ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНЫЕ ЗАЯВКИ
Настоящая заявка испрашивает преимущество по заявке на патент США №62/083,005, поданной 21 ноября 2014 г., заявке на патент США №62/182,314, поданной 19 июня 2015 г., и заявке на патент США №62/211,421, поданной 28 августа 2015 г., каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
ССЫЛКА НА ПЕРЕДАННЫЙ ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ КАК ТЕКСТОВЫЙ ФАЙЛ ЧЕРЕЗ EFS WEB
Перечень последовательностей, записанный в файл 472225SEQLIST.txt с размером 32,7 Кб, созданный 20 ноября 2015 г., включен в настоящий документ путем ссылки.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Хотя имеется определенный прогресс в нацеливании на различные геномные локусы, все еще остается множество типов геномных локусов, нацеливание на которые невозможно провести эффективно, или геномных модификаций, которые невозможно провести эффективно или надлежащим образом с использованием традиционных стратегий нацеливания. Например, возникают сложности при попытке создать нацеленные геномные делеции или другие крупные нацеленные генетические модификации, в особенности в эукариотических клетках и организмах.
В частности, с использованием традиционных стратегий нацеливания сложно эффективно получать клетки или животных, которые являются гомозиготными или компаунд-гетерозиготными (например, гемизиготными) по большой нацеленной геномной делеции или другой геномной модификации. Например, хотя посредством традиционных стратегий нацеливания можно получить мышей поколения F0, гетерозиготных по большой нацеленной геномной делеции, необходимо последующее скрещивание таких гетерозиготных мышей для получения мышей поколения F2, гомозиготных по делеции. Эти дополнительные этапы скрещивания являются дорогостоящими и требуют времени.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Предложены способы и композиции для модификации генома в клетке. В одном аспекте в изобретении предложены способы внесения модификации в геном в клетке, включающие в себя приведение генома в контакт с: (а) первым белком Cas; (b) первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (с) второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (d) тракрРНК; и (е) нацеливающим вектором, содержащим полинуклеотидную вставку, фланкированную 5' гомологичным плечом, гибридизующимся с целевой 5'-последовательностью, и 3' гомологичным плечом, гибридизующимся с целевой 3'-последовательностью, при условии, что если клетка представляет собой эмбрион на стадии единственной клетки, нацеливающий вектор имеет длину не более 5 т.п.н.; причем геном содержит пару из первой и второй гомологичных хромосом, содержащих целевой геномный локус; и при этом первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва в по меньшей мере одной из первой и второй гомологичных хромосом. В одном аспекте в изобретении предложены способы внесения биаллельной модификации в геном в клетке, включающие в себя приведение генома в контакт с: (а) первым белком Cas; (b) первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (с) второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (d) тракрРНК; и (е) нацеливающим вектором, содержащим полинуклеотидную вставку, фланкированную 5' гомологичным плечом, гибридизующимся с целевой 5'-последовательностью, и 3' гомологичным плечом, гибридизующимся с целевой 3'-последовательностью, при условии, что если клетка представляет собой эмбрион на стадии единственной клетки, нацеливающий вектор имеет длину не более 5 т.п.н.; причем геном содержит пару из первой и второй гомологичных хромосом, содержащих целевой геномный локус; и при этом первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва в по меньшей мере одной из первой и второй гомологичных хромосом.
Способы могут дополнительно включать в себя идентификацию клетки, содержащей модифицированный геном. В некоторых способах полинуклеотидная вставка содержит кассету селекции, примыкающую к первому гомологичному плечу, которое гибридизуется с первой целевой последовательностью, причем первое гомологичное плечо представляет собой 5' гомологичное плечо, а первая целевая последовательность представляет собой целевую 5'-последовательность, или причем первое гомологичное плечо представляет собой 3' гомологичное плечо, а первая целевая последовательность представляет собой целевую 3'-последовательность, при этом идентификация включает в себя: (а) получение ДНК из клетки; (b) воздействие на ДНК клетки зондом, связывающимся в пределах первой целевой последовательности, зондом, связывающимся в пределах полинуклеотидной вставки, и зондом, связывающимся в пределах эталонного гена, имеющего известное число копий, при этом каждый зонд генерирует обнаруживаемый сигнал при связывании; (с) обнаружение сигналов, обусловленных связыванием каждого из зондов; и (d) сравнение сигнала от зонда эталонного гена с сигналом от зонда первой целевой последовательности для определения числа копий первой целевой последовательности и сравнение сигнала от зонда эталонного гена с сигналом от зонда полинуклеотидной вставки для определения числа копий полинуклеотидной вставки, при этом число копий полинуклеотидной вставки, равное одной или двум, и число копий первой целевой последовательности, равное двум, указывает на нацеленную вставку полинуклеотидной вставки в целевой геномный локус, и при этом число копий полинуклеотидной вставки, равное одной или более, и число копий первой целевой последовательности, равное трем или более, указывает на случайную вставку полинуклеотидной вставки в геномный локус, отличный от целевого геномного локуса.
В некоторых способах первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR в каждой из первой и второй гомологичных хромосом с получением по меньшей мере одного двухцепочечного разрыва в каждой из первой и второй гомологичных хромосом. В некоторых способах первый белок Cas расщепляет одну из первой и второй последовательностей распознавания РНК CRISPR в по меньшей мере одной из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом.
Некоторые способы дополнительно включают в себя приведение генома в контакт с: третьей РНК CRISPR, гибридизующейся с третьей последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; и четвертой РНК CRISPR, гибридизующейся с четвертой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса. Необязательно первая последовательность распознавания РНК CRISPR и третья последовательность распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 6 т.п.н., от приблизительно 6 т.п.н. до приблизительно 7 т.п.н., от приблизительно 7 т.п.н. до приблизительно 8 т.п.н., от приблизительно 8 т.п.н. до приблизительно 9 т.п.н., от приблизительно 9 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н. или от приблизительно 90 т.п.н. до приблизительно 100 т.п.н. Необязательно вторая последовательность распознавания РНК CRISPR и четвертая последовательность распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 6 т.п.н., от приблизительно 6 т.п.н. до приблизительно 7 т.п.н., от приблизительно 7 т.п.н. до приблизительно 8 т.п.н., от приблизительно 8 т.п.н. до приблизительно 9 т.п.н., от приблизительно 9 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н. или от приблизительно 90 т.п.н. до приблизительно 100 т.п.н. Необязательно первая и третья последовательности распознавания РНК CRISPR представляют собой первую пару последовательностей распознавания РНК CRISPR, а вторая и четвертая последовательности распознавания РНК CRISPR представляют собой вторую пару последовательностей распознавания РНК CRISPR, причем первая пара и вторая пара разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н., от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н.
В некоторых способах первый белок Cas расщепляет по меньшей мере две из первой, второй, третьей и четвертой последовательностей распознавания РНК CRISPR с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом. В некоторых способах первый белок Cas расщепляет по меньшей мере две из первой, второй, третьей и четвертой последовательностей распознавания РНК CRISPR с получением по меньшей мере двух двухцепочечных разрывов как в первой, так и во второй гомологичных хромосомах.
В некоторых способах полинуклеотидную вставку вставляют между целевыми 5'- и 3'-последовательностями. Необязательно целевые 5'- и 3'-последовательности находятся в пределах целевого геномного локуса. Необязательно клетка представляет собой не эмбрион на стадии единственной клетки, а нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC), содержащий по меньшей мере 10 т.п.н.
В некоторых способах приведение генома в контакт как с первой, так и со второй РНК CRISPR приводит к повышению эффективности биаллельной модификации по сравнению с приведением генома в контакт или только с первой РНК CRISPR, или только со второй РНК CRISPR. В некоторых способах клетка является диплоидной, и биаллельная модификация приводит к гомозиготности или компаунд-гетерозиготности в целевом геномном локусе. Необязательно компаунд-гетерозиготность представляет собой гемизиготность. В некоторых способах биаллельная модификация включает в себя делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме. В некоторых способах биаллельная модификация включает в себя делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах. В некоторых способах биаллельная модификация дополнительно включает в себя вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями как в первой, так и во второй гомологичных хромосомах. В некоторых способах биаллельная модификация содержит: (1) делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах; и (2) вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями в первой гомологичной хромосоме, но не во второй гомологичной хромосоме. В некоторых способах биаллельная модификация содержит: (1) делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме; и (2) нарушение локуса между первой и второй последовательностями распознавания РНК CRISPR во второй гомологичной хромосоме. В некоторых способах биаллельная модификация включает в себя: (1) делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме; (2) вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями в первой гомологичной хромосоме; и (3) нарушение локуса между целевыми 5'- и 3'-последовательностями во второй гомологичной хромосоме. В некоторых способах биаллельная модификация содержит: (1) делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме; и (2) вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями в первой гомологичной хромосоме, причем последовательность полинуклеотидной вставки гомологична или ортологична удаленной последовательности.
В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н. или от приблизительно 900 п.н. до приблизительно 1 т.п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н.
В некоторых способах каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии по меньшей мере 50 п.н., по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 900 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н. или по меньшей мере 100 т.п.н. как от целевой 5'-последовательности, так и от целевой 3'-последовательности. В некоторых способах каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 200 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н. или от приблизительно 50 т.п.н. до приблизительно 100 т.п.н. как от целевой 5'-последовательности, так и от целевой 3'-последовательности. В некоторых способах каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии более чем 50 п.н., более чем 100 п.н., более чем 200 п.н., более чем 300 п.н., более чем 400 п.н., более чем 500 п.н., более чем 600 п.н., более чем 700 п.н., более чем 800 п.н., более чем 900 п.н., более чем 1 т.п.н., более чем 2 т.п.н., более чем 3 т.п.н., более чем 4 т.п.н., более чем 5 т.п.н., более чем 6 т.п.н., более чем 7 т.п.н., более чем 8 т.п.н., более чем 9 т.п.н., более чем 10 т.п.н., более чем 20 т.п.н., более чем 30 т.п.н., более чем 40 т.п.н., более чем 50 т.п.н., более чем 60 т.п.н., более чем 70 т.п.н., более чем 80 т.п.н., более чем 90 т.п.н. или более чем 100 т.п.н. как от целевой 5'-последовательности, так и от целевой 3'-последовательности.
В некоторых способах удаленная нуклеиновая кислота составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. В некоторых способах удаленная нуклеиновая кислота составляет по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. Необязательно удаленная нуклеиновая кислота составляет по меньшей мере 550 т.п.н., по меньшей мере 600 т.п.н., по меньшей мере 650 т.п.н., по меньшей мере 700 т.п.н., по меньшей мере 750 т.п.н., по меньшей мере 800 т.п.н., по меньшей мере 850 т.п.н., по меньшей мере 900 т.п.н., по меньшей мере 950 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 1,5 млн п.н. или по меньшей мере 2 млн п.н.
В некоторых способах нацеливающий вектор имеет линейную форму. Необязательно нацеливающий вектор является одноцепочечным или двухцепочечным. В некоторых способах клетка представляет собой не эмбрион на стадии единственной клетки, а нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC), содержащий по меньшей мере 10 т.п.н. В некоторых способах клетка представляет собой не эмбрион на стадии единственной клетки, а нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC), причем общая сумма длин 5' и 3' гомологичных плеч LTVEC составляет по меньшей мере 10 т.п.н. Необязательно LTVEC составляет от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н. или от приблизительно 275 т.п.н. до приблизительно 300 т.п.н. Необязательно общая длина 5' и 3' гомологичных плеч LTVEC составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н.
В некоторых способах клетка представляет собой эукариотическую клетку. Необязательно эукариотическая клетка представляет собой клетку млекопитающего, человеческую клетку, нечеловеческую клетку, клетку грызуна, клетку мыши или клетку крысы. Необязательно эукариотическая клетка представляет собой плюрипотентную клетку, неплюрипотентную клетку, нечеловеческую плюрипотентную клетку, человеческую плюрипотентную клетку, плюрипотентную клетку грызуна, мышиную плюрипотентную клетку, крысиную плюрипотентную клетку, мышиную эмбриональную стволовую (ЭС) клетку, крысиную ЭС-клетку, человеческую ЭС-клетку, стволовую клетку взрослого человека, ограниченную в развитии человеческую клетку-предшественник, человеческую индуцированную плюрипотентную стволовую клетку (ИПС). Необязательно эукариотическая клетка представляет собой эмбрион на стадии единственной клетки. Необязательно эукариотическая клетка представляет собой эмбрион на стадии единственной клетки, а нацеливающий вектор имеет длину от приблизительно 50 нуклеотидов до приблизительно 5 т.п.н. Необязательно эукариотическая клетка представляет собой эмбрион на стадии единственной клетки, а нацеливающий вектор представляет собой одноцепочечную ДНК и имеет длину от приблизительно 60 до приблизительно 200 нуклеотидов.
В некоторых способах первый белок Cas представляет собой Cas9. В некоторых способах первый белок Cas имеет нуклеазную активность в отношении обеих цепей двухцепочечной ДНК.
В некоторых способах первый белок Cas представляет собой никазу. Некоторые способы дополнительно включают в себя приведение генома в контакт с: (f) вторым белком Cas, представляющим собой никазу; (g) третьей РНК CRISPR, гибридизующейся с третьей последовательностью распознавания РНК CRISPR; и (h) четвертой РНК CRISPR, гибридизующейся с четвертой последовательностью распознавания РНК CRISPR; причем первый белок Cas расщепляет первую цепь геномной ДНК в пределах первой последовательности распознавания РНК CRISPR и в пределах второй последовательности распознавания РНК CRISPR, а второй белок Cas расщепляет вторую цепь геномной ДНК в пределах третьей последовательности распознавания РНК CRISPR и в пределах четвертой последовательности распознавания РНК CRISPR.
В некоторых способах первая РНК CRISPR и тракрРНК слиты вместе в первую гидовую РНК (гРНК) и/или вторая РНК CRISPR и тракрРНК слиты вместе во вторую гРНК. В некоторых способах первая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК и/или вторая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК.
В некоторых способах приведение в контакт включает в себя введение первого белка Cas, первой и второй РНК CRISP и тракрРНК в клетку. В некоторых способах (а) первый белок Cas вводят в клетку в виде белка, матричной РНК (мРНК), кодирующей первый белок Cas, или ДНК, кодирующей первый белок Cas; (b) первую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей первую РНК CRISPR; (с) вторую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей вторую РНК CRISPR; и/или (d) тракрРНК вводят в клетку в виде РНК или в виде ДНК, кодирующей тракрРНК. В некоторых способах первый белок Cas, первую РНК CRISPR и тракрРНК вводят в клетку в виде первого комплекса белок-РНК и/или первый белок Cas, вторую РНК CRISPR и тракрРНК вводят в клетку в виде второго комплекса белок-РНК. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) ДНК, кодирующая первую РНК CRISPR, функционально связана со вторым промотором во втором экспрессионном конструкте; (с) ДНК, кодирующая вторую РНК CRISPR, функционально связана с третьим промотором в третьем экспрессионном конструкте; и/или (d) ДНК, кодирующая тракрРНК, функционально связана с четвертым промотором в четвертом экспрессионном конструкте; причем первый, второй, третий и четвертый промоторы активны в клетке. Необязательно первый, второй, третий и/или четвертый экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) молекулы ДНК, кодирующие первую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую первую гидовую РНК (гРНК), и функционально связаны со вторым промотором во втором экспрессионном конструкте; и/или (с) молекулы ДНК, кодирующие вторую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую вторую гРНК, и функционально связаны с третьим промотором в третьем экспрессионном конструкте; причем первый, второй и третий промоторы активны в клетке. Необязательно первый, второй и/или третий экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты.
В некоторых способах клетка модифицирована для снижения негомологичного соединения концов (NHEJ) и/или для повышения генной конверсии или направляемой гомологией репарации (HDR). Необязательно клетка модифицирована для снижения экспрессии или активности ДНК-зависимой протенкиназы (DNA-PK) и/или для снижения экспрессии или активности PARP1. Необязательно клетка модифицирована для снижения экспрессии или активности лигазы IV. Необязательно снижение экспрессии или активности является индуцируемым, обратимым, время-специфическим и/или пространственно-специфическим.
В некоторых способах (1) клетка представляет собой не эмбрион на стадии единственной клетки, а нацеливающий вектор представляет собой большой нацеливающий вектор, причем общая сумма длин 5' и 3' гомологичных плеч составляет по меньшей мере 10 т.п.н.; (2) каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии более чем 200 п.н., более чем 300 п.н., более чем 400 п.н., более чем 500 п.н., более чем 600 п.н., более чем 700 п.н., более чем 800 п.н., более чем 900 п.н., более чем 1 т.п.н., более чем 2 т.п.н., более чем 3 т.п.н., более чем 4 т.п.н., более чем 5 т.п.н., более чем 6 т.п.н., более чем 7 т.п.н., более чем 8 т.п.н., более чем 9 т.п.н., более чем 10 т.п.н., более чем 20 т.п.н., более чем 30 т.п.н., более чем 40 т.п.н., более чем 50 т.п.н., более чем 60 т.п.н., более чем 70 т.п.н., более чем 80 т.п.н., более чем 90 т.п.н. или более чем 100 т.п.н. как от целевой 5'-последовательности, так и от целевой 3'-последовательности; (3) первый белок Cas расщепляет первую и вторую последовательности распознавания РНК CRISPR в по меньшей мере одной из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом; и (4) биаллельная модификация включает в себя делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями в первой гомологичной хромосоме, причем последовательность полинуклеотидной вставки гомологична или ортологична удаленной последовательности.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена любым из вышеуказанных способов; и (b) вынашивание нечеловеческого эмбриона-хозяина в теле суррогатной матери; при этом суррогатная мать дает потомство не относящегося к человеку животного поколения F0, имеющее биаллельную модификацию. Некоторые способы включают в себя: (а) приведение генома нечеловеческой ЭС-клетки в контакт с: (i) первым белком Cas; (ii) первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (iii) второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; (iv) тракрРНК; и (v) нацеливающим вектором, содержащим полинуклеотидную вставку, фланкированную 5' гомологичным плечом и 3' гомологичным плечом, причем геном содержит пару из первой и второй гомологичных хромосом, содержащих целевой геномный локус; и при этом первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательности распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва в по меньшей мере одной из первой и второй гомологичных хромосом; (b) идентификацию нечеловеческой ЭС-клетки, содержащей биаллельную модификацию; (с) введение нечеловеческой ЭС-клетки, содержащей биаллельную модификацию, в тело нечеловеческого эмбриона-хозяина; и (d) вынашивание нечеловеческого эмбриона-хозяина в теле суррогатной матери; при этом суррогатная мать дает потомство не относящегося к человеку животного поколения F0, имеющее биаллельную модификацию.
В некоторых способах первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR в каждой из первой и второй гомологичных хромосом с получением по меньшей мере одного двухцепочечного разрыва в каждой из первой и второй гомологичных хромосом. В некоторых способах первый белок Cas расщепляет одну из первой и второй последовательностей распознавания РНК CRISPR в по меньшей мере одной из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом.
В некоторых способах не относящееся к человеку животное представляет собой мышь, нечеловеческая ЭС-клетка представляет собой мышиную ЭС-клетку, а нечеловеческий эмбрион-хозяин представляет собой мышиного эмбриона-хозяина. В некоторых способах не относящееся к человеку животное представляет собой крысу, нечеловеческая ЭС-клетка представляет собой крысиную ЭС-клетку, а нечеловеческий эмбрион-хозяин представляет собой крысиного эмбриона-хозяина.
В некоторых способах биаллельная модификация приводит к гомозиготности или компаунд-гетерозиготности в целевом геномном локусе. Необязательно компаунд-гетерозиготность представляет собой гемизиготность.
В некоторых способах первая РНК CRISPR и тракрРНК слиты вместе в первую гидовую РНК (гРНК) и/или вторая РНК CRISPR и тракрРНК слиты вместе во вторую гРНК. В некоторых способах первая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК и/или вторая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК.
В некоторых способах приведение в контакт включает в себя введение первого белка Cas, первой и второй РНК CRISP и тракрРНК в клетку. В некоторых способах (а) первый белок Cas вводят в клетку в виде белка, матричной РНК (мРНК), кодирующей первый белок Cas, или ДНК, кодирующей первый белок Cas; (b) первую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей первую РНК CRISPR; (с) вторую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей вторую РНК CRISPR; и/или (d) тракрРНК вводят в клетку в виде РНК или в виде ДНК, кодирующей тракрРНК. В некоторых способах первый белок Cas, первую РНК CRISPR и тракрРНК вводят в клетку в виде первого комплекса белок-РНК и/или первый белок Cas, вторую РНК CRISPR и тракрРНК вводят в клетку в виде второго комплекса белок-РНК. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) ДНК, кодирующая первую РНК CRISPR, функционально связана со вторым промотором во втором экспрессионном конструкте; (с) ДНК, кодирующая вторую РНК CRISPR, функционально связана с третьим промотором в третьем экспрессионном конструкте; и/или (d) ДНК, кодирующая тракрРНК, функционально связана с четвертым промотором в четвертом экспрессионном конструкте; причем первый, второй, третий и четвертый промоторы активны в клетке. Необязательно первый, второй, третий и/или четвертый экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) молекулы ДНК, кодирующие первую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую первую гидовую РНК (гРНК), и функционально связаны со вторым промотором во втором экспрессионном конструкте; и/или (с) молекулы ДНК, кодирующие вторую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую вторую гРНК, и функционально связаны с третьим промотором в третьем экспрессионном конструкте; причем первый, второй и третий промоторы активны в клетке. Необязательно первый, второй и/или третий экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты.
В некоторых способах первый белок Cas представляет собой Cas9. В некоторых способах первый белок Cas имеет нуклеазную активность в отношении обеих цепей двухцепочечной ДНК.
В некоторых способах первый белок Cas представляет собой никазу. Некоторые способы дополнительно включают в себя приведение генома в контакт с: (f) вторым белком Cas, представляющим собой никазу; (g) третьей РНК CRISPR, гибридизующейся с третьей последовательностью распознавания РНК CRISPR; и (h) четвертой РНК CRISPR, гибридизующейся с четвертой последовательностью распознавания РНК CRISPR; причем первый белок Cas расщепляет первую цепь геномной ДНК в пределах первой последовательности распознавания РНК CRISPR и в пределах второй последовательности распознавания РНК CRISPR, а второй белок Cas расщепляет вторую цепь геномной ДНК в пределах третьей последовательности распознавания РНК CRISPR и в пределах четвертой последовательности распознавания РНК CRISPR.
В некоторых способах клетка модифицирована для снижения негомологичного соединения концов (NHEJ) и/или для повышения генной конверсии или направляемой гомологией репарации (HDR). Необязательно клетка модифицирована для снижения экспрессии или активности ДНК-зависимой протенкиназы (DNA-PK) и/или для снижения экспрессии или активности PARP1. Необязательно клетка модифицирована для снижения экспрессии или активности лигазы IV. Необязательно снижение экспрессии или активности является индуцируемым, обратимым, время-специфическим и/или пространственно-специфическим.
В изобретении также предложены способы получения не относящегося к человеку животного поколения F0, включающие в себя имплантацию генетически модифицированного эмбриона на стадии единственной клетки, полученного любым из вышеописанных способов, в тело суррогатной матери; при этом суррогатная мать дает потомство не относящегося к человеку животного поколения F0, имеющее биаллельную модификацию.
В изобретении также предложены способы модификации генома в клетке, являющейся гетерозиготной по первому аллелю, включающие в себя приведение генома в контакт с: (а) первым белком Cas; (b) тракрРНК; (с) первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах второго аллеля, причем первый аллель находится на первой гомологичной хромосоме, а второй аллель находится в соответствующем локусе на второй гомологичной хромосоме; и (d) второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах второго аллеля; при этом первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва и концевых последовательностей, подвергающихся рекомбинации, при этом рекомбинация осуществляется между первым и вторым аллелями с получением модифицированного генома, гомозиготного по первому аллелю. Некоторые способы дополнительно включают в себя идентификацию клетки, гомозиготной по первому аллелю.
В некоторых способах первый белок Cas расщепляет первую последовательность распознавания РНК CRISPR и вторую последовательность распознавания РНК CRISPR. В некоторых способах первый белок Cas расщепляет первую последовательность распознавания РНК CRISPR и вторую последовательность распознавания РНК CRISPR с получением по меньшей мере двух двухцепочечных разрывов и концевых последовательностей, подвергающихся рекомбинации. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR расположены в пределах второго аллеля, но не первого аллеля. В некоторых способах белок Cas и первая РНК CRISPR в природных условиях вместе не встречаются.
В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н. или от приблизительно 900 п.н. до приблизительно 1 т.п.н. В некоторых способах первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н.
В некоторых способах различия в последовательности между первым аллелем и вторым аллелем охватывают от приблизительно 100 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. В некоторых способах различия в последовательности между первым аллелем и вторым аллелем охватывают по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 800 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н.
В некоторых способах первый аллель содержит нацеленную модификацию, а второй аллель представляет собой аллель дикого типа. В некоторых способах первый аллель представляет собой аллель дикого типа, а второй аллель содержит вызывающую заболевание мутацию.
В некоторых способах рекомбинация включает в себя генную конверсию. В некоторых способах рекомбинация включает в себя потерю гетерозиготности (LOH).
В некоторых способах клетка представляет собой эукариотическую клетку. Необязательно эукариотическая клетка представляет собой клетку млекопитающего, человеческую клетку, нечеловеческую клетку, клетку грызуна, клетку мыши или клетку крысы. Необязательно эукариотическая клетка представляет собой плюрипотентную клетку, неплюрипотентную клетку, нечеловеческую плюрипотентную клетку, человеческую плюрипотентную клетку, плюрипотентную клетку грызуна, мышиную плюрипотентную клетку, крысиную плюрипотентную клетку, мышиную эмбриональную стволовую (ЭС) клетку, крысиную ЭС-клетку, человеческую ЭС-клетку, стволовую клетку взрослого человека, ограниченную в развитии человеческую клетку-предшественник, человеческую индуцированную плюрипотентную стволовую клетку (ИПС).
В некоторых способах первый белок Cas представляет собой Cas9. В некоторых способах первый белок Cas имеет нуклеазную активность в отношении обеих цепей двухцепочечной ДНК.
В некоторых способах первый белок Cas представляет собой никазу. Некоторые способы дополнительно включают в себя приведение генома в контакт с: (f) вторым белком Cas, представляющим собой никазу; (g) третьей РНК CRISPR, гибридизующейся с третьей последовательностью распознавания РНК CRISPR; и (h) четвертой РНК CRISPR, гибридизующейся с четвертой последовательностью распознавания РНК CRISPR; причем первый белок Cas расщепляет первую цепь геномной ДНК в пределах первой последовательности распознавания РНК CRISPR и в пределах второй последовательности распознавания РНК CRISPR, а второй белок Cas расщепляет вторую цепь геномной ДНК в пределах третьей последовательности распознавания РНК CRISPR и в пределах четвертой последовательности распознавания РНК CRISPR.
В некоторых способах первая РНК CRISPR и тракрРНК слиты вместе в первую гидовую РНК (гРНК), и/или вторая РНК CRISPR и тракрРНК слиты вместе во вторую гРНК. В некоторых способах первая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК, и/или вторая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК.
В некоторых способах приведение в контакт включает в себя введение первого белка Cas, первой и второй РНК CRISP и тракрРНК в клетку. В некоторых способах (а) первый белок Cas вводят в клетку в виде белка, матричной РНК (мРНК), кодирующей первый белок Cas, или ДНК, кодирующей первый белок Cas; (b) первую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей первую РНК CRISPR; (с) вторую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей вторую РНК CRISPR; и/или (d) тракрРНК вводят в клетку в виде РНК или в виде ДНК, кодирующей тракрРНК. В некоторых способах первый белок Cas, первую РНК CRISPR и тракрРНК вводят в клетку в виде первого комплекса белок-РНК и/или первый белок Cas, вторую РНК CRISPR и тракрРНК вводят в клетку в виде второго комплекса белок-РНК. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) ДНК, кодирующая первую РНК CRISPR, функционально связана со вторым промотором во втором экспрессионном конструкте; (с) ДНК, кодирующая вторую РНК CRISPR, функционально связана с третьим промотором в третьем экспрессионном конструкте; и/или (d) ДНК, кодирующая тракрРНК, функционально связана с четвертым промотором в четвертом экспрессионном конструкте; причем первый, второй, третий и четвертый промоторы активны в клетке. Необязательно первый, второй, третий и/или четвертый экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты. В некоторых способах (а) ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; (b) молекулы ДНК, кодирующие первую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую первую гидовую РНК (гРНК), и функционально связаны со вторым промотором во втором экспрессионном конструкте; и/или (с) молекулы ДНК, кодирующие вторую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую вторую гРНК, и функционально связаны с третьим промотором в третьем экспрессионном конструкте; причем первый, второй и третий промоторы активны в клетке. Необязательно первый, второй и/или третий экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты.
В некоторых способах клетка модифицирована для снижения негомологичного соединения концов (NHEJ) и/или для повышения генной конверсии или направляемой гомологией репарации (HDR). Необязательно клетка модифицирована для снижения экспрессии или активности ДНК-зависимой протенкиназы (DNA-PK) и/или для снижения экспрессии или активности PARP1. Необязательно клетка модифицирована для снижения экспрессии или активности лигазы IV. Необязательно снижение экспрессии или активности является индуцируемым, обратимым, время-специфическим и/или пространственно-специфическим.
В изобретении также предложены способы модификации генома в клетке, являющейся гетерозиготной по первому аллелю, включающие в себя приведение генома в контакт с: (а) первым белком Cas; (b) тракрРНК; и (с) первой РНК CRISPR, которая гибридизуется с первой не аллель-специфической последовательностью распознавания РНК CRISPR, причем первый аллель находится на первой гомологичной хромосоме, и последовательность распознавания РНК CRISPR располагается к центромере от локуса, соответствующего первому аллелю на второй гомологичной хромосоме; и при этом первый белок Cas расщепляет первую последовательность распознавания РНК CRISPR с получением двухцепочечного разрыва, а клетку модифицируют так, чтобы она стала гомозиготной по первому аллелю. Некоторые способы дополнительно включают в себя идентификацию клетки, гомозиготной по первому аллелю. В некоторых способах белок Cas и первая РНК CRISPR в природных условиях вместе не встречаются.
Такие способы могут дополнительно включать в себя приведение генома в контакт со второй РНК CRISPR, гибридизующейся со второй не аллель-специфической последовательностью распознавания РНК CRISPR, центромерной относительно локуса, соответствующего первому аллелю на второй гомологичной хромосоме, причем первый белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва. В некоторых способах первый белок Cas расщепляет первую последовательность распознавания РНК CRISPR и вторую последовательность распознавания РНК CRISPR.
В некоторых способах потеря гетерозиготности происходит теломерно от двухцепочечного разрыва.
В некоторых способах первая и вторая последовательности распознавания РНК CRISPR расположены на второй гомологичной хромосоме, но не на первой гомологичной хромосоме. В некоторых способах первый сайт распознавания РНК CRISPR находится на расстоянии от приблизительно 100 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. от центромеры. В некоторых способах первый аллель находится на расстоянии от приблизительно 100 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. от первого сайта распознавания РНК CRISPR. В некоторых способах область второй гомологичной хромосомы, заменяемая при потере гетерозиготности, находится на расстоянии от приблизительно 100 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н.
В некоторых способах клетка представляет собой эукариотическую клетку. Необязательно эукариотическая клетка представляет собой клетку млекопитающего, человеческую клетку, нечеловеческую клетку, клетку грызуна, мышиную клетку, крысиную клетку, плюрипотентную клетку, неплюрипотентную клетку, нечеловеческую плюрипотентную клетку, человеческую плюрипотентную клетку, плюрипотентную клетку грызуна, мышиную плюрипотентную клетку, крысиную плюрипотентную клетку, мышиную эмбриональную стволовую (ЭС) клетку, крысиную ЭС-клетку, человеческую ЭС-клетку, стволовую клетку взрослого человека, ограниченную в развитии человеческую клетку-предшественник, человеческую индуцированную плюрипотентную стволовую клетку (ИПС) или эмбрион на стадии единственной клетки.
В некоторых способах первый белок Cas представляет собой Cas9. В некоторых способах первый белок Cas имеет нуклеазную активность в отношении обеих цепей двухцепочечной ДНК. В некоторых способах первый белок Cas представляет собой никазу. Необязательно первый белок Cas представляет собой никазу, и причем способ дополнительно включает в себя приведение генома в контакт со вторым белком Cas, представляющим собой никазу, с третьей РНК CRISPR, гибридизующейся с третьей последовательностью распознавания РНК CRISPR, и с четвертой РНК CRISPR, гибридизующейся с четвертой последовательностью распознавания РНК CRISPR, при этом первый белок Cas расщепляет первую цепь геномной ДНК в пределах первой последовательности распознавания РНК CRISPR и в пределах второй последовательности распознавания РНК CRISPR, а второй белок Cas расщепляет вторую цепь геномной ДНК в пределах третьей последовательности распознавания РНК CRISPR и в пределах четвертой последовательности распознавания РНК CRISPR.
В некоторых способах первая РНК CRISPR и тракрРНК слиты вместе в первую гидовую РНК (гРНК), и/или вторая РНК CRISPR и тракрРНК слиты вместе во вторую гРНК. В некоторых способах первая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК, и/или вторая РНК CRISPR и тракрРНК представляют собой отдельные молекулы РНК.
В некоторых способах приведение в контакт включает в себя введение первого белка Cas, первой и второй РНК CRISP и тракрРНК в клетку. В некоторых способах первый белок Cas вводят в клетку в виде белка, матричной РНК (мРНК), кодирующей первый белок Cas, или ДНК, кодирующей первый белок Cas. Необязательно ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте, причем первый промотор активен в клетке. В некоторых способах первую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей первую РНК CRISPR. Необязательно ДНК, кодирующая первую РНК CRISPR, функционально связана со вторым промотором во втором экспрессионном конструкте, причем второй промотор активен в клетке. В некоторых способах вторую РНК CRISPR вводят в клетку в виде РНК или в виде ДНК, кодирующей вторую РНК CRISPR. Необязательно ДНК, кодирующая вторую РНК CRISPR, функционально связана с третьим промотором в третьем экспрессионном конструкте, причем третий промотор активен в клетке. В некоторых способах тракрРНК вводят в клетку в виде РНК или в виде ДНК, кодирующей тракрРНК. Необязательно ДНК, кодирующая тракрРНК, функционально связана с четвертым промотором в четвертом экспрессионном конструкте, причем четвертый промотор активен в клетке. Необязательно первый, второй, третий и/или четвертый экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты.
Необязательно ДНК, кодирующая первый белок Cas, функционально связана с первым промотором в первом экспрессионном конструкте; молекулы ДНК, кодирующие первую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую первую гидовую РНК (гРНК), и функционально связаны со вторым промотором во втором экспрессионном конструкте, и/или молекулы ДНК, кодирующие вторую РНК CRISPR и тракрРНК, слиты вместе в ДНК, кодирующую вторую гРНК, и функционально связаны с третьим промотором в третьем экспрессионном конструкте; причем первый, второй и третий промоторы активны в клетке. Необязательно первый, второй и/или третий экспрессионные конструкты являются компонентами одной молекулы нуклеиновой кислоты.
Необязательно первый белок Cas, первую РНК CRISPR и тракрРНК вводят в клетку в виде первого комплекса белок-РНК и/или первый белок Cas, вторую РНК CRISPR и тракрРНК вводят в клетку в виде второго комплекса белок-РНК.
В некоторых способах клетка модифицирована для снижения негомологичного соединения концов (NHEJ) и/или для повышения генной конверсии или направляемой гомологией репарации (HDR). Необязательно клетка модифицирована для снижения экспрессии или активности одного или более из следующих: DNA-PK, PARP1 и лигазы IV. Необязательно снижение экспрессии или активности является индуцируемым, обратимым, время-специфическим и/или пространственно-специфическим.
В некоторых способах первый аллель содержит мутацию. Необязательно мутация представляет собой нацеленную модификацию. В некоторых способах первый аллель представляет собой аллель дикого типа, а соответствующий локус на второй гомологичной хромосоме содержит мутацию.
В изобретении также предложены способы идентификации нацеленной вставки полинуклеотидной вставки в целевой геномный локус в диплоидной клетке, не являющейся эмбрионом на стадии единственной клетки, включающие в себя: (а) получение ДНК из клетки, причем клетку приводили в контакт с большим нацеливающим вектором (LTVEC), содержащим полинуклеотидную вставку, фланкированную первым гомологичным плечом, гибридизующимся с первой целевой последовательностью, и вторым гомологичным плечом, гибридизующимся со второй целевой последовательностью, при этом полинуклеотидная вставка содержит кассету селекции, прилегающую к первому гомологичному плечу; (b) воздействие на ДНК клетки зондом, связывающимся в пределах первой целевой последовательности, зондом, связывающимся в пределах полинуклеотидной вставки, и зондом, связывающимся в пределах эталонного гена, имеющего известное число копий, при этом каждый зонд генерирует обнаруживаемый сигнал при связывании; (c) обнаружение сигналов, обусловленных связыванием каждого из зондов; и (d) сравнение сигнала от зонда эталонного гена с сигналом от зонда первой целевой последовательности для определения числа копий первой целевой последовательности и сравнение сигнала от зонда эталонного гена с сигналом от зонда полинуклеотидной вставки для определения числа копий полинуклеотидной вставки, при этом число копий полинуклеотидной вставки, равное одной или двум, и число копий первой целевой последовательности, равное двум, указывает на нацеленную вставку полинуклеотидной вставки в целевой геномный локус, и при этом число копий полинуклеотидной вставки, равное одной или более, и число копий первой целевой последовательности, равное трем или более, указывает на случайную вставку полинуклеотидной вставки в геномный локус, отличный от целевого геномного локуса.
В некоторых способах сигнал, обусловленный связыванием зонда первой целевой последовательности, используют для определения значения порогового цикла (Ct) для первой целевой последовательности, а сигнал, обусловленный связыванием зонда эталонного гена, используют для определения значения порогового цикла (Ct) для эталонного гена, и число копий первой целевой последовательности определяют путем сравнения значения Ct первой целевой последовательности и значения Ct эталонного гена. В некоторых способах сигнал, обусловленный связыванием зонда полинуклеотидной вставки, используют для определения значения порогового цикла (Ct) для полинуклеотидной вставки, и число копий полинуклеотидной вставки определяют путем сравнения значения Ct первой целевой последовательности и значения Ct эталонного гена.
В некоторых способах кассета селекции содержит ген резистентности к лекарственному средству.
В некоторых способах полинуклеотидная вставка может составлять по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. В некоторых способах расстояние между последовательностями, с которыми связываются зонды в первой целевой последовательности, и кассетой селекции составляет не более чем 100 нуклеотидов, 200 нуклеотидов, 300 нуклеотидов, 400 нуклеотидов, 500 нуклеотидов, 600 нуклеотидов, 700 нуклеотидов, 800 нуклеотидов, 900 нуклеотидов, 1 т.п.н., 1,5 т.п.н., 2 т.п.н., 2,5 т.п.н., 3 т.п.н., 3,5 т.п.н., 4 т.п.н., 4,5 т.п.н. или 5 т.п.н.
Некоторые способы дополнительно включают в себя определение числа копий второй целевой последовательности. Необязательно этап (b) дополнительно включает в себя воздействие на ДНК клетки зондом, связывающимся со второй целевой последовательностью, этап (с) дополнительно включает в себя обнаружение сигнала, обусловленного связыванием зонда второй целевой последовательности, а этап (d) дополнительно включает в себя сравнение сигнала от зонда эталонного гена с сигналом от зонда второй целевой последовательности для определения числа копий второй целевой последовательности.
Некоторые способы дополнительно содержат определение числа копий одной или более дополнительных последовательностей в пределах полинуклеотидной вставки. Необязательно этап (b) дополнительно включает в себя воздействие на ДНК клетки одним или более дополнительными зондами, связывающимися с полинуклеотидной вставкой, этап (с) дополнительно включает в себя обнаружение сигнала, обусловленного связыванием одного или более дополнительных зондов, а этап (d) дополнительно включает в себя сравнение сигнала от зонда эталонного гена с сигналом от одного или более дополнительных зондов полинуклеотидной вставки для определения числа копий одной или более дополнительных последовательностей в пределах полинуклеотидной вставки. Необязательно одна или более дополнительных последовательностей в пределах полинуклеотидной вставки содержит последовательность, примыкающую ко второй целевой последовательности.
В некоторых способах LTVEC выполнен с возможностью удаления эндогенной последовательности из целевого геномного локуса, или клетку дополнительно привели в контакт с белком Cas, первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, и тракрРНК. Необязательно такие способы дополнительно включают в себя определение числа копий эндогенных последовательностей в целевом геномном локусе. Необязательно этап (b) дополнительно включает в себя воздействие на ДНК клетки зондом, связывающимся с эндогенной последовательностью в целевом геномном локусе, этап (с) дополнительно включает в себя обнаружение сигнала, обусловленного связыванием зонда эндогенной последовательности, а этап (d) дополнительно включает в себя сравнение сигнала от зонда эталонного гена с сигналом от зонда эндогенной последовательности для определения числа копий эндогенной последовательности.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фиг. 1 представлена схема одновременной делеции мышиного эктодомена Lrp5 и замены соответствующей человеческой версией LRP5 с использованием LTVEC и либо одной, либо двух гРНК 5'-области (А, В, В2), средней области (С, D) и 3'-области (Е2, Е, F). LTVEC представлен в верхней части фигуры, а мышиный генный локус Lrp5 представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого восемью гидовыми РНК, обозначены вертикальными стрелками под мышиной генной последовательностью. Горизонтальными стрелками представлены ПЦР-праймеры для мышиных и человеческих последовательностей.
На Фиг. 2А представлена общая схема одновременной делеции мышиного гена и замещения соответствующей человеческой версией с использованием LTVEC и двух гидовых РНК (гидовые РНК А и В). LTVEC представлен в верхней части Фиг. 2А, а мышиный генный локус представлен в нижней части Фиг. 2А. Положения сайтов расщепления Cas9, направляемого двумя гидовыми РНК, обозначены стрелками под мышиной генной последовательностью.
На Фиг. 2 В-Е представлены уникальные биаллельные модификации (аллельные типы), возникающие с большей частотой при использовании двух гидовых РНК. Толстые линии с диагональной штриховкой указывают на мышиный ген, точечные линии указывают на делеции в мышином гене, а толстые черные линии указывают на вставку человеческого гена. На Фиг. 2 В показаны гомозиготные коллапсированные аллели (большая делеция, индуцированная CRISPR). На Фиг. 2С показаны гомозиготные нацеленные аллели. На Фиг. 2D показаны гемизиготные нацеленные аллели. На Фиг. 2Е показаны компаунд-гетерозиготные аллели.
На Фиг. 3А и 3В представлены ПЦР-тесты, подтверждающие генотипы выбранных клонов. На Фиг. 3А представлены результаты ПЦР-тестов длинных фрагментов для выбранных ЭС-клонов с использованием праймеров m-lr-f и m-5'-r, которые устанавливают связь между человеческой вставкой и последовательностями за пределами последовательностей, гомологичных 5' гомологичному плечу, таким образом подтверждая правильное нацеливание. На Фиг. ЗВ представлены результаты ПЦР-тестов на 5' Del J, 5' Ins J, Del А+F и Del A+E2. 5' Del J показывает ПЦР-продукты с использованием праймеров m-5'-f и m-5-r, где амплифицируется последовательность дикого типа, окружающая сайт расщепления гРНК А, для определения сохранения или утраты этой последовательности. 5' Ins J показывает ПЦР-продукты с использованием праймеров m-5'-f и h-5'-r, где устанавливается связь между человеческой вставкой и мышиным геномом. Тест даст положительный результат для клонов как с нацеленной, так и со случайной интеграцией. Del А+F демонстрирует ожидаемый размер ампликона (359 п.н.) и реальные полосы для большой делеции, обусловленной двойным расщеплением с гРНК А и F в клонах ВО-F10 и AW-A8. Del А+Е2 демонстрирует эту же идею применительно к клону ВА-А7. NT означает отсутствие матрицы, +/+ показывает контроль дикого типа - родительскую гибридную ЭС клетку VGF1, Н/+ означает гетерозиготный гуманизированный генотип, НУД означает гемизиготный гуманизированный генотип, H/H означает гомозиготный гуманизированный генотип, а А/А означает гомозиготный генотип с делецией.
На Фиг. 4А-С представлен анализ методом флуоресцентной гибридизации in situ (FISH) мышиных клонов ЭС-клеток AW-D9 (Фиг. 4А) и BA-D5 (Фиг. 4С), на которые нацеливали LTVEC для гуманизации Lrp5 в комбинации с Cas9 и двумя гРНК, а также клона BS-C4 (Фиг. 4 В), на который нацеливали только LTVEC. Стрелки показывают положения сигналов гибридизации на полосе В хромосомы 19. Красный сигнал означает гибридизацию только с мышиным зондом (пунктирная стрелка, Фиг. 4В). Сигнал смешанного желтого цвета означает гибридизацию как с красным мышиным зондом, так и с зеленым человеческим зондом. Полоса В одной хромосомы 19, имеющая красный сигнал (пунктирная стрелка), и полоса В другой хромосомы 19, имеющая желтый сигнал (сплошная стрелка), подтверждают нацеливание на правильный локус и гетерозиготный генотип клона BS-C4 (Фиг. 4В). Наличие желтого сигнала на полосах В обеих хромосом 19 (сплошные стрелки, Фиг. 4А и 4С) подтверждают нацеливание на правильный локус и гомозиготные генотипы клонов AW-D9 и BS-C4.
На Фиг. 5 представлена схема хромосомы 19 и тесты, которые определяют события генной конверсии или митотической рекомбинации, обусловленные двумя гидовыми РНК, посредством анализа на потерю гетерозиготности (LOH) в гибридных ЭС-клетках VGF1. Стрелками показаны приблизительные положения тестов на число копий хромосом (CCN) методом количественной ПЦР TaqMan®. Приблизительные положения ПЦР-тестов на полиморфизм структурных вариантов (SV) показаны значками-шевронами с расстояниями (в млн п.н.) от указанного выше локуса Lrp5. Приблизительные положения тестов на аллельную дискриминацию однонуклеотидных вариантов (SNV) TaqMan® показаны значками-треугольниками с указанием расстояний (в млн п.н.) от вышеуказанного локуса Lrp5. Положения сайтов распознавания гРНК для F, Е2, D, В2 и А показаны диагональными стрелками над изображением гена Lrp5.
На Фиг. 6 представлена схема одновременной делеции области от экзона 2 до стоп-кодона мышиного гена С5 (Hc) и замены соответствующей человеческой версией С5 с использованием LTVEC и либо одной, либо двух гРНК 5'-области (А, В), средней области (С, D) и 3'-области (Е, Е2). LTVEC представлен в верхней части фигуры, а мышиный генный локус С5 (Hc) представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого шестью гидовыми РНК, обозначены стрелками под мышиной генной последовательностью.
На Фиг. 7А и 7В представлен анализ методом флуоресцентной гибридизации in situ (FISH) мышиных клонов ЭС-клеток Q-E9 (Фиг. 7А) и О-Е3 (Фиг. 7В), на которые выполняли нацеливание LTVEC для гуманизации Hc в комбинации с Cas9 и двумя гРНК. Стрелки показывают положения сигналов гибридизации на полосе В хромосомы 2. Красный сигнал означает гибридизацию только с мышиным зондом (пунктирная стрелка, Фиг. 7А). Сигнал смешанного желтого цвета означает гибридизацию как с красным мышиным зондом, так и с зеленым человеческим зондом (сплошная стрелка). Полоса В одной хромосомы 2, имеющая красный сигнал (пунктирная стрелка), и полоса В другой хромосомы 2, имеющая желтый сигнал (сплошная стрелка), подтверждают нацеливание на правильный локус и гетерозиготный генотип клона Q-E9 (Фиг. 7А). Наличие желтого сигнала на полосах В обеих хромосом 2 (сплошные стрелки, Фиг. 7В) подтверждает нацеливание на нужный локус и гомозиготный генотип клона АО-Е3.
На Фиг. 8 представлена схема одновременной делеции мышиного гена Ror1 и замены соответствующей человеческой версией ROR1 с использованием LTVEC и либо одной, либо двух гРНК 5'-области (А, В), средней области (D, С) и 3'-области (Е, F). LTVEC представлен в верхней части фигуры, а мышиный генный локус Ror1 представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого шестью гидовыми РНК, обозначены стрелками под мышиной генной последовательностью.
На Фиг. 9 представлена схема одновременной делеции мышиного гена Trpa1 и замены соответствующей человеческой версией TRPA1 с использованием LTVEC и либо одной, либо двух гРНК 5'-области (А, А2, В), средней области (С, D) и 3'-области (Е2, Е, F). LTVEC представлен в верхней части фигуры, а мышиный генный локус Trpa1 представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого восемью гидовыми РНК, обозначены стрелками под мышиной генной последовательностью.
На Фиг. 10А-Е представлены результаты тестов на структурные вариации (SV) клонов BR-B4, BP-G7, BO-G11, BO-F10, В0-А8 и ВС-Н9 с использованием ДНК VGF1 (F1H4), 129 и В6 в качестве контролей. Тесты проводили на следующих расстояниях теломерно относительно локуса Lrp5: 13,7 млн п.н. (Фиг. 10А), 20,0 млн п.н. (Фиг. 10В), 36,9 млн п.н. (Фиг. 10С), 48,3 млн п.н. (Фиг. 10D) и 56,7 млн п.н. (Фиг. 10Е). Положения ПЦР-продуктов для аллелей В6 и 129 показаны стрелками.
На Фиг. 11А-С показаны графики аллельной дискриминации для 0,32 млн п.н. к центромере от Lrp5 (Фиг. 11А), 1,2 млн п.н. к теломере от Lrp5 (Фиг. 11В) и 57,2 млн п.н. к теломере от Lrp5 (Фиг. 11С). Значения на каждой оси представляют относительную интенсивность флуоресценции. На графиках представлены по четыре копии каждого образца, показанные закрашенными точками (аллель В6), незакрашенными точками (аллель 129) и точками с диагональными линиями (оба аллеля В6/129).
На Фиг. 12А-С представлена схема, демонстрирующая возможный механизм митотической рекомбинации на фазе G2 клеточного цикла, способный создавать гомозиготные события и широкоохватную генную конверсию, обнаруживаемые по потере гетерозиготности. На Фиг. 12А показаны реплицированные гомологичные хромосомы с двумя хроматидами в гибридной ЭС-клетке 129/В6, гетерозиготной по нацеленной гуманизации в гомологе 129. Двунаправленные стрелки показывают потенциальные двухцепочечные разрывы, создаваемые при расщеплении белком Cas9, направляемым двумя гРНК, что приводит к взаимному обмену между хроматидами на гомологичных хромосомах путем гомологичной рекомбинации, показанной крестом с ближней к центромере стороны от нацеленного аллеля, что приводит к образованию гомологичных хроматид, показанных на Фиг. 12В. На Фиг. 12С показано, что после митоза и деления клетки возможно четыре типа распределения хромосом по дочерним клеткам. Два типа с сохранением гетерозиготности, гетерозигота родительского типа (Hum/+, вверху слева) и гетерозигота с равным обменом (Hum/+, вверху справа), которые невозможно различить по тестам LOH. В двух других типах наблюдается потеря гетерозиготности, гуманизированная гомозигота (Hum/Hum, например клон ВО-А8, внизу слева) с потерей теломерных аллелей В6 и гомозигота дикого типа (+/+, внизу справа) с потерей теломерных аллелей 129. Этот последний тип будет потерян, поскольку он не сохраняет кассету резистентности к лекарственному средству, имеющуюся в гуманизированном аллеле.
На Фиг. 13 представлена схема одновременной делеции области от экзона 2 до стоп-кодона мышиного гена С5 (Hc) и замены соответствующей человеческой версией С5 с использованием нацеливающего вектора с размерами гомологичных плеч 35 т.п.н. и 31 т.п.н. (LTVEC) или нацеливающего вектора с размерами гомологичных плеч 5 т.п.н. каждый (sTVEC) и либо одной, либо двух гРНК 5'-области (А, В), средней области (С, D) и 3'-области (Е, Е2). Два нацеливающих вектора представлены в верхней части фигуры, а мышиный генный локус С5 (Hc) представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого шестью гидовыми РНК, обозначены вертикальными стрелками под мышиной генной последовательностью, а праймеры, использованные для скрининга, обозначены горизонтальными стрелками. Положения тестов на приобретение аллели (GOA), в которых оценивается число копий вставки, и тестов на потерю аллели (LOA), в которых количественно оценивается мышиная последовательность, нацеленная для делеции, обозначены треугольниками.
На Фиг. 14 показана схема одновременной делеции первых пяти экзонов мышиного гена Cmah и замены на репортер lacZ и кассету селекции гигромицином с использованием LTVEC и двух гРНК 5'-области (А, В). LTVEC представлен в верхней части фигуры, а мышиный генный локус Cmah представлен в нижней части фигуры. Положения сайтов расщепления Cas9, направляемого двумя гидовыми РНК, обозначены вертикальными стрелками под мышиной генной последовательностью, а положения тестов GOA, оценивающих число копий вставки, и тестов LOA, количественно оценивающих мышиную последовательность, нацеленную для делеции, обозначены треугольниками.
На Фиг. 15 представлена схема событий расщепления и полученный при расщеплении продукт (SEQ ID NO: 112) при нацеливании на мышиный локус гена Cmah (SEQ ID NO: 109) двумя гРНК 5'-области (А и В; SEQ ID NO: 107 и 108 соответственно). Последовательности гРНК, гибридизованные с локусом гена Cmah, выделены жирным шрифтом, белки Cas9 показаны пятнистыми овалами, сайты расщепления Cas9 обозначены вертикальными стрелками, а мотивы, прилежащие к протоспейсеру (РАМ), заключены в прямоугольник. Приблизительные положения прямого праймера, зонда и обратного праймера для LOA-теста TaqMan® обозначены горизонтальными полосами и стрелками в верхней части фигуры. 5'- и 3'-фрагменты, полученные после расщепления и удаления, приведены в SEQ ID NO: 110 и 111 соответственно.
На Фиг. 16А-Е представлены возможные механизмы, объясняющие наблюдаемые результаты, в том числе потерю гетерозиготности (LOH), в выполняемых посредством CRISPR/Cas9 экспериментах по гуманизации в ЭС-клетках гибридных мышей F1, имеющих один гаплоидный набор хромосом, происходящий от линии мышей 129S6/SvEvTac, и один гаплоидный набор хромосом, происходящий от линии мышей C57BL/6NTac (В6). На Фиг. 16А представлен взаимный хроматидный обмен в результате митотического кроссинговера, при котором происходит гетерозиготная модификация на хромосоме 129 до репликации генома или после репликации генома, с последующей генной конверсией между сестринскими хроматидами. На Фиг. 16В представлен взаимный хроматидный обмен в результате митотического кроссинговера, при котором одна хроматида 129 модифицируется после репликации генома. На Фиг. 16С представлен взаимный хроматидный обмен в результате митотического кроссинговера, при котором нацеливание LTVEC не произошло, а расщепление Cas9 произошло или на хромосоме 129, или на хромосоме В6 (показано расщепление В6). На Фиг. 16D представлено копирование хроматиды посредством индуцированной разрывом репликации, при котором гетерозиготная модификация происходит на хромосоме 129 до репликации генома или после репликации генома, с последующей генной конверсией между сестринскими хроматидами. На Фиг. 16Е представлено копирование хроматиды посредством индуцированной разрывом репликации, при котором одна хроматида 129 модифицируется после репликации генома. На Фиг. 16F представлено копирование хроматиды посредством индуцированной разрывом репликации, при котором нацеливание LTVEC не произошло, но произошло расщепление Cas9 или на хромосоме 129, или на хромосоме В6 (показано расщепление В6).
На Фиг. 17А-С представлены стратегии скрининга для нацеленных модификаций. На Фиг. 17А представлена стандартная стратегия скрининга модификации аллеля (МОА) для обнаружения гетерозиготного нацеливания большим нацеливающим вектором (LTVEC), при котором эндогенную последовательность в мышиной хромосоме удаляют и заменяют на вставку Neo-SDC. В этой стратегии используют зонды TaqMan® mTU и mTD к областям ближе к 5'-концу и ближе к 3'-концу от эндогенной последовательности, нацеленной для делеции. На Фиг. 17В представлены тесты TaqMan® на сохранение (зонды retU и retD) в комбинации с тестами на модификацию аллеля (МОА) (зонды mTGU, mTM и mTGD для теста на потерю аллеля (LOA) и зонды hTU и hTD для теста на приобретение аллеля (GOA)), применяемые для скрининга на выполняемую посредством CRISPR/Cas9 гуманизацию. На Фиг. 17С представлено использование тестов на сохранение TaqMan® (зонды retU и retD) в комбинации с тестами на потерю аллеля (LOA) (зонды mTGU, mTM и mTGD), применяемыми для скрининга на выполняемые посредством CRISPR/Cas9 делеции с использованием парных гидовых РНК (gU и gD).
На Фиг. 18 представлены схемы (не в масштабе) области из приблизительно 900 т.п.н. локуса тяжелой цепи мышиного иммуноглобулина с сегментами гена вариабельной области, замененными на человеческие варианты (треугольники), и нацеливающий вектор со вставкой Pgk-Neo (промотор фосфоглицераткиназы I, функционально соединенный с геном неомицинфосфотрансферазы), фланкированной сайтами loxP. Две гРНК применяют для расщепления локуса тяжелой цепи мышиного иммуноглобулина на 5'-конце и две гРНК применяют для расщепления этого локуса на 3'-конце, а нацеливающий вектор удаляет и заменяет локус тяжелой цепи мышиного иммуноглобулина на вставку Pgk-Neo. Положения сайтов расщепления Cas9, направляемого четырьмя гидовыми РНК, обозначены вертикальными стрелками под целевым локусом. Заключенные в круг горизонтальные линии означают зонды TaqMan® для тестов на модификацию аллеля (MOA) (hIgH31, hIgH1, mIgHA1, mIgHA7 и hIgH9) и тестов на сохранение (5' IgH Arm 1,5' IgH Arm 2, mIgM-398 и mIgM-1045).
ОПРЕДЕЛЕНИЯ
Термины «белок», «полипептид» и «пептид», используемые в настоящем документе на взаимозаменяемой основе, включают полимерные формы аминокислот любой длины, в том числе кодированные и некодированные аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают полимеры, которые подверглись модификации, например полипептиды с модифицированными основными цепями пептидов.
Термины «нуклеиновая кислота» и «полинуклеотид», используемые в настоящем документе на взаимозаменяемой основе, включают полимерные формы нуклеотидов любой длины, в том числе рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные формы. Эти термины включают одно-, двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
Термин «оптимизация по кодонам» по существу включает в себя процесс модификации нуклеотидной последовательности для повышения экспрессии в конкретных клетках-хозяевах путем замещения по меньшей мере одного кодона нативной последовательности кодоном, который чаще или наиболее часто используется в генах клетки-хозяина при сохранении нативной аминокислотной последовательности. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована так, чтобы использовать в качестве замены кодоны, имеющие более высокую частоту использования в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, нечеловеческую клетку, клетку млекопитающих, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина, по сравнению с встречающейся в природе нуклеотидной последовательностью. Таблицы использования кодонов являются общедоступными, например, в базе данных Codon Usage Database. Эти таблицы можно адаптировать несколькими способами. См. Nakamura et al. (2000) Nucleic Acids Research 28: 292, которая для всех целей полностью включена в настоящий документ путем ссылки. Также имеются компьютерные алгоритмы для оптимизации по кодонам с конкретной последовательностью для экспрессии в конкретном хозяине (см., например, Gene Forge).
Термин «функциональная связь» или «функционально связанный» относится к смежному размещению двух или более компонентов (например, промотора и другого элемента последовательности) таким образом, чтобы оба компонента нормально функционировали и обеспечивалась возможность того, чтобы по меньшей мере один из компонентов мог опосредовать функцию, осуществляемую в отношении по меньшей мере одного из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции.
Термин «комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты вследствие ориентации нуклеотидных оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. В ДНК, как правило, комплементарны основания А и Т, а также Ц и Г. В РНК это, как правило, Ц и Г, а также У и А. Комплементарность может быть идеальной или существенной/достаточной. Идеальная комплементарность двух нуклеиновых кислот означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание связано с комплементарным основанием, образуя пары Уотсона-Крика. Термин «существенная» или «достаточная» комплементарность означает, что последовательность одной цепи не полностью и/или не идеально комплементарна последовательности противоположной цепи, но между основаниями на двух цепях создается достаточная связь, чтобы образовывался стабильный гибридный комплекс при некотором наборе условий гибридизации (например, концентрации солей и температуре). Такие условия можно спрогнозировать, используя последовательности и стандартные математические расчеты для предсказания Tm (температуры плавления) гибридизованных цепей, или путем эмпирического определения Tm традиционными способами. Tm относится к температуре, при которой популяция гибридизационных комплексов, образовавшихся между двумя цепями нуклеиновых кислот, денатурируется на 50%. При температуре ниже Tm преимущество имеет образование гибридизационного комплекса, а при температуре выше Tm преимущество имеет плавление или разделение цепей гибридизационного комплекса. Tm можно оценить для нуклеиновой кислоты с известным содержанием Г+Ц в водном 1 М растворе NaCl с использованием, например, выражения Tm=81,5+0,41(% Г+Ц), хотя в других известных расчетах Tm принимаются во внимание структурные характеристики нуклеиновых кислот.
Термин «условие гибридизации» означает совокупную среду, в которой одна цепь нуклеиновой кислоты связывается со второй цепью нуклеиновой кислоты в результате комплементарного взаимодействия цепей и водородного связывания с образованием гибридизационного комплекса. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие агенты, формамид) в водном или органическом растворе, содержащем нуклеиновые кислоты, и температуру смеси. Свой вклад в среду могут вносить другие факторы, такие как длительность инкубации или размеры инкубационной камеры. См., например, публикацию Sambrook et al., Molecular Cloning, A Laboratory Manual, 2.sup.nd ed., pp. 1.90-1.91, 9.47-9.51, 11.47-11.57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989), которая для всех целей полностью включена в настоящий документ путем ссылки.
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементации, и переменные известны специалистам в данной области. Чем больше степень комплементации между двумя нуклеотидными последовательностями, тем выше температура плавления (Tm) гибридов нуклеиновых кислот, имеющих эти последовательности. При гибридизации нуклеиновых кислот с короткими участками комплементарности (например, с комплементарностью более 35 нуклеотидов или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее) становится важным положение несовпадающих оснований (см. выше Sambrook et al., 11.7-11.8). Как правило, длина пригодной для гибридизации нуклеиновой кислоты составляет по меньшей мере приблизительно 10 нуклеотидов. Значения иллюстративной минимальной длины пригодных для гибридизации нуклеиновых кислот включают по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 22 нуклеотидов, по меньшей мере приблизительно 25 нуклеотидов и по меньшей мере приблизительно 30 нуклеотидов. Кроме того, температуру и концентрацию солей в промывном растворе можно регулировать по мере необходимости в зависимости от таких факторов, как длина области комплементации и степень комплементации.
Для обеспечения специфической гибридизации последовательность полинуклеотида необязательно должна быть на 100% комплементарна целевой нуклеиновой кислоте. Более того, полинуклеотид может гибридизоваться на одном или более сегментах, так что промежуточные или прилегающие сегменты не участвуют в событии гибридизации (например, структура петли или шпильки). Полинуклеотид (например, гРНК) может иметь по меньшей мере 70%-ю, по меньшей мере 80%-ю, по меньшей мере 90%-ю, по меньшей мере 95%-ю, по меньшей мере 99%-ю или 100%-ю комплементарность последовательности относительно целевой области в пределах целевой нуклеотидной последовательности, на которую он нацелен. Например, гРНК, в которой 18 из 20 нуклеотидов комплементарны целевой области и, следовательно, могут специфически гибридизоваться, будет иметь 90% комплементарности. В этом примере остальные некомплементарные нуклеотиды могут быть кластеризованы или рассеяны среди комплементарных нуклеотидов и не должны быть обязательно смежными друг с другом или с комплементарными нуклеотидами.
Процентную долю комплементарности между конкретными участками нуклеотидных последовательностей в пределах нуклеиновых кислот можно определять традиционными способами, используя программы BLAST (средства поиска основных локальных выравниваний) и PowerBLAST, известные в данной области (Altschul et al. (1990) J. Mol. Biol. 215: 403-410; Zhang and Madden (1997) Genome Res. 7: 649-656), или с помощью программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, г. Мэдисон, штат Висконсин, США) с использованием заданных по умолчанию параметров, где используется алгоритм Смита - Ватермана (Adv. Appl. Math., 1981, 2, 482-489).
В предложенных в настоящем документе способах и композициях используется множество разных компонентов. В описании признается, что некоторые компоненты могут иметь активные варианты и фрагменты. Такие компоненты включают в себя, например, белки Cas, РНК CRISPR, тракрРНК и гидовые РНК. Биологическая активность каждого из этих компонентов описана в других разделах настоящего документа.
Термин «идентичность последовательности» или «идентичность» в контексте двух полинуклеотидов или полипептидных последовательностей означает ссылку на остатки, которые одинаковы в двух последовательностях при выравнивании для максимального соответствия в установленном окне сравнения. Если термин «процентная доля идентичности последовательности» применяется в отношении белков, следует понимать, что положения с остатками, не являющимися идентичными, зачастую отличаются консервативными заменами аминокислот, где аминокислотные остатки заменяются другими аминокислотными остатками с аналогичными химическими свойствами (например, зарядом или гидрофобностью) и поэтому не изменяют функциональные свойства молекулы. Если последовательности отличаются консервативными заменами, процентную долю идентичности последовательностей можно скорректировать в сторону увеличения, чтобы учесть консервативный характер замены. Последовательности, отличающиеся такими консервативными заменами, считаются имеющими «сходство последовательностей» или «сходство». Способы внесения данной корректировки хорошо известны специалистам в данной области. Как правило, они включают в себя оценку консервативной замены как частичного, а не полного несовпадения, в результате чего повышается процентное значение идентичности последовательности. Таким образом, например, если идентичной аминокислоте присваивают оценку 1, то неконсервативной замене присваивают оценку, равную нулю, а консервативная замена получает оценку от нуля до 1. Вычисляют оценку консервативных замен, например, в соответствии с реализацией в программе PC/GENE (Intelligenetics, г. Маунтин-Выо, штат Калифорния, США).
Термин «процентная доля идентичности последовательности» означает значение, которое определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения, причем участок полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) в сравнении с эталонной последовательностью (которая не содержит добавлений или делеции) для оптимального выравнивания двух последовательностей. Процентное значение рассчитывают путем определения числа положений, в которых в обеих последовательностях встречаются идентичные нуклеиновые основания или аминокислотные остатки, деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 с получением процентного значения идентичности последовательности.
Если не указано иное, значения идентичности/сходства последовательности включают значение, полученное с использованием программного обеспечения GAP версии 10 с использованием следующих параметров: % идентичности и % сходства нуклеотидной последовательности при использовании значения GAP Weight (штраф за начало гэпа) = 50 и Length Weight (штраф за удлинение) = 3 и матрица замен nwsgapdna. cmp; % идентичности и % сходства аминокислотной последовательности при использовании значения GAP Weight (штраф за начало гэпа) = 8 и Length Weight (штраф за удлинение) = 2 и матрица замен BLOSUM62; или любой эквивалентной программы. Термин «эквивалентная программа» включает в себя любую программу для сравнения последовательностей, которая для любых двух рассматриваемых последовательностей создает выравнивание, обладающее идентичными соответствиями нуклеотидов или аминокислотных остатков и идентичным процентом идентичности последовательности при сравнении с соответствующим выравниванием, получаемым посредством GAP версии 10.
Термин «in vitro» включает искусственные среды и процессы или реакции, возникающие в искусственной среде (например, пробирке). Термин «in vivo» включает естественные среды (например, клетку, или организм, или тело) и процессы или реакции, протекающие в естественной среде. Термин «ех vivo» включает клетки, которые были извлечены из тела индивида, а также процессы или реакции, происходящие внутри таких клеток.
Композиции или способы, «содержащие» или «включающие в себя» один или более упомянутых элементов, могут включать в себя другие конкретно не упомянутые элементы. Например, композиция, которая «содержит» или «включает в себя» белок, может содержать белок отдельно или в комбинации с другими ингредиентами.
Обозначение диапазона значений включает в себя все целые числа, входящие в диапазон или образующие его, а также все поддиапазоны, образованные целыми числами, входящими в диапазон.
Если иное не очевидно из контекста, термин «приблизительно» охватывает значения в пределах стандартных границ погрешности измерения (например, стандартной погрешности среднего (СПС)) указанного значения.
Все элементы, указанные в единственном числе, также могут использоваться и во множественном, если контекст явно не диктует иное. Например, термин «белок Cas» или «по меньшей мере один белок Cas» может включать в себя множество белков Cas, в том числе их смеси.
ПОДРОБНОЕ ОПИСАНИЕ
I. Общее описание
Предложены способы и композиции для модификации генома в клетке. В способах и композициях применяются системы CRISPR/Cas, в которых используются две гидовые РНК (гРНК), нацеленные на разные сайты в пределах одного целевого геномного локуса. Например, в способах и композициях могут применяться системы CRISPR/Cas, в которых используются две гидовые РНК (гРНК) для создания парных двухцепочечных разрывов в разных сайтах в пределах одного целевого геномного локуса. Альтернативно в способах и композициях могут применяться системы CRISPR/Cas, в которых используются две гидовые РНК (гРНК) для создания парных одноцепочечных разрывов в разных сайтах в пределах одного целевого геномного локуса. В некоторых способах две или более гидовых РНК (например, три или четыре) можно использовать, например, для создания двух или более одноцепочечных разрывов или двухцепочечных разрывов в разных сайтах в пределах одного целевого геномного локуса.
Некоторые способы способствуют биаллельным геномным модификациям и включают в себя коллапсирование генома, при котором большую нуклеотидную последовательность удаляют из хромосомы между двумя сайтами расщепления.
Другие способы способствуют биаллельным геномным модификациям и включают в себя одновременную делецию нуклеотидной последовательности внутри клетки и замену экзогенной нуклеотидной последовательностью. Как более подробно описано ниже, эти способы с использованием двух гРНК повышают эффективность получения клеток или животных с биаллельными нацеленными генетическими модификациями посредством способствования созданию таких клеток и животных за один этап нацеливания. Следовательно, сокращается число животных и скрещиваний, необходимое для получения животного с биаллельной нацеленной генетической модификацией.
Другие способы включают в себя генную конверсию или потерю гетерозиготности, при которой геном, являющийся гетерозиготным по аллелю, модифицируют так, чтобы он стал гомозиготным по аллелю, посредством расщепления в сайтах, определяемых двумя гРНК, в соответствующем аллеле на соответствующей гомологичной хромосоме. Как более подробно описано ниже, использование двух гРНК в этих способах повышает частоту генной конверсии и обеспечивает генную конверсию по более крупным участкам хромосомной ДНК.
II. Системы CRISPR/Cas
В способах и композициях, описанных в настоящем документе, для модификации генома внутри клетки могут использоваться системы коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/cиcтемы, ассоциированные с CRISPR (Cas), или компоненты таких систем. Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии или управлении активностью генов Cas. Система CRISPR/Cas может представлять собой систему I типа, II типа или III типа. В способах и композициях, описанных в настоящем документе, для сайт-направленного расщепления нуклеиновых кислот используются системы CRISPR/Cas посредством использования комплексов CRISPR (содержащих гидовую РНК (гРНК), образующую комплекс с белком Cas).
Некоторые системы CRISPR/Cas, используемые в способах, описанных в настоящем документе, не встречаются в природе. «Не встречающаяся в природе» система имеет признаки вмешательства человека, например, один или более компонентов системы изменены или мутированы по сравнению с их природным состоянием, при этом они по меньшей мере по существу не содержат по меньшей мере один другой компонент, с которым они естественно ассоциированы в природе, или они ассоциированы по меньшей мере с другим компонентом, с которым они не ассоциированы в природе. Например, в некоторых системах CRISPR/Cas используются не встречающиеся в природе комплексы CRISPR, которые содержат гРНК и белок Cas, вместе не встречающиеся в природе.
А. Эндонуклеазы, направляемые РНК Cas
Белки Cas обычно содержат по меньшей мере один домен распознавания или связывания РНК. Такие домены могут взаимодействовать с гидовыми РНК (гРНК, более подробно описанными ниже). Белки Cas могут также содержать нуклеазные домены (например, домены ДНКазы или РНКазы), ДНК-связывающие домены, геликазные домены, домены межбелковых взаимодействий, домены димеризации и другие домены. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновых кислот. Расщепление включает разрушение ковалентных связей молекулы нуклеиновой кислоты. В результате расщепления могут образовываться тупые концы или ступенчатые концы, и расщепление может быть одноцепочечным или двухцепочечным.
Примеры белков Cas включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966 и их гомологи или модифицированные варианты.
В некоторых случаях белок Cas относится к системе CRISPR/Cas типа II. Например, белок Cas может являться белком Cas9 или может быть получен из белка Cas9. Белки Cas9 обычно имеют четыре общих основных мотива с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Белок Cas9 может быть, например, получен из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, AlicyclobacHlus acidocaldarius, Bacilluspseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospiraplatensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus или Acaryochloris marina. Дополнительные примеры представителей семейства Cas9 включают описанные в публикации WO 2014/131833, которая полностью включена в настоящий документ путем ссылки. В одном конкретном примере белок Cas9 представляет собой белок Cas9 из S. pyogenes или его производное. Аминокислотную последовательность белка Cas9 из S. pyogenes можно найти, например, в базе данных SwissProt под номером доступа Q99ZW2.
Белки Cas могут быть белками дикого типа (т.е. встречающимися в природе), модифицированными белками Cas (т.е. вариантами белков Cas) или фрагментами белков Cas дикого типа или модифицированных белков Cas. Белки Cas могут также быть активными вариантами или фрагментами белков Cas дикого типа или модифицированных белков Cas. Идентичность последовательности таких активных вариантов или фрагментов белку Cas дикого типа или модифицированному белку Cas или его части может составлять по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют способность разрезания на требуемом сайте расщепления и, следовательно, сохраняют активность, индуцирующую одно- или двухцепочечный разрыв. Тесты для определения активности, индуцирующей одно- или двухцепочечный разрыв, известны и, как правило, измеряют общую активность и специфичность белка Cas на субстратах ДНК, содержащих сайт расщепления.
Белки Cas могут быть модифицированы для увеличения или уменьшения аффинности связывания нуклеиновых кислот, специфичности связывания нуклеиновых кислот и/или ферментативной активности. Белки Cas также могут быть модифицированы для изменения любого другого вида активности или свойства белка, например стабильности. Например, один или более нуклеазных доменов белка Cas могут быть изменены, удалены или инактивированы, или белок Cas может быть усечен так, чтобы удалить домены, которые не являются необходимыми для функционирования белка, или для оптимизации (например, повышения или снижения) активности белка Cas.
Некоторые белки Cas содержат по меньшей мере два нуклеазных домена, таких как домены ДНКазы. Например, белок Cas9 может содержать RuvC-подобный нуклеазный домен и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать отличную цепь двухцепочечной ДНК, чтобы выполнить двухцепочечный разрыв в ДНК. См., например, публикацию Jinek et al. (2012) Science 337: 816-821, которая для всех целей полностью включена в настоящий документ путем ссылки.
Один или оба нуклеазных домена могут быть удалены или подвергнуты мутации таким образом, чтобы прекратить их функционирование или снизить нуклеазную активность. При удалении или мутации одного из нуклеазных доменов полученный белок Cas (например, Cas9) может называться «никазой» и может производить одноцепочечный разрыв на последовательности распознавания РНК CRISPR в пределах двухцепочечной ДНК, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или некомплементарную цепь, но не обе). При удалении или мутации обоих нуклеазных доменов полученный белок Cas (например, Cas9) будет иметь сниженную способность расщеплять обе цепи двухцепочечной ДНК. Примером мутации, превращающей Cas9 в никазу, является мутация D10A (замена аспартата на аланин в положении 10 Cas9) в домене RuvC белка Cas9 из S. pyogenes. Аналогичным образом, мутация Н939А (замена гистидина на аланин в положении аминокислоты 839) или Н840А (замена гистидина на аланин в положении аминокислоты 840) в домене HNH Cas9 из S. pyogenes может превращать Cas9 в никазу. Другие примеры мутаций, превращающих Cas9 в никазу, включают соответствующие мутации Cas9 из S. thermophilics. См., например, публикации Sapranauskas et al. (2011) Nucleic Acids Research 39: 9275-9282 и WO 2013/141680, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки. Такие мутации могут быть созданы с использованием таких известных способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или полный синтез гена. Примеры других мутаций, создающих никазы, можно найти, например, в публикациях WO/2013/176772 A1 и WO/2013/142578 A1, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
Белки Cas также могут представлять собой слитые белки. Например, белок Cas может быть слит с доменом расщепления, доменом эпигенетических модификаций, доменом транскрипционной активации или доменом транскрипционного репрессора. См. публикацию WO 2014/089290, которая для всех целей полностью включена в настоящий документ путем ссылки. Белки Cas также могут быть слиты с гетерологичным полипептидом, обеспечивающим повышенную или сниженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, на С-конце или может находиться внутри белка Cas.
Одним примером слитого белка Cas является белок Cas, слитый с гетерологичным полипептидом, который обеспечивает субклеточную локализацию. Такие последовательности могут включать в себя, например, сигнал ядерной локализации (NLS), например NLS SV40, для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал удержания ER и т.п. См., например, Lange et al. (2007) J. Biol. Chem. 282: 5101-5105, для всех целей полностью включенную в настоящий документ путем ссылки. Например, белки Cas могут быть слиты с одним или более сигналами ядерной локализации (например, двумя или тремя сигналами ядерной локализации). Такие сигналы субклеточной локализации могут быть расположены на N-конце, на С-конце или в других местах внутри белка Cas. NLS может содержать участок из основных аминокислот и может быть представлен одинарной последовательностью или двойной последовательностью.
Белки Cas могут также содержать проникающий в клетку домен. Например, проникающий в клетку домен может быть получен из белка TAT HIV-1, проникающего в клетку мотива TLM из вируса гепатита В человека, MPG, Рер-1, VP22, проникающего в клетку пептида из вируса простого герпеса или из последовательности пептида полиаргинина. См., например, публикацию WO 2014/089290, которая для всех целей полностью включена в настоящий документ путем ссылки. Проникающий в клетку домен может быть расположен на N-конце, на С-конце или в других местах внутри белка Cas.
Белки Cas также могут содержать гетерологичный полипептид для простоты отслеживания или очистки, например флуоресцентный белок, метку очистки или эпитопную метку. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP, ZsYellowl), синие флуоресцентные белки (например, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire, T-sapphire), голубые флуоресцентные белки (например, eCFP, Cerulean, CyPet, AmCyanl, Midoriishi-Cyan), красные флуоресцентные белки (mKate, mKate2, mPlum, мономерный DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-monomer, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry, Jred), оранжевые флуоресцентные белки (mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine, tdTomato) и любой другой подходящий флуоресцентный белок. Примеры меток включают глутатион-S-трансферазу (GST), хитин-связывающий белок (СВР), мальтоза-связывающий белок, тиоредоксин (TRX), поли(NANP), метку тандемной аффинной очистки (ТАР), myc, AcV5, AU1, AU5, Е, ECS, Е2, FLAG, гемагглютинин (НА), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, S1, T7, V5, VSV-G, гистидин (His), биотин-карбоксил-переносящий белок (BCCP) и кальмодулин.
Белки Cas могут быть обеспечены в любом виде. Например, белок Cas может быть обеспечен в виде белка, такого как белок Cas, образующий комплекс с гРНК. Альтернативно белок Cas может быть обеспечен, в виде нуклеиновой кислоты, кодирующей белок Cas, например РНК (например, матричной РНК (мРНК)) или ДНК. Необязательно нуклеиновая кислота, кодирующая белок Cas, может быть оптимизированной по кодонам для эффективной трансляции в белок в конкретной клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована так, чтобы заменить кодоны, имеющие более высокую частоту использования в бактериальной клетке, дрожжевой клетке, человеческой клетке, не относящейся к человеку клетке, клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или любой другой интересующей клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью. Если в клетку вводят нуклеиновую кислоту, кодирующую белок Cas, то белок Cas может экспрессироваться внутри клетки временно, условно или конститутивно.
Нуклеиновые кислоты, кодирующие белки Cas, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. Альтернативно нуклеиновые кислоты, кодирующие белки Cas, могут быть функционально связаны с промотором в экспрессионном конструкте. Экспрессионные конструкты включают в себя любые нуклеотидные конструкты, которые способны управлять экспрессией гена или другой интересующей нуклеотидной последовательности (например, гена Cas) и которые могут переносить такую представляющую интерес нуклеотидную последовательность в клетку-мишень. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в нацеливающем векторе, содержащем полинуклеотидную вставку, и/или в векторе, содержащем ДНК, кодирующую гРНК. Альтернативно она может находиться в векторе или плазмиде, отдельной от нацеливающего вектора и содержащей полинуклеотидную вставку и/или отдельной от вектора, содержащего ДНК, кодирующую гРНК. Промоторы, которые могут использоваться в экспрессионном конструкте, включают, например, промоторы, активные в плюрипотентной клетке крысы, эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Также могут использоваться промоторы, активные в эмбрионе на стадии единственной клетки. Такие промоторы могут представлять собой, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. В других разделах настоящего документа описаны примеры других промоторов.
В. Гидовые РНК (гРНК)
«Гидовая РНК», или «гРНК», включает в себя молекулу РНК, которая связывается с белком Cas и нацеливает белок Cas в специфическое местоположение внутри целевой ДНК. Гидовые РНК могут содержать два сегмента: «сегмент ДНК-нацеливания» и «белок-связывающий сегмент». «Сегмент» представляет собой сегмент, часть или область молекулы, например непрерывный участок из нуклеотидов в РНК. Некоторые гРНК содержат две отдельные молекулы РНК: «РНК-активатор» и «РНК-нацеливатель». Другие гРНК представляют собой одинарные молекулы РНК (одинарный полинуклеотид РНК), которые также можно назвать «одномолекулярной гРНК», «одногидовой РНК» или «огРНК». См., например, публикации WO/2013/176772 A1, WO/2014/065596 A1, WO/2014/089290 A1, WO/2014/093622 А2, WO/2014/099750 A2, WO/2013142578 A1 и WO 2014/131833 А1, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки. Термины «гидовая РНК» и «гРНК» являются включающими и включают как двухмолекулярные гРНК, так и одномолекулярные гРНК.
Примеры двухмолекулярных гРНК включают крРНК-подобную молекулу («РНК CRISPR», или «РНК-нацеливатель», или «крРНК», или «повтор крРНК») и соответствующую тракрРНК-подобную молекулу («транс-действующую РНК CRISPR», или «РНК-активатор», или «тракрРНК», или «каркас»). КрРНК содержит оба из сегмента (одноцепочечные) ДНК-нацеливания гРНК и участка нуклеотидов, который образует одну половину дуплекса дцРНК белок-связывающего сегмента гРНК.
Соответствующая тракрРНК (РНК-активатор) содержит участок нуклеотидов, который образует другую половину дуплекса дцРНК белок-связывающего сегмента гРНК. Участок нуклеотидов крРНК комплементарен участку нуклеотидов тракрРНК и гибридизуется с ним с образованием дуплекса дцРНК белок-связывающего домена гРНК. Таким образом, можно сказать, что каждая крРНК имеет соответствующую тракрРНК. ТракрРНК могут быть любого вида (например, полноразмерные тракрРНК или активные частичные тракрРНК) и различной длины. Формы тракрРНК могут включать в себя первичные транскрипты или подвергнутые процессингу формы. Например, в S. pyogenes к разным формам тракрРНК относятся 171-нуклеотидная, 89-нуклеотидная, 75-нуклеотидная и 65-нуклеотидная версии. См., например, публикацию Deltcheva et al. (2011) Nature 471: 602-607 и WO 2014/093661, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
КрРНК и соответствующая тракрРНК гибридизуются с образованием гРНК. КрРНК дополнительно обеспечивает одноцепочечный сегмент ДНК-нацеливания, который гибридизуется с последовательностью распознавания РНК CRISPR. При использовании для модификации внутри клетки может быть разработана точная последовательность данной молекулы крРНК или тракрРНК, которая будет специфичной для тех видов, в которых будут использоваться молекулы РНК. См., например, публикации Mali et al. (2013) Science 339: 823-826; Jinek et al. (2012) Science 337: 816-821; Hwang et al. (2013) Nat. Biotechnol. 31: 227-229; Jiang et al. (2013) Nat. Biotechnol. 31: 233-239; и Cong et al. (2013) Science 339: 819-823, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
Сегмент ДНК-нацеливания (крРНК) данной гРНК содержит нуклеотидную последовательность, которая является комплементарной последовательности в целевой ДНК. Сегмент ДНК-нацеливания гРНК взаимодействует с целевой ДНК последовательность-специфическим образом посредством гибридизации (т.е. спаривания нуклеотидов). По существу нуклеотидная последовательность сегмента ДНК-нацеливания может изменяться и определяет местоположение внутри целевой ДНК, с которым будут взаимодействовать гРНК и целевая ДНК. Сегмент ДНК-нацеливания обсуждаемой гРНК может быть модифицирован для гибридизации с любой заданной последовательностью в пределах целевой ДНК. Встречающиеся в природе крРНК различаются в зависимости от системы Cas9 и организма, но часто содержат нацеливающий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, публикацию WO 2014/131833, для всех целей полностью включенную в настоящий документ путем ссылки). В случае S. pyogenes длина DR составляет 36 нуклеотидов, а длина нацеливающего сегмента составляет 30 нуклеотидов. DR, расположенный на 3'-конце, является комплементарным и гибридизуется с соответствующей тракрРНК, которая, в свою очередь, связывается с белком Cas9.
Сегмент ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов до приблизительно 100 нуклеотидов. Например, сегмент ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов (нт) до приблизительно 80 нт, от приблизительно 12 нт до приблизительно 50 нт, от приблизительно 12 нт до приблизительно 40 нт, от приблизительно 12 нт до приблизительно 30 нт, от приблизительно 12 нт до приблизительно 25 нт, от приблизительно 12 нт до приблизительно 20 нт или от приблизительно 12 нт до приблизительно 19 нт. Альтернативно сегмент ДНК-нацеливания может иметь длину от приблизительно 19 нт до приблизительно 20 нт, от приблизительно 19 нт до приблизительно 25 нт, от приблизительно 19 нт до приблизительно 30 нт, от приблизительно 19 нт до приблизительно 35 нт, от приблизительно 19 нт до приблизительно 40 нт, от приблизительно 19 нт до приблизительно 45 нт, от приблизительно 19 нт до приблизительно 50 нт, от приблизительно 19 нт до приблизительно 60 нт, от приблизительно 19 нт до приблизительно 70 нт, от приблизительно 19 нт до приблизительно 80 нт, от приблизительно 19 нт до приблизительно 90 нт, от приблизительно 19 нт до приблизительно 100 нт, от приблизительно 20 нт до приблизительно 25 нт, от приблизительно 20 нт до приблизительно 30 нт, от приблизительно 20 нт до приблизительно 35 нт, от приблизительно 20 нт до приблизительно 40 нт, от приблизительно 20 нт до приблизительно 45 нт, от приблизительно 20 нт до приблизительно 50 нт, от приблизительно 20 нт до приблизительно 60 нт, от приблизительно 20 нт до приблизительно 70 нт, от приблизительно 20 нт до приблизительно 80 нт, от приблизительно 20 нт до приблизительно 90 нт или от приблизительно 20 нт до приблизительно 100 нт.
Нуклеотидная последовательность сегмента ДНК-нацеливания, являющаяся комплементарной нуклеотидной последовательности (последовательности распознавания РНК CRISPR) целевой ДНК, может иметь длину по меньшей мере приблизительно 12 нт. Например, последовательность ДНК-нацеливания (т.е. последовательность в пределах сегмента ДНК-нацеливания, являющаяся комплементарной последовательности распознавания РНК CRISPR в пределах целевой ДНК) может иметь длину по меньшей мере приблизительно 12 нт, по меньшей мере приблизительно 15 нт, по меньшей мере приблизительно 18 нт, по меньшей мере приблизительно 19 нт, по меньшей мере приблизительно 20 нт, по меньшей мере приблизительно 25 нт, по меньшей мере приблизительно 30 нт, по меньшей мере приблизительно 35 нт или по меньшей мере приблизительно 40 нт. Альтернативно последовательность ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов (нт) до приблизительно 80 нт, от приблизительно 12 нт до приблизительно 50 нт, от приблизительно 12 нт до приблизительно 45 нт, от приблизительно 12 нт до приблизительно 40 нт, от приблизительно 12 нт до приблизительно 35 нт, от приблизительно 12 нт до приблизительно 30 нт, от приблизительно 12 нт до приблизительно 25 нт, от приблизительно 12 нт до приблизительно 20 нт, от приблизительно 12 нт до приблизительно 19 нт, от приблизительно 19 нт до приблизительно 20 нт, от приблизительно 19 нт до приблизительно 25 нт, от приблизительно 19 нт до приблизительно 30 нт, от приблизительно 19 нт до приблизительно 35 нт, от приблизительно 19 нт до приблизительно 40 нт, от приблизительно 19 нт до приблизительно 45 нт, от приблизительно 19 нт до приблизительно 50 нт, от приблизительно 19 нт до приблизительно 60 нт, от приблизительно 20 нт до приблизительно 25 нт, от приблизительно 20 нт до приблизительно 30 нт, от приблизительно 20 нт до приблизительно 35 нт, от приблизительно 20 нт до приблизительно 40 нт, от приблизительно 20 нт до приблизительно 45 нт, от приблизительно 20 нт до приблизительно 50 нт или от приблизительно 20 нт до приблизительно 60 нт. В некоторых случаях последовательность ДНК-нацеливания может иметь длину приблизительно 20 нт.
ТракрРНК могут быть любого вида (например, полноразмерные тракрРНК или активные частичные тракрРНК) и различной длины. Они могут включать в себя первичные транскрипты или подвергнутые процессингу формы. Например, тракрРНК (в составе одногидовой РНК или в виде отдельной молекулы в составе двухмолекулярной гРНК) может содержать всю или часть последовательности тракрРНК дикого типа или состоять из всей или части последовательности тракрРНК дикого типа (например, приблизительно или более чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности тракрРНК дикого типа). Примеры последовательностей тракрРНК дикого типа из S. pyogenes включают варианты из 171 нуклеотида, 89 нуклеотидов, 75 нуклеотидов и 65 нуклеотидов. См., например, публикацию Deltcheva et al. (2011) Nature 471: 602-607; WO 2014/093661, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки. Примеры тракрРНК в пределах одногидовых РНК (огРНК) включают сегменты тракрРНК, встречающиеся в пределах вариантов огРНК +48, +54, +67 и +85, где «+n» означает, что в огРНК включено до+n нуклеотидов тракрРНК дикого типа. См. патент США 8,697,359, который для всех целей полностью включен в настоящий документ путем ссылки.
Процентное значение комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). В некоторых случаях процентное значение комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет по меньшей мере 60% на протяжении приблизительно 20 смежных нуклеотидов. В одном примере процентное значение комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет 100% на протяжении 14 непрерывных нуклеотидов на 5'-конце последовательности распознавания РНК CRISPR в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 14 нуклеотидов. В другом примере процентное значение комплементарности между последовательностью ДНК-нацеливания и последовательностью распознавания РНК CRISPR в пределах целевой ДНК составляет 100% на протяжении семи непрерывных нуклеотидов на 5'-конце последовательности распознавания РНК CRISPR в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 7 нуклеотидов.
Белок-связывающий сегмент гРНК может содержать два участка нуклеотидов, которые комплементарны друг другу. Комплементарные нуклеотиды белок-связывающего сегмента гибридизуются с образованием дуплекса двухцепочечной РНК (дцРНК). Белок-связывающий сегмент обсуждаемой гРНК взаимодействует с белком Cas, а гРНК управляет связыванием белка Cas со специфической нуклеотидной последовательностью в пределах ДНК с помощью сегмента ДНК-нацеливания.
Гидовые РНК могут содержать модификации или последовательности, обеспечивающие дополнительные желаемые свойства (например, измененную или регулируемую стабильность; субклеточное нацеливание; отслеживание по флуоресцентной метке; сайт связывания для белка или белкового комплекса и т.п.). К примерам таких модификаций относятся, например, 5'-кэп (например, 7-метилгуанилатный кэп (m7G)); 3'-полиаденилированный хвост (т.е. 3'-поли(А) хвост); последовательность-рибопереключатель (например, для обеспечения регулируемой стабильности и/или регулируемой доступности белками и/или белковыми комплексами); последовательность контроля стабильности; последовательность, образующая дуплекс дцРНК (т.е. шпилька); модификация или последовательность, нацеливающая РНК в субклеточное местоположение (например, ядро, митохондрии, хлоропласты и т.п.); модификация или последовательность, обеспечивающая отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который способствует флуоресцентной детекции, последовательность, обеспечивающая флуоресцентную детекцию, и т.д.); модификация или последовательность, обеспечивающая сайт связывания для белков (например, белков, действующих на ДНК, включая активаторы транскрипции, репрессоры транскрипции, ДНК-метилтрансферазы, ДНК-деметилазы, гистонацетилтрансферазы, гистондеацетилазы и т.п.); и их комбинации.
гРНК может содержать нуклеотидную последовательность, кодирующую крРНК и тракрРНК. Например, гРНК может содержать: (а) химерную РНК, имеющую нуклеотидную последовательность 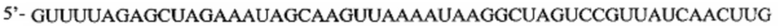
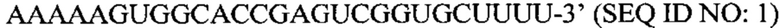 ; или (b) химерную РНК, имеющую нуклеотидную последовательность
; или (b) химерную РНК, имеющую нуклеотидную последовательность 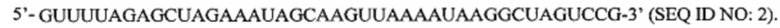
В некоторых случаях крРНК содержит  ;
; 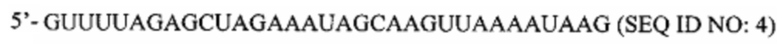 ; или
; или 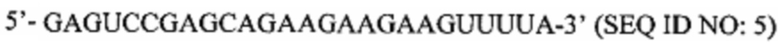
В некоторых случаях тракрРНК содержит 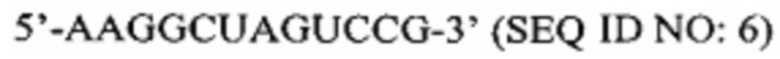 или
или 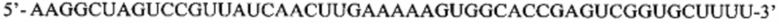
Гидовые РНК могут быть обеспечены в любом виде. Например, гРНК может быть обеспечена в виде РНК, или как две молекулы (отдельно крРНК и тракрРНК), или как одна молекула (огРНК), и необязательно в виде комплекса с белком Cas. гРНК может быть обеспечена также в виде ДНК, кодирующей РНК. ДНК, кодирующая гРНК, может кодировать одну молекулу РНК (огРНК) или отдельные молекулы РНК (например, отдельно крРНК и тракрРНК). В последнем случае ДНК, кодирующая гРНК, может быть обеспечена как отдельные молекулы ДНК, кодирующие соответственно крРНК и тракрРНК.
Если в клетку вводят ДНК, кодирующую гРНК, то гРНК может экспрессироваться внутри клетки временно, условно или конститутивно. ДНК, кодирующие гРНК, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. Альтернативно ДНК, кодирующие гРНК, могут быть функционально связаны с промотором в экспрессионном конструкте. Например, ДНК, кодирующая гРНК, может находиться в нацеливающем векторе, содержащем полинуклеотидную вставку, и/или в векторе, содержащем нуклеиновую кислоту, кодирующую белок Cas. Альтернативно она может находиться в векторе или плазмиде, отдельной от нацеливающего вектора и содержащей полинуклеотидную вставку и/или отдельной от вектора, содержащего нуклеиновую кислоту, кодирующую белок Cas. Промоторы, которые могут использоваться в таких экспрессионных конструктах, включают в себя промоторы, например, активные в плюрипотентной клетке крысы, клетке эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Также могут использоваться промоторы, активные в эмбрионе на стадии единственной клетки. Такие промоторы могут представлять собой, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. В некоторых случаях промотор представляет собой промотор РНК-полимеразы III, такой как промотор U6 человека, промотор U6 полимеразы III крысы или промотор U6 полимеразы III мыши. В других разделах настоящего документа описаны примеры других промоторов.
Альтернативно гРНК можно получать с помощью различных других способов. Например, гРНК можно получать посредством транскрипции in vitro с использованием, например, Т7 РНК-полимеразы (см., например, публикации WO 2014/089290 и WO 2014/065596, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки). Гидовые РНК также могут быть синтетическими молекулами, полученными путем химического синтеза.
С. Последовательности распознавания РНК CRISPR
Термин «последовательность распознавания РНК CRISPR» включает нуклеотидные последовательности, присутствующие в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК при условии существования достаточных условий для связывания. Например, последовательности распознавания РНК CRISPR включают последовательности, которым должна быть комплементарна сконструированная гидовая РНК, где гибридизация между последовательностью распознавания РНК CRISPR и последовательностью ДНК-нацеливания способствует образованию комплекса CRISPR. Полная комплементарность не является обязательным требованием при условии наличия достаточной комплементарности, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Последовательности распознавания РНК CRISPR также включают сайты расщепления для белков Cas, более подробно описанные ниже. Последовательность распознавания РНК CRISPR может содержать любой полинуклеотид, который может быть расположен, например, в ядре или цитоплазме клетки или внутри органеллы клетки, такой как митохондрия или хлоропласт.
На последовательность распознавания РНК CRISPR в пределах целевой ДНК может быть нацелен (т.е. может связываться, или гибридизоваться, или быть комплементарным) белок Cas или гРНК. Подходящие условия связывания ДНК/РНК включают физиологические условия, обычно существующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001), для всех целей полностью включенную в настоящий документ путем ссылки). Цепь целевой ДНК, которая является комплементарной и гибридизуется с белком Cas или гРНК, можно назвать «комплементарной цепью», а цепь целевой ДНК, которая комплементарна «комплементарной цепи» (и, таким образом, не комплементарна белку Cas или гРНК), можно назвать «некомплементарной цепью» или «матричной цепью».
Белок Cas может расщеплять нуклеиновую кислоту в сайте в пределах или за пределами нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК. «Сайт расщепления» включает положение нуклеиновой кислоты, в котором белок Cas производит одноцепочечный разрыв или двухцепочечный разрыв. Например, образование комплекса CRISPR (содержащего гРНК, гибридизованную с последовательностью распознавания РНК CRISPR и образующую комплексы с белком Cas) может приводить к расщеплению одной или обеих цепей внутри или около (например, на расстоянии в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар нуклеотидов от) нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК. Если сайт расщепления находится за пределами нуклеотидной последовательности, с которой будет связываться сегмент ДНК-нацеливания гРНК, то сайт расщепления все еще считается находящимся в пределах «последовательности распознавания РНК CRISPR». Сайт расщепления может находиться только на одной цепи или на обеих цепях нуклеиновой кислоты. Сайты расщепления могут находиться в одном и том же положении на обеих цепях нуклеиновой кислоты (образуя тупые концы) или могут находиться на разных сайтах на каждой цепи (образуя ступенчатые концы). Ступенчатые концы можно получать, например, с использованием двух белков Cas, каждый из которых производит одноцепочечный разрыв на отличающемся сайте расщепления на каждой цепи, создавая таким образом двухцепочечный разрыв. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК таким образом, что образуются нависающие последовательности. В некоторых случаях последовательность распознавания РНК CRISPR никазы на первой цепи разделена с последовательностью распознавания РНК CRISPR никазы на второй цепи по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 пар нуклеотидов.
Сайт-специфическое расщепление целевой ДНК белком Cas9 может происходить в местоположениях, определенных как (i) комплементарностью спариваемых нуклеотидов между гРНК и целевой ДНК, так и (ii) наличием в целевой ДНК короткого мотива, называемого мотивом, прилежащим к протоспейсеру (РАМ). РАМ может фланкировать последовательность распознавания РНК CRISPR. Необязательно последовательность распознавания РНК CRISPR может быть фланкирована РАМ на 3'-конце. Например, сайт расщепления Cas9 может находиться на расстоянии от приблизительно 1 до приблизительно 10 или от приблизительно 2 до приблизительно 5 пар нуклеотидов (например, 3 пар нуклеотидов) ближе к 5'-концу или ближе к 3'-концу от последовательности РАМ. В некоторых случаях (например, когда используется Cas9 из S. pyogenes или близкородственный Cas9) последовательность РАМ некомплементарной цепи может представлять собой 5'-N1GG-3', где N1 представляет собой любой нуклеотид ДНК и находится непосредственно на 3'-конце последовательности распознавания РНК CRISPR некомплементарной цепи целевой ДНК. По существу последовательность РАМ комплементарной цепи будет представлять собой 5'-CCN2-3', где N2 представляет собой нуклеотид ДНК и находится непосредственно на 5'-конце последовательности распознавания РНК CRISPR комплементарной цепи целевой ДНК. В некоторых таких случаях N1 и N2 могут быть комплементарны, а пара нуклеотидов N1-N2 может представлять собой любую пару нуклеотидов (например, N1=Ц и N2=Г; N1=Г и N2=Ц; N1=А и N2=Т или N1=Т и N2=А).
Примеры последовательностей распознавания РНК CRISPR включают последовательность ДНК, комплементарную сегменту ДНК-нацеливания гРНК, или такую последовательность ДНК в дополнение к последовательности РАМ. Например, целевой мотив может представлять собой последовательность ДНК длиной 20 нуклеотидов, непосредственно предшествующую мотиву NGG, распознаваемому белком Cas, например GN19NGG (SEQ ID NO: 8) или N20NGG (SEQ ID NO: 9) (см., например, публикацию WO 2014/165825, которая для всех целей полностью включена в настоящий документ путем ссылки). Гуанин на 5'-конце может способствовать транскрипции РНК-полимеразы в клетках. Другие примеры последовательностей распознавания РНК CRISPR могут включать два гуаниннуклеотида на 5'-конце (например, GGN20NGG; SEQ ID NO: 10) для облегчения эффективной транскрипции Т7-полимеразой in vitro. См., например, публикацию WO 2014/065596, которая для всех целей полностью включена в настоящий документ путем ссылки. Другие последовательности распознавания РНК CRISPR могут иметь длину 4-22 нуклеотида с SEQ ID NO: 8-10, включая 5' G или GG и 3' GG или NGG. Другие последовательности распознавания РНК CRISPR могут иметь длину 14-20 нуклеотидов с SEQ ID NO: 8-10. К конкретным примерам последовательностей распознавания РНК CRISPR относятся последовательности ДНК, комплементарные нуклеиновым кислотам, содержащим любую из SEQ ID NO: 11-38.
Последовательность распознавания РНК CRISPR может представлять собой любую нуклеотидную последовательность, эндогенную или экзогенную по отношению к клетке. Последовательность распознавания РНК CRISPR может представлять собой последовательность, кодирующую генный продукт (например, белок), или некодирующую последовательность (например, регуляторную последовательность), или может включать в себя обе последовательности. В некоторых случаях последовательность распознавания РНК CRISPR может находиться в пределах ассоциированного с заболеванием гена или нуклеиновой кислоты и/или в пределах ассоциированного с сигнальным путем гена или нуклеиновой кислоты. Ассоциированный с заболеванием ген или нуклеиновая кислота включает в себя любой ген или нуклеиновую кислоту, дающую в клетках, полученных из затронутых заболеванием тканей, продукты транскрипции или трансляции в аномальной концентрации или в аномальной форме по сравнению с контрольными тканями или клетками, не имеющими заболевания. Например, ассоциированный с заболеванием ген может нести одну или несколько мутаций или генетических вариаций, которые непосредственно ответственны за этиологию заболевания или находятся в неравновесном сцеплении с одним или более генами, ответственными за этиологию заболевания. Продукты транскрипции или трансляции могут быть известными или неизвестными и могут иметь нормальный или аномальный уровень. Примеры ассоциированных с заболеванием генов и нуклеиновых кислот можно получить на Интернет-сайтах организаций McKusick-Nathans Institute of Genetic Medicine, Johns Hopkins University (г.Балтимор, штат Мэриленд, США) и National Center for Biotechnology Information, National Library of Medicine (г. Бетесда, штат Мэриленд, США). Дополнительные примеры ассоциированных с заболеваниями генов и нуклеиновых кислот представлены в патенте США №8,697,359, который для всех целей полностью включен в настоящий документ путем ссылки.
Мутации в вызывающих заболевания генах могут быть рецессивными мутациями или доминантными мутациями. Диплоидные организмы (т.е. организмы, имеющие две копии каждой хромосомы), как правило, имеют две копии каждого гена. Если две копии у индивида идентичны, то индивид является гомозиготным по гену. Если копии представляют собой разные аллели, индивид является гетерозиготным по гену. Термин «генотип» включает то, несет ли индивид мутации в конкретном гене (или генах), а термин «фенотип» включает физические и функциональные последствия этого генотипа. К рецессивным мутациям относятся мутации, в которых оба аллеля должны быть мутантными, чтобы наблюдался мутантный фенотип (т.е. организм должен быть гомозиготным по мутантному аллелю, чтобы проявился мутантный фенотип). Рецессивные мутации могут, например, инактивировать затронутый ген и приводить к утрате функции. Например, рецессивная мутация может полностью или частично удалять ген из хромосомы, нарушать экспрессию гена или изменять структуру закодированного белка, таким образом изменяя его функцию. Напротив, к доминантным мутациям относятся мутации, при которых доминантный фенотип наблюдается в организме, гетерозиготном по этой мутации (т.е. организм несет один мутантный аллель и один аллель дикого типа). Доминантная мутация может, например, приводить к приобретению функции. Например, доминантная мутация может приводить к повышению активности продукта данного гена, придавать новую активность продукту гена или приводить к ненадлежащей по месту или времени экспрессии. Доминантная мутация также может быть связана с утратой функции. В некоторых случаях, если для нормального функционирования требуются две копии гена, удаление одной копии может приводить к мутантному фенотипу. Такие гены называют гаплонедостаточными. В других случаях мутации в одном аллеле могут приводить к структурному изменению в белке, что приводит к нарушению функции белка дикого типа, кодируемого другим аллелем. Такие мутации называют доминантными негативными мутациями. Некоторые аллели могут быть связаны как с рецессивным, так и с доминантным фенотипом.
Некоторые последовательности распознавания РНК CRISPR находятся в пределах гена или нуклеотидной последовательности, содержащей мутацию. Мутация может представлять собой, например, доминантную мутацию или рецессивную мутацию. В некоторых случаях доминантная мутация находится в пределах клетки, гетерозиготной по доминантной мутации (т.е. клетка содержит аллель дикого типа и мутантный аллель, содержащий доминантную мутацию). В некоторых таких случаях последовательность распознавания РНК CRISPR может находиться в пределах мутантного аллеля, но не аллеля дикого типа. Альтернативно последовательность распознавания РНК CRISPR может находиться в пределах аллеля дикого типа, но не мутантного аллеля.
III. Нацеливающие векторы и полинуклеотидные вставки
В способах и композициях, описанных в настоящем документе, также могут применяться нацеливающие векторы, содержащие полинуклеотидные вставки и гомологичные плечи для модификации генома в клетке. В таких способах полинуклеотидная вставка интегрирована в целевой геномный локус, определяемый гомологичными плечами, в результате события гомологичной рекомбинации. В способах, предложенных в настоящем документе, могут использоваться преимущества нуклеазных агентов (например, белков Cas) в комбинации с событием гомологичной рекомбинации. В таких способах применяется одноцепочечный или двухцепочечный разрыв, создаваемый нуклеазным агентом в сайте расщепления нуклеазой, в комбинации с гомологичной рекомбинацией для упрощения интеграции полинуклеотидной вставки в целевой геномный локус.
А. Нацеливающие векторы и полинуклеотидные вставки для клеток, отличных от эмбрионов на стадии единственной клетки
(I) Полинуклеотидные вставки
В способах, описанных в настоящем документе, можно использовать одну или более отдельных полинуклеотидных вставок, и их можно вводить в клетку при помощи отдельных нацеливающих векторов или одного нацеливающего вектора. Полинуклеотидные вставки содержат сегменты ДНК, предназначенные для интеграции в целевые геномные локусы. Интеграция полинуклеотидной вставки в целевой локус может приводить к добавлению интересующей нуклеотидной последовательности, делеции интересующей нуклеотидной последовательности в целевом локусе и/или замене интересующей нуклеотидной последовательности в целевом локусе (т.е. делеции или вставке).
Полинуклеотидная вставка или соответствующая замещаемая нуклеиновая кислота в целевом локусе может представлять собой кодирующую область, интрон, экзон, нетранслируемую область, регуляторную область, промотор, энхансер или любую их комбинацию. Более того, полинуклеотидная вставка или соответствующая заменяемая нуклеиновая кислота в целевом локусе может иметь любую желаемую длину, например, длину 10-100 нуклеотидов, длину 100-500 нуклеотидов, длину 500 нуклеотидов-1 т.п.н., длину от 1 т.п.н. до 1,5 т.п.н., длину от 1,5 т.п.н. до 2 т.п.н., длину от 2 т.п.н. до 2,5 т.п.н., длину от 2,5 т.п.н. до 3 т.п.н., длину от 3 т.п.н. до 5 т.п.н., длину от 5 т.п.н. до 8 т.п.н., длину от 8 т.п.н. до 10 т.п.н. или более. В других случаях длина может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 800 т.п.н., от приблизительно 800 т.п.н. до 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 2,8 млн п.н., от приблизительно 2,8 млн п.н. до приблизительно 3 млн п.н. В других случаях длина может составлять по меньшей мере 100, 200, 300, 400, 500, 600, 700, 800 или 900 нуклеотидов или по меньшей мере 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 11 т.п.н., 12 т.п.н., 13 т.п.н., 14 т.п.н., 15 т.п.н., 16 т.п.н. или более. Некоторые полинуклеотидные вставки могут быть еще меньше. Например, вставку длиной от приблизительно 4 нуклеотидов до приблизительно 12 нуклеотидов можно вставить для создания сайта расщепления рестрикционным ферментом.
В некоторых нацеливающих векторах полинуклеотидная вставка может составлять от приблизительно 5 т.п.н. до приблизительно 200 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н., от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. Альтернативно полинуклеотидная вставка может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н.
В некоторых случаях замещение нуклеиновой кислоты в целевом локусе приводит к делеции нуклеотидной последовательности длиной в диапазоне от приблизительно 1 т.п.н. до приблизительно 200 т.п.н., от приблизительно 2 т.п.н. до приблизительно 20 т.п.н. или от приблизительно 0,5 т.п.н. до приблизительно 3 млн п.н. В некоторых случаях величина делеции превышает общую длину 5' гомологичного плеча и 3' гомологичного плеча.
В некоторых случаях величина делеции нуклеотидной последовательности находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н., от приблизительно 190 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 800 т.п.н., от приблизительно 800 т.п.н. до 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н., до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 2,8 млн п.н., от приблизительно 2,8 млн п.н. до приблизительно 3 млн п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н.
В других случаях полинуклеотидная вставка или соответствующая заменяемая нуклеиновая кислота в целевом локусе может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
Полинуклеотидная вставка может содержать геномную ДНК или любой другой тип ДНК. Например, полинуклеотидная вставка может происходить из прокариота, эукариота, дрожжей, птицы (например, курицы), не относящегося к человеку млекопитающего, грызуна, человека, крысы, мыши, хомяка, кролика, свиньи, коровы, оленя, овцы, козы, кошки, собаки, хорька, примата (например, мартышки, макаки-резус), домашнего млекопитающего животного, сельскохозяйственного млекопитающего животного или любого другого представляющего интерес организма.
Полинуклеотидная вставка и/или нуклеиновая кислота в целевом локусе может содержать кодирующую последовательность или некодирующую последовательность, например регуляторный элемент (например, промотор, энхансер или элемент, связывающийся с репрессором транскрипции). Например, полинуклеотидная вставка может содержать нокин-аллель по меньшей мере одного экзона эндогенного гена или нокин-аллель всего эндогенного гена (т.е. «нокин с обменом гена»).
Например, полинуклеотидная вставка может быть гомологична или ортологична последовательности, нацеливаемой для делеции, в целевом геномном локусе. Гомологичная или ортологичная полинуклеотидная вставка может замещать последовательность, нацеливаемую для делеции, в представляющем интерес геномном локусе. Гомологичная последовательность включает нуклеотидную последовательность, идентичную или по существу аналогичную известной эталонной последовательности, так что она по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична известной эталонной последовательности. Ортологичная последовательность включает нуклеотидную последовательность от одного вида, которая функционально эквивалентна известной эталонной последовательности от другого вида. Результатом может быть гуманизация локуса, если вставка полинуклеотидной вставки приводит к замещению нечеловеческой нуклеотидной последовательности на гомологичную или ортологичную человеческую полинуклеотидную последовательность (т.е. полинуклеотидная вставка вставляется вместо соответствующей нечеловеческой последовательности ДНК в ее эндогенный геномный локус).
Полинуклеотидная вставка также может содержать условный аллель. Условный аллель может быть многофункциональным аллелем, как описано в публикации US 2011/0104799, которая для всех целей полностью включена в настоящий документ путем ссылки. Например, условный аллель может содержать: (а) активирующую последовательность в смысловой ориентации по отношению к транскрипции целевого гена; (b) кассету селекции лекарственными средствами (DSC) в смысловой или антисмысловой ориентации; (с) находящуюся в антисмысловой ориентации представляющую интерес нуклеотидную последовательность (NSI); и (d) условный по инверсии модуль (COIN, в котором используется разделяющий экзон интрон и инвертируемый модуль, подобный генной ловушке) в обратной ориентации. См., например, публикацию US 2011/0104799, которая для всех целей полностью включена в настоящий документ путем ссылки. Условный аллель может дополнительно содержать рекомбинируемые блоки, которые рекомбинируются при воздействии первой рекомбиназой с образованием условного аллеля, который (i) не имеет активирующей последовательности и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См. публикацию US 2011/0104799, которая для всех целей полностью включена в настоящий документ путем ссылки.
Некоторые полинуклеотидные вставки содержат полинуклеотид, кодирующий селективный маркер. Селективный маркер может входить в кассету селекции. Такие селективные маркеры включают, без ограничений, неомицинфосфотрансферазу (neor), гигромицин-В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин-S-дезаминазу (bsrr), ксантин/гуанинфосфорибозилтрансферазу (gpt) или тимидинкиназу вируса простого герпеса (HSV-k) или их комбинацию. Полинуклеотид, кодирующий селективный маркер, может быть функционально связан с промотором, активным в клетке, на которую выполняется нацеливание. Примеры промоторов описаны в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит репортерный ген. К примерам репортерных генов относятся гены, кодирующие люциферазу, β-галактозидазу, зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (eGFP), голубой флуоресцентный белок (CFP), желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (eYFP), синий флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (eBFP), DsRed, ZsGreen, MmGFP, mPlum, mCherry, tdTomato, mStrawberry, J-Red, mOrange, mKO, mCitrine, Venus, YPet, Emerald, CyPet, Cerulean, T-Sapphire, щелочную фосфатазу и любую их комбинацию. Такие репортерные гены могут быть функционально связаны с промотором, активным в клетке, на которую выполняется нацеливание. Примеры промоторов описаны в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит одну или более экспрессионных кассет или кассет делеции. Данная кассета может содержать представляющую интерес последовательность полинуклеотидов, нуклеиновую кислоту, кодирующую селективный маркер, и/или репортерный ген наряду с различными регуляторными компонентами, влияющими на экспрессию. Примеры пригодных для включения селективных маркеров и репортерных генов описаны более подробно в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит нуклеиновую кислоту, фланкированную целевыми последовательностями сайт-специфической рекомбинации. Хотя вся полинуклеотидная вставка может быть фланкирована такими целевыми последовательностями сайт-специфической рекомбинации, любая область или отдельный представляющий интерес полинуклеотид в пределах полинуклеотидной вставки также может быть фланкирован такими сайтами. Целевые последовательности сайт-специфической рекомбинации, которые могут фланкировать полинуклеотидную вставку или любой представляющий интерес полинуклеотид в полинуклеотидной вставке, могут включать в себя, например, последовательности loxP, lox511, lox2272, lox66, lox71, loxM2, lox5171, FRT, FRT11, FRT71, attp, att, FRT, rox и их комбинацию. В одном примере сайты сайт-специфической рекомбинации фланкируют полинуклеотид, кодирующий селективный маркер и/или репортерный ген, содержащийся в пределах полинуклеотидной вставки. После интеграции полинуклеотидной вставки в целевой локус последовательности между сайтами сайт-специфической рекомбинации можно удалить.
(2) Нацеливающие векторы
Нацеливающие векторы можно использовать для внедрения полинуклеотидной вставки в целевой геномный локус, и они могут содержать полинуклеотидную вставку и гомологичные плечи, фланкирующие полинуклеотидную вставку. Нацеливающие векторы могут иметь линейную форму или форму окружности и могут быть одноцепочечными или двухцепочечными. Нацеливающие векторы могут представлять собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). Для удобства ссылки термины «гомологичные плечи» в настоящем документе относятся к гомологичным плечам 5' и 3' (т.е. ближе к 5'-концу и ближе к 3'-концу). Эта терминология относится к относительному положению гомологичных плеч к полинуклеотидной вставке в пределах нацеливающего вектора. 5' и 3' гомологичные плечи соответствуют областям в пределах целевого локуса, которые в настоящем документе называются «целевой 5'-последовательностью» и «целевой 3'-последовательностью» соответственно. Некоторые нацеливающие векторы содержат 5' и 3' гомологичные плечи без полинуклеотидной вставки. Такие нацеливающие векторы могут функционировать с возможностью удаления последовательности между целевыми 5'- и 3'-последовательностями без вставки полинуклеотидной вставки.
Гомологичное плечо и целевая последовательность «соответствуют» или являются «соответствующими» друг другу, если две области имеют достаточную степень идентичности последовательности друг с другом, чтобы действовать как субстраты для реакции гомологичной рекомбинации. Термин «гомологичные» включает последовательности ДНК, которые или являются идентичными, или имеют идентичность последовательности с соответствующей последовательностью. Идентичность последовательности между данной целевой последовательностью и соответствующим гомологичным плечом, находящимся на нацеливающем векторе, может иметь любую степень идентичности последовательности, которая обеспечивает возникновение гомологичной рекомбинации. Например, степень идентичности последовательности между гомологичным плечом нацеливающего вектора (или его фрагмента) и целевой последовательностью (или ее фрагментом) может составлять по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности таким образом, что последовательности подвергаются гомологичной рекомбинации. Более того, соответствующая область гомологии между гомологичным плечом и соответствующей целевой последовательностью может иметь любую длину, которая является достаточной для стимуляции гомологичной рекомбинации в расщепленном сайте распознавания. Например, данное гомологичное плечо и/или соответствующая целевая последовательность может содержать соответствующие области гомологии, длина которых составляет по меньшей мере приблизительно 5-10 т.п.н., 5-15 т.п.н., 5-20 т.п.н., 5-25 т.п.н., 5-30 т.п.н., 5-35 т.п.н., 5-40 т.п.н., 5-45 т.п.н., 5-50 т.п.н., 5-55 т.п.н., 5-60 т.п.н., 5-65 т.п.н., 5-70 т.п.н., 5-75 т.п.н., 5-80 т.п.н., 5-85 т.п.н., 5-90 т.п.н., 5-95 т.п.н., 5-100 т.п.н., 100-200 т.п.н. или 200-300 т.п.н. тысяч пар нуклеотидов или более (как описано в других разделах настоящего документа для векторов LTVEC) таким образом, что гомологичное плечо имеет достаточную гомологию, чтобы подвергаться гомологичной рекомбинации с соответствующими целевыми последовательностями в пределах генома клетки.
Гомологичные плечи могут соответствовать локусу, который является нативным по отношению к клетке (например, целевому локусу), или альтернативно они могут соответствовать области гетерологичного или экзогенного сегмента ДНК, который был интегрирован в геном клетки, включая, например, трансгены, экспрессионные кассеты или гетерологичные или экзогенные области ДНК. Альтернативно гомологичные плечи нацеливающего вектора могут соответствовать области дрожжевой искусственной хромосомы (YAC), бактериальной искусственной хромосомы (ВАС), человеческой искусственной хромосомы или любой другой сконструированной области, содержащейся в соответствующей клетке-хозяине. Дополнительно гомологичные плечи нацеливающего вектора могут соответствовать или могут быть получены из области из библиотеки ВАС, библиотеки космид или библиотеки фага Р1. В некоторых случаях гомологичные плечи нацеливающего вектора соответствуют локусу, который является нативным, гетерологичным или экзогенным в отношении прокариота, дрожжей, птицы (например, курицы), не относящегося к человеку млекопитающего, грызуна, человека, крысы, мыши, хомяка, кролика, свиньи, коровы, оленя, овцы, козы, кошки, собаки, хорька, примата (например, мартышки, макаки-резус), домашнего млекопитающего животного, сельскохозяйственного млекопитающего животного или любого другого представляющего интерес организма. В некоторых случаях при отсутствии одно- или двухцепочечного разрыва, индуцированного нуклеазным агентом (например, белком Cas), гомологичные плечи соответствуют локусу клетки, на который нельзя выполнить нацеливание с использованием стандартных способов или нацеливание на который может быть только неверным или иметь очень низкую эффективность. В некоторых случаях гомологичные плечи получены из синтетической ДНК.
В некоторых нацеливающих векторах 5' и 3' гомологичные плечи соответствуют целевому геному. Альтернативно гомологичные плечи могут быть получены из родственного генома. Например, целевой геном представляет собой геном мыши первой линии, а нацеливающие плечи получены из генома мыши второй линии, причем первая линия и вторая линия являются разными. В некоторых случаях гомологичные плечи получены из генома одного и того же животного или получены из генома одной линии, например, целевой геном представляет собой геном мыши первой линии, и нацеливающие плечи получены из генома той же мыши или мыши той же линии.
Гомологичное плечо нацеливающего вектора может иметь любую длину, достаточную для стимулирования события гомологичной рекомбинации с соответствующей целевой последовательностью, включая, например, длину по меньшей мере 5-10 т.п.н., 5-15 т.п.н., 5-20 т.п.н., 5-25 т.п.н., 5-30 т.п.н., 5-35 т.п.н., 5-40 т.п.н., 5-45 т.п.н., 5-50 т.п.н., 5-55 т.п.н., 5-60 т.п.н., 5-65 т.п.н., 5-70 т.п.н., 5-75 т.п.н., 5-80 т.п.н., 5-85 т.п.н., 5-90 т.п.н., 5-95 т.п.н., 5-100 т.п.н., 100-200 т.п.н. или 200-300 т.п.н. или более. Как более подробно описано ниже, в больших нацеливающих векторах могут использоваться нацеливающие плечи с большей длиной.
Нуклеазные агенты (например, системы CRISPR/Cas) можно применять в комбинации с нацеливающими векторами для обеспечения модификации целевого локуса. Такие нуклеазные агенты могут обеспечивать гомологичную рекомбинацию между нацеливающим вектором и целевым локусом. При использовании нуклеазных агентов в комбинации с нацеливающим вектором нацеливающий вектор может содержать 5' и 3' гомологичные плечи, соответствующие целевым 5'- и 3'-последовательностям, расположенным достаточно близко к сайту расщепления нуклеазой, чтобы обеспечить событие гомологичной рекомбинации между целевыми последовательностями и гомологичными плечами при одноцепочечном или двухцепочечном разрыве в сайте расщепления нуклеазой. Термин «сайт расщепления нуклеазой» включает последовательность ДНК, в которой нуклеазный агент (например, сайт расщепления Cas9) индуцирует одно- или двухцепочечный разрыв. Целевые последовательности в целевом локусе, соответствующие 5' и 3' гомологичным плечам нацеливающего вектора, «расположены в достаточной близости» к сайту расщепления нуклеазой, если расстояние является таким, чтобы способствовать возникновению события гомологичной рекомбинации между целевыми 5'- и 3'-последовательностями и гомологичными плечами при одно- или двухцепочечном разрыве в сайте распознавания. Таким образом, в конкретных случаях целевые последовательности, соответствующие 5' и/или 3' гомологичным плечам нацеливающего вектора, находятся в пределах по меньшей мере 1 нуклеотида от данного сайта распознавания или в пределах по меньшей мере от 10 нуклеотидов до приблизительно 14 т.п.н. от данного сайта распознавания. В некоторых случаях сайт нуклеазного расщепления непосредственно прилегает к по меньшей мере одной или обеим целевым последовательностям.
Пространственное отношение целевых последовательностей, соответствующих гомологичным плечам нацеливающего вектора и сайту расщепления нуклеазой, может различаться. Например, целевые последовательности могут быть расположены в направлении 5'-конца от сайта нуклеазного расщепления, оба целевых сайта могут быть расположены в направлении 3'-конца от сайта расщепления нуклеазой или целевые последовательности могут фланкировать сайт расщепления нуклеазой.
Совместное использование нацеливающего вектора (включая, например, большой нацеливающий вектор) с нуклеазным агентом может привести к повышению эффективности нацеливания по сравнению с использованием одного только нацеливающего вектора. Например, когда нацеливающий вектор применяют вместе с нуклеазным агентом, эффективность нацеливания нацеливающего вектора можно повысить по меньшей мере в два раза, по меньшей мере в три раза, по меньшей мере в 4 раза или по меньшей мере в 10 раз по сравнению с использованием одного только нацеливающего вектора.
(3) Большие нацеливающие векторы
Некоторые нацеливающие векторы представляют собой «большие нацеливающие векторы» или «LTVEC», к которым относятся нацеливающие векторы, содержащие гомологичные плечи, соответствующие или производные от нуклеотидных последовательностей большего размера, чем те, которые обычно применяют в других подходах, предназначенных для проведения гомологичной рекомбинации в клетках. К LTVEC также относятся нацеливающие векторы, содержащие полинуклеотидные вставки, имеющие нуклеотидные последовательности большей длины, чем у обычно применяемых при других подходах, предназначенных для проведения гомологичной рекомбинации в клетках. Например, LTVEC позволяют выполнять модификацию больших локусов, которую нельзя выполнить с использованием традиционных плазмидных нацеливающих векторов из-за размерных ограничений. Например, целевой локус может представлять собой локус клетки (т.е. 5' и 3' гомологичные плечи могут соответствовать ему), на который в отсутствие одно- или двухцепочечного разрыва, индуцированного нуклеазным агентом (например, белком Cas), нельзя выполнить нацеливание с использованием стандартных способов, или нацеливание может быть только неверным или иметь только очень низкую эффективность.
Примеры LTVEC включают векторы, полученные из бактериальной искусственной хромосомы (ВАС), человеческой искусственной хромосомы или дрожжевой искусственной хромосомы (YAC). Не имеющие ограничительного характера примеры LTVEC и способы их получения описаны, например, в патенте США №6,586,251; патенте США №6,596,541; патенте США 7,105,348; и WO 2002/036789 (PCT/US01/45375), каждый из которых полностью включен в настоящий документ путем ссылки. LTVEC могут иметь линейную форму или кольцевую форму.
LTVEC могут иметь любую длину, включая, например, от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н. или от приблизительно 275 т.п.н. до приблизительно 300 т.п.н. Альтернативно LTVEC может составлять по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более. Размер LTVEC может быть слишком большим, чтобы существовала возможность скрининга событий нацеливания с помощью обычных тестов, например саузерн-блоттинга и ПЦР для амплификации протяженных участков (например, от 1 т.п.н. до 5 т.п.н.).
В некоторых случаях LTVEC включает полинуклеотидную вставку, которая может составлять от приблизительно 5 т.п.н. до приблизительно 200 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. В других случаях полинуклеотидная вставка может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н.
В некоторых LTVEC общая длина 5' гомологичного плеча и 3' гомологичного плеча составляет по меньшей мере приблизительно 10 т.п.н. В других LTVEC длина 5' гомологичного плеча находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 100 т.п.н. и/или длина 3' гомологичного плеча находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 100 т.п.н. Каждое гомологичное плечо может составлять, например, от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. Общая длина 5' и 3' гомологичных плеч может составлять, например, от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. Альтернативно каждое гомологичное плечо может составлять по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н. или по меньшей мере 200 т.п.н. Аналогичным образом, общая длина 5' и 3' гомологичных плеч может составлять по меньшей мере 5 т.п.н., 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н. или по меньшей мере 200 т.п.н.
В некоторых случаях LTVEC и полинуклеотидная вставка сконструированы так, чтобы обеспечить в целевом локусе делецию от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. Альтернативно делеция может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
В других случаях LTVEC и полинуклеотидная вставка предназначены для того, чтобы обеспечить вставку в целевой локус экзогенной полинуклеотидной последовательности длиной в диапазоне от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н. Альтернативно вставка может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
В других случаях длина полинуклеотидной вставки и/или удаляемой области эндогенного локуса составляет по меньшей мере 100, 200, 300, 400, 500, 600, 700, 800 или 900 нуклеотидов или по меньшей мере 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 11 т.п.н., 12 т.п.н., 13 т.п.н., 14 т.п.н., 15 т.п.н., 16 т.п.н. или более.
В. Нацеливающие векторы и полинуклеотидные вставки для эмбрионов на стадии единственной клетки
Нацеливающие векторы для использования в эмбрионах на стадии единственной клетки имеют длину не более чем 5 т.п.н. и могут представлять собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), могут быть одноцепочечными или двухцепочечными и могут иметь кольцевую или линейную форму. Пример нацеливающего вектора для использования в эмбрионах на стадии единственной клетки имеет длину от приблизительно 50 нуклеотидов до приблизительно 5 т.п.н. Например, нацеливающий вектор для использования в эмбрионах на стадии единственной клетки может иметь длину от приблизительно 50 до приблизительно 100, от приблизительно 100 до приблизительно 200, от приблизительно 200 до приблизительно 300, от приблизительно 300 до приблизительно 400, от приблизительно 400 до приблизительно 500, от приблизительно 500 до приблизительно 600, от приблизительно 600 до приблизительно 700, от приблизительно 700 до приблизительно 800, от приблизительно 800 до приблизительно 900 или от приблизительно 900 до приблизительно 1000 нуклеотидов. Альтернативно нацеливающий вектор для использования в эмбрионах на стадии единственной клетки может иметь длину от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н., от приблизительно 2,5 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 3,5 т.п.н., от приблизительно 3,5 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 4,5 т.п.н. или от приблизительно 4,5 т.п.н. до приблизительно 5 т.п.н. Альтернативно нацеливающий вектор для использования в эмбрионах на стадии единственной клетки может иметь длину, например, не более чем 5 т.п.н., 4,5 т.п.н., 4 т.п.н., 3,5 т.п.н., 3 т.п.н., 2,5 т.п.н., 2 т.п.н., 1,5 т.п.н., 1 т.п.н., 900 нуклеотидов, 800 нуклеотидов, 700 нуклеотидов, 600 нуклеотидов, 500 нуклеотидов, 400 нуклеотидов, 300 нуклеотидов, 200 нуклеотидов, 100 нуклеотидов или 50 нуклеотидов. В случае доноров одноцепочечной ДНК примеры нацеливающих векторов могут составлять от приблизительно 60 нуклеотидов до приблизительно 200 нуклеотидов (например, от приблизительно 60 нуклеотидов до приблизительно 80 нуклеотидов, от приблизительно 80 нуклеотидов до приблизительно 100 нуклеотидов, от приблизительно 100 нуклеотидов до приблизительно 120 нуклеотидов, от приблизительно 120 нуклеотидов до приблизительно 140 нуклеотидов, от приблизительно 140 нуклеотидов до приблизительно 160 нуклеотидов, от приблизительно 160 нуклеотидов до приблизительно 180 нуклеотидов или от приблизительно 180 нуклеотидов до приблизительно 200 нуклеотидов).
Такие нацеливающие векторы содержат 5' и 3' гомологичные плечи, соответствующие областям в пределах целевого локуса (целевой 5'-последовательности и целевой 3'-последовательности соответственно). Необязательно нацеливающий вектор содержит полинуклеотидную вставку (например, сегмент ДНК для интеграции в целевой геномный локус), фланкированную 5' и 3' гомологичными плечами. Интеграция полинуклеотидной вставки в целевой локус может приводить к добавлению интересующей нуклеотидной последовательности, делеции интересующей нуклеотидной последовательности в целевом локусе или замене интересующей нуклеотидной последовательности в целевом локусе (т.е. делеции или вставке).
Соответствующая область гомологии между гомологичным плечом и соответствующей целевой последовательностью может иметь любую длину, которая достаточна для стимулирования гомологичной рекомбинации. Примеры гомологичных плеч для использования в эмбрионах на стадии единственной клетки имеют длину от приблизительно 20 нуклеотидов до приблизительно 2,5 т.п.н. (например, длину от приблизительно 30 нуклеотидов до приблизительно 100 нуклеотидов). Например, данное гомологичное плечо и/или соответствующая целевая последовательность может содержать области гомологии длиной от приблизительно 20 до приблизительно 30, от приблизительно 30 до приблизительно 40, от приблизительно 40 до приблизительно 50, от приблизительно 50 до приблизительно 60, от приблизительно 60 до приблизительно 70, от приблизительно 70 до приблизительно 80, от приблизительно 80 до приблизительно 90, от приблизительно 90 до приблизительно 100, от приблизительно 100 до приблизительно 150, от приблизительно 150 до приблизительно 200, от приблизительно 200 до приблизительно 250, от приблизительно 250 до приблизительно 300, от приблизительно 300 до приблизительно 350, от приблизительно 350 до приблизительно 400, от приблизительно 400 до приблизительно 450 или от приблизительно 450 до приблизительно 500 нуклеотидов, так чтобы гомологичные плечи имели достаточную гомологию для того, чтобы проходить гомологичную рекомбинацию с соответствующими целевыми последовательностями в пределах генома клетки. Альтернативно данное гомологичное плечо и/или соответствующая целевая последовательность может содержать соответствующие области гомологии, имеющие длину от приблизительно 0,5 т.п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н. или от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н. В случае одноцепочечных доноров ДНК примеры гомологичных плеч могут иметь длину от приблизительно 30 нуклеотидов до приблизительно 60 нуклеотидов (например, от приблизительно 30 до приблизительно 40 нуклеотидов, от приблизительно 40 нуклеотидов до приблизительно 50 нуклеотидов или от приблизительно 50 нуклеотидов до приблизительно 60 нуклеотидов).
Как описано выше, гомологичные плечи могут соответствовать локусу, нативному для клетки (например, целевому локусу), или альтернативно они могут соответствовать области гетерологичного или экзогенного сегмента ДНК, который был интегрирован в геном клетки. Как описано выше, целевые 5'- и 3'-последовательности предпочтительно расположены достаточно близко к сайту расщепления Cas, чтобы способствовать возникновению события гомологичной рекомбинации между целевыми последовательностями и гомологичными плечами при одноцепочечном разрыве или двухцепочечном разрыве в сайте расщепления Cas.
Полинуклеотидная вставка или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом целевом локусе может иметь разные длины. Пример полинуклеотидной вставки или соответствующей нуклеиновой кислоты в целевом удаляемом и/или заменяемом локусе имеет длину от приблизительно 10 нуклеотидов до приблизительно 5 т.п.н. Например, полинуклеотидная вставка или соответствующая нуклеиновая кислота в целевом удаляемом и/или заменяемом локусе может иметь длину от приблизительно 1 до приблизительно 10, от приблизительно 10 до приблизительно 20, от приблизительно 20 до приблизительно 30, от приблизительно 30 до приблизительно 40, от приблизительно 40 до приблизительно 50, от приблизительно 50 до приблизительно 60, от приблизительно 60 до приблизительно 70, от приблизительно 70 до приблизительно 80, от приблизительно 80 до приблизительно 90, от приблизительно 90 до приблизительно 100, от приблизительно 100 до приблизительно 110, от приблизительно 110 до приблизительно 120, от приблизительно 120 до приблизительно 130, от приблизительно 130 до приблизительно 140, от приблизительно 140 до приблизительно 150, от приблизительно 150 до приблизительно 160, от приблизительно 160 до приблизительно 170, от приблизительно 170 до приблизительно 180, от приблизительно 180 до приблизительно 190, от приблизительно 190 до приблизительно 200, от приблизительно 200 до приблизительно 300, от приблизительно 300 до приблизительно 400, от приблизительно 400 до приблизительно 500, от приблизительно 500 до приблизительно 600, от приблизительно 600 до приблизительно 700, от приблизительно 700 до приблизительно 800, от приблизительно 800 до приблизительно 900 или от приблизительно 900 до приблизительно 1000 нуклеотидов. Например, вставку длиной от приблизительно 4 нуклеотидов до приблизительно 12 нуклеотидов можно вставить для создания сайта расщепления рестрикционным ферментом. Аналогичным образом, полинуклеотидная вставка или соответствующая нуклеиновая кислота в целевом удаляемом и/или заменяемом локусе может иметь длину от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н., от приблизительно 2,5 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 3,5 т.п.н., от приблизительно 3,5 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 4,5 т.п.н. или от приблизительно 4,5 т.п.н. до приблизительно 5 т.п.н. Удаляемая из целевого геномного локуса полинуклеотидная вставка может находиться в диапазоне от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 200 т.п.н., от приблизительно т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 800 т.п.н., от приблизительно 800 т.п.н. до приблизительно 900 т.п.н., от приблизительно 900 т.п.н. до приблизительно 1 млн п.н. или более. Альтернативно нуклеиновая кислота, удаляемая из целевого геномного локуса, может иметь длину от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н., от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н.
Как описано выше, полинуклеотидная вставка может содержать геномную ДНК или любой другой тип ДНК, полинуклеотидная вставка или соответствующая нуклеиновая кислота в целевом удаляемом и/или заменяемом локусе может представлять собой кодирующую область или некодирующую область, и полинуклеотидная вставка может быть гомологична или ортологична последовательности, нацеливаемой для делеции в целевом геномном локусе. Полинуклеотидная вставка может также содержать условный аллель, полинуклеотид, кодирующий маркер селекции, репортерный ген, одну или более экспрессионных кассет, одну или более делеционных кассет или полинуклеотидную вставку, фланкированную целевыми последовательностями сайт-специфической рекомбинации, как описано выше.
С. Промоторы
Различные полинуклеотидные последовательности, описанные в настоящем документе, могут быть функционально связаны с промоторами. Такие промоторы могут быть активны, например, в плюрипотентной клетке крысы, эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Также можно использовать промотор, активный в эмбрионе на стадии единственной клетки. Промотор может представлять собой, например, конститутивно активный промотор, условный промотор, индуцибельный промотор, ограниченный по времени промотор (например, регулируемый стадией развития промотор) или пространственно-ограниченный промотор (например, промотор, специфичный для клеток или тканей). Примеры промоторов можно найти, например, в публикации WO 2013/176772, которая полностью включена в настоящий документ путем ссылки.
К примерам индуцибельных промоторов относятся, например, промоторы с химической регуляцией и промоторы с физической регуляцией. К промоторам с химической регуляцией относятся, например, промоторы с регуляцией спиртом (например, промотор гена алкогольдегидрогеназы (alcA)), промоторы с регуляцией тетрациклином (например, отвечающий на тетрациклин промотор, тетрациклиновая операторная последовательность (tetO), промотор tet-On или промотор tet-Off), промоторы с регуляцией стероидами (например, крысиный глюкокортикоидный рецептор, промотор рецептора эстрогена, промотор рецептора экдизона) или промоторы с регуляцией металлами (например, металлопротеиновый промотор). К промоторам с физической регуляцией относятся, например, промоторы, регулируемые температурой (например, промотор теплового шока), и промоторы, регулируемые светом (например, индуцируемый светом промотор и подавляемый светом промотор).
Тканеспецифические промоторы могут представлять собой, например, нейрон-специфические промоторы, глиальноспецифические промоторы, промоторы, специфичные для мышечных клеток, промоторы, специфичные для сердечных клеток, промоторы, специфичные для почечных клеток, промоторы, специфичные для костных клеток, промоторы, специфичные для эндотелиальных клеток, или промоторы, специфичные для иммунных клеток (например, В-клеточный промотор или Т-клеточный промотор).
К регулируемым стадией развития промоторам относятся, например, промоторы, активные только на эмбриональной стадии развития или только во взрослой клетке.
Также промотор можно выбирать в зависимости от типа клетки. Например, различные известные промоторы используются в эукариотической клетке, клетке млекопитающего, клетке млекопитающего, не относящегося к человеку, плюрипотентной клетке, нечеловеческой плюрипотентной клетке, человеческой плюрипотентной клетке, человеческой ЭС-клетке, стволовой клетке взрослого человека, ограниченной стадией развития человеческой клетке-предшественнице, человеческой ИПС-клетке, человеческой клетке, клетке грызуна, клетке крысы, клетке мыши, клетке хомяка, фибробласте или клетке СНО.
IV. Способы модификации геномов и получения генетически модифицированных не относящихся к человеку животных
А. Способы модификации генома
Предложены различные способы модификации генома в клетке путем использования двух гидовых РНК для нацеливания на разные области в пределах одного целевого геномного локуса. Также предложены способы использования двух или более гидовых РНК (например, трех гидовых РНК или четырех гидовых РНК) для нацеливания на разные области в пределах одного целевого геномного локуса. Способы могут осуществляться in vitro, ex vivo или in vivo. Такие способы способствуют созданию биаллельных генетических модификаций и могут включать в себя коллапсирование генома или другие нацеленные модификации, такие как одновременную делецию нуклеотидной последовательности в геноме и замену экзогенной нуклеотидной последовательностью.
Нацеленная генная модификация путем гомологичной рекомбинации между нацеливающим вектором и целевым локусом может быть очень неэффективной, особенно в типах клеток, отличных от эмбриональных стволовых клеток грызунов. Использование нацеливающего вектора в комбинации с направленным нуклеазой двухцепочечным разрывом ДНК в целевом локусе может существенно повысить эффективность гетерозиготного нацеливания при простых модификациях, таких как малые делеции или вставки.
Комбинирование нацеливающего вектора с нуклеазой CRISPR/Cas9, направляемой одной гидовой РНК (гРНК), также может повышать эффективность гетерозиготного нацеливания при очень больших и низкоэффективных генных модификациях, например делеции мышиного гена с одновременной заменой человеческой версией (гуманизация). Такие модификации могут быть связаны с очень большими (например, >50 т.п.н.) делениями и вставками (см. нацеливание на Lrp5, С5 (Hc), Ror1 и Trpa1 в примере 1).
При направляемой гомологией репарации одного или более двухцепочечных разрывов, созданных в целевом геномном локусе нуклеазой, такой как Cas9, один или более разрывов в целевом геномном локусе сначала подвергаются обработке для получения 3'-одноцепочечного нависания путем усечения 5'-конца. Затем Rad51 полимеризуется на одноцепочечной ДНК с целью поиска гомологичной последовательности, происходит инвазия цепочки в неповрежденную гомологичную матричную дуплексную ДНК (например, в нацеливающий вектор) и образуется промежуточная D-петлевая структура, способствующая репарации одного или более двухцепочечных разрывов с использованием неповрежденной гомологичной ДНК (например, нацеливающего вектора) в качестве шаблона. После этого хромосомные последовательности замещаются полинуклеотидной вставкой из нацеливающего вектора в результате события двойного кроссинговера с участием фланкирующих гомологичных областей. На правильное протекание этого процесса влияют несколько факторов, таких как размер полинуклеотидной вставки, длина областей, гомологичных гомологичным плечам нацеливающего вектора, и положения областей, гомологичных гомологичным плечам нацеливающего вектора (например, относительно одного или более двухцепочечных разрывов).
При увеличении размера полинуклеотидной вставки или удаляемой последовательности в целевом геномном локусе процесс усечения становится более непредсказуемым, стабильность промежуточной D-петлевой структуры снижается и становится более непредсказуемой, а успешность процесса рекомбинации в целом снижается и становится более непредсказуемой. Например, при увеличении размера нацеленной модификации повышается риск внутренней рекомбинации, в особенности если существует сходство между заменяемой и вставляемой последовательностями. При такой внутренней рекомбинации гомологичный рекомбинационный обмен происходит внутри нужной целевой области, и полинуклеотидная вставка не внедряется полностью в целевой геномный локус. Кроме того, традиционно считается, что эффективность опосредованных гомологичной рекомбинацией вставок снижается с увеличением расстояния между двухцепочечным разрывом и сайтом мутации или вставки (например, более 100 п.н. или 200 п.н.). См. Beumer et al. (2013) Genes\Genomes\Genetics 3: 657-664; Elliott et al. (1998) Mol. Cell. Biol. 18: 93-101 и Byrne et al. (2015) Nucleic Acids Research 43 (3): e21, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
Для получения нацеленной генной модификации, создающей большую делецию в целевом геномном локусе с одновременной вставкой большого фрагмента чужеродной ДНК, необходимо формирование двойной омега-структуры в качестве промежуточного продукта рекомбинации. Чем больше модификация, тем ниже стабильность структуры. В типах клеток, не относящихся к эмбрионам на стадии единственной клетки, можно использовать LTVEC, имеющие в сумме общую гомологию 10 т.п.н. или более. LTVEC с гомологичными плечами с общей гомологией 10 т.п.н. или более повышают стабильность промежуточной рекомбинационной двойной омега-структуры и облегчают опосредованную нуклеазой большую делецию и замену большой полинуклеотидной вставкой и дополнительно повышают эффективность нацеливания не только при двухцепочечных разрывах, прилегающих к гомологичным областям, но и эффективность нацеливания при двухцепочечных разрывах вдали от гомологичных областей.
При генных модификациях, связанных с очень большими гуманизациями, комбинирование нацеливающего вектора с нуклеазной системой CRISPR/Cas9, направляемой двумя гРНК, может дополнительно повышать эффективность нацеливания в большей степени, чем при одной гРНК (см. гуманизации Lrp5, С5 (Hc), Ror1 и Trpa1 в примере 1). Использование двух гРНК дает в этом отношении неожиданные результаты. По сравнению с нацеливанием с использованием одной гРНК, которое дает биаллельные модификации с низкой частотой или не дает их вовсе, нацеливание двумя гРНК приводит к созданию клеток с выполненным гомозиготным нацеливанием и компаунд-гетерозиготным нацеливанием (включая клетки с гемизиготным нацеливанием) с существенно более высокой частотой.
Способ создания трех аллельных типов - с выполненным гомозиготным нацеливанием, с гомозиготной делецией и с компаунд-гетерозиготным нацеливанием (в частности, с гемизиготным нацеливанием) - в одном эксперименте с нацеливанием дает новые возможности и повышает эффективность нацеленных генетических модификаций. В случае простой генетической модификации, например нацеленной делеции гена в мышиных ЭС-клетках с его заменой последовательностью, кодирующей белок, который сигнализирует об экспрессии гена (например, β-галактозидазу или флуоресцентный белок), комбинирование нацеливающего вектора с системой CRJSPR/Cas9, направляемой двумя гРНК, повышает продукцию ЭС-клеток с выполненным гетерозиготным нацеливанием, которые затем можно использовать для получения происходящих полностью от ЭС-клеток мышей F0 по способу VelociMouse®. См. Poueymirou et al. (2007) Nat. Biotech. 25: 91-99, для всех целей полностью включенную в настоящий документ путем ссылки. Этих мышей можно использовать для изучения тканеспецифической экспрессии генов с использованием репортерного нокин-аллеля. Клоны ЭС-клеток с выполненным гомозиготным нацеливанием, полученные в том же эксперименте, можно преобразовать в мышей VelociMice с выполненной гомозиготной нацеленной делецией, на которых можно изучать фенотипические последствия нокаута гена, а также экспрессию гена с репортером. Получение мышей VelociMice из ЭС-клеток, имеющих гомозиготную CRISPR-индуцированную делецию целевого гена, позволяет провести верификацию нокаутного фенотипа, наблюдаемого у мышей с выполненным гомозиготным нацеливанием, и позволяет выявить фенотипические различия между чистой делецией гена и делецией, сопровождающейся вставкой репортера и кассеты селекции лекарственным средством. Компаунд-гетерозиготные (и, в частности, гемизиготные) клоны ЭС-клеток, несущие как аллель с нацеленной делецией-вставкой, так и CRISPR-индуцированную делецию, позволяют получить мышей VelociMice с такими же возможностями для исследований, как и у тех, которые получены от клонов с гомозиготным нацеливанием и гомозиготной делецией. Кроме того, этих мышей можно скрещивать для получения из одного клона ЭС-клеток мутантных линий мышей как с нацеливанием, так и с делецией.
Эти преимущества имеют дополнительное значение, если распространить их на случай гуманизации. Важной областью применения гуманизации мышиного гена является создание животной модели для тестирования специфичных для человека лекарственных средств. Чтобы гуманизация стала эффективной моделью, мышиный ген необходимо удалить или инактивировать во избежание взаимодействий между продуктами мышиного и человеческого гена, которые могут нарушить биологическую функцию или надлежащее взаимодействие с лекарственным средством. В то же время человеческий ген должен быть способен заместить биологические функции мышиного варианта. Эти требования можно проверить путем комбинирования нуклеазы Cas9, направляемой двумя гРНК, с нацеливающим вектором, предназначенным для одновременной делеции мышиного гена и замены его на человеческий ген. Мышей VelociMice, полученных из ЭС-клеток, имеющих выполненную гомозиготную гуманизацию, можно сравнить с мышами VelociMice, полученными из ЭС-клеток с гомозиготной CRISPR-индуцированной делецией мышиного гена. Если нокаутная делеция приводит к заметному мутантному фенотипу, и гуманизированные мыши не проявляют этот фенотип, а являются нормальными, то человеческий ген может заместить биологические функции мышиного гена. В качестве животных моделей для изучения механизмов действия и эффективности специфичных для человека лекарственных средств можно использовать или гомозиготных гуманизированных мышей, или мышей, несущих компаунд-гетерозиготную (например, гемизиготную) комбинацию гуманизированного аллеля и аллеля с CRISPR-индуцированной делецией. Компаунд-гетерозиготных (например, гемизиготных) мышей VelociMice можно использовать для получения как гуманизированных, так и имеющих делецию нокаутных мышей посредством традиционного скрещивания. Таким образом, из одного эксперимента с нацеливанием, в котором комбинируют CRISPR-систему, направляемую двумя гРНК, с нацеливающим вектором, получают генетически модифицированных мышей, которые позволяют строить полезные мышиные модели для доклинического исследования лекарственных препаратов и нокаутные линии для изучения биологической функции мышиного гомолога человеческой мишени для лекарственных средств.
(1) Способы получения, стимулирования или повышения частоты биаллельных генетических модификаций
В настоящем документе предложены способы получения биаллельных модификаций в геноме клетки или стимуляции или повышения частоты биаллельнх модификаций генома в клетке. Такие способы могут приводить, например, к коллапсированию генома и удалению крупного фрагмента геномной ДНК между двумя последовательностями геномной ДНК, которые впоследствии рекомбинируют. Такие способы также могут приводить к вставке полинуклеотидной вставки или делеции крупного фрагмента геномной ДНК и ее замене полинуклеотидной вставкой.
Предложенные в настоящем документе способы модификации генома клетки включают в себя приведение генома в контакт с первым белком Cas, первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, и тракрРНК. Необязательно геном можно дополнительно привести в контакт с дополнительными РНК CRISPR, которые гибридизуются с дополнительными последовательностями распознавания РНК CRISPR в пределах целевого геномного локуса, например третьей РНК CRISPR, которая гибридизуется с третьей последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, и/или четвертой РНК CRISPR, которая гибридизуется с четвертой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса. Биаллельные модификации можно получить путем приведения генома в контакт с первым белком Cas, первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса, и тракрРНК. Как более подробно описано ниже, белок Cas, РНК CRISPR и тракрРНК можно вводить в клетку в любом виде и любыми средствами. Аналогичным образом, все или некоторые из белка Cas, РНК CRISPR и тракрРНК можно вводить одновременно или последовательно в любой комбинации. Приведение генома в контакт может осуществляться напрямую (т.е. компонент напрямую контактирует с самим геномом) или косвенно (т.е. компонент взаимодействует с другим компонентом, который напрямую контактирует с геномом).
Геном может содержать пару из первой и второй гомологичных хромосом, содержащих целевой геномный локус. Первый белок Cas может расщеплять одну или обе из этих хромосом в пределах одной или обеих из первой и второй последовательностей распознавания РНК CRISPR (т.е. в первом сайте расщепления в пределах первой последовательности распознавания РНК CRISPR и/или во втором сайте расщепления в пределах второй последовательности распознавания РНК CRISPR). Если также используются третья и/или четвертая РНК CRISPR, то первый белок Cas может расщеплять одну или обе из этих хромосом в пределах одной или обеих из третьей и/или четвертой последовательностей распознавания РНК CRISPR (т.е. в третьем сайте расщепления в пределах третьей последовательности распознавания РНК CRISPR и/или в четвертом сайте расщепления в пределах четвертой последовательности распознавания РНК CRISPR). Затем эти события расщепления могут создавать по меньшей мере один двухцепочечный разрыв в одной или обеих хромосомах. События расщепления также могут создавать по меньшей мере два двухцепочечных разрыва в одной или обеих хромосомах. Если используются никазы Cas, то события расщепления могут создавать по меньшей мере один одноцепочечный разрыв в одной или обеих хромосомах или по меньшей мере два одноцепочечных разрыва в одной или обеих хромосомах. Если используются третья и/или четвертая РНК CRISPR, то события расщепления могут создавать по меньшей мере три или четыре одноцепочечных или двухцепочечных разрыва в одной или обеих хромосомах. Далее концевые последовательности, созданные двухцепочечными разрывами, могут подвергаться рекомбинации, или концевые последовательности, созданные одноцепочечными разрывами, могут подвергаться рекомбинации. После этого можно идентифицировать клетку, имеющую модифицированный геном, содержащий биаллельную модификацию.
Например, первый белок Cas может расщеплять геном в первом сайте расщепления в пределах первой последовательности распознавания РНК CRISPR в первой и второй гомологичных хромосомах и во втором сайте расщепления в пределах второй последовательности распознавания РНК CRISPR по меньшей мере в первой гомологичной хромосоме, таким образом создавая концевые последовательности в первой и второй гомологичных хромосомах. После этого концевые последовательности могут подвергаться рекомбинации с образованием генома с биаллельной модификацией, содержащей нацеленную модификацию. Нацеленная модификация может включать в себя делецию между первой и второй последовательностями распознавания РНК CRISPR в по меньшей мере первой хромосоме.
Первая и вторая последовательности распознавания РНК CRISPR могут находиться в любом месте в пределах целевого геномного локуса. Первая и вторая последовательности распознавания РНК CRISPR могут фланкировать любую интересующую геномную область. Например, первая и вторая последовательности распознавания РНК CRISPR могут фланкировать полную или частичную кодирующую последовательность гена, например локус Lrp5, локус С5 (Hc), локус Ror1 или локус Trpa1. Первая и вторая последовательности распознавания РНК CRISPR также могут фланкировать полную или частичную кодирующую последовательность гена Cmah. Альтернативно первая и вторая последовательности распознавания РНК CRISPR могут фланкировать некодирующую последовательность, такую как регуляторный элемент (например, промотор), или как кодирующую, так и некодирующую последовательности. Третья и четвертая последовательности распознавания РНК CRISPR могут находиться, например, в любом месте в пределах интересующей геномной области, фланкированной первой и второй последовательностями распознавания РНК CRISPR.
Например, третья последовательность распознавания РНК CRISPR может прилегать к первой последовательности распознавания РНК CRISPR, а четвертая последовательность распознавания РНК CRISPR может прилегать ко второй последовательности распознавания РНК CRISPR. Таким образом, первая и третья последовательности распознавания РНК CRISPR могут представлять собой первую пару последовательностей распознавания РНК CRISPR, а вторая и четвертая последовательности распознавания РНК CRISPR могут представлять собой вторую пару последовательностей распознавания РНК CRISPR. Например, первая и третья последовательности распознавания РНК CRISPR (и/или вторая и четвертая последовательности распознавания РНК CRISPR) могут быть разделены на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н., от приблизительно 2,5 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 3,5 т.п.н., от приблизительно 3,5 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 4,5 т.п.н., от приблизительно 4,5 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 6 т.п.н., от приблизительно 6 т.п.н. до приблизительно 7 т.п.н., от приблизительно 7 т.п.н. до приблизительно 8 т.п.н., от приблизительно 8 т.п.н. до приблизительно 9 т.п.н. или от приблизительно 9 т.п.н. до приблизительно 10 т.п.н. Например, первая и третья последовательности распознавания РНК CRISPR (и/или вторая и четвертая последовательности распознавания РНК CRISPR) могут быть разделены на расстояние от приблизительно 100 п.н. до приблизительно 1 т.п.н. Альтернативно первая и третья последовательности распознавания РНК CRISPR (и/или вторая и четвертая последовательности распознавания РНК CRISPR) могут перекрываться.
Первая пара последовательностей распознавания РНК CRISPR может быть расположена вблизи 5'-конца целевого геномного локуса, а вторая пара может быть расположена вблизи 3'-конца целевого геномного локуса. Альтернативно обе из первой и второй пар могут быть расположены вблизи 5'-конца целевого геномного локуса или обе могут быть расположены вблизи 3'-конца целевого локуса. Альтернативно одна или обе пары могут быть расположены внутри в пределах целевого геномного локуса. Например, первая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н. от 5'-конца целевого геномного локуса. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н. от 3'-конца целевого геномного локуса.
Альтернативно первая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н. от 5'-конца целевого геномного локуса. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н. от 3'-конца целевого геномного локуса.
Альтернативно первая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии, например, от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 6 т.п.н., от приблизительно 6 т.п.н. до приблизительно 7 т.п.н., от приблизительно 7 т.п.н. до приблизительно 8 т.п.н., от приблизительно 8 т.п.н. до приблизительно 9 т.п.н., от приблизительно 9 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н. или от приблизительно 90 т.п.н. до приблизительно 100 т.п.н. от 5'-конца целевого геномного локуса. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или первая пара последовательностей распознавания РНК CRISPR может находиться на расстоянии, например, от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 6 т.п.н., от приблизительно 6 т.п.н. до приблизительно 7 т.п.н., от приблизительно 7 т.п.н. до приблизительно 8 т.п.н., от приблизительно 8 т.п.н. до приблизительно 9 т.п.н., от приблизительно 9 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н. или от приблизительно 90 т.п.н. до приблизительно 100 т.п.н. от 3'-конца целевого геномного локуса.
Первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR могут находиться на расстоянии, например, от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. Первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR также могут быть разделены на расстояние, например, от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. Первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR также могут быть разделены, например, на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н. или от приблизительно 900 п.н. до приблизительно 1 т.п.н. Аналогичным образом, первая пара последовательностей распознавания РНК CRISPR может быть разделена от второй пары последовательностей распознавания РНК CMSPR, например, на расстояние от приблизительно 25 п.н. до приблизительно 50 п.н., от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 150 п.н., от приблизительно 150 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 250 п.н., от приблизительно 250 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 350 п.н., от приблизительно 350 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 450 п.н., от приблизительно 450 п.н. до приблизительно 500 п.н., от приблизительно 500 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н., от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н.
Альтернативно первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR могут быть разделены, например, на расстояние по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более. Аналогичным образом, первая пара последовательностей распознавания РНК CRISPR может быть разделена от второй пары последовательностей распознавания РНК CRISPR, например, на расстояние по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
Альтернативно первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR могут быть разделены на расстояние менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н. Аналогичным образом, первая пара последовательностей распознавания РНК CRISPR может быть разделена от второй пары последовательностей распознавания РНК CRISPR, например, на расстояние менее чем 25 п.н., менее чем 50 п.н., менее чем 100 п.н., менее чем 150 п.н., менее чем 200 п.н., менее чем 250 п.н., менее чем 300 п.н., менее чем 350 п.н., менее чем 400 п.н., менее чем 450 п.н., менее чем 500 п.н., менее чем 600 п.н., менее чем 700 п.н., менее чем 800 п.н., менее чем 900 п.н., менее чем 1 т.п.н., менее чем 2 т.п.н., менее чем 3 т.п.н., менее чем 4 т.п.н., менее чем 5 т.п.н. или менее чем 10 т.п.н.
Концевые последовательности, созданные в результате расщепления генома в первом и/или втором сайтах расщепления, могут представлять собой тупые концы или ступенчатые концы, а делеция между первой и второй последовательностями распознавания РНК CRISPR может полностью или частично включать в себя нуклеотидную последовательность между первой и второй последовательностями распознавания РНК CRISPR, включая эти последовательности. Аналогичным образом, концевые последовательности, созданные в результате расщепления генома в третьем и/или четвертом сайтах расщепления, могут представлять собой тупые концы или ступенчатые концы. Например, делеция может включать в себя лишь участок нуклеотидной последовательности между первой и второй последовательностями распознавания РНК CRISPR и/или лишь участок первой последовательности распознавания РНК CRISPR и/или второй последовательности распознавания РНК CRISPR. Альтернативно делеция между первой и второй последовательностями распознавания РНК CRISPR может содержать всю нуклеотидную последовательность между первой и второй последовательностями распознавания РНК CRISPR. Аналогичным образом, делеция может включать в себя первую последовательность распознавания РНК CRISPR и/или вторую последовательность распознавания РНК CRISPR или их участки. В некоторых случаях делеция дополнительно содержит последовательности, расположенные за пределами первой и второй последовательностей распознавания РНК CRISPR (т.е. последовательности, не включающие в себя и не расположенные между первой и второй последовательностями распознавания РНК CRISPR).
Делеция между первой и второй последовательностями распознавания РНК CRISPR может иметь любую длину. Например, удаленная нуклеиновая кислота может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., или от приблизительно 150 т.п.н. до приблизительно 200 т.п.п., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н.
Альтернативно удаленная нуклеиновая кислота может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.п., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более. В некоторых случаях удаленная нуклеиновая кислота может составлять по меньшей мере 550 т.п.н., по меньшей мере 600 т.п.н., по меньшей мере 650 т.п.н., по меньшей мере 700 т.п.н., по меньшей мере 750 т.п.н., по меньшей мере 800 т.п.н., по меньшей мере 850 т.п.н., по меньшей мере 900 т.п.н., по меньшей мере 950 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 1,5 млн п.н., по меньшей мере 2 млн п.н., по меньшей мере 2,5 млн п.н., по меньшей мере 3 млн п.н., по меньшей мере 4 млн п.н., по меньшей мере 5 млн п.н., по меньшей мере 10 млн п.н., по меньшей мере 20 млн п.н., по меньшей мере 30 млн п.н., по меньшей мере 40 млн п.н., по меньшей мере 50 млн п.н., по меньшей мере 60 млн п.н., по меньшей мере 70 млн п.н., по меньшей мере 80 млн п.н., по меньшей мере 90 млн п.н. или по меньшей мере 100 млн п.н. (например, большую часть хромосомы).
Делеция между первой и второй последовательностями распознавания РНК CRISPR может представлять собой точную делецию, при которой удаленная нуклеиновая кислота содержит только нуклеотидную последовательность между первым и вторым сайтом расщепления белком Cas, так что в модифицированном целевом геномном локусе нет дополнительных делеций или вставок. Делеция между первой и второй последовательностями распознавания РНК CRISPR также может представлять собой неточную делецию, выходящую за пределы первого и второго сайтов расщепления белком Cas, в соответствии с неточной репарацией посредством негомологичного соединения концов (NHEJ), что приводит к дополнительным делециям и/или вставкам в модифицированный геномный локус. Например, делеция может на 1 п.н., приблизительно 2 п.н., приблизительно 3 п.н., приблизительно 4 п.н., приблизительно 5 п.н., приблизительно 10 п.н., приблизительно 20 п.н., приблизительно 30 п.н., приблизительно 40 п.н., приблизительно 50 п.н., приблизительно 100 п.н., приблизительно 200 п.н., приблизительно 300 п.н., приблизительно 400 п.н., приблизительно 500 п.н. или более выходить за пределы первого и второго сайтов расщепления белком Cas. Аналогичным образом, модифицированный геномный локус может содержать дополнительные вставки в соответствии с неточной репарацией по методу NHEJ, например вставку приблизительно 1 п.н., приблизительно 2 п.н., приблизительно 3 п.н., приблизительно 4 п.н., приблизительно 5 п.н., приблизительно 10 п.н., приблизительно 20 п.н., приблизительно 30 п.н., приблизительно 40 п.н., приблизительно 50 п.н., приблизительно 100 п.н., приблизительно 200 п.н., приблизительно 300 п.н., приблизительно 400 п.н., приблизительно 500 п.н. или более.
Приведение в контакт может происходить в отсутствие экзогенной донорной последовательности или в присутствии экзогенной донорной последовательности, при условии, что, если клетка представляет собой эмбрион на стадии единственной клетки, то экзогенная последовательность имеет длину не более 5 т.п.н. К экзогенным молекулам или последовательностям относятся молекулы или последовательности, в норме не присутствующие в клетке. К присутствию в норме относится присутствие в соответствии с конкретной стадией развития клетки и условиями окружающей среды. Экзогенная молекула или последовательность, например, может включать в себя мутантную версию соответствующей эндогенной последовательности внутри клетки, например невулканизированную версию эндогенной последовательности. Напротив, к эндогенным молекулам или последовательностям относятся молекулы или последовательности, в норме присутствующие в конкретной клетке на конкретной стадии развития и в конкретных условиях окружающей среды.
Экзогенная донорная последовательность может находиться внутри нацеливающего вектора и может содержать полинуклеотидную вставку, фланкированную 5' и 3' гомологичными плечами, соответствующими 5'- и 3'-целевым последовательностям в пределах генома, при условии, что, если клетка представляет собой эмбрион на стадии единственной клетки, нацеливающий вектор имеет длину не более чем 5 т.п.н. В типах клеток, отличных от эмбриона на стадии единственной клетки, нацеливающий вектор может быть длиннее. В типах клеток, отличных от эмбриона на стадии единственной клетки, нацеливающий вектор может представлять собой, например, большой нацеливающий вектор (LTVEC), описанный в настоящем документе, и может иметь длину по меньшей мере 10 т.п.н. Таким образом, в некоторых способах геном дополнительно приводят в контакт с нацеливающим вектором, и полинуклеотидная вставка вставляется между целевыми 5'- и 3'-последовательностями.
Альтернативно экзогенная донорная последовательность может содержать 5' и 3' гомологичные плечи без полинуклеотидной вставки. Такие нацеливающие векторы без полинуклеотидной вставки могут упрощать точные делеции между целевыми 5'- и 3'-последовательностями в пределах генома. Такие точные делеции могут составлять, например, по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н., по меньшей мере 500 т.п.н., по меньшей мере 550 т.п.н., по меньшей мере 600 т.п.н., по меньшей мере 650 т.п.н., по меньшей мере 700 т.п.н., по меньшей мере 750 т.п.н., по меньшей мере 800 т.п.н., по меньшей мере 850 т.п.н., по меньшей мере 900 т.п.н., по меньшей мере 950 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 1,5 млн п.н. или по меньшей мере 2 млн п.н. или более.
В некоторых таких способах 5' и 3' гомологичные плечи соответствуют целевым 5'- и 3'-последовательностям в целевом геномном локусе, содержащем первую последовательность распознавания РНК CRISPR первой РНК CRISPR и/или вторую последовательность распознавания РНК CRISPR второй РНК CRISPR. Первая и вторая последовательности распознавания РНК CRISPR или первый и второй сайты расщепления могут прилегать к целевой 5'-последовательности, прилегать к целевой 3'-последовательности или не прилегать ни к целевым 5'-последовательностям, ни к целевой 3'-последовательности. Альтернативно первая последовательность распознавания РНК CRISPR или первый сайт расщепления могут прилегать к целевой 5'-последовательности, а вторая последовательность распознавания РНК CRISPR или второй сайт расщепления могут прилегать к целевой 3'-последовательности. Альтернативно первая последовательность распознавания РНК CRISPR или первый сайт расщепления могут прилегать или к целевой 5'-последовательности, или к целевой 3'-последовательности, а вторая последовательность распознавания РНК CRISPR или второй сайт расщепления могут не прилегать ни к целевой 5'-последовательности, ни к целевой 3'-последовательности.
Например, первая и/или вторая последовательности распознавания РНК CRISPR могут быть расположены между целевыми 5'- и 3'-последовательностями, или могут прилегать к целевой 5'-последовательности и/или целевой 3'-последовательности, или находиться в непосредственной близости от них, например, в пределах 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 250 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н. или 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Например, первый и/или второй сайт расщепления могут быть расположены между целевыми 5'- и 3'-последовательностями, или могут прилегать к целевой 5'-последовательности и/или целевой 3'-последовательности, или находиться в непосредственной близости от них, например, в пределах 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 250 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н. или 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Например, первая последовательность распознавания РНК CRISPR или первый сайт расщепления могут находиться в пределах 50 п.н., 100 п.н., 200 п.н., 300 п.н., 400 п.н., 500 п.н., 600 п.н., 700 п.н., 800 п.н., 900 п.н., 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 250 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н. или 500 т.п.н. от целевой 5'-последовательности или обеих целевых 5'- и/или 3'-последовательностей. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или второй сайт расщепления могут находиться в пределах 50 п.н., 100 п.н., 200 п.н., 300 п.н., 400 п.н., 500 п.н., 600 п.н., 700 п.н., 800 п.н., 900 п.н., 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.п., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 250 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н. или 500 т.п.н. от целевой 3'-последовательности или обеих целевых 5'-и/или 3'-последовательностей.
Альтернативно первая и/или вторая последовательности распознавания РНК CRISPR могут быть расположены на расстоянии по меньшей мере 50 п.н., по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 900 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Аналогичным образом, первый и/или второй сайты расщепления могут быть расположены на расстоянии по меньшей мере 50 п.н., по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 900 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Например, первая последовательность распознавания РНК CRISPR или первый сайт расщепления могут быть расположены на расстоянии по меньшей мере 50 п.н., по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 900 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.п., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. от целевой 5'-последовательности или обеих целевых 5'- и/или 3'-последовательностей. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или второй сайт расщепления могут быть расположены на расстоянии по меньшей мере 50 п.н., по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 900 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. от целевой 3'-последовательности или обеих целевых 5'- и/или 3'-последовательностей.
Например, первая и/или вторая последовательность распознавания РНК CRISPR может быть расположена от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 200 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 500 п.н., от приблизительно 500 до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н. или от приблизительно 400 т.п.н. до приблизительно 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Аналогичным образом, первый и/или второй сайт расщепления может быть расположен от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 200 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 500 п.н., от приблизительно 500 до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н. или от приблизительно 400 т.п.н. до приблизительно 500 т.п.н. от целевых 5'- и/или 3'-последовательностей. Например, первая последовательность распознавания РНК CRISPR или первый сайт расщепления может быть расположен от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 200 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 500 п.н., от приблизительно 500 до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п. и., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н. или от приблизительно 400 т.п.н. до приблизительно 500 т.п.н. от целевой 5'-последовательности или обеих целевых 5'-и/или 3'-последовательностей. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или второй сайт расщепления может быть расположен от приблизительно 50 п.н. до приблизительно 100 п.н., от приблизительно 200 п.н. до приблизительно 300 п.н., от приблизительно 300 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 500 п.н., от приблизительно 500 до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 700 п.н., от приблизительно 700 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 900 п.н., от приблизительно 900 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н. или от приблизительно 400 т.п.н. до приблизительно 500 т.п.н. от целевой 3'-последовательности или обеих целевых 5'- и/или 3'-последовательностей.
Альтернативно первая и/или вторая последовательности распознавания РНК CRISPR или первый и/или второй сайты расщепления могут быть расположены на расстоянии более чем 50 п.н., более чем 100 п.н., более чем 200 п.н., более чем 300 п.н., более чем 400 п.н., более чем 500 п.н., более чем 600 п.н., более чем 700 п.н., более чем 800 п.н., более чем 900 п.н., более чем 1 т.п.н., более чем 2 т.п.н., более чем 3 т.п.н., более чем 4 т.п.н., более чем 5 т.п.н., более чем 6 т.п.н., более чем 7 т.п.н., более чем 8 т.п.н., более чем 9 т.п.н., более чем 10 т.п.н., более чем 20 т.п.н., более чем 30 т.п.н., более чем 40 т.п.н., более чем 50 т.п.н., более чем 60 т.п.н., более чем 70 т.п.н., более чем 80 т.п.н., более чем 90 т.п.н. или более чем 100 т.п.н. от целевых 5'- и/или 3'-последовательностей. Например, первая последовательность распознавания РНК CRISPR или первый сайт расщепления может быть расположен на расстоянии более чем 50 п.н., более чем 100 п.н., более чем 200 п.н., более чем 300 п.н., более чем 400 п.н., более чем 500 п.н., более чем 600 п.н., более чем 700 п.н., более чем 800 п.н., более чем 900 п.н., более чем 1 т.п.н., более чем 2 т.п.н., более чем 3 т.п.н., более чем 4 т.п.н., более чем 5 т.п.н., более чем 6 т.п.н., более чем 7 т.п.н., более чем 8 т.п.н., более чем 9 т.п.н., более чем 10 т.п.н., более чем 20 т.п.н., более чем 30 т.п.н., более чем 40 т.п.н., более чем 50 т.п.н., более чем 60 т.п.н., более чем 70 т.п.н., более чем 80 т.п.н., более чем 90 т.п.н. или более чем 100 т.п.н. от целевой 5'-последовательности или обеих целевых 5'- и/или 3'-последовательностей. Аналогичным образом, вторая последовательность распознавания РНК CRISPR или второй сайт расщепления может быть расположен на расстоянии более чем 50 п.н., более чем 100 п.н., более чем 200 п.н., более чем 300 п.н., более чем 400 п.н., более чем 500 п.н., более чем 600 п.н., более чем 700 п.н., более чем 800 п.н., более чем 900 п.н., более чем 1 т.п.н., более чем 2 т.п.н., более чем 3 т.п.н., более чем 4 т.п.н., более чем 5 т.п.н., более чем 6 т.п.н., более чем 7 т.п.н., более чем 8 т.п.н., более чем 9 т.п.н., более чем 10 т.п.н., более чем 20 т.п.н., более чем 30 т.п.н., более чем 40 т.п.н., более чем 50 т.п.н., более чем 60 т.п.н., более чем 70 т.п.н., более чем 80 т.п.н., более чем 90 т.п.н. или более чем 100 т.п.н. от целевой 3'-последовательности или обеих целевых 5'- и/или 3'-последовательностей.
Описанные в настоящем документе способы стимулируют и повышают частоту биаллельных модификаций. В частности, при приведении генома в контакт как с первой, так и с второй РНК CRISPR эффективность получения биаллельных модификаций может повышаться по сравнению с приведением генома в контакт только с первой РНК CRISPR или только со второй РНК CRISPR. Эффективность получения биаллельных модификаций также можно повысить посредством приведения генома в контакт с первой, второй и третьей РНК CRISPR или с первой, второй, третьей и четвертой РНК CRISPR. Биаллельные модификации включают события, когда одна и та же модификация осуществляется в одном и том же локусе на соответствующих гомологичных хромосомах (например, в диплоидной клетке) или когда различные модификации осуществляются в одном и том же локусе на соответствующих гомологичных хромосомах. Гомологичные хромосомы представляют собой хромосомы, которые имеют одинаковые гены в одном и том же локусе, но, вероятно, различные аллели (например, хромосомы, которые спариваются во время мейоза). Термин «аллель» включает одну или более из любых альтернативных форм генетической последовательности. В диплоидной клетке или организме два аллеля с конкретной последовательностью, как правило, занимают соответствующие локусы в паре гомологичных хромосом.
Биаллельная модификация может приводить к гомозиготности по нацеленной модификации или компаунд-гетерозиготности (например, гемизиготности) по нацеленной модификации. В эксперименте с одним нацеливанием на популяции клеток могут быть получены клетки, гомозиготные по нацеленной модификации (например, гуманизации локуса), клетки, которые являются компаунд-гетерозиготными по этой нацеленной модификации (включая клетки, которые являются гемизиготными по нацеленной модификации), и клетки с последовательностями, гомозиготно коллапсированными между первой и второй последовательностями распознавания РНК CRISPR (т.е. большая последовательность нуклеиновой кислоты между двумя последовательностями распознавания РНК CRISPR удалена). Гомозиготность включает ситуации, когда оба аллеля целевого локуса (т.е. соответствующие аллели на обеих гомологичных хромосомах) имеют нацеленную модификацию. Компаунд-гетерозиготность включает ситуации, когда оба аллеля целевого локуса (т.е. аллели на обеих гомологичных хромосомах) были модифицированы, но были модифицированы по-разному (например, нацеленная модификация одного аллеля и инактивация или разрыв другого аллеля). Нарушение эндогенной нуклеотидной последовательности может возникать, например, когда двухцепочечный разрыв, созданный белком Cas, восстанавливается путем репарации ДНК, опосредованной негомологичным соединением концов (NHEJ), вследствие чего получают мутантный аллель, содержащий вставку или делецию в нуклеотидной последовательности, и, таким образом, геномный локус нарушается. К примерам нарушения относится изменение регуляторного элемента (например, промотора или энхансера), миссенс-мутация, несмысловая мутация, мутация сдвига рамки считывания, мутация укорочения, нуль-мутация или вставка или делеция небольшого количества нуклеотидов (например, приводящая к мутации сдвига рамки считывания). Нарушение может приводить к деактивации (т.е. утрате функции) или потере аллеля.
Например, биаллельная модификация может приводить к компаунд-гетерозиготности, если в клетке есть один аллель с нацеленной модификацией и другой аллель, который не способен экспрессироваться или не является функциональным иным образом. Компаунд-гетерозиготность включает гемизиготность. Гемизиготность включает ситуации, когда присутствует только один аллель (т.е. аллель на одной из двух гомологичных хромосом) целевого локуса. Например, биаллельная модификация может приводить к гемизиготности по целевой модификации, если целевая модификация происходит в одном аллеле с соответствующей потерей или делецией другого аллеля.
В конкретном примере биаллельная модификация может содержать делецию между первой и второй последовательностями распознавания РНК CRISPR в паре первых и вторых гомологичных хромосом. Делеции могут происходить одновременно или делеция может происходить сначала в первой гомологичной хромосоме, причем после этого гомозиготность достигается клеткой с использованием первой гомологичной хромосомы в качестве донорной последовательности для репарации одного или более двухцепочечных разрывов во второй гомологичной хромосоме посредством гомологичной рекомбинации, такой как генная конверсия. Удаленная последовательность нуклеиновой кислоты в первой и второй гомологичных хромосомах может быть одинаковой, частично совпадающей или различной. Альтернативно биаллельная модификация может содержать делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и потерей соответствующего аллеля или локуса во второй гомологичной хромосоме. Альтернативно биаллельная модификация может содержать делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и инактивацию или разрыв соответствующего аллеля или локуса между первой и второй последовательностями распознавания РНК CRISPR во второй гомологичной хромосоме.
Если используется донорная последовательность, биаллельная модификация может включать делецию между первой и второй последовательностями распознавания РНК CMSPR, а также вставку полинуклеотидной вставки между целевыми 5'- и 3'-последовательностями в паре первой и второй гомологичных хромосом, что приводит к получению гомозиготного модифицированного генома. Делеция и вставка могут происходить одновременно в обеих хромосомах или делеция и вставка могут сначала происходить в первой гомологичной хромосоме, причем после этого гомозиготность достигается клеткой с использованием первой гомологичной хромосомы в качестве донорной последовательности для репарации двухцепочечного(-ых) разрыва(-ов) во второй гомологичной хромосоме посредством гомологичной рекомбинации, такой как генная конверсия. Например, без ограничения какой-либо конкретной теорией авторы полагают, что вставка полинуклеотидной вставки может происходить в первой гомологичной хромосоме (с расщеплением белком Cas или без него), а затем вторая гомологичная хромосома может модифицироваться посредством события генной конверсии, которая стимулируется расщеплением белком Cas на второй гомологичной хромосоме.
Альтернативно биаллельная модификация может приводить к образованию компаунд-гетерозиготного модифицированного генома. Например, нацеленная модификация может включать делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах, а также вставку полинуклеотидной вставки в первую гомологичную хромосому, но не во вторую гомологичную хромосому. Альтернативно нацеленная модификация может содержать делецию между первой и второй последовательностями распознавания РНК CRISPR, а также вставку полинуклеотидной вставки в первой гомологичной хромосоме и инактивацию или разрыв соответствующего аллеля или локуса во второй гомологичной хромосоме. Альтернативно биаллельная модификация может приводить к гемизиготному модифицированному геному, в котором нацеленная модификация может содержать делецию между первой и второй последовательностями распознавания РНК CRISPR, а также вставку полинуклеотидной вставки в первой гомологичной хромосоме и инактивацию или разрыв соответствующего аллеля или локуса во второй гомологичной хромосоме.
Гомозиготные и компаунд-гетерозиготные (в частности, гемизиготные) нацеленные генетические модификации имеют преимущество, поскольку они могут сделать процесс создания генетически модифицированных животных с этими модификациями (более подробно описанный ниже) более эффективным и менее трудоемким. Во многих ситуациях, таких как удаление гена для изучения эффекта его отсутствия, просто гетерозиготности по нацеленной генетической модификации (т.е. по модификации одного аллеля и отсутствию изменения в другом аллеле) недостаточно. При использовании традиционных стратегий нацеливания могут быть получены животные поколения F0, которые являются гетерозиготными по большой целевой геномной делеции, но для получения животных поколения F1, гомозиготных по делеции, требуется последующее скрещивание этих гетерозиготных животных. Эти дополнительные этапы скрещивания являются дорогостоящими и требуют времени. Благодаря возможности создавать генетически модифицированных животных поколения F0, которые являются гомозиготными или компаунд-гетерозиготными (в частности, гемизиготными) по нацеленной генетической модификации, значительно повышается эффективность и экономия времени, поскольку требуется меньшее число стадий размножения.
(2) Генная конверсия или потеря гетерозиготности
В некоторых способах модифицируемый геном находится в клетке, гетерозиготной по первому аллелю, и ген модифицируется так, что он становится гомозиготным по первому аллелю. Термин «гетерозиготный» включает ситуации, когда геном содержит разные аллели в одном или более соответствующих локусах хромосомы (например, разные аллели в соответствующих локусах на гомологичных хромосомах). Термин «гомозиготный» включает ситуации, когда геном содержит один и тот же аллель в соответствующих локусах хромосом (например, на соответствующих гомологичных хромосомах). В некоторых таких способах гомозиготность может быть достигнута клеткой с использованием первого аллеля в качестве донорной последовательности для репарации двухцепочечного разрыва в соответствующем втором аллеле посредством гомологичной рекомбинации, такой как генная конверсия. Обычно степень генной конверсии ограничена несколькими сотнями пар оснований. См., например, публикацию Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897, которая для всех целей полностью включена в настоящий документ путем ссылки. Однако использование парных гидовых РНК, которые направляют расщепление в разных участках расщепления в пределах одного локуса, может стимулировать и усиливать возможности генной конверсии на более длинных участках.
Такие способы могут быть пригодными в нескольких контекстах. Первый аллель может содержать мутацию. В некоторых способах, например, первый аллель содержит желаемую нацеленную генетическую модификацию. Достижение гомозиготности по этой нацеленной генетической модификации может привести к значительной экономии времени и затрат, если, например, целью является создание не относящегося к человеку животного, гомозиготного по этой модификации. В других способах первый аллель представляет собой аллель дикого типа гена, который соответствует второму вызывающему заболевание аллелю этого гена. Альтернативно второй аллель может содержать любую мутацию. Затем способы можно использовать для достижения конечной цели генной терапии - замены аллеля, вызывающего заболевание, на аллель дикого типа в его природном хромосомном локусе.
В некоторых таких способах для модификации гетерозиготного по первому аллелю генома, в результате которой он становится гомозиготным по первому аллелю, геном приводят в контакт с белком Cas, тракрРНК и первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах второго аллеля, и второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах второго аллеля, причем первый аллель находится на первой гомологичной хромосоме, а второй аллель находится в соответствующем локусе на второй гомологичной хромосоме (т.е. первый аллель и второй аллель могут быть соответствующими аллелями в паре первой и второй гомологичных хромосом). Необязательно геном можно приводить в контакт с дополнительными РНК CRISPR (например, с третьей РНК CRISPR или третьей и четвертой РНК CRISPR), которые гибридизуются с последовательностями распознавания РНК CRISPR в пределах второго аллеля. Белок Cas может расщеплять одну или обе из первой и второй последовательностей распознавания РНК CRISPR (т.е. в первом сайте расщепления в пределах первой последовательности распознавания РНК CRISPR и/или во втором сайте расщепления в пределах второй последовательности распознавания РНК CRISPR). Расщепление генома в первом и/или втором сайтах расщепления может создавать тупые концы или ступенчатые концы в геномной ДНК. После этого сайты расщепления могут пройти репарацию посредством рекомбинации между первым и вторым аллелями, что приводит к получению модифицированного генома, который гомозиготен по первому аллелю. Затем можно идентифицировать клетку, содержащую модифицированный геном.
В некоторых способах первая и/или вторая последовательности распознавания РНК CRISPR расположены в пределах второго аллеля, но не в пределах первого аллеля. Первый и/или второй аллели могут являться аллелями дикого типа или могут содержать нацеленные модификации или другие отклонения от аллеля дикого типа. Например, первый аллель может содержать желаемую нацеленную модификацию, а второй аллель может являться аллелем дикого типа. Альтернативно первый аллель может являться аллелем дикого типа, а второй аллель может содержать нежелательную модификацию, такую как вызывающая заболевание мутация. В некоторых таких способах происходит нацеленная репарация гена или нацеленная коррекция гена, так что вызывающая заболевание мутация во втором аллеле корректируется посредством рекомбинации с использованием первого аллеля в качестве донорной последовательности.
Первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR могут находиться на расстоянии, например, от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н.
Альтернативно первый и второй сайты расщепления или первая и вторая последовательности распознавания РНК CRISPR могут быть разделены, например, по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н.
В некоторых способах различия в последовательности между первым аллелем и вторым аллелем охватывают от приблизительно 100 п.н. до приблизительно 200 п.н., от приблизительно 200 п.н. до приблизительно 400 п.н., от приблизительно 400 п.н. до приблизительно 600 п.н., от приблизительно 600 п.н. до приблизительно 800 п.н., от приблизительно 800 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 4 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н.
В других способах различия между первым аллелем и вторым аллелем охватывают по меньшей мере 100 п.н., по меньшей мере 200 п.н., по меньшей мере 300 п.н., по меньшей мере 400 п.н., по меньшей мере 500 п.н., по меньшей мере 600 п.н., по меньшей мере 700 п.н., по меньшей мере 800 п.н., по меньшей мере 800 п.н., по меньшей мере 1 т.п.н., по меньшей мере 2 т.п.н., по меньшей мере 3 т.п.н., по меньшей мере 4 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 6 т.п.н., по меньшей мере 7 т.п.н., по меньшей мере 8 т.п.н., по меньшей мере 9 т.п.н., по меньшей мере 10 т.п.н., 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н.
В других таких способах для модификации генома в гетерозиготной по первому аллелю клетке, в результате которой он становится гомозиготным по первому аллелю, геном приводят в контакт с белком Cas, тракрРНК и первой не аллель-специфической РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR. Первый аллель находится на первой гомологичной хромосоме, а последовательность распознавания РНК CRISPR является центромерной (т.е. расположена ближе к центромере) по отношению к локусу, соответствующему первому аллелю на второй гомологичной хромосоме. Белок Cas может расщеплять первую последовательность распознавания РНК CRISPR с образованием двухцепочечного разрыва. Затем может произойти рекомбинация с модификацией клетки, в результате которой она становится гомозиготной по первому аллелю.
Необязательно клетка гетерозиготна по одному или более дополнительным аллелям, причем первая последовательность распознавания РНК CRISPR является центромерной по отношению к локусам, соответствующим одному или более дополнительным аллелям на второй гомологичной хромосоме, и рекомбинация модифицирует клетку, в результате чего она становится гомозиготной по одному или более дополнительным аллелям.
Необязательно способ может дополнительно включать приведение генома в контакт со второй не аллель-специфической РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR, центромерной по отношению к локусу, соответствующему первому аллелю во второй гомологичной хромосоме, причем белок Cas расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с образованием по меньшей мере одного двухцепочечного разрыва. Необязательно способ может дополнительно включать приведение генома в контакт с дополнительными не аллель-специфическими РНК CRISPR (например, третьей РНК CRISPR или третьей и четвертой РНК CRISPR), которые гибридизуются с последовательностями распознавания РНК CRISPR, центромерными по отношению к локусу, соответствующему первому аллелю на второй гомологичной хромосоме. Затем можно идентифицировать клетку, содержащую модифицированный геном.
В некоторых способах первая (или вторая, третья, или четвертая) последовательности распознавания РНК CRISPR расположены на второй гомологичной хромосоме, но не на первой гомологичной хромосоме. Первый (или второй, третий, или четвертый) сайт распознавания РНК CRISPR может составлять от приблизительно 100 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. от центромеры.
Первый аллель и/или один или более дополнительных аллелей могут содержать мутацию, такую как нацеленная модификация. Альтернативно первый аллель и/или один или более дополнительных аллелей могут являться аллелем дикого типа, а соответствующие локусы на второй гомологичной хромосоме могут содержать мутации, такие как вызывающая заболевание мутация. Первый аллель может находиться на расстоянии от приблизительно 100 п.н. до приблизительно 1 п.н., от приблизительно 1 до приблизительно 10 п.н., от приблизительно 10 до приблизительно 100 п.н., от приблизительно 100 п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от 80 млн п.н. до 90 млн п.н. или от 90 млн п.н. до 100 млн п.н. от первого сайта распознавания РНК CRISPR. Альтернативно первый аллель может составлять по меньшей мере 100 п.н., по меньшей мере 1 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 10 млн п.н., по меньшей мере 20 млн п.н., по меньшей мере 30 млн п.н., по меньшей мере 40 млн п.н., по меньшей мере 50 млн п.н., по меньшей мере 60 млн п.н., по меньшей мере 70 млн п.н., по меньшей мере 80 млн п.н., по меньшей мере 90 млн п.н. или по меньшей мере 100 млн п.н. или более из первого сайта распознавания РНК CRISPR.
Белок Cas может являться белком Cas9. Он может обладать нуклеазной активностью на обеих цепях двухцепочечной ДНК или он может являться никазой. В некоторых способах белок Cas и первая РНК CRISPR в природных условиях вместе не встречаются.
Рекомбинация может включать потерю гетерозиготности теломерного (т.е. в направлении к теломере) двухцепочечного разрыва (например, полярную или направленную генную конверсию или потерю гетерозиготности). Область второй гомологичной хромосомы, замещаемая потерей гетерозиготности, может составлять от приблизительно 100 п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. Альтернативно заменяемая область второй гомологичной хромосомы может составлять по меньшей мере 100 п.н., по меньшей мере 1 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 10 млн п.н., по меньшей мере 20 млн п.н., по меньшей мере 30 млн п.н., по меньшей мере 40 млн п.н., по меньшей мере 50 млн п.н., по меньшей мере 60 млн п.н., по меньшей мере 70 млн п.н., по меньшей мере 80 млн п.н., по меньшей мере 90 млн п.н. или по меньшей мере 100 млн п.н. или более. Например, может заменяться большая часть хромосомы.
В. Способы создания генетически модифицированного не относящегося к человеку животного
С помощью различных способов, описанных в настоящем документе, можно создавать генетически модифицированных не относящихся к человеку животных. В некоторых случаях способ получения генетически модифицированного не относящегося к человеку животного включает: (1) модификацию генома плюрипотентной клетки с использованием способов, описанных выше; (2) выбор генетически модифицированной плюрипотентной клетки; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион-хозяин и (4) имплантацию эмбриона-хозяина, содержащего генетически модифицированную плюрипотентную клетку, в тело суррогатной матери. Получают клетки-потомки генетически модифицированной плюрипотентной клетки. Донорскую клетку можно вводить в эмбрион-хозяин на любой стадии, такой как стадия бластоцисты или стадия преморулы (т.е. стадия 4 клеток или стадия 8 клеток). Получают клетки-потомки, которые способны передавать генетическую модификацию через зародышевую линию. Плюрипотентная клетка может являться эмбриональной стволовой (ЭС) клеткой (например, ЭС-клеткой мыши или крысы), как описано в других разделах настоящего документа. См., например, патент США №7,294,754, который для всех целей полностью включен в настоящее описание путем ссылки.
Альтернативно способ получения генетически модифицированного не относящегося к человеку животного может включать: (1) модификацию генома эмбриона на стадии единственной клетки с использованием способов, описанных выше; (2) выбор генетически модифицированного эмбриона и (3) имплантацию генетически модифицированного эмбриона в тело суррогатной матери. Получают клетки-потомки, которые способны передавать генетическую модификацию через зародышевую линию.
Для создания не относящегося к человеку млекопитающего животного можно также использовать методики ядерного переноса. В кратком изложении методы ядерного переноса могут включать этапы, на которых происходит: (1) энуклеирование ооцита или обеспечение энуклеированного ооцита; (2) выделение или обеспечение донорской клетки или ядра, которые должны быть объединены с энуклеированным ооцитом; (3) вставка клетки или ядра в энуклеированный ооцит с образованием восстановленной клетки; (4) имплантация восстановленной клетки в матку животного для формирования эмбриона и (5) обеспечение развития эмбриона. В таких способах ооциты обычно извлекают из умерших животных, хотя их можно выделять также из яйцеводов и/или яичников живых животных. До энуклеации ооциты могут созревать в различных средах, известных обычным специалистам в данной области. Энуклеация ооцитов может быть выполнена с помощью ряда способов, хорошо известных обычным специалистам в данной области. Вставку донорской клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки можно проводить перед слиянием посредством микроинъекции донорской клетки под вителлиновый слой. Слияние можно индуцировать с применением электрического импульса постоянного тока в плоскости контакта/слияния (электрослияние), путем воздействия на клетки химическими веществами, стимулирующими слияние, такими как полиэтиленгликоль, или путем использования инактивированного вируса, например вируса Сендай. Реконструированную клетку можно активировать с использованием электричества и/или неэлектрического средства до, во время и/или после слияния ядра донора и ооцита реципиента. Способы активации включают в себя электрические импульсы, химически индуцированный шок, пенетрацию с помощью сперматозоида, повышение уровней двухвалентных катионов в ооцитах, а также снижение фосфорилирования клеточных белков (например, с помощью ингибиторов киназы) в ооците. Активированные реконструированные клетки или эмбрионы можно культивировать в среде, хорошо известной обычным специалистам в данной области, после чего их переносят в матку животного. См., например, US 20080092249, WO/1999/005266 A2, US 20040177390, WO/2008/017234 A1 и патент США №7,612,250, каждый из которых для всех целей полностью включен в настоящий документ путем ссылки.
Некоторые способы получения генетически модифицированного не относящегося к человеку животного включают способы получения не относящегося к человеку животного поколения F0. Такие способы могут включать приведение генома в нечеловеческой ЭС-клетке в контакт с белком Cas, первой РНК CRISPR, которая гибридизуется с первой последовательностью распознавания РНК CRISPR, второй РНК CRISPR, которая гибридизуется со второй последовательностью распознавания РНК CRISPR, и тракрРНК. Белок Cas может расщеплять геном в пределах первой и второй последовательностей распознавания РНК CRISPR с получением концевых последовательностей. Концевые последовательности могут подвергаться рекомбинации с образованием генома с нацеленной модификацией, и нацеленная модификация может включать делецию между первой и второй последовательностями распознавания РНК CRISPR.
Способы могут дополнительно включать: (1) идентификацию нечеловеческой ЭС-клетки, содержащей нацеленную модификацию; (2) введение нечеловеческой ЭС-клетки, содержащей нацеленную модификацию, в нечеловеческий эмбрион-хозяин и (3) гестацию нечеловеческого эмбриона-хозяина в теле суррогатной матери. Затем суррогатная мать может произвести не относящееся к человеку животное поколения F0, содержащее нацеленную модификацию. Эмбрион-хозяин, содержащий генетически модифицированную плюрипотентную или тотипотентную клетку (например, нечеловеческую ЭС-клетку), можно инкубировать до стадии бластоцисты, а затем имплантировать в тело суррогатной матери для получения животного F0. Животные, несущие генетически модифицированный геномный локус, могут быть идентифицированы посредством теста на модификацию аллеля (МОА), как описано в настоящем документе.
Различные способы, представленные в настоящем документе, позволяют получить генетически модифицированное не относящееся к человеку животное F0, причем клетки генетически модифицированного животного F0 содержат нацеленную модификацию. Признано, что в зависимости от способа, используемого для получения животного F0, количество клеток внутри организма животного F0, которые имеют представляющую интерес нуклеотидную последовательность и не содержат рекомбинантную кассету и/или кассету селекции, различается. Введение донорских ЭС-клеток в эмбрион на стадии до морулы из соответствующего организма (например, мышиного эмбриона на стадии 8 клеток) посредством, например, способа VELOCIMOUSE®, позволяет получить более высокий процент клеточной популяции животного F0, содержащей клетки с представляющей интерес нуклеотидной последовательностью с нацеленной генетической модификацией. В конкретных случаях по меньшей мере 50%, 60%, 65% 70%, 75%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада у не относящегося к человеку животного F0 содержит клеточную популяцию, имеющую нацеленную модификацию. В других случаях по меньшей мере одна или более зародышевых клеток животного F0 имеют нацеленную модификацию.
В некоторых случаях клетки генетически модифицированного животного F0 являются гетерозиготными или компаунд-гетерозиготными по отношению к нацеленной модификации. Например, клетки генетически модифицированного животного F0 могут быть гемизиготными по нацеленной модификации. В других случаях клетки генетически модифицированного животного F0 гомозиготны по нацеленной модификации.
В некоторых случаях из животного F0, полученного с помощью способов и композиций, описанных в настоящем документе, может быть выведено животное дикого типа с получением поколения F1, которое гетерозиготно по нацеленной модификации. Затем животных F1 можно скрещивать друг с другом с получением животного F2, гомозиготного по нацеленной модификации. Потомство F1 можно генотипировать с использованием специфических праймеров и/или зондов для определения наличия нацеленной генетической модификации.
С. Геномы и целевые геномные локусы
Геном или целевой геномный локус, модифицированный описанными в настоящем документе способами, может включать в себя любой сегмент или область ДНК внутри клетки. Геном или целевой геномный локус может быть нативным для клетки, может быть гетерологичным или экзогенным сегментом ДНК, встроенной в геном клетки, или может являться их комбинацией. Такие гетерологичные или экзогенные сегменты ДНК могут включать в себя трансгены, экспрессионные кассеты, полинуклеотиды, кодирующие селективные маркеры, или гетерологичные или экзогенные области геномной ДНК.
Геном или целевой геномный локус может также включать в себя внехромосомную ДНК внутри клетки, такую как дрожжевая искусственная хромосома (YAC), бактериальная искусственная хромосома (ВАС), искусственная хромосома человека, или любую другую сконструированную геномную область, содержащуюся в соответствующей клетке-хозяине.
D. Формы Cas9 и гидовой РНК
В некоторых способах приведение генома в контакт включает введение одного или более белков Cas, одной или более РНК CRISPR и одной или более тракрРНК в клетку. Введение может быть реализовано любыми средствами, и один или более компонентов (например, два компонента или все компоненты) можно ввести в клетку одновременно или последовательно в любой комбинации.
РНК CRISPR и тракрРНК могут быть слиты вместе в качестве гидовой РНК (гРНК) для введения в клетку. Альтернативно РНК CRISPR и тракрРНК могут являться разными молекулами РНК. РНК CRISPR можно ввести в клетку в виде РНК или в виде ДНК, кодирующей РНК CRISPR. Аналогичным образом, тракрРНК можно ввести в клетку в виде РНК или в виде ДНК, кодирующей тракрРНК, и гРНК можно ввести в клетку в виде РНК или в виде гРНК, кодирующей ДНК.
Белок Cas можно ввести в клетку в виде белка, информационной РНК (иРНК), кодирующей белок Cas, или ДНК, кодирующей белок Cas. В некоторых способах белок Cas, РНК CRISPR и тракрРНК можно ввести в клетку в виде комплекса белок-РНК. Аналогичным образом, белок Cas и гРНК можно ввести в клетку в виде комплекса белок-РНК. Белок Cas может являться способным к проникновению в клетку белком Cas (например, белком Cas с доменом проникновения в клетку).
ДНК, кодирующая белок Cas, РНК CRISPR или тракрРНК может быть функционально связана с промотором, активным в клетке. Такие ДНК могут находиться в одном или более экспрессионных конструктах. В некоторых способах один или более таких экспрессионных конструктов могут являться компонентами одной молекулы нуклеиновой кислоты. Например, ДНК, кодирующие один или более белков Cas, ДНК, кодирующие одну или более РНК CRISPR, и ДНК, кодирующие одну или более тракрРНК, могут являться компонентами одной молекулы нуклеиновой кислоты. Альтернативно они могут быть разделены в любой комбинации среди двух, трех, четырех или более молекул нуклеиновой кислоты.
Аналогичным образом, ДНК, кодирующая белок Cas, или ДНК, кодирующая гРНК, может быть функционально связана с промотором, активным в клетке. Такие ДНК также могут находиться в одном или более экспрессионных конструктах. В некоторых способах один или более таких экспрессионных конструктов могут являться компонентами одной молекулы нуклеиновой кислоты. Например, ДНК, кодирующие один или более белков Cas, и ДНК, кодирующие одну или более гРНК, могут являться компонентами одной молекулы нуклеиновой кислоты. Альтернативно они могут быть разделены в любой комбинации среди двух, трех, четырех или более молекул нуклеиновой кислоты.
В некоторых способах белок Cas и РНК CRISPR и/или тракрРНК в природных условиях вместе не встречаются. В некоторых способах, например, белок Cas и первая РНК CRISPR в природных условиях вместе не встречаются, белок Cas и вторая CRISPR-РНК в природных условиях вместе не встречаются и/или белок Cas и тракрРНК в природных условиях вместе не встречаются.
В некоторых способах белок Cas представляет собой белок Cas9. Белок Cas может быть слит с гетерологичным полипептидом, таким как сигнал ядерной локализации (NLS). Белок Cas может обладать полной расщепляющей активностью и создавать двухцепочечные разрывы внутри геномной ДНК (например, двухцепочечные разрывы с тупыми концами) или может представлять собой никазу, которая может расщеплять только цепь геномной ДНК.
В некоторых способах используются парные никазы. Например, геном можно приводить в контакт с первой и второй никазами, которые расщепляют противоположные цепи ДНК, благодаря чему геном модифицируется посредством двойного никования. Первая никаза может расщеплять первую цепь геномной ДНК (т.е. комплементарную цепь), а вторая никаза может расщеплять вторую цепь геномной ДНК (т.е. некомплементарную цепь). Альтернативно обе никазы могут расщеплять одну и ту же цепь. Первую и вторую никазы можно создавать, например, посредством мутации каталитического остатка в домене RuvC (например, мутации D10A, описанной в другом разделе настоящего документа) первой никазы и мутации каталитического остатка в домене HNH (например, мутации Н840А, описанной в другом разделе настоящего документа) второй никазы. Альтернативно первая никаза может использоваться для создания обоих одноцепочечных разрывов.
В некоторых таких способах двойное никование можно использовать для создания одного или более двухцепочечных разрывов, имеющих ступенчатые концы. Например, двойное никование используется для создания ступенчатых концов в первом и втором сайтах расщепления. Первая никаза может расщеплять первую цепь ДНК в пределах первой и второй последовательностей распознавания РНК CRISPR, к которым гибридизуются первая и вторая РНК CRISPR, а вторая никаза может расщеплять вторую цепь ДНК в пределах третьей и четвертой целевых последовательностей распознавания РНК CRISPR, к которым гибридизуются третья и четвертая РНК CRISPR. Альтернативно первая никаза может использоваться для никования первой, второй, третьей и четвертой последовательностей распознавания РНК CRISPR. Первая и третья целевые последовательности распознавания РНК CRISPR могут быть расположены таким образом, чтобы создать первый сайт расщепления, так что одноцепочечные разрывы, созданные первой и второй никазами на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. первый сайт расщепления содержит одноцепочечные разрывы в пределах первой и третьей последовательностей распознавания РНК CRISPR). Аналогичным образом, вторая и четвертая последовательности распознавания РНК CRISPR могут быть расположены таким образом, чтобы создать второй сайт расщепления, так что одноцепочечные разрывы, созданные первой и второй никазами на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. второй сайт расщепления содержит одноцепочечные разрывы в пределах второй и четвертой последовательностей распознавания РНК CRISPR). В некоторых случаях одноцепочечные разрывы в пределах первой и третьей последовательностей распознавания РНК CRISPR и/или второй и четвертой последовательностей распознавания РНК CRISPR могут являться смещенными одноцепочечными разрывами. Окно смещения может составлять, например, по меньшей мере приблизительно 5 т.п.н., 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н. или более. См. публикации Ran et al. (2013) Cell 154: 1380-1389; Mali et al. (2013) Nat. Biotech. 31: 833-838 и Shen et al. (2014) Nat. Methods 11: 399-404, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
Е. Способы введения нуклеиновых кислот и белков в клетки
В настоящем документе описаны различные способы и композиции, позволяющие вводить нуклеиновую кислоту в клетку. В некоторых случаях система, применяемая для введения нуклеиновой кислоты, позволяет осуществлять нацеленную интеграцию в конкретном геномном локусе. В таких системах применяются разнообразные компоненты, и для простоты ссылки термин «система нацеленной геномной интеграции» в общем случае включает в себя все компоненты, необходимые для события интеграции (например, один или более нуклеазных агентов, сайтов расщепления нуклеазами, вставок ДНК-полинуклеотидов, нацеливающих векторов, целевых геномных локусов и представляющих интерес полинуклеотидов).
Предложенные в настоящем документе способы могут включать введение в клетку одного или более полинуклеотидов или полипептидных конструктов, содержащих один или более компонентов системы нацеленной геномной интеграции. Введение включает передачу в клетку последовательности (полипептидной или полинуклеотидной) таким образом, что последовательность получает доступ к внутренней части клетки. Предложенные в настоящем документы способы не зависят от конкретного способа введения нуклеиновой кислоты или белка в клетку, а только от того, что нуклеиновая кислота или белок получает доступ к внутренней части по меньшей мере одной клетки. Способы введения нуклеиновых кислот и белков в различные типы клеток известны в данной области и включают в себя, например, способы стабильной трансфекции, способы временной трансфекции и вирус-опосредованные способы.
В некоторых случаях клетки, используемые в способах и композициях, имеют стабильно встроенный в их геном ДНК-конструкт. Например, клетка, применяемая в способах, описанных в настоящем документе, может иметь уже существующий ген, кодирующий Cas, стабильно включенный в ее геном (т.е. клетка со встроенным Cas). Термин «стабильно встроенный» или «стабильно введенный» включает в себя введение полинуклеотида в клетку таким образом, что нуклеотидная последовательность интегрируется в геном клетки и способна передаваться по наследству ее потомству. Для стабильного встраивания ДНК-конструктов или различных компонентов системы нацеленной геномной интеграции может использоваться любой протокол.
Протоколы трансфекции, а также протоколы введения полипептидов или полинуклеотидных последовательностей в клетки могут быть разными. Не имеющие ограничительного характера способы трансфекции включают химические способы трансфекции с использованием липосом; наночастиц; фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc Natl Acad Sci USA 74 (4): 1590-4 и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W.H. Freeman and Company, pp. 96-97); дендримеров или катионных полимеров, таких как ДЭАЭ-декстран или полиэтиленимин. Нехимические способы включают в себя электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция с использованием частиц включает в себя применение «генной пушки» или магнитной трансфекции (Bertram, (2006) Current Pharmaceutical Biotechnology 7, 277-28). Для трансфекции также можно использовать вирусные способы.
В некоторых случаях введение нуклеиновых кислот или белков в клетку опосредуется электропорацией, интрацитоплазматической инъекцией, вирусной инфекцией, аденовирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или Nucleofection™.
Введение нуклеиновых кислот или белков в клетку (например, эмбрион на стадии единственной клетки) также может быть выполнено посредством микроинъекции. В эмбрионах на стадии единственной клетки микроинъекция может быть выполнена в материнский и/или отцовский пронуклеус или в цитоплазму. Если микроинъекцию выполняют только в один пронуклеус, отцовский пронуклеус является предпочтительным в связи с его большим размером. Микроинъекция мРНК выполняется предпочтительно в цитоплазму (например, для доставки мРНК непосредственно в механизмы трансляции), а микроинъекцию белка Cas или кодирующей молекулу нуклеиновой кислоты РНК предпочтительно выполнять в ядро/пронуклеус. Альтернативно микроинъекция может быть осуществлена посредством инъекции как в ядро/пронуклеус, так и в цитоплазму: иглу можно сначала ввести в ядро/пронуклеус и ввести первое количество, а при удалении иглы из эмбриона на стадии единственной клетки в цитоплазму можно ввести второе количество. Если белок Cas вводят в цитоплазму, белок Cas предпочтительно содержит сигнал ядерной локализации, чтобы обеспечить доставку к ядру/пронуклеусу. Способы выполнения микроинъекции хорошо известны. См., например, публикации Nagy et al. (Nagy A, Gertsenstein M, Vintersten K, Behringer R., 2003, Manipulating the Mouse Embryo. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Meyer et al. (2010) Pros Natl Acad Sci USA 107: 15022-15026 и Meyer et al. (2012) Proc Natl Acad Sci USA 109: 9354-9359, каждая из которых для всех целей полностью включена в настоящее описание путем ссылки.
Введение нуклеиновых кислот или белков в клетку может быть выполнено один или множество раз в течение периода времени. Например, введение может выполняться по меньшей мере два раза в течение периода времени, по меньшей мере три раза в течение периода времени, по меньшей мере четыре раза в течение периода времени, по меньшей мере пять раз в течение периода времени, по меньшей мере шесть раз в течение периода времени, по меньшей мере семь раз в течение периода времени, по меньшей мере восемь раз в течение периода времени, по меньшей мере девять раз в течение периода времени, по меньшей мере десять раз в течение периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение периода времени, по меньшей мере тринадцать раз в течение периода времени, по меньшей мере четырнадцать раз в течение периода времени, по меньшей мере пятнадцать раз в течение периода времени, по меньшей мере шестнадцать раз в течение периода времени, по меньшей мере семнадцать раз в течение периода времени, по меньшей мере восемнадцать раз в течение периода времени, по меньшей мере девятнадцать раз в течение периода времени или по меньшей мере двадцать раз в течение периода времени.
Когда в клетку вводят как нуклеазные агенты, так и нацеливающие векторы (например, большой нацеливающий вектор (LTVEC) для клеток, отличных от эмбрионов на стадии единственной клетки), их можно вводить одновременно. Альтернативно нуклеазный агент можно вводить отдельно от нацеливающего вектора. Например, нуклеазный агент можно вводить до введения нацеливающего вектора или его можно вводить после введения нацеливающего вектора.
F. Механизмы рекомбинации и способы изменения распространенности негомологичного соединения концов, генной конверсии или гомологичной рекомбинации
Рекомбинация включает в себя любой процесс обмена генетической информацией между двумя полинуклеотидами и может осуществляться посредством любого механизма. Рекомбинация в ответ на двухцепочечные разрывы (DSB) происходит главным образом посредством двух консервативных путей репарации ДНК: негомологичного соединения концов (NHEJ) и гомологичной рекомбинации (HR). См. публикацию Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897, которая для всех целей полностью включена в настоящий документ путем ссылки. NHEJ включает в себя репарацию двухцепочечных разрывов нуклеиновой кислоты путем прямого сшивания концов разрыва друг с другом без необходимости в гомологичном шаблоне. Сшивание несмежных последовательностей методом NHEJ часто может приводить к делециям, вставкам или перемещениям вблизи области двухцепочечного разрыва. Рекомбинация также может происходить посредством направляемой гомологией репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включает в себя форму репарации нуклеиновых кислот, для которой может потребоваться гомология нуклеотидной последовательности, и в ней используется «донорская» молекула для репарации по ее шаблону «целевой» молекулы (т.е. молекулы, подвергшейся двухцепочечному разрыву), и она приводит к переносу генетической информации от донора к мишени. Без ограничения какой-либо конкретной теорией такой перенос может быть связан с коррекцией несовпадений гетеродуплексной ДНК, сформированной расщепленной мишенью и донором, и/или синтез-зависимым отжигом цепей, при котором донор используется для повторного синтеза генетической информации, которая становится частью мишени, и/или родственными процессами. В некоторых случаях полинуклеотид-донор, участок полинуклеотида-донора, копия полинуклеотида-донора или участок копии полинуклеотида-донора встраивается в целевую ДНК.
В контексте модификации генома клетки, являющейся гетерозиготной по аллелю, чтобы сделать ее гомозиготной по этому аллелю, рекомбинация может включать любые средства, с помощью которых гомозиготные клетки получают из гетерозиготных клеток. Такие средства могут включать в себя, например, потерю гетерозиготности (LOH), генную конверсию или события кроссинговера, происходящие посредством любого известного механизма рекомбинации. Без ограничения какой-либо конкретной теорией авторы полагают, что LOH может происходить, например, при митотической рекомбинации с генной конверсией или без нее или при потере и удвоении хромосом. См., например, публикацию Lefebvre et al. (2001) Nat. Genet. 27: 257-258, которая для всех целей полностью включена в настоящий документ путем ссылки. Генная конверсия в этом контексте может включать в себя однонаправленную передачу генетического материала от донорной последовательности в высокогомологичный акцептор (т.е. нереципрокный обмен генетической информацией от одной молекулы к ее гомологу). Генная конверсия включает в себя любые средства копирования аллеля с помощью любого известного механизма рекомбинации. Например, генная конверсия может включать в себя нереципрокную передачу генетической информации от интактной последовательности в гомологичную область, содержащую двухцепочечный разрыв, и она может возникать между сестринскими хроматидами, гомологичными хромосомами или гомологичными последовательностями или на одной хроматиде, или на разных хромосомах. См., например, публикацию Chen et al. (2007) Nat. Rev. Genet. 8: 762-775, которая для всех целей полностью включена в настоящий документ путем ссылки. В конкретных случаях генная конверсия является непосредственным результатом гомологичной рекомбинации в результате копирования генетической информации из гомологичной хромосомы. Это может привести к локализованной потере гетерозиготности (LOH), когда гомологичные последовательности не являются идентичными.
В качестве примера LOH может происходить посредством взаимного обмена хроматид с помощью митотического кроссинговера или посредством копирования хроматид с помощью индуцированной разрывом репликации. В любом случае может произойти гетерозиготная модификация, в которой на одну хромосому воздействуют до репликации генома. Альтернативно можно воздействовать на одну хроматиду после репликации генома с последующей межхроматидной генной конверсией.
В любом из описанных в настоящем документе способов клетка может представлять собой клетку, которая была модифицирована для повышения или снижения активности NHEJ. Аналогичным образом, клетка может представлять собой клетку, которая была модифицирована для повышения генной конверсии или активности HDR. Такие модификации могут включать в себя модификации в экспрессии или активности генов, участвующих в регуляции NHEJ, генной конверсии и/или HDR. Например, снижение активности NHEJ и/или повышение активности HDR может способствовать биаллельному соединению геномных областей между последовательностями распознавания РНК CRISPR, соответствующими двум гРНК. Без ограничения какой-либо конкретной теорией авторы полагают, что одним механизмом, посредством которого может произойти биаллельное геномное соединение, является NHEJ-опосредованная репарация или HDR-опосредованная репарация в пределах первого аллеля и создание идентичного второго аллеля с помощью механизмов HDR, таких как генная конверсия (см. пример 1). Таким образом, стимулирование HDR-опосредованных путей (например, за счет снижения активности NHEJ или за счет повышения активности HDR) может также способствовать биаллельному коллапсированию геномных областей. Аналогичным образом, без ограничения какой-либо конкретной теорией авторы полагают, что превращение гетерозиготной клетки в гомозиготную клетку с использованием парных гидовых РНК, которые нацелены на один локус, можно стимулировать, если активность NHEJ снижается, а активность HDR (например, активность генной конверсии) соответственно повышается.
Ингибиторы можно использовать для повышения или снижения активности NHEJ или для повышения или снижения активности HDR. Такими ингибиторами могут являться, например, небольшие молекулы или ингибирующие нуклеиновые кислоты, такие как короткие интерферирующие нуклеиновые кислоты (например, малая интерферирующая РНК (миРНК), двухцепочечная РНК (дцРНК), микроРНК (мкРНК) и короткая шпилечная РНК (кшРНК)) или антисмысловые олигонуклеотиды, специфичные для транскрипта гена. Ингибиторы могут быть направлены на ферменты, участвующие в NHEJ или HDR, или на их регуляцию выше участка транскрипции посредством посттрансляционной модификации, например, посредством фосфорилирования, убиквитилирования и сумоилирования.
В клетках млекопитающих NHEJ является преобладающим механизмом репарации DSB и активен в ходе всего клеточного цикла. У позвоночных «канонический», или «классический», путь NHEJ (C-NHEJ) требует ряда ключевых факторов, в том числе ДНК-PK, Ku70-80, Artemis, лигазу IV (Lig4), XRCC4, CLF и Pol μ, для репарации DSB. См. публикацию Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897, которая для всех целей полностью включена в настоящий документ путем ссылки. Во время NHEJ концы ДНК связаны очень обильным защищающим концы белком Ku, который функционирует как док-станция для загрузки других компонентов NHEJ.
Таким образом, в некоторых из описанных в настоящем документе способов клетка была модифицирована для снижения или устранения или для повышения экспрессии или активности факторов, участвующих в C-NHEJ. Например, в некоторых способах клетка была модифицирована для снижения или устранения экспрессии или активности ДНК-PK, Ku70-80, Artemis, лигазы IV (Lig4), XRCC4, CLF и/или Pol. В конкретных способах клетка была модифицирована для снижения или устранения экспрессии или активности ДНК-PK или для повышения экспрессии или активности ДНК-PK (например, экспрессии или активности ДНК-PKcs; примерная последовательность UniProt, обозначенная Р97313). Примеры ингибиторов ДНК-PKcs включают, например, NU7026 и NU7441. См., например, патент США №6,974,867, который для всех целей полностью включен в настоящее описание путем ссылки. В конкретных способах клетка была модифицирована для снижения или устранения экспрессии или активности лигазы IV или для повышения экспрессии или активности лигазы IV. Примером ингибитора лигазы IV является SCR7.
Ингибиторы, нацеленные на белки контрольных точек клеточного цикла, такие как ATM (например, KU55933), CHK1/CHK2 (например, KLD1162 или CHIR-124) и ATR (например, VE 821), могут также использоваться для синергического усиления эффектов специфических ингибиторов репарации ДНК или для предотвращения непреднамеренных побочных эффектов, таких как остановка клеточного цикла и/или апоптоз (см. публикацию Ciccia et al. (2010) Mol Cell 40: 179, которая для всех целей полностью включена в настоящий документ путем ссылки).
Нарушение C-NHEJ может повышать уровни аномального присоединения, опосредованного «альтернативными» путями NHEJ (A-NHEJ), и может также увеличивать восстановление HR. Пути A-NHEJ демонстрируют смещение к микрогомологическим опосредованным соединениям и имеют более медленную кинетику, чем C-NHEJ. Были сделаны предположения о том, что в этом принимает участие ряд факторов, в том числе комплекс MRN (MRE11, RAD50, NBS1), CtIP, XRCC1, PARP, Lig1 и Lig3. См. публикации Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897 и Claybon et al. (2010) Nucleic Acids Res. 38 (21): 7538-7545, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки.
Таким образом, в некоторых из описанных в настоящем документе способов клетка была модифицирована для снижения или устранения или для повышения экспрессии или активности факторов, участвующих в A-NHEJ. Например, в некоторых способах клетка была модифицирована для снижения или устранения экспрессии или активности MRE11, RAD50, NBS1, CtIP, XRCC1, PARP (например, PARP1), Lig1 и/или Lig3. В других способах клетка была модифицирована для повышения экспрессии или активности MRE11, RAD50, NBS1, CtIP, XRCC1, PARP (например, PARP1), Lig1 и/или Lig3. В конкретных способах клетка была модифицирована для снижения или устранения экспрессии или активности PARP1 или для повышения экспрессии или активности PARP1 (примерная последовательность UniProt, обозначенная Р11103). Примеры ингибиторов PARP (например, NU1025, инипариб, олапариб) включают в себя никотинамиды; изохинолиноны и дигидроизохинолиноны; бензимидазолы и индолы; фталазин-1(2Н)-оны и хиназолиноны; изоиндолиноны и их аналоги и производные; фенантридины и фенантридиноны; бензопироны и их аналоги и производные; производные ненасыщенной гидроксамовой кислоты и их аналоги и производные; пиридазины, включая конденсированные пиридазины и их аналоги и производные; и/или другие соединения, такие как кофеин, теофиллин и тимидин, а также их аналоги и производные. См., например, патент США 8,071,579, который для всех целей полностью включен в настоящий документ путем ссылки.
C-NHEJ также демонстрирует конкурентные отношения с HR, так что нарушение C-NHEJ также может привести к увеличению репарации HR. Такая конкуренция между NHEJ и HR может использоваться, так как нарушение NHEJ может приводить к усиленному нацеливанию на ген за счет снижения случайной интеграции и, возможно, повышения нацеленной интеграции посредством гомологичной рекомбинации.
Существует ряд видов репарации посредством гомологичной рекомбинации, в том числе одноцепочечный отжиг, генная конверсия, кроссинговеры и индуцированная разрывом репликация. Одноцепочечный отжиг является незначительным видом репарации HR, при которой гомологичные одноцепочечные последовательности соединяют комплементарные цепи ДНК по обе стороны отрезанного DSB, что приводит к восстановлению хромосомы. Одноцепочечный отжиг выполняет делеции различного размера в зависимости от расстояния, разделяющего две области гомологии последовательностей. Генная конверсия включает нереципрокный обмен генетической информацией от одной молекулы к ее гомологу, что происходит непосредственно при HR в результате копирования генетической информации из гомологичной хромосомы. Это может приводить к локализованной LOH, когда гомологичные последовательности не являются идентичными. Как правило, степень генной конверсии ограничена несколькими сотнями пар оснований. Тем не менее при некоторых генетических состояниях, включая дефицит RAD51C, была зарегистрирована генная конверсия по длинному участку. См. публикацию Nagaraju et al. (2006) Mol. Cell. Biol. 26: 8075-8086, которая для всех целей полностью включена в настоящий документ путем ссылки. Кроссинговеры могут происходить, например, между гомологичными хромосомами, и могут приводить к реципрокным транслокациям, если они происходят в G1, или нереципрокным транслокациям и распространению LOH от места разрыва до дистальной теломеры, если они происходят в G2. Индуцированная разрывом репликация представляет собой вариант HR, при котором после инвазии цепей репликация ДНК продолжается до конца хромосомы. Таким образом, существует много механизмов, посредством которых HR может стимулировать LOH.
Таким образом, в некоторых из описанных в настоящем документе способов клетка была модифицирована для снижения или устранения или для повышения экспрессии или активности факторов, участвующих в HR. Например, в некоторых способах клетка была модифицирована для повышения экспрессии или активности RAD51, RAD52, RAD54, RAD55, RAD51C, BRCA1 и/или BRCA2. В других способах клетка была модифицирована для снижения или устранения экспрессии или активности RAD51, RAD52, RAD54, RAD55, RAD51C, BRCA1 и/или BRCA2.
В некоторых способах экспрессия или активность других белков, участвующих в регуляции NHEJ и/или HR, может быть изменена. Например, в некоторых способах клетка была модифицирована для снижения или устранения экспрессии или активности Chk2, снижения или устранения экспрессии или активности Clspn, снижения или устранения экспрессии или активности Setd2, повышения экспрессии или активности Kat2a и/или повышения экспрессии или активности Rad51. В других способах клетка была модифицирована для повышения экспрессии или активности Chk2, повышения экспрессии или активности Clspn, повышения экспрессии или активности Setd2, снижения или устранения экспрессии или активности Kat2a и/или снижения или устранения экспрессии или активности Rad51.
Chk2 (также известный как Chek2 и Rad53; гомолог у S. pombe - Cds1) представляет собой сериновую/треониновую протеинкиназу, необходимую для опосредованной контрольной точкой остановки клеточного цикла, активации репарации ДНК и апоптоза в ответ на присутствие разрывов в двухцепочечной ДНК. См. публикацию Blaikley et al. (2014) Nucleic Acids Research 42: 5644-5656, которая для всех целей полностью включена в настоящий документ путем ссылки. Clspn (также известный как Claspin; гомолог у S. pombe - Mrc1) представляет собой белок, необходимый для опосредованной контрольной точкой остановки клеточного цикла в ответ на повреждение ДНК. Сообщается, что делеция гомологов Chk2 или Clspn у S. pombe приводит к гиперрекомбинантному фенотипу, демонстрирующему значительно повышенные уровни индуцированной разрывом генной конверсии по сравнению с диким типом. В частности, сообщается, что уровни генной конверсии значительно повышались, а уровни негомологичного соединения концов (NHEJ), конверсии сестринских хроматид (SCC) и потери гетерозиготности (LOH) были снижены. См. публикацию Blaikley et al. (2014) Nucleic Acids Research 42: 5644-5656.
Kat2a (также известен как Gcn5 и Gcn512) представляет собой универсальную гистоновую ацетилтрансферазу, которая способствует активации транскрипции и, по сообщениям, связана с репарацией двухцепочечного разрыва. Kat2a-зависимое ацетилирование лизина 36 гистона Н3 (H3K36) повышает доступность хроматина, улучшает отсечение и способствует гомологичной рекомбинации, одновременно подавляя негомологичное соединение концов. См. публикацию Pai et al. (2014) Nat. Commun. 5: 4091, которая для всех целей полностью включена в настоящий документ путем ссылки. Setd2 (также известный как Kiaa1732, Kmt3a и Set2) представляет собой гистонметилтрансферазу, которая специфически триметилирует лизин 36 гистона H3 (H3K36me3), используя деметилированный лизин 36 (H3K36me2) в качестве субстрата. Setd2-зависимое метилирование H3K36 снижает доступность хроматина, снижает отсечение и стимулирует NHEJ. См. публикацию Pai et al. (2014) Nat. Commun. 5: 4091.
Rad 51 (также известный как Reca, Rad51A и гомолог 1 белка репарации ДНК Rad51) представляет собой белок, который функционирует с Rad52 и другими белками для осуществления обмена цепями при гомологичной рекомбинации, образуя гетеродуплексную ДНК, которая корректируется посредством исправления ошибок спаривания с получением участка генной конверсии. Сообщается, что в клетках млекопитающих повышенная экспрессия Rad51 и Rad52 повышает частоту гомологичной рекомбинации и генной конверсии. См. публикации Yanez & Porter (1999) Gene Ther. 6: 1282-1290 и Lambert & Lopez (2000) EMBO J. 19: 3090-3099, которые для всех целей полностью включены в настоящий документ путем ссылки.
Модификации экспрессии или активности генов, участвующих в регуляции NHEJ, генной конверсии и/или направляемой гомологией репарации, могут быть специфичными с пространственной или временной точек зрения, а также могут быть индуцибельными или временными и обратимыми. Например, можно конструировать различные виды кассет, предусматривающие делецию в конкретных типах клеток и тканей, на конкретных стадиях развития или при индукции. В таких кассетах может использоваться система рекомбиназы, в которой кассета с обеих сторон фланкирована сайтами распознавания рекомбиназы и может быть удалена рекомбиназой, экспрессированной в заданном типе клеток, экспрессированной на заданной стадии развития или экспрессированной или активированной в результате индукции. Такие кассеты дополнительно могут быть сконструированы с включением в них набора из пар сайтов распознавания разными рекомбиназами, которые расположены таким образом, чтобы можно было генерировать нулевые, условные или комбинированные условные/нулевые аллели, как описано в публикации US 2011/0104799, для всех целей полностью включенной в настоящий документ путем ссылки. Регуляцию генов рекомбиназы можно контролировать различными способами, например посредством функционального связывания гена рекомбиназы с клеточноспецифичным, тканеспецифичным или регулируемым в зависимости от стадии развития промотором (или другим регуляторным элементом) или посредством функционального связывания гена рекомбиназы с нетранслируемой областью 3'-UTR, содержащей сайт распознавания для миРНК, активной только в конкретных типах клеток, типах тканей или на стадиях развития. Рекомбиназу также можно регулировать, например, путем использования слитного белка, ставящего рекомбиназу под контроль эффектора или метаболита (например, CreERT2, активность которой положительно контролируется тамоксифеном), или путем постановки рекомбиназы под контроль индуцируемого промотора (например, промотора, активность которого контролируется доксициклином и TetR или вариантами TetR). Примеры различных видов кассет и средств регуляции генов рекомбиназы приведены, например, в патентах US 8,518,392; US 8,354,389 и US 8,697,851, каждый из которых полностью включен в настоящий документ путем ссылки.
G. Клетки и животные
В различных композициях и способах, предложенных в настоящем документе, применяются клетки, такие как клетки от животного. Такие клетки могут быть получены от не относящегося к человеку животного. Такие клетки могут являться эукариотическими клетками, в том числе, например, клетками грибов (например, дрожжей), клетками растений, клетками животных, клетками млекопитающих и клетками человека. Клеткой млекопитающих может быть, например, клетка не относящегося к человеку млекопитающего, клетка человека, клетка грызуна, клетка крысы, клетка мыши, клетка хомяка, фибробласт или клетка СНО. Эукариотическая клетка может быть тотипотентной клеткой, плюрипотентной клеткой, такой как нечеловеческая плюрипотентная клетка (например, эмбрионального стволовая (ЭС) клетка мыши или ЭС-клетка крысы) или плюрипотентная клетка человека, или неплюрипотентной клеткой. Тотипотентные клетки включают недифференцированные клетки, которые могут давать начало любому типу клеток, а плюрипотентные клетки включают недифференцированные клетки, которые обладают способностью развиваться в более чем один дифференцированный тип клеток. Такие плюрипотентные и/или тотипотентные клетки могут представлять собой, например, эмбриональные стволовые (ЭС) клетки или подобные ЭС клетки, такие как индуцированные плюрипотентные стволовые (ИПС) клетки. Эмбриональная стволовая клетка включает в себя полученные из эмбриона тотипотентные или плюрипотентные клетки, которые при введении в эмбрион могут участвовать в развитии любой ткани развивающегося эмбриона. ЭС-клетки могут быть получены из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых листков позвоночных (энтодермы, эктодермы и мезодермы).
Эукариотическая клетка также может являться клеткой, которая не является первичной соматической клеткой. Соматические клетки могут включать в себя любую клетку, которая не является гаметой, зародышевой клеткой, гаметоцитом или недифференцированной стволовой клеткой.
Эукариотические клетки также включают в себя первичные клетки. Первичные клетки включают в себя клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают в себя клетки, которые не являются ни трансформированными, ни бессмертными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не была пассирована в тканевой культуре или была ранее пассирована в тканевой культуре, но не может быть в течение неопределенно длительного времени пассирована в тканевой культуре. Такие клетки могут быть выделены обычными способами и включают в себя, например, соматические клетки, гемопоэтические клетки, эндотелиальные клетки, эпителиальные клетки, фибробласты, мезенхимальные клетки, кератиноциты, меланоциты, моноциты, мононуклеарные клетки, адипоциты, преадипоциты, нейроны, глиальные клетки, гепатоциты, скелетные миобласты и гладкомышечные клетки. Например, первичные клетки могут быть получены из соединительных тканей, мышечных тканей, тканей нервной системы или эпителиальных тканей.
Эукариотические клетки также включают в себя иммортализованные клетки. Иммортализованные клетки включают в себя клетки из многоклеточного организма, которые обычно не размножаются в течение неопределенно длительного времени, но вследствие мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать деление. Такие мутации или изменения могут происходить естественным образом или могут быть намеренно индуцированными. Примерами иммортализованных клеток являются клетки яичника китайского хомячка (СНО), эмбриональные клетки почки человека (например, клетки HEK 293) и мышиные эмбриональные фибробластные клетки (например, клетки 3Т3). Множество типов иммортализованных клеток хорошо известны в данной области.
Иммортализованные или первичные клетки включают в себя клетки, которые обычно используются для культивирования клеток или для экспрессии рекомбинантных генов или белков.
Эукариотические клетки могут также включать в себя эмбрионы на стадии единственной клетки (т.е. оплодотворенные ооциты или зиготы). Такие эмбрионы на стадии единственной клетки могут быть получены из любого генетического фона (например, BALB/c, C57BL/6, 129 или их комбинации), могут быть свежими или замороженными и могут быть получены в результате естественного разведения или экстракорпорального оплодотворения.
Термин «животное» по отношению к клеткам, плюрипотентным и/или тотипотентным клеткам, ЭС-клеткам, донорским клеткам и/или эмбриону-хозяину включает в себя млекопитающих, рыб и птиц. Млекопитающие включают в себя, например, людей, не относящихся к человеку приматов, обезьян, человекообразных обезьян, кошек, лошадей, быков, оленей, бизонов, овец, грызунов (например, мышей, крыс, хомячков, морских свинок), домашний скот (например, жвачных животных, таких как коровы, волы и т.д.; животных семейства полорогих, таких как овцы, козы и т.д.; и животных семейства свиней, такие как свиньи и вепри). Птицы включают в себя, например, цыплят, индеек, страуса, гусей, уток и т.п. Также включены домашние животные и сельскохозяйственные животные. Термин «не относящееся к человеку животное» исключает людей.
Мышиные плюрипотентные и/или тотипотентные клетки могут быть получены из линии 129, линии C57BL/6, смеси линий 129 и C57BL/6, линии BALB/c или линии Swiss Webster. Примеры линий 129 включают в себя 129Р1, 129Р2, 129Р3, 129Х1, 129S1 (например, 129S1/SV, 12981/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1 и 129Т2. См., например, публикацию Festing et al. (1999) Mammalian Genome 10: 836), которая для всех целей полностью включена в настоящий документ путем ссылки. Примеры линий C57BL включают в себя C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal_wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Мышиные плюрипотентные и/или тотипотентные клетки также могут быть получены из смеси вышеупомянутой линии 129 и вышеупомянутого штамма C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогичным образом, мышиные плюрипотентные и/или тотипотентные клетки могут быть получены из смеси вышеупомянутых линий 129 или смеси вышеупомянутых линий BL/6 (например, линии 129S6 (129/SvEvTac)). Конкретным примером мышиной ЭС-клетки является мышиная ЭС-клетка VGF1. См., например, публикацию Auerbach et al. (2000) Biotechniques 29, 1024-1028, 1030, 1032, которая для всех целей полностью включена в настоящий документ путем ссылки.
Плюрипотентная и/или тотипотентная клетка крысы может быть получена из любой линии крыс, включая, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, такую как Fisher F344 или Fisher F6. Плюрипотентные и/или тотипотентные клетки крысы можно также получить из линии, происходящей от смеси двух или более вышеупомянутых линий. Например, плюрипотентная и/или тотипотентная клетки крысы могут быть получены от линии DA или ACI. Линия крыс ACI характеризуется наличием окраски «черный агути» с белым брюхом и лапами и гаплотипом 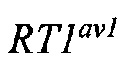 . Такие линии доступны из различных источников, включая Harlan Laboratories. Примером линии крысиных ЭС-клеток от крысы ACI является ЭС-клетка крыс ACI.G1. Линия крысы «темный агути» (DA) характеризуется наличием окраса «агути» и гаплотипа . Такие крысы доступны из различных источников, включая Charles River и Harlan Laboratories. Примерами линии крысиных ЭС-клеток от крысы линии DA является линия крысиных ЭС-клеток DA.2B и крысиных ЭС-клеток DA.2C. В некоторых случаях плюрипотентные и/или тотипотентные клетки крысы получены от инбредной линии крыс. См., например, U.S. 2014/0235933 А1, поданный 20 февраля 2014 г. и для всех целей полностью включенный в настоящий документ путем ссылки.
. Такие линии доступны из различных источников, включая Harlan Laboratories. Примером линии крысиных ЭС-клеток от крысы ACI является ЭС-клетка крыс ACI.G1. Линия крысы «темный агути» (DA) характеризуется наличием окраса «агути» и гаплотипа . Такие крысы доступны из различных источников, включая Charles River и Harlan Laboratories. Примерами линии крысиных ЭС-клеток от крысы линии DA является линия крысиных ЭС-клеток DA.2B и крысиных ЭС-клеток DA.2C. В некоторых случаях плюрипотентные и/или тотипотентные клетки крысы получены от инбредной линии крыс. См., например, U.S. 2014/0235933 А1, поданный 20 февраля 2014 г. и для всех целей полностью включенный в настоящий документ путем ссылки.
Примеры плюрипотентных клеток человека включают в себя ЭС-клетки человека, стволовые клетки взрослого человека, ограниченные в развитии клетки-предшественники человека и индуцированные плюрипотентные стволовые (ИПС) клетки человека, такие как праймированные ИПС-клетки человека и интактные ИПС-клетки человека. Индуцированные плюрипотентные стволовые клетки включают в себя плюрипотентные стволовые клетки, которые могут быть получены непосредственно из дифференцированной взрослой клетки. Человеческие ИПС-клетки можно получать посредством введения в клетку конкретных наборов репрограммирующих факторов, которые могут включать в себя, например, Oct3/4, транскрипционные факторы семейства Sox (например, Sox1, Sox2, Sox3, Sox15), транскрипционные факторы семейства Myc (например, c-Myc, l-Myc, n-Myc), транскрипционные факторы из семейства  (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glis1. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например, эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (2006) Cell 126: 663-676, для всех целей полностью включенную в настоящий документ путем ссылки. Праймированные ЭС-клетки человека и праймированные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам постимплантационных эпибластных клеток, и коммитированы для спецификации линии дифференцировки и дифференцировки. Интактные ЭС-клетки человека и интактные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам ЭС-клеток внутренней клеточной массы предимплантационного эмбриона, и не коммитированы для спецификации линии дифференцировки. См., например, публикацию Nichols and Smith (2009) Cell Stem Cell 4: 487-492, для всех целей полностью включенную в настоящий документ путем ссылки.
(KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glis1. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например, эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (2006) Cell 126: 663-676, для всех целей полностью включенную в настоящий документ путем ссылки. Праймированные ЭС-клетки человека и праймированные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам постимплантационных эпибластных клеток, и коммитированы для спецификации линии дифференцировки и дифференцировки. Интактные ЭС-клетки человека и интактные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам ЭС-клеток внутренней клеточной массы предимплантационного эмбриона, и не коммитированы для спецификации линии дифференцировки. См., например, публикацию Nichols and Smith (2009) Cell Stem Cell 4: 487-492, для всех целей полностью включенную в настоящий документ путем ссылки.
Клетки, которые были имплантированы в эмбрион-хозяин, в настоящем документе могут называться «донорскими клетками». Генетически модифицированная плюрипотентная и/или тотипотентная клетка может быть получена из той же линии, что и эмбрион-хозяин, или из другой линии. Аналогичным образом, суррогатная мать может быть получена из той же линии, что и генетически модифицированная плюрипотентная и/или тотипотентная клетка и/или эмбрион-хозяин, или суррогатная мать может быть получена из линии, отличной от генетически модифицированной плюрипотентной и/или тотипотентной клетки и/или эмбриона-хозяина.
В способах и композициях, описанных в настоящем документе, можно использовать различных эмбрионов-хозяев. Например, плюрипотентные и/или тотипотентные клетки, имеющие нацеленную генетическую модификацию, могут быть введены в эмбрион на стадии до морулы (например, эмбрион на стадии 8 клеток) из соответствующего организма. См., например, публикации US 7,576,259, US 7,659,442, US 7,294,754 и US 2008/0078000 A1, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки. В других случаях донорские ЭС-клетки можно имплантировать в эмбрион-хозяин на стадии 2 клеток, стадии 4 клеток, стадии 8 клеток, стадии 16 клеток, стадии 32 клеток или стадии 64 клеток. Эмбрион-хозяин также может являться бластоцистой или может являться эмбрионом на стадии до бластоцисты, эмбрионом на стадии предморулы, эмбрионом на стадии морулы, эмбрионом на стадии некомпактной морулы или эмбрионом на стадии компактной морулы. При использовании эмбриона мыши стадия эмбриона-хозяина может являться стадией Тейлера 1 (TS1), TS2, TS3, TS4, TS5 и TS6 со ссылкой на стадии Тейлера, описанные в публикации Theiler (1989) The House Mouse: Atlas of Mouse Development, Springer-Verlag, New York, для всех целей полностью включенной в настоящий документ путем ссылки. Например, стадия Тейлера может быть выбрана из TS1, TS2, TS3 и TS4. В некоторых случаях эмбрион-хозяин содержит внеклеточую оболочку ооцита, а донорская клетка представляет собой ЭС-клетку, которую вводят в эмбрион-хозяин через отверстие во внеклеточной оболочке ооцита. В других случаях эмбрион-хозяин представляет собой эмбрион, не содержащий внеклеточной оболочки ооцита. В других случаях стадия морулы эмбриона-хозяина является агрегированной.
Н. Способы идентификации клеток с модифицированными геномами
Некоторые из вышеуказанных способов дополнительно содержат идентификацию клетки, имеющей модифицированный геном. Для идентификации клеток, имеющих нацеленную модификацию, такую как делеция или вставка, могут использоваться различные способы. Такие способы могут включать идентификацию одной клетки, имеющей нацеленную модификацию в целевом локусе (например, между первой и второй последовательностями распознавания РНК CRISPR). Может быть проведен скрининг для идентификации таких клеток с модифицированными геномными локусами.
Этап скрининга может включать количественный тест для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный тест может быть выполнен посредством количественной ПЦР, такой как ПЦР в реальном времени (кПЦР). Во время ПЦР в реальном времени может использоваться первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров содержит флюоресцентный зонд, который распознает амплифицированную последовательность.
Этап скрининга может также включать тест на сохранение, который представляет собой тест, используемый для различения правильно нацеленных вставок полинуклеотидной вставки в целевой геномный локус от случайных трансгенных вставок полинуклеотидной вставки в геномные положения вне целевого геномного локуса. В традиционных тестах на скрининг на нацеленные модификации, таких как ПЦР длинных фрагментов или саузерн-блоттинг, вставленный нацеливающий вектор связывается с целевым локусом. Однако из-за большого размера гомологичного плеча большой нацеливающий вектор (LTVEC) не позволяет проводить скрининг с помощью таких традиционных тестов. Для скрининга нацеливания с помощью большого нацеливающего вектора (LTVEC) можно использовать тесты на модификацию аллеля (МОА), включая тесты на потерю аллеля (LOA) и приобретение аллеля (GOA) (см., например, патент США 2014/0178879 и Frendewey et al. (2010) Methods Enzymol. 476: 295-307, для всех целей полностью включенные в настоящий документ путем ссылки. Тест на потерю аллеля (LOA) инвертирует логику традиционного скрининга и позволяет количественно определять число копий нативного локуса, на который была направлена мутация. В правильно нацеленном клеточном клоне тест LOA позволяет определить один из двух нативных аллелей (для генов не на хромосоме Х или Y), а другой аллель разрывается нацеленной модификацией. Этот же принцип может применяться в обратном направлении в тесте на приобретение аллеля (GOA) для количественного определения числа копий вставленного нацеливающего вектора. Например, комбинированное использование тестов GOA и LOA позволяет выявлять, что правильно нацеленный гетерозиготный клон потерял одну копию нативного целевого гена и приобрел одну копию гена резистентности к лекарственному средству или другого вставленного маркера.
В качестве примера, количественную полимеразную цепную реакцию (кПЦР) можно использовать как способ количественного определения аллелей, но любой способ, который позволяет достоверно различать разницу между отсутствием, одной и двумя копиями целевого гена или между отсутствием, одной и двумя копиями полинуклеотидной вставки, может использоваться для разработки теста МОА. Например, для количественного определения числа копий шаблона ДНК в образце геномной ДНК можно использовать TaqMan®, в особенности при сравнении с эталонным геном (см., например, US 6,596,541, для всех целей полностью включенный в настоящий документ путем ссылки). Контрольный ген количественно определяют в той же геномной ДНК, что и целевой(-ые) ген(-ы) или локус(-ы). Поэтому выполняются две амплификации TaqMan® (каждая со своим соответствующим зондом). Один зонд TaqMan® определяет «Ct» (пороговый цикл) эталонного гена, а другой зонд определяет Ct области целевого(-ых) гена(-ов) или локуса(-ов), которая замещается с помощью успешного нацеливания (т.е. теста LOA). Ct представляет собой значение, которое отражает количество исходной ДНК для каждого из зондов TaqMan®, т.е. менее распространенная последовательность требует большего числа циклов ПЦР для достижения порогового цикла. Снижение числа копий последовательности шаблонов наполовину для реакции TaqMan® приведет к увеличению Ct приблизительно на одну единицу. Реакции TaqMan® в клетках, где один аллель целевого(-ых) гена(-ов) или локуса(-ов) замещен гомологичной рекомбинацией, приводят к увеличению одного Ct для целевой реакции TaqMan® без увеличения Ct для эталонного гена по сравнению с ДНК из нецелевых клеток. Для теста GOA можно использовать другой зонд TaqMan® для определения Ct полинуклеотидной вставки, которая замещает целевой(-ые) ген(-ы) или локус(-ы) путем успешного нацеливания.
Поскольку парные гРНК могут создавать большие Cas-опосредованные делеции в целевом геномном локусе, может быть полезно дополнить стандартные тесты LOA и GOA для проверки правильного нацеливания большим нацеливающим вектором (LTVEC) (т.е. в клетках, отличных от эмбрионов на стадии единственной клетки). Например, тесты LOA и GOA сами по себе не могут различать правильно нацеленные клеточные клоны от клонов, в которых большая Cas-индуцированная делеция целевого геномного локуса совпадает со случайной интеграцией большого нацеливающего вектора (LTVEC) в другом месте генома, в особенности если в тесте GOA используется зонд против кассеты селекции в пределах вставки LTVEC. Поскольку давление отбора в целевой клетке основано на кассете селекции, случайная трансгенная интеграция LTVEC в другом месте генома обычно включает кассету селекции и прилегающие области LTVEC, но исключает более дистальные области LTVEC. Например, если участок LTVEC случайно встраивается в геном, а LTVEC содержит полинуклеотидную вставку длиной приблизительно 5 т.п.н. или более с кассетой селекции, прилегающей к 3' гомологичному плечу, по существу 3' гомологичное плечо, а не 5' гомологичное плечо будет трансгенно интегрировано с кассетой селекции. Альтернативно, если кассета селекции прилегает к 5' гомологичному плечу, по существу 5' гомологичное плечо, а не 3' гомологичное плечо будет трансгенно интегрировано с кассетой селекции. В качестве примера, если тесты LOA и GOA используются для оценки нацеленной интеграции LTVEC, а в тесте GOA используются зонды против кассеты селекции, гетерозиготная делеция в целевом локусе гена в комбинации со случайной трансгенной интеграцией LTVEC даст такое же считывание данных, как и гетерозиготная нацеленная интеграция LTVEC в целевой геномный локус. Чтобы проверить правильное нацеливание с помощью LTVEC, тесты на сохранение можно использовать по отдельности или в сочетании с тестами LOA и/или GOA.
Тесты на сохранение определяют число копий шаблона ДНК в целевой 5'-последовательности (соответствующей 5' гомологичному плечу LTVEC) и/или целевой 3'-последовательности (соответствующей 3' гомологичному плечу LTVEC). В частности, полезно определить число копий шаблона ДНК в целевой последовательности, соответствующей гомологичному плечу, которое прилегает к кассете селекции. В диплоидных клетках число копий больше двух обычно указывает на трансгенную интеграцию LTVEC случайно за пределами целевого геномного локуса, а не в целевой геномный локус, что нежелательно. Правильно нацеленные клоны сохраняют число копий, равное двум. Кроме того, число копий менее двух в таких тестах на сохранение по существу указывает на большие Cas-опосредованные делеции, выходящие за пределы области, нацеливаемой для делеции, что также нежелательно.
В примере теста на сохранение для идентификации нацеленной вставки полинуклеотидной вставки в целевом геномном локусе диплоидной клетки ДНК сначала получают из клетки, которую приводят в контакт с большим нацеливающим вектором (LTVEC), содержащим полинуклеотидную вставку, фланкированную с помощью первого гомологичного плеча, которое гибридизуется с первой целевой последовательностью, и второго гомологичного плеча, которое гибридизуется со второй целевой последовательностью, причем полинуклеотидная вставка содержит кассету селекции, прилегающую к первому гомологичному плечу. Необязательно кассета селекции может содержать ген резистентности к лекарственному средству. Затем на ДНК воздействуют зондом, который связывается в пределах первой целевой последовательности, зондом, который связывается в пределах полинуклеотидной вставки, и зондом, который связывается в пределах эталонного гена, имеющего известное число копий, причем каждый зонд генерирует обнаруживаемый сигнал после связывания. Затем обнаруживают сигналы от связывания каждого из зондов. Сигнал от зонда эталонного гена сравнивают с сигналом от зонда первой целевой последовательности для определения числа копий первой целевой последовательности, и сигнал от зонда эталонного гена сравнивают с сигналом от зонда полинуклеотидной вставки для определения числа копий полинуклеотидной вставки. Число копий полинуклеотидной вставки, равное одному или двум, и число копий первой целевой последовательности, равное двум, как правило, указывает на целевую вставку полинуклеотидной вставки в целевом геномном локусе, а число копий полинуклеотидной вставки, равное одному или более, и число копий первой целевой последовательности, равное трем или более, по существу указывает на случайную вставку полинуклеотидной вставки в геномном локусе, отличном от целевого геномного локуса.
Сигнал от связывания зонда первой целевой последовательности можно использовать для определения значения порогового цикла (Ct) для первой целевой последовательности, сигнал от связывания зонда эталонного гена можно использовать для определения значения порогового цикла (Ct) для эталонного гена, и число копий первой целевой последовательности можно определить посредством сравнения значения Ct первой целевой последовательности и значения Ct эталонного гена. Аналогичным образом, сигнал от связывания зонда полинуклеотидной вставки можно использовать для определения значения порогового цикла (Ct) для полинуклеотидной вставки, а число копий полинуклеотидной вставки можно определить посредством сравнения значения Ct первой целевой последовательности и значения Ct эталонного гена.
Полинуклеотидная вставка в LTVEC может составлять, например, по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более. Расстояние между последовательностями, с которыми связываются зонды в первой целевой последовательности и кассете селекции, может составлять, например, не более 100 нуклеотидов, 200 нуклеотидов, 300 нуклеотидов, 400 нуклеотидов, 500 нуклеотидов, 600 нуклеотидов, 700 нуклеотидов, 800 нуклеотидов, 900 нуклеотидов, 1 т.п.н., 1,5 т.п.н., 2 т.п.н., 2,5 т.п.н., 3 т.п.н., 3,5 т.п.н., 4 т.п.н., 4,5 т.п.н. или 5 т.п.н.
Такие способы могут дополнительно включать в себя дополнительные тесты на сохранение для определения числа копий второй целевой последовательности. Например, такие способы могут дополнительно включать в себя воздействие на ДНК клетки зондом, который связывается со второй целевой последовательностью, обнаруживая сигнал от связывания зонда второй целевой последовательности и сравнивая сигнал от зонда эталонного гена с сигналом от второй целевой последовательности, чтобы определить число копий для второй целевой последовательности.
Аналогичным образом, такие способы могут дополнительно включать в себя дополнительные тесты GOA для определения числа копий одной или более дополнительных последовательностей в пределах полинуклеотидной вставки. Например, такие способы могут дополнительно включать в себя воздействие на ДНК клетки одним или более дополнительными зондами, которые связываются с полинуклеотидной вставкой, обнаружение сигнала от связывания одного или более дополнительных зондов и сравнение сигнала от зонда эталонного гена с сигналом от одного или более дополнительных зондов для полинуклеотидной вставки для определения числа копий для одной или более дополнительных последовательностей в пределах полинуклеотидной вставки.
Аналогичным образом, когда LTVEC выполнен с возможностью удаления эндогенной последовательности из целевого геномного локуса или когда используются парные гРНК (например, для создания спаренных двухцепочечных разрывов в разных местах в пределах одного целевого геномного локуса и удаления промежуточной эндогенной последовательности), такие способы могут дополнительно включать в себя тест LOA для определения числа копий эндогенных последовательностей в целевом геномном локусе. Например, такие способы могут дополнительно включать в себя воздействие на ДНК клетки зондом, который связывается с эндогенной последовательностью в целевом геномном локусе, обнаружение сигнала от связывания зонда эндогенной последовательности и сравнение сигнала от зонда эталонного гена с сигналом от зонда эндогенной последовательности для обнаружения числа копий эндогенной последовательности.
Другие примеры подходящих количественных тестов включают в себя флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(-и) зондом(-ами), Invader Probes®, MMP assays®, молекулярный маяк TaqMan® или Eclipse™ (см., например, US 2005/0144655, для всех целей полностью включенный в настоящий документ путем ссылки).
Для нацеленных генетических модификаций, получаемых без использования LTVEC, можно использовать традиционные тесты для скрининга на нацеленные модификации, такие как ПЦР длинных фрагментов, саузерн-блоттинг или секвенирование по Сенгеру. Такие тесты обычно используют для получения доказательства связи между вставленным нацеливающим вектором и целевым геномным локусом. Например, при ПЦР длинных фрагментов один праймер может распознавать последовательность в пределах вставленной ДНК, а другой распознает последовательность целевого локуса за пределами концов гомологичных плеч нацеливающего вектора.
Все патентные заявки, веб-сайты, другие публикации, номера доступа и т.п., упомянутые выше или ниже, для любых целей полностью включены в настоящий документ путем ссылки в равной мере, как если бы было указано, что каждая индивидуальная позиция отдельно и в индивидуальном порядке включена таким образом путем ссылки на нее. Если в различные моменты времени с номером доступа связаны различные версии последовательности, подразумевается версия, связанная с номером доступа на действительную дату подачи настоящей заявки. Под действительной датой подачи подразумевается наступающая ранее из фактической даты подачи или даты подачи приоритетной заявки, где приведена ссылка на номер доступа (если применимо). Аналогичным образом, если в различные моменты времени издаются различные редакции публикации, веб-сайта и т.п., то, если не указано иное, подразумевается наиболее поздняя изданная версия на действительную дату подачи заявки. Если специально не указано иное, любое свойство, стадия, элемент, вариант осуществления или аспект настоящего изобретения может использоваться в комбинации с любым другим из перечисленного. Хотя настоящее изобретение было подробно описано посредством иллюстрации и примера в целях четкости и лучшего понимания, будет очевидно, что некоторые изменения и модификации могут быть реализованы в рамках объема приложенной формулы изобретения.
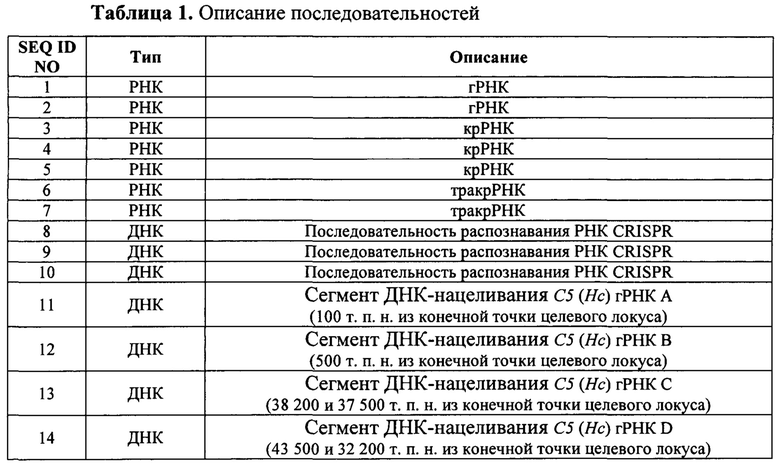
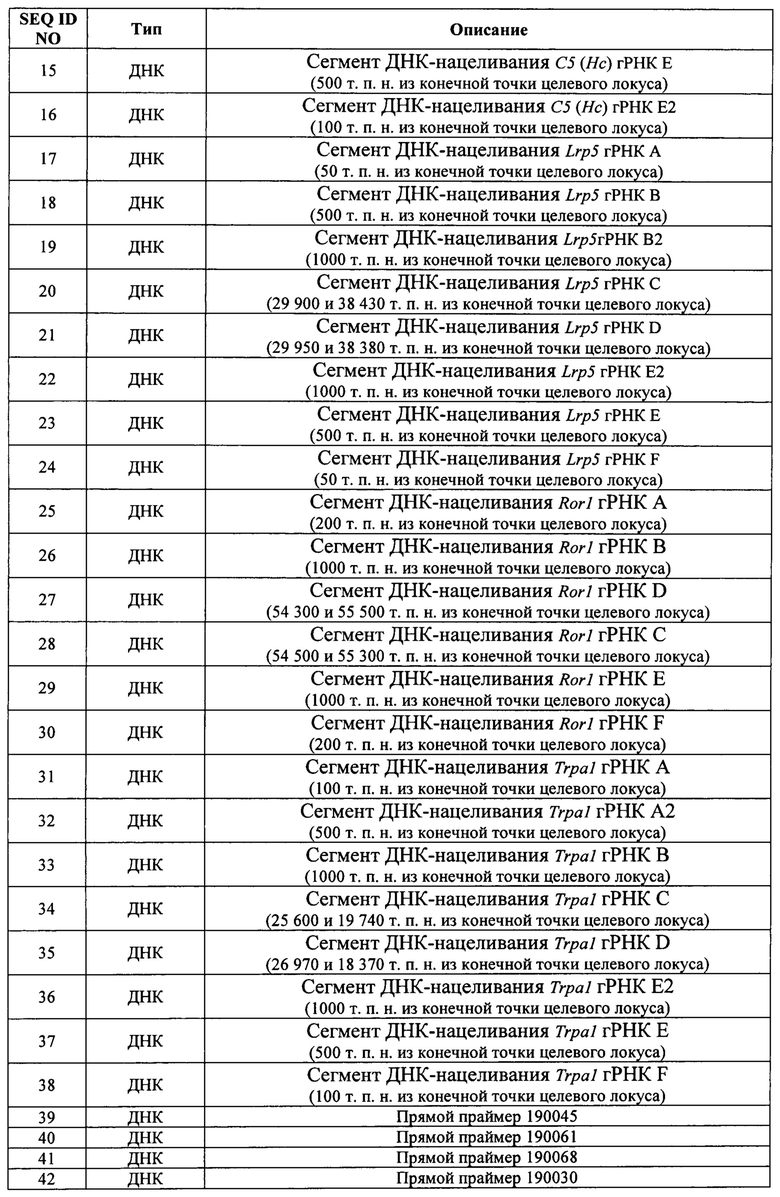
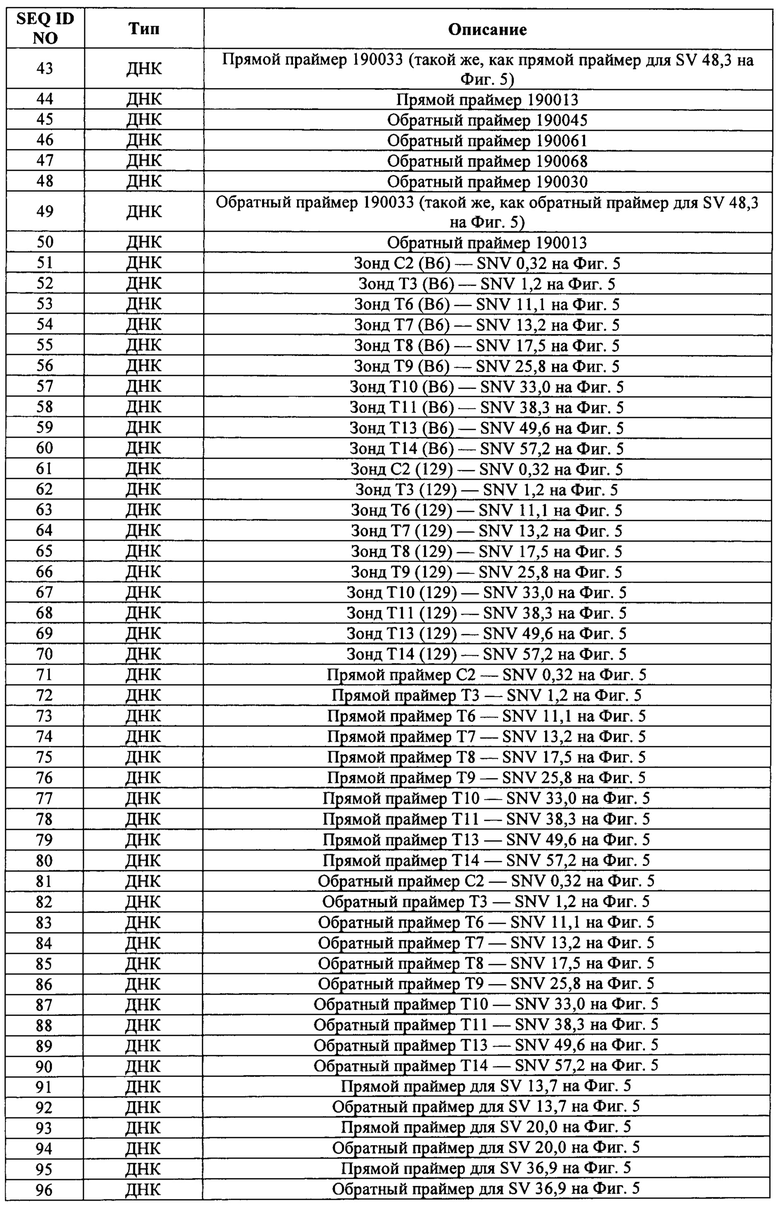
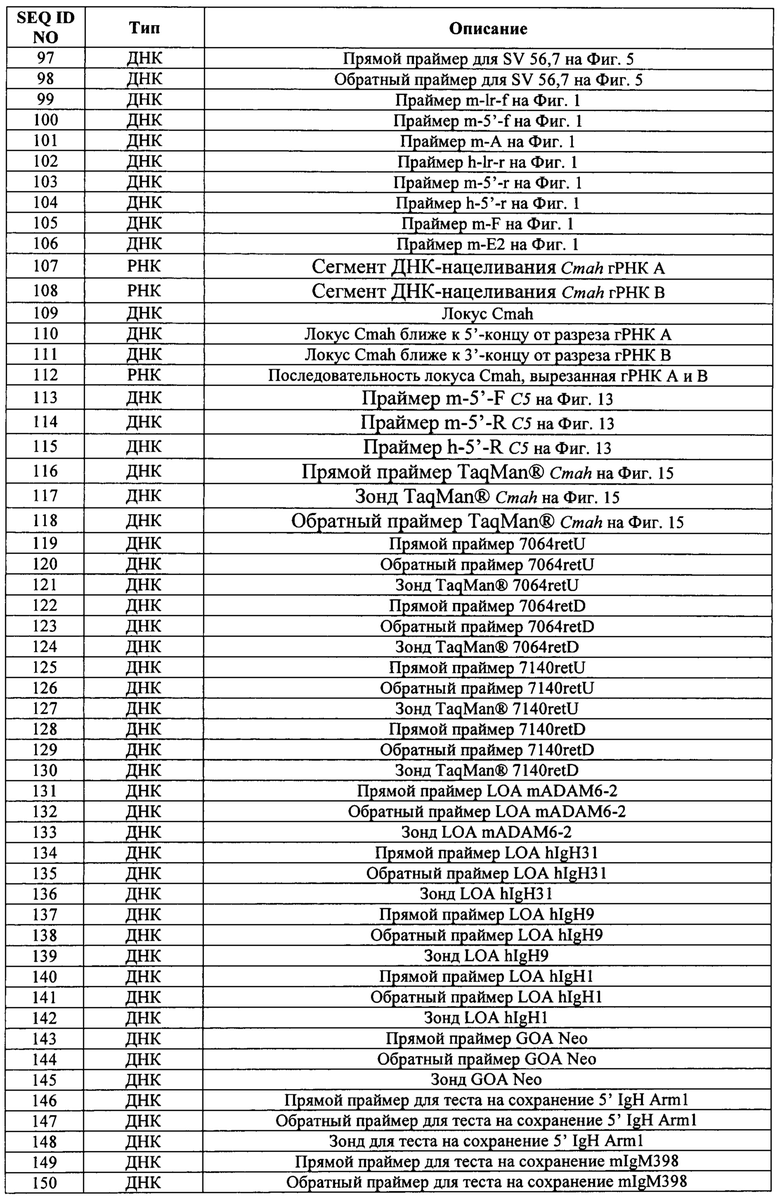
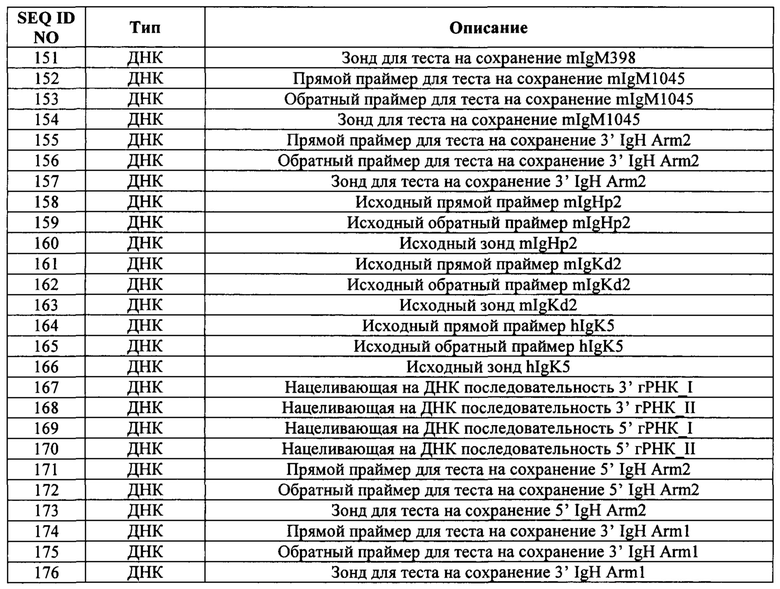
ПРИМЕРЫ
Пример 1. CRISPR/Cas9-опосредованное нацеливание с использованием одной гидовой РНК или двух гидовых РНК
Материалы и способы
Культура ЭС-клеток, скрининг и электропорация
Эксперименты, описанные в настоящем документе, были выполнены с VGF1, нашей XY-гибридной клеточной линией ЭС-клеток C57BL6NTac/129S6SvEvF1 (Poueymirou et al. (2007) Nat. Biotechnol. 25: 91-99; Valenzuela et al. (2003) Nat. Biotechnol. 21: 652-659). ЭС-клетки культивировали так, как описано ранее (Matise et al. (2000) в публикации Joyner, A.L. ed. Gene Targeting: a practical approach, pp. 100-132, Oxford University Press, New York).
Электропорацию (ЕР) проводили с использованием 7,5 млн клеток в кювете с щелью 2 мм в конечном объеме 0,12 мл. Электрические условия ЕР составляли 700 В, сопротивление 400 Ом и емкость 25 мкФ с использованием системы электропорации ВТХ ЕСМ 630 (Harvard Apparatus, г. Холлистон, штат Массачусетс, США). Количество LTVEC на ЕР составляло 0,0015 мг, экспрессирующей Cas9 плазмиды - 0,005 мг, а экспрессирующей огРНК-плазмиды - 0,010 мг. Некоторые процедуры ЕР проводили с добавлением 100 нг плазмиды, придающей резистентность к пуромицину, чтобы обеспечить селекцию клонов без селекции на резистентность к неомицину, экспрессируемую LTVEC. После ЕР клетки помещали на две желатинизированные чашки диаметром 15 сантиметров и ежедневно меняли среду. Отбор сред, содержащих или 100 мкг/мл сульфата G-418, или пуромицин 0,0015 мг/мл, начинали через 48 часов после ЕР и продолжали в течение до 10 дней после ЕР. Колонии собирали в фосфатно-солевом буфере (PBS), добавляли в 96-луночный планшет, содержащий 0,05% трипсина, и оставляли для диссоциации на 15 минут, нейтрализовали средой и использовали для выделения ДНК для скрининга.
Способ модификации аллеля (Frendewey et al. (2010) Methods Enzymol. 476: 295-307) использовали для идентификации правильно нацеленных ЭС-клеточных клонов и определения генотипов аллелей мыши.
Разработка гидовых последовательностей
Приблизительно 200 т.п.н. ДНК, окружающих положение 50 т.п.н., 100 т.п.н., 500 т.п.н. или 1 т.п.н. внутри удаленного участка Lrp5 или других генов, на которые было выполнено нацеливание, как ближе к 5'-концу, так и ближе к 3'-концу, вводили в инструмент разработки CRISPR (crispr.mit.edu) для получения возможных последовательностей гРНК. После этого потенциальные последовательности гРНК фильтровали, чтобы гарантировать, что они позволяют разрезать только эндогенную ДНК, а не гуманизирующую вставку в LTVEC.
Клонирование одногидовой РНК
ОгРНК или клонировали в виде дуплексных олигонуклеотидов (IDT) в рМВ_огРНК (промотор U6) в сайтах BsmbI, слитых с каркасом 77 п.н. для экспрессии бесшовной РНК, или приобретали в виде плазмид с подтвержденной экспрессией в компании GeneCopoeia (гиды LRP5 А, В, В2, Е2, Е и F). Полученные нами плазмиды были подтверждены посредством ПЦР и секвенирования по Сенгеру.
Шаблон ДНК для подтверждения генотипа
ДНК очищали из ЭС-клетки, клоны получали из ЭС-клеток, которые были электропорированы с использованием нацеливающего вектора и плазмиды, экспрессирующей Cas9, и плазмиды, экспрессирующей одну из нескольких гидовых РНК (гРНК), или двух плазмид, экспрессирующих различные комбинации гРНК. Для последующих стандартных ПЦР-тестов были отобраны клоны, идентифицированные с помощью количественных ПЦР-анализов на модификацию аллеля (т.е. потерю аллеля или приобретение аллеля) как имеющие целевую делецию целевого локуса мыши и вставку нацеливающего вектора или имеющие делеции, индуцированные Cas9/гРНК.
Создание олигонуклеотидов
Для каждой комбинации гРНК было разработано по два ПЦР-теста. Первая ПЦР представляла собой тест на делецию для обнаружения коллапсирования между последовательностями распознавания РНК CRISPR различных комбинаций гРНК. Второй ПЦР-тест, который представляет собой тест на 5', включал два ПЦР-теста. Первым был тест на 5' человека на гуманизированные аллели, проводимый через сайт соединения мыши и человека. Вторым был тест на 5' мыши на эндогенные аллели мыши, проводимый через сайт соединения нацеленной 5'-делеции.
Реакция ПЦР и планирование ТОРО
Для амплификации шаблона ДНК ЭС-клеток использовали TaKaRa LA Taq ДНК-полимеразу (кат. № RR002M). Каждую реакционную смесь для теста ПЦР выполняли с отрицательным контролем водой. Контрольные смеси содержали следующее: шаблон ДНК ЭС-клеток - 0,005 мл; буферный раствор 1X LA PCR II (Mg2+plus); 0,01 мМ смесь dNTP; 0,0075 мМ Forward Oligo (каждый); 0,0075 мМ Reverse Oligo (каждый); 5000 ЕД/мл полимеразы LA Taq и дважды дистиллированную воду до 0,025 мл.
Программа термоциклирования ПЦР включала 94°C в течение одной минуты; затем 35 циклов при 94°C в течение 30 секунд, градиент отжига при 60°C в течение 30 секунд и 68°C в течение одной минуты на 1 амплифицируемую т.п.н. с последующей полимеризацией при 72°C в течение 10 минут.
ПЦР-продукты фракционировали электрофорезом на 2%-м агарозном геле с помощью маркера длины ДНК Invitrogen 1 kb plus DNA ladder (кат. №10787-018) и/или маркера длины ДНК Invitrogen 50 bp DNA Ladder (кат. №10416-014). Оставшиеся ПЦР-продукты клонировали в pCR4-TOPO Vector, следуя инструкциям из набора клонирования ТОРО ТА от Invitrogen (кат. № K4575-02) для секвенирования. Реакции клонирования химически трансформировали в клетки One Shot Top 10 и высевали на чашки с агаром, содержащим 0,06 мг/мл X-gal и 0,025 мг/мл канамицина.
Секвенирование
Белые колонии инокулировали в LB, содержащем 0,025 мг/мл канамицина, и инкубировали в течение ночи при встряхивании при 37°C. Каждая колония представляла один ампликон из популяции тестируемых продуктов. ДНК экстрагировали из каждой бактериальной культуры с использованием мини-препаративного набора для выделения плазмид QIAGEN (кат. №12123). ДНК-последовательность вставок определяли в реакционной смеси для секвенирования, которая включала 0,002 мл клонированной ТОРО ПЦР, 1 × раствор энхансера для ПЦР (кол-во 10 шт.) (кат. X11495-017), 0,0075 мМ олиго (M13F или M13R) и дважды дистиллированной Н2О до 0,015 мл.
Секвенирование
В результатах секвенирования обрезали неопределенную последовательность и векторную последовательность pCR4-TOPO, выделяя последовательность вставки, полученной ПЦР. Затем фрагменты последовательности выравнивали по эталону и анализировали вариации.
Секвенирование коллапсированных клонов
ПЦР-продукты из коллапсированных положительных клонов клонировали в вектор pCR4-TOPO по инструкциям производителя (Invitrogen, кат. № K4575-02), затем химически трансформировали в клетки One Shot Top 10 и высевали на чашки с агаром, содержащим 0,060 мг/мл X-gal и 0,025 мг/мл канамицина. ДНК экстрагировали из бактериальных культур с использованием мини-препаративного набора для выделения плазмид QIAGEN (кат. №12123). После этого результаты секвенирования вставки выравнивали по эталону прогнозируемого соединения и анализировали вариации вставочно-делеционной мутации. Прогнозировалось, что Cas9 будет расщеплять 3 пары оснований из РАМ в последовательность, распознаваемую гРНК. Последовательность в пределах прогнозируемого расщепления удаляли из эталона, а оставшуюся часть использовали для выравнивания с результатами.
Аллельные дискриминантные тесты TaqMan® для однонуклеотидных вариантов (SNV)
Объем аллельной дискриминантной реакции TaqMan® составлял 0,008 мл, содержащий геномную ДНК, специфические зонды/праймеры для каждого полиморфизма и смесь TaqMan® Gene Expression PCR Master Mix. Зонды были приобретены в компании Life Technologies (Thermo), а праймеры - в компании IDT. Зонд для аллеля 129 был помечен красителем VIC; зонд для аллеля В6 был помечен красителем FAM. Каждый аллельный тест TaqMan® проводили в четырех повторностях на 384-луночном планшете и анализировали на платформе Applied BioSystems ViiA 7. Программа циклирования ПЦР SNV была следующей: 95°C в течение 10 минут со следующими 40 циклами: 95°C в течение 15 секунд, 60°C в течение 60 секунд и 60°C в течение 30 секунд. Анализ цикла и оценку результатов проводили с использованием программного обеспечения ViiA 7 версии 1.1.
Анализ FISH
Отобранные ЭС-клеточные клоны анализировали в Cell Line Genetics (г. Мэдисон, штат Висконсин, США) или Van Andel Institute (г. Гранд-Рапидс, штат Мичиган, США) с использованием флуоресцентной гибридизации in situ (FISH) по их стандартным процедурам. В качестве зондов для 2-цветного анализа мы использовали ВАС мыши и человека.
Усиленное коллапсирование генома и/или гуманизация целевых локусов
Для осуществления точной одноступенчатой делеции всего или части гена грызуна и необязательно одновременной замены всего или части его человеческого гомолога мы вводили путем электропорации в ЭС-клетки грызунов следующие молекулы нуклеиновых кислот: (1) LTVEC; (2) плазмиду или мРНК, кодирующую эндонуклеазу Cas9; и (3) одну или более плазмид, кодирующих одну или более одногидовых РНК (гРНК) CRISPR или сами гРНК. В каждом эксперименте LTVEC подвергали линеаризации. В некоторых экспериментах LTVEC содержал весь ген человека или часть гена человека, который кодирует генный продукт (белок или РНК), фланкированный с помощью гомологичного плеча ДНК грызуна, предназначенного для направления события гомологичной рекомбинации, которое удаляет ген грызуна и вставляет ген человека. В других экспериментах LTVEC был выполнен с возможностью нацеливания на отдельный локус, такой как локус Ch25h. В любом случае LTVEC также содержал кассету селекции лекарственными средствами, которая направляет экспрессию фермента (например, неомицинфосфотрансферазы), придающую резистентность к антибактериальному лекарственному средству (например, G418).
ЭС-клетки, захватившие LTVEC и внедрившие его в свои геномы, были способны расти и образовывать колонии на чашках Петри в питательной среде, содержащей антибактериальное лекарственное средство. Поскольку мы внедрили в 500-1000 раз больше молекул нуклеиновых кислот, кодирующих CRISPR/Cas9 и гРНК, чем молекул LTVEC, большинство LTVEC-содержащих резистентных к лекарственному средству колоний также по меньшей мере временно содержали компоненты CRISPR/Cas9. Мы собрали резистентные к лекарственному средству колонии и провели скрининг по способу модификации аллеля (Valenzuela et al. (2003) Nat. Biotech. 21: 652-660; Frendewey et al. (2010) Methods Enzymol. 476: 295-307; полностью включены в настоящий документ путем ссылки) с целью идентификации клонов, имеющих гуманизированный аллель с правильным нацеливанием. Кроме того, для проверки правильного нацеливания LTVEC на геном мыши использовали ПЦР-тесты в реальном времени, распознающие последовательности в гомологичных плечах LTVEC, называемые тестами на сохранение. Определение числа копий в этих тестах на сохранение позволяло получать дополнительную информацию, чтобы помочь отличить правильно направленные ЭС-клоны, которые сохранили число копий, равное двум, от клонов, в которых большая Cas9-индуцированная делеция целевого локуса мыши совпадает со случайной интеграцией LTVEC в другом месте в геноме, и в этом случае тесты на сохранение имели число копий, равное трем (или более). Способность парных гРНК создавать крупные Cas9-опосредованные делеции в целевом локусе мыши означала, что стандартные тесты LOA и GOA, как описано ранее, могут быть дополнены тестом на сохранение для обеспечения дополнительного уточнения и проверки правильного нацеливания. Поэтому тесты на сохранение были разработаны и использованы в сочетании с тестами LOA и GOA.
В каждом эксперименте использовали или одну, или две гРНК. гРНК использовали однонаправленное расщепление Cas9 вблизи 5'-конца целевого локуса (т.е. делецию целевого гена мыши), середины целевого локуса или 3'-конца целевого локуса. При использовании двух гРНК одна гРНК направляла расщепление Cas9 около 5'-конца целевого локуса, а другая гРНК направляла расщепление Cas9 в середине целевого локуса или вблизи 3'-конца целевого локуса.
Локус Lrp5
В одной серии экспериментов LTVEC был выполнен с возможностью создания делеции 68 т.п.н. участка гена Lrp5 мыши (гена белка, связанного с рецептором липопротеинов низкой плотности, 5), кодирующего эктодомен, и одновременную замену фрагментом 91 т.п.н. гомологичной последовательности гена LRP5 человека (Фиг. 1). LTVEC содержал фрагмент 91 т.п.н. гена. LRP5 человека, фланкированного гомологичными плечами, содержащими 7 т.п.н. и 33 т.п.н. геномной ДНК, полученной из частей локуса Lrp5 мыши, фланкирующих последовательность 68 т.п.н. гена Lrp5 мыши, предназначенного для делеции. В отдельных экспериментах LTVEC, гуманизирующий Lrp5, комбинировали с плазмидой, кодирующей Cas9, и второй плазмидой, кодирующей одну из восьми гРНК (А, В, В2, С, D, E2, Е, F), предназначенных для создания двухцепочечных разрывов в пределах области гена Lrp5 мыши, на который была нацелена делеция. гРНК разработаны так, чтобы избежать распознавания любой последовательности во вставленном участке гена LRP5 человека. В других экспериментах мы комбинировали LTVEC и кодирующую Cas9 плазмиду с плазмидами, кодирующими две разных гРНК, которые были нацелены на различные сайты в пределах области гена Lrp5 мыши, на который была нацелена делеция.
Проводили скрининг резистентных к лекарственному средству клонов ЭС-клеток на нацеленные гуманизации посредством тестов на модификацию аллеля (Valenzuela et al. (2003) Nat. Biotechnol. 21: 652-659; Frendewey et al. (2010) Methods Enzymol. 476: 295-307) для выявления последовательностей внутри делеций, последовательностей внутри кассеты селекции лекарственными средствами и вставок человеческого гена. Клоны считали получившими правильное нацеливание, если они теряли одну из двух эндогенных мышиных генных последовательностей, приобретали одну копию человеческой вставки, а также сохраняли две копии сохраняемых последовательностей (расположенных в гомологичном плече LTVEC). Два теста на сохранение, использованные в этом скрининге, представляли собой тесты TaqMan® с применением следующих праймеров и зондов: прямой праймер 7064retU,  ; обратный праймер 7064retU,
; обратный праймер 7064retU, 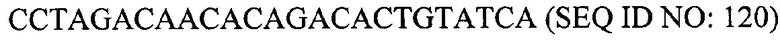 ; зонд 7064retU TaqMan®,
; зонд 7064retU TaqMan®, 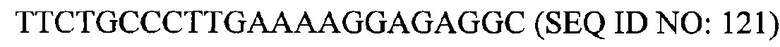 ; прямой праймер 7064retD,
; прямой праймер 7064retD, 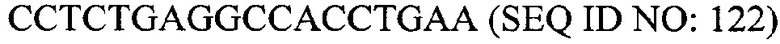 ; обратный праймер 7064retD,
; обратный праймер 7064retD, 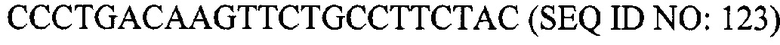 ; зонд 7064retD TaqMan®,
; зонд 7064retD TaqMan®, 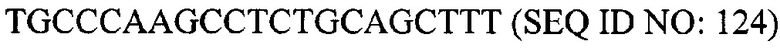 .
.
Результаты выполняемой посредством CRISPR/Cas9 гуманизации гена Lrp5 представлены в таблице 2. Когда в ЭС-клетки вводили только LTVEC, 1,9% прошедших скрининг резистентных к лекарственному средству клонов несли правильно нацеленный гетерозиготный гуманизированный аллель (см. столбец «Гетер. нацел.» в таблице 2, куда входят клоны, в которых ненацеленный аллель совсем не имел мутации или имел небольшую CRISPR-индуцированную мутацию, например, небольшую делецию, вызванную NHEJ). В отличие от этого комбинация LTVEC с эндонуклеазами Cas9, направляемыми семью из восьми протестированных гРНК (А, В, В2, С, D, Е2, Е и F; см. таблицу 1), давала правильно нацеленные моноаллельные гетерозиготные мутации со значениями эффективности в диапазоне от 2,1 до 7,8%. В случае направляемого Cas9 расщепления В2 и D в дополнение к моноаллельному нацеливанию обнаруживалась биаллельная гомозиготная гуманизация с частотой 1,0-2,1%. Нам ни разу не удалось обнаружить биаллельное нацеливание при использовании одного только LTVEC даже в небольших аллелях с простой делецией. Гомозиготные по гуманизированному Lrp5 ЭС-клетки можно преобразовать по способу VELOCIMOUSE® (публикация Poueymirou et al. (2007) Nat. Biotech. 25: 91-99, полностью включенная в настоящий документ путем ссылки) непосредственно в мышей, полностью полученных из ЭС-клеток и готовых к фенотипическим исследованиям и исследованиям эффективности лекарственных средств.
Тесты МОА, разработанные для обнаружения гРНК/Cas9-индуцированных мутаций NHEJ в прогнозируемых сайтах расщепления или поблизости от них, продемонстрировали мутационную активность у всех протестированных гРНК (данные не показаны). Доля моноаллельных или биаллельных гРНК-индуцированных мутаций, обнаруженных среди всех протестированных клонов, варьировала в зависимости от локуса и местоположения. Не наблюдалось четкой корреляции между мутационной активностью гРНК и нацеливанием LTVEC, но самые низкие значения эффективности нацеливания часто были связаны с гРНК, которые имели самые низкие значения частоты мутаций.
Комбинирование двух гРНК, распознающих разные концы нацеленной для делеции области гена Lrp5, повышало общую эффективность нацеленной гуманизации преимущественно посредством повышения частоты событий гомозиготного нацеливания в трех из пяти протестированных комбинаций (таблица 2). Поскольку комбинация гРНК имеет потенциал к созданию больших делеций между сайтами расщепления Cas9, запрограммированными гРНК, мы также наблюдали гемизиготные клоны ЭС-клеток, которые несли нацеленную гуманизацию на одном аллеле Lrp5 и большую CRISPR-индуцированную делецию на другом аллеле (комбинация гРНК A+F, таблица 2). Кроме того, при двух комбинациях гРНК (A+F и А+Е2) мы выявили клоны ЭС-клеток с уникальным генотипом: большие CRISPR-опосредованные делеции на обоих аллелях Lrp5.
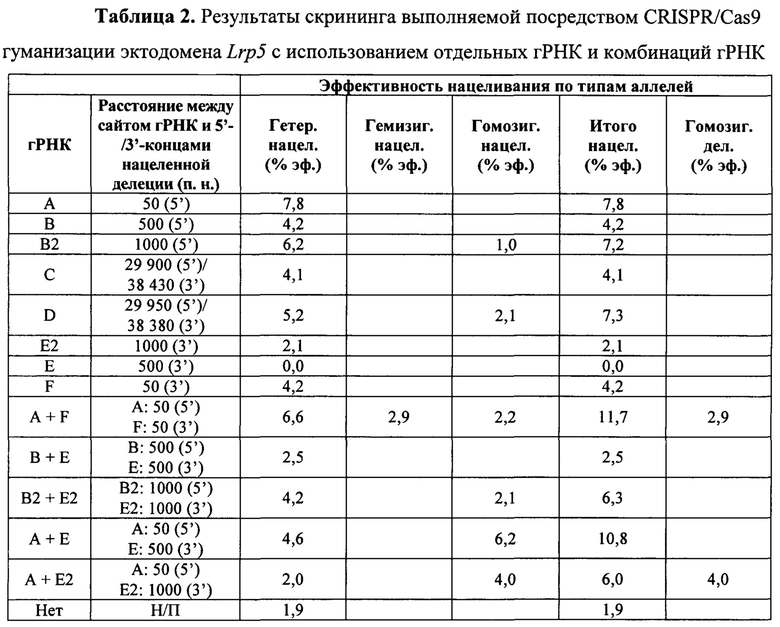
Как показано в таблице 2, существенное увеличение процентной доли клонов, имевших биаллельное нацеливание, наблюдалось при использовании двух гРНК, нацеленных на один локус, по сравнению с одной гРНК (см. Фиг. 2А), и это указывает на то, что использование комбинаций гРНК способствует биаллельным модификациям. На Фиг. 2А представлена общая схема одновременной делеции мышиного гена и замены на соответствующую человеческую версию с использованием LTVEC и двух гидовых РНК (А и В). К уникальным мутантным типам аллелей, которые наблюдались с гораздо более высокой частотой при использовании двух гРНК, относятся аллели с гомозиготным коллапсированием (Фиг. 2B; Δ/Δ), аллели с гомозиготным нацеливанием (Фиг. 2C; Hum/Hum), аллели с гемизиготным нацеливанием (Фиг. 2D; (Hum/Δ)), а также другие аллели с компаунд-гетерозиготным нацеливанием (например, один аллель имеет гуманизацию, нацеленную с помощью LTVEC, а другой аллель имеет CRISPR-индуцированную мутацию, например небольшую делецию) (Фиг. 2Е).
Было проведено несколько ПЦР-тестов для подтверждения и проверки генотипов по тестам МОА. Праймеры показаны на Фиг. 1, и их можно найти в таблице 1. Вектор Lrp5 LTVEC имел 5' гомологичное плечо, которое достаточно короткое (6,9 т.п.н.), чтобы проверить нацеливание посредством ПЦР-теста на физическое соединение между человеческой вставкой и прилегающей мышиной геномной последовательностью (Фиг. 1). Мы наблюдали ожидаемый ПЦР-продукт размером 7,5 т.п.н. при использовании ДНК из клонов, считающихся гетерозиготными, гемизиготными или гомозиготными, но не ДНК из родительской линии ЭС-клеток или из клонов, которые считаются имеющими большие биаллельные делеции (Фиг. 3А), что, таким образом, доказывает нацеливание посредством скрининга МОА (т.е. LOA и GOA) и подтверждает предполагаемые большие биаллельные делеции. ПЦР-тест 5'-Del-J, в котором исследовали последовательности на стыках делеций и вставок (Фиг. 3B), позволил получить продукт из 330 п.н. при использовании ДНК из родительской линии ЭС-клеток и из большинства гетерозиготных гуманизированных клонов (данные не показаны). В случае гетерозиготного клона AW-C3 тест 5'-Del-J давал продукт с размером меньше ожидаемого (Фиг. 3B), и это указывает на то, что расщепление гРНК A/Cas9 индуцировало небольшую делеционную мутацию на не подвергшемся нацеливанию аллеле, которая также была обнаружена в тесте МОА с расщеплением, направляемым гРНК А (данные не показаны). Как ожидалось, тест 5'-Del-J был отрицательным в случае клонов с гемизиготными, гомозиготными и биаллельными делеционными аллелями. ПЦР-тест 5'-Ins-J (Фиг. 3B), в котором исследовали последовательности на стыке между 5'-концом вставки человеческой ДНК и прилегающей мышиной фланкирующей последовательностью, позволил получить продукт из 478 п.н. в гетерозиготных, гемизиготных и гомозиготных клонах, поскольку они имели по меньшей мере один аллель с нацеленной гуманизацией. ПЦР-тест 5'-Ins-J не позволял получать продукты в случае клонов с большими биаллельными делециями (Фиг. 3B). Для подтверждения больших делеций в гемизиготных и биаллельных делеционных клонах мы выполняли ПЦР с праймерами, распознающими последовательности за пределами целевых сайтов двух гРНК. ПЦР Del(A+F), в котором анализировали делецию между сайтами А и F гРНК (Фиг. 1), позволил получить один продукт из приблизительно 360 п.н. при использовании ДНК из клонов AW-A8 и BO-F10 (Фиг. 3B), и это подтверждает, что по меньшей мере один из аллелей Lrp5 имел большую делецию. Аналогичным образом, ПЦР Del(A+E2), во время которого тестировали на большую делецию между сайтами А и Е2 гРНК, позволил получить один продукт из приблизительно 250 п.н. при использовании ДНК из клона ВА-А7. ПЦР делеций в сочетании с тестами на стыки, LOA и GOA подтверждают генотип с большой биаллельной делецией. Результаты теста, представленные на Фиг. 3A и 3B, являются характерными примерами аналогичных тестов, которые мы проводили в дополнение к флуоресцентной гибридизации in situ (FISH; Фиг. 4А-С) для подтверждения биаллельных генотипов, представленных в таблице 2.
Флуоресцентную гибридизацию in situ (FISH) использовали для подтверждения гомозиготной нацеленной гуманизации гена Lrp5. Клоны ЭС-клеток, которые по данным количественного и традиционного ПЦР-тестов были сочтены прошедшими гомозиготное нацеливание в результате экспериментов по нацеливанию, в которых LTVEC для гуманизации Lrp5 (Фиг. 1) комбинировали с Cas9 и двумя гРНК (А+F или А+Е2), направили в коммерческую цитологическую службу для проведения FISH и анализа кариотипа. Искусственную бактериальную хромосому (ВАС), несущую мышиный ген Lrp5, метили красным флуоресцентным маркером и использовали в качестве зонда для идентификации эндогенных локусов Lrp5, а ВАС, несущую человеческий ген LRP5, метили зеленым флуоресцентным красителем и использовали в качестве зонда для идентификации хроматид с проведенным нацеливанием человеческой вставкой. Меченные ВАС-зонды гибридизовали с метафазными пластинками из клонов с проведенным нацеливанием и визуализировали посредством флуоресцентной микроскопии. Хромосомы в метафазных пластинках визуализировали посредством окрашивания DAPI (4',6-диамидино-2-фенилиндол), а отдельные кариотипы в каждом клоне определяли посредством окрашивания по Гимза. Типичный результат представлен на Фиг. 4А для клона AW-D9, который оказался имеющим нормальный кариотип 40XY (не показан). На составной фотографии на Фиг. 4А показано, что как сигнал красного мышиного ВАС-зонда, так и сигнал зеленого человеческого ВАС-зонда колокализовались в цитологической полосе В на обеих копиях мышиной хромосомы 19, т.е. в известном местоположении гена Lrp5. Составная фотография на Фиг. 4С демонстрирует такое же гомозиготное нацеливание в случае другого клона (BA-D5). Эти результаты подтверждают, что фрагмент из 91 т.п.н. человеческого гена LRP5 в LTVEC для гуманизации (Фиг. 1) был успешно вставлен в намеченный локус мышиного Lrp5 на обоих гомологах хромосомы 19 в клонах AW-D9 и BA-D5. В отличие от этого, составная фотография на Фиг. 4B демонстрирует, что как сигнал красного мышиного ВАС-зонда, так и сигнал зеленого человеческого ВАС-зонда колокализовались в цитологической полосе В на одной копии мышиной хромосомы 19 (сплошная стрелка), а в цитологической полосе В на другой копии мышиной хромосомы 19 локализовался только сигнал красного мышиного ВАС-зонда. Эти результаты подтверждают, что фрагмент из 91 т.п.н. человеческого гена LRP5 в LTVEC для гуманизации (Фиг. 1) был успешно вставлен в намеченный локус мышиного Lrp5 только на одной копии хромосомы 19 (гетерозиготное нацеливание). Также они показывают (вместе с другими не показанными контролями), что человеческий ВАС-зонд не гибридизуется перекрестно с мышиным локусом Lrp5, а распознает только человеческую вставку LRP5.
Присутствие в некоторых клонах идентичных CRISPR-индуцированных вставочно-делеционных мутаций, созданных на обоих аллелях посредством очевидной репарации негомологичным соединением концов, свидетельствует о наличии событий генной конверсии в гибридных клетках F1H4 (состоящих на 50% из линии 129SvS6 и на 50% из линии C57BL/6N). Чтобы получить представление о механизме, лежащем в основе улучшенного биаллельного нацеливания при использовании двух гРНК, провели скрининг семи клонов, которые имели или нацеленные гомозиготные гуманизации, или гомозиготные CRISPR-индуцированные крупные делеции после нацеливания с помощью LTVEC и комбинаций гРНК А плюс F или плюс Е2.
На Фиг. 5 представлены примеры тестов, выполненных с возможностью исследования событий генной конверсии, опосредованных двумя гидовыми РНК. В частности, возможную генную конверсию исследовали посредством анализа на потерю гетерозиготности (LOH) в ЭС-клетках гибрида F1H4 (который состоит на 50% из линии 129 SvS6 и на 50% из линии C57BL/6N). Генную конверсию можно продемонстрировать по потере гетерозиготности в известных полиморфизмах между 129SvS6 (129) и C57BL/6N (В6), и, следовательно, были разработаны ПЦР-тесты для дифференцирования этих двух типов аллелей. Полиморфизмы структурных вариантов (SV) тестировали традиционными ПЦР, выполненными с возможностью обнаружения различий между аллелями 129 и В6. Хотя на Фиг. 5 показан только один из SV-тестов, принцип одинаков во всех случаях. На основе структурных вариантов (SV), различающихся у линий мышей В6 и 129, были разработаны праймеры, показанные в таблице 1. Условия создания праймеров были ограничены идентификацией SV размером ~25 п.н. и получения ПЦР-продуктов размером ~300 п.н.; эти условия были подобраны так, чтобы любые изменения можно было видеть на гель-электрофорезе.
Перед проведением ПЦР на клонах провели валидацию и оптимизацию тестов по ДНК ЭС-клеток дикого типа из линий В6, 129 и из ЭС-клеток линии F1H4. Для тестирования на клонах были подобраны наборы праймеров, которые давали различимые полосы ПЦР, специфичные или для аллеля В6, или для аллеля 129, и устойчиво давали такие же две полосы при использовании ДНК F1H4. Для хромосомы 19 (местоположение гена Lrp5) подобрали шесть наборов праймеров - с идентификаторами 190045, 190061, 190068, 190030, 190033, 190013 - для использования на клонах с гуманизированным Lrp5, которые в тестах на модификацию аллеля (МОА) и в традиционном ПЦР были генотипированы как «гомозиготные нацеленные» или «гомозиготные коллапсированные». ПЦР-тесты на SV имели по длине хромосомы 19 от локуса гена Lrp5 в сторону теломерного конца хромосомы протяженность от ~13,7 до ~56,2 млн п.н. от локуса Lrp5. Приблизительные расстояния (в млн п.н.) для SV-тестов на хромосоме 19 от локуса Lrp5 были следующими: 13,7 для теста 190045, 19,0 для теста 190061, 35,0 для теста 190068, 37,4 для теста 190030, 48,3 для теста 190033 и 56,2 для теста 190013. На Фиг. 5 представлен только тест 190033 (обозначен как SV 48,3), а праймеры для тестов 190045, 190061, 190068, 190030, 190033 и 190013 приведены в таблице 1.
ПЦР проводили с ДНК из этих клонов, а также с контрольной ДНК F1H4, контрольной ДНК 129 и контрольной ДНК В6. ПЦР-продукты фракционировали посредством гель-электрофореза на 6%-ных полиакриламидных гелях, которые затем окрашивали GelRed. Клоны, дающие две полосы, соответствовали контролю F1H4, причем предшествующая оптимизация показала, что верхняя полоса специфична для аллеля 129, а нижняя полоса специфична для аллеля В6. Клоны, дающие только одну полосу, демонстрировали или только полосу В6, или только полосу 129. Клоны AW-А7, AW-F10, BA-D5, BA-F2, ВС-Н9 и BR-B4 продемонстрировали только полосу В6 во всех шести тестах, а клон ВО-А8 продемонстрировал только полосу 129 во всех шести тестах. Как было упомянуто ранее, эти клоны были генотипированы как гомозиготные нацеленные или как гомозиготные коллапсированные при МОА и/или ПЦР и включали различные комбинации гРНК (А плюс F, А плюс Е2, В2 и D). Наличие всего одной аллельной полосы свидетельствовало о том, что имеет место событие генной конверсии. Если бы конверсии не было, то должны были присутствовать обе полосы, как в контроле F1H4.
Кроме того, однонуклеотидные вариации (SNV) между аллелями 129 и В6 тестировали посредством тестов TaqMan® на аллельную дискриминацию. Приблизительные местоположения SNV-тестов на хромосоме 19 показаны на Фиг. 5 значками-треугольниками с указанием под ними расстояния (в млн п.н.) от локуса Lrp5. Расстояния (в млн п.н.) от локуса Lrp5 были следующими: 0,32 к центромере от Lrp5 (С2), 1,2 к теломере от Lrp5 (Т3), 11,1 к теломере от Lrp5 (Т6), 13,2 к теломере от Lrp5 (Т7), 17,5 к теломере от Lrp5 (Т8), 25,8 к теломере от Lrp5 (Т9), 33,0 к теломере от Lrp5 (Т10), 38,3 к теломере от Lrp5 (Т11), 49,6 к теломере от Lrp5 (Т13) и 57,2 к теломере от Lrp5 (Т14). 129-специфические и В6-специфические зонды и пары праймеров показаны в таблице 1.
В таблице 3 представлены семь примеров клонов ЭС-клеток, показавших очевидные события генной конверсии по длинному плечу хромосомы 19 в направлении к теломере от целевого локуса Lrp5 по данным LOH для обоих аллелей - SV и SNV. Эти клоны ЭС-клеток были получены из независимых экспериментов по нацеливанию, в которых комбинировали LTVEC для гуманизации Lrp5 (Фиг. 1) с одной или двумя гРНК, как указано. Положения сайтов распознавания гРНК показаны на Фиг. 5 над изображением гена Lrp5 (толстая стрелка, направленная влево). Тесты с генотипированием показали, что шесть из семи клонов имели гомозиготные нацеленные гуманизации гена Lrp5, а один имел гомозиготное коллапсирование (большую делецию между сайтами гРНК). В шести из семи клонов были потеряны аллели 129 и остались только аллели В6. В одном клоне были потеряны аллели В6 и остались только аллели 129. Все клоны остались гетерозиготными по аллелям, протестированным с центромерной стороны от локуса Lrp5 (т.е. все клоны были гетерозиготами В6/129 в тесте SNV C2). Потеря гетерозиготности (LOH), отмеченная в семи клонах, указывает на то, что одним механизмом получения гомозиготных генетически модифицированных аллелей при комбинировании LTVEC с одной или, что чаще, двумя гРНК, является первая нацеленная генетическая модификация на одном аллеле с последующим событием генной конверсии посредством направляемой гомологией рекомбинации, которая копирует нацеленную генетическую модификацию из одной хромосомы в ее гомолог.
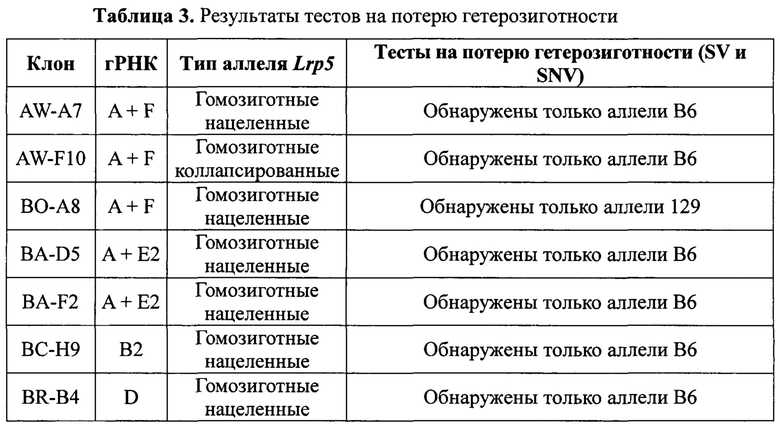
Локус С5 (Hc)
В другой серии экспериментов был разработан LTVEC для создания делеции из 76 т.п.н. мышиного гена компонента 5 системы комплемента (С5 или Hc (гемолитический комплемент)) с одновременной заменой фрагментом из 97 т.п.н. гомологичного человеческого гена С5 (Фиг. 6). Целевой локус содержал экзон 2 до стоп-кодона гена С5 (Hc). LTVEC содержал фрагмент из 97 т.п.н. человеческого гена С5, фланкированный гомологичными плечами, содержащими 35 т.п.н. и 31 т.п.н. геномной ДНК, происходящей из частей мышиного локуса С5 (Hc), фланкирующих последовательность из 76 т.п.н. мышиного гена С5 (Hc), предназначенного для делеции. В отдельных экспериментах LTVEC, гуманизирующий С5 (Hc), комбинировали с плазмидой, кодирующей Cas9, и второй плазмидой, кодирующей одну из шести гРНК (А, В, С, D, Е и Е2; см. таблицу 1), предназначенных для создания двухцепочечных разрывов в области мышиного гена С5 (Hc), нацеленного для делеции. гРНК были созданы так, чтобы избегать распознавания любой последовательности во вставляемом участке человеческого гена С5. В других экспериментах комбинировали LTVEC и кодирующую Cas9 плазмиду с плазмидами, кодирующими две разные гРНК, нацеленные на разные сайты в пределах области мышиного гена С5 (Hc), нацеленного для делеции. В некоторых экспериментах вместо LTVEC, гуманизирующего С5 (Hc), использовали контрольный LTVEC, нацеливающийся на локус Ch25h. Контрольный LTVEC, выполненный с возможностью удаления всей кодирующей последовательности Ch25h (~1 т.п.н.) и вставки кассет селекции пуромицином и неомицином в локус Ch25h, использовали как средство селекции резистентных к лекарственному средству клонов, не нацеленных на гомологичную рекомбинацию в локусе С 5 (Hc).
Результаты выполняемой посредством CRISPR/Cas9 гуманизации гена С5 (Hc) представлены в таблице 4. Они аналогичны результатам, полученными при выполняемой посредством CRISPR/Cas9 гуманизации гена Lrp5. Эффективность нацеливания с помощью одного только LTVEC была выше (6,1%) для гуманизации С5 (Hc), чем для гуманизации Lrp5, но добавление Cas9 и гРНК повышало эффективность нацеливания для четырех из шести протестированных гРНК. Как и в случае с Lrp5, комбинация гРНК (т.е. использование двух гРНК) для гуманизации С5 (Hc) дополнительно повышало общую эффективность нацеливания преимущественно посредством повышения частоты событий гемизиготного и гомозиготного нацеливания. Мы также обнаружили клоны ЭС-клеток с большими CRISPR-индуцированными делениями на обоих аллелях (наблюдавшиеся с частотами 1,8-3,6%). Кроме того, при использовании LTVEC, нацеленного на локус Ch25h, в комбинации с двумя гРНК С5 (Hc) клоны с гомозиготными аллелями, коллапсированными между двумя гРНК с последовательностями распознавания РНК CRISPR, наблюдались с частотами 1,2-6%, и это показывает, что события коллапсирования происходят независимо от событий гомологичной рекомбинации в целевом локусе. Как и в случае с Lrp5, использовали тесты на сохранение для подтверждения клонов с правильным нацеливанием. Два теста на сохранение, использованные в этом скрининге, представляли собой тесты TaqMan® с применением следующих праймеров и зондов: прямой праймер 7140retU, 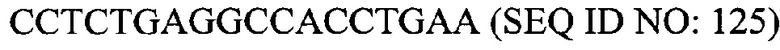 ; обратный праймер 7140retU,
; обратный праймер 7140retU, 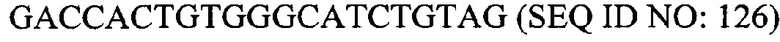 ; зонд 7140retU TaqMan®,
; зонд 7140retU TaqMan®,  ; прямой праймер 7140retD,
; прямой праймер 7140retD, 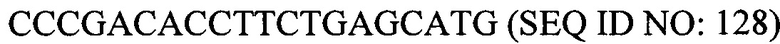 ; обратный праймер 7140retD,
; обратный праймер 7140retD, 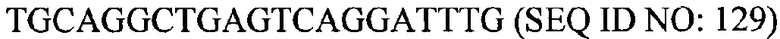 ; зонд 7140retD TaqMan®,
; зонд 7140retD TaqMan®, 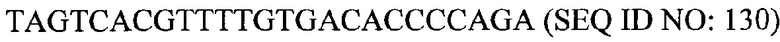 .
.
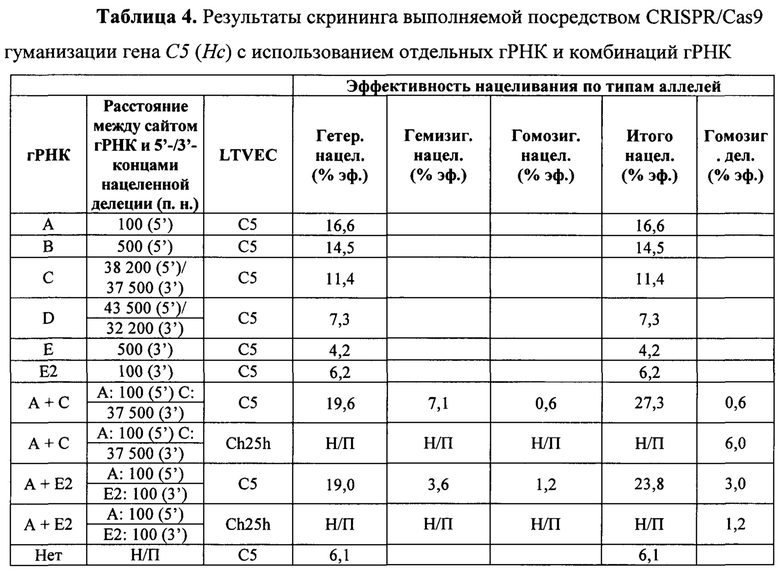
Флуоресцентную гибридизацию in situ (FISH) использовали для подтверждения гомозиготной нацеленной гуманизации гена С5 (Hc). Клоны ЭС-клеток, которые по данным количественного и традиционного ПЦР-тестов были сочтены прошедшими гомозиготное нацеливание в результате экспериментов по нацеливанию, в которых LTVEC для гуманизации С5 (Hc) (Фиг. 6) комбинировали с Cas9 и двумя гРНК, направили в коммерческую цитологическую службу для проведения FISH и анализа кариотипа. Искусственную бактериальную хромосому (ВАС), несущую мышиный ген С5 (Hc), метили красным флуоресцентным маркером и использовали в качестве зонда для идентификации эндогенных локусов, а ВАС, несущую человеческий ген С5, метили зеленым флуоресцентным красителем и использовали в качестве зонда для идентификации хроматид с проведенным нацеливанием человеческой вставкой. Меченные ВАС-зонды гибридизовали с метафазными пластинками из клонов с проведенным нацеливанием и визуализировали посредством флуоресцентной микроскопии. Хромосомы в метафазных пластинках визуализировали посредством окрашивания DAPI (4',6-диамидино-2-фенилиндол), а отдельные кариотипы в каждом клоне определяли посредством окрашивания по Гимза. Типичный результат представлен на Фиг. 7B для клона O-E. На составной фотографии на Фиг. 7B показано, что как сигнал красного мышиного ВАС-зонда, так и сигнал зеленого человеческого ВАС-зонда колокализовались в локусе С5 (Hc) на обеих копиях мышиной хромосомы 2, в известном местоположении гена С5 (Hc). Результаты подтверждают, что фрагмент из 97 т.п.н. человеческого гена С5 в LTVEC для гуманизации (Фиг. 6) был успешно вставлен в намеченный локус мышиного С5 (Hc) на обоих гомологах хромосомы 2 в клоне O-E3. В отличие от этого, на составной фотографии на Фиг. 7A показано, что как сигнал красного мышиного ВАС-зонда, так и сигнал зеленого человеческого ВАС-зонда колокализовались на одной копии мышиной хромосомы 2 (сплошная стрелка), а сигнал красного мышиного ВАС-зонда локализовался только в локусе С5 (Hc) на другой копии мышиной хромосомы 2. Результаты подтверждают, что фрагмент из 97 т.п.н. человеческого гена С5 в LTVEC для гуманизации (Фиг. 6) был успешно вставлен в намеченный локус мышиного С5 (Hc) только на одной копии хромосомы 2 (гетерозиготное нацеливание) в клоне Q-E9.
Локус Ror1
В другой серии экспериментов LTVEC был разработан для создания 110 т.п.н. делеции в мышином гене Ror1 (тирозин-протеинкиназный трансмембранный рецептор ROR1) с одновременной заменой 134 т.п.н. фрагментом гомологичного человеческого гена ROR1 (Фиг. 8). LTVEC содержал фрагмент из 134 т.п.н. человеческого гена ROR1, фланкированный гомологичными плечами, содержащими 41,8 т.п.н. и 96,4 т.п.н. геномной ДНК, полученной из частей мышиного локуса Ror1, фланкирующих последовательность из 110 т.п.н. мышиного гена Ror1, предназначенного для делеции. В отдельных экспериментах LTVEC, гуманизирующий Ror1, комбинировали с плазмидой, кодирующей Cas9, и второй плазмидой, кодирующей одну из шести гРНК (А, В, С, D, Е и F; см. таблицу 1), предназначенных для создания двухцепочечных разрывов в области мышиного гена Ror1, нацеленного для делеции. гРНК были созданы так, чтобы избегать распознавания любой последовательности во вставляемом участке человеческого гена. ROR1. В других экспериментах комбинировали LTVEC и кодирующую Cas9 плазмиду с плазмидами, кодирующими две разные гРНК, нацеленные на разные сайты в пределах гена Ror1, предназначенного для делеции.
Результаты выполняемой посредством CRISPR/Cas9 гуманизации гена Ror1 представлены в таблице 5. Они аналогичны результатам, полученным для выполняемой посредством CRISPR/Cas9 гуманизации генов Lrp5 и С5 (Hc). Эффективность нацеливания с использованием одного только LTVEC составила 0,3%, а добавление Cas9 и гРНК несколько повышало эффективность нацеливания в случае двух из шести протестированных гРНК. Комбинирование гРНК А и F повысило общую эффективность нацеливания на Ror1 до 6,3% путем повышения частоты как гетерозиготного, так и гемизиготного нацеливания. Мы также обнаружили клоны ЭС-клеток с большими CRISPR-индуцированными делениями на обоих аллелях (наблюдавшиеся с частотой 1,6%).
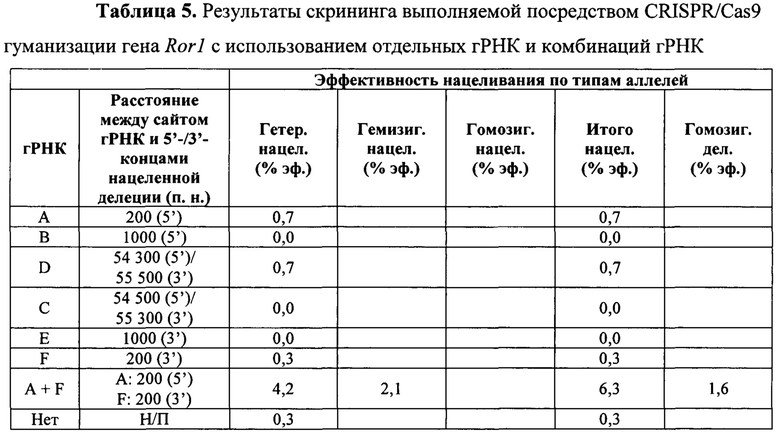
Локус Trpa1
В еще одной серии экспериментов LTVEC был разработан для создания делеции из 45,3 т.п.н. в мышином гене Trpa1 (катионный канал с транзиторным рецепторным потенциалом, подсемейство А, член 1) с одновременной заменой фрагментом из 54,5 т.п.н. гомологичного человеческого гена TRPA1 (Фиг. 9). LTVEC содержал фрагмент из 54,5 т.п.н. человеческого гена TRPA1, фланкированный гомологичными плечами, содержащими 41,0 т.п.н. и 58,0 т.п.н. геномной ДНК, полученной из частей мышиного локуса Trpa1, фланкирующих последовательность из 45,3 т.п.н. мышиного гена Trpa1, предназначенного для делеции. В отдельных экспериментах LTVEC, гуманизирующий Trpa1, комбинировали с плазмидой, кодирующей Cas9, и второй плазмидой, кодирующей одну из восьми гРНК (А, А2, В, С, D, E2, Е и F; см. таблицу 1), предназначенных для создания двухцепочечных разрывов в области мышиного гена Trpa1, нацеленного для делеции. гРНК были созданы так, чтобы избегать распознавания любой последовательности во вставляемом участке человеческого гена TRPA1. В других экспериментах комбинировали LTVEC и кодирующую Cas9 плазмиду с плазмидами, кодирующими две разные гРНК, нацеленные на разные сайты в пределах гена Trpa1, предназначенного для делеции.
Результаты выполняемой посредством CRISPR/Cas9 гуманизации гена Trpa1 представлены в таблице 6. Они аналогичны результатам, полученным для выполняемой посредством CRISPR/Cas9 гуманизации генов Lrp5 и С5 (Hc). Эффективность нацеливания с использованием только LTVEC составила 0,3%, а добавление Cas9 и гРНК повышало эффективность нацеливания в случае шести из восьми протестированных гРНК. Комбинирование гРНК B и F повышало общую эффективность нацеливания на Trpa1 до 3,4% путем повышения частоты событий гетерозиготного, гемизиготного и гомозиготного нацеливания. Мы также обнаружили клоны ЭС-клеток с большими CRISPR-индуцированными делециями на обоих аллелях (наблюдавшиеся с частотой 0,3%).
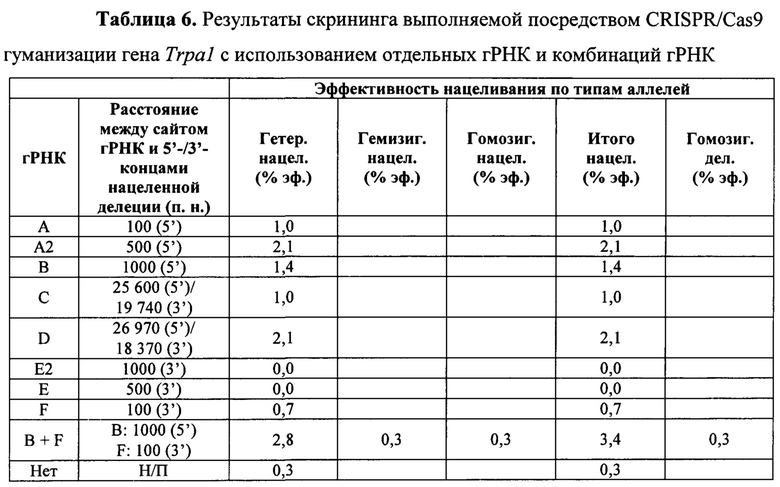
Как показывают эти примеры, использование двух гидовых РНК на далеко расположенных сайтах повысило эффективность гетерозиготной гуманизации в сравнении с использованием отдельных гРНК. Кроме того, использование двух гидовых РНК способствовало биаллельным событиям в сравнении с использованием отдельных гРНК. В отличие от нацеливания при помощи одной гРНК, нацеливание двумя гРНК приводит к созданию клеток с гомозиготным нацеливанием (Hum/Hum), в которых оба аллеля имеют нацеленную гуманизацию, клеток с гомозиготной делецией (Δ/Δ), в которых ни один аллель не подвергся нацеливанию гуманизирующим LTVEC, но оба аллеля имеют крупные делеции, а также клеток с гемизиготным нацеливанием (Hum/Δ), в которых один аллель имеет нацеленную гуманизацию, а другой - индуцированную двумя гРНК/Cas9 большую делецию. Во-первых, мы обнаружили клоны с правильным нацеливанием, имевшие точные и идентичные очень крупные гуманизации в обоих целевых аллелях (например, клетки, гомозиготные по нацеленной генетической модификации). Хотя клетки с гомозиготным нацеливанием также наблюдались при использовании одной гРНК для достижения гуманизации Lrp5, они появлялись с гораздо более низкой частотой, чем при использовании двух гРНК (см. таблицу 2). Аналогичным образом, гомозиготное нацеливание не наблюдалось при использовании одной гРНК для получения гуманизации С5 (Hc) или гуманизации Trpa1, но гомозиготное нацеливание наблюдалось при использовании двух гРНК с нацеливающим вектором (см. таблицы 4 и 6). Аналогичным образом, были обнаружены правильно нацеленные клоны, гемизиготные по генной модификации (т.е. они имели точно нацеленную гуманизацию на одном аллеле и очень большую, иногда разрушающую ген делецию на другом аллеле) при нацеливании на Lrp5, С5 (Hc), Ror1 или Trpa1. Такие модификации совсем не наблюдались при использовании одной гРНК для получения гуманизации Lrp5, С5 (Hc), Ror1 или Trpa1 (см. таблицы 2, 4, 5 и 6 соответственно).
Во-вторых, мы обнаружили клоны, имевшие идентичные очень большие делеции (>45 т.п.н.), индуцированные событиями расщепления посредством Cas9, направляемыми обеими гРНК на обоих нацеленных аллелях (т.е. клетки были гомозиготными по большой, иногда разрушающей ген делеции в целевом локусе). Мутации этого типа не требуют нацеливающего вектора, направленного на тот же ген. Например, как показано в таблице 4, мы наблюдали ЭС-клетки с гомозиготными CRISPR-индуцированными делециями, комбинируя Cas9 и две гРНК с нацеливающим вектором, направленным на другой ген, не связанный с нацеливаемым при помощи гРНК. Следовательно, нуклеаза Cas9, направляемая двумя гРНК, может индуцировать большую делецию в клетках без добавления нацеливающего вектора. В таких случаях временная или устойчивая селекция лекарственным средством, которую обеспечивает вектор, экспрессирующий ген резистентности к лекарственному средству, может облегчить выделение редких клонов с гомозиготной делецией, обогащая популяцию ЭС-клетками, захватившими ДНК.
Пример 2. Анализ больших делеций, вызванных комбинацией гРНК
Аллелъные структуры больших делеций, вызванных комбинацией гРНК
Провели дополнительный анализ последовательностей с клонами, содержащими большие делеций, индуцированные событиями расщепления нуклеазой Cas9, направляемыми двумя гРНК (см. таблицу 7). Эти большие делеций оказались независящими от направляемых LTVEC событий гомологичной рекомбинации в том же локусе в том смысле, что мы получали большие делеции в клоне Lrp5 приблизительно с одинаковой частотой при комбинировании гРНК или с Lrp5 LTVEC, или с LTVEC, нацеливающимся на локус гена Ch25h, расположенный на расстоянии почти 30 млн п.н. (данные не показаны). Чтобы охарактеризовать большие делеции, мы провели ПЦР протяженности делеции на 37 клонах, 15 гемизиготных и 22 с биаллельными большими делециями из четырех гуманизаций и секвенировали отдельные клоны ПЦР-продуктов. Секвенирования подтвердили большие делеции, которые имели диапазон от 38 т.п.н. до 109 т.п.н. Два клона ЭС-клеток (клоны Lrp5 AW-A8 и BP-D3) имели идеально репарированные точные делеции (68,2 т.п.н.) между предсказанными сайтами расщепления Cas9, а один клон (клон Hc Р-В12) имел вставку одной пары оснований в дополнение к делеции 38,1 т.п.н. Двадцать семь клонов ЭС-клеток имели делеции, выходящие за пределы сайтов расщепления Cas9, согласующиеся с неточной репарацией посредством негомологичного соединения концов (NHEJ). Остальные семь клонов ЭС-клеток имели мутации, сочетавшие в себе очевидные NHEJ-индуцированные делеции и вставки (например, клон Lrp5 BP-F6 и клон Hc O-E4), четыре из которых имели вставки более 200 п.н., которые мы смогли проследить до их исходных геномных локусов (данные не показаны). Вставка 210 п.н. в клон Lrp5 ВО-Е9 имела обратную ориентацию относительно идентичной последовательности, лежащей приблизительно на 2600 п.н. за пределами целевого сайта гРНК F в направлении центромеры (хромосома 19+, 3589138-3589347). Эта последовательность присутствовала в длинном 3' гомологичном плече Lrp5 LTVEC. LrpS-клоны BP-F6 и BP-G7 были получены в эксперименте, в котором мы комбинировали Lrp5 гРНК А и F с Cas9 и LTVEC, нацеленным на ген Ch25h, находящийся на расстоянии 30 млн п.н. от Lrp5 в направлении теломеры. Клон BP-F6 имел вставку 266 п.н., которая, по-видимому, происходила из одного конца Ch25h LTVEC в том смысле, что состояла из фрагмента 103 п.н., идентичного части каркаса вектора, соединенного с фрагментом 163 п.н., идентичным последовательности, расположенной вблизи Ch25h и также присутствующей в длинном плече LTVEC (хромосома 19+, 34478136-34478298); этот фрагмент был вставлен в место делеции в обратной ориентации относительно эндогенной хромосомной последовательности. Нс-клон O-E4 имел вставку 254 п.н., имеющую обратную ориентацию относительно идентичной последовательности, обнаруженной в последовательности делеции на расстоянии приблизительно 3,1 т.п.н. от сайта распознавания гРНК А. Вставка 1304 п.н. в Hc-клон S-D5 состояла из двух фрагментов: 1238 п.н. фрагмента, находящегося в той же ориентации, что и идентичная последовательность, обнаруженная в последовательности делеции на расстоянии приблизительно 1,4 т.п.н. от прогнозируемого направляемого гРНК Е2 сайта расщепления Cas9, и фрагмента 66 п.н., представлявшего собой последовательность, удвоенную и имеющую обратную ориентацию относительно идентичной последовательности 25 п.н. за пределами сайта расщепления гРНК Е2.
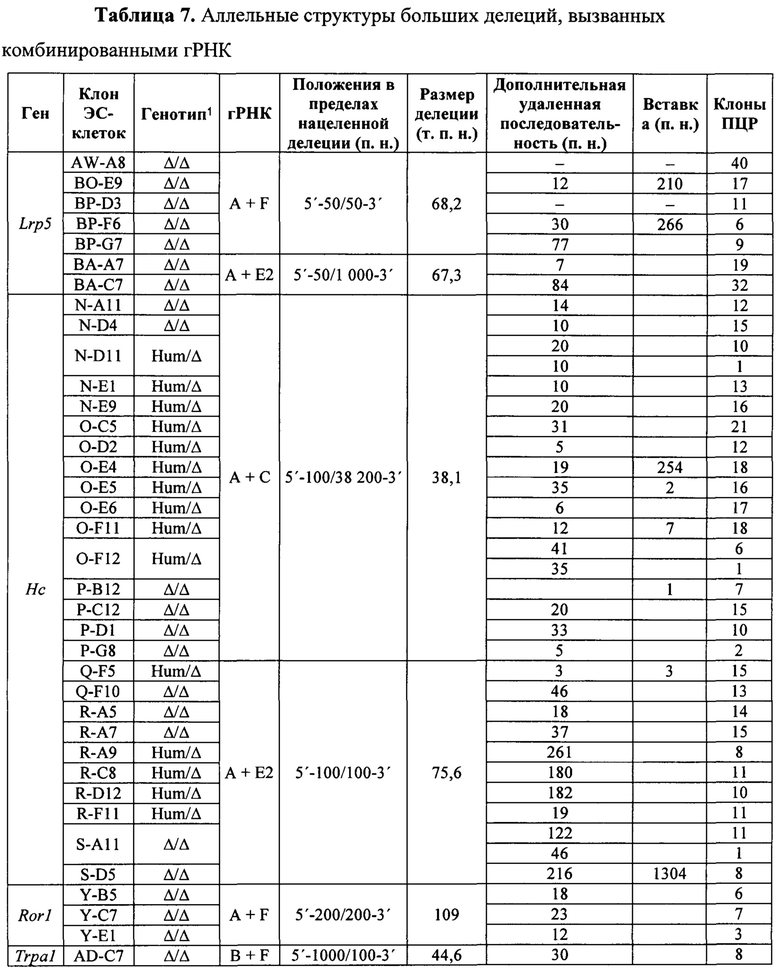
1 Hum/+, нацеленная гуманизация одного из двух нативных аллелей, приводящая к гетерозиготному генотипу; Hum/Δ, биаллельная модификация, в которой один аллель имеет нацеленную гуманизацию, а другой аллель имеет крупную Cas9-гРНК-индуцированную делецию, что приводит к гемизиготному генотипу; Hum/Hum, биаллельная модификация, в которой оба аллеля имеют нацеленную гуманизацию, что приводит к гомозиготному генотипу; Δ/Δ, биаллельная модификация, в которой оба аллеля имеют крупную Cas9-гРНК-индуцированную делецию.
Доказательство генной конверсии в гомозиготных аллелях
Двадцать один из 22 клонов ЭС-клеток с большими биаллельными делениями имели только одну уникальную последовательность (таблица 7), и это показывает, что они являлись гомозиготными аллелями. В случае Нс-клона. S-A11 мы обнаружили ту же последовательность в 11 из 12 ПЦР-клонов. Один клон с другой последовательностью, возможно, имеет два разных делеционных аллеля, но мы также наблюдали этот же результат для двух гемизиготных Hc-клонов - N-D11 и O-F12. Разные гомозиготные делеционные аллели во множестве клонов свидетельствуют о том, что они могли появиться в результате механизма генной конверсии, в котором делеция на одной хромосоме послужила шаблоном для репарации расщеплений Cas9 посредством гомологичной рекомбинации на гомологичной хромосоме. Мы воспользовались линией ЭС-клеток VGF1, представлявшей собой гибрид F1 из линий 129S6SvEvTac (129) и C57BL/6NTac (B6) (Poueymirou et al. (2007) Nat. Biotechnol. 25: 91-99; Valenzuela et al. (2003) Nat. Biotechnol. 21: 652-659), для теста на генную конверсию в виде потери гетерозиготности (Lefebvre et al. (2001) Nat. Genet. 27: 257-258) для структурных (SV) и однонуклеотидных (SNV) вариаций между линиями вокруг локуса Lrp5 на хромосоме 19 (см. Фиг. 5, где приведены пять SV-тестов и десять SNV-тестов, которые использованы ниже) и локуса Hc на хромосоме 2 (не показан). Для подтверждения того, что любая потеря гетерозиготности не была результатом утраты целой хромосомы, были проведены тесты на число копий хромосом (CCN) в сайтах, идентичных для линий 129 и B6. Для гуманизированных или делеционных аллелей Lrp5 мы протестировали множество SV и SNV, расположенных на расстоянии от 1,2 млн п.н. от Lrp5 в направлении теломеры до конца длинного плеча хромосомы 19 (Фиг. 5). Поскольку Lrp5 расположен вблизи центромеры, мы не обнаружили SV и обнаружили только одну SNV с центромерной стороны от гена. В случае Не нам удалось протестировать множество SV и SNV с обеих сторон от гена на хромосоме 2 (не показана). Результаты для шести клонов Lrp5 представлены на Фиг. 10А-Е и 11А-С.
На Фиг. 10А-Е представлены результаты для пяти SV-тестов, положения которых находились в диапазоне от 13,7 млн п.н. от Lrp5 до 56,7 млн п.н. у теломерного конца длинного плеча. Пять SV-тестов дали два продукта разного размера для аллеля 129 (больше) и аллеля B6 (меньше) в контролях 129, B6 и VGF1. Приблизительные положения SV-тестов на карте хромосомы 19 показаны на Фиг. 5 (см. тест SV 13,7, тест SV 20,0, тест SV 36,9, тест SV 48,3 и тест SV 56,7). Номер теста представляет собой число млн п.н. в сторону теломеры от Lrp5. Праймеры для этих тестов представлены в таблице 1, а результаты показаны на Фиг. 10А-Е. Два из клонов, ВС-Н9 (Lrp5Hum/Hum, гРНК B2) и BR-B4 (Lrp5Hum/Hum, гРНК D), продемонстрировали потерю гетерозиготности, при которой сохранились все аллели В6 SV, а третий клон, В0-А8 (Lrp5Hum/Hum, гРНК A+F), сохранил все аллели 129. Три других клона, BO-F10 (+Lrp5Hum/Hum, гРНК A+F), BO-G11 (Lrp5Hum/Hum, гРНК А+F) и BP-G7 (Lrp5Δ/Δ, гРНК А+F), остались гетерозиготными.
Кроме того, однонуклеотидные вариации (SNV) между аллелями 129 и В6 тестировали посредством тестов TaqMan® на аллельную дискриминацию. Приблизительные положения SNV-тестов на карте хромосомы 19 показаны на Фиг. 5 значками-треугольниками с указанием под ними номера теста и ниже приведены расстояния (в млн п.н.) от локуса Lrp5. Расстояния (в млн п.н.) от локуса Lrp5 были следующими: 0,32 к центромере от Lrp5 (С2), 1,2 к теломере от Lrp5 (Т3), 11,1 к теломере от Lrp5 (Т6), 13,2 к теломере от Lrp5 (Т7), 17,5 к теломере от Lrp5 (Т8), 25,8 к теломере от Lrp5 (Т9), 33,0 к теломере от Lrp5 (Т10), 38,3 к теломере от Lrp5 (Т11), 49,6 к теломере от Lrp5 (Т13) и 57,2 к теломере от Lrp5 (Т14). 129-специфические и В6-специфические зонды и пары праймеров показаны в таблице 1. Результаты для трех клонов (ВС-Н9, ВО-А8 и BR-B4), продемонстрировавших с теломерной стороны потерю гетерозиготности (LOH) по данным SV-тестов, представлены на Фиг. 11A-C. SNV-тесты (Фиг. 11A-C и не показанные данные) подтвердили события генной конверсии по длинному плечу хромосомы 19 с теломерной стороны от Lrp5 (SNV 1,2 и SNV 57,2; см. Фиг. 11B и Фиг. 11C соответственно), но тест SNV 0,32 (см. Фиг. 11A) продемонстрировал, что все клоны остались гетерозиготными по аллелю 320 т.п.н. от Lrp5 с центромерной стороны. Из 24 протестированных клонов Lrp5Hum/Hum или Lrp5Δ/Δ мы обнаружили шесть, которые имели признаки потери гетерозиготности по всей длине длинного плеча хромосомы 19 с теломерной стороны от Lrp5. Пять из этих клонов (четыре Lrp5Hum/Hum и один Lrp5Δ/Δ) превратились из гетерозиготных в гомозиготные В6, а шесть клонов (Lrp5Hum/Hum) превратились в гомозиготные 129. CCN-тесты продемонстрировали сохранение двух копий хромосомы 19. Аналогичные тесты на потерю гетерозиготности для 21 гомозиготного Hc-клона. позволили выявить, что два - R-E2 (HcHum/Hum, гРНК А+F) и R-E8 (НсΔ/Δ, гРНК A+F) - показали потерю гетерозиготности, превратившись в гомозиготные 129 по всем SV и SNV с теломерной стороны от гена Hc, но сохранили гетерозиготность по всем аллелям с центромерной стороны. CCN-тесты не показали потери хромосомы 2.
Наши результаты впервые демонстрируют, что CRISPR/Cas9 может улучшить направляемую гомологией репарацию крупных одностадийных гуманизации длиной более 100 т.п.н., что расширяет возможности крупномасштабной генной инженерии. Наиболее заметным и неожиданным преимуществом комбинирования LTVEC и гРНК/Cas9 была их способность стимулировать гомозиготные нацеленные гуманизации. Хотя биаллельные мутации и события гомозиготного нацеливания были отмечены в других экспериментах с CRISPR/Cas9, большинство этих генных модификаций и вставок были на порядки величин меньше, чем в наших гуманизированных аллелях. До использования CRISPR/Cas9 мы никогда не обнаруживали гомозиготного нацеливания посредством LTVEC, а также не наблюдали одновременного нацеливания на более чем один ген при комбинировании множества LTVEC, нацеливающихся на отдельные гены. Учитывая этот опыт, при гРНК/Cas9-индуцированном гомозиготном нацеливании предполагается, что не два LTVEC по отдельности нацеливаются на оба аллеля, а исходное событие нацеливания на один аллель служит шаблоном для гомологичной конверсии другого, чему способствует одно или более расщеплений Cas9. Открытие того, что индуцированные двумя гРНК/Cas9 большие биаллельные делеции также были гомозиготными (таблица 7), дополнительно подтверждает механизм генной конверсии.
Тесты на потерю гетерозиготности (Фиг. 5) продемонстрировали, что крупномасштабная генная конверсия множества аллелей, охватывающая большой фрагмент хромосомы с теломерной стороны целевого гена, ответственна за некоторые гомозиготные гуманизации и большие делеции. Такой тип широкоохватной направленной генной конверсии соответствует митотической рекомбинации между реплицированными хроматидами гомологичных хромосом на фазе G2 клеточного цикла (Lefebvre et al. (2001) Nat. Genet. 27: 257-258) (Фиг. 12). Хотя этот механизм объясняет лишь небольшую часть гомозиготных событий, он может обеспечивать средство, с помощью которого можно использовать гРНК/Cas9-расщепление для стимулирования крупномасштабной конверсии из гетерозиготного в гомозиготное состояние множества аллелей на большом участке хромосомы. Однако большая часть гомозиготных событий, по-видимому, является результатом локальной генной конверсии, механизм которой требует дальнейшего исследования.
Дополнительное свидетельство крупноохватной направленной генной конверсии было получено в результате анализа трех клонов, полученных после электропорации гибридных ЭС-клеток F1H4 (которые на 50% состоят из линии 129SvS6 и на 50% - из линии C57BL/6N) плазмидами, кодирующими Lrp5 гРНК А и F, плазмидой, кодирующей Cas9, и LTVEC, нацеленным на ген Ch25h, находящийся на расстоянии 30 млн п.н. от Lrp5 в теломерном направлении. Три клона были изначально отнесены к первому типу после первичного скрининга с использованием тестов TaqMan® в пределах прогнозируемой делеции между 2 гРНК (500 п.н. на 5'-конце и 2 т.п.н. на 3'-конце), но последующие тесты TaqMan® на аллельную дискриминацию с оценкой однонуклеотидных вариантов (SNV) между аллелями 129 и В6 неожиданно выявили потерю гетерозиготности. К использованным SNV-теетам относились один центромерный тест (SNV 0,32) и два теломерных теста (SNV 1,2 и SNV 57,2) (см. Фиг. 5). Как показано в таблице 8, центромерный SNV-тест (0,32 млн п.н.) подтвердил сохранение гетерозиготности во всех трех клонах. Однако оба теломерных SNV-теста показали, что клоны ВР-Е7 и ВР-Н4 были гомозиготны по аллелю 129, и оба теломерных SNV-теста показали, что клон ВР-Е6 был гомозиготен по аллелю В6. Все три клона продемонстрировали сохранение двух копий хромосомы 19, и все три клона были трансгенными по нацеливанию LTVEC (т.е. было выполнено нацеливание на локус Ch25h). Эти результаты открывают возможность для форсирования гомозиготности с использованием нацеленного расщепления CRISPR/Cas9.
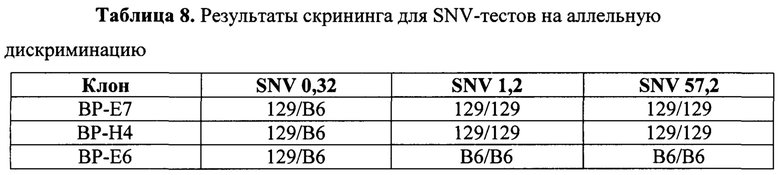
Результаты, наблюдаемые при экспериментах с выполняемой посредством CRISPR/Cas9 гуманизации с LTVEC на мышиных ЭС-клетках гибрида F1H4 (которые на 50% состоят из линии 129SvS6 и на 50% - из линии C57BL/6N) (см. Фиг. 16A-F), можно объяснить с использованием нескольких возможных механизмов. Такие механизмы могут быть связаны с взаимным хроматидным обменом посредством митотического кроссинговера (см. Фиг. 16A-C) или с копированием хроматиды в результате индуцированной разрывом репликации (см. Фиг. 16D-E). В любом случае гетерозиготная модификация может происходить в хромосоме 129 или хромосоме В6, на которую выполнено нацеливание LTVEC перед репликацией генома (см. Фиг. 16A и 16D). Альтернативно нацеливание LTVEC на одну хроматиду 129 или одну хроматиду В6 могло быть выполнено после репликации генома, после чего следовала межхроматидная генная конверсия (см. Фиг. 16B и 16Е). Альтернативно нацеливание LTVEC в целевом геномном локусе может отсутствовать, но может происходить расщепление Cas9 на хромосоме 129 или хромосоме В6 (см. Фиг. 16C и 16F). Эта последняя возможность может объяснять результаты, наблюдаемые в клонах ВР-Е7, ВР-Н4 и ВР-Е6. Возможные исходы представлены на Фиг. 16A-F. В случае Фиг. 16F также можно наблюдать потерю гетерозиготности (LOH) с сохранением аллелей В6, если Cas9 расщепляет хроматиду 129. В вышеописанных экспериментах наблюдались события потери гетерозиготности, приводящие к тому, что оба аллеля оказывались нацеленными (Hum/Hum) или оба аллеля оказывались аллелями дикого типа (+/+).
Пример 3. Влияние размеров гомологичных плеч LTVEC на эффективность нацеливания
Для определения влияния размера гомологичных плеч на эффективность нацеливания было проведено сравнение двух LTVEC для создания делеции из 76 т.п.н. мышиного гена компонента 5 системы комплемента (С5 или Hc (гемолитический комплемент)) с одновременной заменой на фрагмент из 97 т.п.н. гомологичного человеческого гена C5 (Фиг. 13). Целевой локус содержал экзон 2 до стоп-кодона гена С 5 (Hc). Первый LTVEC содержал фрагмент из 97 т.п.н. человеческого гена С5, фланкированный гомологичными плечами, содержащими 35 т.п.н. и 31 т.п.н. геномной ДНК, происходящей из частей мышиного локуса С5 (Hc), фланкирующих последовательность из 76 т.п.н. мышиного гена С5 (Hc), предназначенного для делеции (см. нацеливающий вектор, обозначенный LTVEC на Фиг. 13). Второй LTVEC содержал фрагмент из 97 т.п.н. человеческого гена С5, фланкированный гомологичными плечами, содержащими по 5 т.п.н. геномной ДНК, происходящей из участков мышиного локуса С5 (Hc), фланкирующих последовательность из 76 т.п.н. мышиного гена С5 (Hc), предназначенного для делеции (см. нацеливающий вектор, обозначенный sTVEC на Фиг. 13).
В отдельных экспериментах LTVEC, гуманизирующие С5 (Hc), комбинировали с плазмидой, кодирующей Cas9, и второй плазмидой, кодирующей одну или две из шести гРНК (А, В, С, D, Е и Е2; см. таблицу 1), предназначенных для создания двухцепочечных разрывов в области мышиного гена С5 (Hc), нацеленного для делеции. гРНК были созданы так, чтобы избегать распознавания любой последовательности во вставляемом участке человеческого гена С5.
Результаты выполняемой посредством CRISPR/Cas9 гуманизации гена С5 (Hc) представлены в таблице 9. Эффективность нацеливания одного только первого LTVEC (гомологичные плечи 35 т.п.н. и 31 т.п.н.) была выше эффективности нацеливания одного только второго LTVEC (гомологичные плечи 5 т.п.н. и 5 т.п.н.). Однако значения общей эффективности нацеливания каждого LTVEC в комбинации с гРНК А и Е2 были практически идентичными (см. таблицу 9), и это указывает на то, что размеры гомологичных плеч 5 т.п.н. (т.е. общая сумма 10 т.п.н.) являются достаточными для повышения эффективности нацеливания, наблюдаемого при нацеливании на локус С5 (Hc) при помощи CRISPR/Cas9 в комбинации с нацеливанием LTVEC.
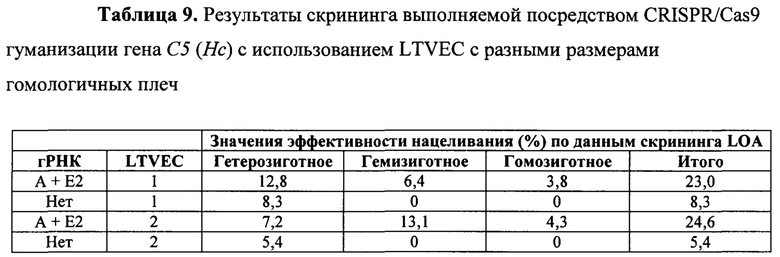
Пример 4. Влияние более коротких расстояний между последовательностями распознавания РНК CRISPR на эффективность нацеливания
Для определения влияния более коротких расстояний между последовательностями распознавания РНК CRISPR и сайтами расщепления на эффективность нацеливания был разработан LTVEC для создания делеции из 18,2 т.п.н. мышиного гена гидролазы цитидинмонофосфат-N-ацетилнейраминовой кислоты (Cmah) и одновременной замены вставкой, содержащей репортер lacZ и кассету селекции по резистентности к гигромицину. Использовали LTVEC с двумя близко расположенными нацеливающими последовательностями гРНК (Фиг. 14). Целевой локус содержал первые пять экзонов гена Cmah. LTVEC содержал вставку 8,8 т.п.н. lacZ-hygr, фланкированную гомологичными плечами, содержащими 120 т.п.н. и 57 т.п.н. геномной ДНК из частей мышиного локуса Cmah, фланкирующих последовательность из 18,2 т.п.н. мышиного гена Cmah, предназначенного для делеции. LTVEC комбинировали с плазмидами, кодирующими Cas9 и две гРНК (А и В), предназначенные для создания двухцепочечных разрывов вблизи 5'-конца области мышиного гена Cmah, нацеленного для делеции. Две гРНК нацеливались на близко расположенные последовательности вблизи ATG на 5'-конце последовательности, предназначенной для делеции, причем сайты расщепления были разнесены на 27 п.н. (см. Фиг. 15). Расщепление при помощи Cas9, направляемое этими двумя гРНК, позволяло получать удаленную последовательность 27 п.н с тупыми концами. В качестве контроля использовали один только LTVEC.
Проводили скрининг резистентных к лекарственному средству клонов ЭС-клеток на нацеленные гуманизации посредством тестов на модификацию аллеля (Valenzuela et al. (2003) Nat. Biotechnol. 21: 652-659; Frendewey et al. (2010) Methods Enzymol. 476: 295-307) для выявления последовательностей внутри делеций, последовательностей внутри кассеты селекции лекарственными средствами и вставок человеческого гена. Клоны считали получившими правильное нацеливание, если они теряли одну из двух эндогенных последовательностей мышиного гена и приобретали одну копию вставки lacZ-hygr. Кроме того, для проверки правильного нацеливания LTVEC на геном мыши использовали ПЦР-тесты в реальном времени, распознающие последовательности в гомологичных плечах LTVEC, называемые тестами на сохранение. Определение числа копий в этих тестах на сохранение позволяло получать дополнительную информацию, чтобы помочь отличить правильно направленные ЭС-клоны, которые сохранили число копий, равное двум, от клонов, в которых большая Cas9-индуцированная делеция целевого локуса мыши совпадает со случайной интеграцией LTVEC в другом месте в геноме, и в этом случае тесты на сохранение имели число копий, равное трем (или более). Способность парных гРНК создавать крупные Cas9-опосредованные делеции в целевом локусе мыши означала, что стандартные тесты LOA и GOA, как описано ранее, могут быть дополнены тестом на сохранение для обеспечения дополнительного уточнения и проверки правильного нацеливания. Поэтому тесты на сохранение были разработаны и использованы в сочетании с тестами LOA и GOA.
Результаты эксперимента с нацеливанием на Cmah представлены в таблице 10. В контрольном эксперименте с нацеливанием одним только вектором LTVEC 5,4% (3/56) прошедших скрининг клонов имели гетерозиготную (Het) делеционно-заместительную мутацию; 95% клонов сохранили дикий тип (WT) в локусе Cmah. В эксперименте с нацеливанием с использованием CRISPR наблюдали пять разных мутантных аллельных типов в дополнение к нескольким клонам дикого типа. Мы наблюдали три типа получивших LTVEC-нацеливание аллелей: (1) Het; (2) Нот (гомозиготная делеция-замена) и (3) Hemi (делеция-замена на одном аллеле и гРНК/Cas9-индуцированная мутация на другом аллеле). Эти три типа составляли 43,5% (106/244) всех прошедших скрининг клонов. По сравнению с использованием одного только LTVEC мы наблюдали 8-кратное улучшение нацеливания на ген Cmah по меньшей мере в одном получившем нацеливание аллеле. Мы также наблюдали два типа аллелей, несущих только вызванные гРНК/Cas9 вставочно-делеционные мутации: (1) Het, в котором мы обнаружили вставку-делецию в одном из двух аллелей дикого типа; и (2) биаллельные вставочно-делеционные мутации, которые могли быть или гомозиготными (Hom), или гетерозиготными (Hemi). Лишь 3,7% исследованных клонов сохранили дикий тип без обнаруживаемых мутаций в локусе Cmah. В целом более 94% клонов имели Cas9-индуцированные мутации при использовании комбинации гРНК А и В.
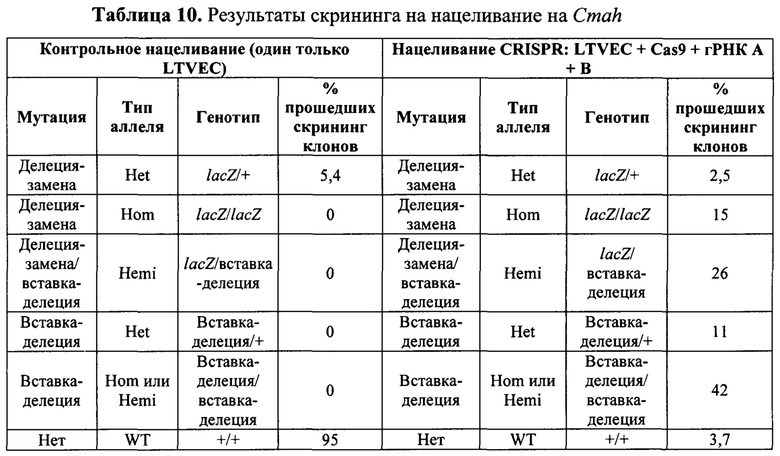
Пример 5. Большое коллапсирование с использованием пары гРНК в эмбрионах на стадии единственной клетки
Для получения большой нацеленной делеции в эмбрионах на стадии единственной клетки был разработан эксперимент по созданию делеции из 68 т.п.н. участка мышиного гена Lrp5 (родственный рецептору липопротеина низкой плотности белок 5), кодирующего эктодомен, с одновременной вставкой 4-нуклеотидной вставки посредством использования одноцепочечной донорной последовательности ДНК (длина 124 нуклеотида), где 4-нуклеотидная вставка фланкирована двумя 60-нуклеотидными гомологичными плечами. При помещении в локус 4-нуклеотидная вставка создает сайт рестрикционного фермента. В отдельных экспериментах белок Cas9 в виде белка доставляли посредством цитоплазматической инъекции (CI) или Cas9 в виде мРНК доставляли посредством инъекции в пронуклеус (PNI) или электропорации (ЕР). Cas9 комбинировали с двумя гРНК (A+F), разработанными для создания двухцепочечных разрывов в пределах области мышиного гена Lrp5, намеченного для делеции, и необязательно с донором гомологичной рекомбинации. гРНК вводили в виде РНК. После этого оценивали частоту получаемых моноаллельных и биаллельных мутаций.
Результаты представлены в таблице 11, включая опосредованную негомологичным соединением концов (NHEJ) делецию между целевыми сайтами двух гидовых РНК или делецию, выполненную с помощью направляемой гомологией репарации с донорной оцДНК. Биаллельные мутации наблюдались, когда парные гидовые РНК и Cas9 вводили вместе с донорной оцДНК посредством цитоплазматической инъекции. В каждой наблюдавшейся биаллельной мутации одна хромосома была модифицирована при помощи NHEJ-опосредованной делеции, а одна хромосома была модифицирована посредством делеции с направляемой гомологией репарацией (HDR). Эти результаты указывают на то, что цитоплазматические пьезоинъекции мРНК приводят к устойчивой направляемой гомологией репарации с потенциальной гомозиготной рекомбинацией с донором.
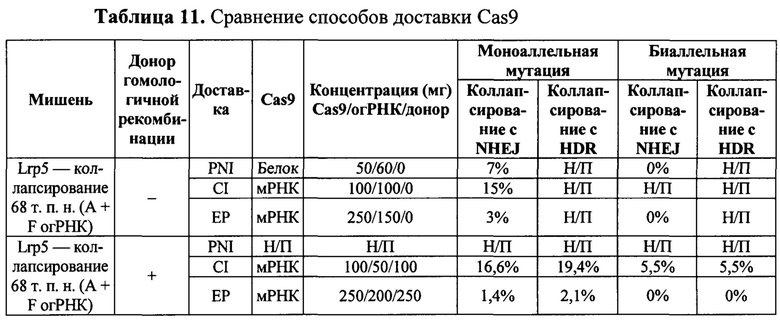
Пример 6. Тесты на сохранение для различения нацеленных вставок от трансгенных вставок и нацеленных делеций от делеций, выходящих за пределы целевой области
Стандартные стратегии скрининга на модификацию аллеля (МОА) (см., например. Фиг. 17A) определяют число копий зондов TaqMan® путем сравнения среднего по значениям Ct четырех биологических копий каждого образца с медианным Ct по всем образцам. Для потери аллеля используют зонды TaqMan® к областям ближе к 5'-концу (mTU) и ближе к 3'-концу (mTD) от области целевого геномного локуса, нацеленного для делеции. Для приобретения аллеля используют зонды TaqMan® к кассете резистентности к неомицину. Однако такие зонды можно разработать к любой области полинуклеотидной вставки. Для диплоидного клона с гетерозиготным нацеливанием число копий TaqMan® для каждого из зондов mTU, mTD и Neo должно быть равно единице. Для диплоидных клонов с гомозиготным нацеливанием число копий TaqMan® для каждого из mTU и mTD должно быть равно нулю, а число копий TaqMan® для Neo должно быть равно двум. Аналогичным образом, для диплоидных клонов без нацеливания число копий TaqMan® для каждого из mTU и mTD должно быть равно двум, а число копий TaqMan® для Neo должно быть равно нулю. Для диплоидных гетерозиготных коллапсированных клонов число копий TaqMan® для mTU и mTD должно быть равно единице, а число копий для Neo должно быть равно нулю. Для диплоидных гомозиготных коллапсированных клонов число копий TaqMan® для каждого из зондов mTU, mTD и Neo должно быть равно нулю.
Однако поскольку парные гРНК могут создавать большие Cas-опосредованные делеции в целевом геномном локусе, они могут быть полезны для улучшения стандартных тестов LOA и GOA для проверки правильности нацеливания посредством LTVEC. Например, тесты LOA и GOA сами по себе могут не различать имеющие правильное нацеливание клеточные клоны от клонов, в которых большая Cas-индуцированная делеция целевого геномного локуса совпадает со случайной интеграцией LTVEC в другое место генома. Поскольку давление отбора в целевой клетке основано на кассете селекции, случайная трансгенная интеграция LTVEC в другом месте генома обычно включает кассету селекции и прилегающие области LTVEC, но исключает более дистальные области LTVEC. Например, если тесты LOA и GOA используются для оценки нацеленной интеграции LTVEC и в тесте GOA применяются зонды к кассете селекции, то гетерозиготная делеция в целевом геномном локусе в комбинации со случайной трансгенной интеграцией LTVEC даст те же показатели, что и гетерозиготная нацеленная интеграция LTVEC в целевой геномный локус. Чтобы проверить правильное нацеливание с помощью LTVEC, тесты на сохранение можно использовать по отдельности или в сочетании с тестами LOA и/или GOA.
В тестах на сохранение TaqMan® используются зонды ближе к 5'-концу и ближе к 3'-концу, соответствующие целевой 5'-последовательности для 5' гомологичного плеча (зонд retU) и целевой 3'-последовательности для 3' гомологичного плеча (зонд retD) (см. Фиг. 17B, где показано использование тестов на сохранение TaqMan® в комбинации с тестами GOA и LOA при скрининге выполняемой посредством CRISPR/Cas9 гуманизации с применением селекции неомицином). На Фиг. 17B также показано, как разные зонды в полинуклеотидной вставке можно использовать для тестов GOA (см. зонд hTU ближе к 5'-концу и зонд hTD ближе к 3'-концу). Результаты тестов GOA, LOA и теста на сохранение для разных типов нацеленных модификаций и трансгенных вставок показаны в таблице 12.
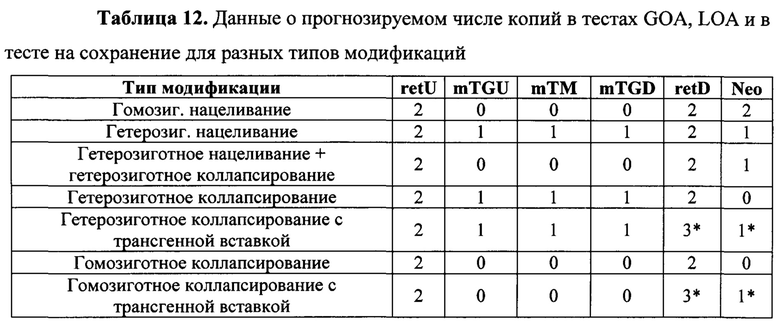
Тесты TaqMan на сохранение также можно использовать в комбинации с тестами LOA для скрининга выполняемых посредством CRISPR/Cas9 делеций с применением парных гРНК (см. Фиг. 17C). В таких тестах число копий retU и retD во всех случаях должно оставаться равным двум. Число копий менее двух указывает на большие Cas9-опосредованные делеции, выходящие за пределы области, нацеленной для делеции. Результаты теста LOA и теста на сохранение для разных типов модификаций, связанных с коллапсированием, показаны в таблице 13.
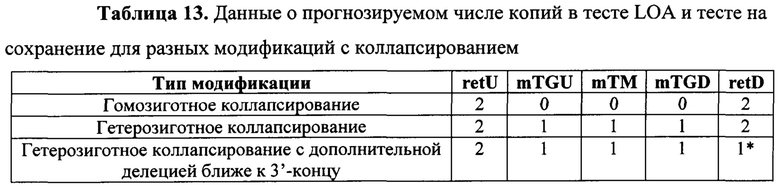
Пример 7. CRISPR/Cas9-опосредованное нацеливание с использованием четырех гидовых РНК
Для выполнения точной одноэтапной делеции области из приблизительно 900 т.п.н. модифицированного мышиного локуса тяжелой цепи иммуноглобулина и одновременной замены на вставку Pgk-Neo (промотор фосфоглицераткиназы I, функционально связанный с геном неомицинфосфотрансферазы), фланкированную сайтами loxP, мы ввели в мышиные ЭС-клетки методом электропорации следующие молекулы нуклеиновых кислот: (1) LTVEC, (2) плазмиду, кодирующую эндонуклеазу Cas9; и (3) одну или более плазмид, кодирующих четыре одногидовых CRISPR РНК (гРНК). В каждом эксперименте LTVEC подвергали линеаризации. Локус, нацеленный для модификации, представлял собой область из приблизительно 900 т.п.н. мышиного локуса тяжелой цепи иммуноглобулина, в которой генные сегменты вариабельных областей (VH, DH, JH) были заменены человеческими аналогами (см. Фиг. 18). Был создан LTVEC, который содержал вставку Pgk-Neo длиной приблизительно 2 т.п.н., фланкированную 19 т.п.н. 5' гомологичного плеча и 13 т.п.н. 3' гомологичного плеча, чтобы направлять событие гомологичной рекомбинации, делегирующее область из приблизительно 900 т.п.н. из целевого локуса и вставляющее кассету селекции лекарственным средством, которая направляет экспрессию неомицинфосфотрансферазы, придающую резистентность к G418.
Из четырех использованных гРНК две направляли расщепление Cas9 вблизи 5'-конца целевого локуса (5' гРНК_I и 5' гРНК_II на Фиг. 18) и две направляли расщепление Cas9 вблизи 3'-конца целевого локуса (3' гРНК_I и 3' гРНК_II на Фиг. 18). Целевые последовательности 5' гРНК_I и 5' гРНК_II находились на расстоянии приблизительно 150 п.н. друг от друга, а целевые последовательности 3' гРНК_I и 3' гРНК_II перекрывались, причем целевой сайт 3' гРНК_II был смещен на 1 п.н. относительно целевого сайта 3' гРНК_I.
ЭС-клетки, захватившие LTVEC и внедрившие его в свои геномы, были способны расти и образовывать колонии на чашках Петри в питательной среде, содержащей антибактериальное лекарственное средство. Мы собрали резистентные к лекарственному средству колонии и провели скрининг по способу модификации аллеля (Valenzuela et al. (2003) Nat. Biotech. 21: 652-660; Frendewey et al. (2010) Methods Enzymol. 476: 295-307; полностью включенные в настоящий документ путем ссылки) с целью идентификации клонов, имеющих гуманизированный аллель с правильным нацеливанием (см. таблицу 14 ниже). Кроме того, использовали ПЦР-тесты в реальном времени, распознающие последовательности в гомологичных плечах LTVEC, называемые тестами на сохранение, чтобы проверить правильность нацеливания LTVEC на геном мыши (см. таблицу 14 ниже).
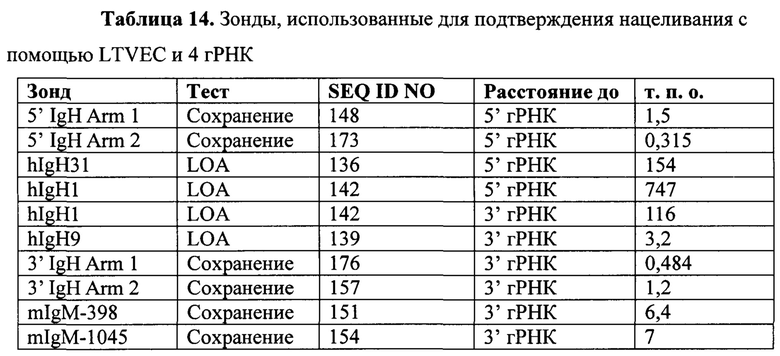
В полученных ЭС-клетках с нацеливанием область из приблизительно 900 т.п.н. была удалена и заменена вставкой Pgk-Neo в обоих аллелях (см. Фиг. 18). Эти большие делеция и замена достигались с неожиданно высокой частотой (эффективность приблизительно 1,2% для биаллельной делеции).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>Frendewey, David
Lai, Ka-Man Venus
Auerbach, Wojtek
Lee, JeffreyD.
Mujica, Alexander O.
Droguett, Gustavo
Trzaska, Sean
Hunt, Charleen
Gagliardi, Anthony
Valenzuela, David M.
Voronina, Vera
Macdonald, Lynn
Murphy, Andrew J.
Yancopoulos, George D.
<120>СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С
ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК
<130> 57766-472225
<150> US 62/083,005
<151> 2014-11-21
<150> US 62/182,314
<151> 2015-06-19
<150> US 62/211,421
<151> 2015-08-28
<160> 176
<170>FastSEQ для Windows, версия 4.0
<210> 1
<211> 80
<212>РНК
<213>Искусственнаяпоследовательность
<220>
<223>гРНК
<400> 1
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcuuuu 80
<210> 2
<211> 42
<212>РНК
<213>Искусственная последовательность
<220>
<223>гРНК
<400> 2
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cg 42
<210> 3
<211> 30
<212>РНК
<213>Искусственная последовательность
<220>
<223>крРНК
<400> 3
guuuuagagc uagaaauagc aaguuaaaau 30
<210> 4
<211> 33
<212>РНК
<213>Искусственная последовательность
<220>
<223>крРНК
<400> 4
guuuuagagc uagaaauagc aaguuaaaau aag 33
<210> 5
<211> 26
<212>РНК
<213>Искусственная последовательность
<220>
<223>крРНК
<400> 5
gaguccgagc agaagaagaa guuuua 26
<210> 6
<211> 12
<212>РНК
<213>Искусственная последовательность
<220>
<223>тракрРНК
<400> 6
aaggcuaguc cg 12
<210> 7
<211> 50
<212>РНК
<213>Искусственная последовательность
<220>
<223>тракрРНК
<400> 7
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugcuuuu 50
<210> 8
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность распознавания РНК CRISPR
<220>
<221>misc_feature
<222> (2)...(21)
<223>n = а, т, ц или г
<400> 8
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 9
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность распознавания РНК CRISPR
<220>
<221>misc_feature
<222> (1)...(21)
<223>n = а, т, ц или г
<400> 9
nnnnnnnnnn nnnnnnnnnn ngg 23
<210> 10
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность распознавания РНК CRISPR
<220>
<221>misc_feature
<222> (3)...(23)
<223>n = а, т, ц или г
<400> 10
ggnnnnnnnn nnnnnnnnnn nnngg 25
<210> 11
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК A
<400> 11
atcacaaacc agttaaccgg 20
<210> 12
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК B
<400> 12
tttcagacga gccgacccgg 20
<210> 13
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК C
<400> 13
tgtgtgtcat agcgatgtcg 20
<210> 14
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК D
<400> 14
aacaggtacc ctatcctcac 20
<210> 15
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК E
<400> 15
ggcccggacc tagtctctct 20
<210> 16
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания C5 (Hc) гРНК E2
<400> 16
tcgtggttgc atgcgcactg 20
<210> 17
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК A
<400> 17
gggaacccac agcatactcc 20
<210> 18
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК B
<400> 18
gaatcatgca cggctacccc 20
<210> 19
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК B2
<400> 19
tgctcctatg gggaggcgcg 20
<210> 20
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК C
<400> 20
actgagatca atgaccccga 20
<210> 21
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК D
<400> 21
gggtcgcccg gaacctctac 20
<210> 22
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК E2
<400> 22
cttggataac attgataccc 20
<210> 23
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК E
<400> 23
ggggcagagc ccttatatca 20
<210> 24
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Lrp5 гРНК F
<400> 24
tcgctcacat taatccctag 20
<210> 25
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК А
<400> 25
tgtgggcctt tgctgatcac 20
<210> 26
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК B
<400> 26
aatctatgat cctatggcct 20
<210> 27
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК D
<400> 27
tgccaatagc agtgacttga 20
<210> 28
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК C
<400> 28
gggaagaatg ggctattgtc 20
<210> 29
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК E
<400> 29
ggttgtttgt gctgatgacg 20
<210> 30
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Ror1 гРНК F
<400> 30
ccgtcctagg ccttctacgt 20
<210> 31
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК А
<400> 31
gtactgggga atcggtggtc 20
<210> 32
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223> Сегмент ДНК-нацеливания Trpa1 гРНК A2
<400> 32
cacgcactcc aaatttatcc 20
<210> 33
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК B
<400> 33
ctaagtgtgt atcagtacat 20
<210> 34
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК C
<400> 34
tgccctgcac aataagcgca 20
<210> 35
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК D
<400> 35
actcattgaa acgttatggc 20
<210> 36
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223> Сегмент ДНК-нацеливания Trpa1 гРНК E2
<400> 36
agtaagggtg gattaaattc 20
<210> 37
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК E
<400> 37
gccatctaga ttcatgtaac 20
<210> 38
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Trpa1 гРНК F
<400> 38
gactagaaat gttctgcacc 20
<210> 39
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 190045
<400> 39
gagctcatag ccaacagctt g 21
<210> 40
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223> 190061 прямой праймер
<400> 40
atgcatcaga tcacgctcag 20
<210> 41
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 190068
<400> 41
gtccttgtgg catttccaac 20
<210> 42
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 190030
<400> 42
ccagtatggt gtcagttaat agcg 24
<210> 43
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 190033
<400> 43
ctgtgcagaa agcagcctc 19
<210> 44
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 190013
<400> 44
cctctccctc taggcacctg 20
<210> 45
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190045
<400> 45
tctttaaggg ctccgttgtc 20
<210> 46
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190061
<400> 46
aagaccaacc attcacccag 20
<210> 47
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190068
<400> 47
ttcccagtcc aagtcaaagg 20
<210> 48
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190030
<400> 48
ctgttatctg caaggcaccc 20
<210> 49
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190033
<400> 49
acaactggat cctgattcgc 20
<210> 50
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 190013
<400> 50
taagagggca tgggtgagac 20
<210> 51
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд C2 (B6)
<400> 51
aattcagaag acctatcgta 20
<210> 52
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T3 (B6)
<400> 52
tatgtgtata ggtgtttgga t 21
<210> 53
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T6 (B6)
<400> 53
tacattgcta aatgaaacc 19
<210> 54
<211> 16
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T7 (B6)
<400> 54
cgcagtcatg cacata 16
<210> 55
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T8 (B6)
<400> 55
ttataaagcc cagtatgtac 20
<210> 56
<211> 14
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T9 (B6)
<400> 56
tgctgcataa tcag 14
<210> 57
<211> 18
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T10 (B6)
<400> 57
tcaggagtga attggata 18
<210> 58
<211> 16
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T11 (B6)
<400> 58
ctgctactta cctttg 16
<210> 59
<211> 13
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T13 (B6)
<400> 59
aggaggaaaa cgc 13
<210> 60
<211> 17
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T14 (B6)
<400> 60
cctttgttcc tcataag 17
<210> 61
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд C2 (129)
<400> 61
aattcagaag acctattgta 20
<210> 62
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T3 (129)
<400> 62
tatgtgtataggtgtttgcat 21
<210> 63
<211> 16
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T6 (129)
<400> 63
cattgctaca tgaaac 16
<210> 64
<211> 16
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T7 (129)
<400> 64
cgcagtcatg cacgta 16
<210> 65
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T8 (129)
<400> 65
tgagaattta taaagcccaa tat 23
<210> 66
<211> 14
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T9 (129)
<400> 66
tgctgcatga tcag 14
<210> 67
<211> 15
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T10 (129)
<400> 67
tcaggagtga atcgg 15
<210> 68
<211> 16
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T11 (129)
<400> 68
ctgctagtta cctttg 16
<210> 69
<211> 15
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T13 (129)
<400> 69
aggaggaaga cgcag 15
<210> 70
<211> 17
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд T14 (129)
<400> 70
ctttgttctt cataagc 17
<210> 71
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер C2
<400> 71
atgagggatt tccttaatca gacaa 25
<210> 72
<211> 29
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T3
<400> 72
tggtatgttt attcttactc aaggttttg 29
<210> 73
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T6
<400> 73
gggcaactga tggaaagaac tc 22
<210> 74
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T7
<400> 74
gactgacgca caaacttgtc ctt 23
<210> 75
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T8
<400> 75
cccaaagcat ataacaagaa caaatg 26
<210> 76
<211> 18
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T9
<400> 76
gcaggacgca ggcgttta 18
<210> 77
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T10
<400> 77
gcatcctcat ggcagtctac atc 23
<210> 78
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T11
<400> 78
cctgcccctt gatgagtgtt 20
<210> 79
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T13
<400> 79
ccctctttga tatgctcgtg tgt 23
<210> 80
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер T14
<400> 80
tcccacaggt ccatgtcttt aa 22
<210> 81
<211> 28
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер C2
<400> 81
agactacaatgagctaccatcataaggt 28
<210> 82
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T3
<400> 82
caaccatcta aaactccagt tcca 24
<210> 83
<211> 28
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T6
<400> 83
tgtgtaacag gacagttgaa tgtagaga 28
<210> 84
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T7
<400> 84
cttaaaaccc gccctgcat 19
<210> 85
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T8
<400> 85
ctacaggaga tgtggctgtt ctatgt 26
<210> 86
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T9
<400> 86
tcagcgtgat tcgcttgtag tc 22
<210> 87
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T10
<400> 87
tgcatagctg tttgaataat gacaag 26
<210> 88
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T11
<400> 88
tgcagcatct ctgtcaagca a 21
<210> 89
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T13
<400> 89
gcaacaacat aacccacagc ataa 24
<210> 90
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер T14
<400> 90
gctaagcgtt tggaagaaat tcc 23
<210> 91
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер SV 13.7
<400> 91
taggctctaa ggatgctggc 20
<210> 92
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер SV 13.7
<400> 92
aagcagcttc aaaccctctg 20
<210> 93
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер SV 20.0
<400> 93
ttacttggcc ttggaactgc 20
<210> 94
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер SV 20.0
<400> 94
tgattcgtaa tcgtcactgc c 21
<210> 95
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер SV 36.9
<400> 95
tcctgtcccg agaaactgtc 20
<210> 96
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер SV 36.9
<400> 96
agctggcttt cagagagctg 20
<210> 97
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер SV 56.7
<400> 97
ttagaaagtg ccaaccaggc 20
<210> 98
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер SV 56.7
<400> 98
ctctggctag gaacaatggc 20
<210> 99
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-lr-f
<400> 99
gttaggtgca gggtctactc agctg 25
<210> 100
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-5’-f
<400> 100
ggaggagagg agaagcagcc 20
<210> 101
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-A
<400> 101
ggaggagagg agaagcagcc 20
<210> 102
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер h-lr-r
<400> 102
gcaaacagcc ttcttcccac attcgg 26
<210> 103
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-5’-r
<400> 103
ttgctttcag tagttcaggt gtgc 24
<210> 104
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер h-5'-r
<400> 104
ggcgttgtca ggaagttgcc 20
<210> 105
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-F
<400> 105
tgaagttgag aggcacatga gg 22
<210> 106
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>праймер m-E2
<400> 106
tagagtagcc acaggcagca aagc 24
<210> 107
<211> 21
<212>РНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Cmah гРНК A
<400> 107
gugacaggaa ggcuucucac c 21
<210> 108
<211> 21
<212>РНК
<213>Искусственная последовательность
<220>
<223>Сегмент ДНК-нацеливания Cmah гРНК B
<400> 108
gcuuacaagc aauuugcuga c 21
<210> 109
<211> 108
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Локус Cmah
<400> 109
ttgccagtga ccctgtttgc agttagagtt gacaggaagg cttctcaccc gggacatttt 60
aaatgaccca gtcagcaaat tgcttgtaag ttttggtgtt ctttcatt 108
<210> 110
<211> 46
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Локус Cmah ближе к 5’ концу от разреза гРНК А
<400> 110
ttgccagtga ccctgtttgc agttagagtt gacaggaagg cttctc 46
<210> 111
<211> 35
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Локус Cmah ближе к 3’ концу от разреза гРНК В
<400> 111
agcaaattgc ttgtaagttt tggtgttctt tcatt 35
<210> 112
<211> 27
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность локуса Cmah, вырезанная гРНК А и В
<400> 112
acccgggaca ttttaaatga cccagtc 27
<210> 113
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>C5 праймер m-5’-F
<400> 113
accccagcat ctgacgacac c 21
<210> 114
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>C5 праймер m-5’-R
<400> 114
agaaagaccg cagtggaacc 20
<210> 115
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>C5 праймер h-5’-R
<400> 115
tccccacatg cctagtagga g 21
<210> 116
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер TaqMan Cmah
<400> 116
gtgaccctgt ttgcagttag ag 22
<210> 117
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд TaqMan Cmah
<400> 117
acaggaaggc ttctcacccg gga 23
<210> 118
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер TaqMan Cmah
<400> 118
tgaagtcagg aaactgttcc aatg 24
<210> 119
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 7064retU
<400> 119
cctcctgagc tttcctttgc ag 22
<210> 120
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 7064retU
<400> 120
cctagacaac acagacactg tatca 25
<210> 121
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд TaqMan 7064retU
<400> 121
ttctgccctt gaaaaggaga ggc 23
<210> 122
<211> 18
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 7064retD
<400> 122
cctctgaggc cacctgaa 18
<210> 123
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 7064retD
<400> 123
ccctgacaag ttctgccttc tac 23
<210> 124
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд TaqMan 7064retD
<400> 124
tgcccaagcc tctgcagctt t 21
<210> 125
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 7140retU
<400> 125
cccagcatct gacgacacc 19
<210> 126
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 7140retU
<400> 126
gaccactgtg ggcatctgta g 21
<210> 127
<211> 27
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд TaqMan 7140retU
<400> 127
ccgagtctgc tgttactgtt agcatca 27
<210> 128
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер 7140retD
<400> 128
cccgacacct tctgagcatg 20
<210> 129
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер 7140retD
<400> 129
tgcaggctga gtcaggattt g 21
<210> 130
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд TaqMan 7140retD
<400> 130
tagtcacgtt ttgtgacacc ccaga 25
<210> 131
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер LOA mADAM6-2
<400> 131
agggctgagg gagaacatat ac 22
<210> 132
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер LOA mADAM6-2
<400> 132
aggcctgatg caggagctat 20
<210> 133
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд LOA mADAM6-2
<400> 133
tcctctcagc tggattaaca gcatca 26
<210> 134
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер LOA hIgH31
<400> 134
atcacactca tcccatcccc 20
<210> 135
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер LOA hIgH31
<400> 135
cacagggaag caggaactgc 20
<210> 136
<211> 29
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд LOA hIgH31
<400> 136
cccttcccta agtaccacag agtgggctc 29
<210> 137
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер LOA hIgH9
<400> 137
tcctccaacg acaggtccc 19
<210> 138
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер LOA hIgH9
<400> 138
gatgaactga cgggcacagg 20
<210> 139
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд LOA hIgH9
<400> 139
tccctggaac tctgccccga caca 24
<210> 140
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер LOA hIgH1
<400> 140
cagtcccgtt gatccagcc 19
<210> 141
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер LOA hIgH1
<400> 141
ggatatgcag cactgtgcca c 21
<210> 142
<211> 30
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд LOA hIgH1
<400> 142
cccatcaggg attttgtatc tctgtggacg 30
<210> 143
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер GOA Neo
<400> 143
ggtggagagg ctattcggc 19
<210> 144
<211> 17
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер GOA Neo
<400> 144
gaacacggcg gcatcag 17
<210> 145
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд GOA Neo
<400> 145
tgggcacaac agacaatcgg ctg 23
<210> 146
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>5’ IgH Arm1, прямой праймер для теста на сохранение
<400> 146
aggatgctgg gaaacagac 19
<210> 147
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>5’ IgH Arm1, обратный праймер для теста на сохранение
<400> 147
gaccactctc aggactctca 20
<210> 148
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>5’ IgH Arm1, зонд для теста на сохранение
<400> 148
tggaaggtcc caaaggaaac caca 24
<210> 149
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер для теста на сохранение mIgM398
<400> 149
gagctcacac cttgaccttt ca 22
<210> 150
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер для теста на сохранение mIgM398
<400> 150
tggtgggacg aacacattta ca 22
<210> 151
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд для теста на сохранение mIgM398
<400> 151
ccagctgtcg cagagatgaa cccc 24
<210> 152
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер для теста на сохранение mIgM1045
<400> 152
tccctccaca gacatcctaa cc 22
<210> 153
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер для теста на сохранение mIgM1045
<400> 153
gttagcggac ttgctgagga a 21
<210> 154
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд для теста на сохранение mIgM1045
<400> 154
tcaccatccc cccctccttt gc 22
<210> 155
<211> 22
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер для теста на сохранение 3’ IgH Arm2
<400> 155
ggtcatgtgg caaggctatt tg 22
<210> 156
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223> 3' обратный праймер для теста на сохранение IgH Arm2
<400> 156
agcctggact ttcggtttgg t 21
<210> 157
<211> 30
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд для теста на сохранение 3’ IgH Arm2
<400> 157
ccactaggta aacttgtagc tgtggtttga 30
<210> 158
<211> 17
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный прямой праймер mIgHp2
<400> 158
gccatgcaag gccaagc 17
<210> 159
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный обратный праймер mIgHp2
<400> 159
agttcttgag ccttagggtg ctag 24
<210> 160
<211> 24
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный зонд mIgHp2
<400> 160
ccaggaaaat gctgccagag cctg 24
<210> 161
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный прямой праймер mIgKd2
<400> 161
gcaaacaaaa accactggcc 20
<210> 162
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный обратный праймер mIgKd2
<400> 162
ggccacattc catgggttc 19
<210> 163
<211> 37
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный зонд mIgKd2
<400> 163
ctgttcctct aaaactggac tccacagtaa atggaaa 37
<210> 164
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный прямой праймер hIgK5
<400> 164
ccccgtcctc ctcctttttc 20
<210> 165
<211> 19
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный обратный праймер hIgK5
<400> 165
tgcaagtgct gccagcaag 19
<210> 166
<211> 32
<212>ДНК
<213>Искусственная последовательность
<220>
<223>исходный зонд hIgK5
<400> 166
tcatgtccat taacccattt accttttgcc ca 32
<210> 167
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность ДНК-нацеливания 3’ гРНК_I
<400> 167
gactactacg gtatggacgt c 21
<210> 168
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность ДНК-нацеливания 3’ гРНК_II
<400> 168
gctactacgg tatggacgtc t 21
<210> 169
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность ДНК-нацеливания 5’ гРНК_I
<400> 169
gaagctgact agtttacgca 20
<210> 170
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>Последовательность ДНК-нацеливания 5’ гРНК_II
<400> 170
gtagcattct tacacctagc a 21
<210> 171
<211> 23
<212>ДНК
<213>Искусственная последовательность
<220>
<223>прямой праймер для теста на сохранение 5’ IgH Arm2
<400> 171
ctgagcatac tgctgcctaa cac 23
<210> 172
<211> 25
<212>ДНК
<213>Искусственная последовательность
<220>
<223>обратный праймер для теста на сохранение 5’ IgH Arm2
<400> 172
gagcagtgca tttcttagtt aagga 25
<210> 173
<211> 26
<212>ДНК
<213>Искусственная последовательность
<220>
<223>зонд для теста на сохранение 5’ IgH Arm2
<400> 173
tgaaatggca gttcttctcc agctgg 26
<210> 174
<211> 20
<212>ДНК
<213>Искусственная последовательность
<220>
<223>3’ IgH Arm1, прямой праймер для теста на сохранение
<400> 174
ggtggagtcc ctggatgatg 20
<210> 175
<211> 21
<212>ДНК
<213>Искусственная последовательность
<220>
<223>3’ IgH Arm1, обратный праймер для теста на сохранение
<400> 175
atccctccag ccataggatt g 21
<210> 176
<211> 28
<212>ДНК
<213>Искусственная последовательность
<220>
<223>3’ IgH Arm1, зонд для теста на сохранение
<400> 176
ctttggaggc tcatttgagg gagatgct 28
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2771532C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ ПОСРЕДСТВОМ ОДНОЭТАПНОГО МНОЖЕСТВЕННОГО НАЦЕЛИВАНИЯ | 2015 |
|
RU2707137C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2711740C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ ИЛИ ПОДДЕРЖАНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2015 |
|
RU2706965C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2745563C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2829155C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАПРАВЛЕННОЙ МОДИФИКАЦИИ ГЕНОМА | 2014 |
|
RU2725520C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАПРАВЛЕННОЙ МОДИФИКАЦИИ ГЕНОМА | 2020 |
|
RU2751237C1 |
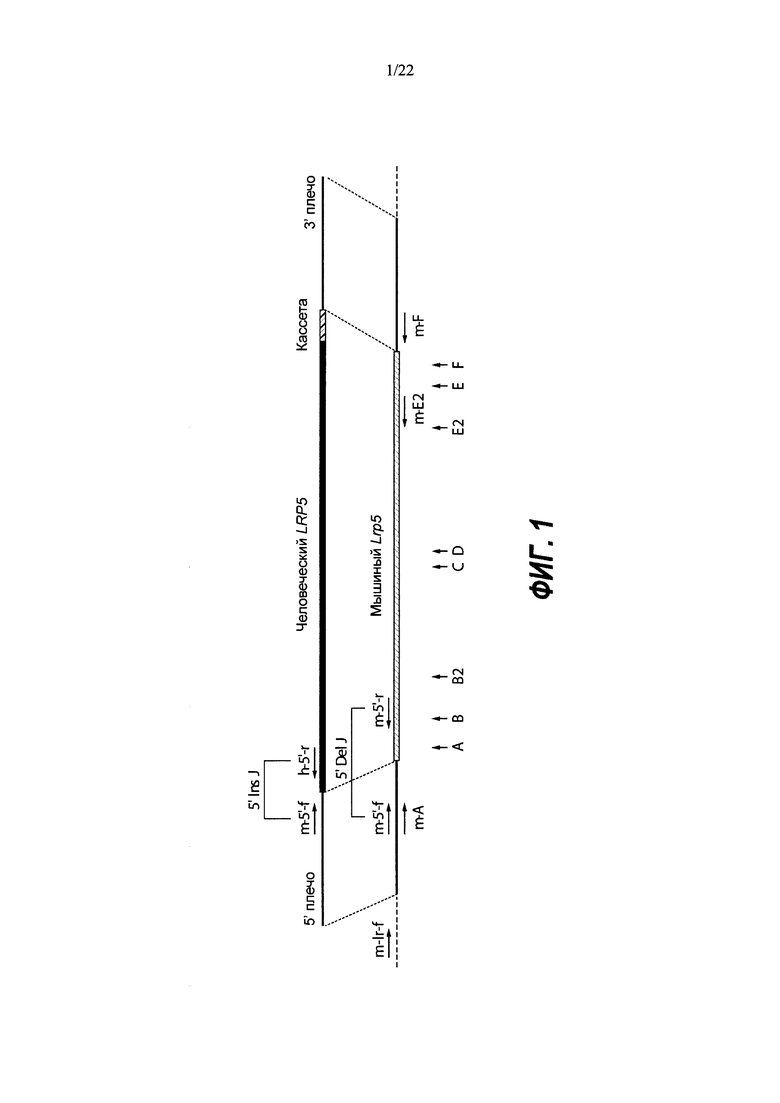
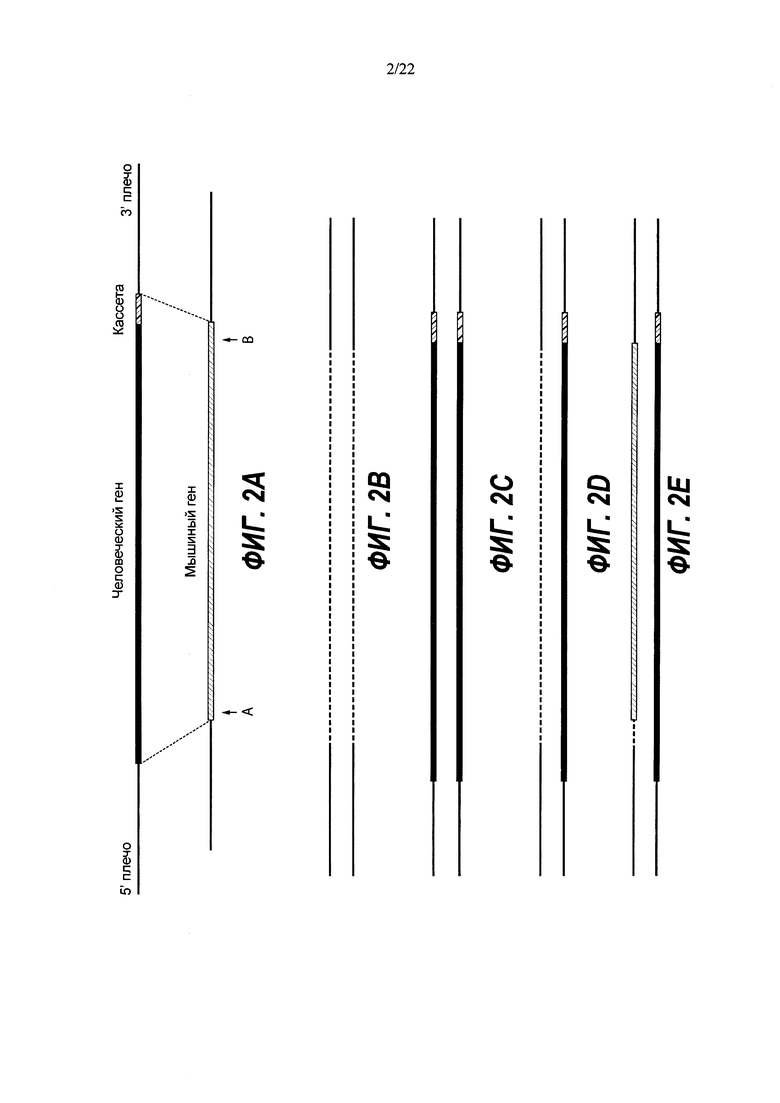
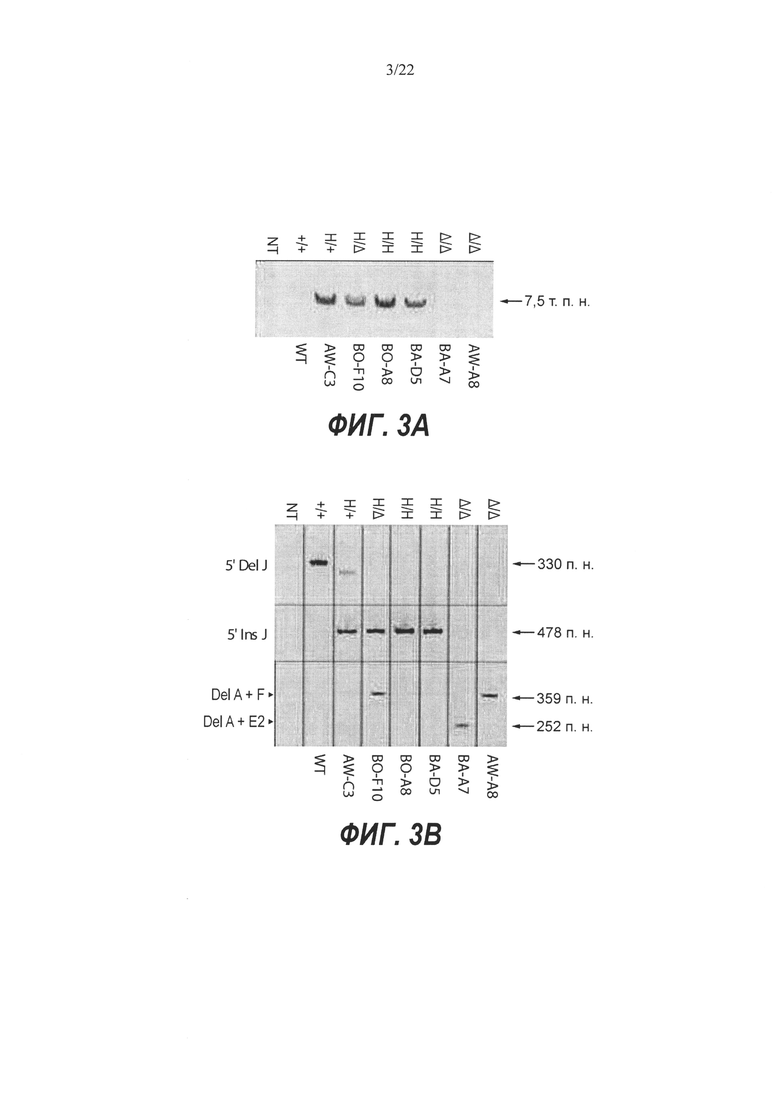
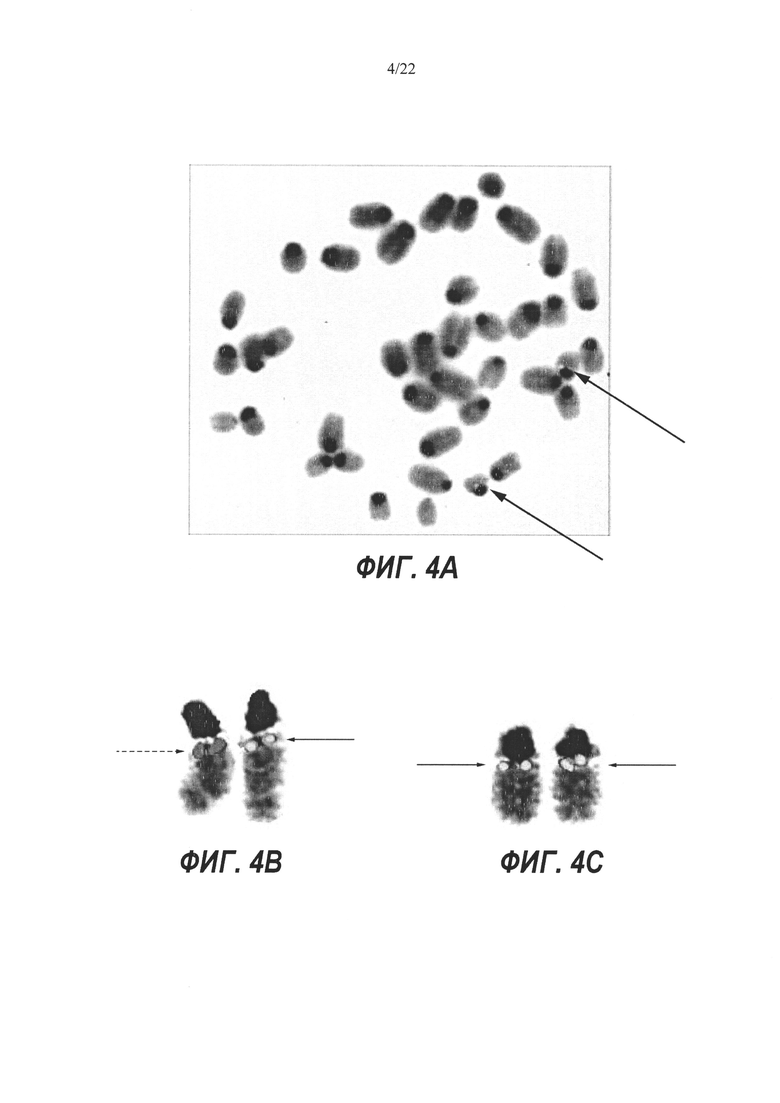
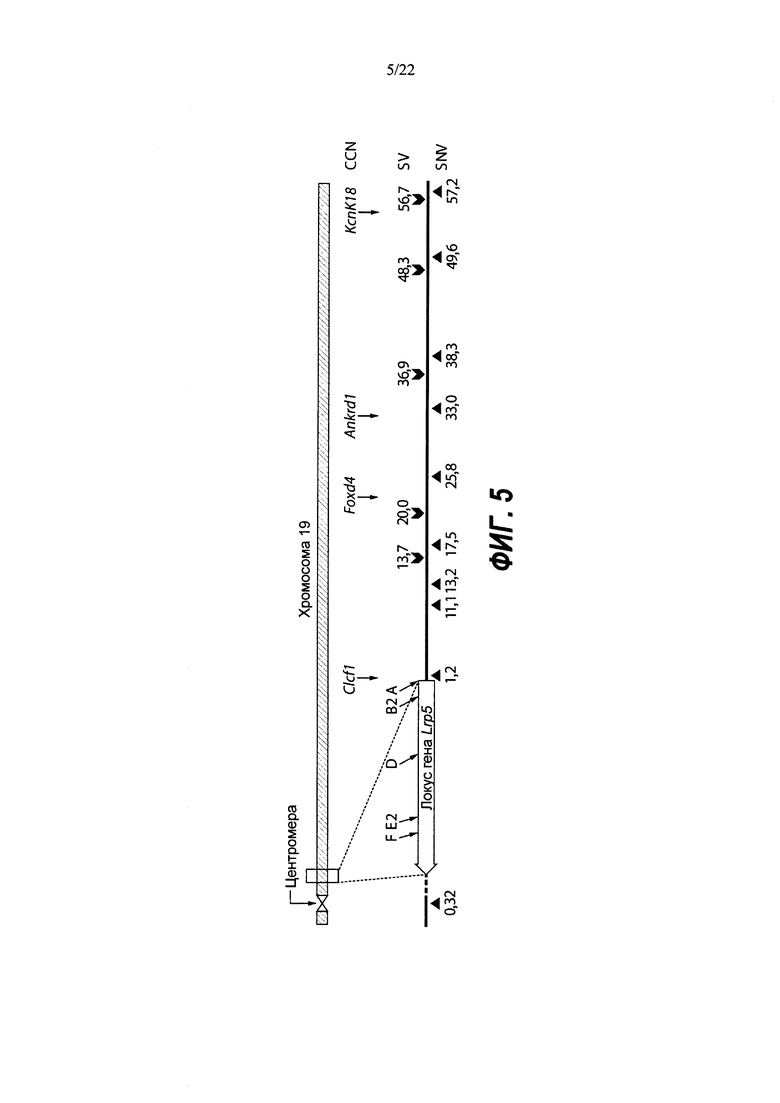
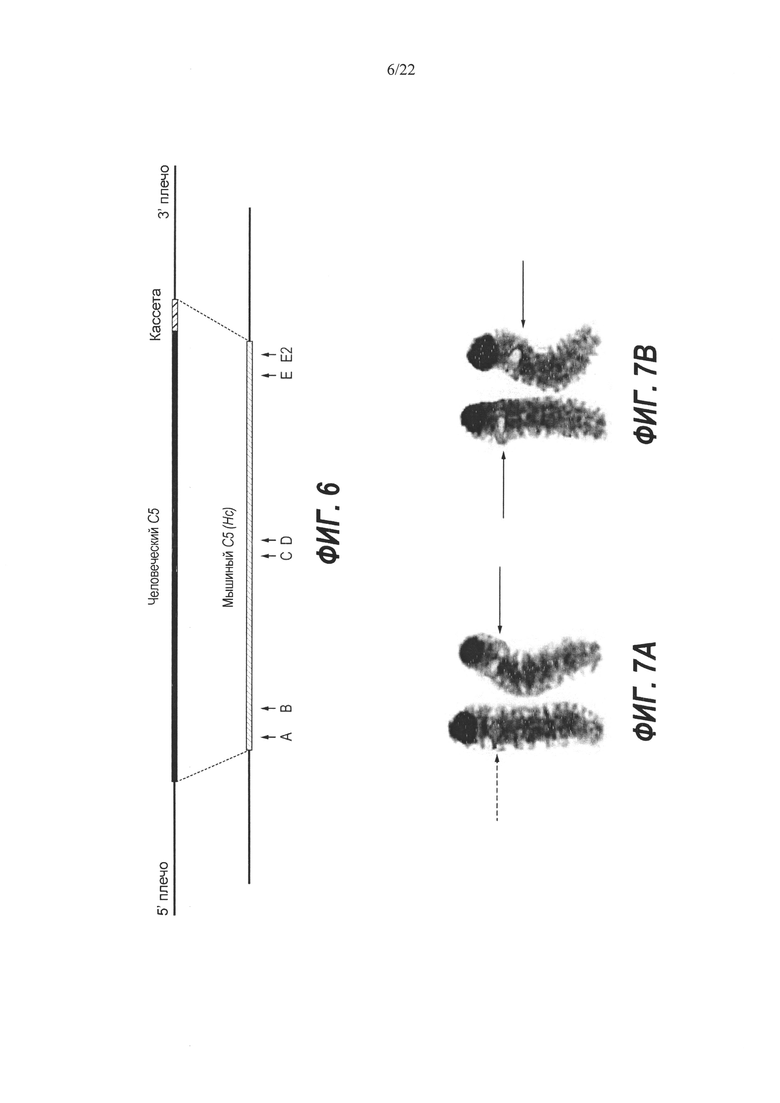
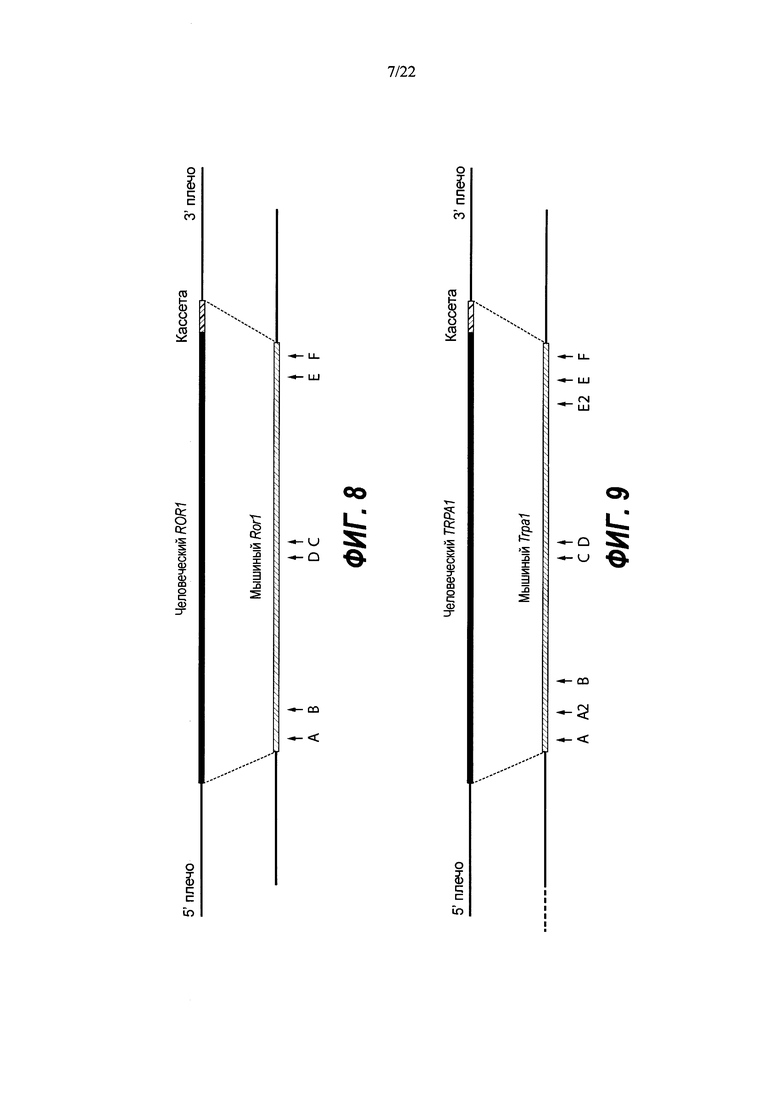
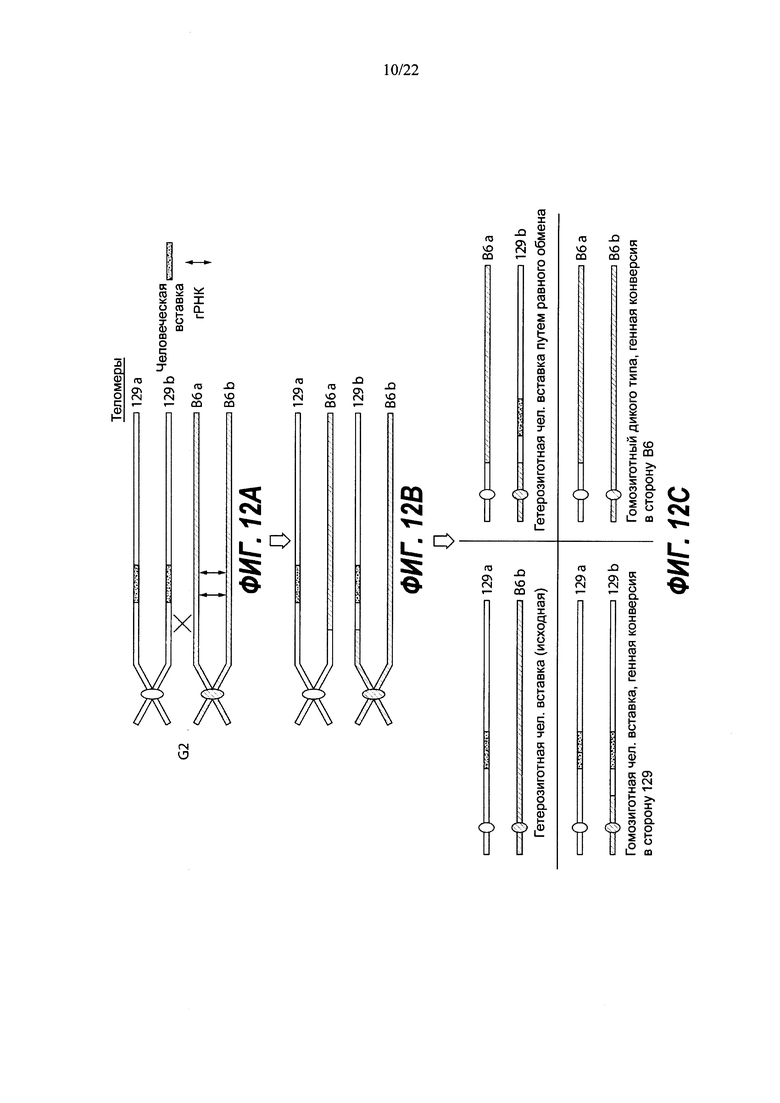
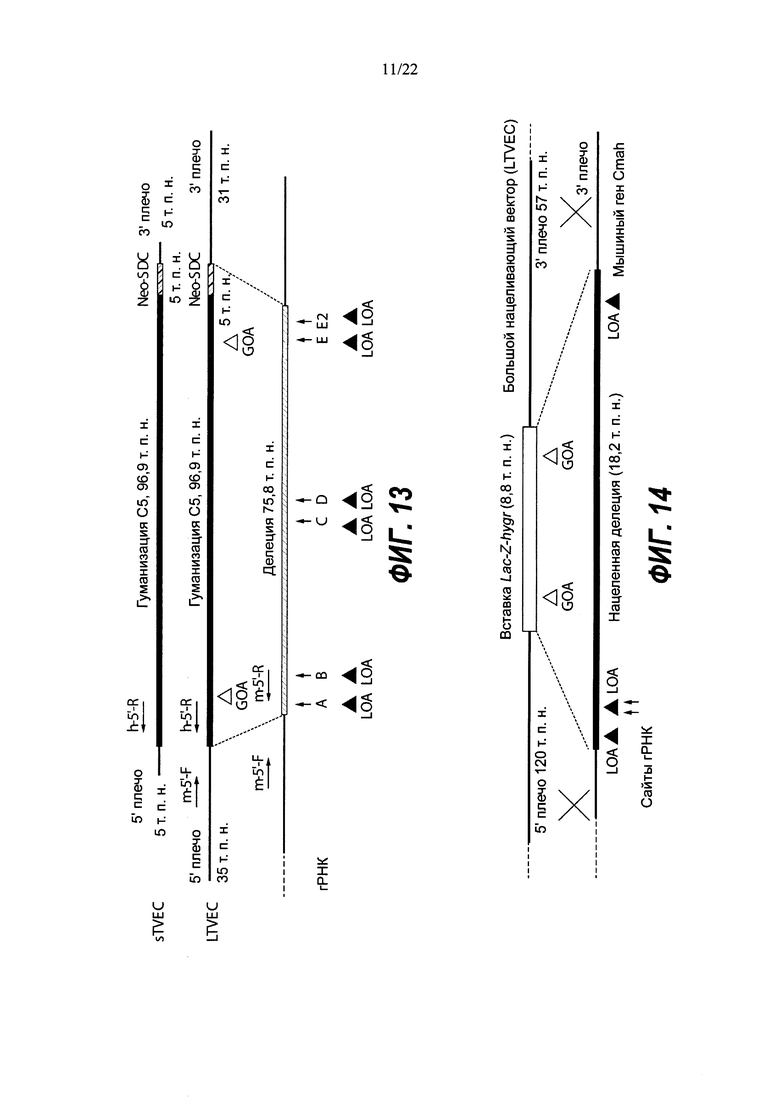
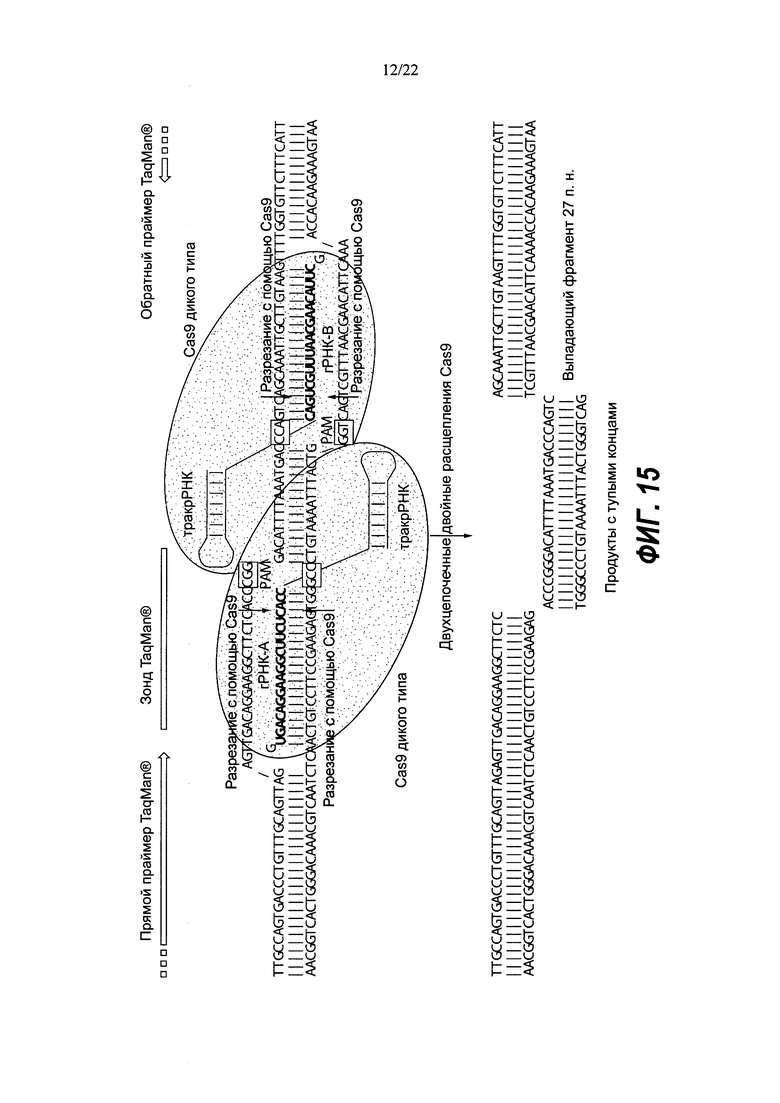
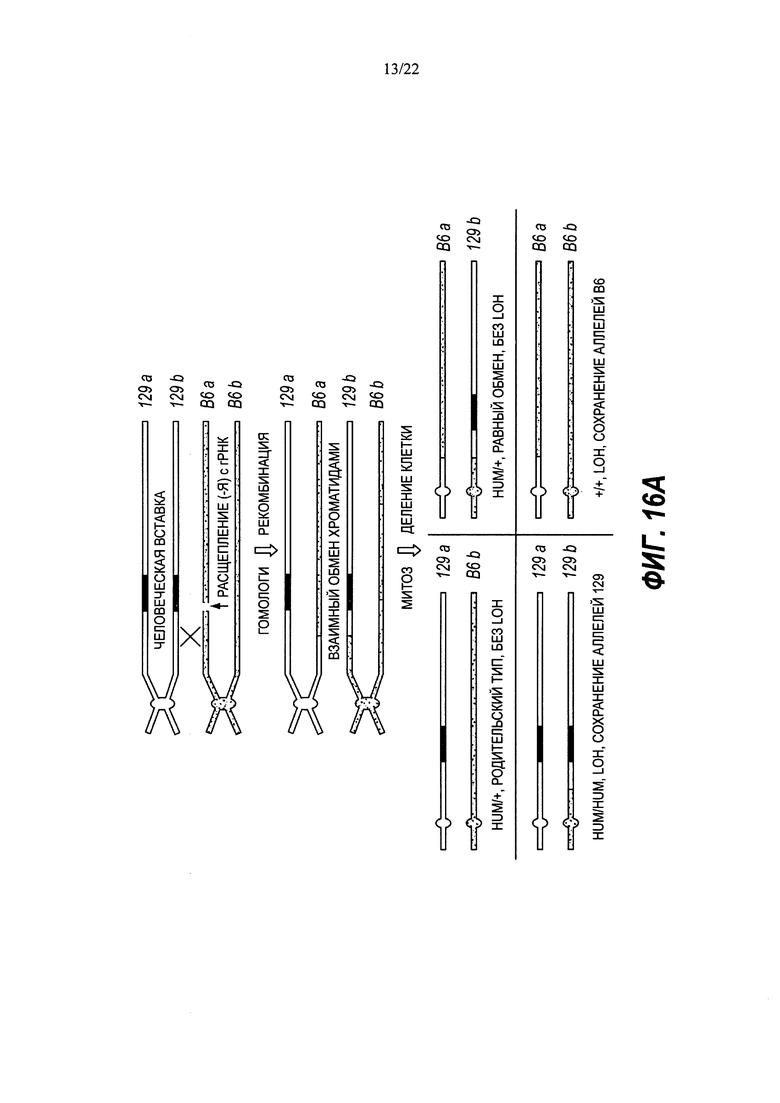
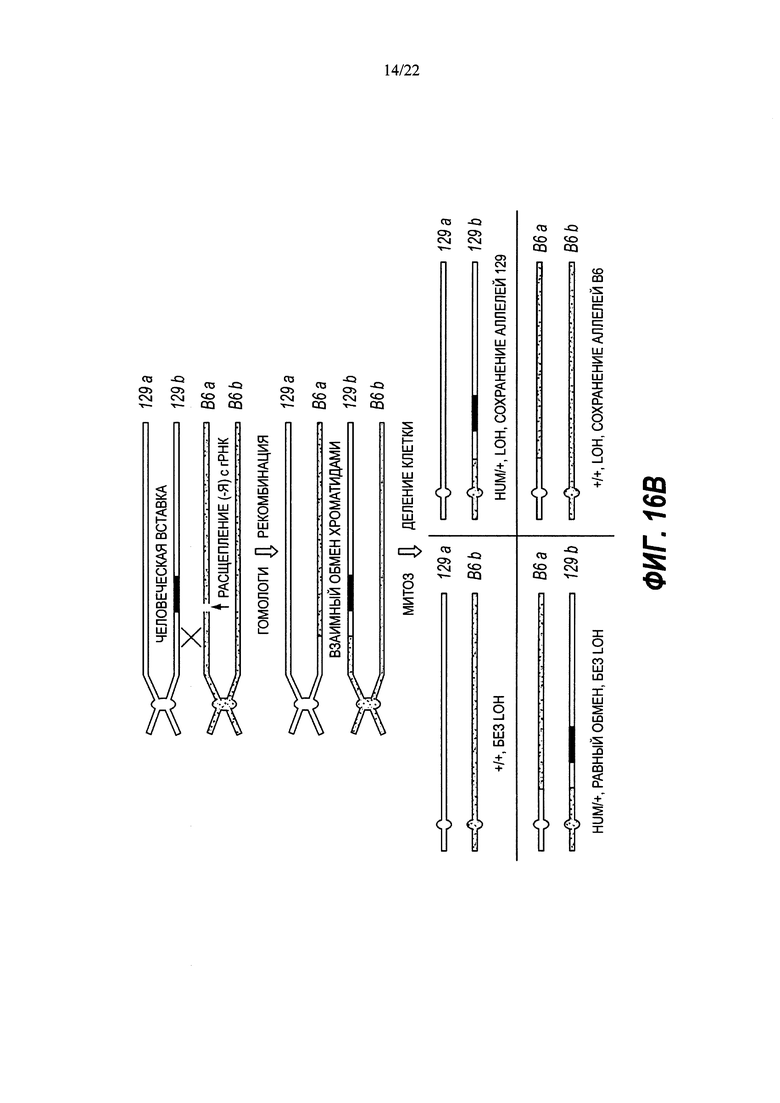
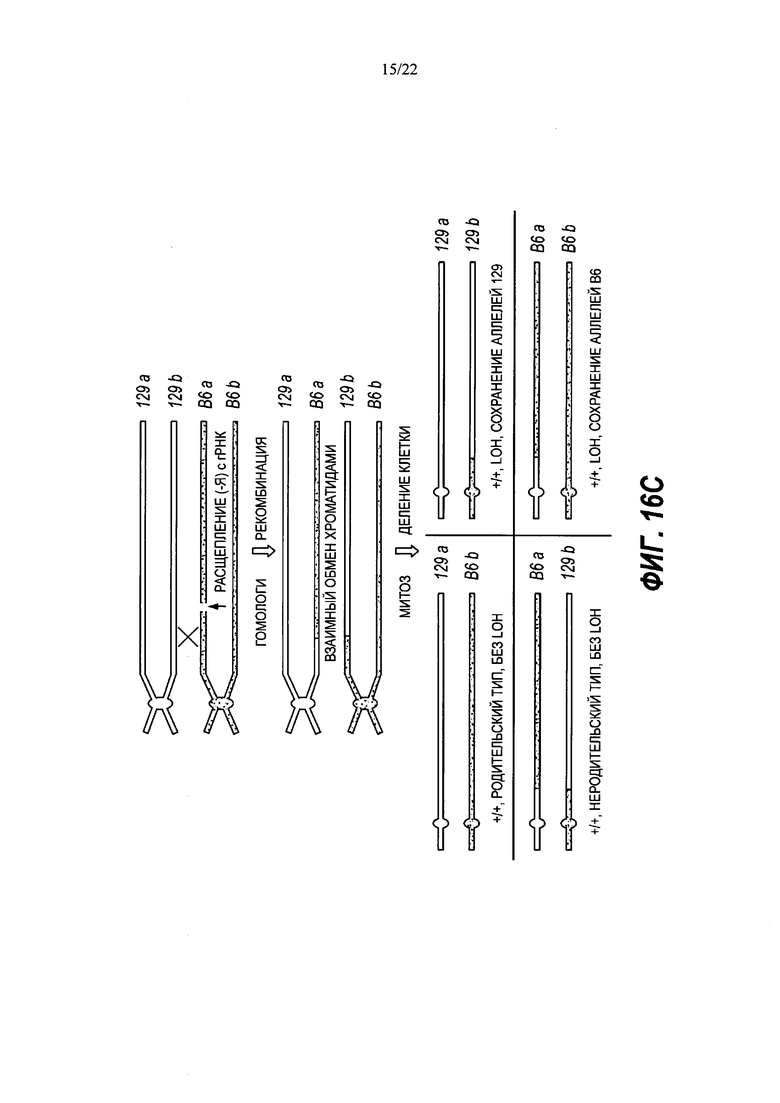
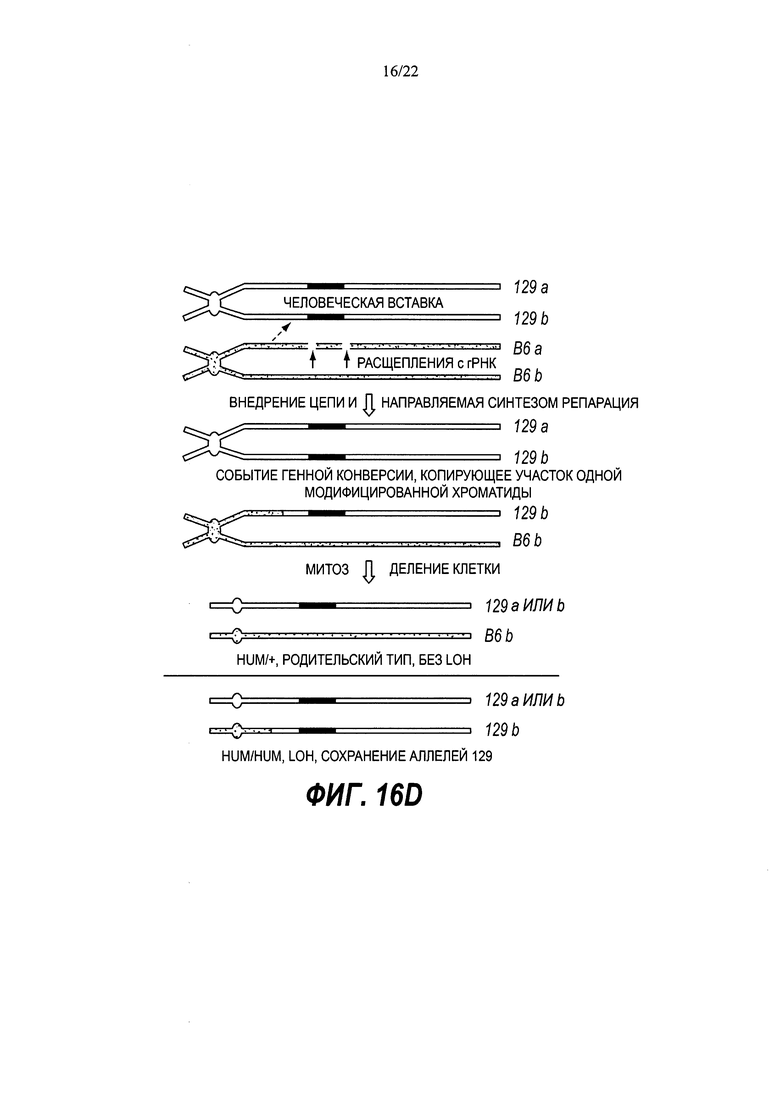
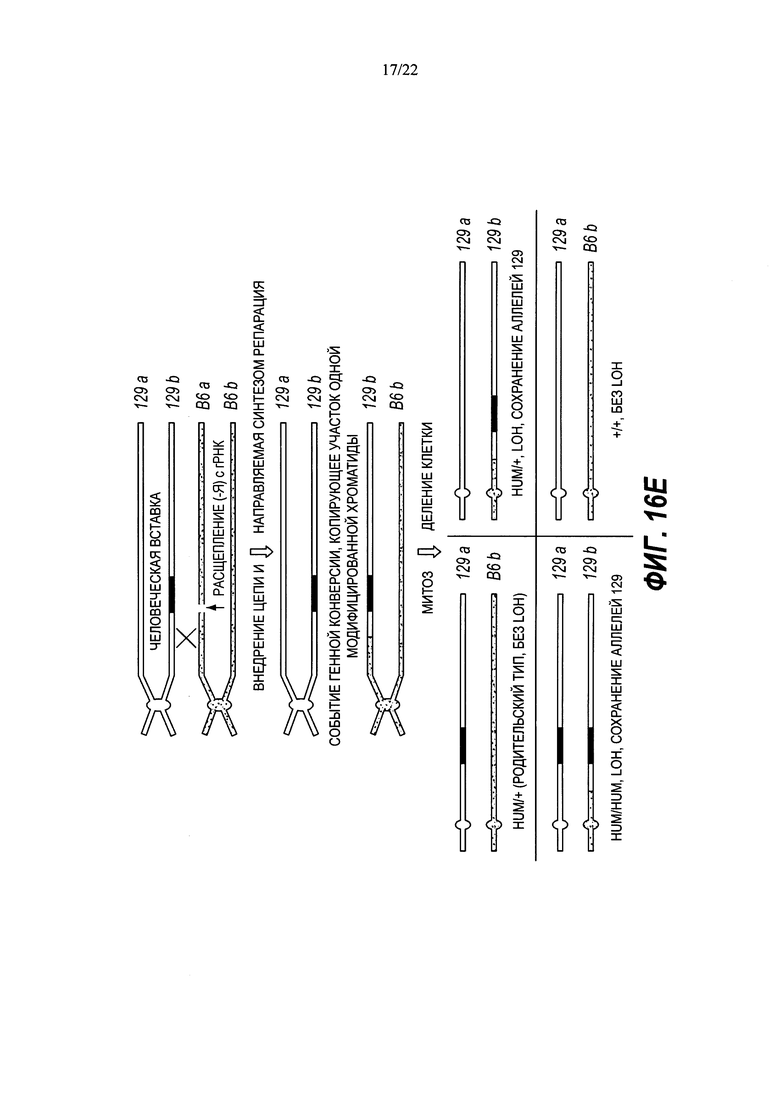
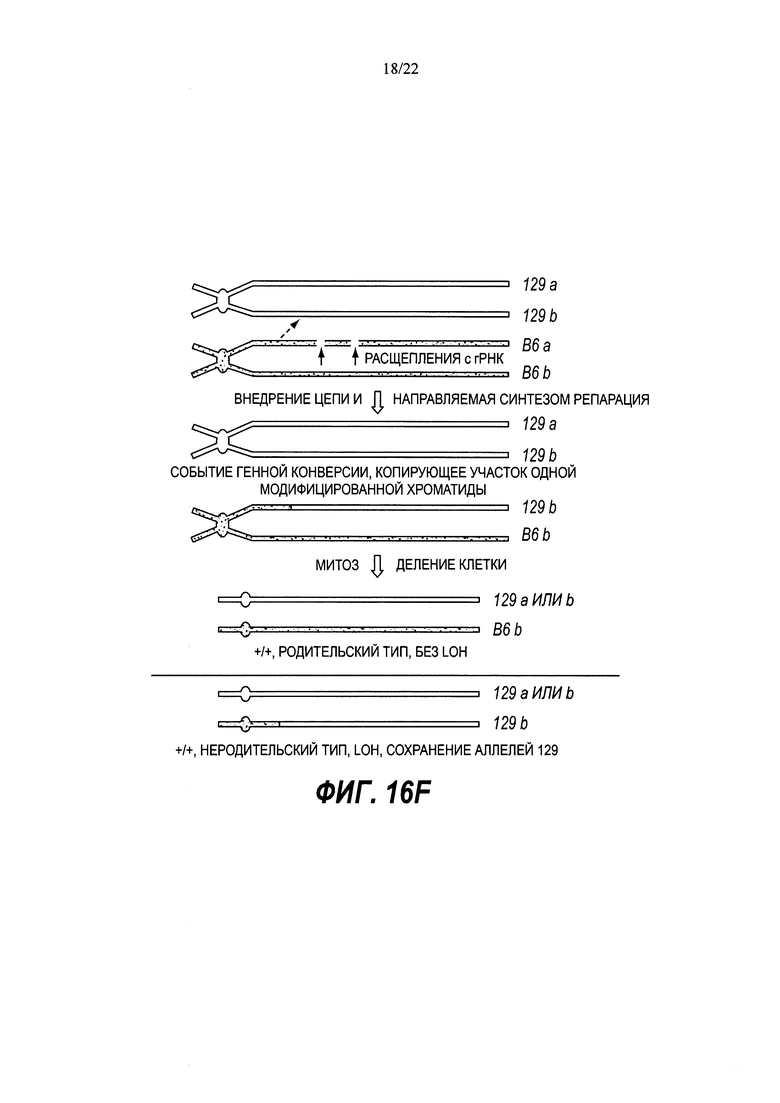
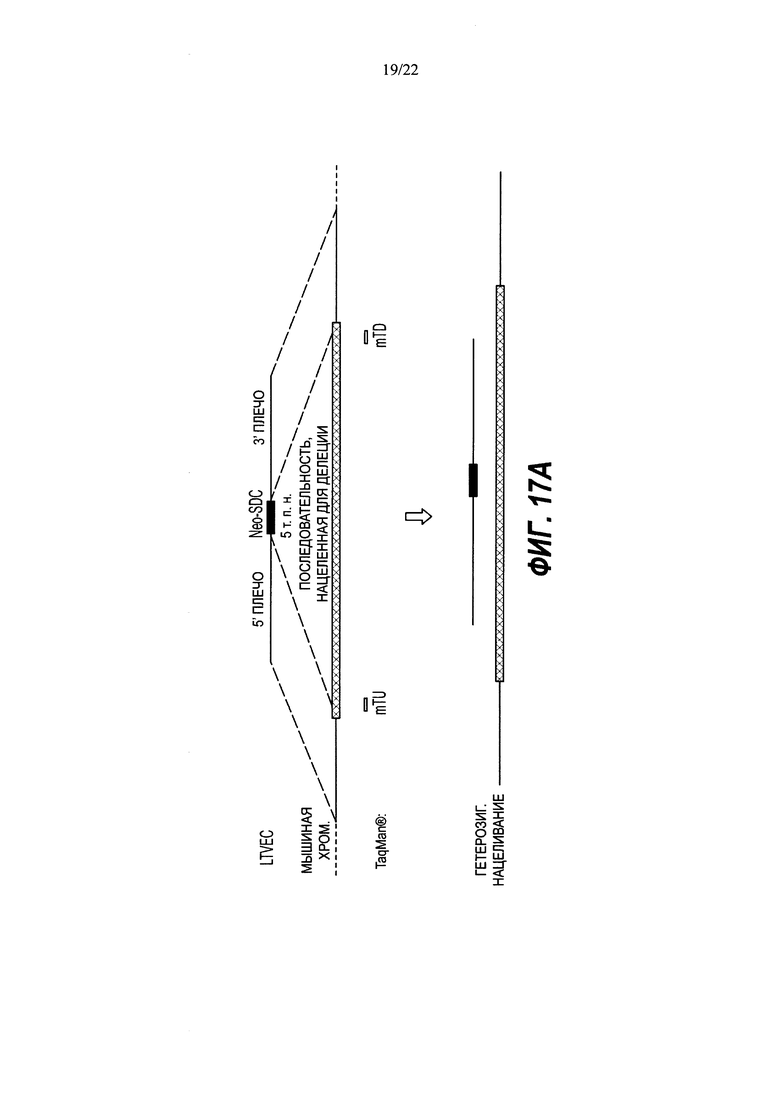
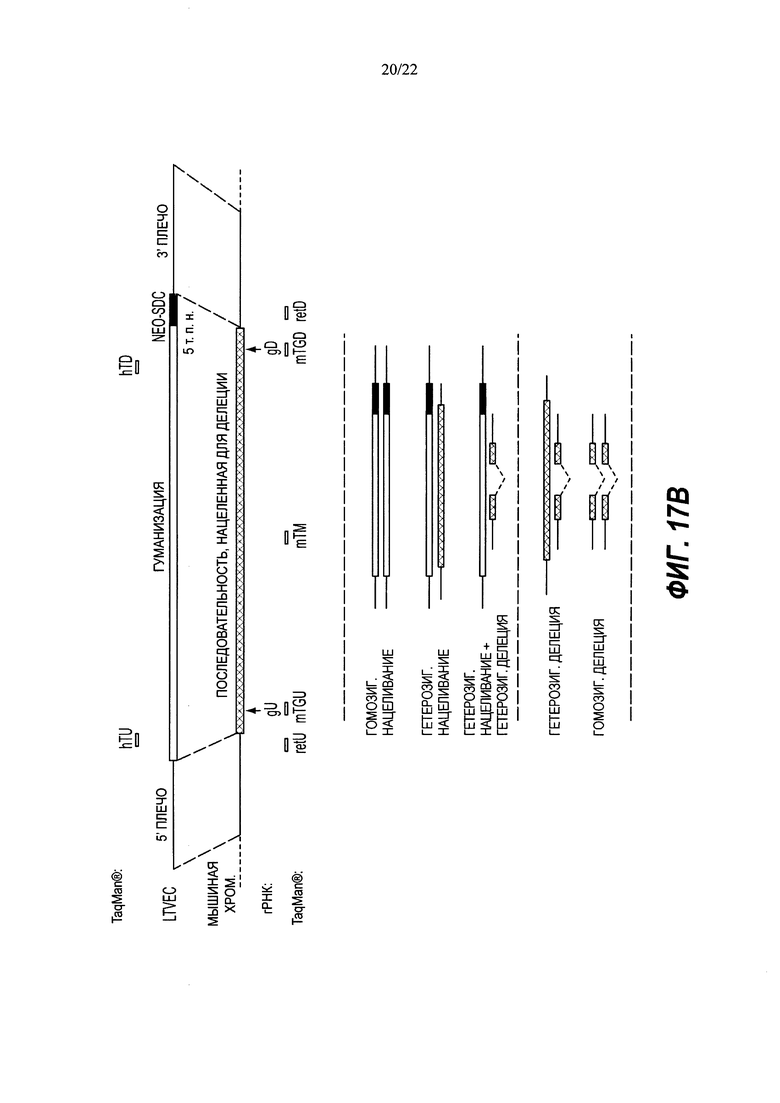
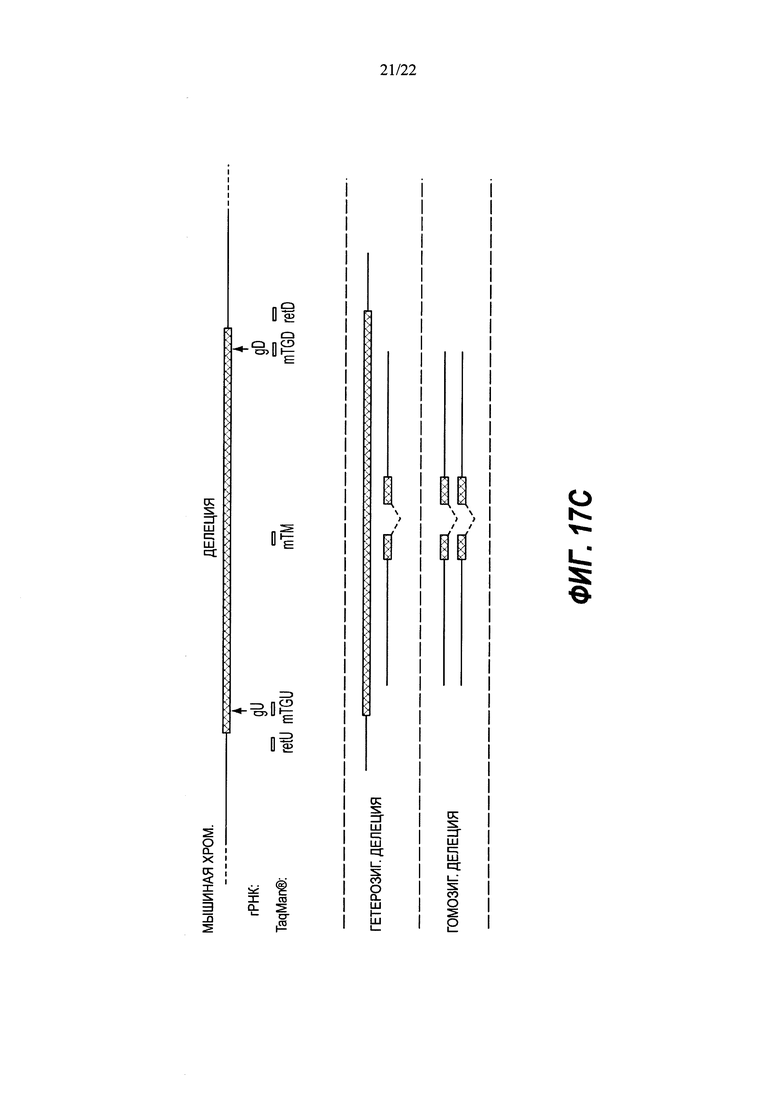
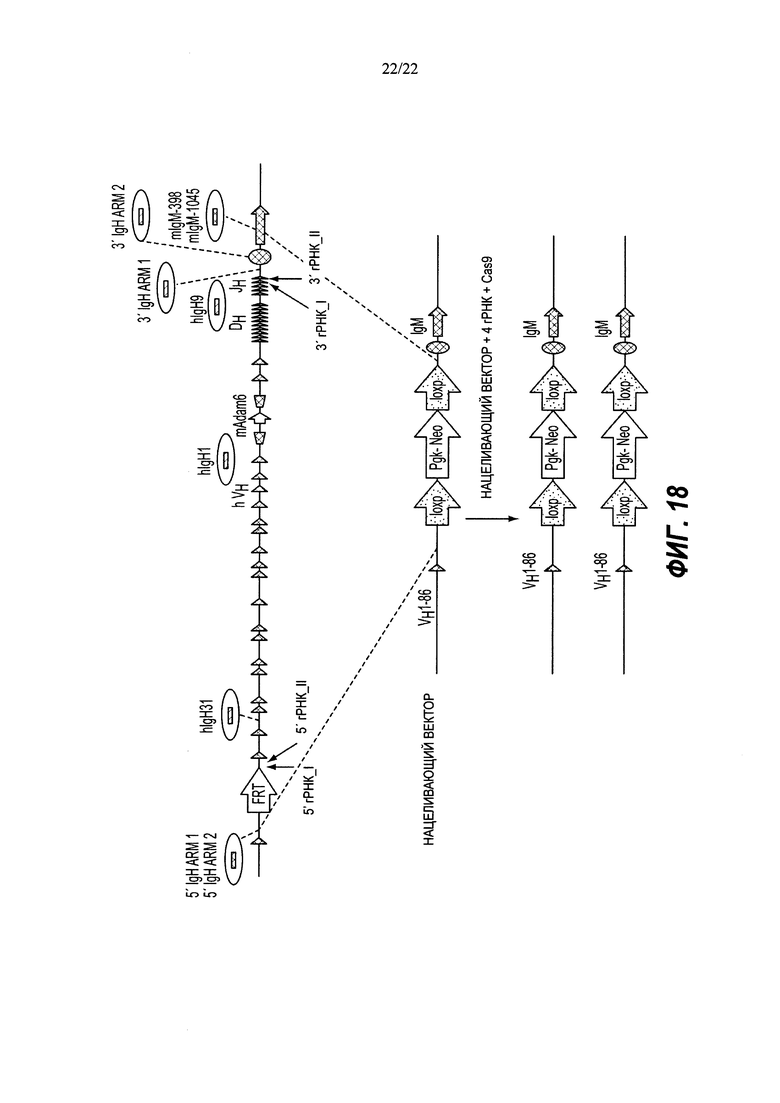
Изобретение относится к генной инженерии. Предложен способ введения биаллельной модификации в целевой геномный локус с использованием системы CRISPR/Cas9, включающий введение Cas9, двух гидовых РНК и нацеливающего вектора, содержащего полинуклеотидную вставку, фланкированную 5' гомологичным плечом, которое гибридизуется с целевой 5'-последовательностью в пределах целевого геномного локуса, и 3' гомологичным плечом, которое гибридизуется с целевой 3'-последовательностью в пределах целевого геномного локуса. Настоящий способ также включает проведение количественного теста для оценки модификации аллеля и теста на сохранение для определения числа копий в образце геномной ДНК области целевой 5'- и/или 3'-последовательности, что позволяет различить правильную нацеленную вставку полинуклеотидной вставки от случайных трансгенных вставок и/или правильные нацеленные делеции от делеций, выходящих за пределы области целевого геномного локуса. 31 з.п. ф-лы, 18 ил., 14 табл., 7 пр.
1. Способ введения биаллельной модификации в целевой геномный локус в геноме клетки, включающий:
(I) введение в популяцию клеток:
(a) белка Cas9 или нуклеиновой кислоты, кодирующей белок Cas9;
(b) первой гидовой РНК или ДНК, кодирующей первую гидовую РНК, причем первая гидовая РНК гибридизуется с первой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса;
(c) второй гидовой РНК или ДНК, кодирующей вторую гидовую РНК, причем вторая гидовая РНК гибридизуется со второй последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; и
(d) нацеливающего вектора, содержащего полинуклеотидную вставку, фланкированную 5’ гомологичным плечом, которое гибридизуется с целевой 5’-последовательностью в пределах целевого геномного локуса, и 3’ гомологичным плечом, которое гибридизуется с целевой 3’-последовательностью в пределах целевого геномного локуса, причем если популяция клеток представляет собой популяцию эмбрионов на стадии единственной клетки, то нацеливающий вектор имеет длину не более 5 т. п. н.,
причем клетки являются нечеловеческими клетками, неплюрипотентными человеческими клетками, стволовыми клетками взрослого человека, ограниченными в развитии человеческими клетками-предшественниками или человеческими индуцированными плюрипотентными клетками, полученными непосредственно из дифференцированной взрослой клетки,
причем геном содержит пару первой и второй гомологичных хромосом, содержащих целевой геномный локус, и
причем белок Cas9 расщепляет по меньшей мере одну из первой и второй последовательностей распознавания РНК CRISPR с получением по меньшей мере одного двухцепочечного разрыва в каждой из первой и второй гомологичных хромосом; и
(II) идентификацию клетки, содержащей модифицированный целевой геномный локус, содержащий биаллельную модификацию, содержащую делецию и/или вставку,
причем идентификация включает проведение количественного теста для оценки модификации аллеля и теста на сохранение,
причем тест для оценки модификации аллеля включает:
(a) тест на приобретение аллеля для определения числа копий области полинуклеотидной вставки в образце геномной ДНК из клетки; и/или
(b) тест на потерю аллеля для определения числа копий в образце геномной ДНК области целевого геномного локуса, нацеленной для делеции, и
причем тест на сохранение определяет число копий в образце геномной ДНК области целевой 5’-последовательности, с которой гибридизуется 5’ гомологичное плечо, и/или определяет число копий в образце геномной ДНК области целевой 3’-последовательности, с которой гибридизуется 3’ гомологичное плечо, и
причем комбинация теста для оценки модификации аллеля и теста на сохранение позволяет различить правильную нацеленную вставку полинуклеотидной вставки в целевой геномный локус от случайных трансгенных вставок полинуклеотидной вставки в геномные местоположения, выходящие за пределы целевого геномного локуса, и/или позволяет различить правильные нацеленные делеции от делеций, выходящих за пределы области целевого геномного локуса, нацеливаемой для делеции.
2. Способ по п. 1, в котором тест для оценки модификации аллеля включает тест на приобретение аллеля, причем тест на приобретение аллеля определяет число копий множества областей полинуклеотидной вставки в образце геномной ДНК.
3. Способ по п. 1 или 2, в котором тест для оценки модификации аллеля включает тест на потерю аллеля, причем тест на потерю аллеля определяет число копий в образце геномной ДНК множества областей в области целевого геномного локуса, нацеленной для делеции.
4. Способ по любому из пп. 1-3, в котором полинуклеотидная вставка содержит кассету селекции и в котором:
(1) целевая 5’-последовательность примыкает к кассете селекции, а тест на сохранение определяет число копий области целевой 5’-последовательности; или
(2) целевая 3’-последовательность примыкает к кассете селекции, а тест на сохранение определяет число копий области целевой 3’-последовательности.
5. Способ по любому из пп. 1-4, в котором этап (II) включает идентификацию клетки, содержащей модифицированный целевой геномный локус, содержащий делецию, причем этап (II) включает тест на сохранение и тест на потерю аллеля и причем число копий менее двух в тесте на сохранение указывает на Cas-опосредованную делецию, выходящую за пределы области целевого геномного локуса, нацеленной для делеции.
6. Способ по любому из пп. 1-5, в котором тест на сохранение определяет число копий области целевой 5’-последовательности и определяет число копий области целевой 3’- последовательности в образце геномной ДНК.
7. Способ по п. 6, в котором тест для оценки модификации аллеля включает тест на потерю аллеля и тест на приобретение аллеля.
8. Способ по любому из пп. 1-7, в котором тест для оценки модификации аллеля включает тест на потерю аллеля и тест на приобретение аллеля и в котором:
(1) тест на приобретение аллеля определяет число копий области полинуклеотидной вставки в образце геномной ДНК путем сравнения с эталонным геном;
(2) тест на потерю аллеля определяет число копий в образце геномной ДНК области целевого геномного локуса, нацеленной для делеции, путем сравнения с эталонным геном, и
(3) тест на сохранение определяет число копий в образце геномной ДНК области целевой 5’-последовательности, с которой гибридизуется 5’ гомологичное плечо, и/или определяет число копий в образце геномной ДНК области целевой 3’-последовательности, с которой гибридизуется 3’ гомологичное плечо, путем сравнения с эталонным геном.
9. Способ по любому из пп. 1-8, в котором полинуклеотидная вставка содержит кассету селекции, примыкающую к первому гомологичному плечу, которое гибридизуется с первой целевой последовательностью,
причем первое гомологичное плечо представляет собой 5’ гомологичное плечо, а первая целевая последовательность представляет собой целевую 5’-последовательность, или причем первое гомологичное плечо представляет собой 3’ гомологичное плечо, а первая целевая последовательность представляет собой целевую 3’-последовательность,
причем идентификация на этапе (II) включает тест на сохранение и тест на приобретение аллеля и включает:
(a) получение ДНК из клетки;
(b) воздействие на ДНК клетки зондом, связывающимся в пределах первой целевой последовательности, зондом, связывающимся в пределах полинуклеотидной вставки, и зондом, связывающимся в пределах эталонного гена, имеющего известное число копий, при этом каждый зонд генерирует обнаруживаемый сигнал при связывании;
(c) обнаружение сигналов, обусловленных связыванием каждого из зондов; и
(d) сравнение сигнала от зонда эталонного гена с сигналом от зонда первой целевой последовательности для определения числа копий первой целевой последовательности и сравнение сигнала от зонда эталонного гена с сигналом от зонда полинуклеотидной вставки для определения числа копий полинуклеотидной вставки,
при этом число копий полинуклеотидной вставки, равное одной или двум в тесте на приобретение аллеля, и число копий первой целевой последовательности, равное двум в тесте на сохранение, указывает на нацеленную вставку полинуклеотидной вставки в целевой геномный локус, и
при этом число копий полинуклеотидной вставки, равное одной или более в тесте на приобретение аллеля, и число копий первой целевой последовательности, равное трем или более в тесте на сохранение, указывает на случайную вставку полинуклеотидной вставки в геномный локус, отличный от целевого геномного локуса.
10. Способ по любому из пп. 1-9, в котором идентификация на этапе (II) включает скрининг на потерю гетерозиготности теломерно целевого геномного локуса.
11. Способ по п. 10, в котором скрининг включает проведение ПЦР-теста на полиморфизм структурных вариантов и/или теста на аллельную дискриминацию однонуклеотидных вариантов.
12. Способ по п. 11, в котором скрининг включает проведение ПЦР-теста на множественный полиморфизм структурных вариантов и/или теста на аллельную дискриминацию множества однонуклеотидных вариантов в разных положениях.
13. Способ по любому из пп. 1-12, в котором идентификация на этапе (II) включает проведение теста на приобретение аллеля, теста на потерю аллеля и теста на сохранение и скрининга на потерю гетерозиготности теломерно целевого геномного локуса.
14. Способ по любому из пп. 1-13, в котором первая и вторая последовательности распознавания РНК CRISPR фланкируют полную или частичную кодирующую последовательность гена.
15. Способ по любому из пп. 1-14, в котором введение как первой, так и второй гидовых РНК или ДНК, кодирующих первую и вторую гидовые РНК, приводит к повышенной эффективности биаллельной модификации по сравнению с введением или только первой гидовой РНК или ДНК, кодирующей первую гидовую РНК, или только второй гидовой РНК или ДНК, кодирующей вторую гидовую РНК.
16. Способ по любому из пп. 1-15, в котором:
(a) белок Cas9 расщепляет первую и вторую последовательности распознавания РНК CRISPR в каждой из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в каждой из первой и второй гомологичных хромосом;
или
(b) белок Cas9 расщепляет первую и вторую последовательности распознавания РНК CRISPR в по меньшей мере одной из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом.
17. Способ по любому из пп. 1-16, в котором этап (I) дополнительно включает введение в популяцию клеток:
(e) третьей гидовой РНК или ДНК, кодирующей третью гидовую РНК, причем третья гидовая РНК гибридизуется с третьей последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса; и
(f) четвертой гидовой РНК или ДНК, кодирующей четвертую гидовую РНК, причем четвертая гидовая РНК гибридизуется с четвертой последовательностью распознавания РНК CRISPR в пределах целевого геномного локуса,
необязательно причем первая и третья последовательности распознавания РНК CRISPR образуют первую пару последовательностей распознавания РНК CRISPR, а вторая и четвертая последовательности распознавания РНК CRISPR образуют вторую пару последовательностей распознавания РНК CRISPR,
необязательно причем первая последовательность распознавания РНК CRISPR примыкает к третьей последовательности распознавания РНК CRISPR, а вторая последовательность распознавания РНК CRISPR примыкает к четвертой последовательности распознавания РНК CRISPR, и
необязательно причем первая пара последовательностей распознавания РНК CRISPR расположена вблизи 5’-конца целевого геномного локуса, а вторая пара последовательностей распознавания РНК CRISPR расположена вблизи 3’-конца целевого геномного локуса.
18. Способ по п. 17, отличающийся тем, что:
(a) первая последовательность распознавания РНК CRISPR и третья последовательность распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п. н. до приблизительно 50 п. н., от приблизительно 50 п. н. до приблизительно 100 п. н., от приблизительно 100 п. н. до приблизительно 150 п. н., от приблизительно 150 п. н. до приблизительно 200 п. н., от приблизительно 200 п. н. до приблизительно 250 п. н., от приблизительно 250 п. н. до приблизительно 300 п. н., от приблизительно 300 п. н. до приблизительно 350 п. н., от приблизительно 350 п. н. до приблизительно 400 п. н., от приблизительно 400 п. н. до приблизительно 450 п. н., от приблизительно 450 п. н. до приблизительно 500 п. н., от приблизительно 500 п. н. до приблизительно 600 п. н., от приблизительно 600 п. н. до приблизительно 700 п. н., от приблизительно 700 п. н. до приблизительно 800 п. н., от приблизительно 800 п. н. до приблизительно 900 п. н., от приблизительно 900 п. н. до приблизительно 1 т. п. н., от приблизительно 1 т. п. н. до приблизительно 2 т. п. н., от приблизительно 2 т. п. н. до приблизительно 3 т. п. н., от приблизительно 3 т. п. н. до приблизительно 4 т. п. н., от приблизительно 4 т. п. н. до приблизительно 5 т. п. н., от приблизительно 5 т. п. н. до приблизительно 6 т. п. н., от приблизительно 6 т. п. н. до приблизительно 7 т. п. н., от приблизительно 7 т. п. н. до приблизительно 8 т. п. н., от приблизительно 8 т. п. н. до приблизительно 9 т. п. н., от приблизительно 9 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 30 т. п. н., от приблизительно 30 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 50 т. п. н., от приблизительно 50 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 70 т. п. н., от приблизительно 70 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 90 т. п. н. или от приблизительно 90 т. п. н. до приблизительно 100 т. п. н.; и/или
(b) вторая последовательность распознавания РНК CR.ISPR и четвертая последовательность распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п. н. до приблизительно 50 п. н., от приблизительно 50 п. н. до приблизительно 100 п. н., от приблизительно 100 п. н. до приблизительно 150 п. н., от приблизительно 150 п. н. до приблизительно 200 п. н., от приблизительно 200 п. н. до приблизительно 250 п. н., от приблизительно 250 п. н. до приблизительно 300 п. н., от приблизительно 300 п. н. до приблизительно 350 п. н., от приблизительно 350 п. н. до приблизительно 400 п. н., от приблизительно 400 п. н. до приблизительно 450 п. н., от приблизительно 450 п. н. до приблизительно 500 п. н., от приблизительно 500 п. н. до приблизительно 600 п. н., от приблизительно 600 п. н. до приблизительно 700 п. н., от приблизительно 700 п. н. до приблизительно 800 п. н., от приблизительно 800 п. н. до приблизительно 900 п. н., от приблизительно 900 п. н. до приблизительно 1 т. п. н., от приблизительно 1 т. п. н. до приблизительно 2 т. п. н., от приблизительно 2 т. п. н. до приблизительно 3 т. п. н., от приблизительно 3 т. п. н. до приблизительно 4 т. п. н., от приблизительно 4 т. п. н. до приблизительно 5 т. п. н., от приблизительно 5 т. п. н. до приблизительно 6 т. п. н., от приблизительно 6 т. п. н. до приблизительно 7 т. п. н., от приблизительно 7 т. п. н. до приблизительно 8 т. п. н., от приблизительно 8 т. п. н. до приблизительно 9 т. п. н., от приблизительно 9 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 30 т. п. н., от приблизительно 30 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 50 т. п. н., от приблизительно 50 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 70 т. п. н., от приблизительно 70 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 90 т. п. н. или от приблизительно 90 т. п. н. до приблизительно 100 т. п. н; и/или
(c) первая и третья последовательности распознавания РНК CRISPR представляют собой первую пару последовательностей распознавания РНК CRISPR, а вторая и четвертая последовательности распознавания РНК CRISPR представляют собой вторую пару последовательностей распознавания РНК CRISPR, причем первая пара и вторая пара разделены на расстояние от приблизительно 25 п. н. до приблизительно 50 п. н., от приблизительно 50 п. н. до приблизительно 100 п. н., от приблизительно 100 п. н. до приблизительно 150 п. н., от приблизительно 150 п. н. до приблизительно 200 п. н., от приблизительно 200 п. н. до приблизительно 250 п. н., от приблизительно 250 п. н. до приблизительно 300 п. н., от приблизительно 300 п. н. до приблизительно 350 п. н., от приблизительно 350 п. н. до приблизительно 400 п. н., от приблизительно 400 п. н. до приблизительно 450 п. н., от приблизительно 450 п. н. до приблизительно 500 п. н., от приблизительно 500 п. н. до приблизительно 600 п. н., от приблизительно 600 п. н. до приблизительно 700 п. н., от приблизительно 700 п. н. до приблизительно 800 п. н., от приблизительно 800 п. н. до приблизительно 900 п. н., от приблизительно 900 п. н. до приблизительно 1 т. п. н., от приблизительно 1 т. п. н. до приблизительно 5 т. п. н., от приблизительно 5 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 100 т. п. н., от приблизительно 100 т. п. н. до приблизительно 150 т. п. н., от приблизительно 150 т. п. н. до приблизительно 200 т. п. н., от приблизительно 200 т. п. н. до приблизительно 300 т. п. н., от приблизительно 300 т. п. н. до приблизительно 400 т. п. н., от приблизительно 400 т. п. н. до приблизительно 500 т. п. н., от приблизительно 500 т. п. н. до приблизительно 1 млн п. н., от приблизительно 1 млн п. н. до приблизительно 1,5 млн п. н., от приблизительно 1,5 млн п. н. до приблизительно 2 млн п. н., от приблизительно 2 млн п. н. до приблизительно 2,5 млн п. н., от приблизительно 2,5 млн п. н. до приблизительно 3 млн п. н., от приблизительно 3 млн п. н. до приблизительно 4 млн п. н., от приблизительно 4 млн п. н. до приблизительно 5 млн п. н., от приблизительно 5 млн п. н. до приблизительно 10 млн п. н., от приблизительно 10 млн п. н. до приблизительно 20 млн п. н., от приблизительно 20 млн п. н. до приблизительно 30 млн п. н., от приблизительно 30 млн п. н. до приблизительно 40 млн п. н., от приблизительно 40 млн п. н. до приблизительно 50 млн п. н., от приблизительно 50 млн п. н. до приблизительно 60 млн п. н., от приблизительно 60 млн п. н. до приблизительно 70 млн п. н., от приблизительно 70 млн п. н. до приблизительно 80 млн п. н., от приблизительно 80 млн п. н. до приблизительно 90 млн п. н. или от приблизительно 90 млн п. н. до приблизительно 100 млн п. н., необязательно причем белок Cas9 расщепляет по меньшей мере две из первой, второй, третьей и четвертой последовательностей распознавания РНК CRISPR с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом, необязательно причем белок Cas9 расщепляет по меньшей мере две из первой, второй, третьей и четвертой последовательностей распознавания РНК CRISPR с получением по меньшей мере двух двухцепочечных разрывов как в первой, так и во второй гомологичных хромосомах.
19. Способ по любому из пп. 1-18, в котором полинуклеотидную вставку вставляют между целевыми 5’- и 3’-последовательностями.
20. Способ по любому из пп. 1-19, в котором клетка, идентифицированная на этапе (II), является диплоидной, а биаллельная модификация приводит к гомозиготности, компаунд- гетерозиготности или гемизиготности в целевом геномном локусе.
21. Способ по любому из пп. 1-20, в котором биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме, необязательно причем:
(a) биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах; необязательно причем биаллельная модификация дополнительно включает вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями как в первой, так и во второй гомологичных хромосомах;
(b) биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах и вставку полинуклеотидной вставки между целевыми 5’- и 3’- последовательностями в первой гомологичной хромосоме, но не во второй гомологичной хромосоме;
(c) биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и нарушение целевого геномного локуса во второй гомологичной хромосоме, причем нарушение возникает из-за опосредованной негомологичным соединением концов (NHEJ) репарации по меньшей мере одного двухцепочечного разрыва;
(d) биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме, вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями в первой гомологичной хромосоме и нарушение целевого геномного локуса во второй гомологичной хромосоме, причем нарушение возникает из-за опосредованной негомологичным соединением концов (NHEJ) репарации по меньшей мере одного двухцепочечного разрыва;
(e) биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями в первой гомологичной хромосоме, причем последовательность полинуклеотидной вставки гомологична или ортологична удаленной последовательности;
(f) удаленная нуклеиновая кислота составляет от приблизительно 5 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 100 т. п. н., от приблизительно 100 т. п. н. до приблизительно 150 т. п. н., от приблизительно 150 т. п. н. до приблизительно 200 т. п. н., от приблизительно 200 т. п. н. до приблизительно 300 т. п. н., от приблизительно 300 т. п. н. до приблизительно 400 т. п. н., от приблизительно 400 т. п. н. до приблизительно 500 т. п. н., от приблизительно 500 т. п. н. до приблизительно 1 млн п. н., от приблизительно 1 млн п. н. до приблизительно 1,5 млн п. н., от приблизительно 1,5 млн п. н. до приблизительно 2 млн п. н., от приблизительно 2 млн п. н. до приблизительно 2,5 млн п. н. или от приблизительно 2,5 млн п. н. до приблизительно 3 млн п. н.; или
(g) удаленная нуклеиновая кислота составляет по меньшей мере 20 т. п. н., по меньшей мере 30 т. п. н., по меньшей мере 40 т. п. н., по меньшей мере 50 т. п. н., по меньшей мере 60 т. п. н., по меньшей мере 70 т. п. н., по меньшей мере 80 т. п. н., по меньшей мере 90 т. п. н., по меньшей мере 100 т. п. н., по меньшей мере 110 т. п. н., по меньшей мере 120 т. п. н., по меньшей мере 130 т. п. н., по меньшей мере 140 т. п. н., по меньшей мере 150 т. п. н., по меньшей мере 160 т. п. н., по меньшей мере 170 т. п. н., по меньшей мере 180 т. п. н., по меньшей мере 190 т. п. н., по меньшей мере 200 т. п. н., по меньшей мере 250 т. п. н., по меньшей мере 300 т. п. н., по меньшей мере 350 т. п. н., по меньшей мере 400 т. п. н., по меньшей мере 450 т. п. н. или по меньшей мере 500 т. п. н., по меньшей мере 550 т. п. н., по меньшей мере 600 т. п. н., по меньшей мере 650 т. п. н., по меньшей мере 700 т. п. н., по меньшей мере 750 т. п. н., по меньшей мере 800 т. п. н., по меньшей мере 850 т. п. н., по меньшей мере 900 т. п. н., по меньшей мере 950 т. п. н., по меньшей мере 1 млн п. н., по меньшей мере 1,5 млн п. н. или по меньшей мере 2 млн п. н.
22. Способ по любому из пп. 1-21, в котором:
(a) каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии по меньшей мере 50 п. н., по меньшей мере 100 п. н., по меньшей мере 200 п. н., по меньшей мере 300 п. н., по меньшей мере 400 п. н., по меньшей мере 500 п. н., по меньшей мере 600 п. н., по меньшей мере 700 п. н., по меньшей мере 800 п. н., по меньшей мере 900 п. н., по меньшей мере 1 т. п. н., по меньшей мере 2 т. п. н., по меньшей мере 3 т. п. н., по меньшей мере 4 т. п. н., по меньшей мере 5 т. п. н., по меньшей мере 6 т. п. н., по меньшей мере 7 т. п. н., по меньшей мере 8 т. п. н., по меньшей мере 9 т. п. н., по меньшей мере 10 т. п. н., по меньшей мере 20 т. п. н., по меньшей мере 30 т. п. н., по меньшей мере 40 т. п. н., по меньшей мере 50 т. п. н., по меньшей мере 60 т. п. н., по меньшей мере 70 т. п. н., по меньшей мере 80 т. п. н., по меньшей мере 90 т. п. н. или по меньшей мере 100 т. п. н. как от целевой 5’-последовательности, так и от целевой 3’-последовательности;
(b) каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии более чем 50 п. н., более чем 100 п. н., более чем 200 п. н., более чем 300 п. н., более чем 400 п. н., более чем 500 п. н., более чем 600 п. н., более чем 700 п. н., более чем 800 п. н., более чем 900 п. н., более чем 1 т. п. н., более чем 2 т. п. н., более чем 3 т. п. н., более чем 4 т. п. н., более чем 5 т. п. н., более чем 6 т. п. н., более чем 7 т. п. н., более чем 8 т. п. н., более чем 9 т. п. н., более чем 10 т. п. н., более чем 20 т. п. н., более чем 30 т. п. н., более чем 40 т. п. н., более чем 50 т. п. н., более чем 60 т. п. н., более чем 70 т. п. н., более чем 80 т. п. н., более чем 90 т. п. н. или более чем 100 т. п. н. как от целевой 5’-последовательности, так и от целевой 3’-последовательности; или
(c) каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии от приблизительно 50 п. н. до приблизительно 100 п. н., от приблизительно 200 п. н. до приблизительно 300 п. н., от приблизительно 300 п. н. до приблизительно 400 п. н., от приблизительно 400 п. н. до приблизительно 500 п. н., от приблизительно 500 п. н. до приблизительно 600 п. н., от приблизительно 600 п. н. до приблизительно 700 п. н., от приблизительно 700 п. н. до приблизительно 800 п. н., от приблизительно 800 п. н. до приблизительно 900 п. н., от приблизительно 900 п. н. до приблизительно 1 т. п. н., от приблизительно 1 т. п. н. до приблизительно 2 т. п. н., от приблизительно 2 т. п. н. до приблизительно 3 т. п. н., от приблизительно 3 т. п. н. до приблизительно 4 т. п. н., от приблизительно 4 т. п. н. до приблизительно 5 т. п. н., от приблизительно 5 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 30 т. п. н., от приблизительно 30 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 50 т. п. н. или от приблизительно 50 т. п. н. до приблизительно 100 т. п. н. как от целевой 5’-последовательности, так и от целевой 3’-последовательности;
(d) первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 1 т. п. н. до приблизительно 5 т. п. н., от приблизительно 5 т. п. н. до приблизительно 10 т. п. н., от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 100 т. п. н., от приблизительно 100 т. п. н. до приблизительно 150 т. п. н., от приблизительно 150 т. п. н. до приблизительно 200 т. п. н., от приблизительно 200 т. п. н. до приблизительно 300 т. п. н., от приблизительно 300 т. п. н. до приблизительно 400 т. п. н., от приблизительно 400 т. п. н. до приблизительно 500 т. п. н., от приблизительно 500 т. п. н. до приблизительно 1 млн п. н., от приблизительно 1 млн п. н. до приблизительно 1,5 млн п. н., от приблизительно 1,5 млн п. н. до приблизительно 2 млн п. н., от приблизительно 2 млн п. н. до приблизительно 2,5 млн п. н. или от приблизительно 2,5 млн п. н. до приблизительно 3 млн п. н.;
(e) первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние по меньшей мере 1 т. п. н., по меньшей мере 2 т. п. н., по меньшей мере 3 т. п. н., по меньшей мере 4 т. п. н., по меньшей мере 5 т. п. н., по меньшей мере 10 т. п. н., по меньшей мере 20 т. п. н., по меньшей мере 30 т. п. н., по меньшей мере 40 т. п. н., по меньшей мере 50 т. п. н., по меньшей мере 60 т. п. н., по меньшей мере 70 т. п. н., по меньшей мере 80 т. п. н., по меньшей мере 90 т. п. н., по меньшей мере 100 т. п. н., по меньшей мере 110 т. п. н., по меньшей мере 120 т. п. н., по меньшей мере 130 т. п. н., по меньшей мере 140 т. п. н., по меньшей мере 150 т. п. н., по меньшей мере 160 т. п. н., по меньшей мере 170 т. п. н., по меньшей мере 180 т. п. н., по меньшей мере 190 т. п. н., по меньшей мере 200 т. п. н., по меньшей мере 250 т. п. н., по меньшей мере 300 т. п. н., по меньшей мере 350 т. п. н., по меньшей мере 400 т. п. н., по меньшей мере 450 т. п. н. или по меньшей мере 500 т. п. н.;
(f) первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние от приблизительно 25 п. н. до приблизительно 50 п. н., от приблизительно 50 п. н. до приблизительно 100 п. н., от приблизительно 100 п. н. до приблизительно 150 п. н., от приблизительно 150 п. н. до приблизительно 200 п. н., от приблизительно 200 п. н. до приблизительно 250 п. н., от приблизительно 250 п. н. до приблизительно 300 п. н., от приблизительно 300 п. н. до приблизительно 350 п. н., от приблизительно 350 п. н. до приблизительно 400 п. н., от приблизительно 400 п. н. до приблизительно 450 п. н., от приблизительно 450 п. н. до приблизительно 500 п. н., от приблизительно 500 п. н. до приблизительно 600 п. н., от приблизительно 600 п. н. до приблизительно 700 п. н., от приблизительно 700 п. н. до приблизительно 800 п. н., от приблизительно 800 п. н. до приблизительно 900 п. н. или от приблизительно 900 п. н. до приблизительно 1 т. п. н.;
(g) первая и вторая последовательности распознавания РНК CRISPR разделены на расстояние менее чем 25 п. н., менее чем 50 п. н., менее чем 100 п. н., менее чем 150 п. н., менее чем 200 п. н., менее чем 250 п. н., менее чем 300 п.н., менее чем 350 п. н., менее чем 400 п. н., менее чем 450 п. н., менее чем 500 п. н., менее чем 600 п. н., менее чем 700 п. н., менее чем 800 п. н., менее чем 900 п. н., менее чем 1 т. п. н., менее чем 2 т. п. н., менее чем 3 т. п. н., менее чем 4 т. п. н., менее чем 5 т. п. н. или менее чем 10 т. п. н.;
(h) при этом каждая из первой и второй последовательностей распознавания РНК CRISPR расположена между целевыми 5’- и 3’-последовательностями; или
(i) первая последовательность распознавания РНК CRISPR находится в пределах 50 п. н., 100 п. н. или 200 п. н. целевой 5’-последовательности, а вторая последовательность распознавания РНК CRISPR находится в пределах 50 п. н., 100 п. н. или 200 п. н. целевой 3’- последовательности.
23. Способ по любому из пп. 1-22, в котором нацеливающий вектор имеет линейную форму и/или в котором нацеливающий вектор является одноцепочечным или двухцепочечным.
24. Способ по любому из пп. 1-23, в котором клетки представляют собой нечеловеческие эукариотические клетки, необязательно причем нечеловеческие эукариотические клетки представляют собой клетки млекопитающего, клетки грызуна, мышиные клетки, крысиные клетки, плюрипотентные клетки, неплюрипотентные клетки, плюрипотентные клетки грызуна, мышиные плюрипотентные клетки, крысиные плюрипотентные клетки, мышиные эмбриональные стволовые (ЭС) клетки, крысиные ЭС-клетки, нечеловеческие ЭС-клетки млекопитающего, нечеловеческие эмбрионы млекопитающего на стадии единственной клетки или мышиные эмбрионы на стадии единственной клетки.
25. Способ по п. 24, в котором клетки представляют собой нечеловеческие эмбрионы млекопитающего на стадии единственной клетки и где:
(a) нацеливающий вектор имеет длину от приблизительно 50 нуклеотидов до приблизительно 5 т. п. н.;
(b) нацеливающий вектор представляет собой одноцепочечную ДНК и имеет длину от приблизительно 60 до приблизительно 200 нуклеотидов;
(c) каждое из белка Cas9 или нуклеиновой кислоты, кодирующей белок Cas9, первой гидовой РНК или ДНК, кодирующей первую гидовую РНК, и второй гидовой РНК или ДНК, кодирующей вторую гидовую РНК, вводят в популяцию эмбрионов на стадии единственной клетки посредством микроинъекции; или
(d) причем этап (I) включает введение нуклеиновой кислоты, кодирующей белок Cas, первой гидовой РНК и второй гидовой РНК в популяцию эмбрионов на стадии единственной клетки, причем нуклеиновая кислота, кодирующая белок Cas, включает РНК и причем каждую из нуклеиновой кислоты, кодирующей белок Cas, первой гидовой РНК и второй гидовой РНК вводят в популяцию эмбрионов на стадии единственной клетки посредством микроинъекции в цитоплазму.
26. Способ по п. 24, в котором клетки не являются эмбрионами на стадии единственной клетки и где:
(a) нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC) длиной по меньшей мере 10 т. п. н., необязательно причем LTVEC составляет от приблизительно 50 т. п. н. до приблизительно 75 т. п. н., от приблизительно 75 т. п. н. до приблизительно 100 т. п. н., от приблизительно 100 т. п. н. до 125 т. п. н., от приблизительно 125 т. п. н. до приблизительно 150 т. п. н., от приблизительно 150 т. п. н. до приблизительно 175 т. п. н., от приблизительно 175 т. п. н. до приблизительно 200 т. п. н., от приблизительно 200 т. п. н. до приблизительно 225 т. п. н., от приблизительно 225 т. п. н. до приблизительно 250 т. п. н., от приблизительно 250 т. п. н. до приблизительно 275 т. п. н. или от приблизительно 275 т. п. н. до приблизительно 300 т. п. н.; или
(b) нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC), в котором общая длина 5’ и 3’ гомологичных плеч LTVEC составляет по меньшей мере 10 т. п. н., необязательно причем общая длина 5’ и 3’ гомологичных плеч LTVEC составляет от приблизительно 10 т. п. н. до приблизительно 20 т. п. н., от приблизительно 20 т. п. н. до приблизительно 30 т. п. н., от приблизительно 30 т. п. н. до приблизительно 40 т. п. н., от приблизительно 40 т. п. н. до приблизительно 50 т. п. н., от приблизительно 50 т. п. н. до приблизительно 60 т. п. н., от приблизительно 60 т. п. н. до приблизительно 70 т. п. н., от приблизительно 70 т. п. н. до приблизительно 80 т. п. н., от приблизительно 80 т. п. н. до приблизительно 90 т. п. н., от приблизительно 90 т. п. н. до приблизительно 100 т. п. н., от приблизительно 100 т. п. н. до приблизительно 110 т. п. н., от приблизительно 110 т. п. н. до приблизительно 120 т. п. н., от приблизительно 120 т. п. н. до приблизительно 130 т. п. н., от приблизительно 130 т. п. н. до приблизительно 140 т. п. н., от приблизительно 140 т. п. н. до приблизительно 150 т. п. н., от приблизительно 150 т. п. н. до приблизительно 160 т. п. н., от приблизительно 160 т. п. н. до приблизительно 170 т. п. н., от приблизительно 170 т. п. н. до приблизительно 180 т. п. н., от приблизительно 180 т. п. н. до приблизительно 190 т. п. н. или от приблизительно 190 т. п. н. до приблизительно 200 т. п. н.
27. Способ по любому из пп. 1-24 и 26, в котором клетки не являются эмбрионами на стадии единственной клетки,
причем нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC), при этом общая сумма длин 5’ и 3’ гомологичных плеч составляет по меньшей мере 10 т. п. н.;
при этом каждая из первой и второй последовательностей распознавания РНК CRISPR расположена на расстоянии более чем 200 п. н., более чем 300 п. н., более чем 400 п. н., более чем 500 п. н., более чем 600 п. н., более чем 700 п. н., более чем 800 п. н., более чем 900 п. н., более чем 1 т. п. н., более чем 2 т. п. н., более чем 3 т. п. н., более чем 4 т. п. н., более чем 5 т. п. н., более чем 6 т. п. н., более чем 7 т. п. н., более чем 8 т. п. н., более чем 9 т. п. н., более чем 10 т. п. н., более чем 20 т. п. н., более чем 30 т. п. н., более чем 40 т. п. н., более чем 50 т. п. н., более чем 60 т. п. н., более чем 70 т. п. н., более чем 80 т. п. н., более чем 90 т. п. н. или более чем 100 т. п.н. как от целевой 5’-последовательности, так и от целевой 3’-последовательности;
при этом белок Cas9 расщепляет первую и вторую последовательности распознавания РНК CRISPR в по меньшей мере одной из первой и второй гомологичных хромосом с получением по меньшей мере двух двухцепочечных разрывов в по меньшей мере одной из первой и второй гомологичных хромосом; и
при этом биаллельная модификация включает делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями в первой гомологичной хромосоме, при этом последовательность полинуклеотидной вставки гомологична или ортологична удаленной последовательности.
28. Способ по любому из пп. 1-27, в котором белок Cas9 имеет нуклеазную активность в отношении обеих цепей двухцепочечной ДНК или причем белок Cas9 представляет собой никазу и при этом этап (I) дополнительно включает введение в популяцию клеток:
(e) третьей гидовой РНК или ДНК, кодирующей третью гидовую РНК, причем третья гидовая РНК гибридизуется с третьей последовательностью распознавания РНК CRISPR; и
(f) четвертой гидовой РНК или ДНК, кодирующей четвертую гидовую РНК, причем четвертая гидовая РНК гибридизуется с четвертой последовательностью распознавания РНК CRISPR;
причем белок Cas9 производит никование первой цепи геномной ДНК в пределах первой и второй последовательностях распознавания РНК CRISPR и белок Cas9 производит никование второй цепи геномной ДНК в пределах третьей и четвертой последовательностях распознавания РНК CRISPR, причем разрывы в пределах первой и третьей последовательностей распознавания РНК CRISPR представляют собой смещенные разрывы для создания первого двухцепочечного разрыва, имеющего ступенчатые концы, и разрывы в пределах второй и четвертой последовательностей распознавания РНК CRISPR представляют собой смещенные разрывы для создания второго двухцепочечного разрыва, имеющего ступенчатые концы.
29. Способ по любому из пп. 1-28, в котором:
(a) первая гидовая РНК содержит первую РНК CRISPR и первую тракрРНК, слитые вместе, и/или вторая гидовая РНК содержит вторую РНК CRISPR и вторую тракрРНК, слитые вместе;
(b) первая гидовая РНК содержит первую РНК CRISPR и первую тракрРНК, причем первая РНК CRISPR и первая тракрРНК представляют собой отдельные молекулы РНК, и/или вторая гидовая РНК содержит вторую РНК CRISPR и вторую тракрРНК, причем вторая РНК CRISPR и вторая тракрРНК представляют собой отдельные молекулы РНК;
(c) этап (I) включает введение белка Cas9 и первой гидовой РНК в популяцию клеток в виде первого комплекса белок-РНК и/или этап (I) включает введение белка Cas9 и второй гидовой РНК в популяцию клеток в виде второго комплекса белок-РНК;
(d) этап (I) включает введение нуклеиновой кислоты, кодирующей белок Cas9, ДНК, кодирующей первую гидовую РНК и ДНК, кодирующей вторую гидовую РНК, в популяцию клеток, где нуклеиновая кислота, кодирующая белок Cas9, содержит ДНК; или
(e) этап (I) включает введение нуклеиновой кислоты, кодирующей белок Cas9, первой гидовой РНК и второй гидовой РНК в популяцию клеток, где нуклеиновая кислота, кодирующая белок Cas9, содержит РНК.
30. Способ по любому из пп. 1-29, в котором популяция клеток модифицирована для снижения негомологичного соединения концов (NHEJ) и/или для повышения генной конверсии или направляемой гомологией репарации (HDR), необязательно причем популяция клеток модифицирована для снижения экспрессии или активности одной или более из: DNA-РК, PARP1 и лигазы IV, и необязательно причем снижение экспрессии или активности является индуцируемым, обратимым, время-специфическим и/или пространственно-специфическим.
31. Способ по любому из пп. 1-30, причем с помощью способа получают:
(a) первую субпопуляцию клеток, содержащих делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах и вставку полинуклеотидной вставки между целевыми 5’- и 3’- последовательностями в первой гомологичной хромосоме, но не во второй гомологичной хромосоме;
(b) вторую субпопуляцию клеток, содержащих делецию между первой и второй последовательностями распознавания РНК CRISPR и вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями как в первой, так и во второй гомологичных хромосомах;
(c) третью субпопуляцию клеток, содержащих делецию между первой и второй последовательностями распознавания РНК CRISPR как в первой, так и во второй гомологичных хромосомах, но не вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями;
(d) четвертую субпопуляцию клеток, содержащих делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме и нарушение целевого геномного локуса во второй гомологичной хромосоме, причем нарушение возникает из-за опосредованной негомологичным соединением концов (NHEJ) репарации по меньшей мере одного двухцепочечного разрыва; или
(e) пятую субпопуляцию клеток, содержащих делецию между первой и второй последовательностями распознавания РНК CRISPR в первой гомологичной хромосоме, вставку полинуклеотидной вставки между целевыми 5’- и 3’-последовательностями в первой гомологичной хромосоме и нарушение целевого геномного локуса во второй гомологичной хромосоме, причем нарушение возникает из-за опосредованной негомологичным соединением концов (NHEJ) репарации по меньшей мере одного двухцепочечного разрыва,
необязательно причем с помощью способа получают первую субпопуляцию клеток, вторую субпопуляцию клеток, третью субпопуляцию клеток, четвертую субпопуляцию клеток и пятую субпопуляцию клеток.
32. Способ по любому из пп. 1-24 и 26-31, в котором клетки представляют собой мышиные или крысиные эмбриональные стволовые (ЭС) клетки и при этом способ дополнительно включает введение мышиной или крысиной ЭС-клетки, идентифицированной на этапе (II), в эмбрион-хозяин.
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| YOSHIMI, K., et al | |||
| "Allele-specific genome editing and correction of disease-associated phenotypes in rats using the CRISPR-Cas platform." Nature communications, 2014 (Published 26 Jun 2014), 5: 4240 | |||
| FUJII, WATARU, et al | |||
| "Efficient generation of genome-modified mice via offset-nicking by CRISPR/Cas system." | |||

