Настоящее изобретение относится к гонадотропинам для применения в лечении бесплодия. В частности, оно относится к фолликулостимулирующему гормону (ФСГ, FSH).
Гонадотропины представляют собой группу гетеродимерных гликопротеиновых гормонов, регулирующих функции гонад в мужском и женском организмах. Они включают фолликулостимулирующий гормон (ФСГ, FSH), лютеинизирующий гормон (ЛГ, LH) и хорионический гонадотропин (ХГ, CG).
Естественная секреция ФСГ происходит в передней доле гипофиза, и функция ФСГ включает поддержание развития фолликула и овуляции. ФСГ содержит альфа-субъединицу из 92 аминокислот, также характерную для других гликопротеиновых гормонов ЛГ и ХГ, и бета-субъединицу из 111 аминокислот, уникальную для ФСГ, обеспечивающую биологическую специфичность гормона (Pierce and Parsons, 1981). Каждая субъединица проходит посттрансляционную модификацию с присоединением сложных углеводных остатков. Обе субъединицы имеют 2 сайта для присоединения N-связанного гликана, альфа-субъединица - по аминокислотам 52 и 78, и бета субъединица - по аминокислотным остаткам 7 и 24 (Rathnam and Saxena, 1975, Saxena and Rathnam, 1976). Таким образом, ФСГ гликозилирован приблизительно на 30% по массе (Dias and Van Roey. 2001. Fox et al. 2001).
ФСГ, очищенный из мочи людей в постменопаузе, в течение многих лет использовали для лечения бесплодия, как для стимуляции овуляции при естественном воспроизведении, так и для обеспечения ооцитов для вспомогательных репродуктивных технологий. В середине 1990-х гг. стали доступны два рекомбинантных варианта ФСГ, Gonal-F (Serono) и Puregon (Organon). Оба эти варианта экспрессируют в клетках яичника китайского хомячка (CHO) (Howles, 1996).
Препараты ФСГ в значительной степени гетерогенны, что связано с различиями в количестве разных изоформ, присутствующих в препарате. Отдельные изоформы ФСГ демонстрируют идентичные аминокислотные последовательности, но различаются по степени их посттрансляционной модификации; конкретные изоформы отличаются гетерогенностью разветвленных углеводных структур и разными количествами присоединенной сиаловой кислоты (концевого сахара), при этом оба параметра, по-видимому, влияют на специфическую биологическую активность изоформы.
Гликозилирование природного ФСГ очень сложно. Гликаны в гипофизарном ФСГ естественного происхождения могут содержать широкий спектр структур, которые могут включать комбинации би-, три- и тетра-разветвленных гликанов (Pierce and Parsons, 1981. Ryan et al., 1987. Baenziger and Green, 1988). Гликаны могут иметь другие модификации: центральное фукозилирование, разделенный пополам глюкозамин, цепи, расширенные ацетиллактозамином, частичное или полное сиалирование, сиалирование с α2,3- и α2,6-связями и замещение галактозы сульфатированным галактозамином (Dalpathado et al., 2006). Кроме того, существуют различия между распределениями гликановых структур в отдельных сайтах гликозилирования. Сопоставимая степень сложности гликановых структур была обнаружена у ФСГ, имеющего происхождение из сыворотки индивидов и из мочи женщин в постменопаузе (Wide et al., 2007).
Гликозилирование продуктов рекомбинантного ФСГ отражает спектр гликозилтрансфераз, представленных в линии клеток-хозяев. Существующие продукты рекомбинантного ФСГ (рФСГ, rFSH) имеют происхождение из сконструированных клеток яичника китайского хомячка (клеток CHO). Спектр гликановых модификаций в рФСГ, имеющем происхождение из клеток CHO, уже спектра, обнаруживаемого у естественных продуктов, имеющих происхождение либо из экстрактов гипофиза, либо из мочи. Примеры меньшей гетерогенности гликанов, обнаруживаемой в рФСГ, имеющем происхождение из клеток CHO, включают отсутствие разделенных пополам глюкозаминов и меньший процент центрального фукозилирования и ацетиллактозаминовых расширений (Hard et al., 1990). В дополнение, в клетках CHO присоединение сиаловой кислоты возможно только через α2,3-связь (Kagawa et al, 1988, Takeuchi et al, 1988, Svensson et al., 1990). Это отличается от продуцируемого естественным образом ФСГ, содержащего гликаны со смесью сиаловых кислот, связанных α2,3- и α2,6-связями.
Было продемонстрировано, что препарат рекомбинантного ФСГ (Organon) отличается по количествам ФСГ с изоэлектрической точкой (pI) ниже 4 (рассматриваемого как кислые изоформы) при сравнении с ФСГ гипофиза, сыворотки или мочи при постменопаузе (Ulloa-Aguirre et al. 1995). Количество кислых изоформ в препаратах, полученных из мочи, было намного больше по сравнению с рекомбинантными продуктами Gonal-f (Serono) и Puregon (Organon) (Andersen et al. 2004). Это, должно быть, отражает меньшее молярное содержание сиаловой кислоты в рФСГ, поскольку содержание отрицательно заряженного гликана, модифицированного сульфатом, в ФСГ невелико. Меньшее содержание сиаловой кислоты по сравнению с природным ФСГ характерно для обоих имеющихся в продаже продуктов ФСГ и, следовательно, должно отражать ограничение технологического процесса (Bassett and Driebergen, 2005).
Существует множество научных работ, в которых предпринят анализ и попытка объяснения отличий гликозилирования ФСГ у разных индивидов и его изменений в течение овуляторного цикла. Одна из основных дискуссий относится к тому наблюдению, что как концентрация ФСГ, так и содержание сиаловой кислоты, снижаются в предовуляторной фазе цикла. Сниженное содержание сиаловой кислоты приводит к образованию более основного ФСГ, как подверженного более быстрому клиренсу, так и более активного в отношении рецептора мишени по меньшей мере in vitro (Zambrano et al. 1996). Вопрос о биологическом значении этих изменений и о том, как они могут быть вовлечены в селекцию доминантного фолликула, остается нерешенным (обзор Ulloa-Aguirre, 2003).
Продолжительность нахождения ФСГ в кровообращении была документирована для веществ из множества источников. Некоторые из этих веществ были фракционированы на основании общего заряда молекулы, что отражено в их pI, где большая кислотность соответствует большему отрицательному заряду. Как определено выше, основной вклад в общий заряд молекулы вносит общее содержание сиаловой кислоты в каждой молекуле ФСГ. Например, рФСГ (Organon) имеет содержание сиаловой кислоты приблизительно 8 моль/моль, в то время как ФСГ, имеющий происхождение из мочи, имеет более высокое содержание сиаловой кислоты (de Leeuw et al. 1996). Соответствующие скорости клиренса из плазмы у крыс составляют 0,34 и 0,14 мл/мин (Ulloa-Aguirre et al. 2003). В другом примере, где образец рекомбинантного ФСГ разделяли на фракции с высокой и низкой pI, активность фракции с высокой pI (c меньшим содержанием сиаловой кислоты) in vivo была снижена, и эта фракция имела более короткий период полувыведения из плазмы (D'Antonio et al. 1999). Также было сообщено, что более основной ФСГ, циркулирующий на поздних стадиях овуляторного цикла, обусловлен понижающей регуляцией α2,3-сиалилтрансферазы в аденогипофизе, что вызвано повышенными уровнями эстрадиола (Damian-Matsumara et al. 1999. Ulloa-Aguirre et al. 2001). О результатах для α2,6-сиалилтрансферазы сообщено не было.
Непосредственное сравнение общего содержания сиаловой кислоты в ФСГ и рФСГ невозможно, поскольку связывание сиаловых кислот обычно происходит двумя путями. ФСГ гипофиза/сыворотки/мочи обычно содержит сиаловую кислоту, связанную как α2,3-, так и α2,6-связями, с преобладанием первых. Однако, рекомбинантные формы, имеющие происхождение из клеток CHO, содержат только α2,3-связи (Kagawa et al, 1988, Takeuchi et al, 1988, Svensson et al., 1990). Это еще одно отличие между природными и современными рекомбинантными продуктами, в дополнение к меньшему общему содержанию сиаловой кислоты в последних.
Для получения фармацевтических человеческих рекомбинантных белков обычно используют клетки CHO. При структурном анализе выявлено, что присоединение сиаловой кислоты происходит исключительно через α2,3-связь. (Kagawa et al, 1988, Takeuchi et al, 1988, Svensson et al., 1990). Многие человеческие гликопротеины содержат как α2,3-, так и α2,6-связи. Таким образом, рекомбинантные белки, экспрессированные с использованием системы CHO, будут отличаться от их природных аналогов по их типу связей с концевыми сиаловыми кислотами. Это является важным соображением при получении биологических агентов для фармацевтического применения, поскольку углеводные группировки могут влиять на фармакологические свойства молекулы.
Желательно иметь продукт рФСГ, более точно повторяющий или имитирующий физиохимический и фармакокинетический профиль продукта, полученного из человеческой мочи. Желательно иметь продукт рФСГ, имеющий улучшенное фармакокинетическое свойство или свойства по сравнению с известным рекомбинантным продуктом.
Согласно настоящему изобретению предложен рекомбинантный ФСГ («рФСГ», «rFSH» или «рекФСГ», «recFSH»), включающий α2,3-сиалирование и α2,6-сиалирование, и, возможно, α2,8-сиалирование. В рФСГ (или препарате рФСГ) по изобретению 10% или более общего сиалирования может представлять собой α2,3-сиалирование, например, 65-85% общего сиалирования может представлять собой α2,3-сиалирование. В рФСГ (или препарате рФСГ) по изобретению 50% или менее общего сиалирования может представлять собой α2,6-сиалирование, например, 15-35% общего сиалирования может представлять собой α2,6-сиалирование. В рФСГ (или препарате рФСГ) по изобретению 5% или менее общего сиалирования может представлять собой α2,8-сиалирование, например, 0,1-4% общего сиалирования может представлять собой α2,8-сиалирование. рФСГ (или препарат рФСГ) по изобретению может иметь содержание сиаловой кислоты (выраженное как отношение молей сиаловой кислоты к молям белка) 6 моль/моль или более, например, от 6 моль/моль до 15 моль/моль.
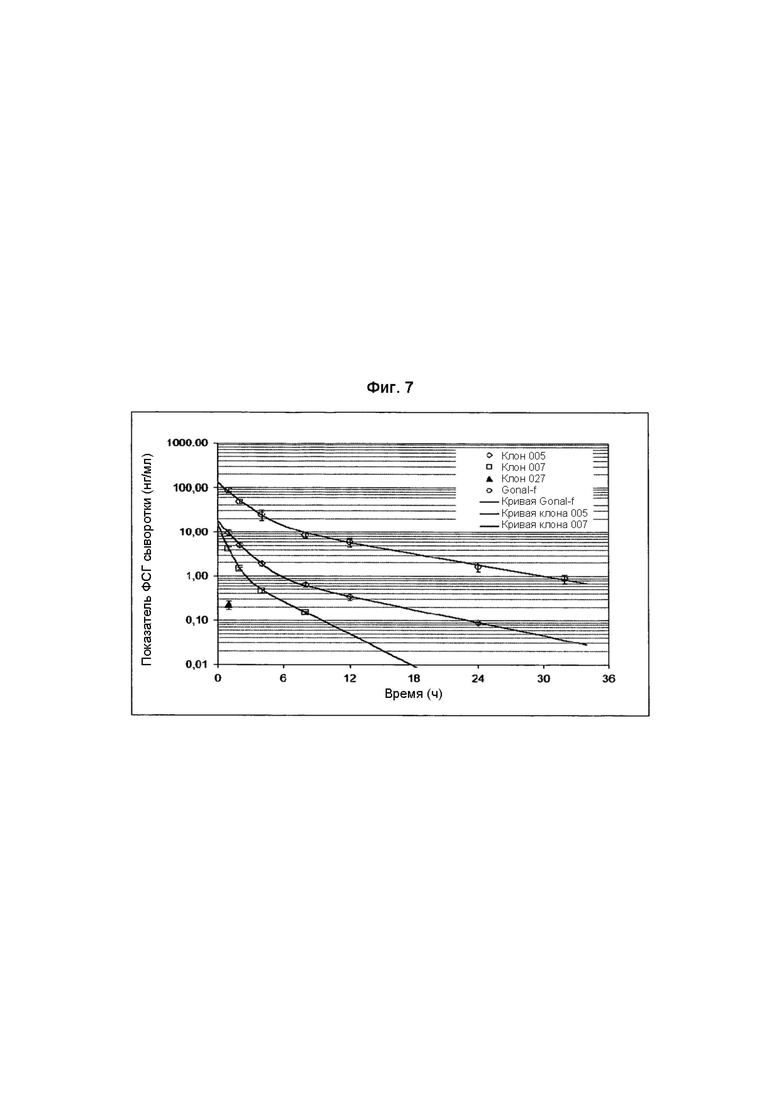
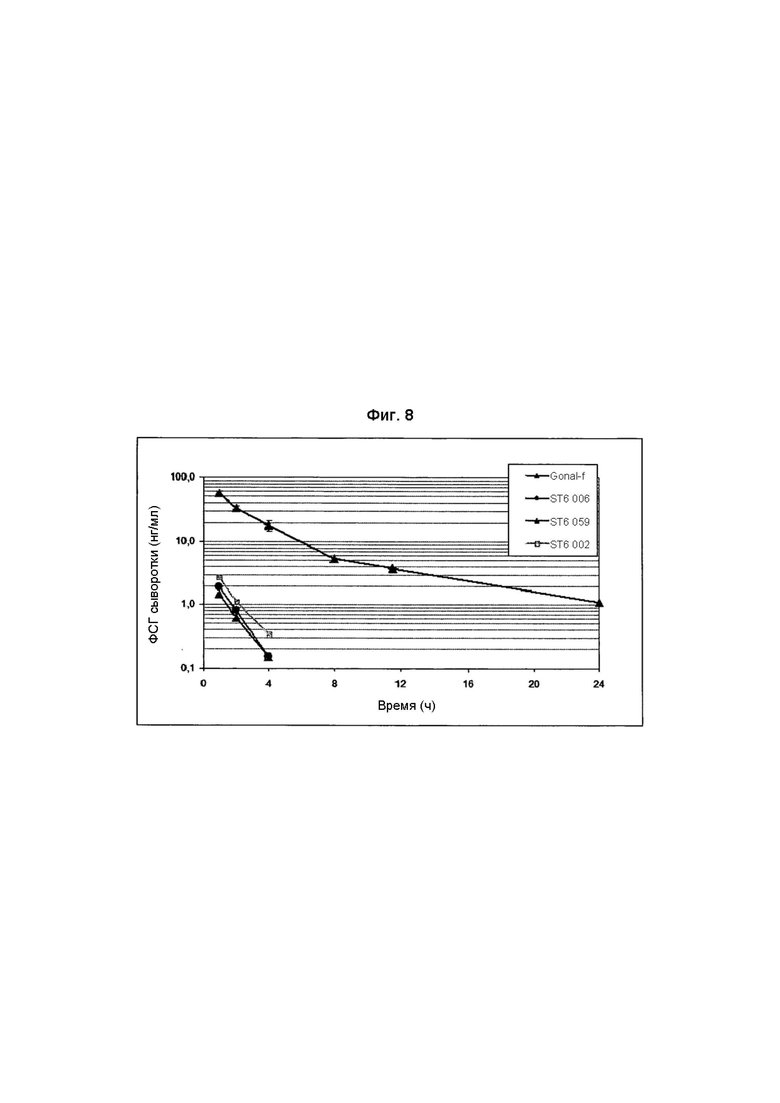
Авторы настоящего изобретения обнаружили, что тип связи с сиаловой кислотой, α2,3- или α2,6-, может оказывать сильное влияние на биологический клиренс ФСГ. В человеческих клеточных линиях, в отличие от клеточных линий CHO, возможна экспрессия рекомбинантного ФСГ с сиаловыми кислотами, присоединенными как α2,3-, так и α2,6-связями. В Примере 4 была получена клеточная линия, образующая рекомбинантный ФСГ, которая экспрессировала ФСГ, содержащий гликаны с низкими уровнями как сиаловой кислоты, связанной α2,3-связями, так и сиаловой кислоты, связанной α2,6-связями (Фиг. 6). Как и можно было предполагать, это основное вещество с ограниченным содержанием сиаловой кислоты (Фиг. 4) было подвержено очень быстрому клиренсу из кровообращения у крыс (Фиг. 7). Клеточную линию затем подвергали второй стадии конструирования с добавлением гена, кодирующего α2,6-сиалилтрансферазу (Пример 5). Полученный рФСГ был сильно сиалирован и демонстрировал содержание сиаловой кислоты и распределение pI, сопоставимые с ФСГ мочи (Фиг. 5). Однако, это вещество было подвержено очень быстрому клиренсу из кровообращения у крыс со скоростью, сопоставимой с исходным веществом, имевшим низкое содержание сиаловой кислоты (Фиг. 8). Это наблюдение было неожиданным, поскольку известно, что часть сиаловой кислоты в природном и биологически активном ФСГ связана α2,6-связями. Было обнаружено, что клиренс α2,6-сиалированного рФСГ опосредован асиалогликопротеиновым рецептором (ASGP-рецептором), обнаруженным в печени (Пример 9). Это было продемонстрировано временной блокадой ASGP-рецепторов с использованием избытка другого субстрата этого рецептора. При блокаде этого рецептора асиалофетуином ожидаемый клиренс высоко сиалированного вещества был восстановлен (Фиг. 9). Это продолжалось в течение нескольких часов, пока блокада не была преодолена и не был восстановлен быстрый клиренс высоко сиалированного рФСГ с α2,6-связями.
Рекомбинантный ФСГ со смесью как сиаловой кислоты, связанной α2,3-связями, так и сиаловой кислоты, связанной α2,6-связями, был получен конструированием человеческой клеточной линии для экспрессии как рФСГ, так и α2,3-сиалилтрансферазы (Пример 4 и 5). Экспрессируемый продукт является сильно кислым и имеет смесь как сиаловых кислот, связанных α2,3-связями, так и сиаловых кислот, связанных α2,6-связями; последние обусловлены активностью эндогенной сиалилтрансферазы (Фиг. 6). Это имеет два преимущества по сравнению с рФСГ, экспрессированным в обычных клетках CHO: во-первых, вещество более высоко сиалировано вследствие комбинированных активностей двух сиалилтрансфераз; и, во-вторых, вещество в большей степени напоминает природный ФСГ. Вероятно, оно является биологически более подходящим, чем рекомбинантные продукты, имеющие происхождение из клеток CHO, продуцирующие сиаловую кислоту, связанную только α2,3-связями (Kagawa et al, 1988, Takeuchi et al, 1988, Svensson et al., 1990), и имеющие сниженное содержание сиаловой кислоты (Ulloa-Aguirre et al. 1995., Andersen et al. 2004).
Авторы настоящего изобретения неожиданно обнаружили, что рФСГ по изобретению может более точно, чем другие рекомбинантные продукты, повторять или имитировать физиохимический и фармакокинетический профиль естественного продукта, полученного из человеческой мочи. Иными словами, рФСГ по изобретению может быть более близким «естественному» ФСГ. Это может иметь существенные преимущества в отношении дозирования и тому подобного. Кроме того, более «естественный» или более «человеческий» продукт может быть более желательным для пациента, который может желать, чтобы терапия, пусть по сути дела искусственная, была по возможности «естественной». У рекомбинантного продукта, имеющего структуру углеводов (например, гликанов), более близкую к природному ФСГ (например, ФСГ мочи человека), чем другие рекомбинантные продукты, могут быть и другие преимущества (например, фармакокинетические преимущества).
Таким образом, согласно изобретению предложен рекомбинантный вариант ФСГ, имеющий смесь сиаловой кислоты, связанной α2,3-связями, и сиаловой кислоты, связанной α2,6-связями, и, следовательно, в большей степени напоминающий природный ФСГ. Ожидают, что применение этого соединения для контролируемой стимуляции яичников в методиках экстракорпорального оплодотворения (ЭКО) и индукции овуляции будет приводить к более естественной стимуляции яичника по сравнению с существующими рекомбинантными продуктами.
Согласно настоящему изобретению предложен рекомбинантный ФСГ («рФСГ», «rFSH» или «рекФСГ», «recFSH») (и/или препарат рекомбинантного ФСГ), включающий α2,3-сиалирование и α2,6-сиалирование. рФСГ или препарат рФСГ может, возможно, дополнительно включать α2,8-сиалирование.
Здесь термин «препарат рекомбинантного ФСГ» включает препарат, для, например, фармацевтического применения, содержащий рекомбинантный ФСГ. В воплощениях данного изобретения рФСГ может присутствовать в виде одной изоформы или как смесь изоформ.
рФСГ (или препарат рФСГ) по изобретению может иметь содержание сиаловой кислоты (выраженное как отношение молей сиаловой кислоты к молям белка) 6 моль/моль или более (Пример 8), например, от 6 моль/моль до 15 моль/моль, например, от 8 моль/моль до 14 моль/моль, например, от 10 моль/моль до 14 моль/моль, например, от 11 моль/моль до 14 моль/моль, например, от 12 моль/моль до 14 моль/моль, например, от 12 моль/моль до 13 моль/моль. рФСГ по изобретению может быть получен или экспрессирован в человеческой клеточной линии.
В рФСГ (или препарате рФСГ) по изобретению 10% или более общего сиалирования может представлять собой α2,3-сиалирование. Например, 20, 30, 40, 50, 60, 70, 80 или 90% или более общего сиалирования может представлять собой α2,3-сиалирование. рФСГ (или препарат рФСГ) может включать α2,3-сиалирование в количестве, составляющем от 65 до 85% общего сиалирования, например, от 70 до 80% общего сиалирования, например, от 71 до 79% общего сиалирования. В рФСГ (или препарате рФСГ) по изобретению 50% или менее общего сиалирования может представлять собой α2,6-сиалирование. Например, 40, 30, 20, 10 или 5% или менее общего сиалирования может представлять собой α2,6-сиалирование. рФСГ (или препарат рФСГ) может включать α2,6-сиалирование в количестве, составляющем от 15 до 35% общего сиалирования, например, от 20 до 30% общего сиалирования, например, от 21 до 29% общего сиалирования. В рФСГ (или препарате рФСГ) по изобретению 5% или менее общего сиалирования может представлять собой α2,8-сиалирование. Например, 2,5% или менее общего сиалирования может представлять собой α2,8-сиалирование. рФСГ (или препарат рФСГ) может включать α2,8-сиалирование в количестве, составляющем от 0,1 до 4% общего сиалирования, например, от 0,5 до 3% общего сиалирования, например, от 0,5 до 2,5% общего сиалирования. Под сиалированием понимают количество остатков сиаловой кислоты, присутствующих в углеводных структурах ФСГ. α2,3-Сиалирование означает сиалирование по 2,3-положению (как хорошо известно в данной области техники), и α2,6-сиалирование означает сиалирование по 2,6-положению (как хорошо известно в данной области техники). Таким образом, «% общего сиалирования, который может представлять собой α2,3-сиалирование» относится к % общего числа остатков сиаловой кислоты, присутствующих в ФСГ, сиалированных по 2,3-положению. Термин «% общего сиалирования, представляющий собой α2,6-сиалирование» относится к % общего числа остатков сиаловой кислоты, присутствующих в ФСГ, сиалированных по 2,6-положению.
рФСГ (или препарат рФСГ) по изобретению может иметь содержание сиаловой кислоты (количество сиалирования на молекулу ФСГ) (исходя из массы белка, а не массы белка с углеводом) 6% или более (например, от 6% до 15%, например, от 7% до 13%, например, от 8% до 12%, например, от 11% до 15%, например, от 12% до 14%) по массе.
Рекомбинантный ФСГ, экспрессированный в клетках яичника китайского хомячка (CHO), содержит исключительно α2,3-сиалирование (Kagawa et al, 1988, Takeuchi et al. 1988, Svensson et al. 1990).
рФСГ по изобретению может быть получен или экспрессирован в человеческой клеточной линии. Это может упростить (и сделать более эффективным) способ получения, поскольку манипулирование и контроль, например, питательной среды для клеток для сохранения сиалирования могут быть менее критичными, чем в известных способах. Способ может также быть более эффективным ввиду меньшего образования основного рФСГ по сравнению с получением известных продуктов рФСГ; происходит образование более кислого рФСГ, и отделение/удаление основного ФСГ менее проблематично. рФСГ может быть получен или экспрессирован в клеточной линии Per.C6, клеточной линии, имеющей происхождение из Per.C6, или модифицированной клеточной линии Per.C6. Клеточная линия может быть модифицирована с использованием α2,3-сиалилтрансферазы. Клеточная линия может быть модифицирована с использованием α2,6-сиалилтрансферазы. Альтернативно или дополнительно, рФСГ может содержать сиаловые кислоты, связанные α2,6-связями (α2,6-сиалирование), обеспеченные активностью эндогенной сиалилтрансферазы (клеточной линии).
рФСГ может быть получен с использованием α2,3- и/или α2,6-сиалилтрансферазы. рФСГ может быть получен с использованием α2,3-сиалилтрансферазы. рФСГ может содержать сиаловые кислоты, связанные α2,6-связями (α2,6-сиалирование), обеспеченные активностью эндогенной сиалилтрансферазы.
В другом аспекте согласно настоящему изобретению предложен способ получения рФСГ и/или препарата рФСГ, как описано здесь (согласно аспектам изобретения), включающий стадию продуцирования или экспрессии рФСГ в человеческой клеточной линии, например, клеточной линии Per.C6, клеточной линии, имеющей происхождение из Per.C6, или модифицированной клеточной линии Per.C6, например, клеточной линии, которая была модифицирована с использованием α2,3-сиалилтрансферазы.
Структура рФСГ содержит гликановые группировки. Возможны разветвления, приводящие к тому, что гликан имеет 1, 2, 3, 4 или более концевых сахарных остатков или «антенн», как хорошо известно в данной области техники. рФСГ по изобретению может содержать гликаны с сиалированием, присутствующим в структурах с одним концевым остатком, и/или с двумя концевыми остатками, и/или с тремя концевыми остатками, и/или с четырьмя концевыми остатками. рФСГ может предпочтительно включать моносиалированные, дисиалированные, трисиалированные и тетрасиалированные гликановые структуры со следующими относительными количествами: 9-15% моносиалированных, 27-30% дисиалированных, 30-36% трисиалированных и 25-29% тетрасиалированных (например, как показано анализом со слабыми анионообменниками (WAX-анализом) заряженных гликанов, как изложено в Примере 8в).
В другом аспекте согласно настоящему изобретению предложен рФСГ, полученный (например, экспрессированный) в человеческой клеточной линии. рФСГ может включать α2,3- и α2,6-сиалирование. рФСГ может быть получен или экспрессирован в клеточной линии Per.C6, клеточной линии, имеющей происхождение из Per.C6, или модифицированной клеточной линии Per.C6. Клеточная линия может быть модифицирована с использованием α2,3-сиалилтрансферазы. Клеточная линия может быть модифицирована с использованием α2,6-сиалилтрансферазы. Альтернативно или дополнительно рФСГ может содержать сиаловые кислоты, связанные α2,6-связями (α2,6-сиалирование), обеспеченные активностью эндогенной сиалилтрансферазы (клеточной линии). В рФСГ (или препарате рФСГ) 10% или более общего сиалирования может представлять собой α2,3-сиалирование, например, 65-85% общего сиалирования может представлять собой α2,3-сиалирование. В рФСГ (или препарате рФСГ) по изобретению 50% или менее общего сиалирования может представлять собой α2,6-сиалирование, например, 15-35% общего сиалирования может представлять собой α2,6-сиалирование. В рФСГ (или препарате рФСГ) по изобретению 5% или менее общего сиалирования может представлять собой α2,8-сиалирование, например, 0,5-4% общего сиалирования может представлять собой α2,8-сиалирование. рФСГ может иметь содержание сиаловой кислоты (выраженное как отношение молей сиаловой кислоты к молям белка) 6 моль/моль или более, например, от 6 моль/моль до 15 моль/моль.
В другом аспекте согласно настоящему изобретению предложена фармацевтическая композиция, содержащая рФСГ, включающий α2,3-сиалирование и α2,6-сиалирование (например, как изложено выше). Фармацевтическая композиция может дополнительно содержать хорионический гонадотропин человека (ХГЧ, hCG) и/или ЛГ.
ХГЧ может быть получен любым способом, известным в данной области техники. При использовании здесь ХГЧ включает ХГЧ, имеющий происхождение из человека, и рекомбинантный ХГЧ. ХГЧ, имеющий происхождение из человека, может быть очищен из любого подходящего источника (например, мочи и плаценты) любым способом, известным в данной области техники. Способы экспрессии и очистки рекомбинантного ХГЧ хорошо известны в данной области техники.
ЛГ может быть получен любым способом, известным в данной области техники. При использовании здесь ЛГ включает ЛГ, имеющий происхождение из человека, и рекомбинантный ЛГ. ЛГ, имеющий происхождение из человека, может быть очищен из любого подходящего источника (например, мочи) любым способом, известным в данной области техники. Способы экспрессии и очистки рекомбинантного ЛГ известны в данной области техники.
Фармацевтическая композиция может быть предназначена для лечения бесплодия, например, для применения, например, во вспомогательных репродуктивных технологиях (ВРТ), индукции овуляции или внутриматочном осеменении (ВО). Фармацевтическая композиция может быть использована, например, по медицинским показаниям, при которых используют известные препараты ФСГ. Согласно настоящему изобретению также предложено применение рФСГ и/или препарата рФСГ, описанного здесь (согласно аспектам изобретения), для изготовления или в изготовлении лекарственного средства для лечения бесплодия. Фармацевтические композиции по настоящему изобретению могут быть изготовлены в форме хорошо известных композиций для любого пути введения лекарственного средства, например, перорального, ректального, парентерального, чрескожного (например, пластырная технология), внутривенного, внутримышечного, подкожного, внутригрудинного, интравагинального, внутрибрюшинного, местного (порошки, мази или капли) или в форме буккального или назального спрея. Типичная композиция содержит фармацевтически приемлемый носитель, такой как водный раствор, нетоксичные эксципиенты, включая соли и консерванты, буферы и тому подобное, как описано, среди прочего, в Remington's Pharmaceutical Sciences fifteenth edition (Matt Publishing Company, 1975), на стр. 1405-1412 и 1461-87, и Национальном фармацевтическом формуляре XIV, четырнадцатого издания (American Pharmaceutical Association, 1975).
Примеры подходящих водных и неводных фармацевтических носителей, разбавителей, растворителей или наполнителей включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное), карбоксиметилцеллюлозу и ее подходящие смеси, растительные масла (такие как оливковое масло) и органические сложные эфиры для инъекций, такие как этилолеат.
Композиции по настоящему изобретению также могут содержать добавки, такие как, без ограничения, консерванты, увлажняющие агенты, эмульгаторы и диспергирующие агенты. Для предотвращения роста микробов могут быть включены антибактериальные и противогрибковые агенты, и они включают, например, парабен, хлорбутанол, фенол, сорбиновую кислоту и тому подобное. Более того, может быть желательным включение изотонических агентов, таких как сахара, хлорид натрия и тому подобное.
В некоторых случаях для обеспечения длительного действия желательно замедлить всасывание ФСГ (и других активных ингредиентов, если они присутствуют) после подкожной или внутримышечной инъекции. Это может быть осуществлено с использованием жидкой суспензии кристаллического или аморфного вещества, плохо растворимого в воде. В этом случае скорость всасывания ФСГ зависит от скорости его растворения, которая, в свою очередь, может зависеть от размера кристаллов и кристаллической формы. Альтернативно, замедленного всасывания парентерально вводимой формы комбинированного ФСГ достигают растворением или суспендированием комбинированного ФСГ в масляном наполнителе.
Депо-формы для инъекций могут быть получены путем изготовления микроинкапсулированных матриц ФСГ (и других агентов, если они присутствуют) в биодеградируемых полимерах, таких как полилактид-полигликолид. В зависимости от соотношения ФСГ и полимера и природы конкретного используемого полимера можно регулировать скорость высвобождения ФСГ. Примеры других биодеградируемых полимеров включают поливинилпирролидон, поли(ортоэфиры), поли(ангидриды) и тому подобное. Депо-композиции для инъекций также изготавливают путем захвата ФСГ в липосомы или микроэмульсии, совместимые с тканями организма.
Композиции для инъекций можно стерилизовать, например, фильтрацией через фильтр, задерживающий бактерии, или включением стерилизующих агентов в форму стерильных твердых композиций, которые могут быть растворены или диспергированы в стерильной воде или другой стерильной среде для инъекций непосредственно перед применением. Возможна доставка композиций для инъекций в любом подходящем контейнере, например, флаконе, предварительно заполненном шприце, инъекционных картриджах и тому подобном.
Возможна доставка композиций для инъекций в виде продукта, включающего фармацевтические композиции, содержащие ФСГ (возможно с ХГЧ, ЛГ и тому подобным). При наличии более чем одного активного ингредиента (то есть, ФСГ и, например, ХГЧ или ЛГ), они могут быть подходящими для раздельного или совместного введения. При раздельном введении, оно может быть последовательным. Возможна доставка продукта в любой подходящей упаковке. Например, продукт может включать несколько предварительно заполненных шприцев, содержащих либо ФСГ или ХГЧ по отдельности, либо комбинацию ФСГ и ХГЧ, упакованных в блистерную упаковку или другое устройство для сохранения стерильности. Продукт может возможно включать инструкции по применению композиций ФСГ и ХГЧ.
pH и точная концентрация различных компонентов фармацевтической композиции корректируют в соответствии с обычной практикой в данной области техники. См. GOODMAN and GILMAN, THE PHARMACOLOGICAL BASIS FOR THERAPEUTICES, 7th ed. В предпочтительном воплощении композиции по изобретению доставляют как композиции для парентерального введения. Общие способы изготовления композиций для парентерального введения известны в данной области техники и описаны в REMINGTON; THE SCIENCE AND PRACTICE OF PHARMACY, supra, на стр. 780-820. Возможна доставка композиций для парентерального введения в виде жидкой композиции или твердого вещества, которое будут смешивать со стерильной средой для инъекций непосредственно перед введением. В особо предпочтительном воплощении композиции для парентерального введения доставляют в стандартной лекарственной форме для простоты введения и единообразия дозирования.
Подробное описание изобретения
Настоящее изобретение будет теперь описано более подробно со ссылкой на следующие Примеры и приложенные графические материалы, описанные ниже.
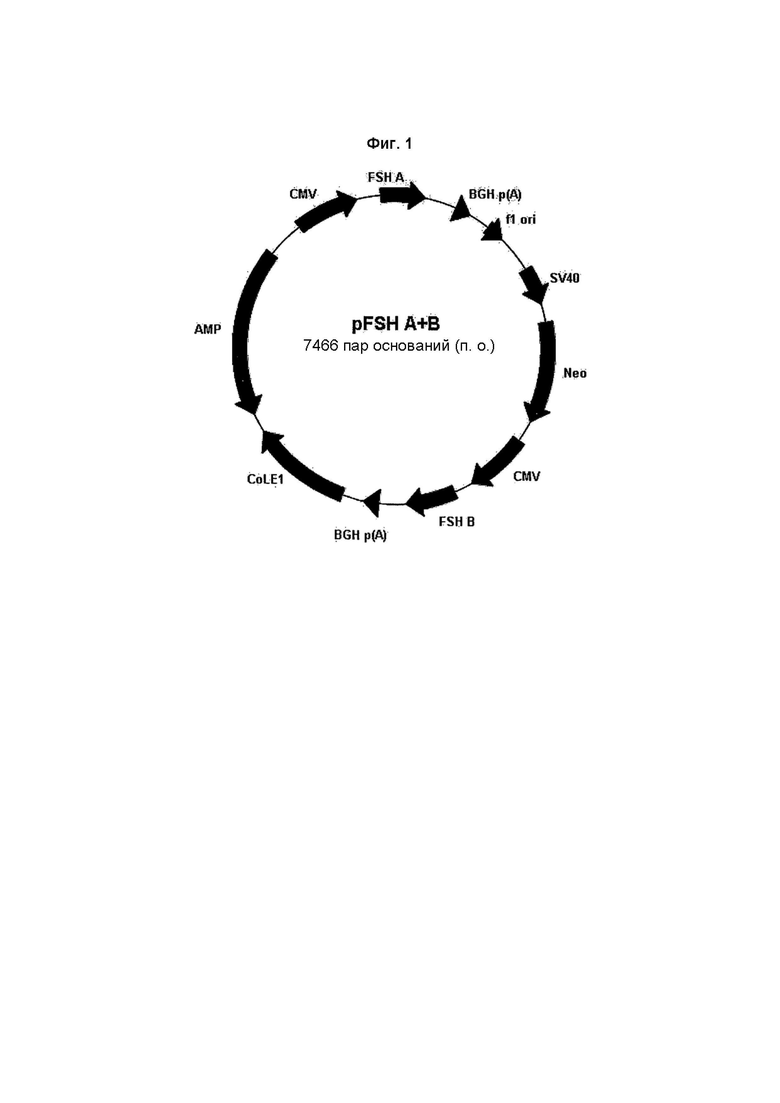
На Фиг. 1 показана плазмидная карта вектора экспрессии pFSHalpha/beta.
CMV обозначает цитомегаловирусный промотор, BGHp(A) обозначает последовательность полиаденилирования бычьего гормона роста, fl ori обозначает репликатор fl, SV40 обозначает промотор вируса обезьян 40, Neo обозначает маркер устойчивости к неомицину, Hyg обозначает маркер устойчивости к гигромицину, SV40 p(A) обозначает последовательность полиаденилирования вируса обезьян 40, FSH A обозначает альфа-полипептид фолликулостимулирующего гормона, FSH В обозначает бета-полипептид фолликулостимулирующего гормона, ColEI обозначает репликатор ColEI, Amp обозначает маркер устойчивости к ампициллину.
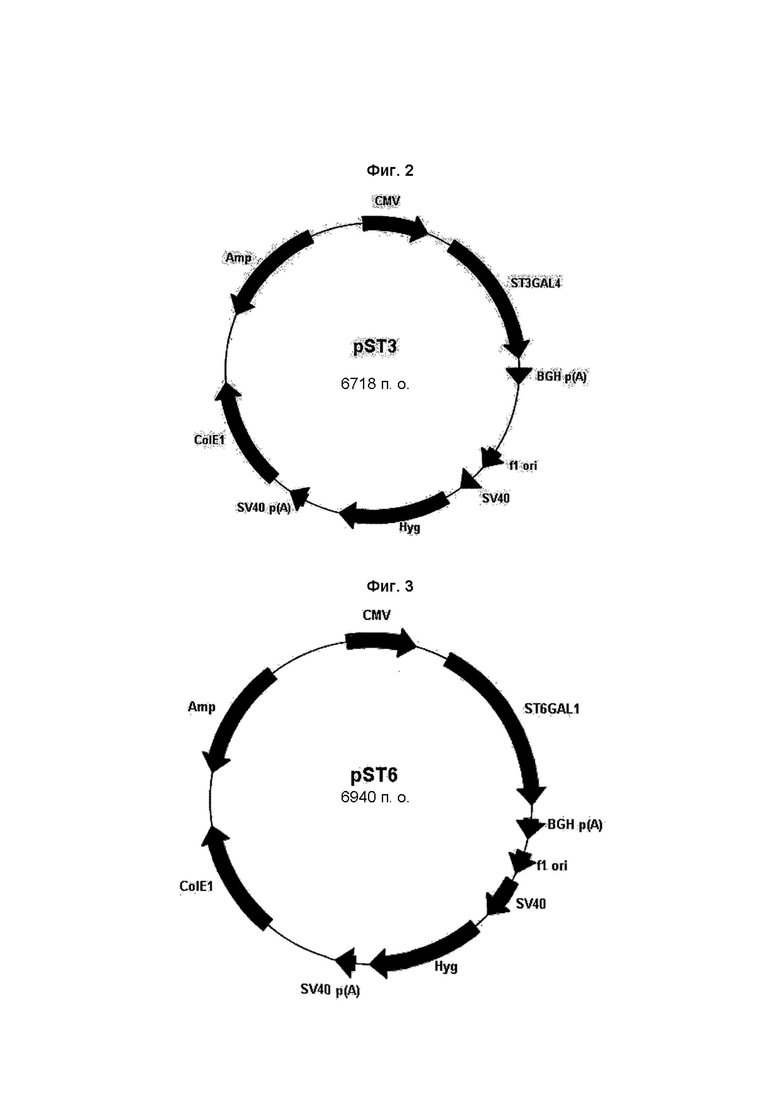
На Фиг. 2 показана плазмидная карта вектора экспрессии α2,3-сиалилтрансферазы pST3 (ST3GAL4). ST3GAL4 обозначает α2,3-сиалилтрансферазу.
На Фиг. 3 показана плазмидная карта вектора экспрессии α2,6-сиалилтрансферазы pST6 (ST6GAL1). ST6GAL1 обозначает α2,6-сиалилтрансферазу.
На Фиг. 4 показано изоэлектрическое фокусирование рекомбинантного ФСГ, продуцированного клетками Per.C6, стабильно экспрессирующими ФСГ.
Супернатанты клеточных культур разделяли в нативных условиях в градиенте pH 3,0-7,0. Клон 005 является типичным из пяти клонов, взятых далее для конструирования с использованием сиалилтрансферазы. Клоны, содержащие менее кислые изоформы, были забракованы.
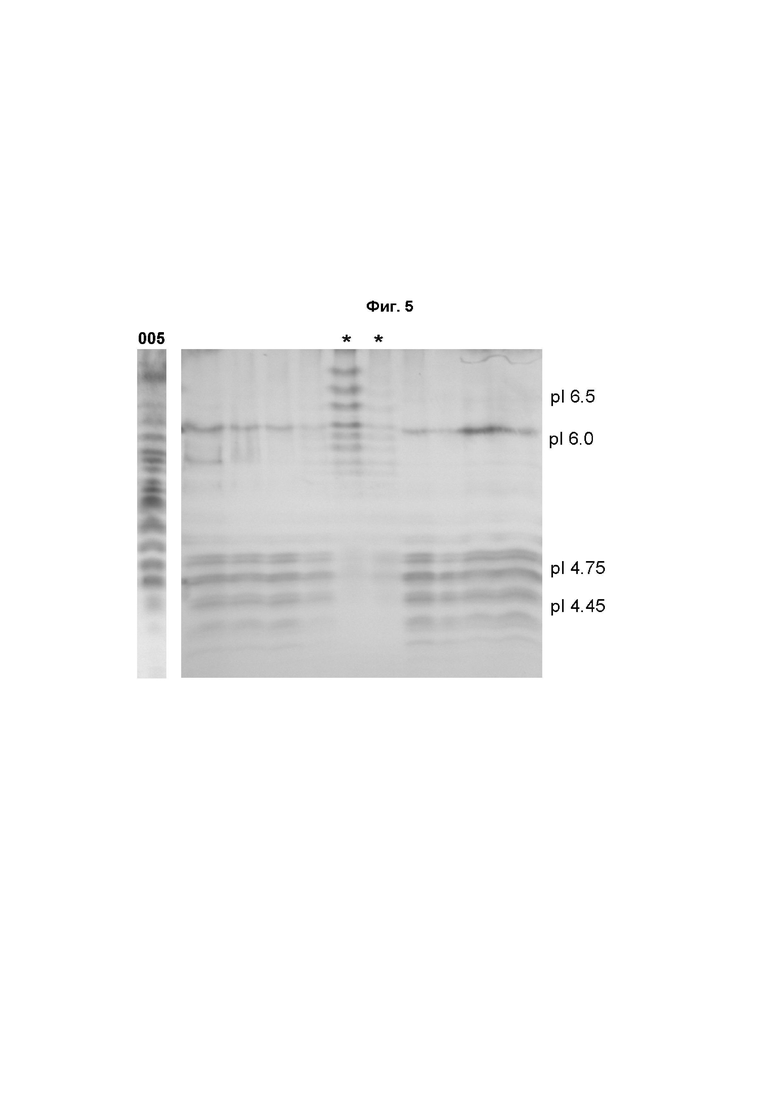
На Фиг. 5 показан пример клонов, анализированных изоэлектрическим фокусированием рекомбинантного ФСГ, продуцированного клетками Per.C6, стабильно экспрессирующими ФСГ, после конструирования с использованием α2,3- или α2,6-сиалилтрансферазы.
Супернатанты клеточных культур разделяли в нативных условиях в градиенте pH 3,0-7,0. Клон 005 является исходной клеточной линией Per.C6, продуцирующей ФСГ. Работы с клонами, демонстрирующими основные или смешанные профили, были прекращены (*). Оставшиеся клоны демонстрируют успешное конструирование с использованием сиалилтрансферазы с увеличением числа молекул сиаловой кислоты на ФСГ.
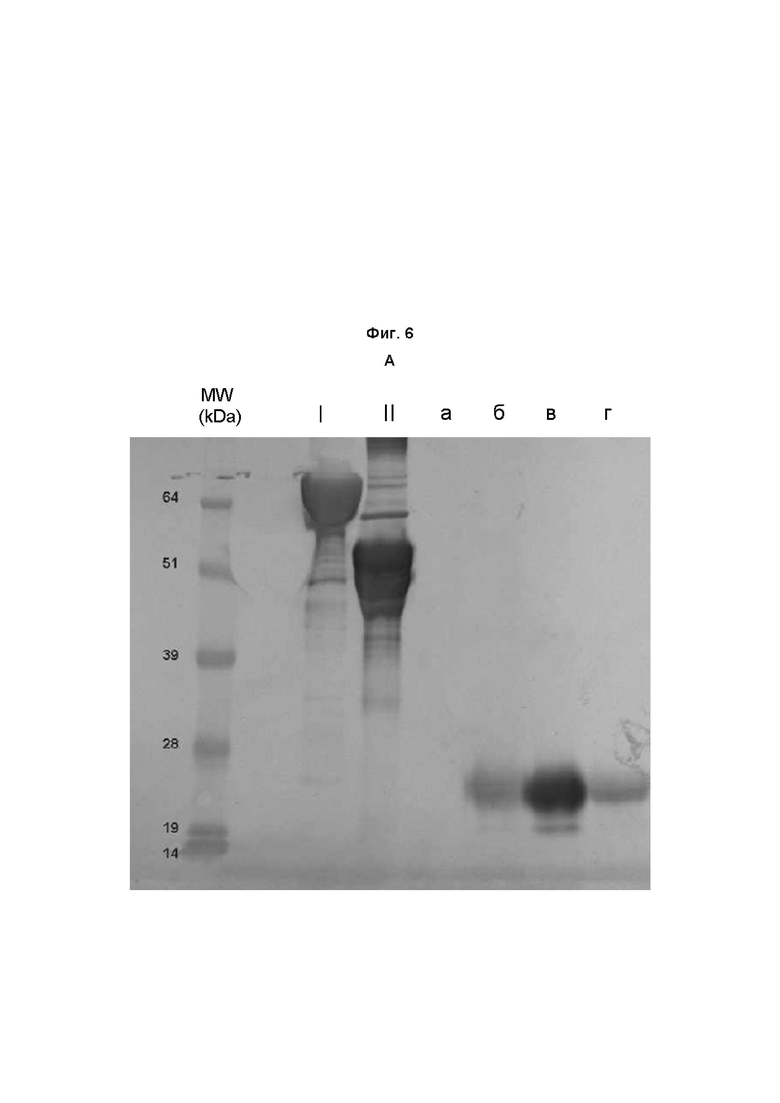
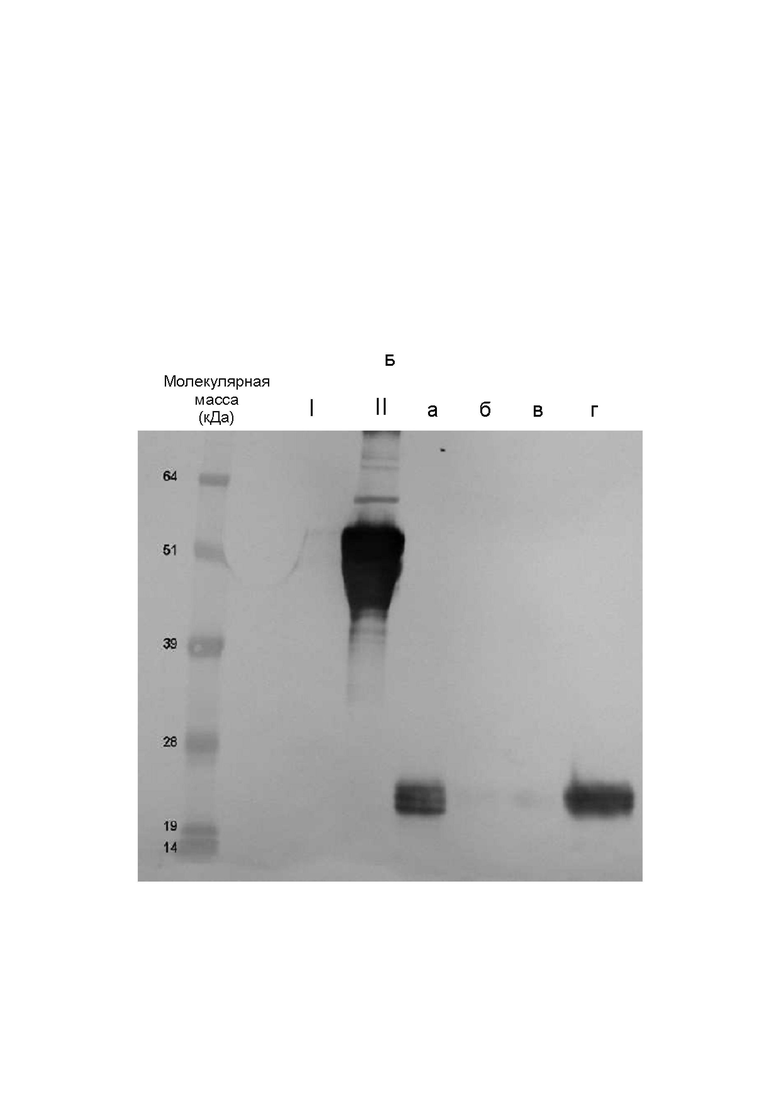
На Фиг. 6 показан анализ связей сиаловых кислот ФСГ из Per.C6.
Очищенный ФСГ из Per.C6 разделяли посредством электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) на двух гелях, переносили на нитроцеллюлозу и визуализировали с использованием набора DIG Glycan Differentiation Kit (номер по каталогу 11 210 238 001, Roche), следуя инструкциям изготовителя. Положительные реакции с агглютинином Sambucus nigra (SNA) указывали на наличие терминально связанной (2-6) сиаловой кислоты (А). Положительные реакции с агглютинином Maackia amurensis (MAA) указывали на наличие терминально связанной (2-3) сиаловой кислоты (Б). Дорожка I представляет собой контроль изготовителя, содержащий только α2,6-связи. Дорожка II представляет собой контроль изготовителя, содержащий α2,6- и α2,3-связи. Образец а представляет собой коммерческий рекомбинантный ФСГ, имеющий происхождение из клеток CHO (Gonal-f, Serono). Образец б представляет собой исходный рФСГ из Per.C6 без конструирования с использованием сиалилтрансферазы. Образец в представляет собой рФСГ из Per.C6 с конструированием с использованием α2,6-сиалилтрансферазы. Образец г представляет собой рФСГ из Per.C6 с конструированием с использованием α2,3-сиалилтрансферазы.
На Фиг. 7 показаны скорости метаболического клиренса (MCR) образцов ФСГ из Per.C6.
В момент времени «ноль» самкам крыс (3 животных на клон) производили болюсную инъекцию рФСГ (1-10 мкг на крысу) в хвостовую вену. Содержание ФСГ в образцах крови, полученных в определенные моменты времени, анализировали посредством твердофазного иммуноферментного анализа (ELISA).
На Фиг. 8 показаны MCR образцов ФСГ из Per.C6, сконструированных с использованием α2,6-сиалилтрансферазы.
В момент времени «ноль» самкам крыс (3 животных на клон) производили болюсную инъекцию рФСГ (1-10 мкг на крысу) в хвостовую вену. Содержание ФСГ в образцах крови, полученных в определенные моменты времени, анализировали посредством ELISA.
На Фиг. 9 показаны скорости метаболического клиренса образцов ФСГ из Per.C6, сконструированных с использованием α2,6-сиалилтрансферазы, при одновременном введении 7500-кратного молярного избытка асиалофетуина для насыщения асиалогликопротеинового рецептора (ASGP-R) в течение 1-2 часов.
На Фиг. 10 показаны скорости метаболического клиренса образцов ФСГ из Per.C6, сконструированных с использованием α2,3-сиалилтрансферазы. Образцы были выбраны по содержанию в них сиаловой кислоты на основании их профиля изоэлектрического фокусирования (IEF-профиля).
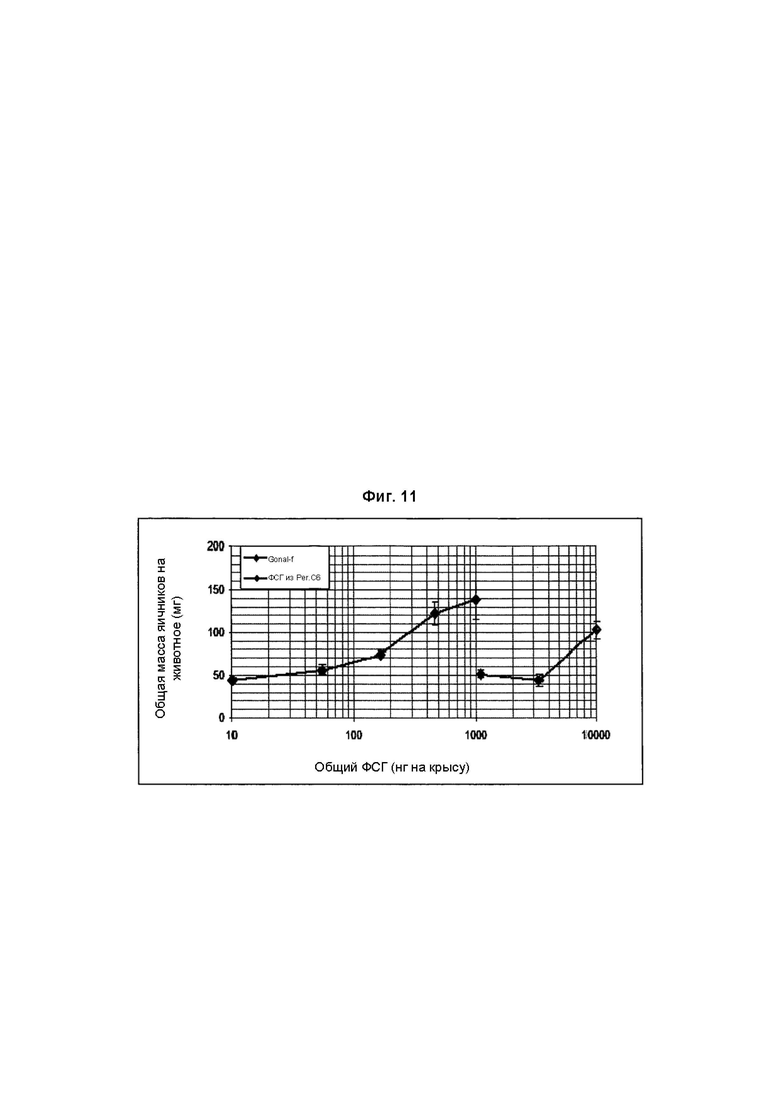
На Фиг. 11 показано увеличение массы яичников клонами рФСГ из Per.C6 от исходных Per.C6, продуцирующих рФСГ, согласно способу Steelman и Pohley (1953).
рФСГ из Per.C6 и стандарты (рФСГ Gonal-f) исследовали в разных дозах (3 крысы на дозу).
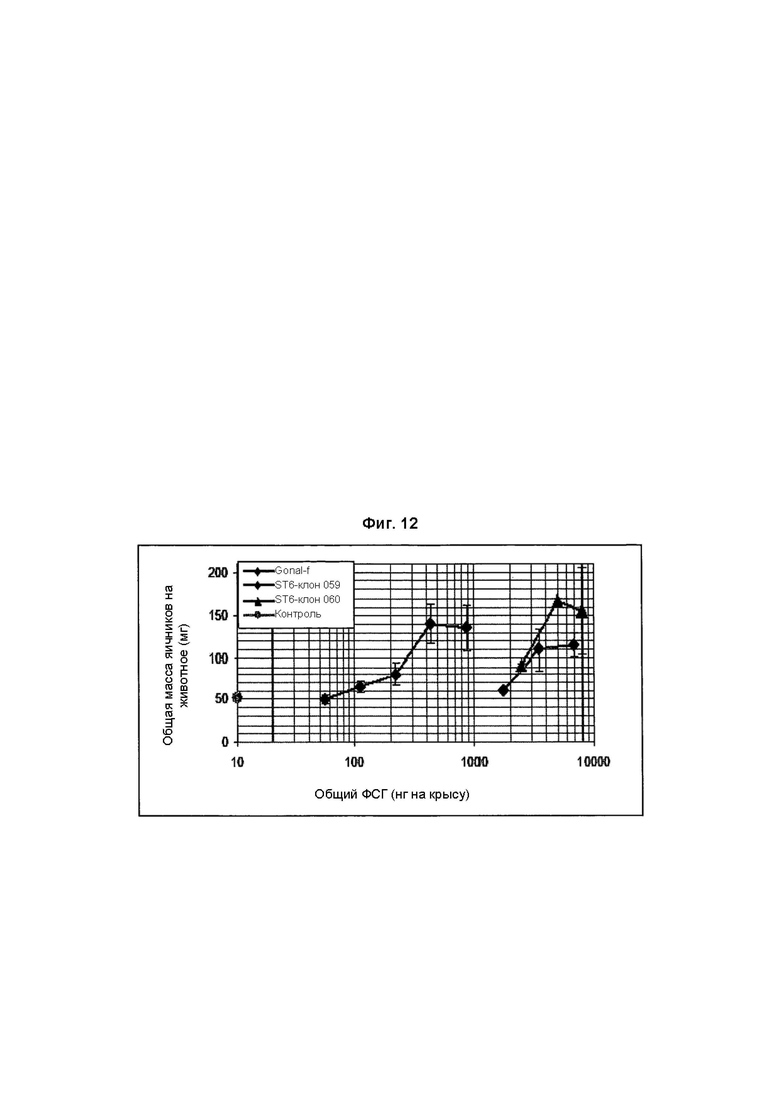
На Фиг. 12 показано увеличение массы яичников клонами рФСГ из Per.C6 от сконструированных (α2,6-сиалилтрансфераза) Per.C6, продуцирующих рФСГ, согласно способу Steelman и Pohley (1953).
рФСГ из Per.C6, сконструированных с использованием α2,6-сиалилтрансферазы, и стандарты (рФСГ Gonal-f) исследовали в разных дозах (3 крысы на дозу).
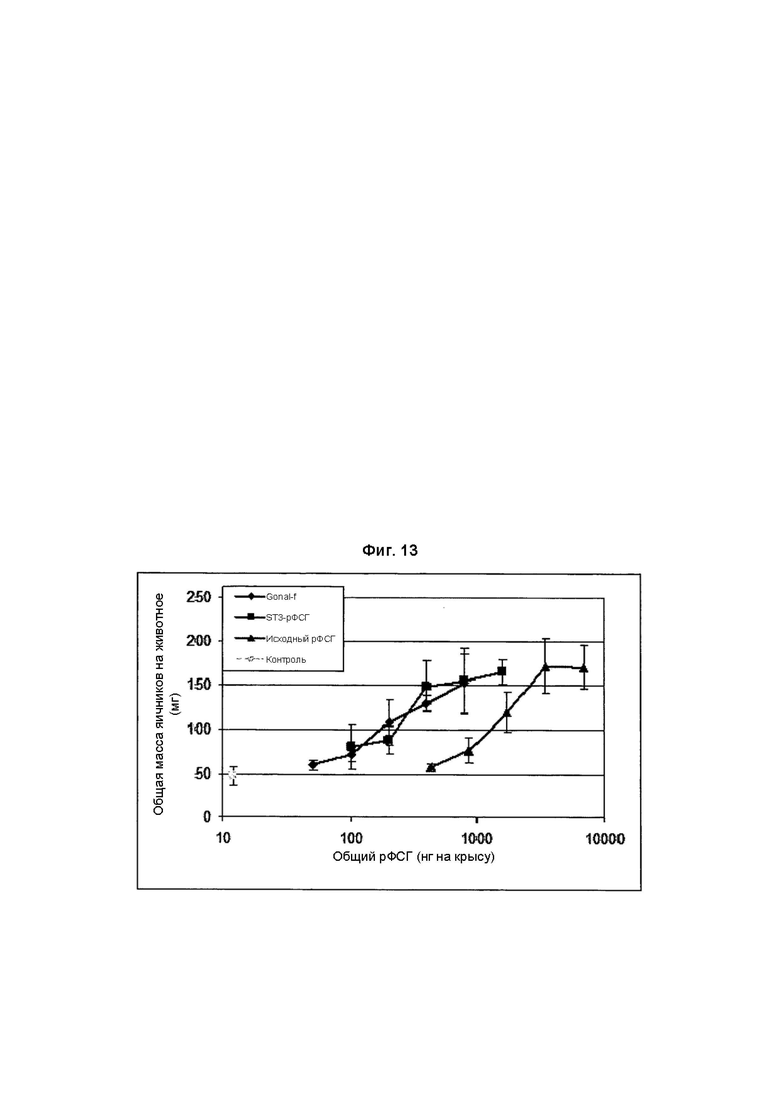
На Фиг. 13 показано увеличение массы яичников клонами рФСГ из Per.C6 от сконструированных (α2,3-сиалилтрансфераза) Per.C6, продуцирующих рФСГ, согласно способу Steelman и Pohley (1953).
Исходный рФСГ из Per.C6, рФСГ из Per.C6, сконструированных с использованием α2,3-сиалилтрансферазы, и стандарты (рФСГ Gonal-f) исследовали в пяти разных дозах (3 крысы на дозу).
Выбор последовательностей
Человеческий ФСГ
Использовали кодирующую область гена альфа-полипептида ФСГ согласно Fiddes and Goodman (1981). Данную последовательность хранят под обозначением AH007338, и на момент конструирования у данной белковой последовательности не было других вариантов. Здесь эта последовательность названа SEQ ID NO: 1.
Использовали кодирующую область гена бета-полипептида ФСГ согласно Keene et al (1989). Данную последовательность хранят под обозначением NM_000510, и на момент конструирования у данной белковой последовательности не было других вариантов. Здесь эта последовательность названа SEQ ID NO: 2.
Сиалилтрансфераза
α2,3-Сиалилтрансфераза. Использовали кодирующую область гена бета-галактозид-альфа-2,3-сиалилтрансферазы 4 (α2,3-сиалилтрансферазы, ST3GAL4) согласно Kitagawa and Paulson (1994). Данную последовательность хранят под обозначением L23767, и здесь она названа SEQ ID NO: 3.
α2,6-Сиалилтрансфераза. Использовали кодирующую область гена бета-галактозамид-альфа-2,6-сиалилтрансферазы 1 (α2,6-сиалилтрансферазы, ST6GAL1) согласно Grundmann et al. (1990). Данную последовательность хранят под обозначением NM_003032, и здесь она названа SEQ ID NO: 4.
ПРИМЕРЫ
Пример 1. Конструирование вектора экспрессии ФСГ
Кодирующую последовательность альфа-полипептида ФСГ (AH007338, SEQ ID NO: 1) и бета-полипептида ФСГ (NM_003032, SEQ ID NO: 2) амплифицировали посредством полимеразной цепной реакции (ПЦР) с использованием комбинаций праймеров FSHa-прямой и FSHa-обратный, и FSHb-прямой и FSHb-обратный, соответственно.
FSHa-прямой 5'-CCAGGATCCGCCACCATGGATTACTACAGAAAAATATGC-3'.
FSHa-обратный 5'-GGATGGCTAGCTTAAGATTTGTGATAATAAC-3'.
FSHb-прямой 5'-CCAGGCGCGCCACCATGAAGACACTCCAGTTTTTC-3'.
FSHb-обратный 5'-CCGGGTTAACTTATTATTCTTTCATTTCACCAAAGG-3'.
Полученную амплифицированную ДНК ФСГ-бета расщепляли рестриктазами AscI и HpaI и вводили в сайты AscI и HpaI в векторе экспрессии млекопитающих под контролем CMV, несущем неомициновый селектируемый маркер. Сходным образом, ДНК ФСГ-альфа расщепляли BamHI и NheI и вводили в сайты BamHI и NheI в векторе экспрессии, уже содержащем ДНК бета-полипептида ФСГ.
Векторную ДНК использовали для трансформации штамма E. coli DH5α. Для амплификации были отобраны шестьдесят колоний, и пятьдесят семь содержали вектор, содержащий как ФСГ-альфа, так и ФСГ-бета. Двадцать из них были выбраны для секвенирования, и все они содержали нужные последовательности согласно SEQ ID NO: 1 и SEQ ID NO: 2. Для трансфекции была выбрана плазмида pFSH A+B#17 (Фиг. 1).
Пример 2. Конструирование вектора экспрессии ST3
Кодирующую последовательность бета-галактозид-альфа-2,3-сиалилтрансферазы 4 (ST3, L23767, SEQ ID NO: 3) амплифицировали посредством ПЦР с использованием комбинации праймеров 2,3ST-прямого и 2,3ST-обратного.
2,3ST-прямой 5'-CCAGGATCCGCCACCATGTGTCCTGCAGGCTGGAAGC 3'.
2,3ST-обратный 5'-TTTTTTTCTTAAGTCAGAAGGACGTGAGGTTCTTG-3'.
Полученную амплифицированную ДНК ST3 расщепляли рестриктазами BamHI и AflII и вводили в сайты BamHI и AflII в векторе экспрессии млекопитающих под контролем CMV, несущем маркер устойчивости к гигромицину. Вектор амплифицировали, как описано выше, и секвенировали. Клон pST3#1 (Фиг. 2) содержал нужную последовательность согласно SEQ ID NO: 3, и он был выбран для трансфекции.
Пример 3. Конструирование вектора экспрессии ST6
Кодирующую последовательность бета-галактозамид-альфа-2,6-сиалилтрансферазы 1 (ST6, NM_003032, SEQ ID NO: 4) амплифицировали посредством ПЦР с использованием комбинации праймеров 2,6ST-прямого и 2,6ST-обратного.
2,6ST-прямой 5'-CCAGGATCCGCCACCATGATTCACACCAACCTGAAG-3'.
2,6ST-обратный 5'-TTTTTTTCTTAAGTTAGCAGTGAATGGTCCGG-3'.
Полученную амплифицированную ДНК ST6 расщепляли рестриктазами BamHI и AflII и вводили в сайты BamHI и AflII в векторе экспрессии млекопитающих под контролем CMV, несущем маркер устойчивости к гигромицину. Вектор амплифицировали, как описано выше, и секвенировали. Клон pST6#11 (Фиг. 3) содержал нужную последовательность согласно SEQ ID NO: 4 и был выбран для трансфекции.
Пример 4. Стабильная экспрессия pFSH A+B в клетках PER.C6 Трансфекция, выделение и скрининг клонов
Клоны Per.C6, продуцирующие ФСГ, получали экспрессией обеих полипептидных цепей ФСГ с одной плазмиды (см. Пример 1).
Для получения стабильных клонов использовали агент для трансфекции на основе липосом совместно с конструкцией pFSH A+B. Стабильные клоны отбирали на среде VPRO, дополненной 10% фетальной телячьей сывороткой (FCS) и содержащей G418. Через три недели после трансфекции вырастали клоны, устойчивые к G418. Для выделения были отобраны в общей сложности 250 клонов. Выделенные клоны культивировали в селекционной среде до 70-80% конфлюэнтности. Супернатанты анализировали на предмет содержания белка ФСГ с применением ФСГ-селективного ELISA и на предмет фармакологичекой активности в отношении рецептора ФСГ в клонированной клеточной линии с применением анализа накопления циклического аденозинмонофосфата (цАМФ). Клоны (98), экспрессирующие функциональный белок, размножали в культурах далее в 24-луночных и 6-луночных планшетах и колбах T80.
Исследования по определению продуктивности и качества вещества от семи клонов проводили в колбах T80 для получения достаточного количества вещества. Клетки культивировали в обогащенных средах, как описано выше, в течение 7 суток и собирали супернатант. Продуктивность определяли с применением ФСГ-селективного ELISA. Определяли изоэлектрический профиль вещества (Пример 6). Типичные образцы показаны на Фиг. 4. Информация, полученная при IEF, была использована для выбора клонов для анализа скорости метаболического клиренса (Пример 9). Клоны (005, 104, 179, 223, 144) с достаточной продуктивностью и качеством были выбраны для конструирования с использованием сиалилтрансферазы.
Пример 5. Степень сиалирования повышена в клетках, сверхэкспрессирующих α2,3- или α2,6-сиалилтрансферазу. Стабильная экспрессия pST3 или pST6 в клетках PER.C6, экспрессирующих ФСГ; трансфекция, выделение и скрининг клонов
Клоны Per.C6, продуцирующие высоко сиалированный ФСГ, получали экспрессией α2,3-сиалилтрансферазы или α2,6-сиалилтрансферазы с отдельных плазмид (см. Примеры 2 и 3) в клетках Per.C6, уже экспрессирующих обе полипептидные цепи ФСГ (см. Пример 4). Четыре клона, полученные из клеток PER.C6®, как изложено в Примере 4, были выбраны на основании их характеристик, включая продуктивность, хорошие ростовые качества, продукцию функционального белка, и они продуцировали ФСГ, включавший некоторое сиалирование.
Стабильные клоны получали, как описано выше в Примере 4. Проводили выделение, размножение и анализ в общей сложности 202 клонов из программы с α2,3-сиалилтрансферазой и 210 клонов из программы с α2,6-сиалилтрансферазой. Конечное число клонов для исследования с α2,3-сиалилтрансферазой составило 12, а для исследования с α2,6-сиалилтрансферазой - 30.
Клоны с α2,3-сиалилтрансферазой адаптировали для свободных от сыворотки сред и суспензионных условий.
Как описано выше, клоны анализировали с применением ФСГ-селективного ELISA, функционального ответа в клеточной линии с рецептором ФСГ, IEF (Пример 6), скорости метаболического клиренса (Пример 9) и анализа Steelman-Pohley (Пример 10). Результаты сравнивали с имеющимся в продаже рекомбинантным ФСГ (Gonal-f, Serono) и исходным ФСГ из клеточных линий Per.C6. Типичные образцы показаны на Фиг. 5. Некоторые клоны не демонстрировали увеличение сиалирования, однако можно наблюдать, что ФСГ, продуцируемый большинством клонов, имеет значительно улучшенное сиалирование (то есть, в среднем больше изоформ ФСГ с большим количеством сиаловых кислот) по сравнению с ФСГ, экспрессированным без α2,3- или α2,6- сиалилтрансферазы.
В заключение, экспрессия ФСГ совместно с сиалилтрансферазой в клетках Per.C6 приводит к повышенным уровням сиалированного ФСГ по сравнению с клетками, экспрессирующими только ФСГ.
Пример 6. Анализ pI изоформ ФСГ, продуцированных Per.C6, изоэлектрическим фокусированием
Электрофорез определяют как транспорт заряженных молекул через растворитель в электрическом поле. Подвижность биологической молекулы в электрическом поле будет зависеть от напряженности поля, общего заряда молекулы, размера и формы молекулы, ионной силы и свойств среды, через которую движутся молекулы.
Изоэлектрическое фокусирование (IEF) является электрофоретической методикой разделения белков на основании их pI. pI представляет собой pH, при котором общий заряд белка равен нулю, и белок не будет двигаться в электрическом поле. Содержание сиаловой кислоты в изоформах ФСГ незначительно смещает точку pI для каждой изоформы, что можно использовать с применением данной методики для визуального отличия изоформ ФСГ из Per.C6 друг от друга.
Изоэлектрические точки изоформ ФСГ, продуцированных в Per.C6, анализировали в супернатантах клеточных культур с применением изоэлектрического фокусирования. Среды клеточных культур от клонов Per.C6, продуцирующих ФСГ, получали, как описано в Примере 4 и 5.
Образцы ФСГ из Per.C6 разделяли в гелях для изоэлектрического фокусирования Novex®, содержащих 5% полиакриламид, в нативных условиях с градиентом pH 3,0-7,0 в растворе амфотерного электролита, pH 3,0-7,0.
Белки переносили на нитроцеллюлозную мембрану и визуализировали с применением первичного моноклонального антитела против ФСГα, вторичного антитела против мышиного IgG, конъюгированного с щелочной фосфатазой, и реагента из 5-бром-4-хлор-3-индолилфосфата (BCIP) и нитросинего тетразолия (NBT) для визуализации полос.
Как указано на Фиг. 4 и 5, полосы отражают изоформы ФСГ, содержащие различные количества молекул сиаловой кислоты.
С применением данного способа были идентифицированы клоны, продуцирующие изоформы ФСГ с большим количеством молекул сиаловой кислоты. Конструирование с использованием α2,3- или α2,6-сиалилтрансферазы приводило к получению клонов с большим содержанием сиаловой кислоты и более низкой pI.
Пример 7. Анализ связей сиаловых кислот в ФСГ из Per.C6
Гликоконъюгаты анализировали с применением способа различения гликанов на основе лектина. Этот способ позволяет описывать гликопротеины и гликоконъюгаты, связанные с нитроцеллюлозой. Лектины селективно распознают определенную группировку, например, сиаловую кислоту, связанную через α2,3-связь. Используемые лектины конъюгированы со стероидным гаптеном дигоксигенином, что делает возможным иммунологическое выявление связанных лектинов.
Очищенный ФСГ из Per.C6 от исходного клона (без дополнительной сиалилтрансферазы), клона, сконструированного с использованием α2,3-сиалилтрансферазы, и клона, сконструированного с использованием α2,6-сиалилтрансферазы, разделяли с применением стандартных методик SDS-PAGE. Имеющийся в продаже рекомбинантный ФСГ (Gonal-f, Serono) использовали в качестве стандарта.
Сиаловую кислоту анализировали с использованием набора DIG Glycan Differentiation Kit (номер по каталогу 11 210 238 001, Roche), следуя инструкциям изготовителя. Положительные реакции с агглютинином Sambucus nigra (SNA) указывали на наличие терминально связанной (2-6) сиаловой кислоты (А). Положительные реакции с агглютинином Maackia amurensis (MAA) указывали на наличие терминально связанной (α2-3) сиаловой кислоты (Б).
В заключение, исходный клон 005 содержал низкие уровни как α2,3-, так и α2,6-сиаловой кислоты. Клоны, сконструированные с использованием α2,3-сиалилтрансферазы, содержали высокие уровни α2,3-связей сиаловых кислот и низкие уровни α2,6-связей сиаловых кислот. Клоны, сконструированные с использованием α2,6-сиалилтрансферазы, содержали высокие уровни α2,6-связей сиаловых кислот и низкие уровни α2,3-связей сиаловых кислот. Стандартный контроль Gonal-f содержит только α2,3-связи сиаловых кислот. Это согласуется с тем, что известно о рекомбинантных белках, полученных в клетках яичника китайского хомячка (CHO) (Kagawa et al, 1988, Takeuchi et al, 1988, Svensson et al., 1990).
В заключение, конструирование клеток Per.C6, продуцирующих ФСГ, с использованием α2,3- или α2,6-сиалилтрансферазы успешно увеличивало число молекул сиаловой кислоты, конъюгированных с ФСГ, в образце.
Пример 8а. Количественное определение общей сиаловой кислоты
Сиаловая кислота представляет собой связанный с белками углевод, который считают моносахаридом, и она встречается в комбинации с другими моносахаридами, такими как галактоза, манноза, глюкозамин, галактозамин и фукоза.
Общую сиаловую кислоту в очищенном рФСГ (Пример 11) измеряли с использованием набора для ферментативного количественного определения сиаловой кислоты, следуя протоколу изготовителя (Sigma, Sialic-Q). Кратко, N-ацетилнейраминовая кислота-альдолаза катализирует превращение сиаловой кислоты в N-ацетилманнозин и пировиноградную кислоту. Пировиноградная кислота может быть восстановлена до молочной кислоты восстановленным β-никотинамидадениндинуклеотидом (β-NADH) и лактатдегидрогеназой. Окисление β-NADH может быть точно измерено спектрофотометрией.
Концентрацию белка измеряли в титрационных микропланшетах с использованием коммерческого набора для анализа с бицинхониновой кислотой (BCA-анализа) (Sigma, В 9643) на основании способа Lowry (Lowry et al, 1951).
Было измерено общее содержание сиаловой кислоты в ФСГ из Per.C6, и было обнаружено, что оно превышало 6 моль/моль.
Пример 8б. Количественное определение относительных количеств α2,3-, α2,6- и α2,8-сиаловой кислоты
Относительные процентные количества α2,3-, α2,6- и α2,8-сиаловой кислоты в очищенном рФСГ (Пример 11) измеряли с применением известных методик.
Каждый образец рФСГ иммобилизовали (гелевый блок), промывали, восстанавливали, алкилировали и расщепляли пептид-N-гликаназой F (PNGase F) в течение ночи. N-гликаны затем экстрагировали и подвергали дальнейшей обработке. Проводили мечение N-гликанов флуорофором 2AB для нормально-фазовой высокоэффективной жидкостной хроматографии (NP-HPLC) и высокоэффективной жидкостной хроматографии со слабыми анионообменниками (WAX-HPLC), как подробно описано Royle et al. N-гликаны анализировали нормально-фазовой (NP) HPLC на амидной колонке TSK (как подробно описано Royle et al) и время удерживания выражали в глюкозных единицах (GU).
Образцы экстрагированных объединенных гликанов (экстрагированных, как описано выше) расщепляли различными сиалидазами для определения связей. NAN 1 (рекомбинантная сиалидаза) высвобождает связанные α2,3-связями невосстанавливающие концевые сиаловые кислоты (N-ацетилнейраминовую кислоту (NeuNAc) и N-гликолилнейраминовую кислоту (NeuNGc)), ABS (сиалидаза Arthrobacter ureafaciens) высвобождает связанные α2,3-, α2,6- и α2,8-связями невосстанавливающие концевые сиаловые кислоты (NeuNAc и NeuNGc). Образцы анализировали посредством NP-HPLC для сравнения нерасщепленного образца с образцом, расщепленным NAN1, и образцом, расщепленным ABS. Сравнение трех записей NP-HPLC (нерасщепленный, расщепленный NAN1, расщепленный ABS) демонстрирует, что расщепление ABS и NAN1 дает разные результаты. Это указывает на то, что образцы содержат сиаловые кислоты с α2,3-, α2,6- и α2,8-связями. Для структур, присутствующих в пулах нерасщепленных гликанов, были вычислены относительные процентные количества, и было обнаружено, что они составляют 65%-85% (например, 77,75%) для α2,3-сиалирования, 15-35% (например, 21,46%) для α2,6-сиалирования и 0,1-3% для α2,8-сиалирования.
Пример 8в. Количественное определение относительных количеств моно-, ди-, три- и тетраразветвленных сиалированных структур
Относительные процентные количества моно-, ди-, три- и тетрасиалированных структур в гликанах, экстрагированных из очищенного рФСГ (Пример 11), измеряли с применением известных методик.
Каждый образец рФСГ иммобилизовали (гелевый блок), промывали, восстанавливали, алкилировали и расщепляли пептид-N-гликаназой F (PNGase F) в течение ночи. N-гликаны затем экстрагировали и подвергали дальнейшей обработке. Проводили мечение N-гликанов флуорофором 2AB для NP-HPLC и WAX-HPLC, как подробно описано Royle et al.
HPLC со слабыми анионообменниками (WAX) для разделения N-гликанов по заряду (Пример 8б) проводили, как изложено в Royle et al, с фетуиновым N-гликановым стандартом в качестве контроля. Элюирование гликанов проводили согласно числу содержавшихся в них сиаловых кислот. Все образцы включали моно- (1S), ди- (2S), три- (3S) и тетра- (4S) сиалированные структуры. Было обнаружено, что относительные количества сиалированных структур были в пределах следующих соотношений (1S : 2S : 4S : 4S): 9-15% : 27-30% : 30-36% : 25-29% (например, 10,24 : 28,65 : 35,49 : 25,62).
Пример 9. Определение скоростей метаболического клиренса рФСГ
Для определения скорости метаболического клиренса (MCR) образцов ФСГ из Per.C6 самкам крыс (3 животных на клон), находящимся в сознании, в момент времени «ноль» производили болюсную инъекцию рФСГ (1-10 мкг на крысу, на основании количественной оценки образцов посредством ELISA, DRG EIA 1288) в хвостовую вену. Образцы крови (400 мкл) отбирали из кончика хвоста на 1, 2, 4, 8, 12, 24 и 32 часы после инъекции исследуемого образца. Сыворотку собирали центрифугированием и анализировали в ней содержание ФСГ посредством ELISA (DRG EIA 1288).
Асиалогликопротеиновый рецептор (ASGP-R) распознает десиалированные гликопротеины (с концевой галактозой), такие как асиалофетуин (ASF). (Pricer and Ashwell, 1971. Van Lenten and Ashwell, 1972). Рецептор ASGP и связанный десиалированный гликопротеин поступают внутрь клетки, где происходят регенерация рецептора и деградация лиганда (Regoeczi et al, 1978, Steer and Ashwell, 1980).
Для исследования роли этого механизма в клиренсе вещества ФСГ из Per.C6 ASGP-R насыщали асиалофетуином. Скорость метаболического клиренса исходного вещества или вещества из клеток, сконструированных с использованием α2,6- или α2,3-сиалилтрансферазы, определяли, как описано, с одновременным введением по меньшей мере 7500-кратного молярного избытка асиалофетуина для насыщения ASGP-R в течение 1-2 часов.
Вещество, полученное из исходных продуцирующих ФСГ клонов Per.C6, содержало некоторое количество вещества с небольшой MCR, но, большей частью, было подвержено быстрому клиренсу (Фиг. 7). Наиболее удачный клон 005, содержавший наиболее сиалированное вещество, конструировали с использованием α2,6- или α2,3-сиалилтрансферазы (Пример 5). Несмотря на то, что клоны, сконструированные с использованием α2,6-сиалилтрансферазы, демонстрировали повышенное сиалирование (Фиг. 5), MCR не была улучшена (Фиг. 7). Блокада ASGR приводила к восстановлению MCR α2,6-вещества до уровня стандарта, демонстрируя, что даже с увеличенными α2,6-связями вещество подвержено быстрому клиренсу (Фиг. 8). Конструирование с использованием α2,3-сиалилтрансферазы приводило к получению клонов с MCR, сопоставимой со стандартом (Фиг. 9), и изменение содержания сиаловой кислоты согласовалось с тем, что известно об изоформах ФСГ (Фиг. 10).
Пример 10. Анализ Steelman-Pohley in vivo
Для демонстрации того, что повышение содержания сиаловой кислоты в ФСГ приводит к увеличению биологического эффекта, исследовали увеличение массы яичников у крыс высоко сиалированным ФСГ, таким как полученный в Примере 5.
Увеличение массы яичников, обусловленное рФСГ, продуцированным клонами Per.C6, анализировали согласно способу Steelman and Pohley (1953). рФСГ из Per.C6 из образцов фильтрованной клеточной среды количественно оценивали посредством ELISA (DRG, EIA-1288). Образцы (рФСГ из Per.C6) и стандарты (рФСГ Gonal-f) исследовали в пяти разных дозах (3 животных на дозу). Gonal-f вводили в дозах 50, 100, 200, 400 и 800 нг на крысу. Дозы образцов рассчитывали с использованием их значений площади под кривой (AUC) относительно Gonal-f, обычно 0,05-10 мкг на крысу.
В заключение, недостаточно сиалированное вещество, продуцированное исходными клонами Per.C6, продуцирующими ФСГ, (Фиг. 11) было менее активно в анализе увеличения массы яичников, чем имеющийся в продаже рФСГ. Конструирование с использованием сиалилтрансферазы с добавлением дополнительных α2,6-связей увеличивало содержание сиаловой кислоты, но не увеличивало активность в анализе in vivo (Фиг. 12). Однако, дополнительные α2,3-связи значительно увеличивали активность (Фиг. 13), и два препарата рекомбинантного ФСГ (имеющие происхождение из Per.C6 и CHO) демонстрируют очень похожие профили в данном анализе.
Пример 11. Контроль получения и очистки
Был разработан способ получения ФСГ в клетках PER.C6, культивируемых в суспензии в свободной от сыворотки среде. Этот способ описан ниже, и он был использован с некоторыми клеточными линиями PER.C6, продуцирующими ФСГ.
ФСГ от исходного клона 005, α2,3-клона 007 и α2,6-клона 059 получали с использованием модификации способа, описанного Lowry et al. (1976).
Для получения ФСГ из PER.C6 клеточные линии адаптировали к свободной от сыворотки среде, то есть, Excell 525 (JRH Biosciences). Сначала клетки культивировали до образования 70%-90% конфлюэнтного монослоя в культуральной колбе T80. При пересеве клетки ресуспендировали в свободной от сыворотки среде Excell 525 + 4 мМ L-глутамина до концентрации клеток 0,3×106 клеток/мл. 25 мл суспензии клеток помещали в 250 мл колбу для шейкера и встряхивали при 100 об/мин при 37°C и 5% CO2. После достижения концентрации клеток более 1×106 клеток/мл, клетки субкультивировали до концентрации клеток 0,2 или 0,3×106 клеток/мл и культивировали далее в колбах для шейкера при 37°C, 5% CO2 и 100 об/мин.
Для получения ФСГ клетки переносили в свободную от сыворотки производственную питательную среду, то есть, VPRO (JRH Biosciences), обеспечивающую поддержание роста клеток PER.C6 до очень высоких концентраций клеток (обычно более 107 клеток/мл в культуре клеток, выращиваемой в замкнутом объеме). Сначала клетки культивировали до концентрации более 1×106 клеток/мл в Excell 525, затем осаждали центрифугированием в течение 5 минут при 1000 об/мин и затем суспендировали в среде VPRO + 6 мМ L-глутамина до концентрации 1×106 клеток/мл. Клетки затем культивировали в колбе для шейкера в течение 7-10 дней при 37°C, 5% CO2 и 100 об/мин. В течение этого периода клетки вырастали до концентрации более 107 клеток/мл. Культуральную среду собирали после начала снижения жизнеспособности клеток. Клетки осаждали центрифугированием в течение 5 минут при 1000 об/мин и супернатант использовали для количественной оценки и очистки ФСГ. Концентрацию ФСГ определяли с использованием ELISA (DRG EIA 1288).
Затем проводили очистку ФСГ с использованием модификации способа, описанного Lowry et al. (1976). Это осуществляли хроматографией на диэтиламиноэтилцеллюлозе (DEAE-целлюлозе), гель-фильтрацией на Sephadex G100, адсорбционной хроматографией на гидроксиапатите и препаративным электрофорезом в полиакриламидном геле.
Во всех хроматографических способах присутствие иммунореактивного ФСГ подтверждали радиоиммунным анализом (RIA) (DRG EIA 1288) и IEF (Пример 6).
Ссылки
Andersen CY, Westergaard LG, and van Wely M. (2004). FSH isoform composition of commercial gonadotrophin preparations: a neglected aspect? Reprod Biomed Online. 9(2), 231-236.
Arey BJ, Stevis PE, Deecher DC, Shen ES, Frail DE, Negro-Vilar A, and Lopez FJ. (1997) Induction of promiscuous G protein coupling of the follicle-stimulating hormone (FSH) receptor: a novel mechanism for transducing pleiotropic actions of FSH isoforms. Mol Endocrinol. 11(5), 517-526.
Baenziger JU and Green ED. (1988). Pituitary glycoprotein hormone oligosaccharides: structure, synthesis and function of the asparagine-linked oligosaccharides on lutropin, follitropin and thyrotropin. Biochim Biophys Acta. 947(2), 287-306.
Bassett RM, and Driebergen R. (2005). Continued improvements in the quality and consistency of follitropin alfa, recombinant human FSH. Reprod Biomed Online. 10(2), 169-177.
 P, Zaga V, Maldonado A,
P, Zaga V, Maldonado A,  C, Timossi C, and Ulloa-Aguirre A. (1999). Oestrogens regulate pituitary alphα2,3-sialyltransferase messenger ribonucleic acid levels in the female rat. J Mol Endocrinol. 23(2), 153-165.
C, Timossi C, and Ulloa-Aguirre A. (1999). Oestrogens regulate pituitary alphα2,3-sialyltransferase messenger ribonucleic acid levels in the female rat. J Mol Endocrinol. 23(2), 153-165.
D'Antonio M., Borrelli F., Datola A., Bucci R. , Mascia M., Polletta P., Piscitelli D., and Papoian R. (1999) Biological characterization of recombinant human follicle stimulating hormone isoforms. Human Reproduction 14, 1160-1167.
Dalpathado DS, Irungu J, Go EP, Butnev VY, Norton K, Bousfield GR, and Desaire H. (2006). Comparative glycomics of the glycoprotein follicle stimulating hormone: glycopeptide analysis of isolates from two mammalian species. Biochemistry. 45(28), 8665-8673. No copy
Dias JA, Van Roey P. (2001). Structural biology of human follitropin and its receptor. Arch Med Res. 32(6), 510-519.
Fiddes, J. C. and Goodman, H. M. (1979) Isolation, cloning and sequence analysis of the cDNA for the alpha-subunit of human chorionic gonadotropin. Nature, 281, 351-356.
Flack, M.R., Bennet, A.P., Froehlich, J. Anasti, JN and Nisula, B. (1994). Increased biological activity due to basic isoforms in recombinant human follicle-stimulating hormone produced in a human cell line. J. Clin. Endocrinol. Metab., 79, 756-760.
Fox KM, Dias JA, and Van Roey P. (2001). Three-dimensional structure of human follicle-stimulating hormone. Mol Endocrinol. 15(3), 378-89.
Grabenhorst E, Hoffmann A, Nimtz M, Zettlmeissl G, and Conradt HS. (1995). Construction of stable BHK-21 cells coexpressing human secretory glycoproteins and human Gal(beta 1-4)GlcNAc-R alpha 2,6-sialyltransferase alpha 2,6-linked NeuAc is preferentially attached to the Gal(beta 1-4)GlcNAc(beta 1-2)Man(alpha 1-3)-branch of diantennary oligosaccharides from secreted recombinant beta-trace protein. Eur J Biochem. 232(3), 718-25.
Green ED and Baenziger JU. (1988). Asparagine-linked oligosaccharides on lutropin, follitropin, and thyrotropin. II. Distributions of sulfated and sialylated oligosaccharides on bovine, ovine, and human pituitary glycoprotein hormones. J Biol Chem. 263(1), 36-44.
Grundmann.U., Nerlich.C, Rein.T. and Zettlmeissl, G. (1990). Complete cDNA sequence encoding human beta-galactoside alpha-2,6-sialyltransferase. G Nucleic Acids Res. 18(3), 667.
Howles, CM. (1996). Genetic engineering of human FSH (Gonal-F). Hum Reprod. Update, 2, 172-191.
Kagawa Y, Takasaki S, Utsumi J, Hosoi K, Shimizu H, Kochibe N, and Kobata A. (1988). Comparative study of the asparagine-linked sugar chains of natural human interferon-beta 1 and recombinant human interferon-beta 1 produced by three different mammalian cells. J Biol Chem. 263(33), 17508-17515.
Keene, J.L., Matzuk, M.M., Otani, Т., Fauser, B,C,J,M., Galway, A.B., Hsueh, A.J.W. and Boime, I. (1989). Expression of Biologically active Human Follitropin in Chinese Hamster Ovary Cells. The Journal of Biological Chemistry, 264(9), 4769-4775.
Kitagawa,H. and Paulson,J.C (1994) Cloning of a novel alpha 2,3-sialyltransferase that sialylates glycoprotein and glycolipid carbohydrate groups. J. Biol. Chem. 269(2), 1394-1401.
Lee EU, Roth J, and Paulson JC (1989) Alteration of terminal glycosylation sequences on N-linked oligosaccharides of Chinese hamster ovary cells by expression of beta-galactoside alpha 2,6-sialyltransferase. J Biol Chem. 264(23), 13848-13855.
de Leeuw, R., Mulders, J., Voortman, G. Rombout, F. Damm, J. and Kloosterboer, L. (1996) Structure-function relationship of recombinant follicle stimulating hormone (Puregon). Mol. Hum. Reprod., 2, 361-369.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. (1951) Protein measurement with the Folin phenol reagent. J Biol Chem. 193(1), 265-75.
Lowry, PJ, McLean, C, Jones RL and Satgunasingam N. (1976) Purification of anterior pituitary and hypothalamic hormones Clin Pathol Suppl (Assoc Clin Pathol). 7, 16-21.
Pierce JG, and Parsons TF (1981) Glycoprotein hormones: structure and function Annu Rev Biochem. 50, 465-495.
Pricer WE Jr, and Ashwell G. (1971). The binding of desialylated glycoproteins by plasma membranes of rat liver. J Biol Chem. 246(15), 4825-33.
Rathnam P, and Saxena BB. (1975). Primary amino acid sequence of follicle-stimulating hormone from human pituitary glands. I. alpha subunit. J Biol Chem.; 250(17):6735-6746.
Regoeczi E, Debanne MT, Hatton MC, and Koj A. (1978) Elimination of asialofetuin and asialoorosomucoid by the intact rat. Quantitative aspects of the hepatic clearance mechanism. Biochim Biophys Acta. 541(3), 372-84.
Royle L, Radcliffe CM, Dwek RA and Rudd PM (2006) Methods in Molecular Biology, ed I Brockhausen-Schutzbach (Humana Press), 347: Glycobiology protocols, 125-144.
Ryan RJ, Keutmann HT, Charlesworth MC, McCormick DJ, Milius RP, Calvo FO and Vutyavanich T. (1987). Structure-function relationships of gonadotropins. Recent Prog Horm Res.;43,:383-429.
Saxena BB and Rathnam P. (1976) Amino acid sequence of the beta subunit of follicle-stimulating hormone from human pituitary glands. J Biol Chem. 251(4), 993-1005.
Steelman SL, and Pohley FM. (1953) Assay of the follicle stimulating hormone based on the augmentation with human chorionic gonadotropin. Endocrinology. 53(6), 604-616.
Steer CJ, and Ashwell G. (1980) Studies on a mammalian hepatic binding protein specific for asialoglycoproteins. Evidence for receptor recycling in isolated rat hepatocytes. J Biol Chem. 255(7), 3008-13.
Svensson EC, Soreghan B, and Paulson JC. (1990) Organization of the beta-galactoside alpha 2,6-sialyltransferase gene. Evidence for the transcriptional regulation of terminal glycosylation. J Biol Chem. 265(34):20863-20868.
Takeuchi M, Takasaki S, Miyazaki H, Kato T, Hoshi S, Kochibe N, and Kobata A (1988). Comparative study of the asparagine-linked sugar chains of human erythropoietins purified from urine and the culture medium of recombinant Chinese hamster ovary cells. J Biol Chem. 263(8), 3657-3663.
Timossi CM, Barrios de Tomasi J, Zambrano E,  R, Ulloa-Aguirre A. (1998). A naturally occurring basically charged human follicle-stimulating hormone (FSH) variant inhibits FSH-induced androgen aromatization and tissue-type plasminogen activator enzyme activity in vitro. Neuroendocrinology. 67(3), 153-163.
R, Ulloa-Aguirre A. (1998). A naturally occurring basically charged human follicle-stimulating hormone (FSH) variant inhibits FSH-induced androgen aromatization and tissue-type plasminogen activator enzyme activity in vitro. Neuroendocrinology. 67(3), 153-163.
Timossi CM, Barrios-de-Tomasi J,  R, Arranz MC, Padmanabhan V, Conn PM, and Ulloa-Aguirre A. (2000). Differential effects of the charge variants of human follicle-stimulating hormone. J Endocrinol. 165(2), 193-205.
R, Arranz MC, Padmanabhan V, Conn PM, and Ulloa-Aguirre A. (2000). Differential effects of the charge variants of human follicle-stimulating hormone. J Endocrinol. 165(2), 193-205.
Ulloa-Aguirre, A., Espinoza, R., Damian-Matsumura, P. and Chappel, S.C. (1988) Immunological and biological potencies of the different molecular species of gonadotrophins. Hum. Reprod. 3, 491-501.
Ulloa-Aguirre, A., Cravioto, A.,  P. Jimenez, M, Zambrano, E and Diaz-Sanchez, V. (1992) Biological characterization of the naturally occurring analogues of intrapituitary human follicle stimulating hormone. Hum. Reprod. 7, 23-30.
P. Jimenez, M, Zambrano, E and Diaz-Sanchez, V. (1992) Biological characterization of the naturally occurring analogues of intrapituitary human follicle stimulating hormone. Hum. Reprod. 7, 23-30.
Ulloa-Aguirre A, Midgley AR Jr, Beitins IZ, and Padmanabhan V. (1995). Follicle-stimulating isohormones: characterization and physiological relevance. Endocr Rev. 16(6), 765-787.
Ulloa-Aguirre A, Maldonado A,  P, and Timossi С (2001). Endocrine regulation of gonadotropin glycosylation. Arch Med Res. 32(6), 520-532.
P, and Timossi С (2001). Endocrine regulation of gonadotropin glycosylation. Arch Med Res. 32(6), 520-532.
Ulloa-Aguirre A, Timossi C, Barrios-de-Tomasi J, Maldonado A, and Nayudu P. (2003). Impact of carbohydrate heterogeneity in function of follicle-stimulating hormone: studies derived from in vitro and in vivo models. Biol Reprod. 69(2), 379-389.
Van Lenten L, and Ashwell G. (1972) The binding of desialylated glycoproteins by plasma membranes of rat liver. Development of a quantitative inhibition assay. J Biol Chem. 247(14), 4633-40.
Wide, L. and Albertsson-Wikland, K. (1990) Change in electrophoretic mobility of human follicle-stimulating hormone in serum after administration of gonadotropin-releasing hormone. J. Clin. Endocrinol. Metab. 70, 271-276.
Wide, L. and Bakos, O. (1993). More basic forms of both human follicle-stimulating hormone and luteinizing hormone in serum at midcycle compared with the follicular or luteal phase. J. Clin. Endocrinol. Metab., 76, 885-889.
Wide L,  T,
T,  I, Eriksson K. (2007) Sulfonation and sialylation of gonadotropins in women during the menstrual cycle, after menopause, and with polycystic ovarian syndrome and in men. J Clin Endocrinol Metab.; 92(11), 4410-4417.
I, Eriksson K. (2007) Sulfonation and sialylation of gonadotropins in women during the menstrual cycle, after menopause, and with polycystic ovarian syndrome and in men. J Clin Endocrinol Metab.; 92(11), 4410-4417.
Zambrano E,  T, Olivares A, Barrios-de-Tomasi J, and Ulloa-Aguirre A. (1999). Receptor binding activity and in vitro biological activity of the human FSH charge isoforms as disclosed by heterologous and homologous assay systems: implications for the structure-function relationship of the FSH variants. Endocrine. 10(2), 113-121.
T, Olivares A, Barrios-de-Tomasi J, and Ulloa-Aguirre A. (1999). Receptor binding activity and in vitro biological activity of the human FSH charge isoforms as disclosed by heterologous and homologous assay systems: implications for the structure-function relationship of the FSH variants. Endocrine. 10(2), 113-121.
Zhang X, Lok SH, and Kon OL (1998) Stable expression of human alpha-2,6-sialyltransferase in Chinese hamster ovary cells: functional consequences for human erythropoietin expression and bioactivity. Biochim Biophys Acta. 1425(3), 441-452.
SEQ ID NO: 1
Альфа-полипептид фолликулостимулирующего гормона
Номер доступа AH007338
Нуклеотидная последовательность ФСГ-альфа

Белковая последовательность ФСГ-альфа

SEQ ID NO: 2
Бета-полипептид фолликулостимулирующего гормона
Номер доступа NM_000510
Нуклеотидная последовательность ФСГ-бета


Белковая последовательность ФСГ-бета

SEQ ID NO: 3
Бета-галактозид-альфа-2,3-сиалилтрансфераза 4
Номер доступа L23767
Нуклеотидная последовательность ST3GAL4


Белковая последовательность ST3GAL4

SEQ ID NO: 4
Бета-галактозамид-альфа-2,6-сиалилтрансфераза 1
Номер доступа NM_003032
Нуклеотидная последовательность ST6GAL1

Белковая последовательность ST6GAL1

| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный фолликулостимулирующий гормон (ФСГ), включающий альфа-2,3- и альфа-2,6-сиалирование | 2009 |
|
RU2682270C2 |
| РЕКОМБИНАНТНЫЙ ФОЛЛИКУЛОСТИМУЛИРУЮЩИЙ ГОРМОН (ФСГ), ВКЛЮЧАЮЩИЙ АЛЬФА-2, 3- И АЛЬФА-2, 6-СИАЛИРОВАНИЕ | 2009 |
|
RU2537268C2 |
| КОМПОЗИЦИЯ ДЛЯ КОНТРОЛИРУЕМОЙ СТИМУЛЯЦИИ ЯИЧНИКОВ | 2012 |
|
RU2613324C2 |
| Композиция для контролируемой стимуляции яичников | 2012 |
|
RU2739037C2 |
| ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, СОДЕРЖАЩИЙ РЕКОМБИНАНТНЫЙ ХОРИОНИЧЕСКИЙ ГОНАДОТРОПНЫЙ ГОРМОН ЧЕЛОВЕКА | 2010 |
|
RU2588650C2 |
| Фармацевтический препарат, содержащий рекомбинантный HCG (хорионический гонадотропный гормон человека) | 2010 |
|
RU2724528C2 |
| Композиция для лечения бесплодия | 2016 |
|
RU2704252C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ САХАРНОЙ ЦЕПИ, СОДЕРЖАЩЕЙ СИАЛОВУЮ КИСЛОТУ | 2012 |
|
RU2597975C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2011 |
|
RU2604811C2 |
| КЛЕТОЧНАЯ ЛИНИЯ huFSHIK, СЕКРЕТИРУЮЩАЯ РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ ФСГ | 2012 |
|
RU2502798C2 |



Изобретение относится к области биотехнологии, конкретно к получению рекомбинантного ФСГ (фолликулостимулирующий гормон), включающего α-2,3- и α-2,6-сиалирование, и может быть использовано в медицине. Полученный ФСГ, где 80% или более общего сиалирования ФСГ представляет собой α-2,3-сиалирование и от 5% до 20% общего сиалирования ФСГ представляет собой α-2,6-сиалирование, может быть эффективно использован для контролируемой стимуляции яичников и индукции овуляции в методиках экстракорпорального оплодотворения (ЭКО). 2 н. и 4 з.п. ф-лы, 13 ил., 11 пр.
1. Рекомбинантный ФСГ (фолликулостимулирующий гормон), включающий α-2,3- и α-2,6-сиалирование, где 80% или более общего сиалирования рекомбинантного ФСГ представляет собой α-2,3-сиалирование и от 5% до 20% общего сиалирования рекомбинантного ФСГ представляет собой α-2,6-сиалирование.
2. Рекомбинантный фолликулостимулирующий гормон (ФСГ), включающий α-2,3- и α-2,6-сиалирование, где от 5% до 20% общего сиалирования рекомбинантного ФСГ представляет собой α-2,6-сиалирование.
3. Рекомбинантный ФСГ по п. 2, продуцированный или экспрессированный в человеческой клеточной линии.
4. Рекомбинантный ФСГ по п. 3, продуцированный или экспрессированный в клеточной линии Per.C6, клеточной линии, имеющей происхождение из Per.C6, или модифицированной клеточной линии Per.C6.
5. Рекомбинантный ФСГ по п. 3, где человеческая клеточная линия представляет собой человеческую клеточную линию, которая модифицирована с использованием α-2,3-сиалилтрансферазы.
6. Рекомбинантный ФСГ по п. 3, содержащий связанное α-2,6-связями сиалирование, обеспеченное активностью эндогенной сиалилтрансферазы.
| FLACK M | |||
| R | |||
| et al | |||
| Increased biological activity due to basic isoforms in recombinant human follicle-stimulating hormone produced in a human cell line | |||
| The Journal of Clinical Endocrinology & Metabolism, 1994, V | |||
| Цилиндрический сушильный шкаф с двойными стенками | 0 |
|
SU79A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| FOX K | |||
| M | |||
| et al | |||
| Three-dimensional structure of human follicle-stimulating hormone | |||
| Molecular endocrinology, 2001, | |||

