Настоящее изобретение относится к антителу против LAG-3. Более конкретно, настоящее изобретение относится к антителу против LAG-3, содержащему вариабельную область, содержащую определяющие комплементарность области (CDR) крысиного антитела против бычьего LAG-3, и константную область антитела животного, отличного от крысы.
УРОВЕНЬ ТЕХНИКИ
Ген активации 3 лимфоцитов (LAG-3), иммуноингибирующий рецептор, был идентифицирован как молекула, тесно связанная с CD4 (непатентный документ №1: Triebel F, Jitsukawa S, Baixeras E, Roman-Roman S, Genevee C, Viegas -Pequignot E, Hercend TJ Exp. Med., 171(5):1393-1405; May 1, 1990). Недавно выяснилось, что эта молекула участвует в иммуносупрессии при хронических инфекциях и опухолях (непатентный документ №2: Blackburn SD, Shin H, Haining WN, Zou T, Workman CJ, Polley A, Betts MR, Freeman GJ, Vignali DA, Wherry EJ. Nat. Immunol., 10(1):29-37; Nov. 30, 2008; Непатентный домент 3: Woo S-R, Turnis ME, Goldberg M V., Bankoti J, Selby M, Nirschl CJ, Bettini ML, Gravano DM, Vogel P, Liu CL, Tangsombatvisit S, Grosso JF, Netto G, Smeltzer MP, Chaux A, Utz PJ, Workman CJ, Pardoll DM, Korman AJ, Drake CG, Vignali DAA. Cancer Res., 72(4):917-927; Feb. 15, 2012). В области здравоохранения в качестве иммунотерапевтического лекарственного средства против опухолей был разработан препарат антител, который ингибирует активность LAG-3, и в настоящее время проводится фаза I его клинического испытания (название антитела: BMS-986016; Bristle-Myers Squibb and Ono Pharmaceutical Co., Ltd.)
На сегодняшний день авторы настоящего изобретения разработали иммунотерапию для лечения резистентных болезней животных, нацеленную на LAG-3, и было обнаружено, что эта новая иммунотерапия может использоваться в отношении многочисленных заболеваний и у множества животных. (Непатентный документ 4: Shirai T, Konnai S, Ikebuchi R, Okagawa T, Suzuki S, Sunden Y, Onuma M, Murata S, Ohashi K. Vet. Immunol. Immunopathol., 144(3-4):462-467; Dec. 15, 2011; Непатентный документ №5: Konnai S, Suzuki S, Shirai T, Ikebuchi R, Okagawa T, Sunden Y, Mingala CN, Onuma M, Murata S, Ohashi K. Comp. Immunol. Microbiol. Infect. Dis., 36(1):63-69; Jan. 2013; Непатентный документ №6: Okagawa T, Konnai S, Nishimori A, Ikebuchi R, Mizorogi S, Nagata R, Kawaji S, Tanaka S, Kagawa Y, Murata S, Mori Y, Ohashi K. Infect. Immun. 84(1):77-89; Oct. 19, 2015.
Однако антитела, которые были получены авторам настоящего изобретения до настоящего момента, являются антителами крысы, и поэтому их невозможно многократно вводить животным, отличным от крысы.
ЛИТЕРАТУРА УРОВНЯ ТЕХНИКИ
Непатентные документы
Непатентный документ №1: Triebel F, Jitsukawa S, Baixeras E, Roman-Roman S, Genevee C, Viegas-Pequignot E, Hercend TJ Exp. Med., 171(5):1393-1405; May 1, 1990.
Непатентный документ №2: Blackburn SD, Shin H, Haining WN, Zou T, Workman CJ, Polley A, Betts MR, Freeman GJ, Vignali DA, Wherry EJ. Nat. Immunol., 10(1):29-37; Nov. 30, 2008.
Непатентный домент №3: Woo S-R, Turnis ME, Goldberg M V., Bankoti J, Selby M, Nirschl CJ, Bettini ML, Gravano DM, Vogel P, Liu CL, Tangsombatvisit S, Grosso JF, Netto G, Smeltzer MP, Chaux A, Utz PJ, Workman CJ, Pardoll DM, Korman AJ, Drake CG, Vignali DAA. Cancer Res., 72(4):917-927; Feb. 15, 2012.
Непатентный документ №4: Shirai T, Konnai S, Ikebuchi R, Okagawa T, Suzuki S, Sunden Y, Onuma M, Murata S, Ohashi K. Vet. Immunol. Immunopathol., 144(3-4):462-467; Dec. 15, 2011.
Непатентный документ №5: Konnai S, Suzuki S, Shirai T, Ikebuchi R, Okagawa T, Sunden Y, Mingala CN, Onuma M, Murata S, Ohashi K. Comp. Immunol. Microbiol. Infect. Dis., 36(1):63-69; Jan. 2013.
Непатентный документ №6: Okagawa T, Konnai S, Nishimori A, Ikebuchi R, Mizorogi S, Nagata R, Kawaji S, Tanaka S, Kagawa Y, Murata S, Mori Y, Ohashi K. Infect. Immun. 84(1):77-89; Oct. 19, 2015.
ЗАДАЧА, РЕШАЕМАЯ ИЗОБРЕТЕНИЕМ
Задачей настоящего изобретения является создание антитела против LAG-3, способного к многократному введению животным, отличным от крысы.
СПОСОБЫ РЕШЕНИЯ ПРОБЛЕМЫ
Авторы настоящего изобретения определили вариабельные области крысиного моноклонального антитела (2D8) против бычьего LAG-3, связывающегося с клетками Cos-7, экспрессирующими бычий LAG-3, и затем объединили гены, кодирующие полученные вариабельные области, с генами, кодирующими константные области бычьего иммуноглобулина (бычий IgG1 с мутациями, введенными в предполагаемые сайты связывания c рецепторам Fcγ в домене CH2 для ингибирования активности ADCC; см. фиг. 1 для аминокислотных положений и мутаций: 247 E→P, 248 L→V, 249 P→A, 250 G→делеция, 344 A→S, 345 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology 2014 Aug; 142(4):551-561), таким образом, получив ген химерного антитела. Этот ген был введен в клетки яичника китайского хомяка (клетки СНО). Посредством культивирования/пролиферации полученных клеток авторам настоящего изобретения удалось получить химерное "крысино-бычье" антитело против бычьего LAG-3. Кроме того, авторы настоящего изобретения определили CDR вариабельной области крысиного моноклонального антитела (2D8) против бычьего LAG-3. Настоящее изобретение было сделано на основе этих обнаружений.
Сущность настоящего изобретения описана ниже.
(1) Антитело против LAG-3, содержащее (а) легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 c аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17) и константную область легкой цепи антитела животного, отличного от крысы; и (b) тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO: 20) и константную область тяжелой цепи антитела животного, отличного от крысы.
(2) Антитело по (1) выше, в котором вариабельная область легкой цепи и вариабельная область тяжелой цепи получены из крысы.
(3) Антитело по (2) выше, в котором вариабельная область легкой цепи представляет собой вариабельную область легкой цепи крысиного антитела против бычьего LAG-3, и вариабельная область тяжелой цепи представляет собой вариабельную область тяжелой цепи крысиного антитела против бычьего LAG-3.
(4) Антитело по (3) выше, в котором вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 1, и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 2.
(5) Антитело по любому из (1)-(4) выше, в котором константная область легкой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области лямбда-цепи или каппа-цепи.
(6) Антитело по любому из (1)-(5) выше, в котором константная область тяжелой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека или указанную аминокислотную последовательность с введенными мутациями, которые снижают активность ADCC и/или активность CDC.
(7) Антитело по (6) выше, в котором животное, отличное от крысы, является особью крупного рогатого скота; константная область легкой цепи бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи; и константная область тяжелой цепи бычьего антитела имеет введенные в нее мутации, которые уменьшают активность ADCC и/или активность CDC.
(8) Антитело по (7) выше, в котором константная область легкой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO: 3, и константная область тяжелой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO: 4.
(9) Антитело по любому из (1)-(8) выше, которое имеет четырехцепочечную структуру, содержащую две легкие цепи и две тяжелые цепи.
(10) Фармацевтическая композиция, содержащая антитело по любому из (1)-(9) выше в качестве активного ингредиента.
(11) Композиция по (10) выше для профилактики и/или лечения злокачественных новообразований и/или воспаления.
(12) Композиция по (11) выше, причем злокачественное новообразование и/или воспаление выбраны из группы, состоящей из опухолевых заболеваний, лейкоза, болезни Джона, анаплазмоза, бактериального мастита, микотического мастита, микоплазменных инфекций (таких как микоплазменный мастит, микоплазменная пневмония или тому подобное), туберкулеза, инфекции Theileria orientalis, криптоспоридиоза, кокцидиоза, трипаносомоза и лейшманиоза.
(13) Искусственная генетическая ДНК, содержащая (а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17) и константную область легкой цепи антитела животного, отличного от крысы; и (b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO: 20) и константную область тяжелой цепи антитела животного, отличного от крысы.
(14) Вектор, содержащий искусственную генетическую ДНК по (13) выше.
(15) Клетка-хозяин, трансформированная вектором (14) выше.
(16) Способ получения антитела, включающий культивирование клетки-хозяина по (15) выше и сбор антитела против LAG-3 из полученной культуры.
(17) ДНК, кодирующая легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17), и константную области легкой цепи антитела животного, отличного от крысы.
(18) ДНК, кодирующая тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 c аминокислотной последовательностью NREDFDY (SEQ ID NO: 20), и константную область тяжелой цепи антитела животного, отличного от крысы.
Настоящее описание охватывает содержание описаний и/или чертежей Японских патентных заявок №2016-159091, на основании которых по настоящей патентной заявке испрашивается приоритет.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Согласно настоящему изобретению было получено новое антитело против LAG-3. Это антитело можно использовать даже в отношении животных, которые не являются крысами.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[Фиг. 1] Аминокислотная последовательность химерного "крысино-бычьего" антитела ch2DD8 против LAG-3. Показаны области CDR1, CDR2 и CDR3 в вариабельной области легкой цепи и вариабельной области тяжелой цепи крысиного антитела 2D8 против бычьего LAG-3. Кроме того, также показаны аминокислоты, введенные как мутации, в бычий IgG1 (домен CH2) (аминокислотные положения и мутации: 247 E→P, 248 L→V, 249 P→A, 250 G→делеция, 344 A→S, 345 P→S).
[Фиг. 2] Схематическое изображение вектора pDC6 и химерного "крысино-бычьего" антитела ch2D8 против LAG-3.
[Фиг. 3] Чистота очищенного химерного "крысино-бычьего" антитела ch2DD8 против LAG-3.
[Фиг. 4] Специфичность связывания химерного "крысино-бычьего" антитела ch2D8 против LAG-3.
[Фиг. 5] Ингибиторная активность химерного "крысино-бычьего" антитела ch2D8 против бычьего LAG-3 в отношении связывания бычьего LAG-3/MHC II
[Фиг. 6] Изменения в ответе IFN-γ под действием химерного "крысино-бычьего" антитела ch2D8 против бычьего LAG-3.
[Фиг. 7] Перекрестная реактивность крысиного антитела 2D8 против бычьего LAG-3 в отношении буйволиного LAG-3
[Фиг. 8] Перекрестная реактивность крысиного антитела 2D8 против бычьего LAG-3 в отношении овечьих Т-клеток
НАИЛУЧШИЕ СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже настоящее изобретение будет описано подробно.
Настоящее изобретение относится к антителу против LAG-3, содержащему (а) легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 c аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17) и константную область легкой цепи антитела животного, отличного от крысы; и (b) тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO: 20) и константную область тяжелой цепи антитела животного, отличного от крысы.
CDR1, CDR2 и CDR3 вариабельной области легкой цепи (VL) крысиного антитела LAG-3 против бычьего PD-L3 представляют собой область, состоящую из аминокислотной последовательности QSLLDSDGNTY (SEQ ID NO: 16), область, состоящую из аминокислотной последовательности SVS, и область, состоящую из аминокислотной последовательности MQATHVPFT (SEQ ID NO: 17), соответственно (см. фиг. 1).
Кроме того, CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи (VH) крысиного антитела 2D8 против бычьего LAG-3 представляют собой область, состоящую из аминокислотной последовательности GFDFDTYP (SEQ ID NO: 18), область, состоящую из аминокислотной последовательности ITIKTHNYAT (SEQ ID NO: 19), и область, состоящую из аминокислотной последовательности NREDFDY (SEQ ID NO: 20), соответственно (см. фиг. 1).
В аминокислотных последовательностях QSLLDSDGNTY (SEQ ID NO: 16), SVS и MQATHVPFT (SEQ ID NO: 17), а также в аминокислотных последовательностях GFDFDTYP (SEQ ID NO: 18), ITIKTHNYAT (SEQ ID NO: 19) и NREDFDY (SEQ ID NO: 20), одна, две, три, четыре или пять аминокислот могут быть удалены, замещены или добавлены. Даже если были введены такие мутации, полученные аминокислотные последовательности способны выполнять функцию CDR VL или CDR VH антитела LAG-3.
Как используется в настоящем документе, термин «антитело» представляет собой концепцию, охватывающую не только полноразмерные антитела, но также и антитела меньшего молекулярного размера, такие как Fab, F(ab)'2, ScFv, диатело, VH, VL, Sc(Fv)2, биспецифичный sc(Fv)2, минитело, мономер scFv-Fc и димер scFv-Fc.
В антителе против LAG-3 по настоящему изобретению, VL и VH могут быть крысиными. Например, VL может быть VL крысиного антитела против бычьего LAG-3, а VH может быть VH крысиного антитела против бычьего LAG-3.
Аминокислотные последовательности VL и аминокислотная последовательность VH крысиного антитела против бычьего LAG-3 показаны как SEQ ID NO: 1 и 2, соответственно. Аминокислотные последовательности SEQ ID NO: 1 и 2 могут иметь делецию(и), замену(и) или добавление(я) в одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если были введены такие мутации, полученные аминокислотные последовательности способны выполнять функцию VL или VH антитела LAG-3.
Существует два типа легкой цепи иммуноглобулина, которые называются каппа-цепью (κ) и лямбда-цепью (λ). В антителе против LAG-3 по настоящему изобретению константная область легкой цепи (CL) антитела животного, отличного от крысы, может иметь аминокислотную последовательность константной области либо каппа-цепи, либо лямбда-цепи. При этом, относительное содержание лямбда-цепи выше у крупного рогатого скота, овец, кошек, собак и лошадей, а каппа-цепи выше у мышей, крыс, людей и свиней. Поскольку цепь с более высоким относительным содержанием считается предпочтительной, то антитело крупного рогатого скота, овцы, кошки, собаки или лошади предпочтительно имеет аминокислотную последовательность константной области лямбда-цепи, а антитело мыши, крысы, человека или свиньи предпочтительно имеет аминокислотную последовательность константной области каппа-цепи (κ).
Константная область тяжелой цепи (СН) антитела животного, отличного от крысы, может иметь аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека. Тяжелая цепь иммуноглобулина классифицируется на γ-цепь, μ-цепь, α-цепь, δ-цепь и ε-цепь в зависимости от разницы в константной области. В зависимости от типа присутствующей тяжелой цепи образуются пять классов (изотипов) иммуноглобулина; ими являются IgG, IgM, IgA, IgD и IgE.
Иммуноглобулин G (IgG) составляет 70-75% иммуноглобулинов человека и является наиболее распространенным мономерным антителом в плазме. IgG имеет четырехцепочечную структуру, состоящую из двух легких цепей и двух тяжелых цепей. IgG1, IgG2 и IgG4 человека имеют молекулярную массу приблизительно 146000, тогда как IgG3 человека имеет длинную шарнирную область, которая соединяет Fab-область и Fc-область, и имеет большую молекулярную массу, равную 170000. IgG1 человека составляет около 65%, IgG2 человека - около 25%, IgG3 человека - около 7% и IgG4 человека - около 3% IgG человека. Они равномерно распределены внутри и снаружи кровеносных сосудов. Обладая сильным сродством к рецепторам Fc и факторам комплемента на поверхности эффекторных клеток, IgG1 человека индуцирует антителозависимую клеточную цитотоксичность (ADCC), а также активирует комплемент, индуцируя комплемент-зависимую клеточную цитотоксичность (CDC). IgG2 и IgG4 человека имеют низкую активность ADCC и CDC, поскольку их сродство к Fc-рецепторам и факторам комплемента низкое.
Иммуноглобулин М (IgM), который составляет приблизительно 10% иммуноглобулинов человека, представляет собой пентамерное антитело, состоящее из пяти основных четырехцепочечных структур, соединенных вместе. Он имеет молекулярный вес 970000. Обычно встречающийся только в крови, IgM продуцируется против инфекционных микроорганизмов и отвечает за иммунитет на ранней стадии.
Иммуноглобулин А (IgA) составляет 10-15% иммуноглобулинов человека. Он имеет молекулярный вес 160000. Секретируемый IgA представляет собой димерное антитело, состоящее из двух молекул IgA, соединенных вместе. IgA1 обнаружен в сыворотке, выделениях из носа, слюне и грудном молоке. В большом количестве IgA2 обнаружен в желудочном соке.
Иммуноглобулин D (IgD) представляет собой мономерное антитело, составляющее не более 1% иммуноглобулинов человека. IgD обнаружен на поверхности В-клеток и участвует в индукции продукции антител.
Иммуноглобулин E (IgE) представляет собой мономерное антитело, которое встречается в очень небольшом количестве, составляя лишь 0,001% или менее иммуноглобулинов человека. Считается, что иммуноглобулин Е вовлечен в иммунный ответ на паразитов, но в развитых странах, где паразиты встречаются редко, IgE, главным образом, вовлечен в бронхиальную астму и аллергию, среди прочего.
Что касается собак, то последовательности IgG-A (эквивалентен IgG2 человека), IgG-B (эквивалентен IgG1 человека), IgG-C (эквивалентен IgG3 человека) и IgG-D (эквивалентен IgG4 человека) были идентифицированы как тяжелая цепь IgG. В антителе по настоящему изобретению константная область тяжелой цепи IgG, не обладающая ни активностью ADCC, ни активностью CDC, является предпочтительной (IgG4 у человека). В случае, когда константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, можно использовать константную область, которая потеряла как активность ADCC, так и активность CDC в результате введения мутаций в соответствующую область иммуноглобулина, эквивалентного IgG4 человека.
Что касается крупного рогатого скота, то последовательности IgG1, IgG2 и IgG3 были идентифицированы как тяжелая цепь IgG. В антителе по настоящему изобретению константная область тяжелой цепи IgG, не обладающая ни активностью ADCC, ни активностью CDC, является предпочтительной (IgG4 у человека). Хотя константная область нативного IgG1 человека обладает активностью ADCC и активностью CDC, известно, что эти типы активности можно уменьшить путем введения аминокислотных замен или делеций в конкретные сайты. У крупного рогатого скота константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, поэтому в соответствующую область иммуноглобулина, эквивалентную IgG1 человека, можно ввести мутации, а затем использовать полученную константную область. В качестве примера, аминокислотная последовательность СН бычьего антитела (цепь IgG1, GenBank: X62916) с мутациями, введенными в домен СН2, и нуклеотидная последовательность для такой аминокислотной последовательности (после оптимизации кодонов) показаны как SEQ ID NO: 4 и 8, соответственно.
Если животным, отличным от крысы, является собака, то антитело против LAG-3 является более предпочтительным, причем (i) CL собачьего антитела имеет аминокислотную последовательность константной области лямбда-цепи и (ii) CH собачьего антитела имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентную IgG4 человека.
Если животным, отличным от крысы, является представитель крупного рогатого скота, то антитело против LAG-3 является более предпочтительным, причем (i) CL бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи и (ii) CH бычьего антитело имеет мутации, которые снижают активность ADCC и/или активность CDC.
Антитело против LAG-3 по настоящему изобретению включает химерные "крысино-бычьи" антитела, бовинизированные антитела и полные антитела бычьего типа. Тем не менее, животные не ограничиваются особями крупного рогатого скота, и могут быть представлены людьми, собаками, свиньями, обезьянами, мышами, кошками, лошадьми, козами, овцами, буйволами, кроликами, хомячками, морскими свинками и тому подобное.
Например, антитело против LAG-3 по настоящему изобретению может представлять собой антитело против LAG-3, в котором CL бычьего антитела имеет аминокислотную последовательность SEQ ID NO: 3, и СН бычьего антитела имеет аминокислотную последовательность SEQ ID NO: 4.
Аминокислотные последовательности SEQ ID NO: 3 и 4 могут иметь делецию(и), замену(и) или добавление(я) в одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если были введены такие мутации, полученные аминокислотные последовательности способны выполнять функцию CL или CH антитела LAG-3.
Антитело против LAG-3 по настоящему изобретению может иметь четырехцепочечную структуру, содержащую две легкие цепи и две тяжелые цепи.
Антитело против LAG-3 по настоящему изобретению может быть получено, как описано ниже. Вкратце, синтезирован искусственный ген, который содержит (i) идентифицированные последовательности вариабельной области крысиного антитела против бычьего LAG-3 и (ii) последовательности константной области антитела животного, отличного от крысы (например, быка) (предпочтительно, иммуноглобулин, эквивалентный IgG1 человека, в котором в соответствующую область введены мутации для снижения активности ADCC и/или активности CDC). Полученный ген встраивают в вектор (например, плазмиду), который затем вводят в клетку-хозяин (например, клетку млекопитающего, такую как клетка CHO). Клетку-хозяин культивирует, и из полученной культуры собирают интересующее антитело.
Аминокислотная последовательность и нуклеотидная последовательность VL крысиного антитела против бычьего LAG-3, идентифицированные авторами изобретения, показаны как SEQ ID NO: 1 и 5, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO: 11.
Аминокислотная последовательность VL и аминокислотная последовательность VH крысиного антитела против бычьего LAG-3, идентифицированные авторами изобретения, показаны как SEQ ID NO: 2 и 6, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO: 12.
Аминокислотная последовательность и нуклеотидная последовательность CL (лямбда-цепь, GenBank: X62917) бычьего антитела показаны как SEQ ID NO: 3 и 7, соответственно. Кроме того, нуклеотидная последовательность после оптимизации кодонов показана как SEQ ID NO: 13.
Аминокислотная последовательность и нуклеотидная последовательность (после оптимизации кодонов) CH (цепь IgG1, модифицированная из GenBank: X62916) бычьего антитела показаны как SEQ ID NO: 4 и 8, соответственно.
Кроме того, SEQ ID NO: 9 показывает аминокислотную последовательность химерной легкой цепи, состоящей из VL крысиного антитела против бычьего LAG-3 и CL (лямбда цепь, GenBank: X62917) бычьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной легкой цепи, состоящей из VL крысиного антитела против бычьего LAG-3 и CL (лямба цепь, GenBank: X62917) бычьего антитела, показана как SEQ ID NO: 14.
SEQ ID NO: 10 показывает аминокислотную последовательность химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего LAG-3 и CL (IgG1 цепь, модифицированная из GenBank: X62916) бычьего антитела. Нуклеотидная последовательность (после оптимизации кодонов) химерной тяжелой цепи, состоящей из VH крысиного антитела против бычьего LAG-3 и CH (IgG1 цепь, модифицированная из GenBank: X62916) бычьего антитела, показана как SEQ ID NO: 15.
Аминокислотные последовательности и нуклеотидные последовательности CL и CH различных животных, отличных от крысы, могут быть получены из известных баз данных для использования в настоящем изобретении.
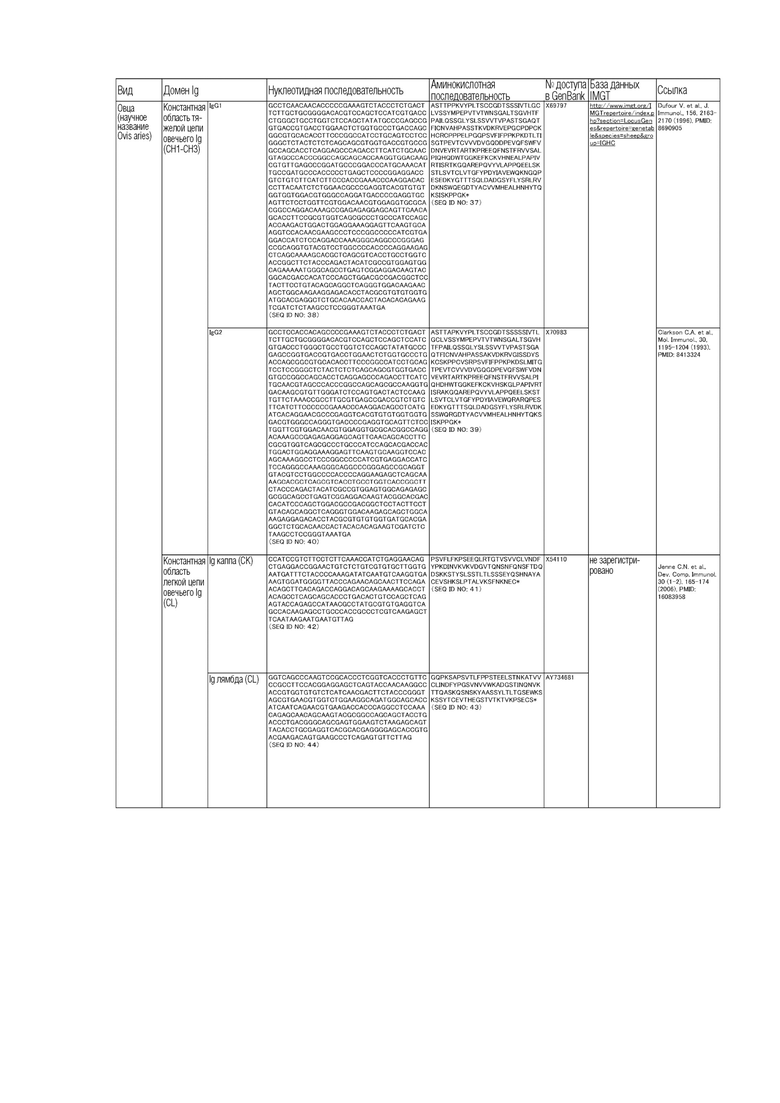
Аминокислотные последовательности и нуклеотидные последовательности бычьих CL и CH приведены в таблице ниже.
Таблица.
(Таблица)


Аминокислотные последовательности и нуклеотидные последовательности CL и CH овец, буйвола и человека приведены в таблице ниже.
(Таблица)


Аминокислотные последовательности SEQ ID NO: 3, 21-28, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57 и 59 могут иметь делецию(и), замену(ы) или добавление(я) в одной или нескольких (например, до пяти, максимально примерно 10) аминокислот. Даже если такие мутации были введены, то полученные аминокислотные последовательности способны выполнять функцию константной области тяжелой цепи или легкой цепи Ig.
Хотя константная область нативного IgG1 человека обладает активностью ADCC и активностью CDC, известно, что эти типы активности можно уменьшить путем введения аминокислотных замен и делеций в конкретные сайты. В случае животных, отличных от человека, где константная область иммуноглобулина, эквивалентного IgG4 человека, не была идентифицирована, мутации могут быть введены в соответствующую область иммуноглобулина, эквивалентного IgG1 человека, так что может быть использована полученная константная область с пониженной активностью ADCC и активностью CDC.
Настоящее изобретение относится к искусственной генетической ДНК, содержащей (а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи (VL), содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17) и константную область легкой цепи (CL) антитела животного, отличного от крысы; и (b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи (VH), содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO: 20) и константную область тяжелой цепи (CH) антитела животного, отличного от крысы. Настоящее изобретение относится к ДНК, кодирующей легкую цепь, содержащую VL, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17), и CL антитела животного, отличного от крысы. Настоящее изобретение также относится к ДНК, кодирующей тяжелую цепь, содержащую VH, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 c аминокислотной последовательностью NREDFDY (SEQ ID NO: 20), и CH антитела животного, отличного от крысы.
В случае (а) легкой цепи, содержащей вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO: 16), CDR2 с аминокислотной последовательностью SVS, и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO: 17) и константную область легкой цепи антитела животного, отличной от крысы; и (b) тяжелой цепи, содержащей вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO: 18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO: 19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO: 20) и константную область тяжелой цепи антитела животного, отличной от крысы, следует обратиться к описанию выше. ДНК (а') представляет собой ДНК (ген), кодирующую легкую цепь (а); и ДНК (b') представляет собой ДНК (ген), кодирующую тяжелую цепь (b). Искусственная генетическая ДНК, содержащая ДНК (a') и ДНК (b'), может быть синтезирована на коммерческом синтезаторе. К искусственной генетической ДНК могут быть добавлены сайты распознавания ферментов рестрикции, последовательности KOZAK, последовательность сигнала присоединения поли-А, последовательности промоторов, последовательности интронов и тому подобное.
Настоящее изобретение также относится к вектору, содержащему вышеуказанную искусственную генетическую ДНК.
В качестве вектора можно использовать плазмиды на основе Escherichia coli (например, pBR322, pBR325, pUC12 или pUC13); плазмиды на основе Bacillus subtilis (например, pUB110, pTP5 или pC194), плазмиды на основе дрожжей (например, pSH19 или pSH15); бактериофаги, такие как фаг λ; вирусы животных, такие как ретровирус или вирус коровьей оспы; или вирусы патогенов насекомых, такие как бакуловирус. В описанных ниже примерах использовали pDC6 (патент Японии №5704753, патент США 9096878, патент ЕС 2385115, патент Гонконга (Китай) HK 1163739 и патент Австралии 2009331326).
Вектор также может содержать промоторы, энхансеры, сигналы сплайсинга, сигналы присоединения поли-А, последовательности интронов, маркеры селекции, точки начала репликации SV40 и так далее.
Настоящее изобретение также относится к клетке-хозяину, трансформированной указанным выше вектором. Можно получить антитело против LAG-3 по изобретению путем культивирования клетки-хозяина и сбора представляющего интерес антитела из полученной культуры. Следовательно, настоящее изобретение также относится к способу получения антитела, включающему культивирование описанной выше клетки-хозяина и сбор антитела против LAG-3 по изобретению из культуры. В способе по настоящему изобретению для получения антитела вектор, содержащий искусственную генетическую ДНК, содержащую ДНК, кодирующую легкую цепь, и ДНК, кодирующую тяжелую цепь, может быть трансфицирован в клетку-хозяина. Альтернативно, вектор, в который введена ДНК, кодирующая легкую цепь, и вектор, в который введена ДНК, кодирующая тяжелую цепь, могут быть совместно трансфицированы в клетку-хозяина.
Примеры клеток-хозяев включают, но не ограничиваются, бактериальные клетки (такие как бактерии Escherichia, бактерии Bacillus или Bacillus subtilis), клетки грибов (такие как дрожжи или Aspergillus), клетки насекомых (такие как клетки S2 или клетки Sf), клетки животных (такие как клетки CHO, клетки COS, клетки HeLa, клетки C127, клетки 3T3, клетки BHK или клетки HEK 293) и клетки растений. Среди них предпочтительной является клетка CHO-DG44 (CHO-DG44 (dfhr-/-)), которая является клеткой с дефицитом дигидрофолатредуктазы.
Введение рекомбинантного вектора в клетку-хозяина может быть осуществлено способами, раскрытыми в Molecular Cloning 2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989 (например, метод на основе фосфата кальция, метод с DEAE-декстраном, трансфекция, микроинъекция, липофекция, электропорация, трансдукция, введение при соскабливании, метод дробовика и тому подобное) или путем инфекции.
Полученный трансформант может быть культивирован в среде с последующим сбором антитела против LAG-3 по настоящему изобретению из культуры. Если антитело секретируется в среду, то среда может быть восстановлена с последующим выделением и очисткой антитела из среды. Если антитело продуцируется внутри трансформированных клеток, то клетки можно лизировать с последующим выделением и очисткой антитела из клеточного лизата.
Примеры сред включают, но не ограничиваются, среду OptiCHO, среду Dynamis, среду CD CHO, среду ActiCHO, среду FortiCHO, среду Ex-Cell CD CHO, среду BalanCD CHO, среду ProCHO 5 и среду Cellvento CHO-100.
рН среды изменяется в зависимости от культивируемой клетки. Обычно используется диапазон рН от 6,8 до 7,6; в основном, подходит диапазон рН от 7,0 до 7,4.
Если культивируемая клетка представляет собой клетки СНО, то культивирование может быть выполнено способами, известными специалистам в данной области. Например, обычно можно проводить культивирование в газофазной атмосфере с концентрацией CO2, равной 0-40%, предпочтительно 2-10%, при 30-39°C, предпочтительно при приблизительно 37°C.
Подходящий период культивирования обычно составляет от одного дня до трех месяцев, предпочтительно от одного дня до трех недель.
Выделение и очистка антитела могут быть выполнены известными способами. Известные способы выделения/очистки, которые можно использовать в настоящем изобретении, включают, но не ограничиваются, способы, использующие разницу в растворимости (такие как высаливание или осаждение растворителем); методы с использованием различий в молекулярной массе (такие как диализ, ультрафильтрация, гель-фильтрация или электрофорез в SDS-полиакриламидном геле); методы, использующие разницу в электрическом заряде (такие как ионообменная хроматография); методы с использованием специфической аффинности (такие как аффинная хроматография); методы, использующие различия в гидрофобности (такие как высокоэффективная жидкостная хроматография с обращенной фазой); и методы, использующие различие в изоэлектрической точке (например, изоэлектрическое фокусирование).
Антитело против LAG-3 по настоящему изобретению можно использовать в качестве лекарственного средства для животных или человека. Следовательно, настоящее изобретение относится к фармацевтической композиции, содержащей вышеописанное антитело против LAG-3 в качестве активного ингредиента.
Фармацевтическая композиция по настоящему изобретению может быть использована для профилактики и/или лечения злокачественных новообразований и/или инфекций. Примеры злокачественных новообразований и/или воспаления включают, но ими не ограничены, опухолевые заболевания (например, меланому, рак легкого, рак желудка, рак почек, рак молочной железы, рак мочевого пузыря, рак пищевода, рак яичников и тому подобное), лейкоз, болезнь Джона, анаплазмоз, бактериальный мастит, микотический мастит, микоплазменные инфекции (такие как микоплазменный мастит, микоплазменная пневмония или тому подобное), туберкулез, инфекцию Theileria orientalis, криптоспоридиоз, кокцидиоз, трипаносомоз и лейшманиоз.
Антитело против LAG-3 по настоящему изобретению может быть растворено в буферах, таких как PBS, физиологический солевой раствор или стерильная вода, необязательно, фильтр-стерилизованная с помощью фильтра или тому подобного, а затем введено животным (включая человека) путем инъекции. К раствору антитела могут быть добавлены добавки (такие как красящие агенты, эмульгаторы, суспендирующие агенты, поверхностно-активные вещества, солюбилизаторы, стабилизаторы, консерванты, антиоксиданты, буферы, изотонизирующие агенты, регуляторы рН и тому подобное). В качестве путей введения могут быть выбраны внутривенное, внутримышечное, внутрибрюшинное, подкожное или внутрикожное введение и тому подобное. Трансназальное или пероральное введение также может быть использовано.
Доза, количество и частота введения антитела против LAG-3 по настоящему изобретению могут изменяться в зависимости от симптомов, возраста и массы тела животного, способа введения, лекарственной формы и тому подобное. Например, 0,1-100 мг/кг массы тела, предпочтительно 1-10 мг/кг массы тела, на взрослое животное обычно можно вводить по меньшей мере один раз с такой частотой, которая позволяет подтвердить желаемый эффект.
Хотя фармацевтическую композицию по настоящему изобретению можно использовать самостоятельно, ее можно применять и в комбинации с хирургическими операциями, лучевой терапией, другими иммунотерапевтическими средствами, такими как противораковая вакцина, или лекарственными средствами с молекулярной мишенью. При таких комбинациях можно ожидать синергетический эффект.
ПРИМЕРЫ
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако настоящее изобретение не ограничивается этими примерами.
[Пример 1] Получение химерного "крысино-бычьего" антитела против бычьего LAG-3
1. Введение
Ген активации 3 лимфоцитов (LAG-3), иммуноингибирующий рецептор, был идентифицирован как молекула, тесно связанная с CD4. Недавно было обнаружено, что LAG-3 вовлечен в иммуносупрессию при хронических инфекциях и опухолях. В этом примере с целью создания нового средства для лечения инфекций у крупного рогатого скота, авторы изобретения получили ген химерного антитела путем связывания гена вариабельной области крысиного моноклонального антитела 2D8 против бычьего LAG-3, способного ингибировать связывание бычьего LAG-3 и MHC класса II с генами, кодирующими константные области бычьего иммуноглобулина (бычий IgG1 и Igλ; однако, были введены мутации в предполагаемые сайты связывания c рецепторам Fcγ в домене CH2 бычьего IgG1 для ингибирования активности ADCC; см. фиг. 1 для аминокислотных положений и мутаций: 247 E→P, 248 L→V, 249 P→A, 250 G→делеция, 344 A→S, 345 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology, 142(4):551-561; Aug. 2014). Этот ген химерного антитела был введен в клетки яичника китайского хомяка (клетки СНО). Посредством культивирования/пролиферации полученных клеток авторам настоящего изобретения удалось получить химерное "крысино-бычье" антитело против бычьего LAG-3 и подтвердить его ингибиторную активность и биологические активности.
2. Материалы, методы и экспериментальные результаты.
2.1 Конструирование клеток, экспрессирующих бычий LAG-3
Была определена нуклеотидная последовательность полноразмерной молекулы кДНК бычьего гена LAG-3 (номер доступа GenBank AB608099; Shirai T, Konnai S, Ikebuchi R, Okagawa T, Sunden Y, Onuma M, Murata S, Ohashi K. Vet. Immunol. Immunopathol, 144(3-4):462-467; Dec. 15, 2011). На основании полученной генетической информации были получены клетки, экспрессирующие бычий LAG-3. Сначала для получения плазмиды, экспрессирующей бычий lAG-3, осуществляли ПЦР, используя синтезированную кДНК, полученную из клеток периферической крови (PBMC) крупного рогатого скота, в качестве матрицы и обработанные ферментами праймеры с сайтами распознавания BglII и EcoRI на 5'конце (boLAG-3-EGFP F и R). Продукты ПЦР расщепляли с помощью EcolRI (Takara) и SmaI (Takara), очищали набором FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в вектор pEGFP-N2 (Clontech), обработанный ферментами рестрикции аналогичным образом. Интересующую полученную плазмиду экспрессии экстрагировали набором QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученная таким образом плазмида обозначена как pEGFP-N2-boLAG-3.
Праймер (boLAG-3-EGFP F): GAAAGATCTATGCTGTGGGAGGCTTGGTT (SEQ ID NO: 61)
Праймер (boLAG-3-EGFP R): CCGGAATTCGGGTTGCTCTGGCTGCAGCT (SEQ ID NO: 62)
Клетки, экспрессирующие бычий LAG-3, получали в соответствии с методиками, описанными ниже. Сначала клетки COS-7 субкультивировали с плотностью 5×104клеток/см2 в 6-луночных планшетах, а затем культивировали в течение ночи в среде RPMI 1640 (Sigma-Aldrich), содержащей 10% инактивированную фетальной бычьей сыворотки (Cell Culture Technologies), пенициллин 200 Ед/мл, стрептомицин 200 мкл/мл и 0,01% L-глютамин (Life Technologies) при 37°С в присутствии 5% СО2. Затем, pEGFP-N2-boLAG-3 или pEGFP-N2 (отрицательный контроль) вводили в клетки COS-7 при 0,4 мкг/см2, используя Lipofectamine 2000 (Invitrogen). Клетки культивировали в течение 48 часов (клетки, экспрессирующие бычий LAG-3-EGFP). Для подтверждения экспрессии бычьего LAG-3 в полученном, таким образом, экспрессирующих клеток, внутриклеточную локализацию EGFP визуализировали с помощью инвертированного конфокального лазерного микроскопа LSM700 (ZEISS).
2.2 Конструирование растворимого бычьего LAG-3
Плазмиду, экспрессирующую бычий LAG-3-Ig, конструировали в соответствии с методиками, описанными ниже. Для амплификации сигнального пептида и внеклеточной области бычьего LAG-3 (номер доступа GenBank AB608099) были сконструированы праймеры с сайтами распознавания NheI и NsiI на 5' конце (boLAG-3-Ig F и R). ПЦР проводили с использованием синтезированной кДНК, полученной из бычьих PBMC, в качестве матрицы. Продукты ПЦР расщепляли NheI (Takara) и NsiI (Takara), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в вектор pCXN2.1-кроличий IgG1 Fc (Niwa H, Yamamura K, Miyazaki J. Gene, 108(2):193-199; Dec. 15, 1991; предоставлен доктором T. Yokomizo, Juntendo University Graduate School of Medicine и модифицирован в лаборатории изобретателей), обработанный ферментами рестрикции аналогичным образом. Плазмиды экспрессии очищали с использованием набора FastGene Xpress Plasmid PLUS (NIPPON Genetics) и хранили при -30°C перед применением в экспериментах. В настоящем документе полученная таким образом плазмида экспрессии обозначена как pCXN2.1-boLAG-3-Ig.
Праймер (boLAG-3-Ig F): CTAGCTAGCCGCCCACCATGCTGTGGGAGGCTTGGTT (SEQ ID NO: 63)
Праймер (boLAG-3-Ig R): TGCATGCATCAGAACAGCTAGGTTGTACG (SEQ ID NO: 64)
Клетки, экспрессирующие растворимый бычий LAG-3-Ig, получали в соответствии с методиками, описанными ниже. Вкратце, 30 мкг pCXN2.1-boLAG-3-Ig вводили в 7,5×107 клеток Expi293F (Life Technologies), используя Expifectamine (Life Technologies). После 7-и дневного культивирования при встряхивании культуральный супернатант собирали. Рекомбинантный Fc-слитый белок очищали от супернатанта с использованием Ab-Capcher Extra (ProteNova). После очистки буфер меняли на PBS (pH 7,4), используя обессоливающую колонку PD-10 (GE Healthcare). Полученный белок хранили при -30°С перед использованием в экспериментах (бычий LAG-3-Ig). Концентрацию очищенного бычьего LAG-3-Ig измеряли с использованием набора Rabbit IgG ELISA Quantitation (Bethyl). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
2.3 Получение клеток, продуцирующих крысиные моноклональные антитела против бычьих LAG-3
Синтезировали пептидную цепь из части внеклеточной области бычьего LAG-3 (аминокислоты с номерами от 71 до 99; аминокислотная последовательность: GSAAPTPRGPGPRRYTVLRLAPGGLRIGK (SEQ ID NO: 72)) путем добавления остатка цистеина к NH2-концу, и связывали с белком гемоцианина молюска (белок-носитель). Получали эмульсию этой пептидной цепи и адъюванта TiterMax Gold (Sigma-Aldrich). Крысу иммунизировали в подушечку лапы полученной эмульсией. После этого, гибридомы получали из подвздошных лимфатических узлов, получая, таким образом, гибридому 2D8, продуцирующую крысиные моноклональные антитела против бычьих LAG-3. Что касается способа конструирования крысиного моноклонального антитела против бычьего LAG-3, то подробное описание приведено в следующем непатентном документе (Okagawa T, Konnai S, Nishimori A, Ikebuchi R, Mizorogi S, Nagata R, Kawaji S, Tanaka S, Kagawa Y, Murata S, Mori Y, Ohashi K. Infect. Immun., 84(1):77-89; Oct. 19, 2015).
2.4 Получение вектора, экспрессирующего химерное "крысино-бычьего" антитело против LAG-3
Химерное "крысино-бычьего" антитело ch2D8 против LAG-3 было получено путем слияния константных областей антитела бычьего IgG1 и Igλ с крысиным антителом против бычьего LAG-3 2D8, используемым в качестве вариабельной области антитела.
Сначала, гены вариабельных областей тяжелой цепи и легкой цепи были идентифицированы с помощью RACE-способа в гибридоме, которая продуцирует крысиное антитело 2D8 против бычьего LAG-3. Затем получали последовательность гена, в котором вариабельные области тяжелой цепи и легкой цепи крысиного антитела 2D8 против бычьего LAG-3 связывали с известными константными областями бычьего IgG1 (тяжелая цепь; модифицирована; номер доступа в GenBank X62916) и бычьего Igλ (легкая цепь; номер доступа в GenBank X62917), соответственно, и была проведена оптимизация кодонов [SEQ ID NO: 9 и 10 (аминокислотные последовательности), SEQ ID NO: 14 и 15 (нуклеотидные последовательности после оптимизации кодонов)]. Следует отметить, что для ингибирования активности ADCC бычьего IgG1 были добавлены мутации в предполагаемые сайты связывания с рецептором Fcγ в домене CH2 (см. фиг. 1, где указаны аминокислотные положения и мутации: 247 E→P, 248 L→V, 249 P→A, 250 G→делеция, 344 A→S, 345 P→S; Ikebuchi R, Konnai S, Okagawa T, Yokoyama K, Nakajima C, Suzuki Y, Murata S, Ohashi K. Immunology, 142(4):551-561; Aug. 2014). Затем ген был искусственно синтезирован таким образом, что последовательность распознавания NotI, последовательность KOZAK, последовательность легкой цепи химерного антитела, последовательность сигнала присоединения поли-A (PABGH), последовательность промотора (PCMV), последовательность распознавания SacI, последовательность интрона (INRBG), последовательность KOZAK, последовательность тяжелой цепи химерного антитела и последовательность распознавания XbaI были расположены в указанном порядке. Синтезированную цепь гена расщепляли с помощью NotI (Takara) и XbaI (Takara), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в сайт клонирования (последовательности распознавания ферментами NotI и XbaI ниже PCMV и между INRBG и PABGH) плазмиды экспрессии pDC6 (любезно предоставлен профессором S. Suzuki, Hokkaido University Research Center for Zoonosis Control), обработанной фрагментами рестрикции таким же образом (фиг. 2). Полученную плазмиду экстрагировали набором QIAGEN Plasmid Midi (Qiagen) и хранили при -30°C перед использованием в экспериментах. В настоящем документе полученная таким образом плазмида обозначена как pDC6-boLAG-3ch2D8.
2.5 Экспрессия химерного "крысино-бычьего" антитела против бычьего LAG-3
Полученную выше pDC6-boLAG-3ch2D8 трансфицировали в клетки CHO-DG44 (CHO-DG44 (dfhr-/-)), которые представляли собой клетки с дефицитом дигидрофолатредуктазы. Через сорок восемь часов среду заменяли средой CD OptiCHO (Life Technologies), содержащей 20 мМ добавки GlutaMAX (Life Technologies). После культивирования в течение 3 недель клетки подвергали селекции и клонированию с помощью лимитирующего разведения. Затем концентрации химерного антитела в культуральных супернатантах измеряли методом дот-блоттинга и ELISA с использованием кроличьего поликлонального антитела против бычьего IgG F(c) (Rockland), проводя таким образом селекцию клонов с высокой экспрессией. Полученный таким образом клон клеток, стабильно экспрессирующий химерное "крысино-бычьего" антитело против бычьего LAG-3, переносили в среду CD Opti-CHO и культивировали при встряхивании в течение 14 дней (125 об/мин, 37°C, 5% CO2). Продукцию химерных антител в культуральном супернатанте измеряли с помощью ELISA с использованием поликлональных антител кролика против бычьего IgG F(c) (Rockland). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric). Культуральный супернатант на 14-й день центрифугировали при 10000 об/мин в течение 10 минут для удаления клеток, и супернатант пропускали через фильтр с размером пор 0,22 мкм Steritop-GP (Millipore) для стерилизации и затем хранили при 4°С перед очисткой.
2.6 Очистка химерного "крысино-бычьего" антитела против бычьего LAG-3
Из культурального супернатанта, полученного, как описано выше, каждое химерное антитело очищали с использованием Ab Capcher Extra (ProteNova). Для связывания со смолой использовали способ с открытой колонкой; PBS pH 7,4 использовали в качестве уравновешивающего буфера и промывочного буфера. В качестве элюирующего буфера использовали 0,1 М глицин-HCl (рН 2,8). В качестве нейтрализующего буфера использовали 1М Трис (рН 9,0). В очищенном антителе заменяли буфер на PBS (pH 7,4), используя обессоливающую колонки PD-10 (GE Healthcare) и концентрировали с использованием Amicon Ultra-15 (50 кДа, Millipore). Очищенное таким образом химерное антитело пропускали через шприцевой фильтр с размером пор 0,22 мкм (Millipore) для стерилизации и хранили при 4°С перед применением в экспериментах.
2.7 Подтверждение чистоты очищенного химерного "крысино-бычьего" антитела против бычьего LAG-3 (фиг. 3).
Чтобы подтвердить чистоту очищенного химерного "крысино-бычьего" антитела против бычьего LAG-3, белки антител обнаруживали с помощью SDS-PAGE и окрашивания CBB. Очищенное химерное "крысино-бычье" антитело ch2D8 против бычьего LAG-3 суспендировали в буфере для образцов Laemmli Sample Buffer (Bio-Rad) и денатурировали при 95°C в течение 5 минут в восстанавливающих условиях (восстановление 2-меркаптоэтанолом; Sigma-Aldrich) или в невосстанавливающих условиях. Используя 10% акриламидный гель, полученные образцы подвергали электрофорезу. В качестве маркеров молекулярной массы использовали Precision Plus Protein All Blue Standards (Bio-Rad). После электрофореза гель окрашивали с помощью Quick-CBB (Wako) и обесцвечивали в дистиллированной воде.
Результаты показаны на фиг. 3. Полосы химерного "крысино-бычьего" антитела против бычьего LAG-3 наблюдали в спрогнозированных положениях, то есть при 25 кДа (легкая цепь) и 50 кДа (тяжелая цепь) в восстанавливающих условиях и при 150-250 кДа в невосстанавливающих условиях.
2.8 Специфичность связывания химерного "крысино-бычьего" антитела против бычьего LAG-3 (фиг. 4).
С помощью проточной цитометрии было подтверждено, что химерное "крысино-бычьего" антитело против бычьего LAG-3 специфически связывается с клетками, экспрессирующими бычье LAG-3 (описано выше). Сначала крысиное антитело 2D8 против бычьего LAG-3 или химерное "крысино-бычьего" антитело ch2D8 против LAG-3 подвергали взаимодействию с клетками, экспрессирующими бычий LAG-3, при комнатной температуре в течение 30 минут. После промывки аллофикоцианин (APC)-меченное козлиное антитело против крысиного Ig (Southern Biotech) или Alexa Fluor 647-меченый козлиный F(ab')2 против бычьего IgG (H+L) (Jackson ImmunoResearch) взаимодействовали при комнатной температуре в течение 30 мин. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG2a (κ) крысы (BD Bioscience) или бычьим антителом IgG1 (Bethyl). После промывки каждое крысиное антитело или химерное "крысино-бычье" антитело, связанное с клеточными поверхностями, детектировали с помощью FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 4. Было обнаружено, что химерное "крысино-бычьего" ch2D8 антитело против бычьего LAG-3 связывается с клетками экспрессирующими бычьими LAG-3 так же как и крысиное антитело 2D8 против бычьего LAG-3.
2.9 Ингибиторная активность химерного "крысино-бычьего" антитела против бычьего LAG-3 в отношении связывания бычьего LAG-3/MHC класса II (фиг. 5)
Используя клеточный клон BL3.1 (клеточный клон, полученный из В-клеточной лимфомы крупного рогатого скота, который высоко экспрессирует MHC класса II) и бычий LAG-3-Ig (описанный выше), тестировали ингибирование связывания бычьего LAG-3/MHC класса II с помощью антител против LAG-3. Сначала, крысиное антитело 2D8 против бычьего LAG-3 или химерное "крысино-бычье" ch2D8 против бычьего LAG-3 в конечной концентрации 0, 1,56, 3,12, 6,25, 12,5 или 25 мкг/мл и бычий LAG-3-Ig в конечной концентрации 3,3 мкг/мл смешивали в 96-луночных планшетах, где они взаимодействовали при 37°С в течение 30 мин. Затем клеточный клон BL3.1 (1×105 клеток) блокировали PBS c добавлением 10% инактивированной козьей сывороткой (Life Technologies) при комнатной температуре в течение 15 минут и проводили реакцию с вышеуказанной реакционной смесью при комнатной температуре в течение 30 минут. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG1 (κ) крысы (BD Bioscience) или бычьим антителом IgG1 (Bethyl). После промывки Alexa Fluor 647-меченный козий F(ab')2 против кроличьего IgG (H+L) (Life Technologies), который заранее подвергали абсорбционной обработки (37°C, 30 мин) IgG, полученным из крысиной сыворотки (Sigma-Aldrich) и IgG, полученный из крысиной сыворотки (Sigma-Aldrich) взаимодействовал при комнатной температуре в течение 30 минут, таким образом обнаруживая бычий LAG-3-Ig, связанный с клеточными поверхностями. Для анализа использовали FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали PBS с добавлением 1% бычьего сывороточного альбумина (Sigma-Aldrich). Принимая долю клеток, связанных с LAG-3-Ig без добавления антител, за 100%, доля клеток, связанных с LAG-3-Ig, для каждой концентрации антител была показана как относительная величина.
Экспериментальные результаты приведены на фиг. 5. Было обнаружено, что химерное "крысино-бычье" антитела ch2D8 против бычьего LAG-3 ингибировало связывание LAG-3-Ig с клетками, экспрессирующими LAG-3, на том же уровне с крысиным антителом 2D8 против бычьего LAG-3.
2.10 Тест биологической активности с использованием химерного "крысино-бычьего" антитела против бычьего LAG-3 (фиг. 6)
Для подтверждения того факта, что ингибирование связывания бычьего LAG-3/MHC класса II химерным "крысино-бычьим" антителом против бычьего LAG-3 активирует лимфоциты, проводили тест биологической активности с использованием IFN-γ в качестве индикатора. Вкратце, РВМС, выделенные из бычьей периферической крови, суспендировали в среде RPMI 1640 (Sigma-Aldrich), содержащей 10% инактивированной фетальной бычьей сыворотки (Cell Culture Technologies), пенициллин 200 Ед/мл, стрептомицин 200 мкг/мл и 0,01% L-глютамин ( Life Technologies) до концентрации 2×106 клеток/мл. К РВМС добавляли 10 мкг/мл крысиного антитела 2D8 против бычьего LAG-3 или химерного "крысино-бычьего" антитела ch2D8 против LAG-3. Затем проводили культивирование при 37°С и 5% СО2 в течение 2 дней. В качестве контрольных антител использовали IgG, полученный из сыворотки крысы (Sigma-Aldrich) и IgG, полученный из бычьей сыворотки (Sigma-Aldrich). Через два дня культивирования собирали культуральный супернатант и измеряли продукцию IFN-γ, используя набор Bovine IFN-γ ELISA (BETYL). Для каждой операции промывки в ELISA использовали Auto Plate Washer BIO WASHER 50 (DS Pharma Biomedical). Поглощение измеряли с помощью микропланшетного ридера MTP-650FA (Corona Electric).
Экспериментальные результаты приведены на фиг. 6. Было обнаружено, что химерное "крысино-бычьего" ch2D8 антитело против бычьего LAG-3 повышает ответ IFN-γ бычьих PBMC так же как и крысиное антитело 2D8 против бычьего LAG-3.
2.11 Анализ CDR крысиного антитела против бычьего LAG-3
Определяющие комплементарность области (CDR) крысиного антитела 2D8 против бычьего LAG-3 определяли с использованием NCBI IGBLAST (http://www.ncbi.nlm.nih.gov/igblast/). Результаты показаны на фиг. 1.
[Пример 2] Применение антитела против LAG-3 у других видов животных
1. Материалы, методы и экспериментальные результаты.
1.1 Идентификация генов овечьего и буйволиного LAG-3
Для определения полноразмерных кодирующих последовательностей (CDS) буйвола (Bubalus bubalis: азиатский буйвол) и кДНК овечье LAG-3, сначала обрабатывали праймеры для амплификации полноразмерных CDS из нуклеотидных последовательностей буйволиного и овечьего LAG-3 (номер доступа GenBank AB608099 и XM_012129455) (buLAG-3 CDS F и R, ovLAG-3 CDS F и R), а затем проводили ПЦР, используя в качестве матрицы синтезированную кДНК, полученную из буйволиного или овечьего PBMC. Для полученных амплифицированных продуктов определяли нуклеотидные последовательности с помощью капиллярного секвенатора согласно обычным методам.
Праймер (buLAG-3 CDS F): ATGCTGTGGGAGGCTTGGTTC (SEQ ID NO: 65)
Праймер (buLAG-3 CDS R): TCAGGGATGCTCTGGCTGCA (SEQ ID NO: 66)
Праймер (ovLAG-3 CDS F): ATGCTGTGGGAGGCTCAGTTCCAGG (SEQ ID NO: 67)
Праймер (ovLAG-3 CDS R): TCAGGGTTGCTCCGGCTGCA (SEQ ID NO: 68)
1.2. Конструирование клеток COS-7, экспрессирующих буйволиный LAG-3
Для получения плазмиды, экспрессирующей буйволиный LAG-3, проводили ПЦР, используя в качестве матрицы синтезированную кДНК, полученную буйволиного LAG-3, и праймеров, сконструированных путем добавления сайтов распознавания SacI и EcoRI на 5' конец (buLAG-3-EGFP F и R). Полученные продукты ПЦР расщепляли с помощью SacI (Takara) и EcoRI (Takara), очищали с помощью набора FastGene Gel/PCR Extraction (NIPPON Genetics) и клонировали в вектор pEGFP-N2 (Clontech), обработанный ферментами рестрикции аналогичным образом. Интересующую плазмиду экспрессии экстрагировали с использованием набора FastGene Xpress Plasmid PLUS (NIPPON Genetics) и хранили при -30°C перед применением в экспериментах. В настоящем документе полученная таким образом плазмида обозначена как pEGFP-N2-buLAG-3.
Праймер (buLAG-3-EGFP F): ATTGAGCTCATGCTGTGGGAGGCTTGGTT (SEQ ID NO: 69)
Праймер (buLAG-3-EGFP R): AATGAATTCGGGATGCTCTGGCTGCAGC (SEQ ID NO: 70)
Клетки COS-7 субкультивировали с плотностью 5 ×104 клеток/см2 в 6-луночных планшетах, а затем культивировали в течение ночи в среде RPMI 1640, содержащей 10% инактивированную фетальной бычьей сыворотки (Invitrogen) и 0,01% L-глютамин (Life Technologies) при 37°С в присутствии 5% СО2. pEGFP-N2-buLAG-3 или pEGFP-N2 (отрицательный контроль) вводили в клетки COS-7 при 0,4 мкг/см2, используя Lipofectamine 2000 (Invitrogen). Клетки культивировали в течение 48 часов (клетки, экспрессирующие бычий LAG-3-EGFP). Для подтверждения экспрессии буйволиного LAG-3 в полученных таким образом экспрессирующих клетках, внутриклеточную локализацию EGFP визуализировали с помощью универсального флуоресцентного микроскопа BZ-9000 (KEYENCE).
1.3. Реакционная способность крысиного антитела 2D8 против буйволиного LAG-3 в отношении буйволиного LAG-3 (фиг. 7)
Методом проточной цитометрии было подтверждено, что крысиное моноклональное антитело против LAG-3 перекрестно реагирует с буйволиным LAG-3. Клетки COS, экспрессирующие буйволиный LAG-3-EGFP, блокировали PBS с добавлением 10% инактивированной козьей сыворотки (Invitrogen) при комнатной температуре в течение 15 минут и проводили реакцию с 10 мкг/мл крысиного антитела 2D8 против бычьего LAG-3 при комнатной температуре в течение 30 минут. После промывки клетки взаимодействовали с меченным APC козьим антителом против крысиного Ig (Beckman Coulter) при комнатной температуре в течение 30 мин. В качестве отрицательного контрольного антитела использовали контроль с изотипом IgG1 (κ) крысы (BD Bioscience). Для анализа использовали FACS Verse (BD Bioscience). Для каждой операции промывки и разбавления антител использовали 1% бычий сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 7. Было подтверждено, что крысиное антитело 2D8 против бычьего LAG-3 связывается с клетками, экспрессирующими буйволиный LAG-3.
1.4. Реакционная способность крысиного антитела 2D8 против LAG-3 с овечьими лимфоцитами (фиг. 8)
Мононуклеарные клетки периферической крови (РВМС) выделяли из периферической крови овец центрифугированием в градиенте плотности с использованием Percoll (GE Healthcare). Выделенные PBMC буйвола суспендировали в среде RPMI 1640 (Sigma-Aldrich), содержащей 10% инактивированную фетальную телячью сыворотку (Invitrogen), пенициллин 200 Ед/мл, стрептомицин 200 мкг/мл и 0,01% L-глютамин (Life Technologies). Плотность клеток доводили до 2×106 клеток/мл. К этим PBMC добавляли 20 нг/мл форбол-12-миристат ацетата (PMA) и 1 мкг/мл йономицина (Sigma-Aldrich), а затем культивировали в течение ночи при 37°C в атмосфере 5% CO2. Культивированные РВМС собирали и блокировали с помощью PBS с добавление 10% инактивированной козьей сыворотки (Invitrogen) при комнатной температуре в течение 15 минут. Затем крысиное антитело 2D8 против бычьего LAG-3 взаимодействовало при комнатной температуре в течение 30 минут. В качестве отрицательного контроля использовали IgG, полученный из крысиной сыворотки (Sigma-Aldrich). После промывки проводили мечение с помощью меченого АРС козьего антитела против крысиного Ig (Beckman Coulter) при комнатной температуре в течение 30 мин. Затем проводили реакцию с мышиным антителом против овечьего CD8 (38.65, AbD Serotec) при комнатной температуре в течение 30 минут. После промывки маркировку осуществляли с использованием меченного PerCP/Cy5.5 козьего антитела против мышиного IgG2a (SAnta Cruz) при комнатной температуре в течение 30 минут. После дополнительной промывки проводили реакцию с мышиным антителом против овечьего CD21, меченным Alexa Flour 488 (GB25A, VMRD), при комнатной температуре в течение 30 минут. Для мечения GB25A использовали набор Zenon Labeling Kit (Life Technologies). Для анализа использовали FACS Verse (BD Biosciences). Для каждой операции промывки и разбавления антител использовали 1% бычий сывороточного альбумина (Sigma-Aldrich).
Экспериментальные результаты приведены на фиг. 8. Крысиное антитело 2D8 против бычьего LAG-3, прочно связывается с CD8+ овечьими T-клетками (CD21-CD8+ клетки) и CD8- T-клетками (CD21-CD8- клетки; т.е. клеточная популяция, содержащая CD4+ T-клетки и γ δ T-клетки), оба активированные стимуляцией PMA/иономицином.
Все публикации, патенты и заявки на патент, которые приводятся в данном документе, тем самым включены в своем полном объеме посредством ссылки.
Промышленная применимость
Антитело против LAG-3 по настоящему изобретению можно использовать для профилактики и/или лечения злокачественного новообразования и инфекций у животных.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> НЭШНЛ ЮНИВЕРСИТИ КОРПОРЕЙШН ХОККАЙДО ЮНИВЕРСИТИ
<120> АНТИТЕЛА ПРОТИВ LAG-3
<130> FP-223PCT
<150> JP P2016-159091
<151> 2016-08-15
<160> 72
<170> PatentIn version 3.5
<210> 1
<211> 132
<212> Белок
<213> Rattus norvegicus
<400> 1
Met Met Ser Pro Val Gln Ser Leu Phe Leu Leu Leu Leu Trp Ile Leu
1 5 10 15
Gly Thr Asn Gly Asp Val Val Leu Thr Gln Thr Pro Pro Thr Leu Ser
20 25 30
Ala Thr Ile Gly Gln Ser Val Ser Ile Ser Cys Arg Ser Ser Gln Ser
35 40 45
Leu Leu Asp Ser Asp Gly Asn Thr Tyr Leu Asn Trp Leu Leu Gln Arg
50 55 60
Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr Ser Val Ser Asn Leu Glu
65 70 75 80
Ser Gly Val Pro Asn Arg Phe Ser Gly Ser Gly Ser Glu Thr Asp Phe
85 90 95
Thr Leu Lys Ile Ser Gly Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr
100 105 110
Cys Met Gln Ala Thr His Val Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys
130
<210> 2
<211> 134
<212> Белок
<213> Rattus norvegicus
<400> 2
Met Val Leu Leu Glu Leu Val Ser Val Ile Ala Leu Phe Gln Gly Val
1 5 10 15
His Cys Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Lys Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asp
35 40 45
Thr Tyr Pro Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp
50 55 60
Trp Val Ala Ser Ile Thr Ile Lys Thr His Asn Tyr Ala Thr Leu Tyr
65 70 75 80
Ala Ala Ser Val Lys Glu Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln
85 90 95
Ser Met Val Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala
100 105 110
Leu Tyr Tyr Cys Asn Arg Glu Asp Phe Asp Tyr Trp Gly Gln Gly Val
115 120 125
Met Val Thr Val Ser Ser
130
<210> 3
<211> 105
<212> Белок
<213> Bos taurus
<400> 3
Gln Pro Lys Ser Pro Pro Ser Val Thr Leu Phe Pro Pro Ser Thr Glu
1 5 10 15
Glu Leu Asn Gly Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ser Val Thr Val Val Trp Lys Ala Asp Gly Ser Thr Ile
35 40 45
Thr Arg Asn Val Glu Thr Thr Arg Ala Ser Lys Gln Ser Asn Ser Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Ser Ser Asp Trp Lys Ser
65 70 75 80
Lys Gly Ser Tyr Ser Cys Glu Val Thr His Glu Gly Ser Thr Val Thr
85 90 95
Lys Thr Val Lys Pro Ser Glu Cys Ser
100 105
<210> 4
<211> 328
<212> Белок
<213> Bos taurus
<400> 4
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Gly Ser Thr Ser Gly Gln Thr Phe
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Ala
85 90 95
Val Asp Pro Thr Cys Lys Pro Ser Pro Cys Asp Cys Cys Pro Pro Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Gly His Asp Asp Pro Glu Val Lys Phe Ser Trp Phe Val Asp Asp
145 150 155 160
Val Glu Val Asn Thr Ala Thr Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Tyr Arg Val Val Ser Ala Leu Arg Ile Gln His Gln Asp Trp
180 185 190
Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Asn Glu Gly Leu Pro
195 200 205
Ser Ser Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Pro Ala Arg Glu
210 215 220
Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys Ser
225 230 235 240
Thr Val Ser Leu Thr Cys Met Val Thr Ser Phe Tyr Pro Asp Tyr Ile
245 250 255
Ala Val Glu Trp Gln Arg Asn Gly Gln Pro Glu Ser Glu Asp Lys Tyr
260 265 270
Gly Thr Thr Pro Pro Gln Leu Asp Ala Asp Ser Ser Tyr Phe Leu Tyr
275 280 285
Ser Lys Leu Arg Val Asp Arg Asn Ser Trp Gln Glu Gly Asp Thr Tyr
290 295 300
Thr Cys Val Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
305 310 315 320
Ser Thr Ser Lys Ser Ala Gly Lys
325
<210> 5
<211> 396
<212> ДНК
<213> Rattus norvegicus
<400> 5
atgatgagtc ctgtccaatc cctgtttttg ttattgcttt ggattctggg aaccaatggt 60
gatgttgtgc tgacccagac tccacccact ttatcggcta ccattggaca atcggtctcc 120
atctcttgca ggtcaagtca gagtctctta gatagtgatg gaaataccta tttaaattgg 180
ttgctacaga ggccaggcca atctccacag cttctaattt attcggtatc caacctggaa 240
tctggggtcc ccaacaggtt cagtggcagt gggtcagaaa cagatttcac actcaaaatc 300
agtggagtgg aggctgaaga tttgggagtt tattactgca tgcaagctac ccatgttcca 360
ttcacgttcg gctcagggac gaagttggaa ataaaa 396
<210> 6
<211> 396
<212> ДНК
<213> Rattus norvegicus
<400> 6
atgatgtctc ccgtccaaag cttgttcctg cttctcctct ggattctggg cacaaacgga 60
gatgtggttc tcacccagac cccccctact ctgtctgcca ccatcggcca gagcgtgtcc 120
atatcctgtc gcagctccca aagcctgctg gactccgatg ggaatactta cctgaattgg 180
ctgttgcagc ggcctggcca gtccccccag ctgttgatct acagcgttag caatctggaa 240
agcggggtcc ccaaccgatt ctccggaagc ggctccgaga ccgattttac cctcaagatc 300
tccggcgtgg aagccgagga cctgggagtg tattattgca tgcaggccac ccatgtgccc 360
ttcaccttcg gtagcggtac caagttggag atcaag 396
<210> 7
<211> 318
<212> ДНК
<213> Bos taurus
<400> 7
cagcccaagt ccccaccctc ggtcaccctg ttcccgccct ccacggagga gctcaacggc 60
aacaaggcca ccctggtgtg tctcatcagc gacttctacc cgggtagcgt gaccgtggtc 120
tggaaggcag acggcagcac catcacccgc aacgtggaga ccacccgggc ctccaaacag 180
agcaacagca agtacgcggc cagcagctac ctgagcctga cgagcagcga ctggaaatcg 240
aaaggcagtt acagctgcga ggtcacgcac gaggggagca ccgtgacgaa gacagtgaag 300
ccctcagagt gttcttag 318
<210> 8
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 8
gctagcacca cagcacctaa agtttaccct ctgtcttcct gctgcggcga caagtcttca 60
tcaactgtta ctcttggatg cctggtctca agttacatgc ccgagcccgt gacagtgacc 120
tggaactcag gcgctctgaa gtctggagtg cacacatttc cagctgtgct tcagtctagc 180
ggcctgtatt ccctcagctc tatggttact gtacctggta gcaccagcgg acagactttc 240
acctgtaatg ttgcccatcc cgcatcttct accaaggtcg ataaagccgt tgaccccact 300
tgcaaaccat ccccttgtga ttgttgtcca ccccctccag tggctggccc ttccgtcttc 360
attttccctc ctaaacctaa ggatactctg accatctcag ggacacccga ggtcacctgt 420
gtcgtcgtgg acgtgggaca tgacgaccca gaagtcaagt tctcatggtt cgtggacgat 480
gtggaggtga acacagcaac aacaaagccc agagaagaac agtttaacag cacatatcgg 540
gtggtcagcg ccttgcgtat tcagcaccag gactggactg gtggcaagga gtttaagtgc 600
aaggtgcata acgaaggtct gccctcttct atagtgagaa ctatctcccg aactaagggc 660
cccgctcggg agccccaggt ttacgtcctt gctccccctc aggaggaact gagtaaatca 720
accgtgagtc tcacctgtat ggttacctca ttttacccag actacatcgc cgtagagtgg 780
cagaggaatg gacagccaga gtctgaggac aaatacggca ctactcctcc ccaactggat 840
gccgactctt cctacttcct ctactccaaa ttgcgagttg accggaactc atggcaggag 900
ggggacacat acacatgcgt cgttatgcac gaggccctgc acaaccatta cacccagaag 960
tccacatcta aaagtgcagg taagtaa 987
<210> 9
<211> 237
<212> Белок
<213> Искусственная последовательность
<220>
<223> химерная L-цепь
<400> 9
Met Met Ser Pro Val Gln Ser Leu Phe Leu Leu Leu Leu Trp Ile Leu
1 5 10 15
Gly Thr Asn Gly Asp Val Val Leu Thr Gln Thr Pro Pro Thr Leu Ser
20 25 30
Ala Thr Ile Gly Gln Ser Val Ser Ile Ser Cys Arg Ser Ser Gln Ser
35 40 45
Leu Leu Asp Ser Asp Gly Asn Thr Tyr Leu Asn Trp Leu Leu Gln Arg
50 55 60
Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr Ser Val Ser Asn Leu Glu
65 70 75 80
Ser Gly Val Pro Asn Arg Phe Ser Gly Ser Gly Ser Glu Thr Asp Phe
85 90 95
Thr Leu Lys Ile Ser Gly Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr
100 105 110
Cys Met Gln Ala Thr His Val Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gln Pro Lys Ser Pro Pro Ser Val Thr Leu Phe Pro
130 135 140
Pro Ser Thr Glu Glu Leu Asn Gly Asn Lys Ala Thr Leu Val Cys Leu
145 150 155 160
Ile Ser Asp Phe Tyr Pro Gly Ser Val Thr Val Val Trp Lys Ala Asp
165 170 175
Gly Ser Thr Ile Thr Arg Asn Val Glu Thr Thr Arg Ala Ser Lys Gln
180 185 190
Ser Asn Ser Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Ser Ser
195 200 205
Asp Trp Lys Ser Lys Gly Ser Tyr Ser Cys Glu Val Thr His Glu Gly
210 215 220
Ser Thr Val Thr Lys Thr Val Lys Pro Ser Glu Cys Ser
225 230 235
<210> 10
<211> 462
<212> Белок
<213> Искусственная последовательность
<220>
<223> химерная Н-цепь
<400> 10
Met Val Leu Leu Glu Leu Val Ser Val Ile Ala Leu Phe Gln Gly Val
1 5 10 15
His Cys Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Lys Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asp
35 40 45
Thr Tyr Pro Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp
50 55 60
Trp Val Ala Ser Ile Thr Ile Lys Thr His Asn Tyr Ala Thr Leu Tyr
65 70 75 80
Ala Ala Ser Val Lys Glu Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln
85 90 95
Ser Met Val Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala
100 105 110
Leu Tyr Tyr Cys Asn Arg Glu Asp Phe Asp Tyr Trp Gly Gln Gly Val
115 120 125
Met Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro
130 135 140
Leu Ser Ser Cys Cys Gly Asp Lys Ser Ser Ser Thr Val Thr Leu Gly
145 150 155 160
Cys Leu Val Ser Ser Tyr Met Pro Glu Pro Val Thr Val Thr Trp Asn
165 170 175
Ser Gly Ala Leu Lys Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
180 185 190
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Gly Ser
195 200 205
Thr Ser Gly Gln Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser
210 215 220
Thr Lys Val Asp Lys Ala Val Asp Pro Thr Cys Lys Pro Ser Pro Cys
225 230 235 240
Asp Cys Cys Pro Pro Pro Pro Val Ala Gly Pro Ser Val Phe Ile Phe
245 250 255
Pro Pro Lys Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val
260 265 270
Thr Cys Val Val Val Asp Val Gly His Asp Asp Pro Glu Val Lys Phe
275 280 285
Ser Trp Phe Val Asp Asp Val Glu Val Asn Thr Ala Thr Thr Lys Pro
290 295 300
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Ala Leu Arg
305 310 315 320
Ile Gln His Gln Asp Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val
325 330 335
His Asn Glu Gly Leu Pro Ser Ser Ile Val Arg Thr Ile Ser Arg Thr
340 345 350
Lys Gly Pro Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln
355 360 365
Glu Glu Leu Ser Lys Ser Thr Val Ser Leu Thr Cys Met Val Thr Ser
370 375 380
Phe Tyr Pro Asp Tyr Ile Ala Val Glu Trp Gln Arg Asn Gly Gln Pro
385 390 395 400
Glu Ser Glu Asp Lys Tyr Gly Thr Thr Pro Pro Gln Leu Asp Ala Asp
405 410 415
Ser Ser Tyr Phe Leu Tyr Ser Lys Leu Arg Val Asp Arg Asn Ser Trp
420 425 430
Gln Glu Gly Asp Thr Tyr Thr Cys Val Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Thr Ser Lys Ser Ala Gly Lys
450 455 460
<210> 11
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 11
atgatgtctc ccgtccaaag cttgttcctg cttctcctct ggattctggg cacaaacgga 60
gatgtggttc tcacccagac cccccctact ctgtctgcca ccatcggcca gagcgtgtcc 120
atatcctgtc gcagctccca aagcctgctg gactccgatg ggaatactta cctgaattgg 180
ctgttgcagc ggcctggcca gtccccccag ctgttgatct acagcgttag caatctggaa 240
agcggggtcc ccaaccgatt ctccggaagc ggctccgaga ccgattttac cctcaagatc 300
tccggcgtgg aagccgagga cctgggagtg tattattgca tgcaggccac ccatgtgccc 360
ttcaccttcg gtagcggtac caagttggag atcaag 396
<210> 12
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 12
atggtgcttc tcgagctggt cagcgtgatt gctctgtttc agggcgtgca ctgcgaagtg 60
cagctggtgg agagtggtgg tgggctcgtg caaccaaaag gcagtctcag gctgagttgt 120
gccgcctccg gattcgattt cgacacctac ccaatgagct gggtcaggca agccccaggg 180
aaaggactcg attgggtggc aagcattacc atcaagacac acaattatgc taccctgtat 240
gccgcaagcg taaaggaacg ctttaccatc tcccgcgatg atagccagtc catggtatat 300
ttgcaaatga ataatttgaa gacagaagat accgctttgt attattgcaa cagagaagat 360
tttgattatt gggggcaggg ggtgatggta accgtgtcca gc 402
<210> 13
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 13
cagcctaagt cccctccttc agtcaccctg tttccaccat ctaccgaaga actcaacggg 60
aataaagcaa cactggtgtg ccttatttct gatttttacc cagggtctgt gacagtggtt 120
tggaaagctg acggttcaac aattacaaga aacgtggaga caacaagggc ttctaagcag 180
tcaaactcta agtatgctgc aagttcttac ctttctctta caagtagtga ctggaaaagt 240
aagggcagtt attcatgcga ggtcactcac gagggaagta ctgtaactaa aactgtaaaa 300
ccatcagagt gttcatag 318
<210> 14
<211> 714
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 14
atgatgtctc ccgtccaaag cttgttcctg cttctcctct ggattctggg cacaaacgga 60
gatgtggttc tcacccagac cccccctact ctgtctgcca ccatcggcca gagcgtgtcc 120
atatcctgtc gcagctccca aagcctgctg gactccgatg ggaatactta cctgaattgg 180
ctgttgcagc ggcctggcca gtccccccag ctgttgatct acagcgttag caatctggaa 240
agcggggtcc ccaaccgatt ctccggaagc ggctccgaga ccgattttac cctcaagatc 300
tccggcgtgg aagccgagga cctgggagtg tattattgca tgcaggccac ccatgtgccc 360
ttcaccttcg gtagcggtac caagttggag atcaagcagc ctaagtcccc tccttcagtc 420
accctgtttc caccatctac cgaagaactc aacgggaata aagcaacact ggtgtgcctt 480
atttctgatt tttacccagg gtctgtgaca gtggtttgga aagctgacgg ttcaacaatt 540
acaagaaacg tggagacaac aagggcttct aagcagtcaa actctaagta tgctgcaagt 600
tcttaccttt ctcttacaag tagtgactgg aaaagtaagg gcagttattc atgcgaggtc 660
actcacgagg gaagtactgt aactaaaact gtaaaaccat cagagtgttc atag 714
<210> 15
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность
<400> 15
atggtgcttc tcgagctggt cagcgtgatt gctctgtttc agggcgtgca ctgcgaagtg 60
cagctggtgg agagtggtgg tgggctcgtg caaccaaaag gcagtctcag gctgagttgt 120
gccgcctccg gattcgattt cgacacctac ccaatgagct gggtcaggca agccccaggg 180
aaaggactcg attgggtggc aagcattacc atcaagacac acaattatgc taccctgtat 240
gccgcaagcg taaaggaacg ctttaccatc tcccgcgatg atagccagtc catggtatat 300
ttgcaaatga ataatttgaa gacagaagat accgctttgt attattgcaa cagagaagat 360
tttgattatt gggggcaggg ggtgatggta accgtgtcca gcgctagcac cacagcacct 420
aaagtttacc ctctgtcttc ctgctgcggc gacaagtctt catcaactgt tactcttgga 480
tgcctggtct caagttacat gcccgagccc gtgacagtga cctggaactc aggcgctctg 540
aagtctggag tgcacacatt tccagctgtg cttcagtcta gcggcctgta ttccctcagc 600
tctatggtta ctgtacctgg tagcaccagc ggacagactt tcacctgtaa tgttgcccat 660
cccgcatctt ctaccaaggt cgataaagcc gttgacccca cttgcaaacc atccccttgt 720
gattgttgtc caccccctcc agtggctggc ccttccgtct tcattttccc tcctaaacct 780
aaggatactc tgaccatctc agggacaccc gaggtcacct gtgtcgtcgt ggacgtggga 840
catgacgacc cagaagtcaa gttctcatgg ttcgtggacg atgtggaggt gaacacagca 900
acaacaaagc ccagagaaga acagtttaac agcacatatc gggtggtcag cgccttgcgt 960
attcagcacc aggactggac tggtggcaag gagtttaagt gcaaggtgca taacgaaggt 1020
ctgccctctt ctatagtgag aactatctcc cgaactaagg gccccgctcg ggagccccag 1080
gtttacgtcc ttgctccccc tcaggaggaa ctgagtaaat caaccgtgag tctcacctgt 1140
atggttacct cattttaccc agactacatc gccgtagagt ggcagaggaa tggacagcca 1200
gagtctgagg acaaatacgg cactactcct ccccaactgg atgccgactc ttcctacttc 1260
ctctactcca aattgcgagt tgaccggaac tcatggcagg agggggacac atacacatgc 1320
gtcgttatgc acgaggccct gcacaaccat tacacccaga agtccacatc taaaagtgca 1380
ggtaagtaa 1389
<210> 16
<211> 11
<212> Белок
<213> Rattus norvegicus
<400> 16
Gln Ser Leu Leu Asp Ser Asp Gly Asn Thr Tyr
1 5 10
<210> 17
<211> 9
<212> Белок
<213> Rattus norvegicus
<400> 17
Met Gln Ala Thr His Val Pro Phe Thr
1 5
<210> 18
<211> 8
<212> Белок
<213> Rattus norvegicus
<400> 18
Gly Phe Asp Phe Asp Thr Tyr Pro
1 5
<210> 19
<211> 10
<212> Белок
<213> Rattus norvegicus
<400> 19
Ile Thr Ile Lys Thr His Asn Tyr Ala Thr
1 5 10
<210> 20
<211> 7
<212> Белок
<213> Rattus norvegicus
<400> 20
Asn Arg Glu Asp Phe Asp Tyr
1 5
<210> 21
<211> 329
<212> Белок
<213> Bos taurus
<400> 21
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Gly Ser Thr Ser Gly Gln Thr Phe
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Ala
85 90 95
Val Asp Pro Thr Cys Lys Pro Ser Pro Cys Asp Cys Cys Pro Pro Pro
100 105 110
Glu Leu Pro Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Gly His Asp Asp Pro Glu Val Lys Phe Ser Trp Phe Val Asp
145 150 155 160
Asp Val Glu Val Asn Thr Ala Thr Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Ala Leu Arg Ile Gln His Gln Asp
180 185 190
Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Asn Glu Gly Leu
195 200 205
Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Pro Ala Arg
210 215 220
Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys
225 230 235 240
Ser Thr Val Ser Leu Thr Cys Met Val Thr Ser Phe Tyr Pro Asp Tyr
245 250 255
Ile Ala Val Glu Trp Gln Arg Asn Gly Gln Pro Glu Ser Glu Asp Lys
260 265 270
Tyr Gly Thr Thr Pro Pro Gln Leu Asp Ala Asp Ser Ser Tyr Phe Leu
275 280 285
Tyr Ser Lys Leu Arg Val Asp Arg Asn Ser Trp Gln Glu Gly Asp Thr
290 295 300
Tyr Thr Cys Val Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Thr Ser Lys Ser Ala Gly Lys
325
<210> 22
<211> 329
<212> Белок
<213> Bos taurus
<400> 22
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Gly Ser Thr Ser Gly Gln Thr Phe
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Ala
85 90 95
Val Asp Pro Thr Cys Lys Pro Ser Pro Cys Asp Cys Cys Pro Pro Pro
100 105 110
Glu Leu Pro Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Gly His Asp Asp Pro Glu Val Lys Phe Ser Trp Phe Val Asp
145 150 155 160
Asp Val Glu Val Asn Thr Ala Thr Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Ala Leu Arg Ile Gln His Gln Asp
180 185 190
Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Asn Glu Gly Leu
195 200 205
Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Pro Ala Arg
210 215 220
Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys
225 230 235 240
Ser Thr Val Ser Leu Thr Cys Met Val Thr Ser Phe Tyr Pro Asp Tyr
245 250 255
Ile Ala Val Glu Trp Gln Arg Asn Gly Gln Pro Glu Ser Glu Asp Lys
260 265 270
Tyr Gly Thr Thr Pro Pro Gln Leu Asp Ala Asp Ser Ser Tyr Phe Leu
275 280 285
Tyr Ser Lys Leu Arg Val Asp Arg Asn Ser Trp Gln Glu Gly Asp Thr
290 295 300
Tyr Thr Cys Val Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Thr Ser Lys Ser Ala Gly Lys
325
<210> 23
<211> 329
<212> Белок
<213> Bos taurus
<400> 23
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Gly Ser Thr Ser Gly Thr Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
85 90 95
Ala Val Asp Pro Arg Cys Lys Thr Thr Cys Asp Cys Cys Pro Pro Pro
100 105 110
Glu Leu Pro Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Gly His Asp Asp Pro Glu Val Lys Phe Ser Trp Phe Val Asp
145 150 155 160
Asp Val Glu Val Asn Thr Ala Thr Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Ala Leu Arg Ile Gln His Gln Asp
180 185 190
Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Asn Glu Gly Leu
195 200 205
Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Pro Ala Arg
210 215 220
Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys
225 230 235 240
Ser Thr Val Ser Leu Thr Cys Met Val Thr Ser Phe Tyr Pro Asp Tyr
245 250 255
Ile Ala Val Glu Trp Gln Arg Asn Gly Gln Pro Glu Ser Glu Asp Lys
260 265 270
Tyr Gly Thr Thr Pro Pro Gln Leu Asp Ala Asp Gly Ser Tyr Phe Leu
275 280 285
Tyr Ser Arg Leu Arg Val Asp Arg Asn Ser Trp Gln Glu Gly Asp Thr
290 295 300
Tyr Thr Cys Val Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Thr Ser Lys Ser Ala Gly Lys
325
<210> 24
<211> 326
<212> Белок
<213> Bos taurus
<400> 24
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ala Ser Ser Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ala Ser Ser Ser Gly Gln Thr Phe
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Ala
85 90 95
Val Gly Val Ser Ile Asp Cys Ser Lys Cys His Asn Gln Pro Cys Val
100 105 110
Arg Glu Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
115 120 125
Met Ile Thr Gly Thr Pro Glu Val Thr Cys Val Val Val Asn Val Gly
130 135 140
His Asp Asn Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Arg Ser Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Tyr Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Thr Gly
180 185 190
Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Gly Leu Ser Ala Pro
195 200 205
Ile Val Arg Ile Ile Ser Arg Ser Lys Gly Pro Ala Arg Glu Pro Gln
210 215 220
Val Tyr Val Leu Asp Pro Pro Lys Glu Glu Leu Ser Lys Ser Thr Leu
225 230 235 240
Ser Val Thr Cys Met Val Thr Gly Phe Tyr Pro Glu Asp Val Ala Val
245 250 255
Glu Trp Gln Arg Asn Arg Gln Thr Glu Ser Glu Asp Lys Tyr Arg Thr
260 265 270
Thr Pro Pro Gln Leu Asp Thr Asp Arg Ser Tyr Phe Leu Tyr Ser Lys
275 280 285
Leu Arg Val Asp Arg Asn Ser Trp Gln Glu Gly Asp Ala Tyr Thr Cys
290 295 300
Val Val Met His Glu Ala Leu His Asn His Tyr Met Gln Lys Ser Thr
305 310 315 320
Ser Lys Ser Ala Gly Lys
325
<210> 25
<211> 326
<212> Белок
<213> Bos taurus
<400> 25
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Gly Ser Thr Ser Gly Gln Thr Phe
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Ala
85 90 95
Val Gly Val Ser Ser Asp Cys Ser Lys Pro Asn Asn Gln His Cys Val
100 105 110
Arg Glu Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
115 120 125
Met Ile Thr Gly Thr Pro Glu Val Thr Cys Val Val Val Asn Val Gly
130 135 140
His Asp Asn Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Arg Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Tyr Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Thr Gly
180 185 190
Gly Lys Glu Phe Lys Cys Lys Val Asn Ile Lys Gly Leu Ser Ala Ser
195 200 205
Ile Val Arg Ile Ile Ser Arg Ser Lys Gly Pro Ala Arg Glu Pro Gln
210 215 220
Val Tyr Val Leu Asp Pro Pro Lys Glu Glu Leu Ser Lys Ser Thr Val
225 230 235 240
Ser Val Thr Cys Met Val Ile Gly Phe Tyr Pro Glu Asp Val Asp Val
245 250 255
Glu Trp Gln Arg Asp Arg Gln Thr Glu Ser Glu Asp Lys Tyr Arg Thr
260 265 270
Thr Pro Pro Gln Leu Asp Ala Asp Arg Ser Tyr Phe Leu Tyr Ser Lys
275 280 285
Leu Arg Val Asp Arg Asn Ser Trp Gln Arg Gly Asp Thr Tyr Thr Cys
290 295 300
Val Val Met His Glu Ala Leu His Asn His Tyr Met Gln Lys Ser Thr
305 310 315 320
Ser Lys Ser Ala Gly Lys
325
<210> 26
<211> 327
<212> Белок
<213> Bos taurus
<400> 26
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ser Ser Cys Cys Gly
1 5 10 15
Asp Lys Ser Ser Ser Gly Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ala Ser Ser Ser Gly Thr Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
85 90 95
Ala Val Gly Val Ser Ser Asp Cys Ser Lys Pro Asn Asn Gln His Cys
100 105 110
Val Arg Glu Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
115 120 125
Leu Met Ile Thr Gly Thr Pro Glu Val Thr Cys Val Val Val Asn Val
130 135 140
Gly His Asp Asn Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val
145 150 155 160
Glu Val His Thr Ala Arg Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
165 170 175
Thr Tyr Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Thr
180 185 190
Gly Gly Lys Glu Phe Lys Cys Lys Val Asn Ile Lys Gly Leu Ser Ala
195 200 205
Ser Ile Val Arg Ile Ile Ser Arg Ser Lys Gly Pro Ala Arg Glu Pro
210 215 220
Gln Val Tyr Val Leu Asp Pro Pro Lys Glu Glu Leu Ser Lys Ser Thr
225 230 235 240
Val Ser Leu Thr Cys Met Val Ile Gly Phe Tyr Pro Glu Asp Val Asp
245 250 255
Val Glu Trp Gln Arg Asp Arg Gln Thr Glu Ser Glu Asp Lys Tyr Arg
260 265 270
Thr Thr Pro Pro Gln Leu Asp Ala Asp Arg Ser Tyr Phe Leu Tyr Ser
275 280 285
Lys Leu Arg Val Asp Arg Asn Ser Trp Gln Arg Gly Asp Thr Tyr Thr
290 295 300
Cys Val Val Met His Glu Ala Leu His Asn His Tyr Met Gln Lys Ser
305 310 315 320
Thr Ser Lys Ser Ala Gly Lys
325
<210> 27
<211> 352
<212> Белок
<213> Bos taurus
<400> 27
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ala Ser Ser Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Arg Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ala Ser Ser Ser Glu Thr Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
85 90 95
Ala Val Thr Ala Arg Arg Pro Val Pro Thr Thr Pro Lys Thr Thr Ile
100 105 110
Pro Pro Gly Lys Pro Thr Thr Pro Lys Ser Glu Val Glu Lys Thr Pro
115 120 125
Cys Gln Cys Ser Lys Cys Pro Glu Pro Leu Gly Gly Leu Ser Val Phe
130 135 140
Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Gly Gln Asp Asp Pro Glu Val
165 170 175
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Arg Thr
180 185 190
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Ala
195 200 205
Leu Arg Ile Gln His Gln Asp Trp Leu Gln Gly Lys Glu Phe Lys Cys
210 215 220
Lys Val Asn Asn Lys Gly Leu Pro Ala Pro Ile Val Arg Thr Ile Ser
225 230 235 240
Arg Thr Lys Gly Gln Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro
245 250 255
Pro Arg Glu Glu Leu Ser Lys Ser Thr Leu Ser Leu Thr Cys Leu Ile
260 265 270
Thr Gly Phe Tyr Pro Glu Glu Ile Asp Val Glu Trp Gln Arg Asn Gly
275 280 285
Gln Pro Glu Ser Glu Asp Lys Tyr His Thr Thr Ala Pro Gln Leu Asp
290 295 300
Ala Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Arg Val Asn Lys Ser
305 310 315 320
Ser Trp Gln Glu Gly Asp His Tyr Thr Cys Ala Val Met His Glu Ala
325 330 335
Leu Arg Asn His Tyr Lys Glu Lys Ser Ile Ser Arg Ser Pro Gly Lys
340 345 350
<210> 28
<211> 352
<212> Белок
<213> Bos taurus
<400> 28
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ala Ser Arg Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ala Ser Thr Ser Glu Thr Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
85 90 95
Ala Val Thr Ala Arg Arg Pro Val Pro Thr Thr Pro Lys Thr Thr Ile
100 105 110
Pro Pro Gly Lys Pro Thr Thr Gln Glu Ser Glu Val Glu Lys Thr Pro
115 120 125
Cys Gln Cys Ser Lys Cys Pro Glu Pro Leu Gly Gly Leu Ser Val Phe
130 135 140
Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Gly Gln Asp Asp Pro Glu Val
165 170 175
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Arg Thr
180 185 190
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Ala
195 200 205
Leu Arg Ile Gln His Gln Asp Trp Leu Gln Gly Lys Glu Phe Lys Cys
210 215 220
Lys Val Asn Asn Lys Gly Leu Pro Ala Pro Ile Val Arg Thr Ile Ser
225 230 235 240
Arg Thr Lys Gly Gln Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro
245 250 255
Pro Arg Glu Glu Leu Ser Lys Ser Thr Leu Ser Leu Thr Cys Leu Ile
260 265 270
Thr Gly Phe Tyr Pro Glu Glu Ile Asp Val Glu Trp Gln Arg Asn Gly
275 280 285
Gln Pro Glu Ser Glu Asp Lys Tyr His Thr Thr Ala Pro Gln Leu Asp
290 295 300
Ala Asp Gly Ser Tyr Phe Leu Tyr Ser Arg Leu Arg Val Asn Lys Ser
305 310 315 320
Ser Trp Gln Glu Gly Asp His Tyr Thr Cys Ala Val Met His Glu Ala
325 330 335
Leu Arg Asn His Tyr Lys Glu Lys Ser Ile Ser Arg Ser Pro Gly Lys
340 345 350
<210> 29
<211> 990
<212> ДНК
<213> Bos taurus
<400> 29
gcctccacca cagccccgaa agtctaccct ctgagttctt gctgcgggga caagtccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggctgtcct tcagtcctcc 180
gggctgtact ctctcagcag catggtgacc gtgcccggca gcacctcagg acagaccttc 240
acctgcaacg tagcccaccc ggccagcagc accaaggtgg acaaggctgt tgatcccaca 300
tgcaaaccat caccctgtga ctgttgccca ccccctgagc tccccggagg accctctgtc 360
ttcatcttcc caccgaaacc caaggacacc ctcacaatct cgggaacgcc cgaggtcacg 420
tgtgtggtgg tggacgtggg ccacgatgac cccgaggtga agttctcctg gttcgtggac 480
gacgtggagg taaacacagc cacgacgaag ccgagagagg agcagttcaa cagcacctac 540
cgcgtggtca gcgccctgcg catccagcac caggactgga ctggaggaaa ggagttcaag 600
tgcaaggtcc acaacgaagg cctcccggcc cccatcgtga ggaccatctc caggaccaaa 660
gggccggccc gggagccgca ggtgtatgtc ctggccccac cccaggaaga gctcagcaaa 720
agcacggtca gcctcacctg catggtcacc agcttctacc cagactacat cgccgtggag 780
tggcagagaa acgggcagcc tgagtcggag gacaagtacg gcacgacccc gccccagctg 840
gacgccgaca gctcctactt cctgtacagc aagctcaggg tggacaggaa cagctggcag 900
gaaggagaca cctacacgtg tgtggtgatg cacgaggccc tgcacaatca ctacacgcag 960
aagtccacct ctaagtctgc gggtaaatga 990
<210> 30
<211> 990
<212> ДНК
<213> Bos taurus
<400> 30
gcctccacca cagccccgaa agtctaccct ctgagttctt gctgcgggga caagtccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtcct tcagtcctcc 180
gggctgtact ctctcagcag catggtgacc gtgcccggca gcacctcagg acagaccttc 240
acctgcaacg tagcccaccc ggccagcagc accaaggtgg acaaggctgt tgatcccaca 300
tgcaaaccat caccctgtga ctgttgccca ccccctgagc tccccggagg accctctgtc 360
ttcatcttcc caccgaaacc caaggacacc ctcacaatct cgggaacgcc cgaggtcacg 420
tgtgtggtgg tggacgtggg ccacgatgac cccgaggtga agttctcctg gttcgtggac 480
gacgtggagg taaacacagc cacgacgaag ccgagagagg agcagttcaa cagcacctac 540
cgcgtggtca gcgccctgcg catccagcac caggactgga ctggaggaaa ggagttcaag 600
tgcaaggtcc acaacgaagg cctcccggcc cccatcgtga ggaccatctc caggaccaaa 660
gggccggccc gggagccgca ggtgtatgtc ctggccccac cccaggaaga gctcagcaaa 720
agcacggtca gcctcacctg catggtcacc agcttctacc cagactacat cgccgtggag 780
tggcagagaa acgggcagcc tgagtcggag gacaagtacg gcacgacccc gccccagctg 840
gacgccgaca gctcctactt cctgtacagc aagctcaggg tggacaggaa cagctggcag 900
gaaggagaca cctacacgtg tgtggtgatg cacgaggccc tgcacaatca ctacacgcag 960
aagtccacct ctaagtctgc gggtaaatga 990
<210> 31
<211> 990
<212> ДНК
<213> Bos taurus
<400> 31
gcctccacca cagccccgaa agtctaccct ctgagttctt gctgcgggga caagtccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtcct tcagtcctcc 180
gggctctact ctctcagcag catggtgacc gtgcccggca gcacctcagg aacccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacaaggc tgttgatccc 300
agatgcaaaa caacctgtga ctgttgccca ccgcctgagc tccctggagg accctctgtc 360
ttcatcttcc caccgaaacc caaggacacc ctcacaatct cgggaacgcc cgaggtcacg 420
tgtgtggtgg tggacgtggg ccacgatgac cccgaggtga agttctcctg gttcgtggac 480
gacgtggagg taaacacagc cacgacgaag ccgagagagg agcagttcaa cagcacctac 540
cgcgtggtca gcgccctgcg catccagcac caggactgga ctggaggaaa ggagttcaag 600
tgcaaggtcc acaacgaagg cctcccagcc cccatcgtga ggaccatctc caggaccaaa 660
gggccggccc gggagccgca ggtgtatgtc ctggccccac cccaggaaga gctcagcaaa 720
agcacggtca gcctcacctg catggtcacc agcttctacc cagactacat cgccgtggag 780
tggcagagaa atgggcagcc tgagtcagag gacaagtacg gcacgacccc tccccagctg 840
gacgccgacg gctcctactt cctgtacagc aggctcaggg tggacaggaa cagctggcag 900
gaaggagaca cctacacgtg tgtggtgatg cacgaggccc tgcacaatca ctacacgcag 960
aagtccacct ctaagtctgc gggtaaatga 990
<210> 32
<211> 981
<212> ДНК
<213> Bos taurus
<400> 32
gcctccacca cagccccgaa agtctaccct ctggcatcca gctgcggaga cacatccagc 60
tccaccgtga ccctgggctg cctggtgtcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggctgtcct tcagtcctcc 180
gggctctact ctctcagcag catggtgacc gtgcccgcca gcagctcagg acagaccttc 240
acctgcaacg tagcccaccc ggccagcagc accaaggtgg acaaggctgt tggggtctcc 300
attgactgct ccaagtgtca taaccagcct tgcgtgaggg aaccatctgt cttcatcttc 360
ccaccgaaac ccaaagacac cctgatgatc acaggaacgc ccgaggtcac gtgtgtggtg 420
gtgaacgtgg gccacgataa ccccgaggtg cagttctcct ggttcgtgga tgacgtggag 480
gtgcacacgg ccaggtcgaa gccaagagag gagcagttca acagcacgta ccgcgtggtc 540
agcgccctgc ccatccagca ccaggactgg actggaggaa aggagttcaa gtgcaaggtc 600
aacaacaaag gcctctcggc ccccatcgtg aggatcatct ccaggagcaa agggccggcc 660
cgggagccgc aggtgtatgt cctggaccca cccaaggaag agctcagcaa aagcacgctc 720
agcgtcacct gcatggtcac cggcttctac ccagaagatg tagccgtgga gtggcagaga 780
aaccggcaga ctgagtcgga ggacaagtac cgcacgaccc cgccccagct ggacaccgac 840
cgctcctact tcctgtacag caagctcagg gtggacagga acagctggca ggaaggagac 900
gcctacacgt gtgtggtgat gcacgaggcc ctgcacaatc actacatgca gaagtccacc 960
tctaagtctg cgggtaaatg a 981
<210> 33
<211> 981
<212> ДНК
<213> Bos taurus
<400> 33
gcctccacca cagccccgaa agtctaccct ctgagttctt gctgcgggga caagtccagc 60
tccaccgtga ccctgggctg cctggtgtcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtcct tcagtcctcc 180
gggctctact ctctcagcag catggtgacc gtgcccggca gcacctcagg acagaccttc 240
acctgcaacg tagcccaccc ggccagcagc accaaggtgg acaaggctgt tggggtctcc 300
agtgactgct ccaagcctaa taaccagcat tgcgtgaggg aaccatctgt cttcatcttc 360
ccaccgaaac ccaaagacac cctgatgatc acaggaacgc ccgaggtcac gtgtgtggtg 420
gtgaacgtgg gccacgataa ccccgaggtg cagttctcct ggttcgtgga cgacgtggag 480
gtgcacacgg ccaggacgaa gccgagagag gagcagttca acagcacgta ccgcgtggtc 540
agcgccctgc ccatccagca ccaggactgg actggaggaa aggagttcaa gtgcaaggtc 600
aacatcaaag gcctctcggc ctccatcgtg aggatcatct ccaggagcaa agggccggcc 660
cgggagccgc aggtgtatgt cctggaccca cccaaggaag agctcagcaa aagcacggtc 720
agcgtcacct gcatggtcat cggcttctac ccagaagatg tagacgtgga gtggcagaga 780
gaccggcaga ctgagtcgga ggacaagtac cgcacgaccc cgccccagct ggacgccgac 840
cgctcctact tcctgtacag caagctcagg gtggacagga acagctggca gagaggagac 900
acctacacgt gtgtggtgat gcacgaggcc ctgcacaatc actacatgca gaagtccacc 960
tctaagtctg cgggtaaatg a 981
<210> 34
<211> 984
<212> ДНК
<213> Bos taurus
<400> 34
gcctccacca cagccccgaa agtctaccct ctgagttctt gctgcgggga caagtccagc 60
tcgggggtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtcct tcagtcctcc 180
gggctctact ctctcagcag catggtgacc gtgcccgcca gcagctcagg aacccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacaaggc tgttggggtc 300
tccagtgact gctccaagcc taataaccag cattgcgtga gggaaccatc tgtcttcatc 360
ttcccaccga aacccaaaga caccctgatg atcacaggaa cgcccgaggt cacgtgtgtg 420
gtggtgaacg tgggccacga taaccccgag gtgcagttct cctggttcgt ggacgacgtg 480
gaggtgcaca cggccaggac gaagccgaga gaggagcagt tcaacagcac gtaccgcgtg 540
gtcagcgccc tgcccatcca gcaccaggac tggactggag gaaaggagtt caagtgcaag 600
gtcaacatca aaggcctctc ggcctccatc gtgaggatca tctccaggag caaagggccg 660
gcccgggagc cgcaggtgta tgtcctggac ccacccaagg aagagctcag caaaagcacg 720
gtcagcctca cctgcatggt catcggcttc tacccagaag atgtagacgt ggagtggcag 780
agagaccggc agactgagtc ggaggacaag taccgcacga ccccgcccca gctggacgcc 840
gaccgctcct acttcctgta cagcaagctc agggtggaca ggaacagctg gcagagagga 900
gacacctaca cgtgtgtggt gatgcacgag gccctgcaca atcactacat gcagaagtcc 960
acctctaagt ctgcgggtaa atga 984
<210> 35
<211> 1059
<212> ДНК
<213> Bos taurus
<400> 35
gcctccacca cagccccgaa agtctaccct ctggcatcca gctgcggaga cacatccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtccg gcagtcctct 180
gggctgtact ctctcagcag catggtgact gtgcccgcca gcagctcaga aacccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacaaggc tgtcactgca 300
aggcgtccag tcccgacgac gccaaagaca actatccctc ctggaaaacc cacaacccca 360
aagtctgaag ttgaaaagac accctgccag tgttccaaat gcccagaacc tctgggagga 420
ctgtctgtct tcatcttccc accgaaaccc aaggacaccc tcacaatctc gggaacgccc 480
gaggtcacgt gtgtggtggt ggacgtgggc caggatgacc ccgaggtgca gttctcctgg 540
ttcgtggacg acgtggaggt gcacacggcc aggacgaagc cgagagagga gcagttcaac 600
agcacctacc gcgtggtcag cgccctgcgc atccagcacc aggactggct gcagggaaag 660
gagttcaagt gcaaggtcaa caacaaaggc ctcccggccc ccattgtgag gaccatctcc 720
aggaccaaag ggcaggcccg ggagccgcag gtgtatgtcc tggccccacc ccgggaagag 780
ctcagcaaaa gcacgctcag cctcacctgc ctgatcaccg gtttctaccc agaagagata 840
gacgtggagt ggcagagaaa tgggcagcct gagtcggagg acaagtacca cacgaccgca 900
ccccagctgg atgctgacgg ctcctacttc ctgtacagca agctcagggt gaacaagagc 960
agctggcagg aaggagacca ctacacgtgt gcagtgatgc acgaagcttt acggaatcac 1020
tacaaagaga agtccatctc gaggtctccg ggtaaatga 1059
<210> 36
<211> 1059
<212> ДНК
<213> Bos taurus
<400> 36
gcctccacca cagccccgaa agtctaccct ctggcatccc gctgcggaga cacatccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagtggcgtg cacaccttcc cggccgtcct tcagtcctcc 180
gggctgtact ctctcagcag catggtgacc gtgcccgcca gcacctcaga aacccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacaaggc tgtcactgca 300
aggcgtccag tcccgacgac gccaaagaca accatccctc ctggaaaacc cacaacccag 360
gagtctgaag ttgaaaagac accctgccag tgttccaaat gcccagaacc tctgggagga 420
ctgtctgtct tcatcttccc accgaaaccc aaggacaccc tcacaatctc gggaacgccc 480
gaggtcacgt gtgtggtggt ggacgtgggc caggatgacc ccgaggtgca gttctcctgg 540
ttcgtggacg acgtggaggt gcacacggcc aggacgaagc cgagagagga gcagttcaac 600
agcacctacc gcgtggtcag cgccctgcgc atccagcacc aggactggct gcagggaaag 660
gagttcaagt gcaaggtcaa caacaaaggc ctcccggccc ccattgtgag gaccatctcc 720
aggaccaaag ggcaggcccg ggagccgcag gtgtatgtcc tggccccacc ccgggaagag 780
ctcagcaaaa gcacgctcag cctcacctgc ctgatcaccg gtttctaccc agaagagata 840
gacgtggagt ggcagagaaa tgggcagcct gagtcggagg acaagtacca cacgaccgca 900
ccccagctgg atgctgacgg ctcctacttc ctgtacagca ggctcagggt gaacaagagc 960
agctggcagg aaggagacca ctacacgtgt gcagtgatgc atgaagcttt acggaatcac 1020
tacaaagaga agtccatctc gaggtctccg ggtaaatga 1059
<210> 37
<211> 331
<212> Белок
<213> Ovis aries
<400> 37
Ala Ser Thr Thr Pro Pro Lys Val Tyr Pro Leu Thr Ser Cys Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Ile Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Ile Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ala Ser Thr Ser Gly Ala Gln Thr
65 70 75 80
Phe Ile Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Gly Cys Pro Asp Pro Cys Lys His Cys Arg Cys Pro
100 105 110
Pro Pro Glu Leu Pro Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys
115 120 125
Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val
130 135 140
Val Val Asp Val Gly Gln Asp Asp Pro Glu Val Gln Phe Ser Trp Phe
145 150 155 160
Val Asp Asn Val Glu Val Arg Thr Ala Arg Thr Lys Pro Arg Glu Glu
165 170 175
Gln Phe Asn Ser Thr Phe Arg Val Val Ser Ala Leu Pro Ile Gln His
180 185 190
Gln Asp Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Asn Glu
195 200 205
Ala Leu Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Gln
210 215 220
Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu
225 230 235 240
Ser Lys Ser Thr Leu Ser Val Thr Cys Leu Val Thr Gly Phe Tyr Pro
245 250 255
Asp Tyr Ile Ala Val Glu Trp Gln Lys Asn Gly Gln Pro Glu Ser Glu
260 265 270
Asp Lys Tyr Gly Thr Thr Thr Ser Gln Leu Asp Ala Asp Gly Ser Tyr
275 280 285
Phe Leu Tyr Ser Arg Leu Arg Val Asp Lys Asn Ser Trp Gln Glu Gly
290 295 300
Asp Thr Tyr Ala Cys Val Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Ile Ser Lys Pro Pro Gly Lys
325 330
<210> 38
<211> 996
<212> ДНК
<213> Ovis aries
<400> 38
gcctcaacaa cacccccgaa agtctaccct ctgacttctt gctgcgggga cacgtccagc 60
tccatcgtga ccctgggctg cctggtctcc agctatatgc ccgagccggt gaccgtgacc 120
tggaactctg gtgccctgac cagcggcgtg cacaccttcc cggccatcct gcagtcctcc 180
gggctctact ctctcagcag cgtggtgacc gtgccggcca gcacctcagg agcccagacc 240
ttcatctgca acgtagccca cccggccagc agcaccaagg tggacaagcg tgttgagccc 300
ggatgcccgg acccatgcaa acattgccga tgcccacccc ctgagctccc cggaggaccg 360
tctgtcttca tcttcccacc gaaacccaag gacaccctta caatctctgg aacgcccgag 420
gtcacgtgtg tggtggtgga cgtgggccag gatgaccccg aggtgcagtt ctcctggttc 480
gtggacaacg tggaggtgcg cacggccagg acaaagccga gagaggagca gttcaacagc 540
accttccgcg tggtcagcgc cctgcccatc cagcaccaag actggactgg aggaaaggag 600
ttcaagtgca aggtccacaa cgaagccctc ccggccccca tcgtgaggac catctccagg 660
accaaagggc aggcccggga gccgcaggtg tacgtcctgg ccccacccca ggaagagctc 720
agcaaaagca cgctcagcgt cacctgcctg gtcaccggct tctacccaga ctacatcgcc 780
gtggagtggc agaaaaatgg gcagcctgag tcggaggaca agtacggcac gaccacatcc 840
cagctggacg ccgacggctc ctacttcctg tacagcaggc tcagggtgga caagaacagc 900
tggcaagaag gagacaccta cgcgtgtgtg gtgatgcacg aggctctgca caaccactac 960
acacagaagt cgatctctaa gcctccgggt aaatga 996
<210> 39
<211> 329
<212> Белок
<213> Ovis aries
<400> 39
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Thr Ser Cys Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Ser Ser Ile Val Thr Leu Gly Cys Leu Val Ser
20 25 30
Ser Tyr Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu
35 40 45
Thr Ser Gly Val His Thr Phe Pro Ala Ile Leu Gln Ser Ser Gly Leu
50 55 60
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ala Ser Thr Ser Gly Ala
65 70 75 80
Gln Thr Phe Ile Cys Asn Val Ala His Pro Ala Ser Ser Ala Lys Val
85 90 95
Asp Lys Arg Val Gly Ile Ser Ser Asp Tyr Ser Lys Cys Ser Lys Pro
100 105 110
Pro Cys Val Ser Arg Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
115 120 125
Asp Ser Leu Met Ile Thr Gly Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Gly Gln Gly Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
145 150 155 160
Asn Val Glu Val Arg Thr Ala Arg Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Phe Arg Val Val Ser Ala Leu Pro Ile Gln His Asp His
180 185 190
Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val His Ser Lys Gly Leu
195 200 205
Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Ala Lys Gly Gln Ala Arg
210 215 220
Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys
225 230 235 240
Ser Thr Leu Ser Val Thr Cys Leu Val Thr Gly Phe Tyr Pro Asp Tyr
245 250 255
Ile Ala Val Glu Trp Gln Arg Ala Arg Gln Pro Glu Ser Glu Asp Lys
260 265 270
Tyr Gly Thr Thr Thr Ser Gln Leu Asp Ala Asp Gly Ser Tyr Phe Leu
275 280 285
Tyr Ser Arg Leu Arg Val Asp Lys Ser Ser Trp Gln Arg Gly Asp Thr
290 295 300
Tyr Ala Cys Val Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Ile Ser Lys Pro Pro Gly Lys
325
<210> 40
<211> 990
<212> ДНК
<213> Ovis aries
<400> 40
gcctccacca cagccccgaa agtctaccct ctgacttctt gctgcgggga cacgtccagc 60
tccagctcca tcgtgaccct gggctgcctg gtctccagct atatgcccga gccggtgacc 120
gtgacctgga actctggtgc cctgaccagc ggcgtgcaca ccttcccggc catcctgcag 180
tcctccgggc tctactctct cagcagcgtg gtgaccgtgc cggccagcac ctcaggagcc 240
cagaccttca tctgcaacgt agcccacccg gccagcagcg ccaaggtgga caagcgtgtt 300
gggatctcca gtgactactc caagtgttct aaaccgcctt gcgtgagccg accgtctgtc 360
ttcatcttcc ccccgaaacc caaggacagc ctcatgatca caggaacgcc cgaggtcacg 420
tgtgtggtgg tggacgtggg ccagggtgac cccgaggtgc agttctcctg gttcgtggac 480
aacgtggagg tgcgcacggc caggacaaag ccgagagagg agcagttcaa cagcaccttc 540
cgcgtggtca gcgccctgcc catccagcac gaccactgga ctggaggaaa ggagttcaag 600
tgcaaggtcc acagcaaagg cctcccggcc cccatcgtga ggaccatctc cagggccaaa 660
gggcaggccc gggagccgca ggtgtacgtc ctggccccac cccaggaaga gctcagcaaa 720
agcacgctca gcgtcacctg cctggtcacc ggcttctacc cagactacat cgccgtggag 780
tggcagagag cgcggcagcc tgagtcggag gacaagtacg gcacgaccac atcccagctg 840
gacgccgacg gctcctactt cctgtacagc aggctcaggg tggacaagag cagctggcaa 900
agaggagaca cctacgcgtg tgtggtgatg cacgaggctc tgcacaacca ctacacacag 960
aagtcgatct ctaagcctcc gggtaaatga 990
<210> 41
<211> 102
<212> Белок
<213> Ovis aries
<400> 41
Pro Ser Val Phe Leu Phe Lys Pro Ser Glu Glu Gln Leu Arg Thr Gly
1 5 10 15
Thr Val Ser Val Val Cys Leu Val Asn Asp Phe Tyr Pro Lys Asp Ile
20 25 30
Asn Val Lys Val Lys Val Asp Gly Val Thr Gln Asn Ser Asn Phe Gln
35 40 45
Asn Ser Phe Thr Asp Gln Asp Ser Lys Lys Ser Thr Tyr Ser Leu Ser
50 55 60
Ser Thr Leu Thr Leu Ser Ser Ser Glu Tyr Gln Ser His Asn Ala Tyr
65 70 75 80
Ala Cys Glu Val Ser His Lys Ser Leu Pro Thr Ala Leu Val Lys Ser
85 90 95
Phe Asn Lys Asn Glu Cys
100
<210> 42
<211> 309
<212> ДНК
<213> Ovis aries
<400> 42
ccatccgtct tcctcttcaa accatctgag gaacagctga ggaccggaac tgtctctgtc 60
gtgtgcttgg tgaatgattt ctaccccaaa gatatcaatg tcaaggtgaa agtggatggg 120
gttacccaga acagcaactt ccagaacagc ttcacagacc aggacagcaa gaaaagcacc 180
tacagcctca gcagcaccct gacactgtcc agctcagagt accagagcca taacgcctat 240
gcgtgtgagg tcagccacaa gagcctgccc accgccctcg tcaagagctt caataagaat 300
gaatgttag 309
<210> 43
<211> 106
<212> Белок
<213> Ovis aries
<400> 43
Gly Gln Pro Lys Ser Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Thr
1 5 10 15
Glu Glu Leu Ser Thr Asn Lys Ala Thr Val Val Cys Leu Ile Asn Asp
20 25 30
Phe Tyr Pro Gly Ser Val Asn Val Val Trp Lys Ala Asp Gly Ser Thr
35 40 45
Ile Asn Gln Asn Val Lys Thr Thr Gln Ala Ser Lys Gln Ser Asn Ser
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Thr Leu Thr Gly Ser Glu Trp Lys
65 70 75 80
Ser Lys Ser Ser Tyr Thr Cys Glu Val Thr His Glu Gly Ser Thr Val
85 90 95
Thr Lys Thr Val Lys Pro Ser Glu Cys Ser
100 105
<210> 44
<211> 321
<212> ДНК
<213> Ovis aries
<400> 44
ggtcagccca agtccgcacc ctcggtcacc ctgttcccgc cttccacgga ggagctcagt 60
accaacaagg ccaccgtggt gtgtctcatc aacgacttct acccgggtag cgtgaacgtg 120
gtctggaagg cagatggcag caccatcaat cagaacgtga agaccaccca ggcctccaaa 180
cagagcaaca gcaagtacgc ggccagcagc tacctgaccc tgacgggcag cgagtggaag 240
tctaagagca gttacacctg cgaggtcacg cacgagggga gcaccgtgac gaagacagtg 300
aagccctcag agtgttctta g 321
<210> 45
<211> 266
<212> Белок
<213> Bubalus bulalis
<400> 45
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
1 5 10 15
Ser Leu Ser Ser Thr Val Thr Ala Pro Ala Ser Ala Thr Lys Ser Gln
20 25 30
Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp
35 40 45
Lys Ala Val Val Pro Pro Cys Arg Pro Lys Pro Cys Asp Cys Cys Pro
50 55 60
Pro Pro Glu Leu Pro Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys
65 70 75 80
Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro Glu Val Thr Cys Val
85 90 95
Val Val Asp Val Gly His Asp Asp Pro Glu Val Lys Phe Ser Trp Phe
100 105 110
Val Asp Asp Val Glu Val Asn Thr Ala Arg Thr Lys Pro Arg Glu Glu
115 120 125
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Ala Leu Pro Ile Gln His
130 135 140
Asn Asp Trp Thr Gly Gly Lys Glu Phe Lys Cys Lys Val Tyr Asn Glu
145 150 155 160
Gly Leu Pro Ala Pro Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Gln
165 170 175
Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro Pro Gln Asp Glu Leu
180 185 190
Ser Lys Ser Thr Val Ser Ile Thr Cys Met Val Thr Gly Phe Tyr Pro
195 200 205
Asp Tyr Ile Ala Val Glu Trp Gln Lys Asp Gly Gln Pro Glu Ser Glu
210 215 220
Asp Lys Tyr Gly Thr Thr Pro Pro Gln Leu Asp Ser Asp Gly Ser Tyr
225 230 235 240
Phe Leu Tyr Ser Arg Leu Arg Val Asn Lys Asn Ser Trp Gln Glu Gly
245 250 255
Gly Ala Tyr Thr Cys Val Val Met His Glu
260 265
<210> 46
<211> 801
<212> ДНК
<213> Bubalus bubalis
<400> 46
gagcggcgtg cacaccttcc cggccgtcct tcagtcctcc gggctctact ctctcagcag 60
cacggtgacc gcgcccgcca gcgccacaaa aagccagacc ttcacctgca acgtagccca 120
cccggccagc agcaccaagg tggacaaggc tgttgttccc ccatgcagac cgaaaccctg 180
tgattgctgc ccaccccctg agctccccgg aggaccctct gtcttcatct tcccaccaaa 240
acccaaggac accctcacaa tctctggaac tcctgaggtc acgtgtgtgg tggtggacgt 300
gggccacgat gaccccgagg tgaagttctc ctggttcgtg gacgatgtgg aggtaaacac 360
agccaggacg aagccaagag aggagcagtt caacagcacc taccgcgtgg tcagcgccct 420
gcccatccag cacaacgact ggactggagg aaaggagttc aagtgcaagg tctacaatga 480
aggcctccca gcccccatcg tgaggaccat ctccaggacc aaagggcagg cccgggagcc 540
gcaggtgtac gtcctggccc caccccagga cgagctcagc aaaagcacgg tcagcatcac 600
ttgcatggtc actggcttct acccagacta catcgccgta gagtggcaga aagatgggca 660
gcctgagtca gaggacaaat atggcacgac cccgccccag ctggacagcg atggctccta 720
cttcctgtac agcaggctca gggtgaacaa gaacagctgg caagaaggag gcgcctacac 780
gtgtgtagtg atgcatgagg c 801
<210> 47
<211> 309
<212> Белок
<213> Bubalus buballis
<400> 47
Ala Ser Ile Thr Ala Pro Lys Val Tyr Pro Leu Thr Ser Cys Arg Gly
1 5 10 15
Glu Thr Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Thr Val Thr Ala Pro Ala Ser Ala Thr Lys Ser Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Thr
85 90 95
Ala Val Gly Phe Ser Ser Asp Cys Cys Lys Phe Pro Lys Pro Cys Val
100 105 110
Arg Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
115 120 125
Met Ile Thr Gly Asn Pro Glu Val Thr Cys Val Val Val Asp Val Gly
130 135 140
Arg Asp Asn Pro Glu Val Gln Phe Ser Trp Phe Val Gly Asp Val Glu
145 150 155 160
Val His Thr Gly Arg Ser Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Tyr Arg Val Val Ser Thr Leu Pro Ile Gln His Asn Asp Trp Thr Gly
180 185 190
Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Gly Leu Pro Ala Pro
195 200 205
Ile Val Arg Thr Ile Ser Arg Thr Lys Gly Gln Ala Arg Glu Pro Gln
210 215 220
Val Tyr Val Leu Ala Pro Pro Gln Glu Glu Leu Ser Lys Ser Thr Val
225 230 235 240
Ser Val Thr Cys Met Val Thr Gly Phe Tyr Pro Asp Tyr Ile Ala Val
245 250 255
Glu Trp His Arg Asp Arg Gln Ala Glu Ser Glu Asp Lys Tyr Arg Thr
260 265 270
Thr Pro Pro Gln Leu Asp Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Arg
275 280 285
Leu Lys Val Asn Lys Asn Ser Trp Gln Glu Gly Gly Ala Tyr Thr Cys
290 295 300
Val Val Met His Glu
305
<210> 48
<211> 929
<212> ДНК
<213> Bubalus bubalis
<400> 48
gcctccatca cagccccgaa agtctaccct ctgacttctt gccgcgggga aacgtccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gagcggcgtg cacaccttcc cggccgtcct tcagtcctct 180
gggctctact ctctcagcag cacggtgacc gcgcccgcca gcgccacaaa aagccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacacggc tgttgggttc 300
tccagtgact gctgcaagtt tcctaagcct tgtgtgaggg gaccatctgt cttcatcttc 360
ccgccgaaac ccaaagacac cctgatgatc acaggaaatc ccgaggtcac atgtgtggtg 420
gtggacgtgg gccgggataa ccccgaggtg cagttctcct ggttcgtggg tgatgtggag 480
gtgcacacgg gcaggtcgaa gccgagagag gagcagttca acagcaccta ccgcgtggtc 540
agcaccctgc ccatccagca caatgactgg actggaggaa aggagttcaa gtgcaaggtc 600
aacaacaaag gcctcccagc ccccatcgtg aggaccatct ccaggaccaa agggcaggcc 660
cgggagccgc aggtgtacgt cctggcccca ccccaggaag agctcagcaa aagcacggtc 720
agcgtcactt gcatggtcac tggcttctac ccagactaca tcgccgtaga gtggcataga 780
gaccggcagg ctgagtcgga ggacaagtac cgcacgaccc cgccccagct ggacagcgat 840
ggctcctact tcctgtacag caggctcaag gtgaacaaga acagctggca agaaggaggc 900
gcctacacgt gtgtagtgat gcatgaggc 929
<210> 49
<211> 352
<212> Белок
<213> Bubalus bubalis
<400> 49
Ala Ser Thr Thr Ala Pro Lys Val Tyr Pro Leu Ala Ser Ser Cys Gly
1 5 10 15
Asp Thr Ser Ser Ser Thr Val Thr Leu Gly Cys Leu Val Ser Ser Tyr
20 25 30
Met Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ala Leu Lys Asn
35 40 45
Gly Val His Thr Phe Pro Ala Val Arg Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Met Pro Thr Ser Thr Ala Gly Thr Gln Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Thr
85 90 95
Ala Val Thr Ala Arg His Pro Val Pro Lys Thr Pro Glu Thr Pro Ile
100 105 110
His Pro Val Lys Pro Pro Thr Gln Glu Pro Arg Asp Glu Lys Thr Pro
115 120 125
Cys Gln Cys Pro Lys Cys Pro Glu Pro Leu Gly Gly Leu Ser Val Phe
130 135 140
Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Thr Ile Ser Gly Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Gly Gln Asp Asp Pro Glu Val
165 170 175
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Arg Met
180 185 190
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Ala
195 200 205
Leu Pro Ile Gln His Gln Asp Trp Leu Arg Glu Lys Glu Phe Lys Cys
210 215 220
Lys Val Asn Asn Lys Gly Leu Pro Ala Pro Ile Val Arg Thr Ile Ser
225 230 235 240
Arg Thr Lys Gly Gln Ala Arg Glu Pro Gln Val Tyr Val Leu Ala Pro
245 250 255
Pro Arg Glu Glu Leu Ser Lys Ser Thr Leu Ser Leu Thr Cys Leu Ile
260 265 270
Thr Gly Phe Tyr Pro Glu Glu Val Asp Val Glu Trp Gln Arg Asn Gly
275 280 285
Gln Pro Glu Ser Glu Asp Lys Tyr His Thr Thr Pro Pro Gln Leu Asp
290 295 300
Ala Asp Gly Ser Tyr Phe Leu Tyr Ser Arg Leu Arg Val Asn Arg Ser
305 310 315 320
Ser Trp Gln Glu Gly Asp His Tyr Thr Cys Ala Val Met His Glu Ala
325 330 335
Leu Arg Asn His Tyr Lys Glu Lys Pro Ile Ser Arg Ser Pro Gly Lys
340 345 350
<210> 50
<211> 1059
<212> ДНК
<213> Bubalus bubalis
<400> 50
gcctccacca cagccccgaa agtctaccct ctggcatcca gctgcgggga cacgtccagc 60
tccaccgtga ccctgggctg cctggtctcc agctacatgc ccgagccggt gaccgtgacc 120
tggaactcgg gtgccctgaa gaacggcgtg cacaccttcc cggccgtccg gcagtcctcc 180
gggctctact ctctcagcag catggtgacc atgcccacca gcaccgcagg aacccagacc 240
ttcacctgca acgtagccca cccggccagc agcaccaagg tggacacggc tgtcactgca 300
aggcatccgg tcccgaagac accagagaca cctatccatc ctgtaaaacc cccaacccag 360
gagcccagag atgaaaagac accctgccag tgtcccaaat gcccagaacc tctgggagga 420
ctgtctgtct tcatcttccc accgaaaccc aaggacaccc tcacaatctc tggaacgccc 480
gaggtcacgt gtgtggtggt ggacgtgggc caggatgacc ccgaagtgca gttctcctgg 540
ttcgtggatg acgtggaggt gcacacagcc aggatgaagc caagagagga gcagttcaac 600
agcacctacc gcgtggtcag cgccctgccc atccagcacc aggactggct gcgggaaaag 660
gagttcaagt gcaaggtcaa caacaaaggc ctcccggccc ccatcgtgag gaccatctcc 720
aggaccaaag ggcaggcccg ggagccacag gtgtatgtcc tggccccacc ccgggaagag 780
ctcagcaaaa gcacgctcag cctcacctgc ctaatcaccg gcttctaccc agaagaggta 840
gacgtggagt ggcagagaaa tgggcagcct gagtcagagg acaagtacca cacgacccca 900
ccccagctgg acgctgacgg ctcctacttc ctgtacagca ggctcagggt gaacaggagc 960
agctggcagg aaggagacca ctacacgtgt gcagtgatgc atgaagcttt acggaatcac 1020
tacaaagaga agcccatctc gaggtctccg ggtaaatga 1059
<210> 51
<211> 105
<212> Белок
<213> Bubalus bubalis
<400> 51
Gln Pro Lys Ser Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Thr Glu
1 5 10 15
Glu Leu Ser Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ser Met Thr Val Ala Arg Lys Ala Asp Gly Ser Thr Ile
35 40 45
Thr Arg Asn Val Glu Thr Thr Arg Ala Ser Lys Gln Ser Asn Ser Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Gly Ser Glu Trp Lys Ser
65 70 75 80
Lys Gly Ser Tyr Ser Cys Glu Val Thr His Glu Gly Ser Thr Val Thr
85 90 95
Lys Thr Val Lys Pro Ser Glu Cys Ser
100 105
<210> 52
<211> 318
<212> ДНК
<213> Bubalus bubalis
<400> 52
cagcccaagt ccgcaccctc agtcaccctg ttcccaccct ccacggagga gctcagcgcc 60
aacaaggcca ccctggtgtg tctcatcagc gacttctacc cgggtagcat gaccgtggcc 120
aggaaggcag acggcagcac catcacccgg aacgtggaga ccacccgggc ctccaaacag 180
agcaacagca agtacgcggc cagcagctac ctgagcctga cgggcagcga gtggaaatcg 240
aaaggcagtt acagctgcga ggtcacgcac gaggggagca ccgtgacaaa gacagtgaag 300
ccctcagagt gttcttag 318
<210> 53
<211> 229
<212> Белок
<213> Homo sapiens
<400> 53
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 54
<211> 690
<212> ДНК
<213> Homo sapiens
<400> 54
gagtccaaat atggtccccc atgcccatca tgcccagcac ctgagttcct ggggggacca 60
tcagtcttcc tgttcccccc aaaacccaag gacactctca tgatctcccg gacccctgag 120
gtcacgtgcg tggtggtgga cgtgagccag gaagaccccg aggtccagtt caactggtac 180
gtggatggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gttcaacagc 240
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa cggcaaggag 300
tacaagtgca aggtctccaa caaaggcctc ccgtcctcca tcgagaaaac catctccaaa 360
gccaaagggc agccccgaga gccacaggtg tacaccctgc ccccatccca ggaggagatg 420
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctaccccag cgacatcgcc 480
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 540
gactccgacg gctccttctt cctctacagc aggctaaccg tggacaagag caggtggcag 600
gaggggaatg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacacag 660
aagagcctct ccctgtctct gggtaaatga 690
<210> 55
<211> 229
<212> Белок
<213> Homo sapiens
<400> 55
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 56
<211> 690
<212> ДНК
<213> Homo sapiens
<400> 56
gagtccaaat atggtccccc gtgcccatca tgcccagcac ctgagttcct ggggggacca 60
tcagtcttcc tgttcccccc aaaacccaag gacactctca tgatctcccg gacccctgag 120
gtcacgtgcg tggtggtgga cgtgagccag gaagaccccg aggtccagtt caactggtac 180
gtggatggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gttcaacagc 240
acgtaccgtg tggtcagcgt cctcaccgtc gtgcaccagg actggctgaa cggcaaggag 300
tacaagtgca aggtctccaa caaaggcctc ccgtcctcca tcgagaaaac catctccaaa 360
gccaaagggc agccccgaga gccacaggtg tacaccctgc ccccatccca ggaggagatg 420
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctaccccag cgacatcgcc 480
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 540
gactccgacg gctccttctt cctctacagc aggctaaccg tggacaagag caggtggcag 600
gaggggaatg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 660
aagagcctct ccctgtctct gggtaaatga 690
<210> 57
<211> 217
<212> Белок
<213> Homo sapiens
<400> 57
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 58
<211> 654
<212> ДНК
<213> Homo sapiens
<400> 58
gcacctgagt tcctgggggg accatcagtc ttcctgttcc ccccaaaacc caaggacact 60
ctcatgatct cccggacccc tgaggtcacg tgcgtggtgg tggacgtgag ccaggaagac 120
cccgaggtcc agttcaactg gtacgtggat ggcgtggagg tgcataatgc caagacaaag 180
ccgcgggagg agcagttcaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac 240
caggactggc tgaacggcaa ggagtacaag tgcaaggtct ccaacaaagg cctcccgtcc 300
tccatcgaga aaaccatctc caaagccaaa gggcagcccc gagagccaca ggtgtacacc 360
ctgcccccat cccaggagga gatgaccaag aaccaggtca gcctgacctg cctggtcaaa 420
ggcttctacc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac 480
tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaagctc 540
accgtggaca agagcaggtg gcaggagggg aacgtcttct catgctccgt gatgcatgag 600
gctctgcaca accactacac gcagaagagc ctctccctgt ctctgggtaa atga 654
<210> 59
<211> 106
<212> Белок
<213> Homo sapiens
<400> 59
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 60
<211> 321
<212> ДНК
<213> Homo sapiens
<400> 60
actgtggctg caccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga 60
actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg 120
aaggtggata acgccctcca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc 180
aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa 240
cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc 300
ttcaacaggg gagagtgtta g 321
<210> 61
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 61
gaaagatcta tgctgtggga ggcttggtt 29
<210> 62
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 62
ccggaattcg ggttgctctg gctgcagct 29
<210> 63
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 63
ctagctagcc gcccaccatg ctgtgggagg cttggtt 37
<210> 64
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 64
tgcatgcatc agaacagcta ggttgtacg 29
<210> 65
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 65
atgctgtggg aggcttggtt c 21
<210> 66
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 66
tcagggatgc tctggctgca 20
<210> 67
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 67
atgctgtggg aggctcagtt ccagg 25
<210> 68
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 68
tcagggttgc tccggctgca 20
<210> 69
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 69
attgagctca tgctgtggga ggcttggtt 29
<210> 70
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 70
aatgaattcg ggatgctctg gctgcagc 28
<210> 71
<211> 516
<212> Белок
<213> Bos taurus
<400> 71
Met Leu Trp Glu Ala Trp Phe Gln Val Trp Leu Phe Leu Gln Leu Leu
1 5 10 15
Trp Ala Ala Ala Val Glu Ala Pro Glu Pro Gly Ala Glu Val Pro Val
20 25 30
Val Trp Ala Gln Glu Gly Ala Pro Ala Gln Leu Pro Cys Ser Pro Thr
35 40 45
Ile Pro Leu Gln Asp Leu Ser Leu Pro Arg Thr Arg Gln Val Thr Trp
50 55 60
Gln His Val Pro Glu Ser Gly Ser Ala Ala Pro Thr Pro Arg Gly Pro
65 70 75 80
Gly Pro Arg Arg Tyr Thr Val Leu Arg Leu Ala Pro Gly Gly Leu Arg
85 90 95
Ile Gly Lys Leu Pro Leu Gln Pro Arg Val Gln Leu Glu Glu Met Gly
100 105 110
Leu Gln Arg Gly Asp Phe Ser Leu Trp Leu Arg Pro Ala Arg Arg Ala
115 120 125
Asp Ala Gly Glu Tyr His Ala Ala Val Arg Phe Gly Asn Arg Ala Leu
130 135 140
Ala Cys Arg Leu Arg Leu Arg Val Gly Gln Ala Ala Val Thr Ala Ser
145 150 155 160
Pro Pro Gly Pro Leu Trp Thr Ser Ser Trp Val Val Leu Asn Cys Ser
165 170 175
Phe Ser Arg Pro Asp Leu Pro Ala Ser Val His Trp Phe Arg Gly Pro
180 185 190
Gly Arg Val Pro Val Gln Glu Ser Pro His His His Leu Val Gly Asn
195 200 205
Phe Leu Phe Leu Pro Gln Val Ser Ser Leu Asp Ser Gly Thr Trp Gly
210 215 220
Cys Ser Leu Thr Tyr Arg Asp Gly Phe Asn Val Ser Ile Thr Tyr Asn
225 230 235 240
Leu Ala Val Leu Gly Leu Glu Pro Arg Ala Thr Leu Thr Val Tyr Ala
245 250 255
Gly Ala Gly Ser Lys Val Glu Leu Pro Cys Arg Leu Pro Pro Gly Val
260 265 270
Gly Ile Gln Ser Ser Leu Thr Ala Met Trp Thr Pro Pro Gly Glu Gly
275 280 285
Pro Asp Leu Leu Val Ala Gly Asp Arg Asn Asn Phe Thr Leu Arg Leu
290 295 300
Glu Ala Val Gly Gln Ala Gln Ala Gly Thr Tyr Thr Cys Arg Val His
305 310 315 320
Leu Gln Gly Arg Gln Leu Ser Ala Thr Val Thr Leu Ala Val Ile Thr
325 330 335
Val Thr Pro Lys Pro Tyr Gly Ser Ser Gly Ser Leu Arg Lys Pro Phe
340 345 350
Cys Glu Val Thr Pro Ala Ser Gly Gln Glu Arg Phe Val Trp Ser Pro
355 360 365
Leu Asp Lys Arg Ser Gln Arg Arg Ser Pro Gly Pro Trp Leu Leu Thr
370 375 380
Pro Asp Ala Arg Pro Leu Ser Gln Pro Trp Gln Cys His Leu Tyr Gln
385 390 395 400
Gly Glu Arg Leu Leu Gly Thr Ala Val Tyr Leu Thr Glu Leu Ser His
405 410 415
Pro Gly Ala Gln Arg Ser Gly Arg Ala Leu Gly Ala Gly Arg Thr Ala
420 425 430
His Leu Pro Leu Leu Ile Leu Gly Leu Leu Phe Leu Leu Leu Leu Val
435 440 445
Thr Gly Ala Ser Ser Phe His Leu Trp Arg Arg Gln Trp Arg Pro Arg
450 455 460
Arg Phe Ser Ala Leu Glu His Gly Thr His Pro Ser Gln Ala Ser Ser
465 470 475 480
Lys Thr Gly Glu Leu Glu Pro Glu Leu Glu Pro Glu Pro Asp Pro Glu
485 490 495
Val Glu Pro Glu Pro Glu Pro Glu Pro Glu Ser Gln Pro Gln Leu Gln
500 505 510
Pro Glu Gln Pro
515
<210> 72
<211> 29
<212> Белок
<213> Bos taurus
<400> 72
Gly Ser Ala Ala Pro Thr Pro Arg Gly Pro Gly Pro Arg Arg Tyr Thr
1 5 10 15
Val Leu Arg Leu Ala Pro Gly Gly Leu Arg Ile Gly Lys
20 25
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-PD-L1 АНТИТЕЛО ДЛЯ ДЕТЕКЦИИ PD-L1 | 2018 |
|
RU2758723C2 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| СВЯЗЫВАЮЩИЙ RGMa БЕЛОК И ЕГО ИСПОЛЬЗОВАНИЕ | 2016 |
|
RU2809500C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ PD-L1 и LAG-3 | 2017 |
|
RU2784388C2 |
| АНТИТЕЛО ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2781148C2 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| НЕВИРУСНЫЙ ИММУНОТАРГЕТИНГ | 2020 |
|
RU2833379C2 |
| Прогнозирование риска развития нежелательной реакции, связанной с введением антитела к ALK2, и способности отвечать на лечение антителом к ALK2 | 2019 |
|
RU2783762C2 |
| АНТИТЕЛО ПРОТИВ ЧЕЛОВЕЧЕСКОГО TLR7 | 2019 |
|
RU2808123C2 |

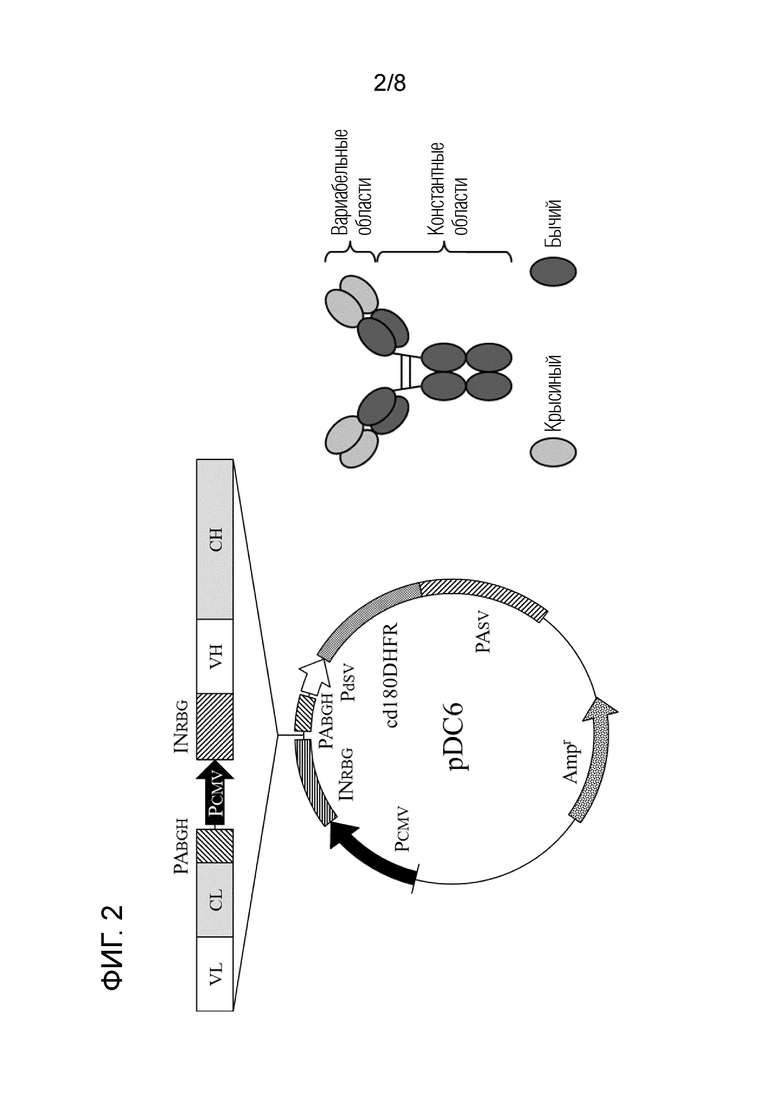

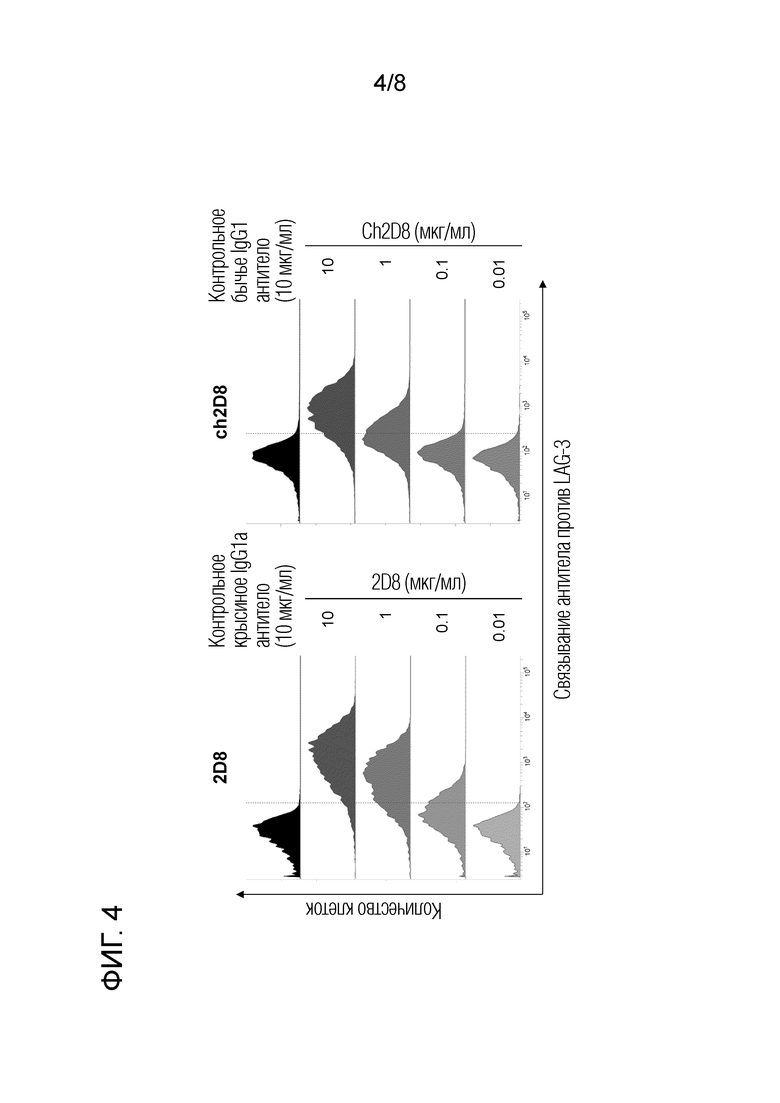
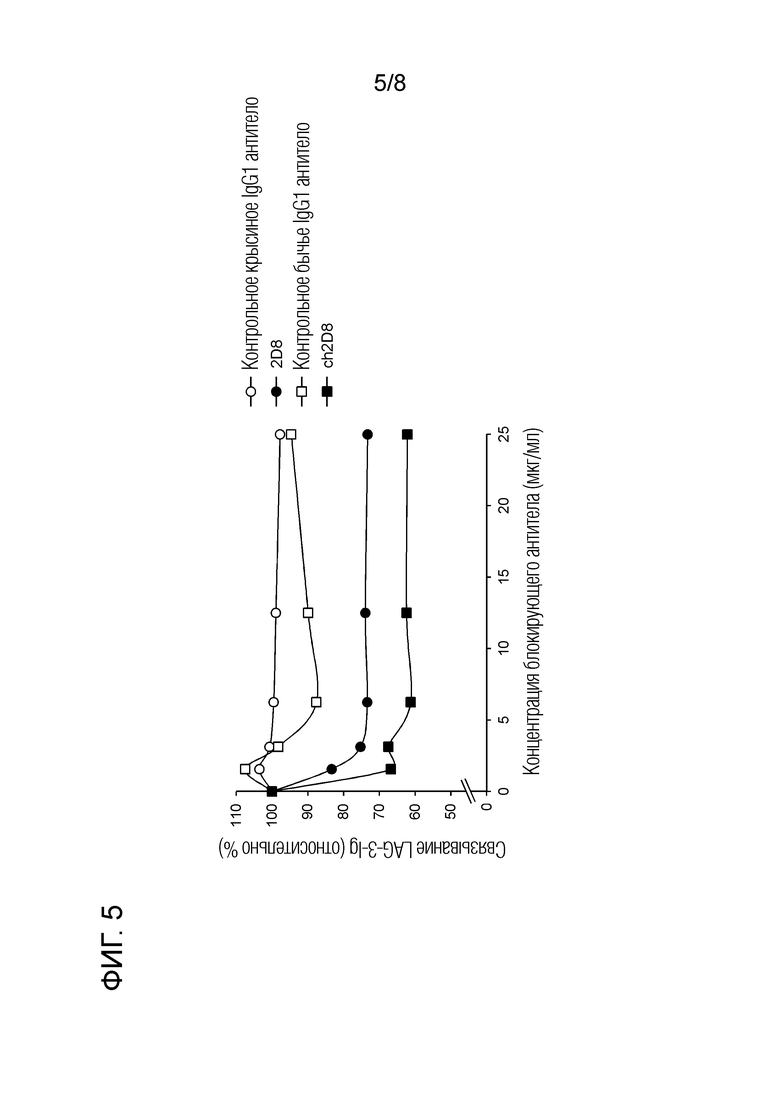
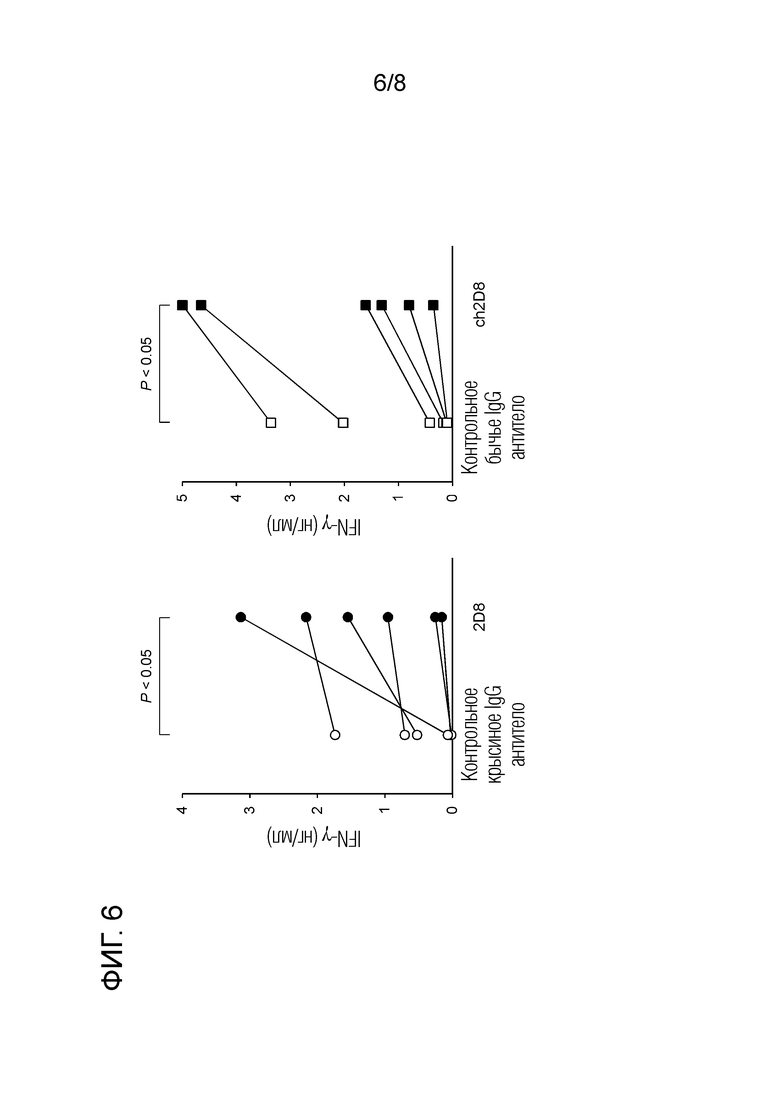
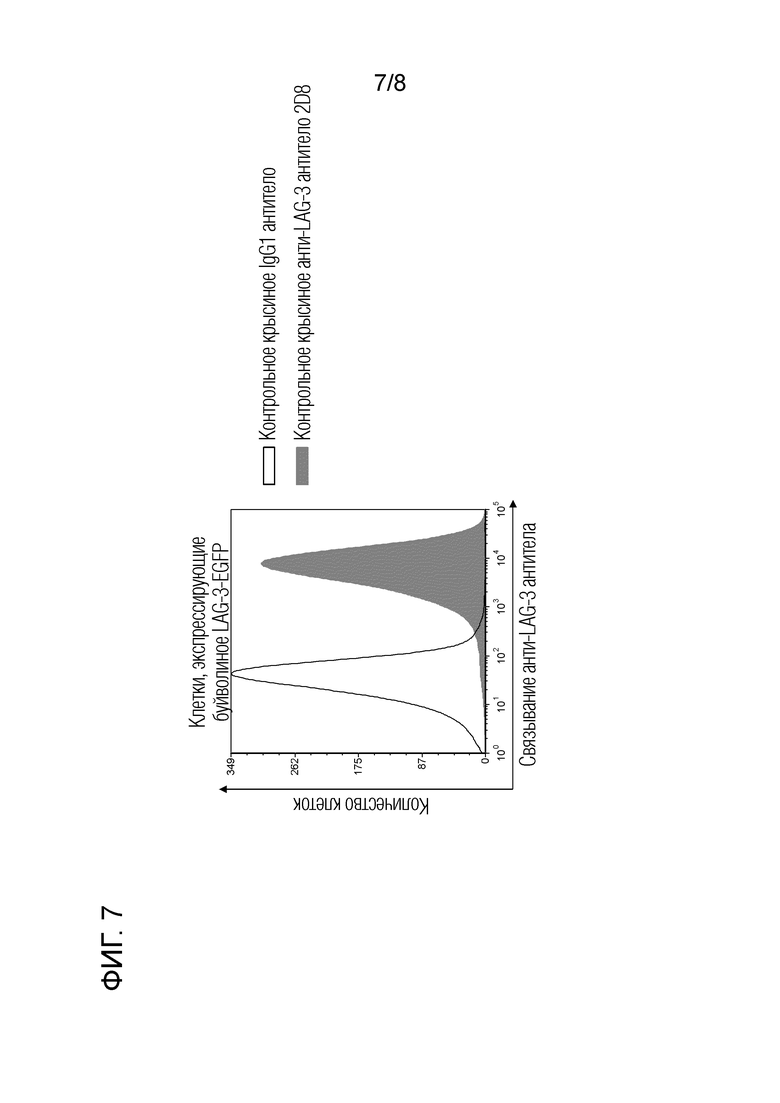
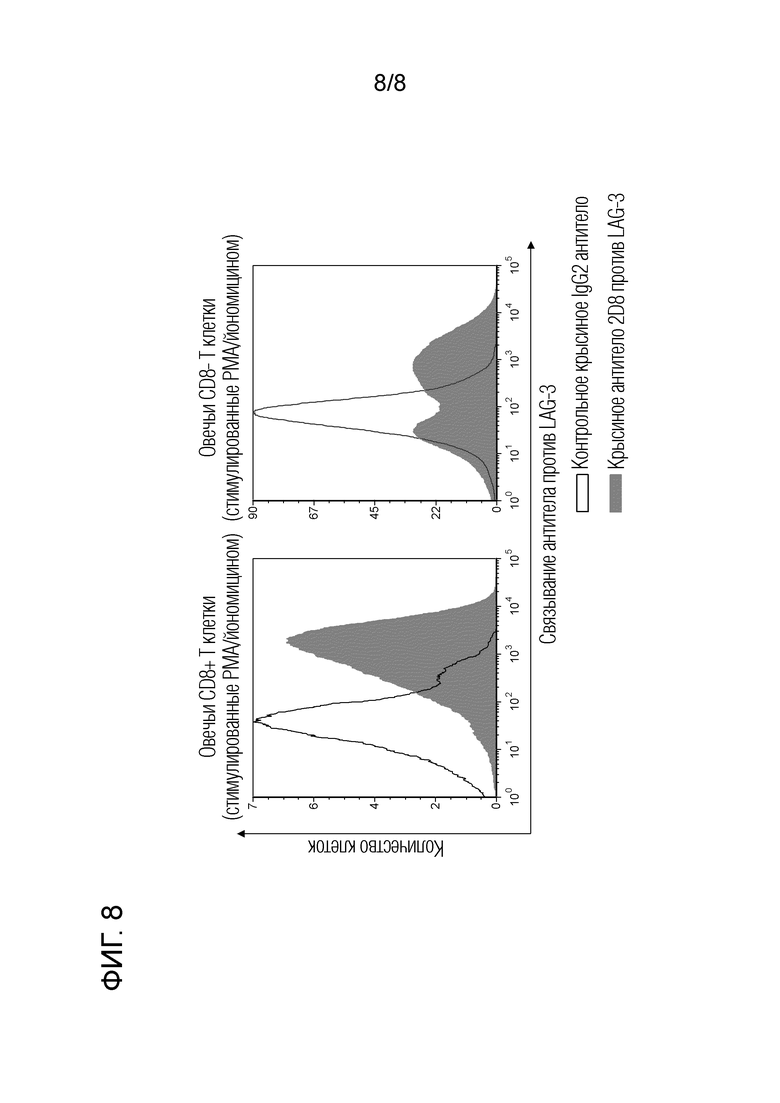
Изобретение относится к области биотехнологии, в частности к антителу против LAG-3, а также к содержащей его фармацевтической композиции. Также раскрыта искусственная генетическая ДНК, кодирующая вышеуказанное антитело или его легкую или тяжелую цепь, а также вектор и клетка, ее содержащие. Изобретение также относится к способу получения антитела против LAG-3, предусматривающему культивирование вышеуказанной клетки-хозяина. Изобретение позволяет эффективно осуществлять профилактику и/или лечение инфекции или злокачественного новообразования. 9 н. и 10 з.п. ф-лы, 8 ил., 2 пр.
1. Антитело против LAG-3, содержащее
(а) легкую цепь, содержащую
вариабельную область легкой цепи, содержащую CDR1 c аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO:16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO:17), и
константную область легкой цепи антитела животного, отличного от крысы; и
(b) тяжелую цепь, содержащую
вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO:18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO:19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO:20), и
константную область тяжелой цепи антитела животного, отличного от крысы.
2. Антитело по п.1, где вариабельная область легкой цепи и вариабельная область тяжелой цепи получены из крысы.
3. Антитело по п.2, где вариабельная область легкой цепи представляет собой вариабельную область легкой цепи крысиного антитела против бычьего LAG-3, вариабельная область тяжелой цепи представляет собой вариабельную область тяжелой цепи крысиного антитела против бычьего LAG-3.
4. Антитело по п.3, где вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO:1 и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:2.
5. Антитело по любому из пп.1-4, где константная область легкой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области лямбда-цепи или каппа-цепи.
6. Антитело по любому из пп.1-5, где константная область тяжелой цепи антитела животного, отличного от крысы, имеет аминокислотную последовательность константной области иммуноглобулина, эквивалентного IgG4 человека, или указанная аминокислотная последовательность имеет введенные мутации, которые снижают активность ADCC и/или активность CDC.
7. Антитело по п.5, где животное, отличное от крысы, является особью крупного рогатого скота; константная область легкой цепи бычьего антитела имеет аминокислотную последовательность константной области лямбда-цепи и константная область тяжелой цепи бычьего антитела имеет введенные в нее мутации, которые уменьшают активность ADCC и/или активность CDC.
8. Антитело по п.7, где константная область легкой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:3 и константная область тяжелой цепи бычьего антитела имеет аминокислотную последовательность SEQ ID NO:4.
9. Антитело по любому из пп.1-8, которое имеет четырехцепочечную структуру, содержащую две легкие цепи и две тяжелые цепи.
10. Фармацевтическая композиция для профилактики и/или лечения злокачественного новообразования, где злокачественное новообразование выбрано из группы, состоящей из опухолевых заболеваний и лейкоза, характеризующаяся тем, что в качестве активного ингредиента содержит антитело по любому из пп.1-9.
11. Композиция по п.10, где опухолевое заболевание выбрано из группы, состоящей из злокачественной меланомы, рака легких, рака желудка, рака почек, рака молочной железы, рака мочевого пузыря, рака пищевода и рака яичников.
12. Фармацевтическая композиция для профилактики и/или лечения инфекции, где инфекция выбрана из группы, состоящей из болезни Джона, анаплазмоза, бактериального мастита, микотического мастита, микоплазменных инфекций, туберкулеза, инфекции Theileria orientalis, криптоспоридиоза, кокцидиоза, трипаносомоза и лейшманиоза, отличающаяся тем, что содержит в качестве активного ингредиента антитело по любому из пп.1-9.
13. Композиция по п.12, где микоплазменная инфекция представляет собой микоплазменный мастит или микоплазменную пневмонию.
14. Искусственная генетическая ДНК, кодирующая антитело по любому из пп.1-9, где ДНК содержит
(а') ДНК, кодирующую легкую цепь, содержащую вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO:16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO:17), и константную область легкой цепи антитела животного, отличного от крысы; и
(b') ДНК, кодирующую тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO:18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO:19) и CDR3 с аминокислотной последовательностью NREDFDY (SEQ ID NO:20), и константную область тяжелой цепи антитела животного, отличного от крысы.
15. Вектор экспрессии, содержащий искусственную генетическую ДНК по п.14.
16. Клетка-хозяин для экспрессии антитела по любому из пп.1-9, где клетка трансформирована вектором по п.15.
17. Способ получения антитела против LAG-3, включающий культивирование клетки-хозяина по п.15 и сбор анти-LAG-3 антитела из полученной культуры.
18. ДНК, кодирующая легкую цепь антитела по любому из пп.1-9, где легкая цепь содержит вариабельную область легкой цепи, содержащую CDR1 с аминокислотной последовательностью QSLLDSDGNTY (SEQ ID NO:16), CDR2 с аминокислотной последовательностью SVS и CDR3 с аминокислотной последовательностью MQATHVPFT (SEQ ID NO:17).
19. ДНК, кодирующая тяжелую цепь антитела по любому из пп.1-9, где тяжелая цепь антитела содержит вариабельную область тяжелой цепи, содержащую CDR1 с аминокислотной последовательностью GFDFDTYP (SEQ ID NO:18), CDR2 с аминокислотной последовательностью ITIKTHNYAT (SEQ ID NO:19) и CDR3 c аминокислотной последовательностью NREDFDY (SEQ ID NO:20).
| TOMOHIRO OKAGAWA et al., Bovine Immunoinhibitory Receptors Contribute to Suppression of Mycobacterium avium subsp | |||
| paratuberculosis-Specific T-CellResponses, Infection and Immunity, January 2016, Vol.84, N.1, pp.77-89 | |||
| TOMOHIRO OKAGAWA, Hokokusho Ushi no Men'eki Yokusei Juyotai no Kino Kaiseki Oyobi Nanchisei Shippei no Shinki Seigyoho eno Oyo |

