо аминокислоты в положении 289;
Ala вместо аминокислоты в положении 297;
Ala вместо аминокислоты в положении 303;
Ala вместо аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr вместо аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg вместо аминокислоты в положении 309;
Ala, His или Ile вместо аминокислоты в положении 311;
Ala или His вместо аминокислоты в положении 312;
Lys или Arg вместо аминокислоты в положении 314;
Ala, Asp или His вместо аминокислоты в положении 315;
Ala вместо аминокислоты в положении 317;
Val вместо аминокислоты в положении 332;
Leu вместо аминокислоты в положении 334;
His вместо аминокислоты в положении 360;
Ala вместо аминокислоты в положении 376;
Ala вместо аминокислоты в положении 380;
Ala вместо аминокислоты в положении 382;
Ala вместо аминокислоты в положении 384;
Asp или His вместо аминокислоты в положении 385;
Pro вместо аминокислоты а в положении 386;
Glu for вместо аминокислоты в положении 387;
Ala или Ser вместо аминокислоты в положении 389;
Ala вместо аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 428;
Lys вместо аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr вместо аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val вместо аминокислоты в положении 436 в системе нумерации EU. Между тем, количество измененных аминокислот конкретно не ограничено; такие аминокислотные изменения включают единичное изменение аминокислоты и изменение аминокислот в двух или более участков. Комбинации изменений аминокислот в двух или более участков включают, например, комбинации изменений, описанные в таблицах с 5-1 по 5-33.
В дополнение к Fc-области IgG1 (SEQ ID NO:13), IgG2 (SEQ ID NO:14), IgG3 (SEQ ID NO:15) или IgG4 (SEQ ID NO:16) человека, Fc-область с модифицированным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, можно соответствующим образом использовать в качестве Fc-области, включенной в рамках настоящего изобретения. В рамках настоящего изобретения, "Fc-область нативного IgG человека" относится к Fc-области, в которой цепь сахаров, связанная с положением 297 (нумерация EU) Fc-области IgG1, IgG2, IgG3 или IgG4 человека, представленной в SEQ ID NO:13, 14, 15 или 16, представляет собой содержащую фукозу цепь сахаров. Такие Fc-области с модифицированным связыванием FcγR можно получать путем модификации аминокислот Fc-области нативного IgG человека. То, является ли активность связывания Fc-области с модифицированным связыванием FcγR с FcγR более высокой, чем у Fc-области нативного IgG человека, можно определять соответствующим образом с использованием способов, описанных в упомянутом выше разделе, касающемся активности связывания.
В рамках настоящего изобретения "модификация аминокислот" или "аминокислотная модификация" Fc-области включает модификацию до аминокислотной последовательности, которая отличается от исходной Fc-области. Исходная Fc-область может представлять собой любую Fc-область, при условии, что варианты, полученные путем модификации исходной Fc-области, могут связываться с FcRn человека в диапазоне нейтральных значений pH. Более того, также в качестве измененной Fc-области по настоящему изобретению предпочтительно можно использовать Fc-область, модифицированную из исходной Fc-области, которая уже модифицирована. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, что в качестве исходной Fc-области можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или модифицированных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности с их исходной Fc-областью, меньшим чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с системой нумерации EU.
Fc-область с модифицированным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, в которой цепь сахаров, связанная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров, содержащуюся в антигенсвязывающих молекулах по настоящему изобретению, можно получать любым способом. В частности, Fc-область с модифицированным связыванием FcγR можно получать путем модификации аминокислот в иммуноглобулине человека IgG-типа, и использовать в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типа для модификации включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3, IgG4 и их варианты).
Аминокислоты в любых положениях можно модифицировать на другие аминокислоты при условии, что активность связывания с Fcγ-рецептором является более высокой, чем у Fc-области нативного IgG человека. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, она предпочтительно содержит модификацию, которая обеспечивает эффект более высокой активности связывания Fcγ-рецептора, чем у Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров. Такие аминокислотные модификации описаны, например, в международных публикациях, таких как WO2007/024249, WO2007/021841, WO2006/031370, WO2000/042072, WO2004/029207, WO2004/099249, WO2006/105338, WO2007/041635, WO2008/092117, WO2005/070963, WO2006/020114, WO2006/116260 и WO2006/023403.
Примеры таких аминокислот, которые можно модифицировать, включают по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU). Путем модификации этих аминокислот можно получить Fc-области (Fc-области с модифицированным связыванием FcγR) с более высокой активностью связывания с Fcγ-рецептором, чем активность связывания с Fcγ-рецептором у Fc-области нативного IgG человека, в которой цепь сахаров, присоединенная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров.
Примеры особенно предпочтительных модификаций для применения в рамках настоящего изобретения включают по меньшей мере одну или более аминокислотных модификаций в Fc-области, выбранных из группы:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU. Более того, количество аминокислот, подлежащих модификации, конкретно не ограничено, и можно модифицировать аминокислоту только в одном участке или можно модифицировать аминокислоты в двух или более участков. Примеры комбинаций для аминокислотных модификаций в двух или более участков включают комбинации, описанные в таблице 6 (таблицы с 6-1 по 6-3).
Среди Fc-областей, пригодных для применения в рамках настоящего изобретения, пригодным примером Fc-области, которая обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором (т.е. имеющей активность селективного связывания с ингибиторным Fcγ-рецептором), которую используют в качестве неограничивающего варианта осуществления Fc-области со свойством наличия более высокой активности связывания со специфическим Fcγ-рецептором, чем с другими Fcγ-рецепторами (т.е. Fc-область, имеющая селективную активность связывания с Fcγ-рецептором), представляет собой Fc-область с одной или несколькими из следующих модификаций аминокислот (указанных посредством нумерации EU) упомянутой выше Fc-области: аминокислота в положении 238 модифицирована на Asp и аминокислота в положении 328 модифицирована на Glu. Fc-области и модификации, описанные в US2009/0136485, можно соответствующим образом выбирать в качестве Fc-области, имеющей активность селективного связывания с ингибиторным Fcγ-рецептором.
В неограничивающем варианте осуществления настоящего изобретения подходящим примером является Fc-область, в которой одна или более аминокислот, указанных посредством нумерации EU в положениях 238 и 328 в соответствии с нумерацией EU, соответственно модифицированы на Asp или Glu в упомянутой выше Fc-области.
Более того, в неограничивающем варианте осуществления настоящего изобретения подходящими примерами Fc-областей являются Fc-области с заменой Pro на Asp в положении 238 (нумерация EU), и одной или несколькими модификациями, выбранными из Trp в качестве аминокислоты в положении 237, Phe в качестве аминокислоты в положении 237, Val в качестве аминокислоты в положении 267, Gln в качестве аминокислоты в положении 267, Asn в качестве аминокислоты в положении 268, Gly в качестве аминокислоты в положении 271, Leu в качестве аминокислоты в положении 326, Gln в качестве аминокислоты в положении 326, Glu в качестве аминокислоты в положении 326, Met в качестве аминокислоты в положении 326, Asp в качестве аминокислоты в положении 239, Ala в качестве аминокислоты в положении 267, Trp в качестве аминокислоты в положении 234, Tyr в качестве аминокислоты в положении 234, Ala в качестве аминокислоты в положении 237, Asp в качестве аминокислоты в положении 237, Glu в качестве аминокислоты в положении 237, Leu в качестве аминокислоты в положении 237, Met в качестве аминокислоты в положении 237, Tyr в качестве аминокислоты в положении 237, Lys в качестве аминокислоты в положении 330, Arg в качестве аминокислоты в положении 330, Asp в качестве аминокислоты в положении 233, Asp в качестве аминокислоты в положении 268, Glu в качестве аминокислоты в положении 268, Asp в качестве аминокислоты в положении 326, Ser в качестве аминокислоты в положении 326, Thr в качестве аминокислоты в положении 326, Ile в качестве аминокислоты в положении 323, Leu в качестве аминокислоты в положении 323, Met в качестве аминокислоты в положении 323, Asp в качестве аминокислоты в положении 296, Ala в качестве аминокислоты в положении 326, Asn в качестве аминокислоты в положении 326, и Met в качестве аминокислоты в положении 330, в соответствии с нумерацией EU.
Для модификации аминокислот антигенсвязывающих доменов можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР с удлинением. Более того, различные известные способы также можно использовать в качестве способа модификации аминокислот для замены аминокислот на аминокислоты, отличные от природных аминокислот (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, можно соответствующим образом использовать бесклеточную систему трансляции (Clover Direct (Protein Express)), содержащую тРНК, в которых амбер-супрессорная тРНК, которая комплементарна кодону UAG (амбер-кодон), который является стоп-кодоном, связана с неприродной аминокислотой.
В одном варианте осуществления вариантов по настоящему изобретению полинуклеотиды, кодирующие антигенсвязывающие молекулы, которые имеют тяжелую цепь, где полинуклеотид, кодирующий Fc-область, модифицированный так, чтобы он имел мутацию аминокислоты, как описано выше, связанную в рамке считывания с полинуклеотидом, кодирующим описанную выше антигенсвязывающую молекулу, активность связывания которой варьирует, в зависимости от выбранных условий.
Настоящее изобретение относится к способам получения антигенсвязывающих молекул, включающим сбор антигенсвязывающих молекул из культуральных сред клеток, в которые введены векторы, в которых полинуклеотид, кодирующий Fc-область, функционально связан в рамке считывания с полинуклеотидом, кодирующим антигенсвязывающий домен, активность связывания которого варьирует в зависимости от условий концентрации ионов. Более того, настоящее изобретение также относится к способам получения антигенсвязывающих молекул, включающим сбор антигенсвязывающих молекул из культуральных сред клеток, в которые введены векторы, сконструированные путем функционального связывания полинуклеотида, кодирующего антигенсвязывающий домен, активность связывания которого варьирует в зависимости от условий концентрации ионов, с полинуклеотидом, кодирующим Fc-область, который предварительно функционально связан с вектором.
в способах получения по настоящему изобретению после получения "антигенсвязывающего домена, для которого подтверждено, что он образует иммунный комплекс" можно добавлять подходящие модификации к домену, при условии, что обеспечивается образование иммунного комплекса. Более того, также предусматривается способ получения антигенсвязывающей молекулы, который включает сбор антигенсвязывающей молекулы из культурального раствора с клетками, содержащими вектор, который содержит полинуклеотид, кодирующий антигенсвязывающую молекулу, которая обладает тяжелой цепью, в которой полинуклеотид, кодирующий антигенсвязывающий домен, способный образовывать иммунный комплекс и модифицированный таким путем, связан в рамке считывания с полинуклеотидом, кодирующим вариант Fc-области, в который добавлены упомянутые выше аминокислотные мутации.
Фармацевтические композиции
Когда вводят общепринятое нейтрализующее антитело против растворимого антигена, ожидается, что удержание антигена в плазме будет пролонгированным вследствие связывания с антителом. Как правило, антитела имеют длительное время полужизни (от одной недели до трех недель), в то время как время полужизни антигена обычно является коротким (одни сутки или менее). Между тем связанные с антителом антигены имеют значительно более длительное время полужизни в плазме по сравнению с тем, когда антигены присутствуют отдельно. По этой причине введение существующего нейтрализующего антитела приводит к увеличению концентрации антигена в плазме. Такие случаи описаны для различных нейтрализующих антител, которые нацелены на растворимые антигены, включая, например, IL-6 (J. Immunotoxicol. (2005) 3, 131-139), амилоид-бета (mAbs (2010) 2 (5), 1-13), MCP-1 (ARTHRITIS & RHEUMATISM (2006) 54, 2387-2392), гепцидин (AAPS J. (2010) 4, 646-657) и рецептор sIL-6 (Blood (2008) 112 (10), 3959-64). Описано, что введение существующих нейтрализующих антител увеличивает общую концентрацию антигена в плазме приблизительно в 10-1000 раз (уровень увеличения варьирует, в зависимости от антигена) от исходного уровня. В рамках настоящего изобретения общая концентрация антигена в плазме относится к концентрации в качестве общего количества антигена в плазме, т.е. сумме концентраций связанного с антителом и не связанного с антителом антигена. Увеличение общей концентрации антигена в плазме является нежелательным для таких фармацевтических средств на основе антител, которые нацелены на растворимый антиген. Причина этого состоит в том, что концентрация антитела должна быть более высокой, чем по меньшей мере общая концентрация антигена в плазме, для нейтрализации растворимого антигена. В частности, "общая концентрация антигена в плазме увеличивается в 10-1000 раз" означает, что для нейтрализации антигена концентрация антитела в плазме (т.е. доза антитела) должна быть в 10-1000 раз более высокой по сравнению с тем, когда увеличение общей концентрации антигена в плазме не происходит. Напротив, если общая концентрация антигена в плазме может быть снижена в 10-1000 раз по сравнению с существующим нейтрализующим антителом, дозу антитела также можно снижать в аналогичной степени. Таким образом, антитела, способные снижать общую концентрацию антигена в плазме путем элиминации растворимого антигена из плазмы, в высокой степени пригодны по сравнению с существующими нейтрализующими антителами.
Без ограничения конкретной теорией, следующий механизм является примером механизма, который может обеспечивать элиминацию антигенных элементов связывания из плазмы антигенсвязывающей молекулой по настоящему изобретению, которая содержит (i) Fc-область и (ii) антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий концентрации ионов, и где антигенсвязывающая молекула может образовывать иммунный комплекс, содержащий две или более антигенсвязывающих молекул и антигены, имеющие два или более антигенных элементов связывания. Когда антигенный элемент связывания представляет собой один элемент (т.е. гомомономер), такой как в sIL-6R, две молекулы ((т.е. два антигенных элемента связывания) антигенов связываются с одной молекулой антитела, содержащей двухвалентный антигенсвязывающий домен, и образуется комплекс одной молекулы антитела против sIL-6R и двух молекул антигена, содержащих два антигенных элемента связывания. Таким образом, этот тип комплекса антиген-антитело имеет только одну Fc-область (нативную Fc-область IgG1), как показано на фиг. 1. Поскольку этот комплекс связывается с двумя молекулами FcRn или одной молекулой FcγR через одну Fc-область, аффинность в отношении этих рецепторов является такой же, как и аффинность обычного IgG-антитела, и полагают, что захват в клетки происходит по большей части неспецифически.
С другой стороны, когда антигенный элемент связывания представляет собой два элемента, как в IgA человека, который является димером гетерокомплекса тяжелых цепей и легких цепей, существует два элемента эпитопов, с которыми антигенсвязывающие домены будут связываться в антигенном элементе связывания. Однако когда двухвалентное антитело против IgA (т.е. антигенсвязывающие домены, содержащиеся в одной молекуле антитела против IgA связываются с тем же эпитопом) связывается с его антигеном IgA, полагают, что связывание каждого из двухвалентных антигенсвязывающих доменов, содержащихся в одной молекуле антитела против IgA с каждым из двух элементов эпитопов, присутствующих на одной молекуле IgA, является затрудненным ввиду расположения эпитопов. В результате, полагают, что отдельные молекулы антитела против IgA связываются с двумя антигенными элементами связывания, присутствующими в двух молекулах IgA, которые связываются с двухвалентными антигенсвязывающими доменами, присутствующими в одной молекуле антитела против IgA, и, следовательно, образуются комплексы антиген-антитело (иммунные комплексы), содержащие по меньшей мере четыре молекулы (т.е. две молекулы IgA, которые представляют собой молекулу антигена, и две молекулы антитела против IgA, которые являются антигенсвязывающими молекулами).
Когда антигенсвязывающая молекула, такая как антитело, связывающееся с молекулой антигена, который содержит два или более антигенных элементов связывания, образует крупный иммунный комплекс, который является по меньшей мере тетрамером, иммунный комплекс может связываться прочно с авидностью через по меньшей мере две или более поливалентных Fc-области с FcγR, FcRn, рецепторами комплемента и т.п. Таким образом, как показано на фиг. 7, комплекс захватывается в клетки, экспрессирующие эти рецепторы, с более высокой эффективностью, чем нативный IgG1. С другой стороны, поскольку опосредуемая Fc-областью аффинность в отношении этих рецепторов у иммунных комплексов, образованных из молекул антигена и антигенсвязывающих молекул, которые, например, связываются с (мономерными) молекулами антигена, содержащими один антигенный элемент связывания, является недостаточной, как упоминалось выше, иммунные комплексы захватываются по большей неспецифически (менее эффективно по сравнению с захватом, опосредуемым связыванием с авидностью) в клетки, экспрессирующие эти рецепторы, как показано на фиг. 1. Таким образом, захват является в большей степени неэффективным, чем захват, опосредуемый связыванием с авидностью.
Когда антигенсвязывающая молекула, такая как антитело, которое связывается с молекулой антигена, содержащей два или более антигенных элементов связывания, представляет собой антитело, которое содержит антигенсвязывающие домены, у которых активность связывания с антигеном варьирует в зависимости от условий концентрации ионов, как например, в случае pH- или Ca-зависимого связывания, и которые образуют комплекс антиген-антитело (иммунный комплекс), содержащий по меньшей мере четыре молекулы (две молекулы антигена и две молекулы антитела) в плазме, после захвата иммунного комплекса в клетки антигены диссоциируют от антител в эндосомах, где условия концентрации ионов отличаются от условий концентрации ионов в плазме. Таким образом, образованный иммунный комплекс распадается в эндосомах клеток, которые захватили иммунные комплексы. Поскольку диссоциированные антигены не могут связываться с FcRn в эндосомах, они деградируют после перемещения в лизосомы. С другой стороны, полагают, что антитела после диссоциации антигена рециклируют в плазму после связывания с FcRn в эндосомах (фиг. 7).
Как описано выше, если антитело, которое содержит нативную константную область IgG1-типа, против мультимерного антигена, содержащего два или более антигенных элементов связывания, и демонстрирует pH- или Ca-зависимое связывание, может образовывать крупный иммунный комплекс и связывается с FcγR, FcRn, рецепторами комплемента и т.п. с авидностью, полагают, что элиминация антигена отдельно может селективно и в значительной степени ускоряться. Полагают, что, когда вводят GA2-IgG1, которое связывается с IgA человека, образуются такие крупные иммунные комплексы. Действительно, как показано в примере 3, GA2-IgG1-FcγR(-), полученное путем внесения в GA2-IgG1 модификаций, которые ухудшают связывание с FcγR мыши, по существу не могут ускорять элиминацию IgA человека подобно GA2-IgG1 по сравнению с IgA человека отдельно, и они продемонстрировали эквивалентный уровень элиминации с IgA человека отдельно. Таким образом, причиной, по которой GA2-IgG1 могло ускорять элиминацию IgA человека, является то, что иммунный комплекс, содержащий GA2-IgG1 и IgA человека, который является мультимерным антигеном, содержащим два или более антигенных элементов связывания, связывается с авидностью с FcγR и быстро захватывается в клетки, экспрессирующие FcγR. IgA, который диссоциирует от иммунного комплекса в эндосомах клеток, которые захватили иммунный комплекс, деградируется в лизосомах. В то же время, диссоциировавшее от IgA антитело, которое связалось с FcRn в эндосомах, впоследствии рециклирует в плазму и может вновь связываться с IgA в плазме. Полагают, что элиминация IgA в плазме значительно ускоряется таким образом. Способ использования Fc-области с вариантом аминокислоты, которая связывается с FcRn в диапазоне нейтральных значений pH, описан в WO2011/122011 в качестве способа ускорения элиминации антигенов из плазмы. Настоящее изобретение является пригодным в качестве способа ускорения элиминации из плазмы мультимерных антигенов, содержащих два или более антигенных элементов связывания без использования упомянутых выше вариантов, и как показано с использованием GA2-N434W, может далее ускорять элиминацию мультимерных антигенов, содержащих два или более антигенных элементов связывания из плазмы посредством комбинирования с упомянутыми выше вариантами. Более того, мультимерные антигены, содержащие два или более антигенных элементов связывания, могут быть элиминированы, помимо плазмы, из интерстициальной жидкости, синовиальной жидкости, периотнеальной жидкости, плевральной жидкости и перикардиальной жидкости при условии, что клетки, контактирующие с интерстициальной жидкостью, синовиальной жидкостью, перитонеальной жидкостью, плевральной жидкостью или перикардиальной жидкостью экспрессируют FcγR или FcRn. Неограничивающий вариант осуществления таких клеток включает клетки и т.п., присутствующие в интерстициальной жидкости, синовиальной жидкости, перитонеальной жидкости, плевральной жидкости и перикардиальной жидкости.
В частности, настоящее изобретение также относится к фармацевтическим композициям, содержащим антигенсвязывающие молекулы по настоящему изобретению, антигенсвязывающим молекулам, полученным способами изменения по настоящему изобретению, или антигенсвязывающим молекулам, полученным способами получения по настоящему изобретению. Антигенсвязывающие молекулы по настоящему изобретению или антигенсвязывающие молекулы, полученные способами получения по настоящему изобретению, пригодны в качестве фармацевтических композиций, поскольку они при введении обладают мощным эффектом снижения концентрации антигена в плазме по сравнению с типичными антигенсвязывающими молекулами, и проявляют улучшенный иммунный ответ in vivo, фармакокинетику и т.п. у животных, которым вводят молекулы. Фармацевтические композиции по настоящему изобретению могут содержать фармацевтически приемлемые носители.
В рамках настоящего изобретения фармацевтические композиции, как правило, относятся к средствам для лечения или профилактики или исследования и диагностики заболеваний.
Фармацевтические композиции по настоящему изобретению могут быть составлены способами, известными специалистам в данной области. Например, их можно использовать парентерально в форме инъекций стерильных растворов или суспензий, включающих воду или другую фармацевтически приемлемую жидкость. Например, такие композиции могут быть составлены путем смешения в форме единичной дозы, требуемой повсеместно одобренной практикой производства лекарственных средств, путем комбинирования надлежащим образом с фармакологически приемлемыми носителями или средами, в частности, со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, поверхностно-активным веществом, стабилизатором, вкусовой добавкой, эксципиентом, носителем, консервантом, связующим веществом и т.п. В таких составах количество активного ингредиента корректируют так, чтобы получить соответствующее количество в заданном диапазоне.
Стерильные композиции для инъекций могут быть составлены с использованием носителей, таких как дистиллированная вода для инъекций, в соответствии со стандартной практикой составления.
Водные растворы для инъекций включают, например, физиологический солевой раствор и изотонические растворы, содержащие декстрозу или другие адъюванты (например, D-сорбит, D-маннозу, D-маннит и хлорид натрия). Также их можно использовать в комбинации с соответствующими солюбилизаторами, например, спиртами (этанол и т.п.), полиспиртами (пропиленгликоль, полиэтиленгликоль и т.п.), неионными поверхностно-активными веществами (полисорбат 80(TM), HCO-50 и т.п.).
Масла включают кунжутное масло и соевые масла. В комбинации с солюбилизаторами можно использовать бензилбензоат и/или бензиловый спирт. Также можно комбинировать буферы (например, фосфатный буфер натрий-ацетатный буфер), успокаивающие средства (например, прокаина гидрохлорид), стабилизаторы (например, бензиловый спирт и фенол) и/или антиоксиданты. Полученными инъекционными препаратами заполняют подходящие ампулы.
Фармацевтические композиции по настоящему изобретению предпочтительно вводят парентерально. Например, вводят композиции в дозированной форме для инъекций, трансназального введения, введения через легкие или чрескожного введения. Например, их можно вводить системно или локально путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции и т.п.
Способы введения можно надлежащим образом выбирать, учитывая возраст и симптомы пациента. Доза фармацевтической композиции, содержащей антигенсвязывающую молекулу, может составлять, например, от 0,0001 до 1,000 мг/кг для каждого введения. Альтернативно доза может составлять, например, от 0,001 до 100000 мг на пациента. Однако настоящее изобретение не ограничивается числовыми величинами, описанными выше. Дозы и способы введения варьируют, в зависимости от массы тела пациента, возраста, симптомов и т.п. Специалисты в данной области могут установить подходящие дозы и способы введения, учитывая факторы, описанные выше.
Более того, настоящее изобретение относится к наборам для применения в способах по настоящему изобретению, которые содержат по меньшей мере антигенсвязывающую молекулу по настоящему изобретению. В дополнение к описанному выше, в наборы могут быть упакованы фармацевтически приемлемые носители, среды, инструкции, в которых описано применение способа, и т.п.
Более того, настоящее изобретение относится к фармацевтическим средствам для элиминации из плазмы комплексов, содержащих два или более антигенных элементов связывания и две или более антигенсвязывающих молекул, присутствующих в плазме, которые содержат в качестве активного ингредиента антигенсвязывающие молекулы по настоящему изобретению или антигенсвязывающие молекулы, полученные способами получения по настоящему изобретению.
Настоящее изобретение относится к способам лечения заболевания, которые включают введение индивидуумам (пациентам, людям и т.д.) антигенсвязывающих молекул по настоящему изобретению или антигенсвязывающих молекул, полученных способами получения по настоящему изобретению. Неограничивающий пример заболевания включает злокачественную опухоль и воспалительные заболевания.
Настоящее изобретение также относится к применению антигенсвязывающих молекул по настоящему изобретению или антигенсвязывающих молекул, полученных способами получения по настоящему изобретению, для изготовления фармацевтического средства для элиминации из плазмы комплексов, содержащих два или более антигенных элементов связывания и две или более антигенсвязывающие молекулы, присутствующих в плазме.
Кроме того, настоящее изобретение относится к применению антигенсвязывающих молекул по настоящему изобретению или антигенсвязывающих молекул, полученных способами получения по настоящему изобретению, для элиминации из плазмы комплексов, содержащих два или более антигенных элементов связывания и две или более антигенсвязывающих молекул, присутствующих в плазме.
Кроме того, настоящее изобретение относится к антигенсвязывающим молекулам по настоящему изобретению и антигенсвязывающим молекулам, полученным способами получения по настоящему изобретению, для применения в способах по настоящему изобретению.
Аминокислоты, содержащиеся в аминокислотных последовательностях по настоящему изобретению, могут быть посттрансляционно модифицированными (например, модификация N-концевого глутамина в пироглутаминовую кислоту путем пироглутамилирования хорошо известна специалистам в данной области). Естественно, такие посттрансляционно модифицированные аминокислоты включены в аминокислотные последовательности по настоящему изобретению.
Все цитированные в описании документы уровня техники включены в настоящее описание в качестве ссылок.
Примеры
Ниже настоящее изобретение конкретно описано с помощью примеров, однако его не следует считать ограниченным ими.
[Пример 1] Получение антител, которые связываются с IgA человека кальций-зависимым образом
1-1. Получение IgA человека (hIgA)
IgA человека (далее сокращенно обозначаемый как "hIgA") получали в качестве антигена с использованием следующих рекомбинантных способов. hIgA экспрессировали путем культивирования клеток, содержащих рекомбинантные векторы, в которые встроены H (WT)-IgA1 (SEQ ID NO:49) и L (WT) (SEQ ID NO:50), и очищали способом, известным специалистам в данной области, с использованием ионообменной хроматографии и гель-фильтрационной хроматографии.
1-2. Антитела с кальций-зависимым связыванием
H54/L28-IgG1, описанное в международной публикации № WO 2009/125825, представляет собой гуманизированное антитело против рецептора IL-6. Fv4-IgG1 представляет собой гуманизированное антитело против рецептора IL-6, которое получено путем сообщения H54/L28-IgG1 свойства связывания с растворимым рецептором IL-6 человека pH-зависимым образом (т.е. связывания в нейтральных условиях и диссоциации в кислых условиях). Исследование in vivo, описанное в международной публикации № WO 2009/125825 с использованием мышей, продемонстрировало, что элиминация растворимого рецептора IL-6 человека значительно ускорялась в группе, в которой вводили смесь Fv4-IgG1 и растворимого рецептора IL-6 человека в качестве антигена, по сравнению с группой, в которой вводили смесь H54/L28-IgG1 и растворимого рецептора IL-6 человека в качестве антигена.
Растворимый рецептор IL-6 человека, связанный с обычным антителом, которое связывается с растворимым рецептором IL-6 человека, рециклирует в плазму вместе с антителом через FcRn. Между тем, антитело, которое связывается с растворимым рецептором IL-6 человека pH-зависимым образом, диссоциирует в кислых условиях в эндосоме от растворимого рецептора IL-6 человека, который связался с антителом. Диссоциировавший растворимый рецептор IL-6 человека деградируется в лизосоме. Таким образом, это может значительно ускорять элиминацию растворимого рецептора IL-6 человека из плазмы. Более того, антитело, которое связывается с растворимым рецептором IL-6 человека pH-зависимым образом, рециклирует в плазму через FcRn после диссоциации от растворимого рецептора IL-6 человека, так что рециклировавшее антитело вновь может связываться с растворимым рецептором IL-6 человека. Путем повторения этого цикла (антитело, которое связалось с антигенами, захватывается в клетки > антигены диссоциируют от антитела > антигены деградируются и антитело рециклирует обратно в плазму), одна молекула антитела повторно связываться с растворимыми рецепторами IL-6 человека несколько раз (фиг. 1).
Между тем, как описано в международной публикации № WO 2009/125825, H54/L28-IgG1 представляет собой гуманизированное антитело против рецептора IL-6 и Fv4-IgG1 представляет собой гуманизированное антитело против рецептора IL-6, полученное путем сообщения H54/L28-IgG1 свойства связывания с растворимым рецептором IL-6 человека pH-зависимым образом (т.е. связывания в нейтральных условиях и диссоциация в кислых условиях). Fv4-IgG1-v2 представляет собой гуманизированное антитело против рецептора IL-6, в котором связывание FcRn увеличено по сравнению с Fv4-IgG1 в нейтральных условиях. Исследование in vivo, описанное в международной публикации № WO 2011/122011 с использованием мышей, продемонстрировало, что элиминация растворимого рецептора IL-6 человека значительно ускорялась в группе, в которой вводили смесь Fv4-IgG1-v2 и растворимого рецептора IL-6 человека в качестве антигена, по сравнению с группой, в которой вводили смесь Fv4-IgG1 и растворимого рецептора IL-6 человека в качестве антигена. Таким образом, сообщалось, что путем усиления связывания с FcRn в нейтральных условиях (pH 7,4) антитела, которое связывается с антигенами pH-зависимым образом, эффект усиленного модифицированного антитела в отношении многократного связывания с антигенами и эффект ускорения элиминации антигенов из плазмы может быть далее улучшен, и путем введения антитела можно элиминировать антигены из плазмы, фиг. 2.
В действии антител, которые связываются с антигенами pH-зависимым образом, представленных на фиг. 1 и 2, используют свойство антител прочно связываться с антигенами в плазме и диссоциировать от антигенов в эндосомах на основе различий условий в плазме и эндосомах, т.е. различий pH (pH 7,4 в плазме; pH 6,0 в эндосомах). Свойства средовых факторов в плазме и эндосомах, а также степень их различий, являются важными для использования таких различий для антигенсвязывающей способности антитела, которое связывается pH-зависимым образом в плазме и эндосомах. Различие в pH соответствует различию в концентрации протонов. В частности, концентрация протонов в плазме (pH 7,4) составляет приблизительно 40 нМ, в то время как концентрация протонов в эндосоме (pH 6,0) составляет приблизительно 1000 нМ; таким образом, концентрация протонов, которая считается средовым фактором в плазме и эндосоме, отличается приблизительно в 25 раз.
Более того, авторы настоящего изобретения предположили, что для достижения действия, проиллюстрированного на фиг. 1 и 2 с помощью различных вариантов осуществления или для достижения этих вариантов осуществления в комбинации, было бы полезным использовать антитело, которое связывается с антигенами в зависимости от средового фактора с большим различием в плазме и эндосоме, отличным от различий в концентрации протонов. Таким образом, авторы изобретения провели поиск средового фактора, концентрация которого значительно различается в плазме и эндосоме, и в результате обнаружили кальций. Концентрация ионизированного кальция составляет приблизительно от 1,1 мМ до 1,3 мМ в плазме и приблизительно 3 мкМ в эндосоме; таким образом, различие в концентрации ионов кальция, которое считается средовым фактором в плазме и в эндосоме, составляет приблизительно 400 раз, и было обнаружено, что оно превышает различие в концентрации протонов (в 25 раз). Таким образом, было сделано заключение, что с использованием антитела, которое связывается с антигеном в условиях высокой концентрации ионов кальция (от 1,1 мМ до 1,3 мМ) и диссоциирует от антигена в условиях низкой концентрации ионов кальция (3 мкМ), антитело может диссоциировать от антигена в эндосоме на эквивалентном или более высоком уровне по сравнению с антителом, которое связывается с антигеном pH-зависимым образом.
1-3. Экспрессия и очистка антител, которые связываются с hIgA
GA1-IgG1 (тяжелая цепь SEQ ID NO:37; легкая цепь SEQ ID NO:38) и GA2-IgG1 (тяжелая цепь SEQ ID NO:39; легкая цепь SEQ ID NO:40) представляют собой антитела, которые связываются с hIgA. Последовательности ДНК, кодирующие GA1-IgG1 (тяжелая цепь SEQ ID NO:37; легкая цепь SEQ ID NO:38) и GA2-IgG1 (тяжелая цепь SEQ ID NO:39; легкая цепь SEQ ID NO:40) встраивали в экспрессирующие плазмиды клеток животных способом, известным специалистам в данной области. Антитела экспрессировали способом, описанным ниже. Клетки линии, происходящей из клеток почки эмбриона человека, FreeStyle 293-F (Invitrogen) суспендировали в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen). Суспензию клеток высевали в планшет с 6 ячейками (3 мл/ячейка) при плотности клеток 1,33×106 клеток/мл. Затем сконструированные плазмиды вводили в клетки способом липофекции. Клетки культивировали в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об/мин). Антитела очищали из выделенных культуральных супернатантов способом, известным специалистам в данной области, с использованием rProtein A SepharoseTM Fast Flow (Amersham Biosciences). Поглощение (длина волны: 280 нм) растворов очищенного антитела измеряли с использованием спектрофотометра. Концентрации антитела определяли из измеренных величин с использованием коэффициента поглощения, вычисленного способом PACE (Protein Science (1995) 4, 2411-2423).
1-4. Оценка полученных антител в отношении кальций-зависимой активности связывания hIgA
Антитела, выделенные, как описано в 1-3, оценивали в отношении их активности связывания hIgA (константа диссоциации KD (M)) с использованием Biacore T200 (GE Healthcare). Подвижные буферы, использованные для измерения, представляли собой:
0,05% tween 20/20 ммоль/л ACES/150 ммоль/л NaCl (pH 7,4 или 5,8), содержащий 3 мкМ или 1,2 мМ CaCl2; и
0,05% tween 20/20 ммоль/л ACES/150 ммоль/л NaCl (pH 8,0), содержащий 0,1 мкМ или 10 мМ CaCl2.
Антителу позволяли связываться с сенсорным чипом CM5 (GE Healthcare), на котором было иммобилизовано подходящее количество рекомбинантного белка A/G (Thermo Scientific) способом присоединения аминов. Затем соответствующую концентрацию hIgA (описанного в 1-1) инжектировали в качестве анализируемого соединения для обеспечения взаимодействия с антителом на сенсорном чипе. Измерение проводили при 37°C. После измерения инжектировали 10 ммоль/л глицин-HCl (pH 1,5) для регенерации сенсорного чипа. Константу диссоциации KD (M) вычисляли из результата измерения путем анализа с аппроксимацией кривой и анализа параметров равновесия с использованием программного обеспечения Biacore T200 Evaluation Software (GE Healthcare). Результат и полученные сенсограммы представлены в таблице 7 и на фиг. 3, соответственно. Было выявлено, что GA2-IgG1 прочно связывалось с hIgA при концентрации Ca2+ 1,2 мМ, в то время как антитело слабо связывалось с hIgA при концентрации Ca2+ 3 мкМ. Более того, было показано, что при концентрации Ca2+ 1,2 мМ, GA2-IgG1 прочно связывалось с IgA человека при pH 7,4, но слабо при pH 5,8. Более конкретно, было выявлено, что GA2-IgG1 связывается с IgA человека pH- и кальций-зависимым образом.

[Пример 2] Получение модифицированных антител, которые связываются с hIgA кальций-зависимым образом
Затем, чтобы далее усилить элиминацию антигена (hIgA) и плазмы, конструировали GA2-N434W (тяжелая цепь SEQ ID NO:41; легкая цепь SEQ ID NO:40) путем внесения аминокислотной замены N434W в GA2-IgG1, которое связывается с hIgA кальций-зависимым образом, чтобы усилить связывание с FcRn мыши при pH 7,4. Более того, конструировали GA2-FcγR(-) (тяжелая цепь SEQ ID NO:42; легкая цепь SEQ ID NO:40) путем внесения аминокислотных замен L235R и S239K в GA2-IgG1 для устранения аффинности связывания FcγR. Модифицированные антитела экспрессировали способом, описанным выше, с использованием плазмид для экспрессии у животных, в которые были встроены последовательности ДНК, кодирующие GA2-N434W (тяжелая цепь SEQ ID NO:41; легкая цепь SEQ ID NO:40) и GA2-FcγR(-) (тяжелая цепь SEQ ID NO:42; легкая цепь SEQ ID NO:40) способом, известным специалистам в данной области. После очистки определяли концентрации антитела. GA2-FcγR(-) оценивали в отношении его связывания с различными FcγR мыши (mFcγRI, mFcγRII, mFcγRIII и mFcγRIV). Результат показал, что GA2-FcγR(-) не связывалось ни с какими из рецепторов.
[Пример 3] Оценка эффекта Ca-зависимых связывающих hIgA антител на время удержания антигена в плазме с использованием нормальных мышей
3-1. Испытание in vivo с использованием нормальных мышей
Кинетику hIgA и антитела против hIgA in vivo оценивали после введения hIgA (IgA человека; полученный, как описано в примере 1), отдельно или в комбинации с антителом против hIgA нормальным мышам (мыши C57BL/6J; Charles River Japan). Раствор hIgA (80 мкг/мл) или смесь hIgA и антитела против hIgA вводили один раз в сутки в дозе 10 мл/кг через хвостовую вену. Использованные антитела против hIgA представляли собой GA1-IgG1, GA2-IgG1, GA2-N434W и GA2-FcγR(-), описанные выше.
В каждой смеси концентрация hIgA составляла 80 мкг/мл. Между тем концентрация антитела против hIgA варьировала, в зависимости от аффинности антитела в отношении hIgA. GA1-IgG1 приготавливали в концентрации 10 мг/мл; GA2-IgG1 в концентрации 2,69 мг/мл; GA2-N434W в концентрации 1 мг/мл; и GA2-FcγR(-) в концентрации 2,69 мг/мл. Согласно прогнозам, в описанных выше условиях большая часть hIgA связывается с антителом, поскольку антитело против hIgA присутствует достаточно в избытке относительно hIgA. Взятие крови от мышей проводили через пять минут, семь часов, одни сутки, двое суток, трое суток и семь суток после введения. Взятую кровь сразу центрифугировали при 12000 об/мин и 4°C в течение 15 минут для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до измерения.
3-2. Определение концентрации антитела против hIgA в плазме у нормальных мышей с помощью ELISA
Концентрации антитела против hIgA в плазме мышей определяли с помощью ELISA. Сначала приготавливали планшеты с иммобилизованным антителом против IgG человека путем распределения аликвотами F(ab')2-фрагмента антитела против IgG человека (специфичного к γ-цепи) (SIGMA) в каждую лунку планшета Nunc-Immuno, MaxiSorp (Nalge nunc International) и позволяя планшетам стоять при 4°C в течение ночи. Образцы антитела против hIgA для стандартной кривой, приготовленные в качестве стандартных растворов плазмы с концентрациями 0,5, 0,25, 0,125, 0,0625, 0,03125, 0,01563 и 0,07813 мкг/мл, и образцы для анализа, приготовленные путем разбавления образцов плазмы мыши в 100 раза или более, распределяли аликвотами в планшеты с иммобилизованным антителом против IgG человека, а затем планшеты инкубировали при 25°C в течение одного часа. Далее в каждую лунку планшетов распределяли аликвотами конъюгат антитела козы против IgG человека (специфичного к γ-цепи) с биотином (BIOT) (Southern Biotechnology Associates Inc.), а затем планшеты инкубировали при 25°C в течение одного часа. Затем в каждую лунку планшетов добавляли стрептавидин-PolyHRP80 (Stereospecific Detection Technologies), после чего планшеты инкубировали при 25°C в течение одного часа. Хромогенную реакцию с использованием в качестве субстрата TMB One Component HRP Microwell Substrate (BioFX Laboratories) завершали с помощью 1 н серной кислоты (Showa Chemical), а затем проводили измерение поглощения реакционной смеси при 450 нм в каждой лунке с использованием устройства для считывания микропланшетов. Концентрацию антитела против hIgA в плазме мыши вычисляли из поглощения на стандартной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Концентрации с течением времени антител GA1-IgG1, GA2-IgG1, GA2-N434W и GA2-FcγR(-) в плазме нормальных мышей, которые были определены способом, описанным выше, представлены на фиг. 4.
3-3. Определение концентрации hIgA в плазме с помощью ELISA
Концентрации hIgA в плазме мыши измеряли с помощью ELISA. Сначала приготавливали планшеты с иммобилизованным антителом против IgA человека путем распределения аликвотами антитела козы против IgA человека (BETHYL) в каждую лунку планшета Nunc-Immuno Plate, MaxiSorp (Nalge nunc International) и позволяя планшетам стоять при 4°C в течение ночи. Образцы hIgA для стандартной кривой, приготовленные в качестве стандартных растворов плазмы с концентрацией 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 и 0,00625 мкг/мл и образцы для анализа, полученные разбавлением образцов плазмы мыши в 100 разили более, распределяли аликвотами в количестве 100 мкл/лунка в планшеты с иммобилизованным антителом против IgA человека, после чего добавляли 500 нг/мл hsIL-6R в количестве 200 мкл/лунка. Полученным планшетам позволяли стоять при комнатной температуре в течение одного часа. Далее после добавления в каждую лунку планшетов биотинилированного антитела против IL-6R человека (R&D) планшеты инкубировали при комнатной температуре в течение одного часа. Затем после распределения аликвотами стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в каждую лунку планшетов, планшеты инкубировали при комнатной температуре в течение одного часа. Хромогенную реакцию с использованием в качестве субстрата TMB One Component HRP Microwell Substrate (BioFX Laboratories) завершали с помощью 1 н серной кислоты (Showa Chemical), а затем проводили измерение поглощения реакционной смеси при 450 нм в каждой лунке с использованием устройства для считывания микропланшетов. Концентрацию в плазме мыши вычисляли из поглощения стандартной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Концентрации hIgA в плазме у нормальных мышей с течением времени после внутривенного введения, при определении описанным выше способом, представлены на фиг. 5.
Результат показал, что элиминация hIgA была замедленной, когда hIgA вводили в комбинации с GA1-IgG1 - антителом, которое проявляет связывание Ca-зависимое связывание (степень зависимости является низкой), по сравнению с тем, когда hIgA вводили отдельно. Напротив, когда hIgA вводили в комбинации с GA2-IgG1, у которого активность Ca-зависимого связывания является в 100 или более раз большей, элиминация hIgA значительно ускорялась по сравнению с тем, когда hIgA вводили отдельно. Концентрацию свободного hIgA в плазме определяли из концентрации антитела в плазме, представленной на фиг. 4, концентрации hIgA в плазме, представленной на фиг. 5 и величины KD для каждого антитела, представленной в таблице 7. Результат представлен на фиг. 6. Как показано на фиг. 6, концентрация свободного антигена (hIgA) в группе, в которой вводили GA2-IgG1, которое связывается с hIgA кальций-зависимым образом, была более низкой, чем в группе, в которой вводили GA1-IgG1. Это демонстрирует, что уровень свободного от антитела антигена (hIgA) (не связанная с антителом форма) может быть снижен путем ускорения элиминации антигена с использованием антитела, которое связывается кальций-зависимым образом. Более того, GA2-N434W с усиленным связыванием FcRn при pH 7,4 ускоряло элиминацию антигена в большей степени, чем GA2-IgG1, и через семь часов после введения концентрация hIgA в плазме была ниже предела обнаружения.
[Пример 4] Получение pH-зависимого антитела против IgE
4-1. Получение антитела против IgE человека
Для получения pH-зависимых антител против IgE человека, IgE человека (тяжелая цепь SEQ ID NO:43; легкая цепь SEQ ID NO:44) (вариабельная область происходит из антитела против глипикана 3 человека) в качестве антигена экспрессировали с использованием FreeStyle293 (Life Technologies). IgE человека получали путем очистки экспрессированного IgE человека с использованием общепринятого способа хроматографии, известного специалистам в данной области.
Из ряда полученных антител отбирали антитело, которое связывается с IgE человека pH-зависимым образом и образует крупный иммунный комплекс, состоящий из двух или более молекул антитела против IgE и двух или более молекул IgE. Отобранное антитело против IgE человека экспрессировали с использованием константной области тяжелой цепи IgG1 человека и константной области легкой цепи человека, а затем очищали. Полученное антитело было названо клоном 278 (тяжелая цепь SEQ ID NO:45; легкая цепь SEQ ID NO:46).
4-2. Оценка антител против IgE человека в отношении их активности связывания и активности pH-зависимого связывания
Антитела, способные диссоциировать от антигенов в эндосоме, можно получать, не только путем конструирования так, чтобы они связывались с антигенами pH-зависимым образом, но также путем конструирования их так, чтобы они связывались с антигенами Ca-зависимым образом. Таким образом, клон 278 и контрольное Xolair (омализумаб; Novartis), у которого активность связывания IgE не зависит от pH/Ca, оценивали в отношении зависимости их активности связывания IgE человека (hIgE) от pH и от pH/Ca.
Более конкретно, активность связывания hIgE (константа диссоциации KD (M)) клона 278 и Xolair оценивали с использованием Biacore T200 (GE Healthcare). Подвижные буферы, используемые в анализе, представляли собой:
1,2 ммоль/л CaCl2/0,05% tween 20, 20 ммоль/л ACES, 150 ммоль/л NaCl, pH 7,4;
1,2 ммоль/л CaCl2/0,05% tween 20, 20 ммоль/л ACES, 150 ммоль/л NaCl, pH 5,8; и
3 мкмоль/л CaCl2/0,05% tween 20, 20 ммоль/л ACES, 150 ммоль/л NaCl, pH 5,8.
Химически синтезированный пептид, имеющий последовательность, происходящую из белка глипикана 3 человека (SEQ ID NO:47), в котором C-концевой Lys биотинилирован (далее сокращенно обозначаемый как "биотинилированный пептид GPC3") добавляли в подходящем количестве и иммобилизовывали на сенсорном чипе SA (GE Healthcare) на основе аффинности между биотином и стрептавидином. IgE человека иммобилизовывали на чипе путем инжектирования его в соответствующей концентрации так, чтобы он улавливался биотинилированным пептидом GPC3. В качестве анализируемого соединения клон 278 инжективровали в соответствующей концентрации и позволяли ему взаимодействовать с IgE человека на сенсорном чипе. Затем инжектровали 10 ммоль/л глицин-HCl (pH 1,5) для регенерации сенсорного чипа. Взаимодействие всегда измеряли при 37°C. Результат измерения анализировали посредством аппроксимации кривой с использованием программного обеспечения для оценки Biacore T200 Evaluation Software (GE Healthcare) для вычисления константы скорости ассоциации ka (1/Mс) и константы скорости диссоциации kd (1/с). Из описанных выше констант вычисляли константу диссоциации KD (M). Более того, вычисляли соотношение KD для каждого антитела в условиях [pH 5,8, 1,2 мМ Ca]-[pH 7,4, 1,2 мМ Ca] для оценки pH-зависимого связывания, в то время как соотношения KD для каждого антитела в условиях [pH 5,8, 3 мкМ Ca]-[pH 7,4, 1,2 мМ Ca] вычисляли для оценки pH/Ca-зависимого связывания. Результат представлен в таблице 8.
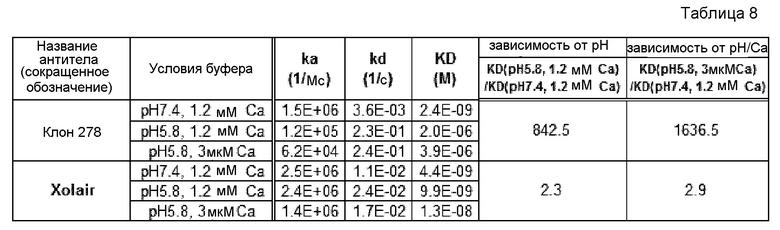
4-3. Оценка образования иммунных комплексов клона 278
То, образует ли клон 278 крупные иммунные комплексы 2:2 или более с IgE человека в нейтральных условиях (pH7,4), и то, диссоциируют ли иммунные комплексы в кислых условиях (pH5,8), оценивали с использованием гель-фильтрации. Клон 278, который был подвергнут диализу против 100 мМ NaCl, разбавляли с использованием буфера для образцов из 20 мМ Tris-HCl, 150 мМ NaCl, 1,2 мМ CaCl2, pH 7,4, в нейтральных условиях и с использованием буфера для образцов из 20 мМ Bis-tris-HCl, 150 мМ NaCl, 3 мкМ CaCl2, pH 5,8, в кислых условиях. Смеси, в которых 100 мкг/мл (0,60 мкМ) hIgE(Asp6), который представляет собой IgE человека (полученный согласно примеру 5), и клон 278 были смешаны в молярном соотношении 1:1 или 1:6, оставляли на два часа или более при комнатной температуре или в автоматическом пробоотборнике при 25°C, а затем анализировали гель-фильтрацией. Подвижную фазу из 20 мМ Tris-HCl, 300 мМ NaCl, 1,2 мМ CaCl2, pH 7,4 использовали в нейтральных условиях, и подвижную фазу из 20 мМ Bis-tris-HCl, 300 мМ NaCl, 3 мкМ CaCl2, pH 5,8 использовали в кислых условиях. Анализы проводили с использованием колонки G4000SWxl (TOSOH) и в условиях скорости потока 0,5 мл/мин и 25°C. Результаты представлены на фиг. 9. Как показано на фиг. 9, было подтверждено, что клон 278 и IgE человека образовывали крупные иммунные комплексы, состоящие из тетрамеров (предполагая, что одна молекула антитела является мономером) или более крупных мультимеров с кажущейся молекулярной массой приблизительно 670 кДа в нейтральных условиях. Более того, такие иммунные комплексы не наблюдали в кислых условиях. Таким образом, было подтверждено, что эти иммунные комплексы диссоциируют pH-зависимым образом аналогично тому, как в описанной выше оценке связывания с использованием Biacore.
Из этих результатов было сделано заключение, что клон 278 был способен ускорять элиминацию IgE человека, аналогично упомянутому выше антителу против IgA GA2-IgG1.
[Пример 5] Оценка in vivo клона 278 и Xolair
5-1. Получение IgE человека (hIgE(Asp6)) для оценки in vivo
hIgE(Asp6) (вариабельная область происходит из антитела против глипикана 3 человека), который представляет собой IgE человека для оценки in vivo, состоящий из тяжелой цепи (SEQ ID NO:48) и легкой цепи (SEQ ID NO:44) получали тем же способом, как описано в примере 1. hIgE(Asp6) представляет собой модифицированную молекулу, поученную путем замены аспарагина на аспарагиновую кислоту в шести участках N-связанного гликозилирования в IgE человека, чтобы на гетерогенность в N-связанной цепи сахаров IgE человека не влияли зависимые от времени изменения концентрации в плазме IgE человека в качестве антигена.
5-2. Оценка клона 278 и Xolair в отношении эффекта ускорения элиминации IgE человека с использованием нормальных мышей
Кинетику hIgE(Asp6) и антитела против IgE человека in vivo оценивали после введения hIgE(Asp6) отдельно или в комбинации с антителом против hIgE (клон 278 и Xolair) мышам C57BL/6J (Charles river Japan). Раствор hIgE(Asp6) (20 мкг/мл) или смесь hIgE(Asp6) и антитела против IgE человека (концентрации представлены в таблице 9) вводили один раз в дозе 10 мл/кг через хвостовую вену. Согласно прогнозам, в описанных выше условиях hIgE(Asp6) связывается практически полностью с антителом, поскольку антитело присутствует достаточно в избытке относительно hIgE(Asp6). Взятие крови от мышей проводили через пять минут, два часа, семь часов, одни сутки, двое суток, четверо или пятеро суток, семь суток, 14 суток, 21 сутки и 28 суток после введения. Взятую кровь сразу центрифугировали при 15000 об/мин и 4°C в течение 5 минут для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°C или ниже до измерения.
5-3. Определение концентрации hIgE(Asp6) в плазме у нормальных мышей
Концентрации hIgA в плазме мыши измеряли с помощью ELISA. Приготавливали образцы для стандартной кривой с концентрацией в плазме 192, 96, 48, 24, 12, 6 и 3 нг/мл. К образцам для стандартной кривой добавляли Xolair (Novartis) в количестве 10 мкг/мл и образцы плазмы мыши для анализа для уравновешивания иммунного комплекса hIgE(Asp6) и антитела против hIgE. После инкубации в течение 30 минут при комнатной температуре, образцы для стандартной кривой и образцы плазмы мыши для анализа распределяли аликвотами в планшеты Immunoplate (MABTECH), на которые было иммобилизовано антитело против IgE человека, или Immunoplate (Nunc F96 MicroWell Plate (Nalge nunc International)), на которые было иммобилизовано антитело против IgE человека (клон 107; MABTECH). Планшетам позволяли стоять при комнатной температуре в течение двух часов или при 4°C в течение ночи. Затем последовательно проводили реакцию с коровым белком GPC3 человека (SEQ ID NO:51), антителом против GPC3, биотинилировнным NHS-PEG4-биотином (Thermo Fisher Scientific) (изготовленным в Chugai pharmaceutical Co., Ltd.), и стрептавидином-PolyHRP80 (Stereospecific Detection Technologies) в каждом случае в течение одного часа. Хромогенную реакцию с использованием в качестве субстрата TMB One Component HRP Microwell Substrate (BioFX Laboratories) завершали 1 н серной кислотой (Showa Chemical), а затем концентрацию в плазме мыши определяли способом, в котором развитие окраски оценивают путем измерения поглощения при 450 нм с использованием устройства для считывания микропланшетов, или способом, в котором проводят люминесцентную реакцию с использованием SuperSignal(r) ELISA Pico Chemiluminescent Substrate (Thermo Fisher Scientific) в качестве субстрата и интенсивность люминесценции измеряют с помощью устройства для считывания микропланшетов. Концентрацию в плазме мыши вычисляли из поглощения или интенсивности люминесценции стандартной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Концентрация (Asp6) в плазме с течением времени после внутривенного введения, которая определена способом, описанным выше, представлена на фиг. 11.
5-4. Определение концентрации антитела против IgE человека в плазме у нормальных мышей
Концентрации антитела против hIgE в плазме мышей определяли с помощью ELISA. Образцы для стандартной кривой приготавливали с концентрации в плазме 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 и 0,00625 мкг/мл. К образцам для стандартной кривой и образцам плазмы мыши для анализа добавляли hIgE(Asp6) в количестве 1 мкг/мл для уравновешивания иммунного комплекса hIgE(Asp6) и антитела против hIgE. После инкубации в течение 30 минут при комнатной температуре образцы для стандартной кривой и образцы плазмы мыши для анализа распределяли аликвотами в Immunoplate (Nunc-Immuno Plate, MaxiSorp (Nalge nunc International)), на которые было иммобилизовано антитело против легкой цепи каппа человека (Bethyl Laboratories). Планшетам позволяли стоять при комнатной температуре в течение двух часов или при 4°C в течение ночи. Затем последовательно проводили реакцию с вторичным антителом против IgG (Fc) человека, конъюгированным с биотином (Pierce Biotechnology) и стрептавидином-PolyHRP80 (Stereospecific Detection Technologies), в каждом случае в течение одного часа. Хромогенную реакцию с использованием в качестве субстрата TMB One Component HRP Microwell Substrate (BioFX Laboratories) завершали 1 н серной кислотой (Showa Chemical), а затем концентрацию в плазме мыши определяли способом, в котором развитие цвета оценивают путем измерения поглощения при 450 нм с помощью устройства для считывания микропланшетов. Концентрацию в плазме мыши вычисляли из поглощения на стандартной кривой с использованием программного обеспечения для анализа SOFTmax PRO (Molecular Devices). Концентрация антитела в плазме с течением времени после внутривенного введения, которая была определена описанным выше способом, представлена на фиг. 10.
Результат показал, что элиминация IgE человека замедлялась, когда IgE человека вводили в комбинации с Xolair, контрольным антителом против IgE, по сравнению с тем, когда IgE человека вводили отдельно. Между тем, элиминация IgE человека значительно ускорялась при введении в комбинации с клоном 278, который имел выраженную активность pH-зависимого связывания IgE человека, по сравнению с тем, когда IgE человека вводили отдельно. В частности, было продемонстрировано, что не только в случае IgA, но также и в случае IgE, элиминация антигена ускорялась путем введения антитела, которое образует крупный иммунный комплекс, по сравнению с тем, когда антиген вводят отдельно.
[Пример 6] Получение вариантов антител, которые демонстрируют кальций-зависимое связывание hIgA
Далее, с целью дальнейшего усиления элиминации антигена (hIgA) из плазмы, Leu в положении 328 (нумерация EU) в GA2-IgG1, которое демонстрирует кальций-зависимое связывание hIgA, заменяли на Tyr для усиления его связывания с FcγR мыши, получив GA2-F1087 (тяжелая цепь SEQ ID NO:52). Последовательность ДНК, кодирующую GA2-F1087 (тяжелая цепь SEQ ID NO:52, и легкая цепь SEQ ID NO:40) встраивали в экспрессирующую плазмиду животных способом, известным специалистам в данной области. Эти варианты антител экспрессировали с использованием плазмиды в соответствии с упомянутым выше способом, и их концентрации определяли после очистки. Антитела, содержащие эту модификацию, продемонстрировали значительно усиленное связывание с FcγR мыши, как показано в справочном примере 5.
[Пример 7] Оценка эффекта на время удержания антигена в плазме у нормальных мышей, которым вводили антитела с Ca-зависимым связыванием hIgA
7-1. Исследование in vivo с использованием нормальных мышей
Нормальным мышам (мышь C57BL/6J; Charles River Japan) вводили hIgA (IgA человека: полученный согласно примеру (1-1)) отдельно или вводили совместно с hIgA и антителом против hIgA, а затем оценивали в отношении динамики hIgA и антитела против hIgA in vivo. Раствор hIgA (80 мкг/мл) или смешанный раствор hIgA и антитела против hIgA вводили один раз в дозе 10 мл/кг в хвостовую вену. Использованные антитела против hIgA представляли собой GA2-IgG1 и GA2-F1087, описанные выше.
Концентрация hIgA составляла 80 мкг/мл во всех случаях и концентрация антитела против hIgA в смешанных растворах составляла 2,69 мг/мл. Антитело против hIgA присутствовало достаточно в избытке относительно hIgA, и, таким образом, практически весь hIgA считался связанным с антителом. В группе введения GA-IgG1 взятие крови проводили у мышей через 5 минут, 7 часов, 1 сутки, 2 суток, 3 суток и 7 суток после введения. В группе введения GA-F1087 взятие крови у мышей проводили через 5 минут, 30 минут, 1 час, 2 часа, 1 сутки, 3 суток и 7 суток после введения. Взятые образцы крови сразу центрифугировали при 4°C и 12000 об/мин в течение 15 минут для получения образцов плазмы. Отделенные образцы плазмы хранили в морозильной камере при -20°C или ниже до измерения.
7-2. Измерение концентрации антитела против hIgA в плазме нормальных мышей способом ELISA
Концентрацию антитела против hIgA в плазме мыши измеряли способом ELISA. Сначала, F(ab’)2-фрагмент антитела против IgG человека (специфичный к γ-цепи) (SIGMA) распределяли в каждую ленку планшетов Nunc-Immuno Plates, MaxiSorp (Nalge nunc International) и позволяли ему стоять в течение ночи при 4°C, получая планшеты с иммобилизованным антителом против IgG человека. Стандартные образцы антител против hIgA для применения в качестве образцов для калибровочной кривой, приготавливали с концентрацией в плазме 0,5, 0,25, 0,125, 0,0625, 0,03125, 0,01563 и 0,007813 мкг/мл; и образцы для анализа плазмы мыши, разбавленные в 100 раз или более, приготавливали и распределяли аликвотами в упомянутые выше планшеты с иммобилизованным антителом против IgG человека, а затем планшеты инкубировали при 25°C в течение одного часа. Затем конъюгат антитела козы против IgG человека (специфичный к γ-цепи) с биотином (BIOT) (Southern Biotechnology Associats Inc.) распределяли в каждую лунку упомянутых выше планшетов, а затем планшеты инкубировали, позволяя реакции протекать при 25°C в течение одного часа. Затем в каждую лунку упомянутых выше планшетов распределяли стрептавидин-PolyHRP80 (Stereospecific Detection Technologies), а затем планшеты инкубировали, позволяя реакции протекать при 25°C в течение одного часа. Развитие окраски осуществляли с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н серной кислотой (Showa Chemical), измеряли поглощение реакционного раствора в каждой лунке при 450 нм на устройстве для считывания микропланшетов. Концентрацию антитела против hIgA в плазме мыши вычисляли на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения в течение времени концентрации в плазме антител GA2-IgG1 и GA2-F1087 у нормальных мышей, определенные этим способом после внутривенного введения, представлены на фиг. 12. Результаты подтвердили, что концентрация антитела в плазме в случае клона GA2-IgG1, которое имеет выраженную pH- и Ca-зависимую активность связывания hIgA, не снижается в значительной степени, даже когда связывание FcγR усиливается.
7-3. Определение концентрации hIgA в плазме с помощью ELISA
Концентрации hIgA в плазме мыши измеряли с помощью ELISA. Сначала антитело козы против IgA человека (BETHYL) распределяли аликвотами в каждую лунку планшета Nunc-Immuno Plate, MaxiSorp (Nalge nunc International). Планшет оставляли стоять при 4°C в течение ночи, получая планшет с иммобилизованным антителом против IgA человека. Стандартные образцы hIgA для применения в качестве образцов для калибровочной кривой, приготавливали при концентрациях в плазме 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 и 0,00625 мкг/мл. Образцы плазмы мыши для анализа приготавливали в разведении в 100-раз или более. 200 мкл hsIL-6R в концентрации 500 нг/мл добавляли к 100 мкл каждого из образцов для калибровочной кривой и образцов плазмы. Полученным смесям позволяли стоять при комнатной температуре в течение одного часа, а затем смешанные растворы распределяли аликвотами в количестве 100 мкл в упомянутый выше планшет с иммобилизованным IgA человека; и их оставляли стоять при комнатной температуре в течение одного часа. Далее, биотинилированное антитело против IL-6R человека (R&D) распределяли аликвотами в каждую лунку упомянутого выше планшета, а затем планшет инкубировали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. Кроме того, стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) распределяли аликвотами в каждую лунку упомянутого выше планшета, и планшет инкубировали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. Развитие окраски осуществляли с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После завершения реакции 1 н серной кислотой (Showa Chemical), измеряли поглощение реакционных растворов в каждой лунке при 450 с использованием устройства для считывания микропланшетов. Концентрации в плазме мыши вычисляли на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентрации hIgA в плазме с течением времени, определенные этим способом у нормальных мышей, которых подвергали внутривенному введению, представлены на фиг. 13.
Результаты показали, что, когда hIgA вводили совместно мышам с GA2-IgG1, которое проявляет в 100 раз или более увеличенную активность Ca-зависимого связывания, элиминация hIgA ускорялась по сравнению с тем, когда hIgA вводили отдельно. Более того, когда hIgA и GA2-F1087, которое имеет усиленное связывание с FcγR, вводили мышам, концентрация hIgA в плазме снижалась ниже диапазона измерения (0,006 мкг/мл или более) через одни сутки после введения, и элиминация hIgA из плазмы была значительно ускорена по сравнению с тем, когда мышам вводили GA-IgG1. Описанные выше результаты указывают на то, что у мышей, которым вводили антитело против hIgA и hIgA, образующие иммунный комплекс, эффект антитела с усиленным связыванием FcγR в отношении удаления антигена (hIgA) из плазмы был усилен по сравнению эффектом удаления антигена (hIgA) антителом, служащим в качестве источника антитела с усиленным связыванием FcγR.
[Пример 8] Получение вариантов антител, которые демонстрируют pH-зависимое связывание IgE человека
Далее, с целью дальнейшего усиления элиминации антигена (IgE человека) из плазмы Leu в положении 328 (нумерация EU) в 278-IgG1, которое демонстрирует pH-зависимое связывание IgE человека, заменяли на Tyr для усиления его связывания с FcγR мыши, с получением 278-F1087 (тяжелая цепь SEQ ID NO:53 и легкая цепь SEQ ID NO:46). Последовательность ДНК, кодирующую 278-F1087, встраивали в экспрессирующую плазмиду животных способом, известным специалистам в данной области. Варианты антител экспрессировали в соответствии упомянутым выше способом с использованием клеток животных, в которые была введения плазмиды, и после очистки определяли их концентрации.
[Пример 9] Оценка 278-IgG1 in vivo
9-1. Получение IgE(hIgE(Asp6)) человека для оценки in vivo
Способ, сходный со способом, описанным в примере 5-1, использовали для получения hIgE (Asp6) (в котором вариабельная область представляет собой вариабельную область антитела против глипикана 3 человека), который представляет собой IgE человека для применения в оценке in vivo. IgE (Asp6) человека представляет собой молекулу, в которой аспарагин модифицирован на аспарагиновую кислоту в шести участках N-гликозилирования IgE человека, так чтобы на гетерогенность N-гликозидов в IgE человека не влияло изменение концентрации в плазме антигена IgE человека.
9-2. Подтверждение эффекта ускорения удаления hIgE человека из плазмы нормальной мыши, которой вводили клон 278
В примере 7 продемонстрировано, что концентрация антигена в плазме была значительно снижена у мышей, которым вводили молекулу с усиленным связыванием с FcγR мыши через pH-зависимое связывание с антигеном IgA человека. Когда связывание FcγR мыши усилено, для дальнейшего подтверждения того, что эффект элиминации растворимого антигена из плазмы может аналогичным образом наблюдаться в организме, в который вводили антитело, которое демонстрирует усиленное связывание с FcγR мыши и связывается pH-зависимым образом с антигенами, отличными от IgA человека, также проводили испытания с использованием антител против антигена IgE человека.
Мышам C57BL/6J (Charles River Japan) вводили hIgE (Asp6) отдельно или совместно вводили hIgE (Asp6) и антитело против hIgE (278-IgG1 или 278-F1087), а затем проводили оценку в отношении динамики hIgE (Asp6) и антитела против IgE человека in vivo. Раствор hIgE (Asp6) (20 мкг/мл) или смешанный раствор hIgE (Asp6) и антитела против IgE человека (все концентрации антитела доводили до концентраций, показанных в таблице 10) вводили один раз в дозе 10 мл/кг в хвостовую вену. В этом случае, поскольку каждое антитело присутствовало достаточно в избытке относительно hIgE (Asp6), считалось, что практически весь hIgE (Asp6) связался с антителом. Взятие крови от мышей проводили через 5 минут, 2 часа, 7 часов, 1 сутки, 2 суток, 4 суток, 5 суток, 7 суток, 14 суток и 21 сутки после введения в группе введения клона 278 (278-IgG1). Взятие крови проводили от мышей через 5 минут, 30 минут, 1 час, 2 часа, 1 сутки, 3 суток, 7 суток, 14 суток и 21 сутки после введения в группе введения 278-F1087. Собранные образцы крови сразу центрифугировали при 4°C и 15000 об/мин в течение 5 минут, получая образцы плазмы. Отделенные образцы плазмы хранили в морозильной камере, установленной на -20°C ниже до проведения анализа.
9-3. Определение концентрации антитела против IgE человека в плазме нормальных мышей
Концентрацию антитела против hIgE определяли с помощью ELISA. Образцы для калибровочной кривой приготавливали с концентрацией в плазме 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 и 0,00625 мкг/мл. Чтобы сделать иммунный комплекс, образовавшийся между hIgE(Asp6) и антителом против hIgE гомогенным, hIgE(Asp6) добавляли в количестве 1 мкг/мл к образцам для калибровочной кривой и образцам плазмы мыши для анализа; и образцы группы введения 278-hIgG1 и соответствующие образцы для калибровочной кривой оставляли стоять при комнатной температуре в течение 30 минут. Более того, образцы группы введения 278-F1087 и соответствующие образцы для калибровочной кривой перемешивали в течение ночи при 37°C. Образцы для калибровочной кривой и образцы плазмы мыши для анализа, которые оставляли стоять или перемешивали, распределяли аликвотами в планшет (Nunc-Immuno Plate, MaxiSorp (Nalge nunc International)) с иммобилизованным антителом против легкой цепи каппа человека (Bethyl Laboratories) и оставляли стоять/перемешиваться при комнатной температуре в течение двух часов (образцы группы введения 278-F1087 и образцы для калибровочной кривой 278-F1087) или оставляли стоять в течение ночи при 4°C (образцы группы введения 278-hIgG1 и образцы 278-hIgG1 для калибровочной кривой). Затем последовательно проводили реакцию с каждым из вторичного антитела кролика против IgG (Fc) человека, конъюгированного с биотином (Pierce Biotechnology) и стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в течение одного часа. Развитие окраски осуществляли с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н серной кислотой (Showa Chemical) концентрацию в плазме мыши определяли по развитию окраски путем измерения поглощения при 450 нм на устройстве для считывания планшета. Концентрации в плазме мыши вычисляли на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентраций антител в плазме с течением времени после внутривенного введения, определенные этим способом, представлены на фиг. 14. Результаты подтвердили, что у мышей, которым водили вариант, полученный путем усиления связывания FcγR у 278-IgG1, которое обладает выраженной активностью pH-зависимого связывания с IgE человека, концентрации антитела в сыворотке этих мышей снижались незначительно, даже когда их сравнивали с концентрациями 278-IgG1.
9-4. Определение концентрации hIgE(Asp6) в плазме нормальной мыши
Концентрацию hIgE(Asp6) в плазме мыши определяли способом ELISA. Образцы для калибровочной кривой приготавливали с концентрацией в плазме 192, 96, 48, 24, 12, 6 и 3 нг/мл. Чтобы сделать иммунный комплекс, образованный между hIgE(Asp6) и антителом против hIgE, гомогенным, к образцам для калибровочной кривой и образцам плазмы мыши для анализа для группы введения 278-hIgG1 добавляли Xolair (Novartis) в количестве 10 мкг/мл, и образцы оставляли стоять при комнатной температуре в течение 30 минут. В группе введения 278-F1087 добавляли 278-F1022 (тяжелая цепь SEQ ID NO:54 и легкая цепь SEQ ID NO:46, полученное аналогично примеру 8) или 278-F760 (тяжелая цепь SEQ ID NO:55 и легкая цепь SEQ ID NO:46, полученное аналогично примеру 8) в количестве 20 мкг/мл, а затем перемешивали при 37°C в течение 60 часов. Образцы плазмы мыши для анализа распределяли аликвотами в Immunoplate с иммобилизованным антителом против IgE человека (MABTECH) или Immunoplate с иммобилизованным антителом против IgE человека (клон 107, MABTECH) (Nunc F96 MicroWell Plate (Nalge nunc International)); и образцы оставляли стоять или перемешивали при комнатной температуре в течение двух часов, или их оставляли стоять в течение ночи при 4°C. Затем последовательно проводили реакцию с коровым белком GPC3 человека (SEQ ID NO:51), антителом против GPC3 (полученным в собственной лаборатории), биотинилированным NHS-PEG4-биотином (Thermo Fisher Scientific), и стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в течение одного часа. Концентрацию в плазме мыши определяли способом, состоящим в реакции развития окраски с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата, остановке реакции 1 н серной кислотой (Showa Chemical), а затем измерении развития окраски по поглощению при 450 нм на устройстве для считывания микропланшетов; или способом, состоящим в осуществлении реакции развития окраски с использованием хемилюминесцентного субстрата SuperSignal(r) ELISA Pico Chemiluminescent Substrate (Thermo Fisher Scientific) в качестве субстрата, а затем измерении интенсивности люминесценции на устройстве для считывания микропланшетов. Концентрации в плазме мыши вычисляли из интенсивности люминесценции или на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентрации hIgE(Asp6) в плазме с течением времени после внутривенного введения, определенные этим способом, представлены на фиг. 15.
В результате, в противоположность элиминации в случае IgE человека отдельно, элиминация IgE человека была ускорена у мышей, которым вводили IgE человека и 278-IgG1, которое обладает выраженной активностью pH-зависимого связывания, по сравнению с IgE человека отдельно. Более того, элиминация IgE человека значительно ускорялась у мышей, которым вводили IgE человека и 278-F1087, которое получено путем усиления связывания 278-IgG1 с FcγR, по сравнению с мышами, которым вводили IgE человека отдельно, и мышами, которым совместно вводили IgE человека и 278-IgG1. Следовательно, было показано, что элиминация антигена ускоряется не только у мышей, которым вводили антитела против IgA с усиленным связыванием FcγR, рассматриваемые до настоящего времени, но также и у мышей, которым вводили антитела против IgE с усиленным связыванием FcγR. Указанные выше результаты показали, что элиминация антигена может быть далее ускорена путем усиления связывания FcγR в каждой из образующих иммунный комплекс пары hIgA-антитело против hIgA и пары hIgE-антитело против hIgE.
[Пример 10] Получение вариантов антител, которые демонстрируют кальций-зависимое связывание hIgA
Далее, с целью усиления элиминации антигена (hIgA) из плазмы, получали варианты с усиленным связыванием с FcRn мыши из GA2-IgG1, которое демонстрирует кальций-зависимое связывание hIgA. Сначала с целью снижения связывания Fc-области с FcγR получали GA2-F760 (тяжелая цепь SEQ ID NO:57) путем замены Leu на Arg в положении 235 и Ser на Lys в положении 239, как указано в соответствии с нумерацией EU в GA2-IgG1. Более того, GA2-F1331, вариант, демонстрирующий более прочное связывание с FcRn при pH 7,4, чем у GA2-F760, получали путем внесения следующих замен в GA2-F760: Gly на Arg в положении 236, Met на Tyr в положении 252, Ser на Thr в положении 254, Thr на Glu в положении 256, Asn на Tyr в положении 434, Tyr на Val в положении 436, Gln на Arg в положении 438, и Ser на Glu в положении 440, как указано в соответствии с нумерацией EU. Последовательности ДНК, кодирующие GA2-F760 (тяжелая цепь SEQ ID NO:57, и легкая цепь SEQ ID NO:40) и GA2-F1331 (тяжелая цепь SEQ ID NO:56, и легкая цепь SEQ ID NO:40) встраивали в экспрессирующие плазмиды для животных способом, известным специалистам в данной области. Эти плазмиды использовали для экспрессии способом, описанным выше, и после очистки определяли концентрации этих вариантов антител. Определяли активность связывания GA2-F760 с FcγR мыши (mFcγRI, mFcγRII, mFcγRIII и mFcγRIV). В результате GA2-F760 не демонстрировало значительного связывания с FcγR мыши.
[Пример 11] Оценка эффекта на время удержания антигена в плазме у мышей с трансгенным FcRn человека, которым вводили антитела с Ca-зависимым связыванием hIgA
11-1. Исследование in vivo с использованием мышей с трансгенным FcRn человека
Мышам с трансгенным FcRn человека (мышь B6.mFcRn-/-.hFcRn Tg line 32 +/+, Jackson Laboratories; Methods Mol Biol. (2010) 602, 93-104) вводили hIgA (IgA человека: полученный согласно примеру (1-1)) отдельно или совместно вводили hIgA и антитело против hIgA; а затем оценивали динамику hIgA и антитела против hIgA in vivo. Раствор hIgA (80 мкг/мл) или смешанный раствор hIgA и антитела против hIgA вводили один раз в дозе 10 мл/кг в хвостовую вену. В качестве антитела против hIgA для введения использовали любое из упомянутых выше GA2-IgG1, GA2-F760 и GA2-F1331.
Концентрация hIgA в смешанном растворе во всех случаях составляла 80 мкг/мл, и концентрация антитела против hIgA составляла 2,69 мг/мл. Поскольку антитело против hIgA присутствовало достаточно в избытке относительно hIgA, считалось, что большая часть hIgA была связана с антителом. Взятие крови от мышей проводили через 15 минут, 1 час, 2 часа, 7 часов, 1 сутки, 3 суток, 7 суток и 14 суток после введения антитела. Взятые образцы крови сразу центрифугировали при 4°C и 12000 об./мин. в течение 15 минут для получения образцов плазмы. Отделенные образцы плазмы хранили в морозильной камере при -20°C или ниже до измерения.
11-2. Определение способом ELISA концентрация антитела против hIgA в плазме мышей с трансгенным FcRn
Концентрацию антитела против hIgA в плазме мыши измеряли способом ELISA. Сначала, F(ab’)2-фрагмент антитела против IgG человека (специфичный к γ-цепи) (SIGMA) распределяли в каждую ленку планшетов Nunc-Immuno Plates, MaxiSorp (Nalge nunc International) и позволяли ему стоять в течение ночи при 4°C, получая планшеты с иммобилизованным антителом против IgG человека. Стандартные образцы антител против hIgA для применения в качестве образцов для калибровочной кривой, приготавливали с концентрацией в плазме 0,5, 0,25, 0,125, 0,0625, 0,03125, 0,01563 и 0,007813 мкг/мл; и образцы для анализа плазмы мыши, разбавленные в 100 раз или более, приготавливали и распределяли аликвотами в упомянутые выше планшеты с иммобилизованным антителом против IgG человека, а затем планшеты инкубировали при 25°C в течение одного часа. Затем конъюгат антитела козы против IgG человека (специфичный к γ-цепи) с биотином (BIOT) (Southern Biotechnology Associats Inc.) распределяли в каждую лунку упомянутых выше планшетов, а затем планшеты инкубировали, позволяя реакции протекать при 25°C в течение одного часа. Затем в каждую лунку упомянутых выше планшетов распределяли стрептавидин-PolyHRP80 (Stereospecific Detection Technologies), а затем планшеты инкубировали, позволяя реакции протекать при 25°C в течение одного часа. Развитие окраски осуществляли с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н серной кислотой (Showa Chemical) измеряли поглощение реакционного раствора в каждой лунке при 450 нм на устройстве для считывания микропланшетов. Концентрацию антитела против hIgA в плазме мыши вычисляли на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения в течение времени концентрации в плазме антител GA2-F1331 и GA2-F760 с трансгенным FcRn человека, определенные этим способом после внутривенного введения, представлены на фиг. 16.
11-3. Определение концентрации hIgA в плазме способом ELISA
Концентрации hIgA в плазме мыши измеряли с помощью ELISA. Сначала антитело козы против IgA человека (BETHYL) распределяли аликвотами в каждую лунку планшета Nunc-Immuno Plate, MaxiSorp (Nalge nunc International). Планшет оставляли стоять в течение ночи при 4°C, получая планшет с иммобилизованным антителом против IgA человека. Стандартные образцы hIgA для применения в качестве образцов для калибровочной кривой, приготавливали при концентрациях в плазме 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 и 0,00625 мкг/мл. Образцы плазмы мыши для анализа приготавливали в разведении в 100-раз или более. 200 мкл hsIL-6R в концентрации 500 нг/мл добавляли к 100 мкл каждого из образцов для калибровочной кривой и образцов плазмы. Полученным смесям позволяли стоять при комнатной температуре в течение одного часа, а затем смешанные растворы распределяли аликвотами в количестве 100 мкл в упомянутый выше планшет с иммобилизованным IgA человека; и их оставляли стоять при комнатной температуре в течение одного часа. Далее, биотинилированное антитело против IL-6R человека (R&D) распределяли аликвотами в каждую лунку упомянутого выше планшета, а затем планшет инкубировали при к.т. в течение одного часа, чтобы позволить реакции протекать. Кроме того, стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) распределяли аликвотами в каждую лунку упомянутого выше планшета, и планшет инкубировали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. Развитие окраски осуществляли с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После завершения реакции 1 н серной кислотой (Showa Chemical), измеряли поглощение реакционных растворов в каждой лунке при 450 с использованием устройства для считывания микропланшетов. Концентрации в плазме мыши вычисляли на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения в плазме концентрации hIgA с течением времени у мышей с трансгенным FcRn человека, определенные этим способом, после внутривенного введения, представлены на фиг. 17.
Результаты показали, что элиминация hIgA из плазмы была значительно ускорена у мышей, которым совместно вводили hIgA и GA2-F1331, которое обладает усиленным связыванием FcRn, по сравнению с элиминацией hIgA из плазмы у мышей, которым вводили hIgA и GA2-F760, которое обладает низкой активностью связывания FcRn человека.
[Пример 12] Получение вариантов антител, которые демонстрируют pH-зависимое связывание IgE человека
Далее, с целью усиления элиминации антигена (IgE человека) из плазмы получали варианты с усиленным связыванием с FcRn мыши из 278-IgG1, которое демонстрирует pH-зависимое связывание IgE человека. Сначала, с целью снижения связывания FcγR мыши, получили 278-F760 (SEQ ID NO:55) путем замены Leu на Arg в положении 235 и Ser на Lys в положении 239, как указано в соответствии с нумерацией EU, в 278-IgG1. Более того, 278-F1331 - вариант, демонстрирующий более прочное связывание FcRn при pH 7,4, чем 278-F760, получали путем внесения следующих замен в 278-F760: Gly на Arg в положении 236, Met на Tyr в положении 252, Ser на Thr в положении 254, Thr на Glu в положении 256, Asn на Tyr в положении 434, Tyr на Val в положении 436, Gln на Arg в положении 438, и Ser на Glu в положении 440, как указано в соответствии с нумерацией EU. Последовательность ДНК, кодирующую 278-F1331 (тяжелая цепь SEQ ID NO:58, и легкая цепь SEQ ID NO:46) или 278-F760 (тяжелая цепь SEQ ID NO:55, и легкая цепь SEQ ID NO:46) встраивали в экспрессирующую плазмиду животных способом, известным специалистам в данной области. Эти варианты антител экспрессировали упомянутым выше способом с использованием клеток животных, в которые была введена плазмида, и после очистки определяли их концентрации.
[Пример 13] Оценка эффекта на время удержания антигена в плазме у мышей с трансгенным FcRn человека, которым вводили антитело с pH-зависимым связыванием hIgE
13-1. Исследование in vivo с использованием трансгенных мышей с FcRn человека
Мышам с трансгенным FcRn человека (мышь B6.mFcRn-/-.hFcRn Tg line 32 +/+, Jackson Laboratories; Methods Mol Biol. (2010) 602, 93-104) совместно вводили hIgE(Asp6) (IgE(Asp6) человека: полученный согласно примеру (5-1)), антитело против hIgE (278-F760 или 278-F1331), и Sanglopor (нормальный иммуноглобулин человека, CSL Behring); а затем оценивали динамику hIgE(Asp6) и антитела против hIgE in vivo. Смешанный раствор hIgE(Asp6), антитела против hIgE и Sanglopor (концентрации представлены в таблице 11) вводили один раз в дозе 10 мл/кг в хвостовую вену. В качестве антитела против hIgE для введения использовали упомянутые выше 278-F760 или 278-F1331.
Поскольку антитело против hIgE присутствовало достаточно в избытке относительно hIgE(Asp6), большую часть hIgE(Asp6) считали связанной антителом. Взятие крови от мышей проводили через 5 минут, 2 часа, 7 часов, 1 сутки, 2 суток, 4 суток, 5 суток, 7 суток, 14 суток, 21 сутки и 28 суток после введения антитела. Взятые образцы крови сразу центрифугировали при 4°C и 12000 об/мин в течение 15 минут с получением образцов плазмы. Отделенные образцы плазмы хранили в морозильной камере при -20°C или ниже до измерения.
13-2. Определение концентрации антитела против hIgE в плазме мышей с трансгенным FcRn человека способом ELISA
Концентрацию антитела против hIgE в плазме мыши определяли с помощью анализа электрохемилюминесценции (ECL). Стандартные образцы антител против hIgE для применения в качестве образцов для калибровочной кривой, приготавливали с концентрацией в плазме 32, 16, 8, 4, 2, 1, 0,5 и 0,25 мкг/мл. Каждый из образцов для калибровочной кривой и образцов плазмы мыши распределяли аликвотами в каждую лунку планшета для ECL с иммобилизованным hIgE(Asp6), а затем планшет инкубировали при 4°C в течение одного часа/в течение ночи, чтобы позволить реакции протекать. Затем антитело козы против IgG человека (специфичное к γ-цепи), конъюгированное с биотином (BIOT) (Southern Biotechnology Associates Inc.) распределяли в каждую лунку упомянутого выше планшета, а затем планшет инкубировали при 25°C в течение одного часа, чтобы позволить реакции протекать. Более того, в каждую лунку упомянутого выше планшета распределяли стрептавидин-PolyHRP80 (Stereospecific Detection Technologies), а затем планшет инкубировали при 25°C в течение одного часа, чтобы позволить реакции протекать. Далее каждый реакционный раствор в планшете подвергали реакции с SULFO-меченым антителом козы против антител кролика (Meso Scale Discovery) при комнатной температуре в течение одного часа. Наконец, в каждый реакционный раствор распределяли буфер для считывания T (x4) (Meso Scale Discovery), и сразу после этого поводили измерение люминесценции реакционного раствора с использованием устройства для считывания Sector Imager 2400 Reader (Meso Scale Discovery). Концентрацию антитела против hIgE в плазме мыши вычисляли на основе ответа из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентраций в плазме антител 278-F1331 и 278-F760 с течением времени у мышей с трансгенным FcRn человека, определенные этим способом, после внутривенного введения, представлены на фиг. 18.
13-3. Определение концентрации hIgE(Asp6) в плазме у мышей с трансгенным FcRn человека
Концентрацию hIgE(Asp6) в плазме мыши определяли способом ELISA. Образцы для калибровочной кривой приготавливали с концентрацией в плазме 192, 96, 48, 24, 12, 6 и 3 нг/мл. Чтобы сделать иммунный комплекс, образованный между hIgE(Asp6) и антителом против hIgE, гомогенным, образцы для калибровочной кривой и образцы плазмы мыши для анализа для группы введения 278-hIgG1 приготавливали путем добавления Xolair (Novartis) в количестве 10 мкг/мл, и образцы оставляли стоять при комнатной температуре в течение 30 минут. Образцы плазмы мыши для анализа распределяли аликвотами в Immunoplate с иммобилизованным антителом против IgE человека (MABTECH) или Immunoplate с иммобилизованным антителом против IgE человека (клон 107, MABTECH) (Nunc F96 MicroWell Plate (Nalge nunc International)); и образцы оставляли стоять или перемешивали при комнатной температуре в течение двух часов, или их оставляли стоять в течение ночи при 4°C. Затем последовательно проводили реакцию с коровым белком GPC3 человека (SEQ ID NO:51), антителом против GPC3 (полученным в собственной лаборатории), биотинилированным NHS-PEG4-биотином (Thermo Fisher Scientific), и стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в течение одного часа. Концентрацию в плазме мыши определяли способом, состоящим в реакции развития окраски с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата, остановке реакции 1 н серной кислотой (Showa Chemical), а затем измерении развития окраски по поглощению при 450 нм на устройстве для считывания микропланшетов; или способом, состоящим в осуществлении реакции развития окраски с использованием хемилюминесцентного субстрата SuperSignal(r) ELISA Pico Chemiluminescent Substrate (Thermo Fisher Scientific) в качестве субстрата, а затем измерении интенсивности люминесценции на устройстве для считывания микропланшетов. Концентрации в плазме мыши вычисляли из интенсивности люминесценции или на основе поглощения из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентрации hIgE(Asp6) в плазме с течением времени после внутривенного введения, определенные этим способом, представлены на фиг. 19.
Результаты показали, что элиминация hIgE из плазмы мыши была значительно ускорена у мышей, которым совместно вводили hIgE и 278-F1331, которое обладает усиленным связыванием FcRn человека, по сравнению с элиминацией hIgE из плазмы у мышей, которым совместно вводили hIgE и 278-F760, которое обладает низкой активностью связывания FcγR мыши. Следовательно, было показано, что элиминация антигена была ускоренной не только у мышей, которым вводили антитела против IgE, обладающие усиленным связыванием FcRn, рассматриваемые до настоящего момента, но также у мышей, которым вводили антитела против IgE, обладающие усиленным связыванием FcRn. Упомянутые выше результаты показали, что элиминация антигена может быть дополнительно ускорена путем усиления связывания FcRn в каждой из образующей иммунный комплекс пары hIgA-антитело против hIgA и пары hIgE-антитело против hIgE.
[Пример 14] Оценка эффекта на время удержания в плазме антигена у нормальных мышей, которым вводили два типа антител против рецептора IL6 человека
14-1. Получение двух типов антител против IL6R
Fv4-IgG1 (тяжелая цепь SEQ ID NO:59 и легкая цепь SEQ ID NO:60) представляет собой антитело против рецептора IL6 человека, обладающее свойством связывания с IL6R человека pH-зависимым образом (связывается в нейтральных условиях и диссоциирует в кислых условиях), как описано в WO 2011/122011. PHX-IgG1 (тяжелая цепь SEQ ID NO:61 и легкая цепь SEQ ID NO:62) представляет собой связывающее IL6R человека антитело. Последовательность ДНК, кодирующую Fv4-IgG1 (тяжела цепь SEQ ID NO:59 и легкая цепь SEQ ID NO:60), PHX-IgG1 (тяжелая цепь SEQ ID NO:61 и легкая цепь SEQ ID NO:62) или PHX-F29 (тяжелая цепь SEQ ID NO:63 и легкая цепь SEQ ID NO:62), полученную путем внесения аминокислотных модификаций в константную область тяжелой цепи PHX-IgG1, встраивали экспрессирующую плазмиду животных способом, известным специалистам в данной области. Эти варианты антитела экспрессировали упомянутым выше способом (описанным в примере 1) с использованием плазмид, и после очистки определяли их концентрации.
14-2. Оценка связывания PHX-IgG1 с рецептором IL6 человека
Взаимодействие между IL-6R и PHX-IgG1, полученным согласно 14-1, анализировали с использованием Biacore T200 (GE Healthcare) для вычисления константы диссоциации (KD). В качестве буфера для анализа взаимодействий использовали буфер из 10 мМ ACES, 150 мМ NaCl, и 0,05% Tween 20, pH 7,4, при 37°C. Белок A/G (Thermo Scientific) иммобилизовывали на сенсорном чипе CM4 Series S (GE Healthcare) способом присоединения аминов, и чипу позволяли улавливать представляющее интерес антитело. Затем подвижный буфер и IL-6R, который был разбавлен с использованием подвижного буфера до 800, 400, 200, 100, 50, 25 и 12,5 нМ, пропускали над чипом с уловленным антителом при скорости потока 2 мкл/мин, чтобы позволить взаимодействию протекать в течение 15 минут. Антитела, уловленные на чипах, промывали путем реакции с 10 мМ глицин-HCl при pH 1,5; и промытые чипы регенерировали и повторно использовали для анализов взаимодействия.
Константу диссоциации KD (моль/л) PHX-IgG1 в отношении IL-6R вычисляли путем проведения стационарного анализа аффинности на сенсограммах, полученных в результате измерения Biacore, с использованием программного обеспечения Biacore Evaluation Software. Константа диссоциации (KD) PHX-IgG1 в отношении IL-6R при pH 7,4, вычисленная этим способом, составила 1,4 E-7 (M).
Далее оценивали зависимость от pH связывания PHX-IgG1 с hIL-6R с использованием Biacore T100. В качестве подвижных буферов для определения связывания PHX-IgG1 с hIL-6R использовали буфер 10 мМ ACES, 150 мМ NaCl, 0,05% Tween 20, pH 7,4, и буфер 10 мМ ACES, 150 мМ NaCl, 0,05% Tween 20, pH 6,0, при 37°C в условиях pH 7,4 и pH6,0, соответственно. Белок A/G (Thermo Scientific) иммобилизовывали на сенсорном чипе CM4 Series S (GE Healthcare) способом присоединения аминов, и чипу позволяли улавливать представляющее интерес антитело. Затем подвижному буферу и IL-6R, который был разбавлен с использованием подвижного буфера до 1000, 250 и 62,5 нМ, позволяли взаимодействовать с чипом с уловленным антителом.
Сенсограммы, полученные путем измерения, проведенного этим способом при pH 7,4 и pH 6,0, представлены на фиг. 20. На фиг. 20 показана связанная с hIL-6R фаза и диссоциировавшая от hIL-6R фаза PHX-IgG1, когда количество уловленных антител, нормализовано к 100 RU. Сравнение результатов, представленных на фиг. 20, указывает на то, что связывание PHX-IgG1 с hIl-6R снижено при pH 6,0 по сравнению с его связыванием при pH 7,4.
14-3. Оценка свойства двух типов антител одновременно связываться с одним и тем же антигеном способом электрохемилюминесценции
То, могут ли два типа антител связываться с одним антигеном одновременно, оценивали способом электрохемилюминесценции. Сначала, Fv4-IgG1 (1 мкг/мл, 100 мкл), биотинилированное с использованием EZ-Link Sulfo-NHS-Biotin (Thermo SCIENTIFIC) добавляли в 96-луночный планшет MULTI-ARRAY(r) Streptavidin Gold Plate, и его инкубировали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. После промывания добавляли 100 мкл растворов hsIL-6R, приготовленных в концентрации 0, 0,2, 1, 5 и 25 мкг/мл, и им позволяли реагировать при комнатной температуре в течение одного часа. После другого промывания добавляли PHX-F29, меченное рутением с использованием SULFO-TAG NHS Ester (Meso Scale Discovery) (0, 0,2, 1, 5, 25 и 125 мкг/мл; 100 мкл), или Fv4-IgG1, меченное рутением тем же способом (5 мкг/мл; 100 мкл), и позволяли им реагировать при комнатной температуре в течение одного часа. После промывания в каждую лунку распределяли 150 мкл буфера для считывания T (x4) (Meso Scale Discovery) и сразу после этого проводили измерение хемилюминесценции с использованием устройства для считывания SECTOR IMAGER 2400 Reader (Meso Scale Discovery).
Результаты представлены на фиг. 21. Когда позволяли реагировать меченному рутением Fv4-IgG1, реакции не наблюдали, независимо от концентрации антигена. С другой стороны, когда позволяли реагировать меченному рутением PHX-F29, наблюдали реакцию, зависимую от концентрации антигена (IL6R). Эти результаты демонстрируют, что PHX-F29 и Fv4-IgG1 связываются одновременно с IL6R. Таким образом, это указывает на то, что эпитоп IL6R, распознаваемый PHX-F29, и эпитоп, распознаваемый Fv4-IgG1, отличаются; и при использовании антител, содержащих вариабельную область PHX-F29 и вариабельную область Fv4-IgG1, соответственно, две антигенсвязывающие молекулы могут связываться с одной молекулой IL6R.
14-4. Испытания in vivo с использованием нормальных мышей
Нормальным мышам (мышь C57BL/6J; Charles River Japan) вводили hsIL-6R (растворимый рецептор IL-6 человека: полученный согласно справочному примеру 1) отдельно или совместно вводили hsIL-6R и антитела против рецептора IL-6 человека, а затем оценивали в отношении динамики hsIL-6R и антител против рецептора IL-6 человека in vivo. Раствор hsIL-6R (5 мкг/мл) или смешанный раствор hsIL-6R и антитела против рецептора IL-6 человека вводили один раз в дозе 10 мл/кг в хвостовую вену. Использованные антитела против рецептора IL-6 человека представляли собой Fv4-IgG1 и PHX-IgG1.
Концентрация hsIL-6R в смешанном растворе в каждом случае составляла 5 мкг/мл, и концентрации антитела против рецептора IL-6 человека отличались для каждой группы введения, как показано в таблице 12. Поскольку антитело против рецептора IL-6 человека присутствовало достаточно в избытке относительно hsIL-6R, считалось, что большая часть hsIL-6R была связана антителом. Взятие крови от мышей проводили через 5 минут, 7 часов, 1 сутки, 2 суток, 3 суток, 7 суток, 14 суток и 21 сутки после введения. Взятие крови от группы введения Fv4-IgG1 (1 мг/кг) (#1) проводили через 5 минут, 7 часов, 1 сутки, 2 суток, 3 суток, 7 суток, 15 суток и 22 суток после введения антитела. Собранные образцы крови сразу центрифугировали при 4°C и 12000 об./мин. в течение 15 минут с получением образцов плазмы. Отделенные образцы плазмы хранили в морозильной камере при -20°C или ниже до измерения.
14-5. Определение концентрации антитела против рецептора IL-6 человека в плазме нормальных мышей способом электрохемилюминесценции
Концентрацию антитела против рецептора IL-6 человека в плазме мыши определяли способом электрохемилюминесценции. Сначала, антитело для улавливания против каппа-цепи (Antibody Solutions) распределяли в 96-луночные планшеты Multi-ARRAY (Meso Scale Discovery) и перемешивали при комнатной температуре в течение одного часа, а затем использовали раствор PBS-Tween, содержащий 5% BSA (масс./об.), для блокирования при комнатной температуре в течение двух часов, получая планшеты с иммобилизованным антителом против IgG человека. Образцы для калибровочной кривой, приготовленные при концентрации в плазме 40,0, 13,3, 4,44, 1,48, 0,494, 0,165 и 0,0549 мкг/мл; и образцы плазмы мыши для анализа, приготовленные в разведении в 500 раз или более, распределяли аликвотами в каждую лунку планшетов с иммобилизованным антителом против IgG человека, а затем планшеты перемешивали в течение одного часа при комнатной температуре. Затем в каждую лунку упомянутых выше планшетов распределяли антитело для улавливания против каппа-цепи человека, конъюгированное с биотином (Антитело Solutions), а затем планшеты перемешивали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. Более того, в каждую лунку планшетов распределяли аликовотами стрептавидин, меченный SULFO-TAG (Meso Scale Discovery), а затем планшеты перемешивали при комнатной температуре в течение одного часа, чтобы позволить реакции протекать. После промывания каждой лунки в лунки распределяли буфер для считывания T (x1) (Meso Scale Discovery) и сразу измеряли хемилюминесценцию реакционных растворов с использованием устройства для считывания Sector Imager 2400 Reader (Meso Scale Discovery). Концентрации в плазме мыши вычисляли на основе ответа из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Изменения концентрации антитела против IL-6R человека с течением времени представлены на фиг. 22.
14-6. Определение концентрации hsIL-6R в плазме способом электрохемилюминесценции
Концентрацию hsIL-6R в плазме мыши определяли способом электрохемилюминесценции. Образцы hsIL-6R для калибровочной кривой приготавливали с концентрацией в плазме 12,5, 6,25, 3,13, 1,56, 0,781, 0,391 и 0,195 нг/мл. Образцы плазмы мыши для анализа приготавливали при разведении в 50 раз или более. Смешанному раствору образца hsIL-6R или образца плазмы мыши для анализа, моноклональному антителу против IL-6R человека (R&D), которое было меченным рутением с использованием SULFO-TAG NHS Ester (Meso Scale Discovery), биотинилированному антителу против IL-6R человека (R&D) и тоцилизумабу (тяжелая цепь SEQ ID NO:64, легкая цепь SEQ ID NO:65) позволяли реагировать в течение ночи при 37°C. Затем раствор PBS-Tween, содержащий 0,5% BSA (масс./об.), использовали для блокирования планшета Streptavidin Gold Multi-ARRAY Plate (Meso Scale Discovery) путем инкубации при 5°C в течение ночи, и смешанный раствор распределяли аликвотами в каждую лунку этого планшета. После инкубации этого планшета в течение дополнительных двух часов при комнатной температуре, чтобы позволить реакции протекать, каждую лунку планшета промывали, а затем в каждую лунку распределяли буфер для считывания T (x2) (Meso Scale Discovery); и сразу измеряли хемилюминесценцию реакционных растворов использованием устройства для считывания SECTOR Imager 2400 Reader (Meso Scale Discovery). Концентрации hsIL-6R вычисляли на основе ответа из калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices).
Вычисленные изменения концентрации IL6R человека (который также описывается как hsIL6R или hsIL-6R, и это относится к одному и тому же белку) с течением времени представлены на фиг. 23. Элиминация IL6R из плазмы была ускорена у мышей, которым вводили PHX-IgG1 вместе с Fv4-IgG1, по сравнению с мышами, которым вводили Fv4-IgG1 отдельно.
Не ограничиваясь конкретной теорией, может быть дано следующее объяснение упомянутому выше ускорению элиминации антигена из плазмы. Даже при введении отдельно, Fv4-IgG1 присутствовало в достаточном количестве относительно hsIL6R, и, таким образом, считалось, что большая часть hsIL6R связана с Fv4-IgG1 в плазме. Когда вводят PHX-IgG1, которое имеет отличающийся эпитоп от эпитопа Fv4-IgG1 и может связываться с hsIL6R вместе с Fv4-IgG1, образуется иммунный комплекс, содержащий Fv4-IgG1 и PHX-IgG1 на одну молекулу hsIL6R. В результате, путем усиленного связывания с Fcγ-рецептором и/или FcR захват в клетки ускорялся, и это могло привести к ускорению элиминации hsIL6R из плазмы. В частности, образование крупных иммунных комплексов, содержащих два или более антитела и два или более антигенных элементов связывания (мономерные антигены) мультиспецифическими антителами, содержащими соответствующие вариабельные области, которые связываются с эпитопами, которые отличаются друг от друга и присутствуют на мономерном антигене, или в неогранчивающем варианте осуществления мультипаратопными антителами, которые представляют собой антитела, вариабельные области которых демонстрируют pH- или Ca-зависимое связывание (биспецифические антитела или бипаратопные антитела, содержащие вариабельную область правого плеча, которая распознает эпитоп A, и вариабельную область левого плеча, которая распознает эпитоп B, как показано на фиг. 8), обеспечивает ускорение элиминации антигена.
[Пример 15] Оценка связывания иммунного комплекса с FcγR с использованием Biacore
15-1. Что касается связывания иммунного комплекса с FcγR
В качестве способов для оценки образования иммунных комплексов, содержащих антигенсвязывающую молекулу и антиген, использовали гель-фильтрацию (эксклюзионную хроматорафию) согласно примеру 4, и использовали способ электрохемиллюминесценции (ECL) согласно примеру 14. Когда антиген или антигенсвязывающая молекула, содержащиеся в иммунном комплексе, содержат FcγR-связывающий домен, такой как константная область иммуноглобулина, иммунный комплекс связывается с FcγR с авидностью. Таким образом, образование иммунного комплекса, содержащего FcγR-связывающий домен, может быть подтверждено способом, в котором используется свойство более прочного связывания FcγR (в частности, свойство более медленной диссоциации), относительно антигенсвязывающей молекулы отдельно или антигена отдельно (The Journal of Biological Chemistry (2001) 276(9), 6591-6604, mAbs (2009) 1 (5), 491-504).
15-2. Получение антител и антигенов для оценки и меченного гистидином FcγRIIIaV человека
Xolair (Novartis), клон 278-IgG1 (полученное согласно примеру 4), GA2-IgG1 (полученное согласно примеру 1), Fv4-IgG1 (полученное согласно примеру 14) и PHX-IgG1 (полученное согласно примеру 14) для применения для оценки образования иммунного комплекса получали упомянутыми выше способами.
hIgE (hIgE(Asp6); полученное согласно примеру 5) и IL6R (справочный пример 1) для применения для оценки получали упомянутыми выше способам, соответственно. Рекомбинантный вариант IgA человека, hIgA-v2 (GC-hIgA), получали способом, описанным ниже. Фрагмент гена, кодирующий GC-hIgA-MYC (тяжелая цепь SEQ ID NO:66 и легкая цепь SEQ ID NO:67), встраивали в экспрессирующий вектор клеток животных. Сконструированный плазмидный вектор вводили в FreeStyle 293 (Invitrogen) с использованием 293Fectin (Invitrogen) вместе с экспрессирующим EBNA1 геном. Затем трансфицированные клетки культивировали при 37°C в 8% CO2 в течение шести суток, и белок GC-hIgA секретировался в культуральный супернатант. Клеточную культуру, содержащую GC-hIgA-MYC, фильтровали через 0,22-мкм фильтра на горловину флакона с получением культурального супернатанта. Очищенный GC-hIgA-MYC получали с использованием ионообменной хроматографии и гель-фильтрации в соответствии со способами, известными специалистам в данной области.
Меченный гистидином FcγRIIIaV человека получали способом справочного примера 2.
15-3. Оценка связывания иммунного комплекса с FcγR
Связывание иммунного комплекса антиген-антитело с FcγR оценивали с использованием Biacore T200 (GE Healthcare).
Подходящее количество антитела с пятью остатками His (QIAGEN) иммобилизовывали на сенсорном чипе CM5 (GE Healthcare) способом присоединения аминов; и инжектировали подходящую концентрацию his-меченного FcγRIIIaV человека. FcγRIIIaV человека иммобилизовывали на чипе, позволяя антителу с пятью остатками His, иммобилизованному на чипе, улавливать FcγRIIIaV человека. Раствор, содержащий 200 нМ антитело или смешанный раствор, содержащий 200 нМ антитело и 200 нМ антиген, инжектировали в качестве анализируемого соединения; и ему позволяли взаимодействовать с FcγRIIIaV человека, иммобилизованном на сенсорном чипе. Затем, инжектировали 10 ммоль/л раствора глицин-HCl, pH 2,5, для регенерации сенсорного чипа. В качестве подвижного буфера использовали 1,2 ммоль/л CaCl2/0,05% tween 20, 20 ммоль/л ACES, 150 ммоль/л NaCl, pH 7,4, и измеряли связывание между иммунным комплексом антиген-антитело и FcγR при 25°C.
Результаты анализов связывания в реакции содержащего антитело раствора или смешанного раствора антитело-антиген с FcγRIIIaV человека представлены на фиг. 24. Чтобы сфокусироваться на диссоциации, на фиг. 24 представлены сенсограммы, на которых уровни связывания нормализованы к 100. Хотя Fv4-IgG1 связывается с IL6R, оно не образует иммунный комплекс. Таким образом, не было выявлено отличий между диссоциацией в реакции содержащего антитело раствора и FcγR и диссоциацией, когда использовали смесь антиген-антитело. Между тем, когда смешанному раствору hIgE и Xolair (J. Pharmacol. Exp. Ther. (1996) 279 (2) 1000-1008), которые, как описано, образуют иммунный комплекс, позволяли связываться с FcγR, наблюдали, что диссоциация от FcγR была более медленной, чем когда Xolair отдельно позволяли связываться с FcγR. Более того, что касается клона 278-IgG1, для которого было подтверждено, что он образует иммунный комплекс, с использованием эксклюзионной хроматографии (гель-фильтрации) в примере 4, когда смешанному раствору клона 278-IgG1 и антигена (hIgE) позволяли связываться с FcγR, диссоциация от FcγR становилась более медленной, чем когда клону 278-IgG1 отдельно позволяли связываться с FcγR. Более того, когда смешанному раствору GA2-IgG1 и hIgA позволяли связываться с FcγR, диссоциация от FcγR также становилась более медленной, чем когда GA2-IgG1 отдельно позволяли связываться с FcγR. Кроме того, когда смешанному раствору Fv4-IgG1, PHX-IgG1 и антигена (IL6R) позволяли связываться с FcγR, диссоциация от FcγR также становилась более медленной, чем когда Fv4-IgG1 или PHX-IgG1 позволяли связываться с FcγR.
Таким образом, когда антиген или антигенсвязывающая молекула, содержащиеся в иммунном комплексе, содержат FcγR-связывающий домен, такой как константная область иммуноглобулина, иммунный комплекс связывается с FcγR с авидностью. Таким образом, было обнаружено, что способ оценки замедления ассоциации от FcγR, когда смешанному раствору антигенсвязывающей молекулы, содержащей FcγR-связывающий домен и т.п., и ее антигена позволяют связываться с FcγR или когда смешанному раствору антигена, содержащему FcγR-связывающий домен и антигенсвязывающую молекулу, которая связывается с этим антигеном, позволяют связываться с FcγR, по сравнению с тем, когда молекуле, содержащей FcγR-связывающий домен, такой как константная область иммуноглобулина, позволяют связываться самой по себе с FcγR, является эффективным способом оценки образования иммунных комплексов, содержащих антигенсвязывающую молекулу и антиген. Было показано, что упомянутая выше оценка иммунных комплексов с использованием диссоциации от FcγR в качестве критерия, является эффективной в случае иммунных комплексов, образованных антителом против антигена, связанного множеством мономеров, и также в случае иммунных комплексов, образованных мономерных антигеном и множеством антител, связывающихся с различными эпитопами.
[Пример 16] Оценка связывания иммунного комплекса с FcRn с использованием Biacore
16-1. Что касается связывания иммунного комплекса с FcRn
В способе оценки образования иммунных комплексов, содержащих антигенсвязывающую молекулу и антиген, когда антиген или антигенсвязывающая молекула, содержащиеся в иммунном комплексе, содержат FcRn-связывающий домен, такой как константная область иммуноглобулина, иммунный комплекс связывается с FcRn с авидностью. Таким образом, образование иммунного комплекса, содержащего FcRn-связывающий домен, может быть подтверждено способом, в котором используется свойство более прочного связывания FcRn (в частности, свойство более медленной диссоциации), чем в случае антигенсвязывающей молекулы отдельно или антигена отдельно (Journal of Immunology (2010) 184 (4) 1968-1976). Однако в описанных до настоящего времени способах связывание между FcRn и FcRn-связывающим доменом оценивали в условиях pH 6,0. Эти условия не только отличались от условий in vivo, но также не пригодны для оценки образцов, авидность которых изменяется при изменении pH. Такие условия можно было использовать, поскольку связывание pH 7,4 нельзя было оценить вследствие низкой авидности между FcRn человека и IgG1 человека. Таким образом, исследовали, является ли возможной оценка иммунных комплексов в условиях pH7,4, которые являются более близкими к условиям in vivo.
16-2. Получение антител и антигенов для оценки и FcRn
Fv4-F22, используемое для оценки, представляет собой вариант константной области тяжелой цепи Fv4-IgG1 (описанный в примере 14). Xolair-F22 представляет собой вариант константной области тяжелой цепи Xolair (Novartis). Последовательность ДНК, кодирующую Fv4-F22 (тяжелая цепь SEQ ID NO:68 и легкая цепь SEQ ID NO:60) и последовательность ДНК, кодирующую Xolair-F22 (тяжелая цепь SEQ ID NO:69 и легкая цепь SEQ ID NO:70) встраивали в экспрессирующую плазмиду клеток животных способом, известным специалистам в данной области. Варианты антител экспрессировали в соответствии с описанным выше способом (описанным в примере 1) с использованием клеток животных, в которые введена плазмида, и после очистки определяли их концентрации. Клон 278-IgG1 (полученный согласно примеру 4), Fv4-IgG1 (полученный согласно примеру 14) и PHX-IgG1 (полученный согласно примеру 14) получали упомянутыми выше способами.
hIgE (hIgE(Asp6), полученный согласно примеру 5) и IL6R (полученный согласно справочному примеру 1), использованные для оценки, получали соответствующими упомянутыми выше способами.
FcRn человека и FcRn мыши получали способами, описанными в справочных примерах 3 и 4, соответственно.
16-3. Оценка связывания иммунного комплекса с FcRn человека
Связывание иммунного комплекса антиген-антитело с FcRn человека оценивали с использованием Biacore T100 (GE Healthcare).
Раствор, содержащий антитело отдельно или смешанный раствор антитело-антиген, представленный в таблице 13, инжектировали в качестве анализируемого соединения на сенсорный чип CM4 (GE Healthcare), на который было иммобилизовано подходящее количество FcRn человека (hFcRn) способом присоединения аминов и взаимодействию позволяли протекать с hFcRn на сенсорном чипе. Затем инжектировали раствор 20 мМ Tris-HCl, 150 мМ NaCl, pH 9,0 или pH 9,5, для регенерации сенсорного чипа. В качестве подвижного буфера использовали буфер 50 ммоль/л Na-фосфат (фосфорная кислота), 150 ммоль/л NaCl, 0,05% Tween 20, pH 7,4, и связывание между иммунным комплексом антиген-антитело и FcRn измеряли при 25°C.

Результаты анализов связывания, когда содержащему антитело раствору или смешанному раствору антитело-антиген позволяли реагировать с hFcRn, представлены на фиг. 25. Чтобы сфокусироваться на диссоциации, на фиг. 25 представлены сенсограммы, на которых уровни связывания нормализованы к 100. Хотя как Fv4-F22 связывается с IL6R, оно не образует иммунный комплекс, содержащий множество молекул, которые содержат константные области иммуноглобулина; и, таким образом, не было выявлено отличий между диссоциацией в реакции содержащего антитело раствора и FcRn, и диссоциацией, когда использовали смешанный раствор антиген-антитело. Между тем, когда смешанному раствору hIgE и Xolair-F22, содержащему вариабельную область Xolair, которое, как было описано, образует иммунный комплекс с hIgE (J. Pharmacol. Exp. Ther. (1996) 279 (2) 1000-1008) позволяли связываться с FcRn, наблюдали, что диссоциация от FcRn является более медленной, чем диссоциация от FcRn, когда Xolair-F22 отдельно было связано с FcRn. Указанное выше показало, что даже в условиях pH 7,4, связывание hFcRn можно было обнаружить с использованием антител с усиленным связыванием с hFcRn. Когда антигенсвязывающая молекула или антиген, содержащиеся в иммунном комплексе, содержат FcRn-связывающий домен, такой как константная область иммуноглобулина, иммунный комплекс более прочно связывается с FcRn. Таким образом, было выявлено, что способ оценки замедления диссоциации от FcRn, когда смешанному раствору антигенсвязывающей молекулы, содержащей FcRn-связывающий домен и т.п., и ее антигена, позволяют связываться с FcRn, или когда смешанному раствору антигена, содержащего FcRn-связывающий домен, и антигенсвязывающей молекулы, которая связывается с этим антигеном, позволяют связываться с FcRn, по сравнению с тем, когда молекуле, содержащей FcRn-связывающий домен, такой как константная область иммуноглобулина и т.п., позволяют связываться самой с FcRn, является эффективным способом оценки образования иммунных комплексов, содержащих антигенсвязывающую молекулу и антиген.
16-4. Оценка связывания иммунного комплекса с FcRn с использованием Biacore
В 16-3 оценивали связывание с FcRn иммунных комплексов, содержащих варианты константных областей иммуноглобулина с усиленным связыванием hFcRn. Поскольку нативный IgG человека может более прочно связываться с FcRn человека, чем с FcRn мыши (Int. Immunol. (2001) 13 (12), 1551-1559), оценивали связывание антитела, содержащего нативную константную область IgG человека с FcRn мыши для исследования того, возможно ли образование иммунного комплекса без модификации константной области иммуноглобулина.
Связывание иммунного комплекса антиген-антитело с FcRn оценивали с использованием Biacore T100 (GE Healthcare). Раствор, содержащий только антитела, или смешанный раствор антитело-антиген, как показано в таблице 14, инжектировали в качестве анализируемого соединения на сенсорный чип CM4 (GE Healthcare), на который было иммобилизовано подходящее количество FcRn мыши (mFcRn) способом присоединения аминов, и позволяли протекать взаимодействию с mFcRn на сенсорном чипе. Затем инжектировали раствор 20 мМ Tris-HCl, 150 мМ NaCl, pH 9,0 или pH 9,5, для регенерации сенсорного чипа. Буфер 50 ммоль/л Na-фосфат (фосфорная кислота), 150 ммоль/л NaCl, 0,05% Tween 20, pH 7,4, использовали в качестве подвижного буфера, и связывание между иммунным комплексом антиген-антитело и FcRn измеряли при 25°C.
Результаты анализов связывания, когда раствору, содержащему только антитела, или смешанному раствору антитело-антиген позволяли реагировать с mFcRn, представлены на фиг. 26 и 27. Чтобы сфокусироваться на диссоциации, на фиг. 26 и 27 представлены сенсограммы, на которых уровни связывания были нормализованы к 100. Когда смешанному раствору антигена (hIgE) и клона 278-IgG1, для которого было подтверждено, что он образует иммунный комплекс с помощью эксклюзионной хроматографии (гель-фильтрации) в примере 4, позволяли связываться с mFcRn, диссоциация от mFcRn также становилась более медленной, чем когда клону 278-IgG1 отдельно позволяли связываться с mFcRn. Более того, когда смешанному раствору hIgE и Xolair, для которого было описано, что оно образует иммунный комплекс (J. Pharmacol. Exp. Ther. (1996) 279 (2) 1000-1008), позволяли связываться с mFcRn, наблюдали, что диссоциация от mFcRn становилась более медленной, чем когда Xolair отдельно позволяли связываться с mFcRn (фиг. 26). С другой стороны, отличий не наблюдали между диссоциацией от mFcRn, когда раствору, содержащему Fv4-IgG1, позволяли связываться с mFcRn, и диссоциацией от mFcRn, когда смешанному раствору Fv4-IgG1 и IL6R позволяли связываться с mFcRn. Более того, не наблюдали отличий между диссоциацией от mFcRn, когда раствору, содержащему PHX-IgG1, позволяли связываться с mFcRn, и диссоциацией от mFcRn, когда смешанному раствору PHX-IgG1 и IL6R позволяли связываться с mFcRn (фиг. 27). Это может быть следствием того, что имеется один антигенный элемент связывания в IL6R при связывании с вариабельной областью, содержащейся в Fv4-IgG1; и когда Fv4-IgG1 отдельно используют в качестве антигенсвязывающей молекулы для IL6R, иммунный комплекс, содержащий две или более молекулы Fv4-IgG1 на одну молекулу IL6R, не образуется. Поскольку в IL6R также имеется один антигенный элемент связывания при связывании с вариабельной областью в PHX-IgG1, существует только один эпитоп в IL6R; и, таким образом, полагают, что, когда PHX-IgG1 отдельно используют в качестве антигенсвязывающей молекулы, нацеленной на IL6R, иммунный комплекс, содержащий две или более молекулы PHX-IgG1 на одну молекулу IL6R, не образуется. В настоящем примере, когда смешанному раствору IL6R, и Fv4-IgG1, и PHX-IgG1, для которых в примере 14 показано, что они имеют эпитопы, которые отличаются друг от друга, позволяли связываться с mFcRn, диссоциация от mFcRn становилась более медленной, чем когда раствору, содержащему антитело Fv4-IgG1 и антитело PHX-IgG1 (только антитела) позволяли связываться с mFcRn. Такое явление более медленной диссоциации является сходным с явлением, наблюдаемым для клона 278. Это указывает на то, что, когда один тип антигенсвязывающей молекулы используют для антигенсвязывающей молекулы, которая связывается с антигеном-мишенью, даже если имеется один антигенный элемент связывания, антигенный элемент связывания антигена может быть заменен двумя или более антигенными элементами связывания с использованием комбинации антигенсвязывающих молекул, которые имеют эпитопы, отличающиеся друг от друга, в качестве антигенсвязывающих молекул, нацеленных на антиген; и в результате может образовываться иммунный комплекс, содержащий две или более антигенсвязывающих молекул.
Вышеуказанное показало, что с использованием FcRn мыши, даже нативную молекулу IgG1 человека без модификации, которая может повлиять на взаимодействие с FcRn, можно оценивать в отношении связывания FcRn в условиях pH 7,4, которые являются такими же, как и условия in vivo. Более того, когда антигенсвязывающая молекула или антиген содержат FcRn-связывающий домен, такой как константная область иммуноглобулина, иммунный комплекс связывается прочно с FcRn; таким образом, образование иммунного комплекса может быть подтверждено путем оценки замедления диссоциации от FcRn иммунного комплекса, содержащего множество молекул, содержащих FcRn-связывающий домен, такой как константная область иммуноглобулина, по сравнению с диссоциацией от FcRn молекулы, содержащей FcRn-связывающий домен константной области иммуноглобулина. Было подтверждено, что упомянутую выше оценку образования иммунного комплекса с использованием FcRn можно использовать для подтверждения образования как иммунного комплекса, образованного из антигена, содержащего множество связанных мономеров, и антитела, которое связывается с этим антигеном, так и из иммунного комплекса, образованного из мономерного антигена и множества антител, обладающих связыванием отличающихся друг от друга эпитопов, которые связываются с этим антигеном.
[Справочный пример 1] Получение растворимого рецептора IL-6 человека (hsIL-6R)
Рекомбинантную форму рецептора IL-6 человека, который является антигеном, получали следующим образом. Конструировали линию CHO, которая постоянно экспрессирует растворимый рецептор IL-6 человека (далее обозначаемый как hsIL-6R), имеющий аминокислотную последовательность от первой до 357-ой аминокислоты с N-конца, как описано в J. Immunol. (1994) 152, 4958-4968, с использованием способа, известного специалистам в данной области. hsIL-6R экспрессировали путем культивирования этой линии CHO. hsIL-6R очищали из культурального супернатанта полученной линии CHO посредством двухстадийного процесса, вовлекающего колоночную хроматографию на колонке Blue Sepharose 6 FF и гель-фильтрационную колоночную хроматографию. Фракцию, которая элюировалась в качестве основного пика на последней стадии, использовали в качестве конечного очищенного продукта.
[Справочный пример 2] Получение меченного гистидином FcγRIIIaV человека
Меченный гистидином FcγRIIIaV человека получали следующим способом. Сначала синтезировали ген внеклеточного домена FcγR способом, хорошо известным специалистам в данной области. В это время получали последовательность каждого FcγR на основе информации, зарегистрированной в NCBI. В частности, FcγRIIIa получали на основе последовательности с номером доступа NCBI № NM_001127593.1, и к C-концу присоединяли His-метку. Более того, известен полиморфизм для FcγRIIIa; однако получение FcγRIIIa осуществляли, ссылаясь на J. Clin. Invest., 1997, 100 (5): 1059-1070.
Полученные фрагменты генов встраивали в экспрессирующий вектор клеток животных, и получали экспрессирующие векторы. Полученные экспрессирующие векторы временно вводили в клетки линии, происходящей из клеток рака почки эмбриона человека FreeStyle293 (Invitrogen) для экспрессии представляющих интерес белков. Клетку культивировали и после сбора полученного культурального супернатанта его пропускали через 0,22-мкм фильтр с получением культурального супернатанта. В принципе, полученные культуральные супернатанты очищали с помощью следующих четырех стадий. Проведенные стадии представляли собой катионообменную колоночную хроматографию (SP Sepharose FF) на стадии 1, аффинную колоночную хроматографию (HisTrap HP) для His-метки на стадии 2, гель-фильтрационную колоночную хроматографию (Superdex200) на стадии 3, и асептическую хроматографию на стадии 4. Очищенные белки подвергали измерению поглощения при 280 нм с использованием спектрофотометра; и из полученных значений вычисляли концентрации очищенных белков с использованием коэффициента поглощения, вычисленного с использованием способов, таких как PACE (Protein Science 1995; 4: 2411-2423).
[Справочный пример 3] Получение растворимого FcRn человека (hFcRn)
FcRn человека образует комплекс с β2-микроглобулином. Праймеры олиго-ДНК конструировали на основе описанной последовательности гена FcRn человека (J Exp Med. 1994 Dec 1; 180(6): 2377-81). кДНК амплифицировали способом ПЦР из кДНК человека (Human Placenta Marathon-Ready cDNA, Clontech) с использованием сконструированных праймеров. Амплифицированную кДНК, кодирующую внеклеточный домен, содержащий сигнальную последовательность (Met1-Leu290), встраивали в экспрессирующий вектор клеток животных. Аналогично, конструировали праймеры для β2-микроглобулина на основе описанной последовательности β2-микроглобулина человека, и кДНК амплифицировали способом ПЦР. Фрагмент кДНК Met1-Met119, содержащий сигнальную последовательность, амплифицировали и встраивали в экспрессирующий вектор клеток животных.
Растворимый FcRn человека (называемый hFcRn) получали по описанной ниже методике. Вектор для экспрессии FcRn человека (SEQ ID NO:71) и вектор для экспрессии β2-микроглобулина (SEQ ID NO:72) трансдуцировали в клетки HEK293H (Invitrogen) способом липофекции с использованием PEI (Polyscience). Трансдуцированные клетки культивировали и культуральный раствор собирали. FcRn человека очищали из собранной культуры каждым из хроматографических способов: IgG Sepharose 6 Fast Flow (Amersham Biosciences) и HiTrap Q HP (GE Healthcare) (J Immunol. 2002 Nov 1; 169(9): 5171-80).
[Справочный пример 4] Получение растворимого FcRn мыши (mFcRn)
Растворимый FcRn мыши получали аналогично растворимому FcRn человека. Сначала ген, кодирующий внеклеточный домен FcRn мыши, и ген, кодирующий β2-микроглобулин мыши, синтезировали способами, известными специалистам в данной области. Для этого последовательности FcRn мыши и β2-микроглобулина мыши получали на основе информации, зарегистрированной в NCBI. В частности, для FcRn мыши фрагмент гена, кодирующий аминокислоты в положениях 1-290, содержащий сигнальную последовательность, получали в качестве внеклеточного домена на основе последовательности с номером доступа NCBI № NP_034319.2 (SEQ ID NO:73). Более того, для β2-микроглобулина фрагмент гена получали на основе последовательности с номером доступа NCBI № NP_033865 (SEQ ID NO:74). Экспрессирующий вектор получали путем встраивания полученных фрагментов генов в экспрессирующий вектор клеток животных. Полученный экспрессирующий вектор временно вводили в клетки, происходящие из клеток карциномы почки эмбриона человека, FreeStyle 293 (Invitrogen) для экспрессии представляющего интерес белка. Трансдуцированные клетки культивировали и культуральный раствор собирали. FcRn мыши очищали из собранной культуры каждым из хроматографических способов: IgG Sepharose 6 Fast Flow (Amersham Biosciences) и Superdex 200 (GE Healthcare).
[Справочный пример 5] Получение антигенсвязывающей молекулы с более высокой активностью связывания FcγR, чем у нативной Fc-области IgG человека и усиленной активностью связывания FcRn человека в условиях диапазона кислых значений pH
(5-1) Получение антитела против рецептора IL-6 человека с усиленным связыванием с FcγR мыши
VH3-IgG1-F1087 (SEQ ID NO:75), которое получено путем замены Lys на Asp в положении 326 (нумерация EU) в VH3-IgG1, и VH3-IgG1-F1182 (SEQ ID NO:76), которое получено путем замены Ser на Asp в положении 239, и Ile на Glu в положении 332 (нумерация EU) в VH3-IgG1, получали в качестве антигенсвязывающих молекул с усиленным связыванием с FcγR мыши. Fv4-IgG1-F1087, содержащее VH3-IgG1-F1087 в качестве тяжелой цепи и VL3-CK (SEQ ID NO:77) в качестве легкой цепи, и Fv4-IgG1-F1182, содержащее VH3-IgG1-F1182 в качестве тяжелой цепи и VL3-в качестве легкой цепи, получали с использованием способа справочного примера 2.
(5-2) Подтверждение активности связывания FcγR мыши
Получали VH3/L (WT)-IgG1-F1087 и VH3/L (WT)-IgG1-F1182, которые содержат VH3-IgG1-F1087 и VH3-IgG1-F1182, соответственно, в качестве тяжелой цепи, и L (WT)-CK (SEQ ID NO:78) в качестве легкой цепи. Оценивали активность связывания этих антител с FcγR мыши и VH3/L (WT)-IgG1-F1022, и результаты представлены в таблице 15. Кратность усиления активности связывания FcγR мыши для каждого из вариантов по сравнению с IgG1 до модификации представлена в таблице 16.
Промышленная применимость
Антигенсвязывающие молекулы по настоящему изобретению обеспечивают ускорение элиминации антигенов с двумя более антигенных элементов связывания из плазмы вследствие наделения их признаком образования крупных иммунных комплексов, содержащих антигены с двумя или более антигенных элементов связывания (эпитопов) и двумя или более антигенсвязывающих молекул (например, антител). Антигенсвязывающие молекулы по настоящему изобретению также обеспечивают дальнейшее ускорение элиминации антигенов вследствие наделения их зависимой от ионов антигенсвязывающей активностью в дополнение к упомянутым выше признакам. Вследствие наделения их такими техническими признаками, антигенсвязывающие молекулы по настоящему изобретению становятся в высокой степени пригодными в качестве фармацевтических препаратов для лечения заболеваний или симптомов (таких как злокачественная опухоль и воспалительное заболевание).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> СОДЕРЖАЩИЙ ЛЕКАРСТВЕННОЕ СРЕДСТВО ПЕРЕНОСЧИК В КЛЕТКУ ДЛЯ
ФОРМИРОВАНИЯ ИММУННОГО КОМПЛЕКСА
<130> C1-A1009Y1PY1P
<150> PCT/JP2011/077619
<151> 2011-11-30
<150> JP 2012-123773
<151> 2012-05-30
<160> 78
<170> PatentIn version 3.4
<210> 1
<211> 468
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Leu Ala Val Gly Cys Ala Leu Leu Ala Ala Leu Leu Ala Ala Pro
1 5 10 15
Gly Ala Ala Leu Ala Pro Arg Arg Cys Pro Ala Gln Glu Val Ala Arg
20 25 30
Gly Val Leu Thr Ser Leu Pro Gly Asp Ser Val Thr Leu Thr Cys Pro
35 40 45
Gly Val Glu Pro Glu Asp Asn Ala Thr Val His Trp Val Leu Arg Lys
50 55 60
Pro Ala Ala Gly Ser His Pro Ser Arg Trp Ala Gly Met Gly Arg Arg
65 70 75 80
Leu Leu Leu Arg Ser Val Gln Leu His Asp Ser Gly Asn Tyr Ser Cys
85 90 95
Tyr Arg Ala Gly Arg Pro Ala Gly Thr Val His Leu Leu Val Asp Val
100 105 110
Pro Pro Glu Glu Pro Gln Leu Ser Cys Phe Arg Lys Ser Pro Leu Ser
115 120 125
Asn Val Val Cys Glu Trp Gly Pro Arg Ser Thr Pro Ser Leu Thr Thr
130 135 140
Lys Ala Val Leu Leu Val Arg Lys Phe Gln Asn Ser Pro Ala Glu Asp
145 150 155 160
Phe Gln Glu Pro Cys Gln Tyr Ser Gln Glu Ser Gln Lys Phe Ser Cys
165 170 175
Gln Leu Ala Val Pro Glu Gly Asp Ser Ser Phe Tyr Ile Val Ser Met
180 185 190
Cys Val Ala Ser Ser Val Gly Ser Lys Phe Ser Lys Thr Gln Thr Phe
195 200 205
Gln Gly Cys Gly Ile Leu Gln Pro Asp Pro Pro Ala Asn Ile Thr Val
210 215 220
Thr Ala Val Ala Arg Asn Pro Arg Trp Leu Ser Val Thr Trp Gln Asp
225 230 235 240
Pro His Ser Trp Asn Ser Ser Phe Tyr Arg Leu Arg Phe Glu Leu Arg
245 250 255
Tyr Arg Ala Glu Arg Ser Lys Thr Phe Thr Thr Trp Met Val Lys Asp
260 265 270
Leu Gln His His Cys Val Ile His Asp Ala Trp Ser Gly Leu Arg His
275 280 285
Val Val Gln Leu Arg Ala Gln Glu Glu Phe Gly Gln Gly Glu Trp Ser
290 295 300
Glu Trp Ser Pro Glu Ala Met Gly Thr Pro Trp Thr Glu Ser Arg Ser
305 310 315 320
Pro Pro Ala Glu Asn Glu Val Ser Thr Pro Met Gln Ala Leu Thr Thr
325 330 335
Asn Lys Asp Asp Asp Asn Ile Leu Phe Arg Asp Ser Ala Asn Ala Thr
340 345 350
Ser Leu Pro Val Gln Asp Ser Ser Ser Val Pro Leu Pro Thr Phe Leu
355 360 365
Val Ala Gly Gly Ser Leu Ala Phe Gly Thr Leu Leu Cys Ile Ala Ile
370 375 380
Val Leu Arg Phe Lys Lys Thr Trp Lys Leu Arg Ala Leu Lys Glu Gly
385 390 395 400
Lys Thr Ser Met His Pro Pro Tyr Ser Leu Gly Gln Leu Val Pro Glu
405 410 415
Arg Pro Arg Pro Thr Pro Val Leu Val Pro Leu Ile Ser Pro Pro Val
420 425 430
Ser Pro Ser Ser Leu Gly Ser Asp Asn Thr Ser Ser His Asn Arg Pro
435 440 445
Asp Ala Arg Asp Pro Arg Ser Pro Tyr Asp Ile Ser Asn Thr Asp Tyr
450 455 460
Phe Phe Pro Arg
465
<210> 2
<211> 428
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Ala Ser Thr Gln Ser Pro Ser Val Phe Pro Leu Thr Arg Cys Cys Lys
1 5 10 15
Asn Ile Pro Ser Asn Ala Thr Ser Val Thr Leu Gly Cys Leu Ala Thr
20 25 30
Gly Tyr Phe Pro Glu Pro Val Met Val Thr Trp Asp Thr Gly Ser Leu
35 40 45
Asn Gly Thr Thr Met Thr Leu Pro Ala Thr Thr Leu Thr Leu Ser Gly
50 55 60
His Tyr Ala Thr Ile Ser Leu Leu Thr Val Ser Gly Ala Trp Ala Lys
65 70 75 80
Gln Met Phe Thr Cys Arg Val Ala His Thr Pro Ser Ser Thr Asp Trp
85 90 95
Val Asp Asn Lys Thr Phe Ser Val Cys Ser Arg Asp Phe Thr Pro Pro
100 105 110
Thr Val Lys Ile Leu Gln Ser Ser Cys Asp Gly Gly Gly His Phe Pro
115 120 125
Pro Thr Ile Gln Leu Leu Cys Leu Val Ser Gly Tyr Thr Pro Gly Thr
130 135 140
Ile Asn Ile Thr Trp Leu Glu Asp Gly Gln Val Met Asp Val Asp Leu
145 150 155 160
Ser Thr Ala Ser Thr Thr Gln Glu Gly Glu Leu Ala Ser Thr Gln Ser
165 170 175
Glu Leu Thr Leu Ser Gln Lys His Trp Leu Ser Asp Arg Thr Tyr Thr
180 185 190
Cys Gln Val Thr Tyr Gln Gly His Thr Phe Glu Asp Ser Thr Lys Lys
195 200 205
Cys Ala Asp Ser Asn Pro Arg Gly Val Ser Ala Tyr Leu Ser Arg Pro
210 215 220
Ser Pro Phe Asp Leu Phe Ile Arg Lys Ser Pro Thr Ile Thr Cys Leu
225 230 235 240
Val Val Asp Leu Ala Pro Ser Lys Gly Thr Val Asn Leu Thr Trp Ser
245 250 255
Arg Ala Ser Gly Lys Pro Val Asn His Ser Thr Arg Lys Glu Glu Lys
260 265 270
Gln Arg Asn Gly Thr Leu Thr Val Thr Ser Thr Leu Pro Val Gly Thr
275 280 285
Arg Asp Trp Ile Glu Gly Glu Thr Tyr Gln Cys Arg Val Thr His Pro
290 295 300
His Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr Ser Gly Pro
305 310 315 320
Arg Ala Ala Pro Glu Val Tyr Ala Phe Ala Thr Pro Glu Trp Pro Gly
325 330 335
Ser Arg Asp Lys Arg Thr Leu Ala Cys Leu Ile Gln Asn Phe Met Pro
340 345 350
Glu Asp Ile Ser Val Gln Trp Leu His Asn Glu Val Gln Leu Pro Asp
355 360 365
Ala Arg His Ser Thr Thr Gln Pro Arg Lys Thr Lys Gly Ser Gly Phe
370 375 380
Phe Val Phe Ser Arg Leu Glu Val Thr Arg Ala Glu Trp Glu Gln Lys
385 390 395 400
Asp Glu Phe Ile Cys Arg Ala Val His Glu Ala Ala Ser Pro Ser Gln
405 410 415
Thr Val Gln Arg Ala Val Ser Val Asn Pro Gly Lys
420 425
<210> 3
<211> 1482
<212> ДНК
<213> Homo sapiens
<400> 3
gcctccacac agagcccatc cgtcttcccc ttgacccgct gctgcaaaaa cattccctcc 60
aatgccacct ccgtgactct gggctgcctg gccacgggct acttcccgga gccggtgatg 120
gtgacctgcg acacaggctc cctcaacggg acaactatga ccttaccagc caccaccctc 180
acgctctctg gtcactatgc caccatcagc ttgctgaccg tctcgggtgc gtgggccaag 240
cagatgttca cctgccgtgt ggcacacact ccatcgtcca cagactgggt cgacaacaaa 300
accttcagcg tctgctccag ggacttcacc ccgcccaccg tgaagatctt acagtcgtcc 360
tgcgacggcg gcgggcactt ccccccgacc atccagctcc tgtgcctcgt ctctgggtac 420
accccaggga ctatcaacat cacctggctg gaggacgggc aggtcatgga cgtggacttg 480
tccaccgcct ctaccacgca ggagggtgag ctggcctcca cacaaagcga gctcaccctc 540
agccagaagc actggctgtc agaccgcacc tacacctgcc aggtcaccta tcaaggtcac 600
acctttgagg acagcaccaa gaagtgtgca gattccaacc cgagaggggt gagcgcctac 660
ctaagccggc ccagcccgtt cgacctgttc atccgcaagt cgcccacgat cacctgtctg 720
gtggtggacc tggcacccag caaggggacc gtgaacctga cctggtcccg ggccagtggg 780
aagcctgtga accactccac cagaaaggag gagaagcagc gcaatggcac gttaaccgtc 840
acgtccaccc tgccggtggg cacccgagac tggatcgagg gggagaccta ccagtgcagg 900
gtgacccacc cccacctgcc cagggccctc atgcggtcca cgaccaagac cagcggcccg 960
cgtgctgccc cggaagtcta tgcgtttgcg acgccggagt ggccggggag ccgggacaag 1020
cgcaccctcg cctgcctgat ccagaacttc atgcctgagg acatctcggt gcagtggctg 1080
cacaacgagg tgcagctccc ggacgcccgg cacagcacga cgcagccccg caagaccaag 1140
ggctccggct tcttcgtctt cagccgcctg gaggtgacca gggccgaatg ggagcagaaa 1200
gatgagttca tctgccgtgc agtccatgag gcagcgagcc cctcacagac cgtccagcga 1260
gcggtgtctg taaatcccga gctggacgtg tgcgtggagg aggccgaggg cgaggcgccg 1320
tggacgtgga ccggcctctg catcttcgcc gcactcttcc tgctcagcgt gagctacagc 1380
gccgccctca cgctcctcat ggtgcagcgg ttcctctcag ccacgcggca ggggaggccc 1440
cagacctccc tcgactacac caacgtcctc cagccccacg cc 1482
<210> 4
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 4
Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
1 5 10 15
<210> 5
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 5
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 6
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 6
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Ser Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 7
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 8
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 8
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Asp Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Val
85 90 95
Leu Arg Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Gln
100 105 110
<210> 9
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 9
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 10
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 10
Asp Ile Val Met Thr Gln Ser Pro Glu Ser Leu Val Leu Ser Leu Gly
1 5 10 15
Gly Thr Ala Thr Ile Asn Cys Arg Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Thr Leu Leu Phe Ser Trp Ala Ser Ile Arg Asp Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Ala Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
65 70 75 80
Ile Ser Asp Leu Gln Ala Glu Asp Ala Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Arg Ala Pro Ser Phe Gly Gln Gly Thr Lys Leu Gln Ile Lys
100 105 110
<210> 11
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asp Pro Gly Gly Gly Glu Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 12
<211> 126
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Thr Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 14
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 15
<211> 377
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 16
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 16
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 17
<211> 365
<212> БЕЛОК
<213> Homo sapiens
<400> 17
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 18
<211> 119
<212> БЕЛОК
<213> Homo sapiens
<400> 18
Met Ser Arg Ser Val Ala Leu Ala Val Leu Ala Leu Leu Ser Leu Ser
1 5 10 15
Gly Leu Glu Ala Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Ala Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser
35 40 45
Gly Phe His Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu
50 55 60
Arg Ile Glu Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp
85 90 95
Glu Tyr Ala Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile
100 105 110
Val Lys Trp Asp Arg Asp Met
115
<210> 19
<211> 1125
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(1125)
<400> 19
atg tgg ttc ttg aca act ctg ctc ctt tgg gtt cca gtt gat ggg caa 48
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
gtg gac acc aca aag gca gtg atc act ttg cag cct cca tgg gtc agc 96
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
gtg ttc caa gag gaa acc gta acc ttg cac tgt gag gtg ctc cat ctg 144
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
cct ggg agc agc tct aca cag tgg ttt ctc aat ggc aca gcc act cag 192
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
acc tcg acc ccc agc tac aga atc acc tct gcc agt gtc aat gac agt 240
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
ggt gaa tac agg tgc cag aga ggt ctc tca ggg cga agt gac ccc ata 288
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
cag ctg gaa atc cac aga ggc tgg cta cta ctg cag gtc tcc agc aga 336
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
gtc ttc acg gaa gga gaa cct ctg gcc ttg agg tgt cat gcg tgg aag 384
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
gat aag ctg gtg tac aat gtg ctt tac tat cga aat ggc aaa gcc ttt 432
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
aag ttt ttc cac tgg aat tct aac ctc acc att ctg aaa acc aac ata 480
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
agt cac aat ggc acc tac cat tgc tca ggc atg gga aag cat cgc tac 528
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
aca tca gca gga ata tct gtc act gtg aaa gag cta ttt cca gct cca 576
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
gtg ctg aat gca tct gtg aca tcc cca ctc ctg gag ggg aat ctg gtc 624
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
acc ctg agc tgt gaa aca aag ttg ctc ttg cag agg cct ggt ttg cag 672
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
ctt tac ttc tcc ttc tac atg ggc agc aag acc ctg cga ggc agg aac 720
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
aca tcc tct gaa tac caa ata cta act gct aga aga gaa gac tct ggg 768
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
tta tac tgg tgc gag gct gcc aca gag gat gga aat gtc ctt aag cgc 816
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
agc cct gag ttg gag ctt caa gtg ctt ggc ctc cag tta cca act cct 864
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
gtc tgg ttt cat gtc ctt ttc tat ctg gca gtg gga ata atg ttt tta 912
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
gtg aac act gtt ctc tgg gtg aca ata cgt aaa gaa ctg aaa aga aag 960
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
aaa aag tgg gat tta gaa atc tct ttg gat tct ggt cat gag aag aag 1008
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
gta att tcc agc ctt caa gaa gac aga cat tta gaa gaa gag ctg aaa 1056
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
tgt cag gaa caa aaa gaa gaa cag ctg cag gaa ggg gtg cac cgg aag 1104
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
gag ccc cag ggg gcc acg tag 1125
Glu Pro Gln Gly Ala Thr
370
<210> 20
<211> 374
<212> БЕЛОК
<213> Homo sapiens
<400> 20
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
Glu Pro Gln Gly Ala Thr
370
<210> 21
<211> 951
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(951)
<400> 21
atg act atg gag acc caa atg tct cag aat gta tgt ccc aga aac ctg 48
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
tgg ctg ctt caa cca ttg aca gtt ttg ctg ctg ctg gct tct gca gac 96
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
agt caa gct gct ccc cca aag gct gtg ctg aaa ctt gag ccc ccg tgg 144
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
atc aac gtg ctc cag gag gac tct gtg act ctg aca tgc cag ggg gct 192
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
cgc agc cct gag agc gac tcc att cag tgg ttc cac aat ggg aat ctc 240
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
att ccc acc cac acg cag ccc agc tac agg ttc aag gcc aac aac aat 288
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
gac agc ggg gag tac acg tgc cag act ggc cag acc agc ctc agc gac 336
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
cct gtg cat ctg act gtg ctt tcc gaa tgg ctg gtg ctc cag acc cct 384
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
cac ctg gag ttc cag gag gga gaa acc atc atg ctg agg tgc cac agc 432
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
tgg aag gac aag cct ctg gtc aag gtc aca ttc ttc cag aat gga aaa 480
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
tcc cag aaa ttc tcc cat ttg gat ccc acc ttc tcc atc cca caa gca 528
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
aac cac agt cac agt ggt gat tac cac tgc aca gga aac ata ggc tac 576
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
acg ctg ttc tca tcc aag cct gtg acc atc act gtc caa gtg ccc agc 624
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
atg ggc agc tct tca cca atg ggg gtc att gtg gct gtg gtc att gcg 672
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
act gct gta gca gcc att gtt gct gct gta gtg gcc ttg atc tac tgc 720
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
agg aaa aag cgg att tca gcc aat tcc act gat cct gtg aag gct gcc 768
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
caa ttt gag cca cct gga cgt caa atg att gcc atc aga aag aga caa 816
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
ctt gaa gaa acc aac aat gac tat gaa aca gct gac ggc ggc tac atg 864
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
act ctg aac ccc agg gca cct act gac gat gat aaa aac atc tac ctg 912
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
act ctt cct ccc aac gac cat gtc aac agt aat aac taa 951
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 22
<211> 316
<212> БЕЛОК
<213> Homo sapiens
<400> 22
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 23
<211> 876
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(876)
<400> 23
atg gga atc ctg tca ttc tta cct gtc ctt gcc act gag agt gac tgg 48
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
gct gac tgc aag tcc ccc cag cct tgg ggt cat atg ctt ctg tgg aca 96
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
gct gtg cta ttc ctg gct cct gtt gct ggg aca cct gca gct ccc cca 144
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
aag gct gtg ctg aaa ctc gag ccc cag tgg atc aac gtg ctc cag gag 192
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
gac tct gtg act ctg aca tgc cgg ggg act cac agc cct gag agc gac 240
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
tcc att cag tgg ttc cac aat ggg aat ctc att ccc acc cac acg cag 288
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
ccc agc tac agg ttc aag gcc aac aac aat gac agc ggg gag tac acg 336
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
tgc cag act ggc cag acc agc ctc agc gac cct gtg cat ctg act gtg 384
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
ctt tct gag tgg ctg gtg ctc cag acc cct cac ctg gag ttc cag gag 432
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
gga gaa acc atc gtg ctg agg tgc cac agc tgg aag gac aag cct ctg 480
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
gtc aag gtc aca ttc ttc cag aat gga aaa tcc aag aaa ttt tcc cgt 528
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
tcg gat ccc aac ttc tcc atc cca caa gca aac cac agt cac agt ggt 576
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
gat tac cac tgc aca gga aac ata ggc tac acg ctg tac tca tcc aag 624
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
cct gtg acc atc act gtc caa gct ccc agc tct tca ccg atg ggg atc 672
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
att gtg gct gtg gtc act ggg att gct gta gcg gcc att gtt gct gct 720
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
gta gtg gcc ttg atc tac tgc agg aaa aag cgg att tca gcc aat ccc 768
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
act aat cct gat gag gct gac aaa gtt ggg gct gag aac aca atc acc 816
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
tat tca ctt ctc atg cac ccg gat gct ctg gaa gag cct gat gac cag 864
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
aac cgt att tag 876
Asn Arg Ile
290
<210> 24
<211> 291
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
Asn Arg Ile
290
<210> 25
<211> 765
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(765)
<400> 25
atg tgg cag ctg ctc ctc cca act gct ctg cta ctt cta gtt tca gct 48
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
ggc atg cgg act gaa gat ctc cca aag gct gtg gtg ttc ctg gag cct 96
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
caa tgg tac agg gtg ctc gag aag gac agt gtg act ctg aag tgc cag 144
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
gga gcc tac tcc cct gag gac aat tcc aca cag tgg ttt cac aat gag 192
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
agc ctc atc tca agc cag gcc tcg agc tac ttc att gac gct gcc aca 240
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
gtt gac gac agt gga gag tac agg tgc cag aca aac ctc tcc acc ctc 288
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
agt gac ccg gtg cag cta gaa gtc cat atc ggc tgg ctg ttg ctc cag 336
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
gcc cct cgg tgg gtg ttc aag gag gaa gac cct att cac ctg agg tgt 384
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
cac agc tgg aag aac act gct ctg cat aag gtc aca tat tta cag aat 432
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
ggc aaa ggc agg aag tat ttt cat cat aat tct gac ttc tac att cca 480
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
aaa gcc aca ctc aaa gac agc ggc tcc tac ttc tgc agg ggg ctt gtt 528
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
ggg agt aaa aat gtg tct tca gag act gtg aac atc acc atc act caa 576
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
ggt ttg tca gtg tca acc atc tca tca ttc ttt cca cct ggg tac caa 624
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
gtc tct ttc tgc ttg gtg atg gta ctc ctt ttt gca gtg gac aca gga 672
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
cta tat ttc tct gtg aag aca aac att cga agc tca aca aga gac tgg 720
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
aag gac cat aaa ttt aaa tgg aga aag gac cct caa gac aaa tga 765
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 26
<211> 254
<212> БЕЛОК
<213> Homo sapiens
<400> 26
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 27
<211> 702
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(702)
<400> 27
atg tgg cag ctg ctc ctc cca act gct ctg cta ctt cta gtt tca gct 48
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
ggc atg cgg act gaa gat ctc cca aag gct gtg gtg ttc ctg gag cct 96
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
caa tgg tac agc gtg ctt gag aag gac agt gtg act ctg aag tgc cag 144
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
gga gcc tac tcc cct gag gac aat tcc aca cag tgg ttt cac aat gag 192
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
agc ctc atc tca agc cag gcc tcg agc tac ttc att gac gct gcc aca 240
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
gtc aac gac agt gga gag tac agg tgc cag aca aac ctc tcc acc ctc 288
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
agt gac ccg gtg cag cta gaa gtc cat atc ggc tgg ctg ttg ctc cag 336
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
gcc cct cgg tgg gtg ttc aag gag gaa gac cct att cac ctg agg tgt 384
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
cac agc tgg aag aac act gct ctg cat aag gtc aca tat tta cag aat 432
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
ggc aaa gac agg aag tat ttt cat cat aat tct gac ttc cac att cca 480
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
aaa gcc aca ctc aaa gat agc ggc tcc tac ttc tgc agg ggg ctt gtt 528
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
ggg agt aaa aat gtg tct tca gag act gtg aac atc acc atc act caa 576
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
ggt ttg gca gtg tca acc atc tca tca ttc tct cca cct ggg tac caa 624
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
gtc tct ttc tgc ttg gtg atg gta ctc ctt ttt gca gtg gac aca gga 672
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
cta tat ttc tct gtg aag aca aac att tga 702
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 28
<211> 233
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 29
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 29
Gly Gly Gly Ser
1
<210> 30
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 30
Ser Gly Gly Gly
1
<210> 31
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 31
Gly Gly Gly Gly Ser
1 5
<210> 32
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 32
Ser Gly Gly Gly Gly
1 5
<210> 33
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 33
Gly Gly Gly Gly Gly Ser
1 5
<210> 34
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 34
Ser Gly Gly Gly Gly Gly
1 5
<210> 35
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 35
Gly Gly Gly Gly Gly Gly Ser
1 5
<210> 36
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 36
Ser Gly Gly Gly Gly Gly Gly
1 5
<210> 37
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 37
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Ala Pro Gly Asn Trp Gly Ser Pro Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 38
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Glu Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Asp Ser Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 39
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 39
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 40
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Asp Asp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Ser Ser Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 41
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 41
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 42
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 42
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 43
<211> 543
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 43
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Gln Ser Pro Ser Val Phe Pro Leu Thr Arg
115 120 125
Cys Cys Lys Asn Ile Pro Ser Asn Ala Thr Ser Val Thr Leu Gly Cys
130 135 140
Leu Ala Thr Gly Tyr Phe Pro Glu Pro Val Met Val Thr Trp Asp Thr
145 150 155 160
Gly Ser Leu Asn Gly Thr Thr Met Thr Leu Pro Ala Thr Thr Leu Thr
165 170 175
Leu Ser Gly His Tyr Ala Thr Ile Ser Leu Leu Thr Val Ser Gly Ala
180 185 190
Trp Ala Lys Gln Met Phe Thr Cys Arg Val Ala His Thr Pro Ser Ser
195 200 205
Thr Asp Trp Val Asp Asn Lys Thr Phe Ser Val Cys Ser Arg Asp Phe
210 215 220
Thr Pro Pro Thr Val Lys Ile Leu Gln Ser Ser Cys Asp Gly Gly Gly
225 230 235 240
His Phe Pro Pro Thr Ile Gln Leu Leu Cys Leu Val Ser Gly Tyr Thr
245 250 255
Pro Gly Thr Ile Asn Ile Thr Trp Leu Glu Asp Gly Gln Val Met Asp
260 265 270
Val Asp Leu Ser Thr Ala Ser Thr Thr Gln Glu Gly Glu Leu Ala Ser
275 280 285
Thr Gln Ser Glu Leu Thr Leu Ser Gln Lys His Trp Leu Ser Asp Arg
290 295 300
Thr Tyr Thr Cys Gln Val Thr Tyr Gln Gly His Thr Phe Glu Asp Ser
305 310 315 320
Thr Lys Lys Cys Ala Asp Ser Asn Pro Arg Gly Val Ser Ala Tyr Leu
325 330 335
Ser Arg Pro Ser Pro Phe Asp Leu Phe Ile Arg Lys Ser Pro Thr Ile
340 345 350
Thr Cys Leu Val Val Asp Leu Ala Pro Ser Lys Gly Thr Val Asn Leu
355 360 365
Thr Trp Ser Arg Ala Ser Gly Lys Pro Val Asn His Ser Thr Arg Lys
370 375 380
Glu Glu Lys Gln Arg Asn Gly Thr Leu Thr Val Thr Ser Thr Leu Pro
385 390 395 400
Val Gly Thr Arg Asp Trp Ile Glu Gly Glu Thr Tyr Gln Cys Arg Val
405 410 415
Thr His Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr
420 425 430
Ser Gly Pro Arg Ala Ala Pro Glu Val Tyr Ala Phe Ala Thr Pro Glu
435 440 445
Trp Pro Gly Ser Arg Asp Lys Arg Thr Leu Ala Cys Leu Ile Gln Asn
450 455 460
Phe Met Pro Glu Asp Ile Ser Val Gln Trp Leu His Asn Glu Val Gln
465 470 475 480
Leu Pro Asp Ala Arg His Ser Thr Thr Gln Pro Arg Lys Thr Lys Gly
485 490 495
Ser Gly Phe Phe Val Phe Ser Arg Leu Glu Val Thr Arg Ala Glu Trp
500 505 510
Glu Gln Lys Asp Glu Phe Ile Cys Arg Ala Val His Glu Ala Ala Ser
515 520 525
Pro Ser Gln Thr Val Gln Arg Ala Val Ser Val Asn Pro Gly Lys
530 535 540
<210> 44
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 44
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 45
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 45
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr His
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Asn Ser Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Asn Leu Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Val
85 90 95
Phe Ser Ser Gly Ser His Asp Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 46
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 46
Ala Tyr Asp Met Thr Gln Thr Pro Ala Ser Val Glu Val Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Glu Leu Ile
35 40 45
Tyr Gly Thr Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ile Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Gly Val Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Ser Glu Asp Asn
85 90 95
Ile Asp Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 47
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 47
Val Asp Asp Ala Pro Gly Asn Ser Gln Gln Ala Thr Pro Lys Asp Asn
1 5 10 15
Glu Ile Ser Thr Phe His Asn Leu Gly Asn Val His Ser Pro Leu Lys
20 25 30
<210> 48
<211> 543
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 48
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Gln Ser Pro Ser Val Phe Pro Leu Thr Arg
115 120 125
Cys Cys Lys Asn Ile Pro Ser Asp Ala Thr Ser Val Thr Leu Gly Cys
130 135 140
Leu Ala Thr Gly Tyr Phe Pro Glu Pro Val Met Val Thr Trp Asp Thr
145 150 155 160
Gly Ser Leu Asp Gly Thr Thr Met Thr Leu Pro Ala Thr Thr Leu Thr
165 170 175
Leu Ser Gly His Tyr Ala Thr Ile Ser Leu Leu Thr Val Ser Gly Ala
180 185 190
Trp Ala Lys Gln Met Phe Thr Cys Arg Val Ala His Thr Pro Ser Ser
195 200 205
Thr Asp Trp Val Asp Asp Lys Thr Phe Ser Val Cys Ser Arg Asp Phe
210 215 220
Thr Pro Pro Thr Val Lys Ile Leu Gln Ser Ser Cys Asp Gly Gly Gly
225 230 235 240
His Phe Pro Pro Thr Ile Gln Leu Leu Cys Leu Val Ser Gly Tyr Thr
245 250 255
Pro Gly Thr Ile Asp Ile Thr Trp Leu Glu Asp Gly Gln Val Met Asp
260 265 270
Val Asp Leu Ser Thr Ala Ser Thr Thr Gln Glu Gly Glu Leu Ala Ser
275 280 285
Thr Gln Ser Glu Leu Thr Leu Ser Gln Lys His Trp Leu Ser Asp Arg
290 295 300
Thr Tyr Thr Cys Gln Val Thr Tyr Gln Gly His Thr Phe Glu Asp Ser
305 310 315 320
Thr Lys Lys Cys Ala Asp Ser Asn Pro Arg Gly Val Ser Ala Tyr Leu
325 330 335
Ser Arg Pro Ser Pro Phe Asp Leu Phe Ile Arg Lys Ser Pro Thr Ile
340 345 350
Thr Cys Leu Val Val Asp Leu Ala Pro Ser Lys Gly Thr Val Asp Leu
355 360 365
Thr Trp Ser Arg Ala Ser Gly Lys Pro Val Asp His Ser Thr Arg Lys
370 375 380
Glu Glu Lys Gln Arg Asn Gly Thr Leu Thr Val Thr Ser Thr Leu Pro
385 390 395 400
Val Gly Thr Arg Asp Trp Ile Glu Gly Glu Thr Tyr Gln Cys Arg Val
405 410 415
Thr His Pro His Leu Pro Arg Ala Leu Met Arg Ser Thr Thr Lys Thr
420 425 430
Ser Gly Pro Arg Ala Ala Pro Glu Val Tyr Ala Phe Ala Thr Pro Glu
435 440 445
Trp Pro Gly Ser Arg Asp Lys Arg Thr Leu Ala Cys Leu Ile Gln Asn
450 455 460
Phe Met Pro Glu Asp Ile Ser Val Gln Trp Leu His Asn Glu Val Gln
465 470 475 480
Leu Pro Asp Ala Arg His Ser Thr Thr Gln Pro Arg Lys Thr Lys Gly
485 490 495
Ser Gly Phe Phe Val Phe Ser Arg Leu Glu Val Thr Arg Ala Glu Trp
500 505 510
Glu Gln Lys Asp Glu Phe Ile Cys Arg Ala Val His Glu Ala Ala Ser
515 520 525
Pro Ser Gln Thr Val Gln Arg Ala Val Ser Val Asn Pro Gly Lys
530 535 540
<210> 49
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 49
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Pro Thr Ser Pro Lys Val Phe
115 120 125
Pro Leu Ser Leu Cys Ser Thr Gln Pro Asp Gly Asn Val Val Ile Ala
130 135 140
Cys Leu Val Gln Gly Phe Phe Pro Gln Glu Pro Leu Ser Val Thr Trp
145 150 155 160
Ser Glu Ser Gly Gln Gly Val Thr Ala Arg Asn Phe Pro Pro Ser Gln
165 170 175
Asp Ala Ser Gly Asp Leu Tyr Thr Thr Ser Ser Gln Leu Thr Leu Pro
180 185 190
Ala Thr Gln Cys Leu Ala Gly Lys Ser Val Thr Cys His Val Lys His
195 200 205
Tyr Thr Asn Pro Ser Gln Asp Val Thr Val Pro Cys Pro Val Pro Ser
210 215 220
Thr Pro Pro Thr Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
225 230 235 240
Cys Cys His Pro Arg Leu Ser Leu His Arg Pro Ala Leu Glu Asp Leu
245 250 255
Leu Leu Gly Ser Glu Ala Asn Leu Thr Cys Thr Leu Thr Gly Leu Arg
260 265 270
Asp Ala Ser Gly Val Thr Phe Thr Trp Thr Pro Ser Ser Gly Lys Ser
275 280 285
Ala Val Gln Gly Pro Pro Glu Arg Asp Leu Cys Gly Cys Tyr Ser Val
290 295 300
Ser Ser Val Leu Pro Gly Cys Ala Glu Pro Trp Asn His Gly Lys Thr
305 310 315 320
Phe Thr Cys Thr Ala Ala Tyr Pro Glu Ser Lys Thr Pro Leu Thr Ala
325 330 335
Thr Leu Ser Lys Ser Gly Asn Thr Phe Arg Pro Glu Val His Leu Leu
340 345 350
Pro Pro Pro Ser Glu Glu Leu Ala Leu Asn Glu Leu Val Thr Leu Thr
355 360 365
Cys Leu Ala Arg Gly Phe Ser Pro Lys Asp Val Leu Val Arg Trp Leu
370 375 380
Gln Gly Ser Gln Glu Leu Pro Arg Glu Lys Tyr Leu Thr Trp Ala Ser
385 390 395 400
Arg Gln Glu Pro Ser Gln Gly Thr Thr Thr Phe Ala Val Thr Ser Ile
405 410 415
Leu Arg Val Ala Ala Glu Asp Trp Lys Lys Gly Asp Thr Phe Ser Cys
420 425 430
Met Val Gly His Glu Ala Leu Pro Leu Ala Phe Thr Gln Lys Thr Ile
435 440 445
Asp Arg Leu Ala Gly Lys Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
450 455 460
<210> 50
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 50
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 51
<211> 545
<212> БЕЛОК
<213> Homo sapiens
<400> 51
Gln Pro Pro Pro Pro Pro Pro Asp Ala Thr Cys His Gln Val Arg Ser
1 5 10 15
Phe Phe Gln Arg Leu Gln Pro Gly Leu Lys Trp Val Pro Glu Thr Pro
20 25 30
Val Pro Gly Ser Asp Leu Gln Val Cys Leu Pro Lys Gly Pro Thr Cys
35 40 45
Cys Ser Arg Lys Met Glu Glu Lys Tyr Gln Leu Thr Ala Arg Leu Asn
50 55 60
Met Glu Gln Leu Leu Gln Ser Ala Ser Met Glu Leu Lys Phe Leu Ile
65 70 75 80
Ile Gln Asn Ala Ala Val Phe Gln Glu Ala Phe Glu Ile Val Val Arg
85 90 95
His Ala Lys Asn Tyr Thr Asn Ala Met Phe Lys Asn Asn Tyr Pro Ser
100 105 110
Leu Thr Pro Gln Ala Phe Glu Phe Val Gly Glu Phe Phe Thr Asp Val
115 120 125
Ser Leu Tyr Ile Leu Gly Ser Asp Ile Asn Val Asp Asp Met Val Asn
130 135 140
Glu Leu Phe Asp Ser Leu Phe Pro Val Ile Tyr Thr Gln Leu Met Asn
145 150 155 160
Pro Gly Leu Pro Asp Ser Ala Leu Asp Ile Asn Glu Cys Leu Arg Gly
165 170 175
Ala Arg Arg Asp Leu Lys Val Phe Gly Asn Phe Pro Lys Leu Ile Met
180 185 190
Thr Gln Val Ser Lys Ser Leu Gln Val Thr Arg Ile Phe Leu Gln Ala
195 200 205
Leu Asn Leu Gly Ile Glu Val Ile Asn Thr Thr Asp His Leu Lys Phe
210 215 220
Ser Lys Asp Cys Gly Arg Met Leu Thr Arg Met Trp Tyr Cys Ser Tyr
225 230 235 240
Cys Gln Gly Leu Met Met Val Lys Pro Cys Gly Gly Tyr Cys Asn Val
245 250 255
Val Met Gln Gly Cys Met Ala Gly Val Val Glu Ile Asp Lys Tyr Trp
260 265 270
Arg Glu Tyr Ile Leu Ser Leu Glu Glu Leu Val Asn Gly Met Tyr Arg
275 280 285
Ile Tyr Asp Met Glu Asn Val Leu Leu Gly Leu Phe Ser Thr Ile His
290 295 300
Asp Ser Ile Gln Tyr Val Gln Lys Asn Ala Gly Lys Leu Thr Thr Thr
305 310 315 320
Ile Gly Lys Leu Cys Ala His Ser Gln Gln Arg Gln Tyr Arg Ser Ala
325 330 335
Tyr Tyr Pro Glu Asp Leu Phe Ile Asp Lys Lys Val Leu Lys Val Ala
340 345 350
His Val Glu His Glu Glu Thr Leu Ser Ser Arg Arg Arg Glu Leu Ile
355 360 365
Gln Lys Leu Lys Ser Phe Ile Ser Phe Tyr Ser Ala Leu Pro Gly Tyr
370 375 380
Ile Cys Ser His Ser Pro Val Ala Glu Asn Asp Thr Leu Cys Trp Asn
385 390 395 400
Gly Gln Glu Leu Val Glu Arg Tyr Ser Gln Lys Ala Ala Arg Asn Gly
405 410 415
Met Lys Asn Gln Phe Asn Leu His Glu Leu Lys Met Lys Gly Pro Glu
420 425 430
Pro Val Val Ser Gln Ile Ile Asp Lys Leu Lys His Ile Asn Gln Leu
435 440 445
Leu Arg Thr Met Ser Met Pro Lys Gly Arg Val Leu Asp Lys Asn Leu
450 455 460
Asp Glu Glu Gly Phe Glu Ala Gly Asp Cys Gly Asp Asp Glu Asp Glu
465 470 475 480
Cys Ile Gly Gly Ala Gly Asp Gly Met Ile Lys Val Lys Asn Gln Leu
485 490 495
Arg Phe Leu Ala Glu Leu Ala Tyr Asp Leu Asp Val Asp Asp Ala Pro
500 505 510
Gly Asn Ser Gln Gln Ala Thr Pro Lys Asp Asn Glu Ile Ser Thr Phe
515 520 525
His Asn Leu Gly Asn Val His Ser Pro Leu Lys His His His His His
530 535 540
His
545
<210> 52
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Tyr Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 53
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 53
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr His
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Asn Ser Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Asn Leu Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Val
85 90 95
Phe Ser Ser Gly Ser His Asp Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Tyr Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 54
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 54
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr His
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Asn Ser Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Asn Leu Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Val
85 90 95
Phe Ser Ser Gly Ser His Asp Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Asp Ala Tyr Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 55
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 55
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr His
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Asn Ser Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Asn Leu Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Val
85 90 95
Phe Ser Ser Gly Ser His Asp Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Arg Gly Gly Pro Lys Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 56
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 56
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Arg Arg Gly Pro Lys Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Tyr His Val Thr Arg Lys Glu Leu Ser
435 440 445
Leu Ser Pro
450
<210> 57
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 57
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Arg Trp Glu Thr Ala Ile Ser Ser Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 58
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 58
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr His
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Asn Ser Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Asn Leu Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Val
85 90 95
Phe Ser Ser Gly Ser His Asp Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Arg Arg Gly Pro Lys Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Tyr
420 425 430
His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440
<210> 59
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 59
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 60
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 60
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 61
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Pro Val Pro Gly Val Tyr Tyr Tyr Tyr Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro
450
<210> 62
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 62
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ala Asn His
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Ser Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Thr Arg Ala Thr Asp Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Ser Ile Ser Gly Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Asp Trp Pro Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Phe Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Pro Val Pro Gly Val Tyr Tyr Tyr Tyr Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Val Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Tyr His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro
450
<210> 64
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 64
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 65
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 65
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 66
<211> 460
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 66
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Pro Thr Ser Pro Lys Val Phe Pro Leu Ser Leu
115 120 125
Cys Ser Thr Gln Pro Asp Gly Asn Val Val Ile Ala Cys Leu Val Gln
130 135 140
Gly Phe Phe Pro Gln Glu Pro Leu Ser Val Thr Trp Ser Glu Ser Gly
145 150 155 160
Gln Gly Val Thr Ala Arg Asn Phe Pro Pro Ser Gln Asp Ala Ser Gly
165 170 175
Asp Leu Tyr Thr Thr Ser Ser Gln Leu Thr Leu Pro Ala Thr Gln Cys
180 185 190
Leu Ala Gly Lys Ser Val Thr Cys His Val Lys His Tyr Thr Asn Pro
195 200 205
Ser Gln Asp Val Thr Val Pro Cys Pro Val Pro Ser Thr Pro Pro Thr
210 215 220
Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser Cys Cys His Pro
225 230 235 240
Arg Leu Ser Leu His Arg Pro Ala Leu Glu Asp Leu Leu Leu Gly Ser
245 250 255
Glu Ala Asn Leu Thr Cys Thr Leu Thr Gly Leu Arg Asp Ala Ser Gly
260 265 270
Val Thr Phe Thr Trp Thr Pro Ser Ser Gly Lys Ser Ala Val Gln Gly
275 280 285
Pro Pro Glu Arg Asp Leu Cys Gly Cys Tyr Ser Val Ser Ser Val Leu
290 295 300
Pro Gly Cys Ala Glu Pro Trp Asn His Gly Lys Thr Phe Thr Cys Thr
305 310 315 320
Ala Ala Tyr Pro Glu Ser Lys Thr Pro Leu Thr Ala Thr Leu Ser Lys
325 330 335
Ser Gly Asn Thr Phe Arg Pro Glu Val His Leu Leu Pro Pro Pro Ser
340 345 350
Glu Glu Leu Ala Leu Asn Glu Leu Val Thr Leu Thr Cys Leu Ala Arg
355 360 365
Gly Phe Ser Pro Lys Asp Val Leu Val Arg Trp Leu Gln Gly Ser Gln
370 375 380
Glu Leu Pro Arg Glu Lys Tyr Leu Thr Trp Ala Ser Arg Gln Glu Pro
385 390 395 400
Ser Gln Gly Thr Thr Thr Phe Ala Val Thr Ser Ile Leu Arg Val Ala
405 410 415
Ala Glu Asp Trp Lys Lys Gly Asp Thr Phe Ser Cys Met Val Gly His
420 425 430
Glu Ala Leu Pro Leu Ala Phe Thr Gln Lys Thr Ile Asp Arg Leu Ala
435 440 445
Gly Lys Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
450 455 460
<210> 67
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 67
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 68
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 68
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 69
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu
420 425 430
His Glu Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 70
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 70
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 71
<211> 365
<212> БЕЛОК
<213> Homo sapiens
<400> 71
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 72
<211> 119
<212> БЕЛОК
<213> Homo sapiens
<400> 72
Met Ser Arg Ser Val Ala Leu Ala Val Leu Ala Leu Leu Ser Leu Ser
1 5 10 15
Gly Leu Glu Ala Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Ala Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser
35 40 45
Gly Phe His Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu
50 55 60
Arg Ile Glu Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp
85 90 95
Glu Tyr Ala Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile
100 105 110
Val Lys Trp Asp Arg Asp Met
115
<210> 73
<211> 290
<212> БЕЛОК
<213> Mus musculus
<400> 73
Met Gly Met Pro Leu Pro Trp Ala Leu Ser Leu Leu Leu Val Leu Leu
1 5 10 15
Pro Gln Thr Trp Gly Ser Glu Thr Arg Pro Pro Leu Met Tyr His Leu
20 25 30
Thr Ala Val Ser Asn Pro Ser Thr Gly Leu Pro Ser Phe Trp Ala Thr
35 40 45
Gly Trp Leu Gly Pro Gln Gln Tyr Leu Thr Tyr Asn Ser Leu Arg Gln
50 55 60
Glu Ala Asp Pro Cys Gly Ala Trp Met Trp Glu Asn Gln Val Ser Trp
65 70 75 80
Tyr Trp Glu Lys Glu Thr Thr Asp Leu Lys Ser Lys Glu Gln Leu Phe
85 90 95
Leu Glu Ala Leu Lys Thr Leu Glu Lys Ile Leu Asn Gly Thr Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Ala Ser Asp Asn Ser Ser Val
115 120 125
Pro Thr Ala Val Phe Ala Leu Asn Gly Glu Glu Phe Met Lys Phe Asn
130 135 140
Pro Arg Ile Gly Asn Trp Thr Gly Glu Trp Pro Glu Thr Glu Ile Val
145 150 155 160
Ala Asn Leu Trp Met Lys Gln Pro Asp Ala Ala Arg Lys Glu Ser Glu
165 170 175
Phe Leu Leu Asn Ser Cys Pro Glu Arg Leu Leu Gly His Leu Glu Arg
180 185 190
Gly Arg Arg Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Gly Asn Ser Gly Ser Ser Val Leu Thr Cys Ala Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Lys Phe Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ser Gly Ser Gly Asn Cys Ser Thr Gly Pro Asn Gly Asp Gly Ser
245 250 255
Phe His Ala Trp Ser Leu Leu Glu Val Lys Arg Gly Asp Glu His His
260 265 270
Tyr Gln Cys Gln Val Glu His Glu Gly Leu Ala Gln Pro Leu Thr Val
275 280 285
Asp Leu
290
<210> 74
<211> 119
<212> БЕЛОК
<213> Mus musculus
<400> 74
Met Ala Arg Ser Val Thr Leu Val Phe Leu Val Leu Val Ser Leu Thr
1 5 10 15
Gly Leu Tyr Ala Ile Gln Lys Thr Pro Gln Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Pro Glu Asn Gly Lys Pro Asn Ile Leu Asn Cys Tyr Val Thr
35 40 45
Gln Phe His Pro Pro His Ile Glu Ile Gln Met Leu Lys Asn Gly Lys
50 55 60
Lys Ile Pro Lys Val Glu Met Ser Asp Met Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Ile Leu Ala His Thr Glu Phe Thr Pro Thr Glu Thr Asp
85 90 95
Thr Tyr Ala Cys Arg Val Lys His Ala Ser Met Ala Glu Pro Lys Thr
100 105 110
Val Tyr Trp Asp Arg Asp Met
115
<210> 75
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 75
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Tyr Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 76
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 76
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Glu Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 77
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 77
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 78
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА АНТИГЕН-МИШЕНЬ | 2012 |
|
RU2722829C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| СПЕЦИФИЧНАЯ К ТКАНИ-МИШЕНИ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2013 |
|
RU2743463C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ МНОГОКРАТНО СВЯЗЫВАТЬСЯ С МНОЖЕСТВОМ АНТИГЕННЫХ МОЛЕКУЛ | 2018 |
|
RU2757786C2 |
| ВАРИАНТ Fc-ОБЛАСТИ | 2014 |
|
RU2757124C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
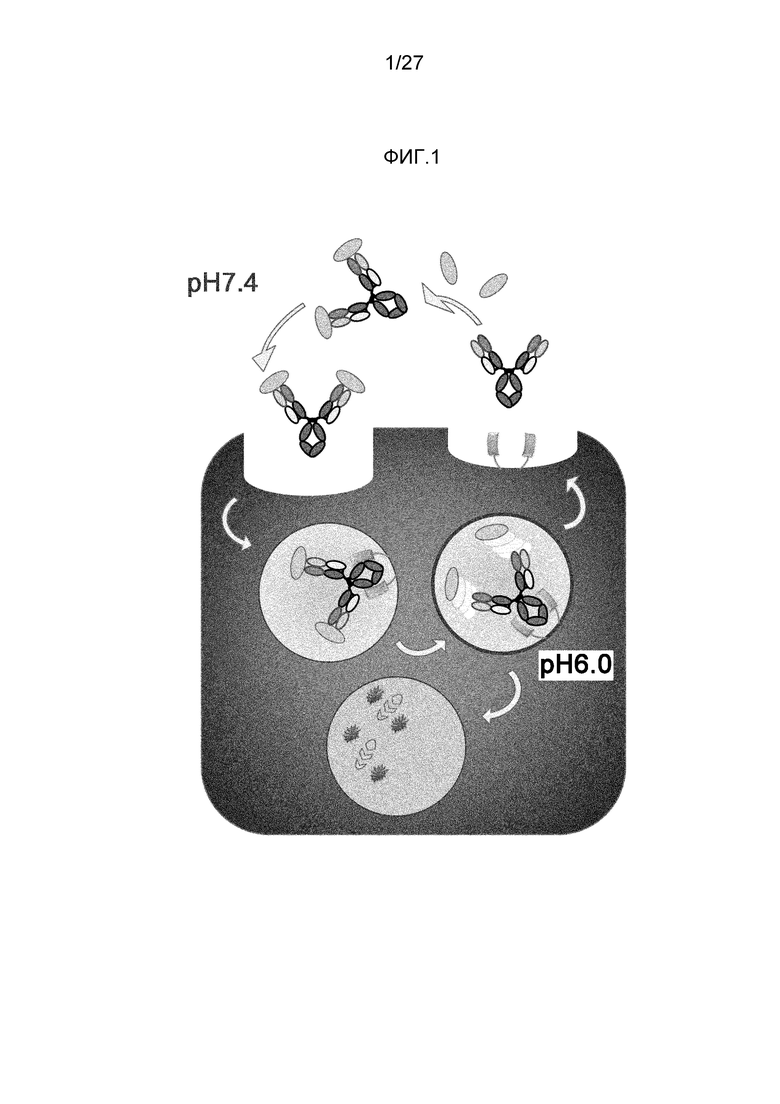
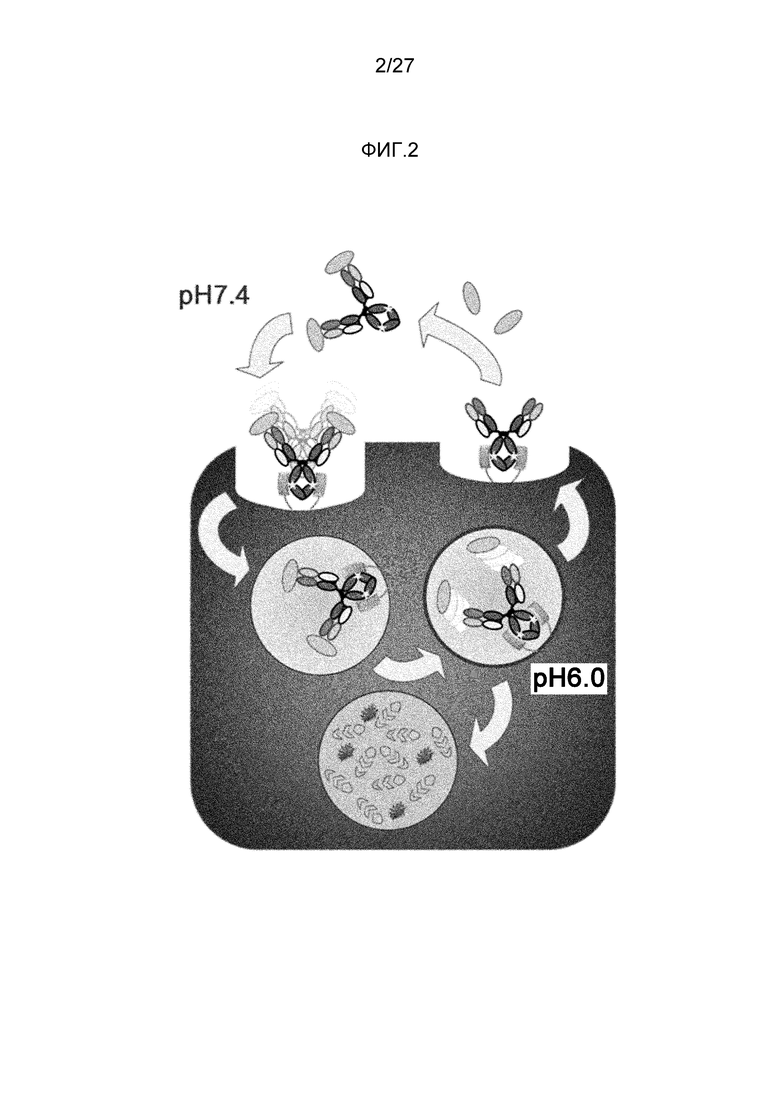
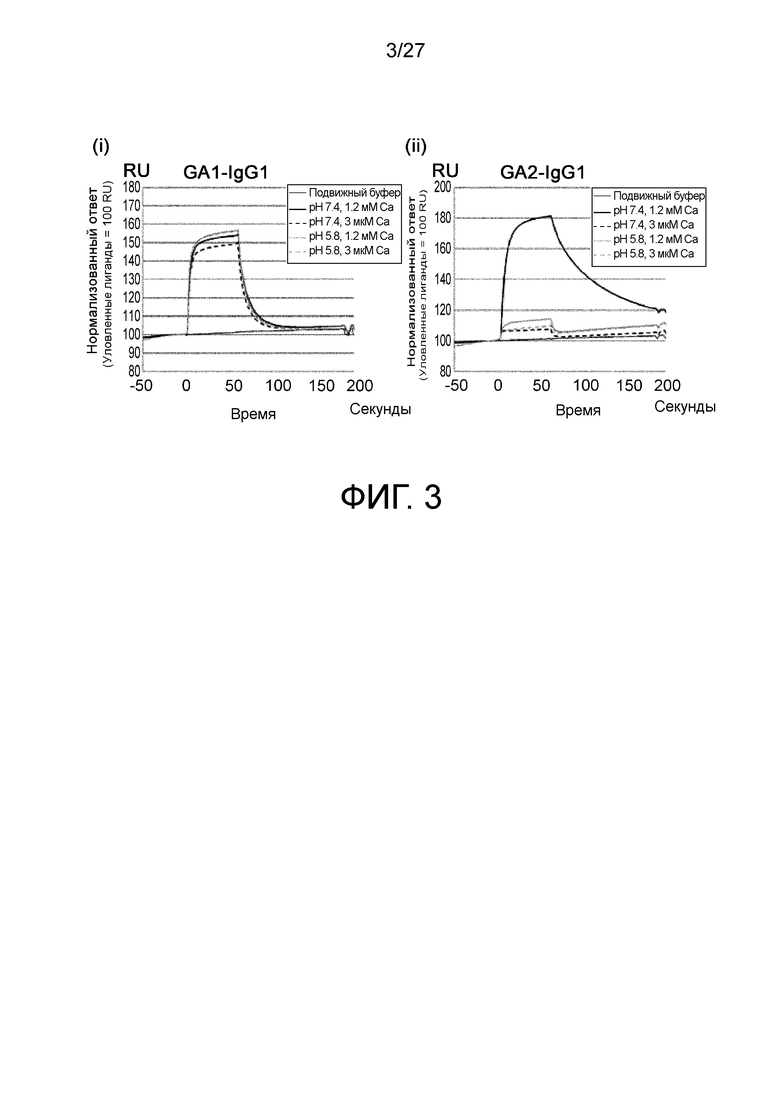

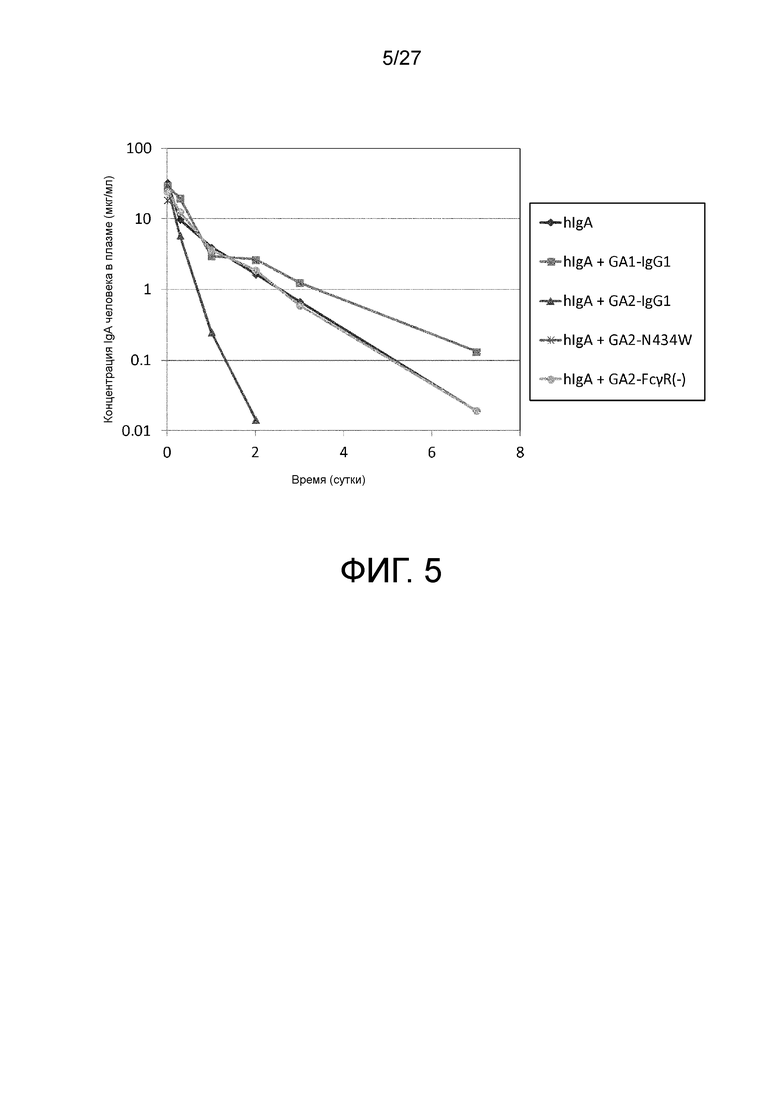
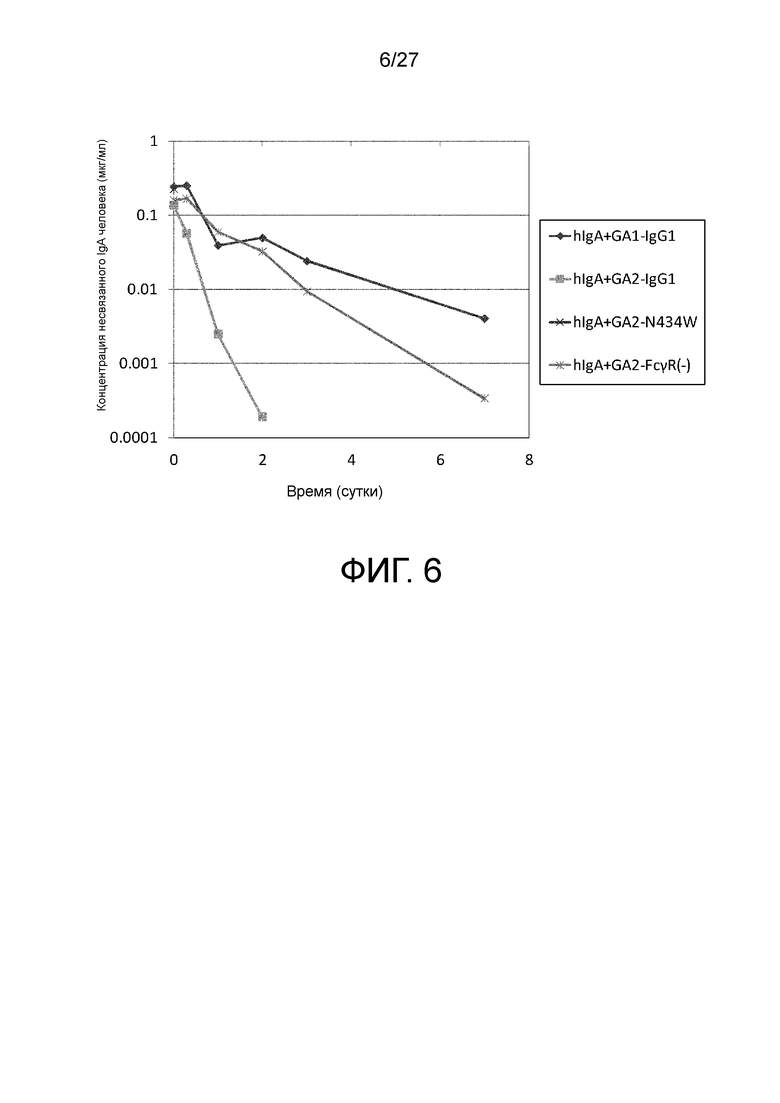
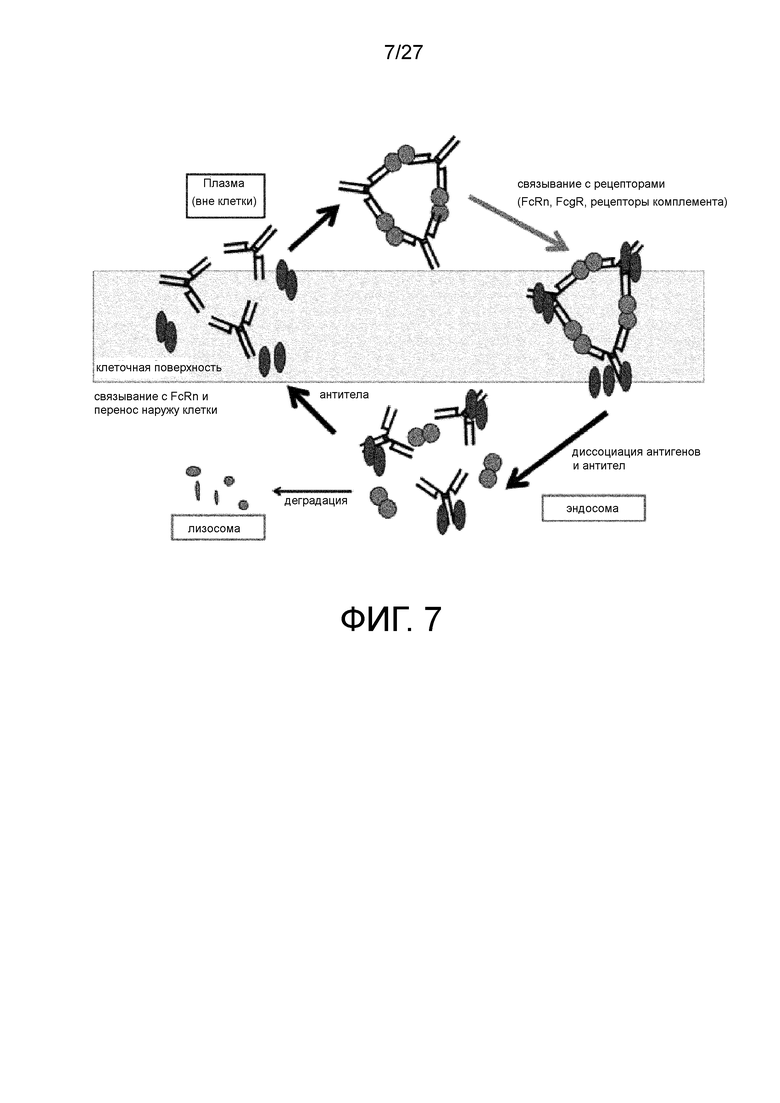

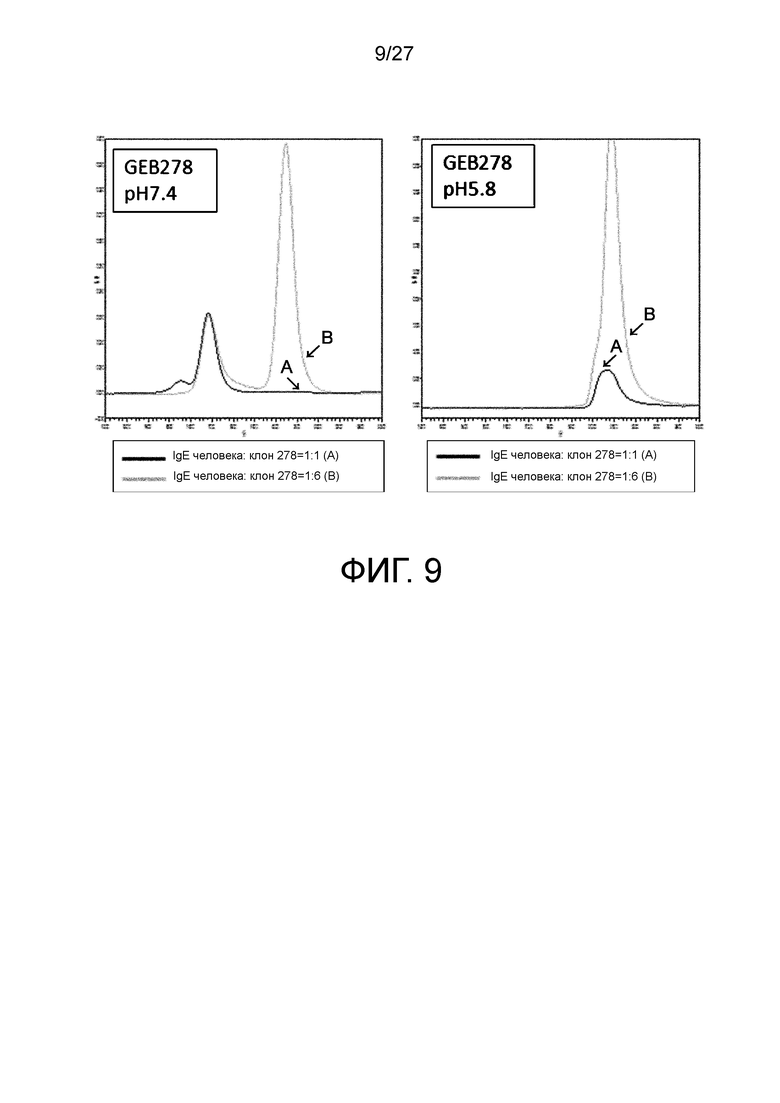
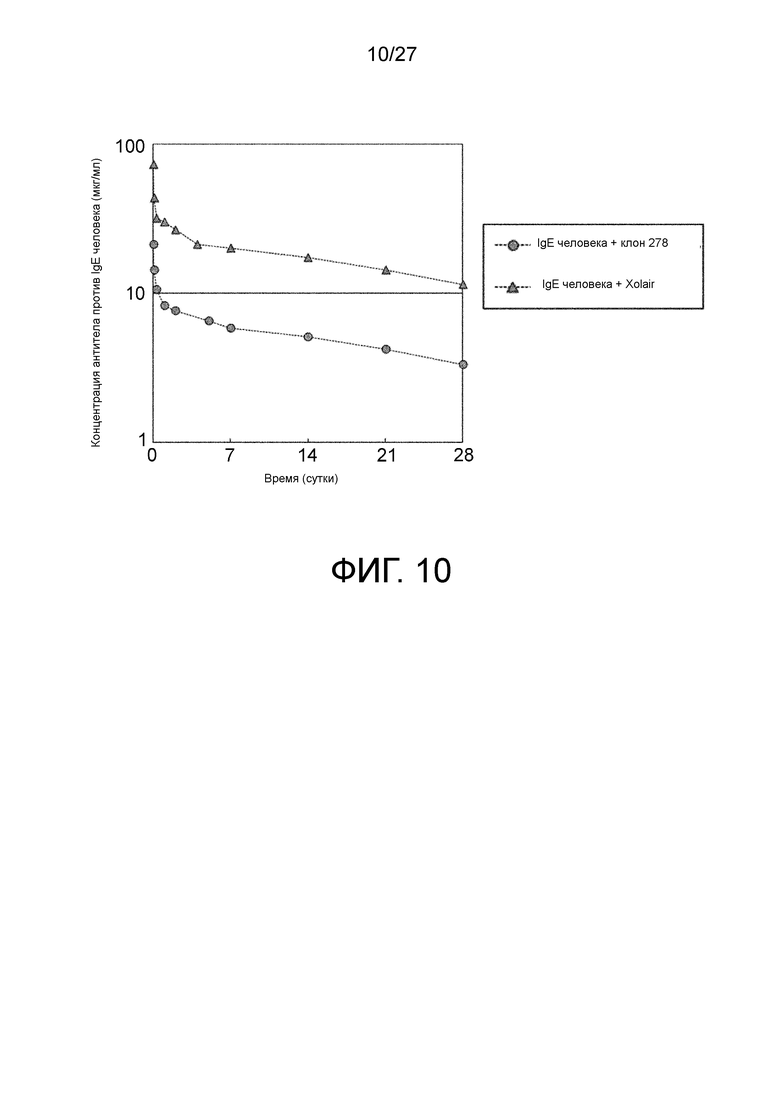
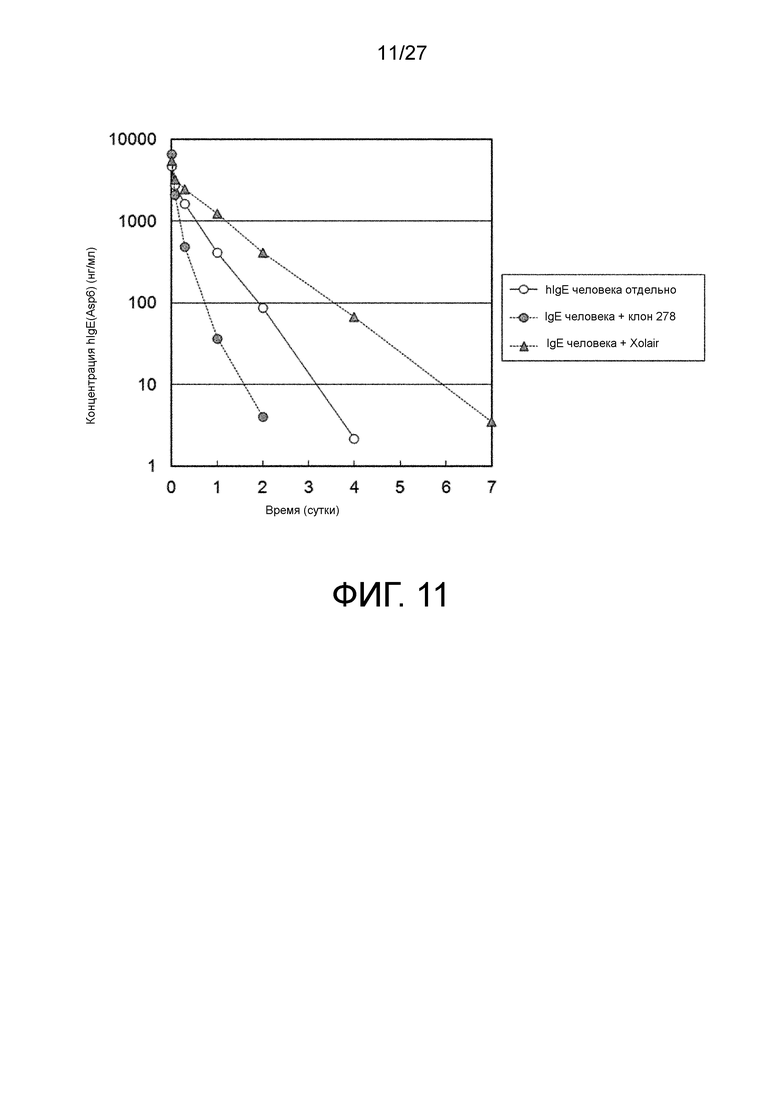
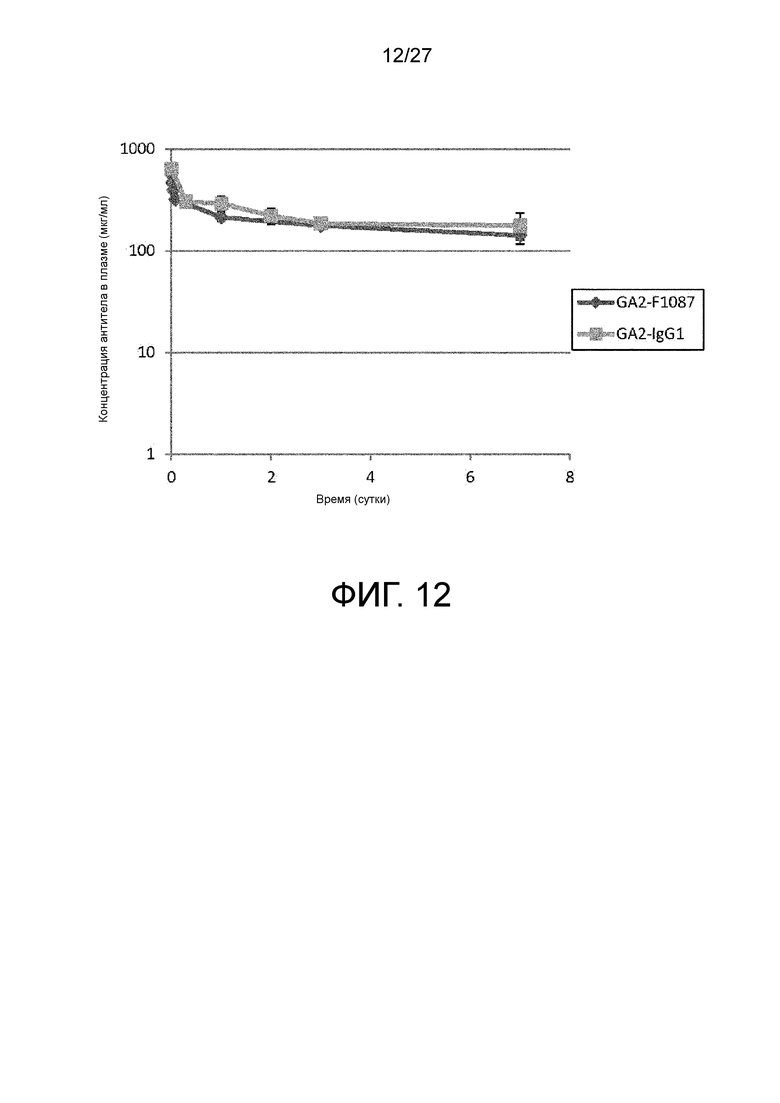
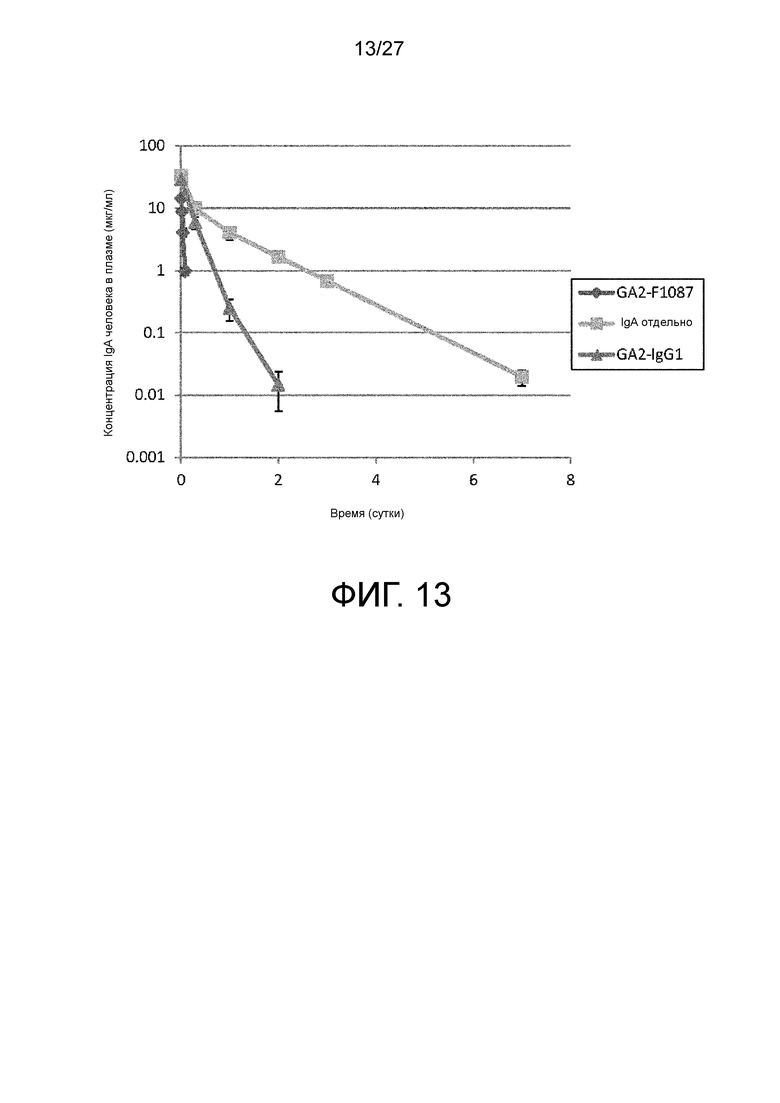
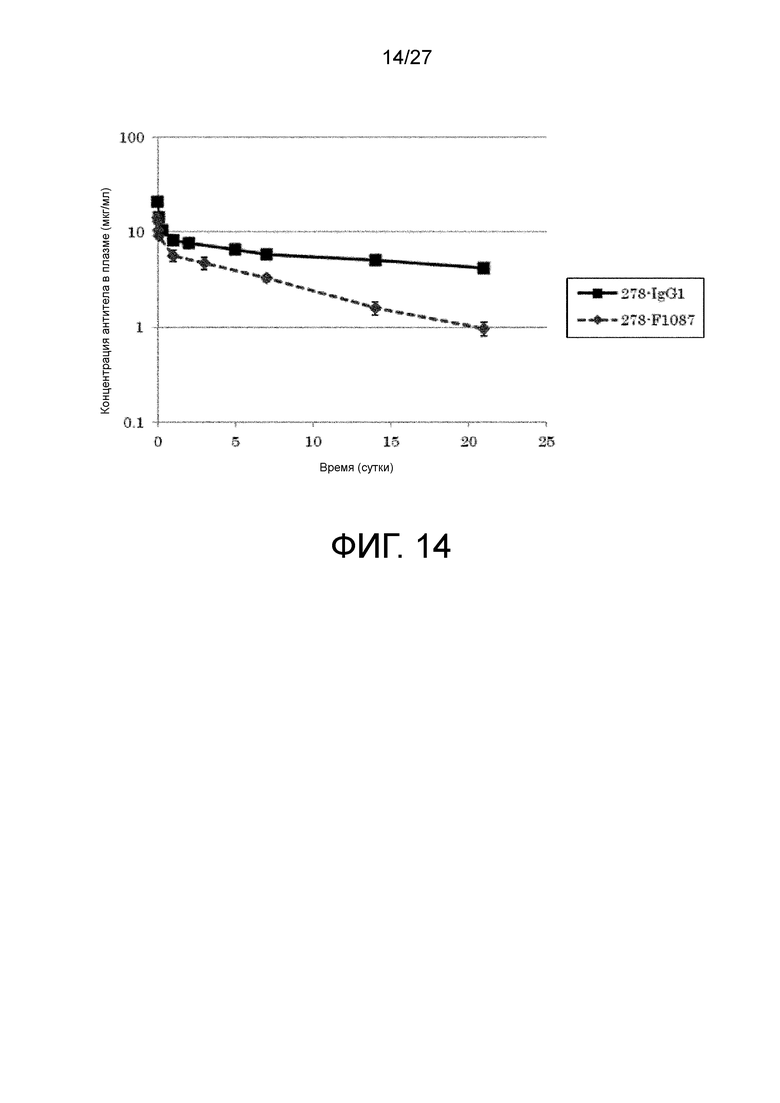
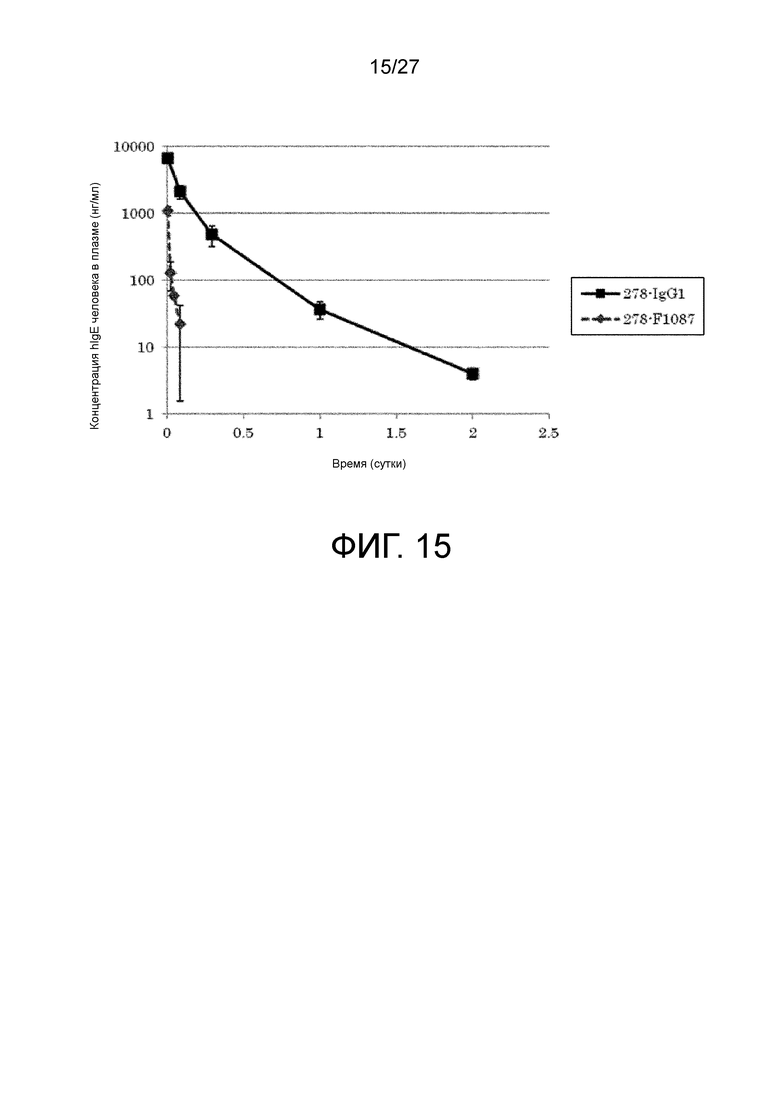
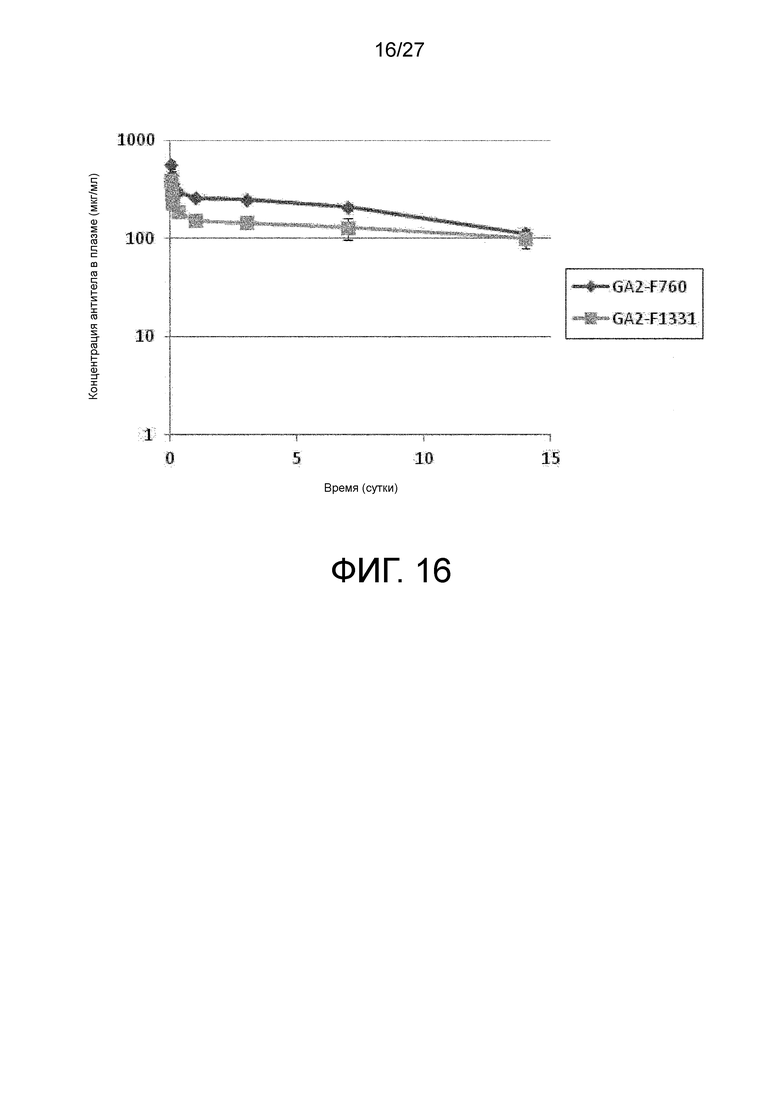
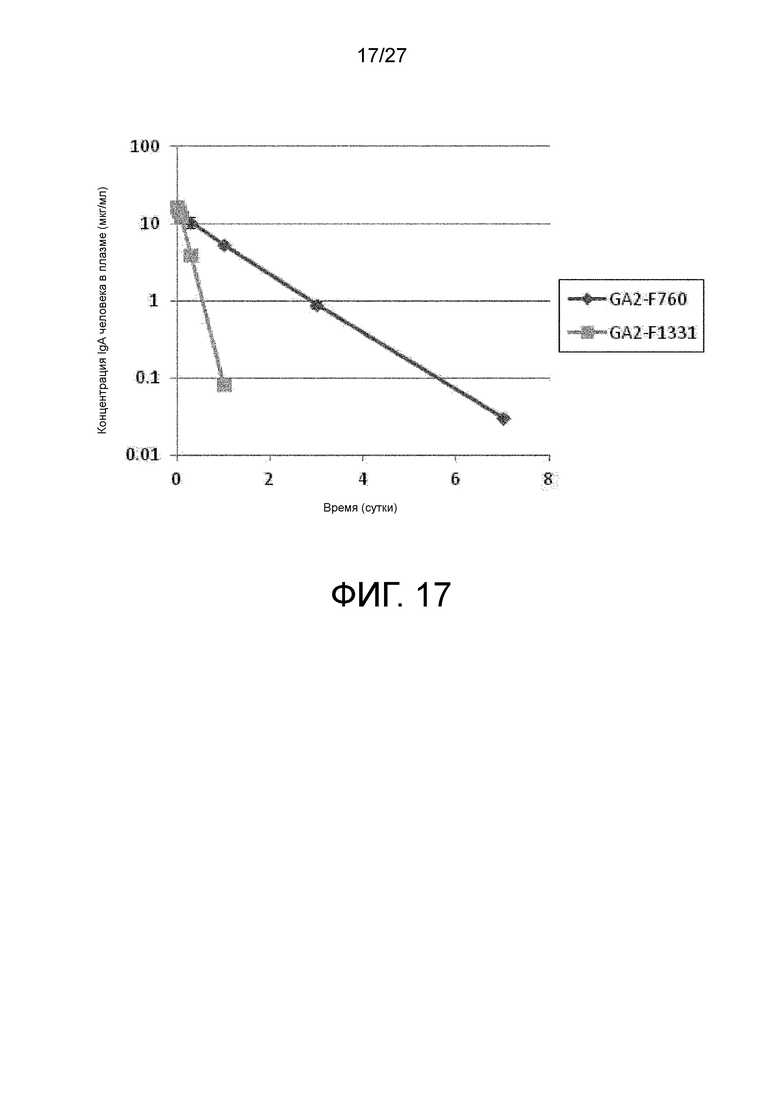
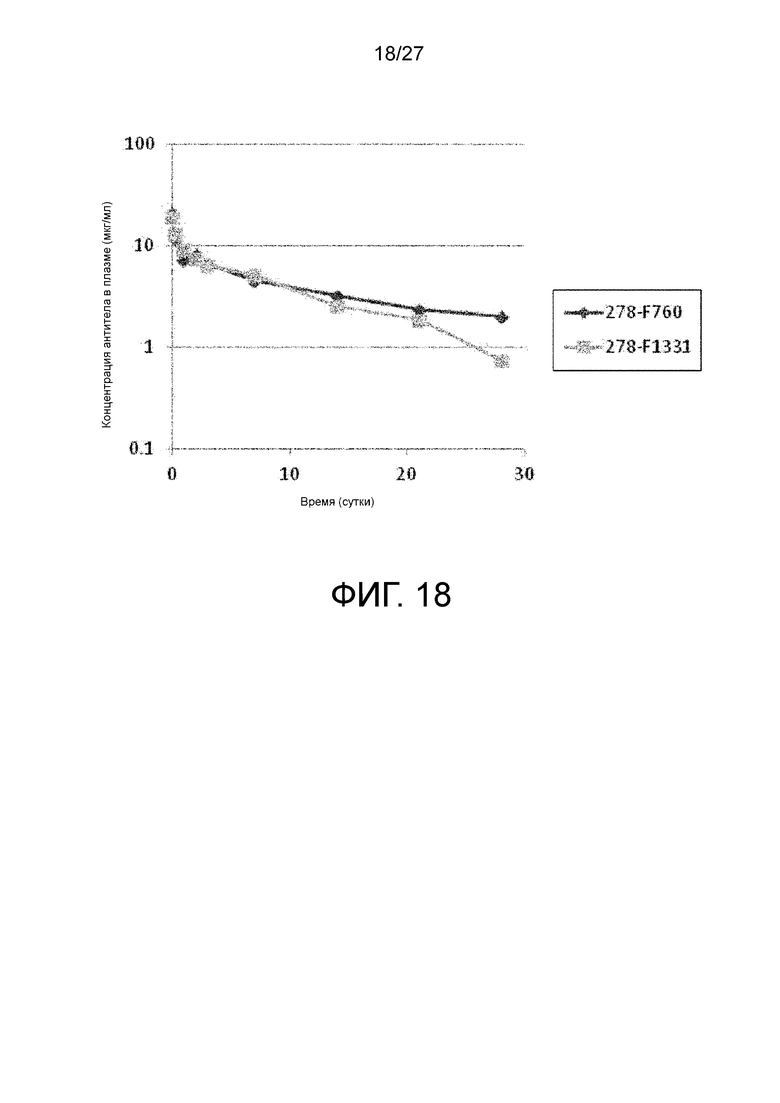
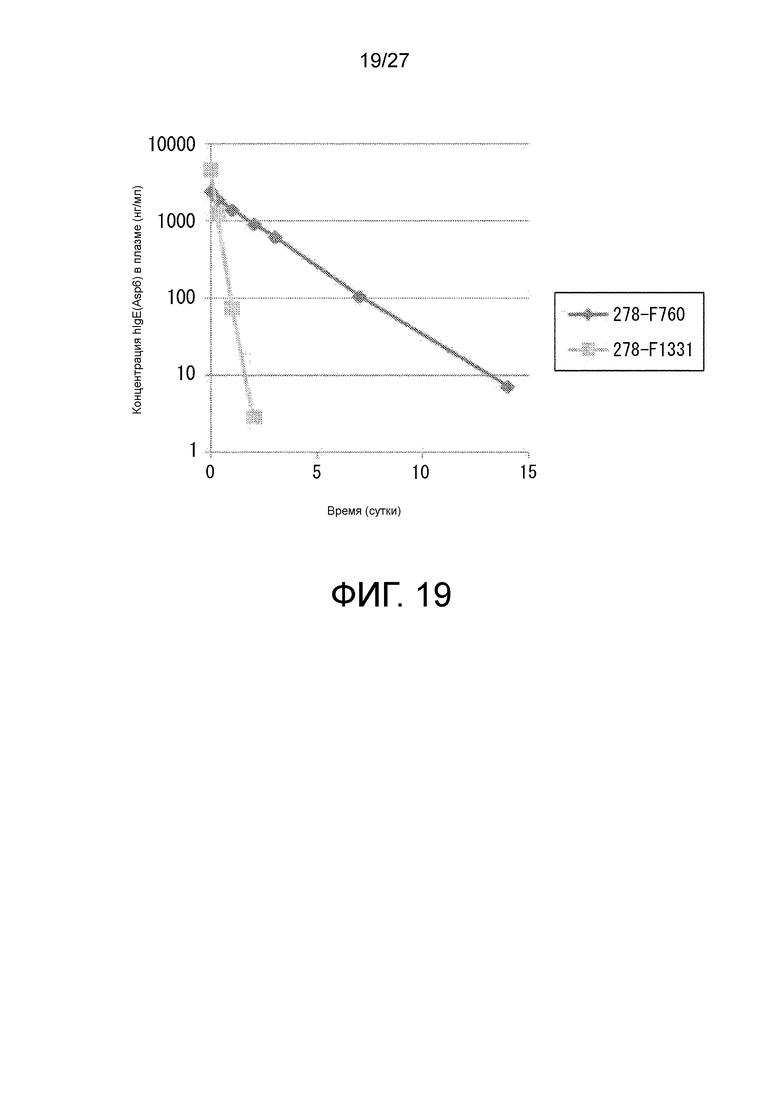
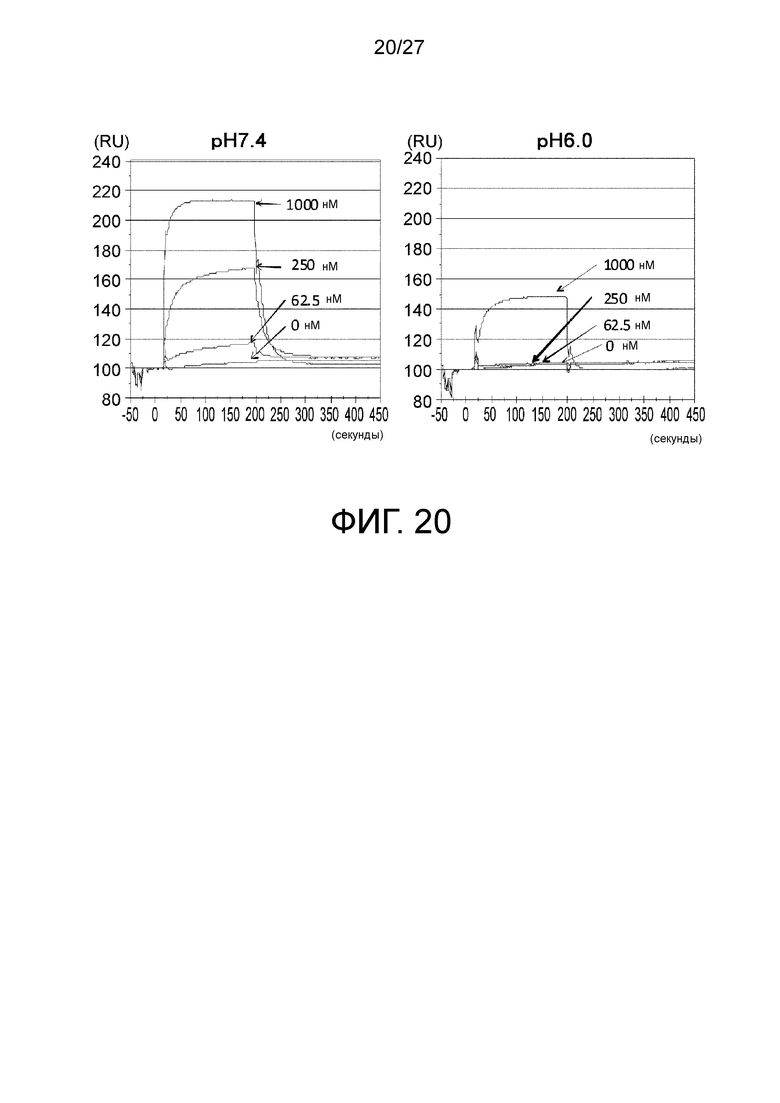
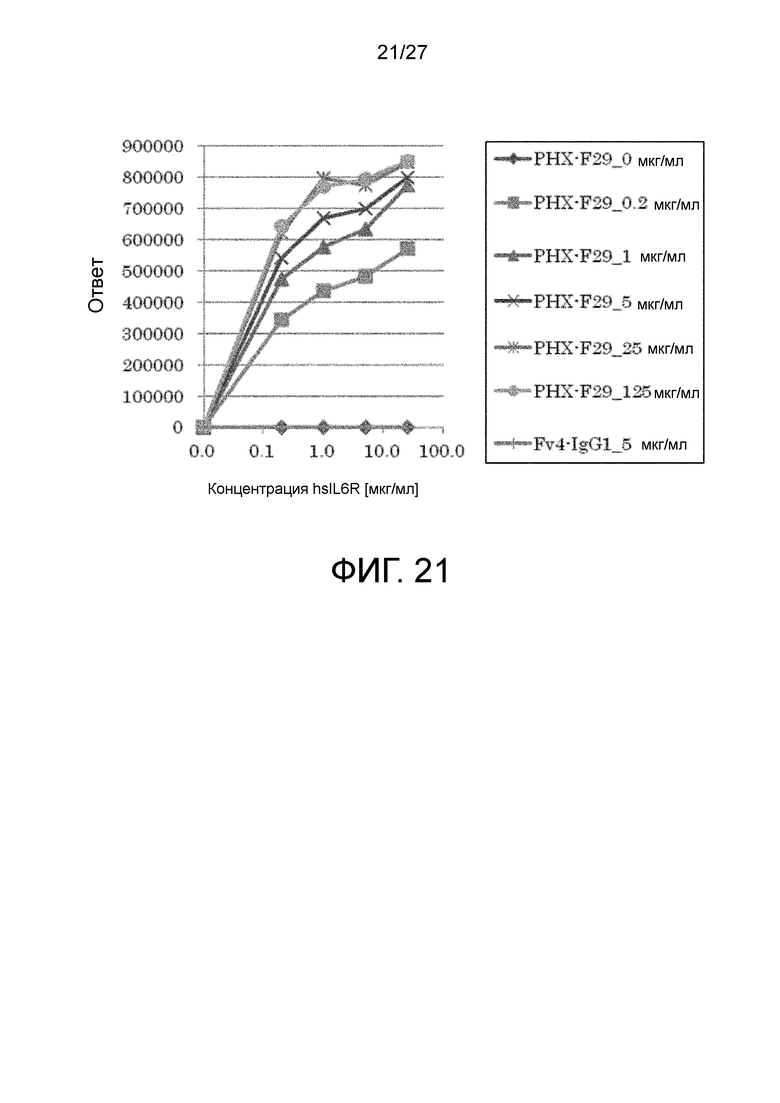
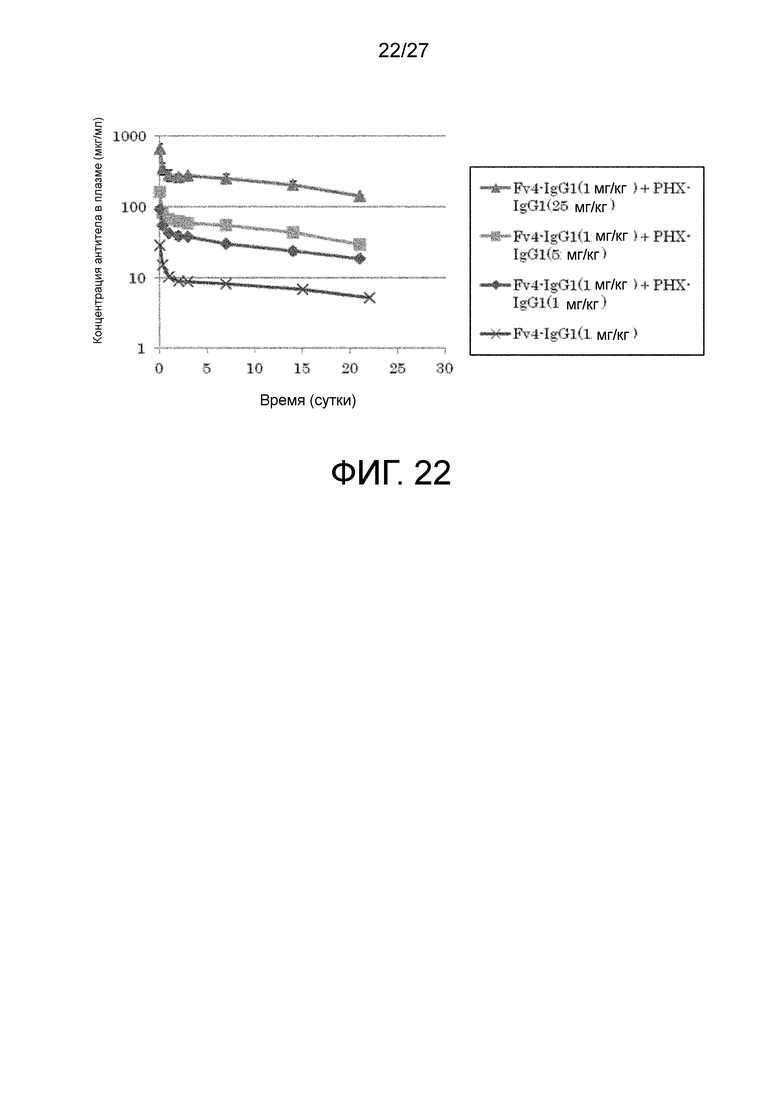
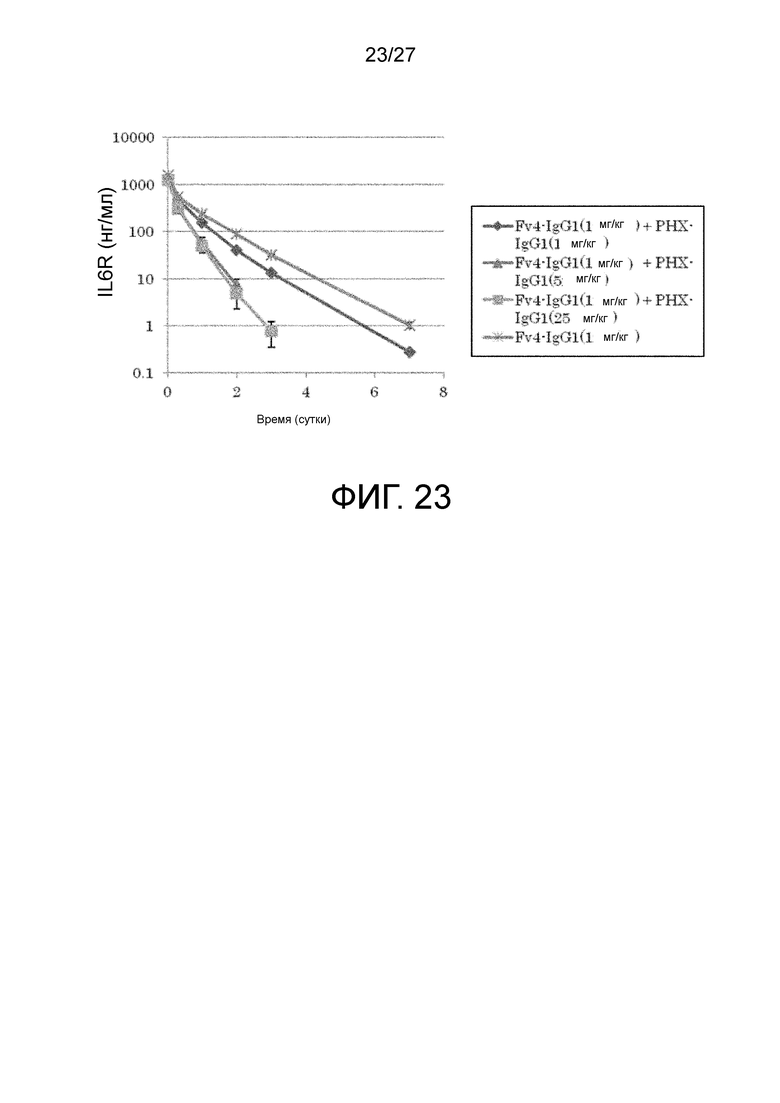
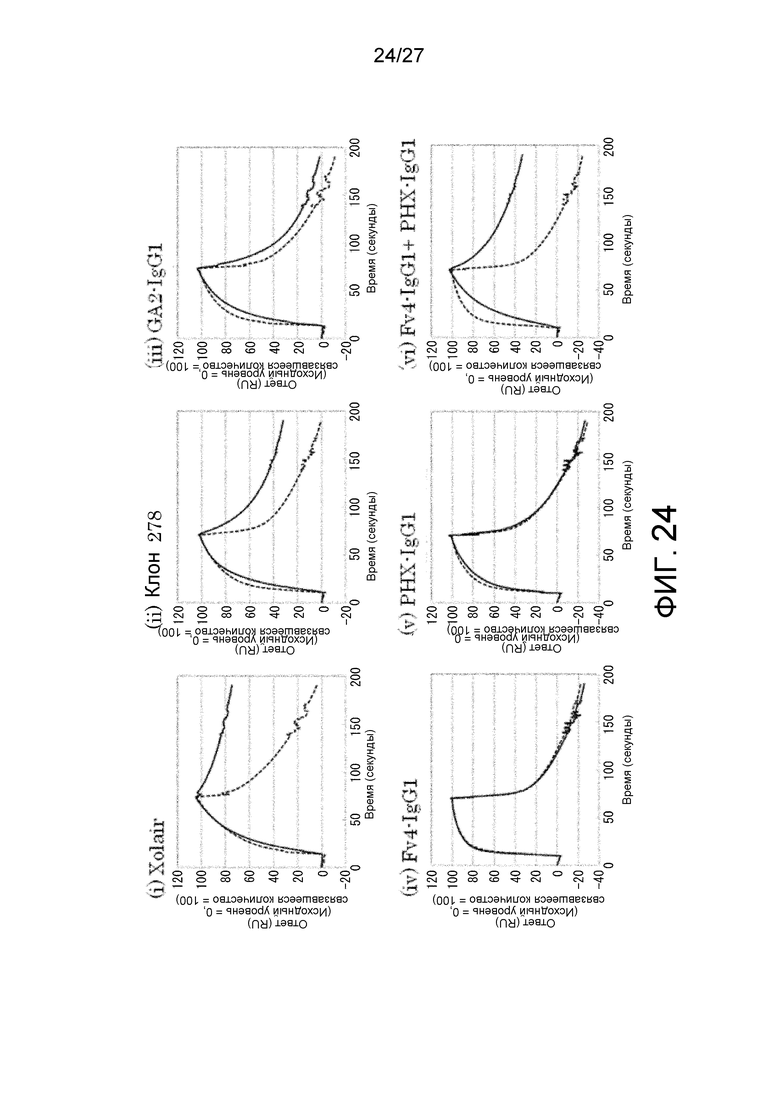
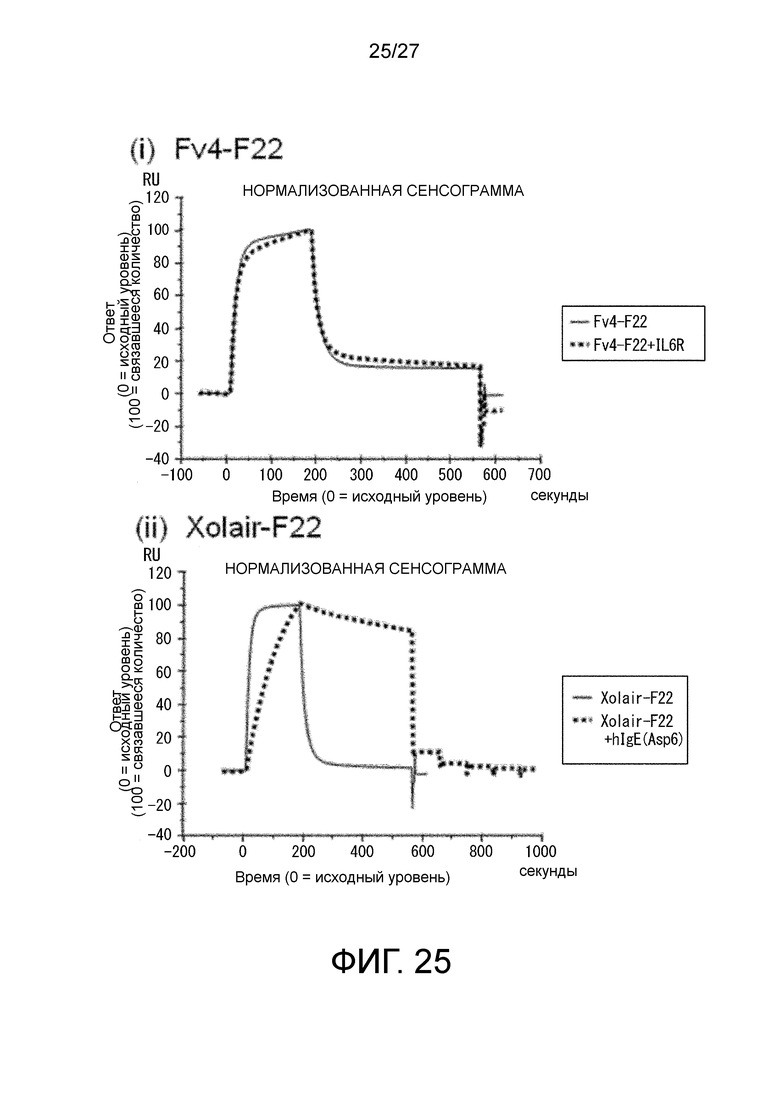
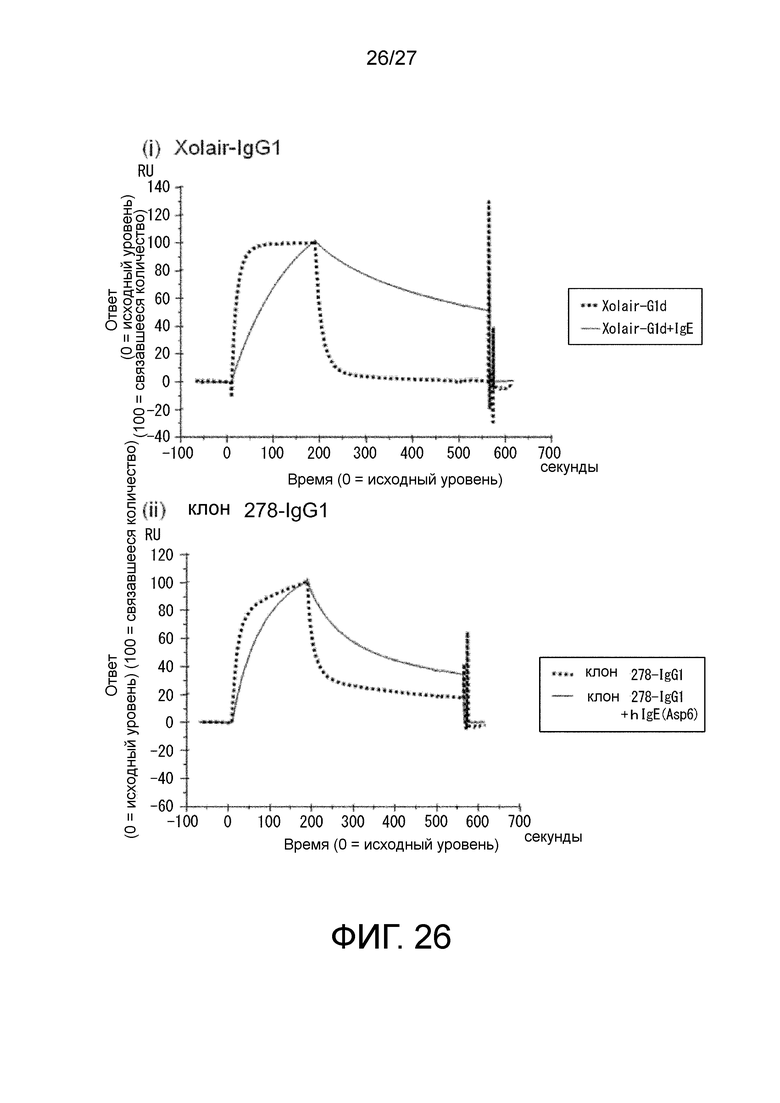
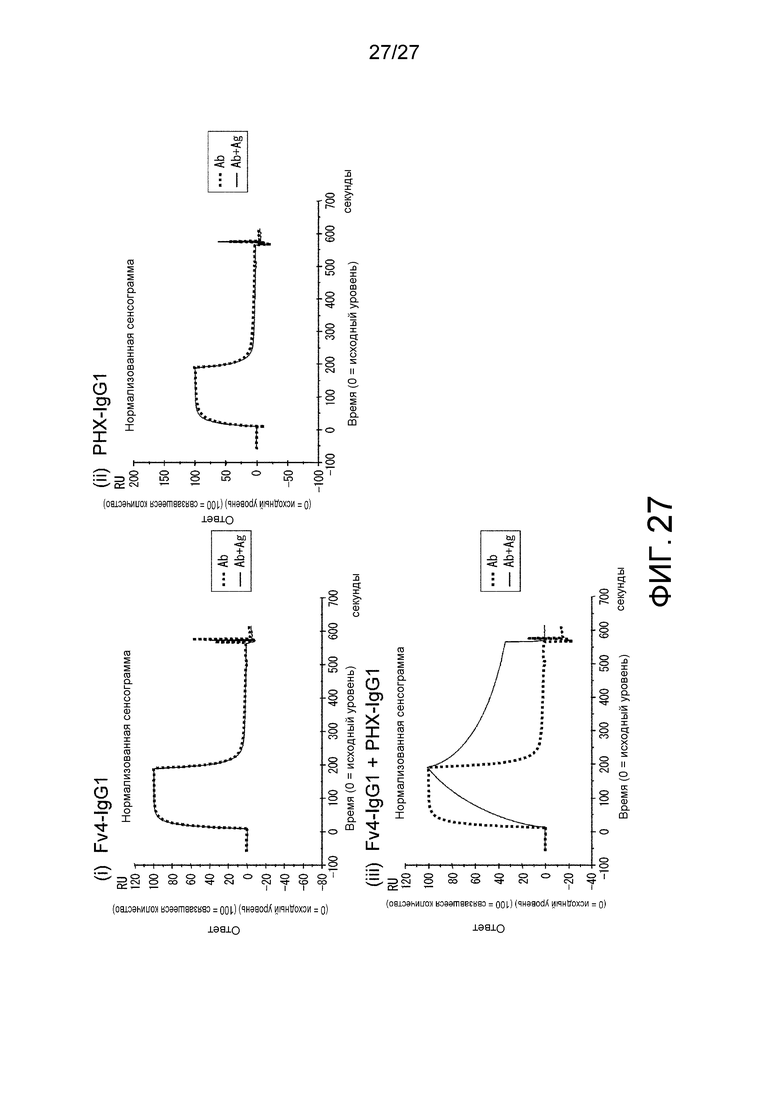
Изобретение относится к области биохимии, в частности к применению антитела, содержащего Fc-область человеческого IgG и два антигенсвязывающих домена для элиминации антигенов из плазмы. Также раскрыт способ скрининга антитела, обладающего функцией элиминации антигена из плазмы, включающий контактирование антигена с антигенсвязывающим доменом при рН 5.8 и рН 7.4 соответственно, получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий рН и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более. Изобретение также относится к способу получения антитела, обладающего функцией элиминации антигена из плазмы, предусматривающему получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий рН, и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более. Изобретение позволяет эффективно осуществлять получение антитела, обладающего функцией элиминации антигена. 5 н. и 18 з.п. ф-лы, 16 табл., 16 пр., 27 ил.
1. Применение антитела, содержащего (i) Fc-область человеческого IgG, представленную в любой из последовательностей SEQ ID NO:13, 14, 15 и 16 или ее вариант, где ее вариант обладает по крайней мере одним из следующих свойств:
(1) Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(2) Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(3) Fc-область с высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, и
(ii) два антигенсвязывающих доменов,
где по меньшей мере один из доменов представляет собой антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий рН, и где антитело образует иммунный комплекс, который содержит
(a) два или более антитела и
(b) два или более антигенов, где антигены содержат два или более антигенных элементов связывания, и являются гомомультимерами,
для элиминации антигенов из плазмы,
где антигенсвязывающий домен, антигенсвязывающая активность которого изменяется в зависимости от условий рН, включает гистидин, замещающий одну или более аминокислоту в CDRs антигенсвязывающего домена или включенный в него, и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более.
2. Применение по п.1, где антиген представляет собой любой из GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), TNF, TNF-альфа, TNF-альфа-бета, TNF-бета-2, TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд ODF TRANCE/RANK, лиганд OPG), TNFSF12 (лиганд Apo-3 TWEAK, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд AITR лиганда GITR, TL6), TNFSF1A (TNF-a коннектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд gp34 OX40, TXGP1), TNFSF5 (лиганд CD154 CD40, gp39, HIGM1, IMD3, TRAP), TNFSF6 (лиганд Apo-1 лиганда Fas, лиганд APT1), TNFSF7 (лиганд CD70 CD27), TNFSF8 (лиганд CD153 CD30), TNFSF9 (лиганд CD137 лиганда 4-1BB), VEGF, IgE, IgA, IgG, IgM, RANKL, TGF-альфа, TGF-бета, панспецифического TGF-бета и IL-8.
3. Применение по п. 1 , где антитело представляет собой мультиспецифическое или мультипаратопное антитело.
4. Применение по любому из пп.1-3, где Fc-область соответствует любой из SEQ ID NO:13, 14, 15 и 16.
5. Применение по любому из пп.1-4, где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16.
6. Применение по п.5, где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442 и 447 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16.
7. Применение по п.6, где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442; Ile, Pro или
Thr в качестве аминокислоты в положении 308;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16.
8. Применение по любому из пп.1-3, где Fc-область представляет собой Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16.
9. Применение по п.8, где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из положений 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16.
10. Применение по п.9, где Fc-область содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Met в качестве аминокислоты в положении 237;
Ile в качестве аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 250;
Phe, Trp или Tyr в качестве аминокислоты в положении 252;
Thr в качестве аминокислоты в положении 254;
Glu в качестве аминокислоты в положении 255;
Asp, Asn, Glu или Gln в качестве аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в качестве аминокислоты в положении 257;
His в качестве аминокислоты в положении 258;
Ala в качестве аминокислоты в положении 265;
Ala или Glu в качестве аминокислоты в положении 286;
His в качестве аминокислоты в положении 289;
Ala в качестве аминокислоты в положении 297;
Ala в качестве аминокислоты в положении 303;
Ala в качестве аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в качестве аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg в качестве аминокислоты в положении 309;
Ala, His или Ile в качестве аминокислоты в положении 311;
Ala или His в качестве аминокислоты в положении 312;
Lys или Arg в качестве аминокислоты в положении 314;
Ala, Asp или His в качестве аминокислоты в положении 315;
Ala в качестве аминокислоты в положении 317;
Val в качестве аминокислоты в положении 332;
Leu в качестве аминокислоты в положении 334;
His в качестве аминокислоты в положении 360;
Ala в качестве аминокислоты в положении 376;
Ala в качестве аминокислоты в положении 380;
Ala в качестве аминокислоты в положении 382;
Ala в качестве аминокислоты в положении 384;
Asp или His в качестве аминокислоты в положении 385;
Pro в качестве аминокислоты в положении 386;
Glu в качестве аминокислоты в положении 387;
Ala или Ser в качестве аминокислоты в положении 389;
Ala в качестве аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 428;
Lys в качестве аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr в качестве аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val в качестве аминокислоты в положении 436;
как указано в соответствии с нумерацией EU в аминокислотной последовательности Fc-области, соответствующей любой из SEQ ID NO:13, 14, 15 и 16.
11. Применение по любому из пп.1-3, 6 или 7, где Fc-область включает Fc-область, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека.
12. Применение по п.11, где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, которые отличаются от аминокислот нативной Fc-области IgG человека, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU).
13. Применение по п.12, где Fc-область содержит в ее аминокислотной последовательности по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или
Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440;
как указано в соответствии с нумерацией EU.
14. Применение по п.1, где Fc-область обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором.
15. Применение по п.14, где ингибиторный Fcγ-рецептор представляет собой FcγRIIb человека.
16. Применение по п.14, где активирующий Fcγ-рецептор представляет собой FcγRIa человека, FcγRIIa (R) человека, FcγRIIa (H) человека, FcγRIIIa (V) человека или FcγRIIIa (F) человека.
17. Применение по любому из пп.14-16, где аминокислота в положении 238 или 328 (нумерация EU) в Fc-области отличается от аминокислоты в нативной Fc-области IgG человека.
18. Применение по п.17, где аминокислота в положении 238 Fc-области представляет собой Asp, или аминокислота в положении 328 Fc-области представляет собой Glu, как указано в соответствии с нумерацией EU.
19. Применение по п.17, где аминокислотная последовательность Fc-области содержит по меньшей мере одну или более аминокислот, выбранных из группы, состоящей из:
Asp в качестве аминокислоты в положении 233;
Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Leu, Met, Phe, Trp или Tyr в качестве аминокислоты в положении 237;
Asp в качестве аминокислоты в положении 239;
Ala, Gln или Val в качестве аминокислоты в положении 267;
Asn, Asp или Glu в качестве аминокислоты в положении 268;
Gly в качестве аминокислоты в положении 271;
Ala, Asn, Asp, Gln, Glu, Leu, Met, Ser или Thr в качестве аминокислоты в положении 326;
Arg, Lys или Met в качестве аминокислоты в положении 330;
Ile, Leu или Met в качестве аминокислоты в положении 323; и
Asp в качестве аминокислоты в положении 296;
как указано в соответствии с нумерацией EU.
20. Способ скрининга антитела, обладающего функцией элиминации антигена из плазмы, где способ включает:
(a) контактирование антигена с антигенсвязывающим доменом при рН 5.8 и рН 7.4 соответственно;
(b) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий рН и имеет значение KD (pH 5.8)/KD (pH 7.4),
определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более;
(с) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (b) выше;
(d) функциональное связывание гена, полученного согласно (с) выше, с геном, кодирующим Fc-область человеческого IgG, представленную в любой из последовательностей SEQ ID NO:13,14,15 и 16 или ее вариант, где ее вариант обладает по крайней мере одним из следующих свойств:
(1) Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(2) Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной любой из SEQ ID NO:13, 14, 15 и 16;
(3) Fc-область с высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG;
(e) культивирование клетки, содержащей гены, функционально связанные согласно (d) выше;
(f) выделение антитела из культурального раствора, полученного согласно (е) выше;
(g) контактирование антитела, полученного согласно (f) выше, с антигеном; и
(h) оценку образования иммунного комплекса, (i) содержащего два или более антитела и (ii) два или более антигена, где антигены включают две или более антигенсвязывающие единицы и являются гомомультимерами;
(i) отбор целевого антитела, по результатам оценки образования иммунного комплекса на стадии (h).
21. Способ получения антитела, обладающего функцией элиминации антигена из плазмы, где способ включает:
(a) контактирование антигена с антителом, содержащим Fc-область человеческого IgG, представленную в любой из последовательностей SEQ ID NO:13, 14, 15 и 16 или ее вариант, где ее вариант обладает по крайней мере одним из следующих свойств:
(1) Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(2) Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(3) Fc-область с высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человек;
и два или более антигенсвязывающих доменов, где по меньшей мере один из антигенсвязывающих доменов обладает антигенсвязывающей активностью, которая изменяется в зависимости от условий рН и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более;
(b) оценку образования иммунного комплекса, содержащего (i) два или более антитела и (ii) два или более антигена, где антигены включают две или более антигенсвязывающие единицы;
(c) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антитело, для которой подтверждено, что она образует иммунный комплекс, согласно (b) выше; и
(d) выделение антитела из культурального раствора, полученного согласно (c) выше.
22. Способ получения антитела, обладающего функцией элиминации антигена из плазмы, где способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий рН и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область человеческого IgG, представленную в любой из последовательностей SEQ ID NO:13, 14, 15 и 16 или ее вариант, где ее вариант обладает по крайней мере одним из следующих свойств:
(1) Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;(2) Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(3) Fc-область с высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека и (d) культивирование клетки, содержащей гены, функционально
связанные согласно (c) выше;
(e) выделение антитела культурального раствора, полученного согласно (d) выше;
(f) контактирование антитела, полученного согласно (e) выше, с антигеном;
(g) оценку образования иммунного комплекса, содержащего (i) два или более антитела и (ii) два или более антигена, где антигены включают две или более антигенсвязывающие единицы;
(h) культивирование клетки-хозяина, содержащей вектор, который содержит ген, кодирующий антитело, для которой подтверждено, что она образует иммунный комплекс, согласно (g) выше; и
(i) выделение антитела из культурального раствора, полученного согласно (h) выше.
23. Способ получения антитела, обладающего функцией элиминации антигена из плазмы, где способ включает:
(a) получение антигенсвязывающего домена, антигенсвязывающая активность которого изменяется в зависимости от условий рН, и имеет значение KD (pH 5.8)/KD (pH 7.4), определенное как соотношение KD для данного антигена при pH 5.8 и KD для данного антигена при pH 7.4, равное 2 или более;
(b) получение гена, кодирующего антигенсвязывающий домен, выбранный согласно (a) выше;
(c) функциональное связывание гена, полученного согласно (b) выше, с геном, кодирующим Fc-область человеческого IgG, представленную в любой из последовательностей SEQ ID NO:13, 14, 15 и 16 или ее вариант, где ее вариант обладает по крайней мере одним из следующих свойств:
(1) Fc-область с усиленной активностью связывания FcRn в условиях диапазона кислых значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(2) Fc-область с усиленной активностью связывания FcRn в условиях диапазона нейтральных значений pH по сравнению с Fc-областью, представленной в любой из SEQ ID NO:13, 14, 15 и 16;
(3) Fc-область с высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека;
(d) культивирование клетки, содержащей гены, функционально связанные согласно (c) выше; и
(e) выделение антитела из культурального раствора, полученного согласно (d) выше; и где способ дополнительно включает контактирование антитела, полученного способом получения, с антигеном, и оценку образования иммунного комплекса, содержащего два или более антитела и
(ii) два или более антигена, где антигены включают две или более антигенсвязывающие единицы.
| МНОГОПОЛЯРНАЯ ЭЛЕКТРОЛИЗНАЯ ВАННА ДЛЯ ПОЛУЧЕНИЯ ЖИДКИХ МЕТАЛЛОВ ЭЛЕКТРОЛИЗОМ РАСПЛАВОВ И СПОСОБ УСТАНОВКИ ЭЛЕКТРОЛИЗНЫХ ВАНН | 2004 |
|
RU2275443C2 |
| WO2011122011 A2, 06.10.2011 | |||
| US2007009523 A1, 11.01.2007 | |||
| WO2006053301 A2, 18.05.2006 | |||
| АНТИТЕЛА К Aβ-ПЕПТИДУ И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2341533C2 |

