ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНЫЕ ЗАЯВКИ
Настоящая заявка относится к заявке США №62/094,104, поданной 19 декабря 2014 г., заявке США №62/167,408, поданной 28 мая 2015 г., и заявке США №62/205,524, поданной 14 августа 2015 г., каждая из которых для любых целей полностью включена в настоящий документ путем ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПОДАННЫЙ В ВИДЕ ТЕКСТОВОГО ФАЙЛА ЧЕРЕЗ СЕТЬ EFS-WEB
Перечень последовательностей, записанный в файл 472224SEQLIST.txt с размером 16,7 кб, созданный 16 декабря 2015 г., включен в настоящий документ путем ссылки.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Гомологичная рекомбинация с использованием нацеливающих векторов, выполненных с возможностью добавления, удаления или замещения конкретной нуклеотидной последовательности в геномном локусе, является популярным подходом для достижения требуемой геномной модификации не относящихся к человеку животных.
Хотя модификация генома посредством гомологичной рекомбинации была значительно усовершенствована за последние два десятка лет, все еще сохраняются сложности с достижением приемлемой частоты нацеливания с применением очень больших нацеливающих векторов (LTVEC) во многих обстоятельствах, например, когда большой участок генома грызуна замещают большим геномным фрагментом человека или при нацеливании на определенные типы клеток, например фибробласты или другие соматические клетки.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Предложены способы и композиции для модификации целевого геномного локуса в клетке посредством системы нацеливания, в которой применяются два или более нацеливающих векторов, способных рекомбинировать друг с другом с образованием одного сегмента непрерывной нуклеиновой кислоты. Необязательно нацеливающие векторы представляют собой большие нацеливающие векторы (LTVEC). Необязательно каждый LTVEC имеет размер по меньшей мере 10 т.п.н.
В изобретении предложены способы модификации целевого геномного локуса в клетке, включающие в себя: (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение в клетку первого большого нацеливающего вектора (LTVEC), содержащего первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, содержащего вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3'-гомологичным плечом, причем первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, а первое 3' гомологичное плечо первого LTVEC и второе 5' плечо второго LTVEC гомологичны друг другу или соответственно дополнительным 5' и 3' гомологичным плечам одного или более дополнительных LTVEC, при этом каждый содержит дополнительную полинуклеотидную вставку, фланкированную дополнительным 5' гомологичным плечом и дополнительным 3' гомологичным плечом, при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки, одной или более дополнительных полинуклеотидных вставок одного или более дополнительных LTVEC, если они имеются, и второй полинуклеотидной вставки между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, одну или более дополнительных полинуклеотидных вставок, если они имеются, и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус. Необязательно размер каждого из первого LTVEC, второго LTVEC и одного или более дополнительных LTVEC составляет по меньшей мере 10 т.п.н. В некоторых таких способах дополнительные LTVEC представляют собой один или более из других LTVEC, которые, если они есть, вставлены между первым LTVEC и вторым LTVEC.
В изобретении также предложены способы двойного нацеливания для модификации целевого геномного локуса в клетке, включающие в себя (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение в клетку первого большого нацеливающего вектора (LTVEC), который имеет размер по меньшей мере 10 т.п.н. и содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус.
Необязательно первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки в целевой геномный локус.
В некоторых таких способах клетка представляет собой человеческую клетку. В некоторых таких способах клетка представляет собой нечеловеческую клетку. В некоторых таких способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В некоторых таких способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В некоторых из вышеупомянутых способов нуклеазный агент представляет собой нуклеазу с «цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), или мегануклеазу. В некоторых из вышеупомянутых способов нуклеазный агент содержит белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК). Необязательно белок Cas представляет собой Cas9.
В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты.
В некоторых способах суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет от приблизительно 50 т.п.н. до приблизительно 500 т.п.н., от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н. или от приблизительно 450 т.п.н. до приблизительно 500 т.п.н. Необязательно суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет от приблизительно 100 т.п.н. до приблизительно 500 т.п.н. Необязательно суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет приблизительно 300 т.п.н.
В некоторых способах клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую полинуклеотидную вставку и вторую полинуклеотидную вставку, которые имеют суммарный размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н.
В некоторых способах первая перекрывающаяся последовательность первого LTVEC по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична первой перекрывающейся последовательности второго LTVEC. В некоторых способах размер первой перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В некоторых способах размер первой перекрывающейся последовательности составляет по меньшей мере 10 т.п.н. или по меньшей мере 20 т.п.н.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. Необязательно делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах комбинированное использование первого LTVEC и второго LTVEC приводит к повышению эффективности нацеливания по сравнению с использованием одиночного LTVEC. Необязательно повышение эффективности нацеливания составляет по меньшей мере 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC или второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC или второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена любым из вышеуказанных способов; и (b) гестацию не относящегося к человеку эмбриона-хозяина в теле суррогатной матери, при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
В изобретении также предложены способы тройного нацеливания для модификации целевого геномного локуса в клетке, включающие в себя: (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение в клетку первого большого нацеливающего вектора (LTVEC), который имеет длину по меньшей мере 10 т.п.н. и содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, второго LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, и третьего LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит третью полинуклеотидную вставку, фланкированную третьим 5' гомологичным плечом и третьим 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, второе 3' гомологичное плечо второго LTVEC имеет вторую перекрывающуюся последовательность, гомологичную третьему 5' гомологичному плечу третьего LTVEC, и первое 5' гомологичное плечо первого LTVEC и третье 3' гомологичное плечо третьего LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, интегрированные в целевой геномный локус.
Необязательно первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, а вторая полинуклеотидная вставка и второе 3' гомологичное плечо и третья полинуклеотидная вставка и третье 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки в целевой геномный локус.
В некоторых таких способах клетка представляет собой человеческую клетку. В некоторых таких способах клетка представляет собой нечеловеческую клетку. В некоторых таких способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В некоторых таких способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В некоторых таких способах нуклеазный агент представляет собой нуклеазу с «цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), или мегануклеазу. В некоторых таких способах нуклеазный агент содержит белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК). Необязательно белок Cas представляет собой Cas9.
В некоторых таких способах одна или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки получены от вида, который отличается от вида, к которому относится клетка. В некоторых таких способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка и третья полинуклеотидная вставка представляют собой человеческие нуклеиновые кислоты.
В некоторых таких способах суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет от приблизительно 50 т.п.н. до приблизительно 700 т.п.н., от приблизительно 50 т.п.н. до приблизительно 500 т.п.н., от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н. или от приблизительно 650 т.п.н. до приблизительно 700 т.п.н. Необязательно суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет от приблизительно 100 т.п.н. до приблизительно 700 т.п.н. Необязательно суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет приблизительно 400 т.п.н.
В некоторых таких способах клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, которые имеют суммарный размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 700 т.п.н.
В некоторых таких способах первая перекрывающаяся последовательность первого LTVEC по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична первой перекрывающейся последовательности второго LTVEC и/или вторая перекрывающаяся последовательность второго LTVEC по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична второй перекрывающейся последовательности третьего LTVEC. В некоторых таких способах размер первой перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. и/или размер второй перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В некоторых таких способах размер первой перекрывающейся последовательности составляет по меньшей мере 10 т.п.н. или по меньшей мере 20 т.п.н. и/или размер второй перекрывающейся последовательности составляет по меньшей мере 10 т.п.н. или по меньшей мере 20 т.п.н.
В некоторых таких способах интеграция одной или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки или третьей полинуклеотидной вставки в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе;
(b) делеции эндогенной последовательности в целевом геномном локусе; или
(c) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. Необязательно делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC, второго LTVEC или третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н., общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых таких способах общая длина 5' и 3' гомологичных плеч первого LTVEC, второго LTVEC или третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н.
до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н; общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена любым из вышеуказанных способов; и (b) вынашивание нечеловеческого эмбриона-хозяина в теле суррогатной матери; при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
В изобретении также предложены способы усиления гомологичной рекомбинации в целевом геномном локусе в клетке, включающие в себя введение в клетку первой нуклеиновой кислоты и второй нуклеиновой кислоты, при этом первая и вторая нуклеиновые кислоты содержат перекрывающуюся нуклеотидную последовательность. В некоторых таких способах гомологичная рекомбинация усилена по сравнению со способами, в которых в клетку вводят только одиночную нуклеиновую кислоту.
В некоторых таких способах гомологичная рекомбинация усиливается в целевом геномном локусе без использования нуклеазного агента. Некоторые такие способы дополнительно включают в себя введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в целевом геномном локусе или вблизи него. В некоторых таких способах нуклеазный агент представляет собой нуклеазу с «цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), или мегануклеазу. В некоторых таких способах нуклеазный агент содержит белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК). Необязательно белок Cas представляет собой Cas9.
В некоторых таких способах способ усиливает гомологичную рекомбинацию первой нуклеиновой кислоты, второй нуклеиновой кислоты или обеих в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых первую нуклеиновую кислоту вводят без второй нуклеиновой кислоты. Некоторые такие способы усиливают гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых вторую нуклеиновую кислоту вводят без первой нуклеиновой кислоты. Необязательно усиление гомологичной рекомбинации составляет по меньшей мере 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В некоторых таких способах перекрывающаяся последовательность первой нуклеиновой кислоты по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична перекрывающейся последовательности второй нуклеиновой кислоты. В некоторых таких способах перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. Необязательно перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 15 т.п.н., от приблизительно 15 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 25 т.п.н., от приблизительно 25 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 35 т.п.н., от приблизительно 35 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 45 т.п.н., от приблизительно 45 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 55 т.п.н., от приблизительно 55 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 65 т.п.н. или от приблизительно 65 т.п.н. до приблизительно 70 т.п.н. В некоторых таких способах перекрывающаяся последовательность составляет по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 25 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 35 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 45 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 55 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 65 т.п.н. или по меньшей мере 70 т.п.н. Необязательно перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н.
В некоторых таких способах первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной целевому геномному локусу, за исключением перекрывающейся последовательности. Необязательно первый нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. или от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. Необязательно первый нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC). Необязательно первый LTVEC имеет длину по меньшей мере 10 т.п.н. Необязательно первый нацеливающий вектор представляет собой первый большой нацеливающий вектор (LTVEC) в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. Необязательно общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н.
В некоторых таких способах первая нуклеиновая кислота представляет собой первый нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом. Необязательно первый нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. или от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. и/или второй нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. или от приблизительно 5 т.п.н.
до приблизительно 10 т.п.н. Необязательно первый нацеливающий вектор представляет собой первый большой нацеливающий вектор (LTVEC) и/или второй нацеливающий вектор представляет собой второй большой нацеливающий вектор (LTVEC). Необязательно первый LTVEC имеет длину по меньшей мере 10 т.п.н. и/или второй LTVEC имеет длину по меньшей мере 10 т.п.н. Необязательно первый нацеливающий вектор представляет собой первый большой нацеливающий вектор (LTVEC) в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. и/или второй нацеливающий вектор представляет собой второй большой нацеливающий вектор (LTVEC) в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. Необязательно первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. и/или второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. Необязательно общая длина 5' и 3' гомологичных плеч первого LTVEC или второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. Необязательно общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н., а общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н.
В некоторых способах перекрывающаяся последовательность расположена на 3'-конце первой нуклеиновой кислоты и 5'-конце второй нуклеотидной последовательности. В некоторых способах перекрывающаяся нуклеотидная последовательность облегчает рекрутирование механизмов рекомбинации в целевом геномном локусе.
В некоторых таких способах клетка представляет собой человеческую клетку. В некоторых таких способах клетка представляет собой нечеловеческую клетку. В некоторых таких способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В некоторых таких способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена любым из вышеуказанных способов; и (b) вынашивание нечеловеческого эмбриона-хозяина в теле суррогатной матери; при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
Предложены способы и композиции для модификации целевого геномного локуса в клетке посредством системы нацеливания, в которой применяются два или более нацеливающих векторов, способных рекомбинировать друг с другом с образованием одного сегмента непрерывной нуклеиновой кислоты. В различных вариантах осуществления нацеливающие векторы представляют собой большие нацеливающие векторы (LTVEC). Необязательно каждый LTVEC имеет размер по меньшей мере 10 т.п.н.
В одном варианте осуществления предложен способ модификации целевого геномного локуса в клетке. Такой способ включает в себя введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса, введение первого большого нацеливающего вектора (LTVEC), содержащего первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, содержащего вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим сегментам в пределах целевого геномного локуса, и первое 3' гомологичное плечо первого LTVEC и второе 5' плечо второго LTVEC гомологичны друг другу или соответственно дополнительным 5' и 3' гомологичным плечам одного или более дополнительных LTVEC, каждый из которых содержит дополнительную вставку, фланкированную дополнительным 5' гомологичным плечом и дополнительным 3' гомологичным плечом, при этом целевой геномный локус модифицирован путем интеграции первой вставки, одной или более дополнительных вставок одного или более дополнительных LTVEC, если они имеются, и второй полинуклеотидной вставки между соответствующими геномными сегментами. Необязательно размер каждого из первого LTVEC, второго LTVEC и одного или более дополнительных LTVEC составляет по меньшей мере 10 т.п.н. Способ дополнительно включает в себя отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, одну или более дополнительных полинуклеотидных вставок, если они имеются, и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус. В таких способах дополнительные LTVEC представляют собой один или более других LTVEC, которые, если они имеются, вставлены между первым LTVEC и вторым LTVEC.
В другом варианте осуществления предложен способ двойного нацеливания для модификации целевого геномного локуса в клетке. Такой способ включает в себя введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса, введение первого большого нацеливающего вектора (LTVEC), содержащего первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, содержащего вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом. Необязательно первый LTVEC имеет размер по меньшей мере 10 т.п.н. и второй LTVEC имеет размер по меньшей мере 10 т.п.н. В таком способе первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого локуса, так что целевой геномный локус модифицирован путем интеграции первой и второй полинуклеотидных вставок между соответствующими геномными сегментами. Способ дополнительно включает в себя отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус.
В некоторых таких способах первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки в целевой геномный локус.
В другом варианте осуществления предложен способ тройного нацеливания для модификации целевого геномного локуса в клетке. Такой способ включает в себя введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса, введение первого большого нацеливающего вектора (LTVEC), содержащего первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, второго LTVEC, содержащего вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, и третьего LTVEC, содержащего третью полинуклеотидную вставку, фланкированную третьим 5' гомологичным плечом и третьим 3' гомологичным плечом Необязательно первый LTVEC имеет размер по меньшей мере 10 т.п.н., второй LTVEC имеет размер по меньшей мере 10 т.п.н. и третий LTVEC имеет размер по меньшей мере 10 т.п.н. В таком способе первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, второе 3' гомологичное плечо второго LTVEC имеет вторую перекрывающуюся последовательность, гомологичную третьему 5' гомологичному плечу третьего LTVEC, и первое 5' гомологичное плечо первого LTVEC и третье 3' гомологичное плечо третьего LTVEC гомологичны соответствующим геномным сегментам в пределах целевого локуса, так что целевой геномный локус модифицирован путем интеграции первой, второй и третьей полинуклеотидных вставок между соответствующими геномными сегментами. Способ дополнительно включает в себя отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, интегрированные в целевой геномный локус.
В некоторых таких способах первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, а вторая полинуклеотидная вставка и второе 3' гомологичное плечо и третья полинуклеотидная вставка и третье 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки в целевой геномный локус.
В одном варианте осуществления клетка представляет собой плюрипотентную клетку. В другом варианте осуществления плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку. В некоторых вариантах осуществления плюрипотентная клетка представляет собой гемопоэтическую стволовую клетку или нейрональную стволовую клетку. В другом варианте осуществления клетка представляет собой индуцированную плюрипотентную стволовую (ИПС) клетку.
В одном варианте осуществления целевой геномный локус находится в геноме клетки. В другом варианте осуществления целевой геномный локус находится на внехромосомной ДНК внутри клетки.
В одном варианте осуществления клетка представляет собой фибробластную клетку.
В некоторых способах клетка представляет собой нечеловеческую клетку. В других способах клетка получена от человека. В некоторых вариантах осуществления клетка представляет собой клетку млекопитающих. В другом варианте осуществления клетка млекопитающих получена от грызуна. В некоторых случаях грызун представляет собой мышь, крысу или хомяка.
В некоторых из вышеуказанных способов нуклеазный агент экспрессируется из экспрессионного конструкта, содержащего нуклеотидную последовательность, кодирующую нуклеазу, и причем нуклеиновая кислота функционально связана с активным в клетке промотором. В других способах нуклеазный агент экспрессируется из мРНК, кодирующей нуклеазу. В некоторых таких способах нуклеаза представляет собой нуклеазу с «цинковыми пальцами» (ZFN). В других таких способах нуклеаза представляет собой эффекторную нуклеазу, подобную активатору транскрипции (TALEN). В других способах нуклеаза представляет собой мегануклеазу.
В некоторых из вышеупомянутых способов нуклеазный агент содержит белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК). В некоторых таких способах белок Cas представляет собой Cas9.
В некоторых из вышеуказанных способов первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка. В одном варианте осуществления первая полинуклеотидная вставка, вторая полинуклеотидная вставка и/или третья полинуклеотидная вставка получены от разных видов. В некоторых способах одна или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки получены от вида, который отличается от вида, к которому относится клетка. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты. В другом варианте осуществления первая полинуклеотидная вставка, вторая полинуклеотидная вставка и/или третья полинуклеотидная вставка представляют собой человеческие нуклеиновые кислоты. В некоторых способах одна или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки представляют собой человеческие нуклеиновые кислоты.
В одном варианте осуществления суммарный размер первой и второй полинуклеотидных вставок составляет от приблизительно 50 т.п.н. до приблизительно 500 т.п.н., от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н. или от приблизительно 450 т.п.н. до приблизительно 500 т.п.н. В другом варианте осуществления суммарный размер первой и второй полинуклеотидных вставок составляет от приблизительно 100 т.п.н. до приблизительно 500 т.п.н. В другом варианте осуществления суммарный размер первой и второй полинуклеотидных вставок составляет приблизительно 300 т.п.н.
В некоторых вариантах осуществления клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, длина которых находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н.
В одном варианте осуществления суммарный размер первой, второй и третьей полинуклеотидных вставок составляет от приблизительно 50 т.п.н. до приблизительно 700 т.п.н., от приблизительно 50 т.п.н. до приблизительно 500 т.п.н., от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н. или от приблизительно 650 т.п.н. до приблизительно 700 т.п.н.
В некоторых вариантах осуществления клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую, вторую и третью полинуклеотидные вставки, которые имеют размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 700 т.п.н. Необязательно суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет от приблизительно 100 т.п.н. до приблизительно 700 т.п.н. В некоторых вариантах осуществления суммарный размер первой, второй и третьей полинуклеотидных вставок составляет приблизительно 400 т.п.н.
В некоторых из вышеуказанных способов первая перекрывающаяся последовательность первого LTVEC по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична первой перекрывающейся последовательности второго LTVEC. В некоторых из вышеуказанных способов вторая перекрывающаяся последовательность второго LTVEC по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична второй перекрывающейся последовательности третьего LTVEC. В любом из вышеуказанных способов перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В конкретном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. В другом конкретном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. В некоторых из вышеуказанных способов первая перекрывающаяся последовательность и/или вторая перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В некоторых способах первая перекрывающаяся последовательность и/или вторая перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. или по меньшей мере 20 т.п.н.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. В некоторых способах интеграция одной или более из первой, второй и третьей полинуклеотидных вставок в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к добавлению экзогенной последовательности в целевой геномный локус. В одном варианте осуществления интеграция первой, второй и/или третьей полинуклеотидной вставки в целевой геномный локус приводит к добавлению экзогенной последовательности в целевой геномный локус.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к делеции эндогенной последовательности в целевом геномном локусе. В другом варианте осуществления интеграция первой, второй и/или третьей полинуклеотидных вставок в целевой геномный локус приводит к делеции эндогенной последовательности в целевом геномном локусе. В некоторых таких способах делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к нокину, нокауту, точечной мутации, перестановке доменов, перестановке экзонов, перестановке интронов, перестановке регуляторных последовательностей, перестановке генов или их комбинации. В еще одном варианте осуществления интеграция первой, второй и/или третьей полинуклеотидных вставок в целевой геномный локус приводит к нокину, нокауту, точечной мутации, перестановке доменов, перестановке экзонов, перестановке интронов, перестановке регуляторных последовательностей, перестановке генов или их комбинации.
В некоторых из вышеуказанных способов комбинированное использование первого LTVEC и второго LTVEC приводит к повышению эффективности нацеливания по сравнению с использованием одиночного LTVEC. Необязательно повышение эффективности нацеливания составляет по меньшей мере 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC или второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых вариантах осуществления общая длина 5' и 3' гомологичных плеч первого LTVEC, второго LTVEC или третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н., общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. и общая длина 5' и 3' гомологичных плеч третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 150 т.п.н. В других вариантах осуществления общая длина 5' и 3' гомологичных плеч первого LTVEC, второго LTVEC или третьего LTVEC составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до 150 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н.; общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до 150 т.п.н. и общая длина 5' и 3' гомологичных плеч третьего LTVEC составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до 150 т.п.н.
Дополнительно предложен способ получения поколения F0 не относящегося к человеку животного. Такой способ включает в себя введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческую ЭС-клетку получали любым из вышеуказанных способов, и вынашивание не относящегося к человеку эмбриона-хозяина в теле суррогатной матери, так что суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
В изобретении также предложены способы усиления гомологичной рекомбинации в целевом геномном локусе в клетке, включающие в себя введение в клетку первой нуклеиновой кислоты и второй нуклеиновой кислоты, при этом первая и вторая нуклеиновые кислоты содержат перекрывающуюся нуклеотидную последовательность. В некоторых таких способах гомологичная рекомбинация усилена по сравнению со способами, в которых в клетку вводят только одиночную нуклеиновую кислоту. В некоторых таких способах гомологичная рекомбинация усиливается в целевом геномном локусе без использования нуклеазного агента. Другие такие способы дополнительно включают в себя введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в целевом геномном локусе или вблизи него.
В одном аспекте предложен способ усиления гомологичной рекомбинации в геномном локусе в клетке без использования нуклеазного агента, включающий в себя введение в клетку первой нуклеиновой кислоты и второй нуклеиновой кислоты, причем первая и вторая нуклеиновые кислоты содержат перекрывающуюся нуклеотидную последовательность.
В одном варианте осуществления способ усиливает гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых первую нуклеиновую кислоту вводят без второй нуклеиновой кислоты. В одном варианте осуществления способ усиливает гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых вторую нуклеиновую кислоту вводят без первой нуклеиновой кислоты. В одном варианте осуществления способ усиливает гомологичную рекомбинацию первой и второй нуклеиновых кислот в целевом геномном локусе.
В одном варианте осуществления усиление гомологичной рекомбинации составляет по меньшей мере 0,5 раза, 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты гомологична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты на 100% идентична перекрывающейся последовательности второй нуклеиновой кислоты.
В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В некоторых способах перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 10 т.п.н. до приблизительно 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 15 т.п.н. до приблизительно 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 20 т.п.н. до приблизительно 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 25 т.п.н. до приблизительно 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 30 т.п.н. до приблизительно 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 35 т.п.н. до приблизительно 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 40 т.п.н. до приблизительно 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 45 т.п.н. до приблизительно 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 50 т.п.н. до приблизительно 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 55 т.п.н. до приблизительно 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 60 т.п.н. до приблизительно 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 65 т.п.н. до приблизительно 70 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 70 т.п.н.
В одном варианте осуществления первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, а первая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления первая нуклеиновая кислота представляет собой первый нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом. В одном варианте осуществления первая полинуклеотидная вставка и вторая полинуклеотидная вставка представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты.
В одном варианте осуществления нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н.
В одном варианте осуществления нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC). В некоторых способах нацеливающий вектор представляет собой LTVEC в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый нацеливающий вектор представляет собой первый LTVEC в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. и/или второй нацеливающий вектор представляет собой второй LTVEC в диапазоне от приблизительно 20 т.п.н. до приблизительно 200 т.п.н. В одном варианте осуществления LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый нацеливающий вектор представляет собой первый LTVEC и/или второй нацеливающий вектор представляет собой второй LTVEC. В некоторых способах первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. В некоторых способах второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. и второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления общая длина 5' и 3' гомологичных плеч LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность расположена на 3'-конце первой нуклеиновой кислоты и 5'-конце второй нуклеотидной последовательности. В одном варианте осуществления перекрывающаяся последовательность расположена на 5'-конце первой нуклеиновой кислоты и 3'-конце второй нуклеотидной последовательности.
В одном варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка получены от разных видов. В другом варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка представляют собой человеческие нуклеиновые кислоты. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. В некоторых способах интеграция одной или более из первой, второй и третьей полинуклеотидных вставок в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
В одном варианте осуществления интеграция первой и/или второй вставок в геномный локус приводит к добавлению экзогенной последовательности в геномном локусе.
В некоторых вариантах осуществления клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, длина которых находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н. В некоторых способах клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, которые имеют суммарный размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н.
В другом варианте осуществления интеграция первой и/или второй полинуклеотидных вставок в геномный локус приводит к делеции эндогенной последовательности в целевом геномном локусе. В одном варианте осуществления делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах клетка представляет собой человеческую клетку. В других способах клетка представляет собой нечеловеческую клетку. В некоторых способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В других способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В одном варианте осуществления клетка представляет собой плюрипотентную клетку. В другом варианте осуществления плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку. В некоторых вариантах осуществления плюрипотентная клетка представляет собой гемопоэтическую стволовую клетку или нейрональную стволовую клетку. В другом варианте осуществления клетка представляет собой индуцированную плюрипотентную стволовую (ИПС) клетку.
В одном варианте осуществления целевой геномный локус находится в геноме клетки. В другом варианте осуществления целевой геномный локус находится на внехромосомной ДНК внутри клетки.
В одном варианте осуществления клетка представляет собой фибробластную клетку.
В некоторых способах клетка представляет собой нечеловеческую клетку. В других способах клетка получена от человека. В некоторых вариантах осуществления клетка представляет собой клетку млекопитающих. В другом варианте осуществления клетка млекопитающих получена от грызуна. В некоторых случаях грызун представляет собой мышь, крысу или хомяка.
В некоторых из вышеуказанных способов перекрывающаяся нуклеотидная последовательность облегчает рекрутирование механизмов рекомбинации в целевой геномный локус.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена вышеуказанными способами; и (b) гестацию не относящегося к человеку эмбриона-хозяина в теле суррогатной матери, при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
В другом аспекте предложен способ усиления гомологичной рекомбинации в целевом геномном локусе в клетке с помощью нуклеазного агента, включающий в себя введение в клетку: (i) первой нуклеиновой кислоты и второй нуклеиновой кислоты, причем первая и вторая нуклеиновые кислоты содержат перекрывающуюся нуклеотидную последовательность; и (ii) нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в геномном локусе или вблизи него.
В одном варианте осуществления способ усиливает гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых первую нуклеиновую кислоту вводят без второй нуклеиновой кислоты. В одном варианте осуществления способ усиливает гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых вторую нуклеиновую кислоту вводят без первой нуклеиновой кислоты. В одном варианте осуществления способ усиливает гомологичную рекомбинацию первой и второй нуклеиновых кислот в целевом геномном локусе. В одном варианте осуществления усиление гомологичной рекомбинации составляет по меньшей мере 0,5 раза, 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты гомологична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты на 100% идентична перекрывающейся последовательности второй нуклеиновой кислоты.
В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 10 т.п.н. до приблизительно 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 15 т.п.н. до приблизительно 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 20 т.п.н. до приблизительно 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 25 т.п.н. до приблизительно 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 30 т.п.н. до приблизительно 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 35 т.п.н. до приблизительно 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 40 т.п.н. до приблизительно 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 45 т.п.н. до приблизительно 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 50 т.п.н. до приблизительно 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 55 т.п.н. до приблизительно 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 60 т.п.н. до приблизительно 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 65 т.п.н. до приблизительно 70 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 70 т.п.н.
В одном варианте осуществления первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, а первая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной целевому геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления первая нуклеиновая кислота представляет собой первый нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом. В одном варианте осуществления первая полинуклеотидная вставка и вторая полинуклеотидная вставка представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты.
В одном варианте осуществления нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н.
В одном варианте осуществления нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC). В некоторых способах нацеливающий вектор представляет собой большой нацеливающий вектор в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый нацеливающий вектор представляет собой первый LTVEC и/или второй нацеливающий вектор представляет собой второй LTVEC. В некоторых способах первый нацеливающий вектор представляет собой первый большой нацеливающий вектор в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. и/или второй нацеливающий вектор представляет собой второй большой нацеливающий вектор в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. В одном варианте осуществления LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. Необязательно первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. Необязательно второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. и второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления общая длина 5' и 3' гомологичных плеч LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность расположена на 3'-конце первой нуклеиновой кислоты и 5'-конце второй нуклеотидной последовательности. В одном варианте осуществления перекрывающаяся последовательность расположена на 5'-конце первой нуклеиновой кислоты и 3'-конце второй нуклеотидной последовательности.
В одном варианте осуществления нуклеазный агент экспрессируется из экспрессионного конструкта, содержащего нуклеотидную последовательность, кодирующую нуклеазу, и причем нуклеиновая кислота функционально связана с активным в клетке промотором. В одном варианте осуществления нуклеазный агент экспрессируется из мРНК, кодирующей нуклеазу. В одном варианте осуществления нуклеаза представляет собой нуклеазу с «цинковыми пальцами» (ZFN). В одном варианте осуществления нуклеаза представляет собой эффекторную нуклеазу, подобную активатору транскрипции (TALEN). В одном варианте осуществления нуклеаза представляет собой мегануклеазу.
В одном варианте осуществления нуклеазный агент содержит белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК). В одном варианте осуществления белок Cas представляет собой Cas9.
В одном варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка получены от разных видов. В другом варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка представляют собой человеческие нуклеиновые кислоты. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. В некоторых способах интеграция одной или более из первой, второй и третьей полинуклеотидных вставок в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
В одном варианте осуществления интеграция первой и/или второй вставок в геномный локус приводит к добавлению экзогенной последовательности в геномном локусе.
В некоторых вариантах осуществления клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, длина которых находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н. В некоторых способах клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, которые имеют суммарный размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н.
В другом варианте осуществления интеграция первой и/или второй полинуклеотидных вставок в геномный локус приводит к делеции эндогенной последовательности в целевом геномном локусе. В одном варианте осуществления делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах клетка представляет собой человеческую клетку. В других способах клетка представляет собой нечеловеческую клетку. В некоторых способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В других способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В одном варианте осуществления клетка представляет собой плюрипотентную клетку. В другом варианте осуществления плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку. В некоторых вариантах осуществления плюрипотентная клетка представляет собой гемопоэтическую стволовую клетку или нейрональную стволовую клетку. В другом варианте осуществления клетка представляет собой индуцированную плюрипотентную стволовую (ИПС) клетку.
В одном варианте осуществления целевой геномный локус находится в геноме клетки. В другом варианте осуществления целевой геномный локус находится на внехромосомной ДНК внутри клетки.
В одном варианте осуществления клетка представляет собой фибробластную клетку.
В некоторых способах клетка представляет собой нечеловеческую клетку. В других способах клетка получена от человека. В некоторых вариантах осуществления клетка представляет собой клетку млекопитающих. В другом варианте осуществления клетка млекопитающих получена от грызуна. В некоторых случаях грызун представляет собой мышь, крысу или хомяка.
В некоторых из вышеуказанных способов перекрывающаяся нуклеотидная последовательность облегчает рекрутирование механизмов рекомбинации в целевой геномный локус.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена вышеуказанными способами; и (b) гестацию не относящегося к человеку эмбриона-хозяина в теле суррогатной матери, при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
В другом аспекте предложен способ усиления гомологичной рекомбинации в целевом геномном локусе в клетке путем загрузки механизмов рекомбинации на нацеливающий вектор, включающий в себя введение в клетку первой нуклеиновой кислоты и второй нуклеиновой кислоты, причем первая и вторая нуклеиновые кислоты содержат перекрывающуюся нуклеотидную последовательность, и при этом перекрывающаяся нуклеотидная последовательность облегчает рекрутирование механизмов рекомбинации в целевой геномный локус.
В одном варианте осуществления способ усиливает гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых первую нуклеиновую кислоту вводят без второй нуклеиновой кислоты. В одном варианте осуществления способ усиливает гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе. Некоторые такие способы усиливают гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе по сравнению со способами, в которых вторую нуклеиновую кислоту вводят без первой нуклеиновой кислоты. Некоторые такие способы усиливают гомологичную рекомбинацию первой и второй нуклеиновых кислот в целевом геномном локусе. В одном варианте осуществления усиление гомологичной рекомбинации составляет по меньшей мере 0,5 раза, 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз.
В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты гомологична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична перекрывающейся последовательности второй нуклеиновой кислоты. В одном варианте осуществления перекрывающаяся последовательность первой нуклеиновой кислоты на 100% идентична перекрывающейся последовательности второй нуклеиновой кислоты.
В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 10 т.п.н. до приблизительно 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 15 т.п.н. до приблизительно 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 20 т.п.н. до приблизительно 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 25 т.п.н. до приблизительно 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 30 т.п.н. до приблизительно 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 35 т.п.н. до приблизительно 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 40 т.п.н. до приблизительно 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 45 т.п.н. до приблизительно 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 50 т.п.н. до приблизительно 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 55 т.п.н. до приблизительно 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 60 т.п.н. до приблизительно 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 65 т.п.н. до приблизительно 70 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 15 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 25 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 30 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 35 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 40 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 45 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 55 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 60 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 65 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 70 т.п.н.
В одном варианте осуществления первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, а первая нуклеиновая кислота не содержит нуклеотидной последовательности, гомологичной геномному локусу, за исключением перекрывающейся последовательности.
В одном варианте осуществления первая нуклеиновая кислота представляет собой первый нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом. В одном варианте осуществления первая полинуклеотидная вставка и вторая полинуклеотидная вставка представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты.
В одном варианте осуществления нацеливающий вектор составляет от приблизительно 1 т.п.н. до приблизительно 2 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. В одном варианте осуществления нацеливающий вектор составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н.
В одном варианте осуществления нацеливающий вектор представляет собой большой нацеливающий вектор (LTVEC). В некоторых способах нацеливающий вектор представляет собой LTVEC в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый нацеливающий вектор представляет собой первый LTVEC и/или второй нацеливающий вектор представляет собой второй LTVEC. В некоторых способах первый нацеливающий вектор представляет собой первый LTVEC в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. и/или второй нацеливающий вектор представляет собой второй LTVEC в диапазоне от приблизительно 10 т.п.н. до приблизительно 200 т.п.н. В одном варианте осуществления LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. Необязательно первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. Необязательно второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. В некоторых способах первый LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н. и второй LTVEC составляет от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н. или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления общая длина 5' и 3' гомологичных плеч LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. В некоторых способах общая длина 5' и 3' гомологичных плеч первого LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н. и общая длина 5' и 3' гомологичных плеч второго LTVEC составляет от 10 т.п.н. до приблизительно 200 т.п.н.
В одном варианте осуществления перекрывающаяся последовательность расположена на 3'-конце первой нуклеиновой кислоты и 5'-конце второй нуклеотидной последовательности. В одном варианте осуществления перекрывающаяся последовательность расположена на 5'-конце первой нуклеиновой кислоты и 3'-конце второй нуклеотидной последовательности.
В одном варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка получены от разных видов. В другом варианте осуществления первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка представляют собой человеческие нуклеиновые кислоты. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка. В некоторых способах первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты.
В некоторых способах интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации. В некоторых способах интеграция одной или более из первой, второй и третьей полинуклеотидных вставок в целевой геномный локус приводит к одному или более из: (а) добавления экзогенной последовательности в целевом геномном локусе; (b) делеции эндогенной последовательности в целевом геномном локусе; или (с) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
В одном варианте осуществления интеграция первой и/или второй вставок в геномный локус приводит к добавлению экзогенной последовательности в геномном локусе.
В некоторых вариантах осуществления клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую и вторую полинуклеотидные вставки, длина которых находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н. В некоторых способах клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую полинуклеотидную вставку и вторую полинуклеотидную вставку, которые имеют суммарный размер в диапазоне от приблизительно 5 т.п.н. до приблизительно 500 т.п.н.
В другом варианте осуществления интеграция первой и/или второй полинуклеотидных вставок в геномный локус приводит к делеции эндогенной последовательности в целевом геномном локусе. В одном варианте осуществления делеция эндогенной последовательности в целевом геномном локусе составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах клетка представляет собой человеческую клетку. В других способах клетка представляет собой нечеловеческую клетку. В некоторых способах клетка представляет собой плюрипотентную клетку, гемопоэтическую стволовую клетку, нейрональную стволовую клетку или фибробластную клетку. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку. В других способах клетка представляет собой клетку млекопитающих. Необязательно клетка млекопитающих представляет собой клетку грызуна. Необязательно клетка грызуна представляет собой клетку мыши или клетку крысы.
В одном варианте осуществления клетка представляет собой плюрипотентную клетку. В другом варианте осуществления плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку. В некоторых вариантах осуществления плюрипотентная клетка представляет собой гемопоэтическую стволовую клетку или нейрональную стволовую клетку. В другом варианте осуществления клетка представляет собой индуцированную плюрипотентную стволовую (ИПС) клетку.
В одном варианте осуществления целевой геномный локус находится в геноме клетки. В другом варианте осуществления целевой геномный локус находится на внехромосомной ДНК внутри клетки.
В одном варианте осуществления клетка представляет собой фибробластную клетку.
В одном варианте осуществления клетка представляет собой нечеловеческую клетку. В другом варианте осуществления клетка получена от человека. В другом варианте осуществления клетка представляет собой клетку млекопитающих. В другом варианте осуществления клетка млекопитающих получена от грызуна. В другом варианте осуществления грызун представляет собой мышь, крысу или хомяка.
В изобретении также предложены способы получения не относящихся к человеку животных поколения F0, включающие в себя: (а) введение нечеловеческой ЭС-клетки в нечеловеческий эмбрион-хозяин, причем нечеловеческая ЭС-клетка получена вышеуказанными способами; и (b) гестацию не относящегося к человеку эмбриона-хозяина в теле суррогатной матери, при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модификацию. Необязательно не относящееся к человеку животное представляет собой мышь или крысу.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фиг. 1 представлена схема события двойного геномного нацеливания, во время которого выполняют нацеливание на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащей кассету селекции гигромицином. Кассета селекции гигромицином расщепляется нуклеазой с «цинковыми пальцами» (ZFN) или комплексом CRISPR/Cas, и нацеливание выполняется двумя большими нацеливающими векторами, содержащими кассету селекции неомицином и более 280 т.п.н. сегментов вариабельного гена каппа-цепи иммуноглобулина человека. Каждый большой нацеливающий вектор содержит перекрывающуюся последовательность из приблизительно 20 т.п.н., что обеспечивает гомологичную рекомбинацию между большими нацеливающими векторами. Событие нацеливания точно вставляло сегменты вариабельного гена каппа-цепи иммуноглобулина человека из обоих нацеливающих векторов за один этап нацеливания. Местоположения различных зондов, используемых для подтверждения события нацеливания, показаны в виде заключенных в окружности прямоугольников. Мышиная последовательность представлена диагональной штриховкой снизу вверх, человеческая последовательность представлена без штриховки, а сайты рекомбинации и кассеты селекции представлены диагональной пунктирной штриховкой сверху вниз. Схема выполнена не в масштабе и, например, не отражает фактическое число сегментов вариабельного гена.
На Фиг. 2 представлена схема одиночного события нацеливания, в котором нацеливание на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащей кассету селекции гигромицином, выполняется одним большим нацеливающим вектором, содержащим кассету селекции неомицином и 120 т.п.н. сегментов вариабельного гена каппа-цепи иммуноглобулина человека. Местоположения различных зондов, используемых для подтверждения события нацеливания, показаны в виде заключенных в окружности прямоугольников. Мышиная последовательность представлена диагональной штриховкой снизу вверх, человеческая последовательность представлена без штриховки, а сайты рекомбинации и кассеты селекции представлены диагональной пунктирной штриховкой сверху вниз. Схема выполнена не в масштабе и, например, не отражает фактическое число сегментов вариабельного гена.
На Фиг. 3 представлена схема нацеливания и разрушения кассеты селекции гигромицином с использованием системы CRISPR/Cas9 и иллюстрируется положение в пределах гена гигромицина сайтов распознавания CRISPR для различных гидовых РНК (гРНК), которые нацелены на различные последовательности в гене гигромицина. Схема выполнена не в масштабе.
На Фиг. 4 представлена схема события тройного геномного нацеливания, в котором выполняют нацеливание на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащей кассету селекции гигромицином. Кассета селекции гигромицином расщепляется нуклеазой с «цинковыми пальцами» (ZFN) или комплексом CRISPR/Cas, и нацеливание выполняется тремя большими нацеливающими векторами, содержащими кассету селекции неомицином и приблизительно 370 т.п.н. сегментов вариабельного гена каппа-цепи иммуноглобулина человека. Каждый большой нацеливающий вектор содержит перекрывающуюся последовательность от приблизительно 20 т.п.н. до приблизительно 60 т.п.н., что обеспечивает гомологичную рекомбинацию между большими нацеливающими векторами. Событие нацеливания точно вставляло сегменты вариабельного гена каппа-цепи иммуноглобулина человека из всех трех нацеливающих векторов за один этап нацеливания. Местоположения различных зондов, используемых для подтверждения события нацеливания, показаны в виде заключенных в окружности прямоугольников. Мышиная последовательность представлена диагональной штриховкой снизу вверх, человеческая последовательность представлена без штриховки, а сайты рекомбинации и кассеты селекции представлены диагональной пунктирной штриховкой сверху вниз. Схема выполнена не в масштабе и, например, не отражает фактическое число сегментов вариабельного гена.
ОПРЕДЕЛЕНИЯ
Термины «белок», «полипептид» и «пептид», используемые в настоящем документе на взаимозаменяемой основе, включают полимерные формы аминокислот любой длины, в том числе кодированные и некодированные аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают полимеры, которые подверглись модификации, например полипептиды с модифицированными основными цепями пептидов.
Термины «нуклеиновая кислота» и «полинуклеотид», используемые в настоящем документе на взаимозаменяемой основе, включают полимерные формы нуклеотидов любой длины, в том числе рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные формы. Эти термины включают одно-, двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
Целевой геномный локус означает область генома, которая подлежит модификации путем нацеленной модификации с нацеливающим вектором. Область может быть определена как область в пределах внешних границ сегментов геномной ДНК, соответствующих гомологичным плечам в пределах нацеливающего вектора. Целевой геномный локус может включать в себя любой или весь ген или группу генов, один или более интронов, один или более экзонов, одну или более регуляторных последовательностей и т.п.
Термин «оптимизация по кодонам» по существу включает в себя процесс модификации нуклеотидной последовательности для повышения экспрессии в конкретных клетках-хозяевах путем замещения по меньшей мере одного кодона нативной последовательности кодоном, который чаще или наиболее часто используется в генах клетки-хозяина при сохранении нативной аминокислотной последовательности. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована так, чтобы использовать в качестве замены кодоны, имеющие более высокую частоту использования в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, нечеловеческую клетку, клетку млекопитающих, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина, по сравнению со встречающейся в природе нуклеотидной последовательностью. Таблицы использования кодонов являются общедоступными, например, в базе данных Codon Usage Database. Эти таблицы можно адаптировать несколькими способами. См. Nakamura et al. (2000) Nucleic Acids Research 28:292. Также имеются компьютерные алгоритмы для оптимизации по кодонам с конкретной последовательностью для экспрессии в конкретном хозяине (см., например, Gene Forge).
Термин «функциональная связь» или «функционально связанный» относится к смежному размещению двух или более компонентов (например, промотора и другого элемента последовательности) таким образом, чтобы оба компонента нормально функционировали и обеспечивалась возможность того, чтобы по меньшей мере один из компонентов мог опосредовать функцию, осуществляемую в отношении по меньшей мере одного из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции.
Термин «плюрипотентная клетка» или «плюрипотентная стволовая клетка», используемый в настоящем документе, включает в себя недифференцированную клетку, которая обладает способностью развиваться в более чем один дифференцированный тип клеток. Такие плюрипотентные клетки могут представлять собой, например, эмбриональную стволовую клетку (ЭС-клетку) млекопитающих или индуцированную плюрипотентную стволовую клетку (ИПС-клетку) млекопитающих.
Термин «эмбриональная стволовая клетка» или «ЭС-клетка» включает в себя полученную из эмбриона тотипотентную или плюрипотентную клетку, которая способна к недифференцированной пролиферации in vitro, а при введении в эмбрион может участвовать в развитии любой ткани развивающегося эмбриона.
Термин «индуцированная плюрипотентная стволовая клетка» или «ИПС-клетка» включает в себя плюрипотентную стволовую клетку, которая может быть получена непосредственно из дифференцированной взрослой клетки. Человеческие ИПС-клетки можно получать путем введения в неплюрипотентную клетку конкретных наборов репрограммирующих факторов, в которые могут входить, например, Oct3/4, транскрипционные факторы семейства Sox (например, Sox1, Sox2, Sox3, Sox15), транскрипционные факторы семейства Мус (например, с-Мус, 1-Мус, n-Мус), транскрипционные факторы семейства  -подобных (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glis1. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (Cell (2006) Vol. 126(4), pp. 663-676).
-подобных (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glis1. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (Cell (2006) Vol. 126(4), pp. 663-676).
Термин «зародышевая линия» в отношении нуклеотидной последовательности иммуноглобулина включает в себя нуклеотидную последовательность, которая может быть передана потомству.
Термин «комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты вследствие ориентации нуклеотидных оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. В ДНК, как правило, комплементарны основания А и Т, а также Ц и Г. В РНК это, как правило, Ц и Г, а также У и А. Комплементарность может быть идеальной или существенной/достаточной. Идеальная комплементарность двух нуклеиновых кислот означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание связано с комплементарным основанием, образуя пары Уотсона-Крика. Термин «существенная» или «достаточная» комплементарность означает, что последовательность одной цепи не полностью и/или не идеально комплементарна последовательности противоположной цепи, но между основаниями на двух цепях создается достаточная связь, чтобы образовывался стабильный гибридный комплекс при наборе условий гибридизации (например, концентрации солей и температуре). Такие условия можно спрогнозировать, используя последовательности и стандартные математические расчеты для предсказания Tm (температуры плавления) гибридизованных цепей, или путем эмпирического определения Tm традиционными способами. Tm означает температуру, при которой популяция гибридизационных комплексов, образованных между двумя цепями нуклеиновых кислот, денатурирована на 50% (т.е. популяция молекул двухцепочечной нуклеиновой кислоты наполовину диссоциируется на отдельные цепи). При температуре ниже Tm преимущество имеет образование гибридизационного комплекса, а при температуре выше Tm преимущество имеет плавление или разделение цепей гибридизационного комплекса. Tm можно оценить для нуклеиновой кислоты с известным содержанием Г+Ц в водном 1 М растворе NaCl с использованием, например, выражения Tm = 81,5+0,41 (% Г + Ц), хотя в других известных расчетах Tm принимаются во внимание структурные характеристики нуклеиновых кислот.
Термин «условие гибридизации» включает в себя совокупную среду, в которой одна цепь нуклеиновой кислоты связывается со второй цепью нуклеиновой кислоты в результате взаимодействия комплементарных цепей и водородного связывания с образованием гибридизационного комплекса. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие агенты, формамид) в водном или органическом растворе, содержащем нуклеиновые кислоты, и температуру смеси. Свой вклад в среду могут вносить другие факторы, такие как длительность инкубации или размеры инкубационной камеры. См., например, публикацию Sambrook et al., Molecular Cloning, A Laboratory Manual, 2.sup.nd ed., pp. 1.90-1.91, 9.47-9.51, 1 1.47-11.57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989).
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементации, и переменные известны специалистам в данной области. Чем больше степень комплементации между двумя нуклеотидными последовательностями, тем выше температура плавления (Tm) гибридов нуклеиновых кислот, имеющих эти последовательности. При гибридизации нуклеиновых кислот с короткими участками комплементарности (например, с комплементарностью более 35 нуклеотидов или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее) становится важным положение несовпадающих оснований (см. выше Sambrook et al., 11.7-11.8). Как правило, длина пригодной для гибридизации нуклеиновой кислоты составляет по меньшей мере приблизительно 10 нуклеотидов. Значения иллюстративной минимальной длины пригодных для гибридизации нуклеиновых кислот включают по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 22 нуклеотида, по меньшей мере приблизительно 25 нуклеотидов и по меньшей мере приблизительно 30 нуклеотидов. Кроме того, температуру и концентрацию солей в промывном растворе можно регулировать по мере необходимости в зависимости от таких факторов, как длина области комплементации и степень комплементации.
Для обеспечения специфической гибридизации последовательность полинуклеотида необязательно должна быть на 100% комплементарна целевой нуклеиновой кислоте. Более того, полинуклеотид может гибридизоваться на одном или более сегментах, так что промежуточные или прилегающие сегменты не участвуют в событии гибридизации (например, структура петли или шпильки). Полинуклеотид (например, гРНК) может иметь по меньшей мере 70%-ную, по меньшей мере 80%-ную, по меньшей мере 90%-ную, по меньшей мере 95%-ную, по меньшей мере 99%-ную или 100%-ную комплементарность последовательности относительно целевой области в пределах целевой нуклеотидной последовательности, на которую он нацелен. Например, гРНК, в которой 18 из 20 нуклеотидов комплементарны целевой области и, следовательно, могут специфически гибридизоваться, будет иметь 90% комплементарности. В этом примере остальные некомплементарные нуклеотиды могут быть кластеризованы или рассеяны среди комплементарных нуклеотидов и не должны быть обязательно смежными друг с другом или с комплементарными нуклеотидами.
Процентную долю комплементарности между конкретными участками нуклеотидных последовательностей в пределах нуклеиновых кислот можно определять традиционными способами, используя программы BLAST (средства поиска основных локальных выравниваний) и PowerBLAST, известные в данной области (Altschul et al. (1990) J. Mol. Biol. 215:403-410; Zhang and Madden (1997) Genome Res. 7:649-656), или с помощью программы Gap (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, г. Мэдисон, штат Висконсин, США) с использованием заданных по умолчанию параметров, где используется алгоритм Смита - Ватермана (Adv. Appl. Math., 1981, 2, 482-489).
В предложенных в настоящем документе способах и композициях используется множество разных компонентов. В описании признается, что некоторые компоненты могут иметь активные варианты и фрагменты. Такие компоненты включают в себя, например, нуклеазные агенты, белки Cas, РНК CRISPR, тракрРНК и гидовые РНК. Биологическая активность каждого из этих компонентов описана в других разделах настоящего документа.
Термин «идентичность последовательности» или «идентичность» в контексте двух полинуклеотидов или полипептидных последовательностей означает ссылку на остатки, которые одинаковы в двух последовательностях при выравнивании для максимального соответствия в установленном окне сравнения. Если термин «процентная доля идентичности последовательности» применяется в отношении белков, следует понимать, что положения с остатками, не являющимися идентичными, зачастую отличаются консервативными заменами аминокислот, где аминокислотные остатки заменяются другими аминокислотными остатками с аналогичными химическими свойствами (например, зарядом или гидрофобностью) и поэтому не изменяют функциональные свойства молекулы. Если последовательности отличаются консервативными заменами, процентную долю идентичности последовательностей можно скорректировать в сторону увеличения, чтобы учесть консервативный характер замены. Последовательности, отличающиеся такими консервативными заменами, считаются имеющими «сходство последовательностей» или «сходство». Средства для осуществления этой регулировки хорошо известны. Как правило, они включают в себя оценку консервативной замены как частичного, а не полного несовпадения, в результате чего повышается процентное значение идентичности последовательности. Таким образом, например, если идентичной аминокислоте присваивают оценку 1, то неконсервативной замене присваивают оценку, равную нулю, а консервативная замена получает оценку от нуля до 1. Вычисляют оценку консервативных замен, например, в соответствии с реализацией в программе PC/GENE (Intelligenetics, г. Маунтин-Вью, штат Калифорния, США).
Термин «процентное значение идентичности последовательности» означает значение, которое определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения, причем участок полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) в сравнении с эталонной последовательностью (которая не содержит добавлений или делеции) для оптимального выравнивания двух последовательностей. Процентное значение рассчитывают путем определения числа положений, в которых в обеих последовательностях встречаются идентичные нуклеиновые основания или аминокислотные остатки, деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 с получением процентного значения идентичности последовательности.
Если не указано иное, значения идентичности/сходства последовательности включают значение, полученное с использованием программного обеспечения GAP версии 10 с использованием следующих параметров: % идентичности и % сходства нуклеотидной последовательности при использовании значения GAP Weight (штраф за начало гэпа) = 50 и Length Weight (штраф за удлинение) = 3 и матрица замен nwsgapdna.cmp; % идентичности и % сходства аминокислотной последовательности при использовании значения GAP Weight (штраф за начало гэпа) = 8 и Length Weight (штраф за удлинение) = 2 и матрица замен BLOSUM62; или любой эквивалентной программы. Термин «эквивалентная программа» включает в себя любую программу для сравнения последовательностей, которая для любых двух рассматриваемых последовательностей создает выравнивание, обладающее идентичными соответствиями нуклеотидов или аминокислотных остатков и идентичным процентом идентичности последовательности при сравнении с соответствующим выравниванием, получаемым посредством GAP версии 10.
«Гомологичная» последовательность включает в себя нуклеотидную последовательность, которая либо идентична, либо по существу аналогична известной эталонной последовательности, так что она на по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентична известной эталонной последовательности. «Ортологичная» последовательность включает в себя нуклеотидную последовательность от одного вида, которая функционально эквивалентна известной эталонной последовательности у другого вида.
Термин «in vitro» включает искусственные среды и процессы или реакции, возникающие в искусственной среде (например, пробирке). Термин «in vivo» включает естественные среды (например, клетку, или организм, или тело) и процессы или реакции, протекающие в естественной среде. Термин «ех vivo» включает клетки, которые были извлечены из тела индивида, а также процессы или реакции, происходящие внутри таких клеток.
Композиции или способы, «содержащие» или «включающие в себя» один или более упомянутых элементов, могут включать в себя другие конкретно не упомянутые элементы. Например, композиция, которая «содержит» или «включает в себя» белок, может содержать белок отдельно или в комбинации с другими ингредиентами.
Обозначение диапазона значений включает в себя все целые числа, входящие в диапазон или образующие его, а также все поддиапазоны, образованные целыми числами, входящими в диапазон.
Если иное не очевидно из контекста, термин «приблизительно» охватывает значения в пределах стандартных границ погрешности измерения (например, стандартной погрешности среднего (СПС)) указанного значения.
Все элементы, указанные в единственном числе, также могут использоваться и во множественном, если контекст явно не диктует иное. Например, термин «белок Cas» или «по меньшей мере один белок Cas» может включать в себя множество белков Cas, в том числе их смеси.
ПОДРОБНОЕ ОПИСАНИЕ
I. Модификация геномных локусов с использованием множества нацеливающих векторов
Предложены способы и композиции для модификации целевого геномного локуса в клетке. В таких способах применяется множество больших нацеливающих векторов (LTVEC), которые способны рекомбинировать друг с другом с образованием одного сегмента непрерывной нуклеиновой кислоты. В таких способах могут применяться 1, 2, 3, 4, 5, 6 или более LTVEC за один этап нацеливания. Также предложены способы и композиции для усиления гомологичной рекомбинации в целевом геномном локусе в клетке. В таких способах применяются две или более нуклеиновых кислот, содержащих одну или более перекрывающихся последовательностей. Любой из описанных в настоящем документе способов может осуществляться in vitro, ex vivo или in vivo.
А. Двойное нацеливание
Предложены способы и композиции для модификации целевого геномного локуса в клетке посредством способа двойного нацеливания. В способах и композициях применяются два больших нацеливающих вектора (LTVEC) (например, первый LTVEC и второй LTVEC), которые способны рекомбинировать друг с другом с образованием одного сегмента непрерывной нуклеиновой кислоты. Первый LTVEC содержит первую полинуклеотидную вставку, а второй LTVEC содержит вторую полинуклеотидную вставку. Полинуклеотидные вставки фланкированы 5' и 3' гомологичными плечами. Первая полинуклеотидная вставка и ее 3' гомологичное плечо и вторая полинуклеотидная вставка и ее 5' гомологичное плечо могут являться перекрывающимися фрагментами одной и той же непрерывной нуклеиновой кислоты. 3' гомологичное плечо первого LTVEC и 5' гомологичное плечо второго LTVEC перекрываются (т.е. комплементарны друг другу) и первая и вторая вставки фланкируют перекрывающиеся гомологичные плечи. Такие способы включают в себя три события рекомбинации, которые могут происходить в любом порядке: (1) рекомбинация между 3' гомологичным плечом первого LTVEC и 5' гомологичным плечом второго LTVEC; (2) рекомбинация между 5' гомологичным плечом первого LTVEC и соответствующим сегментом в целевом локусе; и (3) рекомбинация между 3' гомологичным плечом второго LTVEC и соответствующим сегментом в целевом локусе. Эта трехкомпонентная рекомбинация восстанавливает непрерывную нуклеиновую кислоту в целевом локусе с использованием перекрывающейся последовательности гомологичных плеч, расположенных между первой и второй полинуклеотидными вставками.
Каждый из LTVEC также содержит либо 5', либо 3' гомологичное плечо, которое гомологично области ДНК в пределах целевого геномного локуса или вблизи него, что обеспечивает рекомбинацию и интеграцию одного сегмента непрерывной нуклеиновой кислоты. Таким образом, посредством события трехкомпонентной рекомбинации можно выполнить большую модификацию нуклеиновой кислоты (т.е. делецию, вставку и/или замещение) в целевом локусе за один этап нацеливания.
Три события рекомбинации могут происходить в любом порядке. В одном варианте осуществления событие рекомбинации между перекрывающимися последовательностями двух LTVEC происходит перед гомологичной рекомбинацией с целевым локусом. В другом варианте осуществления рекомбинация с целевым локусом происходит до рекомбинации между двумя LTVEC. В еще одном варианте осуществления три события рекомбинации могут происходить одновременно.
В одном варианте осуществления предложен способ модификации целевого геномного локуса в клетке. Такой способ включает в себя введение первого большого нацеливающего вектора (LTVEC), который содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, который содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, при этом целевой геномный локус модифицирован путем интеграции первой и второй полинуклеотидных вставок между соответствующими геномными сегментами. Способ дополнительно включает в себя отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус.
В. Тройное нацеливание
Также предложены способы и композиции для модификации целевого геномного локуса в клетке посредством способов тройного нацеливания. В способах и композициях применяются три больших нацеливающих вектора (LTVEC) (например, первый LTVEC, второй LTVEC и третий LTVEC), которые способны рекомбинировать друг с другом с образованием одного сегмента непрерывной нуклеиновой кислоты. Первый LTVEC содержит первую полинуклеотидную вставку, второй LTVEC содержит вторую полинуклеотидную вставку, а третий LTVEC содержит третью полинуклеотидную вставку. Полинуклеотидные вставки фланкированы 5' и 3' гомологичными плечами. Первая полинуклеотидная вставка и ее 3' гомологичное плечо и вторая полинуклеотидная вставка и ее 5' гомологичное плечо могут являться перекрывающимися фрагментами одной и той же непрерывной нуклеиновой кислоты. Вторая полинуклеотидная вставка и ее 3' гомологичное плечо и третья полинуклеотидная вставка и ее 5' гомологичное плечо могут являться перекрывающимися фрагментами одной и той же непрерывной нуклеиновой кислоты. 3' гомологичное плечо первого LTVEC и 5' гомологичное плечо второго LTVEC перекрываются (т.е. комплементарны друг другу) и первая и вторая вставки фланкируют перекрывающиеся гомологичные плечи. 3' гомологичное плечо второго LTVEC и 5' гомологичное плечо третьего LTVEC перекрываются (т.е. комплементарны друг другу), а вторая и третья вставки фланкируют перекрывающиеся гомологичные плечи.
Такие способы включают в себя четыре события рекомбинации, которые могут происходить в любом порядке: (1) рекомбинация между 3' гомологичным плечом первого LTVEC и 5' гомологичным плечом второго LTVEC; (2) рекомбинация между 3' гомологичным плечом второго LTVEC и 5' гомологичным плечом третьего LTVEC; (3) рекомбинация между 5' гомологичным плечом первого LTVEC и соответствующим сегментом в целевом локусе; и (4) рекомбинация между 3' гомологичным плечом третьего LTVEC и соответствующим сегментом в целевом локусе. Эта четырехкомпонентная рекомбинация восстанавливает непрерывную нуклеиновую кислоту в целевом локусе с использованием перекрывающейся последовательности гомологичных плеч, расположенных между первой и второй полинуклеотидными вставками и между второй и третьей полинуклеотидными вставками.
Первый и третий LTVEC также содержат либо 5', либо 3' гомологичное плечо, которое гомологично области ДНК в пределах целевого геномного локуса или вблизи него, что обеспечивает рекомбинацию и интеграцию одного сегмента непрерывной нуклеиновой кислоты. Таким образом, с помощью события четырехкомпонентной рекомбинации можно осуществить большую модификацию нуклеиновой кислоты (т.е. делецию, вставку и/или замещение) в целевом локусе за один этап нацеливания.
Четыре события рекомбинации могут происходить в любом порядке. В одном варианте осуществления событие рекомбинации между перекрывающимися последовательностями трех LTVEC происходит перед гомологичной рекомбинацией с целевым локусом. В другом варианте осуществления рекомбинация с целевым локусом происходит до рекомбинации между тремя LTVEC. В еще одном варианте осуществления четыре события рекомбинации могут происходить одновременно.
В одном варианте осуществления предложен способ модификации целевого геномного локуса в клетке. Такой способ включает в себя введение первого большого нацеливающего вектора (LTVEC), который содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, второго LTVEC, который содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, и третьего LTVEC, который содержит третью полинуклеотидную вставку, фланкированную третьим 5' гомологичным плечом и третьим 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, второе 3' гомологичное плечо второго LTVEC имеет перекрывающуюся последовательность, гомологичную третьему 5' гомологичному плечу третьего LTVEC, и первое 5' гомологичное плечо первого LTVEC и третье 3' гомологичное плечо третьего LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, при этом целевой геномный локус модифицирован путем интеграции первой, второй и третьей полинуклеотидных вставок между соответствующими геномными сегментами. Способ дополнительно включает в себя отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, интегрированные в целевой геномный локус.
С. Нацеливание множеством LTVEC
Способы нацеливания, предложенные в настоящем документе для создания генетической модификации за один этап нацеливания, дают новые возможности и повышают эффективность нацеленных модификаций гена помимо тех, которые достигаются по способу нацеливания с одиночной LTVEC. Нацеливание двумя, тремя или более LTVEC, которые способны рекомбинировать друг с другом, позволяет модифицировать более крупный сегмент ДНК. События рекомбинации могут происходить в любом порядке. Например, событие рекомбинации между перекрывающимися последовательностями LTVEC может происходить перед гомологичной рекомбинацией с целевым локусом. Альтернативно рекомбинация с целевым локусом может происходить до рекомбинации между LTVEC или события рекомбинации могут происходить одновременно.
Способы нацеливания, описанные в настоящем документе, дают несколько преимуществ по сравнению с существующими способами нацеливания с одиночным LTVEC, в том числе повышенную эффективность нацеливания, увеличение достижимого размера генетической модификации и сокращение числа этапов нацеливания, необходимых для получения больших геномных модификаций, что экономит время и поддерживает плюрипотентность модифицированных эмбриональных стволовых клеток. Это имеет особое значение в случае больших геномных модификаций, так как способы позволяют модифицировать геномный локус с комбинацией полинуклеотидных вставок из двух, трех или более LTVEC за один этап. Таким образом, такие модификации могут обеспечивать очень большие (например, >50 т.п.н.) делеции, замещения и вставки в пределах нацеленного геномного локуса.
Например, время, требуемое для последовательного использования трех LTVEC с целью модификации целевого геномного локуса, скрининга на нацеленную модификацию и ее подтверждение, составляет приблизительно девять месяцев, в то время как такая же модификация может быть выполнена и подтверждена с использованием трех LTVEC одновременно лишь за примерно четыре месяца.
Последовательные модификации также создают более высокий риск потери плюрипотентности и потенциала передачи зародышевой линии при модификации плюрипотентных клеток, таких как эмбриональные стволовые клетки. По мере увеличения числа пассажей в культуре и увеличения числа электропораций накапливаются хромосомные и кариотипические аномалии, что может привести к утрате компетентности зародышевой линии. См., например, публикации Buehr et al. (2008) Cell 135:1287-1298; Li et al. (2008) Cell 135(7): 1299-1310; и Liu et al. (1997) Dev. Dyn. 209:85-91, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки. Нацеливание с одновременным использованием множества LTVEC вместо последовательного сокращает число пассажей и число электропораций и, таким образом, повышает способность выполнять генетические манипуляции в плюрипотентных клетках, таких как эмбриональные стволовые клетки, с сохранением компетентности их зародышевой линии.
В конкретных вариантах осуществления генетическая модификация включает в себя модификацию одной или более эндогенных нуклеиновых кислот, замену одной или более эндогенных нуклеиновых кислот, замещение эндогенной нуклеиновой кислоты на гетерологичную нуклеиновую кислоту, нокаут или нокин. В конкретных примерах генетическую модификацию вводят путем введения по меньшей мере двух больших нацеливающих векторов (LTVEC) в клетку. В другом примере генетическую модификацию вводят путем введения в клетку по меньшей мере трех больших нацеливающих векторов (LTVEC). В таких примерах LTVEC могут содержать ДНК, которая должна быть вставлена в целевой геномный локус клетки.
В некоторых вариантах осуществления способы модификации целевого геномного локуса включают в себя введение генетической модификации в клетки млекопитающих. Аналогичным образом, в изобретении предложены клетки млекопитающих, содержащие генетическую модификацию.
Можно использовать различные способы получения нацеленных генетических модификаций в клетках. Например, как описано выше, для нацеленной генетической модификации применяется система, которая будет создавать нацеленную генетическую модификацию посредством события гомологичной рекомбинации. В других случаях клетку можно модифицировать с использованием нуклеазных агентов, которые осуществляют одно- или двухцепочечный разрыв в нацеленном геномном локусе. Затем одно- или двухцепочечный разрыв восстанавливается по пути негомологичного соединения концов (NHEJ). Примеры способов создания таких нацеленных генетических модификаций подробно описаны в других разделах настоящего документа, включая, например, использование больших нацеливающих векторов. См. также публикации Wang et al. (2013) Cell 153:910-918, Mandalos et al. (2012) PLOS ONE 7:e45768:1-9 и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждая из которых включена в настоящий документ путем ссылки.
Нацеленная генная модификация путем гомологичной рекомбинации между нацеливающим вектором и целевым локусом может быть очень неэффективной, особенно в типах клеток, отличных от эмбриональных стволовых клеток грызунов. Использование нацеливающего вектора в комбинации с направляемым нуклеазой двухцепочечным разрывом ДНК в целевом локусе может значительно повысить эффективность нацеливания для модификаций, таких как делеции или вставки. Аналогичным образом, использование нацеливающего вектора в комбинации с направляемым нуклеазой одноцепочечным разрывом ДНК в целевом локусе может значительно повысить эффективность нацеливания для модификаций.
В некоторых вариантах осуществления LTVEC могут применяться в комбинации с нуклеазными агентами, которые осуществляют одно- или двухцепочечный разрыв в пределах целевого геномного локуса. Такой способ дополнительно включает в себя введение нуклеазного агента в клетку. В одном варианте осуществления нуклеазный агент представляет собой нуклеазу с «цинковыми пальцами» (ZFN). В другом варианте осуществления нуклеазный агент представляет собой систему коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/CRISPR-ассоциированного белка (Cas).
В одном варианте осуществления предложен способ модификации целевого геномного локуса в клетке путем применения множества LTVEC. Такой способ включает в себя: (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение первого большого нацеливающего вектора (LTVEC), содержащего первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, содержащего вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, и первое 3' гомологичное плечо первого LTVEC и второе 5' плечо второго LTVEC гомологичны друг другу или соответственно дополнительным 5' и 3' гомологичным плечам одного или более дополнительных LTVEC, каждый из которых содержит дополнительную полинуклеотидную вставку, фланкированную дополнительным 5' гомологичным плечом и дополнительным 3' гомологичным плечом; при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки, одной или более дополнительных полинуклеотидных вставок одного или более дополнительных LTVEC, если они имеются, и второй полинуклеотидной вставки между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, одну или более дополнительных полинуклеотидных вставок, если они имеются, и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус. В таких способах дополнительные LTVEC представляют собой один или более других LTVEC, которые, если они имеются, вставлены между первым и вторым LTVEC.
В одном варианте осуществления предложен способ двойного нацеливания для модификации целевого геномного локуса в клетке, причем способ включает в себя: (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение первого большого нацеливающего вектора (LTVEC), который содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, который содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса; при этом целевой геномный локус модифицирован путем интеграции первой и второй полинуклеотидных вставок между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус. В таких способах первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки в целевой геномный локус.
В одном варианте осуществления предложен способ тройного нацеливания для модификации целевого геномного локуса в клетке, причем способ включает в себя: (а) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; (b) введение первого большого нацеливающего вектора (LTVEC), который содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, второго LTVEC, который содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, и третьего LTVEC, который содержит третью полинуклеотидную вставку, фланкированную третьим 5' гомологичным плечом и третьим 3' гомологичным плечом, причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, второе 3' гомологичное плечо второго LTVEC имеет перекрывающуюся последовательность, гомологичную третьему 5' гомологичному плечу третьего LTVEC, и первое 5' гомологичное плечо первого LTVEC и третье 3' гомологичное плечо третьего LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса; при этом целевой геномный локус модифицирован путем интеграции первой, второй и третьей полинуклеотидных вставок между соответствующими геномными сегментами; и (с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, интегрированные в целевой геномный локус. В таких способах тройного нацеливания первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, и вторая полинуклеотидная вставка и второе 3' гомологичное плечо и третья полинуклеотидная вставка и третье 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки в целевой геномный локус.
В некоторых случаях два, три или более LTVEC могут быть введены одновременно. Альтернативно два, три или более LTVEC могут быть введены последовательно или могут быть введены в разные моменты времени.
Различные компоненты системы нацеливания могут включать в себя, например, нацеливающие векторы, нуклеазные агенты, целевой геномный локус, полинуклеотидные вставки, представляющие интерес полинуклеотиды и/или другие компоненты, каждый из которых подробно описан в других разделах настоящего документа.
D. Нацеливание с использованием множества перекрывающихся нуклеиновых кислот
Способы нацеливания, предложенные в настоящем документе для создания генетической модификации за один этап нацеливания, дают новые возможности и повышают эффективность нацеленных модификаций гена помимо тех, которые достигаются с помощью одиночной нуклеиновой кислоты. Нацеливание двумя, тремя или более нуклеиновыми кислотами, которые способны рекомбинировать друг с другом, позволяет модифицировать более крупный сегмент ДНК и может обеспечивать повышенную эффективность нацеливания в сравнении с использованием только одиночных нуклеиновых кислот даже при отсутствии нуклеазных агентов. Такие способы без нуклеазных агентов могут иметь преимущество по сравнению с теми, в которых применяются нуклеазные агенты, так как скрининг, необходимый для способов с использованием нуклеаз, является более сложным и требует много времени, включая дополнительные этапы скрининга для подтверждения расщепления и проверки внецелевых эффектов. Нуклеиновые кислоты (например, LTVEC) с перекрывающимися областями достаточной длины могут усиливать гомологичную рекомбинацию в целевом геномном локусе даже при отсутствии нацеленной нуклеазы. Например, использование двух нуклеиновых кислот с перекрывающейся областью достаточной длины может усиливать гомологичную рекомбинацию в целевом геномном локусе по сравнению с использованием одиночной нуклеиновой кислоты. Хотя для практического применения не требуется понимания механизма, считается, что гомологичная рекомбинация в таких условиях усиливается путем загрузки механизмов рекомбинации (например, ExoI, Rad51, BRCA2 и т.д.) на нуклеиновые кислоты (например, LTVEC), что, таким образом, облегчает рекрутирование механизмов рекомбинации в целевой локус.
В настоящем документе предложены способы модификации целевого геномного локуса или усиления гомологичной рекомбинации в целевом геномном локусе в клетке, включающие в себя введение в клетку первой и второй нуклеиновых кислот, причем первая и вторая нуклеиновые кислоты содержат перекрывающуюся последовательность. Первой и второй нуклеиновыми кислотами могут быть, например, линейные нуклеиновые кислоты. Такие способы могут также включать в себя введение в клетку трех или более нуклеиновых кислот, которые способны рекомбинировать друг с другом. Например, первая и вторая нуклеиновые кислоты могут иметь первую перекрывающуюся последовательность, а вторая и третья нуклеиновые кислоты могут иметь вторую перекрывающуюся последовательность. В некоторых способах целевой геномный локус модифицируется или гомологичная рекомбинация в целевом геномном локусе усиливается без помощи нуклеазы. В других способах целевой геномный локус модифицируется или гомологичная рекомбинация в целевом геномном локусе усиливается с помощью нуклеазы, которая осуществляет одно- или двухцепочечный разрыв в целевом геномном локусе или вблизи него, такой как нуклеаза с «цинковыми пальцами», эффекторная нуклеаза, подобная активатору транскрипции (TALEN), мегануклеаза или Cas9 и гидовая РНК.
Способ может усиливать гомологичную рекомбинацию первой нуклеиновой кислоты в целевом геномном локусе, может усиливать гомологичную рекомбинацию второй нуклеиновой кислоты в целевом геномном локусе или может усиливать гомологичную рекомбинацию как первой, так и второй нуклеиновых кислот в целевом геномном локусе. Например, гомологичная рекомбинация первой нуклеиновой кислоты в целевом геномном локусе может быть усилена по сравнению со способами, в которых первую нуклеиновую кислоту вводят без второй нуклеиновой кислоты. Аналогичным образом, гомологичная рекомбинация второй нуклеиновой кислоты в целевом геномном локусе может быть усилена по сравнению со способами, в которых вторую нуклеиновую кислоту вводят без первой нуклеиновой кислоты. Усиление гомологичной рекомбинации может составлять, например, по меньшей мере 1,5 раза, 2 раза, 2,5 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз, 10 раз, 11 раз, 12 раз, 13 раз, 14 раз, 15 раз, 16 раз, 17 раз, 18 раз, 19 раз или 20 раз. В некоторых способах усиление без нуклеазы может быть сопоставимо с усилением с нуклеазой. Например, кратность изменения усиления с помощью нуклеазы может составлять 0,5 раза, 0,6 раза, 0,7 раза, 0,8 раза, 0,9 раза, 1,0 раза, 1,1 раза, 1,2 раза, 1,3 раза, 1,4 раза, 1,5 раза, 1,6 раза, 1,7 раза, 1,8 раза, 1,9 раза, 2 раза, 3 раза, 4 раза, 5 раз, 6 раз, 7 раз, 8 раз, 9 раз или 10 раз по сравнению с усилением без нуклеазы. В некоторых случаях усиление без нуклеазы может быть таким же или большим, чем усиление с нуклеазой.
Перекрывающаяся последовательность первой нуклеиновой кислоты может быть гомологична перекрывающейся последовательности второй нуклеиновой кислоты. Например, перекрывающаяся последовательность первой нуклеиновой кислоты может быть по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,9% идентична перекрывающейся последовательности второй нуклеиновой кислоты. Альтернативно перекрывающаяся последовательность первой нуклеиновой кислоты может быть на 100% идентична перекрывающейся последовательности второй нуклеиновой кислоты.
Перекрывающая последовательность может составлять, например, от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. или более. Например, перекрывающаяся последовательность может составлять от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 15 т.п.н., от приблизительно 15 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 25 т.п.н., от приблизительно 25 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 35 т.п.н., от приблизительно 35 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 45 т.п.н., от приблизительно 45 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 55 т.п.н., от приблизительно 55 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 65 т.п.н., от приблизительно 65 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 220 т.п.н., от приблизительно 220 т.п.н. до приблизительно 240 т.п.н., от приблизительно 240 т.п.н. до приблизительно 260 т.п.н., от приблизительно 260 т.п.н. до приблизительно 280 т.п.н. или от приблизительно 280 т.п.н. до приблизительно 300 т.п.н. Например, перекрывающаяся последовательность может составлять от приблизительно 20 т.п.н. до приблизительно 60 т.п.н. Альтернативно перекрывающаяся последовательность может составлять по меньшей мере 1 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 25 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 35 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 45 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 55 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 65 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 220 т.п.н., по меньшей мере 240 т.п.н., по меньшей мере 260 т.п.н. по меньшей мере 280 т.п.н. или по меньшей мере 300 т.п.н.
Перекрывающаяся последовательность может быть расположена в любом месте в пределах первой и второй нуклеиновых кислот. Например, перекрывающаяся последовательность может быть расположена на 3'-конце первой нуклеиновой кислоты и 5'-конце второй нуклеиновой кислоты. Альтернативно перекрывающаяся последовательность может быть расположена на 5'-конце первой нуклеиновой кислоты и 3'-конце второй нуклеиновой кислоты.
В некоторых способах первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом. Вторая нуклеиновая кислота может представлять собой любую нуклеиновую кислоту, содержащую перекрывающуюся последовательность, такую как плазмида, нацеливающий вектор или большой нацеливающий вектор. В некоторых способах вторая нуклеиновая кислота не содержит нуклеотидную последовательность, которая гомологична целевому геномному локусу, за исключением перекрывающейся последовательности. Например, вторая нуклеиновая кислота может состоять по существу или состоять из перекрывающейся последовательности.
В некоторых способах первая нуклеиновая кислота представляет собой нацеливающий вектор, содержащий первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, а вторая нуклеиновая кислота представляет собой второй нацеливающий вектор, содержащий вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом.
Размер первого нацеливающего вектора может быть любым. Аналогичным образом, размер второго нацеливающего вектора может быть любым. Например, первый и/или второй нацеливающие векторы могут составлять от приблизительно 1 т.п.н. до приблизительно 2 т.п.н., от приблизительно 2 т.п.н. до приблизительно 5 т.п.н. или от приблизительно 5 т.п.н. до приблизительно 10 т.п.н. Первый нацеливающий вектор также может представлять собой большой нацеливающий вектор (LTVEC). Аналогичным образом, второй нацеливающий вектор может представлять собой LTVEC. Примеры размеров LTVEC приведены в других разделах настоящего документа. Например, первый и/или второй LTVEC может составлять от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н., от приблизительно 650 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 750 т.п.н., от приблизительно 750 т.п.н. до приблизительно 800 т.п.н.
В некоторых способах первая нуклеиновая кислота представляет собой LTVEC, а вторая нуклеиновая кислота представляет собой меньшую нуклеиновую кислоту, содержащую перекрывающуюся последовательность, такую как плазмида или нацеливающий вектор. В некоторых способах вторая нуклеиновая кислота не содержит нуклеотидную последовательность, которая гомологична целевому геномному локусу, за исключением перекрывающейся последовательности. Например, вторая нуклеиновая кислота может состоять по существу или состоять из перекрывающейся последовательности.
В некоторых способах первая полинуклеотидная вставка и вторая полинуклеотидная вставка представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты. В некоторых способах первая и/или вторая полинуклеотидные вставки могут быть получены от видов, которые отличаются от вида, к которому относится клетка. Например, первая и/или вторая полинуклеотидные вставки могут представлять собой человеческие нуклеиновые кислоты.
Способы могут приводить к интеграции первой и/или второй полинуклеотидных вставок в целевой геномный локус. Интеграция может приводить к добавлению последовательности в целевом геномном локусе, делеции последовательности в целевом геномном локусе или замещению последовательности в целевом геномном локусе. Например, интеграция может приводить к добавлению экзогенной последовательности в целевом геномном локусе, делеции эндогенной последовательности в целевом геномном локусе или замещению эндогенной последовательности на экзогенную последовательность в целевом геномном локусе. Первая полинуклеотидная вставка, вторая полинуклеотидная вставка или комбинация первой и второй полинуклеотидных вставок, которые вставлены в целевой геномный локус, могут составлять, например, от приблизительно 5 т.п.н. до приблизительно 500 т.п.н. Другие примеры размеров полинуклеотидной вставки и вставок описаны в других разделах настоящего документа. Делеция в целевом геномном локусе может составлять, например, от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н. Другие примеры размеров делений приведены в других разделах настоящего документа.
Клетка с проведенным нацеливанием может относиться к любому из типов клеток, представленных в настоящем документе, а целевой геномный локус может представлять собой любую ДНК в клетке. Например, целевой геномный локус может находиться в геноме клетки или он может находиться на внехромосомной ДНК в клетке.
II. Полинуклеотидные вставки и нацеливающие векторы
А. Полинуклеотидная вставка
В способах, описанных в настоящем документе, можно применять одну или более полинуклеотидных вставок, и их можно вводить в клетку посредством отдельных нацеливающих векторов или одного и того же нацеливающего вектора. Полинуклеотидные вставки содержат сегменты ДНК, предназначенные для интеграции в целевые геномные локусы. Интеграция полинуклеотидной вставки в целевой локус может приводить к добавлению представляющей интерес нуклеотидной последовательности, делеции представляющей интерес нуклеотидной последовательности в целевом локусе и/или замещению представляющей интерес нуклеотидной последовательности в целевом локусе.
Способы обеспечивают модификацию геномного локуса полинуклеотидными вставками, размер которых больше возможно достижимого размера при использовании традиционных методик одиночного нацеливания (т.е. одиночного LTVEC). В таких способах полинуклеотидные вставки включены в два, три или более LTVEC. LTVEC выполнены таким образом, что они способны рекомбинировать друг с другом, образуя один большой сегмент ДНК, содержащий комбинированные полинуклеотидные вставки из двух, трех или более LTVEC.
В таких способах полинуклеотидные вставки фланкированы 5' и 3' гомологичными плечами. 3' гомологичное плечо, фланкирующее первую полинуклеотидную вставку, и 5' гомологичное плечо, фланкирующее вторую полинуклеотидную вставку, представляют собой перекрывающиеся фрагменты одной и той же непрерывной нуклеиновой кислоты, которую затем реформируют путем рекомбинации между перекрывающимися фрагментами гомологичных плеч. В таких способах рекомбинация между двумя LTVEC приводит к непрерывной полинуклеотидной вставке с перекрывающейся последовательностью гомологичных плеч, расположенных между первой и второй полинуклеотидными вставками. Способы тройного нацеливания включают дополнительную рекомбинацию между вторым LTVEC и третьим LTVEC, в котором 3' гомологичное плечо, фланкирующее вторую полинуклеотидную вставку, и 5' гомологичное плечо, фланкирующее третью полинуклеотидную вставку, представляют собой перекрывающиеся фрагменты одной и той же непрерывной нуклеиновой кислоты, которую затем реформируют путем рекомбинации между перекрывающимися фрагментами гомологичных плеч. В таких способах тройного нацеливания результатом рекомбинации между тремя LTVEC является непрерывная полинуклеотидная вставка с перекрывающейся последовательностью гомологичных плеч, расположенных между первой, второй и третьей полинуклеотидными вставками. В одном варианте осуществления перекрывающаяся последовательность гомологичных плеч включает в себя участок полинуклеотидной вставки.
Таким образом, эти способы обеспечивают модификацию геномного локуса полинуклеотидными вставками из двух, трех или более LTVEC за один этап нацеливания, таким образом эффективно увеличивая общий размер полинуклеотидной вставки с одновременным сокращением числа этапов нацеливания.
Полинуклеотидная вставка или соответствующая замещаемая нуклеиновая кислота в целевом локусе может представлять собой кодирующую область, интрон, экзон, нетранслируемую область, регуляторную область, промотор, энхансер или любую их комбинацию. Более того, размер полинуклеотидной вставки (т.е. комбинированных полинуклеотидных вставок из двух, трех или более LTVEC) или соответствующая замещаемая нуклеиновая кислота в целевом локусе может иметь любую требуемую длину, например, длину 10-100 нуклеотидов, длину 100-500 нуклеотидов, длину 500 нуклеотидов - 1 т.п.н., длину от 1 т.п.н. до 1,5 т.п.н., длину от 1,5 т.п.н. до 2 т.п.н., длину от 2 т.п.н. до 2,5 т.п.н., длину от 2,5 т.п.н. до 3 т.п.н., длину от 3 т.п.н. до 5 т.п.н., длину от 5 т.п.н. до 8 т.п.н., длину от 8 т.п.н. до 10 т.п.н. или более. В других случаях длина может составлять от приблизительно 50 т.п.н. до приблизительно 700 т.п.н., от приблизительно 50 т.п.н. до приблизительно 500 т.п.н., от приблизительно 50 т.п.н. до приблизительно 300 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н., от приблизительно 650 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 800 т.п.н., от приблизительно 800 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н., до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 2,8 млн п.н., от приблизительно 2,8 млн п.н. до приблизительно 3 млн п.н. Альтернативно комбинированные полинуклеотидные вставки из двух, трех или более LTVEC или соответствующая замещаемая нуклеиновая кислота в целевом локусе могут составлять от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 6 млн п.н., от приблизительно 6 млн п.н. до приблизительно 7 млн п.н., от приблизительно 7 млн п.н. до приблизительно 8 млн п.н., от приблизительно 8 млн п.н. до приблизительно 9 млн п.н. или от приблизительно 9 млн п.н. до приблизительно 10 млн п.н. В других случаях длина может составлять по меньшей мере 100, 200, 300, 400, 500, 600, 700, 800 или 900 нуклеотидов или по меньшей мере 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 11 т.п.н., 12 т.п.н., 13 т.п.н., 14 т.п.н., 15 т.п.н., 16 т.п.н. или более. Например, комбинированные полинуклеотидные вставки из двух, трех или более LTVEC или соответствующая замещаемая нуклеиновая кислота в целевом локусе могут составлять по меньшей мере 20 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н., по меньшей мере 500 т.п.н., по меньшей мере 550 т.п.н., по меньшей мере 600 т.п.н., по меньшей мере 650 т.п.н., по меньшей мере 700 т.п.н., по меньшей мере 750 т.п.н., по меньшей мере 800 т.п.н., по меньшей мере 850 т.п.н., по меньшей мере 900 т.п.н., по меньшей мере 950 т.п.н., по меньшей мере 1 млн п.н., по меньшей мере 1,5 млн п.н., по меньшей мере 2 млн п.н., по меньшей мере 2,5 млн п.н., по меньшей мере 3 млн п.н., по меньшей мере 4 млн п.н., по меньшей мере 5 млн п.н., по меньшей мере 6 млн п.н., по меньшей мере 7 млн п.н., по меньшей мере 8 млн п.н., по меньшей мере 9 млн п.н., по меньшей мере 10 млн п.н. В одном варианте осуществления размер полинуклеотидной вставки составляет от приблизительно 5 т.п.н. до приблизительно 700 т.п.н. В одном варианте осуществления размер полинуклеотидной вставки составляет от приблизительно 5 т.п.н. до приблизительно 500 т.п.н. В другом варианте осуществления размер полинуклеотидной вставки составляет от приблизительно 100 т.п.н. до приблизительно 700 т.п.н. В другом варианте осуществления размер полинуклеотидной вставки составляет от приблизительно 100 т.п.н. до приблизительно 500 т.п.н. В конкретном варианте осуществления полинуклеотидная вставка составляет приблизительно 140 т.п.н. В другом конкретном варианте осуществления полинуклеотидная вставка составляет приблизительно 370 т.п.н. В другом конкретном варианте осуществления полинуклеотидная вставка составляет приблизительно 300 т.п.н. В другом конкретном варианте осуществления полинуклеотидная вставка составляет приблизительно 400 т.п.н.
В некоторых отдельных нацеливающих векторах (т.е. до рекомбинации с другим нацеливающим вектором) полинуклеотидная вставка может составлять 10-100 нуклеотидов в длину, 100-500 нуклеотидов в длину, 500 нуклеотидов - 1 т.п.н. в длину, от 1 т.п.н. до 1,5 т.п.н. в длину, от 1,5 т.п.н. до 2 т.п.н. в длину, от 2 т.п.н. до 2,5 т.п.н. в длину, от 2,5 т.п.н. до 3 т.п.н. в длину или от 3 т.п.н. до 5 т.п.н. в длину. В других случаях длина может составлять от приблизительно 5 т.п.н. до приблизительно 200 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. Альтернативно полинуклеотидная вставка может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н. Альтернативно полинуклеотидная вставка может составлять от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н., от приблизительно 650 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 750 т.п.н. или от приблизительно 750 т.п.н. до приблизительно 800 т.п.н.
В некоторых случаях замещение нуклеиновой кислоты в целевом локусе приводит к делеции нуклеотидной последовательности длиной в диапазоне от приблизительно 1 т.п.н. до приблизительно 200 т.п.н., от приблизительно 2 т.п.н. до приблизительно 20 т.п.н. или от приблизительно 0,5 т.п.н. до приблизительно 3 млн п. н. В некоторых случаях величина делеции превышает общую длину 5' гомологичного плеча и 3' гомологичного плеча.
В некоторых случаях величина делеции нуклеотидной последовательности находится в диапазоне от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н., от приблизительно 190 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 800 т.п.н., от приблизительно 800 т.п.н. до 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н., до приблизительно 2,5 млн п.н., от приблизительно 2,5 млн п.н. до приблизительно 2,8 млн п.н., от приблизительно 2,8 млн п.н. до приблизительно 3 млн п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н. или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. Альтернативно делеция может составлять от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н.
В других случаях полинуклеотидная вставка или соответствующая замещаемая нуклеиновая кислота в целевом локусе может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н., по меньшей мере 500 т.п.н., по меньшей мере 550 т.п.н., по меньшей мере 600 т.п.н., по меньшей мере 650 т.п.н. или по меньшей мере 700 т.п.н. или более.
Полинуклеотидная вставка может содержать геномную ДНК или любой другой тип ДНК. Например, полинуклеотидная вставка может происходить из прокариота, эукариота, дрожжей, птицы (например, курицы), не относящегося к человеку млекопитающего, грызуна, человека, крысы, мыши, хомяка, кролика, свиньи, коровы, оленя, овцы, козы, кошки, собаки, хорька, примата (например, мартышки, макаки-резус), домашнего млекопитающего животного, сельскохозяйственного млекопитающего животного или любого другого представляющего интерес организма.
Полинуклеотидная вставка и/или нуклеиновая кислота в целевом локусе может содержать кодирующую последовательность или некодирующую последовательность, например регуляторный элемент (например, промотор, энхансер или элемент, связывающийся с репрессором транскрипции). Например, полинуклеотидная вставка может содержать нокин-аллель по меньшей мере одного экзона эндогенного гена или нокин-аллель всего эндогенного гена (т.е. «нокин с обменом гена»). Например, полинуклеотидная вставка может быть гомологична или ортологична последовательности, нацеливаемой для делеции, в целевом геномном локусе. Гомологичная или ортологичная полинуклеотидная вставка может замещать последовательность, нацеливаемую для делеции, в представляющем интерес геномном локусе. Результатом может быть гуманизация локуса, если вставка полинуклеотидной вставки приводит к замещению нечеловеческой нуклеотидной последовательности гомологичной или ортологичной человеческой полинуклеотидной последовательностью (т.е. полинуклеотидная вставка вставляется вместо соответствующей нечеловеческой последовательности ДНК в ее эндогенный геномный локус).
Полинуклеотидная вставка также может содержать условный аллель. Условный аллель может являться многофункциональным аллелем, как описано в публикации US 2011/0104799, которая полностью включена в настоящий документ путем ссылки. Например, условный аллель может содержать: (а) активирующую последовательность в смысловой ориентации по отношению к транскрипции целевого гена; (b) кассету селекции лекарственными средствами (DSC) в смысловой или антисмысловой ориентации; (с) находящуюся в антисмысловой ориентации представляющую интерес нуклеотидную последовательность (NSI); и (d) условный по инверсии модуль (COIN, в котором используется разделяющий экзон интрон и инвертируемый модуль, подобный генной ловушке) в обратной ориентации. См., например, публикацию US 2011/0104799, которая полностью включена в настоящий документ путем ссылки. Условный аллель может дополнительно содержать рекомбинируемые блоки, которые рекомбинируются при воздействии первой рекомбиназой с образованием условного аллеля, который (i) не имеет активирующей последовательности и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См. публикацию US 2011/0104799.
Некоторые полинуклеотидные вставки содержат полинуклеотид, кодирующий селективный маркер. Селективный маркер может входить в кассету селекции. Такие селективные маркеры включают, без ограничений, неомицинфосфотрансферазу (neor), гигромицин-В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин-S-дезаминазу (bsrr), ксантин/гуанинфосфорибозилтрансферазу (gpt) или тимидинкиназу вируса простого герпеса (HSV-k) или их комбинацию. Полинуклеотид, кодирующий селективный маркер, может быть функционально связан с промотором, активным в клетке, на которую выполняется нацеливание. Примеры промоторов описаны в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит репортерный ген. К примерам репортерных генов относятся гены, кодирующие люциферазу, β-галактозидазу, зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (eGFP), голубой флуоресцентный белок (CFP), желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (eYFP), синий флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (eBFP), DsRed, ZsGreen, MmGFP, mPlum, mCherry, tdTomato, mStrawberry, J-Red, mOrange, mKO, mCitrine, Venus, YPet, Emerald, CyPet, Cerulean, T-Sapphire, щелочную фосфатазу и любую их комбинацию. Такие репортерные гены могут быть функционально связаны с промотором, активным в клетке, на которую выполняется нацеливание. Примеры промоторов описаны в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит одну или более экспрессионных кассет или кассет делеции. Данная кассета может содержать представляющую интерес последовательность полинуклеотидов, нуклеиновую кислоту, кодирующую селективный маркер, и/или репортерный ген наряду с различными регуляторными компонентами, влияющими на экспрессию. Примеры пригодных для включения селективных маркеров и репортерных генов описаны более подробно в других разделах настоящего документа.
В некоторых нацеливающих векторах полинуклеотидная вставка содержит нуклеиновую кислоту, фланкированную целевыми последовательностями сайт-специфической рекомбинации. Хотя вся полинуклеотидная вставка может быть фланкирована такими целевыми последовательностями сайт-специфической рекомбинации, любая область или отдельный представляющий интерес полинуклеотид в пределах полинуклеотидной вставки также могут быть фланкированы такими сайтами. Целевые последовательности сайт-специфической рекомбинации, которые могут фланкировать полинуклеотидную вставку или любой представляющий интерес полинуклеотид в полинуклеотидной вставке, могут включать в себя, например, последовательности loxP, lox511, lox2272, lox66, lox71, loxM2, lox5171, FRT, FRT11, FRT71, attp, att, FRT, rox и их комбинацию. В одном примере сайты сайт-специфической рекомбинации фланкируют полинуклеотид, кодирующий селективный маркер и/или репортерный ген, содержащийся в пределах полинуклеотидной вставки. После интеграции полинуклеотидной вставки в целевой локус последовательности между сайтами сайт-специфической рекомбинации можно удалить.
В. Представляющие интерес полинуклеотиды
Любой представляющий интерес полинуклеотид может содержаться в различных полинуклеотидных вставках и, таким образом, может быть интегрирован в целевой геномный локус. Способы, описанные в настоящем документе, обеспечивают интеграцию по меньшей мере 1, 2, 3, 4, 5, 6 или более представляющих интерес полинуклеотидов в целевой геномный локус.
При интеграции в целевой геномный локус представляющий интерес полинуклеотид в пределах полинуклеотидной вставки может вводить одну или более генетических модификаций в клетку. Генетическая модификация может представлять собой делецию эндогенной нуклеотидной последовательности и/или добавление экзогенного, или гетерологичного, или ортологичного полинуклеотида в целевой геномный локус. В одном варианте осуществления генетическая модификация содержит замещение эндогенной нуклеотидной последовательности в целевом геномном локусе представляющим интерес экзогенным полинуклеотидом. Таким образом, в настоящем документе предложены способы, позволяющие выполнять генетическую модификацию, содержащую нокаут, делецию, вставку, замещение (нокин), точечную мутацию, перестановку доменов, перестановку экзонов, перестановку интронов, перестановку регуляторных последовательностей, перестановку генов или их комбинацию в целевом геномном локусе. Такие модификации могут происходить при интеграции в целевой геномный локус первой, второй, третьей, четвертой, пятой, шестой, седьмой или любой последующей полинуклеотидной вставки.
Представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус может содержать последовательность, являющуюся нативной или гомологичной для клетки, в которую его вводят; представляющий интерес полинуклеотид может быть гетерологичным по отношению к клетке, в которую его вводят; представляющий интерес полинуклеотид может быть экзогенным по отношению к клетке, в которую его вводят; представляющий интерес полинуклеотид может быть ортологичным по отношению к клетке, в которую его вводят; или представляющий интерес полинуклеотид может быть получен от вида, отличного от вида клетки, в которую его вводят. «Гомологичный» в отношении последовательности включает в себя последовательность, которая является нативной по отношению к клетке. Термин «гетерологичная» в отношении последовательности включает в себя последовательность, полученную от чужеродного вида, или, если она получена от того же вида, по существу модифицированную относительно ее нативной формы в композиции и/или в геномном локусе в результате целенаправленного вмешательства человека. Термин «экзогенная» в отношении последовательности включает в себя последовательность, которая получена от чужеродного вида. Термин «ортологичный» включает в себя полинуклеотид от одного вида, который функционально эквивалентен известной эталонной последовательности у другого вида (т.е. видовой вариант). Представляющий интерес полинуклеотид может быть получен от любого представляющего интерес организма, в том числе, без ограничений, не относящегося к человеку животного, грызуна, хомяка, мыши, крысы, человека, обезьяны, птицы, сельскохозяйственного млекопитающего животного или несельскохозяйственного млекопитающего животного. Представляющий интерес полинуклеотид может дополнительно содержать кодирующую область, некодирующую область, регуляторную область или геномную ДНК. Таким образом, 1, 2, 3, 4, 5, 6, 7-я и/или любая из последующих полинуклеотидных вставок может содержать такие последовательности.
В одном варианте осуществления представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус гомологичен человеческой нуклеиновой кислоте. В других дополнительных вариантах осуществления представляющий интерес полинуклеотид, интегрированный в целевой локус, представляет собой фрагмент геномной нуклеиновой кислоты. В одном варианте осуществления геномная нуклеиновая кислота представляет собой геномную нуклеиновую кислоту мыши, геномную нуклеиновую кислоту человека, нуклеиновую кислоту не относящегося к человеку животного, нуклеиновую кислоту грызуна, нуклеиновую кислоту крысы, нуклеиновую кислоту хомяка, нуклеиновую кислоту обезьяны, нуклеиновую кислоту сельскохозяйственного млекопитающего животного или нуклеиновую кислоту несельскохозяйственного млекопитающего животного или их комбинацию.
В одном варианте осуществления представляющий интерес полинуклеотид может составлять от приблизительно 500 нуклеотидов до приблизительно 200 т.п.н., как описано выше. Представляющий интерес полинуклеотид может составлять от приблизительно 500 нуклеотидов до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 200 т.п.н., от приблизительно 5 т.п.н. до приблизительно 700 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н, от приблизительно 190 т.п.н до приблизительно 200 т.п.н, от приблизительно 200 т.п.н до приблизительно 300 т.п.н, от приблизительно 300 т.п.н до приблизительно 400 т.п.н., от приблизительно 400 т.п.н до приблизительно 500 т.п.н., от приблизительно 500 т.п.н до приблизительно 600 т.п.н. или от приблизительно 600 т.п.н. до приблизительно 700 т.п.н.
Представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или вставленный в целевой геномный локус может кодировать полипептид, может кодировать миРНК, может кодировать длинную некодирующую РНК или может содержать любые регуляторные области или представляющие интерес некодирующие области, в том числе, например, регуляторную последовательность, последовательность промотора, последовательность энхансера, последовательность, связывающую транскрипционный репрессор, или делецию некодирующей белок последовательности, но не содержит делецию кодирующей белок последовательности. Кроме того, представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или вставленный в целевой геномный локус может кодировать белок, который экспрессируется в нервной системе, костной системе, пищеварительной системе, кровеносной системе, мышечной системе, дыхательной системе, сердечно-сосудистой системе, лимфатической системе, эндокринной системе, мочевыделительной системе, репродуктивной системе или их комбинации.
Представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус может содержать генетическую модификацию в кодирующей последовательности. Такие генетические модификации включают в себя, без ограничений, делеционную мутацию кодирующей последовательности или слияние двух кодирующих последовательностей.
Представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус может содержать полинуклеотид, кодирующий мутантный белок. В одном варианте осуществления мутантный белок характеризуется измененной характеристикой связывания, измененной локализацией, измененной экспрессией и/или измененным характером экспрессии. В одном варианте осуществления представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус содержит по меньшей мере один аллель заболевания. В таких случаях аллель заболевания может являться доминантным аллелем или аллель заболевания является рецессивным аллелем. Более того, аллель заболевания может представлять собой аллель однонуклеотидного полиморфизма (SNP). Представляющий интерес полинуклеотид, кодирующий мутантный белок, может быть получен от любого организма, включая, без ограничений, кодирующий мутантный белок полинуклеотид от млекопитающего, не относящегося к человеку млекопитающего, грызуна, мыши, крысы, человека, обезьяны, сельскохозяйственного млекопитающего животного или домашнего млекопитающего животного.
Представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус может также содержать регуляторную последовательность, включая, например, последовательность промотора, последовательность энхансера, последовательность, связывающую транскрипционный репрессор, или последовательность терминатора транскрипции. В конкретных вариантах осуществления представляющий интерес полинуклеотид в пределах полинуклеотидной вставки и/или интегрированный в целевой геномный локус содержит полинуклеотид, имеющий делецию некодирующей белок последовательности, но не содержит делецию кодирующей белок последовательности. В одном варианте осуществления делеция некодирующей белок последовательности содержит делецию регуляторной последовательности. В другом варианте осуществления делеция регуляторного элемента содержит делецию последовательности промотора. В одном варианте осуществления делеция регуляторного элемента содержит делецию последовательности энхансера. Такой представляющий интерес полинуклеотид может быть получен от любого организма, включая, без ограничений, кодирующий мутантный белок полинуклеотид от млекопитающего, не относящегося к человеку млекопитающего, грызуна, мыши, крысы, человека, обезьяны, сельскохозяйственного млекопитающего животного или домашнего млекопитающего животного.
Нацеленная генетическая модификация может включать в себя нацеленное изменение представляющего интерес полинуклеотида. Такие нацеленные модификации включают в себя, без ограничений, добавление одного или более нуклеотидов, делеции одного или более нуклеотидов, замены одного или более нуклеотидов, нокаут представляющего интерес полинуклеотида или его участка, нокин представляющего интерес полинуклеотида или его участка, замещение эндогенной нуклеотидной последовательности гетерологичной нуклеотидной последовательностью или их комбинацию. В конкретных вариантах осуществления по меньшей мере 1, 2, 3, 4, 5, 7, 8, 9, 10, 100, 500 или более нуклеотидов или по меньшей мере от 10 т.п.н. до 500 т.п.н. или более изменяют таким образом, чтобы образовать нацеленную геномную модификацию.
С. Нацеливающие векторы
Нацеливающие векторы можно использовать для внедрения полинуклеотидной вставки в целевой геномный локус, и они могут содержать полинуклеотидную вставку и гомологичные плечи, фланкирующие полинуклеотидную вставку. Нацеливающие векторы могут иметь линейную форму или форму окружности, и они могут быть одноцепочечными или двухцепочечными. Нацеливающие векторы могут представлять собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). Для удобства ссылки термины «гомологичные плечи» в настоящем документе относятся к гомологичным плечам 5' и 3' (т.е. ближе к 5' концу и ближе к 3' концу). Эта терминология относится к относительному положению гомологичных плеч к полинуклеотидной вставке в пределах нацеливающего вектора. 5' и 3' гомологичные плечи соответствуют областям в пределах целевого локуса или области в пределах другого нацеливающего вектора, которые в настоящем документе называются «целевой 5'-последовательностью» и «целевой 3'-последовательностью» соответственно. В некоторых случаях гомологичные плечи могут также функционировать в качестве целевой 5'- или 3'-последовательности.
В настоящих способах применяются два, три или более нацеливающих векторов, которые способны рекомбинировать друг с другом. В различных вариантах осуществления нацеливающие векторы представляют собой большие нацеливающие векторы (LTVEC), как описано в других разделах настоящего документа. В таких способах каждый из первого, второго и третьего нацеливающих векторов содержит 5' и 3' гомологичное плечо. 3' гомологичное плечо первого нацеливающего вектора содержит последовательность, которая перекрывается с 5' гомологичным плечом второго нацеливающего вектора (т.е. перекрывающиеся последовательности), что обеспечивает гомологичную рекомбинацию между первым и вторым LTVEC.
В случае способов двойного нацеливания 5' гомологичное плечо первого нацеливающего вектора и 3' гомологичное плечо второго нацеливающего вектора гомологичны соответствующим сегментам в пределах целевого геномного локуса (т.е. целевой последовательности), что способствует гомологичной рекомбинации первого и второго нацеливающих векторов с соответствующими геномными сегментами и модифицирует целевой геномный локус.
В случае способов тройного нацеливания 3' гомологичное плечо второго нацеливающего вектора содержит последовательность, которая перекрывается с 5' гомологичным плечом третьего нацеливающего вектора (т.е. перекрывающиеся последовательности), что обеспечивает гомологичную рекомбинацию между вторым и третьим LTVEC. 5' гомологичное плечо первого нацеливающего вектора и 3' гомологичное плечо третьего нацеливающего вектора гомологичны соответствующим сегментам в пределах целевого геномного локуса (т.е. целевой последовательности), что способствует гомологичной рекомбинации первого и третьего нацеливающих векторов с соответствующими геномными сегментами и модифицирует целевой геномный локус.
Гомологичное плечо и целевая последовательность или два гомологичных плеча «соответствуют» или являются «соответствующими» друг другу, если две области имеют достаточную степень идентичности последовательности друг с другом для того, чтобы они действовали как субстраты для реакции гомологичной рекомбинации. Термин «гомологичные» включает последовательности ДНК, которые или являются идентичными, или имеют идентичность последовательности с соответствующей последовательностью. Идентичность последовательности между данной целевой последовательностью и соответствующим гомологичным плечом, находящимся на нацеливающем векторе (т.е. перекрывающаяся последовательность), или между двумя гомологичными плечами может иметь любую степень идентичности последовательности, которая обеспечивает прохождение гомологичной рекомбинации. Например, значение идентичности последовательности между гомологичным плечом нацеливающего вектора (или его фрагмента) и целевой последовательностью другого нацеливающего вектора или целевой последовательностью целевого геномного локуса (или ее фрагментом) может составлять по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности, так что последовательности подвергаются гомологичной рекомбинации.
Более того, соответствующая область гомологии между гомологичным плечом и соответствующей целевой последовательностью может иметь любую длину, которая достаточна для стимуляции гомологичной рекомбинации в целевом геномном локусе. Например, данное гомологичное плечо и/или соответствующая целевая последовательность может содержать соответствующие области гомологии, длина которых составляет по меньшей мере приблизительно 5-10 т.п.н., 5-15 т.п.н., 5-20 т.п.н., 5-25 т.п.н., 5-30 т.п.н., 5-35 т.п.н., 5-40 т.п.н., 5-45 т.п.н., 5-50 т.п.н., 5-55 т.п.н., 5-60 т.п.н., 5-65 т.п.н., 5-70 т.п.н., 5-75 т.п.н., 5-80 т.п.н., 5-85 т.п.н., 5-90 т.п.н., 5-95 т.п.н., 5-100 т.п.н., 100-200 т.п.н. или 200-300 т.п.н. или более (как описано в других разделах настоящего документа для векторов LTVEC), так что гомологичное плечо имеет достаточную гомологию, чтобы подвергаться гомологичной рекомбинации с соответствующими целевыми последовательностями в пределах целевого геномного локуса или в пределах другого нацеливающего вектора.
Длина перекрывающихся последовательностей 3' гомологичного плеча первого нацеливающего вектора и 5' гомологичного плеча второго нацеливающего вектора или 3' гомологичного плеча второго нацеливающего вектора и 5' гомологичного плеча третьего нацеливающего вектора может быть любой, достаточной для стимуляции гомологичной рекомбинации между нацеливающими векторами. Например, данная перекрывающаяся последовательность гомологичного плеча может содержать соответствующие перекрывающиеся области, длина которых составляет по меньшей мере приблизительно 1-5 т.п.н., 5-10 т.п.н., 5-15 т.п.н., 5-20 т.п.н., 5-25 т.п.н., 5-30 т.п.н., 5-35 т.п.н., 5-40 т.п.н., 5-45 т.п.н., 5-50 т.п.н., 5-55 т.п.н., 5-60 т.п.н., 5-65 т.п.н., 5-70 т.п.н., 5-75 т.п.н., 5-80 т.п.н., 5-85 т.п.н., 5-90 т.п.н., 5-95 т.п.н., 5-100 т.п.н., 100-200 т.п.н. или 200-300 т.п.н. или более, так что перекрывающаяся последовательность гомологичного плеча имеет достаточную гомологию, чтобы подвергаться гомологичной рекомбинации с соответствующей перекрывающейся последовательностью в пределах другого нацеливающего вектора. В одном варианте осуществления перекрывающаяся последовательность составляет 1-5 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет от приблизительно 10 т.п.н. до приблизительно 70 т.п.н. В другом варианте осуществления перекрывающаяся последовательность составляет от приблизительно 10 т.п.н. до приблизительно 50 т.п.н. В одном варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 10 т.п.н. В другом варианте осуществления перекрывающаяся последовательность составляет по меньшей мере 20 т.п.н. Например, перекрывающаяся последовательность может составлять от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 15 т.п.н., от приблизительно 15 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 25 т.п.н., от приблизительно 25 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 35 т.п.н., от приблизительно 35 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 45 т.п.н., от приблизительно 45 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 180 т.п.н, от приблизительно 180 т.п.н до приблизительно 200 т.п.н, от приблизительно 200 т.п.н до приблизительно 220 т.п.н, от приблизительно 220 т.п.н до приблизительно 240 т.п.н., от приблизительно 240 т.п.н. до приблизительно 260 т.п.н., от приблизительно 260 т.п.н до приблизительно 280 т.п.н. или от приблизительно 280 т.п.н. до приблизительно 300 т.п.н. Например, перекрывающаяся последовательность может составлять от приблизительно 20 т.п.н. до приблизительно 60 т.п.н. Альтернативно перекрывающаяся последовательность может составлять по меньшей мере 1 т.п.н., по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 25 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 35 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 45 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 220 т.п.н., по меньшей мере 240 т.п.н., по меньшей мере 260 т.п.н. по меньшей мере 280 т.п.н. или по меньшей мере 300 т.п.н.
Гомологичные плечи могут соответствовать локусу, который является нативным по отношению к клетке (например, целевому локусу), или альтернативно они могут соответствовать области гетерологичного или экзогенного сегмента ДНК, который был интегрирован в геном клетки, включая, например, трансгены, экспрессионные кассеты или гетерологичные или экзогенные области ДНК. Альтернативно гомологичные плечи могут соответствовать области на нацеливающем векторе в клетке. Гомологичные плечи нацеливающего вектора могут соответствовать области дрожжевой искусственной хромосомы (YAC), искусственной бактериальной хромосомы (ВАС), человеческой искусственной хромосомы или любой другой сконструированной области, содержащейся в соответствующей клетке-хозяине. Дополнительно гомологичные плечи нацеливающего вектора могут соответствовать или могут быть получены из области из библиотеки ВАС, библиотеки космид или библиотеки фага Р1. В некоторых случаях гомологичные плечи нацеливающего вектора соответствуют локусу, который является нативным, гетерологичным или экзогенным в отношении прокариота, дрожжей, птицы (например, курицы), не относящегося к человеку млекопитающего, грызуна, человека, крысы, мыши, хомяка, кролика, свиньи, коровы, оленя, овцы, козы, кошки, собаки, хорька, примата (например, мартышки, макаки-резус), домашнего млекопитающего животного, сельскохозяйственного млекопитающего животного или любого другого представляющего интерес организма. В некоторых случаях при отсутствии одно- или двухцепочечного разрыва, индуцированного нуклеазным агентом (например, белком Cas), гомологичные плечи соответствуют локусу клетки, на который нельзя выполнить нацеливание с использованием стандартных способов или нацеливание на который может быть только неверным или иметь очень низкую эффективность. В некоторых случаях гомологичные плечи получены из синтетической ДНК.
В некоторых нацеливающих векторах одно из 5' или 3' гомологичных плеч соответствует целевому геномному локусу, а другое из 5' или 3' гомологичных плеч соответствует области на другом нацеливающем векторе.
В некоторых нацеливающих векторах 5' и 3' гомологичные плечи соответствуют целевому геному. Альтернативно гомологичные плечи могут быть получены из родственного генома. Например, целевой геном представляет собой геном мыши первой линии, а нацеливающие плечи получены из генома мыши второй линии, причем первая линия и вторая линия являются разными. В некоторых случаях гомологичные плечи получены из генома одного и того же животного или получены из генома одной линии, например, целевой геном представляет собой геном мыши первой линии и нацеливающие плечи получены из генома той же мыши или мыши той же линии.
Гомологичное плечо нацеливающего вектора может иметь любую длину, достаточную для стимуляции события гомологичной рекомбинации с соответствующей целевой последовательностью, включая, например, длину по меньшей мере 1-5 т.п.н., 5-10 т.п.н., 5-15 т.п.н., 5-20 т.п.н., 5-25 т.п.н., 5-30 т.п.н., 5-35 т.п.н., 5-40 т.п.н., 5-45 т.п.н., 5-50 т.п.н., 5-55 т.п.н., 5-60 т.п.н., 5-65 т.п.н., 5-70 т.п.н., 5-75 т.п.н., 5-80 т.п.н., 5-85 т.п.н., 5-90 т.п.н., 5-95 т.п.н., 5-100 т.п.н., 100-200 т.п.н. или 200-300 т.п.н. или более. Как более подробно описано ниже, в больших нацеливающих векторах могут использоваться нацеливающие плечи с большей длиной.
Нуклеазные агенты (например, системы CRISPR/Cas) можно применять в комбинации с нацеливающими векторами для обеспечения модификации целевого локуса. Такие нуклеазные агенты могут обеспечивать гомологичную рекомбинацию между нацеливающим вектором и целевым локусом. При использовании нуклеазных агентов в комбинации с нацеливающим вектором нацеливающий вектор может содержать 5' и 3' гомологичные плечи, соответствующие целевым 5'- и 3'-последовательностям, расположенным достаточно близко к сайту расщепления нуклеазой, чтобы обеспечить событие гомологичной рекомбинации между целевыми последовательностями и гомологичными плечами при одноцепочечном или двухцепочечном разрыве в сайте расщепления нуклеазой. Термин «сайт расщепления нуклеазой» включает последовательность ДНК, в которой нуклеазный агент (например, сайт расщепления Cas9) индуцирует одно- или двухцепочечный разрыв. Целевые последовательности в целевом локусе, соответствующие 5' и 3' гомологичным плечам нацеливающего вектора, «расположены в достаточной близости» к сайту расщепления нуклеазой, если расстояние является таким, чтобы способствовать возникновению события гомологичной рекомбинации между целевыми 5'- и 3'-последовательностями и гомологичными плечами при одно- или двухцепочечном разрыве в сайте распознавания. Таким образом, в конкретных случаях целевые последовательности, соответствующие 5' и/или 3' гомологичным плечам нацеливающего вектора, находятся в пределах по меньшей мере 1 нуклеотида от данного сайта распознавания или в пределах по меньшей мере от 10 нуклеотидов до приблизительно 14 т.п.н. от данного сайта распознавания. В некоторых случаях сайт нуклеазного расщепления непосредственно прилегает к по меньшей мере одной или обеим целевым последовательностям.
Пространственное отношение целевых последовательностей, соответствующих гомологичным плечам нацеливающего вектора и сайту расщепления нуклеазой, может различаться. Например, целевые последовательности могут быть расположены в направлении 5'-конца от сайта расщепления нуклеазой, целевые последовательности могут быть расположены в направлении 3'-конца от сайта распознавания или целевые последовательности могут фланкировать сайт расщепления нуклеазой.
Совместное использование нацеливающего вектора (включая, например, большой нацеливающий вектор) с нуклеазным агентом может привести к повышению эффективности нацеливания по сравнению с использованием одного только нацеливающего вектора. Например, когда нацеливающий вектор используют вместе с нуклеазным агентом, эффективность нацеливания нацеливающего вектора можно увеличить по меньшей мере в два раза, по меньшей мере в три раза, по меньшей мере в 4 раза, по меньшей мере в 10 раз или в пределах диапазона, образованного этими целыми числами, например, в 2-10 раз, по сравнению с использованием только нацеливающего вектора.
D. Большие нацеливающие векторы
Некоторые нацеливающие векторы представляют собой «большие нацеливающие векторы» или «LTVEC», к которым относятся нацеливающие векторы, содержащие гомологичные плечи, соответствующие нуклеотидным последовательностям большего размера, чем те, которые обычно применяют в других подходах, предназначенных для проведения гомологичной рекомбинации в клетках, или производные от них. Длина LTVEC может составлять, например, по меньшей мере 10 т.п.н., или общая длина 5' гомологичного плеча и 3' гомологичного плеча может составлять, например, по меньшей мере 10 т.п.н. К LTVEC также относятся нацеливающие векторы, содержащие полинуклеотидные вставки, имеющие нуклеотидные последовательности большей длины, чем у обычно применяемых при других подходах, предназначенных для проведения гомологичной рекомбинации в клетках. Например, LTVEC позволяют выполнять модификацию больших локусов, которую нельзя выполнить с использованием традиционных плазмидных нацеливающих векторов из-за размерных ограничений. Например, целевой локус может представлять собой (т.е. 5' и 3' гомологичные плечи могут соответствовать ему) локус клетки, на который при отсутствии одно- или двухцепочечного разрыва, индуцированного нуклеазным агентом (например, белком Cas), нельзя выполнить нацеливание с использованием традиционного способа, или нацеливание может быть только неверным или может иметь только очень низкую эффективность.
В предложенных в настоящем документе способах применяются два или три LTVEC, которые способны рекомбинировать друг с другом и с целевым геномным локусом в трехкомпонентном или четырехкомпонентном событии рекомбинации, как описано в других разделах настоящего документа. Эти способы позволяют модифицировать большие локусы, чего невозможно достичь с помощью одиночного LTVEC.
Примеры LTVEC включают векторы, полученные из бактериальной искусственной хромосомы (ВАС), человеческой искусственной хромосомы или дрожжевой искусственной хромосомы (YAC). Примеры LTVEC и способов их получения описаны, например, в патенте США №6,586,251; патенте США №6,596,541; патенте США 7,105,348 и WO 2002/036789 (PCT/US01/45375), каждый из которых полностью включен в настоящий документ путем ссылки. LTVEC могут иметь линейную форму или кольцевую форму.
LTVEC могут иметь любую длину, включая, например, от приблизительно 20 т.п.н. до приблизительно 300 т.п.н., от приблизительно 20 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 75 т.п.н., от приблизительно 75 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н. или от приблизительно 275 т.п.н. до приблизительно 300 т.п.н. Альтернативно LTVEC может составлять по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более. Размер LTVEC может быть слишком большим, чтобы существовала возможность скрининга событий нацеливания с помощью обычных тестов, например саузерн-блоттинга и ПЦР для амплификации протяженных участков (например, от 1 т.п.н. до 5 т.п.н.).
В некоторых случаях LTVEC включает полинуклеотидную вставку, которая может составлять от приблизительно 5 т.п.н. до приблизительно 200 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. В других случаях полинуклеотидная вставка может составлять от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н. В некоторых случаях LTVEC содержит полинуклеотидную вставку в диапазоне от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н. от приблизительно 650 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 750 т.п.н. или от приблизительно 750 т.п.н. до приблизительно 800 т.п.н.
В некоторых LTVEC общая длина 5' гомологичного плеча и 3' гомологичного плеча составляет по меньшей мере приблизительно 10 т.п.н. В других LTVEC длина 5' гомологичного плеча находится в диапазоне от приблизительно 1 т.п.н. до приблизительно 100 т.п.н. и/или длина 3' гомологичного плеча находится в диапазоне от приблизительно 1 т.п.н. до приблизительно 100 т.п.н. Общая длина 5' и 3' гомологичных плеч может составлять, например, от приблизительно 1 т.п.н. до приблизительно 5 т.п.н., от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 30 т.п.н., от приблизительно 30 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 50 т.п.н., от приблизительно 50 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 70 т.п.н., от приблизительно 70 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 90 т.п.н., от приблизительно 90 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 110 т.п.н., от приблизительно 110 т.п.н. до приблизительно 120 т.п.н., от приблизительно 120 т.п.н. до приблизительно 130 т.п.н., от приблизительно 130 т.п.н. до приблизительно 140 т.п.н., от приблизительно 140 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 160 т.п.н., от приблизительно 160 т.п.н. до приблизительно 170 т.п.н., от приблизительно 170 т.п.н. до приблизительно 180 т.п.н., от приблизительно 180 т.п.н. до приблизительно 190 т.п.н. или от приблизительно 190 т.п.н. до приблизительно 200 т.п.н. Альтернативно каждое гомологичное плечо может составлять по меньшей мере 5 т.п.н., по меньшей мере 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., по меньшей мере 180 т.п.н., по меньшей мере 190 т.п.н. или по меньшей мере 200 т.п.н. Аналогичным образом, общая длина 5' и 3' гомологичных плеч может составлять по меньшей мере 5 т.п.н., 10 т.п.н., по меньшей мере 15 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 110 т.п.н., по меньшей мере 120 т.п.н., по меньшей мере 130 т.п.н., по меньшей мере 140 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 160 т.п.н., по меньшей мере 170 т.п.н., 180 т.п.н., по меньшей мере 190 т.п.н. или по меньшей мере 200 т.п.н.
В некоторых случаях LTVEC и полинуклеотидная вставка выполнены так, чтобы обеспечивать в целевом локусе делецию эндогенной последовательности от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., или от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 800 т.п.н., от приблизительно 500 т.п.н. до приблизительно 1 млн п.н., от приблизительно 1 млн п.н. до приблизительно 1,5 млн п.н., от приблизительно 1,5 млн п.н. до приблизительно 2 млн п.н., от приблизительно 2 млн п.н. до приблизительно 2,5 млн п.н., или от приблизительно 2,5 млн п.н. до приблизительно 3 млн п.н. Альтернативно деления может составлять от приблизительно 3 млн п.н. до приблизительно 4 млн п.н., от приблизительно 4 млн п.н. до приблизительно 5 млн п.н., от приблизительно 5 млн п.н. до приблизительно 10 млн п.н., от приблизительно 10 млн п.н. до приблизительно 20 млн п.н., от приблизительно 20 млн п.н. до приблизительно 30 млн п.н., от приблизительно 30 млн п.н. до приблизительно 40 млн п.н., от приблизительно 40 млн п.н. до приблизительно 50 млн п.н., от приблизительно 50 млн п.н. до приблизительно 60 млн п.н., от приблизительно 60 млн п.н. до приблизительно 70 млн п.н., от приблизительно 70 млн п.н. до приблизительно 80 млн п.н., от приблизительно 80 млн п.н. до приблизительно 90 млн п.н. или от приблизительно 90 млн п.н. до приблизительно 100 млн п.н. Альтернативно деления может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
В других случаях LTVEC и полинуклеотидная вставка предназначены для того, чтобы обеспечить вставку в целевой локус экзогенной полинуклеотидной последовательности длиной в диапазоне от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н. или от приблизительно 350 т.п.н. до приблизительно 400 т.п.н. Альтернативно вставка может составлять от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н., от приблизительно 650 т.п.н. до приблизительно 700 т.п.н., от приблизительно 700 т.п.н. до приблизительно 750 т.п.н. или от приблизительно 750 т.п.н. до приблизительно 800 т.п.н. Альтернативно вставка может составлять по меньшей мере 10 т.п.н., по меньшей мере 20 т.п.н., по меньшей мере 30 т.п.н., по меньшей мере 40 т.п.н., по меньшей мере 50 т.п.н., по меньшей мере 60 т.п.н., по меньшей мере 70 т.п.н., по меньшей мере 80 т.п.н., по меньшей мере 90 т.п.н., по меньшей мере 100 т.п.н., по меньшей мере 150 т.п.н., по меньшей мере 200 т.п.н., по меньшей мере 250 т.п.н., по меньшей мере 300 т.п.н., по меньшей мере 350 т.п.н., по меньшей мере 400 т.п.н., по меньшей мере 450 т.п.н. или по меньшей мере 500 т.п.н. или более.
В других случаях длина полинуклеотидной вставки и/или удаляемой области эндогенного локуса составляет по меньшей мере 100, 200, 300, 400, 500, 600, 700, 800 или 900 нуклеотидов или по меньшей мере 1 т.п.н., 2 т.п.н., 3 т.п.н., 4 т.п.н., 5 т.п.н., 6 т.п.н., 7 т.п.н., 8 т.п.н., 9 т.п.н., 10 т.п.н., 11 т.п.н., 12 т.п.н., 13 т.п.н., 14 т.п.н., 15 т.п.н., 16 т.п.н. или более.
Е. Геномы и целевые геномные локусы
Геном или целевой геномный локус, модифицированный описанными в настоящем документе способами, может включать в себя любой сегмент или область ДНК внутри клетки. Геном или целевой геномный локус может быть нативным для клетки, может быть гетерологичным или экзогенным сегментом ДНК, встроенной в геном клетки, или может являться их комбинацией. Такие гетерологичные или экзогенные сегменты ДНК могут включать в себя трансгены, экспрессионные кассеты, полинуклеотиды, кодирующие селективные маркеры или гетерологичные или экзогенные области геномной ДНК.
Геном или целевой геномный локус может также включать в себя внехромосомную ДНК внутри клетки, такую как дрожжевая искусственная хромосома (YAC), бактериальная искусственная хромосома (ВАС), искусственная хромосома человека, или любую другую сконструированную геномную область, содержащуюся в соответствующей клетке-хозяине.
III. Нуклеазные агенты
В способах и композициях для модификации целевого геномного локуса, предложенных в настоящем документе, может применяться нуклеазный агент, который индуцирует одно- или двухцепочечный разрыв в требуемом сайте распознавания.
Термин «сайт распознавания для нуклеазного агента» включает в себя последовательность ДНК, в которой нуклеазный агент индуцирует одно- или двухцепочечный разрыв. Сайт распознавания для нуклеазного агента может быть эндогенным (или нативным) по отношению к клетке или сайт распознавания может быть экзогенным по отношению к клетке. В конкретных вариантах осуществления сайт распознавания является экзогенным по отношению к клетке и, таким образом, в природе не встречается в геноме клетки. В других дополнительных вариантах осуществления сайт распознавания является экзогенным по отношению к клетке и представляющим интерес полинуклеотидам, которые требуется расположить в целевом локусе. В дополнительных вариантах осуществления экзогенный или эндогенный сайт распознавания присутствует в геноме клетки-хозяина только один раз. В конкретных вариантах осуществления идентифицирован эндогенный или нативный сайт, который присутствует в геноме только один раз. Впоследствии такой сайт можно использовать для разработки нуклеазных агентов, которые будут создавать в эндогенном сайте распознавания одно- или двухцепочечный разрыв.
Длина сайта распознавания может изменяться и включает в себя, например, сайты распознавания для пары нуклеаз с «цинковыми пальцами» (ZFN) длиной приблизительно 30-36 п.н. (т.е. приблизительно 15-18 п.н. для каждой ZFN), для эффекторной нуклеазы, подобной активатору транскрипции (TALEN), длиной приблизительно 36 п.н. или для гидовой РНК CRISPR/Cas9 длиной приблизительно 20 п.н.
В описанных в настоящем документе способах и композициях может использоваться любой нуклеазный агент, который индуцирует одно- или двухцепочечный разрыв в требуемом сайте распознавания. Встречающийся в природе или нативный нуклеазный агент может применяться при условии, что нуклеазный агент индуцирует в требуемом сайте распознавания одно- или двухцепочечный разрыв. Альтернативно может применяться модифицированный или сконструированный нуклеазный агент. «Сконструированный нуклеазный агент» включает в себя нуклеазу, сконструированную (модифицированную или производную) из ее нативной формы для того, чтобы она специфически распознавала и индуцировала одно- или двухцепочечный разрыв в требуемом сайте распознавания. Таким образом, сконструированный нуклеазный агент может быть получен из нативного встречающегося в природе нуклеазного агента или он может быть создан или синтезирован искусственно. Модификация нуклеазного агента может быть незначительной, например, она может представлять собой модификацию одной аминокислоты в агенте расщепления белка или одного нуклеотида в агенте расщепления нуклеиновой кислоты. В некоторых вариантах осуществления сконструированная нуклеаза индуцирует одно- или двухцепочечный разрыв в сайте распознавания, причем сайт распознавания не является последовательностью, которая могла бы быть распознана нативным (не сконструированным или не модифицированным) нуклеазным агентом. Создание одно- или двухцепочечного разрыва в сайте распознавания или другой ДНК в настоящем документе может называться «разрезанием» или «расщеплением» сайта распознавания или другой ДНК.
Также предложены активные варианты и фрагменты сайтов распознавания, приведенные в качестве примера. Идентичность последовательности таких активных вариантов данному сайту распознавания может составлять по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют биологическую активность и, следовательно, могут распознаваться и расщепляться нуклеазным агентом последовательность-специфическим образом. Тесты для измерения двухцепочечного разрыва сайта распознавания посредством нуклеазного агента известны в данной области (например, из публикации TAQMAN® qPCR assay, Frendewey D. et al., Methods in Enzymology, 2010, 476:295-307, которая полностью включена в настоящий документ путем ссылки).
Сайт распознавания нуклеазного агента может быть расположен в любом месте в целевом локусе или вблизи него. Сайт распознавания может быть расположен в пределах кодирующей области гена или в пределах регуляторных областей, влияющих на экспрессию гена. Сайт распознавания нуклеазного агента может быть расположен в интроне, экзоне, промоторе, энхансере, регуляторной области или любой некодирующей белок области. В конкретных вариантах осуществления сайт распознавания расположен в пределах полинуклеотида, кодирующего селективный маркер. Такое положение может находиться в пределах кодирующей области селективного маркера или в пределах регуляторных областей, влияющих на экспрессию селективного маркера. Таким образом, сайт распознавания нуклеазного агента может быть расположен в интроне селективного маркера, промоторе, энхансере, регуляторной области или любой не кодирующей белок области полинуклеотида, кодирующего селективный маркер. В конкретных вариантах осуществления одно- или двухцепочечный разрыв на сайте распознавания нарушает активность селективного маркера. Известны способы тестирования на наличие или отсутствие функционального селективного маркера.
В одном варианте осуществления нуклеазный агент представляет собой эффекторную нуклеазу, подобную активатору транскрипции (TALEN). TAL-эффекторные нуклеазы представляют собой класс последовательность-специфических нуклеаз, которые можно использовать для выполнения двухцепочечных разрывов в специфических целевых последовательностях в геноме прокариотического или эукариотического организма. TAL-эффекторные нуклеазы создают путем слияния нативного или сконструированного эффектора, подобного активатору транскрипции (TAL), или его функциональной части с каталитическим доменом эндонуклеазы, таким как, например, FokI. Уникальный модульный TAL-эффекторный ДНК-связывающий домен позволяет разрабатывать белки с потенциально любой заданной специфичностью распознавания ДНК. Таким образом, для распознавания целевых сайтов со специфической ДНК могут разрабатываться ДНК-связывающие домены TAL-эффекторных нуклеаз, и, таким образом, их можно использовать для двухцепочечных разрывов в требуемых целевых последовательностях. См. публикации WO 2010/079430; Morbitzer et al. (2010) PNAS 10.1073/pnas.1013133107; Scholze & Boch (2010) Virulence 1:428-432; Christian et al. Genetics (2010) 186:757-761; Li et al. (2010) Nuc. Acids Res. (2010) doi:10.1093/nar/gkq704; и Miller et al. (2011) Nature Biotechnology 29:143-148; все из которых включены в настоящий документ путем ссылки.
Примеры подходящих TAL-нуклеаз и способы получения подходящих TAL-нуклеаз описаны, например, в публикациях US 2011/0239315 A1, US 2011/0269234 A1, US 2011/0145940 A1, US 2003/0232410 A1, US 2005/0208489 A1, US 2005/0026157 A1, US 2005/0064474 A1, US 2006/0188987 A1 и US 2006/0063231 A1 (каждая настоящим включена в настоящий документ путем ссылки). В различных вариантах осуществления сконструированы TAL-эффекторные нуклеазы, которые «выполняют разрез» в целевой нуклеотидной последовательности или вблизи нее, например в представляющем интерес локусе или представляющем интерес геномном локусе, в котором целевая нуклеотидная последовательность находится на последовательности, подлежащей модификации посредством нацеливающего вектора, или вблизи нее. TAL-нуклеазы, подходящие для использования с различными способами и композициями, предложенными в настоящем документе, включают в себя те, которые специально разработаны для связывания в целевой нуклеотидной последовательности, подлежащей модификации посредством нацеливающих векторов, или вблизи нее, как описано в настоящем документе.
В одном варианте осуществления каждый мономер TALEN содержит 33-35 повторов TAL, которые распознают одну пару нуклеотидов посредством двух гипервариабельных остатков. В одном варианте осуществления нуклеазный агент представляет собой химерный белок, содержащий ДНК-связывающий домен на основе TAL-повторов, функционально связанный с независимой нуклеазой. В одном варианте осуществления независимая нуклеаза представляет собой эндонуклеазу FokI. В одном варианте осуществления нуклеазный агент содержит первый ДНК-связывающий домен на основе TAL-повторов и второй ДНК-связывающий домен на основе TAL-повторов, причем каждый из первого и второго ДНК-связывающих доменов на основе TAL-повторов функционально связан с нуклеазой FokI, при этом первый и второй ДНК-связывающие домены на основе TAL-повторов распознают две непрерывные целевые последовательности ДНК в каждой цепи целевой последовательности ДНК, разделенные последовательностью спейсера различной длины (12-20 п.н.), и при этом субъединицы нуклеазы FokI димеризуются для создания активной нуклеазы, которая выполняет двухцепочечный разрыв в целевой последовательности.
Нуклеазный агент, применяемый в различных способах и композициях, описанных в настоящем документе, может дополнительно содержать нуклеазу с «цинковыми пальцами» (ZFN). В одном варианте осуществления каждый мономер ZFN содержит 3 или более ДНК-связывающих доменов на основе «цинковых пальцев», причем каждый ДНК-связывающий домен на основе «цинковых пальцев» связывается с субсайтом из 3 п.н. В других вариантах осуществления ZFN представляет собой химерный белок, содержащий ДНК-связывающий домен на основе «цинковых пальцев», функционально связанный с независимой нуклеазой. В одном варианте осуществления независимая эндонуклеаза представляет собой эндонуклеазу FokI. В одном варианте осуществления нуклеазный агент содержит первую ZFN и вторую ZFN, причем каждая из первой ZFN и второй ZFN функционально связана с субъединицей нуклеазы FokI, при этом первая и вторая ZFN распознают две непрерывные целевые последовательности ДНК в каждой цепи целевой последовательности ДНК, разделенные спейсером из приблизительно 5-7 п.н., и при этом субъединицы нуклеазы FokI димеризуются для создания активной нуклеазы, которая выполняет двухцепочечный разрыв. См., например, публикации US 20060246567; US 20080182332; US 20020081614; US 20030021776; WO/2002/057308A2; US 20130123484; US 20100291048; WO/2011/017293 А2 и Gaj et al. (2013) Trends in Biotechnology, 31 (7):397-405, каждая из которых полностью включена в настоящий документ путем ссылки.
В еще одном варианте осуществления нуклеазный агент представляет собой мегануклеазу. Мегануклеазы были разделены на четыре семейства на основе консервативных мотивов последовательностей, семейства представляют собой LAGLIDADG, GIY-YIG, H-N-H и His-Cys box. Эти мотивы участвуют в координации ионов металлов и гидролизе фосфодиэфирных связей. Мегануклеазы отличаются своими длинными сайтами распознавания и допуском некоторых видов полиморфизма последовательностей в их ДНК-субстратах. Домены мегануклеаз, структура и функции известны; см., например, публикации Guhan and Muniyappa (2003) Crit Rev Biochem Mol Biol 38:199-248; Lucas et al., (2001) Nucleic Acids Res 29:960-9; Jurica and Stoddard, (1999) Cell Mol Life Sci 55:1304-26; Stoddard, (2006) Q Rev Biophys 38:49-95; и Moure et al., (2002) Nat Struct Biol 9:764. В некоторых примерах используют встречающийся в природе вариант и/или сконструированное производное мегануклеазы. Известны способы модификации кинетики, взаимодействия кофакторов, экспрессии, оптимальных условий и/или специфичности сайтов распознавания и скрининга по активности, см., например, публикации Epinat et al., (2003) Nucleic Acids Res 31:2952-62; Chevalier et al., (2002) Mol Cell 10:895-905; Gimble et al., (2003) Mol Biol 334:993-1008; Seligman et al., (2002) Nucleic Acids Res 30:3870-9; Sussman et al., (2004) J Mol Biol 342:31-41; Rosen et al., (2006) Nucleic Acids Res 34:4791-800; Chames et al., (2005) Nucleic Acids Res 33:e178; Smith et al., (2006) Nucleic Acids Res 34:e149; Gruen et al., (2002) Nucleic Acids Res 30:e29; Chen and Zhao, (2005) Nucleic Acids Res 33:e154; WO 2005105989; WO 2003078619; WO 2006097854; WO 2006097853; WO 2006097784 и WO 2004031346.
Здесь может использоваться любая мегануклеаза, включая, без ограничений, I-SceI, I-SceII, I-SceIII, I-SceIV, I-SceV, I-SceVI, I-SceVII, I-CeuI, I-CeuAIIP, I-CreI, I-CrepsbIP, I-CrepsbIIP, I-CrepsbIIIP, I-CrepsbIVP, I-TliI, I-PpoI, PI-PspI, F-SceI, F-SceII, F-SuvI, F-TevI, F-TevII, I-AmaI, I-AniI, I-ChuI, I-CmoeI, I-CpaI, I-CpaII, I-CsmI, I-CvuI, I-CvuAIP, I-DdiI, I-DdiII, I-DirI, I-DmoI, I-HmuI, I-HmuII, I-HsNIP, I-LlaI, I-MsoI, I-NaaI, I-NanI, I-NcIIP, I-NgrIP, I-NitI, I-NjaI, I-Nsp236IP, I-PakI, I-PboIP, I-PcuIP, I-PcuAI, I-PcuVI, I-PgrIP, I-PobIP, I-PorI, I-PorIIP, I-PbpIP, I-SpBetaIP, I-ScaI, I-SexIP, I-SneIP, I-SpomI, I-SpomCP, I-SpomIP, I-SpomIIP, I-SquIP, I-Ssp6803I, I-SthPhiJP, I-SthPhiST3P, I-SthPhiSTe3bP, I-TdeIP, I-TevI, I-TevII, I-TevIII, I-UarAP, I-UarHGPAIP, I-UarHGPA13P, I-VinIP, I-ZbiIP, PI-MtuI, PI-MtuHIP PI-MtuHIIP, PI-PfuI, PI-PfuII, PI-PkoI, PI-PkoII, PI-Rma43812IP, PI-SpBetaIP, PI-SceI, PI-TfuI, PI-TfuII, PI-ThyI, PI-TliI, PI-TliII или любые активные варианты или их фрагменты.
В одном варианте осуществления мегануклеаза распознает двухцепочечные последовательности ДНК из 12-40 пар нуклеотидов. В одном варианте осуществления мегануклеаза распознает одну идеально соответствующую целевую последовательность в геноме. В одном варианте осуществления мегануклеаза представляет собой самонаводящуюся нуклеазу. В одном варианте осуществления самонаводящаяся нуклеаза принадлежит к семейству самонаводящихся нуклеаз LAGLIDADG. В одном варианте осуществления семейство самонаводящихся нуклеаз LAGLIDADG выбирают из I-SceI, I-CreI и I-Dmol.
Нуклеазные агенты могут дополнительно содержать рестрикционные эндонуклеазы, которые включают в себя эндонуклеазы I типа, II типа, III типа и IV типа. Рестрикционные эндонуклеазы I типа и III типа распознают специфические сайты распознавания, но, как правило, осуществляют расщепление в разных положениях на расстоянии от сайта связывания нуклеазы, которые могут находиться на расстоянии сотен пар нуклеотидов от сайта расщепления (сайта распознавания). В системах II типа рестрикционная активность не зависит от какой-либо метилазной активности и расщепление обычно происходит в специфических сайтах в пределах сайта связывания или вблизи него. Большинство ферментов II типа разрезают палиндромные последовательности, однако ферменты IIа типа распознают непалиндромные сайты распознавания и осуществляют расщепление за пределами сайта распознавания, ферменты IIb типа разрезают последовательности дважды на двух сайтах за пределами сайта распознавания, а ферменты IIs типа распознают асимметричный сайт распознавания и осуществляют расщепление с одной стороны и на определенном расстоянии от сайта распознавания, составляющем приблизительно 1-20 нуклеотидов. Рестрикционные ферменты IV типа нацелены на метилированную ДНК. Рестрикционные ферменты дополнительно описаны и классифицированы, например, в базе данных REBASE (веб-страница по адресу rebase.neb.com; Roberts et al., (2003) Nucleic Acids Res 31:418-20), Roberts et al., (2003) Nucleic Acids Res 31:1805-12 и Belfort et al., (2002) in Mobile DNA II, pp. 761-783, Eds. Craigie et al. (ASM Press, Washington, DC).
Нуклеазный агент, применяемый в различных способах и композициях, может также содержать систему коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/CRISPR-ассоциированного белка (Cas) или компоненты такой системы. Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии или управлении активностью генов Cas. Система CRISPR/Cas может представлять собой систему I типа, II типа или III типа. В способах и композициях, описанных в настоящем документе, для сайт-направленного расщепления нуклеиновых кислот используются системы CRISPR/Cas посредством использования комплексов CRISPR (содержащих гидовую РНК (гРНК), образующую комплекс с белком Cas).
Некоторые системы CRISPR/Cas, используемые в способах, описанных в настоящем документе, не встречаются в природе. «Не встречающаяся в природе» система имеет признаки вмешательства человека, например, один или более компонентов системы изменены или мутированы по сравнению с их природным состоянием, при этом они по меньшей мере по существу не содержат по меньшей мере один другой компонент, с которым они естественно ассоциированы в природе, или они ассоциированы по меньшей мере с другим компонентом, с которым они не ассоциированы в природе. Например, в некоторых системах CRISPR/Cas используются не встречающиеся в природе комплексы CRISPR, которые содержат гРНК и белок Cas, вместе не встречающиеся в природе.
Белки Cas обычно содержат по меньшей мере один домен распознавания или связывания РНК. Такие домены могут взаимодействовать с гидовыми РНК (гРНК, более подробно описанными ниже). Белки Cas могут также содержать нуклеазные домены (например, домены ДНКазы или РНКазы), ДНК-связывающие домены, геликазные домены, домены межбелковых взаимодействий, домены димеризации и другие домены. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновых кислот. Расщепление включает разрушение ковалентных связей молекулы нуклеиновой кислоты. В результате расщепления могут образовываться тупые концы или ступенчатые концы, и расщепление может быть одноцепочечным или двухцепочечным. Белок Cas может обладать полной активностью расщепления и создавать двухцепочечный разрыв в целевом геномном локусе (например, двухцепочечный разрыв с тупыми концами) или он может быть никазой, которая создает одноцепочечный разрыв в целевом геномном локусе.
Примеры белков Cas включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966 и их гомологи или модифицированные варианты.
В некоторых случаях белок Cas относится к системе CRISPR/Cas типа II. Например, белок Cas может являться белком Cas9 или может быть получен из белка Cas9. Белки Cas9 обычно имеют четыре общих основных мотива с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Белок Cas9 может быть получен из, например, Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, AlicyclobacHlus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus или Acaryochloris marina. Белки Cas9 также могут быть получены из Staphylococcus aureus. Дополнительные примеры представителей семейства Cas9 включают описанные в публикации WO 2014/131833, которая полностью включена в настоящий документ путем ссылки. В одном конкретном примере белок Cas9 представляет собой белок Cas9 из S. pyogenes или его производное. Аминокислотную последовательность белка Cas9 из S. pyogenes можно найти, например, в базе данных SwissProt под номером доступа Q99ZW2.
Белки Cas могут быть белками дикого типа (т.е. встречающимися в природе), модифицированными белками Cas (т.е. вариантами белков Cas) или фрагментами белков Cas дикого типа или модифицированных белков Cas. Белки Cas могут также быть активными вариантами или фрагментами белков Cas дикого типа или модифицированных белков Cas. Идентичность последовательности таких активных вариантов или фрагментов белку Cas дикого типа или модифицированному белку Cas или его части может составлять по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют способность разрезания на требуемом сайте расщепления и, следовательно, сохраняют активность, индуцирующую одно- или двухцепочечный разрыв. Тесты для определения активности, индуцирующей одно- или двухцепочечный разрыв, известны и, как правило, измеряют общую активность и специфичность белка Cas на субстратах ДНК, содержащих сайт расщепления.
Белки Cas могут быть модифицированы для увеличения или уменьшения аффинности связывания нуклеиновых кислот, специфичности связывания нуклеиновых кислот и/или ферментативной активности. Белки Cas также могут быть модифицированы для изменения любого другого вида активности или свойства белка, например стабильности. Например, один или более нуклеазных доменов белка Cas могут быть изменены, удалены или инактивированы, или белок Cas может быть усечен так, чтобы удалить домены, которые не являются необходимыми для функционирования белка, или для оптимизации (например, повышения или снижения) активности белка Cas.
Некоторые белки Cas содержат по меньшей мере два нуклеазных домена, таких как домены ДНКазы. Например, белок Cas9 может содержать RuvC-подобный нуклеазный домен и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать отличную цепь двухцепочечной ДНК, чтобы выполнить двухцепочечный разрыв в ДНК. См., например, публикацию Jinek et al. (2012) Science 337:816-821, которая полностью включена в настоящий документ путем ссылки.
Один или оба нуклеазных домена могут быть удалены или подвергнуты мутации таким образом, чтобы прекратить их функционирование или снизить нуклеазную активность. При делеции или мутировании одного из нуклеазных доменов полученный белок Cas (например, Cas9) может называться «никаза» и может осуществлять одноцепочечный разрыв на целевой последовательности в пределах двухцепочечной ДНК, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или некомплементарную цепь, но не обе). При делеции или мутировании обоих нуклеазных доменов полученный белок Cas (например, Cas9) будет иметь сниженную способность к расщеплению обеих цепей двухцепочечной ДНК (например, нуклеаза-пустой белок Cas). Примером мутации, превращающей Cas9 в никазу, является мутация D10A (замена аспартата на аланин в положении 10 Cas9) в домене RuvC белка Cas9 из S. pyogenes. Аналогичным образом, мутация Н939А (замена гистидина на аланин в положении аминокислоты 839) или Н840А (замена гистидина на аланин в положении аминокислоты 840) в домене HNH Cas9 из S. pyogenes может превращать Cas9 в никазу. Другие примеры мутаций, превращающих Cas9 в никазу, включают соответствующие мутации Cas9 из S. thermophilus. См., например, публикации Sapranauskas et al. (2011) Nucleic Acids Research 39:9275-9282 и WO 2013/141680, каждая из которых полностью включена в настоящий документ путем ссылки. Такие мутации могут быть созданы с использованием таких известных способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или полный синтез гена. Примеры других мутаций, создающих никазы, можно найти, например, в публикациях WO/2013/176772 A1 и WO/2013/142578 A1, каждая из которых включена в настоящий документ путем ссылки.
Белки Cas также могут представлять собой слитые белки. Например, белок Cas может быть слит с доменом расщепления, доменом эпигенетических модификаций, доменом транскрипционной активации или доменом транскрипционного репрессора. См. публикацию WO 2014/089290, которая полностью включена в настоящий документ путем ссылки. Белки Cas также могут быть слиты с гетерологичным полипептидом, обеспечивающим повышенную или сниженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, на С-конце или может находиться внутри белка Cas.
Одним примером слитого белка Cas является белок Cas, слитый с гетерологичным полипептидом, который обеспечивает субклеточную локализацию. Такие последовательности могут включать в себя, например, сигнал ядерной локализации (NLS), например NLS SV40, для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал удержания ER и т.п. См., например, Lange et al. (2007) J. Biol. Chem. 282:5101-5105. Белок Cas может содержать, например, один или более сигналов ядерной локализации (например, два сигнала ядерной локализации). Такие сигналы субклеточной локализации могут быть расположены на N-конце, на С-конце или в других местах внутри белка Cas. NLS может содержать участок из основных аминокислот и может быть представлен одинарной последовательностью или двойной последовательностью.
Белки Cas могут также содержать проникающий в клетку домен. Например, проникающий в клетку домен может быть получен из белка TAT HIV-1, проникающего в клетку мотива TLM из вируса гепатита В человека, MPG, Рер-1, VP22, проникающего в клетку пептида из вируса простого герпеса или из последовательности пептида полиаргинина. См., например, публикацию WO 2014/089290, которая полностью включена в настоящий документ путем ссылки. Проникающий в клетку домен может быть расположен на N-конце, на С-конце или в других местах внутри белка Cas.
Белки Cas также могут содержать гетерологичный полипептид для простоты отслеживания или очистки, например флуоресцентный белок, метку очистки или эпитопную метку. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP, ZsYellowl), синие флуоресцентные белки (например, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire, T-sapphire), голубые флуоресцентные белки (например, eCFP, Cerulean, CyPet, AmCyanl, Midoriishi-Cyan), красные флуоресцентные белки (mKate, mKate2, mPlum, мономерный DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-monomer, HcRed-Tandem, HcRed1, AsRed2, eqFP611, mRaspberry, mStrawberry, Jred), оранжевые флуоресцентные белки (mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine, tdTomato) и любой другой подходящий флуоресцентный белок. Примеры меток включают глутатион-S-трансферазу (GST), хитин-связывающий белок (СВР), мальтоза-связывающий белок, тиоредоксин (TRX), поли(NANP), метку тандемной аффинной очистки (ТАР), myc, AcV5, AU1, AU5, Е, ECS, Е2, FLAG, гемагглютинин (НА), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, S1, T7, V5, VSV-G, гистидин (His), биотин-карбоксил-переносящий белок (BCCP) и кальмодулин.
Белки Cas могут быть обеспечены в любом виде. Например, белок Cas может быть обеспечен в виде белка, такого как белок Cas, образующий комплекс с гРНК. Альтернативно белок Cas может быть обеспечен в виде нуклеиновой кислоты, кодирующей белок Cas, например РНК (например, матричной РНК (мРНК)) или ДНК. Необязательно нуклеиновая кислота, кодирующая белок Cas, может быть оптимизированной по кодонам для эффективной трансляции в белок в конкретной клетке или организме. Если в клетку вводят нуклеиновую кислоту, кодирующую белок Cas, то белок Cas может экспрессироваться в клетке временно, условно или конститутивно.
Нуклеиновые кислоты, кодирующие белки Cas, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. Альтернативно нуклеиновые кислоты, кодирующие белки Cas, могут быть функционально связаны с промотором в экспрессионном конструкте. Экспрессионные конструкты включают в себя любые нуклеотидные конструкты, которые способны управлять экспрессией гена или другой представляющей интерес нуклеотидной последовательности (например, гена Cas) и которые могут переносить такую представляющую интерес нуклеотидную последовательность в клетку-мишень. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в векторе, содержащем ДНК, кодирующую гРНК. Альтернативно она может находиться в векторе или плазмиде, отдельной от вектора, содержащего ДНК, кодирующую гРНК. Промоторы, которые могут использоваться в экспрессионном конструкте, включают, например, промоторы, активные в плюрипотентной клетке крысы, эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. В других разделах настоящего документа описаны примеры других промоторов.
«Гидовая РНК», или «гРНК», включает в себя молекулу РНК, которая связывается с белком Cas и нацеливает белок Cas в специфическое местоположение внутри целевой ДНК. Гидовые РНК могут содержать два сегмента: «сегмент ДНК-нацеливания» и «белок-связывающий сегмент». «Сегмент» представляет собой сегмент, часть или область молекулы, например непрерывный участок из нуклеотидов в РНК. Некоторые гРНК содержат две отдельные молекулы РНК: «РНК-активатор» и «РНК-нацеливатель». Другие гРНК представляют собой одинарные молекулы РНК (одинарный полинуклеотид РНК), которые также можно назвать «одномолекулярной гРНК», «одногидовой РНК» или «огРНК». См., например, публикации WO/2013/176772 A1, WO/2014/065596 A1, WO/2014/089290 A1, WO/2014/093622 A2, WO/2014/099750 A2, WO/2013142578 A1 и WO 2014/131833 А1, каждая из которых включена в настоящий документ путем ссылки. Термины «гидовая РНК» и «гРНК» являются включающими и включают как двухмолекулярные гРНК, так и одномолекулярные гРНК.
Примеры двухмолекулярных гРНК включают крРНК-подобную молекулу («РНК CRISPR», или «РНК-нацеливатель», или «крРНК», или «повтор крРНК») и соответствующую тракрРНК-подобную молекулу («транс-действующую РНК CRISPR», или «РНК-активатор», или «тракрРНК», или «каркас»). КрРНК содержит как сегмент (одноцепочечный) ДНК-нацеливания гРНК, так и участок нуклеотидов, который образует одну половину дуплекса дцРНК белок-связывающего сегмента гРНК.
Соответствующая тракрРНК (РНК-активатор) содержит участок нуклеотидов, который образует другую половину дуплекса дцРНК белок-связывающего сегмента гРНК. Участок нуклеотидов крРНК комплементарен участку нуклеотидов тракрРНК и гибридизуется с ним с образованием дуплекса дцРНК белок-связывающего домена гРНК. Таким образом, можно сказать, что каждая крРНК имеет соответствующую тракрРНК.
КрРНК и соответствующая тракрРНК гибридизуются с образованием гРНК. крРНК дополнительно обеспечивает одноцепочечный сегмент ДНК-нацеливания, который гибридизуется с целевой последовательностью. При использовании для модификации внутри клетки может быть разработана точная последовательность данной молекулы крРНК или тракрРНК, которая будет специфичной для тех видов, в которых будут использоваться молекулы РНК. См., например, публикации Mali et al. (2013) Science 339:823-826; Jinek et al. (2012) Science 337:816-821; Hwang et al. (2013) Nat. Biotechnol. 31:227-229; Jiang et al. (2013) Nat. Biotechnol. 31:233-239; и Cong et al. (2013) Science 339:819-823, каждая из которых включена в настоящий документ путем ссылки.
Сегмент ДНК-нацеливания (крРНК) данной гРНК содержит нуклеотидную последовательность, которая является комплементарной последовательности в целевой ДНК. Сегмент ДНК-нацеливания гРНК взаимодействует с целевой ДНК последовательность-специфическим образом посредством гибридизации (т.е. спаривания нуклеотидов). По существу нуклеотидная последовательность сегмента ДНК-нацеливания может изменяться и определяет местоположение внутри целевой ДНК, с которым будут взаимодействовать гРНК и целевая ДНК. Сегмент ДНК-нацеливания обсуждаемой гРНК может быть модифицирован для гибридизации с любой заданной последовательностью в пределах целевой ДНК. Встречающиеся в природе крРНК различаются в зависимости от системы Cas9 и организма, но часто содержат нацеливающий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, WO 2014/131833). В случае S. pyogenes длина DR составляет 36 нуклеотидов, а длина нацеливающего сегмента составляет 30 нуклеотидов. DR, расположенный на 3'-конце, является комплементарным и гибридизуется с соответствующей тракрРНК, которая, в свою очередь, связывается с белком Cas9.
Сегмент ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов до приблизительно 100 нуклеотидов. Например, сегмент ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов (нт) до приблизительно 80 нт, от приблизительно 12 нт до приблизительно 50 нт, от приблизительно 12 нт до приблизительно 40 нт, от приблизительно 12 нт до приблизительно 30 нт, от приблизительно 12 нт до приблизительно 25 нт, от приблизительно 12 нт до приблизительно 20 нт или от приблизительно 12 нт до приблизительно 19 нт. Альтернативно сегмент ДНК-нацеливания может иметь длину от приблизительно 19 нт до приблизительно 20 нт, от приблизительно 19 нт до приблизительно 25 нт, от приблизительно 19 нт до приблизительно 30 нт, от приблизительно 19 нт до приблизительно 35 нт, от приблизительно 19 нт до приблизительно 40 нт, от приблизительно 19 нт до приблизительно 45 нт, от приблизительно 19 нт до приблизительно 50 нт, от приблизительно 19 нт до приблизительно 60 нт, от приблизительно 19 нт до приблизительно 70 нт, от приблизительно 19 нт до приблизительно 80 нт, от приблизительно 19 нт до приблизительно 90 нт, от приблизительно 19 нт до приблизительно 100 нт, от приблизительно 20 нт до приблизительно 25 нт, от приблизительно 20 нт до приблизительно 30 нт, от приблизительно 20 нт до приблизительно 35 нт, от приблизительно 20 нт до приблизительно 40 нт, от приблизительно 20 нт до приблизительно 45 нт, от приблизительно 20 нт до приблизительно 50 нт, от приблизительно 20 нт до приблизительно 60 нт, от приблизительно 20 нт до приблизительно 70 нт, от приблизительно 20 нт до приблизительно 80 нт, от приблизительно 20 нт до приблизительно 90 нт или от приблизительно 20 нт до приблизительно 100 нт.
Нуклеотидная последовательность сегмента ДНК-нацеливания, комплементарная нуклеотидной последовательности (целевой последовательности) целевой ДНК, может иметь длину по меньшей мере приблизительно 12 нт. Например, последовательность ДНК-нацеливания (т.е. последовательность в пределах сегмента ДНК-нацеливания, комплементарная целевой последовательности в пределах целевой ДНК) может иметь длину по меньшей мере приблизительно 12 нт, по меньшей мере приблизительно 15 нт, по меньшей мере приблизительно 18 нт, по меньшей мере приблизительно 19 нт, по меньшей мере приблизительно 20 нт, по меньшей мере приблизительно 25 нт, по меньшей мере приблизительно 30 нт, по меньшей мере приблизительно 35 нт или по меньшей мере приблизительно 40 нт. Альтернативно последовательность ДНК-нацеливания может иметь длину от приблизительно 12 нуклеотидов (нт) до приблизительно 80 нт, от приблизительно 12 нт до приблизительно 50 нт, от приблизительно 12 нт до приблизительно 45 нт, от приблизительно 12 нт до приблизительно 40 нт, от приблизительно 12 нт до приблизительно 35 нт, от приблизительно 12 нт до приблизительно 30 нт, от приблизительно 12 нт до приблизительно 25 нт, от приблизительно 12 нт до приблизительно 20 нт, от приблизительно 12 нт до приблизительно 19 нт, от приблизительно 19 нт до приблизительно 20 нт, от приблизительно 19 нт до приблизительно 25 нт, от приблизительно 19 нт до приблизительно 30 нт, от приблизительно 19 нт до приблизительно 35 нт, от приблизительно 19 нт до приблизительно 40 нт, от приблизительно 19 нт до приблизительно 45 нт, от приблизительно 19 нт до приблизительно 50 нт, от приблизительно 19 нт до приблизительно 60 нт, от приблизительно 20 нт до приблизительно 25 нт, от приблизительно 20 нт до приблизительно 30 нт, от приблизительно 20 нт до приблизительно 35 нт, от приблизительно 20 нт до приблизительно 40 нт, от приблизительно 20 нт до приблизительно 45 нт, от приблизительно 20 нт до приблизительно 50 нт или от приблизительно 20 нт до приблизительно 60 нт. В некоторых случаях последовательность ДНК-нацеливания может иметь длину приблизительно 20 нт.
ТракрРНК могут быть любого вида (например, полноразмерные тракрРНК или активные частичные тракрРНК) и различной длины. Они могут включать в себя первичные транскрипты или подвергнутые процессингу формы. Например, тракрРНК (в составе одногидовой РНК или в виде отдельной молекулы в составе двухмолекулярной гРНК) может содержать всю последовательность тракрРНК дикого типа или ее часть или состоять из всей или части последовательности тракрРНК дикого типа (например, приблизительно или более чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности тракрРНК дикого типа). Примеры последовательностей тракрРНК дикого типа из S. pyogenes включают варианты из 171 нуклеотида, 89 нуклеотидов, 75 нуклеотидов и 65 нуклеотидов. См., например, публикацию Deltcheva et al. (2011) Nature 471:602-607; WO 2014/093661, каждая из которых полностью включена в настоящий документ путем ссылки. Примеры тракрРНК в пределах одногидовых РНК (огРНК) включают сегменты тракрРНК, встречающиеся в пределах вариантов огРНК +48, +54, +67 и +85, где «+n» означает, что в огРНК включено до +n нуклеотидов тракрРНК дикого типа. См. публикацию US 8,697,359, полностью включенную в настоящий документ путем ссылки.
Процентное значение комплементарности между последовательностью ДНК-нацеливания и целевой последовательностью в пределах целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). В некоторых случаях процентное значение комплементарности между последовательностью ДНК-нацеливания и целевой последовательностью в пределах целевой ДНК составляет по меньшей мере 60% на протяжении приблизительно 20 смежных нуклеотидов. В одном примере процентное значение комплементарности между последовательностью ДНК-нацеливания и целевой последовательностью в пределах целевой ДНК составляет 100% на протяжении 14 смежных нуклеотидов на 5'-конце целевой последовательности в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 14 нуклеотидов. В другом примере процентное значение комплементарности между последовательностью ДНК-нацеливания и целевой последовательностью в пределах целевой ДНК составляет 100% на протяжении семи смежных нуклеотидов на 5'-конце целевой последовательности в пределах комплементарной цепи целевой ДНК и всего 0% в оставшейся части. В таком случае можно считать, что длина последовательности ДНК-нацеливания составляет 7 нуклеотидов.
Белок-связывающий сегмент гРНК может содержать два участка нуклеотидов, которые комплементарны друг другу. Комплементарные нуклеотиды белок-связывающего сегмента гибридизуются с образованием дуплекса двухцепочечной РНК (дцРНК). Белок-связывающий сегмент обсуждаемой гРНК взаимодействует с белком Cas, а гРНК управляет связыванием белка Cas со специфической нуклеотидной последовательностью в пределах ДНК с помощью сегмента ДНК-нацеливания.
Гидовые РНК могут содержать модификации или последовательности, обеспечивающие дополнительные желаемые свойства (например, измененную или регулируемую стабильность; субклеточное нацеливание; отслеживание по флуоресцентной метке; сайт связывания для белка или белкового комплекса и т.п.). К примерам таких модификаций относятся, например, 5'-кэп (например, 7-метилгуанилатный кэп (m7G)); 3'-полиаденилированный хвост (т.е. 3'-поли(А) хвост); последовательность-рибопереключатель (например, для обеспечения регулируемой стабильности и/или регулируемой доступности белками и/или белковыми комплексами); последовательность контроля стабильности; последовательность, образующая дуплекс дцРНК (т.е. шпилька)); модификация или последовательность, нацеливающая РНК в субклеточное местоположение (например, ядро, митохондрии, хлоропласты и т.п.); модификация или последовательность, обеспечивающая отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который способствует флуоресцентной детекции, последовательность, обеспечивающая флуоресцентную детекцию, и т.д.); модификация или последовательность, обеспечивающая сайт связывания для белков (например, белков, действующих на ДНК, включая активаторы транскрипции, репрессоры транскрипции, ДНК-метилтрансферазы, ДНК-деметилазы, гистонацетилтрансферазы, гистондеацетилазы и т.п.); и их комбинации.
Гидовые РНК могут быть обеспечены в любом виде. Например, гРНК может быть обеспечена в виде РНК, или как две молекулы (отдельно крРНК и тракрРНК), или как одна молекула (огРНК), и необязательно в виде комплекса с белком Cas. гРНК также может быть представлена в виде ДНК, кодирующей гРНК. ДНК, кодирующая гРНК, может кодировать одну молекулу РНК (огРНК) или отдельные молекулы РНК (например, отдельно крРНК и тракрРНК). В последнем случае ДНК, кодирующая гРНК, может быть обеспечена как отдельные молекулы ДНК, кодирующие соответственно крРНК и тракрРНК. Альтернативно ДНК, кодирующая гРНК, может быть представлена в виде одной молекулы ДНК.
Если в клетку вводят ДНК, кодирующую гРНК, то гРНК может экспрессироваться в клетке временно, условно или конститутивно. ДНК, кодирующие гРНК, могут быть стабильно интегрированы в геном клетки и функционально связаны с промотором, активным в клетке. Альтернативно ДНК, кодирующие гРНК, могут быть функционально связаны с промотором в экспрессионном конструкте. Например, ДНК, кодирующая гРНК, может находиться в векторе, содержащем нуклеиновую кислоту, кодирующую белок Cas. Альтернативно она может находиться в векторе или плазмиде, отдельной от вектора, содержащего нуклеиновую кислоту, кодирующую белок Cas. Промоторы, которые могут использоваться в таких экспрессионных конструктах, включают в себя промоторы, например, активные в плюрипотентной клетке крысы, клетке эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Такие промоторы могут представлять собой, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. В некоторых случаях промотор представляет собой промотор РНК-полимеразы III, например человеческий промотор U6.
Альтернативно гРНК можно получать различными другими способами. Например, гРНК могут быть получены с помощью транскрипции in vitro, например с помощью РНК-полимеразы Т7 (см., например, публикации WO 2014/089290 и WO 2014/065596, каждая из которых для всех целей полностью включена в настоящий документ путем ссылки). Гидовые РНК также могут быть синтетическими молекулами, полученными путем химического синтеза.
Целевая последовательность для системы CRISPR/Cas включает в себя нуклеотидные последовательности, присутствующие в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК при условии существования достаточных условий для связывания. Например, целевые последовательности включают в себя последовательности, которым должна быть комплементарна сконструированная гидовая РНК, где гибридизация между целевой последовательностью и последовательностью ДНК-нацеливания способствует образованию комплекса CRISPR. Полная комплементарность не является обязательным требованием при условии наличия достаточной комплементарности, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Целевые последовательности также включают в себя сайты расщепления для белков Cas, более подробно описанные ниже. Целевая последовательность может содержать любой полинуклеотид, который может быть расположен, например, в ядре или цитоплазме клетки или внутри органеллы клетки, такой как митохондрия или хлоропласт.
На целевую последовательность в пределах целевой ДНК может быть нацелен (т.е. может связывать, или гибридизовать, или быть комплементарным) белок Cas или гРНК. Подходящие условия связывания ДНК/РНК включают физиологические условия, обычно существующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001)). Цепь целевой ДНК, которая комплементарна белку Cas или гРНК и гибридизуется с ними, можно назвать «комплементарной цепью», а цепь целевой ДНК, которая комплементарна «комплементарной цепи» (и, таким образом, не комплементарна белку Cas или гРНК), можно назвать «некомплементарной цепью» или «матричной цепью».
Белок Cas может расщеплять нуклеиновую кислоту в сайте в пределах или за пределами нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК. «Сайт расщепления» включает положение нуклеиновой кислоты, в котором белок Cas производит одноцепочечный разрыв или двухцепочечный разрыв. Например, образование комплекса CRISPR (содержащего гРНК, гибридизованную с целевой последовательностью и образующую комплексы с белком Cas) может приводить к расщеплению одной или обеих цепей в нуклеотидной последовательности, присутствующей в целевой ДНК, с которой будет связываться сегмент ДНК-нацеливания гРНК, или вблизи нее (например, на расстоянии в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар нуклеотидов от нее). Если сайт расщепления находится за пределами имеющейся в ДНК нуклеотидной последовательности, с которой будет связываться сегмент ДНК-нацеливания гРНК, то сайт расщепления все еще считается находящимся в пределах «целевой последовательности». Сайт расщепления может находиться только на одной цепи или на обеих цепях нуклеиновой кислоты. Сайты расщепления могут находиться в одном и том же положении на обеих цепях нуклеиновой кислоты (образуя тупые концы) или могут находиться на разных сайтах на каждой цепи (образуя ступенчатые концы (например, нависания)). Ступенчатые концы можно получать, например, с использованием двух белков Cas, которые выполняют одноцепочечный разрыв в разных сайтах расщепления на разных цепях, таким образом выполняя двухцепочечный разрыв. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК таким образом, что образуются нависающие последовательности. В некоторых случаях целевая последовательность никазы на первой цепи разделена с целевой последовательностью никазы на второй цепи по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 пар нуклеотидов.
Сайт-специфическое расщепление целевой ДНК белком Cas9 может происходить в местоположениях, определенных как (i) комплементарностью спариваемых нуклеотидов между гРНК и целевой ДНК, так и (ii) наличием в целевой ДНК короткого мотива, называемого мотивом, прилежащим к протоспейсеру (РАМ). РАМ может фланкировать целевую последовательность. Необязательно целевая последовательность может быть фланкирована РАМ на 3'-конце. Например, сайт расщепления Cas9 может находиться на расстоянии от приблизительно 1 до приблизительно 10 или от приблизительно 2 до приблизительно 5 пар нуклеотидов (например, 3 пар нуклеотидов) ближе к 5' концу или ближе к 3' концу от последовательности РАМ. В некоторых случаях (например, когда используется Cas9 из S. pyogenes или близкородственный Cas9) последовательность РАМ некомплементарной цепи может представлять собой 5'-N1GG-3', где N1 является любым нуклеотидом ДНК и находится непосредственно на 3'-конце целевой последовательности некомплементарной цепи целевой ДНК. По существу последовательность РАМ комплементарной цепи будет представлять собой 5'-СС N2-3', где N2 является любым нуклеотидом ДНК и находится непосредственно на 5'-конце целевой последовательности комплементарной цепи целевой ДНК. В некоторых таких случаях N1 и N2 могут быть комплементарны, а пара нуклеотидов N1-N2 может представлять собой любую пару нуклеотидов (например, N1 = Ц и N2 = Г; N1 = Г и N2 = Ц; N1 = А и N2 = Т или N1 = Т и N2 = А).
Примеры целевых последовательностей включают в себя последовательность ДНК, комплементарную сегменту ДНК-нацеливания гРНК, или такую последовательность ДНК в дополнение к последовательности РАМ. Один пример целевой последовательности содержит нуклеотидную последовательность GNNNNNNNNNNNNNNNNNNNNGG (GN1-20 GG; SEQ ID NO: 1). Гуанин на 5'-конце может способствовать транскрипции РНК-полимеразы в клетках. Другие примеры целевых последовательностей могут включать в себя два гуанин-нуклеотида на 5'-конце для облегчения эффективной транскрипции полимеразой Т7 in vitro. См., например, публикацию WO 2014/065596, которая для всех целей полностью включена в настоящий документ путем ссылки. Другие целевые последовательности могут иметь длину 4-22 нуклеотида SEQ ID NO: 1, включая 5' G и 3' GG. Другие целевые последовательности могут иметь длину от 14 до 20 нуклеотидов SEQ ID NO: 1.
Целевая последовательность может представлять собой любую нуклеотидную последовательность, эндогенную или экзогенную по отношению к клетке. Целевая последовательность может представлять собой последовательность, кодирующую генный продукт (например, белок), или некодирующую последовательность (например, регуляторную последовательность или избыточную ДНК), или может включать в себя обе.
Также предложены активные варианты и фрагменты нуклеазных агентов (т.е. сконструированный нуклеазный агент). Идентичность последовательности таких активных вариантов нативному нуклеазному агенту может составлять по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, причем активные варианты сохраняют способность разрезать требуемый сайт распознавания и, таким образом, сохраняют активность для индуцирования одно- или двухцепочечного разрыва. Например, любой из нуклеазных агентов, описанных в настоящем документе, может быть модифицирован из нативной последовательности эндонуклеазы и выполнен с возможностью распознавания и индуцирования одно- или двухцепочечного разрыва в сайте распознавания, который не распознавался нативным нуклеазным агентом. Таким образом, в некоторых вариантах осуществления сконструированная нуклеаза обладает специфической способностью индуцировать одно- или двухцепочечный разрыв в сайте распознавания, который отличается от сайта распознавания соответствующего нативного нуклеазного агента. Тесты на определение активности, индуцирующей одно- или двухцепочечный разрыв, известны и по существу измеряют общую активность и специфичность эндонуклеазы на субстратах ДНК, содержащих сайт распознавания.
Нуклеазный агент может быть введен в плюрипотентную клетку любыми способами, известными в данной области. Полипептид, кодирующий нуклеазный агент, может быть непосредственно введен в клетку. Альтернативно в клетку может быть введен полинуклеотид, кодирующий нуклеазный агент. Если в клетку вводят полинуклеотид, кодирующий нуклеазный агент, то нуклеазный агент может экспрессироваться в клетке временно, условно или конститутивно. Таким образом, полинуклеотид, кодирующий нуклеазный агент, может содержаться в экспрессионной кассете и может быть функционально связан с условным промотором, индуцируемым промотором, конститутивным промотором или тканеспецифическим промотором. Альтернативно нуклеазный агент вводят в клетку в виде мРНК, кодирующей нуклеазный агент.
В конкретных вариантах осуществления полинуклеотид, кодирующий нуклеазный агент, стабильно интегрирован в геном клетки и функционально связан с активным в клетке промотором. В других вариантах осуществления полинуклеотид, кодирующий нуклеазный агент, находится в том же нацеливающем векторе, который содержит полинуклеотидную вставку, а в других случаях полинуклеотид, кодирующий нуклеазный агент, находится в векторе или плазмиде, которая отделена от нацеливающего вектора, содержащего полинуклеотидную вставку.
Если нуклеазный агент вводят в клетку посредством введения полинуклеотида, кодирующего нуклеазный агент, то такой полинуклеотид, кодирующий нуклеазный агент, может быть модифицирован так, чтобы заменить кодоны, имеющие более высокую частоту использования в представляющей интерес клетке по сравнению со встречающейся в природе полинуклеотидной последовательностью, кодирующей нуклеазный агент. Например, полинуклеотид, кодирующий нуклеазный агент, может быть модифицирован так, чтобы использовать в качестве замены кодоны, имеющие более высокую частоту использования в прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, нечеловеческую клетку, клетку млекопитающих, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина, по сравнению со встречающейся в природе полинуклеотидной последовательностью.
Различные изложенные выше способы могут последовательно повторяться, чтобы обеспечить нацеленную интеграцию любого числа полинуклеотидных вставок в данный целевой геномный локус на хромосоме. Таким образом, различные способы обеспечивают вставку по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более полинуклеотидных вставок в целевой геномный локус на хромосоме. В некоторых вариантах осуществления такие способы последовательного мозаичного расположения позволяют проводить реконструкцию больших геномных областей из клетки животного или из клетки млекопитающих (т.е. человека, не относящегося к человеку животного, грызуна, мыши, обезьяны, крысы, хомяка, домашнего млекопитающего животного или сельскохозяйственного животного) в целевом геномном локусе на хромосоме. В таких случаях перенос и реконструкция геномных областей, которые включают в себя как кодирующие, так и некодирующие области, позволяет сохранить сложность данной области путем по меньшей мере частичного сохранения кодирующих областей, некодирующих областей и вариантов числа копий в пределах нативной геномной области. Таким образом, в различных способах предложены, например, способы создания «гетерологичных» или «экзогенных» геномных областей в любой клетке.
IV. Селективные маркеры
В различных способах и композициях, предложенных в настоящем документе, могут применяться нуклеазные агенты и их соответствующие сайты распознавания в комбинации с селективными маркерами. Как описано в настоящем документе, положение сайта распознавания в полинуклеотиде, кодирующем селективный маркер, обеспечивает эффективный способ, которым идентифицируют события интеграции в целевом локусе. Более того, в настоящем документе предложены различные способы, в которых чередующиеся селективные маркеры, имеющие сайт распознавания нуклеазы, применяются для повышения эффективности и продуктивности интеграции множества представляющих интерес полинуклеотидов в данный нацеленный локус.
В способах и композициях, описанных в настоящем документе, могут использоваться различные селективные маркеры. Такие селективные маркеры могут, например, придавать резистентность к антибиотику, такому как G418, гигромицин, бластицидин, неомицин или пуромицин. Такие селективные маркеры включают в себя неомицинфосфотрансферазу (neor), гигромицин-В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror) и бластицидин-S-дезаминазу (bsrr). В других вариантах осуществления селективный маркер функционально связан с индуцируемым промотором, и экспрессия селективного маркера является токсичной для клетки. Не имеющие ограничительного характера примеры таких селективных маркеров включают в себя ксантин/гуанинфосфорибозилтрансферазу (gpt), гипоксантингуанинфосфорибозилтрансферазу (HGPRT) или тимидинкиназу вируса простого герпеса (HSV-TK).
В одном варианте осуществления сайт распознавания нуклеазы расположен в пределах гена, кодирующего селективный маркер. В конкретном варианте осуществления сайт распознавания нуклеазы расположен в пределах гена гигромицина.
Полинуклеотид, кодирующий селективные маркеры, функционально связан с активным в клетке промотором. Такие экспрессионные кассеты и их различные регуляторные компоненты более подробно описаны в других разделах настоящего документа.
V. Промоторы
Различные полинуклеотидные последовательности, описанные в настоящем документе, могут быть функционально связаны с промоторами. Такие промоторы могут быть активны, например, в плюрипотентной клетке эукариота, млекопитающего, не относящегося к человеку млекопитающего, человека, грызуна, мыши или хомяка. Промотор может представлять собой, например, конститутивно активный промотор, условный промотор, индуцибельный промотор, ограниченный по времени промотор (например, регулируемый стадией развития промотор) или пространственно-ограниченный промотор (например, промотор, специфичный для клеток или тканей). Примеры промоторов можно найти, например, в публикации WO 2013/176772, которая полностью включена в настоящий документ путем ссылки.
К примерам индуцибельных промоторов относятся, например, промоторы с химической регуляцией и промоторы с физической регуляцией. К промоторам с химической регуляцией относятся, например, промоторы с регуляцией спиртом (например, промотор гена алкогольдегидрогеназы (alcA)), промоторы с регуляцией тетрациклином (например, отвечающий на тетрациклин промотор, тетрациклиновая операторная последовательность (tetO), промотор tet-On или промотор tet-Off), промоторы с регуляцией стероидами (например, крысиный глюкокортикоидный рецептор, промотор рецептора эстрогена, промотор рецептора экдизона) или промоторы с регуляцией металлами (например, металлопротеиновый промотор). К промоторам с физической регуляцией относятся, например, промоторы, регулируемые температурой (например, промотор теплового шока), и промоторы, регулируемые светом (например, индуцируемый светом промотор и подавляемый светом промотор).
Тканеспецифические промоторы могут представлять собой, например, нейрон-специфические промоторы, глиальноспецифические промоторы, промоторы, специфичные для мышечных клеток, промоторы, специфичные для сердечных клеток, промоторы, специфичные для почечных клеток, промоторы, специфичные для костных клеток, промоторы, специфичные для эндотелиальных клеток, или промоторы, специфичные для иммунных клеток (например, В-клеточный промотор или Т-клеточный промотор).
К регулируемым стадией развития промоторам относятся, например, промоторы, активные только на эмбриональной стадии развития или только во взрослой клетке.
Также промотор можно выбирать в зависимости от типа клетки. Например, различные известные промоторы находят применение в эукариотической клетке, клетке млекопитающих, нечеловеческой клетке, клетке млекопитающих, не относящихся к человеку, плюрипотентной клетке, нечеловеческой плюрипотентной клетке, плюрипотентной клетке человека, ЭС-клетке человека, стволовой клетке взрослого человека, онтогенетически ограниченной человеческой клетке-предшественнику, ИПС-клетке человека, клетке человека, клетке грызуна, клетке крысы, клетке мыши, клетке хомяка, фибробласту или клетке СНО.
VI. Экспрессионные кассеты
В настоящем документе предложены полинуклеотиды или молекулы нуклеиновой кислоты, содержащие различные компоненты системы нацеливания, предложенной в настоящем документе (т.е. нуклеазные агенты, сайты распознавания, нуклеотидные вставки, представляющие интерес полинуклеотиды, нацеливающие векторы (т.е. LTVEC), селективные маркеры и другие компоненты).
Дополнительно предложены рекомбинантные полинуклеотиды, содержащие различные компоненты системы нацеливания. Термины «рекомбинантный полинуклеотид» и «рекомбинантный ДНК-конструкт» используются в настоящем документе на взаимозаменяемой основе. Рекомбинантный конструкт содержит искусственную или гетерологичную комбинацию нуклеотидных последовательностей, например, регуляторной и кодирующей последовательностей, вместе не встречающихся в природе. В других вариантах осуществления рекомбинантный конструкт может содержать регуляторные последовательности и кодирующие последовательности, полученные из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но размещенные образом, отличным от встречающегося в природе. Такой конструкт может использоваться сам по себе или может использоваться в сочетании с вектором. Если используется вектор, то выбор вектора зависит от способа, который используется для преобразования клеток-хозяев, как хорошо известно специалистам в данной области. Например, можно использовать плазмидный вектор. В настоящем документе предложены генетические элементы, необходимые для успешного преобразования, выбора и размножения клетки-хозяина, которые содержат любой из выделенных фрагментов нуклеиновых кислот. Скрининг можно проводить, среди прочего, с помощью саузерн-блот-анализа ДНК, нозерн-блот-анализа экспрессии мРНК, анализа иммуноблоттинга экспрессии белка или фенотипического анализа.
В конкретных вариантах осуществления один или более компонентов системы нацеливания, описанной в настоящем документе, может быть обеспечен в экспрессионной кассете для экспрессии в прокариотической клетке, эукариотической клетке, бактериальной клетке, дрожжевой клетке, клетке млекопитающих или другом представляющем интерес организме или типе клетки. Кассета может включать в себя 5'- и 3'-регуляторные последовательности, функционально связанные с полинуклеотидом, представленным в настоящем документе. Используемый для обозначения соединения двух кодирующих белок областей термин «функционально связанный» означает, что кодирующие области находятся в одной и той же рамке считывания. В другом случае нуклеотидная последовательность, кодирующая белок, может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.п), чтобы сохранить соответствующее регулирование транскрипции.
Кассета может дополнительно содержать по меньшей мере один дополнительный представляющий интерес полинуклеотид, предназначенный для одновременного введения в организм. Альтернативно дополнительный представляющий интерес полинуклеотид может быть обеспечен на множестве экспрессионных кассет. Такая экспрессионная кассета снабжена множеством сайтов рестрикции и/или сайтов рекомбинации для вставки рекомбинантного полинуклеотида, который должен находиться под транскрипционным контролем регуляторных областей. Экспрессионная кассета может дополнительно содержать гены селективных маркеров.
Экспрессионная кассета может включать в себя в направлении транскрипции 5'-3' область инициации транскрипции и трансляции (т.е. промотор), рекомбинантный полинуклеотид, предложенный в настоящем документе, и область терминации транскрипции и трансляции (т.е. область терминации), действующие в представляющей интерес клетке млекопитающих или клетке-хозяине. Регуляторные области (т.е. промоторы, регуляторные области транскрипции и области терминации трансляции) и/или полинуклеотид, предложенный в настоящем документе, могут быть нативными/аналогичными клетке-хозяину или друг другу. Альтернативно регуляторные области и/или полинуклеотид, предложенный в настоящем документе, могут быть гетерологичны клетке-хозяину или друг другу. Например, промотор, функционально связанный с гетерологичным полинуклеотидом, получен от вида, отличного от вида, от которого был получен полинуклеотид, или, если он получен от того же/аналогичного вида, один или оба по существу модифицированы по сравнению с их изначальной формой и/или локусом, или промотор не является нативным промотором, функционально связанным с полинуклеотидом. Альтернативно регуляторные области и/или рекомбинантный полинуклеотид, предложенный в настоящем документе, могут быть полностью синтетическими.
Область терминации может быть нативной с областью инициации транскрипции, может быть нативной с функционально связанным рекомбинантным полинуклеотидом, может быть нативной с клеткой-хозяином или может быть получена из другого источника (т.е. чужеродного или гетерологичного) по отношению к промотору, рекомбинантному полинуклеотиду, клетке-хозяину или любой их комбинации.
При получении экспрессионной кассеты могут быть проведены манипуляции с различными фрагментами ДНК, чтобы обеспечить подходящую ориентацию последовательностей ДНК. С этой целью для соединения фрагментов ДНК могут применяться адаптеры или линкеры или могут быть проведены другие манипуляции, чтобы обеспечить удобные сайты рестрикции, удалить лишние ДНК, удалить сайты рестрикции или т.п. Для этой цели можно использовать мутагенез in vitro, восстановление праймера, рестрикцию, отжиг, повторные замены, например переходы и трансверсии.
В экспрессионных кассетах, предложенных в настоящем документе, можно использовать некоторое число промоторов. Промоторы можно выбирать на основе требуемого результата. Признано, что различные варианты применения можно расширить за счет использования в экспрессионных кассетах различных промоторов для модуляции времени, места и/или уровня экспрессии представляющего интерес полинуклеотида. Такие экспрессионные конструкты также при необходимости могут содержать промоторную регуляторную область (например, область, придающую индуцибельную или конститутивную, зависящую от окружающей среды или стадии развития, или клеточно- или тканеспецифичную/селективную экспрессию), инициирующий транскрипцию начальный сайт, сайт связывания рибосомы, сигнал РНК-процессинга, сайт терминации транскрипции и/или сигнал полиаденилирования.
Экспрессионная кассета, содержащая полинуклеотиды, предложенные в настоящем документе, также может содержать ген селективного маркера для селекции трансформированных клеток. Гены селективного маркера используются для селекции трансформированных клеток или тканей.
Если применимо, последовательности, применяемые в способах и композициях (т.е. представляющий интерес полинуклеотид, нуклеазный агент и т.п.), могут быть оптимизированы для повышения экспрессии в клетке. Это означает, что гены могут быть синтезированы с использованием кодонов, предпочтительных для данной представляющей интерес клетки, включая, например, кодоны, предпочтительные у млекопитающего, кодоны, предпочтительные у человека, кодоны, предпочтительные у грызуна, кодоны, предпочтительные у мыши, кодоны, предпочтительные у крысы и т.п., для улучшенной экспрессии.
В одном варианте осуществления нуклеазный агент экспрессируется из экспрессионного конструкта, содержащего нуклеотидную последовательность, кодирующую нуклеазу, и нуклеиновая кислота функционально связана с активным в клетке промотором.
VII. Способы создания генетически модифицированных не относящихся к человеку животных
С помощью различных способов, описанных в настоящем документе, можно создавать генетически модифицированных не относящихся к человеку животных. В некоторых случаях способ получения генетически модифицированного не относящегося к человеку животного включает в себя: (1) модификацию генома плюрипотентной клетки с использованием описанных в настоящем документе способов; (2) выбор генетически модифицированной плюрипотентной клетки; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион-хозяин; и (4) имплантацию эмбриона-хозяина, содержащего генетически модифицированную плюрипотентную клетку, в тело суррогатной матери. Получают клетки-потомки генетически модифицированной плюрипотентной клетки. Донорскую клетку можно вводить в эмбрион-хозяин на любой стадии, такой как стадия бластоцисты или стадия преморулы (т.е. стадия 4 клеток или стадия 8 клеток). Получают клетки-потомки, которые способны передавать генетическую модификацию через зародышевую линию. Плюрипотентная клетка может являться эмбриональной стволовой (ЭС) клеткой (например, ЭС-клеткой мыши или крысы), как описано в других разделах настоящего документа. См., например, патент США №7,294,754, который включен в настоящий документ путем ссылки.
Для создания не относящегося к человеку млекопитающего животного можно также использовать методики ядерного переноса. В кратком изложении методы ядерного переноса могут включать следующие этапы: (1) энуклеирование ооцита или обеспечение энуклеированного ооцита; (2) выделение или обеспечение донорской клетки или ядра, которые должны быть объединены с энуклеированным ооцитом; (3) вставка клетки или ядра в энуклеированный ооцит с образованием восстановленной клетки; (4) имплантация восстановленной клетки в матку животного для формирования эмбриона; и (5) обеспечение развития эмбриона. В таких способах ооциты обычно извлекают из умерших животных, хотя их можно выделять также из яйцеводов и/или яичников живых животных. До энуклеации ооциты могут созревать в различных средах, известных обычным специалистам в данной области. Энуклеация ооцитов может быть выполнена с помощью ряда способов, хорошо известных обычным специалистам в данной области. Вставку донорской клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки можно проводить перед слиянием посредством микроинъекции донорской клетки под вителлиновый слой. Слияние можно индуцировать с применением электрического импульса постоянного тока в плоскости контакта/слияния (электрослияние), путем воздействия на клетки химическими веществами, стимулирующими слияние, такими как полиэтиленгликоль, или путем использования инактивированного вируса, например вируса Сендай. Реконструированную клетку можно активировать с использованием электричества и/или неэлектрического средства до, во время и/или после слияния ядра донора и ооцита реципиента. Способы активации включают в себя электрические импульсы, химически индуцированный шок, пенетрацию с помощью сперматозоида, повышение уровней двухвалентных катионов в ооцитах, а также снижение фосфорилирования клеточных белков (например, с помощью ингибиторов киназы) в ооците. Активированные реконструированные клетки или эмбрионы можно культивировать в среде, хорошо известной обычным специалистам в данной области, после чего их переносят в матку животного. См., например, публикации US 20080092249, WO/1999/005266 A2, US 20040177390, WO/2008/017234 A1 и патент США №7,612,250, каждый из которых полностью включен в настоящий документ путем ссылки.
Способы могут дополнительно включать в себя способы получения поколения F0 не относящегося к человеку животного, включающие в себя: (1) идентификацию нечеловеческой ЭС-клетки, содержащей нацеленную модификацию; (2) введение нечеловеческой ЭС-клетки, содержащей нацеленную модификацию, в нечеловеческий эмбрион-хозяин и (3) гестацию нечеловеческого эмбриона-хозяина в теле суррогатной матери. Затем суррогатная мать может произвести не относящееся к человеку животное поколения F0, содержащее нацеленную модификацию. Эмбрион-хозяин, содержащий генетически модифицированную плюрипотентную или тотипотентную клетку (например, нечеловеческую ЭС-клетку), можно инкубировать до стадии бластоцисты, а затем имплантировать в тело суррогатной матери для получения животного F0. Животные, несущие генетически модифицированный геномный локус, могут быть идентифицированы посредством теста на модификацию аллеля (МОА), как описано в настоящем документе.
Различные способы, представленные в настоящем документе, позволяют получить генетически модифицированное не относящееся к человеку животное F0, причем клетки генетически модифицированного животного F0 содержат нацеленную модификацию. Признано, что в зависимости от способа, используемого для получения животного F0, количество клеток в животном F0, которые имеют нацеленную генетическую модификацию, будет варьировать. Введение донорских ЭС-клеток в эмбрион на стадии до морулы из соответствующего организма (например, мышиного эмбриона на стадии 8 клеток), например, по способу VELOCIMOUSE®, обеспечивает то, что большая доля популяции животного F0 будет содержать нацеленную генетическую модификацию. В конкретных случаях по меньшей мере 50%, 60%, 65%, 70%, 75%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада не относящегося к человеку животного F0 содержит клеточную популяцию, имеющую нацеленную модификацию. В других случаях по меньшей мере одна или более зародышевых клеток животного F0 имеют нацеленную модификацию.
В некоторых случаях клетки генетически модифицированного животного F0 являются гетерозиготными или компаунд-гетерозиготными по отношению к нацеленной модификации. Например, клетки генетически модифицированного животного F0 могут быть гемизиготными по нацеленной модификации. В других случаях клетки генетически модифицированного животного F0 гомозиготны по нацеленной модификации.
В некоторых случаях из животного F0, полученного с помощью способов и композиций, описанных в настоящем документе, может быть выведено животное дикого типа с получением поколения F1, которое гетерозиготно по нацеленной модификации. Затем животных F1 можно скрещивать друг с другом с получением животного F2, гомозиготного по нацеленной модификации. Потомство F1 можно генотипировать с использованием специфических праймеров и/или зондов для определения наличия нацеленной генетической модификации.
VIII. Способы введения нуклеиновых кислот и белков в клетки
В настоящем документе описаны различные способы и композиции, позволяющие вводить нуклеиновую кислоту в клетку. В некоторых случаях система, применяемая для введения нуклеиновой кислоты, позволяет осуществлять нацеленную интеграцию в конкретном геномном локусе. В таких системах применяются различные компоненты, и для простоты ссылки в настоящем документе термин «система нацеленной геномной интеграции» по существу относится ко всем компонентам, необходимым для события интеграции (например, один или более нуклеазных агентов, сайты расщепления нуклеазами, полинуклеотидные вставки, нацеливающие векторы, нацеленные геномные локусы и представляющие интерес полинуклеотиды).
Предложенные в настоящем документе способы могут включать введение в клетку одного или более полинуклеотидов или полипептидных конструктов, содержащих один или более компонентов системы нацеленной геномной интеграции. Термин «введение» включает в себя передачу в клетку последовательности (полипептидной или полинуклеотидной) таким образом, что последовательность получает доступ к внутренней части клетки. Предложенные в настоящем документы способы не зависят от конкретного способа введения нуклеиновой кислоты или белка в клетку, а только от того, что нуклеиновая кислота или белок получает доступ к внутренней части по меньшей мере одной клетки. Способы введения нуклеиновых кислот и белков в различные типы клеток известны в данной области и включают в себя, например, способы стабильной трансфекции, способы временной трансфекции и вирус-опосредованные способы.
В некоторых случаях клетки, используемые в способах и композициях, имеют стабильно встроенный в их геном ДНК-конструкт. Термин «стабильно встроенный» или «стабильно введенный» включает в себя введение полинуклеотида в клетку таким образом, что нуклеотидная последовательность интегрируется в геном клетки и способна передаваться по наследству ее потомству. Для стабильного встраивания ДНК-конструктов или различных компонентов системы нацеленной геномной интеграции может использоваться любой протокол.
Протоколы трансфекции, а также протоколы введения полипептидов или полинуклеотидных последовательностей в клетки могут быть разными. Способы трансфекции включают в себя химические способы трансфекции с использованием липосом; наночастиц; фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc Natl Acad Sci USA 74 (4): 1590-4 и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W.H. Freeman and Company, pp. 96-97); дендримеров или катионных полимеров, таких как ДЭАЭ-декстран или полиэтиленимин. Нехимические способы включают в себя электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция с использованием частиц включает в себя применение «генной пушки» или магнитной трансфекции (Bertram, (2006) Current Pharmaceutical Biotechnology 7, 277-28). Для трансфекции также можно использовать вирусные способы.
В некоторых случаях введение нуклеиновых кислот или белков в клетку опосредуется электропорацией, интрацитоплазматической инъекцией, вирусной инфекцией, аденовирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или Nucleofection™.
Введение нуклеиновых кислот или белков в клетку может быть выполнено один или множество раз в течение периода времени. Например, введение может выполняться по меньшей мере два раза в течение периода времени, по меньшей мере три раза в течение периода времени, по меньшей мере четыре раза в течение периода времени, по меньшей мере пять раз в течение периода времени, по меньшей мере шесть раз в течение периода времени, по меньшей мере семь раз в течение периода времени, по меньшей мере восемь раз в течение периода времени, по меньшей мере девять раз в течение периода времени, по меньшей мере десять раз в течение периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение периода времени, по меньшей мере тринадцать раз в течение периода времени, по меньшей мере четырнадцать раз в течение периода времени, по меньшей мере пятнадцать раз в течение периода времени, по меньшей мере шестнадцать раз в течение периода времени, по меньшей мере семнадцать раз в течение периода времени, по меньшей мере восемнадцать раз в течение периода времени, по меньшей мере девятнадцать раз в течение периода времени или по меньшей мере двадцать раз в течение периода времени.
Когда в клетку вводят как нуклеазные агенты, так и нацеливающие векторы (например, LTVEC), они могут вводиться одновременно. Альтернативно нуклеазный агент можно вводить отдельно от нацеливающих векторов. Например, нуклеазный агент можно вводить до введения нацеливающих векторов или его можно вводить после введения нацеливающих векторов. Когда в клетку вводят два или более LTVEC, они могут вводиться одновременно или альтернативно они могут вводиться отдельно.
IX. Клетки и животные
В различных композициях и способах, предложенных в настоящем документе, применяются клетки, такие как клетки от животного. Такими клетками могут быть нечеловеческие клетки, которые могут быть получены от не относящегося к человеку животного. Такие клетки могут являться эукариотическими клетками, в том числе, например, клетками грибов (например, дрожжей), клетками растений, клетками животных, клетками млекопитающих и клетками человека. Клеткой млекопитающих может быть, например, клетка млекопитающих, не относящихся к человеку, клетка человека, клетка грызуна, клетка крысы, клетка мыши, клетка хомяка, фибробласт или клетка СНО. Эукариотическая клетка может быть тотипотентной клеткой, плюрипотентной клеткой, такой как нечеловеческая плюрипотентная клетка (например, эмбриональная стволовая (ЭС) клетка мыши или ЭС-клетка крысы) или плюрипотентная клетка человека, или неплюрипотентной клеткой. Тотипотентные клетки включают в себя недифференцированные клетки, которые могут давать начало любому типу клеток, а плюрипотентные клетки включают в себя недифференцированные клетки, которые обладают способностью развиваться в более чем один дифференцированный тип клеток. Такие плюрипотентные и/или тотипотентные клетки могут представлять собой, например, эмбриональные стволовые (ЭС) клетки или подобные ЭС клетки, такие как индуцированные плюрипотентные стволовые (ИПС) клетки. Эмбриональная стволовая клетка включает в себя полученные из эмбриона тотипотентные или плюрипотентные клетки, которые при введении в эмбрион могут участвовать в развитии любой ткани развивающегося эмбриона. ЭС-клетки могут быть получены из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых листков позвоночных (энтодерма, эктодерма и мезодерма). Такими клетками также могут являться гемопоэтические стволовые клетки или нейрональные стволовые клетки.
Эукариотическая клетка также может являться клеткой, которая не является первичной соматической клеткой. Соматические клетки могут включать в себя любую клетку, которая не является гаметой, зародышевой клеткой, гаметоцитом или недифференцированной стволовой клеткой.
Эукариотические клетки также включают в себя первичные клетки. Первичные клетки включают в себя клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают в себя клетки, которые не являются ни трансформированными, ни бессмертными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не была пассирована в тканевой культуре или была ранее пассирована в тканевой культуре, но не может быть в течение неопределенно длительного времени пассирована в тканевой культуре. Такие клетки могут быть выделены обычными способами и включают в себя, например, соматические клетки, гемопоэтические клетки, эндотелиальные клетки, эпителиальные клетки, фибробласты, мезенхимальные клетки, кератиноциты, меланоциты, моноциты, мононуклеарные клетки, адипоциты, преадипоциты, нейроны, глиальные клетки, гепатоциты, скелетные миобласты и гладкомышечные клетки. Например, первичные клетки могут быть получены из соединительных тканей, мышечных тканей, тканей нервной системы или эпителиальных тканей.
Эукариотические клетки также включают в себя иммортализованные клетки. Иммортализованные клетки включают в себя клетки из многоклеточного организма, которые обычно не размножаются в течение неопределенно длительного времени, но вследствие мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать деление. Такие мутации или изменения могут происходить естественным образом или могут быть намеренно индуцированными. Примерами иммортализованных клеток являются клетки яичника китайского хомячка (СНО), эмбриональные клетки почки человека (например, клетки HEK 293) и мышиные эмбриональные фибробластные клетки (например, клетки 3Т3). Множество типов иммортализованных клеток хорошо известны в данной области.
Иммортализованные или первичные клетки включают в себя клетки, которые обычно используются для культивирования клеток или для экспрессии рекомбинантных генов или белков.
Термин «животное» по отношению к клеткам, плюрипотентным и/или тотипотентным клеткам, ЭС-клеткам, донорским клеткам и/или эмбрионам-хозяевам включает в себя млекопитающих, рыб и птиц. Млекопитающие включают в себя, например, людей, не относящихся к человеку приматов, обезьян, человекообразных обезьян, кошек, лошадей, быков, оленей, бизонов, овец, грызунов (например, мышей, крыс, хомячков, морских свинок), домашний скот (например, жвачных животных, таких как коровы, волы и т.д.; животных семейства полорогих, таких как овцы, козы и т.д.; и животных семейства свиней, таких как свиньи и вепри). Птицы включают в себя, например, цыплят, индеек, страуса, гусей, уток и т.п. Также включены домашние животные и сельскохозяйственные животные. Термин «не относящееся к человеку животное» исключает людей.
Мышиные плюрипотентные и/или тотипотентные клетки могут быть получены из линии 129, линии C57BL/6, смеси линий 129 и C57BL/6, линии BALB/c или линии Swiss Webster. Примеры линий 129 включают в себя 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1 и 129T2. См., например, публикацию Festing et al. (1999) Mammalian Genome 10:836). Примеры линий C57BL включают в себя C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal_wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Мышиные плюрипотентные и/или тотипотентные клетки также могут быть получены из смеси вышеупомянутой линии 129 и вышеупомянутого штамма C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогичным образом, мышиные плюрипотентные и/или тотипотентные клетки могут быть получены из смеси вышеупомянутой линии 129 или смеси вышеупомянутых линий BL/6 (например, линии 129S6 (129/SvEvTac)). Конкретным примером мышиной ЭС-клетки является мышиная ЭС-клетка VGF1. См., например, публикацию Auerbach et al. (2000) Biotechniques 29, 1024-1028, 1030, 1032, которая полностью включена в настоящий документ путем ссылки.
Плюрипотентная и/или тотипотентная клетка крысы может быть получена из любой линии крыс, включая, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, такую как Fisher F344 или Fisher F6. Плюрипотентные и/или тотипотентные клетки крысы можно также получить из линии, происходящей от смеси двух или более вышеупомянутых линий. Например, плюрипотентная и/или тотипотентная клетки крысы могут быть получены от линии DA или ACI. Линия крыс ACI характеризуется наличием окраски «черный агути» с белым брюхом и лапами и гаплотипом RTlavl. Такие линии доступны из различных источников, включая Harlan Laboratories. Примером линии крысиных ЭС-клеток от крысы ACI является ЭС-клетка крыс ACI.G1. Линия крысы «темный агути» (DA) характеризуется наличием окраса «агути» и гаплотипа RTlavl. Такие крысы доступны из различных источников, включая Charles River и Harlan Laboratories. Примерами линии крысиных ЭС-клеток от крысы линии DA является линия крысиных ЭС-клеток DA.2B и крысиных ЭС-клеток DA.2C. В некоторых случаях плюрипотентные и/или тотипотентные клетки крысы получены от инбредной линии крыс. См., например, документ U.S. 2014/0235933 A1, поданный 20 февраля 2014 г., и U.S. 2014/0310828 A1, поданный 16 апреля 2014 г., оба из которых полностью включены в настоящий документ путем ссылки.
Примеры плюрипотентных клеток человека включают в себя ЭС-клетки человека, стволовые клетки взрослого человека, ограниченные в развитии клетки-предшественники человека и индуцированные плюрипотентные стволовые (ИПС) клетки человека, такие как праймированные ИПС-клетки человека и интактные ИПС-клетки человека. См., например, заявку на патент США №14/515,503, поданную 15 октября 2014 г. и полностью включенную в настоящий документ путем ссылки. Индуцированные плюрипотентные стволовые клетки включают в себя плюрипотентные стволовые клетки, которые могут быть получены непосредственно из дифференцированной взрослой клетки. Человеческие ИПС-клетки можно получать посредством введения в клетку конкретных наборов репрограммирующих факторов, которые могут включать в себя, например, Oct3/4, транскрипционные факторы семейства Sox (например, Sox1, Sox2, Sox3, Sox15), транскрипционные факторы семейства Мус (например, с-Мус, 1-Мус, n-Мус), транскрипционные факторы семейства  -подобных (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glisl. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (2006) Cell 126:663-676. Праймированные ЭС-клетки человека и праймированные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам постимплантационных эпибластных клеток, и коммутированы для спецификации линии дифференцировки и дифференцировки. Интактные ЭС-клетки человека и интактные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам ЭС-клеток внутренней клеточной массы предимплантационного эмбриона, и не коммитированы для спецификации линии дифференцировки. См., например, публикацию Nichols and Smith (2009) Cell Stem Cell 4:487-492.
-подобных (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственных транскрипционных факторов, таких как NANOG, LIN28 и/или Glisl. Человеческие ИПС-клетки также можно получить, например, при помощи миРНК - небольших молекул, имитирующих действие транскрипционных факторов или спецификаторов линий дифференцировки. Человеческие ИПС-клетки характеризуются способностью дифференцироваться в любую клетку трех зародышевых листков позвоночных, например эндодермы, эктодермы или мезодермы. Человеческие ИПС-клетки также характеризуются способностью к неограниченному размножению при подходящих условиях культивирования in vitro. См., например, публикацию Takahashi and Yamanaka (2006) Cell 126:663-676. Праймированные ЭС-клетки человека и праймированные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам постимплантационных эпибластных клеток, и коммутированы для спецификации линии дифференцировки и дифференцировки. Интактные ЭС-клетки человека и интактные ИПС-клетки человека включают в себя клетки, которые демонстрируют характеристики, аналогичные характеристикам ЭС-клеток внутренней клеточной массы предимплантационного эмбриона, и не коммитированы для спецификации линии дифференцировки. См., например, публикацию Nichols and Smith (2009) Cell Stem Cell 4:487-492.
Клетки, которые были имплантированы в эмбрион-хозяин, в настоящем документе могут называться «донорскими клетками». Генетически модифицированная плюрипотентная и/или тотипотентная клетка может быть получена из той же линии, что и эмбрион-хозяин, или из другой линии. Аналогичным образом, суррогатная мать может быть получена из той же линии, что и генетически модифицированная плюрипотентная и/или тотипотентная клетка и/или эмбрион-хозяин, или суррогатная мать может быть получена из линии, отличной ПЕРЕЧЕНЬ от генетически модифицированной плюрипотентной и/или тотипотентной клетки и/или эмбриона-хозяина.
В способах и композициях, описанных в настоящем документе, можно использовать различных эмбрионов-хозяев. Например, плюрипотентные и/или тотипотентные клетки, имеющие нацеленную генетическую модификацию, могут быть введены в эмбрион на стадии до морулы (например, эмбрион на стадии 8 клеток) из соответствующего организма. См., например, публикации US 7,576,259, US 7,659,442, US 7,294,754, и US 2008/0078000 A1, все из которых полностью включены в настоящий документ путем ссылки. В других случаях донорские ЭС-клетки можно имплантировать в эмбрион-хозяин на стадии до образования морулы, на стадии 2 клеток, стадии 4 клеток, стадии 8 клеток, стадии 16 клеток, стадии 32 клеток или стадии 64 клеток. Эмбрион-хозяин также может являться бластоцистой или может являться эмбрионом на стадии до бластоцисты, эмбрионом на стадии предморулы, эмбрионом на стадии морулы, эмбрионом на стадии некомпактной морулы или эмбрионом на стадии компактной морулы. При применении эмбриона мыши стадия эмбриона-хозяина может быть стадией Тейлера 1 (TS1), TS2, TS3, TS4, TS5 и TS6 со ссылкой на стадии Тейлера, описанные в публикации Theiler (1989) "The House Mouse: Atlas of Mouse Development," Springer-Verlag, New York. Например, стадия Тейлера может быть выбрана из TS1, TS2, TS3 и TS4. В некоторых случаях эмбрион-хозяин содержит внеклеточую оболочку ооцита, а донорская клетка представляет собой ЭС-клетку, которую вводят в эмбрион-хозяин через отверстие во внеклеточной оболочке ооцита. В других случаях эмбрион-хозяин представляет собой эмбрион, не содержащий внеклеточной оболочки ооцита. В других случаях стадия морулы эмбриона-хозяина является агрегированной.
X. Способы идентификации клеток с модифицированным целевым геномным локусом
Некоторые из вышеуказанных способов дополнительно включают в себя идентификацию клетки, имеющей модифицированный целевой геномный локус (например, модифицированный геном). Для идентификации клеток, имеющих нацеленную модификацию, такую как делеция или вставка, могут использоваться различные способы. Такие способы могут включать в себя идентификацию одной клетки, имеющей нацеленную модификацию в целевом локусе. Может быть проведен скрининг для идентификации таких клеток с модифицированными геномными локусами.
Этап скрининга может включать количественный тест для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный тест может быть выполнен посредством количественной ПЦР, такой как ПЦР в реальном времени (кПЦР). Во время ПЦР в реальном времени может использоваться первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров содержит флюоресцентный зонд, который распознает амплифицированную последовательность.
В других случаях клетки, имеющие нацеленную генетическую модификацию, выбирают с использованием способов, которые включают в себя, например, анализ саузерн-блот, секвенирование ДНК, анализ ПЦР или фенотипический анализ. Затем такие клетки применяют в различных способах и композициях, описанных в настоящем документе.
К другим примерам подходящих количественных тестов относятся флуоресцентная гибридизация in situ (FISH), сравнительная геномная гибридизация, изотермическая амплификация ДНК, количественная гибридизация с иммобилизованным(-и) зондом(-ами), Invader Probes®, ММР assays®, TAQMAN® Molecular Beacon или Eclipse™ (см., например, US 2005/0144655, который полностью включен в настоящий документ путем ссылки).
Этап скрининга по существу также включает в себя тесты, специфичные для плеч, которые используются для того, чтобы различать правильные нацеленные вставки полинуклеотидных вставок в целевой геномный локус от случайных трансгенных вставок полинуклеотидных вставок в местоположения генома за пределами целевого геномного локуса, а также используются для обнаружения правильной сборки двух или более перекрывающихся LTVEC в один конструкт. В традиционных тестах на скрининг на нацеленные модификации, таких как ПЦР длинных фрагментов или саузерн-блоттинг, вставленный нацеливающий вектор связывается с целевым локусом. Однако из-за большого размера гомологичного плеча большой нацеливающий вектор (LTVEC) не позволяет проводить скрининг с помощью таких традиционных тестов. Для скрининга нацеливания с помощью большого нацеливающего вектора (LTVEC) можно использовать тесты на модификацию аллеля (МОА), включая тесты на потерю аллеля (LOA) и приобретение аллеля (GOA) (см., например, патент США 2014/0178879 и публикацию Frendewey et al. (2010) Methods Enzymol. 476:295-307, для всех целей полностью включенную в настоящий документ путем ссылки. Тест на потерю аллеля (LOA) инвертирует логику традиционного скрининга и позволяет количественно определять число копий нативного локуса, на который была направлена мутация. В правильно нацеленном клеточном клоне тест LOA позволяет определить один из двух нативных аллелей (для генов не на хромосоме X или Y), а другой аллель разрывается нацеленной модификацией. Этот же принцип может применяться в обратном направлении в тесте на приобретение аллеля (GOA) для количественного определения числа копий вставленного нацеливающего вектора. Например, комбинированное использование тестов GOA и LOA позволяет выявлять, что правильно нацеленный гетерозиготный клон потерял одну копию нативного целевого гена и приобрел одну копию гена резистентности к лекарственному средству или другого вставленного маркера.
В качестве примера, количественную полимеразную цепную реакцию (кПЦР) можно использовать как способ количественного определения аллелей, но любой способ, который позволяет достоверно определять разницу между отсутствием, одной и двумя копиями целевого гена или между отсутствием, одной и двумя копиями полинуклеотидной вставки, может использоваться для разработки теста МОА. Например, для количественного определения числа копий шаблона ДНК в образце геномной ДНК можно использовать TAQMAN®, в особенности при сравнении с эталонным геном (см., например, US 6,596,541, для всех целей полностью включенный в настоящий документ путем ссылки). Контрольный ген количественно определяют в той же геномной ДНК, что и целевой(-ые) ген(-ы) или локус(-ы). Поэтому выполняются две амплификации TAQMAN® (каждая со своим соответствующим зондом). Один зонд TAQMAN® определяет «Ct» (пороговый цикл) эталонного гена, а другой зонд определяет Ct той области нацеленного гена(-ов) или локуса(-ов), которая замещается с помощью успешного нацеливания (т.е. тест LOA). Ct представляет собой значение, которое отражает количество исходной ДНК для каждого из зондов TAQMAN®, т.е. менее распространенная последовательность требует большего числа циклов ПЦР для достижения порогового цикла. Снижение числа копий последовательности шаблонов наполовину для реакции TAQMAN® приведет к увеличению Ct приблизительно на одну единицу. Реакции TAQMAN® в клетках, где один аллель целевого(-ых) гена(-ов) или локуса(-ов) замещен путем гомологичной рекомбинации, приводят к увеличению одного Ct для целевой реакции TAQMAN® без увеличения Ct для эталонного гена по сравнению с ДНК из клеток без проведенного нацеливания. Для теста GOA можно использовать другой зонд TAQMAN® с целью определения Ct полинуклеотидной вставки, которая замещает целевой(-ые) ген(-ы) или локус(-ы) путем успешного нацеливания.
Может быть полезно расширить стандартные тесты LOA и GOA для проверки правильного нацеливания с помощью LTVEC. Например, тесты LOA и GOA сами по себе могут не различать правильно нацеленные клеточные клоны от клонов, в которых Cas-индуцированная делеция целевого геномного локуса совпадает со случайной интеграцией LTVEC в другое место генома. Поскольку давление отбора в клетке с проведенным нацеливанием основано на кассете селекции, случайная трансгенная интеграция LTVEC в другом месте генома будет по существу включать кассету селекции и прилегающие области LTVEC, но может исключать более дистальные области LTVEC. Например, если участок LTVEC случайно интегрируется в геном, a LTVEC содержит полинуклеотидную вставку длиной приблизительно 5 т.п.н. или более с кассетой селекции, прилегающей к 3' гомологичному плечу, в некоторых случаях 3' гомологичное плечо, а не 5' гомологичное плечо будет трансгенно интегрировано с кассетой селекции. Альтернативно, если кассета селекции прилегает к 5' гомологичному плечу, в некоторых случаях 5' гомологичное плечо, а не 3' гомологичное плечо будет трансгенно интегрировано с кассетой селекции. В качестве примера, если тесты LOA и GOA используются для оценки нацеленной интеграции LTVEC, а в тесте GOA используются зонды против кассеты селекции или любой другой уникальной (неплечевой) области LTVEC, гетерозиготная делеция в целевом геномном локусе в комбинации со случайной трансгенной интеграцией LTVEC даст такое же считывание данных, что и гетерозиготная нацеленная интеграция LTVEC в целевом геномном локусе. Чтобы проверить правильное нацеливание с помощью LTVEC, специфичные для плеч тесты можно использовать в сочетании с тестами LOA и/или GOA.
Специфичные для плеч тесты определяют число копий шаблона ДНК в гомологичных плечах LTVEC. Такие гомологичные плечи могут включать в себя гомологичное плечо LTVEC, которое не перекрывается с другим LTVEC, но соответствует целевой последовательности в клетке (например, гомологичное плечо, перекрывающееся с геномной целевой последовательностью в мышиной клетке (mArm)). Такие гомологичные плечи могут также включать в себя перекрывающееся гомологичное плечо, присутствующее в двух перекрывающихся LTVEC (например, перекрывающаяся человеческая последовательность в 3' гомологичном плече первого LTVEC и 5' гомологичном плече второго LTVEC (hArm)). В экспериментах, в которых в клетку вводят множество перекрывающихся LTVEC, скрининг по существу включает в себя тесты LOA, тесты GOA для всех уникальных вставленных последовательностей и специфичные для плеч тесты для всех областей гомологии (т.е. между LTVEC и целевой последовательностью в клетке и между двумя различными перекрывающимися LTVEC). В качестве примера, в случае трех перекрывающихся LTVEC, введенных в клетку мыши для гуманизации целевого локуса мыши дикого типа, ожидаемое число копий для гетерозиготной нацеленной вставки будет следующим: 2 копии 5' mArm (гомологичное плечо, перекрывающееся с целевой 5'-последовательностью мыши), 1 копия hArm1 (перекрывающаяся последовательность между LTVEC 1 и 2), 1 копия hArm2 (перекрывающаяся последовательность между LTVEC 2 и 3) и 2 копии 3' mArm (гомологичное плечо, перекрывающееся с целевой 3'-последовательностью мыши). В примере выше число копий mArm более двух по существу указывает на трансгенную случайную интеграцию LTVEC за пределами целевого геномного локуса, а не в целевой геномный локус, что нежелательно. Правильно нацеленные клоны сохранили бы число копий mArm, равное двум. Кроме того, число копий mArm менее двух в таких специфичных для плеч тестах по существу указывает на большие Cas-опосредованные делеции, выходящие за пределы области, нацеленной для делеции, что также нежелательно. Аналогичным образом, для гетерозиготных нацеленных модификаций число копий, равное 1, для hArm1 и hArm2 по существу указывает на то, что все три LTVEC были собраны в один конструкт.
Все патентные заявки, веб-сайты, другие публикации, номера доступа и т.п., упомянутые выше или ниже, для любых целей полностью включены в настоящий документ путем ссылки в равной мере, как если бы было указано, что каждая индивидуальная позиция отдельно и в индивидуальном порядке включена таким образом путем ссылки на нее. Если в различные моменты времени с номером доступа связаны различные версии последовательности, подразумевается версия, связанная с номером доступа на действительную дату подачи настоящей заявки. Под действительной датой подачи подразумевается наступающая ранее из фактической даты подачи или даты подачи приоритетной заявки, где приведена ссылка на номер доступа (если применимо). Аналогичным образом, если в различные моменты времени издаются различные редакции публикации, веб-сайта и т.п., то, если не указано иное, подразумевается наиболее поздняя изданная версия на действительную дату подачи заявки. Если специально не указано иное, любое свойство, стадия, элемент, вариант осуществления или аспект настоящего изобретения может использоваться в комбинации с любым другим из перечисленного. Хотя настоящее изобретение было подробно описано посредством иллюстрации и примера в целях четкости и лучшего понимания, будет очевидно, что некоторые изменения и модификации могут быть реализованы в рамках объема приложенной формулы изобретения.
ПРИМЕРЫ
Представленные ниже примеры приведены с целью предоставления специалисту в данной области полного изложения и описания того, как выполнять и применять настоящее изобретение, не должны ограничивать объем того, что авторы изобретения считают своим изобретением, и не предназначены для представления того, что приведенные ниже эксперименты являются всеми или только лишь проведенными экспериментами. Были приложены усилия для обеспечения точности использованных чисел (например, количеств, температуры и т.п.), но следует учитывать некоторые погрешности и отклонения в экспериментах. При отсутствии особых указаний части представляют собой массовые части, молекулярная масса представляет собой средневесовую молекулярную массу, температура указана в градусах по шкале Цельсия, а давление является атмосферным или близким к атмосферному.
Пример 1. Нацеливание локуса TCR-альфа двумя LTVEC в комбинации с нуклеазами с «цинковыми пальцами»
Система двойного нацеливания была разработана таким образом, что два больших нацеливающих вектора (LTVEC) модифицируют геномный локус за один этап нацеливания. Как показано на ФИГ. 1, на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащую кассету селекции гигромицином, было проведено нацеливание системой двойного нацеливания для получения ЭС-клеток, содержащих дополнительные сегменты Igκ гена вариабельного участка.
Данный подход с использованием двойного нацеливания, представленный на ФИГ. 1, включает двойное нацеливание или совместную электропорацию двух разных больших нацеливающих векторов (LTVEC) вместе с нуклеотидными последовательностями, которые кодируют эндонуклеазу (например, нуклеазу с «цинковыми пальцами»), создающую двухцепочечный разрыв в целевом локусе или вблизи него в ЭС-клетках.
В рамках этого подхода первый большой нацеливающий вектор (обозначенный как MAID 1710) содержал 3'-гомологичное плечо из 30 т.п.н., которое включало в себя последовательность человеческих сегментов гена Vκ1-5 и Vκ1-6, последовательность из 120 т.п.н., которая содержала человеческие сегменты гена от Vκ3-7 до Vκ3-15, и 5'-область из 20 т.п.н. («перекрывающаяся область»), которая содержала человеческий сегмент гена Vκ1-16. Второй большой нацеливающий вектор (обозначенный как MAID 6600) содержал 3'-перекрывающуюся область из 20 т.п.н. (область, содержащую человеческий сегмент гена Vκ1-16, такой же, как в первом векторе), последовательность из 140 т.п.н., содержащую человеческие сегменты гена от Vκ1-17 до Vκ2-30, кассету селекции FRT-Ub-Neo-FRT и мышиное 3' гомологичное плечо TCR А из 15,5 т.п.н.
Были разработаны нуклеазы с «цинковыми пальцами» (ZFN), которые распознают и расщепляют целевую последовательность в пределах гена резистентности к гигромицину, чтобы стимулировать гомологичную рекомбинацию двух LTVEC в целевом локусе TCR А. ЭС-клетки, созданные на ФИГ. 1 (MAID 6548, гетерозиготный по всем Jκ сегментам человека и четырем функциональным сегментам Vκ гена человека), подвергали электропорации двумя большими нацеливающими векторами (MAID6600 и MAID1700-обрезанный), описанными выше, и двумя плазмидами, экспрессирующими каждую половину ZFN (1/2), которые связываются с последовательностями распознавания в гене резистентности к гигромицину и катализируют двухцепочечный разрыв в целевом сайте
(TGCGATCGCTGCGGCCGAtcttagCCAGACGAGCGGGTTCGG (SEQ ID NO: 2); при этом сайтом расщепления указан строчными буквами) (см. таблицу 1). Были разработаны две дополнительных ZFN для нацеливания на гигромицин: ZFN(3/4), который нацеливает ген гигромицина на нуклеотидную последовательность CGCTGCGGCCGATCTtagccaGACGAGCGGGTTCGG (SEQ ID NO: 3); и ZFN(5/6), который нацеливает ген гигромицина на нуклеотидную последовательность AGCGTGTCCGACCTGATGcagctcTCGGAGGGCGAAGAA (SEQ ID NO: 4) (см. таблицу 1).

Два больших нацеливающих вектора были вставлены путем гомологичной рекомбинации в последовательность ДНК, замещая область, содержащую и окружающую кассету селекции Hyg. Полученные ЭС-клетки содержали в эндогенном локусе TCR А человеческий вариабельный домен иммуноглобулина, содержащий человеческие сегменты гена от Jκ1 до Jκ5 и от Vκ4-1 до Vκ2-30. Успешное встраивание двух больших нацеливающих векторов было подтверждено с использованием описанных выше тестов TAQMAN® (публикация Lie and Petropoulos, 1998. Curr. Opin. Biotechnology 9:43-48, включенная в настоящий документ путем ссылки) с помощью зондов и праймеров, указанных на ФИГ. 1 и приведенных в таблице 2 ниже (GOA = приобретение аллеля; LOA = потеря аллеля; число копий = проверка числа копий последовательности для отслеживания трансгенной интеграции в сравнении с нацеленной интеграцией; hArm1 = 3' гомологичное плечо из 30 т.п.н. первого большого нацеливающего вектора (MAID 1710); hArm2 = перекрывающаяся область из 20 т.п.н. первого (MAID 1710) и второго (MAID 6600) больших нацеливающих векторов, mArm = 5' гомологичное плечо из 15,5 т.п.н. второго нацеливающего вектора (MAID 6600), контроль (мышь дикого типа) - последовательности, присутствующие в мышином локусе TCR А). Для проверки правильного нацеливания LTVEC на геном мыши использовали тесты ПЦР в реальном времени, распознающие последовательности в гомологичных плечах LTVEC, называемые специфичными для плеч тестами. Определение числа копий этих специфичных для плеч тестов дало дополнительное разъяснение, которое помогает отличать правильно нацеленные ЭС-клоны, которые сохраняют, например, число копий mArm, равное двум, от клонов, у которых Cas9-индуцированная делеция целевого локуса мыши совпадает со случайной интеграцией LTVEC в другом месте генома, и в этом случае число копий mArm будет равно трем (или более).
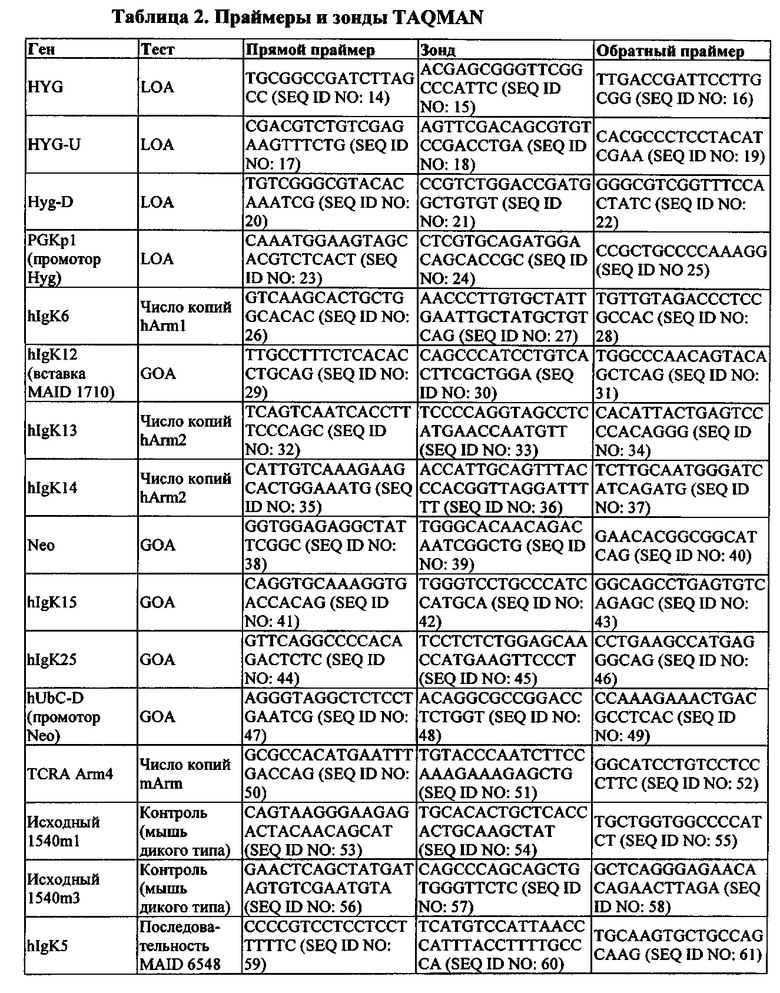
Полученный целевой локус в ЭС-клетках имел следующие соединительные последовательности, в которых мышиные последовательности приведены в скобках, человеческие последовательности представлены обычным шрифтом, сайты множественного клонирования представлены жирным шрифтом, а последовательности Frt представлены курсивом (таблица 3).

Скрининг на модификацию аллеля (МОА) выделенных колоний ЭС-клеток позволил идентифицировать 27 правильно нацеленных клонов среди 960 прошедших скрининг колоний, при этом эффективность нацеливания составила 2,81%.
Альтернативная стратегия получения локуса TCR А, содержащего дополнительные сегменты гена вариабельного участка иммуноглобулина, включает последовательное нацеливание следующими друг за другом большими нацеливающими векторами (см., например, ФИГ. 2). Таким образом, ЭС-клетки, гетерозиготные по всем сегментам гена Jκ человека и четырем функциональным сегментам гена Vκ человека (MAID 6548), подвергали электропорации с большим нацеливающим вектором, содержащим от 5' к 3': мышиное 5' гомологичное плечо из 15,5 т.п.н., кассету селекции Frt-Ub-Neo-Frt, фрагмент из 120 т.п.н., содержащий сегменты гена от Vκ3-7 до Vκ3-15, и человеческое 3' гомологичное плечо из 30 т.п.н., содержащее сегменты гена Vκ1-5 и Vκ1-6 (также присутствуют в последовательности MAID 6548). Успешное встраивание подтверждали с помощью описанных выше тестов TAQMAN® с использованием праймеров и зондов, которые перечислены в таблице 2 выше и указаны на ФИГ. 2: Hyg, hIgK5, hIgK6, hIgK12, Neo, исходный 1540m3, исходный 1540m1. В частности, зонды TCRA Arm4 и hIgK6 использовали в качестве специфичных для плеч зондов для проверки правильного геномного нацеливания LTVEC. Дополнительный набор праймеров и зондов hIgK10 также можно использовать для подтверждения успешного встраивания: прямой праймер - CGATTATGACTGGTTAGGTAGAAAGGTG (SEQ ID NO: 65); зонд - GCCACTGGTTTCTCCAAATGTTTTCAATCCAT (SEQ ID NO: 66); обратный праймер - GGGAGTACTTGGAGATCCCTAAGC (SEQ ID NO: 67).
Полученный целевой локус в ЭС-клетках имел следующие соединительные последовательности, в которых мышиные последовательности приведены в скобках, человеческие последовательности представлены обычным шрифтом, сайты множественного клонирования представлены жирным шрифтом, а последовательности Fit представлены курсивом (таблица 4).

Скрининг МОА выделенных колоний позволил идентифицировать 5 правильно нацеленных клонов среди 440 прошедших скрининг колоний (только LTVEC), при этом эффективность нацеливания составила 1,1%. Результаты скрининга выделенных колоний, нацеленных с использованием LTVEC + ZFN или LTVEC + CRISPR-Cas9, показаны в таблице 9.
По завершении одиночного нацеливания, показанного на ФИГ. 2, на ЭС-клетки может быть проведено последовательное нацеливание большими нацеливающими векторами, содержащими дополнительные Vκ, чтобы суммировать весь репертуар функциональных сегментов гена Vκ иммуноглобулина человека.
В других альтернативных стратегиях двойное или одиночное нацеливание последовательных дополнительных сегментов гена Ig Vκ в локус может быть выполнено с использованием схем двойного (два больших нацеливающих вектора) или одиночного (один большой нацеливающий вектор) нацеливания, которые включают разрушение кассеты (кассет) селекции (например, гигромицином), опосредованное нуклеазой с «цинковыми пальцами» или CRISPR.
ЭС-клетки с выполненным нацеливанием, описанные выше, использовали в качестве донорских ЭС-клеток и вводили в мышиный эмбрион на стадии до морулы, например в мышиный эмбрион на стадии 8 клеток, по способу VELOCIMOUSE® (см., например, US 7,576,259, US 7,659,442, US 7,294,754 и US 2008-0078000 A1). Мышиный эмбрион, содержащий донорские ЭС-клетки, инкубируют до стадии бластоцисты и затем имплантируют в тело суррогатной матери для получения мыши F0, полученной полностью из донорских ЭС-клеток. Мышей F0, полученных полностью из донорской ЭС-клетки, независимо несущих химерный ген человеческого IgK и мышиного Tcra С, идентифицируют посредством генотипирования с помощью теста на модификацию аллеля, который обнаруживает присутствие уникальных последовательностей генов.
Пример 2. Нацеливание гена гигромицина двумя LTVEC в комбинации с системой CRISPR/Cas
Способы двойного нацеливания, описанные в примере 1, в которых используются нуклеазы с «цинковыми пальцами», также применяли с системой CRISPR/Cas9.
Для распознавания различных целевых последовательностей в гене резистентности к гигромицину (последовательность распознавания CRISPR) были разработаны различные гидовые РНК (гРНК). Последовательности распознавания CRISPR в пределах гена гигромицина были следующими: гРНК №1:
ACGAGCGGGTTCGGCCCATTCGG (SEQ ID NO: 70); гРНК №6: CTTAGCCAGACGAGCGGGTTCGG (SEQ ID NO: 71); гРНК №10: GCCGATCTTAGCCAGACGAGCGG (SEQ ID NO: 72) и гРНК №16: CGACCTGATGCAGCTCTCGGAGG (SEQ ID NO: 73). Местоположения последовательностей распознавания в пределах гена гигромицина показаны на ФИГ. 3, на котором изображено CRISPR/Cas-опосредованное разрушение гигромицина в нацеливающем векторе MAID 1545. гРНК №1, гРНК №6, гРНК №10 и гРНК №16 проходили скрининг и были подтверждены как специфически нацеленные на ген гигромицина (см. ФИГ. 3). Результаты первичного скрининга с использованием различных специфичных для гигромицина гРНК приведены в таблице 5.

ЭС-клетки, например, ЭС-клетку, полученную на ФИГ. 1 (MAID 6548, гетерозиготная по всем Jκ сегментам человека и четырем функциональным сегментам гена Vκ человека), подвергали электропорации с двумя большими нацеливающими векторами (описанными в примере 1) вместе с одним вектором или с множеством векторов, кодирующих Cas9 и гРНК (например, гРНК №1, гРНК №6, гРНК №10 или гРНК №16), которые распознают и расщепляют целевой сайт в пределах гена резистентности к гигромицину.
Два больших нацеливающих вектора были введены путем гомологичной рекомбинации в последовательность ДНК, замещая область, содержащую и окружающую кассету селекции Hyg. Успешное встраивание двух больших нацеливающих векторов было подтверждено с помощью тестов TAQMAN®.
ЭС-клетки с проведенным нацеливанием, описанные выше, будут использоваться в качестве донорских ЭС-клеток и вводиться в эмбрион на стадии до морулы, например мышиный эмбрион на стадии 8 клеток, по способу VELOCIMOUSE® (см., например, US 7,576,259, US 7,659,442, US 7,294,754 и US 2008-0078000 A1). Мышиный эмбрион, содержащий генетически модифицированные ЭС-клетки, инкубируют до стадии бластоцисты и затем имплантируют в тело суррогатной матери для получения мыши F0, полученной полностью из донорских ЭС-клеток. Мыши F0, полученные полностью из донорских ЭС-клеток, будут идентифицироваться посредством генотипирования с помощью теста на модификацию аллеля, который обнаруживает присутствие уникальных последовательностей гена.
Пример 3. Нацеливание локуса TCR-альфа тремя LTVEC в комбинации с нуклеазами с «цинковыми пальцами»
Система тройного нацеливания была разработана таким образом, что три больших нацеливающих вектора (LTVEC) модифицируют геномный локус за один этап нацеливания. Как показано на ФИГ. 4, на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащую кассету селекции гигромицином, было проведено нацеливание с использованием системы тройного нацеливания для получения ЭС-клеток, содержащих дополнительные сегменты IgK гена вариабельного участка.
Данный подход с использованием тройного нацеливания, представленный на ФИГ. 4, включает тройное нацеливание или совместную электропорацию трех разных больших нацеливающих векторов (LTVEC) (MAID 6647, MAID 6600 и MAID 1710) вместе с нуклеотидными последовательностями, которые кодируют эндонуклеазу (например, нуклеазу с «цинковыми пальцами» или Cas9 и гРНК), создающую двухцепочечный разрыв в пределах целевого локуса или вблизи него в ЭС-клетках.
В рамках этого подхода первый большой нацеливающий вектор (обозначенный как MAID 1710) содержал 3' гомологичное плечо из 30 т.п.н., которое включает последовательность человеческих сегментов гена Vκ1-5 и Vκ1-6, последовательность из 120 т.п.н., которая содержала человеческие сегменты гена от Vκ3-7 до Vκ3-15, и 5'-область из 20 т.п.н. («перекрывающаяся область»), которая содержала человеческий сегмент гена Vκ1-16. Второй большой нацеливающий вектор (обозначенный как MAID 6600) содержал перекрывающуюся 3'-область из 20 т.п.н. (область, содержащую человеческий сегмент гена Vκ1-16, такую же, как в первом векторе), последовательность из 140 т.п.н., содержащую человеческие сегменты гена от Vκ1-17 до Vκ2-24, и 5'-область из 60 т.п.н. («перекрывающаяся область»), которая содержала человеческий сегмент гена от Vκ3-25 до Vκ2-30. Третий большой нацеливающий вектор (обозначенный как MAID 6647) содержал перекрывающуюся 3'-область из 60 т.п.н. (область, содержащую Vκ3-25-Vκ2-30 человека, такую же, как и во втором векторе), последовательность из 90 т.п.н., содержащую Vκ3-31-Vκ2-40 человека, кассету селекции FRT-Ub-Neo-FRT и 5' гомологичное плечо мышиной TCR А из 15,5 т.п.н.
Были разработаны нуклеазы с «цинковыми пальцами» (ZFN), которые распознают и расщепляют целевую последовательность в пределах гена резистентности к гигромицину, чтобы стимулировать гомологичную рекомбинацию трех LTVEC в целевом локусе TCR А. ЭС-клетки, полученные на ФИГ. 4 (MAID 6548, гетерозиготный по всем Jκ сегментам человека и четырем функциональным сегментам Vκ гена человека), подвергали электропораций тремя большими нацеливающими векторами (MAID6600-обрезанный, MAID1700-обрезанный и MAID6647), описанными выше, и двумя плазмидами, экспрессирующими каждую половину ZFN (1/2), которые связываются с последовательностями распознавания в гене резистентности к гигромицину и катализируют двухцепочечный разрыв в целевом сайте (TGCGATCGCTGCGGCCGAtcttagCCAGACGAGCGGGTTCGG (SEQ ID NO: 2); при этом сайт расщепления указан строчными буквами) (см. таблицу 1).
Три больших нацеливающих вектора вставляли путем гомологичной рекомбинации в последовательность ДНК, замещая область, содержащую и окружающую кассету селекции Hyg. Полученные ЭС-клетки содержали в эндогенном локусе TCR А человеческий вариабельный домен иммуноглобулина, содержащий человеческие сегменты гена от Jκ1 до Jκ5 и от Vκ4-1 до Vκ2-40. Успешное встраивание трех больших нацеливающих векторов было подтверждено с использованием описанных выше тестов TAQMAN® (публикация Lie and Petropoulos, 1998. Curr. Opin. Biotechnology 9:43-48, включенная в настоящий документ путем ссылки) с помощью зондов и праймеров, указанных на ФИГ. 4 и приведенных в таблице 2 выше и таблице 6 ниже (GOA = приобретение аллели; LOA = потеря аллеля; число копий = проверка числа копий последовательности для отслеживания трансгенной интеграции в сравнении с нацеленной интеграцией; hArm1 = 3' гомологичное плечо из 30 т.п.н. первого большого нацеливающего вектора (MAID 1710); hArm2 = перекрывающаяся область из 20 т.п.н. первого (MAID 1710) и второго (MAID 6600) больших нацеливающих векторов, hArm3 = перекрывающаяся область из 60 т.п.н. второго (MAID 6600) и третьего (MAID6647) нацеливающих векторов, mArm=5' гомологичное плечо из 15,5 т.п.н. третьего нацеливающего вектора (MAID 6647), контроль (мышь дикого типа) - последовательности, присутствующие в мышином локусе TCR А). Для проверки правильного нацеливания LTVEC на геном мыши использовали тесты ПЦР в реальном времени, распознающие последовательности в гомологичных плечах LTVEC, называемые специфичными для плеч тестами. Определение числа копий этих специфичных для плеч тестов обеспечило дополнительное разъяснение, которое помогает отличать правильно нацеленные ЭС-клоны, которые сохранили число копий, равное двум, для мышиного зонда (mArm) и число копий, равное одному, для человеческого зонда (hArm1), от клонов, в которых Са89-индуцированная делеция целевого локуса мыши совпадает со случайной интеграцией LTVEC в другом месте генома, и в этом случае будет число копий, равное трем (или более), для мышиного зонда (mArm), и число копий, равное двум (или более), для человеческого зонда (hArm1). Для того чтобы определить правильную сборку трех LTVEC путем гомологичной рекомбинации в требуемый локус, мы использовали специфичный для плеч тест TAQMAN®. Ожидаемое число копий, равное 1, для hArm2 и hArm3 указывает на то, что все три LTVEC были собраны в один конструкт.
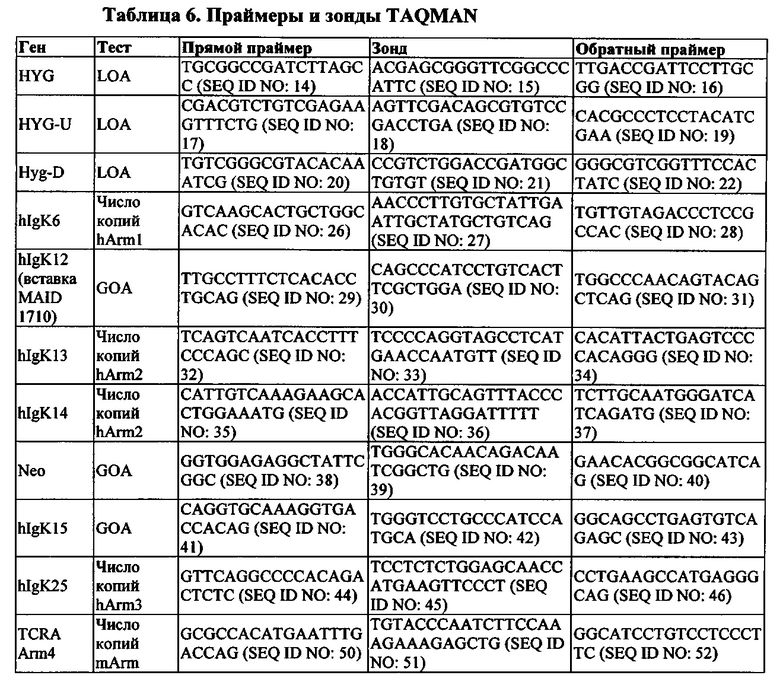

Полученный целевой локус в ЭС-клетках имел следующие соединительные последовательности, в которых мышиные последовательности приведены в скобках, человеческие последовательности представлены обычным шрифтом, сайты множественного клонирования представлены жирным шрифтом, а последовательности Fit представлены курсивом (таблица 7).

Скрининг на модификацию аллеля (МОА) выделенных колоний ЭС-клеток показал эффективность нацеливания, равную 0,4% (см. таблицу 8).

ЭС-клетки с выполненным нацеливанием, описанные выше, использовали в качестве донорских ЭС-клеток и вводили в мышиный эмбрион на стадии до морулы, например мышиный эмбрион на стадии 8 клеток, по способу VELOCIMOUSE® (см., например, US 7,576,259, US 7,659,442, US 7,294,754 и US 2008-0078000 A1). Мышиный эмбрион, содержащий донорские ЭС-клетки, инкубируют до стадии бластоцисты и затем имплантируют в тело суррогатной матери для получения мыши F0, полученной полностью из донорских ЭС-клеток. Мыши F0, полученные полностью из донорской ЭС-клетки, независимо несущие химерный ген человеческого IgK и мышиного Tcra С, идентифицируют посредством генотипирования с помощью теста на модификацию аллеля, который обнаруживает присутствие уникальных последовательностей генов.
Пример 4. Нацеливание гена гигромицина тремя LTVEC в комбинации с системой CRISPR/Cas
Способы тройного нацеливания, описанные в примере 3, в которых используются нуклеазы с «цинковыми пальцами», также применяли с системой CRISPR/Cas9.
Для распознавания различных целевых последовательностей в гене резистентности к гигромицину (последовательность распознавания CRISPR) были разработаны различные гидовые РНК (гРНК). Последовательности распознавания CRISPR в пределах гена гигромицина были следующими: гРНК №1: ACGAGCGGGTTCGGCCCATTCGG (SEQ ID NO: 70); гРНК №6: CTTAGCCAGACGAGCGGGTTCGG (SEQ ID NO: 71); гРНК №10: GCCGATCTTAGCCAGACGAGCGG (SEQ ID NO: 72) и гРНК №16: CGACCTGATGCAGCTCTCGGAGG (SEQ ID NO: 73). Местоположения последовательностей распознавания в пределах гена гигромицина показаны на ФИГ. 3. гРНК №1, гРНК №6, гРНК №10 и гРНК №16 проходили скрининг и были подтверждены как специфически нацеленные на ген гигромицина (см. ФИГ. 3 и таблицу 5).
ЭС-клетки MAID 6548 (гетерозиготные по всем человеческим сегментам Jκ и четырем функциональным сегментам гена Vκ человека) подвергали электропорации с тремя большими нацеливающими векторами, как описано в примере 3, вместе с векторами, кодирующими Cas9 и гРНК №16, которые распознают и расщепляют целевой сайт в пределах гена резистентности к гигромицину.
Три больших нацеливающих вектора были вставлены посредством гомологичной рекомбинации в последовательность ДНК, замещая область, содержащую и окружающую кассету селекции Hyg. Успешное встраивание трех больших нацеливающих векторов было подтверждено с использованием тестов TAQMAN®, описанных в примере 3.
Полученный целевой локус в ЭС-клетках имеет соединительные последовательности, показанные в таблице 7, в которых мышиные последовательности приведены в скобках, человеческие последовательности представлены обычным шрифтом, сайты множественного клонирования представлены жирным шрифтом, а последовательности Fit представлены курсивом.
Скрининг на модификацию аллеля (МОА) выделенных колоний ЭС-клеток показал эффективность нацеливания, равную 0,4% (см. таблицу 8).
ЭС-клетки с проведенным нацеливанием, описанные выше, будут использоваться в качестве донорских ЭС-клеток и вводиться в эмбрион на стадии до морулы, например мышиный эмбрион на стадии 8 клеток, по способу VELOCIMOUSE® (см., например, US 7,576,259, US 7,659,442, US 7,294,754 и US 2008-0078000 A1). Мышиный эмбрион, содержащий генетически модифицированные ЭС-клетки, инкубируют до стадии бластоцисты и затем имплантируют в тело суррогатной матери для получения мыши F0, полученной полностью из донорских ЭС-клеток. Мыши F0, полученные полностью из донорских ЭС-клеток, будут идентифицироваться посредством генотипирования с помощью теста на модификацию аллеля, который обнаруживает присутствие уникальных последовательностей гена.
Пример 5. Усиление нацеливания LTVEC посредством перекрывающихся последовательностей между двумя LTVEC
Систему двойного нацеливания, описанную в примере 1, применяли для модификации геномного локуса за один этап нацеливания с использованием двух больших нацеливающих векторов (LTVEC). Как показано на ФИГ. 1, на клетку, имеющую гетерозиготную модификацию локуса TCR-альфа на мышиной хромосоме 14, содержащую кассету селекции гигромицином, было проведено нацеливание системой двойного нацеливания для получения ЭС-клеток, содержащих дополнительные сегменты Igκ гена вариабельного участка. Два разных LTVEC вместе подвергали совместной электропорации в мышиные эмбриональные стволовые (ЭС) клетки. Необязательно нуклеиновую кислоту, кодирующую эндонуклеазу (либо нуклеазу с «цинковыми пальцами» (ZFN), либо CRISPR-Cas9), подвергали совместной электропорации для создания двухцепочечного разрыва в целевом локусе или вблизи него.
Как показано в примере 1, LTVEC (обозначенный как MAID 1710) содержал 3' гомологичное плечо из 30 т.п.н., которое включало последовательность человеческих сегментов гена Vκ1-5 и Vκ1-6, последовательность из 120 т.п.н., которая включала сегменты гена Vκ3-7 в Vκ3-15 человека, и 5'-область из 20 т.п.н. («перекрывающаяся область»), которая содержала человеческий сегмент гена Vκ1-16. Второй LTVEC (обозначенный как MAID 6600) содержал перекрывающуюся 3'-область из 20 т.п.н. (область, включающая сегмент гена Vκ1-16 человека, такая же, как и в первом векторе), последовательность из 140 т.п.н., содержащую Vκ1-17-Vκ2-30 человека, кассету селекции FRT-Ub-Neo-FRT и 3' гомологичное плечо мышиной TCR А из 15,5 т.п.н.
Успешное нацеливание привело к вставке двух LTVEC посредством гомологичной рекомбинации в последовательность ДНК, замещая область, содержащую и окружающую кассету селекции Hyg. Полученные ЭС-клетки содержали в эндогенном локусе TCR А человеческий вариабельный домен иммуноглобулина, содержащий человеческие сегменты гена от Jκ1 до Jκ5 и от Vκ4-1 до Vκ2-30. Успешное встраивание двух больших нацеливающих векторов было подтверждено с использованием описанных выше тестов TAQMAN (публикация Lie and Petropoulos, 1998. Curr. Opin. Biotechnology 9:43-48, включенная в настоящий документ путем ссылки) с помощью зондов и праймеров, указанных на ФИГ. 1 и в таблице 2.
Для сравнения, система с одиночным LTVEC, описанная в примере 1, также применялась для модификации одного и того же геномного локуса с использованием одиночного LTVEC либо отдельно, либо в комбинации с ZFN или CRISPR-Cas9 (см. ФИГ. 2). Успешное встраивание подтверждали с помощью описанных выше тестов TAQMAN с использованием праймеров и зондов, которые перечислены в таблице 2 выше и указаны на ФИГ. 2.
В таблице 9 представлено сравнение эффективностей нацеливания в экспериментах по нацеливанию с использованием одиночного LTVEC (отдельно, с ZFN или с Cas9), с использованием двух LTVEC одновременно (отдельно, с ZFN или с Cas9) или с использованием двух LTVEC плюс третий LTVEC одновременно (отдельно, с ZFN или с Cas9). Эффективности нацеливания, представленные в таблице 9, представляют собой процентное значение прошедших скрининг ESC-клонов, которые, как было определено, правильно нацелены при начальном скрининге, при скрининге для подтверждения и при скрининге для повторного подтверждения с использованием праймеров и зондов TAQMAN в таблице 2. Нацеливание одним только одиночным LTVEC позволило получить 1,1% правильно нацеленных клонов. Расщепление с помощью ZFN повышало эффективность нацеливания одиночным LTVEC до 4,4%, а расщепление с помощью CRISPR-Cas9 повышало эффективность нацеливания одиночным LTVEC до 5,5%. Неожиданно было обнаружено, что нацеливание 2 LTVEC, имеющими 20 т.п.н. перекрывающейся последовательности, привело к эффективности нацеливания 1,4% даже при отсутствии использования нуклеазы. Эффективность нацеливания повысилась до 2,81% при использовании ZFN и до 1,6% при использовании Cas9.
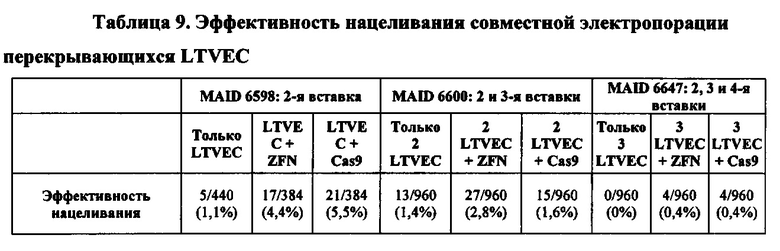
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК | 2015 |
|
RU2734770C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2771532C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ ИЛИ ПОДДЕРЖАНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2015 |
|
RU2706965C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2711740C2 |
| ОПОСРЕДОВАННОЕ НУКЛЕАЗОЙ НАЦЕЛИВАНИЕ С БОЛЬШИМИ НАЦЕЛИВАЮЩИМИ ВЕКТОРАМИ | 2013 |
|
RU2645475C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2829155C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2745563C2 |
| НАПРАВЛЕННАЯ МОДИФИКАЦИЯ ГЕНОМА КРЫСЫ | 2014 |
|
RU2676708C2 |






















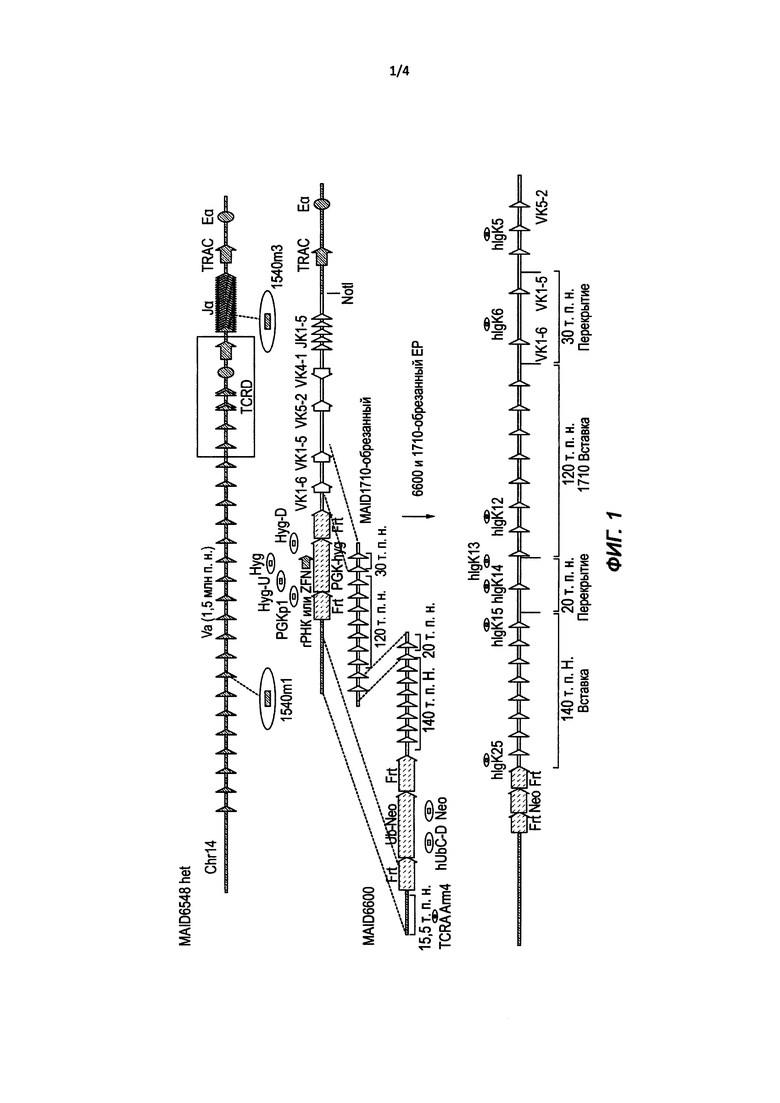

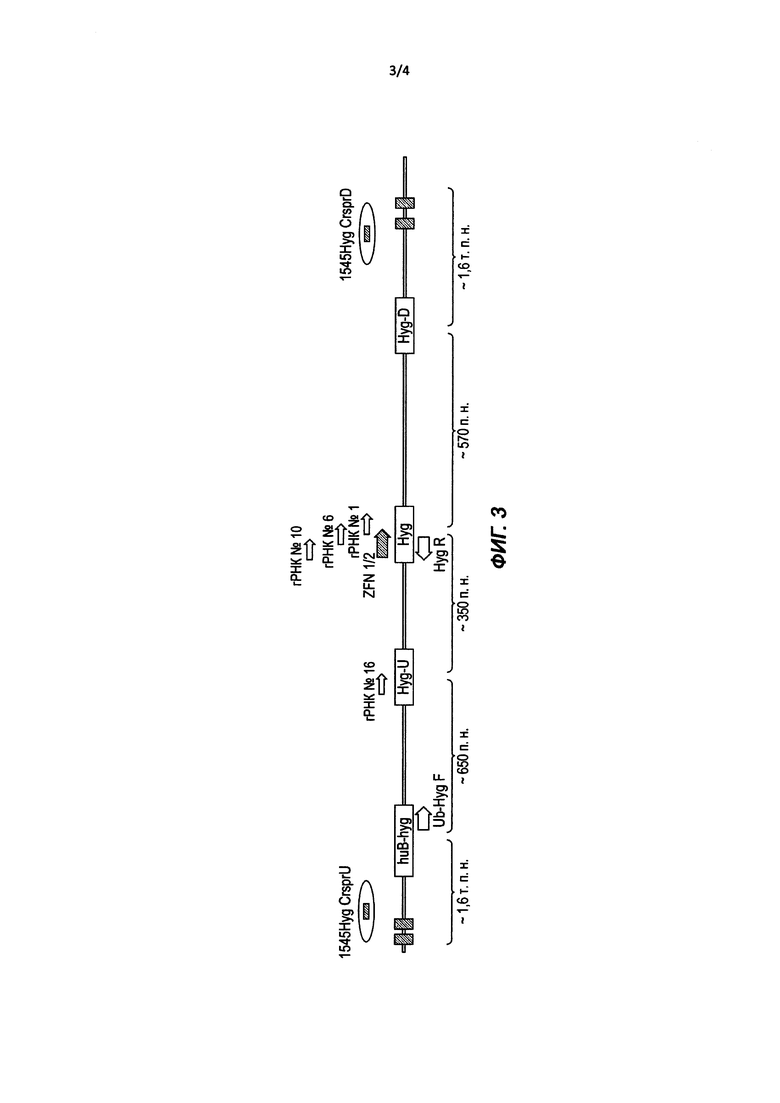

Изобретение относится к биотехнологии. Описан способ модификации целевого геномного локуса в клетке млекопитающего, включающий в себя: введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса; введение в клетку первого большого нацеливающего вектора (LTVEC), который имеет длину по меньшей мере 10 т.п.н. и содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, причем суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет по меньшей мере 100 т.п.н., причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса, при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки между соответствующими геномными сегментами; и отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус. Изобретение позволяет модифицировать целевой геномный локус. 24 з.п. ф-лы, 4 ил., 9 табл., 5 пр.
1. Способ модификации целевого геномного локуса в клетке млекопитающего, включающий в себя:
(a) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса;
(b) введение в клетку первого большого нацеливающего вектора (LTVEC), который имеет длину по меньшей мере 10 т.п.н. и содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, и второго LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом,
причем суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет по меньшей мере 100 т.п.н.,
причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, и первое 5' гомологичное плечо первого LTVEC и второе 3' гомологичное плечо второго LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса,
при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки между соответствующими геномными сегментами; и
(c) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку и вторую полинуклеотидную вставку, интегрированные в целевой геномный локус.
2. Способ по п. 1, в котором первая перекрывающаяся последовательность составляет по меньшей мере 1 т.п.н.
3. Способ по п. 1 или 2, в котором эффективность нацеливания составляет по меньшей мере 1,6%.
4. Способ по любому из пп. 1-3, в котором первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки и второй полинуклеотидной вставки в целевой геномный локус.
5. Способ по любому из пп. 1-4, в котором первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка содержит геномную ДНК,
необязательно причем первая полинуклеотидная вставка и/или вторая полинуклеотидная вставка содержит условный аллель, полинуклеотид, кодирующий селективный маркер, репортерный ген, одну или более экспрессионных кассет или нуклеиновую кислоту, фланкированную целевыми последовательностями сайт-специфической рекомбинации,
необязательно причем геномная ДНК гомологична или ортологична последовательности, нацеливаемой для делеции, в целевом геномном локусе, и
необязательно причем вставка первой полинуклеотидной вставки и второй полинуклеотидной вставки приводит к замещению нечеловеческой нуклеотидной последовательности гомологичной или ортологичной человеческой полинуклеотидной последовательностью.
6. Способ по любому из пп. 1-5, в котором:
(I) первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе получены от вида, который отличается от вида, к которому относится клетка; и/или
(II) первая полинуклеотидная вставка, вторая полинуклеотидная вставка или обе представляют собой человеческие нуклеиновые кислоты.
7. Способ по любому из пп. 1-6, в котором:
(I) суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет от 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н. или от приблизительно 450 т.п.н. до приблизительно 500 т.п.н.;
(II) суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет по меньшей мере 200 т.п.н., необязательно причем суммарный размер первой полинуклеотидной вставки и второй полинуклеотидной вставки составляет приблизительно 300 т.п.н.; или
(III) клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую полинуклеотидную вставку и вторую полинуклеотидную вставку, которые имеют суммарный размер в диапазоне от 100 т.п.н. до приблизительно 500 т.п.н.
8. Способ по любому из пп. 1-7, в котором:
(I) первая перекрывающаяся последовательность первого LTVEC идентична первой перекрывающейся последовательности второго LTVEC; и/или
(II) размер первой перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н.; и/или
(III) размер первой перекрывающейся последовательности составляет по меньшей мере 10 т.п.н., необязательно причем размер первой перекрывающейся последовательности составляет по меньшей мере 20 т.п.н.
9. Способ по любому из пп. 1-8, в котором интеграция первой полинуклеотидной вставки, второй полинуклеотидной вставки или обеих в целевой геномный локус приводит к одному или более из:
(I) добавления экзогенной последовательности в целевом геномном локусе;
(II) делеции эндогенной последовательности в целевом геномном локусе, необязательно причем делеция составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.; и
(III) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
10. Способ по любому из пп. 1-9, в котором:
(I) общая длина 5' и 3' гомологичных плеч первого LTVEC или второго LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н.; и/или
(II) первый LTVEC составляет по меньшей мере 50 т.п.н., и второй LTVEC составляет по меньшей мере 50 т.п.н., необязательно причем первый LTVEC составляет по меньшей мере 100 т.п.н., и второй LTVEC составляет по меньшей мере 100 т.п.н.
11. Способ модификации целевого геномного локуса в клетке млекопитающего, включающий в себя:
(a) введение в клетку нуклеазного агента, который осуществляет одно- или двухцепочечный разрыв в пределах целевого геномного локуса;
(b) введение в клетку первого большого нацеливающего вектора (LTVEC), который имеет длину по меньшей мере 10 т.п.н. и содержит первую полинуклеотидную вставку, фланкированную первым 5' гомологичным плечом и первым 3' гомологичным плечом, второго LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит вторую полинуклеотидную вставку, фланкированную вторым 5' гомологичным плечом и вторым 3' гомологичным плечом, и третьего LTVEC, который имеет длину по меньшей мере 10 т.п.н. и содержит третью полинуклеотидную вставку, фланкированную третьим 5' гомологичным плечом и третьим 3' гомологичным плечом,
причем суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет по меньшей мере 100 т.п.н.,
причем первое 3' гомологичное плечо первого LTVEC имеет первую перекрывающуюся последовательность, гомологичную второму 5' гомологичному плечу второго LTVEC, второе 3' гомологичное плечо второго LTVEC имеет вторую перекрывающуюся последовательность, гомологичную третьему 5' гомологичному плечу третьего LTVEC, и первое 5' гомологичное плечо первого LTVEC и третье 3' гомологичное плечо третьего LTVEC гомологичны соответствующим геномным сегментам в пределах целевого геномного локуса,
при этом целевой геномный локус модифицирован путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки между соответствующими геномными сегментами; и
(с) отбор клетки с проведенным нацеливанием, содержащей первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, интегрированные в целевой геномный локус.
12. Способ по п. 11, в котором первая перекрывающаяся последовательность составляет по меньшей мере 1 т.п.н., и вторая перекрывающаяся последовательность составляет по меньшей мере 1 т.п.н.
13. Способ по п. 11 или 12, в котором эффективность нацеливания составляет по меньшей мере 0,4%.
14. Способ по любому из пп. 11-13, в котором первая полинуклеотидная вставка и первое 3' гомологичное плечо и вторая полинуклеотидная вставка и второе 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, а вторая полинуклеотидная вставка и второе 3' гомологичное плечо и третья полинуклеотидная вставка и третье 5' гомологичное плечо представляют собой перекрывающиеся фрагменты непрерывной нуклеиновой кислоты, которая подвергается реформированию путем интеграции первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки в целевой геномный локус.
15. Способ по любому из пп. 11-14, в котором первая полинуклеотидная вставка, и/или вторая полинуклеотидная вставка, и/или третья полинуклеотидная вставка содержит геномную ДНК,
необязательно причем первая полинуклеотидная вставка, и/или вторая полинуклеотидная вставка, и/или третья полинуклеотидная вставка содержит условный аллель, полинуклеотид, кодирующий селективный маркер, репортерный ген, одну или более экспрессионных кассет или нуклеиновую кислоту, фланкированную целевыми последовательностями сайт-специфической рекомбинации,
необязательно причем геномная ДНК гомологична или ортологична последовательности, нацеливаемой для делеции, в целевом геномном локусе, и
необязательно причем вставка первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки приводит к замещению нечеловеческой нуклеотидной последовательности гомологичной или ортологичной человеческой полинуклеотидной последовательностью.
16. Способ по любому из пп. 11-15, в котором:
(I) одна или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки получены от вида, который отличается от вида, к которому относится клетка; и/или
(II) одна или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки представляют собой человеческие нуклеиновые кислоты.
17. Способ по любому из пп. 11-16, в котором:
(I) суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет от 100 т.п.н. до 125 т.п.н., от приблизительно 125 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 175 т.п.н., от приблизительно 175 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 225 т.п.н., от приблизительно 225 т.п.н. до приблизительно 250 т.п.н., от приблизительно 250 т.п.н. до приблизительно 275 т.п.н., от приблизительно 275 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 350 т.п.н., от приблизительно 350 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 450 т.п.н., от приблизительно 450 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 550 т.п.н., от приблизительно 550 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 650 т.п.н. или от приблизительно 650 т.п.н. до приблизительно 700 т.п.н.;
(II) суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет по меньшей мере 200 т.п.н., необязательно причем суммарный размер первой полинуклеотидной вставки, второй полинуклеотидной вставки и третьей полинуклеотидной вставки составляет приблизительно 400 т.п.н.; или
(III) клетка с проведенным нацеливанием содержит геномную ДНК, содержащую вместе первую полинуклеотидную вставку, вторую полинуклеотидную вставку и третью полинуклеотидную вставку, которые имеют суммарный размер в диапазоне от 100 т.п.н. до приблизительно 700 т.п.н.
18. Способ по любому из пп. 11-17, в котором:
(I) первая перекрывающаяся последовательность первого LTVEC идентична первой перекрывающейся последовательности второго LTVEC, и/или вторая перекрывающаяся последовательность второго LTVEC идентична второй перекрывающейся последовательности третьего LTVEC; и/или
(II) размер первой перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н., и/или размер второй перекрывающейся последовательности составляет от приблизительно 1 т.п.н. до приблизительно 70 т.п.н.; и/или
(III) размер первой перекрывающейся последовательности составляет по меньшей мере 10 т.п.н., и/или размер второй перекрывающейся последовательности составляет по меньшей мере 10 т.п.н., необязательно причем размер первой перекрывающейся последовательности составляет по меньшей мере 20 т.п.н., и/или размер второй перекрывающейся последовательности составляет по меньшей мере 20 т.п.н.
19. Способ по любому из пп. 11 -18, в котором интеграция одной или более из первой полинуклеотидной вставки, второй полинуклеотидной вставки или третьей полинуклеотидной вставки в целевой геномный локус приводит к одному или более из:
(I) добавления экзогенной последовательности в целевом геномном локусе;
(II) делеции эндогенной последовательности в целевом геномном локусе, необязательно причем делеция составляет от приблизительно 5 т.п.н. до приблизительно 10 т.п.н., от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 150 т.п.н., от приблизительно 150 т.п.н. до приблизительно 200 т.п.н., от приблизительно 200 т.п.н. до приблизительно 300 т.п.н., от приблизительно 300 т.п.н. до приблизительно 400 т.п.н., от приблизительно 400 т.п.н. до приблизительно 500 т.п.н., от приблизительно 500 т.п.н. до приблизительно 600 т.п.н., от приблизительно 600 т.п.н. до приблизительно 700 т.п.н. или от приблизительно 700 т.п.н. до приблизительно 800 т.п.н.; или
(III) нокина, нокаута, точечной мутации, перестановки доменов, перестановки экзонов, перестановки интронов, перестановки регуляторных последовательностей, перестановки генов или их комбинации.
20. Способ по любому из пп. 11-19, в котором:
(I) общая длина 5' и 3' гомологичных плеч первого LTVEC, второго LTVEC или третьего LTVEC составляет от приблизительно 10 т.п.н. до приблизительно 20 т.п.н., от приблизительно 20 т.п.н. до приблизительно 40 т.п.н., от приблизительно 40 т.п.н. до приблизительно 60 т.п.н., от приблизительно 60 т.п.н. до приблизительно 80 т.п.н., от приблизительно 80 т.п.н. до приблизительно 100 т.п.н., от приблизительно 100 т.п.н. до приблизительно 120 т.п.н. или от приблизительно 120 т.п.н. до приблизительно 150 т.п.н.; и/или
(II) первый LTVEC составляет по меньшей мере 50 т.п.н., второй LTVEC составляет по меньшей мере 50 т.п.н., и третий LTVEC составляет по меньшей мере 50 т.п.н., необязательно причем первый LTVEC составляет по меньшей мере 100 т.п.н., второй LTVEC составляет по меньшей мере 100 т.п.н., и третий LTVEC составляет по меньшей мере 100 т.п.н.
21. Способ по любому из пп. 1-20, в котором нуклеазный агент представляет собой нуклеазу с «цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), или мегануклеазу.
22. Способ по любому из пп. 1-21, в котором нуклеазный агент представляет собой белок (Cas), ассоциированный с короткими палиндромными повторами, регулярно расположенными группами (CRISPR), и гидовую РНК (гРНК), необязательно причем белок Cas представляет собой Cas9.
23. Способ по любому из пп. 1-22, в котором клетка представляет собой:
(I) человеческую клетку;
(II) нечеловеческую клетку;
(III) клетку грызуна, необязательно причем грызун представляет собой мышь или крысу;
(IV) плюрипотентную клетку, необязательно причем плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку или индуцированную плюрипотентную стволовую (ИПС) клетку;
(V) гемопоэтическую стволовую клетку;
(VI) нейрональную стволовую клетку; или
(VII) фибробластную клетку.
24. Способ по п. 23, в котором клетка представляет собой мышиную ЭС-клетку или крысиную ЭС-клетку, необязательно дополнительно включающий введение мышиной ЭС-клетки или крысиной ЭС-клетки в эмбрион-хозяин.
25. Способ по п. 24, дополнительно включающий вынашивание эмбриона-хозяина в теле суррогатной матери, при этом суррогатная мать производит поколение F0 не относящегося к человеку животного, содержащего модифицированный целевой геномный локус.
| Jallepalli P | |||
| V | |||
| et al | |||
| Securin is required for chromosomal stability in human cells //Cell | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| - Vol | |||
| Транспортер для перевозки товарных вагонов по трамвайным путям | 1919 |
|
SU105A1 |
| - n | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| - pp | |||
| ПРИСПОСОБЛЕНИЕ ДЛЯ АВТОМАТИЧЕСКОЙ БОКОВОЙ СТАБИЛИЗАЦИИ | 1921 |
|
SU445A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Kuijpers N | |||
| G | |||
| A | |||
| et al | |||
| One-step assembly and targeted integration of multigene constructs assisted by the I-SceI meganuclease in Saccharomyces cerevisiae //FEMS yeast research | |||
| - | |||

