Перечень последовательностей
Настоящая заявка подается вместе с перечнем последовательностей в электронном формате. Перечень последовательностей представлен в виде файла с названием BIOL0211WOSEQ.txt, созданного 14 октября 2013 г., размером 184 Кб. Данные в электронном формате для перечня последовательностей включены в данный документ в полном объеме посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Предложены композиции и способы снижения экспрессии мРНК и белка C9ORF72 у животных. Такие способы пригодны для лечения, предупреждения или облегчения протекания нейродегенеративных заболеваний, включающих амиотрофический латеральный склероз (АЛС), лобно-височную деменцию (ЛВД), синдром кортикобазальной дегенерации (КБД), атипичный синдром паркинсонизма и оливопонтоцеребеллярную дегенерацию (ОПЦД).
УРОВЕНЬ ТЕХНИКИ
Амиотрофический латеральный склероз (АЛС) представляет собой нейродегенеративное заболевание, заканчивающееся смертельным исходом, с характерной клинической картиной прогрессирующего паралича, приводящего к смерти от остановки дыхания, как правило, в период от двух до трех лет после появления симптомов (Rowland and Shneider, N. Engl. J. Med., 2001, 344, 1688-1700). АЛС является третьим наиболее распространенным нейродегенеративным заболеванием западного мира (Hirtz et al., Neurology, 2007, 68, 326-337) и в настоящее время для него отсутствуют эффективные способы лечения. Приблизительно 10% случаев в природе являются наследственными, тогда как подавляющее количество пациентов с данным диагностированным заболеванием классифицируются как спорадически заболевшие, поскольку заболевание у них возникает случайным образом в пределах всей популяции (Chio et al., Neurology, 2008, 70, 533-537). На основе клинических, генетических и эпидемиологических данных становится понятно, что АЛС и лобно-височная деменция (ЛВД) представляют собой перекрывающееся пространство общего заболевания, характеризующегося патологическим процессом из-за наличия положительных включений TDP-43 во всей центральной нервной системе (Lillo and Hodges, J. Clin. Neurosci., 2009, 16, 1131-1135; Neumann et al., Science, 2006, 314, 130-133).
К настоящему времени обнаружили несколько генов, являющихся причиной классического наследственного АЛС, например, SOD1, TARDBP, FUS, OPTN и VCP (Johnson et al., Neuron, 2010,68, 857-864; Kwiatkowski et al., Science, 2009, 323, 1205-1208; Maruyama et al., Nature, 2010, 465, 223-226; Rosen et al., Nature, 1993, 362, 59-62; Sreedharan et al., Science, 2008, 319, 1668-1672; Vance et al., Brain, 2009, 129, 868-876). Проведенный недавно анализ сцепления между генными локусами родственников с многочисленными случаями заболеваний АЛС, ЛВД и АЛС-ЛВД показал, что существует важный локус этого заболевания на коротком плече хромосомы 9 (Boxer et al., J. Neurol. Neurosurg. Psychiatry, 2011, 82, 196-203; Morita et al., Neurology, 2006, 66, 839-844; Pearson et al. J. Nerol., 2011, 258, 647-655; Vance et al., Brain, 2006, 129, 868-876). Это локус хромосомы 9р21АЛС-ЛВД расположен в последнем главном аутосомально-доминантном гене, мутация которого вызывает АЛС. Мутация, вызывающая АЛС-ЛВД, представляет собой большую экспансию гексануклеотидного (GGGGCC) повтора в первом интроне гена C9ORF72 (Renton et al., Neuron, 2011, 72, 257-268; DeJesus-Hernandez et al., Neuron, 2011, 72, 245-256). Основной гаплотип, включающий ген C9ORF72, представлен в большинстве случаев мутаций, связанных с этим участком (Renton et al., Neuron, 2011, 72, 257-268). Этот локус на хромосоме 9р21 отвечает почти за половину случаев наследственного АЛС и почти за одну четверть всех случаев возникновения АЛС в группе из 405 пациентов из Финляндии (Laaksovirta et al, Lancet Neurol., 2010, 9, 978-985).
Основной гаплотип, включающий ген C9ORF72, присутствует в большинстве случаев мутаций, связанных с эти участком.
В настоящее время не существует эффективных способов терапии для лечения таких нейродегенеративных заболеваний, поэтому целью настоящей заявки является предоставление композиций и способов для лечения таких нейродегенеративных заболеваний.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В данном документе представлены композиции и способы модуляции уровней мРНК и белка C9ORF72 в клетках, тканях и в организмах животных. В некоторых вариантах реализации изобретения специфические ингибиторы C9ORF72 модулируют экспрессию мРНК и белка C9ORF72. В некоторых вариантах реализации изобретения специфические ингибиторы C9ORF72 являются нуклеиновыми кислотами, белками или малыми молекулами.
В некоторых вариантах реализации изобретения модуляция может происходить в клетке или ткани. В некоторых вариантах реализации изобретения эта клетка или ткань находится в организме животного. В некоторых вариантах реализации изобретения это животное является человеком. В некоторых вариантах реализации изобретения снижаются уровни мРНК C9ORF72. В некоторых вариантах реализации изобретения снижаются уровни белка C9ORF72. В некоторых вариантах реализации изобретения преимущественно снижаются уровни определенных вариантов мРНК C9ORF72. В некоторых вариантах реализации изобретения преимущественно снижаемые уровни вариантов мРНК C9ORF72 являются вариантами, содержащими интрон 1. В некоторых вариантах реализации изобретения интрон 1 содержит экспансию гексануклеотидного повтора. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора связана с заболеванием, ассоциированным с белком C9ORF72. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора связана с заболеванием, ассоциированным с экспансией гексануклеотидного повтора C9ORF72. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора содержит по меньшей мере 30 повторов GGGGCC. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора связана с заболеванием, ассоциированным с фокусами ядер клеток. В некоторых вариантах реализации изобретения композиции и способы, описанные в данном документе, пригодны для снижения уровней мРНК C9ORF72, уровней белка C9ORF72 и уменьшения фокусов ядер клеток. Такое снижение может происходить времязависимым способом или дозозависимым способом.
Предложены также способы, пригодные для предупреждения, лечения и ослабления протекания заболеваний, нарушений и состояний, ассоциированных с белком C9ORF72. В некоторых вариантах реализации изобретения такие заболевание, нарушения и состояния, ассоциированные с белком C9ORF72, являются нейродегенеративными заболеваниями. В некоторых вариантах реализации изобретения нейродегенеративное заболевание представляет собой амиотрофический латеральный склероз (АЛС), лобно-височную деменцию (ЛВД), синдром кортикобазальной дегенерации (КБД), атипичный синдром паркинсонизма и оливопонтоцеребеллярную дегенерацию (ОПЦД).
Такие заболевания, нарушения и состояния могут в целом сопровождаться одним или более факторами риска, причинами и исходами заболевания. Определенные факторы риска и причины развития нейродегенеративного заболевания и, в частности, АЛС и ЛВД, включают генетическую предрасположенность и пожилой возраст.
В некоторых вариантах реализации изобретения способы лечения включают введение индивиду, нуждающемуся в этом, специфического ингибитора C9ORF72. В некоторых вариантах реализации изобретения специфический ингибитор C9ORF72 является нуклеиновой кислотой. В некоторых вариантах реализации изобретения нуклеиновая кислота является антисмысловым соединением. В некоторых вариантах реализации изобретения антисмысловое соединение является одноцепочечным антисмысловым олигонуклеотидом. В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид комплементарен нуклеиновой кислоте C9ORF72.
ПОДРОБНОЕ ОПИСАНИЕ
Должно быть понятно, что как предыдущее общее описание, так и последующее подробное описание сущности изобретения приведены только в качестве примеров и объяснений и не ограничивают данное изобретение, указанное в формуле. В данном документе использование единственного числа включает множественное, если прямо не указано что-либо другое. В данном документе считается, что использование слова «или» обозначает «и/или», если не указано что-либо другое. Кроме того, в данном документе принимается во внимание, что использование слова «и» обозначает «и/или», если не указано что-либо другое. Более того, использование термина «включающий», а также других форм, таких как «включает» и «включенный», не является ограничивающим. Также такие термины, как «элемент» или «компонент» охватывают как элементы, так и компоненты, содержащие одну составляющую, и элементы и компоненты, содержащие более одной подсоставляющей, если прямо не указано что-либо другое.
Заголовки разделов, использованные в данном документе, применены только с целью упорядочивания текста и не должны истолковываться как ограничивающие описанный объект изобретения. Все документы или части документов, цитируемые в этом описании, включая без ограничения патенты, заявки на патенты, опубликованные заявки на патенты, статьи, книги, исследования, номера доступа к базе данных GENBANK и связанную информацию о последовательностях, которую можно получить из баз данных, таких какНациональный центр биотехнологической информации (NCBI) и других данных, относящихся ко всему описанию данного документа, намеренно включены посредством ссылок для частей обсуждаемого здесь документа, а также в их полном объеме.
Определения
Если конкретные определения не предусмотрены, то данная номенклатура используется в связи с, а также в пределах процедур и методик аналитической химии, синтетической органической химии, медицинской и фармацевтической химии, описанных в данном документе, является хорошо известной и обычно используемой в данной области техники. Для химического синтеза и химического анализа могут быть использованы стандартные методики.
Если не указано иное, то указанные ниже термины имеют следующие значения:
«2'-O-метоксиэтильная группа» (также обозначаемая 2'-МОЕ и 2'-ОСН2СН2-ОСН3 и МОЕ) относится к О-метоксиэтил-модификации положения 2' фуранозильного кольца. Модифицированный 2'-O-метоксиэтильный сахар является модифицированным сахаром.
«2'-МОЕ нуклеозид» (также обозначаемый 2'-O-метоксиэтилнуклеозидом) обозначает нуклеозид, содержащий 2'-O-метоксиэтильную группу.
«5-метилцитозин» обозначает цитозин, модифицированный метальной группой, присоединенной к положению 5'. 5-метилцитозин является модифицированным нуклеотидным основанием.
«Около» обозначает нахождение в пределах значения ±7%. Например, если утверждается, что «соединение приводит к по меньшей мере около 70% ингибированию C9ORF72», следовательно это подразумевает, что уровни C9ORF72 ингибированы в пределах диапазона от 63 до 77%.
«Совместное введение» относится к совместному введению двух фармацевтических агентов любым способом, при котором фармацевтическое действие обоих агентов проявляется у пациента одновременно. Совместное введение не требует, чтобы оба фармацевтических агента вводились в одной фармацевтической композиции, в одной и той же лекарственной форме или одинаковым путем. Не требуется, чтобы действие обоих фармацевтических агентов проявляло себя в одно и то же время. Эффекты должно быть перекрывающимися только на некоторый период времени и не должны быть одинаковыми по продолжительности.
«Введение» обозначает поступление фармацевтического агента животному и включает, без ограничения, введение медицинским работником или самостоятельное введение.
Термины «облегчение», или «облегчать», или «облегчающий», относятся к уменьшению по меньшей мере одного показателя, признака или симптома заболевания, нарушения или состояния. Тяжесть показателей может определяться путем субъективных или объективных определений, которые известны специалистам в данной области.
«Животное» относится к человеку или животному, не являющемуся человеком, включая, без ограничения: мышей, крыс, кроликов, собак, кошек, свиней и приматов кроме человека, включая без ограничения обезьян и шимпанзе.
«Антитело» относится к молекуле, для которой характерна способность определенным образом специфически взаимодействовать с антигеном, при этом антитело и антиген определяются относительно друг друга. Обозначение «антитело» может относиться к полной молекуле антитела или к его любому фрагменту или к участку, такому, как тяжелая цепь, легкая цепь, Fab-участок или Fc-участок.
«Антисмысловая активность» обозначает любую доступную для обнаружения или измерения активность, которую можно отнести к гибридизации антисмыслового соединения с его нуклеиновой кислотой-мишенью. В некоторых вариантах реализации изобретения антисмысловая активность представляет снижение количества или экспрессии нуклеиновой кислоты-мишени или белка, кодируемого такой нуклеиновой кислотой-мишенью.
«Антисмысловое вещество» обозначает олигомерное соединение, которое способно претерпевать гибридизацию с нуклеиновой кислотой-мишенью с помощью водородных связей. Примеры антисмысловых соединений включают одноцепочечные и двухцепочечные соединения, такие как антисмысловые олигонуклеотиды, миРНК, мшРНК, оцРНК и вещества, действующие на основе захватывания. Антисмысловые механизмы включают, без ограничения, опосредованный РНКазой H антисмысловой механизм; механизмы интерференции РНК (RNAi), которые используют пути RISC и включают, без ограничения, механизмы с участием миРНК, оцРНК и микроРНК; и механизмы на основе захватывания, включающие, без ограничения, однотипно модифицированные олигонуклеотиды. Определенные антисмысловые соединения могут действовать посредством более, чем одного такого механизма и/или посредством дополнительных механизмов.
«Антисмысловое ингибирование» обозначает снижение уровней нуклеиновой кислоты-мишени или уровней белка-мишени в присутствии антисмыслового соединения комплементарного нуклеиновой кислоте-мишени по сравнению с уровнями нуклеиновой кислоты-мишени или уровнями белка-мишени в отсутствии антисмыслового соединения. Ингибирование может быть обеспечено любыми способами, включая деградацию РНКазой Н, такую как с помощью гэпмера, и стерическую блокировку, такую как с помощью однотипно модифицированного олигонуклеотида.
«Антисмысловой олигонуклеотид» обозначает одноцепочечный олигонуклеотид с последовательностью нуклеотидных оснований, которая допускает гибридизацию с соответствующим сегментом нуклеиновой кислоты-мишени.
«Бициклический сахар» обозначает фуранозильное кольцо, модифицированное образованием мостика между двумя атомами. Бициклический сахар является модифицированным сахаром.
«Бициклический нуклеозид» (также называемый BNA - бициклической нуклеиновой кислотой) обозначает нуклеозид, имеющий сахарный фрагмент, содержащий мостиковое соединение двух атомов углерода сахарного кольца, благодаря чему образуется бициклическая кольцевая система. В некоторых вариантах реализации изобретения этот мостик соединяет 4'-углерод и 2'-углерод сахарного кольца.
«Ассоциированное с C9ORF72 заболевание» обозначает любое заболевание, ассоциированное с любой нуклеиновой кислотой C9ORF72 или продуктом ее экспрессии. Такие заболевания могут включать нейродегенеративное заболевание. Такие нейродегенеративные заболевания могут включать АЛС и ЛВД.
«Ассоциированное с экспансией гексануклеотидного повтора C9ORF72 заболевание» обозначает любое заболевание, связанное нуклеиновой кислотой C9ORF72, содержащей экспансию гексануклеотидного повтора. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора может содержать последовательности GGGGCC, GGGGGG, GGGGGC или GGGGCG, повторяющиеся по меньшей мере 30 раз. Такие заболевания могут включать нейродегенеративное заболевание. Такие нейродегенеративные заболевания могут включать АЛС и ЛВД.
«Нуклеиновая кислота C9ORF72» означает любую нуклеиновую кислоту, кодирующую последовательность C9ORF72. Например, в некоторых вариантах реализации изобретения нуклеиновая кислота C9ORF72 содержит последовательность ДНК, кодирующую C9ORF72, последовательность РНК, транскрибирующуюся с ДНК, кодирующей C9ORF72 (включая геномную ДНК, содержащую интроны и экзоны), и последовательность мРНК, кодирующую C9ORF72. «мРНК C9ORF72» означает мРНК, кодирующую белок C9ORF72.
«Специфический ингибитор C9ORF72» относится к любому агенту, способному специфически ингибировать экспрессию мРНК C9ORF72 и/или белка C9ORF72 на молекулярном уровне. Например, специфические ингибиторы C9ORF72 включают нуклеиновые кислоты (включая антисмысловые соединения), миРНК, аптамеры, антитела, пептиды, малые молекулы и другие агенты, способные ингибировать экспрессию мРНК C9ORF72 и/или белка C9ORF72. Аналогичным образом в некоторых вариантах реализации изобретения специфические ингибиторы C9ORF72 могут влиять на другие молекулярные процессы в организме животного.
«Кэп-структура», или «концевой кэп-фрагмент», обозначает химические модификации, которые были внедрены в какой-либо из концов антисмыслового соединения.
«cEt», или «стерически затрудненный этил» обозначает бициклический нуклеозид, несущий сахарный фрагмент, содержащий мостиковое соединение 4'-углерода и 2'-углерода, при этом данный мостик имеет формулу: 4'-СН(CH3)-O-2'.
«Стерически затрудненный этилнуклеозид» (также обозначаемый cEt-нуклеозид) означает нуклеозид, содержащий бициклический сахарный фрагмент, включающий мостик 4'-СН(СН3)-O-2'.
«Химически отличающийся участок» относится к участку антисмыслового соединения, который определенным образом химически отличается от другого участка того же антисмыслового соединения. Например, участок, несущий 2'-O-метоксиэтилнуклеозиды, химически отличается от участка, несущего нуклеозиды без 2'-O-метоксиэтил модификаций.
«Химерное антисмысловое соединение» обозначает антисмысловое соединение, которое имеет по меньшей мере два химически отличающихся участка.
«Совместное введение» обозначает введение индивиду двух и более фармацевтических агентов. Эти два или более фармацевтических агентов могут быть представлены в виде одной фармацевтической композиции или могут быть представлены в виде отдельных фармацевтических композиций. Каждый из двух и более фармацевтических агентов могут вводиться посредством одного пути или разными путями введения. Совместное введение охватывает параллельное или последовательное введение.
«Комплементарность» обозначает способность к спариванию между нуклеотидными основаниями первой нуклеиновой кислоты и второй нуклеиновой кислоты.
Термин «последовательные нуклеотидные основания» обозначает нуклеотидные основания, непосредственно соседствующие друг с другом.
«Растворитель» обозначает ингредиент композиции, который не обладает фармакологической активностью, но является фармацевтически необходимым или желательным. В качестве примера, растворитель в композиции для инъекций может быть жидкостью, например, солевой раствор.
«Доза» обозначает установленное количество фармацевтического агента, доставляемого за одно введение или за установленный период времени. В некоторых вариантах реализации изобретения доза может быть введена с одним, двумя или более болюсными введениями, таблетками или инъекциями. Например, в некоторых вариантах реализации изобретения, когда требуется подкожное введение, а нужная доза должна быть в таком объеме, который не может быть легко достигнут посредством одной инъекции, в таком случае могут использоваться две или более инъекций для достижения необходимых доз. В некоторых вариантах реализации изобретения данный фармацевтический агент вводят путем инфузии в течение продолжительного периода времени или непрерывно. Дозы могут быть заданы как количество фармацевтического агента за час, день, неделю или месяц.
«Эффективное количество» обозначает количество фармацевтического агента, достаточное для того, чтобы вызвать требуемый физиологический результат у индивида, нуждающегося в данном фармацевтическом агенте. Эффективное количество может отличаться у индивидов в зависимости от здоровья и физического состояния индивида, требующего лечения, таксономической группы индивидов, требующих лечения, лекарственной формы композиции, оценки медицинского состояния индивида и других сопутствующих факторов.
«Экспрессия» означает преобразование информации гена C9ORF72 в мРНК посредством транскрипции и затем в белок посредством трансляции. Экспрессия может привести к фенотипическому проявлению гена C9ORF72.
«Полная комплементарность», или «100% комплементарность», обозначает, что каждое нуклеотидное основание первой нуклеиновой кислоты имеет комплементарное нуклеотидное основание на второй нуклеиновой кислоте. В некоторых вариантах реализации изобретения первая нуклеиновая кислота является антисмысловым соединением, а нуклеиновая кислота-мишень является второй нуклеиновой кислотой.
«Гэпмер» обозначает химерное антисмысловое соединение, в котором внутренний участок, содержащий множество нуклеозидов, что обеспечивает расщепление РНКазой Н, расположено между внешними участками, содержащими один или более нуклеозидов, причем эти нуклеозиды, содержащие данный внутренний участок, химически отличаются от нуклеозида или нуклеозидов, содержащихся во внешних участках. Внутренний участок может называться «гэп», а внешние участки могут называться «крылья».
«Уменьшенный гэп» обозначает химерное антисмысловое соединение, содержащее гэп-сегмент из 9 или меньшего количества последовательных 2'-дезоксирибонуклеотидов расположенных между и непосредственно соседствующих с 5'- и 3'-сегментами крыльев, содержащими от 1 до 6 нуклеозидов.
«Расширенный гэп» означает химерное антисмысловое соединение, содержащее гэп-сегмент из 12 или большего количества последовательно 2'-дезоксирибонуклеотидов расположенных между и непосредственно соседствующих с 5'- и 3'-сегментами крыльев, содержащими от 1 до 6 нуклеозидов.
«Экспансия гексануклеотидного повтора» означает наборы из шести оснований (например, GGGGCC, GGGGGG, GGGGCG или GGGGGC), повторяющиеся по меньшей мере дважды. В некоторых вариантах реализации изобретения такая экспансия гексануклеотидного повтора может быть расположено на интроне 1 нуклеиновой кислоты C9ORF72. В некоторых вариантах реализации изобретения патогенная экспансия гексануклеотидного повтора включает по меньшей мере 30 повторов из GGGGCC, GGGGGG, GGGGCG или GGGGGC в нуклеиновой кислоте C9ORF72 и связанное с заболеванием. В некоторых вариантах реализации изобретения эти повторы являются последовательными. В некоторых вариантах реализации изобретения эти повторы прерываются 1 или более нуклеотидными основаниями. В некоторых вариантах реализации изобретения экспансия гексануклеотидного повтора дикого типа включает 23 или меньшее количество повторов из GGGGCC, GGGGGG, GGGGCG или GGGGGC в нуклеиновой кислоте C9ORF72. В некоторых вариантах реализации изобретения эти повторы являются последовательными. В некоторых вариантах реализации изобретения эти повторы прерываются 1 или более количеством нуклеотидными основаниями.
«Гибридизация» обозначает ренатурацию (соединение) молекул комплементарных нуклеиновых кислот. В некоторых вариантах реализации изобретения молекулы комплементарных нуклеиновых кислот включают антисмысловое соединение и нуклеиновую кислоту-мишень.
«Идентификация животного, пораженного ассоциированным с C9ORF72 заболеванием» указывает на идентификацию животного с диагностированным ассоциированным с белком C9ORF72 заболеванием или предрасположенного к развитию ассоциированного с белком C9ORF72 заболевания. Индивиды, предрасположенные к развитию ассоциированного с C9ORF72 заболевания, включают тех, которые обладают одним и более факторами риска для развития ассоциированного с C9ORF72 заболевания, включая обладающих личной или семейной историей или генетической предрасположенностью к одному или большему количеству ассоциированных с C9ORF72 заболеваний. Такая идентификация может выполняться любым способом, включающим оценку истории болезни индивида и стандартные клинические анализы или обследования, такие как генетическое исследование.
«Непосредственно соседствующий» означает, что между непосредственно соседствующими элементами отсутствуют промежуточные элементы.
«Индивид» указывает на человека или животное кроме человека, выбранное для лечения или оказания терапии.
«Ингибирование C9ORF72» указывает на снижение уровней экспрессии мРНК и/или белка C9ORF72 в присутствии специфического ингибитора C9ORF72, включая антисмысловой олигонуклеотид C9ORF72, по сравнению с уровнями экспрессии мРНК и/или белка C9ORF72 в отсутствии специфического ингибитора C9ORF72, такого как антисмысловой олигонуклеотид C9ORF72.
Термин «межнуклеозидная связь» относится к химической связи между нуклеозидами.
«Связанные нуклеозиды» обозначают соседствующие нуклеозиды, связанные вместе.
Термины «мисмэтч», или «некомплементарное нуклеотидное основание», относятся к случаю, когда нуклеотидное основание первой нуклеиновой кислоты не способно спариваться с соответствующим нуклеотидным основанием второй нуклеиновой кислоты или нуклеиновой кислоты-мишени.
«Модифицированная межнуклеозидная связь» относится к замещению или любому изменению встречающейся в природе межнуклеозидной связи (т.е. фосфодиэфирной межнуклеозидной связи).
«Модифицированное нуклеотидное основание» относится к любому нуклеотидному основанию, отличающемуся от аденина, цитозина, гуанина, тимидина и урацила. «Немодифицированное нуклеотидное основание» означает пуриновые основания: аденин (А) и гуанин (G), и пиримидиновые основания: тимин (Т), цитозин (С) и урацил (U).
«Модифицированный нуклеотид» обозначает нуклеотид, независимо содержащий модифицированный сахарный фрагмент, модифицированную межнуклеозидную связь или модифицированное нуклеотидное основание. «Модифицированный нуклеозид» означает нуклеозид, независимо содержащий модифицированный сахарный фрагмент и модифицированное нуклеотидное основание.
«Модифицированный олигонуклеотид» обозначает олигонуклеотид, содержащий модифицированную межнуклеозидную связь, модифицированный сахар или модифицированное нуклеотидное основание.
Термин «модифицированный сахар» относится к замещению или изменению в природном сахаре.
«Мотив» указывает на набор химически отличающихся участков в антисмысловом соединении.
«Встречающаяся в природе межнуклеозидная связь» обозначает 3' и 5'-фосфодиэфирную связь.
«Природный сахарный фрагмент» обозначает сахар, обнаруженный в ДНК (2'-Н) или РНК (2'-ОН).
«Нуклеиновая кислота» относится к молекуле, состоящей из мономерных нуклеотидов. Нуклеиновая кислота включает рибонуклеиновые кислоты (РНК), дезоксирибонуклеиновые кислоты (ДНК), одноцепочечные нуклеиновые кислоты, двухцепочечные нуклеиновые кислоты, малые интерферирующие рибонуклеиновые кислоты (миРНК) и микроРНК (микроРНК).
«Нуклеотидное основание» означает гетероциклический фрагмент, способный спариваться с основанием другой нуклеиновой кислоты.
«Последовательность нуклеотидных оснований» указывает на порядок последовательных нуклеотидных оснований, независимый от любого сахара, связи или модификации нуклеотидных оснований.
«Нуклеозид» обозначает нуклеотидное основание, связанное с сахаром.
«Нуклеозидный миметик» включает такие структуры, которые используются для замещения сахара или сахара и основания и не обязательно связи в одном или более положений олигомерного соединения, такие, как, например, нуклеозидные миметики, содержащие морфолин, циклогексенил, циклогексил, тетрагидропиранил, бицикло- и трициклосахарные миметики, например, нефуранозные сахарные фрагменты. Нуклеотидный миметик включает такие структуры, которые используются для замещения нуклеозида и связи в одном или более положений олигомерного соединения, такие как, например, пептид-нуклеиновые кислоты или морфолины (морфолины связаны с помощью -N(H)-C(=O)-O- или другой нефосфодиэфирной связи). Заместители сахаров перекрываются немного более широким термином нуклеозидного миметика, но подразумевают только указание на замещение сахарного фрагмента (фуранозного кольца). Тетрагидропиранильные кольца, представленные в данном документе, являются иллюстрациями примера заместителя сахара, в котором фуранозная сахарная группа была замещена системой тетрагидропиранильного кольца.
«Нуклеотид» обозначает нуклеозид, несущий фосфатную группу, ковалентно связанную с сахарной частью данного нуклеозида.
«Олигомерное соединение», или «олигомер», обозначает полимер из связанных субъединиц мономеров, который способен гибридизироваться с по меньшей мере одним участком молекулы нуклеиновой кислоты.
«Олигонуклеотид» означает полимер из связанных нуклеозидов, каждый из которых может быть модифицированным или немодифицированным независимо друг от друга.
«Парентеральное введение» обозначает введение с помощью инъекции или инфузии. Парентеральное введение включает подкожное введение, внутривенное введение, внутримышечное введение, внутриартериальное введение, внутрибрюшинное введение или внутричерепное введение, например, интратекальное или интрацеребровентрикальное введение.
«Пептид» обозначает молекулу, образованную соединением по меньшей мере двух аминокислот амидными связями. Пептид относится к полипептидам и белкам.
«Фармацевтически агент» обозначает субстанцию или субстанции фармацевтической композиции, которая обеспечивает терапевтическую пользу при введении индивиду. В некоторых вариантах реализации изобретения фармацевтическим агентом является антисмысловой олигонуклеотид, нацеленный на последовательность C9ORF72.
«Фармацевтическая композиция» означает смесь субстанций, пригодных для введения индивиду. Например, фармацевтическая композиция может содержать один или более фармацевтических агентов и стерильный водный раствор.
«Фармацевтически приемлемое производное» охватывает фармацевтически приемлемые соли, конъюгаты, пролекарства или изомеры соединений, описанных в данном документе.
Термин «фармацевтически приемлемые соли» охватывает физиологически и фармацевтически приемлемые соли антисмысловых соединений, т.е. соли, сохраняющие требуемую биологическую активность исходного олигонуклеотида и не обладают в дополнение к ней нежелательным токсикологическим действием.
«Фосфоротиоатная связь» указывает на соединение между нуклеозидами, в котором фосфодиэфирная связь модифицирована замещением одного из немостиковых атомов кислорода атомом серы. Фосфоротиоатная связь (P=S) представляет собой модифицированную межнуклеозидную связь.
«Часть» обозначает определенное количество последовательных (т.е., связанных) нуклеотидных оснований нуклеиновой кислоты. В некоторых вариантах реализации изобретения часть представляет собой определенное количество последовательных нуклеотидных оснований нуклеиновой кислоты-мишени. В некоторых вариантах реализации изобретения часть представляет собой определенное количество последовательных нуклеотидных оснований антисмыслового соединения.
«Предупреждать», или «предупреждение», относится к замедлению или упреждению проявления или развития заболевания, нарушения или состояния в течение периода времени - от минут до бесконечности. Предупреждение также означает снижение риска развития заболевания, нарушения или состояния.
«Пролекарство» обозначает, что терапевтический агент приготовлен в виде неактивной формы, которая превращается в свою активную форму в организме или в клетках под воздействием эндогенных ферментов или других химических веществ или условий.
Термин «побочные явления» обозначает физиологические реакции, обусловленные лечением, отличные от требуемого действия. В некоторых вариантах реализации изобретения побочные явления включают реакции в месте инъекции, исследуемые нарушения функционирования печени и почек, токсичность для печени и почек, нарушения центральной нервной системы, миопатии и дискомфорт.
«Одноцепочечный олигонуклеотид» указывает на олигонуклеотид, который не гибридизируется с комплементарной цепью.
Термин «способный специфически гибридизироваться» относится к антисмысловому соединению, имеющему достаточную степень комплементарности между антисмысловым олигонуклеотидом и нуклеиновой кислотой-мишенью для индуцирования требуемого действия, при этом проявляя минимальное или отсутствие влияния на нецелевые нуклеиновые кислоты в условиях, требующих достичь специфического связывания, т.е. в физиологических условиях в случае проведения анализов in vivo и при терапевтических воздействиях.
«Нацеливание», или «целевой», обозначает процесс конструирования и выбора антисмыслового соединения, которое будет специфически гибридизироваться с нуклеиновой кислотой-мишенью и индуцировать требуемое действие.
Термины «нуклеиновая кислота-мишень», «РНК-мишень» и «транскрипт РНК-мишени» все относятся к нуклеиновым кислотам, способным быть мишенями для антисмысловых соединений.
«Сегмент-мишень» означает последовательность нуклеотидов нуклеиновых кислот-мишеней, на которую нацелено антисмысловое соединение. «Сайт-мишень 5'» относится к самому крайнему нуклеотиду с 5'-конца сегмента-мишени. «Сайт-мишень 3'» относится к самому крайнему нуклеотиду с 3'-конца сегмента-мишени.
«Терапевтически эффективное количество» обозначает количество фармацевтического агента, которое обеспечивает терапевтическую пользу для индивида.
Термин «лечить», или «лечение», относится к введению фармацевтической композиции с целью вызвать изменение или улучшение течения заболевания, нарушения или состояния.
«Немодифицированный нуклеотид» обозначает нуклеотид, который состоит из встречающихся в природе нуклеотидных оснований, сахарных фрагментов и межнуклеозидных связей. В некоторых вариантах реализации изобретения немодифицированный нуклеотид является нуклеотидом РНК (т.е. β-D-рибонуклеозиды) или нуклеотидом ДНК (т.е.β-D-дезоксирибонуклеозиды).
Некоторые варианты реализации изобретения
Некоторые варианты реализации изобретения предлагают способы снижения экспрессии мРНК и белка C9ORF72.
Некоторые варианты реализации изобретения предлагают способы лечения, предупреждения или облегчения заболеваний, нарушений и состояний, ассоциированных с C9ORF72, у индивида, нуждающегося в таком воздействии. Подразумеваются также способы получения лекарственного средства для лечения, предупреждения или облегчения заболевания, нарушения и состояния, ассоциированного с C9ORF72. Ассоциированные с белком C9ORF72 заболевания, нарушения и условия включают нейродегенеративные заболевания. В некоторых вариантах реализации изобретения нейродегенеративное заболевание может быть АЛС или ЛВД. В некоторых вариантах реализации изобретения нейродегенеративное заболевание может быть наследственным или спорадическим.
Некоторые варианты реализации изобретения предлагают применение специфического ингибитора C9ORF72 для лечения, предупреждения или облегчения ассоциированного с белком C9ORF72 заболевания. Некоторые варианты реализации изобретения предлагают применение специфического ингибитора C9ORF72 для лечения, предупреждения или облегчения ассоциированного с экспансией гексануклеотидного повтора C9ORF72 заболевания. В некоторых вариантах реализации изобретения эта экспансия гексануклеотидного повтора может содержать последовательности GGGGCC, GGGGGG, GGGGGC или GGGGCG. В некоторых вариантах реализации изобретения специфические ингибиторы C9ORF72 представляют собой нуклеиновые кислоты (включая антисмысловые соединения), пептиды, антитела, малые молекулы и другие агенты, способные ингибировать экспрессию мРНК C9ORF72 и/или белка C9ORF72.
Описанные в данном документе ингибиторы представляют собой соединения, содержащие одноцепочечные антисмысловые олигонуклеотиды комплементарные нуклеиновой кислоте C9ORF72 или гомологу нуклеиновой кислоты C9ORF72.
В некоторых вариантах реализации изобретения нуклеиновая кислота C9ORF72 является нуклеиновой кислотой C9ORF72 человека.
В некоторых вариантах реализации изобретения нуклеиновая кислота C9ORF72 содержит экспансию гексануклеотидного повтора.
В некоторых вариантах реализации изобретения нуклеиновая кислота C9ORF72 не содержит экспансию гексануклеотидного повтора.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид способен специфически гибридизироваться с нуклеиновой кислотой C9ORF72 человека.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид является по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или на 100% комплементарным части нуклеиновой кислоты C9ORF72 человека равной длины.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид является комплементарным любому экзону, интрону, 5'-нетраслируемому участку, 3'-нетраслируемому участку, участку повтора, точке сплайсинга, точке сплайсинга экзон: экзон, сайленсеру экзонного сплайсинга (ESS), энхансеру экзонного сплайсинга (ESE), экзону 1а, экзону 1b, экзону 1с, экзону 1d, экзону 1e, экзону 2, экзону 3, экзону 4, экзону 5, экзону 6, экзону 7, экзону 8, экзону 9, экзону 10, экзону 11, интрону 1, интрону 2, интрону 3, интрону 4, интрону 5, интрону 6, интрону 7, интрону 8, интрону 9 или интрону 10 нуклеиновой кислоты C9ORF72 человека.
Описанные в данном документе олигонуклеотиды представляют собой соединения, которые содержат одноцепочечный антисмысловой олигонуклеотид, состоящий из 12-30 связанных нуклеозидов и содержащий последовательность нуклеотидных оснований, включающих по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19 или по меньшей мере 20 последовательных нуклеотидных оснований последовательностей SEQ ID NO: 30-369.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере одну модификацию.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере одну модифицированную межнуклеозидную связь.
В некоторых вариантах реализации изобретения каждая межнуклеозидная связь одноцепочечного антисмыслового олигонуклеотида является модифицированной межнуклеозидной связью.
В некоторых вариантах реализации изобретения модифицированная межнуклеозидная связь является фосфоротиоатной межнуклеозидной связью.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеозид.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеозид, несущий модифицированный сахар.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеозид, содержащий бициклический сахар.
В некоторых вариантах реализации изобретения бициклический сахар содержит мостик 4'-2', выбранный из: мостика 4'-(СН2)n-O-2', где n равно 1 или 2; и 4'-СН2-O-СН2-2'.
В некоторых вариантах реализации изобретения бициклический сахар содержит мостик 4'-СН(CH3)-O-2'.
В некоторых вариантах реализации изобретения по меньшей мере один модифицированный нуклеозид, несущий модифицированный сахар, содержит небициклический 2'-модифицированный сахарный фрагмент.
В некоторых вариантах реализации изобретения 2'-модифицированный сахарный фрагмент содержит 2'-O-метоксиэтильную группу.
В некоторых вариантах реализации изобретения 2'-модифицированный сахарный фрагмент содержит 2'-O-метильную группу.
В некоторых вариантах реализации изобретения по меньшей мере один модифицированный нуклеозид, несущий модифицированный сахар, содержит заместитель сахара.
В некоторых вариантах реализации изобретения заместитель сахара является морфолином.
В некоторых вариантах реализации изобретения заместитель сахара является пептид-нуклеиновой кислотой.
В некоторых вариантах реализации изобретения каждый нуклеозид является модифицированным.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит по меньшей мере одно модифицированное нуклеотидное основание.
В некоторых вариантах реализации изобретения модифицированное нуклеотидное основание является 5'-метилцитозином.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит:
гэп-сегмент, состоящий из связанных дезоксинуклеозидов;
сегмент 5'-крыла, состоящий из связанных нуклеозидов;
сегмент 3'-крыла, состоящий из связанных нуклеозидов;
при этом данный гэп-сегмент расположен непосредственно соседствующим с или расположен между сегментом 5'-крыла и сегментом 3'-крыла, причем каждый нуклеозид каждого сегмента крыла содержит модифицированный сахар.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид содержит:
гэп-сегмент, состоящий из десяти связанных дезоксинуклеозидов;
сегмент 5'-крыла, состоящий из пяти связанных нуклеозидов;
сегмент 3'-крыла, состоящий из пяти связанных нуклеозидов;
при этом данный гэп-сегмент расположен непосредственно соседствующим с или расположен между сегментом 5'-крыла и сегментом 3'-крыла, причем каждый нуклеозид каждого сегмента крыла содержит 2'-O-метоксиэтильный сахар; и в котором каждая межнуклеозидная связь является фосфоротиоатной связью.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 15 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 16 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 17 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 18 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 19 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 20 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 21 связанного нуклеозида.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 22 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 23 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 24 связанных нуклеозидов.
В некоторых вариантах реализации изобретения одноцепочечный антисмысловой олигонуклеотид состоит из 25 связанных нуклеозидов.
В настоящем документе описаны применения данного соединения в производстве лекарственного средства для лечения нейродегенеративного заболевания.
В данном документе предложены способы преимущественного ингибирования экспрессии транскриптов мРНК, содержащих экспансию гексануклеотидного повтора, путем обеспечения контакта клетки с антисмысловым олигонуклеотидом, нацеленным на экзон 1B в обратном направлении считывания.
Антисмысловые соединения
Олигомерные соединения включают, без ограничения: олигонуклеотиды, олигонуклеозиды, аналоги олигонуклеотидов, миметики олигонуклеотидов, антисмысловые соединения, антисмысловые олигонуклеотиды и миРНК. Олигомерное соединение может быть антисмысловым к нуклеиновой кислоте-мишени, что указывает на то, что оно способно претерпевать гибридизацию с нуклеиновой кислотой-мишенью с помощью водородных связей.
В некоторых вариантах реализации изобретения антисмысловое соединение имеет такую последовательность нуклеотидных оснований, которая записана в направлении 5'-3' и содержит обратно комплементарный фрагмент к сегменту-мишени нуклеиновой кислоты-мишени, на которую это соединение нацелено. В некоторых таких вариантах реализации изобретения антисмысловой олигонуклеотид имеет такую последовательность нуклеотидных оснований, которая записана в направлении 5'-3' и содержит обратно комплементарный фрагмент к сегменту-мишени нуклеиновой кислоты-мишени, на которую этот олигонуклеотид нацелен.
В некоторых вариантах реализации изобретения антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, состоит в длину из 12-30 субъединиц. Другими словами, такие антисмысловые соединения состоят из 12-30 связанных субъединиц. В некоторых вариантах реализации изобретения данное антисмысловое соединение состоит из 8-80, 12-50, 15-30, 18-24, 19-22 или 20 связанных субъединиц. В некоторых вариантах реализации изобретения антисмысловые соединения состоят в длину из 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79 или 80 связанных субъединиц и имеют диапазон длин, определенных любыми двумя из указанных выше значений. В некоторых вариантах реализации изобретения антисмысловое соединение является антисмысловым олигонуклеотидом, а связанные субъединицы представляют собой нуклеозиды.
В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, нацеленные на нуклеиновую кислоту C9ORF72, могут быть укороченные или усеченные. Например, может быть удалена одна субъединица с 5'-конца (5'-усечение) или в альтернативном варианте - с 3'-конца (3'-усечение). Укороченное или усеченное антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, может иметь две субъединицы, удаленные с 5'-конца антисмыслового соединения или в альтернативном варианте иметь две субъединицы, удаленные с 3'-конца. В альтернативном варианте удаленные нуклеозиды могут быть распределены по всему антисмысловому соединению, например, в антисмысловом соединении имеется один удаленный нуклеозид с 5'-конца и один удаленный нуклеозид с 3'-конца.
Когда одна дополнительная субъединица присутствует в удлиненном антисмысловом соединении, то эта дополнительная субъединица может быть расположена на 5'- или 3'-конце антисмыслового соединения. Когда присутствуют две или более субъединиц, то эти добавленные субъединицы могут соседствовать друг с другом, например в антисмысловом соединении, имеющем две субъединицы, добавленные к 5'-концу (5'-добавление) или в альтернативном варианте- к 3'-концу (3'-добавление) данного антисмыслового соединения.
В альтернативном вариантедобавленные субъединицы могут быть распределены по всему антисмысловому соединению, например в антисмысловом соединении имеется одна добавленная субъединица к 5'-концу и одна добавленная субъединица к 3'-концу.
Можно увеличить или уменьшить длину антисмыслового соединения, такого как антисмысловой олигонуклеотид, и/или внедрить некомплементарные основания без исчезновения активности. Так, в статье Woolf et al. (Proc. Natl. Acad. Sci. USA 89: 7305-7309, 1992) наборы антисмысловых олигонуклеотидов длиной 13-25 нуклеотидных оснований исследовали на их способность индуцировать расщепление РНК-мишени в модели с инъецированием ооцитов. Антисмысловые олигонуклеотиды длиной 25 нуклеотидных оснований с 8 или 11 некомплементарными основаниями возле концов этих антисмысловых олигонуклеотидов были способны непосредственно специфически расщеплять мРНК-мишень, хоть и в меньшей степени, чем антисмысловые олигонуклеотиды, которые не содержали некомплементарные основания. Аналогичным образомспецифическое расщепление мишени было достигнуто с использованием антисмысловых олигонуклеотидов размером 13 нуклеотидных оснований, включая те, которые несли 1 или 3 некомплементарные основания.
В статье Gautschi et al (J. Natl. Cancerlnst. 93: 463-471, March 2001) продемонстрирована способность олигонуклеотида, имеющего 100% комплементарность к мРНК bcl-2 и несущего 3 некомплементарных основания к мРНК bcl-xL, снижать экспрессию как гена bcl-2, так и bcl-xL в условиях in vitro и in vivo. Более того, этот олигонуклеотид продемонстрировал сильную противоопухолевую активность in vivo.
Авторами Maher и Dolnick (Nuc. Acid. Res. 16: 3341-3358, 1988) проверены наборы тандемных антисмысловых олигонуклеотидов из 14 нуклеотидных оснований и антисмысловых олигонуклеотидов из 28 и 42 нуклеотидных оснований, содержащих последовательность из двух или трех тандемных антисмысловых олигонуклеотидов, соответственно, относительно их способности подавлять трансляцию дегидрофолатредуктазы (DHFR) человека при анализе на ретикулоцитах кролика. Каждый из этих трех антисмысловых олигонуклеотидов размером 14 нуклеотидных оснований сам по себе был способен ингибировать трансляцию, хоть и в более ограниченной степени, чем антисмысловые олигонуклеотиды размером 28 и 42 нуклеотидных оснований.
Мотивы антисмысловых соединений
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, несут химически модифицированные субъединицы, сгруппированные в наборы, или мотивы, для того, чтобы придать антисмысловым соединениям свойства, такие как усиленная ингибирующая активность, повышенная аффинность связывания нуклеиновой кислоты-мишени или устойчивость к деградации нуклеазами in vivo.
Химерные антисмысловые соединения, как правило, содержат по меньшей мере один участок, модифицированный так, чтобы придать повышенную устойчивость к деградации нуклеазами, повышенное поглощение клетками, повышенную аффинность связывания нуклеиновой кислоты-мишени и/или повышенную ингибирующую активность. Второй участок химерного антисмыслового соединения произвольно может служить субстратом для клеточной эндонуклеазы - РНКазы Н, которая расщепляет нить РНК в дуплексе РНК:ДНК.
Антисмысловые соединения, содержащие гэпмер-мотив, считают химерными антисмысловыми соединениями. В гэпмере внутренний участок, несущий множество нуклеотидов, которые обеспечивают расщепление РНКазой Н, расположен между внешними участками, имеющими множество нуклеотидов, которые химически отличаются от нуклеозидов внутреннего участка. В том случае, если антисмысловой олигонуклеотид несет гэпмер-мотив, то гэп-сегмент, как правило, служит в качестве субстрата для расщепления эндонуклеазами, тогда как сегменты-крылья содержат модифицированные нуклеозиды. В некоторых вариантах реализации изобретения участки гэпмера дифференциируются по типам сахарных фрагментов, каждый из которых содержит отличающийся участок. Типы сахарных фрагментов, которые используются для дифференциации участков гэпмера, в некоторых вариантах реализации могут включать β-D-рибонуклеозиды, β-D-дезоксирибонуклеозиды, 2'-модифицированные нуклеозиды (2'-модифицированные нуклеозиды могут включать среди прочих варианты 2'-МОЕ и 2'-O-CH3) и модифицированные бициклическим сахаром нуклеозиды (такие модифицированные бициклическим сахаром нуклеозиды могут включать те, которые содержат мостик 4'-(CH2)n-O-2', где n=1 или n=2, и 4'-СН2-O-СН2-2'). Предпочтительно, чтобы каждый отдельный участок содержал однотипные сахарные фрагменты. Мотив крыло-гэп-крыло часто описывают как «X-Y-Z», где X представляет длину участка 5'-крыла, Y представляет длину гэп-участка, a Ζ представляет длину участка 3'-крыла. В данном документе считается, что гэпмер, описываемый как «Х-Y-Z», содержит в себе такую конфигурацию, что гэп-сегмент расположен непосредственно соседствующим с каждым сегментом 5'-крыла и сегментом 3'-крыла. Таким образом, не существует промежуточных нуклеотидов между сегментом 5'-крыла и гэп-сегментом или гэп-сегментом и сегментом 3'-крыла. Любое из антисмысловых соединений, описанных в данном документе, может содержать гэпмер-мотив. В некоторых вариантах реализации изобретения значения X и Ζ равны, в других вариантах они разные. В предпочтительном варианте реализации значение Y находится между 8 и 15 нуклеотидами. Значения Χ, Y или Ζ могут быть любыми из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30 или более нуклеотидов. Поэтому гэпмеры, описанные в данном документе, включают, без ограничения, соотношения, например: 5-10-5, 5-10-4, 4-10-4, 4-10-3, 3-10-3, 2-10-2, 5-9-5, 5-9-4, 4-9-5, 5-8-5, 5-8-4, 4-8-5, 5-7-5, 4-7-5, 5-7-4 или 4-7-4.
В некоторых вариантах реализации изобретения антисмысловое соединение имеет мотив «крыломер» с конфигурацией крыло-гэп или гэп-крыло, т.е. конфигурацию Χ-Υ или Υ-Ζ, как описано выше для конфигурации гэпмера. Таким образом, конфигурации гэпмера, описанные в данном документе, включают, без ограничения, например, соотношения: 5-10, 8-4, 4-12, 12-4, 3-14, 16-2, 18-1, 10-3, 2-10, 1-10, 8-2, 2-13, 5-13, 5-8 или 6-8.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат мотив гэпмера 5-10-5.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат мотив гэпмера 5-10-4.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат мотив гэпмера 4-10-4.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат мотив гэпмера 4-10-3.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат мотив гэпмера 5-9-5.
В некоторых вариантах реализации изобретения антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, имеет мотив с уменьшенным гэпом. В некоторых вариантах реализации изобретения антисмысловой олигонуклеотид с уменьшенным гэпом, нацеленный на нуклеиновую кислоту C9ORF72, несет гэп-сегмент из 9, 8, 7 или 6 2'-дезоксирибонуклеотидов, расположенных непосредственно с соседствующими с или между сегментами-крыльями из 5, 4, 3, 2 или 1 химически модифицированного нуклеозида. В некоторых вариантах реализации изобретения химическая модификация включает бициклический сахар. В некоторых вариантах реализации изобретения бициклический сахар содержит мостик 4'-2', выбранный из: мостика 4'-(СН2)n-O-2', где n равен 1 или 2, и 4'-СН2-O-СН2-2'. В некоторых вариантах реализации изобретения бициклический сахар содержит мостик 4'-СН(CH3)-O-2'. В некоторых вариантах реализации изобретения химическая модификация содержит небициклический 2'-модифицированный сахарный фрагмент. В некоторых вариантах реализации изобретения небициклический 2'-модифицированный сахарный фрагмент содержит 2'-O-метилэтильную группу или 2'-O-метильную группу.
В некоторых вариантах реализации изобретения антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, модифицировано однотипным способом. В некоторых вариантах реализации изобретения антисмысловое соединение содержит 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеозидов. В некоторых вариантах реализации изобретения каждый из нуклеозидов является химически модифицированным. В некоторых вариантах реализации изобретения химическая модификация содержит небициклический 2'-модифицированный сахарный фрагмент. В некоторых вариантах реализации изобретения 2'-модифицированный сахарный фрагмент содержит 2'-O-метоксиэтильную группу. В некоторых вариантах реализации изобретения 2'-модифицированный сахарный фрагмент содержит 2'-O-метильную группу. В некоторых вариантах реализации изобретения однотипно модифицированные антисмысловые соединения могут нацеливаться на C9ORF72 или на ее любую часть, такую как экспансия гексануклеотидного повтора. В некоторых вариантах реализации изобретения, нацеливание однотипно модифицированного антисмыслового соединением на экспансию гексануклеотидного повтора снижает синтез повторов РНК путем блокирования взаимодействия с белками, связывающими РНК. В некоторых вариантах реализации изобретения это приводит к отсутствию токсичной РНК в фокусах ядер, которая вместо этого разрушается.
Нуклеиновые кислоты-мишени, участки-мишени и нуклеотидные последовательности
Нуклеотидные последовательности, которые кодируют белок C9ORF72, включают, без ограничения, следующие: комплементарные последовательности с № доступа в базе данных GENBANK NM 001256054.1 (включена в данный документ как SEQ ID NO: 1), № доступа в базе данных GENBANK NT 008413.18, усеченная от нуклеотидного основания 27535000 до 27565000 (включена в данный документ как SEQ ID NO: 2), № доступа в базе данных GENBANK BQ068108.1 (включена в данный документ как SEQ ID NO: 3), № доступа в базе данных GENBANK NM 018325.3 (включена в данный документ как SEQ ID NO: 4), № доступа в базе данных GENBANK DN993522.1 (включена в данный документ как SEQ ID NO: 5), № доступа в базе данных GENBANK NM 145005.5 (включена в данный документ как SEQ ID NO: 6), № доступа в базе данных GENBANK DB079375.1 (включена в данный документ как SEQ ID NO: 7), № доступа в базе данных GENBANK BU 194591.1 (включена в данный документ как SEQ ID NO: 8), идентификатор последовательности 4141014 А (включена в данный документ как SEQ ID NO: 9) и идентификатор последовательность 4008 73 А (включена в данный документ как SEQ ID NO: 10).
Последовательность, описанная под каждым из SEQ ID NO в Примерах, содержащихся в данном документе, не зависит от любой модификации сахарного фрагмента, межнуклеозидной связи или нуклеотидного основания. Как таковое антисмысловое соединение, определенное SEQ ID NO, независимо может содержать одну или более модификаций сахарного фрагмент, межнуклеозидной связи или нуклеотидного основания. Антисмысловые соединения, описанные под номером Isis (№ Isis - нумерация компании Isis Pharmaceutical), указывают на комбинацию последовательности нуклеотидных оснований и мотива.
В некоторых вариантах реализации изобретения участок-мишень представляет собой структурно определенный участок нуклеиновой кислоты-мишени. Например, участок-мишень может охватывать 3'-нетранслируемый участок, 5'-нетранслируемый участок, экзон, интрон, соединение экзона/интрона, кодирующий участок, участок инициации трансляции, участок терминации трансляции или другой определенный участок нуклеиновой кислоты. Структурно определенные участки C9ORF72 могут быть получены по номеру доступа из баз данных последовательностей, таких как NCBI, и такая информация внесена в данный документ посредством ссылки. В некоторых вариантах реализации изобретения участок-мишень может охватывать последовательность от сайта-мишени 5' одного сегмента-мишени в пределах участка-мишени до сайта-мишени 3' другого сегмента-мишени в пределах такого же участка-мишени.
Нацеливание включает определение по меньшей мере одного сегмента-мишени, с которым гибридизируется антисмысловое соединение, вследствие чего проявляется требуемое действие. В некоторых вариантах реализации изобретения требуемое действие заключается в снижении уровней мРНК нуклеиновой кислоты-мишени. В некоторых вариантах реализации изобретения требуемое действие заключается в снижении уровней белка, кодируемого нуклеиновой кислотой-мишенью, или фенотипическом изменении, ассоциированным с нуклеиновой кислотой-мишенью.
Участок-мишень может содержать один или более сегментов-мишеней. Множество сегментов-мишеней в пределах участка-мишени могут быть перекрывающимися. В альтернативном варианте они могут быть неперекрывающимися. В некоторых вариантах реализации изобретения сегменты-мишени в пределах участка-мишени разделены не более чем около 300 нуклеотидами. В некоторых вариантах реализации изобретения сегменты-мишеней в пределах участка-мишени разделены количеством нуклеотидов, которое равно, равно около, равно не более чем, равно не более чем около 250, 200, 150, 100, 90, 80, 70, 60, 50, 40, 30, 20 или 10 нуклеотидам на нуклеиновой кислоте-мишени или является диапазоном, определенным любым из двух предыдущих значений. В некоторых вариантах реализации изобретения сегменты-мишени в пределах участка-мишени разделены не более чем или не более чем около 5 нуклеотидами на нуклеиновой кислоте-мишени. В некоторых вариантах реализации изобретения сегменты-мишени являются последовательными. Подразумеваются участки-мишени, определенные диапазоном, содержащим инициирующую нуклеиновую кислоту, которая является любым из сайтов-мишеней 5' или сайтов-мишеней 3', перечисленных в данном документе.
Подходящие сегменты-мишени могут быть найдены в пределах 5'-нетранслируемого участка, 3'-нетранслируемого участка, интрона, экзона, соединения экзона/интрона. Сегменты-мишени, содержащие инициирующий ко дон или терминирующий ко дон также являются подходящими в качестве сегментов-мишеней. Подходящий сегмент-мишень может специально исключать обозначенный структурно определенный участок, такой как инициирующий кодон или терминирующий кодон.
Установление подходящих сегментов-мишеней предполагает сравнение последовательности нуклеиновой-кислоты мишени с другими последовательностями всего генома. Например, алгоритм BLAST может использоваться для идентификации участков сходства среди разных нуклеиновых кислот. Это сравнение может предотвратить выбор последовательностей антисмысловых соединений, которые могут неспецифическим путем гибридизироваться с последовательностями, отличающимися от выбранной нуклеиновой кислоты-мишени (т.е. нецелевыми или нечетко целевыми последовательностями).
Существуют вариации активности (например, которые определяют по проценту снижения уровней нуклеиновых кислот-мишеней) антисмысловых соединений в пределах участка-мишени. В некоторых вариантах реализации изобретения снижения уровней мРНК C9ORF72 указывают на ингибирование экспрессии C9ORF72. Уменьшение уровней белка C9ORF72 указывает также на ингибирование экспрессии мРНК-мишени. Уменьшение фокусов в присутствии удлиненной РНК C9ORF72 указывает на ингибирование экспрессии C9ORF72. Дополнительно на ингибирование экспрессии C9ORF72 указывают фенотипические изменения. Например, улучшенные двигательная функция и дыхание могут указывать на ингибирование экспрессии C9ORF72.
Гибридизация
В некоторых вариантах реализации изобретения гибридизация происходит между антисмысловым соединением, описанным в данном документе, и нуклеиновой кислотой C9ORF72. Наиболее распространенный механизм гибридизации вовлекает водородные связи (например, водородные связи по Уотсону-Крику, Хугстину или обращенные связи по Хугстину) между комплементарными нуклеотидными основаниями молекул нуклеиновых кислот.
Гибридизация может проходить в различных условиях. Жесткие условия зависят от последовательности и определяются природой и составом молекул нуклеиновых кислот, требующих гибридизации.
Способы определения того, способна ли последовательность специфически гибридизироваться с нуклеиновой кислотой-мишенью, хорошо известны в данной области. В некоторых вариантах реализации изобретения антисмысловые соединения, предложенные в данном документе, способны специфически гибридизироваться с нуклеиновой кислотой C9ORF72.
Комплементарность
Антисмысловое соединение и нуклеиновая кислота-мишень комплементарны друг другу, если достаточное количество нуклеотидных оснований антисмыслового соединения может образовать водородную связь с соответствующими нуклеотидными основаниями нуклеиновой кислоты-мишени так, что будет проявляться требуемое действие (например, антисмысловое ингибирование нуклеиновой кислоты-мишени, такой, как нуклеиновой кислоты C9ORF72).
Некомплементарные нуклеотидные основания между антисмысловым соединением и нуклеиновой кислотой C9ORF72 могут быть приемлемыми при условии, что данное антисмысловое соединение остается способным специфически гибридизироваться с нуклеиновой кислотой-мишени. Более того, антисмысловое соединение может гибридизироваться на протяжении одного или более сегментов нуклеиновой кислоты C9ORF72 так, что промежуточные или соседствующие сегменты не вовлекаются в это гибридизационное событие (например, петлевая структура, некомплементарная или шпильковая структура).
В некоторых вариантах реализации изобретения антисмысловые соединения, представленные в данном документе, или их определенная часть, являются или по меньшей мере являются на 70, 80, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% комплементарными нуклеиновой кислоте C9ORF72, сегменту-мишени или их определенной части. Процент комплементарности антисмыслового соединения с нуклеиновой кислотой-мишенью может быть определен с использованием традиционных способов.
Например, антисмысловое соединение, в котором 18 из 20 нуклеотидных оснований антисмыслового соединения являются комплементарными участку-мишени, и поэтому может специфически гибридизироваться, будет представлять комплементарность на 90%. В этом примере оставшиеся некомплементарные нуклеотидные основания могут быть собраны в кластеры или рассеяны с комплементарными нуклеотидными основаниями и не требуют последовательного расположения друг к другу или к комплементарным нуклеотидным основаниям. Как таковоеантисмысловое соединение, которое содержит в длину 18 нуклеотидных оснований, имеющее 4 (четыре) некомплементарных нуклеотидных оснований, которые фланкированы двумя участками с полной комплементарностью к нуклеиновой кислоте-мишени, может иметь 77,8% общей комплементарности с нуклеиновой кислотой-мишенью, и поэтому должно попадать в пределы объема настоящего изобретения. Процент комплементарности антисмыслового соединения с участком нуклеиновой кислоты-мишени может быть определен традиционным способом с использованием программ BLAST (основные инструменты исследования для локального выравнивания) и программ Power BLAST, известных в данной области (Altschul et al., J. Mol. Biol., 1990, 215,403 410; Zhang and Madden, Genome Res., 1997, 7, 649 656). Процент гомологии, идентичность последовательностей или комплементарность, могут быть определены, например, программой Gap (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, Madison Wis.), используя установки по умолчанию, которая применяет алгоритм Смита и Уотермана (Adv. Appl. Math., 1981, 2,482489).
В некоторых вариантах реализации изобретения антисмысловые соединения, представленные в данном документе, или их определенные части, являются полностью комплементарными (т.е. на 100% комплементарными) нуклеиновой кислоте-мишени или ее определенной части. Например, антисмысловое соединение может быть полностью комплементарным нуклеиновой кислоте C9ORF72, или участку-мишени, или сегменту-мишени или ее последовательности-мишени. В данном документе считается, что «полностью комплементарный» обозначает, что каждое нуклеотидное основание антисмыслового соединения способно к точному спариванию оснований с соответствующими нуклеотидными основаниями нуклеиновой кислоты-мишени. Например, антисмысловое соединение размером 20 нуклеотидных оснований является полностью комплементарным последовательности-мишени, которая в длину составляет 400 нуклеотидных оснований, поскольку имеется соответствующая часть из 20 нуклеотидных оснований нуклеиновой кислоты-мишени, которая полностью комплементарна данному антисмысловому соединению. Полная комплементарность также может быть использована в сравнении с определенной частью первой и/или второй нуклеиновой кислоте. Например, часть размером 20 нуклеотидных оснований антисмыслового соединение размером 30 нуклеотидных оснований может быть полностью комплементарной последовательности-мишени, которая в длину составляет 400 нуклеотидных оснований. Часть размером 20 нуклеотидных оснований антисмыслового соединение размером 30 нуклеотидных оснований полностью комплементарна последовательности-мишени, если последовательность-мишень имеет соответствующую часть размером 20 нуклеотидных оснований, в которой каждое нуклеотидное основание является комплементарным части размером 20 нуклеотидных оснований этого антисмыслового соединения. В то же время целое антисмысловое соединение размером 30 нуклеотидных оснований может или не может быть полностью комплементарным последовательности-мишени, в зависимости от того, комплементарны ли также оставшиеся 10 нуклеотидных оснований антисмыслового соединения последовательности-мишени.
Размещение некомплементарного нуклеотидного основания может находиться на 5'-конце или 3'-конце данного антисмыслового соединения. В альтернативном варианте некомплементарное нуклеотидное основание или нуклеотидные основания могут находиться в антисмысловом соединении во внутреннем положении. Когда присутствуют два или более некомплементарных нуклеотидных оснований, они могут быть последовательными (т.е. связанными) или непоследовательными. В одном варианте реализации изобретения некомплементарное нуклеотидное основание расположено на сегменте-крыле гэпмера антисмыслового олигонуклеотида.
В некоторых вариантах реализации изобретения антисмысловые соединения, которые в длину составлены из или составлены из вплоть до 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидных оснований, содержат не более 4, не более 3, не более 2 или не более 1 некомплементного(ых) нуклеотидного(ых) основания(ий) по сравнению с нуклеиновой кислотой-мишенью, такой, как нуклеиновая кислота C9ORF72, или ее определенной частью.
В некоторых вариантах реализации изобретения антисмысловые соединения, которые в длину составлены из или составлены из вплоть до 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидных оснований, содержат не более 6, не более 5, не более 4, не более 3, не более 2 или не более 1 некомплементного(ых) нуклеотидного(ых) основания(ий) по сравнению с нуклеиновой кислотой-мишенью, такой, как нуклеиновая кислота C9ORF72, или ее определенной частью.
Антисмысловые соединения, предложенные в данном документе, также содержат те, которые комплементарны части нуклеиновой кислоты-мишени. В данном документе считается, что «часть» относится к определенному количеству последовательных (т.е. связанных) нуклеотидных оснований в пределах участка или сегмента нуклеиновой кислоты-мишени. «Часть» может также относиться к определенному количеству последовательных нуклеотидных оснований антисмыслового соединения. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 8 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 9 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 10 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 11 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 12 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 13 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 14 нуклеотидных оснований. В некоторых вариантах реализации изобретения антисмысловые соединения являются комплементарными по меньшей мере части сегмента-мишени размером 15 нуклеотидных оснований. Подразумеваются также антисмысловые соединения, которые комплементарны части из по меньшей мере 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более нуклеотидных оснований сегменту-мишени или из диапазона, определенного любыми двумя из этих значений.
Идентичность
Антисмысловые соединения, предложенные в данном документе, также могут иметь определенный процент идентичности с конкретной нуклеотидной последовательностью, SEQ ID NO, или соединением, представленным специальным номером Isis, или его частью. В данном документе считается, что антисмысловое соединение является идентичным последовательности, описанной в данном документе, если оно имеет такую же способность к спариванию нуклеотидных оснований. Например, РНК, которая в описанной последовательности ДНК содержит урацил вместо тимидина, будет считаться идентичной последовательности ДНК, поскольку и урацил и тимидин образуют пару с аденином. Подразумеваются также укороченные и удлиненные версии антисмысловых соединений, описанные в данном документе, а также соединения, имеющие неидентичные основания по сравнению с антисмысловыми соединениями, описанными в данном документе. Неидентичные основания могут соседствовать друг с другом или быть распределенными по всему антисмысловому соединению. Процент идентичности антисмыслового соединения рассчитывают в соответствии с количеством соединений, которые имеют идентичное спаривание оснований по сравнению с последовательностью, с которой оно сопоставляется.
В некоторых вариантах реализации изобретения антисмысловые соединения или их части являются или по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98, 99 или 100% идентичными одному или большему количеству антисмысловых соединений или последовательностям SEQ ID NO, или их частям, описанным в данном документе.
В некоторых вариантах реализации изобретения часть антисмыслового соединения сравнивается с частью равной длины нуклеиновой кислоты-мишени. В некоторых вариантах реализации изобретения часть размером 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидных оснований сравнивается с частью равной длины нуклеиновой кислоты-мишени.
В некоторых вариантах реализации изобретения часть антисмыслового олигонуклеотида сравнивается с частью равной длины нуклеиновой кислоты-мишени. В некоторых вариантах реализации изобретения часть размером 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидных оснований сравнивается с частью равной длины нуклеиновой кислоты-мишени.
Модификации
Нуклеозид представляет собой комбинацию основания-сахара. Часть нуклеотидного основания (известного как основание) нуклеозида обычно является фрагментом гетероциклического основания. Нуклеотиды являются нуклеозидами, которые дополнительно содержатфосфатную группу, ковалентно связанную с сахарной частью нуклеозида. Для тех нуклеозидов, которые включают пентофуранозильный сахар, фосфатная группа может быть присоединена к 2', 3' или 5'-гидроксильному фрагменту сахара. Олигонуклеотиды образуются посредством ковалентного связывания соседствующих друг с другом нуклеозидов с образованием линейного полимерного олигонуклеотида. В пределах олигонуклеотидной структуры фосфатные группы обычно считаются образующими межнуклеозидные связи данного олигонуклеотида.
Модификации антисмысловых соединений охватывают замены или изменения межнуклеозидных связей, сахарных фрагментов или нуклеотидных оснований. Модифицированные антисмысловые соединения часто преобладают над природными формами, потому что обладают требуемыми свойствами, такими, как, например, усиленное поглощение клетками, усиленная аффинность к нуклеиновой кислоте-мишени, повышенная стабильность в присутствии нуклеаз или повышенная ингибиторная активность.
Химически модифицированные нуклеозиды могут применяться и для увеличения аффинности связывания укороченного или усеченного антисмыслового олигонуклеотида с нуклеиновой кислотой-мишенью. Следовательно, сопоставимые результаты часто могут быть получены с более короткими антисмысловыми соединениями, которые несут такие химически модифицированные нуклеозиды.
Модифицированные межнуклеозидные связи
Встречающаяся в природе межнуклеозидная связь РНК и ДНК представляет собой 3' 5' фосфодиэфирную связь. Антисмысловые соединения, содержащие одну или более модифицированных, такие, что не встречаются в природе, межнуклеозидных связей чаще выбираются для антисмысловых соединений, содержащих встречающиеся в природе межнуклеозидные связи, потому что обладают требуемыми свойствами, такими,как, например, усиленное поглощение клетками, усиленная аффинность к нуклеиновым кислотам-мишеням и повышенная стабильность в присутствии нуклеаз.
Олигонуклеотиды, имеющие модифицированные межнуклеозидные связи, включают межнуклеозидные связи, которые сохраняют атом фосфора, а также межнуклеозидные связи, которые не содержат атом фосфора. Межнуклеозидные связи, содержащие типичный фосфор, включают без ограничения, фосфодиэфиры, фосфотриэфиры, метилфосфонаты, фосфорамидат и фосфоротиоаты. Способы получения содержащих фосфор и несодержащих фосфор связей хорошо известны.
В некоторых вариантах реализации изобретения антисмысловые соединения, нацеленные на нуклеиновую кислоту C9ORF72, содержат одну или более модифицированных межнуклеозидных связей. В некоторых вариантах реализации изобретения модифицированные межнуклеозидные связи являются рассеянными по всему антисмысловому соединению. В некоторых вариантах реализации изобретения модифицированные межнуклеозидные связи являются фосфоротиоатными связями. В некоторых вариантах реализации изобретения каждая межнуклеозидная связь антисмыслового соединения является фосфоротиоатной межнуклеозидной связью.
Модифицированные сахарные фрагменты
Антисмысловые соединения выборочно могут содержать один или более нуклеозидов, в которых каждая сахарная группа была модифицирована. Такие модифицированные сахаром нуклеозиды могут придавать антисмысловым соединениям повышенную стабильность к нуклеазам, увеличенную аффинность связывания или некоторые другие полезные биологические свойства. В некоторых вариантах реализации изобретения нуклеозиды содержат химически модифицированные фрагменты рибофуранозных колец. Примеры химически модифицированных рибофуранозных колец включают, без ограничения, добавление замещающих групп (включая 5'- и 2'-замещающие группы, образование мостиков из негеминальных атомов колец с образованием бициклических нуклеиновых кислот (BNA), замещение атома кислорода рибозильного кольца S, N(R) или C(R1)(R2) (R, R1 и R2 - независимо друг от друга являются Н, C1-С12 алкильной или защитной группой) или их комбинациями. Примеры химически модифицированных сахаров включают 2'-F-5'-метилзамещенный нуклеозид (см. международную заявку PCTWO 2008/101157, опубликованную 21 августа 2008 г. для других описанных 5', 2'-бисзамещенных нуклеозидов) или замещение атома кислорода рибозильного кольца атомом S с последующим замещением положения 2' (см. опубликованную патентную заявку США US 2005-0130923, опубликованную 16 июня 2005 г.) или в альтернативном варианте 5'-замещение BNA (см. международную заявку PCTWO 2007/134181, опубликованную 22 ноября 2007 г., в которой LNA (блокированная нуклеиновая кислота) замещена, например, 5'-метильной или 5'-винильной группой).
Примеры нуклеозидов, содержащих модифицированные сахарные фрагменты, включают, без ограничения, нуклеозиды, содержащие 5'-винил, 5'-метил (R или S), 4'-S, 2-F, 2'-ОСН3, 2'-ОСН2СН3, 2'-OCH2CH2F и 2'-O(СН2)2ОСН3 замещающие группы. Заместитель в положении 2' также может быть выбран из: аллил, амино, азидо, тио, О-аллил, O-C1-С10 алкил, OCF3, OCH2F, O(CH2)2SCH3, O(CH2)2-O-N(Rm)(Rn), O-CH2-C(=O)-N(Rm)(Rn), и O-CH2-C(=O)-N(R1)-(CH2)2-N(Rm)(Rn), где каждый радикал R1, Rm и Rn является независимо H или замещенным или незамещенным C1-C10 алкилом.
В данном документе считается, что термин «бициклический нуклеозид» относится к модифицированному нуклеозиду, содержащему бициклический сахарный фрагмент. Примеры бициклических нуклеозидов включают, без ограничения, нуклеозиды, содержащие мостик между атомами 4' и 2' рибозильного кольца. В некоторых вариантах реализации изобретения антисмысловые соединения, предложенные в данном документе, включают один или более бициклических нуклеозидов, содержащих мостик между атомами 4'-2'. Примеры таких связанных мостиками 4'-2' бициклических нуклеозидов, включают без ограничения одну из формул: 4'-(СН2)-O-2' (LNA); 4'-(CH2)-S-2'; 4*-(СН2)2-O-2' (ΕΝΑ - нуклеиновые кислоты, связанные 4'-этиленовым мостиком); 4'-СН(СН3)-O-2' и 4'-СН(CH2OCH3)-O-2' (и их аналоги, см. патент США 7399845, выданный 15 июля 2008 г.); 4'-С(CH3)(CH3)-O-2' (и ее аналоги, см. опубликованную международную заявку WO/2009/006478, опубликованную 8 января 2009 г.); 4'-СН2-N(ОСН3)-2' (и ее аналоги, см. международную заявку WO/2008/150729, опубликованную 11 декабря 2008 г.); 4'-CH2-O-N(CH3)-2' (см. заявку на патент США US 2004-0171570, опубликованную 2 сентября 2004 г.); 4'-CH2-N(R)-O-2', где R является Н, С1-С12-алкилом или защитной группой (см. патент США 7427672, выданный 23 сентября 2008 г.); 4'-СН2-С(Н)(СН3)-2' (см. статью Chattopadhyaya et al, J. Org. Chem., 2009, 74, 118-134) и 4'-CH2-C(=CH2)-2' (и ее аналоги см. международную заявку WO 2008/154401, опубликованную 8 декабря 2008 г.).
Дополнительные отчеты, касающиеся бициклических нуклеозидов также могут быть найдены в опубликованной литературе (см., например, статьи: Singh et al., Chem. Commun., 1998, 4, 455-456; Koshkin et al, Tetrahedron, 1998, 54, 3607-3630; Wahlestedt et al, Proc. Nati Acad. Sci. U.S.Α., 2000, 97, 5633-5638; Kumar et al, Bioorg. Med. Chem. Lett., 1998, 8, 2219-2222; Singh et al, J. Org. Chem., 1998, 63, 10035-10039; Srivastava et al, J. Am. Chem. Soc, 2007, 129(26) 8362-8379; Elayadi etal, Curr. Opinion Invest. Drugs, 2001, 2, 558-561; Braasch et al, Chem. Biol, 2001, 8, 1-7; и Orum et al, Curr. Opinion Mol Ther., 2001, 3, 239-243; патенты США №№6268490; 6525191; 6670461; 6770748; 6794499; 7034133; 7053207; 7399845; 7547684 и 7696345; публикации патента США № US 2008-0039618; US 2009-0012281; серии патентов США №№60/989574; 61/026995; 61/026998; 61/056564; 61/086231; 61/097787 и 61/099844; опубликованные международные заявки РСТ WO 1994/014226; WO 2004/106356; WO 2005/021570; WO 2007/134181; WO 2008/150729; WO 2008/154401 и WO 2009/006478. Каждый из предыдущих бициклических нуклеозидов может быть получен с содержанием одного или более конфигураций стереохимических сахаров, включая, например, α-L-рибофуранозу и β-D-рибофуранозу (см. международную заявку РСТ - PCT/DK98/00393, опубликованную 25 марта 1999 г. как заявка WO 99/14226).
В некоторых вариантах реализации изобретения бициклические сахарные фрагменты нуклеозидов BNA включают без ограничений, соединения, содержащие по меньшей мере один мостик между положениями 4' и 2' пентофуранозного сахарного фрагмента, в котором такие мостики независимо содержат 1 или от 2 до 4 связанных групп, независимо выбранных из -[C(Ra)(Rb)]n-, -C(Ra)=C(Rb)-, -C(Ra)=N-, -С(=O)-, -C(=NRa)-, -C(=S)-, -O-, -Si(Ra)2-, -S(=O)x- и -N(Ra)-;
где
x равен 0, 1 или 2;
n равен 1, 2, 3 или 4;
каждый радикал Ra и Rb независимо является Н, защитной группой, гидроксилом, C1-С12-алкилом, замещенным С1-С12-алкилом, С2-С12-алкенилом, замещенным С2-С12-алкенилом, С2-С12-алкинилом, замещенным С2-С12-алкинилом, С5-С20-арилом, замещенным С5-С20-арилом, гетероциклическим радикалом, замещенным гетероциклическим радикалом, гетероарилом, замещенным гетероарилом, С5-С7-алициклическим радикалом, замещенным С5-С7-алициклическим радикалом, галогеном, OJ1, NJ1J2, SJ1, N3, COOJ1, ацилом (C(=O)-H), замещенным ацилом, CN, сульфонилом (S(=O)2-J1) или сульфоксилом (S(=O)-J1) и
каждый заместитель J1 и J2 независимо является Н, C1-С12-алкилом, замещенным C1-С12-алкилом, С2-С12-алкенилом, замещенным С2-С12-алкенилом, С2-С12-алкинилом, замещенным С2-С12-алкинилом, С5-С20-арилом, замещенным C5-C20-арилом, ацилом (С(=O)-Н), замещенным ацилом, гетероциклическим радикалом, замещенным гетероциклическим радикалом, С1-С12-аминоалкилом, замещенным С1-С12-аминоалкилом или защитной группой.
В некоторых вариантах реализации изобретения мостик бициклического сахарного фрагмента является структурой -[C(Ra)(Rb)]n-, -[C(Ra)(Rb)]n-O-, -C(RaRb)-N(R)-O- или -C(RaRb)-O-N(R)-. В некоторых вариантах реализации изобретения этот мостик представляет собой структуры 4'-СН2-2', 4'-(СН2)2-2', 4'-(СН2)3-2', 4'-СН2-O-2', 4'-(СН2)2-O-2', 4'-CH2-O-N(R)-2' и 4'-CH2-N(R)-O-2'-, где каждый R независимо является Н, защитной группой или C1-С12-алкилом.
В некоторых вариантах реализации изобретения бициклические нуклеозиды являются дополнительно определенные их изомерной конфигурацией. К примеру, нуклеозид, содержащий 4'-2'-метиленокси мостик, может быть в конфигурации α-L или β-D. Ранее, α-L-метиленокси (4'-СН2-O-2') BNA были включены в антисмысловые олигонуклеотиды, которые показали антисмысловую активность (Frieden et al, Nucleic Acids Research, 2003, 21, 6365-6372).
В некоторых вариантах реализации изобретения бициклические нуклеозиды включают, без ограничения, (А) α-L-метиленокси(4'-CH2-O-2') BNA, (В) β-D-метиленокси (4'-СН2-O-2') BNA, (С) этиленокси(4'-(СН2)2-O-2') BNA, (D) аминокси (4'-CH2-O-N(R)-2') BNA, (Ε) оксиамино (4'-CH2-N(R)-O-2') BNA и (F) метил(метиленокси) (4'-CH(CH3)-O-2') BNA, (G) метилентио (4'-CH2-S-2') BNA, (H) метиленамино (4'-CH2-N(R)-2') BNA, (I) метилкарбоцикло (4'-СН2-СН(СН3)-2') BNA и (J) пропиленкарбоцикло (4'-(СН2)3-2') BNA, как изображено ниже.

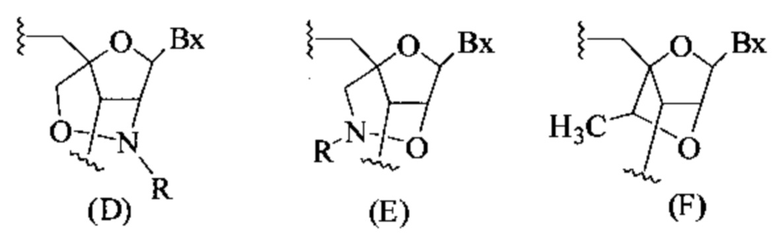

где Вх является фрагментом основания, a R независимо является Н, защитной группой или С1-С12-алкилом.
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу I:
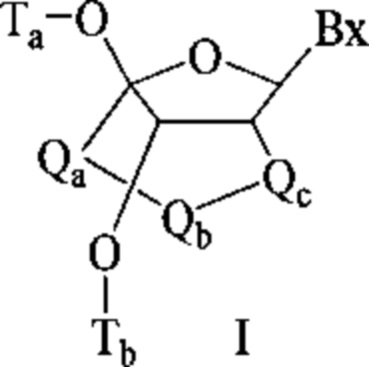
где
Bx является фрагментом гетероциклического основания;
-Qa-Qb-Qc- представляют структуры -CH2-N(Rc)-CH2-, -C(=O)-N(Rc)-CH2-, -CH2-O-N(Rc), -CH2-N(Rc)-O- или -N(Rc)-O-CH2;
Rc является C1-C12-алкилом или аминозащитной группой; и
заместители Та и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу II:

где
Bx является фрагментом гетероциклического основания;
Та и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
Za является C1-С6-алкилом, С2-С6-алкенилом, С2-С6-алкинилом, замещенным C1-С6-алкилом, замещенным С2-С6-алкенилом, замещенным С2-С6-алкинилом, ацилом, замещенным ацилом, замещенным амидом, тиолом или замещенным тиолом.
В одном варианте реализации изобретения каждая из замещающих групп независимо является моно- или полизамещенной с замещающими группами, независимо выбранными из галоген, оксо, гидроксил, OJc, NJcJd, SJc, N3, OC(=X)Jc и NJeC(=X)NJcJd групп, где каждый заместитель Jc, Jd и Je независимо является Н, C1-С6-алкилом или замещенным C1-С6-алкилом, а X является О или NJc.
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу III:
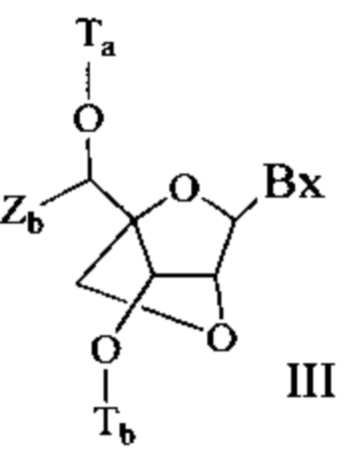
где
Bx является фрагментом гетероциклического основания;
Та и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
Zb является C1-С6-алкилом, С2-С6-алкенилом, С2-С6-алкинилом, замещенным C1-С6-алкилом, замещенным С2-С6-алкенилом, замещенным С2-С6-алкинилом или замещенным ацилом (С(=O)-).
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу IV:
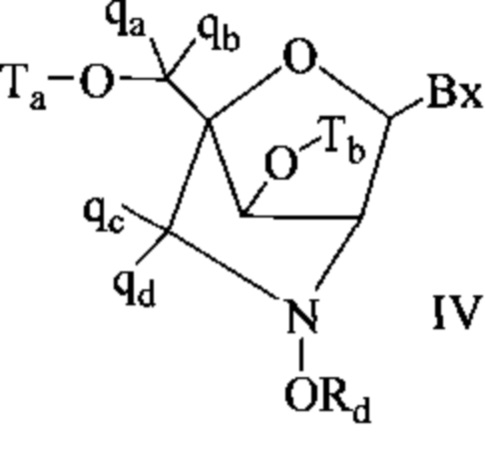
где
Вх является фрагментом гетероциклического основания;
Та и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
Rd является C1-С6-алкилом, замещенным C1-С6-алкилом, С2-С6-алкенилом, замещенным С2-С6-алкенилом, С2-С6-алкинилом или замещенным С2-С6-алкинилом;
каждый заместитель qa, qb, qc и qd является независимо H, галогеном, C1-С6-алкилом, замещенным C1-С6-алкилом, С2-С6-алкенилом, замещенным С2-С6-алкенилом, С2-С6-алкинилом или замещенным С2-С6-алкинилом, C1-С6-алкоксилом, замещенным C1-С6-алкоксилом, ацилом, замещенным ацилом, C1-С6-аминоалкилом или замещенным C1-С6-аминоалкилом.
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу V:
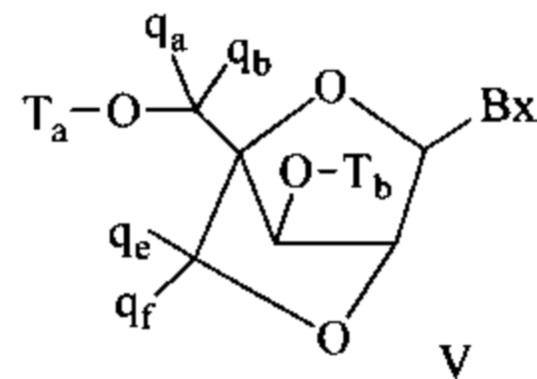
где
Bx является фрагментом гетероциклического основания;
Ta и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
заместители qa, qb, qe и qf являются каждый независимо водородом, галогеном, С1-С12-алкилом, замещенным C2-С12-алкилом, С2-С12-алкенилом, замещенным C2-C12-алкенилом, С2-С12-алкинилом, замещенным С2-С12-алкинилом, С1-С12-алкокси, замещенной С1-С12-алкокси, OJj, SJj, SOJj, SO2Jj, NJjJk, N3, CN, C(=O)OJj, C(=O)NJjJk, C(=O)Jj, O-C(=O)NJjJk, N(H)C(=NH)NJjJk, N(H)C(=O)NJjJk или N(H)C(=S)NJjJk группами;
или qe и qf вместе являются =C(qg)(qh);
qg и qh являются каждый независимо H, галогеном, С1-C12-алкилом или замещенным C1-C12-алкилом.
Были описаны синтез и получение метиленокси-(4'-СН2-O-2')-мономеров BNA аденина, цитозина, гуанина, 5-метилцитозина, тимина и урацила, вместе с их олигомеризацией, и свойства распознавания нуклеиновых кислот (Koshkin et al, Tetrahedron, 1998, 54, 3607-3630). BNA и их получение также описаны в заявках WO 98/39352 и WO 99/14226.
Также были получены аналоги метиленокси-(4'-СН2-O-2') BNA и 2'-тио-BNA (Kumar et al., Bioorg. Med. Chem. Lett., 1998, 8, 2219-2222). Также было описано получение блокированных нуклеозидных аналогов, содержащих олигодезоксирибонуклеотидные дуплексы в качестве субстратов для полимераз нуклеиновых кислот (Wengel et al., WO 99/14226). Более того, в данной области был описан синтез 2'-амино-BNA, нового конформационно ограниченного высокоаффинного олигонуклеотидного аналога (Singh et al, J. Org. Chem., 1998, 63, 10035-10039). В дополнение были получены 2'-амино- и 2'-метиламино-BNA, а ранее сообщили о термической стабильности таких дуплексов с комплементарными цепями РНК и ДНК.
В некоторых вариантах реализации изобретения предложенные бициклические нуклеозиды имеют Формулу VI:

где
Bx является фрагментом гетероциклического основания;
Ta и Tb являются каждый независимо Н, гидроксилзащитной группой, конъюгирующей группой, реакционно-способной фосфорной группой, фосфорным фрагментом или ковалентным соединением с веществом подложки.
каждый qi, qj, qk и ql является независимо Н, галогеном, С1-С12-алкилом, замещенным С1-С12-алкилом, С2-С12-алкенилом, замещенным С2-С12-алкенилом, С2-С12-алкинилом, замещенным С2-С12-алкинилом, С1-С12-алкоксилом, замещенным С1-С12-алкоксилом, OJj, SJj, SOJj, SO2Jj, NJjJk, N3, CN, C(=O)OJj, C(-O)NJjJk, C(=O)Jj, O-C(=O)NJjJk, N(H)C(=NH)NJjJk, N(H)C(=O)NJjJk или N(H)C(=S)NJjJk группами; и
заместители qi и qj или ql и qk вместе являются =C(qg)(qh), где qg и qh являются каждый независимо Н, галогеном, С1-С12-алкилом или замещенным С1-С12-алкилом.
Был описан один карбоциклический бициклический нуклеозид, содержащий мостик 4'-(СН2)3-2' и мостик алкенильного аналога 4'-СН=СН-СН2-2' (Freier et al, Nucleic Acids Research, 1997, 25(22), 4429-4443 и Albaek et al, J. Org. Chem., 2006, 71, 7731-7740). Были также описаны синтез и получение карбоциклических бициклических нуклеозидов вместе с их олигомеризацией и биохимическими исследованиями (Srivastava et al, J. Am. Chem. Soc, 2007, 129(26), 8362-8379).
В данном документе считается, что термин «4'-2' бициклическеский нуклеозид», или «бициклический нуклеозид 4'-2'» относится к бициклическому нуклеозиду, содержащему фуранозное кольцо, включающее мостик, соединяющий два углеродных атома фуранозного кольца, который соединяет углеродный атом 2' с углеродным атомом 4' сахарного кольца.
В данном документе считается, что «моноциклический нуклеозид» относится к нуклеозидам, содержащим модифицированные сахарные фрагменты, которые не являются бициклическими сахарными фрагментами. В некоторых вариантах реализации изобретения сахарный фрагмент или аналог сахарного фрагмента нуклеозида может быть модифицирован или замещен в любом положении.
В данном документе считается, что «2'-модифицированный сахар» указывает на фуранозильный сахар, модифицированный в положении 2'. В некоторых вариантах реализации изобретения такие модификации включают заместители, выбранные изгалогенида, включая без ограничения, замещенную и незамещенную алкокси группу, замещенный и незамещенный тиоалкил, замещенный и незамещенный аминоалкил, замещенный и незамещенный алкил, замещенный и незамещенный аллил и замещенный и незамещенный алкинил. В некоторых вариантах реализации изобретения 2'-модификации выбраны из заместителей, включающих без ограничения: O[(CH2)nO]mCH3, O(CH2)nNH2, O(СН2)nCH3, O(CH2)nF, O(CH2)nONH2, OCH2C(=O)N(H)CH3, и O(CH2)nON[(CH2)nCH3]2, где n и m равны от 1 до около 10. Другие 2'-замещающие группы могут также выбираться из: С1-С12-алкила, замещенного алкила, алкенила, алкинила, алкарила, аралкила, O-алкарила или O-аралкила, SH, SCH3, OCN, Cl, Br, CN, F, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2, гетероциклоалкила, гетероциклоалкарила, аминоалкамино группы, полиалкиламино группы, замещенного силила, расщепляющей РНК группы, репортерной группы, интеркалятора, улучшающей фармакокинетические свойства группы или улучшающей фармакодинамические свойства группы антисмыслового соединения и другие заместители, обладающие сходными свойствами. В некоторых вариантах реализации изобретения модифицированные нуклеозиды содержат боковую цепь 2'-МОЕ (Baker et al, J. Biol. Chem., 1997, 272, 11944-12000). Такое замещение 2'-MOE было описано как обладающее улучшенной аффинностью связывания по сравнению с немодифицированными нуклеозидами и другими модифицированными нуклеозидами, такими как 2'-O-метил, O-пропил и O-аминопропил. Было также показано, что олигонуклеотиды, содержащие заместитель 2'-МОЕ, являются антисмысловыми ингибиторами экспрессии генов с перспективными свойствами для использования in vivo (Martin, Helv. Chim. Acta, 1995,78, 486-504; Altmann et al., Chimia, 1996, 50, 168-176; Altmann et al, Biochem. Soc. Trans., 1996, 24, 630-637 и Altmann et al, Nucleosides Nucleotides, 1997, 76, 917-926).
В данном документе считается, что «модифицированный тетрагидропирановый нуклеозид», или «модифицированный ТГП нуклеозид», обозначает нуклеозид, содержащий шестичленный тетрагидропирановый сахар, замещенный в обычных нуклеозидах пентофуранозидным остатком (сахарный заменитель). Модифицированный ТГП нуклеозид включает, без ограничения, то, что называется в данной области гекситной нуклеиновой кислотой (HNA), анитной нуклеиновой кислотой (ANA), манитной нуклеиновой кислотой (MNA) (см. статью Leumann, Bioorg. Med. Chem., 2002, 10, 841-854), фтор-ΗΝΑ (F-HNA) или те соединения, которые имеют Формулу VII:
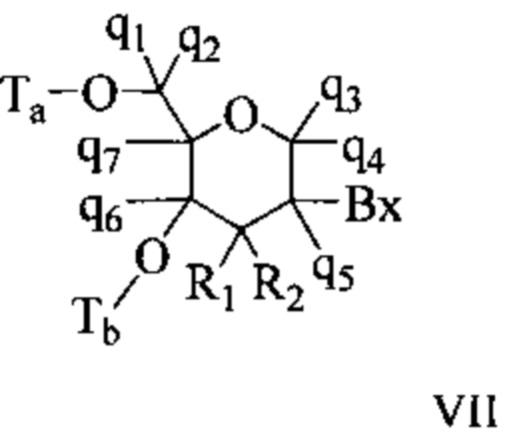
где независимо каждый указанный по меньшей мере один аналог тетрагидропиранового нуклеозида имеет Формулу VII:
Bx является фрагментом гетероциклического основания;
группы Ta и Tb являются каждая независимо межнуклеозидной связывающей группой, соединяющей аналог тетрагидропиранового нуклеозида с антисмысловым соединением или одна из групп Ta и Tb является межнуклеозидной связывающей группой, соединяющей аналог тетрагидропиранового нуклеозида с антисмысловым соединением, а другая группа из Ta и Tb является Н, гидроксилзащитной группой, группой конъюгирующего связывания или 5'- или 3'-концевой группой;
заместители q1, q2, q3, q4, q5, q6 и q7 являются каждый независимо H, C1-С6-алкилом, замещенным C1-С6-алкилом, С2-С6-алкенилом, замещенным С2-С6-алкенилом, С2-С6-алкинилом или замещенным С2-С6-алкинилом; а каждый из радикалов R1 и R2 выбраны из водорода, гидроксила, галогена, замещенной или незамещенной алкокси группы, NJ1J2, SJ1, N3, OC(=X)J1, OC(=X)NJ1J2, NJ3C(=X)NJ1J2 и CN, где X является О, S или NJ1 и каждый заместитель J1, J2 и J3 является независимо H или С1-С6-алкилом.
В некоторых вариантах реализации изобретения предлагаются модифицированные ТГП нуклеозиды Формулы VII, в которых заместители q1, q2, q3, q4, q5, q6 и q7 каждый является H. В некоторых вариантах реализации изобретения по меньшей мере один из заместителей q1, q2, q3, q4, q5, q6 и q7 отличается от Н. В некоторых вариантах реализации изобретения по меньшей мере один из заместителей q1, q2, q3, q4, q5, q6 и q7 является метилом. В некоторых вариантах реализации изобретения предлагаются ТГП нуклеозиды Формулы VII, в которых один из радикалов R1 и R2 является фторогруппой. В некоторых вариантах реализации изобретения R1 является фторогруппой, a R2 является H; R1 является метокси группой, a R2 является Н, и R1 является H, a R2 является метоксиэтокси группой.
В данном документе считается, что термин «2'-модифицированный», или «2'-замещенный», относится к нуклеозиду, содержащему сахар, несущий заместители в положении 2', отличные от Η или ОН. 2'-модифицированные нуклеозиды включают, без ограничения: бициклические нуклеозиды, в которых мостиковое соединение двух углеродных атомов сахарного кольца соединяет углерод 2' и другой углерод сахарного кольца; и нуклеозиды с немостиковыми 2'-заместителями, такими, как аллил, амино, азидо, тио, О-аллил, O-C1-С10-алкил, -OCF3, O-(CH2)2-O-CH3, 2'-O(CH2)2SCH3, O-(CH2)2-O-N(Rm)(Rn) или O-CH2-C(=O)-N(Rm)(Rn), где каждый радикал Rm и Rn независимо является Η или замещенным или незамещенным C1-С10-алкилом. 2'-модифицированые нуклеозиды могут дополнительно содержать другие модификации, например, в других положениях данного сахара и/или в данном нуклеотидном основании.
В данном документе считается, что обозначение «2'-F» относится к нуклеозиду, содержащему сахар, несущий фторогруппу в положении 2'.
В данном документе считается, что каждое обозначение «2'-ОМе», или «2'-ОСН3», или «2'-O-метил»,относится к нуклеозиду, содержащему сахар, несущий -ОСН3 группу в положении 2' сахарного кольца.
В данном документе считается, что каждое обозначение «МОЕ», или «2'-МОЕ», или «2'-ОСН2СН2ОСН3», или «2'-O-метоксиэтил» относится к нуклеозиду, содержащему сахар, несущий -ОСН2СН2ОСН3 группу в положении 2' сахарного кольца.
В данном документе считается, что «олигонуклеотид» относится к соединению, содержащему множество связанных нуклеозидов. В некоторых вариантах реализации изобретения один или более из этого множества нуклеозидов является модифицированным. В некоторых вариантах реализации изобретения олигонуклеотид содержит один и более рибонуклеозидов (РНК) и/или дезоксирибонуклеозидов (ДНК).
В данной области также известны многие другие бицикло- и трициклосахарзаместительные системы колец, которые могут быть использованы для модификации нуклеозидов для введения в антисмысловые соединения (см., например, обзорную статью: Leumann, Bioorg. Med. Chem., 2002, 10, 841-854). Такие системы колец могут претерпевать различные дополнительные замещения для усиления активности.
Способы получения модифицированных сахаров хорошо известны специалистам в данной области.
У нуклеотидов, содержащих модифицированные сахарные фрагменты, фрагменты нуклеотидных оснований (природных, модифицированных или их комбинации) сохраняются для проведения гибридизации с соответствующей нуклеиновой кислотой-мишенью.
В некоторых вариантах реализации изобретения антисмысловые соединения содержат один или более нуклеозидов, несущих модифицированные сахарные фрагменты. В некоторых вариантах реализации изобретения модифицированный сахарный фрагмент является 2'-МОЕ группой. В некоторых вариантах реализации изобретения модифицированные 2'-МОЕ группой нуклеозиды сгруппированы в гэпмер-мотив. В некоторых вариантах реализации изобретения модифицированный сахарный фрагмент является бициклическим нуклеозидом, содержащим мостиковую группу (4'-СН(CH3)-O-2'). В некоторых вариантах реализации изобретения модифицированные (4'-СН(CH3)-O-2') нуклеозиды сгруппированы на протяжении крыльев гэпмер-мотива.
Композиции и способы составления фармацевтических композиций
Антисмысловые олигонуклеотиды могут быть смешаны с фармацевтически приемлемыми действующими или неактивными субстанциями для получения фармацевтических композиций или лекарственных форм. Композиции и способы получения лекарственных форм фармацевтических композиций зависят от ряда критериев, включая, без ограничения, путь введения, степень заболевания и требуемой дозы введения.
Антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, может использоваться в фармацевтических композициях путем сочетания антисмыслового соединения с подходящим фармацевтически приемлемым растворителем или носителем. Фармацевтически приемлемый растворитель включает фосфатно-солевой буферный раствор (ФСБР). ФСБР представляет собой растворитель, пригодный для применения в композициях, которые требуют парентеральной доставки. Соответственно, в одном варианте реализации изобретения, примененном в способах, описанных в данном документе, используется фармацевтическая композиция, содержащая антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72 и фармацевтически приемлемый растворитель. В некоторых вариантах реализации фармацевтически приемлемый растворитель является ФСБР. В некоторых вариантах реализации изобретения это антисмысловое соединение является антисмысловым олигонуклеотидом.
Фармацевтические композиции, содержащие антисмысловые соединения, охватывают любые фармацевтически приемлемые соли, сложные эфиры или соли таких сложных эфиров или любой другой олигонуклеотид, который при введении животному, включая человека, способен обеспечить (прямо или опосредованно) получение биологически активного метаболита или его остатка. Соответственно, например, данное раскрытие также направлено на фармацевтически приемлемые соли антисмысловых соединений, пролекарста, фармацевтически приемлемые соли таких пролекарств и другие биоэквивалентные вещества. Подходящие фармацевтически приемлемые соли включают, без ограничения, соли натрия и калия.
Пролекарство может включать введение дополнительных нуклеозидов в один или оба конца антисмыслового соединения, которое расщепляется эндогенными нуклеазами организма с образованием активного антисмыслового соединения.
Конъюгированные антисмысловые соединения
Антисмысловые соединения могут быть ковалентно связанными с одним или более фрагментами или конъюгатами, которые усиливают активность, распределение в клетках или потребление клетками получающихся антисмысловых олигонуклеотидов. Типичные конъюгирующие группы включают холестериновые фрагменты и липидные фрагменты. Дополнительные конъюгирующие группы включают углеводы, фосфолипиды, биотин, феназин, фолат, фенантридин, антрахинон, акридин, флуоресцеины, родамины, кумарины и красители.
Антисмысловые соединения могут быть модифицированы таким образом, чтобы содержать одну и более стабилизирующих групп, которые обычно присоединяются к одному или обоим концам антисмысловых соединений для усиления свойств, таких, как, например, стабильность к нуклеазам, включая кэп-структуры в составе стабилизирующих групп. Эти концевые модификации защищают антисмысловое соединение, несущее концевую нуклеиновую кислоту, от экзонуклеазной деградации и могут помочь при доставке и/или локализации соединения в клетке. Кэп может быть расположен на 5'-конце (5'-кэп) или на 3'-конце (3'-кэп) или может быть расположен на обоих концах. Кэп-структуры хорошо известны в данной области и включают, например, обращенные дезоксикэпы без азотистых оснований.
Дополнительно 3' и 5'-стабилизирующие группы, которые могут быть использованы на кэпе одного или обоих концов антисмыслового соединения для придания ему стабильности от нуклеаз, включают те, которые описаны в заявке WO 03/004602, опубликованной 16 января 2003 г.
Клеточная культура и обработка антисмысловыми соединениями
Действие антисмысловых соединений на уровень, активность или экспрессию нуклеиновых кислот C9ORF72 может быть протестировано in vitro на различных типах клеток. Типы клеток, используемые в таких анализах, могут быть получены у коммерческих поставщиков (например, Американская коллекция типовых культур, Manassus, VA; Zen-Bio, Inc., Research Triangle Park, NC; Clonetics Corporation, Walkersville, MD) и культивированы в соответствии с инструкциями поставщиков с использованием коммерчески доступных реактивов (например, In vitro gen Life Technologies, Carlsbad, CA). Показательные типы клеток включают, без ограничения, клетки HepG2, клетки Hep3B и первичные гепатоциты.
Испытание антисмысловых олигонуклеотидов in vitro
В данном документе описаны способы для обработки клеток антисмысловыми олигонуклеотидами, которые могут быть надлежащим образом модифицированы для обработки другими антисмысловыми соединениями.
В целом, клетки обрабатывают антисмысловыми олигонуклеотидами, когда клетки достигают в культуре приблизительно 60-80% заселенности.
Один реактив, обычно используемый для введения антисмысловых олигонуклеотидов в культивируемые клетки, включает катионный липидный реактив для трансфекции LIPOFECTIN (Invitrogen, Carlsbad, СА). Антисмысловые олигонуклеотиды смешивают с реактивом LIPOFECTIN в среде OPTI-MEM 1 (Invitrogen, Carlsbad, СА) до достижения требуемой конечной концентрации антисмыслового олигонуклеотида и концентрации LIPOFECTIN, которая обычно лежит в диапазоне 2-12 мкг/мл на 100 нМ антисмыслового олигонуклеотида.
Другой реактив, используемый для введения антисмысловых олигонуклеотидов в культивируемые клетки, включает реактив LIPOFECTAMINE (Invitrogen, Carlsbad, СА). Антисмысловой олигонуклеотид смешивают с реактивом LIPOFECTAMINE в среде с пониженным содержанием сыворотки OPTI-MEM 1 (Invitrogen, Carlsbad, СА) до достижения требуемой концентрации антисмыслового олигонуклеотида и концентрации LIPOFECTAMINE, которая обычно лежит в диапазоне 2-12 мкг/мл на 100 нМ антисмыслового олигонуклеотида.
Другая методика, используемая для введения антисмысловых олигонуклеотидов в культивируемые клетки, включает электропорацию.
Клетки обрабатывают антисмысловыми олигонуклеотидами традиционными способами. Клетки после обработки антисмысловыми олигонуклеотидами, как правило, выращивают 16-24 часов, в это время измеряют уровни РНК или белка нуклеиновой кислоты-мишени методами, известными в данной области и описанные в данном документе. В общем, когда обработки выполняют в многократных повторах, то данные представляют как средние значения повторных обработок.
Концентрация антисмыслового олигонуклеотида, как правило, варьирует для разных клеточных линий. Методы определения оптимальной концентрации антисмыслового олигонуклеотида для конкретной клеточной линии хорошо известны в данной области. Антисмысловые олигонуклеотиды при трансфекции с помощью реактива LIPOFECTAMINE часто используют в концентрациях в диапазонах от 1 нМ до 300 нМ. Антисмысловые олигонуклеотиды при трансфекции с помощью электропорации используют в более высоких концентрациях в диапазоне от 625 до 20000 нМ.
Выделение РНК
Анализ РНК может выполняться на суммарной клеточной РНК или поли(А)+ мРНК. Методы выделения РНК хорошо известны в данной области. РНК получают, используя методы хорошо известные в данной области, например, с применением реактива TRIZOL (Invitrogen, Carlsbad, СА) в соответствии с рекомендованными протоколами производителя.
Анализ ингибирования уровней или экспрессии мишеней
Ингибирование уровней или экспрессии нуклеиновой кислоты C9ORF72 может быть проанализировано разнообразными способами, известными в данной области. Например, нуклеиновая кислота-мишень можно количественно определить, например, путем Нозерн-блоттинг анализа, конкурентной полимеразной цепной реакции (ПЦР) или количественной ПЦР в режиме реального времени. Анализ РНК может выполняться на суммарной клеточной РНК или поли(А)+ мРНК. Методы выделения РНК хорошо известны в данной области.
Анализ Нозерн-блоттинга также является традиционным в этой области. Количественная ПЦР в режиме реального времени может быть беспрепятственно выполнена с использованием коммерчески доступной системы для детекции последовательностей ABI PRISM 7600, 7700 или 7900, поставляемой компанией PE-AppliedBiosystems, FosterCity, СА и используемой согласно инструкций производителя.
Анализ уровней РНК-мишеней методом количественной ПЦР в режиме реального времени Количественное определение уровней РНК-мишеней может выполняться методом количественной ПЦР в режиме реального времени, используя систему для детекции последовательностей ABI PRISM 7600, 7700 или 7900 (PE-AppliedBiosystems, FosterCity, СА) согласно инструкций производителя. Методы количественного определения ПЦР в режиме реального времени хорошо известны в данной области.
Перед проведением ПЦР в режиме реального времени выделенную РНК подвергают реакции с обратной транскриптазой (ОТ), которая синтезирует комплементарную ДНК (кДНК), что потом используется в качестве субстрата для амплификации в ПЦР в режиме реального времени. Реакции ПЦР ОТ и ПЦР в режиме реального времени выполняются последовательно в одной и том же лунке с образцом. Реактивы для ПЦР ОТ и ПЦР в режиме реального времени получают в компании Invitrogen (Carlsbad, СА). Реакции ПЦР-ОТ и ПЦР в режиме реального времени проводят методами, хорошо известными специалистам в данной области.
Количества гена (или РНК)-мишени, полученные методом ПЦР в режиме реального времени нормализируют, используя любой уровень экспрессии гена, экспрессия которого является постоянной, такой, как циклофилин А, или путем количественного определения суммарной РНК, используя краситель RIBOGREEN (Invitrogen, Inc. Carlsbad, СА). Экспрессию циклофилина А количественно определяют ПЦР в режиме реального времени, одновременно проводя анализ с мишенью путем мультиплексирования или анализируя по отдельности. Суммарную РНК количественно определяют, используя реактив для количественного определения РНК RIBOGREEN (Invetrogen, Inc. Eugene, OR). Методы количественного определения РНК с помощью красителя RIBOGREEN изложены в статье Jones, L.J., et al, (Analytical Biochemistry, 1998, 265, 368-374). Для измерения флуоресценции RIBOGREEN используют прибор CYTOFLUOR 4000 (РЕ Applied Biosystems).
Зонды и праймеры разработаны для гибридизации с нуклеиновой кислотой C9ORF72. Способы для конструирования зондов и праймеров для ПЦР в режиме реального времени хорошо известны в данной области и могут включать применение программного обеспечения, такого, как программное обеспечение PRIM EREXPRESS (AppliedBiosystems, FosterCity, СА).
Анализ уровней белков
Антисмысловое ингибирование нуклеиновой кислоты C9ORF72 может оцениваться путем измерения уровней белка C9ORF72. Уровни белка C9ORF72 могут быть оценены или количественно определены различными способами, хорошо известными в данной области, такими как иммунопреципитация, Вестерн-блоттинг анализ (иммуноблоттинг), твердофазный иммуноферментный анализ (ELISA), анализы количественного определения белков, анализы активности белков (например, анализы активности каспазы), иммуногистохимические методы, иммуноцитохимические методы или сортировка флуоресцентно-активированных клеток (FACS). Антитела, направленные на мишень, могут быть идентифицированы и получены из различных источников, таких как каталог антител MSRS (Aerie Corporation, Birmingham, MI), или могут быть получены посредством традиционных методов синтеза моноклональных или поликлональных антител, хорошо известных в данной области. Коммерчески доступные антитела пригодны для обнаружения белка C9ORF72 мыши, крысы, обезьяны и человека.
Испытание антисмысловых соединений in vivo
Антисмысловые соединения, например антисмысловые олигонуклеотиды, испытывают на животных для оценки их способности ингибировать экспрессию белка C9ORF72 и вызывать фенотипические изменения, такие как улучшение двигательной функции и дыхания. В некоторых вариантах реализации изобретения двигательную функцию измеряют с помощью теста с вращающимся стержнем, по силе захвата, лазанию по шесту, активности в тесте «открытое поле», в тесте с балансиром, испытанию по отпечатку задней лапы животного. В некоторых вариантах реализации изобретения интенсивность дыхания измеряют плетисмографией всего организма, устойчивостью к заражению и соответствующими измерениями на животном. Испытание может выполняться на здоровых животных или на экспериментальных моделях заболеваний. Для введения животным антисмысловые олигонуклеотиды изготавливают в виде лекарственной формы с фармацевтически приемлемым растворителем, таким как фосфатно-солевой буферный раствор. Введение включает парентеральные пути введения, такие как внутрибрюшинный, внутривенный и подкожный. Расчет дозы антисмысловых олигонуклеотидов и частоты дозирования находятся в пределах квалификации специалистов в данной области и зависит от таких факторов, как путь введения и масса тела животного. В последующем периоде времени после обработки антисмысловыми олигонуклеотидами выделяют РНК из ткани ЦНС или СМЖ и измеряют изменения экспрессии нуклеиновой кислоты C9ORF72.
Нацеливание на C9ORF72
Антисмысловые олигонуклеотиды, описанные в данном документе, могут гибридизироваться с нуклеиновой кислотой C9ORF72 на любой стадии процессинга РНК. Например, описанные в данном документе предложенные антисмысловые олигонуклеотиды являются комплементарными пре-мРНК или зрелой мРНК. Дополнительно, антисмысловые олигонуклеотиды, описанные в данном документе, могут гибридизироваться с любым элементом нуклеиновой кислоты C9ORF72. Например, описанные в данном документе предложенные антисмысловые олигонуклеотиды являются комплементарными экзону, интрону, 5'-нетраслируемому участку, 3'-нетраслируемому участку, участку повтора, экспансии гексануклеотидного повтора, точке сплайсинга, точке сплайсинга экзонгэкзон, сайленсеру экзонного сплайсинга (ESS), энхансеру экзонного сплайсинга (ESE), экзону 1а, экзону 1b, экзону 1с, экзону 1d, экзону 1e, экзону 2, экзону 3, экзону 4, экзону 5, экзону 6, экзону 7, экзону 8, экзону 9, экзону 10, экзону 11, интрону 1, интрону 2, интрону 3, интрону 4, интрону 5, интрону 6, интрону 7, интрону 8, интрону 9 или интрону 10 нуклеиновой кислоты C9ORF72.
В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, гибридизируются со всеми вариантами C9ORF72. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, гибридизируются с определенными вариантами C9ORF72. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, избирательно гибридизируются с вариантами C9ORF72, содержащими экспансии гексануклеотидного повтора. В некоторых вариантах реализации изобретения такие варианты C9ORF72, содержащие экспансии гексануклеотидного повтора, включают последовательности SEQ ID NO: 1-3 и 6-10. В некоторых вариантах реализации изобретения такая экспансия гексануклеотидного повтора включает по меньшей мере 30 повторов из любых наборов GGGGCC, GGGGGG, GGGGCG или GGGGGC.
В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, ингибируют экспрессию всех вариантов C9ORF72. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, ингибируют экспрессию всех вариантов C9ORF72 в равной степени. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, преимущественно ингибируют экспрессию определенных вариантов C9ORF72. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, преимущественно ингибируют экспрессию вариантов C9ORF72, содержащих экспансию гексануклеотидного повтора. В некоторых вариантах реализации изобретения такие варианты C9ORF72, содержащие экспансию гексануклеотидного повтора, включают последовательности SEQ ID NO: 1-3 и 6-10. В некоторых вариантах реализации изобретения такая экспансия гексануклеотидного повтора включает по меньшей мере 30 повторов из любых наборов GGGGCC, GGGGGG, GGGGCG или GGGGGC. В некоторых вариантах реализации изобретения такая экспансия гексануклеотидного повтора образует фокусы в ядрах клеток. В некоторых вариантах реализации изобретения антисмысловые олигонуклеотиды, описанные в данном документе, пригодны для уменьшения фокусов в ядрах клеток. Фокусы ядер клеток могут быть уменьшены по показателю процента клеток с фокусами, а также количества фокусов на клетку.
На основании ранних исследований, направленных на изучение экспансий повторов, невозможно предсказать, смогут ли антисмысловые олигонуклеотиды, нацеленные на последовательность C9ORF72 за пределами экспансии повтора, успешно ингибировать экспрессию C9ORF72 по двум причинам. Первое - экспансия повтора C9ORF72 расположена в интроне и не известно, содержит ли РНК в фокусах только повторы или также и фланкирующую интронную последовательность. Например, в ранних исследованиях миотонической дистрофии 2 типа (ДМ2), которая является заболеванием, вызываемым мутацией экспансии CCTG в интроне 1 гена ZNF9, определили, что большие экспансии при ДМ2 не предотвращают аллельноспецифический сплайсинг пре-мРНК, ядерный экспорт этих транскриптов или установившиеся уровни мРНК. Данное исследование дополнительно продемонстрировало, что рибонуклеотидные ядерные включения, обнаруженные в связи с этим заболеванием, обогащены экспансией CCUG, но не фланкирующими интронными последовательностями. Благодаря этим данным предположили, что последующие молекулярные эффекты при мутации ДМ2 могут быть инициированы накоплением единственного участка повтора CCUG. Поэтому это исследование подразумевает, что нацеливание на единственную экспансию повтора CCUG может привести к облегчению протекания данного заболевания, поскольку нацеливание на фланкирующие последовательности, особенно на следующий в прямом направлении считывания участок этой экспансии повтора, не влияет на образование рибонуклеотидных ядерных включений (Margolis et al. Hum. Mol. Genet., 2006, 15: 1808-1815). Второе - не известно, насколько быстро интрон 1 гена C9ORF72, который содержит данные повторы, отщепляется и накапливается в фокусах. Поэтому невозможно предсказать, приведет ли нацеливание на пре-мРНК к удалению повторов РНК и фокусов.
Особенности C9OFF72
Антисмысловые олигонуклеотиды, описанные в данном документе, могут гибридизироваться с любым вариантом последовательности C9ORF72 в любом состоянии процессинга в пределах любого элемента гена C9ORF72. Например, антисмысловые олигонуклеотиды, описанные в данном документе, могут гибридизироваться с экзоном, интроном, 5'-нетраслируемым участком, 3'-нетраслируемым участком, участком повтора, экспансией гексануклеотидного повтора, точкой сплайсинга, экзоном: точкой сплайсинга экзона, сайленсером экзонного сплайсинга (ESS), энхансером экзонного сплайсинга (ESE), экзоном 1а, экзоном 1b, экзоном 1с, экзоном 1d, экзоном 1e, экзоном 2, экзоном 3, экзоном 4, экзоном 5, экзоном 6, экзоном 7, экзоном 8, экзоном 9, экзоном 10, экзоном 11, интроном 1, интроном 2, интроном 3, интроном 4, интроном 5, интроном 6, интроном 7, интроном 8, интроном 9 или интроном 10. К примеру, антисмысловые олигонуклеотиды могут нацеливаться на любой из экзонов, описанных в табл.1-5 различных вариантов последовательностей C9ORF72. Антисмысловые олигонуклеотиды, описанные в данном документе, также могут нацеливаться на варианты не описанные ниже, такие, как описанные в базе данных GENBANK. Более того, антисмысловые олигонуклеотиды, описанные в данном документе, также могут нацеливаться на элементы отличные от экзонов, и такие элементы описаны в базе данных GENBANK.
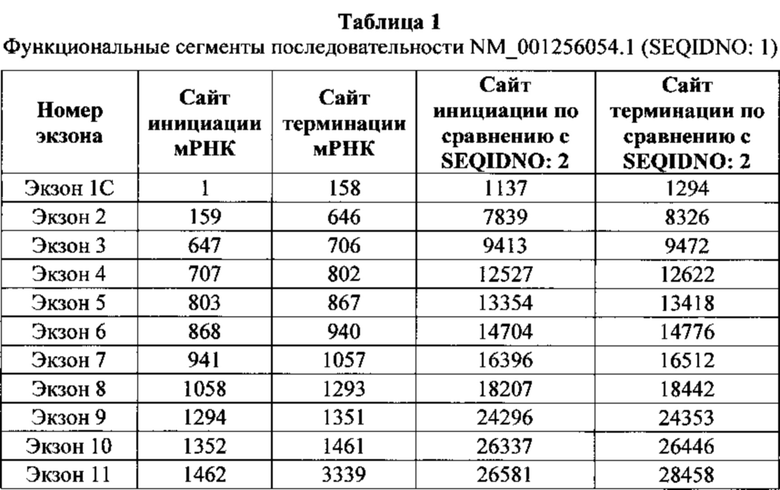





Некоторые показания для применения
В некоторых вариантах реализации изобретения в данном документе предложены способы лечения индивидов, включающие введение одной или более фармацевтических композиций, описанных в данном документе. В некоторых вариантах реализации изобретения индивид страдает нейродегенеративным заболеванием. В некоторых вариантах реализации изобретения у индивида существует риск развития нейродегенеративного заболевания, включая, без ограничения, АЛС или ЛВД. В некоторых вариантах реализации изобретения индивида идентифицировали как страдающего ассоциированным с белком C9ORF72 заболеванием. В некоторых вариантах реализации изобретения индивида идентифицировали как страдающего ассоциированным с экспансией гексануклеотидного повтора C9ORF72 заболеванием. В некоторых вариантах реализации изобретения в данном документе предложены способы для профилактического снижения у индивида экспрессии C9ORF72. Некоторые варианты реализации изобретения включают нуждающегося в лечении индивида путем введения ему терапевтически эффективного количества антисмыслового соединения, нацеленного на нуклеиновую кислоту C9ORF72.
В одном варианте реализации введение терапевтически эффективного количества антисмыслового соединения, нацеленного на нуклеиновую кислоту C9ORF72, сопровождается мониторингом у индивида уровней C9ORF72 для определения ответа индивида на введение данного антисмыслового соединения. Ответ индивида на введение антисмыслового соединения может использоваться врачом для определения количества и длительности терапевтического воздействия.
В некоторых вариантах реализации изобретения введение антисмыслового соединения, нацеленного на нуклеиновую кислоту C9ORF72, приводит к снижению экспрессии C9ORF72 на по меньшей мере 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99%, или на диапазон, определенный любыми двумя из этих значений. В некоторых вариантах реализации изобретения антисмысловое соединение, нацеленное на нуклеиновую кислоту C9ORF72, приводит к улучшению двигательной функции и дыхания животного. В некоторых вариантах реализации изобретения введение антисмыслового соединения к C9ORF72 улучшает двигательную функцию и дыхание на по меньшей мере 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99%, или на диапазон, определенный любыми двумя из этих значений.
В некоторых вариантах реализации изобретения фармацевтические композиции, содержащие антисмысловое соединение, нацеленное на C9ORF72, используют для получения лекарственного средства для лечения пациента, страдающего или склонного к нейродегенеративному заболеванию, включая АЛС и ЛВД.
Определенные виды комбинированной терапии
В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, совместно вводят с одним или большим количеством других фармацевтических агентов. В некоторых вариантах реализации изобретения такая одна или более других фармацевтических композиций разработаны для лечения того же заболевания, нарушения или состояния, что и одна или более фармацевтических композиций, описанных в данном документе. В некоторых вариантах реализации изобретения такая одна или более других фармацевтических композиций разработаны для лечения разных заболеваний, нарушений или состояний, что и одна или более фармацевтических композиций, описанных в данном документе. В некоторых вариантах реализации изобретения такая одна или более других фармацевтических композиций разработаны для лечения нежелательной побочной реакции одной или более фармацевтических композиций, описанных в данном документе. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, совместно вводят с другим фармацевтическим агентом для лечения нежелательной реакции, вызванной другим фармацевтическим агентом. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, совместно вводят с другим фармацевтическим агентом для создания комбинированного действия. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, совместно вводят с другим фармацевтическим агентом для создания синергического действия.
В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, и один или более других фармацевтических агентов вводятся в одно и то же время. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, и один или более других фармацевтических агентов вводятся в разное время. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, и один или более других фармацевтических агентов изготавливают вместе в виде одной лекарственной формы. В некоторых вариантах реализации изобретения одна или более фармацевтических композиций, описанных в данном документе, и один или более других фармацевтических агентов изготавливают по отдельности.
В некоторых вариантах реализации изобретения фармацевтические агенты, которые могут совместно вводиться с фармацевтической композицией, описанной в данном документе, включают препараты Рилузол (Рилутек), Лиоресал (Лиоресал) и Декспрамипексол.
В некоторых вариантах реализации изобретения фармацевтические агенты, которые могут совместно вводиться со специфическим ингибитором C9ORF72, описанным в данном документе, включая без ограничения дополнительный ингибитор C9ORF72. В некоторых вариантах реализации изобретения данный совместно вводимый фармацевтический агент вводится перед введением фармацевтической композиции, описанной в данном документе. В некоторых вариантах реализации изобретения данный совместно вводимый фармацевтический агент вводится после введения фармацевтической композиции, описанной в данном документе. В некоторых вариантах реализации изобретения данный совместно вводимый фармацевтический агент вводится в то же время, что и фармацевтическая композиция, описанная в данном документе. В некоторых вариантах реализации изобретения доза совместно вводимого фармацевтического агента является такой же, что и доза, которая могла бы вводиться, если совместно вводимый фармацевтический агент вводился бы самостоятельно. В некоторых вариантах реализации изобретения доза совместно вводимого фармацевтического агента является более низкой, чем доза, которая могла бы вводиться, если совместно вводимый фармацевтический агент вводился бы самостоятельно. В некоторых вариантах реализации изобретения доза совместно вводимого фармацевтического агента является более высокой, чем доза, которая могла бы вводиться, если совместно вводимый фармацевтический агент вводился бы самостоятельно.
В некоторых вариантах реализации изобретения совместное введение второго соединения усиливает действие первого соединения так, что совместное введение данных соединений приводит к действию, которое превышает действие вводимого самостоятельно первого соединения. В других вариантах реализации изобретения данное совместное введение приводит к действию, которое является аддитивным в отношении действия соединений, назначаемых самостоятельно. В некоторых вариантах реализации изобретения данное совместное введение приводит к действию, которое является сверхаддитивным в отношении действия соединений, назначаемых самостоятельно. В некоторых вариантах реализации изобретения первое соединение является антисмысловым. В некоторых вариантах реализации изобретения второе соединение является антисмысловым.
ПРИМЕРЫ
Примеры, не ограничивающие раскрытие и данные, включенные посредством ссылки.
В то время как определенные соединения, композиции и способы, описанные в данном документе, были описаны с характеристикой в соответствии с определенными вариантами реализации, следующие примеры только иллюстрируют соединения, описанные в данном документе, и не подразумевают ограничение указанных объектов. Каждая из ссылок, перечисленных в настоящей заявке, включена в данный документ в полном объеме посредством ссылки.
Пример 1. Антисмысловое ингибирование гена C9ORF72 человека в клетках HepG2
Антисмысловые олигонуклеотиды были сконструированы для нацеливания на нуклеиновую кислоту C9ORF72 и их действие было испытано на мРНК C9ORF72 in vitro. Данные антисмысловые олигонуклеотиды испытывали в сериях экспериментов, которые обеспечивали сходные культуральные условия. Результаты каждого эксперимента представлены в отдельных таблицах, показанных ниже. Культивированные до плотности 20000 клеток на лунку клетки HepG2 трансфецировали с применением электропорации с помощью антисмыслового олигонуклеотида в концентрации 7000 нМ. По прошествии периода обработки в течение приблизительно 24 часов, РНК выделили из клеток и измерили уровни мРНК C9ORF72 методом количественной ПЦР в режиме реального времени. Для измерения уровней мРНК использовали набор RTS3750 праймеров-зондов человека (прямая последовательность TGTGACAGTTGGAATGCAGTGA, обозначенная в данном документе как SEQ ID NO: 15; обратная последовательность GCCACTTAAAGCAATCTCTGTCTTG, обозначенная в данном документе как SEQ ID NO: 16; последовательность зонда TCGACTCTTTGCCCACCGCCA, обозначенная в данном документе как SEQ ID NO: 17). Уровни мРНК C9ORF72 скорректировали в соответствии с суммарным содержанием РНК, измеренным с помощью красителя RIBOGREEN®. Результаты представлены в виде процента ингибирования C9ORF72 по сравнению с необработанными контрольными клетками.
Антисмысловые олигонуклеотиды в табл. 6-10 были сконструированы в виде 5-10-5 МОЕ-гэпмеров. Данные гэпмеры составлены из 20 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый включает сегмент по 5 нуклеозидов. Каждый нуклеозид на сегменте-крыле 5' и каждый нуклеозид на сегменте-крыле 3' несут модификацию МОЕ. Межнуклеозидные связи на всем протяжении каждого гэпмера являются фосфоротиоатными связями. Все цитозиновые остатки на всем протяжении каждого гэпмера являются 5-метилцитозинами. «Сайт инициации» указывает на самый крайний 5'-нуклеозид последовательности гена человека, на который нацелен антисмысловой олигонуклеотид. «Сайт терминации» указывает на самый крайний 3'-нуклеозид последовательности гена человека, на который нацелен антисмысловой олигонуклеотид. Каждый антисмысловой олигонуклеотид, перечисленный в табл. 6-9, нацелен на одну из последовательностей м РНК C9ORF72 человека, обозначенных в данном документе как SEQ ID NO: 1 (№ доступа в базе данных GENBANK NM 001256054.1) или геномную последовательность C9ORF72 человека, обозначенную в данном документе как SEQ ID NO: 2 (комплементарная последовательность № доступа в базе данных GENBANK NT 008413.18, усеченная от нуклеозида 27535000 до 27565000), или обе последовательности. Сокращение «н/д» указывает на то, что данный антисмысловой олигонуклеотид не был нацелен на эту конкретную последовательность гена. Антисмысловые олигонуклеотиды из табл. 10 нацелены на одну из последовательностей SEQ ID NO: 3 (№ доступа в базе данных GENBANK BQ068108.1) или SEQ ID NO: 4 (№ доступа в базе данных GENBANK NM_018325.3).
Как показано в табл. 6-10, некоторые из олигонуклеотидов, нацеленных на SEQ ID NO: 1, проявляют по меньшей мере 50% ингибирование, включая те, которые нацелены на нуклеотидные основания 90-647, 728-1541, 1598-1863, 1935-2146, 2232-2251, 2429-2576, 2632-2743, 2788-2807, 2860-2879, 2949-2968, 3062-3081, 3132-3151 и 3250-3269 SEQ ID NO 1. Они включают последовательности SEQ ID NO: 32, 33, 34, 35, 36, 37, 38, 40, 41, 42, 43, 44, 45, 46, 47, 50, 51, 53, 55, 56, 57, 61, 62, 64, 66, 67, 72, 73, 75, 76, 81, 82, 85, 89, 90, 91, 92, 93, 94, 96, 97, 100, 102, 103, 109, 111, 112, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 130, 131, 132, 133, 137, 139, 140, 141, 145, 146, 149, 150, 151, 152, 153, 154, 165, 166, 168, 169, 170, 171, 174, 179, 181, 182, 183, 185, 186, 187, 188, 190, 192, 195, 197, 199, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331 и 332. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 70% ингибирование, включая те, которые нацелены на нуклеотидные основания 90-359, 430-479, 550-569, 617-647, 940-959, 1013-1033, 1446-1465, 1687-1706, 1844-1863, 1935-2007 и 2679-2698 SEQ ID NO 1. Они включают последовательности SEQ ID NO: 32, 33, 34, 35, 36, 40, 41, 42, 43, 44, 47, 66, 67, 85, 96, 103, 117, 119, 154, 165, 168, 186, 320, 321, 324, 327, 328 и 331. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 80% ингибирование, включая те, которые нацелены на нуклеотидные основания 90-265 и 310-329. Они включают последовательности SEQ ID NO: 32, 33, 35, 40, 42 и 321. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 90% ингибирование, включая те, которые нацелены на нуклеотидные основания 190-209 и 310-329 SEQ ID NO 1. Они включают последовательности SEQ ID NO: 40 и 321.
Как показано в табл. 6-20, некоторые из олигонуклеотидов, нацеленных на SEQ ID NO: 2, проявляют по меньшей мере 50% ингибирование, включая те, которые нацелены на нуклеотидные основания 1552-1572, 2187-2238, 2728-2779, 3452-2471, 3752-3771, 5025-5044, 5656-5675, 6200-6219, 7594-7613, 7840-8328, 9415-9434, 12526-12545, 13357-13524, 13642-13661, 13790-14130, 14243-14335, 14699-14777, 15587-15606, 16395-16488, 18233-18373, 24306-24340, 24472-24491, 24565-24676, 26400-26424, 26606-26982, 27054-27265, 27351-27370, 27548-27998, 28068-28087, 28181-28270 и 28369-28388 SEQ ID NO 2. Они включают последовательности SEQ ID NO: 32, 33, 34, 35, 36, 37, 38, 40, 41, 42, 43, 44, 45, 46, 47, 50, 51, 53, 55, 56, 57, 64, 67, 72, 73, 75, 76, 81, 82, 85, 89, 90, 91, 92, 93, 94, 96, 97, 100, 102, 103, 111, 112, 115, 117, 118, 119, 121, 122, 123, 124, 125, 126, 130, 131, 132, 133, 137, 139, 140, 141, 145, 146, 149, 150, 151, 152, 153, 154, 165, 166, 168, 169, 170, 171, 174, 179, 181, 182, 183, 185, 186, 187, 188, 190, 192, 195, 197, 199, 205, 206, 208, 211, 212, 224, 226, 230, 231, 250, 251, 252, 256, 300, 301, 304, 306, 307, 310, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331 и 332. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 70% ингибирование, включая те, которые нацелены на нуклеотидные основания 3452-2471, 7840-8159, 8230-8249, 12526-12545, 13642-13661, 14075-14094, 14316-14335, 14758-14777, 16395-16414, 16469, 16488, 24655-24674, 26963, 26982, 27054-27126 и 27798-27817 SEQ ID NO 2. Они включают последовательности SEQ ID NO: 32, 33, 34, 35, 36, 40, 41, 42, 43, 44, 47, 67, 85, 96, 103, 117, 119, 154, 165, 168, 186, 251, 306, 320, 321, 324, 327, 328 и 331. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 80% ингибирование, включая те, которые нацелены на нуклеотидные основания 7848-8023 SEQ ID NO 2. Они включают последовательности SEQ ID NO: 32, 33, 35, 40, 42 и 321. Некоторые из этих олигонуклеотидов проявляют по меньшей мере 90% ингибирование, включая те, которые нацелены на нуклеотидные основания 7870-7889 и 7990-8009 SEQ ID NO 2. Они включают последовательности SEQ ID NO: 40 и 321.



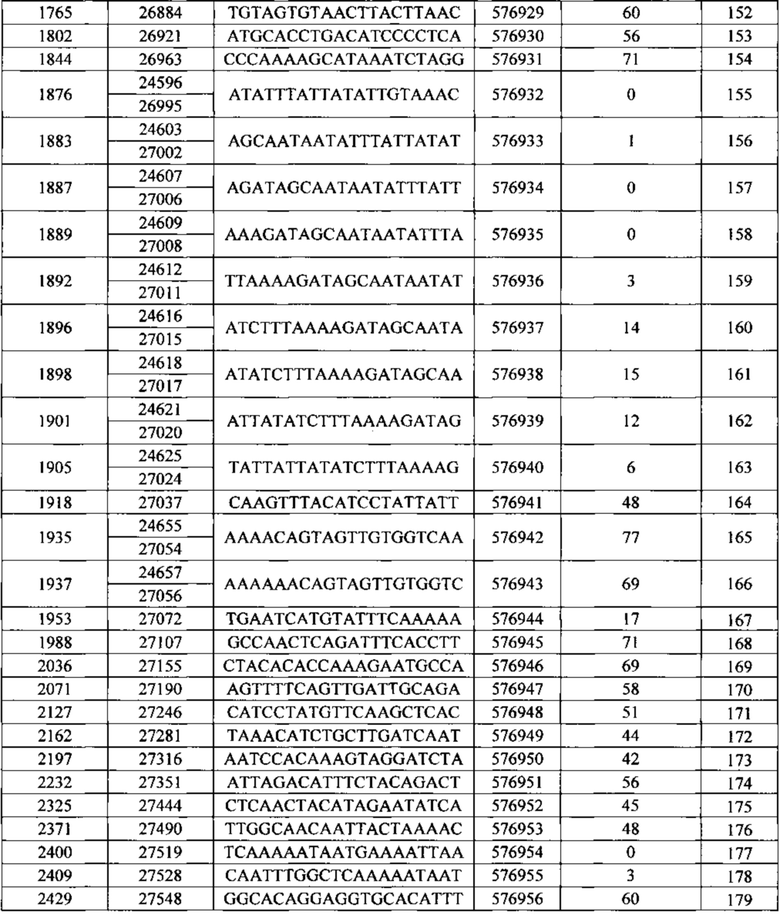

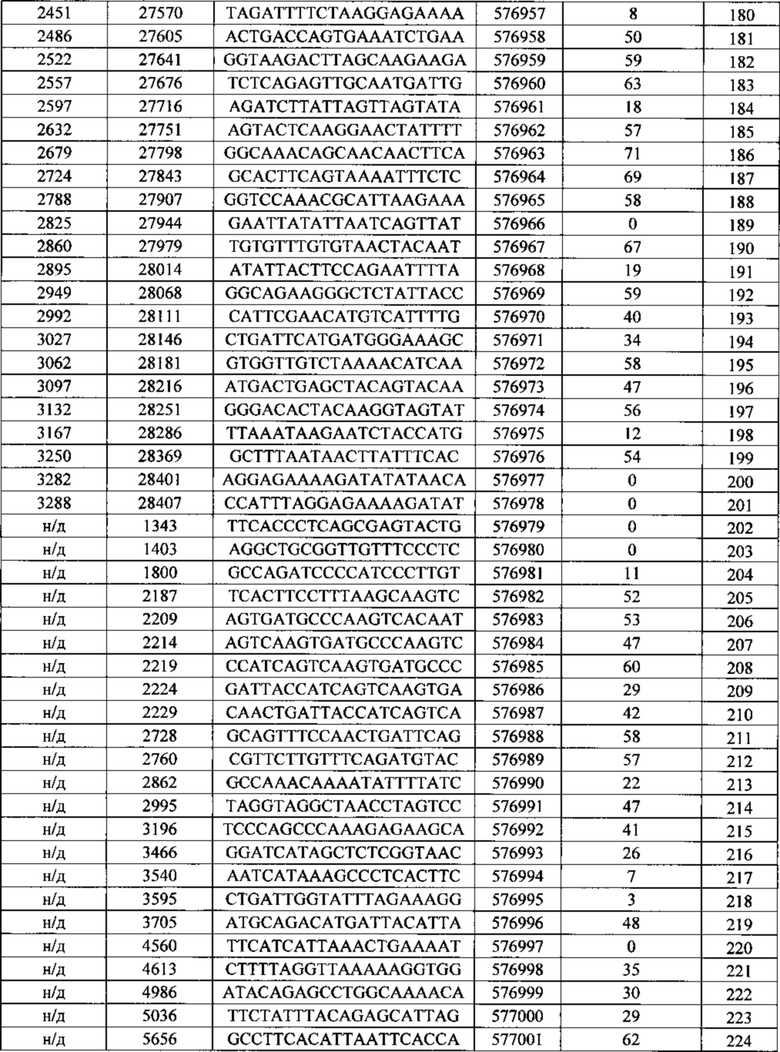


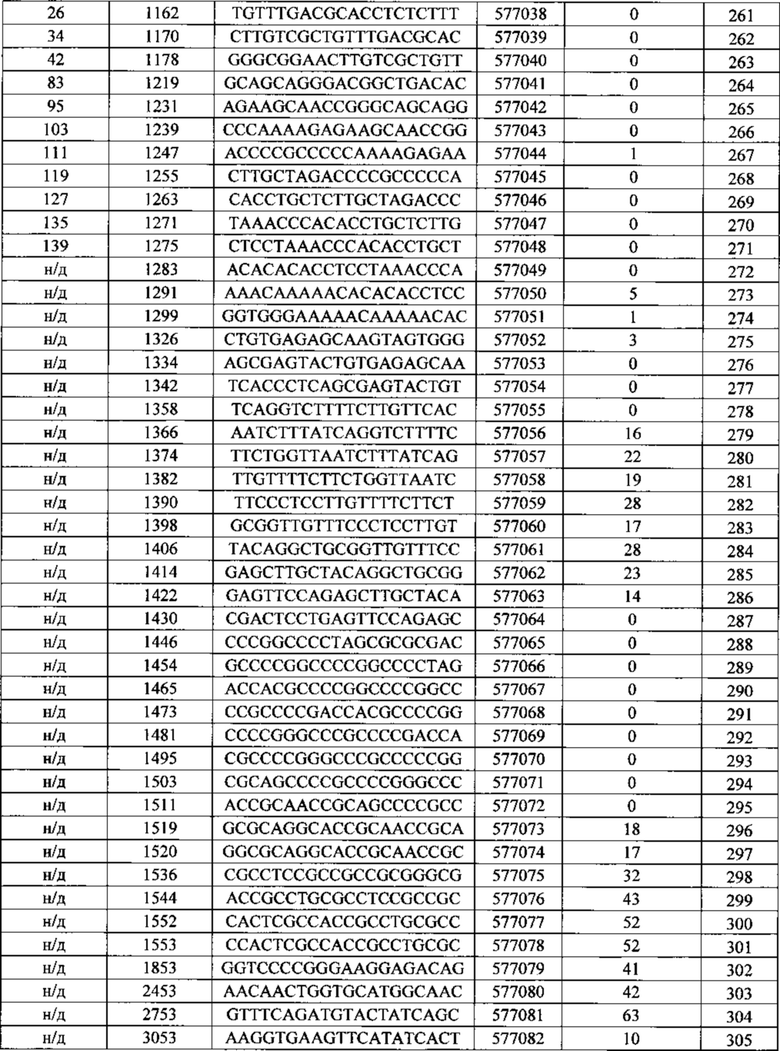
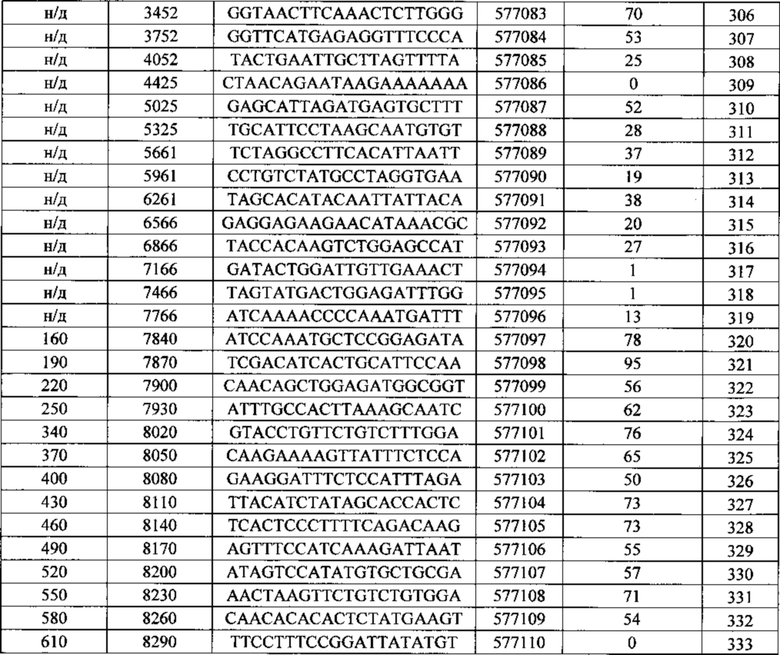

Пример 2. Дозозависимое антисмысловое ингибирование гена C9ORF72 человека в 5 клетках HepG2
Антисмысловые олигонуклеотиды из исследования, описанного выше, проявившие значительное ингибирование мРНК C9ORF72 in vitro, были выбраны и испытаны в различных дозах на клетках HepG2. Данные антисмысловые олигонуклеотиды испытывали в сериях экспериментов, которые обеспечивали сходные культуральные условия. Результаты каждого эксперимента представлены в отдельных таблицах, показанных ниже. Клетки высадили на планшеты в плотности 20000 клеток на лунку и трансфецировали с применением электропорации с помощью концентраций антисмыслового олигонуклеотида 82,3 нМ; 246,9 нМ; 740,7 нМ; 2222,2 нМ; 6666,7 нМ или 20000 нМ. По прошествии периода обработки в течение приблизительно 16 часов РНК выделили из клеток и измерили уровни мРНК C9ORF72 методом количественной ПЦР в режиме реального времени. Для измерения уровней мРНК использовали набор RTS3750 праймеров-зондов для последовательности C9ORF72 человека. Уровни мРНК C9ORF72 скорректировали в соответствии с суммарным содержанием РНК, измеренным с помощью красителя RIBOGREEN®. Результаты представлены в виде процента ингибирования C9ORF72 по сравнению с необработанными контрольными клетками.
Концентрация полумаксимального ингибирования (IC50) для каждого олигонуклеотида также представлена в табл. 11-13. Как показано, уровни мРНК C9ORF72 были снижены дозозависимым способом в обработанных антисмысловыми олигонуклеотидами клетках.
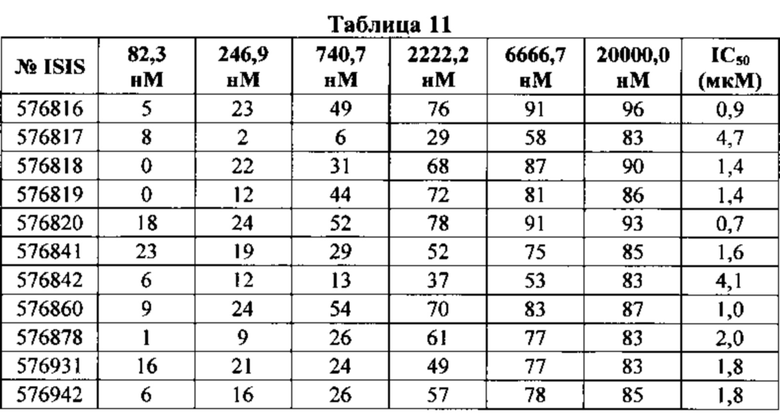

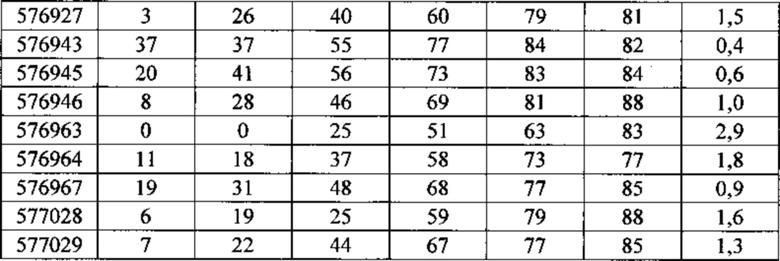
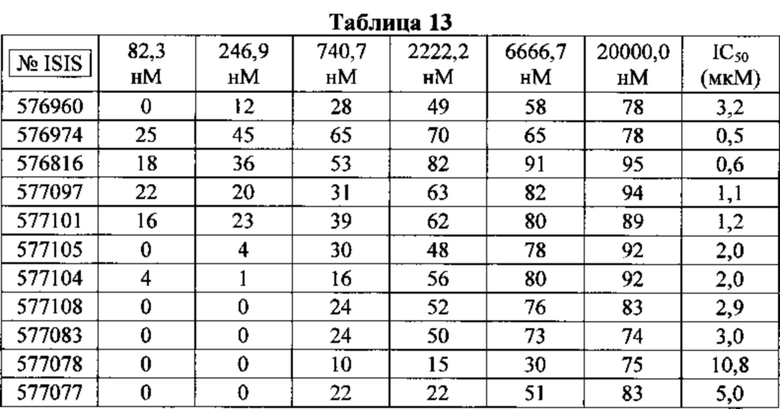
Пример 3. Дозозависимое антисмысловое ингибирование гена C9ORF72 человека в клетках HepG2
Антисмысловые олигонуклеотиды из исследования, описанного выше, проявившие значительное ингибирование мРНК C9ORF72 in vitro, были выбраны и испытаны в различных дозах на клетках HepG2. Данные антисмысловые олигонуклеотиды испытывали в сериях экспериментов, которые обеспечивали сходные культуральные условия. Результаты каждого эксперимента представлены в отдельных таблицах, показанных ниже. Клетки высадили на планшеты в плотности 20000 клеток на лунку и трансфецировали с применением электропорации с помощью концентраций антисмыслового олигонуклеотида 246,9 нМ; 740,7 нМ; 2222,2 нМ; 6666,7 нМ или 20000 нМ. По прошествии периода обработки в течение приблизительно 16 часов РНК выделили из клеток и измерили уровни суммарной мРНК C9ORF72, а также уровни мРНК транскрипта экзона 1, методом количественной ПЦР в режиме реального времени. Для измерения уровней суммарной мРНК C9ORF72 использовали набор RTS3750 праймеров-зондов для последовательности C9ORF72 человека.
Для измерения уровня информационного транскрипта экзона 1 использовали набор RTS3905 праймеров-зондов (прямая последовательность GGGTCTAGCAAGAGCAGGTG, обозначенная в данном документе как SEQ ID NO: 18; обратная последовательность GTCTTGGCAACAGCTGGAGAT, обозначенная в данном документе как SEQ ID NO: 19; последовательность зонда TGATGTCGACTCTTTGCCCACCGC, обозначенная в данном документе как SEQ ID NO: 20). Уровни мРНК C9ORF72 скорректировали в соответствии с суммарным содержанием РНК, измеренным с помощью красителя RIBOGREEN®. Результаты представлены в виде процента ингибирования C9ORF72 по сравнению с необработанными контрольными клетками.
Концентрация полумаксимального ингибирования (IC50) для каждого олигонуклеотида также представлена в табл. 14 и 15. Как показано, уровни мРНК C9ORF72 были снижены дозозависимым способом в обработанных антисмысловыми олигонуклеотидами клетках. Сокращение «н.д.» указывает на то, что для этой конкретной дозы данные отсутствуют.
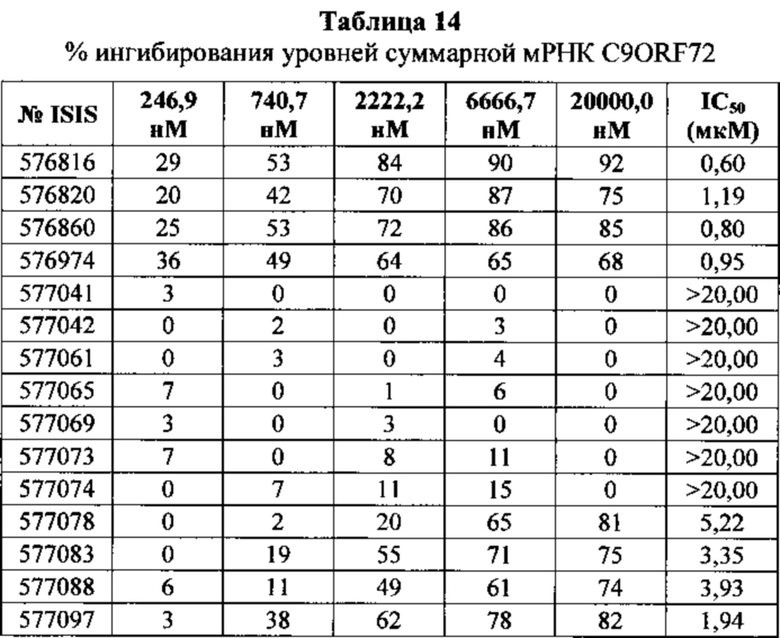

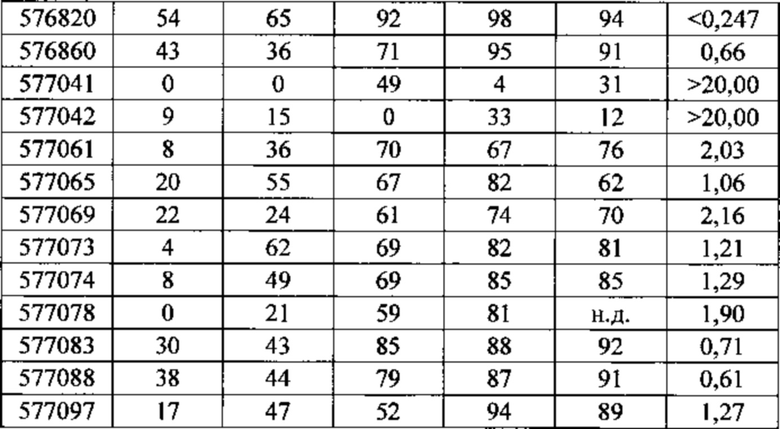
Пример 4. Антисмысловое ингибирование гена C90RF72 человека в клетках HepG2
Антисмысловые олигонуклеотиды были сконструированы для нацеливания на экспансию гексануклеотидного повтора нуклеиновой кислоты C9ORF72 и их действие было испытано на мРНК C9ORF72 in vitro. Данные антисмысловые олигонуклеотиды испытывали в сериях экспериментов, которые обеспечивали сходные культуральные условия. Результаты каждого эксперимента представлены в отдельных таблицах, показанных ниже. Последовательности ISIS 576816 и ISIS 577065 были включены в данные исследования для сравнения. Культивированные до плотности 35000 клеток на лунку фибробласты C9ORF72 трансфецировали с применением электропорации с помощью антисмыслового олигонуклеотида в концентрации 7000 нМ. По прошествии периода обработки в течение приблизительно 24 часов РНК выделили из клеток и измерили уровни мРНК C9ORF72 методом количественной ПЦР в режиме реального времени. Для измерения уровней мРНК использовали наборы праймеров-зондов человека RTS3750, RTS 3905 или RTS4097 (прямая последовательность CAAGCCACCGTCTCACTCAA, обозначенная в данном документе как SEQ ID NO: 24; обратная последовательность GTAGTGCTGTCTACTCCAGAGAGTTACC, обозначенная в данном документе как SEQ ID NO: 25; последовательность зонда CTTGGCTTCCCTCAAAAGACTGGCTAATGT, обозначенная в данном документе как SEQ ID NO: 26). Зонд набора RTS3750 нацелен на экзон 2 транскрипта мРНК, и поэтому измеряет суммарное количество транскриптов мРНК. Зонд набора RTS3905 нацелен на экспансию гексануклеотидного повтора, содержащее транскрипт,, поэтому измеряет только транскрипты мРНК, которые содержат экспансию гексануклеотидного повтора. Зонд набора RTS4097 нацелен на последовательность гена на сайте 3' экспансии гексануклеотидного повтора. Уровни мРНК скорректировали в соответствии с суммарным содержанием РНК, измеренным с помощью красителя RIBOGREEN®. Результаты представлены в виде процента ингибирования C9ORF72 по сравнению с необработанными контрольными клетками. Сокращение «н.д.» указывает на то, что этого конкретного антисмыслового олигонуклеотида отсутствуют данные.
Антисмысловые олигонуклеотиды в табл.16 были сконструированы в виде однотипных МОЕ-олигонуклеотидов или гэпмеров 3-10-3 МОЕ, 4-10-3 МОЕ, 4-10-4 МОЕ, 5-10-4 МОЕ или 5-10-5 МОЕ. Однотипные МОЕ-олигонуклеотиды составлены из 20 нуклеозидов в длину, причем каждый нуклеозид содержит группу 2'-МОЕ. Гэпмеры 3-10-3 МОЕ составлены из 16 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый сегмент включает по три нуклеозида. Гэпмеры 4-10-3 МОЕ составлены из 17 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, включающими по четыре или три нуклеозида, соответственно. Гэпмеры 4-10-4 МОЕ составлены из 18 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый сегмент включает по четыре нуклеозида. Гэпмеры 5-10-4 МОЕ составлены из 19 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, включающими по пять или четыре нуклеозида, соответственно. Гэпмеры 5-10-5 составлены из 20 нуклеозидов в длину, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый сегмент включает по 5 нуклеозидов. Каждый нуклеозид на сегменте-крыле 5' и каждый нуклеозид на сегменте-крыле 3' содержит группу 2'-МОЕ. Межнуклеозидные связи на всем протяжении каждого олигонуклеотида являются фосфоротиоатными связями. Все цитозиновые остатки на всем протяжении каждого олигонуклеотида являются 5-метилцитозинами. «Сайт инициации» указывает на самый крайний 5'-нуклеозид последовательности гена человека, на который нацелен антисмысловой олигонуклеотид. «Сайт терминации» указывает на самый крайний 3'-нуклеозид последовательности гена человека, на который нацелен антисмысловой олигонуклеотид. Каждый антисмысловой олигонуклеотид, перечисленный в табл. 16, нацелен на геномную последовательность C9ORF72 человека, обозначенную в данном документе как SEQ ID NO: 2 (комплементарная последовательность в базе данных GENBANK № доступа NT 008413.18, усеченная от нуклеозида 27535000 до 27565000) или SEQ ID NO: 13, которая является удлиненным вариантом гексануклеотидного повтора от интрона 1 гена C9ORF72.
Полученные данные указывают на то, что определенные антисмысловые олигонуклеотиды преимущественно ингибируют уровни транскриптов мРНК C9ORF72, которые содержат данный гексануклеотидный повтор.
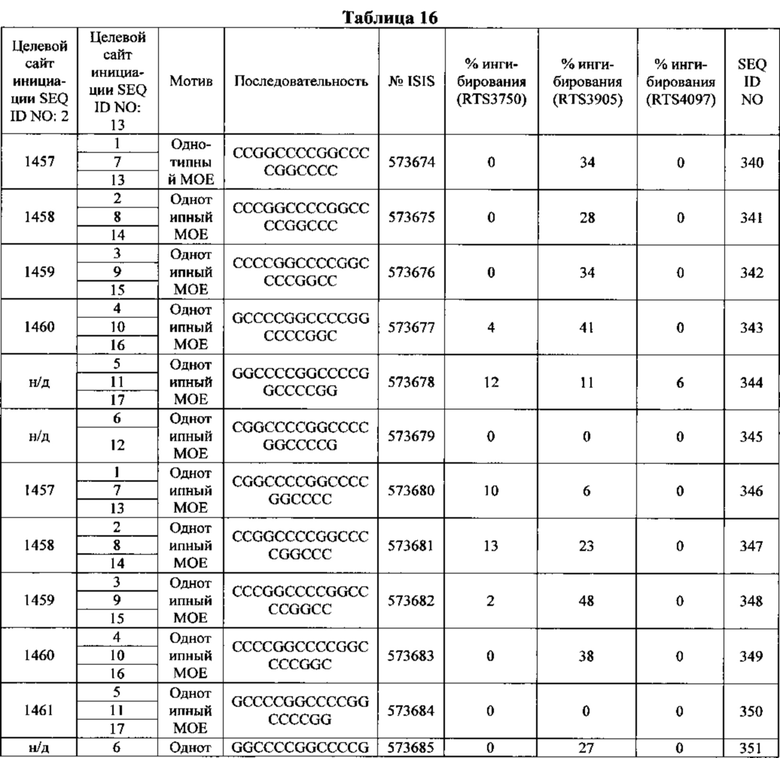
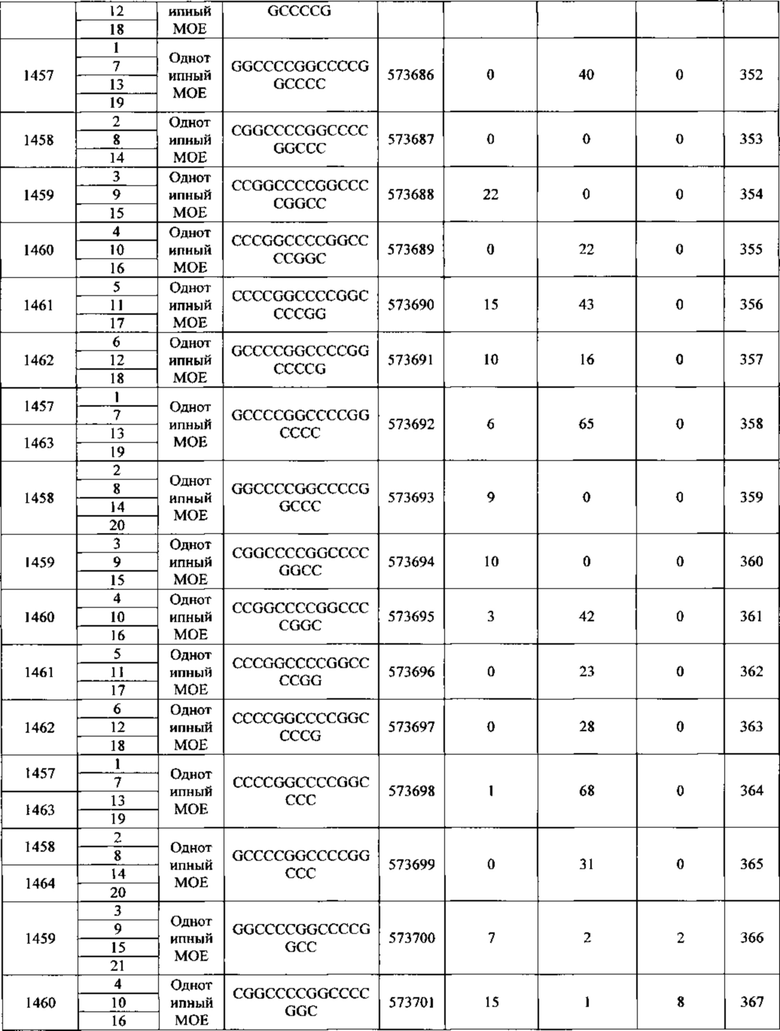

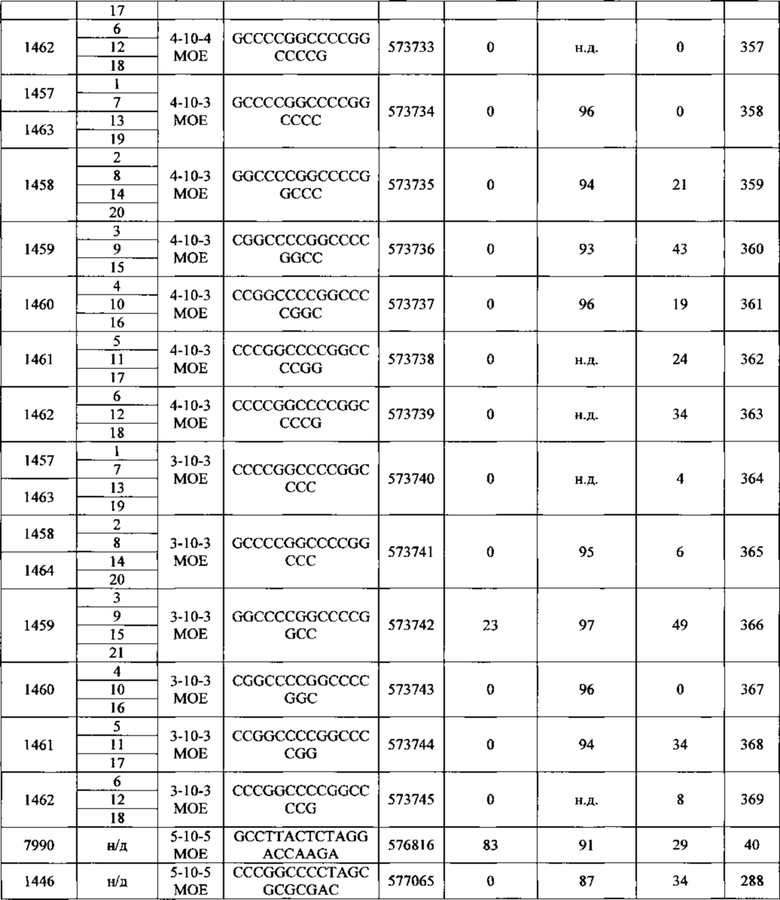
Пример 5. Ингибирование и переносимость при лечении антисмысловыми олигонуклеотидами C9ORF72 грызунов in vivo
Для оценки переносимости ингибирования экспрессии C9ORF72 in vivo сконструировали и оценили на моделях мышей и крыс антисмысловые олигонуклеотиды, нацеленные на нуклеиновую кислоту C9ORF72 мышевидных грызунов.
Последовательность ISIS 571883 сконструировали в виде гэпмера 5-10-5 МОЕ, состоящего в длину из 20 нуклеозидов, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый сегмент включает по пять нуклеозидов. Каждый нуклеозид на сегменте-крыле 5' и каждый нуклеозид на сегменте-крыле 3' несут модификацию МОЕ. Межнуклеозидные связи представляют собой фосфоротиоатные связи. Все цитозиновые остатки на всем протяжении данного гэпмера являются 5-метилцитозинами. Последовательность ISIS 571883 имеет целевой сайт инициации из нуклеозида 33704 геномной последовательности C9ORF72 мышевидных грызунов, обозначенную в данном документе как SEQ ID NO: 11 (комплементарная последовательность из базы данных GENBANK № доступа NT_166289.1, усеченная от нуклеозида 3587000 до 3625000).
Последовательность ISIS 603538 сконструировали в виде гэпмера 5-10-5 МОЕ, состоящего в длину из 20 нуклеозидов, причем центральный гэп-сегмент содержит десять 2'-дезоксирибонуклеотидов и фланкирован сегментами-крыльями как на 5'-конце, так и на 3'-конце, каждый сегмент включает по пять нуклеозидов. Каждый нуклеозид на сегменте-крыле 5' и каждый нуклеозид на сегменте-крыле 3' несут модификацию МОЕ. Межнуклеозидные связи представляют собой или фосфоротиоатные связи или фосфатэфирные связи (Gs Ао Со Со Gs Cs Ts Ts Gs As Gs Ts Ts Ts Gs Co Co Ao Cs А, в которых «s» обозначает фосфоротиоатную межнуклеозидную связь, «о» обозначает фосфатэфирную связь, a A, G, С, Τ - соответствующие нуклеозиды). Все цитозиновые остатки на всем протяжении данного гэпмера являются 5-метилцитозинами. Последовательность ISIS 603538 имеет целевой сайт инициации из нуклеозида 2872 последовательности мРНК C9ORF72 крысы, обозначенную в данном документе как SEQ ID NO: 12 (№ доступа в базе данных GENBANK NM_001007702.1).
Эксперимент на мышах 1
Каждой мыши в группе из 4 мышей линии C57BL/6 инъекционно ввели 50, 100, 300, 500 или 700 мкг последовательности ISIS 571883, введение провели посредством интрацеребровентрикулярной болюсной инъекции. Контрольной группе из четырех мышей линии C57/BL6 сходным образом ввели ФСБР. Животным провели анестезию с помощью 3% изофлурана и поместили на стереотаксическую рамку. После стерилизации места хирургического вмешательства каждой мыши инъецировали на -0,2 мм фасно от брегмы (Международная анатомическая номенклатура) в глубину на 3 мм дорсовентрально к брегме вышеупомянутые дозы ISIS 571883 с использованием шприца Гамильтона. Полученный разрез был закрыт наложением шва. Мышам дали возможность восстановиться в течение 14 дней, после чего провели эвтаназию животных в соответствии с гуманным протоколом, утвержденным Институциональным комитетом по содержанию и использованию животных. Ткани головного и спинного мозга собрали и быстро заморозили в жидком азоте. Перед замораживанием сделали пять поперечных срезов тканей, используя матрицу мозга мыши.
Анализ РНК
Для анализа экспрессии мРНК C9ORF72 экстрагировали РНК из среза головного мозга размером 2-3 мм за местом введения из фронтального кортекса головного мозга и из люмбального среза ткани спинного мозга. Экспрессию мРНК C9ORF72 измерили методом ПЦР в режиме реального времени. Полученные данные представлены в табл. 17. Результаты указывают, что лечение увеличивающимися дозами ISIS 571883 привели к дозозависимому ингибированию экспрессии мРНК C9ORF72.
Оценивали также индукцию микроглиального маркера AIF-1 путем измерения проявления токсичности на ЦНС. Полученные данные представлены в табл. 18. Результаты указывают, что лечение увеличивающимися дозами ISIS 571883 не привели к статистически значимому увеличению экспрессии мРНК AIF-1. Следовательно, инъекция ISIS 571883 на этой модели была признана переносимой.


Эксперимент на мышах 2
Каждой мыши в группе из 4 мышей линии C57BL/6 инъекционно ввели 500 мкг последовательности ISIS 571883, введение провели посредством интрацеребровентрикулярной (ИЦВ) болюсной инъекции по методике, сходной с описанной выше. Контрольной группе из четырех мышей линии C57/BL6 сходным образом ввели ФСБР. Мышей исследовали в равноудаленные временные точки после ИЦВ введения.
Анализ поведения
Применили два стандартных исследования для оценки двигательного поведения: исследование с вращающимся стержнем и анализ силы сжатия. В случае исследований с вращающимся стержнем измеряли время задержки падения. Полученные данные этих исследований представлены в табл. 19 и 20. Результаты указывают, что не было отмечено статистически значимых изменений двигательного поведения мышей по причине антисмыслового ингибирования последовательностью ISIS 571883 или из-за ИЦВ введения. Следовательно, антисмысловое ингибирование гена C9ORF72 на этой модели было признано переносимым.

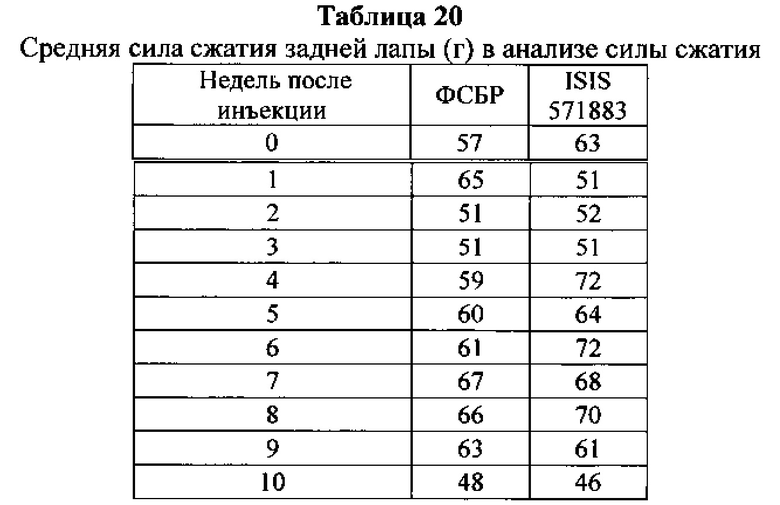
Эксперимент на крысах
Каждой крысе в группе из 4 крыс линии Спрег-Доули инъекционно ввели 700, 1000 или 3000 мкг последовательности ISIS 603538, введение провели посредством интратекальной болюсной инъекции. Контрольной группе из четырех крыс линии Спрег-Доули сходным образом ввели ФСБР. Животным провели анестезию с помощью 3% изофлурана и поместили на стереотаксическую рамку. После стерилизации места хирургического вмешательства каждой крысе инъецировали 30 мкл раствора антисмыслового олигонуклеотида (ACO), вводимого через интратекальный катетер длиной 8 см на глубину 2 см в спинномозговой канал с промывкой 50 мкл. Крысам дали возможность восстановиться в течение 4 недель, после чего провели эвтаназию животных в соответствии с гуманным протоколом, утвержденным Институциональным комитетом по содержанию и использованию животных.
Анализ РНК
Для анализа экспрессии мРНК C9ORF72 экстрагировали из среза головного мозга размером 2-3 мм за местом введения из фронтального кортекса головного мозга и из цервикального и люмбального срезов ткани спинного мозга. Экспрессию мРНК C9ORF72 измерили методом ПЦР в режиме реального времени. Полученные данные представлены в табл. 21. Результаты указывают, что лечение увеличивающимися дозами ISIS 603538 привели к дозозависимому ингибированию экспрессии мРНК C9ORF72.
Оценивали также индукцию микроглиального маркера AIF-1 путем измерения проявления токсичности на ЦНС. Полученные данные представлены в табл. 22. Результаты указывают, что лечение увеличивающимися дозами ISIS 603538 не привели к статистически значимому увеличению экспрессии мРНК AIF-1. Следовательно, инъекция ISIS 603538 на этой модели была признана переносимой.

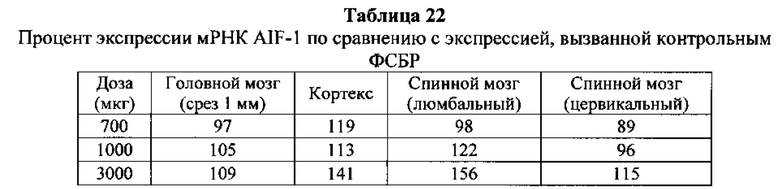
Анализ массы тела
Массу тела крыс измерили во временные точки через равноудаленные интервалы. Полученные данные представлены в табл. 23. Результаты указывают, что лечение увеличивающимися дозами ISIS 603538 не привели к каким-либо статистически значимым изменениям массы тела крыс.
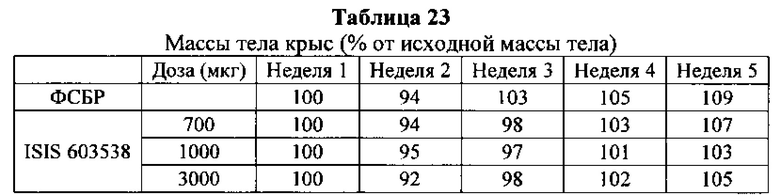
Пример 6. Преимущественное ингибирование экспрессии гена C9ORF72 человека в линиях фибробластов двух пациентов
Две разные клеточные клинии пациентов-людей (F09-152 и F09-229) проанализировали с антисмысловыми олигонуклеотидами, которые нацелены на последовательность пре-мРНК C9ORF72 перед экзоном 1В, т.е. антисмысловые олигонуклеотиды нацелены на экспансию гексануклеотидного повтора, содержащего транскрипт, и антисмысловые олигонуклеотиды нацелены в прямом направлении считывания экзона 1. Целевые сайты инициации и терминации и участки-мишени в сравнении с SEQ ID NO: 1 и 2 для каждого олигонуклеотида представлены в табл.24. Последовательности ISIS 577061 и ISIS 577065 нацелены на последовательность C9ORF72 в обратном направлении считывания экзона 1В и непосредственно в обратном направлении гексануклеотидного повтора. Остальные олигонуклеотиды ISIS из табл. 24 нацелены в прямом направлении считывания последовательности C9ORF72 экзона 1В и гексануклеотидного повтора.

Клетки высадили на планшеты в плотности 20000 клеток на лунку и трансфецировали с применением электропорации с помощью концентраций антисмыслового олигонуклеотида 246,9 нМ; 740,7 нМ; 2222,2 нМ; 6666,7 нМ и 20000 нМ. По прошествии периода обработки в течение приблизительно 16 часов РНК выделили из клеток и измерили уровни мРНК C9ORF72 методом количественной ПЦР в режиме реального времени. Использовали два набора праймеров-зондов: (1) набор RTS3750 праймеров-зондов для гена C9ORF72 человека, который измеряет суммарные уровни мРНК, и (2) набор RTS3905, который нацелен на экспансию гексануклеотидного набора, содержащего транскрипт, что измеряет только транскрипты мРНК, которые содержат экспансию гексануклеотидного набора. Уровни мРНК C9ORF72 скорректировали в соответствии с суммарным содержанием РНК, измеренным с помощью красителя RIBOGREEN®. Результаты представлены в виде процента ингибирования C9ORF72 по сравнению с необработанными контрольными клетками.
Как показано в табл. 25, два олигонуклеотида, которые нацелены в обратном направлении считывания экзона 1В и поэтому направлены на транскрипты мРНК, содержащие экспансию гексануклеотидного повтора (ISIS 577061 и ISIS 577065), не ингибируют уровни суммарной мРНК C9ORF72 (при измерении с помощью набора RTS3750), также, как и ISIS 576974, 576816 и 577083, которые нацелены в прямом направлении экзона 1В и поэтому не направлены на транскрипт мРНК, содержащий экспансию гексануклеотидного повтора. Уровни экспрессии транскрипта мРНК C9ORF72, содержащего экспансию гексануклеотидного повтора, являются низкими (около 10% от суммарной экспрессии продуктов гена C9ORF72), поэтому, олигонуклеотиды, нацеленные на транскрипт мРНК, содержащий экспансию гексануклеотидного повтора, несильно ингибируют суммарную мРНК C9ORF72 (при измерении набором RTS3905), как приведено в табл. 25. Таким образом, последовательности ISIS 577061 и ISIS 577065 преимущественно ингибируют экспрессию транскриптов мРНК, содержащих экспансию гексануклеотидного повтора.
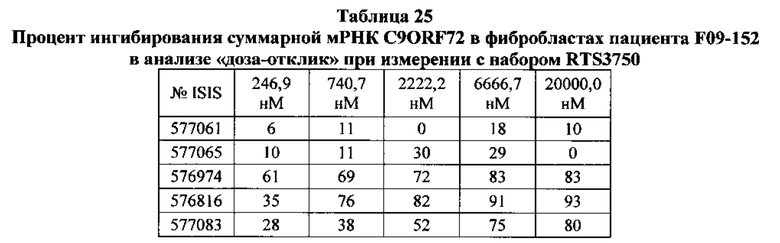

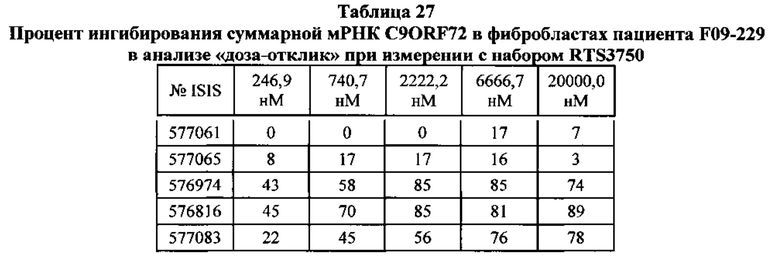
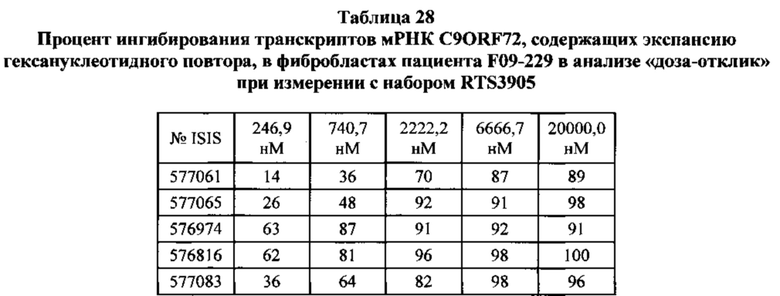
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Isis Pharmaceuticals, Inc.
<120> КОМПОЗИЦИИ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА C9ORF72
<130> BIOL0211WO
<140> PCT/US2013/065073
<141> 2013-10-15
<150> 61/714,132
<151> 2012-10-15
<160> 369
<170> Версия PatentIn 3.5
<210> 1
<211> 3339
<212> ДНК
<213> Homo sapiens
<220>
<221> КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<222> (203)..(1648)
<400> 1
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg gtgtgtcagc cgtccctgct gcccggttgc ttctcttttg 120
ggggcggggt ctagcaagag caggtgtggg tttaggagat atctccggag catttggata 180
atgtgacagt tggaatgcag tg atg tcg act ctt tgc cca ccg cca tct cca 232
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro
1 5 10
gct gtt gcc aag aca gag att gct tta agt ggc aaa tca cct tta tta 280
Ala Val Ala Lys Thr Glu Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu
15 20 25
gca gct act ttt gct tac tgg gac aat att ctt ggt cct aga gta agg 328
Ala Ala Thr Phe Ala Tyr Trp Asp Asn Ile Leu Gly Pro Arg Val Arg
30 35 40
cac att tgg gct cca aag aca gaa cag gta ctt ctc agt gat gga gaa 376
His Ile Trp Ala Pro Lys Thr Glu Gln Val Leu Leu Ser Asp Gly Glu
45 50 55
ata act ttt ctt gcc aac cac act cta aat gga gaa atc ctt cga aat 424
Ile Thr Phe Leu Ala Asn His Thr Leu Asn Gly Glu Ile Leu Arg Asn
60 65 70
gca gag agt ggt gct ata gat gta aag ttt ttt gtc ttg tct gaa aag 472
Ala Glu Ser Gly Ala Ile Asp Val Lys Phe Phe Val Leu Ser Glu Lys
75 80 85 90
gga gtg att att gtt tca tta atc ttt gat gga aac tgg aat ggg gat 520
Gly Val Ile Ile Val Ser Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp
95 100 105
cgc agc aca tat gga cta tca att ata ctt cca cag aca gaa ctt agt 568
Arg Ser Thr Tyr Gly Leu Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser
110 115 120
ttc tac ctc cca ctt cat aga gtg tgt gtt gat aga tta aca cat ata 616
Phe Tyr Leu Pro Leu His Arg Val Cys Val Asp Arg Leu Thr His Ile
125 130 135
atc cgg aaa gga aga ata tgg atg cat aag gaa aga caa gaa aat gtc 664
Ile Arg Lys Gly Arg Ile Trp Met His Lys Glu Arg Gln Glu Asn Val
140 145 150
cag aag att atc tta gaa ggc aca gag aga atg gaa gat cag ggt cag 712
Gln Lys Ile Ile Leu Glu Gly Thr Glu Arg Met Glu Asp Gln Gly Gln
155 160 165 170
agt att att cca atg ctt act gga gaa gtg att cct gta atg gaa ctg 760
Ser Ile Ile Pro Met Leu Thr Gly Glu Val Ile Pro Val Met Glu Leu
175 180 185
ctt tca tct atg aaa tca cac agt gtt cct gaa gaa ata gat ata gct 808
Leu Ser Ser Met Lys Ser His Ser Val Pro Glu Glu Ile Asp Ile Ala
190 195 200
gat aca gta ctc aat gat gat gat att ggt gac agc tgt cat gaa ggc 856
Asp Thr Val Leu Asn Asp Asp Asp Ile Gly Asp Ser Cys His Glu Gly
205 210 215
ttt ctt ctc aat gcc atc agc tca cac ttg caa acc tgt ggc tgt tcc 904
Phe Leu Leu Asn Ala Ile Ser Ser His Leu Gln Thr Cys Gly Cys Ser
220 225 230
gtt gta gta ggt agc agt gca gag aaa gta aat aag ata gtc aga aca 952
Val Val Val Gly Ser Ser Ala Glu Lys Val Asn Lys Ile Val Arg Thr
235 240 245 250
tta tgc ctt ttt ctg act cca gca gag aga aaa tgc tcc agg tta tgt 1000
Leu Cys Leu Phe Leu Thr Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys
255 260 265
gaa gca gaa tca tca ttt aaa tat gag tca ggg ctc ttt gta caa ggc 1048
Glu Ala Glu Ser Ser Phe Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly
270 275 280
ctg cta aag gat tca act gga agc ttt gtg ctg cct ttc cgg caa gtc 1096
Leu Leu Lys Asp Ser Thr Gly Ser Phe Val Leu Pro Phe Arg Gln Val
285 290 295
atg tat gct cca tat ccc acc aca cac ata gat gtg gat gtc aat act 1144
Met Tyr Ala Pro Tyr Pro Thr Thr His Ile Asp Val Asp Val Asn Thr
300 305 310
gtg aag cag atg cca ccc tgt cat gaa cat att tat aat cag cgt aga 1192
Val Lys Gln Met Pro Pro Cys His Glu His Ile Tyr Asn Gln Arg Arg
315 320 325 330
tac atg aga tcc gag ctg aca gcc ttc tgg aga gcc act tca gaa gaa 1240
Tyr Met Arg Ser Glu Leu Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu
335 340 345
gac atg gct cag gat acg atc atc tac act gac gaa agc ttt act cct 1288
Asp Met Ala Gln Asp Thr Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro
350 355 360
gat ttg aat att ttt caa gat gtc tta cac aga gac act cta gtg aaa 1336
Asp Leu Asn Ile Phe Gln Asp Val Leu His Arg Asp Thr Leu Val Lys
365 370 375
gcc ttc ctg gat cag gtc ttt cag ctg aaa cct ggc tta tct ctc aga 1384
Ala Phe Leu Asp Gln Val Phe Gln Leu Lys Pro Gly Leu Ser Leu Arg
380 385 390
agt act ttc ctt gca cag ttt cta ctt gtc ctt cac aga aaa gcc ttg 1432
Ser Thr Phe Leu Ala Gln Phe Leu Leu Val Leu His Arg Lys Ala Leu
395 400 405 410
aca cta ata aaa tat ata gaa gac gat acg cag aag gga aaa aag ccc 1480
Thr Leu Ile Lys Tyr Ile Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro
415 420 425
ttt aaa tct ctt cgg aac ctg aag ata gac ctt gat tta aca gca gag 1528
Phe Lys Ser Leu Arg Asn Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu
430 435 440
ggc gat ctt aac ata ata atg gct ctg gct gag aaa att aaa cca ggc 1576
Gly Asp Leu Asn Ile Ile Met Ala Leu Ala Glu Lys Ile Lys Pro Gly
445 450 455
cta cac tct ttt atc ttt gga aga cct ttc tac act agt gtg caa gaa 1624
Leu His Ser Phe Ile Phe Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu
460 465 470
cga gat gtt cta atg act ttt taa atgtgtaact taataagcct attccatcac 1678
Arg Asp Val Leu Met Thr Phe
475 480
aatcatgatc gctggtaaag tagctcagtg gtgtggggaa acgttcccct ggatcatact 1738
ccagaattct gctctcagca attgcagtta agtaagttac actacagttc tcacaagagc 1798
ctgtgagggg atgtcaggtg catcattaca ttgggtgtct cttttcctag atttatgctt 1858
ttgggataca gacctatgtt tacaatataa taaatattat tgctatcttt taaagatata 1918
ataataggat gtaaacttga ccacaactac tgtttttttg aaatacatga ttcatggttt 1978
acatgtgtca aggtgaaatc tgagttggct tttacagata gttgactttc tatcttttgg 2038
cattctttgg tgtgtagaat tactgtaata cttctgcaat caactgaaaa ctagagcctt 2098
taaatgattt caattccaca gaaagaaagt gagcttgaac ataggatgag ctttagaaag 2158
aaaattgatc aagcagatgt ttaattggaa ttgattatta gatcctactt tgtggattta 2218
gtccctggga ttcagtctgt agaaatgtct aatagttctc tatagtcctt gttcctggtg 2278
aaccacagtt agggtgtttt gtttatttta ttgttcttgc tattgttgat attctatgta 2338
gttgagctct gtaaaaggaa attgtatttt atgttttagt aattgttgcc aactttttaa 2398
attaattttc attatttttg agccaaattg aaatgtgcac ctcctgtgcc ttttttctcc 2458
ttagaaaatc taattacttg gaacaagttc agatttcact ggtcagtcat tttcatcttg 2518
ttttcttctt gctaagtctt accatgtacc tgctttggca atcattgcaa ctctgagatt 2578
ataaaatgcc ttagagaata tactaactaa taagatcttt ttttcagaaa cagaaaatag 2638
ttccttgagt acttccttct tgcatttctg cctatgtttt tgaagttgtt gctgtttgcc 2698
tgcaataggc tataaggaat agcaggagaa attttactga agtgctgttt tcctaggtgc 2758
tactttggca gagctaagtt atcttttgtt ttcttaatgc gtttggacca ttttgctggc 2818
tataaaataa ctgattaata taattctaac acaatgttga cattgtagtt acacaaacac 2878
aaataaatat tttatttaaa attctggaag taatataaaa gggaaaatat atttataaga 2938
aagggataaa ggtaatagag cccttctgcc ccccacccac caaatttaca caacaaaatg 2998
acatgttcga atgtgaaagg tcataatagc tttcccatca tgaatcagaa agatgtggac 3058
agcttgatgt tttagacaac cactgaacta gatgactgtt gtactgtagc tcagtcattt 3118
aaaaaatata taaatactac cttgtagtgt cccatactgt gttttttaca tggtagattc 3178
ttatttaagt gctaactggt tattttcttt ggctggttta ttgtactgtt atacagaatg 3238
taagttgtac agtgaaataa gttattaaag catgtgtaaa cattgttata tatcttttct 3298
cctaaatgga gaattttgaa taaaatatat ttgaaatttt g 3339
<210> 2
<211> 30001
<212> ДНК
<213> Homo sapiens
<400> 2
caaagaaaag ggggaggttt tgttaaaaaa gagaaatgtt acatagtgct ctttgagaaa 60
attcattggc actattaagg atctgaggag ctggtgagtt tcaactggtg agtgatggtg 120
gtagataaaa ttagagctgc agcaggtcat tttagcaact attagataaa actggtctca 180
ggtcacaacg ggcagttgca gcagctggac ttggagagaa ttacactgtg ggagcagtgt 240
catttgtcct aagtgctttt ctacccccta cccccactat tttagttggg tataaaaaga 300
atgacccaat ttgtatgatc aactttcaca aagcatagaa cagtaggaaa agggtctgtt 360
tctgcagaag gtgtagacgt tgagagccat tttgtgtatt tattcctccc tttcttcctc 420
ggtgaatgat taaaacgttc tgtgtgattt ttagtgatga aaaagattaa atgctactca 480
ctgtagtaag tgccatctca cacttgcaga tcaaaaggca cacagtttaa aaaacctttg 540
tttttttaca catctgagtg gtgtaaatgc tactcatctg tagtaagtgg aatctataca 600
cctgcagacc aaaagacgca aggtttcaaa aatctttgtg ttttttacac atcaaacaga 660
atggtacgtt tttcaaaagt taaaaaaaaa caactcatcc acatattgca actagcaaaa 720
atgacattcc ccagtgtgaa aatcatgctt gagagaattc ttacatgtaa aggcaaaatt 780
gcgatgactt tgcaggggac cgtgggattc ccgcccgcag tgccggagct gtcccctacc 840
agggtttgca gtggagtttt gaatgcactt aacagtgtct tacggtaaaa acaaaatttc 900
atccaccaat tatgtgttga gcgcccactg cctaccaagc acaaacaaaa ccattcaaaa 960
ccacgaaatc gtcttcactt tctccagatc cagcagcctc ccctattaag gttcgcacac 1020
gctattgcgc caacgctcct ccagagcggg tcttaagata aaagaacagg acaagttgcc 1080
ccgccccatt tcgctagcct cgtgagaaaa cgtcatcgca catagaaaac agacagacgt 1140
aacctacggt gtcccgctag gaaagagagg tgcgtcaaac agcgacaagt tccgcccacg 1200
taaaagatga cgcttggtgt gtcagccgtc cctgctgccc ggttgcttct cttttggggg 1260
cggggtctag caagagcagg tgtgggttta ggaggtgtgt gtttttgttt ttcccaccct 1320
ctctccccac tacttgctct cacagtactc gctgagggtg aacaagaaaa gacctgataa 1380
agattaacca gaagaaaaca aggagggaaa caaccgcagc ctgtagcaag ctctggaact 1440
caggagtcgc gcgctagggg ccggggccgg ggccggggcg tggtcggggc gggcccgggg 1500
gcgggcccgg ggcggggctg cggttgcggt gcctgcgccc gcggcggcgg aggcgcaggc 1560
ggtggcgagt gggtgagtga ggaggcggca tcctggcggg tggctgtttg gggttcggct 1620
gccgggaaga ggcgcgggta gaagcggggg ctctcctcag agctcgacgc atttttactt 1680
tccctctcat ttctctgacc gaagctgggt gtcgggcttt cgcctctagc gactggtgga 1740
attgcctgca tccgggcccc gggcttcccg gcggcggcgg cggcggcggc ggcgcaggga 1800
caagggatgg ggatctggcc tcttccttgc tttcccgccc tcagtacccg agctgtctcc 1860
ttcccgggga cccgctggga gcgctgccgc tgcgggctcg agaaaaggga gcctcgggta 1920
ctgagaggcc tcgcctgggg gaaggccgga gggtgggcgg cgcgcggctt ctgcggacca 1980
agtcggggtt cgctaggaac ccgagacggt ccctgccggc gaggagatca tgcgggatga 2040
gatgggggtg tggagacgcc tgcacaattt cagcccaagc ttctagagag tggtgatgac 2100
ttgcatatga gggcagcaat gcaagtcggt gtgctcccca ttctgtggga catgacctgg 2160
ttgcttcaca gctccgagat gacacagact tgcttaaagg aagtgactat tgtgacttgg 2220
gcatcacttg actgatggta atcagttgtc taaagaagtg cacagattac atgtccgtgt 2280
gctcattggg tctatctggc cgcgttgaac accaccaggc tttgtattca gaaacaggag 2340
ggaggtcctg cactttccca ggaggggtgg ccctttcaga tgcaatcgag attgttaggc 2400
tctgggagag tagttgcctg gttgtggcag ttggtaaatt tctattcaaa cagttgccat 2460
gcaccagttg ttcacaacaa gggtacgtaa tctgtctggc attacttcta cttttgtaca 2520
aaggatcaaa aaaaaaaaag atactgttaa gatatgattt ttctcagact ttgggaaact 2580
tttaacataa tctgtgaata tcacagaaac aagactatca tataggggat attaataacc 2640
tggagtcaga atacttgaaa tacggtgtca tttgacacgg gcattgttgt caccacctct 2700
gccaaggcct gccactttag gaaaaccctg aatcagttgg aaactgctac atgctgatag 2760
tacatctgaa acaagaacga gagtaattac cacattccag attgttcact aagccagcat 2820
ttacctgctc caggaaaaaa ttacaagcac cttatgaagt tgataaaata ttttgtttgg 2880
ctatgttggc actccacaat ttgctttcag agaaacaaag taaaccaagg aggacttctg 2940
tttttcaagt ctgccctcgg gttctattct acgttaatta gatagttccc aggaggacta 3000
ggttagccta cctattgtct gagaaacttg gaactgtgag aaatggccag atagtgatat 3060
gaacttcacc ttccagtctt ccctgatgtt gaagattgag aaagtgttgt gaactttctg 3120
gtactgtaaa cagttcactg tccttgaagt ggtcctgggc agctcctgtt gtggaaagtg 3180
gacggtttag gatcctgctt ctctttgggc tgggagaaaa taaacagcat ggttacaagt 3240
attgagagcc aggttggaga aggtggctta cacctgtaat gccagagctt tgggaggcgg 3300
aggcaagagg atcacttgaa gccaggagtt caagctcaac ctgggcaacg tagaccctgt 3360
ctctacaaaa aattaaaaac ttagccgggc gtggtgatgt gcacctgtag tcctagctac 3420
ttgggaggct gaggcaggag ggtcatttga gcccaagagt ttgaagttac cgagagctat 3480
gatcctgcca gtgcattcca gcctggatga caaaacgaga ccctgtctct aaaaaacaag 3540
aagtgagggc tttatgattg tagaattttc actacaatag cagtggacca accacctttc 3600
taaataccaa tcagggaaga gatggttgat tttttaacag acgtttaaag aaaaagcaaa 3660
acctcaaact tagcactcta ctaacagttt tagcagatgt taattaatgt aatcatgtct 3720
gcatgtatgg gattatttcc agaaagtgta ttgggaaacc tctcatgaac cctgtgagca 3780
agccaccgtc tcactcaatt tgaatcttgg cttccctcaa aagactggct aatgtttggt 3840
aactctctgg agtagacagc actacatgta cgtaagatag gtacataaac aactattggt 3900
tttgagctga tttttttcag ctgcatttgc atgtatggat ttttctcacc aaagacgatg 3960
acttcaagta ttagtaaaat aattgtacag ctctcctgat tatacttctc tgtgacattt 4020
catttcccag gctatttctt ttggtaggat ttaaaactaa gcaattcagt atgatctttg 4080
tccttcattt tctttcttat tctttttgtt tgtttgtttg tttgtttttt tcttgaggca 4140
gagtctctct ctgtcgccca ggctggagtg cagtggcgcc atctcagctc attgcaacct 4200
ctgccacctc cgggttcaag agattctcct gcctcagcct cccgagtagc tgggattaca 4260
ggtgtccacc accacacccg gctaattttt tgtattttta gtagaggtgg ggtttcacca 4320
tgttggccag gctggtcttg agctcctgac ctcaggtgat ccacctgcct cggcctacca 4380
aagagctggg ataacaggtg tgacccacca tgcccggccc attttttttt tcttattctg 4440
ttaggagtga gagtgtaact agcagtataa tagttcaatt ttcacaacgt ggtaaaagtt 4500
tccctataat tcaatcagat tttgctccag ggttcagttc tgttttagga aatactttta 4560
ttttcagttt aatgatgaaa tattagagtt gtaatattgc ctttatgatt atccaccttt 4620
ttaacctaaa agaatgaaag aaaaatatgt ttgcaatata attttatggt tgtatgttaa 4680
cttaattcat tatgttggcc tccagtttgc tgttgttagt tatgacagca gtagtgtcat 4740
taccatttca attcagatta cattcctata tttgatcatt gtaaactgac tgcttacatt 4800
gtattaaaaa cagtggatat tttaaagaag ctgtacggct tatatctagt gctgtctctt 4860
aagactatta aattgataca acatatttaa aagtaaatat tacctaaatg aatttttgaa 4920
attacaaata cacgtgttaa aactgtcgtt gtgttcaacc atttctgtac atacttagag 4980
ttaactgttt tgccaggctc tgtatgccta ctcataatat gataaaagca ctcatctaat 5040
gctctgtaaa tagaagtcag tgctttccat cagactgaac tctcttgaca agatgtggat 5100
gaaattcttt aagtaaaatt gtttactttg tcatacattt acagatcaaa tgttagctcc 5160
caaagcaatc atatggcaaa gataggtata tcatagtttg cctattagct gctttgtatt 5220
gctattatta taaatagact tcacagtttt agacttgctt aggtgaaatt gcaattcttt 5280
ttactttcag tcttagataa caagtcttca attatagtac aatcacacat tgcttaggaa 5340
tgcatcatta ggcgattttg tcattatgca aacatcatag agtgtactta cacaaaccta 5400
gatagtatag cctttatgta cctaggccgt atggtatagt ctgttgctcc taggccacaa 5460
acctgtacaa ctgttactgt actgaatact atagacagtt gtaacacagt ggtaaatatt 5520
tatctaaata tatgcaaaca gagaaaaggt acagtaaaag tatggtataa aagataatgg 5580
tatacctgtg taggccactt accacgaatg gagcttgcag gactagaagt tgctctgggt 5640
gagtcagtga gtgagtggtg aattaatgtg aaggcctaga acactgtaca ccactgtaga 5700
ctataaacac agtacgctga agctacacca aatttatctt aacagttttt cttcaataaa 5760
aaattataac tttttaactt tgtaaacttt ttaatttttt aacttttaaa atacttagct 5820
tgaaacacaa atacattgta tagctataca aaaatatttt ttctttgtat ccttattcta 5880
gaagcttttt tctattttct attttaaatt ttttttttta cttgttagtc gtttttgtta 5940
aaaactaaaa cacacacact ttcacctagg catagacagg attaggatca tcagtatcac 6000
tcccttccac ctcactgcct tccacctcca catcttgtcc cactggaagg tttttagggg 6060
caataacaca catgtagctg tcacctatga taacagtgct ttctgttgaa tacctcctga 6120
aggacttgcc tgaggctgtt ttacatttaa cttaaaaaaa aaaaaagtag aaggagtgca 6180
ctctaaaata acaataaaag gcatagtata gtgaatacat aaaccagcaa tgtagtagtt 6240
tattatcaag tgttgtacac tgtaataatt gtatgtgcta tactttaaat aacttgcaaa 6300
atagtactaa gaccttatga tggttacagt gtcactaagg caatagcata ttttcaggtc 6360
cattgtaatc taatgggact accatcatat atgcagtcta ccattgactg aaacgttaca 6420
tggcacataa ctgtatttgc aagaatgatt tgttttacat taatatcaca taggatgtac 6480
ctttttagag tggtatgttt atgtggatta agatgtacaa gttgagcaag gggaccaaga 6540
gccctgggtt ctgtcttgga tgtgagcgtt tatgttcttc tcctcatgtc tgttttctca 6600
ttaaattcaa aggcttgaac gggccctatt tagcccttct gttttctacg tgttctaaat 6660
aactaaagct tttaaattct agccatttag tgtagaactc tctttgcagt gatgaaatgc 6720
tgtattggtt tcttggctag catattaaat atttttatct ttgtcttgat acttcaatgt 6780
cgttttaaac atcaggatcg ggcttcagta ttctcataac cagagagttc actgaggata 6840
caggactgtt tgcccatttt ttgttatggc tccagacttg tggtatttcc atgtcttttt 6900
tttttttttt ttttttgacc ttttagcggc tttaaagtat ttctgttgtt aggtgttgta 6960
ttacttttct aagattactt aacaaagcac cacaaactga gtggctttaa acaacagcaa 7020
tttattctct cacaattcta gaagctagaa gtccgaaatc aaagtgttga caggggcatg 7080
atcttcaaga gagaagactc tttccttgcc tcttcctggc ttctggtggt taccagcaat 7140
cctgagtgtt cctttcttgc cttgtagttt caacaatcca gtatctgcct tttgtcttca 7200
catggctgtc taccatttgt ctctgtgtct ccaaatctct ctccttataa acacagcagt 7260
tattggatta ggccccactc taatccagta tgaccccatt ttaacatgat tacacttatt 7320
tctagataag gtcacattca cgtacaccaa gggttaggaa ttgaacatat ctttttgggg 7380
gacacaattc aacccacaag tgtcagtctc tagctgagcc tttcccttcc tgtttttctc 7440
ctttttagtt gctatgggtt aggggccaaa tctccagtca tactagaatt gcacatggac 7500
tggatatttg ggaatactgc gggtctattc tatgagcttt agtatgtaac atttaatatc 7560
agtgtaaaga agcccttttt taagttattt ctttgaattt ctaaatgtat gccctgaata 7620
taagtaacaa gttaccatgt cttgtaaaat gatcatatca acaaacattt aatgtgcacc 7680
tactgtgcta gttgaatgtc tttatcctga taggagataa caggattcca catctttgac 7740
ttaagaggac aaaccaaata tgtctaaatc atttggggtt ttgatggata tctttaaatt 7800
gctgaaccta atcattggtt tcatatgtca ttgtttagat atctccggag catttggata 7860
atgtgacagt tggaatgcag tgatgtcgac tctttgccca ccgccatctc cagctgttgc 7920
caagacagag attgctttaa gtggcaaatc acctttatta gcagctactt ttgcttactg 7980
ggacaatatt cttggtccta gagtaaggca catttgggct ccaaagacag aacaggtact 8040
tctcagtgat ggagaaataa cttttcttgc caaccacact ctaaatggag aaatccttcg 8100
aaatgcagag agtggtgcta tagatgtaaa gttttttgtc ttgtctgaaa agggagtgat 8160
tattgtttca ttaatctttg atggaaactg gaatggggat cgcagcacat atggactatc 8220
aattatactt ccacagacag aacttagttt ctacctccca cttcatagag tgtgtgttga 8280
tagattaaca catataatcc ggaaaggaag aatatggatg cataaggtaa gtgatttttc 8340
agcttattaa tcatgttaac ctatctgttg aaagcttatt ttctggtaca tataaatctt 8400
atttttttaa ttatatgcag tgaacatcaa acaataaatg ttatttattt tgcatttacc 8460
ctattagata caaatacatc tggtctgata cctgtcatct tcatattaac tgtggaaggt 8520
acgaaatggt agctccacat tatagatgaa aagctaaagc ttagacaaat aaagaaactt 8580
ttagaccctg gattcttctt gggagccttt gactctaata ccttttgttt ccctttcatt 8640
gcacaattct gtcttttgct tactactatg tgtaagtata acagttcaaa gtaatagttt 8700
cataagctgt tggtcatgta gcctttggtc tctttaacct ctttgccaag ttcccaggtt 8760
cataaaatga ggaggttgaa tggaatggtt cccaagagaa ttccttttaa tcttacagaa 8820
attattgttt tcctaaatcc tgtagttgaa tatataatgc tatttacatt tcagtatagt 8880
tttgatgtat ctaaagaaca cattgaattc tccttcctgt gttccagttt gatactaacc 8940
tgaaagtcca ttaagcatta ccagttttaa aaggcttttg cccaatagta aggaaaaata 9000
atatctttta aaagaataat tttttactat gtttgcaggc ttacttcctt ttttctcaca 9060
ttatgaaact cttaaaatca ggagaatctt ttaaacaaca tcataatgtt taatttgaaa 9120
agtgcaagtc attcttttcc tttttgaaac tatgcagatg ttacattgac tgttttctgt 9180
gaagttatct ttttttcact gcagaataaa ggttgttttg attttatttt gtattgttta 9240
tgagaacatg catttgttgg gttaatttcc tacccctgcc cccatttttt ccctaaagta 9300
gaaagtattt ttcttgtgaa ctaaattact acacaagaac atgtctattg aaaaataagc 9360
aagtatcaaa atgttgtggg ttgttttttt aaataaattt tctcttgctc aggaaagaca 9420
agaaaatgtc cagaagatta tcttagaagg cacagagaga atggaagatc aggtatatgc 9480
aaattgcata ctgtcaaatg tttttctcac agcatgtatc tgtataaggt tgatggctac 9540
atttgtcaag gccttggaga catacgaata agcctttaat ggagctttta tggaggtgta 9600
cagaataaac tggaggaaga tttccatatc ttaaacccaa agagttaaat cagtaaacaa 9660
aggaaaatag taattgcatc tacaaattaa tatttgctcc cttttttttt ctgtttgccc 9720
agaataaatt ttggataact tgttcatagt aaaaataaaa aaaattgtct ctgatatgtt 9780
ctttaaggta ctacttctcg aacctttccc tagaagtagc tgtaacagaa ggagagcata 9840
tgtacccctg aggtatctgt ctggggtgta ggcccaggtc cacacaatat ttcttctaag 9900
tcttatgttg tatcgttaag actcatgcaa tttacatttt attccataac tattttagta 9960
ttaaaatttg tcagtgatat ttcttaccct ctcctctagg aaaatgtgcc atgtttatcc 10020
cttggctttg aatgcccctc aggaacagac actaagagtt tgagaagcat ggttacaagg 10080
gtgtggcttc ccctgcggaa actaagtaca gactatttca ctgtaaagca gagaagttct 10140
tttgaaggag aatctccagt gaagaaagag ttcttcactt ttacttccat ttcctcttgt 10200
gggtgaccct caatgctcct tgtaaaactc caatatttta aacatggctg ttttgccttt 10260
ctttgcttct ttttagcatg aatgagacag atgatacttt aaaaaagtaa ttaaaaaaaa 10320
aaacttgtga aaatacatgg ccataataca gaacccaata caatgatctc ctttaccaaa 10380
ttgttatgtt tgtacttttg tagatagctt tccaattcag agacagttat tctgtgtaaa 10440
ggtctgactt aacaagaaaa gatttccctt tacccaaaga atcccagtcc ttatttgctg 10500
gtcaataagc agggtcccca ggaatggggt aactttcagc accctctaac ccactagtta 10560
ttagtagact aattaagtaa acttatcgca agttgaggaa acttagaacc aactaaaatt 10620
ctgcttttac tgggattttg ttttttcaaa ccagaaacct ttacttaagt tgactactat 10680
taatgaattt tggtctctct tttaagtgct cttcttaaaa atgttatctt actgctgaga 10740
agttcaagtt tgggaagtac aaggaggaat agaaacttaa gagattttct tttagagcct 10800
cttctgtatt tagccctgta ggattttttt tttttttttt ttttttggtg ttgttgagct 10860
tcagtgaggc tattcattca cttatactga taatgtctga gatactgtga atgaaatact 10920
atgtatgctt aaacctaaga ggaaatattt tcccaaaatt attcttcccg aaaaggagga 10980
gttgcctttt gattgagttc ttgcaaatct cacaacgact ttattttgaa caatactgtt 11040
tggggatgat gcattagttt gaaacaactt cagttgtagc tgtcatctga taaaattgct 11100
tcacagggaa ggaaatttaa cacggatcta gtcattattc ttgttagatt gaatgtgtga 11160
attgtaattg taaacaggca tgataattat tactttaaaa actaaaaaca gtgaatagtt 11220
agttgtggag gttactaaag gatggttttt ttttaaataa aactttcagc attatgcaaa 11280
tgggcatatg gcttaggata aaacttccag aagtagcatc acatttaaat tctcaagcaa 11340
cttaataata tggggctctg aaaaactggt taaggttact ccaaaaatgg ccctgggtct 11400
gacaaagatt ctaacttaaa gatgcttatg aagactttga gtaaaatcat ttcataaaat 11460
aagtgaggaa aaacaactag tattaaattc atcttaaata atgtatgatt taaaaaatat 11520
gtttagctaa aaatgcatag tcatttgaca atttcattta tatctcaaaa aatttactta 11580
accaagttgg tcacaaaact gatgagactg gtggtggtag tgaataaatg agggaccatc 11640
catatttgag acactttaca tttgtgatgt gttatactga attttcagtt tgattctata 11700
gactacaaat ttcaaaatta caatttcaag atgtaataag tagtaatatc ttgaaatagc 11760
tctaaaggga atttttctgt tttattgatt cttaaaatat atgtgctgat tttgatttgc 11820
atttgggtag attatacttt tatgagtatg gaggttaggt attgattcaa gttttcctta 11880
cctatttggt aaggatttca aagtcttttt gtgcttggtt ttcctcattt ttaaatatga 11940
aatatattga tgacctttaa caaatttttt ttatctcaaa ttttaaagga gatcttttct 12000
aaaagaggca tgatgactta atcattgcat gtaacagtaa acgataaacc aatgattcca 12060
tactctctaa agaataaaag tgagctttag ggccgggcat ggtcagaaat ttgacaccaa 12120
cctggccaac atggcgaaac cccgtctcta ctaaaaatac aaaaatcagc cgggcatggt 12180
ggcggcacct atagtcccag ctacttggga ggatgagaca ggagagtcac ttgaacctgg 12240
gaggagaggt tgcagtgagc tgagatcacg ccattgcact ccagcctgag caatgaaagc 12300
aaaactccat ctcaaaaaaa aaaaaagaaa agaaagaata aaagtgagct ttggattgca 12360
tataaatcct ttagacatgt agtagacttg tttgatactg tgtttgaaca aattacgaag 12420
tattttcatc aaagaatgtt attgtttgat gttattttta ttttttattg cccagcttct 12480
ctcatattac gtgattttct tcacttcatg tcactttatt gtgcagggtc agagtattat 12540
tccaatgctt actggagaag tgattcctgt aatggaactg ctttcatcta tgaaatcaca 12600
cagtgttcct gaagaaatag atgtaagttt aaatgagagc aattatacac tttatgagtt 12660
ttttggggtt atagtattat tatgtatatt attaatattc taattttaat agtaaggact 12720
ttgtcataca tactattcac atacagtatt agccacttta gcaaataagc acacacaaaa 12780
tcctggattt tatggcaaaa cagaggcatt tttgatcagt gatgacaaaa ttaaattcat 12840
tttgtttatt tcattacttt tataattcct aaaagtggga ggatcccagc tcttatagga 12900
gcaattaata tttaatgtag tgtcttttga aacaaaactg tgtgccaaag tagtaaccat 12960
taatggaagt ttacttgtag tcacaaattt agtttcctta atcatttgtt gaggacgttt 13020
tgaatcacac actatgagtg ttaagagata cctttaggaa actattcttg ttgttttctg 13080
attttgtcat ttaggttagt ctcctgattc tgacagctca gaagaggaag ttgttcttgt 13140
aaaaattgtt taacctgctt gaccagcttt cacatttgtt cttctgaagt ttatggtagt 13200
gcacagagat tgttttttgg ggagtcttga ttctcggaaa tgaaggcagt gtgttatatt 13260
gaatccagac ttccgaaaac ttgtatatta aaagtgttat ttcaacacta tgttacagcc 13320
agactaattt ttttattttt tgatgcattt tagatagctg atacagtact caatgatgat 13380
gatattggtg acagctgtca tgaaggcttt cttctcaagt aagaattttt cttttcataa 13440
aagctggatg aagcagatac catcttatgc tcacctatga caagatttgg aagaaagaaa 13500
ataacagact gtctacttag attgttctag ggacattacg tatttgaact gttgcttaaa 13560
tttgtgttat ttttcactca ttatatttct atatatattt ggtgttattc catttgctat 13620
ttaaagaaac cgagtttcca tcccagacaa gaaatcatgg ccccttgctt gattctggtt 13680
tcttgtttta cttctcatta aagctaacag aatcctttca tattaagttg tactgtagat 13740
gaacttaagt tatttaggcg tagaacaaaa ttattcatat ttatactgat ctttttccat 13800
ccagcagtgg agtttagtac ttaagagttt gtgcccttaa accagactcc ctggattaat 13860
gctgtgtacc cgtgggcaag gtgcctgaat tctctataca cctatttcct catctgtaaa 13920
atggcaataa tagtaatagt acctaatgtg tagggttgtt ataagcattg agtaagataa 13980
ataatataaa gcacttagaa cagtgcctgg aacataaaaa cacttaataa tagctcatag 14040
ctaacatttc ctatttacat ttcttctaga aatagccagt atttgttgag tgcctacatg 14100
ttagttcctt tactagttgc tttacatgta ttatcttata ttctgtttta aagtttcttc 14160
acagttacag attttcatga aattttactt ttaataaaag agaagtaaaa gtataaagta 14220
ttcactttta tgttcacagt cttttccttt aggctcatga tggagtatca gaggcatgag 14280
tgtgtttaac ctaagagcct taatggcttg aatcagaagc actttagtcc tgtatctgtt 14340
cagtgtcagc ctttcataca tcattttaaa tcccatttga ctttaagtaa gtcacttaat 14400
ctctctacat gtcaatttct tcagctataa aatgatggta tttcaataaa taaatacatt 14460
aattaaatga tattatactg actaattggg ctgttttaag gctcaataag aaaatttctg 14520
tgaaaggtct ctagaaaatg taggttccta tacaaataaa agataacatt gtgcttatag 14580
cttcggtgtt tatcatataa agctattctg agttatttga agagctcacc tacttttttt 14640
tgtttttagt ttgttaaatt gttttatagg caatgttttt aatctgtttt ctttaactta 14700
cagtgccatc agctcacact tgcaaacctg tggctgttcc gttgtagtag gtagcagtgc 14760
agagaaagta aataaggtag tttattttat aatctagcaa atgatttgac tctttaagac 14820
tgatgatata tcatggattg tcatttaaat ggtaggttgc aattaaaatg atctagtagt 14880
ataaggaggc aatgtaatct catcaaattg ctaagacacc ttgtggcaac agtgagtttg 14940
aaataaactg agtaagaatc atttatcagt ttattttgat agctcggaaa taccagtgtc 15000
agtagtgtat aaatggtttt gagaatatat taaaatcaga tatataaaaa aaattactct 15060
tctatttccc aatgttatct ttaacaaatc tgaagatagt catgtacttt tggtagtagt 15120
tccaaagaaa tgttatttgt ttattcatct tgatttcatt gtcttcgctt tccttctaaa 15180
tctgtccctt ctagggagct attgggatta agtggtcatt gattattata ctttattcag 15240
taatgtttct gaccctttcc ttcagtgcta cttgagttaa ttaaggatta atgaacagtt 15300
acatttccaa gcattagcta ataaactaaa ggattttgca cttttcttca ctgaccatta 15360
gttagaaaga gttcagagat aagtatgtgt atctttcaat ttcagcaaac ctaatttttt 15420
aaaaaaagtt ttacatagga aatatgttgg aaatgatact ttacaaagat attcataatt 15480
tttttttgta atcagctact ttgtatattt acatgagcct taatttatat ttctcatata 15540
accatttatg agagcttagt atacctgtgt cattatattg catctacgaa ctagtgacct 15600
tattccttct gttacctcaa acaggtggct ttccatctgt gatctccaaa gccttaggtt 15660
gcacagagtg actgccgagc tgctttatga agggagaaag gctccatagt tggagtgttt 15720
tttttttttt ttttaaacat ttttcccatc ctccatcctc ttgagggaga atagcttacc 15780
ttttatcttg ttttaatttg agaaagaagt tgccaccact ctaggttgaa aaccactcct 15840
ttaacataat aactgtggat atggtttgaa tttcaagata gttacatgcc tttttatttt 15900
tcctaataga gctgtaggtc aaatattatt agaatcagat ttctaaatcc cacccaatga 15960
cctgcttatt ttaaatcaaa ttcaataatt aattctcttc tttttggagg atctggacat 16020
tctttgatat ttcttacaac gaatttcatg tgtagaccca ctaaacagaa gctataaaag 16080
ttgcatggtc aaataagtct gagaaagtct gcagatgata taattcacct gaagagtcac 16140
agtatgtagc caaatgttaa aggttttgag atgccataca gtaaatttac caagcatttt 16200
ctaaatttat ttgaccacag aatccctatt ttaagcaaca actgttacat cccatggatt 16260
ccaggtgact aaagaatact tatttcttag gatatgtttt attgataata acaattaaaa 16320
tttcagatat ctttcataag caaatcagtg gtctttttac ttcatgtttt aatgctaaaa 16380
tattttcttt tatagatagt cagaacatta tgcctttttc tgactccagc agagagaaaa 16440
tgctccaggt tatgtgaagc agaatcatca tttaaatatg agtcagggct ctttgtacaa 16500
ggcctgctaa aggtatagtt tctagttatc acaagtgaaa ccacttttct aaaatcattt 16560
ttgagactct ttatagacaa atcttaaata ttagcattta atgtatctca tattgacatg 16620
cccagagact gacttccttt acacagttct gcacatagac tatatgtctt atggatttat 16680
agttagtatc atcagtgaaa caccatagaa taccctttgt gttccaggtg ggtccctgtt 16740
cctacatgtc tagcctcagg actttttttt ttttaacaca tgcttaaatc aggttgcaca 16800
tcaaaaataa gatcatttct ttttaactaa atagatttga attttattga aaaaaaattt 16860
taaacatctt taagaagctt ataggattta agcaattcct atgtatgtgt actaaaatat 16920
atatatttct atatataata tatattagaa aaaaattgta tttttctttt atttgagtct 16980
actgtcaagg agcaaaacag agaaatgtaa attagcaatt atttataata cttaaaggga 17040
agaaagttgt tcaccttgtt gaatctatta ttgttatttc aattatagtc ccaagacgtg 17100
aagaaatagc tttcctaatg gttatgtgat tgtctcatag tgactacttt cttgaggatg 17160
tagccacggc aaaatgaaat aaaaaaattt aaaaattgtt gcaaatacaa gttatattag 17220
gcttttgtgc attttcaata atgtgctgct atgaactcag aatgatagta tttaaatata 17280
gaaactagtt aaaggaaacg tagtttctat ttgagttata catatctgta aattagaact 17340
tctcctgtta aaggcataat aaagtgctta atacttttgt ttcctcagca ccctctcatt 17400
taattatata attttagttc tgaaagggac ctataccaga tgcctagagg aaatttcaaa 17460
actatgatct aatgaaaaaa tatttaatag ttctccatgc aaatacaaat catatagttt 17520
tccagaaaat acctttgaca ttatacaaag atgattatca cagcattata atagtaaaaa 17580
aatggaaata gcctctttct tctgttctgt tcatagcaca gtgcctcata cgcagtaggt 17640
tattattaca tggtaactgg ctaccccaac tgattaggaa agaagtaaat ttgttttata 17700
aaaatacata ctcattgagg tgcatagaat aattaagaaa ttaaaagaca cttgtaattt 17760
tgaatccagt gaatacccac tgttaatatt tggtatatct ctttctagtc tttttttccc 17820
ttttgcatgt attttcttta agactcccac ccccactgga tcatctctgc atgttctaat 17880
ctgctttttt cacagcagat tctaagcctc tttgaatatc aacacaaact tcaacaactt 17940
catctataga tgccaaataa taaattcatt tttatttact taaccacttc ctttggatgc 18000
ttaggtcatt ctgatgtttt gctattgaaa ccaatgctat actgaacact tctgtcacta 18060
aaactttgca cacactcatg aatagcttct taggataaat ttttagagat ggatttgcta 18120
aatcagagac cattttttaa aattaaaaaa caattattca tatcgtttgg catgtaagac 18180
agtaaatttt ccttttattt tgacaggatt caactggaag ctttgtgctg cctttccggc 18240
aagtcatgta tgctccatat cccaccacac acatagatgt ggatgtcaat actgtgaagc 18300
agatgccacc ctgtcatgaa catatttata atcagcgtag atacatgaga tccgagctga 18360
cagccttctg gagagccact tcagaagaag acatggctca ggatacgatc atctacactg 18420
acgaaagctt tactcctgat ttgtacgtaa tgctctgcct gctggtactg tagtcaagca 18480
atatgaaatt gtgtctttta cgaataaaaa caaaacagaa gttgcattta aaaagaaaga 18540
aatattacca gcagaattat gcttgaagaa acatttaatc aagcattttt ttcttaaatg 18600
ttcttctttt tccatacaat tgtgtttacc ctaaaatagg taagattaac ccttaaagta 18660
aatatttaac tatttgttta ataaatatat attgagctcc taggcactgt tctaggtacc 18720
gggcttaata gtggccaacc agacagcccc agccccagcc cctacattgt gtatagtcta 18780
ttatgtaaca gttattgaat ggacttatta acaaaaccaa agaagtaatt ctaagtcttt 18840
tttttcttga catatgaata taaaatacag caaaactgtt aaaatatatt aatggaacat 18900
ttttttactt tgcattttat attgttattc acttcttatt tttttttaaa aaaaaaagcc 18960
tgaacagtaa attcaaaagg aaaagtaatg ataattaatt gttgagcatg gacccaactt 19020
gaaaaaaaaa atgatgatga taaatctata atcctaaaac cctaagtaaa cacttaaaag 19080
atgttctgaa atcaggaaaa gaattatagt atacttttgt gtttctcttt tatcagttga 19140
aaaaaggcac agtagctcat gcctgtaaga acagagcttt gggagtgcaa ggcaggcgga 19200
tcacttgagg ccaggagttc cagaccagcc tgggcaacat agtgaaaccc catctctaca 19260
aaaaataaaa aagaattatt ggaatgtgtt tctgtgtgcc tgtaatccta gctattccga 19320
aagctgaggc aggaggatct tttgagccca ggagtttgag gttacaggga gttatgatgt 19380
gccagtgtac tccagcctgg ggaacaccga gactctgtct tatttaaaaa aaaaaaaaaa 19440
aaaatgcttg caataatgcc tggcacatag aaggtaacag taagtgttaa ctgtaataac 19500
ccaggtctaa gtgtgtaagg caatagaaaa attggggcaa ataagcctga cctatgtatc 19560
tacagaatca gtttgagctt aggtaacaga cctgtggagc accagtaatt acacagtaag 19620
tgttaaccaa aagcatagaa taggaatatc ttgttcaagg gacccccagc cttatacatc 19680
tcaaggtgca gaaagatgac ttaatatagg acccattttt tcctagttct ccagagtttt 19740
tattggttct tgagaaagta gtaggggaat gttttagaaa atgaattggt ccaactgaaa 19800
ttacatgtca gtaagttttt atatattggt aaattttagt agacatgtag aagttttcta 19860
attaatctgt gccttgaaac attttctttt ttcctaaagt gcttagtatt ttttccgttt 19920
tttgattggt tacttgggag cttttttgag gaaatttagt gaactgcaga atgggtttgc 19980
aaccatttgg tatttttgtt ttgtttttta gaggatgtat gtgtatttta acatttctta 20040
atcattttta gccagctatg tttgttttgc tgatttgaca aactacagtt agacagctat 20100
tctcattttg ctgatcatga caaaataata tcctgaattt ttaaattttg catccagctc 20160
taaattttct aaacataaaa ttgtccaaaa aatagtattt tcagccacta gattgtgtgt 20220
taagtctatt gtcacagagt cattttactt ttaagtatat gtttttacat gttaattatg 20280
tttgttattt ttaattttaa ctttttaaaa taattccagt cactgccaat acatgaaaaa 20340
ttggtcactg gaattttttt tttgactttt attttaggtt catgtgtaca tgtgcaggtg 20400
tgttatacag gtaaattgcg tgtcatgagg gtttggtgta caggtgattt cattacccag 20460
gtaataagca tagtacccaa taggtagttt tttgatcctc acccttctcc caccctcaag 20520
taggccctgg tgttgctgtt tccttctttg tgtccatgta tactcagtgt ttagctccca 20580
cttagaagtg agaacatgcg gtagttggtt ttctgttcct ggattagttc acttaggata 20640
atgacctcta gctccatctg gtttttatgg ctgcatagta ttccatggtg tatatgtatc 20700
acattttctt tatccagtct accattgata ggcatttagg ttgattccct gtctttgtta 20760
tcatgaatag tgctgtgatg aacatacaca tgcatgtgtc tttatggtag aaaaatttgt 20820
attcctttag gtacatatag aataatgggg ttgctagggt gaatggtagt tctattttca 20880
gttatttgag aaatcttcaa actgcttttc ataatagcta aactaattta cagtcccgcc 20940
agcagtgtat aagtgttccc ttttctccac aaccttgcca acatctgtga ttttttgact 21000
ttttaataat agccattcct agagaattga tttgcaattc tctattagtg atattaagca 21060
ttttttcata tgctttttag ctgtctgtat atattcttct gaaaaatttt catgtccttt 21120
gcccagtttg tagtggggtg ggttgttttt tgcttgttaa ttagttttaa gttccttcca 21180
gattctgcat atccctttgt tggatacatg gtttgcagat atttttctcc cattgtgtag 21240
gttgtctttt actctgttga tagtttcttt tgccatgcag gagctcgtta ggtcccattt 21300
gtgtttgttt ttgttgcagt tgcttttggc gtcttcatca taaaatctgt gccagggcct 21360
atgtccagaa tggtatttcc taggttgtct tccagggttt ttacaatttt agattttacg 21420
tttatgtctt taatccatct tgagttgatt tttgtatatg gcacaaggaa ggggtccagt 21480
ttcactccaa ttcctatggc tagcaattat cccagcacca tttattgaat acggagtcct 21540
ttccccattg cttgtttttt gtcaactttg ttgaagatca gatggttgta agtgtgtggc 21600
tttatttctt ggctctctat tctccattgg tctatgtgtc tgtttttata acagtaccct 21660
gctgttcagg ttcctatagc cttttagtat aaaatcggct aatgtgatgc ctccagcttt 21720
gttctttttg cttaggattg ctttggctat ttgggctcct ttttgggtcc atattaattt 21780
taaaacagtt ttttctggtt ttgtgaagga tatcattggt agtttatagg aatagcattg 21840
aatctgtaga ttgctttggg cagtatggcc attttaacaa tattaattct tcctatctat 21900
gaatatggaa tgtttttcca tgtgtttgtg tcatctcttt atacctgatg tataaagaaa 21960
agctggtatt attcctactc aatctgttcc aaaaaattga ggaggaggaa ctcttcccta 22020
atgaggccag catcattctg ataccaaaac ctggcagaga cacaacagaa aaaagaaaac 22080
ttcaggccaa tatccttgat gaatatagat gcaaaaatcc tcaacaaaat actagcaaac 22140
caaatccagc agcacatcaa aaagctgatc tactttgatc aagtaggctt tatccctggg 22200
atgcaaggtt ggttcaacat acacaaatca ataagtgtga ttcatcacat aaacagagct 22260
aaaaacaaaa accacaagat tatctcaata ggtagagaaa aggttgtcaa taaaatttaa 22320
catcctccat gttaaaaacc ttcagtaggt caggtgtagt gactcacacc tgtaatccca 22380
gcactttggg aggccaaggc gggcatatct cttaagccca ggagttcaag acgagcctag 22440
gcagcatggt gaaaccccat ctctacaaaa aaaaaaaaaa aaaaaaatta gcttggtatg 22500
gtgacatgca cctatagtcc cagctattca ggaggttgag gtgggaggat tgtttgagcc 22560
cgggaggcag aggttggcag cgagctgaga tcatgccacc gcactccagc ctgggcaacg 22620
gagtgagacc ctgtctcaaa aaagaaaaat cacaaacaat cctaaacaaa ctaggcattg 22680
aaggaacatg cctcaaaaaa ataagaacca tctatgacag acccatagcc aatatcttac 22740
caaatgggca aaagctggaa gtattctcct tgagaaccgt aacaagacaa ggatgtccac 22800
tctcaccact ccttttcagc atagttctgg aagtcctagc cagagcaatc aggaaagaga 22860
aagaaagaaa gacattcaga taggaagaga agaagtcaaa ctatttctgt ttgcaggcag 22920
tataattctg tacctagaaa atctcatagt ctctgcccag aaactcctaa atctgttaaa 22980
aatttcagca aagttttggc attctctata ctccaacacc ttccaaagtg agagcaaaat 23040
caagaacaca gtcccattca caatagccgc aaaacgaata aaatacctag gaatccagct 23100
aaccagggag gtgaaagatc tctatgagaa ttacaaaaca ctgctgaaag aaatcagaga 23160
tgacacaaac aaatggaaat gttctttttt aacaccttgc tttatctaat tcacttatga 23220
tgaagatact cattcagtgg aacaggtata ataagtccac tcgattaaat ataagcctta 23280
ttctctttcc agagcccaag aaggggcact atcagtgccc agtcaataat gacgaaatgc 23340
taatattttt cccctttacg gtttctttct tctgtagtgt ggtacactcg tttcttaaga 23400
taaggaaact tgaactacct tcctgtttgc ttctacacat acccattctc tttttttgcc 23460
actctggtca ggtataggat gatccctacc actttcagtt aaaaactcct cctcttacta 23520
aatgttctct taccctctgg cctgagtaga acctagggaa aatggaagag aaaaagatga 23580
aagggaggtg gggcctggga agggaataag tagtcctgtt tgtttgtgtg tttgctttag 23640
cacctgctat atcctaggtg ctgtgttagg cacacattat tttaagtggc cattatatta 23700
ctactactca ctctggtcgt tgccaaggta ggtagtactt tcttggatag ttggttcatg 23760
ttacttacag atggtgggct tgttgaggca aacccagtgg ataatcatcg gagtgtgttc 23820
tctaatctca ctcaaatttt tcttcacatt ttttggtttg ttttggtttt tgatggtagt 23880
ggcttatttt tgttgctggt ttgttttttg tttttttttg agatggcaag aattggtagt 23940
tttatttatt aattgcctaa gggtctctac tttttttaaa agatgagagt agtaaaatag 24000
attgatagat acatacatac ccttactggg gactgcttat attctttaga gaaaaaatta 24060
catattagcc tgacaaacac cagtaaaatg taaatatatc cttgagtaaa taaatgaatg 24120
tatattttgt gtctccaaat atatatatct atattcttac aaatgtgttt atatgtaata 24180
tcaatttata agaacttaaa atgttggctc aagtgaggga ttgtggaagg tagcattata 24240
tggccatttc aacatttgaa cttttttctt ttcttcattt tcttcttttc ttcaggaata 24300
tttttcaaga tgtcttacac agagacactc tagtgaaagc cttcctggat caggtaaatg 24360
ttgaacttga gattgtcaga gtgaatgata tgacatgttt tcttttttaa tatatcctac 24420
aatgcctgtt ctatatattt atattcccct ggatcatgcc ccagagttct gctcagcaat 24480
tgcagttaag ttagttacac tacagttctc agaagagtct gtgagggcat gtcaagtgca 24540
tcattacatt ggttgcctct tgtcctagat ttatgcttcg ggaattcaga cctttgttta 24600
caatataata aatattattg ctatctttta aagatataat aataagatat aaagttgacc 24660
acaactactg ttttttgaaa catagaattc ctggtttaca tgtatcaaag tgaaatctga 24720
cttagctttt acagatataa tatatacata tatatatcct gcaatgcttg tactatatat 24780
gtagtacaag tatatatata tgtttgtgtg tgtatatata tatagtacga gcatatatac 24840
atattaccag cattgtagga tatatatatg tttatatatt aaaaaaaagt tataaactta 24900
aaaccctatt atgttatgta gagtatatgt tatatatgat atgtaaaata tataacatat 24960
actctatgat agagtgtaat atatttttta tatatatttt aacatttata aaatgataga 25020
attaagaatt gagtcctaat ctgttttatt aggtgctttt tgtagtgtct ggtctttcta 25080
aagtgtctaa atgatttttc cttttgactt attaatgggg aagagcctgt atattaacaa 25140
ttaagagtgc agcattccat acgtcaaaca acaaacattt taattcaagc attaacctat 25200
aacaagtaag tttttttttt ttttttgaga aagggaggtt gtttatttgc ctgaaatgac 25260
tcaaaaatat ttttgaaaca tagtgtactt atttaaataa catctttatt gtttcattct 25320
tttaaaaaat atctacttaa ttacacagtt gaaggaaatc gtagattata tggaacttat 25380
ttcttaatat attacagttt gttataataa cattctgggg atcaggccag gaaactgtgt 25440
catagataaa gctttgaaat aatgagatcc ttatgtttac tagaaatttt ggattgagat 25500
ctatgaggtc tgtgacatat tgcgaagttc aaggaaaatt cgtaggcctg gaatttcatg 25560
cttctcaagc tgacataaaa tccctcccac tctccacctc atcatatgca cacattctac 25620
tcctacccac ccactccacc ccctgcaaaa gtacaggtat atgaatgtct caaaaccata 25680
ggctcatctt ctaggagctt caatgttatt tgaagatttg ggcagaaaaa attaagtaat 25740
acgaaataac ttatgtatga gttttaaaag tgaagtaaac atggatgtat tctgaagtag 25800
aatgcaaaat ttgaatgcat ttttaaagat aaattagaaa acttctaaaa actgtcagat 25860
tgtctgggcc tggtggctta tgcctgtaat cccagcactt tgggagtccg aggtgggtgg 25920
atcacaaggt caggagatcg agaccatcct gccaacatgg tgaaaccccg tctctactaa 25980
gtatacaaaa attagctggg cgtggcagcg tgtgcctgta atcccagcta cctgggaggc 26040
tgaggcagga gaatcgcttg aacccaggag gtgtaggttg cagtgagtca agatcgcgcc 26100
actgcacttt agcctggtga cagagctaga ctccgtctca aaaaaaaaaa aaaatatcag 26160
attgttccta cacctagtgc ttctatacca cactcctgtt agggggcatc agtggaaatg 26220
gttaaggaga tgtttagtgt gtattgtctg ccaagcactg tcaacactgt catagaaact 26280
tctgtacgag tagaatgtga gcaaattatg tgttgaaatg gttcctctcc ctgcaggtct 26340
ttcagctgaa acctggctta tctctcagaa gtactttcct tgcacagttt ctacttgtcc 26400
ttcacagaaa agccttgaca ctaataaaat atatagaaga cgatacgtga gtaaaactcc 26460
tacacggaag aaaaaccttt gtacattgtt tttttgtttt gtttcctttg tacattttct 26520
atatcataat ttttgcgctt cttttttttt tttttttttt tttttttcca ttatttttag 26580
gcagaaggga aaaaagccct ttaaatctct tcggaacctg aagatagacc ttgatttaac 26640
agcagagggc gatcttaaca taataatggc tctggctgag aaaattaaac caggcctaca 26700
ctcttttatc tttggaagac ctttctacac tagtgtgcaa gaacgagatg ttctaatgac 26760
tttttaaatg tgtaacttaa taagcctatt ccatcacaat catgatcgct ggtaaagtag 26820
ctcagtggtg tggggaaacg ttcccctgga tcatactcca gaattctgct ctcagcaatt 26880
gcagttaagt aagttacact acagttctca caagagcctg tgaggggatg tcaggtgcat 26940
cattacattg ggtgtctctt ttcctagatt tatgcttttg ggatacagac ctatgtttac 27000
aatataataa atattattgc tatcttttaa agatataata ataggatgta aacttgacca 27060
caactactgt ttttttgaaa tacatgattc atggtttaca tgtgtcaagg tgaaatctga 27120
gttggctttt acagatagtt gactttctat cttttggcat tctttggtgt gtagaattac 27180
tgtaatactt ctgcaatcaa ctgaaaacta gagcctttaa atgatttcaa ttccacagaa 27240
agaaagtgag cttgaacata ggatgagctt tagaaagaaa attgatcaag cagatgttta 27300
attggaattg attattagat cctactttgt ggatttagtc cctgggattc agtctgtaga 27360
aatgtctaat agttctctat agtccttgtt cctggtgaac cacagttagg gtgttttgtt 27420
tattttattg ttcttgctat tgttgatatt ctatgtagtt gagctctgta aaaggaaatt 27480
gtattttatg ttttagtaat tgttgccaac tttttaaatt aattttcatt atttttgagc 27540
caaattgaaa tgtgcacctc ctgtgccttt tttctcctta gaaaatctaa ttacttggaa 27600
caagttcaga tttcactggt cagtcatttt catcttgttt tcttcttgct aagtcttacc 27660
atgtacctgc tttggcaatc attgcaactc tgagattata aaatgcctta gagaatatac 27720
taactaataa gatctttttt tcagaaacag aaaatagttc cttgagtact tccttcttgc 27780
atttctgcct atgtttttga agttgttgct gtttgcctgc aataggctat aaggaatagc 27840
aggagaaatt ttactgaagt gctgttttcc taggtgctac tttggcagag ctaagttatc 27900
ttttgttttc ttaatgcgtt tggaccattt tgctggctat aaaataactg attaatataa 27960
ttctaacaca atgttgacat tgtagttaca caaacacaaa taaatatttt atttaaaatt 28020
ctggaagtaa tataaaaggg aaaatatatt tataagaaag ggataaaggt aatagagccc 28080
ttctgccccc cacccaccaa atttacacaa caaaatgaca tgttcgaatg tgaaaggtca 28140
taatagcttt cccatcatga atcagaaaga tgtggacagc ttgatgtttt agacaaccac 28200
tgaactagat gactgttgta ctgtagctca gtcatttaaa aaatatataa atactacctt 28260
gtagtgtccc atactgtgtt ttttacatgg tagattctta tttaagtgct aactggttat 28320
tttctttggc tggtttattg tactgttata cagaatgtaa gttgtacagt gaaataagtt 28380
attaaagcat gtgtaaacat tgttatatat cttttctcct aaatggagaa ttttgaataa 28440
aatatatttg aaattttgcc tctttcagtt gttcattcag aaaaaaatac tatgatattt 28500
gaagactgat cagcttctgt tcagctgaca gtcatgctgg atctaaactt tttttaaaat 28560
taattttgtc ttttcaaaga aaaaatattt aaagaagctt tataatataa tcttatgtta 28620
aaaaaacttt ctgcttaact ctctggattt cattttgatt tttcaaatta tatattaata 28680
tttcaaatgt aaaatactat ttagataaat tgtttttaaa cattcttatt attataatat 28740
taatataacc taaactgaag ttattcatcc caggtatcta atacatgtat ccaaagtaaa 28800
aatccaagga atctgaacac tttcatctgc aaagctagga ataggtttga cattttcact 28860
ccaagaaaaa gttttttttt gaaaatagaa tagttgggat gagaggtttc tttaaaagaa 28920
gactaactga tcacattact atgattctca aagaagaaac caaaacttca tataatacta 28980
taaagtaaat ataaaatagt tccttctata gtatatttct ataatgctac agtttaaaca 29040
gatcactctt atataatact attttgattt tgatgtagaa ttgcacaaat tgatatttct 29100
cctatgatct gcagggtata gcttaaagta acaaaaacag tcaaccacct ccatttaaca 29160
cacagtaaca ctatgggact agttttatta cttccatttt acaaatgagg aaactaaagc 29220
ttaaagatgt gtaatacacc gcccaaggtc acacagctgg taaaggtgga tttcatccca 29280
gacagttaca gtcattgcca tgggcacagc tcctaactta gtaactccat gtaactggta 29340
ctcagtgtag ctgaattgaa aggagagtaa ggaagcaggt tttacaggtc tacttgcact 29400
attcagagcc cgagtgtgaa tccctgctgt gctgcttgga gaagttactt aacctatgca 29460
aggttcattt tgtaaatatt ggaaatggag tgataatacg tacttcacca gaggatttaa 29520
tgagacctta tacgatcctt agttcagtac ctgactagtg cttcataaat gctttttcat 29580
ccaatctgac aatctccagc ttgtaattgg ggcatttaga acatttaata tgattattgg 29640
catggtaggt taaagctgtc atcttgctgt tttctatttg ttctttttgt tttctcctta 29700
cttttggatt tttttattct actatgtctt ttctattgtc ttattaacta tactctttga 29760
tttattttag tggttgtttt agggttatac ctctttctaa tttaccagtt tataaccagt 29820
ttatatacta cttgacatat agcttaagaa acttactgtt gttgtctttt tgctgttatg 29880
gtcttaacgt ttttatttct acaaacatta taaactccac actttattgt tttttaattt 29940
tacttataca gtcaattatc ttttaaagat atttaaatat aaacattcaa aacaccccaa 30000
t 30001
<210> 3
<211> 1031
<212> ДНК
<213> Homo sapiens
<400> 3
attcccggga tacgtaacct acggtgtccc gctaggaaag agaggtgcgt caaacagcga 60
caagttccgc ccacgtaaaa gatgacgctt ggtgtgtcag ccgtccctgc tgcccggttg 120
cttctctttt gggggcgggg tctagcaaga gcaggtgtgg gtttaggaga tatctccgga 180
gcatttggat aatgtgacag ttggaatgca gtgatgtcga ctctttgccc accgccatct 240
ccagctgttg ccaagacaga gattgcttta agtggcaaat cacctttatt agcagctact 300
tttgcttact gggacaatat tcttggtcct agagtaaggc acatttgggc tccaaagaca 360
gaacaggtac ttctcagtga tggagaaata acttttcttg ccaaccacac tctaaatgga 420
gaaatccttc gaaatgcaga gagtggtgct atagatgtaa agttttttgt cttgtctgaa 480
aagggagtga ttattgtttc attaatcttt gatggaaact ggaatgggga tcgcagcaca 540
tatggactat caattatact tccacagaca gaacttagtt tctacctccc acttcataga 600
gtgtgtgttg atagattaac acatataatc cggaaaggaa gaatatggat gcataaggaa 660
agacaagaaa aatgtccaga agattatctt agaaggcaca gagagaatgg aagatcaggg 720
tcagagtatt attccaatgc ttactggaga agtgattcct gtaatggaaa ctgctttcct 780
ctatgaaatt cccccgggtt cctggaggaa atagatatag gctgatacag ttacccaatg 840
atggatgaat attgggggac cgcctggtca ttgaaaggct ttcttttctc caggaaagaa 900
atttttttcc ttttccataa aaagcttggg aatggaagac aacaattccc attctttttt 960
tgcgttccac ccctatgtga caacagaaat ttttggggaa acaacaacga aaaaatttta 1020
tcccgcgcgc a 1031
<210> 4
<211> 3244
<212> ДНК
<213> Homo sapiens
<220>
<221> КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<222> (108)..(1553)
<400> 4
gggcggggct gcggttgcgg tgcctgcgcc cgcggcggcg gaggcgcagg cggtggcgag 60
tggatatctc cggagcattt ggataatgtg acagttggaa tgcagtg atg tcg act 116
Met Ser Thr
1
ctt tgc cca ccg cca tct cca gct gtt gcc aag aca gag att gct tta 164
Leu Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu Ile Ala Leu
5 10 15
agt ggc aaa tca cct tta tta gca gct act ttt gct tac tgg gac aat 212
Ser Gly Lys Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr Trp Asp Asn
20 25 30 35
att ctt ggt cct aga gta agg cac att tgg gct cca aag aca gaa cag 260
Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys Thr Glu Gln
40 45 50
gta ctt ctc agt gat gga gaa ata act ttt ctt gcc aac cac act cta 308
Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn His Thr Leu
55 60 65
aat gga gaa atc ctt cga aat gca gag agt ggt gct ata gat gta aag 356
Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile Asp Val Lys
70 75 80
ttt ttt gtc ttg tct gaa aag gga gtg att att gtt tca tta atc ttt 404
Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser Leu Ile Phe
85 90 95
gat gga aac tgg aat ggg gat cgc agc aca tat gga cta tca att ata 452
Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu Ser Ile Ile
100 105 110 115
ctt cca cag aca gaa ctt agt ttc tac ctc cca ctt cat aga gtg tgt 500
Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His Arg Val Cys
120 125 130
gtt gat aga tta aca cat ata atc cgg aaa gga aga ata tgg atg cat 548
Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile Trp Met His
135 140 145
aag gaa aga caa gaa aat gtc cag aag att atc tta gaa ggc aca gag 596
Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Ile Leu Glu Gly Thr Glu
150 155 160
aga atg gaa gat cag ggt cag agt att att cca atg ctt act gga gaa 644
Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu Thr Gly Glu
165 170 175
gtg att cct gta atg gaa ctg ctt tca tct atg aaa tca cac agt gtt 692
Val Ile Pro Val Met Glu Leu Leu Ser Ser Met Lys Ser His Ser Val
180 185 190 195
cct gaa gaa ata gat ata gct gat aca gta ctc aat gat gat gat att 740
Pro Glu Glu Ile Asp Ile Ala Asp Thr Val Leu Asn Asp Asp Asp Ile
200 205 210
ggt gac agc tgt cat gaa ggc ttt ctt ctc aat gcc atc agc tca cac 788
Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn Ala Ile Ser Ser His
215 220 225
ttg caa acc tgt ggc tgt tcc gtt gta gta ggt agc agt gca gag aaa 836
Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly Ser Ser Ala Glu Lys
230 235 240
gta aat aag ata gtc aga aca tta tgc ctt ttt ctg act cca gca gag 884
Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe Leu Thr Pro Ala Glu
245 250 255
aga aaa tgc tcc agg tta tgt gaa gca gaa tca tca ttt aaa tat gag 932
Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser Ser Phe Lys Tyr Glu
260 265 270 275
tca ggg ctc ttt gta caa ggc ctg cta aag gat tca act gga agc ttt 980
Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp Ser Thr Gly Ser Phe
280 285 290
gtg ctg cct ttc cgg caa gtc atg tat gct cca tat ccc acc aca cac 1028
Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro Tyr Pro Thr Thr His
295 300 305
ata gat gtg gat gtc aat act gtg aag cag atg cca ccc tgt cat gaa 1076
Ile Asp Val Asp Val Asn Thr Val Lys Gln Met Pro Pro Cys His Glu
310 315 320
cat att tat aat cag cgt aga tac atg aga tcc gag ctg aca gcc ttc 1124
His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser Glu Leu Thr Ala Phe
325 330 335
tgg aga gcc act tca gaa gaa gac atg gct cag gat acg atc atc tac 1172
Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln Asp Thr Ile Ile Tyr
340 345 350 355
act gac gaa agc ttt act cct gat ttg aat att ttt caa gat gtc tta 1220
Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile Phe Gln Asp Val Leu
360 365 370
cac aga gac act cta gtg aaa gcc ttc ctg gat cag gtc ttt cag ctg 1268
His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp Gln Val Phe Gln Leu
375 380 385
aaa cct ggc tta tct ctc aga agt act ttc ctt gca cag ttt cta ctt 1316
Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu Ala Gln Phe Leu Leu
390 395 400
gtc ctt cac aga aaa gcc ttg aca cta ata aaa tat ata gaa gac gat 1364
Val Leu His Arg Lys Ala Leu Thr Leu Ile Lys Tyr Ile Glu Asp Asp
405 410 415
acg cag aag gga aaa aag ccc ttt aaa tct ctt cgg aac ctg aag ata 1412
Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu Arg Asn Leu Lys Ile
420 425 430 435
gac ctt gat tta aca gca gag ggc gat ctt aac ata ata atg gct ctg 1460
Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn Ile Ile Met Ala Leu
440 445 450
gct gag aaa att aaa cca ggc cta cac tct ttt atc ttt gga aga cct 1508
Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe Ile Phe Gly Arg Pro
455 460 465
ttc tac act agt gtg caa gaa cga gat gtt cta atg act ttt taa 1553
Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu Met Thr Phe
470 475 480
atgtgtaact taataagcct attccatcac aatcatgatc gctggtaaag tagctcagtg 1613
gtgtggggaa acgttcccct ggatcatact ccagaattct gctctcagca attgcagtta 1673
agtaagttac actacagttc tcacaagagc ctgtgagggg atgtcaggtg catcattaca 1733
ttgggtgtct cttttcctag atttatgctt ttgggataca gacctatgtt tacaatataa 1793
taaatattat tgctatcttt taaagatata ataataggat gtaaacttga ccacaactac 1853
tgtttttttg aaatacatga ttcatggttt acatgtgtca aggtgaaatc tgagttggct 1913
tttacagata gttgactttc tatcttttgg cattctttgg tgtgtagaat tactgtaata 1973
cttctgcaat caactgaaaa ctagagcctt taaatgattt caattccaca gaaagaaagt 2033
gagcttgaac ataggatgag ctttagaaag aaaattgatc aagcagatgt ttaattggaa 2093
ttgattatta gatcctactt tgtggattta gtccctggga ttcagtctgt agaaatgtct 2153
aatagttctc tatagtcctt gttcctggtg aaccacagtt agggtgtttt gtttatttta 2213
ttgttcttgc tattgttgat attctatgta gttgagctct gtaaaaggaa attgtatttt 2273
atgttttagt aattgttgcc aactttttaa attaattttc attatttttg agccaaattg 2333
aaatgtgcac ctcctgtgcc ttttttctcc ttagaaaatc taattacttg gaacaagttc 2393
agatttcact ggtcagtcat tttcatcttg ttttcttctt gctaagtctt accatgtacc 2453
tgctttggca atcattgcaa ctctgagatt ataaaatgcc ttagagaata tactaactaa 2513
taagatcttt ttttcagaaa cagaaaatag ttccttgagt acttccttct tgcatttctg 2573
cctatgtttt tgaagttgtt gctgtttgcc tgcaataggc tataaggaat agcaggagaa 2633
attttactga agtgctgttt tcctaggtgc tactttggca gagctaagtt atcttttgtt 2693
ttcttaatgc gtttggacca ttttgctggc tataaaataa ctgattaata taattctaac 2753
acaatgttga cattgtagtt acacaaacac aaataaatat tttatttaaa attctggaag 2813
taatataaaa gggaaaatat atttataaga aagggataaa ggtaatagag cccttctgcc 2873
ccccacccac caaatttaca caacaaaatg acatgttcga atgtgaaagg tcataatagc 2933
tttcccatca tgaatcagaa agatgtggac agcttgatgt tttagacaac cactgaacta 2993
gatgactgtt gtactgtagc tcagtcattt aaaaaatata taaatactac cttgtagtgt 3053
cccatactgt gttttttaca tggtagattc ttatttaagt gctaactggt tattttcttt 3113
ggctggttta ttgtactgtt atacagaatg taagttgtac agtgaaataa gttattaaag 3173
catgtgtaaa cattgttata tatcttttct cctaaatgga gaattttgaa taaaatatat 3233
ttgaaatttt g 3244
<210> 5
<211> 761
<212> ДНК
<213> Homo sapiens
<220>
<221> misc_feature
<222> (693)..(693)
<223> n является a, c, g или t
<220>
<221> misc_feature
<222> (722)..(722)
<223> n является a, c, g или t
<400> 5
cacgaggctt tgatatttct tacaacgaat ttcatgtgta gacccactaa acagaagcta 60
taaaagttgc atggtcaaat aagtctgaga aagtctgcag atgatataat tcacctgaag 120
agtcacagta tgtagccaaa tgttaaaggt tttgagatgc catacagtaa atttaccaag 180
cattttctaa atttatttga ccacagaatc cctattttaa gcaacaactg ttacatccca 240
tggattccag gtgactaaag aatacttatt tcttaggata tgttttattg ataataacaa 300
ttaaaatttc agatatcttt cataagcaaa tcagtggtct ttttacttca tgttttaatg 360
ctaaaatatt ttcttttata gatagtcaga acattatgcc tttttctgac tccagcagag 420
agaaaatgct ccaggttatg tgaagcagaa tcatcattta aatatgagtc agggctcttt 480
gtacaaggcc tgctaaagga ttcaactgga agctttgtgc tgcctttccg gcaagtcatg 540
tatgctccat atcccaccac acacatagat gtggatgtca atactgtgaa gcagatgcca 600
ccctgtcatg aacatattta taatcagcgt agatacatga gatccgagct gacagccttc 660
tggagagcca cttcagaaga agacatggct cangatacga tcatctacac tgacgaaagc 720
tntactcctg atttgaatat ttttcaagat gtcttacaca g 761
<210> 6
<211> 1901
<212> ДНК
<213> Homo sapiens
<220>
<221> КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<222> (125)..(793)
<400> 6
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg atatctccgg agcatttgga taatgtgaca gttggaatgc 120
agtg atg tcg act ctt tgc cca ccg cca tct cca gct gtt gcc aag aca 169
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr
1 5 10 15
gag att gct tta agt ggc aaa tca cct tta tta gca gct act ttt gct 217
Glu Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu Ala Ala Thr Phe Ala
20 25 30
tac tgg gac aat att ctt ggt cct aga gta agg cac att tgg gct cca 265
Tyr Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro
35 40 45
aag aca gaa cag gta ctt ctc agt gat gga gaa ata act ttt ctt gcc 313
Lys Thr Glu Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala
50 55 60
aac cac act cta aat gga gaa atc ctt cga aat gca gag agt ggt gct 361
Asn His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala
65 70 75
ata gat gta aag ttt ttt gtc ttg tct gaa aag gga gtg att att gtt 409
Ile Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val
80 85 90 95
tca tta atc ttt gat gga aac tgg aat ggg gat cgc agc aca tat gga 457
Ser Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly
100 105 110
cta tca att ata ctt cca cag aca gaa ctt agt ttc tac ctc cca ctt 505
Leu Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu
115 120 125
cat aga gtg tgt gtt gat aga tta aca cat ata atc cgg aaa gga aga 553
His Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg
130 135 140
ata tgg atg cat aag gaa aga caa gaa aat gtc cag aag att atc tta 601
Ile Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Ile Leu
145 150 155
gaa ggc aca gag aga atg gaa gat cag ggt cag agt att att cca atg 649
Glu Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met
160 165 170 175
ctt act gga gaa gtg att cct gta atg gaa ctg ctt tca tct atg aaa 697
Leu Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ser Ser Met Lys
180 185 190
tca cac agt gtt cct gaa gaa ata gat ata gct gat aca gta ctc aat 745
Ser His Ser Val Pro Glu Glu Ile Asp Ile Ala Asp Thr Val Leu Asn
195 200 205
gat gat gat att ggt gac agc tgt cat gaa ggc ttt ctt ctc aag taa 793
Asp Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Lys
210 215 220
gaatttttct tttcataaaa gctggatgaa gcagatacca tcttatgctc acctatgaca 853
agatttggaa gaaagaaaat aacagactgt ctacttagat tgttctaggg acattacgta 913
tttgaactgt tgcttaaatt tgtgttattt ttcactcatt atatttctat atatatttgg 973
tgttattcca tttgctattt aaagaaaccg agtttccatc ccagacaaga aatcatggcc 1033
ccttgcttga ttctggtttc ttgttttact tctcattaaa gctaacagaa tcctttcata 1093
ttaagttgta ctgtagatga acttaagtta tttaggcgta gaacaaaatt attcatattt 1153
atactgatct ttttccatcc agcagtggag tttagtactt aagagtttgt gcccttaaac 1213
cagactccct ggattaatgc tgtgtacccg tgggcaaggt gcctgaattc tctatacacc 1273
tatttcctca tctgtaaaat ggcaataata gtaatagtac ctaatgtgta gggttgttat 1333
aagcattgag taagataaat aatataaagc acttagaaca gtgcctggaa cataaaaaca 1393
cttaataata gctcatagct aacatttcct atttacattt cttctagaaa tagccagtat 1453
ttgttgagtg cctacatgtt agttccttta ctagttgctt tacatgtatt atcttatatt 1513
ctgttttaaa gtttcttcac agttacagat tttcatgaaa ttttactttt aataaaagag 1573
aagtaaaagt ataaagtatt cacttttatg ttcacagtct tttcctttag gctcatgatg 1633
gagtatcaga ggcatgagtg tgtttaacct aagagcctta atggcttgaa tcagaagcac 1693
tttagtcctg tatctgttca gtgtcagcct ttcatacatc attttaaatc ccatttgact 1753
ttaagtaagt cacttaatct ctctacatgt caatttcttc agctataaaa tgatggtatt 1813
tcaataaata aatacattaa ttaaatgata ttatactgac taattgggct gttttaaggc 1873
aaaaaaaaaa aaaaaaaaaa aaaaaaaa 1901
<210> 7
<211> 562
<212> ДНК
<213> Homo sapiens
<220>
<221> misc_feature
<222> (166)..(166)
<223> n является a, c, g или t
<400> 7
agacgtaacc tacggtgtcc cgctaggaaa gagagatatc tccggagcat ttggataatg 60
tgacagttgg aatgcagtga tgtcgactct ttgcccaccg ccatctccag ctgttgccaa 120
gacagagatt gctttaagtg gcaaatcacc tttattagca gctacntttt gcttactggg 180
acaatattct tggtcctaga gtaaggcaca tttgggctcc aaagacagaa caggtacttc 240
tcagtgatgg agaaataact tttcttgcca accacactct aaatggagaa atccttcgaa 300
atgcagagag tggtgctata gatgtaaagt tttttgtctt gtctgaaaag ggagtgatta 360
ttgtttcatt aatctttgat ggaaactgga atggggatcg cagcacatat ggactatcaa 420
ttatacttcc acagacagaa cttagtttct acctcccact tcatagagtg tgtgttgata 480
gattaacaca tataatccgg aaaggaagaa tatggatgca taaggaaaga caagaaaatg 540
tccagaagat tatcttagaa gg 562
<210> 8
<211> 798
<212> ДНК
<213> Homo sapiens
<220>
<221> КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<222> (81)..(590)
<400> 8
gggctctctt ttgggggcgg ggtctagcaa gagcagatat ctccggagca tttggataat 60
gtgacagttg gaatgcagtg atg tcg act ctt tgc cca ccg cca tct cca gct 113
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro Ala
1 5 10
gtt gcc aag aca gag att gct tta agt ggc aaa tca cct tta tta gca 161
Val Ala Lys Thr Glu Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu Ala
15 20 25
gct act ttt gct tac tgg gac aat att ctt ggt cct aga gta agg cac 209
Ala Thr Phe Ala Tyr Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His
30 35 40
att tgg gct cca aag aca gaa cag gta ctt ctc agt gat gga gaa ata 257
Ile Trp Ala Pro Lys Thr Glu Gln Val Leu Leu Ser Asp Gly Glu Ile
45 50 55
act ttt ctt gcc aac cac act cta aat gga gaa atc ctt cga aat gca 305
Thr Phe Leu Ala Asn His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala
60 65 70 75
gag agt ggt gct ata gat gta aag ttt ttt gtc ttg tct gaa aag gga 353
Glu Ser Gly Ala Ile Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly
80 85 90
gtg att att gtt tca tta atc ttt gat gga aac tgg aat ggg gat cgc 401
Val Ile Ile Val Ser Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg
95 100 105
agc aca tat gga cta tca att ata ctt cca cag aca gaa ctt agt ttc 449
Ser Thr Tyr Gly Leu Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe
110 115 120
tac ctc cca ctt cat aga gtg tgt gtt gat aga tta aca cat ata atc 497
Tyr Leu Pro Leu His Arg Val Cys Val Asp Arg Leu Thr His Ile Ile
125 130 135
cgg aaa gga aga ata tgg atg cat aag gaa aga caa gaa aat gtc cag 545
Arg Lys Gly Arg Ile Trp Met His Lys Glu Arg Gln Glu Asn Val Gln
140 145 150 155
aag att atc tta gaa ggc aca gag aga atg gaa gat cag ggt cag 590
Lys Ile Ile Leu Glu Gly Thr Glu Arg Met Glu Asp Gln Gly Gln
160 165 170
agtattattc caatgcttac tggagaagtg attcctgtaa tgggactgct ttcatctatg 650
aaatcacaca gtgttcctga agaaatagat atagctgata cagtactcca tgatgatgat 710
atttggtgac agctgtcatg aaaggctttc ttctcaagta ggaatttttt cttttcataa 770
aagctgggat gaagccagat tcccatct 798
<210> 9
<211> 169
<212> ДНК
<213> Homo sapiens
<400> 9
aaacagcgac aagttccgcc cacgtaaaag atgatgcttg gtgtgtcagc cgtccctgct 60
gcccggttgc ttctcttttg ggggcggggt ctagcaagag cagatatctc cggagcattt 120
ggataatgtg acagttggaa tgcggtgatg tcgactcttt gcccaccgc 169
<210> 10
<211> 176
<212> ДНК
<213> Homo sapiens
<400> 10
aaaacgtcat cgcacataga aaacagacag acgtaaccta cggtgtcccg ctaggaaaga 60
gaggtgcgtc aaacagcgac aagttccgcc cacgtaaaag atgacgcttg atatctccgg 120
agcatttgga taatgtgaca gttggaatgc agtgatgtcg actctttgcc caccgc 176
<210> 11
<211> 38001
<212> ДНК
<213> Mus musculus
<400> 11
caaacacaca cacacacaca cacacacaca cacacacaca cacacacaca cacacactgg 60
catatcaagt ctctgttagg ctaggcgcat cctctcccac tgaggtcaga caagactgcc 120
cagctagaag aacatatccc acggacaggc aacagctttt gggacagcca cgctccagtt 180
gtttgggact cataaaagac taaactcaca cctgctacaa aagtgcaggg aggcctaggt 240
ccagcctgtg tgtgctcttt gcttagtggt gtctctgaga gccctaagga tcaaagtttg 300
ttgactctgt tggtcttcct gtggagttcc tatccccttc gtccccctcc ccaccctgca 360
atctttccct caactcttct ttaggcctgg tggtggtggt gtggtggcat ggaggaggtg 420
gtggaggggg tgggggtctt taatcccggt tcttgtgaga ccgaagcagg gacgatttcg 480
gagctctgtg agtttgaggc cagcctggtc tatagatcta gttccaggac agtcagagct 540
acatagagaa accctgcccc gagggggggg gggcgcgggg aatggttaaa gattattgca 600
ggacccagct gatctgtgga agaggtaacg ggtgtttatg tttttcgaaa ctcattgaac 660
aatgcacttc aattgtgcgc actttagaaa tataaagcca ccacgcgaaa agctgcgccc 720
caacttaaag gcaatttcca aggtacttct gggtccttgc ggttcagtgg ctgtctaggt 780
tcagaaacga aactggatcc ccgccccgcc cccccgcccc ccccctcccc agcgccctga 840
ggcagtttcg atttcctatg gccagccggc taggcagctt tttcatcggg actccttgga 900
aagtccccac ttcgttcatc tctggcggat ttgcgggagc cagggcgctc atcgatcgcc 960
tggagccaca gaatgacagc ggagcagcgg cagaatctgc aagcattcag agactatatc 1020
aagaagattc tggaccccac ctacatcctc agctacatga gttcctggct cgaggatggt 1080
gagtggtccc caactggggc tctcaggctc tccaccttag cgaggggaaa acatcactca 1140
gatcagaaac aattgaaggc tctgcccccc cccctccccc gcgctgtcct taagttaatt 1200
tgtgtaaccc ggtgtatgtg agactcccag gccatattag agtagacagc atagggattt 1260
gatggtcagg aacaaaattc ctgcaagctg tagtaacttg cataaggatg ccactctttt 1320
ctttctttca atgctgggga aatagtttgt ttctcttatt tacaccttct agactgctgt 1380
gtgcctccct ttgtcctgtc atgagaaact gagaaatcag aatgcgcccg cccctcctta 1440
gattcctgta cagagcaaag agcaaggctt tgggctcggg ccaaaggtgg aggtgggggc 1500
cgcaggaagc aagaggactg actgacacgc acatttctgt caaaggatgt tgctcacagg 1560
aagtccgtgg aagaaaactt tctccagact ccgtgtgttc agagtttaac acagttgttc 1620
atatctagct ttggggattt gattggtgga taatagactc tttgtaaatt gcactgggtg 1680
tttccacctg agcaaacaga cctccccacc tcacccccac ccccagggag aagggagagg 1740
gcgtttgaag gggtgaccga gggcgtgcgg cagctacttt tcattttgcc agttaaagcc 1800
tagatgtctt tcctggcgtt ggacgacggt ggcaactgca ggttaattct gactctcttg 1860
agttccgaag cctaacaggc tatgcagaga ggagtaaaag agcactaccc agggctaccc 1920
acatcccggt tgtgttagag agaagcagca aaaaagccct aatgattggg ggcggggtct 1980
gaggagagga aacccaccca agaggtttct taacaccagg gtcacttgcc tttcaatcct 2040
ttaatctgat ctttagtcat ttacattagc atacaaagta actagtttca atactgaaac 2100
aaagtaacta gtttgttcag ccattcctgc cattgctctt tgttcttatt taattgcctc 2160
ttctgtggct cttccacccc ctttacctgt ccctctctgg atgccctccc ccccaaatgg 2220
taccccgttc tgctttctta taacatgagg ttcatcacac tccctccctc cctccctccc 2280
ccatttaaag tatcatcctt tcctctcagg gtgcctgttt tagtttcatg aattttaggg 2340
ttttggtttt ttgtctgttt agttatgaga tttttttaaa aatgtggatt atgttgaatt 2400
tgtagattgt tcttggtgct agaggccttt ttatagtatt atttccaccc atcttgggag 2460
atctttctga aatcttccag tgtcttcaag aatttttttt tcccactgcc ttagaagttt 2520
gcattgtagc tatcgttcac ctctttggtt agggtttgtt gttatttgtt tgtttgaggc 2580
tattgtgaat agaactccct ccttccccca tatctttctg ggccaggttg ttcttagtat 2640
gtaagtaagc tactggtttc tgtatgttta tttagaaccc tgcctcttgg ttgacttttt 2700
atgagggctg agagtttgtg gtagtctttg gggggtcttt ataggattat ataagaatca 2760
tttgactcat tcctttccta tttgtctaac ttttgtttgt ttgtttgttt gtttttttga 2820
gacagggttt ctctgtatag ccctggcagt cctggaactc actttgtaga ccaggctggc 2880
ctcgaactca gaaatctgcc tgcctctgcc tcccgagtgc tgggattaaa ggcgtgcacc 2940
accacacccg gccatcattt ccaagttaaa gatttgatct acattagacg ccgccacgca 3000
gaaaaccttg agacttggtg gaaaggccaa aggccattaa aataaatttt cttttttctt 3060
tcttccattc tttcctttat tccttccttc ctttcttttt gttttctttc ttttcttttt 3120
cctttttcct gagacagggt ttctctgtat agccctggca tcctggaact cactctgtag 3180
accaggctgg cctcgaactc agaaatccac cagcctttgc ctcccaagtg ttgggattaa 3240
aggcattcgc caccactgcc caaatatttt atttatttat ttatttattt atttttatat 3300
atgtgatgag tacactggaa attccatcaa aaagagcagg tttgactggt gtcactagat 3360
ttactattga tagggatccc taaaggagag ctaaggtaaa gggctctccc tctcctaggt 3420
cttctgcata ccttccttga gtgttctggg ccagatctcc taagctctaa gaatgtgctg 3480
aaaacacact gggaactggc tccctccttg ggaatttgta ctccctctgc tgtgggaaac 3540
ttggatataa gaggctacag gaggacagtg agttataccc caggcacaga gttagcgtgt 3600
acattcaaaa cgcataccat tttgaaagta gcagctgcta gcatttcctg tcacctggtc 3660
aacctggtct ctttagctgc cccacccctt ccacttttct gctgtgtttc ttttactctc 3720
ttagcaaaaa ttggaatgaa agaccacaaa tgtatttgta attcaaaatg cttgctgcat 3780
cagctatact cgttactgtt gccatagggc gttcattccc acccaccccc aaccccttag 3840
tccagcagtt gcttcagagt tttgaagaag aggaggaagc ctttcttctt ccatgtgaca 3900
ccctccactg cgacttctgc ttactgtggg gaacttgagt ggaggacggg agtgtgcata 3960
gatgaaagag tggaggacgg gagtgtgcat agatgaagga gtggaggacg ggagtgtgca 4020
tacatgaagg agtggaggac gggagtgtgc atacatgaag gagtggagga cgggagtgtg 4080
catacatgaa ggagtggagg acgggagtgt gcatacatga aggagtggag gatgggagtg 4140
tgcatacatg aaggagtgga ggacgggagt gtgcatacat gaaggagtgg aggacgggag 4200
tgtgcataca tgaaggagag gaggacggga gtgtgcatag atgaaggaga ggaggacggg 4260
agtgtgcata gatgaaggag aggaggacgg gagtgtgcat agatgaagga gaggaggacg 4320
ggagtgtgca tagatgaagg agaggaggac gggagtgtgc atagatgaag gagtggagga 4380
cgggagtgtg catacatgaa ggagtggagg acgggagtgt gcatacatga aggagtggag 4440
gacgggagtg tgcatacatg aaggagtgga ggacgggagt gtgcatacat gaaggagtgg 4500
aggacgggag tgtgcataca tgaaggagag gaggacggga gtgtgcatag atgaaggaga 4560
ggaggacggg agtgtgcata gatgaaggag aggaggacgg gagtgtgcat agatgaagga 4620
gaggaggacg ggagtgtgca tacatgaagg aaaggaggac gggagtgtgc atagatgaag 4680
gagtggagga cgggagtgtg catacatgaa ggagtggagg acgggagtgt gcatacatga 4740
aggagtggag gacgggagtg tgcatatgaa ggagtggagg acgggagtgt gcatacatga 4800
aggagtggag gacgggagtg tgcatagatg aaggagagga ggacgggagt gtgcatagat 4860
gaaggagagg aggacgggag agtgcataga tgaaggagtg gaggacggga gtgtgcatac 4920
atgaaggagt ggaggacggg agtgtgcata catgaaggag tggaggacgg gagtgtgcat 4980
agatgaagga gaggaggacg ggagtgtgca tagatgaagg agaggaggac gggagagtgc 5040
atagatgaag gagtggagga cgggagtgtg catagatgaa ggagtggagg acgagagtgt 5100
gcatacatga aggagtggag gacgggagtg tgcgaggatg gatgagtgga gtctgctgcc 5160
tctcaaaggt cttctggttc catgagttgt tatgactccc agacccacat gggaaggtct 5220
ggtctgttat cttccagtga ctagtgcttc tgcaggctac tcacttgccc ttgcttctgt 5280
ttgcagagga ggtgcagtac attcaggctg agaagaacaa caagggccca atggaagctg 5340
cctcactctt cctccagtac ctgttgaagc tgcagtcaga gggctggttc caggcctttt 5400
tggatgccct gtaccatgca ggttggttcc ttcttcttcc tcacagttca gagtacttca 5460
ctctgctgcc tcagaaggct gaggggagaa aagtgactcg ttctgtgaca tctgtgtgtg 5520
gcttctgcct caggcgggaa atgtaaagac tattttgaat cagataagag aatggtttat 5580
accagaaata tccaaagcaa tctacagagt tgtaactact aggagaggtg acaatattag 5640
tagcatgccg gtatctttca agaggagaac gagtaaataa atcggtttta taatgtttac 5700
agtgctccat tatactgcaa tgaagcgtgt ggacatgtct gtaaatgaca acccagctga 5760
actgtaggca cgcagcattt aaatttgaat atcataaact ataatagcta taaagttcca 5820
catgagtcaa actaaacata tagggaagga aactggatct tgggcgaccc tggctgacca 5880
gtcctgggga gtaagcttaa taaactctcc ctgtctgact gagatcggtg tcctgtggtt 5940
tgtgaggcaa ttcctggact ctaacactta ggcaattaca tttcttgccc ctctgccact 6000
ctagcttatt cactggtgaa agaaggagaa tactttagtg ttaccaacaa tggggggggg 6060
ggggcggggg atgggaaatg ggaaaaagca ggcccggccc agtgtagtaa gaaagaaaca 6120
ccaaagaaag ccaagggctc ctgttgcttt cattgtattg gagtgtttgt cagtcggctg 6180
ggggatgggg tggggggtgg ggaagcacac ctttaatccc agctctgggg aggcagaggc 6240
aggcagatcc ctgtgagctc caccagtcat ggctggcctc agcaagaact gtatccatca 6300
catcctaaca caggtgtgtt gaattaacat ggtactgtta aagcaaacac gctgccttcc 6360
tcgggtgctg cggtccctag gaagccacac attggcagca tgttggcagc agttgtataa 6420
aaactaatgc ttttttttcc ttttcttttt aattcggtaa aagggtttaa atgtcatttg 6480
ttataaaact tggtttcctg ctatttccag gattaacaat tgacttattc tttctatttc 6540
ctgctttata gaccatcatt ttgatacatt atctatttgc atctcagtga tacatgctta 6600
tcttaccctt ttatttcgtt ttaagaattg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 6660
tgtgtgtgtg tgtgtgtgtg tgtctgagag tgggcatgca tttatgagtg cattgcctag 6720
aggtcagaca ttcccctgga gctggagtta atggcagttg tgagggactg acgtgggtgc 6780
tgggatctga ccccagtccc ctgcaagaac acgatgaacc ttacttgcta agccatctcc 6840
ccagccctta gctgttgcag ttactctcca ttccaaataa gccctggcaa tgaaaacaag 6900
acttaattca tatgaataca tgctgtgcac ctagattggg cagatctacc gctacactac 6960
catcttctcc atctatgaga ctcccccttt tttttttctt tttttctttt ttgtggtttt 7020
ggttttttga gacagggttt ctctgtatag ccctggctgt cctggaactc actttgtaga 7080
ccaggctggc ctcgaactca gaaatccgcc tgcctctgcc tcccaagtgc tgggaattaa 7140
ggcgtgcgcc accaggcggt ttctccaggc tgtgtgcttc tgctccactt ttcttcctcc 7200
tcctctgtgg tatcctctcc ctcttcctct ttctccttct ctcttcccac cttcctctcc 7260
aacttccctt tatcagccca atcaccagct ctcctttatt ttactaattg aggtgggaag 7320
caggtttaca ggaaatcacc ggagtgctga ctcattcctt gttcgcagcc actcaatgca 7380
gaatggaatt accatcaaat ataattagcc ccagggctat ccacaacact tacctagcac 7440
atcaaatggc ccagcagggg atcaagagaa aaggaaactc aacttctgct tattttcctc 7500
atctcttatg tagccccatc agagaagctg ttgttttcct tttgtgggct ctaactaatt 7560
tgaatattat atttaagatc tattctctta agtaaaaatg gcacagctaa ctttaactgt 7620
aaaattatat gaggtttact aggaaaagtc ttgagtttaa gcaagaaagg gaattttaaa 7680
acatttgtat tggaacataa gtgctggaac atctctcttt gcaagtgagg tgctttgtgt 7740
gtacaaccct aagagtttct tttttttttt tttttaattt atttacttca tatttcagac 7800
agatctcatt tcaggtggtt gtgagccacc atgtggttgc tgggatttga actcaggacc 7860
tctggaagag cagtcagtgc tcctctgcgc tgagccatct ctccagcccc cctaagagtt 7920
tcataaagga atagtctgca ttaataaatt cagaaaaggc tcagaatata aagccaatat 7980
catcaagtag gtttccagtt tatgtattaa caaataatgc aaaaaagatt ttaagcaagc 8040
gattcctttt ataacagcac caagaacaat aaagttagga atgccagtgc tcctaactgc 8100
tgaaccacct cagcaatgca aacttttaca tcttagcact aagtccagct cctaattcgt 8160
gaatgtaaga tgtcattcat gtccgtgtcc ctatggttcg tttcagaagt ggtttatggt 8220
cttcgggtca taggtctttc ccctgctcag ctttgcttat tcctaacttt atttaaagtt 8280
ctcactgttg ttataaaagg aatcacttgg ggctggcgag atggctcagt gggtaagagc 8340
acccgactgc tcttccgaag gtcctgagtt caaatcccag caaccacatg gtggctcaca 8400
accatccgta acgagatctg actccctctt ctggagtgtc ggaagacagc tacagtgtat 8460
ttacatataa taaataaata aataaataaa tcttcaaaaa aattctaaaa aaatatggaa 8520
aaaaaaggaa tcacttagtt aaaaatctca ttcctagccg gttgtggtgg cacatacctt 8580
taatcccagc acttgggtgg cagaggcagg cggatttcta agtttgaggt ctacaaagtg 8640
agttccaggt ctctgaaaac cacaaaaaaa aaaaaaaaaa atagcactgg ctgctcttcc 8700
gaaggttctg agttcaaatc cctccaacca catagtggct cacaaccata tgtaatggga 8760
tctgatgccc tcttctggtg tgtctgaaga cagtaacagt gtacttacat ataataaata 8820
aataaatctt tgggtgggag tgagcggggc tggagagaga aggaaaagta tctgaagaca 8880
actacagtgt acttacatat aatagataaa taaatcttta aaaaaatcaa taaatgaaag 8940
atgccaatat tacccagagt tggcacagtg atacctttca taatgccaaa ttttggtggc 9000
aggattgttt gtttattaaa caggaataga aaaatttact ctcaaatttg tatgaaatct 9060
taaatggtca aaatattgga aagagaaact cacttggaaa ccttggggga cttatacttc 9120
ctggtgtcaa aacagtacag aacctccata aagccagata attagaccat tagcccagaa 9180
gtaaactctg aaggatatgg ccaaaggttc ttcaacaagg gtaccatgac cacccaaaag 9240
ggggaaaaaa aacccagtcc ctttaatatg aaaatacatt ggggtaagtg ggtatacata 9300
tgggaatgag aggtatcagg cataatcttg tgctatgatg tgaatttgaa atgttctcat 9360
cacattcatg ttttgagtgc ttggtcccca gcttgtgggg ttttaggagg tagagcctag 9420
ctagccaaag taacacgtgt atacgttcat gcatgtgtgc acacaggtat gagggtactt 9480
gtgtgtagcc cagaggctga cattcagtgt cttccttagg agctctccac cgtatgtttt 9540
tgggaatgga tctctcatta gacccagaat ttaccctctc gggctcgact ggctgggatt 9600
atagggtcat gctgctacac ttggcttttt acatgatagc tggggacagg aactcaggtc 9660
ccagccttgt gtggtgagca cttttctact aagcaccttc ttggtcctgg agctattttg 9720
attgttttag ttttttgggt ctataggggg gagaaaaaaa aaaaaaccac attgtcttcc 9780
cagggccttg aatgaagtaa atgagggtct gagaggcagg cacgcctggt ggatctgtcc 9840
aaaaacccca gagtacggca ttcttggatt cttttagtca gaagtcattt tccttctcca 9900
tttgcccatt gacttaatct tttcttggaa tggtgtggaa ggaaacactt ttcaagggca 9960
ggatgtaaga tttgtatttc ctctggtctt ctttactgtt tcctcttgag aagataaaca 10020
tgatgaattt gactaattta aaagtaaatt gagatgacaa agagatggct ctgtgattaa 10080
gagcacttgc tgatcttgca gaggacccag gtttggttcc tagacttaca tggtagctca 10140
caaccatctg taacttcagt tccaggaatc tgaccctctc ttctgctctc caaagatacc 10200
agacacactc acgatacaca gacacatgca aagtaaaata gaaataaata ttaaaaaaaa 10260
atatattgtg ggttgttgtt aaagtgcgtg aggggcattt tgaagatttt attctaaggt 10320
caaatacaag gcctcatatc tgtccttagg acttgaccct gaaagataat gaattttagg 10380
agacctaaac tgttgggtac caaaaatgag tattactccc attttggaaa atcatgaata 10440
gctgtattag ttgactttaa ctactgtgag taaatgcccg aggaaataaa aagcagaaaa 10500
atgagagcca agaggatctg tagcttctgc acccgtgcgt ggtgaggcag ggcaccatgg 10560
cgggagcatg tgggaaaagc cgcactttct ggtagacagg tggcaaaggg tgccgggcag 10620
gggccctgga caagatgcat tcttcaaagc acatctccag tgactcactt ctcccagaga 10680
gggtacagtt ctcagttgct ccttctgtat gaactcaatg tgctgccagt ggtgacgcca 10740
ggactcgaag gatgtggtca ccaggtcccg cagggtgtgg tcacctctaa agactgtcac 10800
cagctggtgt ccaagcctgc aacctgtcag cctcatggtc tggctcccca gactgtccag 10860
tgactgaggc catttgcaga tggttttcag ttcccttgcc actgatttga acaggattcc 10920
catgattttg acttcaaagc atttttatgt tggatttgct taagaaatcc ccatttctct 10980
tttctttttc aggttactgt ggactttgtg aagccatcga aagttgggac tttcaaaaaa 11040
ttgaaaagtt agaggaacac agattacttt taagacgttt agaaccagaa tttaaggcca 11100
cagttgatcc aaatgatatc ctttctgaac tatccgaatg tttgattaat caggaatgtg 11160
aagaaatcag acaggtaaac caatgccagg tactaaattt gaagaaaaat gcagagacat 11220
tggaaatgcc catttttctg tcttgtttta ggcccaagga taattgaaac ccataaaagc 11280
tctcatctag cagatataat gactagaata gaatttttaa agtgaatggg gtaatttttg 11340
tgctagacta ttagaaaatt atttaaccta tttgcagtta aagttgcccc cttactttaa 11400
aaaaaatagt ggtttatgca taatgcaaat cacaccaaac agtgcaacaa ttaaaaggaa 11460
aaatatgtca ggctcttggg catagataca tttattacag tctcgcagtc acttaactag 11520
tgatgtgatg ccaggcagtt ctctaagcat ctgtggggtt tttgttgttg ttgttgttgt 11580
tgtttgtttg tttgtttttc atgtctaaag taagaaaatt tatcttttgt tttttgtttt 11640
tttgtttttt tagatttctt tattttattt tattatatgt gagtacactg tagctgtctt 11700
cagacactcc agaagagggc gtcagatctt gttacggatg gttgtgagcc actatgtggt 11760
tgctgggatt tgaactcagg accttcggaa gaacagtcgg tgctctcaac ccctgagcca 11820
tctctccagc ccccttttgt ttttgttttt gtttttgttt ttgttttttt gtttgtttgg 11880
tttttttgtc gttgttgttg tttgtttgtt ttgttttttc gagatagggt ttctctgtgt 11940
agccctggct gtcctggaac tcactctgta gaccaggctg gccttgaact cagaaatccg 12000
cctgcctctg cctcccgagt actgggatta aaggcatgca ccaccacgcc cgacgaaaat 12060
gtaccttatt agcactcttt tagggctaaa tgagaggtca tgcacaaaat gtgtatgtca 12120
gcttgatgca tagcagtcta tgcacaatgc atttcagtta tcattagaaa gaaaagtcat 12180
agaacatctg cttagaaaag agacctgctg ctgtgctgtt aggcatttcc aaatggctct 12240
gtgtgccgat acatccttag ggtgaatggt tagcgtctgg gttaacgctt ttaccccagg 12300
attgctcttg gtcagggata taaggattca gaagatgaga acatttgcct tggcatattg 12360
ataacacatt ataaaggaca aaggtgaaga aaggaatatc ttaaaagcta gtgctggaca 12420
gggcaaaaag atgatgctaa ctaagcccta ctcaactata cttcacagtg atttcaatca 12480
gataccgctt ccacaaaagc ttgccagagg aaaggctgag ctgcctgatc agtgtgctgc 12540
atttgtctcc cccagatccg agacactaaa gggagaatgg caggtgcgga gaagatggcc 12600
gaatgtctta tcagatccga caaggaaaac tggccaaagg tcttgcaact tgctttggag 12660
aaagacaaca gcaagtttag tgaattgtgg attgttgata aaggtggggt gctccaagaa 12720
agaaccctgg accctgctgc gctcctccca gttctcccca ctttactttc catcagaggc 12780
gctgttcact tcagatacca aaggctatat ccctaggata caagcagtgg aaagctgaat 12840
tctgggagga agggaactac atggcatgga attaacccga ccaggtcaaa gaatctaggg 12900
aaggcttcca gccccaattt gttatcagag aaatagcttg agaattctag acctaaaggt 12960
tcaaactgca agacttacct ccctatcaga gcagaggctg agtgttgggg gtgatagcta 13020
tggactggtg ctcttgcccg gaagccatct ggactccgac agagcaagag taaacgaaga 13080
ttttctgtgt ttaagccaac ctcatttggc ttccggaaac tcacttcttg ctttaaacag 13140
accttgataa atacctgagt ttctagtttc ctttctcacc tagatttcct tagaacataa 13200
attattccag aaactctcta catcgttggt cagagatgga atcctgtctc tttagtgtgc 13260
tcaggaatga cgcccctgcg ttattggcgt gagttccgga gtggggaggg gctccggatg 13320
caaactgctg agagccccgg gttccacact tggagtcgcg tagttccaga tgaaactgga 13380
attcaattgc caagttgagc ttcaaactca gaataatcct tgcagttgtt ttaagccgtc 13440
aaagtggggc tctctagatg gctcagtgga taaggttcct gccactgatc ctgaagaccc 13500
aaattcaacg tccagggcct acatgataga accaatcccc aaacagtgtc ctcatccctc 13560
ggcacactca ctgtgtcgtg tgtgacacac acagtaaaca aatccatttc aaaaataaat 13620
aaaatgttaa gaaagtgcaa gaccgtgatt gtaagagctc aacggaaatt tagatgttta 13680
gtgttagtgt taggactttt tgggacttcc ccaaccaaaa ccataatcac attgcgcatg 13740
cttttaatcc cagcactcag gaggcagagg caggtggatt tctgagttcg aggccagcct 13800
ggtctacaga gtgagttcca ggacagccag ggctatacag agaaaccctg cctcgaccac 13860
cacccccttc caaaaaaaaa aaaaaaagat tctaagctgt aagctgttat ttgtgtttat 13920
gattgtttgc ttgcctgttt atcacaaagg tttcaaaagg gctgaaagca aggctgatga 13980
ggatgatgga gcggaggcgt ccagcatcca gattttcatt caggaagagc cagagtgtca 14040
gaatctcagt cagaatcccg ggcctccttc aggtaccaag catcgtttgc tctcatccat 14100
gatggtgtcc cccagcactt tgatgccctt tgaaaaaaag tctttttaaa ggatgattaa 14160
gaaaagaaag aatttgtggg gcaataggga cttcataatt agaatccctg ctcctgtctt 14220
ccatggcctc tgcatggcct tcaaccctcc ccctcctctc ccctcccctc cccctccctc 14280
cagtatgtat gccttcatct gtaccgtgtt cccagaactt cagtgtccat gacttctcaa 14340
agcagccttg ctctctaaag aacacttctg ctcactaagc aatggctttg agaatctggg 14400
ctgacagctg gttttcctcg gctgtttttg atgatctgtt cttactttgt tccaagtggc 14460
tttgttttga attaggccat tcttgctgtc ctttttcttg ataaagtttc cacgattaag 14520
aaagaattca tggggctgga gagatagatg gttcagcgtt taagagcacc gactgttctt 14580
tcagagatcc cgagttcaat tcctagcaac cacatagtga ctccagcgtc tgggttaatg 14640
tttttacccc atctgtaatg ggctctggtg tactcttctg gtgtgtcaga ggacagcgac 14700
aatgtgtata ttcatataca ttaaataata aataaatctt caaagagaaa agaaggaagg 14760
aagaagtaac agagagagag agagagagag agagagagag agagagagag agagagagag 14820
agagaacaca ctttggccaa gatcccaaac ctcaaacagg ggcattgttg ctagagtcag 14880
aactcatgtc cactgaatgg cagttgcacc atgattcctt gtagcatgaa cccttcgata 14940
actttgtccc ctctatatta cagaagcgtc ttctaataat ttacacagcc cattgaaacc 15000
aagaaattac caactggagc ttgccctgcc tgccaagaaa gggaaaaata caataatatg 15060
tgcccctact ggtaagtcag ttgctgtcac tcacagaact ctctggcttc gctttttctt 15120
ccccctttgg gggctgtaaa aggaggagtt ttccccgtgg cccatgctgc ccatgggaga 15180
gctggtctag cagcttaagg aacctggaca gcgataagga gggagataag tgtcttcttt 15240
agtttgcttt tggttcttgc tacctgagtg cacgttactt aggaagtagc ttggcacttt 15300
tcagccattg tttaaactgt cattgttagt gcggaggagg gattattagt ttatttgtat 15360
cccagtggtc atagagaagc caaaataagt accattctgg aaaaacagct aacacaggtt 15420
atctgttggt ttttttttct tttctttttt tttctttttt cttccctact aaaaggttgt 15480
ggaaaaacct ttgtgtcgct tcttatatgt gaacaccatc ttaaaaaatt cccatgtgga 15540
caaaaaggga aagtggtctt cttcgctaac caaattcctg tctatgagca gcaggcaact 15600
gtgttctcac gatattttga aagacttggg tatgtactac tacaatcaat ctaactgctt 15660
tgatttttgg tttttgtttt gtttatgttt gtattttaaa ttctagcccc tttggctggt 15720
tttgggggct ttgttgtgtc tggttttggg ggctttgttg tgtctggttt tgggggcttt 15780
gttgtgtctg gttttggggg ctttgttgtg tgttttctga gacagtgtat cacgtagcct 15840
tgagttgtct ccaacccact gtgtagctga ggttggccta gaagagatga tcttcttgct 15900
tctacaagtg ctgagattac agtgtgcact ggcatgcctg gctgttctct gattttcttt 15960
cttttttttt tttttaagat ttatttattt attatatgta agtacactgt agctgtcttc 16020
agacactcca gaagagggca tcagatctca ttatgggtgg ttgtgagcca ccatgtggtt 16080
gctgggattt gaacttcgga ccttcggaag agcagtcagg tgctcttacc cactgagcca 16140
tctcaccagc cctctctgat tttcaaagct atgattaaag gaaaatcgcc atggacttaa 16200
cttttagagg tagttccttt gtgcaataac atttttggtt taactttacc agaaatgcta 16260
agccctcatg tcatgctctg acagttaatg aacttggtgg ccaaatttaa catgtaggcg 16320
atacacaggt catccttaat gatgttatac ttgattggct attactcttt tcaaaatcat 16380
ttctctctta atgacttgaa agaataaata cactgtgatc agctataacc tcttgcattt 16440
cctgactccc cggctttgtg tcaggcctgt gagaaagttc aaggtactac ccagttgtac 16500
tcttttgggc ttgggctgac ttctttaatt gctgctctga cctagacttc tactttgtct 16560
ccttgttcat tcacatcaag gttgatgata agggatttct gtcattcccc aggtacaaca 16620
ttgcgagcat ttctggggca acatctgata gcgtctcagt gcagcacatc attgaagaca 16680
atgatatcat catcctgaca ccccagattc ttgtgaacaa tctcaacaac ggagccatcc 16740
cctcgttgtc tgtcttcact ctgatgatat ttgatgagtg tcataacact agcaaaaacc 16800
acccatacaa tcagatcatg ttcagatacc tagaccacaa acttggagag tcacgggacc 16860
cactgcctca ggtatttcca atcttctaag aagaaccaca gtttttcaga gtcccactta 16920
gttgctcttt tgtagccaca tttgagcttg ccctcctcgg ggtctcagtc catcggtaca 16980
actcagtggt caatgttggt tcattcattt gaccaacagt tgtcccttgg tgtccagggt 17040
agatgccctt cacaaaaaac aaaatctagg ctgcttaagt ctcttgtatg agatgacatt 17100
gcatttatat ataatctaca cactttcctc ttgcatactt taaatcttct ctagattact 17160
gatattgtac agtatgatga aaattttata taaatagttc tagtactgta atttttaggg 17220
aataaaggtg ggaaattcat acatgtttag tacttatgaa gttttaaaaa atatttttga 17280
tccatggttg tatgaattca catttatata acctctggat ttggagggcc agctgtataa 17340
accatgggct tccatggacc ttgtgcattg ttctaggctc tgggacacca atacagaaga 17400
tatagtcctg gctctcatac agttaagttt gcagggagcc aggaacatag tagtcacagc 17460
ttatcatgag gtatgctgca gagacaggta aagggtgttg tcagaacata aggggtaata 17520
aggcatagaa atgaagggaa ctgacagggg cttgccaggt agatagcttc tggtttctag 17580
taagagggtg ctgtgtgtcc aggggctcag ggaaatggaa gggcctgaca tgccccagaa 17640
ccttaaaact ctacagtatc acttgagggt agagtgtgaa gcagggaaca acagtcaagc 17700
tgattgttat gacagatgac ccagagacac caggagggca gagcgtcagt gggcaagtgg 17760
atggcttagc acagggaaca agcagcagcc ttctgatgtc atatgagaag agtcacttca 17820
gagtcattct tacatgtgac aagaggagta caaattctcc ttctgtctat cataggagag 17880
ggggtgcttt gctgaaagtc aacgatatag aaacaggagg gggctagaga ggatgaggac 17940
ggtttgactc aggcactgat agatgcaata aagaatgacg gtagtgtatc tatcaggggc 18000
cccaagaagc tgtaaaccat gaattatata caattctttt gctccaacaa taaccttttt 18060
aggacgtgca ggttaaagga catttagtac aggacccaca gtttgttatt ctcgagtatc 18120
gttgctagga agcagatttc ttaccgtcca gctaatcatt taggtgaatg cttactgaag 18180
ggtgttatca tactgaatct acacagctct cttgtacacg actcactgat tgttgaaggt 18240
atttgtccag gcgcacaaaa tgcatgtgat atgaatgagc ctggaatgga ctttttcttc 18300
ccattgtgat gtttagtaag agactggggg ataaaaaaaa cagggtagcc ctgcctggaa 18360
aggtttcctc tctgttctgg atgacacgct agatttattt ccgagctttg ctccaggggg 18420
gtctttgtgc tggagaatgt cagagagcca gtggtggggt gctccttaca ggtcgttggg 18480
ctgactgcct ccgtcggcgt tggagatgct aagaccgcgg aggaagccat gcaacatatc 18540
tgtaaactct gtgccgccct ggatgcctcc gtgattgcca cagtcagaga caacgttgca 18600
gaactggaac aggtcgttta taagccccag aaaagtaagt ggaggtcagc agcccacacc 18660
tcgcgacttt gtaaccttct gtcccctctg cgtcagagac agtggatgaa gtttgatgct 18720
gtatttgttt ggtaaaagca tagtggttac attgcctatc tttctcccta gtcaacctct 18780
tctccctagc gacgcatgag tctcaaaggt agccagaaag ggacaaacat ccctactctt 18840
taccagcagc tgagtgaagg aggcagtggg aagattcaag cattttgaaa gcctcaatag 18900
ctagtggcgg aatcaggtct ctgtgctccg ggccctaggc aggggctatg tggccatctt 18960
gttcttgtat gtatctgatc attgtagtgg catgacccga atcatgacag ttcaaaaggc 19020
cagaacatgt ttttaaaatg agcttcatta gaagatggtt attacttatt aactacctgt 19080
gtaagcaggg aggtaccgta gttacccacg gctggatctt ggcctgagca ctcgttctgt 19140
gagttgacag caggatcaat ggcagggtca atggcaggat gagcaatggg ggggtggggg 19200
ttgggatggc acaaccctgg ttctttctga gagtcccccg tggagagtgt gaagaaggtg 19260
cctccccacc cacgcccacc ccttagcaac actcaagggt ttttctacag tttgagccct 19320
tggagcttag tctacttcaa agtcattttg tgtcactttc tccgtctatg caaaccctct 19380
acgagctatt ctgagggtgt gtcccagctc ctgcgcgcct tcctttttcc cttattattc 19440
atcttgcggc agcttccccg gagagaatga ggtttcctcc cctctttgag agatgccttc 19500
ctggcctgca cctgcttccc agggctctga tgggcgggtt taggagcaca cctttgtttc 19560
ctttaaggag tgggtgggtt ggggagcagg gggagggggg agggagggga tagggggttt 19620
ttggagggga aaccaggaaa gggaataaca tttgaaatgt aaataacgaa aatatcttta 19680
aagaaaagaa aagaaaagga aagaaagaaa gaacctgcct tctgtgcagc atagggtagc 19740
tcttgtcagc tctctgtcac tgaaacagga catgtgacag gcagttcttt ttctgccaaa 19800
agtacacaaa tgtgaacgat aagctcaatg ggggcactct tgggggctcg gaggtgcgca 19860
ggagaatagg aaatcaggaa aacggggctg gagtatggta tttgccgaaa ccagaaggct 19920
gccagacctg ccacagtaga ggcaccagga aagctgactg agacgctggc ttagactaga 19980
ccaggagaga cactagaatc agaagcagtt ccgaggtcag aggcttctga ccgcctgctg 20040
tgatttgggc cacgtgagct tggagcctgt ggctttaaag gacttaccca ggatggagca 20100
gcttcgggaa atggctgcat aggacctggg tttccttcag cttactcaca tgcctttgac 20160
cccagtttcc aggaaagtgg catcccggac ttcgaacacg tttaaatgca tcatctctca 20220
gctgatgaag gagacagaga agctagccaa ggatgtctcc gaggaacttg gtaagcctgt 20280
gccaagtcct ggagagagaa atctcatgtt tcctgtccct tccatttaga ggtactcatg 20340
gattgctcgt tagtgtcttc agttttgggt gagattatac tcagaggtgg actgacttat 20400
ttattcacac atatttcttt ctgtctctgt atcttcttta tctcttcatt cttttttgcc 20460
atcatttttt tctccattcc tttttaaaaa gatttatttt tatttgatat gtgtagatgt 20520
ttttgtctgc atgtatgtat gtatatcaca tgtatcagat acccgggaac tggagttaca 20580
gacagttgtg agctaccaca tctgctggga atcgaaccca tgtcctctag aagggtagct 20640
ggtgcgcata accactgagg agcccccatt tctctagctg tttttaagac aaggtttttt 20700
tccctgtgtc cctggttggc ctgaaacttg ctatgaagac aaggctggct ttgaacttgc 20760
aggggtcccc ttgcttcagc ttctgagtcc tggggtctct ggcaagcgcc accatacctg 20820
gctcagatat agactttctt aatcctaggt tgtttaggaa ccttatagga gttctttaat 20880
tctctcttgc ctttttcttt ttaaatacaa aacacatcca cctggacata catacctgag 20940
aaatactgtc tttaaatcat cttctaaatt tcctttcttc ctttttttcc cctcttgaga 21000
tagaatctct gtgttcagtg taggctggcc ttgaactggg aactctgccc ctcctcctcc 21060
tcctcctcct cccaagatgt gcatcatcac tgagctgcca ttagagtgcc attgtccctt 21120
ccaagagcag ttcccccagt gacctaacac tctctcactg tcctcagctc ttgaaagtgt 21180
caccacctcc taacctcaca cactgaggac caaccagcct tttgccacat gagcatccag 21240
aaggcactta gacagtagct aaggcacagc actgggggag gagtttgaat aatgaatcca 21300
ctatgggtcc taaagtagta gggtagcaag catgctctct cctctagagt tttggaaact 21360
ctctgtaagg taaagagtaa agagaccagg tagtcagtac atggctcacc taggaacaag 21420
ataacatggt ctgactaaag tggtggatgg acagacggac aggaatagag ttgtatgact 21480
tacttttttg ttttgttttt gtttttaaaa cagtctgtct gtatagctct gactgtcctg 21540
gaactctctt tgctggcctc aaactcacag agatctgcct gcctctgcct ccggaggcat 21600
tcacacttta gaatcttttc ccacctcctc acattgagta tctgtcaata gctgcctcac 21660
ttcttctgga acttggacgt ttttcattgt gaactgggtg tggtggcacc atctctactc 21720
ccagcagctg ggagcctgag tctgaggcca gcttaggcta catactgagc tcctgtcctt 21780
ggggcggagg ctgggaagaa cttgtcactg tttcttgttg gtacccgtcc tgtgttctgt 21840
tattgcaaat gtgagggaag ccatttaaca cacaaatgca tttcacttct ttgaactgta 21900
ctgtgcttgt ctcaagaagc ccaggacaca aaacaataga gcaagcatct ggggctgttc 21960
ccacttcgcc tttccccccc tacccacacc aatcttcccc tgagtctgaa tcgctgtgaa 22020
tcccacagta gaaccaagca gtcaagacat gcacatgcgc acacagatgc ttccgggata 22080
actgtgtttg actccgcctt gtggttggtg ctgcaagtgc tgctctgaga tcaggtgttt 22140
gggcttcata gcaacataga gcatgctggg aagggtcctg gtgctcccat ttttatataa 22200
ctgtctccga tgaagctctt gagacgtgct actctaatgg tatcttcatt ttgaaaggca 22260
aagtgtgtcc ctccttctct tcctcctcct ccttcttcct tacccctctt cttcgttctc 22320
tgttatttct gaactacttt ggctgtcagc cccttaagcc tgcagagcat agacaccaca 22380
gagctaggct tgaattcttg cctcacccac acaatatgag ctttatgaca ttgggggtaa 22440
attagttttc cttttataga agatttattt acttaaaaaa aaattatgtg catgtgcatg 22500
tgcgggatgg tgttgttgcc tccaggggtc agaagagggc gctgaatgcc ctggaactgg 22560
atttacaggt cgttggaagc cacccaatgt gagtgcaggg aactgaactt gggtcctcta 22620
caagggccta actattgagc caccacttct gctccttact caatctttct gaatctgttt 22680
cctctttttt ttttttaaag atttatttat ttattatatg taagtacact gtagctaagc 22740
tatcttcaga cactccagaa gagggagtcg gatcttgtta cggatggttg tgaaccacca 22800
tgtggttgct gggacttgaa ctcaggacct ttggaagaac agtcggtgtt cttatccact 22860
gagccatctc tccagcctgt ttcctcttta aaaaaaaaaa ttaaataatg acctcatgaa 22920
attagaaaat ttcaatgcaa ttatgaagct tgattttggg tcatttagta aatagttatt 22980
ttacacactc ctccccccca ccccccgcgc acgcacacag gcacacacac acacacacac 23040
acacacacac acacacacac acatagctta agacccagtc tacttcagga taaacatctt 23100
tcttataatg aataagaaag aaaatcagag gaccggtgct tgcaaatctt ttatttatct 23160
atttatgttc ttaccctgta ggaaagcttt ttcaaattca aaacagagaa ttcggcaccc 23220
agaaatatga acagtggatt gtcggcgtcc acaaagcgtg ctcagtgttt cagatggcag 23280
acaaagagga ggagagccgg gtctgcaaag cgctcttcct gtacacatca catttgcggg 23340
tacattgctg ctctccaggg cttattctca tcaccgcgcc tcctgggatc tgtactgagg 23400
cagctgagag aacatcagcg tctcaagtct aagagcttag tgaggaactt ttcccgaaag 23460
tcatcactaa ccttatttgt tttctgaaac ttatcatcaa gtctccaaaa actggattaa 23520
aggctcagag tctatgccac acctccctcc agcttgtgac tggtgaccac catctaactg 23580
agctcaaaaa agtggctcct gtggccatat cctgaagctt tcgtggtctt aattttgtta 23640
taaagtcata tattagaatc tcaggggctc tggttaacac agagggaagg agtaactgta 23700
agagccctca gctctgtttg ctatgctctg ggaactattt aaagacttac tccacaccat 23760
gggattgtgg gatctaacgc ttaatggact ttcagcatag gtggtaaggg ccatcgttat 23820
gcaaggccca tgtacacttt aagtatgact tggaatttaa ggggaatgtc aaagctaact 23880
tgcttttgtt attgtttctc aagatatgct gtttctcctc tcccaaggtg gagttttata 23940
atccaaagtg aatctacttt taattttcta gctgagccaa aaatagaagc cagcttttgg 24000
ttcagaggtt tttattgtag acccactaag ggccattcgc cattaaaccc tcagctgtac 24060
tgtatgagaa agattttctg caaaccagtt ttgtgctaaa tacagcgagt tgaacttgag 24120
tgtagtgacc atatgcgacc tcagaaatgt attgagaatc acttttcatt tcaaacagaa 24180
atacaacgat gcactcatca tcagtgagga tgcacagatg acagacgctc taaattacct 24240
caaagccttc ttccacgatg tccgagaagc agcattcgat gagaccgagc gagagcttac 24300
tcggaggttt gaaggtgagg gagatttctg aagtcaggag tccctggggt ctggtggctt 24360
ttgtggcagt gtgcacatcg tagttagcat acgtagccat catgttgggt ttaaggtgag 24420
atttgtaggg gctgtgacgg agcatgacct tagcatggct gaaatcccca gcactaaaaa 24480
acgaacctat gctgaaactt tagagccaac caaccgacaa caggagggtt tggcttcaga 24540
gaaatctaat gcctgtggat ggatctgatg cttgccccac ttttcacttg ggaaaatggg 24600
aaacagtggg atttggaaag ggtgcttcct ctaggtggta ggtagtgcta ttctgattaa 24660
ctcagtaatt cagaaggttt aataacaaca gctcgtgtct gatggtgtca agattgtgct 24720
gtatgtatgt ccttccttcc ttccttcctt ccttccttcc ttccttcctt ccttccttcc 24780
ttccttcctt cctccctccc tccctccctc cctcccccct ttctcttctc ttctcttctc 24840
ttctcttctc ttctcttctc ttctcttctc ttctcttctc ttctcttctc ttctcttctc 24900
cttctcttct tcctcctcct tcatcttctt ctttcttcct cctcttcctt ctccttcttc 24960
ttcaaaacac agtctgtata gccccggctg tcctgaaact cactttgtac accaggctgg 25020
cctcgaactc agaaatctgc ctgcctctgc ctcctgagtg ctgggattaa aggcgtgtgc 25080
caccatgccc agtttgtgca catatatgtg catatgttta ttataaattt tactgataca 25140
ggagatggca tagcacaaaa cacacaaata ataaacacgg agttcatgtt ccacagaatg 25200
cctttggagg tcttttcagt acccttgtgt ccagagccaa ccagagacag caattaccca 25260
atatggagtt ctgaaatgaa agtcagtttt atttcctgtt aatggcagaa ataagaacaa 25320
aacgaaacag cagaagcatt ttggaagctt gcttgtttct cagtgatggg agcaacattt 25380
ttctgagcca gataatagtt tttcaaacac gggtgggaca tttctgcatt tttacgtgat 25440
gcataaacag tagctaaatt taatccccat tatatactta gcactttaca aagtctagcc 25500
agacaataaa ggatgaagca agtgctatct tcatttccat ggtatgggta cttctaggat 25560
caccaatctc caaccatcac catgttgctg aacttgtgta aaattgagca gtaacacaca 25620
ctgacatttc taccattcat acactacagg taagtacaca ctcaagagcg tagataatag 25680
taaactgtaa taaaatgagt taggaaatta ataagcgtgg ctatttgtta catttgtttt 25740
tagtcattga gctgcaagca taaagagttg aaattttaat aatagttata tttaaaacca 25800
ggtccacaag tctgaagaac ttaataactg accataatct ggtttgatct ggttctatct 25860
agtacaccac cagtgtgtgt gtgcgtgtgt gctcctatgc atacttatac attaaaaaaa 25920
aaaaagatat cctatgcttc aatttttaac ataaaataac cttctgacag ctgggtggtg 25980
gtggtggtgc atgtctttaa ttgcagcact caggaggcag gggcaggtgg gtctctgggt 26040
tcaaggccag tctggtttac agagccagtt ctaggacagc cagggctaca cagagaaacc 26100
ttgtttcaag acaaaacgaa acaaaacaac cacaaataaa aaatatatct ttttgatgtt 26160
tccaaatcag caggtgtata taactcttta actttaatag taacagtgta tttacctcag 26220
tttggtagcc tgggatccat tgagctgttt ctcactaagc agtgttttgg ttgttggttt 26280
tttttttttt tttttttttt ttttgtattt agttcatagt ttcaacactg attgtccttg 26340
gaatcttctt cagagttttt tttttttttt ttttttaaag atttatttat ttattatttt 26400
atttaagtac actttagctg acttcagaca caccagaaga gggtgtcaga tctcattacg 26460
gatgttagtg agccaccatg tggttgctgg gatttgaact ctggaccttc ggaagaacag 26520
tcggtgctct taaccactga gccatctctc cagccccaga gttttctaaa tagaactatg 26580
agtcaattcc tatctgtgga ttgctgtatc aaagaacatg tgagttttgt attgctgcgc 26640
tgcttttcta aaggggattc ctgatgaaac gagtgtttac tgctctgatc tctggtgaac 26700
agtggaaagg ttaaccgaaa tagaaggcca ctgtttgttc taaagcttta acatttgtaa 26760
gccttttgca aaatgctctc tatttgcaga aaaactagag gaattagaaa aagtttccag 26820
ggatcccagc aatgagaatc ctaaactaag agacctctac ttggtcttac aagaagagta 26880
ccacttaaag ccagagacca agaccattct cttcgtgaag accagagcac tcgtggatgt 26940
aagtgtgtgt gtttacagat tagctctagt ttattgaaaa ggttgcccgt tcttcactgc 27000
cttataatca agtatccata catgtgtgga cctgttctga tgatttgttc ttacaccaat 27060
tgtcattgtt tgtattgacc cacagttata agtcttggtc ctatagagga agccctgcat 27120
cctttttaaa aatttaaaat ttccacttcc agtcatcctg taggttttga ttaatgacta 27180
atgtgtctta tatacctcac agttatcttc atatcatctt ttaaaaataa tttactcaga 27240
ttttaaaaac cagttttaaa attgggcaat gggctggaga tacagctcaa tggtcaaatg 27300
cttgttcagc atgcatgact ttaatcccca gttctggaaa aagatagata tctctctgtt 27360
tatgtagaat gccttgagtc tggccacagc gcctccctct gtttatgtag aatgcctgca 27420
ttgtttctgc tgagtagtag attacccata gagccagagg cagaaaaagt caagctttat 27480
tattttatga gatccgtgga tcaggatctg gaaaggactg gatacttatg cctcaaggtc 27540
tcctgaggcc acagtcagct cggcactcaa ggctgacctc tcggctcctt ttgcaggttg 27600
ttggcaggct tgtgaagatg agcctccaac atggcagctg cccctgccta cagtgagatg 27660
agagactgag gagaagaggg cctagtagac agacactgcc attttataaa gtccatcttg 27720
gacctgatgt cccaccacat ctcccatatt tcagagataa actacagatt atttttagaa 27780
tataggatgt agaagtcatt aagggtcgct tgtcatgtga tctttgctgt cttcttttgt 27840
taatgaatgt gggtgtttac catgtgcgtg tcgtgcccac agagtccagg aagggggcat 27900
gacatgccct ggaactggag ttaacagaca gttgtaagtt gccatgtagg tgctgggaat 27960
tgaactcagg tcctctggaa gaacaaccag tgctcttaac caccgagcca tctctccagc 28020
cccctttgct gtgttttatt agcattttgt catttttagt atagaggtcc tgcatacatt 28080
ttgttagatt catacctagg tattaatttt agtgttgtca ttccgaaatt gtactttcaa 28140
atatttctca ctgtgggcta ggaagacatc tcagtaaagt gtctaaagta caggcatcag 28200
gacctggctt ccagcaacct ggtaaaaaag ccgagtacag tagagtactc ttgtaatccc 28260
agccctgggg acagagataa gcacaaccct aactggcaat gcccaggtcc cagggagtta 28320
ctcattactc agtcttaggc agaacgaagg tgggtggctg ttaagaaatg atacctaggg 28380
ctggtgagat ggctcagtgg gtaagagcac ctgactgctc ttccgaaggt ccagagttca 28440
aatcccagca accacatggt ggctcacaac catccgtaac gagatctgac tccctcttct 28500
ggtgtgtctg aagacagcta cagtgtactt acatataata aataaataaa tctttaaaaa 28560
aaaaaaaaaa aaaaaagaaa tgatacctga ggttgacctc cacatgcatg tacacacaca 28620
cacacacaca cacatgcgtg cgtggacata ctcccctcca acacagtcag ccatgtacac 28680
ctccacacaa cacacagttc ttccaattgc agctgtctgc tgatatttac tgtgtaatta 28740
atttacatgg attgatcttt caccttaaag ccttgctaaa tttcacttac tctatgtctg 28800
aagcttgtct ttttaatcac ttaaaatatc tcctacatta agccataatg aggcagagtt 28860
ctatatcact agcatcaatt gttgtttgga atttaggatt tgccagtctg aaatccattt 28920
ttatctttag ttgtattctc tttttgcata tacatccatt atatcaaatt gatgtgaggt 28980
ttaaagttta caagtggtgt ctaactggcc gttgcttttc acttttaggc tctgaagaaa 29040
tggattgaag aaaatcctgc actaagcttt ctaaagcctg gcatactgac tgggcgtggc 29100
agaacaaacc gggcaacagg tatttatgtc tattgaatta gatttagtat actatgtata 29160
taaaatgtat aaacactaca ttgttttagt gtttctatca gtcagagctc aaccagagaa 29220
ataaagctac tagattatgc atatgtatgt ggtatatatg taagtatgta tgcttatttg 29280
tttgtttgtt tatttatgta cttagagata gggtttctct gtgtaaccct tgctctcctg 29340
gaactcactc tgtggaccag gctggccttg aattcagaaa tccgcctgcc tctgcctccc 29400
gagtgctggg attaaaggtg tgcgccacca ctgcctggcc tatactgtta ttctttaccc 29460
agtagatttt ttttccccat ctactgcctc tttaatagtt ttaaaaaaac agtccaggca 29520
atcctgaact ctagctagtg tggtctaggg aggaaggtta tcatttccca taagaaccct 29580
atgtggctag ctctcatcac agctacgtcc caagtcatat ctcacgactg tatgacctgc 29640
cctcgctgtt cttcctgcca gtgttgttta cactaaacaa gtctccaccc cttctctctc 29700
cgtcctcagg cattgctctt gtacttttcc attgtggaat ttcccgatct cataaacata 29760
gaatggactc aagtgttgaa tgtgtggttt cgagtctaac actaccctaa tgtggctgga 29820
ttttcaaagt tctttgccat ctctccaaca tgaatccaac ttgattttca agctttgcta 29880
ctgacatata aatcgagcct tgaataatat tttgtgtgct catccatgca tgcatggatc 29940
catggatcca atcatgtgtc caaccactca tccacccatc cgtgcatgca tccatctttc 30000
cgtcatgcat ttagacctta ctcagctcct gcttctgtga agaagcagcc atccctgtca 30060
tctaacaccc gaggtgcccc tccccccgcc acgttcccat cttacagatc tcaccccact 30120
tcctccacaa tggcttcctg ctcatcatcc ttatagataa agatggaatc tttaagcgtc 30180
atatttctac ctgctcagcc ataacccata attgctgacc gagtgttgga tggatgaatg 30240
aattggttag gatgattctg ctattgttgt tttctggatg attctttctt gttttatagc 30300
taacctggga aaaaaggtgg acttttacaa aaagccacag gttgcttggt gtttggacat 30360
tttcagattc ccttatctgt agcattttta cttcctactt tgagacacat gttgtaatgt 30420
ttatgcctta ctatcttcat ctgtcaaatg gaacaataaa tagttgtccc cagctcatag 30480
gttaacgaga atggtgaaac ctgagctttt tttttttttt tttttttttt tgtgaaaatg 30540
actttggcac ttttagatgg ttcaaaatta gtagccagtt tatgagtgag ttgtacagtg 30600
acctctttat ccaacacagg aatgacgctc ccggcacaga agtgtgtgct ggaggcattc 30660
agagccagcg gagataacaa tattctgatt gctacctcgg tcgctgatga aggcattgac 30720
attgctgagt gcaatctcgt cattctctat gagtacgtgg gcaacgtcat caagatgatc 30780
caaaccagag gtgagagcgg ctgatgtcat tcccgccccg cacccgcttt tctcctttcc 30840
tcagctgtac catgtgattg acagcacagc tgactctggt actcgaaatc taaaagctga 30900
ctgccttggt caggattggg tggttatagg tttacccata atactccatt gcaactctcc 30960
acaatggtac tgcaatttta cccagcgttc aatggcatag tcgtgaaaat atcatatcca 31020
ctaggccaga ggctttgcca gtcggcaagt agacctttga tgggtgtggt gagtagctct 31080
ctgtactcca gagtctggtc ccacctgaac cagagtctga cttcctttcc ctttcttgtt 31140
tccccaagaa cagcccccac attccctttc cggaataacg tctctgtgcc tgtcactcat 31200
cagtcacatc catttttcgt cctcctccac cgcttactgt gcggttcagc cagccagact 31260
ccgcttcctg ctcgtccagt ttctcagata ctgtcgctct acatgttagg tcctatctct 31320
gtctctgcca cacacaacct aattcttcta cctagaacaa gcactccttt aaatgcccac 31380
taccgtttat atctgtctct gcaagagcat gacaattgca ttcctttctc cgcattgcag 31440
aagggtcagg tgcgcgtgca cggtgccact gctgcgggct catgccagat tatctgtaaa 31500
ttagtgttgc tggcagtgca gagcaatcag actatgccat tggagacccc atgaaaactg 31560
ccagagatgg cttatctgtg tgctgagcac actggctaga acctgcattt gagtctactc 31620
ttcggttcag cttccctaga aagtaggatg cagtgaatca aagttgaact cgagaaatac 31680
tcctcacatc tctttccagt aacctcagag tttgacatta acacacaaag aaaacgtttt 31740
ctgcaggccg aggaagagca cgagatagca agtgcttcct cctgaccagc agcgctgacg 31800
tgattgaaaa agaaaaggcg aacatgatca aggaaaaaat aatgaatgaa tccatcttaa 31860
gactgcagac atgggatgaa atgaaatttg gaaagacggt aagtctcttt ttctgtgcta 31920
ctcttatgga atctgactag aaataacaaa tgaccatggt tggtcctgag tgtgtgtgtg 31980
tgtgtgtgtg tggtatgtgt ttgtggccat gtgcatttat ttatctttgt gtgctagttt 32040
tggccattca aataaccttt ctgttcgcat gtaggttcac cgcatacagg tgaatgaaaa 32100
actcctcaga gacagtcagc acaaaccaca acctgttcct gacaaagaaa acaagaaact 32160
gctgtgtgga aagtgcaaga attttgcgtg ctacacagct gacattcgag tggttgaggt 32220
gagtggccct ggtgatttag caccggttaa atcttaccat cttccggaga aatggttgta 32280
gcaagaacac tatgttgtgg ggtttcgagt gttgaccatg gtcctgtatt aagaaataaa 32340
atcctgctag gtggtggtgg cacacgcctt taatcccagt acttgggagg cagaggcagg 32400
cggatttctg agttcgaggc cagcctggtc tacagggtga gttccaggac agccagggct 32460
acacagagaa accctgtctt ggaaaaacca aaaaaaaaaa aaaaaaaaag aaataaaatc 32520
ctgcttctat gtgggaacca gaaaggctga tgttatttaa gtccaaaaca gaaaatggtg 32580
cttaacggcg agaagaggag gggggtctaa ttgtagctgc cccagacagt caggcaggat 32640
ggataaggtt tcccgttcca ctgcacagca gggtgaatac tgcttatagt ttctgattca 32700
ttacaactct tacaaagaat tagacgagag gaattcatag cttcagacat aaagagatga 32760
aaactgtcca ggcagaaggg aatgctaatt actctggcga gatcattaga tacttagaaa 32820
ttatcacact gcacacccta agtgaggaca actgtgtgct ttgaaagaca gtctcactca 32880
gaccaggctg gcttcaactt gagattcttc tgtgactcag cctccccagt gcagggactc 32940
taggcatgca ccaccactct ctccaaagag atagtttttc ccagtgcagg gatagaaaat 33000
gaaggctctg ctagatacag tgttatgtcc ttggttagtt ccagggagga ggaagggcta 33060
ggcataaaaa tctgtcattg atttcttagt tttaacaaat gtgcaactgc attcaaaatg 33120
gaactgggct aaaggcattt gcaaatcttc tgaggcatct ctgtaacttt actgtatgtc 33180
aaagattatg ccaaagaaat gttaaggctc tgattttgaa gtgtacatgg ttctagtata 33240
aacctgccag caaatgaatg gtaaagtggg aaaatactat gaatatgaaa ttataaagat 33300
gcttttgtta tggctacata caacatgagc agtgatatct ttgtcatgac caatgtgggt 33360
ccacctttcc taaaggggaa aaaaggctaa tatataaaaa tgacatattc tgctagtgaa 33420
ttctctcttc ctgttttgtt ttctaaactt ccttattgga acagagaatg cttttataat 33480
gaaaacaaaa cacctcattt taaaaaatat aacacttgta ggttagcttt ctactttttc 33540
accccttaaa cttttttttt ttttttttga gacaggatct ctctctatag cactgggtat 33600
cttgaaactc attatatgga gattcccttg tccctgcctc ccaaacgctg ggacgtaaag 33660
gcatgtacca ttacacaagg tcttcttaaa acttttaact aaggcaaaaa acctccagag 33720
acgaatcttg cagtcatcat tcctgctacc gatggcgggg gcagatggct gtgctagcta 33780
ggggaggggt acagtcctta ttatgagtga catccacttt ctgagtctga ttctttagat 33840
gcagaaggtc ctttcagctc caaattcaag gcttctcctt cccagtggcc tggagaacga 33900
gatgcttggt ctgcgcttgc gtctgtaggg tacactcttt tttttttaat tggatatttt 33960
ctttatttac ataaagaaca ccttcagttc ccctacactg gggcatctaa gccttcatag 34020
gaccaaggac ctctcttccc attgatgcat gacaaggcca tcctctgcaa cgcacgcagc 34080
tggagccatg tgtactcctt tgtggatggc ttagtcgctg ggagctcttg ggggactggt 34140
tggttgatat tgtttttctc cctgtggggt tccttcagtt gtagggtact ctcttaagtc 34200
aagtctaagt ggggtctgtg gacagcatgg ctcctgagcc tgttcacaca cataccctgt 34260
gacccctggg tgtcaccagc cctgtgcttt ggtgccttcc tggcctctgg tgaacttgaa 34320
ctttgtatgt gaactcctct ttgcttctga gttggaaagc tgggtttcct cctttctcag 34380
gtgccagacg cccaggaaat gggtcctaat cggcctgggg aagactgtct atatagtttt 34440
tttttcttcc aacttgtaaa ataaatggga tcccactcat tcctgacttt tagctactga 34500
gtggcttcta agtcattttc agacccgttt ctaacattgt gcgctccatc tcttcctcct 34560
agacgtccca ctacactgtc cttggagacg cttttaagga gcgctttgtg tgtaagccac 34620
accctaaacc aaagatctat gacaattttg agaagaaagc aaagatattc tgcgccaaac 34680
agaactgtag ccacgactgg ggaatttttg tgagatacaa gacgttcgag attccagtca 34740
taaaaattga aagtttcgtc gtggaagata ttgtgagcgg agttcagaac cggcactcaa 34800
agtggaagga ctttcatttt gaaaggatac agttcgatcc tgcagaaatg tccgtatgac 34860
ctcaggcttc tccgtctcgt gccgcaggga gccgtgcctt aagcatggag ttgatgagcc 34920
aatgctttct tacccaagct tgcacaatcc tttcttacac aagcctgcac tgtgttgaat 34980
gccagataac ctgactggtt ggtttcaagc tggtgctgtc cacacaaagc acacacgcct 35040
gaactgcggc gccgaatagt ttcttcacca ataactcata gcgtagccct tggccatggt 35100
ggggaggggt taaacttgtc ccttttacac ttttcagaac tgcccgacag ggaacgtgca 35160
gccactcggt acaccgagac gcatgatggc tggcgtgctg gaagggttcc cgttctctgt 35220
ctgctcgatc tgctgtaagc tgcctttgcc cttaatgaca gtgcccttaa gaacagtgac 35280
ttagttcttt ttcaggccac cagactgact gccagatccc ttctgtccct tctgtccctt 35340
gcactgattc ctttccggat ttgaccctgc caccctgtca ccctctgcag agtctcctgg 35400
tttctgtctc ttccttggtt tctttgctga ctcaaatttg gtagttgcaa ggttcagtat 35460
gcacacatat atatttaaaa tgacatataa tttaaaatgt aaaagactat agttgacagc 35520
tatgcttact gagatggtat ttctgttctg ttcattacta tacatcttac ccttgctctc 35580
atctgttctt ttaacttggg ccatttcccg tctttgaata gacatctcaa accctgtctg 35640
tatgtctgtc tgtttcccac ctgtttgaga cagggtctct ctgtagacca gaaactccat 35700
atgtagacta tgctggcctt gaactcacag atccccctgc ctctcaagtg ttgaaattaa 35760
aatctttcac catgcctggc tctagctatt ttcaataaag gctcatgttt aaagtttgaa 35820
ctacttccaa ttcattccct gacgtggctt gttgttgttg tttacttttg gagacactgt 35880
tcctctctgt agccctggct gtgtagacca ggctgccctt ggtcacagag atctgccagc 35940
ttctgcctcc ggagcactga gattgaaggc ctgcatcacc atgcctggct tgccctttct 36000
tcttaaacat tatatattca aatggcattt ccgtgtttct tctcaaggtg tgccagtgct 36060
tcagagagct tagtttgggg ttcttcagat caagagacaa gtgtctgagc gctgttactg 36120
ccaacagagc aaagtactct tcagttcagg gaaggaacag tgctggtttt gtaggcagta 36180
cagtggtttt aacaccttcc tagaacttac ttgtaattca tcagttgtag accatcaatg 36240
gcctaaacca aactgcagag atcatctgac cacataactc cccttccagg acatttacat 36300
ttgaagacta tcccaagccc acccagagca cagtgggtta cccaaactcc ccaggtcaac 36360
cctggaggtc aacagtatga catgggatag cacaccactt ctcacagatg cctagagaaa 36420
ttacccagca acataactct ttggggaaaa aacacctata gggattaggc ttttaattga 36480
tagaataggt agaaaaaaag atatgtagta gttcttgata gtggttactg gtaaaattct 36540
tagtgcaata aaatgaattt gccgagagct gactttcttt tttttcttgt ttttttgaga 36600
cagggtttct ctgtatagct ttggctgtcc tggaatgcac tctgtagtcc aggctggcct 36660
tcgaactcag aaatccgcct tcctctgcct cccaagtgtt gggattaaag gcgtgcgcca 36720
ccacgcctgg ctgagagctg actttcattt atgtctttta gtctatgttg cctttctttg 36780
ctgctacagt ttaagaactc tacagcttgt ataagatacc tactggaaat tatttgagaa 36840
aaaaaacttg taaacattac aataatttaa ttaattaaaa atttatgtat tttatgtata 36900
tgggtgtttt tttctgcaag tctgtgtgca cattagaaga gggcattgga tcctctctat 36960
tgttatagtt ttatgctgct gctgttgttg ctgtgtgtgt gtgtgtgtgt gtgtgctatg 37020
aattgaactc aggacctctg gaagagcagt cagtgctctt aactgctgag ctatctctcc 37080
agtcctggca atgataaatc agttgaagtg aaatagtcct cccccccctt tttttttttt 37140
gccaatgggg aaaagcagac taaatctgag accaaatgaa gttttgagtt gtacactgac 37200
ttaagccact gccaagcata ccctggaatg gagcaaaccc tgggttacta agtactgaat 37260
gaatacaaca ggaaggtttt gagagatggg aaaatgcttg tctttggact tccctgatgg 37320
aagttgcatc tggactctcc catgagcaca tcaccagtcc ccactagagt cctcacaggt 37380
tgccatccat gtgtcctttt tgaggctgag atacaacttg ttctgcaacc acagaccttg 37440
ctgttttgtg gtcagtattg gtatcatagc attttcatcc tgacctggag ccttcagtca 37500
aaggcctcat tgtgcagtaa ggacgctgga ctcctgactc ctatacttaa aacagacttg 37560
gtaatttcaa acaagtcaac cagatgccag tatttctgca tgcatgtctt gtgggatggt 37620
gttgtgaggt cccctgacag atgcactgag tggccaggga gacttttgta cccttttcca 37680
ttttaacagc ccacgggtca ctgtgttgct tccatcatat taacatcaac ttgaaccagt 37740
ggttcctgaa acacttcagt tcattggacc ttgctaatta gcatcctgta aaaacccacc 37800
aacaaatatc aactagacag gtagaatcca agtgaactgt acactcctgg atcatgccag 37860
taactgtttt aataatacac cataaaatat aactacgact tcattttaca aatctgtgtt 37920
taataaacag gtacaggctt gttgggtgcg aacttttaaa actcctaata aaaatgccag 37980
ctatgattat ctttgtttat g 38001
<210> 12
<211> 3435
<212> ДНК
<213> Rattus norvegicus
<220>
<221> КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<222> (370)..(1815)
<400> 12
cgtttgtagt gtcagccatc ccaattgcct gttccttctc tgtgggagtg gtgtctagac 60
agtccaggca gggtatgcta ggcaggtgcg ttttggttgc ctcagatcgc aacttgactc 120
cataacggtg accaaagaca aaagaaggaa accagattaa aaagaaccgg acacagaccc 180
ctgcagaatc tggagcggcc gtggttgggg gcggggctac gacggggcgg actcgggggc 240
gtgggagggc ggggccgggg cggggcccgg agccggctgc ggttgcggtc cctgcgccgg 300
cggtgaaggc gcagcggcgg cgagtggcta ttgcaagcgt ttggataatg tgagacctgg 360
gatgcaggg atg tcg act atc tgc ccc cca cca tct cct gct gtt gcc aag 411
Met Ser Thr Ile Cys Pro Pro Pro Ser Pro Ala Val Ala Lys
1 5 10
aca gag att gct tta agt ggt gaa tca ccc ttg ttg gcg gct acc ttt 459
Thr Glu Ile Ala Leu Ser Gly Glu Ser Pro Leu Leu Ala Ala Thr Phe
15 20 25 30
gct tac tgg gat aat att ctt ggt cct aga gta agg cac att tgg gct 507
Ala Tyr Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala
35 40 45
cca aag aca gac caa gta ctc ctc agt gat gga gaa atc act ttt ctt 555
Pro Lys Thr Asp Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu
50 55 60
gcc aac cac act ctg aat gga gaa att ctt cgg aat gcg gag agt ggg 603
Ala Asn His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly
65 70 75
gca ata gat gta aag ttt ttt gtc tta tct gaa aag ggc gtc att att 651
Ala Ile Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile
80 85 90
gtt tca tta atc ttc gac ggg aac tgg aac gga gat cgg agc act tac 699
Val Ser Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr
95 100 105 110
gga cta tca att ata ctg ccg cag acg gag ctg agt ttc tac ctc cca 747
Gly Leu Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro
115 120 125
ctg cac aga gtg tgt gtt gac agg cta acg cac atc att cga aaa gga 795
Leu His Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly
130 135 140
agg ata tgg atg cac aag gaa aga caa gaa aat gtc cag aaa att gtc 843
Arg Ile Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Val
145 150 155
ttg gaa ggc acc gag agg atg gaa gat cag ggt cag agt atc atc cct 891
Leu Glu Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro
160 165 170
atg ctt act ggg gag gtc atc cct gtg atg gag ctg ctt gcg tct atg 939
Met Leu Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ala Ser Met
175 180 185 190
aga tca cac agt gtt cct gaa gac ctc gat ata gct gat aca gta ctc 987
Arg Ser His Ser Val Pro Glu Asp Leu Asp Ile Ala Asp Thr Val Leu
195 200 205
aat gat gat gac att ggt gac agc tgt cat gaa ggc ttt ctt ctc aat 1035
Asn Asp Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn
210 215 220
gcc atc agc tca cat ctg cag acc tgc ggc tgt tct gtg gtg gta ggc 1083
Ala Ile Ser Ser His Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly
225 230 235
agc agt gca gag aaa gta aat aag ata gta aga aca ctg tgc ctt ttt 1131
Ser Ser Ala Glu Lys Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe
240 245 250
ctg aca cca gca gag agg aag tgc tcc agg ctg tgt gaa gcc gaa tcg 1179
Leu Thr Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser
255 260 265 270
tcc ttt aaa tac gaa tct gga ctc ttt gta caa ggc ttg cta aag gat 1227
Ser Phe Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp
275 280 285
gcg act ggc agt ttt gta cta cct ttc cgg caa gtt atg tat gcc cct 1275
Ala Thr Gly Ser Phe Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro
290 295 300
tat ccc acc aca cac atc gat gtg gat gtc aac act gtc aag cag atg 1323
Tyr Pro Thr Thr His Ile Asp Val Asp Val Asn Thr Val Lys Gln Met
305 310 315
cca ccg tgt cat gaa cat att tat aat caa cgc aga tac atg agg tca 1371
Pro Pro Cys His Glu His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser
320 325 330
gag ctg aca gcc ttc tgg agg gca act tca gaa gag gac atg gct cag 1419
Glu Leu Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln
335 340 345 350
gac acc atc atc tac aca gat gag agc ttc act cct gat ttg aat att 1467
Asp Thr Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile
355 360 365
ttc caa gat gtc tta cac aga gac act cta gtg aaa gcc ttt ctg gat 1515
Phe Gln Asp Val Leu His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp
370 375 380
cag gtc ttc cat ttg aag cct ggc ctg tct ctc agg agt act ttc ctt 1563
Gln Val Phe His Leu Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu
385 390 395
gca cag ttc ctc ctc att ctt cac aga aaa gcc ttg aca cta atc aag 1611
Ala Gln Phe Leu Leu Ile Leu His Arg Lys Ala Leu Thr Leu Ile Lys
400 405 410
tac ata gag gat gac acg cag aag ggg aaa aag ccc ttt aag tct ctt 1659
Tyr Ile Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu
415 420 425 430
cgg aac ctg aag ata gat ctt gat tta aca gca gag ggc gac ctt aac 1707
Arg Asn Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn
435 440 445
ata ata atg gct cta gct gag aaa att aag cca ggc cta cac tct ttc 1755
Ile Ile Met Ala Leu Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe
450 455 460
atc ttc ggg aga cct ttc tac act agt gtc caa gaa cgt gat gtt cta 1803
Ile Phe Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu
465 470 475
atg act ttt taa acatgtggtt tgctccgtgt gtctcatgac agtcacactt 1855
Met Thr Phe
480
gctgttacag tgtctcagcg ctttggacac atccttcctc cagggtcctg ccgcaggaca 1915
cgttacacta cacttgtcag tagaggtctg taccagatgt caggtacatc gttgtagtga 1975
atgtctcttt tcctagacta gatgtaccct cgtagggact tatgtttaca accctcctaa 2035
gtactagtgc tgtcttgtaa ggatacgaat gaagggatgt aaacttcacc acaactgctg 2095
gttggttttg ttgtttttgt tttttgaaac ttataattca tggtttacat gcatcacact 2155
gaaaccctag ttagcttttt acaggtaagc tgtgagttga ctgcctgtcc ctgtgttctc 2215
tggcctgtac gatctgtggc gtgtaggatc acttttgcaa caactaaaaa ctaaagcact 2275
ttgtttgcag ttctacagaa agcaacttag tctgtctgca gattcgtttt tgaaagaaga 2335
catgagaaag cggagtttta ggtgaagtca gttgttggat cttcctttat agacttagtc 2395
ctttagatgt ggtctgtata gacatgccca accatcatgc atgggcactg aatatcgtga 2455
actgtggtat gctttttgtt ggtttattgt acttctgtca aagaaagtgg cattggtttt 2515
tataattgtt gccaagtttt aaggttaatt ttcattattt ttgagccaaa ttaaaatgtg 2575
cacctcctgt gcctttccca atcttggaaa atataatttc ttggcagaag gtcagatttc 2635
agggcccagt cactttcgtc tgacttccct ttgcacagtc cgccatgggc ctggcttaga 2695
agttcttgta aactatgcca gagagtacat tcgctgataa aatcttcttt gcagagcagg 2755
agagcttctt gcctctttcc tttcatttct gcctggactt tggtgttctc cacgttccct 2815
gcatcctaag gacagcagga gaactctgac cccagtgcta tttctctagg tgctattgtg 2875
gcaaactcaa gcggtccgtc tctgtccctg taacgttcgt accttgctgg ctgtgaagta 2935
ctgactggta aagctccgtg ctacagcagt gtagggtata cacaaacaca agtaagtgtt 2995
ttatttaaaa ctgtggactt agcataaaaa gggagactat atttattttt tacaaaaggg 3055
ataaaaatgg aaccctttcc tcacccacca gatttagtca gaaaaaaaca ttctattctg 3115
aaaggtcaca gtggttttga catgacacat cagaacaacg cacactgtcc atgatggctt 3175
atgaactcca agtcactcca tcatggtaaa tgggtagatc cctccttcta gtgtgccaca 3235
ccattgcttc ccacagtaga atcttattta agtgctaagt gttgtctctg ctggtttact 3295
ctgttgtttt agagaatgta agttgtatag tgaataagtt attgaagcat gtgtaaacac 3355
tgttatacat cttttctcct agatggggaa tttggaataa aataccttta aaattcaaaa 3415
aaaaaaaaaa aaaaaaaaaa 3435
<210> 13
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 13
ggggccgggg ccggggccgg ggccggggcc ggggcc 36
<210> 14
<400> 14
000
<210> 15
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 15
tgtgacagtt ggaatgcagt ga 22
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 16
gccacttaaa gcaatctctg tcttg 25
<210> 17
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 17
tcgactcttt gcccaccgcc a 21
<210> 18
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 18
gggtctagca agagcaggtg 20
<210> 19
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 19
gtcttggcaa cagctggaga t 21
<210> 20
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 20
tgatgtcgac tctttgccca ccgc 24
<210> 21
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 21
tcctgtaatg gaactgcttt ca 22
<210> 22
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 22
ggtatctgct tcatccagct tt 22
<210> 23
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 23
ccccggcccc ggcccc 16
<210> 24
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 24
caagccaccg tctcactcaa 20
<210> 25
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 25
gtagtgctgt ctactccaga gagttacc 28
<210> 26
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 26
gtagtgctgt ctactccaga gagttacc 28
<210> 27
<400> 27
000
<210> 28
<400> 28
000
<210> 29
<400> 29
000
<210> 30
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 30
agcgggacac cgtaggttac 20
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 31
gtgggcggaa cttgtcgctg 20
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 32
gtcacattat ccaaatgctc 20
<210> 33
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 33
ggtgggcaaa gagtcgacat 20
<210> 34
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 34
atctctgtct tggcaacagc 20
<210> 35
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 35
aagcaatctc tgtcttggca 20
<210> 36
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 36
acttaaagca atctctgtct 20
<210> 37
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 37
ttgccactta aagcaatctc 20
<210> 38
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 38
cccagtaagc aaaagtagct 20
<210> 39
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 39
actctaggac caagaatatt 20
<210> 40
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 40
gccttactct aggaccaaga 20
<210> 41
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 41
ccaaatgtgc cttactctag 20
<210> 42
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 42
tggagcccaa atgtgcctta 20
<210> 43
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 43
tctgtctttg gagcccaaat 20
<210> 44
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 44
ccatcactga gaagtacctg 20
<210> 45
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 45
atttctccat cactgagaag 20
<210> 46
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 46
aaaagttatt tctccatcac 20
<210> 47
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 47
tggcaagaaa agttatttct 20
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 48
gtgtggttgg caagaaaagt 20
<210> 49
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 49
ctccatttag agtgtggttg 20
<210> 50
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 50
tgcatttcga aggatttctc 20
<210> 51
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 51
ccactctctg catttcgaag 20
<210> 52
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 52
acaaaaaact ttacatctat 20
<210> 53
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 53
ccttttcaga caagacaaaa 20
<210> 54
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 54
aagattaatg aaacaataat 20
<210> 55
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 55
gtttccatca aagattaatg 20
<210> 56
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 56
attgatagtc catatgtgct 20
<210> 57
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 57
agtataattg atagtccata 20
<210> 58
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 58
ggaggtagaa actaagttct 20
<210> 59
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 59
atgtgttaat ctatcaacac 20
<210> 60
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 60
tgcatccata ttcttccttt 20
<210> 61
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 61
ttccttatgc atccatattc 20
<210> 62
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 62
cttgtctttc cttatgcatc 20
<210> 63
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 63
acattttctt gtctttcctt 20
<210> 64
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 64
tctggacatt ttcttgtctt 20
<210> 65
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 65
ataatcttct ggacattttc 20
<210> 66
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 66
ctctgaccct gatcttccat 20
<210> 67
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 67
ttggaataat actctgaccc 20
<210> 68
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 68
cagttccatt acaggaatca 20
<210> 69
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 69
cttcaggaac actgtgtgat 20
<210> 70
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 70
atctatttct tcaggaacac 20
<210> 71
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 71
agtactgtat cagctatatc 20
<210> 72
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 72
tcattgagta ctgtatcagc 20
<210> 73
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 73
tcatcatcat tgagtactgt 20
<210> 74
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 74
ccaatatcat catcattgag 20
<210> 75
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 75
tcatgacagc tgtcaccaat 20
<210> 76
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 76
aagccttcat gacagctgtc 20
<210> 77
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 77
agaagaaagc cttcatgaca 20
<210> 78
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 78
tacttgagaa gaaagccttc 20
<210> 79
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 79
attcttactt gagaagaaag 20
<210> 80
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 80
aaaaattctt acttgagaag 20
<210> 81
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 81
agatggtatc tgcttcatcc 20
<210> 82
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 82
caatctaagt agacagtctg 20
<210> 83
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 83
ttaagcaaca gttcaaatac 20
<210> 84
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 84
ctttaaatag caaatggaat 20
<210> 85
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 85
gccatgattt cttgtctggg 20
<210> 86
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 86
gctttaatga gaagtaaaac 20
<210> 87
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 87
tctacagtac aacttaatat 20
<210> 88
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 88
ataattttgt tctacgccta 20
<210> 89
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 89
cactgctgga tggaaaaaga 20
<210> 90
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 90
tggtttaagg gcacaaactc 20
<210> 91
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 91
ttgcccacgg gtacacagca 20
<210> 92
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 92
cagatgagga aataggtgta 20
<210> 93
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 93
acacattagg tactattact 20
<210> 94
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 94
tttttatgtt ccaggcactg 20
<210> 95
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 95
aataggaaat gttagctatg 20
<210> 96
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 96
ggcactcaac aaatactggc 20
<210> 97
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 97
tacatgtaaa gcaactagta 20
<210> 98
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 98
taaaatttca tgaaaatctg 20
<210> 99
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 99
aagtgaatac tttatacttt 20
<210> 100
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 100
catcatgagc ctaaaggaaa 20
<210> 101
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 101
ggctcttagg ttaaacacac 20
<210> 102
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 102
tgcttctgat tcaagccatt 20
<210> 103
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 103
atacaggact aaagtgcttc 20
<210> 104
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 104
caaatgggat ttaaaatgat 20
<210> 105
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 105
tgacatgtag agagattaag 20
<210> 106
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 106
ttattgaaat accatcattt 20
<210> 107
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 107
tagtcagtat aatatcattt 20
<210> 108
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 108
gcattgagaa gaaagccttc 20
<210> 109
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 109
aagacctgat ccaggaaggc 20
<210> 110
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 110
tgagctgatg gcattgagaa 20
<210> 111
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 111
acaacggaac agccacaggt 20
<210> 112
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 112
ttagtgtcaa ggcttttctg 20
<210> 113
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 113
gacggctgac acaccaagcg 20
<210> 114
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 114
tgatggcatt gagaagaaag 20
<210> 115
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 115
tttactttct ctgcactgct 20
<210> 116
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 116
tcttatttac tttctctgca 20
<210> 117
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 117
ggcataatgt tctgactatc 20
<210> 118
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 118
ataacctgga gcattttctc 20
<210> 119
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 119
ccctgactca tatttaaatg 20
<210> 120
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 120
ccagttgaat cctttagcag 20
<210> 121
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 121
catacatgac ttgccggaaa 20
<210> 122
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 122
gacatccaca tctatgtgtg 20
<210> 123
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 123
tgttcatgac agggtggcat 20
<210> 124
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 124
ttataaatat gttcatgaca 20
<210> 125
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 125
cagctcggat ctcatgtatc 20
<210> 126
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 126
ctccagaagg ctgtcagctc 20
<210> 127
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 127
gtatcctgag ccatgtcttc 20
<210> 128
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 128
aatcaggagt aaagctttcg 20
<210> 129
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 129
aaaatattca aatcaggagt 20
<210> 130
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 130
tctctgtgta agacatcttg 20
<210> 131
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 131
gagtgtctct gtgtaagaca 20
<210> 132
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 132
cactagagtg tctctgtgta 20
<210> 133
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 133
gctttcacta gagtgtctct 20
<210> 134
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 134
gatccaggaa ggctttcact 20
<210> 135
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 135
aaagtacttc tgagagataa 20
<210> 136
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 136
aactgtgcaa ggaaagtact 20
<210> 137
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 137
gtcaaggctt ttctgtgaag 20
<210> 138
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 138
agagatttaa agggcttttt 20
<210> 139
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 139
atcttcaggt tccgaagaga 20
<210> 140
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 140
ccctctgctg ttaaatcaag 20
<210> 141
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 141
tgttaagatc gccctctgct 20
<210> 142
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 142
attattatgt taagatcgcc 20
<210> 143
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 143
agagccatta ttatgttaag 20
<210> 144
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 144
ataaaagagt gtaggcctgg 20
<210> 145
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 145
acactagtgt agaaaggtct 20
<210> 146
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 146
gttcttgcac actagtgtag 20
<210> 147
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 147
taaaaagtca ttagaacatc 20
<210> 148
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 148
tattaagtta cacatttaaa 20
<210> 149
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 149
ctttaccagc gatcatgatt 20
<210> 150
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 150
ttctggagta tgatccaggg 20
<210> 151
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 151
acttaactgc aattgctgag 20
<210> 152
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 152
tgtagtgtaa cttacttaac 20
<210> 153
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 153
atgcacctga catcccctca 20
<210> 154
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 154
cccaaaagca taaatctagg 20
<210> 155
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 155
atatttatta tattgtaaac 20
<210> 156
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 156
agcaataata tttattatat 20
<210> 157
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 157
agatagcaat aatatttatt 20
<210> 158
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 158
aaagatagca ataatattta 20
<210> 159
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 159
ttaaaagata gcaataatat 20
<210> 160
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 160
atctttaaaa gatagcaata 20
<210> 161
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 161
atatctttaa aagatagcaa 20
<210> 162
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 162
attatatctt taaaagatag 20
<210> 163
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 163
tattattata tctttaaaag 20
<210> 164
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 164
caagtttaca tcctattatt 20
<210> 165
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 165
aaaacagtag ttgtggtcaa 20
<210> 166
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 166
aaaaaacagt agttgtggtc 20
<210> 167
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 167
tgaatcatgt atttcaaaaa 20
<210> 168
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 168
gccaactcag atttcacctt 20
<210> 169
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 169
ctacacacca aagaatgcca 20
<210> 170
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 170
agttttcagt tgattgcaga 20
<210> 171
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 171
catcctatgt tcaagctcac 20
<210> 172
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 172
taaacatctg cttgatcaat 20
<210> 173
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 173
aatccacaaa gtaggatcta 20
<210> 174
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 174
attagacatt tctacagact 20
<210> 175
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 175
ctcaactaca tagaatatca 20
<210> 176
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 176
ttggcaacaa ttactaaaac 20
<210> 177
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 177
tcaaaaataa tgaaaattaa 20
<210> 178
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 178
caatttggct caaaaataat 20
<210> 179
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 179
ggcacaggag gtgcacattt 20
<210> 180
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 180
tagattttct aaggagaaaa 20
<210> 181
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 181
actgaccagt gaaatctgaa 20
<210> 182
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 182
ggtaagactt agcaagaaga 20
<210> 183
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 183
tctcagagtt gcaatgattg 20
<210> 184
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 184
agatcttatt agttagtata 20
<210> 185
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 185
agtactcaag gaactatttt 20
<210> 186
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 186
ggcaaacagc aacaacttca 20
<210> 187
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 187
gcacttcagt aaaatttctc 20
<210> 188
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 188
ggtccaaacg cattaagaaa 20
<210> 189
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 189
gaattatatt aatcagttat 20
<210> 190
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 190
tgtgtttgtg taactacaat 20
<210> 191
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 191
atattacttc cagaatttta 20
<210> 192
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 192
ggcagaaggg ctctattacc 20
<210> 193
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 193
cattcgaaca tgtcattttg 20
<210> 194
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 194
ctgattcatg atgggaaagc 20
<210> 195
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 195
gtggttgtct aaaacatcaa 20
<210> 196
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 196
atgactgagc tacagtacaa 20
<210> 197
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 197
gggacactac aaggtagtat 20
<210> 198
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 198
ttaaataaga atctaccatg 20
<210> 199
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 199
gctttaataa cttatttcac 20
<210> 200
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 200
aggagaaaag atatataaca 20
<210> 201
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 201
ccatttagga gaaaagatat 20
<210> 202
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 202
ttcaccctca gcgagtactg 20
<210> 203
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 203
aggctgcggt tgtttccctc 20
<210> 204
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 204
gccagatccc catcccttgt 20
<210> 205
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 205
tcacttcctt taagcaagtc 20
<210> 206
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 206
agtgatgccc aagtcacaat 20
<210> 207
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 207
agtcaagtga tgcccaagtc 20
<210> 208
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 208
ccatcagtca agtgatgccc 20
<210> 209
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 209
gattaccatc agtcaagtga 20
<210> 210
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 210
caactgatta ccatcagtca 20
<210> 211
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 211
gcagtttcca actgattcag 20
<210> 212
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 212
cgttcttgtt tcagatgtac 20
<210> 213
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 213
gccaaacaaa atattttatc 20
<210> 214
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 214
taggtaggct aacctagtcc 20
<210> 215
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 215
tcccagccca aagagaagca 20
<210> 216
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 216
ggatcatagc tctcggtaac 20
<210> 217
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 217
aatcataaag ccctcacttc 20
<210> 218
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 218
ctgattggta tttagaaagg 20
<210> 219
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 219
atgcagacat gattacatta 20
<210> 220
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 220
ttcatcatta aactgaaaat 20
<210> 221
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 221
cttttaggtt aaaaaggtgg 20
<210> 222
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 222
atacagagcc tggcaaaaca 20
<210> 223
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 223
ttctatttac agagcattag 20
<210> 224
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 224
gccttcacat taattcacca 20
<210> 225
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 225
tgtgttattg cccctaaaaa 20
<210> 226
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 226
tgtattcact atactatgcc 20
<210> 227
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 227
aagttattta aagtatagca 20
<210> 228
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 228
gacattgaag tatcaagaca 20
<210> 229
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 229
tgttaagtaa tcttagaaaa 20
<210> 230
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 230
ggcatacatt tagaaattca 20
<210> 231
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 231
accttatgca tccatattct 20
<210> 232
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 232
gaattctctt gggaaccatt 20
<210> 233
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 233
atattcaact acaggattta 20
<210> 234
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 234
atgtgttctt tagatacatc 20
<210> 235
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 235
ccttatacag atacatgctg 20
<210> 236
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 236
tagatgcaat tactattttc 20
<210> 237
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 237
tgtacttccc aaacttgaac 20
<210> 238
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 238
ctgaagctca acaacaccaa 20
<210> 239
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 239
gtctatagaa tcaaactgaa 20
<210> 240
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 240
ttgaatcaat acctaacctc 20
<210> 241
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 241
tgcctctttt agaaaagatc 20
<210> 242
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 242
atggaatcat tggtttatcg 20
<210> 243
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 243
aaagctcact tttattcttt 20
<210> 244
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 244
ggtgccgcca ccatgcccgg 20
<210> 245
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 245
gagagaagct gggcaataaa 20
<210> 246
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 246
tctgaccctg cacaataaag 20
<210> 247
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 247
atagtgtgtg attcaaaacg 20
<210> 248
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 248
actgtatcag ctatctaaaa 20
<210> 249
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 249
ttatttgtat aggaacctac 20
<210> 250
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 250
tgtgagctga tggcactgta 20
<210> 251
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 251
ccttatttac tttctctgca 20
<210> 252
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 252
ggaataaggt cactagttcg 20
<210> 253
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 253
atttgcaaca atttttaaat 20
<210> 254
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 254
ataaactacc aatgatatcc 20
<210> 255
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 255
tacctgatcc aggaaggctt 20
<210> 256
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 256
ttcccgaagc ataaatctag 20
<210> 257
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 257
ttgagaagca tgaaattcca 20
<210> 258
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 258
gcgggacacc gtaggttacg 20
<210> 259
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 259
ctttcctagc gggacaccgt 20
<210> 260
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 260
gcacctctct ttcctagcgg 20
<210> 261
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 261
tgtttgacgc acctctcttt 20
<210> 262
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 262
cttgtcgctg tttgacgcac 20
<210> 263
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 263
gggcggaact tgtcgctgtt 20
<210> 264
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 264
gcagcaggga cggctgacac 20
<210> 265
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 265
agaagcaacc gggcagcagg 20
<210> 266
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 266
cccaaaagag aagcaaccgg 20
<210> 267
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 267
accccgcccc caaaagagaa 20
<210> 268
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 268
cttgctagac cccgccccca 20
<210> 269
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 269
cacctgctct tgctagaccc 20
<210> 270
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 270
taaacccaca cctgctcttg 20
<210> 271
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 271
ctcctaaacc cacacctgct 20
<210> 272
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 272
acacacacct cctaaaccca 20
<210> 273
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 273
aaacaaaaac acacacctcc 20
<210> 274
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 274
ggtgggaaaa acaaaaacac 20
<210> 275
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 275
ctgtgagagc aagtagtggg 20
<210> 276
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 276
agcgagtact gtgagagcaa 20
<210> 277
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 277
tcaccctcag cgagtactgt 20
<210> 278
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 278
tcaggtcttt tcttgttcac 20
<210> 279
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 279
aatctttatc aggtcttttc 20
<210> 280
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 280
ttctggttaa tctttatcag 20
<210> 281
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 281
ttgttttctt ctggttaatc 20
<210> 282
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 282
ttccctcctt gttttcttct 20
<210> 283
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 283
gcggttgttt ccctccttgt 20
<210> 284
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 284
tacaggctgc ggttgtttcc 20
<210> 285
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 285
gagcttgcta caggctgcgg 20
<210> 286
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 286
gagttccaga gcttgctaca 20
<210> 287
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 287
cgactcctga gttccagagc 20
<210> 288
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 288
cccggcccct agcgcgcgac 20
<210> 289
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 289
gccccggccc cggcccctag 20
<210> 290
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 290
accacgcccc ggccccggcc 20
<210> 291
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 291
ccgccccgac cacgccccgg 20
<210> 292
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 292
ccccgggccc gccccgacca 20
<210> 293
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 293
cgccccgggc ccgcccccgg 20
<210> 294
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 294
cgcagccccg ccccgggccc 20
<210> 295
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 295
accgcaaccg cagccccgcc 20
<210> 296
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 296
gcgcaggcac cgcaaccgca 20
<210> 297
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 297
ggcgcaggca ccgcaaccgc 20
<210> 298
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 298
cgcctccgcc gccgcgggcg 20
<210> 299
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 299
accgcctgcg cctccgccgc 20
<210> 300
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 300
cactcgccac cgcctgcgcc 20
<210> 301
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 301
ccactcgcca ccgcctgcgc 20
<210> 302
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 302
ggtccccggg aaggagacag 20
<210> 303
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 303
aacaactggt gcatggcaac 20
<210> 304
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 304
gtttcagatg tactatcagc 20
<210> 305
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 305
aaggtgaagt tcatatcact 20
<210> 306
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 306
ggtaacttca aactcttggg 20
<210> 307
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 307
ggttcatgag aggtttccca 20
<210> 308
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 308
tactgaattg cttagtttta 20
<210> 309
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 309
ctaacagaat aagaaaaaaa 20
<210> 310
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 310
gagcattaga tgagtgcttt 20
<210> 311
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 311
tgcattccta agcaatgtgt 20
<210> 312
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 312
tctaggcctt cacattaatt 20
<210> 313
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 313
cctgtctatg cctaggtgaa 20
<210> 314
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 314
tagcacatac aattattaca 20
<210> 315
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 315
gaggagaaga acataaacgc 20
<210> 316
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 316
taccacaagt ctggagccat 20
<210> 317
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 317
gatactggat tgttgaaact 20
<210> 318
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 318
tagtatgact ggagatttgg 20
<210> 319
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 319
atcaaaaccc caaatgattt 20
<210> 320
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 320
atccaaatgc tccggagata 20
<210> 321
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 321
tcgacatcac tgcattccaa 20
<210> 322
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 322
caacagctgg agatggcggt 20
<210> 323
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 323
atttgccact taaagcaatc 20
<210> 324
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 324
gtacctgttc tgtctttgga 20
<210> 325
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 325
caagaaaagt tatttctcca 20
<210> 326
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 326
gaaggatttc tccatttaga 20
<210> 327
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 327
ttacatctat agcaccactc 20
<210> 328
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 328
tcactccctt ttcagacaag 20
<210> 329
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 329
agtttccatc aaagattaat 20
<210> 330
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 330
atagtccata tgtgctgcga 20
<210> 331
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 331
aactaagttc tgtctgtgga 20
<210> 332
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 332
caacacacac tctatgaagt 20
<210> 333
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 333
ttcctttccg gattatatgt 20
<210> 334
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 334
tttccattac aggaatcact 20
<210> 335
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 335
atcagcctat atctatttcc 20
<210> 336
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 336
tcaatgacca ggcggtcccc 20
<210> 337
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 337
ctttttatgg aaaaggaaaa 20
<210> 338
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 338
tgtttcccca aaaatttctg 20
<210> 339
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 339
agatatccac tcgccaccgc 20
<210> 340
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 340
ccggccccgg ccccggcccc 20
<210> 341
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 341
cccggccccg gccccggccc 20
<210> 342
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 342
ccccggcccc ggccccggcc 20
<210> 343
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 343
gccccggccc cggccccggc 20
<210> 344
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 344
ggccccggcc ccggccccgg 20
<210> 345
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 345
cggccccggc cccggccccg 20
<210> 346
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 346
cggccccggc cccggcccc 19
<210> 347
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 347
ccggccccgg ccccggccc 19
<210> 348
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 348
cccggccccg gccccggcc 19
<210> 349
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 349
ccccggcccc ggccccggc 19
<210> 350
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 350
gccccggccc cggccccgg 19
<210> 351
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 351
ggccccggcc ccggccccg 19
<210> 352
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 352
ggccccggcc ccggcccc 18
<210> 353
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 353
cggccccggc cccggccc 18
<210> 354
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 354
ccggccccgg ccccggcc 18
<210> 355
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 355
cccggccccg gccccggc 18
<210> 356
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 356
ccccggcccc ggccccgg 18
<210> 357
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 357
gccccggccc cggccccg 18
<210> 358
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 358
gccccggccc cggcccc 17
<210> 359
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 359
ggccccggcc ccggccc 17
<210> 360
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 360
cggccccggc cccggcc 17
<210> 361
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 361
ccggccccgg ccccggc 17
<210> 362
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 362
cccggccccg gccccgg 17
<210> 363
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 363
ccccggcccc ggccccg 17
<210> 364
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 364
ccccggcccc ggcccc 16
<210> 365
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 365
gccccggccc cggccc 16
<210> 366
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 366
ggccccggcc ccggcc 16
<210> 367
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 367
cggccccggc cccggc 16
<210> 368
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 368
ccggccccgg ccccgg 16
<210> 369
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 369
cccggccccg gccccg 16
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УМЕНЬШЕНИЯ УРОВНЯ мРНК PAPD5 ИЛИ PAPD7 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИОННОГО ГЕПАТИТА В | 2017 |
|
RU2768699C2 |
| КОМПОЗИЦИИ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ C90RF72 | 2014 |
|
RU2748426C2 |
| КОМПОЗИЦИИ ДЛЯ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ ТАУ-БЕЛКА | 2014 |
|
RU2735551C2 |
| КОМПОЗИЦИИ ДЛЯ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ ТАУ-БЕЛКА | 2014 |
|
RU2823361C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2747734C2 |
| МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК) | 2020 |
|
RU2812850C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2777570C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ХАРАКТЕРИЗУЮЩИЕСЯ ЭКСПАНСИЕЙ ГЕКСАНУКЛЕОТИДНЫХ ПОВТОРОВ В ЛОКУСЕ C9ORF72 | 2017 |
|
RU2760877C2 |
| ИНГИБИТОР СИГНАЛА МИОСТАТИНА | 2019 |
|
RU2820270C2 |
| АНТИСМЫСЛОВЫЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2015 |
|
RU2825834C2 |
Группа изобретений относится к соединениям и способам снижения экспрессии мРНК и/или белка гена C9ORF72, содержащих экспансию гексануклеотидного повтора, у животных с помощью специфических ингибиторов C9ORF72. Такие специфические ингибиторы C9ORF72 включают антисмысловые соединения, содержащие модифицированный олигонуклеотид, состоящий из 16-20 связанных нуклеозидов и имеющий последовательность нуклеотидных оснований, содержащую по меньшей мере 12 последовательных нуклеотидных оснований, которые на 100% комплементарны части равной длины из нуклеотидных оснований 1446-1479 SEQ ID NO: 2. Модифицированный олигонуклеотид может содержать по меньшей мере одну модификацию, выбранную из модифицированного сахара и модифицированной межнуклеозидной связи. Соединение, конъюгаты и композиции на его основе пригодны для лечения, предупреждения или облегчения нейродегенеративного заболевания, связанного с нуклеиновой кислотой C9ORF72, содержащей экспансию гексануклеотидного повтора, у нуждающегося в этом индивида. 8 н. и 47 з.п. ф-лы, 28 табл., 6 пр.
1. Соединение, содержащее модифицированный олигонуклеотид, состоящий из 16-20 связанных нуклеозидов и имеющий последовательность нуклеотидных оснований, содержащую по меньшей мере 12 последовательных нуклеотидных оснований, которые на 100% комплементарны части равной длины из нуклеотидных оснований 1446-1479 SEQ ID NO: 2, причем модифицированный олигонуклеотид содержит по меньшей мере одну модификацию, выбранную из модифицированного сахара и модифицированной межнуклеозидной связи, и причем соединение способно снижать уровень экспрессии мРНК и/или белка C9ORF72, содержащих экспансию гексануклеотидного повтора.
2. Соединение, содержащее модифицированный олигонуклеотид, состоящий из 16-20 связанных нуклеозидов и имеющий последовательность нуклеотидных оснований, содержащую по меньшей мере 12 последовательных нуклеотидных оснований из любой из SEQ ID NO: 288, 340-343, 346-349, 352, 355-358, 361-367 и 369, причем модифицированный олигонуклеотид содержит по меньшей мере одну модификацию, выбранную из модифицированного сахара и модифицированной межнуклеозидной связи, и причем соединение способно снижать уровень экспрессии мРНК и/или белка C9ORF72, содержащих экспансию гексануклеотидного повтора.
3. Соединение по п. 2, отличающееся тем, что последовательность нуклеотидных оснований содержит по меньшей мере 12 последовательных нуклеотидных оснований из SEQ ID NO: 288.
4. Соединение по любому из пп. 1-3, отличающееся тем, что последовательность нуклеотидных оснований модифицированного олигонуклеотида по меньшей мере на 90% комплементарна части равной длины SEQ ID NO: 2.
5. Соединение по любому из пп. 1-3, отличающееся тем, что последовательность нуклеотидных оснований модифицированного олигонуклеотида на 100% комплементарна части равной длины SEQ ID NO: 2.
6. Соединение по любому из пп. 1-3, отличающееся тем, что модифицированный олигонуклеотид является одноцепочечным модифицированным олигонуклеотидом.
7. Соединение по п. 6, отличающееся тем, что одноцепочечный модифицированный олигонуклеотид является гэпмером.
8. Соединение по любому из пп. 1-3, отличающееся тем, что по меньшей мере одна межнуклеозидная связь модифицированного олигонуклеотида является модифицированной межнуклеозидной связью.
9. Соединение по п. 8, отличающееся тем, что каждая межнуклеозидная связь модифицированного олигонуклеотида является модифицированной межнуклеозидной связью.
10. Соединение по п. 8, отличающееся тем, что модифицированная межнуклеозидная связь является фосфоротиоатной межнуклеозидной связью.
11. Соединение по п. 9, отличающееся тем, что модифицированная межнуклеозидная связь является фосфоротиоатной межнуклеозидной связью.
12. Соединение по любому из пп. 1-3, отличающееся тем, что по меньшей мере одно нуклеотидное основание модифицированного олигонуклеотида является модифицированным нуклеотидным основанием.
13. Соединение по п. 12, отличающееся тем, что модифицированное нуклеотидное основание является 5'-метилцитозином.
14. Соединение по любому из пп. 1-3, отличающееся тем, что по меньшей мере один нуклеозид модифицированного олигонуклеотида содержит модифицированный сахар.
15. Соединение по п. 14, отличающееся тем, что каждый нуклеозид модифицированного олигонуклеотида содержит модифицированный сахар.
16. Соединение по п. 14, отличающееся тем, что модифицированный сахар является бициклическим сахаром.
17. Соединение по п. 16, отличающееся тем, что бициклический сахар содержит химический мостик между положениями 4' и 2' сахара, причем химический мостик независимо выбран из 4'-CH(R)-O-2' и 4'-(СН2)2-O-2', где R независимо выбран из Н, С1-С6-алкила и C1-С6-алкокси.
18. Соединение по п. 17, отличающееся тем, что химический мостик представляет собой 4'-CH(R)-O-2', и где R является метилом, Н или -СН2-О-СН3.
19. Соединение по п. 15, отличающееся тем, что модифицированный сахар является бициклическим сахаром.
20. Соединение по п. 19, отличающееся тем, что бициклический сахар содержит химический мостик между положениями 4' и 2' сахара, причем химический мостик независимо выбран из 4'-CH(R)-O-2' и 4'-(СН2)2-O-2', где R независимо выбран из Н, C1-С6-алкила и C1-С6-алкокси.
21. Соединение по п. 20, отличающееся тем, что химический мостик представляет собой 4'-CH(R)-O-2', и где R является метилом, Н или -СН2-О-СН3.
22. Соединение по п. 14, отличающееся тем, что модифицированный сахар содержит 2'-O-метоксиэтильную группу.
23. Соединение по п. 14, отличающееся тем, что модифицированный сахар содержит 2'-O-метильную группу.
24. Соединение по любому из пп. 1-3, отличающееся тем, что модифицированный олигонуклеотид имеет последовательность нуклеотидных оснований, которая комплементарна участку C9ORF72, отличному от экспансии гексануклеотидного повтора, причем экспансия гексануклеотидного повтора содержит любую из GGGGCC, GGGGGG, GGGGCG и GGGGGC.
25. Соединение по п. 15, отличающееся тем, что модифицированный сахар содержит 2'-О-метоксиэтильную группу или 2'-O-метильную группу.
26. Соединение по любому из пп. 1-3, отличающееся тем, что по меньшей мере один нуклеозид модифицированного олигонуклеотида содержит заместитель сахара.
27. Соединение по п. 26, отличающееся тем, что заместитель сахара является морфолином или пептид-нуклеиновой кислотой.
28. Конъюгированное антисмысловое соединение, содержащее соединение по любому из пп. 1-3 конъюгированное с любым из холестеринового фрагмента, липидного фрагмента, углевода, фосфолипида, биотина, феназина, фолата, фенантридина, антрахинона, акридина, флуоресцеина, родамина, кумарина и красителя, причем соединение способно снижать уровень экспрессии мРНК и/или белка C9ORF72, содержащих экспансию гексануклеотидного повтора.
29. Композиция, содержащая терапевтически эффективное количество соединения по любому из пп. 1-3 и фармацевтически приемлемый растворитель или носитель, причем композиция предназначена для лечения нейродегенеративного заболевания.
30. Композиция по п. 29, отличающаяся тем, что фармацевтически приемлемый растворитель является фосфатно-солевым буферным раствором (ФСБР), и/или модифицированный олигонуклеотид соединения является натриевой солью.
31. Композиция, содержащая терапевтически эффективное количество соединения по п. 28 и фармацевтически приемлемый растворитель или носитель, причем композиция предназначена для лечения нейродегенеративного заболевания.
32. Композиция по п. 31, отличающаяся тем, что фармацевтически приемлемый растворитель является фосфатно-солевым буферным раствором (ФСБР), и/или модифицированный олигонуклеотид соединения является натриевой солью.
33. Модифицированный олигонуклеотид, состоящий из 16-20 связанных нуклеозидов и имеющий последовательность нуклеотидных оснований, содержащую по меньшей мере 12 последовательных нуклеотидных оснований, которые на 100% комплементарны части равной длины из нуклеотидных оснований 1446-1479 SEQ ID NO: 2, и причем модифицированный олигонуклеотид способен снижать уровень экспрессии мРНК и/или белка C9ORF72, содержащих экспансию гексануклеотидного повтора.
34. Модифицированный олигонуклеотид, состоящий из 16-20 связанных нуклеозидов и имеющий последовательность нуклеотидных оснований, содержащую по меньшей мере 12 последовательных нуклеотидных оснований из любой из SEQ ID NO: 288, 340-343, 346-349, 352, 355-358, 361-367 и 369, и причем модифицированный олигонуклеотид способен снижать уровень экспрессии мРНК и/или белка C9ORF72, содержащих экспансию гексануклеотидного повтора.
35. Модифицированный олигонуклеотид по п. 34, отличающийся тем, что последовательность нуклеотидных оснований содержит по меньшей мере 12 последовательных нуклеотидных оснований из SEQ ID NO: 288.
36. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что последовательность нуклеотидных оснований модифицированного олигонуклеотида по меньшей мере на 90% комплементарна части равной длины SEQ ID NO: 2.
37. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что последовательность нуклеотидных оснований модифицированного олигонуклеотида на 100% комплементарна части равной длины SEQ ID NO: 2.
38. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что по меньшей мере одна межнуклеозидная связь модифицированного олигонуклеотида является модифицированной межнуклеозидной связью.
39. Модифицированный олигонуклеотид по п. 38, отличающийся тем, что каждая межнуклеозидная связь модифицированного олигонуклеотида является модифицированной межнуклеозидной связью.
40. Модифицированный олигонуклеотид по п. 38, отличающийся тем, что модифицированная межнуклеозидная связь является фосфоротиоатной межнуклеозидной связью.
41. Модифицированный олигонуклеотид по п. 39, отличающийся тем, что модифицированная межнуклеозидная связь является фосфоротиоатной межнуклеозидной связью.
42. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что по меньшей мере один нуклеозид модифицированного олигонуклеотида содержит модифицированный сахар.
43. Модифицированный олигонуклеотид по п. 42, отличающийся тем, что каждый нуклеозид модифицированного олигонуклеотида содержит модифицированный сахар.
44. Модифицированный олигонуклеотид по п. 42, отличающийся тем, что модифицированный сахар является бициклическим сахаром.
45. Модифицированный олигонуклеотид по п. 44, отличающийся тем, что бициклический сахар содержит химический мостик между положениями 4' и 2' сахара, причем химический мостик независимо выбран из 4'-CH(R)-O-2' и 4'-(СН2)2-O-2', где R независимо выбран из Н, C1-С6-алкила и C1-С6-алкокси.
46. Модифицированный олигонуклеотид по п. 45, отличающийся тем, что химический мостик представляет собой 4'-CH(R)-O-2', и где R является метилом, Н или -СН2-О-СН3.
47. Модифицированный олигонуклеотид по п. 43, отличающийся тем, что модифицированный сахар является бициклическим сахаром.
48. Модифицированный олигонуклеотид по п. 47, отличающийся тем, что бициклический сахар содержит химический мостик между положениями 4' и 2' сахара, причем химический мостик независимо выбран из 4'-CH(R)-O-2' и 4'-(СН2)2-O-2', где R независимо выбран из Н, С1-С6-алкила и C1-С6-алкокси.
49. Модифицированный олигонуклеотид по п. 48, отличающийся тем, что химический мостик представляет собой 4'-CH(R)-O-2', и где R является метилом, Н или -СН2-О-СН3.
50. Модифицированный олигонуклеотид по п. 42, отличающийся тем, что модифицированный сахар содержит 2'-О-метоксиэтильную группу.
51. Модифицированный олигонуклеотид по п. 42, отличающийся тем, что модифицированный сахар содержит 2'-О-метильную группу.
52. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что модифицированный олигонуклеотид имеет последовательность нуклеотидных оснований, которая комплементарна участку C9ORF72, отличному от экспансии гексануклеотидного повтора, причем экспансия гексануклеотидного повтора содержит любую из GGGGCC, GGGGGG, GGGGCG и GGGGGC.
53. Модифицированный олигонуклеотид по любому из пп. 33-35, отличающийся тем, что модифицированный олигонуклеотид содержит:
сегмент 5'-крыла, имеющий от 1 до 6 нуклеозидов, и причем каждый нуклеозид 5'-крыла содержит модифицированный сахар;
сегмент 3'-крыла, имеющий от 1 до 6 нуклеозидов, и причем каждый нуклеозид 3'-крыла содержит модифицированный сахар; и
гэп-сегмент, имеющий от 8 до 15 нуклеозидов, и причем каждый нуклеозид гэп-сегмента является дезоксинуклеозидом; при этом данный гэп-сегмент расположен непосредственно соседствующим с и между сегментом 5'-крыла и сегментом 3'-крыла.
54. Композиция, содержащая терапевтически эффективное количество модифицированного олигонуклеотид по любому из пп. 33-35 и фармацевтически приемлемый носитель или растворитель, причем композиция предназначена для лечения нейродегенеративного заболевания.
55. Композиция по п. 54, отличающаяся тем, что фармацевтически приемлемый растворитель является фосфатно-солевым буферным раствором (ФСБР), и/или модифицированный олигонуклеотид соединения является натриевой солью.
| WO 2011135396 A1, 03.11.2011 | |||
| US2012149757 A1, 14.06.2012 | |||
| RENTON A | |||
| et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| DEJESUS-HERNANDEZ M | |||
| et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |

