ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящее изобретение заявляет преимущество приоритета предварительной заявки на патент США 61/980506 "Способы, наборы и устройство для размножения популяции клеток" ("Methods, Kits And Apparatus For Expanding A Population Of Cells"), поданной в Ведомство США по патентам и товарным знакам 16 апреля 2014 г., содержание которой включено в данный документ с помощью ссылки во всей своей полноте во всех отношениях.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0002] Настоящее изобретение относится к размножению (пролиферации) популяции клеток, такой как популяция лимфоцитов. Настоящее изобретение в целом обеспечивает новые способы и реагенты для размножения (пролиферации) популяций клеток, которым требуется связывание рецептор-связывающей молекулы (такой как первое средство, описанное в данном документе) с рецепторной молекулой на поверхности клетки, посредством чего происходит передача первичного активационного сигнала клеткам. В настоящем изобретении используется реагент для мультимеризации, на котором иммобилизовано (с которым связано) первое средство, которое передает первичный активационный сигнал клеткам. Такой первичный активационный сигнал может сам по себе быть достаточным, чтобы активировать клетки для размножения/пролиферации. Данное первое средство может быть связано либо обратимо, либо также необратимо с реагентом для мультимеризации. На реагенте для мультимеризации может быть иммобилизовано (с ним может быть связано) также второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток. Второе средство при связывании со вспомогательной молекулой на поверхности клеток может тем самым стимулировать активированные клетки к размножению. Также данное второе средство может быть связано либо обратимо, либо также необратимо с реагентом для мультимеризации. Реагент для мультимеризации может быть иммобилизован на твердой либо растворимой подложке. В одном аспекте способ, раскрытый в данном документе, представляет собой серийное размножение популяции клеток, при котором полную популяцию лимфоцитов стимулируют/размножают, реагенты, необходимые для размножения, затем удаляют с помощью хроматографии в подходящей неподвижной фазе, а размножаемые/стимулированные клетки необязательно трансфицируют, например T-клеточным рецептором или химерным антигенным рецептором (CAR), и подвергают второй стимуляции размножения с помощью другой стимулирующей молекулы, которая связывается с введенным T-клеточным рецептором или химерным антигенным рецептором. Настоящее изобретение относится также к устройству для размножения выбранной популяции клеток.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[0003] Разработка методик размножения популяций T-клеток in vitro была ключевой для многих достижений в понимании распознавания T-клетками антигена и активации T-клеток. Разработка способов культивирования для получения клонов антиген-специфических T-клеток человека была применимой в определении антигенов, экспрессируемых патогенами, и опухолей, которые распознаются T-клетками, для создания способов иммунотерапии для лечения ряда заболеваний человека. Антиген-специфические T-клетки можно размножать in vitro для применения в адоптивной клеточной иммунотерапии или терапии рака, при которых инфузии таких T-клеток проявляют противоопухолевую реактивность у опухоленесущего реципиента. В дополнение, адоптивная иммунотерапия применялась также для лечения вирусных инфекций у индивидуумов с ослабленным иммунитетом.
[0004] Способ размножения T-клеток человека in vitro в отсутствие экзогенного фактора роста и вспомогательных клеток, созданный в последние годы, описан в патенте США 6352694 B1 и Европейском патенте EP 0 700 430 B1. В данных патентах раскрыт способ индукции пролиферации популяции T-клеток in vitro. Способ включает приведение популяции T-клеток в контакт с поверхностью твердой фазы, на которой непосредственно иммобилизованы: (a) первое средство, которое передает первичный активационный сигнал T-клеткам, активируя тем самым T-клетки; и (b) второе средство, которое стимулирует вспомогательную молекулу на поверхности T-клеток, стимулируя тем самым активированные T-клетки. Связывание первого средства и второго средства с T-клетками индуцирует пролиферацию/размножение T-клеток. Предпочтительное первое средство, описанное в патенте США 6352694 B1 и Европейском патенте EP 0 700 430 B1, представляет собой моноклональное антитело к CD3, которое связывается с комплексом TCR/CD3 (TCR = T-клеточный рецептор) и тем самым стимулирует передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках. Предпочтительное второе средство согласно этим двум патентам представляет собой моноклональное антитело к CD28, которое связывается со вспомогательной молекулой CD28, присутствующей на T-клетках. Связывание данного второго средства со вспомогательной молекулой CD28 обеспечивает необходимый дополнительный стимул, необходимый для размножения/пролиферации активированных T-клеток. В то же время коммерчески доступными для размножения T-клеток являются Dynabeads® CD3/CD28 (Invitrogen). Dynabeads® CD3⁄CD28 CTS™ представляют собой однородные суперпарамагнитные стерильные непирогенные гранулы из полистирола размером 4,5 мкм, покрытые смесью аффинно очищенных моноклональных антител к молекулам клеточной поверхности CD3 и CD28 на T-клетках человека.
[0005] Однако такие магнитные гранулы, например, сложно интегрировать в способ размножения клеток в условиях, требуемых для клинических испытаний или терапевтических целей, поскольку перед введением размножаемых T-клеток пациенту следует убедиться в том, что магнитные гранулы полностью удалены. Таким образом, настоящее изобретение направлено на обеспечение альтернативного способа размножения популяций клеток, таких как регуляторные T-клетки или центральные T-клетки памяти, для исследовательских, диагностических и особенно терапевтических целей. В идеале данный новый способ должен подходить также для интеграции в автоматизированный процесс, который можно применять для быстрого и простого размножения требуемой популяции клеток для терапевтических путей применения.
[0006] Такая цель реализуется с помощью объектов из независимых пунктов формулы изобретения, в том числе способов, наборов, систем и устройств, упоминаемых в независимых пунктах формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0007] Настоящее изобретение предусматривает способы, наборы, системы и устройство для размножения in vitro требуемой популяции клеток, имеющих рецепторную молекулу на своей поверхности, которая при связывании с подходящим средством может передавать популяции клеток первичный активационный сигнал, посредством чего происходит активация размножения (пролиферации) популяции клеток. Таким образом, способы по настоящему изобретению применяют также для индуцирования пролиферации популяции клеток.
[0008] Согласно первому аспекту настоящее изобретение предусматривает способ размножения популяции клеток in vitro, включающий приведение образца, содержащего популяцию клеток, в контакт с реагентом для мультимеризации,
где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) первое средство, которое передает первичный активационный сигнал клеткам;
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
где первое средство связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки.
[0009] Согласно второму аспекту настоящее изобретение предусматривает способ размножения популяции клеток in vitro, включающий приведение образца, содержащего популяцию клеток, в контакт с реагентом для мультимеризации,
где реагент для мультимеризации находится в растворимой форме и на нем иммобилизовано (связано с ним) первое средство, которое передает первичный активационный сигнал клеткам;
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для связывания первого средства,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
где первое средство связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки.
[0010] Согласно третьему аспекту настоящее изобретение предусматривает набор реагентов для размножения популяции клеток, при этом набор содержит
(i) реагент для мультимеризации,
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z для обратимого связывания первого средства,
(ii) первое средство, которое связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
(iii) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток,
где второе средство содержит партнера по связыванию C2, где партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C2 и связывающим участком Z2,
где второе средство связывается со вспомогательной молекулой на поверхности клеток, стимулируя тем самым активированные клетки.
[0011] Согласно четвертому аспекту настоящее изобретение предусматривает набор реагентов для размножения популяции клеток, при этом набор содержит
(i) реагент для мультимеризации,
где реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z для обратимого связывания первого средства,
(ii) первое средство, которое связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1.
[0012] Согласно пятому аспекту настоящее изобретение предусматривает способ серийного размножения популяции лимфоцитов in vitro, где популяция лимфоцитов содержит T-клетки, при этом способ включает:
приведение образца, который содержит популяцию лимфоцитов, содержащую
T-клетки, в контакт с реагентом для мультимеризации,
где реагент для мультимеризации находится в растворимой форме и на нем обратимо иммобилизовано (i) первое средство, которое передает первичный активационный сигнал T-клеткам, и (ii) второе средство, которое стимулирует вспомогательную молекулу на поверхности T-клеток,
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1,
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z2 для обратимого связывания второго средства,
где второе средство содержит по меньшей мере одного партнера по связыванию C2, причем партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C2 и связывающим участком Z2,
где первое средство связывается с рецепторной молекулой на поверхности T-клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым T-клетки,
где второе средство связывается со вспомогательной молекулой на поверхности T-клеток, стимулируя тем самым активированные клетки, и при этом первое средство и второе средство тем самым совместно индуцируют размножение T-клеток.
[0013] Согласно шестому аспекту настоящее изобретение предусматривает систему из биореактора и неподвижной фазы для хроматографии,
где биореактор является подходящим для размножения клеток,
где неподвижная фаза является подходящей для разделения клеток и удаления реагентов, при этом неподвижная фаза представляет собой матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии, где матрица для гель-фильтрации и/или аффинной хроматографии содержит аффинный реагент, где аффинный реагент содержит связывающий участок Z1, с которым специфически связывается партнер по связыванию C1, содержащийся в первом средстве, и/или аффинный реагент содержит связывающий участок Z2, с которым специфически связывается партнер по связыванию C2, содержащийся во втором средстве, и тем самым подходит для иммобилизации на неподвижной фазе первого средства и/или второго средства, первого партнера по связыванию C1 и/или второго партнера по связыванию C2 в свободной форме,
где биореактор и неподвижная фаза находятся в жидкостном соединении.
[0014] Согласно седьмому аспекту настоящее изобретение предусматривает устройство для очистки и размножения популяции клеток, при этом устройство содержит по меньшей мере одну систему из биореактора и неподвижной фазы для хроматографии согласно шестому аспекту.
[0015] Согласно восьмому аспекту настоящее изобретение предусматривает реагент для мультимеризации, способный к обеспечению размножения популяции клеток,
где реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства, которое передает первичный активационный сигнал клеткам,
где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное первое средство, которое передает первичный активационный сигнал клеткам;
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться по меньшей мере с одним связывающим участком Z1 реагента для мультимеризации,
где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1.
[0016] Согласно девятому аспекту настоящее изобретение предусматривает композицию, способную к обеспечению размножения популяции клеток, при этом композиция содержит:
(i) первый реагент для мультимеризации,
где первый реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства, которое передает первичный активационный сигнал клеткам,
где на первом реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное первое средство, которое передает первичный активационный сигнал клеткам;
где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться по меньшей мере с одним связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
(ii) второй реагент для мультимеризации,
где второй реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z2 для обратимого связывания второго средства, которое стимулирует вспомогательную молекулу на поверхности клеток,
где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток,
где второе средство содержит партнера по связыванию C2, где партнер по связыванию C2 способен связываться по меньшей мере с одним связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C2 и связывающим участком Z2.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0017] Настоящее изобретение будет более понятным при ссылке на подробное описание, рассматриваемое во взаимосвязи с неограничивающими примерами и сопровождающими графическими материалами. На фигурах проиллюстрированы варианты осуществления способов по настоящему изобретению. Без ограничения какой-либо теорией, на фигурах содержатся выводы в отношении механизма, лежащего в основе размножения. Выводы приведены только в иллюстративных целях и служат лишь для того, чтобы обеспечить возможность достижения визуализации способа размножения на молекулярном уровне.
[0018] На фигуре 1 изображен вариант осуществления способа размножения in vitro популяции клеток, которые имеют рецептор клеточной поверхности, при связывании которого первым средством может происходить передача активационного сигнала клеткам к размножению.
[0019] Как показано на фиг. 1a, образец, который содержит популяцию клеток (2), несущих поверхностную рецепторную молекулу (30), приводят в контакт с реагентом для мультимеризации (4). Популяция клеток (2) находится в смеси с другими популяциями клеток (22), не имеющих поверхностной рецепторной молекулы (30). На реагенте для мультимеризации (4) обратимо иммобилизовано (связано с ним) первое средство (6), которое передает первичный активационный сигнал клеткам. Реагент для мультимеризации (4) содержит по меньшей мере один связывающий участок Z1 (42) для обратимого связывания первого средства (6), и первое средство (6) содержит по меньшей мере одного партнера по связыванию C1 (6a), где партнер по связыванию C1 (6a) способен обратимо связываться со связывающим участком Z1 (44) реагента для мультимеризации. Таким образом, при иммобилизации первое средство (6) связывается с реагентом для мультимеризации (4) посредством обратимой связи, образующейся между партнером по связыванию C1 (6a) и связывающим участком Z1 (42). В примере, показанном на фиг. 1, реагент для мультимеризации (4) имеет второй связывающий участок Z2 (44), который в данном примере не применяется. Реагент для мультимеризации (4) сам по себе иммобилизован на твердой подложке (10), такой как магнитная гранула, полимерная гранула или поверхность планшета или реактора для культуры клеток. Популяция клеток (2) может представлять собой, например, популяцию лимфоцитов, такую как популяция B-клеток, которые можно активировать посредством рецептора CD40 (см., например, Carpenter et al, Journal of Translational Medicine 2009, 7:93 "Activation of human B cells by the agonist CD40 antibody CP-870,893 and augmentation with simultaneous toll-like receptor 9 stimulation"). В таком случае молекула клеточной поверхности (30) представляет собой CD40, а первый реагент (6) может представлять собой любую молекулу, связывающуюся с CD40, которая передает требуемый активационный сигнал, например, моноклональное антитело CP-870,893 или его связывающий фрагмент антитела, такой как моновалентный Fab-фрагмент. Партнер по связыванию C1 первого средства (6) может представлять собой, например, любой аффинный пептид, слитый или конъюгированный, например с C-концом одной из двух полипептидных цепей (тяжелой или легкой цепи) молекулы антитела. Партнер по связыванию C1 (6a) может представлять собой, например, стрептавидин-связывающий пептид, такой как пептид Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 01), также известный как "Strep-tag®"), описанный например в патенте США 5506121, или стрептавидин-связывающие пептиды, имеющие два или более последовательно расположенных отдельных связывающих модулей, описанные в Международной патентной публикации WO 02/077018 или патенте США 7981632. В случае применения стрептавидин-связывающего пептида в качестве партнера по связыванию C1, реагент для мультимеризации (4) может представлять собой любой мутантный вариант белка стрептавидина, с которым обратимо связывается стрептавидин-связывающий пептид (= первый партнер по связыванию C1 (6a)) посредством его (биотин-) связывающих участков Z1 (42), схематически показанных на фиг. 1. Такой реагент для мультимеризации может представлять собой мутантный вариант белка (аналог) стрептавидина, содержащий аминокислотную последовательность Val44-Thr45-Ala46-Arg47 (SEQ ID NO: 2) в положениях 44-47 последовательности стрептавидина дикого типа, или мутантный вариант белка (аналог) стрептавидина, содержащий аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 (SEQ ID NO: 3) в положениях 44-47 последовательности стрептавидина дикого типа, оба из которых описаны например в патенте США 6103493 и коммерчески доступны под торговой маркой Strep-Tactin®. В примере на фиг. 1 реагент для мультимеризации (4) может дополнительно содержать мультимерные кальмодулин или глутатион-S-трансферазу, оба из которых образуют обратимые связи с кальмодулин-связывающими пептидами или глутатионом. Таким образом, связывающий участок Z2 (44) может быть образован кальмодулином или глутатион-S-трансферазой. Такой белковый конъюгат, например кальмодулина и мутантного варианта белка стрептавидина, можно получать с помощью стандартных методов химии белков, например с помощью применения бифункциональных линкеров.
[0020] Как показано на фиг. 1b, после приведения популяции клеток (2) в контакт с реагентом для мультимеризации (4) и, как правило, инкубирования популяции клеток с реагентом для мультимеризации (4), популяция клеток (2) образует комплексы/связывается со средством для мультимеризации при помощи первого средства (6). Первое средство специфически связывается с рецепторной молекулой клеточной поверхности, такой как CD40 в данном примере, и передает активационный сигнал к размножению клеток, например B-клеток. Другие популяции клеток (22), содержащиеся в исходном образце, которые не имеют специфической молекулы клеточной поверхности (30), не связываются с реагентом для мультимеризации. В данном отношении следует отметить, что клетки в популяции (2) обычно имеют несколько копий молекулы клеточной поверхности (30) на своей поверхности, и для активации обычно необходимо связывание с этими несколькими копиями. Таким образом, реагент для мультимеризации (4) обычно предоставляет более одного связывающего участка Z1 так, чтобы несколько первых средств (6) могли обратимо связываться для достижения "мультимеризации" первого средства, что означает предоставление первого средства в достаточной плотности к популяции клеток (2) (на схеме на фиг. 1 не показано). В этом отношении следует отметить, что средство для мультимеризации, применяемое в данном документе, как таковое может иметь несколько связывающих участков Z1, например, мутантный вариант белка стрептавидина (являющийся гомотетрамером) в своем нативном состоянии имеет четыре таких связывающих участка Z1. Однако также возможно, чтобы в основе реагента для мультимеризации лежало соединение, как таковое имеющее только один связывающий участок Z1 для обратимого связывания партнера по связыванию C1. Таким примером является мультимерный кальмодулин. Кальмодулин как таковой имеет только один связывающий участок для кальмодулин-связывающих пептидов. Однако кальмодулин можно биотинилировать и затем подвергать реакции с олигомерами стрептавидина (см. также ниже) с получением тем самым реагента для мультимеризации, в котором несколько молекул кальмодулина представлены при достаточной плотности на "каркасе", образуя тем самым мультимерный кальмодулин.
[0021] Как показано на фиг. 1c, после инкубирования (которое обычно осуществляют в течение периода времени, подходящего для достижения размножения требуемой популяции клеток) связывание между партнером по связыванию C1 (6a) первого средства (6) и связывающим участком Z1 реагента для мультимеризации (4) нарушают с помощью разрушения соответствующей обратимой связи. Разрушение может быть достигнуто с помощью добавления конкурирующего средства в инкубационную/реакционную смесь, содержащую популяцию клеток (2), связанных с реагентом для мультимеризации. Для конкурентного разрушения (которое можно понимать как являющееся конкурентным элюированием) обратимой связи между партнером по связыванию C1 (6a) первого средства и связывающим участком Z1 (22) реагента для мультимеризации, инкубационную смесь/популяцию клеток можно приводить в контакт с первым партнером по связыванию C1 в свободной форме (20) или аналогом указанного первого партнера по связыванию C, способными к разрушению связи между первым партнером по связыванию C1 (6a) и связывающим участком Z1 (22). В примере, в котором партнер по связыванию C1 представляет собой стрептавидин-связывающий пептид, который связывается с биотин-связывающим участком стрептавидина, первый партнер C1 в свободной форме (20) может представлять собой соответствующий стрептавидин-связывающий пептид в свободной форме или его аналог, осуществляющий конкурентное связывание. Такой аналог может представлять собой, например, биотин или производное биотина, такое как дестиобиотин.
[0022] Как показано на фиг. 1d, добавление первого партнера в свободной форме (20) или его аналога приводит к оттеснению партнера по связыванию C1 (6a) от реагента для мультимеризации (4), и таким образом, поскольку партнер по связыванию C1 содержится в первом средстве (6), к оттеснению первого средства (6) от реагента для мультимеризации (4). Такое оттеснение первого средства (6), в свою очередь, приводит к диссоциации первого средства (6) из связи с рецептором клеточной поверхности (30), в частности, если аффинность связывания для связи между первым средством и рецептором клеточной поверхности (30), которая таким образом также является обратимой, характеризуется константой диссоциации (Kd) в диапазоне от 10-2 M до 10-13 M. Вследствие такой диссоциации стимуляция популяции клеток (2) также прекращается. Таким образом, настоящее изобретение дает преимущество в том, что период времени стимуляции или размножения популяции клеток можно точно контролировать, и таким образом можно строго контролировать также функциональный статус популяции клеток. В данном случае следует отметить, что аффинность связывания молекул антитела с их антигеном, в том числе например с рецепторной молекулой клеточной поверхности, такой как CD40 в данном примере, обычно находится в диапазоне значений аффинности Kd от 10-7 M до 10-13 M. Таким образом, в настоящем изобретении в качестве первого средства (а также, разумеется, в качестве второго средства, как поясняется ниже) можно применять стандартные моноклональные антитела. Во избежание каких-либо нежелательных эффектов авидности, которые приводят к более сильному связыванию, моноклональные антитела также можно применять в форме их моновалентных фрагментов антител, таких как Fab-фрагменты или одноцепочечные Fv-фрагменты.
[0023] В дополнение, вследствие диссоциации первого средства из связи с молекулой клеточной поверхности (30) настоящее изобретение имеет дополнительное преимущество в том, что стимулированная популяция клеток не содержит стимуляторов в конце периода стимуляции, и в том, что все другие применяемые в способе реагенты, а именно первое средство (6), а также первый партнер в свободной форме (20) партнера по связыванию C1 или его аналог, можно легко удалять из стимулированной популяции клеток (2) с помощью "картриджа для удаления", описанного в Международной заявке на патент WO 2013/124474, тогда как реагент для мультимеризации (4), иммобилизованный на твердой подложке, такой как поверхность биореактора или магнитная гранула, остается там. Таким образом, возвращаясь к удалению средства в свободной форме (6) и первого партнера в свободной форме (20), в соответствии с описанием "картриджа для удаления" в WO 2013/124474 (см., например, со ссылкой на фиг. 4 в ней), элюированный образец, полученный на фиг. 1d данного документа, можно загружать во вторую хроматографическую колонку согласно WO 2013/124474. Данная хроматографическая колонка имеет подходящую неподвижную фазу, которая как является матрицей для аффинной хроматографии, так и в то же время может выступать в качестве матрицы для гель-проникающей хроматографии. На такой матрице для аффинной хроматографии иммобилизован аффинный реагент. В случае данного примера аффинный реагент может представлять собой, например, стрептавидин, мутантный вариант белка стрептавидина, авидин, мутантный вариант белка авидина или их смесь. Первое средство (6), первый партнер в свободной форме (20) партнера по связыванию C1 (также называемый в данном документе "конкурирующим реагентом") связываются с аффинным реагентом, иммобилизуясь тем самым на матрице для хроматографии. В результате элюированный образец, содержащий выделенную и размножаемую популяцию клеток (2), обедняется первым средством (6) и конкурирующим реагентом (20). Размножаемая популяция клеток (2), освобожденная от каких-либо реагирующих веществ, теперь находится в состоянии для дальнейшего применения, например для диагностических применений (например, дальнейшей сортировки FACSTM) или для любого терапевтического применения с использованием клеток.
[0024] На фиг. 2 показан дополнительный вариант осуществления способа размножения по настоящему изобретению. Как показано на фиг. 2a, образец содержит популяцию клеток (2), несущих две специфические молекулы клеточной поверхности (30) и (32). Молекула клеточной поверхности (30) участвует в передаче первичного активационного сигнала популяции клеток, тогда как молекула клеточной поверхности (32) представляет собой вспомогательную молекулу на клеточной поверхности, участвующую в передаче стимула клеткам. Популяция клеток может представлять собой, например, популяцию T-клеток, в которой молекула клеточной поверхности (30) представляет собой комплекс TCR/CD3, а молекула клеточной поверхности (32) представляет собой вспомогательную молекулу CD28. Связывание как с комплексом TCR/CD3 в качестве источника первичного активационного сигнала, так и с CD28 в качестве костимулятора необходимо для размножения/пролиферации T-клеток. Популяция T-клеток (2) находится в смеси с другими популяциями клеток (22), не имеющих поверхностных рецепторных молекул (30) и (32). Также в данном варианте осуществления популяцию клеток (2) приводят в контакт с реагентом для мультимеризации (4). На реагенте для мультимеризации (4) обратимо иммобилизовано (связано с ним) первое средство (6), которое передает первичный активационный сигнал клеткам. В дополнение, на средстве для мультимеризации обратимо иммобилизовано (связано с ним) второе средство (8), которое стимулирует CD28 в качестве вспомогательной молекулы на поверхности клеток.
[0025] Реагент для мультимеризации (4) содержит по меньшей мере один связывающий участок Z1 (42) для обратимого связывания первого средства (6), и первое средство (6) содержит по меньшей мере одного партнера по связыванию C1 (6a), где партнер по связыванию C1 (6a) способен обратимо связываться со связывающим участком Z1 (44) реагента для мультимеризации. Таким образом, при иммобилизации первое средство (6) связывается с реагентом для мультимеризации (4) посредством обратимой связи, образующейся между партнером по связыванию C1 (6a) и связывающим участком Z1 (42). В дополнение, в примере, проиллюстрированном на фиг. 2, второе средство (8) содержит партнера по связыванию C2 (8a), где партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 (44) реагента для мультимеризации (4). Второе средство (8) связывается с реагентом для мультимеризации (4) посредством обратимой связи, образующейся между партнером по связыванию C2 (8a) и связывающим участком Z2 (44). В данном примере первое средство (6) может представлять собой моноклональное антитело к CD3 или его антигенсвязывающий фрагмент, такой как Fab-фрагмент. Второе средство (8) может представлять собой моноклональное антитело к CD28 или его антигенсвязывающий фрагмент, такой как Fab-фрагмент. Первый партнер по связыванию (6a) может представлять собой стрептавидин-связывающий пептид (6a), слитый или конъюгированный с антителом к CD3 или фрагментом антитела к CD3. Второй партнер по связыванию (8a) может представлять собой кальмодулин-связывающий пептид, также конъюгированный или слитый с антителом к CD28 или фрагментом антитела, связывающегося с CD28. В данном контексте следует отметить, что моноклональные антитела, например к CD3 или CD28, хорошо известны (см., например, патент США 6352694 B или Европейский патент EP 0 700 430 B1, обсуждаемые выше) и коммерчески доступны от многочисленных поставщиков, таких как Santa Cruz Biotechnology (Санта-Круз, Калифорния, США), Life Technologies (Карлсбад, Калифорния, США), BD Biosciences (Сан-Хосе, Калифорния, США), Biolegend (Сан-Диего, Калифорния, США) или Miltenyi Biotec (Бергиш-Гладбах, Германия), не говоря уже о других. Соответственно, такие моноклональные антитела могут применяться в качестве первого и второго средства и могут, например, быть химически связанными (конъюгированными) с партнером по связыванию C1 или C2. В качестве альтернативы также возможно клонировать гены вариабельных доменов из линии гибридомных клеток либо применять антитело, аминокислотная последовательность которого известна, и получать рекомбинантным путем соответствующий фрагмент антитела, такой как Fab-фрагмент или Fv. При использовании такого подхода, который описан в данном документе в разделе "Примеры", как для линии гибридомных клеток OKT3 (ATCC® CRL-8001™, описанной в патенте США 4361549), которая вырабатывает моноклональное антитело к CD3, так и для антитела к CD28 28.3, описанного в Vanhove et al, BLOOD, 15 июля 2003 г., том 102, № 2, стр. 564-570 и имеющего номер доступа в GenBank AF451974.1, партнеры по связыванию C1 и C2 в целях удобства доставляются с помощью соответствующего вектора экспрессии, используемого для рекомбинантного получения, так что фрагмент антитела несет партнера по связыванию C1 или C2 в виде пептида слияния как C-конец либо легкой, либо тяжелой цепи (в данном контексте аминокислотная последовательность вариабельного домена тяжелой цепи и вариабельного домена легкой цепи антитела OKT3, которые описаны в Arakawa et al J. Biochem. 120, 657-662 (1996), показаны в иллюстративных целях как SEQ ID NOS: 17 и 18 в прилагаемом перечне последовательностей, в то время как аминокислотная последовательность вариабельного домена антитела к CD28 28.3, описанного Vanhove et al, выше, показаны как SEQ ID NOS: 19 (VH) и 20 (VL) в прилагаемом перечне последовательностей). Такая методика клонирования вариабельных доменов молекулы антитела и рекомбинантного получения соответствующего фрагмента антитела также хорошо известна специалисту в данной области, см., например, Skerra, A. (1994) A general vector, pASK84, for cloning, bacterial production, and single-step purification of antibody Fab fragments. Gene 141, 79-84, или Skerra, A. (1993) Bacterial expression of immunoglobulin fragments. Curr Opin Immunol. 5, 256-562. Наконец, также возможно получать молекулы антител или искусственные связывающие молекулы с антителоподобными свойствами, направленные на заданную мишень, такую как CD3 или CD28, как в примере на фиг. 2, с помощью хорошо известных эволютивных способов, таких как фаговый дисплей (обзор которого приведен, например, в Kay, B.K. et al. (1996) Phage Display of Peptides and Proteins – A Laboratory Manual, 1st Ed., Academic Press, New York NY; Lowman, H.B. (1997) Annu. Rev. Biophys. Biomol. Struct. 26, 401–424, или Rodi, D.J., and Makowski, L. (1999) Curr. Opin. Biotechnol. 10, 87–93), рибосомный дисплей (обзор которого приведен в Amstutz, P. et al. (2001) Curr. Opin. Biotechnol. 12, 400-405) или мРНК-дисплей, описанный в Wilson, D.S. et al. (2001) Proc. Natl. Acad. Sci. USA 98, 3750-3755.
[0026] В случае примера, показанного на фиг. 2, реагент для мультимеризации (4) имеет два разных связывающих участка Z1 (42) и Z2 (44). Если партнер по связыванию C1 (6a) представляет собой стрептавидин-связывающий пептид, то связывающий участок Z1 (42) реагента для мультимеризации (4) предоставляется подходящим мутантным вариантом белка стрептавидина, с которым обратимо связывается стрептавидин-связывающий пептид (6a). Поскольку связывающий партнер C2 представляет собой кальмодулин-связывающий пептид, то связывающий участок Z2 (44) реагента для мультимеризации (4) предоставляется мультимерным кальмодулином. Реагент для мультимеризации (4) может представлять собой одиночную молекулу, например конъюгат мультимерного кальмодулина и стрептавидина (данный альтернативный вариант обычно будет применяться в случае растворимой формы реагента для мультимеризации), или включать в себя также две независимые молекулы. Последний возможный вариант является предпочтительным, если реагент для мультимеризации (44) иммобилизован на твердой подложке, как показано на фиг. 2. В таком случае смесь мутантного варианта белка стрептавидина и кальмодулина может быть нанесена на твердую подложку (иммобилизована на ней), например в молярном соотношении 1:1 по отношению к связывающим участкам Z1 и Z2. В данном случае следует отметить, что вследствие иммобилизации кальмодулина на поверхности твердой подложки отсутствует необходимость в поясняемом выше получении мультимерного кальмодулина, а иммобилизация кальмодулина на поверхности является достаточной для предоставления кальмодулина (который, как упоминается выше, имеет только один связывающий участок для кальмодулин-связывающих пептидов) в достаточно высокой плотности для обеспечения связывания популяции клеток (2). Например, в данном случае в качестве второго реагента (8) можно применять бивалентный фрагмент антитела, имеющий два связывающих участка для CD28, или интактное антитело, имеющее само по себе два идентичных связывающих участка.
[0027] Как показано на фиг. 2b, после приведения популяции T-клеток (2) в контакт с реагентом для мультимеризации (4) и, как правило, инкубирования популяции клеток с реагентом для мультимеризации (4), популяция T-клеток (2) образует комплексы/связывается со средством для мультимеризации при помощи первого средства (6) и второго средства (8). Первое средство (6) и второе средство (8) специфически связываются с комплексом TCR/CD3 и вспомогательной молекулой CD28, индуцируя тем самым пролиферацию/размножение T-клеток.
[0028] Как показано на фиг. 2c, после инкубирования (которое обычно осуществляют в течение периода времени, подходящего для достижения размножения требуемой популяции клеток) связывание между партнером по связыванию C1 (6a) первого средства (6) и связывающим участком Z1 реагента для мультимеризации (4) нарушают с помощью разрушения соответствующей обратимой связи. Аналогично, связывание между партнером по связыванию C2 (8a) второго средства (8) и связывающим участком Z2 реагента для мультимеризации (4) нарушают с помощью разрушения соответствующей обратимой связи. Обратимую связь между партнером по связыванию C1 (6a) первого средства (6) и связывающим участком Z1 можно разрушать с помощью биотина (который выступает в качестве аналога (20) первого партнера в свободной форме), а обратимую связь между партнером по связыванию C2 (8a) первого средства (8) и связывающим участком Z2 можно разрушать с помощью добавления металлохелатора (кальциевого хелатора), такого как EDTA или EGTA (которые выступают в качестве аналога (20) второго партнера в свободной форме), поскольку связывание кальмодулина с кальмодулин-связывающими пептидами зависит от ионов кальция (Ca2+). Разумеется, это означает, что приведение в контакт популяции клеток (2) осуществляют в Ca2+-содержащем буфере.
[0029] Как показано на фиг. 2d, добавление аналога (20) первого партнера в свободной форме и второго партнера в свободной форме соответственно приводит к оттеснению партнеров по связыванию C1 (6a) и C2 (8a) от реагента для мультимеризации (4) и, таким образом, к оттеснению первого средства (6) и второго средства (8) от реагента для мультимеризации (4). Такое оттеснение первого средства (6) и второго средства (8), в свою очередь, приводит к диссоциации первого средства (6) и второго средства (8) из комплекса TCR/CD3 и вспомогательной молекулы CD28, посредством чего прекращается стимуляция/размножение популяции клеток (2). Таким образом, как сказано выше, настоящее изобретение предусматривает преимущество в том, что период времени стимуляции или размножения популяции T-клеток можно точно контролировать, и следовательно можно строго контролировать также функциональный статус популяции T-клеток. После элюирования клеток, проиллюстрированного на фиг. 1d, первое средство (6), второй реагент (8), а также аналог (20) первого партнера в свободной форме по связыванию C1 и второго партнера в свободной форме по связыванию C2 можно легко удалять из стимулированной популяции клеток (2) с помощью "картриджа для удаления", описанного в Международной заявке на патент WO 2013/124474. В дополнение и, что важно, в случае если исходный образец представлял собой популяцию лимфоцитов, например в виде PMBC, полученных на градиенте фиколла, популяция T-клеток (2) теперь доступна для серийного размножения, определенного в данном документе. Поскольку размножившуюся (например благодаря первоначальной стимуляции с помощью CD3/CD28) популяцию клеток можно трансфицировать в ходе размножения, например, T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR, также известным как искусственный T-клеточный рецептор), генетически модифицированные клетки можно затем избавлять от первоначального стимула и после этого стимулировать стимулом второго типа, например посредством введенного de novo рецептора. Такие стимулы второго типа могут включать в себя антигенный стимул в форме комплекса пептид/молекула MHC, когнатного (перекрестно связывающегося) лиганда генетически введенного рецептора (например природного лиганда CAR) или любого лиганда (такого как антитело), который непосредственно связывается с каркасной областью нового рецептора (например с помощью распознавания константных областей в рецепторе). Таким образом, популяцию T-клеток, полученную в результате такого серийного размножения, можно применять для адоптивного переноса клеток.
[0030] На фиг. 3 показан дополнительный вариант осуществления способа размножения по настоящему изобретению. Образец, применяемый в данном примере, содержит также популяцию T-клеток (2), которые несут две специфические молекулы клеточной поверхности (30) и (32), при этом молекула клеточной поверхности (30) представляет собой комплекс TCR/CD3, а молекула клеточной поверхности (32) представляет собой вспомогательную молекулу CD28. На фиг. 3a популяция T-клеток (2) показана после контакта с реагентом для мультимеризации (4). В данном примере на реагенте для мультимеризации (4) также обратимо иммобилизованы (связаны с ним) в качестве первого средства (6) антитело к CD3 или его антигенсвязывающий фрагмент, которые передают первичный активационный сигнал T-клеткам, и в качестве второго средства (8) антитело к CD28 или его антигенсвязывающий фрагмент, которые стимулируют CD28 в качестве вспомогательной молекулы.
[0031] Реагент для мультимеризации (4), показанный в примере на фиг. 3, содержит только один тип связывающего участка Z1 (42) для обратимого связывания как первого средства (6), так и второго средства (8). Как первое средство (6), так и второе средство (8) содержат по меньшей мере одного партнера по связыванию C1 (6a, 8a), где как партнер по связыванию C1 (6a), так и партнер по связыванию (8a) способны обратимо связываться со связывающим участком Z1 (44) реагента для мультимеризации. Таким образом, при иммобилизации первое средство (6) и второе средство (8) соответственно связываются с реагентом для мультимеризации (4) посредством обратимой связи, образующейся между партнером по связыванию C1 (6a) и партнером по связыванию C2, а также связывающим участком Z1 (42). Партнеры по связыванию C1 и C2 могут быть различными либо идентичными. Например, партнер по связыванию C1 может представлять собой стрептавидин-связывающий пептид с последовательностью Trp-Ser-His-Pro-Gln-Phe-Glu-Lys ((SEQ ID NO: 01), "Strep-tag®"), тогда как партнер по связыванию C2 может представлять собой стрептавидин-связывающий пептид с последовательностью Trp-Ser-His-Pro-Gln-Phe-Glu-Lys-(GlyGlyGlySer)3-Trp-Ser-His-Pro-Gln-Phe-Glu-Lys ((SEQ ID NO: 04), также известной как "двойная метка-3")) или последовательностью Trp-Ser-His-Pro-Gln-Phe-Glu-Lys-(GlyGlyGlySer)2-Trp-Ser-His-Pro-Gln-Phe-Glu-Lys ((SEQ ID NO: 05), также известной как "двойная метка-2"), описанный в Junttila et al., Proteomics 5 (2005), 1199-1203 или патенте США 7981632. Все эти стрептавидин-связывающие пептиды связываются с одним и тем же связывающим участком, а именно биотин-связывающим участком стрептавидина. Если один или более таких стрептавидин-связывающих пептидов применяют в качестве партнеров по связыванию C1 и C2, то реагент для мультимеризации (4) представляет собой мутантный вариант белка стрептавидина. Как показано на фиг. 3, применяют растворимый реагент для мультимеризации (4). В случае мутантного варианта белка стрептавидина такой растворимый реагент для мультимеризации может представлять собой, например, олигомер или полимер стрептавидина или авидина, или любой мутантный вариант белка (аналог) стрептавидина или авидина. Олигомер может содержать три или более мономеров стрептавидина, авидина или мутантных вариантов данных белков. Олигомер или полимер может быть сшит при помощи полисахарида. Такие олигомеры или полимеры стрептавидина или авидина, или мутантных вариантов белков стрептавидина или авидина, можно на первой стадии получать с помощью введения карбоксильных остатков в полисахарид, например декстран, как по существу описано в "Noguchi, A., Takahashi, T., Yamaguchi, T., Kitamura, K., Takakura, Y., Hashida, M. & Sezaki, H. (1992). Preparation and properties of the immunoconjugate composed of anti-human colon cancer monoclonal antibody and mitomycin C dextran conjugate. Bioconjugate Chemistry 3, 132-137". На второй стадии стрептавидин или авидин, или мутантные варианты данных белков, связывают посредством первичных аминогрупп внутреннего лизинового остатка и/или свободного N-конца с карбоксильными группами в декстрановом остове с помощью традиционных карбодиимидных химических методов. В качестве альтернативы сшитые олигомеры или полимеры стрептавидина или авидина, или любых мутантных вариантов белков стрептавидина или авидина, можно получать также с помощью сшивания бифункциональными линкерами, такими как глутаровый альдегид, или с помощью других способов, описанных в литературе.
[0032] Применение в качестве партнеров по связыванию C1 и C2, компонентов, которые связываются с одним и тем же связывающим участком (42) средства для мультимеризации, имеет преимущество в том, что как показано на фиг. 3b, одного и того же партнера в свободной форме (первого партнера по связыванию C1, а также второго партнера по связыванию C2) или его аналог можно применять для прекращения размножения популяции T-клеток (2) и для удаления средства для мультимеризации из такой популяции T-клеток (2). В примере на фиг. 3 аналог первого и второго партнеров C1 и C2, такой как биотин или производное биотина (иминобиотин или дестиобиотин) можно в целях удобства применять для прекращения размножения и элюирования популяции T-клеток (2).
[0033] Как показано на фиг. 3c, после элюирования клеток, проиллюстрированного на фиг. 1d, первое средство (6), второй реагент (8), а также биотин в качестве аналога (20) первого партнера в свободной форме по связыванию C1 и второго партнера в свободной форме по связыванию C2 можно легко удалять из стимулированной популяции клеток (2) с помощью "картриджа для удаления", описанного в Международной заявке на патент WO 2013/124474. В дополнение, вариант осуществления с применением растворимого реагента для мультимеризации (4) имеет дополнительное преимущество в том, что в нем можно обойтись без какой-либо твердой подложки, такой как магнитные гранулы. Это означат отсутствие риска загрязнения активированных T-клеток такими магнитными гранулами. Это также означает, что способ, соответствующий стандартам GMP, может быть намного более простым в осуществлении по сравнению с известным способом, таким как применение Dynabeads®, в котором следует предпринимать дополнительные меры, чтобы убедиться в том, что конечная размножившаяся популяция T-клеток не содержит магнитных гранул. Кроме того, применение растворимого средства для мультимеризации делает намного более простым его удаление из активированной популяции клеток (T-клеток, B-клеток или также природных клеток-киллеров), поскольку клетки можно легко осаждать центрифугированием, а надосадочную жидкость, содержащую растворимое средство для мультимеризации, можно сливать. В качестве альтернативы растворимое средство для мультимеризации можно удалять из размножившейся популяции клеток в матрице для гель-проникающей хроматографии картриджа для удаления согласно Международной заявке на патент WO 2013/124474. Поскольку твердая фаза (например магнитные гранулы) отсутствует, настоящее изобретение также обеспечивает автоматическую закрытую систему для размножения клеток, которую можно интегрировать в известные системы для размножения клеток, такие как система для размножения клеток Xuri W25 и система WAVE Bioreactor 2/10, доступные от GE Healthcare (Литл-Чалфонт, Бакингемшир, Великобритания), или система для размножения клеток Quantum®, доступная от Terumo BCT, Inc. (Лейквуд, Колорадо, США).
[0034] На фиг. 4 показаны результаты эксперимента, в котором CD3+ иммунореактивные T-клетки подвергали пролиферации после стимуляции in vitro Fab-фрагментами αCD3 и αCD28, которые были обратимо иммобилизованы на гранулах, покрытых мутантным вариантом белка стрептавидина Strep-Tactin®. Фиг. 4A представляет собой гистограмму, на которой показано распределение стимулированных клеток по размерам (прямое рассеяние), на фиг. 4B изображены гистограммы, на которых представлена степень пролиферации соответственно количеству клеток на число клеточных делений, указанное в верхней части фиг. 4B (0 означает неделившиеся клетки; 5 означает клетки, прошедшие по меньшей мере 5 делений), и на фиг. 4C показано изображение чашки для культивирования через 4 дня стимуляции.
[0035] На фиг. 5 показаны результаты дифференциальной мобилизации внутриклеточного кальция в клетках Jurkat, которые мечены либо антителом αCD3 OKT3, либо Fab-фрагментами OKT3, мультимеризованными при помощи Strep-Tactin® (также называемыми в данном документе Fab-мультимерами). Фиг. 5A: клетки Jurkat нагружали кальций-чувствительным красителем Indo-1-AM и высвобождение кальция запускали с помощью инъекции либо mAb αCD3 (черные квадраты), либо Fab-мультимеров αCD3 OKT3 (полученных из исходной линии клеток OKT3) с предварительным нарушением D-биотином или без него (соответственно темно-серые треугольники и светло-серые кружки), в сравнении с инъекцией PBS (перевернутые белые треугольники). Применение иономицина служило в качестве положительного контроля. Изменения внутриклеточной концентрации Ca2+ с временным разрешением отслеживали с помощью проточной цитометрии по изменению соотношения FL6/FL7. Фиг. 5B: клетки Jurkat, меченные Indo-1-AM, активировали различными стимулами αCD3, описанными на фиг. 4a (OKT3: верхний график и Fab-мультимер αCD3: средний график), с последующим дальнейшим (t=140 с) нарушением передачи сигналов от Fab-мультимера αCD3, опосредованным D-биотином. Инъекция PBS (нижний график) и иономицин служили в качестве отрицательного или положительного контроля. Данные представляют три различных эксперимента.
[0036] На фиг. 6 показан результат обратимого окрашивания клеток Fab-мультимерами антитела к CD3 OKT3. Свежевыделенные PBMC окрашивали моноклональным антителом (левая точечная диаграмма, исходный клон для Fab-мультимеры) либо когнатными меченными PE Fab-мультимерами и анализировали до (второй столбец слева) либо после обработки D-биотином (средний столбец). Затем после последующих этапов промывания выявляли оставшиеся Fab-мономеры с помощью свежеприготовленного меченного PE Strep-Tactin® (второй столбец справа). Вторичное окрашивание Fab-мультимером обратимо окрашенных клеток служило в качестве контроля (правый столбец). Показаны только живые (PIотрицательные) клетки. Числа на точечных диаграммах указывают на процентную долю клеток внутри окон.
[0037] На фиг. 7 показано выделение клеток с помощью обратимого связывания с Fab-фрагментами антител к CD28, мультимеризованными с помощью Strep-Tactin®, меченного фикоэритрином в качестве флуоресцентной метки. Клетки CD28+ отбирали/выделяли с помощью магнитного отбора клеток с помощью Fab-мультимера из свежевыделенных PMBC согласно описанному в Международной заявке на патент WO 2013/011011. Перед отбором клетки подвергали контрольному окрашиванию либо когнатными флуоресцентными αCD28-мультимерами (левая точечная диаграмма) либо антителом, направленным на легкую каппа-цепь иммуноглобулина (вторая слева точечная диаграмма, mAb α-Ig-каппа). После отбора клетки обрабатывали D-биотином и после этого промывали для удаления магнитных гранул и Fab-мономеров. После этого отделенные клетки CD28+ (повторно) окрашивали либо Fab-мультимерами CD28 (вторая справа точечная диаграмма), либо mAb α-Ig-каппа (правая точечная диаграмма) для выявления возможных оставшихся Fab-мономеров. Показаны только живые (PIотрицательные) CD3+ клетки. Числа на точечных диаграммах указывают на процентную долю клеток в окнах.
[0038] На фиг. 8 показаны результаты эксперимента, в котором CD3+ иммунореактивные T-клетки подвергали пролиферации после стимуляции in vitro обратимыми Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном Strep-Tactin®, выступающем в качестве растворимого реагента для мультимеризации. Для экспериментов, результаты которых показаны на фиг. 8, 300000 CD3+ иммунореактивных T-клеток (Tresp) метили 2 мкМ сукцинимидилового сложного эфира карбоксифлуоресцеина (CFSE) и стимулировали различными количествами препарата растворимых олигомеров Strep-Tactin, на которых была иммобилизована комбинация Fab-фрагмента αCD3 и Fab αCD28, оба из которых несли на тяжелой цепи Strep-tag в качестве стрептавидин-связывающего пептида. ("1x" соответствует 3 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28; числа указывают на величину кратности "1x"). Клетки Tresp оставляли либо нестимулированными, либо стимулировали холостыми мультимерами Strep-Tactin (без Fab), служившими в качестве отрицательного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты вместе с 300000 CD3-отрицательных аутологичных питающих клеток (облученных 30 Гр) в 1 мл среды для культивирования клеток, дополненной 20 ед/мл интерлейкина-2 (IL-2). Клетки инкубировали при 37°C без замены среды, и пролиферацию анализировали по разведению CFSE через 5 дней с помощью FACS-анализа (фиг. 8B). На фиг. 8A показано распределение клеток по размеру в культуре через 5 дней. На гистограммах показаны живые CD3+ клетки, а на фиг. 8C показаны клетки после культивирования, которые отделяли стимулирующими реагентами после обработки при помощи 1 мМ D-биотина и промывания. Диссоциацию и удаление мономеров Fab-фрагментов анализировали повторным окрашиванием при помощи Strep-Tactin®, меченного фикоэритрином (ST-PE) в качестве флуоресцентной метки, и иллюстративная гистограмма показана. На фиг. 8D показано абсолютное количество живых (отрицательных по трипановому синему) клеток через 5 дней, которое подсчитывали с помощью счетной камеры Нойбауэра, и откладывали на графике в зависимости от соответствующего условия стимуляции. На фиг. 8D показаны медианные значения количества клеток; при этом планки погрешностей указывают на стандартное отклонение (SD). На фиг. 8E показано изображение чашки для культивирования через 5 дней стимуляции.
[0039] На фиг. 9 изображена иллюстрация способа серийного размножения по настоящему изобретению (фиг. 9a), а на фиг. 9b вкратце описаны некоторые характерные особенности и преимущества серийного размножения.
[0040] На фиг. 10 показана система по настоящему изобретению, которую можно применять в сочетании со способами размножения по настоящему изобретению. Данная система (100) включает биореактор (50), первый "картридж для удаления" (70) и второй "картридж для удаления" (90). Биореактор (50) находится в жидкостном соединении с первым сменным картриджем (70), а первый картридж для удаления находится в жидкостном соединении со вторым картриджем для удаления (90). Такая система (100) может быть частью устройства для автоматизированного размножения и очистки клеток, описанных в данном документе.
[0041] В биореакторе (50) осуществляют способ размножения, описанный в данном документе, например способ размножения, проиллюстрированный на фиг. 3, в котором применяется растворимый реагент для мультимеризации. В данном случае, после прекращения активации/размножения популяции клеток (2) с помощью добавления конкурирующего средства (20) (партнера в свободной форме по связыванию C1 или его аналога), выходящие из биореактора реакционные смеси содержат размножившуюся популяцию клеток (2), первое средство (6), второе средство (8), а также растворимый реагент для мультимеризации (4). В данном примере первое средство (6) представляет собой фрагмент антитела, связывающегося с CD3, содержащий стрептавидин-связывающий пептид в качестве партнера по связыванию C1, второе средство (8) представляет собой фрагмент антитела, связывающегося с CD28, содержащий стрептавидин-связывающий пептид в качестве партнера по связыванию C1, а конкурирующее средство (20) (аналог партнера по связыванию C1 в свободной форме) представляет собой биотин. Данную реакционную смесь направляют в первый картридж для удаления (70). Этот первый картридж для удаления (70) представляет собой картридж для удаления, описанный в Международной заявке на патент WO 2013/124474, содержащий хроматографическую колонку с подходящей неподвижной фазой. Неподвижная фаза может служить как в качестве матрицы для аффинной хроматографии, так и может одновременно выступать в качестве матрицы для гель-проникающей хроматографии. На такой матрице для аффинной хроматографии иммобилизован аффинный реагент. Аффинный реагент может, в случае данного примера, представлять собой, например, стрептавидин, мутантный вариант белка стрептавидина, авидин, мутантный вариант белка авидина или их смесь. Таким образом, первое средство (6) и второе средство (8) связываются с аффинным реагентом посредством их стрептавидин-связывающего пептида. Биотин в качестве конкурирующего средства (20) также связывается с аффинным реагентом. Таким образом, все три данных реагента иммобилизованы на матрице для хроматографии первого картриджа для удаления, а размножаемая популяция клеток (2) и растворимый реагент для мультимеризации (4) проходят через неподвижную фазу. Этот "непрерывный поток" затем направляют во второй картридж для удаления (90). Данный второй картридж для удаления (90) также содержит неподвижную фазу. Эта неподвижная фаза содержит на себе второй аффинный реагент, способный связываться со связывающим участком Z1 (42) реагента для мультимеризации (4). Данный аффинный реагент может представлять собой, например, биотин, ковалентно связанный с неподвижной фазой. Такая неподвижная фаза может представлять собой, например, D-биотин SepharoseTM, которую можно получить от Affiland S.A. (Анc, Льеж, Бельгия). Таким образом, растворимый реагент для мультимеризации (4) связывается (задерживается) неподвижной фазой второго картриджа для удаления (90), а размножаемая популяция клеток (2) проходит через неподвижную фазу и освобождается от каких-либо реагирующих веществ. Популяция клеток (2) имеет теперь надлежащее состояние для любого дальнейшего применения, например, для диагностических путей применения (например дальнейшей сортировки FACSTM) или для любого терапевтического применения с использованием клеток. Разумеется, здесь следует отметить, что возможно также изменять порядок расположения первого "картриджа для удаления" (70) и второго "картриджа для удаления" (90) в системе (100) таким образом, чтобы биореактор (50) находился в (непосредственном) жидкостном соединении со вторым сменным картриджем (90), а первый картридж для удаления (70) был расположен позади него и находился в жидкостном соединении со вторым картриджем для удаления (90). В такой системе, из популяции клеток (2) вначале удаляется реагент для мультимеризации (4), а после этого удаляется первое средство (6), второе (8) и, например, конкурирующее средство (20). Такая система также охватывается настоящим изобретением и также может быть частью устройства для автоматизированного размножения и очистки клеток, описанных в данном документе.
[0042] На фиг. 11 показан дополнительный вариант осуществления системы по настоящему изобретению, которую можно применять в сочетании со способами размножения по настоящему изобретению. Данная система (110) включает биореактор (50), первый "картридж для удаления" (70) и второй "картридж для удаления" (90). Биореактор (50) находится в жидкостном соединении с первым сменным картриджем (70), а первый картридж для удаления находится в жидкостном соединении со вторым картриджем для удаления (90). В дополнение, второй картридж для удаления (110) находится в жидкостном соединении с биореактором (50). Такая система (110) также может быть частью устройства для автоматизированного размножения и очистки клеток, описанных в данном документе. При применении, например, в сочетании со способом размножения, в котором используется растворимый реагент для мультимеризации (4), очищенную размножаемую популяцию клеток (2) получают в виде элюата из второго картриджа для удаления (90). Поскольку картридж для удаления (90) находится в жидкостном соединении с биореактором (50), популяцию клеток (2) можно переносить обратно в биореактор (50), например для серийного клонального размножения, описанного в данном документе, с помощью трансфицирования популяции клеток, например, при помощи гена T-клеточного рецептора и последующего дополнительного (второго) размножения с применением способа размножения по настоящему изобретению.
[0043] На фиг. 12 показан дополнительный вариант осуществления системы по настоящему изобретению, которую можно применять в сочетании со способами размножения по настоящему изобретению. Данная система (120) включает биореактор (50), первый "картридж для удаления" (70) и второй "картридж для удаления" (90). Биореактор (50) находится в жидкостном соединении с первым сменным картриджем (70), а первый картридж для удаления находится в жидкостном соединении со вторым картриджем для удаления (90). Аналогично варианту осуществления, показанному на фиг. 11, второй картридж для удаления (110) находится в жидкостном соединении с биореактором (50). Однако "картридж для отбора" (92), описанный в Международной заявке на патент WO 2013/124474, расположен между вторым картриджем для удаления (90) и биореактором (50). Таким образом, субпопуляцию клеток (2a), содержащуюся в популяции клеток (2), можно отбирать/обогащать с помощью данного "картриджа для отбора" (92), описанного в WO 2013/124474. Такую субпопуляцию клеток (2a) можно также переносить в биореактор (50), например, чтобы подвергнуть серийному размножению, описанному в данном документе. В качестве альтернативы (не показано) такую субпопуляцию клеток (2a) можно применять для клеточной терапии. Здесь опять-таки следует отметить, что применение растворимого реагента для мультимеризации, описанного в данном документе, позволяет конструировать автоматические устройства для очистки и размножения клеток, которые являются функционально закрытыми и, таким образом, не подвержены загрязнению. В дополнение, поскольку растворимый реагент для мультимеризации позволяет избегать необходимости в материалах твердой фазы, таких как магнитные гранулы, такие устройства для очистки клеток можно конструировать в виде непрерывно-проточных устройств.
[0044] На фиг. 13 показаны кинетические характеристики размножения пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток (Tresp), которые стимулировали in vitro либо с помощью Fab-фрагментов αCD3/αCD28, либо с помощью αCD3/αCD28/αCD8, которые были обратимо иммобилизованы на двух типах растворимого олигомерного мутантного варианта белка Strep-Tactin®, выступающего в качестве растворимого реагента для мультимеризации. Первый тип олигомерного Strep-Tactin® представлял собой фракцию олигомерного мутантного варианта белка стрептавидина (n≥3), полученную в примере 5 (также называемую в данном документе "традиционным остовом Strep-Tactin®", проиллюстрированным на фиг. 13 символом-треугольником, обращенным вершиной вниз), а второй тип такого олигомерного мутантного варианта белка стрептавидина, применяемого в качестве растворимого реагента для мультимеризации, представлял собой олигомер, полученный с помощью реакции растворимого олигомерного мутантного варианта белка стрептавидина с биотинилированным сывороточным альбумином человека (HSA). Данный растворимый реагент для мультимеризации на основе HSA также называется в данном документе "крупным остовом Strep-Tactin®". В экспериментах на фиг. 13 размножение осуществляли без замены среды. Результаты для CD4+ иммунореактивных T-клеток показаны на фиг. 13A, результаты для CD8+ иммунореактивных T-клеток показаны на фиг. 13B. В данном случае следует отметить, что применяемые в экспериментах растворимые реагенты для мультимеризации, которые были функционализированы обратимо связывающимися первыми средствами, а также необязательно вторыми и третьими средствами, называются на фигурах "мультимерами Streptamer®".
[0045] На фиг. 14 показаны кинетические характеристики размножения пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток (Tresp), которые стимулировали in vitro Fab-фрагментами αCD3/αCD28, обратимо иммобилизованных на двух типах растворимого олигомерного Strep-Tactin®, выступающего в качестве растворимого реагента для мультимеризации. Первый тип олигомерного Strep-Tactin® представлял собой фракцию олигомерного мутантного варианта белка стрептавидина (n≥3), полученную в примере 5 (также называемую в данном документе "традиционным остовом Strep-Tactin®", проиллюстрированным на фиг. 14 символом-треугольником, обращенным вершиной вверх), а второй тип такого олигомерного мутантного варианта белка стрептавидина, применяемого в качестве растворимого реагента для мультимеризации, представлял собой растворимый реагент для мультимеризации на основе HSA, вышеупомянутый "крупный остов Strep-Tactin®". В экспериментах на фиг. 14 размножение осуществляли с заменой среды. Результаты для CD4+ иммунореактивных T-клеток показаны на фиг. 14, результаты для CD8+ иммунореактивных T-клеток показаны на фиг. 14B.
[0046] На фиг. 15 показаны сводные данные по результатам, полученным на фиг. 13 и 14, в отношении кинетических характеристик размножения для пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток, при этом на фиг. 15A отображены результаты для CD4+ T-клеток, а на фиг. 15B отображены результаты для CD8+ T-клеток. Прямые линии используются для культивирования с заменой среды в день 3, а пунктирные линии отображают значения, полученные для степени размножения без замены среды в день 3. Данные, показанные на фиг. 15, нормализованы по исходному количеству клеток. Показаны данные только для Tresp, стимулированных олигомерным мутантным вариантом белка стрептавидина (n≥3), Tresp, стимулированных коммерчески доступными Dynabeads (положительный контроль), и нестимулированных T-клеток (отрицательный контроль), но не данные о реагенте для мультимеризации с "крупным остовом Strep-Tactin®".
[0047] На фиг. 16 показано раннее образование скоплений T-клеток после активации очищенных CD4+ и CD8+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном мутантном варианте белка стрептавидина (n≥3), описанном в примере 5. На фиг. 16A отображены результаты для CD4+ T-клеток, а на фиг. 16B изображены результаты для CD8+ T-клеток. Показаны данные для Tresp, стимулированных растворимым реагентом для мультимеризации (олигомерным мутантным вариантом белка стрептавидина), Tresp, стимулированных коммерчески доступными Dynabeads (положительный контроль), и нестимулированных T-клеток (отрицательный контроль).
[0048] На фиг. 17 показаны кинетические характеристики размножения и фенотип CD3+ центральных T-клеток памяти (Tcm) (CD3+CD62L+CD45RA- Tcm), поликлонально стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном мутантном варианте белка стрептавидина (при n≥3), описанном в примере 5. На графиках, показанных на фиг. 17, представлена степень пролиферации соответственно количеству клеток, собранных на каждый момент времени, при этом на фиг. 17A показана пролиферация в среде, дополненной только IL-2, а на фиг. 17B показана пролиферация в среде, дополненной IL-2 и IL-15. На фиг. 17C показан проточный цитометрический анализ поверхностной экспрессии CD62L и CD127 спустя 14 дней культивирования в этих различных цитокиновых окружениях.
[0049] На фиг. 18 показаны кинетические характеристики избирательного антиген-специфического (Ag-специфического) размножения из общей популяции очищенных CD3+CD62L+CD45RA- иммунореактивных клеток Tcm, которые стимулировали in vitro как комплексом пептид:молекула MHC (который выступает в качестве первого средства, передающего первичный активационный сигнал клеткам), так и Fab-фрагментом αCD28 (который выступает в качестве второго средства, связывающегося со вспомогательной молекулой на поверхности клеток), а также нестимулированных T-клеток (отрицательный контроль). Как комплекс антиген-специфического пептида с молекулой MHC, так и Fab-фрагмент αCD28 были обратимо иммобилизованы на одном и том же растворимом олигомерном мутантном варианте белка стрептавидина (при n≥3), описанном в примере 5. Пептид, применяемый для антиген-специфического размножения на фиг. 18A, представлял собой пептид CRVLCCYVL (SEQ ID NO: 06), аминокислоты 309–317 предраннего белка 1, рестриктированные с помощью молекулы MHC HLA-C702 (описанной в Ameres et al, PLOS Pathogens, май 2013 г., том 9, номер 5, e1003383), представляют собой эпитоп IE-1/HLA-C7, специфичный для цитомегаловируса (CMV). Молекула MHC-I, которая представляет собой пептид, несущий на своем C-конце тяжелой цепи стрептавидин-связывающий пептид (SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 07), коммерчески доступный как "Twin-Strep-tag®" от IBA GmbH, Геттинген, Германия). На фиг. 18A показан приводимый в качестве примера проточный цитометрический анализ части Ag-специфических клеток, подвергнутых пролиферации с помощью комплекса пептид:MHC-I, специфичного к данному эпитопу IE-1/HLA-C7, в качестве первого средства, которое передает первичный активационный сигнал клеткам, обратимо иммобилизованного на растворимом олигомерном мутантном варианте белка стрептавидина. На графиках на фиг. 18B - фиг. 18E проиллюстрированы кинетические характеристики размножения для дополнительных видов Ag-специфичности соответственно количеству собранных специфических клеток, положительных по мультимеру пептид:MHC-I, на каждый момент времени по аналогии с фиг. 18A, с применением различных комплексов антиген-специфического пептида с молекулой MHC-I в качестве первого средства, которое передает первичный активационный сигнал клеткам, обратимо иммобилизованного на растворимом олигомерном мутантном варианте белка стрептавидина. Более подробно на фиг. 18B показано размножение Ag-специфических клеток, которые размножали с помощью комплекса пептид:MHC-I, специфичного к эпитопу pp65 CMV (аминокислоты 341-350 (QYDPVAALF (SEQ ID NO: 08)), рестриктированные с помощью HLA-A2402), на фиг. 18C показано размножение Ag-специфических клеток, которые размножали с помощью другого комплекса пептид:MHC-I, специфичного к эпитопу pp65 CMV (аминокислоты 265-274 RPHERNGFTV (SEQ ID NO: 09)), рестриктированные с помощью HLA-B702), нафиг. 18D показано размножение Ag-специфических клеток, которые подвергали пролиферации с помощью комплекса пептид:MHC-I, специфичного к эпитопу гексона 5 аденовируса (аминокислоты 114-124 (CPYSGTAYNSL (SEQ ID NO: 10)), рестриктированные с помощью HLA-B702), на фиг. 18E показано размножение Ag-специфических клеток, которые подвергали пролиферации с помощью комплекса пептид:MHC-I, специфичного к эпитопу IE-1309-317/HLA-B7 CMV (приводимые в качестве примера данные FACS см. выше на фиг. 18A). Все комплексы пептид:молекула MHC, несущие Twin Strep®-Tag, коммерчески доступны от IBA GmbH. В данном случае аминокислотные последовательности молекул HLA-A*2402, HLA-B*0702 и HLA-C*0702, которые несут "Twin Strep®-Tag" на своем C-конце, показаны как SEQ ID NO: 21, 22 и 23 в прилагаемом перечне последовательностей, а аминокислотная последовательность β2-микроглобулина (который вместе с α-цепью, то есть в качестве молекул, кодируемых HLA, образует соответствующую молекулу MHC-I) показана как SEQ ID NO: 24 в прилагаемом перечне последовательностей. В дополнение, на фиг. 18F показан приводимый в качестве примера проточный цитометрический анализ поверхностной экспрессии CD62L и CD127 спустя 14 дней культивирования стимулируемых/размножаемых с помощью гексона 5114-124/HLA-B7 клеток из фиг. 18D.
[0050] На фиг. 19 показаны кинетические характеристики избирательного Ag-специфического размножения из очищенных CD3+CD62L+CD45RA- иммунореактивных клеток Tcm, которые стимулировали in vitro a) комплексами антиген-специфического пептида MHC-I и b) Fab-фрагментами αCD28, которые были обратимо иммобилизованы в качестве первого и второго средства на растворимых олигомерных мутантных вариантах белка стрептавидина. Для данной цели 500000 CD3+CD62L+CD45RA- иммунореактивных клеток Tcm (Tresp) подвергали Ag-специфической стимуляции с помощью 3 мкл препарата реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг комплексов пептид:молекула MHC класса I, оснащенных стрептавидин-связывающим пептидом (специфический пептид представлен аминокислотами 114-124 (CPYSGTAYNSL, SEQ ID NO: 10) белка гексона 5 аденовируса, рестриктированными с помощью HLA-B0702, см. выше), и 0,5 мкг Fab αCD28. В качестве альтернативы 4,5 мкл препарата реагента для мультимеризации Strep-Tactin нагружали 0,5 мкг этого комплекса пептид:молекула MHC класса I, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Для сравнения проводили поликлональную стимуляцию с помощью 3 мкл препарата реагента для мультимеризации Strep-Tactin (1 мг/мл), также нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Опять-таки, в качестве вышеописанного альтернативного условия стимуляции применяли 4,5 мкл препарата мультимеров Strep-Tactin, нагруженных 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, поликлонально стимулированные при помощи Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2 и 5 нг/мл IL-15. Клетки инкубировали при 37°C с заменой среды каждые 3 дня, и число клеток анализировали через 7 и 14 дней. На фотографиях, показанных на фиг. 19, представлена степень образования скоплений в день 5 для Ag-специфической стимуляции, проиллюстрированной на примере для эпитопа гексона 5/HLA-B7 аденовируса.
[0051] На фиг. 20 показаны урожайность и фенотип размножения очищенных CD8+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на двух типах растворимого олигомерного Strep-Tactin®, выступающего в качестве растворимого реагента для мультимеризации. Первый тип олигомерного Strep-Tactin® представлял собой фракцию олигомерного мутантного варианта белка стрептавидина (n-мерного, полученного в примере 5 (стандартный остов)), второй тип данного олигомерного мутантного варианта белка стрептавидина, применяемого в качестве растворимого реагента для мультимеризации, представлял собой растворимый олигомер, описанный выше и называемый в данном документе "крупным" остовом Strep-Tactin®. В этих экспериментах фракцию олигомерного стандартного мутантного варианта белка стрептавидина (n≥3) применяли также в качестве реагента для мультимеризации, который был функционализирован либо отдельными Fab-фрагментами (третий столбик на фиг. 20A и фиг. 20B), либо комбинацией Fab-фрагментов αCD3 и αCD28. В дополнение к комбинированной стимуляции Fab-фрагментами αCD3/αCD28, также был иммобилизован дополнительный Fab-фрагмент αCD8 (коммерчески доступный от IBA GmbH, Геттинген, Германия) в целях тестирования на наличие возможности преимущественной стимуляции определенной субпопуляции T-клеток. На фиг. 20A показан график со столбиками, представляющими степень пролиферации соответственно количеству клеток, собранных в день 6, по сравнению с отрицательными контролями (нестимулированные очищенные CD8+ иммунореактивные T-клетки), и нормализованная по положительному контролю (очищенные CD8+ иммунореактивные T-клетки, стимулированные коммерчески доступными Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28)). На фиг. 20B показан проточный цитометрический анализ поверхностной экспрессии CD8 и поверхностной молекулы T-клеток CD45RO (свидетельствующей о пролиферации и активации T-клеток) после культивирования клеток. Различные условия стимуляции сравнивали с помощью однофакторного ANOVA, и значимого различия выявлено не было (n.s.).
[0052] На фиг. 21 показаны урожайность и фенотип размножения очищенных CD8+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном Strep-Tactin®, выступающем в качестве растворимого реагента для мультимеризации, который был функционализирован либо отдельными Fab-фрагментами, либо комбинацией Fab-фрагментов (как уже описано выше). В этих экспериментах CD8+ иммунореактивные T-клетки стимулировали растворимым реагентом для мультимеризации (растворимый олигомерный Strep-Tactin® (1 мг/мл) из примера 5), который был функционализирован различными количествами Fab-фрагментов αCD3 и αCD28, необязательно вместе с Fab-фрагментом αCD8, описанным выше. Термин "1x" соответствует 1,5 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг Fab-фрагмента αCD3 в отдельности, и 1,5 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг Fab αCD28 в отдельности, или 3 мкл препарата олигомерного Strep-Tactin, нагруженного 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab αCD28, или 4,5 мкл препарата мультимеров Strep-Tactin, нагруженных 0,5 мкг Fab αCD3 со Strep-tag, 0,5 мкг Fab αCD8 со Strep-tag и 0,5 мкг Fab αCD28 со Strep-tag. Соответственно, термин "2x" соответствует 3,0 мкг мультимеризованного Strep-Tactin, функционализированного 1 мкг Fab-фрагмента αCD3 в отдельности, и 3,0 мкг мультимеризованного Strep-Tactin, функционализированного 1 мкг Fab αCD28 в отдельности; это означает, что применяли вдвое большее количество иммобилизованного Fab-фрагмента αCD3. Необработанные клетки Tresp служили в качестве отрицательного контроля, а очищенные иммунореактивные T-клетки CD8+, стимулированные коммерчески доступными Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. На фиг. 21A показаны графики, столбики на которых представляют степень пролиферации соответственно количеству клеток, собранных в день 5, по сравнению с отрицательными контролями, и нормализованная по положительному контролю. На фиг. 21B показан FACS-анализ поверхностной экспрессии CD8 и CD45RO после культивирования клеток.
[0053] На фиг. 22 показана активация внутриклеточных сигнальных каскадов трансдуцированных клеток Jurkat, которые были модифицированы для экспрессии химерного антигенного рецептора (CAR) αCD19 и которые стимулировали с помощью олигомерного Strep-Tactin® из примера 5 в качестве растворимого реагента для мультимеризации. Специфичность CAR обычно проистекает из scFv-области, собранной из антиген-связывающей области моноклонального антитела (mAb), которая специфически связывается с целевым/опухолеассоциированным антигеном, таким как CD19, и увязывает его со специфичной для T-клеток передачей сигналов (описанной в Hudecek et al, Clin Cancer Res. 2013 June 15; 19(12): 3153–3164. В экспериментах внеклеточный домен (ECD) CD19, который содержит природный лиганд αCD19-CAR, а также F(ab)2-фрагмент поликлонального αIgG, который распознает спейсер IgG4 F(ab)2 антитела осла к иммуноглобулинам человека (коммерчески доступный от Jackson Immuno Research) в αCD19-CAR, также применяли в данном эксперименте в качестве первого средства, которое передает первичный активационный сигнал клеткам Jurkat. Обратимая иммобилизация на растворимом олигомерном мутантном варианте белка стрептавидина обеспечивалась стрептавидин-связывающим пептидом SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 07), который был слит с C-концом ECD CD19 или биотинилированным (Fab)2-фрагментом αIgG (поскольку мутантный вариант белка стрептавидина "m2" связывается с биотином с пониженным сродством, такое связывание является обратимым и может, например, быть устранено путем добавления избытка биотина в свободной форме). В контрольном эксперименте на фиг. 22A 300000 CD3+ иммунореактивных клеток Jurkat (Jresp) стимулировали различными количествами смеси препаратов олигомерного Strep-Tactin (1 мг/мл), который был функционализирован Fab αCD3 и Fab αCD28 ("x1" соответствует 3 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28 – поликлонального мультимера Streptamer). В эксперименте на фиг. 22B применяли 3 мкл препарата олигомерного Strep-Tactin, который был функционализирован 0,5 мкг (x1) или 1 мкг (x2) внеклеточного домена (ECD) CD19, или 3 мкл препарата олигомерного Strep-Tactin, нагруженного 0,5 мкг (x1) или 1 мкг (x2) αIgG, который распознает спейсер IgG4 (оба из которых являются CAR-специфичными мультимерами Streptamer®). Jresp, стимулированные с помощью Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28) или PMA и иономицина, служили в качестве положительных контролей. Клетки Jresp высевали в пробирки Eppendorf объемом 1,5 мл в 200 мкл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C, помещали на лед и лизировали спустя от 0 мин. до 20 мин. стимуляции.
[0054] На фиг. 23 показано размножение очищенных CD3+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном Strep-Tactin® из примера 5, который служил в качестве растворимого реагента для мультимеризации. В одном эксперименте, в дополнение к Fab-фрагментам αCD3/αCD28, на растворимом олигомере мутантного варианта белка стрептавидина также был иммобилизован
Fab-фрагмент αCD8, коммерчески доступный от IBA GmbH, Геттинген, Германия (номер по каталогу 6-8000-203), в целях тестирования на наличие возможности преимущественной стимуляции in vitro субпопуляции T-клеток CD8+ в смешанной культуре CD3+ реагентом для мультимеризации по настоящему изобретению, на котором также обратимо иммобилизован Fab-фрагмент αCD8. Более подробно, 500000 очищенных CD3+ иммунореактивных T-клеток (Tresp) стимулировали 3 мкл препарата олигомерного стрептавидина (1 мг/мл), нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. В качестве альтернативного подхода 4,5 мкл олигомера Strep-Tactin нагружали 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28, описанных выше. Нестимулированные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные с помощью Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля.
[0055] На фиг. 24 изображены приводимые в качестве примера стратегии получения олигомерных мутантных вариантов белка стрептавидина, которые можно применять в качестве растворимого реагента для мультимеризации по настоящему изобретению. На фиг. 24A показано, что на первой стадии мутантный вариант белка стрептавидина "m2" (SAm2), который содержит аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 (SEQ ID NO: 03) в положениях 44-47 последовательности стрептавидина дикого типа, применяют для получения олигомерных мутантных вариантов белка стрептавидина, имеющих "традиционный остов". На второй стадии можно получать олигомерные растворимые мутантные варианты белка стрептавидина, имеющие "крупный остов", с помощью связывания мутантного варианта белка стрептавидина с биотинилированным белком-носителем, таким как сывороточный альбумин человека (HSA), или с помощью связывания мутантных вариантов белка стрептавидина с синтетическими носителями, такими как PEG. Фиг. 24B: биотинилирование человеческого сывороточного альбумина (HSA).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0056] Настоящее изобретение обеспечивает способы, наборы и устройство для размножения популяции клеток или для индуцирования пролиферации популяции T-клеток.
[0057] Термин "популяция клеток", применяемый в данном документе, охватывает все клетки, которые можно размножать с помощью связывания с рецептором клеточной поверхности первого средства, которое передает первичный активационный сигнал клеткам. Также возможно, что для размножения популяции клеток может быть необходимым связывание второго средства со вторым рецептором клеточной поверхности (вспомогательной молекулой) для выработки костимулирующего сигнала, требуемого для размножения клеток. В некоторых вариантах осуществления популяция клеток может представлять собой популяцию лимфоцитов, в том числе без ограничения популяцию B-клеток, популяцию T-клеток или популяцию естественных клеток-киллеров. Иллюстративные примеры популяций клеток представляют собой B-клетки, несущие CD40 или CD137 (обе популяции клеток могут пролиферировать при связывании только первого средства, которое передает активационный сигнал, например лиганд 4-1BB; или молекулы антитела αCD40 или молекулы антитела αCD137 (см. например Zhang et al., 2010, J Immunol, 184:787-795)). Другие иллюстративные примеры средств (либо первого, либо второго), которые можно применять для размножения B-клеток, представляют собой средства, связывающиеся с IgG, CD19, CD28 или CD14, например, молекулы антител αCD19, αIgG, αCD28 или αCD14. Также предусматривается, что первое или второе средства для размножения B-клеток могут включать в себя лиганды для Toll-подобных рецепторов или интерлейкины, такие как IL-21 (см. например Dienz O, et al. 2009. J. Exp. Med. 206:69). Следует отметить, что липополисахарид-зависимая активация B-клеток также охватывается настоящим изобретением, поскольку липополисахарид может применяться также в качестве первого средства и может быть оснащен партнером по связыванию C1, применяемым в данном документе. Другие иллюстративные примеры подходящих популяций клеток включают популяцию T-клеток, размножающуюся после активации с помощью связывания первого средства с TCR/CD3 и связывания второго средства со вспомогательной молекулой на T-клетке, такой как CD28. В таком случае первое средство стимулирует передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках, а второе средство передает вторичный стимул с помощью связывания с CD28 в качестве вспомогательной молекулы. Средства, которые можно применять для размножения T-клеток, могут включать также интерлейкины, такие как IL-2, IL-7, IL-15 или IL-21 (см., например, Cornish et al. 2006, Blood. 108(2):600-8, Bazdar and Sieg, 2007, Journal of Virology, 2007, 81(22):12670-12674, Battalia et al, 2013, Immunology, 139(1):109-120). Другие иллюстративные примеры средств, которые можно применять для размножения T-клеток, представляют собой средства, связывающиеся с CD8, CD45 или CD90, такие как антитела αCD8, αCD45 или αCD90. Иллюстративные примеры популяции T-клеток включают антиген-специфические T-клетки, T-клетки-хелперы, цитотоксические T-клетки, T-клетки памяти (иллюстративным примером T-клеток памяти являются CD62L+ CD8+ специфические центральные T-клетки памяти) или регуляторные T-клетки (иллюстративным примером Treg являются CD4+ CD25+ CD45RA+ клетки Treg). Термин "T-клетка (популяция)", применяемый в данном документе, включает также T-клетки, содержащие химерный антигенный рецептор (CAR), также известный как искусственные T-клеточные рецепторы или химерные T-клеточные рецепторы. Таким образом, популяцию T-клеток, содержащих химерный антигенный рецептор, можно размножать также с помощью способов, реагентов и устройств по настоящему изобретению. См. в данном отношении также пример 15, в котором клетки Jurkat, которые экспрессируют химерный CD19-специфический антигенный рецептор (CAR), стимулировали с помощью растворимого реагента для мультимеризации по настоящему изобретению. Другой иллюстративный пример подходящей популяции клеток включает естественные клетки-киллеры (NK-клетки), которые можно например размножать с помощью средств, связывающихся с CD16 или CD56, таких как например антитела αCD16 или αCD56. В иллюстративном примере такое антитело αCD16 представляет собой антитело 3G8 с последовательностью VH, приведенной под SEQ ID NO: 25, и последовательностью VL, приведенной под SEQ ID NO: 26 (см. например Hoshino et al, Blood. 1991 Dec 15;78(12):3232-40.). Другое средство, которое можно применять для размножения NK-клеток, может представлять собой IL-15 (см. например Vitale et al. 2002. The Anatomical Record. 266:87-92). Еще один иллюстративный пример подходящей популяции клеток включает моноциты, которые можно например размножать с помощью средства, связывающегося с CD14, такого как молекула антитела αCD14. Популяция клеток может происходить от любого млекопитающего, в том числе без ограничения от человека, кролика, морской свинки, белки, хомяка, кошки, собаки, лемура, козы, свиньи, лошади, макака-резуса, макака или шимпанзе.
[0058] Таким образом, в соответствии с вышеизложенным настоящее изобретение относится к способам избирательной индукции ex vivo размножения популяции клеток, таких как B-клетки, T-клетки или естественные клетки-киллеры, в отсутствие экзогенных факторов роста, таких как лимфокины, и вспомогательных клеток. В дополнение, пролиферацию этих клеток, таких как B-клетки или T-клетки, можно индуцировать без необходимости для антигена, с получением таким образом размножившейся популяции клеток, такой как популяция T-клеток, которая является поликлональной в отношении антигенной реактивности. Способы, раскрытые в данном документе, могут обеспечивать длительную пролиферацию выбранной популяции T-клеток, таких как CD4+ или CD8+ T-клетки, в течение продолжительного периода времени с достижением многократного увеличения количества данных клеток по сравнению с исходной популяцией T-клеток. Как правило, в случае (клонального) размножения популяции лимфоцитов, описанного в данном документе, все потомство может обладать такой же специфичностью к антигену, как и популяция клеток, которая была выбрана для размножения.
[0059] Также в соответствии с вышеизложенным настоящее изобретение обеспечивает способы размножения популяции антиген-специфических T-клеток. Для получения популяции антиген-специфических T-клеток, T-клетки приводят в контакт с антигеном в форме, подходящей для запуска передачи первичного активационного сигнала в T-клетке, т. е. антиген презентируется T-клетке таким образом, что в T-клетке посредством комплекса TCR/CD3 запускается передача сигнала. Например, антиген может быть презентирован T-клетке с помощью антигенпрезентирующей клетки в комплексе с молекулой MHC. Антигенпрезентирующую клетку, такую как B-клетка, макрофаг, моноцит, дендритная клетка, клетка Лангерганса или другая клетка, которая может презентировать антиген T-клетке, можно инкубировать с T-клеткой в присутствии антигена (например растворимого антигена) так, чтобы антигенпрезентирующая клетка презентировала антиген T-клетке. В качестве альтернативы с T-клеткой можно инкубировать клетку, экспрессирующую антиген, представляющий интерес. Например, вместе с T-клеткой можно инкубировать опухолевую клетку, экспрессирующую опухолеассоциированные антигены, для индукции опухолеспецифического ответа. Аналогично, с T-клеткой можно инкубировать клетку, инфицированную патогеном, например вирусом, которая презентирует антигены патогена. После антиген-специфической активации популяции T-клеток клетки можно размножать в соответствии со способами по настоящему изобретению. Например, после формирования специфичности к антигену T-клетки можно размножать с помощью культивирования с антителом к CD3 (применяемым в качестве первого средства) и антителом к CD28 (применяемым в качестве второго средства) согласно способам, описанным в данном документе. В другом варианте осуществления первое средство может представлять собой комплекс MHC-I:пептид, который связывается с популяцией антиген-специфических T-клеток. В таком варианте осуществления можно применять любой известный антиген-специфический пептид, который способен образовывать комплекс с соответствующей молекулой MHC-I. См. в этом отношении примеры 11 и 12, в которых избирательное антиген-специфическое размножение иммунореактивных клеток Tcm из общей популяции CD3+ центральных T-клеток памяти проиллюстрировано на примере для четырех различных антиген-специфических клеток. В качестве альтернативы также возможно применять в качестве первого средства природный лиганд рецептора, который запускает размножение клеток. См. в этом отношении пример 15, в котором внеклеточный домен CD19 вызывал активацию внутриклеточных сигнальных каскадов трансдуцированных клеток Jurkat, которые были модифицированы для экспрессии химерного антигенного рецептора (CAR), связывающегося с CD19.
[0060] Образец популяции клеток можно получать из любого подходящего источника, обычно из любого образца ткани организма или биологической жидкости, такой как кровь. В последнем случае образец может представлять собой, например, популяцию мононуклеарных клеток периферической крови (PBMC), которую можно получить с помощью стандартных способов выделения, таких как применение градиента фиколла для клеток крови. Популяция клеток, подлежащая размножению, может однако находиться также в очищенной форме, и может быть выделена с помощью технологии обратимого окрашивания/выделения клеток, описанной в патенте США 7776562, патенте США 8298782, Международной заявке на патент WO 02/054065 или Международной заявке на патент WO 2013/011011. В качестве альтернативы популяцию клеток можно получать также с помощью сортировки клеток посредством отрицательной магнитной иммунной адгезии, описанной в патенте США 6352694 B1 или Европейском патенте EP 0 700 430 B1. Если способ выделения, описанный в данном документе, применяется в фундаментальном исследовании, то образец может представлять собой клетки из экспериментов по культивированию клеток in vitro. Образец обычно получают в форме жидкости, такой как раствор или дисперсия.
[0061] В соответствии с вышеизложенным, в одном варианте осуществления настоящее изобретение обеспечивает способ размножения популяции клеток in vitro, включающий приведение образца, содержащего популяцию клеток, в контакт с реагентом для мультимеризации, На реагенте для мультимеризации обратимо иммобилизовано (связано с ним) первое средство, которое передает первичный активационный сигнал клеткам, где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства. Первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1. Первое средство связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки.
[0062] В другом варианте осуществления настоящее изобретение обеспечивает способ, в котором на средстве для мультимеризации обратимо иммобилизовано (связано с ним) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток. Второе средство содержит партнера по связыванию C2, где партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C2 и связывающим участком Z2. Второе средство связывается со вспомогательной молекулой на поверхности клеток, стимулируя тем самым активированные клетки. В данном варианте осуществления первое средство может стимулировать передачу сигнала, связанного с комплексом TCR/CD3, в T-клетках и может представлять собой связывающее средство, которое специфически связывается с CD3. В данном варианте осуществления вспомогательная молекула на T-клетке может представлять собой CD28, а второе средство, которое связывается со вспомогательной молекулой, представляет собой связывающий реагент, который специфически связывается с CD28. В данном случае первое средство, которое специфически связывается с CD3, может быть выбрано из группы, включающей антитело к CD3, бивалентный фрагмент антитела из антитела к CD3, моновалентный фрагмент антитела из антитела к CD3 и белковую молекулу с антителоподобными свойствами связывания, связывающуюся с CD3. Второе средство, которое специфически связывается с CD28, также может быть выбрано из группы, включающей антитело к CD28, бивалентный фрагмент антитела из антитела к CD28, моновалентный фрагмент антитела из антитела к CD28 и белковую молекулу с антителоподобными свойствами связывания, связывающуюся с CD28. Бивалентный фрагмент антитела может представлять собой (Fab)2’-фрагмент или бивалентный одноцепочечный Fv-фрагмент, а моновалентный фрагмент антитела может быть выбран из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv). Белковая молекула с антителоподобными свойствами связывания, связывающаяся CD3 или CD28, может представлять собой аптамер, мутантный вариант белка на основе полипептида из семейства липокалинов, аналог антитела на основе глутатион-S-трансферазы, белок на основе анкиринового каркаса, белок на основе кристаллического каркаса, аднектин и авимер.
[0063] Как правило, первое и второе средство, применяемые в настоящем изобретении, могут например представлять собой антитело, его фрагмент и белковую связывающую молекулу с антителоподобными функциями. Примеры (рекомбинантных) фрагментов антител представляют собой Fab-фрагменты, Fv-фрагменты, одноцепочечные Fv-фрагменты (scFv), бивалентный фрагмент антитела, такой как (Fab)2'-фрагмент, диатела, триатела (Iliades, P., et al., FEBS Lett (1997) 409, 437-441), декатела (Stone, E., et al., Journal of Immunological Methods (2007) 318, 88–94) и другие доменные антитела (Holt, L.J., et al., Trends Biotechnol. (2003), 21, 11, 484-490). В некоторых вариантах осуществления один или более связывающих участков первого или второго средства могут представлять собой бивалентную белковую искусственную связывающую молекулу, такую как димерный мутантный вариант белка липокалина, также известный как "дуокалин". В некоторых вариантах осуществления рецептор-связывающий реагент может иметь один второй связывающий участок, т. е. он может быть моновалентным. Примеры моновалентных первого или второго средств включают без ограничения моновалентный фрагмент антитела, белковую связывающую молекулу с антителоподобными свойствами связывания или молекулу MHC. Примеры моновалентных фрагментов антител включают без ограничения Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv), в том числе бивалентный одноцепочечный Fv-фрагмент.
[0064] Как упоминается выше, примером белковой связывающей молекулы с антителоподобными функциями является мутантный вариант белка на основе полипептида из семейства липокалинов (см. например WO 03/029462, Beste et al., Proc. Natl. Acad. Sci. U.S.A. (1999) 96, 1898-1903). Липокалины, такие как билин-связывающий белок, желатиназа-ассоциированный липокалин нейтрофилов человека, аполипопротеин D человека или липокалин слезной жидкости человека, обладают связывающими участками для природных лигандов, которые можно модифицировать таким образом, чтобы они связывали заданную мишень. Дополнительные примеры белковой связывающей молекулы с антителоподобными свойствами связывания, которую можно применять в качестве рецептор-связывающего реагента, специфически связывающегося с рецепторной молекулой, включают без ограничения так называемые аналоги антител на основе глутатион-S-трансферазы (см. например международную заявку на патент WO 96/23879), белки на основе анкиринового каркаса (Mosavi, L.K., et al., Protein Science (2004) 13, 6, 1435-1448) или кристаллического каркаса (например из Международной заявки на патент WO 01/04144), белки, описанные в Skerra, J. Mol. Recognit. (2000) 13, 167-187, аднектины, тетранектины и авимеры. Авимеры, в том числе поливалентные авимерные белки, образующиеся с помощью перестановки экзонов в семействе рецепторных доменов человека, содержат так называемые A-домены, встречающиеся в виде цепочек из нескольких доменов в некоторых рецепторах клеточной поверхности (Silverman, J., et al., Nature Biotechnology (2005) 23, 1556-1561). Аднектины, получаемые из домена фибронектина человека, содержат три петли, которые можно сконструировать для иммуноглобулиноподобного связывания с мишенями (Gill, D.S. & Damle, N.K., Current Opinion in Biotechnology (2006) 17, 653-658). Тетранектины, получаемые из соответствующего гомотримерного белка человека, точно так же содержат петлевые области в домене лектина C-типа, которые можно сконструировать для желаемого связывания (там же). Пептоиды, которые могут выступать в качестве лигандов белков, представляют собой олиго-(N-алкил)-глицины, которые отличаются от пептидов тем, что боковая цепь соединена с атомом амидного азота, а не с α-атомом углерода. Пептоиды обычно устойчивы к действию протеаз и других модифицирующих ферментов, и могу характеризоваться намного более высокой клеточной проницаемостью, чем пептиды (см. например Kwon, Y.-U., and Kodadek, T., J. Am. Chem. Soc. (2007) 129, 1508-1509). Еще несколькими дополнительными примерами подходящих белковых связывающих молекул являются EGF-подобный домен, Kringle-домен, домен I типа фибронектина, домен II типа фибронектина, домен III типа фибронектина, PAN-домен, G1a-домен, SRCR-домен, домен ингибитора Кунитца/трипсина поджелудочной железы быка, тендамистат, домен ингибитора сериновых протеаз типа Kazal, трилистниковый домен (P-типа), домен типа C фактора фон Виллебранда, анафилотоксиноподобный домен, CUB-домен, повтор I типа тиреоглобулина, домен класса A рецептора LDL, Sushi-домен, Link-домен, домен I типа тромбоспондина, домен иммуноглобулина или иммуноглобулиноподобный домен (например, доменные антитела или антитела верблюда, содержащие только тяжелые цепи), домен лектина C-типа, MAM-домен, домен типа A фактора фон Виллебранда, домен соматомедина B, коровый домен WAP-типа, стабилизированный четырьмя дисульфидными связями, домен F5/8 типа С, домен гемопексина, SH2-домен, SH3-домен, EGF-подобный домен ламининового типа, C2-домен, "каппа-антитела" (см. Ill. et al., Protein Eng (1997) 10, 949-57), так называемое "минитело" (Martin et al., EMBO J (1994) 13, 5303-5309), диатело (см. Holliger et al., PNAS USA (1993)90, 6444-6448), так называемые "янусины" (см. Traunecker et al., EMBO J (1991) 10, 3655-3659 или Traunecker et al., Int J Cancer (1992) Suppl 7, 51-52), нанотело, микротело, аффилин, аффитело, ноттин, убиквитин, белок с "цинковыми пальцами", аутофлуоресцирующий белок или белок, богатый лейциновыми повторами. Примером молекулы нуклеиновой кислоты с антителоподобными функциями является аптамер. Аптамер сворачивается в определенный трехмерный мотив и демонстрирует высокое сродство к заданной целевой структуре.
[0065] Обратимся теперь к реагенту для мультимеризации; связывающие участки Z1 и Z2 средства для мультимеризации могут быть идентичными (см. также пример на фиг. 3). В таком случае можно применять одно средство для мультимеризации.
[0066] В варианте осуществления, в котором применяются обратимо связывающиеся первое и необязательно второе средство, реагент для мультимеризации может быть иммобилизован на твердой поверхности. Для иммобилизации реагента для мультимеризации можно применять любую твердую поверхность (подложку). Иллюстративные примеры твердых поверхностей, на которых может быть иммобилизован реагент для мультимеризации, включают магнитную гранулу, полимерную гранулу, планшет для культуры клеток, титрационный микропланшет, мембрану или полое волокно. Полые волокна применяют, например, в качестве биореактора в системе для размножения клеток Quantum®, доступной от Terumo BCT, Inc. (Лейквуд, Колорадо, США). Реагент для мультимеризации обычно ковалентно прикреплен к твердой подложке, однако при желании можно применять также нековалентные взаимодействия для иммобилизации, например на пластиковых субстратах. Как более подробно поясняется также ниже, реагент для мультимеризации может, например, представлять собой мутантный вариант белка стрептавидина или авидина, обратимо связывающийся со стрептавидин-связывающим пептидом. Такие мутантные варианты белка стрептавидина могут быть ковалентно прикреплены к любой поверхности, например к смоле (гранулам), применяемым для хроматографической очистки, и коммерчески доступны в такой форме от IBA GmbH, Геттинген, например, в виде Strep-Tactin® Sepharose, Strep-Tactin® Superflow®, Strep-Tactin® Superflow® высокой емкости или Strep-Tactin® MacroPrep®. Другими иллюстративными примерами реагентов для мультимеризации, являющихся коммерчески легкодоступными, являются смолы для аффинной хроматографии на иммобилизованных ионах металлов (IMAC), такие как смолы TALON® (Westburg, Лейсден, Нидерланды), которые можно применять для обратимой иммобилизации меченных олигогистидином (имеющих His-метку) белков в целом, что в данном документе означает обратимое связывание первого или второго средства, несущего в качестве первого партнера по связыванию C1 или второго партнера по связыванию C2 олигогистидиновую метку, такую как пента- или гексагистидиновая метка. Другими примерами реагентов для мультимеризации являются кальмодулин-сефароза, доступная от GE Life Sciences, которую можно применять вместе с первым или вторым средством, содержащим кальмодулин-связывающий пептид в качестве партнера по связыванию C1 или C2, или сефароза, с которой связан глутатион. В таком случае партнер по связыванию C1 или C2 представляет собой глутатион-S-трансферазу.
[0067] В других вариантах осуществления способа по настоящему изобретению реагент для мультимеризации может быть в растворимой форме. В целом можно применять такие же средства для мультимеризации, как и в случае реагента для мультимеризации, иммобилизованного на твердой подложке. Реагент для мультимеризации в растворимой форме может представлять собой, например, олигомер мутантного варианта белка стрептавидина, олигомер кальмодулина, соединение (олигомер), предоставляющее по меньшей мере две хелатные группы K, где по меньшей мере две хелатные группы способны к связыванию с ионом переходного металла, тем самым делая компонент A способным к связыванию с олигогистидиновой аффинной меткой, мультимерной глутатион-S-трансферазой или биотинилированным белком-носителем.
[0068] Как поясняется выше, первое и второе средство имеют, в дополнение к связывающему участку, способному связываться с соответствующей рецепторной молекулой клеточной поверхности, партнера по связыванию C1 или C2 (который в дальнейшем для простоты поиска будет называться "партнером по связыванию C"). Этот партнер по связыванию C способен связываться со связывающим участком Z реагента для мультимеризации (Z означает либо связывающий участок Z1, либо связывающий участок Z2 реагента для мультимеризации) C. Нековалентная связь, образующаяся между партнером по связыванию C, содержащимся в первом или втором средстве, и связывающим(-ми) участком(-ками) Z реагента для мультимеризации может характеризоваться любыми желаемыми прочностью и сродством, поскольку она является разрушаемой или обратимой в условиях, в которых осуществляют способ по настоящему изобретению. Константа диссоциации (KD) связывания между партнером по связыванию C, содержащимся в рецептор-связывающем реагенте, и связывающим участком Z реагента для мультимеризации может иметь значение в диапазоне от приблизительно 10-2 M до приблизительно
10-13 M. Таким образом, такая обратимая связь может, например, характеризоваться KD от приблизительно 10-2 M до приблизительно 10-13 M, или от приблизительно 10-3 M до приблизительно 10-12 M, или от приблизительно 10-4 M до приблизительно 10-11M, или от приблизительно 10-5 M до приблизительно 10-10 M. KD для такой связи, а также показатели KD, koff и kon для связи, образующейся между связывающим участком B рецептор-связывающего реагента и рецепторной молекулой, можно определять с помощью любых подходящих способов, например, с помощью флуоресцентного титрования, равновесного диализа или поверхностного плазмонного резонанса. Реагент, связывающийся с рецепторной молекулой, может содержать по меньшей мере одного, в том числе двух, трех или более вторых партнеров по связыванию C, и аффинный реагент может содержать по меньшей мере два, как например, три, четыре, пять, шесть, семь, восемь или более связывающих участков для партнера по связыванию, содержащегося в связывающем реагенте рецепторной молекулы. Как описано в патенте США 7776562, патенте США 8298782 или Международной заявке на патент WO 2002/054065, можно выбирать любую комбинацию партнера по связыванию C и аффинного средства с одним или более соответствующими связывающими участками Z, поскольку партнер по связыванию C и связывающий участок Z аффинного средства способны обратимо связываться или мультимеризоваться в (поливалентный) комплекс, что обычно сопровождается эффектом авидности.
[0069] Партнер по связыванию, содержащийся в первом или втором средстве, может представлять собой олигопептид, полипептид, белок, нуклеиновую кислоту, липид, сахарид, олигосахарид или полисахарид. Такой партнер по связыванию характеризуется более высоким сродством к связывающему участку реагента для мультимеризации, чем к другому веществу. Примеры соответствующего партнера по связыванию включают без ограничения молекулу иммуноглобулина, ее фрагмент и белковую связывающую молекулу с антителоподобными функциями.
[0070] В некоторых вариантах осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит биотин, и аффинный реагент содержит аналог стрептавидина или аналог авидина, которые обратимо связываются с биотином.
[0071] В некоторых вариантах осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит аналог биотина, который обратимо связывается со стрептавидином или авидином, и аффинный реагент содержит стрептавидин, авидин, аналог стрептавидина или аналог авидина, которые обратимо связываются с соответствующим аналогом биотина.
[0072] В некоторых дополнительных вариантах осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит стрептавидин- или авидин-связывающий пептид, и аффинный реагент содержит стрептавидин, авидин, аналог стрептавидина или аналог авидина, которые обратимо связываются с соответствующим стрептавидин- или авидин-связывающим пептидом.
[0073] В некоторых вариантах осуществления партнер по связыванию, содержащийся в первом или втором средстве, может содержать стрептавидин-связывающий пептид Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 01), и аффинный реагент может содержать мутантный вариант белка (аналог) стрептавидина, содержащий аминокислотную последовательность Va144-Thr45-Ala46-Arg47 (SEQ ID NO: 02) в положениях 44-47 последовательности стрептавидина дикого типа, или мутантный вариант белка (аналог) стрептавидина, содержащий аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 (SEQ ID NO: 03) в положениях 44-47 последовательности стрептавидина дикого типа, оба из которых описаны например в патенте США 6103493 и коммерчески доступны под торговой маркой Strep-Tactin®. Стрептавидин-связывающие пептиды могут, например, представлять собой одиночные пептиды, такие как "Strep-tag®", например описанный в патенте США 5506121, или стрептавидин-связывающие пептиды, имеющие два или более последовательно расположенных отдельных связывающих модулей, описанные в Международной патентной публикации WO 02/077018 или патенте США 7981632.
[0074] В некоторых вариантах осуществления партнер по связыванию C первого или второго средства содержит компонент, известный специалисту в данной области в качестве аффинной метки. В таком варианте осуществления аффинный реагент содержит соответствующего партнера по связыванию, например антитело или фрагмент антитела, для которого известно его связывание с аффинной меткой. В качестве некоторых иллюстративных примеров известных аффинных меток партнер по связыванию, содержащийся в первом или втором средстве, может содержать олигогистидин, домен иммуноглобулина, мальтозосвязывающий белок, глутатион-S-трансферазу (GST), хитин-связывающий белок (CBP) или тиоредоксин, кальмодулин-связывающий пептид (CBP), пептид FLAG’, HA-метку (последовательность: Tyr-Pro-Tyr-Asp-Val-Pro-Asp-Tyr-Ala (SEQ ID NO: 11)), VSV-G-метку (последовательность: Tyr-Thr-Asp-Ile-Glu-Met-Asn-Arg-Leu-Gly-Lys (SEQ ID NO: 12)), HSV-метку (последовательность: Gln-Pro-Glu-Leu-Ala-Pro-Glu-Asp-Pro-Glu-Asp (SEQ ID NO: 13)), эпитоп T7 (Ala-Ser-Met-Thr-Gly-Gly-Gln-Gln-Met-Gly (SEQ ID NO: 14)), мальтозосвязывающий белок (MBP), эпитоп HSV с последовательностью Gln-Pro-Glu-Leu-Ala-Pro-Glu-Asp-Pro-Glu-Asp (SEQ ID NO: 13) гликопротеин D вируса простого герпеса, эпитоп "Myc" фактора транскрипции c-Myc с последовательностью Glu-Gln-Lys-Leu-Ile-Ser-Glu-Glu-Asp-Leu (SEQ ID NO: 15), V5-метку (последовательность: Gly-Lys-Pro-Ile-Pro-Asn-Pro-Leu-Leu-Gly-Leu-Asp-Ser-Thr, SEQ ID NO: 16) или глутатион-S-трансферазу (GST). В таком варианте осуществления комплекс, образующийся между одним или более связывающими участками реагента для мультимеризации, в данном случае антитела или фрагмента антитела, и антигеном, можно разрушать конкурентным путем с помощью добавления антигена в свободной форме, т. е. пептида в свободной форме (эпитопной метки) или белка в свободной форме (такого как MBP или CBP). Аффинная метка также может представлять собой олигонуклеотидную метку. Такую олигонуклеотидную метку можно, например, применять для гибридизации с олигонуклеотидом с комплементарной последовательностью, связанным с аффинным реагентом или содержащимся в нем.
[0075] В некоторых вариантах осуществления связывание между партнером по связыванию C, содержащимся в первом или втором средстве, и одним или более связывающими участками реагента для мультимеризации происходит в присутствии двухвалентного, трехвалентного или четырехвалентного катиона. В связи с этим в некоторых вариантах осуществления реагент для мультимеризации содержит двухвалентный, трехвалентный или четырехвалентный катион, обычно удерживаемый, например связываемый в комплекс, с помощью подходящего хелатора. Партнер по связыванию, содержащийся в рецептор-связывающем реагенте, может в таком варианте осуществления содержать компонент, содержащий, например, комплексы, бивалентный, трехвалентный или четырехвалентный катион. Примеры соответствующего металлохелатора включают без ограничения этилендиамин, этилендиаминтетрауксусную кислоту (EDTA), этиленгликольтетрауксусную кислоту (EGTA), диэтилентриаминпентауксусную кислоту (DTPA), N,N-бис(карбоксиметил)глицин (также называемый нитрилотриуксусной кислотой, NTA) или 1,2-бис(о-аминофенокси)этан-N,N,N',N'-тетрауксусную кислоту (BAPTA). В качестве примера, EDTA образует комплекс с большинством моновалентных, бивалентных, трехвалентных и четырехвалентных ионов металлов, таких как, например, кальций (Ca2+), магний (Mn2+), медь (Cu2+), железо (Fe2+), кобальт (Co3+) и цирконий (Zr4+), а BAPTA является специфичной для Ca2+. В качестве иллюстративного примера, стандартный способ, применяемый в данной области техники, представляет собой образование комплекса между олигогистидиновой меткой и ионами меди (Cu2+), никеля (Ni2+), кобальта (Co2+) или цинка (Zn2+), предоставляемыми с помощью хелатора нитрилотриуксусной кислоты (NTA).
[0076] В некоторых вариантах осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит кальмодулин-связывающий пептид, и аффинный реагент содержит мультимерный кальмодулин, описанный в патенте США 5985658 или описанный в данном документе, например со ссылкой на фигуру 2. В некоторых вариантах осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит пептид FLAG, и аффинный реагент содержит антитело, связывающееся с пептидом FLAG, например с пептидом FLAG, который связывается с моноклональным антителом 4E11, описанным в патенте США 4851341. В одном варианте осуществления партнер по связыванию C, содержащийся в первом или втором средстве, содержит олигогистидиновую метку, и аффинный реагент содержит антитело или ион переходного металла, связывающиеся с олигогистидиновой меткой. Разрушение всех этих образованных в результате связывания комплексов можно осуществлять с помощью хелатирования ионов металлов, например хелатирования кальция, например с помощью добавления EDTA или EGTA (выше). Кальмодулин, антитела, такие как 4E11, или хелатированные ионы металлов, или хелаторы в свободной форме можно мультимеризовать с помощью традиционных способов, например с помощью биотинилирования и образования комплексов со стрептавидином или авидином или их мультимерами, или с помощью введения карбоксильных остатков в полисахарид, например декстран, фактически как описано в Noguchi, A, et al. Bioconjugate Chemistry (1992) 3, 132-137, на первой стадии и связывания кальмодулина, или антител, или хелатированных ионов металлов, или хелаторов в свободной форме посредством первичных аминогрупп с карбоксильными группами в полисахаридном, например декстрановом, остове с помощью традиционных карбодиимидных химических методов на второй стадии. В таких вариантах осуществления связывание между партнером по связыванию C, содержащимся в первом или втором средстве, и одним или более связывающими участками Z реагента для мультимеризации можно нарушать с помощью хелатирования ионов металлов. Хелатирование металлов можно осуществлять, например, с помощью добавления EGTA или EDTA.
[0077] В некоторых вариантах осуществления, в частности если реагент для мультимеризации находится в растворимой форме и в его основе лежит стрептавидин или авидин, он представляет собой олигомер или полимер стрептавидина или авидина, или любой мутантный вариант белка (аналог) стрептавидина или авидина. Связывающий участок Z служит для связывания авидина или стрептавидина с природным биотином. Соответствующий олигомер или полимер может быть сшит при помощи полисахарида. В одном варианте осуществления олигомеры или полимеры стрептавидина или авидина, или мутантных вариантов белков (аналогов) стрептавидина или авидина получают путем введения карбоксильных остатков в полисахарид, например декстран, фактически как описано в Noguchi, A, et al., Bioconjugate Chemistry (1992) 3, 132-137, на первой стадии. Затем стрептавидин или авидин или их аналоги можно связывать посредством первичных аминогрупп внутреннего лизинового остатка и/или свободного N-конца с карбоксильными группами в декстрановом остове с помощью традиционных карбодиимидных химических методов на второй стадии. В дополнение, сшитые олигомеры или полимеры стрептавидина или авидина, или любого мутантного варианта белка (аналога) стрептавидина или авидина можно получать также с помощью сшивания отдельных молекул стрептавидина или авидина (тетрамерный гомодимер стрептавидина или авидина называется в данном документе "отдельной молекулой", или наименьшим структурным элементом соответствующего олигомера или полимера) посредством бифункциональных молекул, служащих в качестве линкера, таких как глутаровый альдегид, или с помощью других способов, описанных в уровне техники. Например, возможно получать олигомеры мутантных вариантов белка стрептавидина путем введения, на первой стадии, тиольных групп в мутантный вариант белка стрептавидина (такое можно выполнять например с помощью реакции мутантного варианта белка стрептавидина с 2-иминотиоланом (реагентом Трота)) и с помощью активации, в ходе отдельной реакции аминогрупп, имеющихся в мутантном варианте белка стрептавидина. Данную активацию аминогрупп можно осуществлять с помощью реакции мутантного варианта белка стрептавидина с коммерчески доступными гетеробифункциональными сшивающими средствами, такими как сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат сульфо-SMCC) или сукцинимидил-6-[(β-малеимидопропионамидо)гексаноат (SMPH). На второй стадии два полученных таким образом продукта реакции смешивают вместе, что приводит к реакции тиольных групп, содержащихся в одной партии модифицированного мутантного варианта белка стрептавидина, с активированными (с помощью малеимидных функциональных групп) аминокислотами в другой партии модифицированного мутантного варианта белка стрептавидина. С помощью данной реакции образуются мультимеры/олигомеры мутантного варианта белка стрептавидина. Такие олигомеры могут иметь любое подходящее количество "отдельных молекул" или "стрептавидиновых структурных элементов", превышающее 3, и степень олигомеризации может изменяться в зависимости от условий реакции (см. фиг. 24). После реакции этих двух партий модифицированного мутантного варианта белка стрептавидина олигомерный растворимый реагент для мультимеризации обычно выделяют посредством эксклюзионной хроматографии, и в качестве реагента для мультимеризации можно применять любую желаемую фракцию. Как правило, олигомеры не имеют (и не должны иметь) единый молекулярный вес, и для них обычно наблюдается статистическое распределение по весу, такое как распределение Гаусса. Любой олигомер, имеющий более трех гомотетрамеры стрептавидина (структурных элементов; (n≥3)), можно применять в качестве растворимого реагента для мультимеризации. Олигомеры могут иметь, например, от 3 до 25 гомотетрамеров мутантного варианта белка стрептавидина. При молекулярном весе мутантных вариантов белка стрептавидина, таких как мутантные варианты белка "m1" или "m2", более подробно описанных ниже, приблизительно 50 кДа, эти растворимые олигомеры имеют молекулярный вес от приблизительно 150 кДа до приблизительно 1250 кДа. Поскольку каждая молекула/мутантный вариант белка стрептавидина имеет четыре биотин-связывающих участка, то в таком реагенте для мультимеризации представлены от 12 до 100 связывающих участков Z1 (и Z2), описанных в данном документе.
[0078] В соответствии с вышеприведенным раскрытием, в дополнение к таким олигомерным реагентам для мультимеризации, которые содержат только сшитые гомотетрамеры стрептавидина, тетрамерные мутантные варианты белка стрептавидина возможно подвергать реакции с носителем с получением реагентов для мультимеризации, применяемых в настоящем изобретении. В дополнение к вышеописанной реакции с полисахаридом, также возможно применять физиологически или фармацевтически приемлемые белки, такие как сывороточный альбумин (например сывороточный альбумин человека (HSA) или бычий сывороточный альбумин (BSA)), в качестве белка-носителя. В таком случае мутантный вариант белка стрептавидина (в виде отдельного гомотетрамера или также в форме олигомеров с n≥3) можно связывать с белком-носителем с помощью нековалентного взаимодействия. Для данной цели биотинилированный BSA (коммерчески доступный от различных поставщиков, таких как Thermo Fisher Scientific, Sigma Aldrich или Vector Labs, не говоря уже о других) можно подвергать реакции с мутантным вариантом белка стрептавидина. Тем самым, некоторые из олигомеров стрептавидина будут нековалентно связываться посредством одного или более биотин-связывающих участков (Z1, Z2) с биотинилированным белком-носителем, оставляя при этом большинство связывающих участков (Z1, Z2) олигомера доступными для связывания средств, таких как первое средство и необязательно второе средство, а также любое дополнительное средство, описанное в данном документе. Таким образом, с помощью такого подхода можно удобным образом получать растворимый реагент для мультимеризации с множеством связывающих участков Z1 (см. фиг. 24). В качестве альтернативы мутантный вариант белка стрептавидина (в виде отдельного гомотетрамера или также в форме олигомеров с n≥3) можно ковалентно связывать с синтетическим носителем, таким как молекула полиэтиленгликоля (PEG). Для данной цели можно применять любую подходящую молекулу PEG, поскольку молекула PEG и соответствующий реагент для мультимеризации являются растворимыми. Как правило, все молекулы PEG, имеющие молекулярный вес до 1000 Да, являются растворимыми в воде или культуральной среде, которую можно применять в настоящем изобретении. Такой реагент для мультимеризации на основе PEG можно легко получать с помощью коммерчески доступных активированных молекул PEG (например производных PEG-NHS, доступных от NOF North America Corporation, Ирвайн, Калифорния, США, или активированных производных PEG, доступных от Creative PEGWorks, Чапел-Хилл, Северная Каролина, США) с аминогруппами мутантного варианта белка стрептавидина.
[0079] Под стрептавидином или стрептавидином дикого типа (wt-стрептавидином) предполагается аминокислотная последовательность, раскрытая в Argarana et al., Nucleic Acids Res. 14 (1986) 1871-1882. Мутантные варианты белка стрептавидина представляют собой полипептиды, которые отличаются по последовательности от стрептавидина дикого типа одной или более заменами, делециями или добавлениями аминокислот и которые сохраняют свойства связывания wt-стрептавидина. Стрептавидиноподобные полипептиды и мутантные варианты белка стрептавидина представляют собой полипептиды, которые фактически являются иммунологически эквивалентными стрептавидину дикого типа и, в частности, способны к связыванию с биотином, производными биотина или аналогами биотина с таким же или другим сродством по сравнению с wt-стрептавидином. Стрептавидиноподобные полипептиды или мутантные варианты белка стрептавидина могут содержать аминокислоты, не являющиеся частью стрептавидина дикого типа, или они могут содержать только часть стрептавидина дикого типа. Стрептавидиноподобные полипептиды также представляют собой полипептиды, не являющиеся идентичными стрептавидину дикого типа, поскольку реципиент не имеет ферментов, требуемых для превращения вырабатываемого реципиентом полипептида в структуру стрептавидина дикого типа. Термин "стрептавидин" включает также тетрамеры стрептавидина и димеры стрептавидина, в частности, гомотетрамеры стрептавидина, гомодимеры стрептавидина, гетеротетрамеры стрептавидина и гетеродимеры стрептавидина. Каждая субъединица обычно имеет связывающий участок для биотина или аналогов биотина или для стрептавидин-связывающих пептидов. Примеры стрептавидинов или мутантных вариантов белка стрептавидина упоминаются, например, в WO 86/02077, DE 19641876 A1, US 6022951, WO 98/40396 или WO 96/24606.
[0080] В предпочтительном варианте осуществления мутантные варианты белка стрептавидина, применяемые в качестве реагента для мультимеризации, представляют собой мутантные варианты белка стрептавидина, которые описаны в патенте США 6103493, а также в DE 196 41 876.3. Такие мутантные варианты белка стрептавидина имеют по меньшей мере одну мутацию внутри области аминокислотных положений 44-53 на основе аминокислотной последовательности стрептавидина дикого типа. Предпочтение отдается мутантным вариантам белка стрептавидина минимального размера, которые начинаются на N-конце в области аминокислот 10-16 стрептавидина дикого типа и заканчиваются на C-конце в области аминокислот 133-142 стрептавидина дикого типа. Примеры таких предпочтительных мутантных вариантов белка стрептавидина имеют гидрофобную алифатическую аминокислоту вместо Glu в положении 44, любую аминокислоту в положении 45, гидрофобную алифатическую аминокислоту в положении 46 и/или основную аминокислоту вместо Val в положении 47. Мутантный вариант белка стрептавидина может представлять собой мутантный вариант белка, содержащий аминокислотную последовательность Val44-Thr45-Ala46-Arg47 (SEQ ID NO: 02) в положениях последовательности 44-47, или мутантный вариант белка стрептавидина (аналог), содержащий аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 (SEQ ID NO: 03) в положениях 44-47 последовательности стрептавидина дикого типа. Такие мутантные варианты белка описаны, например, в патенте США 6103493 и коммерчески доступны от IBA GmbH в форме мутантного варианта белка "m1" и мутантного варианта белка "m2" под торговой маркой Strep-Tactin®.
[0081] Способ согласно настоящему изобретению можно в некоторых вариантах осуществления применять для обеднения образца реагентами, которые ранее применялись для размножения клеток. Первое или второе средство и соответствующий партнер в свободной форме (конкурирующее средство) могут, к примеру, на данный момент содержаться в элюате из вышеописанного способа размножения. С помощью способа согласно настоящему изобретению такие реагенты можно по меньшей мере фактически, в том числе полностью, удалять из образца, например из популяции клеток. В качестве иллюстративного примера, образец можно обеднять первым или вторым средством, определенным выше, до уровней, находящихся ниже предела выявления, например для FACS или вестерн-блоттинга. Конкурирующий реагент (первый или второй партнер по связыванию в свободной форме или его аналог) можно применять в целях прекращения и контроля размножения популяции клеток и удаления из нее средства для мультимеризации. Этот конкурирующий реагент может иметь связывающий участок, способный к специфическому связыванию связывающего участка Z аффинного реагента в "картридже для удаления" из WO 2013/124474. В таком варианте осуществления соответствующий способ по настоящему изобретению также может служить для обеднения первым и вторым средством и конкурирующим реагентом, в том числе для их удаления.
[0082] Способ согласно настоящему изобретению можно осуществлять при любой температуре, при которой жизнеспособность популяции клеток по меньшей мере фактически не нарушается. При упоминании в данном документе условий, по меньшей мере фактически не являющихся пагубными, не являющихся вредными или по меньшей мере фактически не подвергающими риску жизнеспособность условиями называют те, при которых процентная доля подлежащих размножению клеток в популяции, имеющих полную жизнеспособность, составляет по меньшей мере 70%, в том числе по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 99,5%. В некоторых вариантах осуществления способ согласно настоящему изобретению осуществляют при температуре приблизительно 20°C или выше. В зависимости от популяции клеток, подлежащей размножению, подходящий диапазон температур может составлять, например, от приблизительно 20°C до приблизительно 45°C, в том числе от приблизительно 25°C до приблизительно 40°C или от приблизительно 32°C до 37°C. В некоторых вариантах осуществления способ согласно настоящему изобретению осуществляют при постоянном значении температуры или при выбранном значении температуры ± приблизительно 5°C, ± приблизительно 4°C, ± приблизительно 3°C, ± приблизительно 2°C, ± приблизительно 1°C или ± приблизительно 0,5°C. Специалист в данной области способен определить подходящую температуру эмпирическим путем, принимая во внимание природу клеток и условия размножения. Обычно клетки человека размножают при такой температуре, как 37°C.
[0083] В дополнительном варианте осуществления настоящее изобретение обеспечивает способ размножения популяции клеток in vitro, включающий приведение образца, содержащего популяцию клеток, в контакт с реагентом для мультимеризации, где реагент для мультимеризации находится в растворимой форме, и на нем иммобилизовано (связано с ним) первое средство, которое передает первичный активационный сигнал клеткам. Реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для связывания первого средства, где первое средство содержит по меньшей мере одного партнера по связыванию C1, где партнер по связыванию C1 способен связываться со связывающим участком Z1 реагента для мультимеризации. Первое средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и первое средство связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки. Здесь следует четко отметить, что в случае применения растворимого средства для мультимеризации отсутствует необходимость в том, чтобы связь между партнером по связыванию C1 и связывающим участком Z1 была обратимой.
[0084] В варианте осуществления этого второго средства для мультимеризации на нем иммобилизовано (связано с ним) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток, где второе средство содержит партнера по связыванию C2, где партнер по связыванию C2 способен связываться со связывающим участком Z2 реагента для мультимеризации. Второе средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C2 и связывающим участком Z2, где второе средство связывается со вспомогательной молекулой на поверхности клеток, стимулируя тем самым активированные клетки.
[0085] В одном варианте осуществления этого второго способа связь, образующаяся между партнером по связыванию C1 и связывающим участком Z1, может быть необратимой и/или также связь, образующаяся между партнером по связыванию C2 и связывающим участком Z2, может быть необратимой.
[0086] В другом варианте осуществления данного второго способа связь, образующаяся между партнером по связыванию C1 и связывающим участком Z1, может быть обратимой. Связь, образующаяся между партнером по связыванию C2 и связывающим участком Z2, может быть также обратимой. В данном случае константа диссоциации (Kd) для обратимого связывания между указанным связывающим участком Z1 и указанным партнером по связыванию C1 и/или для обратимого связывания между указанным связывающим участком Z2 и указанным партнером по связыванию C2 может находиться в диапазоне от 10-2 M до 10-13 M.
[0087] В данном втором способе, основанном на применении растворимого реагента для мультимеризации, первый и второй реагент, а также реагент для мультимеризации и все другие реагенты и популяции клеток можно в иных отношениях применять таким же образом, как раскрыто выше для способа, в котором применяется обратимая связь между первым или вторым средством и реагентом для мультимеризации.
[0088] Настоящее изобретение дополнительно предусматривает набор реагентов для размножения популяции клеток, при этом набор содержит
(i) реагент для мультимеризации, где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z для обратимого связывания первого средства,
(ii) первое средство, которое связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки, где первое средство содержит по меньшей мере одного партнера по связыванию C1, причем партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
(iii) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток, где второе средство содержит партнера по связыванию C2, причем партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации с помощью связи, образующейся между партнером по связыванию C2 и связывающим участком Z2, где второе средство связывается со вспомогательной молекулой на поверхности клеток, стимулируя тем самым активированные клетки.
[0089] Настоящее изобретение обеспечивает также набор реагентов для размножения популяции клеток, при этом набор содержит
(i) реагент для мультимеризации, где реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z для обратимого связывания первого средства,
(ii) первое средство, которое связывается с рецепторной молекулой на поверхности клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым клетки, где первое средство содержит по меньшей мере одного партнера по связыванию C1, причем партнер по связыванию C1 способен связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1.
[0090] Этот второй набор реагентов может дополнительно содержать (iii) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток, где второе средство содержит партнера по связыванию C2, причем партнер по связыванию C2 способен связываться со связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C2 и связывающим участком Z2.
[0091] Набор, раскрытый в данном документе, в частности применяют в случае, когда популяция клеток представляет собой популяцию лимфоцитов.
[0092] В соответствии с вышеприведенным раскрытием настоящее изобретение также обеспечивает новые реагенты для мультимеризации и новую композицию, содержащую реагенты для мультимеризации, способные к обеспечению размножения популяции клеток. Такой реагент для мультимеризации, способный к обеспечению размножения популяции клеток, представляет собой реагент для мультимеризации, который находится в растворимой форме и содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства, которое передает первичный активационный сигнал клеткам, где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное первое средство, которое передает первичный активационный сигнал клеткам; где первое средство содержит по меньшей мере одного партнера по связыванию C1, причем партнер по связыванию C1 способен обратимо связываться по меньшей мере с одним связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации с помощью обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1. Здесь следует отметить, что на таком средстве для мультимеризации может быть иммобилизовано любое первое средство, описанное в данном документе.
[0093] Реагент для мультимеризации по настоящему изобретению может дополнительно содержать по меньшей мере один связывающий участок Z2 для обратимого связывания второго средства, которое стимулирует вспомогательную молекулу на поверхности клеток, где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток, где второе средство содержит партнера по связыванию C2, причем партнер по связыванию C2 способен связываться по меньшей мере с одним связывающим участком Z2 реагента для мультимеризации. В данном варианте осуществления второе средство связано с реагентом для мультимеризации посредством связи, образующейся между партнером по связыванию C2 и связывающим участком Z2.
[0094] В соответствии с вышеприведенным раскрытием такой реагент для мультимеризации способен также к обеспечению размножения популяции лимфоцитов или субпопуляции, содержащейся в популяции лимфоцитов. Популяция лимфоцитов, подлежащая размножению, может представлять собой любую подходящую популяцию, например, популяцию B-клеток, популяцию T-клеток или популяцию естественных клеток-киллеров. Популяция T-клеток может представлять собой популяцию антиген-специфических T-клеток, популяцию T-клеток-хелперов, популяцию цитотоксических T-клеток, T-клеток памяти, регуляторных T-клеток или естественных T-клеток-киллеров. Соответственно в таких вариантах осуществления реагента для мультимеризации первое средство способно стимулировать передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках. Первое средство, присутствующее в реагенте для мультимеризации, может таким образом представлять собой связывающий реагент, который специфически связывается с CD3, а второе средство, которое связывается со вспомогательной молекулой, может представлять собой связывающее средство, которое специфически связывается с CD28 или CD137.
[0095] В вариантах осуществления реагента для мультимеризации первое средство, которое специфически связывается с CD3, может представлять собой антитело к CD3, бивалентный фрагмент антитела из антитела к CD3, моновалентный фрагмент антитела из антитела к CD3 и/или белковую молекулу с антителоподобными свойствами связывания, связывающуюся с CD3. В данных вариантах осуществления второе средство, которое специфически связывается с CD28 или CD137, может представлять собой антитело к CD28, бивалентный фрагмент антитела из антитела к CD28, моновалентный фрагмент антитела из антитела к CD28, белковую молекулу, связывающуюся с CD28, с антителоподобными свойствами связывания, антитело к CD137, бивалентный фрагмент антитела из антитела к CD137, моновалентный фрагмент антитела из антитела к CD137, белковую молекулу, связывающуюся с CD137, с антителоподобными свойствами связывания, лиганд 4-1BB и любую их смесь. Таким образом, на реагенте для мультимеризации по настоящему изобретению могут, как правило, быть иммобилизованы один тип первого средства и смесь вторых средств, например антитело к CD3 в качестве первого средства, и например антитело к CD28 и лиганд 4-1BB в качестве (объединенных) вторых средств.
[0096] Если реагент для мультимеризации подлежит применению для размножения B-клеток, то первое средство, иммобилизованное на реагенте для мультимеризации, может представлять собой связывающий реагент, который специфически связывается с CD40 или CD137. В соответствии с настоящим раскрытием, приведенным в данном документе, в таких вариантах осуществления первый связывающий реагент, который специфически связывается с CD40 или CD137, может быть выбран из антитела к CD40, бивалентного фрагмента антитела из антитела к CD40, моновалентного фрагмента антитела из антитела к CD40 и белковой молекулы, связывающейся с CD40, с антителоподобными свойствами связывания или антитела к CD137, бивалентного фрагмента антитела из антитела к CD137, моновалентного фрагмента антитела из антитела к CD137, белковой молекулы, связывающейся с CD137, с антителоподобными свойствами связывания и лиганда CD40 (CD154).
[0097] Также в соответствии с общим раскрытием настоящего изобретения в реагенте для мультимеризации, описанном в данном документе, связывающие участки Z1 и Z2 реагента для мультимеризации могут быть идентичными. Как описано выше, такой реагент для мультимеризации может содержать олигомер или полимер стрептавидина, олигомер или полимер авидина, олигомер или полимер аналога стрептавидина, который обратимо связывается с биотином, олигомер или полимер аналога авидина, который обратимо связывается с биотином, реагент, содержащий по меньшей мере две хелатные группы K, где по меньшей мере две хелатные группы способны к связыванию с ионом переходного металла, тем самым делая реагент способным к связыванию с олигогистидиновой аффинной меткой, мультимерной глутатион-S-трансферазой, мультимерным кальмодулином и биотинилированным белком-носителем.
[0098] Новая композиция, обеспечиваемая в данном документе, которая способна обеспечивать размножение популяции клеток, может содержать:
(i) первый реагент для мультимеризации, где первый реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства, которое передает первичный активационный сигнал клеткам, где на первом реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное первое средство, которое передает первичный активационный сигнал клеткам, где первое средство содержит по меньшей мере одного партнера по связыванию C1, причем партнер по связыванию C1 способен обратимо связываться по меньшей мере с одним связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации с помощью обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1, и
(ii) второй реагент для мультимеризации, где второй реагент для мультимеризации находится в растворимой форме и содержит по меньшей мере один связывающий участок Z2 для обратимого связывания второго средства, которое стимулирует вспомогательную молекулу на поверхности клеток, где на реагенте для мультимеризации обратимо иммобилизовано (связано с ним) указанное второе средство, которое стимулирует вспомогательную молекулу на поверхности клеток, где второе средство содержит партнера по связыванию C2, причем партнер по связыванию C2 способен связываться по меньшей мере с одним связывающим участком Z2 реагента для мультимеризации, где второе средство связано с реагентом для мультимеризации с помощью связи, образующейся между партнером по связыванию C2 и связывающим участком Z2.
[0099] Такая новая композиция представляет собой, например, реакционную смесь, применяемую в примере 13, в котором два отдельных реагента для мультимеризации были функционализированы либо Fab-фрагментом αCD3 в отдельности, либо Fab-фрагментом αCD28 в отдельности. В данном случае следует отметить, что в примере 13 было продемонстрировано, что такая композиция характеризуется такой же эффективностью размножения, как и отдельный реагент для мультимеризации, на котором совместно иммобилизованы как первое средство, так и второе средство. Таким образом, комбинированное применение двух или более реагентов для мультимеризации, функционализированных по отдельности только одним типом реагента (например одним первым или одним вторым средством), функционально эквивалентно применению для размножения одного объединенного реагента для мультимеризации, на котором иммобилизованы как первое средство, так и второе средство. В данном случае следует также отметить, что реагент для мультимеризации по настоящему изобретению может быть функционализирован таким количеством средств (например, одним, двумя, тремя, четырьмя или даже более средствами), которое предполагается для применения в размножении выбранной популяции клеток. Третье или четвертое средство может, например, передавать стимул к размножению желаемой субпопуляции клеток. К примеру, см. в данном случае пример 13, в котором растворимые реагенты для мультимеризации были обратимо функционализированы тремя реагентами, а именно, Fab-фрагментом αCD3 в качестве первого реагента, Fab-фрагментом αCD28 в качестве второго реагента и Fab-фрагментом αCD8 в качестве третьего реагента для обогащения субпопуляции CD8+ T-клеток в образце популяции CD3+ T-клеток (лимфоцитов). С помощью применения таких комбинаций средств, все из которых могут быть обратимо иммобилизованы на одном и том же реагенте для мультимеризации, настоящее изобретение предоставляет возможность преимущественного размножения или избирательного обогащения любой желаемой (суб)популяции клеток из образца, который содержит например ряд различных субпопуляций. В данном случае следует отметить, что также возможно, однако, применять для данной цели три различных реагента для мультимеризации, например, первый реагент для мультимеризации, функционализированный только Fab-фрагментом αCD3, второй реагент для мультимеризации, функционализированный Fab-фрагментом αCD28, и третий реагент для мультимеризации, функционализированный Fab-фрагментом αCD8. Аналогично, возможно применять только два различных реагента для мультимеризации, первый реагент для мультимеризации, функционализированный только Fab-фрагментом αCD3, и второй реагент для мультимеризации, функционализированный как Fab-фрагментом αCD28, так и Fab-фрагментом αCD8. Соответственно, настоящее изобретение позволяет конструировать любой тип желаемого реагента для размножения модульным способом.
[00100] Настоящее изобретение обеспечивает также способ серийного размножения популяции лимфоцитов in vitro, где популяция лимфоцитов содержит T-клетки. Данный способ включает
приведение образца, который содержит популяцию лимфоцитов, содержащую T-клетки, в контакт с реагентом для мультимеризации,
где реагент для мультимеризации находится в растворимой форме и на нем обратимо иммобилизовано (i) первое средство, которое передает первичный активационный сигнал T-клеткам, и (ii) второе средство, которое стимулирует вспомогательную молекулу на поверхности T-клеток,
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z1 для обратимого связывания первого средства,
где первое средство содержит по меньшей мере одного партнера по связыванию C1, причем партнер по связыванию C1 способен обратимо связываться со связывающим участком Z1 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C1 и связывающим участком Z1,
где реагент для мультимеризации содержит по меньшей мере один связывающий участок Z2 для обратимого связывания второго средства,
где второе средство содержит по меньшей мере одного партнера по связыванию C2, причем партнер по связыванию C2 способен обратимо связываться со связывающим участком Z2 реагента для мультимеризации, где первое средство связано с реагентом для мультимеризации посредством обратимой связи, образующейся между партнером по связыванию C2 и связывающим участком Z2,
где первое средство связывается с рецепторной молекулой на поверхности T-клеток, передавая тем самым первичный активационный сигнал клеткам и активируя тем самым T-клетки,
где второе средство связывается со вспомогательной молекулой на поверхности T-клеток, стимулируя тем самым активированные клетки, и при этом первое средство и второе средство тем самым совместно индуцируют размножение
T-клеток.
[00101] В данном способе приведение образца, содержащего популяцию лимфоцитов, которая в свою очередь содержит популяцию T-клеток, в контакт с растворимым реагентом для мультимеризации, на котором иммобилизованы первое и второе средство, приводит к специфичному связыванию T-клеток с реагентом для мультимеризации.
[00102] Приведение образца, который содержит популяцию лимфоцитов, содержащую T-клетки, в контакт с реагентом для мультимеризации можно осуществлять в биореакторе, таком как половолоконный биореактор (например половолоконный биореактор системы для размножения клеток Quantum®) или биореактор в виде пластикового мешка (например Cellbag®, применяемый в системе для размножения клеток Xuri W25 от GE Healthcare).
[00103] Данный способ дополнительно включает приведение популяции лимфоцитов (реакционной смеси, содержащей T-клетки, связанные с реагентом для мультимеризации посредством первого средства и второго средства) в контакт с (i) первым партнером по связыванию C1 в свободной форме или его аналогом, способным к разрушению связи между первым партнером по связыванию C1 и связывающим участком Z1, и (ii) вторым партнером по связыванию C2 в свободной форме или его аналогом, способным к разрушению связи между вторым партнером по связыванию C2 и связывающим участком Z2. Тем самым, обратимая связь между указанным партнером по связыванию C1 первого средства и указанными связывающими участками Z1, а также обратимая связь между указанным партнером по связыванию C2 второго средства и указанным связывающим участком Z2 указанного реагента для мультимеризации разрушается с выведением тем самым в виде элюата T-клеток, связанных с реагентом для мультимеризации посредством первого средства и второго средства, и остановкой размножения T-клеток.
[00104] В данном способе элюат (реакционная смесь, в которой реакция размножения была прекращена с помощью добавления первого(-ых) партнера(-ов) в свободной форме или их аналогов), содержащий размножившуюся популяцию T-клеток, можно подвергать хроматографии на подходящей (первой) неподвижной фазе. Неподвижная (первая) фаза может представлять собой матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии, описанную в Международной заявке на патент WO 2013/124474. Такая матрица для гель-фильтрации и/или аффинной хроматографии содержит аффинный реагент, где аффинный реагент содержит связывающий участок Z1 и/или Z2, с которым специфически связывается партнер по связыванию C1 и/или C2, содержащийся в первом средстве или втором средстве. Тем самым, первое средство, второе средство, первый партнер по связыванию C1 и/или второй партнер по связыванию C2 в свободной форме иммобилизуются на неподвижной фазе. В данном способе первая неподвижная фаза находится в жидкостном соединении с биореактором.
[00105] В одном из вариантов осуществления этого серийного размножения связывающие участки Z1 и Z2 средства для мультимеризации являются идентичными. В дополнение, можно применять одно средство для мультимеризации. Если применяют растворимое средство для мультимеризации, то популяцию T-клеток (или размножившуюся популяцию клеток в общем) отделяют от растворимого реагента для мультимеризации. Отделение/удаление можно осуществлять с помощью второй неподвижной фазы. Для данной цели смесь, содержащую T-клетки и растворимый реагент для мультимеризации, до или после нанесения на первую вышеописанную неподвижную фазу подвергают хроматографии на подходящей второй неподвижной фазе. Эта вторая неподвижная фаза может представлять собой матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии, где матрица для гель-фильтрации и/или аффинной хроматографии содержит аффинный реагент. Аффинный реагент, содержащийся в смоле для хроматографии, содержит партнера по связыванию D, который (специфически) связывается со связывающим участком Z1 и/или, при его наличии, со связывающим участком Z2 реагента для мультимеризации с иммобилизацией тем самым реагента для мультимеризации на неподвижной фазе. Если применяется средство для мультимеризации на основе стрептавидина, и при этом как первое, так и второе средства имеют стрептавидин-связывающий пептид в качестве партнера по связыванию C1 или C2, то партнер по связыванию D, содержащийся в аффинном реагенте этой второй неподвижной фазы, может представлять собой биотин. Растворимый олигомер стрептавидина или мутантный вариант белка стрептавидина, применяемый в качестве реагента для мультимеризации, связывается затем с биотином, который обычно ковалентно связан с матрицей для хроматографии, такой как биотин-SepharoseTM, которая является коммерчески доступной.
[00106] В данном способе серийного размножения первое средство может стимулировать передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках, и первое средство может таким образом представлять собой связывающий реагент, который специфически связывается с CD3. В дополнение, вспомогательная молекула на T-клетке может представлять собой CD28. В таком случае второе средство, которое связывается со вспомогательной молекулой, представляет собой связывающий реагент, который специфически связывается с CD28.
[00107] В данном способе серийного размножения T-клетки можно трансфицировать либо в ходе размножения, либо после него, например T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR, также известный как искусственный T-клеточный рецептор). Эту трансфекцию для введения гена желаемого рецептора можно осуществлять, например, с помощью любого подходящего ретровирусного вектора. Генетически модифицированную популяцию клеток можно затем избавлять от первоначального стимула (например стимула CD3/CD28) и после этого стимулировать вторым типом стимула, например посредством введенного de novo рецептора. Этот второй тип стимула может включать в себя антигенный стимул в форме комплекса пептид/молекула MHC, когнатного (перекрестно связывающегося) лиганда введенного генетическим путем рецептора (например природного лиганда CAR) или любого лиганда (такого как антитело), который непосредственно связывается с каркасом нового рецептора (например путем распознавания константных областей в рецепторе). См. в данном отношении Cheadle et al, "Chimeric antigen receptors for T-cell based therapy" Methods Mol Biol. 2012; 907:645-66 или Barrett et al., Chimeric Antigen Receptor Therapy for Cancer, Annual Review of Medicine Vol. 65: 333-347 (2014).
[00108] В данном способе популяция лимфоцитов, содержащая T-клетки, может представлять собой популяцию мононуклеарных клеток периферической крови (PBMC) или популяцию обогащенных либо очищенных T-клеток. Популяция лимфоцитов может быть, например, получена из цельной крови или из продукта афереза немобилизованной крови или замороженного тканевого препарата.
[00109] В данном способе серийного размножения, основанном на применении растворимого реагента для мультимеризации, первый и второй реагент, а также реагент для мультимеризации и все другие реагенты и популяции клеток можно в иных отношениях применять таким же образом, как раскрыто выше для способа, в котором применяется обратимая связь между первым или вторым средством и реагентом для мультимеризации.
[00110] Настоящее изобретение дополнительно относится к системе из биореактора и первой неподвижной фазы для хроматографии. Биореактор подходит для размножения клеток, а неподвижная фаза подходит для разделения клеток и удаления реагентов. Первая неподвижная фаза представляет собой матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии, где матрица для гель-фильтрации и/или аффинной хроматографии содержит аффинный реагент, где аффинный реагент содержит связывающий участок Z1, с которым специфически связывается партнер по связыванию C1, содержащийся в первом средстве, и/или аффинный реагент содержит связывающий участок Z2, с которым специфически связывается партнер по связыванию C2, содержащийся во втором средстве. Таким образом, первая неподвижная фаза является подходящей для иммобилизации на ней первого средства и/или второго средства, первого партнера по связыванию C1 и/или второго партнера по связыванию C2 в свободной форме. В дополнение, биореактор и неподвижная фаза находятся в жидкостном соединении. Такую систему можно применять в серийном размножении, поясняемом выше, и можно интегрировать в известные системы для размножения клеток, такие как система для размножения клеток Quantum® или система для размножения клеток Xuri W25.
[00111] В данной системе первая неподвижная фаза содержится в хроматографической колонке или представляет собой плоскую неподвижную фазу. Система может дополнительно содержать вторую неподвижную фазу, которая находится в жидкостном соединении с первой неподвижной фазой. Вторая неподвижная фаза может представлять собой матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии, где матрица для гель-фильтрации и/или аффинной хроматографии содержит аффинный реагент. Этот аффинный реагент может содержать партнера по связыванию D, который (специфически) связывается со связывающим участком Z1 реагента для мультимеризации и тем самым подходит для иммобилизации реагента для мультимеризации на неподвижной фазе.
[00112] Настоящее изобретение дополнительно относится к устройству для очистки и размножения популяции клеток, при этом устройство содержит по меньшей мере одну систему из биореактора и первой неподвижной фазы или второй неподвижной фазы для хроматографии, как определено выше.
[00113] Устройство может дополнительно содержать множество систем из биореактора и неподвижной фазы, находящихся в последовательном жидкостном соединении.
[00114] Устройство может содержать входной канал для образца, находящийся в жидкостном соединении с биореактором системы из биореактора и неподвижной фазы для хроматографии. Устройство также может содержать выходной канал для образца для очищенных и размножившихся целевых клеток, при этом выходной канал для образца находится в жидкостном соединении с неподвижной фазой последней из по меньшей мере одной системы из биореактора и неподвижной фазы для хроматографии.
[00115] Наконец, устройство может быть сконструировано в виде функционально закрытой системы.
[00116] Как будет без труда понятно среднему специалисту в данной области из раскрытия настоящего изобретения, другие смеси веществ, химически или механически связанных между собой, средства, пути применения, способы или этапы, которые существуют в настоящее время или будут разработаны позже, выполняющие практически ту же функцию или обеспечивающие достижение практически того же результата, что и в соответствующих приводимых в качестве примера вариантах осуществления, описанных в данном документе, можно аналогичным образом использовать в соответствии с настоящим изобретением.
[00117] Экспериментальные примеры
Пример 1. Стимуляция/размножение CD3+ иммунореактивных T-клеток при помощи Fab-фрагментов αCD3/αCD28, которые были обратимо иммобилизованы на гранулах, покрытых мутантным вариантом белка стрептавидина Strep-Tactin®
[00118] 300000 CD3+ CD62L- иммунореактивных T-клеток (Tresp, выделенных с помощью серийного магнитного обогащения из продукта афереза немобилизованной крови донора) метили при помощи 3 мкМ CFSE и стимулировали 5 мкл из 15 мкл препарата гранул Strep-Tactin® (10 мг магнитных частиц/мл, нагруженных 35 мкг гранул Strep-Tactin®/мг), нагруженных 0,5 мкг Fab-фрагмента αCD3 в отдельности, 0,5 мкг Fab-фрагмента αCD28 в отдельности или смесью 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab αCD28.
[00119] Применяемый Fab-фрагмент αCD3 получали из связывающегося с CD3 моноклонального антитела, вырабатываемого линией гибридомных клеток OKT3. Линия гибридомных клеток OKT3 и антитело OKT3 описаны в патенте США 4361549, линия клеток была депонирована в ATCC® под номером доступа CRL-8001™. Применяемый Fab CD28 получали из моноклонального антитела CD28.3 к CD28 человека (Vanhove et al, BLOOD, 15 июля 2003 г., том 102, № 2, стр. 564-570). Нуклеотидная последовательность вариабельных доменов этого антитела CD28.3 была депонирована в GenBank в форме синтетического одноцепочечного конструкта Fv антитела к CD28 человека scFv28.3 под номером доступа в GenBank AF451974.1.
[00120] Оба Fab-фрагмента получали рекомбинантным путем в E. Coli, как описано в Международных заявках на патенты WO 2013/011011 и WO 2013/124474, несущих в качестве константных доменов (CH1 и C-каппа) консенсусную последовательность IgG1. Тяжелая цепь обоих Fab-фрагментов была слита на карбокси-конце с двумя последовательно расположенными стрептавидин-связывающими модулями (SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 07), коммерчески доступными как "Twin-Strep-tag®" от IBA GmbH, Геттинген, Германия). Fab-фрагмент αCD3 применяли в качестве первого средства, при этом стрептавидин-связывающий пептид служил в качестве партнера по связыванию C1, а Fab-фрагмент αCD28 применяли в качестве второго средства, при этом стрептавидин-связывающий пептид служил в качестве партнера по связыванию C2. Мутантный (тетрамерный) вариант белка стрептавидина "Strep-Tactin®" служит в качестве реагента для мультимеризации, на котором были обратимо иммобилизованы оба Fab-фрагмента.
[00121] В эксперименте по размножению стимулированные холостыми гранулами клетки Tresp (без Fab), служили в качестве отрицательного контроля. Клетки Tresp высевали в трех повторностях в 48-луночные планшеты вместе с 300000 CD3 аутологичных питающих клеток (облученных 30 Гр) в 3 мл полной среды для культивирования клеток (RPMI (Gibco), дополненной 10% (объем/объем) фетальной телячьей сывороткой, L-глутамином, b-меркаптоэтанолом, HEPES, пенициллином, стрептомицином и гентамицином), дополненной 10 ед/мл интерлейкина-2 (IL-2). Клетки инкубировали при 37°C без замены среды и анализировали через 4 дня с помощью FACS-анализа. FACS-окрашивание и FACS-анализ проводили спустя 10 мин. инкубирования со 100 мкМ D-биотина. По одному иллюстративному графику на каждое условие показано на фиг. 4. На графиках показаны живые CD3+ клетки, которые окрашивали йодидом пропидия (PI) для установления различий между живыми/мертвыми клетками. Фиг. 4a представляет собой гистограмму, на которой показано распределение стимулированных клеток по размеру (прямое рассеяние). На фиг. 4a показано, что определенную клеточную популяцию клеток Tresp стимулировали и размножали (увеличение размера/количества по сравнению с нестимулированным контролем "только гранулы") при инкубировании в присутствии гранул, на которых была иммобилизована смесь 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab αCD28, после стимуляции in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на гранулах, покрытых мутантным вариантом белка стрептавидина Strep-Tactin®. На фиг. 4B изображены гистограммы разбавления красителя CFSE для оценки пролиферации, на которых представлена степень пролиферации соответственно количеству клеток на число клеточных делений (указанное в верхней части фиг. 4B, 0 означает неделившиеся клетки; 5 означает клетки, прошедшие по меньшей мере 5 делений). Из фиг. 4B можно видеть, что популяция T-клеток, стимулированных гранулами, на которых была иммобилизована смесь 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab αCD28, большей частью прошла три клеточных деления и демонстрирует более однородный паттерн пролиферации, чем с одним стимулом в отдельности (небольшое количество клеток в границах пика для неделившихся клеток "0"). Повышение абсолютной величины пролиферации (большее количество клеток характеризовалось однородной пролиферацией спустя 4 дня стимуляции гранулами, функционализированными при помощи αCD3 и αCD28) также представлено более интенсивным потреблением среды, что изображено изменением цвета индикатора в желтый на фиг. 4C.
Пример 2. Анализ дифференциальной мобилизации внутриклеточного кальция в клетках Jurkat
[00122] Здесь рассматривали проточный цитометрический анализ в режиме реального времени дифференциальной мобилизации внутриклеточного кальция, индуцированной в клетках Jurkat, меченных антителом αCD3 клона OKT3 или Fab-фрагментами OKT3, мультимеризованными при помощи Strep-Tactin®.
[00123] Для данной цели клетки Jurkat нагружали кальций‐чувствительным красителем Indo-1-AM, и высвобождение кальция запускали с помощью инъекции моноклонального антитела αCD3 OKT3 (вырабатываемого линией гибридомных клеток OKT3, см. выше, черные квадраты) либо Fab-фрагментов αCD3 (полученных из исходной линии клеток OKT3), которые были мультимеризованы с помощью обратимого связывания их стрептавидин-связывающего пептида с растворимым Strep-Tactin, конъюгированным с флуоресцентным фикоэритрином. В случае интактных комплексов Fab-мультимера OKT3 со Strep-Tactin высвобождение кальция запускали в течение такого же периода времени, как и в случае с исходным клоном антитела (темно-серые треугольники). Активации клеток можно полностью избежать с помощью инъекции обработанных D-биотином предиссоциированных комплексов Fab-Strep-Tactin (темно-серые кружки), так же как и в случае инъекции отрицательного контроля PBS (перевернутые белые треугольники). Применение иономицина служило в качестве положительного контроля для притока кальция. Изменения внутриклеточной концентрации Ca2+ с временным разрешением отслеживали с помощью проточной цитометрии на основе изменения соотношения FL6/FL7. Из фиг. 5A можно видеть, что как исходное антитело OKT3, так и мультимеризованный моновалентный Fab-фрагмент OKT3 вызывали высвобождение кальция, что означает, что мультимеризованный моновалентный Fab-фрагмент OKT3 является фактически таким же функциональным, как и исходное антитело. Примечательно, что мультимерный Fab-фрагмент OKT3 был неспособен запускать высвобождение кальция, если к Strep-Tactin, на котором был иммобилизован Fab-фрагмент OKT3, добавляли биотин перед добавлением комплекса Fab-фрагмент OKT3-Strep-Tactin. В таком случае биотин разрушал обратимую связь, образующуюся между Strep-Tactin в качестве средства для мультимеризации и Fab-фрагментом OKT3. Таким образом, моновалентный Fab-фрагмент оттеснялся от средства для мультимеризации и после диссоциации был неспособен запускать высвобождение кальция с помощью связывания с CD3 клеток Jurkat.
[00124] В экспериментах, показанных на фиг. 5B, клетки Jurkat, меченные Indo-1-AM, активировали комплексами полученных из OKT3 Fab αCD3 со Strep-Tactin, описанными на фиг. 5a. Инъекция интактных (верхний график) или предиссоциированных комплексов (нижний график) служила соответственно в качестве положительных или отрицательных контролей. В дополнение, стимуляция клеток интактными комплексами Fab-Strep-Tactin с последующей дальнейшей инъекцией D-биотина (вблизи пика активации при t=140 с) приводила к резкому нарушению передачи сигнала от Fab-мультимера αCD3 (средний график). Инъекция иономицина в группе предиссоциированного комплекса Fab служила в качестве положительного контроля. Данные представляют три различных эксперимента. Важно отметить, что на фиг. 5B показано, что добавление D-биотина к образцу вызывает быстрое оттеснение Fab-фрагмента от средства для мультимеризации Strep-Tactin, с помощью чего происходит эффективное прекращение высвобождения кальция даже при продолжающейся стимуляции мобилизации кальция, что демонстрирует, что диссоциированный Fab-фрагмент OKT3 больше не обладает какой-либо биологической активностью. Аналогично, мультимерный Fab-фрагмент OKT3 также был неспособен запускать высвобождение кальция при добавлении биотина к комплексу Fab-мультимера-фрагмента OKT3 со Strep-Tactin перед добавлением образца Fab OKT3-Strep-Tactin к клеткам Jurkat.
Пример 3. Обратимое окрашивание клеток Fab-мультимерами, связывающимися с CD3
[00125] В данном примере рассматривали обратимое окрашивание клеток Fab-мультимерами, связывающимися с CD3. Свежевыделенные PBMC окрашивали либо моноклональным антителом αCD3 клона OKT3 (левая точечная диаграмма, исходный клон для Fab-мультимеров), либо когнатными меченными фикоэритрином (PE) Fab-мультимерами OKT3 и анализировали либо до (второй слева столбец), либо после обработки D-биотином (средний столбец). Затем после последующих этапов промывания выявляли оставшиеся Fab-мономеры с помощью свежеприготовленного меченного PE Strep-Tactin® (второй столбец справа). Вторичное окрашивание Fab-мультимером обратимо окрашенных клеток служило в качестве контроля (правый столбец). На фиг. 6 показаны только живые CD3+ клетки, отрицательные по окрашиванию йодидом пропидия (PI) для установления различий между живыми/мертвыми клетками. Числа на точечных диаграммах указывают на процентную долю клеток внутри окон. Данный эксперимент демонстрирует, что окрашивание CD3+ PBMC Fab-фрагментом антитела к CD3, мультимеризованным при помощи Strep-Tactin в качестве реагента для мультимеризации, является полностью обратимым при добавлении D-биотина, и что моновалентный Fab-фрагмент в отдельности не связывается с молекулой CD3, присутствующей на PBMC.
Пример 4. Обратимое выделение клеток с помощью Fab-мультимеров, связывающихся с CD28
[00126] В данном примере демонстрируется выделение клеток с помощью обратимого связывания Fab-фрагментов антитела к CD28, мультимеризованных при помощи магнитных частиц со Strep-Tactin® (магнитные частицы доступны от IBA GmbH, Геттинген, Германия). Для данной цели применяли Fab-фрагменты, полученные из антитела CD28.3, описанного в примере 1 выше. CD28+ клетки отбирали/выделяли с помощью магнитного отбора клеток при помощи Fab-мультимеров из свежевыделенных PMBC, как фактически описано в Международной заявке на патент WO 2013/011011. Перед отбором клетки подвергали контрольному окрашиванию либо когнатными флуоресцентными αCD28-мультимерами (левая точечная диаграмма), либо антителом, направленным на легкую каппа-цепь иммуноглобулина (вторая слева точечная диаграмма, mAb α-Ig-каппа), в качестве контрольного окрашивания. После отбора CD28+ клетки обрабатывали D-биотином и после этого промывали для удаления магнитных гранул и Fab-мономеров. Отделенные CD28+ клетки после этого (повторно) окрашивали либо Fab-мультимерами CD28 (вторая справа точечная диаграмма), либо mAb α-Ig-каппа (правая точечная диаграмма) для выявления возможных оставшихся Fab-мономеров. Показаны только живые (PI-отрицательные) CD3+ клетки. Числа на точечных диаграммах указывают на процентную долю клеток внутри окон. На фиг. 7 показано, что CD28+ клетки можно выделять из PMBC с помощью такого мультимеризованного Fab-фрагмента антитела к CD28, и что все реагенты для выделения, в том числе мономерные Fab антитела к CD28, после отбора можно удалять.
Пример 5. Стимуляция/размножение CD3+ иммунореактивных T-клеток при помощи Fab-фрагментов αCD3/αCD28, которые были обратимо иммобилизованы на растворимом Strep-Tactin
[00127] В данном примере CD3+ иммунореактивные T-клетки (выделенные с помощью магнитного отбора из образца свежих PBMC, полученных из градиента фиколла) размножали после стимуляции in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном Strep-Tactin®, выступающем в качестве растворимого реагента для мультимеризации. Олигомерный Strep-Tactin® получали путем полимеризации Strep-Tactin® при помощи сульфо-SMCC (сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилата, продукта № 22122 Thermo Scientific) и иминотиолана (продукта № 26101 Thermo Scientific) согласно протоколу производителя (Thermo Scientific). Олигомерный стрептавидин отделяли от мономерного (непрореагировавшего) и димерного мутантного варианта белка стрептавидина с помощью эксклюзионной хроматографии, и полученную таким образом фракцию олигомерного мутантного варианта белка стрептавидина (n≥3) применяли в качестве растворимого реагента для мультимеризации.
[00128] Для размножения in vitro 300000 CD3+ иммунореактивных T-клеток (Tresp) метили 2 мкМ сукцинимидилового сложного эфира карбоксифлуоресцеина (CFSE) и стимулировали различными количествами препарата растворимых олигомеров Strep-Tactin®, на которых была иммобилизована комбинация вышеописанных Fab-фрагмента αCD3 OKT3 и Fab-фрагмента αCD28 из антитела 28.3 (оба из которых несли на тяжелой цепи вышеупомянутый Twin-Strep-tag® в качестве стрептавидин-связывающего пептида). ("1x" соответствует 3 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг мономерного Fab-фрагмента αCD3 и 0,5 мкг мономерного Fab-фрагмента αCD28, числа "0,5x", "2x" и "5x" указывают на соответствующую величину n кратности "1x"). Клетки Tresp оставляли либо нестимулированными, либо стимулировали холостыми мультимерами Strep-Tactin (без Fab), служившими в качестве отрицательных контролей. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты вместе с 300000 CD3-отрицательных аутологичных питающих клеток (облученных 30 Гр) в 1 мл среды для культивирования клеток, дополненной 20 ед/мл IL-2. Клетки инкубировали при 37°C без замены среды, и пролиферацию анализировали по разведению CFSE через 5 дней с помощью FACS-анализа. На фиг. 8A показано расширение распределения пролиферирующих клеток по размеру через 5 дней в культуре по сравнению с отрицательными контролями. На фиг. 8B показано, что CD3+ клетки Tresp подвергались надлежащей стимуляции и интенсивно пролиферировали при инкубировании с растворимым олигомерным Strep-Tactin® (по сравнению с твердыми магнитными частицами Strep-Tactin на фиг. 4), на котором была иммобилизована смесь Fab-фрагментов αCD3 и αCD28. Результаты на фиг. 8a и 8b указывают на то, что в таких условиях in vitro большинство CD3+ иммунореактивных T-клеток делилось (от 2 до 5 клеточных делений) после вовлечения поверхностного CD28 и комплекса TCR/CD3 при помощи Fab-фрагментов αCD3 и αCD28, которые были обратимо иммобилизованы на растворимых олигомерах Strep-Tactin®. После размножения in vitro растворимые стимулирующие реагенты Fab-Strep-Tactin подвергали диссоциации и удаляли после обработки D-биотином. Диссоциацию и удаление Fab-мономеров-фрагментов подвергали проточному цитометрическому анализу с помощью повторного окрашивания клеток при помощи Strep-Tactin®, меченного фикоэритрином (ST-PE). Иллюстративная гистограмма (темно-серая гистограмма) показана в сравнении с соответствующим отрицательным контролем только с ST-PE (светло-серая гистограмма). Из фиг. 8C можно видеть, что оба Fab-фрагмента полностью диссоциировали и в полной мере удалялись из размножившихся клеток. На фиг. 8D показано абсолютное количество (отрицательных по трипановому синему) клеток через 5 дней. Количество подсчитывали с помощью счетной камеры Нойбауэра и откладывали на графике в зависимости от соответствующего условия стимуляции. На фиг. 8D показаны медианные значения количества клеток; планки погрешностей указывают на стандартное отклонение (SD). На фиг. 8D показано, что все смеси Fab-фрагментов αCD3 и Fab-фрагментов αCD28, которые были иммобилизованы на растворимом реагенте для мультимеризации Strep-Tactin, были в равной степени эффективными при размножении CD3+ клеток и обусловливали примерно 4-кратное увеличение абсолютных значений количества клеток.
Пример 6. Кинетические характеристики пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток, стимулированных in vitro обратимыми Fab-мультимерами αCD3/αCD28 Streptamer без замены среды
[00129] В данном примере рассматривали кинетические характеристики размножения для пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток (Tresp), которые стимулировали in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимых олигомерных мутантных вариантах белка стрептавидина. Для данной цели растворимый олигомерный мутантный вариант белка Strep-Tactin® двух различных размеров служил в качестве растворимого реагента для мультимеризации. Первый тип олигомерного Strep-Tactin® представлял собой фракцию олигомерного мутантного варианта белка стрептавидина (n≥3), полученную в примере 5 (также называемую в данном документе "традиционным остовом Strep-Tactin®", проиллюстрированным символом-треугольником, обращенным на фиг. 13 вершиной вверх). Второй тип этого олигомерного мутантного варианта белка стрептавидина, применяемого в качестве растворимого реагента для мультимеризации, представлял собой олигомерный мутантный вариант белка стрептавидина (n≥3), который подвергали реакции с биотинилированным сывороточным альбумином человека (также называемый в данном документе "крупным остовом Strep-Tactin®").
[00130] В данном примере 500000 очищенных CD4+ или CD8+ иммунореактивных T-клеток (Tresp) стимулировали по отдельности этими двумя различными мультимерами Streptamer, как поясняется выше, т. е. либо остовом Strep-Tactin из примера 5 (с применением раствора с концентрацией 1 мг олигомерного мутантного варианта белка стрептавидина/мл), либо крупными остовами Strep-Tactin (0,1 мг/мл). 3 мкл обоих различных остовов также нагружали комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28, применяемых в более ранних примерах, которые несли стрептавидин-связывающий пептид SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 07) на C-конце тяжелой цепи Fab-фрагмента. В дополнение, 4,5 мкл традиционного остова Strep-Tactin нагружали 0,5 мкг Fab-фрагмента αCD3, 0,5 мкг Fab-фрагмента αCD8 (IBA GmbH, Геттинген, который также несет на C-конце Fab-фрагмента стрептавидин-связывающий пептид SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 07)) и 0,5 мкг Fab-фрагмента αCD28. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, стимулированные коммерчески доступными Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток (RPMI 1640 (Gibco), дополненной 10% (объем/объем) фетальной телячьей сывороткой, 0,025% (вес/объем) L-глутамином, 0,025% (вес/объем) L-аргинином, 0,1% (вес/объем) HEPES, 0,001% (вес/объем) гентамицином, 0,002% (вес/объем) стрептомицином, 0,002% (вес/объем) пенициллином), дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C без замены среды, и число клеток анализировали через 1, 3 и 6 дней. В экспериментах на фиг. 13 размножение осуществляли без замены среды. Результаты для CD4+ иммунореактивных T-клеток показаны на фиг. 13A, результаты для CD8+ иммунореактивных T-клеток показаны на фиг. 13B, при этом на графиках представлена степень пролиферации соответственно количеству собранных клеток на каждый момент времени для CD4+ Tresp (фиг. 13A) и для CD8+ Tresp (на фиг. 13B).
[00131] Как можно видеть из фиг. 13A, "меньший" растворимый реагент для мультимеризации, на котором были обратимо иммобилизованы Fab-фрагменты αCD3 и αCD28, обеспечивал такую же степень размножения CD4+ T-клеток, как и Dynabeads (которые на данный момент являются стандартным реагентом для размножения T-клеток), тогда как "более крупный" олигомерный растворимый Strep-Tactin обеспечивал даже лучшее размножение по сравнению с Dynabead. Данное улучшение может быть обусловлено тем, что растворимый "более крупный олигомерный реагент для мультимеризации" способен одновременно связываться с большим количеством T-клеток, чем "меньший" растворимый олигомер, и тем самым способен стимулировать большее количество CD4+ T-клеток, чем "меньший" олигомер.
[00132] Как очевидно из фиг. 13B, при применении растворимых реагентов для мультимеризации по настоящему изобретению, CD8+ T-клетки можно размножать в течение первых 3 дней по меньшей мере так же эффективно, как в случае применения Dynabeads. Примечательно, что в данный период времени эксперимент по размножению, в котором применяли растворимый реагент для мультимеризации, который в дополнение к Fab-фрагментам αCD3 и αCD28 (в качестве первого и второго средства) нес обратимо иммобилизованный на нем Fab-фрагмент αCD8, демонстрировал наилучшую степень размножения в данных условиях культивирования. Это означает, что с помощью применения стимула, специфичного для конкретной субпопуляции клеток (в данном случае Fab-фрагмента αCD8), возможно увеличивать или модулировать избирательность размножения и тем самым иметь возможность получить большие количества клеток желаемой (суб)популяции.
[00133] Таким образом, обобщая вышесказанное, пример 6 демонстрирует, что функциональные возможности применяемого в настоящем изобретении растворимого реагента для мультимеризации в отношении запуска размножения T-клеток сравнимы с существующей стандартной методикой применения Dynabeads для этой цели. Однако, поскольку стимуляцию можно контролировать (и при желании прекращать) с помощью добавления конкурирующего средства, такого как биотин, в случае обратимого взаимодействия между первым и вторым средством и реагентом для мультимеризации на основе стрептавидина, настоящее изобретение обеспечивает значительное преимущество по сравнению с технологией Dynabeads, поскольку условия размножения можно оптимизировать (например, будет возможно остановить стимуляцию в эксперименте на фиг. 13B через 3 дня). В дополнение, поскольку растворимый реагент для мультимеризации можно легко удалять из реакционной смеси (например, путем иммобилизации реагента на колонке с биотином после завершения реакции размножения), способ размножения по настоящему изобретению можно осуществлять и автоматизировать в закрытых системах, для которых, например, необходимо получение клеток для терапевтических целей, соответствующее GMP, не имея дела с удалением гранул, таких как Dynabeads.
Пример 7. Кинетические характеристики пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток, стимулированных in vitro обратимыми Fab-мультимерами αCD3/αCD28 Streptamer с заменой среды
[00134] В данном примере также рассматривали кинетические характеристики размножения для пролиферации очищенных CD4+ и CD8+ иммунореактивных T-клеток (Tresp), которые стимулировали in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимых олигомерных мутантных вариантах белка стрептавидина. Для данной цели растворимый олигомерный мутантный вариант белка Strep-Tactin® двух различных размеров служил в качестве растворимого реагента для мультимеризации. Первый тип олигомерного Strep-Tactin® представлял собой фракцию олигомерного мутантного варианта белка стрептавидина (n≥3), полученную в примере 5 (также называемую в данном документе "традиционным остовом Strep-Tactin®", проиллюстрированным символом-треугольником, обращенным на фиг. 13 вершиной вниз). Второй тип этого олигомерного мутантного варианта белка стрептавидина, применяемого в качестве растворимого реагента для мультимеризации, получали с помощью реакции олигомерного Strep-Tactin (n≥3), полученного в примере 5, с биотинилированным сывороточным альбумином человека. Этот растворимый олигомерный реагент для мультимеризации также называется в данном документе "крупным остовом Strep-Tactin®".
[00135] В данном примере 400000 очищенных CD4+ или CD8+ иммунореактивных T-клеток (Tresp) стимулировали по отдельности этими двумя различными мультимерами Streptamer, как поясняется выше, т. е. либо остовом Strep-Tactin из примера 5 (1,0 мг/мл), либо крупными остовами Strep-Tactin (0,1 мг/мл). 3 мкл обоих различных остовов также нагружали комбинацией 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab-фрагмента αCD28, описанных выше. В дополнение, 4,5 мкл остова Strep-Tactin из примера 5 нагружали 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab-фрагмента αCD28, как описано выше. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, стимулированные с помощью Dynabeads (на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C с заменой среды в день 3, и число клеток анализировали через 1, 3 и 6 дней. Результаты для CD4+ иммунореактивных T-клеток показаны на фиг. 14A, результаты для CD8+ иммунореактивных T-клеток показаны на фиг. 14B, при этом на графиках представлена степень пролиферации соответственно количеству собранных клеток на каждый момент времени для CD4+ Tresp (фиг. 14A) и для CD8+ Tresp (на фиг. 14B).
[00136] Как можно видеть из фиг. 14A, растворимые реагенты для мультимеризации по настоящему изобретению, на которых были обратимо иммобилизованы Fab-фрагменты αCD3 и αCD28, обеспечивали лучшее размножение CD4+ T-клеток, чем Dynabeads.
[00137] Как очевидно из фиг. 14B, при применении растворимых реагентов для мультимеризации по настоящему изобретению, CD8+ T-клетки можно размножать в течение первых 6 дней по меньшей мере так же эффективно, как в случае применения Dynabeads. Примечательно, что в данный период времени эксперимент по размножению, в котором применяли более крупный растворимый реагент для мультимеризации, который нес Fab-фрагменты αCD3 и αCD28 (в качестве первого и второго средства), демонстрировал наилучшую степень размножения в данных условиях культивирования. Это опять-таки может быть обусловлено тем, что растворимый "более крупный олигомерный реагент для мультимеризации" способен одновременно связываться с большим количеством T-клеток, чем "меньший" растворимый олигомер, и тем самым способен стимулировать большее количество CD4+ T-клеток, чем "меньший" олигомер.
Пример 8. Кинетические характеристики размножения очищенных культур CD4+ и CD8+ T-клеток с заменой среды или без нее
[00138] В данном примере сводные данные из примеров 6 и 7 нормализовали по исходному количеству клеток для "меньшего" растворимого реагента для мультимеризации и положительного и отрицательного контроля. Для "более крупного" реагента для мультимеризации нормализованные данные получены не были. Как поясняется в примерах 6 и 7, 400000-500000 CD4+ или CD8+ иммунореактивных T-клеток (Tresp) стимулировали 3 мкл препарата мультимеров Strep-Tactin (1 мг/мл; на которых были иммобилизованы 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab-фрагмента αCD28). Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, стимулированные с помощью Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C с заменой среды (сплошные линии на фиг. 15) или без замены среды (пунктирные линии на фиг. 15) в день 3, и число клеток анализировали через 1, 3 и 6 дней. Как очевидно из нормализованных данных на фиг. 15A, "меньший" растворимый реагент для мультимеризации, на котором были обратимо иммобилизованы Fab-фрагменты αCD3 и αCD28, давал приблизительно 2,5-кратное размножение CD4+ T-клеток, а размножение с помощью Dynabeads давало приблизительно 1,8-кратный показатель размножения. Таким образом, применение растворимого реагента для мультимеризации по настоящему изобретению обеспечивает даже улучшенное размножение CD4+ T-клеток по сравнению с Dynabeads. Аналогично, на фиг. 15B подтверждается, что при применении растворимых реагентов для мультимеризации по настоящему изобретению CD8+ T-клетки можно размножать в течение первых 3 дней по меньшей мере так же эффективно, как в случае применения Dynabeads.
Пример 9. Раннее образование скоплений после активации очищенных CD4+ и CD8+ иммунореактивных T-клеток, стимулированных in vitro обратимыми Fab-мультимерами αCD3/αCD28 Streptamer
[00139] В данном примере 400000 CD4+ или CD8+ иммунореактивных T-клеток (Tresp) стимулировали 3 мкл препарата олигомерного реагента для мультимеризации Strep-Tactin (1 мг/мл), нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, стимулированные с помощью Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C и анализировали при помощи микроскопа через 1 и 2 дня. Стимуляция CD4+ Tresp (фиг. 16A) и CD8+ Tresp (фиг. 16B) показана соответственно для Dynabeads (средний ряд) и мультимеров Streptamer (нижний ряд). На фотографиях представлена степень образования скоплений. Для лучшей видимости приводимые в качестве примера скопления отмечены кружками в случае стимуляции растворимыми олигомерами мутантного варианта белка стрептавидина на фиг. 16A и фиг. 16B. Скопления в пределах стимуляции при помощи Dynabead можно легко увидеть по накоплению темных стимулирующих частиц. Очевидно, что в случае применения способа размножения по настоящему изобретению, в котором используется растворимый олигомерный реагент для мультимеризации, образуются ранние скопления как CD4+, так и CD8+ T-клеток.
Пример 10. Кинетические характеристики размножения и фенотип поликлональной активированной/размножившейся общей популяции CD3+ центральных T-клеток памяти (Tcm)
[00140] В данном примере 500000 CD3+ CD62L+ CD45RA- иммунореактивных клеток Tcm (Tresp) стимулировали 3 мкл препарата растворимого олигомерного Strep-Tactin из примера 5 (1 мг/мл), также нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Кроме того, применяли 4,5 мкл препарата мультимеров Strep-Tactin, нагруженных 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28, в качестве дополнительного условия стимуляции. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, стимулированные с помощью Dynabeads (на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной только 30 ед/мл IL-2 или 30 ед/мл IL-2 и 5 нг/мл IL-15. Клетки инкубировали при 37°C с заменой среды каждые 3 дня, и число клеток анализировали через 7 и 14 дней. На графиках представлена степень пролиферации соответственно количеству собранных клеток на каждый момент времени, на фиг. 17A в среде, дополненной только IL-2, а на фиг. 17B в среде, дополненной IL-2 и IL-15. Как можно видеть как из фиг. 17A, так и из фиг. 17B, растворимый реагент для мультимеризации, с которым обратимо связаны Fab-фрагмент αCD3 и Fab-фрагмент αCD28, дает лучшее размножение клеток, чем Dynabeads. Как дополнительно показано с помощью проточного цитометрического анализа поверхностной экспрессии CD62L и CD127 спустя 14 дней культивирования в различных цитокиновых окружениях на фиг. 17C, в экспериментальных подходах с применением растворимых реагентов для мультимеризации по настоящему изобретению, при обоих типах условий, выбранных в данном случае, сохраняется более высокое содержание долгоживущих T-клеток памяти, экспрессирующих CD127, чем при размножении с помощью Dynabeads. Это иллюстрирует дополнительное преимущество способов по настоящему изобретению.
Пример 11. Избирательное антиген-специфическое размножение иммунореактивных клеток Tcm из общей популяции CD3+ центральных T-клеток памяти (кинетические характеристики и фенотип)
[00141] В данном примере рассматривали кинетические характеристики и фенотип избирательного антиген-специфического (Ag-специфического) размножения из очищенных CD3+ CD62L+ CD45RA- иммунореактивных клеток Tcm.
[00142] Более подробно, CD3+ CD62L+ CD45RA- иммунореактивные клетки Tcm стимулировали in vitro как комплексом пептид:молекула MHC (который выступает в качестве первого средства, передающего первичный активационный сигнал клеткам), так и Fab-фрагментом αCD28 (который выступает в качестве второго реагента, стимулирующего вспомогательную молекулу на поверхности клеток). Как комплекс антиген-специфического пептида с молекулой MHC, так и Fab-фрагмент αCD28 были обратимо иммобилизованы на растворимом олигомерном мутантном варианте белка стрептавидина (при n≥3), описанном в примере 5. Пептид, который применяли для антиген-специфического размножения, представлял собой пептид CRVLCCYVL (SEQ ID NO: 06), аминокислоты 309–317 немедленно-раннего белка 1 (описанного в Ameres et al, PLOS Pathogens, май 2013 г., том 9, номер 5, e1003383), представляющие собой эпитоп IE-1/HLA-C7, специфичный для цитомегаловируса (CMV). Молекула MHC-I, которая представляет собой пептид, несущий на C-конце α-цепи (тяжелой цепи) стрептавидин-связывающий пептид (SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 0GGSAWSHPQFEK7), коммерчески доступный как "Twin-Strep-tag®" от IBA GmbH, Геттинген, Германия).
[00143] Для данной цели 500000 CD3+ CD62L+ CD45RA- иммунореактивных клеток Tcm (Tresp) подвергали Ag-специфической стимуляции с помощью 3 мкл препарата растворимого олигомерного реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг комплексов пептид:молекула MHC класса I, оснащенных стрептавидин-связывающим пептидом, и 0,5 мкг Fab αCD28, описанного выше. В качестве альтернативы 4,5 мкл препарата реагента для мультимеризации Strep-Tactin нагружали 0,5 мкг этих комплексов пептид:молекула MHC класса I, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Для сравнения проводили поликлональную стимуляцию с помощью 3 мкл препарата реагента для мультимеризации Strep-Tactin (1 мг/мл), также нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Опять-таки в качестве описанного выше альтернативного условия стимуляции применяли 4,5 мкл препарата реагента для мультимеризации Strep-Tactin, обратимо нагруженного 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, поликлонально стимулированные с помощью Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2 и 5 нг/мл IL-15. Клетки инкубировали при 37°C с заменой среды каждые 3 дня, и число клеток анализировали через 7 и 14 дней. Приводимый в качестве примера проточный цитометрический анализ фракции Ag-специфических клеток, которую стимулировали/размножали с помощью растворимого олигомера Strep-Tactin, на котором был иммобилизован комплекс пептид:MHC-I для эпитопа IE-1/HLA-C7 (для CMV) (фиг. 18A), демонстрирует, что эти антиген-специфические T-клетки размножались специфически. На графиках на фиг. 18B - фиг. 18E (на которых представлена степень размножения для отдельных видов Ag-специфичности соответственно количеству собранных клеток, положительных по мультимеру пептид:MHC-I, на каждый момент времени по аналогии с экспериментом по размножению, показанным на фиг. 18A) показано, что реагент для мультимеризации, в котором применяется соответствующий комплекс Ag-специфического пептида и молекулы MHC-I, обеспечивал более высокое количество размножившихся клеток (в диапазоне от двадцатикратного увеличения количества клеток в случае Ag-специфических клеток, распознающих эпитоп pp65 CMV (аминокислоты 341-350 (QYDPVAALF (SEQ ID NO: 08)), рестриктированные по HLA-A2402) (см. фиг. 18B) до 98-кратного увеличения количества Ag-специфических клеток, распознающих эпитоп IE-1309-317/HLA-B7 (CRVLCCYVL (SEQ ID NO: 06)) CMV (см. фиг. 18E)), тем самым демонстрируя, что способ размножения по настоящему изобретению в полной мере применим для размножения Ag-специфических клеток. Наконец, приводимый в качестве примера проточный цитометрический анализ поверхностной экспрессии CD62L и CD127 спустя 14 дней культивирования с эпитопом гексона 5/HLA-B7 (для аденовируса), показанный на фиг. 18F, дополнительно подтверждает, что в экспериментальных подходах с применением растворимых реагентов для мультимеризации по настоящему изобретению сохраняется более высокое содержание долгоживущих T-клеток памяти, экспрессирующих CD127, в условиях поликлональной и Ag-специфической стимуляции.
Пример 12. Кинетические характеристики избирательного Ag-специфического размножения и фенотип общей популяции центральных T-клеток памяти
[00144] В данном примере рассматривали кинетические характеристики избирательного Ag-специфического размножения из очищенных CD3+ CD62L+ CD45RA- иммунореактивных клеток Tcm, которые стимулировали in vitro a) комплексами антиген-специфического пептида с MHC-I и b) Fab-фрагментами αCD28, которые были обратимо иммобилизованы в качестве первого и второго средства на растворимых олигомерных мутантных вариантах белка стрептавидина.
[00145] Для данной цели 500000 CD3+CD62L+CD45RA- иммунореактивных клеток Tcm (Tresp) подвергали Ag-специфической стимуляции с помощью 3 мкл препарата реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг комплексов пептид:молекула MHC класса I, оснащенных стрептавидин-связывающим пептидом (специфический пептид представлен аминокислотами 114-124 (CPYSGTAYNSL, SEQ ID NO: 10) белка гексона 5 аденовируса, рестриктированными по HLA-B07), и 0,5 мкг Fab αCD28. В качестве альтернативы 4,5 мкл препарата реагента для мультимеризации Strep-Tactin нагружали 0,5 мкг этого комплекса пептид:молекула MHC класса I, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Для сравнения проводили поликлональную стимуляцию с помощью 3 мкл препарата реагента для мультимеризации Strep-Tactin (1 мг/мл), также нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Опять-таки, в качестве вышеописанного альтернативного условия стимуляции применяли 4,5 мкл препарата мультимеров Strep-Tactin, нагруженных 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28. Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, поликлонально стимулированные с помощью Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2 и 5 нг/мл IL-15. Клетки инкубировали при 37°C с заменой среды каждые 3 дня, и число клеток анализировали через 7 и 14 дней. На изображениях, показанных на фиг. 19, представлена степень образования скоплений в день 5, приводимая в качестве примера Ag-специфическая стимуляция проиллюстрирована для эпитопа гексона 5/HLA-B7 аденовируса. Как можно видеть из фиг. 19, такие клетки, специфичные к антигену аденовируса, можно специфически размножать из исходной популяции CD3+CD62L+CD45RA- иммунореактивных Tcm.
Пример 13. Урожайность и фенотип размножившихся CD8+ T-клеток – разноразмерность растворимого реагента для мультимеризации и добавление αCD8-Fab для стимуляции
[00146] В данном примере рассматривали размножение очищенных CD8+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимых олигомерных мутантных вариантах белка стрептавидина. В дополнение, рассматривали эффект добавления αCD8-Fab к реагенту для мультимеризации для повышения специфичности размножения CD8+ T-клеток.
[00147] Для данной цели 300000 очищенных CD8+ иммунореактивных T-клеток (Tresp) стимулировали по отдельности двумя различными реагентами для мультимеризации на основе Strep-Tactin, а именно, либо небольшим олигомерным реагентом для мультимеризации Strep-Tactin из примера 5 (1 мг/мл), либо более крупными олигомерами Strep-Tactin, описанными выше (0,1 мг/мл). 3 мкл обоих различных реагентов для мультимеризации (остовов) также нагружали комбинацией 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab-фрагмента αCD28, описанных выше. В дополнение, 4,5 мкл меньшего реагента для мультимеризации (остова) Strep-Tactin нагружали 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab-фрагмента αCD28, описанных выше. Кроме того, применяли 3 мкл "меньшего" реагента для мультимеризации (остова) Strep-Tactin, функционализированного только 0,5 мкг Fab-фрагмента αCD3 в отдельности или 0,5 мкг Fab-фрагмента αCD28 в отдельности. Нестимулированные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные с помощью Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C с заменой среды через 3 дня и анализировали через 6 дней. На фиг. 20A изображена степень пролиферации соответственно количеству клеток, собранных в день 6, по сравнению с отрицательными контролями, нормализованная по положительному контролю. На фиг. 20A показано, что размножение CD8+ T-клеток с помощью растворимых реагентов для мультимеризации по настоящему изобретению приводит к более высоким показателям урожайности CD8+ T-клеток, чем размножение с помощью Dynabeads. FACS-анализ поверхностной экспрессии CD8 (фиг. 20B) и поверхностной экспрессии CD45RO (фиг. 20C) после культивирования клеток демонстрирует, что CD8+ T-клетки, которые размножали с помощью либо реагентов для мультимеризации по настоящему изобретению, либо Dynabeads, имели одинаковый фенотип (различные условия стимуляции сравнивали с помощью однофакторного ANOVA, и значимое различие не было выявлено (n.s.)). Улучшенная урожайность CD8+ клеток при применении способов размножения по настоящему изобретению по сравнению с Dynabeads может быть обусловлена тем, что растворимый реагент для мультимеризации может иметь лучший доступ к своим целевым рецепторам на клеточной поверхности, чем антитела, иммобилизованные на Dynabeads. Эта улучшенная урожайность может стать весьма преимущественной при размножении редкой популяции клеток из исходного образца.
[00148] В дополнение, при сравнении урожайности от размножения, достигаемой в случае применения средства для мультимеризации, на котором были совместно иммобилизованы как 0,5 мкг Fab-фрагмента αCD3, так и 0,5 мкг Fab-фрагмента αCD28 (второй слева столбец на фиг. 20B), с урожайностью при применении двух реагентов для мультимеризации, которые были функционализированы только Fab-фрагментом αCD3 в отдельности или Fab-фрагментом αCD28 в отдельности (третий слева столбец на фиг. 20B), можно видеть, что оба эксперимента характеризовались одинаковой эффективностью размножения. Таким образом, эти эксперименты демонстрируют, что применение одного реагента для мультимеризации, на котором совместно иммобилизованы как первое средство, так и второе средство, функционально эквивалентно применению для размножения двух отдельных реагентов для мультимеризации, нагруженных соответственно только первым средством или вторым средством.
Пример 14. Урожайность и фенотип размножившихся CD8+ T-клеток – титрование отдельных растворимых реагентов для мультимеризации с различными соотношениями Fab-фрагментов αCD3 и αCD28, иммобилизованных на них
[00149] В данном примере рассматривали урожайность и фенотип размножившихся CD8+ иммунореактивных T-клеток (Tresp), которые стимулировали in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимых олигомерных мутантных вариантах белка стрептавидина в различных количествах.
[00150] Для данной цели 300000 CD8+ иммунореактивных T-клеток (Tresp) стимулировали различными количествами смеси препаратов "небольшого" олигомерного реагента для мультимеризации Strep-Tactin (1 мг/мл), функционализированного Fab αCD3 в отдельности и Fab αCD28 в отдельности ("1x" соответствует 1,5 мкг реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг Fab αCD3 в отдельности, и 1,5 мкг мультимеризованного Strep-Tactin, функционализированного 0,5 мкг Fab-фрагмента αCD28 в отдельности), или 3 мкл препарата реагента для мультимеризации Strep-Tactin, нагруженного 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28, или 4,5 мкл препарата реагента для мультимеризации Strep-Tactin, нагруженного 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 со Strep-tag и 0,5 мкг Fab αCD28. Необработанные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные с помощью Dynabeads, служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C без замены среды и анализировали через 5 дней. На фиг. 21A отображена степень пролиферации соответственно количеству клеток, собранных в день 5, по сравнению с отрицательными контролями, нормализованная по положительному контролю. На фиг. 21A показано, что размножение CD8+ T-клеток с помощью различных растворимых реагентов для мультимеризации по настоящему изобретению приводит к более высоким показателям урожайности CD8+ T-клеток, чем размножение с помощью Dynabeads (в частности, общее совокупное количество реагента в условии 5x обусловливало оптимальное размножение клеток, в частности, с течением времени/увеличение общего количества клеток благодаря началу клеточного деления). FACS-анализ поверхностной экспрессии CD8 (фиг. 21B) и поверхностной экспрессии CD45RO (фиг. 21C) после культивирования клеток демонстрирует, что CD8+ T-клетки, которые размножали с помощью либо различных реагентов для мультимеризации по настоящему изобретению, либо с помощью коммерчески доступных Dynabeads, имели одинаковый фенотип.
Пример 15. Активация внутриклеточных сигнальных каскадов после стимуляции мультимерами Streptamer клеток Jurkat, трансдуцированных αCD19-CAR
[00151] В данном примере рассматривали активацию внутриклеточных сигнальных каскадов трансдуцированных клеток Jurkat, которые были модифицированы для экспрессии опухолеспецифического химерного антигенного рецептора (CAR), а именно в данном случае CD19, и которые стимулировали с помощью олигомерного Strep-Tactin® из примера 5 в качестве растворимого реагента для мультимеризации.
[00152] Для данной цели 300000 иммунореактивных клеток Jurkat (Jresp) стимулировали (A) различными количествами смеси препаратов реагента для мультимеризации Strep-Tactin (1 мг/мл), функционализированного Fab αCD3 и Fab-фрагментами αCD28, описанными в данном документе ("x1" соответствует 3 мкг реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28 – это обеспечивает наличие "поликлонального реагента для мультимеризации на основе Strep-Tactin"), или (B) 3 мкл препарата реагента для мультимеризации Strep-Tactin, функционализированного 0,5 мкг (x1) или 1 мкг (x2) внеклеточного домена (ECD) CD19 (природный лиганд αCD19-CAR – это обеспечивает наличие "CAR-специфичного реагента для мультимеризации на основе Strep-Tactin"), или 3 мкл препарата реагента для мультимеризации Strep-Tactin, нагруженного 0,5 мкг (x1) или 1 мкг (x2) αIgG (распознающий спейсер IgG4 в αCD19-CAR – это также обеспечивает наличие "CAR-специфичного реагента для мультимеризации на основе мутантного варианта белка стрептавидина"). ECD CD19, оснащенный гексагистидиновой меткой, был получен от Sino Biological/Life technologies (SEQ ID NO: 27) и был функционализирован для связывания с реагентом для мультимеризации на основе стрептавидина с помощью смешивания ECD CD19 с адаптерной молекулой His-STREPPER (IBA GmbH, Германия, номер для заказа 2-0920-005) в молекулярном соотношении 1:1 и инкубирования в течение 15 мин. при комнатной температуре. Адаптерная молекула His-STREPPER содержит хелатную часть, которая связывается с гексагистидиновой меткой и стрептавидин-связывающим пептидом, тем самым временно обеспечивая целевую молекулу, в данном случае ECD CD19, стрептавидин-связывающим пептидом, который может обратимо связываться с реагентом для мультимеризации на основе мутантного варианта белка стрептавидина. Jresp, стимулированные при помощи Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28) или PMA и иономицина, служили в качестве положительных контролей. Клетки Jresp высевали в пробирки Eppendorf объемом 1,5 мл в 200 мкл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C, помещали на лед и лизировали спустя 0 мин. - 20 мин. стимуляции. Выявление фосфорилированной ERK указывает на активную передачу сигналов с помощью MAPK, окрашивание β-актина, ген которого является геном "домашнего хозяйства", указывает на загрузку равных количеств общего белка на каждое условие и момент времени. Как можно видеть из сравнения фиг. 22A, на которой показана активация клеток Jurkat посредством "поликлонального реагента для мультимеризации Strep-Tactin", и фиг. 22B, на которой показана активация клеток Jurkat посредством двух "CAR-специфичных реагентов для мультимеризации на основе Strep-Tactin", клетки Jurkat можно активировать/размножать посредством связывания внеклеточного домена CD19 с CD19-специфичным химерным антигенным рецептором. Поскольку последующую генетическую обработку T-клеток проводят практически исключительно на предварительно выбранных популяциях клеток, неспецифическая активация посредством перекрестного связывания введенных CAR при помощи спейсерного домена IgG4 (который является консервативным в составе разнообразных CAR с различными видами специфичности) расширяет применимость обратимых стимуляции/размножения клеток в таких ситуациях обработки клеток in vitro.
[00153] Таким образом, данный эксперимент демонстрирует, что в принципе любую популяцию клеток, активируемую с помощью связывания средства (лиганда), которое передает первичный активационный сигнал популяции клеток, можно размножать с помощью первого средства, обратимо иммобилизованного на реагенте для мультимеризации, как описано в данном документе.
Пример 16. Урожайность и состав субпопуляции размножившихся CD3+ T-клеток при добавлении Fab αCD8 для стимуляции
Эксперимент демонстрирует размножение очищенных CD3+ иммунореактивных T-клеток, стимулированных in vitro Fab-фрагментами αCD3/αCD28, которые были обратимо иммобилизованы на растворимом олигомерном Strep-Tactin® из примера 5, который служил в качестве растворимого реагента для мультимеризации. В одном эксперименте в дополнение к Fab-фрагментам αCD3/αCD28 на растворимом олигомере мутантного варианта белка стрептавидина был иммобилизован также Fab-фрагмент αCD8, коммерчески доступный от IBA GmbH, Геттинген, Германия (номер по каталогу 6-8000-203), в целях тестирования на наличие возможности преимущественной стимуляции in vitro определенной субпопуляции T-клеток с помощью образованных обратимой связью мультимеров Fab αCD3/αCD28 Streptamer. Более подробно, 500000 очищенных CD3+ иммунореактивных T-клеток (Tresp) стимулировали 3 мкл препарата олигомерного стрептавидина (1 мг/мл), нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. В качестве альтернативного подхода 4,5 мкл олигомерного Strep-Tactin нагружали 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 со Strep-tag и 0,5 мкг Fab αCD28 со Strep-tag. Нестимулированные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные при помощи Dynabeads (гранулы, на которых необратимо иммобилизованы моноклональные антитела αCD3 и αCD28), служили в качестве положительного контроля. Как можно видеть из фиг. 23A, реагент для мультимеризации, обратимо нагруженный Fab-фрагментом αCD3, Fab-фрагментом αCD28, а также Fab-фрагментом αCD8, обеспечивал наиболее высокое количество размножившихся CD3+ T-клеток. При количестве размножившихся клеток 1,1 x 106 урожайность была приблизительно на 30% выше, чем в случае размножения таких T-клеток с помощью коммерчески доступных Dynabeads. В дополнение, и что более важно, как показано на фиг. 23B, в случае применения данного реагента для мультимеризации, несущего Fab-фрагмент αCD3, Fab-фрагмент αCD28 и Fab-фрагмент αCD8, количество CD8+ T-клеток было наиболее высоким по сравнению с размножением как с помощью Dynabeads, так и растворимого реагента для мультимеризации по настоящему изобретению, несущего только Fab-фрагмент αCD3 и Fab-фрагмент αCD28 в качестве первого и второго средства, как описано в данном документе. Таким образом, данный эксперимент также демонстрирует преимущество настоящего изобретения в том, что в дополнение к первому средству, которое обеспечивает первичный активационный сигнал желаемой популяции клеток, и необязательно второму средству, которое обеспечивает костимулирующий сигнал, на реагенте для мультимеризации может быть иммобилизовано дополнительное средство, специфичное для активации желаемой популяции клеток. Таким образом, тем самым настоящее изобретение предоставляет возможность преимущественного размножения или избирательного обогащения любой желаемой (суб)популяции клеток из образца, который например содержит ряд различных субпопуляций.
Пример 17. Параллельное антиген-специфическое размножение иммунореактивных клеток Tcm из единого пула
[00154] В данном примере рассматривали кинетические характеристики параллельного антиген-специфического (Ag-специфического) размножения из единого пула иммунореактивных T-клеток, стимулированных in vitro несколькими образованными обратимой связью мультимерами пептид:MHC/Fab-Streptamer αCH28.
[00155] 500000 CD3+CD62L+CD45RA- иммунореактивных клеток Tcm (Tresp) одновременно стимулировали по нескольким видам Ag-специфичности с применением для каждого вида специфичности 3 мкл мультимеров Strep-Tactin, функционализированных 0,5 мкг соответствующих комплексов пептид:MHC класса I, несущих стрептавидин-связывающий пептид, и 0,5 мкг Fab αCD28, несущего также стрептавидин-связывающий пептид. В качестве альтернативного подхода для каждого вида специфичности применяли 4,5 мкл реагента для мультимеризации на основе Strep-Tactin, функционализированного 0,5 мкг комплексов пептид:MHC класса I, несущих стрептавидин-связывающий пептид, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28, как описано в данном документе. Для сравнения проводили поликлональную стимуляцию с помощью 3 мкл препарата реагента для мультимеризации на основе Strep-Tactin (1 мг/мл), также обратимо нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28. Опять-таки, в качестве вышеописанного альтернативного условия стимуляции можно применять 4,5 мкл препарата реагента для мультимеризации на основе Strep-Tactin, обратимо нагруженного 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28 (каждый из которых несет стрептавидин-связывающий пептид). Необработанные (нестимулированные) клетки Tresp служили в качестве отрицательного контроля, а клетки Tresp, поликлонально стимулированные при помощи Dynabeads (гранулы, покрытые mAb αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2 и 5 нг/мл IL-15. Клетки инкубировали при 37°C с заменой среды каждые 3 дня, и число клеток анализировали через 7 и 14 дней.
Пример 18. Преимущественная пролиферация CD8+ T-клеток среди CD3+ иммунореактивных T-клеток, стимулированных in vitro реагентами для мультимеризации на основе стрептавидина, обратимо функционализированными Fab-фрагментами αCD3/αCD8/αCD28
[00156] 300000 CD3+ иммунореактивных T-клеток (Tresp) стимулировали 3 мкл препарата реагента для мультимеризации Strep-Tactin (1 мг/мл) или препарата реагента для мультимеризации, в котором применяется крупный остов Strep-Tactin (0,1 мг/мл), также нагруженного комбинацией 0,5 мкг Fab αCD3 и 0,5 мкг Fab αCD28, или 4,5 мкл препарата реагента для мультимеризации на основе Strep-Tactin, нагруженного 0,5 мкг Fab αCD3, 0,5 мкг Fab αCD8 и 0,5 мкг Fab αCD28, или 3 мкл смеси препаратов реагента для мультимеризации на основе Strep-Tactin с 0,5 мкг Fab αCD3 в отдельности и 0,5 мкг Fab αCD28 в отдельности (каждый Fab-фрагмент опять-таки несет стрептавидин-связывающий пептид). Необработанные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные при помощи Dynabeads (гранулы, покрытые mAb αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в двух повторностях в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C с заменой среды через 3 дня и анализировали через 6 дней.
Пример 19. Преимущественная пролиферация CD8+ T-клеток среди CD3+ иммунореактивных T-клеток, стимулированных in vitro реагентами для мультимеризации на основе стрептавидина, обратимо функционализированными Fab-фрагментами αCD3 и αCD28
[00157] 300000 CD3+ иммунореактивных T-клеток (Tresp) стимулировали различными количествами смеси препаратов реагента для мультимеризации на основе Strep-Tactin (1 мг/мл), функционализированного Fab-фрагментом αCD3 в отдельности и Fab-фрагментом αCD28 в отдельности (1,5 мкг реагента для мультимеризации на основе Strep-Tactin, функционализированного 0,5 мкг Fab-фрагмента αCD3 в отдельности, и 1,5 мкг реагента для мультимеризации на основе Strep-Tactin, функционализированного 0,5 мкг Fab-фрагмента αCD28 в отдельности), или различными количествами смеси препаратов реагента для мультимеризации на основе Strep-Tactin, функционализированного Fab-фрагментом αCD3 и Fab-фрагментом αCD28 с Fab-фрагментом αCD8 или без него (каждый Fab-фрагмент опять-таки несет стрептавидин-связывающий пептид) (3 мкг реагента для мультимеризации на основе Strep-Tactin, функционализированного 0,5 мкг Fab-фрагмента αCD3 и 0,5 мкг Fab-фрагмента αCD28 – без Fab-фрагмента αCD8, или 4,5 мкл препарата реагента для мультимеризации Strep-Tactin, нагруженного 0,5 мкг Fab-фрагмента αCD3, 0,5 мкг Fab-фрагмента αCD8 и 0,5 мкг Fab-фрагмента αCD28, где Fab-фрагмент опять-таки несет стрептавидин-связывающий пептид). Необработанные клетки Tresp служили в качестве отрицательного контроля, а Tresp, стимулированные при помощи Dynabeads (гранулы, покрытые mAb αCD3 и αCD28), служили в качестве положительного контроля. Клетки Tresp высевали в 48-луночные планшеты в 1 мл среды для культивирования клеток, дополненной 30 ед/мл IL-2. Клетки инкубировали при 37°C с заменой среды через 3 дня и анализировали через 6 дней.
[00158] Указание или обсуждение в настоящем описании документа, опубликованного ранее, не обязательно следует считать признанием того, что данный документ является частью существующего уровня техники или содержит общедоступные сведения.
[00159] Настоящее изобретение, иллюстративно описанное в данном документе, можно подходящим образом осуществлять на практике в отсутствие какого-либо элемента или элементов, ограничения или ограничений, конкретно не раскрытых в данном документе. Таким образом, например, термины "содержащий", "включающий", "включающий в себя" и т. п. следует толковать в расширительном значении и без ограничения. Дополнительно, используемые в данном документе термины и выражения применялись как термины для описания, а не для ограничения, и применение таких терминов и выражений не предназначено для исключения каких-либо эквивалентов показанных и описанных отличительных признаков или их частей, однако авторами признается, что в пределах объема заявляемого изобретения возможны различные модификации. Таким образом следует понимать, что хотя настоящее изобретение было определенным образом раскрыто с помощью приводимых в качестве примера вариантов осуществления и необязательных отличительных признаков, специалисты в данной области могут прибегнуть к модификациям и изменениям, которые являются частью раскрытого в данном документе изобретения, и что такие модификации и изменения считаются находящимися в пределах объема настоящего изобретения.
[00160] Настоящее изобретение было описано в данном документе в широком и общем смысле. Каждый из более узких видовых признаков и субродовых группирований находящихся в пределах общего раскрытия, также образует часть настоящего изобретения. Оно включает общее описание изобретения с условием или отрицательным ограничением, исключающим какой-либо объект изобретения из рода, независимо от того, упоминается ли конкретно в данном документе исключаемый материал.
Другие варианты осуществления находятся в пределах нижеследующей формулы изобретения. В дополнение, если отличительные признаки или аспекты настоящего изобретения описаны в контексте групп Маркуша, то специалистам в данной области будет понятно, что настоящее изобретение, таким образом, также описано в контексте любого отдельного представителя или подгруппы представителей группы Маркуша.
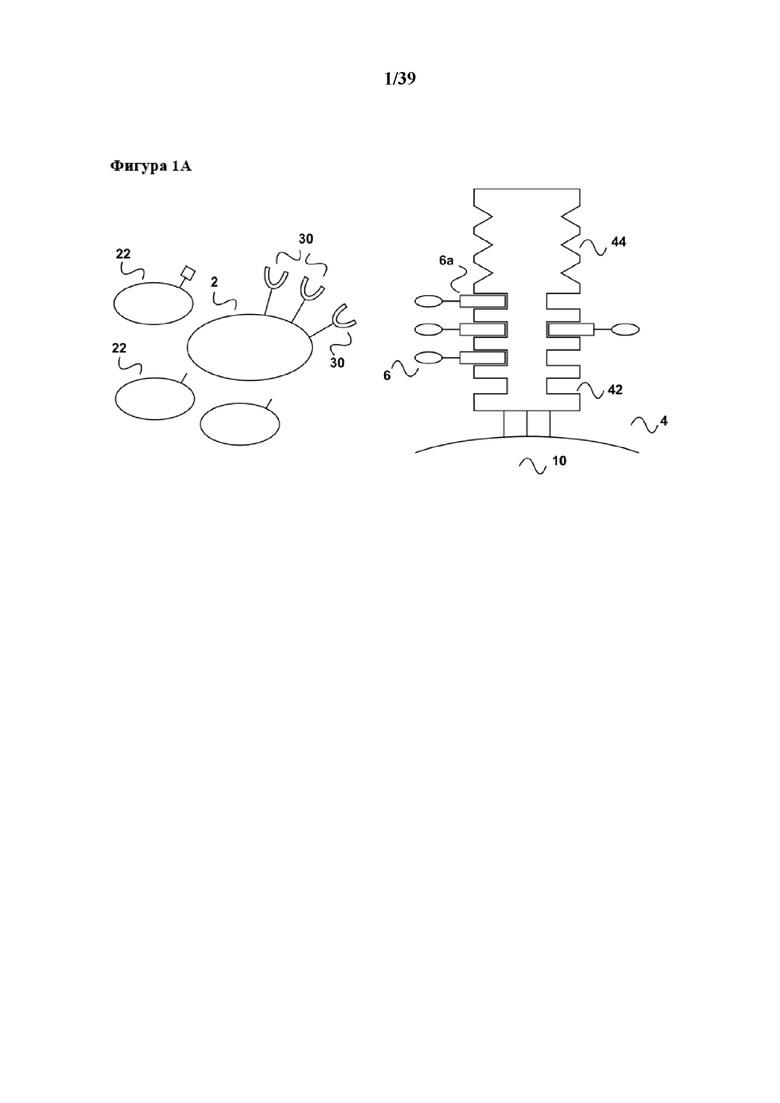
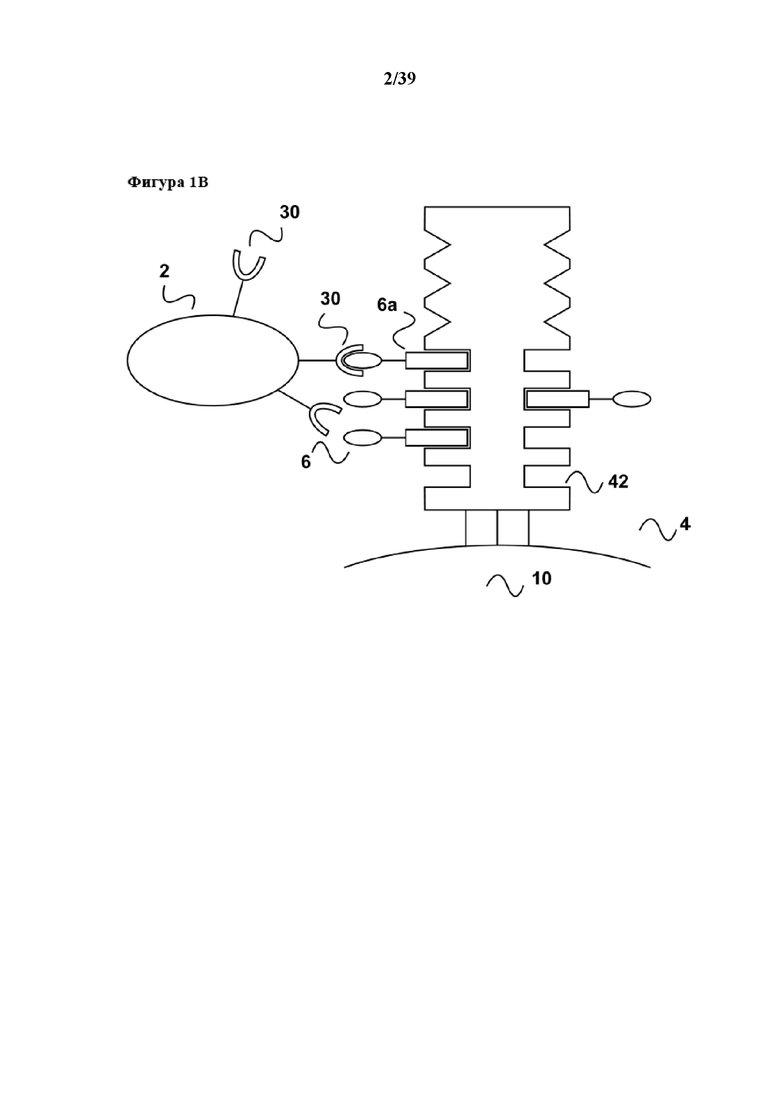
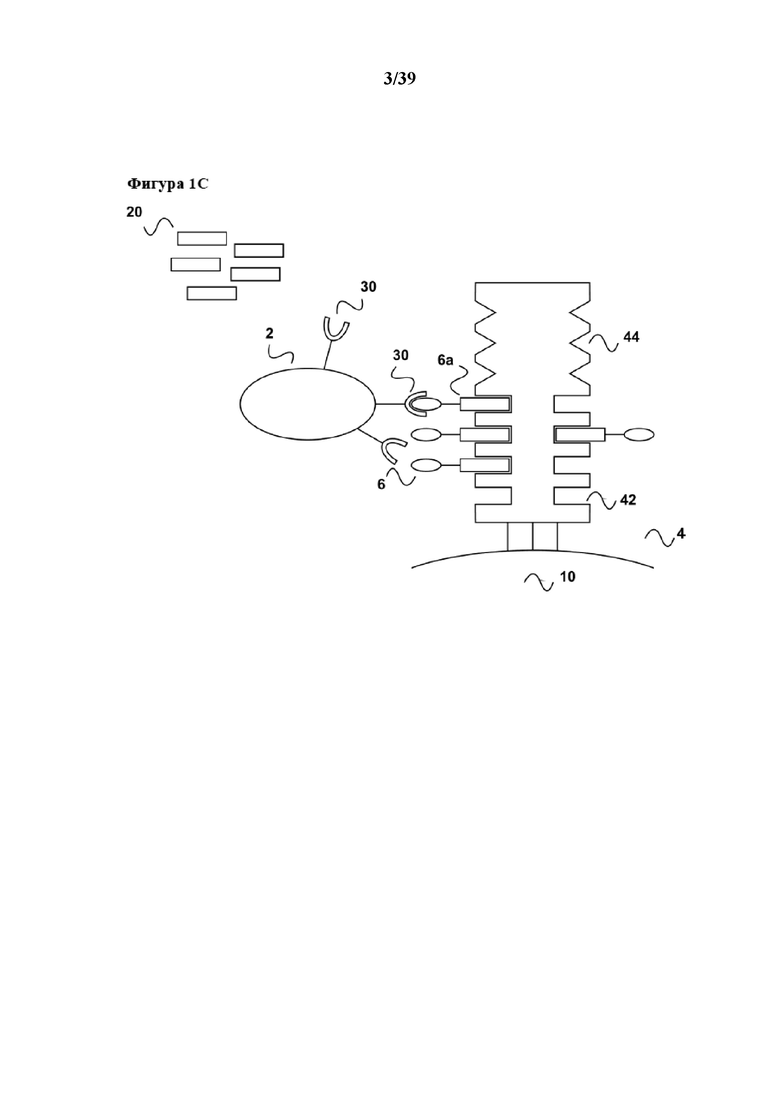
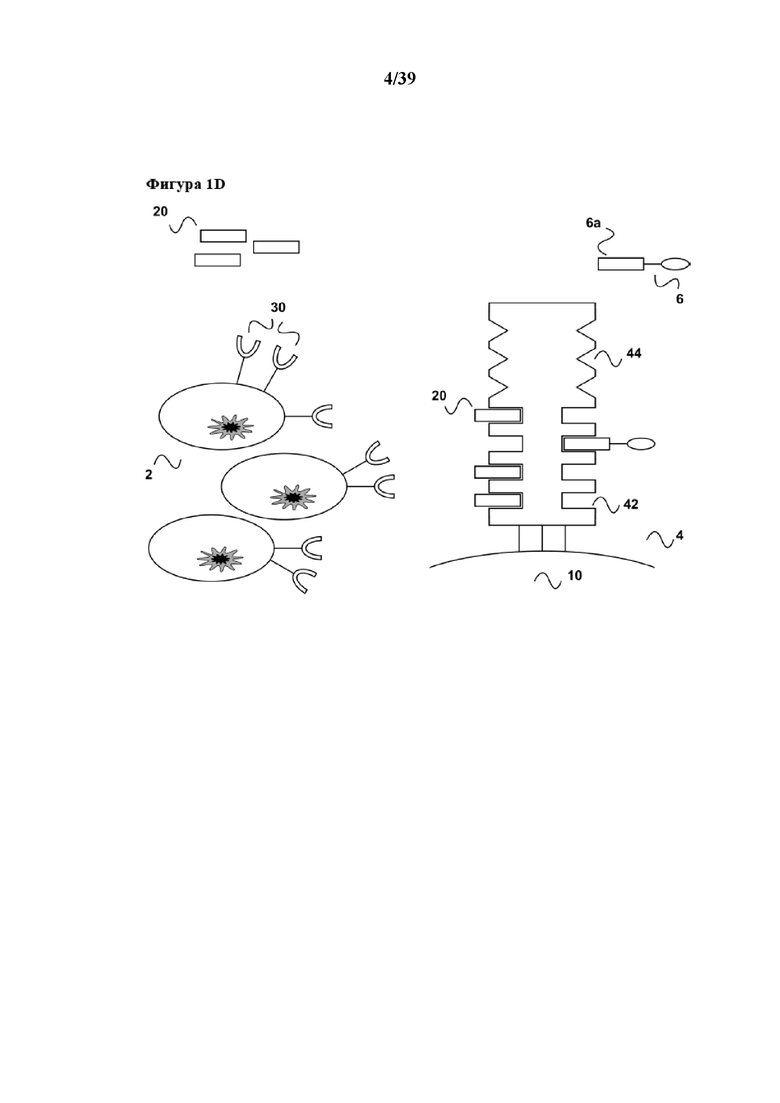
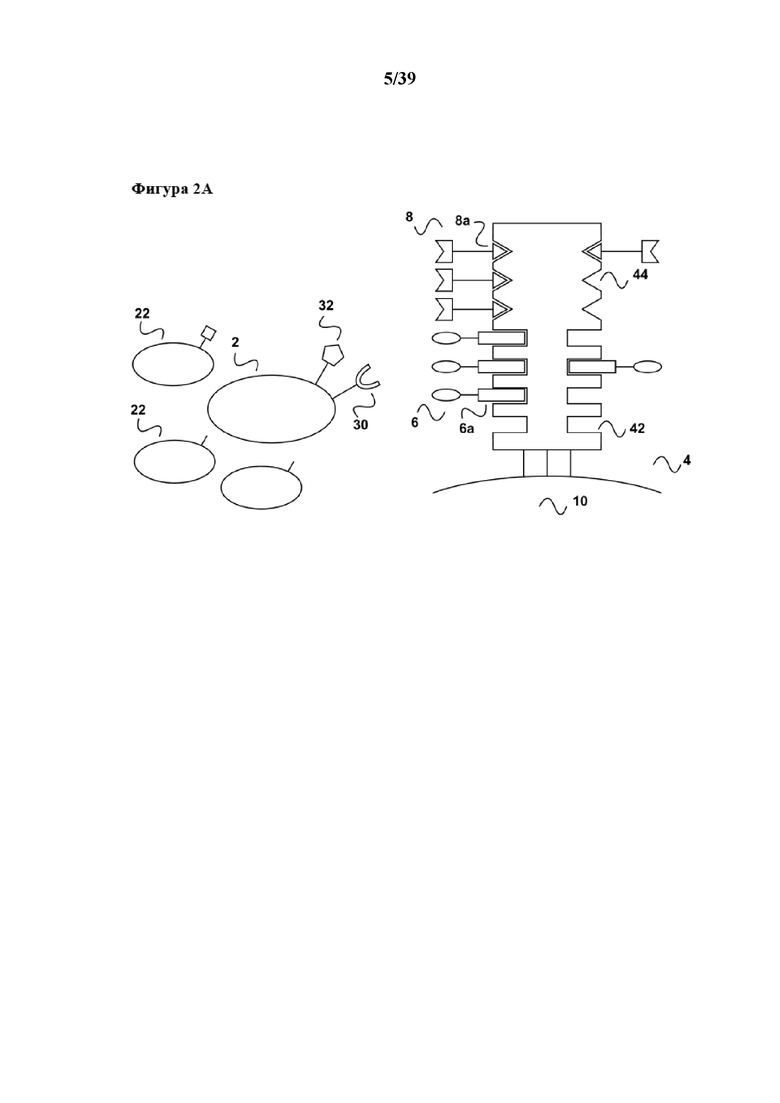
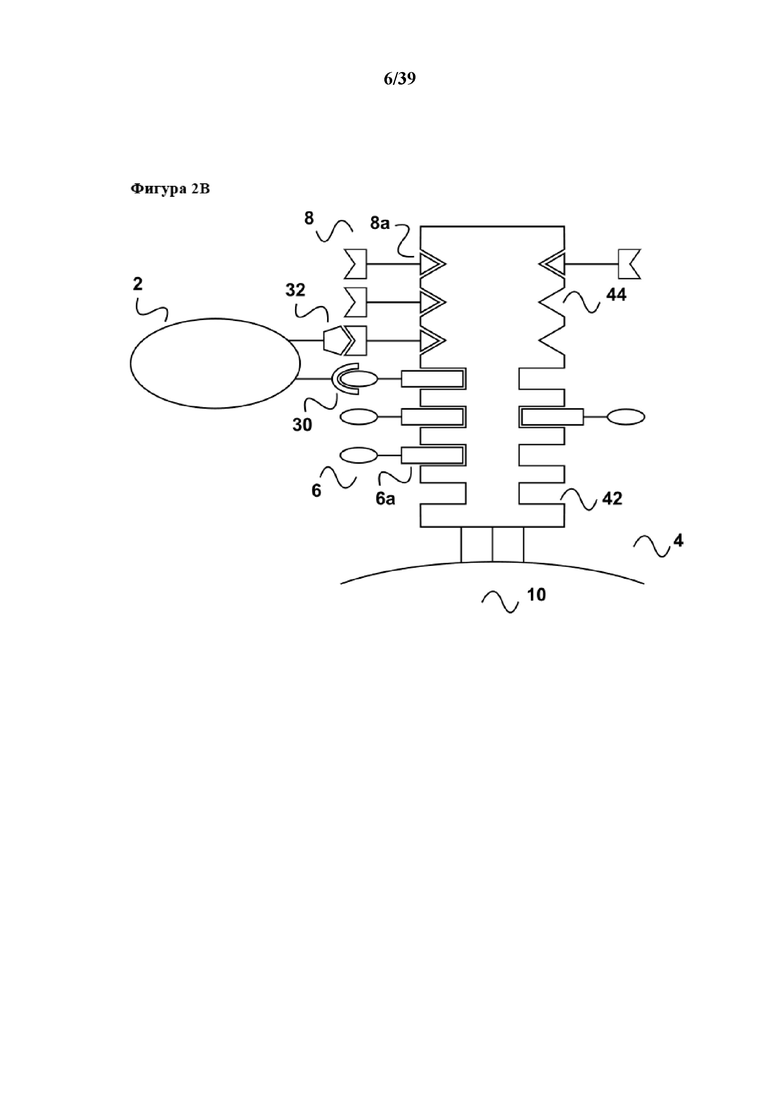
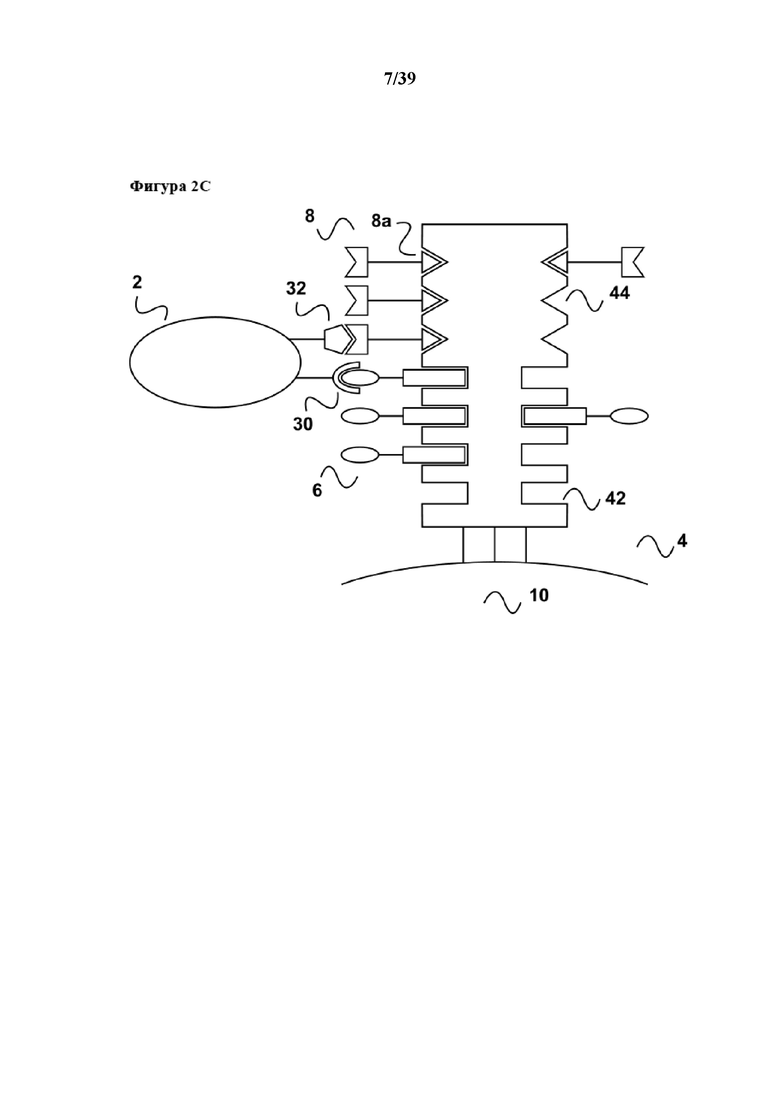
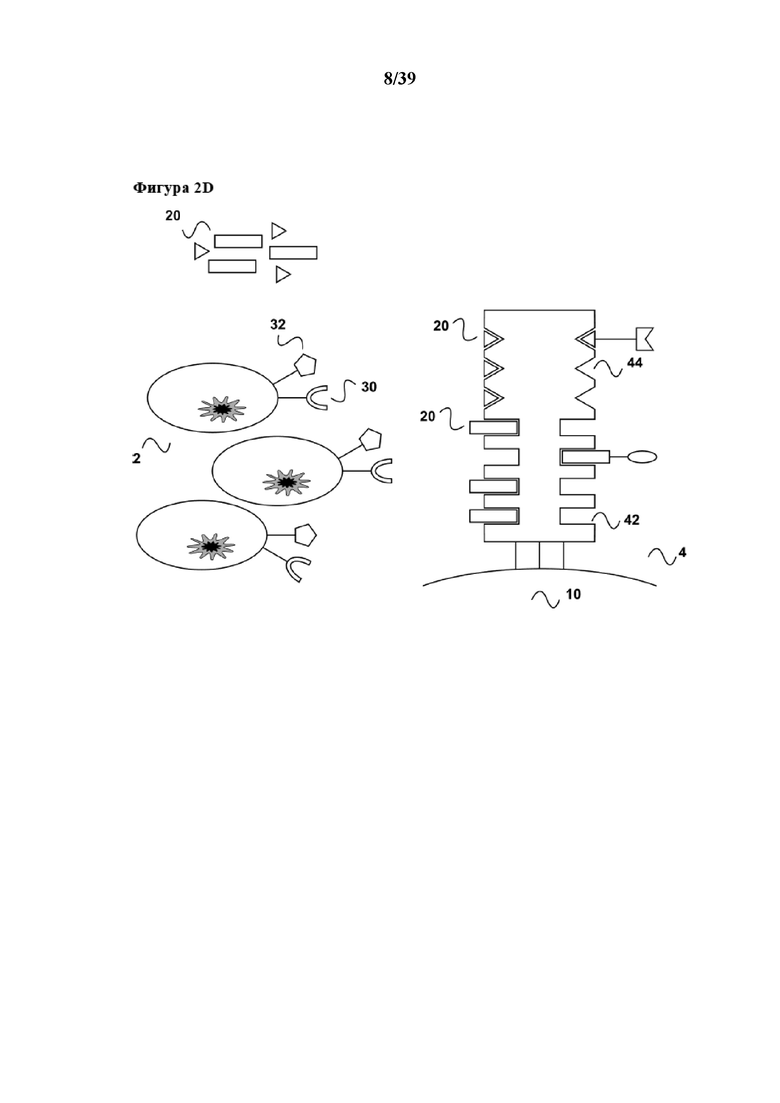
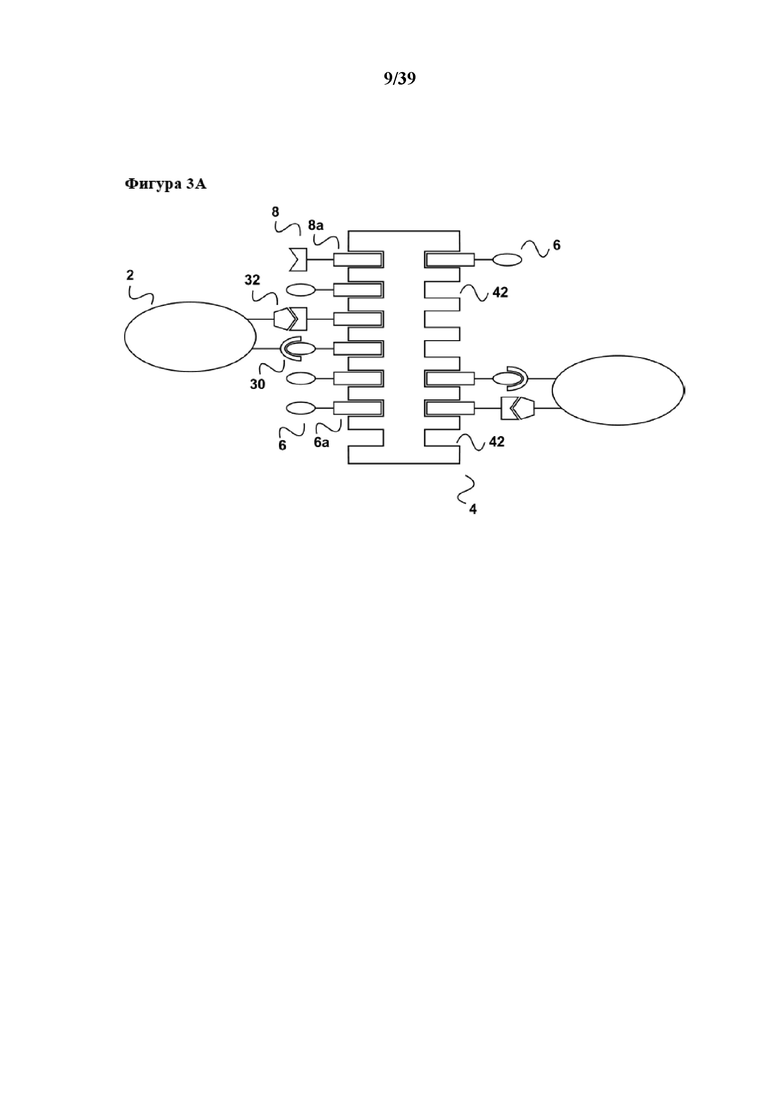
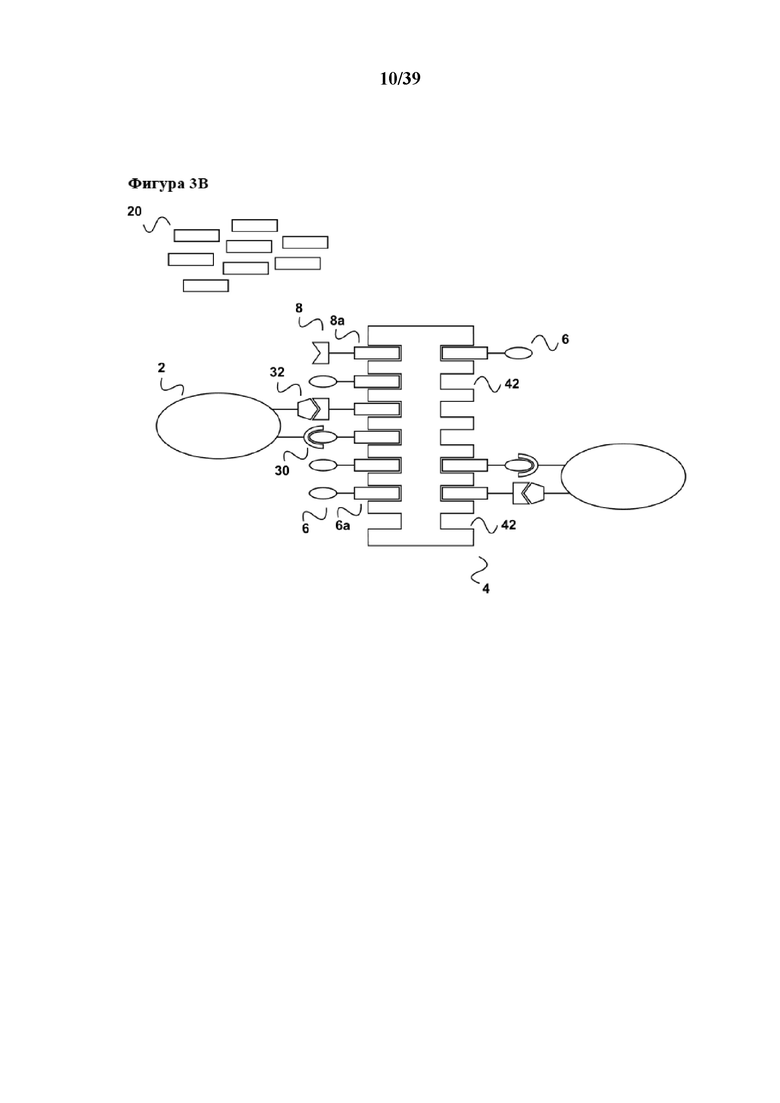
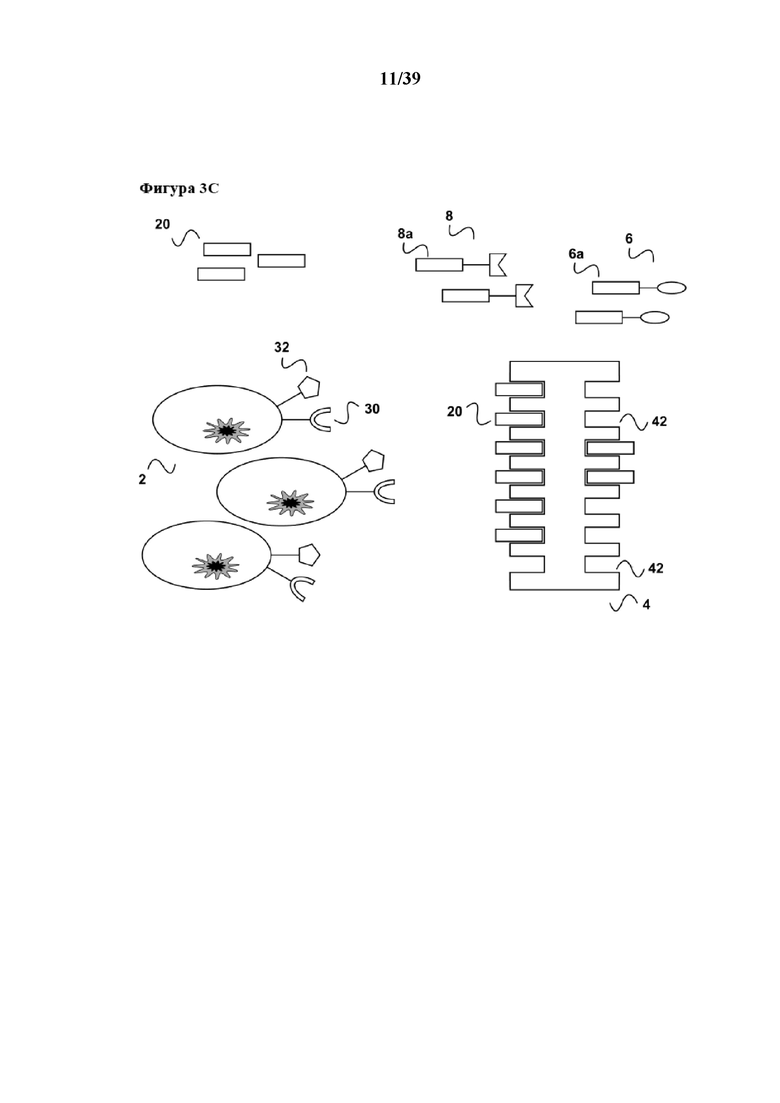
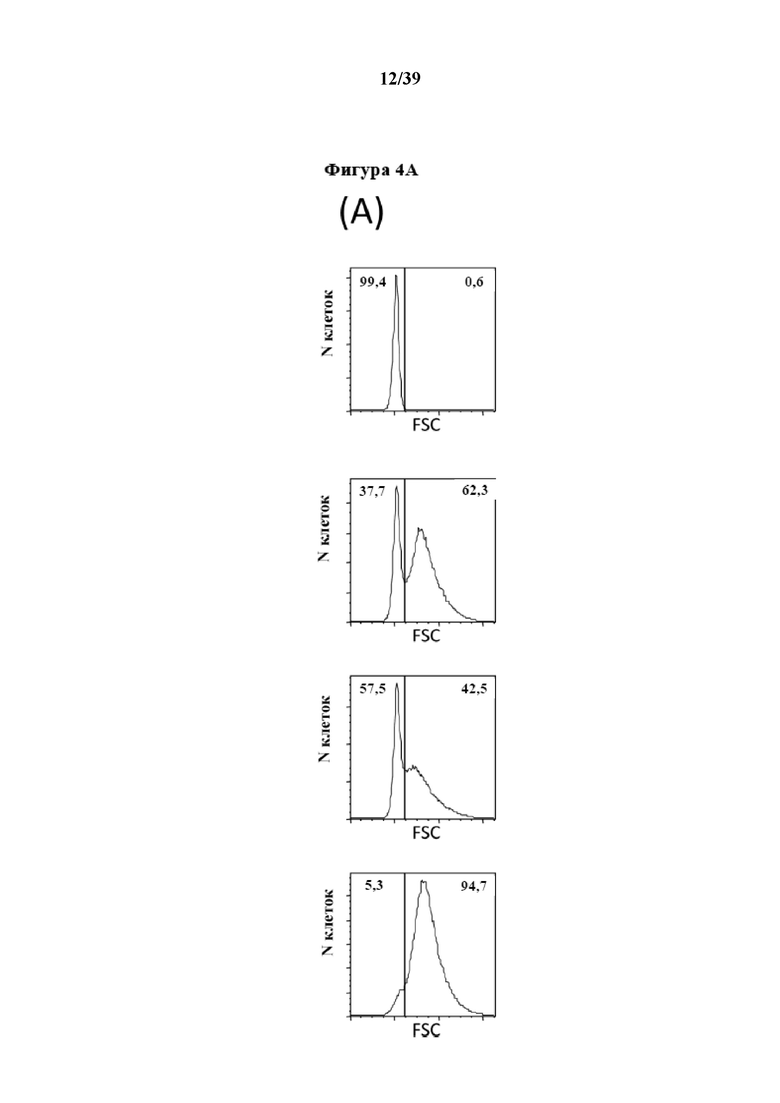
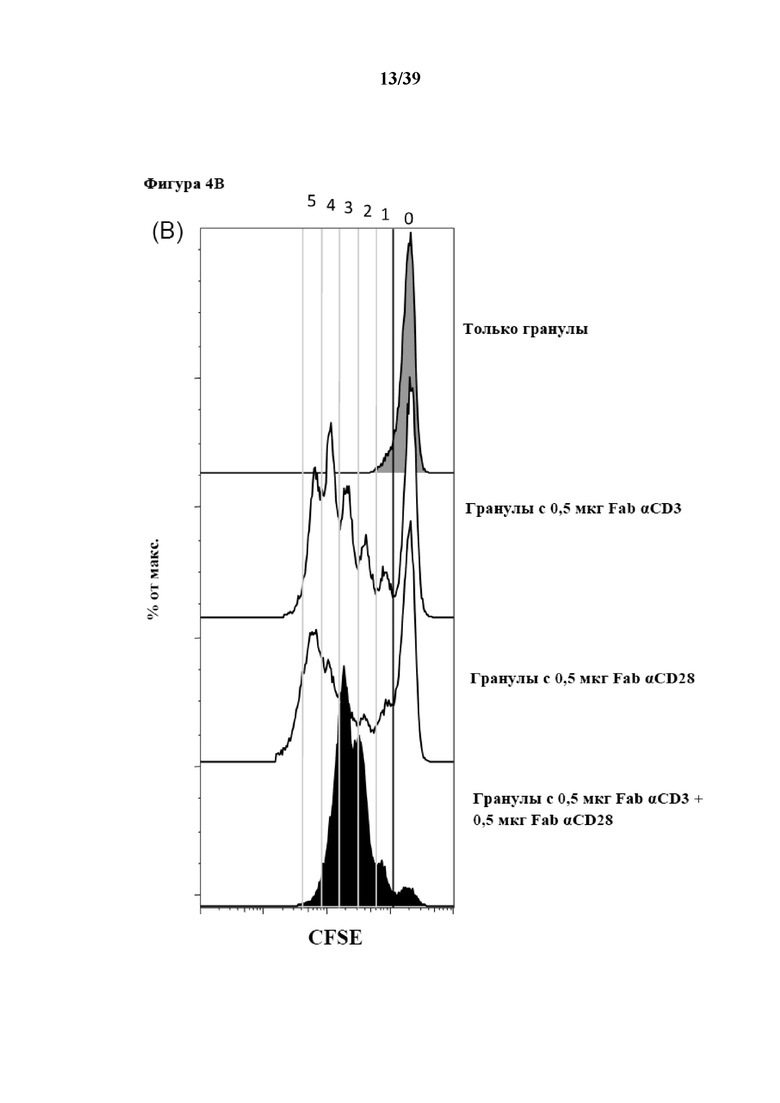

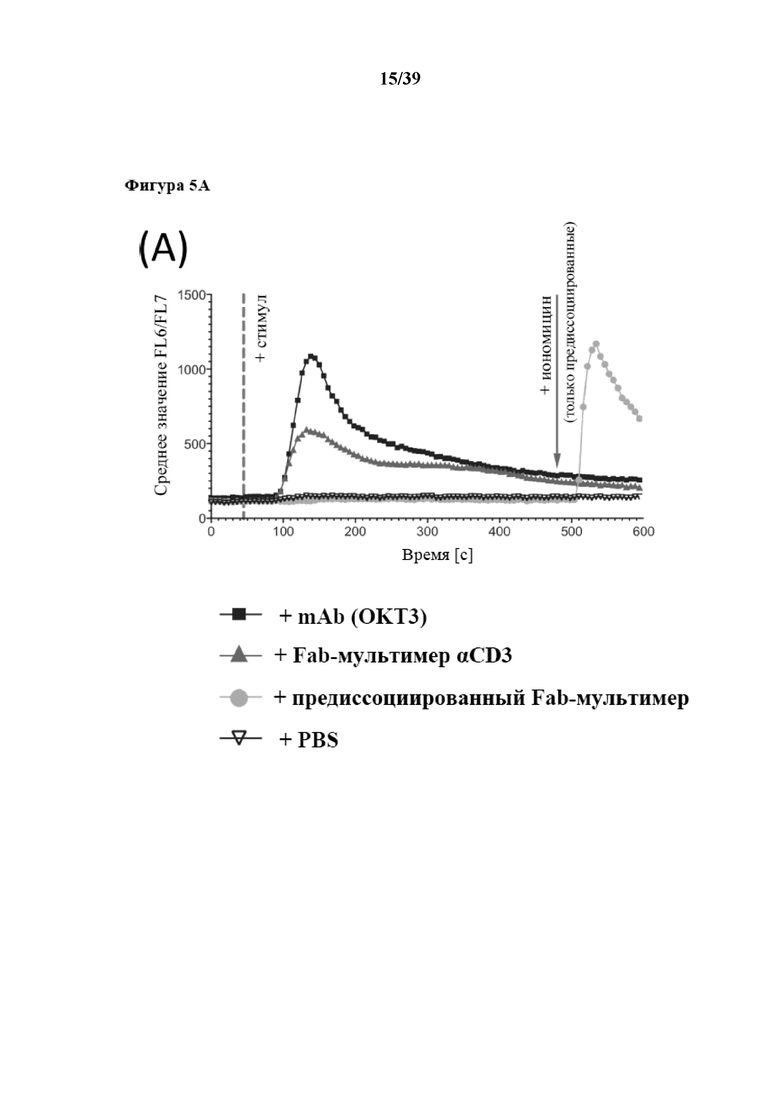
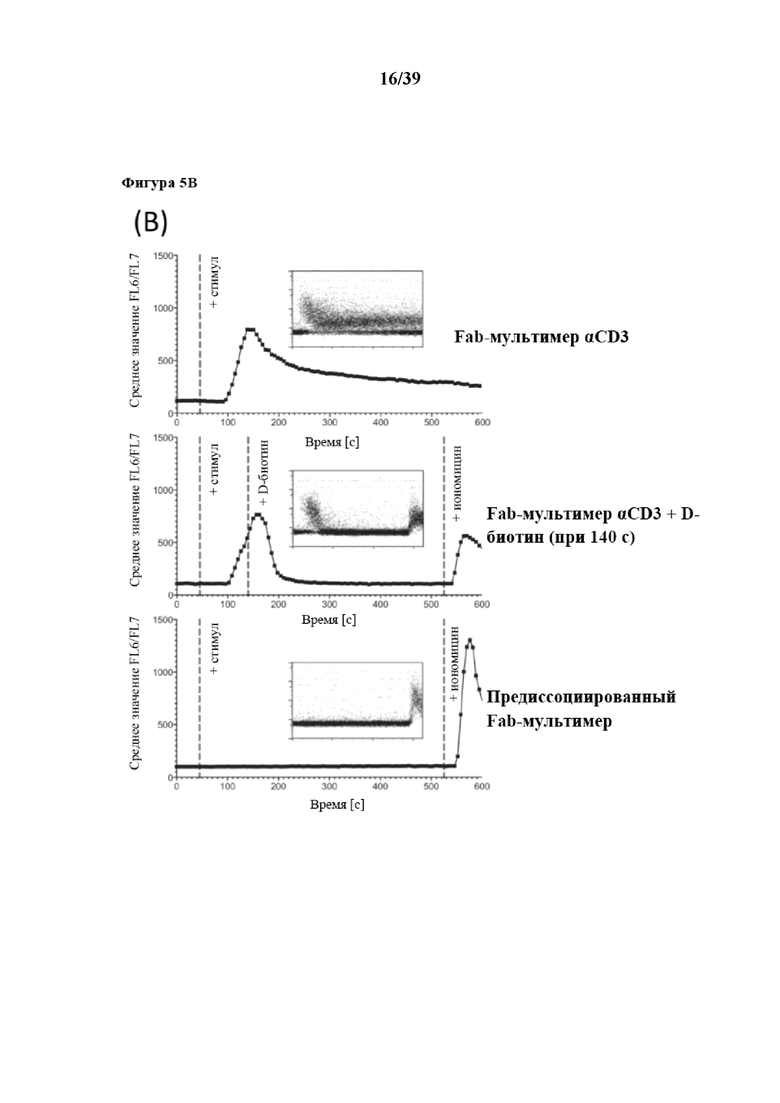
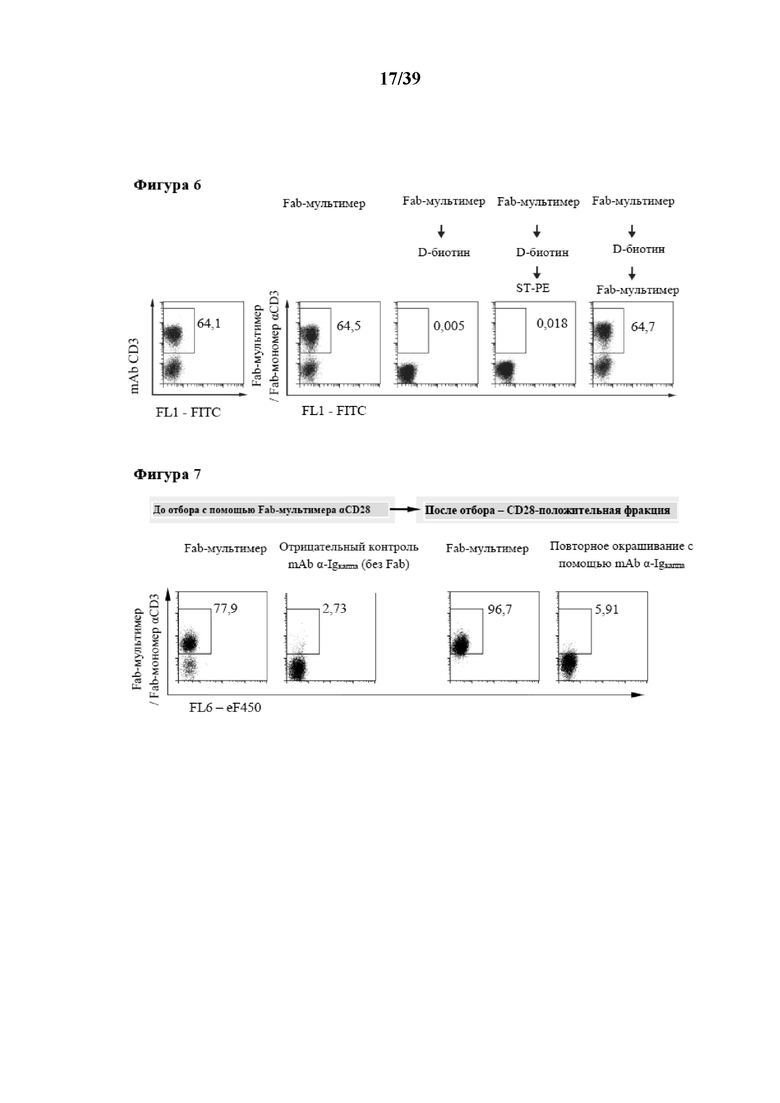
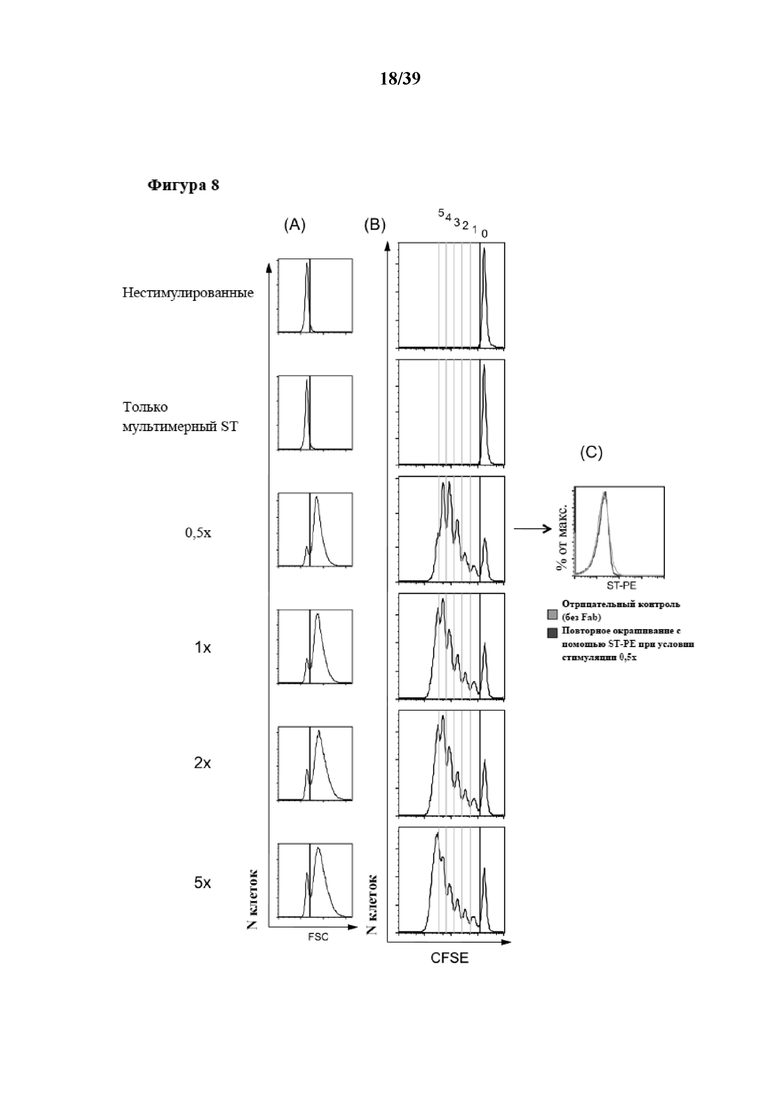
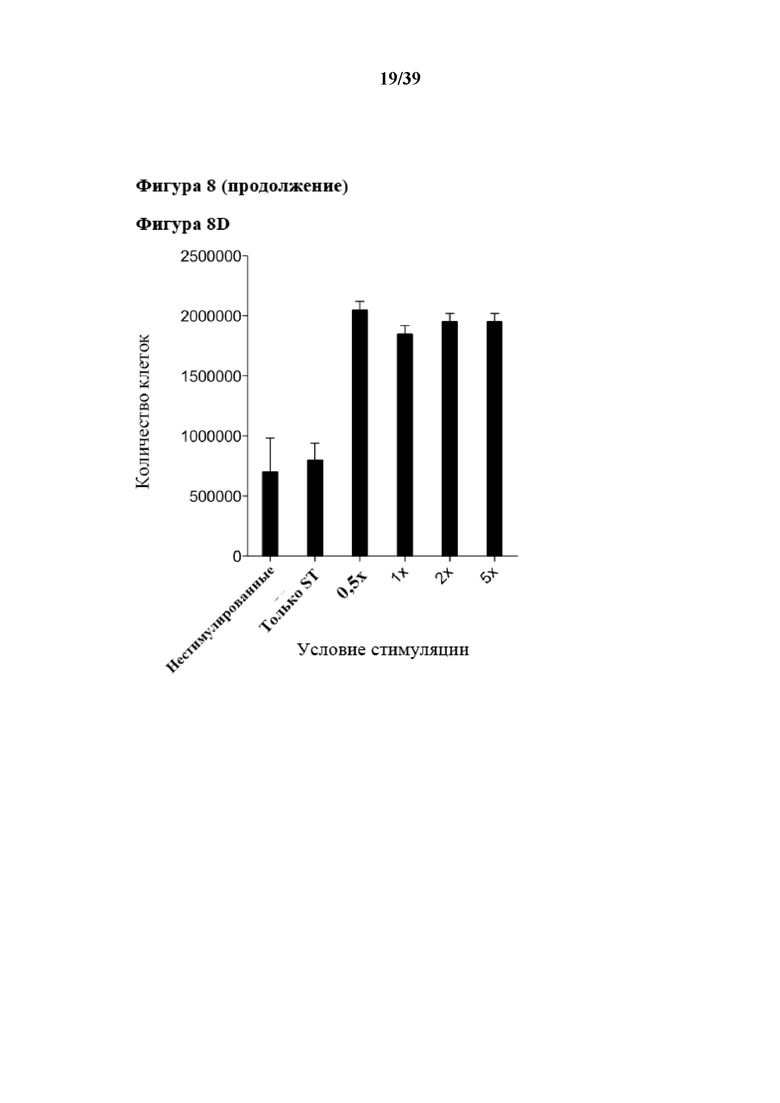
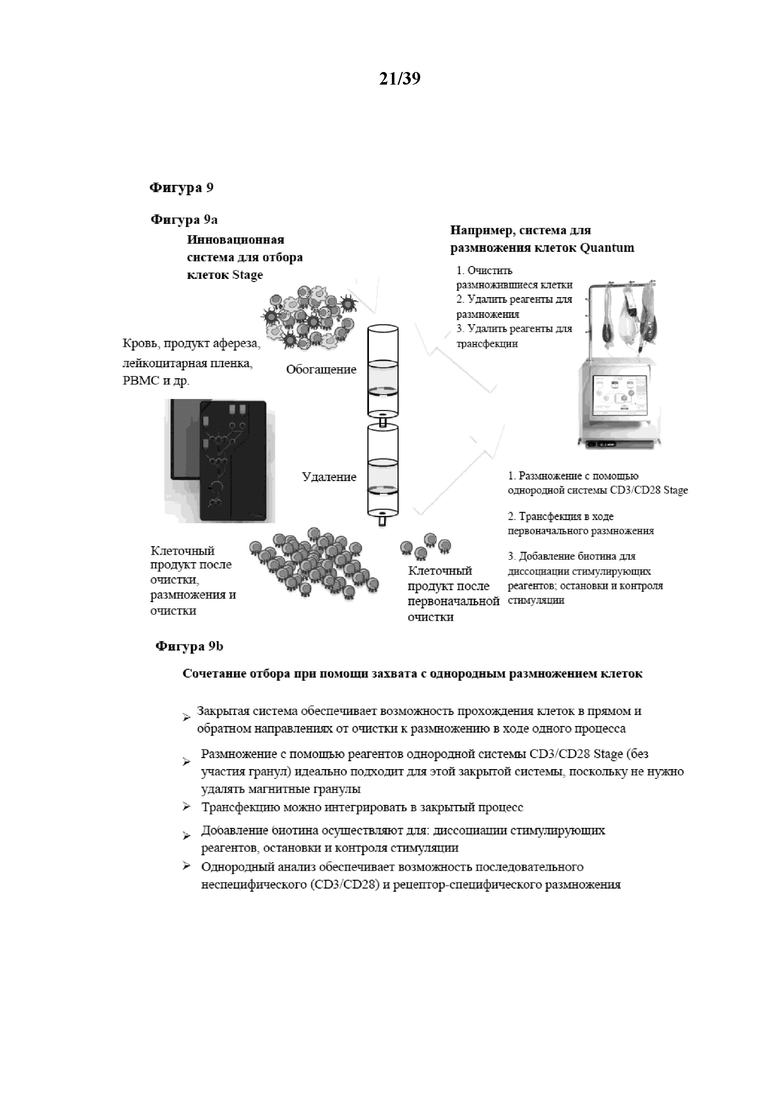
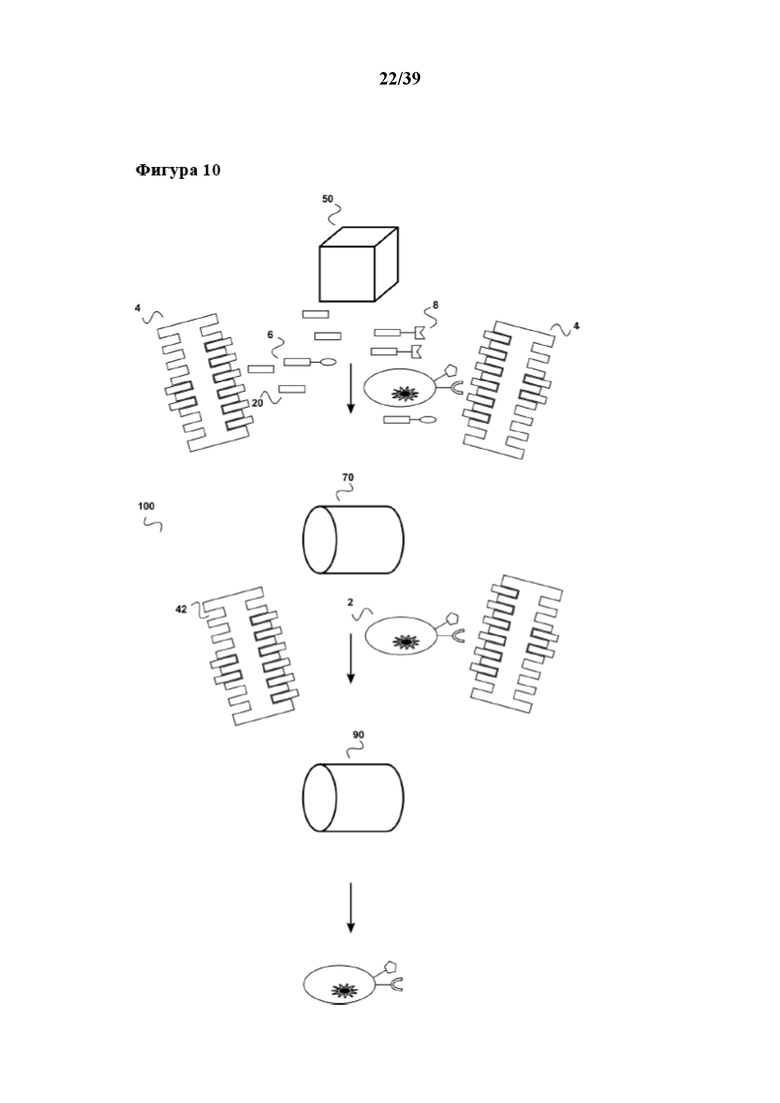
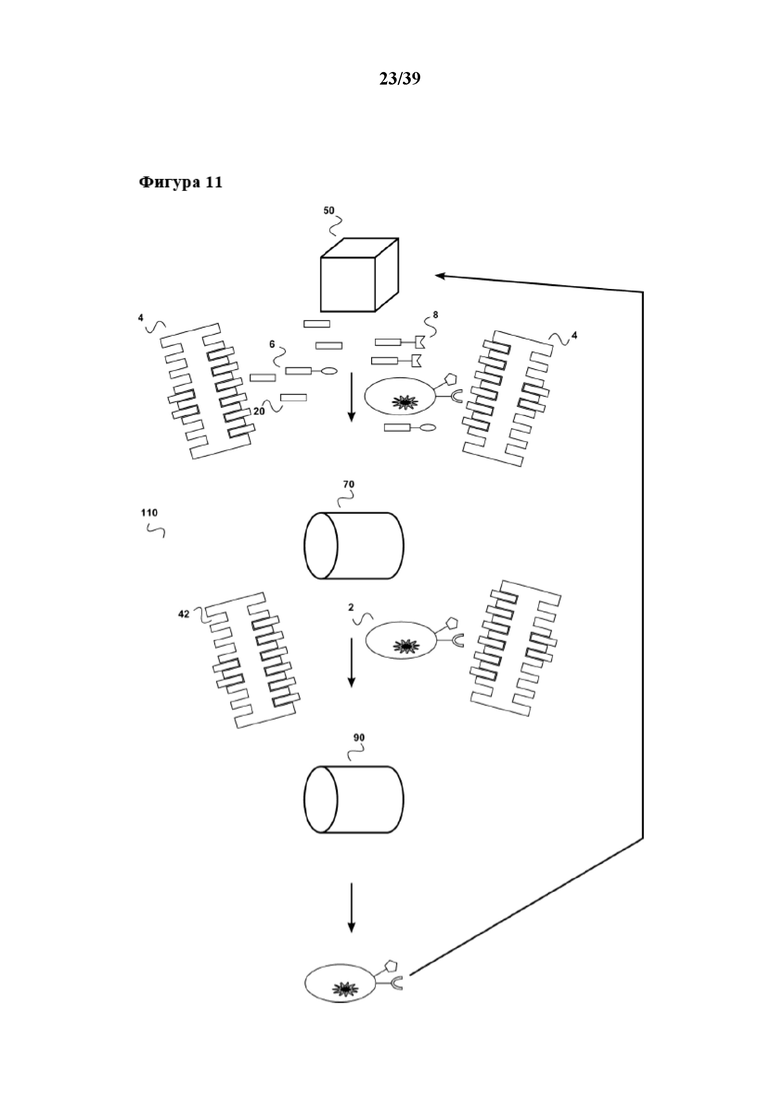
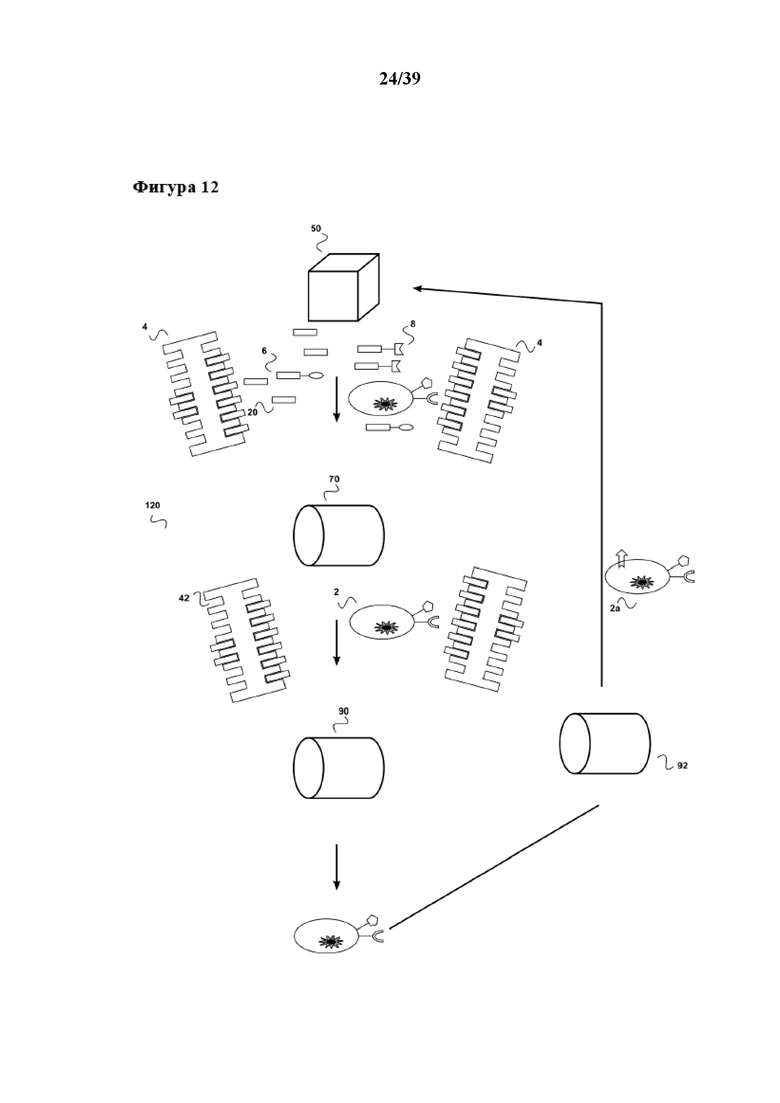
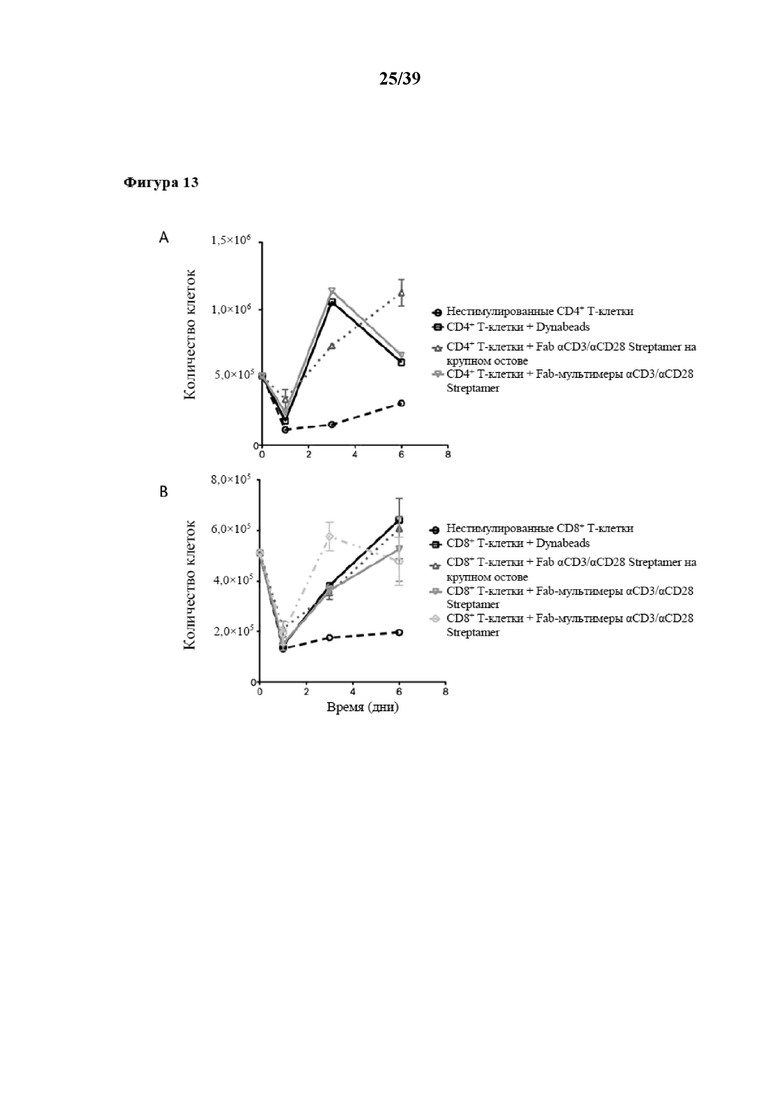
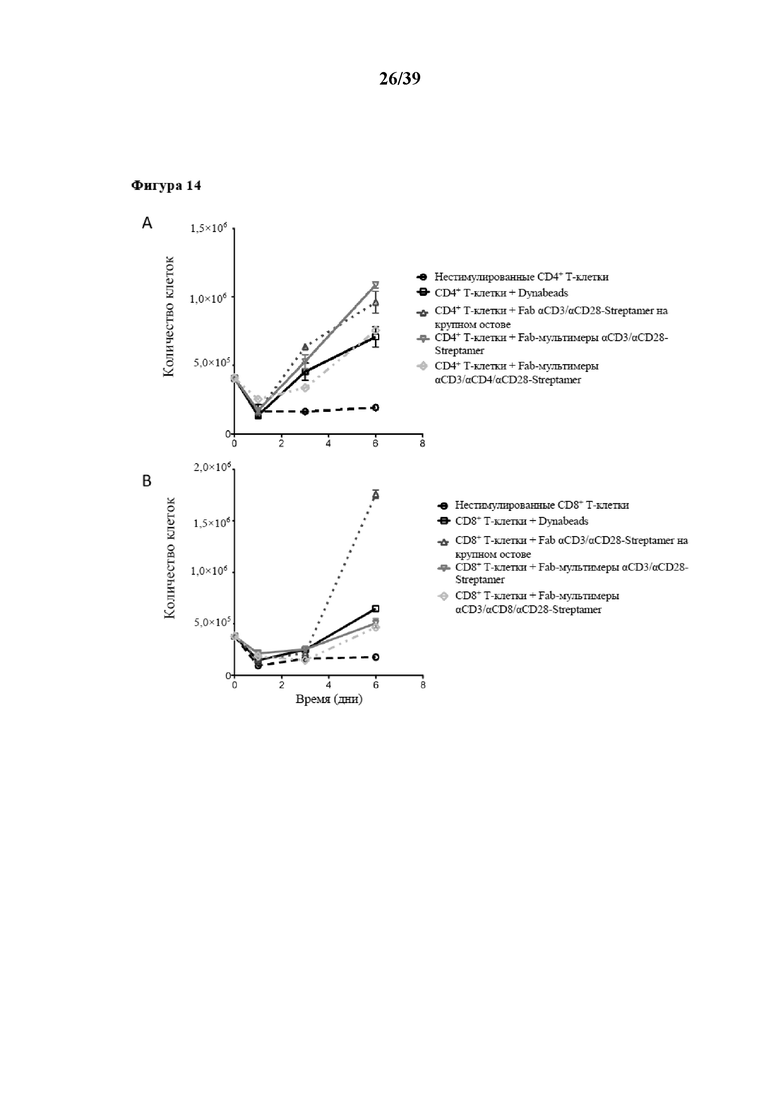
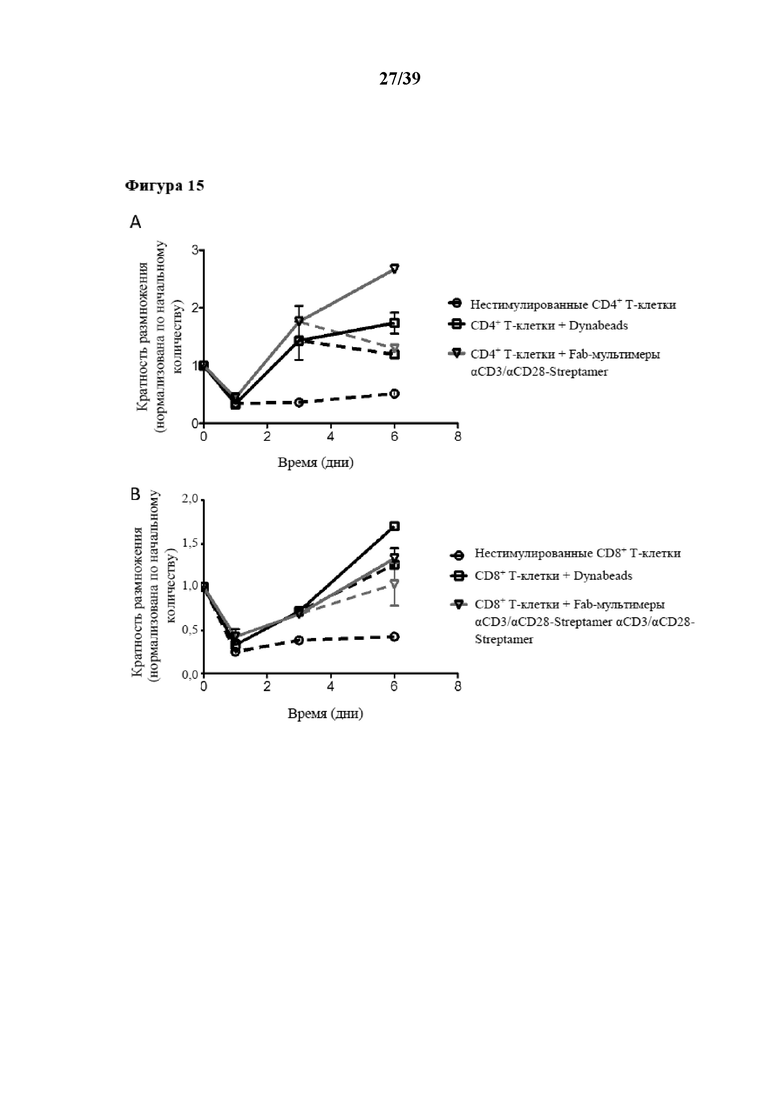
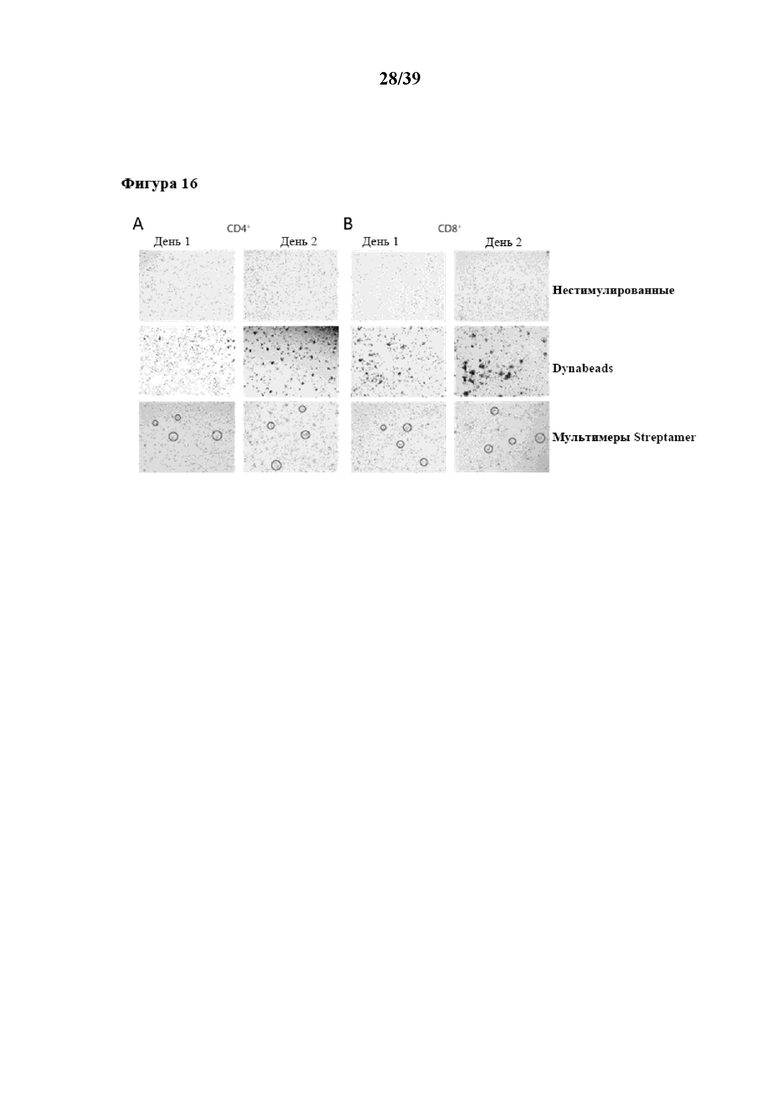
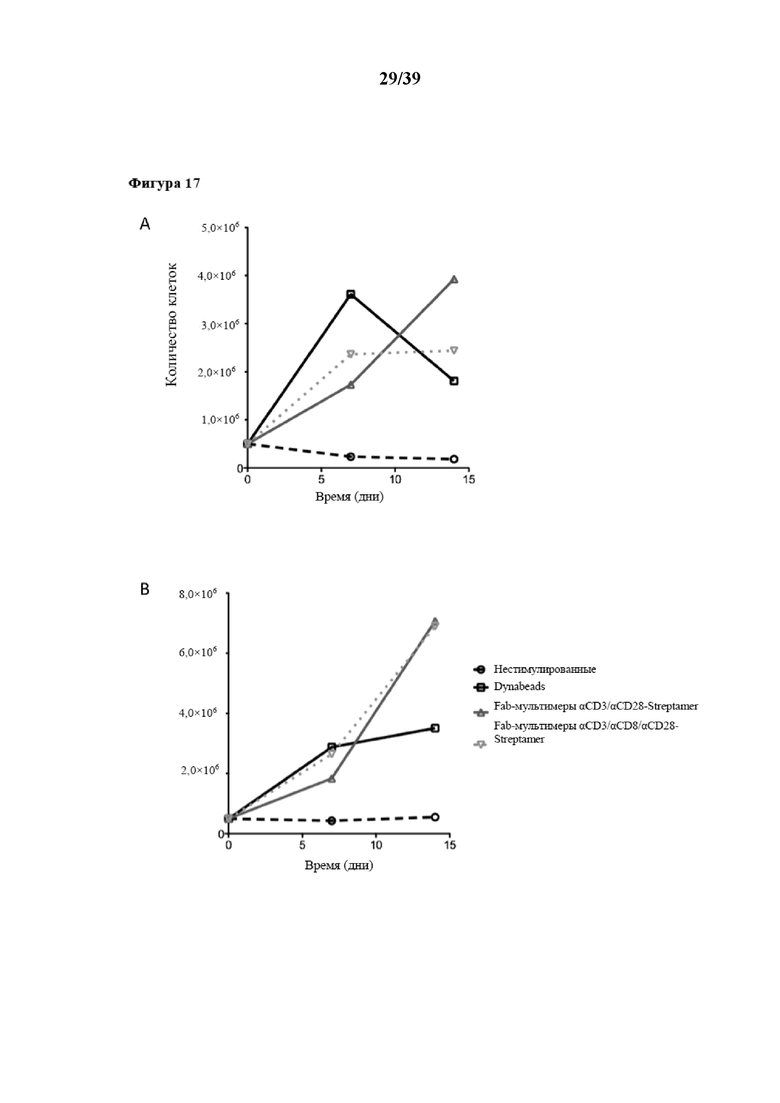
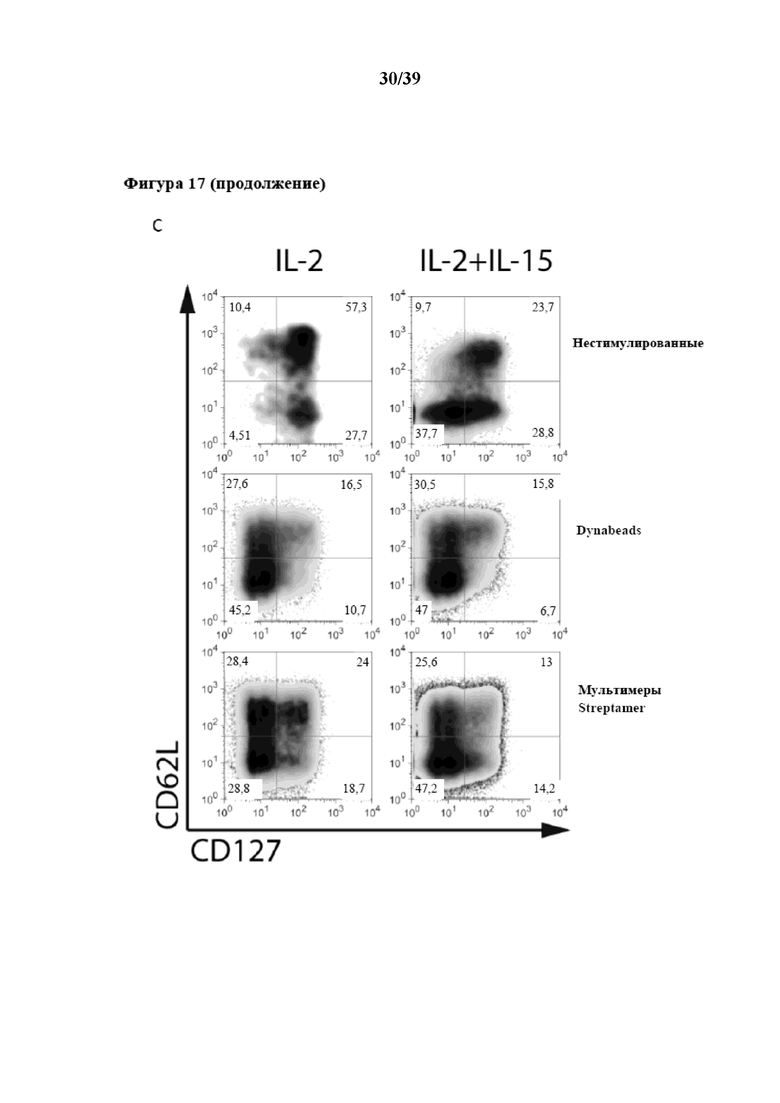
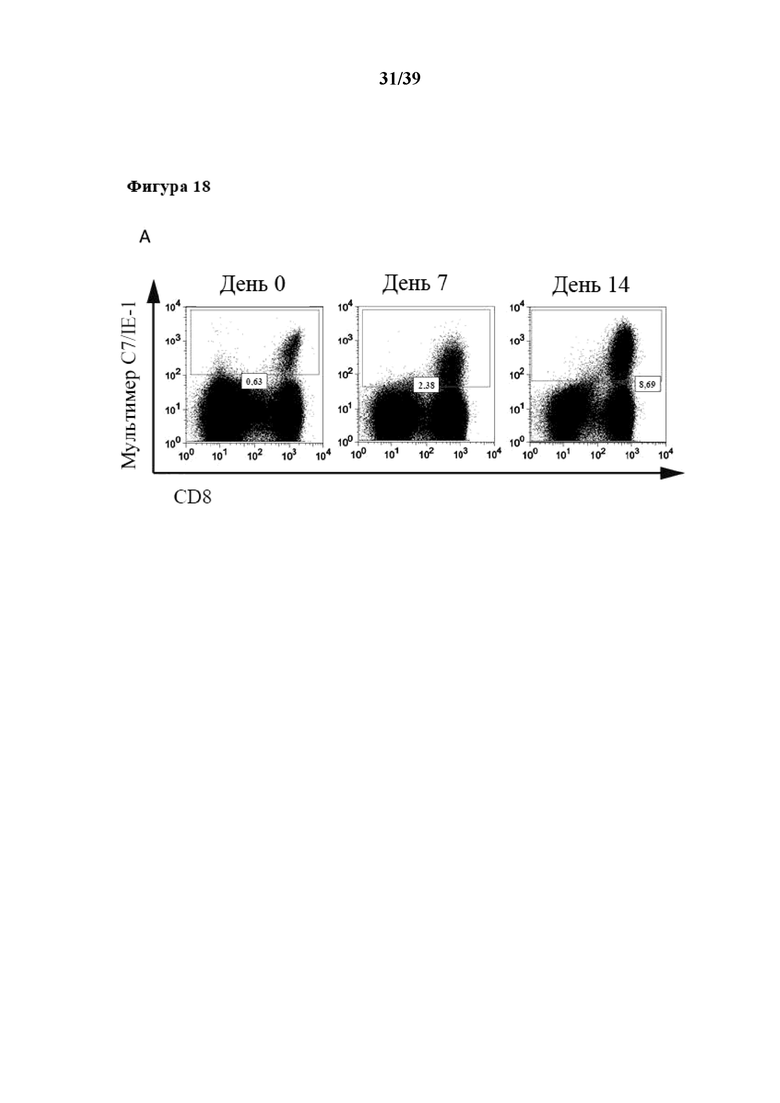
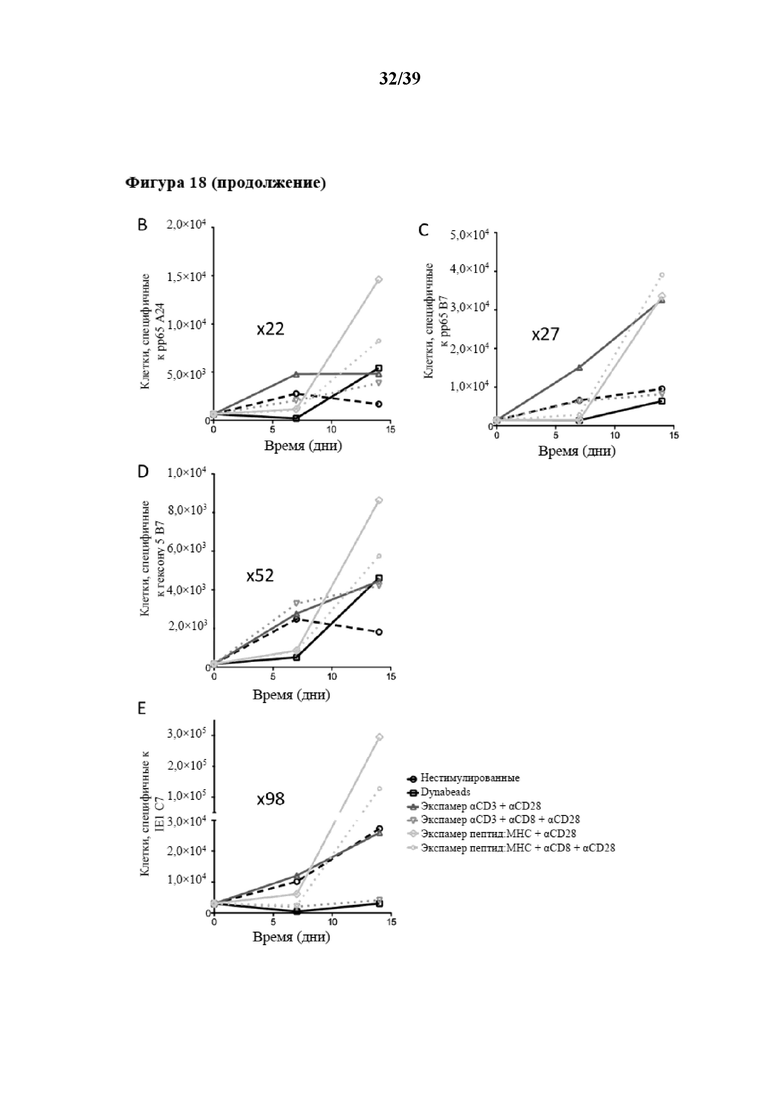
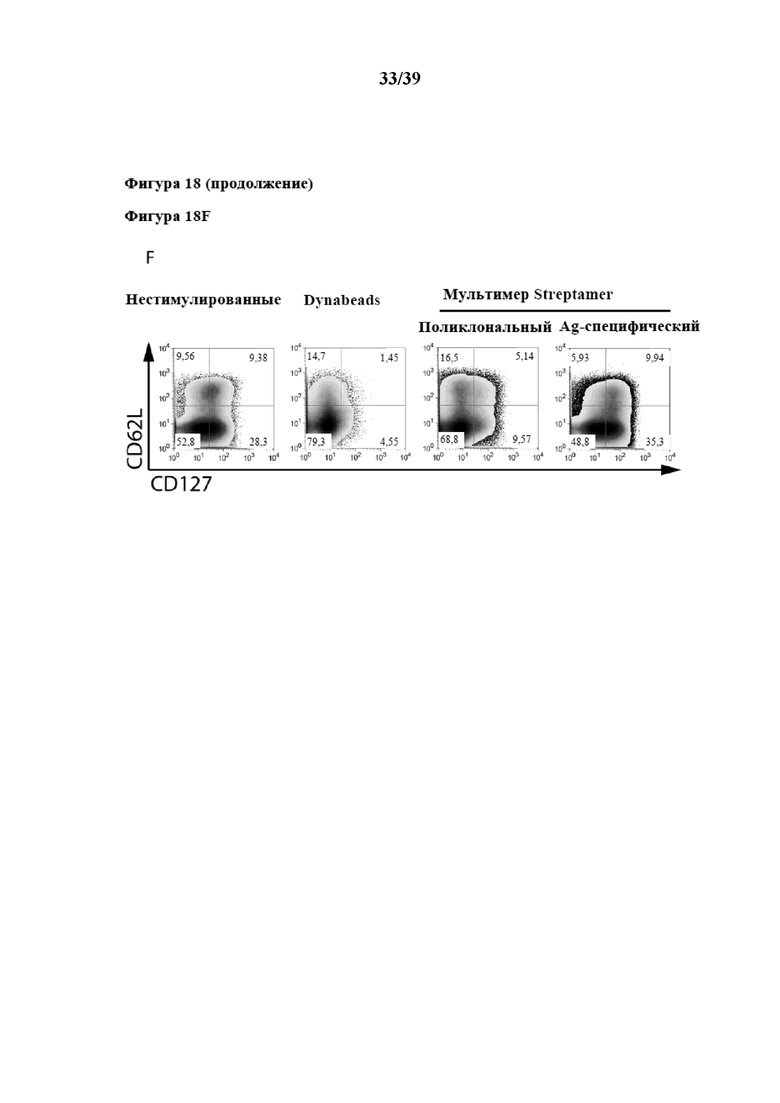
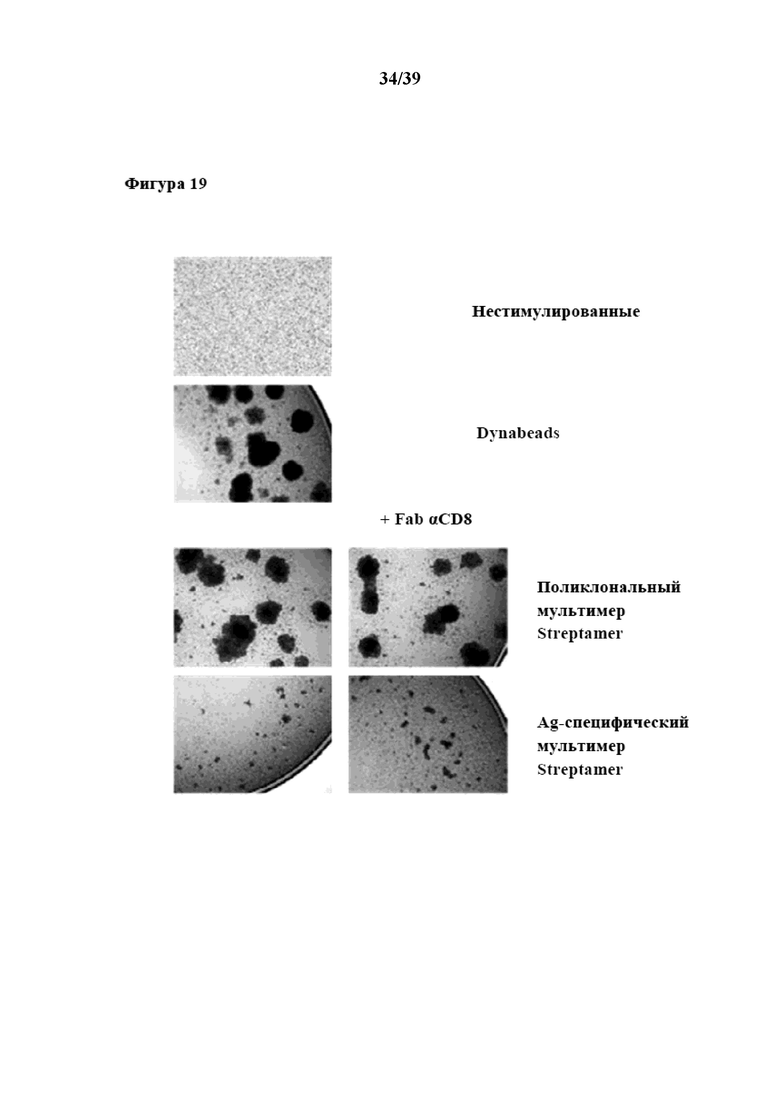
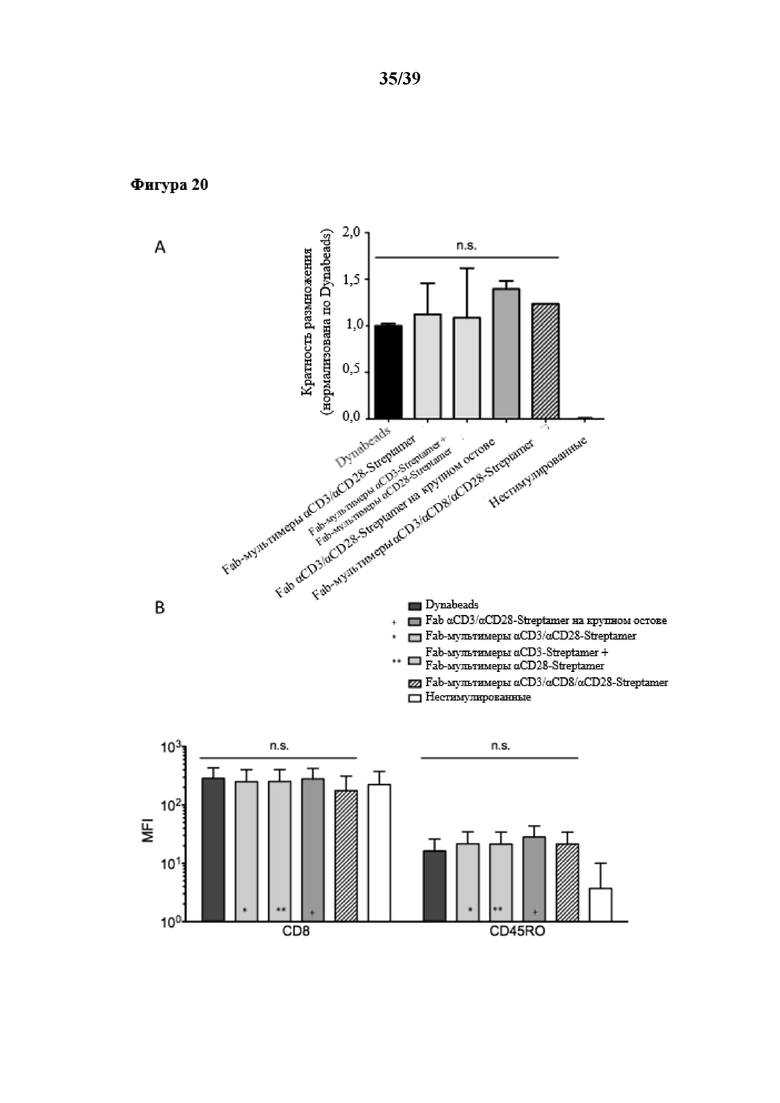
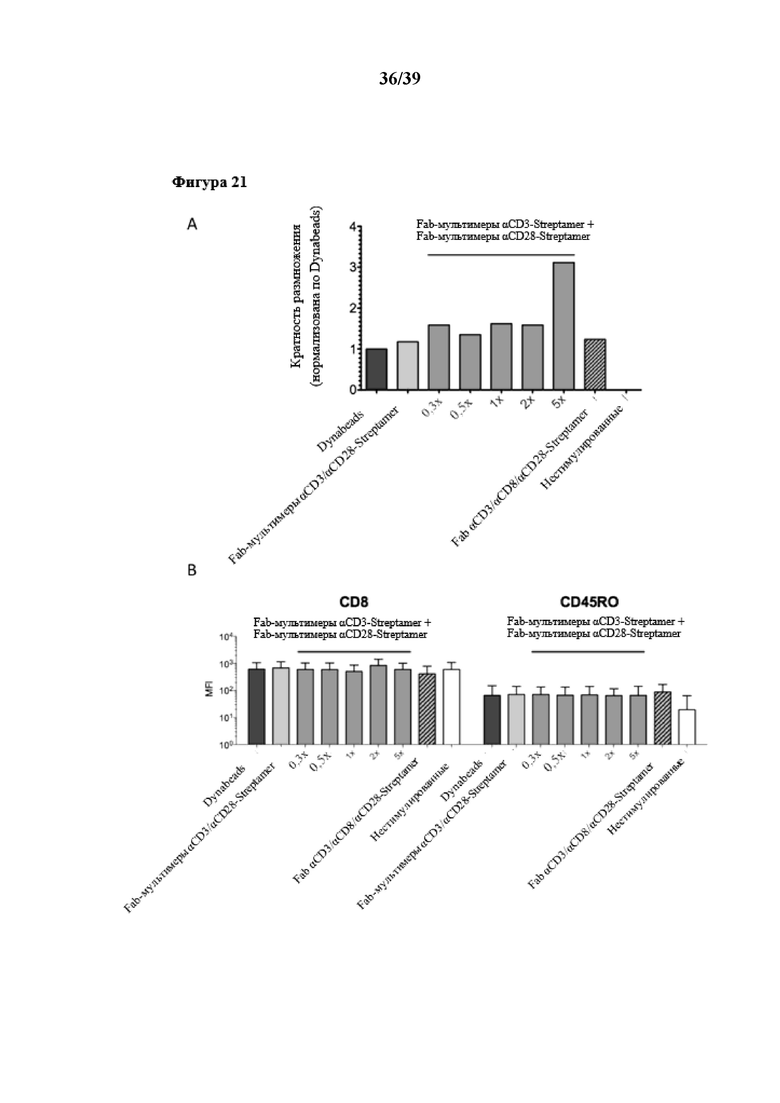

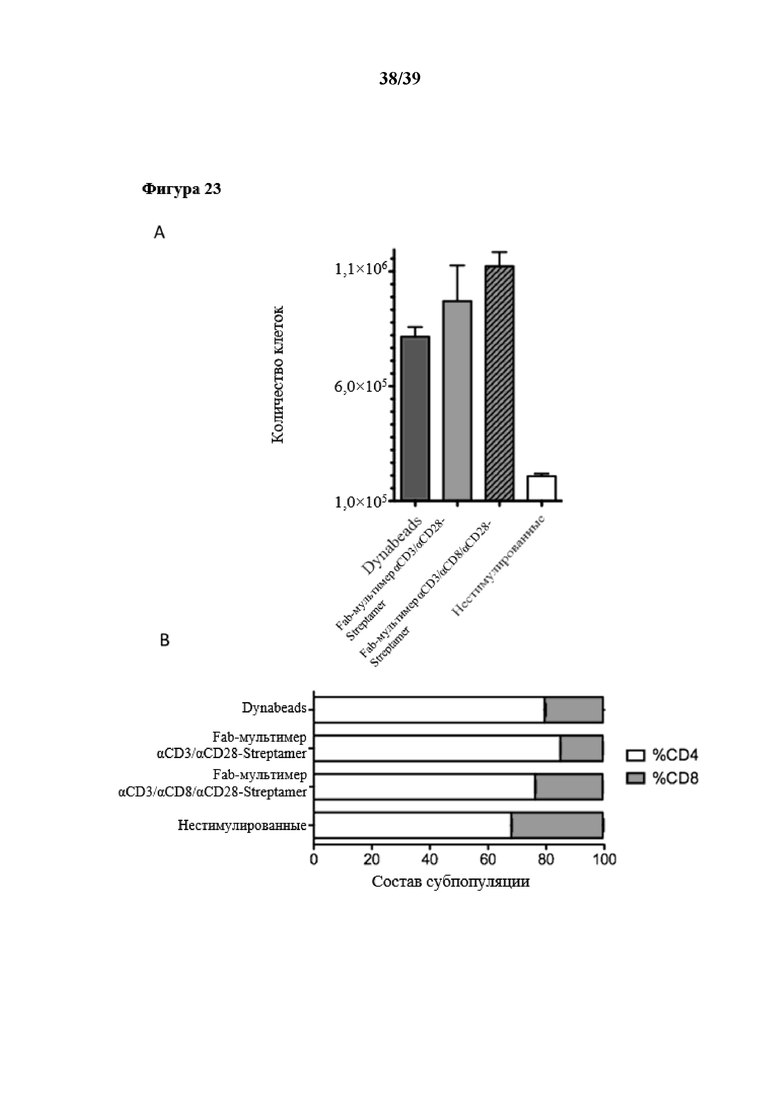
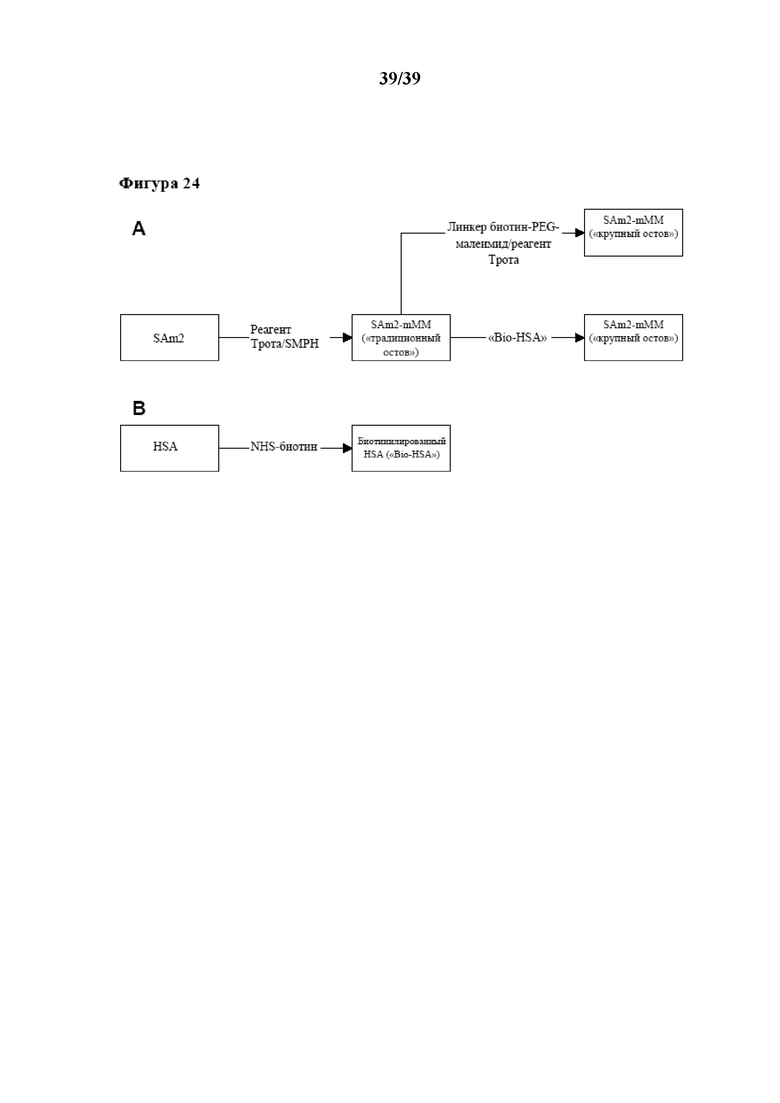
Изобретение относится к области биохимии, в частности к способу размножения популяции лимфоцитов in vitro, включающему инкубирование образца, содержащего популяцию лимфоцитов, с реагентом для мультимеризации, обратимо связанным с первым средством и вторым средством, а также к способу размножения популяции лимфоцитов in vitro, где популяция лимфоцитов содержит Т-клетки, включающему инкубирование образца, содержащего популяцию лимфоцитов, с реагентом для мультимеризации, обратимо связанным с первым средством и вторым средством. Также раскрыты набор реагентов и композиция, предназначенные для размножения популяции лимфоцитов. Изобретение также относится к реагенту для мультимеризации, обратимо связанному с первым средством и вторым средством. Изобретение позволяет эффективно осуществлять размножение популяции лимфоцитов in vitro. 5 н. и 80 з.п. ф-лы, 24 ил., 19 пр.
1. Способ размножения популяции лимфоцитов in vitro, включающий инкубирование образца, содержащего популяцию лимфоцитов, с реагентом для мультимеризации, обратимо связанным с первым средством и вторым средством, где:
реагент для мультимеризации содержит олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина;
первое средство содержит (i) первый стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться с первым связывающим участком реагента для мультимеризации, и (ii) первое антитело или его антигенсвязывающий фрагмент, который связывается с рецепторной молекулой на поверхности лимфоцита в популяции для стимуляции передачи сигнала лимфоциту; и
второе средство содержит (i) второй стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со вторым связывающим участком реагента для мультимеризации, и (ii) второе антитело или его антигенсвязывающий фрагмент, который связывается со вспомогательной молекулой на поверхности лимфоцита для обеспечения костимулирующего сигнала лимфоциту, стимулируя тем самым лимфоциты в популяции.
2. Способ по п. 1, где популяция лимфоцитов содержит B-клетки, T-клетки, естественные клетки-киллеры или их комбинацию.
3. Способ по п. 2, где популяция лимфоцитов содержит T-клетки.
4. Способ по п. 3, где T-клетки содержат антиген-специфические T-клетки, T-клетки-хелперы, цитотоксические T-клетки, T-клетки памяти, регуляторные T-клетки или естественные T-клетки-киллеры.
5. Способ по п. 3 или 4, где первое средство
содержит комплекс MHC-I:пептид или
стимулирует передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках.
6. Способ по любому из пп. 1-5, где первое средство представляет собой связывающий реагент, который специфически связывается с CD3.
7. Способ по любому из пп. 1-6, где вспомогательная молекула на лимфоците представляет собой CD28 или CD137.
8. Способ по любому из пп. 1-7, где второе средство, которое связывается со вспомогательной молекулой, представляет собой связывающий реагент, который специфически связывается с CD28 или CD137.
9. Способ по любому из пп. 1-8, где
указанное первое средство специфически связывается с CD3 и выбрано из группы, включающей антитело к CD3, бивалентный фрагмент антитела из антитела к CD3 и моновалентный фрагмент антитела из антитела к CD3, и/или
второе средство специфически связывается с CD28 или CD137 и выбрано из группы, включающей антитело к CD28, бивалентный фрагмент антитела из антитела к CD28, моновалентный фрагмент антитела из антитела к CD28, антитело к CD137, бивалентный фрагмент антитела из антитела к CD137 и моновалентный фрагмент антитела из антитела к CD137.
10. Способ по п. 9, где бивалентный фрагмент антитела представляет собой F(ab')2-фрагмент или бивалентный одноцепочечный Fv-фрагмент.
11. Способ по п. 9, где моновалентный фрагмент антитела выбран из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv).
12. Способ по п. 2, где популяция лимфоцитов содержит B-клетки и первое средство представляет собой связывающий реагент, который специфически связывается с CD40 или CD137.
13. Способ по п. 12, где первый связывающий реагент, который специфически связывается с CD40 или CD137, выбран из группы, включающей антитело к CD40, бивалентный фрагмент антитела из антитела к CD40, моновалентный фрагмент антитела из антитела к CD40, антитело к CD137, бивалентный фрагмент антитела из антитела к CD137 и моновалентный фрагмент антитела из антитела к CD137.
14. Способ по п. 13, где бивалентный фрагмент антитела представляет собой F(ab')2-фрагмент или бивалентный одноцепочечный Fv-фрагмент.
15. Способ по п. 13, где моновалентный фрагмент антитела выбран из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv).
16. Способ по любому из пп. 12-15, где вспомогательная молекула на B-клетке представляет собой CD40.
17. Способ по любому из пп. 1-16, где реагент для мультимеризации иммобилизован на твердой поверхности.
18. Способ по п. 17, где твердая поверхность выбрана из группы, включающей магнитную гранулу, полимерную гранулу, планшет для культуры клеток, титрационный микропланшет, мембрану, полое волокно и их комбинации.
19. Способ по любому из пп. 1-16, где реагент для мультимеризации находится в растворимой форме.
20. Способ по любому из пп. 1-19, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина сшит при помощи полисахарида или посредством бифункционального линкера.
21. Способ по любому из пп. 1-20, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина содержит три или более мономера стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина.
22. Способ по любому из пп. 1-21, где указанный первый стрептавидин- или авидинсвязывающий пептид и указанный второй стрептавидин- или авидинсвязывающий пептид независимо содержат стрептавидинсвязывающий пептид Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 1) или стрептавидинсвязывающий пептид SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 7).
23. Способ по любому из пп. 1-22, где реагент для мультимеризации содержит мутантный вариант белка стрептавидина, содержащий аминокислотную последовательность Va144-Thr45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа, или мутантный вариант белка стрептавидина, содержащий аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа.
24. Способ по п. 23, где N-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 10-16 аминокислотной последовательности стрептавидина дикого типа и C-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 133-142 аминокислотной последовательности стрептавидина дикого типа.
25. Способ по любому из пп. 1-24, где первый стрептавидин- или авидинсвязывающий пептид и второй стрептавидин- или авидинсвязывающий пептид являются различными.
26. Способ по любому из пп. 1-24, где первый стрептавидин- или авидинсвязывающий пептид и второй стрептавидин- или авидинсвязывающий пептид являются идентичными.
27. Способ по любому из пп. 1-26, дополнительно включающий нарушение связывания между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком указанного реагента для мультимеризации и/или связывания между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком указанного реагента для мультимеризации.
28. Способ по п. 27, где нарушение связывания между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком указанного реагента для мультимеризации и/или нарушение связывания между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком указанного реагента для мультимеризации вызывает прекращение стимуляции лимфоцитов.
29. Способ по любому из пп. 1-28, где константа диссоциации (Kd) для обратимого связывания между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком и/или Kd для обратимого связывания между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком находится в диапазоне от 10-2 до 10-13 M.
30. Способ по любому из пп. 1-29, где указанное обратимое связывание между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком указанного реагента для мультимеризации разрушают с помощью приведения указанной популяции лимфоцитов в контакт с первым стрептавидин- или авидинсвязывающим пептидом в свободной форме или его аналогом, способным к разрушению связывания между первым стрептавидин- или авидинсвязывающим пептидом и первым связывающим участком, и/или
где указанное обратимое связывание между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком указанного реагента для мультимеризации разрушают с помощью приведения указанной популяции лимфоцитов в контакт со вторым стрептавидин- или авидинсвязывающим пептидом в свободной форме или его аналогом, способным к разрушению связывания между вторым стрептавидин- или авидинсвязывающим пептидом и вторым связывающим участком.
31. Способ по любому из пп. 1-30, где обратимое связывание между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком и/или обратимое связывание между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком разрушают с помощью приведения лимфоцитов в контакт с биотином или аналогом биотина.
32. Способ размножения популяции лимфоцитов in vitro, где популяция лимфоцитов содержит Т-клетки, при этом способ включает
инкубирование образца, содержащего популяцию лимфоцитов, с реагентом для мультимеризации, обратимо связанным с первым средством и вторым средством, где:
реагент для мультимеризации находится в растворимой форме и содержит олигомерный или полимерный мутантный вариант белка стрептавидина;
первое средство содержит (i) первый стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться с первым связывающим участком реагента для мультимеризации, и (ii) первое антитело или его антигенсвязывающий фрагмент, который связывается с рецепторной молекулой на поверхности лимфоцита в популяции для стимуляции передачи сигнала лимфоциту; и
второе средство содержит (i) второй стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со вторым связывающим участком реагента для мультимеризации, и (ii) второе антитело или его антигенсвязывающий фрагмент, который связывается со вспомогательной молекулой на поверхности лимфоцита для обеспечения костимулирующего сигнала лимфоциту, стимулируя тем самым лимфоциты в популяции.
33. Способ по п. 32, где рецепторная молекула представляет собой CD3 и первое антитело или его антигенсвязывающий фрагмент специфически связывается с CD3.
34. Способ по п. 33, где вспомогательная молекула представляет собой CD28 и второе антитело или его антигенсвязывающий фрагмент специфически связывается с CD28.
35. Способ по любому из пп. 32-34, где первое и/или второе антитело или его антигенсвязывающий фрагмент выбраны из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv).
36. Способ по любому из пп. 32-35, где обратимое связывание между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком и/или обратимое связывание между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком разрушают с помощью приведения лимфоцитов в контакт с биотином или аналогом биотина.
37. Способ по любому из пп. 1-36, где инкубирование образца с реагентом для мультимеризации приводит к специфичному связыванию Т-клеток с реагентом для мультимеризации.
38. Способ по п. 37, дополнительно содержащий разделение T-клеткок, связанных с реагентом для мультимеризации, и несвязанных клеток популяции лимфоцитов.
39. Способ по любому из пп. 1-38, где инкубирование образца с реагентом для мультимеризации осуществляют в биореакторе, таком как культуральный планшет, половолоконный биореактор или биореактор в виде пластикового мешка.
40. Способ по п. 37, дополнительно содержащий инкубирование Т-клеток, связанных с реагентом для мультимеризации, с помощью первого средства и второго средства с (i) первым стрептавидин- или авидинсвязывающим пептидом в свободной форме или его аналогом, способным к разрушению связывания между первым стрептавидин- или авидинсвязывающим пептидом и первым связывающим участком, и (ii) вторым стрептавидин- или авидинсвязывающим пептидом в свободной форме или его аналогом, способным к разрушению связывания между вторым стрептавидин- или авидинсвязывающим пептидом и вторым связывающим участком,
где указанное обратимое связывание между указанным первым стрептавидин- или авидинсвязывающим пептидом и указанным первым связывающим участком и обратимое связывание между указанным вторым стрептавидин- или авидинсвязывающим пептидом и указанным вторым связывающим участком разрушают с отделением тем самым T-клеток от растворимого реагента для мультимеризации.
41. Способ по п. 40, включающий сбор элюата, который содержит отделенные T-клетки, растворимый реагент для мультимеризации, первое средство и/или второе средство, первый стрептавидин- или авидинсвязывающий пептид в свободной форме или его аналог и второй стрептавидин- или авидинсвязывающий пептид в свободной форме или его аналог.
42. Способ по п. 41, дополнительно содержащий:
проведение в отношении элюата хроматографии на первой неподвижной фазе, содержащей первый аффинный реагент, где первый аффинный реагент содержит один или несколько связывающих участков, способных специфически связываться с первым и/или вторым стрептавидин- или авидинсвязывающим пептидом, с иммобилизацией тем самым первого средства, второго средства, первого стрептавидин- или авидинсвязывающего пептида в свободной форме или его аналога и/или второго стрептавидин- или авидинсвязывающего пептида в свободной форме или его аналога на неподвижной фазе; и/или
проведение в отношении элюата хроматографии на второй неподвижной фазе, содержащей второй аффинный реагент, где второй аффинный реагент содержит один или несколько партнеров по связыванию, способных специфически связываться с первым и/или вторым связывающим участком реагента для мультимеризации, с иммобилизацией тем самым реагента для мультимеризации на неподвижной фазе.
43. Способ по п. 42, где первая и/или вторая неподвижная фаза находится в жидкостном соединении с биореактором и первая и/или вторая неподвижная фаза содержит матрицу для гель-фильтрации и/или матрицу для аффинной хроматографии.
44. Способ по п. 43, где отделяют T-клетки от растворимого реагента для мультимеризации.
45. Способ по любому из пп. 3-11 и 17-44, дополнительно содержащий введение Т-клеточного рецептора или химерного рецептора в Т-клетки, где T-клеточный рецептор или химерный антигенный рецептор вводят в T-клетки после стимуляции или в ходе стимуляции.
46. Способ по п. 45, где T-клетки стимулируют третьим средством, которое связывается с введенным T-клеточным рецептором или химерным антигенным рецептором.
47. Способ по любому из пп. 3-11 и 17-46, где популяция лимфоцитов, содержащая T-клетки, представляет собой популяцию мононуклеарных клеток периферической крови (PBMC) или обогащенную или очищенную популяцию T-клеток.
48. Набор реагентов для размножения популяции лимфоцитов, при этом набор содержит:
(i) реагент для мультимеризации, содержащий олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина, причем реагент для мультимеризации содержит первый связывающий участок и второй связывающий участок,
(ii) первое средство, содержащее (a) первый стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться с первым связывающим участком реагента для мультимеризации, и (b) первое антитело или его антигенсвязывающий фрагмент, который связывается с рецепторной молекулой на поверхности лимфоцита для стимуляции передачи сигнала лимфоциту, и
(iii) второе средство, содержащее (a) второй стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со вторым связывающим участком реагента для мультимеризации, и (b) второе антитело или его антигенсвязывающий фрагмент, который связывается с вспомогательной молекулой на поверхности лимфоцита для обеспечения костимулирующего сигнала лимфоциту.
49. Набор по п. 48, где популяция содержит В-клетки, Т-клетки, естественные клетки-киллеры или их комбинацию.
50. Набор по п. 49, где Т-клетки содержат антиген-специфические T-клетки, T-клетки-хелперы, цитотоксические T-клетки, T-клетки памяти, регуляторные T-клетки или естественные T-клетки-киллеры.
51. Набор по п. 48, где популяция лимфоцитов содержит Т-клетки и первое средство
содержит комплекс MHC-I:пептид или
стимулирует передачу сигнала, связанную с комплексом TCR/CD3, в T-клетках.
52. Набор по п. 51, где первое средство представляет собой связывающий реагент, который специфически связывается с CD3.
53. Набор по п. 51 или 52, где вспомогательная молекула на T-клетках представляет собой CD28.
54. Набор по любому из пп. 51-53, где второе средство, которое связывается со вспомогательной молекулой, представляет собой связывающий реагент, который специфически связывается с CD28.
55. Набор по любому из пп. 48-54, где
указанный первый связывающий реагент специфически связывается с CD3 и выбран из группы, включающей антитело к CD3, бивалентный фрагмент антитела из антитела к CD3 и моновалентный фрагмент антитела из антитела к CD3, и/или
второе средство специфически связывается с CD28 и выбрано из группы, включающей антитело к CD28, бивалентный фрагмент антитела из антитела к CD28 и моновалентный фрагмент антитела из антитела к CD28.
56. Набор по любому из пп. 48-55, где реагент для мультимеризации иммобилизован на твердой поверхности.
57. Набор по п. 56, где твердая поверхность выбрана из группы, включающей магнитную гранулу, полимерную гранулу, планшет для культуры клеток, титрационный микропланшет, мембрану, полое волокно и их комбинации.
58. Набор по любому из пп. 48-55, где реагент для мультимеризации находится в растворимой форме.
59. Набор по любому из пп. 48-58, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина сшит при помощи полисахарида или посредством бифункционального линкера.
60. Набор по любому из пп. 48-59, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина, содержит три или более мономеров стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина.
61. Набор по любому из пп. 48-60, где указанный первый стрептавидин- или авидинсвязывающий пептид или указанный второй стрептавидин- или авидинсвязывающий пептид независимо содержат стрептавидинсвязывающий пептид Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 1), или стрептавидинсвязывающий пептид SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 7).
62. Набор по любому из пп. 48-61, где реагент для мультимеризации содержит мутантный вариант белка стрептавидина, который содержит аминокислотную последовательность Val44-Thr45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа, или мутантный вариант белка стрептавидина, который содержит аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа.
63. Набор по п. 62, где N-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 10-16 аминокислотной последовательности стрептавидина дикого типа и C-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 133-142 аминокислотной последовательности стрептавидина дикого типа.
64. Реагент для мультимеризации, обратимо связанный с первым средством и вторым средством,
где реагент для мультимеризации находится в растворимой форме и содержит олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина;
первое средство содержит (i) первый стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться с первым связывающим участком реагента для мультимеризации, и (ii) первое антитело или его антигенсвязывающий фрагмент, который связывается с рецепторной молекулой на поверхности лимфоцита для стимуляции передачи сигнала лимфоциту; и
второе средство содержит (i) второй стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со вторым связывающим участком реагента для мультимеризации, и (ii) второе антитело или его антигенсвязывающий фрагмент, который связывается со вспомогательной молекулой на клеточной поверхности лимфоцита для обеспечения костимулирующего сигнала лимфоциту, стимулируя тем самым лимфоциты.
65. Реагент для мультимеризации по п. 64, который способен к обеспечению стимулирования популяции лимфоцитов.
66. Реагент для мультимеризации по п. 65, где подлежащая стимулированию популяция лимфоцитов представляет собой популяцию B-клеток, популяцию T-клеток или популяцию естественных клеток-киллеров.
67. Реагент для мультимеризации по п. 66, где популяция T-клеток содержит антиген-специфические T-клетки, T-клетки-хелперы, цитотоксические T-клетки, T-клетки памяти, регуляторные T-клетки или естественные T-клетки-киллеры.
68. Реагент для мультимеризации по п. 67, где первое средство
содержит комплекс MHC-I:пептид
или стимулирует передачу сигнала, связанную с комплексом TCR/CD3, в T-клетке.
69. Реагент для мультимеризации по п. 68, где первое средство специфически связывается с CD3.
70. Реагент для мультимеризации по любому из пп. 64-69, где второе средство, которое связывается со вспомогательной молекулой, представляет собой связывающий реагент, который специфически связывается с CD28 или CD137.
71. Реагент для мультимеризации по любому из пп. 64-70, где
указанное первое средство специфически связывается с CD3 и выбрано из группы, включающей антитело к CD3, бивалентный фрагмент антитела из антитела к CD3 и моновалентный фрагмент антитела из антитела к CD3, и
второе средство специфически связывается с CD28 или CD137 и выбрано из группы, включающей антитело к CD28, бивалентный фрагмент антитела из антитела к CD28, моновалентный фрагмент антитела из антитела к CD28, антитело к CD137, бивалентный фрагмент антитела из антитела к CD137 и моновалентный фрагмент антитела из антитела к CD137.
72. Реагент для мультимеризации по п. 71, где бивалентный фрагмент антитела представляет собой F(ab')2-фрагмент или бивалентный одноцепочечный Fv-фрагмент.
73. Реагент для мультимеризации по п. 71, где моновалентный фрагмент антитела выбран из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv).
74. Реагент для мультимеризации по п. 73, где первое средство представляет собой моновалентный фрагмент антитела из антитела к CD3, который представляет собой Fab, и второе средство представляет собой моновалентный фрагмент антитела из антитела к CD28, который представляет собой Fab.
75. Реагент для мультимеризации по п. 66, где первое средство представляет собой связывающий реагент, который специфически связывается с CD40 или CD137, и где популяция лимфоцитов содержит B-клетки.
76. Реагент для мультимеризации по п. 75, где первый связывающий реагент, который специфически связывается с CD40 или CD137, выбран из группы, включающей антитело к CD40, бивалентный фрагмент антитела из антитела к CD40, моновалентный фрагмент антитела из антитела к CD40, антитело к CD137, бивалентный фрагмент антитела из антитела к CD137 и моновалентный фрагмент антитела из антитела к CD137.
77. Реагент для мультимеризации по п. 76, где бивалентный фрагмент антитела представляет собой F(ab')2-фрагмент или бивалентный одноцепочечный Fv-фрагмент.
78. Реагент для мультимеризации по п. 76, где моновалентный фрагмент антитела выбран из группы, включающей Fab-фрагмент, Fv-фрагмент и одноцепочечный Fv-фрагмент (scFv).
79. Реагент для мультимеризации по п. 66, где вспомогательная молекула на B-клетке представляет собой CD40 или CD137.
80. Реагент для мультимеризации по любому из пп. 64-79, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина сшит при помощи полисахарида или посредством бифункционального линкера.
81. Реагент для мультимеризации по любому из пп. 64-80, где олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина содержит три или более мономеров стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина.
82. Реагент для мультимеризации по любому из пп. 64-81, где указанный первый стрептавидин- или авидинсвязывающий пептид и указанный второй стрептавидин- или авидинсвязывающий пептид независимо содержат стрептавидинсвязывающий пептид Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 1) или стрептавидинсвязывающий пептид SAWSHPQFEK(GGGS)2GGSAWSHPQFEK (SEQ ID NO: 7).
83. Реагент для мультимеризации по любому из пп. 64-82, где реагент для мультимеризации содержит мутантный вариант белка стрептавидина, который содержит аминокислотную последовательность Val44-Thr45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа, или мутантный вариант белка стрептавидина, который содержит аминокислотную последовательность Ile44-Gly45-Ala46-Arg47 в положениях 44-47 последовательности стрептавидина дикого типа.
84. Реагент для мультимеризации по п. 83, где N-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 10-16 аминокислотной последовательности стрептавидина дикого типа и C-концевой аминокислотный остаток мутантного варианта белка стрептавидина находится в области аминокислот 133-142 аминокислотной последовательности стрептавидина дикого типа.
85. Композиция для стимулирования лимфоцитов, содержащая:
(i) первый реагент для мультимеризации, обратимо связанный с первым средством,
где первый реагент для мультимеризации находится в растворимой форме и содержит первый олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина,
где первое средство содержит (a) первый стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со связывающим участком первого реагента для мультимеризации, и (b) первое антитело или его антигенсвязывающий фрагмент, который связывается с рецепторной молекулой на поверхности лимфоцита для стимуляции передачи сигнала лимфоциту, и
(ii) второй реагент для мультимеризации, обратимо связанный со вторым средством,
где второй реагент для мультимеризации находится в растворимой форме и содержит второй олигомер или полимер стрептавидина, авидина, мутантного варианта белка стрептавидина или мутантного варианта белка авидина,
где второе средство содержит (a) второй стрептавидин- или авидинсвязывающий пептид, способный обратимо связываться со связывающим участком второго реагента для мультимеризации, и (b) второе антитело или его антигенсвязывающий фрагмент, который связывается со вспомогательной молекулой на клеточной поверхности лимфоцита для обеспечения костимулирующего сигнала лимфоциту, стимулируя тем самым лимфоциты.
| KNABEL M | |||
| ET AL., Reversible MHC multimer staining for functional isolation of T-cell populations and effective adoptive transfer, NATURE MEDICINE, NATURE PUBLISHING GROUP, vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| НАСАДКА ДЛЯ КАМЕР РЕГЕНЕРАТОРОВ | 1923 |
|
SU631A1 |
| WO2013011011 A2, 24.01.2013 | |||
| STEMBERGER C | |||
| ET AL., Novel Serial Positive Enrichment Technology Enables Clinical Multiparameter Cell Sorting, | |||

