ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В данной заявке содержится список последовательностей, который был представлен в электронном виде в формате ASCII и полностью включен в настоящий документ посредством ссылки. Указанная копия ASCII, созданная 14 июня 2016 года, называется 50474-134WO2_Sequence_Listing_6_14_16_ST25 и имеет размер 133004 байт.
ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к антителам против FcRH5 и способам применения таких антител.
УРОВЕНЬ ТЕХНИКИ
Клеточные пролиферативные расстройства, такие как рак, характеризуются неконтролируемым ростом клеточных популяций. Они являются основной причиной смерти в развитых странах и второй основной причиной смерти в развивающихся странах, при этом каждый год диагностируют 14 миллионов новых случаев рака и регистрируют восемь миллионов смертей. Согласно подсчетам Национального института рака более полумиллиона американцев умрет от рака в 2016 г., что соответствует приблизительно одной из четырех смертей в стране. Так как численность населения старшего возраста увеличилась, одновременно увеличилась частота заболеваемости раком, так как вероятность развития рака после семидесяти возрастает более чем в два раза. Таким образом, рак представляет существенную и постоянно увеличивающуюся социальную нагрузку.
Ген Fc-рецептор-подобного белка 5 (FcRH5, также известный как FcRL5 или IRTA2) принадлежит к семейству из шести недавно идентифицированных генов надсемейства иммуноглобулина (IgSF). Данное семейство генов тесно связано с Fc-рецепторами с консервативной геномной структурой, композицией внеклеточного домена Ig и иммунорецепторным тирозиновым ингибирующим (ITIM) и иммунорецепторным тирозиновым активирующим (ITAM) мотивами (Davis et al., Eur. J. Immunol. 35: 674-80, 2005). Было описано шесть членов семейства рецепторов FcRH/IRTA: FcRH1/IRTA5, FcRH2/IRTA4, FcRH3/IRTA3, FcRH4/IRTA1, FcRH5/IRTA2, и FcRH6 (Polson et al. Int. Immunol. 18(9): 1363-1373, 2006. КДНК FcRH кодируют трансмембранные гликопротеины I типа с множественными Ig-подобными внеклеточными доменами и цитоплазматическими доменами, содержащими консенсусные иммунорецепторные тирозиновые активирующие и/или ингибирующие сигнальные мотивы. Гены FcRH структурно связаны между собой, и их белковые продукты имеют 28-60% внеклеточной идентичности друг с другом. Они также имеют 15-31% идентичность с близкими формами FcR. Существует высокая степень гомологии между различными FcRH.
Лиганд(ы) для FcRH5 неизвестен, но FcRH5 участвует в усиленной пролиферации и снижении экспрессии изотипов во время образования примированных антигеном В-клеток (Dement-Brown et al., J. Leukoc. Biol. 91: 59-67, 2012). Локус FcRH5 имеет три основные изоформы мРНК (FcRH5a, FcRH5b и FcRH5c). Основные изоформы белка FcRH5, кодируемые этими транскриптами, имеют общую аминокислотную последовательность до остатка 560 с общим сигнальным пептидом и шестью внеклеточными Ig-подобными доменами. FcRH5a представляет собой секретируемый гликопротеин с 759 аминокислотами с восемью Ig-подобными доменами, за которыми следуют 13 уникальных, преимущественно полярных аминокислот на его С-конце. FcRH5b расходится с FcRH5a на аминокислотном остатке 560 и простирается на короткий отрезок из 32 дополнительных остатков, гидрофобность которых совместима с ее стыковкой с плазмалеммой через ГФИ-якорь (гликозилфосфатидилинозитол). FcRH5c является самой длинной изоформой, последовательность которой отличается от FcRH5a на аминокислоте 746. FcRH5c кодирует 977-аминокислотный трансмембранный гликопротеин I типа с девятью внеклеточными доменами Ig-типа, несущими восемь потенциальных N-связанных сайтов гликозилирования, 23-аминокислотный трансмембранный домен и 104-аминокислотный цитоплазматический домен тремя консенсусными SH2-связывающими мотивами, имеющими консенсусный ITIM.
Гены FcRH группируются вместе среди классических генов FcR (FcγRI, FcγRII, FcγRIII и FcεRI) в области 1q21-23 хромосомы 1. Эта область содержит одну из наиболее частых вторичных хромосомных аберрациях, связанных со злокачественным фенотипом при гемабластозах, особенно при множественной миеломе (Hatzivassiliou et al. Immunity. 14: 277-89, 2001). FcRH5 экспрессируется только в линии В-клеток, начиная уже с предшественников В-клеток, но не достигает полной экспрессии до стадии зрелой В-клетки. В отличие от большинства известных других поверхностных белков, специфических для В-клеток (например, CD20, CD19 и CD22), FcRH5 продолжает экспрессироваться в плазмоцитах, тогда как другие маркеры, специфические для В-клеток, ослаблены (Polson et al., Int. Immunol. 18: 1363-73, 2006). Кроме того, мРНК FcRH5 сверхэкспрессируется в клеточных линиях множественной миеломы с аномалиями 1q21, обнаруженными при помощи олигонуклеотидных анализов (Inoue Am. J. Pathol. 165: 71-81, 2004). Паттерн экспрессии указывает на то, что FcRH5 может быть мишенью для терапии на основе антител для лечения множественной миеломы. Множественная миелома представляет собой злокачественную опухоль из плазмоцитов, характеризующихся скелетными поражениями, почечной недостаточностью, анемией и гиперкальциемией, и она по существу неизлечима с использованием современных методов лечения. Современные препараты для лечения множественной миеломы включают комбинации ингибитора протеосомы бортезомиба (VELCADE®), иммуномодулятора леналидомида (REVLIMID®) и стероидного дексаметазона.
Терапия на основе моноклональных антител (мкАт) стала важным методом лечения рака. Методы обнаружения и методы лечения на основе FcRH5c-специфических антител могут быть особенно эффективными, поскольку они специально распознают FcRH5, ассоциированный с мембраной целевой клетки, в отличие от антител, которые распознают как растворимые, так и мембранные изоформы FcRH5. Однако только последний Ig-подобный домен FcRH5 (Ig-подобный домен 9) представляет собой уникальную внеклеточную область, которая различает три основные изоформы FcRH5 (например, FcRH5a, FcRH5b и FcRH5c), и существует значительная гомология между Ig-подобными областями внутри FcRH5. Кроме того, последний Ig-подобный домен является высококонсервативным между FcRH1, FcRH2, FcRF3 и FcRH5. Любая терапия на основе антител, специально нацеленная на FcRH5, должна иметь минимальную перекрестную реактивность с другими FcRH, чтобы избежать неблагоприятных побочных эффектов (например, FcRH3 экспрессируется в нормальных NK-клетках).
Принимая во внимание вышесказанное, существует неудовлетворенная потребность в области техники для безопасных и эффективных агентов для использования при лечении клеточных пролиферативных нарушений (например, онкологических заболеваний, например, FcRH5-положительных раковых заболеваний, например, множественной миеломы).
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Данное изобретение относится к антителам против FcRH5 (например, к биспецифическим антителам, например к Т-зависимым биспецифическим антителам (TDB)п против FcRH5), композициям и способам их применения для лечения клеточных пролиферативных нарушений (например, рака, например, FcRH5-позитивных раковых опухолей, например, множественной миеломы).
В первом аспекте изобретение относится к антителу против Fc-рецептор-подобного белка 5 (FcRH5), включая связывающий домен, содержащий следующие шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 2, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 3, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 4, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 5 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 6.
В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23. В некоторых вариантах осуществления изобретения связывающий домен содержит (а) вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 104, (b) вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 105 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие каркасные области вариабельной области тяжелой цепи (FR): (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 104. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 105. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 10, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 14, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 106, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 107 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 53, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осущестления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 106. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 107. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 106, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 107.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 11, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 15 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 20. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 82, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 83 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 82. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 56, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 83. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 82, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 83.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 84, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 85 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 84. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 85. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 84, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 85.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 17 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 22. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 86, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 87 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 86. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 87. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 86, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 87.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 88, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 89 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 88. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 89. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 88, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 89.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 90, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 91 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 90. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 91. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 90, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 91.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 11, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 18 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 22. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 92, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 93 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 92. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 56, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 93. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 92 и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 93.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 11, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 19 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 24. В некоторых вариантах осуществления изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 94, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 95 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 94. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления данного изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 95. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 94 и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 95.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 18 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 25. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 96, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 97 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 53, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 96. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 97. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 96, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 97.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 18 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 25. В некоторых вариантах осуществления изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 98, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 99 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 55, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 98. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 99. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 98, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 99.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 18 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 25. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 100, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 101 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 100. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 101. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 100, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 101.
В других вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 11, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 15 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 20. В некоторых вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 102, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 103 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 102. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 56, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 103. В некоторых вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 102, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 103.
В другом аспекте изобретение отличается антителом против FcRH5, содержащим связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 32, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 33, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 34, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 35, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 36 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 37. В некоторых вариантах осуществления изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 110, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 111 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 66, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 67, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 68 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 69. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 110. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 70, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 71, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 72 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 73. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 111. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 110, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 111.
В другом аспекте изобретение отличается антителом против FcRH5, содержащим связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 38, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 39, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 40, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 41, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 42 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 43. В других вариантах осуществления данного изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 112, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 113 или (с) домен VH, как в (а), и домен VL, как в (b). В других вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 74, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 75, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 76 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 77. В других вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 112. В других вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 78, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 79, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 80 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 81. В других вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 113. В других вариантах осуществления данного изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 112, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 113.
В другом аспекте изобретение отличается антителом против FcRH5, содержащим связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 26, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 27, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 28, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 29, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 30 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 31. В некоторых вариантах осуществления изобретения связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 108, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 109 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 58, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 59, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 60 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 61. В некоторых вариантах осуществления изобретения домен VH содержит аминокислотную последовательность SEQ ID NO: 108. В некоторых вариантах осуществления изобретения антитело дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 62, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 63, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 64 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 65. В некоторых вариантах осуществления изобретения домен VL содержит аминокислотную последовательность SEQ ID NO: 109. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 108, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 109.
В некоторых вариантах осуществления любого из предшествующих аспектов антитело против FcRH5 связано с эпитопом в Ig-подобном домене 9 FcRH5. В некоторых вариантах осуществления изобретения эпитоп содержит часть аминокислот 743-850 SEQ ID NO: 114. В некоторых вариантах осуществления изобретения связывающий домен связывается с FcRH5 человека, FcRH5 яванского макака (cyno) или с обоими. В некоторых вариантах осуществления изобретения связывающий домен специфически не связывается с FcRH1, FcRH2, FcRH3 и/или FcRH4. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 человека с KD около 100 нМ или меньше. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 человека с KD от около 10 пМ до около 100 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 человека с KD от около 100 пМ до около 100 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 человека с KD от около 1 нМ до около 20 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 человека с KD от около 1 нМ до около 10 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 яванского макака с KD около 100 нМ или меньше. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 10 пМ до около 100 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 100 пМ до около 100 нМ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 1 нМ до около 50 нМ.
В других вариантах осуществления изобретения антитело против FcRH5 содержит мутацию сайта агликозилирования. В некоторых вариантах осуществления изобретения мутация сайта агликозилирования является мутацией по типу замены. В некоторых вариантах осуществления изобретения мутация сайта агликозилирования снижает эффекторную функцию антитела против FcRH5. В некоторых вариантах осуществления изобретения мутация по типу замены приходится на аминокислотный остаток N297, L234, L235, D265, и/или Р329 (нумерация ЕС). В некоторых вариантах осуществления изобретения мутация по типу замены выбрана из группы, состоящей из N297G, N297A, L234A, L235A, D265A и P329G. В некоторых вариантах осуществления изобретения мутация по типу замены представляет собой мутацию N297G.
В некоторых вариантах осуществления изобретения антитело против FcRH5 представляет собой антитело IgG.
В некоторых вариантах осуществления изобретения антитело против FcRH5 представляет собой фрагмент антитела, который связывает FcRH5. В некоторых вариантах осуществления изобретения фрагмент антитела выбран из группы, состоящей из фрагментов бис-Fab, Fab, Fab'-SH, Fv, scFv и (Fab')2. В некоторых вариантах осуществления изобретения фрагмент антитела представляет собой фрагмент бис-Fab.
В других вариантах осуществления изобретения антитело против FcRH5 представляет собой полноразмерное антитело.
В некоторых вариантах осуществления изобретения антитело против FcRH5 представляет собой моноспецифическое антитело.
В некоторых вариантах осуществления изобретения антитело против FcRH5 представляет собой мультиспецифическое антитело. В некоторых вариантах осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое антитело. В некоторых вариантах осуществления изобретения биспецифическое антитело содержит второй связывающий домен, который связывает кластер дифференцировки 3 (CD3). В некоторых вариантах осуществления изобретения второй связывающий домен связывается с эпитопом на CD3, содержащим аминокислотный остаток Glu6 CD3. В некоторых вариантах осуществления изобретения эпитоп дополнительно содержит один или более дополнительных аминокислотных остатков, выбранных из группы, состоящей из Gln1, Asp2 и Met7 CD3. В некоторых вариантах осуществления изобретения эпитоп содержит аминокислотные остатки Gln1, Asp2 и Glu6 CD3. В некоторых вариантах осуществления изобретения эпитоп содержит аминокислотные остатки Gln1, Asp2, Glu6 и Met7 CD3. В некоторых вариантах осуществления изобретения эпитоп не содержит аминокислотный остаток Glu5 CD3. В некоторых вариантах осуществления изобретения эпитоп не содержит аминокислотных остатков Gly3 и Glu5 CD3. В некоторых вариантах осуществления изобретения эпитоп состоит из аминокислотных остатков Gln1, Asp2, Glu6 и Met7 CD3. В некоторых вариантах осуществления изобретения второй связывающий домен способен связываться с полипептидом CD3 человека или полипептидом CD3 яванского макака. В некоторых вариантах осуществления изобретения полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3ε человека или полипептид CD3ε яванского макака, соответственно. В некоторых вариантах осуществления изобретения полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3γ человека или полипептид CD3γ яванского макака, соответственно. В некоторых вариантах осуществления изобретения второй связывающий домен связывает полипептид CD3ε человека с KD около 100 нМ или менее. В некоторых вариантах осуществления изобретения второй связывающий домен связывает полипептид CD3ε человека с KD от около 10 пМ до около 100 нМ. В некоторых вариантах осуществления изобретения второй связывающий домен связывает полипептид CD3ε человека с KD от около 100 пМ до около 50 нМ. В некоторых вариантах осуществления изобретения второй связывающий домен связывает полипептид CD3ε человека с KD от около 1 нМ до около 10 нМ.
В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 117, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 120. В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 133, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 134 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 125, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 126, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 127 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 128. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133. В некоторых вариантах осуществления изобретения второй связывающий домен дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 129, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 130, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 131 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 132. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134.
В других вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 124. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 137, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 138 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 125, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 126, (с) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 127 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 128. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137. В некоторых вариантах осуществления изобретения второй связывающий домен дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 129, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 130, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 131 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 132. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138.
В других вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 139, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 140, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 141, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 142, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 143 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 144. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 153, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 154 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 145, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 146, (с) FR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 147 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 148. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153. В некоторых вариантах осуществления изобретения второй связывающий домен дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 149, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 150, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 151 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 152. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154.
В других вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 155, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 156, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 157, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 158, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 159 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 160.
В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 155, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 162, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 157, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 158, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 159 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 160. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 172, (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 173 или (с) домен VH, как в (а), и домен VL, как в (b). В некоторых вариантах осуществления изобретения второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 164, (b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 165, (с) FR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 166 и (d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 167. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172. В некоторых вариантах осуществления изобретения второй связывающий домен дополнительно содержит следующие FR вариабельной области легкой цепи: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 168, (b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 169, (с) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 170 и (d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 171. В некоторых вариантах осуществления изобретения второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173. В некоторых вариантах осуществления изобретения второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173.
В других вариантах осуществления изобретения связывающий домен, который связывает FcRH5, содержит домен VH (VH1), содержащий заряженную область (CR1), и домен VL (VL1), содержащий заряженную область (CR2), причем CR1 в VH1 образует зарядовую пару с CR2 в VL1. В некоторых вариантах осуществления изобретения CR1 содержит основной остаток аминокислоты, a CR2 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR1 содержит мутацию по типу замены Q39K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR1 состоит из мутации по типу замены Q39K. В некоторых вариантах осуществления изобретения CR2 содержит мутацию по типу замены Q38E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR2 состоит из мутации по типу замены Q38E. В некоторых вариантах осуществления изобретения второй связывающий домен, который связывает CD3, содержит домен VH (VH2), содержащий заряженную область (CR3) и домен VL (VL2), содержащий заряженную область (CR4), причем CR4 в VL2 образует зарядовую пару с CR3 в VH2. В некоторых вариантах осуществления изобретения CR4 содержит основной остаток аминокислоты, a CR3 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR4 содержит мутацию по типу замены Q38K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR4 состоит из мутации по типу замены Q38K. В некоторых вариантах осуществления изобретения CR3 содержит мутацию по типу замены Q39E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR3 состоит из мутации по типу замены Q39E. В некоторых вариантах осуществления изобретения домен VL1 связан с константным доменом (CL1) легкой цепи (CL), a VH1 связан с первым константым доменом (СН11) тяжелой цепи (СН1), причем CL1 содержит заряженную область (CR5) и CH11 содержит заряженную область (CR6) и при этом CR5 в CL1 образует зарядовую пару с CR6 в СН11. В некоторых вариантах осуществления изобретения CR5 содержит основной остаток аминокислоты, a CR6 содержит кислотный остаток. В некоторых вариантах осуществления изобретения CR5 содержит мутацию по типу замены V133K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR5 состоит из мутации по типу замены V133K. В некоторых вариантах осуществления изобретения CR6 содержит мутацию по типу замены S183E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR6 состоит из мутации по типу замены S183E.
В других вариантах осуществления изобретения домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), причем CL2 содержит заряженную область (CR7), а СН12 содержит заряженную область (CR8), и при этом CR8 в СН12 образует зарядовую пару с CR7 в CL2. В некоторых вариантах осуществления изобретения CR8 содержит основной остаток аминокислоты, a CR7 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR8 содержит мутацию по типу замены S183K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR8 состоит из мутации по типу замены S183K. В некоторых вариантах осуществления изобретения CR7 содержит мутацию по типу замены V133E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR7 состоит из мутации по типу замены V133E.
В других вариантах осуществления изобретения домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), причем (a) CL2 содержит одну или более мутаций в аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация ЕС), и (b) СН12 содержит одну или более мутаций в аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация ЕС). В некоторых вариантах осуществления изобретения CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F, и/или T178V. В некоторых вариантах осуществления изобретения CL2 содержит следующие мутации по типу замены: F116A, L135V, S174A, S176F, и T178V. В некоторых вариантах осуществления изобретения СН12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A, и/или V185A. В некоторых вариантах осуществления изобретения СН12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A, и V185A.
В других вариантах осуществления изобретения связывающий домен, который связывает FcRH5, содержит домен VH (VH1), содержащий заряженную область (CR1), и домен VL (VL1), содержащий заряженную область (CR2), причем CR2 в VL1 образует зарядовую пару с CR1 в VH1. В некоторых вариантах осуществления изобретения CR2 содержит основной остаток аминокислоты, a CR1 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR2 содержит мутацию по типу замены Q38K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR2 состоит из мутации по типу замены Q38K. В некоторых вариантах осуществления изобретения CR1 содержит мутацию по типу замены Q39E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR1 состоит из мутации по типу замены Q39E. В некоторых вариантах осуществления изобретения второй связывающий домен, который связывает CD3, содержит домен VH (VH2), содержащий заряженную область (CR3) и домен VL (VL2), содержащий заряженную область (CR4), причем CR3 в VH2 образует зарядовую пару с CR4 в VL2. В некоторых вариантах осуществления изобретения CR3 содержит основной остаток аминокислоты, a CR4 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR3 содержит мутацию по типу замены Q39K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR3 состоит из мутации по типу замены Q39K. В некоторых вариантах осуществления изобретения CR4 содержит мутацию по типу замены Q38E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR4 состоит из мутации по типу замены Q38E. В некоторых вариантах осуществления изобретения домен VL1 связан с константным доменом (CL1) легкой цепи (CL), a VH1 связан с первым константым доменом (CH11) тяжелой цепи (СН1), причем CL1 содержит заряженную область (CR5) и СН11 содержит заряженную область (CR6) и при этом CR6 в СН11 образует зарядовую пару с CR5 в CL1. В некоторых вариантах осуществления изобретения CR6 содержит основной остаток аминокислоты, a CR5 содержит кислотный остаток аминокислоты. В некоторых вариантах осуществления изобретения CR6 содержит мутацию по типу замены S183K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR6 состоит из мутации по типу замены S183K. В некоторых вариантах осуществления изобретения CR5 содержит мутацию по типу замены V133E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR5 состоит из мутации по типу замены V133E.
В других вариантах осуществления изобретения домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом CH1 (СН12), причем CL2 содержит заряженную область (CR7), а СН12 содержит заряженную область (CR8), и при этом CR7 в CL2 образует зарядовую пару с CR8 в СН12. В некоторых вариантах осуществления изобретения CR7 содержит основной остаток аминокислоты, a CR8 содержит кислотный остаток. В некоторых вариантах осуществления изобретения CR7 содержит мутацию по типу замены V133K (нумерация ЕС). В некоторых вариантах осуществления изобретения CR7 состоит из мутации по типу замены V133K. В некоторых вариантах осуществления изобретения CR8 содержит мутацию по типу замены S183E (нумерация ЕС). В некоторых вариантах осуществления изобретения CR8 состоит из мутации по типу замены S183E.
В других вариантах осуществления изобретения домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), причем (a) CL2 содержит одну или более мутаций в аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация ЕС), и (b) СН12 содержит одну или более мутаций в аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация ЕС). В некоторых вариантах осуществления изобретения CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F, и/или T178V. В некоторых вариантах осуществления изобретения CL2 содержит следующие мутации по типу замены: F116A, L135V, S174A, S176F, и T178V. В некоторых вариантах осуществления изобретения СН12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A, и/или V185A. В некоторых вариантах осуществления изобретения СН12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A, и V185A. В некоторых вариантах осуществления изобретения антитело против FcRH5 содержит один или более константных доменов тяжелой цепи, причем один или более константных доменов тяжелой цепи выбраны из первого домена СН2 (СН21), первого домена СН3 (СН31), второго домена СН2 (CH22) и второго домена СН3 (СН32). В некоторых вариантах осуществления изобретения по меньшей мере один из одного или более константных доменов тяжелой цепи сопряжен с другим константным доменом тяжелой цепи. В некоторых вариантах осуществления изобретения каждый из СН31 и СН32 содержит выступ (Р1) или впадину (С1), и при этом Р1 или С1 в СН31 могут быть помещены в С1 или Р1 соответственно, в СН32. В некоторых вариантах осуществления изобретения СН3; и СН32 сходятся на поверхности раздела между Р1 и С1. В некоторых вариантах осуществления изобретения каждый из СН21 и СН22 содержит выступ (Р2) или впадину (С2), и при этом Р2 или С2 в СН21 могут быть помещены в С2 или Р2, соответственно, в CH22. В некоторых вариантах осуществления изобретения СН21 и СН22 сходятся на поверхности раздела между Р2 и С2.
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR: (a) HVR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23, и плечо против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123, и при этом каждый из плеча антитела против FcRH5 и плеча антитела против CD3 содержит мутацию по типу замены N297G (нумерация ЕС), и при этом плечо антитела против FcRH5 содержит мутацию по типу замены T366W, а плечо антитела против CD3 содержит замены по типу мутации T366S, L368A и Y407V.
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23, и плечо антитела против CD3, содержащую второй связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L 2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123, и причем плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G, и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G (нумерация ЕС).
В другом аспекте данное изобретение изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133 и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23, и плечо антитела против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123, и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и причем плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G (нумерация ЕС).
В другом аспекте данное изобретение изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и (b) плечо антитела против CD3, содержащую второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (е) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23, и плечо антитела против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119, и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123, и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G, и причем плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом область против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12, (е) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16 и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23, и плечо антитела против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR: (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115, (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116, (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121, (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118, (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119, и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123, и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и причем плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и (б) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, A141I, F170S, S181M, S183A, VI85 А и N297G (нумерация ЕС).
В некоторых вариантах осуществления любого из аспектов данного изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 35 мл/кг/сут. В некоторых вариантах осуществления изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 20 мл/кг/сут у мыши. В некоторых вариантах осуществления изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 12 мл/кг/сут до около 16 мл/кг/сут у мыши. В некоторых вариантах осуществления изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 20 мл/кг/сут до около 40 мл/кг/сут у яванского макака. В некоторых вариантах осуществления изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 25 мл/кг/сут до около 35 мл/кг/сут у яванского макака. В некоторых вариантах осуществления изобретения антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 30 мл/кг/сут до около 35 мл/кг/сут у яванского макака.
В другом аспекте изобретение относится к выделенной нуклеиновой кислоте, кодирующей антитело против FcRH5 по любому из предшествующих аспектов, или его часть, содержащую его связывающий домен, который связывается с FcRH5.
В другом аспекте изобретение относится к вектору, содержащему выделенную нуклеиновую кислоту по предшествующему аспекту.
В другом аспекте изобретение относится к клетке-хозяину, содержащей вектор по предшествующему аспекту. В некоторых вариантах осуществления изобретения клетка-хозяин представляет собой клетку млекопитающего. В некоторых вариантах осуществления изобретения клетка млекопитающего представляет собой клетку яичника китайского хомячка (СНО). В некоторых вариантах осуществления изобретения клетка-хозяина представляет собой прокариотическую клетку. В некоторых вариантах осуществления изобретения прокариотическая клетка представляет собой клетку Е. coli.
В другом аспекте данное изобретение относится к способу получения антитела против FcRH5 по любому из предшествующих аспектов данного изобретения, способ включает культивирование клетки-хозяина по предшествующему аспекту в культуральной среде. В некоторых вариантах осуществления изобретения способ дополнительно включает выделение антитела против FcRH5 из клетки-хозяина или культуральной среды. В некоторых вариантах осуществления изобретения способ дополнительно включает культивирование второй клетки-хозяина, содержащей вторую нуклеиновую кислоту, кодирующую антитело против CD3, которое содержит связывающий домен, который связывает CD3. В некоторых вариантах осуществления изобретения клетки-хозяева совместно культивируют. В некоторых вариантах осуществления изобретения способ дополнительно включает выделение биспецифического антитела против FcRH5 из клеток-хозяев или культуральной среды.
В другом аспекте данное изобретение относится к иммуноконъюгату, содержащему антитело против FcRH5 по любому из предшествующих аспектов и цитотоксическое средство.
В другом аспекте данное изобретение относится к композиции, содержащей антитело против FcRH5 по любому из аспектов данного изобретения. В некоторых вариантах осуществления изобретения композиция дополнительно содержащая фармацевтически приемлемое вспомогательное вещество или разбавитель. В некоторых вариантах осуществления изобретения фармацевтически приемлемое вспомогательное вещество представляет собой буфер, носитель, стабилизатор или консервант. В некоторых вариантах осуществления изобретения композиция представляет собой фармацевтическую композицию. В некоторых вариантах осуществления изобретения композиция дополнительно содержит антагонист, связывающийся с компонентом сигнального пути PD-1, или дополнительный терапевтический агент.
В другом аспекте данное изобретение относится к антителу против FcRH5 по любому из предшествующих аспектов изобретения для применения в качестве лекарственного средства.
В другом аспекте данное изобретение относится к антителу против FcRH5 по любому из предшествующих аспектов изобретения для применения при лечении или замедлении прогрессирования FcRHS-положительного рака у субъекта, нуждающегося в этом.
В другом аспекте данное изобретение относится к антителу против FcRH5 по любому из предшествующих аспектов данного изобретения для применения в повышении иммунной функции у субъекта, страдающего FcRH5-положительным раком. В некоторых вариантах осуществления изобретения FcRH5-положительный рак представляет собой В-клеточный рак. В другом аспекте В-клеточный рак выбирают из группы, состоящей из множественной миеломы (ММ), хронического лимфоцитарного лейкоза (CLL), мантийноклеточной лимфомы (MCL), диффузной В-крупноклеточной лимфомы (DLBCL) и фолликулярной лимфомы (FL). В другом варианте осуществления изобретения В-клеточный рак представляет собой ММ.
В другом аспекте данное изобретение относится к применению антитела против FcRH5 или композиции любого из предшествующих аспектов при изготовлении лекарственного средства для лечения или замедления прогрессирования FcRH5-положительного рака у субъекта. В другом аспекте данное изобретение относится к применению антитела против FcRH5 или композиции любого из предыдущих аспектов при изготовлении лекарственного средства для усиления иммунной функции у субъекта, страдающего FcRH5-положительным раком. В некоторых вариантах осуществления изобретения FcRH5-положительный рак представляет собой В-клеточный рак. В некоторых вариантах осуществления изобретения В-клеточный рак выбран из группы, состоящей из MM, CLL, MCL, DLBCL, и FL. В других вариантах осуществления изобретения В клеточный рак представляет собой ММ.
В другом аспекте данное изобретение относится к способу лечения или замедления прогрессирования FcRH5-положительного рака у субъекта, нуждающегося в этом, причем способ включает введение субъекту антитела против FcRH5 по любому из предшествующих аспектов данного изобретения. В другом аспекте данное изобретение относится к способу усиления иммунной функции у субъекта, страдающего FcRH5-положительным раком, причем способ включает введение субъекту эффективного количества антитела против FcRH5 по любому из предшествующих аспектов данного изобретения. В некоторых вариантах осуществления изобретения FcRH5-положительный рак представляет собой В-клеточный рак. В некоторых вариантах осуществления изобретения В-клеточный рак выбран из группы, состоящей из MM, CLL, MCL, DLBCL, и FL. В других вариантах осуществления изобретения В-клеточный рак представляет собой ММ. В некоторых вариантах осуществления изобретения антитело против FcRH5 связывается с (а) молекулой FcRH5, расположенной на целевой клетке, и (b) молекулой CD3, расположенной на иммунной эффекторной клетке. В некоторых вариантах осуществления изобретения антитело против FcRH5 активирует иммунную эффекторную клетку после связывания с молекулой FcRH5 и молекулой CD3. В некоторых вариантах осуществления изобретения активированная иммунная эффекторная клетка способна оказывать цитотоксическое действие и/или апоптотическое действие на целевую клетку. В некоторых вариантах осуществления изобретения целевая клетка представляет собой плазмоцит. В некоторых вариантах осуществления изобретения плазмоцит представляет собой короткоживущий плазмоцит. В некоторых вариантах осуществления изобретения плазмоцит представляет собой долгоживущую плазмоцит. В некоторых вариантах осуществления изобретения плазмоцит представляет собой миеломную клетку. В некоторых вариантах осуществления изобретения способ включает введение субъекту антитела против FcRH5 в дозе от около 0,01 мг/кг/нед. до около 50 мг/кг/нед. В некоторых вариантах осуществления изобретения способ включает введение субъекту антитела против FcRH5 в дозе от около 0,1 мг/кг/нед. до около 10 мг/кг/нед. В некоторых вариантах осуществления изобретения способ включает введение субъекту антитела против FcRH5 в дозе около 1 мг/кг/нед.
В других вариантах осуществления изобретения способ дополнительно включает введение субъекту антагониста, связывающегося с компонентом сигнального пути PD-1, и/или дополнительного терапевтического агента. В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, или дополнительный терапевтический агент вводят до или после введения антитела против FcRH5. В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, или дополнительный терапевтический агент вводят одновременно с антителом против FcRH5. В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, выбран из группы, состоящей из PD-L1-связывающего антагониста, PD-1-связывающего антагониста и PD-L2-связывающего антагониста. В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой PD-L1-связывающий антагонист. В некоторых вариантах осуществления изобретения PD-L1-связывающий антагонист выбран из группы, состоящей из MPDL3280A (атезолизумаб), YW243.55.S70, MDX-1105, MEDI4736 (дурвалумаб) и MSB0010718C (авелумаб). В некоторых вариантах осуществления изобретения PD-L1-связывающий антагонист представляет собой MPDL3280A (атезолизумаб). В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой PD-1-связывающий антагонист. В некоторых вариантах осуществления изобретения PD-1-связывающий антагонист выбран из группы, состоящей из MDX 1106 (ниволумаб), МK-3475 (пембролизумаб), СТ-011 (пидилизумаб), MEDI-0680 (АМР-514), PDR001, REGN2810, и BGB-108. В некоторых вариантах осуществления изобретения антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой PD-L2-связывающий антагонист.В некоторых вариантах осуществления изобретения PD-L2-связывающий антагонист представляет собой антитело или иммуноадгезин. В некоторых вариантах осуществления изобретения субъект представляет собой стероид, иммуномодулятор (IMiD), ингибитор протеосом (PI) или их комбинацию. В некоторых вариантах осуществления изобретения стероид представляет собой глюкокортикоид. В некоторых вариантах осуществления изобретения глюкокортикоид представляет собой дексаметазон. В некоторых вариантах осуществления изобретения IMiD представляет собой леналидомид. В некоторых вариантах осуществления изобретения PI представляет собой бортезомиб.
В других вариантах осуществления изобретения способ включает введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации по любому из предшествующих аспектов внутривенно, подкожно, внутримышечно, местно, перорально, трансдермально, внутрибрюшинно, внутриорбитально, путем имплантации, путем ингаляции, интратекально, интравентрикулярно или интраназально. В некоторых вариантах осуществления изобретения способ включает внутривенное введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации. В некоторых вариантах осуществления изобретения способ включает подкожное введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации. В некоторых вариантах осуществления любого из предшествующих аспектов субъект представляет собой человека.
В другом аспекте данное изобретение относится к способу обнаружения FcRH5 в биологическом образце у субъекта, причем способ включает: (а) приведение в контакт биологического образца с антителом против FcRH5 по любому из аспектов данного изобретения в условиях, обеспечивающих связывание антитела против FcRH5 с природным FcRH5 в биологическом образце и (b) обнаружение того, образуется ли комплекс между антителом против FcRH5 и природным FcRH5 в биологическом образце. В некоторых вариантах осуществления изобретения биологический образец представляет собой образец крови. В некоторых вариантах осуществления данного аспекта субъект представляет собой человека.
В другом аспекте данное изобретение относится к набору, содержащему антитело против FcRH5 по любому из предшествующих аспектов данного изобретения, и листок-вкладыш в упаковке, содержащий инструкции для применения антитела против FcRH5 для лечения или замедления прогрессирования FcRH5-положительного рака у субъекта. В другом аспекте данное изобретение относится к набору, содержащему антитело против FcRH5 по любому из предшествующих аспектов данного изобретения, и листок-вкладыш в упаковке, содержащий инструкции для применения антитела против FcRH5 для усиления иммунной функции у субъекта, страдающего FcRH5-положительным раком. В некоторых вариантах осуществления данных аспектов субъект представляет собой человека.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1А представлена схематическая диаграмма, демонстрирующая примерную конфигурацию Rational Design (конфигурация 1) TDB против FcRH5, имеющего домены VL, VH, CL и СН1, включая одну или более заряженных областей. Для конфигурации 1 домены VH1, CL1, VL2 и СН12 содержат основные заряженные области, а домены VL1, СН11, VH2 и CL2 содержат кислотные заряженные области.
На фиг. 1В представлена схематическая диаграмма, демонстрирующая примерную конфигурацию Rational Design (конфигурация 2) TDB против FcRH5, имеющего домены VL, VH, CL и СН1, включая одну или более заряженных областей. Для конфигурации 2 домены VH1, CL1, VL2 и СН12 содержат кислотные заряженные области, а домены VL1, CH11, VH2 и CL2 содержат основные заряженные области.
На фиг. 1С представлена схематическая диаграмма, демонстрирующая примерную конфигурацию Rosetta Design (конфигурация 1) TDB против FcRH5, имеющего домены VL, VH, CL и CH1, включая одну или более заряженных областей. Для конфигурации 1 домены VH1, CL1, и VL2 содержат основные заряженные области, а домены VL1, СН11, и VH2 содержат кислотные заряженные области. Кроме того, домен СН12 содержит впадину, а домен CL2 содержит выступ. Впадина и выступ изображены как черный ящик на поверхности раздела CH12/CL2.
На фиг. 1D представлена схематическая диаграмма, демонстрирующая примерную конфигурацию Rosetta Design (конфигурация 2) TDB против FcRH5, имеющего домены VL, VH, CL и СН1, включая одну или более заряженных областей. Для конфигурации 2 домены VH1, CL1, и VL2 содержат кислотные заряженные области, а домены VL1, CH11, и VH2 содержат основные заряженные области. Кроме того, домен СН12 содержит впадину, а домен CL2 содержит выступ. Впадина и выступ изображены в виде черного прямоугольника на поверхности раздела CH12/CL2.
На фиг. 1Е представлена схематическая диаграмма, демонстрирующая примерную альтернативную конфигурацию Rosetta Design (альтернативная конфигурация 1) TDB против FcRH5, имеющего домены VL, VH, CL и СН1, включая одну или более заряженных областей. Для альтернативной конфигурации 1 домены VL1, VH2, и CL2 содержат кислотные заряженные области, а домены VH1, СН12, и VL2 содержат основные заряженные области. Кроме того, домен СН11 содержит впадину, а домен CL1 содержит выступ. Впадина и выступ изображены в виде черного прямоугольника на поверхности раздела CH11/CL1.
На фиг. 1F представлена схематическая диаграмма, демонстрирующая примерную альтернативную конфигурацию Rosetta Design (альтернативная конфигурация 2) TDB против FcRH5, имеющего домены VL, VH, CL и СН1, включая одну или более заряженных областей. Для альтернативной конфигурации 2 домены VL1, VH2, и CL2 содержат основные заряженные области, а домены VH1, СН12, и VL2 содержат кислотные заряженные области. Кроме того, домен СН11 содержит впадину, а домен CL1 содержит выступ. Впадина и выступ изображены в виде черного прямоугольника на поверхности раздела CH11/CL1.
На фиг. 2 представлено выравнивание последовательностей вариабельных доменов легких цепей (VL) выбранных антител против FcRH5. Изменения от 1G7 (см. публикацию патентной заявки США №2015-0098900, которая включена в данное описание посредством ссылки в полном объеме) показаны в черных квадратах. Гипервариабельные области (HVR) обозначены строками выше и/или ниже выравниваний. Эти последовательности доменов VL также представлены как SEQ ID NO: 83, 85, 87, 89, 91, 93, 95, 97, 99, и 101.
На фиг. 3 представлено выравнивание последовательностей вариабельных доменов тяжелых цепей (VH) выбранных антител против FcRH5. Изменения от клона 1G7 (см. публикацию патентной заявки США №2015-0098900) показаны в черных квадратах. Гипервариабельные области (HVR) обозначены строками выше и/или ниже выравниваний. Эти последовательности доменов VH также представлены как SEQ ID NO: 82, 84, 86, 88, 90, 92, 94, 96, 98, и 100.
На фиг. 4 представлена таблица, демонстрирующая влияние на аффинность связывания аминокислотных замен в положении 52 указанных антител против FcRH5.
На фиг. 5А представлено выравнивание последовательностей вариабельных доменов тяжелых цепей (VH) hu1G7.v85 и hu1G7.v93. Изменения в последовательности зародышевой линии человека hIGHV4-59*01 показаны в заштрихованных прямоугольниках. Гипервариабельные области (HVR) обозначены надписями над выравниваниями. Эти последовательности доменов VH также представлены как SEQ ID NO: 104 (hu1G7.v85) и 106 (hu1G7.v93).
На фиг. 5В представлено выравнивание последовательностей вариабельных доменов легких цепей (VL) hu1G7.v85 и hu1G7.v93. Изменения в последовательности зародышевой линии человека hIGKV1-16*01 показаны в заштрихованных прямоугольниках. Гипервариабельные области (HVR) обозначены надписями над выравниваниями. Эти последовательности доменов VL также представлены как SEQ ID NO: 105 (hu1G7.v85) и 107 (hu1G7.v93).
На фиг. 6А представлено выравнивание последовательностей вариабельных доменов тяжелых цепей (VH) hu1G7.v85. 1G7, и консенсусного Н4. Изменения от гуманизированного, с созревшей аффинностью и очищенного клона 1G7 (см. публикацию патентной заявки США №2015-0098900, которая включена в данное описание посредством ссылки в полном объеме) показаны в заштрихованных квадратах. Гипервариабельные области (HVR) обозначены надписями над выравниваниями. Последовательность домена VH hu1G7.v85 раскрыта как SEQ ID NO: 104.
На фиг. 6В представлено выравнивание последовательностей вариабельных доменов легких цепей (VL) hu1G7.v85, 1G7, и консенсусную KI. Изменения от гуманизированного, с созревшей аффинностью и очищенного клона 1G7 показаны в заштрихованных прямоугольниках. Последовательность домена VL hu1G7.v85 раскрыта как SEQ ID NO: 105.
На фиг. 7А показана последовальность вариабельного домена тяжелой цепи (VH) антитела против FcRH5 hu1G7.v93 (SEQ ID NO: 106).
На фиг. 7B показана последовательность вариабельного домена легкой цепи (VL) антитела против FcRH5 hu1G7.v93 (SEQ ID NO: 107).
На фиг. 8 представлен график, демонстрирующий титр антител 1G7.V85 и 1G7.v87 антител.
На фиг. 9А представлено наложение гистограмм, сравнивающих связывание FcRH5/38E4.v1 TDB, имеющих разные области против FcRH5 (т.е. m1G7 («1G7 TDB»), 1G7.v85 («1G7.v85 TDB») и 1G7.v1.4 («1G7.v1.4 TDB»)) к FcRH3-сверхэкспрессирующим клеткам.
На фиг. 9В представлен график, демонстрирующий, что 1G7.v85 TDB не разрушает естественные клетки-киллеры (NK) при концентрациях ≤ 20 мкг/мл, 1G7.v85 TDB имеет медиану ЭК50 в 25 нг/мл на плазмоцитов (PC).
На фиг. 10А представлена серия графиков, сравнивающих способность FcRH5 TDB 1G7.v1.4/38E4.v1 («1G7.v1.4 TDB») связывать FcRH3, FcRH5 человека и FcRH5 яванского макака.
На фиг. 10В представлена серия графиков, сравнивающих способность 1G7.v85 TDB связывать FcRH3, FcRH5 человека и FcRH5 яванского макака.
На фиг. 10С представлена серия графиков, сравнивающих способность FcRH5 TDB 1G7/38E4.v1 («1G7 TDB») связывать FcRH3, FcRH5 человека и FcRH5 яванского макака.
На фиг. 10D представлена серия графиков, сравнивающих способность контрольного антитела связывать FcRH3, FcRH5 человека и FcRH5 яванского макака.
На фиг. 11А показана последовательность последовательности вариабельного домена тяжелой цепи (VH) гуманизированного кроличьего антитела против FcRH5 hu7D8.L1H2. Последовательность домена VH hu7D8.L1H2 раскрыта как SEQ ID NO: 108.
На фиг. 11В показана последовательность последовательности вариабельного домена легкой цепи (VL) гуманизированного кроличьего антитела против FcRH5 hu7D8.L1H2. Последовательность домена VL hu7D8.L1H2 раскрыта как SEQ ID NO: 109.
На фиг. 12А показана последовательность последовательностей вариабельного домена тяжелой цепи (VH) мышиного антитела против FcRH5 17В1. Последовательность домена VH 17В1 раскрыта как SEQ ID NO: 110.
На фиг. 12В показана последовательность последовательности вариабельного домена легкой цепи (VL) мышиного антитела против FcRH5 17В1. Последовательность домена VL 17В1 раскрыта как SEQ ID NO: 111.
На фиг. 13А показана последовательность последовательностей вариабельного домена тяжелой цепи (VH) мышиного антитела против FcRH5 15G8. Последовательность домена VH 15G8 раскрыта как SEQ ID NO: 112.
На фиг. 13В показана последовательность последовательности вариабельного домена легкой цепи (VL) мышиного антитела против FcRH5 15G8. Последовательность домена VL 15G8 раскрыта как SEQ ID NO: 113.
На фиг. 14 представлена серия графиков, демонстрирующих аффинность связывания гуманизированного варианта кроличьих антител против FcRH5 7D8.Rb и h7D8.L1H2 к FcRH5 человека и FcRH5 яванского макака.
На фиг. 15 представлена серия графиков, демонстрирующих, что TDB против FcRH5 1G7.V85 TDB и hu1G7.v93/38E4.v1 («1G7.v93 TDB») связываются со сравнимой аффинностью с FcRH5 человека и FcRH5 яванского макака.
На фиг. 16А-16В представлены графики, демонстрирующие кинетический анализ связывания TDB 1G7.V85 с FcRH5 человека и FcRH5 яванского макака с константами диссоциации (KD) 2,35 нМ и 6,76 нМ, соответственно, как измерено BIACORE® в формате захвата hIgG с использованием модели связывания моновалентной аффинности 1:1.
На фиг. 17А-17В представлены графики, демонстрирующие повышенную FcRH5-индуцированную клеточную токсичность клеток MOLP-2 (т.е. клеток множественной миеломы человека, которые эндогенно экспрессируют FcRH5), с использованием гуманизированных вариантов и вариантов с созревшей аффинностью 1G7, отформатированных в Т-зависимые биспецифические антитела (TDB) с CD3-связывающей областью 38E4.v1 (см. публикацию заявки РСТ № WO 2015-095392 А1, которая включена в данное описание посредством ссылки в полном объеме). На фиг. 17А 1G7 TDB, hu1G7.v1.1/38E4.v1 ("1G7.v1.1 TDB"), hu1G7.v1.2/38E4.v1 ("1G7.v1.2 TDB"), hu1G7.v1.3/38E4.v1 ("1G7.v1.3 TDB"), и 1G7.v1.4 TDB были оценены. На фиг. 17В 1G7. V1.4 TDB, hu1G7.v1.5/38E4.v1 ("1G7.v1.5" TDB), hu1G7.v1.13/38E4.v1 ("1G7.v1.13 TDB"), hu1G7.v1.7/38E4.v1 ("1G7.v1.7 TDB") и hu1G7.v1.13.1/38E4.v1 ("1G7.V1.13.1") были оценены. 1G7.v.1.4 TDB улучшило уничтожение целевых клеток (ЭК50) в 5-13 раз по сравнению с мышиным 1G7 TDB (n=10).
На фиг. 18A-18D представлены графики, сравнивающие способность 1G7.v1.4 TDB и 1G7.v85 TDB активировать Т-клетки (фиг. 18А), уничтожать целевые клетки MOLP-2 (фиг. 18В), уничтожать целевые плазмоциты яванского макака (фиг. 18С) и уничтожать целевые В-клетки яванского макака (фиг. 18D).
На фиг. 19А представлена серия гистограмм, сравнивающих способность 1G7.v85 TDB и hu1G7.v87/38E4.v1 TDB («1G7.v87 TDB») для связывания и перекрестного реагирования с клетками SVT2 мыши, экспрессирующими FcRH5 человека (панель 1), FcRH5 яванского макака (панель 2) и FcRH3 человека (панель 3).
На фиг. 19В-19D представлены графики, сравнивающие способность TDB 1G7.v85 и TDB 1G7.V87 уничтожать целевые клетки MOLP-2 (фиг. 19В), В-клетки человека (фиг. 19С) и В-клетки яванского макака (фиг. 19D).
На фиг. 20A-20D представлены графики, демонстрирующие уменьшенную способность 1G7.v85 TDB связывать FcRH5 после проведения или теста на стресс, вызванный 2,2'-азобис(2-амидопропан)дигидрохлоридом (ААРН) (фиг. 20В), или теста на стресс при избытке освещения (фиг. 20D) по сравнению с бесстрессовыми соответствующими контролями (фиг. 20А и 20С).
На фиг. 21А представлен график, демонстрирующий анализ распределения по размеру 1G7.v85 TDB. 1G7.v85 TDB потерял 0,1% от пика мономера после двух недель стресса в гис-ацетатном растворе при рН 5,5.
На фиг. 21В представлен график, демонстрирующий гетерогенность заряда в 1G7.v85 TDB. 1G7.v85 TDB потерял 7,7% от пика мономера после двух недель стресса в гис-ацетатном растворе при рН 5,5.
На фиг. 22А представлен график, демонстрирующий, что 1G7.v85 TDB не имеет наблюдаемого изменения в уменьшенном массовом профиле легкой цепи после двух недель стресса в гис-ацетатном растворе при рН 5,5.
На фиг. 22В представлен график, демонстрирующий, что 1G7.v85 TDB не имеет наблюдаемого изменения в уменьшенном массовом профиле тяжелой цепи после двух недель стресса в гис-ацетатном растворе при рН 5,5.
На фиг. 23А представлен фосфо-SLP76 вестерн-блот здоровых донорных периферических CD8-клеток, стимулированных 1 мкг/мл 1G7/UCHT1.v9 TDB, 10A8/UCHT1.v9 TDB («10А8 TDB») и TDB против gD/UCHT1.v9 («TDB против gD»), и клеток, экспрессирующих FcRH5 человека с N-концевым маркером экспрессии gD. Для подтверждения равномерной загрузки образца использовали блоттинг для общего SLP76, указывающего на сигналинг TCR.
На фиг. 23В представлен график, демонстрирующий гибель целевых клеток FcRH5 с использованием или TDB 1G7/UCHT1.V9, или TDB против gD и CD8+Т-клетками.
На фиг. 23С представлено схематическое изображение усеченной конструкции FcRH5 с эпитопом gD, теперь мембранно-проксимальным.
На фиг. 23D представлен график, демонстрирующий уничтожение целевых клеток с использованием усеченной конструкции FcRH5 с 1G7 TDB или TDB против gD. Активность проксимального 1G7/UCHT1.v9 TDB увеличилась в 25 раз (ЭК50=20 пМ), и TDB против gD был в состоянии эффективно опосредовать уничтожение клеток (ЭК50=0,19 нМ), если помехи, вызванные ВКД (внеклеточными доменами) были устранены. Усеченную конструкцию экспрессировали в клетках 293.
На фиг. 23Е представлен график, демонстрирующий, что уничтожение целевых клеток для пяти альтернативных TDB против FcRH5, которые распознают мембранно-проксимальный эпитоп (пунктирные линии), эквивалентно уничтожению, опосредованному 1G7/UCHT1.v9 TDB и значительно лучше, чем для 10А8 TDB.
На фиг. 24А представлено наложение гистограмм анализа методом проточной цитометрии SVT2-родительских, gD-FcRH5 полноразмерных и gD-FcRH5-доменных клеток 9.
На фиг. 24В представлен график, демонстрирующий процентное уничтожение целевых клеток при помощи 1G7/UCHT1.v9 TDB, 2Н7/ UCHT1.v9 TDB ("2H7 TDB"), 3G7/ UCHT1.v9 TDB ("3G7 TDB"), 10A8 TDB, и TDB против gD.
На фиг. 25A представлено наложение гистограмм шести клеточных линий (SVT2-вектор, SVT2-FcRH1, SVT2-FCRH2, SVT2-FcRH3, SVT2-FcRH4 и SVT2-FcRH5), демонстрируря, что 1G7.v85 TDB связывается с FcRH5, но не с другими членами семейства.
На фиг. 25В представлено наложение гистограмм трех клеточных линий (293 родительская, 293-FcRH5 полноразмерная и 293-FcRH5-D9-делеция), демонстрируя, что 1G7.v85 TDB связывается с мембранным проксимальным доменом FcRH5.
На фиг. 25С представлено наложение гистограмм трех клеточных линий (SVT2-вектор, SVT2-FcRH5 человека и SVT2-FcRH5 яванского макака), демонстрируя, что TDB 1G7.V85 связывается с FcRH5 яванского макака и FcRH5 человека.
На фиг. 26A-26D представлены наложения гистограмм трех клеточных линий (SVT2-вектор, SVT2-huFcRH5 и SVT2-FcRH5 яванского макака), демонстрирующие связывание 1G7/38E4.v1 TDB («1G7 TDB») с клеточной линией множественной миеломы (ММ) и первичными клетками. Наложенные гистограммы показаны для изотипа-РЕ (фикоэритрин) и 1G7 TDB для клеток MOLP-2 (фиг. 26A), CD20+ В-клеток человека (фиг. 26В), CD38 +CD138+ плазмоцитов (фиг. 26С) и CD38 +CD138+ ММ опухолевых клеток из пунктата костного мозга с ММ (фиг. 26D).
На фиг. 27А представлен график, демонстрирующий дозозависимую активацию CD8+ клеток при стимуляции целевыми клетками (MOLP-2) и 1G7 TDB, обнаруженными методом проточной цитометрии.
На фиг. 27В представлен график, демонстрирующий уничтожение, зависящее от целевой клетки, при помощи 1G7 TDB. Вставка демонстрирует наложение проточной цитометрии родительской линии клеток Fox-NY и клонов, трансфицированных для экспрессии низкого или высокого уровня FcRH5 человека. Планки погрешностей представляют собой стандартное отклонение трех повторностей.
На фиг. 27С-27Е представлены графики, демонстрирующие, что 1G7 TDB индуцирует ответ на пролиферацию CD8 (5 суток), как было обнаружено путем измерения флуоресцентной интесивности CSFE-разведения при помощи проточной цитометрии. Только меченные CFSE CD8+ клетки человека (фиг. 27С), совместное культивирование с MOLP-2 (фиг. 27D) или совместное культивирование с MOLP-2 и 1000 нг/мл 1G7.v85 TDB (фиг. 27Е).
На фиг. 27F представлен график, демонстрирующий уничтожение клеток зависимое от цели, с TDB против FcRH5, содержащими различные плечи антител против CD3 (например, UCHT1.v9, 38E4.v1 и 40G5c) (см. публикацию заявки РСТ № WO 2015/095392 А1, которая включена в данное описание посредством ссылки в полном объеме). Плечи антител против CD3 38E4.v1 (моновалентные KD 0,5 нМ, BIACORE®), UCHT1.v9 (моновалентные KD=2,5 нМ, BIACORE® и Scatchard) и 40G5c (моновалентные KD=51 нМ, BIACORE®) были соединены с плечом антитела против FcRH5 m1G7 (KD=11 нМ, BIACORE®) и тестировали на связывание с очищенными CD8+ клетками человека.
На фиг. 27G представлен график, демонстрирующий зависимое от цели уничтожение клеток с TDB 1G7 и бис-Fab 1G7/38E4.v1 («бис-Fab 1G7»).
На фиг. 28А представлен график, демонстрирующий экспрессию FcRH5 в первичных опухолевых клетках множественной миеломы, здоровых донорных периферических В-клетках и плазмоцитах костного мозга. Экспрессию FcRH5 анализировали с помощью проточной цитометрии и нормировали к экспрессии MOLP-2 внутренного и экспериментального контроля. Относительный уровень FcRH5 рассчитывали следующим образом: (Геометрическое среднее FcRH5 «Х»/геометрическое среднее изотипического контроля «Х»)/(геометрическое среднее FcRH5 MOLP-2/геометрическое среднее изотипического контроля MOLP-2).
На фиг. 28В представлен график, демонстрирующий цитотоксическую активность 1G7.v85 TDB на плазмоцитах человека. Мононуклеарные клетки костного мозга человека (МККМ) культивировали с использованием 1G7.v85 TDB и количество живых CD38+ CD138+ клеток анализировали с помощью проточной цитометрии.
На фиг. 28С представлен график, демонстрирующий цитотоксическую активность 1G7.v85 TDB на разных первичных клетках миеломы, полученных от пациента. МККМ миеломы человека совместно культивировали с CD8+ Т-клетками, выделенными из здорового донора, и 1G7.V85 TBD.
На фиг. 28D представлен график, демонстрирующий, что очень низкая экспрессия FcRH5 в целевых клетках достаточна для сильной активности уничтожения. Число копий FcRH5 на клетку определяли при помощи анализа Скэтчарда.
На фиг. 28Е (сверху) представлен график, демонстрирующий ПЦР в реальном времени уровней мРНК FcRH5 в биопсии костного мозга у пациентов с высоким риском миеломы с увеличением 1q21. Уровень экспрессии мРНК был рассчитан методом дельта Ct (dCt). Статистический анализ проводили с использованием U-критерий Манна-Уитни. (Снизу) Изображения анализа методом FISH (флуоресцентная гибридизация in situ) на первичных биопсиях множественной миеломы демонстрируют нормальный диплоид 1q21 (слева) и смесь приблизительно от трех до шести копий 1q21 (справа). Образец опухоли идентифицировали как увеличение 1q21, если у > 20% опухолевых клеток отмечали три или более копий локуса 1q21.3.
На фиг. 29А представлено наложение гистограмм изотипа-РЕ и клона против FcRH5 1G7-PE, демонстрирующее экспрессию FcRH5 на CD20+ В-клетках яванского макака.
На фиг. 29В представлено наложение гистограмм изотипа-РЕ и клона против FcRH5 1G7-PE, демонстрирующее экспрессию FcRH5 на CD45-CD20-CD38+PC+ плазмоцитов яванского макака.
На фиг. 29C-29D представлены графики, демонстрирующие сравнимую дозозависимую цитотоксическую активность между CD8+ Т-клетками яванского макака и CD8+ Т-клетками человека в анализе уничтожения in vitro с использованием SVT2-FcRH5 яванского макака (фиг. 29С) и MOLP-2 (фиг. 29D) с CD8+ Т-клетками человека или CD8+ Т-клетками яванского макака.
На фиг. 29Е представлен график, демонстрирующий активность уничтожения in vitro 1G7.v85 TDB на CD20+ В-клетках яванского макака (n=14).
На фиг. 29F представлен график, демонстрирующий активность уничтожения in vitro 1G7.v85 TDB на CD45-CD20-CD38+PC+ плазмоцитах из костного мозга яванского макака от восьми разных доноров (n=8).
На фиг. 30А представлен график, демонстрирующий, что Т-клетки селезенки человека, выделенные из селезенки гуманизированных NOD/SCID мышей гамма (NSG) (NOD (диабет без ожирения)/SCID (тяжелый комбинированный иммунодефицит)), имеют сопоставимую активность с периферическими Т-клетками человека от здоровых доноров.
На фиг. 30В представлен график, демонстрирующий, что обработка 1G7 TDB индуцирует регрессию подкожных MOLP-2 ксенотрансплантатных опухолей у гуманизированных мышей NSG. Мыши получали одну внутривенную дозу носителя или 1G7.V85 TDB в 0,5 мг/кг. Указаны средний объем опухоли (черная жирная линия), отдельные объемы опухоли (тонкие линии) и среднее значение контрольной группы (пунктирная линия).
На фиг. 31А представлен график, демонстрирующий концентрацию FcRH5 TDB сыворотке крови, нанесенную на график в течение исследования после однократного введения 1G7.V85 TDB в дозе 1 мг/кг, 2 мг/кг или 4 мг/кг трем животным на группу. Ниже приведена таблица, демонстрирующая фармакодинамические параметры.
На фиг. 31В представлен график, демонстрирующий концентрацию FcRH5 TDB в сыворотке крови (1G7.v85 TDB и 1G7.v87/38E4.v1 TDB («1G7.v87 TDB»)), нанесенную на график в течение исследования после однократного введения анти-gD TDB в дозе 3 мг/кг или FcRH5 TDB в 0,3 мг/кг или 3 мг/кг трем животным на группу. Ниже приведена таблица, демонстрирующая фармакодинамические параметры.
На фиг. 31C-31D представлены графики, демонстрирующие активацию переходных Т-клеток, индуцированных 1G7.v85 TDB, в периферической крови яванского макака после однократного внутривенного введения носителя или 1G7.v85 TDB (1 мг/кг, 2 мг/кг или 4 мг/кг) трем животным на группу.
На фиг. 31Е-31Н представлен график, демонстрирующий абсолютное количество CD20+ В-клеток в периферической крови (фиг. 31Е), селезенке (фиг. 31F), подчелюстном лимфатическом узле (фиг. 31G) и костном мозгу (фиг. 31Н) у яванского макака после однократного внутривенного введения носителя или 1G7.v85 TDB (1 мг/кг, 2 мг/кг или 4 мг/кг) трем животным на группу. На фиг. 31F-31H на график нанесены средние значения для группы и стандартные ошибки среднего значения (СОС).
На фиг. 31I представлен график, демонстрирующий, что 1G7.v85 TDB истощает плазмоциты костного мозга у яванского макака, с насенными на график измеренным СОС для группы.
На фиг. 31J представлен график, демонстрирующий изменение реакции уровня ответа IgG яванского макака на лечение, рассчитанное с использованием формулы {(предварительная доза уровень IgG перед приемом препарата) - (уровень IgG в конце исследования)}/(уровень IgG перед приемом препарата)Х100. Различие между показателями премедикации и после лечения анализируют с помощью критерия Стьюдента для одной выборки. На график нанесены средние значения для группы и стандартные ошибки среднего значения (СОС).
На фиг. 32А-32В представлены графики, демонстрирующие абсолютное количество CD4+ Т-клеток в периферической крови (фиг. 32А) и CD8+ Т-клетки (фиг. 32В) в четырех группах животных после однократного внутривенного введения носителя, 1G7.v85 TDB в дозе 1 мг/кг, 1G7.V85 TDB в 2 мг/кг или 1G7.V85 TDB в 4 мг/кг.
На фиг. 32С представлен график, демонстрирующий уменьшение абсолютного количества CD20+ В-клеток в мезентериальных лимфатических узлах после лечения 1G7.v85 TDB. График изображен в виде отдельных животных и среднего значения группы с СОС.
На фиг. 32D представлен график, демонстрирующий заполнение FcRH5 после однократного внутривенного введения 1G7.v85 TDB в дозе 1 мг/кг, 2 мг/кг или 4 мг/кг.
На фиг. 33A-33F представлены графики, демонстрирующие изменение концентрации цитокинов ИЛ-6 (фиг. 33А), ИЛ-2 (фиг. 33В), ИФН-γ (фиг. 33С), ИЛ-1Rα (фиг. 33D), ИЛ-5 (фиг. 33Е) и МХБ-1 (моноцитарный хемоаттрактный белок) (фиг. 33F) в четырех группах животных после однократного внутривенного введения носителя, 1G7.V85 TDB в дозе 1 мг/кг, 1G7.v85 TDB в 2 мг/кг и 1G7.V85 TDB в 4 мг/кг.
На фиг. 34 представлена серия графиков, демонстрирующих, что обработка 1G7.V85 TDB индуцирует экспрессию PD1 в Т-клетках человека. CD8+ Т-клетки стимулировали в течение 48 часов с использованием 1G7.v85 TDB и MOLP-2 целевых клеток, а затем анализировали при помощи проточной цитометрии.
На фиг. 35А представлен график, демонстрирующий процент PD1+ в CD8+ Т-клетках у яванского макака после однократного внутривенного введения носителя, 1 мг/кг, 2 мг/кг и 4 мг/кг 1G7. v 85 TDB. Обработка 1G7.v85 TDB приводит к индукции PD1 в Т-клетках яванского макака in vivo.
На фиг. 35В представлен график, демонстрирующий процент PD1+в CD4+Т-клетках у яванского макака после однократного внутривенного введения носителя, 1 мг/кг, 2 мг/кг и 4 мг/кг 1G7.V85 TDB.
На фиг. 36A-36D представлены графики, демонстрирующие экспрессию PD-1 в CD4+ Т-клетках из крови (фиг. 36А), CD8+ Т-клетках из селезенки (фиг. 36В), CD8+ Т-клетках из лимфатического узла (фиг. 36С) и CD8+ Т-клетках из костного мозга (фиг. 36D), которые анализировали при помощи проточной цитометрии через семь суток после введения доз 1G7.v85 TDB или носителя.
На фиг. 37А-37В представлены графики, демонстрирующие способность 1G7.V85 TDB перенаправлять активность предварительно стимулированных CD8+ Т-клеток для уничтожения HEK-293Т клеток, экспрессирующих FcRH5 и PD-1 («клетки 293-FcRH5-PD-L1») в присутствии или в отсутствии антитела против PD-L1 или против PD-1. Кривая на фиг. 37А изображена с использованием средних значений и стандартной ошибки среднего (SD) в трех повторностях.
На фиг. 38А представлен график, демонстрирующий уничтожение целевых клеток SVT2-FcRH5 при помощи 1G7.v85 TDB в присутствии и отсутствии дексаметазона (Dex).
На фиг. 38В представлена серия графиков, демострирующих высвобождение цитокинов (то есть ИЛ-2, ИЛ-6, ФНО-α и ИФН-γ) после лечения 1G7.v85 TDB в присутствии и в отсутствии 1 мкМ Dex.
На фиг. 39 представлено схематическое изображение, демонстрирующее получение бис-Fab из человеческого IgG1.
На фиг. 40А представлен график, демонстрирующий связывание FcRH5 с помощью бис-Fab A-D и F (ab')2 А, как определено при помощи ИФА.
На фиг. 40В представлен график, демонстрирующий связывание CD3 с помощью бис-Fab A-D и F (ab')2 А, как определено при помощи ИФА.
На фиг. 41 представлен график, демонстрирующий количество мишень-опосредованной активации Т-клеток при помощи бис-Fab A-D и F(ab')2 А.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. ОПРЕДЕЛЕНИЯ
Термин "около", используемый в данном документе, относится к обычному диапазону ошибок для соответствующего значения и хорошо известный специалисту в данной области техники. В данном документе применение «около» по отношению к величине или параметру включает (и описывает) варианты осуществления, которые относятся непосредственно к этой величине или параметру.
В контексте данного документа "акцепторная каркасная область человека" представляет собой каркасную область, которая содержит аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), которая получена из каркасной области иммуноглобулина человека или консенсусной каркасной области человека, как определено ниже. Акцепторная каркасная область человека, "полученная из" каркасной области иммуноглобулина человека или консенсусной каркасной области человека, может содержать такую же самую аминокислотную последовательность, как и указанная, или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления изобретения количество аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления изобретения последовательность VL-акцепторного каркаса человека идентична последовательности VL-каркаса иммуноглобулина человека или консенсусной последовательности каркаса человека.
Термин «аффинность» относится к силе суммарных общих нековалентных взаимодействий между одиночным сайтом связывания молекулы (например, антитела) и ее партнера по связыванию (например, антигена). Если не указано иное, в данном контексте термин «аффинность связывания» относится к присущему молекуле сродству связывания, отражающему взаимодействие между членами пары связывающихся компонентов (например, антителом и антигеном) при их соотношении 1:1. Аффинность молекулы X к ее партнеру Y в целом можно выразить в виде константы диссоциации (KD). Аффинность можно измерить с помощью общепринятых в данной области техники способов, в том числе тех, которые описаны в данном документе. Конкретные иллюстративные и типичные варианты осуществления данного изобретения, относящиеся к измерению аффинности связывания, описаны далее.
Антитело с «созревшей аффинностью» относится к антителу с одним или более изменениями в одной или более гипервариабельных областях (HVR) по сравнению с исходным антителом, которое не содержит таких изменений, при этом указанные изменения приводят к улучшению аффинности антитела в отношении антигена.
Термин «антитело против FcRH5» или «антитело, связывающееся с FcRH5» относится к антителу, которое способно к связыванию FcRH5 с достаточной аффинностью, так что антитело становится пригодным в качестве диагностического и/или терапевтического средства для таргетинга FcRH5. В одном варианте осуществления изобретения степень связывания антитела против FcRH5 с неродственным не-FcRH5 белком составляет менее чем около 10% связывания антитела с FcRH5 по результатам измерения, например, посредством радиоиммуноанализа (РИА). В некоторых вариантах осуществления изобретения антитело, которое связывается с FcRH5, имеет константу диссоциации (KD) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ, или ≤ 0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В некоторых вариантах осуществления изобретения антитело против FcRH5 связывается с эпитопом FcRH5, который является консервативным среди FcRH5 из различных видов.
Термины «антитело против CD3» и «антитело, которое связывается с CD3» относятся к антителу, которое способно связывать CD3 с достаточной аффинностью, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента для нацеливания на CD3. В одном варианте осуществления изобретения степень связывания антитела против CD3 с неродственным, не являющимся CD3 белком составляет менее около 10% от связывания антитела с CD3 согласно данным, например, радиоиммуноанализа (РИА). В некоторых вариантах осуществления изобретения антитело, которое связывается с CD3, имеет константу диссоциации (KD) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ, или ≤ 0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В определенных вариантах осуществления изобретения антитело против CD3 связывается с эпитопом CD3, который является консервативным для CD3 от разных видов.
Термин «антитело» в данном документе используется в самом широком смысле и охватывает различные структуры антител, включая, но не ограничиваясь ими, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител (например, бис-Fab), при условии, что они проявляют требуемую антигенсвязывающую активность.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются этим, бис-Fab; Fv; Fab; Fab', Fab'-SH; F(ab')2; диатела; линейные антитела; молекулы одноцепочечных антител (например, scFv); и мультиспецифические антитела, образованные из фрагментов антител.
Под «связывающим доменом» подразумевается часть соединения или молекулы, которая специфически связывается с целевым эпитопом, антигеном, лигандом или рецептором. Связывающие домены включают, но не ограничиваются этим, антитела (например, моноклональные, поликлональные, рекомбинантные, гуманизированные и химерные антитела), фрагменты антител или их части (например, фрагменты бис-Fab, фрагменты Fab, F(ab')2, scFv-антитела, SMIP (иммунофармацевтическое средство на основе модульного белка малого размера), домены антител, диатела, мини-антитела, scFv-Fc, аффитела, нанотела и VH- и/или VL- домены антител), рецепторы, лиганды, аптамеры и другие молекулы, имеющие определенного партнера по связыванию.
Используемый в данном документе термин «заряженная область» относится к местоположению полипептида (например, антитела), который включает одну или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) основных или кислых аминокислот, которые способны образовывать зарядовую пару с родственной заряженной областью, имеющей одну или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) или основных, или кислых аминокислот, если заряженная область и ее родственная заряженная область имеют противоположный общий относительный заряд.
Используемый в данном документе термин «зарядовая пара» относится к связи, которая образуется между двумя заряженными областями противоположного общего заряда.
«Химиотерапевтический агент» представляет собой химическое соединение, применяемое в лечении рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа или циклофосфамид (ЦИТОКСАН®); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметиломеламин; ацетогенины (особенно буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, МАРИНОЛ®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включая синтетический аналог топотекан (ГИКАМТИН®)), СРТ-11 (иринотекан, КАМПТОЗАР®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорофосфамид, эстрамустин, ифосфамид мехлорэтамин, мехлорэтамин оксид гидрохлорид, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, особенно калихеамицин гамма 1I и калихеамицин омега I1 (см., например, Nicolaou et al. Angew. Chem Intl. Ed. Engl. 33: 183-186 (1994)); CDP323, оральный ингибитор интегрина альфа-4; динемицин, включая динемицин А; эсперамицин; а также неокарциностатиновый хромофор и родственные хромопротеиновые энедииновые антибиотические хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая АДРИАМИЦИН®-морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролидино-доксорубицин, липосомальный доксорубицин НСl для инъекций (ДОКСИЛ®), липосомальный доксорубицин TLC D-99 (МИОЦЕТ®), пэгилированный липосомальный доксорубицин (КЕЛИКС®) и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, порфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (ГЕМЗАР®), тегафур (УФТОРАЛ®), капецитабин (КСЕЛОДА®), эпотилон и 5-фтороурацил (5-FU); комбретастатин; аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие функции надпочечников, такие как аминоглутетимид, митотан, трилостан; добавку для восполнения дефицита фолиевой кислоты, такую как фролиновая кислота; ацеглатон; альдофосфамид гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; эллиптиниум ацетат; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, Oreg.); разоксан; ризоксин; сизофуран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2', 2'-трихлортриэтиламин; трихотецены (особенно токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин (ЭЛДИЗИН®, ФИЛДЕЗИН®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-С»); тиотепу; таксоид, например, паклитаксел (ТАКСОЛ®, Bristol Myers Squibb Oncology, Princeton, N.J.), альбумин-стабилизированный нанодисперсный паклитаксел (АБРАКСАН™) и доцетаксел (ТАКСОТЕР®, Rhome-Poulene Rorer, Antony, France); хлорамбуцил; 6-тиогуанин; меркаптопурин; метотрексат; платиновые агенты, такие как цисплатин, оксалиплатин (например, ЭЛОКСАТИН®) и карбоплатин; алкалоиды барвинка, которые предотвращают полимеризацию тубулина, приводящую к образованию микротрубочек, включая винбластин (ВЕЛБАН®), винкристин (ОНКОВИН®), виндезин (ЭЛДИЗИН®, ФИЛДЕЗИН®) и винорелбин (НАВЕЛБИН®); этопозид (VP-16); ифосфамид; митоксантрон; лейковорин; новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (ДМФО); ретиноиды, такие как ретиноевая кислота, включая бексаротен (ТАРГРЕТИН®); бифосфонаты, такие как клодронат (например, БОНЕФОС® или ОСТАК®), этидронат (ДИДРОКАЛ®), NE-58095, золедроновую кислоту/золедронат (ЗОМЕТА®), алендронат (ФОСАМАКС®), памидронат (АРЕДИЯ®), тилудронат (СКЕЛИД®) или ризедронат (АКТОНЕЛ®); троксацитабин (аналог 1,3-диоксолан-нуклеозид-цитозина); антисмысловые олигонуклеотиды, в частности те, которые ингибируют экспрессию генов в сигнальных каскадах, вовлеченных в аберрантную клеточную пролиферацию, таких как, например, РКС-альфа, Raf, H-Ras и рецептор эпидермального фактора роста (EGF-R) (например, эрлотиниб (Тарцева™)); и VEGF-A, который снижает клеточную пролиферацию; вакцины, такие как вакцина ТЕРАТОП® и вакцины для генной терапии, например, вакцина АЛЛОВЕКТИН®, вакцина ЛЕУВЕКТИН® и вакцина ВАКСИД®; ингибитор топоизомеразы 1 (например, ЛУРТОТЕКАН®); rmRH (например, АБАРЕЛИКС®); BAY439006 (сорафениб; Bayer); SU-11248 (сунитиниб, СУТЕНТ®, Pfizer); перифозин, ингибитор СОХ-2 (например, целекоксиб или эторикоксиб), ингибитор протеасом (например, PS341); бортезомиб (ВЕЛКЕЙД®); CCI-779; типифарниб (R11577); сорафениб, АВТ510; ингибитор Bcl-2, такой как облимерсен натрий (ГЕНАСЕНС®); пиксантрон; ингибиторы EGFR; ингибиторы тирозинкиназы; ингибиторы серинтреонинкиназы, такие как рапамицин (сиролимус, РАПАМУН®); ингибиторы фарнезилтрансферазы, такие как лонафарниб (SCH 6636, САРАСАР™); и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных соединений; а также комбинации двух или более вышеуказанных соединений, такие как CHOP - сокращение, обозначающее комбинированную терапию циклофосфамидом, доксорубицином, винкристином и преднизолоном; и FOLFOX - сокращение, обозначающее схему лечения оксалиплатином (ЭЛОКСАТИН™) в комбинации с 5-FU и лейковорином, и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных соединений; а также комбинации двух или более вышеуказанных соединений
Химиотерапевтический агент согласно определению данного документа включает «антигормональные агенты» или «эндокринные терапевтические средства», действие которых состоит в регуляции, снижении, блокировании или ингибировании действий гормонов, которые могут стимулировать рост рака. Антиэстрогены и селективные модуляторы рецепторов эстрогена (SERM), включая, например, тамоксифен (включая тамоксифен НОЛВАДЕКС®), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и ФАРЕСТОН (торемифен); ингибиторы ароматазы, которые ингибируют фермент ароматазу, которая регулирует выработку эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, мегестрола ацетат МЕГАС®, эксеместан АРОМАЗИН®, форместанин, фадрозол, ворозол РИВИЗОР®, летрозол ФЕМАРА® и анастрозол АРИМИДЕКС®; и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; а также троксацитабин (аналог 1,3-диоксолан-нуклеозид-цитозина); антисмысловые олигонуклеотиды, в частности те, которые ингибируют экспрессию генов в сигнальных каскадах, вовлеченных в аберрантную клеточную пролиферацию, таких как, например, РКС-альфа, Raf и H-Ras; рибозимы, такие как ингибитор экспрессии VEGF (например, рибозим АНГИОЗИМ®) и ингибитор экспрессии HER2; вакцины, такие как вакцины для генной терапии, например, вакцина АЛЛОВЕКТИН®, вакцина ЛЕУВЕКТИН® и вакцина ВАКСИЛ®; ПРОЛЕЙКИН® rIL-2; ингибитор топоизомеразы 1 ЛУРТОТЕКАН®; rmRH АБАРЕЛИКС®; винорелбин и эсперамицины (см., патент США №4675187) и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных соединений; а также комбинации двух или более вышеуказанных соединений.
Термин «иммуномодулирующее средство» или «IMiD» относится к классу лекарств, который модифицирует ответ иммунной системы или функционирование иммунной системы, такой как стимуляция образования антител и/или ингибирование активности периферических клеток крови и включают, но не ограничиваются ими, талидомид (α-N-фталимидоглутаримид) и его аналоги, РЕВЛИМИД® (леналидомид), ACTI-MID™ (помалидомид), ОТЕЗЛА® (апремистаст) и фармацевтически приемлемые соли или их кислот.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остаток тяжелой и/или легкой цепи получен из другого источника или вида.
Используемый в данном документе термин «FcRH5» относится к любому нативному FcRH5, который является результатом продуцирования белка FcRH5 в клетке. Термин включает FcRH5 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, человек и яванский макак) и грызуны (например, мыши и крысы), если не указано иное. Термин также охватывает встречающиеся в природе варианты FcRH5, например, сплайсинговые варианты или аллельные варианты. Аминокислотная последовательность иллюстративной последовальности белка FcRH5 человека показана в SEQ ID NO: 114. Аминокислотная последовательность иллюстративного белка FcRH5 яванского макака показана в SEQ ID NO: 215.
Употребляемый в данном документе термин «кластер дифференцировки 3» или «CD3» относится, если не указано иное, к любому нативному CD3 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), включая, например, цепи CD3ε, CD3γ, CD3α и CD3β. Данный термин включает «полноразмерный» непроцессированный CD3 (например, непроцессированный или немодифицированный CD3ε или CD3γ), а также любую форму CD3, которая образуется в результате процессинга в клетке. Данный термин также включает варианты CD3 природного происхождения, включая, например, сплайсинговые варианты или аллельные варианты. CD3 включает, например, человеческий белок CD3ε (NCBI RefSeq No. NP_000724), длина которого составляет 207 аминокислот, и человеческий белок CD3γ (NCBI RefSeq No. NP_000064), длина которого составляет 182 аминокислоты.
«Класс» антитела относится к типу константного домена или константной области, содержащейся в его тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно подразделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1, и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Следует понимать, что аспекты и варианты осуществления изобретения, описанные в данном документе, включают «включающий", "состоящий" и "состоящий по существу из" аспектов и вариантов осуществления изобретения.
Термин «цитотоксический агент», используемый в данном документе относится к веществу, которое ингибирует или препятствует фунционированию клеток и/или приводящему к гибели или разрушению клеток. Цитотоксические агенты включают, но не ограничиваются этими, радиоактивные изотопы (например. At211,I131,I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или их производные; и различные противоопухолевые или противораковые агенты, описанные ниже.
«Нарушением» является любое состояние, которое может быть улучшено вследствие лечения, включая, но не ограничиваясь этим, хронические и острые нарушения или заболевания, включая те патологические состояния, которые провоцируют у млекопитающего соответствующее нарушение.
Термины "клеточное пролиферативное растройство" и "пролиферативное растройство" относятся к растройствам, которые связаны с определенной степенью аномальной пролиферации клеток. В одном варианте осуществления изобретения клеточное пролиферативное расстройство представляет собой рак. В одном варианте осуществления изобретения клеточное пролиферативное расстройство представляет собой опухоль.
Термины «рак» и «раковый» относятся к или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым ростом/пролиферацией клеток. Примеры рака включают, но не ограничиваются ими, миелому, карциному, лимфому (например, ходжкинскую и неходжкинскую лимфому), бластому, саркому и лейкемию. В некоторых вариантах осуществления изобретения рак представляет собой FcRH5-положительный рак. Более конкретные примеры таких видов рака включают множественную миелому (ММ), хронический лимфоцитарный лейкоз (CLL), мантийноклеточную лимфому (MCL), диффузную В-крупноклеточную лимфому (DLBCL), фолликулярную лимфому (FL), острый миелоидный лейкоз (AML) миелодиспластический синдром (MDS), хронический миелолейкоз (CML), хронический миеломоноцитарный лейкоз, острый промиелоцитарный лейкоз (APL), хроническое миелопролиферативное расстройство, тромбоцитарный лейкоз, В-клеточный острый лимфобластный лейкоз из клеток-предшественников (pre-B-ALL), Т-клеточный острый лимфобластный лейкоз из клеток-предшественников (pre-T-ALL), мастоцитоз, базофильный лейкоз, тучноклеточную саркому, полиморфно-клеточную саркому, лимфоидный лейкоз и недифференцированный лейкоз. В некоторых вариантах осуществления изобретения рак представляет собой В-клеточный рак. В частности, рак может включать патологические состояния, связанные с избыточным образованием антител, такие как моноклональная гаммапатия, амилоидоз легкой цепи, моноклональная гаммапатия неясного генеза и солитарная плазмацитома, изолированная плазмоцитома и экстрамедуллярная плазмоцитома.
Термин "опухоль", используемый в данном документе, относится ко всем видам роста и пролиферации опухолевой клетки, будь то злокачественный или доброкачественный, и всем предраковым и раковым клеткам и тканям. Термины «рак», «раковый», «расстройство пролиферации клеток» «пролиферативное расстройство» и «опухоль» не являются взаимоисключающими в данном документе.
«Эффекторные функции» относятся к тем видам биологической активности, присущей области Fc антитела, которые изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (КЗЦ); связывание с рецептором Fc; антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ); фагоцитоз; понижающую регуляцию клеточных поверхностных рецепторов (например, В-клеточного рецептора) и активацию В-клетки.
«Эффективное количество» соединения, например, антитела против FcRH5 согласно изобретению или содержащей его композиции (например, фармацевтической композиции), представляет собой по меньшей мере минимальное количество, необходимое для достижения желаемого терапевтического или профилактического результата, такого как определяемое улучшение или предотвращение конкретного нарушения (например, клеточного пролиферативного нарушения, например, рака). Эффективное количество в данном документе может варьироваться в зависимости от таких факторов, как состояние болезни, возраста, пола и массы пациента и от способности антитела вызывать желаемый ответ у индивидуума. Эффективное количество является таким, для которого любые токсические или вредные эффекты лечения перевешиваются терапевтически благоприятными эффектами. Для профилактического применения, благоприятные или желаемые результаты включают результаты, такие как устранение или снижение риска, уменьшения тяжести или задержки начала заболевания, в том числе биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, присутствовавшие при развитии заболевания. Для терапевтического применения, благоприятные или желаемые результаты включают клинические результаты, такие как снижение одного или более симптомов, в результате заболевания, повышения качества жизни людей, страдающих от этой болезни, уменьшение дозы других лекарственных средств, необходимых для лечения заболевания, усиление эффекта другого препарата, среди которых таргетинг, замедление прогрессирования заболевания, и/или увеличения выживаемости индивидуума. В случае рака или опухоли эффективное количество лекарственного средства может иметь эффект в снижении количества раковых клеток; уменьшении размера опухоли; ингибировании (т.е., замедлить до некоторой степени или желательно остановить) инфильтрации раковых клеток в периферические органы; ингибировании (т.е., замедлить до некоторой степени и желательно остановить) метастазирования опухоли; ингибировании до некоторой степени роста опухоли; и/или облегчении до некоторой степени одного или более симптомов, связанных с расстройством. Эффективное количество можно вводить в один или более приемов. Для целей данного изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для достижения профилактического или терапевтического воздействия или непосредственно, или косвенно. Как понимается в клиническом контексте эффективное количество лекарственного средства, соединения или фармацевтической композиции может или не может быть достигнуто в комбинации с другим препаратом, соединением или фармацевтической композицией. Таким образом, "эффективное количество" может рассматриваться в контексте введения одного или более терапевтических агентов, и один агент может рассматриваться как получаемый в эффективном количестве, если в комбинации с одним или более другими агентами желаемый результат может быть достигнут.
В данном контексте термин «область Fc» в данном документе используется для обозначения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере фрагмент константной области. Данный термин включает нативные последовательности областей Fc и варианты областей Fc. В одном варианте осуществления изобретения область Fc тяжелой цепи человеческого IgG находится в пределах от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако, область Fc может содержать или не содержать С-концевой лизин (Lys447). Если в данном документе не указано иное, нумерация аминокислотных остатков в области Fc или константной области соответствует системе нумерации ЕС, также называемой индексом ЕС, как это описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в составе VH (или VL), как правило, определяются в следующей последовательности: FR1-Н1(L1)-FR2-H2(L2)-FR3-Н3(L3)-FR4.
Термин «FcRH5-позитивная клетка» относится к клетке, которая экспрессирует FcRH5 на ее поверхности. В некоторых вариантах осуществления изобретения, FcRH5 представляет собой один или более FcRH5a, FcRH5b, FcRH5c, идентификатор последовательности базы данных UniProt Q96RD9-2, и/или FcRH5d. В некоторых вариантах осуществления изобретения FcRH5 представляет собой FcRH5c.
Термин «FcRH5-положительный рак» относится к раку, содержащему клетки, которые экспрессируют FcRH5 на их поверхности. Для определения того, экспрессирует ли клетка FcRH5 на поверхности, экспрессию мРНК FcRH5 считают коррелирующей с экспрессией FcRH5 на поверхности клетки. В некоторых вариантах осуществления изобретения экспрессию мРНК FcRH5 определяют методом, выбранным из гибридизации in situ и ОТ-ПЦР (включая количественную ОТ-ПЦР). Альтернативно, экспрессия FcRH5 на поверхности клетки может быть определена, например, с использованием антител к FcRH5 в способе, таком как иммуногистохимия, проточная цитометрия и т.д. В некоторых вариантах осуществления изобретения, FcRH5 представляет собой один или более FcRH5a, FcRH5b, FcRH5c, идентификатор последовательности базы данных UniProt Q96RD9-2, и/или FcRH5d. В некоторых вариантах осуществления изобретения FcRH5 представляет собой FcRH5c.
Термины «полноразмерное антитело», «интактное антитело» и «полное антитело» взаимозаменяемо употребляются в данном документе для обозначения антитела, имеющего структуру, в значительной степени сходную со структурой нативного антитела, или содержащего тяжелые цепи, которые содержат область Fc, согласно определению в данном документе.
Термин «гликозилированные формы FcRH5» относится к встречающимся в природе формам FcRH5, которые посттрансляционно модифицируются добавлением углеводных остатков.
Термин "агент, ингибирующий рост", используемый в данном документе, относится к соединению или композиции, которая ингибирует рост клетки или in vitro, или in vivo. В одном варианте осуществления изобретения агент, ингибирующий рост, представляет собой ингибирующее рост антитело, которое предотвращает или снижает пролиферацию клетки, экспрессирующей антиген, с которым связывается антитело. В другом варианте осуществления изобретения ростингибирующий агент может быть один, который значительно снижает процент клеток в S-фазе. Примеры агентов, ингибирующих рост, включают агенты, которые блокируют прохождение по клеточному циклу (в месте, отличном от S-фазы), такие как агенты, которые индуцируют блокировку G1 и блокировку М-фазы. Классические блокаторы М-фазы включают алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Агенты, блокирующие G1, также приводят к остановке S-фазы, например, ДНК-алкилирующие агенты, например, тамоксифен, преднизолон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-С. Дополнительную информацию можно найти в работе The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1 под названием "Cell cycle regulation, oncogenes, and antineoplastic drugs", написанной Murakami. (W.B. Saunders, Philadelphia, 1995), например, с. 13. Таксаны (паклитаксел и доцетаксел) представляют собой противораковые препараты, получаемые из тисового дерева. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из европейского тиса, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки путем предотвращения деполимеризации, что в результате приводит к ингибированию митоза в клетках.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются как взаимозаменяемые и относятся к клеткам, в которые введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформанты» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство вне зависимости от числа пассажей. Потомство может не быть полностью идентичным исходной клетке по содержанию нуклеиновых кислот, и может содержать мутации. Данное описание включает мутантное потомство, обладающее той же функцией или биологической активностью, которую наблюдали во время скрининга или отбора из первоначально трансформированных клеток.
«Антитело человека» представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого организмом человека или клеткой человека, и/или полученного из нечеловеческого источника, в котором используются репертуары антител человека или другие последовательности, кодирующие антитела человека. Из этого определения антитела человека, в частности, исключено гуманизированное антитело, содержащее антигенсвязывающие остатки нечеловеческого происхождения. Человеческие антитела могут быть получены с использованием различных методик известных в данной области техники, включая библиотеки фагового дисплея. Hoogenboom and Winter. J. Mol. Biol. 227:381, 1991; Marks et al. J. Mol. Biol. 222:581,1991. Также доступными для получения моноклональных человеческих антител являются методы, описанные в Cole et al. Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al. J. Immunol., 147(1):86-95, 1991. См. также van Dijk and van de Winkel. Curr. Opin. Pharmacol. 5:368-74, 2001. Антитела человека могут быть получены путем введения антигена трансгенному животному, которое было изменено для получения таких антител в ответ на антигенную стимуляцию, но чьи эндогенные локусы были в неактивном состоянии, например, иммунизированной ксеномыши (см, например, патенты США №№6075181 и 6150584 о технологии XENOMOUSE™). См. также, например, Li et al. Proc. Natl. Acad. Sci. USA. 103:3557-3562, 2006 о человеческих антителах, полученных с помощью гибридомной технологии с человеческими В-клетками..
"Консенсусная каркасная область человека" представляет собой каркасную область, которая содержит наиболее часто встречающиеся аминокислотные остатки при выборе последовательностей каркасной области VL или VH иммуноглобулина человека. Как правило, последовательности VL или VH иммуноглобулина человека выбирают из последовательностей подгруппы вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу по классификации Kabat et al. Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3. В одном варианте осуществления изобретения для VL подгруппа представляет собой подгруппу каппа I, согласно Kabat et al., выше. В одном варианте осуществления изобретения подгруппа VH представляет собой подгруппу III из Kabat et al., выше.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки от нечеловеческих HVR и аминокислотные остатки от FR человека. В некоторых вариантах осуществления изобретения гуманизированное антитело будет содержать по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или, по существу, все HVR (например, CDR) соответствуют таковым из антител нечеловеческого происхождения, и все или по существу все FR соответствуют таковым из человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области человека, полученного из антитела человека. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергли гуманизации.
Используемый в данном документе термин «гипервариабельный участок» или «HVR» относится к каждому из участков вариабельного домена антитела, которые являются гипервариабельными по последовательности («определяющие комплементарность участки» или «CDR») и/или образуют структурно определенные петли («гипервариабельные петли»), и/или содержат контактирующие с антигенами остатки («антигенные контакты»). Как правило, антитела содержат шесть HVR; три в VH (H1, H2, Н3), и три в VL (L1, L2, L3). Типовые HVR в данном документе включают:
(a) гипервариабельные петли, встречающиеся на аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917, 1987);
(b) CDR, встречающиеся на аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (HI), 50-65 (H2) и 95-102 (Н3) (Kabat et al.. Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD(1991));
(c) антигенные контакты, встречающиеся на аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (H2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745, 1996); и
(d) комбинации из (а), (b) и/или (с), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (H2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) нумеруются в данном документе в соответствии с Kabat et al., выше.
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или более гетерологичными молекулами(ой), включая цитотоксический агент, без ограничений.
«Субъект» или «индивид» представляет собой млекопитающее. Млекопитающие включают, но не ограничиваются этими, одомашненных животных (например, коров, овец, кошек, собак, лошадей), приматов (например, людей и приматов, не являющихся человеком, таких, как обезьяны), кроликов и грызунов (например, мышей и крыс). В определенных вариантах осуществления изобретения субъект или индивид является человеком.
«Выделенное» антитело представляет собой антитело, которое было отделено от компонентов его природного окружения. В некоторых вариантах осуществления изобретения антитело является очищенным до более чем 95% или 99% чистоты, определяемой, например, посредством электрофореза (например, ДСН-ПААГ-электрофорез, изоэлектрического фокусирования (ИЭФ), капиллярного электрофореза) или метода хроматографии (например, ионообменной или обращенно-фазовой ВЭЖХ). Обзор методов оценки чистоты антител см., например, в Flatman et al. J. Chromatogr. В 848:79-87, 2007.
"Выделенная" нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, отделенную от компонентов среды, окружающей ее в природе. Выделенная нуклеиновая кислота содержит молекулу нуклеиновой кислоты, которая содержится в клетках, обычно содержащих указанную молекулу нуклеиновой кислоты, однако указанная молекула нуклеиновой кислоты присутствует вне хромосомы или в области хромосомы, отличающейся от ее естественного местоположения в хромосоме.
Выражение «выделенная нуклеиновая кислота, кодирующая антитело против FcRH5» относится к одной или более молекулам нуклеиновой кислоты, кодирующим тяжелые и легкие цепи антитела (или их фрагменты), включая такую(ие) молекулу(ы) нуклеиновых кислот, находящиеся в одном векторе или в отдельных векторах, и такая(ие) молекула(ы) нуклеиновых кислот, присутствующие в одном или более местах клетки-хозяина.
«Выделенная нуклеиновая кислота, кодирующая антитело против CD3» относится к одной или более молекулам нуклеиновых кислот, кодирующим тяжелые и легкие цепи антитела (или их фрагменты), включая молекулы нуклеиновых кислот в одном векторе или отдельных векторах и молекулы нуклеиновых кислот, присутствующие в одной или более местах в клетке-хозяине.
Используемый в данном документе термин «моноклональное антитело» относится к антителу, полученному из популяции в значительной степени гомогенных антител, т.е., отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один эпитоп за исключением возможных вариантных антител, например, содержащих мутации природного происхождения или возникших во время получения препарата моноклональных антител, при этом такие варианты в общем случае присутствуют в незначительном количестве. В отличие от препаратов поликлональных антител, которые обычно содержат различные антитела против различных детерминант (эпитопов), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты антигена. Таким образом, определение "моноклональный" указывает на характеристику антитела, полученного по существу из однородной популяции антител, и не должно быть истолковано как необходимость получения антитела каким-либо конкретным способом. Например, моноклональные антитела для использования согласно данному изобретению можно получить с помощью различных способов, включая, без ограничений, метод гибридомы, методы рекомбинантных ДНК, методы фагового дисплея и способы с использованием трансгенных животных, полностью или частично содержащих локусы человеческого иммуноглобулина, причем такие способы и другие типичные способы получения моноклональных антител описаны в данном документе.
"Неконъюгированное антитело" означает антитело, которое не конъюгировано с гетерологическим компонентом (например, цитотоксическим компонентом) или радиометкой. В фармацевтическом препарате может присутствовать неконъюгированное антитело.
Термин «нативные антитела» относится к природным молекулам иммуноглобулинов, имеющим разные структуры. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой около 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, соединенных дисульфидными связями. В направлении от N- к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, сопровождающийся тремя константными доменами (СН1, СН2 и СН3). Аналогичным образом, в направлении от N- к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, сопровождающимся константным легким (CL) доменом. В зависимости от аминокислотной последовательности своего константного домена легкую цепь антитела можно отнести к одному из двух типов, называемых каппа (κ) и лямбда (λ).
Термин «листок-вкладыш в упаковку» относится к инструкциям, которые обычно вкладывают в коммерческие упаковки терапевтических продуктов для продажи, содержащие информацию о показаниях, использовании, дозах, приеме, комбинированной терапии, противопоказаниях и/или предупреждениях, касающихся использования таких терапевтических продуктов.
Термин "антагонист, связывающийся с компонентом сигнального пути PD-1" относится к молекуле, которая ингибирует взаимодействие партнера, связывающегося с компонентом сигнального пути PD-1, или одного, или более из своих партнеров по связыванию, с тем, чтобы устранить Т-клеточную дисфункцию, которая происходит в следствие передачи сигналов по сигнальному пути PD-1, с результатом в том, чтобы восстановить или усилить Т-клеточную функцию (например, пролиферацию, продукцию цитокинов, и/или уничтожение клеток-мишеней). Используемый в данном документе, антагонист, связывающийся с компонентом сигнального пути PD-1, включает PD-1-связывающий антагонист, PD-L1-связывающий антагонист и PD-L2-связывающий антагонист.
Термин «PD-1-связывающий антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала вследствие взаимодействия PD-1 с одним или более его партнерами по связыванию, такими как PD-L1, PD-L2. В некоторых вариантах осуществления изобретения PD-1-связывающий антагонист является молекулой, которая ингибирует связывание PD-1 с одним или более его партнерами по связыванию. В конкретном варианте осуществления изобретения PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L1 и/или PD-L2. Например, PD-1-связывающие антагонисты включают антитела против PD-1, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или нарушают сигнальную трансдукцию, возникающую из-за взаимодействия PD-1 с PD-L1 и/или PD-L2. В одном варианте осуществления изобретения PD-1-связывающий антагонист снижает отрицательный костимулирующий сигнал, опосредуемый белками клеточной поверхности, экспрессируемыми на Т-лимфоцитах, которые опосредуют сигнализацию через PD-1, чтобы сделать дисфункциональные Т-клетки менее дисфункциональными (например, повышая эффекторный ответ на распознавание антигена). В некоторых вариантах осуществления изобретения PD-1-связывающий антагонист представляет собой антитело против PD-1. В конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой MDX-1106 (ниволумаб). В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой MK-3475 (пембролизумаб). В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой СТ-011 (пидилизумаб). В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой АМР-224. В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой MED1-0680. В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой PDR001. В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой REGN2810. В другом конкретном варианте осуществления изобретения PD-1-связывающий антагонист представляет собой BGB-108.
Термин «PD-L1-связывающий антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала вследствие взаимодействия PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, В7-1. В некоторых вариантах осуществления изобретения PD-L1-связывающий антагонист представляет собой молекулу, которая ингибирует связывание PD-L1 с его партнерами по связыванию. В конкретном варианте осуществления изобретения PD-L1-связывающий антагонист ингибирует связывание PD-L1 с PD-1 и/или В7-1. В некоторых вариантах осуществления изобретения PD-L1-связывающие антагонисты включают антитела против PD-L1, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или нарушают сигнальную трансдукцию, возникающую из-за взаимодействия PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, В7-1. В одном варианте осуществления изобретения PD-L1-связывающий антагонист уменьшает негативный костимулирующий сигнал, опосредованный или с помощью клеточных поверхностных белков, экспрессируемых на Т-лимфоцитах, которые опосредуют сигналинг через PD-L1, так как делают дисфункциональную Т-клетку менее дисфункциональной (например, повышение эффекторного отклика на распознавание антигена). В некоторых вариантах осуществления изобретения PD-L1-связывающий антагонист представляет собой антитело против PD-L1. В еще одном конкретном варианте осуществления изобретения антитело против PD-L1 представляет собой MPDL3280A (атезолизумаб, продаваемый как TECENTRIQ™ с Лекарственный вестник ВОЗ (международные незарегистрированные названия для фармацевтических веществ), рекомендуемый INN (международное непатентованное название): список 74, т. 29, №3, 2015 (см. стр. 387)). В конкретном варианте осуществления изобретения антитело против PD-L1 представляет собой YW 243.55.S70. В другом конкретном варианте осуществления изобретения антитело против PD-L1 представляет собой MDX-1105. В другом конкретном варианте осуществления изобретения антитело против PD-L1 представляет собой MSB0015718C. В еще одном конкретном варианте осуществления изобретения антитело против PD-L1 представляет собой MEDI4736.
Термин «PD-L2-связывающий антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала вследствие взаимодействия PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В некоторых вариантах осуществления изобретения PD-L2-связывающий антагонист является молекулой, которая ингибирует связывание PD-L2 с одним или более его партнерами по связыванию. В конкретном варианте осуществления изобретения PD-L2 -связывающий антагонист ингибирует связывание PD-L2 с PD-1. В некоторых вариантах осуществления изобретения PD-L2-связывающие антагонисты включают антитела анти-PD-L2, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или нарушают сигнальную трансдукцию, возникающую из-за взаимодействия PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В одном варианте осуществления изобретения PD-L2-связывающий антагонист снижает отрицательный костимулирующий сигнал, опосредуемый белками клеточной поверхности, экспрессируемыми на Т-лимфоцитах, которые опосредуют сигнализацию через PD-L2, чтобы сделать дисфункциональные Т-клетки менее дисфункциональными (например, повышая эффекторный ответ на распознавание антигена). В некоторых вариантах осуществления изобретения PD-L2-связывающий антагонист представляет собой иммуноадгезин.
Употребляемый в данном документе термин «белок» относится к любому нативному белку из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Данный термин включает «полноразмерный» непроцессированный белок, а также любую форму белка, которая является результатом процессинга в клетке. Данный термин также включает варианты белка природного происхождения, например, сплайсинговые варианты или аллельные варианты.
«Процент (%) идентичности аминокислотных последовательностей» относительно контрольной полипептидной последовательности определяется как процентная доля аминокислотных остатков в кандидатной последовательности, которые являются идентичными с аминокислотными остатками в контрольной полипептидной последовательности, после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимальной идентичности последовательностей, но без учета каких-либо консервативных замен как части идентичности последовательностей. Выравнивание с целью определения процента идентичности аминокислотных последовательностей может осуществляться различными способами в пределах уровня знаний, доступных специалисту в данной области техники, например, с использованием общедоступного компьютерного программного обеспечения, такого как BLAST, BLAST-2, ALIGN или программного обеспечения Megalign (DNASTAR). Специалисты в данной области могут определить параметры, подходящие для выравнивания последовательностей, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания сравниваемых последовательностей по всей длине. Однако для целей данного изобретения значения % идентичности аминокислотных последовательностей генерируются с использованием компьютерной программы сравнения последовательностей ALIGN-2. Компьютерная программа для выравнивания последовательностей ALIGN-2 была разработана Genentech, Inc., а исходная программа была подана вместе с документацией пользователя в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где она зарегистрирована под № регистрации авторского права США TXU510087. Программа ALIGN-2 находится в свободном доступе в Genentech, Inc., Южный Сан-Франциско, штат Калифорния, или ее можно скомпилировать из исходного кода. Для применения в операционной системе UNIX, включающей цифровую версию UNIX V4.0D, программу ALIGN-2 нужно скомпилировать. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и остаются неизменными.
В тех случаях, когда для сравнения аминокислотных последовательностей используется ALIGN-2, % идентичности аминокислотной последовательности данной аминокислотной последовательности А к, с, или по отношению к данной аминокислотной последовательности В (что в альтернативном варианте может быть сформулировано как данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности к, с или по отношению к данной аминокислотной последовательности В) рассчитывают следующим образом:
100 умножить на соотношение X/Y,
где Х представляет собой число аминокислотных остатков, оцененных программой выравнивания последовательностей ALIGN-2 как идентичные совпадения при программном выравнивании А и В, и где Y представляет общее количество аминокислотных остатков в В. Следует понимать, что там, где длина аминокислотной последовательности А не равняется длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А к В не будет равен % идентичности аминокислотной последовательности В к А. Если специально не указано иное, все значения % идентичности аминокислотной последовательности, которые используется в данном документе, получают, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
Термин «фармацевтический состав» относится к препарату, который находится в такой форме, чтобы делать биологическую активность содержащегося в нем активного ингредиента эффективной, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому вводят состав.
«Фармацевтически приемлемый носитель» относится к ингредиенту фармацевтического препарата, отличающемуся от активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемый носитель включает буфер, вспомогательное вещество, стабилизатор или консервант, без ограничений.
В данном документе термин «лечение» (и его грамматические варианты, например, «лечить») относится к клиническому вмешательству при попытке изменить естественное течение заболевания у индивидуума, подвергаемого лечению, и может осуществляться для профилактики или в процессе клинического проявления патологии. Желаемые эффекты лечения включают, без ограничения, предотвращение возникновения или рецидива заболевания, частичное снятие симптомов, уменьшение каких-либо прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, улучшение или временное облегчение болезненного состояния, и ремиссию или улучшение прогноза. В некоторых вариантах осуществления антитела согласно данному изобретению используют для задержки развития заболевания или замедления прогрессирования заболевания.
В контексте данного документа «замедление прогрессирования» нарушения или заболевания означает отсрочивать, тормозить, замедлять, задерживать, стабилизировать и/или откладывать развитие заболевания или нарушения (например, клеточного пролиферативного нарушения, например, рака). Это замедление может быть разным по продолжительности, в зависимости от истории болезни и/или индивидуума, которого лечат. Для специалиста в данной области техники очевидно, что существенное или значительное замедление может, на практике, включать предотвращение развития заболевания у субъекта. Например, можно замедлить позднюю стадию рака, такую как развитие метастаз.
Термин «эпитоп» относится к конкретному сайту на молекуле антигена, с которой связывается антитело. В некоторых вариантах осуществления изобретения конкретный сайт на молекуле антигена, с которым связывается антитело, определяется путем осаждения гидроксильным радикалом (например, FcRH5-связывающий домен). В некоторых вариантах осуществления изобретения конкретный сайт на молекуле антигена, с которым связывается антитело, определяется при помощи кристаллографии.
Под термином «снижать» или «ингибировать» подразумевается способность приводить к общему снижению, составляющему, например, 20% или более, 50% или более или 75%, 85%, 90%, 95% или более. В определенных вариантах осуществления изобретения термины снижать или ингибировать могут относиться к эффекторной функции антитела, которая опосредуется областью Fc антитела, при этом такие эффекторные функции включают, в частности, комплемент-зависимую цитотоксичность (КЗЦ), антитело-зависимую клеточную цитотоксичность (АЗКЦ) и антитело-зависимый клеточный фагоцитоз (АЗКФ).
Термин «вариабельный участок» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела в общем случае имеют сходные структуры, при этом каждый домен содержит четыре консервативные каркасные области (FR) и три гипервариабельные области (HVR). (См., например, Kindt et al. Kuby Immunology, 6th ed. W.H. Freeman and Co., стр. 91 (2007).) Для придания антигенсвязывающей специфичности может быть достаточно одного домена VH или VL. Кроме того, антитела, которые связывают конкретный антиген, можно выделить, используя домен VH или VL из антитела, которое связывает антиген, для скрининга библиотеки последовательностей, комплементарных доменам VL или VH, соответственно. См., например., Portolano et al. J. Immunol. 150:880-887, 1993; Clarkson et al. Nature 352:624-628, 1991.
"Вариант области Fc" содержит аминокислотную последовательность, которая отличается от таковой у природной последовательности области Fc в силу по меньшей мере одной аминокислотной модификации, предпочтительно одной или более аминокислотных(ой) замен(ы). Предпочтительно вариант области Fc содержит по меньшей мере одну аминокислотную замену по сравнению с природной последовательностью области Fc или с областью Fc исходного полипептида, например, от около одной до около десяти аминокислотных замен, а предпочтительно от около одной до около пяти аминокислотных замен в природной последовательности области Fc или в области Fc исходного полипептида. В данном документе вариант области Fc предпочтительно будет имеет по меньшей мере около 80% гомологии с природной последовательностью области Fc и/или с областью Fc исходного полипептида, и наиболее предпочтительно по меньшей мере около 90% гомологии с ней, более предпочтительно по меньшей мере около 95% гомологии с ней.
Термин «вектор» в контексте данного документа относится к молекуле нуклеиновой кислоты, способной обеспечивать репродукцию другой, связанной с ней нуклеиновой кислоты. Термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, внедренный в геном клетки-хозяина, в которую его внедрили. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы в данном документе называют «экспрессионными векторами».
Используемый в данном документе термин «введение» означает способ введения субъекту дозы соединения (например, антитела против FcRH5 согласно данному изобретению или нуклеиновой кислоты, кодирующей антитело против FcRH5 согласно данному изобретению) или композиции (например, фармацевтической композиции, например, фармацевтической композиции, содержащей антитело против FcRH5 согласно данному изобретению). Композиции, применяемые в описанных в данном документе способах, можно вводить, например, внутримышечно, внутривенно, интрадермально, чрескожно, внутриартериально, внутрибрюшинно, внутриочагово, интракраниально, внутрисуставно, внутрь простаты, внутриплеврально, интратрахеально, интраназально, интравитреально, внутривагинально, ректально, местно, внутриопухолево, перитонеально, подкожно, субконъюнктивально, интравезикулярно, мукозально, интраперикардиально, внутрипуповинно, интраокулярно, перорально, местно, локально, путем ингаляции, путем инъекции, путем инфузии, путем длительной инфузии, путем локализованной перфузии непосредственно через клетки-мишени, при помощи катетера, при помощи лаважа, в кремах или в липидных композициях. Способ введения может изменяться в зависимости от различных факторов (например, соединения или композиции для введения, степени тяжести патологического состояния, заболевания или нарушения, подлежащего лечению).
II. КОМПОЗИЦИИ И МЕТОДЫ
В одном аспекте изобретение частично основано на антителах против FcRH5. В некоторых вариантах осуществления изобретения антитела против FcRH5 являются мультиспецифическими (например, биспецифическими) и связывают в дополнение к FcRH5 или его фрагменту вторую биологическую молекулу (например, антиген клеточной поверхности, например Т-клеточный маркер, например CD3 (например, CD3ε и/или CD3γ)). Антитела согласно изобретению полезны, например, для диагностики и/или лечения, или замедления прогрессирования клеточного пролиферативного нарушения (например, рака, например, FcRH5-положительного рака, например множественной миеломы) у субъекта.
A. Типовые антитела против FcRH5
В одном аспекте изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 2; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 3; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 4; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 5; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 6.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 может иметь вариабельный домен (VH) тяжелой цепи, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85% 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 104, и/или вариабельный домен (VL) легкой цепи, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 105. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может иметь вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO: 104. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105. В конкретном случае антитело против FcRH5 может представлять собой lG7.v85 или его производное или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 10; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 14; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85% 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 106, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 107. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 53, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 106. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 107. В конкретном случае антитело против FcRH5 антитело может представлять собой 1G7.м93 или его производное или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 106, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 107.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 82, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 83. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 82. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 83. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1 или его производное или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 82, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 83.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащий аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 84, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 85. В некоторых случаях антитело против FcRH5 дополнительно содержит каркасные области тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4 SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 84. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 85. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.1 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 84, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 85.
В некоторых случаях антитело против FcRH5 имеет связывающий домен содержащий шесть гипервариабельных областей (HVR) (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 17; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 22. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 86, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 87. В некоторых случаях антитело против FcRH5 дополнительно содержит каркасные области тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащие аминокислотные последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 86. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VL, содержащий аминокислотную последовательность SEQ ID NO: 87. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.2 или его производное или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 86, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 87.
В некоторых случаях антитело против FcRH5, имеющее связывающий домен, содержит шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 88, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 89. В некоторых случаях антитело против FcRH5 дополнительно содержит каркасные области тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащие аминокислотные последовательности SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 88. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 89. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.3 или его производное или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 88, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 89.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 может иметь связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85% 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 90, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 91. В некоторых случаях антитело против FcRH5 дополнительно содержит каркасные области тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4 SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 90. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 91. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.4 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 90, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 91.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 18; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 22. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 92, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 93. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 92. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50 и 51, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 93. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.5 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 92, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 93.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 19; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 24. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 94, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 95. В некоторых случаях антитело против FcRH5 содержит каркасные области тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащие аминокислотные последовательности SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 94. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51, соответственно. В некоторых случаях антитело против FcRH5 содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 95. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.6 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 94, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 95.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 18; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 25. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 96, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 97. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 53, 54, 46, и 47. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 96. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51. В некоторых случаях антитело против FcRH5 содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 97. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1.7 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 96, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 97.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 18; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 25. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 98, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 99. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 55, 46 и 47, соответственно. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 98. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 99. В конкретном случае антитело против FcRH5 может представлять собой 1G7. v1.13 или его производное, или клонально родственное антитело. В некоторых случаях, например, антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 98, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 99.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 18; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 25. В некоторых случаях антитело против FcRH5 содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 100, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 101. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 100. В некоторых случаях антитело против FcRH5 по любому из пп. 79-82, отличающееся тем, что антитело дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 57, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 101. В конкретном случае антитело против FcRH5 может представлять собой 1G7. v1.13. или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 100, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 101.
В некоторых случаях антитело против FcRH5 содержит связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 102, и/или (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 103. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 102. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 103. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v87 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 102, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 103.
В некоторых случаях антитело против FcRH5 может имет связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 32; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 33; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 34; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 35; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 36; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 37. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 110, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 111. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 66, 67, 68 и 69, соответственно. В некоторых случаях антитело против FcRH5 по п. 95, отличающееся тем, что домен VH, содержащий аминокислотную последовательность SEQ ID NO: 110. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 70, 71, 72 и 73. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 111. В конкретном случае антитело против FcRH5 может представлять собой 17В1 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 110, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 111.
В некоторых случаях антитело против FcRH5, имеющий связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 38; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 39; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 40; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 41; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 42; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 43. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 112, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 113. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 74, 75, 76 и 77, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 112. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 78, 79, 80, и 81. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 113. В конкретном случае антитело против FcRH5 может представлять собой 15G8 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 112 и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 113.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 26; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 27; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 28; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 29; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 30; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 31. В некоторых случаях антитело против FcRH5 содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 108, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 109. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 58, 59, 60 и 61, соответственно. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 108. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 62, 63, 64 и 65. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 109. В конкретном случае антитело против FcRH5 может представлять собой 7D8 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, содержащее (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 108, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 109.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 185, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 186. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 179, 54, 46 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 185. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 186. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1A или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 185, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 186.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 187, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 188. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 180, 54, 46 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 187. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 188. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1B или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 187, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 188.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 189, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 190. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 181, 54, 46 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 189. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 190. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1C или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 189, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 190.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 191, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 192. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 23, 54, 46 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 191. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 192. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1D или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 191 и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 192.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 193, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 194. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 182, 46 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 193. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 194. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1E или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 193 и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 194.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 175; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 195, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 196. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 195. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 196. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1F или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 195, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 196.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 176; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 197, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 198. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 197. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 198. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1G или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 197, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 198.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 177; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 199, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 200. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 199. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 200. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1H или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 199, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 200.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 178; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 201, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 202. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 201. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 202. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1I или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 201, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 202.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%. 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 203, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 204. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 183 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 203. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 204. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1J или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 203, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 204.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 205, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 206. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 184 и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 205. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 206. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1K или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 205, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 206.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 7; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 10; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 11; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 15; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 20. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 207, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 208. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 207. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 208. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v1L или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 207, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 208.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 14; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 209, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 210. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 209. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 210. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v86 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 209, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 210.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 14; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 211, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 212. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 53, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 211. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 212. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v191 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 211, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 212.
В нескольких аспектах изобретение относится к антителу против FcRH5, имеющему связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 10; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 14; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23. В некоторых случаях антитело против FcRH5 согласно формуле изобретения может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 213, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 214. В некоторых случаях антитело против FcRH5 содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащих последовательности SEQ ID NO: 52, 54, 46, и 47. В некоторых случаях антитело против FcRH5 может содержать домен VH, содержащий аминокислотную последовательность SEQ ID NO: 213. В некоторых случаях антитело против FcRH5 дополнительно содержит по меньшей мере одну (например, 1,2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 48, 56, 50, и 51. В некоторых случаях антитело против FcRH5 может содержать домен VL, содержащий аминокислотную последовательность SEQ ID NO: 214. В конкретном случае антитело против FcRH5 может представлять собой 1G7.v92 или его производное, или клонально родственное антитело. В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, имеющее связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 213, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 214.
В некоторых вариантах осуществления изобретения антитело, предложенное в данном документе антитело, представляет собой моноклональное, человеческое, гуманизированное или химерное антитело. В некоторых случаях антитело против FcRH5 представляет собой антитело IgG. Антитело против FcRH5 может быть полноразмерным антителом и/или моноспецифическим антителом. В некоторых вариантах осуществления изобретения антитело против FcRH5 может связываться с эпитопом в Ig-подобном домене 9 FcRH5. Например, эпитоп может содержать часть аминокислот 743-850 SEQ ID NO: 114. В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека или FcRH5 яванского макака (cyno) или с обоими. В других случаях связывающий домен специфически не связывается с FcRH1, FcRH2, FcRH3 и/или FcRH4.
В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 45 мл/кг/сут (например, около 1 мл/кг/сут, 5 мл/кг/сут, 10 мл/кг/сут, 11 мл/кг/сут, 12 мл/кг/сут, 13 мл/кг/сут, 14 мл/кг/сут, 15 мл/кг/сут, 16 мл/кг/сут, 17 мл/кг/сут, 18 мл/кг/сут, 19 мл/кг/сут, 20 мл/кг/сут, 21 мл/кг/сут, 22 мл/кг/сут, 23 мл/кг/сут, 24 мл/кг/сут, 25 мл/кг/сут, 26 мл/кг/сут, 27 мл/кг/сут, 28 мл/кг/сут, 29 мл/кг/сут, 30 мл/кг/сут, 31 мл/кг/сут, 32 мл/кг/сут, 33 мл/кг/сут, 34 мл/кг/сут, 35 мл/кг/сут, 36 мл/кг/сут, 37 мл/кг/сут, 38 мл/кг/сут, 39 мл/кг/сут, 40 мл/кг/сут, 41 мл/кг/сут, 42 мл/кг/сут, 43 мл/кг/сут или 44 мл/кг/сут).
В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 1 мл/кг/сут до около 5 мл/кг/сут, от около 6 мл/кг/сут до около 10 мл/кг/сут, от около 11 мл/кг/сут до около 15 мл/кг/сут, от около 16 мл/кг/сут до около 20 мл/кг/сут, от около 21 мл/кг/сут до около 25 мл/кг/сут, от около 26 мл/кг/сут до около 30 мл/кг/сут, от около 31 мл/кг/сут до около 35 мл/кг/сут, от около 36 мл/кг/сут до около 40 мл/кг/сут, от около 41 мл/кг/сут до около 45 мл/кг/сут у мыши. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 35 мл/кг/сут у мыши. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 20 мл/кг/сут у мыши. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 12 мл/кг/сут до около 16 мл/кг/сут у мыши.
В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 1 мл/кг/сут до около 5 мл/кг/сут, от около 6 мл/кг/сут до около 10 мл/кг/сут, от около 11 мл/кг/сут до около 15 мл/кг/сут, от около 16 мл/кг/сут до около 20 мл/кг/сут, от около 21 мл/кг/сут до около 25 мл/кг/сут, от около 26 мл/кг/сут до около 30 мл/кг/сут, от около 31 мл/кг/сут до около 35 мл/кг/сут, от около 36 мл/кг/сут до около 40 мл/кг/сут, от около 41 мл/кг/сут до около 45 мл/кг/сут у яванского макаки. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 20 мл/кг/сут до около 40 мл/кг/сут у яванского макаки. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 25 мл/кг/сут до около 35 мл/кг/сут у яванского макаки. В некоторых случаях антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 30 мл/кг/сут до около 35 мл/кг/сут у яванского макаки.
В дополнительном аспекте антитело против FcRH5 согласно любому из вышеуказанных вариантов осуществления изобретения может включать любые из признаков, по отдельности или в комбинации, как описано ниже в разделах 1-7:
1. Аффинность антитела
В конкретных случаях антитело против FcRH5 связывается с FcRH5 человека и/или яванского макаки с константой диссоциации (KD)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 nM, ≤0,1 нМ, ≤0,01 нМ, или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М, например, от 10-10 до 10-13 М, например, от 10-11 до 10-13 М, например, от 10-12 до 10-13 М). Например, в некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD≤100 нМ (например, ≤90 нМ, ≤80 нМ, ≤70 нМ, ≤60 нМ, ≤50 нМ, ≤40 нМ, ≤30 нМ, ≤20 нМ, ≤10 нМ, ≤5 нМ, ≤1 нМ, ≤750 пМ, ≤500 пМ, ≤250 пМ, ≤100 пМ, ≤50 пМ, ≤25 пМ, ≤10 пМ, ≤5 пМ, или ≤1 пМ) или менее.
В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD от около 1 пМ до около 500 нМ (например, от около 1 пМ до 200 пМ, от 100 пМ до 300 пМ, от 200 пМ до 400 пМ, от 300 пМ до 500 пМ, от 400 пМ до 600 пМ, от 500 пМ до 700 пМ, от 600 пМ до 800 пМ, от 700 пМ до 900 пМ, от 800 пМ до 1 нМ, от 900 пМ до 100 нМ, от 1 нМ до 200 нМ, от 100 нМ до 300 нМ, от 200 нМ до 400 нМ или от 300 нМ до 500 нМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD от около 1 пМ до около 1 нМ (например, от около 1 пМ до 100 пМ, от 50 пМ до 150 пМ, от 100 пМ до 200 пМ, от 150 пМ до 250 пМ, от 200 пМ до 300 пМ, от 250 пМ до 350 пМ, от 300 пМ до 400 пМ, от 350 пМ до 450 пМ, от 400 пМ до 500 пМ, от 450 пМ до 550 пМ, от 500 пМ до 600 пМ, от 550 пМ до 650 пМ, от 600 пМ до 700 пМ, от 650 пМ до 750 пМ, от 700 пМ до 800 пМ, от 750 пМ до 850 пМ, от 800 пМ до 900 пМ, от 850 пМ до 950 пМ или от 900 пМ до 1 нМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD от около 100 пМ до 500 пМ (например, около 100 пМ, 125 пМ, 150 пМ, 175 пМ, 200 пМ, 225 пМ, 250 пМ, 275 пМ, 300 пМ, 325 пМ, 350 пМ, 375 пМ, 400 пМ, 425 пММ, 450 пМ, 475 пМ или 500 пМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD от около 100 пМ до 160 пМ (например, около 100 пМ, 105 пМ, 110 пМ, 115 пМ, 120 пМ, 125 пМ, 130 пМ, 135 пМ, 140 пМ, 145 пМ, 150 пМ, 155 пМ или 160 пМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 человека с KD от около 1 нМ до около 150 нМ (например, около 1 нМ, 2 нМ, 3 нМ, 4 нМ, 5 нМ, 6 нМ, 7 нМ, 8 нМ, 9 нМ, 10 нМ, 11 нМ, 12 нМ, 13 нМ, 14 нМ, 15 нМ, 16 нМ, 17 нМ, 18 нМ, 19 нМ, 20 нМ, 21 нМ, 22 нМ, 23 нМ, 24 нМ, 25 нМ, 26 нМ, 27 нМ, 28 нМ, 29 нМ, 30 нМ, 31 нМ, 32 нМ, 33 нМ, 34 нМ, 35 нМ, 36 нМ, 37 нМ, 38 нМ, 39 нМ, 40 нМ, 41 нМ, 42 нМ, 43 нМ, 44 нМ, 45 нМ, 46 нМ, 47 нМ, 48 нМ, 49 нМ, 50 нМ, 55 нМ, 60 нМ, 65 нМ, 70 нМ, 75 нМ, 80 нМ, 85 нМ, 90 нМ, 95 нМ, 100 нМ, 105 нМ, 110 нМ, 115 нМ, 120 нМ, 125 нМ, 130 нМ, 135 нМ, 140 нМ, 145 нМ или 150 нМ).
В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD ≤100 нМ (например, ≤90 нМ, ≤80 нМ, ≤70 нМ, ≤60 нМ, ≤50 нМ, ≤40 нМ, ≤30 нМ, ≤20 нМ, ≤10 нМ, ≤5 нМ, ≤1 нМ, ≤750 пМ, ≤500 пМ, ≤250 пМ, ≤100 пМ, ≤50 пМ, ≤25 пМ, ≤10 пМ, ≤5 пМ или ≤1 пМ, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М, например, от 10-10 до 10-13 М, например, от 10-11 до 10-13 М, например, от 10-12 до 10-13 М) или менее. В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD от около 1 пМ до около 500 нМ (например, от около 1 пМ до 200 пМ, от 100 пМ до 300 пМ, от 200 пМ до 400 пМ, от 300 пМ до 500 пМ, от 400 пМ до 600 пМ, от 500 пМ до 700 пМ, от 600 пМ до 800 пМ, от 700 пМ до 900 пМ, от 800 пМ до 1 нМ, от 900 пМ до 100 нМ, от 1 нМ до 200 нМ, от 100 нМ до 300 нМ, от 200 нМ до 400 нМ или от 300 нМ до 500 нМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD от около 1 пМ до около 1 нМ (например, от около 1 пМ до 100 пМ, от 50 пМ до 150 пМ, от 100 пМ до 200 пМ, от 150 пМ до 250 пМ, от 200 пМ до 300 пМ, от 250 пМ до 350 пМ, от 300 пМ до 400 пМ, от 350 пМ до 450 пМ, от 400 пМ до 500 пМ, от 450 пМ до 550 пМ, от 500 пМ до 600 пМ, от 550 пМ до 650 пМ, от 600 пМ до 700 пМ, от 650 пМ до 750 пМ, от 700 пМ до 800 пМ, от 750 пМ до 850 пМ, от 800 пМ до 900 пМ, от 850 пМ до 950 пМ или от 900 пМ до 1 нМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD от около 100 пМ до 500 пМ (например, около 100 пМ, 125 пМ, 150 пМ, 175 пМ, 200 пМ, 225 пМ, 250 пМ, 275 пМ, 300 пМ, 325 пМ, 350 пМ, 375 пМ, 400 пМ, 425 пММ, 450 пМ, 475 пМ или 500 пМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD от около 100 пМ до 160 пМ (например, около 100 пМ, 105 пМ, 110 пМ, 115 пМ, 120 пМ, 125 пМ, 130 пМ, 135 пМ, 140 пМ, 145 пМ, 150 пМ, 155 пМ или 160 пМ). В некоторых случаях антитело против FcRH5 связывается с FcRH5 яванского макака с KD от около 1 нМ до около 150 нМ (например, около 1 нМ, 2 нМ, 3 нМ, 4 нМ, 5 нМ, 6 нМ, 7 нМ, 8 нМ, 9 нМ, 10 нМ, 11 нМ, 12 нМ, 13 нМ, 14 нМ, 15 нМ, 16 нМ, 17 нМ, 18 нМ, 19 нМ, 20 нМ, 21 нМ, 22 нМ, 23 нМ, 24 нМ, 25 нМ, 26 нМ, 27 нМ, 28 нМ, 29 нМ, 30 нМ, 31 нМ, 32 нМ, 33 нМ, 34 нМ, 35 нМ, 36 нМ, 37 нМ, 38 нМ, 39 нМ, 40 нМ, 41 нМ, 42 нМ, 43 нМ, 44 нМ, 45 нМ, 46 нМ, 47 нМ, 48 нМ, 49 нМ, 50 нМ, 55 нМ, 60 нМ, 65 нМ, 70 нМ, 75 нМ, 80 нМ, 85 нМ, 90 нМ, 95 нМ, 100 нМ, 105 нМ, 110 нМ, 115 нМ, 120 нМ, 125 нМ, 130 нМ, 135 нМ, 140 нМ, 145 нМ или 150 нМ).
В одном варианте осуществления изобретения KD измеряли анализом связывания антигена, меченного радиоактивным изотопом (РИА). В одном варианте осуществления изобретения РИА выполняли с использованием версии Fab представляющего интерес антитела и его антигена. Например, афинность связывания Fab с антигеном в растворе измеряли путем уравновешивания Fab минимальной концентрацией антигена, меченного (125I)-меткой, в присутствии серий титрований немеченого антигена, и последующего улавливания связанного антигена на планшете, покрытом антителом против Fab (см., например, Chen. et al. J. Mol. Biol. 293:865-881, 1999). Чтобы определить условия анализа, многолуночные планшеты MICROTITER® (Thermo Scientific) покрывали в течение ночи захватывающим антителом против Fab (Cappel Labs) в концентрации 5 мкг/мл в 50 мкМ растворе карбоната натрия (рН 9,6), а затем блокировали 2% (мас./об.) раствором бычьего сывороточного альбумина в PBS в течение от двух до пяти часов при комнатной температуре (около 23°С). В неадсорбентном планшете (Nunc #269620) смешивали 100 пМ или 26 пМ [125I]-антигена с серийными разведениями целевого Fab (например, в соответствии с оценкой антитела против VEGF, Fab-12, в Presta et al.. Cancer Res. 57:4593-4599, 1997). Затем представляющий интерес Fab инкубировали в течение ночи; однако инкубацию можно продолжать в течение более длительного периода (например, около 65 часов), чтобы обеспечить достижение равновесия. Затем смесь переносили на планшет для захвата и инкубировали при комнатной температуре (например, в течение часа). Затем раствор удаляли и планшет промывали восемь раз 0,1% раствором полисорбата 20 (TWEEN-20®) в PBS. Когда планшеты высыхали, добавляли 150 мкл/лунку сцинтиллятора (MICROSCINT-20™; Packard), а планшеты считывали на гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Концентрации каждого Fab, обеспечивавшие связывание, меньшее или равное 20% от максимального, отбирали для использования в конкурентном анализе связывания.
Согласно еще одному варианту осуществления изобретения KD измеряли при помощи метода поверхностного плазменного резонанса с использованием BIACORE®. Например, анализ с использованием BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Пискатауэй, Нью-Джерси) выполняли при 25°С с чипами с иммобилизованным антигеном СМ5 при приблизительно 10 единицах ответа (ЕО). В одном варианте осуществления изобретения биосенсорные чипы на основе карбоксиметилированного декстрана (СМ5, BIACORE, Inc.) активируются N-этил-N'-(3-диметиламинопропил)-карбодиимид гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями изготовителя. Антиген разводят 10 мкМ ацетата натрия, рН 4,8, до 5 мкг/мл (~0,2 мкМ) перед инжекцией при скорости потока 5 мкл/минута для достижения приблизительно 10 единиц ответа (ЕО) сопряженного белка. После введения антигена вводили 1 М раствор этаноламина для блокирования непрореагировавших групп. Для измерения кинетики вводят двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS, содержащий 0,05% полисорбата 20 (TWEEN-20™) в качестве сурфактанта (PBST), при 25°С на скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и диссоциации (koff) рассчитывались как простые однозначные модели связывания Лангмюра (BIACORE® Evaluation Software version 3.2) путем одновременного фитирования сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (KD) рассчитывали как соотношение koff/kon. См., например, Chen et al., Mol. Biol, 293:865-881, 1999. Если скорость ассоциации превышает 106 М-1с-1 согласно данным вышеописанного поверхностного плазменного резонанса, то скорость ассоциации можно определить при помощи метода затухания флуоресценции, при котором измеряют повышение или снижение интенсивности флуоресценции (возбуждение = 295 нм; испускание = 340 нм, 16 нм полоса пропускания) при 25°С для 20 нМ антитела к антигену (Fab-формы) в PBS, рН 7,2, в присутствии увеличивающихся концентраций антигена, измеряемое при помощи спектрометра, такого как спектрофотометр с устройством остановки потока (Aviv Instruments) или спектрофотометр серии 8000 SLM-AMINCO™ (ThermoSpectronic) с перемешивающей кюветой.
2. Фрагменты антител
В определенных вариантах осуществления изобретения антитело, предложенное в данном документе, представляет собой фрагмент антитела. Фрагменты антител включают, без ограничений, фрагменты бис-Fab, Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv и другие фрагменты, описанные ниже. Обзор некоторых фрагментов антител см. в Hudson et al. Nat. Med. 9:129-134, 2003. Для обзора фрагментов scFv см., например,  , in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), стр. 269-315, 1994; см. также WO 93/16185; патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное in vivo время полужизни, см. в патенте США №5869046.
, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), стр. 269-315, 1994; см. также WO 93/16185; патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное in vivo время полужизни, см. в патенте США №5869046.
Фрагменты антитела, в которых два Fab связаны через бисмалеимид, называются в данном документе бисмалеимидо-(тио-Fab)2 или бис-Fab.
Диатела представляют собой фрагменты антител с двумя антигенсвязывающими участками, которые могут быть двухвалентными или биспецифическими. См., например, ЕР 404097; WO 1993/01161; Hudson et al. Med. 9:129-134, 2003; и Hollinger et al. Proc. Natl. Acad. Sci. USA 90:6444-6448, 1993. Триатела и тетратела также описаны в Hudson et al. Nat. Med. 9:129-134, 2003.
Однодоменные антитела представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи антитела или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления изобретения однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Уолтем, Массачусетс; см., например., патент США №6248516 В1).
Фрагменты антител могут быть получены различными способами, включая, без ограничений, протеолитическое расщепление интактного антитела, а также получение при помощи рекомбинантных клеток-хозяев (например, Е. coli или фага), как описано в данном документе.
3. Химерные и гуманизированные антитела
В определенных вариантах осуществления изобретения антитело, предложенное в данном документе, представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в патенте США №4816567 и статье Morrison et al. Natl. Acad. Sci. USA, 81:6851-6855, 1984). В одном примере химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область, полученную от антитела мыши, крысы, хомяка, кролика или отличного от человека примата, например, обезьяны) и константную область человека. В дополнительном примере, химерное антитело представляет собой антитело с "переключенным классом", у которого класс или подкласс был изменен по сравнению с исходным антителом. Термин «химерные антитела» включает антигенсвязывающие фрагменты таких антител.
В некоторых вариантах осуществления изобретения химерное антитело представляет собой гуманизированное антитело. Как правило, антитело нечеловеческого происхождения гуманизируют с целью снижения иммуногенности для человека, сохраняя при этом специфичность и аффинность исходного антитела нечеловеческого происхождения. Как правило, гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR, например, CDR (или их фрагменты) происходят от антитела нечеловеческого происхождения, a FR (или их фрагменты) происходят от последовательностей антитела человека. В некоторых случаях гуманизированное антитело также содержит по меньшей мере фрагмент константной области человека. В некоторых вариантах осуществления изобретения некоторые FR остатки гуманизированного антитела заменены соответствующими остатками из антитела нечеловеческого происхождения (например, антитела, из которого получены остатки HVR), например, для восстановления или улучшения специфичности антитела или аффинности.
Гуманизированные антитела и способы их изготовления рассмотрены, например, в публикации Almagro et al. Front. Biosci. 13:1619-1633, 2008, и дополнительно описаны, например, в Riechmann et al., Nature 332:323-329, 1988; Queen et al., Proc. Natl Acad. Sci. USA 86:10029-10033 (1989); патентах США №5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36:25-34, 2005 (описание прививания определяющей специфичность области (SDR)); Padlan, Mol. Immunol. 28:489-498, 1991 (описание "изменение поверхности");  et al., Methods 36:43-60, 2005 (описание "перетасовки FR"); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260, 2000 (описание подхода "направленной селекции" к перетасовке FR).
et al., Methods 36:43-60, 2005 (описание "перетасовки FR"); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260, 2000 (описание подхода "направленной селекции" к перетасовке FR).
Человеческие каркасные области, которые могут быть использованы для гуманизации, включают, но не ограничиваются этими, каркасные области, выбранные с использованием способа «наилучшего соответствия» (см., например, Sims et al. J. Immunol. 151:2296, 1993); каркасные области, полученные из консенсусной последовательности антител человека конкретной подгруппы вариабельных областей легкой или тяжелой цепи (см., например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285, 1992; и Presta et al. J. Immunol. 151:2623, 1993); зрелые (соматически мутированные) каркасные области человека или каркасные области зародышевой линии человека (см., например, Almagro et al. Front. Biosci. 13:1619-1633, 2008); и каркасные области, полученные в результате скрининга библиотек FR (см., например, Baca et al. J. Biol. Chem. 272:10678-10684, 1997 и Rosok et al., J. Biol. Chem. 271:22611-22618, 1996).
4. Антитела человека
В некоторых вариантах осуществления изобретения антитело, предложенное в данном документе, представляет собой антитело человека. Антитела человека могут быть получены с использованием различных способов, известных в данной области техники. Антитела человека описаны в общих чертах у van Dijk and van de Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) и Lonberg, Curr. Opin. Immunol. 20:450-459, 2008.
Человеческие антитела можно получить путем введения иммуногена трансгенному животному, модифицированному с целью продукции интактных человеческих антител или интактных антител с человеческими вариабельными областями в ответ на антигенную стимуляцию. Такие животные, как правило, содержат все или часть локусов иммуноглобулина человека, которые замещают эндогенные локусы иммуноглобулина или которые находятся во внехромосомном пространстве или случайным образом интегрированы в хромосомы животного. Как правило, у таких трансгенных мышей инактивированы эндогенные локусы иммуноглобулинов. Для обзора способов получения человеческих антител из трансгенных животных, см. Lonberg, Nat. Biotech. 23:1117-1125, 2005. Также см., например, патенты США №№6075181 и 6150584, описывающие технологию XENOMOUSE™; патент США №5770429, описывающий технологию HuMab®; патент США №7041870, описывающий технологию K-М MOUSE®, и публикацию патентной заявки США №US 2007/0061900, описывающую технологию VelociMouse®). Человеческие вариабельные области из интактных антител, образованных такими животными, могут дополнительно модифицироваться, например, путем объединения с разными человеческими консервативными областями.
Антитела человека также можно получать при помощи способов на основе гибридом. Описаны линии клеток миеломы человека и гетеромиеломы мыши-человека для продукции человеческих моноклональных антител (см., например, Kozbor J. Immunol., 133: 3001, 1984; Brodeur et al. Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al. J. Immunol., 147: 86,1991). Антитела человека, полученные с использованием технологии человеческой В-клеточной гибридомы, описаны в Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562, 2006. Дополнительные способы включают способы, описанные, например, в патенте США №7189826 (описывающая продуцирование моноклональных антител IgM человека из гибридомных клеточных линий) и статье Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (описывающая гибридомы человека-человека). Технология с использованием человеческих гибридом (технология Trioma) также описана в Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91, 2005.
Антитела человека также можно получать путем выделения последовательностей вариабельного домена Fv-клона из библиотек фагового дисплея человеческого происхождения. Такие последовательности вариабельного домена можно затем объединять с требуемым константным доменом человека. Методики отбора антител человека из библиотек антител описаны ниже.
5. Антитела, полученные из библиотек
Антитела согласно данному изобретению могут быть выделены путем скрининга комбинаторных библиотек антител с необходимой активностью или свойствами. Например, в данной области техники известно множество способов получения библиотек фагового дисплея и скрининга таких библиотек в отношении антител, обладающих необходимыми связывающими характеристиками. Такие способы рассмотрены, например, в публикации Hoogenboom et al. in Methods in Molecular Biology 178:1-37 ( et al., ed., Human Press, Totowa, NJ, 2001), и дополнительно описаны, например, в статьях McCafferty et al., Nature 348:552-554; Clackson et al, Nature 352: 624-628, 1991; Marks et al. Mol Biol. 222: 581-597 (1992); Marks and Bradbury, in Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310, 2004; Lee et al. J. Mol. Biol. 340(5):1073-1093,2004; Fellouse, Proc. Natl. Acad. Set USA 101(34): 12467-12472, 2004; и Lee et al., J. Immunol. Methods 284(1-2):119-132, 2004.
et al., ed., Human Press, Totowa, NJ, 2001), и дополнительно описаны, например, в статьях McCafferty et al., Nature 348:552-554; Clackson et al, Nature 352: 624-628, 1991; Marks et al. Mol Biol. 222: 581-597 (1992); Marks and Bradbury, in Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310, 2004; Lee et al. J. Mol. Biol. 340(5):1073-1093,2004; Fellouse, Proc. Natl. Acad. Set USA 101(34): 12467-12472, 2004; и Lee et al., J. Immunol. Methods 284(1-2):119-132, 2004.
В некоторых методах фагового дисплея репертуары генов VH и VL отдельно клонируют при помощи полимеразной цепной реакции (ПЦР) и повторно комбинируют случайным образом в фаговых библиотеках, которые затем можно подвергнуть скринингу на антигенсвязывающий фаг, как описано в Winter et al. Ann. Rev. Immunol, 12: 433-455, 1994. Фаг, как правило, представляет фрагменты антител, как одноцепочечные фрагменты Fv (scFv), так и фрагменты Fab. Библиотеки из иммунизированных источников позволяют получить антитела с высокой аффинностью к иммуногену без необходимости конструирования гибридом. В качестве альтернативы, можно клонировать наивный репертуар (например, человека) для получения единого источника антител к широкому спектру чужеродных антигенов и аутоантигенов без иммунизации, как описано в статье Griffiths et al., EMBO J, 12: 725-734, 1993. Наконец, наивные библиотеки можно также получить путем синтеза, клонируя сегменты V-генов стволовых клеток, не подвергавшиеся реаранжировке, и используя ПЦР-праймеры, содержащие случайные последовательности, для кодирования гипервариабельной области CDR3 и осуществления реаранжировки in vitro, как описано в статье Hoogenboom and Winter, J. Mol. Biol, 227: 381-388, 1992. Патентные публикации, в которых описаны фаговые библиотеки антител человека, включают, например: патент США №5750373, и публикации патентов США №№2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936, и 2009/0002360. Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
6. Мультиспецифические антитела, включая FcRH5 Т-зависимые биспецифические (TDB) антитела
В любом из вышеприведенных аспектов предложенное в данном документе антитело против FcRH5 является мультиспецифическим антителом, например, биспецифическим антителом. Мультиспецифические антитела представляют собой моноклональные антитела, которые имеют специфичности связывания в отношении по меньшей мере двух различных сайтов. В некоторых вариантах осуществления изобретения биспецифические антитела могут связываться с разными эпитопами FcRH5.
В некоторых вариантах осуществления изобретения одна из специфичностей связывания направлена на FcRH5, а другая - на любой другой CD3 (например, CD3ε или CD3γ). Такие биспецифические антитела против FcRH5 также упоминаются как FcRH5 Т-зависимые биспецифические (TDB) антитела или TDB против FcRH5. В некоторых случаях второй связывающий домен связывается с эпитопом на CD3, содержащим аминокислотный остаток Glu6 CD3. В некоторых случаях эпитоп дополнительно содержит один или более дополнительных аминокислотных остатков, выбранных из группы, состоящей из Gln1, Asp2 и Met7 CD3. В некоторых случаях эпитоп содержит аминокислотные остатки Gln1, Asp2 и Glu6 CD3. В некоторых случаях эпитоп содержит аминокислотные остатки Glnl, Asp2, Glu6 и Met7 CD3. В некоторых случаях эпитоп не содержит аминокислотный остаток Glu5 CD3. В некоторых случаях эпитоп не содержит аминокислотных остатков Gly3 и Glu5 CD3. В некоторых случаях эпитоп состоит из аминокислотных остатков Gln1, Asp2, Glu6 и Met7 CD3.
В других случаях второй связывающий домен способен связываться с полипептидом CD3 человека или полипептидом CD3 яванского макака. В некоторых случаях полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3ε человека или полипептид CD3ε яванского макака, соответственно. В некоторых случаях полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3γ человека или полипептид CD3γ яванского макака, соответственно.
В конкретных случаях второй связывающий домен связывает полипептид CD3ε человека с константой диссоциации (KD)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например от 10-9 М до 10-13 М). Например, в некоторых случаях второй связывающий домен связывает полипептид CD3ε человека с KD≤100 нМ (например, ≤90 нМ, ≤80 нМ, ≤70 нМ, <60 нМ, ≤50 нМ, ≤40 нМ, ≤30 нМ, ≤20 нМ, ≤10 нМ, ≤5 нМ, ≤1 нМ, ≤750 пМ, ≤500 пМ, ≤250 пМ, ≤100 пМ, ≤50 пМ, ≤25 пМ, ≤10 пМ, ≤5 рМ или ≤1 пМ) или менее.
В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 117; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 120.
В некоторых случаях, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123. В некоторых случаях изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовальности с или последовательность SEQ ID NO: 133, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 134. В некоторых случаях антитело против FcRH5 содержит второй связывающий домен, содержащий по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащий последовательности SEQ ID NO: 125,126, 127 и 128, соответственно. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133. В некоторых случаях второй связывающий домен дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 129, 130, 131 и 132, соответственно. В некоторых случаях второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134. Соответственно, в некоторых случаях вариант полуантитела антитела против FcRH5 согласно изобретению может быть спаривают с вариантом полуантитела антитела против CD3 38E4.v1 с образованием TDB против FcRH5 (то есть антитело против FcRH5/38E4.v1 TDB).
В некоторых случаях, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 122; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123. В некоторых случаях изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовальности с или последовательность SEQ ID NO: 135, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 136. В некоторых случаях антитело против FcRH5 содержит второй связывающий домен, содержащий по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащий последовательности SEQ ID NO: 125,126, 127 и 128, соответственно. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 135. В некоторых случаях второй связывающий домен дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 129, 130, 131 и 132, соответственно. В некоторых случаях второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 136. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 135, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 136. Соответственно, в некоторых случаях вариант полуантитела антитела против FcRH5 согласно изобретению может быть спаривают с вариантом полуантитела антитела против CD3 38E4.v1 с образованием TDB против FcRH5 (то есть антитело против FcRH5/38E4.v1 TDB).
В некоторых случаях, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 124. В некоторых случаях изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовальности с или последовательность SEQ ID NO: 137, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 138. В некоторых случаях антитело против FcRH5 содержит второй связывающий домен, содержащий по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащий последовательности SEQ ID NO: 125, 126, 127 и 128, соответственно. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137. В некоторых случаях второй связывающий домен дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 129, 130, 131 и 132, соответственно. В некоторых случаях второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138. Соответственно, в некоторых случаях вариант полуантитела антитела против FcRH5 согласно изобретению могут быть спарены с вариантом полуантитела антитела против CD3 38E4.v11 с образованием TDB против FcRH5 (то есть антитело против FcRH5/38E4.v11 TDB).
В некоторых случаях, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 139; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 140; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 141; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 142; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 143; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 144. В некоторых случаях изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовальности с или последовательность SEQ ID NO: 153, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 154. В некоторых случаях антитело против FcRH5 содержит второй связывающий домен, содержащий по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащий последовательности SEQ ID NO: 145, 146, 147 и 148, соответственно. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153. В некоторых случаях второй связывающий домен дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 149, 150, 151 и 152, соответственно. В некоторых случаях второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154. Соответственно, в некоторых случаях вариант полуантитела антитела против FcRH5 согласно изобретению могут быть спарены с вариантом полуантитела антитела против CD3 hu40G5c с образованием TDB против FcRH5 (то есть антитело против FcRH5/hu40G5c TDB).
В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 155; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 156; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 157; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 158; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 159; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 160.
В некоторых случаях, изобретение обеспечивает антитело против FcRH5, причем второй связывающий домен содержит по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1 содержащей аминокислотную последовательность SEQ ID NO: 155; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 162; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 157; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 158; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 159; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 160. В некоторых случаях, например, второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 172, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% (например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 173. В некоторых случаях, например, антитело против FcRH5 содержит второй связывающий домен, содержащий по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-Н4, содержащий последовательности SEQ ID NO: 164, 165, 166 и 167, соответственно. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172. В некоторых случаях второй связывающий домен дополнительно содержит по меньшей мере одну (например, 1, 2, 3 или 4) из каркасных областей легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащих последовательности SEQ ID NO: 168, 169, 170 и 171, соответственно. В некоторых случаях второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173. В некоторых случаях второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173. Соответственно, в некоторых случаях вариант полуантитела антитела против FcRH5 согласно изобретению может быть спаривают с вариантом полуантитела антитела против CD3 huUCHT1.v9 с образованием TDB против FcRH5 (то есть антитело против FcRH5/huUCHT1.v9 TDB).
В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, причем связывающий домен, который связывает FcRH5, содержит домен VH (VH1), содержащий заряженную область (CR1) и домен VL (VL1), содержащий заряженную область (CR2), при этом CR2 в VH1 образует зарядовую пару с CR2 в VL1. В некоторых случаях CR1 содержит основной остаток аминокислоты, a CR2 содержит кислотный остаток аминокислоты. В некоторых случаях CR1 содержит мутацию по типу замены Q39K (нумерация ЕС). В некоторых случаях CR1 состоит из мутации по типу замены Q39K. В некоторых случаях CR2 содержит мутацию по типу замены Q38E (нумерация ЕС). В некоторых случаях CR2 состоит из мутации по типу замены Q38E. В некоторых случаях второй связывающий домен, который связывает CD3, содержит домен VH (VH2), содержащий заряженную область (CR3) и домен VL (VL2), содержащий заряженную область (CR4), причем CR4 в VL2 образует зарядовую пару с CR3 в VH2. В некоторых случаях CR4 содержит основной остаток аминокислоты, a CR3 содержит кислотный остаток аминокислоты. В некоторых случаях CR4 содержит мутацию по типу замены Q38K (нумерация ЕС). В некоторых случаях CR4 состоит из мутации по типу замены Q38K. В некоторых случаях CR3 содержит мутацию по типу замены Q39E (нумерация ЕС). В некоторых случаях CR3 состоит из мутации по типу замены Q39E. В некоторых случаях домен VL1 связан с константным доменом (CL1) легкой цепи (CL), и VH1 связан с первым константым доменом (СН11) тяжелой цепи (СН1), причем CL1 содержит заряженную область (CR5) и СН11 содержит заряженную область (CR6) и при этом CR5 в CL1 образует зарядовую пару с CR6 в СН11. В некоторых случаях CR5 содержит основной остаток аминокислоты, и CR6 содержит кислотный остаток. В некоторых случаях CR5 содержит мутацию по типу замены V133K (нумерация ЕС). В некоторых случаях CR5 состоит из мутации по типу замены V133K. В некоторых случаях CR6 содержит мутацию по типу замены S183E (нумерация ЕС). В некоторых случаях CR6 состоит из мутации по типу замены S183E.
В некоторых случаях изобретение обеспечивает антитело против FcRH5, причем домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), при этом CL2 содержит заряженную область (CR7) и СН12 содержит заряженную область (CR8), и при этом CR8 в СН12 образует зарядовую пару с CR7 в CL2. В некоторых случаях CR8 содержит основной остаток аминокислоты, a CR7 содержит кислотный остаток аминокислоты. В некоторых случаях CR8 содержит мутацию по типу замены S183K (нумерация ЕС). В некоторых случаях CR8 состоит из мутации по типу замены S183K. В некоторых случаях CR7 содержит мутацию по типу замены V133E (нумерация ЕС). В некоторых случаях CR7 состоит из мутации по типу замены V133E.
В других случаях, например, изобретение обеспечивает антитело против FcRH5, причем домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), причем CL2 содержит одну или более мутаций в аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация ЕС), и СН12 содержит одну или более мутаций в аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация ЕС). В некоторых вариантах осуществления изобретения CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F, и/или T178V. В некоторых случаях CL2 содержит следующие мутации по типу замены: F116А, L135V, S174A, S176F, и T178V. В некоторых вариантах осуществления изобретения СН12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A, и/или V185A. В некоторых случаях СН12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A, и V185A.
В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, причем связывающий домен, который связывает FcRH5, содержит домен VH (VH1), содержащий заряженную область (CR1) и домен VL (VL1), содержащий заряженную область (CR2), при этом CR2 в VL1 образует зарядовую пару с CR1 в VH1. В некоторых случаях CR2 содержит основной остаток аминокислоты, a CR1 содержит кислотный остаток аминокислоты. В некоторых случаях CR2 содержит мутацию по типу замены Q38K (нумерация ЕС). В некоторых случаях CR2 состоит из мутации по типу замены Q38K. В некоторых случаях CR1 содержит мутацию по типу замены Q39E (нумерация ЕС). В некоторых случаях CR1 состоит из мутации по типу замены Q39E. В некоторых случаях второй связывающий домен, который связывает CD3, содержит домен VH (VH2), содержащий заряженную область (CR3) и домен VL (VL2), содержащий заряженную область (CR4), причем CR3 в VH2 образует зарядовую пару с CR4 в VL2. В некоторых случаях CR3 содержит основной остаток аминокислоты, a CR4 содержит кислотный остаток аминокислоты. В некоторых случаях CR3 содержит мутацию по типу замены Q39K (нумерация ЕС). В некоторых случаях CR3 состоит из мутации по типу замены Q39K. В некоторых случаях CR4 содержит мутацию по типу замены Q38E (нумерация ЕС). В некоторых случаях CR4 состоит из мутации по типу замены Q38E. В некоторых случаях домен VL1 связан с константным доменом (CL1) легкой цепи (CL), и VH1 связан с первым константым доменом (СН11) тяжелой цепи (СН1), причем CL1 содержит заряженную область (CR5) и СН11 содержит заряженную область (CR6) и при этом CR6 в СН11 образует зарядовую пару с CR5 в CL1. В некоторых случаях CR6 содержит основной остаток аминокислоты, a CR5 содержит кислотный остаток аминокислоты. В некоторых случаях CR6 содержит мутацию по типу замены S183K (нумерация ЕС). В некоторых случаях CR6 состоит из мутации по типу замены S183K. В некоторых случаях CR5 содержит мутацию по типу замены V133E (нумерация ЕС). В некоторых случаях CR5 состоит из мутации по типу замены V133E.
В некоторых случаях, например, изобретение обеспечивает антитело против FcRH5, причем домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), при этом CL2 содержит заряженную область (CR7) и СН12 содержит заряженную область (CR8), и при этом CR7 в CL2 образует зарядовую пару с CR8 в СН12. В некоторых случаях CR7 содержит основной остаток аминокислоты, и CR8 содержит кислотный остаток. В некоторых случаях CR7 содержит мутацию по типу замены V133K (нумерация ЕС). В некоторых случаях CR7 состоит из мутации по типу замены V133K. В некоторых случаях CR8 содержит мутацию по типу замены S183E (нумерация ЕС). В некоторых случаях CR8 состоит из мутации по типу замены S183E.
В других случаях, например, изобретение обеспечивает антитело против FcRH5, причем домен VL2 связан с доменом CL (CL2), a VH2 связан с доменом СН1 (СН12), причем CL2 содержит одну или более мутаций в аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация ЕС), и СН12 содержит одну или более мутаций в аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация ЕС). В некоторых вариантах осуществления изобретения CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F, и/или T178V. В некоторых случаях CL2 содержит следующие мутации по типу замены: F116А, L135V, S174A, S176F, и T178V. В некоторых вариантах осуществления изобретения СН12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A, и/или V185A. В некоторых случаях СН12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A, и VI85А. В некоторых случаях антитело против FcRH5 содержит один или более константных доменов тяжелой цепи, причем один или более константных доменов тяжелой цепи выбраны из первого домена СН2 (СН21), первого домена СН3 (СН31), второго домена СН2 (СH22) и второго домена СН3 (СН32). В некоторых случаях по меньшей мере один из одного или более константных доменов тяжелой цепи сопряжен с константным доменом другой тяжелой цепи. В некоторых случаях каждый из СН31 и СН32 содержит выступ (Р1) или впадину (С1), и Р1 или С1 в СН31 могут быть помещены в С1 или Р1, соответственно, в СН32. В некоторых случаях СН31 и СН32 сходятся на поверхности раздела между Р1 и С1. В некоторых случаях каждый из СН21 и СН22 содержит (Р2) или впадину (С2), а Р2 или С2 в CH21 могут быть помещены в С2 или Р2, соответственно, в СН22. В некоторых случаях СН21 и СН22 сходятся на поверхности раздела между Р2 и С2.
В другом аспекте изобретение относится к антителу против против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123; и при этом каждая из области против FcRH5 и плечо антитела против CD3 содержат мутацию по типу замены N297G (нумерация ЕС); и при этом плечо антитела против FcRH5 содержит мутацию по типу замены T366W, а область против CD3 содержит мутации по типу замены T366S, L368A и Y407V. В некоторых случаях антитело против FcRH5 содержит область против FcRH5, содержащую первый связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и при этом плечо антитела против FcRH5 и плечо антитела против CD3, содержащее второй связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и при этом каждый из плеча антитела против FcRH5 и плеча антитела против CD3 содержат мутацию по типу замены N297G (нумерация ЕС); и при этом область против FcRH5 содержит мутацию по типу замены T366W, а плечо антитела против CD3 содержит мутации по типу замены T366S, L368A и Y407V.
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутацию по типу замены Q39E, S183K и N297G (нумерация ЕС). В некоторых случаях антитело против FcRH5 содержит область против FcRH5, содержащую первый связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и при этом область против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G (нумерация ЕС).
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 104 и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 105, причем плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G; и (b) область против против CD3, содержащую второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 133 и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G (нумерация ЕС). В некоторых случаях изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G (нумерация ЕС).
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутацию по типу замены Q39K, S183E и N297G (нумерация ЕС). В некоторых случаях антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 104 и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G (нумерация ЕС). В некоторых случаях изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 104, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G (нумерация ЕС).
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) FTVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС). В некоторых случаях антитело против FcRH5 содержит область против FcRH5, содержащую первый связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183E и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте данное изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 104 и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 105, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, A141I, F170S, S181M, S183A, V185А и N297G (нумерация ЕС). В некоторых случаях изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38E и V133K, и тяжелую цепь, содержащую мутации по типу замены Q39K, S183Е и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38K, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39E, А141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащую второй связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС). В некоторых случаях антитело против FcRH5 содержит область против FcRH5, содержащую первый связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и плечо антитела против CD3, содержащее второй связывающий домен, содержащий шесть гипервариабельных областей (HVR): (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
В другом аспекте изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 104 и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) идентичность последовательности с или последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G; и (b) плечо антитела против CD3, содержащую второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, АНН, F170S, S181M, S183A, V185A и N297G (нумерация ЕС). В некоторых случаях изобретение относится к антителу против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит: (а) плечо антитела против FcRH5, содержащее первый связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105, при этом плечо антитела против FcRH5 содержит легкую цепь, содержащую мутации по типу замены Q38K и V133E, и тяжелую цепь, содержащую мутации по типу замены Q39E, S183K и N297G, и (b) плечо антитела против CD3, содержащее второй связывающий домен, содержащий домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134, при этом плечо антитела против CD3 содержит легкую цепь, содержащую мутации по типу замены Q38E, F116A, L135V, S174A, S176F и T178V, и тяжелую цепь, содержащую мутации по типу замены Q39K, A141I, F170S, S181M, S183A, V185A и N297G (нумерация ЕС).
7. Варианты антител
В некоторых вариантах осуществления изобретения варианты аминокислотной последовательности антитела против FcRH5 согласно изобретению (например, биспецифические антитела против FcRH5 согласно изобретению, которые связываются с FcRH5 и второй биологической молекулой, например CD3, такие как антитела TDB против FcRH5 согласно изобретению или их варианты). Например, может существовать потребность в улучшении аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотных последовательностей антитела можно получить путем внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно проводить любые комбинации делеций, вставок и замен, при условии, что конечная конструкция обладает необходимыми характеристиками, например, антигенсвязывающими.
a. Варианты замены, вставки и делеции
В некоторых вариантах осуществления изобретения предложены варианты антитела, содержащие одну или более аминокислотных замен. Сайты, представляющие интерес для мутагенеза с заменами, включают HVR и FR. Консервативные замены показаны в табл. 1 под заголовком «предпочтительные замены». Более существенные изменения представлены в табл. 1 под заголовком «типовые замены», и как дополнительно описано ниже со ссылкой на классы аминокислотных боковых цепей. Аминокислотные замены можно вносить в представляющее интерес антитело, и проводить скрининг полученных продуктов в отношении необходимой активности, например, сохранения/улучшения связывания антигена, снижения иммуногенности или улучшения АЗКЦ или КЗЦ.
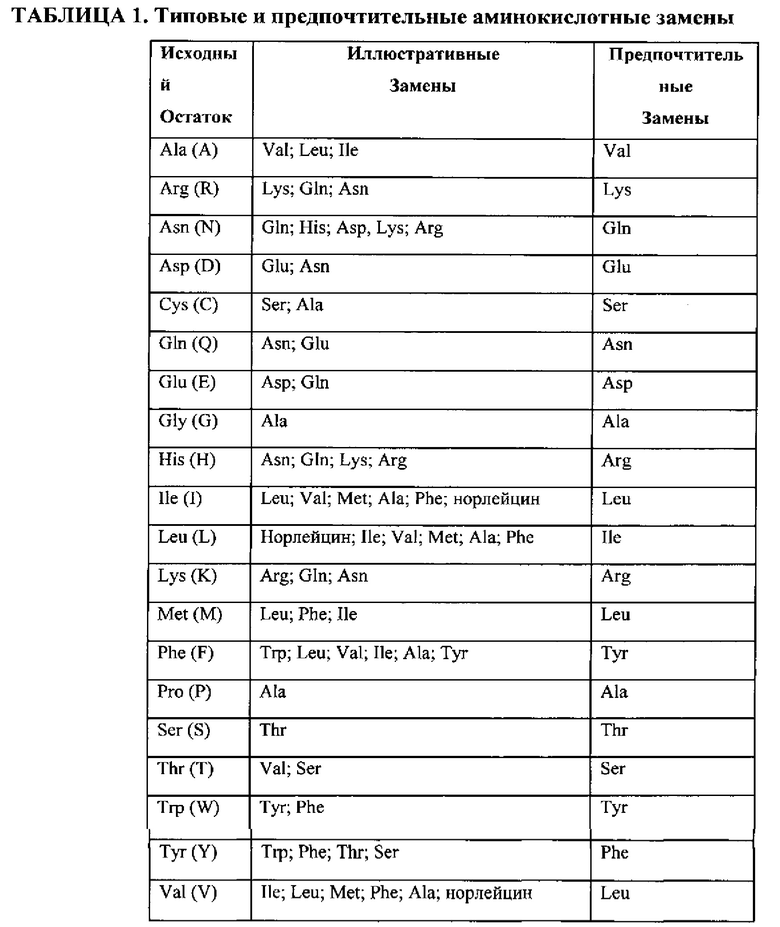
Аминокислоты можно поделить на группы в соответствии с общими свойствами боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены приводят к замене представителя одного из этих классов на представителя другого класса.
Один из вариантов замены включает замену одного или более аминокислотных остатков гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный вариант(ы), выбранный для дальнейшего изучения, будет иметь модификации (например, улучшения) определенных биологических свойств (например, повышенную аффинность, пониженную иммуногенность) по сравнению с родительским антителом, и/или будет иметь по существу сохраненные определенные биологические свойства исходного антитела. Типовой заменяемый вариант представляет собой антитело со зрелой аффинностью, который может быть беспрепятственно получен, например, с использованием методик созревания аффинности на основе фагового дисплея, таких как описанные в данном документе. Вкратце один или более аминокислотных остатков HVR подвергают мутации и вариантные антитела представляют на поверхности фага и проводят скрининг в отношении конкретного вида биологической активности (например, аффинности связывания).
Модификации (например, замены) в HVR можно осуществить, например, с целью улучшения аффинности антитела. Такие изменения могут быть произведены в «горячих точках» HVR, т.е. остатках, кодируемых кодонами, которые подвергаются мутации с высокой частотой во время процесса соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207:179-196, 2008) и/или остатках, которые контактируют с антигеном, с тестированием полученного варианта VH или VL на аффинность связывания. Созревание аффинности путем конструирования и повторного отбора из вторичных библиотек было описано, например, в Methods in Molecular Biology 178:1-37 ( et al., ed., Human Press, Totowa, NJ, (2001).) В некоторых вариантах осуществления изобретения с созреванием аффинности многообразие вводится в вариабельные гены, выбранные для созревания любым из множества способов (например, ПЦР с внесением ошибок, перетасовка цепей или олигонуклеотидспецифический мутагенез). Затем создают вторичную библиотеку. Затем проводят скрининг библиотеки для идентификации любых вариантов антител с требуемой аффинностью. Другой способ для внесения разнообразия включает подходы, направленные на HVR, в которых несколько аминокислотных остатков HVR (4-6 аминокислотных остатков подряд) являются рандомизированными. Остатки HVR, вовлеченные в связывание антигена, могут быть конкретно идентифицированы, например, с использованием сканирующего аланином мутагенеза или моделирования. Мишенями в частности являются CDR-H3 и CDR-L3.
В определенных вариантах осуществления изобретения замены, вставки или делеции могут находиться в одной или более HVR до той меры, пока такие изменения существенно не снижают способность антитела связывать антиген. Например, в HVR можно проводить консервативные замены (например, консервативные замены, предложенные в данном документе), которые не существенно снижают аффинность связывания. Такие изменения могут быть, например, за пределами остатков, контактирующих с антигеном, в HVR. В некоторых вариантах осуществления изобретения в вариантах последовательностей VH и VL, приведенных ранее, каждый HVR или не изменен, или содержит не более одной, двух или трех аминокислотных замен.
Удобный способ выявления остатков или областей антитела, которые могут быть мишенями для мутагенеза, называется «аланин-сканирующим мутагенезом», описанным Cunningham and Wells (1989) Science, 244:1081-1085. В этом способе аминокислотный остаток или группу целевых аминокислотных остатков (например, заряженные аминокислотные остатки, такие как Arg, Asp, His, Lys и Glu) идентифицируют и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) для определения влияния на взаимодействие антитела с антигеном. Дополнительные замены могут быть внесены в те расположения аминокислот, которые проявляли функциональную чувствительность к исходным заменам. В альтернативном или дополнительном варианте используют кристаллическую структуру комплекса антиген-антитело для идентификации точек контакта между антитело и антигеном. Такие контактные остатки и соседние остатки можно использовать в качестве мишеней или удалять из кандидатов для замены. Модификации могут быть скринированы для определения наличия в них желаемых свойств.
Аминокислотные вставки включают амино- и/или карбокси-концевые слияния с диапазоном длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или более аминокислотных остатков внутри последовательности. Примеры концевых вставок включают антитело с N-концевым остатком метионила. Другие инсерционные варианты молекулы антитела включают слияние с N- или С-концом антитела с ферментом (например, для терапии ADEPT) или полипептидом, который увеличивает период полужизни антитела из сыворотки.
b. Гликозилированные варианты
В некоторых вариантах осуществления изобретения антитело против FcRH5 согласно изобретению изменяли с целью увеличения или уменьшения степени гликозилирования антитела. Добавление или удаление участков гликозилирования в антитело против FcRH5 согласно изобретению традиционно можно осуществлять путем изменения аминокислотной последовательности так, чтобы создать или удалить один или более участков гликозилирования. Добавление или удаление сайта гликозилирования может изменять эффекторную функцию антитела, такого как антитело против FcRH5 (например, TDB против FcRH5). В некоторых вариантах осуществления изобретения антитело против FcRH5 (например, TDB против FcRH5) может содержать мутацию участка агликозилирования. В некоторых вариантах осуществления изобретения мутация сайта агликозилирования является мутацией по типу замены. В некоторых вариантах осуществления изобретения мутация участка агликозилирования снижает эффекторную функцию антитела против FcRH5 по меньшей мере на 1% или более (например, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%, или более). В некоторых вариантах осуществления изобретения мутация замещения приходится на аминокислотный остаток N297, L234, L235, D265, и/или Р329 (нумерация ЕС). В некоторых вариантах осуществления изобретения мутация по типу замены выбрана из группы, состоящей из N297G, N297A, L234A, L235A, D265A, и P329G. В некоторых вариантах осуществления изобретения мутация по типу замены представляет собой мутацию N297G.
Если антитело содержит область Fc, можно изменять присоединенный к ней углевод. Нативные антитела, вырабатываемые клетками млекопитающих, как правило, содержат разветвленный, биантеннальный олигосахарид, который в общем случае присоединен посредством N-связи к Asn297 домена СН2 области Fc. См., например, Wright et al. TIBTECH 15:26-32 (1997. Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стволе» биантеннальной олигосахаридной структуры. В некоторых вариантах осуществления изобретения можно выполнить модификации олигосахарида в антителе согласно данному изобретению с целью создания вариантов антител с определенными улучшенными свойствами.
В одном варианте осуществления изобретения предложены варианты антител против FcRH5, содержащие углеводную структуру с недостаточным количеством фукозы, присоединенной (непосредственно или косвенно) к области Fc. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяется путем подсчитывания среднего количества фукозы в сахарной цепи в Asn 297 по отношению к сумме всех гликоструктур, прикрепленных к Asn297 (например, сложные, гибридные и структуры с высоким содержанием маннозы), как измерено с помощью масс-спектрометрии MALDI-TOF, описанной, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному в положении 297 в области Fc (согласно нумерации ЕС остатков области Fc); при этом Asn297 также может быть расположен на около ±3 аминокислот выше или ниже положения 297, т.е. между положениями 294 и 300, вследствие незначительных вариаций последовательностей в антителах. Такие фукозилированные варианты могут обладать улучшенной функцией АЗКЦ. См., например, публикации патентов США №№. US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, связанных с «дефукозилированными» или «фукозодефицитными» вариантами антитела включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki et al. J. Mol. Biol. 336:1239-1249, 2004; Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614, 2004. Примеры клеточных линий, которые способны продуцировать дефукозилированные антитела, включают клетки СНО Lec13, дефектные по фукозилированию белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545,1986; заявка на патент США № US 2003/0157108 A1, Presta, L, и WO 2004/056312 A1, Adams et al., особенно в примере 11), и нокаутные линии клеток, такие как нокаутные клетки СНО по гену альфа-1,6-фукозилтрансферазы, FUT8, (см., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87:614, 2004; Kanda, Y. et al. Biotechnol. Bioeng., 94 (4):680-688, 2006; и WO 2003/085107).
Дополнительно предложены антитела против FcRH5 с разделенными пополам олигосахаридами, например, в которых биантеннальный олигосахарид, присоединенный к области Fc антитела, разделен GlcNAc. Такие варианты антител могут обладать сниженным уровнем фукозилирования и/или улучшенной функцией АЗКЦ. Примеры таких модификаций антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США №6602684 (Umana et al.,); и патенте США №2005/0123546 (Umana et al). Кроме того, предлагаются варианты антител, содержащие по меньшей мере один остаток галактозы в составе олигосахарида, присоединенного к области Fc. Такие варианты антител могут обладать улучшенной функцией КЗЦ. Такие модификации антител описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
c. Варианты доменов VH, VL, CH1 и CL
В некоторых вариантах осуществления изобретения одна или более модификаций аминокислот могут быть введены в вариабельный домен тяжелой цепи (VH, например, VH1 и/или VH2), вариабельный домен легкой цепи (VL, например, VL1 и/или VL2), константный домен тяжелой цепи (СН1, например, СН11 и/или СН12), и/или константный домен легкой цепи (CL, например CL1 и/или CL2) антитела против FcRH5 согласно изобретению (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например, антителом TDB против FcRH5 согласно изобретению или его вариантом). Домены VH, VL, СН1 и/или CL могут иметь аминокислотные модификации (например, замены) в одной или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) положений аминокислот. В конкретных вариантах осуществления изобретения аминокислотные замены включают кислотные и/или основные аминокислотные остатки. Аминокислотные модификации могут образовывать заряженные области в доменах VH, VL, СН1 и/или CL (например, области, содержащие кислотные и/или основные аминокислотные остатки). Заряженные области в доменах VH, VL, СН1 и/или CL могут взаимодействовать со вторыми заряженными областями противоположного общего заряда с образованием зарядовой пары. Например, аминокислотные модификации могут опосредовать формирование зарядовой пары между заряженными областями, присутствующими в доменах VL1 и VH1 плеча FcRH5 TDB. В некоторых случаях аминокислотные модификации могут опосредовать формирование зарядовой пары между заряженными областями, присутствующими в доменах CL1 и СН11 плеча FcRH5 TDB. В некоторых случаях аминокислотные модификации могут опосредовать образование зарядовой пары между заряженными областями, присутствующими в доменах VL2 и VH2 второго плеча TDB (например, плечо CD3 TDB). В некоторых случаях аминокислотные модификации могут опосредовать образование зарядовой пары между заряженными областями, присутствующими в доменах CL2 и СН12 второго плеча TDB (например, область CD3 TDB). Примеры конфигураций FcRH5 TDB, содержащих варианты VH, VL, СН1 и/или CL, представлены на фиг. 1A-1D.
В некоторых вариантах осуществления изобрения антитело против FcRH5 (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например, CD3, такой как антитело против FcRH5 TDB согласно изобретению), содержит одну или более асимметричных модификаций в областях VH, VL, СН1 и/или CL для облегчения корректного спаривания тяжелой и легкой цепей. В других вариантах осуществления изобретения антитело против FcRH5 дополнительно содержит одну или более модификаций в области Fc для облегчения гетеродимеризации двух плеч (например, плечо антитела против FcRH5 и плечо антитела против CD3) антитела против FcRH5 (например, FcRH5 TDB).
В некоторых вариантах осуществления изобретения домен СН11 содержит аминокислотную замену в S183 (нумерация ЕС), a CL1 домен содержит аминокислотную замену в V133 (нумерация ЕС). В других вариантах осуществления изобретения легкая цепь первого плеча (например, FcRH5-связывающее плечо) антитела против FcRH5 представляет собой цепь каппа. В некоторых вариантах осуществления изобретения легкая цепь второго плеча (например, связывающее плечо антитела против CD3) антитела против FcPvH5 представляет собой цепь каппа. В некоторых вариантах осуществления изобретения легкие цепи в обоих областях антитела против FcRH5 (например, FcRH5-связывающее плечо и CD3-связывающее плечо) представляют собой цепи каппа.
В некоторых вариантах осуществления изобретения домен CH11 содержит мутацию S183E, и домен CL1 содержит мутацию V133K. В некоторых вариантах осуществления изобретения домен СН11 содержит мутацию S183K, и домен CL1 содержит мутацию V133Е.
В некоторых вариантах осуществления изобретения домен СН11 содержит мутацию S183K, домен CL1 содержит мутацию V133E, домен СН12 содержит мутацию S183E, и домен CL2 содержит мутацию V133K.
В некоторых вариантах осуществления изобретения антитело против FcRH5 дополнительно содержит мутацию Q39E в домене VH1, мутацию Q38K в домене VL1, мутацию Q39K в домене VH2 и мутацию Q38E в домене VL2.
В некоторых вариантах осуществления изобретения антитело против FcRH5 дополнительно содержит мутацию выступа (например, выпячивание) в домене СН31 и мутацию впадины (например, полость) в домене СН32. В некоторых вариантах осуществления изобретения мутация выступа содержит мутацию T366W (нумерация ЕС). В некоторых вариантах осуществления изобретения мутация впадины содержит по меньшей мере одну, по меньшей мере две или все три мутации T366S, L368A и Y407V (нумерация ЕС). В некоторых вариантах осуществления изобретения антитело против FcRH5 дополнительно содержит мутацию T366W в первой тяжелой цепи и мутации T366S, L368A и Y407V во второй тяжелой цепи.
В некоторых вариантах осуществления изобретения домен CH11 содержит мутации A141I, F170S, S181M, S183A и V185A, а домен CL1 содержит мутации F116A, L135V, S174A, S176F и T178V. В некоторых других вариантах осуществления изобретения домен СН11 содержит мутации A141I, F170S, S181M, S183A и V185A; домен CL1 содержит мутации F116A, L135V, S174A, S176F и T178V; домен СН12 содержит мутацию S183E; и домен CL2 содержит мутацию V133K.
d. Варианты области Fc
В некоторых вариантах осуществления изобретения одна или более аминокислотных модификаций могут быть введены в область Fc антитела против FcRH5 согласно изобретению (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например, CD3, такой как антитело TDB согласно изобретению или его вариантом), получая, таким образом, вариант области Fc (см., например, патент США №2012/0251531, полное содержание которой включено в настоящий документ посредством ссылки). В других вариантах осуществления изобретения антитело против FcRH5 содержит одну или более модификаций в области Fc для облегчения гетеродимеризации двух плеч (например, плечо антитела против FcRH5 и плечо антитела против CD3) антитела против FcRH5 (например, FcRH5 TDB). В некоторых вариантах осуществления изобретения антитело против FcRH5 (например, FcRH5 TDB), имеющее одну или более модификаций Fc, может также иметь одну или более модификаций в доменах VH, VL, СН1 и/или CL, как описано выше. В некоторых вариантах осуществления изобретения антитело против FcRH5 (например, TDB против FcRH5), содержащее модификации Fc, VH, VL, CH1 и/или CL, могут быть получено с помощью метода получения одной клетки антитела, как описано в данном документе. Вариант области Fc может содержать последовательность человеческой области Fc (например, человеческой области Fc IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одной или более аминокислотных положениях.
В некоторых вариантах осуществления в данном изобретении предложен вариант антитела против FcRH5, который обладает некоторыми, но не всеми эффекторными функциями, что делает ее подходящим кандидатом для практических применений, в которых важное значение имеет период полужизни антитела in vivo, хотя отдельные эффекторные функции (такие как комплемент и АЗКЦ) не являются необходимыми или являются вредными. Цитотоксические анализы могут быть проведены in vitro и/или in vivo для подтверждения восстановления/ослабления активности КЗЦ и/или АЗКЦ. Например, анализы связывания с Fc-рецептором (FcR) могут быть проведены с целью подтверждения того, что антитело не может связываться с FcγR (следовательно, вероятно, из-за недостаточности активности АЗКЦ), но при этом сохраняет способность связываться с FcRn. Первичные клетки для опосредования АЗКЦ экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Сводная информация об экспрессии FcR на кроветворных клетках приведена в таблице 3 на странице 464 статьи Ravetch et al. Annu. Rev. Immunol. 9:457-492, 1991. Неограничивающие примеры анализов in vitro для оценки активности АЗКЦ представляющей интерес молекулы, описаны в патенте США №5500362 (см., например, Hellstrom et al. Proc. Natl Acad. Sci. USA 83:7059-7063, 1986) и Hellstrom et al. Proc. Natl Acad. Sci. USA 82:1499-1502, 1985; 5821337 (см. Bruggemann, et al. J. Exp Med. 166:1351-1361, 1987). В альтернативном варианте могут быть использованы нерадиоактивные методы анализа (см., например, ACTI™ нерадиоактивный анализ цитотоксичности для проточной цитометрии (CellTechnology, Inc. Маунтин-Вью, Калифорния; и CytoTox 96® нерадиоактивный анализ цитотоксичности (Promega, Мадисон, Висконсин). Эффекторные клетки, пригодные для такого анализа, включают мононуклеары периферической крови (МКПК) и натуральные клетки-киллеры (NK). В альтернативном варианте, или дополнительно, активность АЗКЦ представляющей интерес молекулы может оцениваться in vivo, например, на животной модели, такой как описано в Clynes et al. Natl Acad. Sci. USA 95:652-656, 1998. Также можно проводить анализ связывания C1q, чтобы подтвердить, что антитело неспособно связывать C1q и, следовательно, у него отсутствует активность КЗЦ. См., например, ннализ связывания C1q и С3с методом ИФА в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента, может быть выполнен анализ КЗЦ (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996); Cragg, M.S. et al., 101:1045-1052, 2003; and Cragg, M.S. and M.J. Glennie Blood. 103:2738-2743, 2004). Определение связывания с FcRn и in vivo клиренса/времени полужизни также может быть выполнено с использованием способов, известных в данной области техники (см., например, Petkova, S.B. et al. Intl. Immunol. 18(12):1759-1769, 2006).
Антитела со сниженной эффекторной функцией включают те, которые содержат замены одного или более остатков области Fc 238, 265, 269, 270, 297, 327 и 329 (патенты США №№6737056 и 8219149). Такие мутанты Fc включают мутантов Fc с заменами в двух или более аминокислотных положениях 265, 269, 270, 297 и 327, в том числе так называемые мутанты Fc "DANA" с заменой остатков 265 и 297 на аланин (патенты США №№7332581 и 8219149).
В определенных вариантах осуществления изобретения пролин в положении 329 человеческой области Fc дикого типа в антителе заменен глицином или аргинином или аминокислотным остатком, достаточно крупным, чтобы разрушить «сэндвич» пролина в области раздела Fc/Fc-гамма-рецептор, который образуется между пролином 329 из Fc и остатками триптофана Trp87 и Trp110 из FcgRIII (Sondermann et al. Nature. 406:267-273, 2000). В определенных вариантах осуществления изобретения антитело содержит по меньшей мере одну дополнительную аминокислотную замену. В одном варианте осуществления изобретения дополнительная аминокислотная замена представляет собой S228P, Е233Р, L234A, L235A, L235E, N297A, N297D или P331S, а в другом варианте осуществления изобретения по меньшей мере одна дополнительная аминокислотная замена представляет собой L234A и L235A области Fc человеческого IgG1 или S228P и L235E области Fc человеческого IgG4 (см., например, US 2012/0251531), и в другом варианте осуществления изобретения по меньшей мере одна дополнительная аминокислотная замена представляет собой L234A и L235A, и P329G области Fc человеческого IgG1.
Описаны некоторые модификации антитела с улучшенным или уменьшенным связыванием с FcR (см., например, патент США №6737056; WO 2004/056312, и Shields et al. J. Biol. Chem. 9(2):6591-6604, 2001).
В некоторых вариантах осуществления изобретения вариант антитела содержит область Fc с одной или более аминокислотными заменами, которые улучшают АЗКЦ, например, с заменами в положениях 298, 333 и/или 334 области Fc (нумерация остатков по системе ЕС).
В некоторых вариантах осуществления изобретения изменения сделаны в области Fc, что привело к изменению (то есть или к улучшению, или к ухудшению) связывания C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в патенте США №6194551, международной патентной заявке WO 99/51642 и публикации Idusogie et al. J. Immunol. 164: 4178-4184, 2000.
Антитела с увеличенным временем полужизни и улучшенным связыванием с неонатальным рецептором Fc (FcRn), который отвечает за перенос материнских IgG к плоду (Guyer et al., J. Immunol. 117:587, 1976; Kim et al. J. Immunol. 24:249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Данные антитела содержат область Fc с одной или более заменами в ней, которые улучшают связывание области Fc с FcRn. Такие варианты Fc включают варианты с заменами одного или более остатков области Fc: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену аминокислотного остатка 434 в области Fc (патент США №7371826).
См. также публикацию Duncan et al. Nature 322:738-40, 1988; патент США №5648260; патент США №5624821; и Международную патентную заявку WO 94/29351 в отношении других примеров вариантов области Fc.
В других вариантах осуществления изобретения антитело против FcRH5 содержит мутацию сайта агликозилирования. Мутация сайта агликозилирования может быть мутацией по типу замены. Мутация сайта агликозилирования может снижать эффекторную функцию антитела против FcRH5 по сравнению с немутированной версией по меньшей мере на 1% или более (например, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%). В некоторых случаях мутация по типу замены приходится на аминокислотный остаток N297, L234, L235, D265, и/или Р329 (нумерация ЕС). Дополнительно, мутация по типу замены может быть выбрана из группы, состоящей из N297G, N297A, L234A, L235A, D265A, и P329G. В конкретных случаях мутация по типу замены представляет собой мутацию N297G.
e. Варианты выступов и впадин
Могут также быть разработаны варианты выступов и впадин антител, в которых модификация аминокислоты (например, замена) одного или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) аминокислотных остатков создают выпячивание (выступ) или полость (впадину). В некоторых случаях выпячивание, расположенное на одном полипептиде антитела, можно разместить в полости, расположенной на втором полипептиде антитела, так что два составных полипептида антитела могут встречаться на границе раздела между выступом и полостью. Выступ может быть образован путем замены одного или более аминокислотных остатков одним или более аминокислотными остатками большего размера. Полость может быть образована путем замемы одного или более аминокислотных остатков одним или более аминокислотными остатками меньшего размера. Выпячивание или полость могут быть введены в любое из плеч антитела против FcRH5 (например, TDB против FcRH5 (например, или в плечо антитела против FcRH5, или в плечо антитела против CD3)). Модификации выступов и впадин (например, домена Fc) особенно полезны для увеличения общего выхода, гомогенности и стабильности биспецифических антител (например, TDB). В некоторых случаях (например, когда пара выпячивание-полость расположена на границе раздела LC/HC (например, VH и VL или CL и СН1)), модификации аминокислот, которые вводят модификации выпячиваний и полостей, могут уменьшать замещение легкой цепи. В некоторых вариантах осуществления изобретения выпячивания формируются путем введения мутации T366W в одно плечо антитела против FcRH5 (например, или плечо антитела против FcRH5, или плечо антитела против CD3). В некоторых вариантах осуществления изобретения мутация T366W находится в плече антитела против FcRH5. В некоторых вариантах осуществления изобретения мутация T366W находится в плече антитела против CD3. В некоторых вариантах осуществления изобретения впадины формируются путем введения мутации T366W, L368A, и/или Y407V в одно плечо антитела против FcRH5 (например, или плечо антитела против FcRH5, или плечо антитела против CD3). В некоторых вариантах осуществления изобретения мутация T366W, L368A и/или Y407V находится в плече антитела против FcRH5. В некоторых вариантах осуществления изобретения мутация T366W, L368A и/или Y407V находится в плече антитела против CD3.
Методики для получения мультиспецифических антител включают, но не ограничиваются рекомбинантной коэкспрессией двух пар тяжелой цепи-легкой цепи иммуноглобулинов, имеющих разную специфичность (см. Milstein and Cuello, Nature 305: 537 (1983)), WO 93/08829, и Traunecker et al., EMBO J. 10: 3655 (1991)), и метод конструирования «выступ-во-впадину» (см., например, патент США №5731168) Метод конструирования мультиспецифических антител «выступ-во-впадину» можно применять для получения первого плеча, содержащего выступ, и второго плеча, содержащего впадину, в которой возможно связывание выступа первого плеча. Например, антитела TDB согласно изобретению могут содержать выступ, расположенный на плече антитела против CD3, и впадину, расположенную на его плече, нацеленном на опухоль. С другой стороны, антитела TDB согласно изобретению могут содержать выступ, расположенный на его плече, нацеленном на опухоль, и впадину, расположенную на плече антитела против CD3. Мультиспецифические антитела также можно конструировать, используя метод кроссинговера иммуноглобулина (также известный как обмен Fab-доменов или формат CrossMab) (см., например, WO 2009/080253; Schaefer et al., Proc. Natl. Acad. Sci. USA, 108:11187-11192 (2011)). Мультиспецифические антитела также могут быть получены путем конструирования с использованием эффекта электростатического взаимодействия для получения молекул Fc-гетеродимерных антител (WO 2009/089004А1); путем перекрестного связывания двух или более антител или фрагментов (см., например, патент США №4676980, и Brennan et al., Science, 229: 81 (1985)); с использованием лейциновых застежек для создания биспецифических антител (см., например, Kostelny et al., J. Immunol., 148 (5): 1547-1553 (1992)); с использованием технологии «диател» для получения фрагментов биспецифических антител (см., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)); и с использованием одноцепочечных Fv (sFv) димеров (см., например, Gruber et al., J. Immunol, 152:5368 (1994)); и получения триспецифических антител, как описано, например, в Tutt et al. J. Immunol. 147: 60 (1991).
f. Варианты антител, сконструированные с использованием цистеина
В некоторых вариантах осуществения изобретения может быть желательно создать антитела, сконструированные с использованием цистеина, например, «thioMAb,» в которых один или более остатков замещены остатками цистеина. В конкретных вариантах осуществления изобретения замещенные остатки расположены в доступных участках антитела. Таким образом, в результате замены таких остатков на цистеин, реактивные тиольные группы располагаются в доступных участках антитела и могут быть использованы для конъюгации антитела с другими компонентами, такими как лекарственные компоненты или линкер-лекарственные компоненты, для создания иммуноконъюгата, как дополнительно описано в данном документе. В некоторых вариантах осуществления изобретения цистеином можно заменить один или более из следующих остатков: V205 (система нумерации по Kabat) в легкой цепи, А118 (система нумерации ЕС) в тяжелой цепи и S400 (система нумерации ЕС) в области Fc тяжелой цепи. Цистеин-сконструированные антитела можно получать так, как описано, например, в патенте США №7521541, который включен в данный документ посредством ссылки в полном объеме.
g. Другие производные антител
В некоторых вариантах осуществления изобретения антитело против FcRH5 согласно изобретению (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например, CD3, такой как антитело TDB согласно изобретению или его вариантом), предложенной в данном документе, может быть дополнительно модифицировано, чтобы содержать дополнительные небелковые компоненты, которые известны в данной области техники и легко доступны. Фрагменты, пригодные для получения производных антитела, включают, без ограничений, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, без ограничений, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1, 3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо статистические сополимеры), декстран или поли(n-винилпирролидон) полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры оксида/этиленоксида, полиоксиэтилированные полнолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущество при производстве вследствие его стабильности в воде. Указанный полимер может обладать любой молекулярной массой и быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может изменяться и, в случае присоединения более одного полимера, они могут представлять собой одинаковые или разные молекулы. В целом, количество и/или тип полимеров, используемых для дериватизации, можно определить с учетом факторов, включающих, без ограничений, конкретные свойства или функции антитела, подлежащие улучшению, независимо от того, будет ли производное антитела использоваться в терапии при определенных условиях и т.д.
В другом варианте осуществления изобретения предложены конъюгаты антитела и небелкового компонента, которые можно избирательно нагревать путем облучения. В одном варианте осуществления изобретения небелковый компонент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102:11600-11605, 2005). Излучение может иметь любую длину волны и включает, без ограничений, длины волн, которые не наносят вред обычным клеткам, но которые нагревают небелковый компонент до температуры, при которой клетки, расположенные близко к антитело-небелковому компоненту, погибают.
В. Рекомбинантные методы и композиции
Антитела против FcRH5 согласно изобретению (например, биспецифические антитела против FcRH5 согласно изобретению, которые связываются с FcRH5 и второй биологической молекулой, например, CD3, такие как антитела TDB согласно изобретению или их варианты) могут быть получены с использованием рекомбинантных методов и композиции, например, как описано в патенте США №4816567. В одном варианте осуществления изобретения предложена выделенная нуклеиновая кислота, кодирующая антитело против FcRH5, описанное в данном документе. В одном варианте осуществления изобретения клетка-хозяин представляет собой эукариотическую клетку, например, клетку яичника китайского хомячка (СНО). В одном варианте осуществления изобретения клетка-хозяин является прокариотической, например, клеткой Е. coli. В одном варианте осуществления изобретения предложен способ получения антитела против FcRH5, причем способ включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующей антитело, как указано выше, в условиях, подходящих для экспрессии антитела, и, в некоторых случаях, выделение антитела из клетки-хозяина (или из среды для культивирования клетки-хозяина). В некоторых вариантах осуществления изобретения способ дополнительно включает культивирование второй клетки-хозяина, содержащей вторую нуклеиновую кислоту, кодирующую антитело против CD3, которое содержит связывающий домен, который связывает CD3. В некоторых вариантах осуществления изобретения клетки-хозяева совместно культивируют. Еще один вариант осуществления изобретения включает выделение биспецифического антитела против FcRH5 из клетки-хозяина или культуральной среды. В некоторых вариантах осуществления изобретения антитела против FcRH5 и антитела против CD3 продуцируются в одной и той же клетке-хозяине (например, одноклеточный подход). В некоторых вариантах осуществления изобретения антитела против FcRH5 и антитела против CD3 продуцируются в отдельных клетках-хозяевах (например, двухклеточной подход).
В одноклеточном и двухклеточном подходах одна или более плазмид, кодирующих TDB против FcRH5 (например, полуантитело против FcRH5 и полуантитело против CD3), вводят в одну или более клеток-хозяев для культивирования и экспрессии TDB. В одном случае одна плазмида может кодировать как полуантитело против FcRH5, так и полуантитело против CD3. Альтернативно, полуантитела могут кодироваться отдельными плазмидами. В другом случае тяжелая цепь каждого полуантитела кодируется на первой плазмиде, тогда как легкая цепь каждого полуантитела кодируется на второй плазмиде. В одноклеточном подходе TDB против FcRH5 продуцируется в одном хозяине. В двухклеточном подходе TDB против FcRH5 получают путем экспрессии полуантител в разных хозяевах (например, отдельных культурах одних и тех же клеток хозяина или отдельных культурах разных клеток-хозяев). В двухклеточном подходе два хозяина можно культивировать в одном сосуде или в разных сосудах. Две культуры хозяина могут быть объединены до лизиса и очистки TDB против FcRH5 или два полуантитела могут быть очищены отдельно.
В некоторых вариантах осуществления изобретения антитело против FcRH5 согласно изобретению (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например CD3, такой как антитело TDB против FcRH5 согласно изобретению), которое имеет (например, мутации доменов VH, VL, CH1, CL и/или Fc, описанные выше) получают с использованием одноклеточного подхода, что приводит к улучшению правильного спариванию тяжелой цепи и легкой цепи и/или повышенному выходу антитела против FcRH5 по сравнению с антителом против FcRH5, которое не было модифицировано с учетом асимметричных модификаций.
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела могут быть получены в бактериях, в частности, когда гликозилирование и эффекторная функция Fc не нужны. Для экспрессии фрагментов антител и полипептидов в бактериях см., например, патенты США №№5648237, 5789199 и 5840523 (см., например, Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), pp. 245-254, описывающие экспрессию фрагментов антител в E.coli). После экспрессии антитело может быть выделено из пасты бактериальных клеток в растворимой фракции и может быть дополнительно очищено.
В дополнение к прокариотам эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, являются подходящими клонирующими или экспрессирующими хозяевами для векторов, кодирующих антитела, включая грибы и дрожжевые штаммы, у которых пути гликозилирования «гуманизированы», что приводит к получению антитела с частично или полностью человеческим профилем гликозилирования. См., Gerngross, Nat. Biotech. 22:1409-1414, 2004, и Li et al. Nat. Biotech. 24:210-215, 2006.
Подходящие клетки-хозяева для экспрессии гликозилированного антитела также получены из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, особенно для трансфекции клеток Spodoptera frugiperda.
Культуры клеток растений также могут быть использованы в качестве хозяев. См., например, патенты США №№5959177, 6040498, 6420548, 7125978 и 6417429 (описывающие технологию PLANTIBODIES™ для производства антител в трансгенных растениях).
Клетки позвоночных могут также использоваться в качестве хозяев. Например, могут быть подходящими клеточные линии млекопитающих, которые приспособлены для роста в суспензии. Другими примерами подходящих клеточных линий клеток млекопитающих являются линия CV1 почки обезьяны, трансформированная при помощи SV40 (COS-7); линия эмбриональных клеток почки человека (293 или 293-клетки, как описано, например, в Graham et al., J. Gen Virol. 36:59, 1977); клетки почки новорожденного хомяка (ВНK); мышиные клетки Сертоли (клетки ТМ4, как описано, например, в Mather Biol. Reprod. 23: 243-251, 1980); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK, клетки печени серой крысы (BRL 3А), клетки легких человека (W138), клетки печени человека (Hep G2), клетки опухоли молочной железы мыши (ММТ 060562), клетки TRI, как описано, например, в Mather et al. Annals NY Acad. Sci. 383:44-68, 1982; клетки MRC-5; и клетки FS4. Другие подходящие линии клеток млекопитающих включают клетки яичника китайского хомячка (СНО), включая клетки DHFR-CHO (Urlaub et al. Proc. Natl. Acad. Sci. USA 77:4216, 1980); и миеломные клеточные линии, такие как Y0, NS0 и Sp2/0. Для обзора некоторых линий клеток-хозяев млекопитающих, подходящих для продуцирования антител, см., например, Yazaki et al. Methods in Molecular Biology, 248:255-268, 2003.
C. Анализы
Антитела против FcRH5 согласно изобретению (например, биспецифические антитела против FcRH5 согласно изобретению, которые связываются с FcRH5 и второй биологической молекулой, например CD3, такие как антитела TDB согласно изобретению или их варианты), могут быть идентифицированы, скринированы или характеризуются их физико-химическими свойствами и/или биологической активностью с помощью различных анализов, известных в данной области техники.
1. Анализы связывания и другие анализы
В одном аспекте антитело против FcRH5 согласно изобретению тестируют на его антигенсвязывающую активность, например, с помощью известных методов, таких как ИФА, вестерн-блоттинг и т.д.
В другом аспекте конкурентные анализы могут быть использованы для идентификации антитела, которое конкурирует с антителом против FcRH5 согласно изобретению за связывание с FcRH5.
В иллюстративном конкурентном анализе иммобилизованный FcRH5 инкубируют в растворе, содержащем первое меченое антитело, которое связывается с FcRH5 и вторым немеченым антителом, которое тестируется на его способность конкурировать с первым антителом за связывания с FcRH5. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный FcRH5 инкубируют в растворе, содержащем первое меченое антитело, но не второе немеченое антитело. После инкубации в условиях, допускающих связывание первого антитела с FcRH5, избыточное несвязанное антитело удаляют и измеряют количество метки, связанной с иммобилизованным FcRH5. Если количество метки, связанной с иммобилизованным FcRH5, существенно уменьшается в тестируемом образце относительно контрольного образца, то это указывает на то, что второе антитело конкурирует с первым антителом за связывание с FcRH5. См., например, Harlow and Lane (1988) Antibodies: A Laboratory Manual. Ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
2. Анализ активности
В одном аспекте представлены анализы для идентификации антител против FcRH5, обладающих биологической активностью. Биологическая активность может включать, например, связывание с FcRH5 или его пептидный фрагментом или in vivo, in vitro, или ex vivo. В случае мультиспецифического (например, биспецифического) антитела против FcRH5 согласно изобретению (например, антитело TDB, имеющее одно плечо антитела против FcRH5 и одно плечо, которое распознает вторую биологическую молекулу, например, CD3), биологическая активность может также включать например, активацию эффекторных клеток (например, активацию Т-клеток (например, CD8+ и/или CD4+ Т-клеток)), расширение популяции эффекторных клеток (т.е. увеличение количества Т-клеток), уменьшение популяции целевых клеток (т.е. уменьшение в популяции клеток, экспрессирующих вторую биологическую молекулу на их клеточных поверхностях) и/или уничтожение целевых клеток. Предложены антитела, обладающие такой биологической активностью in vivo и/или in vitro. В некоторых вариантах осуществления изобретения антитело согласно изобретению тестируют на такую биологическую активность, как подробно описано в приведенных ниже примерах.
В некоторых вариантах осуществления изобретения активность антитела против FcRH5 (например, TDB против FcRH5) включает способность обеспечивать уничтожение В-клеток и/или активацию цитотоксических Т-клеток. В определенных вариантах осуществления изобретения антитело TDB против FcRH5 согласно изобретению исследуют в отношении такого уничтожения В-клеток и/или активации цитотоксического эффекта биологической активности Т-клеток любыми способами, описанными в данном документе, в частности, в примерах. В некоторых вариантах осуществления любого из этих методов анализа активности, МКПК можно выделять из цельной крови здорового донора методом фиколл-сепарации. В частности, человеческую кровь можно набирать в гепаринизированные шприцы и выделять МКПК при помощи Leucosep и Ficoll Paque Plus. В случае необходимости сепарацию Т-клеток CD4+ и Т-клеток CD8+ можно осуществлять при помощи наборов Miltenyi в соответствии с инструкциями производителя.
Кроме того, клетки можно промыть в среде RPMI, содержащей 10% FBS, дополненной GlutaMax, пенициллином и стрептомицином, и добавить ~0,2 миллиона суспендированных клеток в 96-луночный планшет с U-образным дном. Клетки можно культивировать в RPMI1640, дополненной 10% FBS, при 37°С в увлажненном стандартном инкубаторе для культивирования клеток. В случае анализа уничтожения клеток BJAB 20000 клеток BJAB можно инкубировать с эффекторными клетками, человеческими МКПК (huPBMC) или очищенными Т-клетками, в указанном соотношении для каждого анализа, в присутствии различных концентраций антител TDB в течение 24 часов. В случае анализа уничтожения эндогенных В-клеток 200000 человеческих МКПК можно инкубировать с различными концентрациями антител TDB в течение 24 часов.
После культивирования клетки можно промывать буфером FACS (0,5% БСА, 0,05 % азида Na в PBS). Затем клетки можно окрашивать в буфере FACS, промывать буфером FACS и суспендировать в 100 мкл буфера FACS, содержащем 1 мкг/мл пропидиум иодида. Данные можно получать при помощи проточного цитометра FACSCalibur и анализировать при помощи FlowJo. Живые В-клетки могут быть выделены как PI-негативные CD 19+ или PI-негативные CD20+ В-клетки при помощи проточной цитометрии, а абсолютное количество клеток можно получить при помощи гранул FITC, добавленных в реакционную смесь в качестве внутреннего контроля счета. Процент (%) уничтожения клеток можно рассчитать на основании необработанных TDB контрольных образцов. Активированные Т-клетки можно выявлять по поверхностной экспрессии CD69 и CD25, используя анти-СD69-FITC и анти-СD25-РЕ.
D. Иммуноконъюгаты
В данном изобретении также предложены иммуноконъюгаты, которые содержат антитело против FcRH5, например, мультиспецифическое антитело против FcRH5, например, TDB против FcRH5, которое конъюгировано с одним или более цитотоксическими агентами, такими как химиотерапевтические агенты или лекарственные средства, агенты, ингибирующие рост, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном варианте осуществления изобретения иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC), в котором антитело представляет собой конъюгированное с одним или более лекарственными средствами, включая, без ограничений, майтанзиноид (см. патенты США №5208020, 5416064 и Европейский патент ЕР 0425235 В1); ауристатин, такой как фрагменты DE и DF лекарственного средства монометилауристатина (ММАЕ и MMAF) (см. патенты США №5635483 и 5780588, и 7498298); доластатин; калихеамицин или его производные (см. патенты США №5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, и 5877296; Hinmanetal. Cancer Res. 53:3336-3342, 1993; и Lode et al. Cancer Res. 58:2925-2928, 1998); антрациклин, такой как дауномицин или доксорубицин (см. Kratz et al., Current Med. Chem. 13:477-523, 2006; Jeffrey et al. Bioorganic & Med. Chem. Letters 16:358-362, 2006; Torgov et al. Bioconj. Chem. 16:717-721, 2005; Nagy et al. Proc. Natl. Acad. Sci. USA 97:829-834, 2000; Dubowchik et al. Bioorg. & Med. Chem. Letters 12:1529-1532,2002; King et al. J. Med. Chem. 45: 4336-4343, 2002; и патент США №6630579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен и СС1065.
В другом варианте осуществления изобретения иммуноконъюгат включает антитело против FcRH5 (например, мультиспецифическое антитело против FcRH5, например, TDB против FcRH5), описанное в данном документе, конъюгированное с ферментативно активным токсином или его фрагментом, включая, без ограничений, цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модекцина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
В другом варианте осуществления изобретения иммуноконъюгат содержит антитело против FcRH5 (например, мультиспецифическое антитело против FcRH5, например, TDB против FcRH5), описанное в данном документе, конъюгированное с радиоактивным атомом с образованием радиоконъюгата. Для получения радиоконъюгатов доступно множество радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Когда для выявления используют радиоконъюгат, то он может содержать радиоактивный атом для сцинтиграфических исследований, например tc99m или I123, или спиновую метку для визуализации ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как тот же йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента могут быть получены при помощи множества агентов, связывающих бифункциональные белки, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие, как дисукцинимидилсуберат), альдегиды (такие, как глутаральдегид), бис-азидо соединения (такие, как бис(р-азидобензоил)-гександиамин), производные бис-диазония (такие, как бис(р-диазоний бензоил)-этилендиамин), диизоцианаты (такие, как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина может быть получен, как описано в Vitetta et al., Science 238:1098, 1987. 1-изотиоцианатобензил-3-метилдиэтилентриамин пентауксусная кислота (MX-DTPA), меченная углеродом 14, представляет собой типовой хелатирующий агент для конъюгации радионуклеотида с антителом. См. WO 94/11026. Линкер может представлять собой "расщепляемый линкер", который облегчает высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотолабильный линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметильный линкер или дисульфидсодержащий линкер (Chari et al., Cancer Res. 52:127-131, 1992; патент США №5208020).
Иммуноконъюгаты или ADC прямо подразумевают, но не ограничиваются конъюгатами, полученными при помощи перекрестно-сшивающих реагентов, включая, без ограничений, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB, которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Е. Фармацевтические составы
Фармацевтические составы антитела против FcRH5 согласно изобретению (например, биспецифическое антитело против FcRH5 согласно изобретению, которое связывается с FcRH5 и второй биологической молекулой, например, CD3, например, TDB против FcRH5), получают путем смешивания такого антитела, имеющего желаемую степень чистоты с одним или более необязательными фармацевтически приемлемыми носителями, буферами, стабилизаторами и/или консервантами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители являются в общем случае нетоксичными для реципиентов в применяемых дозировках и концентрациях и включают, но не ограничиваются этим, буферные вещества, например, фосфат, цитрат и другие органические соли; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (например, хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензэтония; фенол; бутиловый или бензиловый спирт; алкилпарабены, например, метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и m-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, например, сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, например, поливинилпирролидон; аминокислоты, например, глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, например, ЭДТА; сахара, например, сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, например, натрия; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионогенные сурфактанты, такие как полиэтиленгликоль (ПЭГ). Примеры фармацевтически приемлемых носителей в данном документе дополнительно включают средства, обеспечивающие внутритканевое распределение лекарственных средств, такие как растворимые нейтрально-активные гликобелки, содержащие гиалуронидазу (sHASEGP), например, растворимые гликобелки РН-20 человека, содержащие гиалуронидазу, такие как rHuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые примеры sHASEGP и способы применения, в том числе rHuPH20, описаны в публикациях патентов США №№2005/0260186 и 2006/0104968. В одном аспекте sHASEGP объединяют с одним или более дополнительными глюкозаминогликаназами, такими как хондроитиназы.
Типовые лиофилизированные составы антител описаны в патенте США №6267958. Водные составы антител включают те, которые описаны в патенте США №6171586 и WO 2006/044908, последние составы содержат гис-ацетатный буфер.
Состав согласно данному документу также может содержать более одного активного ингредиента, если это необходимо в случае конкретного показания, лечение которого проводят, предпочтительно те, которые имеют комплементарные активности и не оказывают вредного воздействия друг на друга. Например, может существовать потребность в дополнительном терапевтическом агенте (например, химиотерапевтическом агенте, цитотоксическом агенте, агенте, ингибирующем рост, и/или антигормональном агенте, таком как те, что перечислены выше). Такие активные ингредиенты соответственно присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, при помощи технологий коацервации или путем межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, микросферы альбумина, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методы раскрыты в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
В некоторых случаяъ композиция может содержать антагонист, связывающийся с компонентом сигнального пути PD-1, и/или дополнительный терапевтический агент. Например, антагонист, связывающийся с компонентом сигнального пути PD-1, может быть выбран из группы, состоящей из MPDL3280A (атезолизумаб), YW243.55.S70, MDX-1105, MEDI473 (дурвалумаб) и MSB0010718C (авелумаб), MDX 1106 (ниволумаб), МК-3475 (пембролизумаб), СТ-011 (пидилизумаб), MEDI-0680 (АМР-514), PDR001, REGN2810 и BGB-108. Кроме того, композиция, содержащая антитело против FcRH5 согласно изобретению (например, TDB против FcRH5), может содержать стероид, иммуномодулятор (IMiD), ингибитор протеосом (PI) или их комбинацию.
Могут быть получены препараты замедленного высвобождения. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, при этом матрицы представлены в виде формованных изделий, например, пленок или микрокапсул.
Составы, предназначенные для введения in vivo, обычно являются стерильными. Стерильность может легко достигаться, например, путем фильтрации через мембраны для стерильной фильтрации.
F Терапевтические способы и композиции
Любое из антител против FcRH5 согласно изобретению (например, биспецифические антитела против FcRH5 согласно изобретению, которые связываются с FcRH5 и второй биологической молекулой, например, CD3, например, TDB против FcRH5), могут быть использованы в терапевтических методах.
В одном аспекте предложено антитело против FcRH5 для применения в качестве лекарственного средства. В других аспектах предложено антитело против FcRH5 для применения в лечении или замедлении прогрессирования клеточного пролиферативного расстройства (например, рака, например, FcRH5-положительного рака, например, множественной миеломы (ММ)) и/или усилении иммунной функции у индивидуума. В определенных вариантах осуществления изобретения предложено антитело против FcRH5 для применения в способе лечения. В некоторых вариантах осуществления изобретение обеспечивает антитело против FcRH5 для применения в способе лечения человека, имеющего клеточное пролиферативное расстройство (например, рак, например, FcRH5-положительный рак, например ММ), включающий введение индивидууму эффективного количества антитела против FcRH5. В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже.
В других вариантах осуществления изобретение обеспечивает антитело против FcRH5 для применения в повышении иммунной функции у индивидуума, имеющего клеточное пролиферативное расстройство, такое как FcRH5-положительный рак (например, ММ). В некоторых вариантах осуществления изобретение обеспечивает антитело против FcRH5 для применения в способе усиления иммунной функции у индивидуума, имеющего клеточное пролиферативное расстройство, включающем введение индивидууму эффективного антитела против FcRH5 для активации эффекторных клеток (например, Т-клеток, например, CD8+ и/или CD4+ Т-клеток), способных оказывать цитотоксический и/или апоптотический эффект на целевые клетки (например, клетку, экспрессирующую вторую биологическую молекулу, распознаваемую антителом против FcRH5 согласно изобретению, таким как антитело TDB против FcRH5 согласно изобретению) у индивидуума. Антитело против FcRH5 может связываться как с молекулой FcRH5, расположенной на целевой клетке, так и с молекулой CD3, расположенной на иммунной эффекторной клетке. Целевая клетка может быть плазмоцитом, такой как долго- или короткоживущий плазмоцит. Кроме того, целевая клетка может быть клеткой миеломы. «Индивидуум» согласно любому из вышеописанных вариантов осуществления изобретения может представлять собой человека.
В дополнительном аспекте данное изобретение относится к применению антитела против FcRH5 в производстве или изготовлении лекарственного средства. В одном варианте осуществления изобретения медикамент предназначен для лечения клеточного пролиферативного нарушения (например, рака, например, FcRH5-положительного рака, например, ММ). В дополнительном варианте осуществления изобретения медикамент предназначен для применения в способе лечения клеточного пролиферативного расстройства, включающем введение индивидууму, имеющему клеточное пролиферативное расстройство, эффективного количества лекарственного средства. В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже.
В дополнительном варианте осуществления изобретения лекарственное средство предназначено для активации эффекторных клеток, активирующих эффекторные клетки (например, Т-клетки, например, CD8+ и/или CD4+ Т-клетки), способные оказывать цитотоксическое и/или апоптотическое действие на целевые клетки (например, клетку, экспрессирующую вторую биологическую молекулу, распознаваемую антителом против FcRH5 согласно изобретению, таким как антитело TDB против FcRH5 согласно изобретению) у индивидуума. В дополнительном варианте осуществления изобретения лекарственное средство предназначено для применения в способе усиления иммунной функции у индивидуума, имеющего клеточное пролиферативное расстройство, включающем введение индивидууму количества, эффективного лекарственного средства для активации эффекторных клеток (например, Т-клеток, например CD8+ и/или CD4+ Т-клеток), расширить (увеличить) популяцию эффекторных клеток, уменьшить количество целевых клеток (например, клетку, экспрессирующую вторую биологическую молекулу, распознаваемую антителом против FcRH5 согласно изобретению, таким как антитело TDB против FcRH5 согласно изобретению) и/или убить целевую клетку (например, опухолевую целевую клетку). «Индивидуум» согласно любому из вышеописанных вариантов осуществления изобретения может представлять собой человека.
В дополнительном аспекте изобретение относится к способу лечения клеточного пролиферативного расстройства (например, рака). FcRH5-положительный рак может представлять собой В-клеточный рак. В некоторых случаях В-клеточный рак может представлять собой множественную миелому (ММ), хронический лимфоцитарный лейкоз (CLL), мантийноклеточную лимфому (MCL), диффузную В-крупноклеточную лимфому (DLBCL) и/или фолликулярную лимфому (FL). В конкретных случаях, В-клеточный рак представляет собой ММ. В других случаях FcRH5-положительный рак представляет собой В-клеточный рак. В одном варианте осуществления изобретения способ включает введение индивидууму, имеющему такое клеточное пролиферативное расстройство (например, рак, например, FcRH5-положительный рак, например множественную миелому (ММ)), эффективного количества антитела против FcRH5. В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже. «Индивидуум» согласно любому из вышеописанных вариантов осуществления изобретения может представлять собой человека.
Антитела согласно изобретению могут применяться как самостоятельно, так и в комбинации с другими агентами в терапии. Например, антитело согласно изобретению можно совместно вводить с по меньшей мере одним дополнительным терапевтическим агентом. В некоторых вариантах осуществления изобретения дополнительный терапевтический агент представляет собой антагонист, связывающийся с компонентом сигнального пути PD-1, такой как MPDL3280A (атезолизумаб), YW243.55.S70, MDX-1105, MEDI473 (дурвалумаб) и MSB0010718C (авелумаб), MDX 1106 (ниволумаб), МK-3475 (пембролизумаб), СТ-011 (пидилизумаб), MEDI-0680 (АМР-514), PDR001, REGN2810 и/или BGB-108. Кроме того, субъекту может также вводиться стероид, иммуномодулятор (IMiD), ингибитор протеосом (PI) или их комбинация. В одном варианте осуществления изобретения глюкокортикоид представляет собой дексаметазон. В некоторых вариантах осуществления изобретения IMiD представляет собой леналидомид. В некоторых вариантах осуществления изобретения PI представляет собой бортезомиб.
Такие комбинированные терапии, указанные ранее, охватывают комбинированное введение (в котором два или более терапевтических агента включены в одну и ту же или отдельные композиции), а также раздельное введение, и в этом случае, введение антитела согласно изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического агента или агентов. В одном варианте осуществления изобретения введение антитела против FcRH5 и введение дополнительного терапевтического агента может разделять около одного месяца или около одной, двух или трех недель, или около одной, двух, трех, четырех, пяти или шести суток.
Антитело согласно изобретению (и/или любой дополнительный терапевтический агент) можно вводить любым подходящим способом, в том числе внутривенно, подкожно, внутримышечно, местно, перорально, трансдермально, внутрибрюшинно, внутрикорбитально, путем имплантации, путем ингаляции, интратекально, интравентрикулярно или интраназально. Введение дозы можно проводить любым удобным способом, например, путем инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или постоянным. В данном документе рассматриваются различные схемы дозирования, включая, без ограничений, одноразовое или многоразовые введения через временные промежутки, в виде одноразовой дозы введения и импульсной инфузии.
Антитела согласно изобретению можно включать в состав препарата, дозировать и вводить с помощью способов, согласующихся с надлежащей медицинской практикой. Факторы, которые следует учитывать в данном контексте, включают конкретное расстройство, подлежащее лечению, конкретное млекопитающее, подвергаемое лечению, клиническое состояние отдельно взятого пациента, причину расстройства, место доставки средства, способ введения, схему введения и другие факторы, известные практикующим врачам. Антитело необязательно готовят вместе с одним или более агентами, которые на данный момент используются для профилактики или лечения соответствующего нарушения. Эффективное количество таких других агентов зависит от количества антитела, которое присутствует в составе, типа нарушения или лечения, и других факторов, которые обсуждались ранее. Такие агенты, как правило, вводятся в тех же дозах и с помощью тех же путей, которые описаны в данном документе, или в дозе, составляющей от около 1 до 99% дозы, описанной в данном документе, или в любой дозе и с помощью любого пути, который эмпирически/клинически считается пригодным.
Для профилактики или лечения заболевания ((например, пролиферативное клеточное расстройство, например рак, например, FcRH5-положительный рак, например, ММ)), подходящая доза антитела согласно изобретению (используемого самостоятельно или в комбинации с одним или более другими дополнительными терапевтическими агентами будет зависеть от типа заболевания, подлежащего лечению, типа антитела, тяжести и течения заболевания, того, вводится ли антитело с профилактической или терапевтической целью, предыдущей терапии, истории болезни пациента, чувствительности к антителу и предписания лечащего врача. Антитело удобно вводить пациенту одноразово или в течение серии курсов лечения.
В качестве общей нормы терапевтически эффективное количество антитела против FcRH5, вводимого человеку, соответствует диапазону от около 0,01 до около 100 мг/кг массы тела пациента, вне зависимости от того, осуществляют одно введение или более. В некоторых вариантах осуществления изобретения ежедневно вводимое количество антитела составляет, например, от около 0,01 до около 0,01, до около 55 мг/кг, 50 мг/кг, от 0,01 до около 45 мг/кг, от около 0,01 до около 40 мг/кг, от около 0,01 до около 35 мг/кг, от около 0,01 до около 30 мг/кг, от около 0,01 до около 25 мг/кг, от около 0,01 до около 20 мг/кг, от около 0,01 до около 15 мг/кг, от около 0,01 до около 10 мг/кг, от около 0,01 до около 5 мг/кг или от около 0,01 до около 1 мг/кг. В одном варианте осуществления изобретения антитело против FcRH5, описанное в данном документе, вводят человеку в дозе, составляющей около 100 мг, около 200 мг, около 300 мг, около 400 мг, около 500 мг, около 600 мг, около 700 мг, около 800 мг, около 900 мг, около 1000 мг, около 1100 мг, около 1200 мг, около 1300 мг или около 1400 мг, на 1 сутки 21-суточных циклов. Дозу можно вводить в виде однократной дозы или многократных доз (например, 2 или 3 доз), например, инфузий. В случае повторных введений в течение нескольких суток или более, в зависимости от состояния, лечение, как правило, проводят до достижения желаемой степени подавления симптомов заболевания. Одна типовая доза антитела соответствует диапазону от около 0,05 мг/кг до около 10 мг/кг. Таким образом, пациенту можно вводить одну или более доз, составляющих около 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). В некоторых вариантах осуществления изобретения антитело против FcRH5 вводят в дозе от около 0,01 мг/кг/нед. до около 10 мг/кг/нед. В некоторых вариантах осуществления изобретения антитело против FcRH5 вводят в дозе от около 0,1 мг/кг/нед. до около 10 мг/кг/нед. В некоторых вариантах осуществления изобретения антитело против FcRH5 вводят в дозе 1 мг/кг/нед.
Такие дозы можно вводить с интервалами, например, каждую неделю или каждые три недели (например, чтобы пациент получал от около двух до около двадцати или, например, около шести доз антитела против FcRH5). Можно вводить начальную повышенную нагрузочную дозу, за которой следует одна или более пониженных доз. Эффективность такой терапии легко контролировать при помощи традиционных методик и методов анализа.
В некоторых вариантах осуществления изобретения способы могут дополнительно включать дополнительную терапию. Дополнительная терапия может представлять собой лучевую терапию, хирургию, химиотерапию, генную терапию, ДНК-терапию, вирусную терапию, РНК-терапию, иммунотерапию, трансплантацию костного мозга, нанотерапию, терапию моноклональными антителами или комбинацию вышеперечисленного. Дополнительная терапия может быть в виде адъювантной или неоадъювантной терапии. В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой введение низкомолекулярного ферментативного ингибитора или антиметастатического агента. В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой введение агентов, ограничивающих побочные эффекты, (например, агенты, предназначенные для уменьшения возникновения и/или выраженности побочных эффектов лечения, такие как агенты против тошноты и т.д.). В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой лучевую терапию. В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой хирургическую операцию. В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой комбинацию лучевой терапии и хирургической операции. В некоторых вариантах осуществления изобретения дополнительная терапия представляет собой гамма-облучение. В некоторых вариантах осуществления изобретения дополнительная терапия может представлять собой отдельное введение одного или более из описанных выше терапевтических агентов.
G. Способы и композиции для диагностики и детекции
В некоторых вариантах осуществления изобретения любое из антител против FcRH5 согласно изобретению можно применять для обнаружения наличия FcRH5 в биологическом образце. Термин «обнаружение», используемый в данном документе, охватывает количественное или качественное обнаружение. В определенных вариантах осуществления изобретения биологический образец включает клетку или ткань.
В одном варианте осуществления изобретения предложено антитело против FcRH5 для применения в способе диагностики или обнаружения. В дополнительном аспекте предлагается способ выявления присутствия природного FcRH5 в биологическом образце. В некоторых вариантах осуществления изобретения способ включает приведение в контакт биологического образца с описанным в данном документе антителом против FcRH5 в условиях, обеспечивающих связывание антитела против FcRH5 с природным FcRH5, и выявление того, образуется ли комплекс между антителом против FcRH5 и природным FcRH5. Такой способ может выполнять в условиях in vitro или in vivo. В некоторых случаях биологический образец представляет собой образец крови. В некоторых случаяъ субъект представляет собой человека.
В некоторых вариантах осуществления изобретения предложены меченые антитела против FcRH5. Метки включают, но не ограничиваясь этим, метки или фрагменты, обнаруживаемые прямым способом (например, флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, обнаруживаемые непрямым способом, например, через ферментативную реакцию или молекулярные взаимодействия. Типовые метки включают, без ограничений, радиоизотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например люциферазу светляков и бактериальную люциферазу (патенте США №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (ПХ), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахаридооксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат дегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сопряженные с ферментом, который использует пероксид водорода для окисления предшественника красителя, такого как ПХ, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и тому подобное.
H. Промышленные изделия
В другом аспекте изобретение относится к промышленному изделию, содержащему материалы, применяемые для лечения, профилактики и/или диагностики клеточных пролиферативных расстройств (например, рака, например, FcRH5-положительного рака, например, множественной миеломы (ММ)) у субъекта (например, человека). Промышленное изделие включает контейнер и этикетку или листок-вкладыш в упаковке, нанесенный на или связанный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для растворов для внутривенного введения и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, одну или в комбинации с другой композицией, эффективной в лечении, профилактики и/или диагностировании патологического состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для растворов для внутривенного введения или флакон с пробкой, прокалываемой с помощью иглы для подкожных инъекций). По меньшей мере один активный агент в композиции представляет собой антитело согласно изобретению. На этикетке или листке-вкладыше в упаковке указано то, что композиция используется для выбранного патологического состояния. Кроме того, промышленное изделие может содержать (а) первый контейнер с содержащейся в нем композицией, при этом композиция содержит антитело согласно изобретению; и (б) второй контейнер с содержащейся в нем композицией, при этом композиция содержит дополнительный цитотоксический или другой терапевтический агент. Промышленное изделие в данном варианте осуществления изобретения может дополнительно содержать листок-вкладыш в упаковке с указанием того, что композиции могут быть использованы для лечения конкретного патологического состояния. В альтернативном или дополнительном варианте промышленное изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БВДИ), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Дополнительно могут быть включены другие вещества, желательные с коммерческой и потребительской точки зрения, включая другие буферы, растворители, фильтры, иглы и шприцы. В некоторых случаях набор включает листок-вкладыш в упаковке, содержащий инструкции по применению антитела для лечения или задержки прогрессирования FcRH5-положительного рака (например, ММ) у субъекта. В других случаях набор включает листок-вкладыш в упаковку, содержащий инструкции по применению антитела для усиления иммунной функции у субъекта, имеющего FcRH5-положительный рак (например, ММ).
III ПРИМЕРЫ
Ниже приведены примеры способов и композиций согласно данному изобретению. Следует понимать, что учитывая общее описание, представленное выше, можно осуществить различные другие варианты осуществления данного изобретения.
Пример 1: Генерация антител против FcRH5
Иммунизация и скрининг
Мышей BALB/с (Charles River, Холлистер, Калифорния) иммунизировали 2 мкг, 10 мкг или 100 мкг/инъекция на мышь. Антигены суспендировали в адъюванте монофосфориллипида A (MPL) и трегалозы дикориномиколята (TDM) (Ribi) или адъюванте Фрейнда и вводили в подушечку лапы, брюшину или основание хвоста каждого иммунизированного животного. Мыши получали в общей сложности от 9 до 18 доз и от 1 до 2 предшествующих стимуляции инъекций только PBS, интраперитонеально (и/п) и/или внутрисуставно за 2-4 суток до слияния. В кампании иммунизации А, описанной в публикации патента США №2015/0098900, получили исходное антитело против FcRH5 1G7, которое оптимизировано и дополнительно описано в данном документе.
А. Кампания иммунизации В
Для получения изоформ-специфических антител к мембранно-проксимальному Ig-подобному домену FcRH5, мышей иммунизировали белком Е11 (аминокислоты 745-850 SEQ ID NO: 114) с N-концевой гистидиновой меткой, которая экспрессировалась в клетках СНО. Пять мышей иммунизировали 10 мкг меченного His белка E11 в адъюванте Ribi при помощи и/п введения. Мыши получили 18 доз, за которыми следуют два предшествующих слиянию инъекции в PBS только через и/п введение за семь и четыре сутки до слияния. После 18 доз рекомбинантного белка Е11 сыворотку анализировали на FcRH5-связывающие антитела с использованием проточной цитометрии и титрованием в ИФА с использованием Е11 в качестве антигена. Значительная реактивность была обнаружена в клетках SVT2, которые экспрессировали полноразмерный человеческий FcRH5, полноразмерный белок FcRH5 яванского макака или Е11, но не трансформированные вектором клетки SVT2, что указывает на то, что FcRH5-связывающие антитела присутствуют в сыворотке всех пяти иммунизированных мышей. После 20 доз лимфоциты иммунизированных мышей подвергали электрослитию с клетками миеломы мыши P3X63Ag8U.1.22.
Клоны тестировали на связывание с рекомбинантными белками Е11 человека и яванского макака при помощи ИФА. Восемь клонов, которые были перекрестно-реактивными как для FcRH5 человека, так и для FcRH5 яванского макака в ИФА, тестировали на связывание с клетками SVT2, которые экспрессировали полноразмерный FcRH5 человека, полноразмерный FcRH5 яванского макака или Е11 белок, при помощи проточной цитометрии. Все восемь клонов связывали клетки SVT2, экспрессирующие человеческий домен E11, трансмембранный домен и цитоплазматические домены.
Для тестирования связывания с раковыми клетками, которые эндогенно экспрессировали FcRH5 или которые трансфицировали FcRH5, клетки поднимали с использованием EDTA/PBS, и 1×105 клетки суспендировали в 100 мкл и инкубировали с первичными антителами (1 объем супернатанта количественно оцененного субклона не IgG, 4 мкг/мл супернатанта количественно оцененного субклона IgG или 2 мкг/мкл очищенных моноклональных антител). Клетки дважды промывали буфером FACS (PBS, 1% BSA, 2 мМ EDTA) и инкубировали с разведением 1:1000 козьего антимышиного антитела, помеченного РЕ (фикоэритрин) или 1:100 козьего антимышиного АРС. Клетки промывали дважды буфером FACS, и анализ проточной цитометрии проводили на FACSCalibur. Прямое мечение антител с Xenon выполняли в соответствии с протоколом производителя (Invitrogen), если это указано. Однако только три клона связывались с клетками, которые экспрессировали полноразмерный FcRH5 человека и не связывались с клетками, которые экспрессировали полноразмерный FcRH5 яванского макака (таблица 2).

*-домен E11, трансмембранный домен и цитоплазматический домен FcRH5 человека
В. Кампания С по иммунизации и исследовании мкАт
Для компании иммунизации С, N-терминально His-меченый белок E11 был получен в клетках СНО. Мышей иммунизировали первоначальной инъекцией 100 мкг белка Е11 в полном адъюванте Фрейнда в основание хвоста. Мыши получили в общей сложности 17 доз, за которыми следовала предшествующая слиянию инъекция только в PBS и/п и внутрисуставная инъекция за двое суток до слияния (таблица 3).
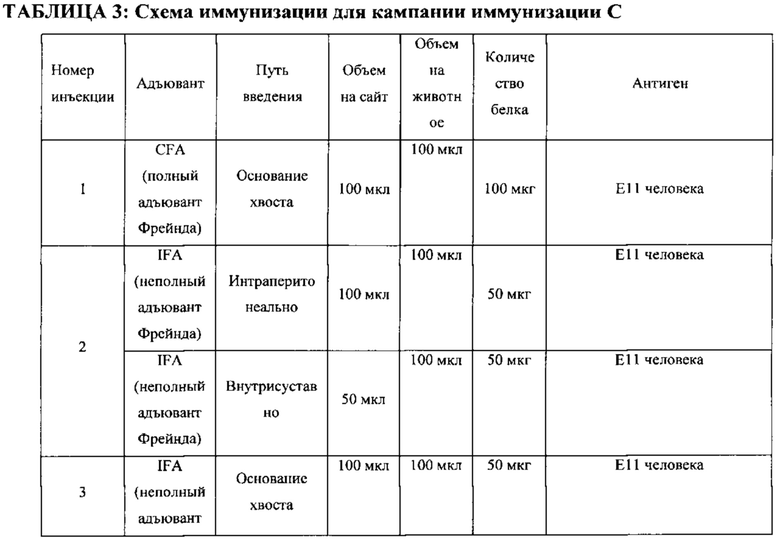
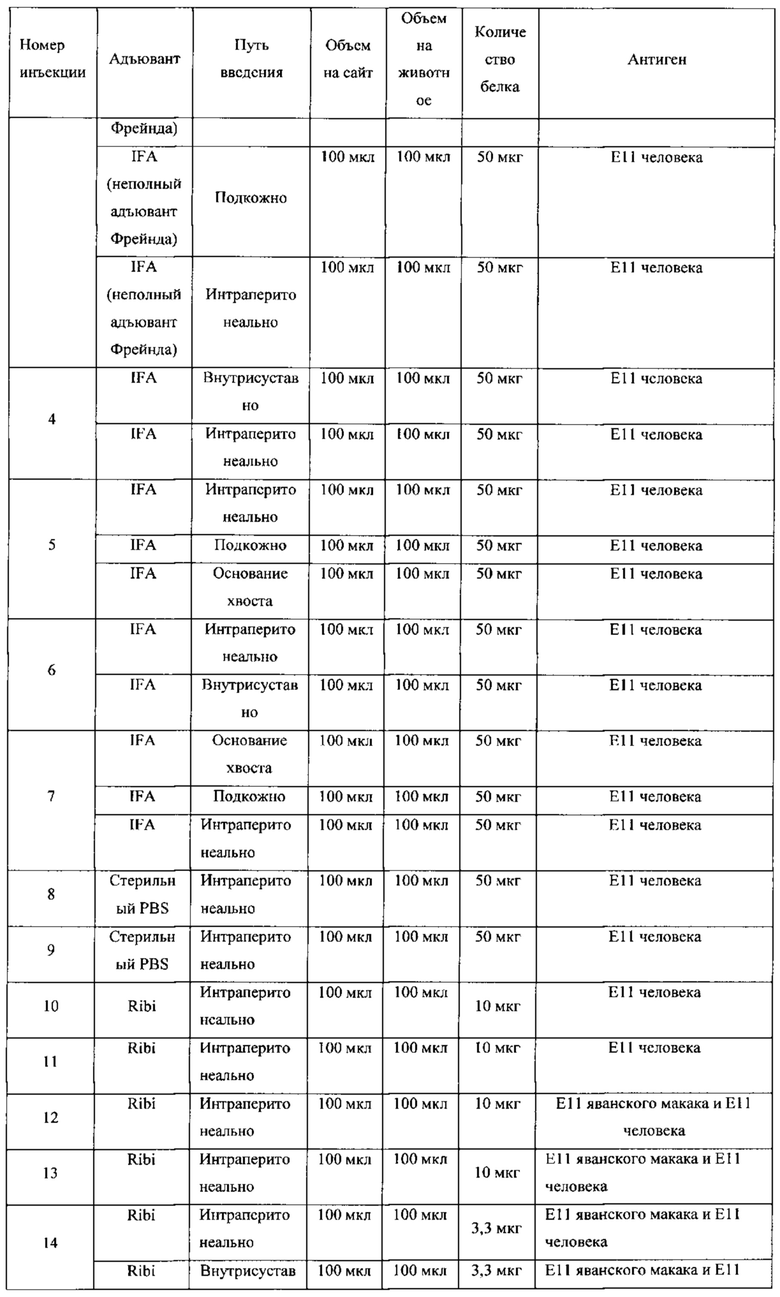
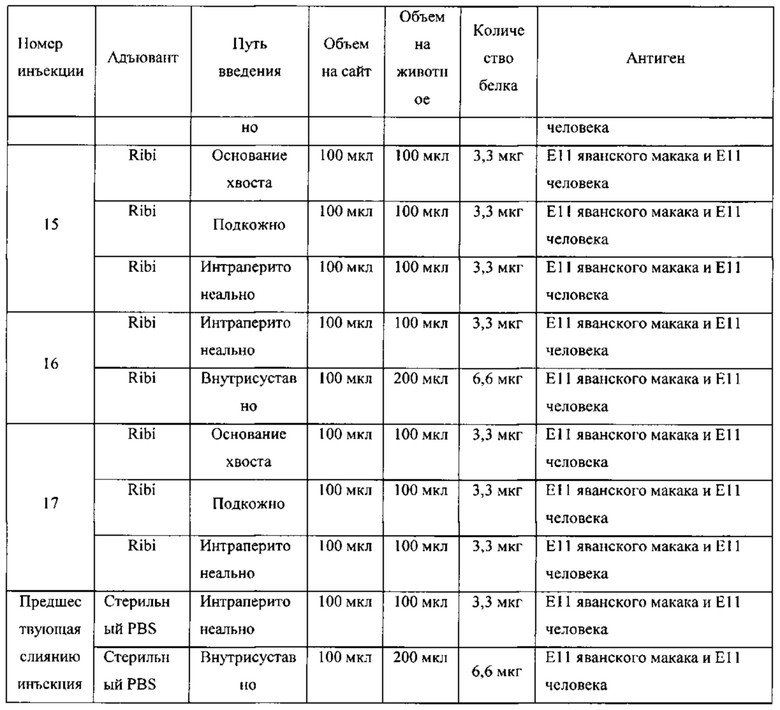
После 15 доз рекомбинантного белка Е11 с последующими совместными инъекциями рекомбинантных белков Е11 человека и яванского макака, сыворотку анализировали на FcRH5-связывающие антитела при помощи проточной цитометриии и титрования в ИФА с использованием белка Е11 в качестве антигена. Значительная реактивность была обнаружена в клетках SVT2, которые экспрессировали полноразмерный человеческий FcRH5, полноразмерный белок FcRH5 яванского макака или домен Е11 человека, трансмембранный домен и цитоплазматический домен, но не трансформированные вектором клетки SVT2, что указывает на то, что FcRH5-связывающие антитела присутствуют в сыворотке всех пяти иммунизированных мышей. После конечной инъекции PBS в лимфоциты иммунизированных мышей подвергали электрослитию с клетками миеломы мыши P3X63Ag8U.1.22.
Клоны тестировали на связывание с рекомбинантными белками E11 человека и яванского макака и связывание с клетками SVT2, которые экспрессировали полноразмерный FcRH5 человека; полноразмерный FcRH5 яванского макака; полноразмерные человеческие FcRH1, FcRH2, FcRH3 или FcRH4; или домена E11 человека, трансмембранный домен и цитоплазматические домены FcRH5 при помощи проточной цитометрии. В ИФА было 44 клона положительных по белку E11 человека и 32 клона положительных по белку E11 яванского макака. Из них было идентифицировано в общей сложности 16 клонов, которые были связаны с клетками, которые экспрессировали FcRH5 человека и клетки, которые экспрессировали FcRH5 яванского макака, но не клетки, которые экспрессировали FcRH1, FcRH2, FcRH3 или FcRH4, что указывает на специфическую реакционную межвидовую реактивность FcRH5 (таблица 4).
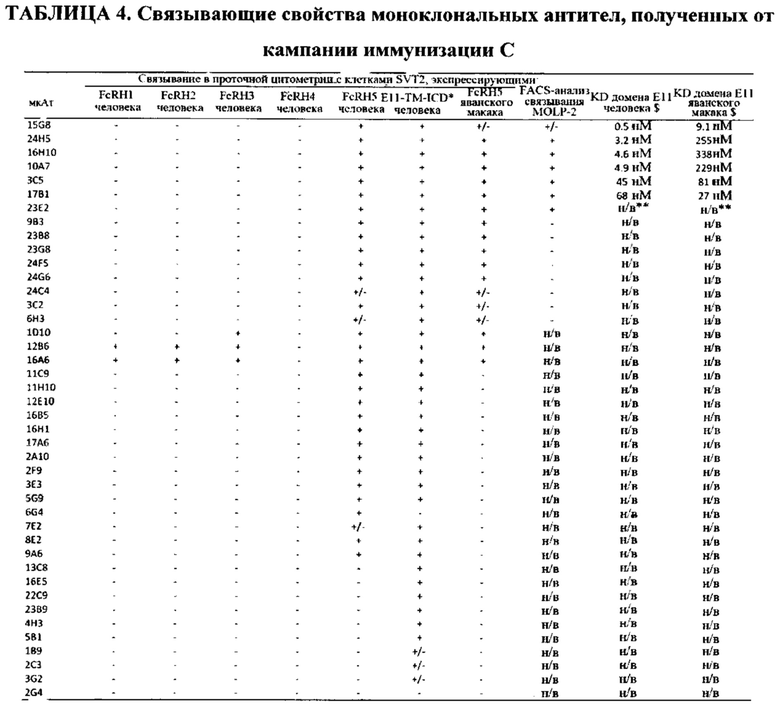
*- домен E11, трансмембранный домен в цитоплазматический домен FcRH5 человека
$ KD. аффинность моновалентного связывания определяли при помошн поверхностного плазменного резонанса в BIACORE®. Эксперименты с доменом Е11 человека выполняли от 1 до 3 раз и высчитывали среднее. Эксперименты с доменом Е11 яванского макака выполняли один раз для каждого антитела.
** Нерекомбинантные клоны получали для антитела 23Е2.
Очищенный IgG из 16 выбранных клонов моноклональных антител (мкАт) дополнительно исследовали на связывание с клетками, которые экспрессируют FcRH5 человека эндогенно (клетки MOLP-2). Было показано, что в общей сложности восемь моноклональных антител связывали клетки миеломы MOLP-2. Основываясь на анализе последовальности было обнаружено, что два из этих восьми клонов мкАт были избыточными, а один из них не был клонирован молекулярно. Шесть молекулярно клонированных MOLP-2-положительных клонов мкАт экспрессировали рекомбинантно в виде мышиного IgG2a и тестировали на аффинность при помощи поверхностного плазмонного резонанса в аппарате BIACORE® Т200 в формате захвата IgG.
Вкратце, тестируемые антитела захватывали на проточных кюветах 2, 3 или 4 с проточной кюветой 1 в качестве стандартного образца на чипе СМ5 серии S белка A (GE Life Sciences, Piscataway, NJ). В качестве аналита использовали фрагмент белка FcRH5 с линейной скоростью потока 30 мкл/мин. Между инъекциями поверхность захвата регенерировали 30-секундной инъекцией 10 мМ глицина, рН 1,5 при линейной скорости потока 10 мкл/мин. Взаимодействие оценивали при 25°С в 10 мМ HEPES, рН 7,4, 150 мМ NaCl, 0,05% Твин-20 (HBSP). Контрольные данные эталонной проточной кюветы и от введения только одного буфера вычитали перед кинетическим анализом. Кинетические параметры рассчитывали путем подгонки данных к модели связывания 1: 1. Вычитание величин стандартных образцов и подгонку данных выполняли с использованием программного обеспечения BIAevaluation (GE Life Sciences, Пискатауэй, Нью-Джерси).
Аффинности моновалентного связывания этих антител к домену Е11 человека варьировались от 0,5 нМ до 68 нМ, тогда как аффинности для домена E11 яванского макака варьировались от 9,1 нМ до 338 нМ (таблица 4). Только один клон имел аффинность к домену Е11 человека выше, чем у антитела 1G7, мкАт 15G8. Однако разница KD между доменами E11 человека и яванского макака составляла около 20 раз, что делает этот клон непригодным для дальнейшего клинического исследования. Кроме того, несмотря на высокую аффинность этого клона антитела к выделенному домену Е11 человека, связывание этого клона антитела с клетками MOLP-2 было очень слабым; оно было слабее, чем клоны с более низкой аффинностью к выделенному домену Е11 человека, что указывает на то, что этот клон не удовлетворительно связывается с нативным белком FcRH5 человека.
Пример 2: Генерация и характеристика вариантов антител против FcRH5 от кампаний иммунизации А и С
А. Гуманизация моноклоналъных антител против FcRH5 от кампании иммунизации А
Антитело против FcRH5 1G7 из кампании иммунизации А, описанное в публикации патента США №2015/0098900, было гуманизировано методом трансплантации HVR, как описано ранее (Presta et al. Cancer Res. 57: 4593-4599, 1997), за исключением того, что консенсусные каркасные области VH4 и Vκ1 (Dennis, Current Trends in Monoclonal Ant. Develop, and Manufac. 9-28, 2010) были использованы в качестве акцепторных каркасных областей. Трансплантат тяжелой цепи также включал мышиные остатки в положениях каркасной области 37, 48, 67, 71, 73, 78, 93 и 94 по Кабат, а трансплантат легкой цепи также включал мышиные остатки в положениях каркасной области 36 и 43 для надлежащего представления HVR и контакт доменов VH/VL. Остатки пронумерованы в соответствии с Kabat et al. (Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD, (1991). Последовательность гуманизированного 1G7, называемая в данном документе hu1G7.v1 или 1G7.v1, показана на фиг. 2 и 3.
В. Созревание аффинности и характеристика вариантов hu1G7.v1
Для усиления эффективности гуманизированное антитело hu1G7.v1 подвергали аффинному созреванию с использованием фагового дисплея. Вкратце, рандомизированные библиотеки фрагмента антитела Fab на поверхности бактериофага М13 отображали и подвергали скринингу по связыванию с биотинилированным фрагментом белка FcRH5. Подходящие мутации были идентифицированы секвенированием ДНК отдельных клонов. Предполагалось, чт подходящие мутации приведут к вариантам с улучшенной аффинностью (Li et al., MAbs. 6: 437-445, 2014). Клоны антител с выбранными мутациями тестировали на аффинность при помощи поверхностного плазмонного резонанса (таблицы 5А и 5В).
Вкратце, поверхность захвата IgG человека была сгенерирована на чипе СМ5 серии S с использованием аминной связи и набора для захвата IgG человека (GE Life Sciences, Пискатауэй, Нью-Джерси). Затем тестируемые антитела захватывали в проточных кюветах 2, 3 или 4, причем проточная кювета 1 использовалась в качестве стандартного образца. В качестве аналита использовали фрагмент белка FcRH5 с линейной скоростью потока >30 мкл/мин. Между инъекциями поверхность захвата регенерировали 30-секундной инъекцией 3 М хлорида магния или 10 мМ глицина, рН 1,5. Взаимодействие оценивали при 25°С в 10 мМ HEPES рН 7,4,150 мМ NaCl и 0,05% Tween 20 (HBSP). Контрольные данные эталонной проточной кюветы и от введения только одного буфера вычитали перед кинетическим анализом. Кинетические параметры рассчитывали путем подгонки данных к модели связывания 1:1. Вычитание величин стандартных образцов и подгонку данных выполняли с использованием программного обеспечения BIAevaluation (GE Life Sciences, Пискатауэй, Нью-Джерси).
Для начальных экспериментов по скринингу аффинности, антитела захватывали непосредственно из очищенных культуральных сред, собранных после временной трансфекции клеток Expi293 (Invitrogen) (таблицы 5А-5В). Способность каждого антитела связывать фрагмент белка FcRH5 оценивали при 100 нМ и 500 нМ (эксперимент 1, таблицы 5А-5В) или 0 нМ, 20 нМ, 100 нМ и 500 нМ (эксперимент 2, таблицы 5А-5В). Ассоциацию и диссоциацию контролировали при линейной скорости потока 30 мкл/мин, в течение 180 с и 600 с, соответственно. Все последовательности, представленные в таблице 5А, раскрыты в SEQ ID NO: 4-6 (т.е. широкие консенсусные последовательности L1, L2, L3 FcRH5). Все последовательности, представленные в таблице 5В, раскрыты в SEQ ID NO: 1-3 (т.е. широкие консенсусные последовательности H1, Н2, Н3 FcRH5). Наблюдали улучшения аффинности в 5,2 раза (таблица 5А). Поскольку каждая отдельная мутация не обеспечивала желаемых улучшений, было решено объединить различные мутации с акцентом на мутации VL S30R, I32L, A51G, S52Y, Y53N, T56S, H91Q, Y92F, S93Q и S94P и мутации VH S28K, L29T, и S56T.
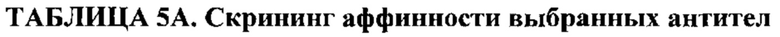
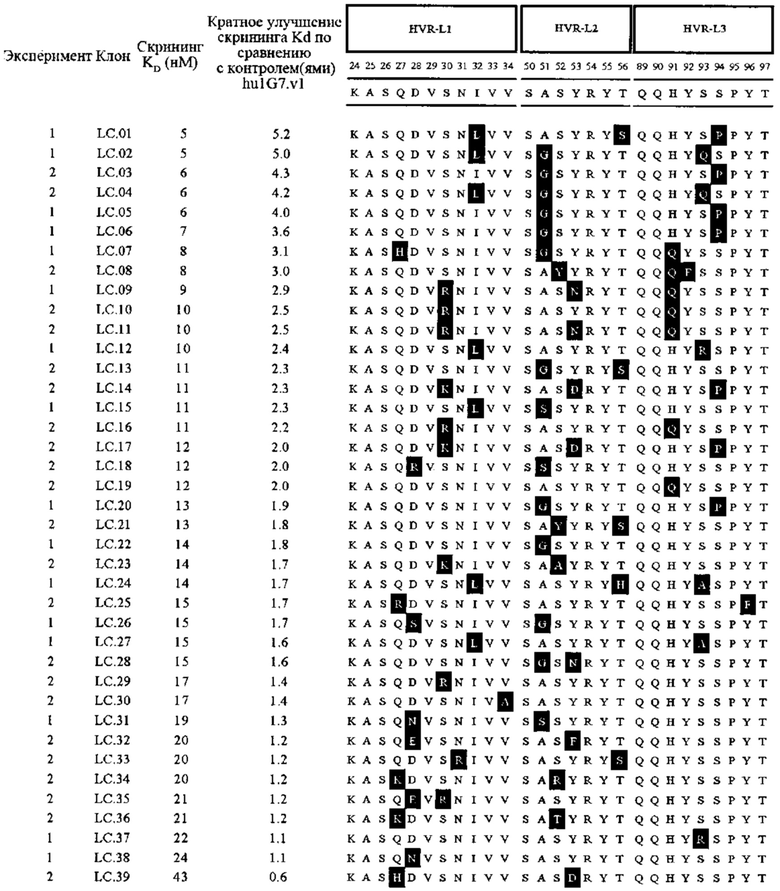
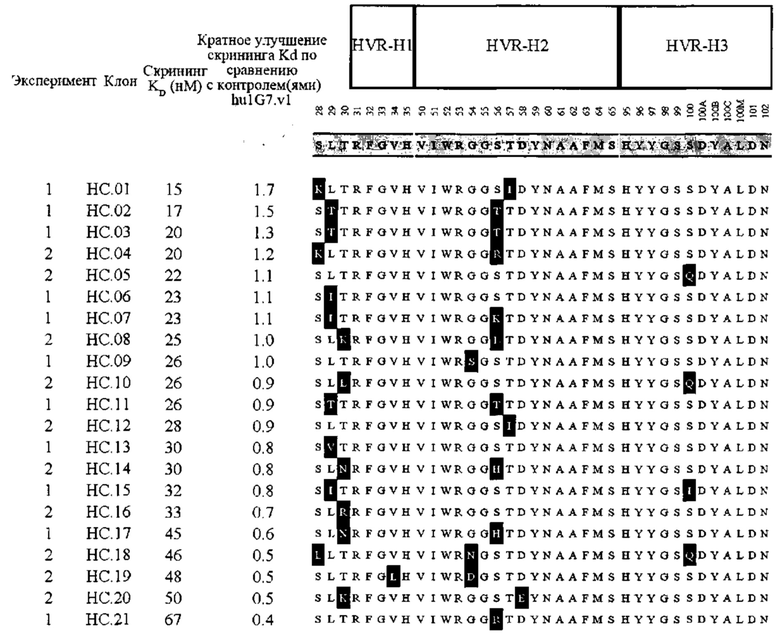
Библиотеки фагового дисплея антител были разработаны с использованием как жесткого, так и мягкого сайт-направленного мутагенеза. Для библиотек L188 и L189 использовали мягкий мутагенез, чтобы подвергнуть мутации остатки VL 27-34, 50-56, 91-94 и 96 (L188) или остатки VH 28-35, 50-58 и 95-100d (L189). В положениях, которые подвергались мутагенезу, олигонуклеотиды мягкого мутагенеза использовали 91% исходного кодирующего нуклеотида и 3% от каждого другого нуклеотида. Для библиотек L190 и L191 использовали жесткий мутагенез (кодоны NNK) для мутации остатков VL 27-34, 50-56 и 89-96 (L190) или остатков VH 28-35, 50-58 и 95-100d (L191), один кодон одновременно с тремя мутациями HVR (Li et al., MAbs. 6: 437-445, 2014).
Было проведено до четырех раундов селекции по связыванию с фрагментом белка FcRH5. Селекцию проводили как в твердой фазе (целевой белок, адсорбированный непосредственно на микропланшет), так и в растворе. Для селекции в растворе использовали биотинилированный целевой белок для захвата целевых комплексов фаг-антитело. Клоны с выбранными мутациями экспрессировались в клетках Expi293 (Thermo Fisher Scientific, Waltham, MA) и супернатанты подвергали скринингу при помощи поверхностного плазмонного резонанса.
Библиотеки фагового дисплея антител были получены при помощи мутагенеза по Кункелю (Kunkel et al. Methods in Enzymology, 154: 367-382, 1987), используя в качестве матричной одноцепочечной ДНК, продуцируемой в бактериях CJ236, и очищали с использованием набора Qiagen QIAprep Spin М13 Kit (QIAGEN, Inc., Валенсия, Калифорния). Матричная ДНК для исходных библиотек созревания аффинности содержала стоп-кодоны в выбранных для мутагенеза HVR; матричная ДНК для последующей «целевой» фаговой библиотеки, описанной ниже, не использовала стоп-кодоны матричной HVR. Если не указано иное, остатки в вариабельных областях антитела нумеруются в соответствии с системой Kabat. Ферменты были приобретены у New England Biolabs (Ипсвич, Массачусетс). Реакционную смесь после мутагенеза по Кункелю очищали с использованием мини спин-колонок QIAprep (QIAGEN, Inc., Валенсия, Калифорния) и трансформировали в XLI Blue Е. coli (Agilent Technologies, Санта-Клара, Калифорния) путем электропорации. После восстановления в среде SOC при 37°С культуры инфицировали вспомогательным фагом и инкубировали при 37°С в течение дополнительных 0,5-2 часов перед добавлением антибиотиков карбенициллина и канамицина. Культуры затем инкубировали при 37°С в течение дополнительных 4-5 часов с последующей дополнительной инкубацией в течение ночи при 30°С. Бактериофаг очищали из остветленной культуральной среды путем осаждения приблизительно 1/6 объема осаждающего реагента (20% ПЭГ, 2,5 М хлорида натрия). Осажденный фаг осаждали центрифугированием и растоворяли в фосфатно-солевом буферном растворе (PBS). Раствор дополнительно осветляли центрифугированием при 14000 g. Очищенный фаг хранили в 50% глицерине в морозильнике -20°С и повторно растворяли в PBS перед использованием.
В подходе рационального дизайна шесть мутаций VL, идентифицированных из экспериментов с фагами (S30R, I32L, A51G, T56S, H91Q и S94P), были объединены с дополнительной гуманизирующей мутацией VL S43A и оценены парами для оценки совместимости (таблица 6). Для оценки совместимости выбранных мутаций VL (таблица 6) антитела были взяты непосредственно из осветленных культуральных сред, собранных после временной трансфекции клеток Expi293. Связывание фрагмента белка FcRH5 оценивали при 6 нМ, 19 нМ, 56 нМ, 167 нМ и 500 нМ при линейной скорости потока 30 мкл/мин. Ассоциацию и диссоциацию наблюдали в течение 300 с и 600 с, соответственно. Последовательности, раскрытые в таблице 6, представляют собой отдельные аминокислоты (то есть число в верхней части колонки представляет собой аминокислотное число в последовательности антитела). Результаты показали, что мутации VL I32L и H91Q, при одновременном улучшении аффинности в отдельности, не приносят преимуществ при объединении. Однако другие тестируемые последовательности дали дополнительные улучшения аффинности до четырех раз по сравнению с контролем hu1G7.v1. На основании этих результатов были разработаны антитела hu1G7.v1.1, hu1G7.v1.2, hu1G7.v1.3 и hu1G7.v1.4 (фиг. 2 и 3). Антитело hu1G7.v1.1 было разработано для включения шести мутаций VL S30R, I32L, A51G, T56S, H91Q и S94P. Антитело hu1G7.v1.2 было разработано для включения десяти мутаций VL S30R, I32L, A51G, S52Y, Y53N, T56S, H91Q, Y92F, S93Q и S94P. Антитела hu1G7.v1.3 и hu1G7.v1.4 были основаны на hu1G7.v1.1 и предназначены для исключения или I32L (hu1G7.v1.3), или H91Q (hu1G7.v1.4).

Также была разработана целевая библиотека фагов для исследования мутаций представляющих интерес (мутации VL S30R, I32L, A51G, S52Y, Y53N, T56S, H91Q, Y92F, S93Q, S94P, мутации VH S28K, L29T, S56T). В целях дальнейшего увеличения сходства с последовательностями антител человека, целевая библиотека фагов также допускала ограниченную вариацию в положениях каркасных областей следующим образом: S43A в LC, L48I в НС и/или N73T в НС. Было получено около 108 трансформантов. Селекцию выполняли с использованием фрагмента белка FcRH5 в качестве «приманки». Сначала проводили селекцию с твердофазным иммобилизованным белком-мишенью, причем выбор фазы раствора использовали на более поздних этапах. Из этой библиотеки были получены антитела hu1G7.v1.5, hu1G7.v1.7, hu1G7.v1.13 и hu1G7.v1.13.1 (фиг. 2 и 3).
Данные секвенирования из экспериментов с целевой библиотекой фагов также поддерживали концепцию (таблица 5А-5 В) отрицательного взаимодействия между мутациями I32L и H91Q VL. После пяти раундов отбора на фаге и последующего анализа 163 VL-последовательностей, частота глутамина в положении 91 VL оказалась значительно выше в присутствии lle32 (49/70 клонов), чем в присутствии Leu32 (5/93 клонов). Этот эффект не наблюдали в невыбранной библиотеке, в которой Gln91 наблюдали в 50% (9/18) анализируемых клонов, которые содержали лейцин в положении 32. Эти результаты предполагали отрицательную селекцию клонов, которые содержали как I32L, так и H91Q, несмотря на положительное обогащение каждой из этих мутаций в других случаях.
Восприимчивость к окислению исследовали путем инкубации антитела с 1 мМ 2,2'-азобис (2-метилпропионамидин) дигидрохлорида (ААРН) в течение 16 часов с последующим расщеплением трипсином для создания пептидов, которые можно было анализировать с использованием жидкостной хроматографии с масс-спектрометрией (LC-MS). Для каждого пептида в образце определяли время удерживания в LC, и высокоточные данные о массах и фрагментации ионов пептида (информация о аминокислотной последовательности) получали в MS. Экстрагированные ионные хроматограммы (XIC) были получены для представляющих интерес пептидов (нативные и модифицированные ионы пептидов) из наборов данных в окне ±10 ppm, а пики были интегрированы для определения области. Относительные проценты модификации были рассчитаны для каждого образца следующим образом: (площадь модифицированного пептида)/(площадь модифицированного пептида плюс площадь нативного пептида)×100. Затем эти относительные проценты сравнивали между контрольным и обработанным ААРН образцами.
После 16-часовой инкубации при 40°С в присутствии 1 мМ ААРН наблюдали увеличение окисления Trp52 и Met64 HVR-H2 (44,5% и 52,7%, соответственно). Кроме того, после стресса при избытке освещения окисление Trp52 увеличилось с 2% до 62%, а мутация Trp-52нс приводила к потере аффинности для вариантов 1 G7.v1.4, измеренных при помощи BIACORE® (фиг. 4). Замены Trp52 фенилаланином, тирозином, лейцином или гистидином и замены Met64 фенилаланином, изолейцином, валином или лейцином были исследованы в связи с легкими цепями из антител hu1G7.v1, hu1G7.v1.3, hu1G7.v1. 4, hu1G7.v1.5, hu1G7.v1.6 и hu1G7.v1.7 (фиг. 2 и 3). В частности, мутация W52 на F, Y, L или Н приводила к значениям KD в 119 нМ, 131 нМ, 92 нМ и 60,4 нМ, соответственно. Хотя мутация М64 на v в 1G7.v85 привела к KD размером 2,5 нМ. Для скрининга аффинности вариантов Trp52 и Met64 HVR-H2 (таблица 7) антитела экспрессировали путем транзиентной трансфекции клеток Expi293 и очищали с использованием наконечников колонок, которые были сделаны на заказ с аффинной смолой MabSelect SuRe (Glygen Corp., Columbia, MD & GE Life Sciences, Пискатауэй, Нью-Джерси). Контрольные антитела hu1G7.v1 и hu1G7.v1.5 экспрессировали с помощью транзиентной трансфекции клеток 293Т и очищали с использованием MabSelect SuRe. Связывания фрагмента белка FcRH5 с захваченными антителами наблюдали при 100 нМ и 500 нМ. Ассоциацию и диссоциацию наблюдали при линейной скорости потока 40 мкл/мин, в течение 180 с и 300 с, соответственно. Результаты, показанные в таблице 7, продемонстрировали, что Met64 HVR-H2 может быть изменен на валин, сохраняя при этом высокую аффинность к FcRH5.
Антитело hu1G7.v1.4 было выбрано на основе аффинности и селективности к FcRH5 по сравнению с другими членами семейства FcRH. Для улучшения устойчивости этого антитела к окислению мутация M64V, описанная в таблице 7, также была включена в область VH. Полученное антитело hu1G7.v1.4.M64V было обозначено как hu1G7.v85 (фиг. 5А-5В) и было получено в виде двухвалентного IgG и в виде Т-клеточно-зависимого биспецифического антитела (TDB). Последовательность hu1G7.v85 представлена на фиг. 6А-6В и фиг. 5А-5В. Кинетический анализ hu1G7.v85 показан в таблице 8. Для сравнения показан анализ hu1G7.v1.4. Различия, наблюдаемые в KD hu1G7.v1.4 (таблицы 7 и 8), находятся в пределах экспериментальной вариации, ожидаемой для этих молекул.
Также была получена версия с не созревшей аффинностью hu1G7.v.l, hu1G7.v87, которая содержит HVR 1G7 мыши в описанной выше гуманизированной каркасной области с мутацией M64V для улучшения устойчивости к окислению.

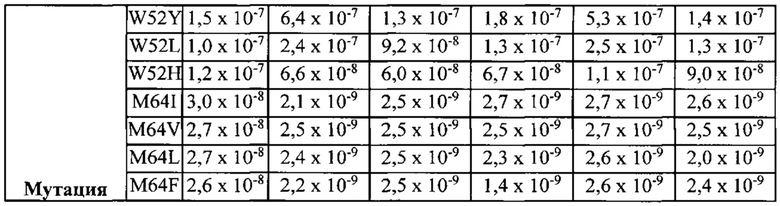
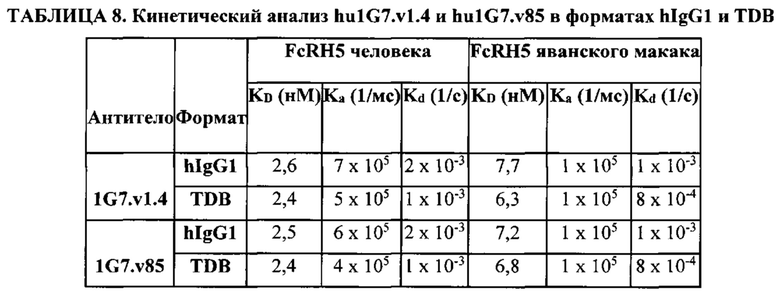
При дополнительном методе созревания аффинности используется секвенирование по Сэнгеру для рандомизации положений HVR тяжелой цепи; однако этим способом не было выявлено никаких желательных мутаций. Поэтому было расширено количество положений, протестированных на варианты и глубину скрининга с использованием секвенирования нового поколения (секвенирование Illumina). Была разработана библиотека, в которой выбранные положения в вариабельной области тяжелой цепи были рандомизированы с помощью кодона NNK, который кодирует любую аминокислоту или янтарный стоп-кодон. Конструкция допускает изменение только одной аминокислоты в вариабельных областях антитела на один клон. Положения были выбраны из HVR и положений каркасных областей в прямом контакте или расположенные близко к HVR. Положения по Kabat 1, 2, 24-40, 43, 45-78, 80-83, 85, 86, 91 и 93-102 были рандомизированы. Библиотека была создана путем синтеза ДНК (GeneWiz), с получением 75 независимых линейных фрагментов ДНК с одним положением в каждом фрагменте, рандомизированном с кодоном NNK. Линейные фрагменты ДНК объединяли и клонировали в один из векторов фагового дисплея моновалентного фрагмента Fab (Lee et al., J. Immunol. Methods 284: 119-132, 2004), содержащий вариабельную область легкой цепи без мутаций созревания аффинности. Продукты для лигирования электропорировали в XL-1 Е. coli, суперинфицировали хелперным фагом М13 KO7 (New England Biolabs) и выращивали, как описано. Связующие вещества были отобраны в трех циклах сортировки с FcRH5 человека или яванского макака на параллельных треках. Для этой селекции планшеты ИФА, покрытые нейтравидином (Pierce), использовали для захвата биотинилированного домена 8 FcRH5 человека или яванского макака с концентрацией 50 нг/мл в PBS. Фаг (1 ОП268/мл) инкубировали с иммобилизованным FcRH5 в течение двух часов при комнатной температуре, а несвязанный фаг удаляли, промывая 10 раз PBS. Библиотеку и выбранный фаг использовали для инфицирования XL-1 E. coli, плазмидную ДНК экстрагировали и вставки амплифицировали с помощью 15-цикловой ПЦР-амплификации с использованием ДНК-полимеразы Phusion (New England Biolabs) с последующей очисткой ампликонов в агарозном геле. ДНК из фага из исходной библиотеки (2 ОП268) экстрагировали с помощью набора для очистки ДНК М13 (Qiagen) и 200 нг ДНК использовали в качестве матрицы для амплификации вставок с использованием тех же условий, что и выше. Ампликон были секвенированы при помощи секвенирования Illumina, как описано ранее (Koenig et al., J. Biol. Chem. 290: 21773-21786, 2015). Последовательности фильтруют и анализируют, как описано выше (Koenig et al. J. Biol. Chem. 290: 21773-21786, 2015), удаляя все последовательности с более чем одной мутацией в вариабельной области, последовательности с мутациями, не соответствующими использованию вырожденного кодона NNK, и последовательности со стоп-кодонами. Обогащение рассчитывали путем деления частоты данной мутации в данном положении в сортированной выборке на частоту той же самой мутации в несортированной выборке (исходной библиотеке), как описано ранее (Fowler et al., Nat. Methods 7: 741-746, 2010), используя следующую формулу: 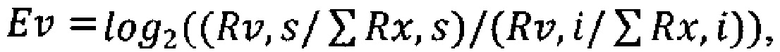 где Eν - обогащение мутанта, Rν,s - число прочтений с мутацией ν в положении s в сортированной совокупности, Rx,s - число прочтений в сортированной совокупности с мутациями в том же положении, что и у варианта Rν,s, Rν,i - число прочтений того же мутанта, что и Rν,s на входе, несортированной совокупности и Rx,i - сумму всех вариантов во входной, несортированной совокупности с мутациями в том же положении, что и вариант Rν,i. Степень Eν для отбора с использованием FcRH5 человека и яванского макака показаны в таблицах 9 и 10, предпочтительно, а серые прямоугольники представляют собой мутации, которые не были идентифицированы в выбранном образце или в библиотеке.
где Eν - обогащение мутанта, Rν,s - число прочтений с мутацией ν в положении s в сортированной совокупности, Rx,s - число прочтений в сортированной совокупности с мутациями в том же положении, что и у варианта Rν,s, Rν,i - число прочтений того же мутанта, что и Rν,s на входе, несортированной совокупности и Rx,i - сумму всех вариантов во входной, несортированной совокупности с мутациями в том же положении, что и вариант Rν,i. Степень Eν для отбора с использованием FcRH5 человека и яванского макака показаны в таблицах 9 и 10, предпочтительно, а серые прямоугольники представляют собой мутации, которые не были идентифицированы в выбранном образце или в библиотеке.
В таблице 11 показаны мутации с оценками по меньшей мере 0,5 в отборе с FcRH5 человека и по меньшей мере 0 в отборе с FcRH5 яванского макака. В таблице 11 мутации, выбранные для дальнейшего анализа (т.е. мутации, которые по меньшей мере не особо предпочтительны для связывания с FcRH5 человека и которые являются нейтральными для связывания FcRH5 яванского макака) выделяются черным или серым цветом, а мутация L29T, идентифицированная при помощи секвенирования по Сэнгеру выделена серым цветом. В общей сложности 11 положений имели мутации, отвечающие этим критериям, с несколькими вариантами в некоторых положениях. Одно положение имело мутацию, которая вводила сайт гликозилирования (G54N) и не была выбрана для дальнейшего изучения. Другой вариант, L29T, был идентифицирован путем скрининга с использованием секвенирования по Сэнгеру, но был идентифицирован только в клонах с мутацией S56T. Этот вариант показал среднее улучшение аффинности только в 1,2 раза. Мутант S56T имел сильный отрицательный коэффициент обогащения -2,1 (таблица 9), что указывает на то, что мутация L29T может иметь благоприятный эффект, который был скрыт из-за присутствия второй мутации S56T у двойного мутанта. Из остальных девяти положений была выбрана мутация с наивысшей оценкой, за исключением положений 1 и 65, где были выбраны две мутации. Эти варианты были получены с помощью синтеза ДНК (GenWiz) и экспрессированы в виде IgG1 человека в комбинации с легкой цепью hu1G7.v1 (без мутаций созревания аффинности) в клетках Expi293 в 1-миллилитровых культурах. Варианты IgG1 человека очищали хроматографией с протеином А и тестировали в BIACORE® на аффинность. Из двух тестируемых мутаций только две, L29T и S100P оказали значительное влияние на аффинность при тестировании в BIACORE® с растворимым FcRH5 человека. Отдельно мутация L29T улучшала аффинность в 2,2 раза по сравнению с мутацией S100P, которая улучшала аффинность в 3 раза. Эти мутации тестировали изолированно и в комбинации в контексте легкой цепи с созревшей аффинностью с мутациями I32L, A51G, T56S и S94P, присутствующими в hu1G7.v86. L29T тяжелой цепи улучшала аффинность hu1G7.v86 примерно в 2 раза, по сравнению с легкой цепью без мутаций. Напротив, S100P, которая по сравнению с легкой цепью без мутаций улучшил аффинность в 3 раза, улучшил аффинность hu1G7.v86 в 1,5 раза. Комбинированные мутации L29T и S100P улучшили аффинность hu1G7.v86 в 2,5 раза. Таким образом, даже используя глубокое исследование с секвенированием нового поколения и тестированием многих положений каркасной области в дополнение к положениям CDR для предпочтительных мутаций в тяжелой цепи, можно добиться лишь 2,5-кратного улучшения аффинности. Результаты показали, что мутации, которые в значительной степени способствовали аффинности при тестировании по отдельности, имели гораздо меньшее влияние на молекулы, у которых уже были другие мутации. Это особенно верно в отношении мутации S100P в тяжелой цепи, которая отдельно улучшает аффинность hu1G7.v1 примерно в 3 раза по сравнению с молекулой с мутациями легкой цепи I32L, A51G, T56S и S94P и мутацией тяжелой цепи L29T имела небольшое дополнительное влияние на аффинность (таблица 12, сравнение последовальности hu1G7.v91 и hu1G7.v93; hu1G7.v93, представленная на фиг. 7А-7В). Таким образом, хотя значительное улучшение аффинности может быть достигнуто путем добавления нескольких мутаций к легкой цепи, относительно небольшое улучшение может быть достигнуто путем введения мутаций в тяжелую цепь.

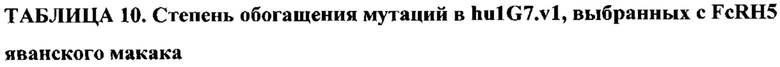


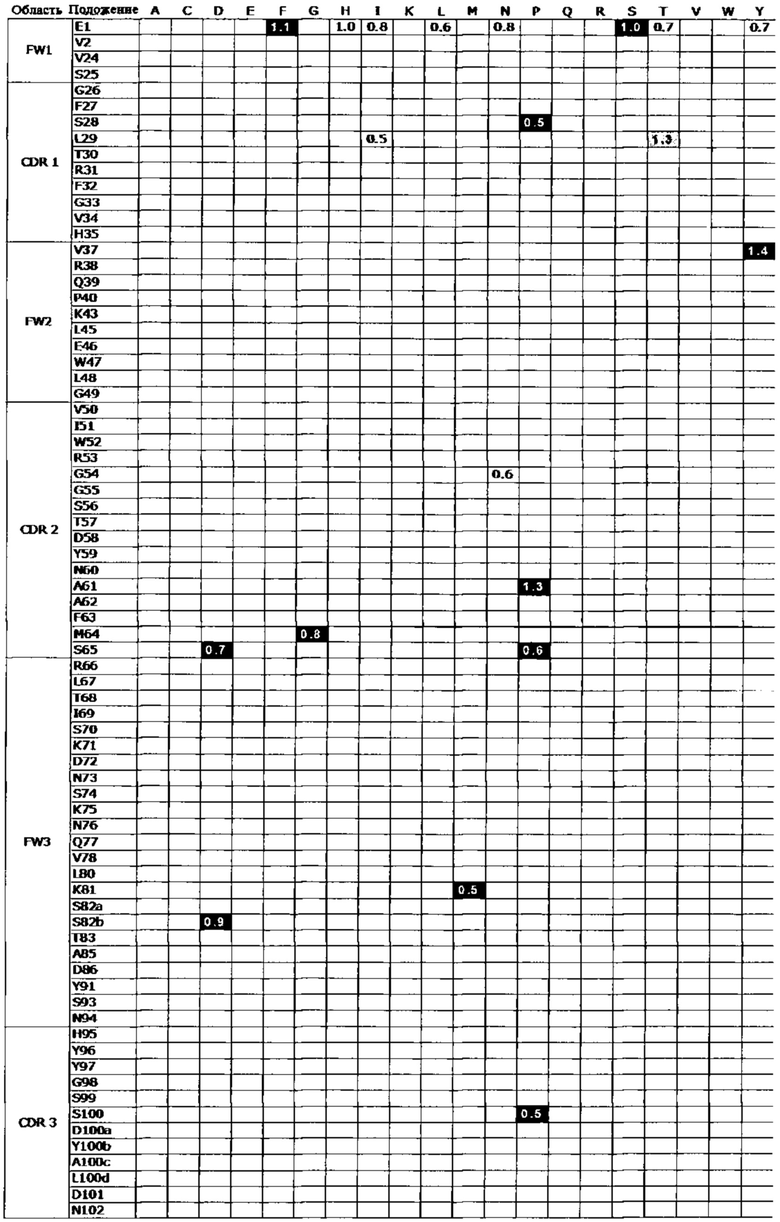

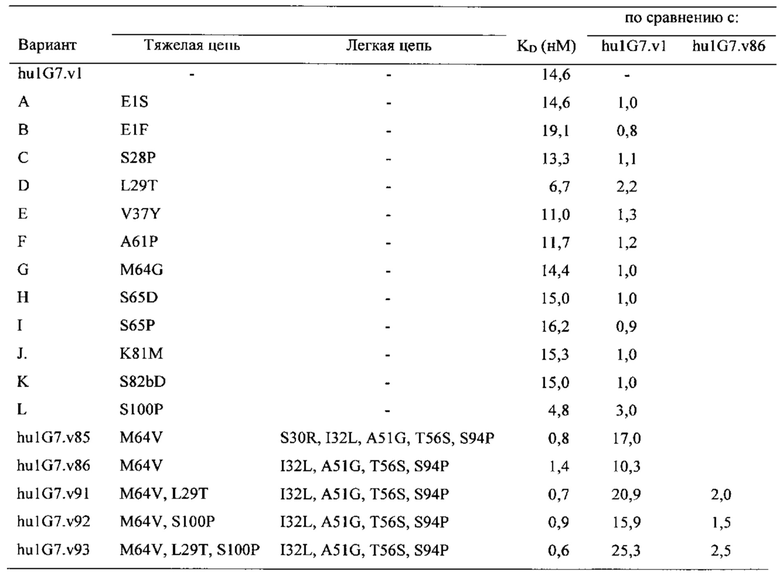
Пример 3: Генерация и исследование in vitro типовых TDB против FcRH5
Антитела против FcRH5, описанные в данном документе, использовались для генерации Т-клеточно-зависимых биспецифических (TDB) антител, содержащих связывающие детерминанты антитела против FcRH5 на одном плече и антитела против CD3ε человека на другом плече. FcRH5-связывающие детерминанты включают гуманизированные и с созревшей аффинностью клоны моноклональных антител 1G7, 1G7.v85 и 1G7.v87. Гуманизированные связывающие детерминанты для антител против CD3ε включают клон высокоаффинного антитела 38E4.v1, высокоаффинный клон 38E4.v11 и низкоаффинный клон 40G5 (ЭК50 для hCD3ε=1,0 нМ, 50 пМ и 13 нМ, соответственно). Клоны антител против CD3 38E4.v1, 38E4.v1l и 40G5c связывают полипипептид CD3ε человека (фрагмент полипептида CD3ε человека, состоящий из аминокислот 1-26 или 1-27 (SEQ ID NO: 174)) и аминокислотный остаток Glu5 CD3ε не требуется для связывания (см. также публикацию заявки РСТ № WO 2015/095392 и публикацию патентной заявки США №2015-0166661, каждая из которых включена в данное описание посредством ссылки в полном объеме).
Иллюстративные FcRH5 TDB были охарактеризованы для оценки их терапевтического потенциала. Было обнаружено, что гуманизированные полноразмерные молекулы FcRH5 TDB уничтожают плазмоциты человека и опухолевые клетки первичной миеломы, полученные из пациентов, в крайне низких (пМ) дозах и вызывают устойчивую пролиферацию Т-клеток, TDB против. FcRH5 были эффективны в подавлении роста ксенотрансплантатов миеломы in vivo и привели к полному истощению В-клеток и плазмоцитов у приматов при хорошо переносимых дозах, которые, как ожидается, будут насыщать мишень. Полное истощение плазмоцитов обеспечило убедительные доказательства эффективности в микроокружении костного мозга. Активность TDB против. FcRH5 коррелирует с экспрессией мишени (например, экспрессией FcRH5), предполагая, что пациенты с высокой степенью риска миеломы с увеличениме числа копий хромосомы 1q могут быть чувствительны к этой иммунотерапии.
Материалы и способы
А. Антитела
а. Продуцирование TDB
1. Метод рационального дизайна
Чтобы оптимизировать экспрессию и увеличение выхода TDB, мутации были сконструированы в доменах Fc, VH, VL, CH1 и CL антител против FcRH5 (например, TDB против. FcRH5), для управления соответствующим образованием мономера антитела. В частности, модификации аминокислот, которые вводили заряженные области в области VH, VL, СН1 и/или CL, обеспечивали механизм для уменьшения ошибочных спариваний тяжелой цепи и легкой цепи. Сопряженные заряженные области, имеющие противоположные общие заряды, управляют и содействуют в соотвествующем спаривании тяжелой цепи и легкой цепи, увеличивая общий выход продукции антитела против FcRH5 (например, TDB против. FcRH5). Иллюстративные конфигурации Rational Design, включая модификации аминокислот доменов Fc, VH, VL, CH1 и CL, представлены на фиг. 1А-1В.
2. Метод моделирования Rosetta
Кроме того, или, альтернативно, для оптимизации экспрессии и увеличения выхода TDB, мутации были сконструированы в доменах Fc, VH, VL, CH1 и CL антител против FcRH5 (например, TDB против. FcRH5), чтобы обусловить соответствующее образование мономера антитела. Метод моделирования Rosetta обеспечил дополнительный механизм управления соотвествующего спаривания тяжелой цепи и легкой цепи в дополнение к мутациям с заряженной областью в форме мутаций в виде выступов и впадин в доменах СН1 и CL. Иллюстративные конфигурации Rosetta Design, включая модификации аминокислот доменов Fc, VH, VL, CH1 и CL, представлены на фиг. 1С-IF.
3. Одноклеточный подход для получения TDB против FcRH5
В одном подходе TDB против FcRH5 могут быть получены путем культивирования клеток-хозяев, которые были совместно трансфицированы двумя плазмидами, каждая из которых кодирует одно из двух плеч TDB против FcRH5 (например, первая плазмида, кодирующая полуантитело против FcRH5 и вторую плазмиду, кодирующую полуантитело против CD3). Трансфекцию клеток-хозяев (например, клеток бактерий, млекопитающих или насекомых) проводили в 96-луночном планшете. Для скрининга продурования TDB против FcRH5 можно отбирать и оценивать примерно от 2000 до 3000 клонов при помощи ИФА и анализа методом гомогенной флуоресценции с временным разрешением (HTRF) интактной IgG на их способность связывать целевой антиген FcRH5. Клоны, продуцирующие TDB против FcRH5, способные связываться с FcRH5, или их фрагментом, были выбраны для размножения и дальнейшего скрининга (например, на связывание с CD3). Лучшие клоны затем могут быть выбраны для дальнейшего анализа на основе процента продуцируемых биспецифических антител (бисАт), титра и эксплутационных свойств (PQ).
Иллюстративный одноклеточный подход, который может быть использован для получения антител TBD против FcRH5 согласно изобретению, описан в международной патентной заявке № PCT/US 16/28850, которая полностью включена в данное описание посредством ссылки.
4. Двухклеточный подход для получения TDB против FcRH5
Альтернативно, TDB против FcRH5 могут быть получены путем культивирования гемимеров антител (например, полуантител) отдельно (то есть в двух разных клеточных линиях) с использованием ферментации с высокой клеточной плотностью и последующим независимым выделением каждого полуантитела при помощи хроматографии с протеином А. Затем очищенные полуантитела можно объединить, например, в молярном соотношении 1: 1 и инкубировать в 50 мМ Трис, рН 8,5, в присутствии 2 мМ ДТТ (дитиотреитол) в течение 4 часов, чтобы обеспечить отжиг и восстановление дисульфидов в шарнирной области. Диализ против того же буфера без ДТТ в течение 24-48 часов приводил к образованию межцепочечных дисульфидных связей.
TDB могут быть альтернативно получены путем трансфекции двух плазмид, каждая из которых кодирует различные плечи TDB, в отдельные клетки-хозяева. Клетки-хозяева могут быть совместно культивированы или культивированы отдельно. Трансфекцию клеток-хозяев можно проводить в 96-луночном планшете. Для скрининга продуцирования TDB отбирают и оценивают от 2000 до 3000 клонов при помощи ИФА и анализа методом гомогенной флуоресценции с временным разрешением (HTRF) интактной IgG на их способность связывать выбранный антиген (например, FcRH5). Клоны, продуцирующие TDB, способные связываться с FcRH5 или их фрагментом, выбирают для размножения и дальнейшего скрининга. Лучшие клоны выбирают для дальнейшего анализа на основе процента продуцируемых биспецифических антител (бисАт), титра и PQ.
5. Иллюстративные способы получения
В одном примере, используя эту стратегию, TDB против FcRH5 были получены с помощью стратегии совместного культивирования с использованием клеток E. coli, экспрессирующих первое полуантитело (впадина), и клеток E. coli, экспрессирующих второе полуантитело (выступ), выращивали вместе в смесительной колбе с заданным соотношением, так что они продуцировали одинаковые количества каждого полуантитела (см. Spiess et al., Nat. Biotechnol. 31 (8): 753-8, 2013; публикацию заявки PCT № WO 2011/069104, полное содержание которой включено в настоящий документ посредством ссылки. Затем собирали бульон совместно культивированных бактерий, клетки разрушали в микрофлюидизаторе и антитела очищали при помощи аффинности к протеину А. Было замечено, что во время микрофлюидизации и захвата белка А два плеча гибридизировались и образовывали шарнирные межцепочечные дисульфидные мостики.
В другом примере полноразмерные биспецифические антитела были получены, как описано ранее (Junttila et al. Cancer Res. 74: 5561-5571, 2014; Sun et al. Science Trans. Med. 7: 287ra270, 2015). Вкратце, два полуантитела (например, антитело против FcRH5 (например, оптимизированные варианты 1G7) и антитела против CD3 (например, 38E4.v1)), содержащие мутации «впадины» или «выступы» в их доменах СН3, были экспрессированы при помощи транзиентной трансфекции клеток СНО и последующей очистки аффинной хроматографией с протеином А. Равные количества двух полуантител инкубировали с 200-молярным избытком восстановленного глутатиона при рН 8,5 в течение ночи при 32°С для образования дисульфидных связей выступ-впадина. Собранное биспецифическое антитело (например, TDB против FcRH5) очищали от загрязнений посредством хроматографии с гидрофобным взаимодействием. Очищенные TDB против FcRH5 исследовали на чистоту с помощью масс-спектрометрии, эксклюзионной хроматографии (ЭХ) и гель-электрофореза.
b. Очистка TDB против FcRH5
TDB против FcRH5 очищали от загрязняющих веществ при помощи хроматографии с гидрофобным взаимодействием (HIC). Полученный в результате материал анализировали в отношении уровней эндотоксинов, используя портативную тест-систему Endosafe и, в случае необходимости, содержание эндотоксинов уменьшали путем промывки белка 0,1% Тритоном Х-114. Молекулярные массы TDB анализировали с помощью масс-спектрометрии (LC-ESI/TCF), как описано ранее (Jackman et al. The Journal of Biological Chemistry. 285: 20850-9, 2010). TDB против FcRH5 также анализировали с помощью аналитической эксклюзионной хроматографии на колонке Zenix SEC-300 (Sepax Technologies USA) с использованием системы ВЭЖХ Agilent 1:100. Присутствие остаточных фрагментов антител определяли количественно при помощи электрофореза с использованием биоанализатора 2100 и чипа Protein 230.
с. Меченые антитела
Все непосредственно меченые антитела для проточной цитометрии, за исключением упомянутых выше, были приобретены у BD Bioscience. Антитела против PD-1 человека былы приобретены у Affymetrix. Козьи антитела против IgG человека и козьи антитела против IgG мыши были приобретены у Jackson Immunoresearch. Антитело против PC-FITC (РС-фосфорилхолин, FITC-флуоресцеин изотиоцианат) (клон Vs38c) было приобретено у DAKO. Антитело SLP-76 для вестерн-блоттинга было приобретено у Cell Signaling Technology. P-SLP76 (Ser376) был создан в Genentech, Inc.
Для обнаружения FcRH5 из множественной миеломы (ММ) и плазмы крови здорового донора и В-клеток при помощи проточной цитометрии, антитело против FcRH5 1G7 было помечено РЕ (фикоэритрином) от Southern Biotec. Для микроскопии TDB были помечены Alexa Fluor 647 с использованием соответствующего набора для мечения белков (ThermoFisher) в соответствии с инструкциями производителя. TDB подвергали диализу в PBS, при рН 7,2 перед мечением и обычно обеспечивали соотношение краситель/белок равное ~4.
В. Анализы на молекулярную стабильность
Для оценки молекулярной стабильности тесты для TDB против FcRH5 на тепловой стресс проводили при температуре от 30°С до 40°С в течение четырех недель. TDB против FcRH5 инкубировали в концентрации 1 мг/мл в 20 мМ гис-ацетата, 240 мМ сахарозы, рН 5,5 и оценивали через 2 недели. TDB были оценены на <5%-ное изменение N-дезаминирования/D-изомеризации, при <2,5%-ном изменении потери мономера по ЭХ и при <16% в основной пиковой нагрузке по IEC на 2 недели. Используемый буфер для ЭХ состоял из 0,25 М KCl, 0,2 М K3PO4, с рН 6,3. LC-MS проводили в восстановленных условиях с использованием ТСЕР (трихлорэтилфосфат) при 60°С в течение 10 минут. Анализ LC-MS/MS проводили путем RCM-триптического пептидного картирования (RCM-восстановление/алкилирование) с восстановлением ДТТ, кэппированием IAA (иодуксусная кислота) и выщелачиванием при рН 8,2.
Анализ окисления ААРН проводили для оценки <35% окисления Trp и <1,1 Met Ox/Met256 (ОХ - окисление). TDB против FcRH5 инкубировали в концентрации 1,0 мг/мл и подвергали стрессу в 1 мМ ААРН в течение 16 часов. Был также проведен анализ фотоокисления для оценки Trp OX/TrpOX_ApoMabW53 >0,5. Для анализа фотоокисления TDB инкубировали в концентрации 1,0 мг/мл в течение 48 часов и подвергали воздействию света в 2,4 миллиона люкс-часов. Окисление оценивали при помощи анализа LC-MS/MS с RCM-триптическим пептидным картированием с уменьшением ДТТ, кэппированием IAA и выщелачиванием при рН 8,2.
C. Клеточная культура и получение стабильной клеточной линии
Клетки HEK-293Т и HEK-Т, экспрессирующие комплекс TCR 1G4 и ключевые сигнальные компоненты TCR (James et al. Nature 487: 64-69, 2012), культивировали в DMEM (Sigma Aldrich). Все другие клеточные линии культивировали в RPMI. Все среды были дополнены 10%-ным FBS, инактивированным нагреванием, (Life Technologies), 1 мМ HEPES (Lonza), 2 мМ глутамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина (Sigma Aldrich).
Для оценки образования иммунологического синапса клетки SVT2 были инфицированы или ретровирусом, кодирующим полноразмерный FcRH5, имеющим N-концевую метку gD, или вирусом, кодирующим усеченный FcRH5 (то есть FcRH5, включая делецию аминокислот 1-744), имеющий N-концевую метку gD. Чтобы оценить целевую зависимость уничтожения TDB против FcRH5, клетки FOX-NY были инфицированы лентивирусом, кодирующим полноразмерный FcRH5. Клоны, полученные из одной клетки, с уровнями дифференциальной экспрессии FcRH5 затем выбирали с помощью 2 мкг/мл пуромицина.
Чтобы оценить влияние сигналинга PD-1/PD-L1 на активность TDB против FcRH5, клетки 293 были инфицированы лентивирусом, кодирующим FcRH5 с последующей трансфекцией плазмиды, кодирующей PD-L1 человека, с использованием липофектамина (Invitrogen).
D. Анализ радиолигандного связывания клеток
Антитело против FcRH5 1G7.v85 иодировали с использованием метод иодирования до удельной эффективной активности 20 мкКи/мкг.Смеси для конкурентной реакции, содержащие фиксированную концентрацию йодированного антитела и уменьшающиеся концентрации серийно разведенного немеченого антитела, помещали в 96-луночные планшеты. Линии клеток, экспрессирующие эндогенный FcRH5 человека (например, MOLP-2, R1-1, KARPAS 620 и KMS21-BM), промывали буфером для связывания, который состоял из среды Игла, модифицированной по способу Дульбекко (DMEM) с 2% фетальной бычьей сывороткой (FBS), 50 мМ HEPES (рН 7,2) и 0,1% азида натрия, а затем добавлли в 96-луночные планшеты. Конкурирующие реакции с клетками анализировали в трех повторностях для каждой концентрации немеченого антитела и инкубировали в течение двух часов при комнатной температуре. Через два часа инкубации смеси для конкурентных реакций переносили в фильтровальный планшет Millipore Multiscreen (г. Биллерика, штат Массачусетс) и промывали четыре раза с помощью буфера для связывания для отделения свободного от связанного йодированного антитела. Активность высушенных воздухом фильтров подсчитывали на гамма-счетчике Wallac Wizard 2470 (PerkinElmer Life and Analytical Sciences Inc.; Уэлсли, Массачусетс) и данные связывания оценивали с использованием программного обеспечения NewLigand (Genentech), в котором для определения аффинности связывания антитела применен алгоритм подгона Мансона и Родбарда (Munson and Anal. Biochem. 107: 22-39, 1980).
E. Векторы и транзиентная трансфекция для микроскопии
FcRH5, имеющий N-концевую метку gD, сливали с флуоресцентным белком mRuby2, сначала вставляя FcRH5 в лентивирусный вектор pHR-SIN, перед лигированием последовательности ДНК mRuby2 в этот вектор, тем самым создавая pHR-FcRH5-mRuby2. Затем промотор SFFV в этом векторе заменяли промотором mHSP, создавая pHRI-FcRH5-mRuby2, который использует более слабый промотор, чем pHR, что дает более физиологические уровни экспрессии FcRH5-Ruby. Векторы, экспрессирующие LCK, ZAP70, CSK/CBP и CD45, были описаны ранее (James et al. Nature. 487: 64-69, 2012). Используемая конструкция CD45 представляла собой изоформу RO или конструкцию, содержащую цитоплазматический домен CD45 с трансмембранным и внеклеточным доменами CD43, который, как известно, имитирует функцию CD45. До трансфекции конструкций клетки HEK высевали примерно до 60% конфлюентности в 6-луночных планшетах. Затем векторы транзиентно трансфецировали при соответствующих соотношениях с использованием GeneJuice (Novagen), следуя инструкциям производителя. Клетки использовали в экспериментах через 24-48 часов после трансфекции.
F. Микроскопия и анализ
Для визуализации клеточных конъюгатов 3x105 клеток каждого типа клеток, которые должны быть отображены, собирали из культуры и ресуспендировали в 100 мкл 20 нМ TDB в RPMI-1640 (без фенолового красного). После 20-30-минутной инкубации, чтобы обеспечить конъюгацию клеток, клетки промывали PBS, ресуспендировали в среде для визуализации DMEMgfp2 (Evrogen) и добавляли к 35-миллиметровым чашкам для визуализации (Mattek). Для визуализации клеток при 37°С использовали систему спиннинг-диск конфокальной микроскопии Andor. Все изображения были проанализированы, и все представленные изображения были обработаны эквивалентным образом с использованием ImageJ. У представленных изображений вычитали фон, и затем обрезали, чтобы сфокусироваться на паре клеток, и оптимизировали контраст.Степень кластеризации и сегрегации белка определяли по интенсивности флуоресцентно меченых белков в плазмалемме. Плазмалемму выбирали вручную, проводя линию, а среднюю интенсивность флуоресценции плазмалеммы в пределах области контакта клетка-клетка делили на среднюю интенсивность флуоресценции плазмалеммы вне области контакта клетка-клетка для расчета степени кластеризации или сегрегации. Чтобы получить изображение области контакта пары клеток, конъюгированных TDB из серии срезов по оси Z, стек изображений сначала восстанавливали из свертки, а затем обрезали, чтобы выделить область контакта с помощью программного обеспечения Huygens.
G. Анализ цитотоксичности in vitro и активации Т-клеток
Целевые клетки метили карбоксифлуоресцеин сукцинимидиловым эфиром (CFSE) в соответствии с протоколом производителя (Life Technology, #С34554). Меченые CFSE целевые клетки и очищенные CD8+ клетки смешивали в соотношении эффекторные клетки к целевым клеткам (Е:Т) 3:1 и инкубировали с TDB в течение 48 часов. В конце инкубации клетки анализировали с помощью проточной цитометрии на FACSCalibur в автоматическом режиме. Число живых целевых клеток подсчитывали путем гейтирования по CFSE+/PI-отрицательным клеткам. Процент цитотоксичности рассчитывали следующим образом: % цитотоксичности (число живых целевых клеток без TDB - количество живых целевых клеток с ТБВ)/(число живых целевых клеток без TDB) X 100.
а. Анализ цитотоксичности in vitro на линиях клеток
Разделение мононуклеарных клеток периферической крови (МКПК) и CD8+, анализы жизнеспособности на основе Cell Titer Glo (Promega) и проточной цитометрии (48 ч) проводили, как описано ранее в Junttila et al. Cancer Res. 74: 5561-5571, 2014. CD8+ клетки использовали в качестве эффекторов в соотношении эффектор-мишень 3:1.
b. Человеческие плазмоциты и образцы первичной множественной миеломы
Пунктат человеческого костного мозга здоровых доноров (ALLCELLS) разводили в PBS, а моноклуеарные клетки костного мозга (МККМ) выделяли обычным градиентным разделением (Lymphoprep, STEMCELL). Анализ жизнеспособности на основе проточной цитометрии использовали для исследования влияния 72-часовой терапии TDB против FcRH5 на плазмоциты МККМ. Замороженные МККМ человека из пациентов с ММ были приобретены у Conversant Bio. Миеломные МККМ смешивали с недавно выделенными CD8+ Т-клетками здорового донора, а совместную культуру клеток обрабатывали TDB против FcRH5 в течение 72 часов. PI-негативные CD38+CD138+ клетки подсчитывали с помощью проточной цитометрии.
Н. Анализ активации и пролиферации Т-клеток
Анализ активации Т-клеток ранее был описан в Junttila et al. Cancer Res. 74:5561-5571, 2014. Свежевыделенные CD8+ Т-клетки метили CFSE и смешивали с целевыми клетками (например, клетки MOLP-2) в соотношении 1: 1 и совместно культивировали с 1 мкг/мл TDB в течение 48 часов или пяти суток. Клетки окрашивали антителами против CD8-APC (BD Bioscience, #555634), антителами против CD69-PE (BD Bioscience, #555531) и/или антителами против CD25-APC (BD Bioscience, №555434) и разведение интенсивности флуоресценции CFSE анализировали с помощью проточной цитометрии.
I. Анализ методом проточной цитометрии для плазмоцитов яванского макака
Пунктат костного мозга яванского макака разводили (1:10) в буфере для лизиса АСК (Life Technology, #А1 0492) дважды. Клетки костного мозга яванского макака окрашивали антителом против CD45, антителом против CD20 и антителом против CD38. После промывки клетки фиксировали и пермеабилизировали с помощью набора IntraStain (DAKO). Затем клетки окрашивали антителом против PC (клон Vs38c). Плазмоциты яванского макака классифицировали при помощи проточной цитометрии как CD45-CD20-CD38+PC+.
J. Анализ уровня IgG яванского макака методом ИФА
Общий IgG сыворотки яванского макака определяли количественно с использованием стандартного колориметрического сэндвич-ИФА. В качестве антитела для захвата и детектирования использовали козье антитело против IgG обезьяны (Bethyl А140-202А) и конъюгированное с пероксидазой хрена (HRP) козье антитело против IgG обезьяны (Bethyl A140-202P), соответственно. IgG яванского макака (Cell Sciences CSI20163A) использовали в качестве стандарта количественной оценки белка.
K. Вестерн-блоттинг
Свежевыделенные человеческие CD8+ Т-клетки и клетки 293T-FcRH5 (соотношение 2: 1) обрабатывали 1 мкг/мл 1G7.v85/38E4.v1 («1G7.v85 TDB»), 10A8/38E4.v1 («10А8 TDB» ») или aHra-gD/38E4.v1 («TDB против gD») TDB и промывали в фосфатно-солевом буферном растворе (PBS) при 4°С и лизировали буфером для лизиса RIP A (Cell Signaling Technology). PSLP76 (Ser376) и SLP76 были обнаружены с использованием стандартного вестерн-блоттинга и антител.
Результаты
А. Аффинность связывания TDB против FcRH5
Среди продуцируемых TDB против FcRH5 были молекулы, содержащие клон 1G7 в качестве FcRH5-связывающего домена и выбранные клоны в качестве CD3-связывающего домена, включая 38E4.v1, 40G5c и 38.E4.v1l. Антитела hu1G7.v1.1 ("1G7.v1.1 TDB"), hu1G7.v1.2 ("1G7.v1.2 TDB"), hu1G7.v1.3 (1G7.v1.3 TDB"), hu1G7. v1.4 («1G7.v1.4 TDB»), hu1G7.v1.5 («1G7.v1.5 TDB»), hu1G7.v1.7 («1G7.v1.7 TDB»), hu1G7.v1. 13 («1G7.v1.13 TDB») и hu1G7.v1.13.1 («1G7.v1.13.1 TDB») генерировали в виде TDB против FcRH5 с плечом антитела против CD3 38E4.v1 и определяли при помощи BIACORE®, как правило, как описано в данном документе, с высокой аффинностью к растворимому фрагменту белка FcRH5 (таблица 13). Исследования экспрессии 1G7.v85 и 1G7.v87 в форматах полуантител, включая мутанты по типу «выступ», дали титр выше нежелательного уровня (фиг. 8).
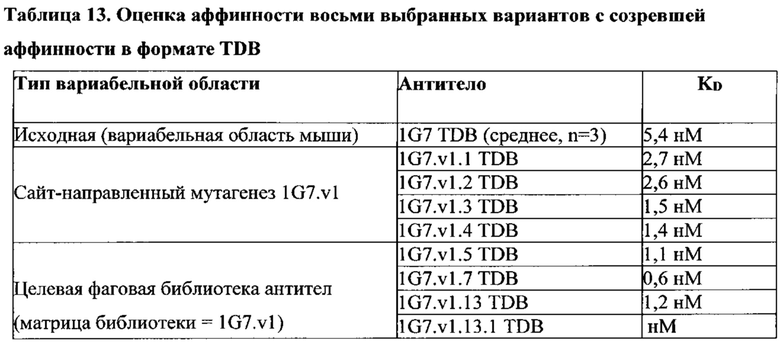
В. Связывание и перекрестная реактивность TDB против FcRH5
Клетки SVT2 мыши, трансфецированные FcRH5 человека, FcRH5 яванского макака, FcPVH1 человека, FcRH2, FcRH3 и FcRH4, использовали для проверки связывания и перекрестной реактивности TDB против FcRH5. Способ выполняли следующим образом. Культивированные клетки SVT2 поднимали с использованием неферментого буфера для диссоциации клеток (Sigma, #С5914). 1×105 клеток суспендировали в 100 мкл и инкубировали с TDB против FcRH5 при 3 мкг/мл. Затем клетки промывали буфером FACS (PBS, 1% BSA, 2 мМ EDTA) и инкубировали с козьим антителом против Fc РЕ (Jackson Immunoresearch, №109-116-170) при разведении 1:100. Клетки промывали дважды буфером FACS до проведения анализа проточной цитометрии на FACSCalibur.
После гуманизации и созревания аффинности мышиного 1G7 было получено восемь созревших вариантов в формате TDB. Оптимизированные 1G7 TDB были созданы в формате «высуп-во-впадину» с 38E4.v1 в качестве плеча антитела против CD3. Все восемь вариантов показали увеличение в 5-15 раз аффинности к мышиному 1G7 TDB при помощи BIACORE® в формате TDB (таблица 13). Все варианты продемонстрировали отрицательное связывание с FcRH1 или FcRH4 с разной степенью положительного связывания с FcRH2 или FcRH3. TDB 1G7.v1.4 показал наименьшую перекрестную реактивность для FcRH2 и FcRH3 (таблица 14). Связывание TDB против FcRH5, содержащих различные плечи антител против FcRH5 (то есть 1G7, 1G7.v85 или 1G7.v1.4) с FcRH3, также оценивали с помощью проточной цитометрии (фиг. 9А).
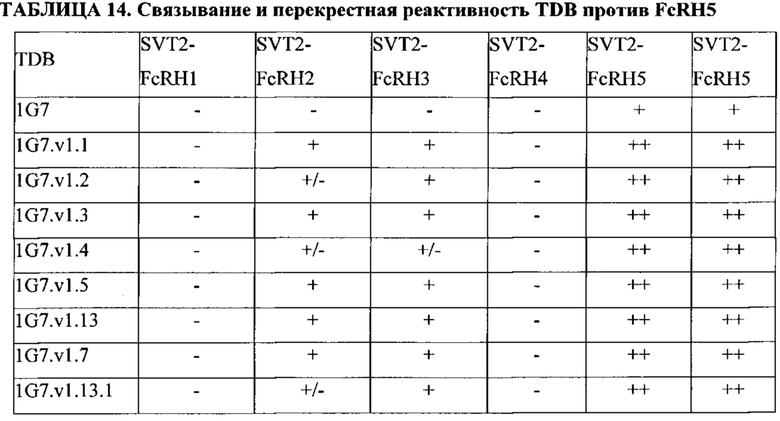
Способности TDB 1G7.v1.4, 1G7.v85 и 1G7 для связывания huFcRH5 и huFcRH3 оценивали при помощи анализа BIACORE® (фиг. 10A-10D). Аффинности TDB 1G7.v85 и TDB 1G7.v1.4 были сопоставимы. 1G7.v1.4 TDB, связанное с FcRH5 человека и FcRH3 человека с KD 2,4 нМ и и ~80 нМ, соответственно и 1G7.v85 TDB, связанное с FcRH5 человека и FcRH3 человека с KD 2,4 нМ и ~90 нМ, соответственно (фиг. 10A-10D). Дополнительные варианты антител против FcRH5 hu7D8.LlH2, 17B1-VH66, 17В1 и 15G8 (последовательности, представленные на фиг. 11А-13В) также были сгенерированы в формате TDB с плечом антитела против CD3 38E4.v1. TDB hu7D8.LlH2, 17В1 и 15G8 были протестированы на связывания с FcRH5 человека и яванского макака. Результаты представлены на фиг. 14 и в таблице 4.
Сравнение связывания 1G7.v85 TDB и 1G7.v93 TDB с FcRH5 человека и FcRH5 яванского макака проводили при помощи анализа BIACORE® (фиг. 15). TDB 1G7.v85 проявило KD равную 2,1 нМ для связывания с FcRH5 человека и KD равную 7,8 нМ для связывания с FcRH5 яванского макака. 1G7.v93 TDB проявило KD равную 1,1 нМ для FcRH5 человека и 8,1 нМ для FcRH5 яванского макака.
Проведены эксперименты по определению кинетических характеристик (см. таблицу 8). В этих экспериментах связывание растворимого фрагмента белка FcRH5 с очищенными антителами в формате антитела TDB человека анализировали при шести разных ненулевых концентрациях (серия разведения 1:3, одна концентрация вводилась дважды как повторность). Скорость потока устанавливали на уровне 40 мкл/мин, и ассоциацию и диссоциацию наблюдали в течение 600 с. KD для FcRH5 человека и FcRH5 яванского макака, полученная при помощи кинетического анализа аффинности моновалентного TDB 1G7.v85 с использованием формата захвата hIgG и модели связывания 1:1, составила 2,4 нМ и 6,8 нМ, соответственно (фиг. 16А-16В). Мутация окисленного Met-64НС на Va1 в TDB 1G7.v85 не влияла на аффинность связывания.
С. Анализы цитотоксичности in vitro
Для оценки функции гуманизированных вариантов TDB против FcRH5 тестировали клеточную цитотоксичность на клетках FcRH5 MOLP-2, которые эндогенно экспрессировали FcRH5. Все варианты показали сходную активность в отношении активности целевых клеток, но значительно увеличивали активность по сравнению с мышиным 1G7 (фиг. 17А-17В). В случае варианта TDB 1G7.v1.4 ЭК50 улучшалась в 5-13 раз по сравнению с мышиным TDB 1G7 (n=10).
Основываясь на своей высокой цитотоксической активности и низкой перекрестной реактивности к другим членам семьи, для дальнейшего анализа был выбран TDB 1G7.v1.4. Чтобы повысить стабильность молекулы TDB, плечо 1G7.v1.4 подвергали мутации на потенциальных участках окисления, создавая, таким образом, очищенную версию 1G7.v85. Две версии, 1G7.v1.4 и 1G7.v85, были оценены в формате TDB в различных анализах, включая активацию Т-клеток, активность уничтожения целевых клеток MOLP-2, истощение В-клеток яванского макака in vitro и истощение плазмоцитов яванского макака (фиг. 18A-18D). Во всех проведенных анализах TDB 1G7.v85 не отличалось от TDB 1G7.v1.4.
Гуманизированное и очищенное TDB 1G7.v87 также было сравнимо с TDB 1G7.v85. В соответствии с его более низкой аффинностью, TDB 1G7.v87 демонстрирует уменьшенное связывание с FcRH5 яванского макака по сравнению с TDB 1G7.v85, как показано при помощи метода проточной цитометрии (фиг. 19А). 1G7.v87 также продемострировал отрицательную перекрестную реактивность к FcRH3.
Цитотоксичность к целевым клеткам TDB 1G7.v87 и TDB 1G7.v85 оценивали на CD8+ клетках MOLP-2 (фиг. 19В) и на МКПК от четырех здоровых доноров (фиг. 19С). Во всех донорах TDB 1G7.v87 был отрицательным по уничтожению В-клеток человека, тогда как TDB 1G7.v85 показал некоторую степень положительной активности уничтожения. Также были протестированы МКПК яванского макака от четырех здоровых доноров (фиг. 19D). Ранжирование ЭК50 трех версий TDB 1G7 согласуется для всех трех доноров. TDB 1G7.v87 сопоставимо с мышиным TDB 1G7, но имеет значительно меньшую активность, чем TDB 1G7.v85. TDB 1G7.v87 продемонстрировал значительно более слабую активность уничтожения, чем TDB 1G7.v85 на В-клетках яванского макака in vitro. TDB 1G7.v85 также оценивали на активность на NK-клетках человека и обнаружили, что оно не имеет активности уничтожения при ≤20 мкг/мл (фиг. 9В). ЭК50 TDB 1G7.v85 была в 5-8 раз лучше, чем у TDB 1G7.v87.
D. Молекулярная оценка устойчивости TDB против FcRH5
Химическая стабильность hu1G7.v1 до созревания аффинности указывает на склонность к окислению в Met-64НС и Trp-52НС. Образцы TDB 1G7.v85 подвергали стрессу посредством тестов с ААРН и стрессу при избытке освещения и оценивали на связывание FcRH5 с помощью BIACORE®. TDB 1G7.v85 продемонстрировал снижение связывания FcRH5 после тестов с ААРН и на стресс при избытке освещения по сравнению с не подвергающимся стрессу контролем (фиг. 20A-20D). Стабильность мономера и гетерогенность заряда в TDB 1G7.v85 оценивали при помощи эксклюзионной хроматографии (ЭХ) и динамического капиллярного изоэлектрофокусирования (icIEF), соответственно, после того, как их подвергали стрессу в буфере с низким рН (гис-ацетат, рН 5,5) в течение 2 недель (фиг. 21А-21В). Наблюдаемое изменение пиковых потерь мономера было небольшим как для стабильности мономера (0,1%), так и для гетерогенности заряда (7,7%) (фиг. 21А-21В). Кроме того, не наблюдали заметное изменение массы или легких цепей, или тяжелых цепей 1G7.v85 TDB после двух недель стресса, обусловленного низким рН (фиг. 22А-22В). После теплового стресса при 30°С в течение 2 недель пиковые потери мономера составляли 0,1% по ЭХ и 8% по icIEF.
Е. Мембранно-проксималъный эпитоп необходим для эффективного сигналинга TCR и активности уничтожения TDB против FcRH5
Чтобы описать молекулярные события, которые приводят к активации рецептора Т-клеток (TCR) при стимуляции TDB против FcRH5, была использована восстановленная система (James et al., Nature. 487: 64-69, 2012) которая позволяет контролировать исходные события, ведущие к активации рецептора, управляемым способом. Используя CD8 клетки здорового донора, TDB 1G7/UCHT1.v9 приводил к очень устойчивому фосфорилированию SLP76, что свидетельствует о сигналинге TCR и опосредованном эффективном уничтожении целевых клеток (фиг. 23А-23В, ЭК50=0,5 нМ). Напротив, TDB против gD, которое нацелено на мембранно-дистальный эпитоп, не приводил к обнаружению сигналинга TCR и не мог опосредовать уничтожение Т-клеток (фиг. 23А-23В). Было также подтверждено, что активность TDB определялась расположением эпитопа и размером внеклеточного домена путем нацеливания на клетки, которые экспрессировали сильно усеченную мишень, которая сохраняла эпитопы 1G7 и gD (фиг. 23С). Активность проксимального 1G7/UCHT1.v9 TDB увеличилась в 25 раз (фиг. 23, ЭК50=20 пМ) и TDB против gD было способно эффективно опосредовать уничтожение клеток (ЭК50=0,19 нМ), если помехи, вызванные ВКД (внеклеточными доменами) были устранены. Возможность уровень дифференциальной экспрессии мишени, являющейся причиной разности активностей между клеточными линиями, была исключена из анализа методом проточной цитометрии (фиг. 24А).
Чтобы продемонстрировать, что различия в активности уничтожения были связаны с эпитопом, а не со свойствами клонов специфических антител, было опробовано всего пять уникальных клонов антител, нацеленных на мембранный проксимальный домен к FcRH5 в формате TDB, и продемонстрировали, что активность каждого клона была в 20 раз выше по сравнению с 10А8 (фиг. 23Е). Уровень эндогенной экспрессии FcRH5 при множественной миеломе является низким (например, от 100 до 2000 копий/клетка) и сопоставим с линией клеток миеломы MOLP-2 (например, ~2200 копий/клетка). Если Т-клетки были перенацелены на уничтожение клеток MOLP-2, только мембранные проксимальные TDB индуцировали уничтожение клеток MOLP-2. Нацеливание на среднюю область FcRH5 с использованием TDB 10А8 не привела к достаточно сильной стимуляции TCR, требуемому для уничтожения клеток миеломы (фиг. 24 В).
F. TDB против FcRH5 индуцирует зависимое от мишени клеточное уничтожение и пролиферацию Т-клеток
На фиг. 18А обнаружено, что 1G7.v1.4 TDB и 1G7.v85 TDB обладают неразличимыми способностями активации Т-клеток. TDB 1G7.v85 оценивали по специфичности эпитопа и перекрестной реактивности 1G7 яванского макака (фиг. 25А-25С). 1G7/38E4.v1 TDB ("1G7 TDB"), связанный с клетками MOLP-2, В-клетками здоровых доноров, клетками костного мозга и первичными опухолевыми клетками миеломы, как ожидалось (фиг. 26A-26D). Доклиническое исследование активности 1G7 TDB было начато с определения механизма действия с использованием CD8 клеток здоровых доноров. Обработка 1G7 TDB клеток, экспрессирующих мишень, приводила к дозозависимой активации Т-клеток (фиг. 27А). Цитотоксическая активность TDB 1G7 наблюдалась только для положительных по мишени клеток и коррелировала с уровнем экспрессии FcRH5 (фиг. 27В). Т-клеточная активация путем стимуляции 1G7 TDB и целевой клеткой приводила к устойчивой пролиферации Т-клеток. Через пять суток 95% CD8 клеток претерпели целых шесть клеточных делений, что было продемонстрировано путем разбавления флуоресцентного красителя CSFE (фиг. 27С-27Е). Был дополнительно оценен вклад плеча антитела против CD3 и домена Fc в цитотоксическую активность TDB, и было обнаружено, что для достижения цитотоксической активности требуется высокоаффинное плечо антитела против CD3, и напротив домен Fc не является необходимым (фиг. 27F-27G).
G. TDB против FcRH5 опосредует эффективное уничтожение нормальных плазмоцитов и первичных миеломных клеток, полученных из пациентов
Экспрессию FcRH5 в CD138+CD38+ клетках множественной миеломы и нормальных плазмоцитах костного мозга изучали с использованием 1G7.v85 TDB и проточной цитометрии. Все опухолевые клетки, полученные из пациентов, и все нормальные плазмоциты экспрессировали FcRH5 во всех образцах, что свидетельствует о 100% уровне распространения миеломы (фиг. 28А). Была обнаружена значительная вариабельность между пациентами в уровне экспрессии в миеломах. Как правило, уровень экспрессии в опухолевых клетках был не значительно повышен по сравнению с нормальными плазмоцитами, что указывает на то, что получение специфического к опухолевым клеткам TDB против FcRH5, щадящего нормальные плазмоциты, вероятно, нецелесообразно. Уровень экспрессии в нормальных В-клетках стабильно и значительно ниже по сравнению с нормальными плазмоцитами и опухолевыми клетками множественной миеломы.
Ген FcRH5 расположен в хромосомной точке разрыва в lq21 (Hatzivassiliou et al. Immunity. 14: 277-289, 2001). Анализ ~20 биопсий первичных множественных миеломы продемонстрировал значительную связь между экспрессией РНК FcRH5 и увеличением lq21 (фиг. 28Е), демонстрируя, что хромосомная транслокация может приводить к сверхэкспрессии FcRH5 у пациентов с высоким риском миеломы.
Способность 1G7.v85 TDB уничтожать плазмоциты анализировали путем нацеливания на мононуклеарные клетки костного мозга (МККМ), выделенные из пунктата костного мозга здоровых доноров (фиг. 28В). 1G7.v85 TDB индуцировал дозозависимое, высокоэффективное и мощное (ЭК50=85-180 пМ) уничтожение плазмоцитов. Подобную устойчивую активность была обнаружена, когда МККМ из нескольких пациентов с миеломой подвергали обработке 1G7.v85 TDB (фиг. 28С). Примерно 100% уничтожение миеломных клеток с высокой эффективностью (ЭК50=60-1200 пМ) было обнаружено независимо от истории лечения в течение жизни.
Поскольку экспрессия FcRH5 является изменчивой в миеломе (фиг. 28А) и активность 1 G7.v85 TDB коррелирует с уровнем экспрессии (фиг. 28D), независимо от того, будут ли пациенты в нижнем конце спектра экспрессии прогнозируемо реагировать на TDB, FcRH5 также будет исследована. Линию клеток миеломы MOLP-2 идентифицировали как контрольную клеточную линию, которая имеет уровень экспрессии FcRH5, который аналогичен средней экспрессии в плазмоцитах и клетках первичной ММ (фиг. 28А). Мы также идентифицировали несколько линий раковых клеток, которые экспрессируют чрезвычайно низкие уровни мишени и определяли количество FcRH5 на клетку с использованием анализа Скэтчарда. Диапазон сайтов связывания FcRH5 в этих клеточных линиях варьировался от 2200 до 160 на клетку. Несмотря на очень низкое число копий мишени, 1G7.v85 TDB индуцировало устойчивое уничтожение всех тестируемых клеточных линий (ЭК50=2-230 пМ). Расчеты занятости показали, что всего лишь 50 молекул TDB (2%-ное заполнение в MOLP-2, ЭК50=58 пМ) были достаточными для индукции активации Т-клеток и апоптоза целевых клеток.
Таким образом, FcRH5 экспрессируется у всех пациентов с миеломой. TDB против FcRH5 могут уничтожать человеческие плазмоциты и первичные опухолевые клетки миеломы, полученные из пациентов, в пМ дозах. Поскольку для перенаправления активности Т-клеток требуется не большое количество TDB, молекула эффективно уничтожает клетки при очень низкой экспрессии мишени. Полученные результаты демонстрирурют, что TDB против FcRH5 могут быть широко активными у пациентов с миеломой и не подтверждают исключение пациентов из терапии на основании уровня экспрессии FcRH5.
Н. Яванский макак (cyno) является подходящей моделью безопасности и эффективности для TDB против FcRH5
Анализ методом проточной цитометрии периферических В-клеток и плазмоцитов костного мозга подтвердил, что FcRH5 экспрессируется во всей В-клеточной линии дифференцировки в яванском макаке, подобно человеческой (фиг. 29А-29В; Poison et al. Int. Immunol. 18: 1363-1373, 2006). Аминокислотные последовательности FcRH5 человека и яванского макака на 89% идентичны, и TDB 1G7.v85 связывается с FcRH5 и CD3 яванского макака со сравнимой аффинностью. Обработка in vitro целевых клеток, экспрессирующих FcRH5 яванского макака, или клеток MOLP-2, экспрессирующих FcRH5 человека, привела к устойчивому уничтожению с использованием периферических Т-клеток из человека или яванского макак с сопоставимой эффективностью (фиг. 29C-29D). Добавление 1G7.v85 TDB к образцам МКПК/МККМ из яванского образца приводила к зависимому от дозы и устойчивому уничтожению В-клеток яванского макака (фиг. 29Е) и плазмоциты костного мозга (фиг. 29F). На фиг. 18С обнаружено, что 1G7.v1.4 TDB и 1G7.v85 TDB обладают неразличимыми способностями к уничтожению плазмоцитов яванского макака. Эти результаты подтверждают яванского макака как подходящую модель безопасности и эффективности для антител против FcRH5/CD3.
Пример 4. Характеристика in vivo типовых TDB против FcRH5
Материалы и способы
A. Исследования эффективности in vivo в мышиных моделях
а. Мышиная модель ксенотрансплантата huNSG/MOLP-2
Самки гуманизированных мышей NOD.Cg-Prkdcscid I12rgtmlWj1/SzJ (NOD/scid gamma, NSG) NOD ((диабет без ожирения)/SCID (тяжелый комбинированный иммунодефицит)) были получены из Лаборатории Джексона. В день инокуляции клеток пяти животных инокулировали 0,2 мл опухолевых клеток MOLP-2 в концентрации 100 миллионов клеток/мл в HBSS/матригель (HBSS - сбалансированный соляной раствор Хэнка), подкожно в правый бок. Как только объемы опухолей достигли объема в диапазоне 100-250 мм3, животные были рандомизированы на две группы: контрольную группу, получающую носитель, и группу лечения, и первые средства лечения были введены в это время (0 сутки). Все процедуры выполняли один раз в неделю путем внутривенной инъекции в хвостовую вену (в/в), в общей сложности ввели четыре дозы. Контрольная группа получала 0,1 мл 20 мМ ацетата гистидина, рН 5,5, 240 мМ сахарозы, 0,02% буфера TW-20. Группа лечения получала 0,1 мл 1G7.v85 TDB в концентрации 0,5 мг/кг.Опухоли измеряли 1-2 раза в неделю с помощью циркуля на протяжении всего исследования, а массу тела животных измеряли не реже одного раза в неделю. В течение всего этого исследования дважды в неделю проводили клинический осмотр, чтобы отслеживать состояние здоровья животных.
В. Токсикологическое исследование у яванских макак
Толерантность, профиль токсичности, фармакокинетика (РK) и фармакодинамика (PD) TDB против FcRH5 оценивали у наивных самцов яванских макак (cyno) в Charles River Laboratories (CRL). Яванские макаки получали разовую дозу носителя, 1, 2 или 4 мг/кг 1G7.v85 TDB при помощи внутривенной инфузии (1 ч) и подвергали вскрытию через семь суток после начала лечения. В течение первых пяти часов и по окончанию исследования для животных тщательно вели подробные клинические наблюдения, контролировали интенсивность дыхания и температуру тела. Информацию о клинических наблюдениях у клетки и массе тела собирали ежедневно. Образцы крови собирали путем венопункции через предварительное исследование бедренной вены и в определенные моменты времени на протяжении всего исследования для общего анализа крови, биохимического анализа сыворотки, анализа коагулирующей активности и ФК (фармакокинетика) общего количества антител) и конечных точек ФД (фармакодинамика). ФД включала количественные измерения цитокинов (ИЛ-1β, ИЛ-1RA, ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-12/23, ИЛ-13, IL-17, Г-КСФ (гранулоцитарный колониестимулирующий фактор), ГМ-КСФ (гранулоцитарно-макрофагальный колониестимулирующий фактор), ИФН-γ, ФНО-α и МХБ-1 (моноцитарный хемотаксический белок 1)), проточной цитометрии Т-лимфоцитов, В-лимфоцитов, NK-клеток, активированных Т-лимфоцитов, PD-1 и циркулирующего IgG яванского макака. Костный мозг отбирали у анестезированных животных путем аспирации с предварительно исследованной плечевой кости и до вскрытия на 8-е сутки для оценки В-лимфоцитов и плазмоцитов методом проточной цитометрии. При вскрытии измеряли массу органов, а органы и ткани тщательно изучали путем макроскопического и микроскопического исследования. Селезеночные, брыжеечные и нижнечелюстные лимфатические узлы оценивали на Т-лимфоциты, В-лимфоциты и NK-клетки. Все процедуры выполняли в соответствии с Законом о благосостоянии животных, Руководством по содержанию и использованию лабораторных животных и Управлением по охране здоровья животных.
С. ФКФД-исследование яванских макак и мышей
Концентрации TDB против FcRH5 в сыворотке определяли с помощью общего ИФА. В качестве реагента для захвата использовали антитело против человеческого IgG овец, а в качестве реагента для выявления использовали овечье антитело против IgG человека, конъюгированное с пероксидазой хрена (HRP). Анализ концентрации в сыворотке крови во времени с доступными образцами анализировали с помощью некомпартментной модели внутривенного болюсного введения (Phoenix™ WinNonlin®, версия 6.3, Pharsight Corporation, Маунтин-Вью, Калифорния). В анализе данных использовали номинальное время сбора проб и концентрацию номинальной дозы. Весь токсикокинетический (ТК) анализ основывался на данных отдельных животных.
Результаты
A. TDB против FcRH5 подавляет рост прижившихся опухолей MOLP-2 у мышей, воссозданных иммунными клетками человека
Моделирование антимиеломной активности TDB против FcRH5 у мышей является сложным, поскольку антитела против CD3 не реагируют перекрестно с мышиным CD3, а ортолог FcRH5 отсутсвует у мыши. Таким образом, модель мыши с восстановленной иммунной системой человека была установлена путем трансплантации CD34+ отобранных человеческих гемопоэтических стволовых клеток облученным мышам (мышам huNSG). CD8+ клетки человека, собранные из селезенки huNSG мышей, смогли уничтожить клетки MOLP2 в условиях in vitro с эффективностью сопоставимой с периферическими CD8+ клетками человека от здоровых доноров (фиг. 30А). Через 20 недель после трансплантации мышей huNSG инокулировали пятью миллионами клеток MOLP-2 подкожно. Мышей с прижившимися опухолями 100-200 мм3 лечили с помощью одной в/в дозы носителя или 0,5 мг/кг TDB против FcRH5. Лечение TDB против FcRH5 приводило к регрессии опухоли у всех животных (фиг. 30В), что указывает на то, что лечение TDB против FcRH5 подавляет рост опухоли in vivo.
В. TDB против FcRH5 имеет длительный период полужизни в сыворотке
Исследование с однократным введением предназначалось для оценки безопасности, эффективности, фармакокинетических (ФК) и фармако динамических (ФД) свойств 1G7.v85 TDB у приматов, не относящихся к человеку. Яванские макаки получали одну дозу носителя или 1-4 мг/кг 1G7.v85 TDB в виде медленной внутривенной инфузии. Животных тщательно контролировали на наличие нежелательных реакций. Образцы крови отбирали через 0, 2, 6 и 24 ч на анализ цитокинов, клиническая лабораторная диагностика и ФК/ФД ответ.Исследование прекратили через семь суток после начала лечения. TDB против FcRH5 продемонстрировал дозозависимую экспозицию (Cmax и AUC) между 1 и 4 мг/кг и выводился 29-33 мл/сут/кг во всех когортах (фиг. 31А), а уровень Cmax при дозе 4 мг/кг составлял 129 мкг/мл. Это примерно в 2000 раз выше, чем требуется для достижения в условиях in vitro ЭК50 уничтожения плазмоцитов человека и MOLP-2. Исследование ФК, которое не является обязательным, также проводили у мышей SCID.bg и было обнаружено, что TDB 1G7.v85 имеет сравнимые скорости выведения, как и у TDB против gD (фиг. 31В). Расчеты заполнения рецепторов дали основания для предположения о взаимодействии FcRH5 близком к насыщенному на В-клетках периферической крови при Cmax при всех уровнях дозы (фиг. 32D). Эти результаты показали, что TDB 1G7.v85 имеет продолжительный период полужизни in vivo и согласовывается со схемой применения препарата раз в неделю или с меньшей частотой.
С. TDB против FcRH5 истощает В-клетки и плазмоциты костного мозга у яванского макака
Анализ методом проточной цитометрии периферической крови продемонстрировал устойчивий фармакологический эффект на всех уровнях доз. Лечение 1G7.v85 TDB приводило к активации Т-клеток и временной лимфопении (ответ в виде синдрома скопления лейкоцитов по краю участка воспаления) в течение 24 часов (фиг. 31C-31D). В-клетки не выявляли в крови через семь суток после введения дозы, что указывает на то, что, что они были истощены TDB 1G7.v85 (фиг. 31Е). Напротив, CD4+ и CD8+ клетки восстанавливаются в конце исследования (фиг. 32А-32В). Все уровни доз приводили к полному истощению В-клеток в селезенке и костном мозге (фиг. 31F и 31Н). Лечение FcRH5 также индуцировало устойчивое дозозависимое истощение В-клеток из лимфатических узлов (фиг. 31G и 32С).
Истощение плазмоцитов костного мозга яванского макака в условиях in vivo является ключевой конечной точкой эффективности в доклиническом исследовании антитела против FcRH5/CD3. Полное истощение плазмоцитов было обнаружено у животных, получавших лечение в дозе 2-4 мг/кг (фиг. 31I). Лечение 1G7.v85 TDB также приводило к дозозависимому снижению IgG яванского макака, ожидаемому вторичному результату, возникающему в результате истощения плазмоцитов. Теоретически, полное истощение плазмоцитов должно снизить уровень IgG до ~30-40% к 7-ым суткам. Измеренное снижение IgG у яванского макака составляло 37% в группе 2 мг/кг и 44% в группе 4 мг/кг (фиг. 31J). Таким образом, TDB 1G7.v85 вызвало устойчивый ФД ответ у яванского макака, соответствующий его механизму действия. Полное истощение плазмоцитов обеспечивает убедительные доказательства эффективности в микроокружении костного мозга.
D. TDB против FcRH5 хорошо переносится яванским макакой
TDB 1G7.v85 хорошо переносится яванским макакой при уровнях дозы ≤4 мг/кг.Легкие/умеренные побочные эффекты, которые были выявлены, были одинаковыми на всех уровнях дозы, и мы не определили четкого дозозависимого эффекта. Клинические наблюдения были ограничены обратимым увеличением температуры тела в пределах 0,4-1,6°С в течение четырех часов после введения дозы. Влияние на общий анализ крови состояло из ожидаемой острой и обратимой лимфопении, связанной с синдромом скопления лейкоцитов по краю участка воспаления. Как и ожидалось, были обнаружены признаки острого и обратимого провоспалительного состояния (увеличение СРБ, фибриногена, протромбинового времени и активированного частичного тромбопластинового времени). Лечение вызывало обратимое увеличение АЛТ, ACT и общего билирубина.
В соответствии с механизмом действия TDB 1G7.v85 индуцировал быстрое, в целом легкое/умеренное высвобождение цитокинов (фиг. 33A-33F). Все уровни дозы индуцировали провоспалительный ответ (включая ИЛ-6, ИЛ-5, ИФН-гамма, ИЛ-2, ИЛ-13, Г-КСФ и МХБ-1) и противовоспалительный ответ для противодействия этому (ИЛ-1R) достигал максимума на 2-6 ч. Все цитокины были восстановлены до нормального уровня в течение 24 часов. Не было обнаружено признаков экстенсивного или длительного высвобождения цитокинов. Обширный гистопатологический анализ, включающий детальный анализ центральной нервной системы (ЦНС), не выявил значительной органотоксичности. Таким образом, максимальная переносимая доза не была достигнута в исследовании. TDB 1G7.v85 хорошо переносится на уровнях дозы, которые, как ожидается, будут насыщать мишень и достаточны для полного истощения В-клеток и плазмоцитов. При нежелательных явлениях не был обнаружен дозозависимый эффект.
Пример 5. Комбинированная терапия FcRH5
Для изучения возможных комбинированных терапий FcRH5, TDB против FcRH5 тестировали в комбинации с каждым из двух иллюстративных антагонистов, связывающихся с компонентом сигнального пути PD-1. Эти эксперименты показали, что, хотя обратная связь сигналинга PD-1/PD-L1 может уменьшить уничтожение, опосредованное TDB против FcRH5, блокада PD-L1 преодолела это ингибирование, что привело к повышению терапевтической эффективности.
Материалы и способы
A. Антитела
Все меченые антитела для проточной цитометрии, за исключением упомянутых выше, были приобретены у BD Bioscience. Используемое антитело против PD-1 представляло собой KEYTRUDA® (пембролизумаб), а антитело против PD-L1 было получено в Genentech, Inc. Козьи антитела против IgG человека и козьи антитела против IgG мыши были приобретены у Jackson Immunoresearch. Антитело против PC-FITC (РС-фосфорилхолин, FITC-флуоресцеин изотиоцианат) (клон Vs38c) было приобретено у DAKO.
B. Анализ индукции и цитотоксичности PD-1 с помощью антитела против PD-L1
Свежевыделенные CD8+ Т-клетки человека смешивали с клетками MOLP-2 в соотношении 1: 1 и совместно культивировали в присутствии 1000 нг/мл 1G7.v85 TDB в течение 48 часов. Клетки окрашивали флуоресцинизотиоцианат (FITC)-меченым антителом против CD8 («анти-CD8-FITC»), фикоэритрин (РЕ)-конъюгированным антителом против CD69 («анти-CD69-PE») и аллоцикоцианин (АРС)-конъюгированным антителом против PD-1 («анти-PD-1-APC») и анализировали методом проточной цитометрии. Анализ цитотоксичности клеток HEK-293Т, экспрессирующих FcRH5 и PD-L1 («клетки 293-FcRH5-PD-L1») проводили, как было описано выше, с или без 10 мг/мл антитела против PD-L1 или антитела против PD-1 и анализировали с помощью проточной цитометрии.
C. Клеточная культура и получение стабильной клеточной линии
Влияние сигналинга PD-1/PD-L1 на активность TDB 1G7.v85 оценивали путем инфицирования клеток HEK-293Т лентивирусом, кодирующим FcRH5, с последующей трансфекцией человеческой PD-L1 -кодирующей плазмиды с использованием липофектамина (Invitrogen).
D. Анализ цитотоксичности in vitro и активации Т-клеток
Целевые клетки метили карбоксифлуоресцеин сукцинимидиловым эфиром (CFSE) в соответствии с протоколом производителя (Life Technology, #С34554). Меченые CFSE целевые клетки и очищенные CD8+ клетки смешивали в соотношении эффекторные клетки к целевым клеткам (Е:Т) 3:1 и инкубировали с 1G7.v85 TDB в течение от 24 до 48 часов. В конце инкубации клетки анализировали с помощью проточной цитометрии на FACSCalibur в автоматическом режиме. Число живых целевых клеток подсчитывали путем гейтирования по CFSE+/PI-отрицательным клеткам. Процент цитотоксичности рассчитывали следующим образом: % цитотоксичности (число живых целевых клеток без TDB - количество живых целевых клеток с TDB)/(число живых целевых клеток без TDB) X 100.
Результаты
А. Блокада PD-1/PD-L1 усиливает активностъ ТDВ против FcRH5
Сильный TCR-стимулирующий сигнал может привести к иммунодепрессивной обратной связи, которая ограничивает активность Т-клеток. Путь PD-1/PD-L1 является критическим компонентом этой обратной связи и терапевтически подтвержденным механизмом ускользания от иммунологического надзора в нескольких опухолевых признаках. PD-L1 часто экспрессируется миеломными опухолевыми клетками (Gorgun et al., Amer. Assoc. for Cancer Res. 21: 4607-4618, 2015), и его сигналинг может ограничивать активность Т-клеток у пациентов с миеломой. PD-1 отсутствует в покоящихся Т-клетках, индуцированных при активации Т-клеток и ограничивающих активность Т-клеток при хронической инфекции (Zou et al. Science Tran. Med. 8: 328rv324, 2016). Стимуляция 1G7.v85 TDB (48 часов) человеческих CD8+клеток здорового донора в присутствии FcRH5-экспрессирующих клеток приводила к значительной индукции PD-1 в Т-клетках (фиг. 34). Сигнал обратной связи также активируется in vivo. Значительное увеличение PD-1-положительных Т-клеток наблюдали в Т-клетках яванского макака на всех уровнях дозы. Индукция PD-1 была обнаружена как в CD8+, так и в CD4+клетках в крови, селезенке, лимфатических узлах и костном мозге (фиг. 35А-35В и 36A-36D).
TDB 1G7.v85-опосредованное уничтожение целевых клеток с помощью примированных CD8+ клеток оценивали в присутствии и отсутствии антагонистов PD-1/PD-L1. Эффективность 1G7.v85 TDB для примирования CD8+Т-клеток с целью уничтожения целевых клеток, экспрессирующих PD-L1, была незначительной (фиг. 37А). Блокирование сигналинга PD-1/PD-L1 с использованием антитела против PD-L1 значительно увеличивало эффективность TDB 1G7.v85-опосредованного уничтожения (фиг. 37А). В конкретном эксперименте примированные CD8+ клетки смешивали с клетками 293-FcRH5-PD-Ll и обрабатывали только 1G7.v85 TDB или в комбинации с антителом против PD-L1, или антителом против PD-1 (пембролизумаб) (фиг. 37В). Комбинированная терапия антителом против PD-L1 или антителом против PD-1 (пембролизумаб) значительно повышала эффективность TDB 1G7.v85. ЭК50 для обоих комбинированных терапий составила 0,4 нг/мл, тогда как ЭК50 для терапии только 1G7.v85 TDB составила 0,63 нг/мл (фиг. 37В).
Эти результаты демострируют, что 1G7.v85 TDB-опосредованная активация Т-клеток приводит к индукции PD-1 в Т-клетках in vitro и in vivo. Эти результаты дополнительно демонстрируют, что сигналинг PD-1/PD-L1 может ограничить TDB против FcRH5-опосредованное уничтожение и что блокада PD-L1 может преодолеть это ингибирование и привести к повышению эффективности. Эти данные подкрепляют применение TDB против FcRH5 в комбинации с антагонистом, связывающимся с компонентом сигнального пути PD-1, таким как антитело против PD-1 или антитело против PD-L1.
В. Дексаметазон снижает цитокиновый ответ первой дозы, не влияя на активность TDB против FcRH5
Уничтожение целевых клеток также оценивали в присутствии и отсутствии дексаметазона (Dex), компонента стандарта лечения миеломы, который оказывает противовоспалительное и иммунодепрессантное действие (фиг. 38А). ИК50 для TDB 1G7.v85 в буфере или в ДМСО составляла 7 пМ и 6 пМ соответственно. В присутствии 0,1 мкМ или 1 мкМ Dex, ИК50 для TDB 1G7.v85 составляла 16 пМ и 25 пМ, соответственно. Комбинированное лечение Dex имело лишь незначительное влияние на эффективность 1G7.v85 TDB и значительно уменьшало уровни ИЛ-2, ИЛ-6, ФНО-α и ИФН-γ. Эти результаты демонстрируют, что дексаметазон можно применять в комбинации с терапией TDB против FcRH5 для смягчения цитокинового ответа первой дозы у пациентов (фиг. 38 В).
Пример 6. Получение и исследование бис-Fab против FcRH5
А. Получение тио-Fab и шарнирных цис-Fab и продуцирование белка
Для получения фрагментов антитела со свободными сульфгидрильными группами, замены цистеина (Cys) вводили в конструкции антител в различных положениях в вариабельных или константных доменах легких цепей или тяжелых цепей с помощью направленного мутагенеза для создания тио-мкАт, как описано ранее в Junutula и другие. J. Immunol Methods 332 (1-2): 41-52, 2008. Тио-Fab генерировали ферментативно из тио-мкАт путем разбавления тио-мкАт до 1 мг/мл в 25 мМ Tris, рН 8,0, с последующим ферментативным расщеплением при 37°С в течение 1 часа с использованием Lys-C (Wako Chemicals USA, Inc., Ричмонд, Виргиния) при соотношении фермента к антителу 1:1000 (масса:масса). Расщепление Lys-C прекращали с помощью 5 мкМ ингибитора протеазы тозил-L-лизинхлорметилкетона (TLCK) (Bachem, Торренс, Калифорния) и очищали при помощи катионо-ионообменной хроматографии на 5 мл колонке Hi-Trap SP FF (GE Healthcare, Пискатауэй, Нью-Джерси) с использованием 50 мМ натрий-ацетатного буфера и градиента концентрации NaCl 0-300 мМ на 10 объемов колонки (CV). Тио-Fab, полученные этим методом, иногда упоминаются как «ферментативные тио-Fab». В другом подходе конструкты ДНК, кодирующие Fab, имеющие сконструированные остатки Cys или конструкты ДНК, кодирующие фрагменты тяжелой цепи, содержащие один нативный остаток Cys в шарнирной области, субклонировали в экспрессионные плазмидные векторы и экспрессировали непосредственно в клетках СНО. Тио-Fab, полученные этим методом, иногда упоминаются как «рекомбинантные тио-Fab». Для антител, не имеющих сконструированного остатка Cys, был использован третий подход, который берет за основу природный остаток Cys, присутствующий в шарнирной области lgG. Этот метод используется для создания «шарнирный цис-Fab» и более подробно описан ниже.
Для получения шарнирних цис-Fab из нативных антител, которые не содержат сконструированного цистеина для использования в реакциях синтеза, была использована следующая ферментативная процедура, как показано на панели 1 фиг. 39. Оба исходных антитела FcRH5 и CD3 расщепляли при помощи обработки пепсином (1% мас./мас.) в натрий-ацетатном буфере при рН 4,5. После расщепления в течение 1 часа F(ab')2 выделяли из смеси для расщепления путем захвата на катионообменной смоле SP-HP и очищали с помощью градиента концетрации соли 0-1 M NaCl на 10 CV. Затем F(ab')2 восстанавливали в буфере, содержащем 25 мМ MES, рН 5,8, 2 мМ ЭДТА и 300 мМ NaCl. После восстановления с помощью 1 мМ ТСЕР Fab окисляли, как показано на панели 2 фиг. 39, путем добавления 5 мМ DHAA (дегидроаскорбиновая кислота) для перестройки дисульфида между тяжелой цепью и легкой цепью. Было отмечено, что обычно, в этих условиях реакции был перестроен только дисульфид между тяжелой цепью и легкой цепью; два остатка цистеина в шарнирной области оставались неокисленными.
Два свободных тиола (остатки Cys) на шарнире, как показано на панели 2 фиг. 39, затем подвергали взаимодействию с 1 эквимолярным количеством N-этилмалеимида (NEM) (Sigma Aldrich, Сент-Луис, Миссури). Полученную смесь, содержащую однократно модифицированные, дважды модифицированные и немодифицированные Fab, затем подвергали взаимодействию, как показано на панели 3 фиг. 39, с избытком бисмалеимидного сшивающего агента.
Эти условия реакции дали три продукта: Fab с одним сшивающим агентом и одним NEM, Fab с двумя NEM, и Fab, содержащими только один сшивающий агент.Было обнаружено, что Fab, содержащие только один сшивающий агент, не имеют свободного цистеина. Таким образом, в этих условиях реакции один сшивающий агент очень эффективно реагировал с обоими цистеинами, с образованием молекулы, в которой цистеины были циклизованы сшивающим агентом. Продукт, содержащий вышеуказанные три продукта реакции, очищали из реакционной смеси (для удаления нежелательных компонентов реакции) с помощью гель-фильтрации и использовали в комбинации с другими шарнирными цис-Fab, как показано на панели 4 фиг. 39, полученными аналогичним образом, или с тио-Fab. Только шарнирные цис-Fab или тио-Fab, полученные, как описано и содержащие один сшивающий агент, один свободный малеимид и один свободный сульфгидрил, могли реагировать в реакциях синтеза бис-Fab, подробно описанных ниже.
B. Экспрессия и очистка белка
Чтобы облегчить очистку, Fab экспрессировали с использованием Flag- или гистидиновой метки. Экспрессию в клетках СНО выполняли при помощи стандартных процедур. Аффинную очистку после культивирования клеток проводили с использованием смолы, содержащей мкАт против Flag, или смолы, содержащей никелевые шарики. Очищенные тио-Fab исследовали при помощи ДСН-ПААГ-электрофореза и масс-спектрометрии. Эти исследования часто демонстрировали повышение массы на 275 Да и 306 Да. Было обнаружено, что эти увеличения массы являются дисульфидными аддуктами на неспаренном цистеине, которые удаляют восстановлением и окислением для получения тио-Fab для сшивания с бисмалеимидом. Восстановление и окисление тио-Fab проводили следующим образом. Во-первых, тио-Fab восстанавливали в течение 24 часов путем добавления 2 мМ трис(2-карбоксиэтил) фосфина HCl (ТСЕР-HCl, также называемого ТСЕР) (Pierce [Thermo Fisher Scientific], Рокфорд, Иллинойс) в буфере, содержащем 25 мМ MES, рН 5,8, 300 мл NaCl и 5 мМ ЭДТА. После восстановления белок окисляли добавлением 5 мМ дегидроаскорбиновой кислоты (DHAA) (Sigma-Aldrich, Сент-Луис, Миссури). Выделенные тио-Fab анализировали с помощью ДСН-ПААГ и масс-спектрометрии для обеспечения надлежащего восстановления и окисления белков.
C. Синтез бис-Fab
Для ковалентного связывания двух Fab можно использовать два разных типа сшивающих агентов: бис-малеимид и пару адаптеров DBCO (азодибензоциклооктин)-ПЭГ-малеимид/бромацетамид-ПЭГ-азид.
Конъюгирование с использованием бисмалеимидных сшивающих агентов
На первом этапе синтеза бис-Fab использовали тио-Fab или шарнирные цис-Fab с неспаренным цистеином. Как правило, тио-Fab или шарнирный цис-Fab находились в том же буфере, в котором было проведено восстановление (фиг. 39, панель 1) и окисление (фиг. 39, панель 2) (MES, рН 5,8, 2 мМ EDTA и 300 мМ NaCl) при концентрации белка 1 мг/мл. На этой стадии были два потенциальных нежелательных продукта реакции: дисульфидные димеры и сшитые димеры. Наличие белка в концентрации 1 мг/мл на этой стадии синтеза было важной особенностью реакции, поскольку димеризация была минимизирована при этой концентрации белка. Кроме того, контроль реакции с использованием буфера с низким рН с помощью ЭДТА помог минимизировать димеризацию.
К реакционной смеси добавляли пятикратный избыток бисмалеимидного сшивающего агента (Quanta BioDesign, Пауэлл, Огайо), как показано на панели 3 фиг. 39. Этот пятикратный избыток сшивающего агента был также полезен для минимизации нежелательной димеризации. Реакционную смесь инкубировали при комнатной температуре (RT) или 37°С в течение четырех часов до завершения. Затем смесь концентрировали до объема, подходящего для гель-фильтрации. Использовали 22-миллиметровую колонку S-200 Tricom (GE Healthcare, Пискатауэй, Нью-Джерси) для синтеза количества от мкг до мг.Эта первая стадия гель-фильтрации позволила удалить неиспользованный сшивающий агент, получая очищенный тио-Fab или шарнирный цис-Fab, конъюгированный с сшивающим агентом. Условия, описанные выше, обычно приводят к по меньшей мере 90% или более целевого продукта. Ни один тио-Fab или шарнирный цис-Fab не сохранял свободные тиолы, так как все они были конъюгированы или с сшивающим агентом, или связаны дисульфидом с другим тио-Fab или шарнирным цис-Fab через неспаренные цистеины. Выделенный и очищенный тио-Fab (или шарнирный цис-Fab) плюс сшивающий агент затем добавляли ко второму тио-Fab (или шарнирному цис-Fab) и концентрировали до 5 мг/мл или выше, обычно до объема, подходящего для гель-фильтрации, как показано на панели 4 фиг. 39. Концентрация белка, составляющая не менее 5 мг/мл на этой стадии синтеза, была важна для завершения реакции. Более низкие концентрации белка приводят к образованию только небольших количеств сшитых димеров бис-Fab. Было установлено, что, не ограничиваясь теорией, стерический эффект или связанная с вязкостью переменная, которая препятствует образованию сшитых димеров бис-Fab, была преодолена за счет увеличения концентрации реагентов. Кроме того, был испытан диапазон концентраций белка до 65 мг/мл включительно. Установлена корреляция между концентрацией белка и временем реакции, заключающаяся в том, что чем выше концентрация белка, тем быстрее реакция завершается. Через 2-24 часа при комнатной температуре или 37°С реакция завершалась, как было определено при помощи масс-спектрометрии. Как правило, один реагент находился в избытке и оставался несвязанным в конечной смеси.
Конъюгирование с использованием DBCO-ПЭГ-малеимидом/бромацетамид-ПЭГ-азидом.
Один из очищенных и деблокированных Fab подвергали взаимодействию в 5-молярном избытке DBCO-PEG-малеимида (№760676, Sigma) в 50 мМ HEPES при рН 8, а другой Fab подвергали взаимодействию с 5-молярным избытком азид-ПЭГ-малеимида (№21097 BroadPharm) в 50 мМ HEPES при рН 8. После одночасовой инкубации при 37°С реакцию проверяли при помощи масс-спектрометрии на завершение реакции. Конъюгированные Fab очищали от избытка сшивающего агента SEC и затем смешивали при соотношении 1:1 и доводили до концентрации выше 5 мг/мл и инкубировали в течение ночи при комнатной температуре.
Независимо от используемого сшивающего агента, продукты завершенной реакции снова очищали при помощи гель-фильтрации; на этот раз был получен пик димеров, который содержал 100 кД бис-Fab, необратимо сшитый через свободную аминокислоту цистеина (в случае тио-Fab) или через неспаренный цистеин, расположенный в шарнирной области (в случае несконструированного шарнирного цис-Fab). Прогресс реакции на обеих стадиях обычно контролировали с помощью масс-спектрометрии, которая четко показала присутствие обоих реагентов и образование продукта бис-Fab. Чистоту целевого продукта после второй гель-фильтрации определяли с помощью масс-спектрометрии и ДСН-ПААГ-электрофореза. При восстановлении и анализе методом ДСН-ПААГ электрофореза, наблюдали необратимая сшивка по наличию 50 кД полосы, представляющей невосстанавливаемые сшитые цепи. Используя описанный выше процесс в небольших масштабах, достигали выхода микрограммовых количеств продукта из микрограммовых количеств исходных веществ. Кроме того, в большем масштабе обычно достигали выхода миллиграммовых количеств продукта из миллиграммовых количеств исходных веществ.
D. Синтез бис-Fab, нацеленных на CD3 и FcRH5
Были созданы биспецифические бис-Fab, полученные из двух разных антител, которые нацелены на CD3 и FcRH5. Исходным антителом против CD3 может быть любое антитело против CD3, такое как 38E4v.1, 38E4.v11 или 40G5. В одном варианте осуществления изобретения бис-Fab использует 38E4.v1 в качестве компонента антитела против CD3 с последовательностью легкой цепи SEQ ID NO: 134 и последовательность тяжелой цепи SEQ ID NO: 133. Используемое исходное антитело против FcRH5 также может быть любым антителом против FcRH5, например, описанными в данном документе антителами, такими как hu1G7.v85 и hu1G7.v87. Конкретно, один примерный бис-Fab включает последовательность легкой цепи SEQ ID NO: 105 и вариабельную последовательность тяжелой цепи SEQ ID NO: 104.
Для каждого из этих антител рекомбинантные тио-Fab были получены в клетках СНО, как описано выше. Затем бис-Fab синтезировали из тио-Fab в комбинаторном формате с использованием матрицы синтеза, начиная примерно с 2 мг каждого тио-Fab. Различные тио-Fab были объединены для синтеза четырех уникальных молекул бис-Fab. Примерно один мг каждого бис-Fab извлекали из синтеза для показанных примеров, но ожидается, что выход продукта будет варьироваться в зависимости от различных тио-Fab. Каждому из бис-Fab был присвоен уникальный идентификатор. Чистоту каждого бис-Fab анализировали с помощью ДСН-ПААГ электрофореза и масс-спектрометрии с использованием стандартных способов, хорошо известных в данной области техники.
Используя описанный выше подход матричной рекомбинации, была синтезирована серия структурных вариантов бис-Fab, полученных из CD3 и FcRH5. Для синтеза бис-Fab были выбраны четыре точки тио-присоединения; одно из положений находилось в тяжелой цепи плеча тио-Fab антитела против CD3 (например, в положении 76 (Cys76HC)), одно положение находилось в легкой цепи плеча тио-Fab анитела против CD3 (например, в положении 22 (Cys22LC)), одно из положений находилось в тяжелой цепи плеча тио-Fab антитела против FcRH5 (например, в положении 114 (Cys114HC)), и одно положение находилось в легкой цепи плеча тио-Fab антитела против FcRH5 (например, в положении 149 (Cys149LC)). Другие положения могут быть использованы для введения необходимых цистеинов. Fab, содержащие точки тио-присоединения, были получены из трех разных источников; (1) тио-мкАт с цистеиновыми заменами, которые были расщеплены лизином-С, чтобы высвободить тио-Fab из антитела, (2) тио-Fab с цистеиновыми заменами, которые были непосредственно экспрессированы и очищены от клеток СНО, и (3) шарнирых цис-Fab, получаемых ферментативным методом, описанным выше для присоединения одного сшивающего агента к шарнирной области несконструированного антитела после расщепления пепсином. Этот подход привел к различным точкам замен в тио-Fab для рекомбинации с другими тио-Fab, что дает структурные варианты (см. таблицу 15).
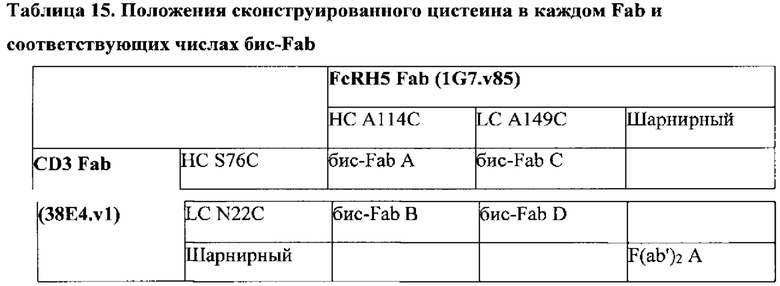
E. Биологическая активность бис-Fab против FcRH5
Затем каждый из структурных вариантов бис-Fab тестировали на их способность связывания с каждым из антигенов (то есть CD3 и FcRH5) с помощью проточной цитометрии. Бис-Fab, полученный путем связывания Cys в шарнирной области, напоминал структуру природного антитела и служил в качестве контроля. Хотя все конструкции бис-Fab имели сходную способность связывания с FcRH5 (фиг. 40А), большая часть бис-Fab продемонстрировала снижение связывания с CD3 по сравнению с эталонным бис-Fab (фиг. 40 В). Биологическую активность оценивали в анализе активации Т-клеток in vitro, который служил заменителем для Т-клеточной активности уничтожения (фиг. 41). В этом анализе использовали клеточную линию Jurkat (JurkatDual, Invivogen), стабильно трансфицированную репортером люциферазы под контролем фактора транскрипции NF-kB, и преимущество использования этого анализа по сравнению с анализом Т-клеточного уничтожения заключалось в том, что количество клеток, которые можно использовать неограничено. В тестах на уничтожение клеток МКПК количество тестов было ограничено количеством клеток, которые можно было получить от одного донора. Анализ выполняли следующим образом. Соответствующую клеточную линию, такую как MOLP-2, выбирали в качестве целевых клеток и совместно культивировали с клетками Jurkat. В каждую лунку добавляли 10000 целевых клеток и 50000 эффекторных клеток (Jurkat) (10000 целевых клеток на лунку, общий объем 200 мкл, с отношением мишень: эффектор=1:5) с или без присутствия бис-Fab. После инкубации в течение ночи 10 мкл супернатанта из различных лунок анализировали на активность люциферазы, используя 50 мкл QUANTI-LUC (Invivogen), и люминесценцию определяли количественно на люминометре Envision (Perkin Elmer).
Интересно, что хотя последовательности HVR каждого варианта бис-Fab, полученного для антитела против CD3 или антитела против FcRH5, не изменялись при генерации самих бис-Fab, максимальную стимуляцию Т-клеток наблюдали для каждого бис-Fab, демонстрировавшего отклонение, одного бис-Fab от другого, просто основанное на положении цистеин-сконструированной поперечной связи. Без привязки к какой-либо теории это может иметь последствия для того, как токсичность и/или эффективность бис-Fab можно модулировать в соответствии с конкретной терапевтической или диагностической потребностью. Другое наблюдение заключалось в том, что некоторые бис-Fab (например, бис-Fab С) имели значительно уменьшенную аффинность к CD3, но активность активации Т-клеток была сравнима с таковой для эталонного F(ab')2 А (фиг. 40 В и 41, таблица 15).
F. Биологическая активность FcRH5 бис-Fab с использованием эндогенных В-клеток человека
Каждый вариант бис-Fab был испытан на его биологическую эффективность в Т-зависимом киллинг-анализе бис-Fab по уничтожению периферических эндогенных В-клеток следующим образом: 200000 МКПК человека, выделенных из каждого из трех здоровых доноров, добавляли в лунку с бис-Fab или без них. После 48-часовой инкубации клетки окрашивали соответствующим антигеном клеточной поверхности линии целевых клеток (В-клетки=CD20) (5 мкл/лунку) и иодид пропидия для оценки жизнеспособности клеток, а затем анализировали с помощью проточной цитометрии. Бис-Fab-зависимую активность уничтожения вычисляли по следующему уравнению: % уничтожения=(1-количество живых клеток с бис-Fab/количество живых клеток без бис-Fab)×l00. В качестве положительного контроля использовали F(ab')2, который был получен из антитела TDB против FcRH5 с «выступом-во-впадину», в котором плечи антитела против CD3 и плечи антитела против FcRH5 каждой Fab имели такую же последовательность, что и у используемого бис-Fab (за исключением не связанных с цистеином точечных мутаций) (см., например, Ridgway et al. Protein Eng., 9: 617-621, 1996). Как правило, результаты демонстрировали воспроизводимость, несмотря на использование разных донорных клеток.
Количество тестируемых в данном документе бис-Fab против FcRH5 для полумаксимального лизиса периферических эндогенных В-клеток человека или значение активности ЭК50, выраженное в нг/мл, рассчитывали для каждого испытанного выше бис-Fab. В целом ожидалось, что закономерности в активности каждой бис-Fab, испытанной в эндогенном анализе В-клеток человека, будут сходиться с результатами, определенными в анализе с использованием метода проточной цитометрии, описанном выше (фиг. 40А-40В).
--->
SEQUENCE LISTING
<110> Genentech, Inc.
F. Hoffmann-La Roche AG
<120> HUMANIZED AND AFFINITY MATURED ANTIBODIES TO FcRH5 AND METHODS OF
USE
<130> 50474-134WO2
<150> US 62/180,459
<151> 2015-06-16
<160> 215
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 1
Arg Phe Gly Val His
1 5
<210> 2
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa is Ala or Pro
<220>
<221> MISC_FEATURE
<222> (15)..(15)
<223> Xaa is Met, Gly, or Val
<220>
<221> MISC_FEATURE
<222> (16)..(16)
<223> Xaa is Ser, Asp, or Pro
<400> 2
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Xaa Ala Phe Xaa Xaa
1 5 10 15
<210> 3
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa is Ser or Pro
<400> 3
His Tyr Tyr Gly Ser Xaa Asp Tyr Ala Leu Asp Asn
1 5 10
<210> 4
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa is Ser or Arg
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa is Ile or Leu
<400> 4
Lys Ala Ser Gln Asp Val Xaa Asn Xaa Val Val
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa is Ala or Gly
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> Xaa is Ser or Tyr
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is Tyr or Asn
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa is Thr or Ser
<400> 5
Ser Xaa Xaa Xaa Arg Tyr Xaa
1 5
<210> 6
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> Xaa is His or Gln
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is Tyr or Phe
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa is Gln or Ser
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa is Ser or Pro
<400> 6
Gln Gln Xaa Xaa Xaa Xaa Pro Tyr Thr
1 5
<210> 7
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 7
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met Ser
1 5 10 15
<210> 8
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 8
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val Ser
1 5 10 15
<210> 9
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 9
His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn
1 5 10
<210> 10
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 10
His Tyr Tyr Gly Ser Pro Asp Tyr Ala Leu Asp Asn
1 5 10
<210> 11
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 11
Lys Ala Ser Gln Asp Val Ser Asn Ile Val Val
1 5 10
<210> 12
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 12
Lys Ala Ser Gln Asp Val Arg Asn Leu Val Val
1 5 10
<210> 13
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 13
Lys Ala Ser Gln Asp Val Arg Asn Ile Val Val
1 5 10
<210> 14
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 14
Lys Ala Ser Gln Asp Val Ser Asn Leu Val Val
1 5 10
<210> 15
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 15
Ser Ala Ser Tyr Arg Tyr Thr
1 5
<210> 16
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 16
Ser Gly Ser Tyr Arg Tyr Ser
1 5
<210> 17
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 17
Ser Gly Tyr Asn Arg Tyr Ser
1 5
<210> 18
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 18
Ser Gly Tyr Tyr Arg Tyr Thr
1 5
<210> 19
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 19
Ser Gly Tyr Tyr Arg Tyr Ser
1 5
<210> 20
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 20
Gln Gln His Tyr Ser Ser Pro Tyr Thr
1 5
<210> 21
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 21
Gln Gln Gln Tyr Ser Pro Pro Tyr Thr
1 5
<210> 22
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 22
Gln Gln Gln Phe Gln Pro Pro Tyr Thr
1 5
<210> 23
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 23
Gln Gln His Tyr Ser Pro Pro Tyr Thr
1 5
<210> 24
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 24
Gln Gln Gln Tyr Gln Pro Pro Tyr Thr
1 5
<210> 25
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 25
Gln Gln His Tyr Gln Pro Pro Tyr Thr
1 5
<210> 26
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 26
Gly Ile Asp Leu Ser Ser Tyr Ala Met Gly
1 5 10
<210> 27
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 27
Thr Ile Gly Thr Gly Gly Thr Pro Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 28
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 28
Ala Arg Asp Met Tyr Thr Gly Ser Ile Tyr Tyr Asp Met
1 5 10
<210> 29
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 29
Gln Ser Ser Gln Ser Val Phe Asn Asn Ala Gln Leu Ser
1 5 10
<210> 30
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 30
Asp Thr Ser Asn Leu Ala Ser
1 5
<210> 31
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 31
Leu Gly Ala Tyr Thr Asp Asp Ala Asp Asn Ala
1 5 10
<210> 32
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 32
Gly Tyr Thr Phe Thr His Tyr Ala Met His
1 5 10
<210> 33
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 33
Met Ile Ser Ser Ser Ser Gly Asn Thr Asn Phe Asn Gln Lys Phe Met
1 5 10 15
Asp
<210> 34
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 34
Ala Arg Trp Ser Tyr Tyr Ala Leu Phe Asp Tyr
1 5 10
<210> 35
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 35
Ser Ala Asp Ser Ser Val Asp Tyr Ile His
1 5 10
<210> 36
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 36
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 37
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 37
Phe Gln Gly Ser Gly Tyr Pro Leu Thr
1 5
<210> 38
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 38
Gly Tyr Thr Phe Thr Asn Tyr Val Ile His
1 5 10
<210> 39
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 39
Tyr Ile Ser Pro Tyr Asn Asp Tyr Thr Lys Tyr Asn Glu Lys Phe Glu
1 5 10 15
Gly
<210> 40
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 40
Ala Arg Arg Asp Tyr Tyr Gly Ser Ser Asp Tyr
1 5 10
<210> 41
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 41
Ser Ala Ser Ser Ser Val Asp Tyr Met His
1 5 10
<210> 42
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 42
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 43
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 43
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 44
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa is Glu, Ser, or Phe
<220>
<221> MISC_FEATURE
<222> (28)..(28)
<223> Xaa is Ser or Pro
<220>
<221> MISC_FEATURE
<222> (29)..(29)
<223> Xaa is Leu or Thr
<400> 44
Xaa Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Xaa Xaa Thr
20 25 30
<210> 45
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa is Val or Tyr
<220>
<221> MISC_FEATURE
<222> (13)..(13)
<223> Xaa is Leu or Ile
<400> 45
Trp Xaa Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Xaa Gly
1 5 10
<210> 46
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 46
Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu Lys
1 5 10 15
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser Asn
20 25 30
<210> 47
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 47
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 48
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 48
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 49
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa is Ala or Ser
<400> 49
Trp Phe Gln Gln Lys Pro Gly Lys Xaa Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 50
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 50
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 51
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 51
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 52
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr
20 25 30
<210> 53
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Thr Thr
20 25 30
<210> 54
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 54
Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu Gly
1 5 10
<210> 55
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 55
Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile Gly
1 5 10
<210> 56
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 56
Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 57
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 57
Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 58
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 58
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser
20 25
<210> 59
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 59
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
1 5 10
<210> 60
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 60
Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Thr Val Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
20 25 30
<210> 61
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 61
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 62
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 62
Asp Ala Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr
20
<210> 63
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 63
Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 64
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 64
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 65
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 65
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 66
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 66
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Gly Ser
20 25
<210> 67
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 67
Trp Val Lys Gln Ser His Thr Lys Ser Leu Glu Trp Ile Gly
1 5 10
<210> 68
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 68
Lys Ala Thr Met Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr Leu Glu
1 5 10 15
Leu Ala Arg Leu Thr Phe Gly Asp Ser Ala Ile Tyr Tyr Cys
20 25 30
<210> 69
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 69
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 70
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 70
Glu Asn Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys
20
<210> 71
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 71
Trp Tyr Gln Gln Lys Ser Asn Thr Ser Pro Lys Leu Trp Ile Tyr
1 5 10 15
<210> 72
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 72
Gly Val Pro Gly Arg Phe Ser Gly Ser Gly Ser Gly Asn Ser Tyr Ser
1 5 10 15
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 73
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 73
Phe Gly Ser Gly Thr Lys Leu Glu Leu Lys
1 5 10
<210> 74
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 74
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser
20 25
<210> 75
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 75
Trp Met Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 76
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 76
Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Asn Thr Ala Tyr Met Glu
1 5 10 15
Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
20 25 30
<210> 77
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 77
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
1 5 10
<210> 78
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 78
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys
20
<210> 79
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 79
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Lys Trp Ile Tyr
1 5 10 15
<210> 80
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 80
Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
1 5 10 15
Leu Thr Ile Gly Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
20 25 30
<210> 81
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 81
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 82
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 82
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 83
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 83
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 84
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 84
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 85
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gln Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 86
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 86
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 87
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Asn Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gln Phe Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 88
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 88
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 89
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 89
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gln Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 90
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 90
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 91
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 91
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 92
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 92
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 93
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gln Phe Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 94
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 94
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 95
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 95
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gln Tyr Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 96
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 96
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Thr Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 97
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 97
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 98
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 98
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 99
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 99
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 100
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 100
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 101
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 101
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Tyr Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Gln Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 102
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 103
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 103
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 104
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 104
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 105
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 105
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Arg Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 106
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Thr Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Pro Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 107
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 107
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 108
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Gly Thr Gly Gly Thr Pro Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg Asp Met Tyr Thr Gly Ser Ile Tyr Tyr Asp Met Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 109
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 109
Asp Ala Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Ser Val Phe Asn Asn
20 25 30
Ala Gln Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Ala Tyr Thr Asp
85 90 95
Asp Ala Asp Asn Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 110
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 110
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Gly Ser Gly Tyr Thr Phe Thr His Tyr
20 25 30
Ala Met His Trp Val Lys Gln Ser His Thr Lys Ser Leu Glu Trp Ile
35 40 45
Gly Met Ile Ser Ser Ser Ser Gly Asn Thr Asn Phe Asn Gln Lys Phe
50 55 60
Met Asp Lys Ala Thr Met Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ala Arg Leu Thr Phe Gly Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Trp Ser Tyr Tyr Ala Leu Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 111
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 111
Glu Asn Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Asp Ser Ser Val Asp Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Ser Asn Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Gly Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Asn Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Phe Gln Gly Ser Gly Tyr Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 112
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 112
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Val Ile His Trp Met Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Pro Tyr Asn Asp Tyr Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Glu Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Tyr Gly Ser Ser Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 113
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 113
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asp Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Lys Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Gly Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 114
<211> 977
<212> PRT
<213> Homo sapiens
<400> 114
Met Leu Leu Trp Val Ile Leu Leu Val Leu Ala Pro Val Ser Gly Gln
1 5 10 15
Phe Ala Arg Thr Pro Arg Pro Ile Ile Phe Leu Gln Pro Pro Trp Thr
20 25 30
Thr Val Phe Gln Gly Glu Arg Val Thr Leu Thr Cys Lys Gly Phe Arg
35 40 45
Phe Tyr Ser Pro Gln Lys Thr Lys Trp Tyr His Arg Tyr Leu Gly Lys
50 55 60
Glu Ile Leu Arg Glu Thr Pro Asp Asn Ile Leu Glu Val Gln Glu Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Ala Gln Gly Ser Pro Leu Ser Ser Pro Val
85 90 95
His Leu Asp Phe Ser Ser Ala Ser Leu Ile Leu Gln Ala Pro Leu Ser
100 105 110
Val Phe Glu Gly Asp Ser Val Val Leu Arg Cys Arg Ala Lys Ala Glu
115 120 125
Val Thr Leu Asn Asn Thr Ile Tyr Lys Asn Asp Asn Val Leu Ala Phe
130 135 140
Leu Asn Lys Arg Thr Asp Phe His Ile Pro His Ala Cys Leu Lys Asp
145 150 155 160
Asn Gly Ala Tyr Arg Cys Thr Gly Tyr Lys Glu Ser Cys Cys Pro Val
165 170 175
Ser Ser Asn Thr Val Lys Ile Gln Val Gln Glu Pro Phe Thr Arg Pro
180 185 190
Val Leu Arg Ala Ser Ser Phe Gln Pro Ile Ser Gly Asn Pro Val Thr
195 200 205
Leu Thr Cys Glu Thr Gln Leu Ser Leu Glu Arg Ser Asp Val Pro Leu
210 215 220
Arg Phe Arg Phe Phe Arg Asp Asp Gln Thr Leu Gly Leu Gly Trp Ser
225 230 235 240
Leu Ser Pro Asn Phe Gln Ile Thr Ala Met Trp Ser Lys Asp Ser Gly
245 250 255
Phe Tyr Trp Cys Lys Ala Ala Thr Met Pro Tyr Ser Val Ile Ser Asp
260 265 270
Ser Pro Arg Ser Trp Ile Gln Val Gln Ile Pro Ala Ser His Pro Val
275 280 285
Leu Thr Leu Ser Pro Glu Lys Ala Leu Asn Phe Glu Gly Thr Lys Val
290 295 300
Thr Leu His Cys Glu Thr Gln Glu Asp Ser Leu Arg Thr Leu Tyr Arg
305 310 315 320
Phe Tyr His Glu Gly Val Pro Leu Arg His Lys Ser Val Arg Cys Glu
325 330 335
Arg Gly Ala Ser Ile Ser Phe Ser Leu Thr Thr Glu Asn Ser Gly Asn
340 345 350
Tyr Tyr Cys Thr Ala Asp Asn Gly Leu Gly Ala Lys Pro Ser Lys Ala
355 360 365
Val Ser Leu Ser Val Thr Val Pro Val Ser His Pro Val Leu Asn Leu
370 375 380
Ser Ser Pro Glu Asp Leu Ile Phe Glu Gly Ala Lys Val Thr Leu His
385 390 395 400
Cys Glu Ala Gln Arg Gly Ser Leu Pro Ile Leu Tyr Gln Phe His His
405 410 415
Glu Gly Ala Ala Leu Glu Arg Arg Ser Ala Asn Ser Ala Gly Gly Val
420 425 430
Ala Ile Ser Phe Ser Leu Thr Ala Glu His Ser Gly Asn Tyr Tyr Cys
435 440 445
Thr Ala Asp Asn Gly Phe Gly Pro Gln Arg Ser Lys Ala Val Ser Leu
450 455 460
Ser Val Thr Val Pro Val Ser His Pro Val Leu Thr Leu Ser Ser Ala
465 470 475 480
Glu Ala Leu Thr Phe Glu Gly Ala Thr Val Thr Leu His Cys Glu Val
485 490 495
Gln Arg Gly Ser Pro Gln Ile Leu Tyr Gln Phe Tyr His Glu Asp Met
500 505 510
Pro Leu Trp Ser Ser Ser Thr Pro Ser Val Gly Arg Val Ser Phe Ser
515 520 525
Phe Ser Leu Thr Glu Gly His Ser Gly Asn Tyr Tyr Cys Thr Ala Asp
530 535 540
Asn Gly Phe Gly Pro Gln Arg Ser Glu Val Val Ser Leu Phe Val Thr
545 550 555 560
Val Pro Val Ser Arg Pro Ile Leu Thr Leu Arg Val Pro Arg Ala Gln
565 570 575
Ala Val Val Gly Asp Leu Leu Glu Leu His Cys Glu Ala Pro Arg Gly
580 585 590
Ser Pro Pro Ile Leu Tyr Trp Phe Tyr His Glu Asp Val Thr Leu Gly
595 600 605
Ser Ser Ser Ala Pro Ser Gly Gly Glu Ala Ser Phe Asn Leu Ser Leu
610 615 620
Thr Ala Glu His Ser Gly Asn Tyr Ser Cys Glu Ala Asn Asn Gly Leu
625 630 635 640
Val Ala Gln His Ser Asp Thr Ile Ser Leu Ser Val Ile Val Pro Val
645 650 655
Ser Arg Pro Ile Leu Thr Phe Arg Ala Pro Arg Ala Gln Ala Val Val
660 665 670
Gly Asp Leu Leu Glu Leu His Cys Glu Ala Leu Arg Gly Ser Ser Pro
675 680 685
Ile Leu Tyr Trp Phe Tyr His Glu Asp Val Thr Leu Gly Lys Ile Ser
690 695 700
Ala Pro Ser Gly Gly Gly Ala Ser Phe Asn Leu Ser Leu Thr Thr Glu
705 710 715 720
His Ser Gly Ile Tyr Ser Cys Glu Ala Asp Asn Gly Leu Glu Ala Gln
725 730 735
Arg Ser Glu Met Val Thr Leu Lys Val Ala Val Pro Val Ser Arg Pro
740 745 750
Val Leu Thr Leu Arg Ala Pro Gly Thr His Ala Ala Val Gly Asp Leu
755 760 765
Leu Glu Leu His Cys Glu Ala Leu Arg Gly Ser Pro Leu Ile Leu Tyr
770 775 780
Arg Phe Phe His Glu Asp Val Thr Leu Gly Asn Arg Ser Ser Pro Ser
785 790 795 800
Gly Gly Ala Ser Leu Asn Leu Ser Leu Thr Ala Glu His Ser Gly Asn
805 810 815
Tyr Ser Cys Glu Ala Asp Asn Gly Leu Gly Ala Gln Arg Ser Glu Thr
820 825 830
Val Thr Leu Tyr Ile Thr Gly Leu Thr Ala Asn Arg Ser Gly Pro Phe
835 840 845
Ala Thr Gly Val Ala Gly Gly Leu Leu Ser Ile Ala Gly Leu Ala Ala
850 855 860
Gly Ala Leu Leu Leu Tyr Cys Trp Leu Ser Arg Lys Ala Gly Arg Lys
865 870 875 880
Pro Ala Ser Asp Pro Ala Arg Ser Pro Ser Asp Ser Asp Ser Gln Glu
885 890 895
Pro Thr Tyr His Asn Val Pro Ala Trp Glu Glu Leu Gln Pro Val Tyr
900 905 910
Thr Asn Ala Asn Pro Arg Gly Glu Asn Val Val Tyr Ser Glu Val Arg
915 920 925
Ile Ile Gln Glu Lys Lys Lys His Ala Val Ala Ser Asp Pro Arg His
930 935 940
Leu Arg Asn Lys Gly Ser Pro Ile Ile Tyr Ser Glu Val Lys Val Ala
945 950 955 960
Ser Thr Pro Val Ser Gly Ser Leu Phe Leu Ala Ser Ser Ala Pro His
965 970 975
Arg
<210> 115
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 115
Ser Tyr Tyr Ile His
1 5
<210> 116
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 116
Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Asp
<210> 117
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa is Tyr or Ala
<400> 117
Asp Gly Tyr Ser Arg Xaa Tyr Phe Asp Tyr
1 5 10
<210> 118
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 118
Lys Ser Ser Gln Ser Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 119
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 119
Trp Thr Ser Thr Arg Lys Ser
1 5
<210> 120
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa is Lys or Thr
<400> 120
Xaa Gln Ser Phe Ile Leu Arg Thr
1 5
<210> 121
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 121
Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr
1 5 10
<210> 122
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 122
Asp Gly Tyr Ser Arg Ala Tyr Phe Asp Tyr
1 5 10
<210> 123
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 123
Lys Gln Ser Phe Ile Leu Arg Thr
1 5
<210> 124
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 124
Thr Gln Ser Phe Ile Leu Arg Thr
1 5
<210> 125
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 125
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr
20 25 30
<210> 126
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 126
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 127
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 127
Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr Leu Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 128
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 128
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 129
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 129
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys
20
<210> 130
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 130
Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 131
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 131
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
20 25 30
<210> 132
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 132
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 133
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 133
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 134
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 134
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 135
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 135
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Ala Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 136
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 136
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 137
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 137
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 138
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 138
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 139
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 139
Asn Tyr Tyr Ile His
1 5
<210> 140
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 140
Trp Ile Tyr Pro Gly Asp Gly Asn Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 141
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 141
Asp Ser Tyr Ser Asn Tyr Tyr Phe Asp Tyr
1 5 10
<210> 142
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 142
Lys Ser Ser Gln Ser Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 143
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 143
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 144
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 144
Thr Gln Ser Phe Ile Leu Arg Thr
1 5
<210> 145
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 145
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
20 25 30
<210> 146
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 146
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 147
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 147
Arg Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr Leu Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 148
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 148
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 149
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 149
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys
20
<210> 150
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 150
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 151
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 151
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
20 25 30
<210> 152
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 152
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 153
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 153
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Tyr Ser Asn Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 154
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 154
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 155
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 155
Gly Tyr Thr Met Asn
1 5
<210> 156
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa is Ser or Thr
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa is Asn or Ala
<220>
<221> MISC_FEATURE
<222> (13)..(13)
<223> Xaa is Gln or Asp
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa is Lys or Ser
<220>
<221> MISC_FEATURE
<222> (15)..(15)
<223> Xaa is Val or Phe
<220>
<221> MISC_FEATURE
<222> (17)..(17)
<223> Xaa is Gly or Asp
<400> 156
Leu Ile Asn Pro Tyr Lys Gly Val Xaa Thr Tyr Xaa Xaa Xaa Xaa Lys
1 5 10 15
Xaa
<210> 157
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 157
Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val
1 5 10
<210> 158
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 158
Arg Ala Ser Gln Asp Ile Arg Asn Tyr Leu Asn
1 5 10
<210> 159
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 159
Tyr Thr Ser Arg Leu Glu Ser
1 5
<210> 160
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 160
Gln Gln Gly Asn Thr Leu Pro Trp Thr
1 5
<210> 161
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 161
Leu Ile Asn Pro Tyr Lys Gly Val Thr Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 162
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 162
Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 163
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 163
Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 164
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 164
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
20 25 30
<210> 165
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 165
Trp Val Arg Gln Ala Pro Gly Lys Asp Leu Glu Trp Val Ala
1 5 10
<210> 166
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 166
Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 167
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 167
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 168
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 168
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 169
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 169
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 170
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 170
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 171
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 171
Phe Gly Gln Gly Thr Lys Leu Glu Leu Lys
1 5 10
<210> 172
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 172
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Asp Leu Glu Trp Val
35 40 45
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 173
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 173
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 174
<211> 28
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 174
Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys
20 25
<210> 175
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 175
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Pro Ala Phe Met Ser
1 5 10 15
<210> 176
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 176
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Gly Ser
1 5 10 15
<210> 177
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 177
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met Asp
1 5 10 15
<210> 178
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 178
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met Pro
1 5 10 15
<210> 179
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 179
Ser Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr
20 25 30
<210> 180
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 180
Phe Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr
20 25 30
<210> 181
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 181
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Pro Leu Thr
20 25 30
<210> 182
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 182
Trp Tyr Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu Gly
1 5 10
<210> 183
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 183
Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu Met
1 5 10 15
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser Asn
20 25 30
<210> 184
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 184
Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu Lys
1 5 10 15
Leu Ser Asp Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser Asn
20 25 30
<210> 185
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 185
Ser Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 186
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 186
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 187
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 187
Phe Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 188
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 188
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 189
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 189
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Pro Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 190
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 190
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 191
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 191
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Thr Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 192
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 192
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 193
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 193
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Tyr Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 194
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 194
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 195
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 195
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Pro Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 196
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 196
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 197
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 197
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Gly
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 198
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 198
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 199
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 199
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Asp Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 200
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 200
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 201
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 201
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Pro Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 202
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 202
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 203
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 203
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Met Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 204
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 204
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 205
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 205
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Asp Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 206
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 206
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 207
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 207
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Pro Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 208
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 208
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Ile
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 209
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 209
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 210
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 210
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 211
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 211
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Thr Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Ser Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 212
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 212
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 213
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 213
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Arg Phe
20 25 30
Gly Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Val
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ser
85 90 95
Asn His Tyr Tyr Gly Ser Pro Asp Tyr Ala Leu Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 214
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct
<400> 214
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Asn Leu
20 25 30
Val Val Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 215
<211> 975
<212> PRT
<213> Macaca fascicularis
<400> 215
Met Leu Leu Trp Val Ile Leu Leu Val Leu Ala Pro Val Ser Gly Gln
1 5 10 15
Phe Val Arg Thr Tyr Lys Ser His Phe Leu Gln Pro Pro Trp Thr Thr
20 25 30
Val Phe Arg Gly Glu Arg Val Asn Leu Thr Cys Lys Gly Phe Gly Phe
35 40 45
Tyr Ser Ser Gln Lys Thr Lys Trp Tyr Tyr Arg His Leu Gly Lys Glu
50 55 60
Ile Ser Arg Glu Thr Gln Lys Asn Thr Leu Glu Val Gln Glu Ser Gly
65 70 75 80
Glu Tyr Arg Cys Gln Ala Gln Gly Ser Pro Leu Ser Ser Pro Val Ser
85 90 95
Leu Asp Phe Ser Ser Ala Ser Leu Ile Leu Gln Ala Pro Leu Ser Val
100 105 110
Phe Glu Gly Asp Ser Val Val Leu Arg Cys Arg Ala Lys Ala Glu Val
115 120 125
Thr Leu Lys Thr Thr Ile Tyr Lys Asn Glu Asn Val Leu Ala Phe Leu
130 135 140
Asn Lys Ser Thr Asp Phe His Ile Ser His Ala Ser Leu Lys Asp Asn
145 150 155 160
Gly Ala Tyr Arg Cys Thr Gly Tyr Lys Glu Thr Cys Cys Leu Val Ser
165 170 175
Ser Asn Thr Val Lys Ile Gln Val Gln Glu Ser Phe Thr Arg Pro Val
180 185 190
Leu Arg Val Ser Ser Phe Gln Pro Ile Ser Gly Ser Pro Val Thr Leu
195 200 205
Thr Cys Glu Thr Gln Leu Ser Leu Glu Arg Ser Asp Val Pro Leu Gln
210 215 220
Phe Cys Phe Phe Arg Asn Asp Gln Met Leu Gly Ser Gly Cys Ser Leu
225 230 235 240
Ser Pro Lys Phe Arg Ile Thr Ala Met Trp Ser Lys Asp Ser Gly Ser
245 250 255
Tyr Trp Cys Lys Ala Ala Thr Met Cys Tyr Asp Thr Thr Ser Asn Ser
260 265 270
Leu Arg Ser Trp Ile Gln Val Leu Ile Pro Ala Ser His Pro Val Leu
275 280 285
Thr Leu Ser Pro Glu Lys Ala Leu Asn Phe Glu Gly Thr Lys Val Lys
290 295 300
Leu His Cys Glu Thr Gln Glu Asp Ser Leu Arg Thr Leu Tyr Lys Phe
305 310 315 320
Tyr His Asp Gly Val Pro Leu Arg Tyr Lys Ser Val Arg Cys Glu Lys
325 330 335
Gly Ala Ser Ile Ser Phe Ser Leu Thr Thr Glu His Ser Gly Asn Tyr
340 345 350
Tyr Cys Thr Ala Asp Asn Gly His Gly Ala Lys Pro Ser Glu Ala Val
355 360 365
Ser Leu Ser Val Thr Val Pro Val Ser Arg Pro Val Leu Thr Leu Ser
370 375 380
Ser Ala Glu Asp Leu Ile Ser Glu Gly Ala Lys Leu Thr Leu His Cys
385 390 395 400
Glu Ala Gln Arg Gly Ser Leu Pro Ile Val Tyr Gln Phe His His Glu
405 410 415
Asn Ala Ser Leu Gly Asn Arg Ser Ala His Ser Ala Gly Gly Val Ala
420 425 430
Ile Ser Phe Ser Leu Thr Ala Asp His Ser Gly Asn Tyr Tyr Cys Thr
435 440 445
Ala Asn Asn Gly Phe Gly Pro Gln Arg Ser Glu Ala Val Ser Leu Ser
450 455 460
Ile Thr Val Pro Val Ser Arg Pro Val Leu Thr Leu Ser Ser Ala Glu
465 470 475 480
Ala Leu Thr Phe Glu Gly Ala Thr Val Thr Leu Tyr Cys Glu Val Gln
485 490 495
Arg Gly Ser Pro Arg Ile Leu Tyr Gln Phe Tyr His Glu Asp Val Pro
500 505 510
Leu Gly Ser Asn Ser Thr Pro Ser Val Gly Lys Val Ser Phe Ser Phe
515 520 525
Ser Leu Thr Ala Ala His Ser Gly Asn Tyr Tyr Cys Thr Ala Asp Asn
530 535 540
Gly Phe Gly Pro Gln Arg Ser Glu Ala Val Ser Leu Phe Val Thr Val
545 550 555 560
Pro Val Ser Arg Pro Ile Leu Thr Leu Arg Val Pro Arg Ala Gln Ala
565 570 575
Val Val Gly Asp Leu Leu Glu Leu Arg Cys Glu Ala Leu Arg Gly Ser
580 585 590
Pro Pro Ile Met Tyr Trp Phe Tyr His Glu Asp Val Thr Leu Gly Ser
595 600 605
Ser Ser Val Pro Ser Gly Gly Glu Ala Ser Phe Asn Leu Ser Leu Thr
610 615 620
Ala Glu His Ser Gly Asn Tyr Ser Cys Glu Ala Asn Asn Gly Leu Val
625 630 635 640
Ala Gln His Ser Asp Thr Ile Ser Leu Ser Val Ile Val Pro Val Ser
645 650 655
Arg Pro Ile Leu Thr Phe Arg Ala Pro Arg Ala Gln Ala Val Val Gly
660 665 670
Asp Leu Leu Glu Leu His Cys Glu Ala Leu Arg Gly Ser Ser Pro Ile
675 680 685
Leu Tyr Trp Phe Tyr His Glu Asp Val Thr Leu Gly Lys Ile Ser Ala
690 695 700
Pro Ser Gly Gly Gly Ala Tyr Phe Asn Leu Ser Leu Thr Thr Glu His
705 710 715 720
Ser Gly Ile Tyr Ser Cys Glu Ala Asp Asn Gly Leu Glu Ala Gln Arg
725 730 735
Ser Glu Met Val Thr Leu Lys Val Ala Val Pro Val Ser Arg Pro Val
740 745 750
Leu Thr Leu Arg Ala Pro Arg Ala Gln Val Ala Val Gly Asp Leu Leu
755 760 765
Glu Leu His Cys Glu Ala Leu Arg Gly Ser Pro Leu Ile Leu Tyr Gln
770 775 780
Phe Tyr His Glu Asp Val Thr Leu Gly Asn Ser Ser Ala Leu Ser Gly
785 790 795 800
Gly Ala Phe Phe Asn Leu Ser Leu Thr Ala Glu His Gly Asn Tyr Ser
805 810 815
Cys Glu Ala Asp Asn Gly Leu Gly Ala Gln Arg Ser Glu Thr Val Thr
820 825 830
Leu Tyr Leu Thr Gly Leu Thr Glu Asn Arg Ser Gly Pro Val Ala Thr
835 840 845
Gly Val Thr Gly Gly Leu Leu Ser Leu Ala Gly Leu Ala Ala Val Ala
850 855 860
Leu Leu Leu Tyr Cys Trp Leu Ser Arg Lys Ala Gly Arg Glu Pro Ala
865 870 875 880
Ser Asp Pro Cys Arg Ser Pro Ser Asp Leu Asp Ser Gln Glu Pro Thr
885 890 895
Tyr His Asn Val Pro Ala Trp Glu Glu Leu Gln Pro Val Tyr Ser Asn
900 905 910
Val Asn Pro Arg Gly Glu Asn Val Val Tyr Ser Glu Val Arg Ile Ile
915 920 925
Arg Glu Lys Lys Lys His Ala Val Ala Ser Asn Pro Arg His Leu Arg
930 935 940
Asn Lys Gly Ser Cys Ile Ile Tyr Ser Glu Val Lys Val Ala Ser Thr
945 950 955 960
Pro Ala Ser Arg Cys Leu Phe Leu Ala Ser Ser Ala Pro His Arg
965 970 975
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-FcRH5 АНТИТЕЛА | 2014 |
|
RU2687132C2 |
| АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2818569C1 |
| АНТИТЕЛА ПРОТИВ НtrА1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2750285C2 |
| БИСПЕЦИФИЧНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2800779C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |
| АНТИТЕЛА К LAG3 | 2018 |
|
RU2778053C2 |
| АНТИТЕЛА К PD1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2746409C1 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2817838C2 |
| АНТИ-TIM3 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723708C2 |
| АНТИТЕЛА ПРОТИВ ТАУ-БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2732122C2 |
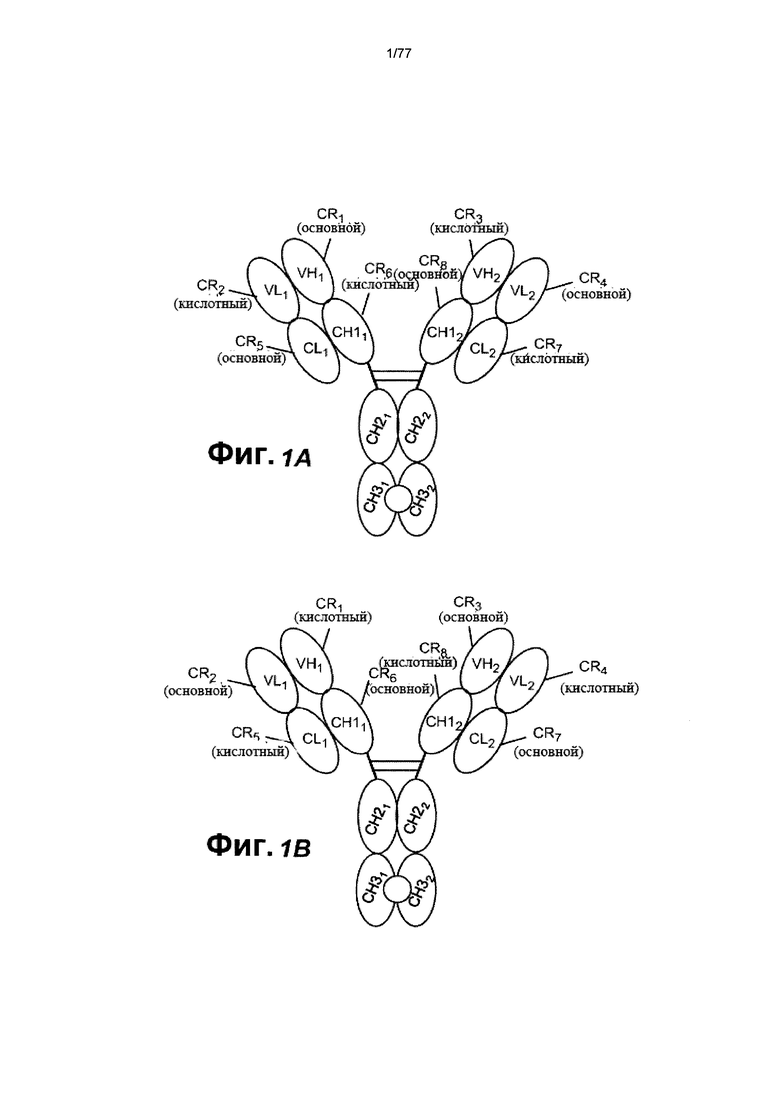
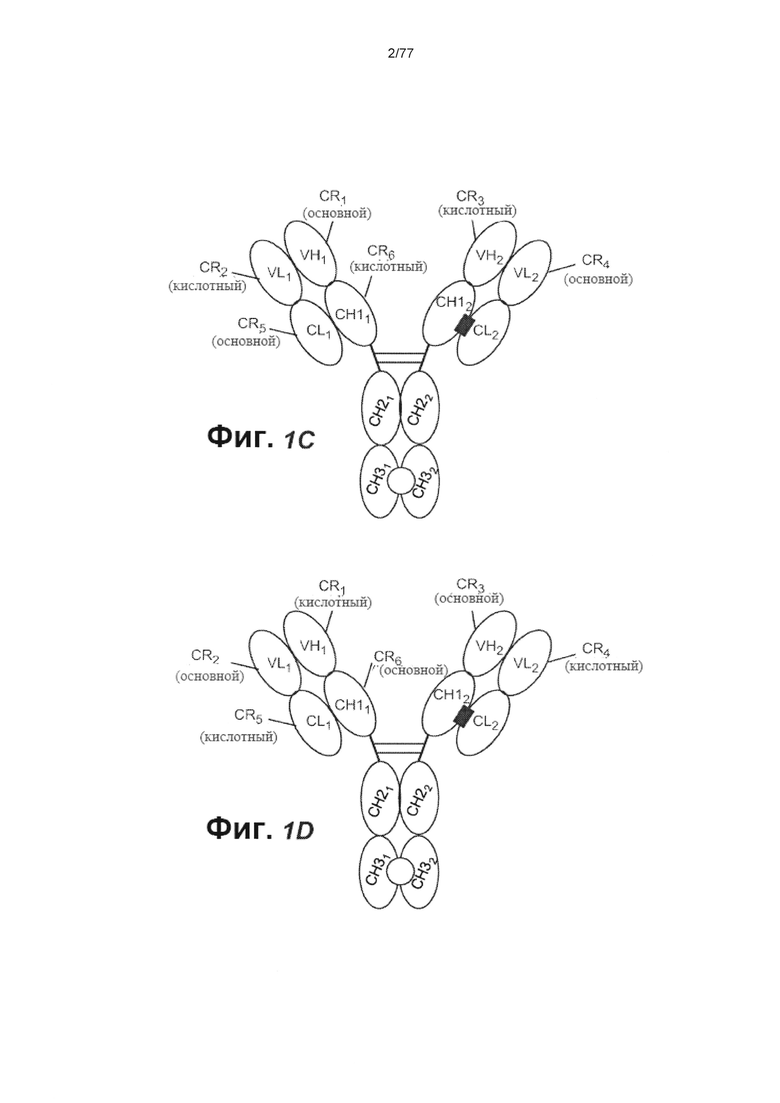
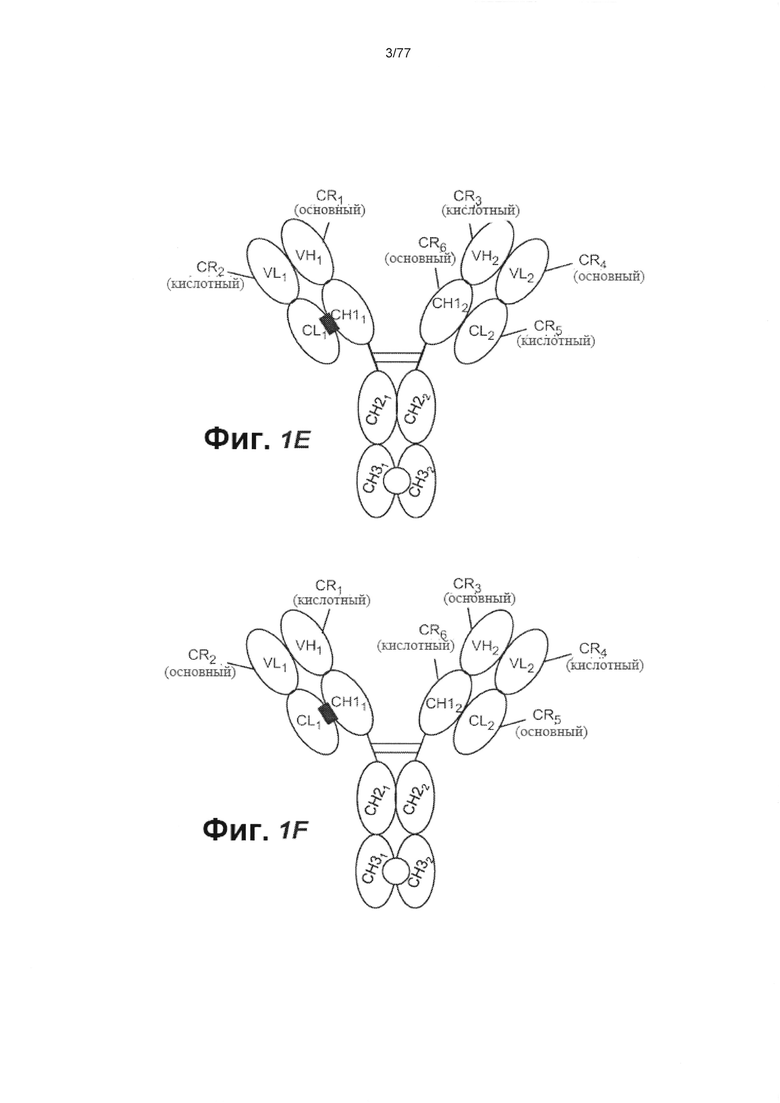

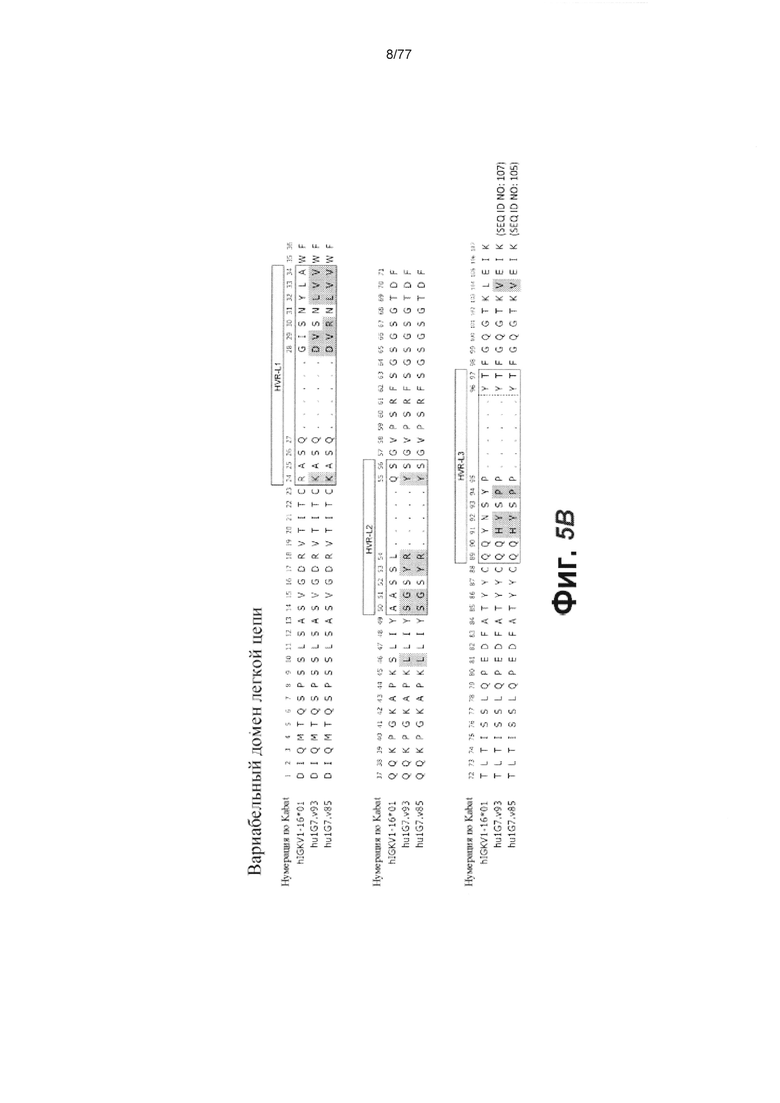
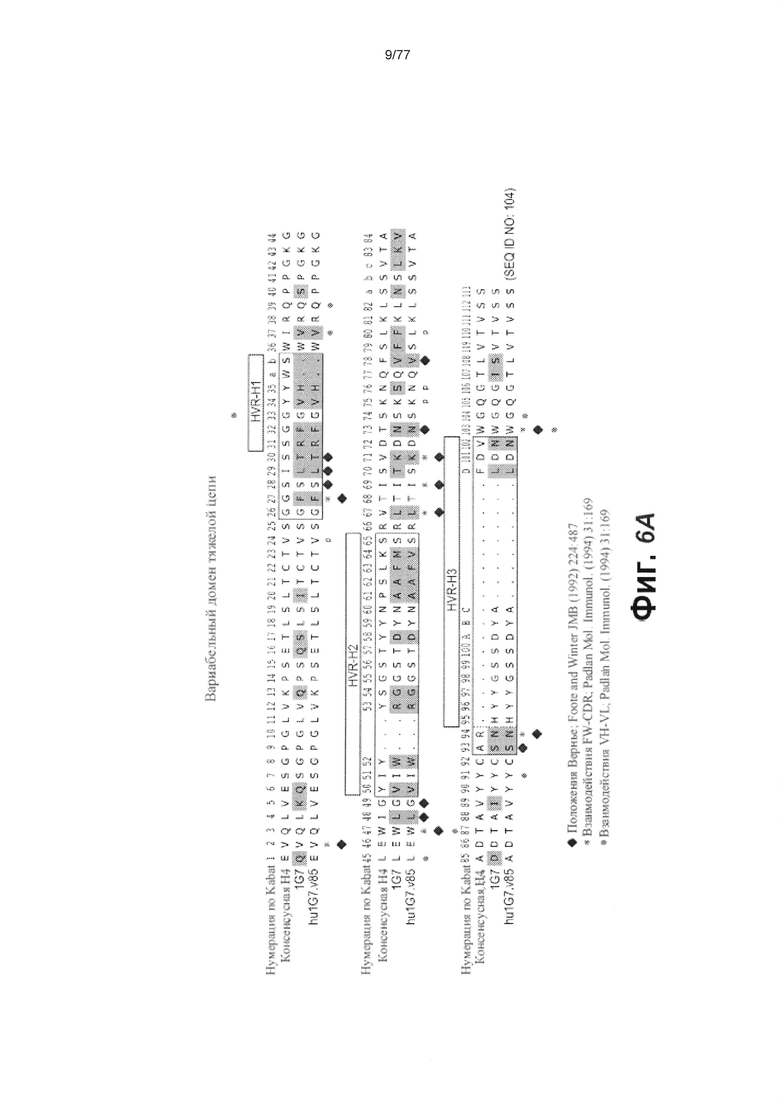
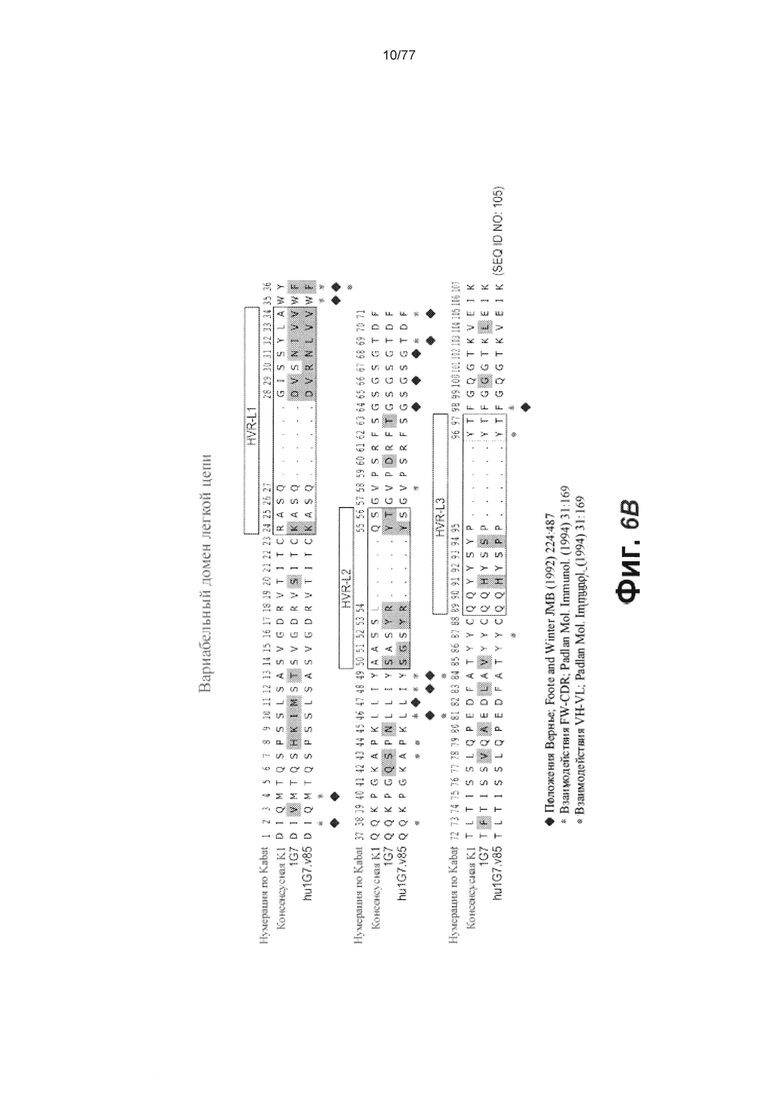
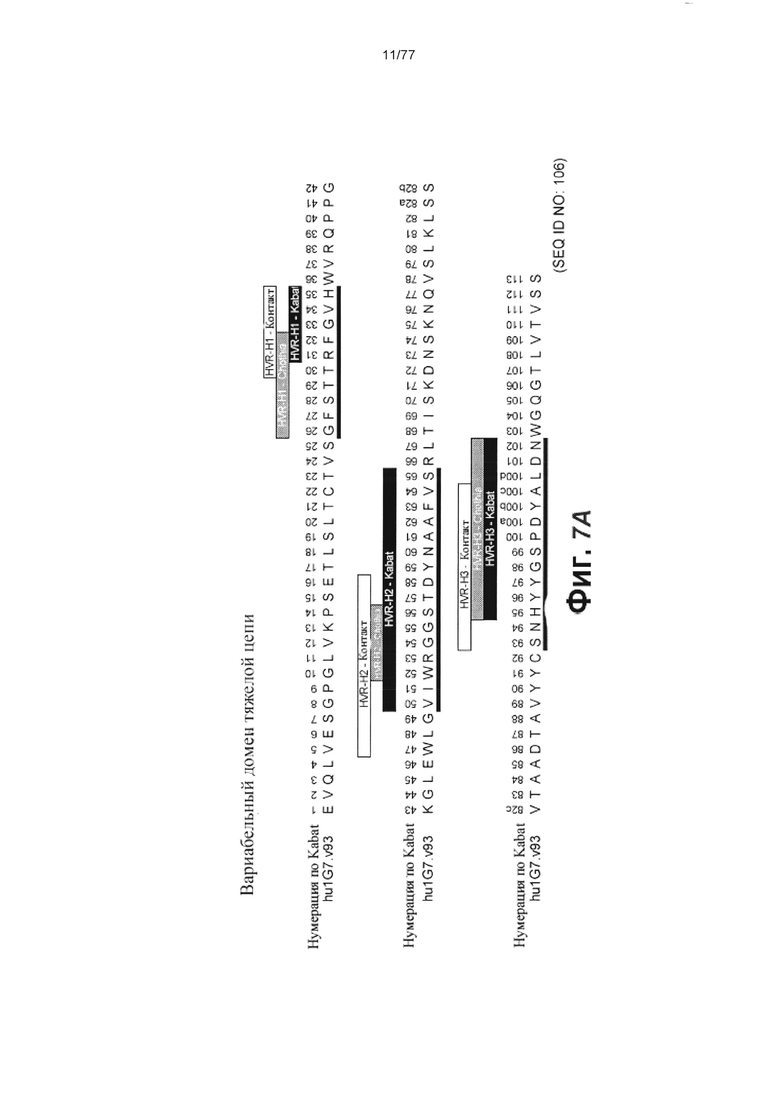
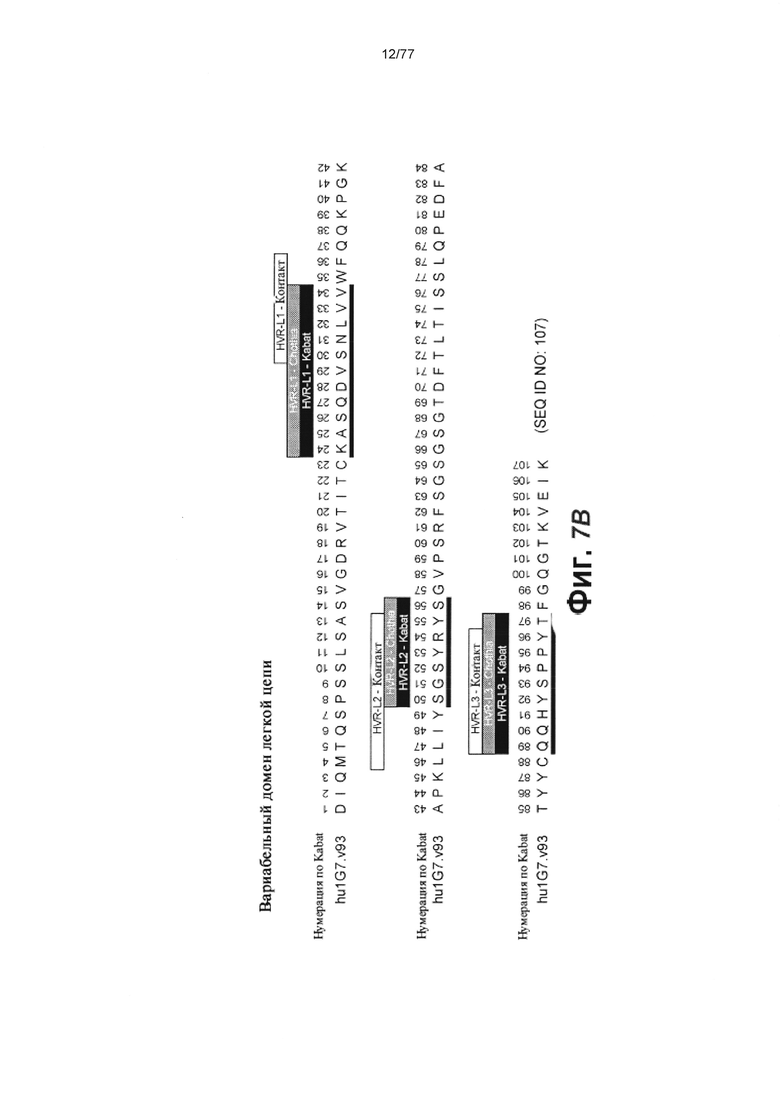
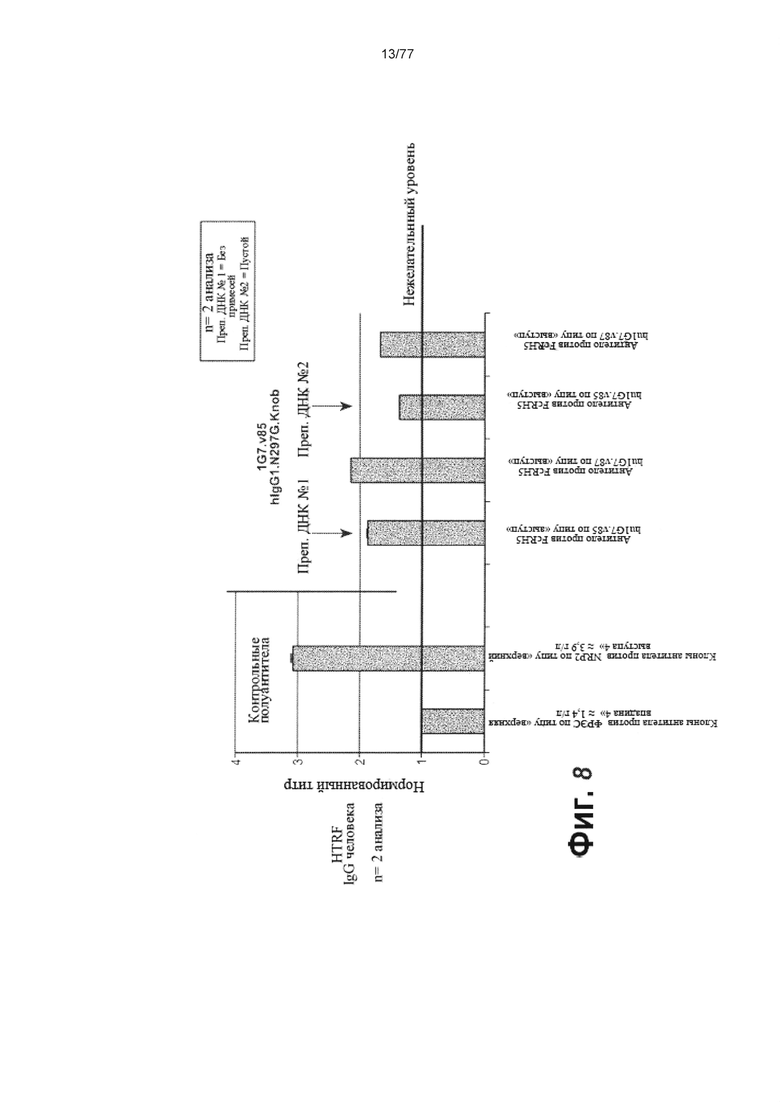
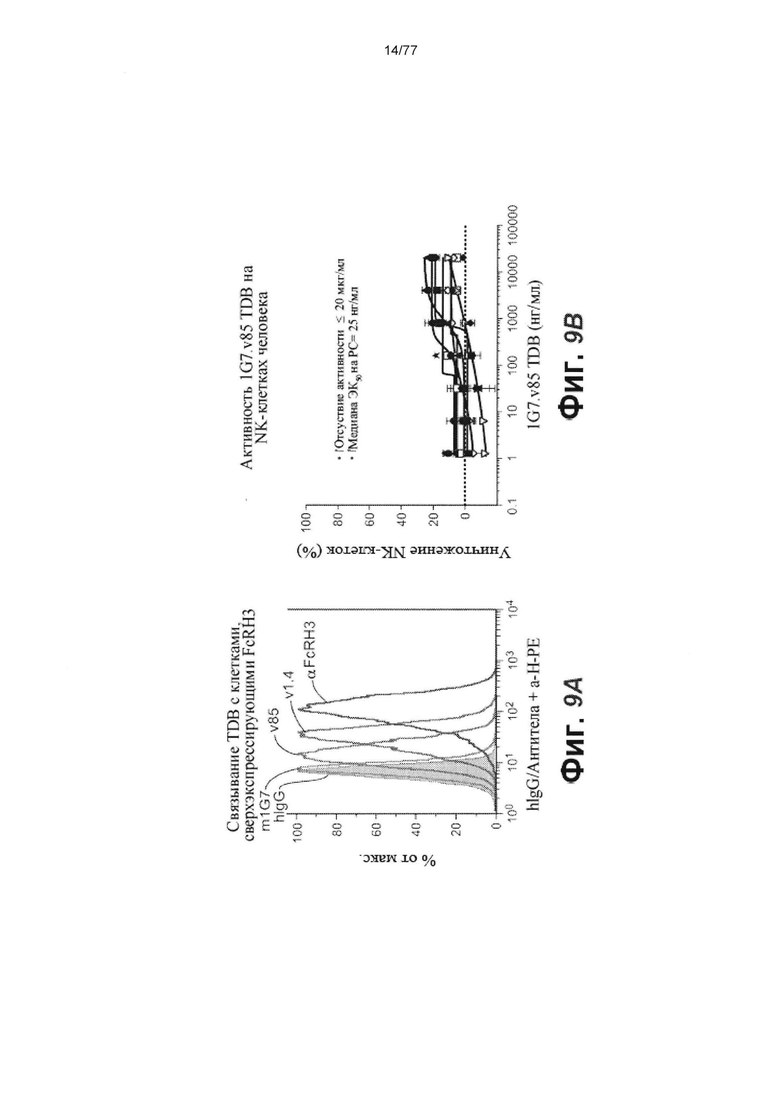
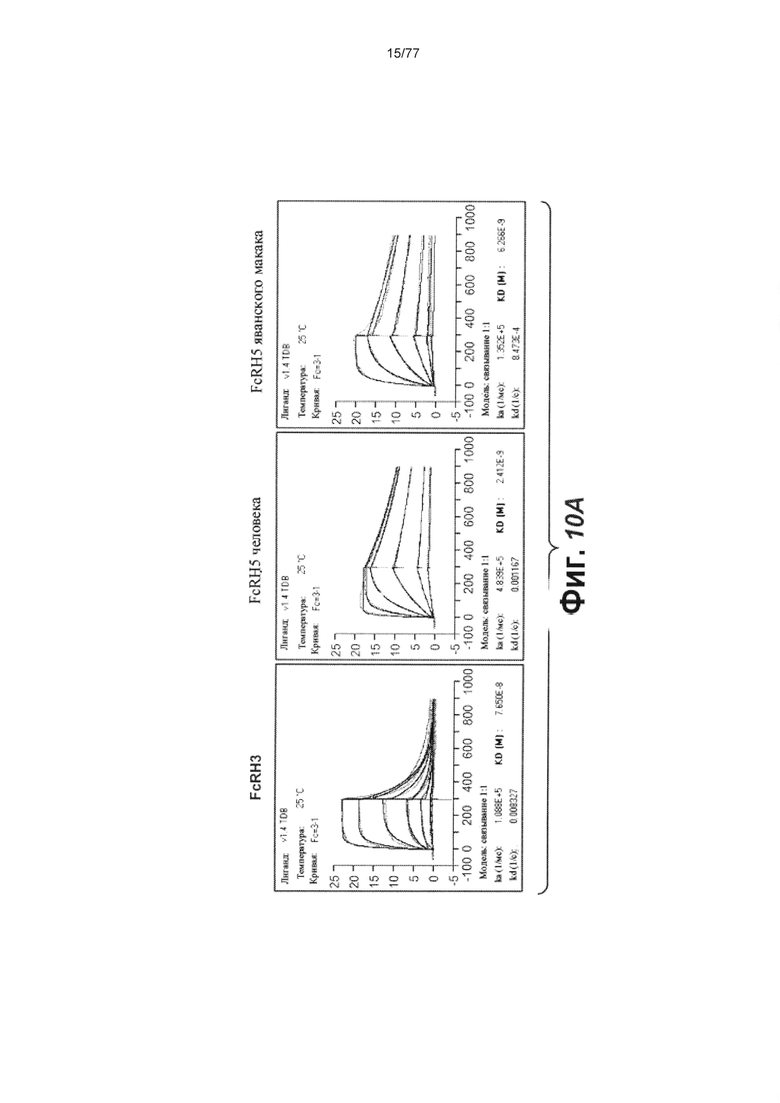
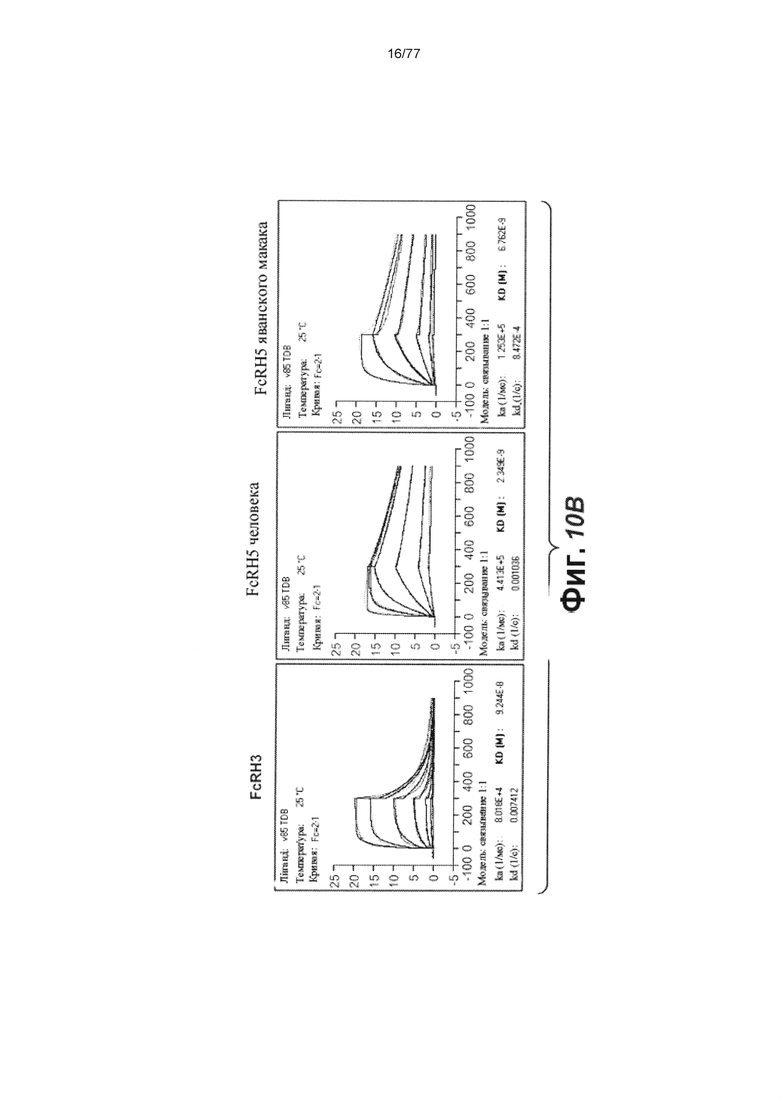
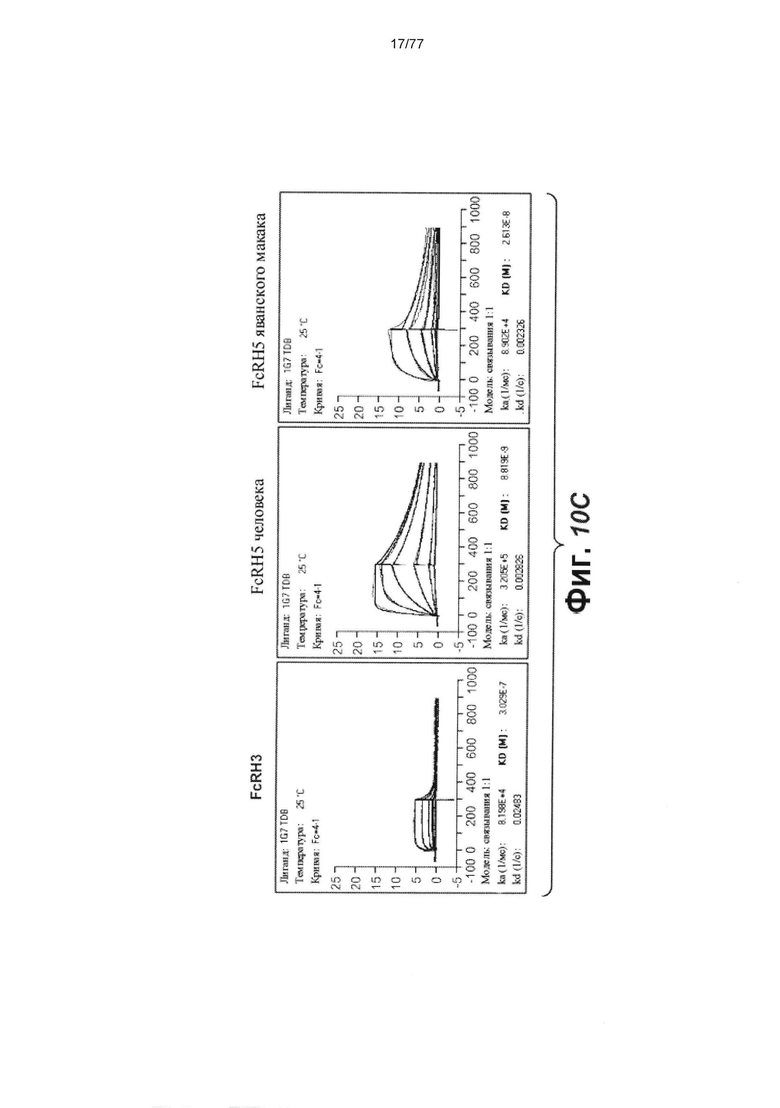

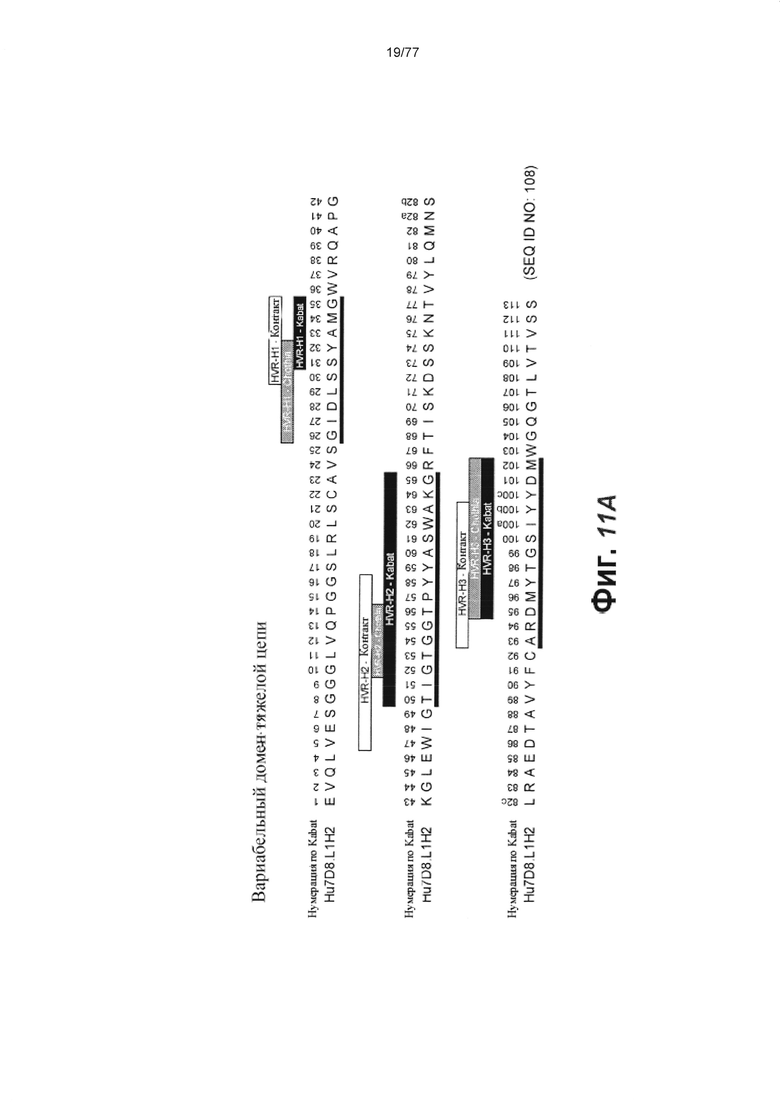
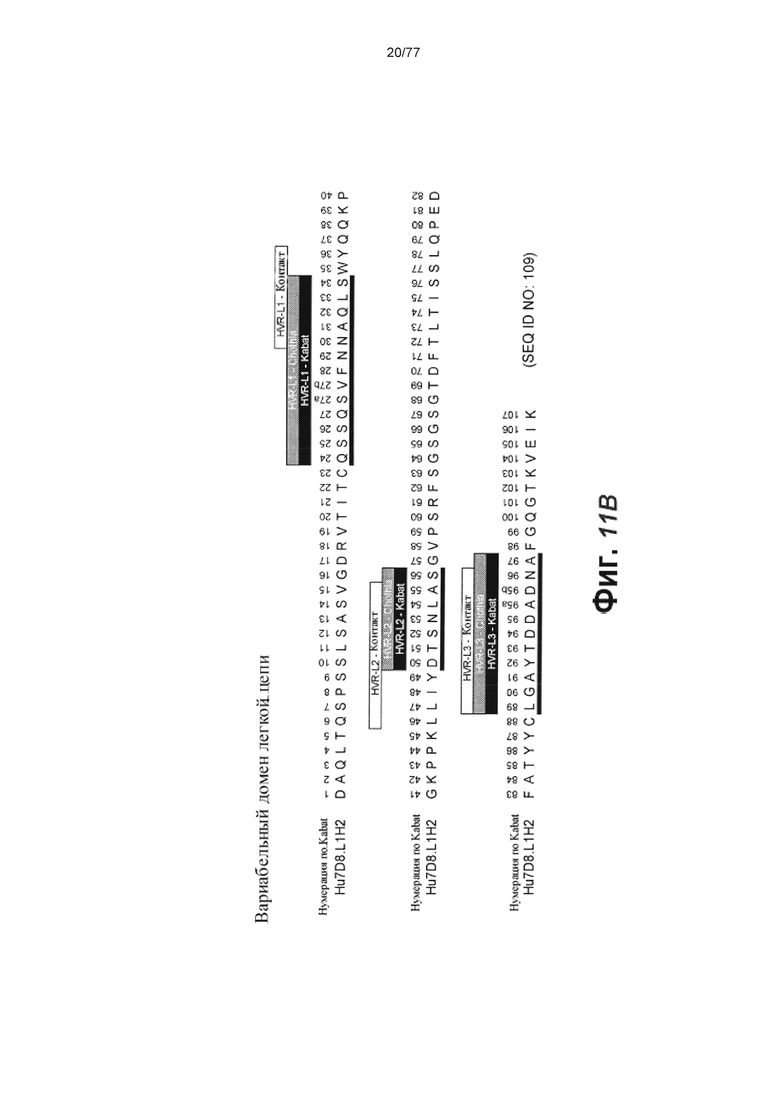
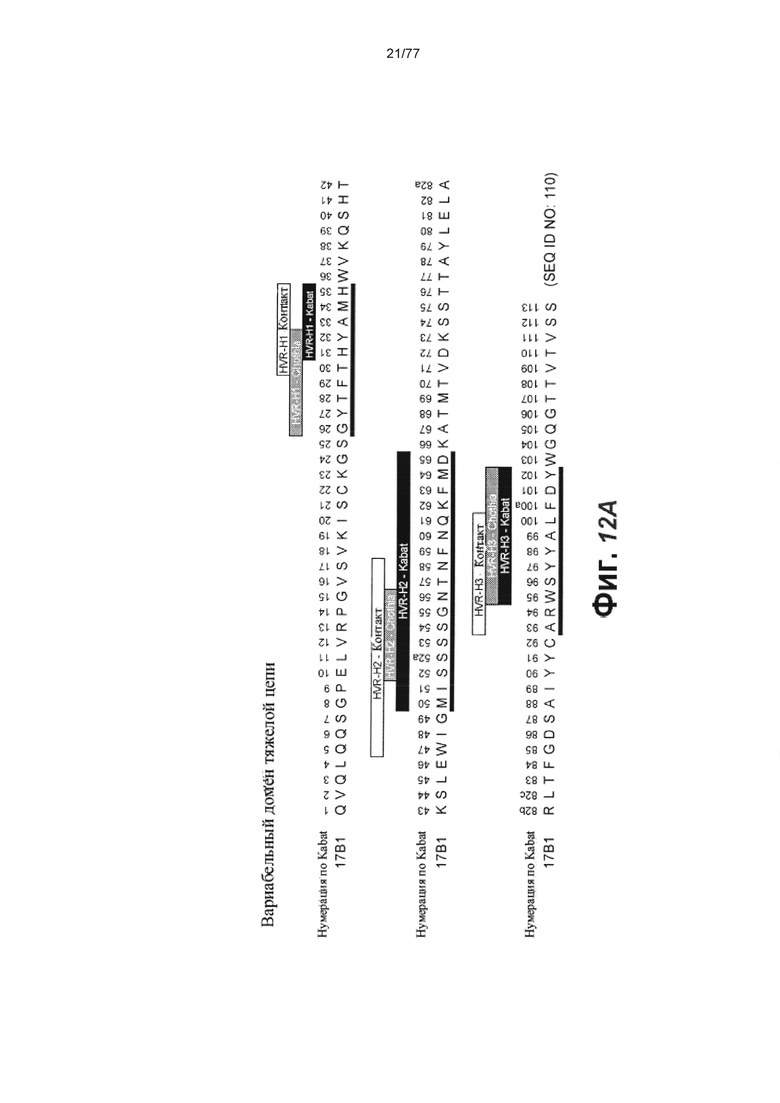
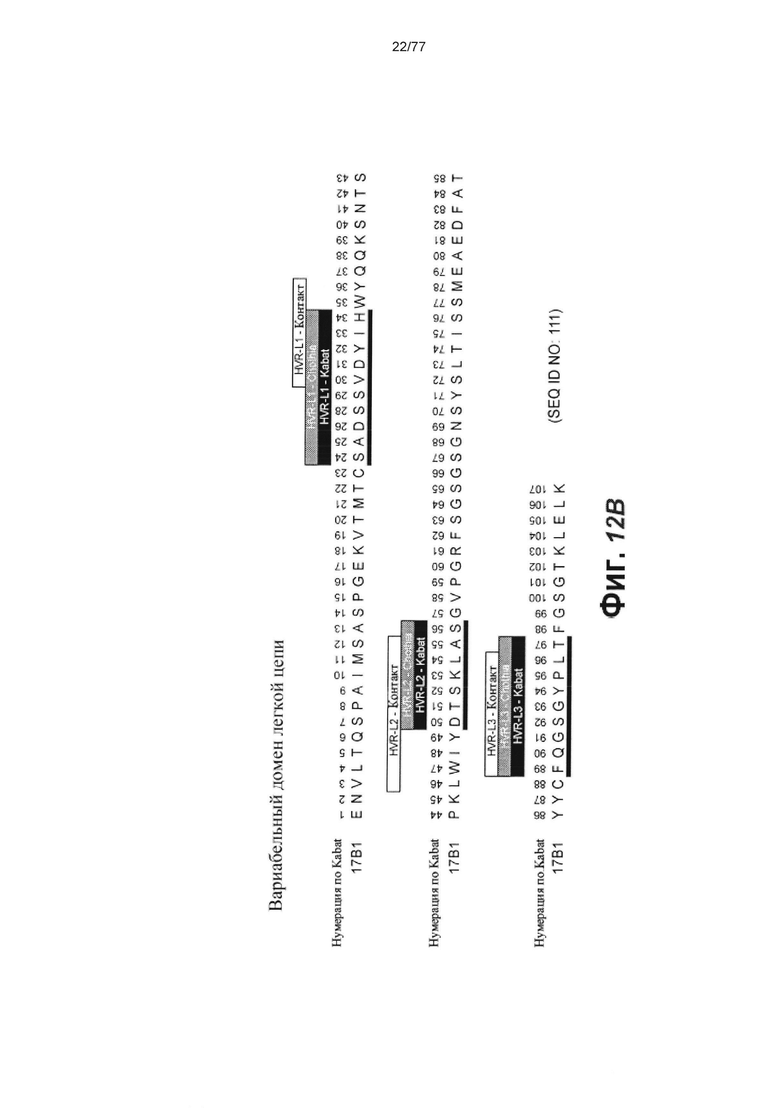
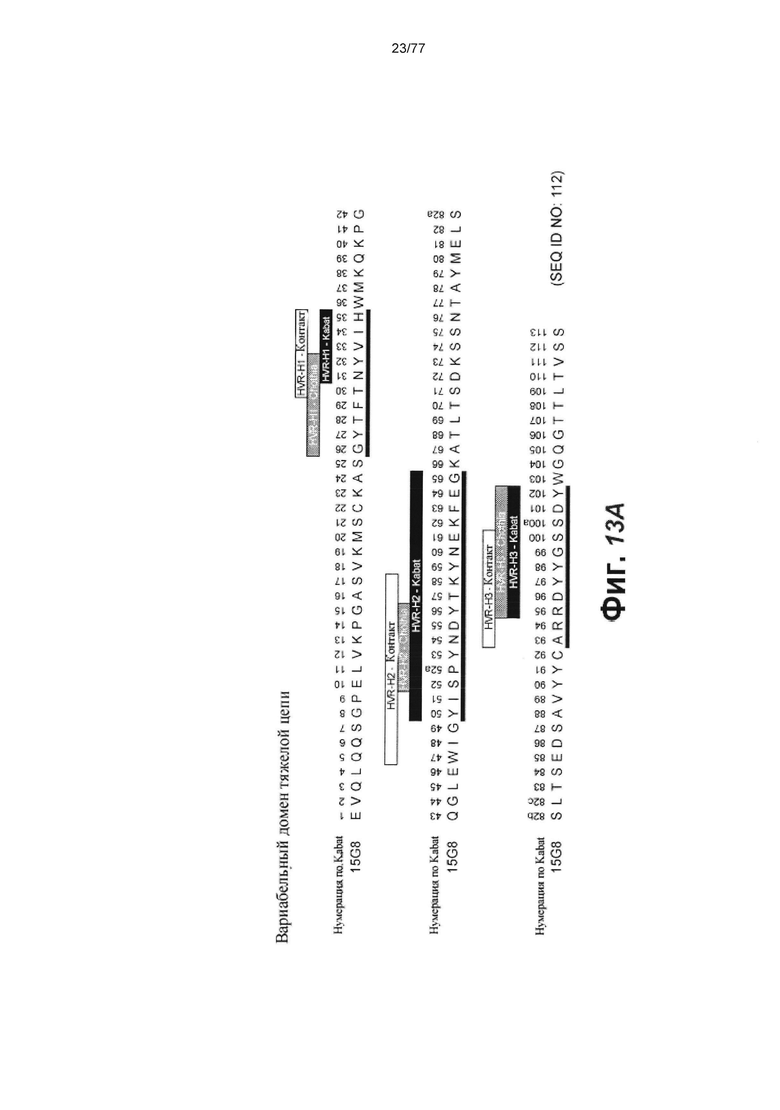
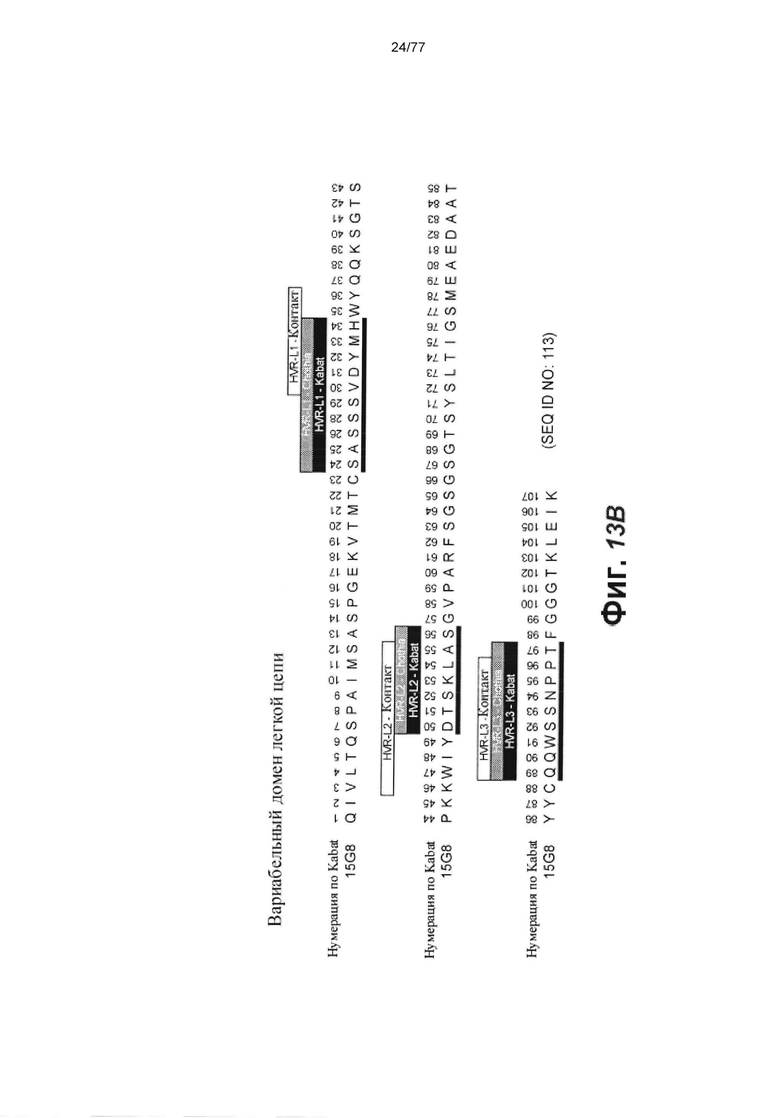
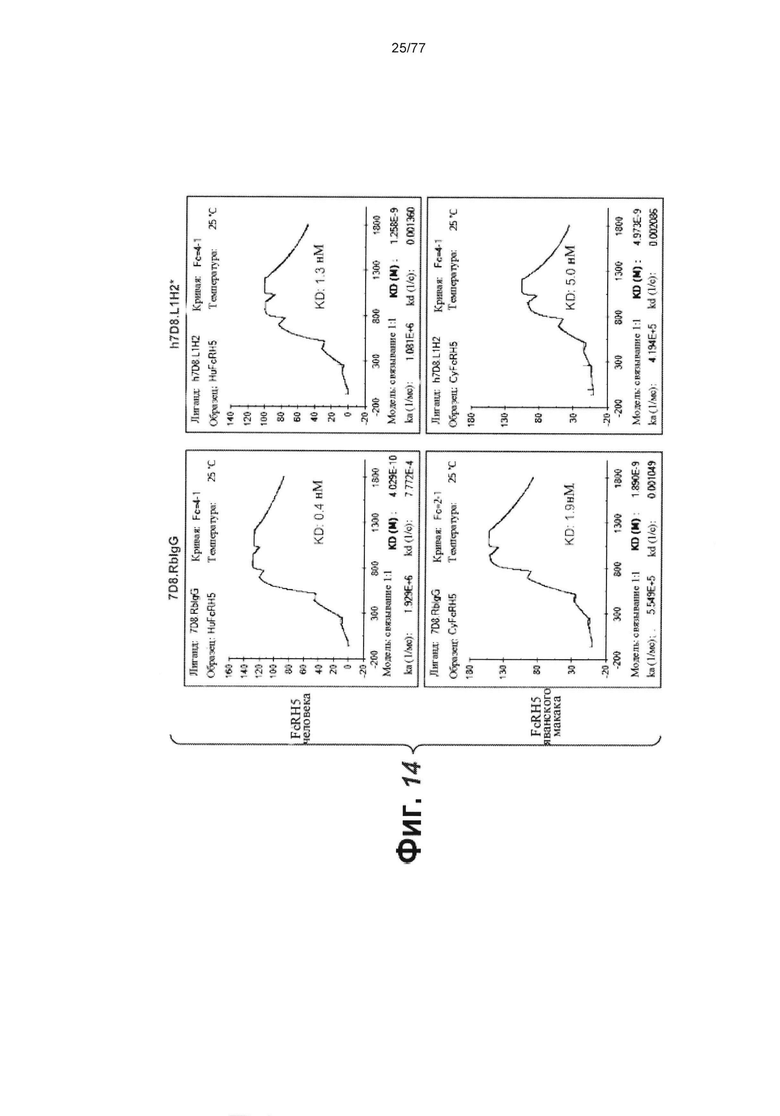
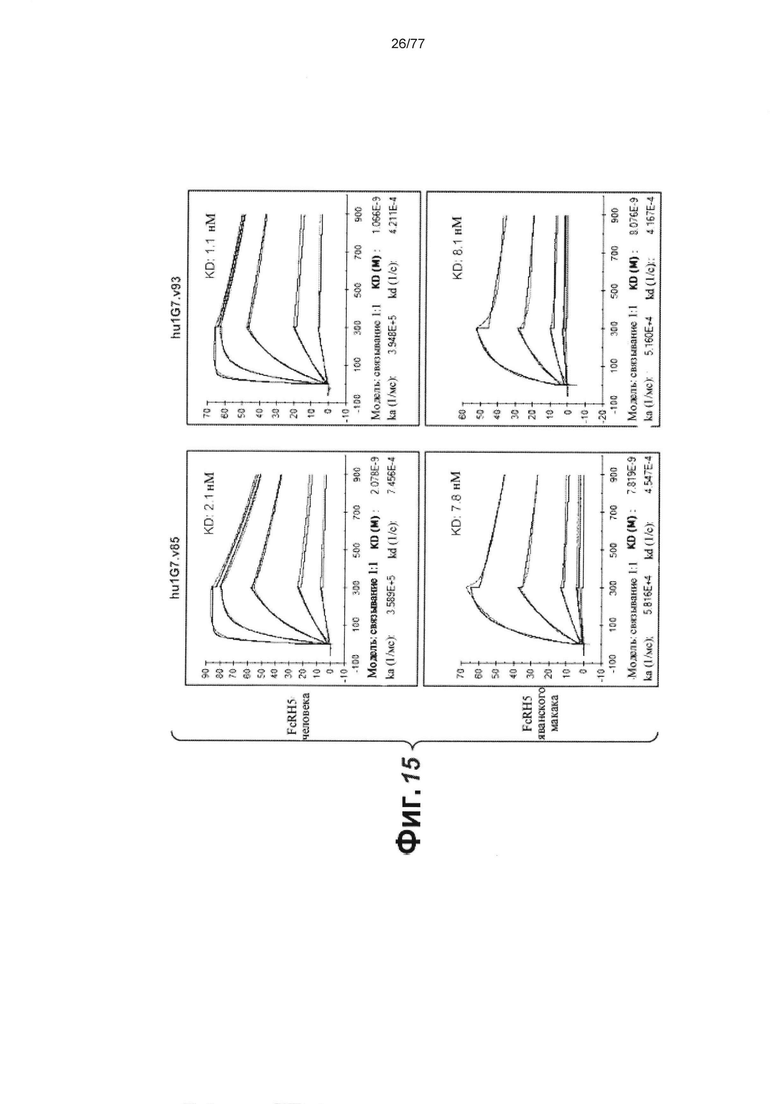
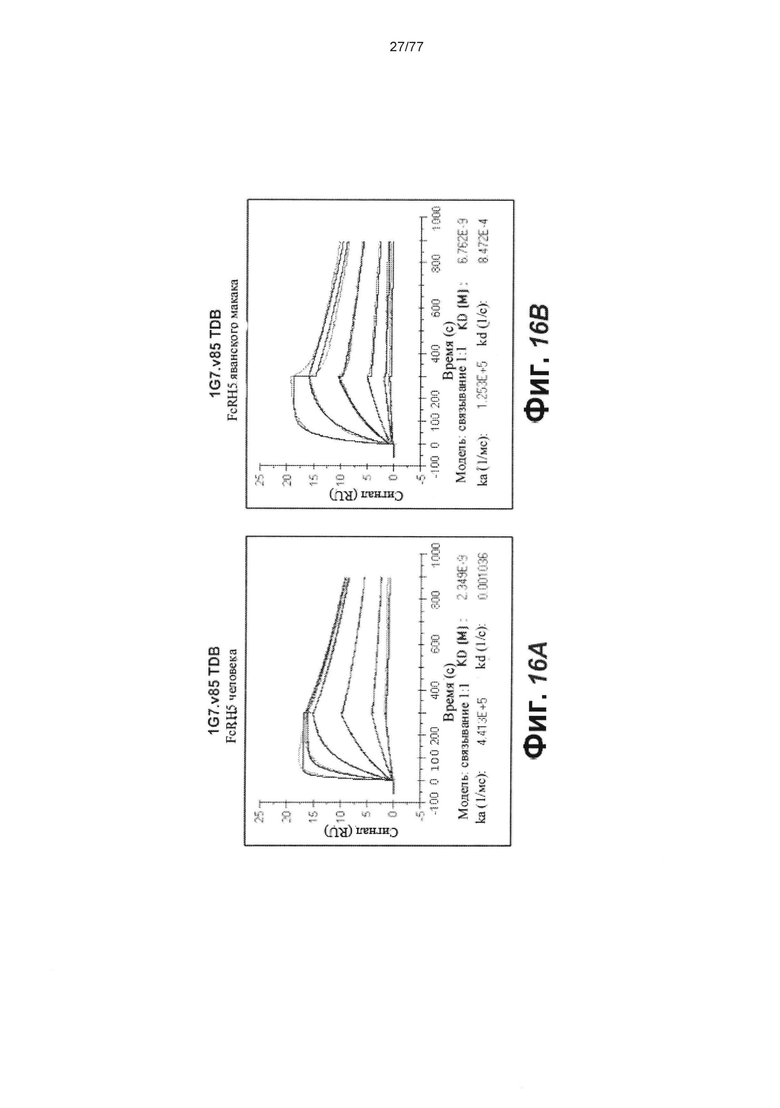
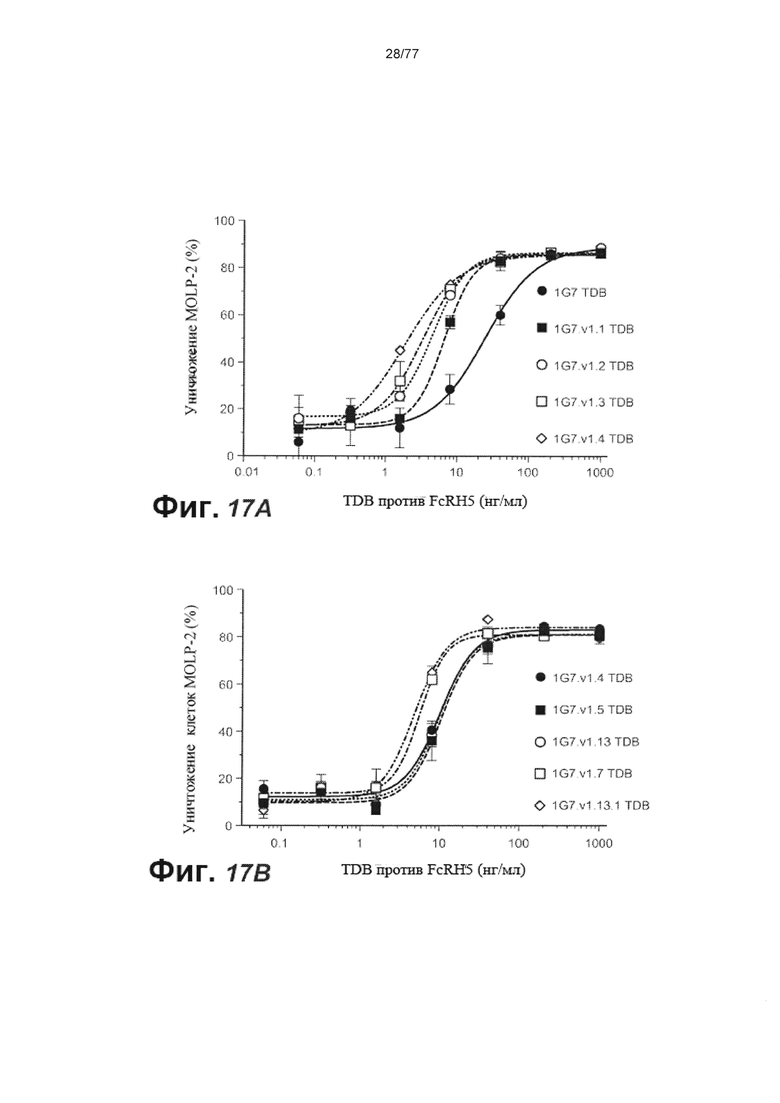
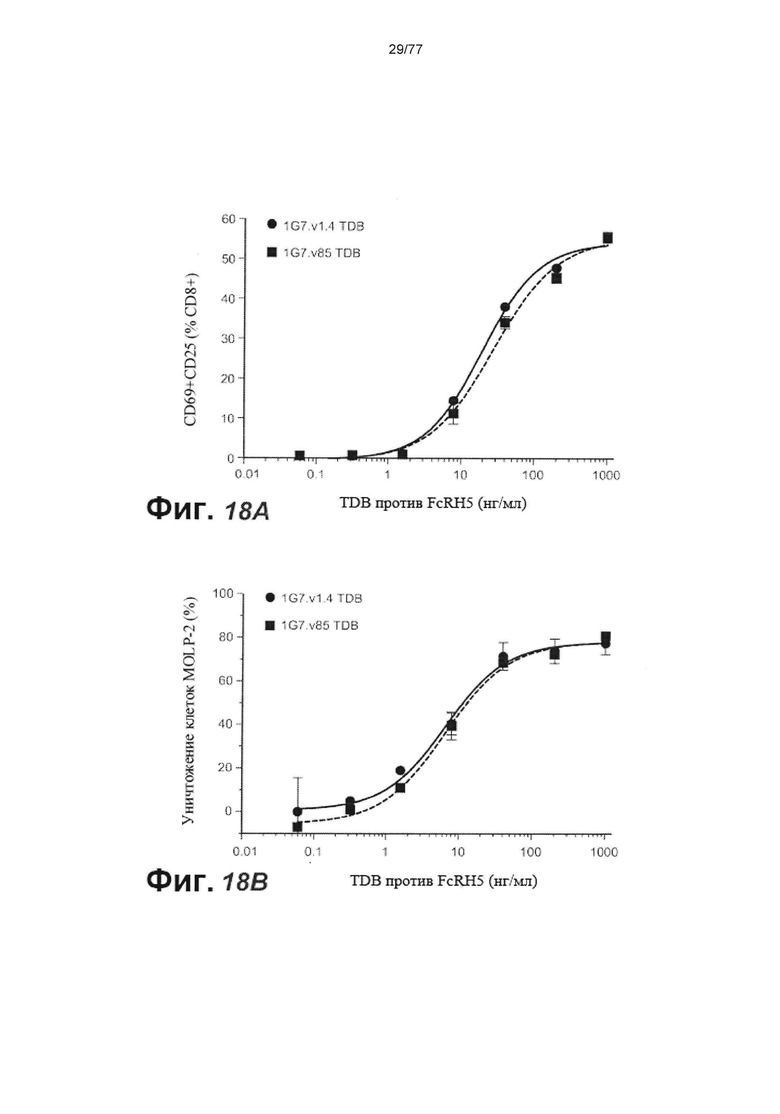
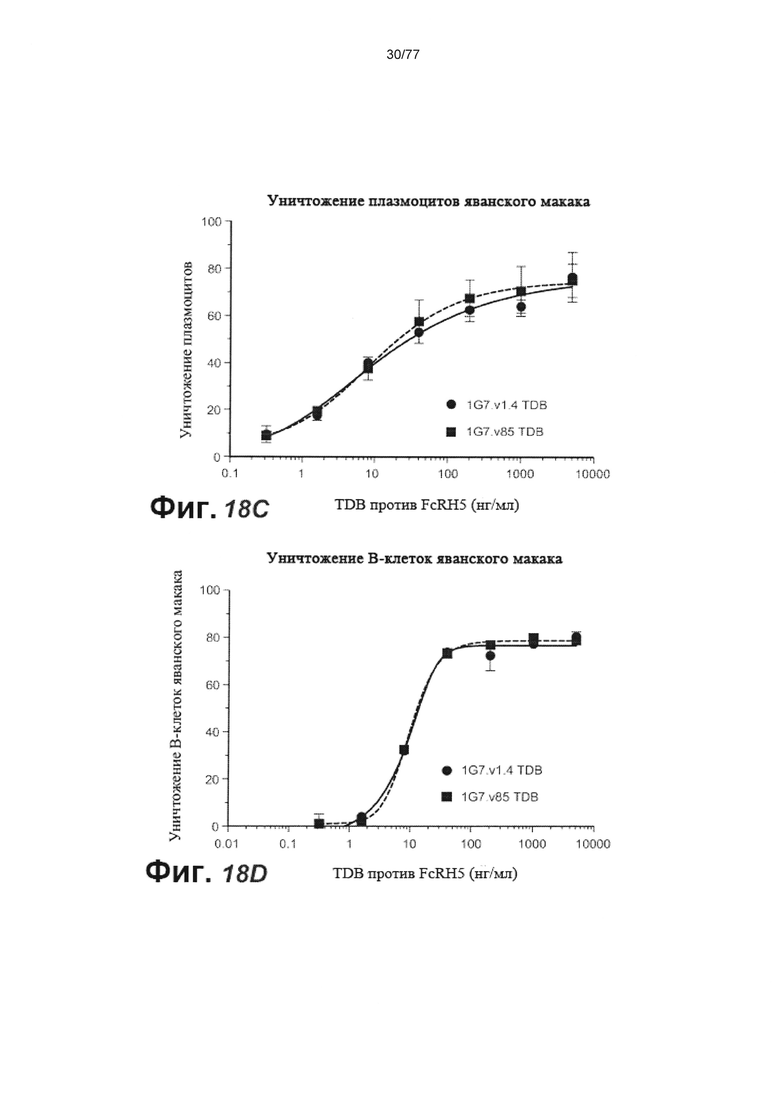
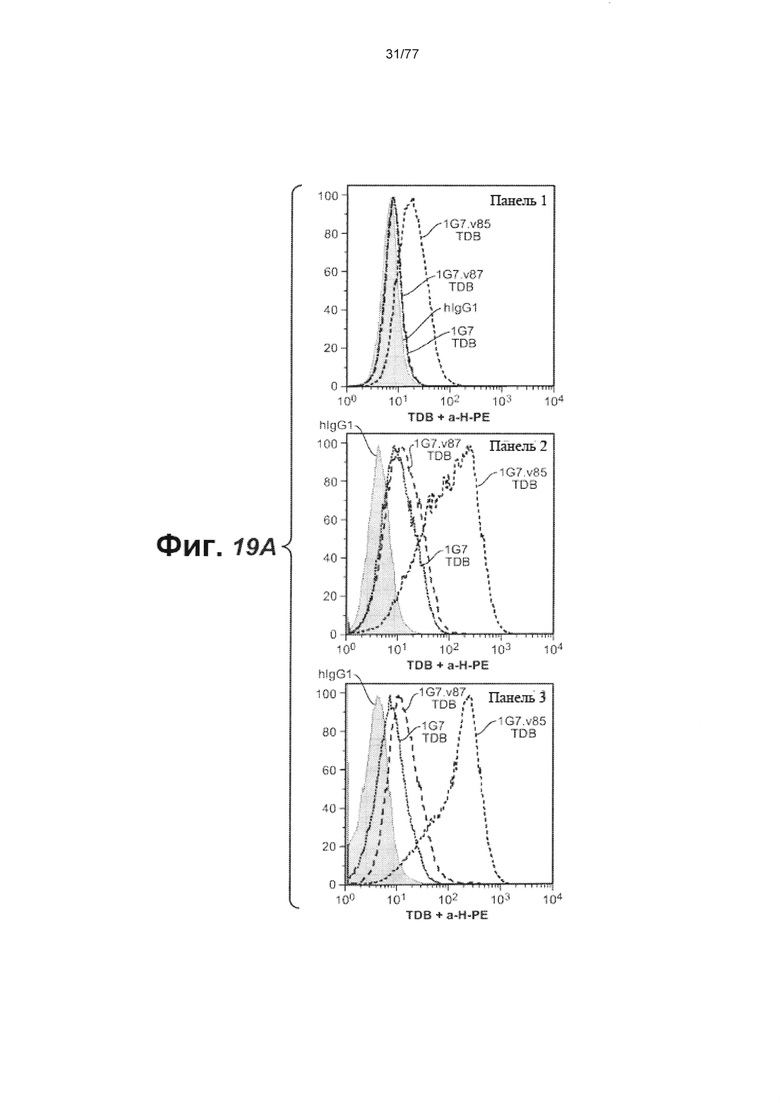
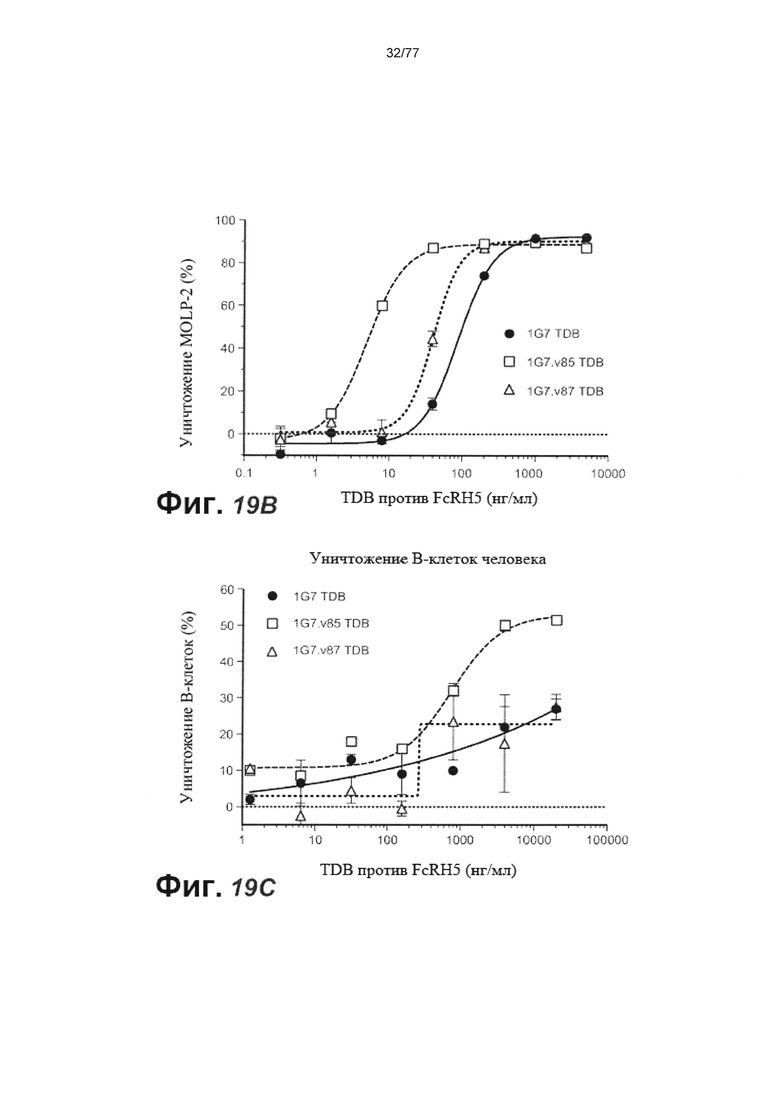
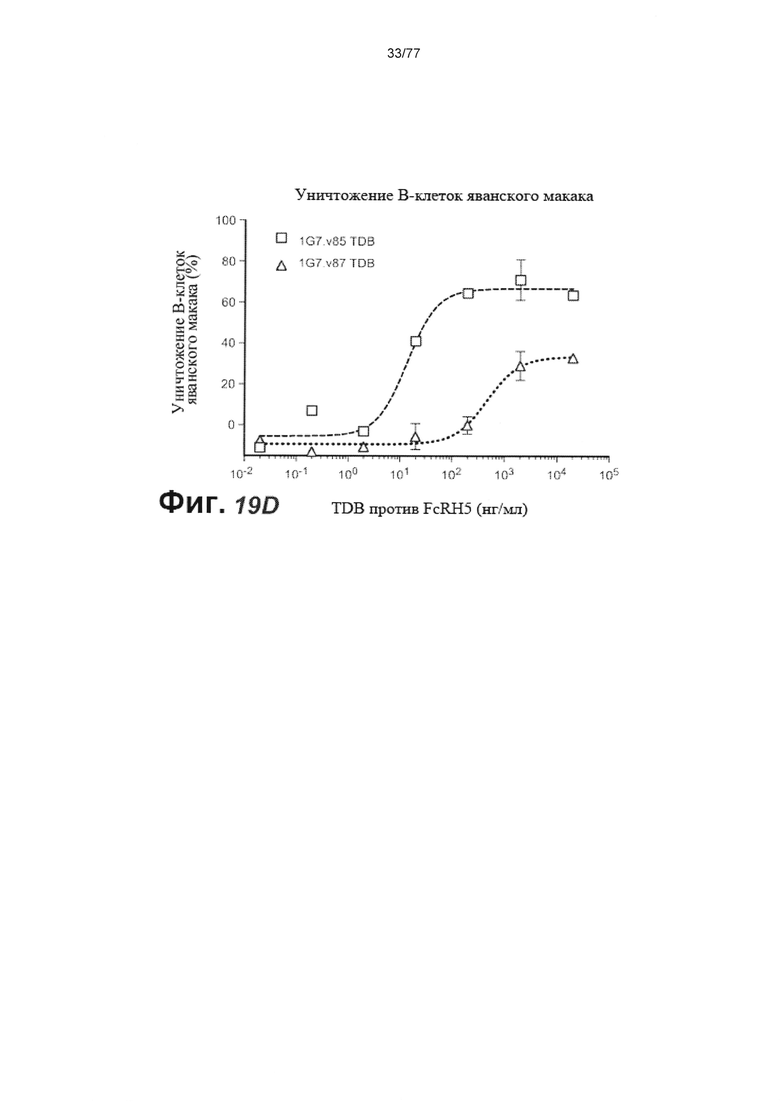
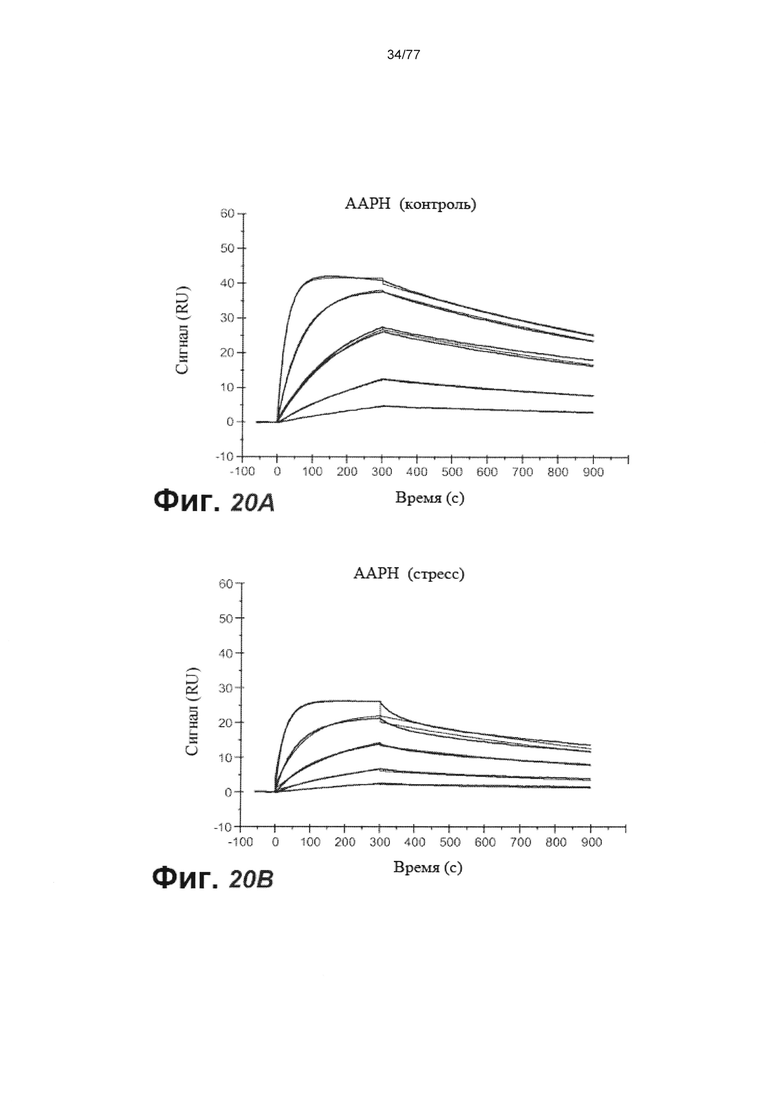
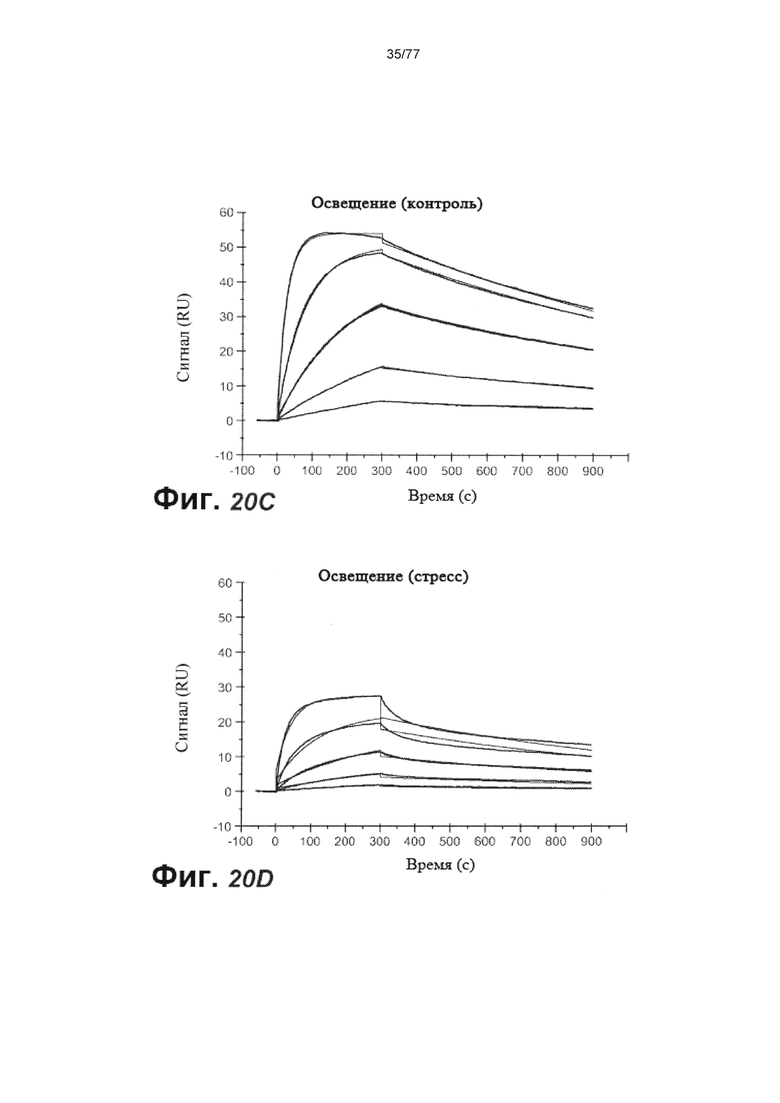
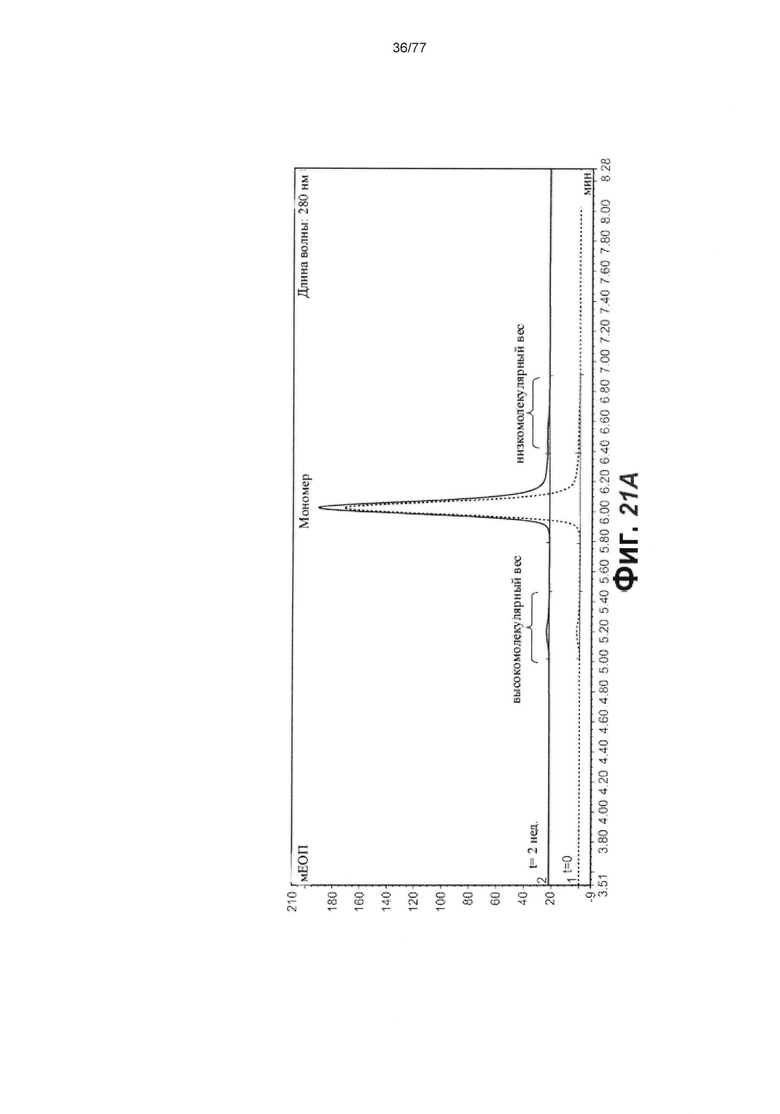
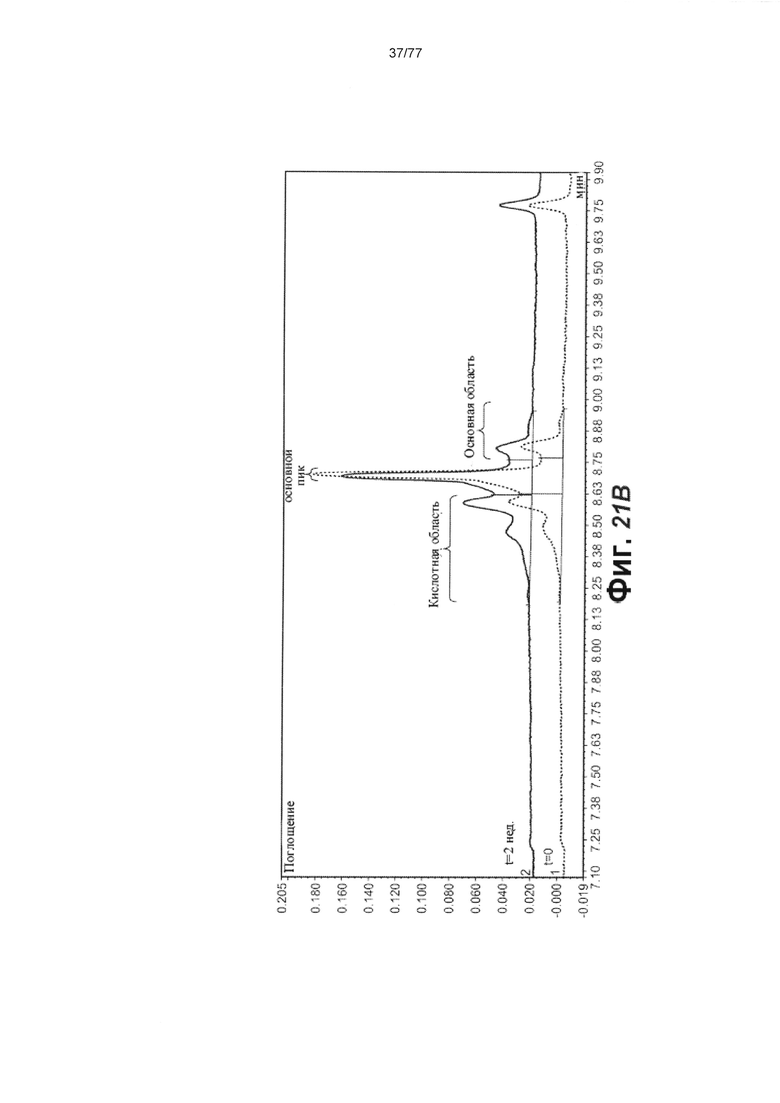
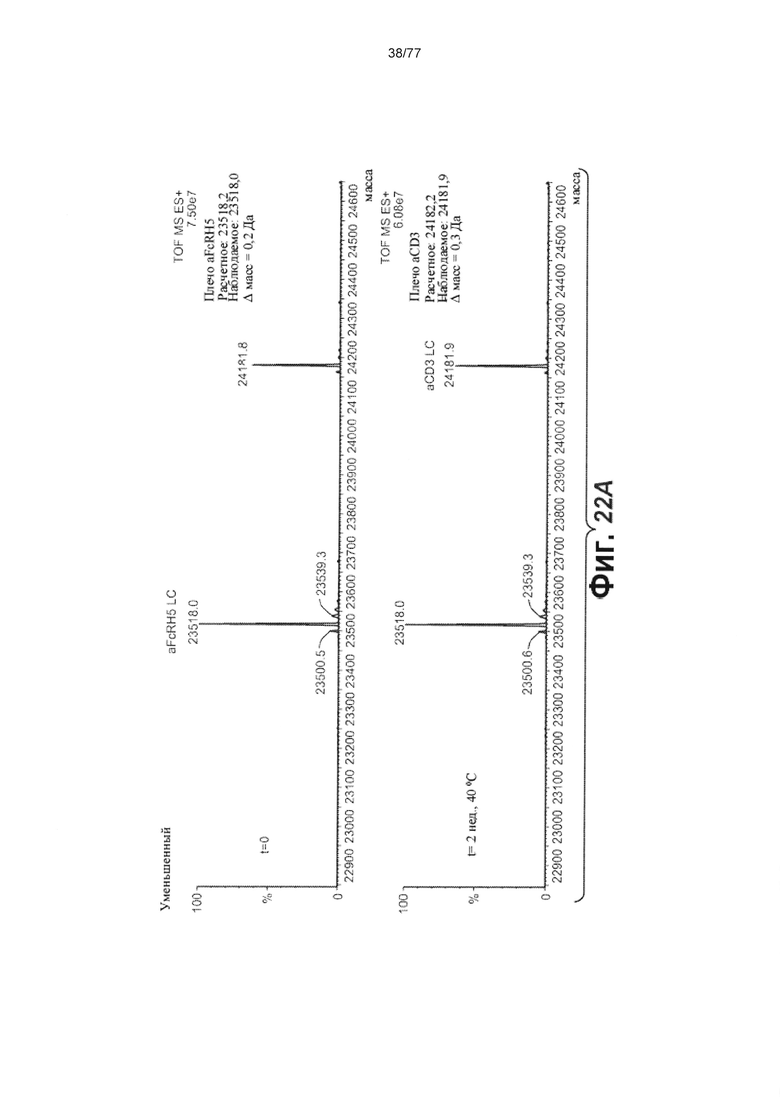

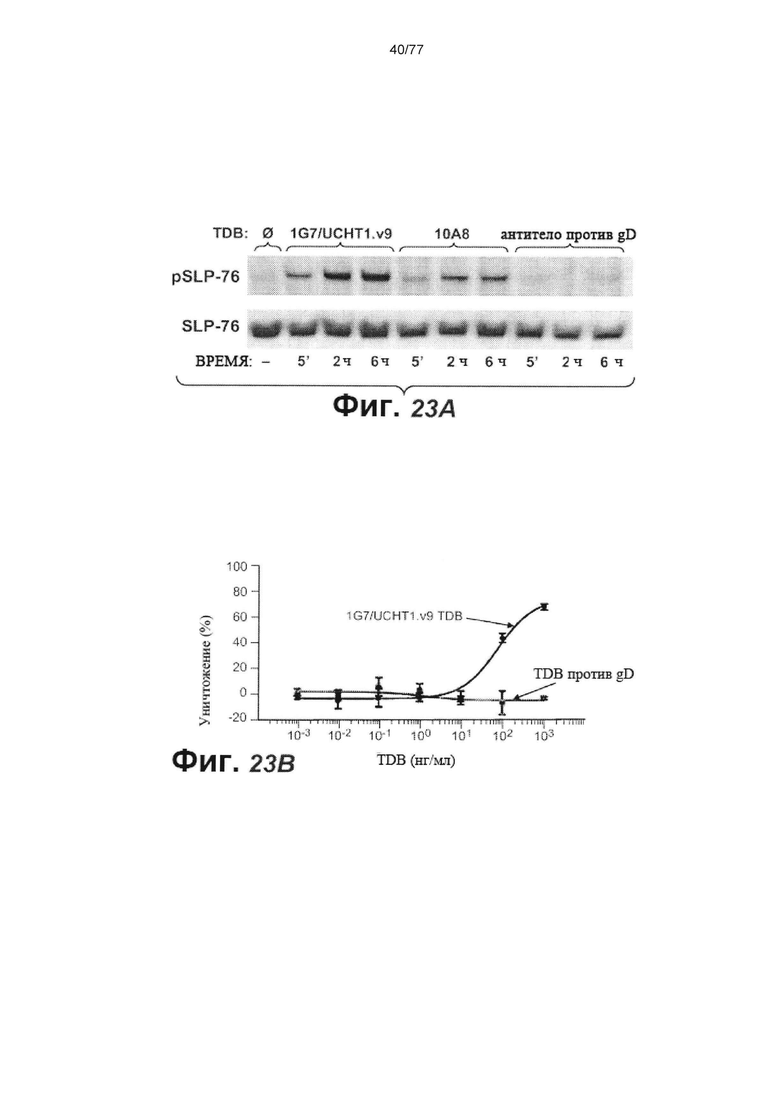
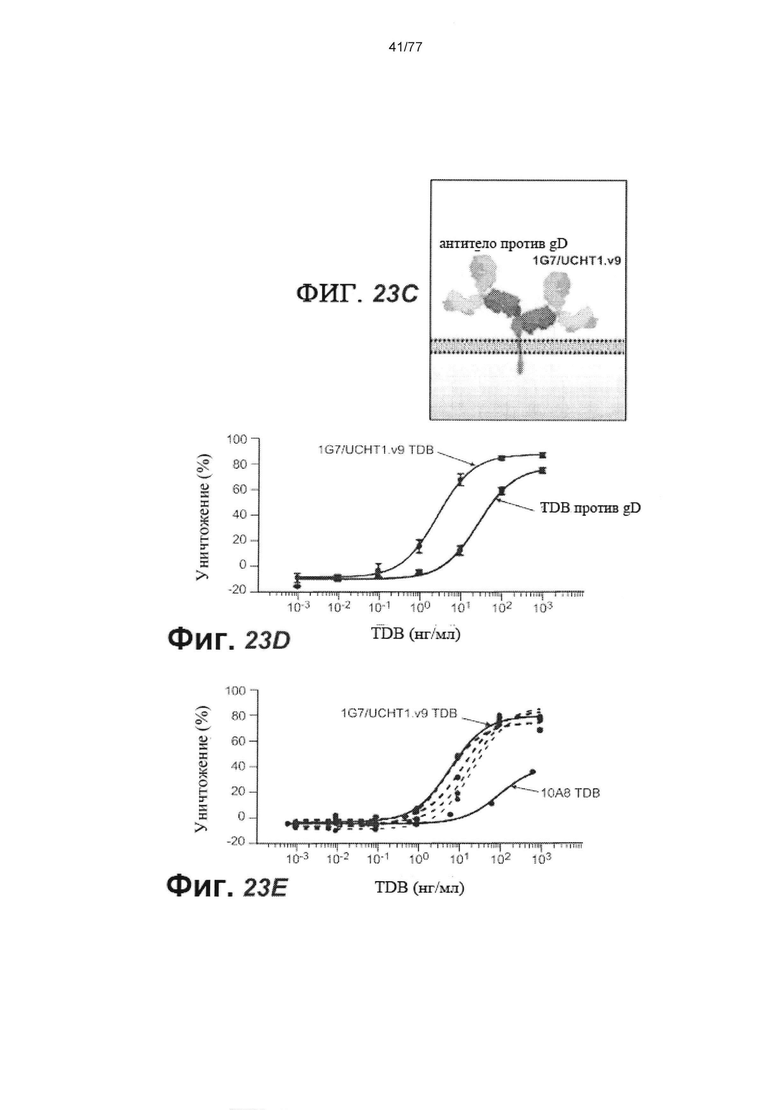

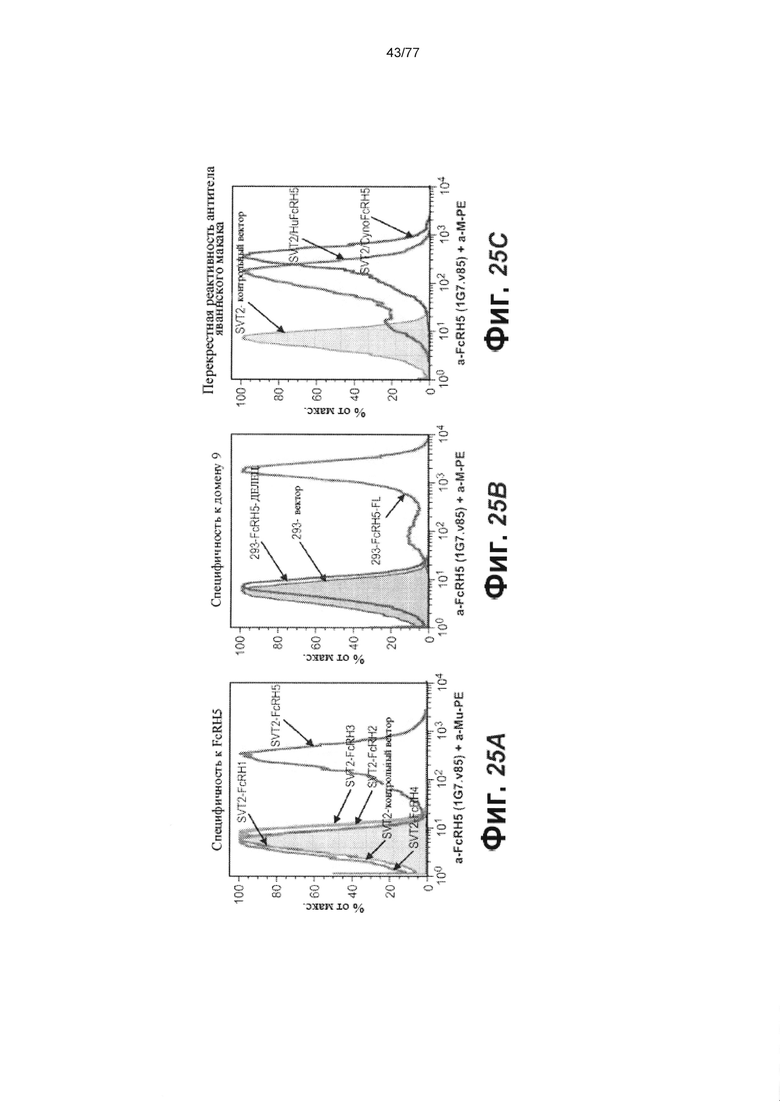
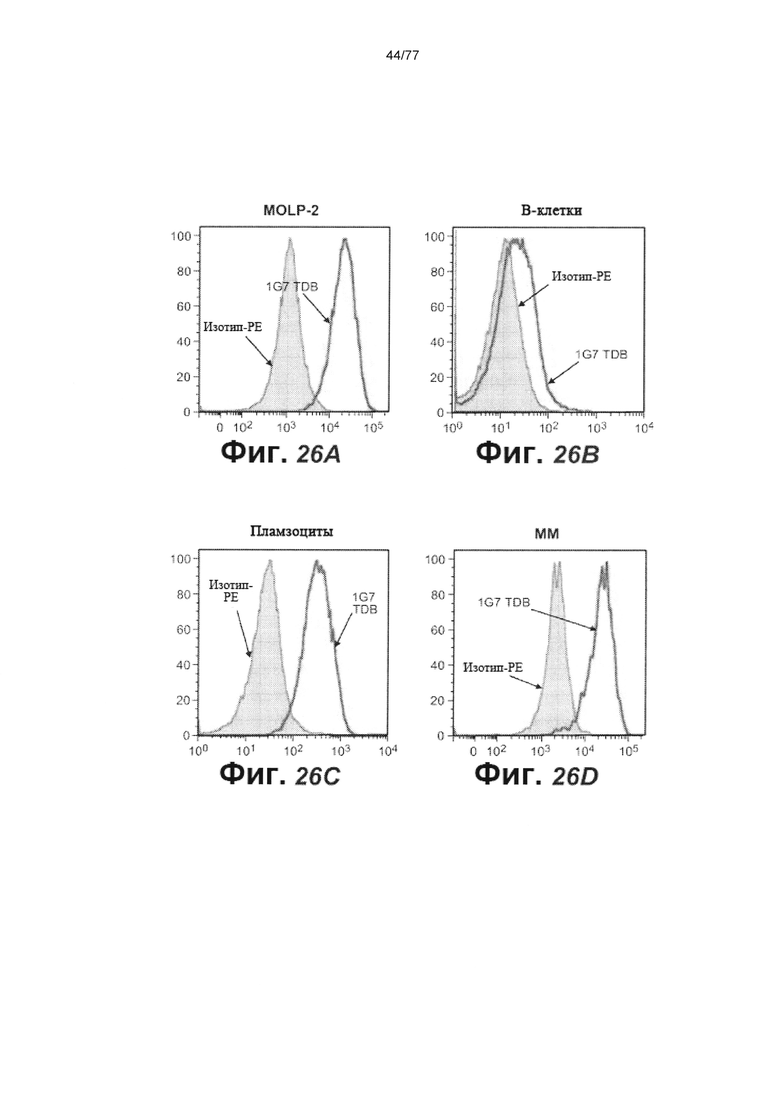
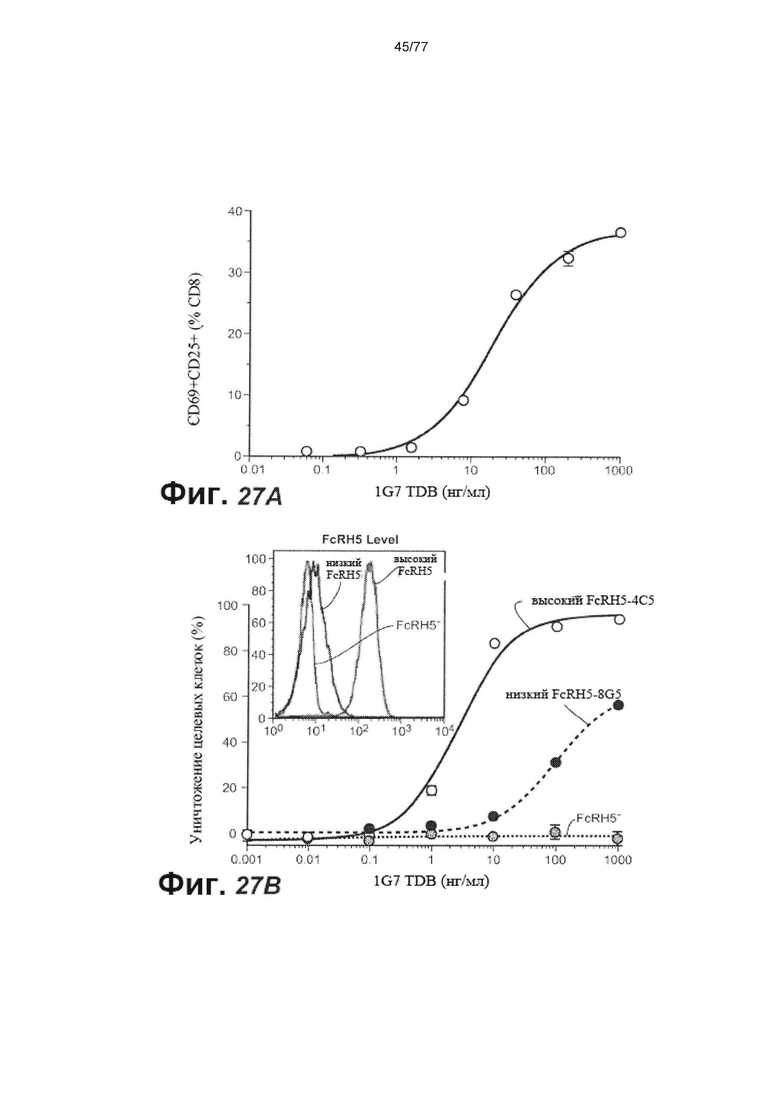
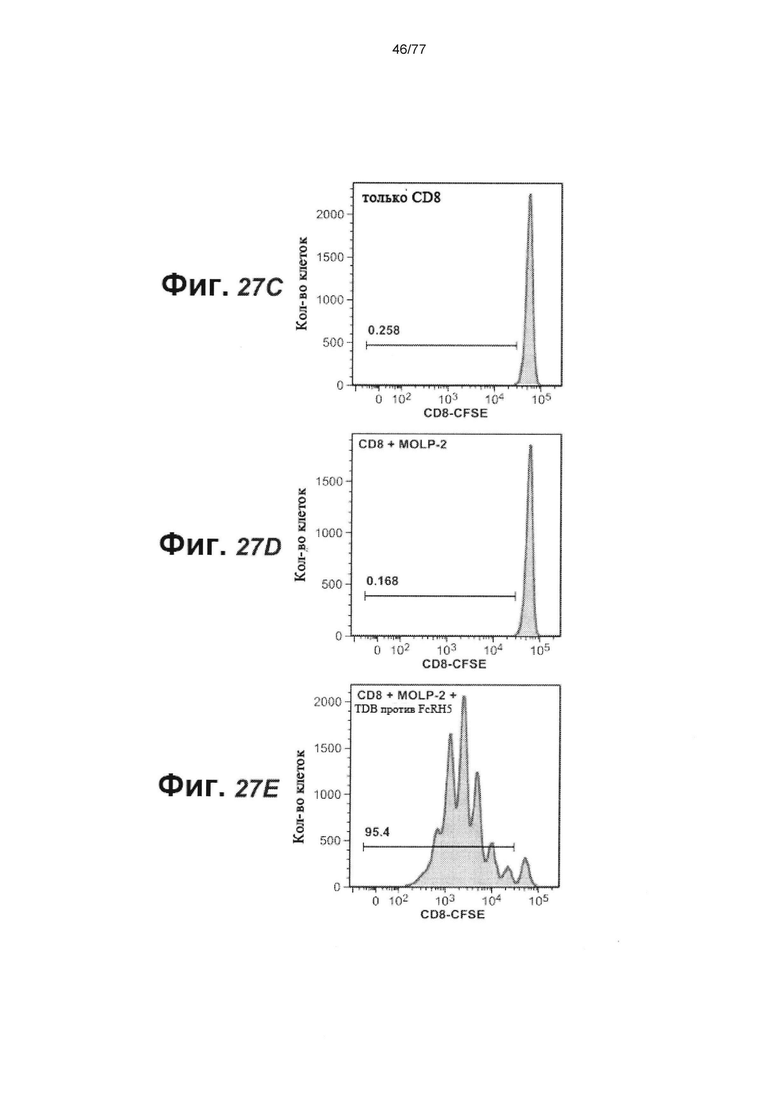
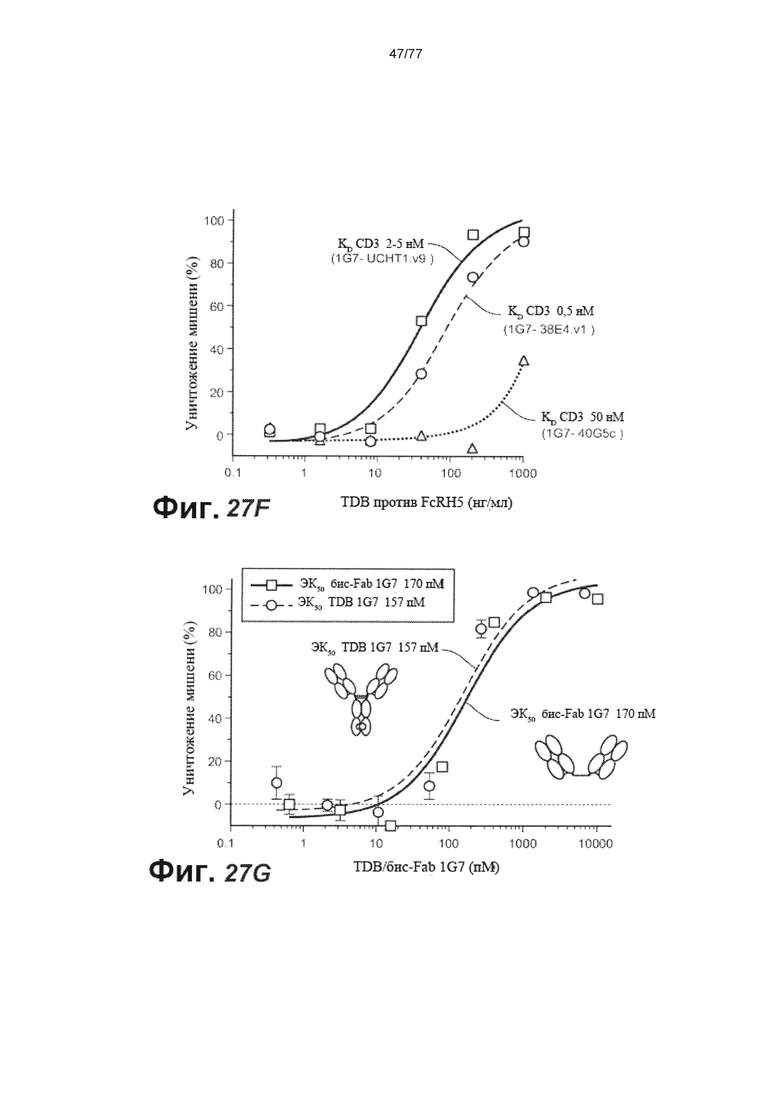
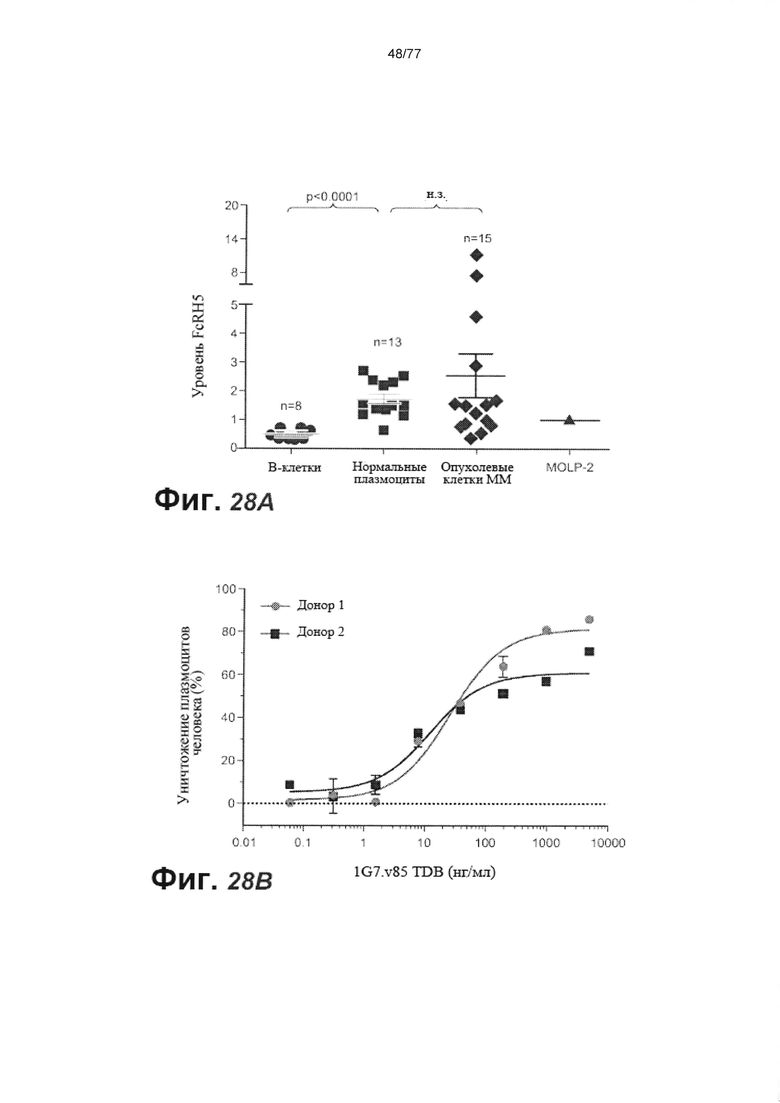
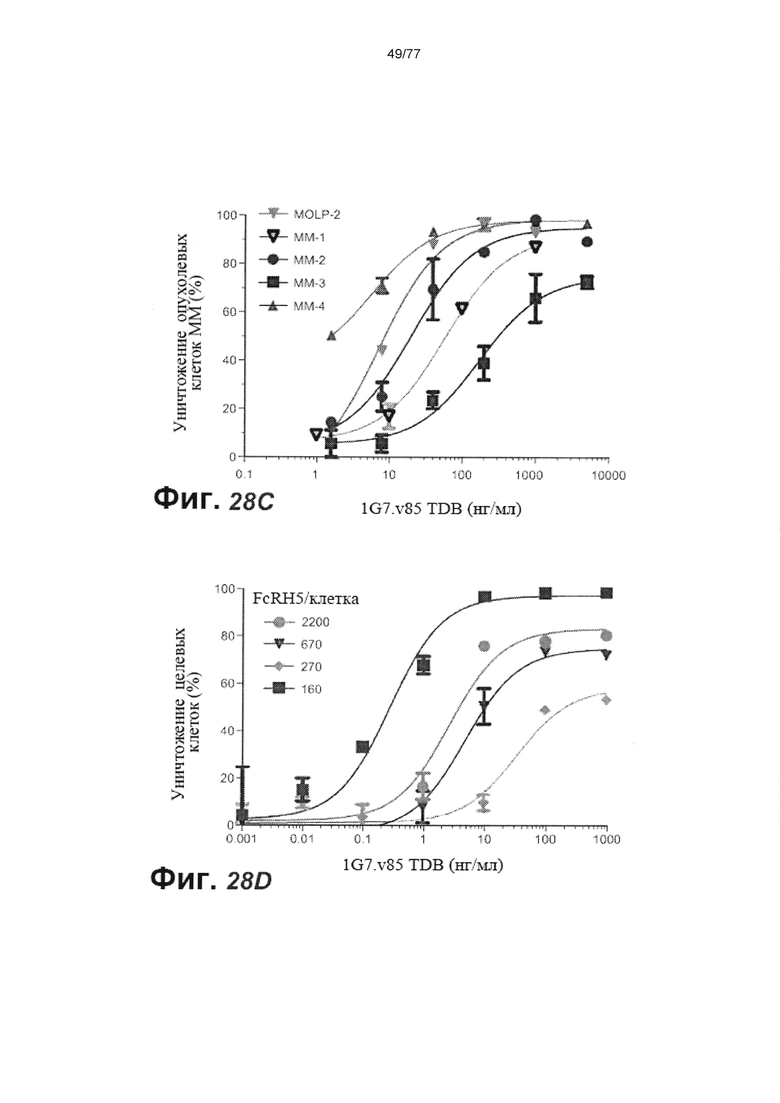
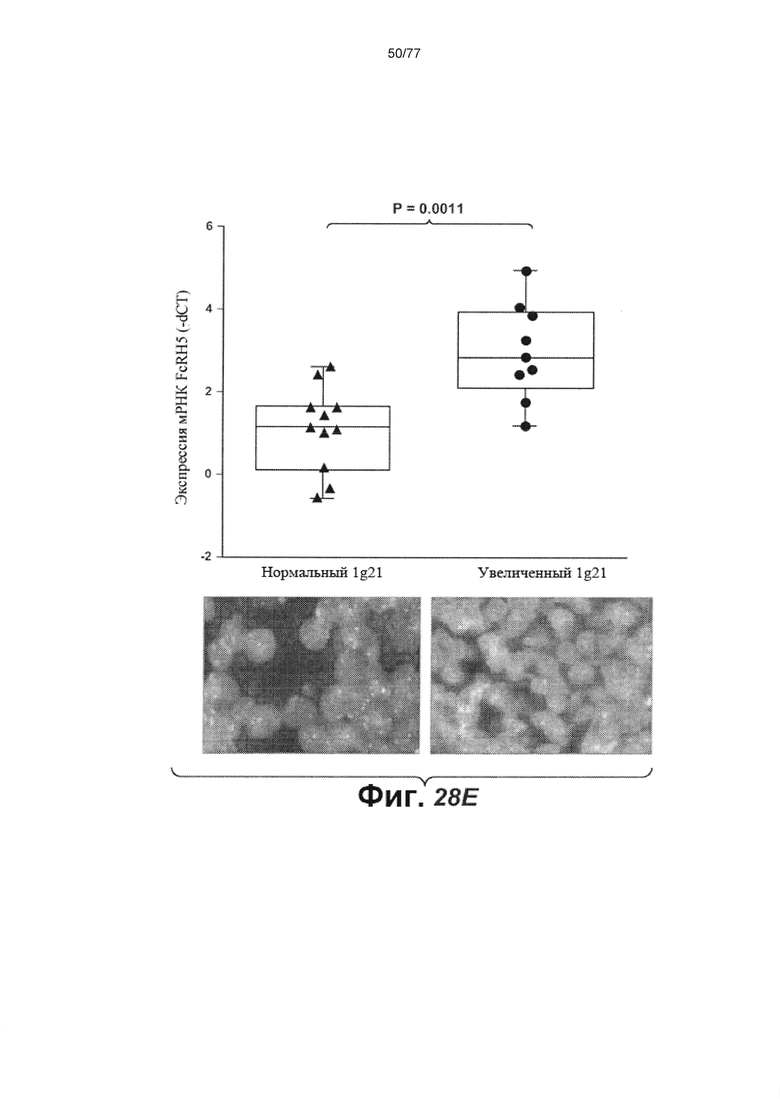
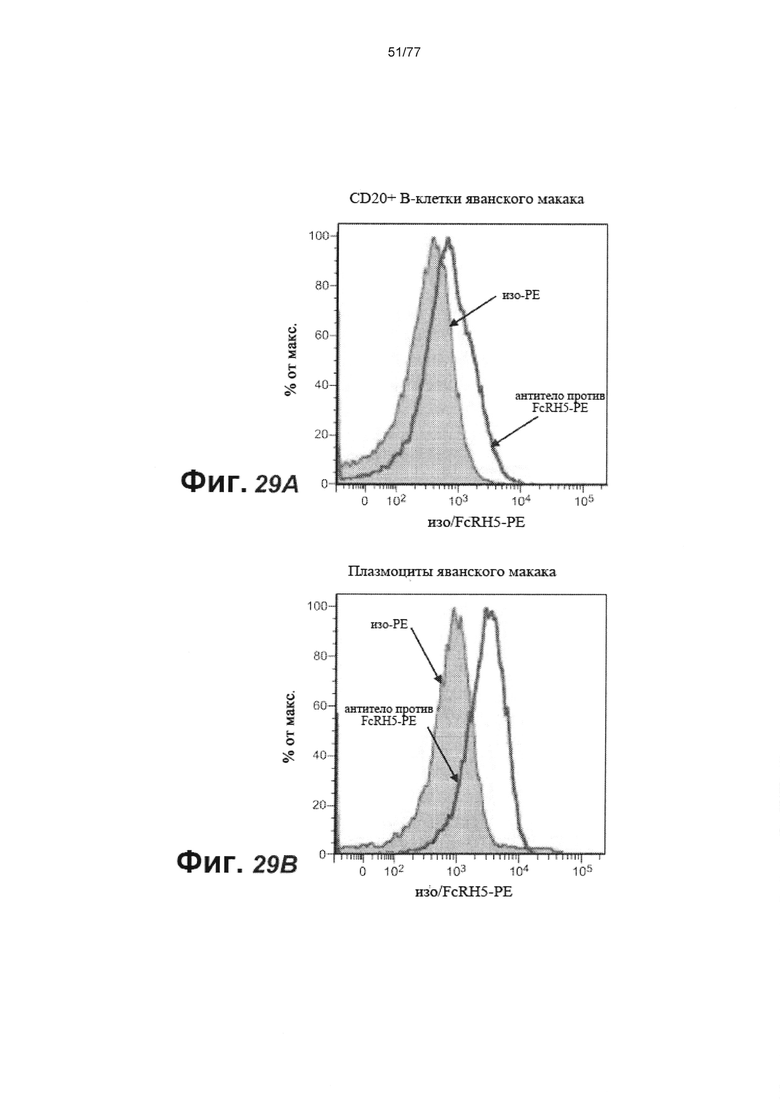
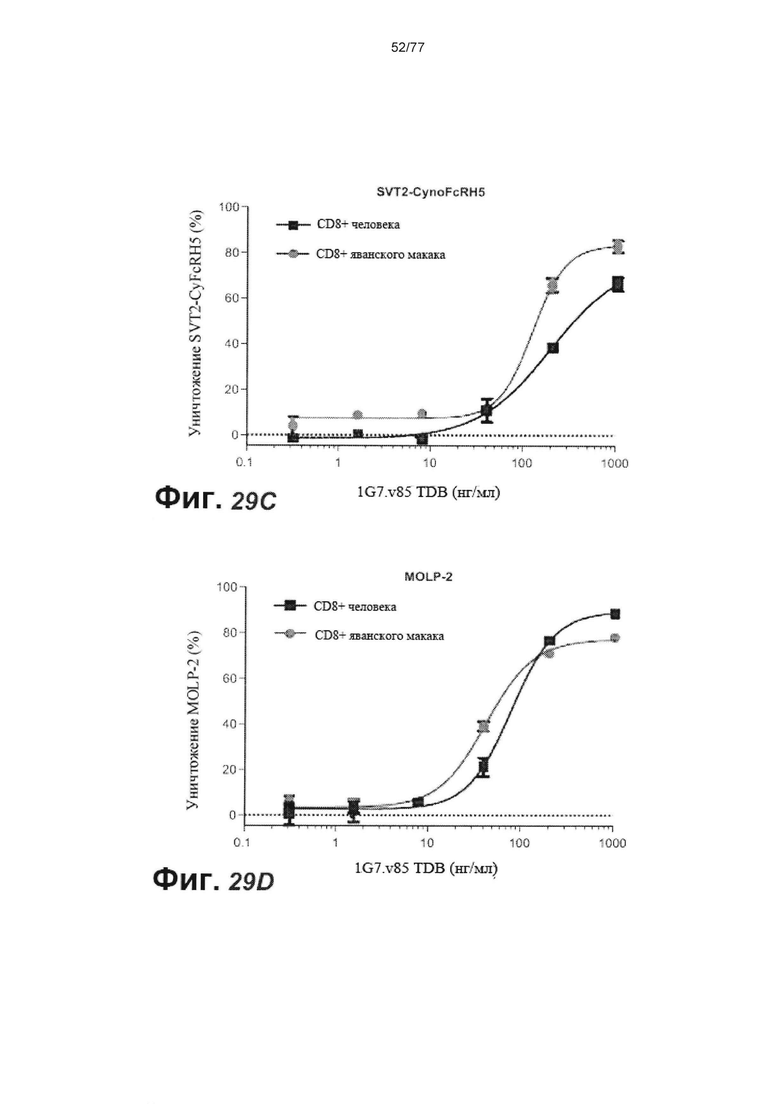
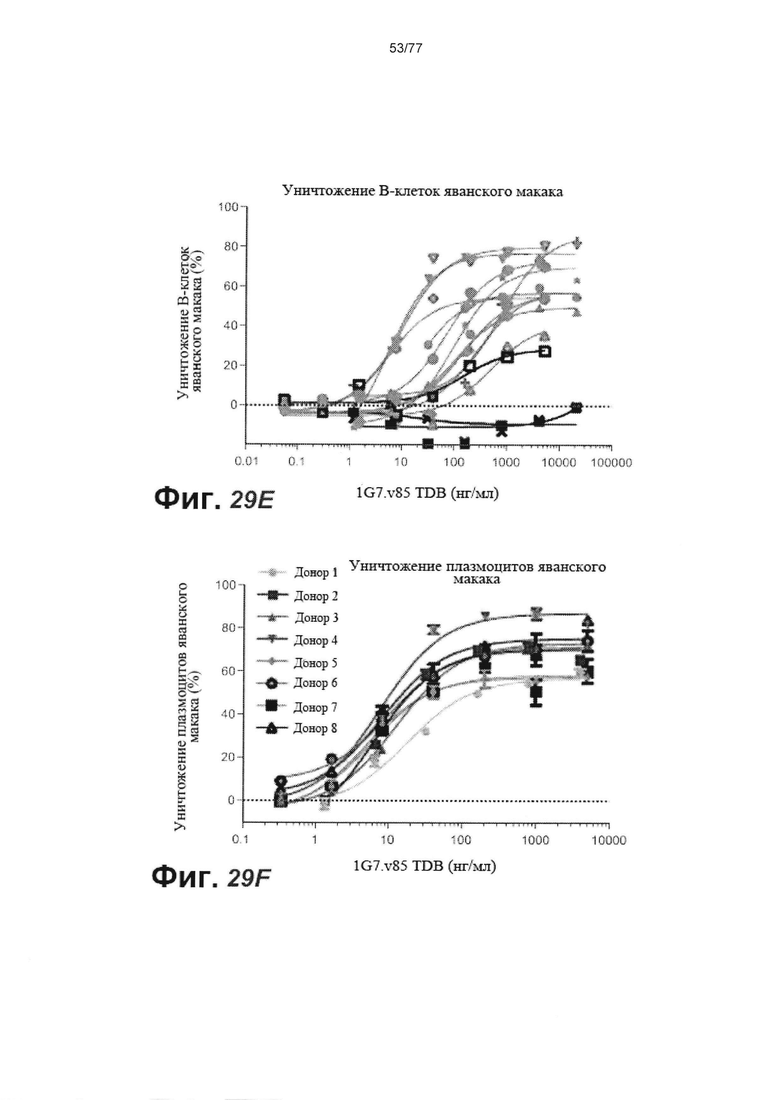
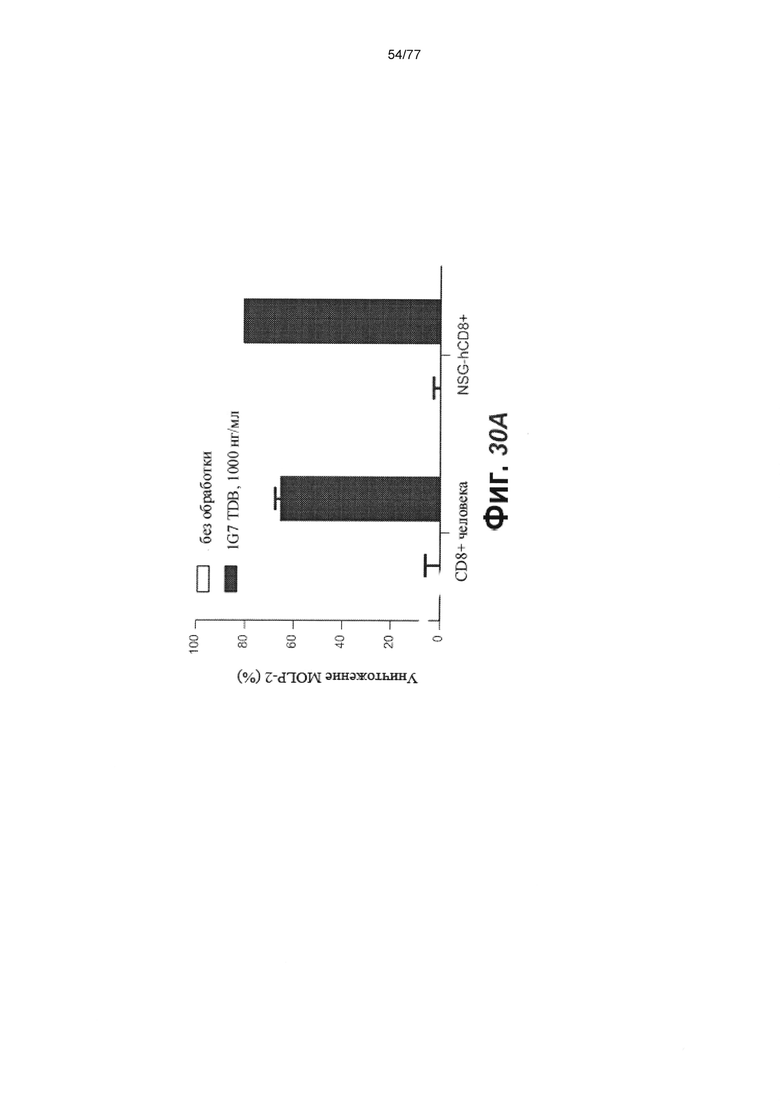

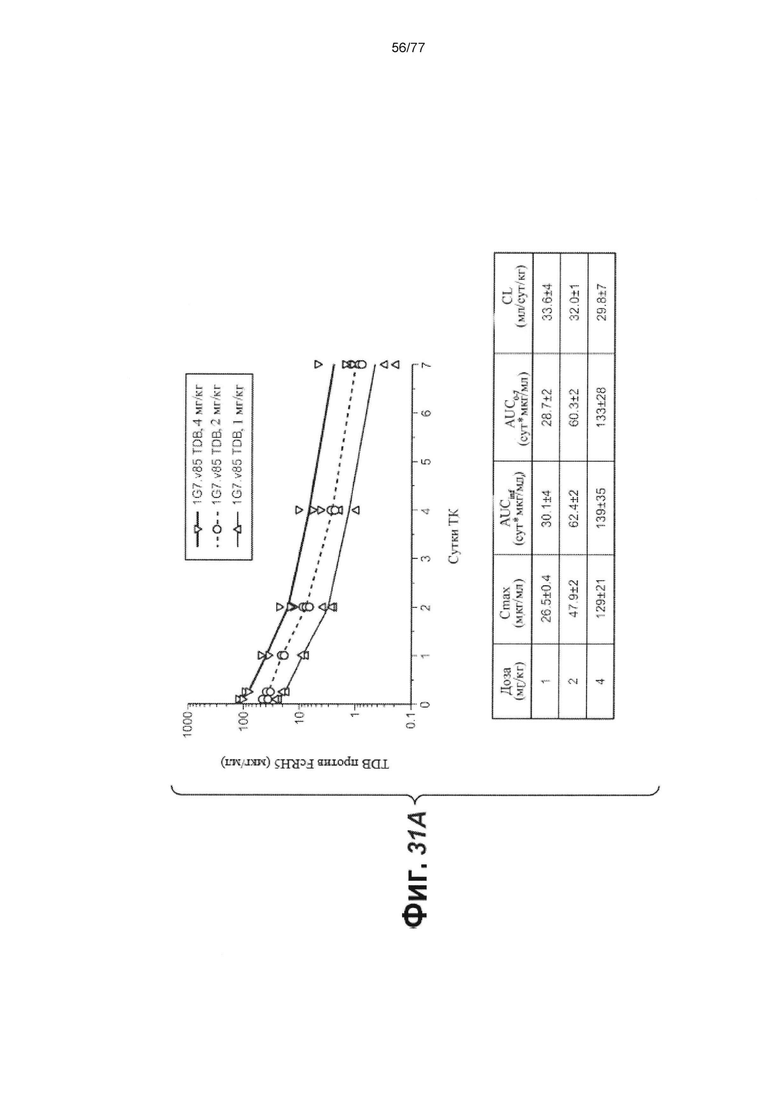
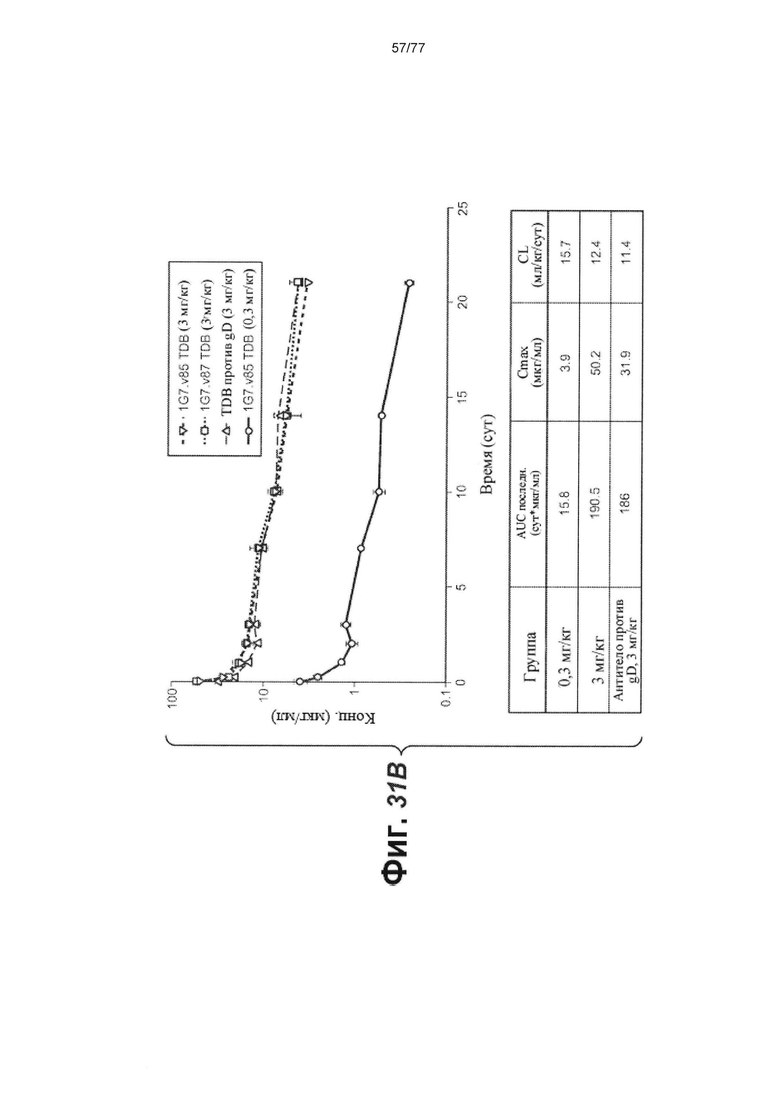
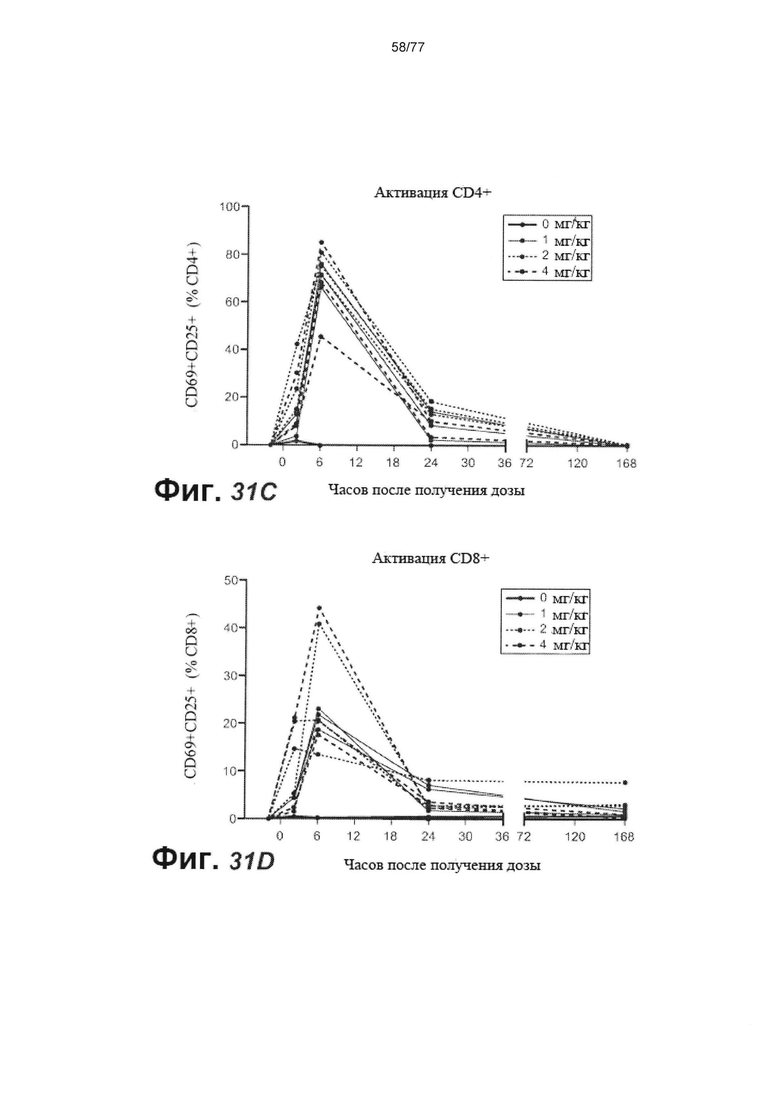
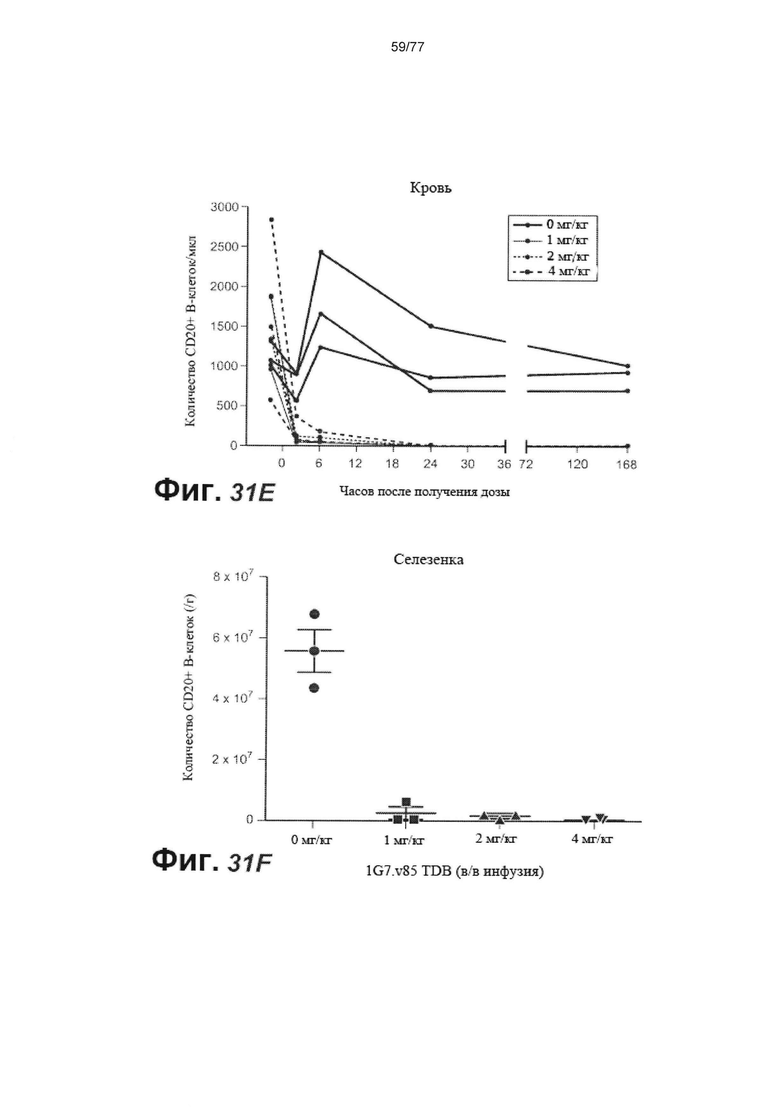
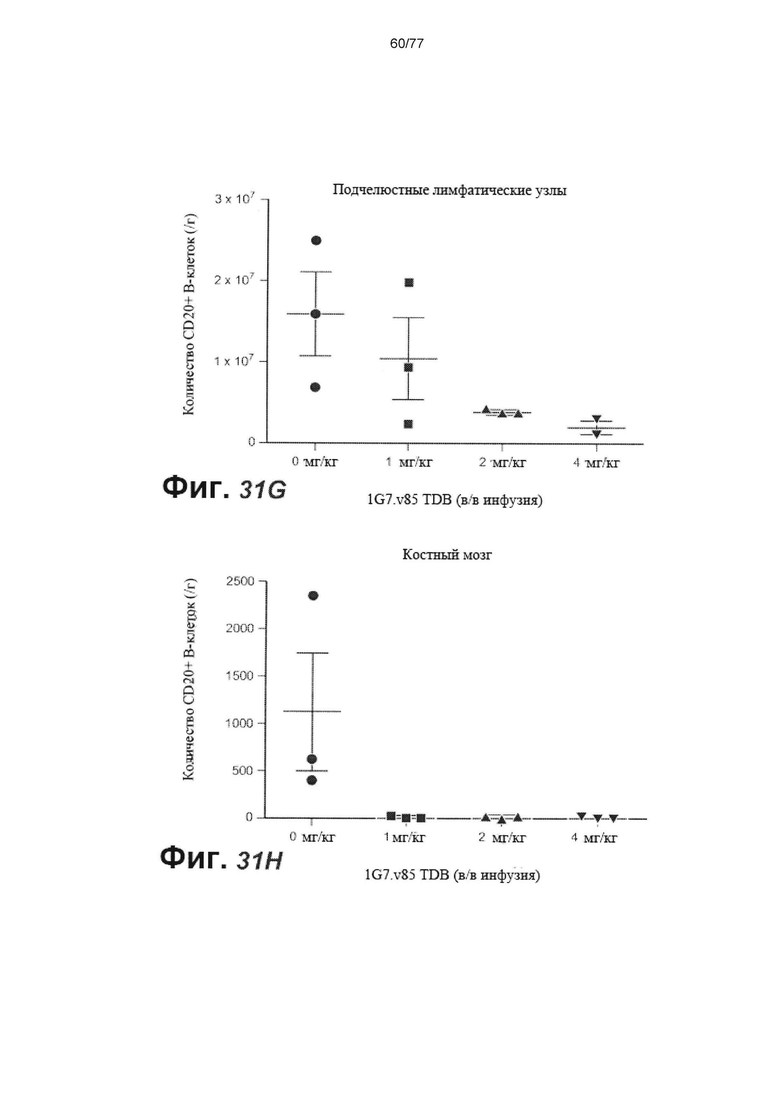

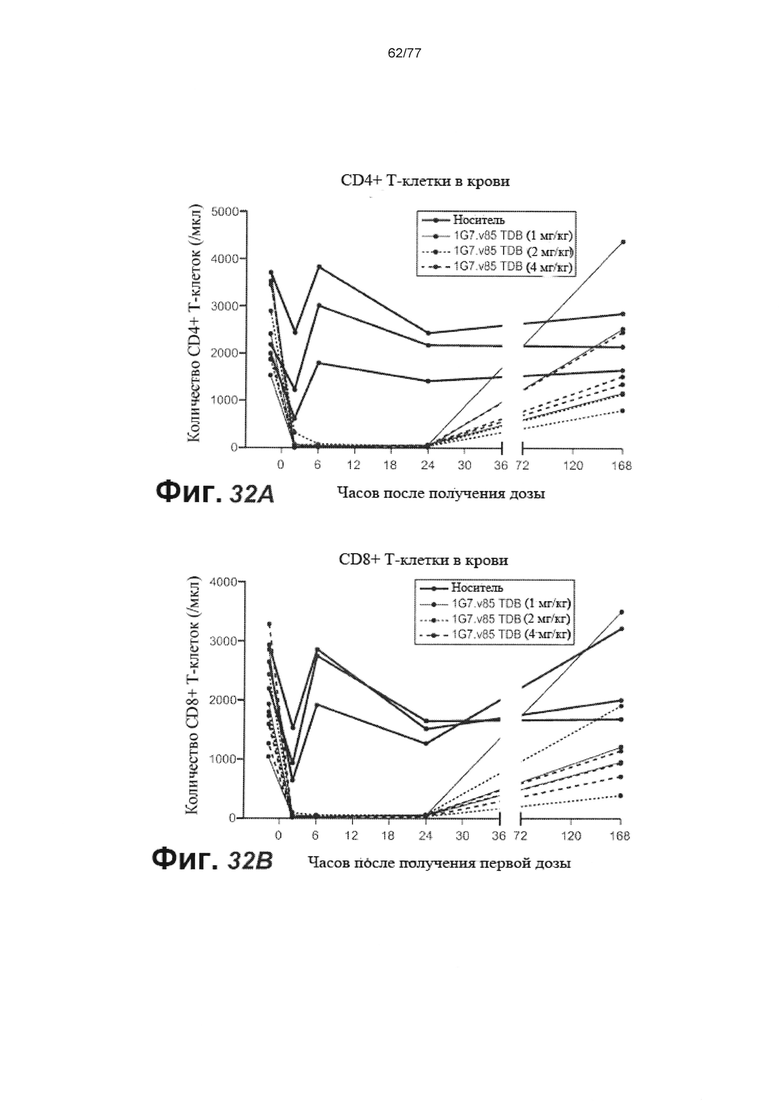
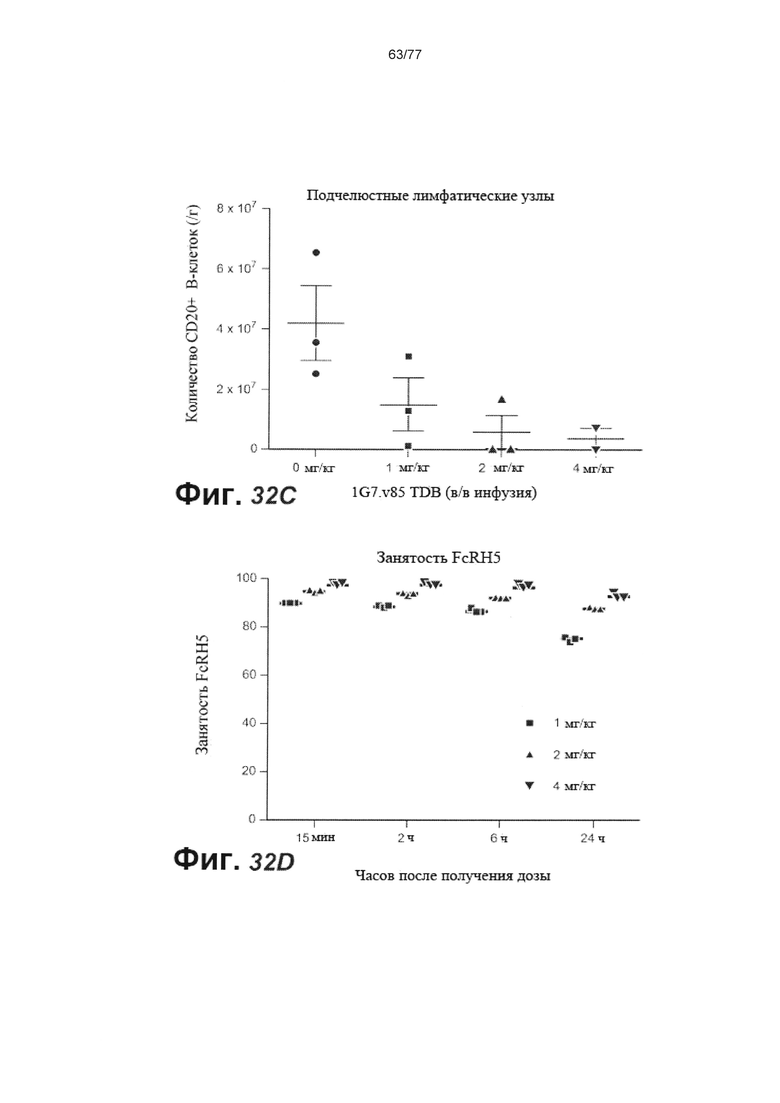
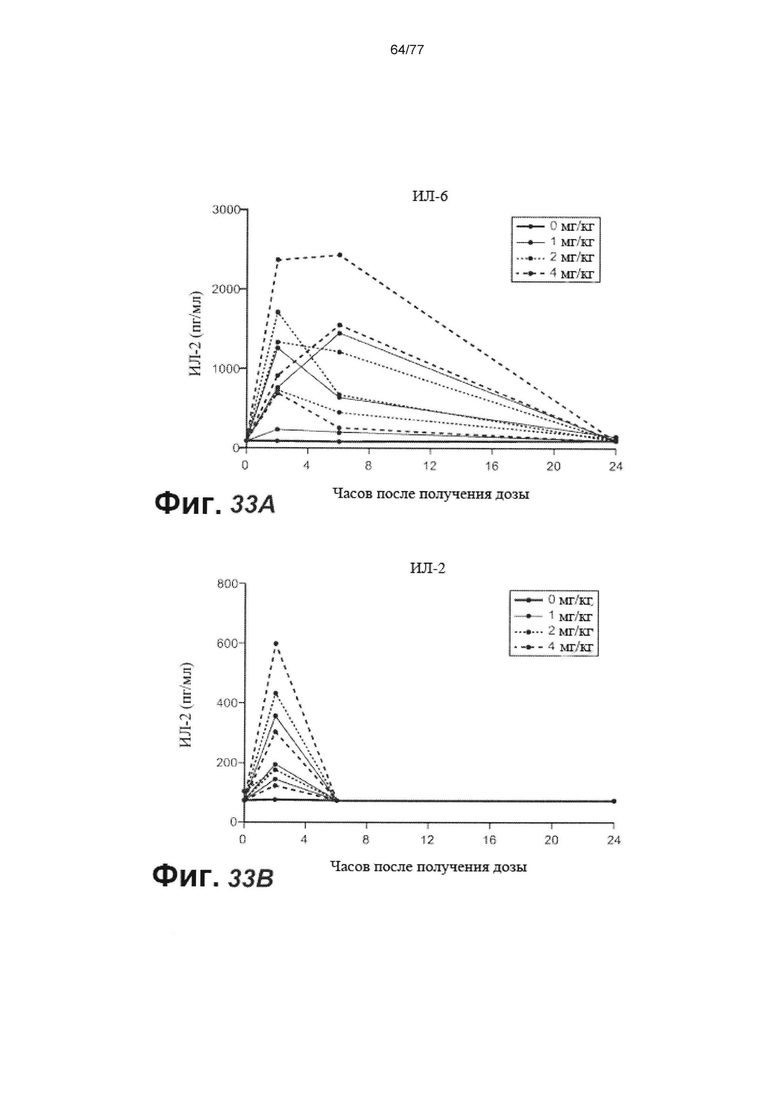

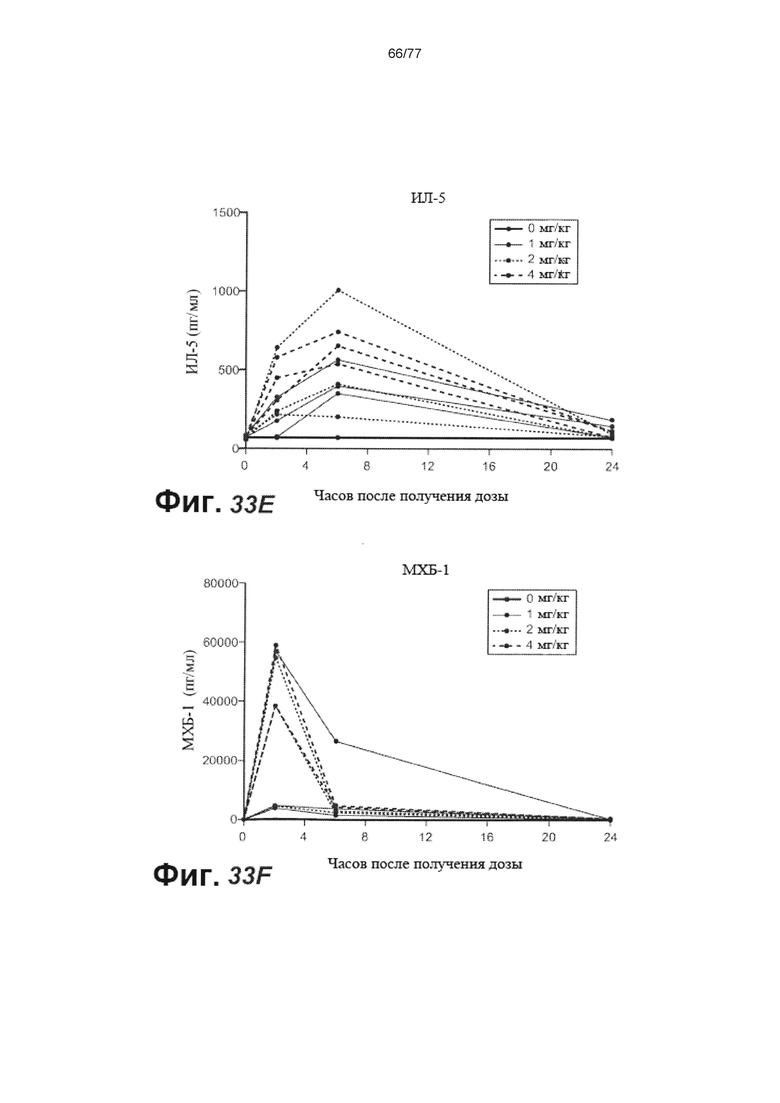
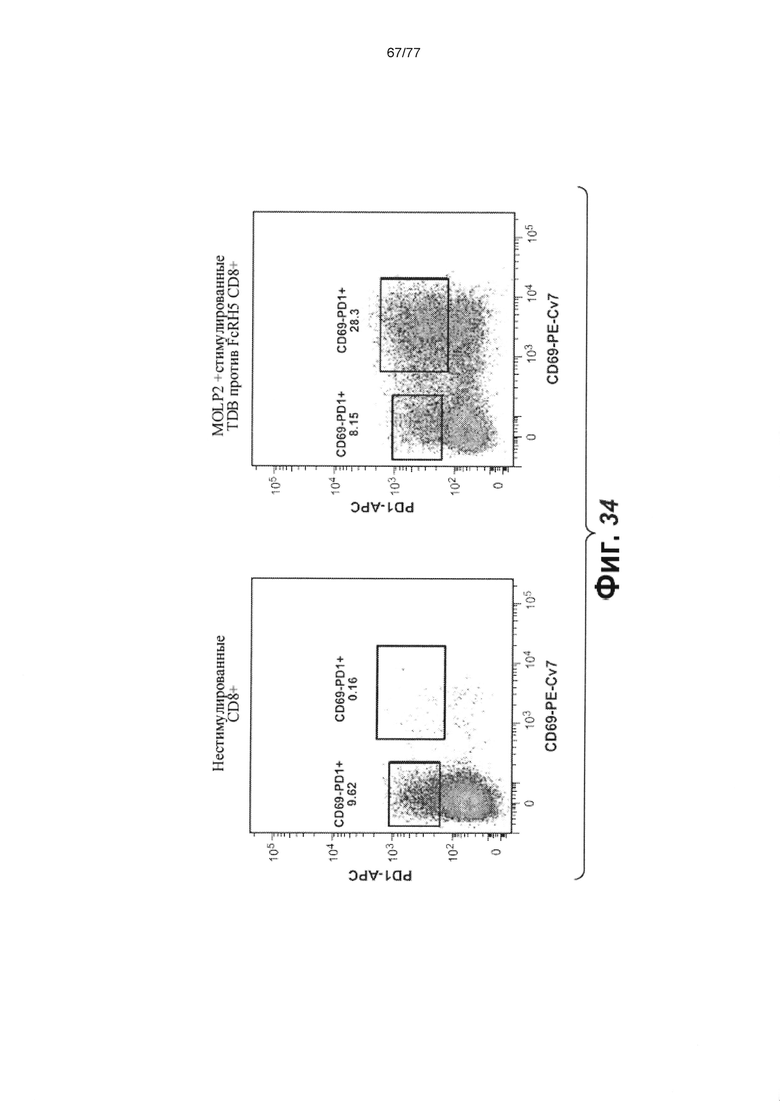
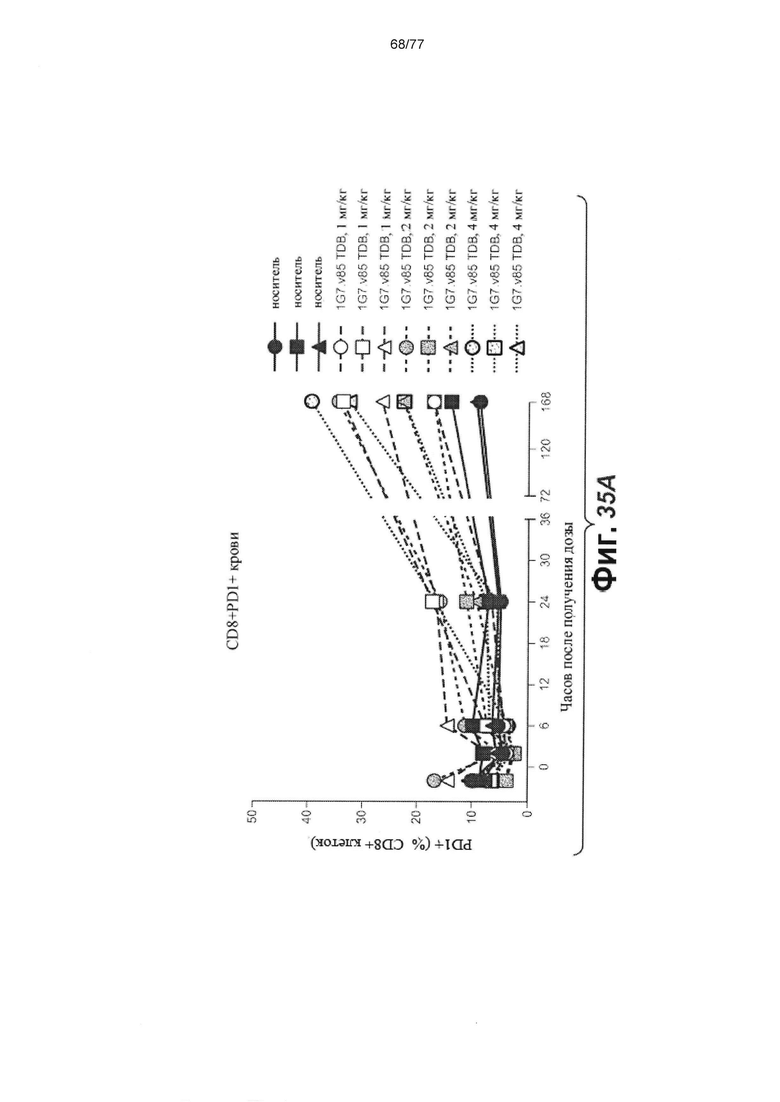
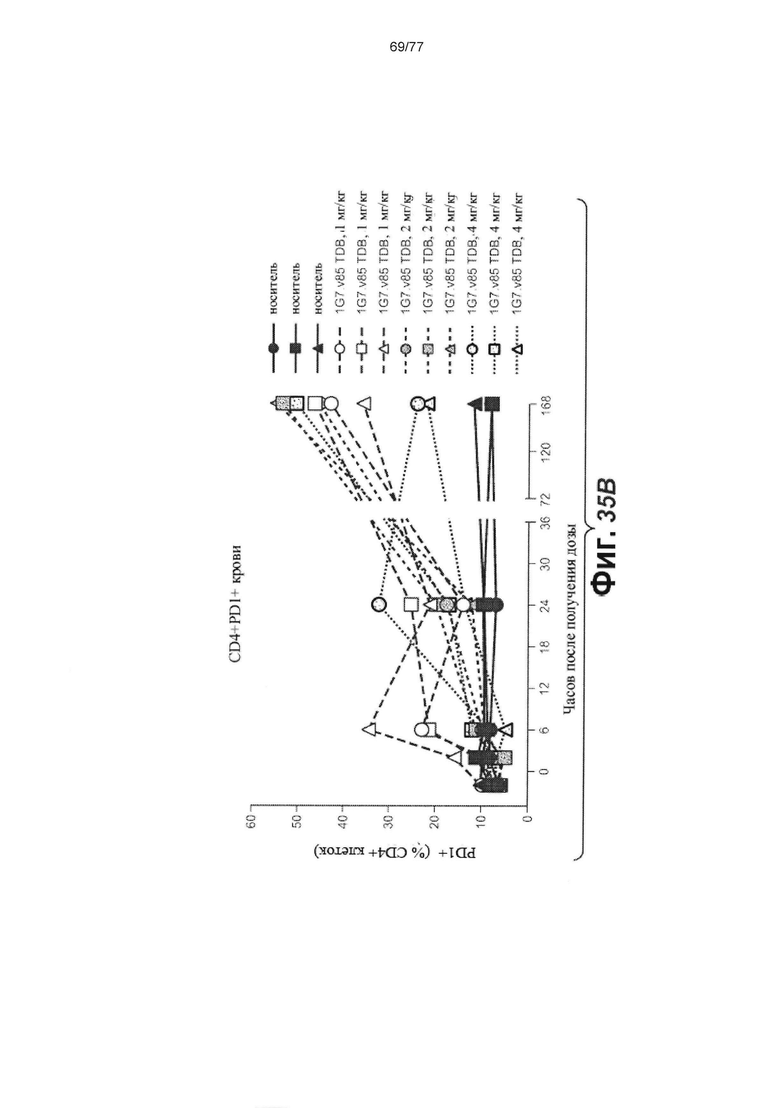
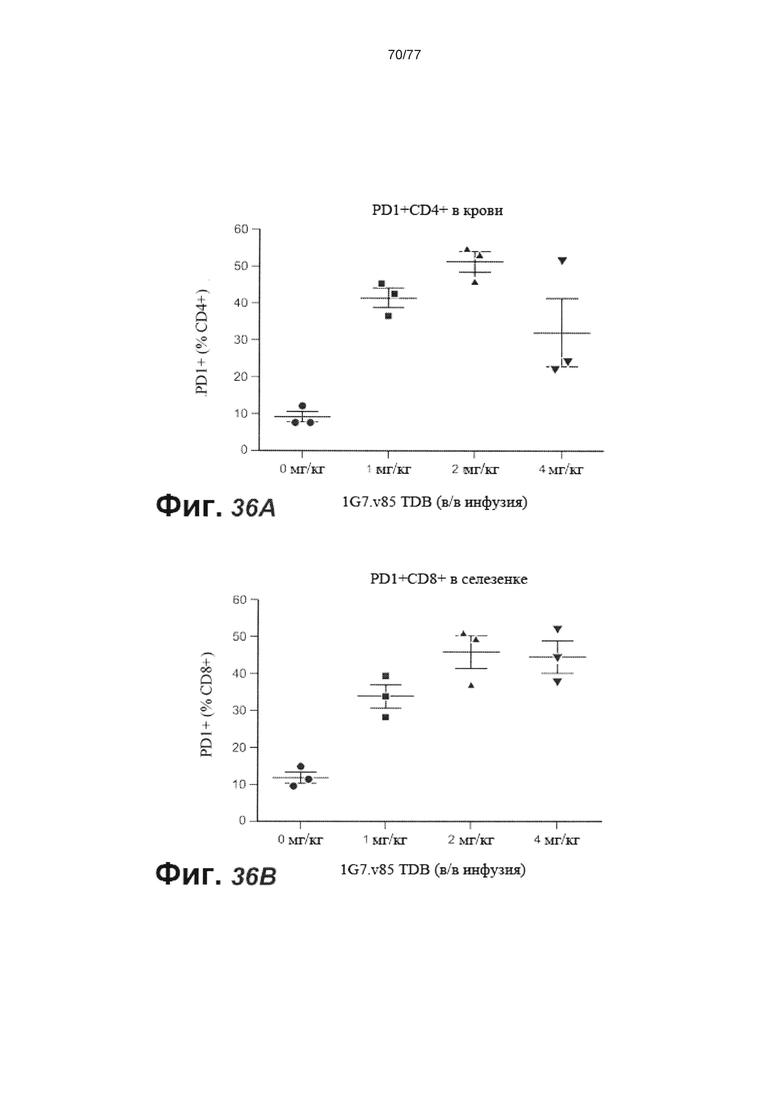
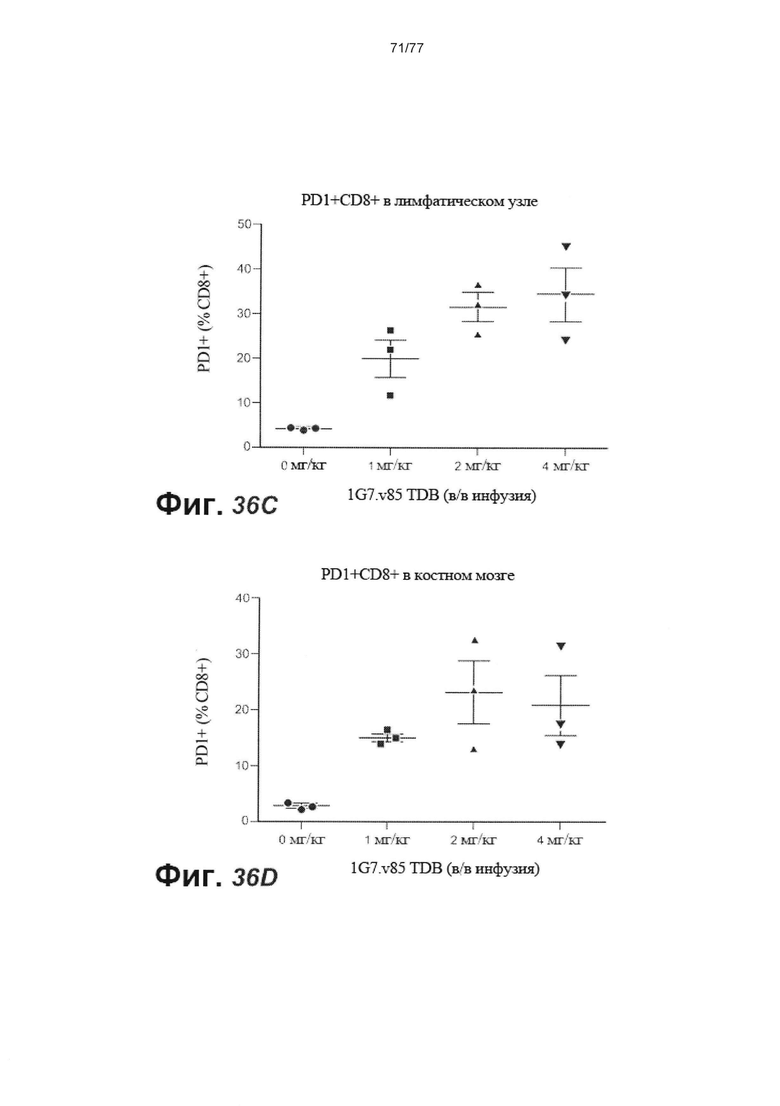

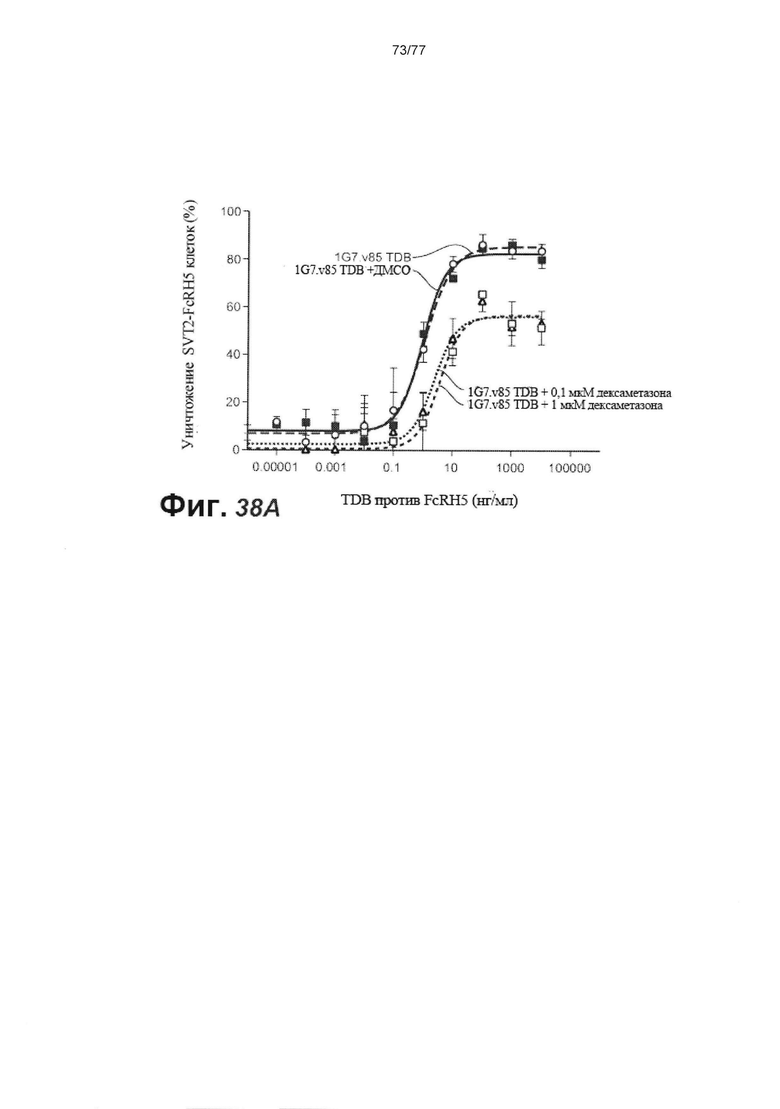
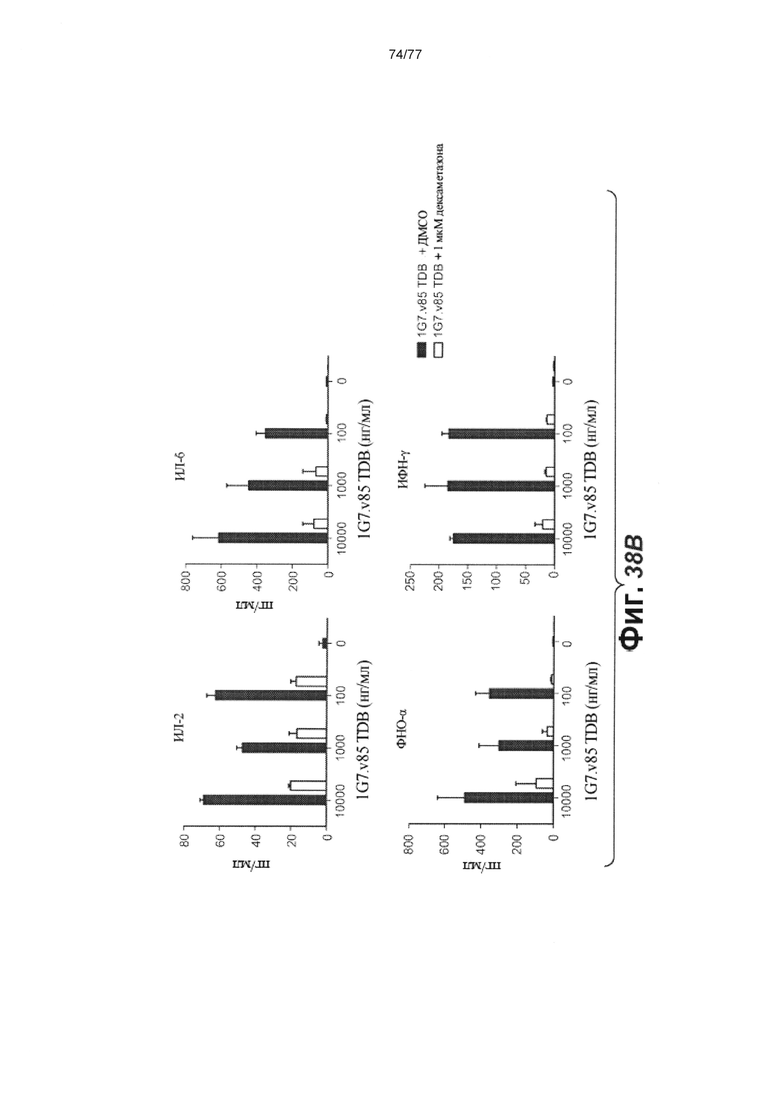
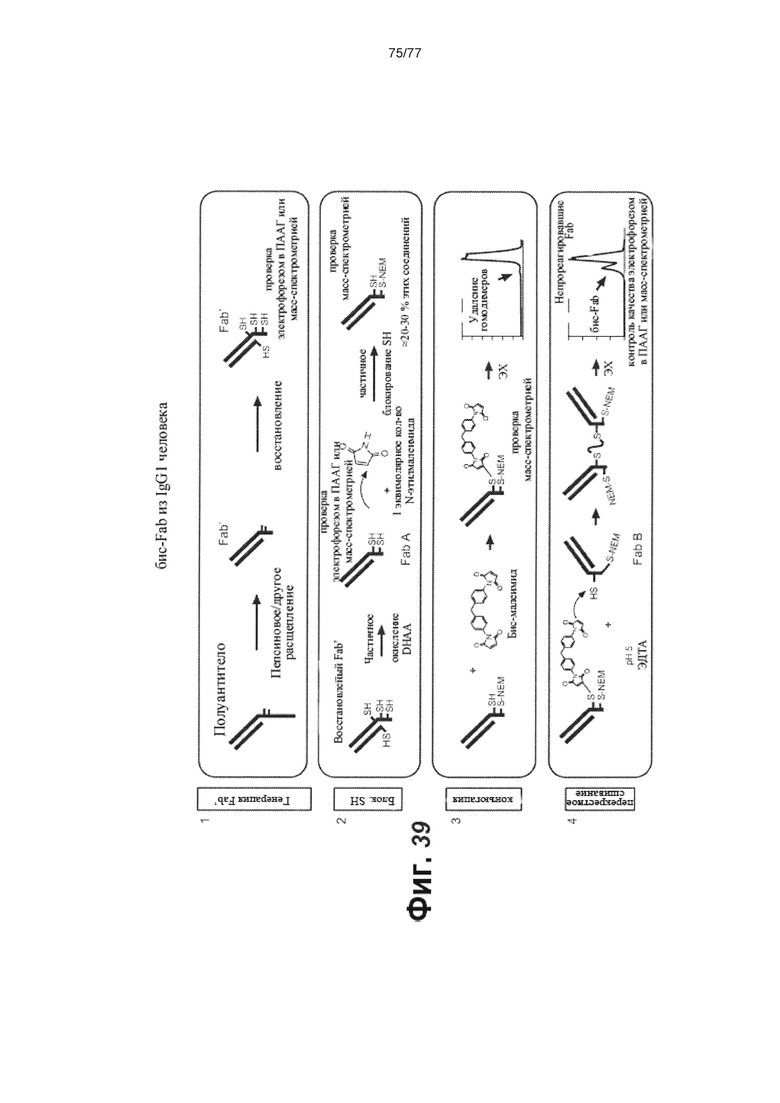
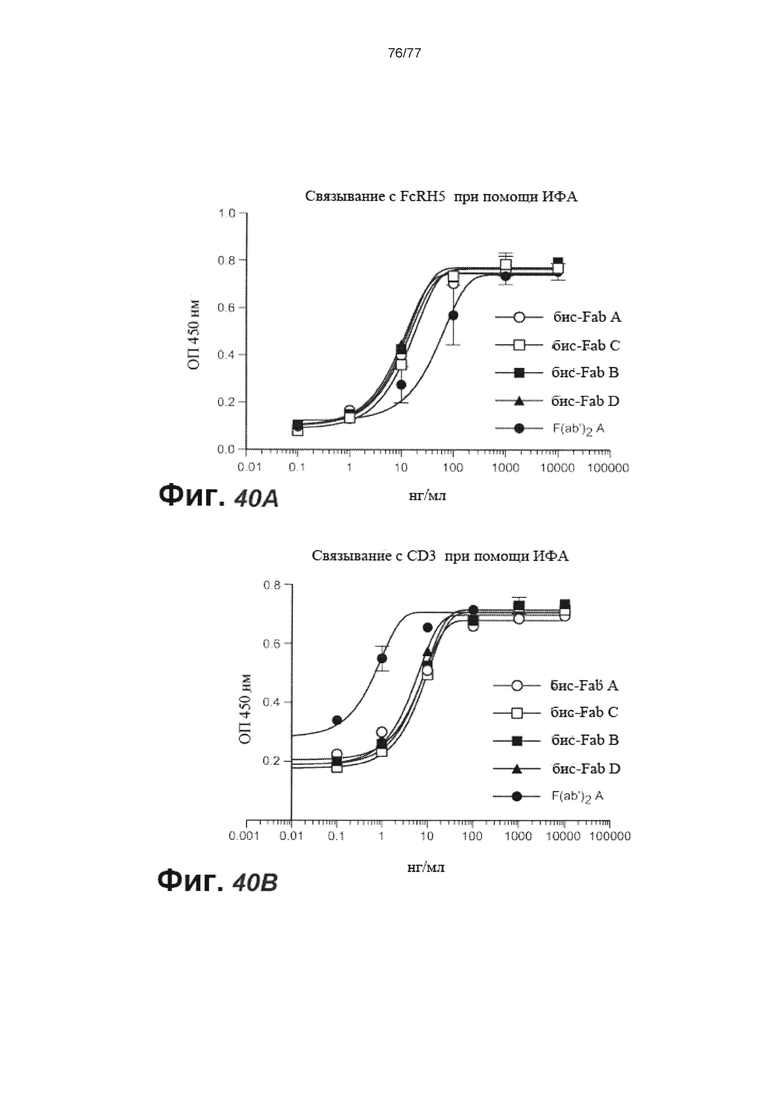
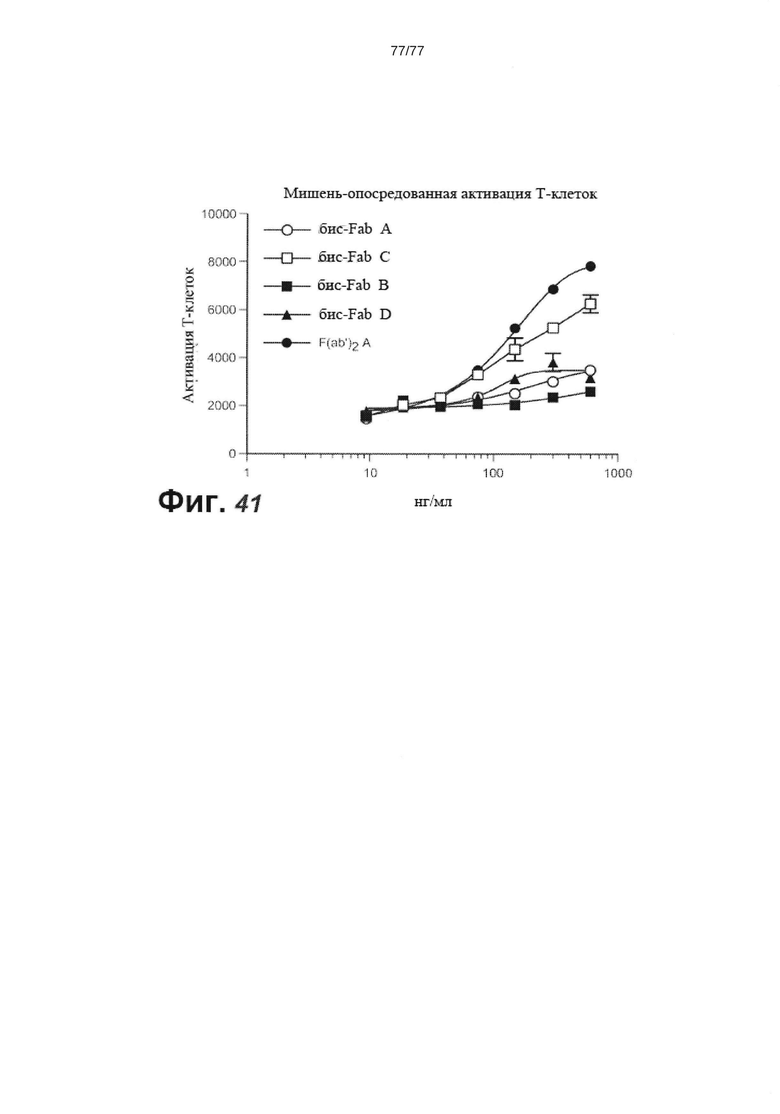
Изобретение относится к области биохимии, в частности к антителу против подобного Fc-рецептору белка 5 (FcRH5). Также раскрыты нуклеиновая кислота, кодирующая указанное антитело, вектор и клетка-хозяин, экспрессирующие указанное антитело, иммуноконъюгат, композиция и набор, содержащие указанное антитело. Раскрыты способы получения указанного антитела, лечения, обнаружения FcRH5 и усиления иммунной функции с помощью указанного антитела, применение указанного антитела для изготовления лекарственного средства для лечения или замедления прогрессирования FcRH5-пoлoжитeльнoгo рака у субъекта, применение указанного антитела для изготовления лекарственного средства для усиления иммунной функции у субъекта, имеющего FcRH5-пoлoжитeльный рак. Изобретение позволяет эффективно лечить рак, ассоциированный с FcRH5. 17 н. и 134 з.п. ф-лы, 41 ил., 15 табл., 6 пр.
1. Антитело против подобного Fc-рецептору белка 5 (FcRH5), отличающееся тем, что антитело против FcRH5 содержит связывающий домен, содержащий следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23.
2. Антитело против FcRH5 по п. 1, отличающееся тем, что связывающий домен содержит
(a) вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 104; (b) вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 105, или (с) домен VH, как в (а), и домен VL, как в (b).
3. Антитело против FcRH5 по п. 1 или 2, отличающееся тем, что антитело дополнительно содержит следующие каркасные области (FR) вариабельной области тяжелой цепи:
(a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 52;
(b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 54;
(c) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 46; и
(d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 47.
4. Антитело против FcRH5 по п. 3, отличающееся тем, что домен VH содержит аминокислотную последовательность SEQ ID NO: 104.
5. Антитело против FcRH5 по любому из пп. 1-4, отличающееся тем, что антитело дополнительно содержит следующие FR-области вариабельной области легкой цепи:
(a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 48;
(b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 57;
(c) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 50; и
(d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 51.
6. Антитело против FcRH5 по п. 5, отличающееся тем, что домен VL содержит аминокислотную последовательность SEQ ID NO: 105.
7. Антитело против FcRH5, причем антитело против FcRH5 содержит связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 104, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 105.
8. Антитело против FcRH5 по любому из пп. 1-7, отличающееся тем, что антитело против FcRH5 связывается с эпитопом в Ig-подобном домене 9 FcRH5.
9. Антитело против FcRH5 по п. 8, отличающееся тем, что эпитоп содержит часть аминокислот 745-850 SEQ ID NO: 114.
10. Антитело против FcRH5 по п. 8 или 9, отличающееся тем, что связывающий домен связывается с FcRH5 человека, или FcRH5 яванского макака (cyno), или с обоими.
11. Антитело против FcRH5 по п. 10, отличающееся тем, что связывающий домен специфически не связывается с FcRHl, FcRH2, FcRH3 и/или FcRH4.
12. Антитело против FcRH5 по п. 10 или 11, отличающееся тем, что антитело против FcRH5 связывает FcRH5 человека с KD около 100 нМ или менее.
13. Антитело против FcRH5 по п. 12, отличающееся тем, что антитело против FcRH5 связывает FcRH5 человека с KD от около 10 пМ до около 100 нМ.
14. Антитело против FcRH5 по п. 13, отличающееся тем, что антитело против FcRH5 связывает FcRH5 человека с KD от около 100 пМ до около 100 нМ.
15. Антитело против FcRH5 по п. 14, отличающееся тем, что антитело против FcRH5 связывает FcRH5 человека с KD от около 1 нМ до около 20 нМ.
16. Антитело против FcRH5 по п. 15, отличающееся тем, что антитело против FcRH5 связывает FcRH5 человека с KD от около 1 нМ до около 10 нМ.
17. Антитело против FcRH5 по любому из пп. 12-16, отличающееся тем, что антитело против FcRH5 связывает FcRH5 яванского макака с KD около 100 нМ или менее.
18. Антитело против FcRH5 по п. 17, отличающееся тем, что антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 10 пМ до около 100 нМ.
19. Антитело против FcRH5 по п. 18, отличающееся тем, что антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 100 пМ до около 100 нМ.
20. Антитело против FcRH5 по п. 19, отличающееся тем, что антитело против FcRH5 связывает FcRH5 яванского макака с KD от около 1 нМ до около 50 нМ.
21. Антитело против FcRH5 по любому из пп. 1-20, отличающееся тем, что антитело против FcRH5 содержит мутацию по типу замены в аминокислотном остатке N297, L234, L235, D265 и/или Р329 (нумерация ЕС).
22. Антитело против FcRH5 по п. 21, отличающееся тем, что мутация по типу замены выбрана из группы, состоящей из N297G, N297A, L234A, L235A, D265A и P329G.
23. Антитело против FcRH5 по п. 22, отличающееся тем, что мутация по типу замены представляет собой мутацию N297G.
24. Антитело против FcRH5 по любому из пп. 1-23, отличающееся тем, что антитело против FcRH5 представляет собой моноклональное, человеческое, гуманизированное или химерное антитело.
25. Антитело против FcRH5 по любому из пп. 1-24, отличающееся тем, что антитело против FcRH5 представляет собой антитело IgG.
26. Антитело против FcRH5 по любому из пп. 1-25, отличающееся тем, что антитело против FcRH5 представляет собой фрагмент антитела, который связывает FcRH5.
27. Антитело против FcRH5 по п. 26, отличающееся тем, что фрагмент антитела выбран из группы, состоящей из фрагментов бис-Fab, Fab, Fab'-SH, Fv, scFv и (Fab')2.
28. Антитело против FcRH5 по п. 27, отличающееся тем, что фрагмент антитела представляет собой фрагмент бис-Fab.
29. Антитело против FcRH5 по любому из пп. 1-25, отличающееся тем, что антитело против FcRH5 представляет собой полноразмерное антитело.
30. Антитело против FcRH5 по любому из пп. 1-29, отличающееся тем, что антитело против FcRH5 представляет собой моноспецифическое антитело.
31. Антитело против FcRH5 по любому из пп. 1-30, отличающееся тем, что антитело против FcRH5 представляет собой биспецифическое антитело, содержащее второй связывающий домен, который связывает кластер дифференцировки 3 (CD3), при этом второй связывающий домен содержит:
(I)
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123;
(II)
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 124;
(III)
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 139;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 140;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 141;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 142;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 143; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 144 или
(IV)
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 155;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 162;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 157;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 158;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 159; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 160.
32. Антитело против FcRH5 по п. 31, отличающееся тем, что второй связывающий домен связывается с эпитопом на CD3, содержащим аминокислотный остаток Glu6 CD3.
33. Антитело против FcRH5 по п. 32, отличающееся тем, что эпитоп дополнительно содержит один или более дополнительных аминокислотных остатков, выбранных из группы, состоящей из Glnl, Asp2 и Met7 CD3.
34. Антитело против FcRH5 по п. 32 или 33, отличающееся тем, что эпитоп содержит аминокислотные остатки Glnl, Asp2 и Glu6 CD3.
35. Антитело против FcRH5 по любому из пп. 32-34, отличающееся тем, что эпитоп содержит аминокислотные остатки Glnl, Asp2, Glu6 и Met7 CD3.
36. Антитело против FcRH5 по любому из пп. 32-35, отличающееся тем, что эпитоп не содержит аминокислотный остаток Glu5 CD3.
37. Антитело против FcRH5 по любому из пп. 32-36, отличающееся тем, что эпитоп не содержит аминокислотные остатки Gly3 и Glu5 CD3.
38. Антитело против FcRH5 по любому из пп. 32-37, отличающееся тем, что эпитоп состоит из аминокислотных остатков Glnl, Asp2, Glu6 и Met7 CD3.
39. Антитело против FcRH5 по любому из пп. 32-38, отличающееся тем, что второй связывающий домен способен связываться с полипептидом CD3 человека или полипептидом CD3 яванского макака.
40. Антитело против FcRH5 по п. 39, отличающееся тем, что полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3ε человека или полипептид CD3ε яванского макака соответственно.
41. Антитело против FcRH5 по п. 39, отличающееся тем, что полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3γ человека или полипептид CD3γ яванского макака соответственно.
42. Антитело против FcRH5 по любому из пп. 31-41, отличающееся тем, что второй связывающий домен связывает полипептид CD3ε человека с KD около 100 нМ или менее.
43. Антитело против FcRH5 по п. 42, отличающееся тем, что второй связывающий домен связывает полипептид CD3ε человека с KD от около 10 пМ до около 100 нМ.
44. Антитело против FcRH5 по п. 43, отличающееся тем, что второй связывающий домен связывает полипептид CD3ε человека с KD от около 100 пМ до около 50 нМ.
45. Антитело против FcRH5 по п. 44, отличающееся тем, что второй связывающий домен связывает полипептид CD3ε человека с KD от около 1 нМ до около 10 нМ.
46. Антитело против FcRH5 по любому из пп. 31-45, отличающееся тем, что второй связывающий домен содержит следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123.
47. Антитело против FcRH5 по п. 46, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 133; (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 134; или (с) домен VH, как в (а), и домен VL, как в (b).
48. Антитело против FcRH5 по п. 46 или 47, отличающееся тем, что второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи:
(a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 125;
(b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 126;
(c) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 127; и
(d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 128.
49. Антитело против FcRH5 по п. 48, отличающееся тем, что второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133.
50. Антитело против FcRH5 по любому из пп. 46-49, отличающееся тем, что второй связывающий домен дополнительно содержит следующие FR-области вариабельной области легкой цепи:
(a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 129;
(b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 130;
(c) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 131; и
(d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 132.
51. Антитело против FcRH5 по п. 50, отличающееся тем, что второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134.
52. Антитело против FcRH5 по любому из пп. 46-51, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 133, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 134.
53. Антитело против FcRH5 по любому из пп. 31-45, отличающееся тем, что второй связывающий домен содержит следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 124.
54. Антитело против FcRH5 по п. 53, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 137; (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 138; или (с) домен VH, как в (а), и домен VL, как в (b).
55. Антитело против FcRH5 по п. 53 или 54, отличающееся тем, что второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи:
(a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 125;
(b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 126;
(c) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 127; и
(d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 128.
56. Антитело против FcRH5 по п. 55, отличающееся тем, что второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137.
57. Антитело против FcRH5 по любому из пп. 53-56, отличающееся тем, что второй связывающий домен дополнительно содержит следующие FR-области вариабельной области легкой цепи:
(a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 129;
(b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 130;
(c) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 131; и
(d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 132.
58. Антитело против FcRH5 по п. 57, отличающееся тем, что второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138.
59. Антитело против FcRH5 по любому из пп. 53-58, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 137, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 138.
60. Антитело против FcRH5 по любому из пп. 31-45, отличающееся тем, что второй связывающий домен содержит следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 139;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 140;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 141;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 142;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 143; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 144.
61. Антитело против FcRH5 по п. 60, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 153; (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 154; или (с) домен VH, как в (а), и домен VL, как в (b).
62. Антитело против FcRH5 по п. 60 или 61, отличающееся тем, что второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи:
(a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 145;
(b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 146;
(c) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 147; и
(d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 148.
63. Антитело против FcRH5 по п. 62, отличающееся тем, что второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153.
64. Антитело против FcRH5 по любому из пп. 60-63, отличающееся тем, что второй связывающий домен дополнительно содержит следующие FR-области вариабельной области легкой цепи:
(a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 149;
(b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 150;
(c) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 151; и
(d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 152.
65. Антитело против FcRH5 по п. 64, отличающееся тем, что второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154.
66. Антитело против FcRH5 по любому из пп. 60-65, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 153, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 154.
67. Антитело против FcRH5 по любому из пп. 31-45, отличающееся тем, что второй связывающий домен содержит следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 155;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 162;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 157;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 158;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 159; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 160.
68. Антитело против FcRH5 по п. 67, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 172; (b) домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95 % идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 173; или (с) домен VH, как в (а), и домен VL, как в (b).
69. Антитело против FcRH5 по п. 67 или 68, отличающееся тем, что второй связывающий домен содержит следующие FR-области вариабельной области тяжелой цепи:
(a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 164;
(b) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 165;
(c) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 166; и
(d) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 167.
70. Антитело против FcRH5 по п. 69, отличающееся тем, что второй связывающий домен содержит домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172.
71. Антитело против FcRH5 по любому из пп. 67-70, отличающееся тем, что второй связывающий домен дополнительно содержит следующие FR-области вариабельной области легкой цепи:
(a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 168;
(b) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 169;
(c) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 170; и
(d) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 171.
72. Антитело против FcRH5 по п. 71, отличающееся тем, что второй связывающий домен содержит домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173.
73. Антитело против FcRH5 по любому из пп. 67-72, отличающееся тем, что второй связывающий домен содержит (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 172, и (b) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 173.
74. Антитело против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и
плечо антитела против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123.
75. Антитело против FcRH5, которое связывается с FcRH5 и CD3, причем антитело против FcRH5 содержит плечо против FcRH5, содержащее первый связывающий домен, содержащий следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 16; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 23; и
плечо антитела против CD3, содержащее второй связывающий домен, содержащий следующие шесть HVR:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 115;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 116;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 121;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 123; и
при этом плечо антитела против FcRH5 и плечо антитела против CD3, каждое, содержат мутацию по типу замены N297G (нумерация ЕС); и
при этом плечо антитела против FcRH5 содержит мутацию по типу замены T366W, и плечо против CD3 содержит мутации по типу замены T366S, L368A и Y407V (нумерация ЕС).
76. Антитело против FcRH5 по любому из пп. 1-52, 74 и 75, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 35 мл/кг/сут.
77. Антитело против FcRH5 по п. 76, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 10 мл/кг/сут до около 20 мл/кг/сут у мыши.
78. Антитело против FcRH5 по п. 77, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 12 мл/кг/сут до около 16 мл/кг/сут у мыши.
79. Антитело против FcRH5 по любому из пп. 76-78, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 20 мл/кг/сут до около 40 мл/кг/сут у яванского макака.
80. Антитело против FcRH5 по п. 79, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 25 мл/кг/сут до около 35 мл/кг/сут у яванского макака.
81. Антитело против FcRH5 по п. 80, отличающееся тем, что антитело против FcRH5 имеет клиренс после внутривенной инъекции от около 30 мл/кг/сут до около 35 мл/кг/сут у яванского макака.
82. Выделенная нуклеиновая кислота, кодирующая антитело против FcRH5 по любому из пп. 1-81, или его часть, содержащая его связывающий домен, который связывается с FcRH5.
83. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 82.
84. Клетка-хозяин для продуцирования антитела против FcRH5, где клетка-хозяин содержит вектор экспрессии по п. 83.
85. Клетка-хозяин по п. 84, отличающаяся тем, что клетка-хозяин представляет собой клетку млекопитающего и при этом клетка млекопитающего не является эмбриональной стволовой клеткой человека.
86. Клетка-хозяин по п. 85, отличающаяся тем, что клетка млекопитающего представляет собой клетку яичника китайского хомячка (СНО).
87. Клетка-хозяин по п. 84, отличающаяся тем, что клетка-хозяин представляет собой прокариотическую клетку.
88. Клетка-хозяин по п. 87, отличающаяся тем, что прокариотическая клетка представляет собой клетку Е. coli.
89. Способ получения антитела против FcRH5 по любому из пп. 1-81, включающий стадию, в которой культивируют клетки-хозяина по п. 84 в культуральной среде.
90. Способ по п. 89, отличающийся тем, что способ дополнительно включает выделение антитела против FcRH5 из клетки-хозяина или культуральной среды.
91. Способ по п. 89, дополнительно включающий культивирование второй клетки-хозяина, содержащей вторую нуклеиновую кислоту, кодирующую антитело против CD3, которое содержит связывающий домен, который связывает CD3.
92. Способ по п. 91, отличающийся тем, что клетки-хозяева совместно культивируют.
93. Способ по п. 91 или 92, отличающийся тем, что способ дополнительно включает выделение биспецифического антитела против FcRH5 из клетки-хозяина или культуральной среды.
94. Иммуноконъюгат для применения в лечении или замедлении прогрессирования FcRH5-положительного рака, содержащий антитело против FcRH5 по любому из пп. 1-81 и цитотоксический агент.
95. Эффективное количество композиции для применения в лечении или замедлении прогрессирования FcRH5-положительного рака, содержащее антитело против FcRH5 по любому из пп. 1-81.
96. Композиция по п. 95, дополнительно содержащая фармацевтически приемлемое вспомогательное вещество или разбавитель.
97. Композиция по п. 96, отличающаяся тем, что фармацевтически приемлемое вспомогательное вещество представляет собой буфер, носитель, стабилизатор или консервант.
98. Композиция по п. 97, отличающаяся тем, что композиция представляет собой фармацевтическую композицию.
99. Композиция по любому из пп. 95-98, отличающаяся тем, что композиция дополнительно содержит антагонист, связывающийся с компонентом сигнального пути PD-1, или дополнительный терапевтический агент.
100. Антитело против FcRH5 по любому из пп. 1-81 для применения в качестве лекарственного средства.
101. Антитело против FcRH5 по любому из пп. 1-81 для применения при лечении или замедлении прогрессирования FcRH5-положительного рака у субъекта, нуждающегося в этом.
102. Антитело против FcRH5 по любому из пп. 1-81 для применения в повышении иммунной функции у субъекта, имеющего FcRH5-положительный рак.
103. Антитело против FcRH5 по п. 101 или 102, отличающееся тем, что FcRH5-положительный рак представляет собой В-клеточный рак.
104. Антитело против FcRH5 по п. 103, отличающееся тем, что В-клеточный рак выбран из группы, состоящей из множественной миеломы (ММ), хронического лимфоцитарного лейкоза (CLL), мантийноклеточной лимфомы (MCL), диффузной В-крупноклеточной лимфомы (DLBCL) и фолликулярной лимфомы (FL).
105. Антитело против FcRH5 по п. 104, отличающееся тем, что В-клеточный рак представляет собой ММ.
106. Применение антитела против FcRH5 по любому из пп. 1-81 или композиции по любому из пп. 95-99 при изготовлении лекарственного средства для лечения или замедления прогрессирования FcRH5-пoлoжитeльнoгo рака у субъекта.
107. Применение антитела против FcRH5 по любому из пп. 1-81 или композиции по любому из пп. 95-99 при изготовлении лекарственного средства для усиления иммунной функции у субъекта, имеющего FcRH5-пoлoжитeльный рак.
108. Применение по п. 106 или 107, отличающееся тем, что FcRH5-положительный рак представляет В-клеточный рак.
109. Применение по п. 108, отличающееся тем, что В-клеточный рак выбран из группы, состоящей из MM, CLL, MCL, DLBCL и FL.
110. Применение по п. 109, отличающееся тем, что В-клеточный рак представляет собой MM.
111. Способ лечения или замедления прогрессирования FcRH5-положительного рака у субъекта, нуждающегося в этом, при этом способ включает введение субъекту антитела против FcRH5 по любому из пп. 1-81.
112. Способ усиления иммунной функции у субъекта, имеющего FcRH5-положительный рак, причем способ включает введение субъекту эффективного количества антитела против FcRH5 по любому из пп. 1-81.
113. Способ по п. 111 или 112, отличающийся тем, что FcRH5-положительный рак представляет В-клеточный рак.
114. Способ по п. 113, отличающийся тем, что В-клеточный рак выбран из группы, состоящей из MM, CLL, MCL, DLBCL и FL.
115. Способ по п. 114, отличающийся тем, что В-клеточный рак представляет собой ММ.
116. Способ по любому из пп. 111-115, отличающийся тем, что антитело против FcRH5 связывается с (а) молекулой FcRH5, расположенной на целевой клетке, и (b) молекулой CD3, расположенной на иммунной эффекторной клетке.
117. Способ по п. 116, отличающийся тем, что антитело против FcRH5 активирует иммунную эффекторную клетку после связывания с молекулой FcRH5 и молекулой CD3.
118. Способ по п. 117, отличающийся тем, что активированная иммунная эффекторная клетка способна оказывать цитотоксическое и/или апоптотическое действие на целевую клетку.
119. Способ по любому из пп. 116-118, отличающийся тем, что целевая клетка представляет собой плазмоцит.
120. Способ по п. 119, отличающийся тем, что плазмоцит представляет собой короткоживущий плазмоцит.
121. Способ по п. 119, отличающийся тем, что плазмоцит представляет собой долгоживущий плазмоцит.
122. Способ по любому из пп. 119-121, отличающийся тем, что плазмоцит представляет собой миеломную клетку.
123. Способ по любому из пп. 111-122, отличающийся тем, что способ включает введение субъекту антитела против FcRH5 в дозе от около 0,01 мг/кг/нед. до около 50 мг/кг/нед.
124. Способ по п. 123, отличающийся тем, что способ включает введение субъекту антитела против FcRH5 в дозе от около 0,1 мг/кг/нед. до около 10 мг/кг/нед.
125. Способ по п. 124, отличающийся тем, что способ включает введение субъекту антитела против FcRH5 в дозе около 1 мг/кг/нед.
126. Способ по любому из пп. 111-125, дополнительно включающий введение субъекту антагониста, связывающегося с компонентом сигнального пути PD-1, и/или дополнительного терапевтического агента.
127. Способ по п. 126, отличающийся тем, что антагонист, связывающийся с компонентом сигнального пути PD-1, или дополнительный терапевтический агент вводят до или после введения антитела против FcRH5.
128. Способ по п. 127, отличающийся тем, что антагонист, связывающийся с компонентом сигнального PD-1, или дополнительный терапевтический агент вводят одновременно с антителом против FcRH5.
129. Способ по любому из пп. 126-128, отличающийся тем, что антагонист, связывающийся с компонентом сигнального пути PD-1, выбран из группы, состоящей из PD- L1-связывающего антагониста, PD-1-связывающего антагониста и РD-L2-связывающего антагониста.
130. Способ по п. 129, отличающийся тем, что антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой PD-L1-связывающий антагонист.
131. Способ по п. 130, отличающийся тем, что PD-L1-связывающий антагонист выбран из группы, состоящей из MPDL3280A (атезолизумаба), MDX-1105, MEDI4736 (дурвалумаба) и MSB0010718C (авелумаба).
132. Способ по п. 131, отличающийся тем, что PD-L1-связывающий антагонист представляет собой MPDL3280A (атезолизумаб).
133. Способ по п. 129, отличающийся тем, что антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой PD-1-связывающий антагонист.
134. Способ по п. 133, отличающийся тем, что PD-1-связывающий антагонист выбран из группы, состоящей из MDX 1106 (ниволумаба), МК-3475 (пембролизумаба), MEDI-0680 (АМР- 514), PDR001, REGN2810 и BGB-108.
135. Способ по п. 129, отличающийся тем, что антагонист, связывающийся с компонентом сигнального пути PD-1, представляет собой РD-L2-связывающий антагонист.
136. Способ по п. 135, отличающийся тем, что РD-L2-связывающий антагонист представляет собой антитело или иммуноадгезин.
137. Способ по любому из пп. 111-136, дополнительно включающий введение субъекту стероида, иммуномодулятора (IMiD), ингибитора протеосом (PI) или их комбинации.
138. Способ по п. 137, отличающийся тем, что стероид представляет собой глюкокортикоид.
139. Способ по п. 138, отличающийся тем, что глюкокортикоид представляет собой дексаметазон.
140. Способ по п. 137, отличающийся тем, что IMiD представляет собой леналидомид.
141. Способ по п. 137, отличающийся тем, что PI представляет собой бортезомиб.
142. Способ по любому из пп. 111-141, отличающийся тем, что способ включает введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации внутривенно, подкожно, внутримышечно, местно, перорально, трансдермально, внутрибрюшинно, внутриорбитально, путем имплантации, путем ингаляции, интратекально, интравентрикулярно или интраназально.
143. Способ по п. 142, отличающийся тем, что способ включает внутривенное введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации.
144. Способ по п. 142, отличающийся тем, что способ включает подкожное введение антитела против FcRH5, антагониста, связывающегося с компонентом сигнального пути PD-1, стероида, IMiD, PI или их комбинации.
145. Способ по любому из пп. 111-144, отличающийся тем, что субъект представляет собой человека.
146. Способ обнаружения FcRH5 в биологическом образце субъекта, причем способ включает:
(а) приведение в контакт биологического образца с антителом против FcRH5 по любому из пп. 1-81 в условиях, обеспечивающих связывание антитела против FcRH5 с природным FcRH5 в биологическом образце, и
(b) обнаружение того, образуется ли комплекс между антителом против FcRH5 и природным FcRH5 в биологическом образце.
147. Способ по п. 146, отличающийся тем, что биологический образец представляет собой образец крови.
148. Способ по п. 146 или 147, отличающийся тем, что субъект представляет собой человека.
149. Набор, содержащий антитело по любому из пп. 1-81 и листок-вкладыш в упаковке, содержащий инструкции по применению антитела для лечения или замедления прогрессирования FcRH5-положительного рака у субъекта.
150. Набор, содержащий антитело по любому из пп. 1-81 и листок-вкладыш в упаковке, содержащий инструкции по применению антитела для усиления иммунной функции у субъекта, имеющего FcRH5-положительный рак.
151. Набор по п. 149 или 150, отличающийся тем, что субъект представляет собой человека.
| Способ понижения термостабильности фенилдиазоамино соединений | 1953 |
|
SU104460A1 |
| СПОСОБЫ ЛЕЧЕНИЯ, ДИАГНОСТИКИ И МОНИТОРИНГА РЕВМАТОИДНОГО АРТРИТА | 2010 |
|
RU2539112C2 |
| WO 2010114940 A1, 07.10.2010. | |||

