Настоящее изобретение относится к полипептиду, содержащему (a) первый набор из шести определяющих комплементарность областей (CDR), скомпонованный для связывания первого антигена; и (b) (ba) второй набор из шести CDR, скомпонованный для связывания второго антигена; или (bb) лиганд, способный связываться со вторым антигеном; где (i) указанный первый антиген выбран из малого поверхностного антигена вируса гепатита B (HBV); среднего поверхностного антигена HBV и большого поверхностного антигена HBV; и (ii) указанный второй антиген выбран из поверхностных антигенов, презентируемых иммунными эффекторными клетками, такими как натуральные киллеры (NK) и цитотоксические T-лимфоциты (CTL).
В настоящем описании цитирован ряд документов, включая патентные заявки и инструкции изготовителей. Содержание этих документов, хотя и не считается связанным с патентоспособностью настоящего изобретения, включено в настоящее описание в качестве ссылки в полном объеме. Более конкретно, все цитированные документы включены в качестве ссылок в той степени, как если бы было конкретно и индивидуально указано, что каждый индивидуальный документ включен в качестве ссылки.
Приблизительно 350 миллионов человек хронически инфицированы вирусом гепатита B (HBV). Инфекция HBV может влечь за собой цирроз печени и печеночно-клеточную карциному (HCC), которые вызывают приблизительно один миллион смертей в год (Ganem et al., Hepatitis B virus infection-natural history and clinical consequences. N Engl J Med; 350:1118-29 (2004)). Инфекции HBV в настоящее время не могут контролироваться приблизительно у 5% взрослых пациентов и приблизительно у 90% новорожденных. В таком случае инфекция HBV становится хронической. Вероятной причиной является недостаточный клеточный иммунный ответ. Доступные в настоящее время противовирусные лекарственные средства, которые используют для лечения инфекции HBV, ингибируют репликацию вируса. Однако ковалентно замкнутая кольцевая ДНК (кзкДНК) остается в ядре инфицированных гепатоцитов и может вызывать реактивацию инфекции HBV после того, как пациент прекращает принимать медикаменты. Таким образом, намереваясь излечить инфекцию полностью, было бы необходимым устранить инфицированные HBV клетки, содержащие указанную кзкДНК (Protzer et al., Nat Immunol Rev 12: 2013-213 (2012)).
Однако такое цитотоксическое устранение инфицированных клеток HBV (будь то цитотоксические T-лимфоциты или натуральные киллеры (NK)) не происходит или не происходит в достаточной степени.
Инфицированные клетки, содержащие кзкДНК HBV, экспонируют на их поверхности поверхностные вирусные белки. Полагают, что это происходит, несмотря на то, что вирус высвобождается во внутриклеточные везикулы, поскольку ряд поверхностных белков HBV остаются встроенными во внутриклеточную мембрану эндоплазматической сети. В ходе процессов везикулярного транспорта указанная внутриклеточная мембрана может сливаться с клеточной мембраной, что приводит к экспонированию поверхностных белков HBV на поверхности инфицированной клетки.
Bohne et al. (T cells redirected against Hepatitis B virus surface proteins eliminate infected hepatocytes. Gastroenterology; 134:239-247 (2008)) и Krebs et al. (T Cells Expressing a Chimeric Antigen Receptor That Binds Hepatitis B Virus Envelope Proteins Control Virus Replication in Mice. Gastroenterology (2013)) описывают химерные рецепторы антигенов, которые при ретровирусной доставке и экспрессии на поверхности T-клеток позволяют первичным T-клеткам человека и мыши распознавать гепатоциты, экспонирующие малый поверхностный антиген HBV и лизировать клетки с реплицирующимся HBV.
Биспецифические антитела, как правило, используют в области онкологии. В качестве примера см. Hartmann et al. (Treatment of refractory Hodgkin disease with an anti-CD 16/CD30 bispecific antibody. Blood; 89:2042-7 (1997)).
В EP 2 524 699 A1 описаны трифункциональные антитела. Эти антитела "обладают функциональной Fc-областью" и "должны состоять из тяжелых цепей иммуноглобулинов различных подклассов". С другой стороны в Hornig und Farber-Schwarz описана в главе 40 "Antibody Engineering" (ed. Patrick Channes, Humane Press, 2012) конструкция scFv, которая лишена Fc-части.
В Liao et al. (Oncology Reports 3, 637-644 (1996)) описаны биспецифические моноклональные антитела, перенацеливающие эффекторные клетки на лизис ксенотрансплантатов гепатомы человека у мышей nude. Описанные биспецифические антитела получают путем слияния двух гибридом с получением гибридомной клеточной линии, экспрессирующей комбинации тяжелая/легкая цепь двух различных антител. Это может приводить спариванию двух различных тяжелых цепей, а также к спариванию идентичных тяжелых цепей, что дает начало случайной смеси моноспецифических исходных антител и биспецифических антител. Биспецифические антитела содержат тяжелую и легкую цепи и димеризуются с образованием молекулы Ig, которая не является единой полипептидной цепью.
Ввиду уровня техники, техническая проблема может состоять в предоставлении альтернативных или усовершенствованных средств и способов для лечения инфекции HBV, а также состояний, вызываемых инфекцией HBV, таких как цирроз печени или печеночно-клеточная карцинома. Выражаясь в терминах клеточной биологии, техническая проблема может состоять в предоставлении средств и способов для устранения клеток, содержащих кзкДНК HBV. Эта техническая проблема решается прилагаемой формулой изобретения.
Таким образом, в первом аспекте настоящее изобретение относится к полипептиду, содержащему (a) первый набор из шести определяющих комплементарность областей (CDR), скомпонованных для связывания первого антигена; и (b) (ba) второй набор из шести CDR, скомпонованных для связывания второго антигена; или (bb) лиганд, способный связываться со вторым антигеном; где (i) указанный первый антиген выбран из малого поверхностного антигена HBV; среднего поверхностного антигена HBV и большого поверхностного антигена HBV; и (ii) указанный второй антиген выбран из поверхностных антигенов, презентируемых иммунными эффекторными клетками, такими как натуральные киллеры (NK) и цитотоксические T-лимфоциты (CTL).
Термин "полипептид" определяет молекулу, которая представляет собой поликонденсат аминокислот, которые образуют одну единую цепь с одним N-концом и одним C-концом. Составляющие аминокислоты включают 20 встречающихся в природе протеиногенных аминокислот. Предпочтительно, указанный полипептид состоит исключительно из указанных встречающихся в природе протеиногенных аминокислот. Вместе с тем, термин распространяется на молекулы, которые, в дополнение к указанным встречающимся в природе протеиногенным аминокислотам, содержат вплоть до 20%, 10%, 5%, 2% или 1% аминокислот, которые выбраны из не встречающихся в природе α-аминокислот, β-аминокислот, D-аминокислот, селеноцистеина, селенометионина, гидроксипролина, пирролизина и орнитина. Более того, понятно, что одна или более, как например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот, могут быть фосфорилированными. Последнее применимо, в частности, к серину, треонину и тирозину. Также могут присутствовать другие посттрансляционные модификации, известные в данной области, включая гликозилирование. Гликозилирование включает N-связанное гликозилирование, как правило, по аспарагину, и O-связанное гликозилирование, как правило, по остаткам серина или треонина. N- и/или C-конец может быть защищенным защитными группами, включая ацетил для N-конца и амин для C-конца. Тип связи между аминокислотами, содержащимися в указанном полипептиде, ограничивается амидными (CONH) связями. Термин "амидная связь" включает пептидные связи, которые соединят α-карбоксилат данной аминокислоты с α-аминогруппой следующей аминокислоты. "Амидная связь" также распространяется на изопептидные связи, которые представляют собой амидную связь, которая не присутствует на основной цепи полипептида. Например, вместо α-аминогруппы может быть вовлечена аминогруппа боковой цепи лизина. Аналогично, вместо α-карбоксильной группы может быть вовлечен карбоксилат боковой цепи глутамата или аспартата. Предусматривается встречаемость одного или более, как например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 изопептидных связей. Хотя предпочтительными являются полипептиды, в которых составляющие аминокислоты связаны друг с другом исключительно пептидными связями.
Как правило, отсутствует верхний предел количества аминокислот в полипептиде. Как можно видеть из иллюстративных полипептидных последовательностей, содержащихся в списке последовательностей, полипептиды по настоящему изобретению обычно содержат несколько сот аминокислот, предпочтительно от 250 до 1000, от 400 до 900 или от 700 до 800 аминокислот. Различить пептиды с одной стороны и полипептид с другой стороны просто: пептиды имеют 30 или менее аминокислот и полипептиды имеют более 30 аминокислот.
Термин "определяющая комплементарность область", сокращенно обозначаемый как "CDR" имеет общепринятое в данной области значение. Она представляет собой короткую подпоследовательность, как правило, в диапазоне от приблизительно 3 до приблизительно 25 аминокислот, которая сообщает антителу способность специфически распознавать эпитоп антигена. Как правило, в вариабельном домене легкой цепи антитела присутствует три CDR и в вариабельном домене тяжелой цепи антитела присутствует три CDR. Хотя CDR обычно являются частью доменов иммуноглобулинов, в соответствии с настоящим изобретением в этом отсутствует необходимость. Достаточной является аминокислотная последовательность, которая содержит указанные CDR, при условии, что указанная аминокислотная последовательность при сворачивании в физиологических условиях презентирует указанные CDR в пространственной близости и сохраняет их способность распознавать их антиген. Упомянутая пространственная близость и способность связывать антиген выражаются термином "скомпонованы для связывания антигена", как используют в описанном выше основном варианте осуществления. Термин "домен иммуноглобулина" известен в данной области и относится к последовательности, как правило из 70-100 аминокислот, предполагая трехмерную структуру 2-слойного сэндвича между 7 и 9 антипараллельными β-цепями.
Каждый из указанного первого набора из шести CDR, а также указанного второго набора из шести CDR определяет участок связывания.
Понятно, что, помимо указанного первого набора и указанного второго набора в полипептиде по изобретению отсутствуют другие CDR.
Термин "антиген" имеет его принятое в данной области значение. Он относится к молекуле, которая специфически распознается и связывается набором из шести CDR, которые, как правило, презентируются доменами иммуноглобулинов. Конкретная часть антигена, распознаваемая и связываемая указанными CDR, также известна как эпитоп.
Термин "лиганд" имеет его принятое в данной области значение. Лиганд представляет собой контрструктуру для рецептора. Более конкретно, лиганд способен связываться, предпочтительно специфически связываться, с распознаваемым им рецептором. В соответствии с изобретением указанный лиганд предпочтительно представляет собой иммунолиганд.
Иммунолиганд представляет собой лиганд, который способен связываться с рецептором, присутствующим на поверхности иммунной эффекторной клетки. Предпочтительными иммунными эффекторными клетками являются, как определено выше, NK-клетки и CTL. Предпочтительными являются иммунолиганды, которые, когда они связаны с распознаваемым ими рецептором на поверхности иммунной эффекторной клетки, проявляют стимулирующий и/или костимулирующий эффект. Термины "активировать" и "стимулировать" используют в этом контексте эквивалентно. Рецепторы, связываемые предпочтительными иммунолигандами, дополнительно указаны ниже.
Поверхностные белки S/M/L HBV представляют собой малые, средние и большие поверхностные белки на наружной оболочке HBV (Stibbe, W., and W. H. Gerlich. Structural relationships between minor and major proteins of hepatitis B surface antigen. J. Virol. 1983 46:626-628).
Три поверхностных антигена HBV транскрибируются и транслируются с одной рамки считывания и отличаются друг от друга длиной N-концевой части. Таким образом, большой поверхностный антиген содержит часть, которая не присутствует ни в среднем, ни в малом поверхностном антигене, и средний поверхностный антиген содержит часть, которая, в то время как содержится в большом антигене, не содержится в малом антигене. Малый антиген состоит из последовательности, которая содержится на C-концевой части как среднего, так и большого антигена.
Большой поверхностный антиген HBV может встраиваться в цитоплазматическую мембрану двумя способами. С внеклеточной стороны может находиться либо N-конец, либо C-конец. Обе конфигурации встречаются в клетках, инфицированных HBV.
Указанный второй антиген представляет собой поверхностный антиген, презентируемый иммунными эффекторными клетками, предпочтительно, специфически презентируемый NK-клетками и/или CTL. Иммунные эффекторные клетки представляют собой клетки, которые перенацеливаются на инфицированные HBV клетки, причем указанные инфицированные HBV клетки презентируют упомянутые поверхностные антигены HBV на их поверхности.
Особенно предпочтительно, чтобы связывание в соответствии с изобретением, в частности, между CDR и антигенами, а также между лигандами и антигенами, было специфическим. Термины "специфически связывается" и "специфически связывающийся" (имеющие то же значение, что и "специфически взаимодействующий") используют в соответствии с настоящим изобретением для обозначения того, что эти связывающие части не реагируют перекрестно или по существу не реагируют перекрестно с эпитопом или структурой, сходной со структурой антигена-мишени. Перекрестную реактивность исследуемой панели молекул можно исследовать, например, путем оценки связывания указанной панели молекул в общепринятых условиях с представляющий интерес эпитопом, а также с рядом более или менее близкородственных (структурно и/или функционально) эпитопов. Специфическими для представляющего интерес эпитопа считаются только те молекулы, которые связываются с представляющим интерес эпитопом в его соответствующем контексте (например, конкретный мотив в структуре белка), но не связываются или по существу не связываются с какими-либо другими эпитопами.
Первый аспект включает варианты осуществления, где положения (a) и (ba) вместе являются единственными связывающими участками, присутствующими на указанном полипептиде, а также варианты осуществления, где положения (a) и (bb) вместе являются единственными связывающими участками, присутствующими на указанном полипептиде.
Хроническая инфекция HBV характеризуется иммунотолерантным статусом. Более конкретно, клетки CTL и NK пациента функционируют так, что полного устранения инфицированных клеток, или полного контроля репликации HBV, или полной элиминации HBV не происходит. Полипептиды по изобретению представляют собой биспецифические молекулы в том отношении, что они специфически распознают поверхностный антиген HBV с одной стороны и поверхностьный антиген иммунных эффекторных клеток с другой стороны. Можно полагать, что такие биспецифические молекулы сообщают искусственную специфичность иммунным эффекторным клеткам. В действительности, клетки CTL и NK перенацеливаются полипептидами по изобретению (также обозначаемые как являющиеся "биспецифическими"), так что они привлекаются к инфицированным HBV клеткам и уничтожают их.
Связывание полипептидов по изобретению с инфицированными HBV клетками с одной стороны и привлечение иммунных эффекторных клеток с другой стороны может происходить в любом порядке или также одновременно.
В частности, предполагается системное применение полипептидов по изобретению либо путем инъекции, либо в качестве перорально применяемой формы, и позволение им связаться с инфицированными HBV или экспрессирующими антиген HBV клетками-мишенями и привлечь указанные иммунные эффекторные клетки к указанным клеткам-мишеням.
Вместе с тем, также предусматривается приведение в контакт полипептидов по изобретению с иммунными эффекторными клетками (или популяцией мононуклеарных клеток периферической крови, содержащей указанные эффекторные клетки), так что указанные эффекторные клетки становятся нагруженными указанными полипептидами. Такие эффекторные клетки (или популяция PBMC, содержащая такие нагруженные эффекторные клетки), которые нагружены in vitro или ex vivo, можно вводить пациенту, страдающему инфекцией HBV или состоянием, ассоциированным с ней и определенным ниже. Такое введение можно осуществлять внутривенно, например, в Arteria hepatica. Иммунная эффекторная клетка с полипептидом в соответствии с настоящим изобретением, связываемым с поверхностным антигеном указанной иммунной эффекторной клетки, также является аспектом настоящего изобретения. Этот аспект дополнительно описан ниже.
Это уничтожение, в частности, в отношении анивирусных иммунных медиаторов (например, цитокинов), секретируемых иммунными клетками, обеспечивает устранение инфекции HBV, или длительный контроль инфекции HBV, или элиминацию опухолевых клеток, экспрессирующих поверхностные антигены HBV. Предпочтительные или иллюстративные биспецифические полипептиды в соответствии с настоящим изобретением обеспечивают удивительно высокие уровни уничтожения инфицированных HBV клеток или опухолевых клеток печени (также известных как клетки гепатомы), реплицирующих HBV или экспрессирующих поверхностные антигены HBV; см. примеры, прилагаемые к настоящему описанию.
Учитывая, что биспецифические полипептиды в соответствии с настоящим изобретением обеспечивают адаптированную специфичность для иммунных эффекторных клеток, природная имманентная специфичность иммунных клеток или презентация им антигенов становятся несущественными. По существу, большой набор эффекторных клеток-кандидатов подвержен перенацеливанию. Более того, полипептиды по изобретению обладают биодоступностью и временем полужизни, которые по меньшей мере сравнимы с биодоступностью и временем полужизни моноклональных антител.
В предпочтительном варианте осуществления (a) указанный первый набор из шести CDR содержится в первом фрагменте scFv; и/или (b) (ba) указанный второй набор из шести CDR содержится во втором фрагменте scFv; или (bb) указанный лиганд представляет собой иммунолиганд, предпочтительно способный связываться с NKG2D/CD314 (такой как лиганды MICA, MICB, ULBP1-6), NKp30/NCR3/CD337 (такой как лиганд B7-H6), 4-1BB/CD137 (такой как лиганд 4-1BB-L/CD137L) или OX40/CD134 (такой как лиганд OX40-L/CD252). Наклонная черта ("/") разделяет альтернативные принятые в данной области обозначения. В скобках представлены предпочтительные репрезентативные представители данного рода антигенов.
Термин "scFv" является общепринятым в данной области. Это сокращение обозначает "одноцепочечный вариабельный фрагмент" антитела и определяет полипептид, способный специфически распознавать и связывать эпитоп антигена. Как отмечалось выше, три CDR презентируются вариабельным доменом легкой цепи (VL) антитела и три CDR презентируются вариабельным доменом тяжелой цепи (VH) антитела. В scFv два вариабельных домена соединены друг с другом пептидным линкером. Полученная слитая конструкция представляет собой единую полипептидную цепь. Это обеспечивает простоту экспрессии молекулы scFv. Схематическое изображение представлено на фиг.1.
Термины "VH-домен" и "VL-домен" используют в соответствии с определениями, установленными в данной области. Таким образом, они относятся к вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) иммуноглобулинов, соответственно. Как правило, каждый из VH- и VL-доменов содержит три определяющих комплементарность области (CDR), где CDR представляют собой высоковариабельные области, в основном ответственные за связывание антигена.
Пептидный линкер предпочтительно используют для связывания либо вариабельных областей scFv, либо для связывания scFv с областью димеризации и/или спейсерной областью, предпочтительно, с Fc. Как правило, пептидные линкеры имеют длину от 3 до 30 аминокислот, предпочтительно от 5 до 25 или от 10 до 20 аминокислот. Предпочтительными являются линкеры, которые не нарушают или по существу не нарушают структуру и/или функцию доменов или полипептидов, которые они связывают (связывание приводит к одной непрерывной полипептидной цепи). Линкеры включают Gly-богатые линкеры, такие как линкер (Gly4Ser)3 (SEQ ID NO: 47), который используют в предпочтительных полипептидах по изобретению для связывания доменов VH/VL scFv, специфичных к CTL или NK, и линкер Yol (SEQ ID NO: 48; AKTTPKLEEGEFSEARV, как описано в Sellrie et al., Journal of Biochemistry and Molecular Biology, Vol. 40, No. 6, November 2007, pp. 875-880), который используют в предпочтительных полипептидах по изобретению для связывания доменов VH/VL scFv, специфичных к поверхностным антигенам HBV. Также линкер (Gly4Ser)4 (SEQ ID NO: 49) можно использовать для связывания доменов VH/VL scFv, специфичных к поверхностным антигенам HBV.
Термин "антитело", как используют в рамках изобретения, имеет его принятое в данной области значение. Предпочтительно, он относится к моноклональному антителу. Моноклональные антитела можно получать, например, способами, первоначально описанными в Kohler and Milstein, Nature 256 (1975), 495, и Galfré, Meth. Enzymol. 73 (1981), 3, которые включают слияние клеток миеломы мыши с клетками селезенки, происходящими из иммунизированных млекопитающих, с модификациями, разработанными в данной области. Более того, антитела или их фрагменты, направленные на упомянутые выше поверхностные белки HBV, можно получать с использованием способов, которые описаны, например, в Harlow and Lane "Antibodies, A Laboratory Manual", CSH Press, Cold Spring Harbor, 1988. Получение химерных антител описано, например, в WO 89/09622. Другим источником антител для применения в соответствии с настоящим изобретением являются так называемые ксеногенные антитела. Общий принцип получения ксеногенных антител, таких как антитела человека, у мышей описан, например, в WO 91/10741, WO 94/02602, WO 96/34096 и WO 96/33735. Антитела для применения в соответствии с изобретением или их соответствующую иммуноглобулиновую цепь(и) можно далее модифицировать с использованием общепринятых способов, известных в данной области, например, с использованием делеции(ий), инсереции(ий), замены(замен), вставки(вставок) и/или рекомбинации(ий) и/или любой другой модификации(ий) аминокислот, известной в данной области, либо отдельно, либо в комбинации. Способы внесения таких модификаций в последовательность ДНК или полипептида, лежащую в основе аминокислотной последовательности цепи иммуноглобулина, хорошо известны специалисту в данной области; см., например, Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989. Модификации полипептидов также включают посттрансляционные модификации, такие как гликозилирование.
В следующем предпочтительном варианте осуществления указанный первый набор из шести CDR связывает эпитоп указанного первого антигена, причем указанный эпитоп расположен (a) в указанном малом поверхностном антигене HBV; или (b) в части указанного большого поверхностного антигена HBV, которая не содержится в указанном малом поверхностном антигене HBV; или (c) в части указанного большого поверхностного антигена HBV, структура которой отличается от указанного малого поверхностного антигена.
Положение (a) относится к эпитопам, присутствующим в малом поверхностном антигене HBV. Вследствие описанной выше взаимосвязи между малым, средним и большим поверхностными антигенами HBV, вся последовательность малого антигена содержится в среднем или большом антигене. Как правило, но не обязательно, трехмерный эпитоп, презентируемый малым поверхностным антигеном, также презентируется средним и/или большим поверхностными антигенами.
В соответствии с положением (b), предпочтительно, чтобы указанная часть указанного большого поверхностного антигена HBV также не содержалась в указанном среднем поверхностном антигене HBV. Что касается положения (c), понятно, что "отличие в структуре" включает эпитопы указанного большого поверхностного антигена HBV, которые содержат или состоят из последовательностей, которые являются частью последовательности указанного малого поверхностного антигена HBV, где указанные эпитопы не присутствуют на указанном малом поверхностном антигене HBV. В соответствии с положением (c), более того, предпочтительно, чтобы указанный эпитоп находился в части указанного большого поверхностного антигена HBV, структура которой отличается от указанного среднего поверхностного антигена HBV.
Указанное положение (a), т.е. что указанным первым антигеном называется малый поверхностный антиген HBV, является особенно предпочтительным в отношении всех аспектов и вариантов осуществления настоящего изобретения.
В соответствии с положениями (b) и (c), полипептид специфически распознает большой поверхностный антиген HBV.
В следующем предпочтительном варианте осуществления указанный поверхностный антиген, презентируемый иммунными эффекторными клетками, выбран из CD3, CD28, 4-1BB, OX40, CD16, CD56, NKG2D и NKp30/NCR3. Таким образом, настоящее изобретение относится к полипептиду, содержащему (a) первый набор из шести определяющих комплементарность областей (CDR), скомпонованный для связывания первого антигена; и (b) (ba) второй набор из шести CDR, скомпонованный для связывания второго антигена; или (bb) лиганд, способный связываться со вторым антигеном; где (i) указанный первый антиген выбран из малого поверхностного антигена вируса гепатита B (HBV); среднего поверхностного антигена HBV; и большого поверхностного антигена HBV; и (ii) указанный второй антиген выбран из поверхностных антигенов, презентируемых иммунными эффекторными клетками, такими как натуральные киллеры (NK) и цитотоксические T-лимфоциты (CTL), где (c) указанный первый набор из шести CDR содержится в первом фрагменте scFv; и (d) (da) указанный второй набор из шести CDR содержится во втором фрагменте scFv; или (db) указанный лиганд представляет собой иммунный лиганд, способный связываться с NKG2D, такой как лиганды MICA, MICB, ULBP1-6; NKp30, такой как лиганд B7-H6, 4-1BB, такой как лиганд 4-1BB-L; или OX40, такой как лиганд OX40-L; и где указанный поверхностный антиген, презентируемый иммунными эффекторными клетками, выбран из CD3, CD28, 4-1BB, OX40, CD16, CD56, NKG2D и NKp30.
CD3 означает эпсилон-цепь CD3, которая является частью CD3-T-клеточного рецепторного комплекса. (Borst, J. et al., The delta- and epsilon-chains of the human T3/T-cell receptor complex are distinct polypeptides. Nature. 1984. 312: 455-458).
CD28 представляет собой главный костимулирующий рецептор T-клеток (Lesslauer, W. et al., T90/44 (9.3 antigen). A cell surface molecule with a function in human T cell activation. Eur. J. Immunol. 1986. 16: 1289-1296).
4-1 BB (CD137) представляет собой костимулирующий рецептор активированных T-клеток и NK-клеток (Kwon, B.S. et al., cDNA sequences of two inducible T-cell genes. Proc. Natl. Acad. Sci. U. S. A. 1989. 86:.1963-1967).
OX40 (CD134) представляет собой вторичный костимулирующий рецептор. (Arch, R. H. et al., Mol. Cell. Biol. 1998. 18: 558-565). 4-1 BB и OX40 являются представителями семейства рецепторов фактора некроза опухоли (TNF), который связывает ассоциированные с рецептором TNF лиганды и активирует ядерный фактор каппа-B.
CD16 (FcγRllla) представляет собой низкоаффинный Fc-рецептор, экспрессируемый NK-клетками, подгруппой активированных цитотоксических T-клеток, а также клеточными типами миеломоноцитарной линии дифференцировки, связывающимися с Fc-доменом молекул IgG. (Lanier, L.L. et al., Functional properties of a unique subset of cytotoxic CD3+ T lymphocytes that express Fc receptors for IgG (CD16/Leu-11 antigen). J. Exp. Med. 1985. 162: 2089-2106).
CD56 (NCAM) представляет собой молекулу адгезии, экспрессируемую NK-клетками. (Lanier, L.L. et al., Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. J. Exp. Med. 1989. 169: 2233-2238).
NKG2D представляет собой активирующий рецептор, экспрессируемый NK-клетками (Houchins, J. et al., DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrabe proteins on human natural killer cells. 1991. J. Exd. Med. 173: 1017-1020V.
NKp30 (NCR3) представляет собой рецептор, экспрессируемый NK-клетками (Pende, D. et al., Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. 2000. J. Exp. Med. 192: 337-346).
CD3, CD28, 4-1BB и OX40 присутствуют на поверхности CTL. Связывание полипептида по изобретению с любым из этих поверхностных антигенов вовлекает стимуляцию или костимуляцию CTL.
CD16, CD56, NKG2D, NKp30/NCR3 и 4-1 BB присутствуют на поверхности NK-клеток. Связывание полипептида по изобретению с любым из этих поверхностных антигенов вовлекает стимуляцию или костимуляцию NK-клеток.
Что касается CTL человека, предпочтительными являются CD3 и CD28. Что касается NK-клеток человека, предпочтительными являются CD16 и CD56.
Упомянутые поверхностные антигены обозначают общепризнанными названиями (также см. Kenneth Murphy, Janeway's Immunobiology, 7th edition, Garland Science; William E. Paul, Fundamental Immunology, 7th edition, Lippincott Williams & Wilkins).
В следующем предпочтительном варианте осуществления указанный полипептид дополнительно содержит область димеризации. Указанная область димеризации может обеспечить ковалентную и/или нековалентную димеризацию.
Посредством димеризации биспецифические двухвалентные антитела преобразуются в биспецифические четырехвалентные (или даже тетраспецифические четырехвалентные, если различные биспецифические антитела коэкспрессируются в продуцирующей клетке). Ожидается, что биспецифические четырехвалентные реагенты, как описано в настоящем описании, будут обладать увеличенной авидностью, сходной с общепринятыми моноспецифическими антителами, поскольку они способны связывать две молекулы антигена одного типа их N-концевой стороной и их C-концевой стороной, соответственно.
В особенно предпочтительном варианте осуществления указанная область димеризации, которая связывает два полипептида по изобретению, состоит из шарнирной области тяжелой цепи IgG или содержит остатки цистеина, ответственные за димеризацию тяжелых цепей антитела. Предпочтительно, указанная область димеризации состоит из подпоследовательности длиной 32 аминокислоты, и так называемая шарнирная область тяжелой цепи (EPKSSDKTHTCPPCPAPEFEGAPSVFLFPPKP, см. SEQ ID NO: 43-46) содержит два остатка цистеина (подчеркнуты в указанной выше последовательности), ответственных за димеризацию тяжелых цепей. Предпочтительно осуществляют мутацию одного остатка цистеина в шарнирной области тяжелой цепи IgG, который опосредует межмолекулярную дисульфидную связь между константными доменами тяжелой и легкой цепей IgG в природном антителе, на серин для предотвращения аберрантных дисульфидных мостиков.
Домены димеризации, пригодные для нековалентной димеризации, известны в данной области и включают лейциновые молнии.
В следующем предпочтительном варианте осуществления указанный полипептид дополнительно содержит спейсерную область, причем указанная спейсерная область предпочтительно содержит CH2-домен и CH3-домен, и указанная спейсерная область расположена между (i) указанным первым фрагментом scFv и (ii) указанным вторым фрагментом scFv или указанным рекомбинантным лигандом в аминокислотной последовательности указанного полипептида.
Преимущественной является спейсерная область, содержащая или состоящая из CH2-домена и CH3-домена, в частности, происходящего из IgG. Их способность связывать белок A обеспечивает эффективную секрецию из продуцирующих клеток и/или последующую очистку от реагентов.
Как указанные домены CH2 и CH3 с одной стороны, так и указанная область димеризации с другой стороны, могут быть предоставлены соответствующей областью молекулы IgG, предпочтительно молекулы IgG1 или IgG2, еще более предпочтительно молекулы IgG1 или IgG2 человека (hIgG1, hIgG2). Предпочтительная подпоследовательность молекулы hIgG1, в которой представлен CH2-домен, CH3-домен и домен димеризации, представлена в последовательностях 43-46. Предпочтительно, и это применимо к упомянутым последовательностям, в часть hIgG1, в частности, указанный домен CH2, внесена мутация во множестве положений для уменьшения или устранения связывания с Fc-рецепторами (указанные полужирным шрифтом и курсивом в подпоследовательностях, приведенных ниже). Более часто, в Fc-область, в частности, в CH2-домен и/или CH3-домен, может быть внесена мутация в одном или более положениях для уменьшения или устранения связывания с Fc-рецепторами. Такая методика известна в данной области и описана, например, в Armour et al., Recombinant human IgG molecules lacking Fcgamma receptor I binding and monocyte triggering activities. Eur. J. Immunol. 1999. 29: 2613-2624 и Lazaret al., Engineered antibody Fc variants with enhanced effector function. Proc. Natl. Acad. Sci. U. S. A. 2006. 103: 4005-4010. Это является преимущественным, поскольку запуск антителозависимой клеточно-опосредуемой цитотоксичности (ADCC) не является предпочтительным в соответствии с изобретением.
Иными словами, Fc-фрагмент антитела можно использовать для внесения спейсерной области и области димеризации. Термин "Fc-фрагмент" известен квалифицированному специалисту и определяет фрагмент IgG, который получен расщеплением папаином и содержит домены CH2 и CH3.
Между указанным первым scFv-фрагментом и указанной спейсерной областью и/или между указанной спейсерной областью и указанным вторым scFv-фрагментом присутствует/присутствуют (a) линкерная последовательность(и). Предпочтительные линкерные последовательности описаны в настоящем описании выше. Как можно видеть из предпочтительных последовательностей, содержащихся в списке последовательностей, в частности, из последовательностей SEQ ID NO: 43-46, такие линкерные последовательности могут состоять из остатков глицина или остатков глицина и серина.
На фиг.2 проиллюстрирована молекулярная архитектура предпочтительных полипептидов по изобретению, которые содержат область димеризации (шарнирная область hIgG), а также область CH2 и CH3, отделяющую два фрагмента scFv друг от друга.
Термины "CH2-домен" и "CH3-домен" имеют их принятое в данной области значение. Они относятся ко второму и третьему константным доменам тяжелых цепей антитела.
Понятно, что особенно предпочтительный вариант осуществления относится к полипептиду, содержащему
(a) первый набор из шести определяющих комплементарность областей (CDR), скомпонованных для связывания первого антигена; и (b) (ba) второй набор из шести CDR, скомпонованных для связывания второго антигена; или (bb) лиганд, способный связываться со вторым антигеном; где (i) указанный первый антиген выбран из малого поверхностного антигена вируса гепатита B (HBV); среднего поверхностного антигена HBV и большого поверхностного антигена HBV; и (ii) указанный второй антиген выбран из поверхностных антигенов, презентируемых иммунными эффекторными клетками, такими как натуральные киллеры (NK) и цитотоксические T-лимфоциты (CTL), где (c) указанный первый набор из шести CDR содержится в первом scFv-фрагменте; и (d) (da) указанный второй набор из шести CDR содержится во втором scFv-фрагменте; или (db) указанный лиганд представляет собой иммунолигнад, предпочтительно способный связываться с NKG2D, такой как лиганды MICA, MICB, ULBP1-6; NKp30, такой как лиганд B7-H6; 4-1BB, такой как лиганд 4-1BB-L; или OX40, такой как лиганд OX40-L, где указанный поверхностный антиген, презентируемый иммунными эффекторными клетками, выбран из CD3, CD28, 4-1BB, OX40, CD16, CD56, NKG2D, NKp30 и 4-1BB, и где указанный полипептид дополнительно содержит область димеризации и спейсерную область, причем указанная область димеризации и указанная спейсерная область предпочтительно являются такими, как определено выше.
В следующем предпочтительном варианте осуществления (a) указанный первый набор из шести CDR имеет последовательности SEQ ID NO: 1-6, 7-12 или 13-18; и/или (b) указанный второй набор из шести CDR имеет последовательности SEQ ID NO: 19-24, 25-30, 31-36 или 37-42.
Как общепринято в данной области и, более того, как очевидно из прилагаемого списка последовательностей, порядок CDR в каждом наборе из шести CDR, как описано выше, является следующим: CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи.
C8, 5F9, 5A19, OKT3, 9.3, A9 и NCAM29.2, как используют в списке последовательностей, обозначают антитело, из которого происходят соответствующие CDR, и относятся к предпочтительному антителу против HBs, ко второму отличающемуся антителу против HBs, к антителу против большого поверхностного антигена HBV, к антителу против CD3 человека, к антителу против CD28 человека, к антителу против CD16 человека и к антителу против CD56 человека, соответственно. "HB" обозначает малый поверхностный антиген HBV.
Особенно предпочтительным является то, что указанный полипептид содержит или состоит из любой аминокислотной последовательности из SEQ ID NO: 43-46 или аминокислотной последовательности, которая обладает по меньшей мере 80% идентичностью с любой из SEQ ID NO: 43-46 при условии, что CDR указанной аминокислотной последовательности, обладающей по меньшей мере 80% идентичностью, являются идентичными CDR, содержащимся в любой из SEQ ID NO: 43-46, соответственно. В SEQ ID NO: 43 последние три остатка "GNS" являются необязательными.
Предпочтительные уровни идентичности последовательностей включают по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% и по меньшей мере 99%. Средства и способы для определения идентичности последовательностей хорошо известны в данной области. Предпочтительным алгоритмом для попарного определения идентичности последовательности является Basic local alignment search tool (BLAST), как описано, например, в McGinnis and Madden (Nucleic Acid Research 32, W20-W25 (2004)).
Положение указанных CDR в данной последовательности, в настоящем случае в последовательностях SEQ ID NO: 43-46, можно определять принятыми в данной области способами, и известные принятые в данной области способы включают системы Chothia, Kabat и LeFranc/IMGT, соответственно. В отсутствие какого либо указания на обратное понятно, что CDR согласно определенным выше особенно предпочтительным вариантам осуществления представляют собой CDR, определенные выше, а именно, первый набор, имеющий последовательности SEQ ID NO: 1-6, 7-12 или 13-18, и второй набор, имеющий последовательности SEQ ID NO: 19-24, 25-30, 31-36 или 37-42. Как можно видеть из последовательностей в прилагаемом списке последовательностей, эти конкретные последовательности CDR (подчеркнуты в последовательностях, приведенных ниже) действительно содержатся в последовательностях SEQ ID NO: 43-46.
Последовательности SEQ ID NO: 1-6 определяют CDR и SEQ ID NO: 37-40 определяют биспецифические полипептиды, способные связывать конкретный эпитоп в малом поверхностном антигене HBV. Этот эпитоп расположен в a-детерминанте, которая экспонируется на поверхности инфицированных клеток и вирионов, соответственно. Термин "a-детерминанта" используют для обозначения области в малом поверхностном антигене HBV, где расположены основные эпитопы для индукции защитного гуморального иммунного ответа. Эти CDR, а также полипептиды SEQ ID NO: 43-46, имеют то преимущество, что их можно использовать для всех серотипов HBV.
Во втором аспекте настоящее изобретение относится нуклеиновой кислоте, кодирующей полипептиды, определенные выше. Предпочтительные варианты осуществления полипептидов дают начало соответствующим предпочтительным вариантам осуществления указанной нуклеиновой кислоты.
Термин "нуклеиновая кислота" имеет его принятое в данной области значение и конкретно не ограничен. Предпочтительными являются ДНК, такая как геномная ДНК или кДНК, а также РНК, такая как мРНК. Хотя это и не является предпочтительным, предусматривается применение нуклеотидных производных, которые включают 2'-дериватизированные нуклеотиды, такие как 2'-метилнуклеотиды; пептидные нуклеотиды, которые встречаются в пептидных нуклеиновых кислотах, и т.п.
В третьем аспекте настоящее изобретение относится ковалентно связанному комплексу, содержащему или состоящему из первого и второго полипептида, где между указанным первым и указанным вторым полипептидами существует по меньшей мере одна ковалентная связь, предпочтительно по меньшей мере один дисульфидный мостик между остатком Cys указанного первого полипептида и остатком Cys указанного второго полипептида, причем указанные первый и второй полипептиды являются такими, как определено в соответствии с изобретением.
Предпочтительными являются две ковалентные связи между указанным первым и указанным вторым полипептидами, предпочтительно, две дисульфидных связи, как представлено на фиг.2.
Также предусматривается комплекс, содержащий или состоящий из первого и второго полипептидов, где указанный первый и указанный второй полипептиды связаны друг с другом нековалентно.
Иллюстративное изображение такого ковалентно связанного комплекса представлено на фиг.2. Предпочтительным является указанный комплекс, являющийся димером.
В четвертом аспекте настоящее изобретение относится к композиции, содержащей или состоящей из одного или более полипептидов по изобретению и/или одного или более комплексов по изобретению при условии, что по меньшей мере два полипептида содержатся в указанной композиции, причем эти два полипептида отличаются друг от друга в отношении первого антигена и/или второго антигена, с которыми они связываются.
В предпочтительном варианте осуществления указанного четвертого аспекта указанные два полипептида представляют собой (a) (i) полипептид, связывающийся с малым или большим поверхностными антигенами HBV и CD3; и (ii) полипептид, связывающийся с малым или большим поверхностными антигенами HBV и CD28; или (b) (i) полипептид, связывающийся с малым или большим поверхностными антигенами HBV и CD16; и (ii) полипептид, связывающийся с малым или большим поверхностными антигенами HBV и CD56.
Как альтернатива (a), так и альтернатива (b), в частности, в той степени, в которой они относятся к полипептидам, связывающимся с малым поверхностным антигеном HBV этого предпочтительного варианта осуществления, обеспечивают неожиданно высокие уровни элиминации вплоть до 95% по сравнению с отрицательным контролем. Ожидается, что это обеспечит полное устранение инфицированных HBV клеток или положительных по антигену HBV опухолевых клеток, особенно после многократного применения в ситуации in vivo.
Было обнаружено, что комбинированное применение биспецифических молекул, связывающихся с двумя различными маркерами CTL или маркерами NK, обеспечивает синергические эффекты. На фиг.3 и 4B показано сравнение специфического лизиса клеток-мишеней при введении биспецифических конструкций.
В особенно предпочтительном варианте осуществления указанные два полипептиды содержат или состоят из последовательностей (a) SEQ ID NO: 43 и 44; или (b) SEQ ID NO: 45 и 46.
Каждая из последовательностей SEQ ID NO: 43-46 позволяет образование двух дисульфидных мостиков, когда образуется гомодимер. Вместе с тем, предусматривается преднамеренное образование также и гетеродимеров. Примером гетеродимера является ковалентно связанный комплекс двух полипептидов по настоящему изобретению, где первый полипептид связывается с поверхностным антигеном HBV и первым маркером, презентируемым иммунной эффекторной клеткой, и второй полипептид связывается с поверхностным антигеном HBV и вторым маркером иммунной эффекторной клетки. Два маркера иммунной эффекторной клетки могут представлять собой, например, CD3 и CD28, или, альтернативно, CD16 и CD56.
В следующем аспекте настоящее изобретение относится к фармацевтической композиции, содержащей или состоящей из одного или более полипептидов по изобретению, одного или более комплексов по изобретению и/или одной или более композиций по изобретению.
Фармацевтическая композиция, кроме того, может содержать фармацевтически приемлемые носители, эксципиенты и/или разбавители. Примеры подходящих фармацевтических носителей, эксципиентов и/или разбавителей хорошо известны в данной области и включают фосфатно-солевые буферы, воду, эмульсии, такие как эмульсии масло/вода, различные типы смачивающих веществ, стерильные растворы и т.д. Композиции, содержащие такие носители, можно составлять хорошо известными общепринятыми способами. Эти фармацевтические композиции можно вводить индивидууму в подходящей дозе. Введение подходящих композиций может осуществляться различными путями, например, посредством внутривенного, подкожного или перорального введения, которые являются предпочтительными вариантами, и, более того, посредством внутрибрюшинного, внутримышечного, местного, внутрикожного, интраназального или внутрибронхиального введения. Составы для перорального введения включают таблетки и сиропы. Особенно предпочтительно, чтобы указанное введение проводили посредством инъекции. Композиции также можно вводить прямо в заданную область, например, путем биолистической доставки во внешнюю или внутреннюю заданную область. Режим дозирования определяется лечащим врачом и клиническими факторами. Как хорошо известно в области медицины, дозировки для любого пациента зависят от множества факторов, включая размер пациента, площадь поверхности тела, возраст, конкретное вводимое соединение, пол, время и путь введения, общее состояние здоровья и другие лекарственные средства, вводимые одновременно. Белковое фармацевтически активное вещество может присутствовать в количествах от 1 нг до 10 мг/кг массы тела на дозу; однако предусматриваются дозы ниже или выше этого иллюстративного диапазона, особенно учитывая вышеупомянутые факторы. Если режим представляет собой непрерывную инфузию, количество также должно находиться в диапазоне от 1 нг до 10 мг на килограмм массы тела в минуту.
Особенно предпочтительным является внутривенное введение.
В следующем аспекте настоящее изобретение относится к одному или более полипептидам по любому из изобретений, к одному или более комплексам по изобретению и/или к одной или более композициям по любому из изобретений для применения в способе лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызываемое указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется одним или более поверхностными антигенами HBV. Предпочтительно, чтобы указанная печеночно-клеточная карцинома характеризовалась экспрессией одного или более из определенных выше поверхностных антигенов HBV.
В следующем аспекте настоящее изобретение относится к способу лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызываемое инфекцией HBV, выбрано из цирроза печени и печеночно-клеточной карциномы, причем указанный способ включает введение терапевтически эффективного количества или профилактического количества, соответственно, одного или более полипептидов по изобретению, одного или более комплексов по изобретению и/или одной или более композиций по изобретению пациенту, нуждающемуся в этом.
Предпочтительно, чтобы указанная фармацевтическая композиция, указанные полипептид/комплекс/композиция для применения в способе лечения и указанный способ лечения, указанные полипептиды, комплексы и/или композиции были единственными фармацевтически активными средствами, содержащимися или используемыми.
Вместе с тем, предусматривается преднамеренное включение одного или более дополнительных фармацевтически активных средств в комбинированную терапию. Такие дополнительные фармацевтически активные средства могут быть выбраны из интерферонов или других иммуномодуляторов (например, таких как интерферон-альфа 2a или 2b, интерферон-лямбда), противовирусных средств прямого действия, таких как аналоги нуклеоз(т)идов (например, такие как ламивудин (Epivir-HBV, Zeffix или Heptodin), адефовир дипивоксил (Hepsera, Preveon), энтекавир (Baraclude, Entaliv), телбувидин (Tyzeka, Sebivo), тенофовир (Viread)), ингибиторы проникновения (например, такие как Myrcludex-B), другие противовирусные средства или цитокины, такие как интерлейкин-2.
В следующем аспекте настоящее изобретение относится к способу уничтожения клеток, инфицированных HBV, in vitro, причем указанный способ включает культивирование указанных клеток, инфицированных HBV, с (i) иммунными эффекторными клетками и (ii) одним или более полипептидами по изобретению, одним или более комплексами по изобретению и/или одной или более композициями по изобретению.
В предпочтительном варианте осуществления способа in vitro указанные иммунные эффекторные клетки (i) содержатся в мононуклеарных клетках периферической крови; или (ii) представляют собой или содержат NK-клетки и/или CTL.
В следующем аспекте, настоящее изобретение относится к иммунной эффекторной клетке in vitro или ex vivo, которая имеет полипептид по изобретению или комплекс в соответствии с изобретением, связанный с поверхностным антигеном указанной иммунной эффекторной клетки. Предпочтительные иммунные эффекторные клетки и предпочтительные поверхностные антигены, презентируемые иммунными эффекторными клетками, являются такими, как определено выше. Такая иммунная эффекторная клетка является пригодной для введения пациенту, страдающему инфекцией HBV, циррозом печени или печеночно-клеточной карциномой. Таким образом, также предусматривается фармацевтическая композиция, содержащая или состоящая из иммунной эффекторной клетки, которая имеет связанный с ее поверхностным антигеном полипептид по изобретению или комплекс в соответствии с изобретением. Также предусматривается иммунная эффекторная клетка, которая имеет связанный с ее поверхностным антигеном полипептид по изобретению или комплекс в соответствии с изобретением для применения в способе лечения или предупреждения инфекции HBV, цирроза печени или печеночно-клеточной карциномы.
Последовательности, описанные в настоящей заявке









Изобретение иллюстрируется чертежами.
Фиг.1:
scFv-фрагменты получают путем слияния двух вариабельных доменов. Слияние вовлекает применение гибкого пептидного линкера, который не нарушает или по существу не нарушает структуру каждого вариабельного домена.
Фиг.2:
Димеризация двух полипептидов по изобретению путем образования дисульфидных связей. Каждый полипептид содержит биспецифическое двухвалентное антитело. Природная димеризация антител в эндоплазматической сети продуцирующих клеток может приводить к образованию биспецифического четырехвалентного антитела, или три- или тетраспецифического четырехвалентного антитела, если два биспецифических двухвалентных антитела коэкспрессируются (не представлено).
Фиг.3:
Сравнение специфической элиминации продуцирующих поверхностный антиген HBV клеток-мишеней гепатомы после введения единичных биспецифических антител и синергические эффекты одновременного введения двух CTL-специфических или двух специфичных к NK-клеткам биспецифических антител. Использован анализ Blue Cell Viability Assay CellTiter.
Фиг.4:
A) Секреция цитокинов как признак активации иммунных эффекторных клеток в присутствии биспецифических антител по настоящему изобретению. Инфицированные HBV клетки HepaRG сокультивировали с PBMC в присутствии или в отсутствие указанных биспецифических антител.
B) Специфическая элиминация инфицированных HBV клеток-мишеней в сокультуре с иммунными клетками и биспецифическими антителами.
Фиг.5:
Вариабельность клеток-мишеней, сокультивируемых с PBMC в присутствии индивидуальных реактивных в отношении HBs биспецифических антител. Единичные биспецифические антитела опосредуют лизис клеток-мишеней. A, C, E: Эффект стимуляции посредством αHBs x αCD3 (A), αHBs × αCD28 (C) или суммарный (E). B, D, F: Эффект стимуляции αHBs × αCD3 [FcΔADCC] (B), αHBs × αCD28 [FcΔADCC] (D) или суммарный (F). Стрелка указывает на добавление PBMC и биспецифических антител. Кривые с точками соответствуют трансфицированным HBs клеткам HuH7-S, кривые с ромбами соответствуют родительским клеткам гепатомы HuH7. Используют анализ цитотоксичности в реальном времени xCELLigence. Время нормализации клеточного индекса: 0 ч.
Фиг.6:
Жизнеспособность клеток-мишеней, сокультивируемых с PBMC в присутствии реактивных к HBs биспецифических антител. Комбинация биспецифических антител опосредует массивное уничтожение клеток-мишеней. A: Эффект стимуляции αHBs × αCD3 и αHBs × αCD28. B: Эффект стимуляции αHBs × αCD3 [FcΔADCC] и αHBs × αCD28 [FcΔADCC]. C, D: Эффект индивидуальных биспецифических антител по сравнению с комбинациями. Стрелка указывает на добавление PBMC и биспецифических антител. Кривые с точками соответствуют клеткам HuH7-S, кривые с ромбами соответствуют клеткам HuH7. Время нормализации клеточного индекса: 0 ч.
Фиг.7:
Жизнеспособность клеток-мишеней, сокультивируемых с PBMC, в присутствии различных концентраций биспецифических антител. Смеси 50 мкл/50 мкл содержащих антитело супернатантов αHBs x αCD3/ αHBs × αCD28 (A), или αHBs × αCD3 [FcΔADCC]/ αHBs x αCD28 [FcΔADCC] (B), индуцировали лизис клеток-мишеней раньше, чем смеси 25 мкл/25 мкл, что указывает на дозозависимые эффекты. Стрелками указано добавление PBMC и биспецифических антител. Кривые с точками соответствуют клеткам HuH7-S, кривые с ромбами соответствуют клеткам HuH7. Время нормализации клеточного индекса: 0 ч.
Фиг.8:
Жизнеспособность клеток-мишеней, сокультивируемых с различными количествами PBMC в присутствии смеси αHBs x αCD3 и αHBs × αCD28. 2×105 PBMC опосредуют значительно более раннюю элиминацию клеток HuH7-S, чем 1×105 PBMC. Стрелками указано добавление PBMC и биспецифических антител. Кривые с точками соответствуют клеткам HuH7-S, кривые с ромбами соответствуют клеткам HuH7. Время нормализации клеточного индекса: 0 ч.
Фиг.9:
Жизнеспособность клеток-мишеней, сокультивируемых с PBMC в присутствии смесей αHBs x αCD3/ αHBs x αCD28 в течение различных периодов времени. Супернатанты, содержавшие биспецифические антитела, извлекали после указанных периодов стимуляции. Стимуляция только в течение 4 ч привела к небольшому снижению жизнеспособности клеток-мишеней (конечная жизнеспособность 78,5%). Стимуляция PBMC биспецифическими антителами в течение 8 ч или дольше индуцировала элиминацию клеток-мишеней. После стимуляции в течение 8 ч и 12 ч уничтожение клеток-мишеней было замедленным по сравнению со стимуляцией в течение 24 ч или 48 ч, что указывает на непрерывную активацию и перенацеливание эффекторных клеток. Однако конечная жизнеспособность HuH7-S через 48 ч была сравнимой: стимуляция в течение 8 ч: 14,7%; стимуляция в течение 12 ч: 11,7%, стимуляция в течение 24 ч: 5,1%, стимуляция в течение 48 ч: 3,2%. Стрелка указывает на добавление PBMC и биспецифических конструкций. Представлена кинетика жизнеспособности для клеток HuH7-S. Время нормализации клеточного индекса: 0 ч.
Фиг.10:
Секреция IL-2, IFN-γ и TNF-α из PBMC после сокультивирования с клетками HuH7-S/HuH7 в присутствии αHBs × αCD3/ αHBs × αCD28 в различные моменты времени. A: Концентрация IL-2 возрастала с течением времени и достигала плато приблизительно через 24 ч с концентрацией приблизительно 1550 пг/мл. B: Секреция IFN-γ начиналась между 8 ч и 12 ч и возрастала вплоть до 12000 пг/мл (48 ч). C: Продукция TNF-α поддавалась обнаружению уже через 4 ч, возрастала непрерывно, достигала пика через 24 ч (1700 пг/мл) и снижалась до 1400 пг/мл после 48 ч. Можно было обнаружить высокую фоновую секрецию TNF-α в отсутствие HBs (клетки HuH7) причем наивысшая концентрация снижалась через 4 ч (~70 пг/мл) до 9 пг/мл после сокультивирования в течение 48 ч.
Фиг.11:
Окрашивание LAMP-1 после сокультивирования PBMC с клетками HuH7-S/HuH7 в присутствии биспецифических антител. Поверхностная экспрессия маркера эндосомальной дегрануляции LAMP-1 обнаруживается на CD4+ (A, B) и CD8+ (C, D) T-клетках после сокультивирования с клетками HuH7-S (черная линия) или HuH7 (серая линия) в присутствии либо αHBs × αCD3/ αHBs x αCD28 (A, C), либо αHBs × αCD3 [FcΔADCC]/ αHBs × αCD28 [FcΔADCC] (B, D).
Фиг.12:
FACS-анализ PBMC, сокультривируемых с клетками HuH7-S или HuH7 в присутствии αHBs x αCD3/ αHBs × αCD28 через 8 ч, 12 ч и 24 ч. A, B; Проценты IFNγ+/IL-2+/TNFa+/CD154* CD4+ T-клеток (A) или IFNγ+/IL-2+/TNFa+/CD154+ CD8+ (B) T-клеток. C, D: Окна булевых комбинаций для IFNγ+, IL-2+ и/или TNFa+ CD4+ (C), или IFNy+, IL-2+ и/или TNFa+ CD8+ (D) T-клеток.
Фиг.13:
FACS-анализ PBMC, сокультивируемых с иммобилизованным или растворимым HBsAg в присутствии αHBs × αCD3 [FcΔADCC]/ αHBs x αCD28 [FcΔADCC] после 24 ч и 48 ч. A, B: Проценты IFNγ+/IL-2+/TNFα+ /CD154+ CD4+ T-клеток (A) или IFNγ+/IL-2+/TNFa+/CD154+ CD8+ (B) T-клеток. C, D: Окна булевых комбинаций для IFNγ+, IL-2+ и/или TNFa+ CD4+ (C), или IFNy+, IL-2+ и/или TNFa+ CD8+ (D) T-клеток.
Фиг.14:
HBsAg в супернатанте клеток HuH7-S (110.8 S/CO), клеток HepG2.2.15 (41.7 S/CO) и инфицированных HBV клеток HepaRG (16.5 S/CO).
Фиг.15:
Жизнеспособность инфицированных HBV/неинфицированных клеток HepaRG, сокультивированных с PBMC, в присутствии биспецифических антител. αHBs × αCD3 (A) и αHBs x αCD3/ αHBs × αCD28 (B) опосредуют значительный лизис клеток-мишеней. Конечная жизнеспособлность необработанных клеток составляет 65,9% (HBV+) и 62,9% (HBV-). Стрелками указано добавление PBMC и биспецифических конструкций. Кривые с точками соответствуют инфицированным HBV клеткам HepaRG, кривые с ромбами соответствуют неинфицированным клеткам HepaRG. Время нормализации клеточного индекса в анализе xCELLigence: 0 ч.
Фиг.16:
Уменьшение размера опухоли у животных, которых лечили биспецифическими антителами. Мышей, имеющих положительные по HBV подкожные опухоли HepG2.2.15, обрабатывали PBMC человека и смесью биспецифических антител αHBs × αCD3 и αHBs × αCD28 в течение четырех последовательных суток. Мышей умерщвляли и размер опухоли анализировали.
Изобретение иллюстрируется примерами.
Пример 1
Материалы и способы для примера 2
Клонирование и продуцирование биспецифических антител
Комплементарные ДНК, кодирующие вариабельные тяжелые и вариабельные легкие цепи антитела против CD3 (OKT3), антитела против CD28 (9.3), антитела против CD16 (A9) и антитела против CD56 (NCAM29.2) получали посредством амплификации способом ПЦР обратно транскрибированных мРНК из соответствующей гибридомы с использованием набора праймеров, охватывающих все подтипы VH и Vκ/Vλ. Продукты ПЦР лигировали в pCR2.1-TOPO (Invitrogen, Life Technologies) и секвенировали. Антитело против HBsAg scFv C8 было предоставлено в кодон-оптимизированной форме в плазмиде pMP71-C8. С использованием праймеров, содержащих соответствующие участки рестрикции на 5' и 3'-фланкирующих областях, кДНК вариабельной области тяжелой цепи и вариабельной области легкой цепи, кодирующие упомянутые выше антитела, собирали с помощью глицин-серинового линкера в scFv. scFv OKT3, 9.3, A9 и NCAM29.2 (с N-концевым удлинением посредством (Gly)3-4) клонировали на 3'-конце кДНК, присутствующей в pBluescript KS II+ (Stratagene), которая кодирует Fc-домен (шарнирная область, CH2, CH3) IgG1 человека, которая удлинена глицин-сериновым линкером GlyAsnSer(Gly4Ser)3AlaSer на 5'-конеце, и последовательность StrepTag (WSHPQFEK) и во второй серии конструкций дополнительный глицин-сериновый линкер (Gly4Ser)3 на 3'-конце. Кодирующую последовательность C8 scFv клонировали на 5'-конце упомянутого 5'-глицин-серинового линкера. Полную последовательность scFv-линкер-hIgG1Fc-линкер-scFv субклонировали в экспрессирующий вектор млекопитающих pcDNA3.1(-) (Invitrogen). Плазмидную ДНК Maxi-prep использовали для трансфекции клеток HEK293 с использованием реагента для трансфекции peqFECT (Peqlab). Стабильные трансфектанты отбирали с использованием 0,8-1,0 мг/мл G418 и увеличивали в количестве. Супернатанты трансфектантов HEK собирали и анализировали способом ELISA в отношении концентрации секретируемых биспецифических антител и вестерн-блоттингом в отношении целостности секретируемых антител с использованием меченных пероксидазой антител козы, специфичных к IgG-Fc человека.
Условия культивирования клеток и инфицирование HBV
Клетки гепатомы HuH7 (Nakabayaski, et al. 1982. Growth of human hepatoma cell lines with differentiated functions in chemically defined medium. Cancer Res. 42: 3858-3863) и клетки HEK293 поддерживали в модифицированной способом Дульбекко среде Игла (DMEM), дополненной 10% эмбриональной телячьей сывороткой (FBS), пенициллином (100 Е/мл), стрептомицином (100 мкг/мл) и L-глутамином (2 ммоль/л) (все от GIBCO, Life Technologies).
Мононуклеарные клетки периферической крови (PBMC) выделяли центрифугированием в градиенте плотности из гепаринизированной цельной крови с использованием среды для разделения лимфоцитов LSM 1077 (PAA). 25 мл крови наслаивали на 13 мл LSM 1077. После центрифугирования при 2000 об/мин в течение 20 мин (без торможения) при комнатной температуре PBMC собирали и культивировали в среде RPMI 1640, дополненной 10% эмбриональной телячьей сывороткой (FBS), пенициллином (100 Е/мл), стрептомицином (100 мкг/мл) и L-глутамином (2 ммоль/л) (все от GIBCO). После стадии покоя в течение ночи PBMC или отсортированные NK-клетки использовали для экспериментов по сокультивированию.
Клетки HepaRG поддерживали в среде Williams E Medium (Invitrogen GmbH, Karlsruhe, Германия), дополненной L-глутамином (5 ммоль/л), глюкозой (0,06% [масс./об.]), HEPES (23 ммоль/л, pH7,4), гентамицином (50 (мкг/мл), пенициллином (501 Е/мл), стрептомицином (50 пг/мл), инозином (37 пмоль/л), гидрокортизоном (4,8 пг/мл) и инсулином (1 мкг/мл). Перед инфицированием клеткам HepaRG позволяли дифференцироваться в течение 4 недель с использованием среды для дифференцировки (среда Williams E Medium (как описано выше), дополненной DMSO (1,75%). Клетки HepaRG инфицировали с использованием исходных культур HBV при конечной m.o.i. 200 и PEG (5%) в среде для дифференцировки. Инфекционный инокулят удаляли после инкубации в течение ночи, и заменяли средой для дифференцировки, и культивировали в течение 6 суток. Для сокультур с перенацеленными T-клетками авторы настоящего изобретения заменили среду для дифференцировки на свободную от гидрокортизона среду за 2 суток до начала сокультивирования, чтобы избежать иммуносупрессии, опосредуемой гидрокортизоном.
Трансфекция плазмидами, кодирующими поверхностный антиген HBV
Клетки HuH-7 трансфицировали плазмидами, кодирующими различные поверхностные антигены, с использованием реагента для трансфекции FuGene (Promega). В 8 лунок 96-луночного планшета добавляли 3 мкл FuGENE, 1 мкг плазмидной ДНК добавляли к 100 мкл OptiMEM (Gibco). Раствор для трансфекции инкубировали в течение 15 мин при комнатной температуре, чтобы FuGENE связался с плазмидной ДНК. Использовали конечный объем 100 мкл на лунку после добавления дополнительного OptiMEM и проводили инкубацию в течение по меньшей мере 24 ч.
Магнитно-активируемая клеточная сортировка (MACS) для NK-клеток
Клетки NK выделяли из PBMC с использованием набора для выделения CD56+CD16+ NK-клеток человека (Miltenyi). На первой стадии отрицательной селекции все клетки, не являющиеся NK-клетками, удаляли с использованием моноклональных антител, направленных против антигенов, не экспрессируемых на поверхности NK-клеток. На второй стадии положительной селекции NK-клетки выделяли с использованием моноклональных антител против CD16, конъюгированных с микрогранулами из оксида железа, и удерживали внутри магнитного поля. После выделения NK-клетки культивировали в среде RPMI-1640, как описано выше.
Сокультура HBV-положительных клеток-мишеней и перенацеленных эффекторных клеток
Клетки-мишени культивировали в 96-луночном планшете при смыкании монослоя. Добавляли 1×105 эффекторнных клеток в объеме среды 100 мкл на лунку. Добавляли 100 мкл на лунку супернатантов HEK, содержавших биспецифические антитела. Для определения синергических эффектов добавляли 50 мкл супернатанта каждого биспецифического антитела на лунку. Необработанные клетки-мишени, инкубированные с 200 мкл среды или с эффекторными клетками отдельно или с биспецифческими антителами отдельно служили в качестве отрицательного контроля.
Твердофазный иммуноферментный анализ (ELISA) активации эффекторных клеток
Секрецию цитокинов вследствие активации эффекторных клеток выявляли с помощью ELISA. Использовали ELISA MAX™ для IFN-γ человека (BioLegend). Поглощение при 450 нм выявляли с использованием программы Magellan6 и lnfiniteF200 (Tecan).
Анализ жизнеспособности клеток-мишеней
Жизнеспособность клеток-мишеней после сокультивирования определяли с использованием анализа жизнеспособности клеток CellTiter-Blue (Promega). Этот анализ основан на способности живых клеток конвертировать окислительно-восстановительный краситель (резазурин) во флуоресцентный конечный продукт (резоруфин) вследствие метаболической активности. Нежизнеспособные клетки быстро утрачивают их метаболическую способность и, таким образом, не генерируют флуоресцентный сигнал. После удаления супернатанта к сокультурам добавляли 100 мкл на лунку бесцветной DMEM, содержавшей 20% реагент CellTiter-Blue и инкубировали при 37°C в течение 2 часов. Сигнал флуоресценции регистрировали при 560 нм с использованием InfiniteF200 (Tecan).
Пример 2
Результаты
В первой линии экспериментов авторы настоящего изобретения оценивали активность биспецифических конструкций антител, направленных на поверхностные антигены CTL CD3 и CD28 и на поверхностные антигены NK-клеток CD16 и CD56. Авторы настоящего изобретения использовали трансфицированные плазмидой клеточные линии гепатомы, продуцирующие поверхностные антигены HBV. После налаживания экспрессии белка HBV эти клетки-мишени сокультивировали вместе с иммунными эффекторными клетками, а именно PBMC и выделенными NK-клетками, и биспецифическими конструкциями антител. PBMC содержат приблизительно 70% T-клеток, но только 7% NK-клеток. Таким образом, авторы настоящего изобретения проводили магнитное выделение CD16+ CD56+ NK-клеток. В качестве отрицательных контролей авторы настоящего изобретения анализировали сокультуры с HBV-отрицательными клетками-мишенями, преинкубированные с супернатантами, содержащими HBV и субвирусные частицы. Этот контроль использовали для исключения активации эффекторных клеток вследствие неспецифического связывания частиц HBV на поверхности HBV-отрицательных клеток-мишеней. Более того, авторы настоящего изобретения сокультивировали HBV-положительные клетки-мишени с иммунными эффекторными клетками в отсутствие биспецифических конструкций для оценки неспецифической фоновой цитотоксичности. Для исключения цитотоксического эффекта биспецифических конструкций авторы настоящего изобретения получили культуры HBV-положительных клеток-мишеней без иммунных эффекторных клеток в присутствии биспецифических конструкций.
Эти эксперименты продемонстрировали специфическую активацию CTL при сокультивировании в присутствии CD3- или CD28-спсецфических конструкций, как определяют по секреции провоспалительного цитокина интерферона-гамма (IFN-γ), составляющей вплоть до 7000 пг/мл. Этот эффект далее усиливался совместным введением CD3- и CD28-специфических конструкций, демонстрирующих синергический эффект.
Более того, биспецифические конструкции опосредовали специфическую цитотоксическую элиминацию HBsAg-продуцирующих клеточных линий гепатомы HuH7 (фиг.3), составляющую вплоть до 90% снижения жизнеспособности клеток-мишеней по сравнению с контролями. Этот цитотоксический ответ наблюдали для сокультур PBMC и HBV-положительных клеток-мишеней вместе с биспецифическими конструкциями, направленными против CD3 и CD28, а также для выделенных NK-клеток с конструкциями, направленными против CD16 и CD56. Совместное введение конструкций, специфичных к CTL и NK-клеткам, далее увеличивало цитотоксический эффект синергично до уровней элиминации выше 95%. Авторы настоящего изобретения наблюдали неспецифическую фоновую цитотоксичность от 15% до 40% для CTL и NK-клеток, соответственно.
Во втором раунде экспериментов авторы настоящего изобретения использовали инфицированные HBV клетки гепатомы HepaRG. Эта клеточная линия позволяет инфицирование HBV после дифференцировки в течение четырех недель и отражает естественную ситуацию для HBV-инфицированных тканей. Как правило, уровни инфицирования для клеток HepaRG никогда не достигают 100%, и эта смесь инфицированных и неинфицированных клеток имитирует ситуацию у HBV-инфицированного индивидуума при противовирусной терапии, имеющего как инфицированные, так и неинфицированные клетки, в присутствии свободных внеклеточных вирусных частиц.
В сокультурах иммунных эффекторных клеток и совместно введенных биспецифических конструкций HBV-инфицированные клетки HepaRG опосредовали эффективную активацию как CTL, так и NK-клеток с впечатляющими уровнями IFN-γ вплоть до 60000 пг/мл (фиг.4A). В этом эксперименте авторы настоящего изобретения не выделяли или не увеличивали в количестве NK-клетки перед сокультивированием.
Более того, биспецифические конструкции антител приводили к цитотоксическому ответу активированных иммунных эффекторных клеток, что приводило к специфической элиминации HBV-инфицированных клеток-мишеней (фиг.4B). Авторы настоящего изобретения наблюдали уровни элиминации от 50% до 70% для NK-клеток и CTL, соответственно. В этих экспериментах неспецифическая фоновая цитотоксичность отсутствовала.
Пример 3
Способы для примера 4
Для анализа терапевтического потенциала биспецифических конструкций антител в отношении успешного перенацеливания T-клеток на HBV-положительные клетки проводили эксперименты по сокультивированию in vitro и проводили их детальный анализ. Авторы настоящего изобретения использовали биспецифические конструкции антител, содержащие одноцепочечные связывающие домены, направленные против CD3 человека (αCD3) и CD28 человека (αCD28) и, кроме того, конструкции, содержавшие направленные мутации в их спейсерном Fc-домене, которые должны устранять антителозависимую клеточную цитотоксичность (ΔADCC), путем препятствования связыванию рецептора Fcγ. Их конструировали в качестве меры безопасности для исключения неспецифической активации натуральных киллеров. С другой стороны все конструкции биспецифических антител содержали специфический связывающий домен C8 S-белка HBV (HBsAg). Мононуклеарные клетки периферической крови (PBMC), выделенные из свежей венозной крови здоровых доноров, сокультивировали с различными клеточными линиями гепатомы человека в качестве заместительных моделей для инфекции HBV. Авторы настоящего изобретения использовали HuH7-S (трансгенный по S-антигену HBV) и в качестве отрицательного контроля родительскую клеточную линию HuH7, инфицированную HBV, или в качестве контроля неинфицированные клетки HepaRG. Клетки HepG2.2.15 (HBV с трансгенным геномом) использовали в качестве контролей для количественного определения маркера HBV. Для предоставления биспецифических конструкций антител добавляли супернатант продуцирующих клеточных линий, содержащих биспецифические антитела. Для визуализации изменений жизнеспособности клеток-мишеней вследствие цитотоксичности, опосредуемой биспецифическими антителами, с течением времени, использовали систему xCELLigence. Этот способ позволяет мониторинг в реальном времени жизнеспособности клеток при культивировании в течение длительных периодов времени. Таким образом, клетки-мишени гепатомы высевали на специально сконструированные микропланшеты для титрования, которые содержали гребенчатыми золотыми микроэлектродами для неинвазивного мониторинга жизнеспособности прикрепляющихся клеток-мишеней с использованием электрического сопротивления в качестве считываемых данных. Цитотоксическая элиминация приводит к изменению сопротивления, которое может быть конвертировано в так называемую величину клеточного индекса (CI), которую используют для мониторинга жизнеспособности клеток.
Сокультивирование с клетками-мишенями
На нулевые сутки высевали 3×104 клеток HuH7-S/HuH7 на лунку в 96-леночном планшете (E-Plate 96). На 1 сутки супернатант удаляли и в соответствующие лунки добавляли 1×105 первичных PBMC человека в 100 мкл среды PBMC или только 100 мкл для контролей. Кроме того, добавляли 100 мкл супернатанта, содержащего биспецифические антитела, по отдельности или в комбинациях. В качестве отрицательного контроля в лунки добавляли 100 мкл среды DMEM до общего объема 200 мкл. Мониторинг культур проводили в течение 48 ч или 72 ч в среде xCELLigence.
Клетки HepaRG выращивали до смыкания монослоя, подвергали дифференцировке в течение 21 суток и инфицировали HBV перед экспериментом по иммунотерапии.
Для инфицирования клеток HepaRG приготавливали исходную культуру вируса в среде для дифференцировки, содержавшей PEG, и 50 мкл добавляли на лунку. Конечная концентрация PEG составляла 5% и MOI исходной культуры вируса была установлена как 200 (7,5×106 вирусных частиц/лунка). Через 16 ч после добавления основной смеси для инфицирования клетки промывали 3 раза PBS для удаления остаточного вируса. Добавляли среду для дифференцировки и среду заменяли каждые 3 суток в течение всего 12 суток. Перед экспериментами по сокультивировани среду заменяли на среду для сокультивирования (обедненную иммунодепрессантом гидрокортизон). Успешное инфицирование HBV клеток HepaRG исследовали путем измерения HBsAg (Axsym) и HBeAg (ВЕР III System) в супернатанте инфицированных клеток.
Получение PBMC
PBMC для экспериментов по сокультивированию выделяли из цельной крови. Гепаринизированную свежую кровь разбавляли 1:1 промывочной средой RPMI. 25 мл разбавленной крови наслаивали на 15 мл Percoll и центрифугировали при 960 g в течение 20 мин без торможения в поворотной центрифуге. PBMC выделяли и переносили в 50 мл среды RPMI. После промывания клетки ресуспендировали в 10 мл среды PBMC и определяли число клеток. Концентрацию доводили до 2×106 клеток/мл для обеспечения оптимальных условий. PBMC оставляли в состоянии покоя в течение ночи 37°C.
Активированная флуоресценцией сортировка клеток (FACS)
Для исследования эффекторных функций перенацеленных PBMC проводили FACS-анализ. Таким образом, анализировали секрецию провоспалительных цитокинов IFN-γ, IL-2 и TNF-α, а также экспрессию маркера активации CD154 (CD40L) и маркера дегрануляции LAMP-1 (CD107a), соответственно. Измерение продукции цитокинов проводили с использованием внутриклеточного окрашивания цитокинов. Таким образом, 0,2 мкг/мл брефелдина A (BFA) наносили на клетки и инкубировали в течение 4 часов при 37°C.
BFA блокирует прямой транспорт между эндоплазматической сетью и аппаратом Гольджи и, вследствие этого, происходит ингибирование экзоцитоза цитокинов. В случае одновременного окрашивания LAMP-1, антитело наносили за 1 ч до добавления BFA (для обеспечения транслокации LAMP-1 на клеточную поверхность). Затем клетки переносили в 96-луночный планшет (круглодонный) и промывали два раза 200 мкл буфера FACS. Для окрашивания жизнеспособных клеток и исключения погибших клеток использовали набор LIVE/DEAD Fixable Near-IR Dead Cell Stain Kit. Для фиксации и повышения проницаемости клетки ресуспендировали в 100 мкл реагента Cytofix/Cytoperm и инкубировали на льду в темноте в течение 20 мин. После промывания клетки ресуспендировали в полученной смеси антител или только окрашивали соответствующими отдельными цветами для систематической компенсации. Окрашивание проводили на льду в темноте в течение 30 мин. После промывания клетки ресуспендировали в 200 мкл буфера FACS и переносили в пробирки FACS для сбора данных. Сбор данных проводили с использованием либо FACSCanto II, либо LSR Fortessa. Для регистрации данных использовали программное обеспечение FACS Diva, анализ проводили с использованием программного обеспечения FlowJo.
Эксперименты на животных
Для первого испытания биспецифических конструкций in vivo проводили эксперименты на иммунодеффицитных мышах Rag2/IL2Rγnull (международная номенклатура: B10;B6-Rag2tm1Fwa II2rgtm1WjI). Авторы настоящего изобретения инъецировали мышам в возрасте 6 недель 5×106 клеток трансгенной по HBV клеточной линии гепатомы человека HepG2.2.15. Клетки инъецировали подкожно в бок животных. Это приводило к образованию опухоли в течение 14 суток. Мониторинг репликации HBV в опухоли проводили путем определения виремии HBV. PBMC человека выделяли из свежей пуповинной крови человека и стимулировали на планшетах, предварительно покрытых антителами против CD3 и CD28 человека в концентрации клеток 0,25×106 PBMC на мл в течение 3 суток. Затем клетки поддерживали в среде для культивирования клеток, содержавшей 300 Е/мл IL-2, в течение 7 суток.
На 14 сутки после индукции опухоли мышам внутрибрюшинно инъецировали 2×107 PBMC на мышь и вводили 100 мкл биспецифических конструкций антител против αCD3/αCD28 в супернатанте продуцирующих клеток HEK в хвостовую вену на животное в течение четырех последовательных суток. Мышей умерщвляли на 18 сутки после индукции опухоли и анализировали в отношении размера опухоли. Затем образцы сыворотки и ткани хранили для дальнейшего анализа.
Пример 4
Биспецифические антитела опосредуют специфическую элиминацию клеток-мишеней, экспрессирующих поверхностный белок HBV (HuH7-S)
Для исследования того, осуществляют ли биспецифические конструкции антител успешное перенацеливание T-клеток на клетки, экспрессирующие HBsAg, и индуцируют ли они лизис клеток-мишеней, выделенные PBMC сокультивировали с клетками HuH7-S в присутствии биспецифических конструкций антитела. Клетки HuH7-S подвергали стабильной трансфекции для экспрессии HBsAg и, таким образом, имитировали HBV-инфицированные гепатоциты. Это обеспечивало продукцию и секрецию субвирусных частиц в супернатант и включение HBsAg в клеточную мембрану. В качестве отрицательного контроля служили нетрансфицированные клетки HuH7.
Индивидуальные биспецифические антитела индуцируют уничтожение клеток-мишеней
Для анализа того, способны ли индивидуальные биспецифические антитела стимулировать активацию T-клеток и опосредовать лизис клеток-мишеней PBMC сокультивировали с клетками HuH7-S/HuH7 в присутствии биспецифических четырехвалентных антител αHBs × αCD3, αHBs × αCD28, αHBs × αCD3 [FcΔADCC] или αHBs x αCD28 [FcΔADCC]. Стимуляция эффекторных клеток единичными биспецифическими антителами приводила к специфическому уничтожению клеток-мишеней, экспрессирующих HBsAg (фиг.5). Биспецифические антитела, направленные против CD3, опосредовали элиминацию клеток-мишеней раньше и сильнее, чем конструкции, направленные против CD28, поскольку конечная жизнеспособность клеток HuH7-S, обработанных αCD3, составляла только 6,4% (αCD3ΔADCC: 15,5%) по сравнению с 44,42% для αCD28 (αCD28ΔADCC: 48,9%). Более того αCD3ΔADCC и αCD28ΔADCC требовали больше времени для индукции лизиса клеток-мишеней по сравнению с αCD3 и αCD28, соответственно. Опосредуемое αCD3ΔADCC уничтожение начиналось приблизительно через 35 ч после начала сокультивирования, в то время как αCD3 приводило к снижению жизнеспособности клеток-мишеней уже через 12 ч. Также можно было наблюдать небольшой временной сдвиг приблизительно 20 ч между лизисом клеток-мишеней, опосредуемым αCD28ΔADCC и αCD28. Стимуляция αCD3 также приводила к поддающемуся обнаружению лизису HBsAg-отрицательных клеток HuH7 с конечной жизнеспособностью 78,1%, что указывает на неспецифическую активацию. Указанное было справедливым для стимуляции αCD3ΔADCC в некоторых экспериментах, даже если это не показано здесь. Жизнеспособность клеток HuH7 в ходе сокультивирования в присутствии других биспецифических конструкций оставалась на уровне 100%.
Эти данные демонстрируют, что стимуляция каждым из индивидуальных биспецифических антител индуцирует элиминацию клеток-мишеней без дальнейшей костимуляции.
Биспецифические антитела опосредуют лизис клеток-мишеней синергичным образом
Для дальнейшего анализа того, приводит ли комбинация биспецифических конструкций к увеличенной активности и, таким образом, цитотоксичности эффекторных клеток, PBMC сокультивировали с клетками HuH7-S/HuH7 в присутствии комбинаций либо αCD3/αCD28, либо αCD3ΔADCC/αCD28ΔADCC. Как показано на фиг.6, комбинация биспецифических конструкций привела к массивному уничтожению экспрессирующих HBsAg клеток-мишеней с остаточной жизнеспособностью 1,2% (αCD3/αCD28) и 4,4% (αCD3ΔADCC/αCD28ΔADCC), в то время клетки HuH7 практически не элиминировались (конечная жизнеспособность клеток HuH7: αCD3/αCD28: 92,4%; αCD3ΔADCC/αCD28ΔADCC: 100,4%).
Вновь, опосредуемый αCD3/αCD28 лизис клеток-мишеней был более быстрым, чем уничтожение, индуцируемое конструкциями с мутантной Fc-областью, даже если уничтожение клеток-мишеней начиналось приблизительно в тот же момент времени через приблизительно 11 ч (фиг. 6A, B). Комбинация биспецифических антител привела к более быстрой элиминации клеток-мишеней по сравнению с лизисом, индуцируемым индивидуальными биспецифическими конструкциями (фиг.6C, D). Это ожидалось, поскольку T-клетки получали не только один сигнал, как в случае присутствия индивидуальных конструкций, но получали как активирующий, так и костимулирующий сигнал, если присутствовали антитела, направленные против CD3 и CD28.
Таким образом, комбинация биспецифических конструкций опосредует специфический лизис поверхностного белка HBV, экспрессирующего клетки-мишени, синергичным образом.
Биспецифические антитела индуцируют элиминацию клеток-мишеней зависимым от концентрации образом
Для исследования того, имело ли количество биспецифических антител эффект на лизис клеток-мишеней для сокультивирования использовали два различных количества биспецифических конструкций. Таким образом, использовали обычное количество антител (100 мкл супернатанта всего  высокое) и его половину (50 мкл супернатант всего низкое). Более низкое количество биспецифических антител также могло индуцировать лизис клеток-мишеней (конечная жизнеспособность клеток HuH7-S: αCD3/αCD28: 12,6%; αCD3ΔADCC/αCD28ΔADCC: 15,9%), в то время как более высокое количество индуцировало более быструю элиминацию клеток-мишеней (фиг.7), причем оставалось только 1,5% (αCD3/αCD28) и 2,1% (αCD3ΔADCC/αCD28ΔADCC) жизнеспособных клеток. Клетки HuH7 оставались неизмененными в любом случае. Комбинация либо αCD3/αCD28, либо αCD3ΔADCC/αCD28ΔADCC индуцировала уничтожение клеток-мишеней зависимым от концентрации образом.
высокое) и его половину (50 мкл супернатант всего низкое). Более низкое количество биспецифических антител также могло индуцировать лизис клеток-мишеней (конечная жизнеспособность клеток HuH7-S: αCD3/αCD28: 12,6%; αCD3ΔADCC/αCD28ΔADCC: 15,9%), в то время как более высокое количество индуцировало более быструю элиминацию клеток-мишеней (фиг.7), причем оставалось только 1,5% (αCD3/αCD28) и 2,1% (αCD3ΔADCC/αCD28ΔADCC) жизнеспособных клеток. Клетки HuH7 оставались неизмененными в любом случае. Комбинация либо αCD3/αCD28, либо αCD3ΔADCC/αCD28ΔADCC индуцировала уничтожение клеток-мишеней зависимым от концентрации образом.
Увеличенные концентрации эффекторных клеток усиливают лизис клеток-мишеней
Кроме того, интерес представляло то, влияет ли количество эффекторных клеток на элиминацию клеток-мишеней. Таким образом, обычное количество PBMC, использованное для сокультивирования (1x105), сравнивали с двойным количеством (2×105). Как продемонстрировано на фиг.8, более высокое количество PBMC индуцировало лизис клеток HuH7-S в присутствии αCD3/αCD28 значительно быстрее с конечной жизнеспособностью 4,5% по сравнению 11,7%, но также уничтожалось больше клеток HuH7, если присутствовало двойное количество PBMC (конечная жизнеспособность клеток HuH7: 2×105 PBMC: 83,8%; 1×105 PBMC: 102,7%).
Эти данные указывают на то, что элиминация клеток-мишеней зависит от количества эффекторных клеток.
Биспецифические антитела опосредуют уничтожение клеток-мишеней после сокультивирования в течение только 8 ч
Для исследования вопроса, как долго биспецифические антитела должны присутствовать в процессе сокультивирования для активации T-клеток и, таким образом, индукции цитотоксичности, супернатант сокультур, содержавших биспецифические антитела, удаляли через различные моменты времени и добавляли новую стандартную среду DMEM. Если супернатант, содержавший αCD3/αCD28, удаляли после 4 ч, PBMC индуцировали только небольшое снижение жизнеспособности клеток-мишеней (78,5%), но не были способны индуцировать лизис всех клеток-мишеней (фиг.9). Если супернатант, содержавший биспецифические антитела, присутствовал в течение 8 ч или дольше, PBMC были способны индуцировать элиминацию клеток-мишеней. Как проиллюстрировано на фиг.10, для PBMC требовалось больше времени для индукции лизиса клеток-мишеней, если стимуляция αCD3/αCD28 длилась 8 ч или 12 ч по сравнению с 24 ч или 48 ч, однако эффект после 48 ч был практически сходным (конечная жизнеспособность HuH7-S: 8 ч: 14,7%; 12 ч: 11,7%, 24 ч: 5,1%, 48 ч: 3,2%).
Биспецифические антитела опосредуют эффекторные функции T-клеток в процессе сокультивирования либо с клетками HBsAa, либо с клетками HuH7-S
Для исследования активации и функциональности T-клеток в процессе экспериментов по сокультивированию секрецию цитокинов исследовали посредством либо ELISA, либо FACS-анализа.
Биспецифические конструкции опосредуют секрецию IFN-γ, TNF-α и IL-2
В хронологическом эксперименте анализировали, когда PBMC начинают секретировать цитокины при контакте с биспецифическими антителами и как динамика развивается с течением времени. Таким образом, супернатант сокультур удаляли через 4 ч, 8 ч, 12 ч, 24 ч и 48 ч после добавления PBMC и αCD3/αCD28. Продукцию цитокинов измеряли способом ELISA для IL-2, IFN-γ и TNF-α. Секреция IL-2 возрастала с течением времени, однако после 4 ч IL-2 практически не поддавался обнаружению, после 8 ч концентрация составляла уже 316 пг/мл и в течение последующих 4 часов, концентрация увеличилась практически в четыре раза (1119 пг/мл). Не происходило дальнейшего повышения между 24 ч и 48 ч и оказалось, что концентрация IL-2 достигла плато при концентрации приблизительно 1550 пг/мл (фиг.10A). Секреция IFN-γ (фиг.10B) требовала больше времени, после 8 ч все еще обнаруживались очень низкие уровни. Между 8 ч и 12 ч T-клетки начинали секретировать IFN-γ, поскольку их концентрация уже составляла 1800 пг/мл после 12 ч. Затем (24 ч) наблюдали увеличение продуцирования IFN-γ, концентрация возрастала приблизительно до 10000 пг/мл. Наиболее высокое количество обнаруживалось после 48 ч (12000 пг/мл). Как для IL-2, так и для IFNγ концентрация отрицательных по HBsAg клеток увеличивалась с течением времени, причем наивысшим было количество после 48 ч (IL-2: 45 пг/мл; IFNγ: 200 пг/мл), которое также соответствует наблюдениям, касающимся жизнеспособности клеток.
Секреция TNF-α (фиг.10C) увеличивалась вплоть до 24 ч, где она достигала максимальной концентрации (1700 пг/мл). Затем она снижалась и составляла только 1400 пг/мл после 48 ч. Напротив, секреция TNF-α начиналась раньше, чем остальных, с приблизительно 100 пг/мл после 4 ч с последующим устойчивым увеличением вплоть до 24 ч. Интересно, что продукция TNF-α в клетках HuH7 происходила в точности противоположным образом. Имея относительно высокую фоновую концентрацию по сравнению с другими цитокинами, она продемонстрировала наиболее высокую концентрацию через 4 ч (~70 пг/мл), которая снижалась в течение времени и составляла только 9 пг/мл после 48 ч. В PBMC индуцируется секреция IL-2, IFN-γ и TNF-α при контакте с αCD3/αCD28 в ходе сокультивирования с клетками, экспрессирующими HBsAg, в то время как среди индивидуальных цитокинов динамика секреции отличается.
Биспецифические конструкции активируют CD8+ T-клетки, а также CD4+ T-клетки
Чтобы проанализировать, демонстрируют ли PBMC также дегрануляцию цитотоксических везикул исследовали транслокацию LMAP-1 (CD107a), маркера дегрануляции. После сокультивирования с клетками HuH7-S/HuH7 в присутствии αCD3/αCD28 или αCD3ΔADCC/αCD28ΔADCC, CD8+ T-клетки продемонстрировали очевидный сдвиг окрашивания LAMP-1, в то время в образцах, стимулированных αCD3ΔADCC/αCD28ΔADCC, сигнал был более сильным по сравнению с αCD3/αCD28 (фиг.11 C, D). Интересно, что то же наблюдение можно сделать для CD4+ T-клеток (фиг.11 A, B). Для αCD3/αCD28 транслокация LAMP-1 была более выраженной в CD8+ T-клетках, для αCD3ΔADCC/αCD28ΔADCC она было в точности обратной.
Эти данные демонстрируют, что не только CD8+ T-клетки, но также CD4+ индуцируются к секреции цитотксических гранул при контакте с биспецифическими антителами и HBsAg.
Для исследования полифункциональности T-клеток после экспериментов по сокультивированию PBMC окрашивали в отношении IFN-γ, IL-2 и TNF-α, а также в отношении маркера активации CD154 (CD40L), который в основном экспрессируется на CD4+ T-клетках, через 8 ч, 12 ч и 24 ч после добавления PBMC и αCD3/αCD28 (фиг.12). CD4+ T-клетки продемонстрировали устойчивое увеличение количества IFN-γ+ T-клеток (9,3% после 24 ч), IL-2+ T-клеток (11,3% после 24 ч), TNF-α+ T-клеток (14,7% после 24 ч) и CD154+ T-клеток (28,0% после 24 ч), в то время как основное увеличение происходило между 12 ч и 24 ч (фиг.12A).
Указанное было справедливым для CD8+ T-клеток, в то время как процент IFN-γ+ и IL-2+ клеток на 18,4% и 11,3% превышал процент CD4+ T-клеток. Количество TNF- α+ и CD154+ CD8+ T-клеток было сниженным на 10,1% и 6,25% по сравнению с CD4+ T-клетками (фиг.12B). PBMC в случае клеток HuH7 продемонстрировали отсутствие активации в любом образце. Для дальнейшего анализа T-клеток, секретирующих цитокины, использовали окна булевых комбинаций (фиг.12C, D). После 24 ч 3,1% CD4+ T-клеток и 2,1% CD8+ T-клеток были IFNγ+, IL-2+ и TNFα+, что указывает на полифункциональность T-клеток. Таким образом, αCD3/αCD28 опосредует активацию PBMC в процессе сокультивирования с клетками HuH7-S/HuH7, что приводит к полифункциональным CD4+ и CD8+ T-клеткам.
Для исключения того, что были обнаружены ложноположительные сигналы вследствие неспецифического связывания с погибшими клетками-мишенями в ходе FACS-анализа, PBMC культивировали в присутствии иммобилизованного HBsAg. Кроме того, исследовали эффект растворимого HBsAg, поскольку инфицированные HBV пациенты обладают высокими количествами HBsAg в их крови. PBMC вновь окрашивали в отношении IFN-γ, IL-2 и TNF-α, а также в отношении CD154, но только через 24 ч и 48 ч после добавления PBMC и αCD3ΔADCC/αCD28ΔADCC (фиг.13). Вновь CD4+ T-клетки продемонстрировали увеличение количества IFN-γ+ T-клеток (3,4% после 24 ч, 6,8% после 48 ч) и CD154+ T-клеток (17,2% после 24 ч, 19,9% после 48 ч). После 48 ч было меньше IL-2+ T-клеток (4,9%) по сравнению с 24 ч (5,5%), количество TNF-α+ T-клеток также снижалось (14,9% после 24 ч, 8,1% после 48 ч) (фиг.13A). CD8+ T-клетки продемонстрировали только снижение количества TNF-α+ T-клеток (12,6% после 24 ч, 7,4% после 48 ч), в то время как это снижение также наблюдали в ELISA (фиг.10). Процент IFN-γ+, IL-2+ и CD154+ CD8+ T-клеток возрастал между 24 ч и 48 ч (IFNγ+: 4,7% после 24 ч, 8,5% после 48 ч, IL-2+: 5,1% после 24 ч, 7,2% после 48 ч, CD154+: 8,3% после 24 ч, 10,4% после 48 ч) (фиг.13B). Вновь, процент IFN-γ+ и IL-2+ CD8+ T-клеток превышал процент CD4+ T-клеток и количество TNF-α+ и CD154+ CD8+ T-клеток было сниженным по сравнению с CD4+ T-клетками. После 48 ч также некоторые T-клетки, по-видимому, активировались растворимым HBsAg, поскольку TNFα+ T-клетки достигали 1,1% (CD4+ T-клетки) и 1,0% (CD8+ T-клетки), CD154+ T-клетки достигали 2,7% (CD4+ T-клетки) и 3,1% (CD8+ T-клетки), IFNγ+ CD8+ T-клетки достигали 1,2% и IL-2+ CD8+ T-клетки достигали 1,4%. Вновь, для дальнейшего анализа T-клеток, секретирующих цитокины использовали булевы окна (фиг.13C, D). 0,35% (после 24 ч) и 0,63% (после 48 ч) CD4+ T-клеток, 0,3% (после 24 ч) и 1,0% (после 48 ч) CD8+ T-клеток были IFN-γ+, IL-2+ и TNF-α+, что указывает на полифункциональность T-клеток. αCD3ADCC/αCD28ΔADCC опосредует активацию PBMC в процессе сокультивирования с иммобилизованными HBsAg-клетками, что приводит к полифункциональным CD4+ и CD8+ T-клеткам. Активация вследствие растворимого HBsAg остается низкой.
Биспецифические антитела опосредуют секрецию IFNγ и уничтожение инфицированных HBV клеток HepaRG
Наконец, представляло интерес то, способны ли биспецифические антитела перенацеливать T-клетки на инфицированные HBV клетки HepaRG. Успех инфицирования исследовали путем измерения уровня HBsAg в супернатанте инфицированных клеток. По сравнению с результатом для клеток HuH7-S или HepG2.2.15, концентрация HBsAg, продуцируемого инфицированными HBV клетками HepaRG, была очень низкой. Кроме того, величины в различных лунках существенно варьировались, на что указывало относительно высокое стандартное отклонение (фиг.14).
Тем не менее, инфицирование было успешным, и проводили сокультивирование PBMC с клетками HepaRG в присутствии биспецифических антител. Как можно видеть на фиг.15, жизнеспособность необработанных клеток снижалась с течением времени до остаточной жизнеспособности 65,9% (HBV+) и 62,9% (HBV-) после 56 ч. По сравнению с этим αCD3 и комбинация αCD3/αCD28 опосредовали специфический лизис инфицированных HBV клеток HepaRG. αCD28 отдельно не могло индуцировать специфическую элиминацию клеток-мишеней. Если αCD3 присутствовало в процессе сокультивирования, жизнеспособность инфицированных HBV клеток снижалась до 25,3%, в то время как неинфицированные клетки HepaRG оставались на уровне 53,5% (фиг.15A). Стимуляция эффекторных клеток посредством αCD3/αCD28 также привела к значительному уничтожению инфицированных HBV клеток HepaRG (фиг.15B), причем 37,5% клеток-мишеней оставались жизнеспособными (не инфицированные клетки HepaRG: 62,4%).
Таким образом, αCD3 или αCD3/αCD28 индуцируют специфический лизис инфицированных HBV клеток HepaRG.
Биспецифические антитела опосредуют уменьшение положительных по HBV опухолей in vivo
Иммунодефицитным мышам, которым была инъецирована трансгенная по HBV клеточная линия гепатомы человека HepG2.2.15 для развития подкожных HBV-положительных опухолей, инъецировали PBMC человека и биспецифические конструкции, направленные против CD3 и CD28 (фиг.16). Это лечение приводило к значительному снижению размера опухолей по сравнению с животными, которым не проводили введения, или с животными, которым проводили имитирующее введение (животные, которым вводили PBMC и PBS). У животных, которым проводили лечение, размер опухоли снижался приблизительно на пятьдесят процентов.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ ГЕПАТИТА В И СВЯЗАННЫХ СОСТОЯНИЙ | 2016 |
|
RU2733496C2 |
| ПОЛИВАЛЕНТНАЯ АНТИГЕН-СВЯЗЫВАЮЩАЯ FV-МОЛЕКУЛА | 2011 |
|
RU2613368C2 |
| НОВОЕ PSMA-СВЯЗЫВАЮЩЕЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2762704C2 |
| АНТИГЕННЫЙ РЕЦЕПТОР | 2020 |
|
RU2832189C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО РЕЦЕПТОРА АНТИГЕНА ПРОТИВ CD19 | 2015 |
|
RU2718542C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С УСТОЙЧИВОСТЬЮ К РАСТВОРИМОМУ СЕА | 2006 |
|
RU2426743C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2535992C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2708311C2 |




























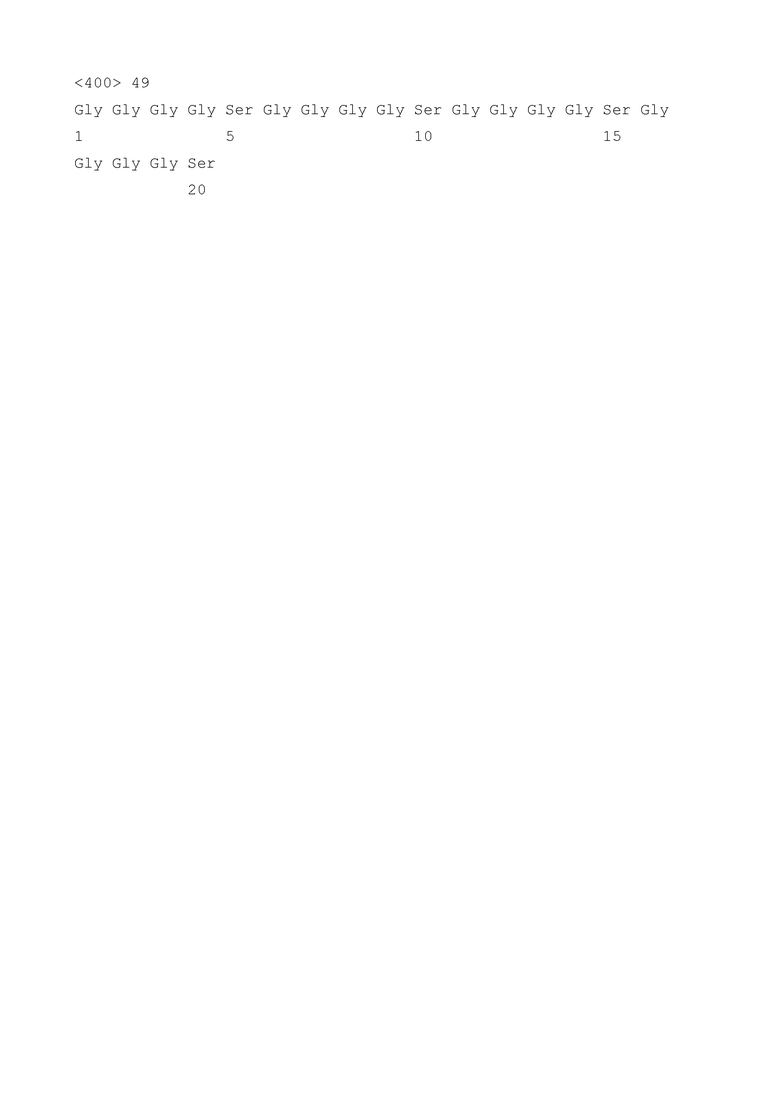
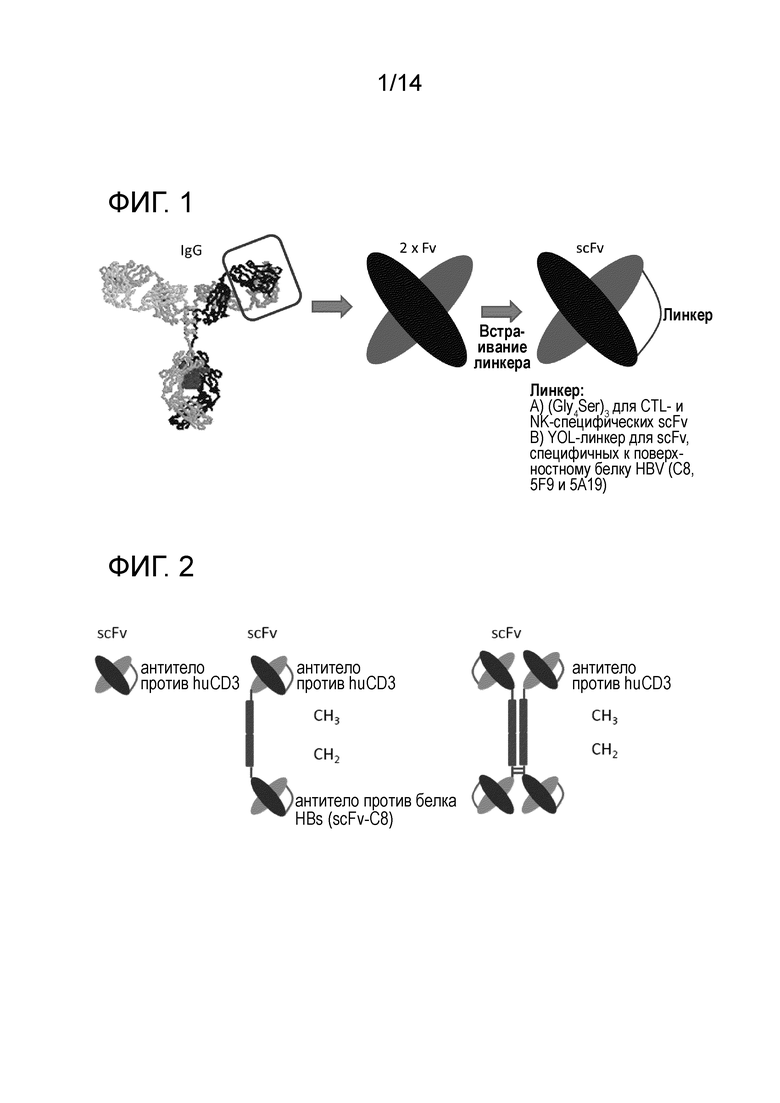
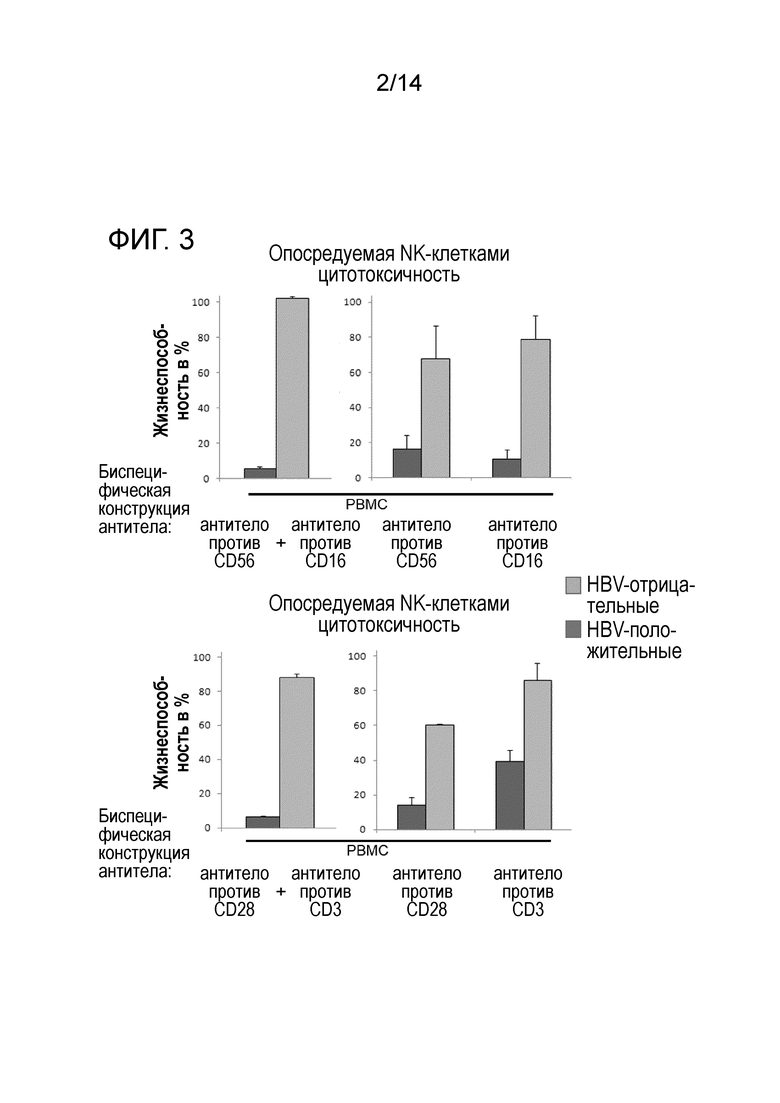
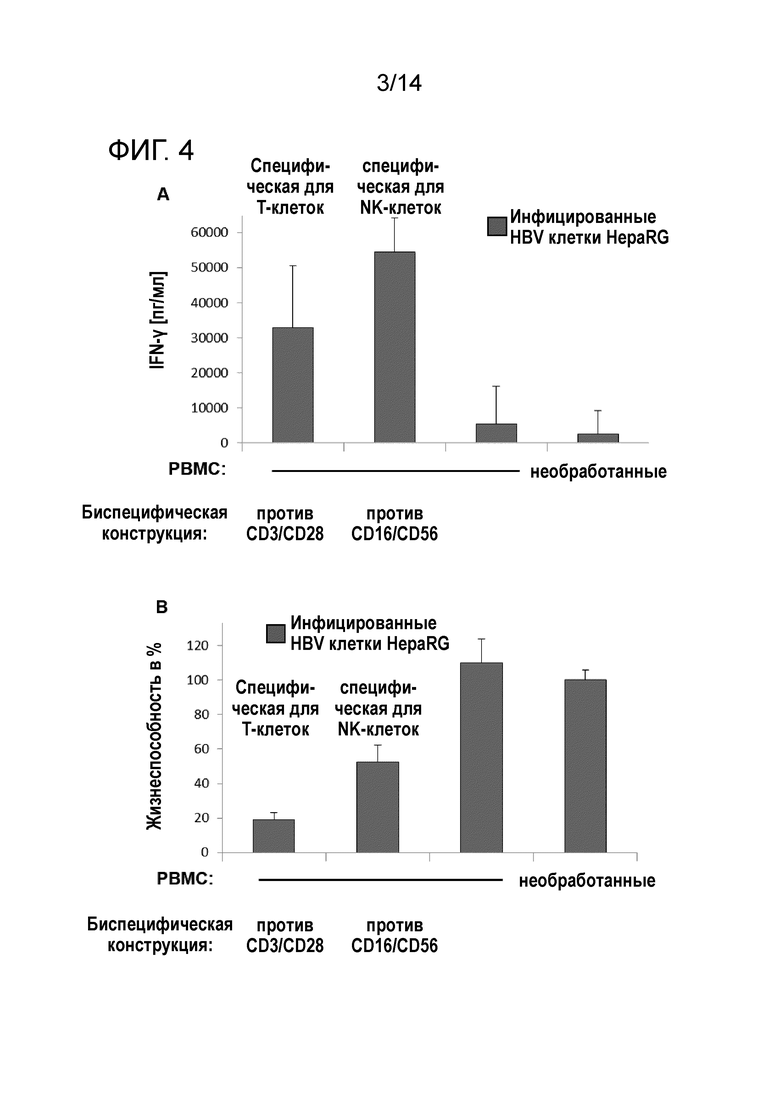
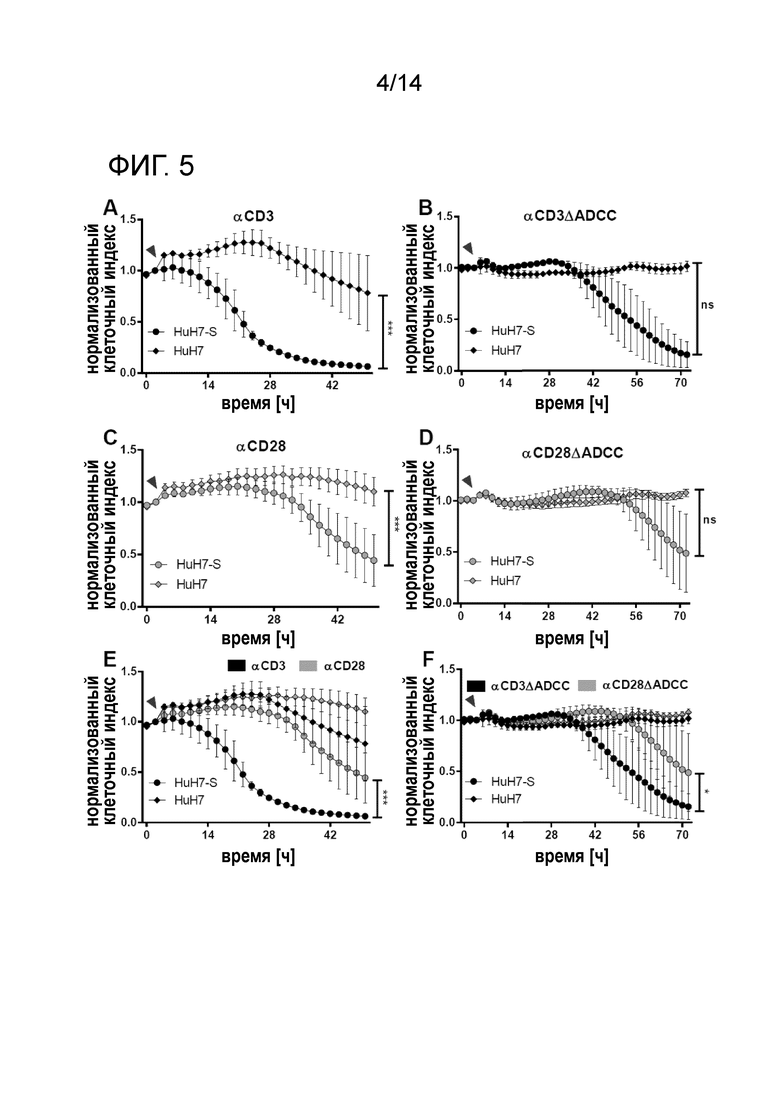
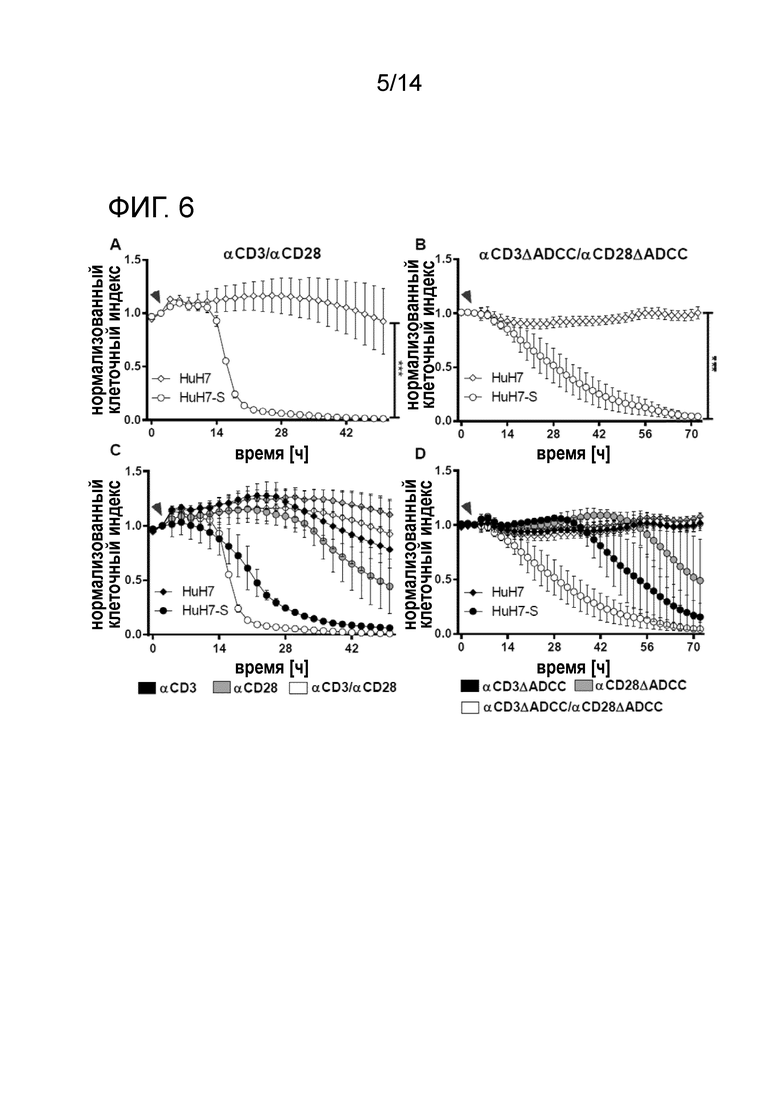
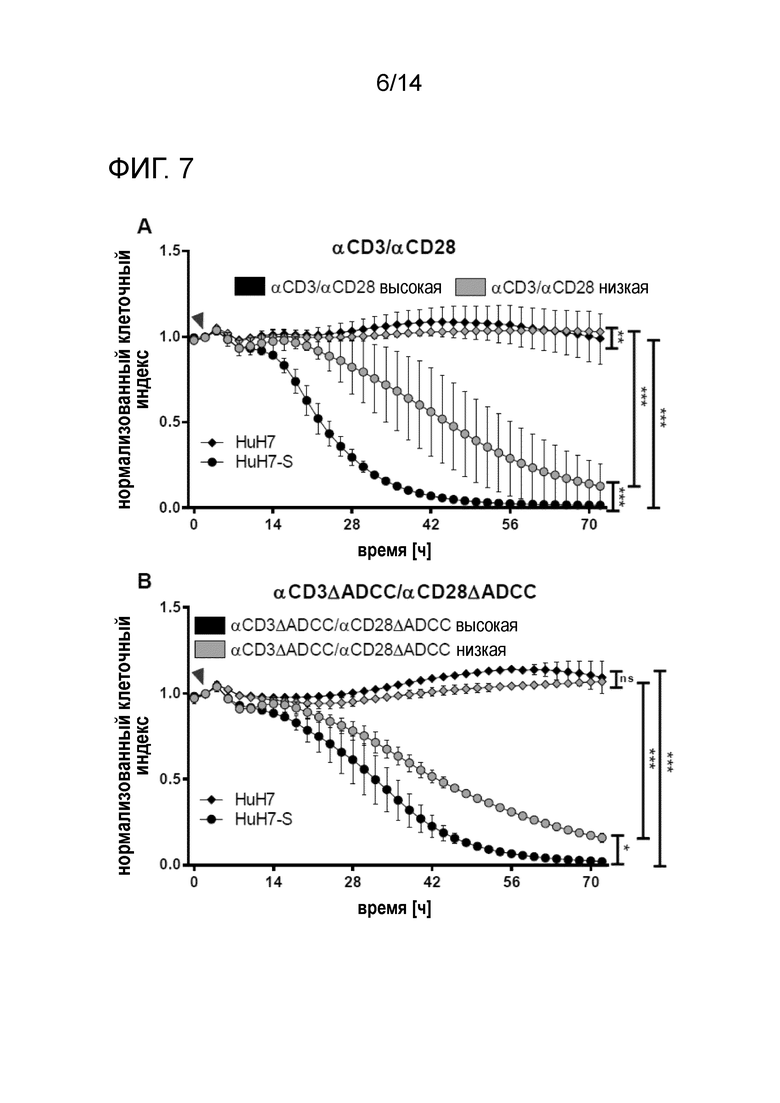
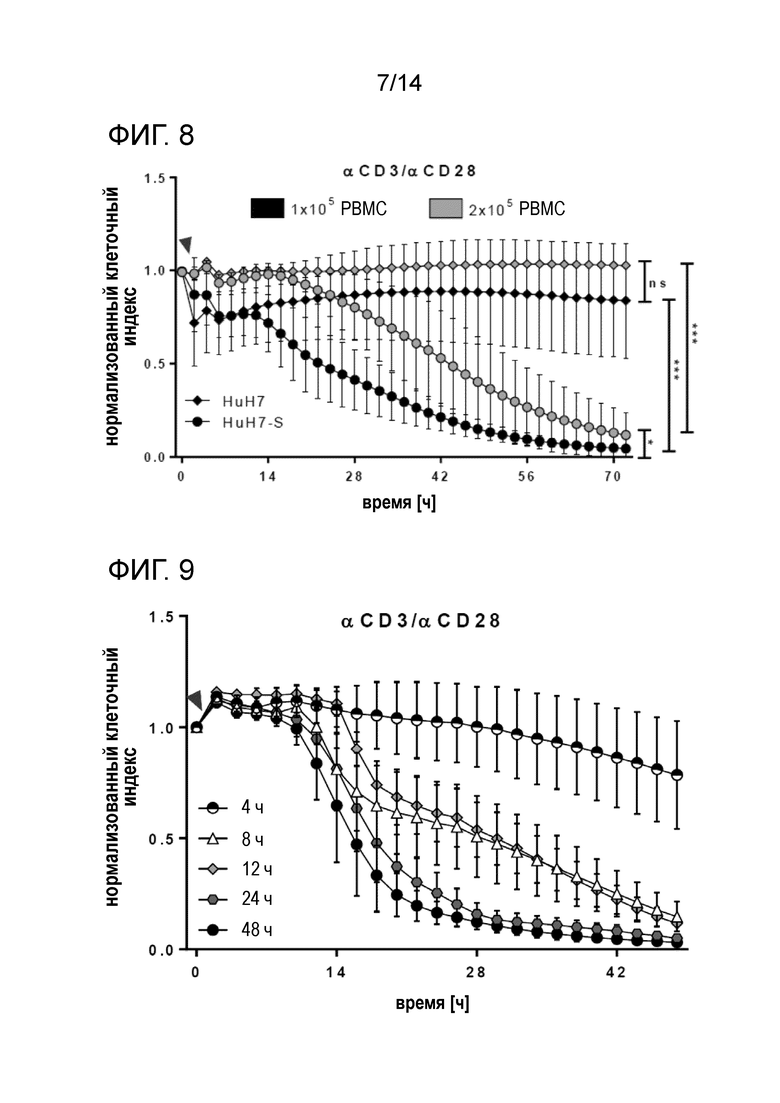
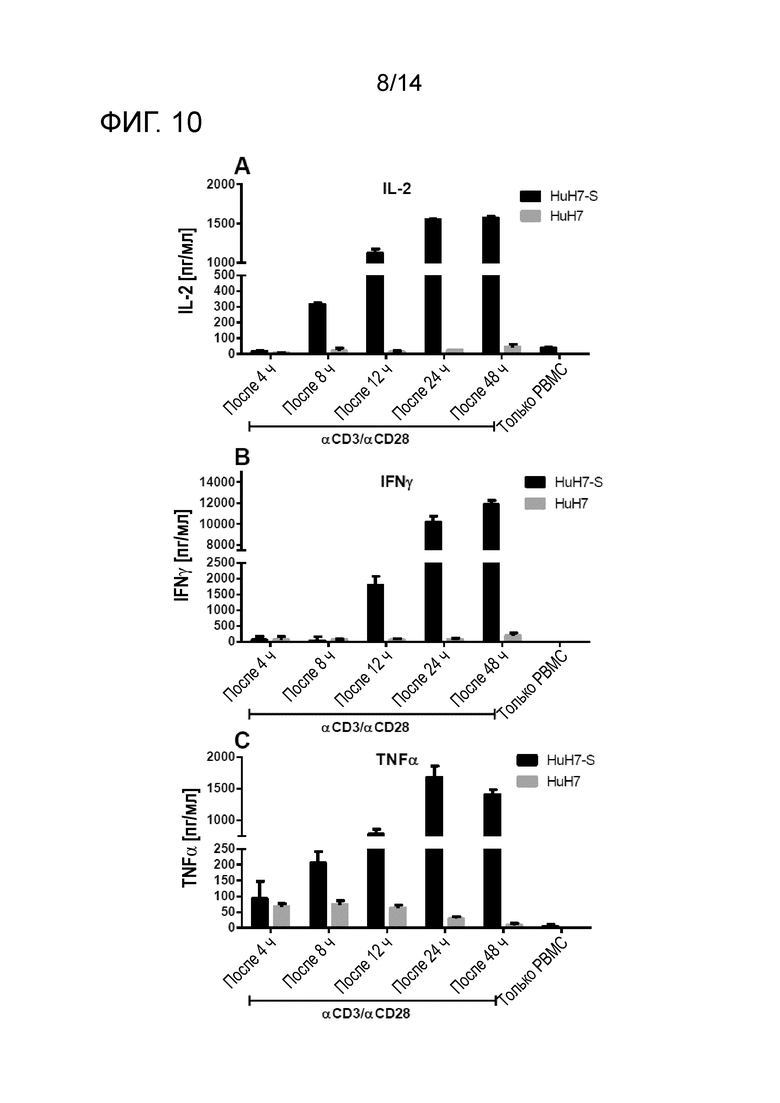
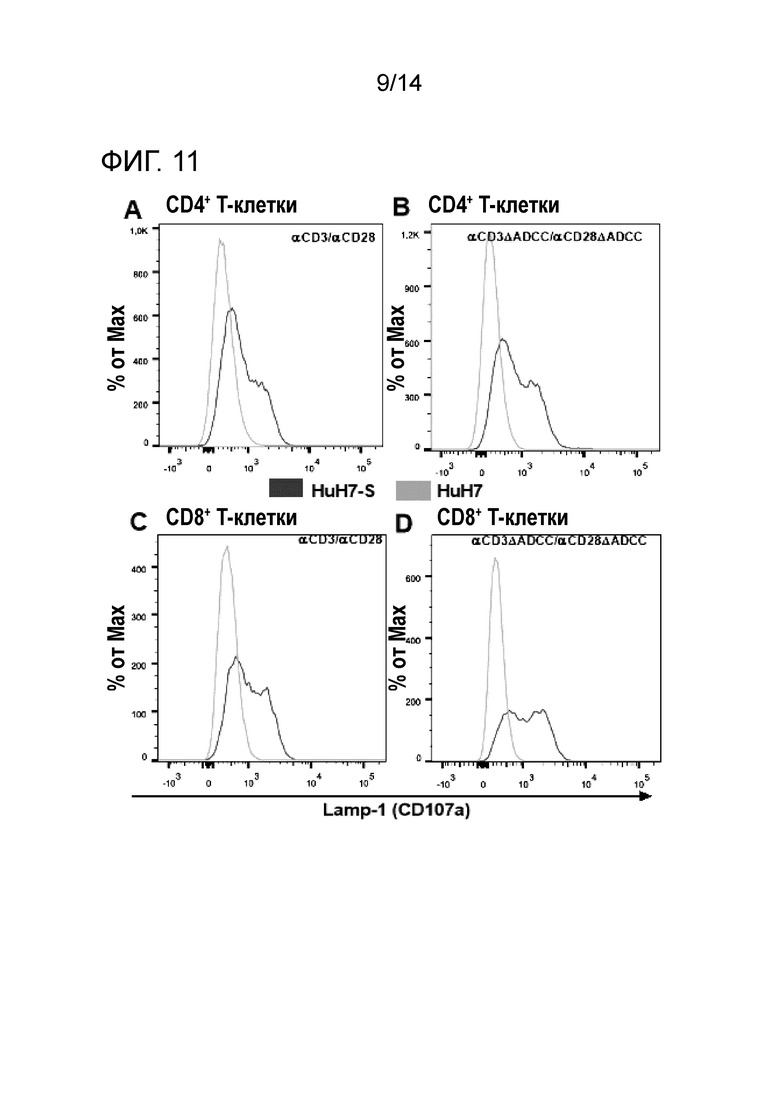
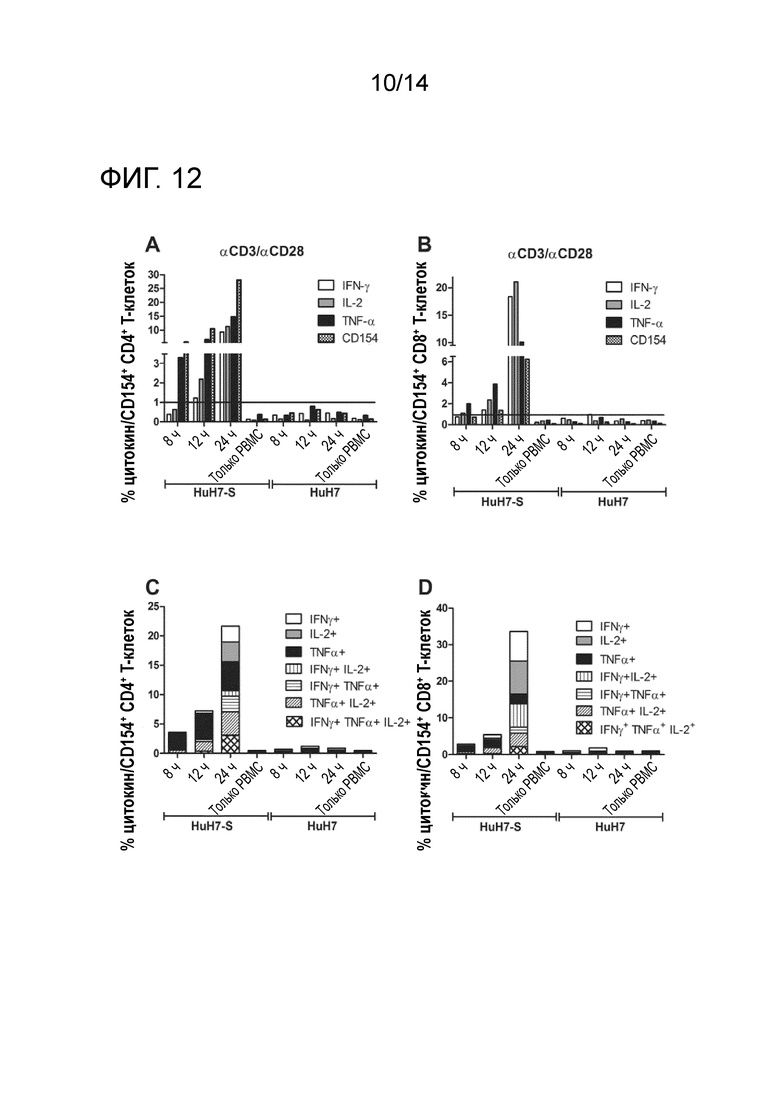
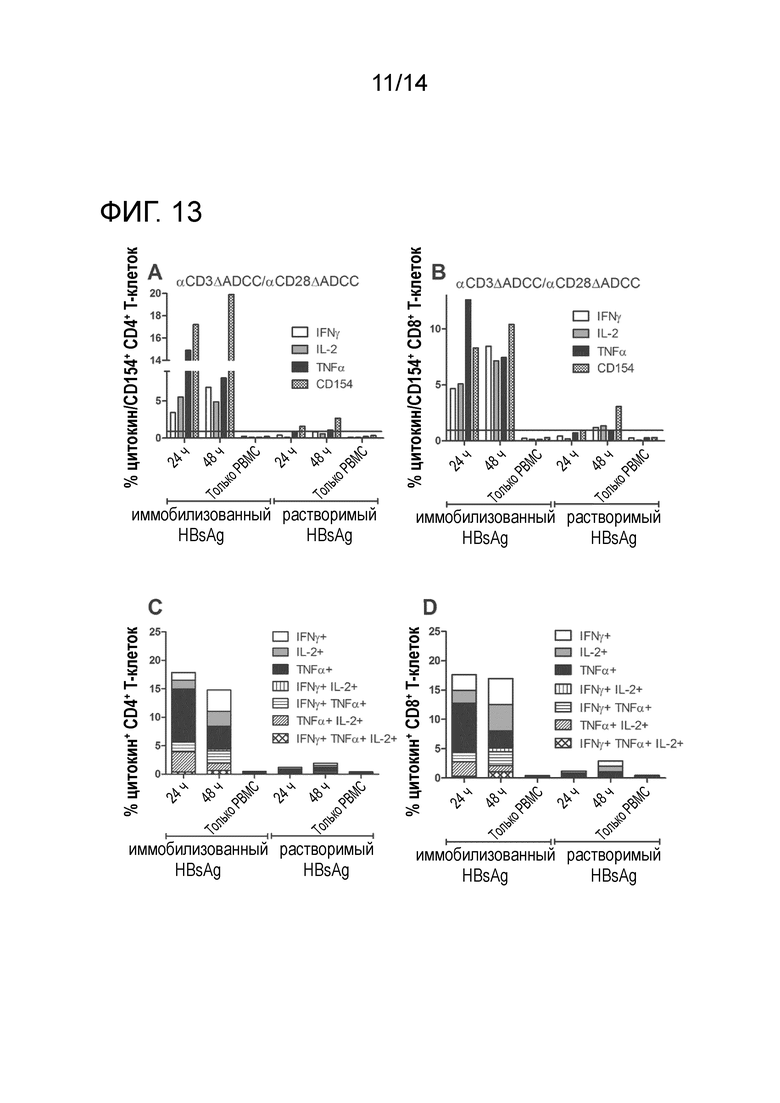
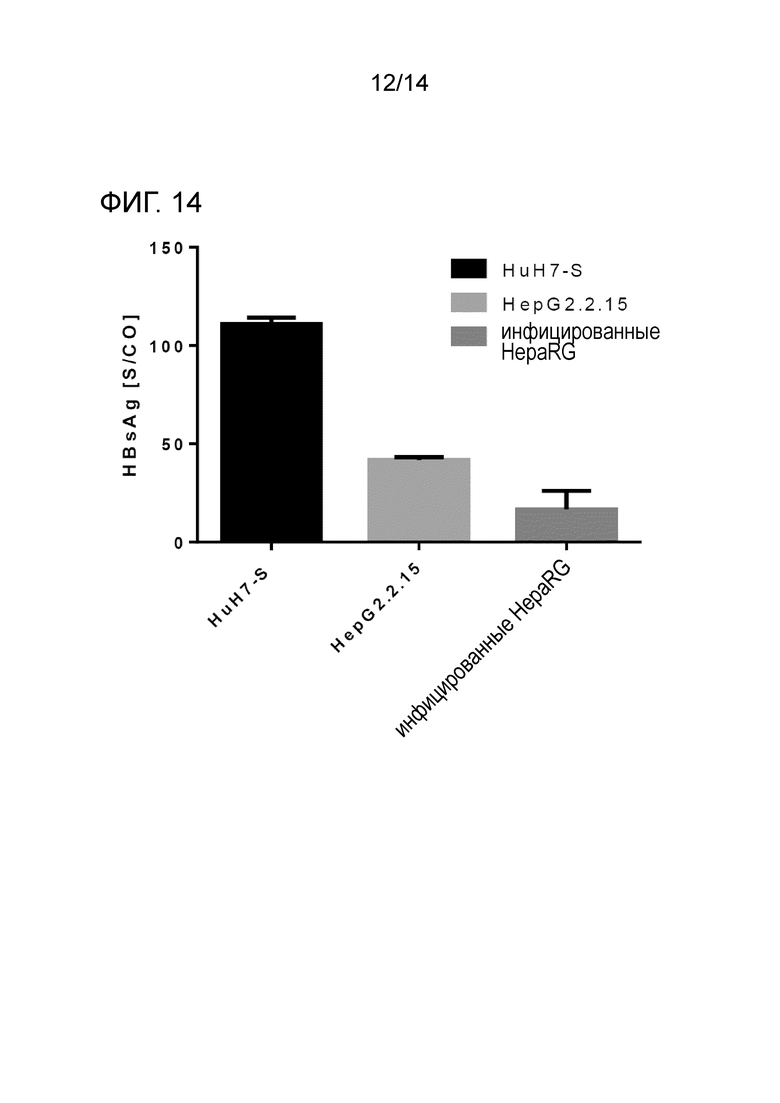
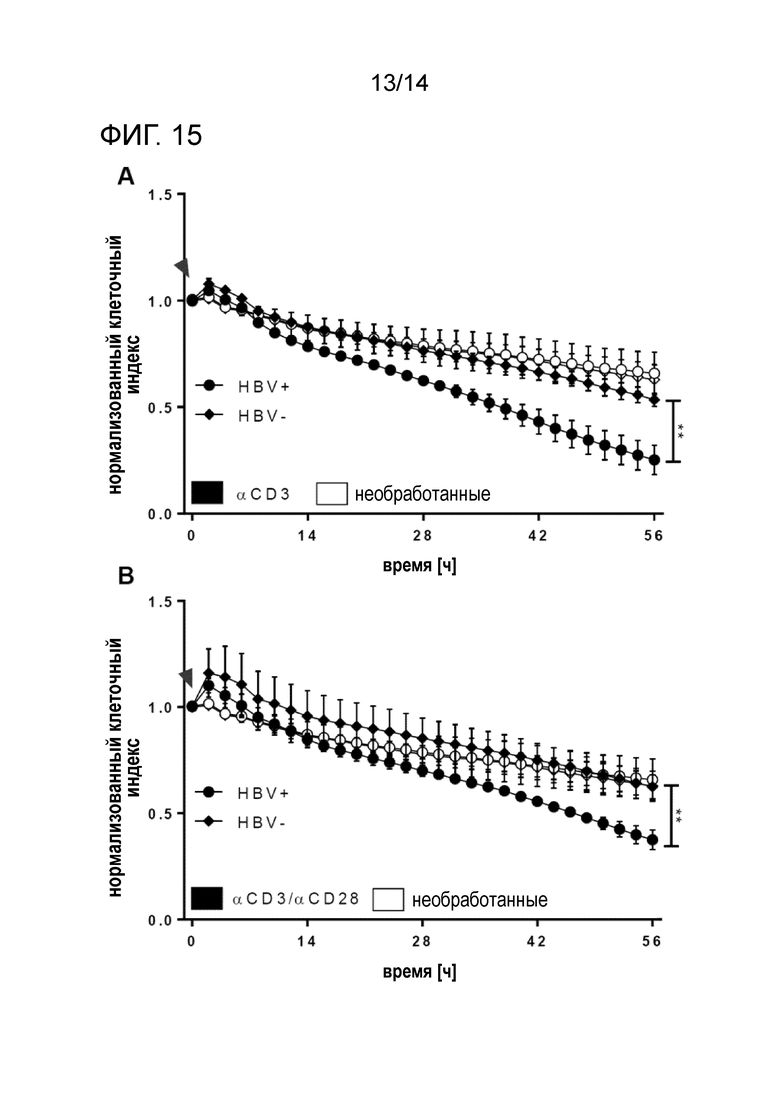
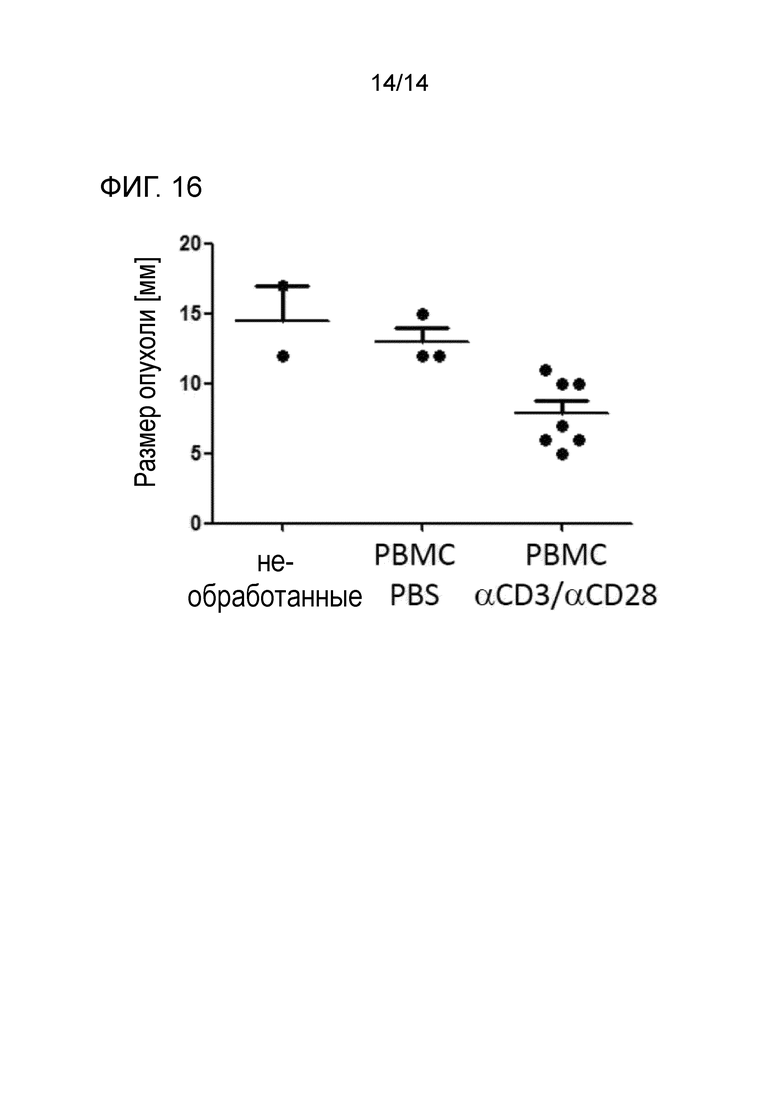
Настоящее изобретение относится к иммунологии. Предложен полипептид, способный связываться с поверхностным антигеном вируса гепатита B (HBV) и поверхностным антигеном, презентируемым иммунными эффекторными клетками. Также рассмотрена нуклеиновая кислота, кодирующая указанный полипептид; комплекс, содержащий два полипептида; иммунная эффекторная клетка и композиции для лечения и предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, в частности цирроза печени, печеночно-клеточной карциномы и рака печени. Полипептид по настоящему изобретению обеспечивает специфический лизис инфицированных HBV клеток и может найти дальнейшее применение в лечении заболеваний, вызванных инфекцией HBV. 7 н. и 9 з.п. ф-лы, 16 ил., 4 пр.
1. Полипептид, способный связываться с первым и вторым антигенами, содержащий
(a) первый набор из шести определяющих комплементарность областей (CDR), связывающих первый антиген; и
(b) (ba) второй набор из шести CDR, связывающих второй антиген; или
(bb) лиганд, способный связываться со вторым антигеном;
где
(i) указанный первый антиген выбран из малого поверхностного антигена вируса гепатита B (HBV); среднего поверхностного антигена HBV и большого поверхностного антигена HBV; и
(ii) указанный второй антиген выбран из поверхностных антигенов, презентируемых иммунными эффекторными клетками, такими как натуральные киллеры (NK) и цитотоксические T-лимфоциты (CTL), где
указанный первый набор из шести CDR имеет последовательности SEQ ID NO: 1-6, 7-12 или 13-18; и
указанный второй набор из шести CDR имеет последовательности SEQ ID NO: 19-24, 25-30, 31-36 или 37-42,
где в каждом наборе из шести CDR порядок CDR является следующим: CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи,
где указанные CDR являются частью иммуноглобулиновых доменов, и где
указанный лиганд представляет собой иммунолиганд, предпочтительно способный связываться с NKG2D, NKp30/NCR3, 4-1 BB или OX40.
2. Полипептид по п.1, где
(a) указанный первый набор из шести CDR содержится в первом scFv-фрагменте; и/или
(b) (ba) указанный второй набор из шести CDR содержится во втором scFv-фрагменте.
3. Полипептид по п.1 или 2, где указанный первый набор из шести CDR связывает эпитоп указанного первого антигена, причем эпитоп расположен
(a) в указанном малом поверхностном антигене HBV;
(b) в части указанного большого поверхностного антигена HBV, которая не содержится в указанном малом поверхностном антигене HBV; или
(c) в части указанного большого поверхностного антигена HBV, структура которой отличается от указанного малого поверхностного антигена HBV.
4. Полипептид по п.1 или 2, где указанный поверхностный антиген, презентируемый иммунными эффекторными клетками, выбран из CD3, CD28, 4-1 BB, OX40, CD16, CD56, NKG2D и NKp30/NCR3.
5. Полипептид по п. 1 или 2, где указанный полипептид дополнительно содержит область димеризации, где указанная область димеризации обеспечивает ковалентную и/или нековалентную димеризацию.
6. Полипептид по п.1 или 2, где указанный полипептид дополнительно содержит спейсерную область, причем указанная спейсерная область предпочтительно содержит CH2-домен и CH3-домен, при этом указанная спейсерная область расположена между
(i) указанным первым scFv-фрагментом и
(ii) указанным вторым scFv-фрагментом или указанным рекомбинантным лигандом в аминокислотной последовательности указанного полипептида,
и в указанный CH2-домен и/или указанный CH3-домен предпочтительно внесены мутации в одном или нескольких положениях для снижения или устранения связывания Fc-рецепторов.
7. Полипептид по п. 1 или 2, где указанный полипептид содержит или состоит из аминокислотной последовательности по любому из SEQ ID NO: 43-46 или аминокислотной последовательности, которая проявляет по меньшей мере 80% идентичность с любой из SEQ ID NO: 43-46 при условии, что CDR указанной аминокислотной последовательности, проявляющей по меньшей мере 80% идентичность, идентичны SEQ ID NO: 1-6, 7-12, 13-18, 19-24, 25-30, 31-36 или 37-42, соответственно.
8. Нуклеиновая кислота, кодирующая полипептид по любому из предшествующих пунктов.
9. Комплекс для лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызванное указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или нескольких поверхностных антигенов HBV, содержащий или состоящий из двух полипептидов, причем указанные полипептиды определены в любом из пп.1-7, где
(a) между указанными полипептидами существует по меньшей мере ковалентная связь, предпочтительно по меньшей мере один дисульфидный мостик между остатками Cys указанных полипептидов; или
(b) указанные полипептиды связаны друг с другом нековалентно.
10. Комплекс по п.9, где указанный комплекс содержит или состоит из четырехвалентного антитела, которое является биспецифическим, триспецифическим или тетраспецифическим.
11. Композиция для лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызванное указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или более поверхностных антигенов HBV, содержащая в эффективном количестве один или более полипептидов по любому из пп.1-7 и/или один или более комплексов по п.9 при условии, что по меньшей мере два полипептида содержатся в указанной композиции, причем эти два полипептида отличаются друг от друга в отношении первого антигена и/или второго антигена, с которым они связываются.
12. Композиция по п.11, где указанные два полипептида представляют собой
(a) (i) полипептид, связывающийся с малым или большим поверхностным антигеном HBV и CD3; и
(ii) полипептид, связывающийся с малым или большим поверхностным антигеном HBV и CD28;
или
(b) (i) полипептид, связывающийся с малым или большим поверхностным антигеном HBV и CD16; и
(ii) полипептид, связывающийся с малым или большим поверхностным антигеном HBV и CD56.
13. Композиция по п.11 или 12, где указанная композиция содержит четырехвалентное антитело, которое является биспецифическим, триспецифическим или тетраспецифическим.
14. Фармацевтическая композиция для лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызванное указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или более поверхностных антигенов HBV, содержащая в эффективном количестве один или более полипептидов по любому из пп.1-7, один или более комплексов по п.9 или 10 и/или один или более композиций по любому из пп.11-13.
15. Применение одного или нескольких полипептидов по любому из пп.1-7, одного или нескольких комплексов по п.9 или 10 и/или одной или нескольких композиций по любому из пп.11-13 для лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызванное указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или нескольких поверхностных антигенов HBV.
16. Иммунная эффекторная клетка in vitro или ex vivo для лечения или предупреждения инфекции HBV и/или состояния, вызываемого указанной инфекцией HBV, причем указанное состояние, вызванное указанной инфекцией HBV, выбрано из цирроза печени, печеночно-клеточной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или более поверхностных антигенов HBV, которая имеет полипептид по любому из пп.1-7, или комплекс по п.9 или 10, связанные с поверхностным антигеном указанной иммунной эффекторной клетки.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| СЫРЬЕВАЯ СМЕСЬ ДЛЯ ИЗГОТОВЛЕНИЯ БЕТОНА | 2013 |
|
RU2524699C1 |
| BOHNE F | |||
| et al., "T Cells Redirected Against Hepatitis B Virus Surface Proteins Eliminate Infected Hepatocytes", Gastroenterology, 2008, 134: 239-247 | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| PIZARRO J.C | |||
| et al., "Structural and functional characterization of a monoclonal antibody specific for the preS1 region of hepatitis B virus", FEBS Lett., 2001, 509(3):463-8 | |||
| Вертикальный двукратный двигатель внутреннего горения | 1928 |
|
SU13564A1 |

