ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к олигонуклеотидам (олигомерам), которые являются комплементарными лиганду-1 запрограммированной смерти (PD-L1), приводящим к понижению экспрессии PD-L1 в печени. Настоящее изобретение также относится к способу облегчения исчерпания Т-клеток, вызванного инфекцией печени или раком в печени. Релевантными инфекциями являются хронические инфекции HBV (вирус гепатита В), HCV (вирус гепатита С) и HDV (вирус гепатита D) и паразитарные инфекции, подобные малярии и токсоплазмозу (например, вызванные протистами Plasmodium, в частности, видами P. vivax, P. malariae и P. falciparum).
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Известно то, что костимулирующий путь, состоящий из рецептора запрограммированной смерти-1 (PD-1) и его лиганда - PD-L1 (или B7-H1, или CD274) - непосредственно способствует исчерпанию Т-клеток, приводящему к отсутствию контроля за вирусами во время хронических инфекций печени. Путь PD-1 также играет роль в автоиммунитете, так как у мышей с нарушениями данного пути развиваются аутоиммунные заболевания.
Было показано то, что антитела, которые блокируют взаимодействие между PD-1 и PD-L1, усиливают ответы Т-клеток, в частности, ответ цитотоксических Т-клеток CD8+ (см. Barber et al 2006 Nature Vol 439 p682 и Maier et al 2007 J. Immunol. Vol 178 p 2714).
В WO 2006/042237 описан способ диагностики рака посредством оценки экспрессии PD-L1 (B7-H1) в опухолях и предложена доставка пациенту агента, который препятствует взаимодействию PD-1/PD-L1. Препятствующими агентами могут быть антитела, фрагменты антител, миРНК (малая интерферирующая РНК) или антисмысловые олигонуклеотиды. Нет ни конкретных примеров таких препятствующих агентов, ни какого-либо упоминания хронических инфекций печени.
Опосредованное РНК-интерференцией ингибирование PD-L1 с использованием двухцепочечных молекул РНК (дцРНК, РНКи или миРНК) также было раскрыто, например, в WO 2005/007855, WO 2007/084865 и US 8507663. Ни в одной из них не описана таргетная доставка в печень.
В Dolina et al. 2013 Molecular Therapy-Nucleic Acids, 2 e72 описана доставка in vivo молекул миРНК, нацеленных на PD-L1, в клетки Купфера, усиливая, посредством этого клиренс клетками NK (природная клетка-киллер) и Т-клетками CD8+ у мышей, инфицированных MCMV (мышиный цитомегаловирус). В данной статье делается заключение о том, что молекулы миРНК, нацеленные на PD-L1, доставленные в гепатоциты, не являются эффективными в отношении усиления эффекторной функции Т-леток CD8+.
Подход с миРНК значительно отличается от подхода с одноцепочечными антисмысловыми олигонуклеотидами, так как биораспределение и механизмы действия являются совершенно разными. Как описано в Xu et al 2003 Biochem. Biophys. Res. Comm. том 306, страницы 712-717, антисмысловые олигонуклеотиды и миРНК имеют разные предпочтения в отношении сайтов-мишеней в мРНК.
В WO 2016/138278 описано ингибирование иммунных контрольных точек, включая PD-L1, с использованием двух или более чем двух одноцепочечных антисмысловых олигонуклеотидов, которые связаны на их 5'-концах. В данной заявке не упоминается HBV или таргетная доставка в печень.
ЦЕЛЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении идентифицированы новые олигонуклеотиды и конъюгаты олигонуклеотидов, которые очень эффективно понижают уровень мРНК PD-L1 в клетках печени - как в паренхимных (например, гепатоциты), так и в непаренхимных клетках, таких как клетки Купфера и синусоидальные эндотелиальные клетки печени (LSEC). Посредством понижения или сайленсинга PD-L1 олигонуклеотиды и конъюгаты олигонуклеотидов уменьшают опосредованное PD-1 ингибирование и, посредством этого, активизируют иммуностимуляцию истощенных Т-клеток. Облегчение исчерпания Т-клеток при хронической патогенной инфекции печени будет приводить к возобновленному иммунному контролю и пониженным уровням вирусных антигенов в крови во время хронической патогенной инфекции печени. Клетки природные киллеры (NK) и Т-клетки природные киллеры (NKТ) также могут активироваться олигонуклеотидами и конъюгатами олигонуклеотидов по настоящему изобретению.
Конъюгаты олигонуклеотидов обеспечивают местное снижение уровня PD-L1 в клетках печени и, следовательно, уменьшают риск аутоиммунных побочных эффектов, таких как пневмонит, невирусный гепатит и колит, ассоциированный с системным исчерпанием PD-L1.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к олигонуклеотидам или их конъюгатам, нацеленным на нуклеиновую кислоту, способным модулировать экспрессию PD-L1 и лечить или предупреждать заболевания, связанные с функционированием PD-L1. Олигонуклеотиды или конъюгаты олигонуклеотидов, в частности, могут использоваться для лечения заболеваний, при которых был истощен иммунный ответ против инфекционного агента.
Соответственно, в первом аспекте изобретения предложены олигонуклеотиды, которые содержат непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью к нуклеиновой кислоте мишени PD-L1. Данный олигонуклеотид может представлять собой антисмысловой олигонуклеотид, предпочтительно с гэпмерной конструкцией. Предпочтительно данный олигонуклеотид способен ингибировать экспрессию PD-L1 посредством расщепления нуклеиновой кислоты-мишени. Расщепление предпочтительно достигается посредством рекрутирования нуклеазы.
В другом аспекте данный олигонуклеотид конъюгирован с по меньшей мере одной конъюгатной группировкой, нацеленной на рецептор асиалогликопротеина, такой как конъюгатная группировка, содержащая по меньшей мере одну N-ацетилгалактозаминную (GalNAc) группировку. Конъюгатная группировка и олигонуклеотид могут быть связаны друг с другом посредством линкера, в частности, биорасщепляемого линкера.
В другом аспекте согласно изобретению предложены фармацевтические композиции, содержащие олигонуклеотиды или конъюгаты олигонуклеотидов по изобретению и фармацевтически приемлемые разбавители, носители, соли и/или адъюванты.
В другом аспекте согласно изобретению предложены способы in vivo или in vitro для понижения экспрессии PD-L1 в клетке-мишени, которая экспрессирует PD-L1, посредством введения в указанную клетку олигонуклеотида или композиции по изобретению в эффективном количестве.
В другом аспекте согласно изобретению предложены олигонуклеотиды, конъюгаты олигонуклеотидов или фармацевтические композиции для применения в восстановлении иммунитета против вируса или паразита.
В другом аспекте согласно изобретению предложены олигонуклеотиды, конъюгаты олигонуклеотидов или фармацевтические композиции для применения в качестве лекарственного средства.
В другом аспекте согласно изобретению предложены способы лечения или предупреждения заболевания, расстройства или дисфункции посредством введения терапевтически или профилактически эффективного количества олигонуклеотида по изобретению субъекту, страдающему от или восприимчивому к заболеванию, расстройству или дисфункции, в частности, к заболеваниям, выбранным из вирусных инфекций печени или паразитарных инфекций.
В другом аспекте олигонуклеотид, конъюгаты олигонуклеотида или фармацевтическая композиция по изобретению используются для лечения или предупреждения вирусных инфекций печени, таких как инфекции HBV, HCV и HDV, или паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, или рака печени, или метастазов в печени.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: иллюстрирует типичные конъюгаты антисмыслового олигонуклеотида, где данный олигонуклеоид либо представлен волнистой линией (А-Г), либо как «олигонуклеотид» (Д-З), либо как Т2 (И), а конъюгатные группировки, нацеленные на рецептор асиалогликопротеина, представляют собой трехвалентные N-ацетилгалактозаминные группировки. Соединения А-Г содержат дилизиновую разветвляющую молекулу, ПЭГ3 спейсер и три концевые GalNAc углеводные группировки. В соединении А и Б олигонуклеотид непосредственно присоединен к конъюгатной группировке, нацеленной на рецептор асиалогликопротеина, без линкера. В соединении В и Г олигонуклеотид непосредственно присоединен к конъюгатной группировке, нацеленной на рецептор асиалогликопротеина, через С6 линкер. Соединения Д-И содержат утраивающую разветвляющую молекулу, спейсеры варьирующей длины и структуры, и три концевые GalNAc углеводные группировки.
Фиг. 2: график, демонстрирующий ЕС50 (полумаксимальная эффективная концентрация) (А) и нокдаун PD-L1 как % от воздействия физиологическим раствором (Б) для соединений, протестированных в Примере 2, в связи с их положением на нуклеиновой кислоте-мишени. Клеточными линиями, в которых тестировали данное соединение, были  и
и 
Фиг. 3: структурная формула трехвалентного GalNAc кластера (GN2). GN2 является полезным в настоящем изобретении в качестве конъюгатной группировки. Волнистая линия иллюстрирует сайт конъюгирования данного кластера, например, с С6 аминолинкером или непосредственно с олигонуклеотидом.
Фиг. 4: структурная формула CMP ID NO 766_2.
Фиг. 5: структурная формула CMP ID NO 767_2.
Фиг. 6: структурная формула CMP ID NO 768_2.
Фиг. 7: структурная формула CMP ID NO 769_2.
Фиг. 8: структурная формула CMP ID NO 770_2.
Фиг. 9: вестерн-блот, выявляющий экспрессию белка PD-L1 в печени от животных, индуцированных поли(IC), после обработки физиологическим раствором и указанными CMP ID NO. Каждый блот показывает голый олигонуклеотид относительно версии того же самого олигонуклеотида, конъюгированной с GalNAc, блот А) CMP ID NO 744_1 и 755_2, Б) CMP ID NO 747_1 и 758_2, В) CMP ID NO 748_1 и 759_2, Г) CMP ID NO 752_1 и 763_2, и Д) CMP ID NO 753_1 и 764_2. Верхняя полоса представляет собой контроль загрузки - винкулин, нижняя полоса представляет собой белок PD-L1. Первая дорожка в каждом блоте демонстрирует мышей, обработанных физиологическим раствором, без индукции поли(IC). Данные мыши экспрессируют очень мало белка PD-L.
Фиг. 10: популяция одноядерных клеток в печени после обработки  носителем (группа 10 и 1),
носителем (группа 10 и 1),  ДНК-вакциной (группа 11 и 2),
ДНК-вакциной (группа 11 и 2),  антителом против PD-L1 (группа 12),
антителом против PD-L1 (группа 12),  голым АСО (антисмысловой олигонуклеотид) против PD-L1 плюс ДНК-вакциной (группа 7) или
голым АСО (антисмысловой олигонуклеотид) против PD-L1 плюс ДНК-вакциной (группа 7) или  АСО против PD-L1, конъюгированным с GalNAc, плюс ДНК-вакциной (группа 8) представлена для каждой группы индивидуальных животных, и среднее значение показано для каждой группы вертикальной линией (см. Таблица 18). Оценивали статистическую значимость между группой ДНК-вакцины и тремя группами обработки, и, при наличии, она показана * между группами (* - Р меньше 0,05; *** - Р меньше 0,001, и **** Р меньше 0,0001). А) представляет число Т-клеток в печени после обработки. Б) представляет долю Т-клеток CD4+, и В) представляет долю Т-клеток CD8+.
АСО против PD-L1, конъюгированным с GalNAc, плюс ДНК-вакциной (группа 8) представлена для каждой группы индивидуальных животных, и среднее значение показано для каждой группы вертикальной линией (см. Таблица 18). Оценивали статистическую значимость между группой ДНК-вакцины и тремя группами обработки, и, при наличии, она показана * между группами (* - Р меньше 0,05; *** - Р меньше 0,001, и **** Р меньше 0,0001). А) представляет число Т-клеток в печени после обработки. Б) представляет долю Т-клеток CD4+, и В) представляет долю Т-клеток CD8+.
Фиг. 11: модуляция PD-L1-позитивных клеток в печени после обработки носителем (группа 10 и 1), ДНК-вакциной (группа 11 и 2), антителом против PD-L1 (группа 12), голым АСО против PD-L1 плюс ДНК-вакциной (группа 7) или АСО против PD-L1, конъюгированным с GalNAc, плюс ДНК-вакциной (группа 8) представлена для каждой группы индивидуальных животных, и среднее значение показано для каждой группы вертикальной линией (см. Таблицу 19). Оценивали статистическую значимость между группой ДНК-вакцины и тремя группами обработки, и, при наличии, она показана * между группами (* - Р меньше 0,05 и **** - Р меньше 0,0001). А) представляет процентную долю Т-клеток CD8+, которые экспрессируют PD-L1 в печени после обработки. Б) представляет процентную долю Т-клеток CD4+, которые экспрессируют PD-L1 в печени после обработки, и В) представляет процентную долю В-клеток, которые экспрессируют PD-L1 в печени после обработки.
Фиг. 12: специфичные в отношении антигена HBV цитокин-секретирующие клетки CD8+ после обработки носителем (группа 10 и 1), ДНК-вакциной (группа 11 и 2), антителом против PD-L1 (группа 12), голым АСО против PD-L1 плюс ДНК-вакциной (группа 7) или АСО против PD-L1, конъюгированным с GalNAc, плюс ДНК-вакциной (группа 8) представлены для каждой группы индивидуальных животных, и среднее значение показано для каждой группы вертикальной линией (см. Таблицу 20). Оценивали статистическую значимость между группой ДНК-вакцины и тремя группами обработки, и, при наличии, она показана * между группами (* - Р меньше 0,05). А) представляет процентную долю секретирующих IFN-γ (интерферон-гамма) Т-клеток CD8+ в печени, которые являются специфичными в отношении антигена PreS2+S HBV, после обработки. Б) представляет процентную долю секретирующих IFN-γ Т-клеток CD8+ в печени, которые являются специфичными в отношении корового антигена HBV, после обработки, и В) представляет процентную долю секретирующих IFN-γ и TNF-α (фактор некроза опухолей-альфа) Т-клеток CD8+ в печени, которые являются специфичными в отношении антигена PreS2+S HBV, после обработки.
Фиг. 13: ДНК HBV, HВsAg и HВeAg у мышей AAV/HBV после обработки антисмысловым CMP NO: 759_2 против PD-L1, конъюгированным с GalNAc  , по сравнению с носителем
, по сравнению с носителем  . Вертикальная линия показывает конец обработки.
. Вертикальная линия показывает конец обработки.
ОПРЕДЕЛЕНИЯ
Олигонуклеотид
Термин «олигонуклеотид» в том виде, в котором он используется в данном документе, определяется таким образом, как обычно понятно специалисту, как молекула, содержащая два или более чем два ковалентно связанных нуклеозида. Такие ковалентно связанные нуклеозиды также могут называться молекулами нуклеиновой кислоты или олигомерами. Олигонуклеотиды обычно получают в лаборатории посредством твердофазного химического синтеза с последующей очисткой. При ссылке на последовательность олигонуклеотида, делается ссылка на последовательность или порядок группировок нуклеиновых оснований или их модификаций ковалентно связанных нуклеотидов или нуклеозидов. Олигонуклеотид по изобретению является искусственным, синтезируется химически и типично очищается или выделяется. Олигонуклеотид по изобретению может содержать один или более чем один модифицированный нуклеозид или нуклеотид.
Антисмысловые олигонуклеотиды
Термин «антисмысловой олигонуклеотид» в том виде, в котором он используется в данном документе, определяется как олигонуклеотид, способный модулировать экспрессию гена-мишени посредством гибридизации с нуклеиновой кислотой-мишенью, в частности с непрерывной последовательностью нуклеиновой кислоты-мишени. Антисмысловые олигонуклеотиды по существу не являются двухцепочечными и, следовательно, не являются миРНК. Предпочтительно антисмысловые олигонуклеотиды по настоящему изобретению являются одноцепочечными.
Непрерывная нуклеотидная последовательность
Термин «непрерывная нуклеотидная последовательность» относится к области олигонуклеотида, которая является комплементарной нуклеиновой кислоте-мишени. Данный термин используется в данном документе взаимозаменяемо с термином «непрерывная последовательность нуклеиновых оснований» и термином «последовательность олигонуклеотидного мотива». В некоторых воплощениях все нуклеотиды данного олигонуклеотида составляют непрерывную нуклеотидную последовательность. В некоторых воплощениях олигонуклеотид содержит непрерывную нуклеотидную последовательность и, возможно, может содержать дополнительный(ные) нуклеотид(ды), например, область нуклеотидного линкера, которая может использоваться для присоединения функциональной группы к непрерывной нуклеотидной последовательности. Область нуклеотидного линкера может быть или может не быть комплементарной нуклеиновой кислоте-мишени.
Нуклеотиды
Нуклеотиды представляют собой структурные элементы олигонуклеотидов и полинуклеотидов и для целей настоящего изобретения включают и встречающиеся в природе, и не встречающиеся в природе нуклеотиды. В природе нуклеотиды, такие как нуклеотиды ДНК и РНК, содержат группировку сахара рибозы, группировку нуклеинового основания и одну или более чем одну фосфатную группу (которая отсутствует в нуклеозидах). Нуклеозиды и нуклеотиды также могут взаимозаменяемо называться «звеньями» или «мономерами».
Модифицированный нуклеозид
Термин «модифицированный нуклеозид» или «модификация нуклеозида» в том виде, в котором он используется в данном документе, относится к нуклеозидам, модифицированным по сравнению с эквивалентным нуклеозидом ДНК или РНК посредством введения одной или более чем одной модификации сахарной группировки или группировки (нуклеинового) основания. В предпочтительном воплощении модифицированный нуклеозид содержит модифицированную сахарную группировку. Термин «модифицированный нуклеозид» также может использоваться в данном документе взаимозаменяемо с термином «аналог нуклеозида» или модифицированные «звенья» или модифицированные «мономеры».
Модифицированная межнуклеозидная связь
Термин «модифицированная межнуклеозидная связь» определяется так, как обычно понятно специалисту: как связи, отличные от фосфодиэфирных (РО) связей, которые ковалентно связывают вместе два нуклеозида. Нуклеотиды с модифицированными межнуклеозидными связями также называются «модифицированными нуклеотидами». В некоторых воплощениях модифицированная межнуклеозидная связь увеличивает устойчивость олигонуклеотида к нуклеазе по сравнению с фосфодиэфирной связью. Для олигонуклеотидов, встречающихся в природе, межнуклеозидная связь включает фосфатные группы, создающие фосфодиэфирную связь между соседними нуклеозидами. Модифицированные межнуклеозидные связи являются особенно полезными в стабилизации олигонуклеотидов для применения in vivo, и они могут служить для защиты против нуклеазного расщепления в областях нуклеозидов ДНК или РНК в олигонуклеотиде по изобретению, например в пределах области гэпа (gap) гэпмерного олигонуклеотида, а также в областях модифицированных нуклеозидов.
В одном воплощении олигонуклеотид содержит одну или более чем одну межнуклеозидную связь, модифицированную от природного фосфодиэфира до связи, которая, например, является более устойчивой к нуклеазной атаке. Устойчивость к нуклеазе можно определять посредством инкубирования олигонуклеотида в сыворотке крови или посредством применения анализа устойчивости к нуклеазе (например, фосфодиэстеразе яда змеи (SVPD)), которые оба хорошо известны в данной области. Межнуклеозидные связи, которые способны усиливать устойчивость олигонуклеотида к нуклеазе, называются нуклеазоустойчивыми межнуклеозидными связями. В некоторых воплощениях по меньшей мере 50% межнуклеозидных связей в олигонуклеотиде или в его непрерывной нуклеотидной последовательности являются модифицированными как, например, по меньшей мере 60%, как, например, по меньшей мере 70%, как, например, по меньшей мере 80% или как, например, по меньшей мере 90% межнуклеозидных связей в олигонуклеотиде или в его непрерывной нуклеотидной последовательности являются модифицированными. В некоторых воплощениях все межнуклеозидные связи олигонуклеотида или его непрерывной нуклеотидной последовательности являются модифицированными. Будет понятно то, что в некоторых воплощениях нуклеозиды, которые связывают олигонуклеотид по изобретению с ненуклеотидной функциональной группой, такой как конъюгат, могут содержать фосфодиэфир. В некоторых воплощениях все межнуклеозидные связи олигонуклеотида или его непрерывной нуклеотидной последовательности являются нуклеазоустойчивыми межнуклеозидными связями.
Модифицированные межнуклеозидные связи могут быть выбраны из группы, содержащей фосфоротиоат, дифосфоротиоат и боранофосфат. В некоторых воплощениях модифицированные межнуклеозидные связи, например, фосфоротиоатная, дифосфоротиоатная или боранофосфатная, являются совместимыми с рекрутированием олигосахаридом по изобретению РНКазы Н.
В некоторых воплощениях межнуклеозидная связь, такая как фосфоротиоатная межнуклеозидная связь, содержит серу (S).
Фосфоротиоатная межнуклеозидная связь является особенно полезной из-за устойчивости к нуклеазе, полезной фармакокинетики и легкости получения. В некоторых воплощениях по меньшей мере 50% межнуклеозидных связей в олигонуклеотиде или его непрерывной нуклеотидной последовательности представляют собой фосфоротиоат, как, например, по меньшей мере 60%, как, например, по меньшей мере 70%, как, например, по меньшей мере 80% или как, например, по меньшей мере 90% межнуклеозидных связей в олигонуклеотиде или его непрерывной нуклеотидной последовательности представляют собой фосфоротиоат. В некоторых воплощениях все межнуклеозидные связи олигонуклеотида или его непрерывной нуклеотидной последовательности представляют собой фосфоротиоат.
В некоторых воплощениях олигонуклеотид содержит одну или более чем одну нейтральную межнуклеозидную связь, в частности, межнуклеозидную связь, выбранную из фосфотриэфира, метилфосфоната, MMI, амида-3, формацеталя или тиоформацеталя.
Дополнительные межнуклеозидные связи раскрываются в WO2009/124238 (включенной в данный документ посредством ссылки). В одном воплощении межнуклеозидная связь выбрана из линкеров, раскрытых в WO2007/031091 (включенной в данный документ посредством ссылки). В частности, межнуклеозидная связь может быть выбрана из -O-P(O)2-O-, -O-P(O,S)-O-, -O-P(S)2-O-, -S-P(O)2-O-, -S-P(O,S)-O-, -S-P(S)2-O-, -O-P(O)2-S-, -O-P(O,S)-S-, -S-P(O)2-S-, -O-PO(RH)-O-, 0-PO(OCH3)-0-, -O-PO(NRH)-O-, -O-PO(OCH2CH2S-R)-O-, -O-PO(BH3)-O-, -O-PO(NHRH)-O-, -O-P(O)2-NRH-, -NRH-P(O)2-O-, -NRH-CO-O-, -NRH-CO-NRH-, и/или межнуклеозидный линкер может быть выбран из группы, состоящей: -O-CO-O-, -O-CO-NRH-, -NRH-CO-CH2-, -O-CH2-CO-NRH-, -O-CH2-CH2-NRH-, -CO-NRH-CH2-, -CH2-NRHCO-, -O-CH2-CH2-S-, -S-CH2-CH2-O-, -S-CH2-CH2-S-, -CH2-SO2-CH2-, -CH2-CO-NRH-, -O-CH2-CH2-NRH-CO -, -CH2-NCH3-O-CH2-, где RH выбран из водорода и C1-4-алкила.
Нуклеазоустойчивые связи, такие как фосфотиоатные связи, являются особенно полезными в областях олигонуклеотидов, способных рекрутировать нуклеазу, при формировании дуплекса с нуклеиновой кислотой-мишенью, таких как область G для гэпмеров или немодифицированная область нуклеозидов хэдмеров (headmer) и тэйлмеров (tailmer). Однако, фосфоротиоатные связи могут быть также полезными в областях, не рекрутирующих нуклеазу, и/или в областях, усиливающих аффинность, таких как области F и F' для гэпмеров, или в области модифицированных нуклеозидов хэдмеров и тэйлмеров.
Каждая из сконструированных областей может, однако, содержать межнуклеозидные связи, отличные от фосфоротиоатных, такие как фосфодиэфирные связи, в частности, в областях, где модифицированные нуклеозиды, такие как LNA (запертые нуклеиновые кислоты), защищают связь от нуклеазной деградации. Включение фосфодиэфирных связей, как, например, одной или двух связей, в частности между или рядом с модифицированными нуклеозидными звеньями (типично в областях, не рекрутирующих нуклеазу), может модифицировать биодоступность и/или биораспределение олигонуклеотида - см. WO2008/113832, включенную в данный документ посредством ссылки.
В одном воплощении все межнуклеозидные связи в олигонуклеотиде представляют собой фосфоротиоатные и/или боранофосфатные связи. Предпочтительно все межнуклеозидные связи в олигонуклеотиде представляют собой фосфоротиоатные связи.
Нуклеиновое основание
Нуклеиновое основание включает пуриновую (например, аденин и гуанин) и пиримидиновую (например, урацил, тимин и цитозин) группировку, присутствующую в нуклеозидах и нуклеотидах, которые образуют водородные связи при гибридизации нуклеиновых кислот. В контексте настоящего изобретения термин нуклеиновое основание также охватывает модифицированные нуклеиновые основания, которые могут отличаться от встречающихся в природе нуклеиновых оснований, но являются функциональными во время гибридизации нуклеиновых кислот. В данном контексте термин «нуклеиновое основание» относится и к встречающимся в природе нуклеиновым основаниям, таким как аденин, гуанин, цитозин, тимин, урацил, ксантин и гипоксантин, также как и к не встречающимся в природе вариантам. Такие варианты, например, описываются в Hirao et al (2012) Accounts of Chemical Research, том 45, страница 2055 и Bergstrom (2009) Current Protocols in Nucleic Acid Chemistry Suppl. 37 1.4.1.
В некоторых воплощениях группировка нуклеинового основания модифицируется заменой пурина или пиримидина на модифицированный пурин или пиримидин, такой как замещенный пурин или замещенный пиримидин, как, например, нуклеиновое основание, выбранное из изоцитозина, псевдоизоцитозина, 5-метилцитозина, 5-тиозолоцитозина, 5-пропинилцитозина, 5-пропинилурацила, 5-бромурацил-5-тиазолоурацила, 2-тиоурацила, 2'-тиотимидина, инозина, диаминопурина, 6-аминопурина, 2-аминопурина, 2,6-диаминопурина и 2-хлор-6-аминопурина.
Группировки нуклеиновых оснований могут быть указаны буквенным кодом для каждого соответствующего нуклеинового основания, например, A, T, G, C или U, где каждая буква возможно может включать модифицированные нуклеиновые основания с эквивалентной функцией. Например, в проиллюстрированных в качестве примеров олигонуклеотидах группировки нуклеиновых оснований выбраны из A, T, G, C и 5-метилцитозина. Возможно для гэпмеров LNA можно использовать 5-метилцитозин нуклеозидов LNA.
Модифицированный олигонуклеотид
Термин «модифицированный олигонуклеотид» описывает олигонуклеотид, содержащий один или более чем один нуклеозид с модифицированным сахаром и/или модифицированными межнуклеозидными связями. Термин «химерный олигонуклеотид» представляет собой термин, который использовали в литературе для описания олигонуклеотидов с модифицированными нуклеозидами.
Комплементарность
Термин «комплементарность» описывает способность нуклеозидов/нуклеотидов к образованию пар оснований по Уотсону-Крику. Парами оснований по Уотсону-Крику являются гуанин (G)-цитозин (С) и аденин (А)-тимин (Т)/урацил (U). Будет понятно то, что олигонуклеотиды могут содержать нуклеозиды с модифицированными нуклеиновыми основаниями, например, 5-метилцитозин часто используется вместо цитозина, и термин «комплементарность» как таковой охватывает образование пар оснований по Уотсону-Крику между немодифицированными и модифицированными нуклеиновыми основаниями (см., например, Hirao et al (2012) Accounts of Chemical Research, том 45, страница 2055 и Bergstrom (2009) Current Protocols in Nucleic Acid Chemistry Suppl. 37 1.4.1).
Термин «% комплементарности» в том виде, в котором он используется в данном документе, относится к числу нуклеотидов непрерывной нуклеотидной последовательности в молекуле нуклеиновой кислоты (например, в олигонуклеотиде) в процентах, которые, в данном положении, являются комплементарными (т.е. образуют пары оснований по Уотсону-Крику) в данном положении непрерывной нуклеотидной последовательности отдельной молекулы нуклеиновой кислоты (например, нуклеиновой кислоты-мишени). Данная процентная доля рассчитывается подсчетом числа выровненных оснований, которые образуют пары между двумя последовательностями (при выравнивании с использованием последовательности-мишени 5'-3' и последовательности олигонуклеотида от 3' к 5'), деля на общее число нуклеотидов в олигонуклеотиде и умножая на 100. При таком сравнении нуклеиновое основание/нуклеотид, который не выравнивается (не образует пару оснований), называется несоответствием.
Термин «полностью комплементарный» относится к 100%-ной комплементарности.
Следующее представляет собой пример олигонуклеотида (SEQ ID NO: 5), который является полностью комплементарным нуклеиновой кислоте-мишени (SEQ ID NO: 772).
5'gcagtagagccaatta3' (SEQ ID NO:772)
3'cgtcatctcggttaat5' (SEQ ID NO: 5)
Идентичность
Термин «идентичность» в том виде, в котором он используется в данном документе, относится к числу нуклеотидов в процентах непрерывной нуклеотидной последовательности в молекуле нуклеиновой кислоты (например, в олигонуклеотиде), которые в данном положении являются идентичными (т.е. по их способности образовать пары оснований по Уотсону-Крику с комплементарным нуклеозидом) непрерывной нуклеотидной последовательности в данном положении отдельной молекулы нуклеиновой кислоты (например, нуклеиновой кислоты-мишени). Данная процентная доля рассчитывается посредством подсчета числа выровненных оснований, которые являются идентичными между двумя последовательностями, включая пробелы, деля на общее число нуклеотидов в олигонуклеотиде и умножая на 100.
Процент идентичности = (Соответствия × 100)/Длину выровненной области (с пробелами).
Гибридизация
Термин «осуществление гибридизации» или «гибридизуется» в том виде, в котором он используется в данном документе, следует понимать как две нити нуклеиновой кислоты (например, олигонуклеотид и нуклеиновая кислота-мишень), образующие водородные связи между парами оснований на противоположных нитях, формируя, посредством этого, дуплекс. Аффинность связывания между двумя нитями нуклеиновой кислоты представляет собой силу гибридизации. Она часто описывается в показателях температуры плавления (Tm), определенной как температура, при которой половина олигонуклеотидов образует дуплекс с нуклеиновой кислотой-мишенью. При физиологических условиях Tm не является строго пропорциональной аффинности (Mergny and Lacroix, 2003, Oligonucleotides 13:515-537). Свободная энергия Гиббса в стандартном состоянии - ΔG° - является более точным представлением аффинности связывания и связана с константой диссоциации (Kd) реакции согласно уравнению ΔG° = -RTln(Kd), где R представляет собой газовую постоянную, и Т представляет собой абсолютную температуру. Следовательно, очень низкая ΔG° реакции между олигонуклеотидом и нуклеиновой кислотой-мишенью отражает сильную гибридизацию между олигонуклеотидом и нуклеиновой кислотой-мишенью. ΔG° представляет собой энергию, связанную с реакцией, где концентрации в воде составляют 1 М, рН составляет 7, и температура составляет 37°С. Гибридизация олигонуклеотидов с нуклеиновой кислотой-мишенью представляет собой спонтанную реакцию, и для спонтанных реакций ΔG° меньше, чем ноль. ΔG° можно измерять экспериментально, например, посредством применения способа изотермической титрационной калориметрии (ITC), как описано в Hansen et al., 1965, Chem. Comm. 36-38 и Holdgate et al., 2005, Drug Discov Today. Специалисту будет известно, что доступно имеющееся в продаже оборудование для измерений ΔG°. ΔG° также можно оценивать численно посредством применения модели ближайшего соседа, как описывается SantaLucia, 1998, Proc Natl Acad Sci USA. 95: 1460-1465, с использованием подходящим образом выведенных термодинамических параметров, описанных Sugimoto et al., 1995, Biochemistry 34:11211-11216 и McTigue et al., 2004, Biochemistry 43:5388-5405. Для того чтобы иметь возможность модулирования их намеченной нуклеиновой кислоты-мишени посредством гибридизации, олигонуклеотиды по настоящему изобретению гибридизуются с нуклеиновой кислотой-мишенью с оценочными значениями ΔG° меньше -41,8 кДж для олигонуклеотидов, которые имеют 10-30 нуклеотидов в длину. В некоторых воплощениях степень или сила гибридизации измеряется посредством свободной энергии Гиббса в стандартном состоянии - ΔG°. Олигонуклеотиды могут гибридизоваться с нуклеиновой кислотой-мишенью с оценочными значениями ΔG° меньше интервала -41,8 кДж, как, например, меньше -62,8 кДж, как, например, меньше -83,7 кДж и как, например, меньше -104,6 кДж для олигонуклеотидов, которые имеют 8-30 нуклеотидов в длину. В некоторых воплощениях олигонуклеотиды гибридизуются с нуклеиновой кислотой-мишенью с оценочным значением ΔG° от -41,8 до -251 кДж, как, например, от -50,2 до -167,4 кДж, как, например, от -62,8 до -125,5 кДж или от -66,9 до 113 кДж, как, например, от -75,3 до -104,6 кДж.
Нуклеиновая кислота-мишень
Согласно настоящему изобретению нуклеиновая кислота-мишень представляет собой нуклеиновую кислоту, которая кодирует PD-L1 млекопитающего и может представлять собой, например, ген, РНК, мРНК и пре-мРНК, зрелую мРНК или последовательность кДНК. Следовательно, данная мишень может называться нуклеиновая кислота-мишень PD-L1. Олигонуклеотид по изобретению, например, может быть нацелен на области экзонов PD-L1 млекопитающего, или, например, может быть нацелен на область интрона пре-мРНК PD-L1 (см. Таблицу 1).
Таблица 1: экзоны и интроны человеческого PD-L1
Подходящим образом, нуклеиновая кислота-мишень кодирует белок PD-L1, в частности PD-L1 млекопитающего, такой как PD-L1 человека (см., например, Таблицы 2 и 3, в которых приводится ссылка на последовательности мРНК и пре-мРНК для человеческого, обезьяньего и мышиного PD-L1). В контексте настоящего изобретения пре-мРНК также считается нуклеиновой кислотой, которая кодирует белок.
В некоторых воплощениях нуклеиновая кислота-мишень выбрана из группы, состоящей из SEQ ID NO: 1, 2 и 3 или их встречающихся в природе вариантов (например, последовательностей, кодирующих белок PD-L1 млекопитающего).
При использовании олигонуклеотида по изобретению в исследовании или диагностике нуклеиновая кислота-мишень может представлять собой кДНК или синтетическую нуклеиновую кислоту, происходящую из ДНК или РНК.
Для применения in vivo или in vitro олигонуклеотид по изобретению типично способен ингибировать экспрессию нуклеиновой кислоты-мишени PD-L1 в клетке, которая экспрессирует нуклеиновую кислоту-мишень PD-L1. Непрерывная последовательность нуклеиновых оснований олигонуклеотида по изобретению типично комплементарна нуклеиновой кислоте-мишени PD-L1 при измерении по длине олигонуклеотида, возможно, за исключением одного или двух несоответствий, и, возможно, исключая линкерные области на основе нуклеотидов, которые могут связывать олигонуклеотид с возможной функциональной группой, такой как конъюгат или другие некомплементарные концевые нуклеотиды (например, область D' или D''). Нуклеиновая кислота-мишень в некоторых воплощениях может представлять собой РНК или ДНК, такую как матричная РНК, как, например, зрелая мРНК или пре-мРНК. В некоторых воплощениях нуклеиновая кислота-мишень представляет собой РНК или ДНК, которая кодирует белок PD-L1 млекопитающего, такой как человеческий PD-L1, например, последовательность пре-мРНК человеческого PD-L1, такую как последовательность, раскрытая как SEQ ID NO 1, или последовательность человеческой мРНК с регистрационным номером NM_014143 NCBI (Национальный центр биотехнологической информации). Дополнительная информация по типичым нуклеиновым кислотам-мишеням приводится в Таблицах 2 и 3.
Таблица 2: информация по геному и сборке для PD-L1 между видами.
CM000671.2
000364345.1
CM001012.2
Прям. - прямая нить. Геномные координаты дают последовательность пре-мРНК (геномная последовательность). Ссылка NCBI дает последовательность мРНК (последовательность кДНК).
* База данных эталонных последовательностей Национального центра биотехнологической информации представляет собой всеохватывающий, интегрированный, неизбыточный, хорошо аннотированный набор эталонных последовательностей, включающих геномные, последовательности транскриптов и белков. Она размещается на www.ncbi.nlm.nih.gov/refseq.
Таблица 3: подробности последовательностей для PD-L1 между видами.
Последовательность-мишень
Термин «последовательность-мишень» в том виде, в котором он используется в данном документе, относится к последовательности нуклеотидов, присутствующей в нуклеиновой кислоте-мишени, которая содержит последовательность нуклеиновых оснований, которая является комплементарной олигонуклеотиду по изобретению. В некоторых воплощениях последовательность-мишень состоит из области на нуклеиновой кислоте-мишени, которая является комплементарной непрерывной нуклеотидной последовательности олигонуклеотида по изобретению. В некоторых воплощениях последовательность-мишень длиннее, чем комплементарная последовательность одного олигонуклеотида, и, например, может представлять предпочтительную область нуклеиновой кислоты-мишени, на которую могут быть нацелены несколько олигонуклеотидов по изобретению.
Последовательность-мишень может представлять собой подпоследовательность нуклеиновой кислоты-мишени.
В некоторых воплощениях подпоследовательность представляет собой последовательность, выбранную из группы, состоящей из а1-а149 (см. Таблицы 4). В некоторых воплощениях подпоследовательность представляет собой последовательность, выбранную из группы, состоящей из экзона мРНК PD-L1, такого как экзон человеческой мРНК PD-L1, выбранный из группы, состоящей из e1, e2, e3, e4, e5, e6 и e7 (см. Таблицу 1 выше).
В некоторых воплощениях подпоследовательность представляет собой последовательность, выбранную из группы, состоящей из интрона мРНК PD-L1, такого как интрон человеческой мРНК PD-L1, выбранный из группы, состоящей из i1, i2, i3, i4, i5 и i6 (см. Таблицу 1 выше).
Олигонуклеотид по изобретению содержит непрерывную нуклеотидную последовательность, которая является комплементарной или гибридизуется с нуклеиновой кислотой-мишенью, такой как подпоследовательность нуклеиновой кислоты-мишени, такой как последовательность-мишень, описанная в данном документе.
Данный олигонуклеотид содержит непрерывную нуклеотидную последовательность из по меньшей мере 8 нуклеотидов, которая является комплементарной или гибридизуется с последовательностью-мишенью, присутствующей в молекуле нуклеиновой кислоты-мишени. Непрерывная нуклеотидная последовательность (и, следовательно, последовательность-мишень) содержит по меньшей мере 8 смежных нуклеотидов, как, например, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 смежных нуклеотидов, как, например, 12-25, как, например, 14-18 смежных нуклеотидов.
Клетка-мишень
Термин «клетка-мишень» в том виде, в котором он используется в данном документе, относится к клетке, которая экспрессирует нуклеиновую кислоту-мишень. В некоторых воплощениях клетка-мишень может находиться в условиях in vivo или in vitro. В некоторых вопощениях клетка-мишень представляет собой клетку млекопитающего, такую как клетка грызуна, как, например, клетка мыши или клетка крысы, или клетку примата, как, например, клетка обезьяны или клетка человека.
В предпочтительных воплощениях клетка-мишень экспрессирует мРНК PD-L1, такую как пре-мРНК PD-L1 или зрелая мРНК PD-L1. Поли А хвост мРНК PD-L1 типично не принимается во внимание для нацеливания антисмыслового олигонуклеотида.
Вариант, встречающийся в природе
Термин «вариант, встречающийся в природе» относится к вариантам гена PD-L1 или транскриптам, которые происходят из тех же самых генетических локусов, что и нуклеиновая кислота-мишень, но могут отличаться, например, за счет вырожденности генетического кода, вызывающей множественность кодонов, кодирующих ту же самую аминокислоту, или благодаря альтернативному сплайсингу пре-мРНК, или присутствию полиморфизмов, таких как однонуклеотидные полиморфизмы и аллельные варианты. На основе присутствия достаточно комплементарной последовательности к олигонуклеотиду, олигонуклеотид по изобретению, следовательно, может быть нацелен на нуклеиновую кислоту-мишень и ее встречающиеся в природе варианты.
В некоторых воплощениях встречающиеся в природе варианты имеют по меньшей мере 95%, как, например, по меньшей мере 98% или по меньшей мере 99% гомологии с нуклеиновой кислотой-мишенью PD-L1 млекопитающего, такой как нуклеиновая кислота-мишень, выбранная из группы, состоящей из SEQ ID NO 1, 2 и 3.
В гене PD-L1 известны многочисленные однонуклеотидные полиморфизмы, например, однонуклеотидные полиморфизмы, раскрытые в следующей таблице (начало/эталонная последовательность человеческой пре-мРНК представляет собой SEQ ID NO 2).
Модуляция экспрессии
Термин «модуляция экспрессии» в том виде, в котором он используется в данном документе, следует понимать как общий термин для способности олигонуклеотида изменять количество PD-L1 по сравнению с количеством PD-L1 до введния данного олигонуклеотида. В качестве альтернативы, модуляцию экспрессии можно определять посредством ссылки на контрольный экспермент. Обычно понятно то, что контроль представляет собой индивида или клетку-мишень, обработанную композицией физиологического раствора, или индивида или клетку-мишень, обработанную ненацеленым олигонуклеотидом (имитация). Однако он также может представлять собой индивида, которого лечили стандартом лечения.
Одним типом модуляции является способность олигонуклеотида ингибировать, осуществлять понижающую регуляцию, уменьшать, подавлять, удалять, останавливать, блокировать, предотвращать, ослаблять, снижать, устранять или прекращать экспрессию PD-L1, например, посредством деградации мРНК или блокировки транскрипции. Другим типом модуляции является способность олигонуклеотида восстанавливать, увеличивать или усиливать экспрессию PD-L1, например, посредством репарации сайтов сплайсинга или предотвращения сплайсинга, или устранения или блокировки ингибирующих механизмов, таких как репрессия микроРНК.
Высокоаффинные модифицированные нуклеозиды
Высокоаффинный модифицированный нуклеозид преставляет собой модифицированный нуклеотид, который, при включении в олигонуклеотид, увеличивает аффинность олигонуклеотида в отношении его комплементарной мишени, например, при измерении по температуре плавления (Tm). Высокоаффинный модифицированный нуклеозид по настоящему изобретению предпочтительно приводит к увеличению температуры плавления от +0,5 до +12°С, более предпочтительно - от +1,5 до +10°С, и наиболее предпочтительно - от +3 до +8°С на модифицированный нуклеозид. В данной области известны многочисленные высокоаффинные модифицированные нуклеозиды, и они включают, например, многие 2'-замещенные нуклеозиды, а также запертые нуклеиновые кислоты (LNA) (см., например, Freier & Altmann; Nucl. Acid Res., 1997, 25, 4429-4443 и Uhlmann; Curr. Opinion in Drug Development, 2000, 3(2), 293-213).
Модификации сахара
Олигомер по изобретению может содержать один или более чем один нуклеозид, который имеет модифицированную сахарную группировку, т.е. модификацию сахарной группировки по сравнению с рибозной сахарной группировкой, находящейся в ДНК и РНК.
Были получены многочисленные нуклеозиды с модификацией рибозной сахарной группировки, главным образом, с целью улучшения определенных свойств олигонуклеотидов, таких как аффинность и/или нуклеазоустойчивость.
Такие модификации включают модификации, где модифицируется структура рибозного кольца, например, посредством замены на гексозное кольцо (HNA) или бициклическое кольцо, которое типично имеет бирадикальный мостик между углеродами С2 и С4 на рибозном кольце (LNA), или на несвязанное рибозное кольцо, у которого типично отсутствует связь между углеродами С2 и С3 (например, UNA). Другие нуклеозиды с модифицированным сахаром включают, например, бициклогексозные нуклеиновые кислоты (WO2011/017521) или трициклические нуклеиновые кислоты (WO 2013/154798). Модифицированные нуклеозиды также включают нуклеозиды, где сахарная группировка заменяется на несахарную группировку, например, в случае пептидных нуклеиновых кислот (PNA) или морфолинонуклеиновых кислот.
Модификации сахаров также включают модификации, сделанные посредством изменения замещающих групп на рибозном кольце на группы, отличные от водорода, или группы 2'-OH, находящейся в природе в нуклеозидах ДНК и РНК. Заместители, например, можно вводить в 2'-, 3'-, 4'- или 5'-положения. Нуклеозиды с модифицированными сахарными группировками также включают 2'-модифицированные нуклеозиды, такие как 2'-замещенные нуклеозиды. В самом деле, значительное внимание было потрачено на разработку 2'-замещенных нуклеозидов, и было обнаружено то, что многочисленные 2'-замещенные нуклеозиды имеют полезные свойства при включении в олигонуклеотиды, такие как повышенная устойчивость нуклеозидов и повышенная аффинность.
2'-Модифицированные нуклеозиды
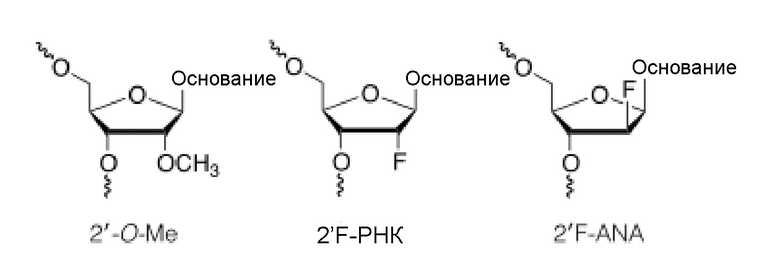
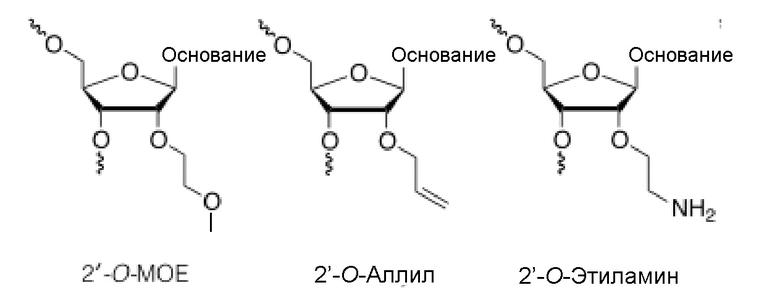
Нуклеозид с 2'-модифицированным сахаром представляет собой нуклеозид, который имеет в положении 2' заместитель, отличный от Н или -ОН (2'-замещенный нуклеозид) или содержит 2'-связанный бирадикал и включает 2'-замещенные нуклеозиды и нуклеозиды LNA (связанные 2'-4' бирадикальным мостиком). Например, 2'-модифицированный сахар может обеспечивать повышенную аффинность связывания и/или повышенную нуклеазоустойчивость олигонуклеотида. Примерами 2'-замещенных модифицированных нуклеозидов являются 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-алкокси-РНК, 2'-O-метоксиэтил-РНК (МОЕ), 2'-амино-ДНК, 2'-фтор-РНК и 2'-фтор-ANA (F-ANA) нуклеозид. Относительно дополнительных примеров, пожалуйста, см., например, Freier & Altmann; Nucl. Acid Res., 1997, 25, 4429-4443; Uhlmann; Curr. Opinion in Drug Development, 2000, 3(2), 293-213 и Deleavey and Damha, Chemistry and Biology 2012, 19, 937. Ниже приводятся иллюстрации некоторых 2'-замещенных модифицированных нуклеозидов.
Нуклеозиды запертых нуклеиновых кислот (LNA)
Нуклеозиды LNA представляют собой модифицированные нуклеозиды, которые содержат линкерную группу (именуемую бирадикал или мостик) между C2' и C4' рибозного сахарного кольца нуклеотида. Данные нуклеозиды также называются в литературе мостиковой нуклеиновой кислотой или бициклической нуклеиновой кислотой (BNA).
В некоторых воплощениях модифицированный нуклеозид или нуклеозиды LNA олигомера по изобретению имеют общую структуру формулы I или II:
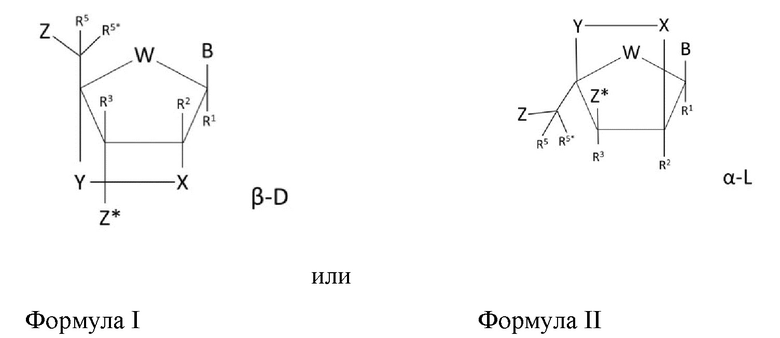
где W выбран из -O-, -S-, -N(Ra)-, -C(RaRb)-, таким образом, что в некоторых воплощениях -О-; В обозначает нуклеиновое основание или модифицированную группировку нуклеинового основания;
Z обозначает межнуклеозидную связь с соседним нуклеозидом или 5'-концевую группу;
Z* обозначает межнуклеозидную связь с соседним нуклеозидом или 3'-концевую группу;
Х обозначает группу, выбранную из списка, состоящего из -C(RaRb)-, -C(Ra)=C(Rb)-, -C(Ra)=N-, -O-, -Si(Ra)2-, -S-, -SO2-, -N(Ra)- и >C=Z.
В некоторых воплощениях Х выбран из группы, состоящей из: -O-, -S-, NH-, NRaRb, -CH2-, CRaRb, -C(=CH2)- и -C(=CRaRb)-.
В некоторых воплощениях Х представляет собой -О-.
Y обозначает группу, выбранную из группы, состоящей из -C(RaRb)-, -C(Ra)=C(Rb)-, -C(Ra)=N-, -O-, -Si(Ra)2-, -S-, -SO2-, -N(Ra)- и >C=Z.
В некоторых воплощениях Y выбран из группы, состоящей из: -CH2-, -C(RaRb)-, -CH2CH2-, -C(RaRb)-C(RaRb)-, -CH2CH2CH2-, -C(RaRb)C(RaRb)C(RaRb)-, -C(Ra)=C(Rb)- и -C(Ra)=N-.
В некоторых воплощениях Y выбран из группы, состоящей из: -CH2-, -CHRa-, -CHCH3-, CRaRb-.
или -Х-Y- совместно обозначают двухвалентную линкерную группу (также именуемую радикал), состоящую из 1, 2, 3 или 4 групп/атомов, выбранных из группы, состоящей из -C(RaRb)-, -C(Ra)=C(Rb)-, -C(Ra)=N-, -O-, -Si(Ra)2-, -S-, -SO2-, -N(Ra)- и >C=Z.
В некоторых воплощениях -Х-Y- обозначает бирадикал, выбранный из групп, состоящих из: -X-CH2-, -X-CRaRb-, -X-CHRa-, -X-C(HCH3)-, -O-Y-, -O-CH2-, -S-CH2-, -NH-CH2-, -O-CHCH3-, -CH2-O-CH2, -O-CH(CH3CH3)-, -O-CH2-CH2-, OCH2-CH2-CH2-,-O-CH2OCH2-, -O-NCH2-, -C(=CH2)-CH2-, -NRa-CH2-, N-O-CH2, -S-CRaRb- и -S-CHRa-.
В некоторых воплощениях -Х-Y- обозначает -O-CH2- или -O-CH(CH3)-,
где Z выбран из -О-, -S- и -N(Ra)-,
и Ra, и, при наличии Rb, каждый независимо выбран из водорода, возможно замещенного С1-6-алкила, возможно замещенного С2-6-алкенила, возможно замещенного С2-6-алкинила, гидрокси, возможно замещенного С1-6-алкокси, С2-6-алкоксиалкила, С2-6-алкенилокси, карбокси, С1-6-алкоксикарбонила, С1-6-алкилкарбонила, формила, арила, арилоксикарбонила, арилокси, арилкарбонила, гетероарила, гетероарилоксикарбонила, гетероарилокси, гетероарилкарбонила, амино, моно- и ди(С1-6-алкил)амино, карбамоила, моно- и ди(С1-6-алкил)амино-карбонила, амино-С1-6-алкил-аминокарбонила, моно- и ди(С1-6-алкил)амино-С1-6-алкил-аминокарбонила, С1-6-алкил-карбониламино, карбамидо, С1-6-алканоилокси, сульфоно, С1-6-алкилсульфонилокси, нитро, азидо, сульфанила, С1-6-алкилтио, галогена, где арил и гетероарил возможно могут быть замещены, и где два геминальных заместителя Ra и Rb могут вместе обозначать возможно замещенный метилен (=СН2), где для всех хиральных центров асимметрические группы могут находиться либо в R, либо в S ориентации,
где R1, R2, R3, R5 и R5* независимо выбраны из группы, состоящей из: водорода, возможно замещенного С1-6-алкила, возможно замещенного С2-6-алкенила, возможно замещенного С2-6-алкинила, гидрокси, С1-6-алкокси, С2-6-алкоксиалкила, С2-6-алкенилокси, карбокси, С1-6-алкоксикарбонила, С1-6-алкилкарбонила, формила, арила, арилокси-карбонила, арилокси, арилкарбонила, гетероарила, гетероарилокси-карбонила, гетероарилокси, гетероарилкарбонила, амино, моно- и ди(С1-6-алкил)амино, карбамоила, моно- и ди(С1-6-алкил)-амино-карбонила, амино-С1-6-алкил-аминокарбонила, моно- и ди(С1-6-алкил)амино-С1-6-алкил-аминокарбонила, С1-6-алкил-карбониламино, карбамидо, С1-6-алканоилокси, сульфоно, С1-6-алкилсульфонилокси, нитро, азидо, сульфанила, С1-6-алкилтио, галогена, где арил и гетероарил возможно могут быть замещены, и где два геминальных заместителя могут вместе обозначать оксо, тиоксо, имино или возможно замещенный метилен.
В некоторых воплощениях R1, R2, R3, R5 и R5* независимо выбраны из С1-6-алкила, такого как метил, и водорода.
В некоторых воплощениях все R1, R2, R3, R5 и R5* представляют собой атомы водорода.
В некоторых воплощениях все R1, R2, R3 представляют собой атомы водорода, и либо R5, либо R5* также представляет собой атом водорода, а другой из R5 и R5* отличается от атома водорода, как, например, представляет собой С1-6-алкил, такой как метил.
В некоторых воплощениях Rа представляет собой либо атом водорода, либо метил. В некоторых воплощениях Rb, при наличии, представляет собой либо атом водорода, либо метил.
В некоторых воплощениях один или оба из Rа и Rb представляют собой атом водорода.
В некоторых воплощениях один из Rа и Rb представляет собой атом водорода, а другой отличается от атома водорода.
В некоторых воплощениях один из Rа и Rb представляет собой метил, а другой представляет собой атом водорода.
В некоторых воплощениях оба из Rа и Rb представляют собой метил.
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-СН2-, W представляет собой О, и все R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды LNA раскрыты в WO 99/014226, WO 00/66604, WO 98/039352 и WO 2004/046160, которые все включены в данный документ посредством ссылки, и они включают то, что обычно известно как нуклеозиды бета-О-окси LNA и альфа-L-окси LNA.
В некоторых воплощениях бирадикал -X-Y- представляет собой -S-СН2-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды тио LNA раскрыты в WO 99/014226 и WO 2004/046160, которые включены тем самым посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -NH-СН2-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды амино LNA раскрыты в WO 99/014226 и WO 2004/046160, которые тем самым включены посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-СН2-СН2- или -О-СН2-СН2-СН2-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды LNA раскрыты в WO 00/047599 и Morita et al, Bioorganic & Med. Chem. Lett. 12 73-76, которые тем самым включены посредством ссылки, и включают то, что обычно известно как нуклеиновые кислоты, связанные 2'-O-4'C-этиленовым мостиком (ENA).
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-СН2-, W представляет собой О, и все из R1, R2, R3 и один из R5 и R5* представляют собой атомы водорода, а другой из R5 и R5* отличается от атома водорода, как, например, С1-6алкил, такой как метил. Такие 5'-замещенные нуклеозиды LNA раскрыты в WO2007/134181, которая тем самым включена посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-CRaRb, где один или оба из Ra и Rb отличаются от атома водорода, как, например, метил, W представляет собой О, и все из R1, R2, R3 и один из R5 и R5* представляют собой атом водорода, а другой из R5 и R5* отличается от атома водорода, как, например, С1-6алкил, такой как метил. Такие бис-модифицированные нуклеозиды LNA раскрыты в WO2010/077578, которая тем самым включена в данный документ посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- обозначает двухвалентную линкерную группу -O-CH(CH2OCH3)- (2'O-метоксиэтилбициклическая нуклеиновая кислота - Seth at al., 2010, J. Org. Chem. Vol 75(5) pp. 1569-81). В некоторых воплощениях бирадикал -X-Y- обозначает двухвалентную линкерную группу -O-CH(CH2CH3) - (2'O-этилбициклическая нуклеиновая кислота - Seth at al., 2010, J. Org. Chem. Vol 75(5) pp. 1569-81). В некоторых воплощениях бирадикал -X-Y- представляет собой -О-CНRa, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие 6'-замещенные нуклеозиды LNA раскрыты в WO 10036698 и WO 07090071, которые обе тем самым включены посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-CН(СН2ОСН3)-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды LNA также известны в данной области как циклические МОЕ (сМОЕ) и раскрыты в WO 07090071.
В некоторых воплощениях бирадикал -X-Y- обозначает двухвалентную линкерную группу -O-CH(CH3)- либо в R-, либо в S-конфигурации. В некоторых воплощениях бирадикал -X-Y- вместе обозначает двухвалентную линкерную группу -O-CH2-O-CH2- (Seth at al., 2010, J. Org. Chem). В некоторых воплощениях бирадикал -X-Y- представляет собой -О-CН(СН3)-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие 6'-метилнуклеозиды LNA также известны в данной области как сЕТ нуклеозиды и могут представлять собой либо (S)cET, либо (R)cET стереоизомеры, как раскрыто в WO 07090071 (бета-D) и WO 2010/036698 (альфа-L), которые оба тем самым включены посредством ссылки).
В некоторых воплощениях бирадикал -X-Y- представляет собой -О-CRaRb-, в котором ни один из Ra или Rb не является атомом водорода, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. В некоторых воплощениях и Ra, и Rb представляют собой метил. Такие 6'-двухзамещенные нуклеозиды LNA раскрыты в WO 2009006478, которая тем самым включена посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -S-CHRa-, W представляет собой О, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие 6'-замещенные нуклеозиды тио LNA раскрыты в WO 11156202, которая тем самым включена посредством ссылки. В некоторых воплощениях 6'-замещенных тио LNA Ra представляет собой метил.
В некоторых воплощениях бирадикал -X-Y- представляет собой -C(=CH2)-C(RaRb)-, как, например, -C(=CH2)-CH2- или -C(=CH2)-CH(CH3)-, W представляет собой O, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. Такие нуклеозиды винилкарбо LNA раскрыты в WO 08154401 и WO 09067647, которые обе тем самым включены посредством ссылки.
В некоторых воплощениях бирадикал -X-Y- представляет собой -N(-ORa)-, W представляет собой O, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. В некоторых воплощениях Ra представляет собой C1-6алкил, такой как метил. Такие нуклеозиды LNA также известны как N-замещенные LNA и раскрываются в WO 2008/150729, которая тем самым включена посредством ссылки. В некоторых воплощениях бирадикал -X-Y- вместе обозначает двухвалентную линкерную группу -O-NRa-CH3- (Seth at al., 2010, J. Org. Chem). В некоторых воплощениях бирадикал -X-Y- представляет собой -N(Ra)-, W представляет собой O, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. В некоторых воплощениях Ra представляет собой C1-6алкил, такой как метил.
В некоторых воплощениях один или оба из R5 и R5* представляют собой атом водорода, и, при замещении, другой из R5 и R5* представляет собой C1-6алкил, такой как метил. В таком воплощении все из R1, R2, R3 могут представлять собой атомы водорода, и бирадикал -X-Y- может быть выбран из -O-CH2- или -O-C(HCRa)-, такого как -O-C(HCH3)-.
В некоторых воплощениях бирадикал представляет собой -CRaRb-O-CRaRb-, такой как CH2-O-CH2-, W представляет собой O, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. В некоторых воплощениях Ra представляет собой C1-6алкил, такой как метил. Такие нуклеозиды LNA также известны как конформационно ограниченные нуклеотиды (CRN) и раскрываются в WO 2013036868, которая тем самым включена посредством ссылки.
В некоторых воплощениях бирадикал представляет собой -O-CRaRb-O-CRaRb-, такой как O-CH2-O-CH2-, W представляет собой O, и все из R1, R2, R3, R5 и R5* представляют собой атомы водорода. В некоторых воплощениях Ra представляет собой C1-6алкил, такой как метил. Такие нуклеозиды LNA также известны как нуклеотиды СОС и раскрываются в Mitsuoka et al., Nucleic Acids Research 2009 37(4), 1225-1238, которая тем самым включена посредством ссылки.
Будет понятно то, что, если не определено, нуклеозиды LNA могут находиться в бета-D или альфа-L стереоизоформе.
Некоторые примеры нуклеозидов LNA представлены на Схеме 1.
Схема 1
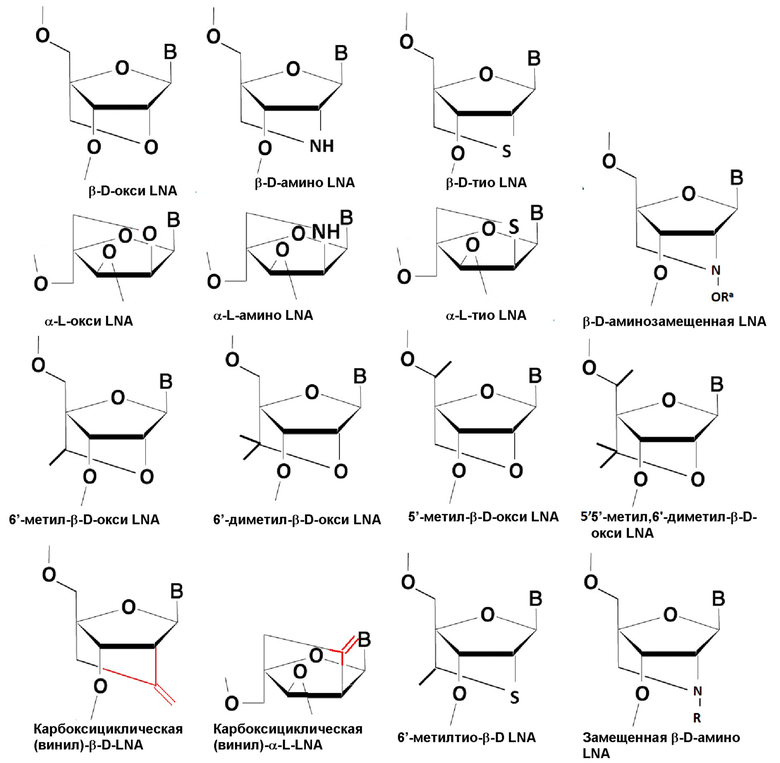
Как проиллюстрировано в Примерах, в предпочтительных воплощениях изобретения нуклеозиды LNA в олигонуклеотидах представляют собой нуклеозиды бета-D-окси-LNA.
Деградация, опосредованная нуклеазой
Термин «деградация, опосредованная нуклеазой» относится к олигонуклеотиду, способному опосредовать деградацию комплементарной нуклеотидной последовательности при образовании дуплекса с такой последовательностью.
В некоторых воплощениях олигонуклеотид может функционировать посредством деградации нуклеиновой кислоты-мишени, опосредованной нуклеазой, где олигонуклеотиды по изобретению способны рекрутировать нуклеазу, в частности, эндонуклеазу, предпочтительно эндорибонуклеазу (РНКазу), такую как РНКаза Н. Примерами конструкций олигонуклеотидов, которые работают посредством механизмов, опосредованных нуклеазой, являются олигонуклеотиды, которые типично содержат область из по меньшей мере 5 или 6 нуклеозидов ДНК и фланкированы на одной стороне или на обеих сторонах нуклеозидами, увеличивающими аффинность, например, гэпмеры, хэдмеры и тэйлмеры.
Активность и рекрутирование РНКазы Н
Термин «активность РНКазы Н антисмыслового олигонуклеотида» относится к его способности при нахождении в дуплексе с комплементарной молекулой РНК рекрутировать РНКазу Н. В WO 01/23613 предложены способы in vitro для определения активности РНКазы Н, которые можно использовать для определения способности рекрутировать РНКазу Н. Типично олигонуклеотид считается способным рекрутировать РНКазу Н, если он, при предоставлении с комплементарной последовательностью нуклеиновой кислоты-мишени, имеет исходную скорость, измеренную в пмоль/л/мин, по меньшей мере 5%, как, например, по меньшей мере 10% или больше, чем 20% от исходной скорости, определенной при использовании олигонуклеотида, имеющего такую же последовательность оснований, что и тестируемый модифицированный олигонуклеотид, но содержащего только мономеры ДНК с фосфоротиоатными связями между всеми мономерами в олигонуклеотиде, и с использованием методологии, предложенной в Примерах 91-95 WO 01/23613 (включенной тем самым посредством ссылки).
Гэпмер
Термин «гэпмер» в том виде, в котором он используется в данном документе, относится к антисмысловому олигонуклеотиду, который содержит область олигонуклеотидов (гэп), рекрутирующую РНКазу Н, которая фланкирована 5' и 3' областями (фланги или крылья), которые содержат один или более чем один модифицированный нуклеозид, усиливающий аффинность. В данном документе описываются разные конструкции гэпмеров, и они характеризуются по их способности рекрутировать РНКазу Н. Хэдмеры и тэйлмеры представляют собой олигонуклеотиды, способные рекрутировать РНКазу Н, где отсутствует один из флангов, т.е. только один из концов олигонуклеотида содержит модифицированные нуклеозиды, усиливающие аффинность. Для хэдмеров отсутстует 3'-фланг (т.е. 5'-фланг содержит модифицированные нуклеозиды, усиливающие аффинность), и для тэйлмеров отсутстует 5'-фланг (т.е. 3'-фланг содержит модифицированные нуклеозиды, усиливающие аффинность).
Гэпмер LNA
Термин «гэпмер LNA» относится к гэпмерному олигонуклеотиду, в котором по меньшей мере один из модифицированных нуклеозидов, усиливающих аффинность, представляет собой нуклеозид LNA.
Гэпмер со смешанными крыльями
Термин «гэпмер со смешанными крыльями» или «гэпмер со смешанными флангами» относится к гэпмеру LNA, в котором по меньшей мере одна фланговая область содержит по меньшей мере один нуклеозид LNA и по меньшей мере один модифицированный нуклеозид, не являющийся LNA, такой как по меньшей мере один 2'-замещенный модифицированный нуклеозид, такой как, например, нуклеозид(ды) 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-O-алкокси-РНК, 2'-O-метоксиэтил-РНК (МОЕ), 2'-амино-ДНК, 2'-фтор-РНК и 2'-F-ANA. В некоторых воплощениях гэпмер со смешанными крыльями имеет один фланг, который содержит только нуклеозиды LNA (например, 5' или 3'), и другой фланг (3' или 5' соответственно) содержит 2'-замещенный(ные) модифицированный(ные) нуклеозид(ды) и возможно нуклеозиды LNA.
Олигонуклеотид с разорванным гэпом
Термин «олигонуклеотид с разорванным гэпом» используется по отношению к гэпмеру, способному поддерживать рекрутирование РНКазы Н, даже несмотря на то, что область гэпа разрывается нуклеозидом, не рекрутирующим РНКазу Н (нуклеозидом, разрывающим гэп, Е), таким образом, что область гэпа содержит меньше, чем 5 последовательных нуклеозидов ДНК. Нуклеозиды, не рекрутирующие РНКазу Н, представляют собой, например, нуклеозиды в 3'-эндоконформации, такие как LNA, где мостик между C2' и C4' рибозного сахарного кольца нуклеозида находится в бета-конформации, как, например, нуклеозид бета-D-окси LNA или ScET. Способность олигонуклеотида с разорванным гэпом рекрутировать РНКазу Н типично является специфичной в отношении последовательности или даже соединения - см. Rukov et al. 2015 Nucl. Acids Res. Vol. 43 pp. 8476-8487, в которой раскрыты «олигонуклеотиды с разорванным гэпом», которые рекрутируют РНКазу Н, которые в некоторых случаях обеспечивает более специфичное расщеплениее РНК-мишени.
В некоторых воплощениях олигонуклеотид по изобретению представляет собой олигонуклеотид с разорванным гэпом. В некоторых воплощениях олигонуклеотид с разорванным гэпом содержит 5'-фланг (F), гэп (G) и 3'-фланг (F'), где гэп разорван нуклеозидом, не рекрутирующим РНКазу Н (нуклеозидом, разрывающим гэп, Е), таким образом, что гэп содержит по меньшей мере 3 или 4 последовательных нуклеозида ДНК. В некоторых воплощениях нуклеозид, разрывающий гэп (Е), представляет собой нуклеозид LNA, где мостик между C2' и C4' рибозного сахарного кольца нуклеозида находится в бета-конформации, и он размещается в пределах области гэпа таким образом, что нуклеозид LNA, разрывающий гэп, фланкирован 5' и 3' по меньшей мере 3(5') и 3(3') или по меньшей мере 3(5') и 4(3'), или по меньшей мере 4(5') и 3(3') нуклеозидами ДНК, и где данный олигонуклеотид способен рекрутировать РНКазу Н.
Олигонуклеотид с разорванным гэпом может быть представлен следующими формулами:
F-G-E-G-F'; в частности F1-7-G3-4-E1-G3-4-F'1-7
D'-F-G-F', в частности D'1-3-F1-7- G3-4-E1-G3-4-F'1-7
F-G-F'-D'', в частности F1-7- G3-4-E1-G3-4-F'1-7-D''1-3
D'-F-G-F'-D'', в частности D'1-3-F1-7- G3-4-E1-G3-4-F'1-7-D''1-3
Где области D' и D'' являются такими, как описано в разделе «Конструкция гэпмера».
В некоторых воплощениях нуклеозид, разрывающий гэп (Е), представляет собой бета-D-окси LNA или ScET, или другие нуклеозиды бета-LNA, показанные на Схеме 1).
Конъюгат
Термин «конъюгат» в том виде, в котором он используется в данном документе, относится к олигонуклеотиду, который ковалентно связан с ненуклеотидной группировкой (конъюгатная группировка или область С, или третья область), также именуемому конъюгатом олигонуклеотида.
Конъюгирование олигонуклеотида по изобретению с одной или более чем одной ненуклеотидной группировкой может улучшать фармакологию данного олигонуклеотида, например, посредством влияния на активность, клеточное распределение, клеточное поглощение или стабильность олигонуклеотида. В некоторых воплощениях конъюгатная группировка нацеливает олигонуклеотид в печень. В то же самое время конъюгат служит для уменьшения активности олигонуклеотида в типах клеток, тканях или органах, не являющихся мишенями, например, активность вне мишени или активность в типах клеток, тканях или органах, не являющихся мишенями. В одном воплощении изобретения конъюгат олигонуклеотида по изобретению демонстрирует улучшенное ингибирование PD-L1 в клетке-мишени по сравнению с неконъюгированным олигонуклеотидом. В другом воплощении конъюгат олигонуклеотида по изобретнию имеет улучшенное клеточное распределение между печенью и другими органами, такими как селезенка или почки (т.е. больше конъюгированного олигонуклеотида поступает в печень, чем в селезенку или в почки), по сравнению с неконъюгированным олигонуклеотидом. В другом воплощении конъюгат олигонуклеотида по изобретнию демонстрирует улучшенное клеточное поглощение в печень конъюгата олигонуклеотида по сравнению с неконъюгированным олигонуклеотидом.
В WO 93/07883 и WO 2013/033230 предложены подходящие конъюгатные группировки, которые тем самым являются включенными посредством ссылки. Дополнительными подходящими конъюгатными группировками являются группировки, способные к связыванию с рецептором асиалогликопротеина (ASGPr). В частности, подходящими для связывания с ASGPr являются трехвалентные N-ацетилгалактозаминные конъюгатные группировки, см., например, WO 2014/076196, WO 2014/207232 и WO 2014/179620 (включенные тем самым посредством ссылки). Конъюгатная группировка является по существу частью конъюгатов антисмысловых олигонуклеотидов, которая не состоит из нуклеиновых кислот.
Конъюгаты олигонуклеотидов и их синтез также были описаны во всеохватывающих обзорах Manoharan в Antisense Drug Technology, Principles, Strategies, and Applications, S.T. Crooke, ed., Ch. 16, Marcel Dekker, Inc., 2001 и Manoharan, Antisense and Nucleic Acid Drug Development, 2002, 12, 103, каждый из которых включен в данный документ посредством ссылки во всей его полноте.
В одном воплощении ненуклеотидная группировка (конъюгатная группировка) выбрана из группы, состоящей из углеводов, лигандов рецептора поверхности клетки, лекарственных веществ, гормонов, липофильных веществ, полимеров, белков, пептидов, токсинов (например, бактериальных токсинов), витаминов, вирусных белков (например, капсидов) или их комбинаций.
Линкеры
Связка или линкер представляет собой соединение между двумя атомами, которое связывает одну химическую группу или интересующий сегмент с другой химической группой или интересующим сегментом посредством одной или более чем одной ковалентной связи. Конъюгатные группировки могут присоединяться к олигонуклеотиду непосредственно или через связывающую группировку (например, линкер или связку). Линкеры служат для ковалентного присоединения третьей области, например, конъюгатной группировки (области С), к первой области, например, олигонуклеотиду или непрерывной нуклеотидной последовательности, комплементарной нуклеиновой кислоте-мишени (область А).
В некоторых воплощениях изобретения конъюгат или конъюгат олигонуклеотида по изобретению возможно может содержать линкерную область (вторая область или область В, и/или область Y), которая располагается между олигонуклеотидом или непрерывной нуклеотидной последовательностью, комплементарной нуклеиновой кислоте-мишени (область А или первая область), и конъюгатной группировкой (область С или третья область).
Область В относится к биорасщепляемым линкерам, содержащим или состоящим из физиологически лабильной связи, которая является расщепляемой при условиях, которые обычно встречаются или являются аналогичными условиям, которые встречаются в организме млекопитающего. Условия, при которых физиологически лабильные линкеры подвергаются химическому превращению (например, расщеплению), включают такие химические условия, как рН, температура, окислительные или восстановительные условия или агенты и концентрация соли, обнаруженные или аналогичные условиям, встречающимся в клетках млекопитающих. Внутриклеточные условия у млекопитающих также включают присутствие ферментативной активности, обычно присутствующей в клетке млекопитающего, как, например, от протеолитических ферментов или гидролитических ферментов, или нуклеаз. В одном воплощении биорасщепляемый линкер чувствителен к расщеплению нуклеазой S1. В предпочтительном воплощении нуклеазочувствительный линкер содержит от 1 до 10 нуклеозидов, как, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеозидов, более предпочтительно - от 2 до 6 нуклеозидов, и наиболее предпочтительно - от 2 до 4 связанных нуклеозидов, содержащих по меньшей мере две последовательные фосфодиэфирные связи, как, например, по меньшей мере 3 или 4, или 5 последовательных фосфодиэфирных связей. Предпочтительно нуклеозидами являются ДНК или РНК. Биорасщепляемые линкеры, содержащие фосфодиэфир, более подробно описываются в WO 2014/076195 (включенной тем самым посредством ссылки).
Область Y относится к линкерам, которые не обязательно являются биорасщепляемыми, но, главным образом, служат для ковалентного присоединения конъюгатной группировки (области С или третьей области) к олигонуклеотиду или непрерывной нуклеотидной последовательности, комплементарной нуклеиновой кислоте-мишени (области А или первой области). Линкеры области Y могут содержать структуру цепи или олигомер из повторяющихся звеньев, такой как этиленгликоль, аминокислотные звенья или аминоалкильные группы. Конъюгаты олигонуклеотидов по настоящему изобретению могут быть сконструированы из следующих элементов областей: A-C, A-B-C, A-B-Y-C, A-Y-B-C или A-Y-C. В некоторых воплощениях линкер (область Y) представляет собой аминоалкил, как, например, С2-С36 аминоалкильную группу, включающую, например, С6-С12 аминоалкильные группы. В предпочтительном воплощении линкер (область Y) представляет собой С6 аминоалкильную группу.
Лечение
Термин «лечение» в том виде, в котором он используется в данном документе, относится как к лечению существующего заболевания (например, заболевания или расстройства, на которые дается ссылка в данном документе), так и к предупреждению заболевания, т.е. профилактике. Следовательно, будет понятно, что лечение в том виде, в котором на него дается ссылка в данном документе, в некоторых воплощениях может быть профилактическим.
Восстановление иммунного ответа против патогенов
Иммунный ответ подразделяется на врожденный и адаптивный иммунный ответ. Врожденная иммунная система обеспечивает немедленный, но неспецифичный ответ. Адаптивный иммунный ответ активируется врожденным иммунным ответом и является высокоспецифичным по отношению к конкретному патогену. При презентации антигена, происходящего из патогена, на поверхности антигенпрезентирующих клеток иммунные клетки адаптивного иммунного ответа (т.е. Т- и В-лимфоциты) активируются посредством их антигенспецифичных рецепторов, приводя к патогенспецифичному иммунному ответу и развитию иммунологической памяти. Хронические вирусные инфекции, такие как HBV и HCV, ассоциированы с исчерпанием Т-клеток, характеризуемым неотвечаемостью вирусоспецифичных Т-клеток. Исчерпание Т-клеток хорошо изучено, относительно обзора см., например, Yi et al 2010 Immunology 129, 474-481. Хронические вирусные инфекции также ассоциированы с пониженной функцией NK-клеток, которые являются врождеными иммунными клетками. Усиление иммунного ответа против вирусов является важным для клиренса хронической инфекции. Восстановление иммунного ответа против патогенов, опосредованного Т-клетками и NK-клетками, можно оценивать посредством измерения пролиферации, секреции цитокинов и цитолитической функции (Dolina et al. 2013 Molecular Therapy-Nucleic Acids, 2 e72 и Пример 6 в данном документе).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к применению антисмысловых олигонуклеотидов, их конъюгатов и содержащих их фармацевтических композиций для восстановления иммунного ответа против патогенов, которые инфицировали животное, в частности человека. Конъюгаты антисмысловых олигонуклеотидов по настоящему изобретению являются особенно полезными против патогенов, которые инфицировали печень, в частности, против хронических инфекций печени, подобных HBV. Данные конъюгаты обеспечивают таргетное распределение олигонуклеотидов и предотвращают системный нокдаун нуклеиновой кислоты-мишени.
Олигонуклеотиды по изобретению
Данное изобретение относится к олигонуклеотидам, способным модулировать экспрессию PD-L1. Модуляция может достигаться гибридизацией с нуклеиновой кислотой-мишенью, кодирующей PD-L1, или с той, которая участвует в регуляции PD-L1. Нуклеиновая кислота-мишень может представлять собой последовательность PD-L1 млекопитающего, такую как последовательность, выбранная из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и/или SEQ ID NO: 3. Нуклеиновая кислота-мишень может представлять собой пре-мРНК, мРНК или любую последовательность РНК, экспрессируемую из клетки млекопитающего, которая поддерживает экспрессию или регуляцию PD-L1.
Олигонуклеотид по изобретению представляет собой антисмысловой олигонуклеотид, который нацелен на PD-L1.
В одном аспекте изобретения олигонуклеотиды по изобретению конъюгированы с конъюгатной группировкой, в частности, с конъюгатной группировкой, нацеленной на рецептор асиалогликопротеина.
В некоторых воплощениях антисмысловой олигонуклеотид по изобретению способен модулировать экспрессию мишени посредством осуществления ее ингибирования или понижающей регуляции. Предпочтительно такая модуляция дает ингибирование экспрессии по меньшей мере 20% по сравнению с нормальным уровнем экспрессии мишени, более предпочтительно по меньшей мере 30%-ное, 40%-ное, 50%-ное, 60%-ное, 70%-ное, 80%-ное или 90%-ное ингибирование по сравнению с нормальным уровнем экспрессии мишени. Предпочтительно такая модуляция дает ингибирование экспрессии по меньшей мере 20% по сравнению с уровнем экспрессии при заражении клетки или организма инфекционным агентом, или при обработке агентом, имитирующим заражение инфекционным агентом (например, поли I:C или LPS (липополисахарид)), более предпочтительно по меньшей мере 30%-ное, 40%-ное, 50%-ное, 60%-ное, 70%-ное, 80%-ное или 90%-ное ингибирование по сравнению с с уровнем экспрессии при заражении клетки или организма инфекционным агентом, или при обработке агентом, имитирующим заражение инфекционным агентом (например, поли I:C или LPS). В некоторых воплощениях олигонуклеотиды по изобретению могут быть способны ингибировать уровни экспрессии мРНК PD-L1 по меньшей мере на 60% или 70% in vitro с использованием клеток KARPAS-299 или ТНР1. В некоторых воплощениях соединения по изобретению могут быть способны ингибировать уровни экспрессии белка PD-L1 по меньшей мере на 50% in vitro с использованием клеток KARPAS-299 или ТНР1. Подходящим образом в примерах предложены анализы, которые можно использовать для измерения РНК PD-L1 (например, Пример 1). Модуляция мишени запускается гибридизацией между непрерывной нуклеотидной последовательностью олигонуклеотида и нуклеиновой кислотой-мишенью. В некоторых воплощениях олигонуклеотид по изобретению содержит несоответствия между данным олигонуклеотидом и нуклеиновой кислотой-мишенью. Несмотря на несоответствия гибридизация с нуклеиновой кислотой-мишенью все еще может быть достаточной для демонстрации желательной модуляции экспрессии PD-L1. Пониженная аффинность связывания, происходящая из-за несоответствий, преимущественно может компенсироваться большим числом нуклеотидов в олигонуклеотиде и/или большим числом модифицированных нуклеозидов, способных увеличивать аффинность связывания с мишенью, таких как 2'-модифицированные нуклеозиды, включая LNA, присутствующих в пределах последовательности олигонуклеотида.
В некоторых воплощениях антисмысловой олигонуклеотид по изобретению способен восстанавливать патогенспецифичные Т-клетки. В некоторых воплощениях олигонуклеотиды по изобретению способны увеличивать число патогенспецифичных Т-клеток по меньшей мере на 40%, 50%, 60% или 70% по сравнению с необработанными контролями или контролями, обработанными стандартом лечения. В одном воплощении антисмысловой олигонуклеотид или конъюгат по изобретению способен увеличивать уровень HBV-специфичных Т-клеток по сравнению с необработанными контролями или контролями, обработанными стандартом лечения. Подходящим образом, в данных примерах предложены анализы, которые можно использовать для измерения HBV-специфичных Т-клеток (например, пролиферация Т-клеток, секреция цитокинов и цитолитическая активность). В другом воплощении антисмысловой олигонуклеотид или конъюгат по изобретению способен увеличивать уровень HСV-специфичных Т-клеток по сравнению с необработанными контролями или контролями, обработанными стандартом лечения. В другом воплощении антисмысловой олигонуклеотид или конъюгат по изобретению способен увеличивать уровень HDV-специфичных Т-клеток по сравнению с необработанными контролями или контролями, обработанными стандартом лечения.
В некоторых воплощениях антисмысловой олигонуклеотид по изобретению способен уменьшать уровни HBsAg у животного или человека. В некоторых воплощениях олигонуклеотиды по изобретению способны уменьшать уровни HВsAg по меньшей мере на 40%, 50%, 60% или 70%, более предпочтительно по меньшей мере на 80%, 90% или 95% по сравнению с уровнем до обработки. Наиболее предпочтительно олигонуклеотиды по изобретению способны достигать сероконверсии HВsAg у животного или человека, иницированного HBV.
Один аспект настоящего изобретения относится к антисмысловому олигонуклеотиду, который содержит непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью с нуклеиновой кислотой-мишенью PD-L1.
В некоторых воплощениях данный олигонуклеотид содержит непрерывную последовательность, которая является по меньшей мере на 90% комплементарной, как, например, по меньшей мере на 91%, как, например, по меньшей мере на 92%, как, например, по меньшей мере на 93%, как, например, по меньшей мере на 94%, как, например, по меньшей мере на 95%, как, например, по меньшей мере на 96%, как, например, по меньшей мере на 97%, как, например, по меньшей мере на 98% или на 100% комплементарной области нуклеиновой кислоты-мишени.
В предпочтительном воплощении олигонуклеотид по изобретению или его непрерывная нуклеотидная последовательность является полностью комплементарным (на 100% комплементарным) области нуклеиновой кислоты-мишени, или в некоторых воплощениях может содержать одно или два несоответствия между олигонуклеотидом и нуклеиновой кислотой-мишенью.
В некоторых воплощениях олигонуклеотид содержит непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью, как, например, с полной (или 100%-ной) комплементарностью с областью нуклеиновой кислоты-мишени, присутствующей в SEQ ID NO: 1 или SEQ ID NO: 2. В некоторых воплощениях последовательность олигонуклеотида является на 100% комплементарной соответствующей области нуклеиновой кислоты-мишени, присутствующей в SEQ ID NO: 1 и SEQ ID NO: 2. В некоторых воплощениях последовательность олигонуклеотида является на 100% комплементарной соответствующей области нуклеиновой кислоты-мишени, присутствующей в SEQ ID NO: 1 или SEQ ID NO: 3.
В некоторых воплощениях олигонуклеотид или конъюгат олигонуклеотида содержит непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью, как, например, 100%-ной комплементарностью с соответствующей областью нуклеиновой кислоты-мишени, где непрерывная нуклеотидная последовательность является комплементарной подпоследовательности нуклеиновой кислоты-мишени, выбранной из группы, состоящей из положения 371-3068, 5467-12107 и 15317-19511 на SEQ ID NO: 1. В другом воплощении подпоследовательность нуклеиновой кислоты-мишени выбрана из группы, состоящей из положения 371-510, 822-1090, 1992-3068, 5467-5606, 6470-12107, 15317-15720, 15317-18083, 18881-19494 и 1881-19494 на SEQ ID NO: 1. В предпочтительном воплощении подпоследовательность нуклеиновой кислоты-мишени выбрана из группы, состоящей из положения 7300-7333, 8028-8072, 9812-9859, 11787-11873 и 15690-15735 на SEQ ID NO: 1.
В некоторых воплощениях олигонуклеотид или конъюгат олигонуклеотида содержит непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью, как, например, 100%-ной комплементарностью с соответствующей областью нуклеиновой кислоты-мишени, присутствующей в SEQ ID NO: 1, где область нуклеиновой кислоты-мишени выбрана из группы, состоящей из областей от а1 до а449 в Таблице 4.
Таблица 4: области SEQ ID NO 1, на которые может осуществляться нацеливание с использованием олигонуклеотида по изобретению
SEQ ID NO 1
SEQ ID NO 1
В некотором воплощении олигонуклеотид или непрерывная нуклеотидная последователность является комплементарным области нуклеиновой кислоты-мишени, где данная область нуклеиновой кислоты-мишени выбрана из группы, состоящей из a7, a26, a43, a119, a142, a159, a160, a163, a169, a178, a179, a180, a189, a201, a202, a204, a214, a221, a224, a226, a243, a254, a258, 269, a274, a350, a360, a364, a365, a370, a372, a381, a383, a386, a389, a400, a427, a435 и a438.
В предпочтительном воплощении олигонуклеотид или непрерывная нуклеотидная последовательность является комплементарной области нуклеиновой кислоты-мишени, где область нуклеиновой кислоты-мишени выбрана из группы, состоящей из a160, a180, a221, a269 и a360.
В некоторых воплощениях олигонуклеотид по изобретению содержит или состоит из 8-35 нуклеотидов в длину, как, например, от 9 до 30, как, например, от 10 до 22, как, например, от 11 до 20, как, например, от 12 до 18, как, например, от 13 до 17 или от 14 до 16 смежных нуклеотидов в длину. В предпочтительном воплощении олигонуклеотид содержит или состоит из 16-20 нуклеотидов в длину. Следует понимать то, что любой интервал, приведенный в данном документе, включает конечные точки интервала. Соответственно, если говорится то, что олигонуклеотид включает от 10 до 30 нуклеотидов, включены и 10, и 30 нуклеотидов.
В некоторых воплощениях непрерывная нуклеотидная последовательность содержит или состоит из 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 смежных нуклеотидов в длину. В предпочтительном воплощении олигонуклеотид содержит или состоит из 16, 17, 18, 19 или 20 нуклеотидов в длину.
В некоторых воплощениях олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из последовательности, выбранной из группы, состоящей из последовательностей, перечисленных в Таблице 5.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5-743 (см. мотивы последовательностей, перечисленные в Таблице 5).
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5-743 и 771.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 6, 8, 9, 13, 41, 42, 58, 77, 92, 111, 128, 151, 164, 166, 169, 171, 222, 233, 245, 246, 250, 251, 252, 256, 272, 273, 287, 292, 303, 314, 318, 320, 324, 336, 342, 343, 344, 345, 346, 349, 359, 360, 374, 408, 409, 415, 417, 424, 429, 430, 458, 464, 466, 474, 490, 493, 512, 519, 519, 529, 533, 534, 547, 566, 567, 578, 582, 601, 619, 620, 636, 637, 638, 640, 645, 650, 651, 652, 653, 658, 659, 660, 665, 678, 679, 680, 682, 683, 684, 687, 694, 706, 716, 728, 733, 734 и 735.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с SEQ ID NO: 287.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с SEQ ID NO: 342.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с SEQ ID NO: 640.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с SEQ ID NO: 466.
В некоторых воплощениях антисмысловой олигонуклеотид или непрерывная нуклеотидная последовательность содержит или состоит из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной идентичностью, предпочтительно со 100%-ной идентичностью с SEQ ID NO: 566.
В воплощениях, где олигонуклеотид длиннее, чем непрерывная нуклеотидная последовательность (которая комплементарна нуклеиновой кислоте-мишени), мотивы последоваельностей в Таблице 5 образуют часть непрерывной нуклеотидной последовательности антисмысловых олигонуклеотидов по изобретению. В некоторых воплощениях последовательность олигонуклеотида является эквивалентной непрерывной нуклеотидной последовательности (например, если не добавляются биорасщепляемые линкеры).
Понятно то, что непрерывные последовательности из нуклеиновых оснований (мотив последовательности) могут быть модифицированы, например, для увеличения устойчивости к нуклеазе и/или аффинности связывания с нуклеиновой кислотой-мишенью. Модификации описываются в определениях и в разделе «Конструкция олигонуклеотидов». В Таблице 5 перечислены предпочтительные конструкции каждого мотива последовательности.
Конструкция олигонуклеотида
Термин «конструкция олигонуклеотида» относится к картине модификаций сахара нуклеозидов в последовательности олигонуклеотида. Олигонуклеотиды по изобретению содержат нуклеозиды с модифицированным сахаром и также могут содержать нуклеозиды ДНК или РНК. В некоторых воплощениях олигонуклеотид содержит нуклеозиды с модифицированным сахаром и нуклеозиды ДНК. Включение модифицированных нуклеозидов в олигонуклеотид по изобретению может увеличивать аффинность олигонуклеотида в отношении нуклеиновой кислоты-мишени. В данном случае модифицированные нуклеозиды можно называть модифицированными нуклеотидами, увеличивающими аффинность, модифицированные нуклеозиды также можно называть звеньями.
В одном воплощении олигонуклеотид содержит по меньшей мере 1 модифицированный нуклеозид, как, например, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15 или по меньшей мере 16 модифицированных нуклеозидов. В одном воплощении олигонуклеотид содержит от 1 до 10 модифицированных нуклеозидов, как, например, от 2 до 8 модифицированных нуклеозидов, как, например, от 3 до 7 модифицированных нуклеозидов, как, например, от 4 до 6 модифицированных нуклеозидов, как, например, 3, 4, 5, 6 или 7 модифицированных нуклеозидов.
В одном воплощении олигонуклеотид содержит один или более чем один нуклеозид с модифицированным сахаром, как, например, нуклеозиды, модифицированные 2' сахаром. Предпочтительно олигонуклеотид по изобретению содержит один или более чем один нуклеозид, модифицированный 2'-сахаром, независимо выбранный из группы, состоящей из 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-O-алкокси-РНК, 2'-O-метоксиэтил-РНК, 2'-амино-ДНК, 2'-фтор-ДНК, арабинонуклеиновой кислоты (ANA), 2'-фтор-ANA и нуклеозидов LNA. Даже более предпочтительно один или более чем один модифицированный нуклеозид представляет собой запертую нуклеиновую кислоту (LNA).
В другом воплощении олигонуклеотид содержит по меньшей мере одну модифицированную межнуклеозидную связь. В предпочтительном воплощении все межнуклеозидные связи в пределах непрерывной нуклеотидной последовательности представляют собой фосфоротиоатные или боранофосфатные межнуклеозидные связи. В некоторых воплощениях все межнуклеозидные связи в непрерывной последовательности олигонуклеотида представляют собой фосфоротиоатные связи.
В некоторых воплощениях олигонуклеотид по изобретению содержит по меньшей мере один нуклеозид LNA, как, например, 1, 2, 3, 4, 5, 6, 7 или 8 нуклеозидов LNA, как, например, от 2 до 6 нуклеозидов LNA, как, например, от 3 до 7 нуклеозидов LNA, от 4 до 6 нуклеозидов LNA или 3, 4, 5, 6 или 7 нуклеозидов LNA. В некоторых воплощениях по меньшей мере 75% модифицированных нуклеозидов в олигонуклеотиде представляют собой нуклеозиды LNA, как, например, 80%, как, например, 85%, как, например, 90% модифицированных нуклеозидов представляют собой нуклеозиды LNA В еще одном другом воплощении все модифицированные нуклеозиды в олигонуклеотиде представляют собой нуклеозиды LNA. В другом воплощении олигонуклеотид может содержать и бета-D-окси-LNA, и один или более чем один из следующих нуклеозидов LNA: тио-LNA, амино-LNA, окси-LNA и/или ENA либо в бета-D, либо в альфа-L конфигурациях, или их комбинации. В другом воплощении все цитозиновые звенья LNA представляют собой 5-метилцитозин. В предпочтительном воплощении олигонуклеотид или непрерывная нуклеотидная последовательность имеет по меньшей мере 1 нуклеозид LNA на 5'-конце и по меньшей мере 2 нуклеозида LNA на 3'-конце нуклеотидной последовательности.
В некоторых воплощениях олигонуклеотид по изобретению содержит по меньшей мере один модифицированный нуклеозид, который представляет собой нуклеозид 2'-MOE-РНК, как, например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеозидов 2'-MOE-РНК. В некоторых воплощениях по меньшей мере один из указанных модифицированных нуклеозидов представляет собой 2'-фтор-ДНК, как, например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеозидов 2'-фтор-ДНК.
В некоторых воплощениях олигонуклеотид по изобретению содержит по меньшей мере один нуклеозид LNA и по меньшей мере один 2'-замещенный модифицированный нуклеозид.
В некоторых воплощениях изобретения олигонуклеотид содержит и нуклеозиды, модифицированные 2'-сахаром, и звенья ДНК. Предпочтительно олигонуклеотид содержит и нуклеотиды (звенья) LNA, и ДНК. Предпочтительно объединенное общее число звеньев LNA и ДНК составляет 8-30, как, например, 10-25, предпочтительно 12-22, как, например, 12-18, даже более предпочтительно 11-16. В некоторых воплощениях изобретения нуклеотидная последовательность олигонуклеотида, такая как непрерывная нуклеотидная последовательность, состоит из по меньшей мере из одного или двух нуклеотидов LNA, и остальные нуклеозилы представляют собой звенья ДНК. В некоторых воплощениях олигонуклеотид содержит только нуклеозиды LNA и встречающиеся в природе нуклеозиды (такие как нуклеозиды РНК или ДНК, наиболее предпочтительно нуклеозиды ДНК), возможно с модифицированными межнуклеозидными связями, такими как фосфоротиоатная.
В одном воплощении изобретения олигонуклеотид по изобретению способен рекрутировать РНКазу Н.
Структурная конструкция олигонуклеотида по изобретению может быть выбрана из гэпмеров, олигонуклеотидов с разорванным гэпом, хэдмеров и тэйлмеров.
Конструкция гэпмера
В предпочтительном воплощении олигонуклеотид по изобретению имеет конструкцию или структуру гэпмера, также именуемую в данном документе просто «гэпмер». В структуре гэпмера олигонуклеотид содержит по меньшей мере три отличные структурные области: 5'-фланг, гэп и 3'-фланг, F-G-F' в ориентации '5 -> 3'. В данной конструкции фланкирующие области F и F' (также именуемые области крыльев) содержат непрерывный отрезок модифицированных нуклеозидов, которые являются комплементарными нуклеиновой кислоте-мишени PD-L1, тогда как область гэпа - G - содержит непрерывный отрезок нуклеотидов, который способен рекрутировать нуклеазу, предпочтительно эндонуклеазу, такую как РНКаза, например, РНКаза Н, когда олигонуклеотид находится в дуплексе с нуклеиновой кислотой-мишенью. Нуклеозиды, которые способны рекрутировать нуклеазу, в частности РНКазу Н, могут быть выбраны из группы, состоящей из ДНК, альфа-L-окси-LNA, 2'-фтор-ANA и UNA. Области F и F', фланкирующие 5'- и 3'-концы области G, предпочительно содержат нуклеозиды, не рекрутирующие нуклеазу (нуклеозиды с 3'-эндоструктурой), более предпочтительно один или более чем один модифицированный нуклеозид, увеличивающий аффинность. В некоторых воплощениях 3'-фланг содержит по меньшей мере один нуклеозид LNA, предпочтительно по меньшей мере 2 нуклеозида LNA. В некоторых воплощениях 5'-фланг содержит по меньшей мере один нуклеозид LNA. В некоторых воплощениях обе 5'- и 3'-фланкирующие области содержат нуклеозид LNA. В некоторых воплощениях все нуклеозиды в данных фланкирующих областях представляют собой нуклеозиды LNA. В других воплощениях фланкирующие области могут содержать и нуклеозиды LNA, и другие нуклеозиды (смешанные фланги), такие как нуклеозиды ДНК и/или модифицированные нуклеозиды, не являющиеся нуклеозидами LNA, такие как 2'-замещенные нуклеозиды. В данном случае гэп определяется как непрерывная последовательность из по меньшей мере 5 нуклеозидов, рекрутирующих РНКазу Н (нуклеозиды с 2'-эндоструктурой, предпочтительно ДНК), фланкированная на 5'- и 3'-конце модифицированным нуклеозидом, увеличивающим аффинность, предпочтительно LNA, таким как бета-D-окси-LNA. Следовательно, нуклеозиды 5'-фланкирующей области и 3'- фланкирующей области, которые являются смежными с областью гэпа, представляют собой модифицированные нуклеозиды, предпочтительно нуклеозиды, не рекрутирующие нуклеазу.
Область F
Область F (5'-фланг или 5'-крыло), присоединенная к '5-концу области G, включает, содержит или состоит из по меньшей мере одного модифицированного нуклеозида, как, например, из по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7 модифицированных нуклеозидов. В одном воплощении область F содержит или состоит из от 1 до 7 модифицированных нуклеозидов, как, например, из 2-6 модифицированных нуклеозидов, как, например, из 2-6 модифицированных нуклеозидов, как, например, из 2-5 модифицированных нуклеозидов, как, например, из 2-4 модифицированных нуклеозидов, как, например, из 1-3 модифицированных нуклеозидов, как, например, из 1, 2, 3 или 4 модифицированных нуклеозидов. Область F определяется наличием по меньшей мере одного модифицированного нуклеозида на 5'-конце и 3'-конце данной области.
В некоторых воплощениях модифицированные нуклеозиды в области F имеют 3'-эндоструктуру.
В одном воплощении один или более чем один модифицированный нуклеозид в области F представляет собой 2'-модифицированный нуклеозид. В одном воплощении все нуклеозиды в области F представляет собой 2'-модифицированные нуклеозиды.
В другом воплощении область F содержит ДНК и/или РНК, помимо 2'-модифицированных нуклеозидов. Фланги, содержащие ДНК и/или РНК, характеризуются наличием 2'-модифицированного нуклеозида на 5'-конце и 3'-конце (рядом с областью G) области F. В одном воплощении область F содержит нуклеозиды ДНК, как, например, от 1 до 3 смежных нуклеозидов ДНК, как, например, от 1 до 3 или от 1 до 2 смежных нуклеозидов ДНК. Данные нуклеозиды ДНК во флангах предпочтительно не должны быть способны рекрутировать РНКазу Н. В некоторых воплощениях 2'-модифицированные нуклеозиды и нуклеозиды ДНК и/или РНК в области F чередуются с 1-3 2'-модифицированными нуклеозидами и 1-3 нуклеозидами ДНК и/или РНК. Такие фланги также могут называться флангами с чередованием. Длина 5'-фланга (области F) в олигонуклеотидах с флангами с чередованием может составлять от 4 до 10 нуклеозидов, как, например, от 4 до 8, как, например, от 4 до 6 нуклеозидов, как, например, 4, 5, 6 или 7 модифицированных нуклеозидов. В некоторых воплощениях только 5'-фланг олигонуклеотида является флангом с чередованием. Конкретными примерами области F с чередующимися нуклеозидами являются:
2'1-3-N'1-4-2'1-3
2'1-2-N'1-2-2'1-2- N'1-2-2'1-2,
где 2' показывает модифицированный нуклеозид, и N' представляет собой РНК или ДНК. В некоторых воплощениях все модифицированные нуклеозиды во флангах с чередованием представляют собой LNA, а N' представляет собой ДНК. В другом воплощении один или более чем один 2'-модифицированный нуклеозид в области F выбран из звеньев 2'-O-алкил-РНК, 2'-O-метил-РНК, звеньев 2'-амино-ДНК, звеньев 2'-фтор-ДНК, 2'-алкокси-РНК, звеньев МОЕ, звеньев LNA, звеньев арабинонуклеиновой кислоты (ANA) и звеньев 2'-фтор-ANA.
В некоторых воплощениях область F содержит и нуклеозид LNA, и 2'-замещенный модифицированный нуклеозид. Они часто называются олигонуклеотидами со смешанными крыльями или со смешанными флангами.
В одном воплощении изобретения все модифицированные нуклеозиды в области F представляют собой нуклеозиды LNA. В другом воплощении все нуклеозиды в области F представляют собой нуклеозиды LNA. В другом воплощении нуклеозиды LNA в области F независимо выбраны из группы, состоящей из окси-LNA, тио-LNA, амино-LNA, cET и/или ENA либо в бета-D, либо в альфа-L конфигурациях, или их комбинации. В предпочтительном воплощении область F содержит по меньшей мере 1 звено бета-D-окси LNA на 5'-конце непрерывной последовательности.
Область G
Область G (область гэпа) предпочтительно включает, содержит или состоит из по меньшей мере 4, как, например, из по меньшей мере 5, как, например, из по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15 или по меньшей мере 16 последовательных нуклеозидов, способных рекрутировать вышеупомянутую нуклеазу, в частности РНКазу Н. В другом воплощении область G включает, содержит или состоит из от 5 до 12 или от 6 до 10, или от 7 до 9, как, например, из 8 последовательных нуклеотидных звеньев, способных рекрутировать вышеупомянутую нуклеазу.
Нуклеозидные звенья в области G, которые способны рекрутировать нуклеазу, в одном воплощении выбраны из группы, состоящей из ДНК, альфа-L-LNA, C4'-алкилированой ДНК (как описано в PCT/EP2009/050349 и Vester et al., Bioorg. Med. Chem. Lett. 18 (2008) 2296 - 2300, которые обе включены в данный документ посредством ссылки), нуклеозидов, происходящих из арабинозы, подобных ANA и 2'F-ANA (Mangos et al. 2003 J. AM. CHEM. SOC. 125, 654-661), UNA (незапертая нуклеиновая кислота) (как описано в Fluiter et al., Mol. Biosyst., 2009, 10, 1039, включенной в данный документе посредством ссылки). UNA представляет собой незапертую нуклеиновую кислоту, типично где связь между С2 и С3 рибозы была удалена, образуя незапертый «сахарный» остаток.
В еще одном другом воплощении по меньшей мере одно нуклеозидное звено в области G представляет собой нуклеозидное звено ДНК, как, например, от 1 до 18 звеньев ДНК, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 звеньев ДНК, предпочтительно от 2 до 17 звеньев ДНК, как, например, от 3 до 16 звеньев ДНК, как, например, от 4 до 15 звеньев ДНК, как, например, от 5 до 14 звеньев ДНК, как, например, от 6 до 13 звеньев ДНК, как, например, от 7 до 12 звеньев ДНК, как, например, от 8 до 11 звеньев ДНК, более предпочтительно от 8 до 17 звеньев ДНК или от 9 до 16 звеньев ДНК, от 10 до 15 звеньев ДНК или от 11 до 13 звеньев ДНК, как, например, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 звеньев ДНК. В некоторых воплощениях область G состоит из 100% звеньев ДНК.
В других воплощениях область G может состоять из смеси ДНК и других нуклеозидов, способных опосредовать расщепление РНКазой Н. Область G может состоять из по меньшей мере 50% ДНК, более предпочтительно - из 60%, 70% или 80% ДНК, и даже более предпочтительно из 90% или 95% ДНК.
В еще одном другом воплощении по меньшей мере одно нуклеозидное звено в области G представляет собой нуклеозидное звено альфа-L-LNA, как, например, по меньшей мере одно звено альфа-L-LNA, как, например, 2, 3, 4, 5, 6, 7, 8 или 9 звеньев альфа-L-LNA. В другом воплощении область G содержит по меньшей мере одно звено альфа-L-LNA, которое представляет собой звено альфа-L-окси-LNA. В другом воплощении область G содержит комбинацию нуклеозидных звеньев ДНК и альфа-L-LNA.
В некоторых воплощениях нуклеозиды в области G имеют 2'-эндоструктуру.
В некоторых воплощениях область G может содержать нуклеозид, разрывающий гэп, приводящий к олигонуклеотиду с разорванным гэпом, который способен рекрутировать РНКазу Н.
Область F'
Область F' (3'-фланг или 3'-крыло), присоединенная к '3-концу области G, включает, содержит или состоит из по меньшей мере одного модифицированного нуклеозида, как, например, из по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7 модифицированных нуклеозидов. В одном воплощении область F' содержит или состоит из от 1 до 7 модифицированных нуклеозидов, как, например, из 2-6 модифицированных нуклеозидов, как, например, из 2-4 модифицированных нуклеозидов, как, например, из 1-3 модифицированных нуклеозидов, как, например, из 1, 2, 3 или 4 модифицированных нуклеозидов. Область F' определяется наличием по меньшей мере одного модифицированного нуклеозида на 5'-конце и на 3'-конце данной области.
В некоторых воплощениях модифицированные нуклеозиды в области F' имеют 3'-эндоструктуру.
В одном воплощении один или более чем один модифицированный нуклеозид в области F' представляет собой 2'-модифицированный нуклеозид. В одном воплощении все нуклеозиды в области F' представляют собой 2'-модифицированные нуклеозиды.
В одном воплощении один или более чем один модифицированный нуклеозид в области F' представляет собой 2'-модифицированный нуклеозид.
В одном воплощении все нуклеозиды в области F' представляет собой 2'-модифицированные нуклеозиды. В другом воплощении область F' содержит ДНК и/или РНК, помимо 2'-модифицированных нуклеозидов. Фланги, содержащие ДНК и/или РНК, характеризуются наличием 2'-модифицированного нуклеозида на 5'-конце (смежном с областью G) и 3'-конце области F'. В одном воплощении область F' содержит нуклеозиды ДНК, как, например, от 1 до 4 смежных нуклеозидов ДНК, как, например, от 1 до 3 или от 1 до 2 смежных нуклеозидов ДНК. Данные нуклеозиды ДНК во флангах предпочтительно не должны быть способны рекрутировать РНКазу Н. В некоторых воплощениях 2'-модифицированные нуклеозиды и нуклеозиды ДНК и/или РНК в области F' чередуются с 1-3 2'-модифицированными нуклеозидами и 1-3 нуклеозидами ДНК и/или РНК, причем такие фланги также могут называться флангами с чередованием. Длина 3'-фланга (области F') в олигонуклеотидах с флангами с чередованием может составлять от 4 до 10 нуклеозидов, как, например, от 4 до 8, как, например, от 4 до 6 нуклеозидов, как, например, 4, 5, 6 или 7 модифицированных нуклеозидов. В некоторых воплощениях только 3'-фланг олигонуклеотида является флангом с чередованием. Конкретными примерами области F' с чередующимися нуклеозидами являются:
2'1-3-N'1-4-2'1-3
2'1-2-N'1-2-2'1-2- N'1-2-2'1-2,
где 2' показывает модифицированный нуклеозид, и N' представляет собой РНК или ДНК. В некоторых воплощениях все модифицированные нуклеозиды во флангах с чередованием представляют собой LNA, а N' представляет собой ДНК. В другом воплощении модифицированные нуклеозиды в области F' выбраны из звеньев 2'-O-алкил-РНК, 2'-O-метил-РНК, звеньев 2'-амино-ДНК, звеньев 2'-фтор-ДНК, 2'-алкокси-РНК, звеньев МОЕ, звеньев LNA, звеньев арабинонуклеиновой кислоты (ANA) и звеньев 2'-фтор-ANA.
В некоторых воплощениях область F' содержит и LNA и 2'-замещенный модифицированный нуклеозид. Они часто называются олигонуклеотидами со смешанными крыльями или со смешанными флангами.
В одном воплощении изобретения все модифицированные нуклеозиды в области F' представляют собой нуклеозиды LNA. В другом воплощении все нуклеозиды в области F' представляют собой нуклеозиды LNA. В другом воплощении нуклеозиды LNA в области F' независимо выбраны из группы, состоящей из окси-LNA, тио-LNA, амино-LNA, cET и/или ENA либо в бета-D, либо в альфа-L конфигурациях, или их комбинации. В предпочтительном воплощении область F' имеет по меньшей мере 2 звена бета-D-окси LNA на 3'-конце непрерывной последовательности.
Область D' и D''
Область D' и D'' может быть присоединена к 5'-концу области F или к 3'-концу области F' соответственно. Область D' или D'' является опцией.
Область D' и D'' может независимо содержать от 0 до 5, как, например, от 1 до 5, как, например, от 2 до 4, как, например, 0, 1, 2, 3, 4 или 5 дополнительных нуклеотидов, которые могут быть комплементарными или некомплементарными нуклеиновой кислоте-мишени. В данном отношении олигонуклеотид по изобретению может в некоторых воплощениях содержать непрерывную нуклеотидную последовательность, способную модулировать мишень, которая фланкирована на 5'- и/или 3'-конце дополнительными нуклеотидами. Такие дополнительные нуклеотиды могут служить в качестве нуклеазочувствительного биорасщепляемого линкера (см. определение линкеров). В некоторых воплощениях дополнительные 5'- и/или 3'-концевые нуклеозиды связаны фосфодиэфирными связями и могут представлять собой ДНК или РНК. В другом воплощении дополнительные 5'- и/или 3'-концевые нуклеозиды представляют собой модифицированные нуклеозиды, которые могут быть включены, например, для увеличения нуклеазоустойчивости или для легкости синтеза. В одном воплощении олигонуклеотид по изобретению содержит область D' и/или D'' на 5'- или 3'-конце непрерывной нуклеотидной последовательности. В другом воплощении область D' и/или D'' состоит из 1-5 нуклеозидов ДНК или РНК, связанных фосфодиэфирными связями, которые не являются комплементарными нуклеиновой кислоте-мишени.
Гэпмерный олигонуклеотид по настоящему изобретению может быть представлен следующими формулами:
3'-F-G-F'-3'; в частности, F1-7-G4-12-F'1-7
5'-D'-F-G-F'-3', в частности, D'1-3-F1-7-G4-12-F'1-7
5'-F-G-F'-D''-3', в частности, F1-7-G4-12-F'1-7-D''1-3
5'-D'-F-G-F'-D'-3'', в частности, D'1-3-F1-7-G4-12-F'1-7-D''1-3
Предпочтительное число и типы нуклеозидов в областях F, G и F', D' и D'' были описаны выше. Конъюгаты олигонуклеотидов по настоящему изобретению имеют область С, ковалентно присоединенную либо к 5'-, либо к 3'-концу олигонуклеотида, в частности гэпмерных олигонуклеотидов, представленных выше.
В одном воплощении конъюгат олигонуклеотида по изобретению содержит олигонуклеотид с формулой 5'-D'-F-G-F'-3' или 5'-F-G-F'-D''-3', где области F и F' независимо содержат 1-7 модифицированных нуклеозидов, G представляет собой область из 6-16 нуклеозидов, которые способны рекрутировать РНКазу Н, и область D' или D'' содержит 1-5 нуклеозидов, связанных фосфодиэфирными связями. Предпочтительно область D' или D'' присутствует в конце олигонуклеотида, где рассматривается конъюгирование с конъюгатной группировкой.
Примеры олигонуклеотидов с флангами с чередованием могут быть представлены следующими формулами:
2'1-3-N'1-4-2'1-3-G6-12-2'1-2-N'1-4-2'1-4
2'1-2-N'1-2-2'1-2-N'1-2-2'1-2-G6-12-2'1-2-N'1-2-2'1-2- N'1-2-2'1-2
F-G6-12-2'1-2-N'1-4-2'1-4
F-G6-12-2'1-2-N'1-2-2'1-2-N'1-2-2'1-2
2'1-3-N'1-4-2'1-3-G6-12-F'
2'1-2-N'1-2-2'1-2-N1-2-2'1-2-G6-12-F'
где фланг указан F или F', если он содержит только 2'-модифицированные нуклеозиды, такие как нуклеозиды LNA. Предпочтительное число и типы нуклеозидов в областях с чередованием и области F, G и F', D' и D'' были описаны выше.
В некоторых воплощениях олигонуклеотид представляет собой гэпмер, состоящий из 16, 17, 18, 19, 20, 21, 22 нуклеотидов в длину, где каждая из областей F и F' независимо состоит из 1, 2, 3 или 4 модифицированных нуклеозных звеньев, комплементарных нуклеиновой кислоте-мишени PD-L1, и область G состоит из 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 нуклеозидных звеньев, способных рекрутировать нуклеазу при нахождении в дуплексе с нуклеиновой кислотой-мишенью PD-L1, и область D' состоит из 2 ДНК, связанных фосфодиэфирной связью.
В других воплощениях олигонуклеотид представляет собой гэпмер, в котором каждая из областей F и F' независимо состоит из 3, 4, 5 или 6 модифицированных нуклеозных звеньев, таких как нуклеозидные звенья, содержащие сахар 2'-O-метоксиэтил-рибозу (2'-MOE), или нуклеозидные звенья, содержащие сахар 2'-фтор-дезоксирибозу, и/или звенья LNA, и область G состоит из 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 нуклеозидных звеньев, таких как звенья ДНК или другие нуклеозиды, рекрутирующие нуклеазу, такие как L-LNA или смесь нуклеозидов ДНК и нуклеозидов, рекрутирующих нуклеазу.
В другом конкретном воплощении олигонуклеотид представляет собой гэпмер, в котором каждая из областей F и F' состоит из двух звеньев LNA, и область G состоит из 12, 13, 14 нуклеозидных звеньев, предпочтительно звеньев ДНК. Конкретные конструкции гэпмеров данной природы включают 2-12-2, 2-13-2 и 2-14-2.
В другом конкретном воплощении олигонуклеотид представляет собой гэпмер, в котором каждая из областей F и F' независимо состоит из трех звеньев LNA, и область G состоит из 8, 9, 10, 11, 12, 13 или 14 нуклеозидных звеньев, предпочтительно звеньев ДНК. Конкретные конструкции гэпмеров данной природы включают 3-8-3, 3-9-3 3-10-3, 3-11-3, 3-12-3, 3-13-3 и 3-14-3.
В другом конкретном воплощении олигонуклеотид представляет собой гэпмер, в котором каждая из областей F и F' состоит из четырех звеньев LNA, и область G состоит из 8 или 9, 10, 11 или 12 нуклеозидных звеньев, предпочтительно звеньев ДНК. Конкретные конструкции гэпмеров данной природы включают 4-8-4, 4-9-4, 4-10-4, 4-11-4 и 4-12-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 6 нуклеозидами и, независимо, из 1-4 модифицированных нуклеозидов в крыльях, включая гэпмеры 1-6-1, 1-6-2, 2-6-1, 1-6-3, 3-6-1, 1-6-4, 4-6-1, 2-6-2, 2-6-3, 3-6-2 2-6-4, 4-6-2, 3-6-3, 3-6-4 и 4-6-3.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 7 нуклеозидами и, независимо, из 1-4 модифицированных нуклеозидов в крыльях, включая гэпмеры 1-7-1, 2-7-1, 1-7-2, 1-7-3, 3-7-1, 1-7-4, 4-7-1, 2-7-2, 2-7-3, 3-7-2, 2-7-4, 4-7-2, 3-7-3, 3-7-4, 4-7-3 и 4-7-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 8 нуклеозидами и, независимо, из 1-4 модифицированных нуклеозидов в крыльях, включая гэпмеры 1-8-1, 1-8-2, 1-8-3, 3-8-1, 1-8-4, 4-8-1,2-8-1, 2-8-2, 2-8-3, 3-8-2, 2-8-4, , 4-8-2, 3-8-3, 3-8-4, 4-8-3 и 4-8-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 9 нуклеозидами и, независимо, из 1-4 модифицированных нуклеозидов в крыльях, включая гэпмеры 1-9-1, 2-9-1, 1-9-2, 1-9-3, 3-9-1, 1-9-4, 4-9-1, 2-9-2, 2-9-3, 3-9-2, 2-9-4, 4-9-2, 3-9-3, 3-9-4, 4-9-3 и 4-9-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 10 нуклеозидами, включая гэпмеры 1-10-1, 2-10-1, 1-10-2, 1-10-3, 3-10-1, 1-10-4, 4-10-1, 2-10-2, 2-10-3, 3-10-2, 2-10-4, 4-10-2, 3-10-3, 3-10-4, 4-10-3 и 4-10-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 11 нуклеозидами, включая гэпмеры 1-11-1, 2-11-1, 1-11-2, 1-11-3, 3-11-1, 1-11-4, 4-11-1, 2-11-2, 2-11-3, 3-11-2, 2-11-4, 4-11-2, 3-11-3, 3-11-4, 4-11-3 и 4-11-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 12 нуклеозидами, включая гэпмеры 1-12-1, 2-12-1, 1-12-2, 1-12-3, 3-12-1, 1-12-4, 4-12-1, 2-12-2, 2-12-3, 3-12-2, 2-12-4, 4-12-2, 3-12-3, 3-12-4, 4-12-3 и 4-12-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 13 нуклеозидами, включая гэпмеры 1-13-1, 2-13-1, 1-13-2, 1-13-3, 3-13-1, 1-13-4, 4-13-1, 2-13-2, 2-13-3, 3-13-2, 2-13-4, 4-13-2, 3-13-3, 3-13-4, 4-13-3 и 4-13-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 14 нуклеозидами, включая гэпмеры 1-14-1, 2-14-1, 1-14-2, 1-14-3, 3-14-1, 1-14-4, 4-14-1, 2-14-2, 2-14-3, 3-14-2, 2-14-4, 4-14-2, 3-14-3, 3-14-4, 4-14-3 и 4-14-4.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 15 нуклеозидами, включая гэпмеры 1-15-1, 2-15-1, 1-15-2, 1-15-3, 3-15-1, 1-15-4, 4-15-1, 2-15-2, 2-15-3, 3-15-2, 2-15-4, 4-15-2 и 3-15-3.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 16 нуклеозидами, включая гэпмеры 1-16-1, 2-16-1, 1-16-2, 1-16-3, 3-16-1, 1-16-4, 4-16-1, 2-16-2, 2-16-3, 3-16-2, 2-16-4, 4-16-2 и 3-16-3.
Конкретные конструкции гэпмеров данной природы включают конструкции F-G-F', выбранные из группы, состоящей из гэпа с 17 нуклеозидами, включая гэпмеры 1-17-1, 2-17-1, 1-17-2, 1-17-3, 3-17-1, 1-17-4, 4-17-1, 2-17-2, 2-17-3 и 3-17-2.
Во всех случаях конструкция F-G-F' может дополнительно включать область D' и/или D'', которая может иметь 1, 2 или 3 нуклеозидных звена, таких как звенья ДНК, таких как 2 звена ДНК, связанных 2-фосфодиэфиром. Предпочтительно нуклеозиды в области F и F' представляют собой модифицированные нуклеозиды, тогда как нуклеотиды в области G предпочтительно представляют собой немодифицированные нуклеозиды.
При каждой конструкции предпочтительным модифицированным нуклеозидом является LNA.
В другом воплощении все межнуклеозидные связи в гэпе в гэпмере представляют собой фосфоротиоатные и/или боранофосфатные связи. В другом воплощении все межнуклеозидные связи во флангах (области F и F') в гэпмере представляют собой фосфоротиоатные и/или боранофосфатные связи. В другом предпочтительном воплощении все межнуклеозидные связи в области D' и D'' в гэпмере представляют собой фосфодиэфирные связи.
Для конкретных гэпмеров, как раскрыто в данном документе, при аннотации остатков цитозина (С) как 5-метил-цитозин в разных воплощениях один или более чем один С, присутствующий в олигонуклеотиде, может представлять собой немодифицированный остаток С.
В конкретном воплощении гэпмер представляет собой так называемый шортмер, как описано в WO2008/113832, включенной в данный документ посредством ссылки.
Дополнительные конструкции гэпмеров раскрыты в WO2004/046160, WO2007/146511 и включены посредством ссылки.
Для некоторых воплощений изобретения олигонуклеотид выбран из группы олигонуклеотидных соединений с CMP-ID-NO от 5_1 до 743_1 и 771_1.
Для некоторых воплощений изобретения олигонуклеотид выбран из группы олигонуклеотидных соединений с CMP-ID-NO 6_1, 8_1, 9_1, 13_1, 41_1, 42_1, 58_1, 77_1, 92_1, 111_1, 128_1, 151_1, 164_1, 166_1, 169_1, 171_1, 222_1, 233_1, 245_1, 246_1, 250_1, 251_1, 252_1, 256_1, 272_1, 273_1, 287_1, 292_1, 303_1, 314_1, 318_1, 320_1, 324_1, 336_1, 342_1, 343_1, 344_1, 345_1, 346_1, 349_1, 359_1, 360_1, 374_1, 408_1, 409_1, 415_1, 417_1, 424_1, 429_1, 430_1, 458_1, 464_1, 466_1, 474_1, 490_1, 493_1, 512_1, 519_1, 519_1, 529_1, 533_1, 534_1, 547_1, 566_1, 567_1, 578_1, 582_1, 601_1, 619_1, 620_1, 636_1, 637_1, 638_1, 640_1, 645_1, 650_1, 651_1, 652_1, 653_1, 658_1, 659_1, 660_1, 665_1, 678_1, 679_1, 680_1, 682_1, 683_1, 684_1, 687_1, 694_1, 706_1, 716_1, 728_1, 733_1, 734_1, and 735_1.
В одном предпочтительном воплощении изобретения олигонуклеотид представляет собой CMP-ID-NO: 287_1.
В другом предпочтительном воплощении изобретения олигонуклеотид представляет собой CMP-ID-NO: 342_1.
В другом предпочтительном воплощении изобретения олигонуклеотид представляет собой CMP-ID-NO: 640_1.
В другом предпочтительном воплощении изобретения олигонуклеотид представляет собой CMP-ID-NO: 466_1.
В другом предпочтительном воплощении изобретения олигонуклеотид представляет собой CMP-ID-NO: 566_1.
В другом воплощении изобретения непрерывная нуклеотидная последовательность олигонуклеотидных мотивов и олигонуклеотидных соединений по изобретению содержит от двух до четырех дополнительных нуклеозидов, связанных фосфодиэфирной связью, на 5'-конце непрерывной нуклеотидной последовательности (например, области D'). В одном воплощении нуклеозиды служат в качестве биорасщепляемого линкера (см. раздел «Биорасщепляемые линкеры»). В предпочтительном воплощении динуклеотид са (цитидин-аденозин) связан с 5'-концом непрерывной нуклеотидной последовательности (т.е. с любым из мотивов последовательностей или олигонуклеотидных соединений, перечисленных в Таблице 5) посредством фосфодиэфирной связи. В предпочтительном воплощении динуклеотид са не является комплементарным последовательности-мишени в положении, где остальная часть непрерывного нуклеотида является комплементарной.
В некоторых воплощениях изобретения олигонуклеотид или непрерывная нуклеотидная последовательность выбрана из группы, состоящей из последовательностей нуклеотидных мотивов с SEQ ID NO: 766, 767, 768, 769 и 770.
В некоторых воплощениях изобретения олигонуклеотид выбран из группы, состоящей из олигонуклеотидных соединений с CMP-ID-NO 766_1, 767_1, 768_1, 769_1 и 770_1.
Углеводные конъюгатные группировки
Углеводные конъюгатные группировки включают галактозу, лактозу, N-ацетилгалактозамин, маннозу и моннозо-6-фосфат, но не ограничиваются ими. Углеводные конъюгаты могут использоваться для увеличения доставки или активности в целом ряде тканей, таких как печень и/или мышцы. См., например, EP1495769, WO99/65925, Yang et al., Bioconjug Chem (2009) 20(2): 213-21. Zatsepin & Oretskaya Chem Biodivers. (2004) 1(10): 1401-17.
В некоторых воплощениях углеводные конъюгатные группировки являются многовалентными, таким образом, что, например, 2, 3 или 4 идентичные или неидентичные углеводные группировки могут быть ковалентно связаны с олигонуклеотидом, возможно через линкер или линкеры. В некоторых воплощениях согласно изобретению предложен конъюгат, содержащий олигонуклеотид по изобретению и углеводную конъюгатную группировку. В некоторых воплощениях конъюгатная группировка представляет собой или может содержать маннозу или маннозо-6-фосфат. Это особенно полезно для нацеливания в мышечные клетки, см., например, US 2012/122801.
Конъюгатные группировки, способные к связыванию с рецептором асиалогликопротеина (ASGPr) являются особенно полезными для нацеливания в гепатоциты в печени. В некоторых воплощениях согласно изобретению предложен конъюгат олигонуклеотида, содержащий олигонуклеотид по изобретению и конъюгатную группировку, нацеленную на рецептор асиалогликопротеина. Конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, содержит одну или более чем одну углеводную группировку, способную к связыванию с рецептором асиалогликопротеина (углеводные группировки, связывающиеся с ASGPr) с аффинностью, равной или большей, чем аффинность галактозы. Аффинности многочисленных производных галактозы в отношении рецептора асиалогликопротеина были исследованы (см., например, Jobst, S.T. and Drickamer, K. JB.C. 1996, 271, 6686) или легко определяются с использованием способов, типичных в данной области.
Одним аспектом настоящего изобретения является конъюгат антисмыслового олигонуклеотида, содержащий а) олигонуклеотид (область А), содержащий непрерывную нуклеотидную последовательность из 10-30 нуклеотидов в длину с по меньшей мере 90%-ной комплементарностью к нуклеиновой кислоте-мишени PD-L1; и б) по меньшей мере одну конъюгатную группировку, нацеленную на рецептор асиалогликопротеина (область С), ковалентно присоединенную к олигонуклеотиду в а). Олигонуклеотид или непрерывная нуклеотидная последовательность может быть такой, как описано в любом из разделов «Олигонуклеотиды по изобретению», «Конструкция олигонуклеотидов» и «Конструкция гэпмеров».
В некоторых воплощениях конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, содержит по меньшей мере одну углеводную группировку, связывающуюся с ASPGr, выбранную из группы, состоящей из галактозы, галактозамина, N-формил-галактозамина, N-ацетилгалактозамина, N-пропионил-галактозамина, N-н-бутаноил-галактозамина и N-изобутаноилгалактозамина. В некоторых воплощениях конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, является одновалентной, двухвалентной, трехвалентной или четырехвалентной (т.е. содержащей 1, 2, 3 или 4 концевые углеводные группировки, способные к связыванию с рецептором асиалогликопротеина). Предпочтительно конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, является двухвалентной, даже более предпочтительно она является трехвалентной. В предпочтительном воплощении конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, содержит 1-3 группировки N-ацетилгалактозамина (GalNAc) (также именуемая GalNAc конъюгат). В некоторых воплощениях конъюгат олигонуклеотида содержит конъюгатную группировку, нацеленную на рецептор асиалогликопротеина, которая является трехвалентной N-ацетилгалактозаминной (GalNAc) группировкой. GalNAc конъюгаты использовали с фосфодиэфирными, метилфосфонатными и PNA антисмысловыми олигонуклеотидами (например, US 5994517 и Hangeland et al., Bioconjug Chem. 1995 Nov-Dec;6(6):695-701, Biessen et al 1999 Biochem J. 340, 783-792, и Maier et al 2003 Bioconjug Chem 14, 18-29 ) и миРНК (например, WO 2009/126933, WO 2012/089352 и WO 2012/083046) и с модифицированными нуклеозидами LNA и 2'-MOE (WO 2014/076196, WO 2014/207232 и WO 2014/179620, включенные тем самым посредством ссылки).
Для получения конъюгатной группировки, нацеленной на рецептор асиалогликопротеина, углеводные группировки, связывающиеся с ASPGr (предпочтительно GalNAc), присоединяются к разветвляющей молекуле через C-1 углероды сахаридов. Углеводные группировки, связывающиеся с ASPGr, предпочтительно связываются с разветвляющей молекулой через спейсеры. Предпочтительным спейсером является гибкий гидрофильный спейсер (патент США 5885968; Biessen et al. J. Med. Chern. 1995 Vol. 39 p. 1538-1546). Предпочтительным гибким гидрофильным спейсером является ПЭГ (полиэтиленгликоль) спейсер. Предпочтительным ПЭГ спейсером является ПЭГ3 спейсер (три этиленовых звена). Разветвляющей молекулой может быть любая маленькая молекула, которая допускает присоединение двух или трех концевых углеводных группировок, связывающихся с ASPGr, и дополнительно обеспечивает присоединение точки разветвления к олигонуклеотиду. Типичной разветвляющей молекулой является дилизин. Молекула дилизина содержит три аминогруппы, посредством которых три углеводные группировки, связывающиеся с ASPGr, могут быть присоединены к карбоксильной реакционноспособной группе, через которую дилизин может быть присоединен к олигонуклеотиду. Альтернативными разветвляющими молекулами могут быть удвоитель или утроитель, такие как поставляемые Glen Research. В некоторых воплощениях разветвляющая молекула может быть выбрана из группы, состоящей из 1,3-бис-[5-(4,4'-диметокситритилокси)пентиламидо]пропил-2-[(2-цианоэтил)-(N,N-диизопропил)] фосфорамидита (каталожный номер Glen Research: 10-1920-xx), три-2,2,2-[3-(4,4'-диметокситритилокси)пропилоксиметил]этил-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита (каталожный номер Glen Research: 10-1922-xx), трис-2,2,2-[3-(4,4'- диметокситритилокси)пропилоксиметил]метиленоксипропил-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита и 1-[5-(4,4'-диметокси-тритилокси)пентиламидо]-3-[5-флюоренометокси-карбонил-окси-пентиламидо]-пропил-2-[(2-цианоэтил)-(N,N-диизопропил)]-фосфорамидита (каталожный номер Glen Research: 10-1925-xx). В WO 2014/179620 и заявке PCT № PCT/EP2015/073331 описано получение разных конъюгатных группировок GalNAc (включенных тем самым посредством ссылки). Между разветвляющей молекулой и олигонуклеотидом может быть вставлен один или более чем один линкер. В предпочтительном воплощении линкер представляет собой биорасщепляемый линкер. Линкер может быть выбран из линкеров, описанных в разделе «Линкеры» и его подразделах.
Конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, в частности, конъюгатная группировка GalNAc, может быть присоединена к 3'- или 5'-концу олигонуклеотида с использованием способов, известных в данной области. В предпочтительных воплощениях конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, связывается с 5'-концом олигонуклеотида.
Модуляторы фармакокинетики в отношении доставки миРНК были описаны в WO 2012/083046 (включенной тем самым посредством ссылки). В некоторых воплощениях углеводная конъюгатная группировка содержит модулятор фармакокинетики, выбранный из группы, состоящей из гидрофобной группы, имеющей 16 или более атомов углерода, гидрофобной группы, имеющей 16-20 атомов углерода, пальмитоила, гексадек-8-еноила, олеила, (9Е,12Е)-октадека-9,12-диеноила, диоктаноила, С16-С20-ацила и холестерина. В предпочтительном воплощении углеводная конъюгатная группировка, содержащая модулятор фармакокинетики, представляет собой GalNAc конъюгат.
Предпочтительные углеводные конъюгатные группировки содержат от одной до трех концевых углеводных группировок, связывающихся с ASPGr, предпочтительно группировку(ки) N-ацетилгалактозамина. В некоторых воплощениях углеводная конъюгатная группировка содержит три углеводные группировки, связывающиеся с ASPGr, предпочтительно N-ацетилгалактозаминные группировки, связанные через спейсер с разветвляющей молекулой. Спейсерная молекула может иметь длину от 8 до 30 атомов. Предпочтительная углеводная конъюгатная группировка содержит три концевые GalNAc группировки, связанные через ПЭГ спейсер с разветвляющей молекулой дилизином. Предпочтительно ПЭГ спейсер представляет собой 3ПЭГ. Подходящие конъюгатные группировки, нацеленные на рецептор асиалогликопротеина, показаны на Фиг. 1. Предпочтительная конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, показана на Фиг. 3.
Другие GalNAc конъюгатные группировки могут включать, например, маленькие пептиды с присоединенными GalNAc группировками, такие как Tyr-Glu-Glu-(аминогексил-GalNAc)3 (YEE(ahGalNAc)3; гликотрипептид, который связывается с рецептором асиалогликопротеина на гепатоцитах, см., например, Duff, et al., Methods Enzymol, 2000, 313, 297); галактозные кластеры на основе лизина (например, L3G4; Biessen, et al., Cardovasc. Med., 1999, 214); и галактозные кластеры на основе холана (например, мотив распознавания углевода для рецептора асиалогликопротеина).
В некоторых воплощениях изобретения конъюгат антисмыслового олигонуклеотида выбран из группы, состоящей из следующих CPM ID NO: 766_2, 767_2, 768_2, 769_2 и 770_2.
В предпочтительном воплощении конъюгат антисмыслового олигонуклеотида соответствует соединению, представленному на Фиг. 4.
В другом предпочтительном воплощении конъюгат антисмыслового олигонуклеотида соответствует соединению, представленному на Фиг. 5.
В другом предпочтительном воплощении конъюгат антисмыслового олигонуклеотида соответствует соединению, представленному на Фиг. 6.
В другом предпочтительном воплощении конъюгат антисмыслового олигонуклеотида соответствует соединению, представленному на Фиг. 7.
В другом предпочтительном воплощении конъюгат антисмыслового олигонуклеотида соответствует соединению, представленному на Фиг. 8.
Линкеры
Биорасщепляемые линкеры (область В)
Применение конъюгата часто ассоциировано с улучшенными фармакокинетическими или фармакодинамическими свойствами. Однако присутствие конъюгатной группировки может препятствовать активности олигонуклеотида против намеченной мишени, например посредством стерического препятствия, предотвращающего гибридизацию или рекрутирование нуклеазы (например, РНКазы Н). Применение физиологически лабильной связи (биорасщепляемого линкера) между олигонуклеотидом (область А или первая область) и конъюгатной группировкой (область С или третья область) обеспечивает улучшенне свойства из-за присутствия конъюгатной группировки при обеспечении того, что при нахождении в ткани-мишени конъюгатная группа не предотвращает эффективную активность олигонуклеотида.
Расщепление физиологически лабильной связи происходит спонтанно при достижении молекулой, содержащей лабильную связь, подходящей внутри- и/или внеклеточной среды. Например, рН-лабильная связь может расщепляться при поступлении молекулы в подкисленную эндосому. Таким образом, рН-лабильная связь может рассматриваться как связь, расщепляемая в эндосоме. Связи, расщепляемые ферментом, могут расщепляться при воздействии ферментов, таких как ферметы, присутствующие в эндосоме или в лизосоме, или в цитоплазме. Дисульфидная связь может расщепляться при поступлении молекулы в более восстанавливающую среду цитоплазмы клетки. Таким образом, дисульфид может рассматриваться как связь, расщепляемая в цитоплазме. рН-лабильная связь в том виде, в котором данный термин используется в данном документе, представляет собой лабильную связь, которая селективно расщепляется при кислотных условиях (рН меньше 7). Такие связи также могут называться эндосомно-лабильными связыми, так как клеточные эндосомы и лизосомы имеют рН меньше, чем 7.
Для биорасщепляемых линкеров, ассоциированных с конъюгатной группировкой для таргетной доставки, предпочтительным является то, что скорость расщепления, наблюдаемая в ткани-мишени (например, в мышце, печени, почке или опухоли), была больше, чем наблюдающаяся в сыворотке крови. Подходящие способы для определения уровня (%) расщепления в ткани-мишени относительно сыворотки или расщепления нуклеазой S1 описываются в разделе «Материаы и методы». В некоторых воплощениях биорасщепляемый линкер (также именуемый физиологически лабильный линкер или нуклеазочувствительный линкер, или область В) в конъюгате по изобретению расщепляется по меньшей мере примерно на 20%, как, например, расщепляется по меньшей мере примерно на 30%, как, например, расщепляется по меньшей мере примерно на 40%, как, например, расщепляется по меньшей мере примерно на 50%, как, например, расщепляется по меньшей мере примерно на 60%, как, например, расщепляется по меньшей мере примерно на 70%, как, например, расщепляется по меньшей мере примерно на 75% по сравнению со стандартом.
В некоторых воплощениях конъюгат олигонуклеотида по изобретению содержит три области: i) первую область (область А), которая содержит 10-25 смежных нуклеотидов, комплементарных нуклеиновой кислоте-мишени; ii) вторую область (область В), которая содержит биорасщепляемый линкер, и iii) третью область (область С), которая содержит конъюгатную группировку, такую как конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, где третья область ковалентно связана со второй областью, которая ковалентно связана с первой областью.
В одном воплощении настоящего изобретения конъюгат олигонуклеотида содержит биорасщепляемый линкер (область В) между непрерывной нуклеотидной последовательностью (область А) и конъюгатной группировкой, нацеленной на рецептор асиалогликопротеина (область С).
В некоторых воплощениях биорасщепляемый линкер может находиться или на 5'-конце и/или на 3'-конце смежных нуклеотидов, комплементарных нуклеиновой кислоте-мишени (область А). В предпочтительном воплощении биорасщепляемый линкер находится на 5'-конце.
В некоторых воплощениях расщепляемый линкер является чувствительным к нуклеазе(зам), которые, например, могут экспрессироваться в клетке-мишени. В некоторых воплощениях биорасщепляемый линкер состоит из 2-5 последовательных фосфодиэфирных связей. Данный линкер может представлять собой короткую область (например, из 1-10, как подробно описано в определении линкеров) нуклеозидов, связанных фосфодиэфирными связями. В некоторых воплощениях нуклеозиды в области В биорасщепляемого линкера (возможно независимо) выбраны из группы, состоящей из нуклеозидов ДНК и РНК или их модификаций, которые не препятствуют нуклеазному расщеплению. Модификации нуклеозидов ДНК и РНК, которые не препятствуют нуклеазному расщеплению, могут представлять собой не встречающиеся в природе нуклеиновые основания. Некоторые нуклеозиды с модифицированным сахаром также могут обеспечивать нуклеазное расщепление, как, например, альфа-L-окси-LNA. В некоторых воплощениях все нуклеозиды области В содержат (возможно независимо) либо сахар 2'-OH-рибозу (РНК), либо 2'-H сахар - т.е. РНК или ДНК. В предпочтительном воплощении по меньшей мере два последовательных нуклеозида области В представляют собой нуклеозиды ДНК или РНК (как, например, по меньшей мере 3 или 4, или 5 последовательных нуклеозидов ДНК или РНК). В даже более предпочтительном воплощении нуклеозиды области В представляют собой нуклеозиды ДНК. Предпочтительно область В состоит из от 1 до 5 или от 1 до 4, как, например, из 2, 3, 4 последовательных нуклеозидов ДНК, связанных фосфодиэфирной связью. В предпочтительных воплощениях область В является такой короткой, что она не рекрутирует РНКазуН. В некоторых воплощениях область В содержит не больше, чем 3 или больше, чем 4 последовательных нуклеозида ДНК и/или РНК, связанных фосфодиэфирной связью (таких как нуклеозиды ДНК).
Когда область В состоит из нуклеозидов, связанных фосфодиэфирной связью, область А и В могут вместе образовать олигонуклеотид, который связан с областью С. В данном контексте область А может отличаться от области В тем, что область А начинается с по меньшей мере одного, предпочтительно по меньшей мере двух модифицированных нуклеозидов с повышенной аффинностью связывания с нуклеиновой кислотой-мишенью (например, LNA или нуклеозиды с 2'-замещенной сахарной группировкой), и область А сама по себе способна к модуляции экспрессии нуклеиновой кислоты-мишени в релевантной линии клеток. Кроме того, если область А содержит нуклеозиды ДНК или РНК, они связаны нуклеазоустойчивой межнуклеозидной связью, такой как фосфоротиоатная или боранофосфатная. Область В, с другой стороны, содержит фосфодиэфирные связи между нуклеозидами ДНК или РНК. В некоторых воплощениях область В не является комплементарной или содержит по меньшей мере 50% несоответствий с нуклеиновой кислотой-мишенью.
В некоторых воплощениях область В не является комплементарной последовательности нуклеиновой кислоты-мишени или смежным нуклеотидам, комплементарным нуклеиновой кислоте-мишени в области А.
В некоторых воплощениях область В является комплементарной последовательности нуклеиновой кислоты-мишени. В данном отношении области А и В вместе могут образовать одну непрерывную последовательность, которая является комплементарной последовательности-мишени.
В некоторых аспектах изобретения межнуклеозидная связь между первой (область А) и второй областью (область В) может рассматриваться как часть второй области.
В некоторых воплощениях последовательность оснований в области В выбрана для обеспечения оптимального сайта расщепления эндонуклеазой на основе преобладающих эндонуклеазных расщепляющих ферментов, присутствующих в ткани-мишени или в клетке-мишени, или в субклеточном компартменте-мишени. В данном отношении посредством выделения клеточных экстрактов из тканей-мишеней и тканей, не являющихся мишенями, могут быть выбраны последовательности расщепления эндонуклеазой для применения в области В на основе предпочтительной расщепляющей активности в желательной клетке-мишени (например, печени/гепатоцитах) по сравнению с клеткой, не являющейся мишенью (например, почки). В данном отношении эффективность соединения в отношении понижающей регуляции мишени может быть оптимизирована для желательной ткани/клетки.
В некоторых воплощениях область В содержит динуклеотид последовательности AA, AT, AC, AG, TA, TT, TC, TG, CA, CT, CC, CG, GA, GT, GC, или GG, в которм С может представлять собой 5-метилцитозин, и/или Т может быть заменен U. Предпочтительно межнуклеозидная связь представляет собой фосфодиэфирную связь. В некоторых воплощениях область В содержит тринуклеотид последовательности AAA, AAT, AAC, AAG, ATA, ATT, ATC, ATG, ACA, ACT, ACC, ACG, AGA, AGT, AGC, AGG, TAA, TAT, TAC, TAG, TTA, TTT, TTC, TAG, TCA, TCT, TCC, TCG, TGA, TGT, TGC, TGG, CAA, CAT, CAC, CAG, CTA, CTG, CTC, CTT, CCA, CCT, CCC, CCG, CGA, CGT, CGC, CGG, GAA, GAT, GAC, CAG, GTA, GTT, GTC, GTG, GCA, GCT, GCC, GCG, GGA, GGT, GGC и GGG, в которм С может представлять собой 5-метилцитозин, и/или Т может быть заменен U. Предпочтительно межнуклеозидные связи представляют собой фосфодиэфирные связи. В некоторых воплощениях область В содержит тетрануклеотид последовательности AAAX, AATX, AACX, AAGX, ATAX, ATTX, ATCX, ATGX, ACAX, ACTX, ACCX, ACGX, AGAX, AGTX, AGCX, AGGX, TAAX, TATX, TACX, TAGX, TTAX, TTTX, TTCX, TAGX, TCAX, TCTX, TCCX, TCGX, TGAX, TGTX, TGCX, TGGX, CAAX, CATX, CACX, CAGX, CTAX, CTGX, CTCX, CTTX, CCAX, CCTX, CCCX, CCGX, CGAX, CGTX, CGCX, CGGX, GAAX, GATX, GACX, CAGX, GTAX, GTTX, GTCX, GTGX, GCAX, GCTX, GCCX, GCGX, GGAX, GGTX, GGCX и GGGX, где Х может быть выбран из группы, состоящей из A, T, U, G, C и их аналогов, где С может представлять собой 5-метилцитозин, и/или Т может быть заменен U. Предпочтительно межнуклеозидные связи представляют собой фосфодиэфирные связи. Будет понятно то, что при ссылке на (встречающиеся в природе) нуклеиновые основания A, T, U, G, C они могут быть заменены аналогами нуклеиновых оснований, которые функционируют как эквивалент природных нуклеиновых оснований (например, пара оснований с комплементарным нуклеозидом).
Другие линкеры (область Y)
Линкер может иметь по меньшей мере две функциональные группы: одну для присоединения олигонуклеотида и другую - для присоединения конъюгатной группировки. Типичные функциональные группы линкера могут быть электрофильными для осуществления реакции с нуклеофильными группами на олигонуклеотиде или конъюгатной группировке или нуклеофильными для осуществления реакции с электрофильными группами. В некоторых воплощениях функциональные группы линкера включают амино, гидроксил, карбоновую кислоту, тиол, фосфорамидат, фосфоротиоат, фосфат, фосфит, ненасыщенные группы (например, с двойными или тройными связями) и тому подобное. Некоторые типичные линкеры (область Y) включают 8-амино-3,6-диоксаоктановую кислоту (ADO), сукцинимидил-4-(N-малеимидометил)циклогексан-l-карбоксилат (SMCC), 6- аминогексановую кислоту (AHEX или AHA), 6-аминогексилокси, 4-аминомасляную кислоту, 4-аминоциклогексилкарбоновую кислоту, сукцинимидил-4-(N-малеимидометил)циклогексан-l-карбокси-(6-амидо-капроат) (LCSMCC), сукцинимидил-м-малеимидо-бензоилат (MBS), сукцинимидил-N-e-малеимидо-капроилат (EMCS), сукцинимидил-6-(бета-малеимидо-пропионамидо)гексаноат (SMPH), сукцинимидил-N-(a-малеимидоацетат) (AMAS), сукцинимидил-4-(п-малеимидофенил)бутират (SMPB), бета-аланин (бета-ALA), фенилглицин (PHG), 4-аминоциклогексановую кислоту (ACHC), бета-(циклопропил)аланин (бета-CYPR), аминододекановую кислоту (ADC), алилендиолы, полиэтиленгликоли, аминокислоты и тому подобное. В некоторых воплощениях линкер (область Y) представляет собой аминоалкил, как, например, С2-С36 аминоалкильную группу, включающую, например, С6-С12 аминоалкильные группы. В предпочтительном воплощении линкер (область Y) представляет собой С6 аминоалкильную группу. Аминоалкильная группа может быть добавлена к олигонуклеотиду (область А или область А-В) в качестве части стандартного синтеза олигонуклеотида, например, с использованием (например, защищенного) аминоалкилфосфорамидита. Связывающая группа между аминоалкилом и олигонуклеотдом может представлять собой, например, фосфоротиоат или фосфодиэфир, или одну из других нуклеозидных связывающих групп, на которые дается ссылка в данном документе. Аминоалкильная группа квалентно связывается с 5' или 3'-концом олигонуклеотида. Имеющиеся в продаже аминоалкильные линкеры представляют собой, например, 3'-аминомодифицирующий реактив для связывания на 3'-конце олигонуклеотида, и для связывания на 5'-конце олигонуклеотида доступен 5'-аминомодификатор С6. Данные реактивы доступны у Glen Research Corporation (Sterling, Va.). Данные соединения или аналогичные соединения были использованы Krieg, et al, Antisense Research and Development 1991, 1, 161 для связывания флюоресцеина с 5'-концом олигонуклеотида. В данной области известен широкий спектр других линкерных групп, и они могут быть полезными при присоединении конъюгатных группировок к олигонуклеотидам. Обзор многих полезных линкерных групп можно найти, например, в Antisense Research and Applications, S. T. Crooke and B. Lebleu, Eds., CRC Press, Boca Raton, Fla., 1993, p. 303-350. Другие соединения, такие как акридин, присоединяли к 3'-концевой фосфатной группе олигонуклеотида посредством полиметиленовой связи (Asseline, et al., Proc. Natl. Acad. Sci. USA 1984, 81, 3297). Любую из приведенных выше групп можно использовать в качестве одиночного линкера (область Y) или в комбинации с одним или более чем одним другим линкером (область Y-Y' или область Y-B, или B-Y).
Линкеры и их применение в получении конъюгатов олигонуклеотидов приводятся во всей данной области, как, например, в WO 96/11205 и WO 98/52614 и в патентах США № 4948882; 5525465; 5541313; 5545730; 5552538; 5580731; 5486603; 5608046; 4587044; 4667025; 5254469; 5245022; 5112963; 5391723; 5510475; 5512667; 5574142; 5684142; 5770716; 6096875; 6335432 и 6335437, WO 2012/083046, каждая из которых включена посредством ссылки во всей ее полноте.
Способ изготовления
В другом аспекте согласно изобретению предложены способы изготовления олигонуклеотидов по изобретению, включающие проведение взаимодействия нуклеотидных звеньев и, посредством этого, образование ковалентно связанных смежных нуклеотидных звеньев, содержащихся в олигонуклеотиде. Предпочтительно в данном способе используется фосфорамидитная химия (см., например, Caruthers et al, 1987, Methods in Enzymology том 154, страницы 287-313). В другом воплощении данный способ дополнительно включает проведение взаимодействия непрерывной нуклеотидной последовательности с конъюгируемой группировкой (лигандом). В другом аспекте предложен способ изготовления композиции по изобретению, включающий смешивание олигонуклеотида или конъюгированного олигонуклеотида по изобретению с фармацевтически приемлемым разбавителем, растворителем, носителем, солью и/или адъювантом.
Фармацевтическая композиция
В другом аспекте согласно изобретению предложена фармацевтическая композиция, содержащая любой из вышеупомянутых олигонуклеотидов и/или конъюгатов олигонуклеотидов и фармацевтически приемлемый разбавитель, растворитель, носитель, соль и/или адъювант. Фармацевтически приемлемый разбавитель включает фосфатно-солевой буферный раствор (PBS), а фармацевтически приемлемые соли включают натриевые и калиевые соли, но не ограничиваются ими. В некоторых воплощениях фармацевтически приемлемый разбавитель представляет собой стерильный фосфатно-солевой буферный раствор. В некоторых воплощениях олигонуклеотид используется в фармацевтически приемлемом разбавителе в виде раствора в концентрации 50-300 мкМ.
Подходящие препараты для применения в настоящем изобретении находятся в Remington's Pharmaceutical Sciences, Mack Publishing Company, Philadelphia, Pa., 17th ed., 1985. Относительно краткого обзора способов доставки лекарственного средства, см., например, Langer (Science 249:1527-1533, 1990). В WO 2007/031091 предложены дполнительные подходящие и предпочтительные примеры фармацевтически приемлемых разбавителей, носителей и адъювантов (включенные тем самым посредством ссылки). Подходящие дозировки, препараты, пути введения, композиции, лекарственные формы, комбинации с другими терапевтическими агентами, препараты пролекарств также приводятся в WO 2007/031091.
Олигонуклеотиды или конъюгаты олигонуклеотидов по изобретению могут быть смешаны с фармацевтически приемлемыми активными или инертными веществами для приготовления фармацевтических композиций или препаратов. Композиции и способы для приготовления фармацевтических композиций зависят от целого ряда критериев, включающих путь введения, степень заболевания или дозу, подлежащую введению, но не ограничивающихся ими.
Данные композиции можно стерилизовать традиционными методиками стерилизации, или они могут подвергаться стерилизующему фильтрованию. Образующиеся водные растворы могут быть упакованы для применения в том виде, в котором они находятся, или лиофилизируются, причем лиофилизированный препарат объединяется со стерильным водным раствором перед введением. рН препаратов типично будет составлять от 3 до 11, более предпочтительно - от 5 до 9 или от 6 до 8, и наиболее предпочтительно - от 7 до 8, как, например, от 7 до 7,5. Образующиеся композиции в твердой форме можно упаковывать в многочисленные единичные стандартные дозы, причем каждая из них содержит фиксированное количество вышеупомянутого агента или агентов, как, например, в запечатанной упаковке или таблетках, или капсулах. Композицию в твердой форме также можно упаковывать в контейнер для гибкого количества, такой как поддающийся выдавливанию тюбик, разработанный для местно наносимого крема или мази.
В некоторых воплощениях олигонуклеотид или конъюгат олигонуклеотида по изобретению представляет собой пролекарство. В частности, в отношении конъюгатов олигонуклеотидов конъюгатная группировка отщепляется от олигонуклеотида, как только пролекарство доставляется к месту действия, например, в клетку-мишень.
Применения
Олигонуклеотиды или конъюгаты олигонуклеотидов по настоящему изобретению могут использоваться в качестве исследовательских реактивов, например, для диагностики, терапии и профилактики.
В исследовании такие олигонуклеотиды или конъюгаты олигонуклеотидов можно использовать для специфичной модуляции синтеза белка PD-L1 в клетках (например, в культурах клеток in vitro) и в экспериментальных животных, облегчая, посредством этого, функциональный анализ мишени или оценку ее полезности в качестве мишени для терапевтического вмешательства. Типично модуляция мишени достигается посредством осуществления деградации или ингибирования мРНК, продуцирующей белок, предотвращая, посредством этого, образование белка, или посредством осуществления деградации или ингибирования модулятора гена или мРНК, продуцирующей белок.
При использовании олигонуклеотида по изобретению в исследовании или диагностике нуклеиновая кислота-мишень может представлять собой кДНК или синтетическую нуклеиновую кислоту, полученную из ДНК или РНК.
Согласно настоящему изобретению предложен способ in vivo или in vitro модуляции экспрессии PD-L1 в клетке-мишени, которая экспрессирует PD-L1, причем указанный способ включает введение в указанную клетку олигонуклеотида или конъюгата олигонуклеотида по изобретению в эффективном количестве.
В некоторых воплощениях клетка-мишень представляет собой клетку млекопитающего, в частности, человеческую клетку. Клетка-мишень может представлять собой культуру клеток in vitro или клетку in vivo, образующую часть ткани млекопитающего. В предпочтительных воплощениях клетка-мишень присутствует в печени. Клетка-мишень печени может быть выбрана из паренхимных клеток (например, гепатоцитов) и непаренхимных клеток, таких как клетки Купфера, LSEC, звездчатые клетки (или клетки Ито), холангиоциты и лейкоциты, ассоциированные с печенью (включающие Т-клетки и NK-клетки). В некоторых воплощениях клетка-мишень представляет собой антигенпрезентирующую клетку. Антигенпрезентирующие клетки подвергают дисплею чужеродные антигены, образующие комплекс с главным комплексом гистосовместимости (МНС) класса I или класса II на их поверхностях. В некоторых воплощениях антигенпрезентирующие клетки экспрессируют МНС класса II (т.е. профессиональные антигенпрезентирующие клетки, такие как дендритные клетки, макрофаги и В-клетки).
При диагностике данные олигонуклеотиды могут быть использованы для выявления и количественного измерения экспрессии PD-L1 в клетке и тканях посредством норзерн-блоттинга, гибридизации in situ или аналогичных методик.
Для терапии олигонуклеотиды или конъюгаты олигонуклеотидов по настоящему изобретению или их фармацевтические копозиции могут вводиться животному или человеку, у которого подозревается наличие заболевания или расстройства, которое может быть облегчено или вылечено посредством уменьшения экспрессии PD-L1, в частности, посредством уменьшения экспрессии PD-L1 в клетках-мишенях печени.
Согласно изобретению предложены способы лечения или предупреждения заболевания, включающие введение терапевтически или профилактически эффективного количества олигонуклеотида, конъюгата олигонуклеотида или фармацевтической композиции по изобретению субъекту, страдающему от или восприимчивому к заболеванию.
Данное изобретение также относится к олигонуклеотиду, конъюгату олигонуклеотида или фармацевтической композиции согласно изобретению для применения в качестве лекарственного средства.
Олигонуклеотид, конъюгат олигонуклеотида или фармацевтическую композицию согласно изобретению типично вводят в эффективном количестве.
Согласно изобретению также предложено применение олигонуклеотида или конъюгата олигонуклеотида, или фармацевтической композиции по изобретению, как описано, для изготовления лекарственного средства для лечения заболевания или расстройства, на которые дается ссылка в данном документе. В одном воплощении данное заболевание выбрано из: а) вирусных инфекций печени, таких как инфекции HBV, HCV и HDV; б) паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, и в) рака печени или метастазов в печени.
В одном воплощении данное изобретение относится к олигонуклеотидам, конъюгатам олигонуклеотидов или фармацевтическим композициям для применения в лечении заболеваний или расстройств, выбранных из вирусных или паразитарных инфекций. В другом воплощении данное заболевание выбрано из: а) вирусных инфекций печени, таких как инфекции HBV, HCV и HDV; б) паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, и в) рака печени или метастазов в печени.
Заболевание или расстройство, на которые дается ссылка в данном документе, ассоциировано с истощением иммунной системы. В частности, данное заболевание или расстройство ассоциировано с истощением вирусоспецифичных ответов Т-клеток. В некоторых воплощениях заболевание или расстройство может быть облегчено или вылечено посредством уменьшения экспрессии PD-L1.
Способы по изобретению предпочтительно используют для лечения или профилактики против заболеваний, ассоциированных с истощением иммунной системы.
В одном воплощении изобретения олигонуклеотид, конъюгат олигонуклеотида или фармацевтические композиции по изобретению используются в восстановлении иммунного ответа против рака печени или метастазов в печени.
В одном воплощении изобретения олигонуклеотид, конъюгат олигонуклеотида или фармацевтические композиции по изобретению используются в восстановлении иммунного ответа против патогена. В некоторых воплощениях патоген может находиться в печени. Данными патогенами могут быть вирус или паразит, в частности, вирусы или паразиты, описанные в данном документе. В предпочтительном воплощении патоген представляет собой HBV.
Данное изобретение дополнительно относится к применению олигонуклеотида, конъюгата олигонуклеотида или фармацевтической композиции, как определено в данном документе, для изготовления лекарственного средства для восстановления иммунитета против вирусной или паразитарной инфекции, как упомянуто в данном документе.
Олигонуклеотиды или конъюгаты олигонуклеотидов, или фармацевтические композиции по настоящему изобретению можно использовать в лечении вирусных инфекций, в частности, вирусных инфекций в печени, где подвержен влиянию путь PD-1 (см., например, Kapoor and Kottilil 2014 Future Virol Vol. 9 pp. 565-585 и Salem and El-Badawy 2015 World J Hepatol Vol. 7 pp. 2449-2458). Вирусные инфекции печени могут быть выбраны из группы, состоящей из вирусов гепатита, в частности HBV, HCV и HDV, в частности, хронических форм данных инфекций. В одном воплощении олигонуклеотиды или конъюгаты олигонуклеотидов, или фармацевтические композиции по настоящему изобретению используются для лечения HBV, в частности, хронического HBV. Показателями хронических инфекций HBV являются высокие уровни вирусной нагрузки (ДНК HBV) и даже более высокие уровни пустых частиц HВsAg (более чем 100-кратный избыток относительно вирионов) в системе кровообращения.
Олигонуклеотиды или конъюгаты олигонуклеотидов по настоящему изобретению также можно использовать для лечения вирусных инфекций печени, которые наблюдаются в качестве сопутствующих инфекций ВИЧ (вирус иммунодефицита человека). Другими вирусными инфекциями, которые можно лечить олигонуклеотидами или конъюгатами олигонуклеотидов, или фармацевтическими композициями по настоящему изобретению, являются lcmv (вирус лимфоцитарного хориоменингита) и ВИЧ в качестве моноинфекции, HSV-1 и -2, и другие герпесвирусы. Данные вирусы не являются гепатотрофическими, однако, они могут быть чувствительными к понижающей регуляции PD-L1.
В некоторых воплощениях восстановление иммунитета или иммунного ответа включает улучшение ответа Т-клеток и/или NK-клеток, и/или ослабление исчерпания Т-клеток, в частности, восстанавливается HBV-специфичный Т-клеточный ответ, HСV-специфичный Т-клеточный ответ и HDV-специфичный Т-клеточный ответ. Улучшение Т-клеточного ответа, например, можно оценивать как увеличение уровня Т-клеток в печени, в частности, увеличение уровня Т-клеток CD8+ и/или CD4+ по сравнению с контролем (т.е. уровнем до обработки или уровнем у субъекта, обработанного носителем). В другом воплощении восстанавливается или возрастает именно уровень вирусоспецифичных Т-клеток CD8+ по сравнению с контролем, в частности, восстанавливается или возрастает уровень HBV-специфичных Т-клеток CD8+ или HСV-специфичных Т-клеток CD8+, или HDV-специфичных Т-клеток CD8+ по сравнению с контролем. В предпочтительном воплощении возрастает уровень Т-клеток CD8+, специфичных в отношении антигена s HBV (HВsAg) и/или Т-клеток CD8+, специфичных в отношении антигена e HBV (HBeAg), и/или Т-клеток CD8+, специфичных в отношении антигена кора HBV (HBсAg), у субъектов, обработанных олигонуклеотидом, конъюгатом олигонуклеотида или фармацевтической композицией по настоящему изобретению по сравнению с контролем. Предпочтительно Т-клетки CD8+, специфичные в отношении антигена HBV, продуцируют один или более чем один цитокин, такой как интерферон-гамма (IFN-γ) или фактор некроза опухолей альфа (TNF-α). Увеличение уровня Т-клеток CD8+, описанных выше, в частности, наблюдается в печени. Описанное в данном документе увеличение должно быть статистически значимым по сравнению с контролем. Предпочтительно данное увеличение составляет по меньшей мере 20%, как, например, 25%, как, например, 50%, как, например, 75% по сравнению с контролем. В другом воплощении олигонуклеотидами или конъюгатами олигонуклеотидов по настоящему изобретению активируются природные клетки-киллеры (NK) и/или природные Т-клетки-киллеры (NKТ).
Олигонуклеотиды или конъюгаты олигонуклеотидов, или фармацевтические композиции по настоящему изобретению можно использовать в лечении паразитарных инфекций, в частности, паразитарных инфекций, где подвергается влиянию путь PD-1 (см., например, Bhadra et al. 2012 J Infect Dis vol 206 pp. 125-134; Bhadra et al. 2011 Proc Natl Acad Sci U S A Vol. 108 pp. 9196-9201; Esch et al. J Immunol vol 191 pp 5542-5550; Freeman and Sharpe 2012 Nat Immunol Vol 13 pp. 113-115; Gutierrez et al. 2011 Infect Immun Vol 79 pp. 1873-1881; Joshi et al. 2009 PLoS Pathog Vol 5 e1000431; Liang et al. 2006 Eur J Immunol Vol. 36 pp 58-64; Wykes et al. 2014 Front Microbiol Vol 5 pp 249). Паразитарные инфекции могут быть выбраны из группы, состоящей из малярии, токсоплазмоза, лейшманиоза и трипаносомоза. Малярийная инфекция вызвана протистом рода Plasmodium, в частности вида P. vivax, P. malariae и P. falciparum. Токсоплазмоз представляет собой паразитарное заболевание, вызванное Toxoplasma gondii. Лейшманиоз представляет собой заболевание, вызванное протозойными паразитами рода Leishmania. Трипаносомоз вызван простейшим рода Trypanosoma. Болезнь Чагаса, которая является тропической формой, вызывается видом Trypanosoma cruzi, а расстройство сна вызывается видом Trypanosoma brucei.
В некоторых воплощениях восстановление иммунитета включает восстановление ответа паразитоспецифичных Т-клеток и NK-клеток, в частности, ответа Plasmodium-специфичных Т-клеток, ответа Toxoplasma gondii-специфичных Т-клеток и NK-клеток, ответа Leishmania-специфичных Т-клеток и NK-клеток, ответа Trypanosoma cruzi-специфичных Т-клеток и NK-клеток или ответа Trypanosoma brucei-специфичных Т-клеток и NK-клеток. В другом воплощении восстанавливается именно ответ паразитоспецифичных Т-клеток CD8+ и NK-клеток.
Введение
Олигонуклеотиды или фармацевтические композиции по настоящему изобретению можно вводить местно (как, например, на кожу, посредством ингаляции, в глаз или в ухо), энтерально (как, например, перорально или через желудочно-кишечный тракт) или парентерально (как, например, внутривенно, подкожно, внутримышечно, интрацеребрально, интрацеребровентрикулярно или подоболочечно).
В предпочтительном воплощении олигонуклеотид или фармацевтические композиции по настоящему изобретению вводятся посредством парентрального пути, включающего внутривенную, внутриартериальную, подкожную, внутрибрюшинную или внутримышечную инъекцию или инфузию, внутриоболочечное или внутричерепное, например, интрацеребральное или интравентрикулярное, интравитреальное введение. В одном воплощении активный олигонуклеотид или конъюгат олигонуклеотида вводится внутривенно. В другом воплощении активный олигонуклеотид или конъюгат олигонуклеотида вводится подкожно.
В некоторых воплощениях олигонуклеотид, конъюгат олигонуклеотида или фармацевтическая композиция по изобретению вводится в дозе 0,1-15 мг/кг, как, например, 0,1-10 мг/кг, как, например, 0,2-10 мг/кг, как, например, 0,25-10 мг/кг, как, например, 0,1-5 мг/кг, как, например, 0,2-5 мг/кг, как, например, 0,25-5 мг/кг. Введение может осуществляться один раз в неделю, каждую 2-ую неделю, каждую третью неделю или даже один раз в месяц.
Комбинированные терапии
В некоторых воплощениях олигонуклеотид, конъюгат олигонуклеотида или фармацевтическая композиция по изобретению предназначены для применения в комбинированном лечении с другим терапевтическим агентом. Данный терапевтический агент может представлять собой, например, стандарт лечения для описанных выше заболеваний или расстройств.
Для лечения хронических инфекций HBV в качестве стандарта лечения рекомендуется комбинация противовирусных лекарственных средств и модуляторов иммунной системы. Эффективными противовирусными лекарственными средствами против HBV являются, например, аналоги нуклеоз(т)идов. Имеется пять аналогов нуклеоз(т)идов, лицензированных для терапии HBV, а именно: ламивудин (эпивир), адефовир (гепсера), тенофовир (виреад), телбивудин (тизека), энтекавир (бараклуд). Они являются эффективными в подавлении вирусной репликации (ДНК HBV), но не имеют влияния на уровни HВsAg. Другие антивирусные лекарственные средства включают рибавирин и терапию антителом против HBV (моноклональным или поликлональным). Модуляторами иммунной системы могут быть, например, интерферон-альфа-2а и ПЭГилированный интерферон-альфа-2а (пегасис) или агонисты TLR7 (например, GS-9620), или терапевтические вакцины. Лечение IFN-α демонстрирует лишь очень умеренный эффект в уменьшении вирусной нагрузки, но приводит к некоторому снижению уровня HВsAg, хотя и очень не эффективно (меньше 10% после 48 недель терапии).
Олигонуклеотид или конъюгаты олигонуклеотида по настоящему изобретению также могут быть объединены с другими противовирусными лекарственными средствами, эффективными против HBV, такими как антисмысловые олигонуклеотиды, описанные в WO2012/145697 и WO 2014/179629, или молекулы миРНК, описанные в WO 2005/014806, WO 2012/024170, WO 2012/2055362, WO 2013/003520 и WO 2013/159109.
При введении олигонуклеотидов или конъюгатов олигонуклеотидов по данному изобретению в комбинированных терапиях с другими агентами, они могут вводиться индивиду последовательно или сопутствующе. В качестве альтернативы, фармацевтические композиции согласно настоящему изобретению могут содержать комбинацию олигонуклеотида или конъюгата олигонуклеотида по настоящему изобретению в ассоциации с фармацевтически приемлемым эксципиентом, как описано в данном документе, и с другим терапевтическим или профилактическим агентом, известным в данной области.
ВОПЛОЩЕНИЯ
Следующие воплощения настоящего изобретения можно использовать в комбинации с любыми другими воплощениями, описанными в данном документе.
1. Антисмысловой олигонуклеотид, который содержит или состоит из непрерывной нуклеотидной последовательности из 10-30 нуклеотидов в длину, способный уменьшать экспрессию PD-L1.
2. Олигонуклеотид по воплощению 1, в котором непрерывная нуклеотидная последовательность является по меньшей мере на 90% комплементарной нуклеиновой кислоте-мишени PD-L1.
3. Олигонуклеотид по воплощению 1 или 2, в котором непрерывная нуклеотидная последовательность комплементарна нуклеиновой кислоте-мишени, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и/или SEQ ID NO: 3.
4. Олигонуклеотид по воплощениям 1-3, в котором непрерывная нуклеотидная последовательность комплементарна области в пределах положения 1 и 15720 на SEQ ID NO: 1.
5. Олигонуклеотид по воплощениям 1-4, где данный олигонуклеотид способен гибридизоваться с нуклеиновой кислотой-мишенью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и/или SEQ ID NO: 3, с ΔG° меньше -41,8 кДж.
6. Олигонуклеотид по воплощениям 1-5, в котором непрерывная нуклеотидная последовательность комплементарна подпоследовательности нуклеиновой кислоты-мишени, где данная подпоследовательность выбрана из группы, состоящей из положения 371-3068, 5467-12107, 15317-15720, 15317-18083, 15317-19511 и 18881-19494 на SEQ ID NO: 1.
7. Олигонуклеотид по воплощению 6, в котором подпоследовательность выбрана из группы, состоящей из положения 7300-7333, 8028-8072, 9812-9859, 11787-11873 и 15690-15735 на SEQ ID NO: 1.
8. Олигонуклеотид по воплощениям 2-7, в котором нуклеиновая кислота-мишень представляет собой РНК.
9. Олигонуклеотид по воплощению 8, где РНК представляет собой мРНК.
10. Олигонуклеотид по воплощению 9, где мРНК представляет собой пре-мРНК или зрелую мРНК.
11. Олигонуклеотид по воплощениям 1-10, в котором непрерывная нуклеотидная последовательность содержит или состоит из по меньшей мере 14 непрерывных нуклеотидов, в частности, из 15, 16, 17, 18, 19, 20, 21, 22, 23 или 24 непрерывных нуклеотидов.
12. Олигонуклеотид по воплощениям 1-10, в котором непрерывная нуклеотидная последовательность содержит или состоит из 16-20 нуклеотидов.
13. Олигонуклеотид по воплощениям 1-10, где данный олигонуклеотид содержит или состоит из 14-35 нуклеотидов в длину.
14. Олигонуклеотид по воплощению 13, где данный олигонуклеотид содержит или состоит из 18-22 нуклеотидов в длину.
15. Олигонуклеотид по воплощениям 1-14, где данный олигонуклеотид или непрерывная нуклеотидная последовательность является одноцепочечной.
16. Олигонуклеотид по воплощениям 1-15, в котором непрерывная нуклеотидная последовательность комплементарна подпоследовательности нуклеиновой кислоты-мишени, где данная подпоследовательность выбрана из группы, состоящей из A7, A26, A43, A119, A142, A159, A160, A163, A169, A178, A179, A180, A189, A201, A202, A204, A214, A221, A224, A226, A243, A254, A258, 269, A274, A350, A360, A364, A365, A370, A372, A381, A383, A386, A389, A400, A427, A435 и A438.
17. Олигонуклеотид по воплощению 16, в котором подпоследовательность выбрана из группы, состоящей из A221, A360, A180, A160 и A269.
18. Олигонуклеотид по воплощениям 1-17, где данный олигонуклеотид не является миРНК и не является комплементарным самому себе.
19. Олигонуклеотид по воплощениям 1-18, в котором непрерывная нуклеотидная последовательность содержит или состоит из последовательности, выбранной из SEQ ID NO: 5-743 или 771.
20. Олигонуклеотид по воплощениям 1-19, в котором непрерывная нуклеотидная последовательность содержит или состоит из последовательности, выбранной из SEQ ID NO: 6, 8, 9, 13, 41, 42, 58, 77, 92, 111, 128, 151, 164, 166, 169, 171, 222, 233, 245, 246, 250, 251, 252, 256, 272, 273, 287, 292, 303, 314, 318, 320, 324, 336, 342, 343, 344, 345, 346, 349, 359, 360, 374, 408, 409, 415, 417, 424, 429, 430, 458, 464, 466, 474, 490, 493, 512, 519, 519, 529, 533, 534, 547, 566, 567, 578, 582, 601, 619, 620, 636, 637, 638, 640, 645, 650, 651, 652, 653, 658, 659, 660, 665, 678, 679, 680, 682, 683, 684, 687, 694, 706, 716, 728, 733, 734 и 735.
21. Олигонуклеотид по воплощениям 1-20, в котором непрерывная нуклеотидная последовательность содержит или состоит из последовательности, выбранной из SEQ ID NO: 466, 640, 342, 287 и 566.
22. Олигонуклеотид по воплощениям 1-21, в котором непрерывная нуклеотидная последовательность имеет от нуля до трех несоответствий по сравнению с нуклеиновой кислотой-мишенью, которой она комплементарна.
23. Олигонуклеотид по воплощению 22, в котором непрерывная нуклеотидная последовательность имеет одно несоответствие по сравнению с нуклеиновой кислотой-мишенью.
24. Олигонуклеотид по воплощению 22, в котором непрерывная нуклеотидная последовательность имеет два несоответствия по сравнению с нуклеиновой кислотой-мишенью.
25. Олигонуклеотид по воплощению 22, в котором непрерывная нуклеотидная последовательность является полностью комплементарной последовательности нуклеиновой кислоты-мишени.
26. Олигонуклеотид по воплощениям 1-25, содержащий один или более чем один модифицированный нуклеозид.
27. Олигонуклеотид по воплощению 26, в котором один или более чем один модифицированный нуклеозид представляет собой высокоаффинные модифицированные нуклеозиды.
28. Олигонуклеотид по воплощению 26 или 27, в котором один или более чем один модифицированный нуклеозид представляет собой нуклеозид, модифицированный 2'-сахаром.
29. Олигонуклеотид по воплощению 28, в котором один или более чем один нуклеозид, модифицированный 2'-сахаром, независимо выбран из группы, состоящей из нуклеозидов 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-алкокси-РНК, 2'-O-метоксиэтил-РНК, 2'-амино-ДНК, 2'-фтор-ДНК, 2'-фтор-ANA и LNA.
30. Олигонуклеотид по воплощению 28, в котором один или более чем один модифицированный нуклеозид представляет собой нуклеозид LNA.
31. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид LNA представляет собой окси-LNA.
32. Олигонуклеотид по воплощению 31, в котором модифицированный нуклеозид представляет собой бета-D-окси-LNA.
33. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид представляет собой тио-LNA.
34. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид представляет собой амино-LNA.
35. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид представляет собой сЕТ.
36. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид представляет собой ENA.
37. Олигонуклеотид по воплощению 30, в котором модифицированный нуклеозид LNA выбран из бета-D-окси-LNA, альфа-L-окси-LNA, бета-D-амино-LNA, альфа-L-амино-LNA, бета-D-тио-LNA, альфа-L-тио-LNA, (S)cET, (R)cET, бета-D-ENA и альфа-L-ENA.
38. Олигонуклеотид по воплощениям 30-37, в котором, помимо модифицированного нуклеозида LNA, имеется по меньшей мере один 2'-замещенный модифицированный нуклеозид.
39. Олигонуклеотид по воплощению 38, в котором 2'-замещенный модифицированный нуклеозид выбран из группы, состоящей из 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-алкокси-РНК, 2'-O-метоксиэтил-РНК (МОЕ), 2'-амино-ДНК, 2'-фтор-ДНК, 2'-фтор-ANA.
40. Олигонуклеотид по любому из воплощений 1-39, где данный олигонуклеотид содержит по меньшей мере одну модифицированную межнуклеозидную связь.
41. Олигонуклеотид по воплощению 40, в котором модифицированная межнуклеозидная связь является устойчивой к нуклеазе.
42. Олигонуклеотид по воплощению 40 или 41, в котором по меньшей мере 50% межнуклеозидных связей в пределах непрерывной нуклеотидной последовательности представляют собой фосфоротиоатные межнуклеозидные связи или боранофосфатные межнуклеозидные связи.
43. Олигонуклеотид по воплощению 40 или 41, в котором все межнуклеозидные связи в пределах непрерывной нуклеотидной последовательности представляют собой фосфоротиоатные межнуклеозидные связи.
44. Олигонуклеотид по воплощениям 1-43, где данный олигонуклеотид способен рекрутировать РНКазу Н.
45. Олигонуклеотид по воплощению 44, где данный олигонуклеотид представляет собой гэпмер.
46. Олигонуклеотид по воплощению 44 или 45, где данный олигонуклеотид представляет собой гэпмер формулы 5'-F-G-F'-3', где области F и F' независимо содержат или состоят из 1-7 модифицированных нуклеозидов, и G представляет собой область из 6-16 нуклеозидов, которые способны рекрутировать РНКазу Н.
47. Олигонуклеотид по воплощению 44 или 45, где гэпмер имеет формулу 5'-D'-F-G-F'-3' или 5'-F-G-F'-D''-3', где области F и F' независимо содержат 1-7 модифицированных нуклеозидов, G представляет собой область из 6-16 нуклеозидов, которые способны рекрутировать РНКазу Н, и области D' или D'' содержат 1-5 нуклеозидов, связанных фосфодиэфирной связью.
48. Олигонуклеотид по воплощению 47, в котором D' или D'' являются опционными.
49. Олигонуклеотид по воплощению 47, в котором область D' состоит из двух нуклеозидов, связанных фосфодиэфирной связью.
50. Олигонуклеотид по воплощению 49, в котором нуклеозиды, связанные фосфодиэфирной связью, представляют собой са (цитидин-аденозин).
51. Олигонуклеотид по воплощению 46 или 47, в котором модифицированный нуклеозид представляет собой нуклеозид, модифицированный 2'-сахаром, независимо выбранный из группы, состоящей из нуклеозидов 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-алкокси-РНК, 2'-O-метоксиэтил-РНК, 2'-амино-ДНК, 2'-фтор-ДНК, арабинонуклеиновой кислоты (ANA), 2'-фтор-ANA и LNA.
52. Олигонуклеотид по воплощениям 46-51, в котором один или более чем один модифицированный нуклеозид в области F и F' представляет собой нуклеозид LNA.
53. Олигонуклеотид по воплощению 52, в котором все модифицированные нуклеозиды в области F и F' представляет собой нуклеозиды LNA.
54. Олигонуклеотид по воплощению 53, в котором область F и F' состоит из нуклеозидов LNA.
55. Олигонуклеотид по воплощениям 52-54, в котором все модифицированные нуклеозиды в области F и F' представляет собой нуклеозиды окси-LNA.
56. Олигонуклеотид по воплощению 52, в котором по меньшей мере одна из областей F и F' дополнительно содержит по меньшей мере один 2'-замещенный модифицированный нуклеозид, независимо выбранный из группы, состоящей из 2'-O-алкил-РНК, 2'-O-метил-РНК, 2'-алкокси-РНК, 2'-O-метоксиэтил-РНК, 2'-амино-ДНК и 2'-фтор-ДНК.
57. Олигонуклеотид по воплощениям 46-56, в котором нуклеозиды, рекрутирующие РНКазу Н, в области G независимо выбраны из ДНК, альфа-L-LNA, C4'-алкилированной ДНК, ANA, 2'-F-ANA и UNA.
58. Олигонуклеотид по воплощению 57, в котором нуклеозиды в области G представляют собой нуклеозиды ДНК и/или альфа-L-LNA.
59. Олигонуклеотид по воплощению 57 или 58, в котором область G состоит из по меньшей мере 75% нуклеозидов ДНК.
60. Олигонуклеотид по воплощениям 1-59, где данный олигонуклеотид выбран из любого олигонуклеотида CMP ID NO: от 5_1 до 743_1 и 771_1 (Таблица 5).
61. Олигонуклеотид по воплощениям 1-60, где данный олигонуклеотид выбран из группы, состоящей из CMP ID NO: 6_1, 8_1, 9_1, 13_1, 41_1, 42_1, 58_1, 77_1, 92_1, 111_1, 128_1, 151_1, 164_1, 166_1, 169_1, 171_1, 222_1, 233_1, 245_1, 246_1, 250_1, 251_1, 252_1, 256_1, 272_1, 273_1, 287_1, 292_1, 303_1, 314_1, 318_1, 320_1, 324_1, 336_1, 342_1, 343_1, 344_1, 345_1, 346_1, 349_1, 359_1, 360_1, 374_1, 408_1, 409_1, 415_1, 417_1, 424_1, 429_1, 430_1, 458_1, 464_1, 466_1, 474_1, 490_1, 493_1, 512_1, 519_1, 519_1, 529_1, 533_1, 534_1, 547_1, 566_1, 567_1, 578_1, 582_1, 601_1, 619_1, 620_1, 636_1, 637_1, 638_1, 640_1, 645_1, 650_1, 651_1, 652_1, 653_1, 658_1, 659_1, 660_1, 665_1, 678_1, 679_1, 680_1, 682_1, 683_1, 684_1, 687_1, 694_1, 706_1, 716_1, 728_1, 733_1, 734_1 и 735_1.
62. Олигонуклеотид по воплощениям 1-61, где данный олигонуклеотид выбран из группы, состоящей из CMP ID NO: 287_1, 342_1, 466_1, 640_1, 566_1, 766_1, 767_1, 768_1, 769_1 и 770_1.
63. Конъюгат антисмыслового олигонуклеотида, содержащий:
а. олигонуклеотид по любому из воплощений 1-62 (область А); и
б. по меньшей мере одну конъюгатную группировку (область С), ковалентно присоединенную к указанному олигонуклеотиду.
64. Конъюгат олигонуклеотида по воплощению 63, в котором конъюгатная группировка выбрана из углеводов, лигандов рецептора поверхности клетки, лекарственных веществ, гормонов, липофильных веществ, полимеров, белков, пептидов, токсинов, витаминов, вирусных белков или их комбинаций.
65. Конъюгат олигонуклеотида по воплощению 63 или 64, в котором конъюгатная группировка представляет собой углеводсодержащую группировку.
66. Конъюгат олигонуклеотида по воплощению 65, в котором углеводная конъюгатная группировка содержит по меньшей мере одну группировку, нацеленную на рецептор асиалогликопротеина, ковалентно присоединенную к олигонуклеотиду по любому из пп. 1-62.
67. Конъюгат олигонуклеотида по воплощению 66, в котором конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, содержит по меньшей мере одну углеводную группировку, выбранную из группы, состоящей из галактозы, галактозамина, N-формил-галактозамина, N-ацетилгалактозамина, N-пропионил-галактозамина, N-н-бутаноил-галактозамина и N-изобутаноилгалактозамина.
68. Конъюгат олигонуклеотида по воплощению 66 или 67, в котором конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, является одновалентной, двухвалентной, трехвалентной или четырехвалентной.
69. Конъюгат олигомера по воплощению 68, в котором конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, состоит из двух-четырех концевых GalNAc группировок, ПЭГ спейсера, связывающего каждую GalNAc группировку с разветвляющей молекулой.
70. Конъюгат олигонуклеотида по воплощениям 66-69, в котором конъюгатная группировка, нацеленная на рецептор асиалогликопротеина, представляет собой трехвалентную N-ацетилгалактозаминную (GalNAc) группировку.
71. Конъюгат олигонуклеотида по воплощениям 66-70, в котором конъюгатная группировка выбрана из одной из трехвалентных GalNAc группировок на Фиг. 1.
72. Конъюгат олигонуклеотида по воплощению 71, в котором конъюгатная группировка представляет собой трехвалентную GalNAc группировку на Фиг. 3.
73. Конъюгат олигонуклеотида по воплощению 63-72, в котором присутствует линкер между олигонуклеотидом или непрерывной олигонуклеотидной последовательностью и конъюгатной группировкой.
74. Конъюгат олигонуклеотида по воплощению 73, в котором линкер представляет собой физиологически лабильный линкер (область В).
75. Конъюгат олигонуклеотида по воплощению 74, в котором физиологически лабильный линкер представляет собой нуклеазочувствительный линкер.
76. Конъюгат олигонуклеотида по воплощению 74 или 75, в котором физиологически лабильный линкер состоит из 2-5 последовательных фосфодиэфирных связей.
77. Конъюгат олигонуклеотида по воплощению 76, в котором физиологически лабильный линкер является эквивалентным области D' или D'', присутствующей в воплощении 47-50.
78. Конъюгат олигонуклеотида по любому из воплощений 63-77, где данный конъюгат олигонуклеотида выбран из CMP ID NO: 766_2, 767_2, 768_2, 769_2 и 770_2.
79. Конъюгат олигонуклеотида по воплощению 78, где данный конъюгат олигонуклеотида выбран из конъюгата олигонуклеотида, представленного на Фиг. 4, 5, 6, 7 и 8.
80. Конъюгат олигонуклеотида по воплощениям 63-76, который демонстрирует улучшенное ингибирование PD-L1 в клетке-мишени или улучшенное клеточное распределение между печенью и селезенкой, или улучшенное клеточное поглощение в печени конъюгата олигонуклеотида по сравнению с неконъюгированным олигонуклеотидом.
81. Фармацевтическая композиция, содержащая олигонуклеотид по воплощениям 1-62 или конъюгат по воплощениям 63-80 и фармацевтически приемлемый разбавитель, носитель, соль и/или адъювант.
82. Способ изготовления олигонуклеотида по воплощению 1-62, включающий проведение взаимодействия нуклеотидных звеньев, образуя, посредством этого, ковалентно связанные смежные нуклеотидные звенья, содержащиеся в олигонуклеотиде.
83. Способ по воплощению 82, дополнительно включающий проведение взаимодействия непрерывной нуклеотидной последовательности с ненуклеотидной конъюгатной группировкой.
84. Способ изготовления композиции по воплощению 81, включающий смешивание олигонуклеотида с фармацевтически приемлемым разбавителем, носителем, солью и/или адъювантом.
85. Способ in vivo или in vitro модулирования экспрессии PD-L1 в клетке-мишени, которая экспресирует PD-L1, причем указанный способ включает введение олигонуклеотида по воплощениям 1-62 или конъюгата по воплощениям 63-80, или фармацевтической композиции по воплощению 81 в эффективном количестве в указанную клетку.
86. Способ лечения или предупреждения заболевания, включающий введение терапевтически или профилактически эффективного количества олигонуклеотида по воплощениям 1-62 или конъюгата по воплощениям 63-80, или фармацевтической композиции по воплощению 81 субъекту, страдающему от или восприимчивому к заболеванию.
87. Способ восстановления иммунитета против вируса или паразита, включающий введение терапевтически или профилактически эффективного количества конъюгата олигонуклеотида по воплощениям 63-80 или олигонуклеотида по воплощениям 1-62, или фармацевтической композиции по воплощению 81 субъекту, инфицированному вирусом или паразитом.
88. Способ по воплощению 87, причем восстановление иммунитета представляет собой увеличение в печени Т-клеток CD8+, специфичных в отношении одного или более чем одного антигена HBV по сравнению с контролем.
89. Олигонуклеотид по воплощениям 1-62 или конъюгат по воплощениям 63-80, или фармацевтическая композиции по воплощению 81 для применения в качестве лекарственного средства для лечения или предупреждения заболевания у субъекта.
90. Применение олигонуклеотида по воплощениям 1-62 или конъюгата по воплощениям 63-80 для получения лекарственного средства для лечения или предупреждения заболевания у субъекта.
91. Олигонуклеотид по воплощениям 1-62 или конъюгат по воплощениям 63-80, или фармацевтическая композиции по воплощению 81 для применения в восстановлении иммунитета против вируса или паразита.
92. Применение по воплощению 91, где восстановление иммунитета представляет собой увеличение в печени Т-клеток CD8+, специфичных в отношении одного или более чем одного антигена HBV по сравнению с контролем.
93. Применение по воплощению 92, где антиген HBV представляет собой HBsAg.
94. Способ, олигонуклеотид или применение по воплощениям 86-93, где заболевание ассоциировано с активностью PD-L1 in vivo.
95. Способ, олигонуклеотид или применение по воплощениям 86-94, где заболевание ассоциировано с повышенной экспрессией PD-L1 в антигенпрезентирующей клетке.
96. Способ, олигонуклеотид или применение по воплощению 95, где экспрессия PD-L1 снижается по меньшей мере на 30% или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 95% по сравнению с экспрессией без обработки или до обработки олигонуклеотидом по воплощениям 1-62 или конъюгатом по воплощениям 63-80, или фармацевтической композицией по воплощению 81.
97. Способ, олигонуклеотид или применение по воплощениям 86-95, где заболевание выбрано из вирусной инфекции печени или паразитарных инфекций.
98. Способ, олигонуклеотид или применение по воплощению 98, где вирусная инфекция представляет собой HBV, HCV или HDV.
99. Способ, олигонуклеотид или применение по воплощениям 86-95, где заболевание представляет собой хроническую инфекцию HBV.
100. Способ, олигонуклеотид или применение по воплощению 98, где паразитарная инфекция представляет собой малярию, токсоплазмоз, лейшманиоз или трипаносомоз.
101. Способ, олигонуклеотид или применение по воплощениям 86-100, где субъект представляет собой млекопитающее.
102. Способ, олигонуклеотид или применение по воплощению 101, где млекопитающее представляет собой человека.
ПРИМЕРЫ
Материалы и методы
Мотивы последовательностей и олигонуклеотидные соединения
Таблица 5: список последовательностей олигонуклеотидных мотивов (указанных по SEQ ID NO), нацеленных на транскрипт человеческого PD-L1 (SEQ ID NO: 1), их конструкции, а также специфичные антисмысловые олигонуклеотидные соединения (указанные по CMP ID NO), разработанные на основе мотива последовательности.
Мотивы последовательностей представляют собой непрерывную последовательность нуклеиновых оснований, присутствующую в олигонуклеотиде.
Конструкции относятся к конструкции гэпмеров - F-G-F' - где каждое число представляет число последовательных модифицированных нуклеозидов, например, 2' модифицированных нуклеозидов (первое число - 5'-фланг), с последующим числом нуклеозидов ДНК (второе число - область гэпа), с последующим числом модифицированных нуклеозидов, например, 2' модифицированных нуклеозидов (третье число - 3'-фланг), которому возможно предшествуют или после которого возможно следуют дополнительные повторяющиеся области ДНК и LNA, которые не являются необходимой частью данной непрерывной последовательности, которая является комплементарной нуклеиновой кислоте-мишени.
Олигонуклеотидные соединения представляют конкретные конструкции мотивов последовательности. Заглавные буквы представляют нуклеозиды бета-D-окси LNA, строчные буквы представляют нуклеозиды ДНК, все C LNA представляют собой 5-метилцитозин, все межнуклеозидные связи представляют собой фосфоротиоатные межнуклеозидные связи.
Таблица 6: олигонуклеотиды, нацеленные на мышиный транскрипт PD-L1 (SEQ ID NO: 4), их конструкция, а также специфичные олигонуклеотидные соединения (указанные по CMP ID NO), разработанные на основе последовательности мотива.
Мотивы последовательностей представляют собой непрерывную последовательность нуклеиновых оснований, присутствующую в олигонуклеотиде.
Конструкции относятся к конструкции гэпмеров - F-G-F' - где каждое число представляет число последовательных модифицированных нуклеозидов, например, 2' модифицированных нуклеозидов (первое число - 5'-фланг), с последующим числом нуклеозидов ДНК (второе число - область гэпа), с последующим числом модифицированных нуклеозидов, например, 2' модифицированных нуклеозидов (третье число - 3'-фланг), которому возможно предшествуют или после которого возможно следуют дополнительные повторяющиеся области ДНК и LNA, которые не являются необходимой частью данной непрерывной последовательности, которая является комплементарной нуклеиновой кислоте-мишени.
Олигонуклеотидные соединения представляют конкретные конструкции мотивов последовательности. Заглавные буквы представляют нуклеозиды бета-D-окси LNA, строчные буквы представляют нуклеозиды ДНК, все C LNA представляют собой 5-метилцитозин, все межнуклеозидные связи представляют собой фосфоротиоатные межнуклеозидные связи.
Таблица 7: последовательности олигонуклеотидных мотивов и антисмысловые соединения с 5'-ca биорасщепляемым линкером.
Заглавные буквы представляют нуклеозиды бета-D-окси LNA, строчные буквы представляют нуклеозиды ДНК, все C LNA представляют собой 5-метилцитозин, подстрочный символ o представляет фосфодиэфирную межнуклеозидную связь, и, если не указано иначе, другие межнуклеозидные связи представляют собой фосфоротиоатные межнуклеозидные связи.
Таблица 8: антисмысловые олигонуклеотидные соединения, конъюгированные с GalNAc
GN2 представляет трехвалентный GalNAc кластер, показанный на Фиг. 3, С6 представляет аминоалкильную группу с 6 атомами углерода, заглавные буквы представляют нуклеозиды бета-D-окси LNA, строчные буквы представляют нуклеозиды ДНК, все C LNA представляют собой 5-метилцитозин, подстрочный символ o представляет фосфодиэфирную нуклеозидную связь, и, если не указано иначе, другие межнуклеозидные связи представляют собой фосфоротиоатные межнуклеозидные связи. Химические формулы, представляющие некоторые из данных молекул, показаны на Фиг. 4-8.
Мышиные модели AAV/HBV
Модель Пастера:
В Институте Пастера создали и разводили трансгенных по HLA-A2.1-/HLA-DR1 нокаутированных по H-2 класса I-/класса II мышей (именуемых в данном документе HLA-A2/DR1). Данные мыши представляют собой экспериментальную модель in vivo для исследований человеческой иммунной функции без какого-либо мешающего влияния ответа мышиного МНС (главный комплекс гистосовместиости) (Pajot et al 2004 Eur J Immunol. 34(11):3060-9.
В данных исследованиях использовали вектор на основе аденосателлитного вируса (AAV) серотипа AAV 2/8, несущий компетентный к репликации ДНК-геном HBV. Вектор AAV-HBV (партия GVPN #6163) разводили в стерильном фосфатно-солевом буферном растворе (PBS) с достижением титра 5×1011 вг (геном вектора)/мл. Мышам внутривенно (в.в.) инъецировали 100 мкл данного разведенного раствора (доза/мышь: 5×1010 вг) в хвостовую вену. В крови мышей-носителей HBV выявляли полные вирусные частицы, содержащие ДНК HBV. HBcAg выявляли в течение вплоть до одного года в печени, наряду с циркулирующими белками HBV - HBeAg и HBsAg - в крови. У всех мышей, трансдуцированных AAV2/8-HBV, HBsAg, HBeAg и ДНК HBV сохранялись в сыворотке в течение по меньшей мере одного года (Dion et al 2013 J Virol 87:5554-5563).
Шанхайская модель:
В данной модели у мышей, инфицированных рекомбинантным аденосателлитным вирусом (AAV), несущим геном HBV (AAV/HBV), поддерживается стабильная виремия и антигенимия в течение больше, чем 30 недель (Dan Yang, et al. 2014 Cellular & Molecular Immunology 11, 71-78).
Самцов мышей C57BL/6 (4-6-недельных), не содержащих специфических патогенов, приобретали в SLAC (Центр животных Шанхайской лаборатории Китайской академии наук) и содержали в виварии в индивидуально вентилируемых клетках. Следовали руководствам по уходу и применению животных, как указывается WuXi IACUC (Институциональный комитет по уходу и применению животных, номер потокола WUXI IACUC R20131126-мышь). Мышам давали акклиматизироваться к новым условиям в течение 3 суток и группировали их согласно схеме экперимента.
Рекомбинантный AAV-HBV разводили в PBS, 200 мкл на инъекцию. Данный рекомбинантный вирус несет 1,3 копии генома HBV (генотип D, серотип ayw).
В сутки 0 всем мышам инъецировали через хвостовую вену 200 мкл AAV-HBV. В сутки 6, 13 и 20 после инъекции AAV всем мышам делали подчелюстное кровопускание (0,1 мл крови/мышь) для отбора сыворотки. В сутки 22 после инъекции мыши со стабильной виремией были готовы для обработки олигонуклеотидами. Данные олигонуклеотиды могут быть неконъюгированными или конъюгированными с GalNAc.
ДНК-вакцина
Плазмидная ДНК не содержала эндотоксинов и была изготовлена Plasmid-Factory (Германия). pCMV-S2.S ayw кодирует домены preS2 и S HBsAg (генотип D), и их экспрессия контролируется немедленным ранним промотором гена цитомегаловируса (Michel et al 1995 Proc Natl Acad Sci U S A 92:5307-5311). pCMV-HBc кодирует капсид HBV, несущий Ag (антиген) кора вируса гепатита (HBc) (Dion et al 2013 J Virol 87:5554-5563).
Обработку ДНК-вакциной проводили, как описано в данном документе. За пять суток до вакцинации в мышцы мышей инъецировали кардиотоксин (CaTx, латоксан реф. L81-02, 50 мкл/мышцу). CaTx деполяризует мышечные волокна с индукцией дегенерации клеток, через 5 суток после инъекции появятся новые мышечные волокна, и они будут получать ДНК-вакцину для лучшей эффективности трансфекции. Каждый из pCMV-S2.S ayw и pCMVCore в концентрации 1 мг/мл смешивали в равном количестве, и каждая мышь получала всего 100 мкг двухсторонней внутримышечной инъекции в обработанные кардиотоксином передние большеберцовые мышцы, как было описано ранее в Michel et al 1995 Proc Natl Acad Sci U S A 92:5307-5311, под анестезией (100 мкл 12,5 мг/мл кетамина, 1,25 мг/мл ксилазина).
Антитело против PD-L1
Это мышиное антитело IgG1 против мышиного PD-L1 клона 6Е11, произведенное внутри Genetech. Оно представляет собой имитирующее антитело, которое перекрестно блокирует атезолизумаб, и имеет аналогичную активность, блокирующую in vitro атезолизумаб, произведенный внутри Roche. Данное антитело вводили внутрибрюшинной инъекцией (в.б.) в дозе 12,5 мкг/г.
Синтез олигонуклеотидов
Синтез олигонуклеотидов обычно известен в данной области. Ниже представлен протокол, который можно применять. Олигонуклеотиды по настоящему изобретению возможно были получены слегка отличными способами в показателях использованных прибора, подложки и концентраций.
Олигонуклеотиды синтезируются на уридиновых универсальных подложках с использованием фосфорамидитного подхода на Oligomaker 48 в масштабе 1 мкмоль. В конце синтеза олигонуклеотиды отщепляются от твердой подложки с использованием водного аммиака в течение 5-16 часов при 60°С. Олигонуклеотиды очищаются посредством ВЭЖХ (высокоэффективная жидкостная хроматография) с обращенной фазой (ВЭЖХ-ОФ) или посредством экстракций твердой фазы и характеризуются посредством СВЭЖХ (сверхэффективная жидкостная хроматография), и далее подтверждается молекулярная масса посредством МС-ЭРИ (масс-спектрометрия с электрораспылительной ионизацией).
Элонгация олигонуклеотида:
Связывание β-цианоэтил-фосфорамидитов (ДНК-А(Bz), ДНК-G(ibu), ДНК-C(Bz), ДНК-Т, LNA-5-метил-C(Bz), LNA-A(Bz), LNA-G(dmf) или LNA-T) проводится посредством применения раствора 0,1 М амидита, защищенного 5'-O-DMT, в ацетонитриле и DCI (4,5-дицианоимидазол) в ацетонитриле (0,25 М) в качестве активатора. Для последнего цикла может быть использован фосфорамидит с желательными модификациями, например, С6-линкер для присоединения конъюгатной группы или конъюгатная группа как таковая. Тиолирование для введения фосфортиоатных связей проводится с использованием ксантангидрида (0,01 М в ацетонитриле/пиридине 9:1). Фосфодиэфирные связи могут вводиться с использованием 0,02 М йода в THF/пиридине/воде 7:2:1. Остальными реактивами являются реактивы, типично используемые для синтеза олигонуклеотидов.
Для конъюгирования после твердофазного синтеза можно использовать имеющийся в продаже С6 аминолинкер фосфорамидит в последнем цикле твердофазного синтеза, и после снятия защиты и отщепления от твердой подложки выделяется связанный аминолинкером олигонуклеотид со снятой защитой. Конъюгаты вводятся через активацию функциональной группы с использованием стандартных способов синтеза.
В качестве альтернативы, конъюгатную группировку можно добавлять к олигонуклеотиду, в то время как он все еще находится на твердой подложке с использованием фосфорамидита с GalNAc или с кластером GalNAc, как описано в PCT/EP2015/073331 или в заявке EP № 15194811.4.
Очистка посредством ВЭЖХ-ОФ:
Неочищенные соединения очищают препаративной ВЭЖХ-ОФ на колонке Phenomenex Jupiter C18 10 мкм 150×10 мм. В качестве буферов используют 0,1 М ацетат аммония, рН 8, и ацетонитрил при скорости тока 5 мл/мин. Отобранные фракции лиофилизируют с получением очищенного соединения - типично в виде белого твердого вещества.
Сокращения:
ВЭЖХ-ОФ: высокоэффективная жидкостная хроматография с обращенной фазой
Анализ Tm
Дуплексы олигонуклеотида и РНК-мишени (связанной фосфатом - РО) разводят до 3 мМ в 500 мл воды, не содержащей РНКазы, и смешивают с 500 мл 2× Tm-буфера (200 мМ NaCl, 0,2 мМ EDTA (этилендиаминтетрауксусная кислота), 20 мМ фосфат Na, pH 7,0). Данный раствор нагревают до 95°С в течение 3 мин и затем дают осуществляться отжигу при комнатной температуре в течение 30 мин. Температуры плавления дуплекса (Tm) измеряют на спектрофотометре Lambda 40, работающем в ультрафиолетовой/видимой области, оснащенном программатором температуры Пелетье РТР6, используя программу PE Templab (Perkin Elmer). Температура повышается от 20°С до 95°С и затем снижается до 25°С, с записью поглощения при 260 нм. Для оценки Tm дуплекса используются первая производная и локальные максимумы как плавления, так и отжига.
Анализ тканеспецифичного расщепления линкера in vitro
Меченные FAM олигонуклеотиды с биорасщепляемым линкером, подлежащие тестированию (например, фосфодиэфирный линкер ДНК (РО линкер)), подвергаются расщеплению in vitro с использованием гомогенатов релевантных тканей (например, печени или почки) и сыворотки.
Образцы ткани и сыворотки отбирают у подходящего животного (например, мыши, обезьяны, свиньи или крысы) и гомогенизируют в гомогенизационном буфере (0,5% Igepal CA-630, 25 мМ Tris, рН 8,0, 100 мМ NaCl, pH 8,0 (доведенный 1 н. NaOH). В гомогенаты ткани и сыворотку впрыскивают олигонуклеотид до концентраций 200 мкг/г ткани. Образцы инкубируются в течение 24 часов при 37°С, и затем данные образцы подвергают экстракции фенолом-хлороформом. Растворы подвергают анализам ВЭЖХ АО (высокоэффективная жидкостная хроматография на анионообменной колонке) на Dionex Ultimate 3000 с использованием колонки Dionex DNApac p-100 и градиента в интервале от 10 мМ до 1 М перхлората натрия при рН 7,5. Содержание расщепленного и нерасщепленного олигонуклеотида определяется относительно стандарта с использованием как флюоресцентного детектора при 615 нм, так и УФ детектора при 260 нм.
Анализ расщепления нуклеазой S1
Меченные FAM олигонуклеотиды с линкерами, чувствительными к нуклеазе S1 (например, фосфодиэфирный ДНК-линкер (РО линкер)), подвергаются расщеплению in vitro в экстракте нуклеазы S1 или сыворотке.
100 мкМ олигонуклеотидов подвергаются расщеплению in vitro нуклеазой S1 в буфере для нуклеазы (60 U на 100 мкл) в течение 20 и 120 минут. Ферментативную активность останавливают добавлением в раствор буфера EDTA. Растворы подвергают анализам ВЭЖХ АО на Dionex Ultimate 3000 с использованием колонки Dionex DNApac p-100 и градиента в интервале от 10 мМ до 1 М перхлората натрия при рН 7,5. Содержание расщепленного и нерасщепленного олигонуклеотида определяется относительно стандарта с использованием как флюоресцентного детектора при 615 нм, так и УФ детектора при 260 нм.
Получение одноядерных клеток печени
Клетки печени от мышей AAV/HBV получали, как описано ниже, и согласно способу, описанному Tupin et al 2006 Methods Enzymol 417:185-201, с небольшими модификациями. После умерщвления мышей осуществляли перфузию печени 10 мл стерильного PBS через портальную вену печени с использованием шприца с иглой калибра 25. Когда орган становлся бледным, данный орган отбирали в сбалансированный солевой раствор Хэнкса (HBSS) (GIBCO® HBSS, 24020) плюс 5 % фетальной телячьей сыворотки с удаленным комплементом (FCS). Отобранную печень аккуратно продавливали через 100 мкм клеточное сито (BD Falcon, 352360), и клетки суспендировали в 30 мл HBSS плюс 5% FCS. Суспензию клеток центрифугировали при 50 g в течение 5 мин. Супернатанты затем центрифугировали при 289 g в течение 10 мин при 4°С. После центрифугирования супернатанты отбрасывали, и осадки ресуспендировали в 15 мл при комнатной температуре в 35%-ном изотоничном растворе перколла (Percoll от GE Healthcare #17-0891-01, разведенный в RPMI 1640 (GIBCO, 31870)) и переносили в 15 мл пробирку. Клетки далее центрифугировали при 1360 g в течение 25 мин при комнатной тмпературе. Супернатант отбрасывали отсасыванием, и осадок, содержащий одноядерные клетки, дважды промывали HBSS плюс 5% FCS.
Клетки культивировали в полной среде (α-минимальная незаменимая среда (Gibco, 22571), дополненная 10% FCS (Hyclone, # SH30066, партия APG21570), 100 U/мл пенициллина плюс 100 мкг/мл стрептомицина плюс 0,3 мг/мл L-глутамина (Gibco, 10378), 1× заменимыми аминокислотами (Gibco, 11140), 10 мМ Hepes (Gibco, 15630), 1 мМ пируватом натрия (Gibco, 11360) и 50 мкM β-меркаптоэтанолом (LKB, 1830)).
Поверхностное мечение клеток
Клетки высевали в 96-луночные планшеты с U-образным дном и промывали PBS для FACS (флуоресцентная сортировка клеток) (PBS, содержащий 1% бычьего сывороточного альбумина и 0,01% азида натрия). Клетки инкубировали с 5 мкл PBS для FACS, содержащего крысиное антитело против мышиного CD16/CD32 и маркер жизнеспособности LD фиксируемый желтый, Thermofisher, L34959, в течение 10 мин в темноте при 4°С. Затем клетки окрашивали в течение 20 мин в темноте при 4°С с использованием 25 мкл PBS для FACS, содержащего моноклональные антитела (Mab) против NK P46 BV421 (крысиное Mab против мышиного NK P46, Biolegend, 137612) и F4/80 (крысиное Mab против мышиного F4/80, конъюгированное с FITC (флюоресцеинизотиоцианат), BD Biolegend, 123108), и также добавляли два дополнительных поверхностных маркера: PD1 (крысиное Mab против мышиного PD1, конъюгированное с PE (фосфоэстераза), BD Biosciences, 551892) и PDL1 (крысиное Mab против мышиного PDL1 BV711, Biolegend, 124319).
Анализ внутриклеточного окрашивания на цитокины (ICS)
Анализы ICS проводили и на спленоцитах, и на одноядерных клетках печени. Клетки высевали в 96-луночные планшеты с U-образным дном. Планшеты с клетками инкубировали в течение ночи при 37° либо в одной полной среде в качестве негативного контроля, либо с пептидами, описанными в Таблице 9, в концентрации 2 мкг/мл. После одного часа инкубации добавляли брефельдин А (Sigma, B6542) в концентрации 2 мкг/мл.
После культуры в течение ночи клетки промывали PBS для FACS и инкубировали с 5 мкл PBS для FACS, содержащего крысиное антитело против мышиного CD16/CD32 и маркер жизнеспособности LD фиксируемый желтый, Thermofisher, L34959, в течение 10 мин в темноте при 4°С. Затем клетки окрашивали в течение 20 мин в темноте при 4°С с использованием 25 мкл PBS для FACS, содержащего Mab. Смесь состояла из моноклональных антител против CD3 (Mab хомяка против мышиного CD3-PerCP, BD Biosciences, 553067), против CD8 (крысиное Mab против мышиного CD8-APC-H7, BD Biosciences, 560182), против CD4 (крысиное Mab против мышиного CD4-РЕ-Cy7, BD Biosciences, 552775) и против NK-клеток (крысиное Mab против мышиных NK P46 BV421, Biolegend, 137612). Клетки фиксировали после нескольких промывок и делали проницаемыми в течение 20 мин в темноте при комнатной температуре с использованием Cytofix/Cytoperm, промывали раствором Perm/Wash (BD Biosciences, 554714) при 4°С.
Внутриклеточное окрашивание на цитокины с использованием антител против IFNγ (интерферон-гамма) (крысиное Mab против мышиного IFNγ-APC, клон XMG1.2, BD Biosciences, 554413) и фактора некроза опухолей-альфа (TNFα) (крысиное Mab против мышиного TNFα-FITC, клон MP6-XT22; 1/250 (BD Biosciences 554418) проводили в течение 30 мин в темноте при 4°С. Перед анализом посредством проточной цитометрии с использованием анализатора MACSQuant клетки промывали Perm/Wash и ресуспендировали в PBS для FACS, содержащем 1% формальдегида.
Живые клетки CD3+CD8+CD4- и CD3+CD8-CD4+ сортировали и представляли на точечном графке. Определяли две области для сортировки позитивных клеток в отношении каждого цитокина. Число событий, обнаруженное при данных сортировках, делили на общее число событий в родительской популяции для получения процентных содержаний отвечающих Т-клеток. Для каждой мыши процентная доля, полученная в одной среде, рассматривалась как фон и вычиталась из процентной доли, полученной при стимулированиях пептидами.
Порог позитивности определяли согласно фону эксперимента, т.е. средней процентной доле окрашенных клеток, полученной для каждой группы в условиях одной среды, плюс два стандартных отклонения. Только процентная доля цитокина, представленного по меньшей мере в 5 событиях, считалась позитивной.
Таблица 9: эпитопы, ограниченные HLA-A2/DR1, содержащиеся в коровом белке HBV и доменах оболочки HВsAg (S2+S).
HLA
(SEQ ID NO: 773)
Gastroenterology 1997;112:193-199
(SEQ ID NO: 774)
Gastroenterology 1997;112:193-199
(SEQ ID NO: 775)
(SEQ ID NO: 776)
(SEQ ID NO: 777)
1994;153:5586-5592.
(SEQ ID NO: 778)
(SEQ ID NO: 779)
(SEQ ID NO: 780)
(SEQ ID NO: 781)
(SEQ ID NO: 782)
(SEQ ID NO: 783)
Пример 1. Тестирование эффективности in vitro
Проводили прогулку по гену по человеческому транскрипту PD-L1, используя, главным образом, 16-20-мерные гэпмеры. Тестирование эффективности проводили в эксперименте in vitro в моноцитарной линии клеток человеческого лейкоза ТНР1 и в линии клеток человеческой неходжкинской лимфомы K (KARPAS-299).
Линии клеток
Линию клеток ТНР1 и Karpas-299 исходно приобретали в Европейской коллекции аутентичных культур клеток (ЕСАСС) и поддерживали, как рекомендовано поставщиком, в увлажненном инкубаторе при 37°С с 5% СО2.
Эффективность олигонуклеотидов
Клетки ТНР-1 (3×104 в RPMI-Glutamax, 10% FBS, 1% пенициллина-стрептомицина (Thermo Fisher Scientific) добавляли к олигонуклеотидам (4-5 мкл) в 96-луночные круглодонные планшеты и культивировали в течение 6 суток в конечном объеме 100 мкл/лунку. Олигонуклеотиды подвергали скринингу в одной единственной концентрации (20 мкМ) и в интервале дозы-концентрациях от 25 мкМ до 0,004 мкМ (разведение 1:3 в воде). Общую мРНК экстрагировали с использованием набора для клеточной РНК в большом объеме MagNA Pure 96 на системе MagNA Pure 96 System (Roche Diagnostics) согласно инструкциям изготовителя. Для анализа экспрессии генов проводили кПЦР-ОТ (количественная полимеразная цепная реакция, сопряженная с обратной транскрипцией) с использованием набора TaqMan RNA-to-ct 1-Step (Thermo Fisher Scientific) на приборе QuantStudio (Applied Biosystems) с предварительно сконструированными праймерами Taqman, нацеленными на человеческий PD-L1 и ACTB, используемый в качестве эндогенного контроля (Thermo Fisher Scientific). Относительный уровень экспрессии мРНК PD-L1 рассчитывали с использованием способа 2(-дельта дельта С(Т)), а процентную долю ингибирования выражали в % по сравнению с контрольным образцом (необработанные клетки).
Клетки Karpas-299 культивировали в RPMI 1640, 2 мМ глутамине и 20% FBS (Sigma). Клетки, посеянные в количестве 10000 клеток/лунку в 96-луночные планшеты, инкубировали в течение 24 часов перед добавлением олигонуклеотидов, растворенных в PBS. Конечная концентрация олигонуклеотидов была в одной дозе 5 мкМ в конечном объеме культуры 100 мкл/лунку, или их добавляли в дозе, варьирующей от 50 мкМ, 15,8 мкМ, 5,0 мкМ, 1,58 мкМ, 0,5 мкМ, 0,158 мкМ, 0,05 мкМ до 0,0158 мкМ в объеме культуры 100 мкл. Клетки отбирали через 3 суток после добавления олигонуклеотидных соединений, и РНК экстрагировали с использованием набора для очистки РНК PureLink Pro 96 (Ambion) согласно инструкциям изготовителя. кДНК синтезировали с использованием обратной транскриптазы M-MLT, случайных декамеров RETROscript, ингибитора РНКазы (Ambion) и набора 100 мM дНТФ (дезоксинуклеотидтрифосфат) (Invitrogen, уровень качества для ПЦР) согласно инструкции изготовителя. Для анализа экспрессии генов проводили кПЦР с использованием мастер-микса (2×) TaqMan Fast Advanced (Ambion) в дуплексе, установленном с праймерами TaqMan для анализов PD-L1 (Applied Biosystems; Hs01125299_m1) и TBP (Applied Biosystems; 4325803). Относительный уровень экспрессии мРНК PD-L1 показан в Таблице 10 как % от контрольного образца (клетки, обработанные PBS).
Таблица 10: эффективность in vitro соединений против PD-L1 в линиях клеток ТНР1 и KARPAS-299 (среднее из экспериментов с n, равным 3). Уровни мРНК PD-L1 нормировали к ТВР в клетках KARPAS-299 или к АСТВ в клетках ТНР1, и они показаны как % от контроля (клетки, обработанные PBS).
5 мкМ CMP
20 мкМ CMP
Пример 2 - тестирование эффективности in vitro в кривой доза-ответ
Набор олигонуклеотидов из Таблицы 10 тестировали в клетках KERPAS-299 с использованием полу-log серийных разведений в PBS (от 50 мкМ, 15,8 мкМ, 5,0 мкМ, 1,58 мкМ, 0,5 мкМ, 0,158 мкМ, 0,05 мкМ до 0,0158 мкМ олигонуклеотида) в анализе эффективности in vitro, описанном в Примере 1. Для данных олигонуклеотидов оценивали IC50 (полумаксимальная ингибирующая концентрация) и максимальное ингибирование (% остаточной экспрессии PD-L1).
Расчеты ЕС50 (полумаксимальная эффективная концентрация) проводили в GraphPad Prism6. IC50 и максимальный уровень нокдауна PD-L1 показаны в Таблице 11 как % от контрольных клеток (обработанных PBS).
Таблица 11: максимальное ингибирование как % от обработки физиологическим раствором и ЕС50 в линии клеток KARPAS-299.
(% остаточной экспрессии PD-L1; % от обработанных физиологическим раствором)
Набор олигонуклеотидов из Таблицы 6 тестировали в клетках ТНР-1 с использованием серийного разведения 1:3 в воде от 25 мкМ до 0,004 мкМ в анализе эффективности in vitro, описанном в Примере 1. Для данных олигонуклеотидов оценивали IC50 и максимальное ингибирование (% остаточной экспрессии PD-L1).
Расчеты ЕС50 проводили в GraphPad Prism6. IC50 и максимальный уровень нокдауна PD-L1 показаны в Таблице 12 как % от контрольных клеток (обработанных PBS).
Таблица 12: максимальное ингибирование как % от обработки физиологическим раствором и ЕС50 в линии клеток ТНР1.
(% остаточной экспрессии PD-L1; % от обработанных физиологическим раствором)
Результаты в Таблицах 7 и 8 также показаны на Фиг. 2 в связи с их положением, где они нацелены на пре-мРНК PD-L1 SEQ ID NO: 1.
Из этого можно видеть то, что почти все из соединений имеют значения ЕС50 меньше 1 мкМ и целевой нокдаун меньше 25% от уровня экспрессии PD-L1 в контрольных клетках (обработанных физиологическим раствором).
Пример 3 - активность и эффективность in vitro и снижение уровня PD-L1 in vivo у мышей, индуцированных поли(I:C), с использованием голых и конъюгированных с GalNAc антисмысловых олигонуклеотидов к PD-L1
Тестирование эффективности и активности проводили в эксперименте in vitro в исследованиях доза-ответ в клетках МСР-11 с использованием олигонуклеотидов в Таблице 6. Те же самые олигонуклеотиды, а также версии, конъюгированные с GalNAc (Таблица 8, CMP ID NO 755_2-765_2), тестировали in vivo у самок мышей C57BL/6J, индуцированных поли(I:C), на их способность уменьшать экспрессию мРНК и белка PD-L1.
Анализ in vitro
Клетки МСР-11 (исходно купленные в АТСС (Американская коллекция типовых клеточных культур)), суспендированные в DMEM (среда Игла, модифицированная по Дульбекко) (кат. № Sigma D0819), дополненной 10% лошадиной сыворотки, 2 мМ L-глутамином, 0,025 мг/мл гентамицином и 1 мМ пируватом натрия, добавляли при плотности 8000 клеток/лунку к олигонуклеотидам (10 мкл) в 96-луночные круглодонные планшеты и культивировали в течение 3 суток в конечном объеме 200 мкл/лунку в увлажненном инкубаторе при 37°С с 5% СО2. Олигонуклеотиды подвергали скринингу в интервале концентраций (50 мкМ, 15,8 мкМ, 5,0 мкМ, 1,58 мкМ, 0,5 мкМ, 0,158 мкМ, 0,05 мкМ до 0,0158 мкМ).
Общую мРНК экстрагировали с использованием набора для очистки РНК PureLink Pro 96 (Ambion) согласно инструкциям изготовителя. кДНК синтезировали с использованием обратной транскриптазы M-MLT, случайных декамеров RETROscript, ингибитора РНКазы (Ambion) и набора 100 мM дНТФ (Invitrogen, уровень качества для ПЦР) согласно инструкции изготовителя. Для анализа экспрессии генов проводили кПЦР с использованием мастер-микса (2×) TaqMan Fast Advanced (Ambion) в дуплексе, установленном с праймерами TaqMan для анализа PD-L1 (Thermo Fisher Scientific; FAM-MGB Mm00452054-m1) и Gusb (Thermo Fisher Scientific; VIC-MGB-PL Mm01197698-m1). Относительный уровень экспрессии мРНК PD-L1 показан в Таблице 9 как % остаточной экспрессии PD-L1 в % от контрольных образцов с PBS (клетки, обработанные PBS). Расчеты ЕС50 проводили в GraphPadPrism6. ЕС50 и максимальный уровень нокдауна PD-L1 показаны в Таблице 13 как % от контрольных (PBS) клеток.
Анализ in vivo
Самкам мышей C57BL/6J (20-23 г; 5 мышей на группу) п.к. инъецировали 5 мг/кг неконъюгированных олигонуклеотидов к мышиному PD-L1 или 2,8 мг/кг конъюгированных с GalNAc олигонуклеотидов к мышиному PD-L1. Через трое суток мышам в.в. инъецировали 10 мг/кг поли(I:C) (LWM, Invivogen). Мышей умерщвляли через 5 ч после инъекции поли(I:C), и образцы печени помещали в RNAlater (Thermo Fisher Scientific) для экстракции РНК или замораживали на сухом льду для экстракции белка.
Общую мРНК экстрагировали из гомогенизированных образцов печени с использованием набора для очистки РНК PureLink Pro 96 (Ambion) согласно инструкциям изготовителя. кДНК синтезировали с использованием обратной транскриптазы M-MLT, случайных декамеров RETROscript, ингибитора РНКазы (Ambion) и набора 100 мM дНТФ (Invitrogen, уровень качества для ПЦР) согласно инструкции изготовителя. Для анализа экспрессии генов проводили кПЦР с использованием мастер-микса (2×) TaqMan® Fast Advanced (Ambion) в дуплексе, установленном с праймерами TaqMan для анализа мРНК PD-L1 (Thermo Fisher Scientific; FAM-MGB Mm00452054-m1) и ТВР (Thermo Fisher Scientific; VIC-MGB-PL Mm00446971_m1). Относительный уровень экспрессии мРНК PD-L1 показан в Таблице 13 как % контрольных образцов от мышей, которым инъецировали физиологический раствор и поли(I:C).
Гомогенаты печени получали посредством проведения гомогенизации образцов печени в 2 мл на 100 мг ткани реактива для эктракции тканевого белка T-PER® (Thermo Fisher Scientific), смешанного с 1× смесью ингибиторов протеаз Halt, не содержащей EDTA (Thermo Fisher Scientific). Концентрации белка в гомогенатах печени измеряли с использованием реактива для анализа кумасси плюс (Бредфорд) (Thermo Scientific) согласно инструкциям изготовителя. Гомогенаты печени (40 мкг белка) разделяли на 4-12%-ных Bis-Tris Plus полиакриламидных гелях (Thermo Fisher Scientific) в 1× электродном буфере MOPS и переносили на нитроцеллюлозные мембраны с использованием системы блоттинга iBLOT Dry (Thermo Fisher Scientific) согласно инструкциям изготовителя. Каждый блот разрезали горизонтально на две части по полосе 64 кДа. После блокирования в TBS, содержащем 5% обезжиренного молока и 0,05% Tween 20, мембраны инкубировали в течение ночи при 4°С с кроличьим моноклональным антителом против винкулина (Abcam, кат. № ab129002), разведенным 1:10000 (верхние мембраны), или с поликлональным антителом козы против mPD-L1 (R&D Systems, кат. № AF1019), разведенным 1:1000 (нижние мембраны) в TBS, содержащем 5% обезжиренного молока и 0,05% Tween 20. Мембраны промывали в TBS, содержащем 0,05% Tween 20, и подвергали воздействию в течение 1 ч при комнатной температуре свиного антитела против IgG кролика, конъюгированного с HRP (пероксидаза хрена) (DAKO), разведенного 1:3000 (верхние мембраны), или кроличьего антитела против IgG козы, конъюгированного с HRP (DAKO), разведенного 1:2000 в TBS, содержащем 5% обезжиренного молока и 0,05% Tween 20. После промывки мембран реактивность выявляли с использованием ECL select (Amersham GE Healthcare). Для каждой группы мышей, обработанных олигонуклеотидами, оценивали интенсивность полос PD-L1 по отношению к полосам винкулина посредством сравнения с интенсивностями полос PD-L1/винкулин мышей, которым инъецировали физиологический раствор и поли(I:C) (контроль). Результаты показаны в Таблице 13, и вестерн-блоты с парами голых и конъюгированных олигонуклеотидов показаны на Фиг. 9А-Д.
Таблица 13: эффективность in vitro и in vivo олигонуклеотидов к мышиному PD-L1.
(% от контроля)
(относительно контроля)
+++: аналогичная интенсивности полосы PD-L1/винкулина контроля; ++: слабее, чем интенсивность полосы PD-L1/винкулина контроля; +: значительно слабее, чем интенсивность полосы PD-L1/винкулина контроля; nd - не определяется.
Из данных Таблицы 13 можно видеть то, что конъюгирование олигонуклеотидов с GalNAc явно улучшает уменьшение уровня PD-L1 in vivo. Уменьшение уровня мРНК обычно коррелирует с уменьшением уровня белка PD-L1. За исключением CMP ID NO: 754_1, низкое значение ЕС50 in vitro обычно отражает хорошее снижение уровня мРНК PD-L1 in vivo, если только олигонуклеотид конъюгирован с GalNAc.
Пример 4 - PK (фармакокинетика)/PD (фармакодинамика) in vivo в сортированных гепатоцитах и непаренхимных клетках от мышей, индуцированных поли(I:C)
Исследовали распределение голых и конъюгированных с GalNAc олигонуклеотидов, а также уменьшение уровня мРНК PD-L1 в гепатоцитах и непаренхимных клетках, выделенных из мышей, индуцированных поли(I:C).
Самкам мышей C57BL/6J (n равно 3 на группу) п.к. (подкожно) инъецировали 5 мг/кг неконъюгированного олигонуклеотида (748_1) или 7 мг/кг олигонуклеотидов, конъюгированных с GalNAc (759_2), нацеленных на мышиную мРНК PD-L1. Через двое суток мышам в.б. (внутрибрюшинно) инъецировали 15 мг/кг поли(I:C) (LWM, Invivogen). Через 18-20 ч после инъекции поли(I:C) мышей анестезировали, и подвергали печень перфузии при скорости тока 7 мл в мин через полую вену с использованием сбалансированного солевого раствора Хэнкса, содержащего 15 мМ Hepes и 0,38 мM EGTA, в течение 5 мин, с последующей перфузией раствором коллагеназы (сбалансированный солевой раствор Хэнкса, содержащий 0,17 мг/мл коллагеназы типа 2 (Worthington 4176), 0,03% BSA, 3,2 мM CaCl2 и 1,6 г/л NaHCO3) в течение 12 мин. После перфузии печень удаляли и открывали капсулу печени, суспензию печени фильтровали через 70 мкм клеточное сито с использованием среды William E, и аликвоту клеточной суспензии (смешанные клетки печени) удаляли для более позднего анализа. Остаток клеточной суспензии центрифугировали в течение 3 мин при 50 g. Супернатант отбирали для более поздней очистки непаренхимных клеток. Осадок ресуспендировали в 25 мл среды William E (Sigma, кат. № W1878, дополненная 1× пенициллином/стрептомицином, 2 мМ L-глутамином и 10% FBS (ATCC #30-2030)), смешивали с 25 мл среды William E, содержащей 90% перколла, и гепатоциты осаждали центрифугированием при 50 g в течение 10 мин. После 2-кратной промывки в среде William E осажденные гепатоциты ресуспендировали в среде William E. Супернатант, содержащий непаренхимные клетки, центрифугировали при 500 g 7 мин, и данные клетки ресуспендировали в 4 мл среды RPMI и центрифугировали через два слоя перколла (25% и 50% перколла) при 1800 g в течение 30 мин. После отбора непаренхимных клеток между двумя слоями перколла, данные клетки промывали и ресуспендировали в среде RPMI.
Общую мРНК экстрагировали из очищенных гепатоцитов, непаренхимных клеток и общей суспензии печени (нефракционированные клетки печени) с использованием набора для очистки РНК PureLink Pro 96 (Ambion) согласно инструкциям изготовителя. кДНК синтезировали с использованием обратной транскриптазы M-MLT, случайных декамеров RETROscript, ингибитора РНКазы (Ambion) и набора 100 мM дНТФ (Invitrogen, уровень качества для ПЦР) согласно инструкции изготовителя. Для анализа экспрессии генов проводили кПЦР с использованием мастер-микса (2×) TaqMan Fast Advanced (Ambion) в дуплексе, установленном с праймерами TaqMan для анализа PD-L1 (Thermo Fisher Scientific; FAM-MGB Mm00452054-m1) и ТВР (Thermo Fisher Scientific; VIC-MGB-PL Mm00446971_m1). Относительный уровень экспрессии мРНК PD-L1 показан в Таблице 10 как % контрольных образцов от мышей, которым инъецировали физиологический раствор и поли(I:C).
Анализ содержания олигонуклеотидов проводили с использованием ELISA (твердофазный иммуноферментный анализ) с использованием биотинилированного захватывающего зонда с последовательностью 5'-TACCGT-s-Bio-3' и выявляющего зонда, конъюгированного с дигоксигенином, с последовательностью 5'- DIG-C12-S1-CCTGTG - 3'. Данные зонды состояли только из LNA с фосфодиэфирной связью. Образцы печени (приблизительно 50 мг) гомогенизировали в 1,4 мл буфера для лизиса MagNa pure (Roche, кат. № 03604721001) в 2 мл пробирке Эппендорфа, содержащей один 5 мм шарик из нержавеющей стали. Образцы гомогенизировали на гомогенизаторе Retsch MM400 (Merck Eurolab), пока не был получен однородный лизат. Образцы инкубировали в течение 30 мин при комнатной температуре. Стандарты получали впрыскиванием неконъюгированного антисмыслового олигонуклеотидного соединения (CMP ID NO 748_1) в определенных концентрациях в образец необработанной печени, и обрабатывали их также, как и образцы. Впрыснутые концентрации выбирали так, чтобы они соответствовали ожидаемому содержанию олигонуклеотидов в образце (в пределах ~10-кратных отличий).
Гомогенизированные образцы разводили минимум в 10 раз в 5 × буфере SSCT (750 мМ NaCl и 75 мМ цитрат натрия, содержащий 0,05% (об./об.) Tween-20, pH 7,0), и делали серию 6 разведений в 2 раза с использованием раствора для захвата-выявления (35 нМ захватывающий зонд и 35 нМ выявляющий зонд в 5 × буфере SSCT) и инкубировали в течение 30 мин при комнатной температуре. Образцы переносили в 96-луночный планшет, покрытый стрептавидином (Nunc, кат. № 436014), по 100 мкл в каждую лунку. Планшеты инкубировали в течение 1 часа при комнатной температуре с легким встряхиванием. В каждой лунке была сделана промывка три раза 2× буфером SSCT, было добавлено 100 мкл фрагмента Fab против DIG-AP (Roche Applied Science, кат. № 11093274910), разведенного 1:4000 в PBST (фосфатно-солевой буферный раствор, содержащий 0,05% (об./об.) Tween-20, pH 7,2, свежеприготовленный), и проведена инкубация в течение 1 часа при комнатной температуре с легким встряхиванием. Была сделана промывка три раза 2× буфером SSCT, и было добавлено 100 мкл раствора субстрата щелочной фосфатазы (АР) (субстрат Blue Phos, KPL, код продукта 50-88-00, свежеприготовленный). Интенсивность окрашивания измеряли спектрофотометрически при 615 нм после 30-минутной инкубации с легким встряхиванием. Необработанные данные экспортировали из ридеров (программа Gen5 2.0) в формат Excel и далее анализировали в Excel. Стандартные кривые получали с использованием программы GraphPadPrism 6 и логистической регрессионной модели 4PL.
Таблица 14: экспрессия PD-L1 и содержание олигонуклеотидов в общей суспензии печени, гепатоцитах и непаренхимных клетках от поли(I:C) мышей, обработанных неконъюгированными и конъюгированными с GalNAc олигонуклеотидами, n равно 3.
(% от физиологического раствора-поли(I:C))
(нг/105 клеток)
Результаты показывают то, что голые (CMP ID NO: 748_1) и конъюгированные (CMP ID NO: 759_2) олигонуклеотиды одинаково хорошо снижают уровень мРНК PD-L1 в общих клетках печени. В выделенных гепатоцитах эффект конъюгированного олигонуклеотида почти в 5 раз сильнее, чем эффект голого олигонуклеотида, тогда как голые олигонуклеотиды демонстрировали в два раза более сильный эффект, чем олигонуклеотиды, конъюгированные с GalNAc, в непаренхимных клетках. В гепатоцитах и непаренхимных клетках уменьшение экспрессии мРНК PD-L1 в некоторой степени коррелирует с содержанием олигонуклеотидов в данных типах клеток.
Пример 5 - нокдаун PD-L1 in vivo у мышей AAV/HBV с использованием голых и конъюгированных с GalNAc антисмысловых олигонуклеотидов к PD-L1
В настоящем исследовании мышей AAV/HBV обрабатывали голыми или конъюгированными с GalNAc антисмысловыми олигонуклеотидами против PD-L1, и оценивали в печени экспрессию мРНК PD-L1 и экспрессию генов HBV.
Самок мышей HLA-A2/DR1 5-8-недельного возраста (5 животных на группу) предобрабатывали в неделю -1 носителем (физиологический раствор), голыми антисмысловыми олигонуклеотидами к PD-L1 (CMP ID NO 752_1 п.к. в дозе 5 мг/кг) и антисмысловыми олигонуклеотидами к PD-L1, конъюгированными с GalNAc (CMP ID NO 763_2 п.к. в дозе 7 мг/кг), данные дозы соответствуют эквимолярным концентрациям олигонуклеотидов. Данных мышей трансдуцировали 5×1010 вг AAV-HBV в неделю 0 (относительно дополнительных подробностей см. описание мышиной модели AAV/HBV в разделе Материалы и методы). С W1 (неделя 1) после трансдукции AAV-HBV до W4 мыши получали 4 дополнительные п.к. инъекции олигонуклеотидов к PD-L1 или носителя (физиологический раствор), которые давали с интервалом в одну неделю.
Образцы крови отбирали за одну неделю до трансдукции и через одну неделю после каждой инъекции.
Мышей умерщвляли через две недели после последних инъекций, и их печень удаляли после перфузии PBS. Печень разрезали на меньшие куски и прямо замораживали.
Для измерения экспрессии генов HBV ДНК экстрагировали из сыворотки с использованием Qiagen Biorobot, используя QIAamb One для всего набора нуклеиновых кислот, кат. № 965672, сыворотку разводили 1:20 в PBS, всего 100 мкл лизировали в 200 мкл буфера AL. ДНК элюировали из набора в 100 мкл.
Для кПЦР в реальном времени использовали мастер-микс для экспрессии генов TaqMan (кат. № 4369016, Applied Biosystems) вместе со смесью праймеров, приготовленной посредством добавления 1:1:0,5 следующих праймеров: F3_core, R3_core, P3_core (Integrated DNA Technologies, все разведены до концентрации 100 мкМ каждый):
Прямой (F3_core): CTG TGC CTT GGG TGG CTT T (SEQ ID NO: 784)
Обратный (R3_core): AAG GAA AGA AGT CAG AAG GCA AAA (SEQ ID NO: 785)
Зонд (P3_core): 56-FAM-AGC TCC AAA/ZEN/TTC TTT ATA AGG GTC GAT GTC CAT G-3IABkFQ (SEQ ID NO: 786)
Получали стандартную кривую с использованием плазмиды HBV (генотип D, GTD), используя 10-кратные разведения, начиная с 1×109 копий/мкл вплоть до 1 копии/мкл, и использовали в 5 мкл на реакцию.
Для каждой реакции добавляли 10 мкл мастер-микса для экспрессии генов, 4,5 мкл воды, 0,5 мкл смеси праймеров и 5 мкл образца или стандарта, и проводили кПЦР.
Для анализа рассчитывали число копий/мл/лунку с использованием стандартной кривой. Результаты показаны в Таблице 15.
Экспрессию мРНК PD-L1 измеряли с использованием кПЦР.
мРНК экстрагировали из замороженных кусков печени, которые добавляли в 2 мл пробирки, содержащие керамические шарики (пробирки Lysing Matrix D, 116913500, mpbio) и 1 мл тризола.
Кусок печени гомогенизировали с использованием дезинтегратора тканей Precellys. В гомогенат добавляли 200 мкл хлороформа, перемешивали на вибромешалке и центрифугировали при 4°С в течение 20 мин при 10000 об./мин. Прозрачную фазу, содержащую РНК (около 500 мкл) переносили в свежую пробирку, и добавляли такой же объем 70%-ного EtOH. После хорошего перемешивания данный раствор переносили на колонку RNeasy spin, и РНК далее экстрагировали, следуя руководству набора RNeasy для мининабора RNeasy, кат. № 74104, Qiagen (включающего расщепление РНК, набор ДНКазы, не содержащий РНКазу, кат. № 79254). Элюирование осуществляли в 50 мкл Н2О. Конечную концентрацию РНК измеряли и доводили до 100 нг/мкл для всех образцов.
кПЦР проводили на 7,5 мкл РНК с использованием набора RNA-to-ct 1-step от Taqman, кат. № 4392938, Thermo Fisher, согласно инструкциям изготовителя. Использованные смешанные праймеры содержали PD-L1_1-3 (номер праймера Mm00452054_m1, Mm03048247_m1 и Mm03048248_m1) и эндогенные контроли (ATCB Mm00607939_s1, CANX Mm00500330_m1, YWHAZ Mm03950126_s1 и GUSB Mm01197698_m1).
Данные анализировали с использованием способа 2^-ddct. Для расчета значений dct использовали среднее всех четырех эндогенных контролей. Экспрессию PD-L1 выражали относительно среднего значения для эндогенных контролей и в % от физиологического раствора.
Таблица 15: экспрессия мРНК PD-L1 и ДНК HBV у мышей AAV/HBV, обработанных неконъюгированными и конъюгированными с GalNAc олигонуклеотидами, n равно 5.
(% от физиологического раствора)
(% от физиологического раствора)
Из данных результатов можно видеть то, что и голые, и конъюгированные с GalNAc олигонуклеотиды способны понижать экспрессию мРНК PD-L1 в печени мыши AAV/HBV, причем олигонуклеотид, конъюгированный с GalNAc, является в некоторой степени лучшим. Оба олигонуклеотида также приводили в некоторому снижению уровня ДНК HBV в сыворотке.
Пример 6 - влияние in vivo на ответ Т-клеток у мышей AAV/HBV
В настоящем исследовании мышей AAV/HBV из модели Пастера обрабатывали антителом или антисмысловыми олигонуклеотидами, нацеленными на PD-L1. Антисмысловые олигонуклеотиды были либо голыми, либо конъюгированными с GalNAc. Во время обработки животные были иммунизированы ДНК-вакциной против антигенов HBs и HBc (см. раздел Материалы и методы) для обеспечения эффективного примирования Т-клеток антигенпрезентирующими клетками. Оценивали, как данная обработка влияла на популяцию клеток в печени и селезенке, а также на экспрессию PD-L1 в данных популяциях и то, мог ли быть идентифицирован HBV-специфичный ответ Т-клеток.
Протокол обработки:
Самок мышей HLA-A2/DR1 обрабатывали согласно протоколам, приведенным ниже. Данное исследование проводили в двух отдельных подисследованиях с небольшими различиями в схемах введения, как показано в Таблице 16 и 17 ниже.
ДНК-вакцину и антитело против PD-L1 вводили, как описано в разделе Материалы и методы. Использованными антисмысловыми олигонуклеотидами были CMP ID NO 748_1 (голый) в дозе 5 мг/кг и CMP ID NO: 759_2 (конъюгированный с GalNAc) в дозе 7 мг/кг, которые оба вводили в виде подкожных инъекций (п.к.).
Таблица 16: протокол обработки мышей AAV/HBV ДНК-вакциной и ДНК-вакциной плюс антителом против PD-L1, 6 мышей в каждой группе
(Группа 10)
(Группа 11)
(Группа 13)
Изотипическое - мышиное контрольное Ab IgG, CaTx - кардиотоксин, ДНК - ДНК-вакцина, Ab - антитело против PD-L1, и * - отбор сыворотки.
Таблица 17: протокол обработки мышей AAV/HBV ДНК-вакциной и ДНК-вакциной плюс голым или конъюгированным олигонуклеотидом к PD-L1 (ASO), 7 мышей в каждой группе
(Группа 1)
(Группа 2)
(Группа 7)
(Группа 8)
ДНК - ДНК-вакцина, CaTx - кардиотоксин, Ab - антитело против PD-L1, ASO - голый олигонуклеотид к PD-L1, GN-ASO - конъюгированный с GalNAc олигонуклеотид к PD-L1 и * - отбор сыворотки.
В момент умерщвления отбирали одноядерные клетки крови, селезенки и печени каждой мыши из каждой группы, и обедняли эритроцитами (лизирующий буфер, BD biosciences, 555899). Одноядерные клетки печени требуют специфического приготовления, как описано в разделе Материалы и методы.
Популяции клеток:
В печени популяцию клеток анализировали поверхностным мечением на одноядерные клетки печени (см. Материалы и методы) с использованием цитометрии.
Не отмечали значимых изменений в частотах NK-клеток в селезенке и печени обработанных мышей по сравнению с контрольными группами (т.е. группами, иммунизированными носителем и ДНК). В Таблице 18 показано то, что в печени группы, обработанные голым олигонуклеотидом к PD-L1 (CMP ID NO 748_1) и конъюгированным с GalNAc олигонуклеотидом к PD-L1 (CMP ID NO: 759_2), имели значимое увеличение числа Т-клеток по сравнению с каждой из контрольных групп (т.е. группами, иммунизированными носителем и ДНК), также представленных на Фиг. 10А. Данное увеличение было обусловлено увеличением обеих популяций Т-клеток: CD4+ и CD8+ (Таблица 18 и Фиг. 10Б и 10В соответственно).
Таблица 18: Т-клетки в печени после обработки, в миллионах клеток
(миллионы)
Ab против PD-L1 (Группа 13)
ASO к PD-L1 (Группа 7)
ASO GN-PD-L1 (Группа 8)
Экспрессия PD-L1:
Экспрессию белка PD-L1 оценивали на макрофагах, В- и Т-клетках из селезенки и печени во время умерщвления. Присутствие антитела против PD-L1 в смеси антител, метящих поверхность (см. Материалы и методы), обеспечивало количественное измерение клеток, экспрессирующих PD-L1, посредством цитометрии.
В селезенке не наблюдали значимого различия между обработками в % макрофагов, В-клеток и Т-клеток CD4+, экспрессирующих PD-L1. % Т-клеток CD8+, экспрессирующих PD-L1, был ниже у мышей, обработанных голым олигонуклеотидом к PD-L1 (CMP ID NO 748_1) и конъюгированным с GalNAc олигонуклеотидом к PD-L1 (CMP ID NO: 759_2), по сравнению с другими обработками (данные не показаны).
В печени PD-L1 экспрессировался, главным образом, на Т-клетках CD8+ со средней частотой 32% и 41% в контрольных группах (две группы носителя и комбинированные группы с ДНК-вакцинацией соответственно, Фиг. 11А). Обработка голым олигонуклеотидом к PD-L1 или конъюгированным с GalNAc олигонуклеотидом к PD-L1 приводила к уменьшению частоты Т-клеток CD8+, экспрессирующих PD-L1 (см. Таблицу 19, Фиг. 11А). Значимые различия в % клеток, экспрессирующих PD-L1, также были отмечены для В-клеток и Т-клеток CD4+ после обработки ASO, хотя данные типы клеток экспрессируют значимо меньше PD-L1, чем Т-клетки CD8+ (см. Таблицу 19 и Фиг. 11Б и В). Обработка Ab против PD-L1 также приводила к кажущемуся снижению в экспрессии PD-L1 во всех типах клеток. Однако возможно то, что данное снижение частично обусловлено блокадой эпитопа PD-L1 антителом против PD-L1, используемым для обработки, таким образом, что предотвращается связывание с PD-L1 антитела, выявляющего PD-L1, в смеси антител для мечения поверхности. Следовательно, то, что проявляется как понижающая регуляция PD-L1 антителом против PD-L1, используемым для обработки, может быть результатом конкуренции за эпитоп между антителом для обработки и выявляющим антителом.
Таблица 19: % популяции клеток печени с экспрессией PD-L1
(Группа 11)
Ab против PD-L1
(Группа 13)
(Группа 1)
(Группа 2)
ASO к PD-L1
(Группа 7)
ASO GN-PD-L1
(Группа 8)
Ответ HBV-специфичных Т-клеток:
NK-клетки и Т-клетки CD4+ и CD8+, продуцирующие провоспалительные цитокины, выявляли с использованием анализов окрашивания на внутриклеточные цитокины (см. раздел Материалы и методы), выявляющих продукцию IFNγ и TNFα.
В селезенке на выявлялись NK-клетки, секретирующие IFNγ и TNFα, и выявлялось мало Т-клеток CD4+, секретирующих IFNγ и TNFα, (частота меньше 0,1%) в момент умерщвления. IFNγ-продуцирующие Т-клетки CD8+, нацеленные на два антигена HBV, выявлялись у мышей, обработанных голым олигонуклеотидом к PD-L1 или конъюгированным с GalNAc олигонуклеотидом к PD-L1, а также у мышей из данного исследования, которые получали только ДНК-вакцину (данные не показаны).
В печени иммунизированных ДНК мышей-носителей HBV не выявляли NK-клеток, продуцирующих IFNγ, в момент умерщвления, тогда как секретирующие IFNγ Т-клетки CD4+, специфичные в отношении кора или S2+S, выявляли в печени немногих иммунизированных ДНК мышей с низкой частотой (меньше 0,4%, данные не показаны). Специфичные в отношении S2+S HBV T-клетки CD8+, продуцирующие IFNγ, выявляли у большинства мышей, иммунизированных ДНК. Частота Т-клеток CD8+, секретирующих IFNγ, возрастала у мышей, обработанных комбинацией ДНК-вакцины и голого олигонуклеотида к PD-L1 или конъюгированного с GalNAc олигонуклеотида к PD-L1, тогда как обработка антителом против PD-L1 не добавляла какого-либо кажущегося дополнительного эффекта к вакцинации ДНК (Фиг. 12). Т-клетки CD8+, продуцирующие IFNγ, нацеленные на антигены оболочки и кора, были выявлены у большинства групп, иммунизированных ДНК (за исключением антитела против PD-L1) (Фиг. 12Б). Большинство S2-S-специфичных Т-клеток продуцировали и IFNγ, и TNFα (Фиг. 12В). Данные результаты также показаны в Таблице 20.
Таблица 20: % Т-клеток CD8+, специфичных в отношении антигена HBV (S2-S или кора) из общей популяции клеток, продуцирующих IFNγ или IFNγ плюс TNFα
(% клеток, секретирующих IFNγ)
(% клеток, секретирующих IFNγ)
(% клеток, секретирующих IFNγ плюс TNFα)
(Группа 10)
(Группа 11)
Ab против-PD-L1
(Группа 1)
(Группа 2)
ASO PD-L1
ASO GN-PD-L1
Пример 7 - влияние in vivo на антиген HBV и ДНК HBV в сыворотке мышей AAV/HBV
В настоящем исследовании мышей AAV/HBV модели Шанхай (см. раздел Материалы и методы) обрабатывали конъюгированным с GalNAc антисмысловым олигонуклеотидом к PD-L1 CMP ID NO 759_2.
Оценивали, как данная обработка влияла на уровни антигенов HBe и HBs, и ДНК HBV в сыворотке по сравнению с животными, обработанными носителем.
Протокол обработки:
В данном исследовании использовали самцов мышей C57BL/6, инфицированных рекомбинантным аденосателлитным вирусом (AAV), несущим геном HBV (AAV/HBV), как описано в Шанхайской модели в разделе Материалы и методы. Мышам (6 мышей на группу) один раз в неделю в течение 8 недель инъецировали антисмысловой олигонуклеотид CMP ID NO: 759_2 в дозе 5 мг/кг или носитель (физиологический раствор), причем и тот, и другой вводили в виде подкожных инъекций (п.к.). Образцы крови отбирали каждую неделю на протяжении обработки, а также 6 недель после обработки. Уровни ДНК HBV, HВsAg и HВeAg измеряли в образцах сыворотки, как описано ниже. Результаты для первых 10 недель показаны в Таблице 21 и на Фиг. 13. Данное исследование все еще продолжалось во время подачи заявки, поэтому данные для остающихся 4 недель не были получены.
Выявление HBsAg и HBeAg:
Сывороточные уровни HBsAg и HBeAg определяли в сыворотке инфицированных мышей AAV-HBV с использованием хемолюминисцентного иммуноанализа HBsAg (CLIA) и набора для CLIA HBеAg (Autobio diagnostics Co. Ltd., Zhengzhou, Китай, кат. № CL0310-2 и CL0312-2 соответственно) согласно протоколу изготовителя. Вкратце, 50 мкл сыворотки переносили в планшет для микротитрования, покрытый соответствующим антителом, и добавляли 50 мкл реактива с конъюгированным ферментом. Планшет инкубировали в течение 60 мин на шейкере при комнатной температуре перед промывкой всех лунок шесть раз промывочным буфером с использованием автоматической промывающей машины. В каждую лунку добавляли 25 мкл субстрата А и затем 25 мкл субстрата Б. Планшет инкубировали в течение 10 мин при RT (комнатная температура) перед измерением люминисценции с использованием люминисцентного ридера Envision. HBsAg приводится в единицах IU (международные единицы)/мл, где 1 нг HBsAg соответствует 1,14 IU. HBeAg приводится в единицах NCU (национальная клиническая единица)/мл сыворотки.
Экстракция ДНК HBV и кПЦР:
Исходную сыворотку мышей разводили в 10 раз (1:10) фосфатно-солевым буферным раствором (PBS). ДНК экстрагировали с использованием робота MagNA Pure 96 (Roche). 50 мкл разведенной сыворотки смешивали в картридже для обработки с 200 мкл внешнего лизирующего буфера MagNA Pure 96 (Roche, кат. № 06374913001) и инкубировали в течение 10 минут. ДНК затем экстрагировали с использованием «Набора для маленького объема ДНК и вирусных нуклеиновых кислот MagNA Pure 96» (Roche, кат. № 06543588001) и протокола «Вирусная NA плазма SV внешний лизис 2.0». Объем элюции ДНК составлял 50 мкл.
Количественное измерение экстрагированной ДНК HBV проводили с использованием прибора для кПЦР Taqman (Vii7, Life technologies). Каждый образец ДНК тестировали в ПЦР в двойной повторности. 5 мкл образца ДНК добавляли к 15 мкл мастер-микса для ПЦР, содержащего 10 мкл мастер-микса для экспрессии генов TaqMan (Applied Biosystems, кат. № 4369016), 0,5 мкл праймера/зонда для кПЦР PrimeTime XL (IDT) и 4,5 мкл дистиллированной воды, в 384-луночный планшет, и ПЦР проводили с использованием следующих установок: инкубация UDG (2 мин, 50°С), ферментативная активация (10 мин, 95°С) и ПЦР (40 циклов по 15 с, 95°С для денатурации и 1 мин, 60°С для отжига и элонгации). Число копий ДНК рассчитывали из значений Ct на основе стандартной кривой плазмидной ДНК HBV, полученной программой ViiA7.
Последовательности праймеров и зондов TaqMan (IDT):
Прямой праймер к кору (F3_core): CTG TGC CTT GGG TGG CTT T (SEQ ID NO: 784)
Обратный праймер (R3_core): AAG GAA AGA AGT CAG AAG GCA AAA (SEQ ID NO: 785)
Зонд Taqman (P3_core): 56-FAM/AGC TCC AAA /ZEN/TTC TTT ATA AGG GTC GAT GTC CAT G/3IABkFQ (SEQ ID NO: 786).
Таблица 21: уровни ДНК HBV, HBsAg и HBeAg в сыворотке от мышей AAV/HBV после обработки антисмысловым олигонуклеотидом к PD-L1, конъюгированным с GalNAc.
Из данного исследования можно видеть то, что конъюгированный с GalNAc антисмысловой олигонуклеотид к PD-L1 CMP NO 759_2 имеет значимое влияние на уменьшение уровней ДНК HBV, HBsAg и HBeAg в сыворотке после 6 недель обработки, и влияние, которое поддерживается в течение по меньшей мере 2 недель после завершения обработки.
Пример 8 - нокдаун PD-L1 in vitro в человеческих первичных гепатоцитах с использованием олигонуклеотидов к PD-L1, конъюгированных с GalNAc
Способность антисмысловых олигонуклеотидных соединений к PD-L1, конъюгированных с GalNAc, уменьшать уровень транскрипта PD-L1 в первичных человеческих гепатоцитах исследовали с использованием геномики.
Культура клеток
Криоконсервированные человеческие гепатоциты суспендировали в WME, дополненной 10% фетальной телячьей сыворотки, пенициллином (100 U/мл), стрептомицином (0,1 мг/мл) и L-глутамином (0,292 мг/мл), при плотности приблизительно 5×106 клеток/мл и высевали в покрытые коллагеном 24-луночные планшеты (Becton Dickinson AG, Allschwil, Швейцария) в плотности 2×105 клеток/лунку. Клетки предкультивировали в течение 4 ч, обеспечивая прикрепление к чашкам для культуры клеток до начала обработки олигонуклеотидами в конечной концентрации 100 мкМ. Использованные олигонуклеотиды показаны в Таблице 21 и Таблице 8, носителем был PBS. Среду для посева заменяли 315 мкл бессывороточной WME (дополненной пенициллином (100 U/мл), стрептомицином (0,1 мг/мл), L-глутамином (0,292 мг/мл)), и в клеточную культуру добавляли 35 мкл 1 мМ маточных растворов олигонуклеотидов в PBS, и оставляли на клетках на 24 часа или 66 часов.
Получение библиотеки
Профилирование экспрессии транскриптов проводили с использованием химии Illumina Stranded mRNA на платформе для секвенирования Illumina с использованием стратегии секвенирования 2×51 п.о. парных концевых считываемых фрагментов и минимальной глубины считывания 30М на образец (Q в квадрате ЕА). Клетки лизировали в лунках посредством добавления 350 мкл буфера RLT от Qiagen, и присваивали им номер доступа по рандомизированной схеме.
мРНК очищали с использованием мининабора Rneasy от Qiagen. мРНК количественно измеряли, и оценивали целостность с использованием биоанализатора Agilent. При исходной оценке качества выделенной РНК наблюдали то, что все образцы удовлетворяли входному показателю качества 100 нг с баллами RIN (число целостности РНК) больше 7,0.
Для всех образцов получали секвенированные библиотеки с использованием препарата библиотеки Illumina TrueSeq Stranded mRNA, начиная со 100 нг общей РНК. Конечные библиотеки кДНК анализировали на распределение размеров с использованием биоанализатора Agilent (набор DNA 1000), количественно измеряли посредством кПЦР (набор KAPA Library Quant) и нормировали к 2 нМ в препарате для секвенирования. Для связывания библиотек кДНК с поверхностью проточной ячейки использовали набор для получения стандартных кластеров v5, а cBot использовали изотермически для амплификации присоединеных конструкций кДНК вплоть до клональных кластеров, каждый из которых состоит из ~1000 копий. Последовательность ДНК определяли технологией секвенирования посредством синтеза с использованием набора TruSeq SBS.
Обработка данных
Считываемые фрагменты секвенирования с парными концами Illumina длиной 2×51 п.о. картировали на человеческом эталонном геноме hg19 с использованием программы для выравнивания коротких считываемых фрагментов GSNAP. Выравнивания в формате SAM превращали в сортированные файлы выравнивания в формате ВАМ с использованием программы SAMTOOLS. Числа считываемых генов оценивали для PD-L1 на основе аннотации экзонов из RefSeq NCBI, определенной соответствующим файлом GTF для hg19. Стадию нормирования, учитывающую разный размер библиотеки каждого образца, применяли с использованием пакета R DESeq2.
Уменьшение уровня транскриптов PD-L1 после инкубирования с антисмысловыми олигонуклеотидными соединениями к PD-L1, конъюгированными с GalNAc, показаны в Таблице 22.
Таблица 22: Уменьшение уровня транскриптов PD-L1 в человеческих первичных гепатоцитах после обработки олигонуклеотидами, конъюгированными с GalNAc, n равно 4.
(числа, скоррекрированные в зависимости от размера библиотеки)
(числа, скоррекрированные в зависимости от размера библиотеки)
Все пять антисмысловых соединений, конъюгированных с GalNAc, демонстрировали значимое уменьшение уровня транскриптов PD-L1 после 24 и 66 часов инкубации по сравнению с образцами, обработанными носителем.
Пример 9 - ЕС50 конъюгированных и голых антисмысловых олигонуклеотидов к PD-L1 в клетках ASGPR-HepaRG, инфицированных HBV
Эффективность двух голых и эквивалентных конъюгированных с GalNAc антисмысловых олигонуклеотидов к PD-L1 сравнивали в клетках ASGPR-HepaRG, инфицированных HBV.
Линия клеток
Клетки HepaRG (Biopredic International, Saint-Gregoire, Франция) культивировали в среде Williams E (дополненной 10% ростовой добавки HepaRG (Biopredic)). Из данной линии клеток получали линию клеток HepaRG, стабильно сверхэкспрессирующую человеческий ASGPR1 и ASGPR2, с использованием лентивирусного способа. Пролиферирующие клетки HepaRG трансдуцировали при MOI (множественность заражения) 300 с использованием лентивируса, произведенного по требованию Sirion biotech (CLV-CMV-ASGPR1-T2a_ASGPR2-IRES-Puro), кодирующего человеческий ASGPR1 и 2 под контролем промотора CMV (цитомегаловирус) и ген устойчивости к пуромицину. Трансдуцированные клетки подвергали селекции с 1 мкг/мл пуромицина в течение 11 суток и затем поддерживали при такой же концентрации антибиотика для обеспечения стабильной экспрессии трансгенов. Сверхэкспрессию ASGPR1/2 подтверждали как на уровне мРНК посредством кПЦР-ОТ (ASGPR1: 8560 раз по сравнению с нетрансдуцированными, ASGPR2: 2389 раз по сравнению с нетрансдуцированными), так и на уровне белка посредством анализа проточной цитометрией.
Клетки дифференцировались с использованием 1,8% DMSO в течение по меньшей мере 2 недель до инфекции. HBV генотипа D получали из супернатанта культуры клеток HepG2.2.15 и концентрировали с использованием осаждения с ПЭГ. Для оценки активности тестируемых соединений против HBV дифференцированные клетки ASGPR-HepaRG в 96-луночных планшетах инфицировали HBV при МОI от 20 до 30 в течение 20 ч до того, как клетки были промыты 4 раза PBS для удаления инокулума HBV.
Эффективность олигонуклеотида
Следующие олигонуклеотиды:
добавляли к клеткам ASGPR-HepaRG, инфицированным HBV, в сутки 7 и сутки 10 после инфекции с использованием серийных разведений от 25 мкМ до 0,4 нМ (разведения 1:4 в PBS). Клетки отбирали в сутки 13 после инфекции.
Общую мРНК экстрагировали с использованием набора для клеточной РНК в большом объеме MagNA Pure 96 на системе MagNA Pure 96 (Roche Diagnostics) согласно инструкциям изготовителя. Для анализа экспрессии генов проводили кПЦР-ОТ, как описано в Примере 5.
Данные анализировали с использованием способа 2^-ddct. Актин В использовали в качестве эндогенного контроля для расчета значений dct. Экспрессия PD-L1 приводится относительно эндгенных контролей и относительно солевого носителя.
Расчеты ЕС50 проводили в GraphPad Prism6, и они показаны в Таблице 23.
Таблица 23: ЕС50 в клетках ASGPR-HepaRG, инфицированных HBV, n равно 4.
Эти данные явно показывают то, что конъюгирование с GalNAc антисмысловых олигонуклеотидов к PD-L1 значительно улучшает значения ЕС50.
Пример 10 - стимулирование функции Т-клеток в РВМС, полученных от пациентов с хронической инфекцией HBV
Исследовали, могли ли голые антисмысловые соединения к PD-L1 увеличивать функцию Т-клеток пациентов, хронически инфицированных HBV (CHB), после стимулирования одноядерных клеток периферической крови (РВМС) ex vivo антигеном HBV.
Замороженные РВМС от трех пациентов, хронически инфицированных HBV, оттаивали и высевали при плотности 200000 клеток/лунку в 100 мкл среды (RPMI1640 плюс GlutaMax плюс 8% человеческой сыворотки плюс 25 мМ Hepes плюс 1% пенициллин/стрептомицин). На следующие сутки клетки стимулировали 1 мкМ белковой смесью большого белка оболочки HBV или 1 мкМ белковой смесью белка кора HBV (см. Таблицу 9) с 5 мкМ CMP ID NO: 466_1 или CMP ID NO: 640_1 или без них в 100 мкл среды, содержащей 100 пг/мл IL-12 (интерлейкин-12) и 5 нг/мл IL-7 (стимулирование конканавалином применяли только в сутки 8). Через четверо суток обработку антисмысловым олигонуклеотидом к PD-L1 обновляли с использованием среды, содержащей 50 IU IL-2. В сутки 8 после первой стимуляции клетки повторно стимулировали белковой смесью или 5 мкг/мл конканавалина А плюс антисмысловой олигонуклеотид к PD-L1 в течение 24 ч. В течение последних 5 ч стимулирования добавляли 0,1 мкл брефельдина А, 0,1 мкл моненсина и 3 мкл антитела против человеческого CD-107 (APC).
Через 24 ч клетки промывали буфером для окрашивания (PBS плюс 1% BSA плюс 0,09% азид натрия плюс EDTA), и применяли окрашивание поверхности в течение 30 мин при 4°С [антитело против человеческого CD3 (BV 605), антитело против человеческого CD4 (FITC), антитело против человеческого CD8 (BV711), антитело против человеческого PDL1 (BV421), антитело против человеческого PD1 (PerCP-Cy5.5) и окрашивание на живые и мертвые клетки (BV510) (BD Biosciences)]. Клетки фиксировали в фиксирующем буфере BD в течение 15 мин при 4°С. На следующее утро клетки пермеабилизировали буфером для пермеабилизации/промывки BD в течение 15 мин при 4°С, и осуществляли внутриклеточное окрашивание в течение 30 мин при 4°С [антитело против человеческого IFNγ (PE)]. После промывки в буфере для пермеабилизации/промывки клетки растворяли в 250 мкл буфера для окрашивания.
Измерения FACS проводили на BD Fortessa (BD Biosciences). Для анализа всю популяцию клеток сначала сортировали на живые клетки (окрашивание на живые и мертвые, BV510) и затем на клетки CD3+ (BV605). Клетки CD3+ затем обозначали на графике как CD107+ (APC) относительно IFNγ+ (PE). Результаты показаны в Таблице 24.
Таблица 24: эффект обработки ASO к PD-L1 на Т-клетки CD3+ из РВМС, выделенных от трех пациентов, хронически инфицированных HBV.
р-р
р-р
р-р
CD107+
CD107+
CD107-
Из этих данных можно видеть то, что сама по себе стимуляция антигеном способна индуцировать активацию Т-клеток (увеличение % клеток CD3+, экспрессирующих IFNγ и/или CD107a) в РВМС пациентов СНВ (n равно 3). Добавление антисмыслового олигонуклеотида к PD-L1 CMP 466_1 или 640_1 приводило к дополнительному увеличению ответа Т-клеток CD3+. Данное увеличение, главным образом, наблюдали в группе, стимулированной оболочкой HBV.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф.Хоффманн-ля Рош АГ
Рош Инновейшен Сентер Копенгаген A/С
<120> ОЛИГОНУКЛЕОТИДЫ ДЛЯ ПОНИЖЕНИЯ ЭКСПРЕССИИ PD-L1
<130> P33358-WO
<160> 786
<170> PatentIn version 3.5
<210> 1
<211> 20064
<212> ДНК
<213> homo sapiens
<400> 1
ggcgcaacgc tgagcagctg gcgcgtcccg cgcggcccca gttctgcgca gcttcccgag 60
gctccgcacc agccgcgctt ctgtccgcct gcaggtaggg agcgttgttc ctccgcgggt 120
gcccacggcc cagtatctct ggctagctcg ctgggcactt taggacggag ggtctctaca 180
ccctttcttt gggatggaga gaggagaagg gaaagggaac gcgatggtct agggggcagt 240
agagccaatt acctgttggg gttaataaga acaggcaatg catctggcct tcctccaggc 300
gcgattcagt tttgctctaa aaataattta tacctctaaa aataaataag ataggtagta 360
taggataggt agtcattctt atgcgactgt gtgttcagaa tatagctctg atgctaggct 420
ggaggtctgg acacgggtcc aagtccaccg ccagctgctt gctagtaaca tgacttgtgt 480
aagttatccc agctgcagca tctaagtaag tctcttcctg cgctaagcag gtccaggatc 540
cctgaacgga atttatttgc tctgtccatt ctgagaaccc aaaggagtcc taaaagagga 600
atggaggagc ctaagaataa aaatagtata ataaaacatt tcttagacac attgaccttg 660
gcctatgtca aagttcagtc tgggtttgtc ttataacaca aggagtaaaa gtaccattgt 720
tctacctctt tttttaatac ttgaaaaaaa tttactgtgg atgcttttct atgaattaaa 780
taaccttcta aaaaatgttt tcattgctgc attcgattag attgggtaac taaatgaaat 840
taattcctca ctgttgggta taaaggttat ttacagtggt tctgtcttag ccattcactg 900
aactcattgc atatatatct ctggaatatt gctgattgtt tccttcaagt aaacttagaa 960
gtgtaactac ttagtcaaag agcctgaata ttttaaaggc cttttgaaga aaactgaaaa 1020
tgctttccag aaaggatgta tcagttgaca atgacagtcg tcaacagtat ttaaggagaa 1080
ctatgatact ctgaagaaaa acttagcctt tctcagtaaa agtaggtagg cagaggccac 1140
atgacagcag ttagagtgtg gtcttcaagg aagtcacaga aatactgtgg ggaattgaaa 1200
ccccatgtgg aaaatgtaca agagtgtctc agtgtgactg agaaggaggt tgggcatggg 1260
gtttcatgga gtttaataaa gtttggtcac ttagtagagg tttaataaat caactgtctt 1320
aatctttgat cctacttaag aatttttttt ttgtttttgt agagatgggg ctcttgttat 1380
gttgcccagg ctgttctcga actcctagcc tcaggcgatc ctccctcctc aggctccaga 1440
agtcctggga ttactggcgg gagccaccat gcaggcctct tgctcctact tttgagaaag 1500
gaagtttaac cggttttttt tgtctttttt tttttttttt tgagacagag tctcactctg 1560
ttgcccatgc tggagtgcag tggtgcaatc tcagctcact gcctcccggg ttcaagtgat 1620
tctcctgcct cagcctcccg agtagctggg actacaggca cctgccacca cgcccagcta 1680
atttttgtat ttttagtaga aatggggttt caccatattg gccaggctga tctcgaactc 1740
ctgacctcag gtgatccgcc tgcctcggcc tcccaaagtg ctgggattac aggcatgagc 1800
cactgctcct ggctgcttaa ctttttctct atctcatcct cctacccatc ctacccttgg 1860
aagatagaga agtagtatta gttccatagt gttatactgg gcttccccca gggacaaacc 1920
cacttcccca acctgaatga gccatcactt cttccccagt ttacatttca ttgctcttta 1980
aatgtctcca ttcggatatg ggaattcaca tatggtcata attcttacct gaagaagatg 2040
tcagtcttct tctcttagac caactgccct gatatgaggt ttagaggtta aagaacatgt 2100
gtgtatttac atgatctttg tattctgcct tttcgtccct cactaatgac agctgcaccc 2160
caaggaaatg gagctgtgga agagagggtt tgataagaaa ttaagtaaat attggatcta 2220
atccatcacc ctccaggaag cctttattac tcctaaaaat ttcaaccaaa ttcattaaag 2280
gacaagaact ccaccagagt aggccataaa cattggcaaa attagttgta atccatgact 2340
agatttaatg tccctttgtt ttattcccat atggttataa tgctttgctt ggcattaggg 2400
gtattttaag ttttcttctg cctagtaagt gaatttgtgt ttataataca ataatcataa 2460
aatatcacat taatatttta taactgtaca gttataaaat attttataag taatatttat 2520
attttataag taatatttta taactgtaca gttaactctg gcccaaggaa aagatagtct 2580
gatagatgct gcagccccat tttagcaaat gtgacctcac aggcctgaat gccatcgcta 2640
ttccacatct acaggataga cggaaaggaa agaaataaaa aaataggtac ctaacactgg 2700
caagaggatg atgactcatg ttatttcact taaccttttt atcttttaac atgaaggact 2760
catacaggtt gataagaaac cagtgacata aacagaccaa aaaatgatca gatctttcaa 2820
attagcaaaa aaataatatt ttttaaacaa tgggtgaaaa tacagtgtaa cagtaccaat 2880
tatcaacatg tgttgagaac cagaaaaatg ttctttttct ttgatcagca acactatttg 2940
ggaaaatcta tcctcagggc ctagcctggg gccctggcac acagtaggca ctcaacgaat 3000
atttgctgaa cacacaaata cttatgatat tttaaaaaat tggcaacaat ctgataccta 3060
acaatagagg gattaaatat tatggaactg ttaaataaga tgcttatgaa taccatgcag 3120
taagatgggc aatatttatg ccataagctt taatgaaaca aatgggtatt aaatgtatga 3180
taaggttata aattactttt taaaagatta cagggaaaaa aattgaaaga tatacactga 3240
aatgtttttt gctcacagtg gtgacaaggt ttctcagcac tggcactgtt gacgttttag 3300
gctgtatgtc tttgctgtgg gaggctggcc tgtgcactgc agggtgtttg gcagcactct 3360
tggcctctgc ccctagatag caatagcagt cctccctcaa ccagcccaat tttgacaacc 3420
aaaaatgttt ccaggcatca ccagatgctc cctgggtgag agtgatgaaa tagtagggga 3480
ttttcccctt cttttcttat tttctgtaat tccattatat tactttaata ataaagaaaa 3540
aaacataaaa aataaacgaa tgttattatt ctacgtcagt ttggatgttt ggactccatt 3600
ttggggttct ttccattata tcacttggtc tgctaaacat tctacggttt ggtaaggtga 3660
agtgattcat gaaattttgg ttttattttt ttcctgatac taaaaataaa acattctttc 3720
acttggaaat ttggacacag aacaccaaaa aaaatccata atctcatctc tctttttctg 3780
tcttttcctt ccttttttcc ctttaaaaac aataaagagt gaaacctacc tgttctccct 3840
ctaatttaat tcctaaatat aatcactgtc aatatcttgg acatttcctg tgtctaaaca 3900
cacacacaca cttttttttt tcagcaaaag tggatttctg ctacatgtag tgttctgcaa 3960
cttactttct atgtgtttac aaaatcagta catgtacata tgctgaattc agtccttaat 4020
ggtattatat tttgtgaata taccaaaatt tgtttaacca cttagacaat ctaggatatt 4080
ctcagtttgc tgttatgagc aatgctcttc ctttacatat acagacatat atatatatat 4140
gtgtgtgtgt gtgtttttgt tttagtagga tagatttcta ggagagggtg aaaggtctta 4200
tgacatccgc atttacgatt gtaataggaa gtatcaaagt gccccctaaa gaaaaaaatc 4260
ctcccattag tgggtaagaa agcctatttg ttcatatctt cacaaacact aaatattaga 4320
aatatttaca attgtggtca agctcataag tgaaaatggt atttcatatc ttatattttt 4380
tattgtgaga ttgaacatct ttcatatgtt tacatgtcac ctgtatttct tattctctga 4440
actatatgtt atgacctttc actttttttc ctcatgggtt atgtgtagtt tgtatagttg 4500
tcttattgat tgttaggagc tatttatata ttaggaacat taatctcctg tcttatatat 4560
acgtggcatc gattagttga tcatttgtga gttcatgtct gtatacaaag attggagagg 4620
cactaagagg gaaaacttac ctctttctta tcaaagtttg taaatatatg tataacagaa 4680
gagggagaaa atattaataa atgcacagat tggctgaaat agagtataaa tcttttactc 4740
ccctacttca acataaactg caaaaggaga gtgacttttc tttcactctg acttccgtat 4800
tcctcatgct taaaatagtg cctagcacag aagaggtgct caatcagtgt ttgctaaacg 4860
aaataattag tcacatttca agcaggatga ctaaatgaag aatagaatct aggcagatac 4920
tctggaagag tggctgtgag tcattcatat cttagtatga attagtcaaa tccaactctc 4980
tccccttccc actccccact gttagtagaa gaatctgttt attgagagaa tagatttata 5040
atttagaata agtgagaggg gcagaagagg agattttgaa ggatggcacc tgaaggagga 5100
ctagcatggc tgagacagtg aagtggaagc cttgaatagc taaagggtaa gatgaaagta 5160
tttagctgta gggggaaaaa gcattgacag gttggaaaag taaaagtcag attctccttg 5220
ctctgaaatt ttgtacaggg caggttctac taggtatgtt acaatgcaga aaaaacatga 5280
aataattgag aggaatttgg tgcaatatta tcttcttggc ttcttttgag tgggcagatt 5340
tttttcacgg cctgtaacta taataaattt gaaacttctc atcttttagt aacttttttc 5400
acttaagttt atgtggctgt gggcaatgga atgaagatat tgaacttcca attccctgtt 5460
gggtttccac aattacaagt caatcatgac tggttattag aagactattt cagttagaac 5520
caccaagtcc catattgtca tattgtatgt ttaattatta agtgaagcag tcttcttttc 5580
gtgttttcca taattagggc attccagaaa gatgaggata tttgctgtct ttatattcat 5640
gacctactgg catttgctga acggtaagac accaaatcct tccattaggt tctatatttt 5700
aaatatttta accatgagtt taaaactaaa atgatcattt aaaatgcatg caattttctt 5760
atagagagaa cattctattc tttcttctac tttacacaat ggcaaagtct tctttctact 5820
ttacgcaatg ataaagttac ctgtgtcatt ttgtaaaaat atagagaata tagacaaatt 5880
gaaagacaca aaataatcta ttacccattt cccagggtta actactgaaa atatctgggg 5940
aaatggcctg tatgtataca tttatttgtt tgctttcaac aaggccaaga tcctttgatc 6000
tttcagtctt ggttgctctg tgacatgcct ttcctgatga ggatacttta aggaagaatt 6060
gtaagataca tggaaaatgt caggctaaca cagtactggc atcaccctgt gctctttcct 6120
gaactccata ccaatgtact tcttgccaga aaactgatca aaagtttagg gaagtaaaaa 6180
gagatgactg ttagaatcta ccattccctc tatgtaggaa gcaaataggt gtcctgtcaa 6240
aggacattct ggggatgtct acatgaaacc aagtctccct ggttgtaagg actccatctc 6300
catataatat ttatacagta atatatgttt ataaattgtg ggggcaactt gtttagctaa 6360
ttttattatt ctgctattgg gacactgtgt ctcagcatga gatatagtgt cccaaaacat 6420
atttcaagcc cattggataa aatatgtgtt tagcaagttc ttaaatataa tgataacata 6480
accgaccaga taaagtgatt tataaacgct gtgccaattt tgtaaatgtt tcgaggaatt 6540
ttcccttttc tgaagattgt ccttctttct ttttagcatt tactgtcacg gttcccaagg 6600
acctatatgt ggtagagtat ggtagcaata tgacaattga atgcaaattc ccagtagaaa 6660
aacaattaga cctggctgca ctaattgtct attgggaaat ggaggataag aacattattc 6720
aatttgtgca tggagaggaa gacctgaagg ttcagcatag tagctacaga cagagggccc 6780
ggctgttgaa ggaccagctc tccctgggaa atgctgcact tcagatcaca gatgtgaaat 6840
tgcaggatgc aggggtgtac cgctgcatga tcagctatgg tggtgccgac tacaagcgaa 6900
ttactgtgaa agtcaatggt aagaattatt atagatgaga ggcctgatct ttattgaaaa 6960
catattccaa gtgttgaaga cttttcattc ttgtaagtcc atacttattt tcaaacagaa 7020
cagcatagtc tgttcattca ttcattcaat tcatgaattc attcacataa ttatccaatt 7080
tcttgagcac ctatttgata gtcactggaa atccagagac aaacaacaca gagccatgtt 7140
ctacagtatg tacagttttc caaaaagaat ttctagtctt tactttttta ttacaaatgg 7200
aatacgtata cttgcaaata attcagatac tgtggaagag atcaaatgaa ttgcaaaagt 7260
gtccctcctc ccttcaccac tatctcccat ggcatgcaga gagagtaacc attatttgtg 7320
tgtccctcca gaaatttttt tattcaacta ctattttttt attttattag gtccgtcagt 7380
tttccttttt tgagcctctc tatatcaaat gcaaataaat atattcagaa caaaccccac 7440
tgtaaggttc acattaaaaa agacttgaag tcaccctatg aagacaaaaa ataatcacat 7500
taagtgtgaa agaacctatt cttccagtac aggataagcc atacttactg ggcatatatt 7560
catcttgaaa atctatactg atgttgtctt ggggaattga aaaggaacta ggagtgttag 7620
ttcctcggta ttgacccaca gttatgttat caggtcactt gagttcaaag ttttgtgttg 7680
gcactagcta agtaaaggaa aacacctctg ctttcattgt tgagtttcac agaattgaga 7740
gctgaaagga tcccaggcag gagcagctaa tccaaactcc cacaaagaac aaaaatcccc 7800
cagaggatct tctgttctta tatttcctgc aatggcgtcc ctgtcatatc ccacaatggc 7860
ctccctgcca tttggatatc ccttccatat cctgttgaaa ttactcccta atagtaagct 7920
gaaatctgcc cctctagttg tagtcttggg attatttcat ttacatgatg accttttaat 7980
atttgactag aattaaatca tctccccttg gtctttccat tcctgggcta actaccatca 8040
atctgagggc taacaataca agtagaaaaa gtatacattt gtcactgatc actgatcaat 8100
tattaatcaa tgatcactga taactataaa ctcaaaaaca aaatcatgtg gggattaaga 8160
gaaatgtatc agttttatgt tgtatttctg gtccctgata ctggctcagg taatgccact 8220
attgtcaaga agataccact tgtaaagtag atttaatttt cattatattt taccatatgc 8280
ttctccattc atgacatctc ttgagatgtt gtggtttata ctttcagttt ttctccagtc 8340
catccgcaaa tatcaggcat ctactgtgtt ccaagatatt aaagaaatca tcatgactta 8400
gcctcatcaa cagcattgct agatctggga tggaaaggaa gagtataatc ctggcagtca 8460
ggaagaaggc agcataaagt ataagtttct gcttccaaaa aaggtctctc atcagcctgt 8520
agggagtgtg tagggaaggg acagctgtcc ttgtagtagg gaagggtttt attcaggtcg 8580
tctgggctcc ataatatccc ttgtgtatct gcagtctcct ttgccatgga tcaacacaat 8640
aggaaatctt ccggcactga tggtttttcc aagggggagt tcttcctgga gcaaagcaaa 8700
tgaccaacca ggtttgagga cctgatttgt ttgacaattc cattttgtat tgtaaattac 8760
ttaattggca ttctactccc aatccatctt gtcatttgca tacagtggtt ttgggattga 8820
gttcagctat accaaaagtc tgaaccttct gcacttagaa caaggcaacc accaagcttc 8880
acttgcactg aggccgtgtc tccaatggaa atgaggcagc tggcttgcag gagcttccca 8940
actcagggaa gtagaactcc tgagtcacct ccatatgcaa atgatttcac agtaatgctg 9000
ttgaacttca cttcccatca cagcaaatgt gtggtaacat agcttcccca caggagttta 9060
ctcaccatgg tattttaaag gtgaaacatt tcaaaactga aatttgaaag aatttagttt 9120
tggattcact caattatcac tatcacttcg ggtgttattg cacctttctt gtttgtgagt 9180
ttaaatgcca gactctcagg ccactaactt tcaattaaaa gtgtttttct ttaatcgctg 9240
aacctaacag cagggaaaac gaaatgttca ttcagacttt cagaaccttc aatgagatta 9300
ggcagctgaa agatcaaagt gttgcatagt tgtcccgata aagctatttg gatcatatgg 9360
accaaatcga ctgctgtcat tccccaccaa ccccatctct ccccaaaatt cccagccctg 9420
tttaagtgtt ctctgtagca tttatctcta tctagtatat tgtgtagcat atcatatcat 9480
acttttctgt tttgtttatt gtctctctcc tcctagaata taaactccac aagcacaaag 9540
atttgggcct gttttataat attgttgcat ccccagggcc tgatatacag cagagtggtg 9600
gtacgaaaag agcacacaaa aaaatatttg ttgagtcaat gaatgaatga tttcctcaaa 9660
taggattagc ctaaaatttt ggaaacatga acagatttgg atatgtgaaa atttatttcc 9720
agactgttca tcaggaactg ttagcagctt ctaaagggta cactggagca gcagtagtaa 9780
aaggaggaag aggagcagct ctgctactgc tactatcgag tactactaca attagcactt 9840
gcttattctg tgtgttaggc cctgtactga acactctgtc taaattagtt catttcctcc 9900
tggaaatgac tctagggggt aagtgcttca tcatgtaaga tgagtatttt tcacattttg 9960
ttgtgtctga aatctgagtg tgtctttcaa tgatggaatc tttgattcca tgataagtgg 10020
tattattccc attttaagga tgaggaaact gaggtccaaa gaaattaagt aatttgccca 10080
aattcaccca gcctagaaaa tgataaagct agttctaaac ccaagcagat tagctctgaa 10140
gtctgggccc ttaataacca ctttttattg cctatatttg tacctctggt gtacgtatca 10200
agttatatgt tgacttcaaa actatcatga ccttttcttg gttttgattg tccaacatta 10260
gtatagtgtt ctgggtctgc aaaaattttg attactcatc tcatctgtaa aacattttga 10320
actcgtgtgt ttgtgcatgc acatttgtgt gtaattataa aaattttact ttctgttaat 10380
atataagttg tatcataaga aactgccgtt tttgaagagc aaaaaaaggt tgaatgttac 10440
cagttacatc tggttcaacc taatagacat ttgtacaaaa acagacattt taagaggttg 10500
aaataaaaat ttaataaaca atattttcag tttttactaa ttgtgatgct tcactatcat 10560
tagctaatat gtcaaggcat aatatacctt agggtgaact ttatcattaa caaaggtgga 10620
tggtgtcaat aatcttgagg tttgtgtttt tttatataac actgcgaggt ctaattaagt 10680
acttactgtt taccacctca tacagtggcc gataaaaagt gtcacttctg ctgtttcctc 10740
tgggttgtgc ttgaattatt agtattatct tcagtcctca gtttctttgt gggaaacttt 10800
ttaattagtt gtttaatttt gtaagatggt tagtttagtc aaaattagat aagagaattt 10860
gaaaatccgt agctacccca aagcaaccta cacataagaa ctattatttt tgtgttttga 10920
aatcataatt ttattgattt ccagtgtttc cactggtagt ggtttcattg atataggagt 10980
atcaaaacat cactcattat ttatttcagt ttcatttgat cctagccgtt ttgtattaac 11040
tctctgtgaa gaaattacct cacaaatcta ttgctgtcct tggtaaagga atggagaatt 11100
aaggctctag atcattagtg gttacactat agtattagaa gtaaaaaaaa gattatacca 11160
acaaaataag aacatgttaa tgtacttgta atgaataaac atgaataaag ctcttatgct 11220
atataggtgc actaaacaat ctactagaat tgtcagcaaa ctacgtatct taatcctgaa 11280
agggtcccaa accaatgatc taaaattgaa tcaaactttc ttccttgagc ataattactt 11340
aaatgattta ttaaaatagc cagcatttaa aagcttaaaa tgtaaatatc ataatgtggt 11400
atcctagata gcatcccaga acagaaaaag gatattaggg aaaaactgga ggaatggaat 11460
aaattatgca gtttagttat taataatgta ctaacgtcct tagttatgac gattgtacca 11520
tggtaatgta agatactaac aatagaggaa accgggtaag gagtatacag taactctata 11580
ctatctttgc aacttttttg taaatttaaa acttctaaaa taaagaacaa atttaaacat 11640
taaaaagtat caccaggaac atatatcact gtttacagat gaaatactat gtattttcat 11700
atctaatttc tgatcattga cttcaaatca gaaaagtgaa tgacacctca aaatcaggtt 11760
ttctgtttac tgaagtctaa gaaaagaaag cataccagct ggagagattc atgtttataa 11820
agacagattt ataacaacaa aaataaaata tccaagaata aatttaagaa gaagcacttt 11880
actgagaaac atatgaaaac ctgaacaaat ggagagggat attttgtatt tgaatagaaa 11940
gacttctggt ttaaagataa ttctctttaa attatttttt gtagaaattt aaggggtaca 12000
agagcagtgt tgtcacatgg atatattaca tagtggtgaa gtctggggtt ttagtgtaaa 12060
ttaatcttta cattttgttt gagcccaata aatgtaccaa catgattttt atagaaagat 12120
agtcattcct attaatccaa acttgtccca actttgaatt gaattgaggc agagctagca 12180
ggtgttcccc acggctgagg catctgaaca ttaagcatat ccctctgaga accagcctgc 12240
attgatactc tttctaatgt ggacagcatc aagctatgta cgtagttctg tgctcagcaa 12300
aagccctgac ttctttttgt ttatgtccta gccccataca acaaaatcaa ccaaagaatt 12360
ttggttgtgg atccagtcac ctctgaacat gaactgacat gtcaggctga gggctacccc 12420
aaggccgaag tcatctggac aagcagtgac catcaagtcc tgagtggtaa gaccaccacc 12480
accaattcca agagagagga gaagcttttc aatgtgacca gcacactgag aatcaacaca 12540
acaactaatg agattttcta ctgcactttt aggagattag atcctgagga aaaccataca 12600
gctgaattgg tcatcccagg taatattctg aatgtgtcca ttaaaatatg tctaacactg 12660
tcccctagca cctagcatga tgtctgccta tcatagtcat tcagtgattg ttgaataaat 12720
gaatgaatga ataacactat gtttacaaaa tatatcctaa ttcctcacct ccattcatcc 12780
aaaccatatt gttacttaat aaacattcag cagatattta tggaatatac cttttgttcc 12840
atgcattgta gtactcattg gatacacata gaataataag actcagttca cactcttcag 12900
gaaacagata aaaaactaag aaacaaacaa aaaacaggca atccaacacc atgtgggaaa 12960
tgctttcata gccgggaaac ctggggaata cctgagagga atactcaatt caggccttgt 13020
ttcaggaatc caaatcctgg cacatcagag ctgcttccct ctttccaggg tggcaggaaa 13080
taaatggaac atatttttct atcttatgcc aaacatgagg gaccctttct ccccggtgcc 13140
tctcccaagg tagtctacaa tatttcaact ctagcagtct gcttagtgca tagaacatga 13200
ggctgtgtgt ccctgggcaa attactagac ttctgtgtgc ttcactttcc ctgtaggatt 13260
ataatctact gagcaagctt attgtaaggg tcagattagc aacagtgtat gaaaatgatt 13320
tgagaccatt gcctgcacaa attcaactat ttttttttat ctcactactc tacagaagta 13380
ggtagggtgg gagacagagt ctgatgagag gctcagaatg tgaaagaaag tgaggcgagt 13440
gagcatgata tttaatataa acacaaagat attctgagaa gagctgctca ctgccccctc 13500
ccccaataca tgttgatagg aaaatgccac gtacttcagc aaaaacaact gaaaaattag 13560
atagaaaagt caatcaatag gaaaagataa tccaggacgg tgttgtgaac agaaagaggg 13620
ggaaaaaact ttagaaaatg atggggatgc tcttactggg gtacgagtcc tcaggtattg 13680
aactggcttt cagtaaaagc tagattagtg ggttcctgcc atttacaagc tgttttatga 13740
caacttactt gttgggtggc ctacagtaac tcacctaact gcactgagtc tgtttcctca 13800
tctgtaaatt ggggattttt ttttaaatac ctggcatgcc taactcataa agttgttctg 13860
aaactgaaat aaaacatacg tgaacaggca ttgtaaactg taagttacgg aaaaagctgg 13920
ctgttgttgt gtctttaaag tttcacctgg gtagtcaaag atggatcatg ggtctcagtg 13980
gagagctgag ccaggcagga gctgactaag ggtgagaggt gggagttagc agcctctgaa 14040
catctgtgta ccatgggacc ccctttcctc ctgcatggta ccccagacaa ggagcctagt 14100
aagagatact aatggcttgt tgtccagaga tgttcaaact gcagagaaag ataagacaac 14160
aagcattggc ctccaatcat gatgacagat aggaggaggt gggagctcct tagcagtgct 14220
ggttggcctt ccatgttcta ctgtgggcca tctctgccat gtactgtagg ctactagctt 14280
ctatattaaa gaatgcaaga ggggccagga gcggaggctc atgcctgtaa tctcagcact 14340
ttgggaggcc aaggtgggca gatcacttga ggtcaggagt ttgtgaccag cctggccaac 14400
atggtgaaac tctgccttta ctaaaaatat aaaaattagc tgggtgtggt ggtgtgcacc 14460
tgtaatccca gctactcggg agactgaggc acaagaattg cttgaacctg ggaggcggaa 14520
gttgcagtga gcccagattg cgccactgca ctccaccctg ggcaacagag aaagactctg 14580
cctcaaaaaa aaaaaaaaaa agcaagagga agtgaaataa tcaaggccgc catttaatag 14640
tgagcagcca ctccatgtgg tactgtgcaa gcacattata aatattagcc tcacaagaaa 14700
tgtattagca tttgtatttt gtacactggt taagtatctt gcccaagacc tcaaaactgg 14760
ttaagggcag cagaatttag ccccagcacc accttttcaa agcctgggct tctcacactt 14820
ctccatgctg ttcccatttt aacacaggta tctcgccatt ccagccactc aaactttggc 14880
atttaagaaa attatcctaa agctaaacta aacttcaagg atgaccattc tcctgacccc 14940
ttcccatcaa aattttatct ttagtcagtt tgttttcgtt ttgttttgtt tttcagaact 15000
acctctggca catcctccaa atgaaaggac tcacttggta attctgggag ccatcttatt 15060
atgccttggt gtagcactga cattcatctt ccgtttaaga aaaggtagta tttccttaat 15120
tgcagtggtc tccactgggg gtgaggaagg ggtgagaatt ggatcatggc tgcaaggaaa 15180
cccgacttaa cctctgcaag gtggtgcaaa ggcattccac tgttcaacag caattatatt 15240
gaagctgagt gggatcactg ggtgaagatg aagcgtaagg ggtgaggggc aggagaatgg 15300
gtatggatgg aggtagaaga tgcagtgtca tacagttttt ttctatcatg aaaataacca 15360
cagacttaca gaagagaaag agctaaaatg cccgtcattt tcagttgcat tttagtcttg 15420
cattagttgc aaccagctgg tttctgggta ccctaagtaa taaaaatagt tcctctgtag 15480
aactgtagta tgtttaccat agagtatttt gcaaaatttt tggtagagga tgttacataa 15540
tttgcatgtg ttcatttctc catttacctg tgggaacaat taaaatccag gaaaatgagt 15600
atattcaaat aatttcctcc catttaagat gagtcagagt aaataattcc tccaatactt 15660
agagaagtat accaagagat ccagtgatgg tatagagttg tctgatgtta aatagggaag 15720
tagaatatgg aaggggattc caatagtcgt tgaaaaattc cccataaccc cttacatggg 15780
ggaaagtagt gttaactgag agagtagaga taagctgttt ccaaaaatta tattcttaac 15840
aggactgaga tagccagaat ataaggatca agtttcaatg acagtaagat cctgagatgg 15900
agttgatttg cacaaagaaa taattgttgc cagcatgcat tttgaatatt tctctggaaa 15960
aaaagattag ttggcagtag aaatggatag aaatcaatag atattaaaat acctcagaat 16020
ttggttcatc tctgggaaaa gatgaaaaat aaaagtgtat actcctcaag aacatctagg 16080
atcaaaagca tgtgccctac actattgaat taattaacct cataagttgg gacctgtgga 16140
ataaggatgt ccaccagact tcctagggat tacaaatgtt tcacagaact tgaaatttaa 16200
acttgggtca ctgtatggga tgtagagctg tgctatatgg aaataaaaat gatttctttt 16260
tctcaaggga gaatgatgga tgtgaaaaaa tgtggcatcc aagatacaaa ctcaaagaag 16320
caaagtggta agaatatcag aaggaattgg gaagtaaaag tcaaaggaaa caaaaagcta 16380
aagcaataac aaagagaaat ccatcagtca taatctcctc tccttttaaa gaatgctggt 16440
tcccctttgc ctcacagcta acacaagaac tcctccaccg tctgaggagg tttaggagca 16500
gggaagggga aggagtcagc ttcatttgct aatcttctgt tgccctgcac cctagcagct 16560
ccttgcagca ggggacaagg atgacttagg tggatggata attaattgat tctaaaatat 16620
tgtgtgtcag tattgtaata ctatgttaat tgcaccatgc acggtatctc atttaatccc 16680
ccaccccttg ccattaccaa agagagagag agagagagag agagaaatac tagaatttat 16740
cctcatttta cagtagagaa aacagagggt caagaagata atgtaaagtg cccaagaaca 16800
cacagctgat cacaaaaatc aagcttgggg gccattagcc taaccacaga cccttactct 16860
taacccatct gcttcaatcc attttgctac aaatgtttac atttataagc agggcagaaa 16920
aacctcatcc aggttattga actaagaaga aagttatatt aaggtttcta atttttttaa 16980
tgtagttaga aaccaaactt aacaatgagc ccaagtttaa agcagtctaa ttaacctgga 17040
caagctcagg caagtttcat tctgtggccc atagcatcat ctgtgttgta aagctaagta 17100
gcaaatgttg tttgggtcat gctgggggac aagccatccc aatttgctca ggactgaggg 17160
gttttccagg atatcatgta aggataattg ggtacaaata taacctgctg ctttctctca 17220
tttcaaattt atcatttatc atatcagcaa ctatgagtta tgttttttat tagatttctt 17280
gttacttttt ccccagacca cttcccatga aattaatata ctattatcac tctccagata 17340
cacatttgga ggagacgtaa tccagcattg gaacttctga tcttcaagca gggattctca 17400
acctgtggtt taggggttca tcggggctga gcgtgacaag aggaaggaat gggcccgtgg 17460
gatgcaggca atgtgggact taaaaggccc aagcactgaa aatggaacct ggcgaaagca 17520
gaggaggaga atgaagaaag atggagtcaa acagggagcc tggagggaga ccttgatact 17580
ttcaaatgcc tgaggggctc atcgacgcct gtgacaggga gaaaggatac ttctgaacaa 17640
ggagcctcca agcaaatcat ccattgctca tcctaggaag acgggttgag aatccctaat 17700
ttgagggtca gttcctgcag aagtgccctt tgcctccact caatgcctca atttgttttc 17760
tgcatgactg agagtctcag tgttggaacg ggacagtatt tatgtatgag tttttcctat 17820
ttattttgag tctgtgaggt cttcttgtca tgtgagtgtg gttgtgaatg atttcttttg 17880
aagatatatt gtagtagatg ttacaatttt gtcgccaaac taaacttgct gcttaatgat 17940
ttgctcacat ctagtaaaac atggagtatt tgtaaggtgc ttggtctcct ctataactac 18000
aagtatacat tggaagcata aagatcaaac cgttggttgc ataggatgtc acctttattt 18060
aacccattaa tactctggtt gacctaatct tattctcaga cctcaagtgt ctgtgcagta 18120
tctgttccat ttaaatatca gctttacaat tatgtggtag cctacacaca taatctcatt 18180
tcatcgctgt aaccaccctg ttgtgataac cactattatt ttacccatcg tacagctgag 18240
gaagcaaaca gattaagtaa cttgcccaaa ccagtaaata gcagacctca gactgccacc 18300
cactgtcctt ttataataca atttacagct atattttact ttaagcaatt cttttattca 18360
aaaaccattt attaagtgcc cttgcaatat caatcgctgt gccaggcatt gaatctacag 18420
atgtgagcaa gacaaagtac ctgtcctcaa ggagctcata gtataatgag gagattaaca 18480
agaaaatgta ttattacaat ttagtccagt gtcatagcat aaggatgatg cgaggggaaa 18540
acccgagcag tgttgccaag aggaggaaat aggccaatgt ggtctgggac ggttggatat 18600
acttaaacat cttaataatc agagtaattt tcatttacaa agagaggtcg gtacttaaaa 18660
taaccctgaa aaataacact ggaattcctt ttctagcatt atatttattc ctgatttgcc 18720
tttgccatat aatctaatgc ttgtttatat agtgtctggt attgtttaac agttctgtct 18780
tttctattta aatgccacta aattttaaat tcataccttt ccatgattca aaattcaaaa 18840
gatcccatgg gagatggttg gaaaatctcc acttcatcct ccaagccatt caagtttcct 18900
ttccagaagc aactgctact gcctttcatt catatgttct tctaaagata gtctacattt 18960
ggaaatgtat gttaaaagca cgtattttta aaattttttt cctaaatagt aacacattgt 19020
atgtctgctg tgtactttgc tatttttatt tattttagtg tttcttatat agcagatgga 19080
atgaatttga agttcccagg gctgaggatc catgccttct ttgtttctaa gttatctttc 19140
ccatagcttt tcattatctt tcatatgatc cagtatatgt taaatatgtc ctacatatac 19200
atttagacaa ccaccatttg ttaagtattt gctctaggac agagtttgga tttgtttatg 19260
tttgctcaaa aggagaccca tgggctctcc agggtgcact gagtcaatct agtcctaaaa 19320
agcaatctta ttattaactc tgtatgacag aatcatgtct ggaacttttg ttttctgctt 19380
tctgtcaagt ataaacttca ctttgatgct gtacttgcaa aatcacattt tctttctgga 19440
aattccggca gtgtaccttg actgctagct accctgtgcc agaaaagcct cattcgttgt 19500
gcttgaaccc ttgaatgcca ccagctgtca tcactacaca gccctcctaa gaggcttcct 19560
ggaggtttcg agattcagat gccctgggag atcccagagt ttcctttccc tcttggccat 19620
attctggtgt caatgacaag gagtaccttg gctttgccac atgtcaaggc tgaagaaaca 19680
gtgtctccaa cagagctcct tgtgttatct gtttgtacat gtgcatttgt acagtaattg 19740
gtgtgacagt gttctttgtg tgaattacag gcaagaattg tggctgagca aggcacatag 19800
tctactcagt ctattcctaa gtcctaactc ctccttgtgg tgttggattt gtaaggcact 19860
ttatcccttt tgtctcatgt ttcatcgtaa atggcatagg cagagatgat acctaattct 19920
gcatttgatt gtcacttttt gtacctgcat taatttaata aaatattctt atttattttg 19980
ttacttggta caccagcatg tccattttct tgtttatttt gtgtttaata aaatgttcag 20040
tttaacatcc cagtggagaa agtt 20064
<210> 2
<211> 20261
<212> ДНК
<213> macaca fascicularis
<400> 2
gtaaaatcaa ggtgcgttca gatgttggct tgttgtaaat ttctgtttat attaataaca 60
taccaaatgt ggatttgttt taatcttcgg aactctttcc ggtgaaaacc tcatttacaa 120
gaaaactgga ctgacaggtt tcactttctg tttcatttct atacatagct ttattcctag 180
gacaccaaca ccactcgcta cccaaactga aagcttcccc gattccgccg aaggtcagga 240
aagtccaatg ccgggcaaac tggatttgct gccttgcgca gaggtgggcg ggaccccgcc 300
tccgggccgg gcgccaagtt gagcagctgg cacgcctcgc gaagccccag tcctgaagcc 360
ccagtcctgc gctgcttccc gaggctccgc accagccgcg cttctctctg cctgcaggta 420
gggagcgttg ttcctccgca ggtgcccacg gcccagcatc tctggctaac tcgctgggca 480
ccttaggacg gaggatctct acaccctttc tttgggatgg agagaggagg agggaaaggg 540
aaggcgatgg tctagggggc agtagagcca attacctgtt ggggttaata agaacaggca 600
atgcatctgg gcttcctcca ggcgcaattc agttttgctc taaaaataat ttatacctct 660
aaaaataaat aaggtgggta gtataggata ggtagtcatt cttatgcgac cgtgtgttca 720
gaatatagct ctgatgctag gctggaagtc tggacacggg tcctagtcca ccgtcagctg 780
cttgctagta atatgacttg tgtaagtcat cccagctgca gcagataagt aagtctcttc 840
ctgcgctaag cacgtccagg acccctgaac ggaatttatt tgctctgtcc attctgaaaa 900
cccaaaggag tcctaaaaga ggaatggagg agcctaagaa taaaaatagt ataataaaac 960
atttcttaga catgttgacc ttggcctatg tcaaagttca gtctgggttt gtcttataac 1020
ataaggagta aaagtaccat tgttctacct ctttttttaa tacttgaaaa aaaatttact 1080
gtagatgctt ttctattaat taaataacct tctaaaaaat gtttttagtg ctgcattcga 1140
ttaggttgga taactaaatg aaattaattc ctcactgttg ggtataaagg ttatttacag 1200
tggttctgtc ttagctattc actgaacatc attacataga tatctctgga atattgctga 1260
ttgtttccgt caataaactt agaagtgtaa ctacttagtc aaagagactg aatattttaa 1320
aggcattttg aagaaaactg aaaatgcttt ccagaaagga tgtatcagtt gacaatgatg 1380
gttgtcaaca gtatttaagg agaactatga tactctgaag aaaaacttag cctttctcag 1440
tagaagcagg taggcagagg ccacatgaca gcagttagag tgtggtcttc aaggaagtca 1500
cagaaatact gtggggaatt gaaaccccaa gtggaaaatg tacaagagtg tctcagtgtg 1560
actgagaagg aggttgggct tggggtttaa cttaagaatt tttttctttt tcttttgtgg 1620
agatagggct tttgttatgt tacccaggct ggtcttgaac tcctagcctc aggcgatcct 1680
cccgcctcag gctgcagaag tcctgggatt actggccgga gccaccatgc aggcctcttg 1740
ctcctacttt tgagaaagga agcttaacct tttttttttg tttgtttgtt tttgtttttt 1800
gttttttttg agacgagtct cactctgttg cccaagctgg agtgcagtgg tgccatctca 1860
gctcacggca acctctgcct cctgggttca agtgattctc ctgcctcagc ctcccgagta 1920
gctgggacta caggcaccca ccaccacgcc cggctaattt ttgtattttt agtagagatg 1980
gggtttcacc atattggtca ggctgatctc gaactcctga ctcaggtgat ccacctgcct 2040
cagcctccca aagtgctggg attacaggcg tgagccaccg cgcctggcca cttgactttt 2100
tctctatctc ttccttctac ccatcctacc cttggaagat agagaagtaa tatcagttcc 2160
atcgtgttat actgggcttc ccccagggac aaacccactt ccccaacctg aatgagccat 2220
cacttcttcc ccagtttaca tttcattgct ctttgaatgt ctccgttcgg atatgggaat 2280
tcacatgtgg tcataattct tacctgaaga agactgacgt cttcttctct tagaccaact 2340
gccctgatgt gaggtttaga ggttaaagaa catgtgtgta tttacatgat ctttgtattc 2400
tgccttttcg tccctcacta atgacagctg aaccccaagg aaatagagct gtggaggaga 2460
gggtttgatg agaaagtagg taaatattgg atctaatcca tcatcttcca ggaaacctcc 2520
attacttcta aaaatttcaa ccaaattcgt taaaggacaa gaactccacc agagtagggc 2580
cataaacatt ggcaaaatta gttgtaatct atgactagat ttaatgtccc tttgttttat 2640
tcacatatgg ttataatgct ttgcttggaa ttaggggtat tttaagtttt cttctgccta 2700
gtaagtgtat ttgtgcttat aatacaataa ttataaaata tcacattaat attttataac 2760
tgtacagtta actctggccc aaggaaaaga tagtccggta gatgctgcag cctgattttg 2820
tatctaaccc tggcaagagg ataatgactc atgttatttc acttaccctt tttatctttt 2880
aacatgaagg gctcatatag gtcaataaga aaccagtgat ataaacagac caaaaaatga 2940
tcagatcttt cacattagca aaaaaaaata ttttttaaac aatacccaac tgggtgaaaa 3000
tacagtgtaa cagtaccaaa tatcaacatg tgtcaagaac cagaaaaatg tttgttttct 3060
ttgatcagca acactatttg aggaaatcta tcctcagggc ctagcctggg gcctggcaca 3120
cagtaggcac tgaacaaata tttgctgaac acacacatac ttatgatatt tttaaaattg 3180
gcaacaatcc aatacccaat aatagaggaa ttaaatatta tagaactgtt caataagatg 3240
cttacgaata tcatgcagta agatgggcaa tatttatatc ataagcttaa atgaaacaaa 3300
tgggtattaa aggtatggta aggttataaa ttacttttta agagattaaa gggaaaagac 3360
tgaaagatat atactgaaat gctcacagtg gtgacaaggt tccccagcct tggcactatt 3420
gacattttgg gctatgtctt tgctgtggga ggctggcctg tgctctgcag gaggtttggc 3480
agcactcttg gtttctaccc ctagatagca gtagcaaccc tccctcaacc agcccaattt 3540
tgacaaccaa aaatgtttcc aggcatcacc agattctccc tgggtgagag tgatgaaaca 3600
gtaggtgatt ttccccttct tttctcattt tctgtaattt tgtcacatta cgttaataat 3660
aaggaaaaaa cataaaaaat agatgaattt attattctac ctcagtttgg atgtttggac 3720
tccctttggg ggttctttcc attatatcac ttggtctgct aaacagtcta tggtttggca 3780
aggtgaaatg attcatgaaa ttttgttttt attttttacc tgatactaaa agtaaaacat 3840
tcattcgctt gaaaatttgg acacagaaca ccaaaaaaaa tccataatct catctctctt 3900
tttctgtctt ttccttcctt ttttcccttt aaaaacaaga gtgaaaacct accggttctc 3960
cctccaattt aattcctaaa tataatcact gttaacatct tggacatttc ctgtgtctaa 4020
acacacatac tcactttttc ttttttttag caaaaagtgg atttctgcta catgtagtgt 4080
tccgcaactt cctacatgtt tacaaaatca gtacatttac atatgctgaa ttcagtcctt 4140
aatggtatta tattttgtga atataacaaa atttatttaa ccacttagac aatctaagat 4200
attctcagtt tgctgttatg agcaatgctc ttcctttaca tatacatata tatgtgtgtg 4260
tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg ttttagtagg atagatttct aggagagggc 4320
gaaatgtcat atgacatcca catttacaat tgtaatagga agtatcaaag tgccccctaa 4380
aaaaaaaaat tcctcccatt agtgtgtgag aaagcctatt tgttcatatc ttcacaaaca 4440
ctaaatatta gaaatattta caattgtgtt caagctaaga agtgaaaaat ggtatttcat 4500
atcttataat ttttgttgtg agattgaaca tatttcctat gtttacatgt cacctgtatt 4560
tcttattctc tgaactatac gttatgacct ttcacttatt ttcctcatgg gttatgtgta 4620
gtttgtgtag ttgtcttatt gattgttagg agctatttat atattaggaa cattaatctc 4680
ctgtcttata tgtatgtggc attgattagt tgatcatttg tgagttcatg tctgtataca 4740
aagattagag aggcagtaag agggaaaact tacctctttc ttatcaaagt ttgtaaatat 4800
atgtataaca gaagagagag aaaacattaa taaatgctga aataaagtat aaatttttta 4860
ctccactact tcaacataaa ctacaaaagg agagtgactt ttctttcatg ctgacttcca 4920
tattccccat gcctaaaata gtgcctagca cagaagaggt gctcaatcag tgtttgctaa 4980
atgaaagaat tagtaacatt tcaagcagga tgactaaatg aagaatagaa tctaggcaga 5040
tactctggaa gagtgactgt gagtcattca tggtcttggt atgaattagt caaatccagc 5100
tttctcccct tcccactccc cactgttagt agaataatct gtttattgag agaatagatt 5160
tataatttag aataagaggg gcagaagagg agattttgaa ggatggcacc tgaaggagga 5220
ctagcatggc tgagatagtg aagtggaagt cttggaaagc taaagggtaa gatgaaagta 5280
tttagccgta gggggaaaaa gcattgacag gttggaaatg taaaagtcag attctccttg 5340
ctttgaaatt ttgtacaggg caggttctac taggtatgtt acaatgcaga aaaaacatga 5400
aatagttgag aggaatttgg tgaaatatta tcttcctggc ttcttttgag tgagcagatt 5460
tttttcaggg cctgtaacta taataaattt gaaacttctc atcttttagt aacttttttc 5520
acttaagttt atgtggctgt gggcaatgga atgaaggtat tgaacttcct attccctgct 5580
gggtttccac aattacaagt caatcatgac tggttattag aagactattt caattagaac 5640
caccaagtcc cataatgtca tattgtgtgt ttaattatta agtaaagcag tcttcttttt 5700
gtgttttcca taattagcac attccagaaa gatgaggata tttgctgtct ttatattcac 5760
gatctactgg catttgctga atggtaagac accaaatcct tccattaggt tctatatttt 5820
aaatatttta accatgagtt taaaactaaa atgataattt aaaatgcatg caattttctt 5880
atagagagaa tattctcttc tttcttctac tttacacaat gacaaagtct ttcttctgct 5940
ttacacaatg ataaagttcc ctgtgtcatt gtgtaaaaat atagagaata tagacaaatt 6000
gaaagacaca aaataatcta ttacccattt cccagggtta actactgaaa atatctgggg 6060
aaatggcctg tatgtatata tttatttgtt tgttttcaac aaggccagga tctttcaatc 6120
tttcaatctt ggttgctctg tgacatgcct ttcctgatga gaatactcta aggaagaatt 6180
gtaggataca tggaaaatgt cagggtaaca cagtactggc accaccctgc ggtcctttct 6240
gaactccata ccaatgtact tcttgccaga aaactgatca aaagtttagg gaagtaaaaa 6300
gagatgttag aatctaccat tccctctatg taggaagcaa ataggtgtcc agtcaaagga 6360
cattctgggg atgtctacat gaaaccaagt ctcctggttg taagtactcc atctccatat 6420
aatatttcta cagtaatata tgtttataaa ttgtgggggc aacttgttta gctaatttta 6480
ttattctgtt attgggacac tatgtctctg catgagacat agtgtcccaa aacatatttc 6540
aagcccattg gataaaatat gtatttagca agttcttaaa tataatgata acataactga 6600
ccagataagg tgatttttaa atgctgtgcc aactttataa atgttttgag gaattttccc 6660
ttttctgaag gttattcttc tttcttttta gcatttactg tcacggttcc caaggaccta 6720
tatgtggtag agtatggcag caatatgaca attgaatgca aattcccagt agaaaaacaa 6780
ttagacctga cttcactaat tgtctattgg gaaatggagg ataagaacat tattcaattt 6840
gtgcatggag aggaagacct gaaggttcag catagtaact acagacagag ggcccagctg 6900
ttgaaggacc agctctccct gggaaatgct gcacttcgga tcacagatgt gaaattgcag 6960
gatgcagggg tttaccgctg catgatcagc tatggtggtg ccgactacaa gcggattacc 7020
gtgaaagtca atggtaagaa ttattataga tgagaggcct gatctttata gaaaacatat 7080
tctaagtgtt gaagactttt cattcttata agtccatact tattttcaaa cagaatagca 7140
tagtctcttc attcattcat tcagttcatg aattcattca cgtaagtctc caattagcat 7200
ttcttgagca cctatatgat agtcattgga aatccagaga caacacagag ccatgttcta 7260
cggtatgtac agttttccaa aaataattcc tagtctttac ttttttatta taaatgtaat 7320
acatatactt gcaaagaatt cagatactat ggaagagatc aaatgaattg caaaagtgtc 7380
cctcctccct tcaccactat ctcccatgag ataaccaaga gacactccaa gagagtaacc 7440
attatttgtg tgtccctcca gaaccttttt tattcaacta ccattttttt attttattag 7500
gtctgtcagt tttccttttt tgagcctctc tatatcaaat gctaataaat atattcagat 7560
caaaccccac tgtaaggttc atattaaaaa agacttgaag tctccctatg aagacaaaaa 7620
ataatcatat taagtgtaaa agaacttatt cttccagtac agtataaact atactcactg 7680
ggcatatatt catcttgaaa atctatactg atgttgtctt ggggaattga agaggaacta 7740
ggagtgtcaa ttcctgggaa ctgacccaca gttatgtcat caggtcactt gagttcgaag 7800
ttttgtgttg gcactagcta agtaaaggaa aacacctctg ctttcattgt tgagtttcat 7860
agaattgaga gctgaaagga tcccaggcag gagcgactaa tccaaactcc cacaaagaac 7920
aaaaatcccc cagaggatct tctgttcata tatttcctgc agtggcatcc ctgtcatatc 7980
ccacaatggc atccctgcca tttggactcc ccttccatat cctgttgaaa ttactcccta 8040
atagtaagct gaaatctgcc cctctagttg tagttttgga attatttcat ttccatgatg 8100
accttttaat atttgactag aattaaatca tctccccttg gtatttccat tcctggacta 8160
actaccatca atctgagggc taacaataca agtagaaaaa gtctacactt gtcattgatc 8220
actgatcaat gattaatcaa tgatcactga taattataaa ctcaaaaaca aaatcatgta 8280
gggattaaga gaaatgtatc agttttatgt tgtatttctg gtccctgatt ctggctcaaa 8340
taatgctact attgtcaaga agatatcact tgtaaagtag atttaatttt cattatattt 8400
taccatgtgc ttctccattc acggcatttc ttgagatgtt gtggtttata ctttcagttt 8460
ttctccagtc catcagcaaa tatcaggcat ctactgtgtt ccaagatatt aaagaaatca 8520
tcatgactta gcctcatcaa cagcattgct agatctggga tggaaaagaa gagtataatc 8580
ctggcagtca ggaagaaggc tgcataaagt ataagtttct gcttccaaag aagatctctc 8640
atcagcctgt agggagtgta tagggagggg acagctgtcc ttgtagtagc aaagggtttt 8700
attcaggtca tctgggctcc ataatatccc ttgtgtatct gcagtctcct ttgccatgga 8760
tcaacacaat aggaaatctt ccggcactga tggtttttcc aagggggagt tcttcaggga 8820
gcaaagcaaa tgaccaacca ggtttgagga cctgatttga caattccatt ttgtatttta 8880
aattagttaa tttgcattct agtcccaatc catcttgtca tttgcagaca gtggttttgg 8940
ggttgagttg agctatacca aaagtctgaa ccttctgcac ttagaacaag gaaggcaacc 9000
accaagcttc acttgcactg aggcagtgtc tccaatggaa acgaggtagc tggcttgcag 9060
aagctttcca actcagggaa gtagaactcc tgagtcacct ccatatgcaa ataatttcac 9120
agtactgctg ttgaacttca cttcccatca cagcaaatgt gtggtaacat agctttgcca 9180
caggagttta ctcaccatgg gattttaaag gtgaaacatt tcaaaactga aatttgaaag 9240
aatttagttt tggattcact caattatcat gatcactttg ggtgttattg cacctttcat 9300
gtttgtgagt ttaaatacca gactctcagg cctctaactt tcaattaaaa gtgtttttct 9360
ttaatcactg aacctaatag tagggaaaac gaaatgttca ttcagacttt caggaccttc 9420
aatgagatga ggcagctgaa agatcaaagt gttgcatagt tatcccagta aagctatttg 9480
gatcgtatgg accagatcaa ctgctgtcat tccccaccaa ccccatcttt ccccaaaatt 9540
cccagccctg tttaagtgtt ctctgtagca tttatctcta tctagtatat tgtgtagcat 9600
atcatatcat acttttctat tttgtttatt gtctctctcc tcctagaata taaactccac 9660
aagcacagag atttgggtct gttttttaat attgttgtat ccccagggct tgacgtaaag 9720
cagagtggta gtatgacaaa agcacacaaa aaaatatttg ttgagtcaat gaatgaatga 9780
tttcctcaaa taggattagc ctaaaatttt ggaaacatga acagacttgg atatatgaaa 9840
atttatttcc aaaactgttc atcaggaact gttagcacct tctaaagggt acactgaagc 9900
agcagtagta aaaggaggag gaggagaagc agctctgcta ctactattat cgagtactac 9960
tacaactatc gagtactact acaactatcg agtactacta caattagcac ttgcttattc 10020
tgtgtgttag gtcctgtact gaacattctg cctaaattag ttcatttcct cctggaaatg 10080
actctgtggg gtaggtgctt catcatgtaa gatgagtatt tttcacactt cactgtctct 10140
gaaatctgag tgtgtctttc aatgatggaa tctttgattt catgataagt ggtattattc 10200
ccattttaag gatgaggaaa ctggggtcca aagaaattaa gtaatttgcc caaattcacc 10260
tagcctcgta aatgataaag ctagttctaa atccaagcag attggctctg aagtctgggc 10320
ccttaataac cacttattgc ctgtatttgc acctctggtg tatgtatcaa gttatatatt 10380
ggcttcaaaa ctatcatgac cttttcttga ttttgattgt tcaatattag tatagtgttc 10440
tagatctagt agaccagggg tctgcaaaaa attttgatta ctcacctcat ctgtaaaaca 10500
ttttaaactt gtgtgtctgt gcaggcacat ttgtgtgtaa ttataaaaaa tttactatct 10560
attaatatat aggttgtacc gtaagaaaaa ttgccatttt tgaagagcaa aaaaggttga 10620
atattaccag tttcatctgg ttcaacctaa tagacatttg tacaaaaaca gacattttaa 10680
gaggatgaaa taaaaattta ataaacaata ttttcaattt ttactaattg tgacgcttca 10740
ctattgttag ctaatatgtc aaggcatgat ataccttagg gtggaattta tcattaacaa 10800
aggtggatag tgtcaataat cttgaggttt gtgttttttt atataacact gtgaggtcta 10860
attaagtact taattgttta caacctcata cagtcgccaa taataagtgt cacttctgct 10920
gtttcctctg ggttgtgctt gaattattag tattatcttc aatcctcagt ttctttgtgg 10980
aaaacttttt aattagttgt ttaattttgt aagatggtta gtttagtcaa aattagataa 11040
gagaatctga aaatccataa ttaccccaaa gcaacccact cataagaact attatttttg 11100
tgttttgaaa tcataatttt attgatttcc agtgtttcca ctggtagtgg tttcattgat 11160
gtaggagtat caaaacatca ctaattattt atttcagttt tgtttgatcc tagctgtttt 11220
gtgttaactt tgaagaaatt acatcacaga tctattgttg tccttggtaa aggaatggag 11280
agttaaggct ctagatcatt agtggttatg ctgtagtatt aggagtaaaa aaaaagatta 11340
tatcaacaaa ataagaacat gttaatgtac ttgtaataga taaacatgaa taaagctctt 11400
atgctatata gatgcactga acaatctact agaattgtca gcaaacggta tcttaatcct 11460
aaaagggtcc caaaccaatg atctaaaatt gaatcaaact ttcttccttg agcataatta 11520
tttaagtgat ttattaaaat agccagcatt taaaagctta aaatataagt atcataatgt 11580
ggtatcctag atagatccca gaacagagaa aggatattag ggaaaaactg gaggaatgga 11640
ataaattatg cagtttagtt attaataatg tactaatgtc cttagttatg accattgtac 11700
catggtaaag taagatacta acaatagagg aaattgggta aggggtatat gtaactctat 11760
actatctttg caattttttt gtaaatttaa aacttctaaa ataaagaaca aatttgaaca 11820
ttaaaaagtg tcgccaggaa catgtatcac tgtttacaga tgaaacagta tgtattttta 11880
tatctaattt ctgatcattg gcttcaaatc agaaaagtga atgacacatc aagatcaggt 11940
tttctgttta ctaaataaag tctaagaaaa caaagcatac cagctggaga gattcatgtt 12000
tataaagaca gatttataac aacaaaaata aaatatccaa gaataaattt aagaagaaat 12060
agggcactat gtaaaaagta tagcacttta ctgagaaaca tatgaaaacc tgaatacatg 12120
gagagaggta ttttatattt gaatagaaag attgctggtt taaagataat tctctttaaa 12180
ttttttttgt agaaatttaa gaggtacaag agcagttttg tcacacggat atattacata 12240
gtggtgaagt ctggggtttt agtgtaaatt aatctttaca ttttgtttga gcccaataaa 12300
tgtaccaaca tgatttttat agcaagatag tcattcctat taacccaaac ttgtcccaac 12360
tttgaactga actgaggcag agctagcagg tgttccccac tgctgaggca tctgaacatt 12420
aagcgtatcc ctctgagaac cagcctgcat tgatcctctt tctaatgtag acagcatcaa 12480
gctatatatc tagttctgtg ctcagcaaaa gccctgactt ctttttgctt atgtcctagc 12540
tccatacaac aaaatcaacc aaagaatttt ggttgtcgat ccagtcacct ctgaacatga 12600
actaacatgt caggctgagg gctaccccaa ggccgaagtc atttggacaa gcagtgacca 12660
tcaagtcctg agtggtaaga ccaccaccac caattccaag agagaggaga agcttttaaa 12720
tgtgaccagc acactgagaa tcaacacaac agctaatgag attttctact gcatttttag 12780
gagattagat cctgaggaaa accatacagc tgaattggtc atcccaggta atattctgaa 12840
tgtgtccatt aaaatatgtc taacactgtc ccctagcacc tagtatgatg tctgcctatc 12900
atagtcattc agtgtttgtt gaataaatga attaatgaat aacattatat ttacaaaatg 12960
tatcctaatt cctcacttcc attcatccaa atcatattgt tacttaataa acattcacca 13020
aatatttatt gaatatgcct tttgttccat gcattgtagt actcatttga cacacataga 13080
ataataagac tcacgttcac actcttcagg aaacagataa aaaacaaatg aacaaacaaa 13140
aaacaggcaa tccaatacca tgtgggaaat gctttcatac catgtgggaa acctggggga 13200
atacctgaga ggaatattca attcaggcca tgtttcagga atccaaatcc tggcacatca 13260
gagccgcctc cttcttacta gggtttctgt ggcaggaaat aaatggaacg tatttttcta 13320
tcttatgcca aacaggaggg accctttctc ccctgtgcct ctcccaaggt agtctacaat 13380
atttcaacgc tagcagtctg tttagtgcac aggacatgag gctgtgtatc cctgggcaaa 13440
ttgctacact tctgtgtgct tcactttctc tgtaggatta taacctactg agcaaggtta 13500
ttgtgggggt caaattagca acagtgtatg aaaatgattt gagaccagtg cctgcacaaa 13560
ttcaactatt tttttttatc tcactactct atagaagtag gtaggatggg agacagagtc 13620
tgatgggagg ctcagaatgt gaaagtaagt gaggtgagtg agcatgatat ttcatataaa 13680
cacaaagata ctctgagaag agcttctcac ttcccccgcc cccaatagat gttgacagga 13740
aaatgccatg tacttcagca aaaacagctg aaaaattaga cataaaagtc aatcaatagg 13800
aaaagataat ccaggatggt cttgtgaaca gaaagaggga aaaaaaaagt ttagaaaatg 13860
atggggatgc tcttactggg gtatgagtcc tcaggtattc aactggcttt cagaaaaagc 13920
tagactagtg ggttcctgcc atttaaaagc tgttttatga caacttactt gttgggtggc 13980
ctacagtaac tcacttaact gtgctgagtc tgtttcctca tctgtaaatt ggggattttt 14040
taaaataact ggcatgccta actcataaag ttgttctgaa actgaaataa aacatatatg 14100
aacaggcatt gtaaactgta agttacggaa aaagctggct gttgttgtgt ctttaaagct 14160
tcacctgggt agttagagat ggatcatggg tctcagtgga gagctgagcc aggcaggagc 14220
tgactaaggg taagaggtgg gagttagcaa tctctgaaca tctgtgtgcc atgggacccc 14280
ttttcctcct gcatggtacc ccagacaagg agcctagtaa gagatactaa tgacttgttg 14340
tccagagatg ttcaaactgc agagaaagat aagacaacaa gcattggcct ccaatcatga 14400
tgacagatag gaggaggtgg gagctcctta gcagtgctgg ttggttttcc atgttctact 14460
gtgggccatc tctgccatgt actgtaggct actaacttct atattaaaaa atgcaagagg 14520
ggccgggagt ggaggctcat gcctgtaatc tcagcacttt gggaggccaa ggtgggcaga 14580
tcacttgagg tcaggagttt gtaaccagcc tggccaacat ggtgaatctc tgcctctact 14640
aaaaatacaa aaattagcca gatgtggtgg cgtgcacccg taatcccagc tactcgggaa 14700
gctgaggcac gagaattgct tgaacctggg aggcggaggt tgcagtgagc caagattgtg 14760
ccactgccct ccagcctggg caacagagaa agactttgcc tcaaaaataa ataaataaat 14820
aaataaataa ataaataaat aaatggaagt gaaataatca aggccaccat ttaatactga 14880
gtagccactc catgtggtac tgtgctaagc acattataaa atattagcct cacaagaaat 14940
gtattagcat ttgtattttg tacactggtt aagtatcttg cccaagacct caaaactggt 15000
taaggggcag cagaatttaa ccccagcgcc accttttcaa agtctgggct tcttacactt 15060
ctccatgctg ttcccatttt aacagatgta tctcgccatt ccagccactc aaactttggc 15120
atttaagaaa attatcctaa agctaaacta aacttcaagg atgactattc tcctgatgac 15180
cccttcccat caaaatttta tctttagtca gtttgttttt gttttgtttt gtttttcaga 15240
actacctctg gcgcttcctc caaatgaaag gactcacttg gtaattctgg gagccatctt 15300
tttactcctt ggtgtagcac tgacattcat cttctattta agaaaaggta gtatttcctt 15360
aattgcagtg gtctccactg gggatgagga gggggtgaga attggatcga tcatggctgc 15420
aaggaaacct gacttaacct ctgcagggtg gtgcaaaggc attccactat tcaacagtaa 15480
ttatattgaa gctgcatggg atcactgggt gaagatgagg tgtaaggggt gagggacagg 15540
agaatgggta tggatggagg tagaagatgc agtgtcatac aatttttttc tatcatgaaa 15600
ataaccacag acttactgta aagaaggagc taaaatgcct gtcattttca gttgcatttt 15660
agttttgcat tagttgcacc cagctggttt ctgggtactc taagtaataa aaatagttcc 15720
tctgtagaac tgtagtattt tcaccataga gtattttgta aaattattgg tagaggatgt 15780
tacataattt gcatgtgttc ctttctccat ttacctgtgg gaacaattaa aatccaggaa 15840
aatgagtata ttcaaataat ttcctcccat ttatgatgat tcagagtaaa taattcctct 15900
gatacttaga gaagtatacc aagagatcca gtgattgtat agagttgtct gatgttaaat 15960
agggaagtag aatatggaag ggaattccaa tagtcgttga aaaattcccc acaacccctt 16020
acatggggga aagtggtgtt aactgagata gtagagataa gctgttacca aaaattatgt 16080
tcttaacagg attgagatag ccagaatata aggatcaagt ttcaatgaca gtaagatcct 16140
gagatgcagt tgatttgcac aaagaaataa ttgttgccag cttgcatttt gaatatttct 16200
ctggaaaaag agattagttg gcagtagaaa tgaatagaaa tcaatagata ttaaaatacc 16260
tcagaatttg attcatctct gggaaaagat gaaaaataaa agtgtatagt cctcaagaaa 16320
atctgggatc aaaagcatgg gccttaccct attgaattaa ttaacctcag aagttgggaa 16380
ctgtggaata aggatgtcca ccagacttcc tagggattac aaatgtttca tagaacttga 16440
aatttaaact tgggtcactg tatgggatgt agagctgtgc tatatggaaa taaaaatgat 16500
ttctttttct caagggagaa tgatggatat gaaaaaatgt ggcattcgag ttacaaactc 16560
aaagaagcaa cgtggtaaga atatcagaag gaattgggaa gtagaaggca aaggaaacaa 16620
aaagctaaag caataacaaa gagaaatcca ttagtcataa tctcctctcc ttttaaagaa 16680
tgctggttcc cctttgcctc acaactaata caagaacttc tccaccatct caggaagttt 16740
agggatggcc ttcaagagta gagagtaggg agcagctctg tggagagagg agaggagcag 16800
ggaaggggaa ggagtcagct tctctttgct aatctgttgc cctgcaccct agcagctccc 16860
tgcagcaggg gacaaggttg acttaggtgg atggataatt aattgattct aaaatattgt 16920
gtgtcagtat tgtatattgt aatactatgt taactgcgcc atgcacggta tctcatttaa 16980
tcccccaccc cttgccatta ccaaaaagag agagagaaaa atactagaat tatcctcatt 17040
ttacagtaga gaaaacagag ggtcaagaag ataaagtaaa gtgcccaaga acacacaact 17100
gatcacaaat atcaagcttg gggtccatta gcctaaccac agacccttac tcttaaccca 17160
tctgcttcaa tccattttgc tacaaatgtt tacatttata tgcagggcag aaaagtctca 17220
tccagtttat tgaactaaga agaaagttat attaaggtgt ctaatttttt ttaatgtagt 17280
tagaaaccaa acttaacaat gagcccaagt ttaaagcagt ctaattaact tgacaagctc 17340
aggcaagttt cattctgtgg cctgtagcat catctgtgtt gtaaagctaa gtagcaaatg 17400
ttatttgggt catcctgggg ggaaagtcat cccaatttgc tcaagactga ggggtttttc 17460
aggatatcat gtaaggataa ttgggtacaa atataacctg cttctttctc tcatttcaaa 17520
tttatcattt atcatctcag caactatgag ttatgttttt tattagattt cttgttactt 17580
tttccccaga ccgctcccca tgaaattaat atactattat cactctccag atacacaatt 17640
ggaggagacg taatccagca ttggaacttc tgatcttcaa gcagggattc tcagcctgtg 17700
gtttgggggt tcgtcagggc tgagcatgac cagaggaatg aatgggcccg tgggatgcat 17760
gcagtatggg acttaaaagg cccaagcact gaaaatggaa cctggcgaaa gcagaggagg 17820
agaatgaaga aaaatggagt tgaacaggga gcgtggaggg agaccttgat actttcaaat 17880
gcctgagggg ctcatcggtg catgtgacag ggagaaagga tacttctgaa caaggagcct 17940
ccaagcaaat catccactgc tcatcttagg aaaacgggtt gagaatccct aatttgaggg 18000
tcagttcctg cagaagtgcc ctttgcctcc actcaatgcc tcaatttgtt ttctgcgtga 18060
ctgagggtcc cagtgttgga acagtattta tgtatgagat tttcctattt attttgagtc 18120
tgtgaggtct tcttgtcatg ggagtgtggt tgtgaatgat ttcttttgaa gatatattgt 18180
agtagatgtt acaattttgt cgccaaacta aacttgatgc ttaatgactt gctcacatct 18240
agtaaaacat ggagtatttg taaggtgctt ggtctcctct ataactacaa gtacacattg 18300
gaagcataaa gatcaaaccg ttgatttgta taggatgtca cctttattta acccattaat 18360
actctgattg acttaatctt attctcagac ctcaagtgtc tgtgcagtat ctgttccatt 18420
taaatatcag ctttataatt atgtggtacc atacacacat aatctccttt catcgctgta 18480
accaccctgt tgtgatgacc actattattt tacccattgt acagctgagg aagcaaacag 18540
attaagtaac ttgccaaaac cagtaaatag cagagctcag actgccaccc actgtccttt 18600
tataatacaa tttacagcta tattttactt taagcaattc atttattcaa aacccattta 18660
ttaagtgccc ttgcaatatc aatcactgta ccaggcattg aatctacaga tgtgagcaag 18720
agaaagtacc tgtcctcaag gagcttggag tataataagg agattaataa gaaaatatat 18780
tattacaatc tagtccagtg tcatagcata aggatgatgt gaggagaaaa gctgagcagt 18840
gttgccaaga ggaggaaata ggccaatgtg gtctgggaca gttgaatgta tttaaacatc 18900
ttaataatca aagtaatttt catttacaaa gagaagtcag tacttaaaat aaccctgaaa 18960
aataacactg gaattccttt tctagcatta tatttatccc tgatttgcct ttgccataca 19020
atctaatgct tgtttatata gtgtctgata ttgtttaaca gttctgtctt ttctattcaa 19080
atgctattaa attttaaatt catacctttc catgattcaa aattcaaaag atcccatggg 19140
agatggtttg aaaatctcca cttcatcctc caagccattc aagtttcctt tccagaagca 19200
actgctactg ccttttattc atatgttctt ctaaagatag tctacatttg gaaatgtatg 19260
ttaaaagcat atatttttaa atttttttcc ctaaatagta acacattata tgtctgctgt 19320
gcactttgct atttttattt attgtagtgt ttcttatgta gcagatggaa tgaatttgaa 19380
gctcccaagg gtcaggacac atgccttctt tgtttctaag ttatctttcc catagctttt 19440
cataatcttt catatgattt agtacatgtt aaatatgtgc tacatataca tttagacaac 19500
cagcatttgt taagtatttg ctctaggact gagtttggat ttatgtttgc tcaaaaggag 19560
acccatgggc tctccagggt gcactgagtc aatctagtcc taaaaagcaa tcttattatt 19620
aactctgtat gacagaatca tatctggaac ttttgttttc tgctttctgt caagtataaa 19680
cttcactttg atgctgtact tgcaaaatca cattttcttt ctggaaattc cagtagtgta 19740
ccttgactgc tagttaccct gtgccagaaa agcctcattc gttgtgcttg aaccctttaa 19800
tgccaccagc tgtcatcact acacaggcct cctaagaggc ttcctggagg ttttgagatt 19860
cagatgccct gagagatccc agagtttcct ttccctcttg gccacattct ggtgtcagtg 19920
acaaggaata ccttcgcttt gccacccgtc aaggttgaag aaacagcgtc tccaacagag 19980
ctccttgtgt tatctgtttg tacatgtgca tttgtacagt aatttgtgtg acagtgttct 20040
ttgtgtgaat tacaggcaag aactgtggct gagcaaggca catagtctac tcagtctatt 20100
cctaactcct ccttttggtg ttggatttgt aaggcacttt atcccttttg tctcatgttt 20160
catcgtaaat ggcataggca gagatgatat ctaattctgc atttgattgt cactttttgt 20220
acctgcatta atttaataaa atatccttat ttattttgtt a 20261
<210> 3
<211> 20340
<212> ДНК
<213> macaca fascicularis
<220>
<221> misc_feature
<222> (1658)..(1881)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (2742)..(2745)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (2747)..(2747)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (2749)..(2749)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (4764)..(4802)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (8775)..(8778)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (8980)..(8984)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (9161)..(9293)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (9344)..(9348)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (9840)..(9874)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (9958)..(9973)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (10437)..(10452)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (10603)..(10603)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (10830)..(10830)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (10989)..(11006)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (11105)..(11122)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (11457)..(11457)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (13710)..(13710)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (14015)..(14049)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (14518)..(14585)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (14689)..(14699)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (14707)..(14767)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (14837)..(14865)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (15703)..(15741)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (17403)..(17415)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (19122)..(19135)
<223> n представляет собой a, c, g или t
<400> 3
ctgaaagctt ccccgattcc gccgaaggtc aggaaagtcc aatgccgggc aaactggatt 60
tgctgccttg cgcagaggtg ggcgggaccc cgcctccggg ccgggcgcca agttgagcag 120
ctggcacgcc tcgcgaagcc ccagtcctga agccccagtc ctgcgctgct tcccgaggct 180
ccgcaccagc cgcgcttctc tctgcctgca ggtagggagc gttgttcctc cgcaggtgcc 240
cacggcccag catctctggc taactcgctg ggcaccttag gacggaggat ctctacaccc 300
tttctttggg atggagagag gaggagggaa agggaaggcg atggtctagg gggcagtaga 360
gccaattacc tgttggggtt aataagaaca ggcaatgcat ctgggcttcc tccagcgcaa 420
ttcagttttg ctctaaaaat aatttatacc tctaaaaata aataaggtgg gtagtatagg 480
ataggtagtc attcttatgc gaccgtgtgt tcagaatata gctctgatgc taggctggaa 540
gtctggacac gggtcctagt ccaccgtcag ctgcttgcta gtaatatgac ttgtgtaagt 600
catcccagct gcagcagata agtaagtctc ttcctgcgct aagcacgtcc aggacccctg 660
aacggaattt atttgctctg tccattctga aaacccaaag gagtcctaaa agaggaatgg 720
aggagcctaa gaataaaaat agtataataa aacatttctt agacaggttg accttggcct 780
atgtcaaagt tcagtctggg tttgtcttat aacataagga gtaaaagtac cattgttcta 840
cctctttttt taatacttga aaaaaaattt actgtagatg cttttctatt aattaaataa 900
ccttctaaaa aatgttttta gtgctgcatt cgattaggtt ggataactaa atgaaattaa 960
ttcctcactg ttgggtataa aggttattta cagtggttct gtcttagcta ttcactgaac 1020
atcattacat agatatctct ggaatattgc tgattgtttc cgtcaataaa cttagaagtg 1080
taactactta gtcaaagaga ctgaatattt taaaggcatt ttgaagaaaa ctgaaaatgc 1140
tttccagaaa ggatgtatca gttgacaatg atggttgtca acagtattta aggagaacta 1200
tgatactctg aagaaaaact tagcctttct cagtagaagc aggtaggcag aggccacatg 1260
acagcagtta gagtgtggtc ttcaaggaag tcacagaaat actgtgggga attgaaaccc 1320
caagtggaaa atgtacaaga gtgtctcagt gtgactgaga aggaggttgg gcttggggtt 1380
taacttaaga attttttctt ttcttttgtg gagatagggc ttttgttatg ttacccaggc 1440
tggtcttgaa ctcctagcct caggcgatcc tcccgcctca ggctgcagaa gtcctgggat 1500
tactggccgg agccaccatg caggcctctt gctcctactt ttgagaaagg aagcttaacc 1560
tttttttttt gtttgttttt gttttttgtt ttttttgtag acgagtctca ctctgttgcc 1620
caagctggag tgcagtggtg ccatctcagc tcacagcnnn nnnnnnnnnn nnnnnnnnnn 1680
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 1740
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 1800
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 1860
nnnnnnnnnn nnnnnnnnnn ngactttttc tctatctctt ccttctaccc atcctaccct 1920
tggaagatag agaagtaata tcagttccat cgtgttatac tgggcttccc ccagggacaa 1980
acccacttcc ccaacctgaa tgagccatca cttcttcccc agtttacatt tcattgctct 2040
ttgaatgtct ccgttcggat atgggaattc acatgtggtc ataattctta cctgaagaag 2100
actgacgtct tcttctctta gaccaactgc cctgatgtga ggtttagagg ttaaagaaca 2160
tgtgtgtatt tacatgatct ttgtattctg ccttttcgtc cctcactaat gacagctgaa 2220
ccccaaggaa atagagctgt ggaggagagg gtttgatgag aaagtaggta aatattggat 2280
ctaatccatc atcttccagg aaacctccat tacttctaaa aatttcaacc aaattcgtta 2340
aaggacaaga actccaccag agtagggcca taaacattgg caaaattagt tgtaatctat 2400
gactagattt aatgtccctt tgttttattc acatatggtt ataatgcttt gcttggaatt 2460
aggggtattt taagttttct tctgcctagt aagtgtattt gtgcttataa tacaataatt 2520
ataaaatatc acattaatat tttataactg tacagttaac tctggcccaa ggaaaagata 2580
gtccggtaga tgctgcagcc tgattttgta tctaaccctg gcaagaggat aatgactcat 2640
gttatttcac ttaccctttt tatcttttaa catgaagggc tcatataggt caataagaaa 2700
ccagtgatat aaacagacca aaaaatgatc agatctttca cnnnnanana aaaaaaatat 2760
ttttaaacaa tacccaactg ggtgaaaata caatgtaaca gtaccaaata tcaacatgtg 2820
tcaagaacca gaaaaatgtt tgttttcttt gatcagcaac actatttgag gaaatctatc 2880
ctcagggcct agcctggggc ctggcacaca gtaggcactg aacaaatatt tgctgaacac 2940
acacatactt atgatatttt taaaattggc aacaatccaa tacccaataa tagaggaatt 3000
aaatattata gaactgttca ataagatgct tacgaatatc atgcagtaag atgggcaata 3060
tttatatcat aagcttaaat gaaacaaatg ggtattaaag gtatggtaag gttataaatt 3120
actttttaag agattaaagg gaaaagactg aaagatatat actgaaatgc tcacagtggt 3180
gacaaggttc cccagccttg gcactattga cattttgggc tatgtctttg ctgtgggagg 3240
ctggcctgtg ctctgcagga ggtttggcag cactcttggt ttctacccct agatagcagt 3300
agcaaccctc cctcaaccag cccaattttg acaaccaaaa atgtttccag gcatcaccag 3360
attctccctg ggtgagagtg atgaaacagt aggtgatttt ccccttcttt tctcattttc 3420
tgtaattttg tcacattacg ttaataataa ggaaaaaaca taaaaaatag atgaatttat 3480
tattctacct cagtttggat gtttggactc cctttggggg ttctttccat tatatcactt 3540
ggtctgctaa acagtctatg gtttggcaag gtgaaatgat tcatgaaatt ttgtttttat 3600
tttttacctg atactaaaag taaaacattc attcgcttga aaatttggac acagaacacc 3660
aaaaaaaatc cataatctca tctctctttt tctgtctttt ccttcctttt ttccctttaa 3720
aaacaagagt gaaaacctac cggttctccc tccaatttaa ttcctaaata taatcactgt 3780
taacatcttg gacatttcct gtgtctaaac acacatactc actttttctt ttttttagca 3840
aaaagtggat ttctgctaca tgtagtgttc cgcaacttcc tacatgttta caaaatcagt 3900
acatttacat acgctgaatt cagtccttaa tggtattata ttttgtgaat ataacaaaat 3960
ttatttaacc acttagacaa tctaagatat tctcagtttg ctgttatgag caatgctctt 4020
cctttacata tacatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtt 4080
ttagtaggat agatttctag gagagggcga aatgtcatat gacatccaca tttacaattg 4140
taataggaag tatcaaagtg ccccctaaaa aaaaaaattc ctcccattag tgtgtgagaa 4200
agcctatttg ttcatatctt cacaaacact aaatattaga aatatttaca attgtgttca 4260
agctaagaag tgaaaaatgg tatttcatat cttataattt ttgttgtgag attgaacata 4320
tttcctatgt ttacatgtca cctgtatttc ttattctctg aactatacgt tatgaccttt 4380
cacttatttt cctcatgggt tatgtgtagt ttgtgtagtt gtcttattga ttgttaggag 4440
ctatttatat attaggaaca ttaatctcct gtcttatatg tatgtggcat tgattagttg 4500
atcatttgtg agttcatgtc tgtatacaaa gattagagag gcagtaagag ggaaaactta 4560
cctctttctt atcaaagttt gtaaatatat gtataacaga agagagagaa aacattaata 4620
aatgctgaaa taaagtataa atttttactc cactacttca acataaacta caaaaggaga 4680
gtgacttttc tttcatgctg acttccatat tccccatgcc taaaatagtg cctagcacag 4740
aagaggtgct caatcagtgt ttgnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 4800
nnaaatgaag aatagaatct aggcagatac tctggaagag tgactgtgag tcattcatgg 4860
tcttggtatg aattagtcaa atccagcttt ctccccttcc cactccccac tgttagtaga 4920
ataatctgtt tattgagaga atagatttat aatttagaat aagaggggca gaagaggaga 4980
ttttgaagga tggcacctga aggaggacta gcatggctga gatagtgaag tggaagtctt 5040
ggaaagctaa agggtaagat gaaagtattt agccgtaggg ggaaaaagca ttgacaggtt 5100
ggaaatgtaa aagtcagatt ctccttgctt tgaaattttg tacagggcag gttctactag 5160
gtatgttaca atgcagaaaa aacatgaaat agttgagagg aatttggtga aatattatct 5220
tcctggcttc ttttgagtga gcagattttt ttcagggcct gtaactataa taaatttgaa 5280
acttctcatc ttttagtaac ttttttcact taagtttatg tggctgtggg caatggaatg 5340
aaggtattga acttcctatt ccctgctggg tttccacaat tacaagtcaa tcatgactgg 5400
ttattagaag actatttcaa ttagaaccac caagtcccat aatgtcatat tgtgtgttta 5460
attattaagt aaagcagtct tctttttgtg ttttccataa ttagcacatt ccagaaagat 5520
gaggatattt gctgtcttta tattcacgat ctactggcat ttgctgaatg gtaagacacc 5580
aaatccttcc attaggttct atattttaaa tattttaacc atgagtttaa aactaaaatg 5640
ataatttaaa atgcatgcaa ttttcttata gagagaatat tctcttcttt cttctacttt 5700
acacaatgac aaagtctttc ttctgcttta cacaatgata aagctccctg tgtcattgtg 5760
taaaaatata gagaatatag acaaattgaa agacacaaaa taatctatta cccatttccc 5820
agggttaact actgaaaata tctggggaaa tggcctgtat gtatatattt atttgtttgt 5880
tttcaacaag gccaggatct ttcaatcttt caatcttggt tgctctgtga catgcctttc 5940
ctgatgagaa tactctaagg aagaattgta ggatacatgg aaaatgtcag ggtaacacag 6000
tactggcacc accctgcggt cctttctgaa ctccatacca atgtacttct tgccagaaaa 6060
ctgatcaaaa gtttagggaa gtaaaaagag atgttagaat ctaccattcc ctctatgtag 6120
gaagcaaata ggtgtccagt caaaggacat tctggggatg tctacatgaa accaagtctc 6180
ctggttgtaa gtactccatc tccatataat atttctacag taatatatgt ttataaattg 6240
tgggggcaac ttgtttagct aattttatta ttctgttatt gggacactat gtctctgcat 6300
gagacatagt gtcccaaaac atatttcaag cccattggat aaaatatgta tttagcaagt 6360
tcttaaatat aatgataaca taactgacca gataaggtga tttttaaatg ctgtgccaac 6420
tttataaatg ttttgaggaa ttttcccttt tctgaaggtt attcttcttt ctttttagca 6480
tttactgtca cggttcccaa ggacctatat gtggtagagt atggcagcaa tatgacaatt 6540
gaatgcaaat tcccagtaga aaaacaatta gacctgactt cactaattgt ctattgggaa 6600
atggaggata agaacattat tcaatttgtg catggagagg aagacctgaa ggttcagcat 6660
agtaactaca gacagagggc ccagctgttg aaggaccagc tctccctggg aaatgctgca 6720
cttcggatca cagatgtgaa attgcaggat gcaggggttt accgctgcat gatcagctat 6780
ggtggtgccg actacaagcg gattaccgtg aaagtcaatg gtaagaatta ttatagatga 6840
gaggcctgat ctttatagaa aacatattct aagtgttgaa gacttttcat tcttataagt 6900
ccatacttat tttcaaacag aatagcatag tctcttcatt cattcattca gttcatgaat 6960
tcattcacgt aagtctccaa ttagcatttc ttgagcacct atatgatagt cattggaaat 7020
ccagagacaa cacagagcca tgttctacgg tatgtacagt tttccaaaaa taattcctag 7080
tctttacttt tttattataa atgtaataca tatacttgca aagaattcag atactatgga 7140
agagatcaaa tgaattgcaa aagtgtccct cctcccttca ccactatctc ccatgagata 7200
accaagagac actccaagag agtaaccatt atttgtgtgt ccctccagaa ccttttttat 7260
tcaactacca tttttttatt ttattaggtc tgtcagtttt ccttttttga gcctctctat 7320
atcaaatgct aataaatata ttcagatcaa accccactgt aaggttcata ttaaaaaaga 7380
cttgaagtct ccctatgaag acaaaaaata atcatattaa gtgtaaaaga acttattctt 7440
ccagtacagt ataaactata ctcactgggc atatattcat cttgaaaatc tatactgatg 7500
ttgtcttggg gaattgaaga ggaactagga gtgtcaattc ctgggaactg acccacagtt 7560
atgtcatcag gtcacttgag ttcgaagttt tgtgttagca ctagctaagt aaaggaaaac 7620
acctctgctt tcattgttga gtttcataga attgagagct gaaaggatcc caggcaggag 7680
cgactaatcc aaactcccac aaagaacaaa aatcccccag aggatcttct gttcatatat 7740
ttcctgcagt ggcatccctg tcatatccca caatggcatc cctgccattt ggactcccct 7800
tccatatcct gttgaaatta ctccctaata gtaagctgaa atctgcccct ctagttgtag 7860
ttttggaatt atttcatttc catgatgacc ttttaatatt tgactagaat taaatcatct 7920
ccccttggta tttccattcc tggactaact accatcaatc tgagggctaa caatacaagt 7980
agaaaaagtc tacacttgtc attgatcact gatcaatgat taatcaatga tcactgataa 8040
ttataaactc aaaaacaaaa tcatgtaggg attaagagaa atgtatcagt tttatgttgt 8100
atttctggtc cctgattctg gctcaaataa tgctactatt gtcaagaaga tatcacttgt 8160
aaagtagatt taattttcat tatattttac catgtgcttc tccattcacg gcatttcttg 8220
agatgttgtg gtttatactt tcagtttttc tccagtccat cagcaaatat caggcatcta 8280
ctgtgttcca agatattaaa gaaatcatca tgacttagcc tcatcaacag cattgctaga 8340
tctgggatgg aaaagaagag tataatcctg gcagtcagga agaaggctgc ataaagtata 8400
agtttctgct tccaaagaag atctctcatc agcctgtagg gagtgtatag ggaggggaca 8460
gctgtccttg tagtagcaaa gggttttatt caggtcatct gggctccata atatcccttg 8520
tgtatctgca gtctcctttg ccatggatca acacaatagg aaatcttccg gcactgatgg 8580
tttttccaag ggggagttct tcagggagca aagcaaatga ccaaccaggt ttgaggacct 8640
gatttgacaa ttccattttg tattttaaat tagttaattt gcattctagt cccaatccat 8700
cttgtcattt gcagacagtg gttttggggt tgagttgagc tataccaaaa gtctgaacct 8760
tctgcactta gaacnnnnaa ggcaaccacc aagcttcact tgcactgagg cagtgtctcc 8820
aatggaaacg aggtagctgg cttgcagaag ctttccaact cagggaagta gaactcctga 8880
gtcacctcca tatgcaaata atttcacagt actgctgttg aacttcactt cccatcacag 8940
caaatgtgtg gtaacatagc tttgccacag gagtttactn nnnnccatgg gattttaaag 9000
gtgaaacatt tcaaaactga aatttgaaag aatttagttt tggattcact caattatcat 9060
gatcactttg ggtgttattg cacctttcat gtttgtgagt ttaaatacca gactctcagg 9120
cctctaactt tcaattaaaa gtgtttttct ttaatcactg nnnnnnnnnn nnnnnnnnnn 9180
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 9240
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnctgtcat 9300
tccccaccaa ccccatcttt ccccaaaatt cccagccctg tttnnnnngt ttttctttaa 9360
tcactgaacc taatagtagg gaaaacgaaa tgttcattca gactttcagg accttcaatg 9420
agatgaggca gctgaaagat caaagtagtt gcatagttat cccagtaaag ctatttggat 9480
cgtatggacc agatcaactg ctgtcattcc ccaccaaccc catctttccc caaaattccc 9540
agccctgttt aagtgttctc tgtagcattt atctctatct agtatattgt gtagcatatc 9600
atatcatact tttctatttt gtttattgtc tctctcctcc tagaatataa actccacaag 9660
cacagagatt tgggtctgtt ttttaatatt gttgtatccc cagggcttga cgtaaagcag 9720
agtggtagta tgacaaaagc acacaaaaaa tatttgttga gtcaatgaat gaatgatttc 9780
ctcaaatagg attagcctaa aatttggaaa catgaacaga cttggatata tgaaaatttn 9840
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnttctaa agggtacact gaagcagcag 9900
tagtaaaagg aggaggagga gaagcagctc tgctactact attatcgagt actactannn 9960
nnnnnnnnnn nnnctacaac tatcgagtac tactacaatt agcacttgct tattctgtgt 10020
gttaggtcct gtactgaaca ttctgcctaa attagttcat ttcctcctgg aaatgactct 10080
gtggggtagg tgcttcatca tgtaagatga gtatttttca cacttcactg tctctgaaat 10140
ctgagtgtgt ctttcaatga tggaatcttt gatttcatga taagtggtat tattcccatt 10200
ttaaggatga ggaaactggg gtccaaagaa attaagtaat ttgcccaaat tcacctagcc 10260
tcgtaaatga taaagctagt tctaaatcca agcagattgg ctctgaagtc tgggccctta 10320
ataaccactt attgcctgta tttgcacctc tggtgtatgt atcaagttat atattggctt 10380
caaaactatc atgacctttt cttgattttg attgttcaat attagtatag tgttctnnnn 10440
nnnnnnnnnn nngggtctgc aaaaaatttt gattactcac ctcatctgta aaacatttta 10500
aacttgtgtg tctgtgcagg cacatttgtg tgtaattata aaaaatttac tatctattaa 10560
tatataggtt gtaccgtaag aaaaattgcc atttttgaag agncaaaaaa ggttgaatat 10620
taccagtttc atctggttca acctaataga catttgtaca aaaacagaca ttttaagagg 10680
atgaaataaa aatttaataa acaatatttt caatttttac taattgtgac gcttcactat 10740
tgttagctaa tatgtcaagg catgatatac cttagggtgg aatttatcat taacaaaggt 10800
ggatagtgtc aataatcttg aggtttgtgn ttttttatat aacactgtga ggtctaatta 10860
agtacttaat tgtttacaac ctcatacagt cgccaataat aagtgtcact tctgctgttt 10920
cctctgggtt gtgcttgaat tattagtatt atcttcaatc ctcagtttct ttgtggaaaa 10980
ctttttaann nnnnnnnnnn nnnnnnaaga tggttagttt agtcaaaatt agataagaga 11040
atctgaaaat ccataattac cccaaagcaa cccactcata agaactatta tttttgtgtt 11100
ttggnnnnnn nnnnnnnnnn nntttccagt gtttccactg gtagtggttt cattgatgta 11160
ggagtatcaa aacatcacta attatttatt tcagttttgt ttgatcctag ctgttttgtg 11220
ttaactttga agaaattaca tcacagatct attgttgtcc ttggtaaagg aatggagagt 11280
taaggctcta gatcattagt ggttatgctg tagtattagg agtaaaaaaa agattatatc 11340
aacaaaataa gaacatgtta atgtacttgt aatagataaa catgaataaa gctcttatgc 11400
tatatagatg cactgaacaa tctactagaa ttgtcagcaa acggtatctt aatcctnaaa 11460
agggtcccaa accaatgatc taaaattgaa tcaaactttc ttccttgagc ataattattt 11520
aagtgattta ttaaaatagc cagcatttaa aagcttaaaa tataagtatc ataatgtggt 11580
atcctagata gatcccagaa cagagaaagg atattaggga aaaactggag gaatggaata 11640
aattatgcag tttagttatt aataatgtac taatgtcctt agttatgacc attgtaccat 11700
ggtaaagtaa gatactaaca atagaggaaa ttgggtaagg ggtatatgta actctatact 11760
atctttgcaa tttttttgta aatttaaaac ttctaaaata aagaacaaat ttgaacatta 11820
aaaagtgtcg ccaggaacat gtatcactgt ttacagatga aacagtatgt atttttatat 11880
ctaatttctg atcattggct tcaaatcaga aaagtgaatg acacatcaag atcaggtttt 11940
ctgtttacta aataaagtct aagaaaacaa agcataccag ctggagagat tcatgtttat 12000
aaagacagat ttataacaac aaaataaaat atccaagaat aaatttaaga agaaataggg 12060
cactatgtaa aaagtatagc actttactga gaaacatatg aaaacctgaa tacatggaga 12120
gaggtatttt atatttgaat agaaagattg ctggtttaaa gataattctc tttaaatttt 12180
ttttgttaga aatttaagag gtacaagagc agttttgtca cacggatata ttacatagtg 12240
gtgaagtctg gggttttagt gtaaattaat ctttacattt tgtttgagcc caataaatgt 12300
accaacatga tttttatagc aagatagtca ttcctattaa cccaaacttg tcccaacttt 12360
gaactgaact gaggcagagc tagcaggtgt tccccactgc tgaggcatct gaacattaag 12420
cgtatccctc tgagaaccag cctgcattga tcctctttct aatgtagaca gcatcaagct 12480
atatatctag ttctgtgctc agcaaaagcc ctgacttctt tttgcttatg tcctagctcc 12540
atacaacaaa atcaaccaaa gaattttggt tgtcgatcca gtcacctctg aacatgaact 12600
aacatgtcag gctgagggct accccaaggc cgaagtcatt tggacaagca gtgaccatca 12660
agtcctgagt ggtaagacca ccaccaccaa ttccaagaga gaggagaagc ttttaaatgt 12720
gaccagcaca ctgagaatca acacaacagc taatgagatt ttctactgca tttttaggag 12780
attagatcct gaggaaaacc atacagctga attggtcatc ccaggtaata ttctgaatgt 12840
gtccattaaa atatgtctaa cactgtcccc tagcacctag tatgatgtct gcctatcata 12900
gtcattcagt gtttgttgaa taaatgaatt aatgaataac attatattta caaaatgtat 12960
cctaattcct cacttccatt catccaaatc atattgttac ttaataaaca ttcaccaaat 13020
atttattgaa tatgcctttt gttccatgca ttgtagtact catttgacac acatagaata 13080
ataagactca cgttcacact cttcaggaaa cagataaaaa acaaatgaac aaacaaaaaa 13140
caggcaatcc aataccatgt gggaaatgct ttcataccat gtgggaaacc tgggggaata 13200
cctgagagga atattcaatt caggccatgt ttcaggaatc caaatcctgg cacatcagag 13260
ccgcctcctt cttactaggg tttctgtggc aggaaataaa tggaacgtat ttttctatct 13320
tatgccaaac aggagggacc ctttctcccc tgtgcctctc ccaaggtagt ctacaatatt 13380
tcaacgctag cagtctgttt agtgcacagg acatgaggct gtgtatccct gggcaaattg 13440
ctacacttct gtgtgcttca ctttctctgt aggattataa cctactgagc aaggttattg 13500
tgggggtcaa attagcaaca gtgtatgaaa atgatttgag accagtgcct gcacaaattc 13560
aactattttt ttttatctca ctactctata gaagtaggta ggatgggaga cagagtctga 13620
tgggaggctc agaatgtgaa agtaagtgag gtgagtgagc atgatatttc atataaacac 13680
aaagatactc tgagaagagc ttctcacttn ccccgccccc aatagatgtt gacaggaaaa 13740
tgccacgtac ttcagcaaaa acagctgaaa aattagacat aaaagtcaat caataggaaa 13800
agataatcca ggatggtctt gtgaacagaa agaggaaaaa aaaagtttag aaaatgatgg 13860
ggatgctctt actggggtat gagtcctcag gtattcaact ggctttcaga aaaagctaga 13920
ctagtgggtt cctgccattt aaaagctgtt ttatgacaac ttacttgttg ggtggcctac 13980
agtaactcac ttaactgtgc tgagtctgtt tcctnnnnnn nnnnnnnnnn nnnnnnnnnn 14040
nnnnnnnnna tgcctaactc ataaagttgt tctgaaactg aaataaaaca tatatgaaca 14100
ggcattgtaa actgtaagtt acggaaaaag ctggctgttg ttgtgtcttt aaagcttcac 14160
ctgggtagtt agagatggat catgggtctc agtggagagc tgagccaggc aggagctgac 14220
taagggtaag aggtgggaat tagcaatctc tgaacatctg tgtgccatgg gacccctttt 14280
cctcctgcat ggtaccccag acaaggagcc tagtaagaga tactaatgac ttgttgtcca 14340
gagatgttca aactgcagag aaagataaga caacaagcat tggcctccaa tcatgatgac 14400
agatagagga ggtgggagct ccttagcagt gctggttggt tttccatgtt ctactgtggg 14460
ccatctctgc catgtactgt aggctactaa cttctatatt aaaaaatgca agaggggnnn 14520
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 14580
nnnnntcagg agtttgtaac cagcctggcc aacatggtga atctctgcct ctactaaaaa 14640
tacaaaaatt agccagatgt ggtggcgtgc acccgtaatc ccagctacnn nnnnnnnnna 14700
ggcacgnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 14760
nnnnnnnagc atgggcaaca gagaaagact ttgcctcaaa aataaataaa aaataaataa 14820
ataaataaat aaataannnn nnnnnnnnnn nnnnnnnnnn nnnnncattt aatactgagt 14880
agccactcca tgtggtactg tgctaagcac attataaaat attagcctca caagaaatgt 14940
attagcattt gtattttgta cactggttaa gtatcttgcc caagacctca aaactggtta 15000
aggggcagca gaatttaacc ccagcgccac cttttcaaag tctgggcttc ttacacttct 15060
ccatgctgtt cccattttaa cagatgtatc tcgccattcc agccactcaa actttggcat 15120
ttaagaaaat tatcctaaag ctaaactaaa cttcaaggat gactattctc ctcatgaccc 15180
cttcccatca aaattttatc tttagtcagt ttgtttttgt tttgttttgt ttttcagaac 15240
tacctctggc gcttcctcca aatgaaagga ctcacttggt aattctggga gccatctttt 15300
tactccttgg tgtagcactg acattcatct tctatttaag aaaaggtagt atttccttaa 15360
ttgcagtggt ctccactggg gatgaggagg gggtgagaat tggatcgatc atggctgcaa 15420
ggaaacctga cttaacctct gcagggtggt gcaaaggcat tccactattc aacagtaatt 15480
atattgaagc tgcatgggat cactgggtga agatggggtg taaggggtga gggacaggag 15540
aatgggtatg gatggaggta gaagatgcag tgtcatacaa tttttttcta tcatgaaaat 15600
aaccacagac ttactgtaaa gaaggagcta aaatgcctgt cattttcagt tgcattttag 15660
ttttgcatta gttgcaccca gctggtttct gggtactcta acnnnnnnnn nnnnnnnnnn 15720
nnnnnnnnnn nnnnnnnnnn nccatagagt attttgtaaa attattggca gaggatgtac 15780
ataatttgca tgtgttcctt tctccattta cctgtgggaa caattaaaat ccaggaaaat 15840
gagtatattc aaataatttc ctcccattta tgatgattca gagtaaataa ttcctctgat 15900
acttagagaa gtataccaag agatccagtg attgtataga gttgtctgat gttaaatagg 15960
gaagtagaat atggaaggga ttccaatagt cgttgaaaaa ttccccacaa ccccttacat 16020
gggggaaagt ggtgttaact gagatagtag agataagctg ttaccaaaaa ttatgttctt 16080
aacaggattg agatagccag aatataagga tcaagtttca atgacagtaa gatcctgaga 16140
tgcagttgat ttgcacaaag aaataattgt tgccagcttg cattttgaat atttctctgg 16200
aaaaagagat tagttggcag tagaaatgaa tagaaatcaa tagatattaa aatacctcag 16260
aatttgattc atctctggga aaagatgaaa aataaaagtg tatagtcctc aagaaaatct 16320
gggatcaaaa gcatgggcct taccctattg aattaattaa cctcagaagt tgggaactgt 16380
ggaataagga tgtccaccag acttcctagg gattacaaat gtttcataga acttgaaatt 16440
taaacttggg tcactgtatg ggatgtagag ctgtgctata tggaaataaa aatgatttct 16500
ttttctcaag ggagaatgat ggatatgaaa aaatgtggca ttcgagttac aaactcaaag 16560
aagcaacgtg gtaagaatat cagaaggaat tgggaagtag aaggcaaagg aaacaaaaag 16620
ctaaagcaat aacaaagaga aatccattag tcataatctc ctctcctttt aaagaatgct 16680
ggttcccctt tgcctcacaa ctaatacaag aacttctcca ccatctcagg aagtttaggg 16740
atggccttca agagtagaga gtagggagca gctctgtgga gagaggagag gagcagggaa 16800
ggggaaggag tcagcttctc tttgctaatc tgttgccctg caccctagca gctccctgca 16860
gcaggggaca aggttgactt aggtggatgg ataattaatt gattctaaaa tattgtgtgt 16920
cagtattgta tattgtaata ctatgttaac tgcgccatgc acggtatctc atttaatccc 16980
ccaccccttg ccattaccaa aaagagagag agaaaaatac tagaattatc ctcattttac 17040
agtagagaaa acagagggtc aagaagataa agtaaagtgc ccaagaacac acaactgatc 17100
acaaatatca agcttggggt ccattagcct aaccacagac ccttactctt aacccatctg 17160
cttcaatcca ttttgctaca aatgtttaca ttatatgcag ggcagaaaag tctcatccag 17220
tttattgaac taagaagaaa gttatattaa ggtgtctaat tttttttaat gtagttagaa 17280
accaaactta acaatgagcc caagtttaaa gcagtctaat taacttgaca agctcaggca 17340
agtttcattc tgtggcctgt agcatcatct gtgttgtaaa gctaagtagc aaatgttatt 17400
tgnnnnnnnn nnnnnggtca tcctgggggg aaagtcatcc caatttgctc aagactgagg 17460
ggtttttcag gatatcatgt aaggataatt gggtacaaat ataacctgct tctttctctc 17520
atttcaaatt tatcatttat catctcagca actatgagtt atgtttttta ttagatttct 17580
tgttactttt tccccagacc gctccccatg aaattaatat actattatca ctctccagat 17640
acacaattgg aggagacgta atccagcatt ggaacttctg atcttcaagc agggattctc 17700
agcctgtggt ttgggggttc gtcagggctg agcatgacca gaggaatgaa tgggcccgtg 17760
ggatgcatgc agtatgggac ttaaaaggcc caagcactga aaatggaacc tggcgaaagc 17820
agaggaggag aatgaagaaa aatggagttg aacagggagc gtggagggag accttgatac 17880
tttcaaatgc ctgaggggct catcggtgca tgtgacaggg agaaaggata cttctgaaca 17940
aggagcctcc aagcaaatca tccactgctc atcttaggaa aacgggttga gaatccctaa 18000
tttgagggtc agttcctgca gaagtgccct ttgcctccac tcaatgcctc aatttgtttt 18060
ctgcgtgact gagggtccca gtgttggaac agtatttatg tatgagattt tcctatttat 18120
tttgagtctg tgaggtcttc ttgtcatggg agtgtggttg tgaatgattt cttttgaaga 18180
tatattgtag tagatgttac aattttgtcg ccaaactaaa cttgatgctt aatgacttgc 18240
tcacatctag taaaacatgg agtatttgta aggtgcttgg tctcctctat aactacaagt 18300
acacattgga agcataaaga tcaaaccgtt gatttgtata ggatgtcacc tttatttaac 18360
ccattaatac tctgattgac ttaatcttat tctcagacct caagtgtctg tgcagtatct 18420
gttccattta aatatcagct ttataattat gtggtaccat acacacataa tctcctttca 18480
tcgctgtaac caccctgttg tgatgaccac tattatttta cccattgtac agctgaggaa 18540
gcaaacagat taagtaactt gccaaaacca gtaaatagca gagctcagac tgccacccac 18600
tgtcctttta taatacaatt tacagctata ttttacttta agcaattcat ttattcaaaa 18660
cccatttatt aagtgccctt gcaatatcaa tcactgtacc aggcattgaa tctacagatg 18720
tgagcaagag aaagtacctg tcctcaagga gcttggagta taataaggag attaataaga 18780
aaatatatta ttacaatcta gtccagtgtc atagcataag gatgatgtga ggagaaaagc 18840
tgagcagtgt tgccaagagg aggaaatagg ccaatgtggt ctgggacagt tgaatgtatt 18900
taaacatctt aataatcaaa gtaattttca tttacaaaga gaagtcagta cttaaaataa 18960
ccctgaaaaa taacactgga attccttttc tagcattata tttatccctg atttgccttt 19020
gccatacaat ctaatgcttg tttatatagt gtctgatatt gtttaacagt tctgtctttt 19080
ctattcaatg ctattaattt taaattcata cctttccatg annnnnnnnn nnnnngatcc 19140
catgggagat ggtttgaaaa tctccacttc atcctccaag ccattcaagt ttcctttcca 19200
gaagcaactg ctactgcctt ttattcatat gttcttctaa agatagtcta catttggaaa 19260
tgtatgttaa aagcatatat ttttaaattt ttttccctaa atagtaacac attatatgtc 19320
tgctgtgcac tttgctattt ttatttattg tagtgtttct tatgtagcag atggaatgaa 19380
tttgaagctc ccaaaggtca ggacacatgc cttctttgtt tctaagttat ctttcccata 19440
gcttttcata atctttcata tgatttagta catgttaaat atgtgctaca tatacattta 19500
gacaaccagc atttgttaag tatttgctct aggactgagt ttggatttat gtttgctcaa 19560
aaggagaccc atgggctctc cagggtgcac tgagtcaatc tagtcctaaa aagcaatctt 19620
attattaact ctgtatgaca gaatcatatc tggaactttt gttttctgct ttctgtcaag 19680
tataaacttc actttgatgc tgtacttgca aaatcacatt ttctttctgg aaattccagt 19740
agtgtacctt gactgctagt taccctgtgc cagaaaagcc tcattcgttg tgcttgaacc 19800
ctttaatgcc accagctgtc atcactacac aggcctccta agaggcttcc tggaggtttt 19860
gagattcaga tgccctgaga gatcccagag tttcctttcc ctcttggcca cattctggtg 19920
tcagtgacaa ggaatacctt cgctttgcca cccgtcaagg ttgaagaaac agcgtctcca 19980
acagagctcc ttgtgttatc tgtttgtaca tgtgcatttg tacagtaatt tgtgtgacag 20040
tgttctttgt gtgaattaca ggcaagaact gtggctgagc aaggcacata gtctactcag 20100
tctattccta actcctcctt ttggtgttgg atttgtaagg cactttatcc cttttgtctc 20160
atgtttcatc gtaaatggca taggcagaga tgatatctaa ttctgcattt gattgtcact 20220
ttttgtacct gcattaattt aataaaatat ccttatttat tttgttactt ggtacaccag 20280
catgtccatt ttcttgttta ttttgtgttt cataaaatgc ttagtttaac atcccggtgg 20340
<210> 4
<211> 20641
<212> ДНК
<213> mus musculus
<400> 4
gcctcatgcc aggctgcact tgcacgtcgc gggccagtct cctcgcctgc aggtaaggga 60
gcatcttctc gcggaatccg cttgcagggc actttaaaga gccagaatcc ctagaccttt 120
ttaggacgga gaagggaacc ggtttcctgg gaaagttaag aactcagaat ccgcagtttt 180
gtgtgtttat ggatcttgtg ggtaggtagc tgggtcagaa gagatgaatt aattggtcct 240
agcgcgactt gactgtttgc tagcaatgac tgggtctttc cacttgaagc atctccggag 300
gtcccttcct ctgtcgaggt ctaggatgct ggagcttaag attttcattc tatctgccca 360
gagaacctaa aggatttttg gaagaaaatg tcccaaacag ttcttagata cagtgaccta 420
ggctatgtta aagtttaggt tggggttgtc ataagaggtg aaagtacaat attctatctt 480
tttttaaaaa taaattatta caggtgcttt cctatgaatt acttagccag ttttttttta 540
attgctatat tccattgtgc tgggttatta aagggcatca ctctcccact gaggaatgca 600
caggttagta atagtgcctt actagctcag tcatccactg accatctgtg tatgtgactg 660
tctggaatat ggctaggtat ttcatttaga taaactctta tatgtataag tacttcatta 720
tgtttgagtg tctgaaaggc ctttgtgtag aaaaaaaaaa aacaacaacc tgagaataca 780
ttccagaaag tatccgcagc tgatgatgat agtgaccaag cggtgtttaa ggagaacttt 840
atacagggaa aatattcacc tttcccagct gagcatcata gataggaaga catgatagtg 900
gttatatgct gctctaagga gggatggaga agggatggat ttgagcacag ggttcaggat 960
catttgatag aatttgaaaa ctgggtaaat tttcactctt ttgagaaagg aagtttgcat 1020
ctgcttcccc tccccgcccc aggcactctt ccccccccca tccccccccg tgcccccccc 1080
ccccatggga agataaagaa gtggcgttcc aagatcttca tctggacttc ccctgggaca 1140
aacctatttc cccatcttga atgagcgtca cttctttact ttaaactctg ttctgtgaaa 1200
ggctctcagc tcttgtctgg gaatttgtga aaggttacag tttcttgccg gagggaggga 1260
gaaggtgacc ctggactctt agcccatgta agaggtgcag gtttagaggt caaaggccat 1320
gctgatttac atggtccctg aatttcccct tttgatctac tgctcacacc tgcaccctga 1380
gagccaggga ggggaggata tgatactgaa gaaggtgact atcagaccca acccatcacc 1440
tttcagactt tatactaagc tttaaagcag actcataagc aatagataac tgttccaact 1500
gagtctgcca tcaacgtagg aaaaattagg tacagtcttc agtggatatc atgtacctca 1560
ttgtgactgt ctagtactcg gcttggccat ttctcccagg gcctcattgc aacagtcact 1620
gtgtcctcac aggcctggat gtcatcctgc ctcatctgct tgggcagatg aactggagag 1680
atatgattaa taatgagtca tttgattttg ctctttttgt aaatgtcctt ggtttggttt 1740
ggtttgttta ttttttgttt tgttgttgtt gtttgttttt attttcatgc tcttgttgtt 1800
gtttgagaca gagtctcact atgtagccca ggctagcctc ttcccccagt ctctcagatg 1860
ctgttgcatc tgcccttttc taacatcaat gattcataat aatcagcaag aaggctatga 1920
ccttaacagt caaaataagt aatcaggtaa ttattgagag ctaaatattt atttaaacaa 1980
atcaagttgc atttttcaag aattgaatgg ggggatctaa aaagtaatgt ttgtgctgca 2040
caaagatgta gtggtatcag ttgccatagt ctttctccct agatccgcag atctccttag 2100
ggaaatctat aattcaagcc taaacttgga cctggtacac aactggtact cgcccaaaca 2160
ttggccaagc acacacatat ttgtgactct gaaaaaaggg acaacacata acagcaaagg 2220
cgctccgtgt taggggcctg ttaaatacaa gacttgggga aattatgcca taatcttaca 2280
ttttaaaaat ggctattaca tgtgtgagac aacgggggct ggagagatac ctcagaggta 2340
ctaggcacac atatggcaag cagatgtaca agcaggcaaa acattcttaa aaaacaaaaa 2400
caaaacccca aaacagattt ctttctatat tatttatatg gggttctgcc tgcgtgtaca 2460
cctgaagggc agaggagagc accaggtctc ataggtggtt ggttgtaagc taccatatgg 2520
ttgctaggaa ttaaactcag gactttggta agagcagcca gtgctcttaa cctctgagcc 2580
atccctccag tcccagacaa aacactctta cacataaaat aaatcaatct ttaaaagtgt 2640
agaggaaaat ggctgaacag aaactattga cagacatgct gagagtggtc accgtttctc 2700
cagcttggca ctggtggtgc tgtaggccat ctcagtgtct gtcaccttgg ggcggggcgg 2760
ggggcgggag ggggggaagg cggcagggca gggtcttggc agcatctcgg gcctctattt 2820
catcacatca gtaacagctc tccttcgtcc caagcaagct ttggcaacca aataaatatg 2880
ttagccaata ttaccaaacg gtccctagag gagaatgtgt gtaatgtgcc tacattcgtt 2940
tttataacaa caccaacaat atccaaccat aaaaataaac tgaaatggtc actctacctc 3000
agtgacaacg tctgagctcc gtttcagggt ccctcacaag aaattcgata ggtgtaatga 3060
tttatttttc ctaataagta aaatgattat tcactcattt gaaaattgga cacacacaca 3120
aaaaaatgaa gaaaaaattt gctatttcat ctctcctttt ctatctcttt ccctttaaaa 3180
aaagtataaa tgtgggctaa agagatggct cagcagttaa gagcactggc tgctcttccg 3240
gaggtactga gttcaaactg caccaaccac atggtggctc acaaccatct gtaatgggat 3300
ctgatgccct cttctggtgt gtctagagac agtgacagtg tactcacata cataaaataa 3360
gtaaaaaaaa aaaaaacaaa actttttttt aagtataaat gtgtaagaac ttacctgtct 3420
tccctataaa agccacaatt acctttgcag ataatcactg tcaggcagtg tcttcaagag 3480
aagtggggtt gttctacgac acaagaatat ttttgtttat atactaccct ctaagctcct 3540
tcctacttct ctgcaaactc atcccctgtg catatactga aatctatccg tcacggtatt 3600
ttgagtatac acagttatcc gtcacggtat tttgagtata cacagtttta ttagctaata 3660
catggggtat ttcaagtctg ggactgttgt tggctgtgtc atctctgtta catatgactt 3720
atattttcta agatgaattt ccaagacagg aatagagaga tcatacgaca tgcacagtta 3780
aagttttaag agattacaaa attgaccttt aagaagctat acaatttcac atcctttcca 3840
ggaaaggtca agaaatccct gtatcttcac taatgctacc tatgaggagt ctgtaatttt 3900
ggtagagcag aaggtccaaa tggtatcaca gatcctccat ggactttctt atcgagagcg 3960
ttctttccgc ttttattgat tatccttcga gctatgtgag actctcatcc gtcttcctct 4020
tgaccgatgt atagttttct cactgagttc agaggctagg atgcagaagc cgtcctcgtc 4080
agctttatac acctttcaga ttgactgtga atgagcacac aggtcagaga gacggtaagg 4140
gaagacctac tctcccatga acgttgtaaa catgcgaata acacagaagc agaaaatgta 4200
aactattaga ctggctggaa tgtagcgctc acctttgagc ccactccttt agacaccggt 4260
ttcatggaag aatggctcct gtttcccacc tgctgcttca gcagtgcccc aaatagtgcc 4320
agattcagag gagatactca gtgtttgccg agtgaattag gaaagtgttc agcggagggg 4380
acactagcat tcagacatcc agaccttaga gtaaccgggc cagatggaag tctccttcct 4440
tcccatagct gttaccactg tactgtcttt actggggaaa agatttatca tgtggaatag 4500
gtgcgaggca gaggtgagat tttaatgggg aggaagcatt gaaaagtgaa agtgaaaatt 4560
ggatgctctt tgctttgaag ctttgcctaa agcaggtttt agctttcaaa tacgtttcaa 4620
tgttgaaaga acgcatgata catatggagg ggggcctggg gggggtcctt ggctgagttt 4680
gaatgtacat taacaatctg gggggctaat aactcaatgt aaagctgctg atcccatcat 4740
actgacttct ttccacttgg ttctacatgg ctttgagtta caaaatgaaa gcattgaatt 4800
ttgaactgtt cagctgtgtt tccacacttg caaatcggtt gttggccagc cctcagaatt 4860
gcttcagtta cagctggctc gtctgctctt tccagactgg cttttagggc ttatgtatat 4920
atgagaagga cacatttact agtgtctcct tgctctgcta ttgaaattaa gcagacctct 4980
ctgtgtttcc cgttactaga tagttcccaa aacatgagga tatttgctgg cattatattc 5040
acagcctgct gtcacttgct acggggtaag tcaccaaatc ttttcagtgg gttctatatt 5100
ttcaatattt tagctatgaa ttaaaaatgg aagtaatttg tggggtgtgt atgtgtgtgt 5160
atatgtgtgt gtagaggggg gtctgtgtgt atgtgcagtt gctaggcaca cataaagcgt 5220
tcataggaca acctagagct tagtcctcac cttctacctt gtttgagaca aggtctctta 5280
tttgttgtac attgctgagt cctgtagttc ggctagctca gaacctcctg ggggctctcc 5340
tgtctccacc tcccagtcca ctgagattgt aggcacatgc tactgcacct ggcttctacc 5400
tggtctctgg ggatttgaac ttgggtccat gggctacaca gcaagtcgtt tacttactgg 5460
gcaatcactc catcccctaa gataattata aggaatatac cttgcttatc caaacacatt 5520
ctcattctcc tttgccataa ataagttact tggcaaatat attgtatgta tttttaataa 5580
ataaataaaa tcttaaaaat aaataaaatt atttgtgaag acaaaaaaaa taagttactt 5640
ggaaaggatg aaggaaaata ctggagcttt gggtgtggtt tagtagtaga acacttggct 5700
gatgtaaaaa aaaagcccta ggtgcaatcc caacaccaga aacaaatgaa ggaatgaaca 5760
acaaccgccc ccacccccca ggggatgaat ataaaaatat caggtaatac agaactaaca 5820
ggtgatccgt ttcctatgaa taactactga acattcccag ggaggtggcc cactgataat 5880
atatttttat ttattggttc cttttaaaca agactgggaa tatattatct agcttgcatc 5940
accaccacca ccccccaccc ccgccccatg aagttatttc aaagaagaat tttagtgttc 6000
atgtgattcc ctaaataaaa tgatagtaac cttttaccca ggttttcaga tgtgtttgga 6060
ggagttttct gtcttctgag ggctggtcct ctttcctttt cagcgtttac tatcacggct 6120
ccaaaggact tgtacgtggt ggagtatggc agcaacgtca cgatggagtg cagattccct 6180
gtagaacggg agctggacct gcttgcgtta gtggtgtact gggaaaagga agatgagcaa 6240
gtgattcagt ttgtggcagg agaggaggac cttaagcctc agcacagcaa cttcaggggg 6300
agagcctcgc tgccaaagga ccagcttttg aagggaaatg ctgcccttca gatcacagac 6360
gtcaagctgc aggacgcagg cgtttactgc tgcataatca gctacggtgg tgcggactac 6420
aagcgaatca cgctgaaagt caatggtaag aattaccctg gatggggaag gcttcatccg 6480
tatttaaaac agctccctaa tgttgagagc tcttcattct tgagagttcg cacgcacttc 6540
tcacagaaca acagcagcct gttcttctcg ctcgtttgtt cattcgttcg ttcacacact 6600
tcaccagtga aaaagcctag cactgtgtgt ttgatagtaa cttgagattc agtaccagat 6660
aatactcagc catgctttgc agtcagtacc atgatcttgc aaaggtgaaa tgccaggtgt 6720
ttgtttctta tcataaatgc aatatataat atattacata gatgtataga tataactgtg 6780
taacatgcaa taagatataa tatgcatata tttcatataa cataatgtat aatatataat 6840
gtataataat atatactaca atatatagtt atatgcatag ttatatattg catttatgat 6900
aaaaagcaaa cacctggcat ttcacttttg caagcttttt gaattacttg taaatatata 6960
tacatgcaaa catacataca cacacatgtt tttttacaag taatttgaat gtcatggaaa 7020
gaaatagaat cataaaaatg tccctcctcc ctaactacca tcttctaagc ataaatatac 7080
agtaactact atttgtacat ccctccatga ctttttgatt ggattactgt ttatatttaa 7140
tctatcaggc ttagcacatt ttctttcctt tgaatacctc catacaaaat tcaatgtgtg 7200
tttatatata tatgtatata tatagttata tcatatcata tatcatacaa agttttatat 7260
atgtatacat atataaacac acatatctac acatacatac acatttttta tatatataca 7320
caatatataa tgtatatgtg tgtgtgtgtg catatacctc tatatctatc tatctatcta 7380
tctatctatc tatctatcta tctatctatc tatagcttct actgtaaggg tcacttttta 7440
aaaaattaag gttaatctat gaaggatgag aagtgaagat cttaagtgta gaagaagccg 7500
ttcttccaca gagatggtac aggctacact cagcaggcat gcattcattt tcagggcctg 7560
catctctggg agtgctgagg aggaactatg agtgtaagtt cctgggtaac gagccacaga 7620
aatgtcatca ggtcccttga gttcacggtt ctgggttgga actaactaag ggaaggaaaa 7680
caccttgcta accacccctg ctttcactgt tgggttgcat agacctgaga gcttcactca 7740
tcgcaggcgt gcacacagcc agcccagttg cccaccagga ctggaaatcc ccaaggctct 7800
cctcttcaca tacctcccac agtggtgtcc ctcgaatcag gtccctttcg tggtctgcgg 7860
aaacttcacc ttgatggaca ctgagccgaa ccccgtcttc tccaggcaga cgttgcagat 7920
ggtctgcctg ctccttttcg gttttcataa taactttaaa actatttggc tagaatcaga 7980
tagtctcctt cgatcttttt actcctaaac tagtgtgaag ctcaatatta tacataggaa 8040
aaattcacat tcctcataag ggatcacggc agttattaac gcaatcttat gtagagatta 8100
ggaaaaatgt gtccattcga cagtttatta atgggtctgt gatggtgggt cgcttagagc 8160
ttatgaagat gccacttgcc ttatttcggt gcttggctct aacttccatt accttttact 8220
tacaattctt catttgtgac actcatgaaa ggagactcgg gatatggttg ttaatatctt 8280
gagcagctgt tagtacatca gcaaatatga ggctcctgtt tggttccaag atttttaaag 8340
acattgtgaa gctggaaggc catgagcagt caaagttgct ggacctgggg cactggaagg 8400
agctcaaggc gggcagtcag gaagaaggga gcagggtgta agcctctgtg acaagaggtc 8460
tctcatggct tgttaggagt aggtgactct ggtaaggagg aatttactct agattatgtg 8520
ctcagtgata tctctgatac caaagacaat cttttggagc gcacggcttc tccacggagg 8580
gacctgtgat gcggcaaagc tggtatccaa gcagtcttga aggcctgctt gactattttg 8640
ttttcttaca gaaatcacta cgtcgatatt ttactcccag ttcaccttgc catttgcatg 8700
cacgttagct ggtctaggcc aagagtctga accttggttg tggctcaaat ctgtaatctc 8760
agctatgtgg gaagctgagg caggagaatc gcgcaattag gacttgtctg agctacgaac 8820
atatagcagg ttcaagagca gcctgggcaa tttggcaaga tccgttctca aagacataca 8880
tacatacata tatttggcaa gatccgttct caaagacata catacataca tacatttggc 8940
aagatccgtt ctcaaagaca tacatacatg catgcatgca taaatacatg tataaatgaa 9000
tataaaatta aaataggaac aggactgagg acatgcccag tgatgtacag gaccctggtt 9060
tcagtttcta gcaccatggt cgctggcatg caaaagcttt ccatgtcaag gaagtagaac 9120
tcctgaattc tgtgctatgc aaatgacttc acagcaaggt tccttcccat cactgcaagt 9180
gtgtggtggt gaagcagccc tacagggagt tcactcgcca tggggtttta aaggtgaatt 9240
atttcaaaac tgaaatctga aagagtgtag ttttggatta actcagtttt tattatctct 9300
ttggataatg ttctaccttc aaaggtttac agaagaccct caggcatcag gcttttaatt 9360
gaaaaatatt cagctttaat cactgacctc aatatcagaa aagaatgaag tgtccattta 9420
gtagtcctcg gaccgtagat ggcattaagc agctgtaaga aacacgctca tcacatggtt 9480
caacaattaa ggattgccat ggtgcacccg atcccccagc tctgtttaaa ccttttctct 9540
gtcatctatg cctatctaag atgtctcagc atatcatgct tattgtctct ttccttctct 9600
tgaacacaag ctctaggggt aggggctttg gtcagtttta ttgttggacc tgttggcctt 9660
gacgtattat gaaggggcag tgtaataaga aaagaaaacc gtgtttgttg atggaatggt 9720
ttaaaaaaaa attagcttaa aatgtatgct atacgaacgg gcatgaatct gggagatttt 9780
agactgttcc ttgtgagccg taggaacttt cttgtttctg tgttcacagt cctccgcagc 9840
tgtgctcatg gtcctccaca tctgcctaac aacagcaggg gtagatgtta taattaacac 9900
tcacagaaca cctgatatgt gagaggttgt gccctgggct tcttgctcat atgaattcat 9960
ttcatcttta acataactct tcgggggagg cacttgccca tgaccatggc ttgttttctt 10020
tcctatcgac tatctttcaa atcggaatgt gtctttccct aggtgacatc tcagattcaa 10080
tgttacacag cactgctgtg cccattttca ggcattaaaa atgcgatccg cctaaattcc 10140
accccgataa aattatgcag ccaggttcgg aacccaacta gcccatctct aaaaatctat 10200
gcccttccta gcctcgcttt gttgcctgtc tttgtacctc tggagtgggt atcaaggaac 10260
agaatggctt taaacatgat ttgaactttt cctggttttt gatcacagca gtaacagagt 10320
gttctagaga taagccagag gtttgaaaac aaacagaact tgtctttgag gtgttataaa 10380
catgtgtgtg attataaagg atatatctgg tgccaatagg tatagcttat attataagaa 10440
atggccattg tgaagatcag aaggagataa ctcactgatt tcaggtggtt gattcaatct 10500
aacacatgat atattctgaa actgtacaaa aaagatattc aaaagaagaa acacatggaa 10560
caaagagtat atggaaattc tgacaactca gaatcagtgt atgttaagat gaagttcatg 10620
attaacaaat ggagatggtc gcagtcatcc tgaagcttat tttagccaat gctttatcac 10680
atccaactca atcttcgttt tttttttttc agatggatct tcccattttt aattaacctt 10740
atcatgatag cccctagtaa ggcatgtcct gacatttctt ttcttttttt ttatagctta 10800
tgtttctttt tttaaaaatt tttttactac atattttcct caattacatt tccaatgcta 10860
tcccaaaagt cccccatacc ctcctccccc actcccctac ccacccactc ccactttttg 10920
gccctggggt tcccctgtac tgggggatat aaagtctgcg tgtccaatgg gcctctcttt 10980
ccagtgatgg cctactaggc catcttttga tacatatgca gcttgagtca agagctccgg 11040
ggtactggtt agttcatagt gttgttccac ctatagggtt gcagatccct ttagttcctt 11100
gggtgctttc tctagcttct ccattgggag ccctgtgatc catccaatag ctgactgtga 11160
gcatccactt ctgtgtttgc taggtcccgg ccaagtctca caagagacag ctatttcagg 11220
gtcctttcag catatgcttg ctagtgtatg caatggtgtc atcgtttgga ggctaattat 11280
gggatggctc cctggatatg gcagtctcta aatggtccat ccttttgtct cagctccaaa 11340
ctttgtctct gtaactcctt tcaatcttcg tttttaatgt ccgaaaatga ctggtaacgt 11400
cactgcttac atttcccttt gctttgtgtt gaattgtgag ctttatcttt gatctggagt 11460
ttctttaaga agactttgta atgtgcttat ttttaaagat ggtgaattca attaaagata 11520
gacaagaggg acgggtctgt atttcatctg gtagagtgct tacctgccat tcatgaagtt 11580
ctggttcgat ccccggcaca gcatagaacc acatgcggtg gcaactcagt tgcctgtgac 11640
cccagcactc aggagctaga ggcaggttac ctttgactac acagcaagtt caaggctagc 11700
ctggtctaca tgcaactctt cctcaaaggg aaaacaaaac aaaaatagac aggagaattg 11760
gaaacatcca tagaaaccca aatatgaatc aatagactcc cctcttcctg tgtcagacat 11820
tcagactcag tgacagccgt caccgtttat gaggcatgac tgattttaca gatcctagag 11880
caagcgtgca caattctggt gtttcctgcc tgcgtcctca cacacacgct caccttaggg 11940
acggaggcag cggaggtcta gtcagtttca gacgaatcca ctcatcagct gctcaccatg 12000
gtcttctagg tgaagcagaa ctctccttcc atcagcaccc gtgaaaaaca aaagtggaaa 12060
gcagatattt cactccgaat gctttatcat tggcagtgat ttcaaattta aaccactaat 12120
ttcccagaag aattaggtca cgggccatgt ggcattagac cctattgtgg tgacattatt 12180
tcctgatgtg cctatttaat aatgtatagt ggtctcatct ttatttagtt ggtcttgaat 12240
gtgataaata caagactggg caagtgttca tcaaagctgt tacttgaaat tgtaattata 12300
cctattttgt tggtggtgtt ttttcttggt tcatggacaa ccaaacacat tacggctttg 12360
ctttgtctta gctactctgt gtcaagttcc tgtgaaaata ccgtattaca gtgctcttat 12420
tacatctttg gtaatgaagt agtcacctaa gcccctatcc tgggctctct atggctcact 12480
aaagaatcgt aagttggata atgattgctg aagaaataga tatgttttat ttacattgag 12540
gtagaaagac ctgtggtttc taggtaacag tttacttaac ttagttgcta catctagttt 12600
gcactcagta aacaccctgg tgtggttttt acaacaacaa ggttgttcct gaagcccaaa 12660
catggcccag atttgaatag aagacctgag gcagactaac actcactccc tgctgccaaa 12720
gtagcagagc cggggacata tttctccaat agccagcccg agttgatgct ctttgtaagc 12780
agacaccaca aagccacacg gctagcccta agatgggaga gccctgacct ctctttgctt 12840
ctgacctagc cccataccgc aaaatcaacc agagaatttc cgtggatcca gccacttctg 12900
agcatgaact aatatgtcag gccgagggtt atccagaagc tgaggtaatc tggacaaaca 12960
gtgaccacca acccgtgagt gggaagagaa gtgtcaccac ttcccggaca gaggggatgc 13020
ttctcaatgt gaccagcagt ctgagggtca acgccacagc gaatgatgtt ttctactgta 13080
cgttttggag atcacagcca gggcaaaacc acacagcgga gctgatcatc ccaggtgagt 13140
tgcctaactc gtccccggat tcctagcacg atggccatcc gccatagtca tttagcagta 13200
gttggccgag tgtatgaata catgaaactt acattggcta gacgtattct agttcctctt 13260
tctcttgcgt gcaagccata atgctacttg gtcaacactc agcaagaatt aagtgtttgc 13320
tcctgacaga cagtatgtta gtcattgagt attcgtggct ggataggaca taagttactc 13380
tgctccagga aacaggtgac agtcgcatgt aggagatgct cttgtaactt gggcaccacg 13440
ggcaacgtga gagcaagctt atgcttcagg aatgtaaacc tttgtcacat caaagctgcc 13500
ctcttccctc ccgggctgct gtagtagaaa gcaaatgcat tgcctttttt tgtatcttat 13560
attcaacagg aagaatcctt tcttcccata ctcctcttca gggcattccc tgatacttca 13620
actccaggaa gtctgcctgt gcatagaaca cgaggctgat gcctatgggc aaccgactag 13680
ctgggttgta actcaccagg ccagggcatt gtagggatag aatgagctgc acgaaaatga 13740
ttgtaggaaa atcgtttgag aattcagctg tctttaaaat tcatttacct tacagaaata 13800
gggccacagg agacagtttg gtgagaggct cagagcatta tatgaatatg aacccagagg 13860
tactcactct gaggagggtt gcttctgcct ccatcgacac ccctttccca atgtatgtca 13920
acagtacatt gatgtctaga gatagctctg tgaagtttag agaagatata taaaactaca 13980
ttaaaaagtc aaccaatatg aaaaatgaag cccaggatgg gcgtggagcc aaaaggagca 14040
cacaagtttg gaaaaatgta aaataaaaag gaagacactc taactagaat tggacttttc 14100
cataaaagct atattgttgg tttcctacca ttaaatgttt tatggggctg gagacgttgt 14160
agctcagttg ggagagtact tgcctagcat ccacaatgct ctgaccccag cgccatataa 14220
acgtatatag ggctcaccta tagtctaaac actttggtgg tggaagccgg agggtcagtt 14280
caaggtcatc ctcagcttca tagcaagttc aaggccagta tggactgact acaagagacc 14340
ctgcctcaaa gacaaaacaa gcaaacatca tcataataaa acagcaaaaa gctgttctct 14400
ggccatttct ttgtaggaca tcctggagta acttaaccac tctgagtgtc tcttctcatc 14460
tgtaacttgg gttcatagtg ggaccaaata ggagctgagt tacaacagga gatggaagtt 14520
agcaatctca gagcatccca gagccctggg acccattttt ctctttctca aaactccagg 14580
cagagaaaca gggagatgtt aatgactgga gtgcccaaga gtctcagaca tggaagaaac 14640
acaacccgca ctggtccaca tttgtgggag acagaagcgg cttagcagtg ccccgctttc 14700
ccatgttcgg gagttggcct tccacaccaa gtcagccatt actgtctata ttaatgagtg 14760
cacaaggaag ttactgcact aaggttacca tttaatattg cagtcacttt ctgtggcaac 14820
atgtcacagg cctggagttc ttctctgtgc tgctcccagt ttcttaggag atctgtcttc 14880
ttcttccaac ccttacctgg acattaactt tcagtgatta agattaccct aagaccaaac 14940
tatatttcag gagtcatcat tcccctgact atggcgcagc aaagccttac cttccatcag 15000
cttctcttta tccttttcca gaactgcctg caacacatcc tccacagaac aggactcact 15060
gggtgcttct gggatccatc ctgttgttcc tcattgtagt gtccacggtc ctcctcttct 15120
tgagaaaaca aggtatttcc tccattgctg tactgcctgc agggaagtgc atgagggtcc 15180
cttcaccatt gccttgagca ggcagtgcca gagatggtac caccctgaca agaacagacc 15240
cacatcacag ggcgtcctac actgcacgct agctgaacct caccaggaaa cactgacttg 15300
aaaggtgctg ccctccattc cctgggcttg ctttcagtta cacagagtaa gttttaacag 15360
tgactctggt gtactgagtc ttccttaagt cctgttcaca ttgttgctct gcgacatctc 15420
ctaccttgga gctcggtgtt ggaccctgtt gtaatatgaa gaggtatcag ttggacaagg 15480
gcgtagggaa gctaaaagac ttagctttca aggcgtgggg gagggggagg gcattgtgag 15540
gttccctgtt cagcggttca gagtaaagct gggcagcaca caagtttaga caacaaggaa 15600
gatgctctaa ctggggttgg aggtcaatgg tgcagggata ggtagcaatg gaggctaaag 15660
aagaaagcca taaggtttct ttctacagac tctgaagaca gagcactgtg gtgcctattg 15720
gtctcaggtg aactcaagtc ttgtgttcct tacaccacct attgagctgg gttctggata 15780
ccctaagtgg aaaaacactt ctgtctaaaa agagctgtgg catctttacc acaaagcact 15840
tttttttttt gcaagatttc tgaagacgaa atgacacagt ttttaaaata atttccatat 15900
atttatttca gtacttaccc ataagaatga ctgaaatcta ggaaaatgaa tatattcaaa 15960
taatgccttc ctattgtaaa gtgacttaga gtaaaccttt tcaccaaata ctttgaaatg 16020
cataccaagt gagccaatga tgcatggact tgtgtgaagc caaatgggga aatagaatat 16080
gggagggaac atcaatcatc agagaggaat tccccatcac ccttttggag ggcgcgtgta 16140
tgggaaaagt gttgtaactg gggcggtagc gaccagctgc ttccaaagaa taagccctga 16200
gcagaaccaa gagggttgcc agggtgtgaa gattaagctt cagagacaca tctggcttag 16260
ggctagccta ggacttgcac aggaccgcag ttgttgccta atggcatctg ggatgattgg 16320
ctgaggatga ggaaagcaga gagtcagtta ggtggtaagc tagctcagga tgtgggacgt 16380
ttctgaggga agggtggaag acaaagtaga cagtcctaaa aaccatctga gaatgaaagt 16440
atgtacccta aattgttgaa taattaactt tgcggcgtgg gatctggaat aaggaagttc 16500
acgggctaaa agcctggtga ttaggggatt gatcggtgtc tcctaaaaac cttaagttga 16560
aaatcgagtc cctgtgggtt gtagtagcat cttgctataa ggaattaaga ttgatttctt 16620
cttctttagt gagaatgcta gatgtggaga aatgtggcgt tgaagataca agctcaaaaa 16680
accgaaatgg taagtgtgag taacgaggga ggggcaagcc gagggaatga gtgggacaga 16740
gcagccaagc agggtcctgc aagggctgca gttccagcgc tctgtaggct gaggcagggg 16800
gatagggagt tccaggccag caaccacaca gccaaaacaa aacaaacaaa acaaaaaaca 16860
aagcaaaagg aagagcagaa aaagctactg ttaccaacaa gagacatact ttagccgtga 16920
tctcctctcc ttccaaagga agctgtttgt gggggctgct ggttcctctg cctaaggatc 16980
gccacatctg agaaaactct gggtttgctg ctgcggtgtc tgtgtggtag ccctgcaacc 17040
agggacgctg ttgacttatt tgaatgggca tttgattaag caaatgattc tgctgtgtgg 17100
ttagctgtta actgtttcat gtgtatgata tggcatttca tttcatcatc ctgcccccaa 17160
tcagataaat atgagaatca tctccattgt atagtaggga aaacagggct gagaagataa 17220
tgcgaggtgc ctaagaacac acagacttgc taccacagac gtccgttctt tgctaggtgc 17280
ttcaatctat gctgctacta atgtctgtat ttataagagt agaaaactct gcccgggcgt 17340
tggtggccca cgcctttaat cccagcactc gggaggcaga ggtagtcgga tttctgagtt 17400
tgaggccagc ctggtctaca gagtgagttc caggacagcc aggctacaca gagaaaccct 17460
gtctcgaaaa aaatcaaaaa agaaaaaaga acaagaaaac tctaactgaa ctatcttgta 17520
gtaaattaca gtgtgttcta attcatttta gtgttgttag aaaccaaaca gtaaggccag 17580
gcagaactaa agtggtttcg ctaacttggg cagacccaag caggttccct cctgtacatt 17640
accatttatt ctgctgagtc accctgagga accggcctgg cttgctcaag actgaggatt 17700
tctcagccat gaacactttc cattttaaaa ctagggcagt cccggacaag caaggatggt 17760
tagtcatcct tcatcatgta agggtagtca gacataggta gtatctgtct ctgtcttacg 17820
gcaacgttac tatctgctct agcaaacatc ccatttgttt atcttacata tcatacgttt 17880
cccccccaca tattccctgc ccctgtgaaa ttattatact atgatcactc tccagataca 17940
caattcgagg agacgtaagc agtgttgaac cctctgatcg tcgattggca gcttgtggtc 18000
tgtgaaagaa agggcccatg ggacatgagt ccaaagactc aagatggaac ctgagggaga 18060
gaaccaagaa agtgttggga gaggagcctg gaacaacgga cattttttcc agggagacac 18120
tgctaagcaa gttgcccatc agtcgtcttg ggaaatggat tgagggttcc tggcttagca 18180
gctggtcctt gcacagtgac cttttcctct gctcagtgcc gggatgagag atggagtcat 18240
gagtgttgaa gaataagtgc cttctattta ttttgagtct gtgtgttctc actttgggca 18300
tgtaattatg actggtgaat tctgacgaca tgatagatct taagatgtag tcaccaaact 18360
caactgctgc ttagcatcct ccgtaactac tgatacaagc agggaacaca gaggtcacct 18420
gcttggtttg acaggctctt gctgtctgac tcaaataatc tttatttttc agtcctcaag 18480
gctcttcgat agcagttgtt ctgtatcagc cttataggtg tcaggtatag cactcaacat 18540
ctcatctcat tacaatagca accctcatca ccatagcaac agctaacctc tgttatcctc 18600
acttcatagc caggaagctg agcgactaag tcacttgccc acagagtatc agctctcaga 18660
tttctgttct tcagccactg tcctttcagg atagaatttg tcgttaagaa attaatttaa 18720
aaactgatta ttgagtagca ttgtatatca atcacaacat gccttgtgca ctgtgctggc 18780
ctctgagcat aaagatgtac gccggagtac cggtcggaca tgtttatgtg tgttaaatac 18840
tcagagaaat gttcattaac aaggagcttg cattttagag acactggaaa gtaactccag 18900
ttcattgtct agcattacat ttacctcatt tgctatcctt gccatacagt ctcttgttct 18960
ccatgaagtg tcatgaatct tgttgaatag ttcttttatt ttttaaatgt ttctatttaa 19020
atgatattga catctgaggc gatagctcag ttggtaaaac cctttcctca caagtgtgaa 19080
accctgagtc ttatccctag aacccacata aaaaacagtt gcgtatgttt gtgcatgctt 19140
ttgatcccag cactagggag gcagaggcag gcagatcctg agctctcatt gaccacccag 19200
cctagcctac atggttagct ccaggcctac aggagctggc agagcctgaa aaacgatgcc 19260
tagacacaca cacacacaca cacacacaca cacacacaca cacacaccat gtactcatag 19320
acctaagtgc accctcctac acatgcacac acatacaatt caaacacaaa tcaacaggga 19380
attgtctcag aatggtcccc aagacaaaga agaagaaaaa caccaaacca gctctattcc 19440
ctcagcctat cctctctact ccttcctaga agcaactact attgtttttg tatataaatt 19500
tacccaacga cagttaatat gtagaatata tattaaagtg tctgtcaata tatattatct 19560
ctttctttct ttcttccttt ctttctttct ttctttcttt ctttctttct ttctttcttt 19620
ctttcttcct tccttccttc cttccttcct tccttccttc ctttctttct ttctttcttt 19680
ttttctgtct atctgtacct aaatggttgc tcactatgca ttttctgtgc tcttcgccct 19740
ttttatttaa tgtatggata tttatgctgc ttccagaatg gatctaaagc tctttgtttc 19800
taggttttct cccccatcct tctaggcatc tctcacactg tctaggccag acaccatgtc 19860
tgctgcctga atctgtagac accatttata aagcacgtac tcaccgagtt tgtatttggc 19920
ttgttctgtg tctgattaaa gggagaccat gagtccccag ggtacactga gttaccccag 19980
taccaagggg gagccttgtt tgtgtctcca tggcagaagc aggcctggag ccattttggt 20040
ttcttccttg acttctctca aacacagacg cctcacttgc tcattacagg ttctcctttg 20100
ggaatgtcag cattgctcct tgactgctgg ctgccctgga aggagcccat tagctctgtg 20160
tgagcccttg acagctactg cctctcctta ccacaggggc ctctaagata ctgttaccta 20220
gaggtcttga ggatctgtgt tctctggggg gaggaaagga ggaggaaccc agaactttct 20280
tacagttttc cttgttctgt cacatgtcaa gactgaagga acaggctggg ctacgtagtg 20340
agatcctgtc tcaaaggaaa gacgagcata gccgaacccc cggtggaacc ccctctgtta 20400
cctgttcaca caagcttatt gatgagtctc atgttaatgt cttgtttgta tgaagtttaa 20460
gaaaatatcg ggttgggcaa cacattctat ttattcattt tatttgaaat cttaatgcca 20520
tctcatggtg ttggattggt gtggcacttt attcttttgt gttgtgtata accataaatt 20580
ttattttgca tcagattgtc aatgtattgc attaatttaa taaatatttt tatttattaa 20640
a 20641
<210> 5
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 5
taattggctc tactgc 16
<210> 6
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 6
tcgcataaga atgact 16
<210> 7
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 7
tgaacacaca gtcgca 16
<210> 8
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 8
ctgaacacac agtcgc 16
<210> 9
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 9
tctgaacaca cagtcg 16
<210> 10
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 10
ttctgaacac acagtc 16
<210> 11
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 11
acaagtcatg ttacta 16
<210> 12
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 12
acacaagtca tgttac 16
<210> 13
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 13
cttacttaga tgctgc 16
<210> 14
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 14
acttacttag atgctg 16
<210> 15
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 15
gacttactta gatgct 16
<210> 16
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 16
agacttactt agatgc 16
<210> 17
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 17
gcaggaagag acttac 16
<210> 18
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 18
aataaattcc gttcagg 17
<210> 19
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 19
gcaaataaat tccgtt 16
<210> 20
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 20
agcaaataaa ttccgt 16
<210> 21
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 21
cagagcaaat aaattcc 17
<210> 22
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 22
tggacagagc aaataaat 18
<210> 23
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 23
atggacagag caaata 16
<210> 24
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 24
cagaatggac agagca 16
<210> 25
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 25
ttctcagaat ggacag 16
<210> 26
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 26
ctgaactttg acatag 16
<210> 27
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 27
aagacaaacc cagactga 18
<210> 28
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 28
tataagacaa acccagac 18
<210> 29
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 29
ttataagaca aacccaga 18
<210> 30
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 30
tgttataaga caaaccc 17
<210> 31
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 31
tagaacaatg gtacttt 17
<210> 32
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 32
gtagaacaat ggtact 16
<210> 33
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 33
aggtagaaca atggta 16
<210> 34
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 34
aagaggtaga acaatgg 17
<210> 35
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 35
gcatccacag taaatt 16
<210> 36
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 36
gaaggttatt taattc 16
<210> 37
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 37
ctaatcgaat gcagca 16
<210> 38
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 38
tacccaatct aatcga 16
<210> 39
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 39
tagttaccca atctaa 16
<210> 40
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 40
catttagtta cccaat 16
<210> 41
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 41
tcatttagtt acccaa 16
<210> 42
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 42
ttcatttagt taccca 16
<210> 43
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 43
gaattaattt catttagt 18
<210> 44
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 44
cagtgaggaa ttaattt 17
<210> 45
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 45
ccaacagtga ggaatt 16
<210> 46
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 46
cccaacagtg aggaat 16
<210> 47
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 47
tatacccaac agtgagg 17
<210> 48
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 48
ttatacccaa cagtgag 17
<210> 49
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 49
tttataccca acagtga 17
<210> 50
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 50
cctttatacc caacag 16
<210> 51
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 51
taacctttat acccaa 16
<210> 52
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 52
aataaccttt ataccca 17
<210> 53
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 53
gtaaataacc tttata 16
<210> 54
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 54
actgtaaata acctttat 18
<210> 55
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 55
atatatatgc aatgag 16
<210> 56
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 56
agatatatat gcaatg 16
<210> 57
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 57
gagatatata tgcaat 16
<210> 58
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 58
ccagagatat atatgc 16
<210> 59
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 59
caatattcca gagatat 17
<210> 60
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 60
gcaatattcc agagata 17
<210> 61
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 61
agcaatattc cagagat 17
<210> 62
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 62
cagcaatatt ccagag 16
<210> 63
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 63
aatcagcaat attccag 17
<210> 64
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 64
acaatcagca atattcc 17
<210> 65
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 65
actaagtagt tacacttct 19
<210> 66
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 66
ctaagtagtt acacttc 17
<210> 67
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 67
gactaagtag ttacactt 18
<210> 68
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 68
tgactaagta gttaca 16
<210> 69
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 69
ctttgactaa gtagtta 17
<210> 70
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 70
ctctttgact aagtag 16
<210> 71
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 71
gctctttgac taagta 16
<210> 72
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 72
ccttaaatac tgttgac 17
<210> 73
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 73
cttaaatact gttgac 16
<210> 74
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 74
tccttaaata ctgttg 16
<210> 75
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 75
tctccttaaa tactgtt 17
<210> 76
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 76
tatcatagtt ctcctt 16
<210> 77
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 77
agtatcatag ttctcc 16
<210> 78
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 78
gagtatcata gttctc 16
<210> 79
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 79
agagtatcat agttct 16
<210> 80
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 80
cagagtatca tagttc 16
<210> 81
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 81
ttcagagtat catagt 16
<210> 82
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 82
cttcagagta tcatag 16
<210> 83
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 83
ttcttcagag tatcata 17
<210> 84
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 84
tttcttcaga gtatcat 17
<210> 85
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 85
gagaaaggct aagttt 16
<210> 86
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 86
gacactcttg tacatt 16
<210> 87
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 87
tgagacactc ttgtaca 17
<210> 88
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 88
tgagacactc ttgtac 16
<210> 89
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 89
ctttattaaa ctccat 16
<210> 90
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 90
accaaacttt attaaa 16
<210> 91
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 91
aaacctctac taagtg 16
<210> 92
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 92
agattaagac agttga 16
<210> 93
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 93
aagtaggagc aagaggc 17
<210> 94
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 94
aaagtaggag caagagg 17
<210> 95
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 95
gttaagcagc caggag 16
<210> 96
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 96
agggtaggat gggtag 16
<210> 97
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 97
aagggtagga tgggta 16
<210> 98
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 98
caagggtagg atgggt 16
<210> 99
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 99
ccaagggtag gatggg 16
<210> 100
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 100
tccaagggta ggatgg 16
<210> 101
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 101
cttccaaggg taggat 16
<210> 102
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 102
atcttccaag ggtagga 17
<210> 103
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 103
agaagtgatg gctcatt 17
<210> 104
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 104
aagaagtgat ggctcat 17
<210> 105
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 105
gaagaagtga tggctca 17
<210> 106
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 106
atgaaatgta aactggg 17
<210> 107
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 107
caatgaaatg taaactgg 18
<210> 108
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 108
gcaatgaaat gtaaactg 18
<210> 109
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 109
agcaatgaaa tgtaaact 18
<210> 110
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 110
gagcaatgaa atgtaaac 18
<210> 111
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 111
tgaattccca tatccga 17
<210> 112
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 112
agaattatga ccatat 16
<210> 113
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 113
aggtaagaat tatgacc 17
<210> 114
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 114
tcaggtaaga attatgac 18
<210> 115
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 115
cttcaggtaa gaattatg 18
<210> 116
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 116
tcttcaggta agaatta 17
<210> 117
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 117
cttcttcagg taagaat 17
<210> 118
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 118
tcttcttcag gtaagaa 17
<210> 119
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 119
tcttcttcag gtaaga 16
<210> 120
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 120
tggtctaaga gaagaag 17
<210> 121
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 121
gttggtctaa gagaag 16
<210> 122
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 122
agttggtcta agagaa 16
<210> 123
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 123
cagttggtct aagagaa 17
<210> 124
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 124
gcagttggtc taagagaa 18
<210> 125
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 125
cagttggtct aagaga 16
<210> 126
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 126
gcagttggtc taagaga 17
<210> 127
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 127
gcagttggtc taagag 16
<210> 128
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 128
ctcatatcag ggcagt 16
<210> 129
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 129
cacacatgtt ctttaac 17
<210> 130
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 130
taaatacaca catgttct 18
<210> 131
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 131
gtaaatacac acatgttc 18
<210> 132
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 132
tgtaaataca cacatgtt 18
<210> 133
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 133
gatcatgtaa atacacac 18
<210> 134
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 134
agatcatgta aatacaca 18
<210> 135
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 135
caaagatcat gtaaatacac 20
<210> 136
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 136
acaaagatca tgtaaataca 20
<210> 137
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 137
gaatacaaag atcatgta 18
<210> 138
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 138
agaatacaaa gatcatgt 18
<210> 139
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 139
cagaatacaa agatcatg 18
<210> 140
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 140
gcagaataca aagatca 17
<210> 141
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 141
aggcagaata caaagat 17
<210> 142
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 142
aaggcagaat acaaaga 17
<210> 143
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 143
attagtgagg gacgaa 16
<210> 144
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 144
cattagtgag ggacga 16
<210> 145
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 145
gagggtgatg gattag 16
<210> 146
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 146
ttaggagtaa taaagg 16
<210> 147
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 147
ttaatgaatt tggttg 16
<210> 148
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 148
ctttaatgaa tttggt 16
<210> 149
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 149
catggattac aactaa 16
<210> 150
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 150
tcatggatta caacta 16
<210> 151
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 151
gtcatggatt acaact 16
<210> 152
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 152
cattaaatct agtcat 16
<210> 153
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 153
gacattaaat ctagtca 17
<210> 154
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 154
agggacatta aatcta 16
<210> 155
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 155
caaagcatta taacca 16
<210> 156
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 156
acttactagg cagaag 16
<210> 157
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 157
cagagttaac tgtaca 16
<210> 158
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 158
ccagagttaa ctgtac 16
<210> 159
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 159
gccagagtta actgta 16
<210> 160
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 160
tgggccagag ttaact 16
<210> 161
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 161
cagcatctat cagact 16
<210> 162
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 162
tgaaataaca tgagtcat 18
<210> 163
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 163
gtgaaataac atgagtc 17
<210> 164
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 164
tctgtttatg tcactg 16
<210> 165
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 165
gtctgtttat gtcact 16
<210> 166
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 166
tggtctgttt atgtca 16
<210> 167
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 167
ttggtctgtt tatgtc 16
<210> 168
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 168
tcacccattg tttaaa 16
<210> 169
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 169
ttcagcaaat attcgt 16
<210> 170
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 170
gtgtgttcag caaatat 17
<210> 171
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 171
tctattgtta ggtatc 16
<210> 172
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 172
attgcccatc ttactg 16
<210> 173
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 173
tattgcccat cttact 16
<210> 174
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 174
aaatattgcc catctt 16
<210> 175
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 175
ataaccttat cataca 16
<210> 176
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 176
tataacctta tcatac 16
<210> 177
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 177
ttataacctt atcata 16
<210> 178
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 178
tttataacct tatcat 16
<210> 179
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 179
actgctattg ctatct 16
<210> 180
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 180
aggactgcta ttgcta 16
<210> 181
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 181
gaggactgct attgct 16
<210> 182
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 182
acgtagaata ataaca 16
<210> 183
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 183
ccaagtgata taatgg 16
<210> 184
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 184
ttagcagacc aagtga 16
<210> 185
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 185
gtttagcaga ccaagt 16
<210> 186
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 186
tgacagtgat tatatt 16
<210> 187
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 187
tgtccaagat attgac 16
<210> 188
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 188
gaatatccta gattgt 16
<210> 189
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 189
caaactgaga atatcc 16
<210> 190
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 190
gcaaactgag aatatc 16
<210> 191
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 191
tcctattaca atcgta 16
<210> 192
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 192
ttcctattac aatcgt 16
<210> 193
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 193
actaatggga ggattt 16
<210> 194
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 194
tagttcagag aataag 16
<210> 195
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 195
taacatatag ttcaga 16
<210> 196
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 196
ataacatata gttcag 16
<210> 197
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 197
cataacatat agttca 16
<210> 198
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 198
tcataacata tagttc 16
<210> 199
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 199
tagctcctaa caatca 16
<210> 200
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 200
ctccaatctt tgtata 16
<210> 201
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 201
tctccaatct ttgtat 16
<210> 202
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 202
tctatttcag ccaatc 16
<210> 203
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 203
cggaagtcag agtgaa 16
<210> 204
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 204
ttaagcatga ggaata 16
<210> 205
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 205
tgattgagca cctctt 16
<210> 206
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 206
gactaattat ttcgtt 16
<210> 207
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 207
tgactaatta tttcgt 16
<210> 208
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 208
gtgactaatt atttcg 16
<210> 209
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 209
ctgcttgaaa tgtgac 16
<210> 210
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 210
cctgcttgaa atgtga 16
<210> 211
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 211
atcctgcttg aaatgt 16
<210> 212
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 212
attataaatc tattct 16
<210> 213
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 213
gctaaatact ttcatc 16
<210> 214
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 214
cattgtaaca taccta 16
<210> 215
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 215
gcattgtaac atacct 16
<210> 216
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 216
taatattgca ccaaat 16
<210> 217
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 217
gataatattg caccaa 16
<210> 218
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 218
agataatatt gcacca 16
<210> 219
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 219
gccaagaaga taatat 16
<210> 220
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 220
cacagccaca taaact 16
<210> 221
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 221
ttgtaattgt ggaaac 16
<210> 222
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 222
tgacttgtaa ttgtgg 16
<210> 223
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 223
tctaactgaa atagtc 16
<210> 224
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 224
gtggttctaa ctgaaa 16
<210> 225
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 225
caatatggga cttggt 16
<210> 226
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 226
atgacaatat gggact 16
<210> 227
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 227
tatgacaata tgggac 16
<210> 228
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 228
atatgacaat atggga 16
<210> 229
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 229
cttcacttaa taatta 16
<210> 230
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 230
ctgcttcact taataa 16
<210> 231
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 231
aagactgctt cactta 16
<210> 232
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 232
gaatgcccta attatg 16
<210> 233
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 233
tggaatgccc taatta 16
<210> 234
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 234
gcaaatgcca gtaggt 16
<210> 235
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 235
ctaatggaag gatttg 16
<210> 236
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 236
aatatagaac ctaatg 16
<210> 237
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 237
gaaagaatag aatgtt 16
<210> 238
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 238
atgggtaata gattat 16
<210> 239
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 239
gaaagagcac agggtg 16
<210> 240
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 240
ctacatagag ggaatg 16
<210> 241
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 241
gcttcctaca tagagg 16
<210> 242
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 242
tgcttcctac atagag 16
<210> 243
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 243
tgggcttgaa atatgt 16
<210> 244
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 244
cattatattt aagaac 16
<210> 245
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 245
tcggttatgt tatcat 16
<210> 246
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 246
cactttatct ggtcgg 16
<210> 247
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 247
aaattggcac agcgtt 16
<210> 248
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 248
accgtgacag taaatg 16
<210> 249
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 249
tgggaaccgt gacagta 17
<210> 250
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 250
ccacatatag gtcctt 16
<210> 251
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 251
catattgcta ccatac 16
<210> 252
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 252
tcatattgct accata 16
<210> 253
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 253
caattgtcat attgct 16
<210> 254
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 254
cattcaattg tcatattg 18
<210> 255
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 255
tttctactgg gaatttg 17
<210> 256
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 256
caattagtgc agccag 16
<210> 257
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 257
gaataatgtt cttatcc 17
<210> 258
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 258
cacaaattga ataatgttct 20
<210> 259
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 259
catgcacaaa ttgaataat 19
<210> 260
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 260
atcctgcaat ttcacat 17
<210> 261
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 261
ccaccatagc tgatca 16
<210> 262
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 262
accaccatag ctgatca 17
<210> 263
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 263
caccaccata gctgatc 17
<210> 264
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 264
tagtcggcac caccat 16
<210> 265
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 265
cttgtagtcg gcaccac 17
<210> 266
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 266
cttgtagtcg gcacca 16
<210> 267
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 267
cgcttgtagt cggcac 16
<210> 268
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 268
tcaataaaga tcaggc 16
<210> 269
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 269
tggacttaca agaatg 16
<210> 270
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 270
atggacttac aagaat 16
<210> 271
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 271
gctcaagaaa ttggat 16
<210> 272
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 272
tactgtagaa catggc 16
<210> 273
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 273
gcaattcatt tgatct 16
<210> 274
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 274
tgaagggagg agggacac 18
<210> 275
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 275
agtggtgaag ggaggag 17
<210> 276
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 276
tagtggtgaa gggaggag 18
<210> 277
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 277
atagtggtga agggaggag 19
<210> 278
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 278
tagtggtgaa gggagga 17
<210> 279
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 279
atagtggtga agggagga 18
<210> 280
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 280
tagtggtgaa gggagg 16
<210> 281
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 281
atagtggtga agggagg 17
<210> 282
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 282
gatagtggtg aagggagg 18
<210> 283
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 283
atagtggtga agggag 16
<210> 284
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 284
gatagtggtg aagggag 17
<210> 285
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 285
gagatagtgg tgaagg 16
<210> 286
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 286
catgggagat agtggt 16
<210> 287
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 287
acaaataatg gttactct 18
<210> 288
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 288
acacacaaat aatggtta 18
<210> 289
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 289
gagggacaca caaataat 18
<210> 290
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 290
atatagagag gctcaa 16
<210> 291
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 291
ttgatataga gaggct 16
<210> 292
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 292
gcatttgata tagaga 16
<210> 293
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 293
tttgcatttg atatag 16
<210> 294
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 294
ctggaagaat aggttc 16
<210> 295
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 295
actggaagaa taggtt 16
<210> 296
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 296
tactggaaga ataggt 16
<210> 297
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 297
tggcttatcc tgtact 16
<210> 298
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 298
atggcttatc ctgtac 16
<210> 299
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 299
tatggcttat cctgta 16
<210> 300
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 300
gtatggctta tcctgt 16
<210> 301
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 301
atgaatatat gcccagt 17
<210> 302
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 302
gatgaatata tgccca 16
<210> 303
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 303
caagatgaat atatgcc 17
<210> 304
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 304
gacaacatca gtataga 17
<210> 305
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 305
caagacaaca tcagta 16
<210> 306
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 306
cactcctagt tccttt 16
<210> 307
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 307
aacactccta gttcct 16
<210> 308
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 308
taacactcct agttcc 16
<210> 309
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 309
ctaacactcc tagttc 16
<210> 310
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 310
tgataacata actgtg 16
<210> 311
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 311
ctgataacat aactgt 16
<210> 312
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 312
tttgaactca agtgac 16
<210> 313
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 313
tcctttactt agctag 16
<210> 314
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 314
gagtttggat tagctg 16
<210> 315
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 315
tgggatatga caggga 16
<210> 316
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 316
tgtgggatat gacagg 16
<210> 317
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 317
atatggaagg gatatc 16
<210> 318
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 318
acaggatatg gaaggg 16
<210> 319
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 319
atttcaacag gatatgg 17
<210> 320
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 320
gagtaatttc aacagg 16
<210> 321
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 321
agggagtaat ttcaaca 17
<210> 322
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 322
attagggagt aatttca 17
<210> 323
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 323
cttactatta gggagt 16
<210> 324
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 324
cagcttacta ttaggg 16
<210> 325
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 325
tcagcttact attagg 16
<210> 326
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 326
atttcagctt actattag 18
<210> 327
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 327
ttcagcttac tattag 16
<210> 328
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 328
cagatttcag cttact 16
<210> 329
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 329
gactacaact agaggg 16
<210> 330
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 330
agactacaac tagagg 16
<210> 331
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 331
aagactacaa ctagag 16
<210> 332
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 332
atgatttaat tctagtcaaa 20
<210> 333
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 333
tttaattcta gtcaaa 16
<210> 334
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 334
gatttaattc tagtca 16
<210> 335
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 335
atgatttaat tctagtca 18
<210> 336
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 336
gatgatttaa ttctagtca 19
<210> 337
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 337
gatttaattc tagtca 16
<210> 338
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 338
gatgatttaa ttctagtc 18
<210> 339
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 339
tgatttaatt ctagtc 16
<210> 340
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 340
gagatgattt aattcta 17
<210> 341
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 341
gagatgattt aattct 16
<210> 342
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 342
cagattgatg gtagtt 16
<210> 343
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 343
ctcagattga tggtag 16
<210> 344
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 344
gttagccctc agattg 16
<210> 345
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 345
tgtattgtta gccctc 16
<210> 346
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 346
acttgtattg ttagcc 16
<210> 347
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 347
agccagtatc agggac 16
<210> 348
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 348
ttgacaatag tggcat 16
<210> 349
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 349
acaagtggta tcttct 16
<210> 350
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 350
aatctacttt acaagt 16
<210> 351
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 351
cacagtagat gcctgata 18
<210> 352
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 352
gaacacagta gatgcc 16
<210> 353
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 353
cttggaacac agtagat 17
<210> 354
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 354
atatcttgga acacag 16
<210> 355
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 355
tctttaatat cttggaac 18
<210> 356
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 356
tgatttcttt aatatcttg 19
<210> 357
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 357
tgatgatttc tttaatatc 19
<210> 358
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 358
aggctaagtc atgatg 16
<210> 359
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 359
ttgatgaggc taagtc 16
<210> 360
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 360
ccaggattat actctt 16
<210> 361
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 361
gccaggatta tactct 16
<210> 362
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 362
ctgccaggat tatact 16
<210> 363
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 363
cagaaactta tactttatg 19
<210> 364
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 364
aagcagaaac ttatact 17
<210> 365
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 365
gaagcagaaa cttatact 18
<210> 366
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 366
tggaagcaga aacttatact 20
<210> 367
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 367
tggaagcaga aacttatac 19
<210> 368
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 368
aagcagaaac ttatac 16
<210> 369
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 369
tggaagcaga aacttata 18
<210> 370
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 370
aagggatatt atggag 16
<210> 371
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 371
tgccggaaga tttcct 16
<210> 372
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 372
atggattggg agtaga 16
<210> 373
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 373
agatggattg ggagta 16
<210> 374
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 374
aagatggatt gggagt 16
<210> 375
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 375
acaagatgga ttggga 16
<210> 376
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 376
agaaggttca gacttt 16
<210> 377
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 377
gcagaaggtt cagact 16
<210> 378
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 378
tgcagaaggt tcagac 16
<210> 379
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 379
agtgcagaag gttcag 16
<210> 380
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 380
aagtgcagaa ggttca 16
<210> 381
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 381
taagtgcaga aggttc 16
<210> 382
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 382
tctaagtgca gaaggt 16
<210> 383
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 383
ctcaggagtt ctacttc 17
<210> 384
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 384
ctcaggagtt ctactt 16
<210> 385
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 385
atggaggtga ctcaggag 18
<210> 386
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 386
atggaggtga ctcagga 17
<210> 387
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 387
atggaggtga ctcagg 16
<210> 388
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 388
tatggaggtg actcagg 17
<210> 389
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 389
atatggaggt gactcagg 18
<210> 390
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 390
tatggaggtg actcag 16
<210> 391
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 391
atatggaggt gactcag 17
<210> 392
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 392
catatggagg tgactcag 18
<210> 393
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 393
atatggaggt gactca 16
<210> 394
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 394
catatggagg tgactca 17
<210> 395
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 395
catatggagg tgactc 16
<210> 396
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 396
gcatatggag gtgactc 17
<210> 397
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 397
tgcatatgga ggtgactc 18
<210> 398
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 398
ttgcatatgg aggtgactc 19
<210> 399
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 399
tttgcatatg gaggtgactc 20
<210> 400
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 400
gcatatggag gtgact 16
<210> 401
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 401
tgcatatgga ggtgact 17
<210> 402
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 402
ttgcatatgg aggtgact 18
<210> 403
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 403
tttgcatatg gaggtgact 19
<210> 404
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 404
tgcatatgga ggtgac 16
<210> 405
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 405
ttgcatatgg aggtgac 17
<210> 406
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 406
tttgcatatg gaggtgac 18
<210> 407
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 407
tttgcatatg gaggtga 17
<210> 408
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 408
tttgcatatg gaggtg 16
<210> 409
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 409
aagtgaagtt caacagc 17
<210> 410
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 410
tgggaagtga agttca 16
<210> 411
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 411
atgggaagtg aagttc 16
<210> 412
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 412
gatgggaagt gaagtt 16
<210> 413
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 413
ctgtgatggg aagtgaa 17
<210> 414
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 414
attgagtgaa tccaaa 16
<210> 415
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 415
aattgagtga atccaa 16
<210> 416
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 416
gataattgag tgaatcc 17
<210> 417
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 417
gtgataattg agtgaa 16
<210> 418
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 418
aagaaaggtg caataa 16
<210> 419
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 419
caagaaaggt gcaata 16
<210> 420
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 420
acaagaaagg tgcaat 16
<210> 421
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 421
atttaaactc acaaac 16
<210> 422
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 422
ctgttaggtt cagcga 16
<210> 423
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 423
tctgaatgaa catttcg 17
<210> 424
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 424
ctcattgaag gttctg 16
<210> 425
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 425
ctaatctcat tgaagg 16
<210> 426
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 426
cctaatctca ttgaag 16
<210> 427
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 427
actttgatct ttcagc 16
<210> 428
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 428
actatgcaac actttg 16
<210> 429
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 429
caaatagctt tatcgg 16
<210> 430
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 430
ccaaatagct ttatcg 16
<210> 431
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 431
tccaaatagc tttatc 16
<210> 432
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 432
gatccaaata gcttta 16
<210> 433
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 433
atgatccaaa tagctt 16
<210> 434
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 434
tatgatccaa atagct 16
<210> 435
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 435
taaacagggc tgggaat 17
<210> 436
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 436
acttaaacag ggctgg 16
<210> 437
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 437
acacttaaac agggct 16
<210> 438
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 438
gaacacttaa acaggg 16
<210> 439
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 439
agagaacact taaacag 17
<210> 440
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 440
ctacagagaa cactta 16
<210> 441
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 441
atgctacaga gaacact 17
<210> 442
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 442
ataaatgcta cagagaaca 19
<210> 443
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 443
agataaatgc tacagaga 18
<210> 444
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 444
tagagataaa tgctaca 17
<210> 445
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 445
tagatagaga taaatgct 18
<210> 446
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 446
caatatacta gatagaga 18
<210> 447
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 447
tacacaatat actagatag 19
<210> 448
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 448
ctacacaata tactag 16
<210> 449
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 449
gctacacaat atacta 16
<210> 450
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 450
atatgctaca caatatac 18
<210> 451
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 451
tgatatgcta cacaat 16
<210> 452
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 452
atgatatgat atgctac 17
<210> 453
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 453
gaggagagag acaataaa 18
<210> 454
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 454
ctaggaggag agagaca 17
<210> 455
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 455
tattctagga ggagaga 17
<210> 456
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 456
ttatattcta ggaggag 17
<210> 457
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 457
gtttatattc taggag 16
<210> 458
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 458
tggagtttat attctagg 18
<210> 459
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 459
cgtaccacca ctctgc 16
<210> 460
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 460
tgaggaaatc attcattc 18
<210> 461
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 461
tttgaggaaa tcattcat 18
<210> 462
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 462
aggctaatcc tatttg 16
<210> 463
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 463
tttaggctaa tcctat 16
<210> 464
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 464
tgctccagtg taccct 16
<210> 465
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 465
tagtagtact cgatag 16
<210> 466
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 466
ctaattgtag tagtactc 18
<210> 467
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 467
tgctaattgt agtagt 16
<210> 468
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 468
agtgctaatt gtagta 16
<210> 469
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 469
gcaagtgcta attgta 16
<210> 470
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 470
gaggaaatga actaattta 19
<210> 471
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 471
caggaggaaa tgaacta 17
<210> 472
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 472
ccctagagtc atttcc 16
<210> 473
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 473
atcttacatg atgaagc 17
<210> 474
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 474
gacacactca gatttcag 18
<210> 475
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 475
agacacactc agatttcag 19
<210> 476
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 476
aagacacact cagatttcag 20
<210> 477
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 477
agacacactc agatttca 18
<210> 478
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 478
aagacacact cagatttca 19
<210> 479
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 479
aaagacacac tcagatttca 20
<210> 480
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 480
gaaagacaca ctcagatttc 20
<210> 481
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 481
aagacacact cagatttc 18
<210> 482
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 482
aaagacacac tcagatttc 19
<210> 483
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 483
tgaaagacac actcagattt 20
<210> 484
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 484
tgaaagacac actcagatt 19
<210> 485
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 485
tgaaagacac actcagat 18
<210> 486
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 486
attgaaagac acactca 17
<210> 487
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 487
tcattgaaag acacact 17
<210> 488
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 488
ttccatcatt gaaaga 16
<210> 489
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 489
ataataccac ttatcat 17
<210> 490
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 490
ttacttaatt tctttgga 18
<210> 491
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 491
ttagaactag ctttatca 18
<210> 492
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 492
gaggtacaaa tatagg 16
<210> 493
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 493
cttatgatac aactta 16
<210> 494
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 494
tcttatgata caactt 16
<210> 495
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 495
ttcttatgat acaact 16
<210> 496
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 496
cagtttctta tgatac 16
<210> 497
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 497
gcagtttctt atgata 16
<210> 498
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 498
tacaaatgtc tattaggtt 19
<210> 499
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 499
tgtacaaatg tctattag 18
<210> 500
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 500
agcatcacaa ttagta 16
<210> 501
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 501
ctaatgatag tgaagc 16
<210> 502
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 502
agctaatgat agtgaa 16
<210> 503
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 503
atgccttgac atatta 16
<210> 504
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 504
ctcaagatta ttgacac 17
<210> 505
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 505
acctcaagat tattga 16
<210> 506
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 506
aacctcaaga ttattg 16
<210> 507
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 507
cacaaacctc aagattatt 19
<210> 508
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 508
gtacttaatt agacct 16
<210> 509
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 509
agtacttaat tagacc 16
<210> 510
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 510
gtatgaggtg gtaaac 16
<210> 511
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 511
aggaaacagc agaagtg 17
<210> 512
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 512
gcacaaccca gaggaa 16
<210> 513
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 513
caagcacaac ccagag 16
<210> 514
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 514
ttcaagcaca acccag 16
<210> 515
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 515
aattcaagca caaccc 16
<210> 516
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 516
taataattca agcacaacc 19
<210> 517
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 517
actaataatt caagcac 17
<210> 518
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 518
ataatactaa taattcaagc 20
<210> 519
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 519
tagatttgtg aggtaa 16
<210> 520
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 520
agccttaatt ctccat 16
<210> 521
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 521
aatgatctag agcctta 17
<210> 522
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 522
ctaatgatct agagcc 16
<210> 523
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 523
actaatgatc tagagc 16
<210> 524
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 524
cattaacatg ttcttatt 18
<210> 525
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 525
acaagtacat taacatgttc 20
<210> 526
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 526
ttacaagtac attaacatg 19
<210> 527
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 527
gctttattca tgtttat 17
<210> 528
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 528
gctttattca tgttta 16
<210> 529
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 529
agagctttat tcatgttt 18
<210> 530
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 530
ataagagctt tattcatg 18
<210> 531
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 531
cataagagct ttattca 17
<210> 532
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 532
agcataagag ctttat 16
<210> 533
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 533
tagattgttt agtgca 16
<210> 534
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 534
gtagattgtt tagtgc 16
<210> 535
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 535
gacaattcta gtagatt 17
<210> 536
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 536
ctgacaattc tagtag 16
<210> 537
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 537
gctgacaatt ctagta 16
<210> 538
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 538
aggattaaga tacgta 16
<210> 539
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 539
caggattaag atacgt 16
<210> 540
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 540
tcaggattaa gatacg 16
<210> 541
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 541
ttcaggatta agatac 16
<210> 542
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 542
aggaagaaag tttgattc 18
<210> 543
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 543
tcaaggaaga aagtttga 18
<210> 544
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 544
ctcaaggaag aaagtttg 18
<210> 545
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 545
tgctcaagga agaaagt 17
<210> 546
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 546
aattatgctc aaggaaga 18
<210> 547
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 547
taggatacca cattatga 18
<210> 548
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 548
cataatttat tccattcctc 20
<210> 549
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 549
tgcataattt attccat 17
<210> 550
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 550
actgcataat ttattcc 17
<210> 551
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 551
ctaaactgca taatttatt 19
<210> 552
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 552
ataactaaac tgcata 16
<210> 553
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 553
ttattaataa ctaaactgc 19
<210> 554
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 554
tagtacatta ttaataact 19
<210> 555
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 555
cataactaag gacgtt 16
<210> 556
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 556
tcataactaa ggacgt 16
<210> 557
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 557
cgtcataact aaggac 16
<210> 558
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 558
tcgtcataac taagga 16
<210> 559
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 559
atcgtcataa ctaagg 16
<210> 560
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 560
gttagtatct tacatt 16
<210> 561
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 561
ctctattgtt agtatc 16
<210> 562
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 562
agtatagagt tactgt 16
<210> 563
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 563
ttcctggtga tacttt 16
<210> 564
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 564
gttcctggtg atactt 16
<210> 565
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 565
tgttcctggt gatact 16
<210> 566
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 566
ataaacatga atctctcc 18
<210> 567
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 567
ctttataaac atgaatctc 19
<210> 568
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 568
ctgtctttat aaacatg 17
<210> 569
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 569
ttgttataaa tctgtctt 18
<210> 570
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 570
ttaaatttat tcttggata 19
<210> 571
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 571
cttaaattta ttcttgga 18
<210> 572
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 572
cttcttaaat ttattcttg 19
<210> 573
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 573
tatgtttctc agtaaag 17
<210> 574
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 574
gaattatctt taaacca 17
<210> 575
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 575
cccttaaatt tctaca 16
<210> 576
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 576
acactgctct tgtacc 16
<210> 577
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 577
tgacaacact gctctt 16
<210> 578
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 578
tacatttatt gggctc 16
<210> 579
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 579
gtacatttat tgggct 16
<210> 580
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 580
ttggtacatt tattgg 16
<210> 581
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 581
catgttggta catttat 17
<210> 582
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 582
aatcatgttg gtacat 16
<210> 583
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 583
aaatcatgtt ggtaca 16
<210> 584
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 584
gacaagtttg gattaa 16
<210> 585
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 585
aatgttcaga tgcctc 16
<210> 586
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 586
gcttaatgtt cagatg 16
<210> 587
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 587
cgtacatagc ttgatg 16
<210> 588
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 588
gtgaggaatt aggata 16
<210> 589
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 589
gtaacaatat ggtttg 16
<210> 590
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 590
gaaatattgt agacta 16
<210> 591
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 591
ttgaaatatt gtagac 16
<210> 592
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 592
aagtctagta atttgc 16
<210> 593
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 593
gctcagtaga ttataa 16
<210> 594
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 594
catacactgt tgctaa 16
<210> 595
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 595
atggtctcaa atcatt 16
<210> 596
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 596
caatggtctc aaatca 16
<210> 597
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 597
ttcctattga ttgact 16
<210> 598
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 598
tttctgttca caacac 16
<210> 599
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 599
aggaacccac taatct 16
<210> 600
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 600
taaatggcag gaaccc 16
<210> 601
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 601
gtaaatggca ggaacc 16
<210> 602
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 602
ttgtaaatgg caggaa 16
<210> 603
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 603
ttatgagtta ggcatg 16
<210> 604
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 604
ccaggtgaaa ctttaa 16
<210> 605
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 605
cccttagtca gctcct 16
<210> 606
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 606
acccttagtc agctcc 16
<210> 607
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 607
cacccttagt cagctc 16
<210> 608
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 608
tctcttacta ggctcc 16
<210> 609
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 609
cctatctgtc atcatg 16
<210> 610
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 610
tcctatctgt catcat 16
<210> 611
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 611
gagaagtgtg agaagc 16
<210> 612
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 612
catccttgaa gtttag 16
<210> 613
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 613
taataagatg gctccc 16
<210> 614
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 614
caaggcataa taagat 16
<210> 615
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 615
ccaaggcata ataaga 16
<210> 616
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 616
tgatccaatt ctcacc 16
<210> 617
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 617
atgatccaat tctcac 16
<210> 618
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 618
cgcttcatct tcaccc 16
<210> 619
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 619
tatgacactg catctt 16
<210> 620
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 620
gtatgacact gcatct 16
<210> 621
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 621
tgtatgacac tgcatc 16
<210> 622
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 622
ttctcttctg taagtc 16
<210> 623
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 623
ttctacagag gaacta 16
<210> 624
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 624
actacagttc tacaga 16
<210> 625
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 625
ttcccacagg taaatg 16
<210> 626
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 626
attatttgaa tatactcatt 20
<210> 627
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 627
tgggaggaaa ttatttg 17
<210> 628
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 628
tgactcatct taaatg 16
<210> 629
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 629
ctgactcatc ttaaat 16
<210> 630
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 630
tttactctga ctcatc 16
<210> 631
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 631
tattggagga attatt 16
<210> 632
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 632
gtattggagg aattat 16
<210> 633
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 633
tggtatactt ctctaagtat 20
<210> 634
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 634
gatctcttgg tatact 16
<210> 635
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 635
cagacaactc tatacc 16
<210> 636
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 636
aacatcagac aactcta 17
<210> 637
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 637
tttaacatca gacaactc 18
<210> 638
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 638
taacatcaga caactc 16
<210> 639
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 639
atttaacatc agacaa 16
<210> 640
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 640
cctatttaac atcagac 17
<210> 641
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 641
tccctattta acatca 16
<210> 642
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 642
tcaacgacta ttggaat 17
<210> 643
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 643
cttatattct ggctat 16
<210> 644
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 644
atccttatat tctggc 16
<210> 645
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 645
gatccttata ttctgg 16
<210> 646
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 646
tgatccttat attctg 16
<210> 647
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 647
attgaaactt gatcct 16
<210> 648
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 648
actgtcattg aaactt 16
<210> 649
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 649
tcttactgtc attgaa 16
<210> 650
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 650
aggatcttac tgtcatt 17
<210> 651
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 651
gcaaatcaac tccatc 16
<210> 652
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 652
gtgcaaatca actcca 16
<210> 653
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 653
caattatttc tttgtgc 17
<210> 654
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 654
tggcaacaat tatttctt 18
<210> 655
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 655
gctggcaaca attatt 16
<210> 656
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 656
atccatttct actgcc 16
<210> 657
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 657
taatatctat tgatttcta 19
<210> 658
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 658
tcaatagtgt agggca 16
<210> 659
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 659
ttcaatagtg tagggc 16
<210> 660
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 660
aggttaatta attcaatag 19
<210> 661
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 661
catttgtaat ccctag 16
<210> 662
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 662
acatttgtaa tcccta 16
<210> 663
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 663
aacatttgta atccct 16
<210> 664
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 664
taaatttcaa gttctg 16
<210> 665
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 665
gtttaaattt caagttct 18
<210> 666
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 666
ccaagtttaa atttcaag 18
<210> 667
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 667
acccaagttt aaatttc 17
<210> 668
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 668
catacagtga cccaagttt 19
<210> 669
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 669
acatcccata cagtga 16
<210> 670
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 670
agcacagctc tacatc 16
<210> 671
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 671
atatagcaca gctcta 16
<210> 672
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 672
tccatatagc acagct 16
<210> 673
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 673
atttccatat agcaca 16
<210> 674
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 674
tttatttcca tatagca 17
<210> 675
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 675
tttatttcca tatagc 16
<210> 676
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 676
aaggagagga gattatg 17
<210> 677
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 677
agttcttgtg ttagct 16
<210> 678
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 678
gagttcttgt gttagc 16
<210> 679
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 679
attaattatc catccac 17
<210> 680
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 680
atcaattaat tatccatc 18
<210> 681
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 681
agaatcaatt aattatcc 18
<210> 682
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 682
tgagataccg tgcatg 16
<210> 683
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 683
aatgagatac cgtgca 16
<210> 684
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 684
ctgtggttag gctaat 16
<210> 685
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 685
aagagtaagg gtctgtggtt 20
<210> 686
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 686
gatgggttaa gagtaa 16
<210> 687
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 687
agcagatggg ttaaga 16
<210> 688
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 688
tgtaaacatt tgtagc 16
<210> 689
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 689
cctgcttata aatgta 16
<210> 690
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 690
tgccctgctt ataaat 16
<210> 691
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 691
tcttcttagt tcaata 16
<210> 692
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 692
tggtttctaa ctacat 16
<210> 693
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 693
agtttggttt ctaacta 17
<210> 694
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 694
gaatgaaact tgcctg 16
<210> 695
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 695
attatcctta catgat 16
<210> 696
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 696
gtacccaatt atcctt 16
<210> 697
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 697
tgtacccaat tatcct 16
<210> 698
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 698
ttgtacccaa ttatcc 16
<210> 699
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 699
tttgtaccca attatc 16
<210> 700
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 700
agcagcaggt tatatt 16
<210> 701
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 701
tgggaagtgg tctggg 16
<210> 702
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 702
ctggagagtg ataata 16
<210> 703
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 703
aatgctggat tacgtc 16
<210> 704
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 704
caatgctgga ttacgt 16
<210> 705
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 705
ttgttcagaa gtatcc 16
<210> 706
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 706
gatgatttgc ttggag 16
<210> 707
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 707
gaaatcattc acaacc 16
<210> 708
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 708
ttgtaacatc tactac 16
<210> 709
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 709
cattaagcag caagtt 16
<210> 710
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 710
ttactagatg tgagca 16
<210> 711
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 711
tttactagat gtgagc 16
<210> 712
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 712
gaccaagcac cttaca 16
<210> 713
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 713
agaccaagca ccttac 16
<210> 714
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 714
atgggttaaa taaagg 16
<210> 715
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 715
tcaaccagag tattaa 16
<210> 716
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 716
gtcaaccaga gtatta 16
<210> 717
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 717
attgtaaagc tgatat 16
<210> 718
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 718
cacataattg taaagc 16
<210> 719
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 719
gaggtctgct atttac 16
<210> 720
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 720
tgtagattca atgcct 16
<210> 721
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 721
cctcattata ctatga 16
<210> 722
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 722
ccttatgcta tgacac 16
<210> 723
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 723
tccttatgct atgaca 16
<210> 724
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 724
aagatgttta agtata 16
<210> 725
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 725
ctgattatta agatgt 16
<210> 726
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 726
tggaaaggta tgaatt 16
<210> 727
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 727
acttgaatgg cttgga 16
<210> 728
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 728
aacttgaatg gcttgg 16
<210> 729
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 729
caatgtgtta ctattt 16
<210> 730
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 730
acaatgtgtt actatt 16
<210> 731
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 731
catctgctat ataaga 16
<210> 732
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 732
cctagagcaa atactt 16
<210> 733
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 733
cagagttaat aataag 16
<210> 734
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 734
gttcaagcac aacgaa 16
<210> 735
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 735
agggttcaag cacaac 16
<210> 736
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 736
tgttggagac actgtt 16
<210> 737
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 737
aaggaggagt taggac 16
<210> 738
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 738
ctatgccatt tacgat 16
<210> 739
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 739
tcaaatgcag aattag 16
<210> 740
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 740
agtgacaatc aaatgc 16
<210> 741
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 741
aagtgacaat caaatg 16
<210> 742
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 742
gtgtaccaag taacaa 16
<210> 743
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 743
tgggatgtta aactga 16
<210> 744
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 744
agtttacatt ttctgc 16
<210> 745
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 745
tatgtgaaga ggagag 16
<210> 746
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 746
cacctttaaa acccca 16
<210> 747
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 747
tcctttataa tcacac 16
<210> 748
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 748
acggtatttt cacagg 16
<210> 749
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 749
gacactacaa tgagga 16
<210> 750
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 750
tggtttttag gactgt 16
<210> 751
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 751
cgacaaattc tatcct 16
<210> 752
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 752
tgatatacaa tgctac 16
<210> 753
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 753
tcgttgggta aattta 16
<210> 754
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 754
tgctttataa atggtg 16
<210> 755
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 755
caagtttaca ttttctgc 18
<210> 756
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 756
catatgtgaa gaggagag 18
<210> 757
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 757
cacctttaaa acccca 16
<210> 758
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 758
catcctttat aatcacac 18
<210> 759
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 759
caacggtatt ttcacagg 18
<210> 760
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 760
cagacactac aatgagga 18
<210> 761
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 761
catggttttt aggactgt 18
<210> 762
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 762
cacgacaaat tctatcct 18
<210> 763
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 763
catgatatac aatgctac 18
<210> 764
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 764
catcgttggg taaattta 18
<210> 765
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 765
catgctttat aaatggtg 18
<210> 766
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 766
caacaaataa tggttactct 20
<210> 767
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 767
cacagattga tggtagtt 18
<210> 768
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 768
cacctattta acatcagac 19
<210> 769
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 769
cactaattgt agtagtactc 20
<210> 770
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 770
caataaacat gaatctctcc 20
<210> 771
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный мотив
<400> 771
tgatttaatt ctagtca 17
<210> 772
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень
<400> 772
gcagtagagc caatta 16
<210> 773
<211> 10
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 773
Phe Leu Pro Ser Asp Phe Phe Pro Ser Val
1 5 10
<210> 774
<211> 15
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 774
Gly Arg Glu Thr Val Leu Glu Tyr Leu Val Ser Phe Gly Val Trp
1 5 10 15
<210> 775
<211> 15
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 775
Thr Thr Phe His Gln Thr Leu Gln Asp Pro Arg Val Arg Gly Leu
1 5 10 15
<210> 776
<211> 16
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 776
Gln Ala Gly Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Pro Gln Ser
1 5 10 15
<210> 777
<211> 9
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 777
Phe Leu Leu Thr Arg Ile Leu Thr Ile
1 5
<210> 778
<211> 15
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 778
Thr Ser Leu Asn Phe Leu Gly Gly Thr Thr Val Cys Leu Gly Gln
1 5 10 15
<210> 779
<211> 9
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 779
Phe Leu Gly Gly Thr Thr Val Cys Leu
1 5
<210> 780
<211> 9
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 780
Trp Leu Ser Leu Leu Val Pro Phe Val
1 5
<210> 781
<211> 21
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 781
Ser Leu Leu Val Pro Phe Val Gln Trp Phe Val Gly Leu Ser Pro Thr
1 5 10 15
Val Trp Leu Ser Val
20
<210> 782
<211> 10
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 782
Gly Leu Ser Pro Thr Val Trp Leu Ser Val
1 5 10
<210> 783
<211> 10
<212> ПРТ
<213> Искусственная
<220>
<223> Эпитоп HBV
<400> 783
Ser Ile Leu Ser Pro Phe Leu Pro Leu Leu
1 5 10
<210> 784
<211> 19
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 784
ctgtgccttg ggtggcttt 19
<210> 785
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 785
aaggaaagaa gtcagaaggc aaaa 24
<210> 786
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 786
ttctttataa gggtcgatgt ccatg 25
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ОЛИГОНУКЛЕОТИДЫ ДЛЯ ПОНИЖЕНИЯ ЭКСПРЕССИИ PD-L1 | 2017 |
|
RU2835926C1 |
| МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УМЕНЬШЕНИЯ УРОВНЯ мРНК PAPD5 ИЛИ PAPD7 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИОННОГО ГЕПАТИТА В | 2017 |
|
RU2768699C2 |
| ОЛИГОНУКЛЕОТИДЫ ДЛЯ ИНДУЦИРОВАНИЯ ОТЦОВСКОЙ ЭКСПРЕССИИ UBE3A | 2016 |
|
RU2742007C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УМЕНЬШЕНИЯ УРОВНЯ мРНК PAPD5 ИЛИ PAPD7 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИОННОГО ГЕПАТИТА В | 2018 |
|
RU2766360C2 |
| КОНЪЮГАТЫ И ИХ ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2018 |
|
RU2795400C2 |
| МОДУЛЯЦИЯ ЭКСПРЕССИИ ПРЕКАЛЛИКРЕИНА (ПКК) | 2014 |
|
RU2712559C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ К АЛЬФА-СИНУКЛЕИНУ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2773197C2 |
| СОЕДИНЕНИЕ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА C9ORF72 И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2730677C2 |
| МОДИФИЦИРОВАННЫЕ ОЛИГОНУКЛЕОТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2802836C2 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ TAU | 2014 |
|
RU2745324C2 |
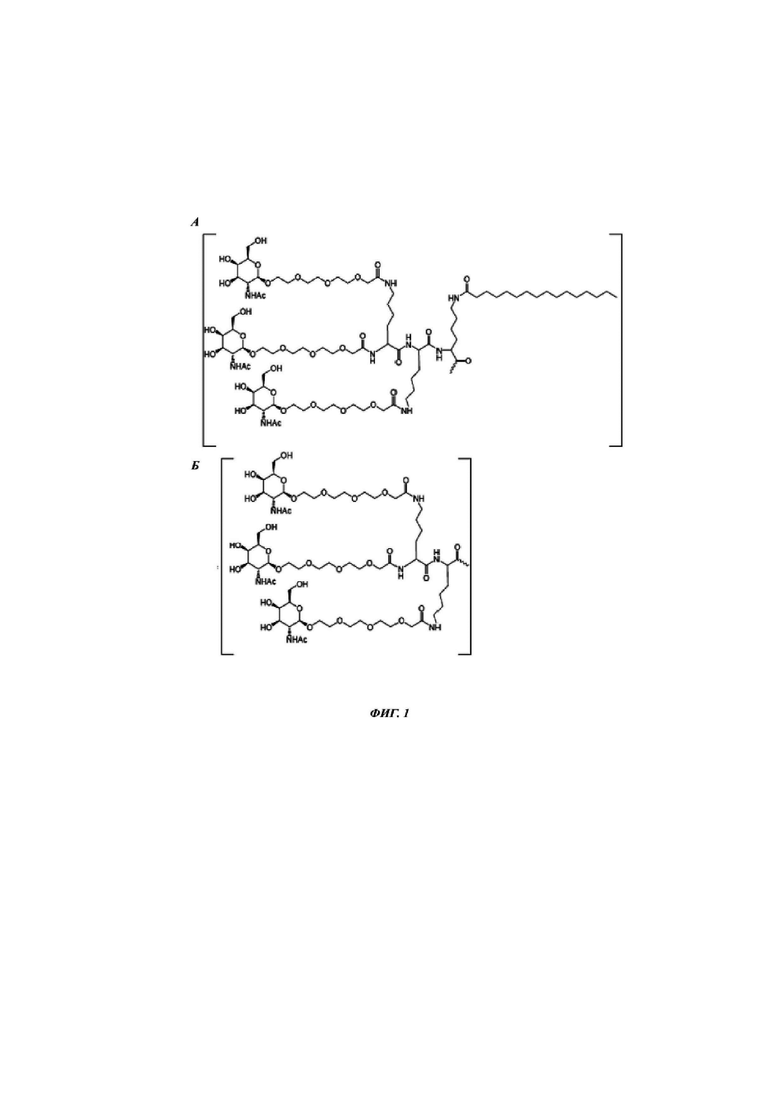
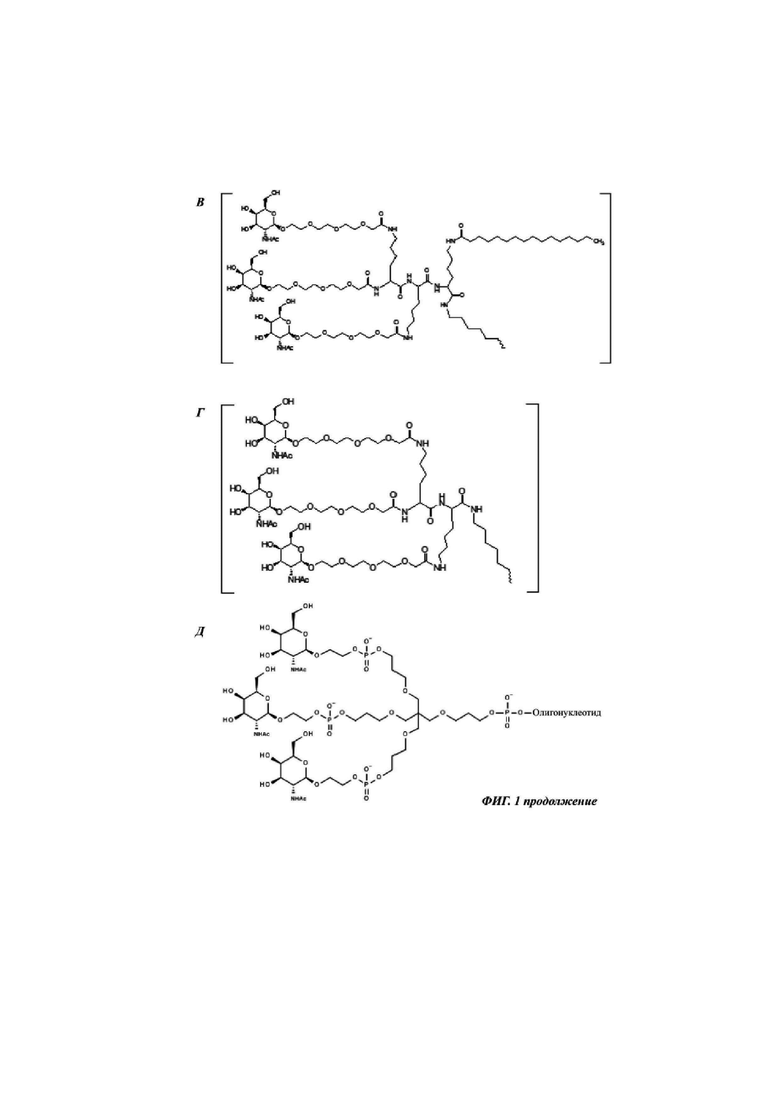
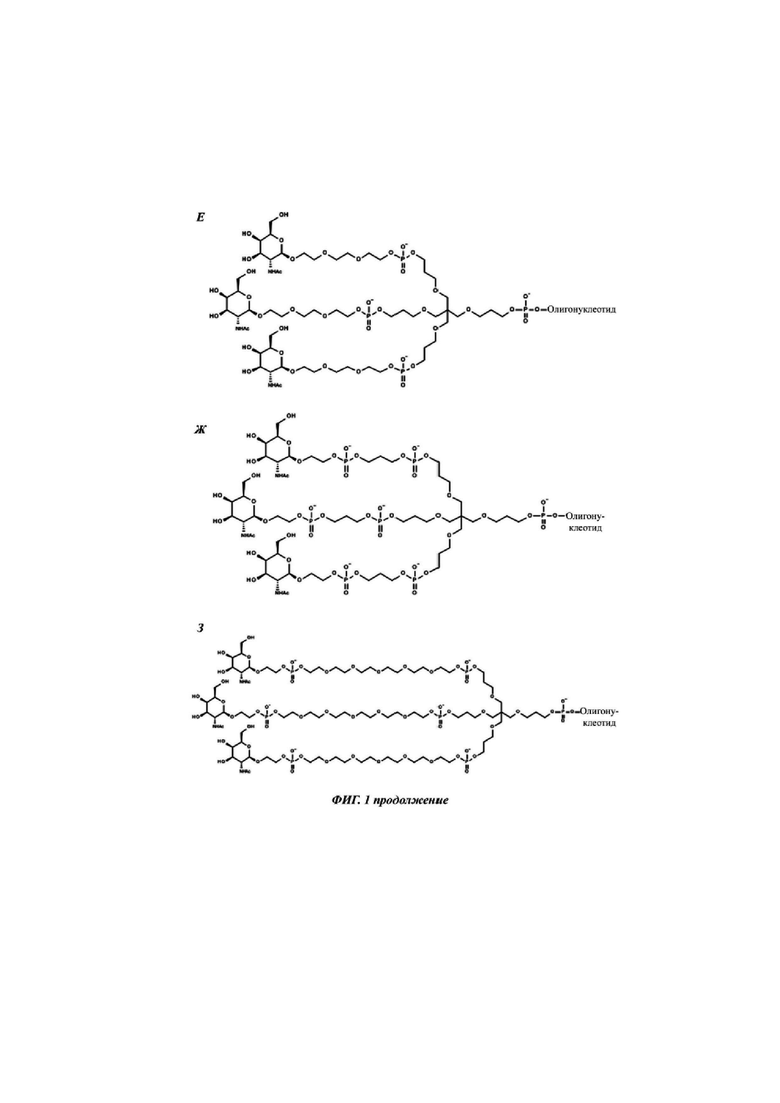
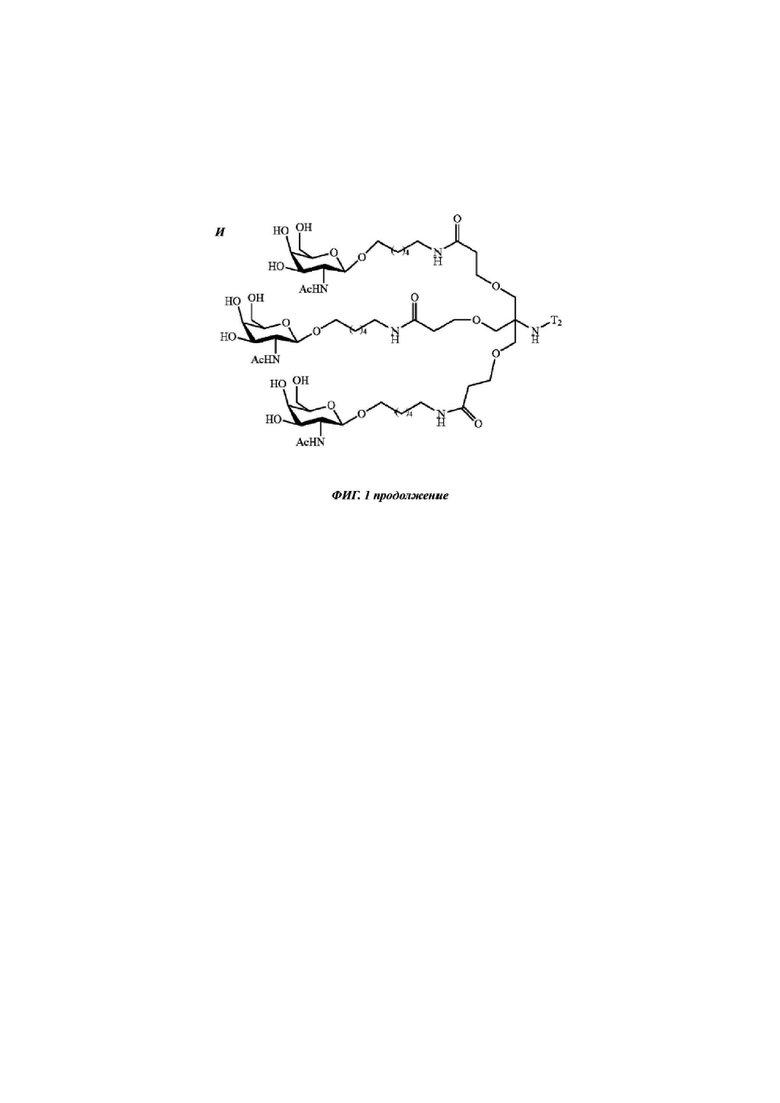

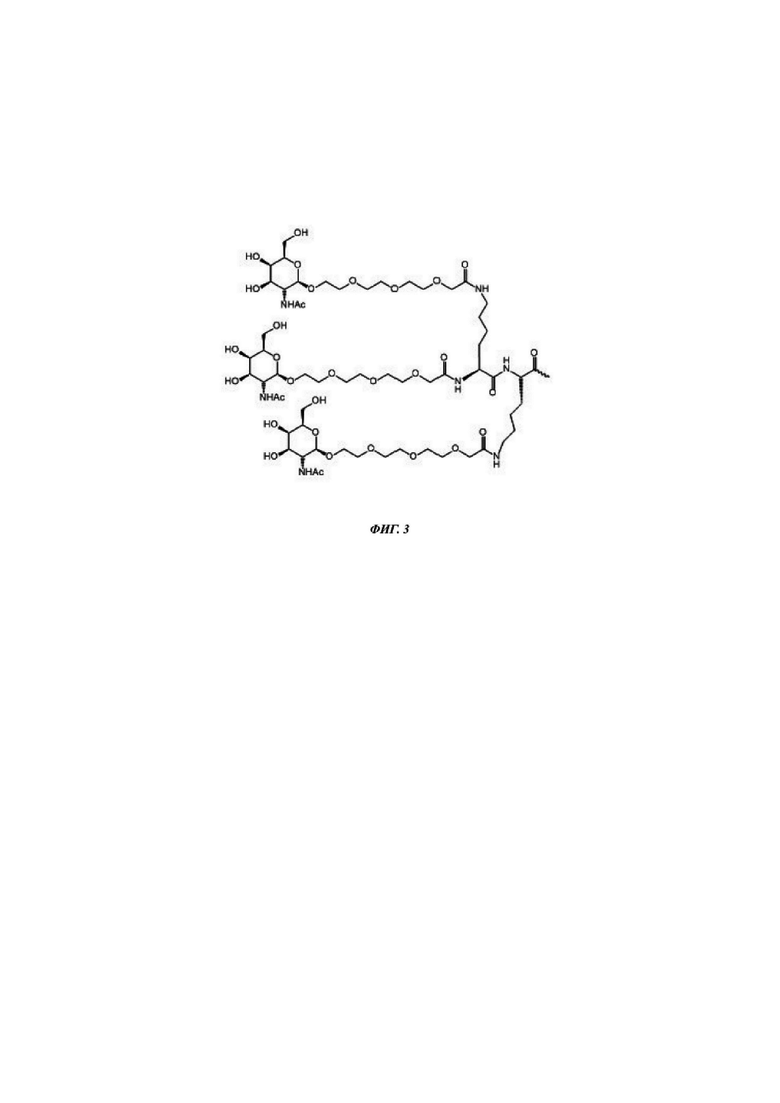
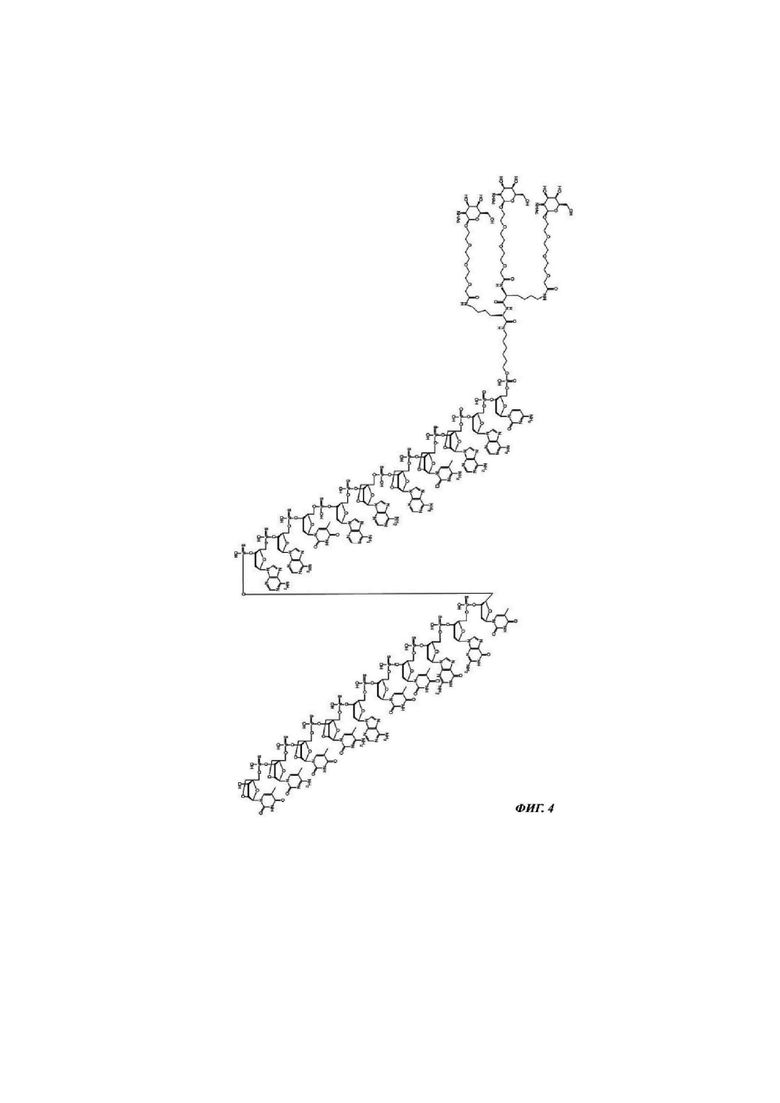
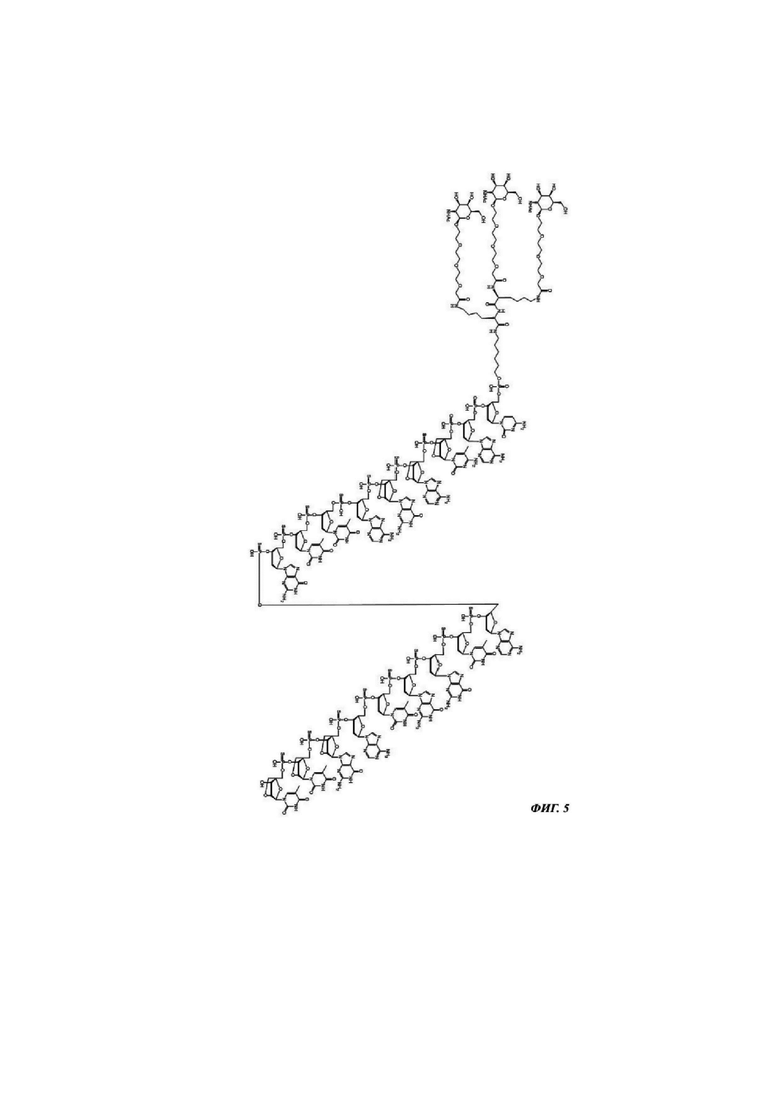
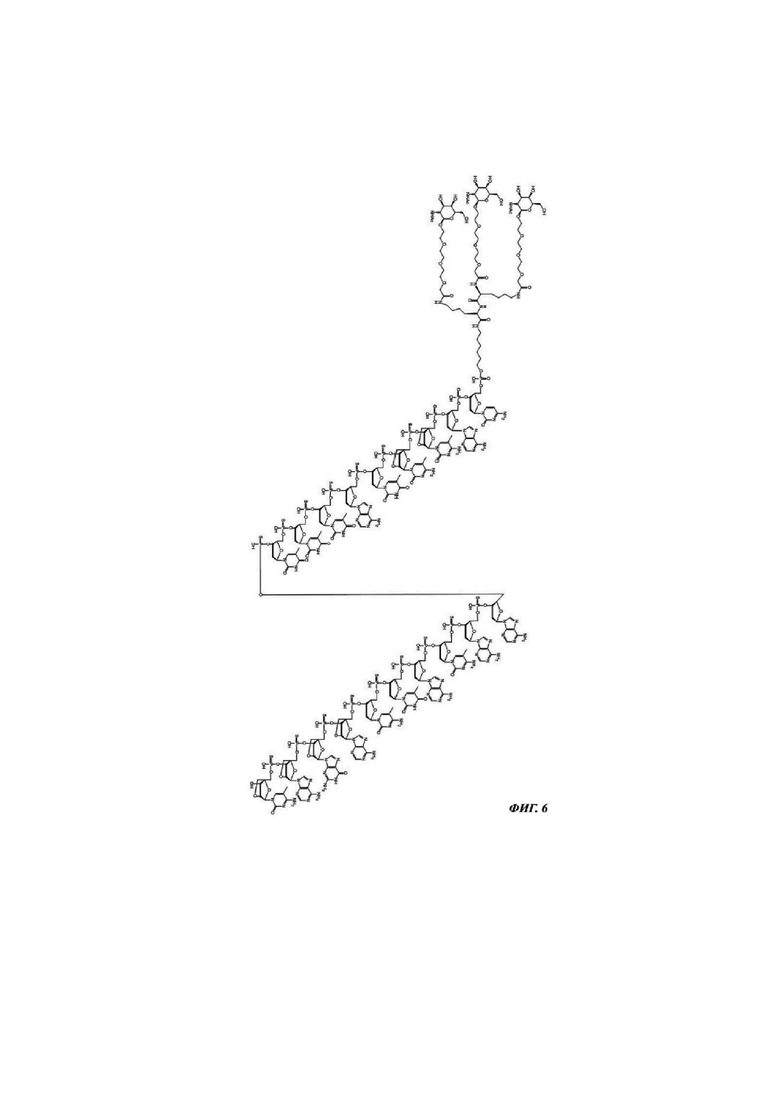
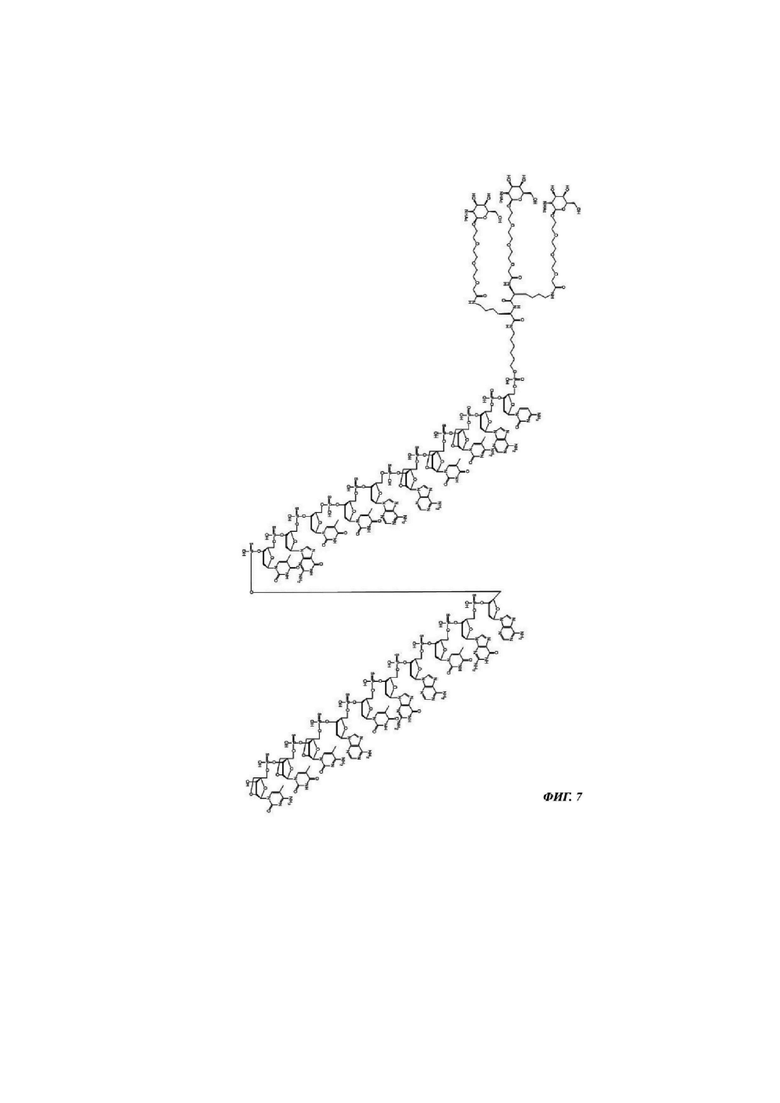
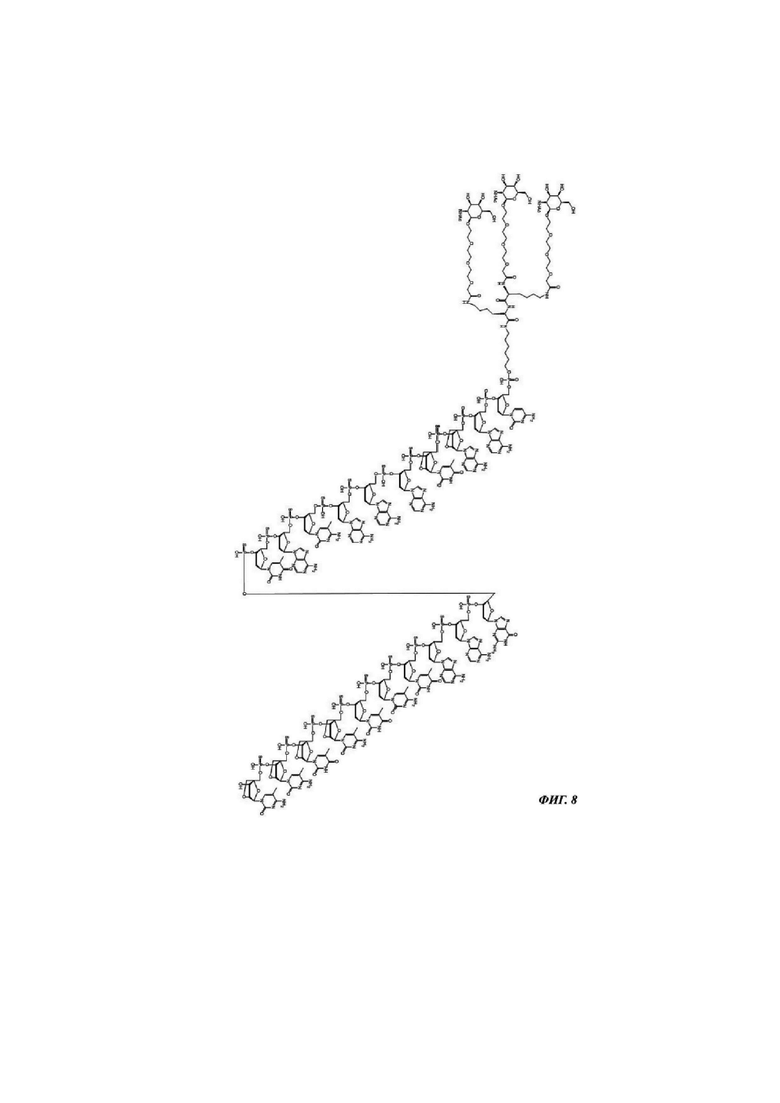
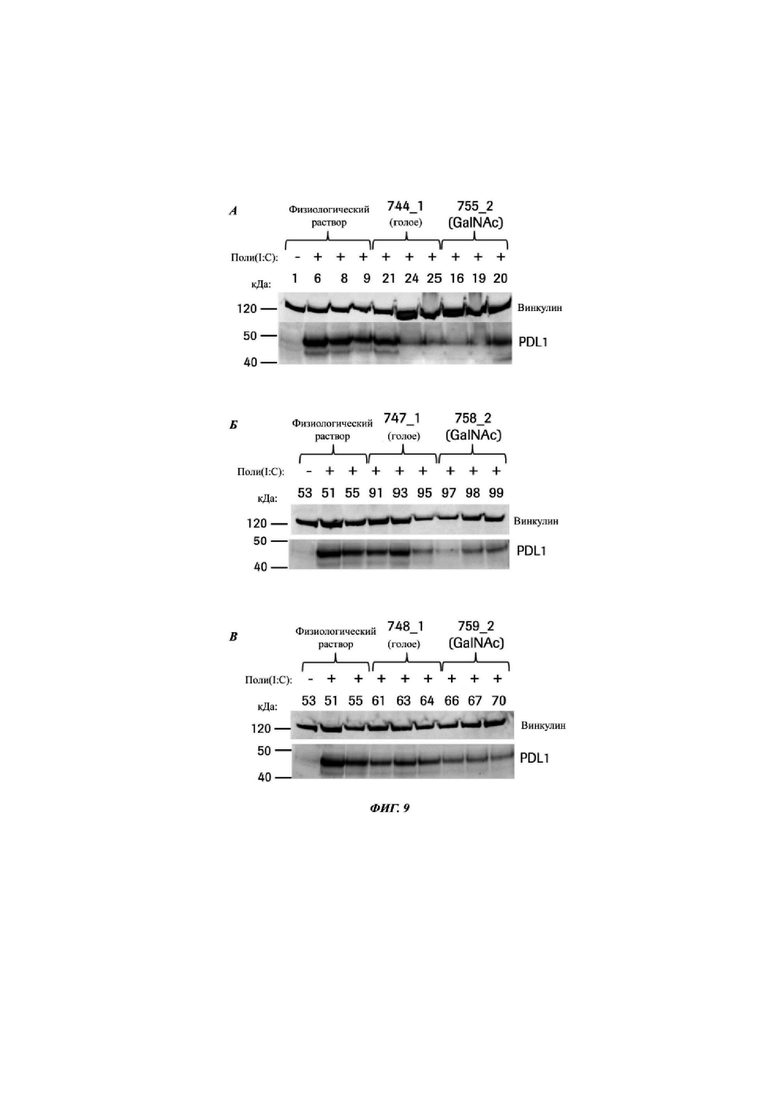
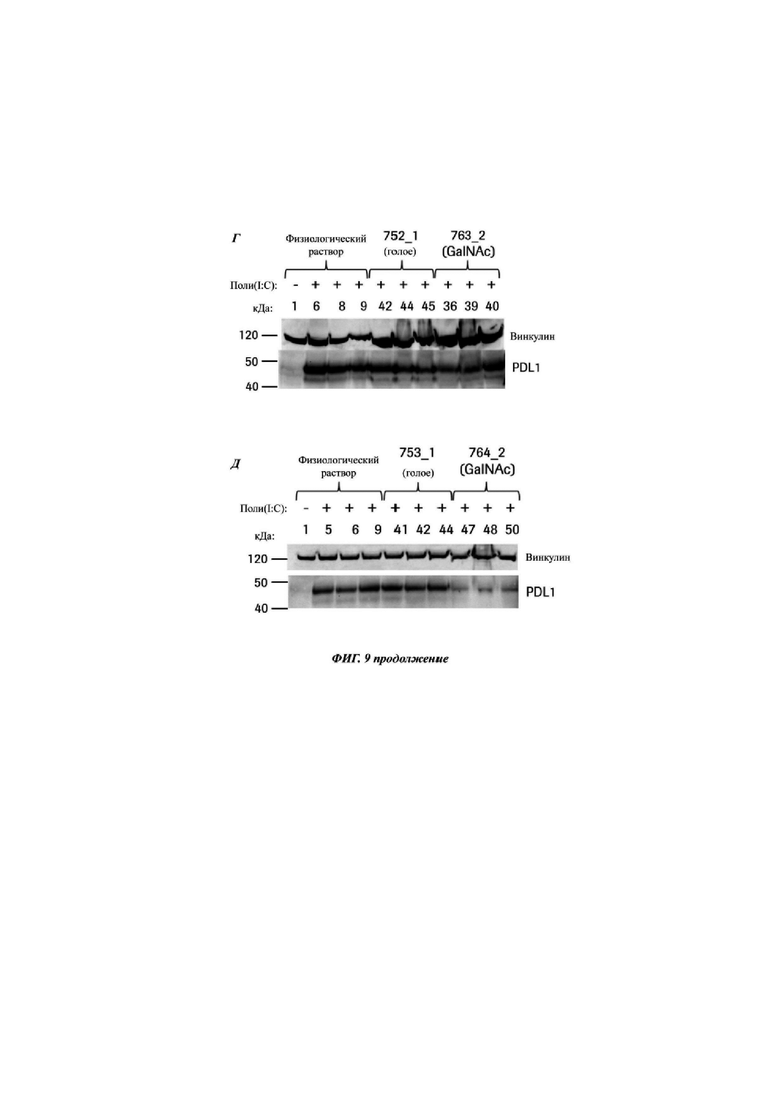
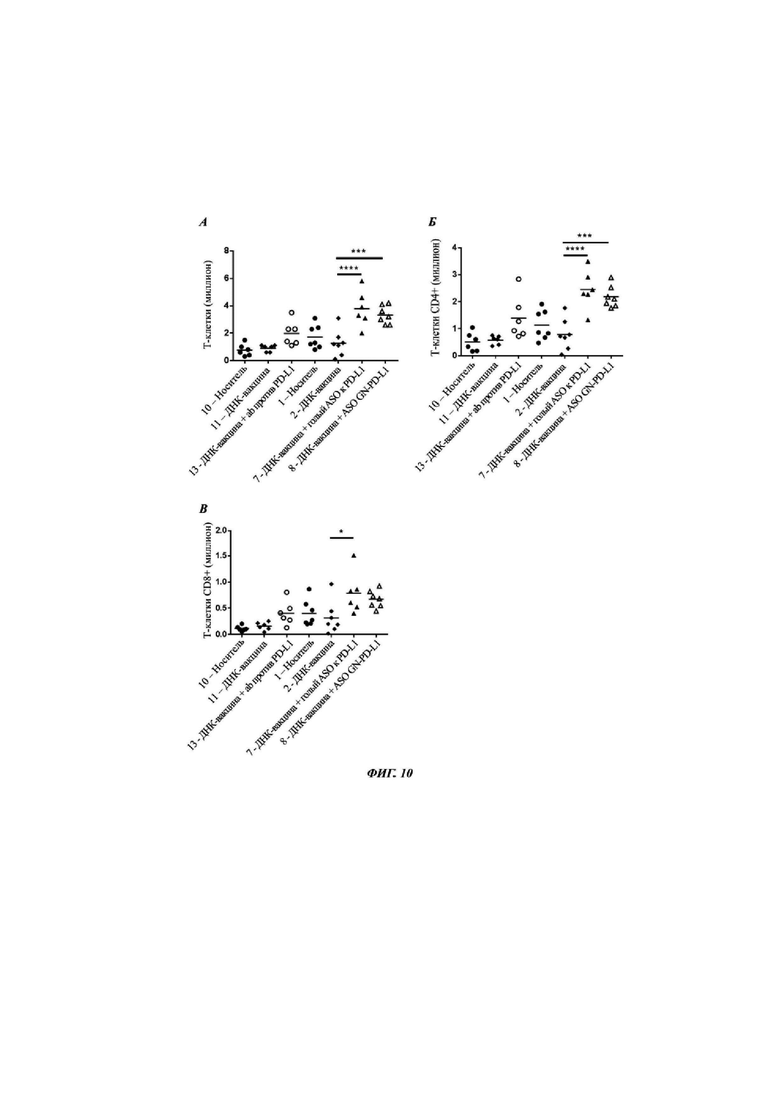
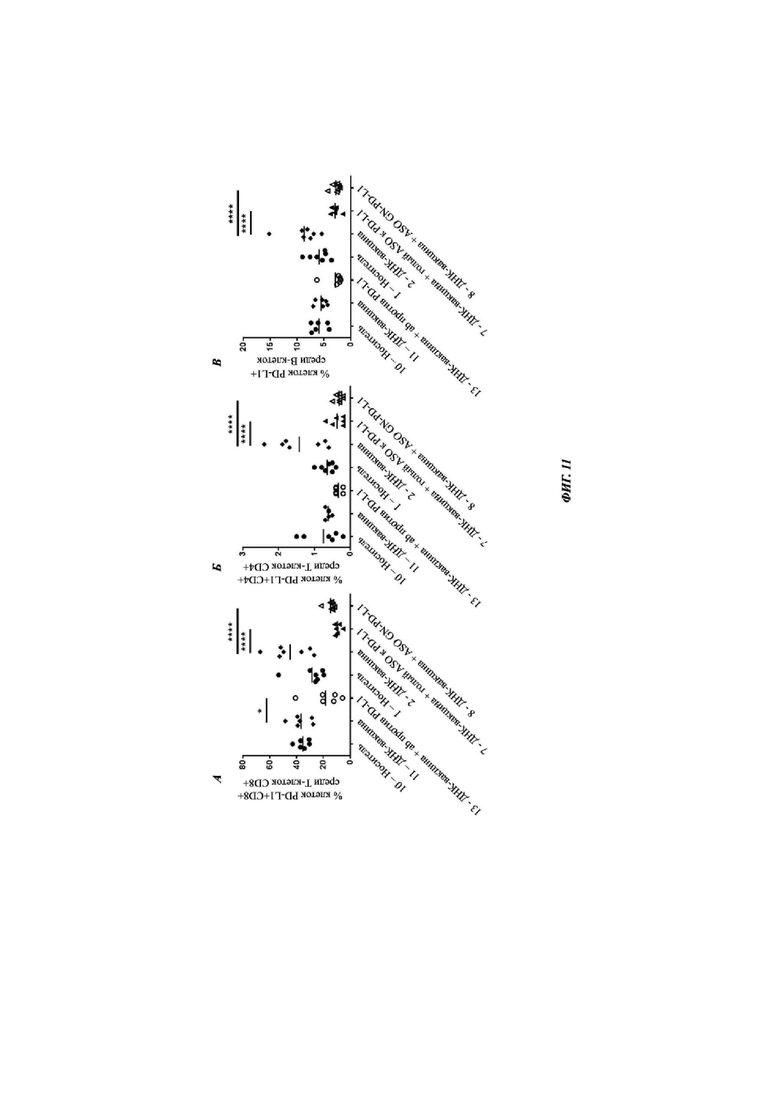
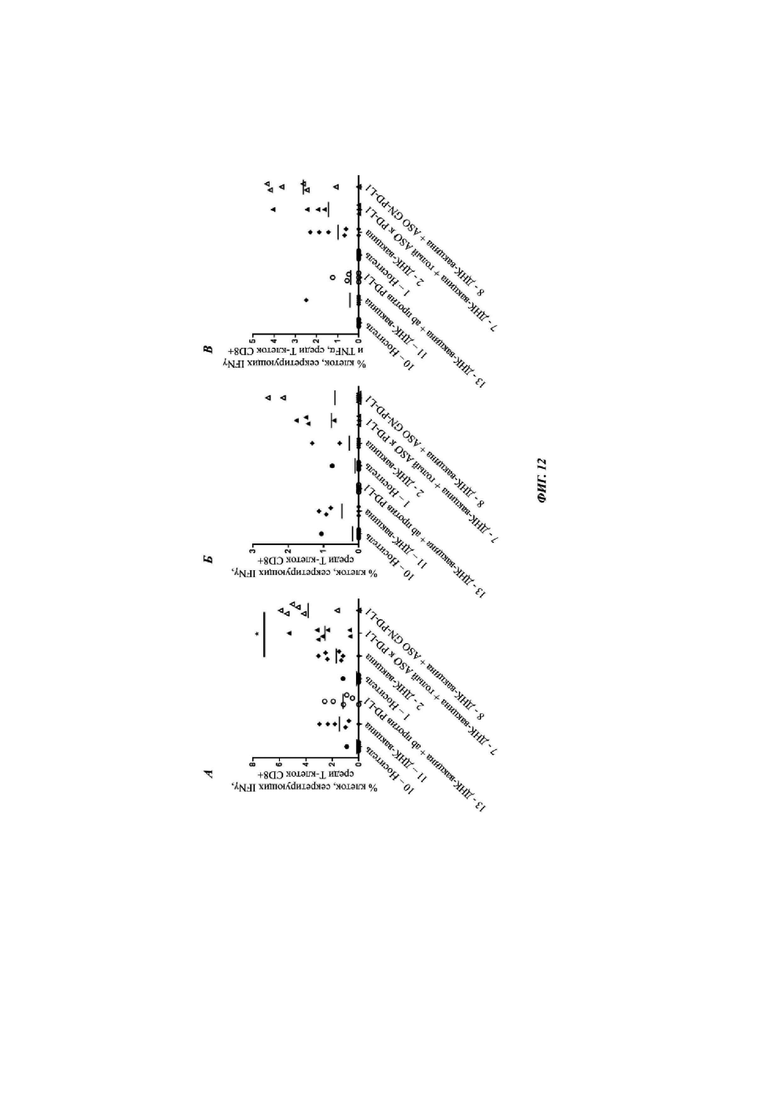
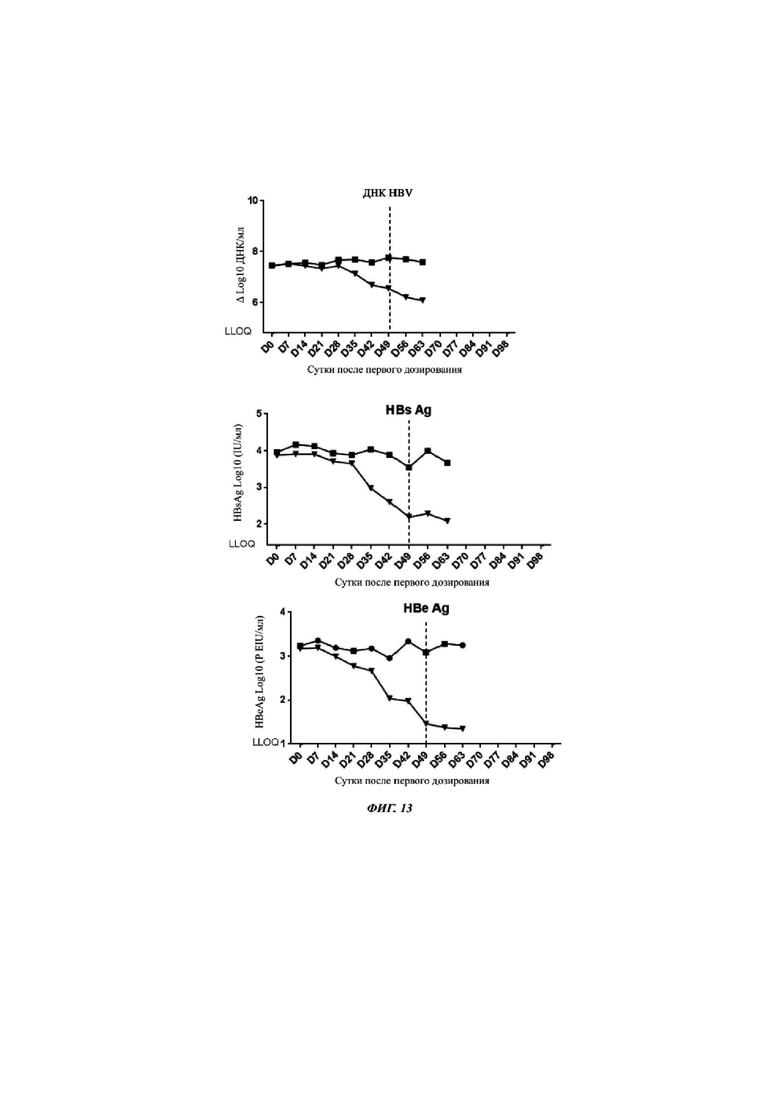
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая антисмысловой олигонуклеотид для снижения экспрессии лиганда-1 запрограммированной смерти (PD-L1), конъюгат вышеуказанного антисмыслового олигонуклеотида, содержащий конъюгатную группировку, нацеленную на рецептор асиалогликопротеина, фармацевтическую композицию для лечения или предупреждения вирусных инфекций печени, таких как инфекции HBV, HCV и HDV, паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, или рака печени или метастазов в печени, способ in vivo или in vitro снижения экспрессии PD-L1 в клетке-мишени, применение антисмыслового олигонуклеотида, конъюгата антисмыслового олигонуклеотида или фармацевтической композиции для получения лекарственного средства для лечения инфекции HBV, способ лечения или предупреждения заболевания, где указанное заболевание представляет собой вирусную инфекцию печени, такую как HBV, HCV и HDV, паразитарную инфекцию, такую как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, или рак печени или метастазы в печени, способ лечения или предупреждения инфекции HBV. Изобретение расширяет арсенал средств для снижения экспрессии PD-L1. 7 н. и 7 з.п. ф-лы, 13 ил., 24 табл., 9 пр.
1. Антисмысловой олигонуклеотид для снижения экспрессии лиганда-1 запрограммированной смерти (PD-L1), где антисмысловой олигонуклеотид имеет формулу CCtatttaacatcAGAC, где заглавные буквы представляют нуклеозиды бета-D-окси LNA, строчные буквы представляют нуклеозиды ДНК, все C LNA представляют собой 5-метилцитозин, а все межнуклеозидные связи представляют собой фосфоротиоатные межнуклеозидные связи.
2. Конъюгат антисмыслового олигонуклеотида для снижения экспрессии PD-L1, содержащий олигонуклеотид по п. 1 и конъюгатную группировку, ковалентно присоединенную к указанному олигонуклеотиду, где конъюгатная группировка представляет собой группировку, нацеленную на рецептор асиалогликопротеина, где группировка, нацеленная на рецептор асиалогликопротеина, представляет собой трехвалентную N-ацетилгалактозаминную (GalNAc) группировку.
3. Конъюгат антисмыслового олигонуклеотида по п. 2, где присутствует линкер между указанным олигонуклеотидом и указанной конъюгатной группировкой, где указанный линкер представляет собой физиологически лабильный, нуклеазочувствительный линкер, содержащий динуклеотид цитидин-аденозин.
4. Фармацевтическая композиция для лечения или предупреждения вирусных инфекций печени, таких как инфекции HBV, HCV и HDV; паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз; или рака печени или метастазов в печени, содержащая антисмысловой олигонуклеотид по п. 1 или конъюгат антисмыслового олигонуклеотида по п. 2 или 3 и фармацевтически приемлемый разбавитель, растворитель, носитель и/или адъювант.
5. Фармацевтическая композиция по п. 4, где фармацевтически приемлемый разбавитель представляет собой стерильный фосфатно-солевой буферный раствор.
6. Способ in vivo или in vitro снижения экспрессии PD-L1 в клетке-мишени, которая экспрессирует PD-L1, причем указанный способ включает введение в указанную клетку антисмыслового олигонуклеотида по п. 1, конъюгата олигонуклеотида по п. 2 или 3 или фармацевтической композиции по п. 4 или 5 в эффективном количестве.
7. Антисмысловой олигонуклеотид по п. 1, конъюгат антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтическая композиция по п. 4 или 5 для применения в восстановлении иммунного ответа против вирусных инфекций печени, таких как HBV, HCV и HDV.
8. Антисмысловой олигонуклеотид, конъюгат антисмыслового олигонуклеотида или фармацевтическая композиция для применения по п. 7, где вирус представляет собой HBV.
9. Антисмысловой олигонуклеотид по п. 1, конъюгат антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтическая композиция по п. 4 или 5 для применения в восстановлении иммунного ответа против паразитарных инфекций, таких как малярия, токсоплазмоз, лейшманиоз и трипаносомоз.
10. Антисмысловой олигонуклеотид, конъюгат антисмыслового олигонуклеотида или фармацевтическая композиция для применения по любому из пп. 7-9, где восстановление иммунного ответа представляет собой увеличение в печени уровня Т-клеток CD8+, специфичных в отношении одного или более чем одного антигена HBV, по сравнению с контролем.
11. Антисмысловой олигонуклеотид по п. 1, конъюгат антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтическая композиция по п. 4 или 5 для применения в лечении инфекции HBV.
12. Применение антисмыслового олигонуклеотида по п. 1, конъюгата антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтической композиции по п. 4 или 5 для получения лекарственного средства для лечения инфекции HBV.
13. Способ лечения или предупреждения заболевания, где указанное заболевание представляет собой вирусную инфекцию печени, такую как HBV, HCV и HDV, паразитарную инфекцию, такую как малярия, токсоплазмоз, лейшманиоз и трипаносомоз, или рак печени или метастазы в печени, включающий введение терапевтически или профилактически эффективного количества антисмыслового олигонуклеотида по п. 1, конъюгата антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтической композиции по п. 4 или 5 субъекту, страдающему от или восприимчивому к заболеванию.
14. Способ лечения или предупреждения инфекции HBV, включающий введение терапевтически или профилактически эффективного количества антисмыслового олигонуклеотида по п. 1, конъюгата антисмыслового олигонуклеотида по п. 2 или 3 или фармацевтической композиции по п. 4 или 5 субъекту, страдающему от или восприимчивому к инфекции HBV.
| WO 2011127180 A1, 13.10.2011 | |||
| WO 2014118272 A1, 07.08.2014 | |||
| WO 2015114146, 06.08.2015 | |||
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВ HCV, ПРИГОДНЫХ ДЛЯ ИСПОЛЬЗОВАНИЯ В ВАКЦИНЕ ИЛИ ИММУНОАНАЛИЗЕ, АСИАЛОГЛИКОПРОТЕИН (ВАРИАНТЫ), КОМПОЗИЦИЯ ДЛЯ ИСПОЛЬЗОВАНИЯ В ВАКЦИНЕ ИЛИ В ИММУНОАНАЛИЗЕ (ВАРИАНТЫ), СПОСОБ ОЧИСТКИ АСИАЛОГЛИКОПРОТЕИНА И СПОСОБ ПОНИЖЕНИЯ СОДЕРЖАНИЯ ИЛИ ЭЛИМИНАЦИИ HCV | 1991 |
|
RU2123528C1 |

