ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к антителу против PD-L1. Более конкретно, настоящее изобретение относится к антителу против PD-L1, содержащему вариабельную область, содержащую определяющие комплементарность области (CDR) крысиного антитела против бычьего PD-L1, и константную область антитела животного, отличного от крысы.
УРОВЕНЬ ТЕХНИКИ
Белок программируемой гибели клеток 1 (PD-1), иммуноингибирующий рецептор и лиганд белка программируемой гибели клеток 1 (PD-L1) представляют собой молекулы, идентифицированные профессором Tasuku Honjo et al., Kyoto University, как факторы, которые ингибируют избыточный иммунный ответ и глубоко связанные с иммунотолерантностью (непатентный документ № 1Ishida Y, Agata Y, Shibahara K, Honjo T The EMB 1992 Nov; 11(11):3887-3895). Недавно было выяснено, что эти молекулы также участвуют в иммуносупрессии опухолей. В области здравоохранения человека был разработан и внедрен в практику препарат антител, который ингибирует эффект PD-1 (Opdivo™, Ono Pharmaceutical Co., Ltd.)
На сегодняшний день авторы настоящего изобретения разработали иммунотерапию для лечения резистентных болезней животных, нацеленную на PD-1 или PD-L1, и было обнаружено, что эта новая иммунотерапия может использоваться в отношении многочисленных заболеваний и у множества животных. (Непатентный документ № 2: Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology. 2014 Aug;142(4):551-61; непатентный документ № 3: Maekawa N, Konnai S, Ikebuchi R, Okagawa T, Adachi M, Takagi S, Kagawa Y, Nakajima C, Suzuki Y, Murata S, Ohashi K. PLoS One. 2014 Jun 10; 9(6):e98415; непатентный документ № 4: Mingala CN, Konnai S, Ikebuchi R, Ohashi K. Comp. Immunol. Microbiol. Infect. Dis. 2011 Jan; 34(1):55-63.)
Однако антитела, которые были получены авторам настоящего изобретения до настоящего момента, являются антителами крысы, и поэтому их невозможно многократно вводить животным, отличным от крысы.
ЛИТЕРАТУРА УРОВНЯ ТЕХНИКИ
Непатентные документы
Непатентный документ № 1: Ishida Y, Agata Y, Shibahara K, Honjo T The EMB 1992 Nov; 11(11):3887-3895.
Непатентный документ № 2: Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology. 2014 Aug; 142(4):551-61.
Непатентный документ № 3: Maekawa N, Konnai S, Ikebuchi R, Okagawa T, Adachi M, Takagi S, Kagawa Y, Nakajima C, Suzuki Y, Murata S, Ohashi K. PLoS One. 2014 Jun 10; 9(6):e98415.
Непатентный документ № 4: Mingala CN, Konnai S, Ikebuchi R, Ohashi K. Comp. Immunol. Microbiol. Infect. Dis. 2011 Jan; 34(1):55-63.
ЗАДАЧА, РЕШАЕМАЯ ИЗОБРЕТЕНИЕМ
Задачей настоящего изобретения является создание антитела против PD-L1, способного к многократному введению животным, отличным от крысы.
СПОСОБЫ РЕШЕНИЯ ПРОБЛЕМЫ
Авторы настоящего изобретения определили вариабельные области крысиного моноклонального антитела (4G12) против бычьего PD-L1, способного ингибировать связывание собачьего PD-1 и PD-L1, и затем объединили гены, кодирующие полученные вариабельные области, с генами, кодирующими константные области собачьего иммуноглобулина (IgG-D, эквивалентен IgG4 человека), тем самым получив ген химерного антитела, который был введен в клетки яичника китайского хомячка (клетки СНО). Посредством культивирования/пролиферации полученных клеток СНО авторам настоящего изобретения удалось получить химерное "крысино-собачье" антитело против PD-L1. Кроме того, авторы настоящего изобретения определили CDR вариабельной области крысиного моноклонального антитела 4G12 против бычьего PD-L1.
Кроме того, авторы настоящего изобретения определили вариабельные области крысиного моноклонального антитела 4G12 против бычьего PD-L1, способного ингибировать связывание бычьего PD-1 и PD-L1, и затем объединили гены, кодирующие полученные вариабельные области, с генами, кодирующими константные области бычьего иммуноглобулина (бычий IgG1 с мутациями, введенными в предполагаемые сайты связывания c рецепторам Fcγ в домене CH2 для ингибирования активности ADCC; см. фиг.19 для аминокислотных положений и мутаций: 250 E→ P, 251 L→V, 252 P→A, 253 G→делеция, 347 A→S, 348 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug; 142(4):551-561), таким образом, получив ген химерного антитела. Этот ген был введен в клетки яичника китайского хомяка (клетки СНО). Посредством культивирования/пролиферации полученных клеток авторам настоящего изобретения удалось получить химерное "крысино-бычье" антитело против PD-L1. Настоящее изобретение было сделано на основе этих обнаружений.
Сущность настоящего изобретения описана ниже.
(1) Антитело против PD-L1, содержащее (а) легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT, и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи антитела животного, отличного от крысы; и (b) тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40), и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы.
(2) Антитело по (1) выше, в котором вариабельная область легкой цепи и вариабельная область тяжелой цепи получены из крысы.
(3) Антитело по (2) выше, в котором вариабельная область легкой цепи представляет собой вариабельную область легкой цепи крысиного антитела против бычьего PD-L1, и вариабельная область тяжелой цепи представляет собой вариабельную область тяжелой цепи крысиного антитела против бычьего PD-L1.
(4) Антитело по (3) выше, в котором вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO:1, и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:2.
(5) Антитело по любому из (1)-(4) выше, в котором константная область легкой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области лямбда-цепи или каппа-цепи.
(6) Антитело по любому из (1)-(5) выше, в котором константная область тяжелой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека.
(7) Антитело по любому из (1)-(5) выше, причем животное, отличное от крысы, представляет собой особь крупного рогатого скота, и константная область тяжелой цепи бычьего антитела имеет введенные в него мутации, которые снижают активность ADCC и/или активность CDC.
(8) Антитело по (6) выше, причем животное, отличное от крысы, является собакой; константная область легкой цепи собачьего антитела имеет аминокислотную последовательность константной области лямбда-цепи; и константная область тяжелой цепи антитела собаки имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека.
(9) Антитело по (7) выше, в котором константная область легкой цепи бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи, а константная область тяжелой цепи бычьего антитела имеет введенные в нее мутации, которые уменьшают активность ADCC и/или активность CDC.
(10) Антитело по (8) выше, в котором константная область легкой цепи собачьего антитела имеет аминокислотную последовательность SEQ ID NO:3, и константная область тяжелой цепи собачьего антитела имеет аминокислотную последовательность SEQ ID NO:4.
(11) Антитело по (9) выше, в котором константная область легкой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:100, и константная область тяжелой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:102.
(12) Антитело по любому из (1)-(11) выше, которое имеет четырехцепочечную структуру, содержащую две легкие цепи и две тяжелые цепи.
(13) Фармацевтическая композиция, содержащая антитело по любому из (1)-(12) выше в качестве активного ингредиента.
(14) Композиция по (13) выше для профилактики и/или лечения злокачественных новообразований и/или воспаления.
(15) Композиция по (14) выше, причем злокачественное новообразование и/или воспаление выбраны из группы, состоящей из опухолевых заболеваний, лейкоза, болезни Джона, анаплазмоза, бактериального мастита, микотического мастита, микоплазменных инфекций (таких как микоплазменный мастит, микоплазменная пневмония или тому подобное), туберкулеза, инфекции Theileria orientalis, криптоспоридиоза, кокцидиоза, трипаносомоза и лейшманиоза.
(16) Искусственная генетическая ДНК, содержащая (а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи антитела животного, отличного от крысы; и (b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы.
(17) Вектор, содержащий искусственную генетическую ДНК по (16) выше.
(18) Клетка-хозяин, трансформированная вектором (17) выше.
(19) Способ получения антитела, включающий культивирование клетки-хозяина по (18) выше и сбор антитела против PD-L1 из полученной культуры.
(20) ДНК, кодирующая легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную области легкой цепи антитела животного, отличного от крысы.
(21) ДНК, кодирующая тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 c аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы.
Настоящее описание охватывает содержание описаний и/или чертежей Японских патентных заявок № 2016-159088, № 2016-159089, № 2017-110723 и № 2017-61454, на основании которых по настоящей патентной заявке испрашивается приоритет.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Согласно настоящему изобретению было получено новое антитело против PD-L1. Это антитело можно использовать даже в отношении животных, которые не являются крысами.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[Фиг.1] Ингибирование связывания рекомбинантного собачьего PD-L1 с рекомбинантным собачьим PD-1. Связывание собачьего PD-L1-Ig с собачьим PD-1-Ig определяли на планшетах для ELISA. Оптическая плотность (O.D.) без добавления антитела была принята за 100%. О.D. при каждой концентрации антитела была показана как относительная величина. Среди крысиных моноклональных антител 4G12 (крысиные IgG2a (κ)), 5A2 (крысиные IgG1 (κ)) и 6G7 (крысиные IgM (κ)) против бычьего PD-L1, которые показали перекрестную реакцию с собачьим PD-L1, клоны 4G12 и 6G7 показали высокую способность ингибировать связывание.
[Фиг.2] Схематическое изображение вектора pDC6 и химерного "крысино-собачьего" антитела против PD-L1.
[Фиг.3] Экспрессия и очистка химерных "крысино-собачьих" антител c4G12 и c6G7 против PD-L1. SDS-PAGE проводили в невосстанавливающих условиях, а затем визуализировали полосы окрашиванием CBB. а: очистка только белком А. b: а+гель-фильтрационная хроматография.
[Фиг.4] Активность ингибирования связывания PD-1/PD-L1 химерных "крысино-собачьих" антител c4G12 и c6G7 против PD-L1.
[Фиг.5] Создание клеточных клонов, способных к высокой экспрессии химерного "крысино-собачьего" антитела c4G12 против PD-L1.
[Фиг.6] Изображения SDS-PAGE химерного "крысино-собачьего" антитела c4G12 против PD-L1. Крысиные антитела 4G12 против бычьего PD-L1 и химерные "крысино-бычьи" антитела c4G12 против PD-L1 подвергали электрофорезу в восстанавливающих и невосстанавливающих условиях, а затем визуализировали полосы путем окрашивания CBB. В восстанавливающих условиях полоса тяжелой цепи антитела была обнаружена при приблизительно 50 кДа, а полоса легкой цепи антитела - при приблизительно 25 кДа. Никаких других полос, кроме интересующих, обнаружено не было.
[Фиг.7] Ингибирующая активность крысиного антитела 4G12 против бычьего PD-L1 и химерного "крысино-собачьего" антитела c4G12 в отношении связывания собачьего PD-L1/PD-L1 и CD80/PD-L1. Крысиное моноклональное антитело 4G12 против бычьего PD-L1 и химерное "крысино-собачье" антитело c4G12 против PD-L1 уменьшало связывание PD-L1-Ig с собачьим PD-1-Ig и CD80-Ig. Никаких изменений из-за химеризации антитела в активности ингибирования связывания не наблюдалось.
[Фиг.8] Эффект активации иммунных клеток собаки под действием химерного "крысино-собачьго" антитела c4G12 против PD-L1. РВМС собак культивировали при стимуляции в течение 3 дней, а затем определяли концентрацию IL-2 и IFN-γ в супернатанте с помощью ELISA. Дополнительно, аналог нуклеиновой кислоты EdU добавляли в культуральную среду на 2-ой день при стимуляции, а затем с помощью проточной цитометрии определяли захват EdU. Химерное "крысино-собачье" антитело c4G12 против PD-L1 повышало продукцию IL-2 и IFN-γ PBMC собак и усиливало пролиферацию CD4+ и CD8+ лимфоцитов.
[Фиг.9] Экспрессия PD-L1 при меланоме полости рта (A) и недифференцированной саркоме (B)
[Фиг. 10] Изображения СТ и визуализация опухоли в тесте с лечение путем введения собаке с меланомой полости рта химерного "крысино-собачьего" антитела c4G12 против PD-L1. (a,d) Перед началом лечения, (b,e) на 10-ой неделе лечения и (c, f) на 34-ой неделе лечения. Заметный противоопухолевый эффект отмечали после пяти введений антитела (на 10-ой неделе после начала лечения). На 34-ой неделе было подтверждено дальнейшее уменьшение опухоли.
[Фиг.11] Зависимые от времени изменения максимального диаметра опухоли у собаки с меланомой полости рта, показанной на фиг.10. Снижение на 30% или более по сравнению с исходным максимальным диаметром было расценено как частичный ответ (PR).
[Фиг. 12] Изображения СТ в тесте с лечением путем введения собаке с недифференцированной саркомой химерного "крысино-собачьего" антитела c4G12 против PD-L1. (a,с) Перед началом лечения, (b,d) на 3-ей неделе лечения. Заметное уменьшение опухоли отмечали после двух введений антитела.
[Фиг. 13] Изображения СТ в тесте с лечением путем введения собаке с меланомой полости рта (и метастазами в легких) химерного "крысино-собачьего" антитела c4G12 против PD-L1. (a,d,g) Перед началом лечения, (b,e,h) на 6-ой неделе лечения и (c,f,i) на 18-ой неделе лечения. Множественные метастатические поражения легких исчезали через девять введений антитела.
[Фиг.14] Зависимые от времени изменения в пропорциях выживаемости собак с меланомой полости рта после появления метастазов в легких. В группе с введением антитела продолжительность выживания была повышена по сравнению с контрольной группой.
[Фиг.15] Проиллюстрированы области CDR1, CDR2 и CDR3 вариабельной области легкой цепи и вариабельной области тяжелой цепи крысиного антитела 4G12 против бычьего PD-L1.
[Фиг.16] Перекрестная реактивность крысиного антитела 4G12 против бычьего PD-L1. Было подтверждено, что крысиное антитело 4G12 против бычьего PD-L1 связывается с овечьим PD-L1 и свиным PD-L1.
[Фиг.17] Реакционная способность крысиного антитела 4G12 против бычьего PD-L1 с лейкоцитами буйвола. Крысиное антитело 4G12 против бычьего PD-L1 прочно связывается с макрофагами крови (CD14+ CD11b+ клетки) буйвола, при этом крысиное антитело 4G12 против бычьего PD-L1 слабо связывается с лимфоцитами (CD14-CD11b- клетки) буйвола. Считается, что это различие в связывании отражает уровни экспрессии PD-L1 в макрофагах и лимфоцитах.
[Фиг.18] Тест на ингибирование крысиного антитела 4G12 против бычьего PD-L1 в отношении связывания овечьего или свиного PD-1/PD-L1. Было показано, что крысиное антитело 4G12 против бычьего PD-L1 способно ингибировать связывание овечьего и свиного PD-1/PD-L1 зависимым от концентрации образом.
[Фиг.19] Аминокислотная последовательность химерного "крысино-бычьего" антитела ch4G12 против PD-L1. Показаны области CDR1, CDR2 и CDR3 в вариабельной области легкой цепи и вариабельной области тяжелой цепи крысиного антитела 4G12 против бычьего PD-L1. Кроме того, также показаны аминокислоты, введенные как мутации, в бычий IgG1 (домен CH2) (аминокислотные положения и мутации: 250 E→P, 251 L→V, 252 P→A, 253 G→делеция, 347 A→S, 348 P→S).
[Фиг.20] Схематическое изображение вектора pDC6 и химерного "крысино-бычьего" антитела ch4G12 против PD-L1.
[Фиг.21] Подтверждение чистоты очищенного химерного "крысино-бычьего" антитела ch4G12 против PD-L1.
[Фиг.22] Специфичность связывания химерного "крысино-бычьего" антитела ch4G12 против PD-L1.
[Фиг.23] Ингибирующая активность химерного "крысино-бычьего" антитела ch4G12 против PD-L1 в отношении связывания бычьего PD-1/PD-L1 (результаты теста на ингибирование связывания клеток, экспрессирующих бычий PD-L1, и растворимого бычьего PD-1).
[Фиг.24] Ингибирующая активность химерного "крысино-бычьего" антитела ch4G12 против PD-L1 в отношении связывания бычьего PD-1/PD-L1 (результаты теста на ингибирование связывания клеток, экспрессирующих бычьи PD-1, и растворимого бычьего PD-L1).
[Фиг.25] Активирующий эффект химерного "крысино-бычьего" антитела ch-GG12 против PD-L1 на ответ бычьих лимфоцитов (в единицах пролиферации клеток).
[Фиг.26] Активирующий эффект химерного "крысино-бычьего" антитела ch-GG12 против PD-L1 на ответ бычьих лимфоцитов на антиген BLV (в единицах продукции IFN-γ).
[Фиг.27] Реакция пролиферации Т-клеток на антиген BLV у теленка, экспериментально инфицированного BLV, путем введения химерного "крысино-бычьего" антитела ch4G12 против бычьего PD-L1.
[Фиг.28] Изменения провирусной нагрузки BLV у теленка, экспериментально инфицированного BLV, при введении химерного "крысино-бычьего" антитела ch4G12 против бычьего PD-L1.
НАИЛУЧШИЕ СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже настоящее изобретение будет описано подробно.
Настоящее изобретение относится к антителу против PD-L1, содержащему (а) легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи антитела животного, отличного от крысы; и (b) тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы.
CDR1, CDR2 и CDR3 вариабельной области легкой цепи (VL) крысиного антитела 4G12 против бычьего PD-L1 представляют собой область, состоящую из аминокислотной последовательности QSLLYSENQKDY (SEQ ID NO:37), область, состоящую из аминокислотной последовательности WAT, и область, состоящую из аминокислотной последовательности GQYLVYPFT (SEQ ID NO:38), соответственно (см. фиг.15).
Кроме того, CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи (VH) крысиного антитела 4G12 против бычьего PD-L1 представляют собой область, состоящую из аминокислотной последовательности GYTFTSNF (SEQ ID NO:39), область, состоящую из аминокислотной последовательности IYPEYGNT (SEQ ID NO:40), и область, состоящую из аминокислотной последовательности ASEEAVISLVY (SEQ ID NO:41), соответственно (см. фиг.15).
В аминокислотных последовательностях QSLLYSENQKDY (SEQ ID NO:37), WAT и GQYLVYPFT (SEQ ID NO:38) и в аминокислотных последовательностях GYTFTSNF (SEQ ID NO:39), IYPEYGNT (SEQ ID NO:40) и ASEEAVISLVY (SEQ ID NO:41), одна, две, три, четыре или пять аминокислот могут быть удалены, замещены или добавлены.
Как используется в настоящем документе, термин «антитело» представляет собой концепцию, охватывающую не только полноразмерные антитела, но также и антитела меньшего молекулярного размера, такие как Fab, F(ab)'2, ScFv, диатело, VH, VL, Sc(Fv)2, биспецифичный sc(Fv)2, минитело, мономер scFv-Fc и димер scFv-Fc.
В антителе против PD-L1 по настоящему изобретению, VL и VH могут быть крысиными. Например, VL может быть VL крысиного антитела против бычьего PD-L1, а VH может быть VH крысиного антитела против бычьего PD-L1.
Аминокислотные последовательности VL и аминокислотная последовательность VH крысиного антитела против бычьего PD-L1 показаны как SEQ ID NO:1 и 2, соответственно. Аминокислотные последовательности SEQ ID NO:1 и 2 могут иметь делецию(и), замену(и) или добавление(я) в одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если были введены такие мутации, полученные аминокислотные последовательности способны выполнять функцию VL или VH антитела PD-L1.
VL и VH антитела животного, отличного от крысы, могут быть получены от животного, которое продуцирует PD-L1, который перекрестно реагирует с крысиным антителом 4G12 против бычьего PD-L1.
Существует два типа легкой цепи иммуноглобулина, которые называются каппа-цепью (κ) и лямбда-цепью (λ). В антителе против PD-L1 по настоящему изобретению константная область легкой цепи (CL) антитела животного, отличного от крысы, может иметь аминокислотную последовательность константной области либо каппа-цепи, либо лямбда-цепи. При этом, относительное содержание лямбда-цепи выше у овец, кошек, собак, лошадей и крупного рогатого скота, а каппа-цепи выше у мышей, крыс, людей и свиней. Поскольку цепь с более высоким относительным содержанием считается предпочтительной, то антитело овцы, кошки, собаки, лошади или быка предпочтительно имеет аминокислотную последовательность константной области лямбда-цепи, а антитело мыши, крысы, человека или свиньи предпочтительно имеет аминокислотную последовательность константной области каппа-цепи.
Константная область тяжелой цепи (СН) антитела животного, отличного от крысы, может иметь аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека. Тяжелая цепь иммуноглобулина классифицируется на γ-цепь, μ-цепь, α-цепь, δ-цепь и ε-цепь в зависимости от разницы в константной области. В зависимости от типа присутствующей тяжелой цепи образуются пять классов (изотипов) иммуноглобулина; ими являются IgG, IgM, IgA, IgD и IgE.
Иммуноглобулин G (IgG) составляет 70-75% иммуноглобулинов человека и является наиболее распространенным мономерным антителом в плазме. IgG имеет четырехцепочечную структуру, состоящую из двух легких цепей и двух тяжелых цепей. IgG1, IgG2 и IgG4 человека имеют молекулярную массу приблизительно 146000, тогда как IgG3 человека имеет длинную шарнирную область, которая соединяет Fab-область и Fc-область, и имеет большую молекулярную массу, равную 170000. IgG1 человека составляет около 65%, IgG2 человека - около 25%, IgG3 человека - около 7% и IgG4 человека - около 3% IgG человека. Они равномерно распределены внутри и снаружи кровеносных сосудов. Обладая сильным сродством к рецепторам Fc и факторам комплемента на поверхности эффекторных клеток, IgG1 человека индуцирует антителозависимую клеточную цитотоксичность (ADCC), а также активирует комплемент, индуцируя комплемент-зависимую клеточную цитотоксичность (CDC). IgG2 и IgG4 человека имеют низкую активность ADCC и CDC, поскольку их сродство к Fc-рецепторам и факторам комплемента низкое.
Иммуноглобулин М (IgM), который составляет приблизительно 10% иммуноглобулинов человека, представляет собой пентамерное антитело, состоящее из пяти основных четырехцепочечных структур, соединенных вместе. Он имеет молекулярный вес 970000. Обычно встречающийся только в крови, IgM продуцируется против инфекционных микроорганизмов и отвечает за иммунитет на ранней стадии.
Иммуноглобулин А (IgA) составляет 10-15% иммуноглобулинов человека. Он имеет молекулярный вес 160000. Секретируемый IgA представляет собой димерное антитело, состоящее из двух молекул IgA, соединенных вместе. IgA1 обнаружен в сыворотке, выделениях из носа, слюне и грудном молоке. В большом количестве IgA2 обнаружен в желудочном соке.
Иммуноглобулин D (IgD) представляет собой мономерное антитело, составляющее не более 1% иммуноглобулинов человека. IgD обнаружен на поверхности В-клеток и участвует в индукции продукции антител.
Иммуноглобулин E (IgE) представляет собой мономерное антитело, которое встречается в очень небольшом количестве, составляя лишь 0,001% или менее иммуноглобулинов человека. Считается, что иммуноглобулин Е вовлечен в иммунный ответ на паразитов, но в развитых странах, где паразиты встречаются редко, IgE, главным образом, вовлечен в бронхиальную астму и аллергию, среди прочего.
Что касается собак, то последовательности IgG-A (эквивалентен IgG2 человека), IgG-B (эквивалентен IgG1 человека), IgG-C (эквивалентен IgG3 человека) и IgG-D (эквивалентен IgG4 человека) были идентифицированы как тяжелая цепь IgG. В антителе по настоящему изобретению константная область тяжелой цепи IgG, не обладающая ни активностью ADCC, ни активностью CDC, является предпочтительной (IgG4 у человека). В случае, когда константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, можно использовать константную область, которая потеряла как активность ADCC, так и активность CDC в результате введения мутаций в соответствующую область иммуноглобулина, эквивалентного IgG4 человека.
Что касается крупного рогатого скота, то последовательности IgG1, IgG2 и IgG3 были идентифицированы как тяжелая цепь IgG. В антителе по настоящему изобретению константная область тяжелой цепи IgG, не обладающая ни активностью ADCC, ни активностью CDC, является предпочтительной (IgG4 у человека). Хотя константная область нативного IgG1 человека обладает активностью ADCC и активностью CDC, известно, что эти типы активности можно уменьшить путем введения аминокислотных замен или делеций в конкретные сайты. У крупного рогатого скота константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, поэтому в соответствующую область иммуноглобулина, эквивалентную IgG1 человека, можно ввести мутации, а затем использовать полученную константную область. В качестве примера, аминокислотная последовательность СН бычьего антитела (цепь IgG1, GenBank: X62916) с мутациями, введенными в домен СН2, и нуклеотидная последовательность для такой аминокислотной последовательности (после оптимизации кодонов) показаны как SEQ ID NO:102 и 102, соответственно.
Если животным, отличным от крысы, является собака, то антитело против PD-L1 является более предпочтительным, причем (i) CL собачьего антитела имеет аминокислотную последовательность константной области лямбда-цепи и (ii) CH собачьего антитела имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентную IgG4 человека.
Если животным, отличным от крысы, является представитель крупного рогатого скота, то антитело против PD-L1 является более предпочтительным, причем (i) CL бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи и (ii) CH бычьего антитело имеет мутации, которые снижают активность ADCC и/или активность CDC.
Антитело против PD-L1 по настоящему изобретению включает химерные "крысино-собачьи" антитела, канинизированные антитела, полные антитела собачьего типа, химерные "крысино-бычьи" антитела, бовинизированные антитела и полные антитела бычьего типа. Тем не менее, животные не ограничиваются собаками и быками и могут быть представлены людьми, свиньями, обезьянами, мышами, кошками, лошадьми, козами, овцами, буйволами, кроликами, хомячками, морскими свинками и тому подобное.
Например, антитело против PD-L1 по настоящему изобретению может представлять собой антитело против PD-L1, в котором CL собачьего антитела имеет аминокислотную последовательность SEQ ID NO:3, и СН собачьего антитела имеет аминокислотную последовательность SEQ ID NO:4.
В качестве дополнительного примера, антитело против PD-L1 по настоящему изобретению может представлять собой антитело против PD-L1, в котором CL бычьего антитела имеет аминокислотную последовательность SEQ ID NO:100, и CH бычьего антитела имеет аминокислотную последовательность SEQ ID NO:102.
Аминокислотные последовательности SEQ ID NO:3 и 4, а также SEQ ID NO:100 и 102, могут иметь делецию(и), замену(и) или добавление(я) в одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если были введены такие мутации, полученные аминокислотные последовательности способны выполнять функцию CL или CH антитела PD-L1.
Антитело против PD-L1 по настоящему изобретению может иметь четырехцепочечную структуру, содержащую две легкие цепи и две тяжелые цепи.
Антитело против PD-L1 по настоящему изобретению может быть получено, как описано ниже. Вкратце, синтезирован искусственный ген, который содержит (i) идентифицированные последовательности вариабельной области крысиного антитела против бычьего PD-L1 и (ii) последовательности константной области антитела животного, отличного от крысы (например, собаки или быка) (предпочтительно, антитело IgG4 человека; антитело, эквивалентное антителу IgG4 человека, или иммуноглобулин, эквивалентный IgG1 человека, в котором в соответствующую область введены мутации для снижения активности ADCC и/или активности CDC). Полученный ген встраивают в вектор (например, плазмиду), который затем вводят в клетку-хозяин (например, клетку млекопитающего, такую как клетка CHO). Клетку-хозяин культивирует, и из полученной культуры собирают интересующее антитело.
Аминокислотная последовательность и нуклеотидная последовательность VL крысиного антитела против бычьего PD-L1, идентифицированные авторами изобретения, показаны как SEQ ID NO:1 и 5, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO:15.
Аминокислотная последовательность VL и аминокислотная последовательность VH крысиного антитела против бычьего PD-L1, идентифицированные авторами изобретения, показаны как SEQ ID NO:2 и 6, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO:16.
Аминокислотная последовательность и нуклеотидная последовательность CL (лямбда-цепь, GenBank: E02824.1) собачьего антитела показаны как SEQ ID NO:3 и 7, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO:17.
Аминокислотная последовательность и нуклеотидная последовательность CL (лямбда-цепь, GenBank: X62917) бычьего антитела показаны как SEQ ID NO:100 и 101, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO:104.
Аминокислотная последовательность и нуклеотидная последовательность CH (цепь IgG-D, GenBank: AF354267.1) собачьего антитела показаны как SEQ ID NO:4 и 8, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO:18.
Аминокислотная последовательность и нуклеотидная последовательность (после оптимизации кодонов) CH (цепь IgG1, модифицированная из GenBank: X62916) бычьего антитела показаны как SEQ ID NO:102 и 103, соответственно.
Кроме того, SEQ ID NO:9 показывает аминокислотную последовательность химерной легкой цепи, состоящей из VL крысиного антитела против бычьего PD-L1 и CL (лямбда цепь, GenBank: E02824.1) собачьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной легкой цепи, состоящей из VL крысиного антитела против бычьего PD-L1 и CL (лямба цепь, GenBank: E02824.1) собачьего антитела, показана как SEQ ID NO:19.
Кроме того, SEQ ID NO:105 показывает аминокислотную последовательность химерной легкой цепи, состоящей из VL крысиного антитела против бычьего PD-L1 и CL (лямбда цепь, GenBank: X62917) бычьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной легкой цепи, состоящей из VL крысиного антитела против бычьего PD-L1 и CL (лямба цепь, GenBank: X62917) бычьего антитела, показана как SEQ ID NO:107.
SEQ ID NO:9 показывает аминокислотную последовательность химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего PD-L1 и CL (IgG-D цепь, GenBank: AF354267.1) собачьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего PD-L1 и CH (IgG-D цепь, GenBank: AF354267.1) собачьего антитела, показана как SEQ ID NO:20.
SEQ ID NO:106 показывает аминокислотную последовательность химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего PD-L1 и CL (IgG1 цепь, модифицированная из GenBank: X62916) бычьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего PD-L1 и CH (IgG1 цепь, модифицированная из GenBank: X62916) бычьего антитела, показана как SEQ ID NO:108.
Аминокислотные последовательности и нуклеотидные последовательности CL и CH различных животных, отличных от указанных выше, могут быть получены из известных баз данных для использования в настоящем изобретении.
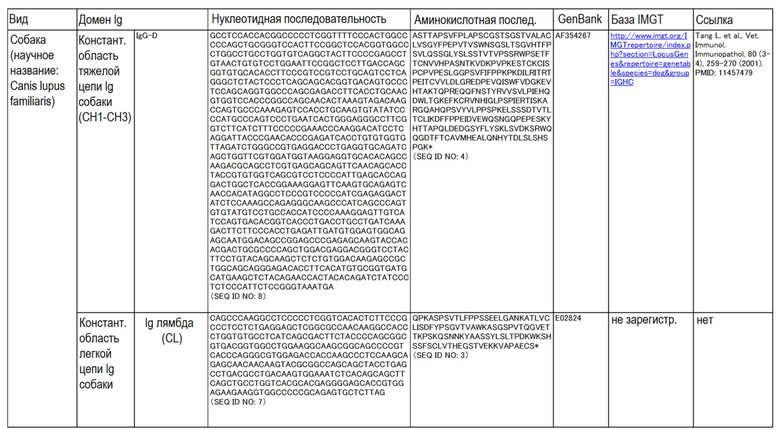
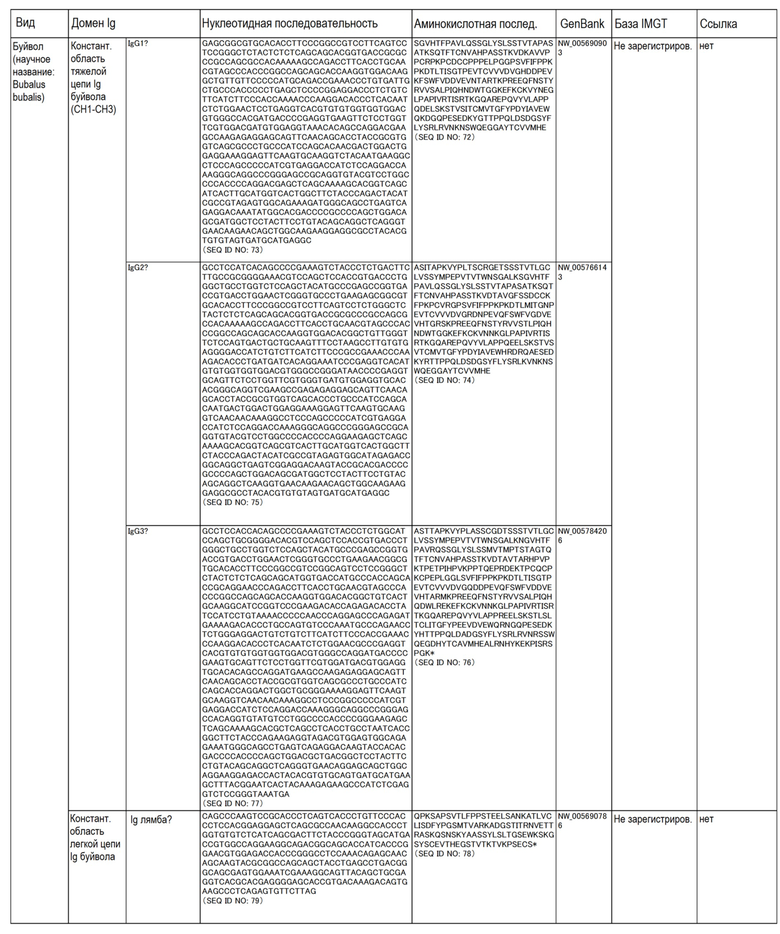
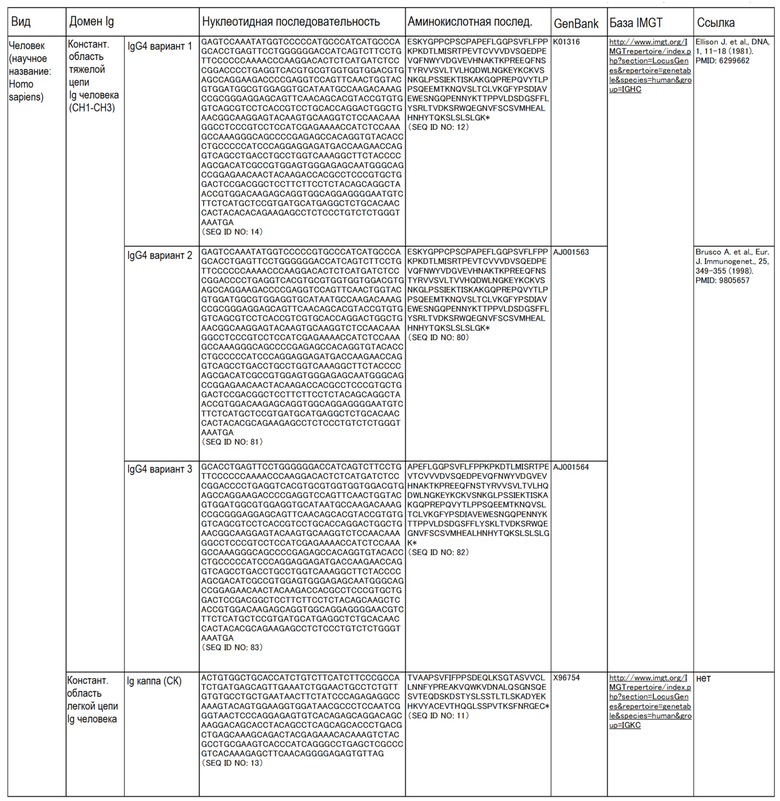
Аминокислотные последовательности и нуклеотидные последовательности CL и CH собак, овец, свиней, буйвола, человека и крупного рогатого скота приведены в таблице ниже.
Таблица.
(Таблица)





Аминокислотные последовательности SEQ ID NO:4, 3, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 78, 12, 80, 82, 84-91, 100, 102 и 11 могут иметь делецию(и), замену(и) или добавление(я) одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если такие мутации были введены, то полученные аминокислотные последовательности способны выполнять функцию константной области тяжелой цепи или легкой цепи Ig.
Хотя константная область нативного IgG1 человека обладает активностью ADCC и активностью CDC, известно, что эти типы активности можно уменьшить путем введения аминокислотных замен и делеций в конкретные сайты. В случае животных, отличных от человека, где константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, мутации могут быть введены в соответствующую область иммуноглобулина, эквивалентного IgG1 человека, так что может быть использована полученная константная область с пониженной активностью ADCC и активностью CDC.
Настоящее изобретение относится к искусственной генетической ДНК, содержащей (а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи (VL), содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT, и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи (CL) антитела животного, отличного от крысы; и (b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи (VH), содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40), и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи (CH) антитела животного, отличного от крысы. Настоящее изобретение также относится к ДНК, кодирующей легкую цепь, содержащую VL, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38) и CL антитела животного, отличного от крысы (то есть, ДНК (а') выше). Кроме того, настоящее изобретение также относится к ДНК, кодирующей тяжелую цепь, содержащую VH, содержащую CDR1 с GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41) и CH антитела животного, отличного от крысы (то есть, ДНК (b') выше).
Для (а) легкой цепи, содержащей вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи антитела животного, отличного от крысы; и для (b) тяжелой цепи, содержащей вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы, следует обратиться к предшествующему описанию. ДНК (а') представляет собой ДНК (ген), кодирующую легкую цепь (а); и ДНК (b') представляет собой ДНК (ген), кодирующую тяжелую цепь (b). Искусственная генетическая ДНК, содержащая ДНК (a') и ДНК (b'), может быть синтезирована на коммерческом синтезаторе. К искусственной генетической ДНК могут быть добавлены сайты распознавания ферментов рестрикции, последовательности KOZAK, последовательность сигнала присоединения поли-А, последовательности промоторов, последовательности интронов и тому подобное.
Настоящее изобретение также относится к вектору, содержащему вышеуказанную искусственную генетическую ДНК.
В качестве вектора можно использовать плазмиды на основе Escherichia coli (например, pBR322, pBR325, pUC12 или pUC13); плазмиды на основе Bacillus subtilis (например, pUB110, pTP5 или pC194), плазмиды на основе дрожжей (например, pSH19 или pSH15); бактериофаги, такие как фаг λ; вирусы животных, такие как ретровирус или вирус коровьей оспы; или вирусы патогенов насекомых, такие как бакуловирус. В описанных ниже примерах использовали pDC6 (патент Японии № 5704753, патент США 9096878, патент ЕС 2385115, патент Гонконга (Китай) HK1163739 и патент Австралии 2009331326).
Вектор также может содержать промоторы, энхансеры, сигналы сплайсинга, сигналы присоединения поли-А, последовательности интронов, маркеры селекции, точки начала репликации SV40 и так далее.
Настоящее изобретение также относится к клетке-хозяину, трансформированной указанным выше вектором. Можно получить антитело против PD-L1 по изобретению путем культивирования клетки-хозяина и сбора представляющего интерес антитела из полученной культуры. Следовательно, настоящее изобретение также относится к способу получения антитела, включающему культивирование описанной выше клетки-хозяина и сбор антитела против PD-L1 по изобретению из культуры. В способе по настоящему изобретению для получения антитела вектор, содержащий искусственную генетическую ДНК, содержащую ДНК, кодирующую легкую цепь, и ДНК, кодирующую тяжелую цепь, может быть трансфицирован в клетку-хозяина. Альтернативно, вектор, в который введена ДНК, кодирующая легкую цепь, и вектор, в который введена ДНК, кодирующая тяжелую цепь, могут быть совместно трансфицированы в клетку-хозяина.
Примеры клеток-хозяев включают, но не ограничиваются, бактериальные клетки (такие как бактерии Escherichia, бактерии Bacillus или Bacillus subtilis), клетки грибов (такие как дрожжи или Aspergillus), клетки насекомых (такие как клетки S2 или клетки Sf), клетки животных (такие как клетки CHO, клетки COS, клетки HeLa, клетки C127, клетки 3T3, клетки BHK или клетки HEK 293) и клетки растений. Среди них предпочтительной является клетка CHO-DG44 (CHO-DG44 (dfhr-/-)), которая является клеткой с дефицитом дигидрофолатредуктазы.
Введение рекомбинантного вектора в клетку-хозяина может быть осуществлено способами, раскрытыми в Molecular Cloning 2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989 (например, метод на основе фосфата кальция, метод с DEAE-декстраном, трансфекция, микроинъекция, липофекция, электропорация, трансдукция, введение при соскабливании, метод дробовика и тому подобное) или путем инфекции.
Полученный трансформант может быть культивирован в среде с последующим сбором антитела против PD-L1 по настоящему изобретению из культуры. Если антитело секретируется в среду, то среда может быть восстановлена с последующим выделением и очисткой антитела из среды. Если антитело продуцируется внутри трансформированных клеток, то клетки можно лизировать с последующим выделением и очисткой антитела из клеточного лизата.
Примеры сред включают, но не ограничиваются, среду OptiCHO, среду Dynamis, среду CD CHO, среду ActiCHO, среду FortiCHO, среду Ex-Cell CD CHO, среду BalanCD CHO, среду ProCHO 5 и среду Cellvento CHO-100.
рН среды изменяется в зависимости от культивируемой клетки. Обычно используется диапазон рН от 6,8 до 7,6; в основном, подходит диапазон рН от 7,0 до 7,4.
Если культивируемая клетка представляет собой клетки СНО, то культивирование может быть выполнено способами, известными специалистам в данной области. Например, обычно можно проводить культивирование в газофазной атмосфере с концентрацией CO2, равной 0-40%, предпочтительно 2-10%, при 30-39°C, предпочтительно при приблизительно 37°C.
Подходящий период культивирования обычно составляет от одного дня до трех месяцев, предпочтительно от одного дня до трех недель.
Выделение и очистка антитела могут быть выполнены известными способами. Известные способы выделения/очистки, которые можно использовать в настоящем изобретении, включают, но не ограничиваются, способы, использующие разницу в растворимости (такие как высаливание или осаждение растворителем); методы с использованием различий в молекулярной массе (такие как диализ, ультрафильтрация, гель-фильтрация или электрофорез в SDS-полиакриламидном геле); методы, использующие разницу в электрическом заряде (такие как ионообменная хроматография); методы с использованием специфической аффинности (такие как аффинная хроматография); методы, использующие различия в гидрофобности (такие как высокоэффективная жидкостная хроматография с обращенной фазой); и методы, использующие различие в изоэлектрической точке (например, изоэлектрическое фокусирование).
Антитело против PD-L1 по настоящему изобретению можно использовать в качестве лекарственного средства для животных или человека. Следовательно, настоящее изобретение относится к фармацевтической композиции, содержащей вышеописанное антитело против PD-L1 в качестве активного ингредиента.
Фармацевтическая композиция по настоящему изобретению может быть использована для профилактики и/или лечения злокачественных новообразований и/или инфекций. Примеры злокачественных новообразований и/или воспаления включают, но ими не ограничены, опухолевые заболевания (например, меланому, рак легкого, рак желудка, рак почек, рак молочной железы, рак мочевого пузыря, рак пищевода, рак яичников и тому подобное), лейкоз, болезнь Джона, анаплазмоз, бактериальный мастит, микотический мастит, микоплазменные инфекции (такие как микоплазменный мастит, микоплазменная пневмония или тому подобное), туберкулез, инфекцию Theileria orientalis, криптоспоридиоз, кокцидиоз, трипаносомоз и лейшманиоз.
Антитело против PD-L1 по настоящему изобретению может быть растворено в буферах, таких как PBS, физиологический солевой раствор или стерильная вода, необязательно, фильтр-стерилизованная с помощью фильтра или тому подобного, а затем введено животным (включая человека) путем инъекции. К раствору антитела могут быть добавлены добавки (такие как красящие агенты, эмульгаторы, суспендирующие агенты, поверхностно-активные вещества, солюбилизаторы, стабилизаторы, консерванты, антиоксиданты, буферы, изотонизирующие агенты, регуляторы рН и тому подобное). В качестве путей введения могут быть выбраны внутривенное, внутримышечное, внутрибрюшинное, подкожное или внутрикожное введение и тому подобное. Трансназальное или пероральное введение также может быть использовано.
Доза, количество и частота введения антитела против PD-L1 по настоящему изобретению могут изменяться в зависимости от симптомов, возраста и массы тела животного, способа введения, лекарственной формы и тому подобное. Например, 0,1-100 мг/кг массы тела, предпочтительно 1-10 мг/кг массы тела, на взрослое животное обычно можно вводить по меньшей мере один раз с такой частотой, которая позволяет подтвердить желаемый эффект.
Хотя фармацевтическую композицию по настоящему изобретению можно использовать самостоятельно, ее можно применять и в комбинации с хирургическими операциями, лучевой терапией, другими иммунотерапевтическими средствами, такими как противораковая вакцина, или лекарственными средствами с молекулярной мишенью. При таких комбинациях можно ожидать синергетический эффект.
ПРИМЕРЫ
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако настоящее изобретение не ограничивается этими примерами.
[Пример 1] Химерное "крысино-собачье" антитело против PD-L1
1. Введение
Белок программируемой гибели клеток 1 (PD-1), иммуноингибирующий рецептор и лиганд белка программируемой гибели клеток 1 (PD-L1) представляют собой молекулы, идентифицированные профессором Tasuku Honjo et al., Kyoto University, как факторы, которые ингибируют избыточный иммунный ответ и глубоко связанные с иммунотолерантностью. Недавно было выяснено, что эти молекулы также участвуют в иммуносупрессии опухолей. В этом примере, с целью получения нового средства для терапии неопластических заболеваний у собак получали ген химерного антитела, в котором ген вариабельной области крысиного моноклонального антитела против бычьего PD-L1 (4G12), способного ингибировать связывание PD-1 собаки с PD-L1, связывали с геном константной области собачьего иммуноглобулина (IgG4). Полученный ген химерного антитела вводили в клетки яичника китайского хомяка (клетки СНО), которые культивировали с получением химерного "крысино-собачье" антитела c4G12 против PD-L1. Эффект этого химерного антитела был подтвержден in vitro и in vivo.
2. Материалы и методы
2.1 Клетки, продуцирующие бычье моноклональное антитело PD-L1
Была идентифицирована нуклеотидная последовательность бычьего PD-L1 (Ikebuchi R, Konnai S, Shirai T, Sunden Y, Murata S, Onuma M, Ohashi K. Vet Res. 2011 Sep 26; 42: 103). На основании информации о последовательности был получен бычий рекомбинантный PD-L1. Крысу иммунизировали этим рекомбинантным белком с получением крысиного антитела против бычьего PD-L1 (Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology. 2014 Aug; 142 (4): 551-61; в этой статье описан клон 4G12, который впоследствии будет служить вариабельной областью интересующего собачьего химерного антитела).
2.2 Идентификация полноразмерных генов собачьего PD-1 и PD-L1
Для определения полноразмерных ДНК собачьего PD-1 и PD-L1 сначала были созданы ПЦР праймеры на основе предполагаемых нуклеотидных последовательностей собачьего PD-1 и PD-L1, уже зарегистрированных в National Center for Biotechnology Information (NCBI) (номер доступа в GenBank; XM_543338 и XM_541302). Вкратце, были созданы праймеры для амплификации внутренней последовательности открытой рамки считывания (ORF) каждого гена (внутренние F и R cPD-1, внутренние F и R cPD-L1) и осуществлена ПЦР. Для амплифицированных продуктов определяли нуклеотидные последовательности с помощью капиллярного секвенатора согласно обычным методам. Кроме того, для определения нуклеотидных последовательностей полноразмерной кДНК PD-1 и PD-L1 были получены праймеры (cPD-1 5' GSP и 3' GSP; cPD-L1 5' GSP и 3' GSP) на основе последовательности кДНК собачьего PD-1 и PD-L1, как определено выше. Затем осуществляли 5'-RACE и 3'-RACE, используя систему 5'-RACE для быстрой амплификации концов кДНК, и систему 3'-RACE для быстрой амплификации концов кДНК (Invitrogen), соответственно. Интересующие полученные фрагменты генов секвенировали, как описано (Maekawa N, Konnai S, Ikebuchi R, Okagawa T, Adachi M, Takagi S, Kagawa Y, Nakajima C, Suzuki Y, Murata S, Ohashi K. PLoS One. 2014 Jun 10; 9(6):e98415.
Праймер (cPD-1 внутренний F): AGGATGGCTCCTAGACTCCC (SEQ ID NO:21)
Праймер (cPD-1 внутренний R): AGACGATGGTGGCATACTCG (SEQ ID NO:22)
Праймер (cPD-L1 внутренний F): ATGAGAATGTTTAGTGTCTT (SEQ ID NO:23)
Праймер (cPD-L1 внутренний R): TTATGTCTCTTCAAATTGTATATC (SEQ ID NO:24)
Праймер (cPD-1 5' GSP): GTTGATCTGTGTGTTG (SEQ ID NO:25)
Праймер (cPD-1 3´ GSP): CGGGACTTCCACATGAGCAT (SEQ ID NO:26)
Праймер (cPD-L1 5´ GSP): TTTTAGACAGAAAGTGA (SEQ ID NO:27)
Праймер (cPD-L1 3´ GSP): GACCAGCTCTTCTTGGGGAA (SEQ ID NO:28)
2.3. Конструирование клеток COS-7, экспрессирующих собачий PD-1 и PD-L1
Для получения плазмид, экспрессирующих собачий PD-1-EGFP и PD-L1-EGFP, осуществляли ПЦР, используя в качестве матрицы синтезированную кДНК, полученную из PBMC бигля, и праймеры, полученные путем добавления сайтов распознавания XhoI и BamHI (PD-1) и сайтов распознавания BglII и EcoRI (PD-L1) на 5' конце (cPD-1-EGFP F и R; cPD-L1-EGFP F и R). Полученные продукты ПЦР расщепляли с помощью XhoI(Takara) и BamHI (Takara) (PD-1) и с помощью BglII (New England Biolabs) и EcoRI (Takara) (PD-L1), а затем очищали набором FastGene Gel/PCR Extraction (NIPPON Genetics) с последующим клонированием в вектор pEGFP-N2 (Clontech), обработанный ферментами рестрикции аналогичным образом. Интересующие полученные плазмиды экспрессии экстрагировали с помощью набора QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученные таким образом плазмиды экспрессии обозначены как pEGFP-N2-cPD-1 и pEGFP-N2-cPD-L1.
Праймер (cPD-1-EGFP F): CCGCTCGAGATGGGGAGCCGGCGGGGGCC (SEQ ID NO:29)
Праймер (cPD-1-EGFP R): CGCGGATCCTGAGGGGCCACAGGCCGGGTC (SEQ ID NO:30)
Праймер (cPD-L1-EGFP F): GAAGATCTATGAGAATGTTTAGTGTC (SEQ ID NO:31)
Праймер (cPD-L1-EGFP R): GGAATTCTGTCTCTTCAAATTGTATATC (SEQ ID NO:32)
Клетки COS-7 субкультивировали с плотностью 5×104 клеток/см2 в 6-луночных планшетах, а затем культивировали в течение ночи в среде RPMI 1640, содержащей 10% инактивированной фетальной бычьей сыворотки и 0,01% L-глютамина, при 37°С в присутствии 5% CO2. pEGFP-N2-cPD-1, pEGFP-N2-cPD-L1 или pEGFP-N2 (отрицательный контроль) вводили в клетки COS-7 при 0,4 мкг/см2, используя Lipofectamine 2000 (Invitrogen). Клетки культивировали в течение 48 часов (cPD-1-EGFP экспрессирующая клетка и cPD-L1-EGFP экспрессирующая клетка). Для подтверждения экспрессии собачьего PD-1 и PD-L1 в полученных таким образом экспрессирующих клетках, внутриклеточную локализацию усиленного зеленого флуоресцентного белка (EGFP) визуализировали с помощью инвертированного конфокального лазерного микроскопа LSM700 (ZEISS) (Maekawa N, Konnai S, Ikebuchi R, Okagawa T, Adachi M, Takagi S, Kagawa Y, Nakajima C, Suzuki Y, Murata S, Ohashi K. PLoS One. 2014 Jun 10; 9(6):e98415.
2.4 Конструирование рекомбинантных собачьих PD-1, PD-L1 и CD80
Для амплификации внеклеточных областей собачьих PD-1, PD-L1 и CD80, оцененных по их предполагаемым аминокислотным последовательностям, были сконструированы праймеры. Вкратце, конструировали праймеры с последовательностью распознавания NheI или EcoRV (PD-1 и PD-L1), добавленной на 5' конце (cPD-1-Ig F и R; cPD-L1-Ig F и R), или с последовательностью распознавания EcoRV или KpnI (CD80), добавленной на 5' конце (cCD80-Ig F и R). ПЦР проводили с использованием в качестве матрицы синтезированной кДНК, полученной из PBMC бигля. Продукты ПЦР расщепляли NheI (Takara) и EcoRV (Takara) или EcoRV (Takara) и KpnI (New England Biolabs) и очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics). Каждую очищенную таким образом ДНК клонировали в вектор pCXN2.1-кроличий Fc IgG (Niwa et al., 1991; Zettlmeissl et al., 1990; любезно предоставлен доктором T. Yokomizo, Juntendo University Graduate School of Medicine, и модифицирован в лаборатории авторами), обработанный ферментами рестрикции аналогичным образом. Плазмиды экспрессии очищали с помощью набора QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C до использования в экспериментах. В настоящем документе полученные таким образом плазмиды экспрессии обозначены как pCXN2.1-cPD-1-Ig, pCXN2.1-cPD-L1-Ig и pCXN2.1-cCD80-Ig, соответственно.
Праймер (cPD-1-Ig F): CGCGGCTAGCATGGGGAGCCGGCGGGGGCC (SEQ ID NO:33)
Праймер (cPD-1-Ig R): CGCGGATATCCAGCCCCTGCAACTGGCCGC (SEQ ID NO:34)
Праймер (cPD-L1-Ig F): CGCGGCTAGCATGAGAATGTTTAGTGTCTT (SEQ ID NO:35)
Праймер (cPD-L1-Ig R): CGCGGATATCAGTCCTCTCACTTGCTGGAA (SEQ ID NO:36)
Праймер (cCD80-Ig F): CGCGGATATCATGGATTACACAGCGAAGTG (SEQ ID NO:129)
Праймер (cCD80-Ig R): CGGGGTACCCCAGAGCTGTTGCTGGTTAT (SEQ ID NO:130)
Каждый вектор экспрессии трансфицировали в клетки Expi293F (Life Technologies) с получением культурального супернатанта, содержащего рекомбинантный Ig-слитый белок. Полученный рекомбинантный белок очищали от супернатанта с помощью Ab Capcher Extra (мутант белка A; ProteNova). После замены буфера на забуференный фосфатом физиологический солевой раствор (PBS; pH 7,4), используя PD-MidiTrap G-25 (GE Healthcare), каждый рекомбинантный белок хранили при -30°C перед использованием в экспериментах (cPD-1-Ig, cPD- L1-Ig и cCD80-Ig). Концентрацию каждого белка измеряли с помощью набора для анализа Pierce BCA Protein Assay (Thermo Fisher Scientific) перед использованием в последующих экспериментах.
2.5. Идентификация крысиного моноклонального антитела против бычьего PD-L1, проявляющего перекрестную реактивность в отношении собачьего PD-L1
Для идентификации крысиного моноклонального антитела против бычьего PD-L1, проявляющего перекрестную реактивность в отношении собачьего PD-L1, проводили проточную цитометрию, используя антитело против бычьего PD-L1, полученное в разделе 2.1 выше. Антитело против бычьего PD-L1 (10 мкг/мл) взаимодействовало с клетками 2×105-1×106 при комнатной температуре в течение 30 мин. После промывки антитело против бычьего PD-L1 детектировали с помощью меченного аллофикоцианином козьего антитела против крысиного Ig (Beckman Coulter). Для анализа использовали FACS Verse (Becton, Dickinson and Company). В качестве отрицательного контроля использовали контроль с изотипом крысиного IgG2a (κ) (BD Biosciences), контроль с изотипом крысиного IgG1 (κ) (BD Biosciences) и контроль с изотипом крысиного IgM (κ) (BD Biosciences). Для каждой стадии промывки и разбавления антител использовали PBS c 10% инактивированной козьей сывороткой (Maekawa N, Konnai S, Ikebuchi R, Okagawa T, Adachi M, Takagi S, Kagawa Y, Nakajima C, Suzuki Y, Murata S, Ohashi K. PLoS One. 2014 Jun 10; 9(6):e98415, в этой статье описано использование трех моноклональных антител против бычьего PD-L1: 4G12 (крысиный IgG2a (κ)), 5A2 (крысиный IgG1 (κ)) и 6G7 (крысиный IgM (κ)).
2.6 Тест на селекцию вариабельной области для определения химерного "крысино-собачьего" антитела
Из 10 клонов крысиного моноклонального антитела против бычьего PD-L1, которые показали перекрестную реактивность в отношении собачьего PD-L1, 4G12 (крысиный IgG2a (κ)), 5A2 (крысиный IgG1 (κ)) и 6G7 (крысиный IgM (к)), были отобраны и оценены на способность антител ингибировать связывание собачьего PD-1/PD-L1. Вкратце, собачий PD-1-Ig (полученный в разделе 2.4 выше) иммобилизовали на 96-луночных планшетах с плоским дном и блокировали 1% BSA и 0,05% PBS, содержащим Tween 20. Собачий PD-L1-Ig (полученный в разделе 2.4 выше) биотинилировали, используя набор для конъюгации Lightning-Link Biotin Conjugation (Innova Bioscience), и проводили реакцию при различных концентрациях (0, 2,5, 5 и 10 мкг/мл) крысиного антитела 4G12, 5A2 и 6G7 против PD-L1 при 37°С в течение 30 мин, а затем добавляли в 96-луночные планшеты. Связывание cPD-L1-Ig с cPD-1-Ig измеряли по цветовой реакции, используя Neutravidin-HRP (Thermo Fisher Scientific) и однокомпонентный субстрат TMB (Bethyl Laboratories). В результате крысиные моноклональные антитела 4G12 и 6G7 против PD-L1 показали хорошую ингибирующую активность в отношении связывания собачьего PD-1/PD-L1, тогда как 5A2 не проявляло ингибирующей активности в отношении связывания (фиг.1).
2.7 Получение вектора экспрессии химерного "крысино-собачьего" антитела против PD-L1 (фиг.2)
Используя в качестве вариабельной области крысиные моноклональные антитела 4G12 и 6G7 против бычьего PD-L1, которые показали хорошую ингибирующую активность в отношении связывания собачьего PD-1/PD-L1 (фиг.1), были созданы два типа химерных "крысино-собачьих" антител против PD-L1.
Вкратце, гены вариабельной области тяжелой цепи и легкой цепи были идентифицированы из гибридом, продуцирующих крысиные моноклональные антитела 4G12 и 6G7 против бычьего PD-L1. Кроме того, гены вариабельной области тяжелой цепи и легкой цепи вышеуказанных крысиных антител связывали с константной областью IgG4 тяжелой цепи и константной областью лямбда легкой цепи известного собачьего антитела, соответственно, с получением нуклеотидных последовательностей с последующей оптимизацией кодонов (SEQ ID NO:9 и 10 (аминокислотные последовательности), SEQ ID NO:19 и 20 (нуклеотидные последовательности после оптимизации кодонов). Затем проводили синтез генов таким образом, чтобы сайт распознавания рестриктазы NotI, последовательность KOZAK, последовательность легкой цепи химерного антитела, последовательность сигнала присоединения поли-A (PABGH), последовательность промотора (PCMV), сайт распознавания рестриктазы SacI, последовательность интрона (INRBG), последовательность KOZAK, последовательность тяжелой цепи химерного антитела и сайт распознавания рестриктазы XbaI были расположены в указанном порядке. Каждую синтезированную генную цепь встраивали в сайт клонирования (последовательности распознавания рестриктаз NotI и XbaI ниже PCMV и между INRBG и PABGH) вектора экспрессии pDC6 (любезно предоставлен профессором S.Suzuki, Hokkaido University Research Center for Zoonosis Control), используя последовательности распознавания ферментов рестрикции так, чтобы перечисленные выше последовательности были расположены в указанном порядке (фиг.2). Таким образом, были сконструированы векторы, экспрессирующие химерное "крысино-собачье" антитело против PD-L1. Каждый вектор экспрессии трансфицировали в клетки Expi293F (Life Technologies) с получением культурального супернатанта, содержащего химерное антитело. Химерное антитело очищали от супернатанта с помощью Ab Capcher Extra (мутант белка A; ProteNova) и дополнительно очищали гель-фильтрационной хроматографией. SDS-PAGE проводили в невосстанавливающих условиях с использованием 10% акриламидного геля. Полосы окрашивали с помощью набора Quick-CBB (Wako Pure Chemical) и обесцвечивали в дистиллированной воде. Хотя загрязняющие белки наблюдали после очистки только белком А, высокоочищенное антитело можно получить с помощью гель-фильтрационной хроматографии (фиг.3). С помощью проточной цитометрии было подтверждено, что полученные очищенные антитела специфически связываются с клетками, экспрессирующими собачий PD-L1 (данные не показаны). Если ингибирующую активность двух химерных антител в отношении связывания собачьего PD-1/PD-L1 исследовали способом, описанным в разделе 2.6 выше, то химерное "крысино-собачье" антитело c4G12 против PD-L1 показало, что активность ингибирования связывания аналогична активности первоначального крысиного моноклонального антитело 4G12 против бычьего PD-L1, и способность ингибировать связывание была явно ослаблена у химерного "крысино-собачьего" антитела c6G7 против PD-L1 (фиг.4). Следовательно, химерное "крысино-собачье" антитело c4G12 против PD-L1 было выбрано в качестве терапевтического антитела, в которое встроены последовательности вариабельной области крысиного моноклонального антитела 4G12 против бычьего PD-L1 (SEQ ID NO:2 и 1 (аминокислотные последовательности) и SEQ ID NO:16 и 15 (нуклеотидные последовательности после оптимизации кодонов)). Аминокислотная последовательность и нуклеотидная последовательность (после оптимизации кодонов) легкой цепи c4G12 показаны в SEQ ID NO:9 и 19, а аминокислотная последовательность и нуклеотидная последовательность (после оптимизации кодонов) тяжелой цепи c4G12 показаны в SEQ ID NO:10 и 20.
2.8 Экспрессия химерного "крысино-собачьего" антитела c4G12 против PD-L1
Вектор pDC6, экспрессирующий химерное "крысино-собачье" антитело c4G12 против PD-L1, использованный в разделе 2.7 выше, трансфицировали в клетки CHO-DG44 (CHO-DG44 (dfhr-/-)), которые представляли собой клетки с дефицитом дигидрофолатредуктазы, и с помощью дот-блоттинга отбирали клоны с высокой экспрессией. Кроме того, амплификацию генов осуществляли путем добавления нагрузки на клетки в среде, содержащей 60 нМ метотрексат (Mtx). Клетки, стабильно экспрессирующие химерное "крысино-собачье" антитело c4G12 против PD-L1 (название клона: 4.3F1) после амплификации генов, переносили в среду Opti-CHO без Mtx и культивировали при встряхивании в течение 14 дней (125 об/мин, 37°C, 5% CO2). Выживаемость клеток рассчитывали путем окрашивания трипановым синим (фиг. 5). Продукцию химерных антител в культуральном супернатанте измеряли с помощью ELISA (фиг. 5). Культуральный супернатант на 14-ый день центрифугировали при 10000 об./мин в течение 10 минут для удаления клеток, затем пропускали через фильтр с размером пор 0,22 мкм, а затем проводили стадии очистки антитела.
Следует отметить, что при замене среды на среду Dynamis и проведении необходимых подкормок, продукция антител улучшалась примерно в два раза по сравнению с обычной продукцией (данные не показаны).
2.9. Очистка химерного "крысино-собачьего" антитела c4G12 против PD-L1
Культуральный супернатант, полученный, как описано выше, очищали с помощью Ab Capcher Extra (ProteNova). Для связывания со смолой использовали способ с открытой колонкой; PBS с pH 7,4 использовали в качестве уравновешивающего буфера и промывочного буфера. В качестве буфера для элюции использовали буфер для элюции IgG Elution Buffer (Thermo Scientific). В качестве нейтрализующего буфера использовали 1 М Трис. Очищенное антитело концентрировали и ультрафильтрацией заменяли буфер PBS, используя Amicon Ultra-15 (50 кДа, Millipore). Полученное антитело пропускали через фильтр с размером пор 0,22 мкм для использования в соответствующих экспериментах.
2.9. Подтверждение очистки химерного "крысино-собачьего" антитела c4G12 против PD-L1 (фиг.6)
Чтобы подтвердить чистоту очищенного антитела, белки антитела обнаруживали с помощью SDS-PAGE и окрашивания CBB. Используя градиентный гель SuperSep Ace 5-20% (Wako), крысиное моноклональное антитело против бычьего PD-L1 4G12 и химерное "крысино-собачье" антитело c4G12 против PD-L1 подвергали электрофорезу в восстанавливающих и невосстанавливающих условиях. Полосы окрашивали с помощью набора Quick-CBB (Wako) и обесцвечивали в дистиллированной воде. Полосы оценивали в положениях молекулярных масс, соответствующих антителам. Полосы загрязняющих белков визуально обнаружены не были.
2.11. Измерение авидности связывания с cPD-L1-His крысиного моноклонального антитела 4G12 против бычьего PDG-L1 и химерного "крысино-собачьего" антитела c4G12 против PD-L1
Для амплификации внеклеточной области собачьего PD-L1, оцененной по ее предполагаемой аминокислотной последовательности, конструировали праймеры. Вкратце, конструировали праймер с последовательностью распознавания NheI, добавленной на 5' конец (cPD-L1-His F), и праймер с последовательностью распознавания EcoRV и последовательностью метки 6xHis, добавленными на 5' конец (cPD-L1-His R). ПЦР проводили с использованием в качестве матрицы синтезированной кДНК, полученной из PBMC бигля. Продукты ПЦР расщепляли NheI(Takara) и EcoRV (Takara) и очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics). Очищенную таким образом ДНК клонировали в вектор pCXN2.1 (Niwa et al., 1991; предоставлен доктором T. Yokomizo, Juntendo University Graduate School of Medicine), обработанный ферментами рестрикции аналогичным образом. Плазмиды экспрессии очищали с помощью набора QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C до использования в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pCXN2.1-cPD-L1-His.
Праймер (cPD-L1-His F): CGCGGCTAGCATGAGAATGTTTAGTGTCTT (SEQ ID NO:131)
Праймер (cPD-L1-His R): CGCGGATATCTTAATGGTGATGGTGATGGTGAGTCCTCTCACTTGCTGG (SEQ ID NO:132)
Вектор экспрессии трансфицировали в клетки Expi293F (Life Technologies) с получением культурального супернатанта, содержащего рекомбинантный слитый белок. Полученный рекомбинантный белок очищали от супернатанта, используя аффинную смолу TALON Metal (Clontech), и буфер заменяли на PBS, используя Amicon Ultra-4 Ultracel-3 (Merck Millipore). Полученный таким образом рекомбинантный белок хранили при 4°С до использования в экспериментах (cPD-L1-His). Концентрацию белка измеряли с помощью набора для анализа Pierce BCA Protein Assay (Thermo Fisher Scientific) для использования в последующих экспериментах.
С использованием анализатора биомолекулярного взаимодействия (Biacore X100) оценивали авидность связывания с cPD-L1-His крысиного моноклонального антитела 4G12 против бычьего PD-L1 и химерного "крысино-собачьего" антитела c4G12 против PD-L1. Вкратце, антигистидиновое антитело фиксировали на цензорном чипе CM5 с последующим захватом cPD-L1-His. Затем, в качестве аналита для наблюдения специфического связывания добавляли моноклональные антитела. Оба антитела проявляли специфическое связывание, и их авидность была почти сопоставима (таблица 1). Кроме того, авидность связывания собачьего PD-1-Ig и CD80-Ig с cPD-L1-His измеряли аналогичным образом и было обнаружено, что она явно ниже, чем у химерного "крысино-собачьего" антитела c4G12 против PD-L1 (таблица 1).
Таблица 1. Авидность связывания каждого антитела и рекомбинантного белка с собачьим PD-L1-His
2.12. Ингибирующая активность химерного "крысино-собачьего" антитела c4G12 против связывания собачьего PD-1/PD-L1 и связывания CD80/PD-L1 (фиг.7)
Используя собачьи PD-1-Ig, PD-L1-Ig и CD80-Ig (описаны выше), антитело против PD-L1 тестировали на способность ингибировать связывание собачьего PD-1/PD-L1 и CD80/PD-L1. Вкратце, собачий PD-1-Ig или CD80-Ig иммобилизовали на 96-луночных планшетах с плоским дном. Собачий PD-L1-Ig взаимодействовал с различными концентрациями (0, 2,5, 5 и 10 мкг/мл) крысиного антитела 4G12 против бычьего PD-L1 или химерного "крысино-собачьего" антитела c4G12 против PD-L1, соответственно, и в соответствии с теми же процедурами, как описано в разделе 2.6 выше, и оценивали связывание собачьего PD-L1-Ig. Никакого изменения в активности ингибирования связывания из-за химеризации антитела не наблюдалось.
2.13. Активация иммунных клеток собак химерным "крысино-собачьим" антителом c4G12 против PD-L1 (фиг. 8)
PBMC собак культивировали при стимуляции супер-антигеном Staphylococcal Enterotoxin B (SEB) в течение трех дней, и изменения в продукции цитокинов при добавлении химерного "крысино-собачьего" антитела c4G12 против PD-L1 измеряли с помощью ELISA с использованием Duoset ELISA на собачьих IL-2 или IFN-γ (R&D системы). Химерное "крысино-собачье" антитело c4G12 против PD-L1 повышало продукцию IL-2 и IFN-γ PBMC собаки. Кроме того, в культуральную среду добавляли аналог нуклеиновой кислоты EdU на 2-ой день при стимуляции SEB. Через два часа измеряли захват EdU с помощью проточной цитометрии с использованием набора для анализа проточной цитометрии Click-iT Plus EdU (Life Technologies). В результате захват EdU в CD4+ и CD8+ лимфоцитах собак повышался при добавлении химерного "крысино-собачьего" антитела c4G12 против PD-L1, что указывает на повышенную способность к пролиферации клеток.
2.14 Отбор собак, пораженных опухолью, для использования в тесте c инокуляцией собак
Поскольку ожидается, что лечение будет более высоко эффективным, если опухоли экспрессируют PD-L1, то анализ экспрессии PD-L1 в опухолевом участке у собак проводили иммуногистохимическим окрашиванием. Вкратце, образцы опухолевой ткани, фиксированные формальдегидом и залитые в парафин, нарезали на срезы толщиной 4 мкм с помощью микротома, прикрепляли и высушивали на предметном стекле, покрытом силаном (стекло Matsunami) и депарафинизировали ксилолом/спиртом. Пока полученные срезы замачивали в цитратном буфере [0,37 г лимонной кислоты (Wako Pure Chemical), 2,4 г дигидрата тринатрийцитрата (Kishida Chemical), 1000 мл дистиллированной воды], проводили обработку антигена в течение 10 минут микроволновым излучением с последующим окрашиванием, используя автоматическое имитирующее устройство Nichirei. В качестве предварительной обработки образцы срезов замачивали в 0,3% метанольном растворе, содержащем пероксид водорода, при комнатной температуре в течение 15 минут и промывали PBS. Затем добавляли моноклональное антитело против бычьего PD-L1 и реакцию проводили при комнатной температуре в течение 30 минут. После промывания PBS добавляли краситель histofine simple stain MAX-PO (Rat) (Nichirei Bioscience) и реакцию осуществляли при комнатной температуре в течение 30 минут с последующим окрашиванием 3,3'-диаминобензидин тетрагидрохолридом и наблюдали с помощью светового микроскопа. Собак с меланомой полости рта или недифференцированной саркомой, у которых опухолевые клетки были PD-L1-позитивными, использовали в следующем тесте с инокуляцией (клиническое испытание). Моноклональное антитело против бычьего PD-L1 получали из крысиной гибридомы, продуцирующей крысиное моноклональное антитело против бычьего PD-L1 (Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology. 2014 Aug;142(4):551-61).
2.15 Тест с инокуляцией у собак
Что касается химерного "крысино-собачьего" антитела c4G12 против PD-L1, которое должно быть инокулировано собакам в ходе клинического испытания, культуральный супернатант, полученный с помощью процедур, описанных в разделе 2.8 выше, очищали аффинной хроматографией с использованием MabSelect SuRe LX (GE Healthcare) и затем проводили хроматографию на гидроксиапатите с использованием колонки BioScale CHT20-I (Bio-Rad) для удаления загрязнений и полимерных белков. Содержащие агрегаты фракции дополнительно очищали с помощью анионообменной хроматографии с использованием предварительно упакованной колонки HiScreen Q-Sepharose HP (GE Healthcare).
(1) Тест на безопасность: полученное химерное "крысино-собачье" антитело c4G12 против PD-L1 вводили внутривенно собаке (бигль, стерилизованная самка, 13 лет, весом около 10 кг) в дозе 2 мг/кг, каждые 2 недели, всего 3 раза. Ни анафилаксии, ни других побочных эффектов, которые могли бы вызвать какие-либо проблемы в клинических испытаниях, не отмечалось. (2) Клиническое испытание 1: Полученное химерное "крысино-собачье" антитело c4G12 против PD-L1 вводили внутривенно PD-L1-позитивной собаке с рецидивирующей меланомой полости рта (фиг.9A) (карликовая такса, кобель, 11 лет) (около 7,5 кг массы тела) в дозе 2 мг/кг или 5 мг/кг каждые 2 недели, всего 22 раза. На 10-й неделе после начала лечения было замечено заметное уменьшение размера опухоли. На 34-й неделе после начала лечения было подтверждено еще большее снижение (фиг.10). В период наблюдения 44 недели метастазов в лимфатические узлы или легкое не наблюдалось. Уменьшение на 30% или более максимального диаметра опухоли по сравнению с исходным уровнем определяли как PR (частичный ответ), критерий PR был достигнут на 16-20-й неделе и на 34-й неделе и позднее (фиг. 11).
(3) Клиническое испытание 2: Химерное "крысино-собачье" антитело c4G12 против PD-L1 вводили внутривенно собаке с недифференцированной саркомой, первичное поражение которой было PD-L1-позитивным (фиг. 9B) и у которого было множество метастатических поражений в мышцах по всему телу (вест-хайленд-уайт-терьер, кастрированный кобель, 12 лет, вес около 8 кг) в дозе 5 мг/кг каждые 2 недели, всего 2 раза. На 3-й неделе от начала лечения была выявлена четкая регрессия опухоли (фиг. 12).
(4) Клиническое испытание 3: Химерное "крысино-собачье" антитело c4G12 против PD-L1 вводили внутривенно собаке с меланомой полости рта, первичное поражение у которой было удалено хирургическим путем (бигль, стерилизованная самка, 11 лет, вес около 10 кг) в дозе 2 мг/кг или 5 мг/кг каждые 2 недели, всего 9 раз. На 18-й неделе после начала лечения множество легочных метастатических поражений исчезло (фиг. 13),
(5) Клиническое испытание 4: Химерное "крысино-собачье" антитело c4G12 против PD-L1 вводили внутривенно 4 собакам с меланомой полости рта с легочными метастазами в дозе 2 мг/кг или 5 мг/кг каждые 2 недели. Не смотря на то, что в период наблюдения не отмечалось явного уменьшения размера опухоли, продолжительность выживания обработанных собак после подтверждения легочного метастазирования, как правило, была больше, чем у контрольной группы (антитело не вводили, предыдущая контрольная группа: n=15) (фиг. 14). Следовательно, продолжительность жизни может быть увеличена путем введения антител.
2.16 Анализ CDR антитела против PD-L1
Определяющие комплементарность области (CDR) крысиного антитела 4G12 против бычьего PD-L1 определяли с использованием NCBI IGBLAST (http://www.ncbi.nlm.nih.gov/igblast/). Результаты показаны на фиг.15.
[Пример 2] Применение антитела против PD-L1 у других видов животных
1.1 Идентификация генов PD-L1 овцы, свиньи и буйвола
Для определения полноразмерных кодирующих последовательностей (CDS) кДНК PD-L1 овцы, свиньи и буйвола, сначало были созданы праймеры для амплификации полноразмерных CDS из нуклеотидных последовательностей генов PD-L1 овцы, свиньи и водяного буйвола (номер доступа GenBank; XM_004004362, NM_001025221 и XM_613366) (ovPD-L1 CDS F и R; poPD-L1 CDS F и R; buPD-L1 CDS F1, R1, F2 и R2), а затем была проведена ПЦР. Для полученных амплифицированных продуктов нуклеотидные последовательности определяли с помощью капиллярного секвенатора в соответствии с общепринятыми методами (Mingala CN, Konnai S, Ikebuchi R, Ohashi K. Comp. Immunol. Microbiol. Infect. Dis. 2011 Jan; 34(1):55-63; в этой статье был идентифицирован ген PD-L1 буйвола).
Праймер (ovPD-L1 CDS F): ATGAGGATATATAGTGTCTTAACAT (SEQ ID NO:109)
Праймер (ovPD-L1 CDS R): TTACGTCTCCTCAAAATGTG (SEQ ID NO:110)
Праймер (poPD-L1 CDS F): ATGAGGATATGTAGTATCTTTACAT (SEQ ID NO:111)
Праймер (poPD-L1 CDS R): TTACGTCTCCTCAAATTGTGT (SEQ ID NO:112)
Праймер (buPD-L1 CDS F1): ATGAGGATATATAGTGTCTT (SEQ ID NO:113)
Праймер (buPD-L1 CDS R1): GCCACTCAGGACTTGGTGAT (SEQ ID NO:114)
Праймер (buPD-L1 CDS F2): GGGGGTTTACTGTTGCTTGA (SEQ ID NO:115)
Праймер (buPD-L1 CDS R2): TTACGTCTCCTCAAATTGT (SEQ ID NO:116)
1.2 Конструирование клеток COS-7, экспрессирующих овечье PD-1, овечье PD-L1, свиной PD-1 и свиной PD-L1
Для получения плазмид, экспрессирующих овечьи PD-1, овечьи PD-L1, свиные PD-1 и свиные PD-L1, проводили ПЦР с использованием синтезированной кДНК, полученной из PBMC овцы или свиньи, в качестве матрицы и праймеров, полученных путем добавления BglII и SmaI (PD-1 овцы), HindIII и SmaI (PD-1 свиньи) или сайты узнавания XhoI и SmaI (PD-L1 овцы и свиньи) на 5' конце (ovPD-1-EGFP F и R; ovPD-L1- EGFP F и R; poPD-1-EGFP F и R или poPD-L1-EGFP F и R). Полученные продукты ПЦР расщепляли с помощью BglII (Takara) и SmaI (Takara) (PD-1 овцы), HindIII (Takara) и SmaI (Takara) (PD-1 свиньи), а также XhoI (Takara) и SmaI (Takara) (PD-L1 овцы и свиньи), затем очищали набором FastGene Gel/PCR Extraction Kit (NIPPON Genetics) и клонировали в вектор pEGFP-N2 (Clontech), обработанный ферментами рестрикции аналогичным образом. Плазмиды экспрессии экстрагировали с использованием набора FastGene Xpress Plasmid PLUS (NIPPON Genetics) и хранили при -30°C перед применением в экспериментах. В настоящем документе полученные таким образом плазмиды обозначены как pEGFP-N2-ovPD-1, pEGFP-N2-ovPD-L1, pEGFP-N2-poPD-1 или pEGFP-N2-poPD-L1.
Праймер (ovPD-1-EGFP F): GAAGATCTATGGGGACCCCGCGGGCGCCG (SEQ ID NO:117)
Праймер (ovPD-1-EGFP R): GACCCGGGGAGGGGCCAGGAGCAGTGTCC (SEQ ID NO:118)
Праймер (ovPD-L1-EGFP F): CCGCTCGAGATGAGGATATATAGTGTCT (SEQ ID NO:119)
Праймер (ovPD-L1-EGFP R): ATCCCGGGCGTCTCCTCAAAATGTGTAG (SEQ ID NO:120)
Праймер (poPD-1-EGFP F): ACTAAGCTTATGGGGACCCCGCGGG (SEQ ID NO:121)
Праймер (poPD-1-EGFP R): ACTCCCGGGGAGGGGCCAAGAGCAGT (SEQ ID NO:122)
Праймер (poPD-L1-EGFP F): CCGCTCGAGATGAGGATATGTAGTATCTT (SEQ ID NO:123)
Праймер (poPD-L1-EGFP R): ATCCCGGGCGTCTCCTCAAATTGTGTATC (SEQ ID NO:124)
Клетки COS-7 субкультивировали с плотностью 5×104 клеток/см2 в 6-луночных планшетах, а затем культивировали в течение ночи в среде RPMI 1640, содержащей 10% инактивированной фетальной бычьей сыворотки и 0,01% L-глютамина, при 37°С в присутствии 5% CO2. pEGFP-N2-ovPD-1, pEGFP-N2-ovPD-L1, pEGFP-N2-poPD-1, pEGFP-N2-poPD-L1 или pEGFP-N2 (отрицательный контроль) вводили в клетки COS-7 при 0,4 мкг/см2, используя Lipofectamine 2000 (Invitrogen). Клетки культивировали в течение 48 часов (экспрессирующая ovPD-1-EGFP клетка, экспрессирующая ovPD-L1-EGFP клетка, экспрессирующая poPD-1-EGFP клетка и экспрессирующая poPD-L1-EGFP клетка). Для подтверждения экспрессии PD-1 овцы, PD-L1 овцы, PD-1 свиньи и PD-L1 свиньи в приготовленных таким образом экспрессирующих клетках, внутриклеточную локализацию EGFP визуализировали с помощью инвертированного конфокального лазерного микроскопа LSM700 (ZEISS) или универсального флуоресцентного микроскопа BZ-9000 (KEYENCE).
1.3 Конструирование рекомбинантного овечьего PD-L1 и свиного PD-L1
Для амплификации внеклеточных областей овечьих PD-L1 и свиного PD-L1, оцененных по их предполагаемым аминокислотным последовательностям, были сконструированы праймеры. Вкратце, были сконструированы праймеры с последовательностью распознавания NheI или EcoRV, добавленную на 5' конец (ovPD-L1-Ig F и R или poPD-L1-Ig F и R). ПЦР проводили с использованием синтезированной кДНК, полученной из PBMC овец и свиней, в качестве матрицы. Продукты ПЦР расщепляли NheI(Takara) и EcoRV (Takara) и очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics). Каждую очищенную таким образом ДНК клонировали в вектор pCXN2.1-кроличий IgG (Niwa et al., 1991; Zettlmeissl et al., 1990; любезно предоставлен доктором T. Yokomizo, Juntendo University Graduate School of Medicine, и модифицирован в лаборатории автором), обработанный ферментами рестрикции аналогичным образом. Плазмиды экспрессии очищали с использованием набора FastGene Xpress Plasmid PLUS (NIPPON Genetics) и хранили при -30°C перед применением в экспериментах. В настоящем документе полученные таким образом плазмиды экспрессии обозначены как pCXN2.1-ovPD-L1-Ig и pCXN2.1-poPD-L1-Ig, соответственно.
Праймер (ovPD-L1-Ig F): GACGCTAGCATGAGGATATATAGTGTCT (SEQ ID NO:125)
Праймер (ovPD-L1-Ig R): GCTCTGATATCCCTCGTTTTGCTGGAT (SEQ ID NO:126)
Праймер (poPD-L1-Ig F): GACGCTAGCATGAGGATATGTAGTATCTT (SEQ ID NO:127)
Праймер (poPD-L1-Ig R): AGCTTGATATCCCTCTTTCTTGCTGGATC (SEQ ID NO:128)
Тридцать микрограммов pCXN2.1-ovPD-L1-Ig или pCXN2.1-poPD-L1-Ig вводили в клетки 7,5×107 Expi293F (Life Technologies) с использованием Expifectamin (Life Technologies). После 6-и дневного встряхивания культуры собирали супернатант культуры. Культуральный супернатант содержал рекомбинантный Fc-слитый белок. Полученный рекомбинантный Fc-слитый белок очищали от супернатанта с использованием Ab-Capcher Extra (ProteNova). После очистки буфер заменяли на PBS (pH 7,4), используя обессоливающую колонку PD-10 (GE Healthcare). Полученный рекомбинантный белок хранили при -30°С перед использованием в эксперименте (овечий PD-L1-Ig). Концентрация очищенного овечьего PD-L1-Ig и свиного PD-L1-Ig измеряли с помощью набора Rabbit IgG ELISA Quantitation (BETHYL). Для каждой операции промывки в ELISA использовали Auto Palte Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
1.4 Реакционная способность крысиного антитела 4G12 против бычьего PD-L1 с овечьим и свиным PD-L1
Методом проточной цитометрии было подтверждено, что крысиное моноклональное антитело против PD-L1 перекрестно реагирует с овечьим и свиным PD-L1. Клетки COS-7, экспрессирующие овечьи или свиные PD-L1-EGFP, блокировали PBS с добавлением 10% инактивированной козьей сыворотки при комнатной температуре в течение 15 минут и проводили реакцию с 10 мкг/мл крысиного антитела 4G12 против бычьего PD-L1 при комнатной температуре в течение 30 минут. После промывки клетки взаимодействовали с меченным аллофикоцианином козьим антителом против крысиного Ig (Beckman Coulter) при комнатной температуре в течение 30 мин. Для анализа использовали FACS Verse (BD Bioscience). В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина.
Экспериментальные результаты приведены на фиг. 16. Было подтверждено, что крысиное антитело 4G12 против бычьего PD-L1 связывается с овечьим и свиным PD-L1.
1.5 Реакционная способность крысиного антитела 4G12 против бычьего PD-L1 с лейкоцитами буйвола.
Периферическая кровь буйвола (Bubalus bubalis; азиатский буйвол) была гемолизирована с буфером ACK для выделения лейкоцитов. После блокирования PBS с добавлением 10% инактивированной козьей сыворотки при комнатной температуре в течение 15 минут, проводили реакцию с крысиным антителом 4G12 против бычьего PD-L1, комплексом перидинин-хлорофилл-белок/цианин 5.5-меченным антителом против бычьего CD14 (мышиный IgG1, CAM36A, VMRD) и антителом против бычьего CD11b (мышиный IgG2b, CC126, AbD Serotec) при комнатной температуре в течение 30 минут. После промывки проводили реакцию с меченным аллофикоцианином козлиным антителом против крысиного IgG (Beckman Coulter) и с меченным флуоресцеином изотиоцианатом козьим антителом против мышиного IgG2 (Beckman Coulter) при комнатной температуре в течение 30 минут. Для анализа использовали FACS Calibur (BD Biosciences). В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 10% инактивированной козьей сывороткой.
Экспериментальные результаты приведены на фиг. 17. Крысиное антитело 4G12 против бычьего PD-L1 прочно связывается с макрофагами крови (CD14+CD11b+клетки) буйвола. С другой стороны, крысиное антитело 4G12 против бычьего PD-L1 слабо связывается с лимфоцитами (CD14-CD11b-клетками) водяного буйвола. Считается, что это различие в свойствах связывания отражает уровни экспрессии PD-L1 в макрофагах и лимфоцитах.
1.6 Тест на ингибирование связывания овечьего или свиного PD-1/PD-L1 с антителом 4G12 против бычьего PD-L1
Используя клетки COS-7, экспрессирующие овечье PD-1-EGFP, и клетки COS-7, экспрессирующие рекомбинантный овечий белок PD-L1-Ig или свиной PD-1-EGFP, и рекомбинантный свиной белок PD-L1-Ig, тестировали ингибирование овечьего или свиного PD-1/PD-L1 с помощью крысиного антитела (4G12) против бычьего PD-L1. Вкратце, крысиное антитело 4G12 против бычьего PD-L1 в различных концентрациях (0, 1, 5, 10, 20, 50 мкг/мл) заранее подвергали взаимодействию с овечьим PD-L1-Ig (конечная концентрация 1 мкг/мл) или свиным PD-L1-Ig (конечная концентрация 5 мкг/мл) при 37°С в течение 30 мин. Затем антитело 4G12 взаимодействовало с 2×105 клетками COS-7, экспрессирующими овечьи PD-1-EGF, или клетками COS-7, экспрессирующими свиного PD-1-EGFP, при 37°C в течение 30 минут. После промывки, проводили детекцию овчьего PD-L1-Ig или свиного PD-L1-Ig на клеточной поверхности с помощью меченного Alexa Fluor 647 козьего F(ab')2 против кроличьего IgG (H+L) (Life Technologies). Для анализа использовали FACS Verse (BD Biosciences). В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience). Принимая долю клеток, связанных с PD-L1-Ig без добавления антител, за 100%, доля клеток, связанных с PD-L1-Ig, для каждой концентрации антител была показана как относительная величина.
Результаты показывают, что крысиное антитело 4G12 против бычьего PD-L1 способно ингибировать связывание PD-1/PD-L1 овцы и PD-1/PD-L1 свиньи зависимым от концентрации образом (фиг.18).
[Пример 3]
1. Введение
Белок программируемой гибели клеток 1 (PD-1), иммуноингибирующий рецептор и лиганд белка программируемой гибели клеток 1 (PD-L1) представляют собой молекулы, идентифицированные профессором Tasuku Honjo et al., Kyoto University, как факторы, которые ингибируют избыточный иммунный ответ и глубоко связанные с иммунотолерантностью. Недавно было выяснено, что эти молекулы также участвуют в иммуносупрессии опухолей. В этом примере с целью создания нового средства для лечения инфекций у крупного рогатого скота, авторы изобретения получили ген химерного антитела путем связывания гена вариабельной области крысиного моноклонального антитела (4G12) против бычьего PD-L1, способного ингибировать связывание бычий PD-1 и PD-L1 с генами, кодирующими константные области бычьего иммуноглобулина (бычий IgG1 с мутациями, введенными в предполагаемые сайты связывания c рецепторам Fcγ в домене CH2 для ингибирования активности ADCC; см. фиг.19 для аминокислотных положений и мутаций: 250 E→ P, 251 L→V, 252 P→A, 253 G→делеция, 347 A→S, 348 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug; 142(4):551-561). Этот ген химерного антитела был введен в клетки яичника китайского хомяка (клетки СНО). Путем культивирования/пролиферации полученных клеток авторы настоящего изобретения получили химерное "крысино-бычьего" антитело (ch4G12) против PD-L1 и подтверждали его действие in vitro и in vivo.
2. Материалы и методы
Конструирование клеток, экспрессирующих бычий PD-1 и PD-L1
Были определены нуклеотидные последовательности полноразмерных кДНК гена бычьего PD-1 (номер доступа в GenBank AB510901; Ikebuchi R, Konnai S, Sunden Y, Onuma M, Ohashi K. Microbiol. Immunol. 2010 May; 54(5):291-298) и ген бычьего PD-L1 (номер доступа GenBank AB510902; Ikebuchi R, Konnai S, Shirai T, Sunden Y, Murata S, Onuma M, Ohashi K. Vet. Res. 2011 Sep. 26; 42:103). На основании полученной генетической информации были получены клетки, экспрессирующие на мембране бычий PD-1 и бычий PD-L1. Сначала для получения плазмиды, экспрессирующей бычий PD-1 или бычий PD-L1, осуществляли ПЦР, используя синтезированную кДНК, полученную из PBMC крупного рогатого скота, в качестве матрицы и обработанные ферментами праймеры с сайтами распознавания NotI и HindIII (бычий PD-1) и сайтами распознавания NheI и XhoI (бычий PD-1) на 5' конце (boPD-1-myc F и R; boPD-L1-EGFP F и R). Продукты ПЦР расщепляли NotI (Takara) и HindIII (Takara; бычий PD-1), NheI (Takara) и XhoI (Takara; бычий PD-L1), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в вектор pCMV-Tag1 (Agilent Technologies; бычий PD-1) или в вектор pEGFP-N2 (Clontech; бычий PD-L1), обработанный фрагментами рестрикции таким же образом. Интересующую полученную плазмиду экспрессии экстрагировали набором QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pCMV-Tag1-boPD-1.
Праймер (boPD-1-myc F): ATATGCGGCCGCATGGGGACCCCGCGGGCGCT (SEQ ID NO:133)
Праймер (boPD-1-myc R): GCGCAAGCTTTCAGAGGGGCCAGGAGCAGT (SEQ ID NO:134)
Праймер (boPD-L1-EGFP F): CTAGCTAGCACCATGAGGATATATAGTGTCTTAAC (SEQ ID NO:135)
Праймер (boPD-L1-EGFP R): CAATCTCGAGTTACAGACAGAAGATGACTGC (SEQ ID NO:136)
Клетки, экспрессирующие на мембране бычий PD-1, получали в соответствии с методиками, описанными ниже. Сначала 2,5 мкг pCMV-Tag1-boPD-1 вводили в 4×106 клеток CHO-DG44 с помощью Lipofectamine LTX (Invitrogen). Через сорок восемь часов среду заменяли средой CD DG44 (Life Technologies), содержащей 800 мкг/мл G418 (Enzo Life Science), 20 мл/л добавки GlutaMAX (Life Technologies) и 18 мл/л 10% Pluronic F-68 (Life Technologies), а затем проводили селекцию. Полученные в результате экспрессирующие клетки взаимодействовали с крысиным антителом против бычьего PD-1 5D2 при комнатной температуре. После промывки клетки дополнительно взаимодействовали с антителом против крысиного IgG, меченного микрогранулами (Miltenyi Biotec), при комнатной температуре. Клетки, экспрессирующие бычий PD-1 на высоких уровнях, выделяли с помощью Auto MACS (Miltenyi Biotec). Затем проводили повторное выделение тем же образом и получали еще более высокую чистоту. Полученные экспрессирующие клетки подвергали клонированию путем лимитирования разведения, тем самым получая клон клеток CHO DG44, экспрессирующий бычий PD-1 на высоком уровне (экспрессирующие бычьи PD-1 клетки).
Клетки, экспрессирующие на мембране бычий PD-L1, получали в соответствии с методиками, описанными ниже. Сначала 2,5 мкг pEGFP-N2-boPD-L1 или pEGFP-N2 (негативный контроль) вводили в 4×106клеток CHO-DG44, используя Lipofectamine LTX (Invitrogen). Через сорок восемь часов среду заменяли средой CD DG44 (Life Technologies), содержащей 800 мкг/мл G418 (Enzo Life Science), 20 мл/л добавки GlutaMAX (Life Technologies) и 18 мл/л 10% Pluronic F-68 (Life Technologies), а затем проводили селекцию и клонирование с помощью лимитированного разведения (клон клеток, эксперссирующих бычий PD-L1). Для подтверждения экспрессии бычьего PD-1 в полученном, таким образом, клон экспрессирующих клеток, внутриклеточную локализацию EGFP визуализировали с помощью инвертированного конфокального лазерного микроскопа LSM700 (ZEISS).
Конструирование растворимого бычьего PD-1 и PD-L1
Плазмиду, экспрессирующую бычий PD-1-Ig, конструировали в соответствии с методиками, описанными ниже. Вкратце, сигнальный пептид и внеклеточная область бычьего PD-1 (номер доступа в GenBank AB510901) связывали с Fc-доменом константной области известного бычьего IgG1 (номер доступа GenBank X62916) с получением последовательности гена. После того, как кодоны были оптимизированы для клеток CHO, синтез генов осуществляли таким образом, чтобы последовательность распознавания NotI, последовательность KOZAK, последовательность сигнального пептида бычьего PD-1, последовательность внеклеточной области гена бычьего PD-1, последовательность Fc-области бычьего IgG1 и последовательность распознавания XbaI находились в указанном порядке. Следует отметить, что бычий IgG1 был мутирован для ингибирования активности ADCC; более конкретно, мутации были введены в предполагаемые сайты связывания домена CH2 Fcγ-рецепторов (сайты мутаций: 185 E→P, 186 L→V, 187 P→A, 189 G→делеция, 281 A→S, 282 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug; 142(4):551-561; аминокислотная последовательность PD-1-Ig и сайты мутации описаны на фиг.2 этой статьи). Цепь синтезированного гена расщепляли с помощью NotI (Takara) и XbaI (Takara), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и встраивали в сайт клонирования (последовательности ферментов рестрикции NotI и XbaI ниже PCMV и между INRBG и PABGH) вектора экспрессии pDN11 (любезно предоставлен профессором S.Suzuki, Hokkaido University Research Center for Zoonosis Control), обработанного ферментами рестрикции таким же образом получая вектор, экспрессирующий PD-1-Ig. Плазмиду экспрессии очищали набором QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C до использования в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pDN11-boPD-1-Ig.
Плазмиду, экспрессирующую бычий PD-L1-Ig, конструировали в соответствии с методиками, описанными ниже. Для амплификации сигнального пептида и внеклеточной области бычьего PD-L1 (номер доступа GenBank AB510902) были сконструированы праймеры с сайтами распознавания NheI и EcoRV с 5'-конца (boPD-L1-Ig F и R). ПЦР проводили с использованием синтезированной кДНК, полученной из бычьих PBMC, в качестве матрицы. Продукты ПЦР обрабатывали ферментами NheI (Takara) и EcoRV (Takara), очищали набором FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в Fc-вектор pCXN2.1-кроличий IgG1 (Niwa et al., 1991; Zettlmeissl et al., 1990; любезно предоставлен доктором T. Yokomizo, Juntendo University Graduate School of Medicine, и модифицирован в лаборатории автором), обработанный ферментами рестрикции аналогичным образом. Плазмиду экспрессии очищали набором QIAGEN Plasmid Midi (Qiagen) или набором FastGene Xpress Plasmid PLUS (NIPPON Genetics) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pCXN2.1-boPD-L1-Ig.
Праймер (boPD-L1-Ig F): GCTAGCATGAGGATATATAGTGTCTTAAC (SEQ ID NO:137)
Праймер (boPD-L1-Ig R): GATATCATTCCTCTTTTTTGCTGGAT (SEQ ID NO:138)
Клетки, экспрессирующие на растворимый бычий PD-1-Ig, получали в соответствии с методиками, описанными ниже. Вкраце, 2,5 мкг pDN11-boPD-1-Ig вводили в 4×106 клеток CHO-DG44 с помощью Lipofectamine LTX (Invitrogen). Через сорок восемь часов среду заменяли средой OptiCHO AGT (Life Technologies), содержащей 800 мкг/мл G418 (Enzo Life Science) и 20 мл/л добавки GlutaMAX (Life Technologies). После культивирования в течение 3 недель клетки подвергали селекции. Вкратце, концентрации рекомбинантного Fc-слитого белка в культуральных супернатантах полученных клонов клеток измеряли с помощью ELISA с использованием кроличьего поликлонального антитела против бычьего IgG F(c) (Rockland), чтобы тем самым отбрая те клеточные клоны, которые экспрессируют рекомбинантный Fc-слитый белок на высоком уровне. Полученный высокоэкспрессирующий клеточный клон переносили в среду, не содержащую G418, и культивировали при встряхивании в течение 14 дней с последующим сбором культурального супернатанта. Культуральный супернатант, содержащий рекомбинантный Fc-слитый белок, подвергали ультрафильтрации с Centricon Plus-70 (Millipore). Затем рекомбинантный Fc-слитый белок очищали с помощью Ab-Capcher Extra (ProteNova). После очистки буфер заменяли на забуференный фосфатом физиологический солевой раствор (PBS; pH 7,4), используя обессоливающую колонку PD-10 (GE Healthcare). Полученный белок хранили при -30°С перед использованием в экспериментах (бычий PD-1-Ig). Концентрацию очищенного бычьего PD-1-Ig измеряли с помощью ELISA с использованием кроличьих IgG F(c) поликлональных антител (Rockland). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
Клетки, экспрессирующие на растворимый бычий PD-L1-Ig, получали в соответствии с методиками, описанными ниже. Вкратце, 30 мкг pCXN2.1-boPD-L1-Ig вводили в 7,5×107 клеток Expi293F (Life Technologies), используя Expifectamine (Life Technologies). После 7-и дневного культивирования при встряхивании культуральный супернатант собирали. Рекомбинантный белок очищали от супернатанта с использованием Ab-Capcher Extra (ProteNova; бычий PD-L1-Ig). После очистки буфер меняли на PBS (pH 7,4), используя обессоливающую колонку PD-10 (GE Healthcare). Полученный белок хранили при -30°С перед использованием в эксперименте (бычий PD-L1-Ig). Концентрацию очищенного бычьего PD-L1-Ig измеряли с использованием набора Rabbit IgG ELISA Quantitation (Bethyl). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
Получение клеток, продуцирующих крысиные моноклональные антитела против бычьих PD-L1
Крысу иммунизировали в подушечку стопы бычьим PD-L1-Ig (Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug.; 142 (4): 551- 561; бычий PD-L1-Ig получали способом, описанным в этой статье, и использовали для иммунизации). Гибридомы получали из подвздошных лимфатических узлов, получая, таким образом, гибридому 4G12, продуцирующую крысиные моноклональные антитела против бычьих PD-L1. Что касается способа получения крысиного моноклонального антитела бычьего PD-L1, то подобное описание приведено в следующем непатентном документе (Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Vet. Res. 2013 Jul. 22; 44:59; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug.; 142(4):551-561).
Получение вектора, экспрессирующего химерное "крысино-бычьего" антитело против PD-L1
Химерное "крысино-бычьего" антитело ch4G12 против PD-L1 было получено путем слияния константных областей антитела бычьего IgG1 и Igλ с крысиным антителом против бычьего PD-L1 4G12, используемым в качестве вариабельной области антитела.
Сначала, гены вариабельных областей тяжелой цепи и легкой цепи были идентифицированы в гибридоме, которая продуцирует крысиное антитело 4G12 против бычьего PD-L1. Затем получали последовательность гена, в котором вариабельные области тяжелой цепи и легкой цепи антитела 4G12 связывали с известными константными областями бычьего IgG1 (тяжелая цепь; модифицирована; номер доступа в GenBank X62916) и бычьего Igλ (легкая цепь; номер доступа в GenBank X62917), соответственно, и была проведена оптимизация кодонов [химерное "крысино-бычьего" антитело ch4G12 против PD-L1: SEQ ID NO:105 и 106 (аминокислотные последовательности), SEQ ID NO:107 и 108 (нуклеотидные последовательности после оптимизации кодонов)]. Следует отметить, что для ингибирования активности ADCC бычьего IgG1 были добавлены мутации в предполагаемые сайты связывания с рецептором Fcγ в домене CH2 (см. фиг. 19, где указаны аминокислотные положения и мутации: 250 E→Р, 251 L→V, 252 P→A, 253 G→делеция, 347 A→S, 348 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug; 142(4):551-561). Затем ген был искусственно синтезирован таким образом, что последовательность распознавания NotI, последовательность KOZAK, последовательность легкой цепи химерного антитела, последовательность сигнала присоединения поли-A (PABGH), последовательность промотора (PCMV), последовательность распознавания SacI, последовательность интрона (INRBG), последовательность KOZAK, последовательность тяжелой цепи химерного антитела и последовательность распознавания XbaI были расположены в указанном порядке. Синтезированную цепь гена расщепляли с помощью NotI (Takara) и XbaI (Takara), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в сайт клонирования (последовательности распознавания ферментами NotI и XbaI ниже PCMV и между INRBG и PABGH) плазмиды экспрессии pDC6 (любезно предоставлен профессором S. Suzuki, Hokkaido University Research Center for Zoonosis Control), обработанной фрагментами рестрикции таким же образом (фиг. 20). Полученную плазмиду экстрагировали набором QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pDC6-boPD-L1ch4G12.
Экспрессия химерного "крысино-бычьего" антитела против бычьего PD-L1
PDC6-boPD-L1ch4G12 трансфицировали в клетки CHO-DG44 (CHO-DG44 (dfhr-/-)), которые представляли собой клетки с дефицитом дигидрофолатредуктазы. Через сорок восемь часов среду заменяли средой OptiCHO AGT (Life Technologies), содержащей 20 мг/мл добавки GlutaMAX (Life Technologies). После культивирования в течение 3 недель клетки подвергали селекции и клонированию с помощью лимитирующего разведения. Затем концентрации химерного антитела в культуральных супернатантах измеряли методом дот-блоттинга и ELISA с использованием кроличьего поликлонального антитела против бычьего IgG F(c) (Rockland), проводя таким образом селекцию клонов с высокой экспрессией. Кроме того, отобранные клоны с высоким уровнем экспрессии химерного "крысино-бычьего" антитела против бычьего PD-L1 подвергали генной амплификации путем добавления нагрузки с 60 нМ метотрексат (Mtx)-содержащей средой. Полученный таким образом клон клеток, стабильно экспрессирующий химерное "крысино-бычьего" антитело против бычьего PD-L1, переносили в среду Opti-CHO AGT без Mtx и культивировали при встряхивании в течение 14 дней (125 об/мин, 37°C, 5% CO2). Продукцию химерных антител в культуральном супернатанте измеряли с помощью ELISA с использованием поликлональных антител кролика против бычьего IgG F(c) (Rockland). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric). Культуральный супернатант на 14-й день центрифугировали при 10000 об/мин в течение 10 минут для удаления клеток, и супернатант пропускали через фильтр с размером пор 0,22 мкм Steritop-GP (Millipore) для стерилизации и затем хранили при 4°С перед очисткой.
Очистка химерного "крысино-бычьего" антитела против бычьего PD-L1
Из культурального супернатанта, полученного, как описано выше, каждое химерное антитело очищали с использованием Ab Capcher Extra (ProteNova). Для связывания со смолой использовали способ с открытой колонкой; PBS pH 7,4 использовали в качестве уравновешивающего буфера и промывочного буфера. В качестве буфера для элюции использовали буфер для элюции IgG Elution Buffer (Thermo Fisher Scientific). В качестве нейтрализующего буфера использовали 1М Трис (рН 9,0). В очищенном антителе заменяли буфер на PBS (pH 7,4), используя обессоливающую колонки PD-10 (GE Healthcare) и концентрировали с использованием Amicon Ultra-15 (50 кДа, Millipore). Очищенное таким образом химерное антитело пропускали через шприцевой фильтр с размером пор 0,22 мкм (Millipore) для стерилизации и хранили при 4°С перед применением в экспериментах.
Подтверждение чистоты очищенного химерного "крысино-бычьего" антитела против бычьего PD-L1 (фиг.21).
Чтобы подтвердить чистоту очищенного химерного "крысино-бычьего" антитела против бычьего PD-L1, белки антител обнаруживали с помощью SDS-PAGE и окрашивания CBB. Используя 10% акриламидный гель, очищенное химерное "крысино-бычьего" антитело подвергали электрофорезу в восстанавливающих условиях (восстановление с 2-меркаптоэтанолом от Sigma-Aldrich) и в невосстанавливающих условиях. Полосы окрашивали с помощью набора Quick-CBB (Wako) и обесцвечивали в дистиллированной воде. Результаты показаны на фиг.21. Полосы отмечали в прогнозированных положениях, то есть при 25 кДа и 50 кДа в восстанавливающих условиях и при 150 кДа в невосстанавливающих условиях.
Специфичность связывания химерного "крысино-бычьего" антитела против бычьего PD-L1 (фиг.22).
С помощью проточной цитометрии было подтверждено, что химерное "крысино-бычьего" антитело против бычьего PD-L1 специфически связывается с клетками, экспрессирующими бычье PD-L1 (описано выше). Сначала крысиное антитело 4G12 против бычьего PD-L1 или химерное "крысино-бычьего" антитело ch4G12 против PD-L1 подвергали взаимодействию с клетками, экспрессирующими бычий PD-L1, при комнатной температуре в течение 30 минут. После промывки APC-меченного козлиного антитела против крысиного Ig (Southern Biotech) или Alexa Fluor 647-меченого козлиного F(ab')2 против бычьего IgG (H+L) (Jackson ImmunoResearch) проводили взаимодействие при комнатной температуре в течение 30 мин. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience) или бычьим антителом IgG1 (Bethyl). После промывки каждое крысиное антитело или химерное "крысино-бычьего" антитело, связанное с клеточными поверхностями, детектировали с помощью FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 22. Было обнаружено, что химерное "крысино-бычьего" ch4G12 антитело против бычьего PD-L1 связывается с клетками экспрессирующими бычьими PD-L1 так же как и крысиное антитело 4G12 против бычьего PD-L1.
Ингибиторная активность химерного "крысино-бычьего" антитела против бычьего PD-L1 в отношении связывания бычьего PD-1/PD-L1.
(1) Тест ингибирования связывания на клетках, экспрессирующих бычий PD-L1, и растворимый бычий PD-1 (фиг. 23)
Используя клетки, экспрессирующие бычьи PD-L1 (как описано выше) и бычий PD-1-Ig (как описано выше), тестировали ингибирование связывания бычьего PD-1/PD-L1 с помощью антитела против бычьего PD-L1. Сначала 2×105клеток, экспрессирующих бычий PD-L1, реагировали с различными концентрациями (0, 0,32, 0,63, 1,25, 2,5, 5 или 10 мкг/мл) крысиного антитела 4G12 против бычьего PD-L1 или химерного "крысино-бычьего" антитела ch4G12 против бычьего PD-L1 при комнатной температуре в течение 30 мин. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience) или бычьим антителом IgG1 (Bethyl). После промывки добавляли бычий PD-1-Ig, меченный биотином, с использованием набора Lightning-Link Type A Biotin Labeling (Innova Bioscience) до конечной концентрации 2 мкг/мл с последующим взаимодействием в течение дополнительных 30 минут при комнатной температуре. Затем, после промывки, бычий PD-1-Ig, связанный с клеточными поверхностями, обнаруживали с помощью APC-меченного стрептавидина (BioLegend). Для анализа использовали FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich). Принимая долю клеток, связанных с PD-1-Ig без добавления антител, за 100%, доля клеток, связанных с PD-1-Ig, для каждой концентрации антител была показана как относительная величина.
Экспериментальные результаты приведены на фиг. 23. Было обнаружено, что подобно крысиному антителу 4G12 против бычьего PD-L1, химерное "крысино-бычьего" антитело ch4G12 против бычьего PD-L1 способно ингибировать связывание бычьего PD-1/PD-L1 зависимым от концентрации образом.
(2) Тест ингибирования связывания на клетках, экспрессирующих бычий PD-1, и растворимый бычий PD-L1 (фиг. 24)
Используя клетки, экспрессирующие бычьи PD-1 (как описано выше) и бычий PD-L1-Ig (как описано выше), тестировали ингибирование связывания бычьего PD-1/PD-L1 с помощью антитела против бычьего PD-L1. Во-первых, крысиное антитело 4G12 против бычьего PD-L1 или химерное "крысиное-бычье" антитело ch4G12 против бычьего PD-L1 с конечной концентрацией 0, 0,32, 0,63, 1,25, 2,5, 5 или 10 мкг/мл и бычьего PD-L1-Ig с конечной концентрацией 1 мкг/мл помещали в 96-луночные планшеты, где они взаимодействовали при комнатной температуре в течение 30 мин. Полученную смесь подвергали взаимодействию с 2×105 клеток, экспрессирующих бычий PD-1, при комнатной температуре в течение 30 минут. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience) или бычьим антителом IgG1 (Bethyl). После промывки Alexa Fluor 647-меченный козий F(ab')2 против козлиного IgG (H+L) (Life Technologies) взаимодействовал при комнатной температуре в течение 30 минут, таким образом обнаруживая бычий PD-L1-Ig, связанный с клеточными поверхностями. Для анализа использовали FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich). Принимая долю клеток, связанных с PD-L1-Ig без добавления антител, за 100%, доля клеток, связанных с PD-L1-Ig, для каждой концентрации антител была показана как относительная величина.
Экспериментальные результаты приведены на фиг. 24. Было обнаружено, что подобно крысиному антителу 4G12 против бычьего PD-L1, химерное "крысино-бычьего" антитело ch4G12 против бычьего PD-L1 способно ингибировать связывание бычьего PD-1/PD-L1 зависимым от концентрации образом.
Тест биологической активности с использованием химерного "крысино-бычьего" антитела против бычьего PD-L1
(1) Эффект на пролиферацию клеток (фиг. 25)
Для подтверждения того факта, что ингибирование связывания бычьего PD-1/PD-L1 химерным "крысино-бычьим" антителом против PD-L1 активирует лимфоциты, проводили тест биологической активности с использованием пролиферации клеток в качестве индикатора. Вкратце, бычьи РВМС, выделенные из периферической крови здоровой особи крупного рогатого скота, суспендировали в PBS до концентрации 10×10 6 клеток/мл и взаимодействовали с сукцинимидиловым эфиром карбоксифлуоресцеина (CFSE) при комнатной температуре в течение 20 минут. После двухкратной промывки средой RPMI 1640 (Sigma-Aldrich), содержащей 10% инактивированную фетальную бычью сыворотку (Cell Culture Technologies), антибиотики (стрептомицин 200 мкг/мл, пенициллин 200 Ед/мл) (Life Technologies) и 0,01% L-глютамин (Life Technologies), PBMC подвергали взаимодействию с мышиным антителом против бычьего CD3 (WSU Monoclonal Antibody Center) при 4°C в течение 30 минут. После промывки РВМС взаимодействовали с микрогранулами с антимышиным IgG1 (Miltenyi Biotec) при 4°С в течение 15 минут, а затем выделяли CD3-позитивные Т-клетки, используя autoMACS™ Pro (Miltenyi Biotec). К выделенным CD3-позитивным Т-клеткам добавляли мышиное антитело против бычьего CD3 (WSU Monoclonal Antibody Center) и мышиное антитело против бычьего CD28 (Bio-Rad). Затем клетки культивировали вместе с клетками, экспрессирующими бычий PD-L1 (CD3-позитивные Т-клетки:клетки, экспрессирующие бычий PD-L1=10:1) в присутствии или в отсутствие 10 мкг/мл химерного "крысино-бычьего" антитела ch4G12 против PD-L1. В качестве контроля антител использовали полученный из сыворотки бычий IgG (Sigma-Aldrich); в качестве контроля для клеток, экспрессирующих PD-L1, использовали клетки, экспрессирующие EGFP, трансфицированные pEGFP-N2. После 6-дневного совместного культивирования клетки собирали и реагировали с мышиным антителом против бычьего CD4 и мышиным антителом против бычьего CD8 (Bio-Rad) при комнатной температуре в течение 30 минут. Для мечения антител использовали наборы Zenon Mouse IgG1 Labeling (Life Technologies) или Lightning-Link (Innova Biosciences). Для анализа использовали FACS Verse (BD Biosciences). Для операции промывки и разбавления антитела использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 25. Пролиферация CD4-позитивных и CD8-позитивных Т-клеток была значительно супрессирована при совместном культивировании с клетками, экспрессирующими бычьи PD-L1. Было обнаружено, что химерное "крысино-бычье" антитело ch4G12 против PD-L1 ингибирует такое супрессирование в CD4-позитивных Т-клетках.
(2) Влияние на продукцию IFN-γ (фиг. 26)
Для подтверждения того факта, что ингибирование связывания бычьего PD-1/PD-L1 химерным "крысино-бычьим" антителом против PD-L1 активирует лимфоциты, проводили тест биологической активности с использованием IFN-γ в качестве индикатора. Вкратце, РВМС, выделенные из периферической крови BLV-инфицированной особи крупного рогатого скота, суспендировали в среде RPMI (Sigma-Aldrich), содержащей 10% инактивированной фетальной бычьей сыворотки (Cell Culture Technologies), антибиотики (стрептомицин 200 мкг/мл, пенициллин 200 Ед/мл) ( Life Technologies) и 0,01% L-глютамина (Life Technologies) с получением концентрации 4×106 клеток/мл. К РВМС добавляли 10 мкг/мл крысиного антитела 4G12 против бычьего PD-L1 или химерного "крысино-бычьего" антитела ch4G12 против бычьего PD-L1 и 2% культурального супернатанта BLV-инфицированных фетальных клеток почки (FLK-BLV); культивирование затем проводили при 37°С и 5% СО2 в течение 6 дней. В качестве контрольных антител использовали сывороточный крысиный IgG (Sigma-Aldrich) и сыворотчный бычий IgG (Sigma-Aldrich). Через 6 дней культивирования собирали культуральный супернатант и измеряли продукцию IFN-γ, используя набор Bovine IFN-γ ELISA (BETYL). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
Экспериментальные результаты приведены на фиг. 26. Было обнаружено, что химерное "крысино-бычье" антитело ch4G12 против бычьего PD-L1 повышает ответ IFN-γ бычьих РВМС на антиген BLV таким же образом, как крысиное антитело 4G12 против бычьего PD-L1 (n=10).
Анализ CDR крысиного антитела против бычьего PD-L1
Определяющие комплементарность области (CDR) крысиного антитела 4G12 против бычьего PD-L1 определяли с использованием NCBI IGBLAST (http://www.ncbi.nlm.nih.gov/igblast/). Результаты показаны на фиг.19.
Тест с инокуляцией крупного рогатого скота
Полученное химерное "крысино-бычье" антитело ch4G12 против PD-L1 (приблизительно 260 мг; 1 мг/кг) вводили внутривенно экспериментально BLV- инфицированному теленку (самец голштинской породы, 7 месяцев, 267 кг). Образцы крови собирали в хронологическом порядке от зараженного теленка и затем выделяли РВМС центрифугированием в градиенте плотности.
(1) Реакция пролиферации клеток Т-клеток на антиген BLV (фиг. 27)
Бычьи РВМС суспендировали в PBS и проводили реакцию с CFSE при комнатной температуре в течение 20 минут. После двухкратной промывки средой RPMI 1640 (Sigma-Aldrich), содержащей 10% инактивированную фетальную бычью сыворотку (Cell Culture Technologies), антибиотики (стрептомицин 200 мкг/мл, пенициллин 200 Ед/мл) (Life Technologies) и 0,01% L-глютамин (Life Technologies), концентрацию клеток доводили до 4×106 клеток/мл, используя ту же среду. Культуральный супернатант 2% BLV-инфицированных фетальных клеток почки (FLK-BLV) добавляли в РВМС, которые затем культивировали при 37°С и 5% СО2 в течение 6 дней. В качестве контроля использовали культуральный супернатант из 2% BLV-неинфицированных фетальных клеток почек (FLK). После 6-дневного культивирования PBMC собирали и подвергали взаимодействию с мышиным антителом против бычьего CD4, мышиным антителом против бычьего CD8 и мышиным антителом против бычьего IgM (Bio-Rad) при 4°C в течение 20 минут. Для мечения антител использовали наборы Zenon Mouse IgG1 Labeling (Life Technologies) или Lightning-Link (Innova Biosciences). Для анализа использовали FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 27. В результате введения антитела BLV-специфический ответ в виде пролиферации клеток CD4-позитивных Т-клеток повышался по сравнению с ответом перед введением.
(2) Изменения вирусной нагрузки BLV (фиг. 28)
ДНК выделяли из выделенных бычьих РВМС с использованием набора для очистки Wizard DNA Purification (Promega). Концентрацию экстрагированной ДНК определяли количественно, принимая оптическую плотность (260 нм), измеренную на спектрофотометре Nanodrop 8000 (Thermo Fisher Scientific), в качестве эталона. Для измерения провирусной нагрузки BLV в РВМС, проводили ПЦР в реальном времени с использованием Cycleave PCR Reaction Mix SP (TaKaRa) и зонд/праймер/положительный контроль для обнаружения вируса лейкоза крупного рогатого скота (TaKaRa). Для измерения использовали Light Cycler 480 System II (Roche Diagnosis).
Экспериментальные результаты приведены на фиг. 28. Провирусная нагрузка BLV значительно снижалась до конца периода тестирования по сравнению с нагрузкой до введения.
Все публикации, патенты и заявки на патент, которые приводятся в данном документе, тем самым включены в своем полном объеме посредством ссылки.
Промышленная применимость
Антитело против PD-L1 по настоящему изобретению можно использовать для профилактики и/или лечения злокачественного новообразования и инфекций у животных.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ PD-L1 | 2017 |
|
RU2744911C2 |
| АНТИТЕЛО ПРОТИВ LAG-3 | 2017 |
|
RU2744866C2 |
| АНТИ-PD-L1 АНТИТЕЛО ДЛЯ ДЕТЕКЦИИ PD-L1 | 2018 |
|
RU2758723C2 |
| АНТИТЕЛО ПРОТИВ ЛИГАНДА 1 ЗАПРОГРАММИРОВАННОЙ ГИБЕЛИ КЛЕТОК (PD-L1), ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2727914C2 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2761663C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ ИНТЕРЛЕЙКИНА-31 | 2012 |
|
RU2588462C2 |
| КАНИНИЗИРОВАННЫЕ МЫШИНЫЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ PD-1 | 2014 |
|
RU2676158C1 |
| СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ВИРУСА БЕШЕНСТВА НЕЙТРАЛИЗУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА И НУКЛЕИНОВЫЕ КИСЛОТЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2001 |
|
RU2272809C2 |
| ОПУХОЛЕСПЕЦИФИЧНОЕ АНТИТЕЛО ПРОТИВ EGFR И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2730605C2 |
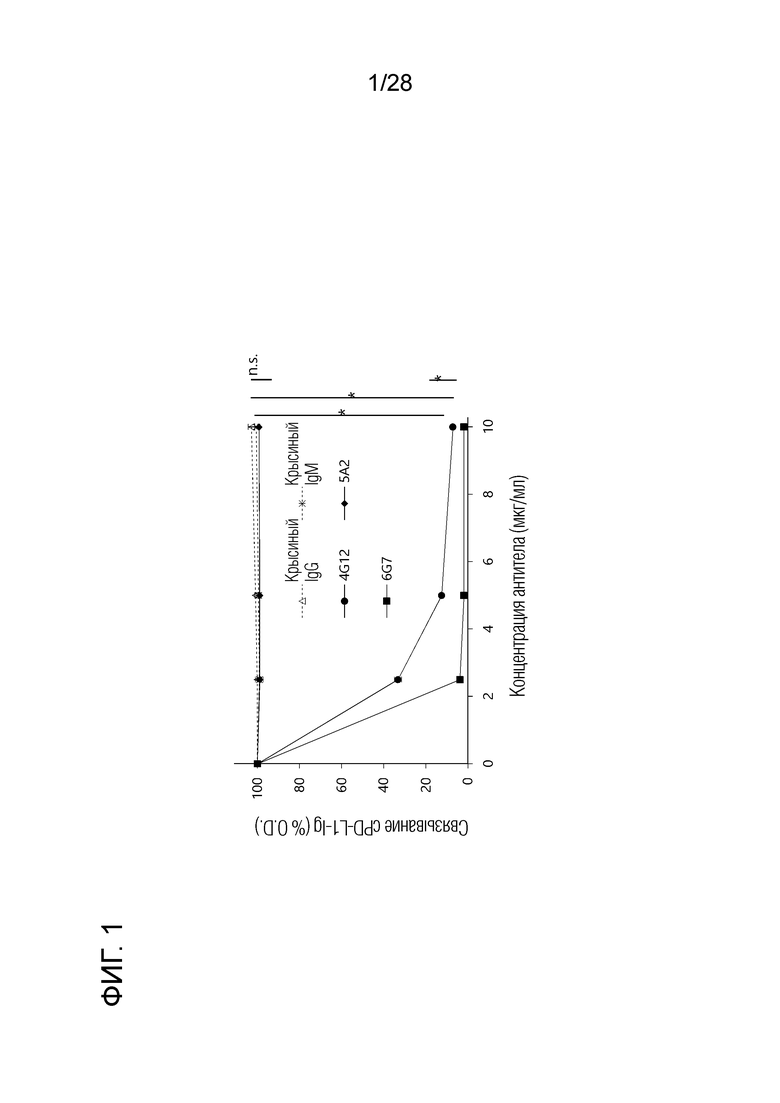
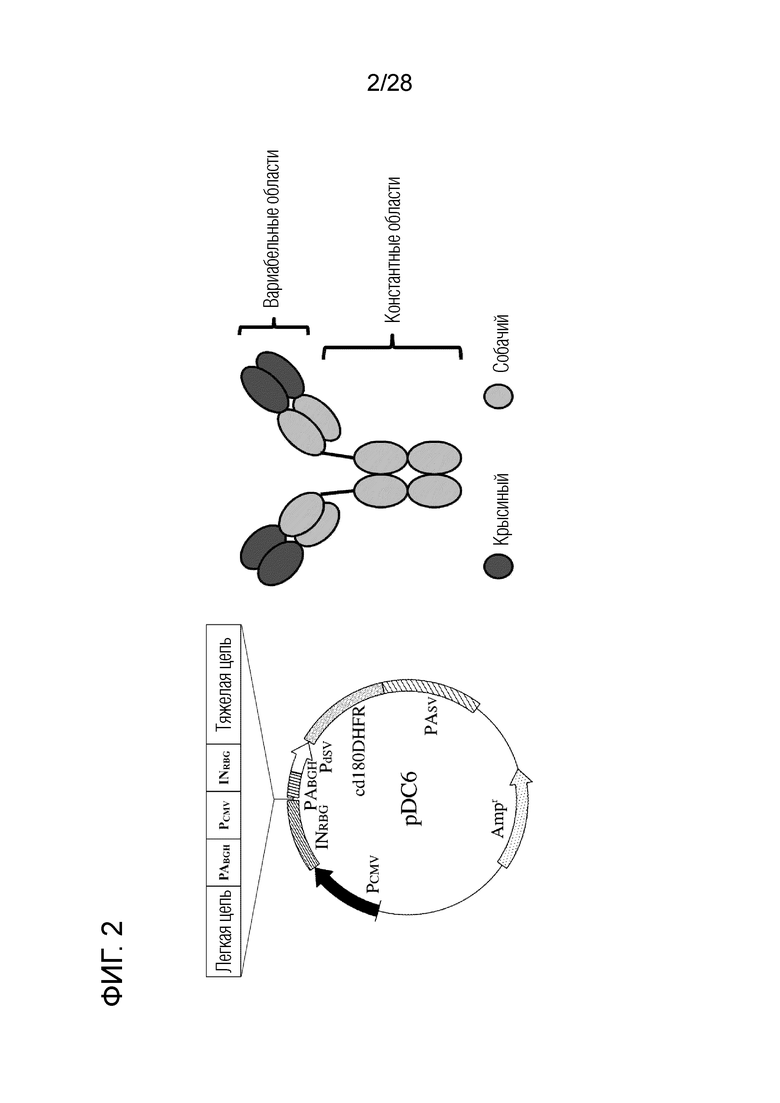

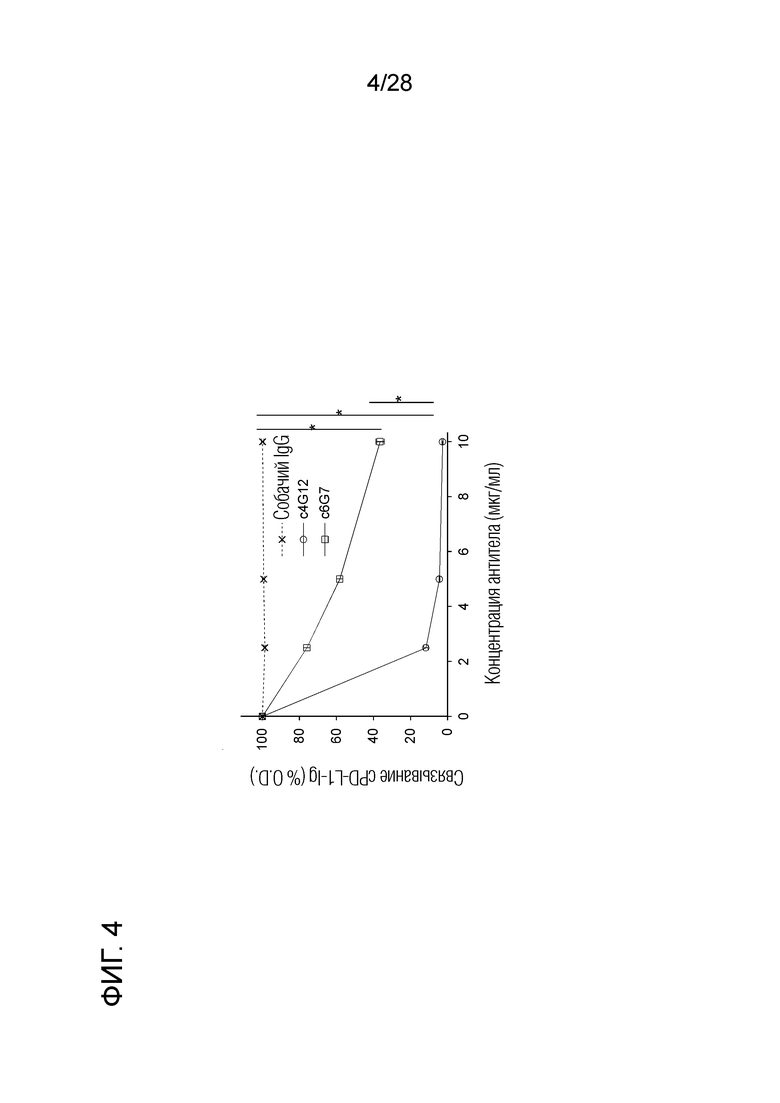
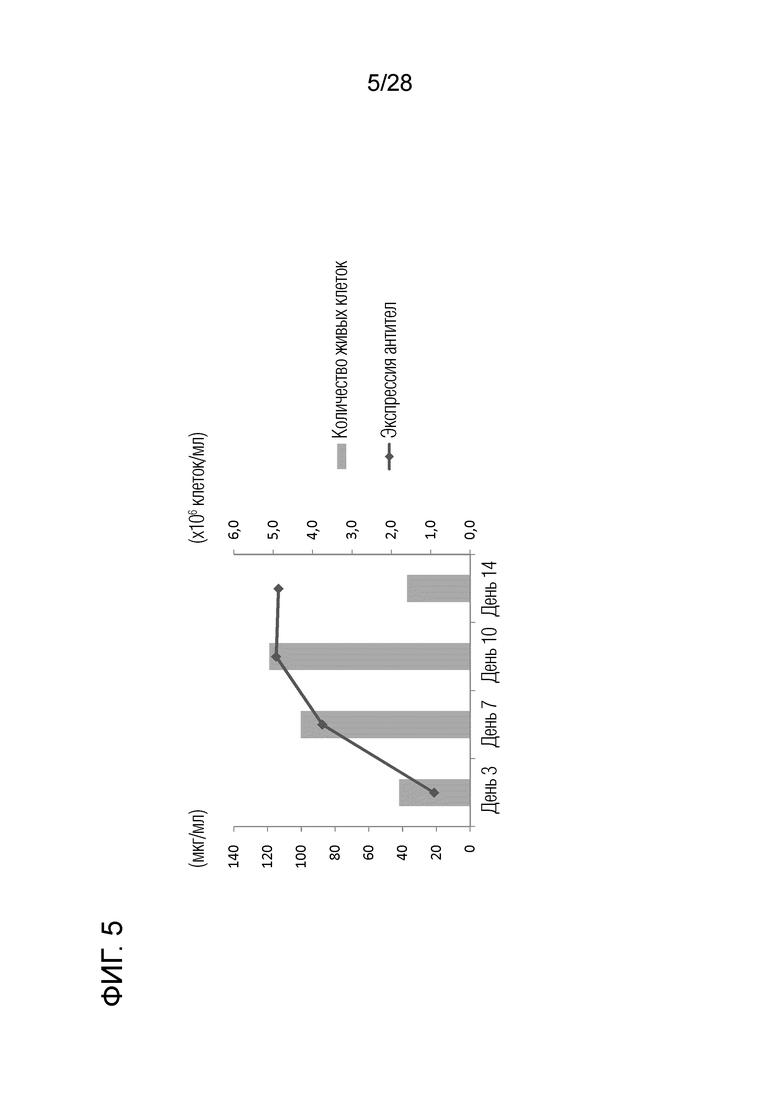

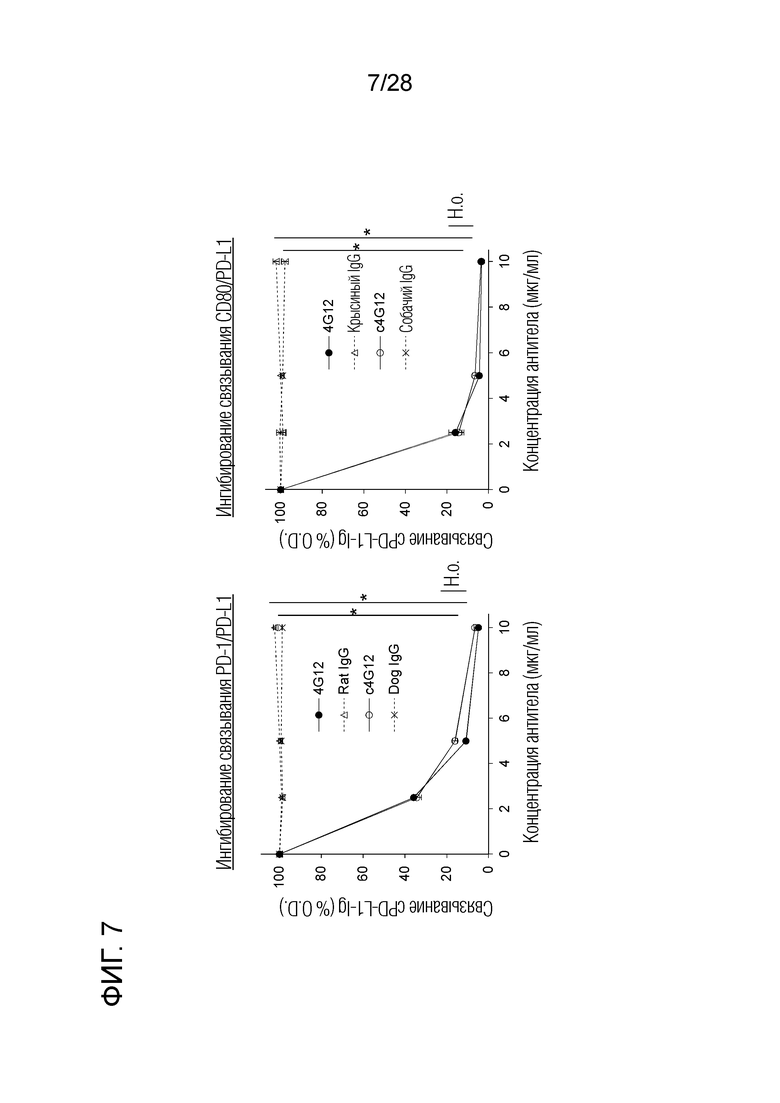
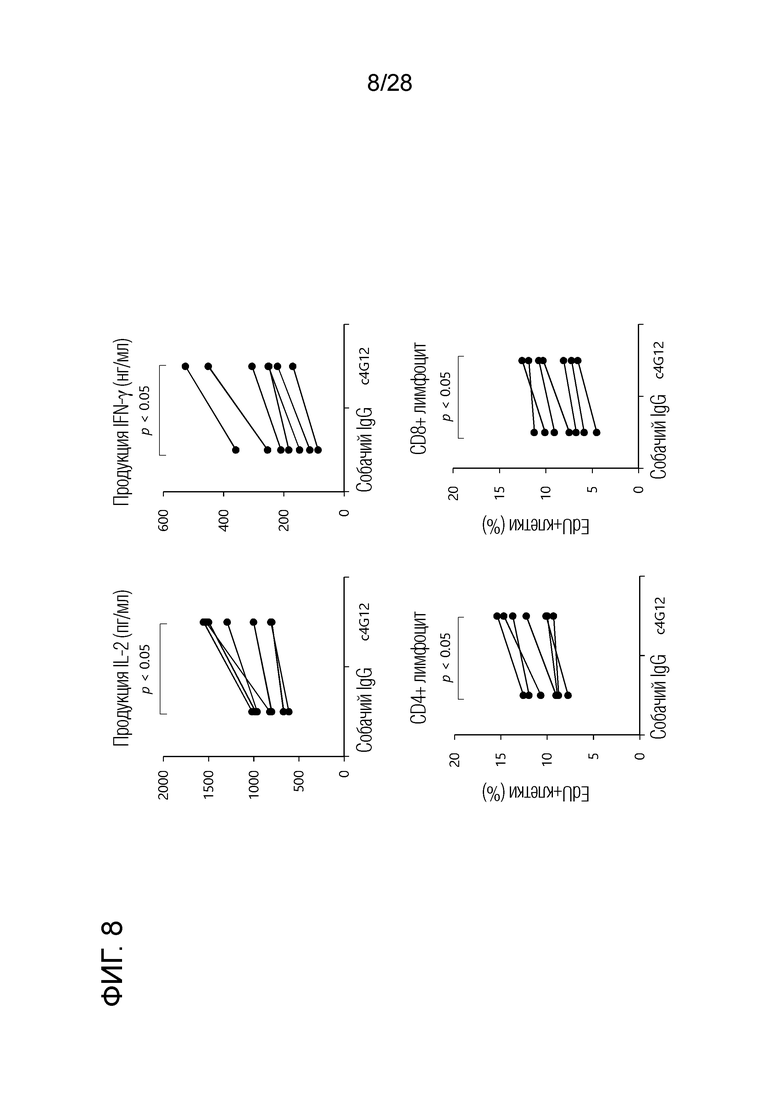

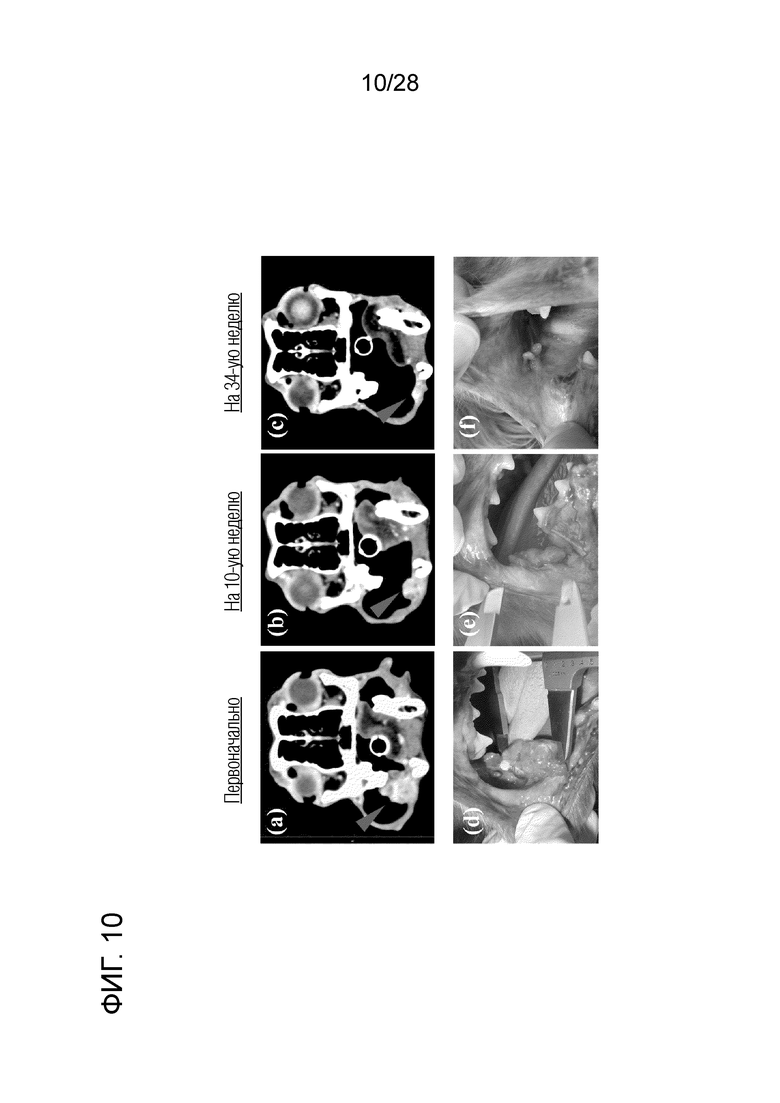
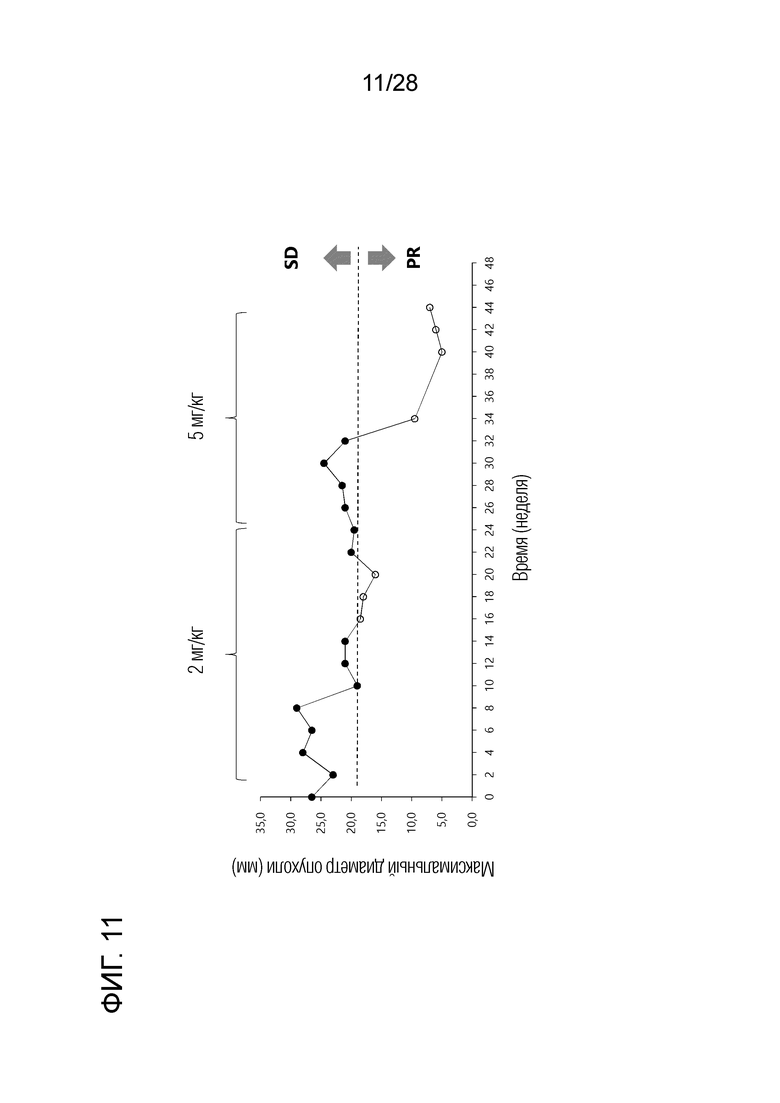
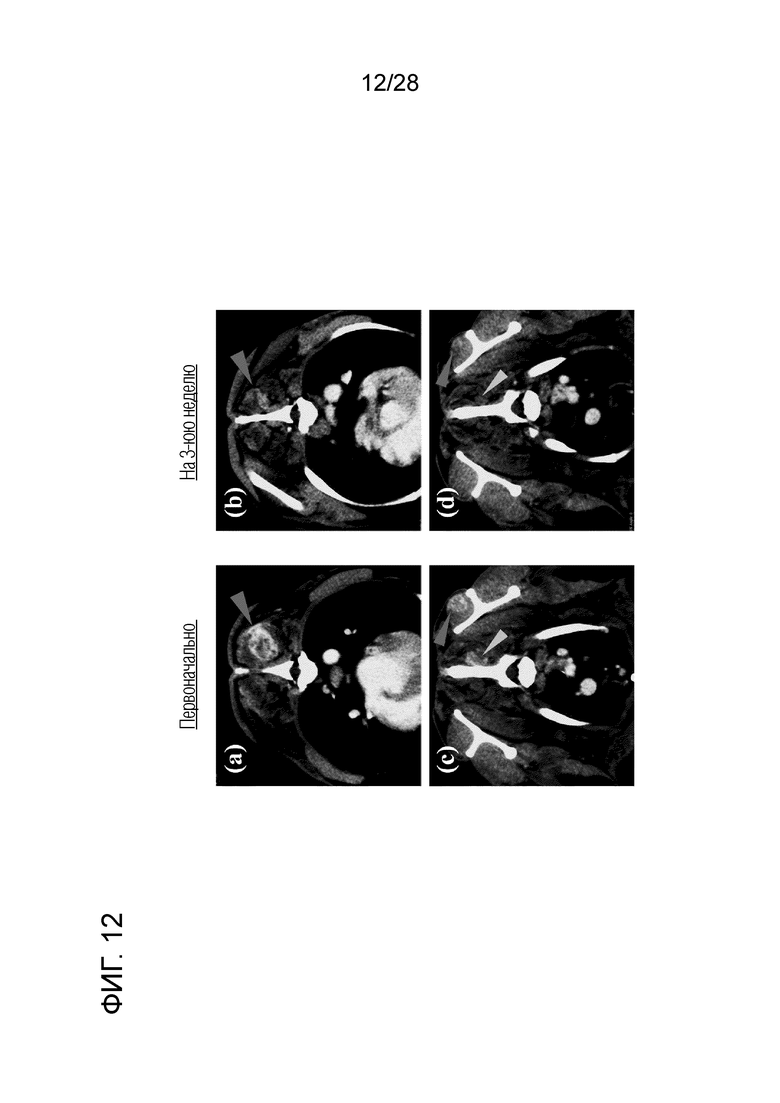
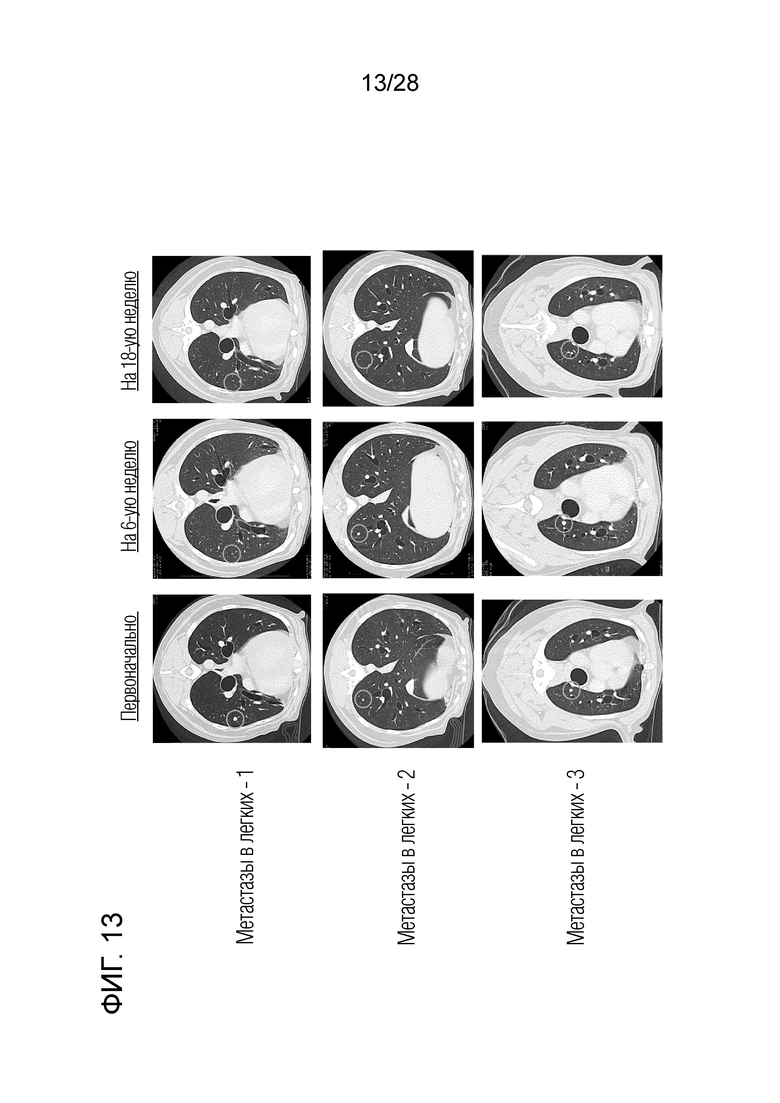
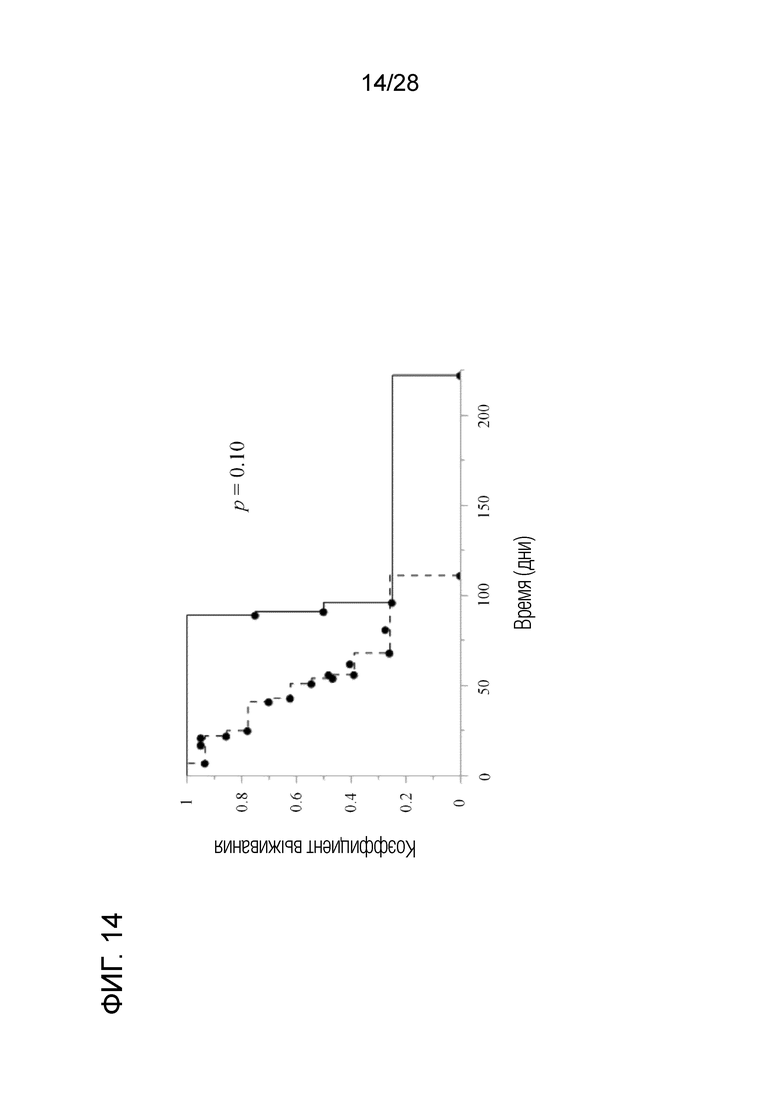

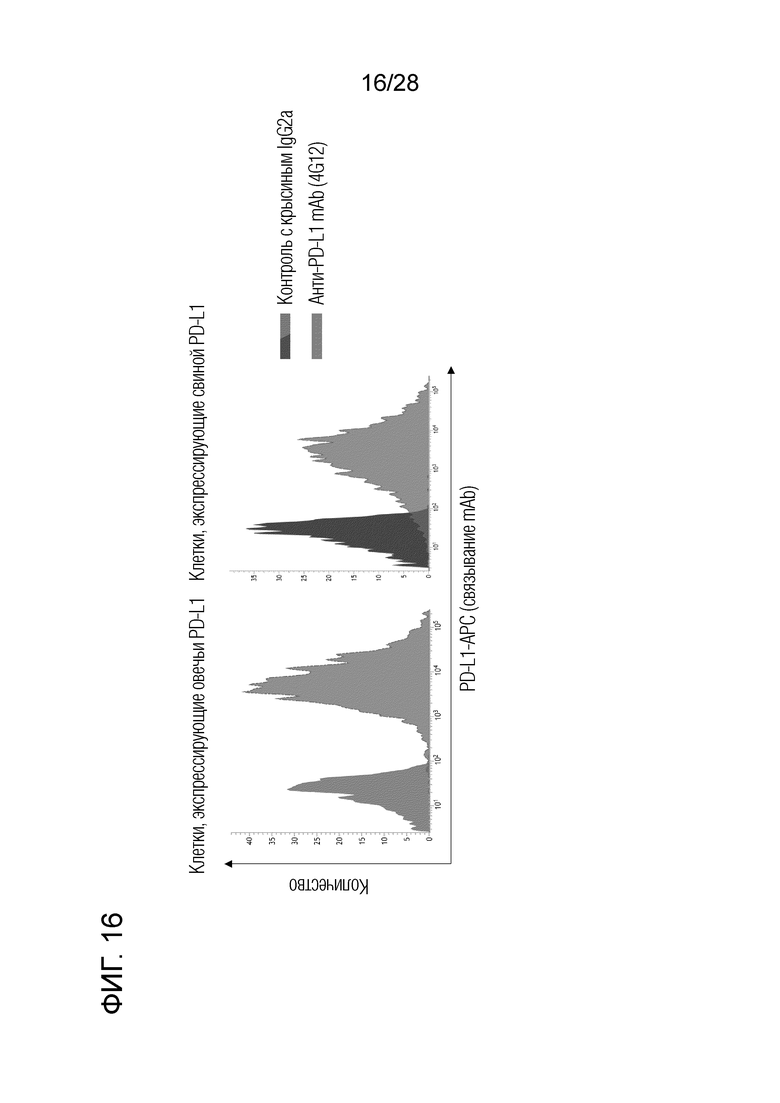
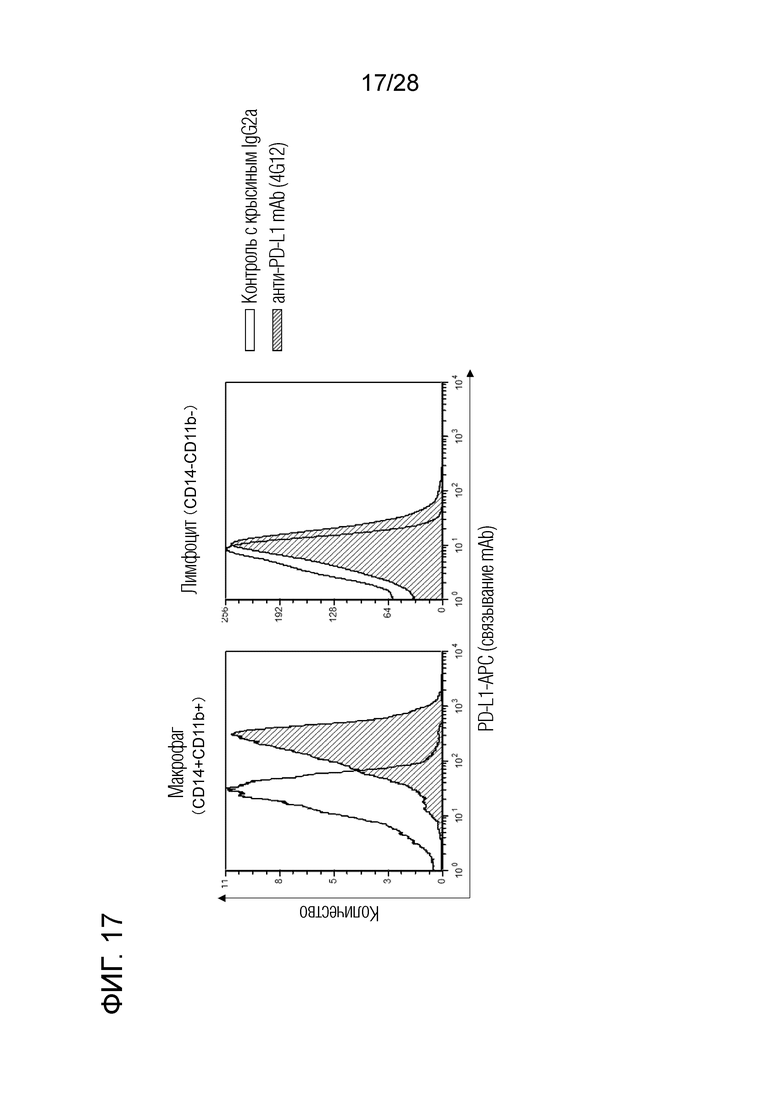
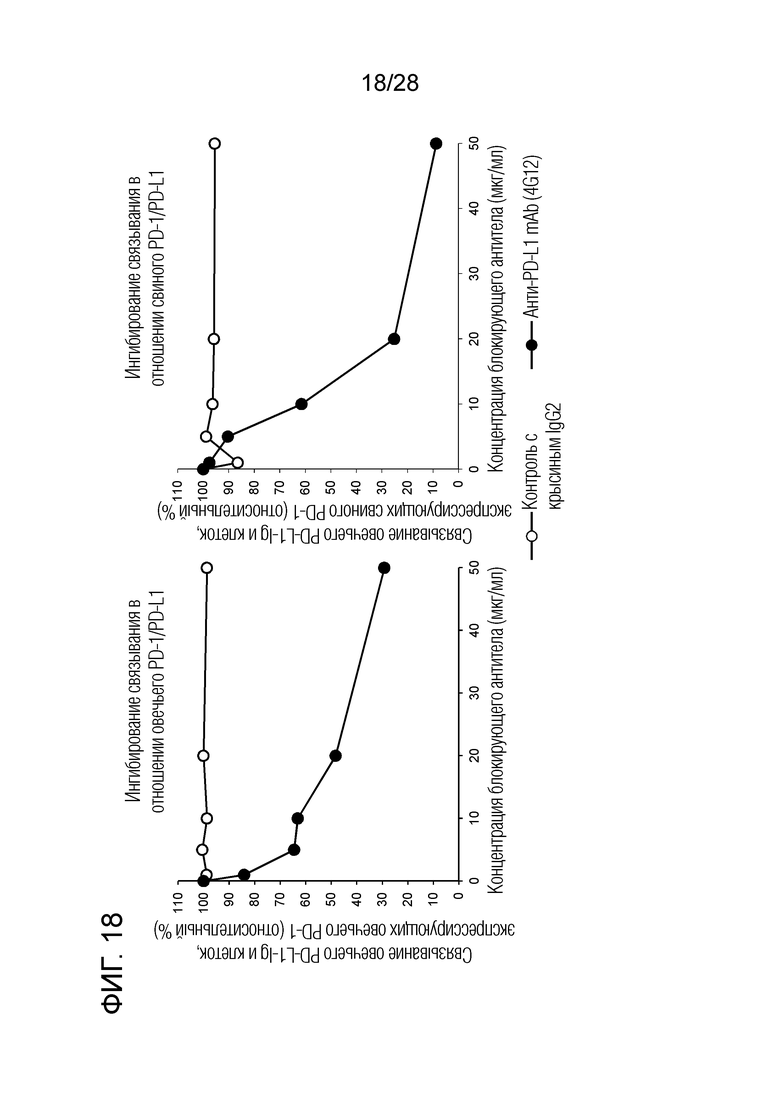

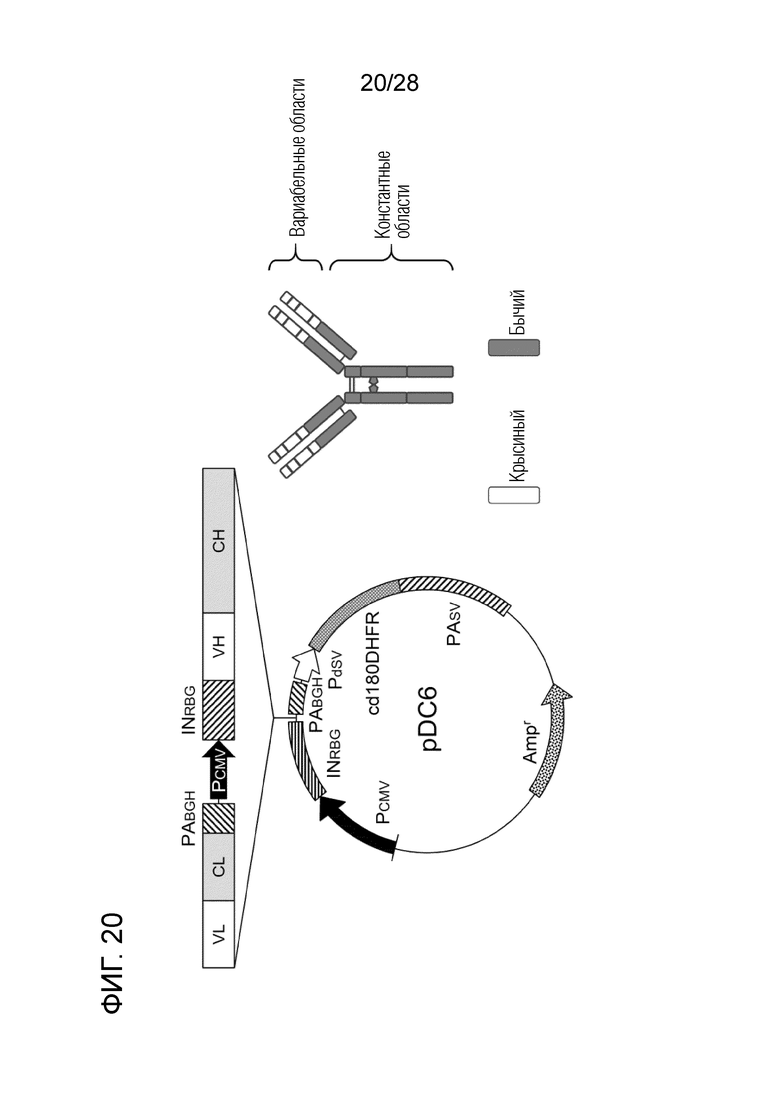
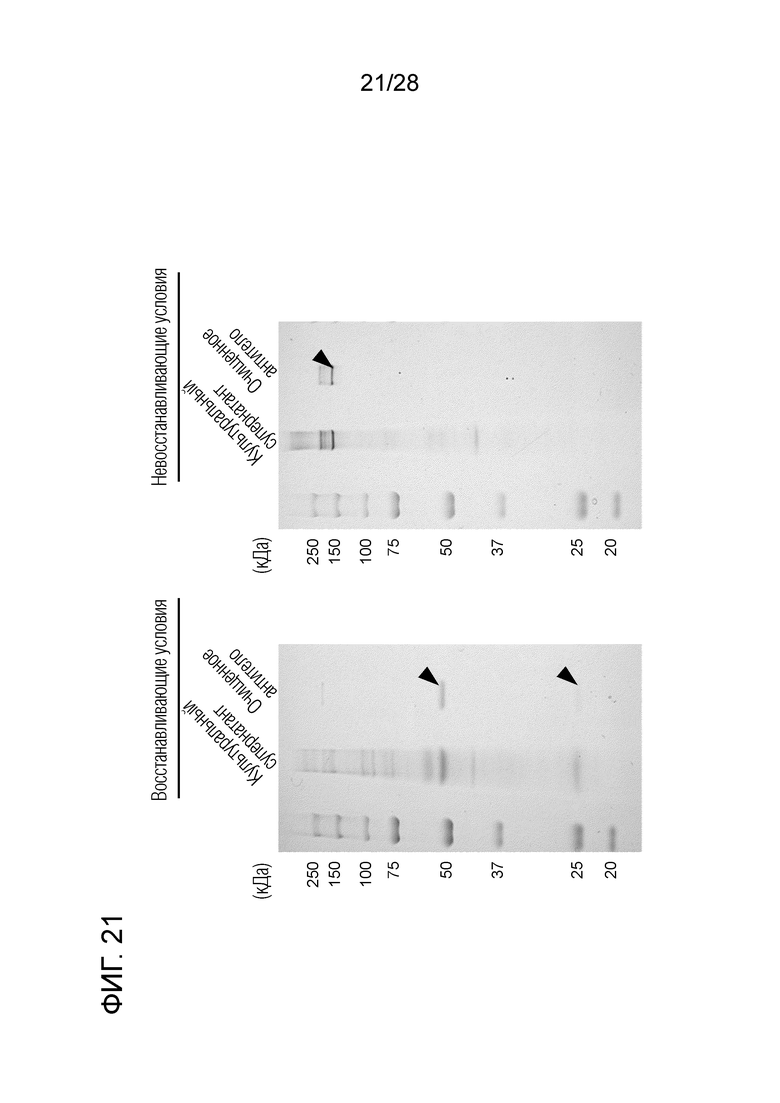
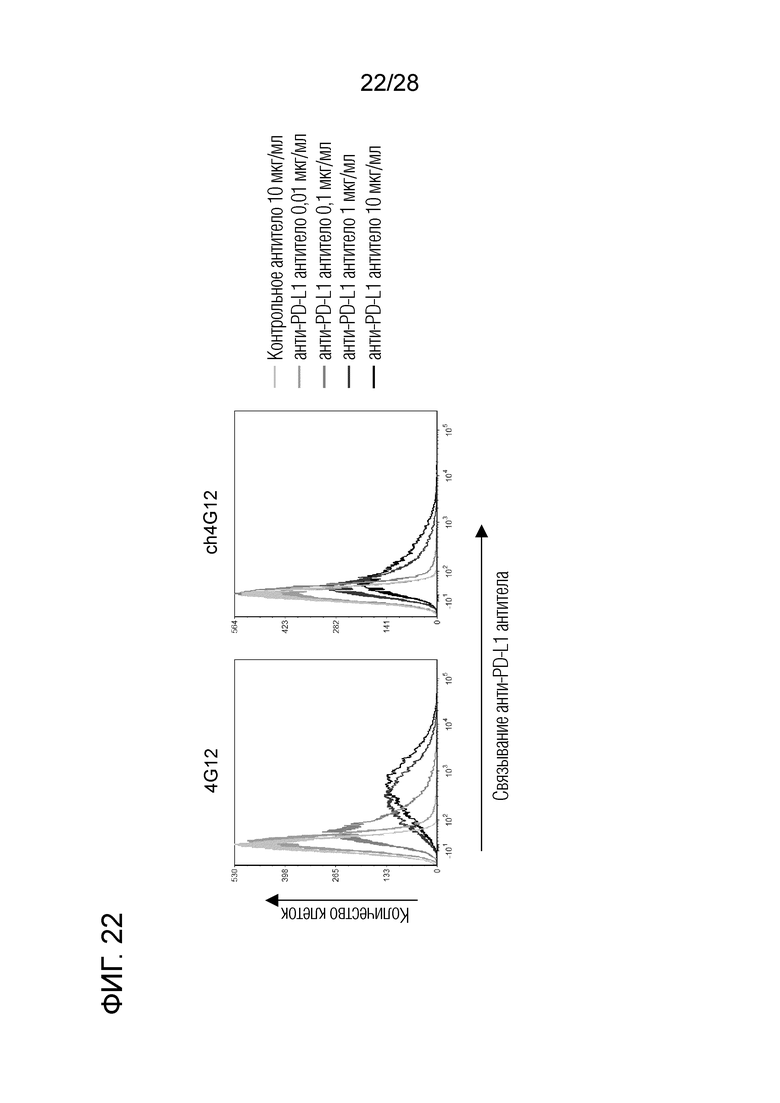
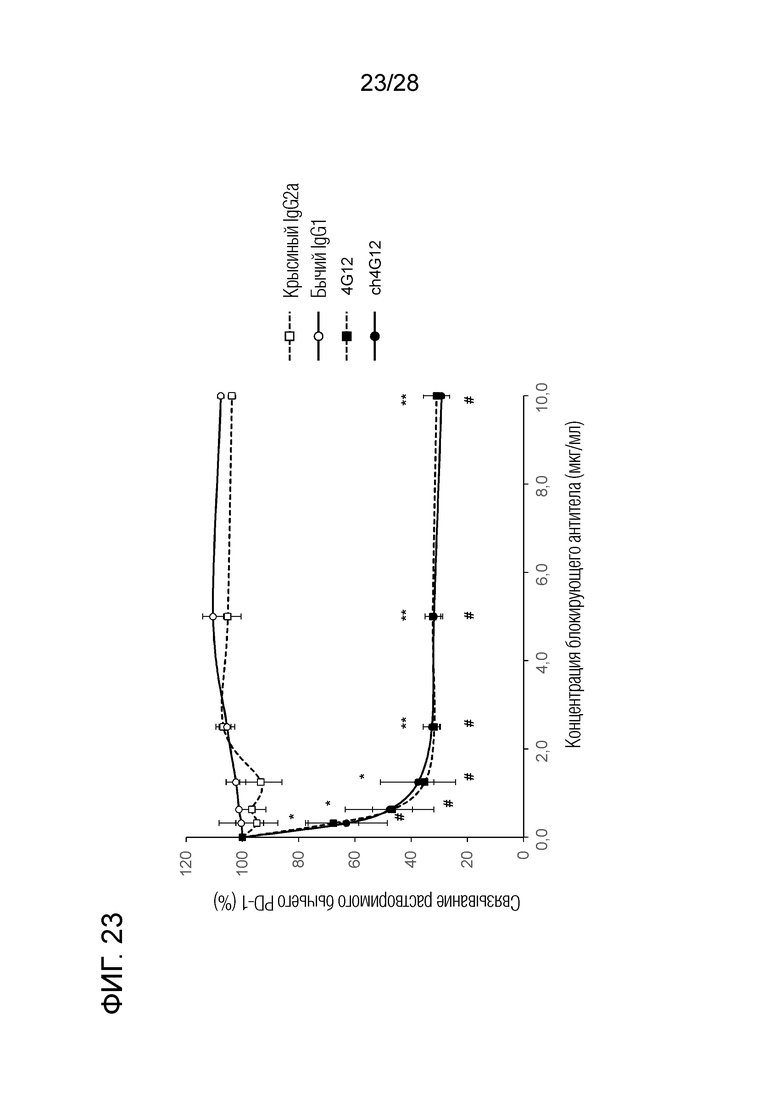
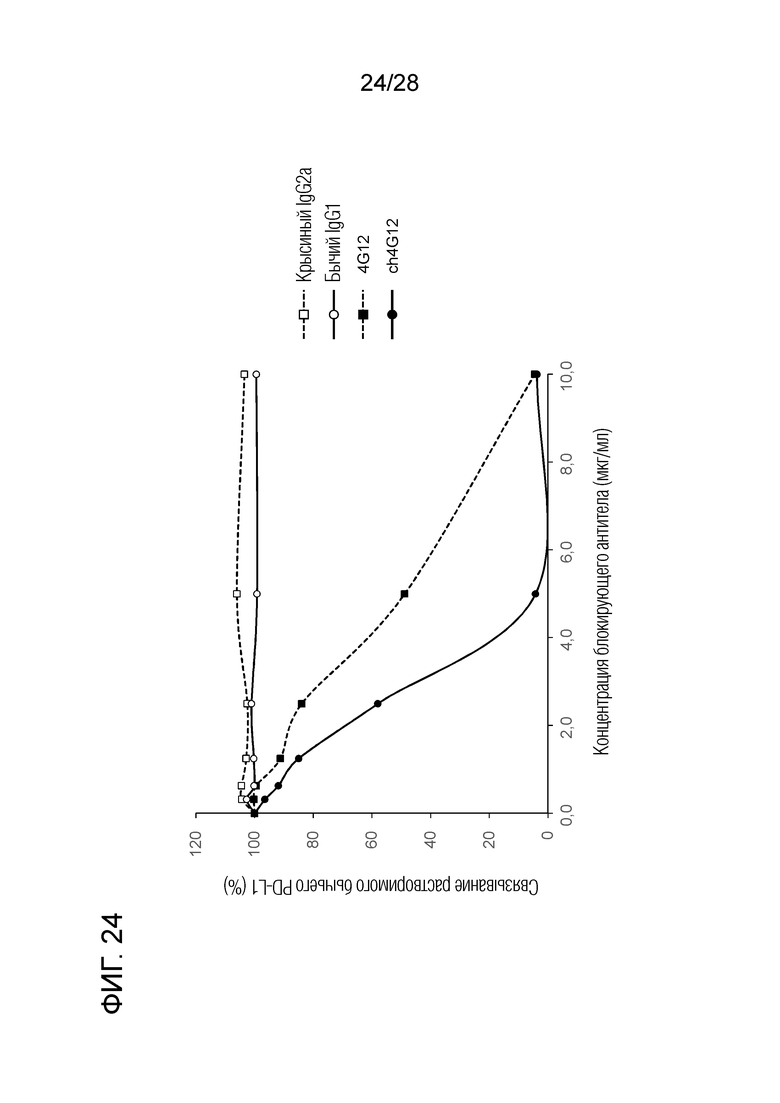
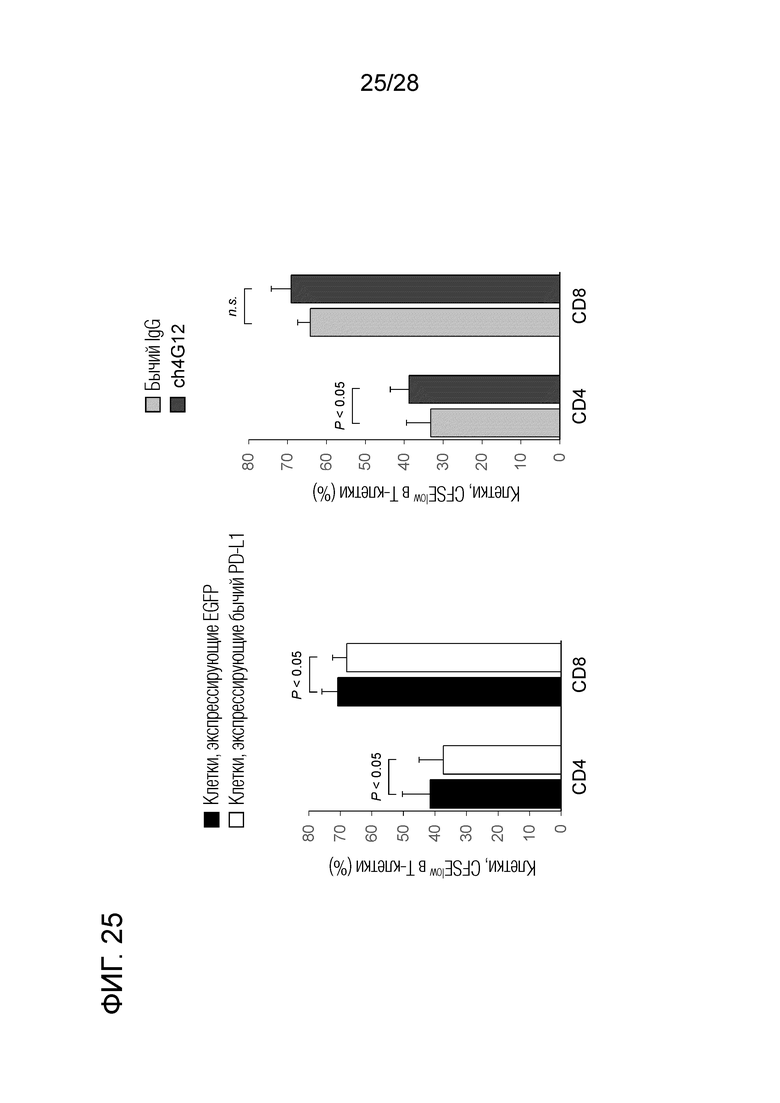
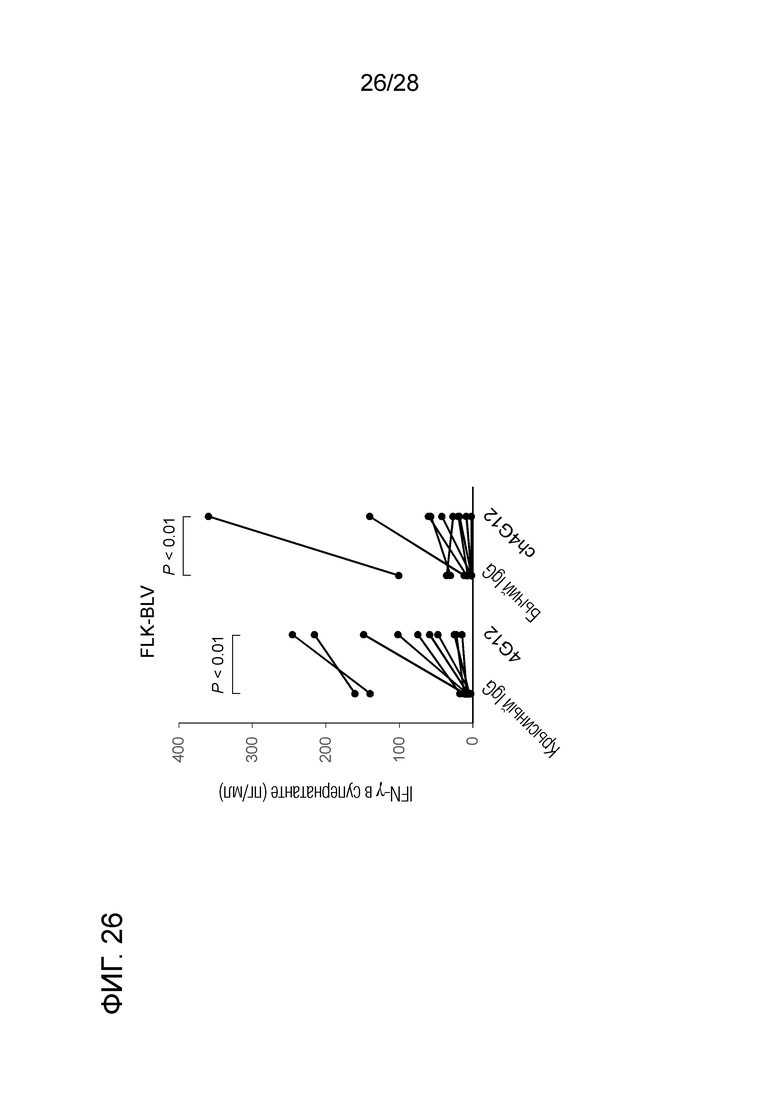
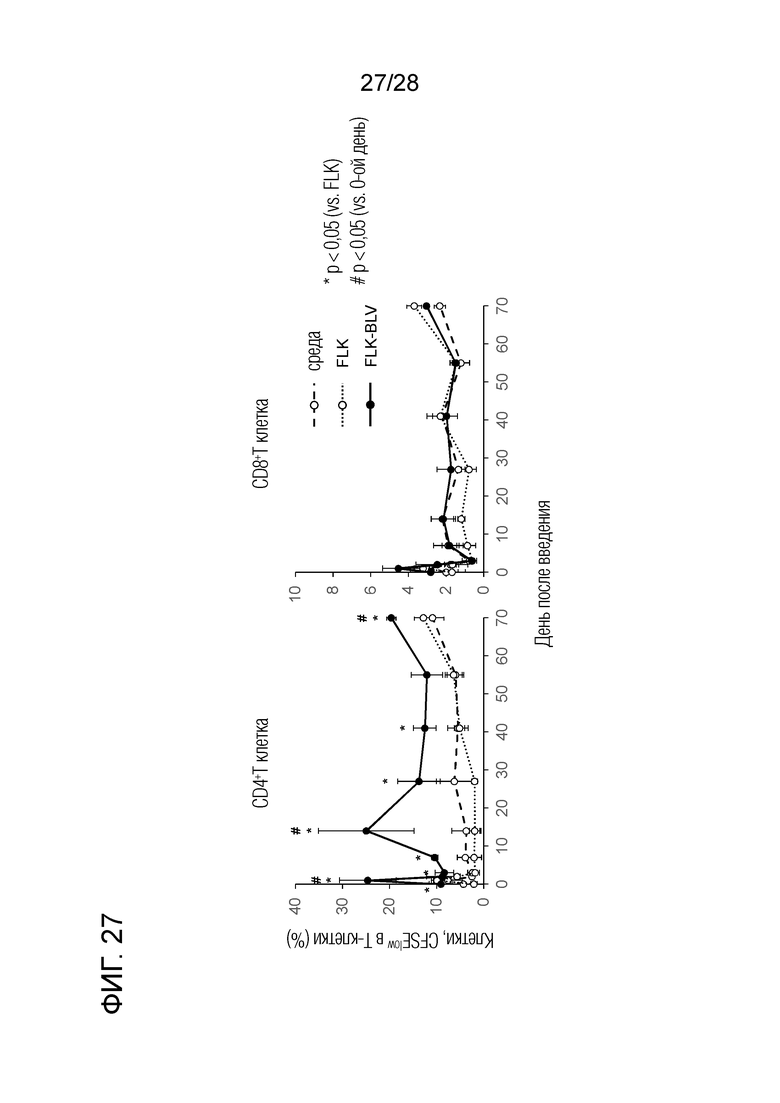
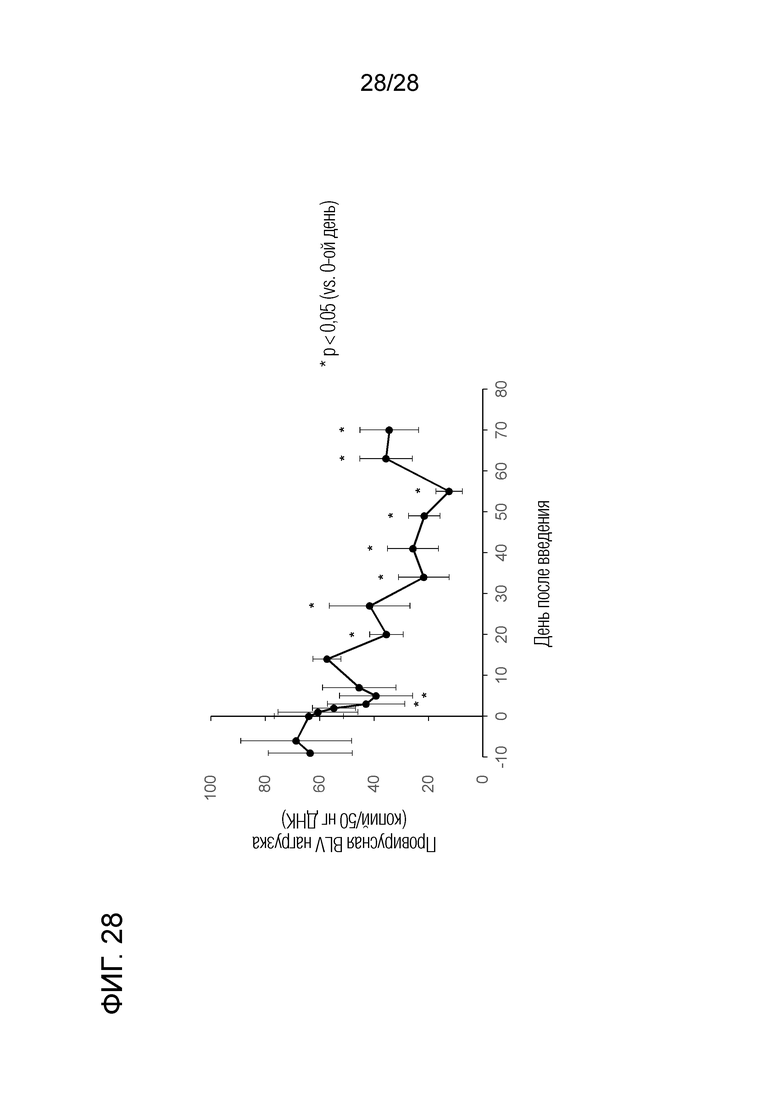
Изобретение относится к области биотехнологии, в частности к антителу против PD-L1, а также к содержащей его фармацевтической композиции. Также раскрыта искусственная генетическая ДНК, кодирующая вышеуказанное антитело или его легкую или тяжелую цепь, а также вектор и клетка, ее содержащие. Изобретение также относится к способу получения антитела против PD-L1, предусматривающему культивирование вышеуказанной клетки-хозяина. Изобретение позволяет эффективно осуществлять профилактику и/или лечение инфекции или злокачественного новообразования. 9 н. и 13 з.п. ф-лы, 1 табл., 3 пр., 28 ил.
1. Антитело против PD-L1, содержащее
(а) легкую цепь, содержащую
вариабельную область легкой цепи, содержащую
CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37),
CDR2 с аминокислотной последовательностью WAT, и
CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и
константную область легкой цепи антитела животного, отличного от крысы; и
(b) тяжелую цепь, содержащую
вариабельную область тяжелой цепи, содержащую
CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39),
CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40), и
CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и
константную область тяжелой цепи антитела животного, отличного от крысы.
2. Антитело по п.1, где вариабельная область легкой цепи и вариабельная область тяжелой цепи получены из крысы.
3. Антитело по п.2, где вариабельная область легкой цепи представляет собой вариабельную область легкой цепи крысиного антитела против бычьего PD-L1, и вариабельная область тяжелой цепи представляет собой вариабельную область тяжелой цепи крысиного антитела против бычьего PD-L1.
4. Антитело по п.3, где вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO:1, и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:2.
5. Антитело по любому из пп.1-4, где константная область легкой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области лямбда-цепи или каппа-цепи.
6. Антитело по любому из пп.1-5, где константная область тяжелой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека.
7. Антитело по любому из пп.1-5, где животное, отличное от крысы, представляет собой быка, и константная область тяжелой цепи бычьего антитела имеет введенные в него мутации, которые снижают активность ADCC и/или активность CDC.
8. Антитело по п.6, где животное, отличное от крысы, является собакой; константная область легкой цепи собачьего антитела имеет аминокислотную последовательность константной области лямбда-цепи; и константная область тяжелой цепи антитела собаки имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека.
9. Антитело по п.7, в котором константная область легкой цепи бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи, а константная область тяжелой цепи бычьего антитела имеет введенные в нее мутации, которые уменьшают активность ADCC и/или активность CDC.
10. Антитело по п.8, в котором константная область легкой цепи собачьего антитела имеет аминокислотную последовательность SEQ ID NO:3, и константная область тяжелой цепи собачьего антитела имеет аминокислотную последовательность SEQ ID NO:4.
11. Антитело по п.9, в котором константная область легкой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:100, и константная область тяжелой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:102.
12. Антитело по любому из пп.1-11, которое имеет четырехцепочечную структуру, включающую две легкие цепи и две тяжелые цепи.
13. Фармацевтическая композиция для профилактики и/или лечения злокачественного новообразования, где злокачественное новообразование выбрано из группы, состоящей из неопластических заболеваний и лейкоза, где композиция отличается тем, что в качестве активного ингредиента содержит антитело по любому из пп.1-12.
14. Композиция по п.13, где неопластическое заболевание представляет собой злокачественную меланому, рак легкого, рак желудка, рак почек, рак молочной железы, рак мочевого пузыря, рак пищевода и рак яичников.
15. Фармацевтическая композиция для профилактики и/или лечения инфекции, где инфекция выбрана из группы, состоящей из болезни Джона, анаплазмоза, бактериального мастита, микотического мастита, микоплазменных инфекций, туберкулеза, инфекции Theileria orientalis, криптоспоридиоза, кокцидиоза, трипаносомоза и лейшманиоза, где композиция отличается тем, что в качестве активного ингредиента содержит антитело по любому из пп.1-12.
16. Композиция по п.13, где микоплазменная инфекция представляет собой микоплазменный мастит или пневмонию, вызванную микоплазмой.
17. Искусственная генетическая ДНК, кодирующая антитело по любому из пп.1-12, где ДНК содержит
(а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT, и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38), и константную область легкой цепи антитела животного, отличного от крысы; и
(b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 с аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41), и константную область тяжелой цепи антитела животного, отличного от крысы.
18. Вектор экспрессии, содержащий искусственную генетическую ДНК по п.17.
19. Клетка-хозяин для экспрессии антитела по любому из пп.1-12, где клетка трансформирована вектором по п.18.
20. Способ получения антитела против PD-L1, включающий культивирование клетки-хозяина по п.19 и сбор антитела против PD-L1 из полученной культуры.
21. ДНК, кодирующая легкую цепь антитела по любому из пп.1-12, где вариабельная область легкой цепи содержит CDR1 с аминокислотной последовательностью QSLLYSENQKDY (SEQ ID NO:37), CDR2 с аминокислотной последовательностью WAT и CDR3 с аминокислотной последовательностью GQYLVYPFT (SEQ ID NO:38).
22. ДНК, кодирующая тяжелую цепь антитела по любому из пп.1-12, где вариабельная область тяжелой цепи содержит CDR1 с аминокислотной последовательностью GYTFTSNF (SEQ ID NO:39), CDR2 с аминокислотной последовательностью IYPEYGNT (SEQ ID NO:40) и CDR3 c аминокислотной последовательностью ASEEAVISLVY (SEQ ID NO:41).
| WO 2015091910 A2, 25.06.2015 | |||
| RYOYO IKEBUCHI et al., Influence of PD-L1 cross-linking on cell death in PD-L1-expressingcell lines and bovine lymphocytes, Immunology, 2014, Vol.142, pp.551-561 | |||
| WO 2015035173 A1, 12.03.2015 | |||
| Резиновая заготовка для подошв | 1929 |
|
SU19344A1 |

