ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В настоящей заявке содержится перечень последовательностей, который был представлен в электронном виде в формате ASCII и полностью включен в настоящий документ посредством ссылки. Указанная копия ASCII, созданная 10 декабря 2020 года, называется 50474-184WO2_Sequence_Listing_12.10.20_ST25 и имеет размер 146127 байт.
ОБЛАСТЬ ТЕХНИКИ
В настоящем документе представлены антитела к Ly6G6D (комплекс лимфоцитарного антигена 6, локус G61) и способы их применения.
УРОВЕНЬ ТЕХНИКИ
Рак остается одной из наиболее сильных смертельных угроз для здоровья человека. В США рак поражает более чем 1,7 миллиона новых пациентов каждый год и является второй главной причиной смерти после заболеваний сердца, что составляет приблизительно одну из четырех смертей. Колоректальный рак (КРР), в частности, является третьей главной причиной смертей от рака в США, и пятилетняя выживаемость является низкой для пациентов с высокой степенью развития КРР. Рак, такой как КРР, представляет значительную и постоянно возрастающую угрозу и нагрузку для общества.
Давнишние подходы к лечению рака включают химиотерапию, лучевую терапию и хирургическое вмешательство для удаления солидных опухолей. В последнее время были разработаны иммунотерапии на основе биспецифических антител. Такие биспецифические антитела способны одновременно связывать антигены клеточной поверхности на цитотоксических клетках и клетках опухолей, чтобы связанная цитотоксическая клетка разрушила связанную клетку опухоли.
Существует неудовлетворенная потребность в данной области в разработке эффективных иммунотерапий на основе биспецифических антител (например, иммунотерапий на основе биспецифических антител к LY6G6D) для применения в лечении рака (например, КРР).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение представляет композиции для лечения рака. Также представлены составы и способы применения.
В первом аспекте настоящее изобретение показывает выделенное антитело, которое связывается с членом G6D семейства лимфоцитарных антигенов 6 (LY6G6D), где антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), где H1 содержит вариабельный (VH) домен (VH1) тяжелой цепи, содержащий следующие определяющие комплементарность области (CDR): (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4 или SEQ ID NO: 111; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5, SEQ ID NO: 112 или SEQ ID NO: 113; и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; и L1 содержит вариабельный (VL) домен легкой цепи (VL1), содержащий следующие CDR: (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3 или любую из SEQ ID NO: 99-107. В некоторых аспектах антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), где H1 содержит вариабельный (VH) домен (VH1) тяжелой цепи, содержащий следующие определяющие комплементарность области (CDR): (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; и L1 содержит вариабельный (VL) домен легкой цепи (VL1), содержащий следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3. В некоторых аспектах антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), где H1 содержит вариабельный (VH) домен (VH1) тяжелой цепи, содержащий следующие определяющие комплементарность области (CDR): (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; и L1 содержит вариабельный (VL) домен легкой цепи (VL1), содержащий следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащую аминокислотную последовательность любой из SEQ ID NO: 99-107. В некоторых аспектах антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), где H1 содержит вариабельный (VH) домен (VH1) тяжелой цепи, содержащий следующие определяющие комплементарность области (CDR): (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 111; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; и L1 содержит вариабельный (VL) домен легкой цепи (VL1), содержащий следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3. В некоторых аспектах антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), где H1 содержит вариабельный (VH) домен (VH1) тяжелой цепи, содержащий следующие определяющие комплементарность области (CDR): (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 112 или SEQ ID NO: 113; и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; и L1 содержит вариабельный (VL) домен легкой цепи (VL1), содержащий следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3.
В некоторых аспектах (a) VH1 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 10; (б) VL1 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 11; или (в) антитело содержит VH1, как в (а), и VL1, как в (б).
В некоторых аспектах VH1 содержит следующие каркасные области (FR): (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 34; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 36; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 37. В некоторых аспектах VH1 содержит аминокислотную последовательность SEQ ID NO: 10.
В некоторых аспектах VH1 содержит следующие FR: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 34; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 58; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 36; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 37. В некоторых аспектах VH1 содержит аминокислотную последовательность SEQ ID NO: 59.
В некоторых аспектах VL1 содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 39; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 41. В некоторых аспектах VL1 содержит аминокислотную последовательность SEQ ID NO: 11.
В некоторых аспектах VL1 содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 61; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 41. В некоторых аспектах VL1 содержит аминокислотную последовательность SEQ ID NO: 60.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с LY6G6D, причем антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), при этом H1 содержит домен VH (VH1), содержащий аминокислотную последовательность SEQ ID NO: 10, и L1 содержит домен VL (VL1), содержащий аминокислотную последовательность SEQ ID NO: 11.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с LY6G6D, причем антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (HI) и полипептид легкой цепи (L1), при этом H1 содержит домен VH (VH1), содержащий аминокислотную последовательность SEQ ID NO: 59, и L1 содержит домен VL (VL1), содержащий аминокислотную последовательность SEQ ID NO: 60.
В некоторых аспектах антитело связывает полипептид LY6G6D человека с KД от приблизительно 100 пМ до 10 нМ при 37°С по результатам измерений с помощью анализа BIAcore. В некоторых аспектах антитело связывает полипептид LY6G6D человека с KД 6,0 нМ или менее; 4 нМ или менее или 2 нМ или менее.
В некоторых аспектах антитело является моноклональным, человеческим, гуманизированным или химерным.
В некоторых аспектах антитело представляет собой фрагмент антитела, который связывает LY6G6D. В некоторых аспектах фрагмент антитела выбран из группы, состоящей из фрагментов Fab, Fab'-SH, Fv, scFv и (Fab')2.
В некоторых аспектах антитело является полноразмерным антителом или антителом IgG.
В некоторых аспектах антитело является моноспецифическим антителом, мультиспецифическим антителом или биспецифическим антителом.
В некоторых аспектах биспецифическое антитело связывается с кластером дифференциации 3 (CD3) и содержит CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), где Н2 содержит домен VH (VH2), и L2 содержит домен VL (VL2).
В некоторых аспектах CD3-связывающий домен способен связываться с полипептидом CD3 человека или полипептидом CD3 яванского макака. В некоторых аспектах полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3ε человека или полипептид CD3ε яванского макака соответственно. В некоторых аспектах полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3γ человека или полипептид CD3γ яванского макака соответственно.
В некоторых аспектах антитело связывает полипептид CD3ε человека с KД от приблизительно 1 нМ до 500 нМ при 37°С по результатам измерений с помощью анализа BIAcore. В некоторых аспектах CD3-связывающий домен связывает полипептид CD3ε человека с KД 250 нМ или менее. В некоторых аспектах CD3-связывающий домен связывает полипептид CD3ε человека с KД 100 нМ или менее. В некоторых аспектах CD3-связывающий домен связывает полипептид CD3ε человека с KД 15 нМ или менее. В некоторых аспектах CD3-связывающий домен связывает полипептид CD3ε человека с KД 10 нМ или менее. В некоторых аспектах CD3-связывающий домен связывает полипептид CD3ε человека с KД 5 нМ или менее.
В некоторых аспектах VH2 содержит следующие CDR: (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (в) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 17; и VL2 содержит следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14.
В некоторых аспектах VH2 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 20; (б) VL2 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 21; или (в) антитело содержит VH2, как в (а), и VL2, как в (б). В некоторых аспектах VH2 содержит аминокислотную последовательность SEQ ID NO: 20. В некоторых аспектах VL2 содержит аминокислотную последовательность SEQ ID NO: 21.
В некоторых аспектах VH2 содержит следующие CDR: (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; и (в) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 17; и VL2 содержит следующие CDR: (г) CDR-L1 содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (e) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51;
В некоторых аспектах (a) VH2 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 20; (б) VL2 содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 55; или (в) антитело содержит VH2, как в (а), и VL2, как в (б). В некоторых аспектах VH2 содержит аминокислотную последовательность SEQ ID NO: 20. В некоторых аспектах VL2 содержит аминокислотную последовательность SEQ ID NO: 55.
В некоторых аспектах VH2 содержит следующие FR: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 43; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 45.
В некоторых аспектах VH2 содержит следующие FR: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 62; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 45.
В некоторых аспектах VL2 содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 47; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах VL2 содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 63; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах каждый H1 и Н2 дополнительно содержит константный домен тяжелой цепи (СН1), и каждый L1 и L2 дополнительно содержит константный домен легкой цепи (CL). В некоторых аспектах СН1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU). В некоторых аспектах СН1 из H1 содержит мутацию S183K, и CL из L1 содержит мутацию V133E. В некоторых аспектах СН1 из Н2 содержит мутацию S183E, и CL из L2 содержит мутацию V133K. В некоторых аспектах СН1 из H1 содержит мутацию S183E, и CL из L1 содержит мутацию V133K. В некоторых аспектах СН1 из Н2 содержит мутацию S183K, и CL из L2 содержит мутацию V133E.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит: LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), причем каждый H1 и Н2 содержит вариабельный домен тяжелой цепи (VH) и константный домен тяжелой цепи (СН1), и каждый L1 и L2 содержит вариабельный домен легкой цепи (VL) и константный домен легкой цепи (CL), где: (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 4 или SEQ ID NO: 111; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5, SEQ ID NO: 112 или SEQ ID NO: 113; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3 или любую из SEQ ID NO: 99-107; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату). В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату).
В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность любой из SEQ ID NO: 99-107; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату).
В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 111; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату). В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 112 или SEQ ID NO: 113; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату).
В другом аспекте настоящее изобретение показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит: LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), причем каждый H1 и Н2 содержит вариабельный домен тяжелой цепи (VH) и константный домен тяжелой цепи (СН1), и каждый L1 и L2 содержит вариабельный домен легкой цепи (VL) и константный домен легкой цепи (CL), где: (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 4 или SEQ ID NO: 111; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5, SEQ ID NO: 112 или SEQ ID NO: 113; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3 или любую из SEQ ID NO: 99-107; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату). В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату). В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность любой из SEQ ID NO: 99-107; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату).
В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 111; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату). В некоторых аспектах (а) LY6G6D-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 111; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 112 или SEQ ID NO: 113; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (iv) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; (б) CD3-связывающий домен содержит следующие шесть CDR: (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51; (в) CH1 из H1 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L1 содержит аминокислотную замену в V133 (нумерация согласно EU), и/или СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU); и (г) VH из H1 содержит аминокислотную замену в положении Q39, и VL из L1 содержит аминокислотные замены в положении Q38, и/или VH из Н2 содержит аминокислотную замену в положении Q39, и VL из L2 содержит аминокислотную замену в положении Q38 (все нумерации по Кабату).
В некоторых аспектах VH из H1 содержит аминокислотную замену в Q39 (нумерация по Кабату), и VL из L1 содержит аминокислотную замену в Q38 (нумерация по Кабату). В некоторых аспектах СН1 из Н2 содержит аминокислотную замену в S183 (нумерация согласно EU), и CL из L2 содержит аминокислотную замену в V133 (нумерация согласно EU). В некоторых аспектах VH из Н2 дополнительно содержит аминокислотную замену в положении Q39 (нумерация по Кабату), и VL из L2 дополнительно содержит аминокислотную замену в положении Q38 (нумерация по Кабату). В некоторых аспектах СН1 из H1 содержит мутацию S183K (нумерация согласно EU), и CL из L1 содержит мутацию V133E (нумерация согласно EU), и СН1 из Н2 содержит мутацию S183E (нумерация согласно EU), и CL из L2 содержит мутацию V133K (нумерация согласно EU). В некоторых аспектах VH из H1 содержит мутацию Q39E (нумерация по Кабату), VL из L1 содержит мутацию Q38K, VH из Н2 содержит мутацию Q39K, и VL из L2 содержит мутацию Q38E (нумерация по Кабату). В некоторых аспектах СН1 из H1 содержит мутацию S183E (нумерация согласно EU), и CL из L1 содержит мутацию V133K (нумерация согласно EU), и СН1 из Н2 содержит мутацию S183K (нумерация согласно EU), и CL из L2 содержит мутацию V133E (нумерация согласно EU). В некоторых аспектах VH из H1 содержит мутацию Q39K (нумерация по Кабату), VL из L1 содержит мутацию Q38E (нумерация по Кабату), VH из Н2 содержит мутацию Q39E (нумерация по Кабату), и VL из L2 содержит мутацию Q38K (нумерация по Кабату).
В некоторых аспектах VH из Н2 содержит аминокислотную последовательность SEQ ID NO: 20, и/или VL из L2 содержит аминокислотную последовательность SEQ ID NO: 21. В некоторых аспектах VH из H1 содержит аминокислотную последовательность SEQ ID NO: 10, и VL из L1 содержит аминокислотную последовательность SEQ ID NO: 11.
В некоторых аспектах VH из Н2 содержит аминокислотную последовательность SEQ ID NO: 20, и/или VL из L2 содержит аминокислотную последовательность SEQ ID NO: 55. В некоторых аспектах VH из H1 содержит аминокислотную последовательность SEQ ID NO: 10, и VL из L1 содержит аминокислотную последовательность SEQ ID NO: 11.
В некоторых аспектах каждый из первого домена СН3 (СН31) Fc-области H1 и второго домена СН3 (СН32) Fc-области Н2 содержит выступ или впадину, и причем выступ или впадина в СН31 может располагаться во впадине или выступе, соответственно, в СН32. В некоторых аспектах СН31 и СН32 сходятся на поверхности между выступом и впадиной.
В некоторых аспектах СН31 Fc-области H1 содержит выступ, и СН32 Fc-области Н2 содержит впадину. В некоторых аспектах (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотной замены T366W (нумерация согласно EU); (б) СН32 Fc-области Н2 содержит впадину, содержащую мутацию аминокислотной замены T366S, L368A или Y407V (нумерация согласно EU), или их комбинацию; или (в) как (а), так и (б).
В некоторых аспектах (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотой замены T366W (нумерация согласно EU);
(б) СН32 Fc-области Н2 содержит впадину, содержащую мутации аминокислотной замены T366S, L368A и Y407V (нумерация согласно EU); или
(в) как (а), так и (б). В некоторых аспектах (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотной замены T366W (нумерация согласно EU), и (б) СН32 Fc-области Н2 содержит впадину, содержащую мутации аминокислотной замены T366S, L368A и Y407V (нумерация согласно EU).
В некоторых аспектах СН31 Fc-области H1 содержит впадину, и СН32 Fc-области Н2 содержит выступ. В некоторых аспектах (а) СН31 Fc-области H1 содержит впадину, содержащую мутацию аминокислотной замены T366S, L368A или Y407V (нумерация согласно EU), или их комбинацию; (б) СН32 Fc-области Н2 содержит выступ, содержащий мутацию аминокислотной замены T366W (нумерация согласно EU); или (в) как (а), так и (б). В некоторых аспектах (а) СН31 Fc-области H1 содержит впадину, содержащую мутации аминокислотной замены T366S, L368A и Y407V (нумерация согласно EU); (б) СН32 Fc-области Н2 содержит выступ, содержащий мутацию аминокислотной замены T366W (нумерация согласно EU); или (в) как (а), так и (б). В некоторых аспектах (а) СН31 Fc-области H1 содержит впадину, содержащую мутации аминокислотной замены T366S, L368A и Y407V (нумерация согласно EU), и (б) СН32 Fc-области второго полипептида тяжелой цепи содержит выступ, содержащий мутацию аминокислотной замены T366W (нумерация согласно EU).
В некоторых аспектах Fc-области являются Fc-областями изотипа IgG человека или их вариантами Fc-области. В некоторых аспектах Fc-области являются вариантами Fc-области изотипа IgG человека. В некоторых аспектах каждый из вариантов Fc-области изотипа IgG человека содержит мутацию в аминокислотном остатке N297 (нумерация согласно EU), что приводит к отсутствию гликозилирования. В некоторых аспектах мутация в аминокислотном остатке N297 представляет собой мутацию по типу замены. В некоторых аспектах мутация в аминокислотном остатке N297 снижает эффекторную функцию Fc-области.
В некоторых аспектах мутация по типу замены представляет собой мутацию N297G или N297A. В некоторых аспектах каждый из вариантов Fc-области изотипа IgG человека содержит мутацию N297G. В некоторых аспектах каждый из вариантов Fc-области изотипа IgG человека содержит мутацию, которая снижает эффекторную функцию Fc-области.
В некоторых аспектах мутация, которая снижает эффекторную функцию Fc-области, представляет собой мутацию по типу замены. В некоторых аспектах мутация по типу замены приходится на аминокислотный остаток Е233, L234, L235, D265 и/или Р329 (нумерация согласно EU). В некоторых аспектах мутация по типу замены представляет собой мутацию Е233Р, L234A, L234V, L235A, D265A или P329G. В некоторых аспектах каждый из вариантов Fc-области изотипа IgG человека содержит мутацию P329G. В некоторых аспектах каждый из вариантов Fc-области изотипа IgG человека содержит мутации N297G и P329G.
В некоторых аспектах варианты Fc-области изотипа IgG человека представляют собой варианты Fc-области изотипов IgG1 или IgG3 человека, причем каждый дополнительно содержит мутацию L234A или L235A. В некоторых аспектах варианты Fc-области изотипа IgG человека представляют собой варианты Fc-области изотипов IgGl или IgG3 человека, причем каждый дополнительно содержит мутации L234A и L235A.
В некоторых аспектах варианты Fc-области изотипа IgG человека представляют собой варианты Fc-области изотипов IgG1 или IgG3 человека, причем каждый дополнительно содержит следующие мутации по типу замены Е233Р, L234V и L235A (нумерация согласно EU) и делецию остатка G236 (нумерация согласно EU). В некоторых аспектах варианты Fc-области изотипа IgG человека представляют собой варианты Fc-области изотипа IgGl человека.
В некоторых аспектах варианты Fc-области изотипа IgG человека представляют собой варианты Fc-области изотипов IgG4 человека, причем каждый дополнительно содержит следующие мутации по типу замены Е233Р, F234V и L235A (нумерация согласно EU) и делецию остатка G236 (нумерация согласно EU).
В некоторых аспектах Fc-области Fc-комплекса являются безэффекторными Fc-областями.
В некоторых аспектах H1 содержит аминокислотную последовательность SEQ ID NO: 7, и L2 содержит аминокислотную последовательность SEQ ID NO: 9. В некоторых аспектах Н2 содержит аминокислотную последовательность SEQ ID NO: 18, и L2 содержит аминокислотную последовательность SEQ ID NO: 19. В некоторых аспектах Н2 содержит аминокислотную последовательность SEQ ID NO: 18, и L2 содержит аминокислотную последовательность SEQ ID NO: 57.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 7; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 9; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 18; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 19.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 64; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 65; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 69; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 70.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 8; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 9; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 67; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 19.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 66; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 65; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 68; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 70.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (a) H1 содержит аминокислотную последовательность SEQ ID NO: 7; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 9; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 18; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 57.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 64; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 65; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 69; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 73.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 8; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 9; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 67; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 57.
В другом аспекте настоящее раскрытие показывает биспецифическое антитело, которое связывается с LY6G6D и CD3, причем биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), и где: (а) H1 содержит аминокислотную последовательность SEQ ID NO: 66; (б) L1 содержит аминокислотную последовательность SEQ ID NO: 65; (в) Н2 содержит аминокислотную последовательность SEQ ID NO: 68; и (г) L2 содержит аминокислотную последовательность SEQ ID NO: 73.
В некоторых аспектах антитело имеет клиренс после внутривенной инъекции от приблизительно 10 мл/кг/сутки до приблизительно 35 мл/кг/сутки.
В другом аспекте настоящее раскрытие показывает одну или более выделенных нуклеиновых кислот, кодирующих антитело по любому из вышеуказанных аспектов или его часть, содержащую связывающий домен, который связывается с LY6G6D.
В другом аспекте настоящее раскрытие показывает один или более векторов, содержащих одну или более выделенных нуклеиновых кислот вышеуказанного аспекта.
В другом аспекте настоящее раскрытие показывает одну или более клеток-хозяев, содержащих один или более векторов вышеуказанного аспекта.
В некоторых аспектах одна или более клеток-хозяев представляют собой одну или более клеток-хозяев млекопитающего. В некоторых аспектах одна или более клеток-хозяев млекопитающего представляют собой одну или более клеток-хозяев яичника китайского хомячка (СНО).
В некоторых аспектах одна или более клеток-хозяев представляют собой одну или более прокариотических клеток-хозяев. В некоторых аспектах одна или более прокариотических клеток-хозяев представляют собой одну или более клеток-хозяев Е. coli.
В другом аспекте настоящее раскрытие показывает способ получения антитела по любому из вышеуказанных аспектов, причем способ включает культивирование одной или более клеток-хозяев вышеуказанного аспекта в культуральной среде. В некоторых аспектах способ дополнительно включает извлечение антитела к LY6G6D из одной или более клеток-хозяев или культуральной среды.
В другом аспекте настоящее раскрытие показывает композицию, содержащую антитело по любому из вышеуказанных аспектов. В некоторых аспектах композиция дополнительно содержит фармацевтически приемлемое вспомогательное вещество или разбавитель. В некоторых аспектах фармацевтически приемлемое вспомогательное вещество представляет собой буфер, носитель, стабилизатор или консервант.В некоторых аспектах композиция представляет собой фармацевтическую композицию.
В другом аспекте настоящее раскрытие показывает антитело по любому из вышеуказанных аспектов для применения в качестве лекарственного средства.
В другом аспекте настоящее раскрытие показывает антитело по любому из вышеуказанных аспектов или композицию по любому из вышеуказанных аспектов для применения в лечении или замедлении прогрессирования LY6G6D-положительного рака у субъекта, нуждающегося в этом. В некоторых аспектах LY6G6D-положительный рак представляет собой колоректальный рак. В некоторых аспектах LY6G6D-положительный рак имеет статус микросателлитной нестабильности (MSI): микросателлитная стабильность (MSS) или низкий уровень микросателлитной нестабильности (MSI-L).
В другом аспекте настоящее раскрытие показывает применение антитела по любому из вышеуказанных аспектов или композиции по любому из вышеуказанных аспектов в производстве лекарственного средства для лечения или замедления прогрессирования LY6G6D-положительного рака у субъекта. В некоторых аспектах LY6G6D-положительный рак представляет собой колоректальный рак. В некоторых аспектах LY6G6D-положительный рак имеет статус MSI: MSS или MSI-L.
В другом аспекте настоящее раскрытие показывает способ лечения или замедления прогрессирования LY6G6D-положительного рака у субъекта, нуждающегося в этом, причем способ включает введение субъекту антитела по любому из вышеуказанных аспектов или композиции по любому из вышеуказанных аспектов. В некоторых аспектах LY6G6D-положительный рак представляет собой колоректальный рак. В некоторых аспектах LY6G6D-положительный рак имеет статус MSI: MSS или MSI-L.
В другом аспекте настоящее раскрытие показывает набор, содержащий антитело по любому из вышеуказанных аспектов и листок-вкладыш в упаковке, содержащий инструкции по применению антитела для лечения или замедления прогрессирования LY6G6D-положительного рака у субъекта. В некоторых аспектах LY6G6D-положительный рак представляет собой колоректальный рак. В некоторых аспектах LY6G6D-положительный рак имеет статус MSI: MSS или MSI-L. В некоторых аспектах субъект представляет собой человека.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с CD3, причем антитело содержит связывающий домен, содержащий следующие CDR: (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и (e) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14. В некоторых аспектах антитело содержит (a) VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 20; (б) VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 21; или (в) VH, как в (а), и VL, как в (б). В некоторых аспектах VH содержит аминокислотную последовательность SEQ ID NO: 20. В некоторых аспектах VL содержит аминокислотную последовательность SEQ ID NO: 21.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с CD3, причем антитело содержит связывающий домен, содержащий следующие CDR: (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 50; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 51. В некоторых аспектах антитело содержит (a) VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 20; (б) VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 55; или (в) VH, как в (а), и VL, как в (б). В некоторых аспектах VH содержит аминокислотную последовательность SEQ ID NO: 20. В некоторых аспектах VL содержит аминокислотную последовательность SEQ ID NO: 55.
В некоторых аспектах VH содержит следующие FR: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 43; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 45.
В некоторых аспектах VH содержит следующие FR: (a) FR-H1, содержащую аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащую аминокислотную последовательность SEQ ID NO: 62; (в) FR-H3, содержащую аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащую аминокислотную последовательность SEQ ID NO: 45.
В некоторых аспектах VL содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 47; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах VL содержит следующие FR: (a) FR-L1, содержащую аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащую аминокислотную последовательность SEQ ID NO: 63; (в) FR-L3, содержащую аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащую аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах антитело связывает полипептид CD3ε человека с KД от приблизительно 1 нМ до 500 нМ при 37°С по результатам измерений с помощью анализа BIAcore. В некоторых аспектах антитело связывает полипептид CD3ε человека с KД 250 нМ или менее; 100 нМ или менее; 15 нМ или менее; 10 нМ или менее или 5 нМ или менее.
В некоторых аспектах антитело является моноклональным, человеческим, гуманизированным или химерным.
В некоторых аспектах антитело представляет собой фрагмент антитела, который связывает CD3. В некоторых аспектах фрагмент антитела выбран из группы, состоящей из фрагментов Fab, Fab'-SH, Fv, scFv и (Fab')2.
В некоторых аспектах антитело представляет собой полноразмерное антитело.
В некоторых аспектах антитело представляет собой антитело IgG.
В некоторых аспектах антитело к CD3 представляет собой моноспецифическое антитело.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с LY6G6D, причем антитело содержит связывающий домен, содержащий следующие определяющие комплементарность области (CDR): (а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 27; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 28; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 29; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 24; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 25; и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 26.
В некоторых аспектах антитело содержит (a) VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 32; (б) VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 33; или (в) VH, как в (а), и VL, как в (б). В некоторых аспектах VH содержит аминокислотную последовательность SEQ ID NO: 32. В некоторых аспектах VL содержит аминокислотную последовательность SEQ ID NO: 33.
В другом аспекте настоящее раскрытие показывает выделенное антитело, которое связывается с LY6G6D, причем антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 30, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 31.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1А представляет собой график, показывающий экспрессию LY6G6D и LY6G6F в нормальной (черный) и опухолевой (красный) тканях в нормализованных считываниях на тысячу пар нуклеотидов на миллион (nRPKM) в Атласе ракового генома (АРГ). LY6G6D значительно сверхэкспресисирован в ткани опухоли толстой кишки.
Фиг. 1Б представляет собой график, показывающий экспрессию LY6G6D и LY6G6F в нормальных тканях в nRPKM в общедоступных данных проекта GTEx.
Фиг. 1В представляет набор коробчатых диаграмм, показывающих экспрессию LY6G6D в nRPKM при видах колоректального рака (КРР), имеющих статус микросателлитной нестабильности (MSI): микросателлитная стабильность (MSS), низкий уровень микросателлитной нестабильности (MSI-L) или высокий уровень микросателлитной нестабильности (MSI-H). Указана связь между статусом MSI для КРР и прогнозом.
Фиг. 2А представляет микрофотографию нормальной ткани толстой кишки, показывая иммуногистохимическое (ИГХ) окрашивание для LY6G6D.
Фиг. 2Б представляет микрофотографию первичной опухоли толстой кишки, показывающую слабое (1+) ИГХ окрашивание для LY6G6D.
Фиг. 2В представляет микрофотографию первичной опухоли толстой кишки, показывающую среднее (2+) ИГХ окрашивание для LY6G6D.
Фиг. 2Г представляет микрофотографию первичной опухоли толстой кишки, показывающую сильное (3+) ИГХ окрашивание для LY6G6D.
Фиг. 3А представляет собой пару графиков, показывающих in vitro уничтожение клеток НТ55 (клеточная линия карциномы толстой кишки человека), дополненное 10Х мононуклеарами периферической крови (МНПК) человека от донора №1 или донора №2 Т-клеточнозависимым биспецифическим антителом LY6G6D (TDB), содержащим LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c. Значения ЕС50 для каждого TDB указаны.
Фиг. 3Б представляет набор графиков, показывающих объем (мм2) опухоли НТ55 ксенотрансплантата у мышей NSG™ после обработки с помощью LY6G6D TDB, содержащих LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 40G5c или 38E4v1. Мышей гуманизировали с помощью МНПК здорового донора. Обработки, содержащие среду доставки и МНПК или содержащие LY6G6D TDB и не содержащие МНПК, представлены в качестве контролей.
Фиг. 3В представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК человека, с помощью LY6G6D TDB, содержащего химерное или гуманизированное LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1, и дополнительную таблицу, показывающую аффинность химерного или гуманизированного плеча 1G4 в анализе BIAcore.
Фиг. 3Г представляет набор графиков, показывающих объем (мм2) опухолей НТ55 ксенотрансплантата у мышей NSG™ после обработки с помощью LY6G6D TDB, содержащего химерное или гуманизированное LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1. Мышей гуманизировали с помощью МНПК здорового донора. Обработки, содержащие среду доставки и МНПК или содержащие LY6G6D TDB и не содержащие МНПК, представлены в качестве контролей.
Фиг. 4А представляет ленточную диаграмму, показывающую место сконструированных сайтов гликозилирования (красные, розовые, зеленые и синие круги) в модели структурной гомологии полипептида LY6G6D. Сайты гликозилирования закодированы цветом на основе их влияния на связывание антитела. Гликозилирование на сайте, отмеченном красным кругом, прерывало связывание 1G4. Гликозилирование на сайте, отмеченном розовым кругом, прерывало связывание 16D7. Гликозилирование на сайте, отмеченном синими кругами, не прерывало связывание 1G4 или 16D7. Зеленые круги отмечают сконструированные сайты гликозилирования в линкере к Fc.
Фиг. 4Б представляет собой схему, показывающую связывание кандидатных антител к LY6G6D с полипептидами LY6G6D, содержащими сконструированные сайты гликозилирования на указанных остатках. Клетки, отмеченные с помощью X, указывают, что связывание не было обнаружено. Клетки, отмеченные с помощью W, указывают, что связывание было значительно снижено.
Фиг. 4В представляет пару диаграмм, показывающих место сконструированных сайтов гликозилирования в модели структурной гомологии полипептида LY6G6D в первой ориентации и второй ориентации, которая была повернута на 180°. Сайты гликозилирования, которые прерывают связывание антител группы 1, 2, 3 и 4 фиг. 4А, указаны зелеными, фиолетовыми, синими и оранжевыми кругами соответственно.
Фиг. 4Г представляет аннотированную последовательность полипептида LY6G6D, в которой аминокислотные остатки, затронутые мутациями гликозилирования, закодированы цветом, как на фиг. 4В, и группы 1, 2, 3 и 4 фиг. 4А указаны путем подчеркивания.
Фиг. 4Д представляет диаграмму, показывающую клоны антитела к LY6G6D кролика, помещенные в четыре отдельные группы эпитопов. Группа 1 содержит три группы последовательностей и включает 20А12, rf.1G4, 6Е10 и 4Н7. Группа 2 содержит шесть групп последовательностей и содержит f.16D7. Каждая из групп 3 и 4 содержит три группы последовательностей.
Фиг. 4Е представляет график, показывающий связывание антител TDB кролика к LY6G6D человека, содержащих указанное LY6G6D-специфичное плечо из группы 1, 2, 3 или 4 фиг. 4Е и CD3-специфичное плечо 40G5c с клетками НТ55 (клеточная линия карциномы толстой кишки человека). Связывание измеряют как среднюю интенсивность флуоресценции (СИФ).
Фиг. 4Ж представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК человека, с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо из группы 1, 2, 3 или 4 фиг. 4Е и CD3-специфичное плечо 40G5c.
Фиг. 4И представляет набор графиков, показывающих связывание 1G4 и антител к LY6G6D кролика 6Е10, 20А12 и 4Н7 с полипетидом Ly6G6D по результатам измерений с помощью анализа BIAcore. Антитела кролика экспрессировались как химерные антигенсвязывающие фрагменты (Fab) с вариабельными доменами кролика и константными доменами человека. Ly6G6D-Fc непосредственно иммобилизировали на чипе, и Fab пропускали при 37°С. КД указана под каждым графиком.
Фиг. 5А представляет диаграмму, показывающую аминокислотную последовательность вариабельной области тяжелой цепи 20A12.QNTv12 (двухклеточная) (SEQ ID NO: 22) согласно системе нумерации Кабата. Определяющие комплементарность области (CDR) CDR H1, CDR Н2 и CDR Н3 указаны. Последовательности CDR согласно системе нумерации Кабата подчеркнуты.
Фиг. 5Б представляет диаграмму, показывающую аминокислотную последовательность вариабельной области легкой цепи 20A12.QNTv12 (двухклеточная) (SEQ ID NO: 23) согласно системе нумерации Кабата. CDR -CDR H1, CDR Н2 и CDR Н3 - указаны. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 5В представляет диаграмму, показывающую аминокислотную последовательность вариабельной области тяжелой цепи 20A12.QNTv12 (одноклеточная), содержащую мутацию аминокислотной замены Q39E (в рамке) в каркасной области (FR) 2 (SEQ ID NO: 10). Эта последовательность вариабельной области тяжелой цепи особенно пригодна для одноклеточного изготовления TDB. Определяющие комплементарность области (CDR) CDR H1, CDR Н2 и CDR Н3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 5Г представляет диаграмму, показывающую аминокислотную последовательность вариабельной области легкой цепи 20A12.QNTv12 (одноклеточная), содержащей мутацию Q38K (в рамке) в FR2 (SEQ ID NO: 11). Эта последовательность вариабельной области легкой цепи особенно пригодна для одноклеточного изготовления TDB. Определяющие комплементарность области (CDR) CDR H1, CDR Н2 и CDR Н3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 6А представляет модель структуры белка, показывающую антигенсвязывающую область фрагмента (Fab) антитела 20A12.QNTv12, связывающегося с полипептидом, содержащим аминокислотные остатки 94-103 LY6G6D (RDCYLGDLCN; SEQ ID NO: 78).
Фиг. 6Б представляет область модели структуры белка, показывающей десять наложенных комплексов, содержащих Fab антитела 20A12.QNTv12, связанного с аминокислотными остатками 94-103 LY6G6D. 20A12.QNTv12 показано как ленточная диаграмма. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма, и аминокислотные остатки маркированы.
Фиг. 6В представляет область модели структуры белка, показывающей тяжелую цепь (НС; розовый) и легкую цепь (LC; зеленый) антитела 20A12.QNTv12, связанного с аминокислотными остатками 94-103 LY6G6D. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма, и аминокислотные остатки маркированы.
Фиг. 6Г представляет область модели структуры белка, показывающей НС (розовый) и LC (зеленый) антитела 20A12.QNTV12, связанного с аминокислотными остатками 94-103 LY6G6D. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма с помощью карты распределения электронной плотности 2Fo-Fc, проведенной при 1,0а (синий).
Фиг. 6Д представляет область модели структуры белка, показывающей антитело 20A12.QNTv12, связанное с аминокислотными остатками 94-103 LY6G6D. CDR H1, CDR Н2 и CDR Н3 (H1, Н2 и Н3, соответственно) вариабельной области тяжелой цепи 20A12.QNTv12 и CDR L1, CDR L2 и CDR L3 (L1, L2 и L3, соответственно) вариабельной области легкой цепи 20A12.QNTv12 маркированы и отмечены цветом в ленточной модели. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма.
Фиг. 6Е представляет область модели структуры белка, показывающей антитело 20A12.QNTv12, связанное с аминокислотными остатками 94-103 LY6G6D. 20A12.QNTv12 показано как пространственная модель. CDR H1, CDR Н2 и CDR НЗ (H1, Н2 и Н3, соответственно) вариабельной области тяжелой цепи 20A12.QNTv12 и CDR L1, CDR L2 и CDR L3 (L1, L2 и L3, соответственно) вариабельной области легкой цепи 20A12.QNTv12 маркированы. Выбранные остатки 20A12.QNTv12 маркированы, и взаимодействия между 20A12.QNTv12 и полипептидом LY6G6D указаны красными пунктирными линиями. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма.
Фиг. 7А представляет область модели структуры белка, показывающей антитело 1G4, связанное с аминокислотными остатками 94-103 LY6G6D. CDR H1, CDR Н2 и CDR Н3 вариабельной области тяжелой цепи 1G4 и CDR L1, CDR L2 и CDR L3 вариабельной области легкой цепи 1G4 маркированы и отмечены цветом в ленточной модели. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма.
Фиг. 7Б представляет область модели структуры белка, показывающей антитело 20A12.QNTv12, связанное с аминокислотными остатками 94-103 LY6G6D. CDR H1, CDR Н2 и CDR Н3 вариабельной области тяжелой цепи 20A12.QNTv12 и CDR L1, CDR L2 и CDR L3 вариабельной области легкой цепи 20A12.QNTv12 маркированы и отмечены цветом в ленточной модели. Полипептид, содержащий остатки 94-103 LY6G6D, показан как палочковая диаграмма.
Фиг. 7В представляет модель структуры белка, показывающую тяжелую цепь (SEQ ID NO: 96) и легкую цепь (SEQ ID NO: 97) антигенсвязывающей области фрагмента (Fab) антитела 20A12.QNTv12, связанного с полипептидом, содержащим аминокислотные остатки 93-104 LY6G6D (HRDCYLGDLCNS; SEQ ID NO: 87), и диаграмму последовательностей остатков 93-104 LY6G6D, показывающую конкретные остатки, с которыми взаимодействует 20A12.QNTv12 (оранжевые и подчеркнутые). Каждый из этих остатков располагается в 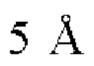 от Fab.
от Fab.
Фиг. 7Г представляет область модели структуры белка, показывающую тяжелую цепь (SEQ ID NO: 96) и легкую цепь (SEQ ID NO: 97) антигенсвязывающей области фрагмента (Fab) антитела 20A12.QNTv12, связывающегося с полипептидом, содержащим аминокислотные остатки 93-104 LY6G6D (HRDCYLGDLCNS; SEQ ID NO: 87). Остатки в 20A12.QNTv12, которые взаимодействуют с полипептидом LY6G6D, маркированы. НС указывает, что остаток находится в тяжелой цепи 20A12.QNTv12; LC указывает, что остаток находится в легкой цепи 20A12.QNTv12.
Фиг. 7Д представляет модель структуры белка, показывающую тяжелую цепь (SEQ ID NO: 94) и легкую цепь (SEQ ID NO: 95) антигенсвязывающей области фрагмента (Fab) антитела 1G4, связанного с поли пептидом, содержащим аминокислотные остатки 93-104 LY6G6D (HRDCYLGDLCNS; SEQ ID NO: 87), и диаграмму последовательностей остатков 93-104 LY6G6D, показывающую конкретные остатки, с которыми взаимодействует 1G4 (оранжевые и подчеркнутые). Каждый из этих остатков располагается в 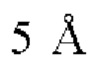 от Fab.
от Fab.
Фиг. 7Е представляет область модели структуры белка, показывающую тяжелую цепь (SEQ ID NO: 94) и легкую цепь (SEQ ID NO: 95) антигенсвязывающей области фрагмента (Fab) антитела 1G4, связывающегося с полипептидом, содержащим аминокислотные остатки 93-104 LY6G6D (HRDCYLGDLCNS; SEQ ID NO: 87). Остатки в 1G4, которые взаимодействует с полипептидом LY6G6D, маркированы. НС указывает, что остаток находится в тяжелой цепи 1G4; LC указывает, что остаток находится в легкой цепи 1G4.
Фиг. 8А представляет схематическую диаграмму, показывающую изготовление LY6G6D TDB, имеющего CD3-специфичное плечо 38E4v1, содержащее Fc-область, имеющую мутации аминокислотных замен T366S, L368A и Y407V, образуя область «впадины», и мутацию N297G, в паре с LY6G6D-специфичным плечом 20A12.QNTv12 (двухклеточная), содержащим Fc-область, имеющую мутацию аминокислотной замены T366W, которая образует область «выступа», и мутацию N297G, где CD3-специфичное плечо и LY6G6D-специфичное плечо образуют полноразмерное TDB IgG1K.
Фиг. 8Б представляет схематическую диаграмму, показывающую технологический процесс для изготовления биспецифического антитела, используя две линии клеток-хозяев (двуклеточная технология). Первое плечо антитела, содержащее область впадины, получается в первой линии клеток-хозяев, а второе плечо антитела, содержащее область выступа, получают во второй линии клеток-хозяев. Плечи антитела очищаются из линий клеток-хозяев и собираются in vitro.
Фиг. 8В представляет схематическую диаграмму, показывающую технологический процесс для изготовления биспецифического антитела, используя одну линию клеток-хозяев (одноклеточная технология). Первое плечо антитела, содержащее область впадины, и второе плечо антитела, содержащее область выступа, получают в и очищают из одной линии клеток-хозяев. Первое плечо и второе плечо антитела содержат мутации аминокислотных замен, как показано на фиг. 8Г или фиг. 8Д.
Фиг. 8Г представляет диаграмму, показывающую биспецифическое антитело, полученное с помощью одной клеточной линии. Мутации аминокислотной замены, вводящие зарядовые пары, указаны. Зарядовые пары содержат мутацию по типу замены Q39K в VH первого плеча и мутацию по типу замены Q38E в VL первого плеча; мутацию по типу замены S183E в СН1 первого плеча и мутацию по типу замены V133K в CL первого плеча; мутацию по типу замены Q39E в VH второго плеча и мутацию по типу замены Q38K в VL второго плеча; и мутацию по типу замены S183K в СН1 второго плеча и мутацию по типу замены V133E в CL второго плеча.
Фиг. 8Д представляет диаграмму, показывающую биспецифическое антитело, полученное с помощью одной клеточной линии. Мутации аминокислотной замены указаны. Мутации аминокислотной замены, вводящие зарядовые пары, указаны. Зарядовые пары содержат мутацию по типу замены Q39E в VH первого плеча и мутацию по типу замены Q38E в VL первого плеча; мутацию по типу замены Q39K в VH второго плеча и мутацию по типу замены Q38E в VL второго плеча; и мутацию по типу замены S183E в СН1 второго плеча и мутацию по типу замены V133K в CL второго плеча. Антитело также содержит мутации Rosetta YT65 и мутации A141I, F170S, S181M, S183A и V185A в СН1 первого плеча и мутации F116A, L135V, S174A, S176F и T178V в CL первого плеча.
Фиг. 9А представляет график, показывающий in vitro уничтожение клеток НТ55 с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®. TDB представлялось в концентрациях от 0,01 до 10000 нг/мл.
Фиг. 9Б представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c. Активацию Т-клеток CD4+ измеряли с помощью активируемой флуоресценцией сортировки клеток (АФСК).
Фиг. 9В представляет график, показывающий in vitro активацию Т-клеток CD8+с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточный) и CD3-специфичное плечо 38E4v1 или 40G5c. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 10А представляет график, показывающий in vitro уничтожение клеток НТ55 с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.v1 и CD3-специфичное плечо 38E4v1 или 40G5c, LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c или LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®. TDB представлялось в концентрациях от 0,01 до 10000 нг/мл.
Фиг. 10Б представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.v1 и CD3-специфичное плечо 38E4v1 или 40G5c, LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c или LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c. Активацию Т-клеток CD4+ измеряли с помощью АФСК.
Фиг. 10В представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.v1 и CD3-специфичное плечо 38E4v1 или 40G5c, LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5 с или LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 10Г представляет график, показывающий in vitro уничтожение клеток НТ55 с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c, LY6G6D-специфичное плечо 20A12.SNVv12 и CD3-специфичное плечо 38E4v1 или 40G5c или LY6G6D-специфичное плечо 6E10.v23 и CD3-специфичное плечо 38E4v1 или 40G5 с. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®.
Фиг. 11А представляет график, показывающий in vitro уничтожение клеток Colo320DM, НТ55 и LS1034 с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®.
Фиг. 11Б представляет набор графиков, показывающих антигенсвязывающую способность LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1, для клеток Colo320DM, НТ55 и LS1034, что измерено с помощью АФСК.
Фиг. 11В представляет набор микрофотографий, показывающих ИГХ окрашивание в дебрисе и в образцах опухолевых ксенотрансплантатов.
Фиг. 11Г представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК человека от здорового донора, с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c или LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c, через 24 часа.
Фиг. 11Д представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК человека от здорового донора, с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 или 40G5c или LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4v1 или 40G5c, через 48 часов. KД для каждого TDB указано в скобках.
Фиг. 11Е представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК человека от десяти доноров, с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1.
Фиг. 11Ж представляет график, показывающий представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 11И представляет таблицу, показывающую значения ЕС50 для уничтожения клеток и активации Т-клеток CD8+ для десяти доноров МНПК.
Фиг. 11К представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 в клетках Colo320DM, НТ55 и LS1034. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 12 представляет график, показывающий объем (мм2) опухолей COLO320DM ксенотрансплантанта у мышей после обработки с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1. Мышей гуманизировали с помощью мононуклеаров периферической крови (МНПК) от здоровых доноров. Обработки, содержащие среду доставки и МНПК или содержащие TDB и не содержащие МНПК, представлены в качестве контролей.
Фиг. 13А представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо rb6E, rb11A11, rb5D3, rb7F2, rb20F12, rb20A12, rb5E4, rb3A4, rb17F11, rb4H7, rb3D3 или гуманизированное 1G4 и CD3-специфичное плечо 40G5c. Активацию Т-клеток CD4+ измеряли с помощью АФСК.
Фиг. 13Б представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо rb6E, rb11A11, rb5D3, rb7F2, rb20F12, rb20A12, rb5E4, rb3A4, rb17F11, rb4H7, rb3D3 или гуманизированное 1G4 и CD3-специфичное плечо 40G5c. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 13В представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК от донора №2, с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо rb6E, rb11A11, rb5D3, rb7F2, rb20F12, rb20A12, rb5E4, rb3A4, rb17F11, rb4H7, rb3D3 или гуманизированное 1G4 и CD3-специфичное плечо 40G5c. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®.
Фиг. 13Г представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо rb6E, rb11A11, rb5D3, rb7F2, rb20F12, rb20A12, rb5E4, rb3A4, rb17F11, rb4H7, rb3D3 или гуманизированное 1G4 и CD3-специфичное плечо 40G5c. Активацию Т-клеток CD4+ измеряли с помощью АФСК.
Фиг. 13Е представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо rb6E, rb11A11, rb5D3, rb7F2, rb20F12, rb20A12, rb5E4, rb3A4, rb17F11, rb4H7, rb3D3 или гуманизированное 1G4 и CD3-специфичное плечо 40G5с. Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 14А представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК от донора №1, с помощью LY6G6D TDB, собранных с помощью двухклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью TDB, собранных с помощью одноклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Конкретные остатки в MD2, MD3 и MD4, которые имеют мутации относительно последовательности 38E4.v1 ДТ, указаны в скобках. Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®.
Фиг. 14Б представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, собранных с помощью двухклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью TDB, собранных с помощью одноклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Активацию Т-клеток CD4+ измеряли с помощью АФСК.
Фиг. 14В представляет график, показывающий in vitro активацию Т-клеток CD8+ с помощью LY6G6D TDB, собранных с помощью двухклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью LY6G6D TDB, собранных с помощью одноклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 15А представляет график, показывающий in vitro уничтожение клеток НТ55, дополненных МНПК от донора №2, с помощью LY6G6D TDB, собранных с помощью двухклеточной систеиы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью TDB, собранных с помощью одноклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CDS-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Уничтожение определяется количественно как % цитотоксичности в анализе CELLTITER-GLO®.
Фиг. 15Б представляет график, показывающий in vitro активацию Т-клеток CD4+ с помощью LY6G6D TDB, собранных с помощью двухклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью LY6G6D TDB, собранных с помощью одноклеточной системы и содержпщих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточное) и CDS-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Активацию Т-клеток CD4+ измеряли с помощью АФСК.
Фиг. 15В представляет график, показывающий in vitro активацию Т-кдеток CD8+ с помощью LY6G6D TDB, собранных с помощью двухклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3 или 38E4.v1 (ДТ), и с помощью LY6G6D TDB, собранных с помощью одноклеточной системы и содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD1, 38E4.v1 MD2, 38E4.v1 MD3, 38E4.v1 MD4 или 38E4.v1 (ДТ). Активацию Т-клеток CD8+ измеряли с помощью АФСК.
Фиг. 16А представляет график, показывающий объем (мм) опухолей LS1034 ксенотрансплантанта у мышей NSG™ после обработки с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 40G5c или 38E4.v1. Мышей гуманизировали с помощью мононуклеаров периферической крови (МНПК) от здоровых доноров. Обработки, содержащие среду доставки и МНПК или содержащие LY6G6D TDB и не содержащие МНПК, представлены в качестве контролей. «3+» указывает оценку ИГХ LY6G6D клеточной линии.
Фиг. 16Б представляет график, показывающий концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 40G5c или 38E4.v1 у мышей LS1034 NSG™ после введения одной дозы TDB.
Фиг. 16В представляет набор графиков, показывающих необработанные данные для анализа объема опухолей, показанного на фиг. 16А.
Фиг. 17А представляет график, показывающий объем (мм2) опухолей НТ55 ксенотрансплантанта у мышей NSG™ после обработки с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 40G5 с или 38E4.v1. Мышей гуманизировали с помощью МНПК здорового донора. Обработки, содержащие среду доставки и МНПК или содержащие TDB и не содержащие МНПК, представлены в качестве контролей. «2+» указывает оценку ИГХ LY6G6D клеточной линии.
Фиг. 17Б представляет набор графиков, показывающих необработанные данные для анализа объема опухолей, показанного на фиг.17А.
Фиг. 17В представляет график, показывающий концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 40G5c или 38E4.v1, у мышей НТТ55 NSG™ после введения одной дозы TDB, что измерено с помощью ИФА для фармакокинетики общего иммуноглобулина (GRIP).
Фиг. 18 представляет график и таблицу, показывающие концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 40G5c или 38E4.v1, и антитела к gD В56 у мышей с тяжелым комбинированным иммунодефицитом (ТКИД) после внутривенного введения одной дозы в 5 мг/кг антитела. Смакс: максимальная концентрация в сыворотке; AUC0-28: площадь под кривой; CL: скорость клиренса; t1/2: время полужизни.
Фиг. 19 представляет схематическую диаграмму и таблицу, показывающие исследование токсичности для LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, у яванских макак (макак).
Фиг. 20А представляет график, показывающий концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, у яванских макак после внутривенного введения одной дозы TDB с указанными дозировками.
Фиг. 20Б представляет график и таблицу, показывающие концентрацию в сыворотке (в мкг/мл) и клиренс (CL) TDB, содержащих различные нацеленные на опухоли плечи, спаренные с CD3-специфичным плечом 38E4v1 или 405Gc, у яванских макак после внутривенного введения одной дозы в 1 мг/кг TDB.
Фиг. 21 представляет набор микрофотографий, показывающих околососудистые/сосудистые инфильтраты мононуклеаров в мозге животного №6003, которому дозировали LY6G6D TDB, содержащее LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в дозе 15 мг/кг. Верхняя левая панель показывает контрольный (нормальный) сосуд оболочки мозга. Нижняя левая панель показывает ненормальный сосуд оболочки мозга животного №6003. Правая панель показывает увеличенный вид ненормального сосуда оболочки мозга.
Фиг. 22А представляет набор графиков, показывающих концентрацию цитокинов G-CSF, IL-IRa, МСР-1, TNF-a, IL-13 и IL-8 (в пг/мл) после обработки яванских макак одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак.
Фиг. 22Б представляет диаграмму разброса, показывающую концентрацию С-реактивного белка (CRP; в пг/мл) после обработки яванских макак одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак.
Фиг. 23А представляет пару графиков, показывающих процент клеток, которые были отрегулированы как экспрессирующие CD3+/CD4+/CD5+ CD25 лимфоциты-Т-хэлперы (Th) (левая панель) и экспрессирующие CD3+/CD8+/CD5+ CD25 Т-цитотоксичные (Тс) лимфоциты (правая панель) в анализе цитометрии в потоке у яванских макак, обработанных одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак. Измерения проводили за 7 дней до обработки (день -7) и в день обработки (день 1 до) и усредняли (средняя преддоза). После окончания инфузии (ОИ) измерения проводили через 2 часа, 6 часов, 24 часов и 168 часов. Пик, показывающий умеренную активацию Т-клеток, отмечали стрелкой.
Фиг. 23Б представляет график, показывающий процент клеток, которые были отрегулированы как Т-лимфоциты CD45+/CD3+ в анализе цитометрии в потоке у яванских макак, обработанных одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак. Пик, показывающий восстановление Т-клеток, отмечали стрелкой.
Фиг. 23В представляет график, показывающий процент клеток, которые были отрегулированы как В-лимфоциты CD45+/CD20+ в анализе цитометрии в потоке у яванских макак, обработанных одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак. Пик, показывающий восстановление В-клеток, отмечали стрелкой.
Фиг. 23Г представляет график, показывающий процент клеток, которые были отрегулированы как естественные клетки-киллеры (NK) CD45+/CD16+ в анализе цитометрии в потоке у яванских макак, обработанных одной дозой LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, в указанных дозировках и у контрольных (необработанных) яванских макак.
Фиг. 24А представляет пару графиков, показывающих связывание LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1, к полипептиду Ly6G6D человека (левая панель) и яванского макак (правая панель), что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 24Б представляет пару графиков, показывающих связывание LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTV12 (двухклеточная) и CD3-специфичное плечо 40G5c, к полипептиду Ly6G6D человека (левая панель) и яванского макак (правая панель), что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 24В представляет пару графиков, показывающих связывание LY6G6D TDB, содержащего LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4.v1, к полипептиду Ly6G6D человека (левая панель) и яванского макак (правая панель), что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 24Г представляет пару графиков, показывающих связывание LY6G6D TDB, содержащего LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 40G5c, к полипептиду Ly6G6D человека (левая панель) и яванского макак (правая панель), что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 25 представляет набор графиков, показывающих связывание LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4.v1 (левая панель), LY6G6D-специфичное плечо 20A12.QNTV12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD1 (центральная панель) или LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4.v1 MD4 (правая панель), к полипептиду Ly6G6D человека, что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 26А представляет график, показывающий объем (мм2) опухолей НТ55 ксенотрансплантанта у мышей NSG™ после обработки с помощью LY6G6D TDB, собранного с помощью одноклеточной системы, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4v1 MD1, 38E4v1 MD4 или 38E4v1 (ДТ), и с помощью LY6G6D TDB, собранного с помощью двухклеточной системы и содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 (ДТ). Обработки, содержащие среду доставки и МНПК или содержащие TDB и не содержащие МНПК, представлены в качестве контролей.
Фиг. 26Б представляет набор графиков, представляющий необработанные данные для анализа объема опухолей, показанного на фиг. 26А.
Фиг. 27 представляет график, показывающий концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, собранного с помощью двухклеточной системы и содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 (ДТ), и LY6G6D TDB, собранные с помощью одноклеточной системы, содержащей LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4v1 MD1 или 38E4v1 MD4.
Фиг. 28 представляет график и таблицу, показывающие концентрацию в сыворотке (в мкг/мл) LY6G6D TDB, собранного с помощью одноклеточной системы, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 (одноклеточная) и CD3-специфичное плечо 38E4v1 MD1 или 38E4v1 MD4, и LY6G6D TDB, собранное с помощью двухклеточной системы и содержащее LY6G6D-специфичное плечо 20A12.QNTv12 (двухклеточная) и CD3-специфичное плечо 38E4v1 (ДТ).
Фиг. 29А представляет выравнивание последовательностей, показывающее аминокислотные последовательности VL клонов к CD3 38E4v1, 40G5c, 38E4v1 MD1 (MD1) и 38E4v1 MD4 (MD4). Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 29Б представляет выравнивание последовательностей, показывающее аминокислотные последовательности VH клонов к CD3 38E4v1, 40G5c, 38E4v1 MD1 (MD1) и 38E4v1 MD4 (MD4). Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 29В представляет график, показывающий результаты анализа получения временной трансфекции, включающие CD3-специфичные плечи 38E4v1, MD1, 38E4v1.S43P, 38E4v1.T51A, 38E4v1.K55E и 38E4v1.K89T.
Фиг. 29Г представляет график, показывающий процент надлежащим образом спаренных биспецифических антител, полученных для LY6G6D TDB, содержащих LY6G6D-специфичное плечо или FcRH5-специфичное плечо и CD3-специфичные плечи 38E4v1 (ДТ), MD1 или вариант 38E4v1, имеющие однократные аминокислотные замены (указанные в скобках) при различных отношениях ДНК легкой цепи плеча-мишени (LC) к ДНК CD3-специфичного плеча (LC плеча-мишени: LC CD3).
Фиг. 29Д представляет график, показывающий концентрацию в сыворотке (в мкг/мл) после введения одной дозы 5 мг/кг моноспецифического двухвалентного антитела к CD3, содержащего CD3-специфичное плечо 38E4v1, 40G5c, MD1, MD4 или 38E4v1 K55E к СВ-17 SCID мышей (n=3 в момент времени). Антитело к gD показано как контроль. Отдельные точки данных (символы) показаны вместе со связанными средними значениями (сплошные линии).
Фиг. 29Е представляет выравнивание последовательностей, показывающее аминокислотные последовательности VL клонов к CD3 38E4v1, 40G5c, MD1 и MD4, содержащих мутацию аминокислотной замены Q38E (в рамке) в каркасной области (FR) 2. Эта последовательность вариабельной области легкой цепи особенно пригодна для одноклеточного изготовления TDB. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 29Ж представляет выравнивание последовательностей, показывающее аминокислотные последовательности VH клонов к CD3 38E4v1, 40G5c, MD1 и MD4, содержащих мутацию аминокислотной замены Q39K (в рамке) в каркасной области (FR) 2. Эта последовательность вариабельной области тяжелой цепи особенно пригодна для одноклеточного изготовления TDB. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны согласно определениям контакта, по Чотия и Кабату. Последовательности CDR, определенные согласно системе нумерации Кабата, подчеркнуты.
Фиг. 30 представляет график, показывающий результаты анализа получения временной трансфекции для двух изготовленных повторов LY6G6D-специфичного плеча 1G4. CD3-специфичное плечо 38E4v1 и GFR1-специфичное плечо представлены как контроли.
Фиг. 31А представляет диаграмму легкой цепи (LC) и тяжелой цепи (НС) клона 20А12 кролика, показывающего сайт гликозилирования в CDR-L3 и дисульфидную связь между двумя цистеиновыми остатками CDR-H1 и CDR-L2.
Фиг. 31Б представляет набор графиков, показывающих связывание вариантов LY6G6D-специфичного плеча 20А12, содержащего CDR 20А12 кролика и каркасные области легкой цепи человека hIGKV.1-5, hIGKV.1-39 и hIGKV.4-1, и каркасные области тяжелой цепи человека hIGHV.3-23 и hIGHV.3-30, что измерено с помощью анализа BIAcore. Показаны процентная идентичность последовательности зародышевой линии человека относительно последовательности rb.20A12 и число положений зон Верньера в каждом варианте.
Фиг. 31В представляет таблицу, показывающую процентную идентичность различных последовательности зародышевой линии VL человека относительно последовательности rb.20A12; число положений зон Верньера в каждой последовательности зародышевой линии человека и преобладание последовательности зародышевой линии у людей.
Фиг. 31Г представляет таблицу, показывающую процентную идентичность различных последовательности зародышевой линии VH человека относительно последовательности rb.20A12; число положений зон Верньера в каждой последовательности зародышевой линии человека и преобладание последовательности зародышевой линии у людей.
Фиг. 32А представляет диаграмму легкой цепи (LC) и тяжелой цепи (НС) клона 20А12 кролика, показывающего сайт гликозилирования в CDR-L3 и дисульфидную связь между двумя цистеиновыми остатками CDR-H1 и CDR-L2.
Фиг. 32Б представляет диаграмму LC и НС гуманизированного варианта 20А12 - 20A12.v1, показывающие сайт гликозилирования CDR-L3 и C35S и мутации аминокислотных замен С50А, которые исключают дисульфидную связь между двумя цистеиновыми остатками CDR-H1 и CDR-L2. 20A12.v1 содержит каркасные области VH hIGHV.3-23 и каркасные области VL hIGKV.1-39. Каркасные области человека были модифицированы в девяти положениях (круги), чтобы содержать остатки Верньера из последовательности 20А12 кролика.
Фиг. 32В представляет диаграмму LC и НС гуманизированного варианта 20А12 тонкой очистки, показывающего сайт гликозилирования в CDR-L3 и мутации аминокислотных замен C35S и С50А, которые исключают дисульфидную связь между двумя цистеиновыми остатками CDR-H1 и CDR-L2. Вариант 20А12 тонкой очистки содержит каркасные области VH hIGHV.3-23 и каркасные области VL hIGKV.1-5. Каркасные области человека были модифицированы в четырех положениях (круги), чтобы содержать остатки Верньера из последовательности 20А12 кролика.
Фиг. 32Г представляет пару диаграмм и таблицу, показывающие КД для rb.20A12 и его различных гуманизированных вариантов. Центральная колонка показывает мутации аминокислотных замен относительно последовательностей тяжелой цепи каркасной области человека (Н) hIGHV.3-23 и последовательностей легкой цепи каркасной области человека (L), которые возвращают положение аминокислоты к остатку Верньера кролика.
Фиг. 33А представляет таблицу, показывающую процентную идентичность различных последовательности зародышевой линии VL человека относительно последовательности rb.6Е10; число положений зон Верньера в каждой последовательности зародышевой линии человека и преобладание последовательности зародышевой линии у людей.
Фиг. 33Б представляет таблицу, показывающую процентную идентичность различных последовательности зародышевой линии VH человека относительно последовательности rb.6Е10; число положений зон Верньера в каждой последовательности зародышевой линии человека и преобладание последовательности зародышевой линии у людей.
Фиг. 34А представляет набор графиков, показывающих связывание rb.20A12 и различные химерные Fab, имеющие вариабельные домены rb.20A12 и константные области человека к полипептиду Ly6G6D, что измерено с помощью анализа BIAcore. Каждый из химерных Fab содержит такие аминокислотные мутации в каждом из С35 CDR-H1 и С50 CDR-H2): C35S-C50A (SA), C35S-C50S (SS), C35I-C50A (IA), C35I-C50S (IS) и C35I-C50I (II). КД для каждого химерного Fab указаны.
Фиг. 34Б представляют диаграмму последовательностей, показывающую сайт гликозилирования на CDR-L3 rb.20A12, имеющей NNT последовательности, и таблицу, показывающую KД вариантов гуманизированной последовательности легкой цепи 20А12 тонкой очистки, описанной выше, с мутациями аминокислотных замен в сайте гликозилирования, что измерено с помощью анализа BIAcore для связывания с LY6G6D.
Фиг. 34В представляет набор графиков, показывающих связывание вариантов Fab 20А12 кролика, причем 20А12 кролика содержит мутации C35I и С50А (IA), и гладкую гуманизированную 20А12, имеющую мутации аминокислотных замен QNT в сайте гликозилирования фиг. 34Б к LY6G6D, что измерено с помощью анализа BIAcore. Ly6G6D-Fc захватывали на чипе белка А, и Fab пропускали при 37°С.
Фиг. 34Г представляет набор графиков, показывающих связывание вариантов Fab гуманизированной последовательности легкой цепи 20А12 тонкой очистки, имеющей мутации аминокислотных замен QNV, SNV, GNT и SNA в сайте гликозилирования фиг. 34Б к полипептиду Ly6G6D, что измерено с помощью анализа BIAcore. Ly6G6D-Fc захватывали на чипе белка А, и Fab пропускали при 37°С.
Фиг. 35А представляет диаграмму, показывающую 20А12 кролика фиг. 32А, и график, показывающий связывание показанного на диаграмме антитела с LY6G6D, что измерено с помощью анализа BIAcore.
Фиг. 35Б представляет диаграмму, показывающую гуманизированный вариант 20А12 20A12.QNTV12 тонкой очистки, и график, показывающий связывание показанного на диаграмме антитела с LY6G6D, что измерено с помощью анализа BIAcore.
Фиг. 36 представляет набор графиков, показывающий связывание 20A12.QNTV12, 6E10.v114, и 1G4 с полипептидами LY6G6D человека и яванского макака, и таблицу, подытоживающую KД для каждого анализа, что измерено с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С.
Фиг. 37 представляет график, показывающий результаты анализа получения временной трансфекции для LY6G6D-специфичных плеч 20A12.QNTV.1 и 20A12.QNTv12. CD3-специфичное плечо 38E4v1 и FGFR1-специфичное плечо представлены как контроли.
Фиг. 38 представляет таблицу, показывающую результаты анализа ИФА бакуловируса (BV) для неспецифичного клиренса для LY6G6D-специфичного плеча 20A12.QNTv12 (20A12.ver1.polished).
Фиг. 39 представляет таблицу, показывающую результаты анализов оценки молекул (ОМ) TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4v1 или 40G5c. Зеленый цвет показывает анализы, для которых не было определено очевидных проблем.
Фиг. 40А представляет выравнивание последовательностей, показывающее аминокислотные последовательности гуманизированных вариантов 20А12, содержащих CDR rb.20A12 и каркасные области VH последовательности зародышевой линии человека hIGHV.3-23 или hIGHV.3-30 и каркасные области VL последовательности зародышевой линии человека hIGHV.1-5, hIGKV.1-39 или hIGKV.4-1, причем каждая имеет остатки Верньера кролика. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 40Б представляет выравнивание последовательностей, показывающее аминокислотные последовательности rb.20A12 и гуманизированных вариантов 20A12.QNTV.1 и 20A12.QNTV12. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 40В представляет выравнивание последовательностей, показывающее аминокислотные последовательности VH последовательности зародышевой линии человека hIGHV.3-23, VL последовательности зародышевой линии человека hIGHV.1-5 и VH и VL гуманизированных вариантов 20А12 20A12.QNTV.1 и 20A12.QNTV12. Остатки Верньера кролика, присутствующие в 20A12.QNTV.1 и 20A12.QNTv12, показаны овалами. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 41 представляет таблицу, показывающую результаты анализов оценки молекул (ОМ) Fab 6E10v1. Зеленый цвет показывает анализы, для которых не было определено очевидных проблем; красный цвет показывает, что проблемы были обнаружены.
Фиг. 42 представляет набор графиков, показывающих связывание вариантов LY6G6D-специфичного плеча 6Е10, содержащего CDR 6Е10 кролика и каркасные области VH последовательности зародышевой линии человека hIGHV.3-53, hIGHV.4-4 или hIGHV.3-48 и каркасные области VL последовательности зародышевой линии человека hIGHV.1-5, hIGKV.3-20 или hIGKV.4-1, что измерено с помощью анаилза BIAcore. Количество положений зон Верньера в каждом варианте показано.
Фиг. 43А представляет выравнивание последовательностей, показывающее аминокислотные последовательности гуманизированных вариантов 6Е10, содержащих CDR rb.6Е10 и каркасные области VH последовательности зародышевой линии человека hIGHV.3-53, hIGHV.4-4 или hIGHV.3-48 и каркасные области VL последовательности зародышевой линии человека hIGHV.1-5, hIGKV.3-20 или hIGKV.4-1. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 43Б представляет выравнивание последовательностей, показывающее аминокислотные последовательности rb.6Е10 и гуманизированных вариантов 6E10.v23 и 6E10.v114. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 43В представляет выравнивание последовательностей, показывающее аминокислотные последовательности VH последовательности зародышевой линии человека hIGHV.3-53*01, VL последовательности зародышевой линии человека hIGHV.3-20*01 и VH и VL rb.6Е10 и гуманизированного варианта 6Е10 6E10.v114. Остатки Верньера кролика, присутствующие в rb.6Е10 и 6E10.v114, показаны овалами. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
Фиг. 43Г представляет выравнивание последовательностей, показывающее аминокислотные последовательности VH последовательности зародышевой линии человека hIGHV.3-48*01, VL последовательности зародышевой линии человека hIGHV. 1-5*01 и VH и VL rb.6Е10 и гуманизированного варианта 6Е10 6E10.v23. Остатки Верньера кролика, присутствующие в rb.6Е10 и 6E10.v23, показаны овалами. Определяющие комплементарность области (CDR) CDR L1, CDR L2 и CDR L3 указаны. Остатки, которые отличаются между последовательностями, выделены.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
I. ОПРЕДЕЛЕНИЯ
В контексте данного документа термин «приблизительно» относится к обычному диапазону погрешностей для соответствующего значения, хорошо известному специалисту в данной области техники. В настоящем документе ссылка на «приблизительно» по отношению к величине или параметру включает (и описывает) аспекты, которые относятся непосредственно к этой величине или параметру.
Следует понимать, что аспекты настоящего изобретения, описанные в настоящем документе, включают «содержащие», «состоящие» и «преимущественно состоящие из» аспектов.
В контексте настоящего документа термин «Ly6G6D» или «комплекс лимфоцитарного антигена 6, локус G61» относится к любому нативному Ly6G6D из любого источника - позвоночного животного, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное, и охватывает «полноразмерное» непроцессированное Ly6G6D, а также любую форму Ly6G6D, которая получается в ходе процессинга в клетке. Данный термин также включает варианты Ly6G6D природного происхождения, включая, например, сплайсинговые варианты или аллельные варианты. Ly6G6D также называется G6D, Ly6-D, C6orf23, транскрипт гена, усиленный мегакариоцитами 1 (MEGT1), и NG25 и раскрыт в патенте США №7951546, который включен ссылкой в настоящий документ во всей своей полноте, как ТАТ201, с аминокислотной последовательностью SEQ ID NO: 75 и нуклеотидной последовательностью, DNA234441, SEQ ID NO: 76. Ly6G6D включает, например, белок Ly6G6D человека (NCBI RefSeq №NP_067079.2), который имеет длину 133 аминокислот.
Термины «антитело к LY6G6D» и «антитело, которое связывается с LY6G6D» относятся к антителу, которое способно связывать LY6G6D с достаточной аффинностью, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента для нацеливания на LY6G6D. В одном варианте осуществления степень связывания антитела к LY6G6D с неродственным, не являющимся LY6G6D белком, составляет менее чем приблизительно 10% от связывания антитела с LY6G6D, что измерено, например, радиоиммуноанализом (РИА). В некоторых вариантах осуществления антитело, которое связывается с LY6G6D, имеет константу диссоциации (KД) ≤1 мкМ, ≤250 нМ, ≤100 нМ, ≤15 нМ, ≤10 нМ, ≤6 нМ, ≤4 нМ, ≤2 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ, или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В определенных вариантах осуществления антитело к LY6G6D связывается с эпитопом LY6G6D, который является консервативным среди LY6G6D от разных видов.
Употребляемый в данном документе термин «кластер дифференцировки 3» или «CD3» относится, если не указано иное, к любому нативному CD3 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), включая, например, цепи CD3ε, CD3γ, CD3α и CD3β. Данный термин включает «полноразмерный» непроцессированный CD3 (например, непроцессированный или немодифицированный CD3ε или CD3γ), а также любую форму CD3, которая образуется в результате процессинга в клетке. Данный термин также включает варианты CD3 природного происхождения, включая, например, сплайсинговые варианты или аллельные варианты. CD3 включает, например, человеческий белок CD3ε (NCBI RefSeq № NP_000724), длина которого составляет 207 аминокислот, и человеческий белок CD3γ (NCBI RefSeq № NP_000064), длина которого составляет 182 аминокислоты.
Термины «антитело к CD3» и «антитело, которое связывается с CD3» относятся к антителу, которое способно связывать CD3 с достаточной аффинностью, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента для нацеливания на CD3. В одном варианте осуществления степень связывания антитела к CD3 с неродственным, не являющимся CD3 белком, составляет менее чем приблизительно 10% от связывания антитела с CD3, что измерено, например, радиоиммуноанализом (РИА). В некоторых вариантах осуществления антитело, которое связывается с CD3, имеет константу диссоциации (KД) ≤1 мкМ, ≤250 нМ, ≤100 нМ, ≤15 нМ, ≤10 нМ, ≤5 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В определенных вариантах осуществления антитело к CD3 связывается с эпитопом CD3, который является консервативным среди CD3 от разных видов.
Термин «антитело» в данном документе используется в самом широком смысле и охватывает различные структуры антител, включая, но не ограничиваясь ими, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител (например, бис-Fab), при условии, что они проявляют требуемую антигенсвязывающую активность.
Термин «аффинность» относится к силе суммарных нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнера по связыванию (например, антигена). Если не указано иное, в контексте данного документа «аффинность связывания» относится к характерной аффинности связывания, которая отражает 1:1 взаимодействие между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y обычно может быть представлена константой диссоциации (KД). Аффинность можно измерять с помощью общепринятых способов, известных в данной области техники, включая описанные в данном документе. Конкретные иллюстративные и типичные аспекты для определения аффинности связывания описаны ниже.
Антитело с «созревшей аффинностью» относится к антителу с одним или более изменениями в одной или более гипервариабельных областях (HVR) по сравнению с родительским антителом, которое не содержит такие изменения, при этом такие изменения приводят к улучшению аффинности антитела в отношении антигена.
В настоящем документе термины «полноразмерное антитело», «интактное антитело» и «целое антитело» используются взаимозаменяемо для обозначения антитела, имеющего структуру, по существу сходную со структурой нативного антитела, или имеющего тяжелые цепи, которые содержат Fc-область, определенную в настоящем документе.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются этим, бис-Fab; Fv; Fab; Fab', Fab'-SH; F(ab')2; диатела; линейные антитела; молекулы одноцепочечных антител (например, scFv, ScFab); и мультиспецифические антитела, образованные из фрагментов антител.
«Однодоменное антитело» относится к фрагменту антитела, содержащему весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных аспектах однодоменное антитело представляет собой человеческое однодоменное антитело (см., например, патент США №6248516 В1). Примеры однодоменных антител включают, помимо прочего, VHH.
«Fab»-фрагмент представляет собой антигенсвязывающий фрагмент, образованный папаиновой ферментацией антител и состоит из целой L-цепи вместе с доменом вариабельной области Н-цепи (VH), и первым константным доменом одной тяжелой цепи (СН1). Папаиновая ферментация антител дает два идентичных Fab-фрагмента. Обработка антитела пепсином приводит к образованию одного крупного фрагмента F(ab')2, который приблизительно соответствует двум связанным дисульфидом Fab-фрагментам, имеющим двухвалентную антигенсвязывающую активность, и, кроме того, способен к перекрестному связыванию антигена. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на карбоксильном конце СН1-домена, содержащих один или более остатков цистеина из шарнирной области антитела. Fab'-SH в контексте данного документа обозначает Fab', в котором остаток(-ки) цистеина константных доменов несет(-ут) свободную тиольную группу. F(ab')2-фрагменты антител первоначально были получены в виде пар Fab'-фрагментов, между которыми находятся шарнирные остатки цистеина. Кроме того, известны другие варианты химического сопряжения фрагментов антител.
«Fv» состоит из димера, состоящего из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочной нековалентной связью. Сворачивание этих двух доменов образует шесть гипервариабельных петель (3 петли в каждой из Н- и L-цепи), которые предоставляют аминокислотные остатки для связывания антигена и обеспечивают специфичность связывания антигена антителом. Однако, даже один вариабельный домен (или половина Fv, содержащая только три HVR, специфичных по отношению к антигену) обладает способностью распознавать и связывать антиген, хотя часто и с более низкой аффинностью, чем целый сайт связывания.
В настоящем документе термин «Fc-область» используется для определения С-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативной последовательностью и варианты Fc-областей. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьироваться, Fc-область тяжелой цепи IgG человека обычно определяют как отрезок от аминокислотного остатка в положении Cys226, или от Pro230, до его карбоксильного конца. С-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области может быть удален, например, в процессе получения или очистки антитела, или при рекомбинантном конструировании нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, композиция интактных антител может включать совокупности антител, в которых удалены все остатки Lys447, совокупности антител, в которых не удалены остатки Lys447, и совокупности антител, содержащие смесь антител, содержащих и не содержащих остаток Lys447.
«Функциональная Fc-область» обладает «эффекторной функцией» Fc-области с нативной последовательностью. Примеры «эффекторных функций» включают связывание C1q; КЗЦ; связывание Fc-рецептора; АЗКЦ; фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, рецептора В-клеток; BCR) и т.д. Для таких эффекторных функций в общем случае необходимо, чтобы Fc-область была объединена со связывающим доменом (например, вариабельным доменом антитела), а оценить их можно, используя различные анализы, описанные, например, в определениях в данном документе.
«Fc-область с нативной последовательностью» включает аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, встречающейся в природе. Fc-области человека с нативной последовательностью включают, без ограничения, Fc-область IgG1 человека с нативной последовательностью (аллотипы не-А и А); Fc-область IgG2 человека с нативной последовательностью; Fc-область IgG3 человека с нативной последовательностью и Fc-область IgG4 человека с нативной последовательностью, а также их встречающиеся в природе варианты.
«Вариант Fc-области» включает аминокислотную последовательность, которая отличается от Fc-области с нативной последовательностью тем, что содержит по меньшей мере одну модификацию аминокислоты, предпочтительно одну или более аминокислотную замену (замен). Предпочтительно вариант Fc-области включает по меньшей мере одну аминокислотную замену по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например, от приблизительно одной до приблизительно десяти аминокислотных замен, и предпочтительно от приблизительно одной до приблизительно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. В данном документе вариант Fc-области предпочтительно обладает по меньшей мере приблизительно 80% гомологией с Fc-областью с нативной последовательностью и/или Fc-областью исходного полипептида, и предпочтительно он обладает по меньшей мере приблизительно 90% гомологией с ними, или предпочтительно он обладает по меньшей мере приблизительно 95% гомологией с ними.
В контексте настоящего документа «Fc-комплекс» относится к доменам СН3 двух Fc-областей, взаимодействующих вместе с образованием димера, или, как в некоторых аспектах, две Fc-области взаимодействуют с образованием димера, причем цистеиновые остатки в шарнирных областях и/или домены СН3 взаимодействуют посредством связей и/или сил {например, сил ван-дер-ваальса, гидрофобных сил, водородных связей, электростатических сил или дисульфидных связей).
В контексте настоящего документа «Fc-компонент» относится к шарнирной области, домену СН2 или домену СН3 Fc-области.
«Шарнирная область» обычно определяется как растяжение от приблизительно остатка 216-230 IgG (нумерация согласно EU), от приблизительно остатка 226-243 IgG (нумерация по Кабату) или от приблизительно остатка 1-15 IgG (уникальная нумерация IMGT).
«Нижняя шарнирная область» Fc-области обычно определяется как растяжение от остатков, непосредственно С-концевыми относительно шарнирной области, т.е. остатков 233-239 Fc-области (нумерация согласно EU).
«Вариант Fc-области» включает аминокислотную последовательность, которая отличается от Fc-области с нативной последовательностью тем, что содержит по меньшей мере одну модификацию аминокислоты, предпочтительно одну или более аминокислотную замену (замен). Предпочтительно вариант Fc-области включает по меньшей мере одну аминокислотную замену по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например, от приблизительно одной до приблизительно десяти аминокислотных замен, и предпочтительно от приблизительно одной до приблизительно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. В данном документе вариант Fc-области предпочтительно обладает по меньшей мере приблизительно 80% гомологией с Fc-областью с нативной последовательностью и/или Fc-областью исходного полипептида, и предпочтительно он обладает по меньшей мере приблизительно 90% гомологией с ними, более предпочтительно он обладает по меньшей мере приблизительно 95% гомологией с ними.
Термины «Fc-рецептор» или «FcR» обозначают рецептор, который связывается с Fc-областью антитела. Предпочтительным FcR является FcR человека с нативной последовательностью. Более того, предпочтительным FcR является FcR, который связывает антитело IgG (гамма-рецептор) и включает в себя рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно-сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые обладают сходными аминокислотными последовательностями, отличающимися, главным образом, своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA в своем цитоплазматическом домене содержит иммунорецепторный тирозиновый активирующий мотив (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене иммунорецепторный тирозиновый ингибирующий мотив (ITIM) (см. обзор М. в  Annu. Rev. Immunol. 15:203-234 (1997)). FcR описаны в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-492 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). К термину «FcR» в данном документе относятся другие FcR, в том числе FcR, которые будут выявлены в будущем. Термин также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994)).
Annu. Rev. Immunol. 15:203-234 (1997)). FcR описаны в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-492 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). К термину «FcR» в данном документе относятся другие FcR, в том числе FcR, которые будут выявлены в будущем. Термин также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994)).
Термин «выступ-во-впадину» или технология «KnH», используемые в настоящем документе, относятся к технологии, направляющей спаривание двух полипептидов вместе in vitro или in vivo путем введения выступа (выпячивания) в один полипептид и полости (впадины) в другой полипептид на поверхности взаимодействия, с помощью которой они взаимодействуют. Например, KnH были встроены в поверхности взаимодействия Fc:Fc, поверхности взаимодействия CL:CH1 или поверхности взаимодействия VH/VL антител (см., например, US 2007/0178552, WO 96/027011, WO 98/050431 и Zhu et al. (1997) Protein Science 6:781-788). Это особенно пригодно в возбуждении спаривания двух различных тяжелых цепей вместе при изготовлении мультиспецифических антител. Например, мультиспецифические антитела, имеющие KnH в своих Fc-областях, могут дополнительно содержать единичные вариабельные домены, связанные с каждой Fc-областью, или дополнительно содержать различные вариабельные домены тяжелой цепи, спаренные с идентичными, одинаковыми или разными вариабельными доменами легкой цепи. Технология KnH может также использоваться для спаривания вместе двух разных внеклеточных доменов рецептора или любых других полипептидных последовательностей, которые содержат различные целевые последовательности распознавания.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно находятся в VH (или VL) в следующем порядке: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
«Область СН1» или «домен СН1» содержит отрезок из остатков от приблизительно остатка 118 до остатка 215 IgG (нумерация согласно EU), от приблизительно остатка 114 до 223 IgG (нумерация по Кабату) или от приблизительно остатка 1,4 до остатка 121 IgG (уникальная нумерация IMGT) (Lefranc М-Р, Giudicelli V, Duroux Р, Jabado-Michaloud J, Folch G, Aouinti S, Carillon E, Duvergey H, Houles A, Paysan-Lafosse T, Hadi-Saljoqi S, Sasorith S, Lefranc G, Kossida S. IMGT®, the international ImMunoGeneTics information system® 25 years on. Nucleic Acids Res. 2015 Jan; 43(Database issue):D413-22). Полипептид ИЛ-15 или полипептид ИЛ-15Rα могут быть ковалентно связаны непосредственно с первым остатком домена СН1 или, альтернативно, могут быть ковалентно связаны с остатком в положении С-конца с первым остатком СН1. В альтернативных аспектах полипептид ИЛ-15 или полипептид ИЛ-15Rα могут быть ковалентно связаны с СН1 посредством линкера, как определено в настоящем документе.
«Домен СН2» Fc-области IgG человека обычно проходит от приблизительно остатков 244 до приблизительно 360 IgG (нумерация по Кабату), от приблизительно остатков 231 до остатков 340 IgG (нумерация согласно EU) или от приблизительно остатков 1,6 до приблизительно 125 IgG (уникальная нумерация IGMT). Домен СН2 является уникальным в том, что он не характеризуется близким спариванием с другим доменом. Точнее, две N-связанные разветвленные углеводные цепи расположены между двумя доменами СН2 интактной нативной молекулы IgG. Было предположено, что этот углевод может заменять домен-доменное спаривание и помогать стабилизировать домен СН2. Burton, Molec. Immunol.22: 161-206 (1985).
«Домен СН3» содержит отрезок из остатков С-конца до домена СН2 в Fc-области (т.е. от приблизительно аминокислотного остатка 361 до приблизительно аминокислотного остатка 478 IgG (нумерация по Кабату), от приблизительно аминокислотного остатка 341 до приблизительно аминокислотного остатка 447 IgG (нумерация согласно EU) или от приблизительно аминокислотного остатка 1,4 до приблизительно аминокислотного остатка 130 IgG (уникальная нумерация IGMT)).
«Домен CL» или «константный домен легкой цепи» содержит отрезок из остатков С-конца до вариабельного домена легкой цепи (VL). Легкая цепь антитела может быть каппа (κ) («Сκ») или лямбда (λ) («Сλ») областью легкой цепи. Область Сκ обычно проходит от приблизительно остатка 108 до остатка 214 IgG (нумерация по Кабату или согласно EU) или от приблизительно остатка 1,4 до остатка 126 IgG (уникальная нумерация IMGT). Остаток Сλ, обычно проходит от приблизительно остатка 107а до остатка 215 (нумерация по Кабату) или от приблизительно остатка 1,5 до остатка 127 (уникальная нумерация IMGT) (Lefranc М-Р, Giudicelli V, Duroux P, Jabado-Michaloud J, Folch G, Aouinti S, Carillon E, Duvergey H, Houles A, Paysan-Lafosse T, Hadi-Saljoqi S, Sasorith S, Lefranc G, Kossida S. IMGT®, the international ImMunoGeneTics information system® 25 years on. Nucleic Acids Res. 2015 Jan; 43(Database issue):D413-22).
Легкая цепь (LC) любого вида позвоночных может быть отнесена к одному из двух четко различающихся типов, называемых каппа и лямбда, на основе аминокислотных последовательностей их константных доменов. В зависимости от аминокислотных последовательностей константного домена их тяжелых цепей (СН), иммуноглобулины можно отнести к различных классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющих тяжелые цепи, обозначаемые α, β, γ, ε и μ соответственно. Классы γ и α далее подразделяются на подклассы на основе относительно небольших различий в последовательности СН и функции, например, у человека экспрессируются следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
В контексте настоящего документа термин «заряженная область» относится к местоположению полипептида (например, антитела), который включает одну или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) основных или кислых аминокислот, которые способны образовывать зарядовую пару с родственной заряженной областью, имеющей одну или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) или основных, или кислых аминокислот, если заряженная область и ее родственная заряженная область имеют противоположный общий относительный заряд.
Используемый в данном документе термин «зарядовая пара» относится к связи, которая образуется между двумя заряженными областями противоположного общего заряда.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из определенного источника или вида, тогда как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида.
«Класс» антитела относится к типу константного домена или константной области, которые содержит его тяжелая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, называются α, β, ε, γ и μ, соответственно.
«Антитело человека» представляет собой антитело, которое имеет аминокислотную последовательность, соответствующую последовательности антитела, вырабатываемого человеком или клеткой человека, или полученного из источника, отличного от человека, в котором используются репертуары антител человека или другие последовательности, кодирующие антитела человека. Это определение антитела человека явным образом исключает гуманизированное антитело, содержащее отличные от человеческих антигенсвязывающие остатки. Антитела человека можно получать, используя различные способы, известные в данной области техники, включая библиотеки фагового дисплея. Hoogenboom and Winter. J. Mol. Biol. 227:381, 1991; Marks et al. J. Mol. Biol. 222:581, 1991. Также для получения моноклональных антител человека доступны способы, описанные в Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol., 147(1):86-95, 1991. См. также van Dijk and van de Winkel. Curr. Opin. Pharmacol. 5:368-74, 2001. Антитела человека можно получить путем введения антигена в организм трансгенного животного, модифицированного с целью выработки таких антител в ответ на антигенную стимуляцию, причем эндогенные локусы указанного животного отключены, например, как в случае иммунизированных ксеномышей (в отношении технологии XENOMOUSE™ смотрите, например, патенты США №№6075181 и 6150584). См. также, например, Li et al. Proc. Natl. Acad. Sci. USA. 103:3557-3562, 2006, об антителах человека, полученных с помощью гибридомной технологии с В-клетками человека.
«Консенсусная последовательность каркасной области человека» представляет собой каркасную область, которая представляет собой наиболее часто встречающиеся аминокислотные остатки при выборе каркасных последовательностей VL или VH иммуноглобулина человека. Обычно выбор последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельных доменов. В общем случае подгруппа последовательностей представляет собой подгруппу согласно Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), тома 1-3. В одном аспекте в случае VL подгруппа представляет собой подгруппу каппа I согласно Кабату и соавт., выше. В одном аспекте для VH подгруппа представляет собой подгруппу III согласно Кабату и соавт., выше.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из отличных от человеческих HVR и аминокислотные остатки из человеческих FR. В определенных аспектах гуманизированное антитело содержит практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют таковым из отличного от человеческого антитела, а все или практически все FR соответствуют таковым из человеческого антитела. В некоторых аспектах, в которых все или по существу все из FR гуманизированного антитела соответствуют таковым антитела человека, любые из FR гуманизированного антитела могут содержать один или более аминокислотных остатков (например, один или более остатков в положениях зон Верньера FR) от отличной от человеческой FR. Гуманизированное антитело, необязательно, может содержать по меньшей мере часть константной области антитела, полученную из человеческого антитела. «Гуманизированная форма» антитела, например, отличного от человеческого антитела, относится к антителу, подвергшемуся гуманизации.
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют сходные структуры, причем каждый домен содержит четыре консервативные каркасные области (FR) и три гипервариабельные области (HVR). (Смотрите, например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., стр. 91 (2007).) Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, можно выделять, используя домен VH или VL из антитела, которое связывает антиген, для скрининга библиотеки комплементарных доменов VL или VH, соответственно. Смотрите, например, Portolano et al., J. Immunol. 150:880-887, 1993; Clarkson et al., Nature 352:624-628, 1991.
В контексте данного документа термин «гипервариабельная область» или «HVR» относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности («определяющие комплементарность области» или «CDR»). В общем случае антитела содержат шесть CDR: три в VH (CDR-H1, CDR-H2, CDR-H3) и три в VL (CDR-L1, CDR-L2, CDR-L3). Типовые CDR по данному документу включают:
(а) CDR, находящиеся в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917, 1987);
(б) CDR, находящиеся в аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)); и
(в) антигенные контакты, находящиеся в аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (НЗ) (MacCallum et al. J. Mol. Biol. 262: 732-745, 1996).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) нумеруются в данном документе согласно Кабату и соавт., выше.
«Одноцепочечные Fv», также сокращаемые как «sFv» или «scFv», представляют собой фрагменты антител, которые содержат VH и VL-домены антитела, соединенные в единую полипептидную цепь. Предпочтительно полипептид scFv дополнительно содержит полипептидный линкер между VH- и VL-доменами, который позволяет scFv образовывать необходимую структуру для связывания антигена. Для обзора фрагментов scFv, см., Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); Malmborg et al., J. Immunol. Methods 183:7-13, 1995.
Под «нацеливающим доменом» подразумевается часть соединения или молекулы, которая специфически связывается с целевым эпитопом, антигеном, лигандом или рецептором. Нацеливающие домены включают, но не ограничиваются этим, антитела (например, моноклональные, поликлональные, рекомбинантные, гуманизированные и химерные антитела), фрагменты антител или их части (например, фрагменты бис-Fab, фрагменты Fab, F(ab')2, scFab, scFv-антитела, SMIP (иммунофармацевтическое средство на основе модульного белка малого размера), антитела с одним доменом, диатела, мини-антитела, scFv-Fc, аффитела, нанотела и VH- и/или VL- домены антител), рецепторы, лиганды, аптамеры, нацеленные на пептиды домены (например, белки цистеинового узла (БЦУ)) и другие молекулы, имеющие определенного партнера по связыванию. Нацеливающий домен может нацеливать, блокировать, агонизировать или антагонизировать антиген, с которым он связывается.
В контексте настоящего документа термин «моноклональное антитело» относится к антителу, полученному из популяции практически гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих мутации природного происхождения или возникающие во время получения препарата моноклональных антител, при этом такие варианты в целом присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклональных антител направлено против одной детерминанты на антигене. Таким образом, определение «моноклональный» указывает на характеристику антитела, как полученного из, по существу, гомогенной популяции антител, и его не следует интерпретировать как требование получения антитела каким-либо конкретным способом. Например, моноклональные антитела для применения в соответствии с настоящим изобретением могут быть получены с помощью различных способов, включая, но не ограничиваясь этим, способ гибридомы, способы рекомбинантной ДНК, способы фагового дисплея и способы с использованием трансгенных животных, содержащих все или часть локусов человеческого иммуноглобулина, причем такие способы и другие типичные способы получения моноклональных антител описаны в данном документе.
Термин «мультиспецифическое антитело» используется в самом широком смысле и, в частности, охватывает антитело, которое имеет полиэпитопную специфичность. В одном аспекте мультиспецифическое антитело связывается с двумя различными мишенями (например, биспецифическое антитело). Такие мультиспецифические антитела включают, помимо прочего, антитело, содержащее вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), где звено VH/VL имеет полиэпитопную специфичность, причем антитела с двумя или более доменами VL и VH, причем каждое звено VH/VL связывается с различным эпитопом, антитела с двумя или более отдельными вариабельными доменами, причем каждый отдельный вариабельный домен связывается с разным эпитопом, полноразмерные антитела, фрагменты антител, такие как Fab, Fv, dsFv, scFv, диатела, биспецифические диатела и триатела, фрагменты антител, которые были ковалентно или нековалентно связаны. «Полиэпитопная специфичность» относится к способности специфически связываться с двумя или более различными эпитопами на одной и той же или различных мишенях. «Моноспецифический» относится к способности связывать только один антиген. В одном аспекте моноспецифическое биэпитопное антитело связывает два различных эпитопа на одной и той же мишени/антигене. В одном аспекте моноспецифическое полиэпитопное антитело связывается с множеством разных эпитопов той же мишени/антигена. Согласно одному аспекту мультиспецифическое антитело представляет собой антитело IgG, которое связывается с каждым эпитопом с аффинностью от 5 мкМ до 0,001 пМ, от 3 мкМ до 0,001 пМ, от 1 мкМ до 0,001 пМ, от 0,5 мкМ до 0,001 пМ или от 0,1 мкМ до 0,001 пМ.
«Оголенное антитело» означает антитело, не конъюгированное с гетерологическим компонентом (например, цитотоксическим компонентом) или радиоактивной меткой. Неконъюгированное антитело может присутствовать в фармацевтическом составе.
«Нативные антитела» относятся к молекулам иммуноглобулинов природного происхождения с различной структурой. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины массой около 150000 Дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидными связями. В направлении от N-конца к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следуют три константных домена (CH1, СН2 и СН3). Аналогичным образом, в направлении от N-конца к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный легкий домен (CL). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
В контексте настоящего документа термин «иммуноадгезин» обозначает молекулы, которые объединяют специфичность связывания гетерологичного белка («адгезина») с эффекторными функциями константных доменов иммуноглобулина. Структурно иммуноадгезины содержат слияние аминокислотной последовательности с желаемой специфичностью связывания, причем эта аминокислотная последовательность отличается от сайта распозначания и связывания антигена антитела (т.е. является «гетерологичной» по сравнению с константной областью антитела) и последовательности константного домена иммуноглобулина (например, последовательности СН2 и/или СН3 IgG). Константные домены адгезина и иммуноглобулина могут быть необязательно разделены аминокислотным спейсером. Типичные последовательности адгезина включают родственные аминокислотные последовательности, которые содержат часть рецептора или лиганда, который связывается с белком, представляющим интерес. Последовательности адгезина могут также быть последовательностями, которые связывают белок, представляющий интерес, а не последовательности рецептора или лиганда (например, последовательности адгезина в пептидных телах). Такие полипептидные последовательности могут быть выбраны или определены различными способами, включают способы фагового дисплея и высокопроизводительные способы сортировки. Последовательность константного домена иммуноглобулина в иммуноадгезине может быть получена из любого иммуноглобулина, такого как подтипы IgG1, IgG2, IgG3 или IgG4, IgA (включая IgA1 и IgA2), IgE, IgD или IgM.
«Химиотерапевтический агент» включает химические соединения, применимые при лечении рака. Примеры химиотерапевтических агентов включают эрлотиниб (Тарцева®, Genentech/OSI Pharm.), бортезомиб (Велкад®, Millennium Pharm.), дисульфирам, галлат эпигаллокатехина, салиноспорамид А, карфилзомиб, 17-AAG (гелданамицин), радицикол, лактатдегидрогеназу А (LDH-A), фулвестрант (Фазлодекс®, AstraZeneca), сунитиб (Сутент®, Pfizer/Sugen), летрозол (Фемара®, Novartis), мезилат иматиниба (Гливек®, Novartis), финасунат (Ваталаниб®, Novartis), оксалиплатин (Элоксатин®, Sanofi), 5-ФУ (5-фторурацил), лейковорин, рапамицин (сиролимус, Парамун®, Wyeth), лапатиниб (Тикерб®, GSK572016, Glaxo Smith Kline), лонафамиб (SCH 66336), сорафениб (Нексавар®, Bayer Labs), гефитиниб (Пресса®, AstraZeneca), AG1478, алкилирующие средства, такие как тиотепа и циклофосфамид Цитоксан®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метилметамины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметиломеламин; ацетогенины (в частности, буллатацин и буллатацинон); камптотецин (включая топотекан и иринотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); адренокортикостероиды (включая преднизон и преднизолон); ацетат ципротерона; 5α-редуктазы, включая финастерид и дутастерид); вориностат, ромидепсин, панобиностат, вальпроевую кислоту, моцетиностат доластатин; альдеслейкин, тальк дуокармицин (включая синтетические аналоги, KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктин; спонгистатин; азотистый аналог иприта, такой как хлорамбуцил, хломафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урацилиприт; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности, калихеамицин γ1I и калихеамицин ω1I (Angew Chem. Intl. Ed. Engl. 1994 33:183-186); динемицин, включая динемицин А; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарзиностатиновый хромофор и родственные хромопротеиновые хромофоры на основе ендииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, каминомицин, карзинофилин, хромомицин, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, Адриамицин® (доксорубицин), морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, порфиромицин, пуромицин, келамицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие функции надпочечников, такие как аминоглютетимид, митотан, трилостан; пополнитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновая кислота; энилурацил; амсакрин; бесстрабуцил; бисантрен; эдатраксат; дефамин; демеколцин; диазиквон; эльфомитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтансиноиды, такие как майтансин и ансамитоцины; митогуазон; митоксантрон; мопидамнол; нитраэрин; пентостатин; фенамет; пирарубицин; лосоксантрон; подофиллиновая кислота; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Юджин, Орегон); разоксан; ризоксин; сизофуран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (в частности, Т-2 токсин, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ара-С»); циклофосфамид; тиотепа; таксоиды, например, таксол (паклитаксел; Bristol-Myers Squibb Oncology, Принстон, Нью-Джерси), Абраксан® (без кремофора), подвергнутые инжинирингу с альбумином составы наночастиц паклитаксела (American Pharmaceutical Partners, Шаумбург, Иллинойс) и Таксотере® (доцетаксел, доксетаксел; Sanofi-Aventis); хлорамбуцил; Гемзар® (гемцитабин); 6-тиогуанин; меркаптопурин; метотрексат; платиновые аналоги, такие как цисплатин и карбоплатин; винбластин; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; Навелбин® (винорелбин); новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; капецитабин (Кселода®); ибандронат; СРТ-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (ДМФО); ретиноиды, такие как ретиноевая кислота; и фармацевтически приемлемые соли, кислоты и производные любых из вышеперечисленного.
Химиотерапевтический агент также включает (i) антигормональные агенты, действие которых заключается в регуляции или ингибировании гормонального влияния на опухоли, такие как антиэстрогены и селективные модуляторы рецепторов эстрогена (SERM), включая, например, тамоксифен (включая Нолвадекс®; тамоксифена цитрат), ралоксифен, дролоксифен, йодоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и Фарестон® (торемифина цитрат); (ii) ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует выработку эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглютетимид, Мегаза® (мегестрола ацетат), Аромазин® (экземестан; Pfizer), форместан, фадрозол, Ривизор® (ворозол), Фемара® (летрозол; Novartis), и Аримидекс® (анастрозол; AstraZeneca); (iii) антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; бусерелин, трипторелин, медроксипрогестерона ацетат, диэтилстильбестрол, премарин, флуоксиместерон, все препараты трансретиноевой кислоты, фенретинид, а также троксацитабин (1,3-диоксолановый аналог нуклеозида цитозина); (iv) ингибиторы протеинкиназы; (v) ингибиторы липид-киназы; (vi) антисмысловые олигонуклеотиды, в частности те, которые ингибируют экспрессию генов в сигнальных путях, участвующих в аберрантной клеточной пролиферации, такие как, например, РКС-альфа, Ralf и H-Ras; (vii) рибозимы, такие как ингибиторы экспрессии VEGF (например, Ангиозим®) и ингибиторы экспрессии HER2; (viii) вакцины, такие как вакцины для генной терапии, например, Алловектин®, Лейвектин® и Ваксид®; Пролейкин®, rIL-2; ингибитор топоизомеразы 1, такой как Луртотекан®; Абареликс® rmRH; и (ix) фармацевтически приемлемые соли, кислоты или производные любого из вышеприведенных агентов.
Химиотерапевтический агент также включает антитела, такие как алемтузумаб (Кампат), бевацизумаб (Авастин®, Genentech); цетуксимаб (Эрбитукс®, Imclone); панитумумаб (Вектибикс®, Amgen), ритуксимаб (Ритуксан®, Genentech/Biogen Idec), пертузумаб (Омнитарг®, 2С4, Genentech), трастузумаб (Герцептин®, Genentech), тозитумомаб (Бексар, Corixia) и конъюгат антитела с лекарственным препаратом, гемтузумаб озогамицин (Милотарг®, Wyeth). Дополнительные гуманизированные моноклональные антитела с терапевтическим потенциалом в качестве агентов в комбинации с соединениями по настоящему изобретению включают: аполизумаб, азелизумаб, атлизумаб, бапинейзумаб, биватузумаб мертанзин, кантузумаб мертанзин, цеделизумаб, цертолизумаб пегол, цидфузитузумаб, цидтузумаб, даклизумаб, экулизумаб, эфализумаб, эпратузумаб, эрлизумаб, фелвизумаб, фонтолизумаб, гемтузумаб озогамицин, инотузумаб озогамицин, ипилимумаб, лабетузумаб, линтузумаб, матузумаб, меполизумаб, мотавизумаб, мотовизумаб, натализумаб, нимотузумаб, ноловизумаб, нумавизумаб, окрелизумаб, омализумаб, паливизумаб, пасколизумаб, пекфузитузумаб, пектузумаб, пекселизумаб, раливизумаб, ранибизумаб, ресливизумаб, реслизумаб, резивизумаб, ровелизумаб, руплизумаб, сибротузумаб, сиплизумаб, сонтузумаб, такатузумаб тетраксетан, тадоцизумаб, тализумаб, тефибазумаб, тоцилизумаб, торализумаб, тукотузумаб целмолейкин, тукуситузумаб, умавизумаб, уртоксазумаб, устекинумаб, визилизумаб и антитело к интерлейкину-12 (ABT-874/J695, Wyeth Research and Abbott Laboratories), которое представляет собой рекомбинантное, имеющее исключительно человеческую последовательность, полноразмерное антитело IgG1 λ, генетически модифицированное для распознавания белка р40 интерлейкина-12.
Химиотерапевтический агент также включает «ингибиторы EGFR», которые относятся к соединениям, которые связываются или иным образом взаимодействуют с EGFR и предотвращают или снижают его сигнальную активность, и, в качестве альтернативного варианта, называются «антагонистами EGFR». Примеры таких агентов включают антитела и малые молекулы, которые связываются с EGFR. Примеры антител, которые связываются с EGFR, включают MAb 579 (АТСС CRL НВ 8506), MAb 455 (АТСС CRL НВ8507), MAb 225 (АТСС CRL 8508), MAb 528 (АТСС CRL 8509) (смотрите патент США №4943533, Mendelsohn et al.) и его варианты, такие как химеризованное антитело 225 (С225 или цетуксимаб; ERBUTIX®) и реконструированное человеческое антитело 225 (Н225) (смотрите WO 96/40210, Imclone Systems Inc.); IMC-11F8, полностью человеческое нацеленное на EGFR антитело (Imclone); антитела, которые связывают мутантный EGFR II типа (патент США №5212290); гуманизированные и химерные антитела, которые связывают EGFR, описанные в патенте США №5891996; и человеческие антитела, которые связывают EGFR, такие как ABX-EGF или панитумумаб (смотрите WO 98/50433, Abgenix/Amgen); EMD 55900 (Stragliotto et al. Eur. J. Cancer 32A:636-640 (1996)); EMD7200 (матузумаб), гуманизированное антитело, направленное против EGFR, которое конкурирует за связывание EGFR как с EGF, так и с TGF-альфа (EMD/Merck); человеческое антитело к EGFR, HuMax-EGFR (GenMab); полностью человеческие антитела, известные как E1.1, Е2.4, Е2.5, Е6.2, Е6.4, E2.11, Е6. 3 и Е7.6. 3 и описанные в US 6235883; MDX-447 (Medarex Inc); и mAb 806 или гуманизированное mAb 806 (Johns et al., J. Biol. Chem. 279(29):30375-30384 (2004)). Антитело к EGFR может быть конъюгировано с цитотоксическим агентом с получением, таким образом, иммуноконъюгата (смотрите, например, ЕР659, 439А2, Merck Patent GmbH). Антагонисты EGFR включают малые молекулы, такие как соединения, описанные в патентах США №№: 5616582, 5457105, 5475001, 5654307, 5679683, 6084095, 6265410, 6455534, 6521620, 6596726, 6713484, 5770599, 6140332, 5866572, 6399602, 6344459, 6602863, 6391874, 6344455, 5760041, 6002008 и 5747498, а также в следующих публикациях РСТ: WO 98/14451, WO 98/50038, WO 99/09016 и WO 99/24037. Конкретные низкомолекулярные антагонисты EGFR включают OSI-774 (СР-358774, эрлотиниб, Тарцева® Genentech/OSI Pharmaceuticals); PD 183805 (CI 1033, 2-пропенамид, N-[4-[(3-хлор-4-фторфенил)амино]-7-[3-(4-морфолинил)пропокси]-6-хиназолинил]-дигидрохлорид, Pfizer Inc.); (ZD1839, гефинитиб (ИРЕССА®) 4-(3'-хлор-4'-фторанилино)-7-метокси-6-(3-морфолинопропокси)хиназолин, AstraZeneca); ZM 105180 ((6-амино-4-(3-метилфениламино)-хиназолин, Zeneca), ВГВХ-1382 (N8-(3-хлор-4-фторфенил)-N2-(1-метилпиперидин-4-ил)-пиримидо[5,4-d]пиримидин-2,8-диамин, Boehringer Ingelheim), PKI-166 ((R)-4-[4-[(1-фенилэтил)амино]-1Н-пирроло[2,3-d]пиримидин-6-ил]фенол), (R)-6-(4-гидроксифенил)-4-[(1-фенилэтил)амино]-7Н-пирроло[2,3-d]пиримидин); CL-387785 (N-[4-[(3-бромфенил)амино]-6-хиназолинил]-2-бутинамид); ЕКВ-569 (N-[4-[(3-хлор-4-фторфенил)амино]-3-циано-7-этокси-6-хинолинил]-4-(диметиламино)-2-бутенамид) (Wyeth); AG 1478 (Pfizer); AG1571 (SU 5271; Pfizer); двойные ингибиторы тирозинкиназы EGFR/HER2, такие как лапатиниб (ТАЙКЕРБ®, GSK572016 или N-[3-хлор-4-[(3-фторфенил)метокси]фенил]-6[5[[[2-метилсульфонил)этил] амино]метил]-2-фуранил]-4-хиназолинамин).
Химиотерапевтические агенты также включают «ингибиторы тирозинкиназы», включая лекарственные препараты, нацеленные на EGFR, указанные в предыдущем параграфе; низкомолекулярный ингибитор тирозинкиназы HER2, такой как ТАK165, доступный от Takeda; CP-724,714, пероральный селективный ингибитор рецепторной тирозинкиназы ErbB2 (Pfizer и OSI); двойные ингибиторы HER, такие как EKB-569 (доступный от Wyeth), который предпочтительно связывает EGFR, но ингибирует как HER2-, так и EGFR-сверхэкспрессирующие клетки; лапатиниб (GSK572016, доступный от Glaxo-SmithKline), пероральный ингибитор тирозинкиназы HER2 и EGFR; PKI-166 (доступный от Novartis); ингибиторы pan-HER, такие как канертиниб (CI-1033; Pharmacia); ингибиторы Raf-1, такие как антисмысловой агент ISIS-5132, доступный от ISIS Pharmaceuticals, который ингибирует сигнализацию Raf-1; ингибиторы ТК, не нацеленные на HER, такие как иматиниба мезилат (Гливек®, доступный от Glaxo SmithKline); многонацеленные ингибиторы тирозинкиназы, такие как сунитиниб (Сутент®, доступный от Pfizer); ингибиторы рецепторной тирозинкиназы VEGF, такие как ваталаниб (PTK787/ZK222584, доступный от Novartis/Schering AG); внеклеточно регулируемый ингибитор киназы I MAPK CI-1040 (доступный от Pharmacia); хиназолины, такие как PD 153035,4-(3-хлоранилино) хиназолин; пиридопиримидины; пиримидопиримидины; пирролопиримидины, такие как CGP 59326, CGP 60261 и CGP 62706; пиразолопиримидины, 4-(фениламино)-7Н-пирроло [2,3-d] пиримидины; куркумин (диферулоилметан, 4,5-бис(4-фторанилино)фталимид); тирфостины, содержащие нитротиофеновые фрагменты; PD-0183805 (Warner-Lamber); антисмысловые молекулы (например, те, которые связываются с HER-кодирующей нуклеиновой кислотой); хиноксалины (патент США №5804396); трифостины (патент США №5804396); ZD6474 (Astra Zeneca); РТК-787 (Novartis/Schering AG); ингибиторы pan-HER, такие как CI-1033 (Pfizer); аффинитак (ISIS 3521; Isis/Lilly); иматиниба мезилат (Гливек®); PKI 166 (Novartis); GW2016 (Glaxo SmithKline); CI-1033 (Pfizer); EKB-569 (Wyeth); семаксиниб (Pfizer); ZD6474 (AstraZeneca); PTK-787 (Novartis/Schering AG); INC-1C11 (Imclone), рапамицин (сиролимус, Рапамун®); или описанные в любой из следующих патентных публикаций: патент США №5804396; WO 1999/09016 (American Cyanamid); WO 1998/43960 (American Cyanamid); WO 1997/38983 (Warner Lambert); WO 1999/06378 (Warner Lambert); WO 1999/06396 (Warner Lambert); WO 1996/30347 (Pfizer, Inc); WO 1996/33978 (Zeneca); WO 1996/3397 (Zeneca) и WO 1996/33980 (Zeneca).
Химиотерапевтические агенты также включают дексаметазон, интерфероны, колхицин, метоприн, циклоспорин, амфотерицин, метронидазол, алемтузумаб, алитретиноин, аллопуринол, амифостин, триоксид мышьяка, аспарагиназу, живую БЦЖ, бевацузимаб, бексаротен, кладрибин, клофарабин, дарбэпоэтин альфа, денилейкин, дексразоксан, эпоэтин альфа, элотиниб, филграстим, гистрелина ацетат, ибритумомаб, интерферон альфа-2а, интерферон альфа-2b, леналидомид, левамизол, месну, метоксален, нандролон, неларабин, нофетумомаб, опрелвекин, палифермин, памидронат, пегадемазу, пегаспаргазу, пегфилграстим, двунатриевый пеметрексед, пликамицин, порфимер натрия, квинакрин, расбуриказу, сарграмостим, темозоломид, VM-26, 6-TG, торемифен, третиноин, ATRA, валрубицин, золедронат и золедроновую кислоту и их фармацевтически приемлемые соли.
Химиотерапевтические агенты также включают гидрокортизон, гидрокортизона ацетат, кортизона ацетат, тиксокортола пивалат, триамцинолона ацетонид, триамцинолоновый спирт, мометазон, амцинонид, будесонид, десонид, флуоцинонид, флуоцинолона ацетонид, бетаметазон, бетам етазон фосфат натрия, дексаметазон, дексаметазон фосфат натрия, флуокортолон, гидрокортизон-17-бутират, гидрокортизон-17-валерат, аклометазона дипропионат, бетаметазона валерат, бетаметазона дипропионат, предникарбат, клобетазон-17-бутират, клобетазон-17-пропионат, флуокортолона капроат, флуокортолона пивалат и флупреднидена ацетат; иммунные селективные противовоспалительные пептиды (ImSAID), такие как фенилаланин-глутамин-глицин (FEG) и его D-изомерная форма (feG) (IMULAN BioTherapeutics, LLC); противоревматические лекарственные препараты, такие как азатиоприн, циклоспорин (циклоспорин А), D-пеницилламин, соли золота, гидроксихлорохин, лефлуномид, миноциклин, сульфасалазин, блокаторы фактора некроза опухоли альфа (TNFa), такие как этанерцепт (Энбрел), инфликсимаб (Ремикад), адалимумаб (Хумира), цертолизумаба пэгол (Симзия), голимумаб (Симпони), блокаторы интерлейкина 1 (IL-1), такие как анакинра (Кинерет), блокаторы костимуляции Т-клеток, такие как абатацепт (Оренсия), блокаторы интерлейкина 6 (IL-6), такие как тоцилизумаб (Актемера®); блокаторы интерлейкина 13 (IL-13), такие как лебрикизумаб; блокаторы интерферона альфа (IFN), такие как ронтализумаб; блокаторы интегрина бета 7, такие как rhuMAb Beta7; блокаторы пути IgE, такие как анти-М1 прайм; блокаторы секретируемого гомотримерного LTa3 и мембраносвязанного гетеротримера LTa1/β2, такие как анти-лимфотоксин альфа (LTa); радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); прочие исследуемые агенты, такие как тиоплатин, PS-341, фенилбутират, ЕТ-18- ОСН3, или ингибиторы фарнесилтрансферазы (L-739749, L-744832); полифенолы, такие как кверцетин, резвератрол, пикеатаннол, эпигаллокатехина галлат, теафлавины, флаванолы, процианидины, бетулиновую кислоту и ее производные; ингибиторы аутофагии, такие как хлорохин; дельта-9-тетрагидроканнабинол (дронабинол, Маринол®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; ацетилкамптотецин, скополектин и 9-аминокамптотецин; подофиллотоксин; тегафур (Уфторал®); бексаротен (Таргретин®); бисфосфонаты, такие как клодронат (например, Бонфос® или Остак®), этидронат (Дидрокал®), NE-58095, золедроновую кислоту/золедронат (Зомета®), алендронат (Фосамакс®), памидронат (Аредиа®), тилудронат (Скелид®) или ризедронат (Актонел®); и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина Тератоп®; перифосин, ингибитор СОХ-2 (например, целекоксиб или эторикоксиб), ингибитор протеосом (например, PS341); CCI-779; типифарниб (R11577); орафениб, АВТ510; ингибитор Вс1-2, такой как облимерсен натрия (Генасенс®); пиксантрон; ингибиторы фарнезилтрансферазы, такие как лонафарниб (SCH 6636, СаразарТМ); и фармацевтически приемлемые соли, кислоты или производные любого из вышеприведенных агентов; а также комбинации двух или более вышеприведенных агентов, такие как CHOP, аббревиатура для комбинированной терапии на основе циклофосфамида, доксорубицина, винкристина и преднизолона; и FOLFOX, аббревиатура для схемы лечения на основе оксалиплатина (Элоксатин™) в комбинации с 5-ФУ и лейковорином.
Химиотерапевтические агенты также включают нестероидные противовоспалительные лекарственные препараты с обезболивающим, жаропонижающим и противовоспалительным эффектами. НПВП включают неселективные ингибиторы фермента циклооксигеназы. Конкретные примеры НПВП включают аспирин, производные пропионовой кислоты, такие как ибупрофен, фенопрофен, кетопрофен, флурбипрофен, оксапрозин и напроксен, производные уксусной кислоты, такие как индометацин, сулиндак, этодолак, диклофенак, производные эноловой кислоты, такие как пироксикам, мелоксикам, теноксикам, дроксикам, лорноксикам и изоксикам, производные фенамовой кислоты, такие как мефенамовая кислота, меклофенамовая кислота, флуфенамовая кислота, толфенамовая кислота, и ингибиторы СОХ-2, такие как целекоксиб, эторикоксиб, люмиракоксиб, парекоксиб, рофекоксиб и валдекоксиб. НПВП могут быть назначены для симптоматического облегчения патологических состояний, таких как ревматоидный артрит, остеоартрит, воспалительные артропатии, анкилозирующий спондилоартрит, псориатический артрит, синдром Рейтера, острая подагра, дисменорея, метастатическая боль в костях, головная боль и мигрень, послеоперационная боль, легкая или умеренная боль вследствие воспаления и поражения тканей, пирексия, непроходимость кишечника и почечная колика.
Термин «цитотоксический агент», используемый в данном документе, относится к веществу, которое ингибирует или препятствует клеточной функции и/или приводит к гибели или разрушению клеток. Цитотоксические агенты включают, без ограничения, радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, Р32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, описанные ниже.
«Нарушением» является любое состояние, которое может быть улучшено вследствие лечения, в том числе, без ограничения, хронические и острые нарушения или заболевания, включая те патологические состояния, которые провоцируют у млекопитающего соответствующее нарушение. В одном аспекте нарушение представляет собой рак, например, колоректальный рак.
Термины «клеточно-пролиферативное нарушение» и «пролиферативное нарушение» относятся к нарушениям, которые связаны с некоторой степенью аномальной клеточной пролиферации. В одном аспекте клеточно-пролиферативное нарушение представляет собой рак. В одном аспекте клеточно-пролиферативное нарушение представляет собой опухоль.
В контексте данного документа термин «опухоль» относится к любым неопластическим клеточным росту и пролиферации, как злокачественным, так и доброкачественным, и к любым предраковым и раковым клеткам и тканям. В контексте данного документа термины «рак», «раковый», «клеточно-пролиферативное нарушение», «пролиферативное нарушение» и «опухоль» не являются взаимоисключающими.
Термины «рак» и «раковый» относятся к или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым ростом/пролиферацией клеток. Аспекты рака включают виды рака с солидными опухолями и виды рака с несолидными опухолями. Солидные раковые опухоли включают, помимо прочего, колоректальный рак, меланому, рак молочной железы, рак легкого, рак головы и шеи, рак мочевого пузыря, рак почек, рак яичников, рак поджелудочной железы или рак простаты, или их метастазирующие формы. Рак может быть LY6G6D-положительным раком.
В некоторых аспектах рак представляет собой колоректальный рак. В контексте настоящего документа термин «колоректальный рак», «КРР», «рак толстой кишки» или «рак кишечника» относится к раку, который развивается из толстого кишечника, например, толстой кишки или прямой кишки. В некоторых аспектах КРР является левосторонней опухолью, т.е. опухолью, возникающей в дистальной части толстой кишки (например, дистальной трети поперечноободочной кишки, левый изгиб нисходящей ободочной кишки, сигмовидная ободочная кишка или прямая кишка). В других аспектах КРР является правосторонней опухолью, т.е. опухолью, возникающей в проксимальной части толстой кишки (например, проксимальных двух третях поперечноободочной кишки, восходящей ободочной кишке и слепой кишке). Правосторонние опухоли могут быть связаны со снижением OS. В некоторых аспектах КРР является метастазирующим. В некоторых аспектах КРР имеет статус микросателлитной нестабильности: «микросателлитная стабильность» (MSS) или «низкий уровень микросателлитной нестабильности» (MSI-L). В других аспектах КРР имеет статус микросателлитной нестабильности «высокий уровень микросателлитной нестабильности» («MSI-H»). В некоторых аспектах КРР является LY6G6D-положительным (LY6G6D+) КРР.
В контексте настоящего документа «статус микросателлитной нестабильности» или «статус MSI» относится к определению характеристики микросателлитной стабильности в опухолевой ткани пациента. Опухолевая ткань пациента может характеризоваться как имеющая «высокий уровень микросателлитной нестабильности» («MSI-Н»), «низкий уровень микросателлитной нестабильности» («MSI-L») или «микросателлитную стабильность» («MSS»). Статус MSI может оцениваться, например, с помощью подхода на основе ПЦР, такого как система анализа MSI (Promega, Мадисон, Висконсин), которая состоит из 5 псевдомономорфных мононуклеотидных повторов (ВАТ-25, ВАТ-26, NR-21, NR-24 и MONO-27) для обнаружения MSI и 2 пентануклеотидных локусов (PentaC и PendaD) для подтверждения идентичности между нормальными и опухолевыми образцами. Размер в основаниях для каждого микросателлитного локуса можно определять, например, с помощью гель-электрофореза, и опухоль можно обозначать MSI-H, если два или более мононуклеотидных локусов изменяются по длине по сравнению с ДНК зародышевой линии. См., например, Le et al. NEJM 372:2509-2520, 2015.
В некоторых аспектах стадия КРР оценивается согласно системе классификации злокачественных опухолей (TNM) Объединенного американского комитета по раку (AJCC)/Международного союза борьбы с раком (UICC). В системе TNM рак обозначается буквой Т (размер опухоли), N (пальпируемые узлы) и/или М (метастазы). T1, Т2, Т3 и Т4 описывают увеличивающийся размер первичного поражения. T1, Т2, Т3 и Т4 могут дополнительно классифицироваться как а или b (например, Т4а или T4b) для обеспечения дополнительной информации о статусе, например, местное распространение, рака. N0, N1, N2, N3 указывает прогрессивно развивающееся вовлечение узлов; а М0 и M1 отражают отсутствие или присутствие отдаленных метастазов. В некоторых аспектах КРР индивидуума представляет КРР стадии I, стадии II или стадии III, например, карциному толстой кишки стадии I, стадии II или стадии III. В некоторых аспектах индивидуум не имеет КРР стадии IV. В некоторых аспектах индивидуум не имеет метастазирующий КРР. В некоторых аспектах КРР индивидуума в эталонной популяции представляет КРР стадии I, стадии II, стадии III мили стадии IV, например, карциному толстой кишки стадии I, стадии II, стадии III или стадии IV.
В некоторых аспектах рак представляет собой рак молочной железы. Дополнительные аспекты рака молочной железы включают положительный в отношении рецепторов гормонов (HR+) рак молочной железы, например, положительный в отношении рецепторов эстрогена (ER+) рак молочной железы, положительный в отношении рецепторов прогестерона (PR+) рак молочной железы или положительный в отношении ER+/PR+ рак молочной железы.
Другие аспекты рака молочной железы включают HER2-положительный (HER2+) рак молочной железы. Еще одни аспекты рака молочной железы включают трижды негативный рак молочной железы (ТНРМЖ). В некоторых аспектах рак молочной железы представляет рак молочной железы ранней стадии. В некоторых аспектах рак представляет собой рак легкого. Дополнительные аспекты рака легкого включают положительный в отношении рецептора эпидермального фактора роста (EGFR+) рак легкого. Другие аспекты рака легкого включают отрицательный в отношении рецептора эпидермального фактора роста (EGFR-) рак легкого. Еще одни аспекты рака легкого включают немелкоклеточный рак легкого, например, сквамозный рак легкого или несквамозный рак легкого. Другие аспекты рака легкого включают мелкоклеточный рак легкого. В некоторых аспектах рак представляет собой рак головы и шеи. Дополнительные аспекты рака головы и шеи включают плоскоклеточную карциному головы и шеи (ПККГШ). В некоторых аспектах рак представляет собой рак мочевого пузыря. Дополнительные аспекты рака мочевого пузыря включают уротеиальный рак мочевого пузыря (УРМП), мышечно-инвазивный рак мочевого пузыря (МИРМП) или немышечно-инвазивный рак мочевого пузыря (НМИРМП). В некоторых аспектах рак представляет собой рак почек. Дополнительные аспекты рака почек включают почечноклеточный рак (ПКР). В некоторых аспектах рак представляет собой рак печени. Дополнительные аспекты рака печени включают гепатоцеллюлярную карциному. В некоторых аспектах рак представляет собой рак простаты. Дополнительные аспекты рака простаты включают кастрационно-резистентный рак простаты (КРРП). В некоторых аспектах рак представляет собой метастазирующую форму солидной опухоли. В некоторых аспектах метастазирующая форма солидной опухоли представляет собой метастазирующую форму меланомы, рака молочной железы, колоректального рака, рака легкого, рака головы и шеи, рака мочевого пузыря, рака почек, рака яичников, рака поджелудочной железы или рака простаты. В одном аспекте рак представляет собой рак с несолидными опухолями. Рак с несолидными опухолями включает, помимо прочего, гематологический рак, например, В-клеточную лимфому. Дополнительные аспекты В-клеточной лимфомы включают, например, хронический лимфолейкоз (ХЛЛ), диффузную В-крупноклеточную лимфому (ДВККЛ), фолликулярную лимфому, миелодиспластический синдром (МДС), неходжикинскую лимфому (НХЛ), острый лимфобластный лейкоз (ОЛЛ), множественную миелому, острый миелолейкоз (ОМЛ) или грибовидный микоз (ГМ).
«Эффекторные функции» относятся к видам биологической активности, характерным для Fc-области антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание C1q и комплементзависимую цитотоксичность (КЗЦ); связывание с Fc-рецептором; антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ); фагоцитоз; понижающую регуляцию клеточных поверхностных рецепторов (например, В-клеточного рецептора) и активацию В-клеток.
«Комплемент-зависимая цитотоксичность» или «CDC (КЗЦ)» относится к лизису клетки-мишени в присутствии комплемента. Активация классического пути комплемента начинается со связывания первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), связанными с распознаваемым им антигеном. Для оценки активации комплемента можно проводить анализ КЗЦ, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
«Антителозависимая клеточно-опосредуемая цитотоксичность» или «АЗКЦ» относится к форме цитотоксичности, при которой секретируемые Ig, связанные с Fc-рецепторами (FcR), находящимися на определенных цитотоксических клетках (например, естественных клетках-киллерах (NK), нейтрофилах и макрофагах), вызывают специфичное для этих цитотоксических эффекторных клеток связывание с несущей антиген клеткой-мишенью, а затем уничтожают клетки-мишени посредством цитотоксических агентов. Антитела являются «оружием» цитотоксических клеток и абсолютно необходимы для такого уничтожения. Первичные клетки для осуществления АЗКЦ, NK-клетки, экспрессируют только FcyRIII, в то время как моноциты экспрессируют FcyRI, FcyRII и FcyRIII. Сводная информация об экспрессии FcR на гемопоэтических клетках приведена в Таблице 3 на странице 464 статьи Ravetch and Kinet. Annu. Rev. Immunol. 9:457-92, 1991. Для оценки активности представляющей интерес молекулы в отношении АЗКЦ, можно проводить анализ АЗКЦ in vitro, такой как анализ, описанный в патенте США №5500362 или 5821337. Эффекторные клетки, подходящие для такого анализа, включают мононуклеарные клетки периферической крови (МКПК) и естественные клетки-киллеры (NK). В альтернативном варианте, или дополнительно, активность АЗКЦ представляющей интерес молекулы может оцениваться in vivo, например, на животной модели, такой как описано в Clynes et al. Proc. Natl. Acad. Sci. USA. 95:652-656, 1998.
«Комплекс» или «образующий комплекс» в контексте настоящего документа относится к связи двух или более молекул, которые взаимодействуют друг с другом посредством связей и/или сил (например, ван-дер-ваальса, гидрофобных, гидрофильных сил), которые не являются пептидными связями. В одном аспекте комплекс является гетеромультимерным. Следует понимать, что термин «белковый комплекс» или «полипептидный комплекс» в контексте настоящего документа включает комплексы, которые имеют небелковое вещество, слитое с белком в белковый комплекс (например, включающий, помимо прочего, химические молекулы, такие как токсин или средство обнаружения).
В контексте данного документа «замедление прогрессирования» нарушения или заболевания означает отсрочивать, тормозить, замедлять, задерживать, стабилизировать и/или откладывать развитие заболевания или нарушения (например, клеточно-пролиферативного нарушения, например, рака). Это замедление может быть разным по продолжительности, в зависимости от истории болезни и/или индивидуума, проходящего лечение. Для специалиста в данной области техники очевидно, что существенное или значительное замедление может, на практике, включать предотвращение развития заболевания у субъекта. Например, можно замедлить позднюю стадию рака, такую как развитие метастазов.
«Эффективное количество» соединения, например, антитела к LY6G6D согласно изобретению или содержащей его композиции (например, фармацевтической композиции), представляет собой по меньшей мере минимальное количество, необходимое для достижения желаемого терапевтического или профилактического результата, такого как определяемое улучшение или предотвращение конкретного нарушения (например, клеточно-пролиферативного нарушения, например, рака). Эффективное количество в данном документе может варьироваться в зависимости от таких факторов, как состояние болезни, возраст, пол и масса пациента и от способности антитела вызывать необходимый ответ у индивидуума. Эффективное количество является таким, для которого любые токсические или вредные эффекты лечения перевешиваются терапевтически благоприятными эффектами. В случае профилактического применения благоприятные или желаемые результаты включают результаты, такие как устранение или снижение риска, уменьшения тяжести или задержка начала заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, присутствовавшие при развитии заболевания. В случае терапевтического применения благоприятные или желаемые результаты включают клинические результаты, такие как уменьшение одного или более симптомов, обусловленных заболеванием, повышение качества жизни страдающих от заболевания, снижение дозы других лекарственных препаратов, необходимых для лечения заболевания, усиление эффекта другого лекарственного препарата, например, за счет нацеливания, замедление прогрессирования заболевания и/или продление выживаемости. В случае рака или опухоли эффективное количество лекарственного препарата может оказывать эффект, состоящий в снижении количества раковых клеток; уменьшении размера опухоли; ингибировании (т.е. замедлении в некоторой степени или, желательно, прекращении) инфильтрации раковых клеток в периферические органы; ингибировании (т.е. замедлении в некоторой степени или, желательно, прекращении) метастазирования опухоли; ингибировании в некоторой степени роста опухоли; и/или облегчении в некоторой степени одного или более симптомов, связанных с нарушением. Эффективное количество можно вводить за один или более приемов. В целях данного изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для обеспечения профилактического или терапевтического лечения прямым или косвенным образом. В понимании в клиническом контексте эффективное количество лекарственного средства, соединения или фармацевтической композиции может или не может быть получено в комбинации с другими лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективное количество» можно рассматривать в контексте введения одного или более терапевтических средств, а одно средство можно рассматривать как вводимое в эффективном количестве, если в комбинации с одним или более другими средствами может быть достигнут или достигается необходимый результат.
Термин «эпитоп» относится к конкретному сайту на молекуле антигена, с которым связывается антитело. В некоторых аспектах конкретный сайт на молекуле антигена, с которым связывается антитело, определяется с помощью футпринтинга гидроксильного радикала. В некоторых аспектах конкретный сайт на молекуле антигена, с которым связывается антитело, определяется при помощи кристаллографии.
В контексте настоящего документа термин «ингибирующий рост агент» относится к соединению или композиции, которые ингибируют рост клетки in vitro, или in vivo. В одном аспекте ингибирующий рост агент представляет собой ингибирующее рост антитело, которое предотвращает или снижает пролиферацию клетки, экспрессирующей антиген, с которым связывается антитело. В другом аспекте ингибирующий рост агент может представлять собой агент, который значительно снижает процент клеток в S-фазе. Аспекты ингибирующих рост агентов включают агенты, которые блокируют развитие клеточного цикла (в месте, отличном от S-фазы), такие как агенты, которые индуцируют блокировку G1 и блокировку М-фазы. Классические блокаторы М-фазы включают алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Агенты, блокирующие G1, также приводят к блокировке S-фазы, например, ДНК-алкилирующие агенты, такие как тамоксифен, преднизолон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-С. Дополнительную информацию можно найти в Mendelsohn and Israel, eds., The Molecular Basis of Cancer, глава 1, по названием "Cell cycle regulation, oncogenes, and antineoplastic drugs" авторства Murakami et al. (W.B. Saunders, Philadelphia, 1995), например, стр. 13. Таксаны (паклитаксел и доцетаксел) представляют собой противораковые лекарственные препараты, получаемые из тисового дерева. Доцетаксел (Таксотер®, Rhone-Poulenc Rorer), получаемый из европейского тиса, представляет собой полусинтетический аналог паклитаксела (Таксол®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки путем предотвращения деполимеризации, что приводит к ингибированию митоза в клетках.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была внесена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформанты» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство вне зависимости от числа пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновой кислоты и может содержать мутации. Сюда включено мутантное потомство, которое имеет такую же функцию или биологическую активность, в отношении которой проводится скрининг или отбор изначально трансформированных клеток.
В настоящем документе термин «вектор» относится к молекуле нуклеиновой кислоты, способной размножать другую нуклеиновую кислоту, с которой она соединена. Этот термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую его внесли. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называются в настоящем документе «векторы экспрессии».
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или более гетерологичными молекулами, включая, но не ограничиваясь этим, цитотоксический агент.
Термин «иммуномодулирующее средство» относится к классу молекул, который модифицирует реакцию иммунной системы или работу иммунной системы. Иммуномодулирующие средства включают, помимо прочего, антагонисты связывания оси PD-1, Т-клеточно-зависимые биспецифические антитела и персонализированные вакцины от рака на основе иРНК, а также талидомид (α-N-фталимидоглутаримид) и его аналоги, Отезла® (апремиласт), Ревлимид® (леналидомид) и Акти-Мид™ (помалидомид) и их фармацевтически приемлемые соли или кислоты.
«Субъект» или «индивидуум» представляет собой млекопитающее. Млекопитающие включают, но не ограничиваются этим, одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и отличных от человека приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В определенных аспектах субъект или индивидуум является человеком.
«Выделенный» белок или пептид представляет собой такой, который было отделен от компонента его природного окружения. В некоторых аспектах белок или пептид очищают до степени чистоты более чем 95% или 99%, что определено, например, электрофорезом (например, ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографическими (например, ионообменная или обращенно-фазовая ВЭЖХ) способами.
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат эту молекулу нуклеиновой кислоты, но при этом молекула нуклеиновой кислоты присутствует внехромосомно или в хромосомном положении, которое отличается от ее природного хромосомного положения.
Термин «связывающий ось PD-1 антагонист» относится к молекуле, которая ингибирует взаимодействие партнера по связыванию оси PD-1 с одним или более его партнерами по связыванию так, чтобы устранить дисфункцию Т-клеток в результате передачи сигнала в сигнальной оси PD-1, что приводит к восстановлению или усилению функции Т-клеток (например, пролиферации, выработке цитокинов, уничтожению целевых клеток). В контексте данного документа связывающий ось PD-1 антагонист включает связывающий PD-1 антагонист, связывающий PD-L1 антагонист и связывающий PD-L2 антагонист.
Термин «связывающий PD-1 антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала в результате взаимодействия PD-1 с одним или более его партнерами по связыванию, такими как PD-L1, PD-L2. В некоторых аспектах связывающий PD-1 антагонист представляет собой молекулу, которая ингибирует связывание PD-1 с одним или более его партнерами по связыванию. В конкретном аспекте связывающий PD-1 антагонист ингибирует связывание PD-1 с PD-L1 и/или PD-L2. Например, связывающие PD-1 антагонисты включают антитела к PD-1, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или препятствует передаче сигнала в результате взаимодействия PD-1 с PD-L1 и/или PD-L2. В одном аспекте связывающий PD-1 антагонист снижает отрицательный костимулирующий сигнал, опосредованный или передаваемый посредством белков клеточной поверхности, экспрессируемых на Т-лимфоцитах, опосредующих сигнал через PD-1, чтобы сделать дисфункциональную Т-клетку менее дисфункциональной (например, повышая эффекторные ответы на распознавание антигена). В некоторых аспектах связывающий PD-1 антагонист представляет собой антитело к PD-1. В конкретном аспекте связывающий PD-1 антагонист представляет собой MDX-1106 (ниволумаб). В другом конкретном аспекте связывающий PD-1 антагонист представляет собой МК-3475 (пембролизумаб). В другом конкретном аспекте связывающий PD-1 антагонист представляет собой АМР-224. В другом конкретном аспекте связывающий PD-1 антагонист представляет собой MED1-0680. В другом конкретном аспекте связывающий ось PD-1 антагонист представляет собой PDR001. В другом конкретном аспекте связывающий ось PD-1 антагонист представляет собой REGN2810. В другом конкретном аспекте связывающий PD-1 антагонист представляет собой BGB-108.
Термин «связывающий PD-L1 антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала в результате взаимодействия PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, В7-1. В некоторых аспектах связывающий PD-L1 антагонист представляет собой молекулу, которая ингибирует связывание PD-L1 с его партнерами по связыванию. В конкретном аспекте связывающий PD-L1 антагонист ингибирует связывание PD-L1 с PD-1 и/или В7-1. В некоторых аспектах связывающие PD-L1 антагонисты включают антитела к PD-L1, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или препятствует передаче сигнала в результате взаимодействия PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, В7-1. В одном аспекте связывающий PD-L1 антагонист снижает негативный костимулирующий сигнал, опосредованный или передаваемый клеточными поверхностными белками, экспрессируемыми на Т-лимфоцитах, которые опосредуют передачу сигнала через PD-L1, так, чтобы сделать дисфункциональную Т-клетку менее дисфункциональной (например, повышая эффекторные ответы на распознавание антигена). В некоторых аспектах связывающий PD-L1 антагонист представляет собой антитело к PD-L1. В еще одном конкретном аспекте антитело к PD-L1 представляет собой MPDL3280A (атезолизумаб, продаваемый как Тецентрик™ с рекомендуемым INN (международное непатентованное название) в соответствии с Лекарственным вестником ВОЗ (международные незарегистрированные названия для фармацевтических веществ): список 74, т. 29, №3, 2015 (см. стр. 387)). В конкретном аспекте антитело к PD-L1 представляет собой YW243.55.S70. В другом конкретном аспекте антитело к PD-L1 представляет собой MDX-1105. В другом конкретном аспекте антитело к PD-L1 представляет собой MSB0015718C. В еще одном конкретном аспекте антитело к PD-L1 представляет собой MEDI4736.
Термин «связывающий PD-L2 антагонист» относится к молекуле, которая снижает, блокирует, ингибирует, подавляет или препятствует передаче сигнала в результате взаимодействия PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В некоторых аспектах связывающий PD-L2 антагонист представляет собой молекулу, которая ингибирует связывание PD-L2 с одним или более его партнерами по связыванию. В конкретном аспекте связывающий PD-L2 антагонист ингибирует связывание PD-L2 с PD-1. В некоторых аспектах связывающие PD-L2 антагонисты включают антитела к PD-L2, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, подавляют или препятствует передаче сигнала в результате взаимодействия PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В одном аспекте связывающий PD-L2 антагонист снижает отрицательный костимулирующий сигнал, опосредованный или передаваемый посредством белков клеточной поверхности, экспрессируемых на Т-лимфоцитах, которые опосредуют передачу сигнала через PD-L2, чтобы сделать дисфункциональную Т-клетку менее дисфункциональной (например, повышая эффекторные ответы на распознавание антигена). В некоторых аспектах связывающий PD-L2 антагонист представляет собой иммуноадгезин.
В контексте настоящего документа термин «белок» относится к любому нативному белку из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Данный термин включает «полноразмерный» непроцессированный белок, а также любую форму белка, которая является результатом процессинга в клетке. Этот термин также охватывает варианты белка природного происхождения, например, сплайс-варианты или аллельные варианты.
«Процент (%) идентичности аминокислотной последовательности» относительно эталонной полипептидной последовательности определяется как процентная доля аминокислотных остатков в кандидатной последовательности, которые являются идентичными с аминокислотными остатками в эталонной полипептидной последовательности, после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен в качестве части идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотной последовательности можно осуществлять различными способами, которые известны в данной области техники, например, используя общедоступное компьютерное программное обеспечение, такое как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. При этом в целях данного документа значения процента (%) идентичности аминокислотных последовательностей получают, используя компьютерную программу для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана компанией Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где он зарегистрирован под номером регистрации авторского права США TXU510087. Программа ALIGN-2 находится в свободном доступе от Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, включая Digital UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не варьируются.
В тех случаях, когда для сравнения аминокислотных последовательностей используют ALIGN-2, % идентичности аминокислотной последовательности заданной аминокислотной последовательности А с или по сравнению с заданной аминокислотной последовательностью В (что в альтернативном варианте можно сформулировать как заданная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности с или по сравнению с заданной аминокислотной последовательностью В) рассчитывают следующим образом:
100 умножить на соотношение X/Y,
где X представляет собой число аминокислотных остатков, оцененных программой выравнивания последовательностей ALIGN-2 как идентичные совпадения при программном выравнивании А и В, и где Y представляет общее количество аминокислотных остатков в В. Следует понимать, что там, где длина аминокислотной последовательности А не равняется длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А к В не будет равен % идентичности аминокислотной последовательности В к А. Если специально не указано иное, все значения % идентичности аминокислотной последовательности, которые используется в данном документе, получают, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
Термин «фармацевтический состав» относится к препарату, который находится в форме, обеспечивающей реализацию биологической активности содержащегося в нем активного ингредиента, и который при этом не содержит дополнительные компоненты, являющиеся неприемлемо токсичными для субъекта, которому будет введен состав.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтическом составе, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но не ограничивается этим, буфер, вспомогательное вещество, стабилизатор или консервант.
Под «лучевой терапией» подразумевается применение направленного гамма-облучения для индукции значительного повреждения клеток так, чтобы ограничить их способность нормально функционировать или разрушить все клетки. Следует понимать, что в данной области техники существует много способов определения дозы и длительности лечения. Типичные варианты лечения включают одноразовое применение, а типичные дозы находятся в диапазоне от 10 до 200 единиц (Грей).
В контексте данного документа термин «лечение» (и его грамматические варианты, такие как «лечить» или «осуществление лечения») относится к клиническому вмешательству с целью изменения естественного течения заболевания индивида, подлежащего лечению, и может проводиться как для профилактики, так и в течении клинической патологии. Необходимые эффекты лечения включают, но не ограничиваются этим, предотвращение появления или повторного появления заболевания, смягчение симптомов, уменьшение каких-либо прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, уменьшение интенсивности или временное облегчение болезненного состояния и ремиссию или улучшение прогноза. В некоторых аспектах антитела по настоящему изобретению (например, антитела к LY6G6D по настоящему изобретению) используют для задержки развития заболевания или для замедления прогрессирования заболевания.
Под термином «снижать» или «ингибировать» подразумевается способность приводить к общему снижению, составляющему, например, 20% или более, 50% или более или 75%, 85%, 90%, 95% или более. В определенных аспектах термины снижать или ингибировать могут относиться к эффекторной функции антитела, которая опосредуется Fc-областью антитела, при этом такие эффекторные функции включают, в частности, комплемент-зависимую цитотоксичность (КЗЦ), антитело-зависимую клеточную цитотоксичность (АЗКЦ) и антитело-зависимый клеточный фагоцитоз (АЗКФ).
Согласно настоящему изобретению, термин «вакцина» относится к фармацевтическому препарату (фармацевтической композиции) или продукту, который при введении вызывает иммунную реакцию, в частности клеточную иммунную реакцию, которая распознает и атакует патоген или болезненные клетки, такие как раковая клетка. Вакцина может использоваться для предотвращения или лечения заболевания. Вакцина может быть противораковой вакциной. В контексте настоящего документа «противораковая вакцина» представляет собой композицию, которая стимулирует иммунную реакцию у субъекта против рака. Противораковые вакцины обычно состоят из источника связанного с раком материала или клеток (антиген), который может быть аутологичным (от самого себя) или аллогенным (от других) для субъекта, вместе с другими компонентами (например, адъювантами), чтобы дополнительно стимулировать и активировать иммунную реакцию против антигена. Противораковые вакцины могут приводить к стимуляции иммунной системы субъекта для продуцирования антител для одного или нескольких специфичных антигенов, и/или для продуцирования Т-клеток-киллеров для борьбы с раковыми клетками, которые имеют такие антигены.
Термин «персонализированная противораковая вакцина» («ППРВ») относится к противораковой вакцине, которая приспособлена к потребностям или конкретными условиям отдельного пациента с раком. В некоторых аспектах ППРВ стимулирует иммунную реакцию против одной или более специфичных для рака соматических мутаций, присутствующих в раковых клетках пациента, как описано, например, в публикациях РСТ №№ WO 2014/082729 и WO 2012/159754. Специфичная для рака соматическая мутация может присутствовать в любой раковой клетке пациента, например, опухолевой клетке, например, эмболической опухолевой клетке. В некоторых аспектах специфичная для рака соматическая мутация обнаруживается с помощью определения последовательности следующего поколения. В некоторых аспектах полипептид, содержащий специфичную для рака соматическую мутацию, или нуклеиновая кислота (например, РНК, например, in vitro транскрибированная РНК), кодирующая полипептид, содержащий специфичную для рака соматическую мутацию, вводят пациенту для стимуляции иммунной реакции пациента.
В контексте настоящего документа «введение» подразумевает способ введения дозы соединения (например, антитела к LY6G6D по настоящему изобретению) субъекту. В некоторых аспектах композиции, используемые в способах в настоящем документе, вводят внутривенно. Композиции, используемые в способах, описанных в настоящем документе, можно вводить, например, внутримышечно, внутривенно, внутрикожно, подкожно, внутриартериально, внутрибрюшинно, в пораженные ткани, внутричерепно, внутрисуставно, внутрипростатически, внутриплеврально, внутритрахеально, интраназально, в стекловидное тело, внутривлагалищно, интраректально, местно, внутриопухолево, перитонеально, подкожно, подконъюктивально, внутрипузырно, в слизистую, внутриперикардиально, внутрипупочно, внутриглазно, перорально, местно, локально, ингаляцией, инъекцией, инфузией, непрерывной инфузией, местной перфузией, омывающей клетки-мишени непосредственно, катетером, зондом, в кремах или в липидных композициях. Способ введения может изменяться в зависимости от различных факторов (например, соединения или композиции для введения, степени тяжести патологического состояния, заболевания или нарушения, подлежащего лечению).
II. КОМПОЗИЦИИ И СПОСОБЫ
В одном аспекте настоящее изобретение основано, отчасти, на антителах к комплексу лимфоцитарного антигена 6, локусу G61 (антитело к LY6G6D). В некоторых вариантах осуществления антитела к LY6G6D являются мультиспецифическими (например, биспецифическими) и связывают в дополнение к LY6G6D или его фрагменту вторую биологическую молекулу (например, поверхностный антиген Т-клетки, например кластер дифференциации 3 (CD3). Антитела по настоящему изобретению пригодны, например, для лечения или замедления развития клеточно-пролиферативного нарушения, например, рака, например, колоректального рака (КРР) (например, LY6G6D-положительного КРР), или для улучшения иммунной функции у субъекта, имеющего такое нарушение.
А. Типовые антитела к LY6G6D
В одном аспекте в изобретении предложены выделенные антитела, которые связываются с LY6G6D. В некоторых аспектах антитело к LY6G6D связывается с полипептидом LY6G6D человека (SEQ ID NO: 75) или полипептидом LY6G6D яванского макака (cyno) (SEQ ID NO: 77). В некоторых аспектах антитело к LY6G6D связывается с эпитопом, содержащим или имеющим аминокислоты 93-104 (SEQ ID NO: 87), 94-103 (SEQ ID NO: 78) или 99-101 (SEQ ID NO: 79) LY6G6D (например, LY6G6D человека). В некоторых аспектах антитело к LY6G6D связывается с одним, двумя, тремя или всеми четырьмя остатками Arg94, Leu 101, Cys102 и Asn103 LY6G6D. В некоторых аспектах антитело к LY6G6D связывается с эпитопом, содержащим остатки Arg94, Asp95, Cys96, Tyr97, Leu98, Gly99, Asp100, Leu101, Cys102 и Asn103 LY6G6D. В некоторых аспектах антитело к LY6G6D связывается с эпитопом, содержащим остатки Arg94, Asp95, Cys96, Tyr97, Leu98, Gly99, Asp100, Leu101, Cys102 и Asn103 LY6G6D.
Эпитоп LY6G6D может быть определен по LY6G6D-связывающему домену антитела к LY6G6D, связывающемуся с фрагментами пептида эпитопа. Эпитоп LY6G6D может также быть определен по сканирующему аланином мутагенезу. В одном варианте осуществления восстановление связывания LY6G6D-связывающего домена с мутированным LY6G6D на 20%, 30%, 50%, 80% или более указывает, что аминокислотный остаток LY6G6D, мутировавший в анализе сканирующего аланином мутагенеза, является эпитопным остатком для LY6G6D-связывающего домена. Альтернативно, эпитоп LY6G6D может быть определен с помощью масс-спектромерии. В некоторых вариантах осуществления эпитоп определяется с помощью кристаллографии (например, способов кристаллографии).
В некоторых вариантах осуществления эпитоп LY6G6D может быть определен способами кристаллографии путем объединения Fab антитела к LY6G6D, растворенного в конкретном условии (например, 0,15 М NaCl, 25 мМ трис, рН 7,5 при 10 мг/мл), с мольным избытком (например, 2-кратным мольным избытком) пептида LY6G6D и исходного скрининга матрицы осадителей в формате диффузии пара в виде сидячей или висячей капли. Оптимизированные кристаллы могут расти, например, из смеси 1:1 с раствором в резервуаре, содержащем 70% об./об. Метилпентандиола и 0,1 М буфера HEPES при рН 7,5. Резервуар можно использовать как криопротектор. Кристаллы можно переносить в криогенную температуру путем быстрого погружения в жидкий азот.
Данные дифракции для кристаллов можно собирать на оси пучка. Записанные дифракции можно интегрировать и масштабировать с помощью программы, такой как HKL2000.
Структура может быть фазированна путем молекулярного замещения (МЗ), например, с помощью программы, такой как Phaser. Например, модель поиска МЗ представляет субъединицу Fab, полученную из кристаллической структуры комплекса HGFA/Fab (код по PDB (Банк данных белков): 2R0L). Пептид LY6G6D встраивается в структуру на основе карты Fo-Fc. Структура может быть затем уточнена с помощью программ REFMAC5 и PHENIX, используя целевые функции наибольшего правдоподобия, способ уточнения анизотропного В-фактора и способ уточнения TLS для достижения схождения.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, имеющее связывающий домен, содержащий по меньшей мере одну, две, три, четыре, пять или шесть гипервариабельных областей (HVR), выбранных из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3 или любую из SEQ ID NO: 99-107.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4 или SEQ ID NO: 111; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5, SEQ ID NO: 112 или SEQ ID NO: 113; (в) a CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (e) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3 или любую из SEQ ID NO: 99-107.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-НЗ, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 99.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 100.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 101.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 102.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 103.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 104.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 105.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 106.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 107.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 111; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-НЗ, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 112; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3.
В некоторых аспектах настоящее изобретение предусматривает антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 113; (в) CDR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3.
В некоторых аспектах антитело к LY6G6D может иметь домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или содержащую последовательность SEQ ID NO: 10, и/или домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или содержащую последовательность SEQ ID NO: 11. В конкретном случае антитело к LY6G6D может быть 20A12.QNTv12 (включая изготовленные из одной клетки и двух клеток варианты) или его производным или клонально родственным антителом. В некоторых аспектах антитело к LY6G6D имеет домен VH, содержащий аминокислотную последовательность SEQ ID NO: 59, или домен VL, содержащий аминокислотную последовательность SEQ ID NO: 60. В некоторых аспектах антитело к LY6G6D имеет домен VH, содержащий аминокислотную последовательность SEQ ID NO: 59, и домен VL, содержащий аминокислотную последовательность SEQ ID NO: 60
В некоторых аспектах антитело к LY6G6D может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 34; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 35; (в) FR-H3, содержащий аминокислотную последовательность SEQ ID NO: 36; и (г) FR-H4, содержащий аминокислотную последовательность SEQ ID NO: 37. В некоторых аспектах антитело к LY6G6D может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 39; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 41.
В некоторых аспектах антитело к LY6G6D содержит все четыре из (a) FR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 34; (б) FR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 35; (в) FR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 36; и (г) FR-Н4, содержащей аминокислотную последовательность SEQ ID NO: 37, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 39; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 41. В некоторых аспектах антитело к LY6G6D может иметь домен VH, содержащий аминокислотную последовательность SEQ ID NO: 10, и/или домен VL, содержащий аминокислотную последовательность SEQ ID NO: 11.
В некоторых аспектах антитело к LY6G6D может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 34; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 58; (в) FR-H3, содержащий аминокислотную последовательность SEQ ID NO: 36; и (г) FR-H4, содержащий аминокислотную последовательность SEQ ID NO: 37. В некоторых аспектах антитело к LY6G6D может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 61; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 41.
В некоторых аспектах антитело к LY6G6D содержит все четыре из (a) FR-Н1, содержащей аминокислотную последовательность SEQ ID NO: 34; (б) FR-Н2, содержащей аминокислотную последовательность SEQ ID NO: 58; (в) FRED, содержащей аминокислотную последовательность SEQ ID NO: 36; и (г) FR-Н4, содержащей аминокислотную последовательность SEQ ID NO: 37, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 38; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 61; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 40; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 41. В некоторых аспектах антитело к LY6G6D может иметь домен VH, содержащий аминокислотную последовательность SEQ ID NO: 59, и/или домен VL, содержащий аминокислотную последовательность SEQ ID NO: 60.
В любом из вышеописанных аспектов антитело к LY6G6D может быть гуманизировано. В одном варианте осуществления антитело к LY6G6D содержит CDR согласно любому из вышеприведенных вариантов осуществления и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область.
В дополнительном аспекте настоящего изобретения антитело к LY6G6D согласно любому из вышеприведенных вариантов осуществления представляет собой моноклональное антитело. В некоторых аспектах антитело к LY6G6D представляет собой химерное или человеческое антитело. В одном аспекте антитело к LY6G6D представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или F(ab')2. В другом аспекте антитело представляет собой полноразмерное антитело, например, интактное антитело IgG1 (например, интактное антитело IgG1), или антитело другого класса или изотипа, как определено в настоящем документе.
В дополнительном аспекте антитело к LY6G6D согласно любому из вышеуказанных вариантов осуществления изобретения может включать любые из признаков, по отдельности или в комбинации, как описано ниже в разделах 1 -8.
Б. Типовые антитела к CD3
В другом аспекте настоящее изобретение предусматривает выделенные антитела, которые связываются с кластером дифференциации 3 (CD3) (например, CD3ε и/или CD3γ) и имеют следующие шесть последовательностей CDR: (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13 или SEQ ID NO: 50; и (e) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14 или SEQ ID NO: 51, последовательность VL, содержащую SEQ ID NO: 21, 55, 90 или 92, последовательности VL и VH, содержащие SEQ ID NO: 21 или 55 и 20, соответственно, или последовательности VL и VH, содержащие SEQ ID NO: 90 или 92 и 89, соответственно. В некоторых случаях антитело к CD3 связывается с полипептидом CD3 человека или полипептидом CD3 яванского макака (супо). В некоторых случаях полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3ε человека (SEQ ID NO: 80) или полипептид CD3ε яванского макака (SEQ ID NO: 81) соответственно. В некоторых случаях полипептид CD3 человека или полипептид CD3 яванского макака представляет собой полипептид CD3γ человека (SEQ ID NO: 82) или полипептид CD3γ яванского макака (SEQ ID NO: 83) соответственно. В некоторых случаях антитело к CD3 связывается с эпитопом в фрагменте CD3 (например, CD3ε человека), состоящем из аминокислот 1-26 (SEQ ID NO: 84) или 1-27 (SEQ ID NO: 85) CD3ε человека.
В некоторых аспектах настоящее изобретение предусматривает антитело к CD3, содержащее связывающий домен, содержащий все шесть из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 13; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 14.
В некоторых аспектах настоящее изобретение предусматривает антитело к CD3, содержащее связывающий домен, содержащий все шесть из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 50; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 51.
В некоторых случаях антитело к CD3 может иметь домен VH, содержащий аминокислотную последовательность с последовательностью SEQ ID NO: 20 или 89, и домен VL, содержащий аминокислотную последовательность с последовательностью SEQ ID NO: 21 или 90. В конкретном случае антитело к CD3 может представлять собой 38E4.v1 MD1 или его производное, или клонально родственное антитело.
В некоторых случаях антитело к CD3 может иметь домен VH, содержащий аминокислотную последовательность с последовательностью SEQ ID NO: 20 или 89, и домен VL, содержащий аминокислотную последовательность с последовательностью SEQ ID NO: 55 или 92. В конкретном случае антитело к CD3 может представлять собой 38E4.v1 MD4 или его производное, или клонально родственное антитело.
В некоторых аспектах антитело к CD3 может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 43 или SEQ ID NO: 62; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 47 или SEQ ID NO: 63; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах антитело к CD3 содержит все четыре из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 43; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 47; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах антитело к CD3 содержит все четыре из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 62; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 63; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49.
В любом из вышеуказанных вариантов осуществления антитело к CD3 может быть гуманизировано. В одном варианте осуществления антитело к CD3 содержит CDR согласно любому из вышеприведенных вариантов осуществления и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область.
В другом аспекте предложено антитело к CD3, причем антитело содержит VH согласно любому из вышеприведенных вариантов осуществления и VL согласно любому из вышеприведенных вариантов осуществления, причем одна или обе из последовательностей вариабельных доменов включают посттрансляционные модификации.
В дополнительном аспекте в изобретении предложено антитело, которое связывается с тем же эпитопом, что и предложенное в данном документе антитело к CD3. Например, в некоторых вариантах осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к CD3, содержащее последовательность VH SEQ ID NO: 20 и последовательность VL SEQ ID NO: 21, или антитело к CD3, содержащее последовательность VH SEQ ID NO: 20 и последовательность VL SEQ ID NO: 55. В некоторых вариантах осуществления представлено антитело, которое связывается с эпитопом во фрагменте CD3 (например, CD3ε человека), состоящем из аминокислот 1-26 (SEQ ID NO: 84) или 1-27 (SEQ ID NO: 85) CD3ε человека.
В дополнительном аспекте настоящего изобретения антитело к CD3 согласно любому из вышеупомянутых вариантов осуществления представляет собой моноклональное антитело. В других вариантах осуществления антитело к CD3 представляет собой химерное или человеческое антитело. В одном варианте осуществления антитело к CD3 представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или F(ab')2. В другом варианте осуществления антитело представляет собой полноразмерное антитело, например, интактное антитело IgG1 (например, интактное антитело IgG1), или антитело другого класса или изотипа, как определено в настоящем документе.
В дополнительном аспекте антитело к CD3 согласно любому из вышеуказанных вариантов осуществления изобретения может включать любые из признаков, по отдельности или в комбинации, как описано ниже в разделах 1-8.
1. Аффинность антитела
В определенных вариантах осуществления предложенное в настоящем документе антитело имеет константу диссоциации (KД) ≤1 мкМ, ≤250 нМ, ≤100 нМ, ≤15 нМ, ≤10 нМ, ≤6 нМ, ≤4 нМ, ≤2 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
В одном варианте осуществления KД измеряется анализом связывания антигена, меченного радиоактивным изотопом (РИА). В одном варианте осуществления РИА проводят с Fab-версией представляющего интерес антитела и его антигеном. Например, аффинность связывания Fab с антигеном в растворе измеряют путем уравновешивания Fab минимальной концентрацией (125I)-меченого антигена в присутствии ряда титрующих концентраций немеченого антигена с последующим захватом связанного антигена на планшете, покрытом антителами к Fab (смотрите, например, Chen et al., J. Mol. Biol. 293:865-881(1999)). Для определения условий анализа многолуночные планшеты MICROTITER® (Thermo Scientific) покрывают на ночь 5 мкг/мл захватывающего антитела к Fab (Cappel Labs) в 50 мМ растворе карбоната натрия (рН 9,6), а затем блокируют 2% (масс./об.) бычьим сывороточным альбумином в ФСБ в течение двух-пяти часов при комнатной температуре (приблизительно 23°С). В неадсорбентном планшете (Nunc №269620) смешивают 100 пМ или 26 пМ [125I]-антигена с серийными разведениями, представляющего интерес Fab (например, в соответствии с оценкой антитела к VEGF, Fab-12, в Presta et al., Cancer Res. 57:4593-4599 (1997)). Затем представляющий интерес Fab инкубируют в течение ночи; при этом инкубацию можно продолжать в течение более длительного периода (например, около 65 часов), чтобы гарантировать достижение равновесия. После этого смеси переносят в планшет для захвата для инкубации при комнатной температуре (например, в течение одного часа). Затем раствор удаляют и восемь раз промывают планшет 0,1% раствором полисорбата 20 (Твин-20) в ФСБ. После высушивания планшетов добавляли 150 мкл/лунку сцинтиллятора (MICROSCINT-20™; Packard) и производили подсчет планшетов на гамма-счетчике TOPCOUNT TM(Packard) в течение десяти минут.Концентрации каждого из Fab, которые обеспечивают связывание, меньшее или равное 20% от максимального связывания, выбирают для применения в конкурентных анализах связывания.
Согласно другому варианту осуществления KД определяют, используя анализ способом поверхностного плазмонного резонанса BIACORE®. Например, анализ с применением BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Пискатауэй, Нью-Джерси) проводят при 37°С с чипами СМ5 с иммобилизованным антигеном при ~10 единицах ответа (ЕО). В одном варианте осуществления биосенсорные чипы на основе карбоксиметилированного декстрана (СМ5, BIACORE, Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимида гидрохлоридом (ЭДК) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят в 10 мМ ацетате натрия, рН 4,8, до 5 мкг/мл (~0,2 мкМ) перед вводом при скорости потока 5 мкл/минута для достижения приблизительно 10 единиц ответа (ЕО) связанного белка. После ввода антигена вводят 1 М этаноламина, чтобы блокировать не вступившие в реакцию группы. Для измерений кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) вводят в ФСБ с содержащим 0,05% полисорбата 20 (Твин-20™) поверхностно-активным веществом (ФСБТ) при 37°С при скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kасс. или ka) и скорости диссоциации (кдисс. или kд) вычисляют, используя простую модель связывания Лэнгмюра «один-к-одному» (программное обеспечение для расчетов BIACORE®, версия 3.2), путем одновременной подгонки сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (KД) рассчитывают как отношение kдисс./kасс. Смотрите, например, Chen et al., J. Mol. Biol. 293:865-881 (1999). Если скорость ассоциации превышает 106 М-1 с-1 согласно данным вышеупомянутого анализа способом поверхностного плазмонного резонанса, то скорость ассоциации можно определить способом гашения флуоресценции, в котором измеряют повышение или снижение интенсивности флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 37°С для 20 нМ антитела к антигену (Fab-формы) в ФСБ, рН 7,2, в присутствии возрастающих концентраций антигена, с измерениями, проводимыми на спектрометре, таком как спектрофотометр с остановкой потока (Aviv Instruments) или спектрофотометр серии 8000 SLM-AMINCO™ (ThermoSpectronic) с кюветой с возможностью перемешивания.
В некоторых вариантах осуществления антитело к LY6G6D, представленное в настоящем документе, связывает полипептид LY6G6D человека с KД от приблизительно 1 нМ до 500 нМ при 37°С, что измерено с помощью анализа BIAcore, например, связывает LY6G6D человека с KД ≤1 мкМ, ≤250 нМ, ≤100 нМ, ≤40 нМ, ≤30 нМ, ≤15 нМ, ≤10 нМ, ≤6 нМ, ≤4 нМ, ≤2 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ. В некоторых вариантах осуществления антитело к LY6G6D, представленное в настоящем документе, связывает полипептид LY6G6D человека с KД от приблизительно 100 нМ до 0,01 нМ; от приблизительно 50 нМ до 5 нМ; от приблизительно 40 нМ до 10 нМ; от приблизительно 35 нМ до 15 нМ или от приблизительно 30 нМ до 20 нМ.
В некоторых вариантах осуществления антитело к CD3, представленное в настоящем документе, связывает полипептид CD3 человека с KД от приблизительно 100 пМ до 10 нМ при 37°С, что измерено с помощью анализа BIAcore, например, связывает CD3 человека с KД ≤1 мкМ, ≤250 нМ, ≤100 нМ, ≤40 нМ, ≤30 нМ, ≤15 нМ, ≤10 нМ, ≤5 нМ, ≤2 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ. В некоторых вариантах осуществления антитело к CD3, представленное в настоящем документе, связывает полипептид CD3 человека с KД от приблизительно 100 нМ до 0,01 нМ; от приблизительно 50 нМ до 5 нМ; от приблизительно 40 нМ до 10 нМ; от приблизительно 35 нМ до 15 нМ или от приблизительно 30 нМ до 20 нМ.
2. Фрагменты антител
В определенных вариантах осуществления антитело, представленное в настоящем документе (например, антитело к LY6G6D или антитело к CD3), представляет собой фрагмент антитела. Фрагменты антител включают, но не ограничиваются этим, фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv и другие фрагменты, описанные ниже. Для обзора некоторых фрагментов антител см. Hudson et al. Nat. Med. 9:129-134 (2003). Обзор фрагментов scFv см., например, в Pluckthun, в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. (Springer-Verlag, New York), pp. 269-315 (1994); также см. WO 93/16185; и патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное время полувыведения in vivo, смотрите в патенте США №5869046.
Диатела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими. См., например, ЕР 404,097; WO 1993/01161; Hudson et al. Nat. Med. 9:129-134, 2003; и Hollinger et al. Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
«Однодоменные антитела» представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах осуществления однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; смотрите, например, патент США №6248516 В1).
Фрагменты антител можно получать различными способами, включая, но не ограничиваясь этим, протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, Е. coli или фагом), как описано в данном документе.
3. Химерные и гуманизированные антитела
В некоторых вариантах осуществления антитело, представленное в настоящем документе (например, антитело к LY6G6D или антитело к CD3), представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в патенте США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В одном примере химерное антитело содержит отличную от человеческой вариабельную область (например, вариабельную область, полученную из антитела мыши, крысы, хомяка, кролика или отличного от человека примата, такого как обезьяна) и человеческую константную область. В дополнительном примере химерное антитело представляет собой антитело с «переключенным классом», класс или подкласс которого был изменен по сравнению с родительским антителом. Химерные антитела включают их антигенсвязывающие фрагменты.
В определенных вариантах осуществления химерное антитело представляет собой гуманизированное антитело. Как правило, отличное от человеческого антитело гуманизируют в целях снижения иммуногенности для человека, сохраняя при этом специфичность и аффинность родительского отличного от человеческого антитела. В общем, гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR (или их части), например, получены из антитела отличного от человеческого происхождения, а FR (или их части) получены из последовательностей человеческого антитела. Гуманизированное антитело, необязательно, также содержит по меньшей мере часть человеческой константной области. В некоторых вариантах осуществления некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками отличного от человеческого антитела (например, антитела, из которого получены остатки HVR), например, с целью восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения рассмотрены, например, в Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008), и дополнительно описаны, например, в Riechmann et al., Nature 332:323-329 (1988); Queen et al., Proc. Nat'l Acad. Sci. USA 86:10029-10033 (1989); патентах США Ms 5821337, 7527791, 6982321 и 7087409; Kashmiri et al, Methods 36:25-34 (2005) (где описано прививание определяющей специфичность области (SDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (где описано «изменение поверхности»); Dall'Acqua et al., Methods 36:43-60 (2005) (где описана «перетасовка FR»); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (где описан подход «направленного отбора» при перетасовке FR).
Человеческие каркасные области, которые можно использовать для гуманизации, включают, но не ограничиваются этим: каркасные области, выбранные способом «наилучшего соответствия» (смотрите, например, Sims et al. J. Immunol. 151:2296 (1993)); каркасные области, полученные из консенсусной последовательности человеческих антител конкретной подгруппы вариабельных областей легкой или тяжелой цепи (смотрите, например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285 (1992); и Presta et al. J. Immunol., 151:2623 (1993)); человеческие зрелые (соматически мутированные) каркасные области или каркасные области зародышевой линии человека (смотрите, например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)); и каркасные области, полученные при скрининге библиотек FR (смотрите, например, Baca et al., J. Biol. Chem. 272:10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
4. Человеческие антитела
В определенных вариантах осуществления антитело, представленное в настоящем документе (например, антитело к LY6G6D или антитело к CD3), представляет собой человеческое антитело. Человеческие антитела могут быть получены с использованием различных технологий, известных в данной области техники. Человеческие антитела в целом описаны в van Dijk and van de Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) и Lonberg, Curr. Opin. Immunol. 20:450-459 (2008).
Человеческие антитела могут быть получены путем введения иммуногена трансгенному животному, модифицированному для выработки интактных человеческих антител или интактных антител с человеческими вариабельными областями в ответ на антигенную стимуляцию. Такие животные, как правило, содержат все или часть локусов человеческого иммуноглобулина, которые замещают эндогенные локусы иммуноглобулина или которые находятся вне хромосом или случайным образом интегрированы в хромосомы животного. Как правило, у таких трансгенных мышей инактивированы эндогенные локусы иммуноглобулина. Обзор способов получения человеческих антител от трансгенных животных смотрите в Lonberg, Nat. Biotech. 23:1117-1125 (2005). Также смотрите, например, патенты США №№6075181 и 6150584, в которых описана технология XENOMOUSE™; патент США №5770429, в котором описана технология HuMab®; патент №7041870, в котором описана технология K-М MOUSE®, и публикацию заявки на патент США № US 2007/0061900, в которой описана технология VELOCIMouse®). Человеческие вариабельные области из интактных антител, вырабатываемых такими животными, можно дополнительно модифицировать, например, путем объединения с отличной человеческой константной областью.
Человеческие антитела можно получать способами на основе гибридом. Были описаны линии клеток человеческой миеломы и мышино-человеческой гетеромиеломы для выработки человеческих моноклональных антител. (Смотрите, например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol, 147: 86 (1991).) Человеческие антитела, полученные посредством технологии человеческой В-клеточной гибридомы, также описаны в Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные способы включают способы, описанные, например, в патенте США №7189826 (в котором описано получение моноклональных человеческих антител IgM из линий клеток гибридомы) и Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (в котором описаны человек-человеческие гибридомы). Технология человеческой гибридомы (технология триомы) описана в Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3):185-91 (2005).
Человеческие антитела также можно получать путем выделения последовательностей вариабельного домена Fv-клона из библиотек фагового дисплея человеческого происхождения. Такие последовательности вариабельного домена затем можно объединять с необходимым человеческим константным доменом, способы отбора человеческих антител из библиотек антител описаны ниже.
5. Антитела из библиотек
Антитела настоящего изобретения (например, антитела к LY6G6D или антитела к CD3) можно выделять путем скрининга комбинаторных библиотек в отношении антител с необходимым видом или видами активности. Например, в данной области техники известны различные способы получения библиотек фагового дисплея и скрининга таких библиотек в отношении антител, обладающих необходимыми характеристиками связывания. Дополнительные способы описаны, например, в Hoogenboom et al. в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001) и дополнительно описаны, например, в McCafferty et al., Nature 348:552-554; Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, в Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2): 119-132(2004).
В некоторых способах на основе фагового дисплея репертуары генов VH и VL по отдельности клонируют с помощью полимеразной цепной реакции (ПЦР) и случайным образом рекомбинируют в фаговых библиотеках, которые затем можно подвергать скринингу в отношении антигенсвязывающего фага, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Как правило, фаг представляет фрагменты антител в виде одноцепочечных Fv-фрагментов (scFv) или Fab-фрагментов. Библиотеки из иммунизированных источников позволяют получать антитела с высокой аффинностью к иммуногену без необходимости конструирования гибридом. В альтернативном варианте можно клонировать наивный репертуар (например, от человека) для получения единого источника антител к широкому спектру не собственных, а также собственных антигенов без какой-либо иммунизации, как описано в Griffiths et al., EMBO J, 12: 725-734 (1993). Наконец, наивные библиотеки также можно получать синтетически, клонируя неперестроенные V-генные сегменты из стволовых клеток и используя ПЦР-праймеры, содержащие случайную последовательность, для кодирования высоковариабельных областей CDR3 и осуществления перестройки in vitro, как описано в Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992). Патентные публикации, в которых описаны фаговые библиотеки человеческих антител, включают, например: патент США №5750373 и патентные публикации США №№2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
6. Мулътиспецифические антитела
В любом из вышеприведенных аспектов предложенное в данном документе антитело к LY6G6D или антитело к CD3 является мультиспецифическим антителом, например, биспецифическим антителом. Мультиспецифические антитела представляют собой антитела (например, моноклональные антитела), которые обладают специфичностями связывания в отношении по меньшей мере двух разных сайтах. В некоторых аспектах биспецифические антитела могут связываться с двумя разными эпитопами LY6G6D. В некоторых аспектах одна из специфичностей связывания направлена на LY6G6D, а другая - на любой другой антиген (например, вторую биологическую молекулу, например, поверхностный антиген Т-клетки, например, CD3). В некоторых аспектах одна из специфичностей связывания направлена на CD3, а другая - на любой другой антиген (например, вторую биологическую молекулу, например, антиген клеточной поверхности, например, опухолевый антиген). В некоторых аспектах одна из специфичностей связывания относится к LY6G6D, а другая - к CD3.
В некоторых аспектах антитело к LY6G6D содержит (а) LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1), содержащий вариабельный (VH) домен (VH1) тяжелой цепи, и полипептид легкой цепи (L1), содержащий вариабельный (VL) домен (VL1) легкой цепи, и (б) CD3-связывающий домен, содержащий полипептид, тяжелой цепи (Н2), содержащий вариабельный (VH) домен (VH2) тяжелой цепи, и полипептид легкой цепи (L2), содержащий вариабельный (VL) домен (VL2) легкой цепи.
В некоторых аспектах антитело к CD3, содержащее первый связывающий домен, содержащий (a) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13 или SEQ ID NO: 50; и (e) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14 или SEQ ID NO: 51, такое как 38E4.v1 MD1 или 38E4.v1 MD4, может иметь второй связывающий домен, который связывается с антигеном клеточной поверхности (например, опухолевым антигеном, например, LY6G6D) на клетке-мишени, отличной от иммунной клетки-эффектора.
В некоторых аспектах антиген клеточной поверхности может экспрессироваться с низким числом копий на клетке-мишени. Например, в некоторых аспектах антиген клеточной поверхности экспрессируется или присутствует в количестве менее чем 35000 копий на клетку-мишень. В некоторых вариантах осуществления антиген клеточной поверхности с низким числом копий составляет от 100 до 35000 копий на клетку-мишень; от 100 до 30000 копий на клетку-мишень; от 100 до 25000 копий на клетку-мишень; от 100 до 20000 копий на клетку-мишень; от 100 до 15000 копий на клетку-мишень; от 100 до 10000 копий на клетку-мишень; от 100 до 5000 копий на клетку-мишень; от 100 до 2000 копий на клетку-мишень; от 100 до 1000 копий на клетку-мишень или от 100 до 500 копий на клетку-мишень. Число копий антигена клеточной поверхности можно определять, например, с помощью стандартного графика Скэтчарда.
Например, в некоторых аспектах антитело к LY6G6D, содержащее связывающий домен, содержащий все шесть из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 2; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 3, может иметь второй связывающий домен, который связывается с CD3. В некоторых аспектах первый связывающий домен, который связывает LY6G6D, содержит по меньшей мере одну (например, 1, 2, 3 или 4) каркасную область тяжелой цепи FR-H1, FR-H2, FR-H3 и FR-H4, содержащую последовательности SEQ ID NO: 34-37, соответственно, и/или по меньшей мере одну (например, 1, 2, 3 или 4) каркасную область легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащую последовательности SEQ ID NO: 38-41, соответственно. В других аспектах первый связывающий домен, который связывает LY6G6D, содержит по меньшей мере одну (например, 1, 2, 3 или 4) каркасную область тяжелой цепи FR-H1, FR-Н2, FR-H3 и FR-H4, содержащую последовательности SEQ ID NO: 34, 58, 36 и 37, соответственно, и/или по меньшей мере одну (например, 1, 2, 3 или 4) каркасной области легкой цепи FR-L1, FR-L2, FR-L3 и FR-L4, содержащую последовательности SEQ ID NO: 38, 61, 40 и 41, соответственно. В некоторых аспектах первый связывающий домен, который связывается с LY6G6D, может, например, содержать (а) домен VH1, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность, SEQ ID NO: 10, и (б) домен VL1, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность, SEQ ID NO: 11, например, как установлено для антитела к LY6G6D 20A12.QNTv12, описанного в настоящем документе. В некоторых аспектах первый связывающий домен, который связывается с LY6G6D, содержит (а) домен VH1, содержащий аминокислотную последовательность, имеющую последовательность SEQ ID NO: 59, и (б) домен VL1, содержащий аминокислотную последовательность, имеющую последовательность SEQ ID NO: 60.
В некоторых аспектах вышеописанного антитела к LY6G6D, имеющего второй связывающий домен, который связывается с CD3, второй домен, связывающийся с CD3, содержит по меньшей мере одну, две, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 13 или SEQ ID NO: 50; и (e) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 14 или SEQ ID NO: 51.
В некоторых аспектах второй домен, связывающийся с CD3, содержит все шесть из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 13; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 14.
В некоторых аспектах второй домен, связывающийся с CD3, содержит связывающийся домен, содержащий все шесть из (a) CDR-H1, содержащей аминокислотную последовательность SEQ ID NO: 15; (б) CDR-H2, содержащей аминокислотную последовательность SEQ ID NO: 16; (в) CDR-H3, содержащей аминокислотную последовательность SEQ ID NO: 17; (г) CDR-L1, содержащей аминокислотную последовательность SEQ ID NO: 12; (д) CDR-L2, содержащей аминокислотную последовательность SEQ ID NO: 50; и (е) CDR-L3, содержащей аминокислотную последовательность SEQ ID NO: 51.
В некоторых случаях второй домен, связывающийся с CD3, содержит домен VH2, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность SEQ ID NO: 20, и/или домен VL2, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность SEQ ID NO: 21. В конкретном случае антитело к CD3 может представлять собой 38E4.v1 MD1 или его производное, или клонально родственное антитело.
В некоторых случаях второй домен, связывающийся с CD3, может иметь домен VH2, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность SEQ ID NO: 20, и/или домен VL2, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности последовательности (например, по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности) с или имеющую последовательность SEQ ID NO: 55. В конкретном случае антитело к CD3 может представлять собой 38E4.v1 MD4 или его производное, или клонально родственное антитело.
В некоторых аспектах второй домен, связывающийся с CD3, может содержать по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 43 или SEQ ID NO: 62; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или по меньшей мере одну (например, 1, 2, 3 или 4) из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 47 или SEQ ID NO: 63; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49.
В некоторых аспектах антитело к CD3 содержит все четыре из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 43; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 47; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49. В некоторых аспектах антитело к LY6G6D может иметь домен VH2, содержащий аминокислотную последовательность SEQ ID NO: 20, и/или домен VL2, содержащий аминокислотную последовательность SEQ ID NO: 21. В других аспектах антитело к LY6G6D может иметь домен VH2, содержащий аминокислотную последовательность SEQ ID NO: 20, и/или домен VL2, содержащий аминокислотную последовательность SEQ ID NO: 55.
В некоторых аспектах антитело к CD3 содержит все четыре из (a) FR-H1, содержащей аминокислотную последовательность SEQ ID NO: 42; (б) FR-H2, содержащей аминокислотную последовательность SEQ ID NO: 62; (в) FR-H3, содержащей аминокислотную последовательность SEQ ID NO: 44; и (г) FR-H4, содержащей аминокислотную последовательность SEQ ID NO: 45, и/или содержит все четыре из (a) FR-L1, содержащей аминокислотную последовательность SEQ ID NO: 46; (б) FR-L2, содержащей аминокислотную последовательность SEQ ID NO: 63; (в) FR-L3, содержащей аминокислотную последовательность SEQ ID NO: 48; и (г) FR-L4, содержащей аминокислотную последовательность SEQ ID NO: 49.
В некоторых вариантах осуществления биспецифическое антитело можно использовать для локализации цитотоксического агента в клетке, которая экспрессирует опухолевый антиген, например, Ly6G6D. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Технологии получения мультиспецифических антител включают, но не ограничиваются этим, рекомбинантную коэкспрессию двух пар тяжелая цепь - легкая цепь иммуноглобулина, имеющих разную специфичность (см. Milstein et al. Nature 305: 537, (1983), WO 93/08829 и Traunecker et al. EMBO J. 10: 3655, (1991)), и конструирование по типу «выступ-во-впадину» (см., например, патент США №5731168). Способ конструирования биспецифических антител «выступ-во-впадину» можно применять для получения первого плеча, содержащего выступ, и второго плеча, содержащего впадину, в которой возможно связывание выступа первого плеча. Выступ мультиспецифических антител по данному изобретению может представлять собой CD3-специфичное плечо в одном варианте осуществления. Альтернативно, выступ мультиспецифических антител по настоящему изобретению может быть плечом к мишени/антигену в одном варианте осуществления. Впадина мультиспецифеских антител по настоящему изобретению может быть CD3-специфичным плечом в одном варианте осуществления. Альтернативно, впадина мультиспецифичных антител по настоящему изобретению может быть плечом к мишени/антигену в одном варианте осуществления. Мультиспецифические антитела также могут быть сконструированы с использованием технологии иммуноглобулина кроссовера (также известный как формат обмена домена Fab или CrossMab) технологии (см., например, в WO 2009/080253; Schaefer et al., Proc. Natl. Acad. Sci. USA, 108:11187-11192 (2011)). Мультиспецифические антитела также можно получать путем конструирования с использованием эффекта электростатического взаимодействия для получения Fc-гетеродимерных молекул антител (WO 2009/089004А1); перекрестного сшивания двух или более антител или фрагментов (см., например, патент США №4676980 и Brennan et al. Science, 229: 81, 1985); использования лейциновых молний для получения биспецифических антител (см., например, Kostelny et al. J. Immunol., 148(5): 1547-1553 (1992)); использования технологии «диател» для получения фрагментов биспецифических антител (см., например, Hollinger et al. Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993)); и использования димеров одноцепочечных Fv (sFv) (см., например, Gruber et al., J. Immunol., 152:5368 (1994)); и получения триспецифических антител, как описано, например, например, в Tutt et al. J. Immunol. 147: 60 (1991).
Сконструированные антитела с тремя или более функциональными антигенсвязывающими сайтами, включая «антитела-осьминоги», также включены в данный документ (см., например, US 2006/0025576 А1).
Антитела или их фрагменты антител могут также включать «FAb двойного действия» или «DAF», содержащие антигенсвязывающий сайт, который связывается с CD3, а также другим отличным антигеном (например, второй биологической молекулой)(смотрите, например, US 2008/0069820).
7. Варианты антител
В некоторых аспектах рассматриваются варианты аминокислотной последовательности антитела к LY6G6D и/или антитела к CD3 по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, например, с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например CD3, такие как антитела TDB по настоящему изобретению или их варианты). Например, может существовать потребность в улучшении аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела можно получать путем внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или с помощью пептидного синтеза. Такие модификации включают, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно проводить любые комбинации делеций, вставок и замен, при условии, что конечная конструкция обладает необходимыми характеристиками, например, антигенсвязывающими.
а. Варианты с заменами, вставками и делециями
В определенных вариантах осуществления предложены варианты антител, содержащие одну или более аминокислотных замен. Сайты, представляющие интерес для заместительного мутагенеза, включают CDR и FR. Консервативные замены приведены в таблице 1 под заголовком «предпочтительные замены». Более существенные изменения приведены в Таблице 1 под заголовком «типовые замены» и дополнительно описаны ниже со ссылкой на классы аминокислотных боковых цепей. Аминокислотные замены можно вносить в представляющее интерес антитело, и проводить скрининг полученных продуктов в отношении необходимой активности, например, сохранения/улучшения связывания антигена, снижения иммуногенности или улучшения АЗКЦ или КЗЦ.
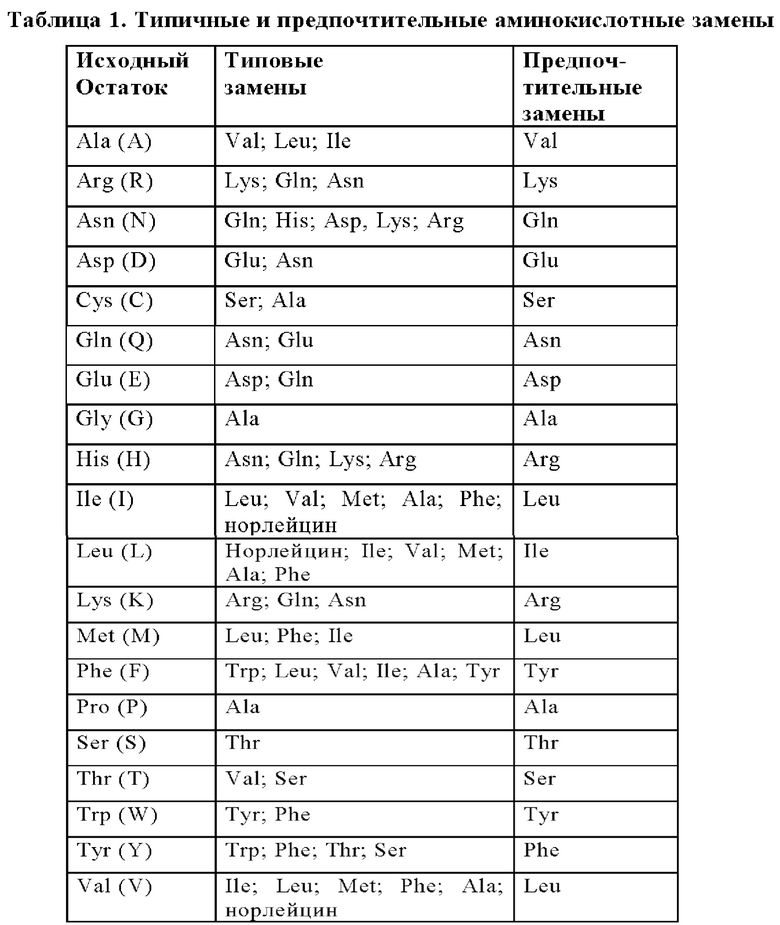
Аминокислоты можно разделить на группы в соответствии с общими свойствами боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены подразумевают замену представителя одного из этих классов представителем другого класса.
Один из типов заместительного варианта включает замену одного или более остатков гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). В общем случае полученные варианты, отобранные для дополнительного исследования, имеют модификации (например, улучшение) определенных биологических свойств (например, повышенную аффинность, сниженную иммуногенность) по сравнению с родительским антителом и/или в значительной степени сохраняют определенные биологические свойства родительского антитела. Типовой заместительный вариант представляет собой антитело с созревшей аффинностью, которое удобно получать, например, используя способы созревания аффинности на основе фагового дисплея, такие как описанные в данном документе. Вкратце, один или более аминокислотных остатков HVR подвергают мутации, а вариантные антитела отображают на поверхности фага и проводят их скрининг в отношении конкретного вида биологической активности (например, аффинности связывания).
Изменения (например, замены) можно осуществлять в CDR, например, чтобы улучшить аффинность антитела. Такие изменения могут быть сделаны в «горячих точках» CDR, т.е. в остатках, кодируемых кодонами, которые с высокой частотой подвергаются мутации во время процесса соматического созревания (смотрите, например, Chowdhury, Methods Mol. Biol. 207:179-196 (2008)), и/или в остатках, которые контактируют с антигеном, а полученный в результате вариант VH или VL исследуют в отношении аффинности связывания. Созревание аффинности путем конструирования и повторного отбора из вторичных библиотек было описано, например, в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, (2001)). В некоторых вариантах осуществления созревания аффинности в вариабельные гены, выбранные для созревания, вносят разнообразие любым из ряда способов (например, ПЦР с внесением ошибок, перетасовки цепей или олигонуклеотид-направленного мутагенеза). Затем создают вторичную библиотеку. После этого библиотеку подвергают скринингу для идентификации любых вариантов антител с необходимой аффинностью. Другой способ внесения разнообразия включает подходы, направленные на CDR, в которых рандомизируют несколько остатков CDR (например, 4-6 остатков за раз). Остатки CDR, вовлеченные в связывание антигена, можно четко идентифицировать, например, используя аланин-сканирующий мутагенез или моделирование. В частности, мишенями часто являются CDR-H3 и CDR-L3.
В некоторых вариантах осуществления замены, вставки или делеции могут находиться в одной или более CDR при условии, что такие изменения по существу не снижают способность антитела связывать антиген. Например, в CDR можно проводить консервативные изменения (например, консервативные замены, предложенные в данном документе), которые существенно не снижают аффинность связывания. Такие изменения могут, например, находиться за пределами контактирующих с антигеном остатков в CDR. В определенных вариантах осуществления вариантных последовательностей VH и VL, представленных выше, каждая CDR остается неизменной или содержит не более одной, двух или трех аминокислотных замен.
Удобный способ идентификации остатков или областей антитела, которые могут быть мишенями для мутагенеза, называется «аланин-сканирующим мутагенезом» и описан Cunningham and Wells (1989) Science, 244:1081-1085. В этом способе идентифицируют остаток или группу целевых остатков (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют их нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), чтобы определить, влияет ли это на взаимодействие антитела с антигеном. Дополнительные замены можно вносить в аминокислотные положения, демонстрирующие функциональную чувствительность к исходным заменам. В альтернативном или дополнительном варианте используют кристаллическую структуру комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут быть нацелены или удалены, как кандидаты для замены. Можно проводить скрининг вариантов для определения наличия у них необходимых свойств.
Вставки аминокислотных последовательностей включают амино- и/или карбокси-концевые слияния с диапазоном длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутрь последовательности одного или множества аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым остатком метионила. Другие инсерционные варианты молекулы антитела включают слияние N- или С-конца антитела с ферментом (например, для терапии ADEPT) или полипептидом, который увеличивает период сывороточного полувыведения антитела.
б. Варианты по гликозилированию
В некоторых вариантах осуществления антитело к LY6G6D и/или антитело к CD3 по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и вторая биологическая молекула, например, CD3) могут быть изменены для повышения или снижения степени, до которой антитело гликозилируется. Добавление или удаление сайтов гликозилирования в антителе к LY6G6D согласно настоящему изобретению традиционно можно осуществлять путем изменения аминокислотной последовательности так, чтобы создать или удалить один или более сайтов гликозилирования.
Если антитело содержит Fc-область, можно изменять присоединенный к ней углевод. Нативные антитела, вырабатываемые клетками млекопитающих, обычно содержат разветвленный биантеннарный олигосахарид, который обычно присоединен посредством N-связи к Asn297 СН2-домена Fc-области. Смотрите, например, Wright et ah TIBTECH 15:26-32 (1997). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стволе» биантеннарной олигосахаридной структуры. В некоторых вариантах осуществления могут быть проведены модификации олигосахарида в антителе, согласно изобретению, для создания вариантов антитела с определенными улучшенными свойствами.
В одном варианте осуществления предложены варианты антител к LY6G6D и/или антител к CD3, содержащие углеводную структуру с недостаточным количеством фукозы, присоединенной (непосредственно или косвенно) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют путем расчета среднего количества фукозы в сахарной цепи в Asn297 относительно суммарного количества всех гликоструктур, присоединенных к Asn 297 (например, сложных, гибридных структур и структур с высоким содержанием маннозы), согласно результатам измерения способом времяпролетной масс-спектрометрии ВП-МАЛДИ, например, как описано в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному приблизительно в положении 297 в Fc-области (нумерация согласно EU остатков Fc-области); при этом Asn297 также может быть расположен на приблизительно ±3 аминокислоты выше или ниже положения 297, т.е. между положениями 294 и 300, вследствие незначительных вариаций последовательностей в антителах. Такие варианты по фукозилированию могут иметь улучшенную функцию АЗКЦ. См., например, публикации заявок на выдачу патента США №№ US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, относящихся к «дефукозилированным» или «фукозодефицитным» вариантам антител, включают: США 2003/0157108; WO 2000/61739; WO 2001/29246; США 2003/0115614; США 2002/0164328; США 2004/0093621; США 2004/0132140; США 2004/0110704; США 2004/0110282; США 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры линий клеток, способных вырабатывать дефукозилированные антитела, включают клетки СНО Lec13, дефицитные по фукозилированию белка (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); заявка на патент США № US 2003/0157108 Al, Presta, L; и WO 2004/056312 A1, Adams et al., в особенности пример 11), и нокаутные линии клеток, такие как клетки СНО с нокаутом гена альфа-1,6-фукозилтрансферазы, FUT8 (смотрите, например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94(4):680-688 (2006); и WO 2003/085107).
Дополнительно предложены варианты антител к LY6G6D и/или антител к CD3 с разделенными пополам олигосахаридами, например, в которых биантеннальный олигосахарид, присоединенный к Fc-области антитела, разделен GlcNAc. Такие варианты антител могут иметь сниженный уровень фукозилирования и/или улучшенную функцию АЗКЦ. Примеры таких вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США №6602684 (Umana et al.,); и патенте США №2005/0123546 (Umana et al). Также предложены варианты антител с по меньшей мере одним остатком галактозы в олигосахариде, присоединенным к Fc-области. Такие варианты антител могут иметь улучшенную функцию КЗЦ. Такие варианты антител описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
в. Варианты Fc-области
В некоторых вариантах осуществления одна или более аминокислотных модификаций могут быть введены в Fc-область антитела к LY6G6D и/или антитела к CD3 настоящего изобретения (например, биспецифического антитела к LY6G6D настоящего изобретения, которое связывается с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например, CD3, при этом образуя вариант Fc-области (см., например, US 2012/0251531). Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одной или более аминокислотных положениях.
В некоторых вариантах осуществления настоящее изобретение рассматривает вариант антитела к LY6G6D и/или антитела к CD3, который обладает некоторыми, но не всеми эффекторными функциями, что делает его подходящим кандидатом для практических применений, в которых важное значение имеет период полувыведения антитела in vivo, хотя отдельные эффекторные функции (такие как комплемент и АЗКЦ) не являются необходимыми или являются вредными. Для подтверждения снижения/уменьшения КЗЦ- и/или АЗКЦ-активности можно проводить анализ цитотоксичности in vitro и/или in vivo. Например, можно проводить анализ связывания Fc-рецептора (FcR), чтобы убедиться в том, что антитело не способно связывать FcγR (следовательно, вероятно, не обладает АЗКЦ-активностью), но сохраняет способность связывать FcRn. Первичные клетки для опосредования АЗКЦ, NK-клетки, экспрессируют только Fc(RIII, тогда как моноциты экспрессируют Fc(RI, Fc(RII и Fc(RIII. Экспрессия FcR на гемопоэтических клетках описана в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol. 9:457-492 (1991). Неограничивающие примеры способов in vitro анализа для оценки АЗКЦ-активности представляющей интерес молекулы, описаны в патенте США №5500362 (смотрите, например, Hellstrom, I. et al. Proc. Nat'I Acad. Sci. USA 83:7059-7063 (1986)) и Hellstrom, I et al., Proc. Nat'l Acad. Sci. USA 82:1499-1502 (1985); 5,821,337 (см. Bruggemann, M. et al., J. Exp.Med. 166:1351-1361 (1987)). В альтернативном варианте можно использовать нерадиоактивные способы анализа (смотрите, например, нерадиоактивный анализ цитотоксичности ACTI™ для проточной цитометрии (CellTechnology, Inc., Маунтин-Вью, Калифорния; и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Мадисон, Висконсин). Эффекторные клетки, подходящие для такого анализа, включают мононуклеарные клетки периферической крови (МКПК) и естественные клетки-киллеры (NK). Альтернативно или дополнительно, АЗКЦ-активность молекулы, представляющей интерес, можно оценить in vivo, например, в модели на животных, такой как та, которая раскрыта в Clynes et al. Proc. Nat'I Acad. Sci. USA 95:652-656 (1998). Также можно проводить анализ связывания C1q, чтобы подтвердить, что антитело не способно связывать C1q и, следовательно, у него отсутствует КЗЦ-активность. Смотрите, например, ИФА-анализ связывания C1q и С3с в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента может быть выполнен анализ КЗЦ (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996); Cragg, M.S. et al. Blood. 101:1045-1052 (2003); и Cragg, M.S. and M.J. Glennie Blood. 103:2738-2743 (2004)). Определение связывания с FcRn и in vivo клиренса/времени полувыведения также может быть осуществлено, используя способы, известные в данной области техники (см., например, Petkova, S.B. et al., Int'l. Immunol. 18(12): 1759-1769 (2006)).
Антитела со сниженной эффекторной функцией включают те, которые содержат замены одного или более остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 (патенты США №№6737056 и 8219149). Такие мутанты Fc включают мутантов Fc с заменами в двух или более аминокислотных положениях 265, 269, 270, 297 и 327, в том числе так называемые мутанты Fc «DANA» с заменой остатков 265 и 297 на аланин (патенты США №№7332581 и 8219149).
В некоторых вариантах осуществления пролин в положении 329 человеческой Fc-области дикого типа в антителе заменен глицином или аргинином или аминокислотным остатком, достаточно крупным, чтобы разрушить «сэндвич» пролина в области раздела Fc/Fc-гамма-рецептор, который образуется между пролином 329 из Fc и остатками триптофана Trp87 и Trp110 из FcgRIII (Sondermann et al. Nature. 406, 267-273, 2000). В определенных вариантах осуществления антитело содержит по меньшей мере одну дополнительную аминокислотную замену. В одном варианте осуществления изобретения дополнительная аминокислотная замена представляет собой S228P, Е233Р, L234A, L235A, L235E, N297A, N297D или P331S, а в другом варианте осуществления по меньшей мере одна дополнительная аминокислотная замена представляет собой L234A и L235A Fc-области человеческого IgG1 или S228P и L235E Fc-области человеческого IgG4 (см., например, US 2012/0251531), и в другом варианте осуществления по меньшей мере одна дополнительная аминокислотная замена представляет собой L234A и L235A, и P329G Fc-области человеческого IgG1.
Описаны некоторые варианты антител с улучшенным или уменьшенным связыванием с FcR. (Смотрите, например, патент США №6737056, WO 2004/056312 и Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001).)
В определенных вариантах осуществления вариант антитела содержит Fc-область с одной или более аминокислотными заменами, которые улучшают АЗКЦ, например, с заменами в положениях 298, 333 и/или 334 Fc-области (нумерация остатков EU).
В некоторых вариантах осуществления в Fc-области проведены изменения, которые приводят к изменению (т.е. к улучшению или снижению) связывания C1q и/или комплементзависимой цитотоксичности (КЗЦ), например как описано в патенте США №6194551, WO 99/51642 и Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Антитела с повышенным временем полувыведения и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Эти антитела содержат Fc-домен с одной или более заменами, которые улучшают связывание Fc-области с FcRn. Такие Fc-варианты включают варианты с заменами в одном или более остатках Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, заменой остатка 434 Fc-области (патент США №7371826).
Смотрите также Duncan & Winter, Nature 322:738-40 (1988); патент США №5648260; патент США №5624821; и WO 94/29351 в отношении других примеров вариантов по Fc-области.
В некоторых аспектах антитело к LY6G6D и/или антитело к CD3 (например, биспецифическое антитело к LY6G6D) содержит Fc-область, содержащую мутацию N297G. В некоторых вариантах осуществления антитело к LY6G6D, содержащее мутацию N297G, содержит LY6G6D-специфичное плечо, содержащее первый связывающий домен, содержащий следующие шесть CDR: (а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1; (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и (e) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3; и CD3-специфичное плечо.
В некоторых вариантах осуществления антитело к LY6G6D, содержащее мутацию N297G, содержит CD3-специфичное плечо, содержащее первый связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 10, и (б) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 11, и CD3-специфичное плечо. В других вариантах осуществления антитело к LY6G6D, содержащее мутацию N297G, содержит CD3-специфичное плечо, содержащее первый связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 59, и (б) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 60, и CD3-специфичное плечо.
В некоторых вариантах осуществления антитело к LY6G6D, содержащее мутацию N297G, содержит один или более константных доменов тяжелой цепи, причем один или более константных доменов тяжелой цепи выбраны из первого домена СН1 (СН11) первого домена СН2 (СН21), первого домена СН3 (СН31), второго домена СН1 (СН12) второго домена СН2 (СН22) и второго домена СН3 (СН32). В некоторых аспектах по меньшей мере один из одного или более константных доменов тяжелой цепи сопряжен с константным доменом другой тяжелой цепи. В некоторых аспектах каждый из доменов СН31 и СН32 содержит выступ или впадину, и причем выступ или впадина в домене СН31 располагается во впадине или выступе, соответственно, в домене СН32. В некоторых аспектах домены СН31 и СН32 сходятся на поверхности между указанным выступом и впадиной. В некоторых аспектах каждый из доменов СН21 и СН22 содержит выступ или впадину, и причем выступ или впадина в домене СН21 располагается во впадине или выступе, соответственно, в домене СН22. В других случаях домены СН21 и СН22 встречаются на поверхности между указанным выступом и впадиной. В некоторых аспектах антитело к LY6G6D представляет собой антитело IgG1.
В некоторых вариантах осуществления антитело к CD3, содержащее мутацию N297G, содержит LY6G6D-специфичное плечо, содержащее первый связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 10 или SEQ ID NO: 59, и (б) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 11 или SEQ ID NO; 60, и CD3-специфичное плечо, где (а) LY6G6D-специфичное плечо содержит мутации по типу замены T366S, L368A, Y407V и N297G, и (б) CD3-специфичное плечо содержит мутации по типу замены T366W и N297G.
В других вариантах осуществления антитело к CD3, содержащее мутацию N297G, содержит LY6G6D-специфичное плечо, содержащее первый связывающий домен, содержащий (а) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 10 или SEQ ID NO: 59, и (б) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 11 или SEQ ID NO; 60, и CD3-специфичное плечо, причем (а) LY6G6D-специфичное плечо содержит мутации по типу замены T366W и N297G, и (б) CD3-специфичное плечо содержит мутации по типу замены T366S, L368A, Y407V и N297G.
г. Варианты антител, сконструированные с использованием цистеина
В некоторых вариантах осуществления может быть желательно создать антитела, сконструированные с использованием цистеина, например, «thioMAb,» в которых один или более остатков замещены остатками цистеина. В конкретных вариантах осуществления, замещенные остатки находятся в доступных сайтах антитела. При замене этих остатков цистеином реакционноспособные тиоловые группы располагаются в доступных сайтах антитела и могут быть использованы для конъюгации антитела с другими фрагментами, такими как фрагменты лекарственного средства или фрагменты линкер-лекарственное средство, для создания иммуноконъюгата, как дополнительно описано в настоящем документе. В определенных вариантах осуществления цистеином может быть замещен любой из следующих остатков: V205 (нумерация по Кабату) легкой цепи; А118 (нумерация EU) тяжелой цепи; и S400 (нумерация EU) Fc-области тяжелой цепи. Антитела, сконструированные с использованием остатков цистеина, можно получить, как описано, например, в патенте США №7521541.
д. Производные антител
В некоторых вариантах осуществления антитело к LY6G6D согласно изобретению (например, биспецифическое антитело к LY6G6D согласно изобретению, которое связывается с LY6G6D, предпочтительно с высокой аффинностью, (например, 20A12.QNTv12) и второй биологической молекулой например, CD3), предложенное в данном документе, может быть дополнительно модифицировано, чтобы содержать дополнительные небелковые компоненты, которые известны в данной области техники и легко доступны. Фрагменты, подходящие для дериватизации антитела, включают, без ограничения, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, но не ограничиваются этим, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или случайные сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры пропропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества при производстве из-за его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество молекул полимера, присоединенных к антителу, может варьироваться, и в случае присоединении более чем одного полимера они могут представлять собой одинаковые или разные молекулы. В общем случае количество и/или тип полимеров, применяемых для дериватизации, можно определить на основании соображений, которые включают, но не ограничиваются этим, конкретные свойства или функции антитела, которые нужно улучшить, то, будут ли производное антитела применять в терапии в определенных условиях и т.д.
В другом варианте осуществления предложены конъюгаты антитела и небелкового фрагмента, который можно избирательно нагревать путем облучения. В одном варианте осуществления небелковый фрагмент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Излучение может иметь любую длину волны и включает, без ограничения, длины волн, которые не вредят обычным клеткам, но которые нагревают небелковый фрагмент до температуры, при которой погибают клетки, расположенные вблизи конъюгата антитело-небелковый фрагмент.
8. Заряженные области
В некоторых аспектах связывающий домен, который связывает LY6G6D или CD3, содержит VH1, содержащий заряженную область (CR1), и VL1, содержащий заряженную область (CR2), где CR1 в VH1 образует зарядовую пару с CR2 в VL1. В некоторых аспектах CR1 содержит остаток основной аминокислоты, a CR2 содержит остаток кислой аминокислоты. В некоторых аспектах CR1 содержит мутацию по типу замены Q39K (нумерация по Кабату). В некоторых аспектах CR1 состоит из мутации по типу замены Q39K. В некоторых аспектах CR2 содержит мутацию по типу замены Q38E (нумерация по Кабату). В некоторых аспектах CR2 состоит из мутации по типу замены Q38E. В некоторых аспектах второй связывающий домен, который связывает CD3, содержит VH2, содержащий заряженную область (CR3), и домен VL2, содержащий заряженную область (CR4), причем CR4 в VL2 образует зарядовую пару с CR3 в VH2. В некоторых аспектах CR4 содержит остаток основной аминокислоты, a CR3 содержит остаток кислой аминокислоты. В некоторых аспектах CR4 содержит мутацию по типу замены Q38K (нумерация по Кабату). В некоторых аспектах CR4 состоит из мутации по типу замены Q38K. В некоторых аспектах CR3 содержит мутацию по типу замены Q39E (нумерация по Кабату). В некоторых аспектах CR3 состоит из мутации по типу замены Q39E. В некоторых аспектах домен VL1 связан с константным доменом (CL1) легкой цепи (CL), и VH1 связан с первым константым доменом (CH1) тяжелой цепи (СН1), причем CL1 содержит заряженную область (CR5), и СН1 содержит заряженную область (CR6), и при этом CR5 в CL1 образует зарядовую пару с CR6 в CH11. В некоторых аспектах CR5 содержит остаток основной аминокислоты, a CR6 содержит остаток кислой аминокислоты. В некоторых аспектах CR5 содержит мутацию по типу замены V133K (нумерация согласно EU). В некоторых аспектах CR5 состоит из мутации по типу замены V133K. В некоторых аспектах CR6 содержит мутацию по типу замены S183E (нумерация согласно EU). В некоторых аспектах CR6 состоит из мутации по типу замены S183E.
В других аспектах домен VL2 связан с доменом CL (CL2), и VH2 связан с доменом СН1 (CH12), причем CL2 содержит заряженную область (CR7), и CH12 содержит заряженную область (CR8), и при этом CR8 в CH12 образует зарядовую пару с CR7 в CL2. В некоторых аспектах CR8 содержит остаток основной аминокислоты, a CR7 содержит остаток кислой аминокислоты. В некоторых аспектах CR8 содержит мутацию по типу замены S183K (нумерация согласно EU). В некоторых аспектах CR8 состоит из мутации по типу замены S183K. В некоторых аспектах CR7 содержит мутацию по типу замены V133E (нумерация согласно EU). В некоторых аспектах CR7 состоит из мутации по типу замены V133E.
В других аспектах домен VL2 связан с доменом CL (CL2), и VH2 связан с доменом CH1 (CH12), причем (a) CL2 содержит одну или более мутаций на аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация согласно EU), и (б) CHl2 содержит одну или более мутаций на аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация согласно EU). В некоторых аспектах CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F и/или T178V. В некоторых аспектах CL2 содержит следующие мутации по типу замены: F116A, L135V, S174A, S176F и T178V. В некоторых аспектах CH12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A и/или V185A. В некоторых аспектах CH12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A и V185A.
В других аспектах связывающий домен, который связывает LY6G6D или CD3, содержит домен VH (VH1), содержащий заряженную область (CR1), и домен VL (VL1), содержащий заряженную область (CR2), причем CR2 в VL1 образует зарядовую пару с CR1 в VH1. В некоторых аспектах CR2 содержит остаток основной аминокислоты, a CR1 содержит остаток кислой аминокислоты. В некоторых аспектах CR2 содержит мутацию по типу замены Q38K (нумерация по Кабату). В некоторых аспектах CR2 состоит из мутации по типу замены Q38K. В некоторых аспектах CR1 содержит мутацию по типу замены Q39E (нумерация по Кабату). В некоторых аспектах CR1 состоит из мутации по типу замены Q39E. В некоторых аспектах второй связывающий домен, который связывает CD3, содержит домен VH (VH2), содержащий заряженную область (CR3), и домен VL (VL2), содержащий заряженную область (CR4), причем CR3 в VH2 образует зарядовую пару с CR4 в VL2. В некоторых аспектах CR3 содержит остаток основной аминокислоты, a CR4 содержит остаток кислой аминокислоты. В некоторых аспектах CR3 содержит мутацию по типу замены Q39K (нумерация по Кабату). В некоторых аспектах CR3 состоит из мутации по типу замены Q39K. В некоторых аспектах CR4 содержит мутацию по типу замены Q38E (нумерация по Кабату). В некоторых аспектах CR4 состоит из мутации по типу замены Q38E. В некоторых аспектах домен VL1 связан с константным доменом (CL1) легкой цепи (CL), и VH1 связан с первым константным доменом (СН11) тяжелой цепи (CH1), причем CL1 содержит заряженную область (CR5), и CH11 содержит заряженную область (CR6) и при этом CR6 в CH11 образует зарядовую пару с CR5 в CL1. В некоторых аспектах CR6 содержит остаток основной аминокислоты, a CR5 содержит остаток кислой аминокислоты. В некоторых аспектах CR6 содержит мутацию по типу замены S183K (нумерация согласно EU). В некоторых аспектах CR6 состоит из мутации по типу замены S183K. В некоторых аспектах CR5 содержит мутацию по типу замены V133E (нумерация согласно EU). В некоторых аспектах CR5 состоит из мутации по типу замены V133E.
В других аспектах домен VL2 связан с доменом CL (CL2), и VH2 связан с доменом CH1 (CH12), причем CL2 содержит заряженную область (CR7), a CH12 содержит заряженную область (CR8), и при этом CR7 в CL2 образует зарядовую пару с CR8 в CH12. В некоторых аспектах CR7 содержит остаток основной аминокислоты, и CR8 содержит кислотный остаток. В некоторых аспектах CR7 содержит мутацию по типу замены V133K (нумерация согласно EU). В некоторых аспектах CR7 состоит из мутации по типу замены V133K. В некоторых аспектах CR8 содержит мутацию по типу замены S183E (нумерация согласно EU). В некоторых аспектах CR8 состоит из мутации по типу замены S183E.
В других аспектах домен VL2 связан с доменом CL (CL2), и VH2 связан с доменом СН1 (CH12), причем (a) CL2 содержит одну или более мутаций на аминокислотных остатках F116, L135, S174, S176 и/или Т178 (нумерация согласно EU), и (б) CH12 содержит одну или более мутаций на аминокислотных остатках А141, F170, S181, S183 и/или V185 (нумерация согласно EU). В некоторых аспектах CL2 содержит одну или более следующих мутаций по типу замены: F116A, L135V, S174A, S176F и/или T178V. В некоторых аспектах CL2 содержит следующие мутации по типу замены: F116A, L135V, S174A, S176F и T178V. В некоторых аспектах CH12 содержит одну или более следующих мутаций по типу замены: A141I, F170S, S181M, S183A и/или V185A. В некоторых аспектах CH12 содержит следующие мутации по типу замены: A141I, F170S, S181M, S183A и V185A. В некоторых аспектах антитело к FcRH5 содержит один или более константных доменов тяжелой цепи, причем один или более константных доменов тяжелой цепи выбраны из первого домена СН2 (СН21), первого домена СН3 (СН31), второго домена СН2 (СН22) и второго домена СН3 (СН32). В некоторых аспектах по меньшей мере один из одного или более константных доменов тяжелой цепи сопряжен с константным доменом другой тяжелой цепи. В некоторых аспектах каждый из СН31 и СН32 содержит выступ (Р1) или впадину (С1), и причем Р1 или С1 в СН31 располагается в С1 или Р1, соответственно, в СН32. В некоторых аспектах СН31 и СН32 сходятся на поверхности раздела между Р1 и С1. В некоторых аспектах каждый из СН21 и СН22 содержит (Р2) или впадину (С2), и причем Р2 или С2 в СН21 располагается в C2 или Р2, соответственно, в СН22. В некоторых аспектах СН21 и СН22 находятся на поверхности раздела между Р2 и С2.
А. Рекомбинантные способы и композиции
Антитела к LY6G6D по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например, CD3) и/или антитела к CD3 по настоящему изобретению (например, 38E4v1 MD1 (MD1) и 38E4v1 MD4 (MD4)) можно получать с помощью рекомбинантных способов и композиций, например, как описано в патенте США №4816567. В одном варианте осуществления предложена выделенная нуклеиновая кислота, кодирующая описанное в данном документе антитело к LY6G6D. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом варианте осуществления предложена выделенная нуклеиновая кислота, кодирующая описанное в данном документе антитело к CD3. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В дополнительном варианте осуществления предложен один или более векторов (например, векторов экспрессии), содержащих такую нуклеиновую кислоту. В дополнительном варианте осуществления предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком варианте осуществления клетка-хозяин содержит (например, была трансформирована при помощи): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном варианте осуществления клетка-хозяин является эукариотической, например, клеткой яичника китайского хомяка (СНО) или лимфоидной клеткой (например, клеткой YO, NSO, Sp20). В одном варианте осуществления предложен способ получения антитела к LY6G6D, причем указанный способ включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую предложенное выше антитело, в условиях, подходящих для экспрессии антитела, и, необязательно, выделение антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантной продукции антитела к LY6G6D и/или антитела к CD3 нуклеиновая кислота, кодирующая антитело, например, как описано выше, является выделенной и вставлена в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть легко выделена и секвенирована с использованием традиционных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
1. Деухклеточные способы для изготовления биспецифических антител
В некоторых аспектах антитело по настоящему изобретению (например, LY6G6D TDB, например, LY6G6D TDB с CD3-специфичным плечом и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)) изготавливают с помощью способа, включающего две линии клеток-хозяев. В некоторых аспектах первое плечо антитела (например, первое плечо, содержащее область впадины) получают в первой линии клеток-хозяев, а второе плечо антитела (например, второе плечо, содержащее область выступа) получают во второй линии клеток-хозяев. Плечи антитела очищают из линий клеток-хозяев и собирают in vitro.
2. Одноклеточные способы для изготовления биспецифических антител
В некоторых аспектах антитело по настоящему изобретению (например, LY6G6D TDB, например, LY6G6D TDB с CD3-специфичным плечом (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)), изготавливают с помощью способаа, включающего одну линию клеток-хозяев. В некоторых аспектах первое плечо антитела (например, первое плечо, содержащее область впадины) и второе плечо антитела (например, второе плечо, содержащее область выступа) получают в и очищают из одной линии клеток-хозяев. Предпочтительно первое плечо и второе плечо экспрессируются на сравнимых уровнях в клетке-хозяине, например, оба экспрессируются на высоком уровне в клетке-хозяине. Подобные уровни экспрессии увеличивают вероятность эффективного получения TDB и снижают вероятность ошибочного спаривания легких цепей (LC) компонентов TDB. Как первое плечо, так и второе плечо антитела могут дополнительно содержать мутации аминокислотных замен, вводящие зарядовые пары, как описано в разделе IIБ (8) в настоящем документе. Зарядовые пары вызывают спаривание родственных пар тяжелой и легкой цепей каждого плеча биспецифического антитела, при этом минимизируя ошибочное спаривание.
3. Клетки-хозяева
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела могут вырабатываться в бактериях, в частности, если не требуется гликозилирование и Fc-эффекторная функция. Экспрессию фрагментов антитела и полипептидов в бактериях см., например, в патентах США №5648237, 5789199 и 5840523. (См. также Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), стр. 245-254, где описана экспрессия фрагментов антитела в Е. coli). После экспрессии антитело можно выделять из бактериальной клеточной пасты в растворимую фракцию и дополнительно очищать.
Помимо прокариот, эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии кодирующих антитела векторов, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что приводит к получению антитела с частично или полностью человеческим профилем гликозилирования. См. Gerngross, Nat. Biotech. 22:1409-1414 (2004), и Li et al., Nat. Biotech. 24:210-215 (2006).
Клетки-хозяева, пригодные для экспрессии гликозилированного антитела, также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
Культуры клеток растений также можно использовать в качестве хозяев. Смотрите, например, патенты США №№5959177, 6040498, 6420548, 7125978 и 6417429 (в которых описана технология PLANTIBODIES™ для получения антител в трансгенных растениях).
Клетки позвоночных также можно использовать в качестве хозяев. Например, можно использовать линии клеток млекопитающих, которые адаптированы для роста в суспензии. Другими примерами полезных линий клеток-хозяев млекопитающих являются линия CV1 почки обезьяны, трансформированная SV40 (COS-7); линия эмбриональных клеток почки человека (293 или клетки 293, как описано, например, в Graham et al., J. Gen Virol. 36:59 (1977)); клетки почек детенышей хомяка (BHK); клетки сертоли мыши (клетки ТМ4, как описано, например, в Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK; клетки печени крысы баффало (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, как описано, например, в Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982); клетки MRC 5; и клетки FS4. Другие подходящие линии клеток-хозяев млекопитающих включают клетки яичника китайского хомяка (СНО), включая DHFR- клетки СНО (Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); и линии клеток миеломы, такие как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для получения антител, см., например, в Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp. 255-268 (2003).
Б. Анализы
Антитела к LY6G6D по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12) и второй биологической молекулой, например CD3, такие как антитела TDB по настоящему изобретению или их варианты), представленные в настоящем документе, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы по их физико-химическим свойствам и/или биологической активности с помощью различных анализов, известных в данной области техники.
1. Анализы связывания и другие анализы
В одном аспекте антитело к LY6G6D и антитело к CD3 по настоящему изобретению тестируют на их антигенсвязывающую активность, например, с помощью известных способов, таких как ИФА, вестерн-блоттинг и т.д.
В другом аспекте конкурентные анализы можно использовать для идентификации антитела, которое конкурирует с антителом к LY6G6D по настоящему изобретению для связывания с LY6G6D, или для идентификации антитела, которое конкурирует с антителом к CD3 по настоящему изобретению для связывания с CD3.
В иллюстративном конкурентном анализе иммобилизованный LY6G6D инкубируют в растворе, содержащем первое меченое антитело, которое связывается с LY6G6D и вторым немеченым антителом, которое тестируется на его способность конкурировать с первым антителом за связывания с LY6G6D. Второе антитело может присутствовать в гибридомном супернатанте. В качестве контроля иммобилизованную LY6G6D инкубируют в растворе, содержащем первое меченное антитело, но не содержащем второе немеченное антитело. После инкубации в условиях, обеспечивающих связывание первого антитела с LY6G6D, удаляют избыток несвязанного антитела и измеряют количество метки, связанной с иммобилизованной LY6G6D. Если количество метки, связанной с иммобилизованной LY6G6D, существенно снижается в исследуемом образце по сравнению с контрольным образцом, то это указывает на то, что второе антитело конкурирует с первым антителом за связывание с LY6G6D. См., например, Harlow and Lane (1988) Antibodies: A Laboratory Manual. Ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Другой иллюстративный конкурентный анализ включает иммобилизованное CD3 и первое меченое антитело, которое связывается с CD3, причем анализ проводят, как описано выше.
2. Анализ активности
В одном аспекте представлены анализы для идентификации их антител к LY6G6D, обладающих биологической активностью. Биологическая активность может включать, например, связывание с LY6G6D (например, LY6G6D на поверхности опухоли) или его пептидным фрагментом или in vivo, in vitro, или ex vivo. В случае мультиспецифического (например, биспецифического) антитела к LY6G6D по настоящему изобретению (например, антитело TDB, имеющее одно LY6G6D-специфичное плечо, например, 20A12.QNTV12, и одно плечо, которое распознает вторую биологическую молекулу, например, антиген клеточной поверхности, например, CD3), биологическая активность может также включать, например, активацию эффекторных клеток (например, активацию Т-клеток (например, Т-клеток CD8+и/или CD4+)), расширение популяции эффекторных клеток (т.е. увеличение количества Т-клеток), уменьшение популяции целевых клеток (т.е. снижение популяции клеток, экспрессирующих LY6G6D на их клеточных поверхностях) и/или уничтожение клеток-мишеней. Предложены антитела, обладающие такой биологической активностью in vivo и/или in vitro. В некоторых вариантах осуществления антитело по настоящему изобретению тестируют на такую биологическую активность, как подробно описано в приведенных ниже примерах.
Кроме того, клетки можно промыть в среде RPMI, содержащей 10% FBS, дополненной GlutaMax, пенициллином и стрептомицином, и добавить ~0,2 миллиона суспендированных клеток в 96-луночный планшет с U-образным дном. Клетки можно культивировать в RPMI1640, дополненной 10% FBS, при 37°С в увлажненном стандартном инкубаторе для культивирования клеток. В случае анализа уничтожения клеток BJAB 20000 клеток BJAB можно инкубировать с эффекторными клетками, или как человеческими МНПК (huPBMC), или очищенными Т-клетками, в указанном соотношении для каждого анализа, в присутствии различных концентраций антител TDB в течение 24 часов.
В. Иммуноконъюгаты
В данном изобретении также предложены иммуноконъюгаты, содержащие предложенное в данном документе антитело к LY6G6D и/или антитело к CD3, конъюгированное с одним или более цитотоксическими агентами, например, химиотерапевтическими агентами или лекарственными препаратами, ингибиторами роста, токсинами (например, белковыми токсинами, ферментативно активными токсинами бактериального, грибкового, растительного или животного происхождения или их фрагментами) или радиоактивными изотопами.
В одном варианте осуществления иммуноконъюгат представляет собой конъюгат антитело - лекарственный препарат (ADC), в котором антитело конъюгировано с одним или более лекарственными препаратами, включая, но не ограничиваясь этим, майтанзиноид (смотрите патенты США №№5208020, 5416064 и европейский патент ЕР 0 425 235 В1); ауристатин, такой как лекарственный фрагменты монометилауристатина DE и DF (ММАЕ и MMAF) (смотрите патенты США №№5635483, 5780588 и 7498298); доластатин; калихеамицин или его производное (смотрите патенты США №№5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman et al., Cancer Res. 53:3336- 3342 (1993); и Lode et al., Cancer Res. 58:2925- 2928 (1998)); антрациклин, такой как дауномицин или доксорубицин (смотрите Kratz et al., Current Med. Chem. 13:477- 523 (2006); Jeffrey et al., Bioorganic & Med. Chem. Letters 16:358- 362 (2006); Torgov et al., Bioconj. Chem. 16:717- 721 (2005); Nagy et al., Proc. Natl. Acad. Sci. USA 97:829- 834 (2000); Dubowchik et al., Bioorg. & Med. Chem. Letters 12:1529- 1532 (2002); King et al., J. Med. Chem. 45:4336- 4343 (2002); и патент США №6,630,579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и СС1065.
В другом варианте осуществления иммуноконъюгат содержит антитело к LY6G6D и/или антитело к CD3, описанное в данном документе, конъюгированное с ферментативно активным токсином или его фрагментом, включая, но не ограничиваясь этим, цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из момордики харантской, курцин, кротин, ингибитор из Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
В другом варианте осуществления иммуноконъюгат содержит антитело к LY6G6D и/или антитело к CD3, описанное в данном документе, конъюгированное с радиоактивным атомом с образованием радиоконъюгата. Для получения радиоконъюгатов доступен ряд радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если радиоконъюгат используют в целях обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, tc99m или I123, или спиновую метку для ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как снова йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента можно создавать, используя ряд бифункциональных агентов, связывающих белок, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидо соединения (такие, как бис(п-азидобензоил)-гександиамин), производные бис-диазония (такие как бис(п-диазоний бензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой типовой хелатирующий агент для конъюгации радионуклида с антителом. Смотрите WO 94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотно-лабильный линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); патент США №5208020).
В контексте данного документа под иммуноконъюгатами или ADC прямо подразумевают, без ограничения конъюгатами, полученными с помощью перекрестно-сшивающих реагентов, помимо прочего, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A).
Г. Способы и композиции для диагностики и выявления
В некоторых вариантах осуществления любое из антитела к LY6G6D и/или антитела к CD3 по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например, CD3) пригодны для обнаружения присутствия LY6G6D и/или CD3 в биологическом образце. В контексте данного документа термин «выявление» включает количественное или качественное выявление. В определенных вариантах осуществления биологический образец включает клетку или ткань.
В одном варианте осуществления предложено антитело к LY6G6D для применения в способе диагностики или обнаружения. В дополнительном аспекте предложен способ обнаружения наличия LY6G6D в биологическом образце. В определенных вариантах осуществления указанный способ включает приведение биологического образца в контакт с описанным в данном документе антителом к LY6G6D в условиях, обеспечивающих возможность связывания антитела к LY6G6D с LY6G6D, и обнаружение того, образовался ли комплекс между антителом к LY6G6D и LY6G6D. Такой способ может быть in vitro или in vivo способом.
В другом варианте осуществления предложено антитело к CD3 для применения в способе диагностики или обнаружения. В дополнительном аспекте предложен способ обнаружения наличия CD3 в биологическом образце. В определенных вариантах осуществления указанный способ включает приведение биологического образца в контакт с описанным в данном документе антителом к CD3 в условиях, обеспечивающих возможность связывания антитела к CD3 с CD3, и обнаружение того, образовался ли комплекс между антителом к CD3 и CD3. Такой способ может быть in vitro или in vivo способом.
В некоторых вариантах осуществления предложены меченые антитела к LY6G6D и/или антитела к CD3. Метки включают, но не ограничиваются этим, метки или фрагменты, выявляемые прямым образом (например, флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, выявляемые непрямым образом, например, посредством ферментативной реакции или молекулярного взаимодействия. Типовые метки включают, но не ограничиваются этим, радиоактивные изотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светляков и бактериальную люциферазу (патент США №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизозим, сахаридооксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат дегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сопряженные с ферментом, который использует пероксид водорода для окисления предшественника красителя, такой как HRP, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
Д. Фармацевтические составы
Фармацевтические составы антитела к LY6G6D и/или антитела к CD3 по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например, CD3) получают путем смешивания такого антитела с желаемой степенью чистоты с одним или более необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в виде лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители в общем случае являются нетоксичными для реципиентов в применяемых дозировках и концентрациях и включают, но не ограничиваются этим: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензил аммония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Типичные фармацевтически приемлемые носители по данному документу дополнительно включают агенты для диспергирования лекарственных препаратов в интерстициальном пространстве, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы РН-20 человека, такие как rHuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые типичные sHASEGP и способы их применения, включая rHuPH20, описаны в патентных публикациях США №№2005/0260186 и 2006/0104968. В одном аспекте sHASEGP комбинируют с одной или более дополнительными гликозаминогликаназами, такими как хондроитиназы.
Типовые лиофилизированные составы антител описаны в патенте США №6267958. Водные составы антител включают описанные в патенте США №6171586 и WO 2006/044908, причем последние составы содержат гистидин-ацетатный буфер.
Состав, представленный в настоящем документе, также может содержать более одного активного ингредиента, если необходимо для конкретного показания, которое лечат, предпочтительно те, которые обладают дополняющими видами активности и не оказывают отрицательного влияния друг на друга. Например, может существовать потребность в дополнительном терапевтическом агенте (например, химиотерапевтическом агенте, цитотоксическом агенте, агенте, ингибирующем рост, и/или антигормональном агенте, таком как те, что перечислены выше). Такие активные ингредиенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты также могут быть заключены в микрокапсулах, полученных, например, с помощью способов коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, при этом матрицы представлены в виде формованных изделий, например, пленок или микрокапсул.
Составы, предназначенные для введения in vivo, в общем случае являются стерильными. Стерильность можно легко обеспечить, например, путем фильтрации через стерильные фильтровальные мембраны.
Е. Терапевтические способы и композиции
Любое из антител к LY6G6D и/или антител к CD3 по настоящему изобретению (например, биспецифические антитела к LY6G6D по настоящему изобретению, которые связываются с LY6G6D, предпочтительно с высокой аффинностью (например, 20A12.QNTv12), и второй биологической молекулой, например, CD3, предпочтительно с высокой аффинностью, например, LY6G6D TDB, имеющие Ly6G6D-специфичное плечо, такие как 20A12.QNTv12, и CD3-специфичное плечо, такие как 38E4.v1 MD1 или 38E4.v1 MD4) можно использовать в терапевтических способаах.
В одном аспекте предложено антитело к LY6G6D для применения в качестве лекарственного средства. В дополнительных аспектах представлено антитело к LY6G6D, например, LY6G6D TDB, имеющее CD3-специфичное плечо (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичное плечо (например, 20A12.QNTv12) для применения в лечении или или замедлении прогрессирования связанного с клеточной пролиферацией нарушения (например, рака, например, колоректального рака). В некоторых вариантах осуществления рак представляет собой LY6G6D-положительный рак (например, LY6G6D-положительный колоректальный рак). В определенных вариантах осуществления изобретения предложено антитело к LY6G6D для применения в способе лечения. В некоторых вариантах осуществления настоящее изобретение представляет антитело к LY6G6D (например, LY6G6D TDB, имеющее CD3-специфичное плечо (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичное плечо (например, 20A12.QNTv12)) для применения в способе лечения индивидуума, имеющего связанное с клеточной пролиферацией нарушение, включающем введение индивидууму эффективного количества антитела к LY6G6D. В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже. В дополнительных вариантах осуществления настоящее изобретение представляет антитело к LY6G6D (например, LY6G6D TDB, имеющее CD3-специфичное плечо (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичное плечо (например, 20A12.QNTv12)) для применения в усилении иммунной функции у индивидуума, имеющего клеточно-пролиферативное нарушение. В некоторых вариантах осуществления настоящее изобретение представляет антитело к LY6G6D для применения в способе усиления иммунной функции у индивидуума, имеющего клеточно-пролиферативное нарушение, включающем введение индивидууму эффективного количества антитела к LY6G6D (например, биспецифического антитела к LY6G6D по настоящему изобретению, которое связывается со второй биологической молекулой, например, CD3) (например, для активации эффекторных клеток (например, Т-клеток, например, Т-клеток CD8+ и/или CD4+), расширения (увеличения) популяции эффекторных клеток, снижения популяции клеток-мишеней (например, клеток, экспрессирующих вторую биологическую молекулу, распознанную антителом к LY6G6D по настоящему изобретению, таким как биспецифическое антитело TDB по настоящему изобретению) и/или уничтожения клеток-мишеней (например, клеток-мишеней опухоли). «Индивидуум» в соответствии с любым из вышеприведенных вариантов осуществления может быть человеком.
В дополнительном аспекте в изобретении предложено применение антитела к LY6G6D в производстве или приготовлении лекарственного средства. В одном варианте осуществления лекарственное средство предназначено для лечения связанного с пролиферацией клеток нарушения (например, рака, например, колоректального рака). В некоторых вариантах осуществления рак представляет собой LY6G6D-положительный рак (например, LY6G6D-положительный колоректальный рак). В дополнительном варианте осуществления лекарственное средство предназначено для применения в способе лечения клеточно-пролиферативного нарушения, включающем введение индивидууму, имеющему клеточно-пролиферативное нарушение, эффективного количества лекарственного средства. В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже. В дополнительном варианте осуществления лекарственное средство предназначено для активации эффекторных клеток (например, Т-клеток, например, Т-клеток CD8+ и/или CD4+), расширения (увеличения) популяции эффекторных клеток, снижения популяции клеток-мишеней (например, популяции клеток, экспрессирующих LY6G6D) и/или уничтожения клеток-мишеней (например, клеток-мишеней опухоли) у индивидуума. В дополнительном варианте осуществления лекарственное средство предназначено для применения в способе усиления иммунной функции у индивидуума, имеющего клеточно-пролиферативное нарушение, включающем введение индивидууму эффективного количества лекарственного средства для активации эффекторных клеток (например, Т-клеток, например, Т-клеток CD8+ и/или CD4+), расширения (увеличения) популяции эффекторных клеток, снижения популяции клеток-мишеней (например, популяции клеток, экспрессирующих LY6G6D) и/или уничтожения клетки-мишени (например, опухолевой клетки-мишени). «Индивидуум» в соответствии с любым из вышеприведенных вариантов осуществления может быть человеком.
В дополнительном аспекте настоящее изобретение представляет способ лечения клеточно-пролиферативного нарушения (например, рака, например, колоректального рака). В некоторых вариантах осуществления рак представляет собой LY6G6D-положительный рак (например, LY6G6D-положительный колоректальный рак). В одном варианте осуществления способ включает введение индивидууму, имеющему такое клеточно-пролиферативное нарушение, эффективного количества антитела к LY6G6D, например, LY6G6D TDB, имеющего CD3-специфичное плечо (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичное плечо (например, 20A12.QNTv12). В одном из таких вариантов осуществления изобретения данный способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже. «Индивидуум» в соответствии с любым из вышеприведенных вариантов осуществления может быть человеком.
В дополнительном аспекте настоящее изобретение представляет способ усиления иммунной функции у индивидуума, имеющему клеточно-пролиферативное нарушение. В одном варианте осуществления способ включает введение индивидууму эффективного количества антитела к LY6G6D (например, LY6G6D TDB, имеющего CD3-специфичное плечо (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичное плечо (например, 20A12.QNTv12)) для активации эффекторных клеток (например, Т-клеток, например, Т-клеток CD8+ и/или CD4+), расширения (увеличения) популяции эффекторных клеток, уменьшения популяции клеток-мишеней (например, популяции клеток, экспрессирующих LY6G6D) и/или уничтожения клеток-мишеней (например, клеток-мишеней опухолей). В одном варианте осуществления «индивидуум» является человеком.
В дополнительном аспекте настоящее изобретение представляет способ лечения колоректального рака, рака пищевода, рака желудка, рака тонкого кишечника, рака толстого кишечника или аденокарциномы (например, колоректальной аденокарциномы, аденокарциномы желудка или аденокарциномы поджелудочной железы), которая может быть метастазирующей аденокарциномой (например, метастазирующей колоректальной аденокарциномой, метастазирующей аденокарциномой желудка или метастазирующей аденокарциномой поджелудочной железы), путем введения эффективного количества антитела к LY6G6D по настоящему изобретению, такого как биспецифическое антитело TDB по настоящему изобретению, такого как нацеливающее TDB к Ly6G6D, такое как LY6G6D TDB с CD3-специфичным плечом высокой аффинности, такое как 38E4.v1 MD1 или 38E4.v1 MD4, и Ly6G6D-специфичное плечо, такое как 20A12.QNTv12. В некоторых аспектах рак имеет статус микросателлитной нестабильности: «микросателлитная стабильность» (MSS) или «низкий уровень микросателлитной нестабильности» (MSI-L). В других аспектах рак имеет статус микросателлитной нестабильности «высокий уровень микросателлитной нестабильности» («MSI-H»). В некоторых аспектах рак является LY6G6D-положительным.
В некоторых аспектах настоящее изобретение представляет способ лечения колоректального рака, например, колоректального рака со статусом микросателлитной нестабильности: «микросателлитная стабильность» (MSS) или «низкий уровень микросателлитной нестабильности» (MSI-L), путем введения эффективного количества антитела к LY6G6D по настоящему изобретению, такого как биспецифическое антитело TDB по настоящему изобретению, такого как нацеленное TDB к Ly6G6D, такое как LY6G6D TDB с CD3-специфичным плечом высокой аффинности, такое как 38E4.v1 MD1 или 38E4.v1 MD4, и Ly6G6D-специфичным плечом, таким как 20A12.QNTv12.
В дополнительном аспекте настоящее изобретение представляет фармацевтические составы, содержащие любое из антител к LY6G6D, представленных в настоящем документе (например, LY6G6D TDB с CD3-специфичным плечом (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)), например, для применения в любом из вышеуказанных терапевтических способов. В одном варианте осуществления фармацевтический состав содержит любое из предложенных в настоящем документе антител к LY6G6D и фармацевтически приемлемый носитель. В другом варианте осуществления фармацевтический состав содержит любое из предложенных в данном документе антител к LY6G6D и по меньшей мере один дополнительный терапевтический агент, например, как описано в настоящем документе.
Антитело по настоящему изобретению (и/или любой дополнительный терапевтический агент) можно вводить любым подходящим способом, включая парентеральное, внутрилегочное и интраназальное, и, если это необходимо для местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. В некоторых вариантах осуществления антитело вводят путем внутривенного введения. В других вариантах осуществления антитело вводят путем подкожного введения. В некоторых вариантах осуществления антитело к LY6G6D, введенное путем подкожной инъекции, проявляет менее токсичный ответ у пациента, чем-то же антитело к LY6G6D, введенное путем внутривенной инъекции. Введение дозы можно проводить любым удобным способом, например, путем инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или постоянным. В данном документе предусмотрены различные схемы введения доз, включая, но не ограничиваясь этим, однократное или многократное введение в различные моменты времени, болезное введение и импульсную инфузию.
Антитела по изобретению составляют, дозируют и вводят способом, соответствующим надлежащей медицинской практике. Факторы, которые необходимо учитывать в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки агента, способ введения, график введения и другие факторы, известные практикующим врачам. Антитело необязательно составляют с одним или более агентами, применяемыми в настоящее время для предотвращения или лечения рассматриваемого нарушения. Эффективное количество таких других средств зависит от количества антитела, присутствующего в составе, типа нарушения или лечения и других факторов, которые обсуждались выше. Другие агенты обычно применяют в таких же дозировках и вводят путями, описанными в данном документе, или в дозировках, составляющих от приблизительно 1 до 99% дозировок, описанных в данном документе, или в любой дозировке и любым путем, которые эмпирически/клинически определены как подходящие.
Для предотвращения или лечения заболевания соответствующая доза антитела по настоящему изобретению (например, антитело к LY6G6D, например, LY6G6D TDB с CD3-специфичным плечом (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)) (при использовании отдельно или в комбинации с одним или более другими дополнительными терапевтическими агентами) будет зависеть от типа заболевания, подлежащего лечению, типа антитела, тяжести и хода заболевания, вводится ли антитело для профилактики или терапевтических целей, предыдущей терапии, истории болезни пациента и реакции на антитело, и выбора лечащего врача. Антитело предпочтительно вводить пациенту за один раз или в течение серии курсов лечения.
В общем положении, терапевтически эффективное количество антитела к LY6G6D (например, LY6G6D TDB с CD3-специфичным плечом (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)), вводимого человеку, будет находиться в диапазоне от приблизительно 0,01 до приблизительно 100 мг/кг веса тела пациента, будь то за одно или несколько введений. В некоторых вариантах осуществления суточное вводимое количество антитела составляет, например, от приблизительно 0,01 до приблизительно 45 мг/кг, от приблизительно 0,01 до приблизительно 40 мг/кг, от приблизительно 0,01 до приблизительно 35 мг/кг, от приблизительно 0,01 до приблизительно 30 мг/кг, от приблизительно 0,01 до приблизительно 25 мг/кг, от приблизительно 0,01 до приблизительно 20 мг/кг, от приблизительно 0,01 до приблизительно 15 мг/кг, от приблизительно 0,01 до приблизительно 10 мг/кг, от приблизительно 0,01 до приблизительно 5 мг/кг или от приблизительно 0,01 до приблизительно 1 мг/кг. В одном варианте осуществления описанное в данном документе антитело к LY6G6D вводят человеку в дозе приблизительно 100 мг, приблизительно 200 мг, приблизительно 300 мг, приблизительно 400 мг, приблизительно 500 мг, приблизительно 600 мг, приблизительно 700 мг, приблизительно 800 мг, приблизительно 900 мг, приблизительно 1000 мг, приблизительно 1100 мг, приблизительно 1200 мг, приблизительно 1300 мг или приблизительно 1400 мг на 1 сутки 21-суточных циклов. Дозу можно вводить в виде однократной дозы или многократных доз (например, 2 или 3 доз), например, инфузий. В случае повторных введений в течение нескольких дней или более, в зависимости от состояния, лечение в общем случае проводят до достижения необходимой степени подавления симптомов заболевания. Одна типовая дозировка антитела находится в диапазоне от приблизительно 0,05 мг/кг до приблизительно 10 мг/кг. Таким образом, пациенту можно вводить одну или более доз, составляющих приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить с интервалами, например, каждую неделю или каждые три недели (например, чтобы пациент получал от приблизительно двух до приблизительно двадцати или, например, приблизительно шести доз антитела к LY6G6D). Можно вводить начальную более высокую нагрузочную дозу, за которой следует одна или более меньших доз. Эффективность такой терапии легко контролировать с помощью традиционных способов и анализов.
Ж. Дополнительные терапевтические агенты
Антитела по данному изобретению можно применять в терапии отдельно или в комбинации с другими агентами. Например, антитело по изобретению можно вводить совместно с по меньшей мере одним дополнительным терапевтическим агентом. В некоторых вариантах осуществления дополнительный терапевтический агент представляет собой химиотерапевтический агент, ингибирующий рост агент, цитотоксический агент, агент, используемый в лучевой терапии, агент против ангиогенеза, апоптический агент, антитубулиновый агент или другой агент, такой как антагонист рецептора эпидермального фактора роста (EGFR) (например, ингибитор тирозинкиназы), ингибитор HER1/EGFR (например, эрлотиниб (Тарцева™), ингибитор тромбоцитарного фактора роста (например, Гливек™ (мезилат иматиниба)), ингибитор СОХ-2 (например, целекоксиб), интерферон, цитокин, антитело, отличное от антитела к CD3 по настоящему изобретению, такое как антитело, которое связывается с одной или более следующими мишенями ErbB2, ErbB3, ErbB4, PDGFR-бета, BlyS, APRIL, ВСМА VEGF или рецептор(ы) VEGF, TRAIL/Аро2, PD-1, PD-L1, PD-L2 или другим биоактивным или органическим химическим агентом.
Такие комбинированные терапии, указанные ранее, охватывают комбинированное введение (в котором два или более терапевтических агента включены в одну и ту же или отдельные композиции), а также раздельное введение, и в этом случае, введение антитела согласно изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического агента или агентов. В одном варианте осуществления введение антитела к LY6G6D и введение дополнительного терапевтического агента может разделять около одного месяца или около одной, двух или трех недель, или около одних, двух, трех, четырех, пяти или шести суток. Антитела к LY6G6D по настоящему изобретению (например, биспецифическое антитело к LY6G6D по настоящему изобретению, которое связывается с второй биологический молекулой, например, CD3) может также использоваться в комбинации с лучевой терапией. В некоторых вариантах осуществления дополнительной терапией может быть хирургическое вмешательство, генная терапия, ДНК-терапия, вирусная терапия, РНК-терапия, иммунотерапия, пересадка костного мозга, нанотерапия, терапия моноклональными антителами или комбинация вышеуказанного. Дополнительная терапия может иметь форму адъювантной или неоадъювантной терапии. В некоторых вариантах осуществления дополнительная терапия представляет собой введение низкомолекулярного ферментативного ингибитора или антиметастатического агента. В некоторых вариантах осуществления дополнительная терапия представляет собой введение агентов, ограничивающих побочные эффекты (например, агентов, предназначенных для снижения возникновения и/или тяжести побочных эффектов лечения, таких как агенты против тошноты и т.д.). В некоторых вариантах осуществления дополнительная терапия представляет собой хирургию. В некоторых вариантах осуществления дополнительная терапия представляет собой комбинацию лучевой терапии и хирургии.
В некоторых вариантах осуществления LY6G6D TDB (например, LY6G6D TDB с CD3-специфичным плечом (например, 38E4.v1 MD1 или 38E4.v1 MD4) и Ly6G6D-специфичным плечом (например, 20A12.QNTv12)) вводят совместно (одновременно, как одну или множество композиций (например, составов)) с одним или более дополнительными терапевтическими агентами, например, любым одним, двумя, тремя, четырьмя, пятью, шестью, семью, восмью, девятью, десятью или всеми одиннадцатью из следующих: Фолфокс (оксалиплатин (Элоксатин™) вместе с 5-фторурацилом и лейковорином), капецитабин (Кселода®), 5-фторурацил (5-ФУ), КапеОкс (Кселокс; капецитабин с оксалиплатином), лейковорин (фолиновая кислота), бевацизумаб (Авастин®), цетуксимаб (Эрбитукс®), панитумумаб (Вектибикс®), регорафениб (Стиварга®), иринотекан (СРТ-11; Камптозар®) и Флокс (5-фторурацил с оксалиплатином). В других вариантах осуществления LY6G6D TDB вводят до одного или более дополнительных терапевтических агентов, например, любого одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти или всех одиннадцати из следующих: Фолфокс (оксалиплатин (Элоксатин™) вместе с 5-фторурацилом и лейковорином), капецитабин (Кселода®), 5-фторурацил (5-ФУ), КапеОкс (Кселокс; капецитабин с оксалиплатином), лейковорин (фолиновая кислота), бевацизумаб (Авастин®), цетуксимаб (Эрбитукс®), панитумумаб (Вектибикс®), регорафениб (Стиварга®), иринотекан (СРТ-11; Камптозар®) и Флокс (5-фторурацил с оксалиплатином). В других вариантах осуществления LY6G6D TDB вводят после одного или более дополнительных терапевтических агентов, например, любого одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти или всех одиннадцати из следующих: Фолфокс (оксалиплатин (Элоксатин™) вместе с 5-фторурацилом и лейковорином), капецитабин (Кселода®), 5-фторурацил (5-ФУ), КапеОкс (Кселокс; капецитабин с оксалиплатином), лейковорин (фолиновая кислота), бевацизумаб (Авастин®), цетуксимаб (Эрбитукс®), панитумумаб (Вектибикс®), регорафениб (Стиварга®), иринотекан (СРТ-11; Камптозар®) и Флокс (5-фторурацил с оксалиплатином).
i. Агенты для ингибирования роста
В некоторых аспектах дополнительый терапевтический агент представляет собой агент для ингибирования роста. Типичные агенты для ингибирования роста включают агенты, которые блокируют прогрессирование клеточного цикла в месте, отличном от S-фазы, например, агенты, которые индуцируют остановку G1 (например, ДНК аликилирующие средства, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил или ара-С) или остановку М-фазы (например, винкристин, винбластин, таксаны (например, паклитаксел и доцетаксел), доксорубицин, эпирубицин, даунорубицин, этопозид или блеомицин).
ii. Виды лучевой терапии
В некоторых аспектах дополнительный терапевтический агент представляет собой лучевую терапию. Под «видами лучевой терапии» подразумевается применение направленного гамма-облучения для индукции значительного повреждения клеток так, чтобы ограничить их способность нормально функционировать или разрушить все клетки. Типичные варианты лечения включают одноразовое применение, а типичные дозы находятся в диапазоне от 10 до 200 единиц (Грей (Гр)).
iii. Цитотоксические агенты
В некоторых аспектах дополнительный терапевтический агент представляет собой цитотоксический агент, например, вещество, которое ингибирует или предотвращает клеточную функцию и/или вызывает некроз или разрушение клеток. Цитотоксические агенты включают, без ограничения, радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты; и противоопухолевые или противораковые агенты.
iv. Иммуномодулиуующие агенты
В некоторых аспектах дополнительный терапевтический агент представляет собой иммуномодулирующий агент, например, антагонист связывания PD-L1, который может быть антагонистом связывания PD-1, антагонистом связывания PD-L1 или антагонистом связывания PD-L2. PD-1 (запрограммированная гибель клетки 1) также называется в данной области «запрограммированная гибель клетки 1», «PDCD1», «CD279» и «SLEB2». Типичный PD-1 человека известен под номером доступа в UniProtKB/Swiss-Prot № Q15116. PD-L1 (лиганд запрограммированной гибели 1) также называется в данной области «лиганд 1 запрограммированной гибели клетки 1», «PDCD1LG1», «CD274», «В7-Н» и «PDL1». Типичный PD-L1 человека известен под номером доступа в UniProtKB/Swiss-Prot № Q9NZQ7.1. PD-L2 (лиганд 2 запрограммированной гибели) также называется в данной области «лиганд 2 запрограммированной гибели клетки 1», «PDCD1LG2», «CD273», «B7-DC», «Btdc» и «PDL2». Типичный PD-L2 человека известен под номером доступа в UniProtKB/Swiss-Prot №Q9BQ51. В некоторых случаях PD-1, PD-L1 и PD-L2 являются человеческими PD-1, PD-L1 и PD-L2.
В некоторых аспектах антагонист связывания PD-1 представляет собой молекулу, которая ингибирует связывание PD-1 с его лигандом-партнером по связыванию. В конкретном аспекте лигандами-партнерами по связыванию PD-1 являются PD-L1 и/или PD-L2. В другом случае антагонист связывания PD-L1 представляет собой молекулу, которая ингибирует связывание PD-L1 с его партнерами по связыванию. В конкретном аспекте партнерами по связыванию PD-L1 являются PD-1 и/или В7-1. В другом случае антагонист связывания PD-L2 представляет собой молекулу, которая ингибирует связывание PD-L2 с его партнерами по связыванию. В конкретном аспекте лигандом-партнером по связыванию PD-L2 является PD-1. Антагонист может представлять собой антитело, его антигенсвязывающий фрагмент, иммуноадгезин, слитый белок или олигопептид.
В некоторых аспектах антагонист связывания PD-1 представляет собой антитело к PD-1 (например, человеческое антитело, гуманизированное антитело или химерное антитело).
В некоторых аспектах антагонист связывания PD-L1 представляет собой антитело к PD-L1, например, как описано ниже. В некоторых аспектах антитело к PD-L1 способно ингибировать связывание между PD-L1 и PD-1 и/или между PD-L1 и В7-1. В некоторых аспектах антитело к PD-L1 представляет собой моноклональное антитело. В некоторых аспектах антитело к PD-L1 представляет собой фрагмент антитела, выбранный из группы, состоящей из фрагментов Fab, Fab'-SH, Fv, scFv и (Fab')2. В некоторых аспектах антитело к PD-L1 представляет собой гуманизированное антитело. В некоторых аспектах антитело к PD-L1 представляет собой человеческое антитело.
В некоторых аспектах ингибитор контрольных точек иммунного ответа представляет собой антагонист, направленный против совместно ингибирующей молекулы (например, антагонист CTLA-4 (например, антитело к CTLA-4), антагонист TIM-3 (например, антитело к TIM-3) или антагонист LAG-3 (например, антитело к LAG-3)), или любую их комбинацию.
В некоторых аспектах ингибитор контрольных точек иммунного ответа представляет собой антагонист, направленный против TIGIT (например, антитело к TIGIT).
В некоторых аспектах дополнительный терапевтический агент представляет собой 5-фторурацил (5-ФУ); иринотекан; капецитабин; оксалиплатин; цетуксимаб; бевацизумаб; панитумумаб; афлиберцепт, регорафениб; рамуцирумаб, TAS-102 (трифлуридин и типирацил); пембролизумаб; ниволумаб; ниволумаб и ипилимумаб; вемурафениб; Фолфоксири и бевацизумаб, терапию против EGFR в комбинации с ингибитором BRAF и/или MEK, необязательно включающий цитотоксический агент; Фолфокс/Фолфири и терапию против EGFR; или Фолфокс/Фолфири/Фолфоксири и бевацизумаб.
З. Готовые изделия
В другом аспекте изобретения предложено готовое изделие, содержащее материалы, применяемые при лечении, предотвращении и/или диагностике вышеописанных нарушений. Готовое изделие содержит контейнер и этикетку или вкладыш в упаковку на нем или связанные с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для в/в раствора и т.д. Контейнеры могут быть выполнены из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией эффективна для лечения, предотвращения и/или диагностики патологического состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с пробкой, прокалываемой гиподермической иглой для инъекций). По меньшей мере один активный агент в композиции представляет собой антитело по изобретению. На этикетке или на вкладыше в упаковку указано, что композицию применяют для лечения выбранного патологического состояния. Кроме того, готовое изделие может содержать (а) первый контейнер с содержащейся в нем композицией, причем указанная композиция содержит антитело по изобретению; и (б) второй контейнер с содержащейся в нем композицией, причем указанная композиция содержит дополнительный цитотоксический или иной терапевтический агент. Готовое изделие в этом варианте осуществления может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного патологического состояния. В альтернативном или дополнительном варианте готовое изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БВДИ), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Оно также может содержать другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
III. ПРИМЕРЫ
Ниже приведены примеры способов и композиций по изобретению. Понятно, что различные другие аспекты могут применяться на практике, учитывая общее описание, представленное выше, и примеры не предназначены для ограничения объема формулы изобретения.
Пример 1. LY6G6D является поверхностный маркером клеток колоректального рака и имеет ограниченную экспрессию в нормальных тканях
Экспрессию члена G6D семейства лимфоцитарного антигена 6 (LY6G6D) (SEQ ID NO: 75) в нормальных и опухолевых тканях человека оценивали с помощью Атласа ракового генома (АРГ) (Grossman et al., New England Journal of Medicine, 375(12): 1109-1112) и данных Genotype-Tissue Expression Project (GTEx) (проект по изучению экспрессии генотипа в разных тканях) (Pierson et al., PLoS Comput Biol, 11, e1004220, 2015) и иммуногистохимии (ИГХ).
А. Экспрессия LY6G6D
Фиг. 1А показывает экспрессию LY6G6D и члена G6D семейства лимфоцитарного антигена 6 (LY6G6F) в нормальных и опухолевых тканях человека в данных АРГ. В опухолевых тканях показание с наивысшей экспрессией LY6G6D давала толстая кишка, a LY6G6D значительно сверхэкспрессировался только в тканях опухоли толстой кишки. Нормальные ткани толстой кишки также показывают некоторую экспрессию LY6G6D, хотя и на намного более низких уровнях. Достойная внимания экспрессия (>1 nRPKM) LY6G6F главным образом обнаруживается в ткани опухоли толстой кишки. Фиг. 1Б показывает экспрессию LY6G6D и LY6G6F в нормальных тканях в данных общедоступного проекта GTEx. LY6G6D больше всего экспрессируется в ткани простаты, яичка, шейки матки и влагалища, и значительная экспрессия обнаруживается в нормальной ткани толстой кишки и поднаборе образцов кожи. LY6G6F больше всего экспрессируется в крови, а затем ткани яичка, селезенки, щитовидной железы и легкого.
Б. Экспрессия LY6G6D в КРР с MSS и MSI-L
LY6G6D больше всего экспрессируется при колоректальном раке (КРР) со статусом микросателлитной нестабильности (MSI): микросателлитная стабильность (MSS) или низкий уровень микросателлитной нестабильности (MSI-L) или высокий уровень микросателлитной нестабильности (MSI-H); КРР с MSS связаны с худшим прогнозом (фиг. 1В).
В. ИГХ окрашивание LY6G6D
Опухоли КРР человека окрашивали по LY6G6D. Приблизительно 20% первичных случаев КРР идентифицировали как LY6G6D-положительные с помощью ИГХ. Оценивали 141 тканевую матрицу (ТМА) первичных опухолей толстой кишки. 21 опухоли показали слабое (1+) ИГХ окрашивание (14%), 5 показали умеренное (2+) окрашивание, и 4 показали сильное (3+) окрашивание (6-7% вместе) (фиг. 2А). На Фиг. 2Б-2Г показано слабое (1+), умеренное (2+) и сильное (3+) ИГХ окрашивание для LY6G6D в ткани первичной опухоли толстой кишки.
Пример 2. Требования к получению для LY6G6D-специфичного плеча 1G4, образования клона антитела к LY6G6D, картирования эпитопов и гуманизации
А. Требования к получению для LY6G6D-специфичного плеча 1G4 TDB к LY6G6D, содержащие химерное LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4.v1, показали in vitro уничтожение клеток НТ55 (линия клеток карциномы толстой кишки человека) (фиг. 3А) и in vivo активность к ксенотрансплантату LS1034 и опухолям НТ55 у мышей NSG™ (фиг. 3Б). 1G4 представляет антитело гибридомы мышей; химерное 1G4 (ch1G4) представляет химерное антитело мыши/человека, в котором вариабельные домены мыши (VH и VL) 1G4 были генетически слиты, соответственно, с константными доменами тяжелой цепи человека (CH1, СН2 и СН3) с мутацией аминокислотной замены N297G в СН2 и которое содержит «область «впадины» и константный домен легкой цепи человека (CL). Гуманизированную версию плеча 1G4 получали и была продемонстрирована in vitro и in vivo эффективность (фиг. 3В и 3Г).
Требования к молекулярной оценке (МО) идентифицировали для аминокислотного остатка W50 CDR2 легкой цепи (LC) гуманизированного 1G4 (WASTRIS; SEQ ID NO: 110). Вкратце, гуманизированное IG4 тестировали на стресс при химических условиях с ААПГ (2,2-азобис(2-амидинопропан)дигидрохлорид), небольшой молекулой, которая, как известно, генерирует свободные радикалы (см., например, Ji et al., J. Pharm. Sci. 98(12):4485-4500, 2009), а также при тепловых условиях при различном рН (двухнедельный тест на тепловой стресс при 40°С, рН 5,5). Остаток W50 легкой цепи идентифицировали как имеющий повышенное окисление (72,0% окисление) после стресса с ААПГ.
Анализы на тепловой стресс имитируют стабильность в течение срока годности продукта. В образцах меняли буфер на 20 мМ ацетат гистидина, 240 мМ сахарозы, рН 5,5 и разводили в концентрации 1 мг/мл. Один мл каждого образца подвергали стрессу при 40°С в течение 2 недель, а второй хранили при -70°С в качестве контроля. Оба образца затем ферментировали с использованием трипсина для получения пептидов, которые могли быть проанализированы с использованием жидкостной хроматографии (ЖХ) - масс-спектрометрии (МС). Для каждого пептида в образце определяли время удержания (что измерено с помощью жидкостной хроматографии), и получали высокоточные данные о массах и фрагментации ионов пептида (информация о аминокислотной последовательности). Извлеченные ионные хроматограммы (XIC) отбирались для искомых пептидов (например, нативные и модифицированные пептидные ионы) из наборов данных в диапазоне +-10 м.д., и пики интегрировались для определения площади. Относительные проценты модификации рассчитывали для каждого образца, взяв (площадь модифицированного пептида), деленную на (площадь модифицированного пептида плюс площадь нативного пептида), умноженную на 100).
Кроме того, плечо 1G4 не проходило анализ на временную трансфекцию для получения (фиг. 30). W50 заменяли на все 18 альтернативные аминокислоты (исключая Cys), но замена, как оказывалось, влияет на аффинность связывания. Наконец, как показано на фиг. 3А, плечо 1G4 эффективно только при спаривании с плечом 38E4.v1 с высокой аффинностью, а не при спаривании с 40G5c с низкой аффинностью. Таким образом, предпринималась новая кампания по обнаружению антител к LY6G6D.
Б. Образование mAb кролика к huLY6G6D
Моноклональные антитела (mAb) к LY6G6D человека (huLY6G6D) генерировали у кроликов. Новозеландских белых кроликов иммунизировали с помощью Ly6G6D человека, и отдельные В-клетки выделяли из иммунизированных кроликов с помощью модифицированного протокола из Offner et al. PLoS ONE, 9(2), 2014. Этот модифицированный процесс включал прямую сортировку АФСК IgG+ huLy6G6D+ В-клеток в отдельные лунки. В-клеточные культуральные супернатанты анализировали способом ИФА в отношении связывания с Ly6G6D человека и нерелевантным контрольным белком. Ly6G6D-специфические В-клетки лизировали и незамедлительно замораживали при -80°С для хранения до проведения молекулярного клонирования. Вариабельные области (VH и VL) каждого моноклонального антитела из В-клеток кролика клонировали в экспрессионные векторы из экстрагированной иРНК, как было описано ранее (Offner et al., PLoS ONE 9(2), 2014). Отдельные рекомбинантные кроличьи антитела экспрессировали в клетках Expi293, а после этого очищали с помощью протеина А. Затем проводили анализ функциональной активности и кинетический скрининг очищенных антител к Ly6G6D. Приблизительно 280 клонов ИФА+ к Ly6G6D получалось. 96 клонов затем группировали в четыре отдельные группы сортировки в зависимости от связываемого эпитопа, как описано ниже.
В. Кинетический анализ и сортировка антител к LY6G6D в зависимости от связываемого эпитопа с помощью гликосконструированного LY6G6D
Систему получения изображений матричной ППР (Carterra ®, США) использовали для тестирования кинетики и сортировки антител в зависимости от связываемого эпитопа панели из 96 моноклональных антител кролика к huLY6G6D, включая 1G4. Для сортировки антител в зависимости от связываемого эпитопа полипептиды LY6G6D подвергали конструированию для введения сайтов гликозилирования на поверхность молекулы (фиг. 4А и 4Б). Сайты выбирали для простоты добавления сайтов гликозилирования с минимальным разрывом относительно природной последовательности. Кандидатные антитела к LY6G6D тестировали на взаимодействие с гликосконструированными полипептидами LY6G6D (фиг. 4Б). Очищенные антитела разводили при 10 мкг/мл в 10 мМ натрий-ацетатном буфере, рН 4,5. Используя аминное сопряжение, антитела напрямую иммобилизовали на чипе SPR sensorprism CMD 200М (XanTec Bioanalytics GmbH, Германия), используя микроспоттер с непрерывным потоком SPRi (Carterra®, США), для создания матрицы из 96 антител. Для анализа использовали IBIS МХ96 SPRi (Carterra®, США), чтобы оценить связывание аналитов с иммобилизованными лигандами. Для кинетического анализа Ly6G6D человека вводили в течение 3 минут от 0 до 300 нМ при 3-кратном разведении с последующим периодом диссоциации 10 минут. Для сортировки антител в зависимости от связываемого эпитопа каждый мутант по гликозилированию Ly6G6D человека сначала вводили в течение 4 минут при 50 нМ, а затем второе 4-минутное введение отдельного моноклонального антитела при 10 мкг/мл. Между циклами поверхность восстанавливали с помощью 10 мМ глицина, рН 1,5. Эксперимент проводили при 25°С в подвижном буфере HBS-T (0,01 М ГЭПЭС, рН 7,4, 0,15 М NaCl, 0,05% поверхностно-активного вещества Р20). Кинетические данные обрабатывали с помощью Scrubber 2.0 (BioLogic Software), и данные сортировки антител в зависимости от связываемого эпитопа обрабатывали с помощью программных средств для сортировки Wasatch (Carterra®, США).
Связывание 1G4 и антитела кролика к LY6G6D 20А12 с полипептидом LY6G6D прерывали в гликосконструированном полипептиде LY6G6D с мутациями аминокислотной замены G99N.L101S (Фиг. 4Б). Связывание других антител кролика к LY6G6D по-разному прерывали мутациями аминокислотных замены P60A.P61S, A73N.H75S, V80S, V80N, T82N или D87N (фиг. 4Б). Клоны антитела кролика и 1G4 помещали в четыре отдельных группы эпитопов на основе результатов анализа гликоконструирования (фиг. 4Д): группа 1 содержит три группы последовательностей и включает 1G4, 20А12, 6Е10 и 4Н7; группа 4 содержит шесть групп последовательностей и содержит f.16D7; и каждая из групп 3 и 4 содержит три группы последовательностей. Аминокислотные остатки, затронутые мутациями гликозилирования, были кодированы цветом, и группы 1, 2, 3 и 4 указаны путем подчеркивания на фиг. 4Г (SEQ ID NO: 88). Антитела к LY6G6D 1G4 и 16D7 связывают эпитопы на противоположных сторонах антигена, как показано на фиг. 4В.
Г. Анализы клеточной цитотоксичности
Для оценки способности новой панели антител кролика нацеливаться на линию клеток Ly6G6D-положительной опухоли, получали биспецифические Т-клеточнозависимые антитела (TDB), содержащие CD3-специфичное плечо 40G5с, спаренное с другими LY6G6D-специфичными плечами кролика. Типичные антитела кролика из каждой группы эпитопа (группа 1, 2, 3 и 4) преобразовывали в половинные антитела с Fc-областью, содержащей область «выступа», химерные вариабельные домены кролика и константные домены человека с мутациями N297G и T366W. После очистки LY6G6D-специфичных плеч «выступа» выравнивали с CD3-специфичным плечом 40G5 с, имеющим Fc-область, содержащую область «выпадины», и анализировали на связывание с и in vitro уничтожение клеток НТ55 (фиг. 4Е, 4Ж и 13А-13Д). Антитела кролика из группы 1 (например, 20А12 и 6Е10), группы, которая включала 1G4, как было обнаружено, являются наиболее эффективными как для связывания, так и уничтожения клеток НТ55.
Д. Кинетический анализ
Аффинность связывания антител к LY6G6D кролика с LY6G6D определяли с помощью машины BIAcore™ Т200 (GE Healthcare Life Sciences). Вкратце, чипы исследовательской категории СМ5 BIAcore™ активировали реагентами 1-этил-3-(3-диметиламинопропил)карбодиимид (ЭДК) и N-гидроксисукцинимид (NHS) в соответствии с инструкциями поставщика. Для измерений кинетики белок Ly6G6D человека связывали с чипами до достижения приблизительно 100 единиц ответа (ЕО) в каждой проточной ячейке. Не вступившие в реакцию связывающие группы блокировали 1 М этаноламином. Антитела кролика экспрессировались как химерные антигенсвязывающие фрагменты (Fab) с вариабельными доменами кролика и константными доменами человека. Десятикратные серийные разведения Fab вводили в буфер HBS-P при 37°С со скоростью потока 30 мкл/мин. Скорости ассоциации (ka) и скорости диссоциации (kд) рассчитывали, используя 1:1 модель связывания Лэнгмюра (оценочное программное обеспечение BIAcore™ Т200 версии 2.0). Равновесную константу диссоциации (KД) рассчитывали как соотношение kд/ka (фиг. 4И).
Е. Гуманизация антител кролика
Моноклональные антитела 20 А12 и 6Е10 кролика из группы 1 гуманизировали, как описано ниже. Номера остатков соответствуют Kabat et al., Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, Md. (1991).
Гуманизация 20A12
Одной проблемой при гуманизации антител кролика является то, что разница между последовательностями кролика и человека больше, чем между последовательностями грызунов и человека. Множество каркасных областей, таким образом, применяли для гуманизации 20А12.
Гипервариабельные области из каждого из кроличьих антител, а именно, положения 24-34 (L1), 50-56 (L2) и 89-97 (L3) в домене VL и положения 26-35 (H1), 50-65 (Н2) и 95-102 (Н3) в домене VH) прививали в две разные акцепторные каркасные области человека. Варианты легкой цепи rb.20A12 получали на основе последовательностей легких цепей зародышевых линий человека hIGHV.1-5, hIGKV.1-39 и hIGKV.4-1, а варианты тяжелой цепи rb.20A12 получали на основе последовательностей тяжелых цепей зародышевых линий человека hIGHV.3-23 и hIGHV.3-30 (фиг. 31Б, 31В и 40А). Эти последовательности зародышевых линий выбирали на основе их высокого преобладания в сыворотке и высокой идентичности последовательности с rb.20A12 (фиг. 31В и 31Г). VL CDR KV1-5*01 и VH CDR HV3-23*01 выбирали для дальнейшего анализа. Последовательность 20А12, содержащая полностью каркасные области человека показана как L2H10 на фиг. 32Г.
Гуманизированные варианты 20А12 оценивали как Fab. Каркасные области зародышевых линий человека (VL и VH) модифицировали во всех предполагаемых положениях Верньера кролика, так что каждое положения Верньера содержало аминокислоту, присутствующую в последовательности антитела кролика (т.е. rb20A12). Положения Верньера кролика идентифицировали на основе известных положений Верньера грызунов. Вариант, в котором все положения Верньера тяжелых цепей и легких цепей были изменены на аминокислоту кролика, называются L1H1 (hu20A12.LlHl) на фиг. 32Г. Вариант, в котором все положения Верньера тяжелых цепей и легких цепей содержат аминокислоты человека, называется L2H10. Для оценки, влияет ли каждое положение Верньера на аффинность связывания антитела или huLY6G6D, положения Верньера кролика по отдельности меняли на аминокислоту соответствующей последовательности человека, т.е. KV1-5*01 или HV3-23*01. Получали один вариант легкой цепи, L2, и восемь дополнительных вариантов тяжелой цепи, Н2-Н9. L2 содержит мутацию аминокислотной замены Р43А относительно последовательности KV 1-5*01. Н2-Н9 содержат, соответственно, мутации аминокислотных замен Q2V, I48V, A49S, K71R, S73N, V78L, F91Y и P105R относительно последовательности HV3-23*01. Аффинность связывания вариантов антител сравнивали с таковой исходного клона, содержащего полностью последовательности каркасных областей человека (L2H10), с помощью анализа BIAcore (фиг. 32Г). Остатки тяжелой цепи кролика S73, Т76, V78 и Р105 определяли как основные остатки Верньера кролика на основе оценки аффинности связывания вариантов антител, описанных выше.
На фиг. 32В показана тонко очищенная версия гуманизированного 20А12, содержащая остатки тяжелой цепи кролика S73, Т76, V78 и Р105 и остатки человека во всех остальных положениях Верньера кролика. Тонко очищенное гуманизированное 20А12 также модифицировали для вмещения аминокислотных замен C35S или C35I и С50А, как описано ниже. Остатки Верньера кролика S73, V78 и Р105, дополнительный остаток Верньера кролика Т76 и аминокислотные замены C35S и С50А также прививали на зародышевую линию человека HV3-30*1 для получения дополнительной гуманизированной тонко очищенной версии rb.20A12.
Приблизительно 20-40% антител кролика содержат дополнительные цистеиновые остатки. Например, клон 20А12 кролика содержит цистеиновую пару в CDR-H1 и CDR-H2 (С35 на CDR-H1 и С50 на CDR-H2) (фиг. 31А; фиг. 40Б). Цистеины в этих двух положениях обычно обнаруживаются и, как считается, образуют дисульфидную связь. Для удаления этой цистеиновой пары, которая может быть необходимой при разработке, С35 и С50 rb.20A12 одновременно мутировали в C35S-C50A, C35S-C50S, C35I-C50A, C35I-C50S, C35I-C50I и C35G-C50T, и варианты оценивали на связывание с LY6G6D. Каждый из вариантов оценивали в виде химерных Fab с вариабельными доменами кролика и константными областями человека. Вариант rb20A12.IA (C35I-C50A), как обнаружили, сохраняет большую часть аффинности исходной молекулы (KД 0,86 нМ) (фиг. 34А и 34В). Мутации C35I-C50A, таким образом, были включены в последовательность тяжелой цепи тонко очищенного гуманизированного 20А12, описанную выше.
Кроме того, последовательность легкой цепи 20А12 кролика содержит мотив гликозилирования (NNT) в CDR3 (фиг. 32А и фиг. 40Б), и этот сайт подтверждали как гликозилированный. Ряд вариантов получали в химерном скелете кролика/человека для удаления этого сайта гликозилирования: NNT заменяли на QNT, QNV, SNA, SNV, ANT, GNT, NNV или NNA (фиг. 34Б). Замены QNT, QNV, SNV, GNT и SNA включали в последовательность легкой цепи тонко очищенного гуманизированного 20А12, описанную выше, и анализировали на связывание с LY6G6D (фиг. 34Б-34Г).
20A12.ONTv12
Вариант 20A12.QNTv12 выбирали как гуманизированное антитело rb.20A12. 20A12.QNTv12
содержит каркасные области VL KV1-5*01; каркасные области VH CDR HV3-23*01, модифицированные аминокислотными заменами S73, Т76, V78 и Р105 (полученные из положений Верньера кролика); и CDR rb.20A12, с заменой цистеиновых остатков в CDR-H1 и CDR-H2 на I и А, соответственно, и мутацией NNT в QNT на сайте гликозилирования в CDR-L3 (фиг. 32А, 40Б и 40В). KД составляет 0,23 нМ для rb.20A12 и 0,14 нМ для 20A12.QNTv 12.
Связывание с LY6G6D
Гуманизированный вариант rb.20A12 - 20A12.QNTv12, гуманизированный вариант rb.6Е10 - 6E10.v114 и LY6G6D-специфичное плечо 1G4 спаривали с CD3-специфичным плечом 38E4v1 или 40G5c как TDB и тестировали на аффинность для LY6G6D человека и яванского макака в анализе BIAcore®. 20А12 кролика, 6Е10 кролика и гуманизированное 20A12.QNTv12 и 20A12.SNVv12 дополнительно анализировали на связывание как антигенсвязывающие фрагменты (Fab). LY6G6D-Fc непосредственно иммобилизирован на чипе, и TDB пропускали при 37°С, за исключением Fab rb6E10, который анализировали при 25°С. Результаты показаны в таблице 2 ниже.
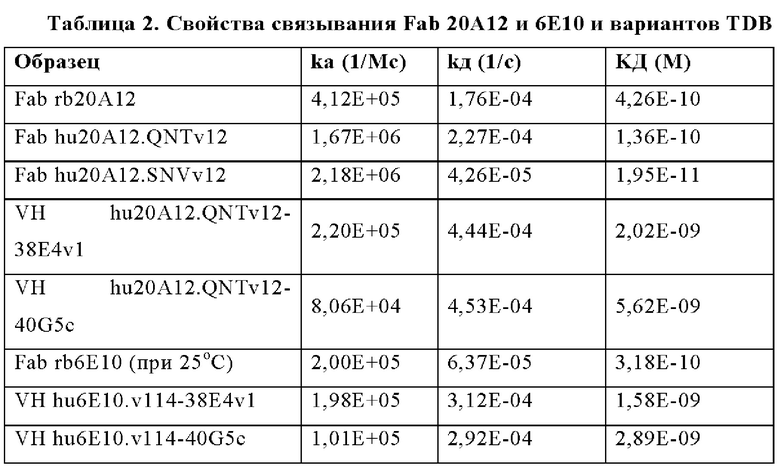
Анализ получения временной трансфекции
Два гуманизированных варианта rb.20A12, 20A12.QNTv1 и 20A12.QNTv12, оценивали в анализе получения временной трансфекции. Через один день после трансфекции в клетки яичника китайского хомячка (СНО) среду роста меняли на патентованную среду для получения. Супернатанты собирали через один день после добавления среды для получения и оценивали на титр антител с помощью Fc-связывающего ИФА. Выход получения нормализовали относительно 38E4v1. aFGFR1.knob предоставляли как контроль. Как 20A12.QNTv.1, так и 20A12.QNTv12 имели приемлемый выход (фиг. 37).
Все из 20A12.V1, 20A12.v1.polished (20A12.QNTv12) и 1G4 показали благоприятные результаты в анализе BV ИФА (тест in vitro на риск атипичного клиренса; Hotzel et al., MAbs, 4: 753-760, 2012) при спаривании со CD3-специфичным плечом 38E4v1 или 40G5c (фиг. 38). Результаты нормализовали к антителу к Lye6E, антитело использовали как контроль для высокого сигнала связывания в анализе BV ИФА.
LY6G6D TDB, содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4v1 или 40G5c, оценивали на требования к оценке молекул с помощью теплового стресса и окислительного стресса с ААПГ. Не идентифицировали никаких требований (фиг. 39).
Гуманизированный вариант rb.20A12 - 20A12.QNTv12, гуманизированный вариант rb.6Е10 - 6E10.v114 и LY6G6D-специфичное плечо 1G4 спаривали с CD3-специфичным плечом 38E4v1 как TDB и тестировали на аффинность для LY6G6D человека и яванского макака в анализе BIAcore®. LY6G6D-Fc был непосредственно иммобилизирован на чипе, и TDB пропускали при 37°С. KД для TDB, содержащих плечо 1G4, была значительно выше, чем или у плеча 20A12.QNTv12, или плеча 6E10.v114 для LY6G6D как человека, так и яванского макака (фиг. 36).
Гуманизация rb.6Е10
Множество каркасных областей применяли для гуманизации 6Е10. CDR VL rb6E10 прививали на последовательности зародышевых линий человека KV1-5*01 и KV3-20*01, a CDR VH прививали на последовательности зародышевых линий человека HV3-53*01 и HV3-48*01 (фиг. 43А). Эти последовательности зародышевых линий выбирали на основе их высокого преобладания в сыворотке и высокой идентичности последовательности с rb.6Е10 (фиг. 33А и 33Б). Как описано выше для 20А12, все положения Верньера VL и VH из антител кролика прививали в соответствующие каркасные области зародышевой линии человека. Привитые компоненты со всеми аминокислотами кролика в положениях Верньера называются версией 1 (hu.6E10.v1a для KV1-5/HV3-53; hu.6E10.v1b для KV1-5/HV3-48 и hu.6E10.v1c для K3-20/HV3-53). Пять дополнительных вариантов легкой цепи получали для каждой зародышевой линии (KV1-5: L2-L6 и KV3-20: L2-L6), и десять дополнительных вариантов тяжелой цепи (Н2-Н11) получали для HV3-53. Для легкой цепи KV1-5 А1а2, Phe36 и Arg43 (L3) были определены как ключевые остатки Верньера кролика на основании оценки аффинности связывания вариантных антител, описанных выше (данные не приведены). Аналогично, для легкой цепи KV3-20 А1а2, Phe36 и Val58 (L4) были определены как ключевые остатки Верньера кролика. Для тяжелой цепи привитый компонент CDR на HV3-53*01 (Н11), как обнаружили, был достаточным для поддержания аффинности к huLy6G6D; не было никаких положений Верньера кролика. Дополнительный привитой компонент CDR получали в зародышевой линии HV3-48*01 (3-48Н2). Тяжелую цепь Н11 спаривали с KV3-20.L4 как 6E10v114. Тяжелую цепь HV3-48.H2 спаривали с KV1-5.L3 как 6E10v23 (фиг. 43А-43Г).
Результаты анализа молекулярной оценки (МО) для варианта 6E10v1 показаны на фиг. 41. D(54)G и D(58)Y в CDR-H2, как было обнаружено, являются нестабильными, с 30,2% увеличением изомеризации за две недели.
Пример 3. Последовательность и кристаллическая структура антитела к LY6G6D hu.20A12.QNTv12
Как описано в примере 2, гуманизированное антитело кролика 20A12.QNTv12, которое имеет перекрывающийся эпитоп с 1G4 (пример 2), определяли как сильное антитело к LY6G6D. 20A12.QNTv12 характеризовался высокой аффинностью связывания относительно Ly6G6D человека как TDB при 37°С (KД приблизительно 2 нМ, относительно приблизительно 16 нМ для 1G4), сравнимой аффинностью связывания с LY6G6D человека и яванского макака, и благоприятными результатами в анализе BV ИФА, тесто на экспрессию и тепловыми и окислительными тестами молекулярной оценки (МО). Аминокислотная последовательность 20A12.QNTv12, содержащая варианты, модифицированные для включения зарядовых пар для одноклеточного изготовления, и кристаллическая структура 20A12.QNTv12, связанного с LY6G6D, описаны ниже.
А. Аминокислотные последовательности 20А12.QNTv12
Аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи 20A12.QNTv12 показаны на фиг. 5А и 5Б (SEQ ID NO: 22 и 23). Аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи вариантов 20A12.QNTv12, модифицированных для включения зарядовых пар для одноклеточного изготовления, показаны на фиг. 5В и 5Г (SEQ ID NO: 10 и 11).
Б. Кристаллические структуры антитела 20А12.QNTv12, связанного с LY6G6D
Для определения кристаллической структуры 20A12.QNTv12, связанного с LY6G6D, полипептид, содержащий аминокислоты 93-104 LY6G6D (SEQ ID NO: 78), совместно кристаллизовали с антигенсвязывающей областью фрагмента (Fab) антитела 20A12.QNTv12 (фиг. 6А-6Е). Кристаллическая структура комплекса 20A12.QNTv12-LY6G6D имела разрешение 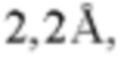 R/Rсвободный 19,9/24,3%; пространственная группа Р1: 82, 138, 139, 68, 75, 90. Fab 20A12.QNTv12 тестировали. Остатки LY6G6D 94-103 были связаны растворенными для всех десяти копий. Полипептид Ly6G6D 94-103 образовывал димер в кристаллической структуре антитела LY6G6D 1G4, и связывался как мономер с 20A12.QNTv12.
R/Rсвободный 19,9/24,3%; пространственная группа Р1: 82, 138, 139, 68, 75, 90. Fab 20A12.QNTv12 тестировали. Остатки LY6G6D 94-103 были связаны растворенными для всех десяти копий. Полипептид Ly6G6D 94-103 образовывал димер в кристаллической структуре антитела LY6G6D 1G4, и связывался как мономер с 20A12.QNTv12.
В. Кристаллическая структура 20А12.QNTv12 относительно 1G4
Кроме того, полипептид, содержащий аминокислоты 93-104 LY6G6D (SEQ ID NO: 87), совместно кристаллизовали с антигенсвязывающей областью фрагмента (Fab) антитела 20A12.QNTv12 (SEQ ID NO: 96-97) или Fab 1G4 (SEQ ID NO: 94 и 95). Конформации пептидного скелета были подобными между структурами 1G4 и 20A12.QNTv12 из-за дисульфидной связи, тогда как боковые цепи показали значительную конформационную подвижность (фиг. 7А и 7Б). 20A12.QNTv12 и 1G4, как было обнаружено, связываются с разными остатками пептида LY6G6D, как показано на фиг. 7В-7Е и в таблицах 3 и 4.
На фиг. 7В и 7Г показано взаимодействие 20A12.QNTv12 с LY6G6D. Остатки в 20A12.QNTv12, которые взаимодействуют с LY6G6D, маркированы на фиг. 7Г. В таблице 3 подытожены остатки поверхности контакта в Fab 20A12.QNTv12: LY6G6D комплексе. Эпитоп Fab 20A12.QNTv12 на LY6G6D человека состоит из остатков Arg94, Asp95, Cys96, Tyr97, Leu98, Gly99, Asp 100, Leu101, Cys102 и Asn103 (RDCYLGDLCN). Каждый из этих остатков располагается в 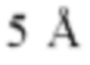 от Fab. Fab 20A12.QNTv12 использует остатки тяжелой цепи Asn31, Asn32, А1а33 и Met34 из CDRH1, Ser52 из CDRH2 и Arg98, Gly99 и Asp 100 из CDRH3 и остатки легкой цепи Thr91, Ser92, Phe93 и Arg94 из CDRL3 для взаимодействия с LY6G6D.
от Fab. Fab 20A12.QNTv12 использует остатки тяжелой цепи Asn31, Asn32, А1а33 и Met34 из CDRH1, Ser52 из CDRH2 и Arg98, Gly99 и Asp 100 из CDRH3 и остатки легкой цепи Thr91, Ser92, Phe93 и Arg94 из CDRL3 для взаимодействия с LY6G6D.
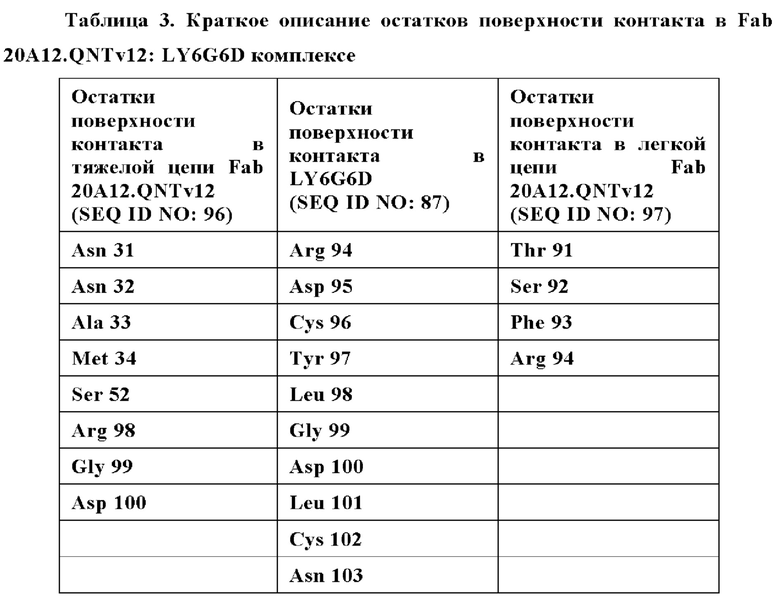
На фиг. 7Д и 7Е показано взаимодействие 1G4 с LY6G6D. Остатки в 1G4, которые взаимодействуют с LY6G6D, маркированы на фиг. 7Е. В таблице 4 подытожены остатки поверхности контакта в Fab 1G4: LY6G6D комплексе. Эпитоп Fab 1G4 на LY6G6D человека состоит из остатков His93, Asp95, Cys96, Tyr97, Leu98, Gly99 и Asp 100; каждый из этих остатков располагается в 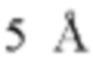 от Fab. В отличие от 20A12.QNTv12, не было обнаружено взаимодействие 1G4 с остатками LY6G6D Arg94, Leu101, Cys102 или Asn103. Таким образом, остатки LY6G6D Arg94, Leu101, Cys102 и Asn103 являются эпитопными остатками, которые уникально связываются 20A12.QNTv12.
от Fab. В отличие от 20A12.QNTv12, не было обнаружено взаимодействие 1G4 с остатками LY6G6D Arg94, Leu101, Cys102 или Asn103. Таким образом, остатки LY6G6D Arg94, Leu101, Cys102 и Asn103 являются эпитопными остатками, которые уникально связываются 20A12.QNTv12.
Fab 1G4 использует остатки тяжелой цепи Thr31, Tyr3 и Val33 из CDRH1 и Arg99 и Asn100 из CDRH3 и остатки легкой цепи Ser97, Tyr98, Ser99 и Ala100 из CDRL3 для взаимодействия с LY6G6D.
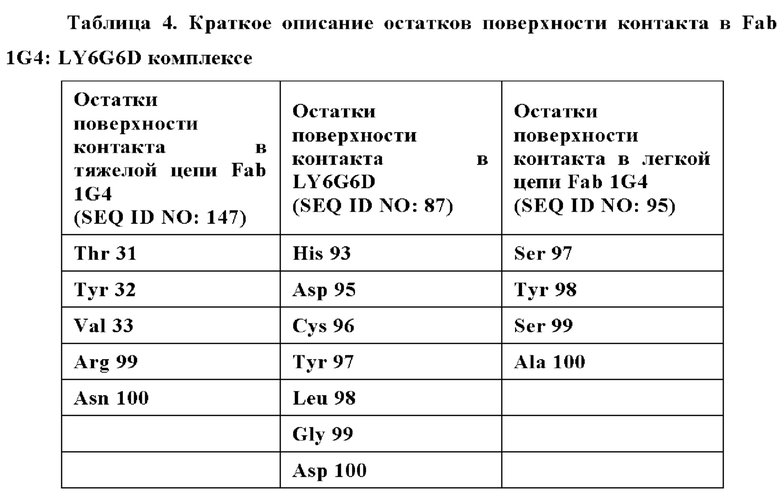
Пример 4. Анализы активности TDB in vitro
Цитотоксичность, связывание клеток, активация Т-клеток и уничтожение клеток
LY6G6D TDB (например, LY6G6D TDB, имеющие LY6G6D-специфичное плечо 20A12.QNTv12 или его вариант), спаренные с CD3-специфичным плечом 38E4v1 или 40G5c, тестировали на активность in vitro, используя клетки НТ55 (линия клеток карциномы толстой кишки человека), которые эндогенно экспрессируют средний уровень LY6G6D (фиг. 9А-9В, 10А-10Г и 11Е-11И). 40G5c и 38E4v1 представляют собой гуманизированные антитела гибридомы, полученные от мыши, иммунизированной ГЛУ (гемоцианином лимфы улитки), конъюгированный пептид, образованный N-концевыми 27 аминокислотами CD3ε человека. 40G5c, как ранее наблюдали, имеет относительно низкую аффинность к CD3, тогда как 38E4v1, как наблюдали, имеет высокую аффинность (патентная публикация США №2015-0166661).
Мононуклеары периферической крови человека (МНПК) выделяли из цельной крови здоровых доноров градиентом фиколла и использовали как эффекторные клетки. Совместную культуру МНПК человека и клеток НТ55 (Е:Т=10:1) инкубировали в присутствии различных концентраций LY6G6D TDB. Нацеленное уничтожение клеток и активацию Т-клеток измеряли через 72 часа инкубации. Нацеленное уничтожение клеток определяли количественно как процент цитотоксичности путем измерения интенсивности люминесценции (RLU) в анализе CellTiter-GLO®. Процент нацеленного уничтожения клеток рассчитывали с помощью следующего уравнения:
% нацеленного уничтожения клеток = {(RLU в необработанной лунке - RLU в обработанной TDB лунке) / (RLU в необработанной лунке)} × 100.
Активацию Т-клеток CD4+ и Т-клеток CD8+ измеряли с помощью активируемой флуоресценцией сортировки клеток (АФСК). Для Т-клеток CD8+ обнаруживали поверхностную экспрессию CD69 и CD25, и процент Т-клеток CD8+, которые были CD69+CD25+, сообщался как активация Т-клеток CD8+.
LY6G6D TDB, содержащее LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4v1, было сильным в индуцировании активации Т-клеток (фиг. 9Б, 9В, 10Б, 10В, 11Ж и 11К) и нацеленном уничтожении клеток (фиг. 9А, 10А, 10Г, 11Г-11Е и 11К) in vitro против НТ55 дозозависимым образом. При спаривании с высокоаффинным CD3-специфичным плечом 38E4v1 20A12.QNTv12 имел сравнимую in vitro силу для уничтожения клеток относительно 1G4. Когда плечо 20A12.QNTv12 спаривали с низкоаффинным CD3-специфичным плечом 40G5c, сила для уничтожения клеток была ниже, чем для плеча 38E4v1; однако, TDB, содержащее плечи 20A12.QNTv12 и 40G5c, имело большую силу для уничтожения клеток, чем TDB, содержащее плечи 1G4 и 40G5c. (фиг. 11Г и 11Д). 20А12 кролика также показало более высокую аффинность связывания с клетками НТ55, чем 1G4 (фиг. 4Е).
В анализе с использованием МНПК от десяти здоровых доноров-людей среднее уничтожение клеток ЕС50 с помощью TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4v1, составляло приблизительно 1,04 нг/мл (7 пМ); средняя активация Т-клеток CD8+ ЕС50 составляла приблизительно 87 нг/мл (583 пМ) (фиг. 11Е-11К). Уничтожение клеток и активация Т-клеток CD8+ также тестировали в клетках Colo320DM (типа С Дюка человека, линия клеток колоректальной аденокарциномы) и LS1034 (типа С Дюка человека, линия клеток колоректальной аденокарциномы) (фиг. 11А), которые экспрессировали различные уровни LY6G6D (фиг. 11Б и 11К).
Пример 5. Анализы активности TDB in vivo и клиренса
А. Объем опухоли
Кандидатные LY6G6D TDB, содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 40G5c или 38E4.v1 тестировали на активность in vivo против ксенотрансплантата LS1034 и опухолей НТ55 у мышей NSG™ (фиг. 16А-16В, 17А-17В). Мышей гуманизировали с помощью мононуклеаров периферической крови (МНПК) от здоровых доноров. Обработки, содержащие среду доставки и МНПК или содержащие TDB и не содержащие МНПК, представлены в качестве контролей. Концентрации TDB в сыворотке измеряли с использованием анализа ИФА Generic Immunoglobulin Pharmacokinetic (GRIP) (Yang et al., J. Immunol. Methods, 8-20, 2008) после введения одной дозы TDB.
Эффективность in vivo LY6G6D TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 40G5c или 38E4.v1 против опухолей LS1034 (LY6G6D ИГХ оценка 3+) и опухолей НТ55 (LY6G6D ИГХ оценка 2+) устанавливали. Для моделей опухолей LS1034 и НТ55 большую эффективность наблюдали для TDB, содержащего высокоаффинное CD3-специфичное плечо 38E4.v1 (1 мг/кг, одна доза, в/в, D0) (фиг. 16А и 17А). Эффективность и ФК были дозозависимыми для LY6G6D TDB, содержащего 38E4.v1, и небольшое отличие в ФК сыворотки между LY6G6D TDB, содержащим 38E4v1, и LY6G6D TDB, содержащим 40G5c, наблюдали (фиг. 16А-16В и 17А-17В). Эффективность in vivo LY6G6D TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4.v1, также тестировали против опухолей Colo320DM (фиг. 12).
Б. Клиренс у мышей SCID
Клиренс LY6G6D TDB, содержащих LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 40G5c или 38E4.v1, измеряли у мышей с тяжелым комбинированным иммунодефицитом (SCID) после внутривенного введения одной 5 мг/кг дозы антитела с помощью анализа ИФА GRIP (фиг. 18). Скорость клиренса (7,67 мл/сутки/кг) была сравнима с другими TDB, спаренными с CD3-специфичным плечом 38E4v1. Скорости клиренса находились в диапазоне от 8,6 до 16 мл/кг/сутки. TDB, содержащее 20A12.QNTv12 и 38E4v1, имело приемлемую фармакокинетику (ФК) у мышей SCID и показало менее чем двукратное отличие в системном CL по сравнению с контрольным антителом к gD 5 В6. Никакие требования для ФК (атипичная ФK) не были очевидными для LY6G6D TDB, содержащего плечи 20A12.QNTV12 и 38E4v1, в несвязанных частицах. Все растворы для дозирования восстанавливали в пределах ±20% номинальной дозы.
Пример 6. Анализ безопасности для яванских макак
А. Исследование токсичности для яванских макак
Исследование токсичности для LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTV12 и CD3-специфичное плечо 38E4.v1, проводили для яванских макак (супо). Исследование имело адаптивную схему с повторяющимися и смещенными группами дозирования (фиг. 19). Группу 2 лечили одной дозой (1 мг/кг) в/в инфузии LY6G6D TDB в 1 день (Д1). Группы 3, 4 и 5 лечили одной дозой (2, 4 и 8 мг/кг соответственно) в/в инфузии, а группу 6 лечили одной дозой (15 мг/кг) в/в инфузии. Окончательную аутопсию проводили на Д8 для оценки гистопатологии (фиг. 19). Целями исследования было снятие риска мишени LY6G6D у здоровых животных и для тестируемых диапазонов доз. ФК/ФД и стандартные конечные значения токсичности включали на основе предыдущего опыта с TDB.
Внутривенную ФК оценивали для всех групп лечения с помощью анализа ИФА GRIP. Результаты показали большую чем доза пропорциональный системное воздействие (нормализованное по дозе AUC0-7). Клиренс (CL) снижался с увеличением дозы TDB (4 мг/кг и выше), что предполагало насыщение опосредованного мишенью распределения лекарственного средства (TMDD) при этих дозах (CD3-опосредованный, в периферической крови) (фиг. 20А; таблица 5). ФК при 1 мг/кг дозе была сравнима с историческими данными для других TDB у яванцев, например, TDB gD/38E4v1, которое показывало CL приблизительно 20 мл/кг/сутки (фиг. 20А и 20Б). Растворы для дозирования восстанавливали в пределах ±20% номинального значения.
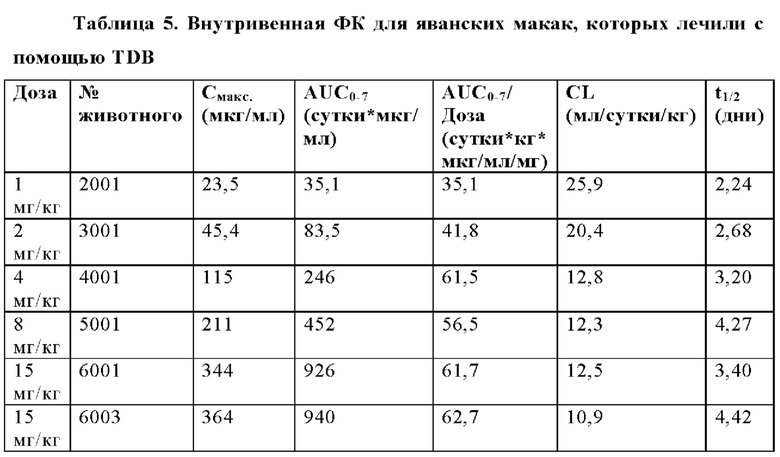
Лечение одной дозой в/в с помощью LY6G6D TDB, содержащего LY6G6D-специфичное плечо 20A12.QNTV12 и CD3-специфичное плечо 38E4.v1, хорошо переносилось при дозах до 15 мг/кг. Клинические наблюдения не включали ветеринарное лечение и влияние на смертность, вес тела или потребление пищи. При дозе 15 мг/кг наблюдали тошноту (умеренную) у животного №6001, и бледность и тремор (незначительный) отмечали у животного №6003 через 4-5 часа после дозы только. Клиническая патология не показывала признаков воспаления или повреждения печени и умеренное повышение С-реактивного белка (CRP) (фиг. 22Б). Анатомическая патология обнаруживала околососудистые/сосудистые мононуклеарные инфильтраты в мозгу одного животного при дозе 15 мг/кг (животное №6003, с заметным незначительным тремором) (фиг. 21).
Концентрацию цитокинов G-CSF, IL-IRa, МСР-1, TNF-a, IL-13, IL-8 и С-реактивного белка (CRP) измеряли после лечения (фиг. 22А и 22Б). Некоторые цитокины показали умеренное повышение концентрации при дозах ≥1 мг/кг. Не было очевидной взаимосвязь доза-реакция. Умеренное увеличение в МСР-1 наблюдали при ≥4 мг/кг (фиг. 22А). Все изменения возвращались на исходный уровень через 24 часа после лечения.
Подсчеты экспрессирующих CD3+/CD4+/CD5+CD25 лимфоцитов-Т-хэлперов (Th), экспрессирующих CD3+/CD8+/CD5+CD25 Т-цитотоксических (Тс) лимфоцитов, Т-лимфоцитов CD45+/CD3+, В-лимфоцитов CD45+/CD20+ и клеток-естественных киллеров CD45+/CD16+ измеряли цитометрией в потоке (фиг. 23А-23Г). Измерения проводили за 7 дней до обработки (день -7) и в день обработки (день 1 до) и усредняли для обеспечения средней предварительной дозы. После окончания инфузии измерения проводили через 2 часа, 6 часов, 24 часов и 168 часов. Наблюдали пики, показывающие умеренную активацию Т-клеток (фиг. 23А), восстановление Т-клеток (фиг. 23Б) и восстановление В-клеток (фиг. 23В).
Пример 7. Анализы BIAcore для аффинности
TDB, содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4.v1 (фиг. 24А) или плечо 40G5C (фиг. 24Б), анализировали на аффинность к полипептиду Ly6G6D человека и яванского макака с помощью анализа BIAcore. Ly6G6D-Fc был непосредственно иммобилизирован на чипе, a TDB пропускали при 37°С. Аналогичные анализы проводили для TDB, содержащих LY6G6D-специфичное плечо 1G4 и CD3-специфичное плечо 38E4.v1 (фиг. 24В) или плечо 40G5C (фиг. 24Г). Результаты этих анализов приведены в Таблице 6.
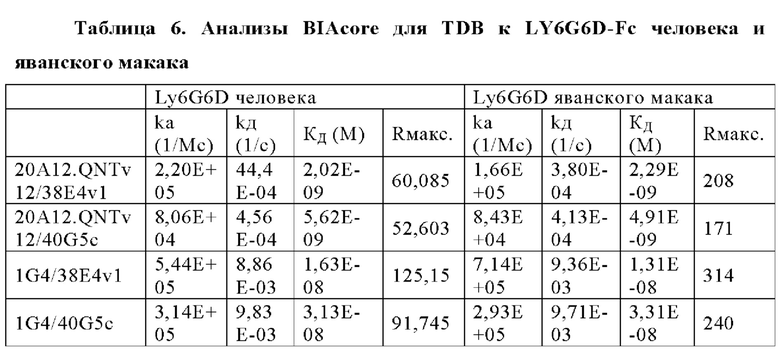
LY6G6D TDB, изготовленное с помощью двухклеточной системы и содержащее LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4.v1, имело высокую аффинность связывания с полипептидами LY6G6D человека и яванского макака и внеклеточным доменом (ECD) человека и яванского макака CD3 (таблица 7).
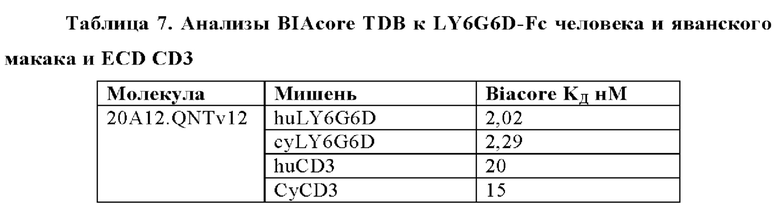
TDB, содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3 38E4.v1 (левая панель; получаемые с помощью двухклеточной системы изготовления), MD1 38E4.v1 (центральная панель; получаемые с помощью одноклеточной системы изготовления), или MD4-специфичное плечо 38E4.v1 (правая панель; получаемые с помощью одноклеточной системы изготовления) анализировали на аффинность к полипептиду Ly6G6D человека (фиг. 25) с помощью анализа BIAcore. Результаты этих анализов приведены в Таблице 8.
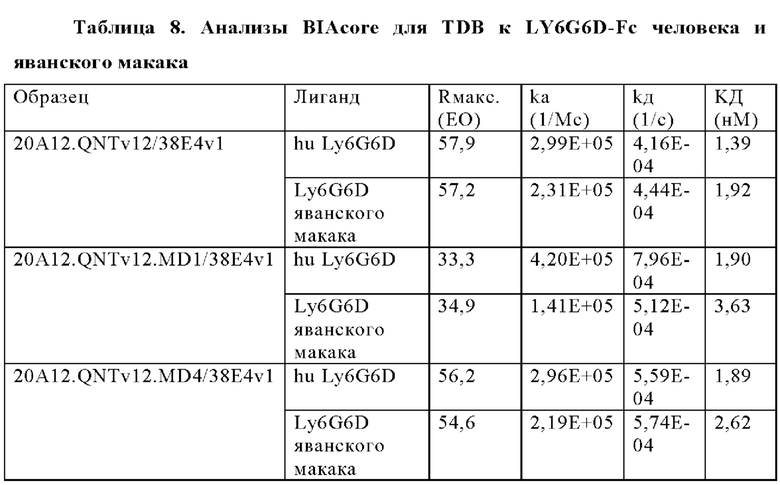
Пример 8. Разработка CD3-специфичных плечей MD1 и MD4 для одноклеточного изготовления
А. Одноклеточная и двухклеточная системы изготовления
Антитела к LY6G6D (например, 20A12.QNTv12) изготавливали как LY6G6D Т-клеточнозависимые биспецифические антитела (LY6G6D TDB) с первым плечом со специфичностью к LY6G6D и вторым плечом со специфичностью к CD3, как показано на фиг. 8А. TDB изготавливали с помощью или двухклеточной системы (фиг. 8Б), или одноклеточной системы (фиг. 8В). Для одноклеточного изготовления важно, чтобы первое плечо и второе плечо экспрессировались на сравнимых уровнях в клетке-хозяине: Большие различия в уровне экспрессии увеличивают вероятность ошибочного спаривания легких цепей (LC) и снижает вероятность правильного спаривания TDB. CD3-специфичное плечо 38E4v1 относительно плохо экспрессируется, тогда как LY6G6D-специфичное плечо 20A12.QNTv12 сильно экспрессируется (фиг. 37). В анализе временной трансфекции для оценки сборки TDB LY6G6D TDB, содержащее плечо 38E4v1 и плечо 20A12.QNTv12, не достигает >80% правильного спаривания, даже когда соотношения ДНК, кодирующих соответствующие плечи, модифицировали в отношении 1:16 (фиг. 29Г).
Два варианта CD3-специфичного плеча 38E4v1, 38E4v1 MD1 (MD1) и 38E4v1 MD4 (MD4), были разработаны как имеющие улучшенную экспрессию относительно 38E4v1 и, таким образом, позволяют улучшенное изготовление одноклеточного формата LY6G6D TDB (например, одноклеточного формата LY6G6D TDB с LY6G6D-специфичным плечом 20A12.QNTv12). MD1 и MD4, как было обнаружено, имеют подходящую биспецифическую к LC-ошибочное спаривание отношение чистоты (т.е. высокую чистоту правильно образованных LY6G6D TDB) и сравнимую in vitro силу, in vivo силу и ФК для 38E4v1 дикого типа.
TDB, изготовленные с помощью одноклеточной системы, были дополнительно модифицированы для содержания мутаций аминокислотных замен, вводящих зарядовые пары, как показано на фиг. 8Г или 8Д, и как описано в настоящем документе.
Б. Одноклеточный вариант 20А12.QNTv12
Для облегчения изготовления в одноклеточной системе последовательности тяжелой цепи и легкой цепи плеча 20A12.QNTv12 модифицировали для содержания мутаций аминокислотных замен, вводящих зарядовые пары (фиг. 5В, 5Г и 8А-8Д).
В. Образование экспрессионных вариантов 38E4v1 MD1 и MD4
Для создания вариантов с улучшенной экспрессией 38E4v1, подходящих для изготовления в одноклеточной системе, последовательность VL 38E4v1 модифицировали остатками из 40G5c, антитела к CD3 с лучшей экспрессией, чем 38E4v1, как описано ниже. Два варианта 38E4v1 - 38E4v1 MD1 (MD1) и 38E4v1 MD4 (MD4) - имеют приемлемое отношение чистоты биспецифического к LC-ошибочное спаривание и сравнимую in vitro силу, in vivo силу и ФК для 38E4v1 дикого типа (ДТ) у мышей.
Варианты вариабельной области легкой цепи (VL) 38E4v1, содержащей одну или более аминокислотных замен, полученных из последовательности легкой цепи низкоаффинного антитела к CD3 40G5c, получали (фиг. 29А). VL MD1 38E4v1 содержит четыре аминокислотные замены относительно последовательности 38E4v1: S43P, Т51А, K55E и K89T (фиг. 29А). VL MD4 содержит только замены S43P и Т51А (фиг. 29А). Варианты, содержащие только S43P, Т51А, K55E или К89Т аминокислотную замену, также получали. Все варианты 38E4v1 содержали последовательность вариабельной области тяжелой цепи (VH) 38E4v1 (фиг. 29Б).
В. Анализ временной трансфекции
Уровни экспрессии 38E4v1, MD1 и вариантов, содержащих только S43P, Т51А, K55E или K89T аминокислотные замены относительно 38E4v1, оценивали в анализе временной трансфекции в 11-9 клетках-хозяевах яичника китайского хомячка (СНО).
Через день после трансфекции среду для роста меняли на среду для получения. Супернатанты собирали через один день после добавления среды для получения и оценивали на титр антител с помощью Fc-связывающего ИФА. Выход получения нормализовали относительно 38E4v1. Легкая цепь 38E4v1, по-видимому, ограничивала выход экспрессии антитела. Неожиданно, замена отдельных положений в легкой цепи 38E4v1 на остатки от низкоаффинного антитела к CD3 40G5c (S43P, Т51А или K89T) приводила к умеренным улучшениям выходов (фиг. 29В), и более значительное увеличение выхода наблюдали, когда эти замены объединяли в варианте MD1, который содержит все из аминокислотных замен S43P, Т51А, K55E, К89Т (фиг. 29А и 29В).
В анализе временной трансфекции для сборки TDB варианты 38E4v1 с улучшенной экспрессией MD1 и MD4 достигали 95% сборки интактных антител в отношении 1:4 ДНК легкой цепи (LC) плеча-мишени (специфичного для LY6G6D) к ДНК CD3-специфичного плеча, тогда как 38E4v1 не достигал >80% надлежащего спаривания даже при отношении 1:16 ДНК LC к ДНК CD3-специфичного плеча (фиг. 29Г). Успешное разработанный клон клеточной линии xFcRH5 представлен как контроль. MD1 и MD4, таким образом, обеспечивают высокий процент надлежаще спаренных биспецифических антител, которые необходимо получить.
Г. Анализ кинетики связывания
Кинетика связывания плечей 38E4v1 MD1 и MD4 показала аффинность к лигандам CD3 человека и яванского макака, сравнимых с высокоаффинным CD3-специфичным плечом 38E4v1 дикого типа (ДТ) (таблица 9). Варианты MD1 и MD4, таким образом, сохраняли высокую аффинность 38E4v1. Анализы проводили с помощью BIAcore Т200: 27mer полипептиды CD3 человека и яванского макака конъюгировали с биотином и иммобилизировали на чипе SA СМ5, и TDB пропускали при 100 мкл/мин. при 37°С.
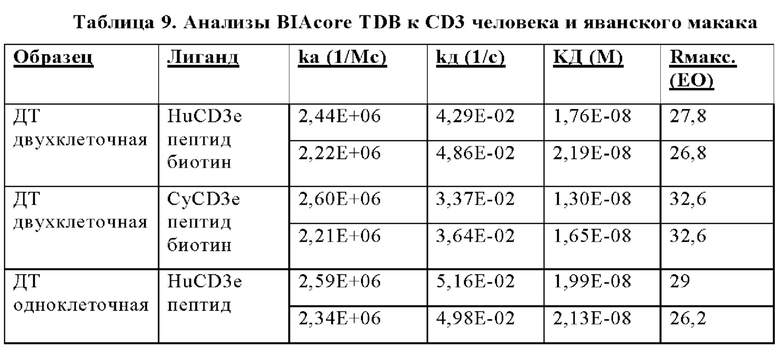
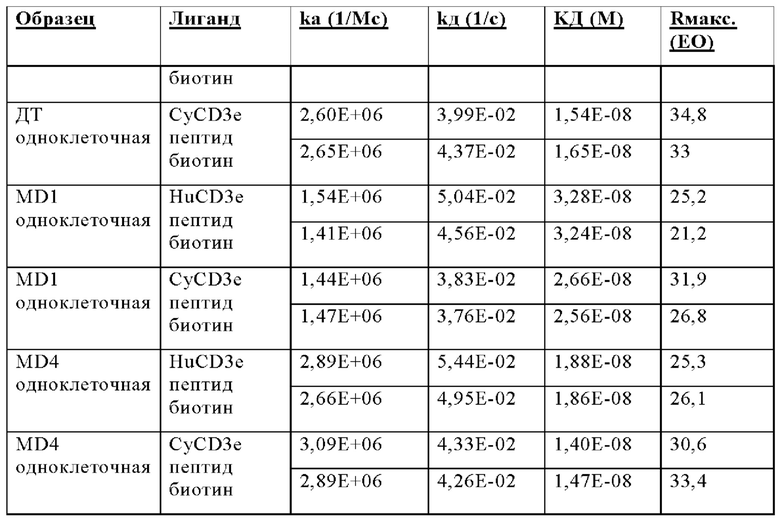
Д. Фармакокипетические анализы
Для оценки фармакокинетики (ФК) различных CD3-специфичных плечей, профили ФК 38E4v1, 40G5c, MD1, MD4 и 38E4v1.K55E измеряли у мышей СВ-17 SCID после введения одной дозы каждого варианта. Антитело к gD использовали как контроль. Антитело к gD является несвязывающим контрольным IgG, нацеленным на эпитоп гликопротеина D вируса простого герпеса. Все антитела тестировали как моноспецифические двухвалентные антитела к CD3, имеющие изотип IgG1 человека с мутацией N297G, для ослабления FcγR-опосредованной эффекторной функции.
Шести группам самок мышей СВ-17.SCID (n=12 на группу; Charles River Laboratories, 251) вводили одну в/в дозу каждого антитела в дозе 5 мг/кг. Самок мышей использовали для удобства. Исторически, мы не наблюдали никаких различий в исследованиях ФК, в которых использовали самцов или самок мышей. Образцы крови собирали из бедренной вены в выбранные моменты времени (3 повтора для каждого момента времени) в течение до 21 дня. Общие концентрации антител в сыворотке определяли с помощью ИФА GRIP (планшет, покрытый антителом к IgG человека и детектируемый с антителом к IgG человека) с пределом обнаружения 15,6 нг/мл и используемый для ФК оценок. Восстановления растворов для дозирования составляли 111%, 110%, 106%, 97,8%, 103% и 106% для антитела к gD, 38E4v1, 40G5c, MD1, MD4 и 38E4v1.K55E соответственно. Восстановления дозирования составляли в пределах 20%; таким образом, номинальные дозы использовали для дальнейшего анализа.
ФК профили собирали от различных мышей в различные моменты времени. Номинальное время отбора образцов и фактические концентрации растворов дозирования использовали в анализе данных. Параметры некомпартментного анализа (НКА) устанавливали с помощью Phoenix WinNonlin® 64 с немногочисленным отбором образцов и введением в/в болюса. Значения стандартных погрешностей обеспечивали. Мышей умерщвляли после введения анестезии с изофлураном (5% изофлуран с 2 л/мин. О2). Все процедуры были ободрены и подтверждены в соответствии с руководствами и принципами, установленными Институциональным комитетом по уходу за животными и их использованию (IACUC) компании Genentech, и проводились на предприятии, аккредитованном Международной ассоциацией оценки и аккредитации лабораторных исследований на животных.
Варианты антител к CD3 не дают перекрестную реакцию с CD3 мышей; таким образом, ФК профили мышей TDB, содержащих варианты антител к CD3, давалидавали возможность сравнивать ФК и неспецифическую скорость выведения вариантов в отсутствие целевого связывания. Профили концентрации в сыворотке-времени вариантов антител к CD3 оценивали, вместе с контрольным антителом к gD (фиг. 29Д). ФК данные каждой группы характеризовали с помощью NCA (таблица 10).
Группа введения антитела к gD имела наивысшее воздействие по сравнению с группами, которым вводили варианты антител к CD3. Группа введения 40G5c имела несколько более низкое воздействие на основе его значения AUCпосл. по сравнению с группой введения антитела к gD и имела наивысшее воздействие по сравнению с остальными вариантами антител к CD3. С другой стороны, 38E4v1 имел самое низкое воздействие на основе его AUCпосл. по сравнению с остальными вариантами антител к CD3.
Среди трех вариантов MD1, MD4 и K55E, MD1 имел наивысшее воздействие на основе его АUСпосл., и его профиль ФК был подобен группе введения 40G5c. Группа введения MD4 имела самое низкое воздействие на основе его AUCпосл. среди трех вариантов, и его ФК профиль был подобен группе введения 38E4v1. Кроме того, MD1 улучшало воздействие 38E4v1 в ~3 раза на основе AUCбеск. у мышей. Кроме того, группы введения антитела к gD, 40G5c и 38E4v1.MD1 имели сравнимые значения Vss, которые составляли приблизительно в 2 раза ниже, чем значения Vss групп введения 38E4v1, MD4 и K55E (таблица 10).
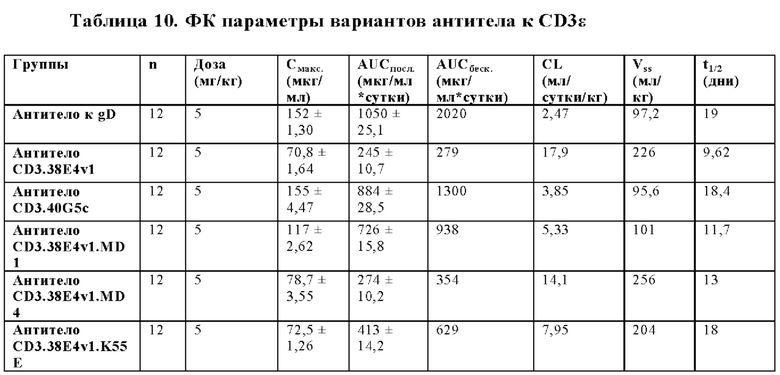
Значения стандартных погрешностей представлены, где применимо. Смакс. = максимальная наблюдаемая концентрация в сыворотке, АUСпосл. = площадь под кривой концентрация в сыворотке-время от времени 0 до последних измеренных моментов времени, день 28. АUСбеск. = площадь под кривой концентрация в сыворотке-время от времени 0, экстраполированное до бесконечности. CL = Клиренс. Vss = Объем распределения при установившемся режиме. t1/2 = конечный период полувыведения.
При условии более низкого воздействия 38E4v1 по сравнению с 40G5c, далее оценивали, имелись ли характеристики CD3-специфичного плеча, которые могут вносить вклад в неспецифичность для наблюдаемого более низкого воздействия. Мы оценивали заряд вариабельной области (Fv) антитела и гидрофобность 38E4v1 и 40G5c с помощью in silico прибора оценки клиренса (iCAT), прибора для расчета на основе последовательности, который дает оценку теоретического риска клиренса антитела у яванских макак (Sharma et al., Proc Natl Acad Sci USA, 111: 18601-6, 2014). Эта оценка основана на параметрах, рассчитанных из последовательности доменов Fv. Оценку iCAT оценивали для двухвалентного антитела к CD3. Рассчитанный заряд Fv 38E4v1 составлял +7,6, что было за пределами заряда для приемлемого in vivo клиренса (приемлемый диапазон включает заряды Fv от ≥ 0 до ≤ +6,2), тогда как рассчитанный заряд Fv 40G5c находился в пределах приемлемого диапазона на уровне +5,6. Рассчитанный заряд Fv для MD1 составлял +4,7, а для MD4 - +7,6. MD1, таким образом, имело улучшенный заряд Fv по сравнению с 38E4v1 и приемлемый теоретический риск для клиренса у яванских макак.
Кроме того, варианты антител к CD3 тестировали с помощью ИФА бакуловируса (BV). ИФА BV представляет собой in vitro прибор, используемый для оценки неспецифического клиренса (Hotzel et al., MAbs, 4: 753-760, 2012). Эта оценка основана на обнаружении ИФА неспецифического связывания с частицами бакуловируса, и может идентифицировать антитела с повышенным риском для быстрого клиренса. Оценка согласно ИФА BV для 38E4v1 составляла 1,15, что было за пределами диапазона прогнозирования приемлемого in vivo клиренса (оценка согласно ИФА BV > 1,0 показывает высокую вероятность быстрого клиренса). Напротив, оценка согласно ИФА BV составляла 0,15 для MD1 и составляла 0,72 для MD4; таким образом, как MD1, так и MD4 были в пределах приемлемого диапазона. И интересно, что, хотя MD4 проходил тест согласно ИФА BV для дополнительного тестирования in vivo, данные in vivo ФК у мышей показали, что MD4 не улучшал воздействие 38E4v1 по сравнению с MD1.
Пример 9. Анализы активности TDB для антител, полученных в одноклеточной системе
Кандидатные LY6G6D TDB, собранные с помощью одноклеточной системы (фиг. 8В-8Д) и содержащие LY6G6D-специфичное плечо 20A12.QNTV12 и CD3-специфичное плечо 38E4v1 MD1 (MD1), 38E4v1 MD4 (MD4) или 38E4v1 (ДТ), тестировали на in vitro активность, in vivo активность и аффинность с полипептидом CD3. TDB, собранное с помощью двухклеточной системы (фиг. 8Б) и содержащее LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо 38E4v1, использовали как контроль.
A. In vitro цитотоксичность, клеточное связывание и активация Т-клеток
LY6G6D TDB, собранные с помощью одноклеточной системы и содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо (ДТ) 38E4v1 MD1, 38E4v1 MD4 или 38E4v1, тестировали на in vitro активность в клетках НТ55, дополненных МНПК от здорового донора, активацию Т-клеток CD4+ и активацию Т-клеток CD8+ (фиг. 14А-14В и 15А-15В). Уничтожение количественно определяли как процент цитотоксичности в анализе CELLTITER-GLO®, и Т-клетки CD4+ и Т-клетки CD8+ измеряли с помощью активированной флуоресценцией сортировки клеток (АФСК), как описано в примере 2. Все TDB дозировали на уровне 1 мг/кг. LY6G6D TDB, содержащие плечи варианта 38E4v1, были сильными в отношении индуцирования активации Т-клеток (фиг. 14Б, 14В, 15Б и 15В) и нацеленного уничтожения клеток (фиг. 14А и 15А) in vitro против НТ55. Варианты MD1 и MD4 таким образом сохраняли высокую силу in vitro 38E4v1, в то же время обеспечивая улучшенную технологичность в одноклеточной системе.
Б. In vivo активность
LY6G6D TDB, содержащие LY6G6D-специфичное плечо 20A12.QNTv12 и CD3-специфичное плечо варианта 38E4v1, собранное с помощью одноклеточной системы, тестировали на in vivo активность против опухолей ксенотрансплантата НТ55 у мышей NSG™ (фиг. 26А и 26Б). Мышей гуманизировали с помощью мононуклеаров периферической крови (МНПК) от здоровых доноров. Обработки, содержащие среду доставки и МНПК или содержащие TDB и не содержащие МНПК, представлены в качестве контролей. Одноклеточные варианты, содержащие CD3-специфичное плечо 38E4v1 MD1 или 38E4v1 MD4, показали сравнимое снижение опухоли относительно двухклеточного LY6G6D TDB.
Клиренс TDB измеряли в мышиной модели опухоли НТ55 и у мышей SCID после внутривенного введения одной дозы антитела с помощью анализа ИФА GRIP (фиг. 27 и 28). ФK одноклеточных вариантов, содержащих CD3-специфичное плечо 38E4v1 MD1 или 38E4v1 MD4, была сравнимой с таковой для двухклеточного TDB. Никаких требований для ФК (например, атипичная ФК) не определяли для одноклеточных вариантов TDB. Растворы дозирования восстанавливали в пределах ±20% номинального значения.
В. Анализы на аффинность к CD3
Кандидатные LY6G6D TDB, собранные с помощью одноклеточной системы и содержащие CD3-специфичное плечо 38E4v1 MD1 или 38E4v1 MD4, показали аффинность к CD3, сравнимую с таковой для TDB, собранных с помощью одноклеточной системы или двухклеточной системы и содержащих CD3-специфичное плечо 38E4v1; MD1 и MD4, такое как 38E4v1, таким образом являются высокоаффинными CD3-специфичными плечами (таблица 11). Анализы проводили с помощью BIAcore Т200. 27Mer полипептиды CD3 человека и яванского макака конъюгировали с биотином и иммобилизовали на чипе SA СМ5. LY6G6D TDB пропускали при 37°С.
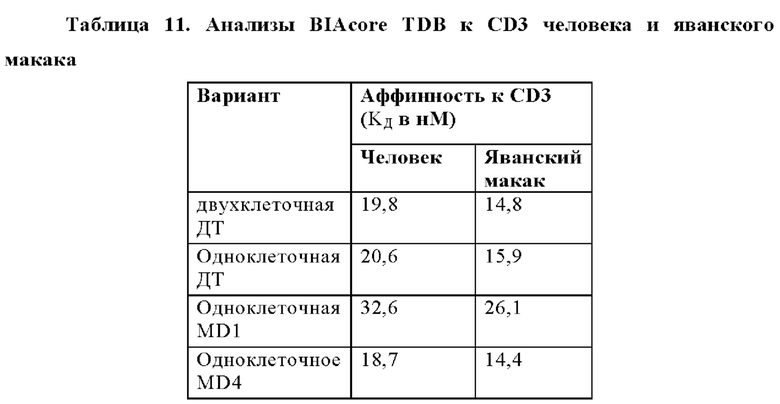
Хотя вышеизложенное изобретение было подробно описано посредством иллюстрации и примеров в целях ясности понимания, описания и примеры не следует воспринимать как ограничивающие объем данного изобретения. Содержание всех патентов и научной литературы, цитируемых в данном документе, в полном объеме и явным образом включено посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЖЕНЕНТЕК, ИНК.
<120> АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ
<130> 50474-184WO2
<150> US 62/948,097
<151> 2019-12-13
<160> 147
<170> PatentIn version 3.5
<210> 1
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 1
Gln Ala Ser Glu Ser Ile Thr Arg Tyr Leu Asn
1 5 10
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 2
Asp Ala Ser Lys Leu Pro Ser
1 5
<210> 3
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 3
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Gln Asn Thr
1 5 10
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 4
Asn Asn Ala Met Ile
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 5
Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala Ser
1 5 10 15
Gly
<210> 6
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 6
Gly Asp Leu
1
<210> 7
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Lys Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 8
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 8
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Lys Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 9
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 9
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Lys Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Glu Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 10
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 10
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 11
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 11
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Lys Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 12
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 12
Lys Ser Ser Gln Ser Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 13
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 13
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 14
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 14
Thr Gln Ser Phe Ile Leu Arg Thr
1 5
<210> 15
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 15
Ser Tyr Tyr Ile His
1 5
<210> 16
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 16
Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Asp
<210> 17
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 17
Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr
1 5 10
<210> 18
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 18
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Glu Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 19
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 19
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Lys Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 20
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 20
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 21
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 21
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 22
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 22
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 23
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 23
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 24
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 24
Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Arg Thr Tyr Leu His
1 5 10 15
<210> 25
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 25
Leu Val Ser Asn Leu Asp Ser
1 5
<210> 26
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 26
Trp Gln Gly Thr His Leu Pro Phe Thr
1 5
<210> 27
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 27
Ser Tyr Trp Ile Gln
1 5
<210> 28
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 28
Glu Ile Leu Pro Gly Ser Asp Asn Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly Lys
<210> 29
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 29
Trp Phe Ala Tyr
1
<210> 30
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 30
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Trp Ile Gln Trp Ile Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Ser Asp Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Lys Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
100 105 110
Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 31
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 31
Asp Ile Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1 5 10 15
His Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Arg Thr Tyr Leu His Trp Leu Phe Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Leu Val Ser Asn Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 32
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 32
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Trp Ile Gln Trp Ile Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Ser Asp Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Lys Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
100 105 110
Ala
<210> 33
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 33
Asp Ile Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1 5 10 15
His Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Arg Thr Tyr Leu His Trp Leu Phe Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Leu Val Ser Asn Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 34
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 34
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val
20 25 30
<210> 35
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 35
Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 36
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 36
Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Met Arg
20 25 30
<210> 37
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 37
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 38
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 39
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 39
Trp Tyr Gln Lys Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 40
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 40
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 41
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 41
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 42
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 42
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr
20 25 30
<210> 43
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 43
Trp Val Arg Glu Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 44
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 44
Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr Leu Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 45
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 45
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 46
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 46
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys
20
<210> 47
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 47
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 48
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 48
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
20 25 30
<210> 49
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 49
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 50
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 50
Trp Ala Ser Thr Arg Lys Ser
1 5
<210> 51
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 51
Lys Gln Ser Phe Ile Leu Arg Thr Phe
1 5
<210> 52
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 52
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 53
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 53
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 54
<400> 54
000
<210> 55
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 55
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 56
<400> 56
000
<210> 57
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 57
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Lys Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 58
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 58
Trp Val Arg Lys Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 59
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 59
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Lys Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 60
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 60
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Glu Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 61
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 61
Trp Tyr Gln Glu Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 62
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 62
Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 63
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 63
Trp Tyr Gln Gln Glu Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 64
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 64
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Lys Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Glu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 65
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 65
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Glu Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Lys Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 66
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 66
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Lys Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Glu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 67
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 67
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Glu Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 68
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 68
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Glu Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Lys Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 69
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 69
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Glu Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Lys Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 70
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 70
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Glu Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 71
<400> 71
000
<210> 72
<400> 72
000
<210> 73
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 73
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Glu Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 74
<400> 74
000
<210> 75
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 75
Met Lys Pro Gln Phe Val Gly Ile Leu Leu Ser Ser Leu Leu Gly Ala
1 5 10 15
Ala Leu Gly Asn Arg Met Arg Cys Tyr Asn Cys Gly Gly Ser Pro Ser
20 25 30
Ser Ser Cys Lys Glu Ala Val Thr Thr Cys Gly Glu Gly Arg Pro Gln
35 40 45
Pro Gly Leu Glu Gln Ile Lys Leu Pro Gly Asn Pro Pro Val Thr Leu
50 55 60
Ile His Gln His Pro Ala Cys Val Ala Ala His His Cys Asn Gln Val
65 70 75 80
Glu Thr Glu Ser Val Gly Asp Val Thr Tyr Pro Ala His Arg Asp Cys
85 90 95
Tyr Leu Gly Asp Leu Cys Asn Ser Ala Val Ala Ser His Val Ala Pro
100 105 110
Ala Gly Ile Leu Ala Ala Ala Ala Thr Ala Leu Thr Cys Leu Leu Pro
115 120 125
Gly Leu Trp Ser Gly
130
<210> 76
<211> 2449
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 76
atgaaacccc agtttgttgg gatcttgctc agctccctgc taggggctgc cttgggtaag
60
gaggcggcca gctagcttct cacacaggcc ttctgccagc cggctccacc gagggcccag
120
gtccagcgcc tcttttctcc tgccaggaaa ccgaatgcgg tgctacaact gtggtggaag
180
ccccagcagt tcttgcaaag aggccgtgac cacctgtggc gagggcagac cccagccagg
240
cctggaacag atcaagctac ctggaaaccg tgagtcctca gtttctccct cttccagcag
300
cctttccctg cctccagccc catgtcaatc cttctggctt ccagaaccct ccaggctcag
360
tctggctctg ggcagatggt gcagctgtta gaggagagca gtctgtaccc cttctggctc
420
ctggcacgga gcccctgaga ggcccacagt ccttgtgccc ccacttcccc acctccttat
480
tctcctaaaa gaatctcata ggcccattag ctcacaaatg aagagctctg gccctgaaag
540
gccaaagtta aaaccaaact tcaaattttc ggcattagtt aaggaccagg gaggggtgtg
600
tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtacatg tttttaatat tttattttaa
660
cataattttg gattgacaga aaagttgcag aaatactcaa cttctcctaa tgctaacatc
720
ttacataacc atagcacaat tatcaaaatc acaaaataac tgatacaata ctactaacta
780
atctacagac tttatttgat ttagcaagat cctacattgc atttagctct catgtcttct
840
tagtctcctc tgatctgtgc cagttctgtt tttctttgtc tttcatgacc ctgacacatt
900
tgaagagccc tgataaatta ttttatacct ggagtttaaa aaattacttt tagggccagt
960
gcagtcactc gcacctgtaa tcccagcact ttaggaggcc aaggtgggag gaccacttga
1020
gcccaagagt tgagaccagc ctgggcaaca tagggagacc ctgtctctac aaaaaacaaa
1080
caaacaaaca aacaaacaga ttaaaaaatt agttgggtgt ggtggcacat gcttgtagtc
1140
ctagctactc agggggctga agagggagga tcgcttgagc ctgggagatt gaagctacaa
1200
tgagccatga tcacgccact acactccagc ctggggaaca aaatgagacc ctgtctcaaa
1260
aataataata ataataattt ttaggctagg cttggtggca cacacttgta atcccagcac
1320
tttgggaggc caaggctgaa gagtcacctg aggtcaggag tttgacacca gcctgggcag
1380
caaagtgaga cccccatctc tacaaaaaat gtttttaaaa aattagccag gcatagtggc
1440
acacacctgt aatctcagtt tcctgagagg ctgaggcagg aggattactt gagcccagga
1500
gtttgaggct atagggaggt atgattgcac caccacactc cagcctgagt gagagagcaa
1560
gatcttttct ctaaaattaa ataaaatcat ttttagatta aacaaaaatt acgtgccgga
1620
tgcagtggct cacgcctgta atcccagcac tttgggaggc caaggcgggt ggataacctg
1680
aggtcgggag ttcaagacca gcctgatcaa tgtggagaaa tctcgtctct actaaaaata
1740
caaaattagc cgggtgtagt ggtgcccgcc tgtaatacca gctactcggg aacctgaggc
1800
aggagaattg cttgaaccca agaggtggag gtcgcggtga gccgagatca caccattgca
1860
ctccagctgg gcaataagag tgaaactccg tctcaaaaaa aaaaaaaaaa ttacagatac
1920
ttgaaatact aaaaattatt ttatagaatg tccctcgata tttatttatc tgatatttgc
1980
catgatgaga ttgaggtcat gcattttaag caagaatact gcagaagtga tgttgcatcc
2040
ttcttgctgc atcacatcag gagtttacaa ggtcaatgca ttaactttga tcacttggtt
2100
tcagggaggt gttttttgag ggggctgaaa atccctttgg gctccttgaa atcacatctg
2160
ctctgcccca gaaggcaagt cctgaagcca ggagtccaac accccagttt cattctctct
2220
ctcagcccca gtgaccttga ttcaccaaca tccagcctgc gtcgcagccc atcattgcaa
2280
tcaagtggag acagagtcgg tgggagacgt gacttatcca gcccacaggg actgctacct
2340
gggagacctg tgcaacagcg ccgtggcaag ccatgtggcc cctgcaggca ttttggctgc
2400
agcagctacc gccctgacct gtctcttgcc aggactgtgg agcggatag
2449
<210> 77
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 77
Met Lys Pro Gln Phe Val Gly Ile Leu Leu Ser Ser Leu Leu Gly Ala
1 5 10 15
Ala Leu Gly Asn Arg Met Arg Cys Tyr Asn Cys Gly Gly Ser Pro Ser
20 25 30
Ser Ser Cys Lys Glu Ala Val Thr Thr Cys Gly Glu Gly Arg Pro Gln
35 40 45
Pro Gly Leu Glu Gln Ile Lys Leu Pro Gly Asn Pro Pro Val Thr Leu
50 55 60
Ile His Gln His Pro Ala Cys Val Ala Ala His His Cys Asn Gln Val
65 70 75 80
Glu Thr Glu Ser Val Gly Asp Val Thr Tyr Pro Ala His Arg Asp Cys
85 90 95
Tyr Leu Gly Asp Leu Cys Asn Ser Ala Val Ala Ser His Val Ala Pro
100 105 110
Ala Gly Ile Leu Ala Ala Ala Ala Thr Ala Leu Thr Cys Leu Leu Pro
115 120 125
Gly Leu Trp Ser Gly
130
<210> 78
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 78
Arg Asp Cys Tyr Leu Gly Asp Leu Cys Asn
1 5 10
<210> 79
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 79
Gly Asp Leu
1
<210> 80
<211> 207
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 80
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 81
<211> 198
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 81
Met Gln Ser Gly Thr Arg Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Ile Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr
20 25 30
Gln Thr Pro Tyr Gln Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Ser Gln His Leu Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys
50 55 60
Asn Lys Glu Asp Ser Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu
65 70 75 80
Met Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro
85 90 95
Glu Asp Ala Ser His His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn
100 105 110
Cys Met Glu Met Asp Val Met Ala Val Ala Thr Ile Val Ile Val Asp
115 120 125
Ile Cys Ile Thr Leu Gly Leu Leu Leu Leu Val Tyr Tyr Trp Ser Lys
130 135 140
Asn Arg Lys Ala Lys Ala Lys Pro Val Thr Arg Gly Ala Gly Ala Gly
145 150 155 160
Gly Arg Gln Arg Gly Gln Asn Lys Glu Arg Pro Pro Pro Val Pro Asn
165 170 175
Pro Asp Tyr Glu Pro Ile Arg Lys Gly Gln Gln Asp Leu Tyr Ser Gly
180 185 190
Leu Asn Gln Arg Arg Ile
195
<210> 82
<211> 182
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 82
Met Glu Gln Gly Lys Gly Leu Ala Val Leu Ile Leu Ala Ile Ile Leu
1 5 10 15
Leu Gln Gly Thr Leu Ala Gln Ser Ile Lys Gly Asn His Leu Val Lys
20 25 30
Val Tyr Asp Tyr Gln Glu Asp Gly Ser Val Leu Leu Thr Cys Asp Ala
35 40 45
Glu Ala Lys Asn Ile Thr Trp Phe Lys Asp Gly Lys Met Ile Gly Phe
50 55 60
Leu Thr Glu Asp Lys Lys Lys Trp Asn Leu Gly Ser Asn Ala Lys Asp
65 70 75 80
Pro Arg Gly Met Tyr Gln Cys Lys Gly Ser Gln Asn Lys Ser Lys Pro
85 90 95
Leu Gln Val Tyr Tyr Arg Met Cys Gln Asn Cys Ile Glu Leu Asn Ala
100 105 110
Ala Thr Ile Ser Gly Phe Leu Phe Ala Glu Ile Val Ser Ile Phe Val
115 120 125
Leu Ala Val Gly Val Tyr Phe Ile Ala Gly Gln Asp Gly Val Arg Gln
130 135 140
Ser Arg Ala Ser Asp Lys Gln Thr Leu Leu Pro Asn Asp Gln Leu Tyr
145 150 155 160
Gln Pro Leu Lys Asp Arg Glu Asp Asp Gln Tyr Ser His Leu Gln Gly
165 170 175
Asn Gln Leu Arg Arg Asn
180
<210> 83
<211> 181
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 83
Met Val Gln Gly Lys Gly Leu Thr Gly Phe Ile Leu Ala Ile Ile Leu
1 5 10 15
Leu Gln Gly Ser Leu Ala Gln Ser Phe Glu Glu Asn Arg Lys Leu Asn
20 25 30
Val Tyr Asn Gln Glu Asp Gly Ser Val Leu Leu Thr Cys His Val Lys
35 40 45
Asn Thr Asn Ile Thr Trp Phe Lys Glu Gly Lys Met Ile Asp Ile Leu
50 55 60
Thr Ala His Lys Asn Lys Trp Asn Leu Gly Ser Asn Thr Lys Asp Pro
65 70 75 80
Arg Gly Val Tyr Gln Cys Lys Gly Ser Lys Asp Lys Ser Lys Thr Leu
85 90 95
Gln Val Tyr Tyr Arg Met Cys Gln Asn Cys Ile Glu Leu Asn Ala Ala
100 105 110
Thr Ile Leu Gly Phe Val Phe Ala Glu Ile Ile Ser Ile Phe Phe Leu
115 120 125
Ala Val Gly Val Tyr Phe Ile Ala Gly Gln Asp Gly Val Arg Gln Ser
130 135 140
Arg Ala Ser Asp Lys Gln Thr Leu Leu Pro Asn Asp Gln Leu Tyr Gln
145 150 155 160
Pro Leu Lys Asp Arg Glu Asp Asp Gln Tyr Ser His Leu Gln Gly Asn
165 170 175
Gln Leu Arg Met Asn
180
<210> 84
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 84
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu
20 25
<210> 85
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 85
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu
20 25
<210> 86
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 86
Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys
1 5 10 15
<210> 87
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 87
His Arg Asp Cys Tyr Leu Gly Asp Leu Cys Asn Ser
1 5 10
<210> 88
<211> 91
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 88
Asn Arg Met Arg Cys Tyr Asn Cys Gly Gly Ser Pro Ser Ser Ser Cys
1 5 10 15
Lys Glu Ala Val Thr Thr Cys Gly Glu Gly Arg Pro Gln Pro Gly Leu
20 25 30
Glu Gln Ile Lys Leu Pro Gly Asn Pro Pro Val Thr Leu Ile His Gln
35 40 45
His Pro Ala Cys Val Ala Ala His His Cys Asn Gln Val Glu Thr Glu
50 55 60
Ser Val Gly Asp Val Thr Tyr Pro Ala His Arg Asp Cys Tyr Leu Gly
65 70 75 80
Asp Leu Cys Asn Ser Ala Gly Asn Ser Val Thr
85 90
<210> 89
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 89
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Glu Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Glu Asn Asp Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 90
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 90
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 91
<400> 91
000
<210> 92
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 92
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 93
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 93
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 94
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 94
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 95
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 95
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Arg Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
<210> 96
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 96
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp
210 215
<210> 97
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 97
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 98
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 98
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Tyr Ser Asn Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 99
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 99
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Asn Asn Thr
1 5 10
<210> 100
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 100
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Gln Asn Val
1 5 10
<210> 101
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 101
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Ser Asn Thr
1 5 10
<210> 102
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 102
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Ser Asn Ala
1 5 10
<210> 103
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 103
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Ala Asn Thr
1 5 10
<210> 104
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 104
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Asn Asn Ala
1 5 10
<210> 105
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 105
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Asn Asn Val
1 5 10
<210> 106
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 106
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Gly Asn Thr
1 5 10
<210> 107
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 107
Gln Ser Thr Ser Phe Arg Gly Arg Ser Tyr Ser Asn Val
1 5 10
<210> 108
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 108
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 109
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 109
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Ile His Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Tyr Ser Asn Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 110
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 110
Trp Ala Ser Thr Arg Ile Ser
1 5
<210> 111
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 111
Asn Asn Ala Met Ser
1 5
<210> 112
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 112
Ser Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala Ser
1 5 10 15
Gly
<210> 113
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 113
Ile Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala Ser
1 5 10 15
Gly
<210> 114
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 114
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 115
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 115
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 116
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 116
Gly Arg Ser Tyr Asn Asn Thr
1 5
<210> 117
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 117
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Asn Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 118
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 118
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Asn Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 119
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 119
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Val Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Asn Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 120
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 120
Glu Gln Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Thr Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 121
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 121
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 122
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 122
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Gln Asn Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 123
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 123
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Ala Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Met Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 124
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 124
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 125
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 125
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 126
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 126
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 127
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 127
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Ile His Trp Val Arg Glu Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Tyr Ser Asn Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 128
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 128
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 129
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 129
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Lys Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Ile Leu Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 130
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 130
Asp Ile Val Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Glu Ser Ile Thr Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Leu Pro Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Thr Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Thr Ser Phe Arg Gly Arg
85 90 95
Ser Tyr Asn Asn Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 131
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 131
Gln Gln Gln Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Asp Phe Val Asn Asn
20 25 30
Ala Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Leu Ser Phe Ala Asp Asn Thr Ala Tyr Tyr Ala Thr Trp Ala
50 55 60
Ser Gly Arg Phe Thr Ile Ser Lys Ile Ser Ser Thr Thr Val Thr Leu
65 70 75 80
Arg Met Thr Ser Leu Thr Gly Ala Asp Thr Ala Thr Tyr Phe Cys Met
85 90 95
Arg Gly Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 132
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 132
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Arg Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 133
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 133
Asp Ala Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Arg Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 134
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 134
Asp Ala Val Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Arg Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 135
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 135
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Ser Leu Ser Thr Asn
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Ser Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 136
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 136
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Pro Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Leu Ser Leu Ser Thr Asn
20 25 30
Ser Ile Asn Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Thr Gln Val Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Ser Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 137
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 137
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Ser Leu Ser Thr Asn
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Ser Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Ser Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 138
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 138
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Ile Thr Ile Asn Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Arg Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Thr Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Arg Gly Thr Ala Val Val Val Lys
100 105 110
<210> 139
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 139
Asp Ala Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Arg Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 140
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 140
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Glu Ser Val Trp His Asn
20 25 30
Asn Trp Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Ser Asp Ala His Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 141
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 141
Gln Ser Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Leu Ser Leu Ser Thr Asn Ser
20 25 30
Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr Leu Arg
65 70 75 80
Met Thr Ser Leu Thr Val Ala Asp Thr Ala Thr Tyr Phe Cys Ala Ser
85 90 95
Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 142
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 142
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Ser Leu Ser Thr Asn
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Arg Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 143
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 143
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Ser Leu Ser Thr Asn
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Tyr His Gly Asp Gly Ile Ala Asp Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Gly Ala Gly Tyr Ser Leu Phe Thr Leu Trp Gly Arg Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 144
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 144
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 145
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 145
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 146
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 146
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 147
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический конструкт
<400> 147
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Leu Ser
20 25 30
Gly Asn Gln Glu Asn Tyr Leu Ala Trp His Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Thr Leu Ile Thr Trp Ala Ser Thr Arg Ile Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Ser Ala Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2822200C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| МОЛЕКУЛЫ АНТИ-SIGLEC-9 АНТИТЕЛА | 2020 |
|
RU2833330C1 |
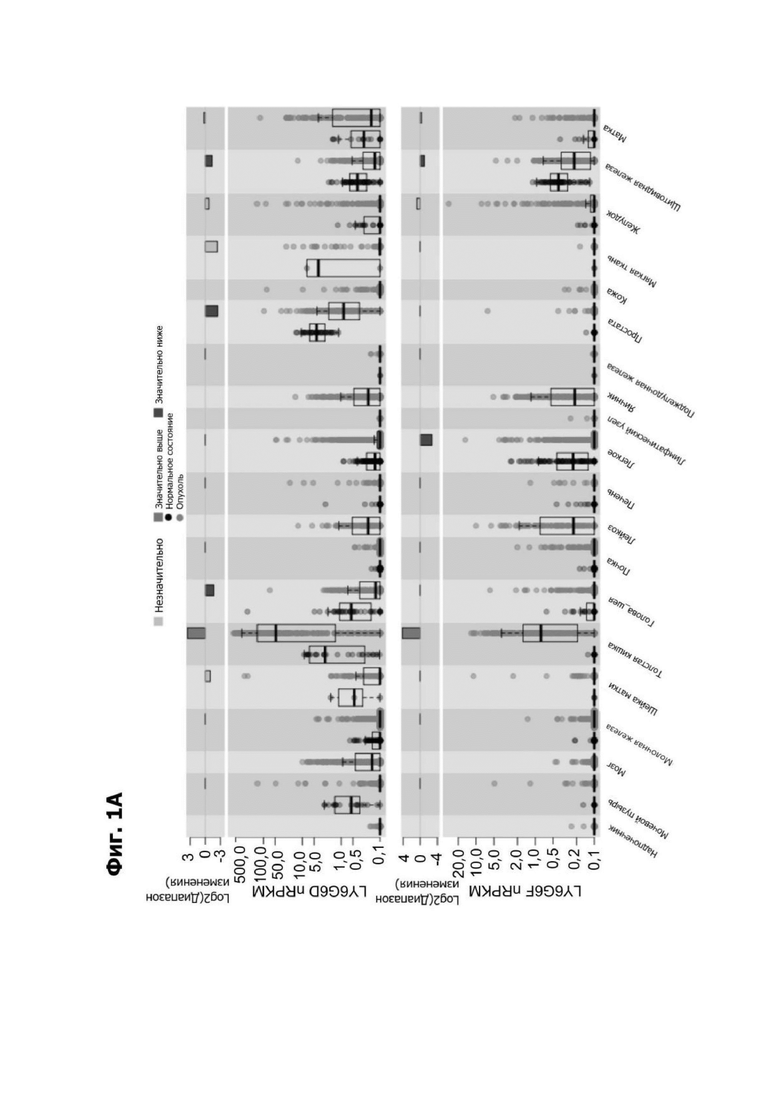
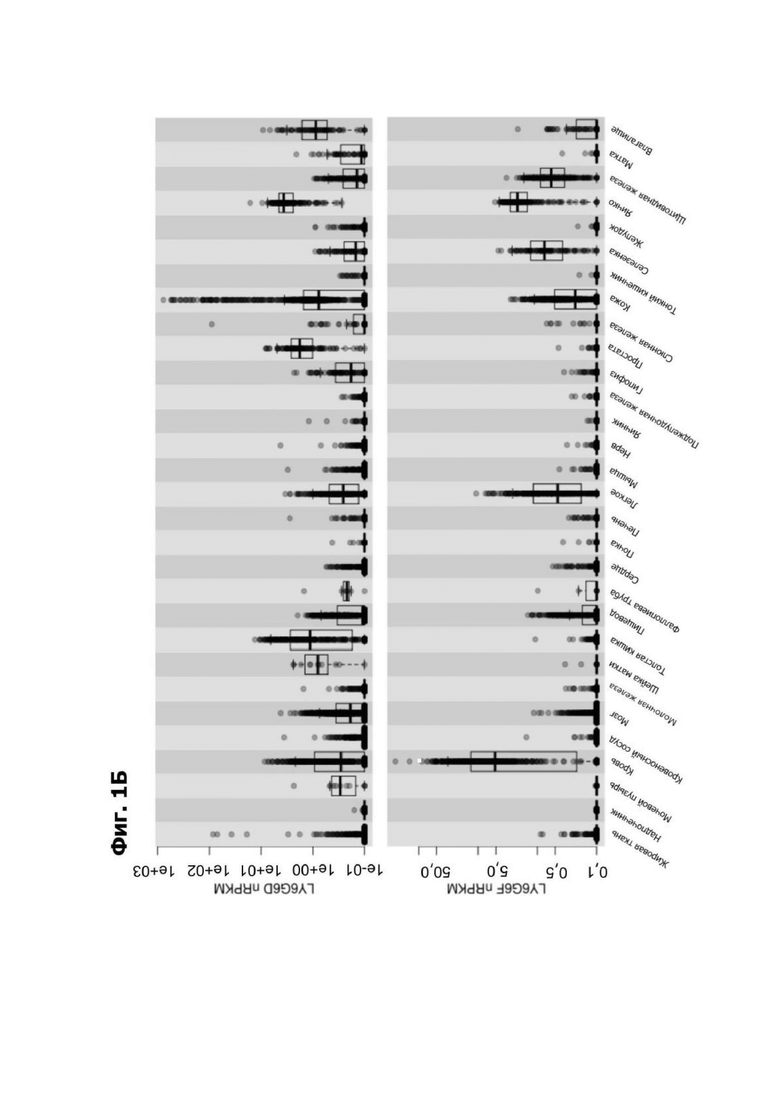
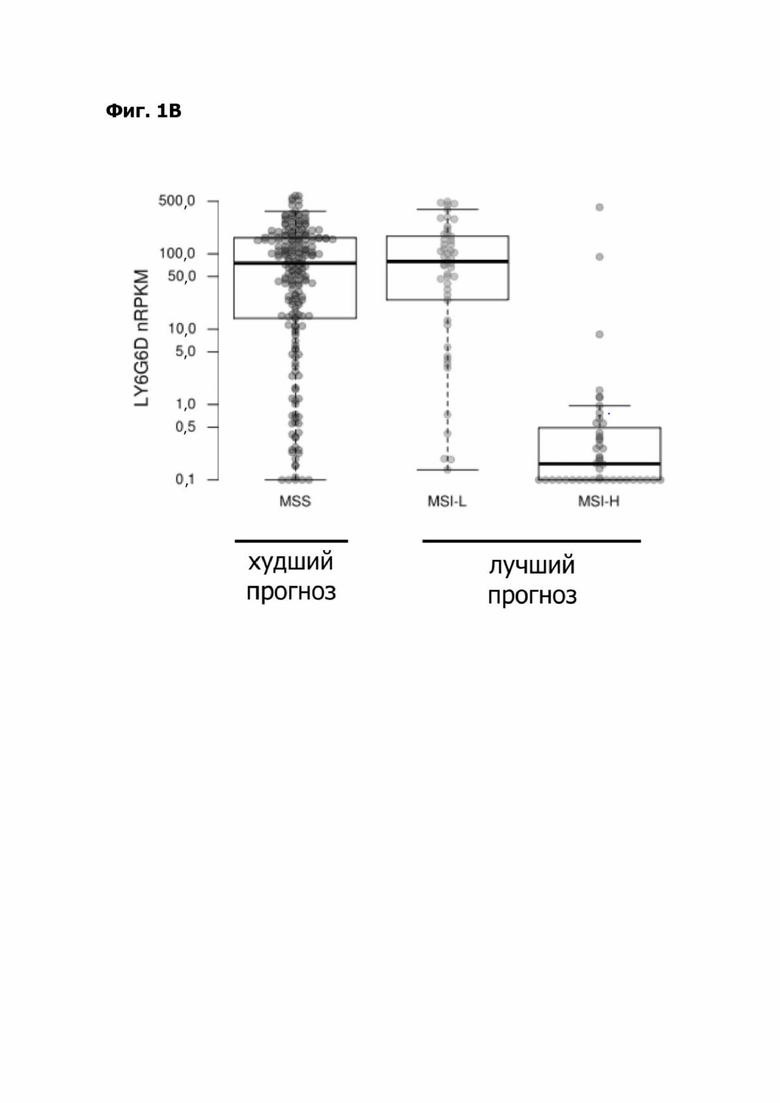
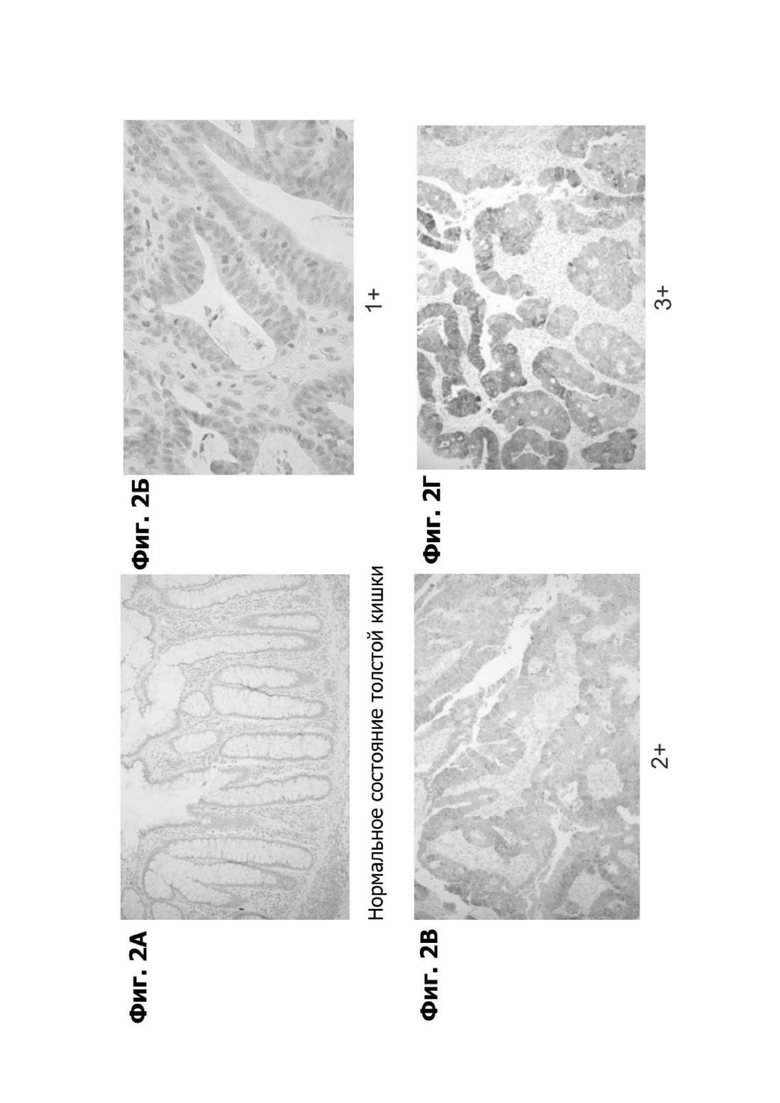
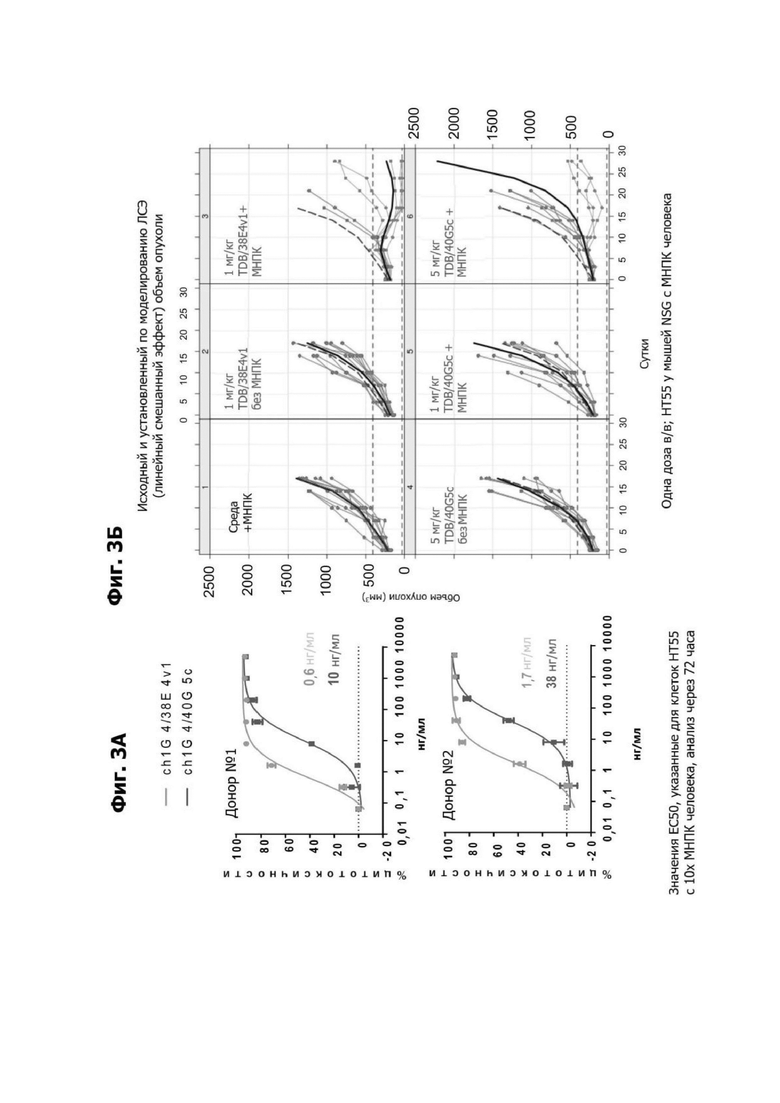
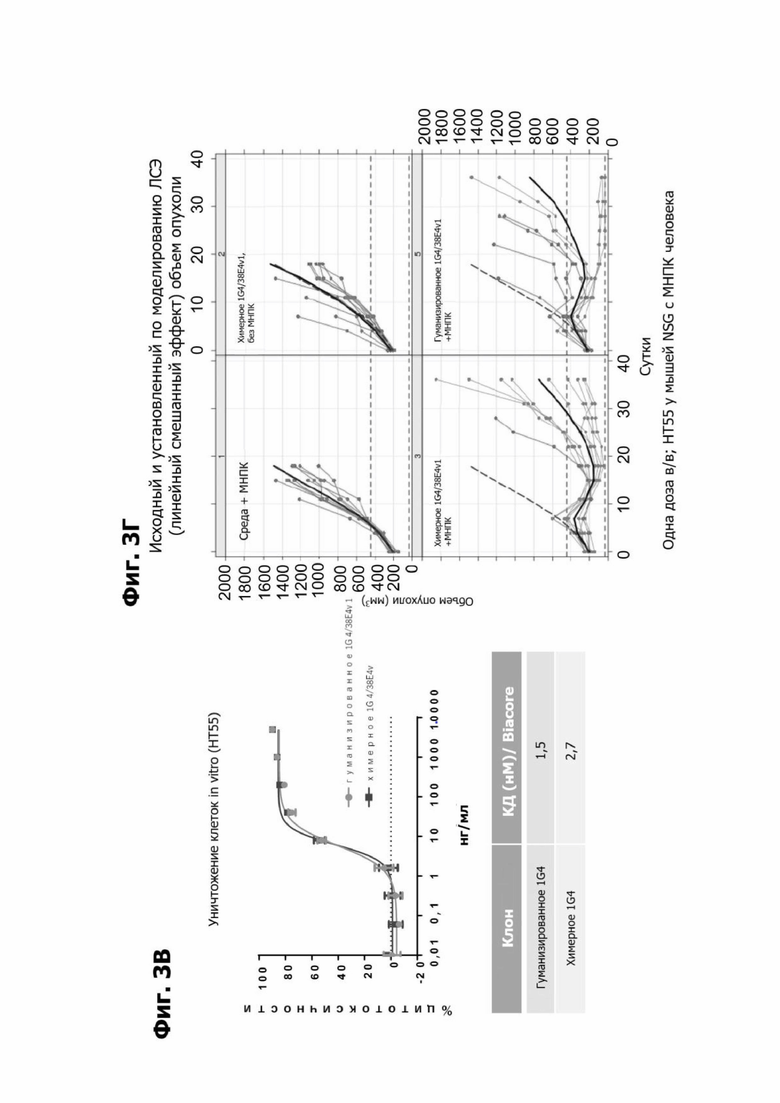
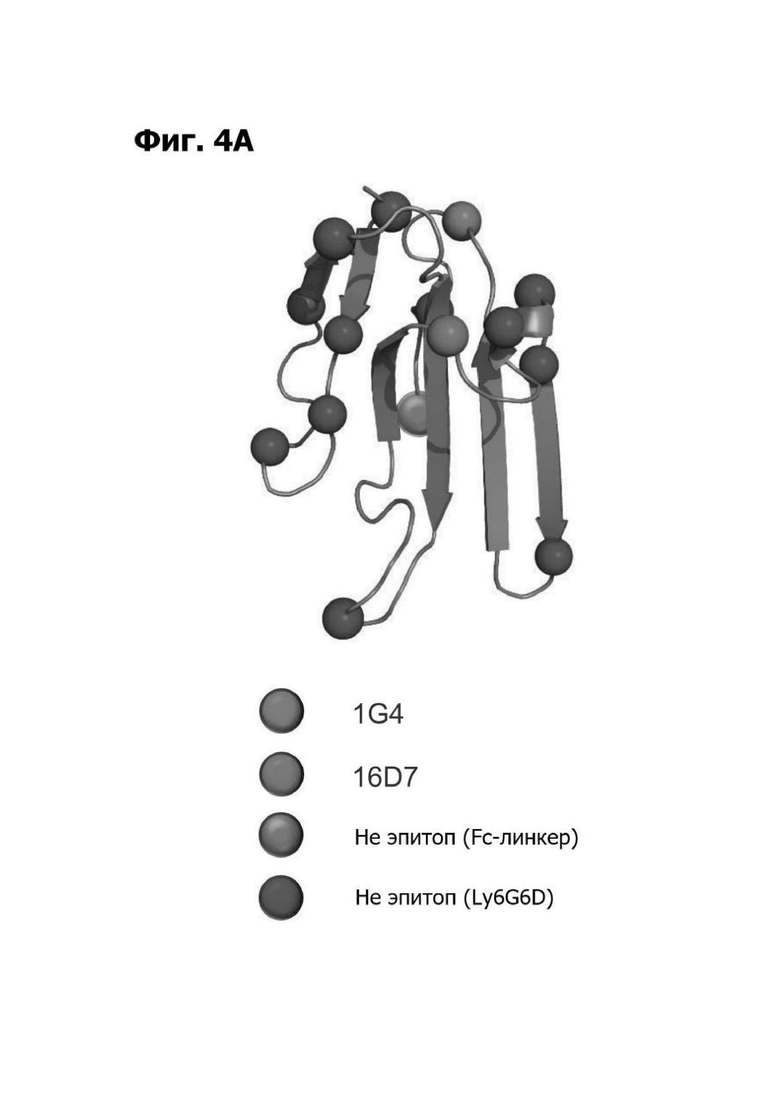
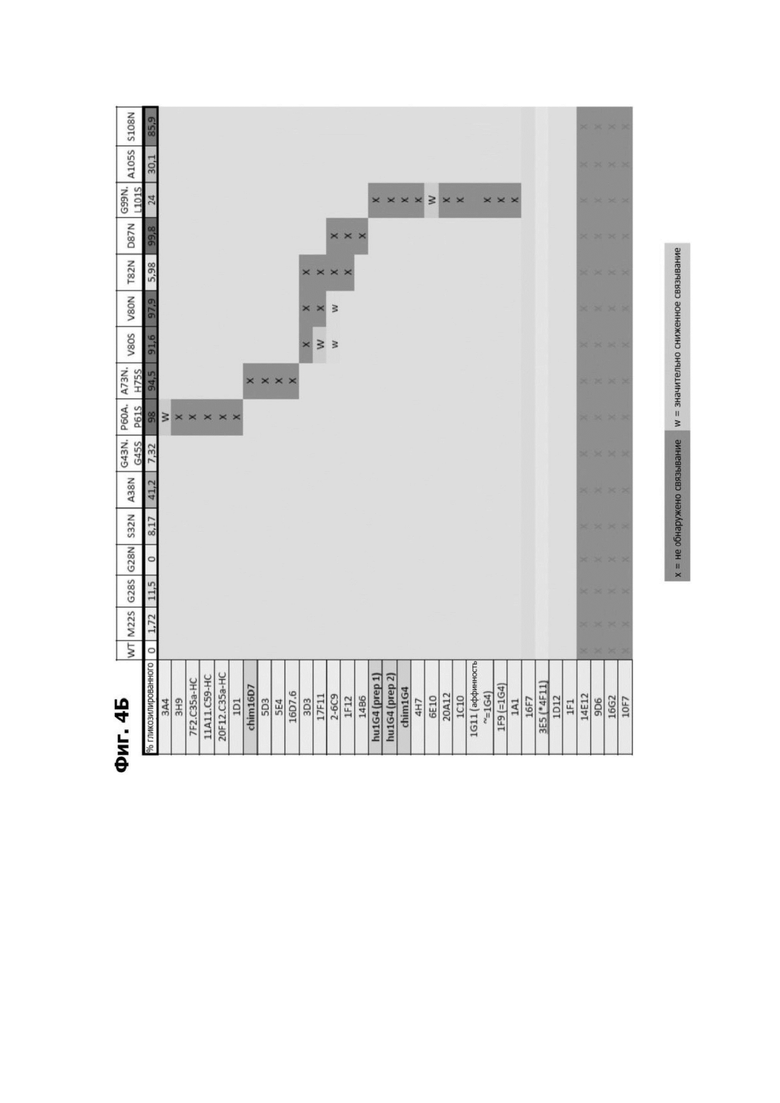

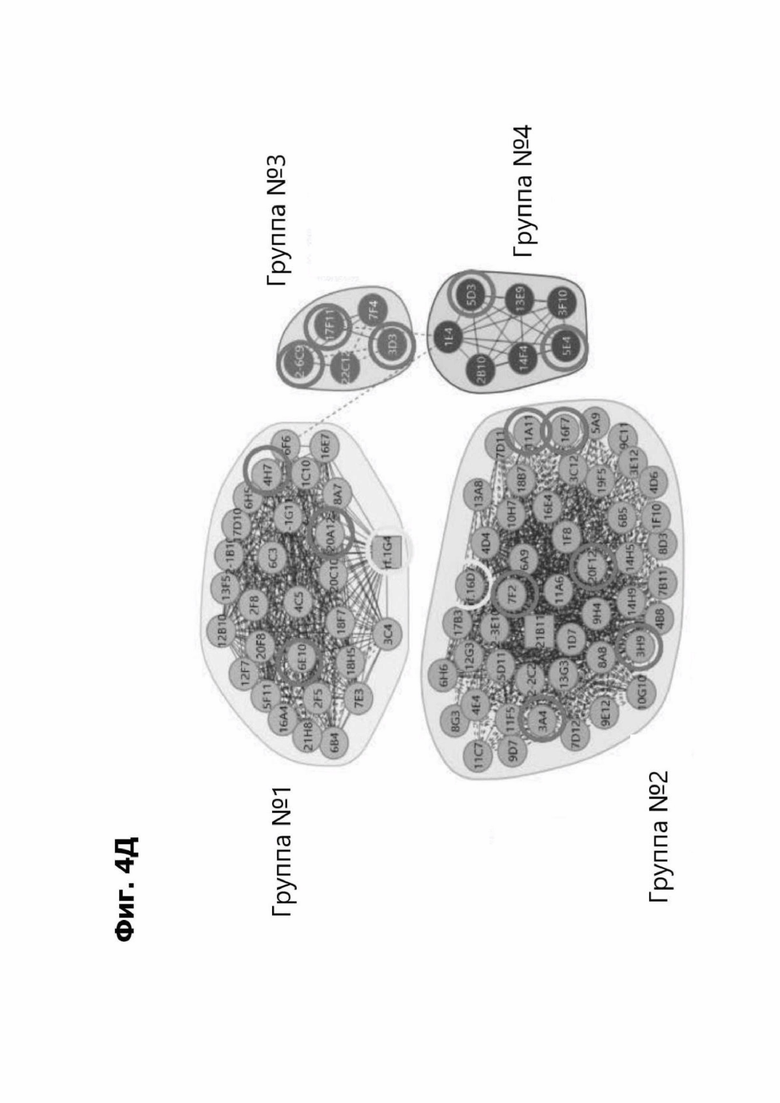
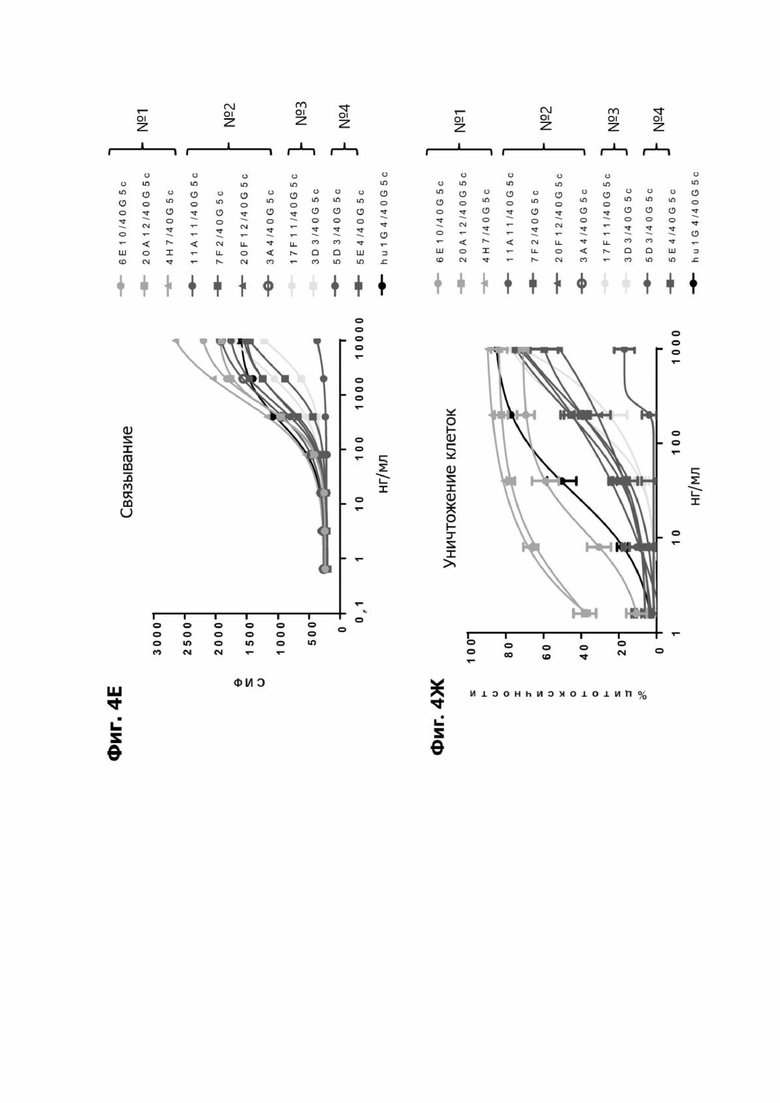
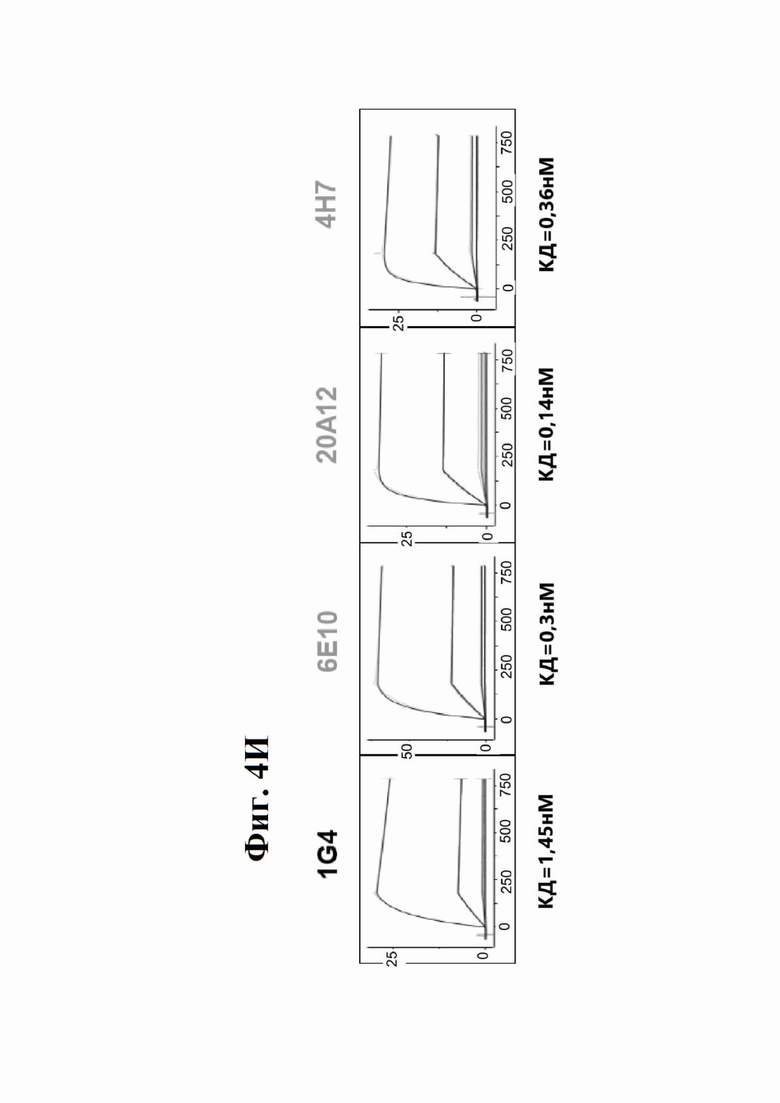
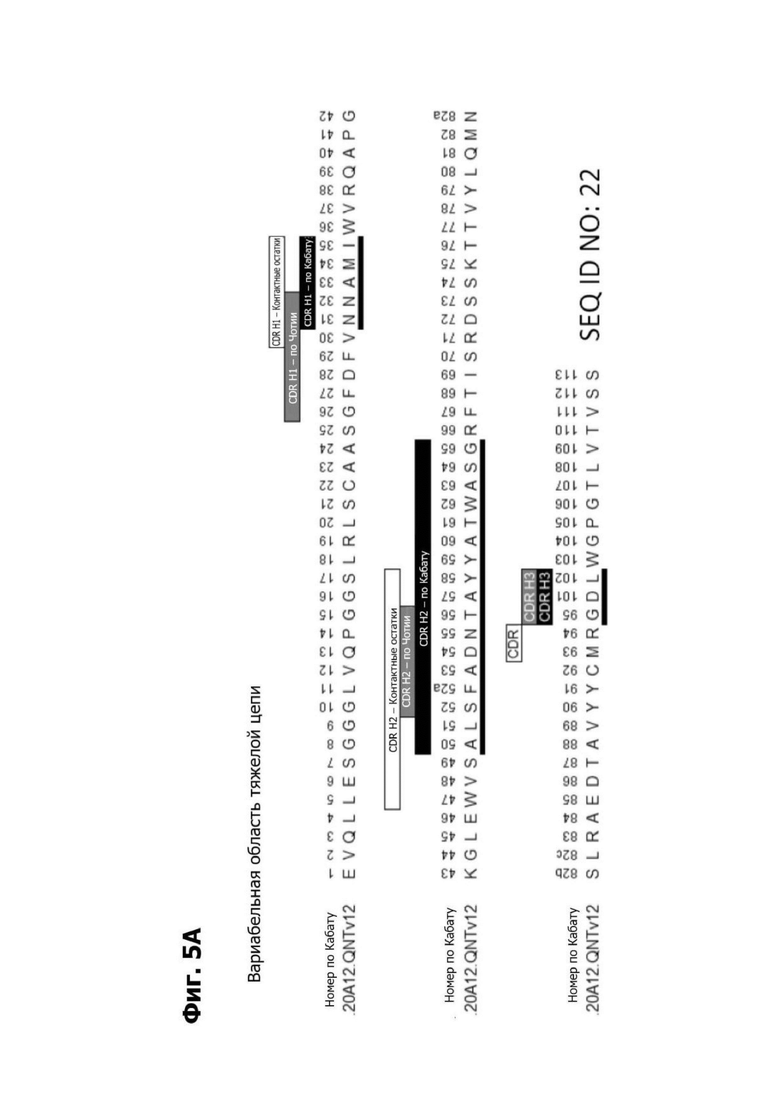
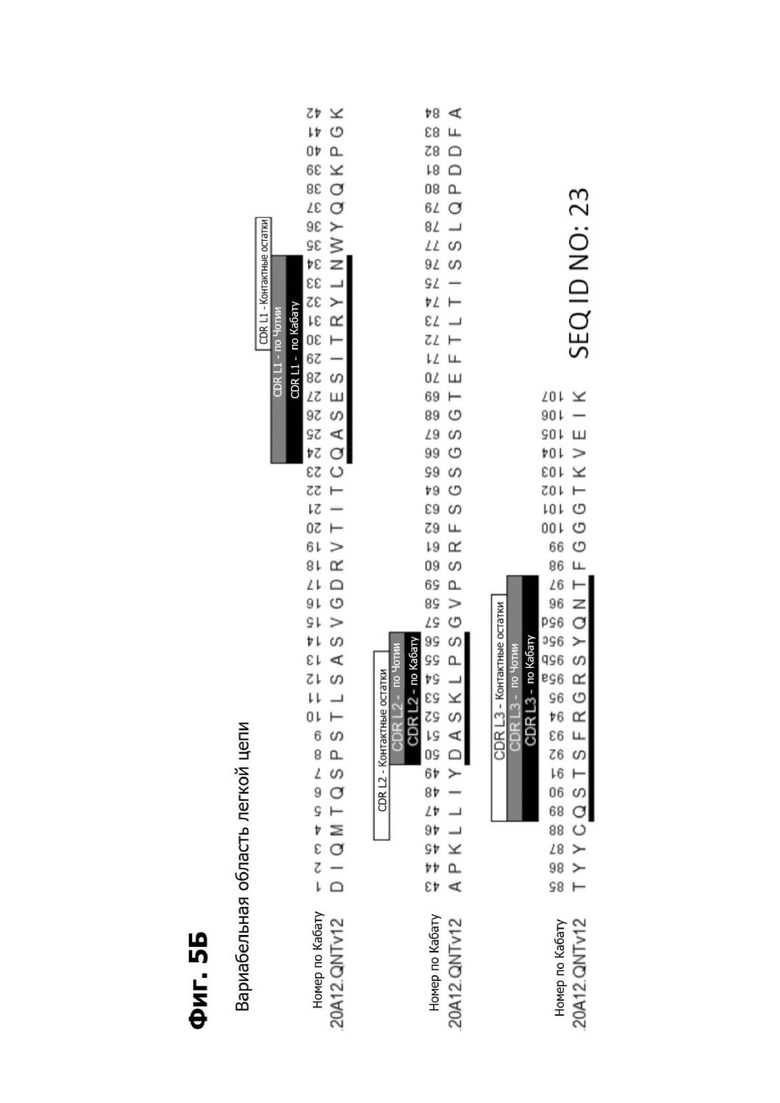
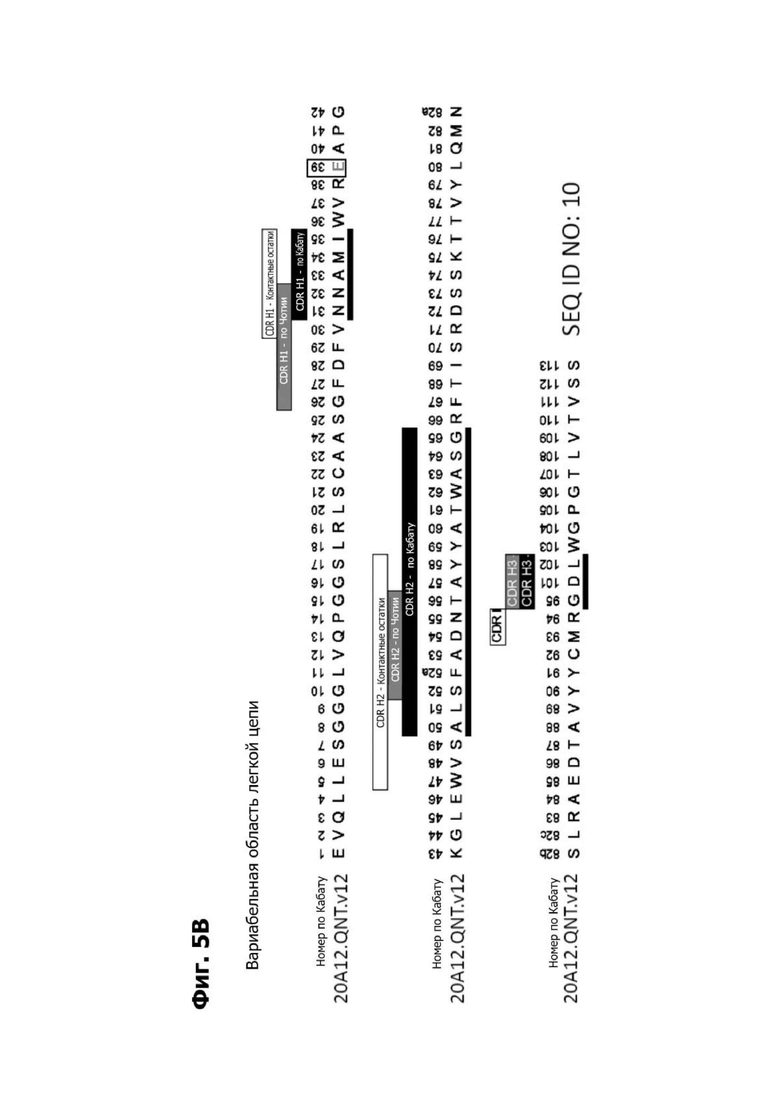
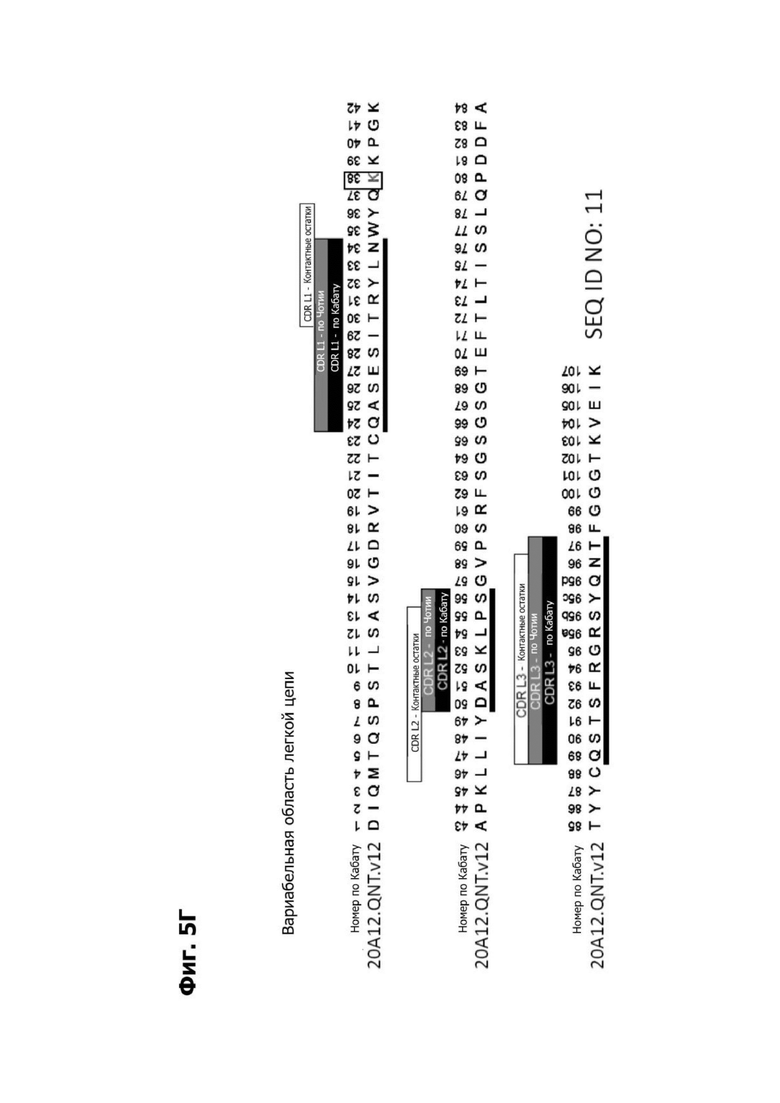
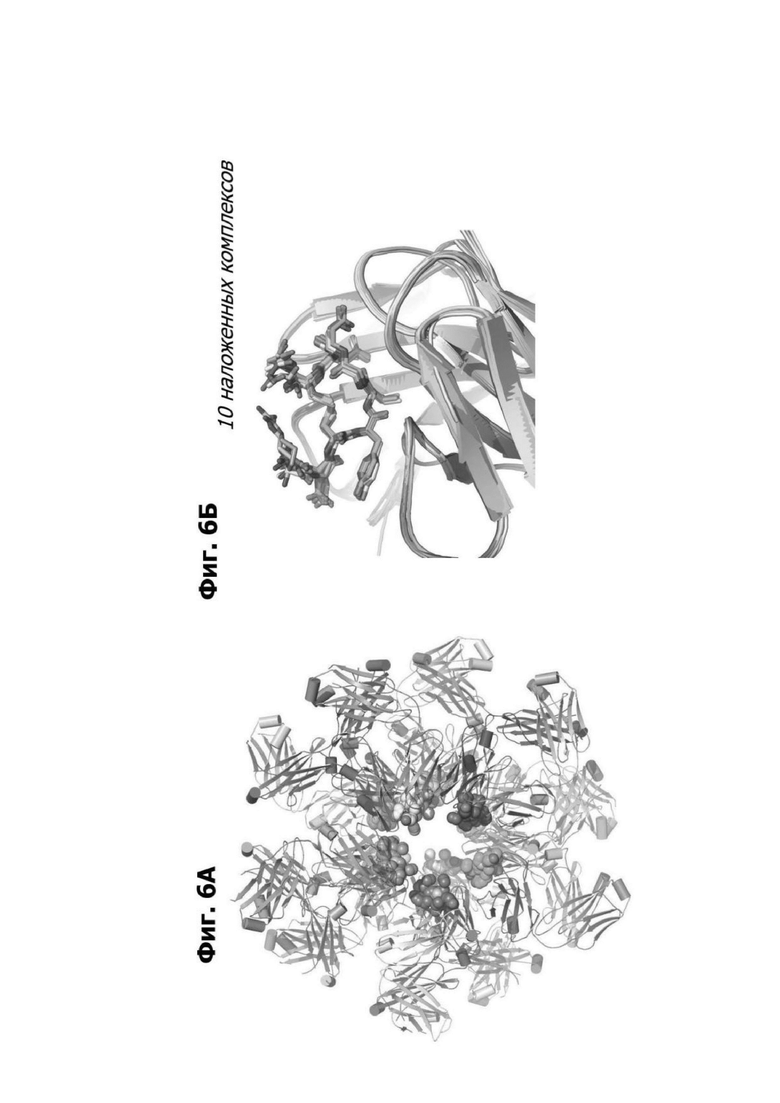
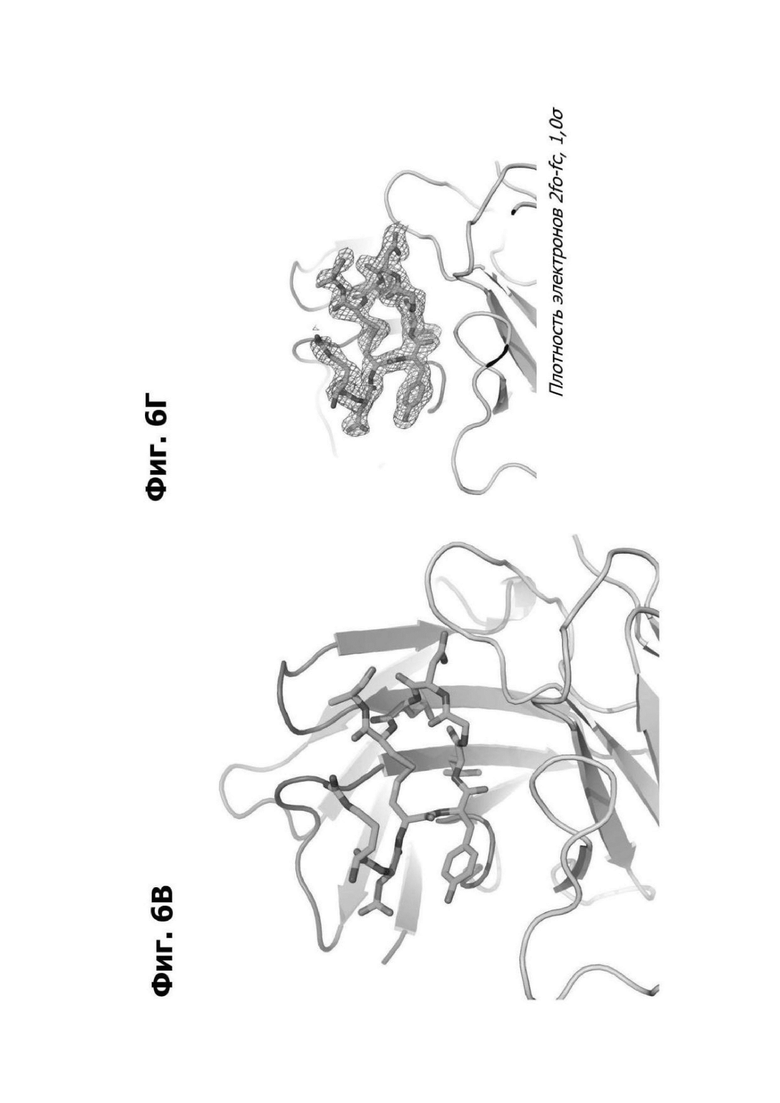
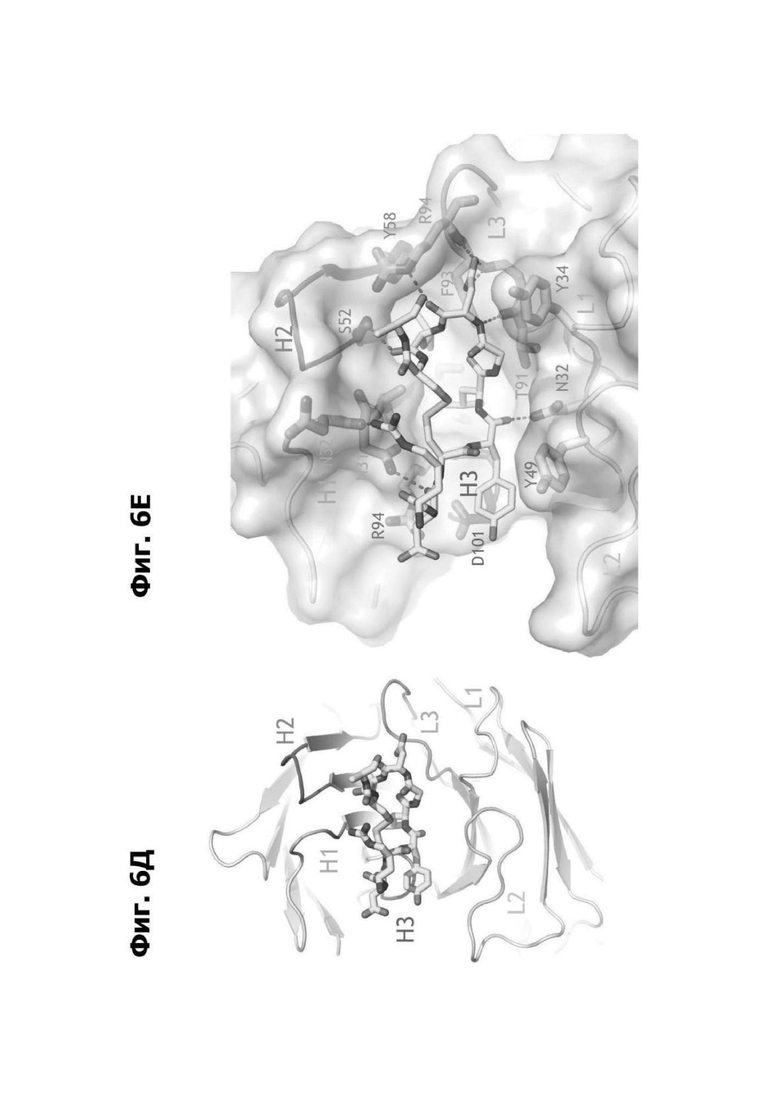
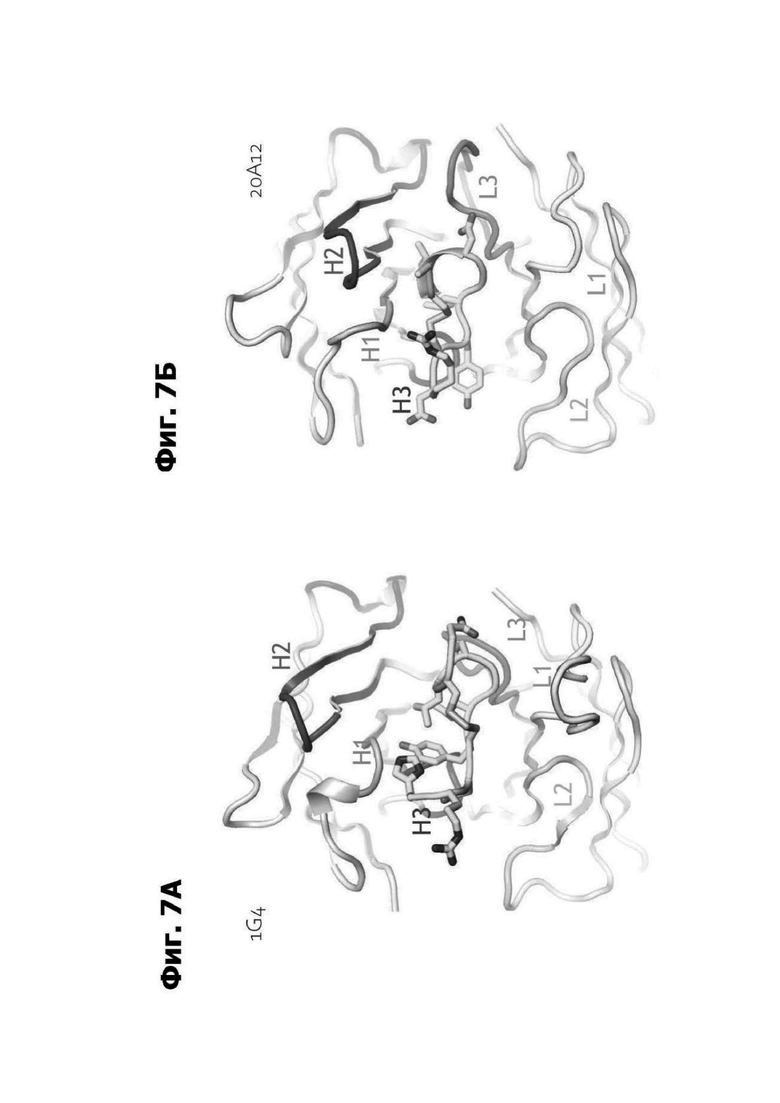
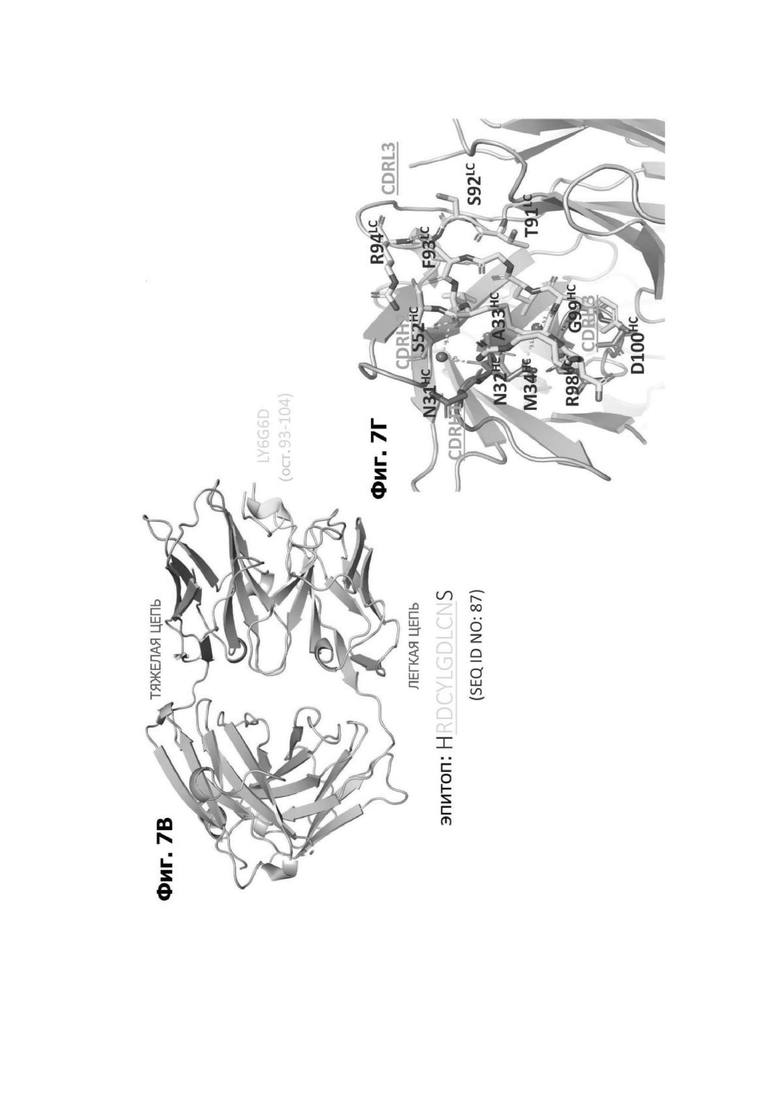
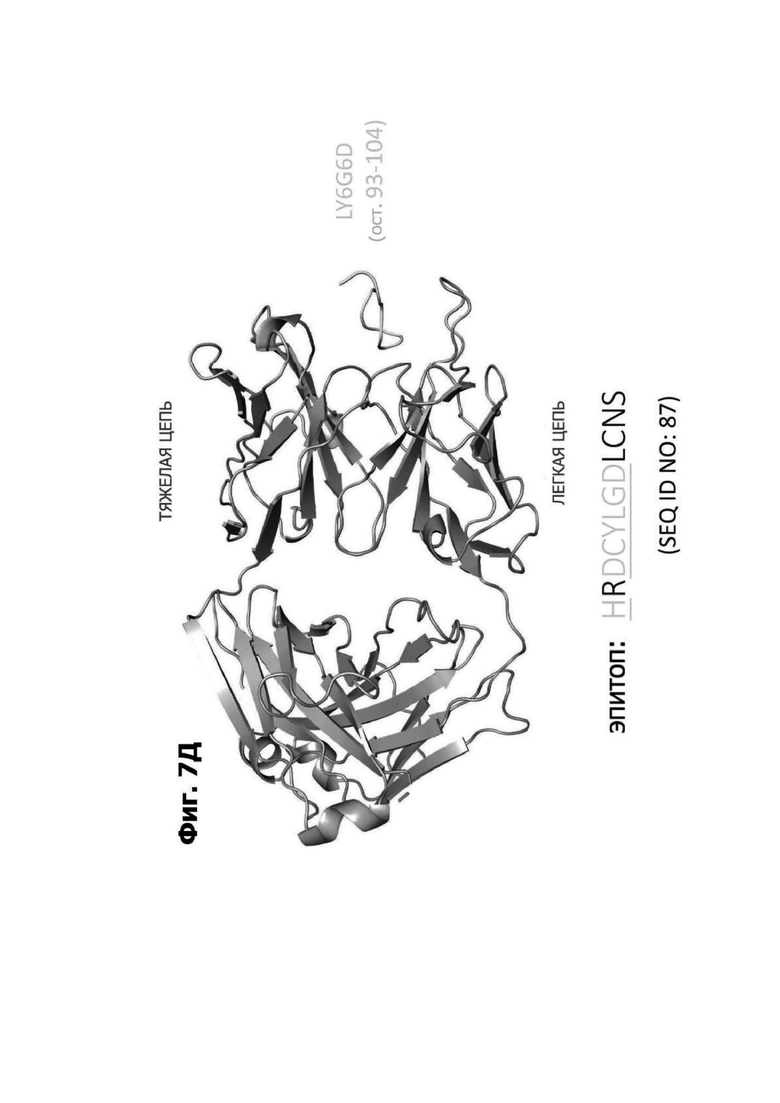
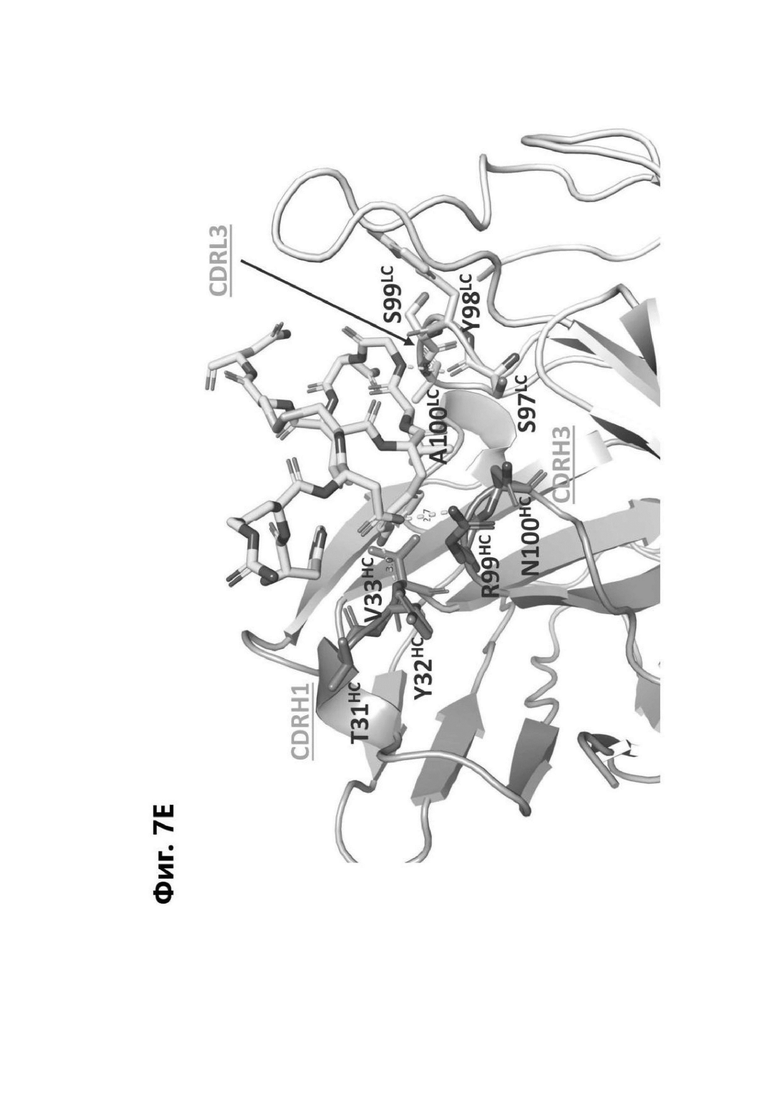
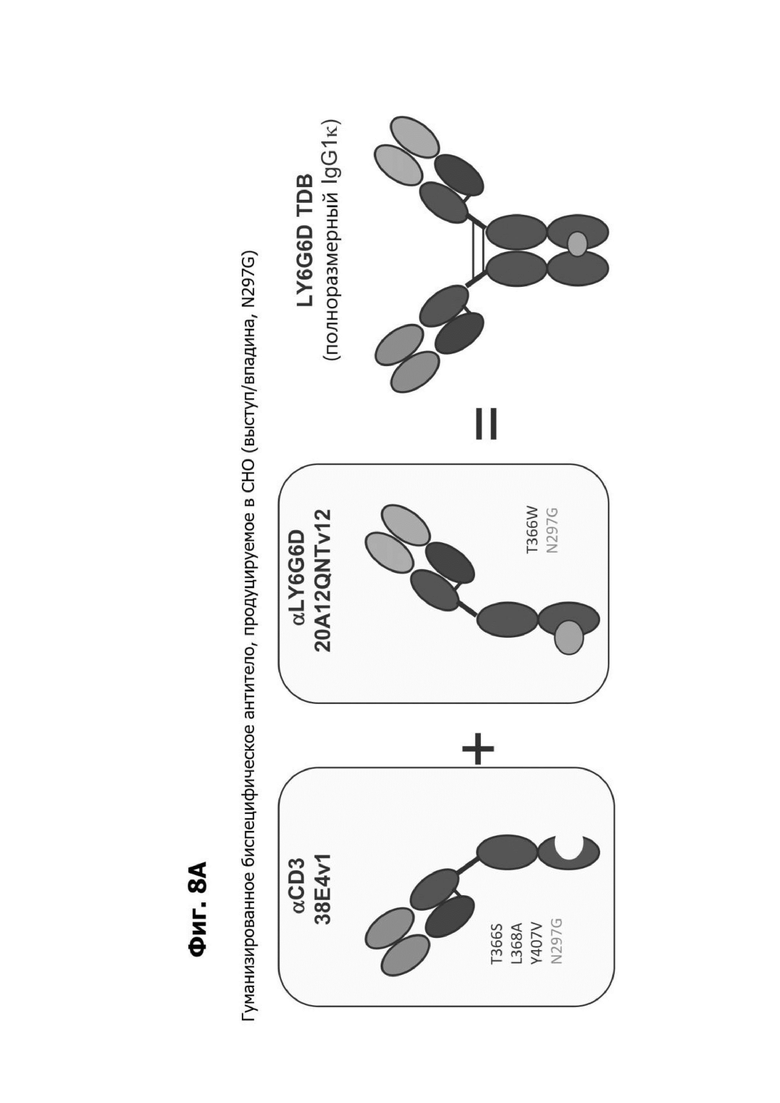
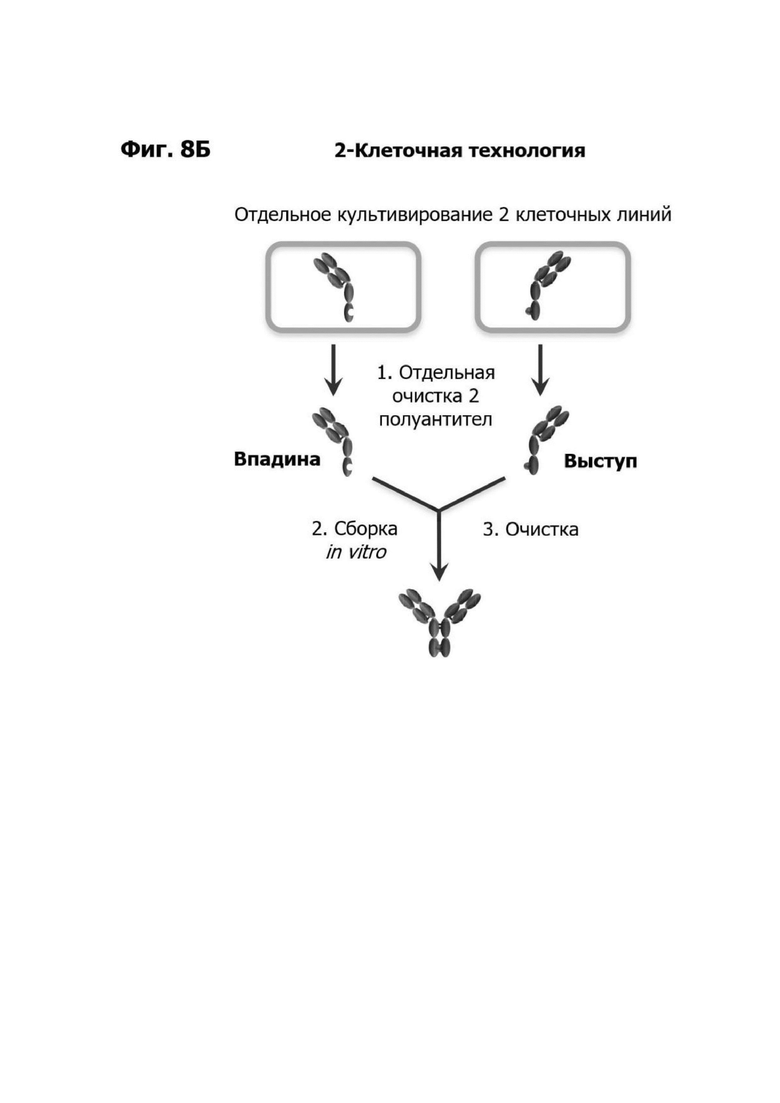
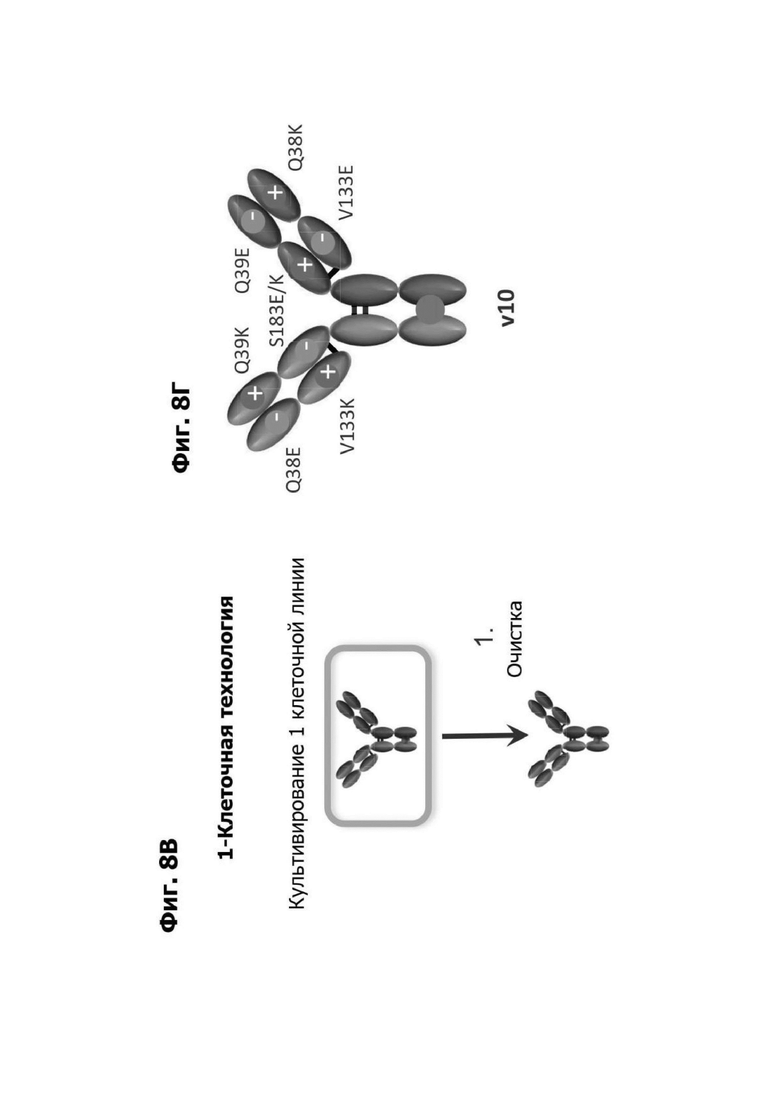
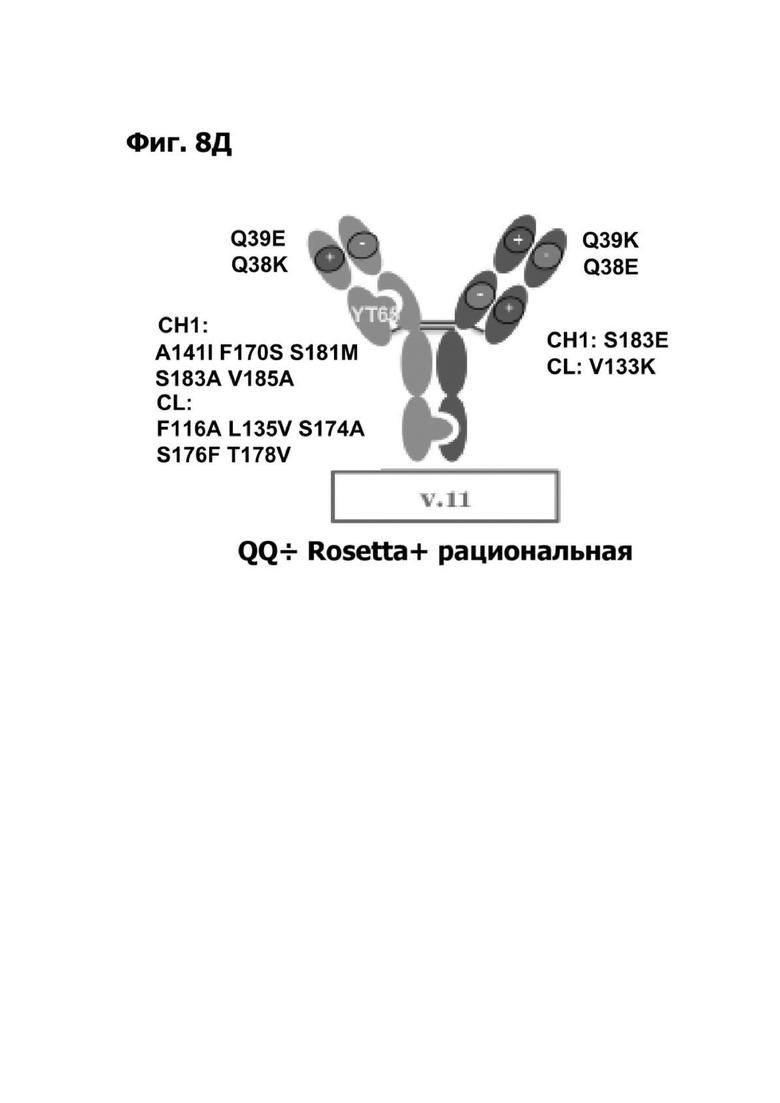
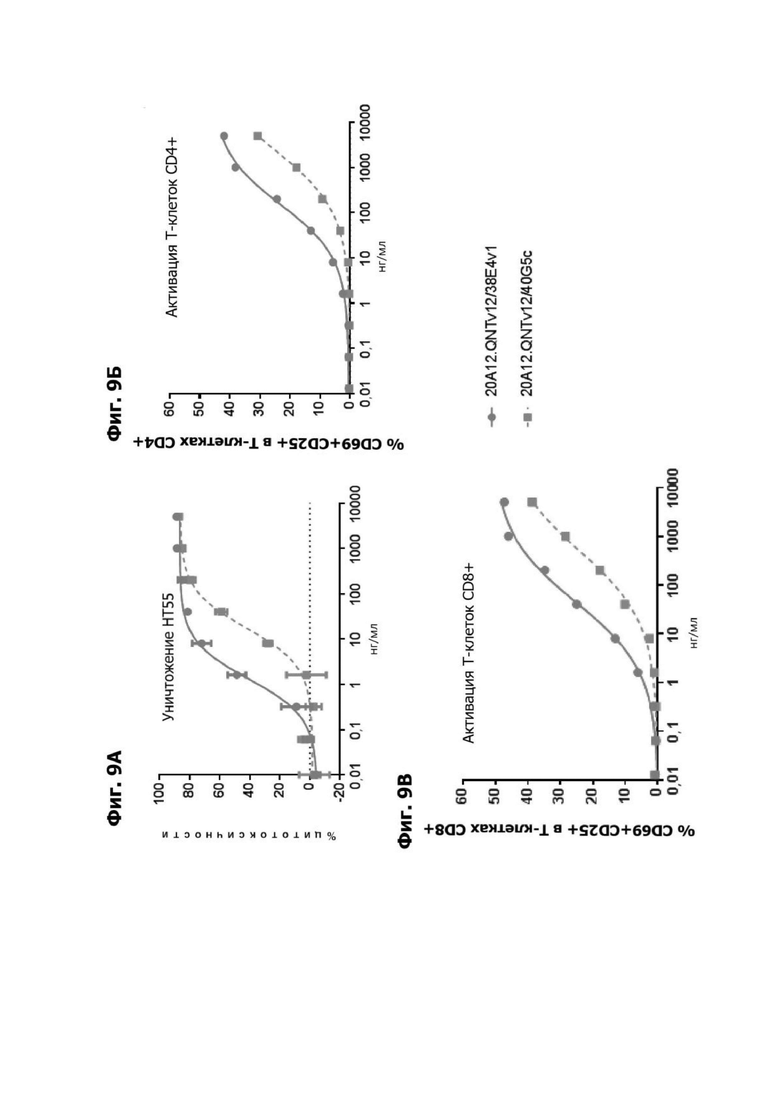
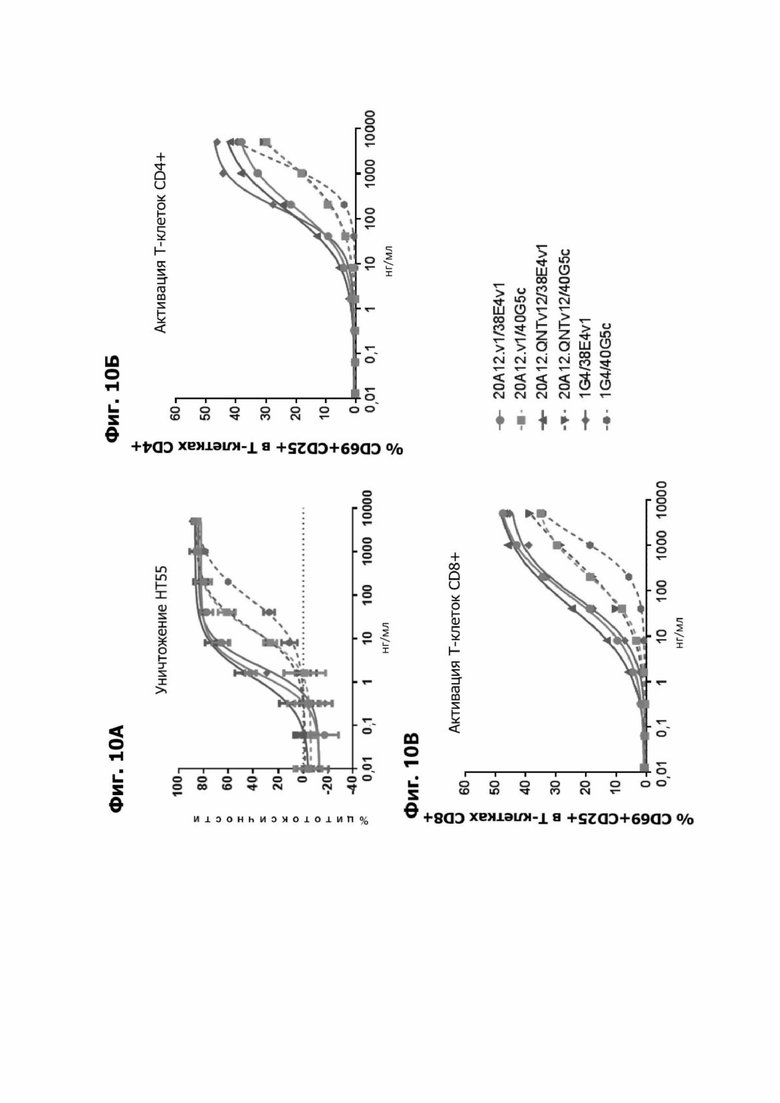
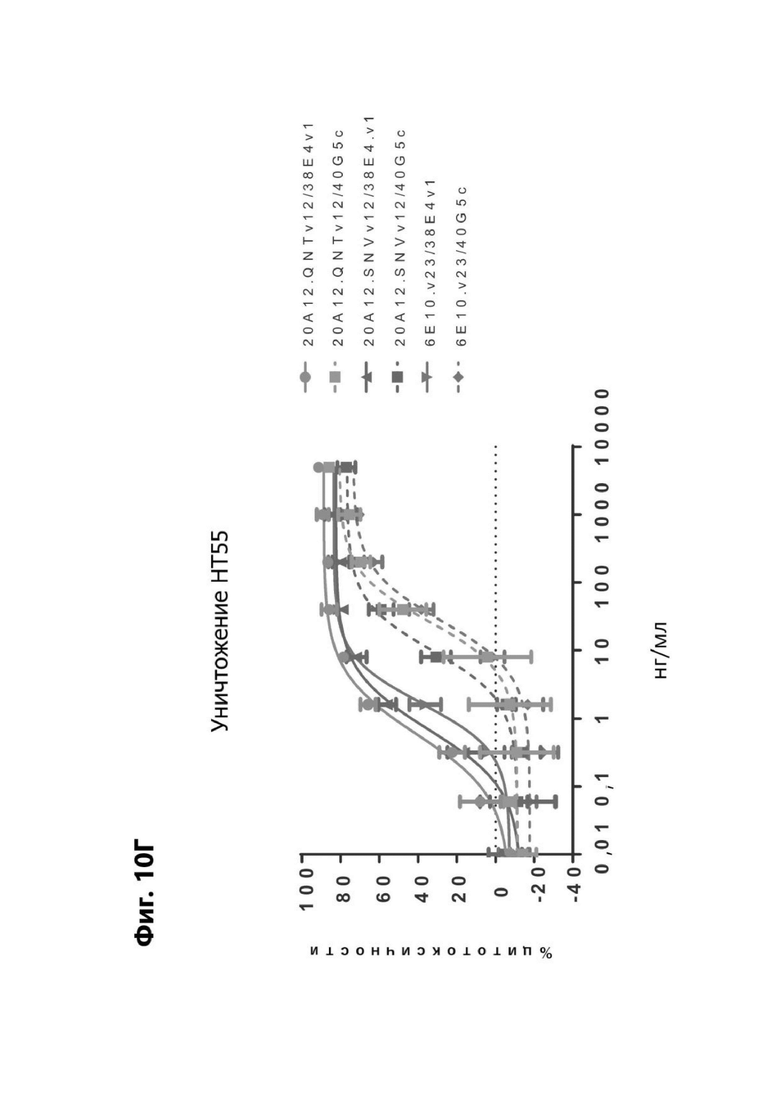
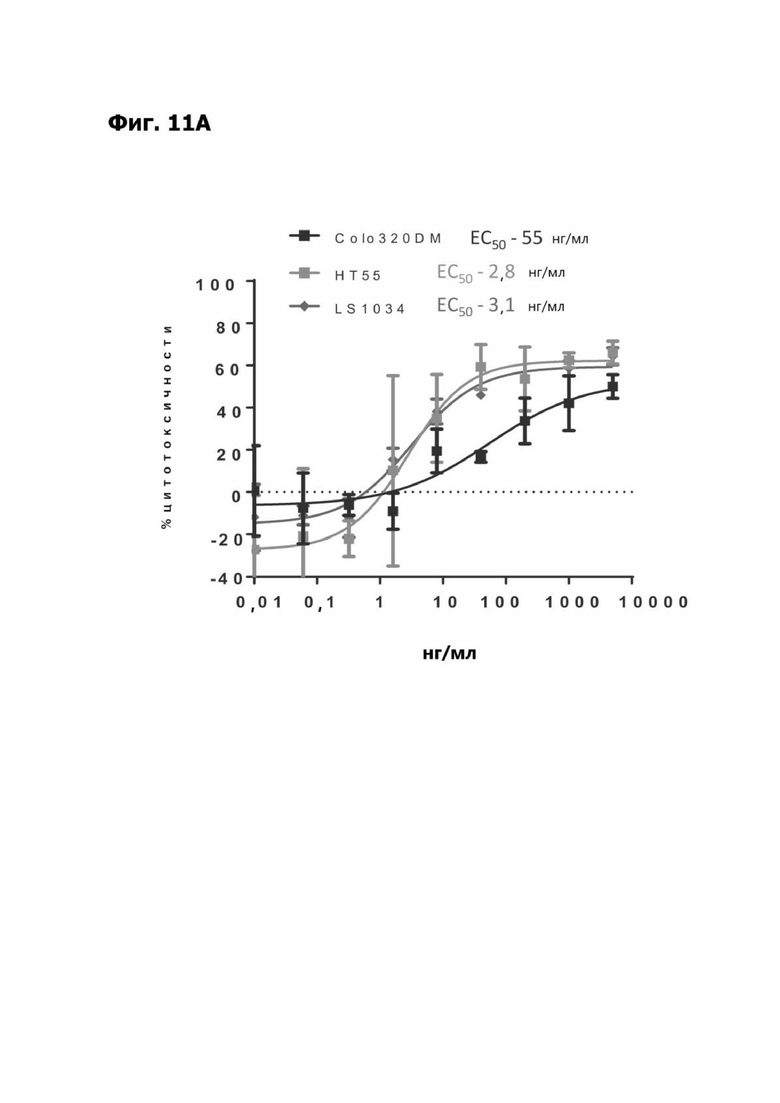
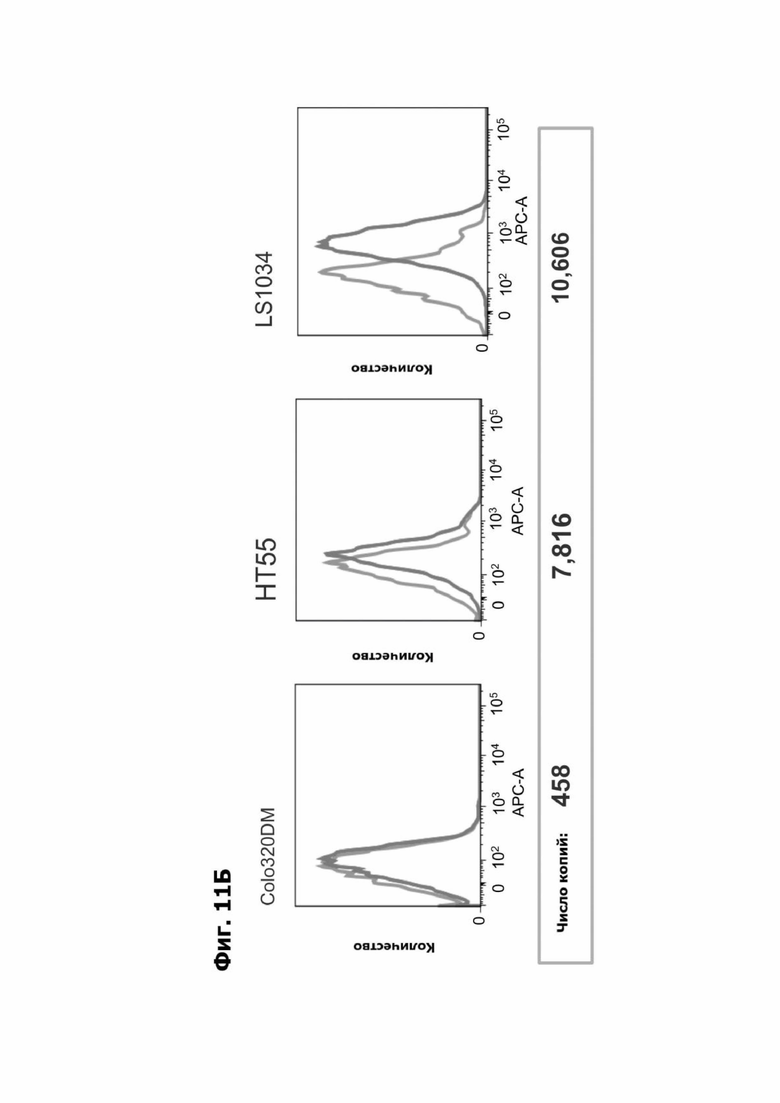
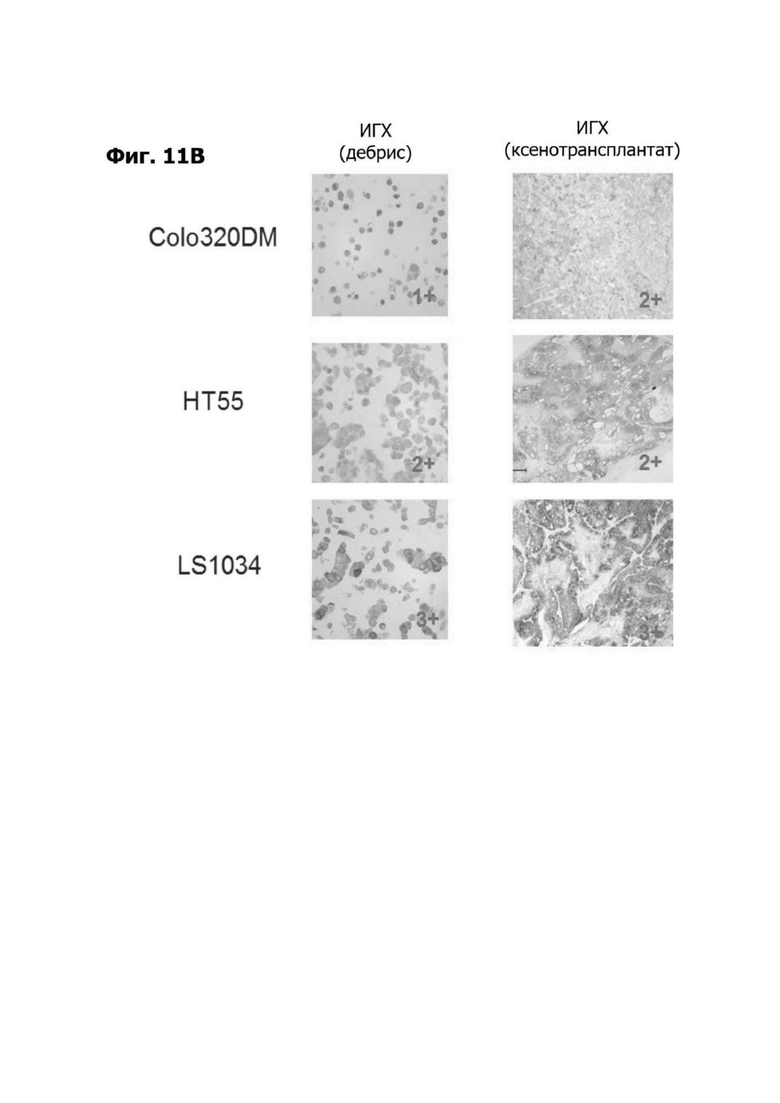
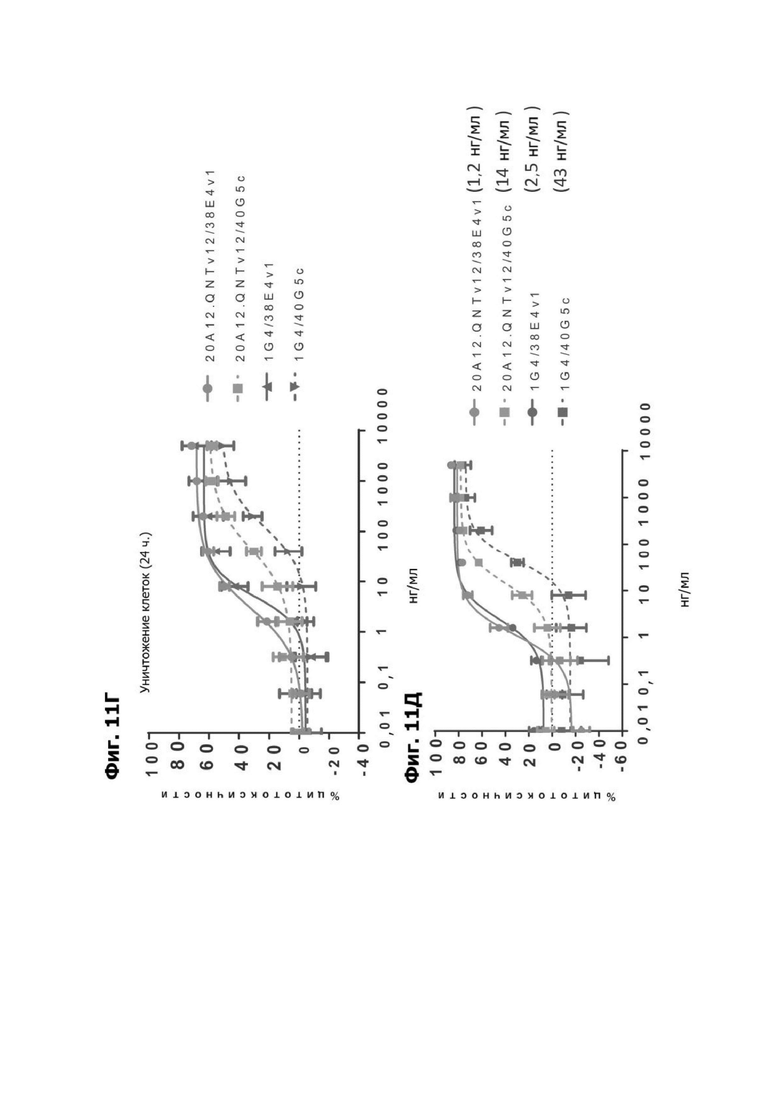

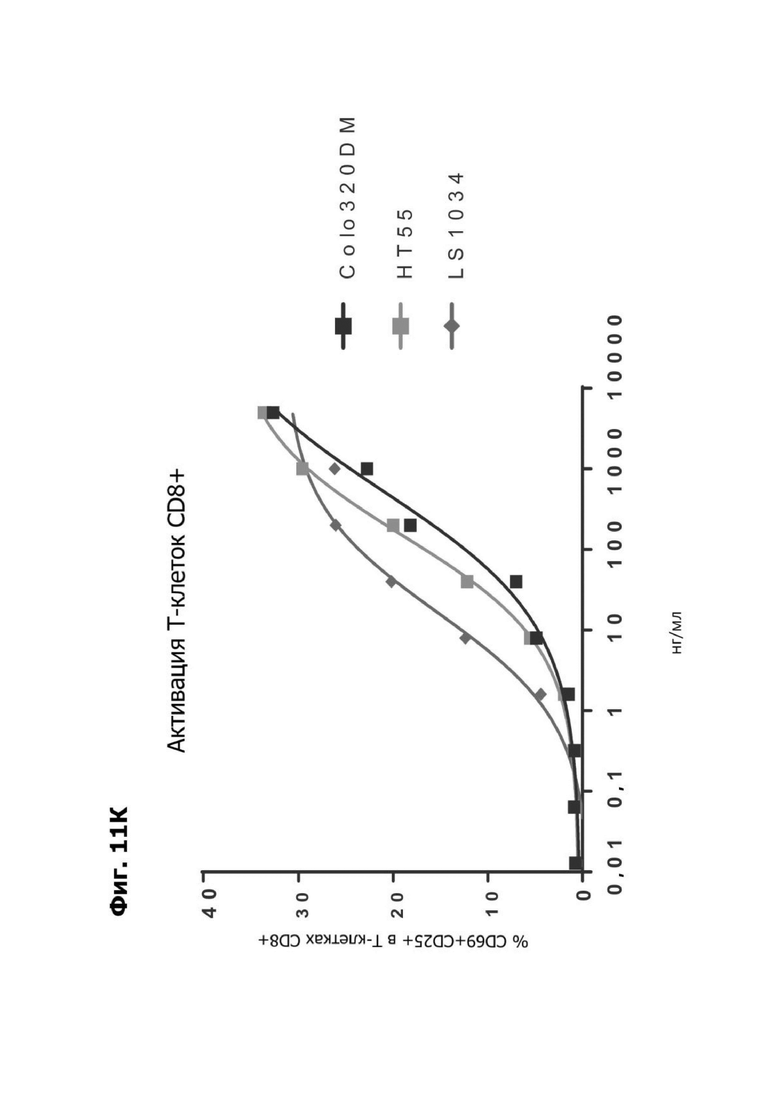
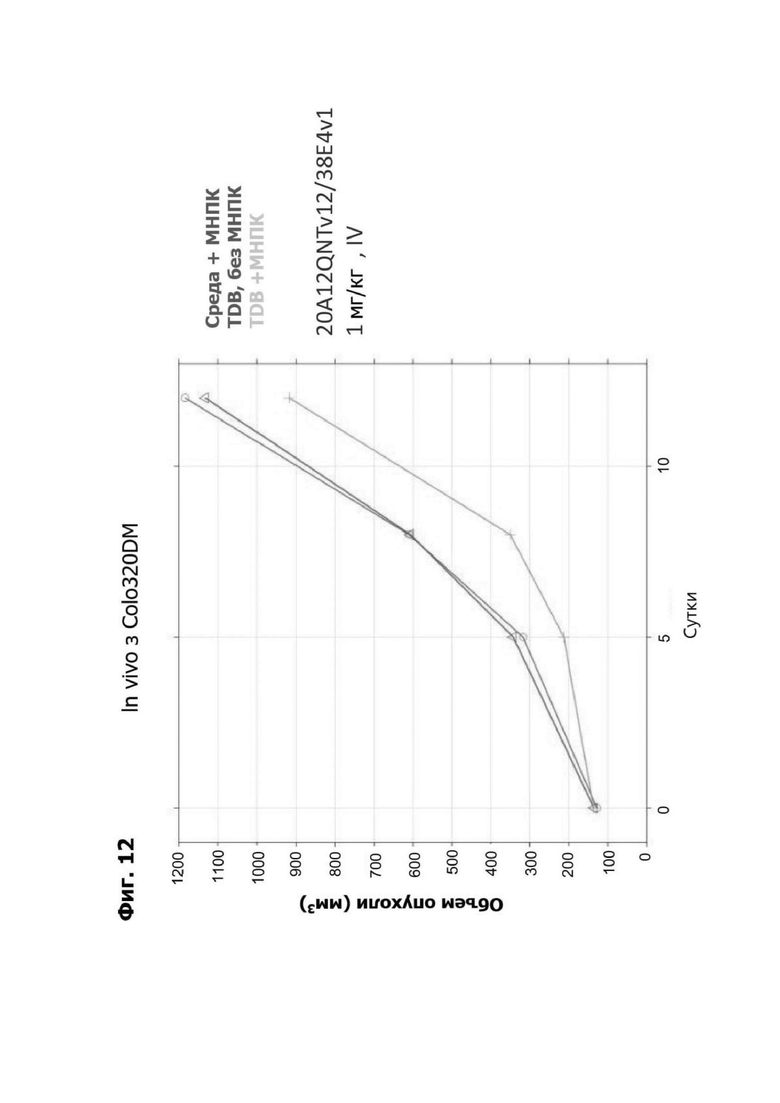
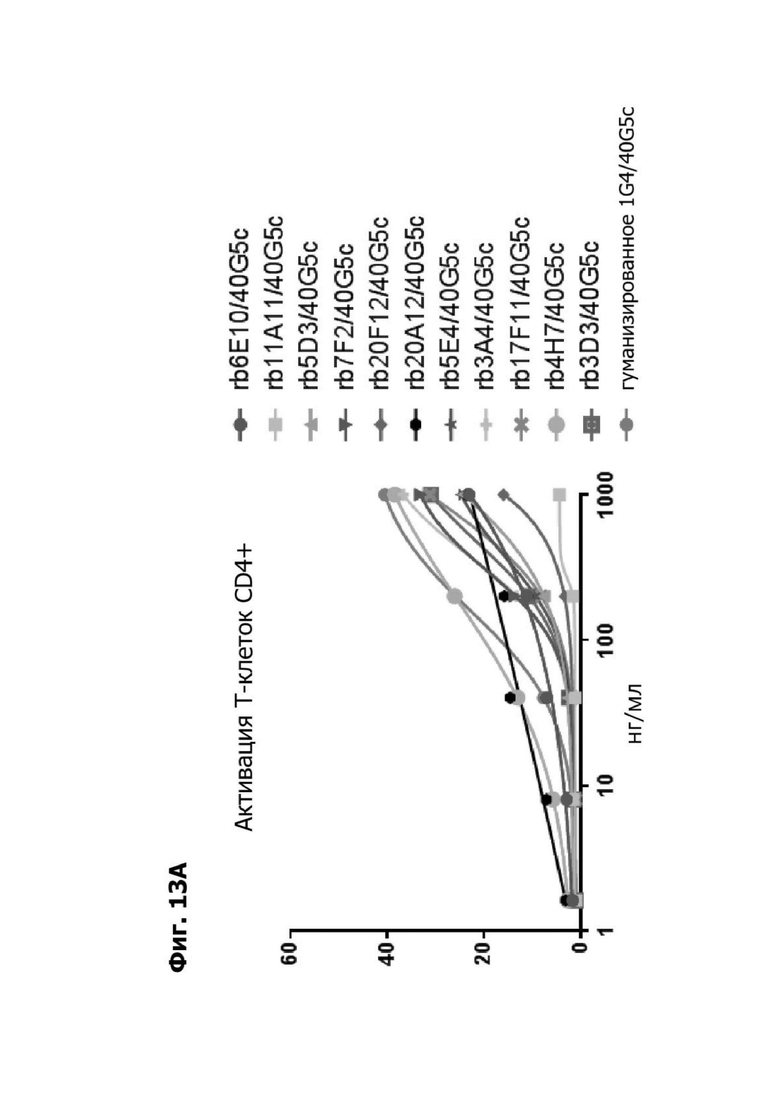
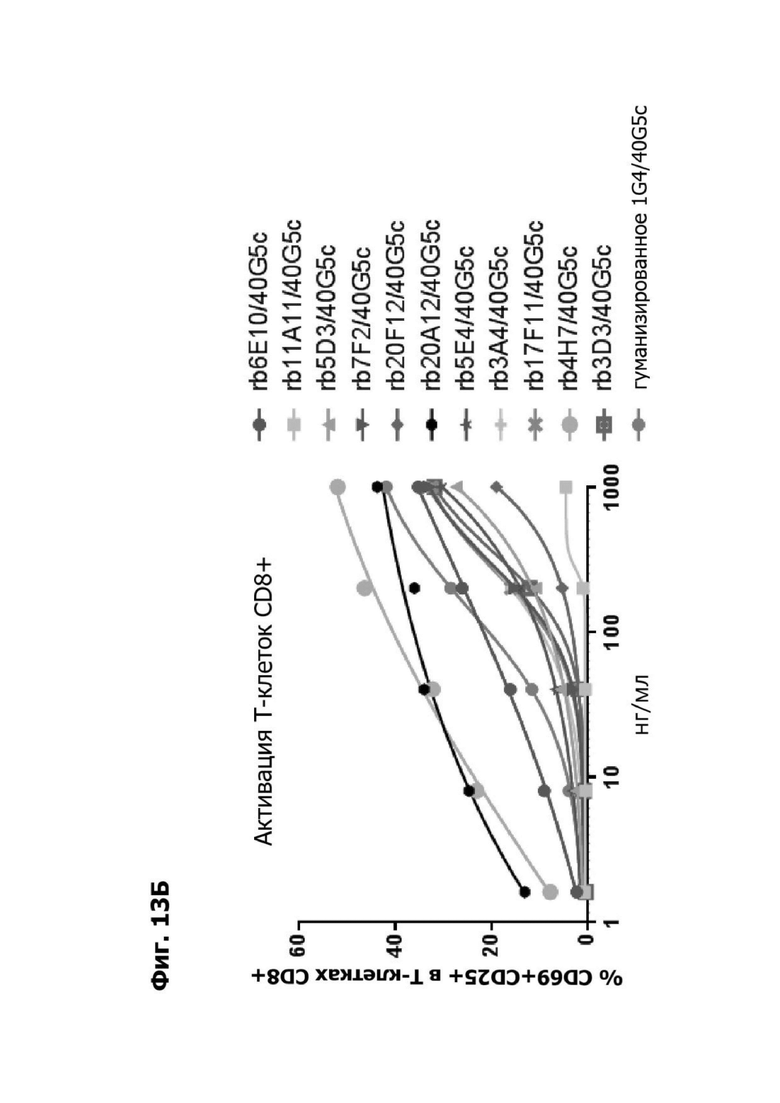
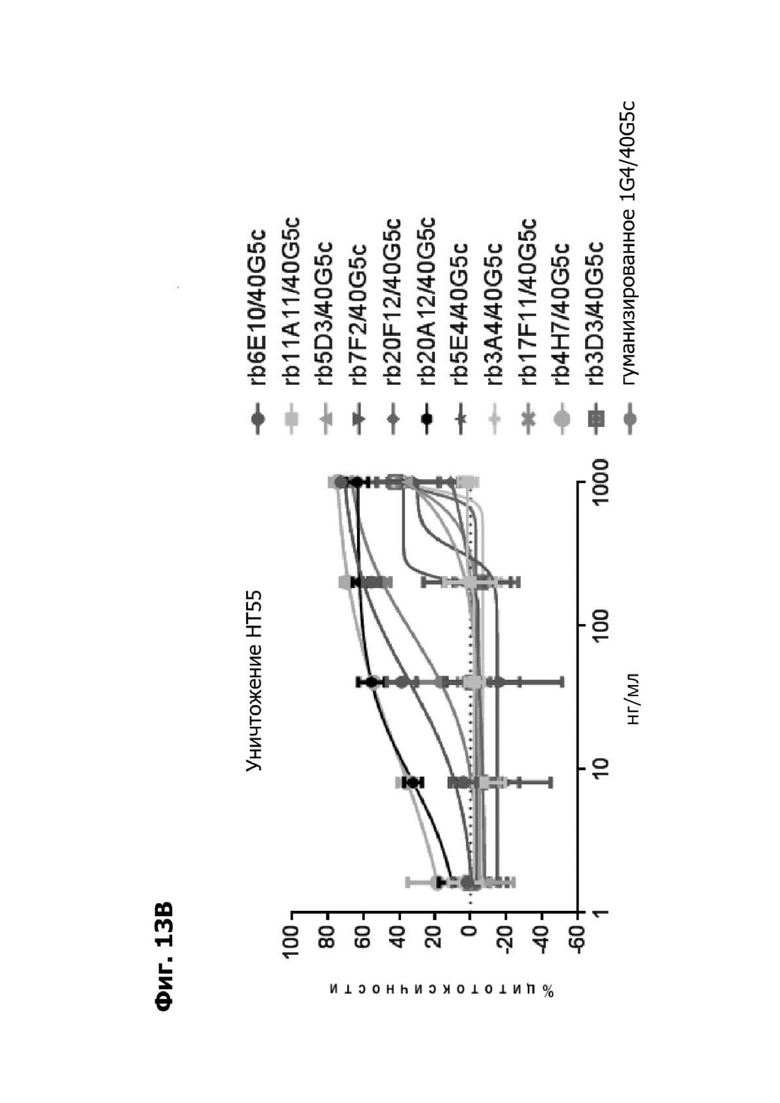
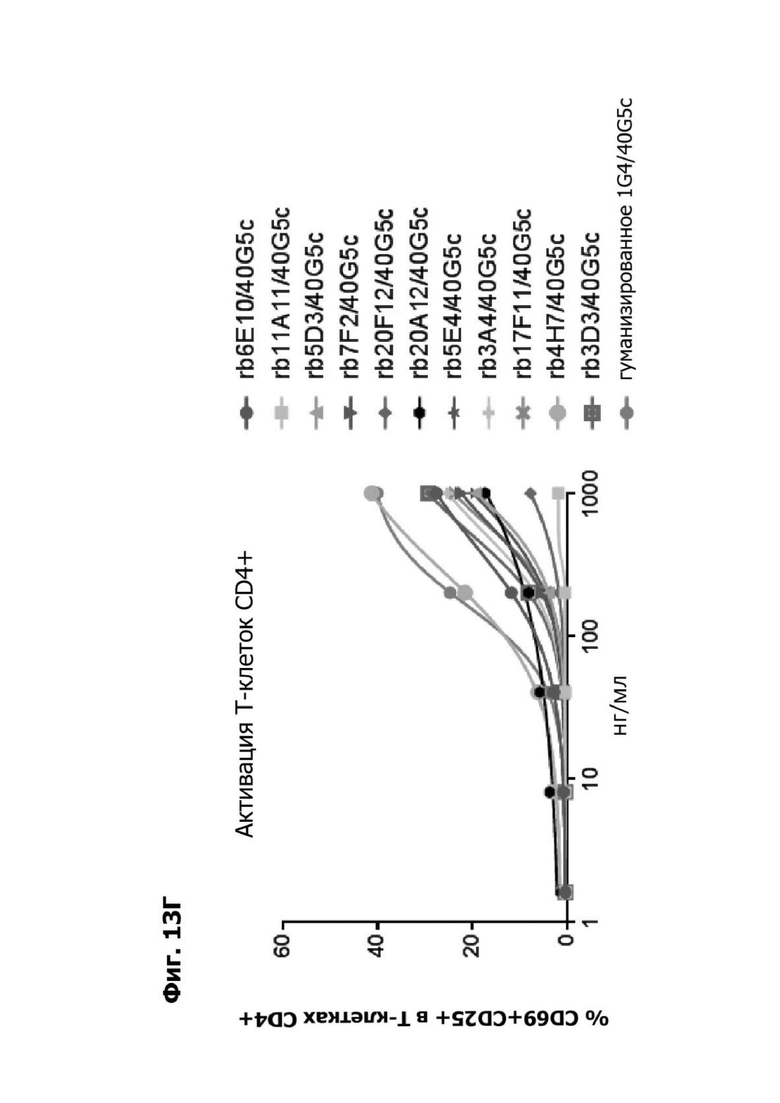
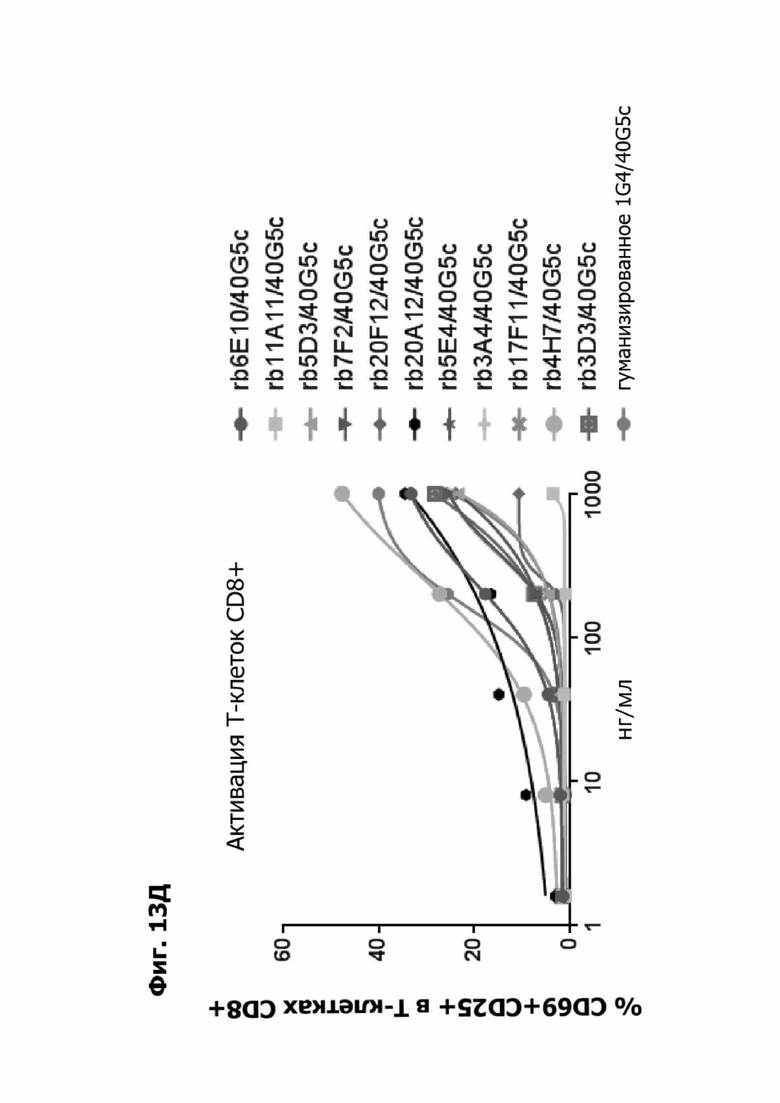
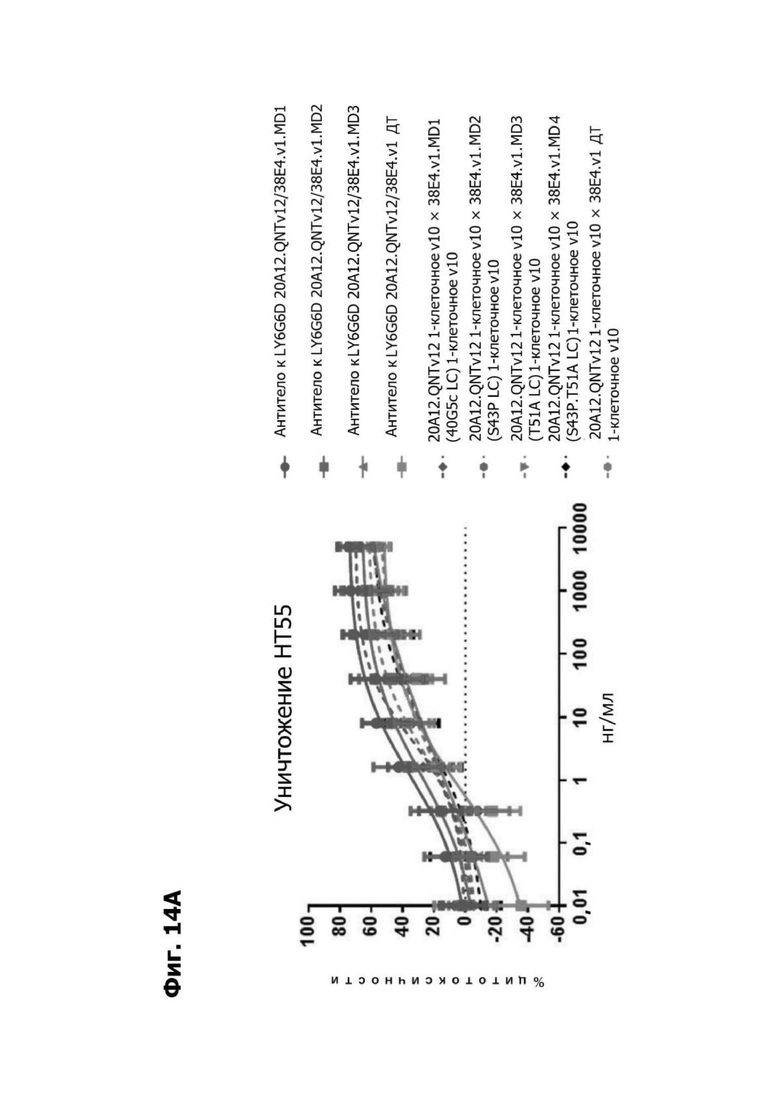
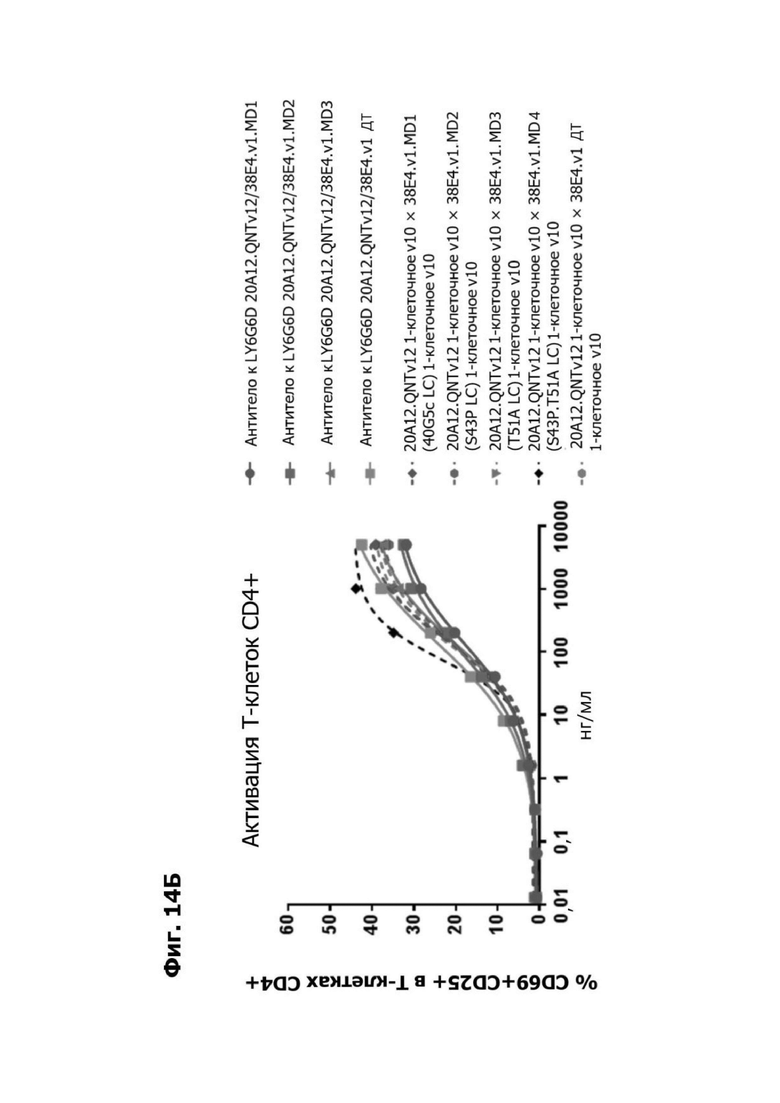
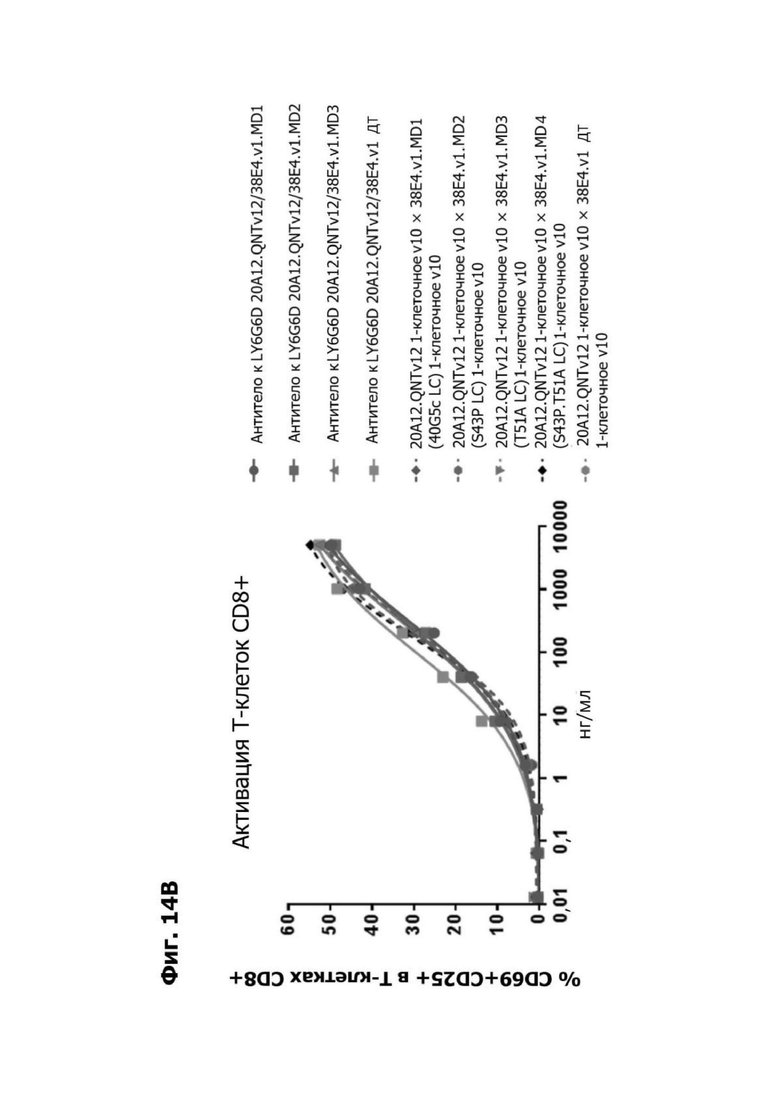
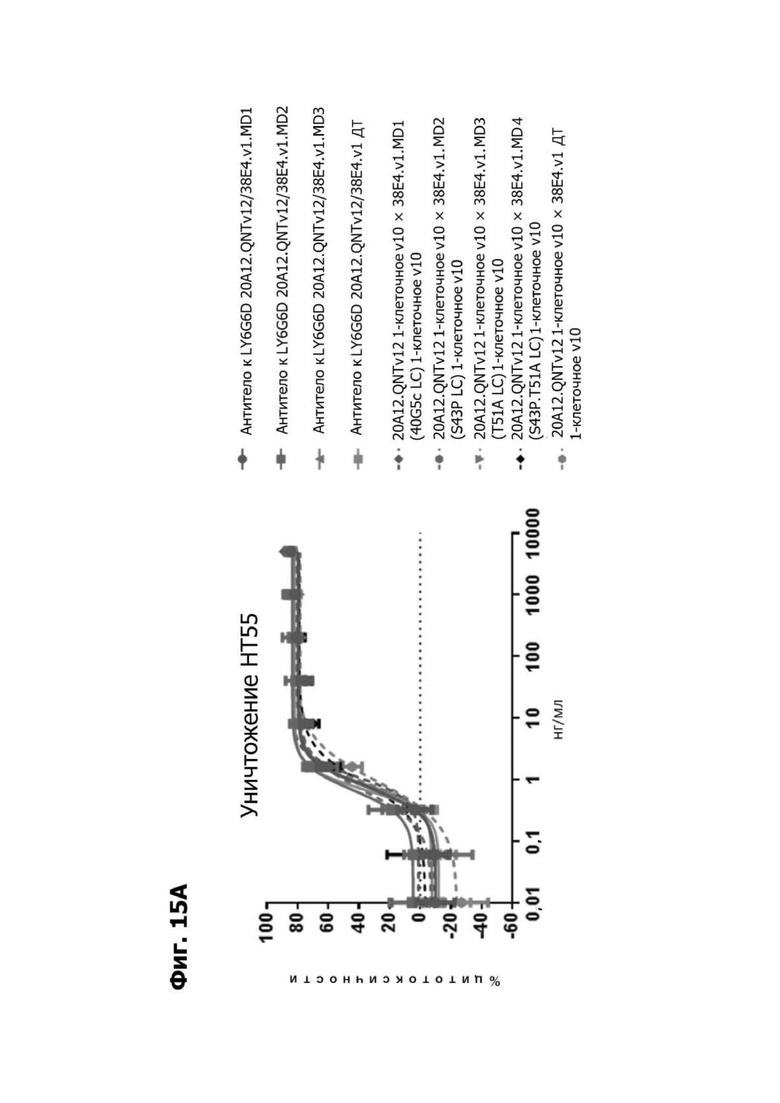
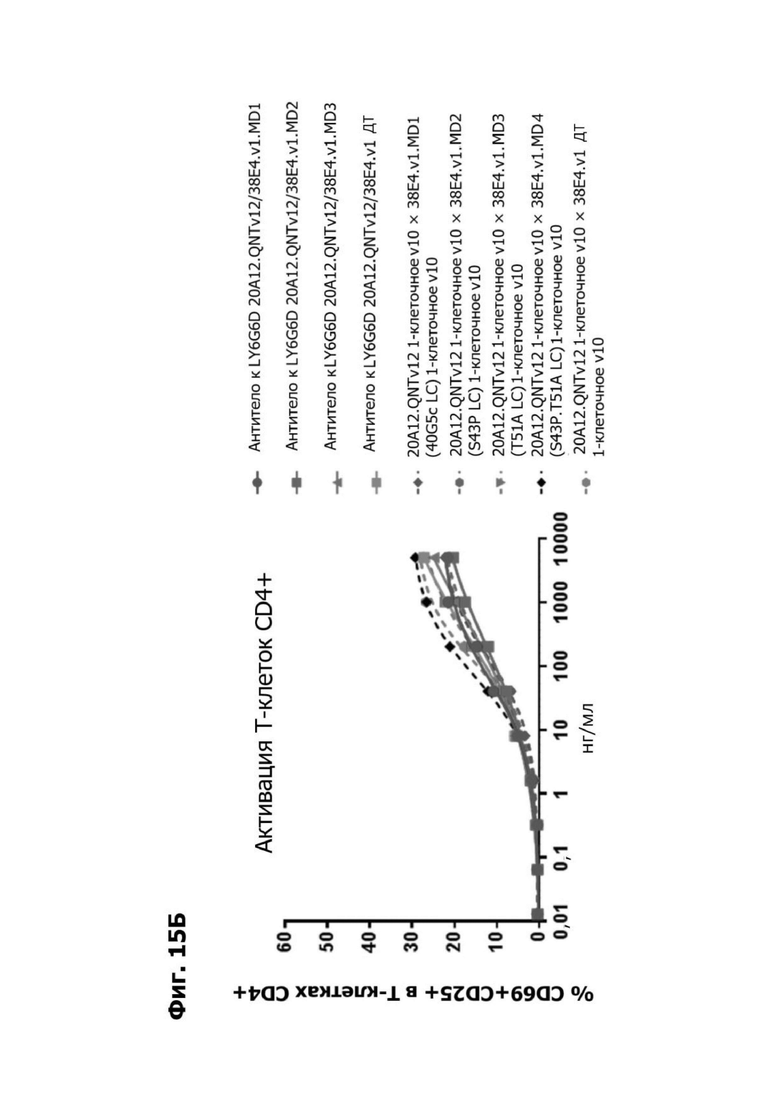
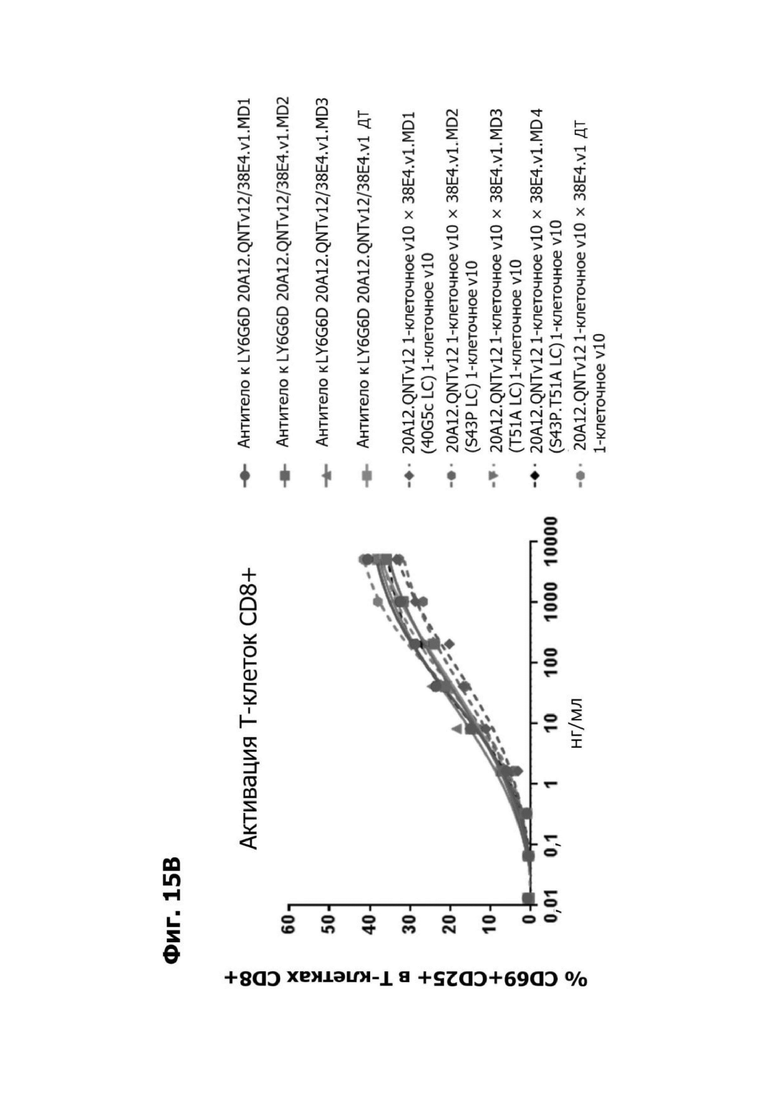
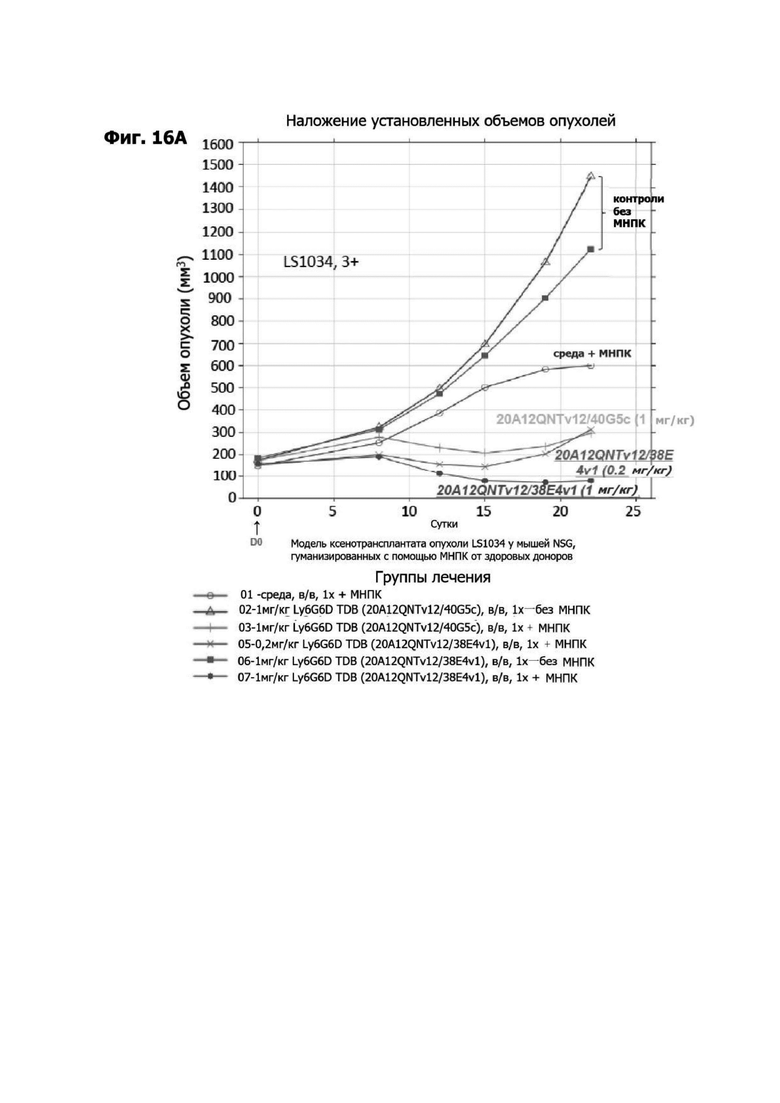
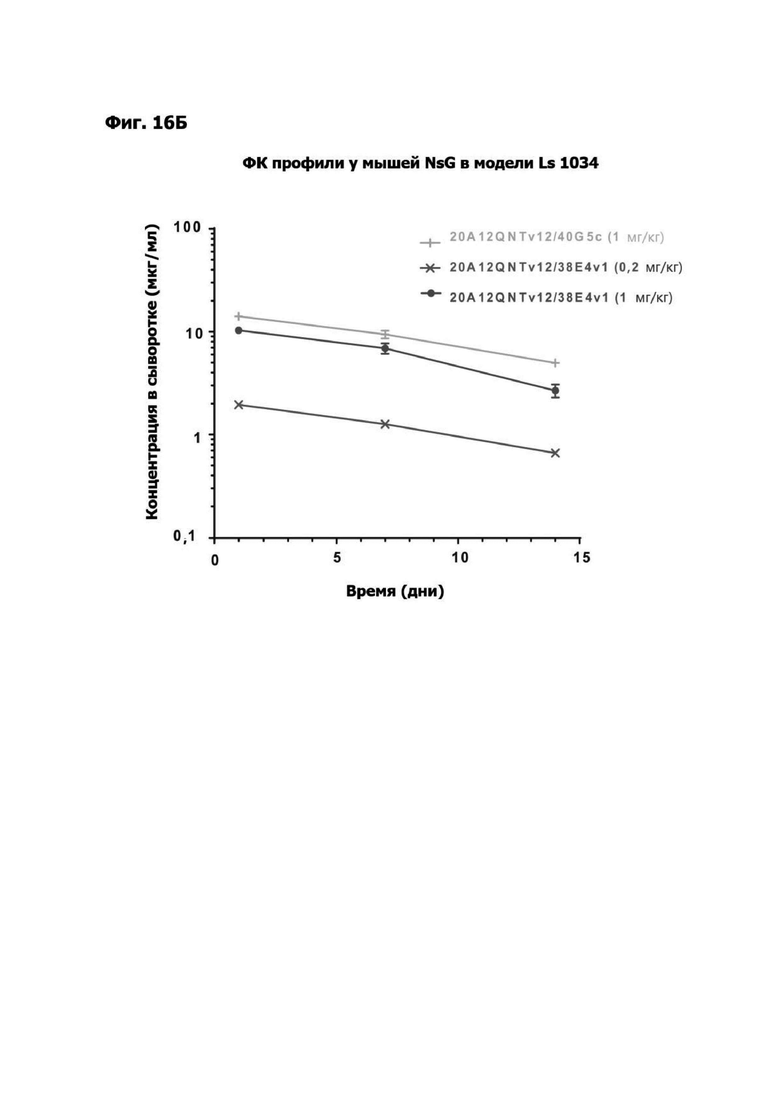
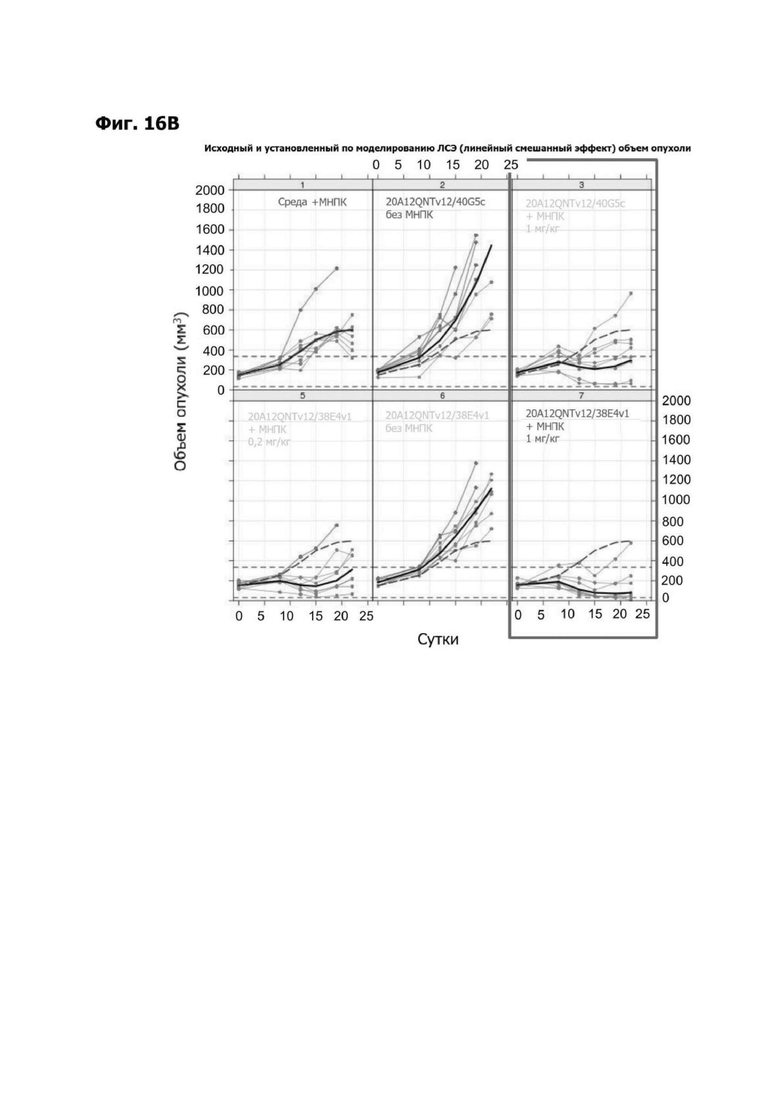
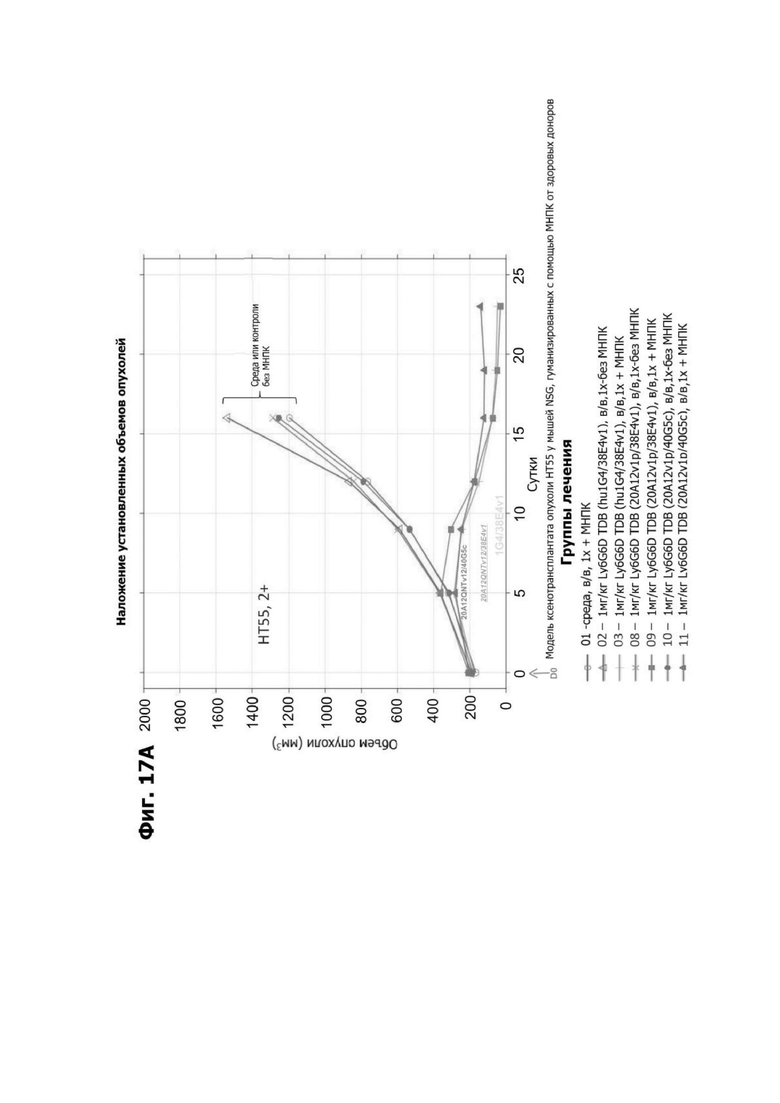
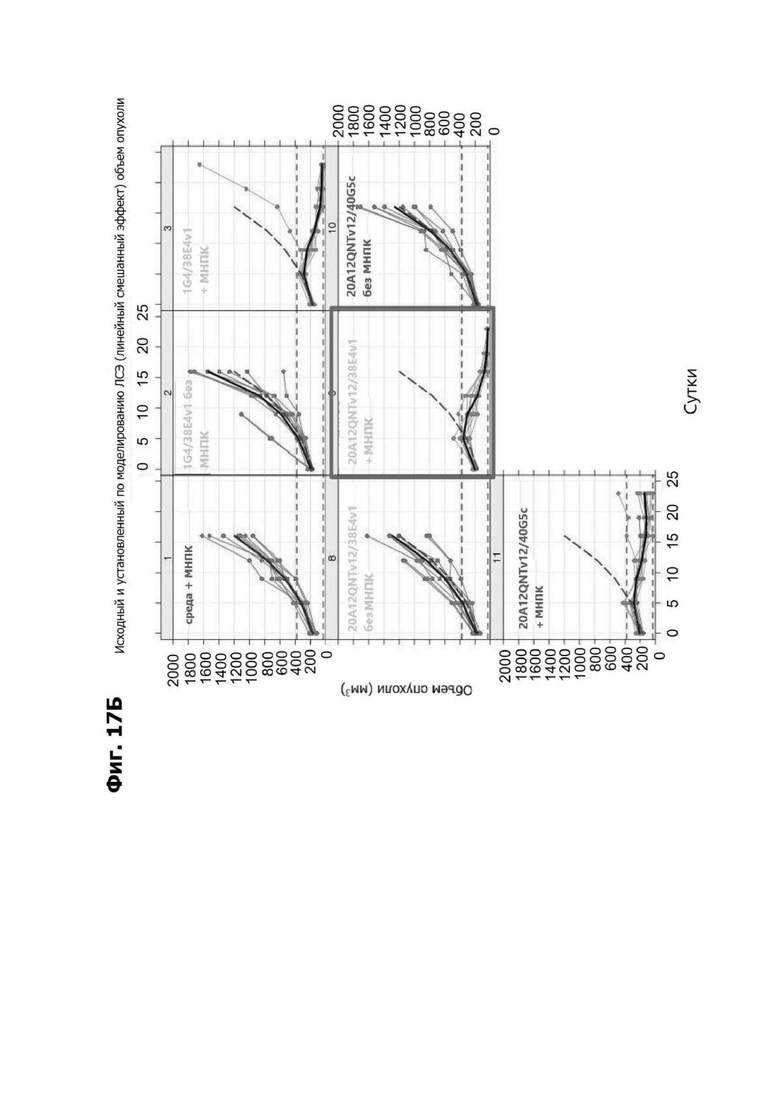
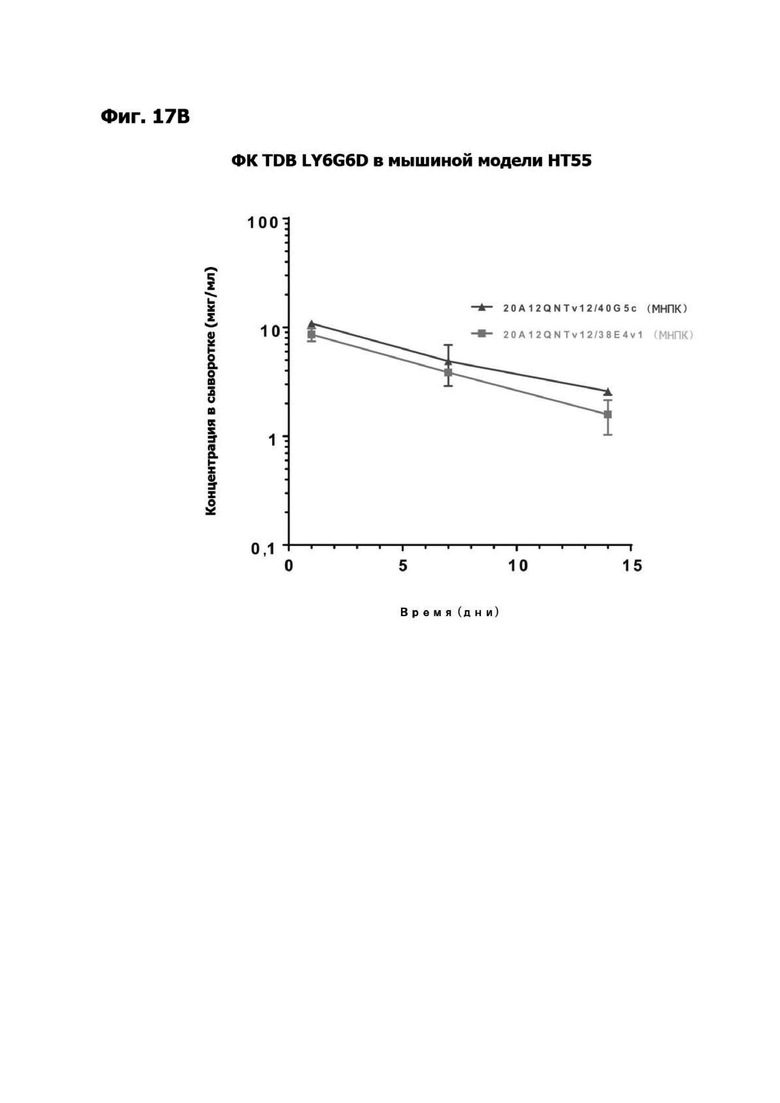
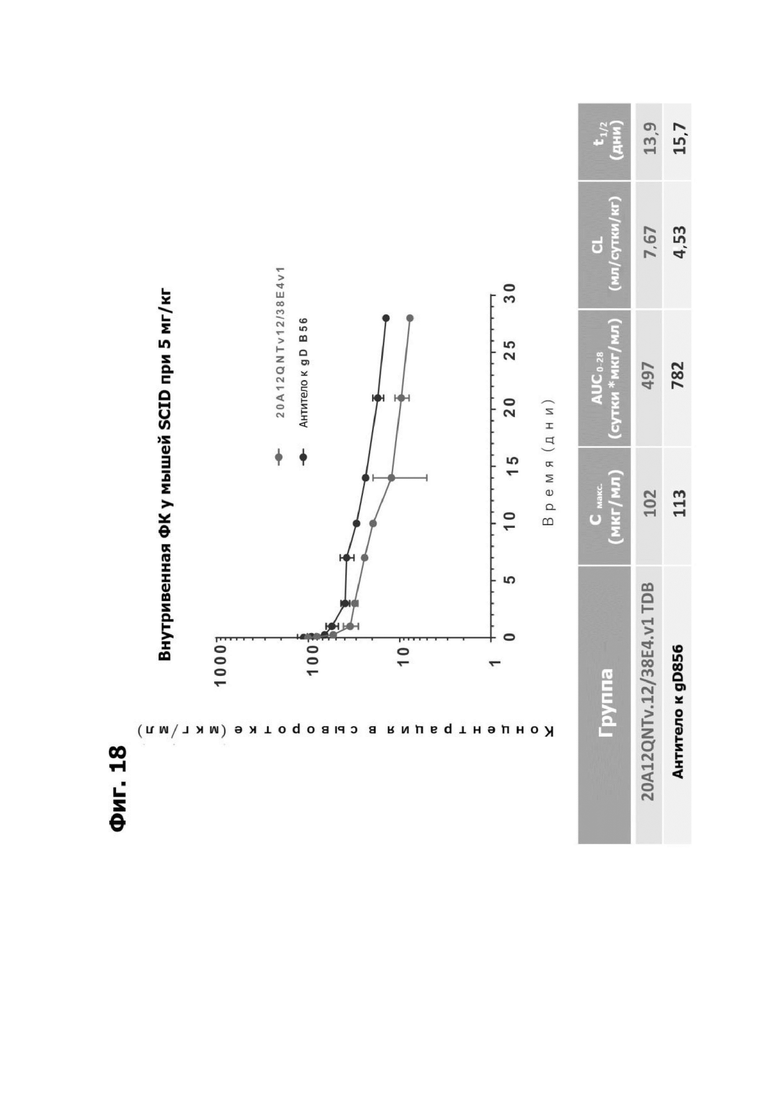
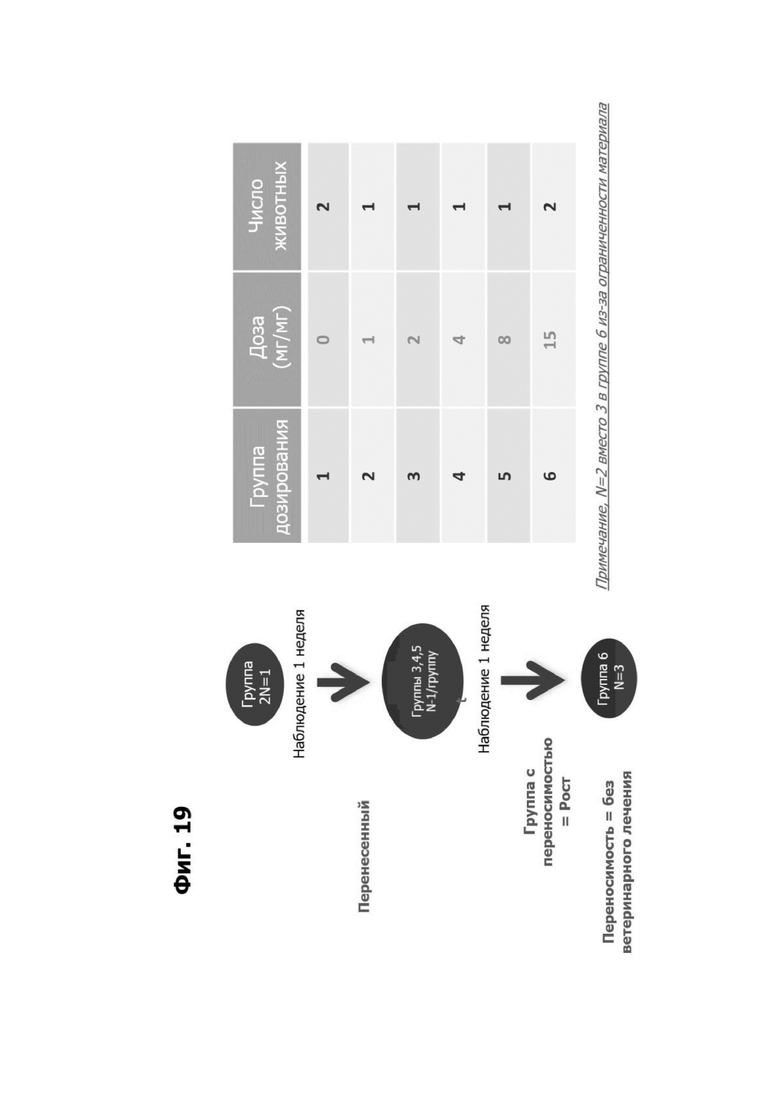
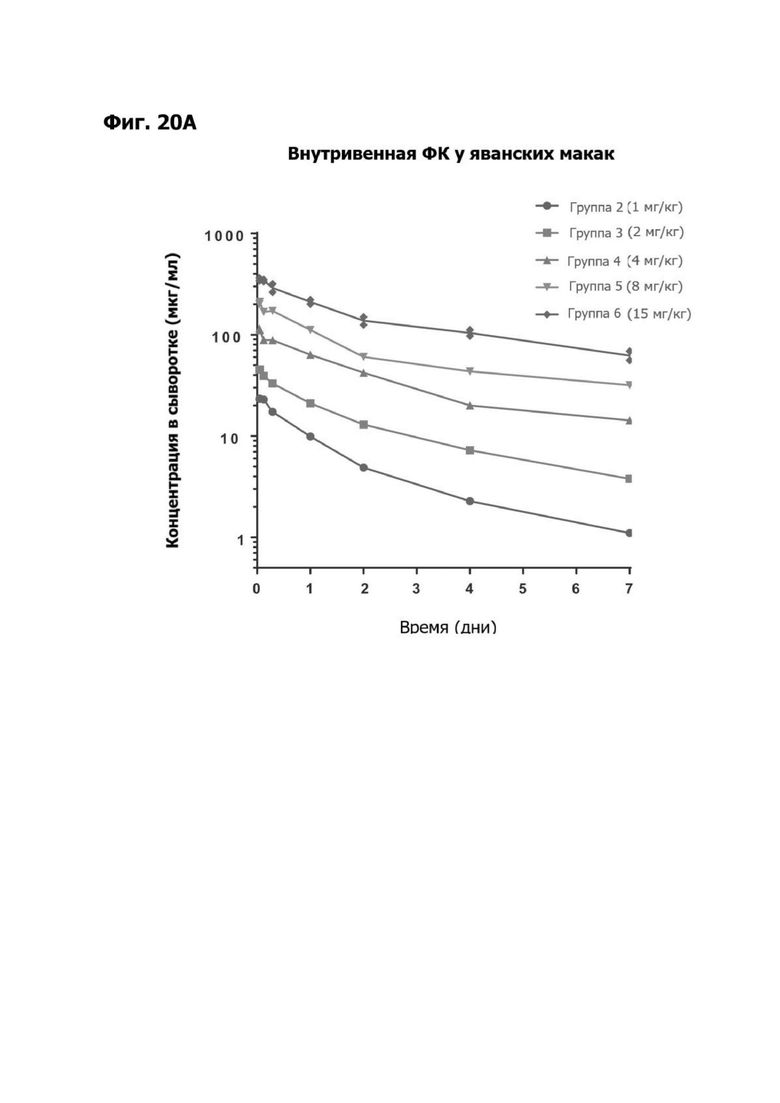
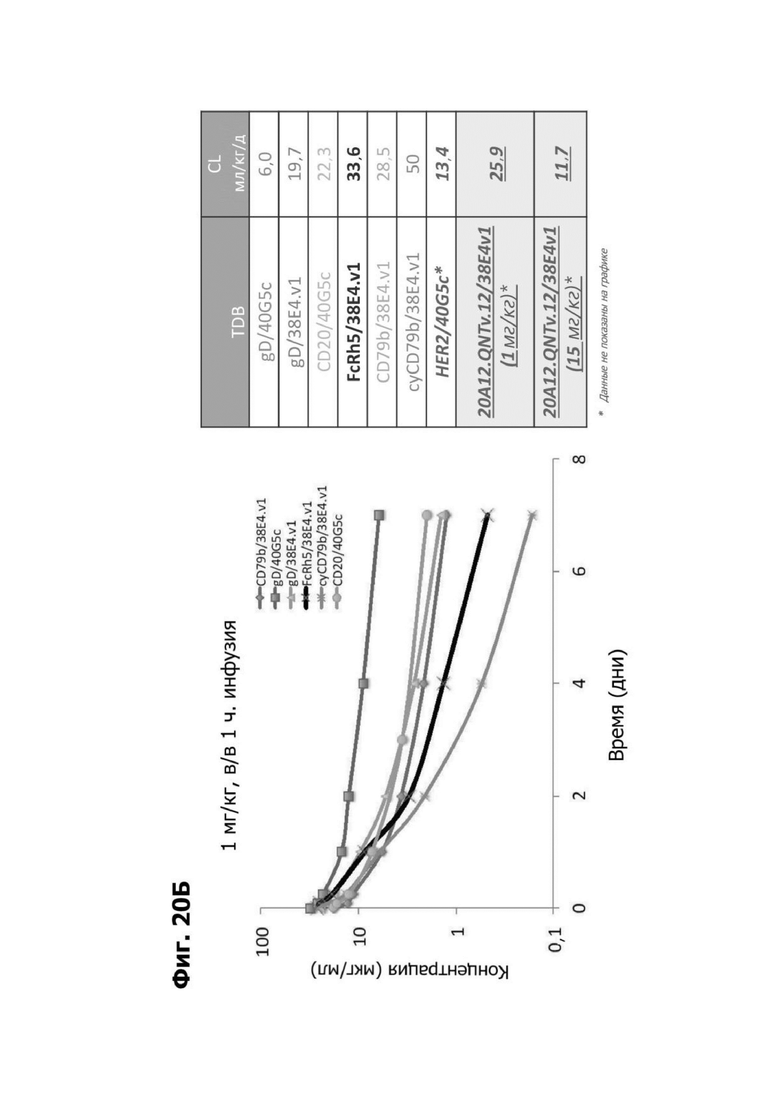
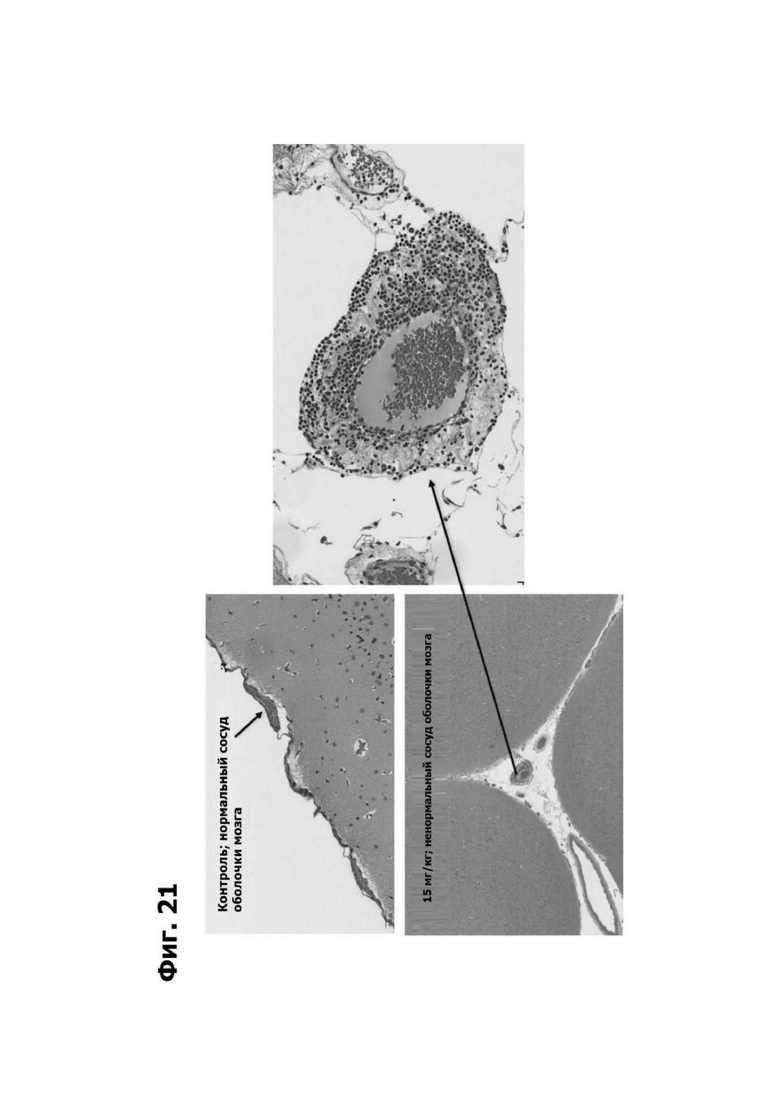
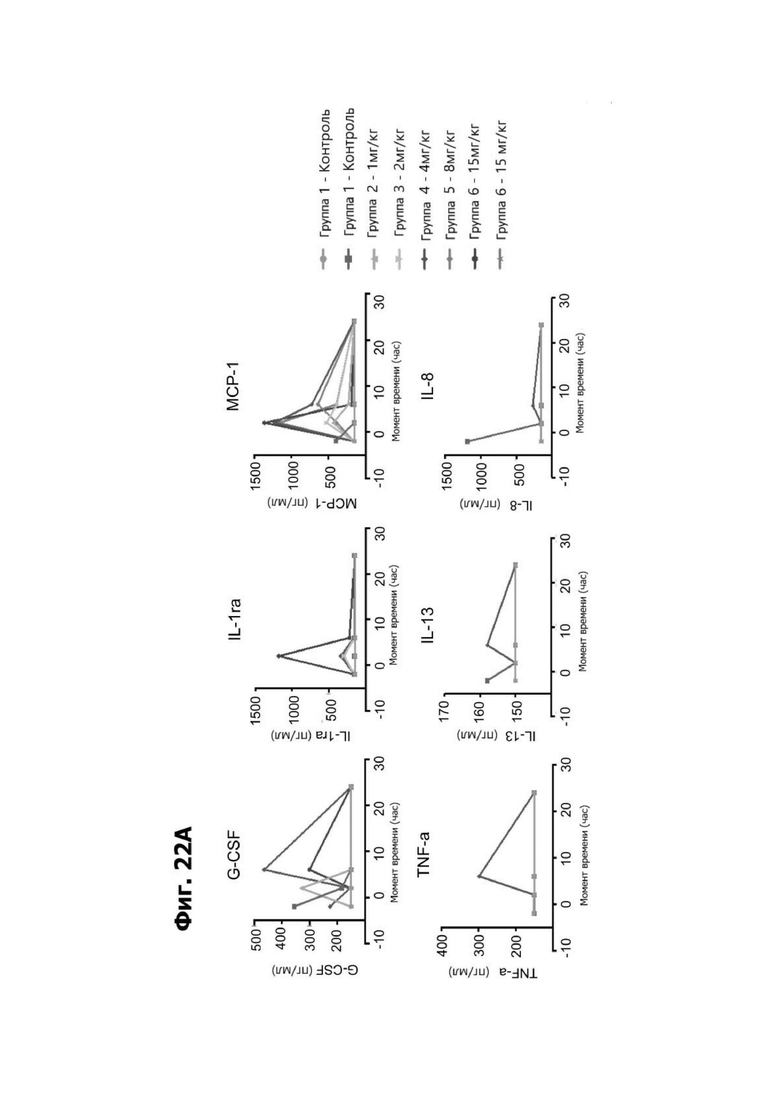
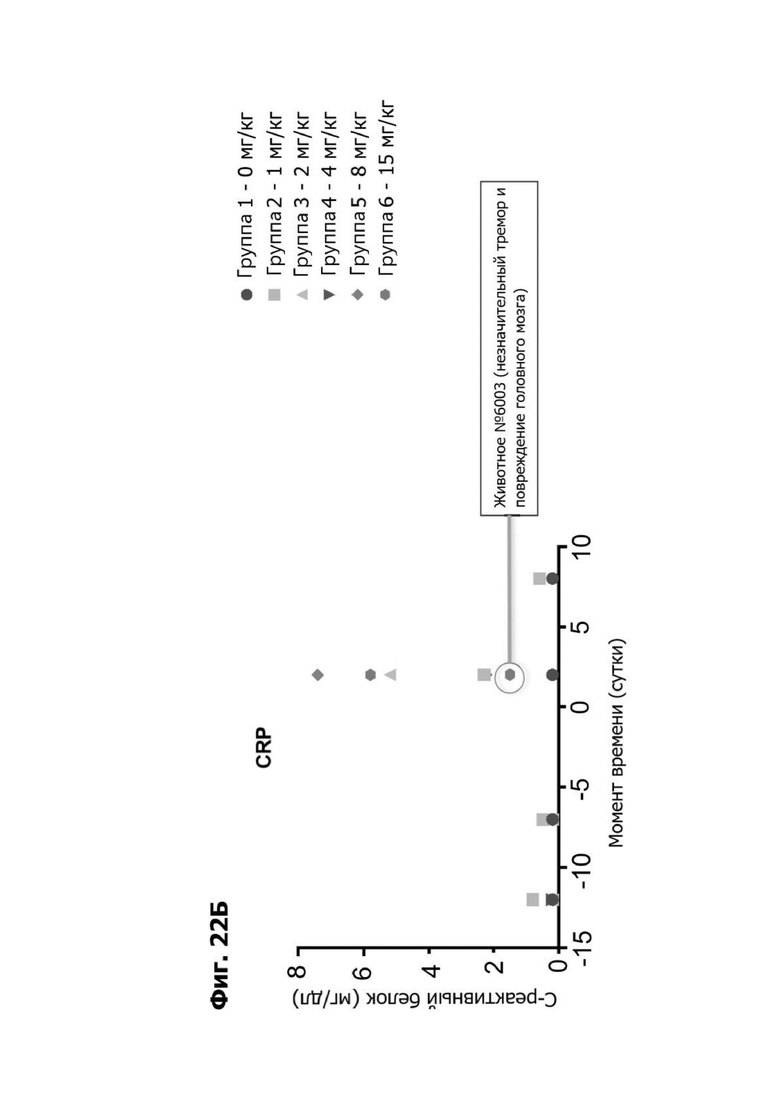
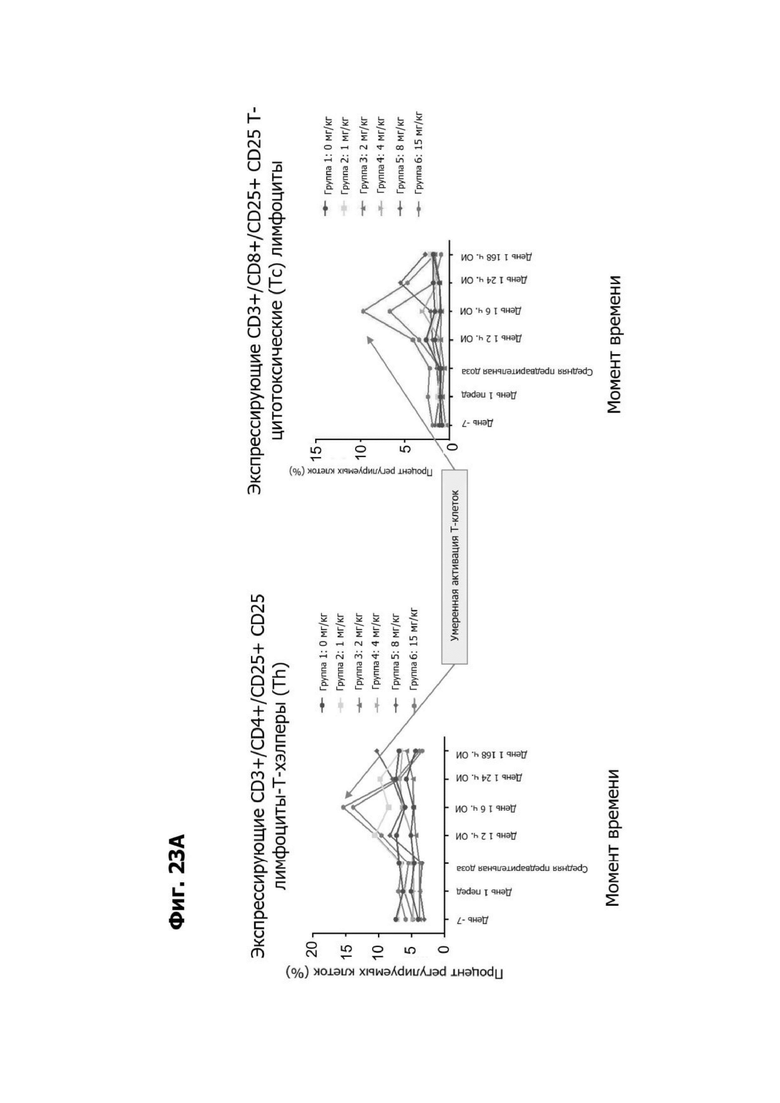
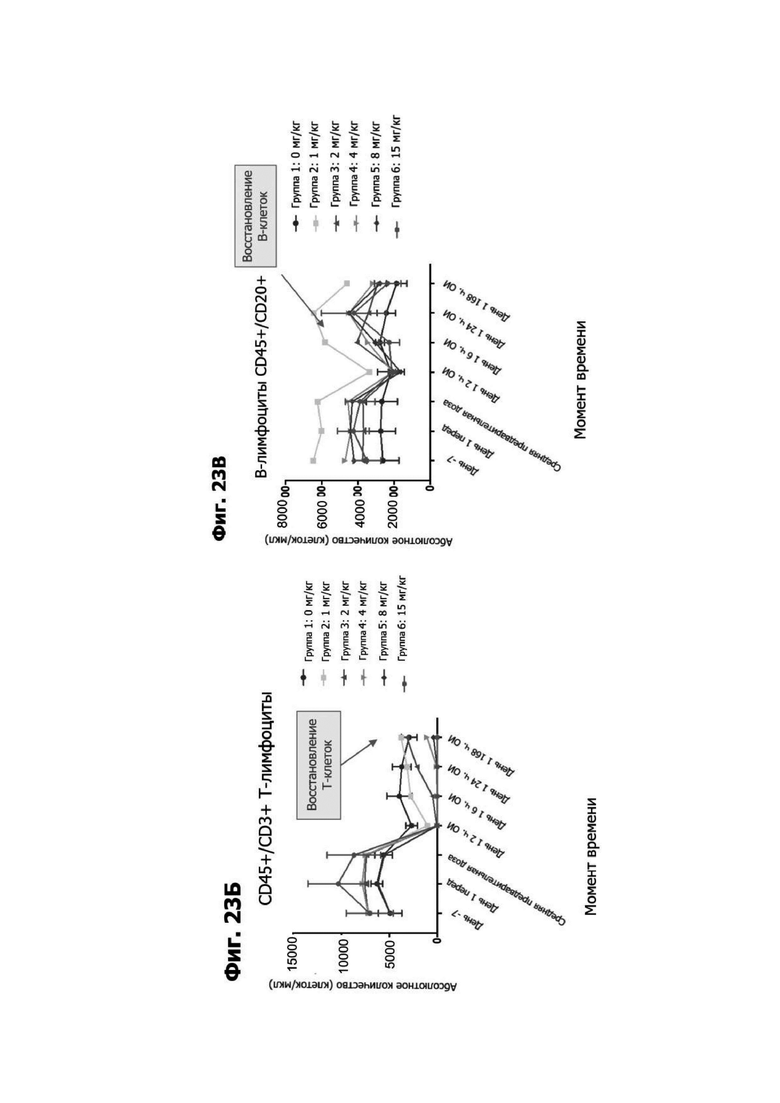
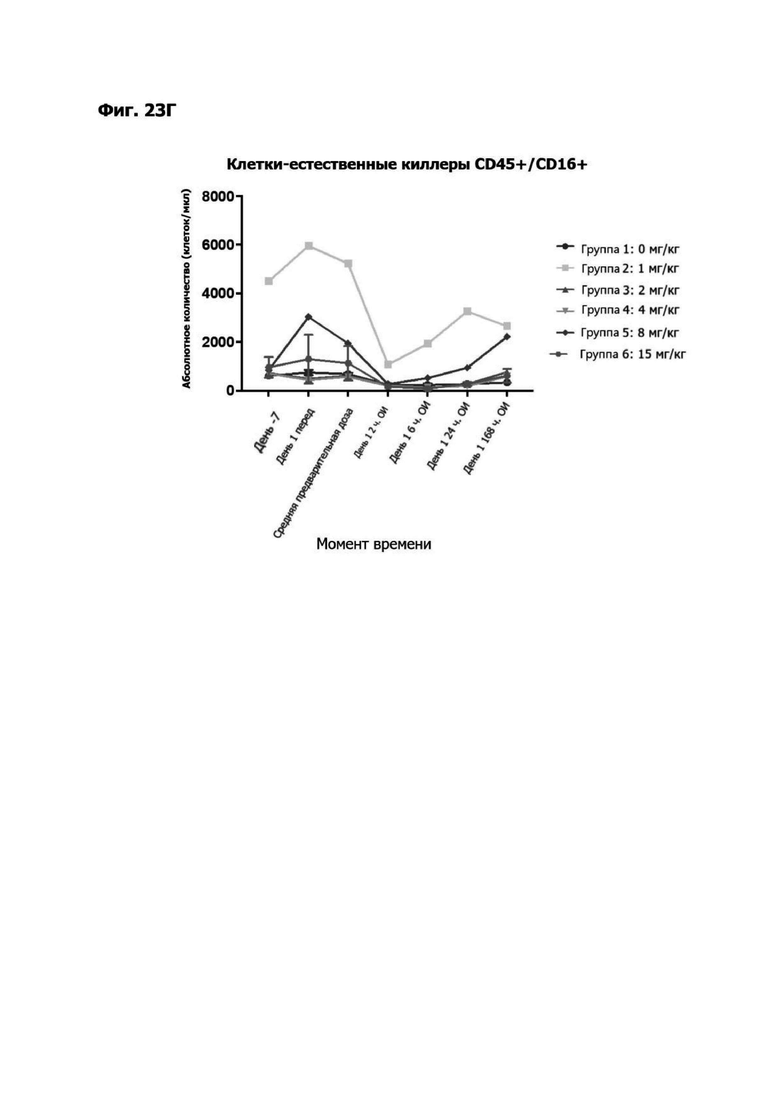
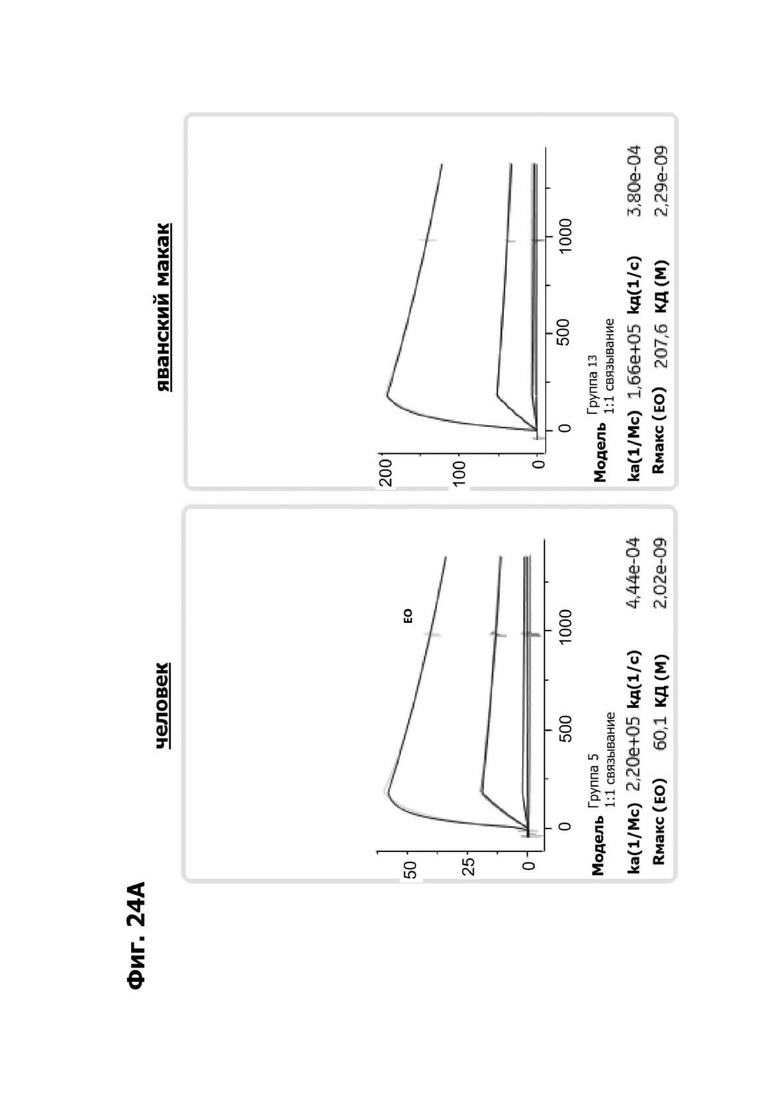

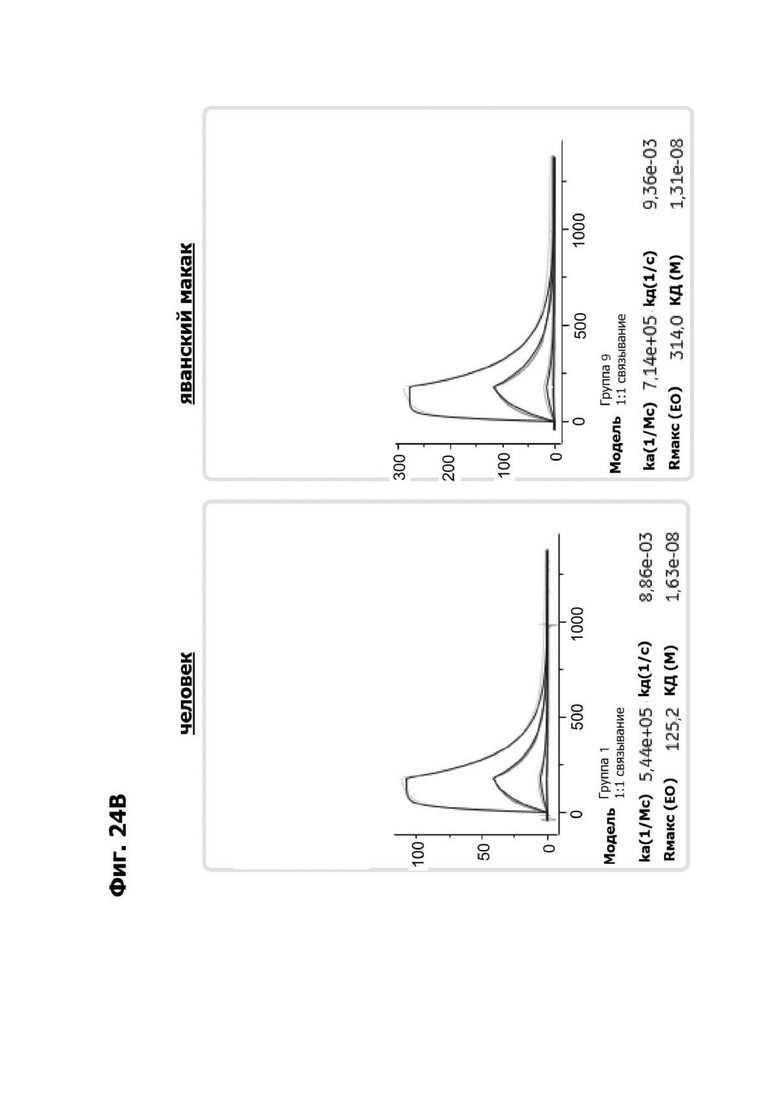
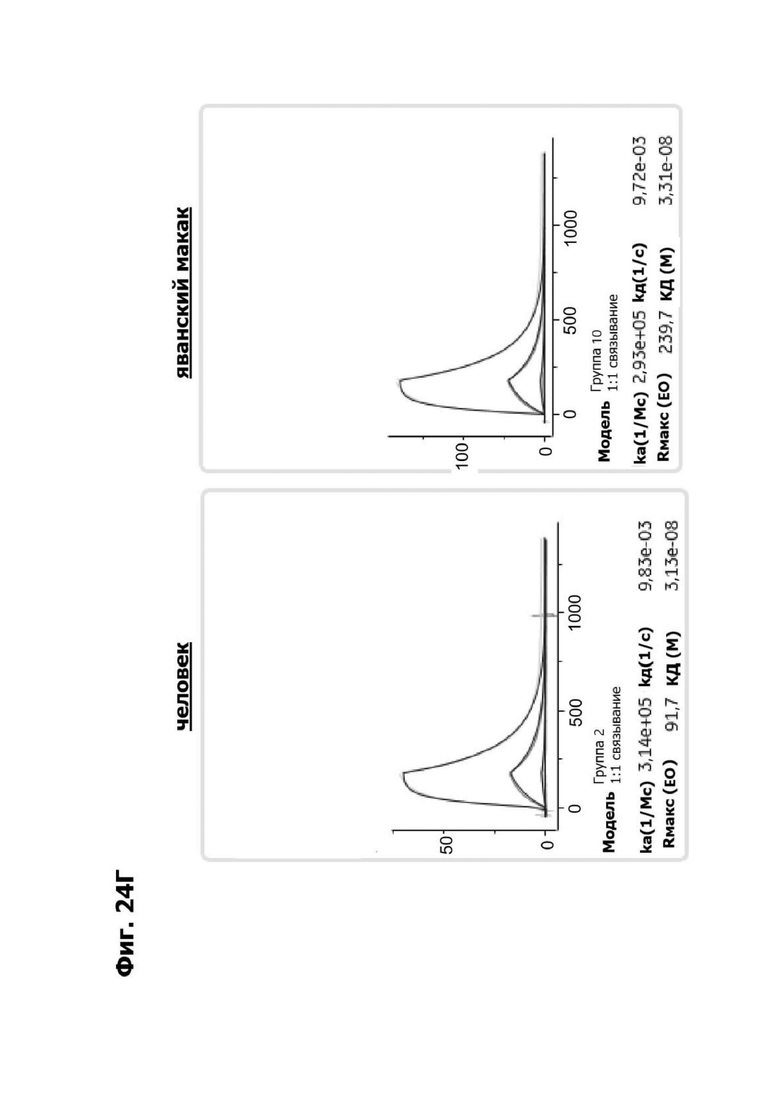
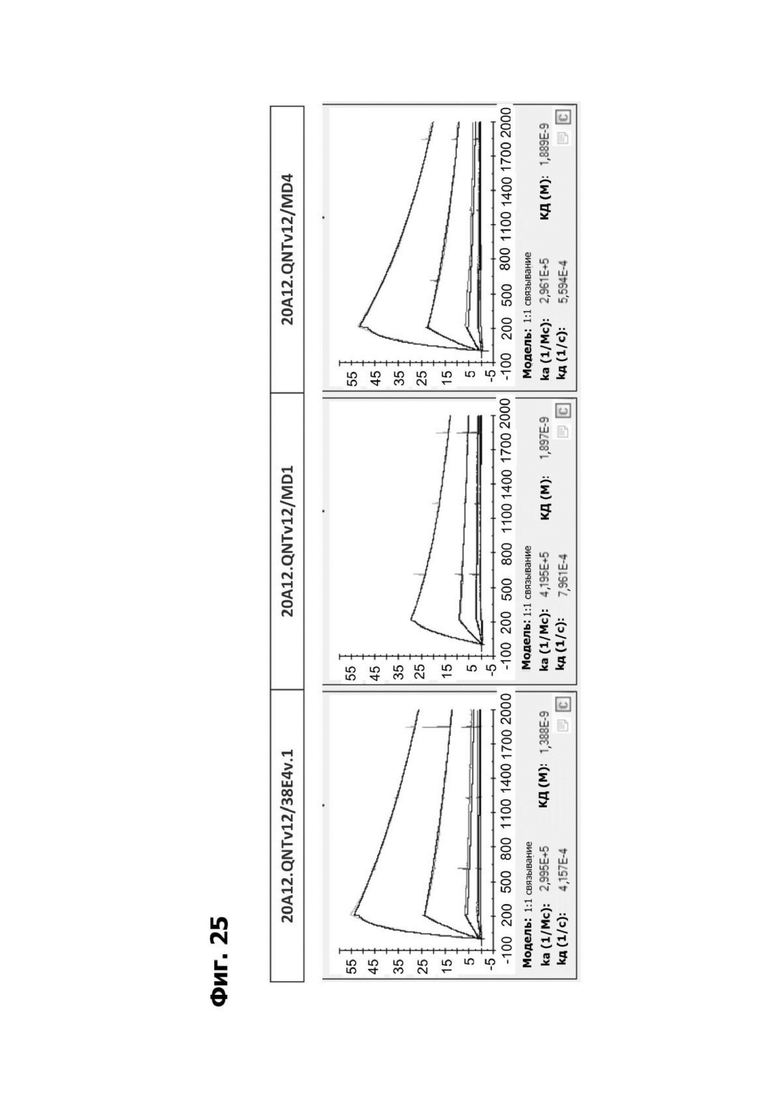
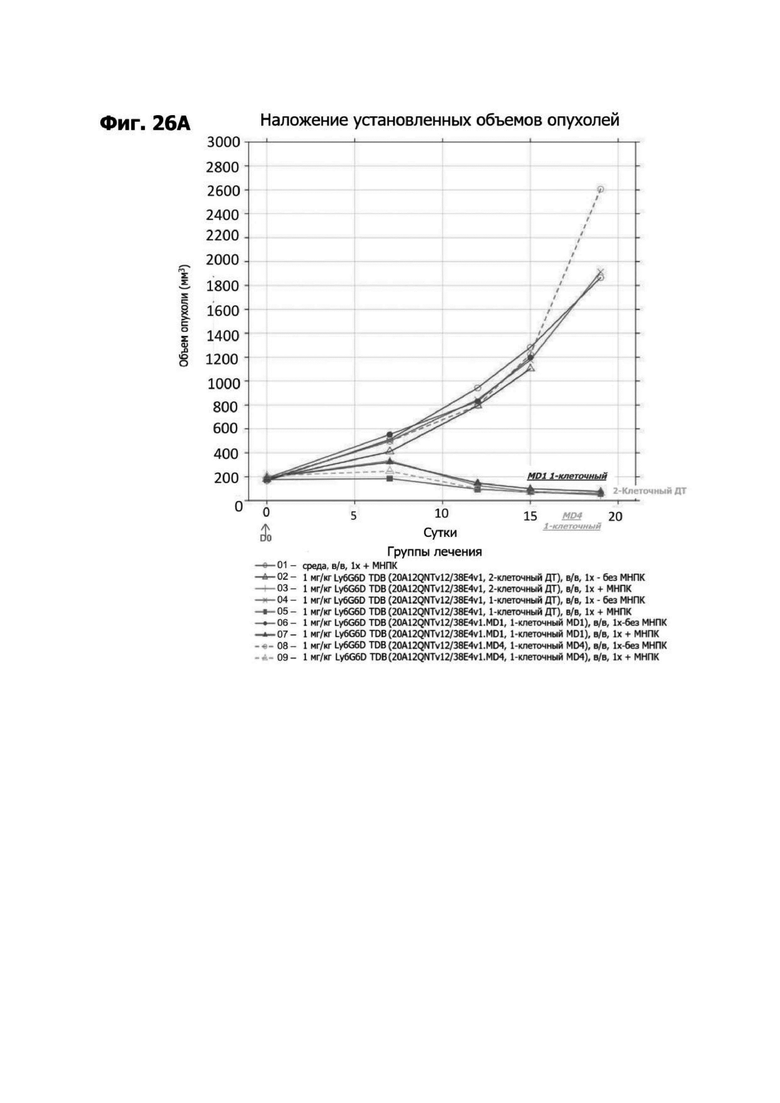
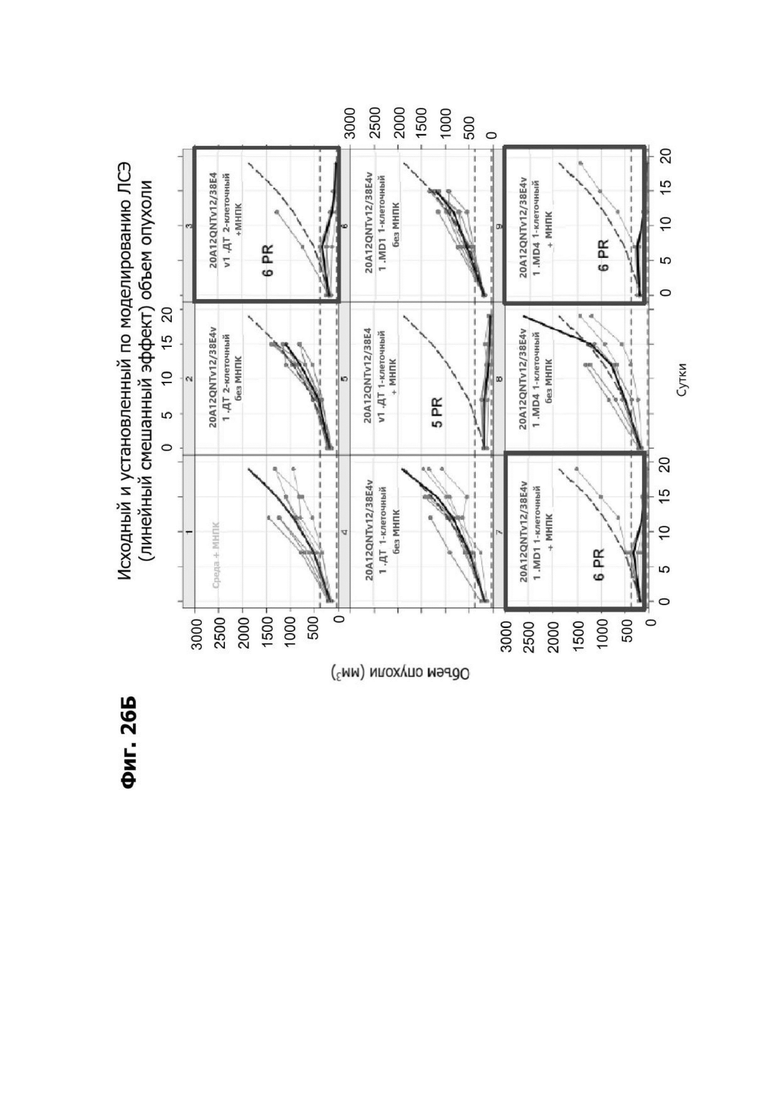
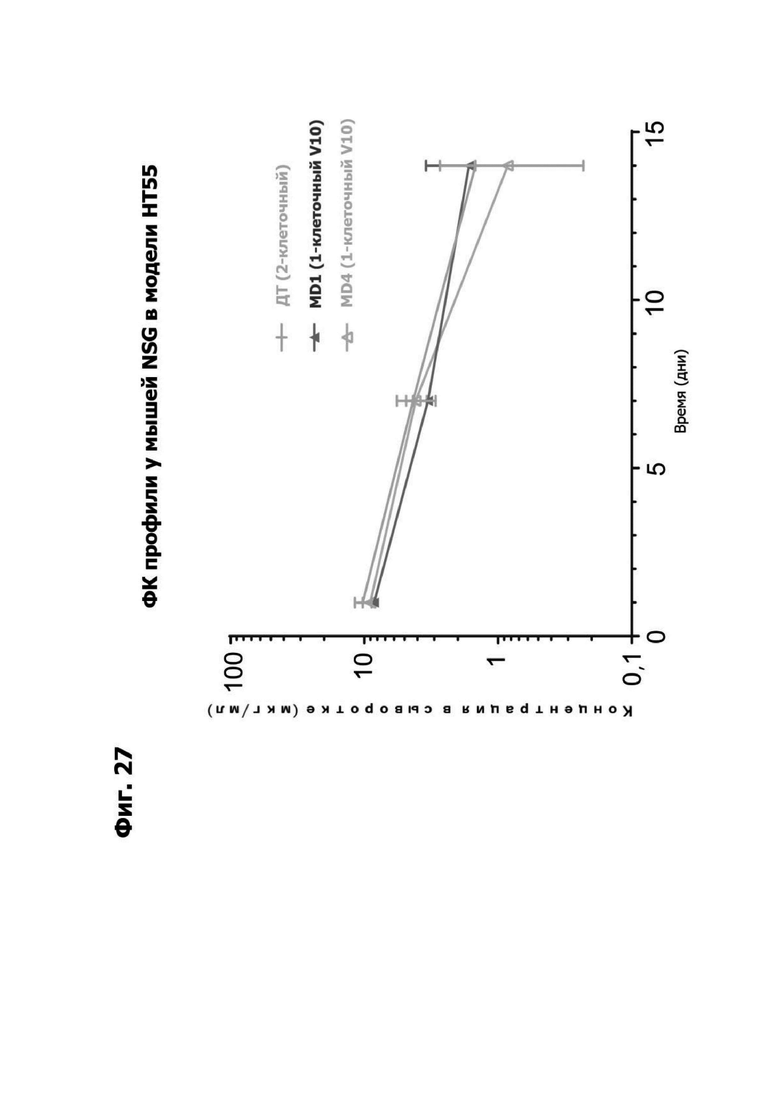

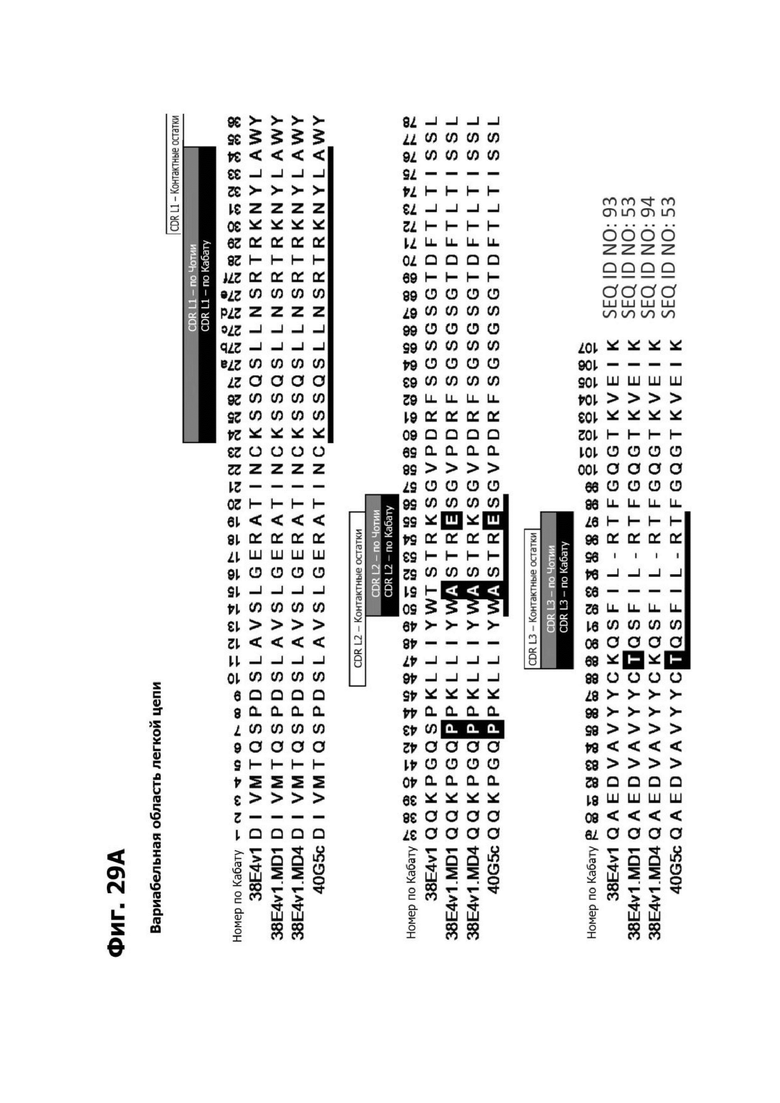
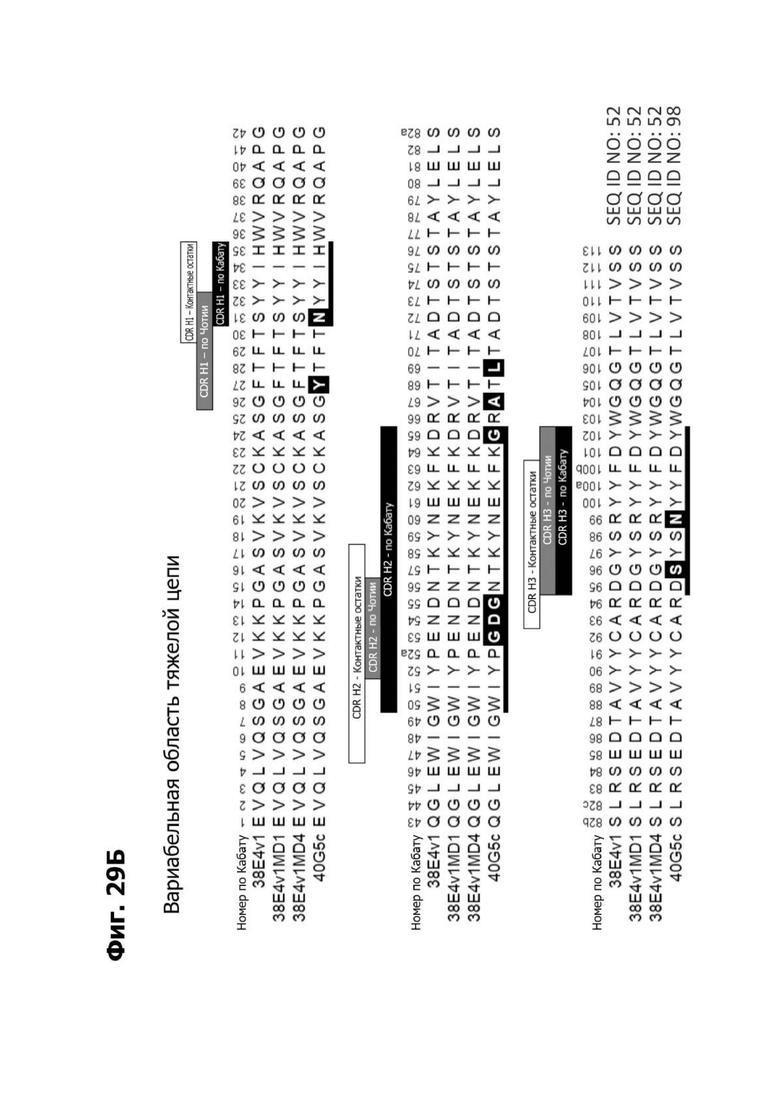
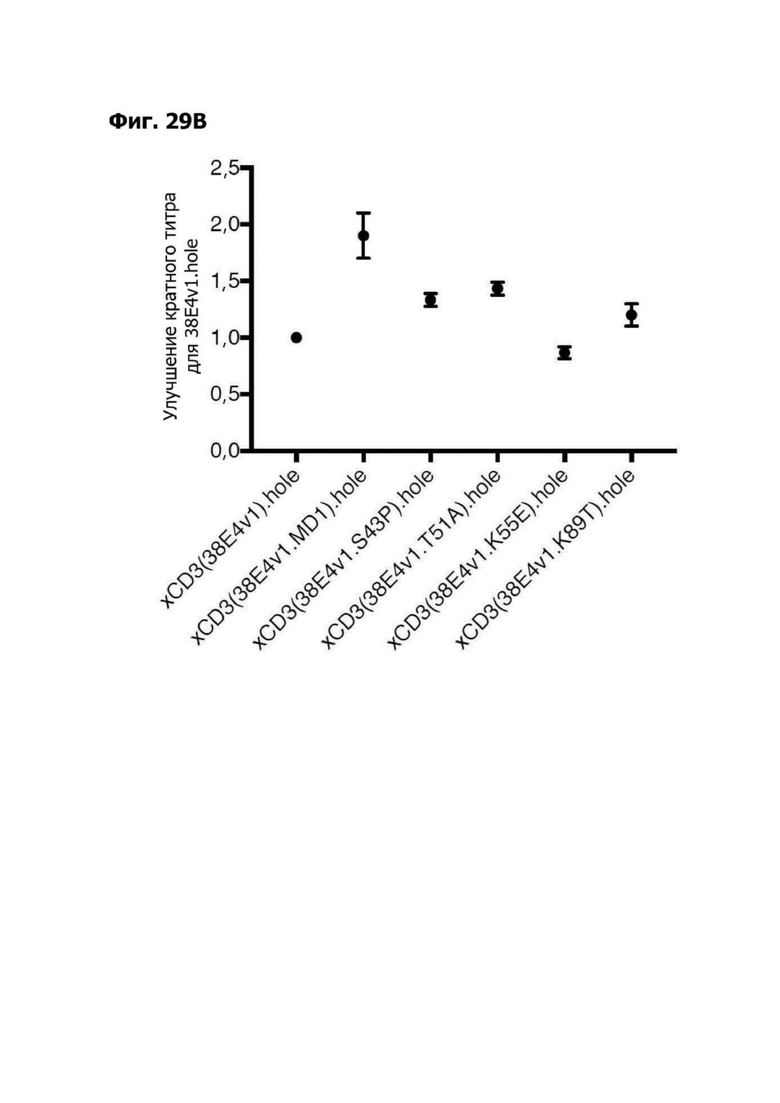
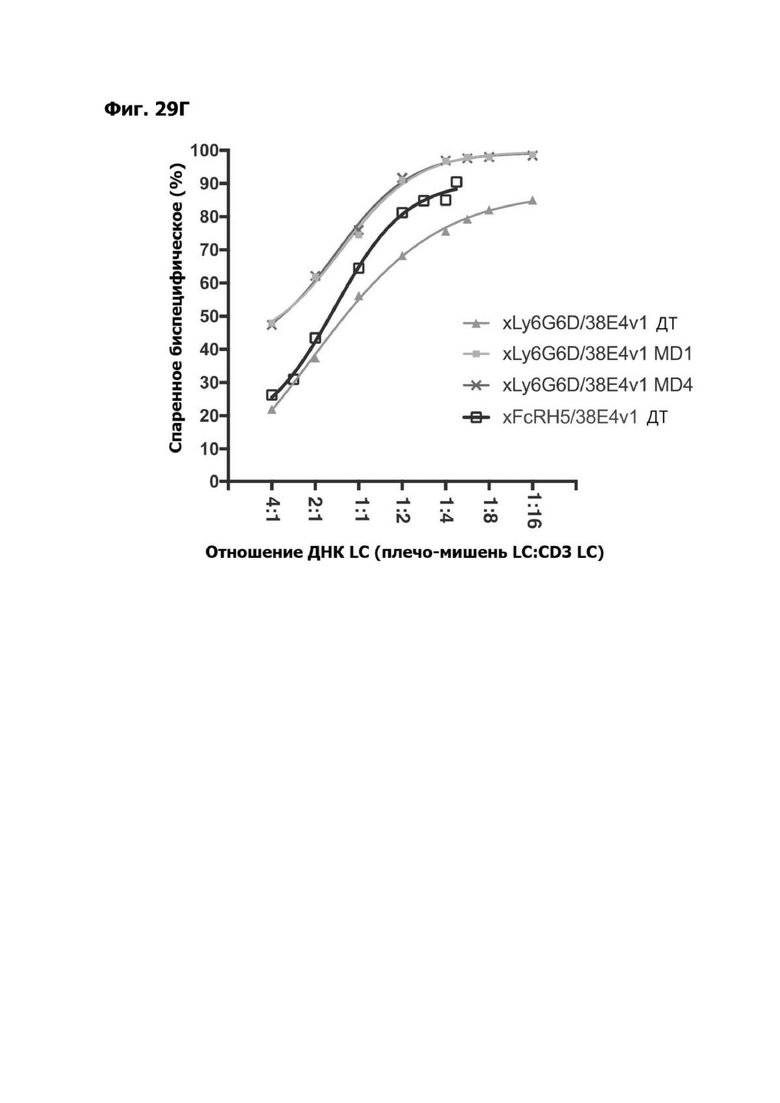
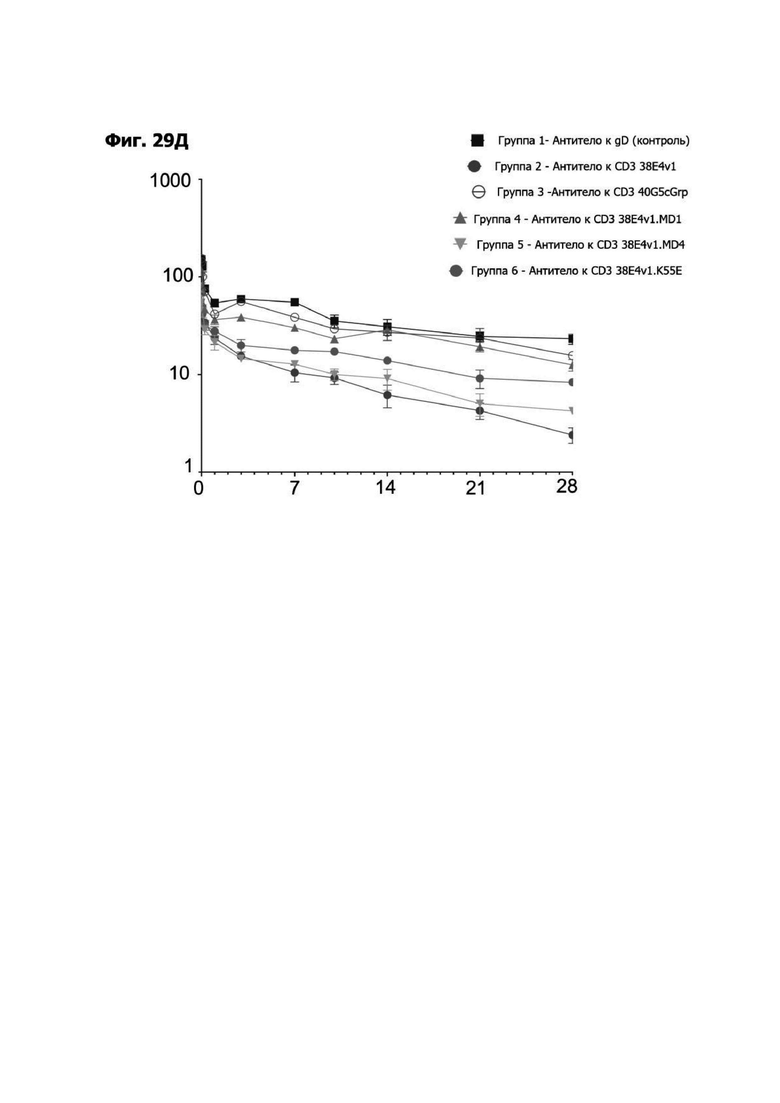

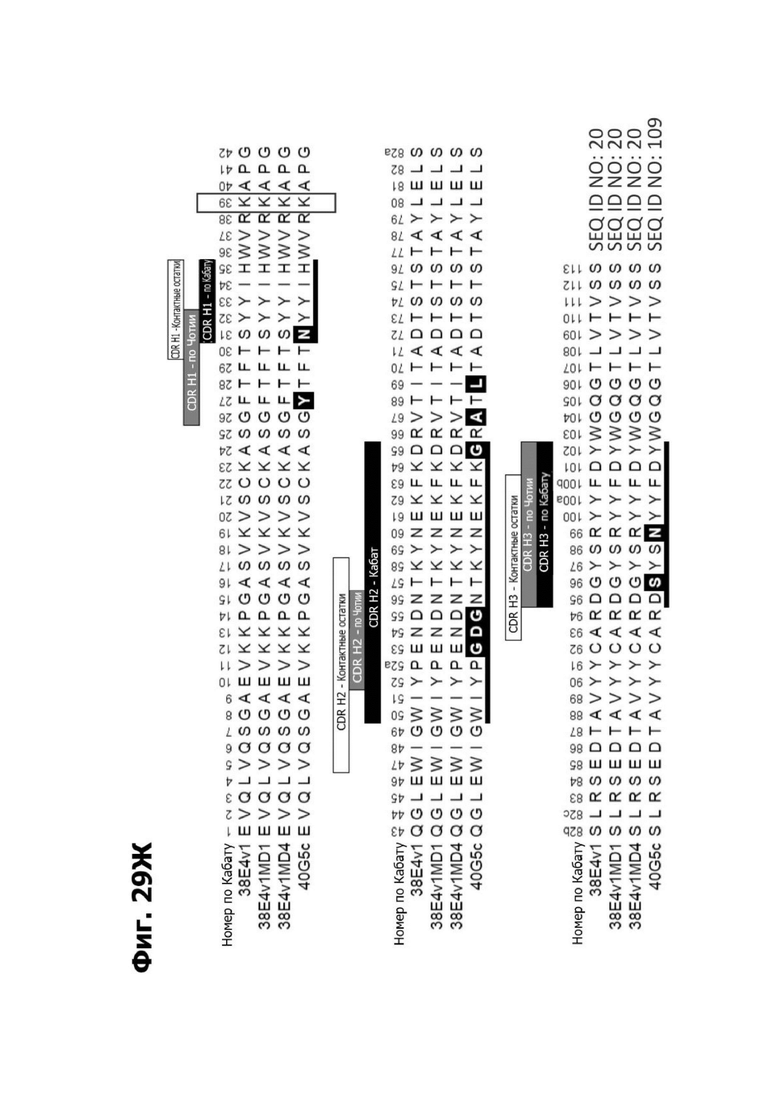
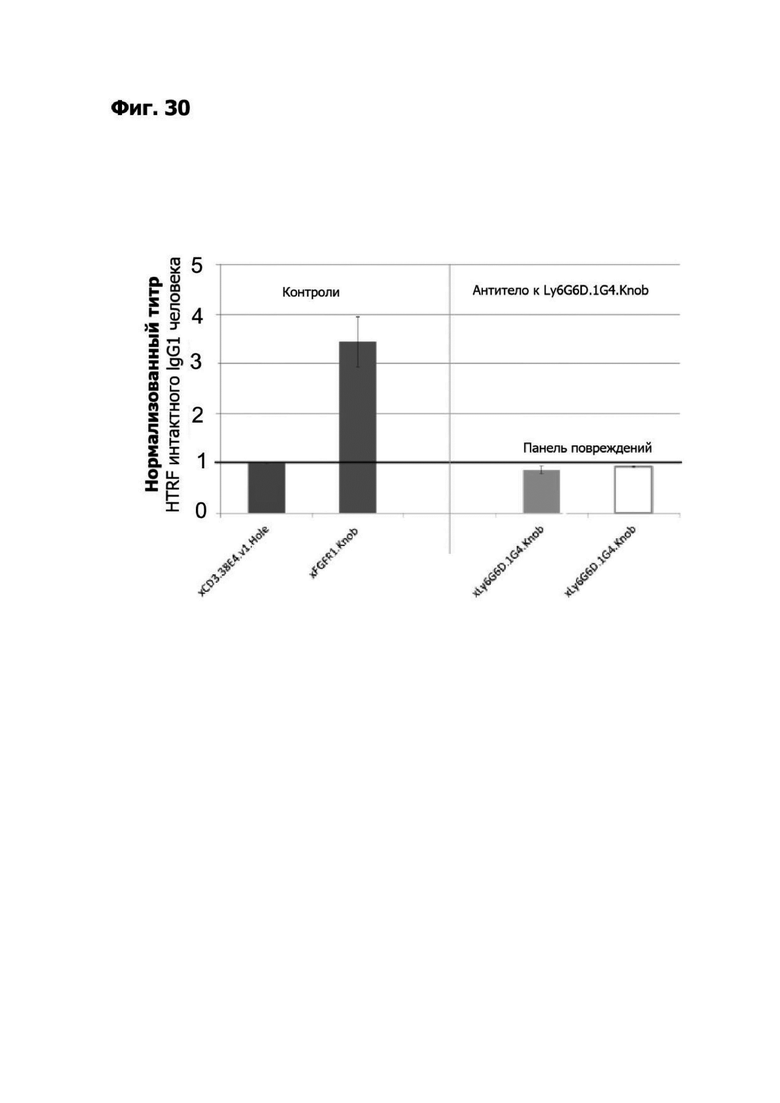
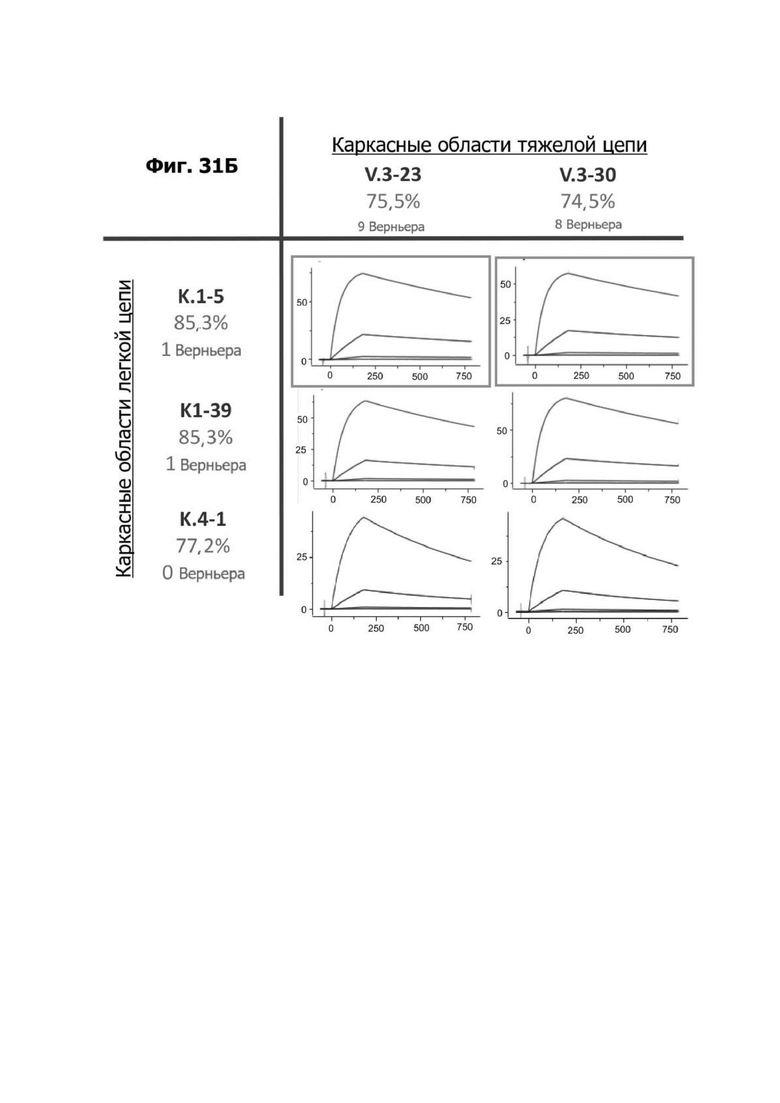
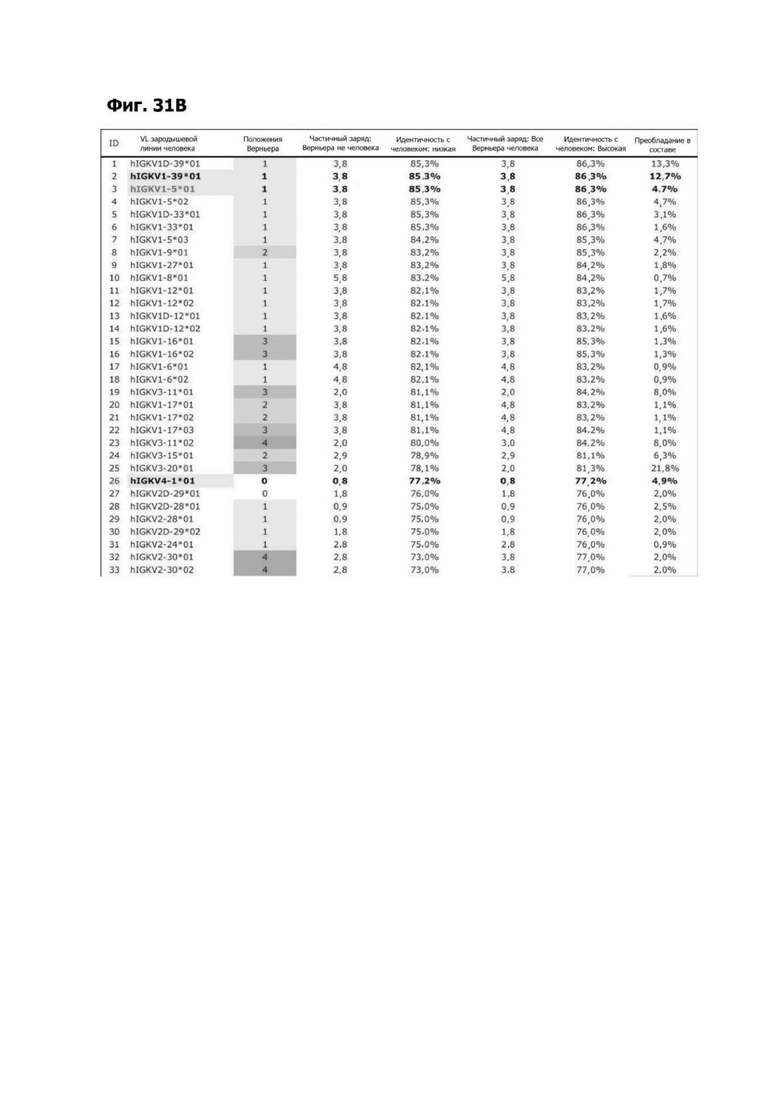
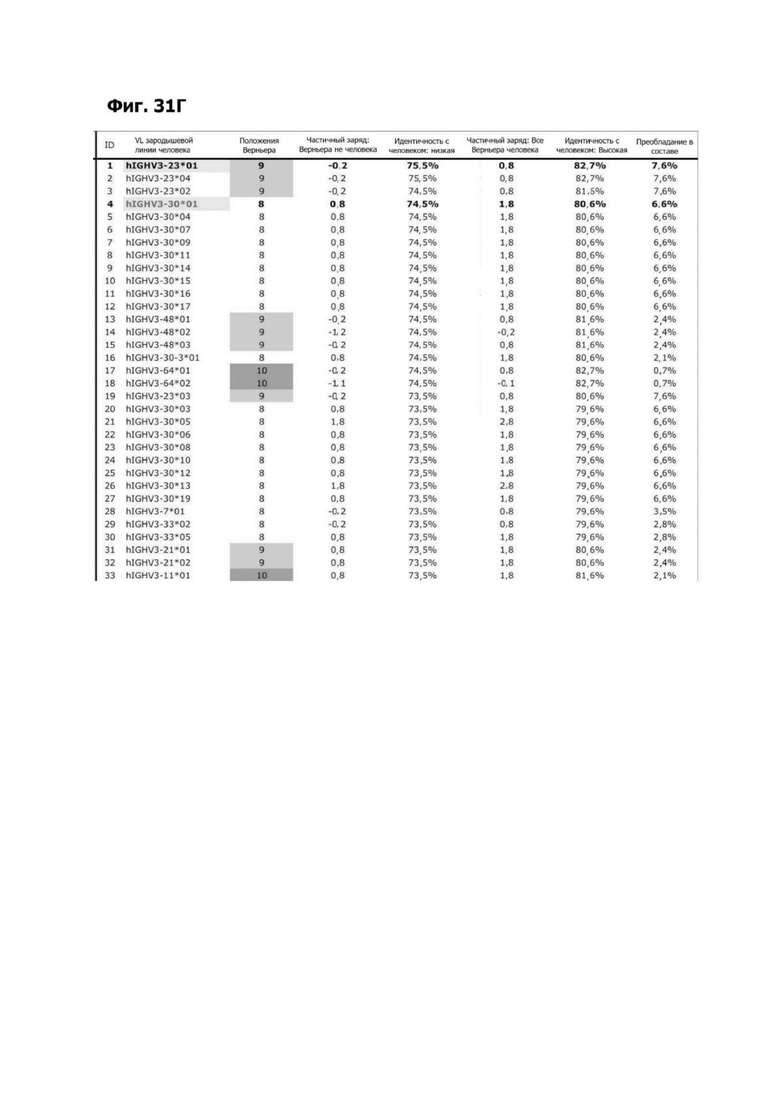
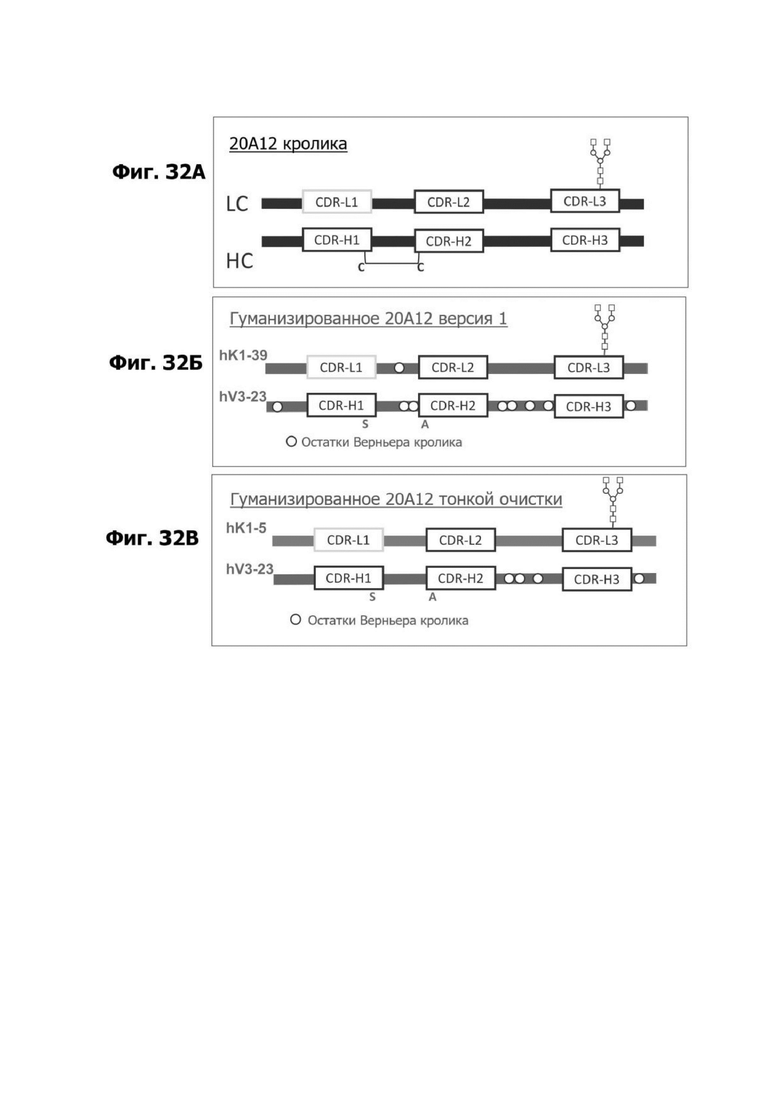
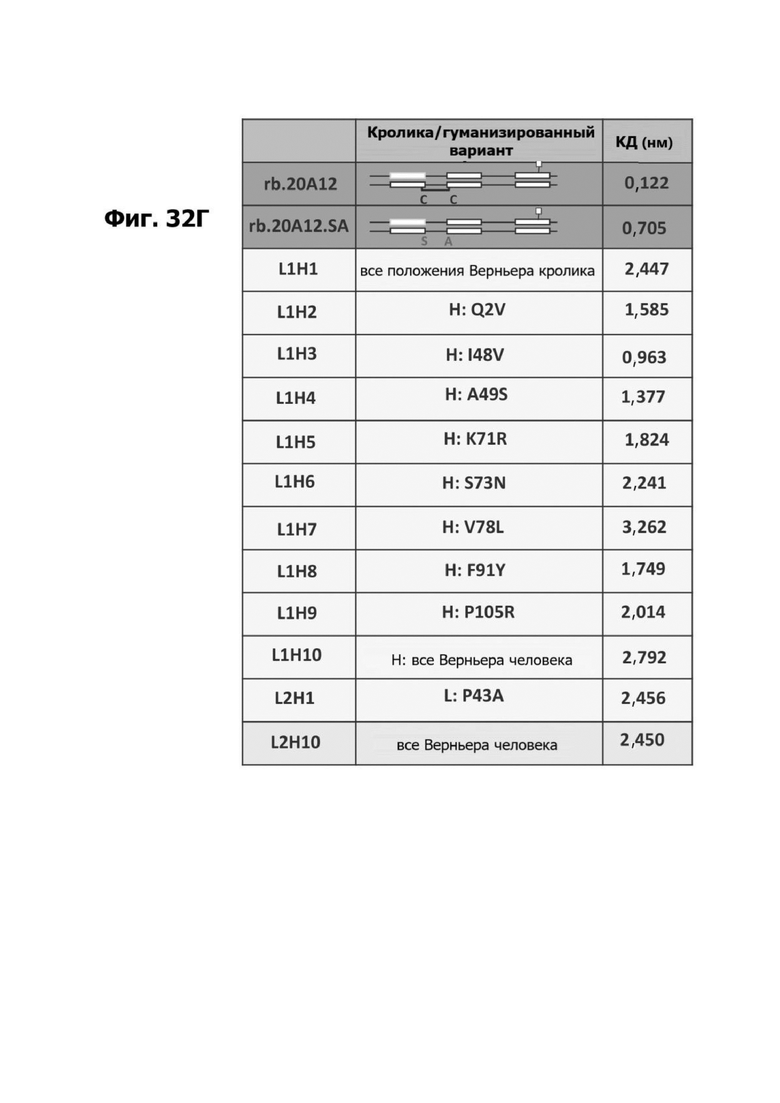
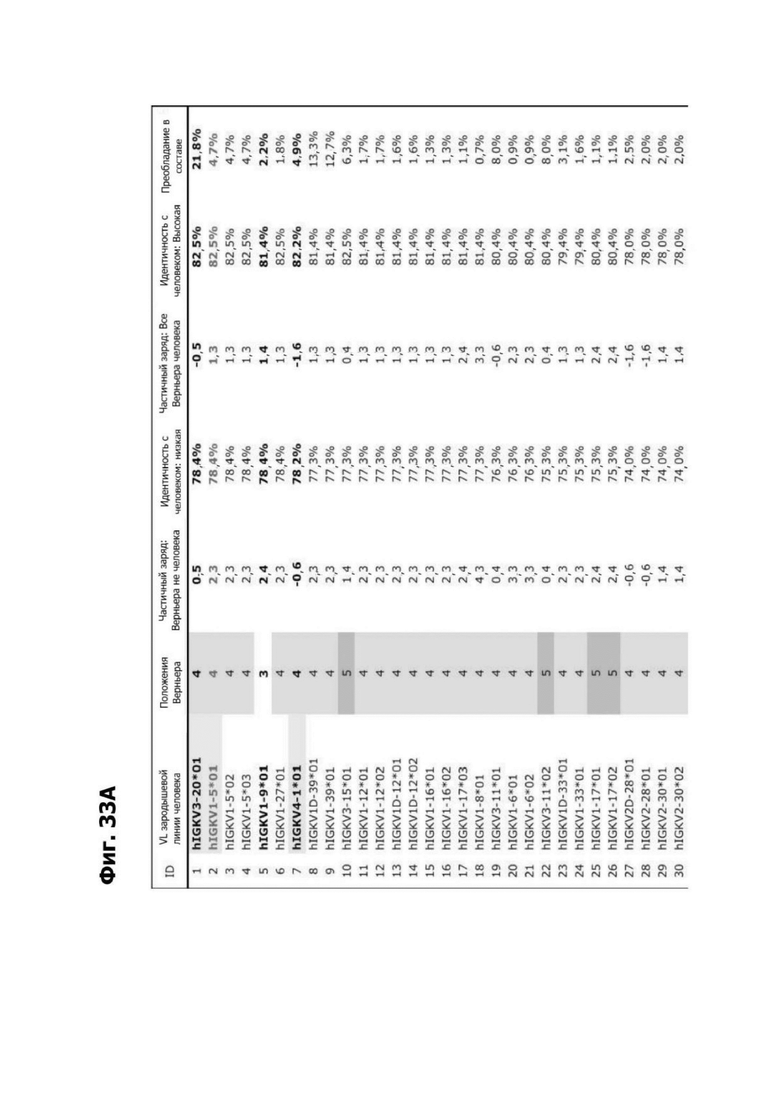

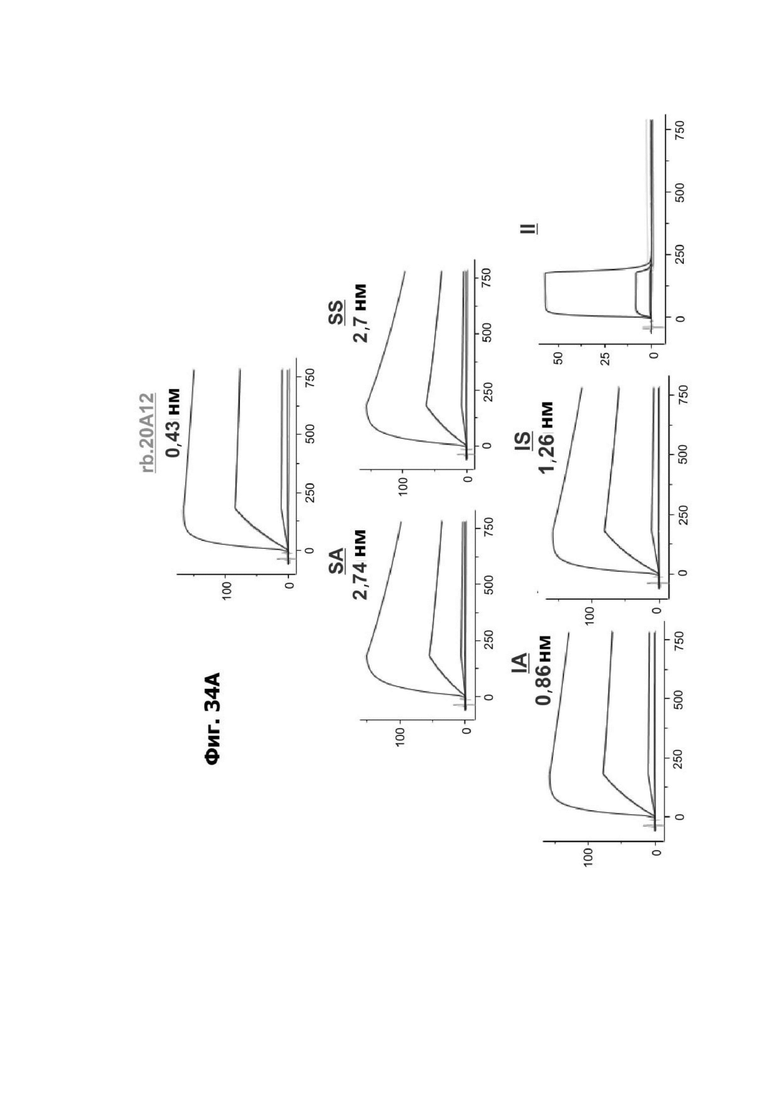
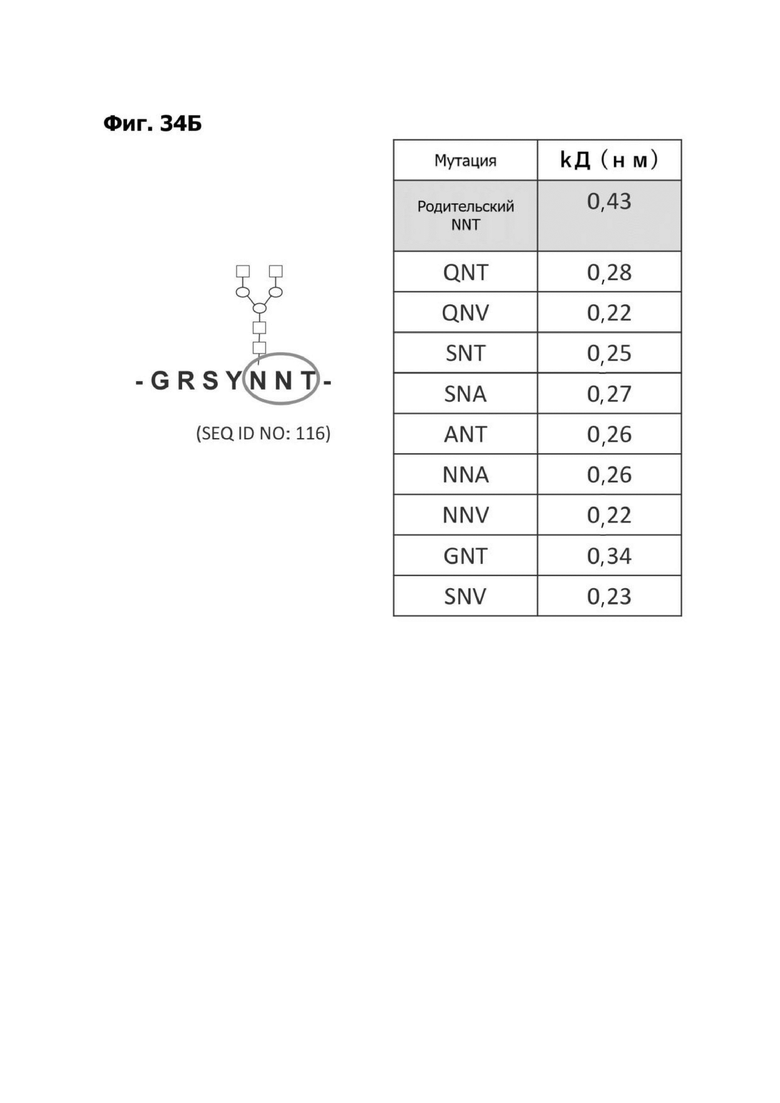
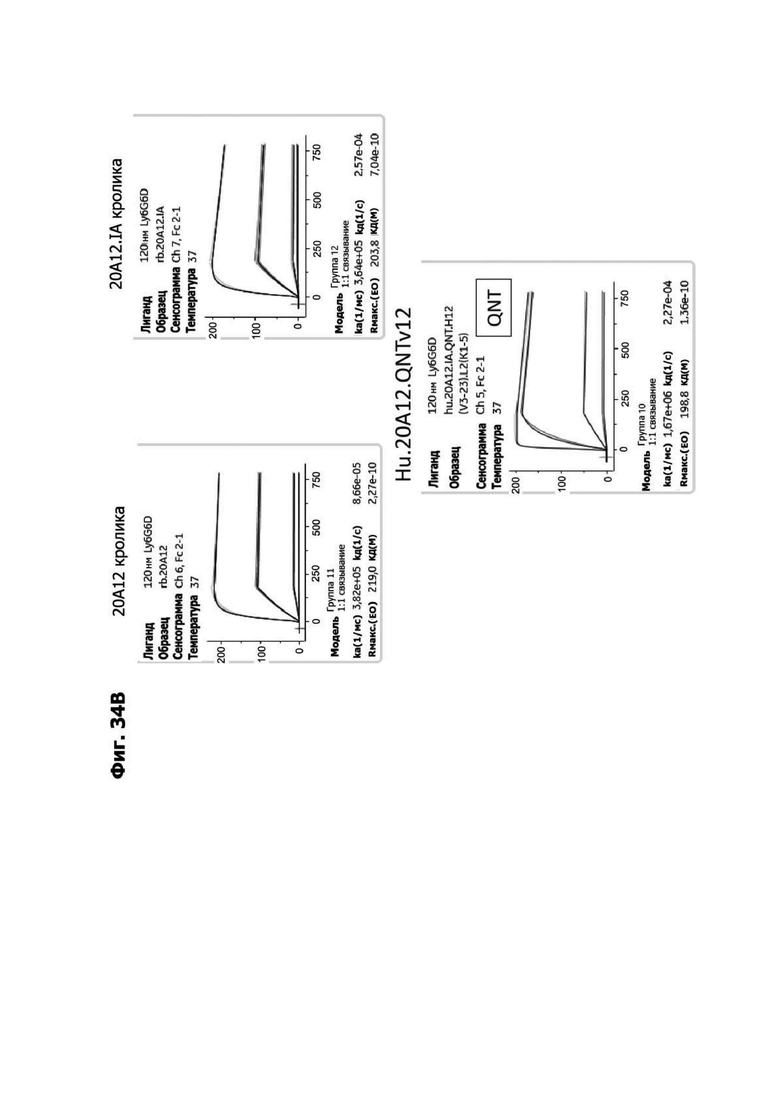
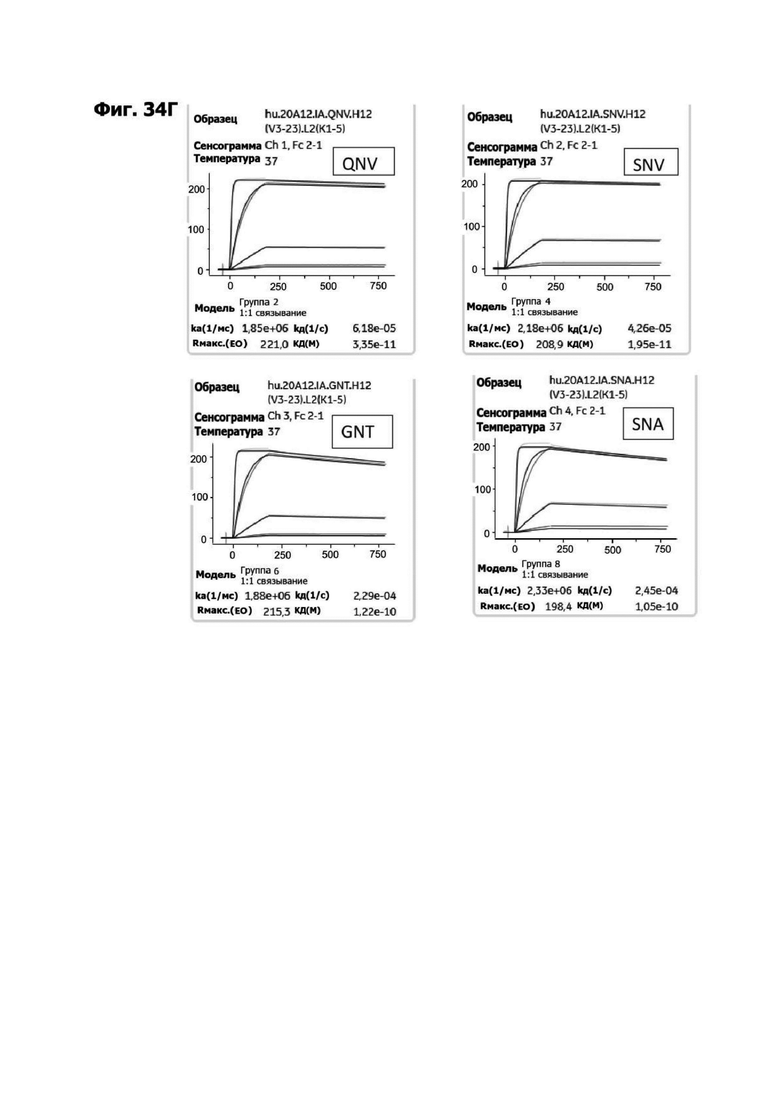
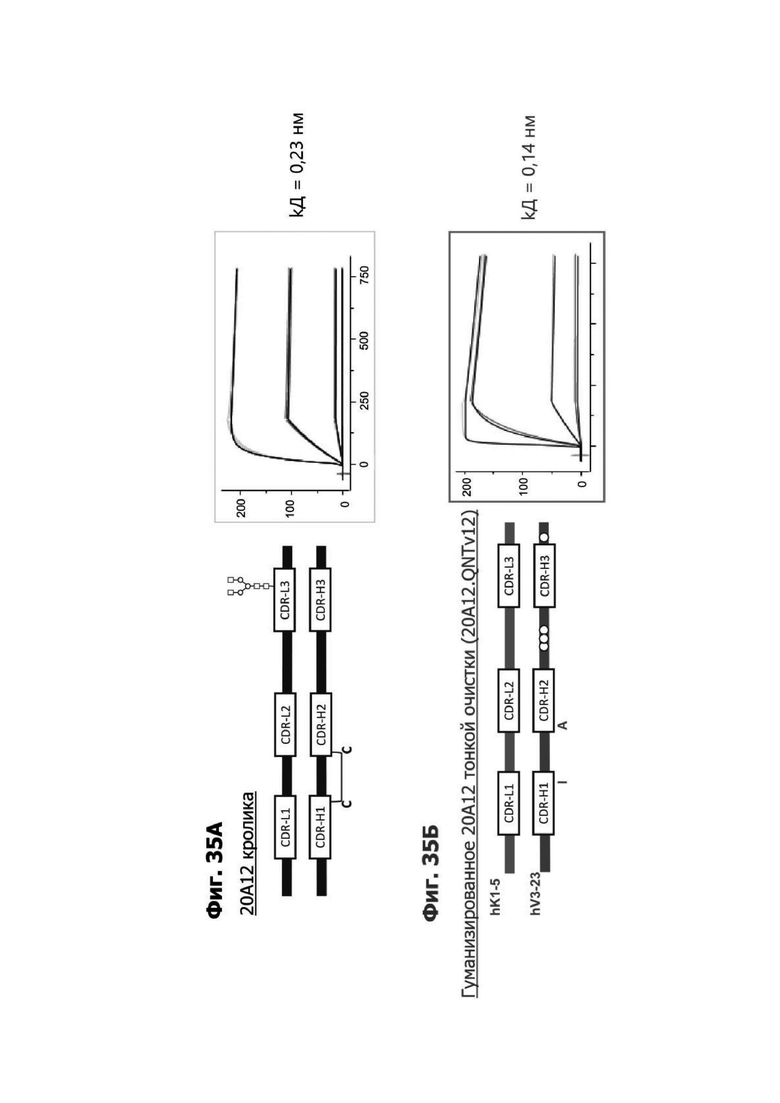
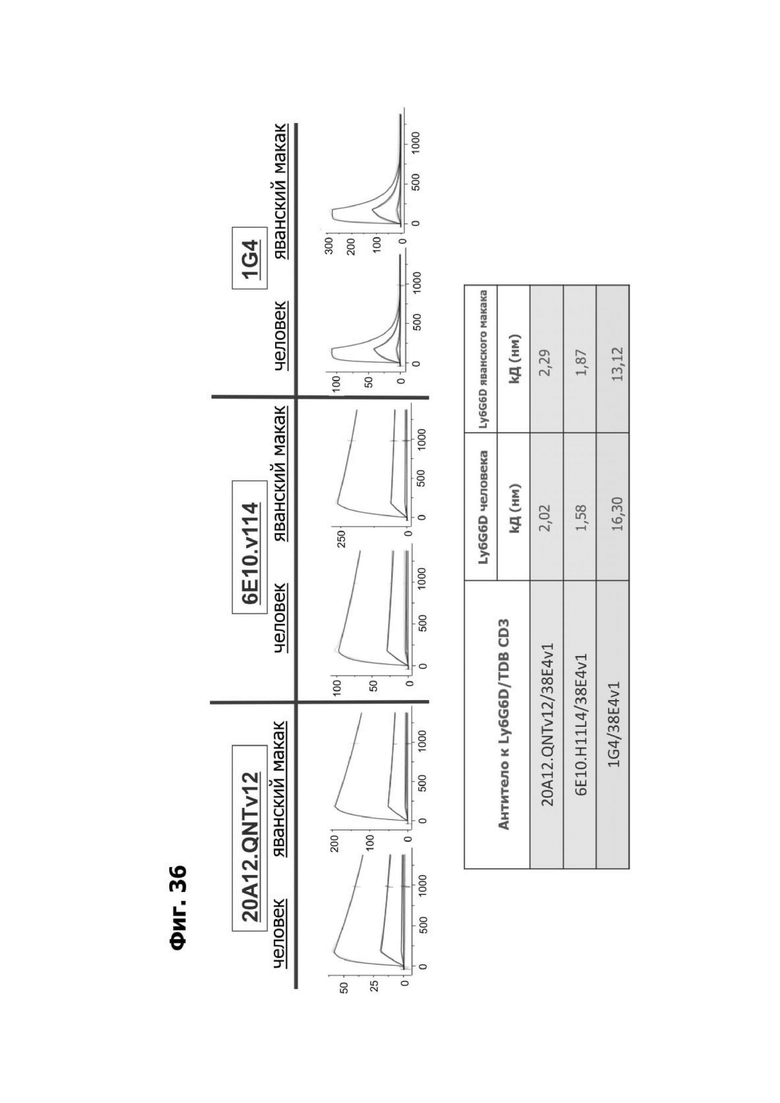
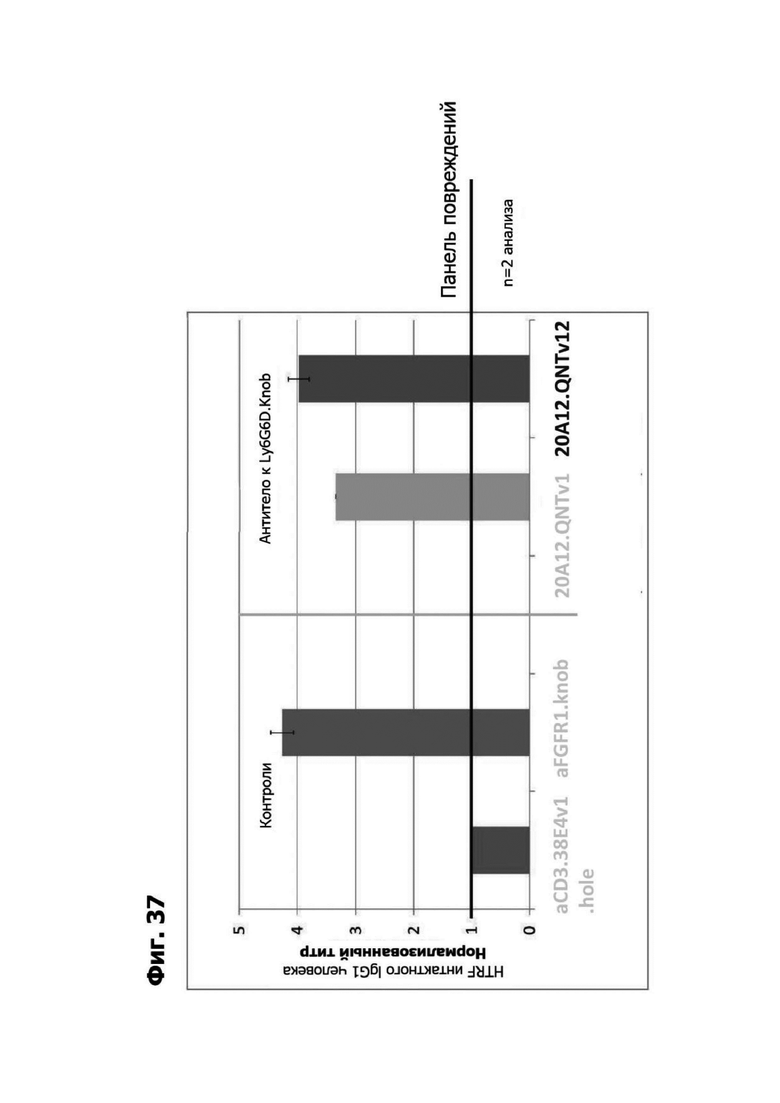


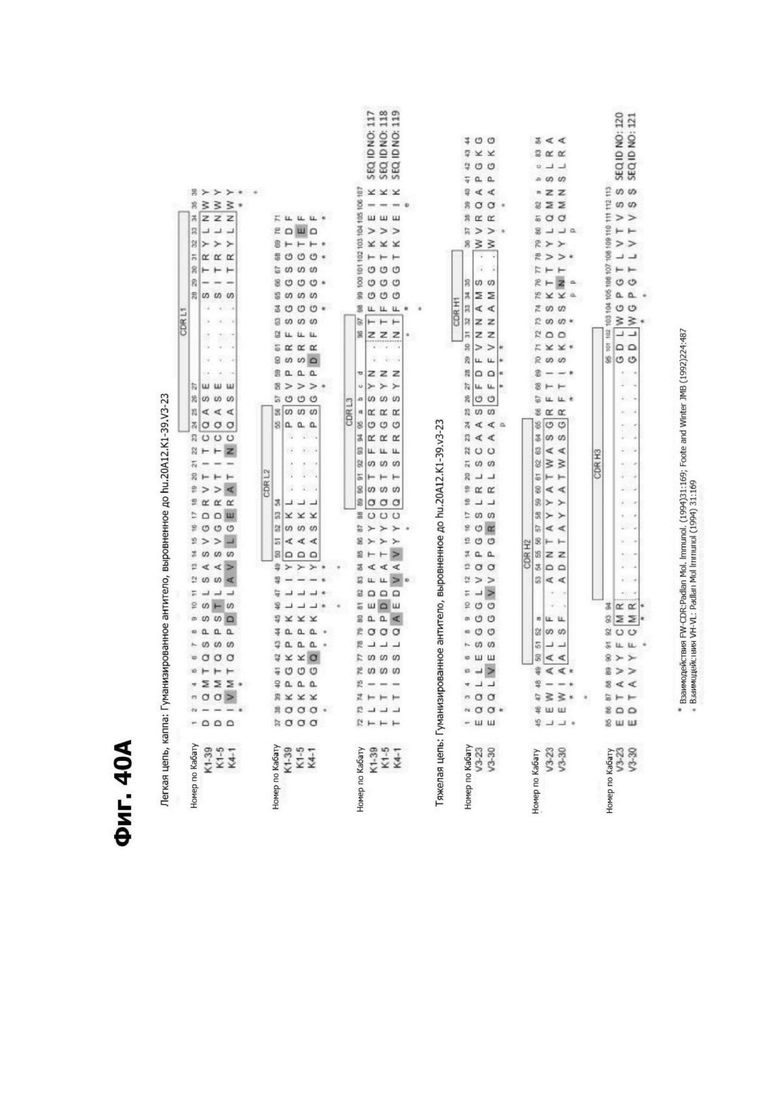
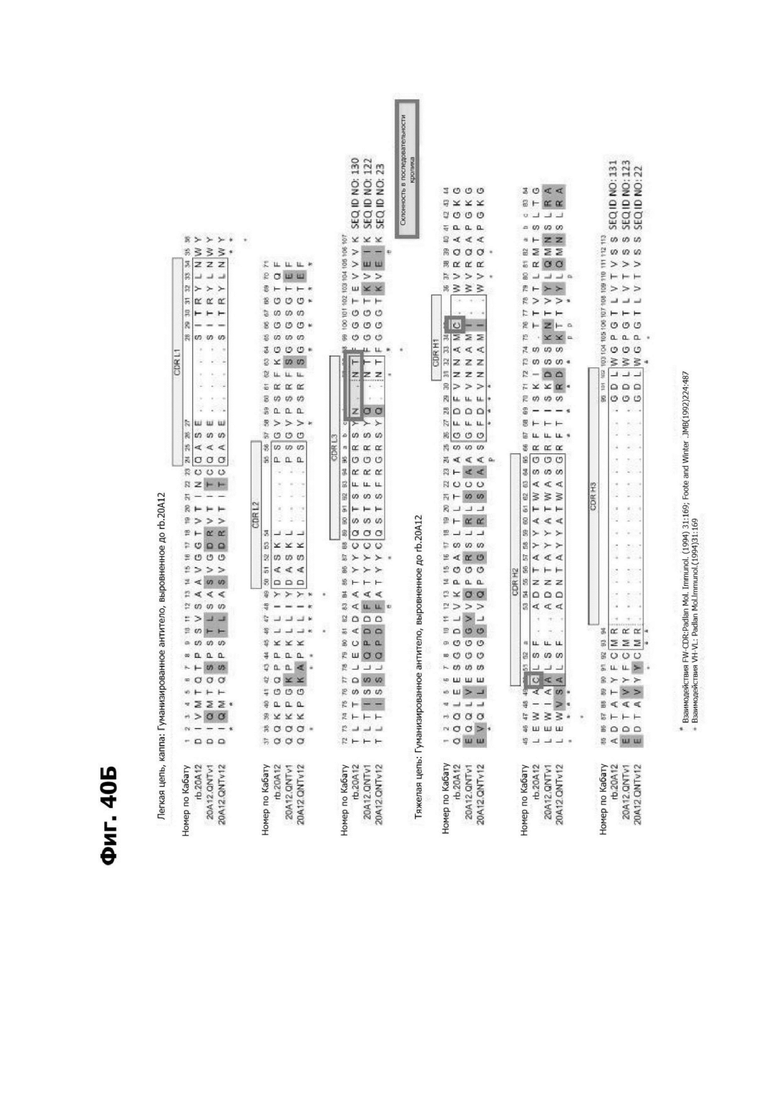
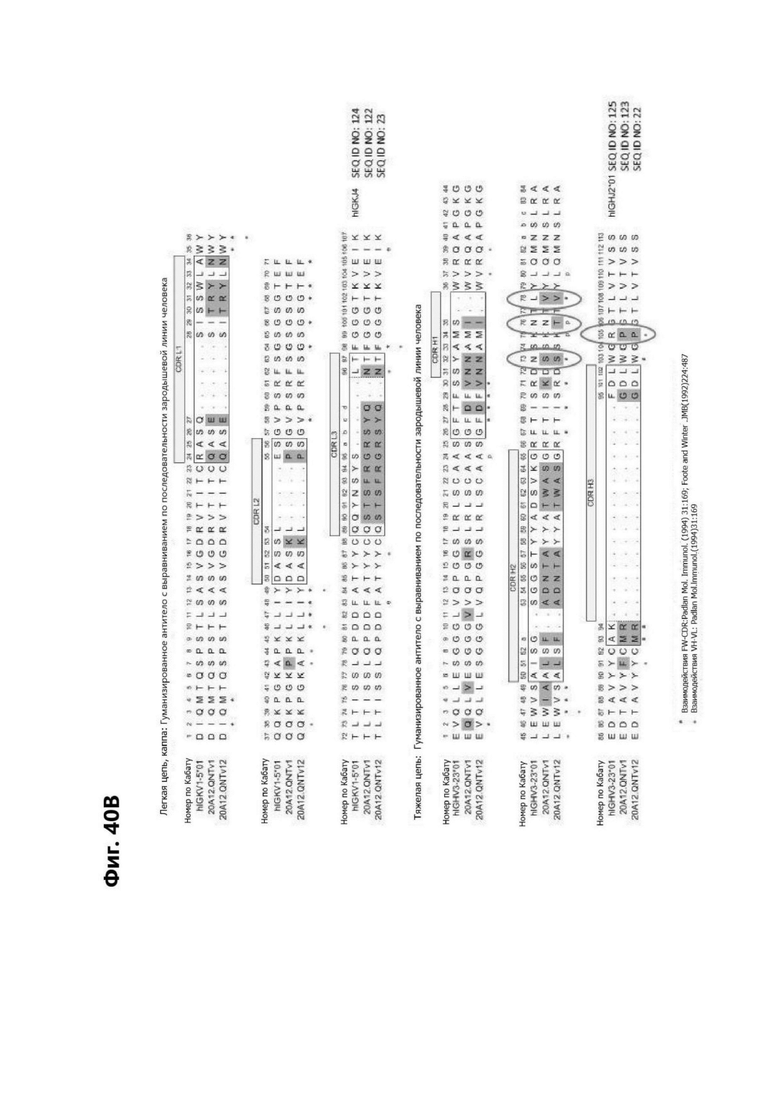

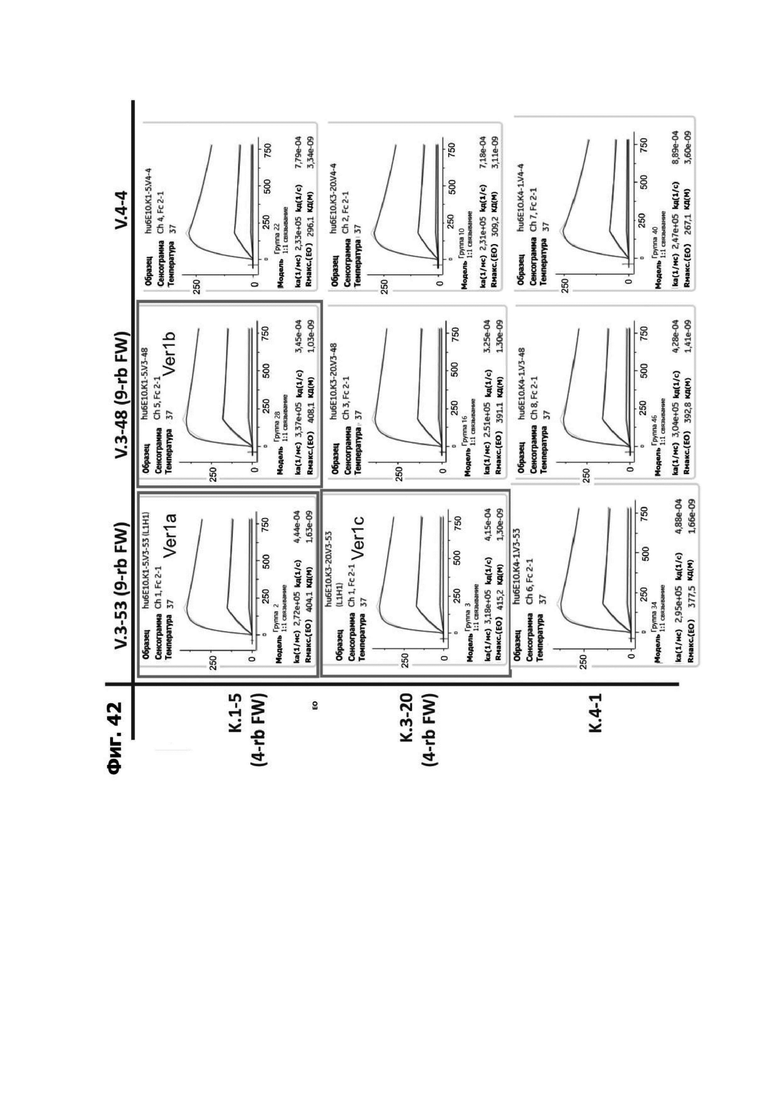

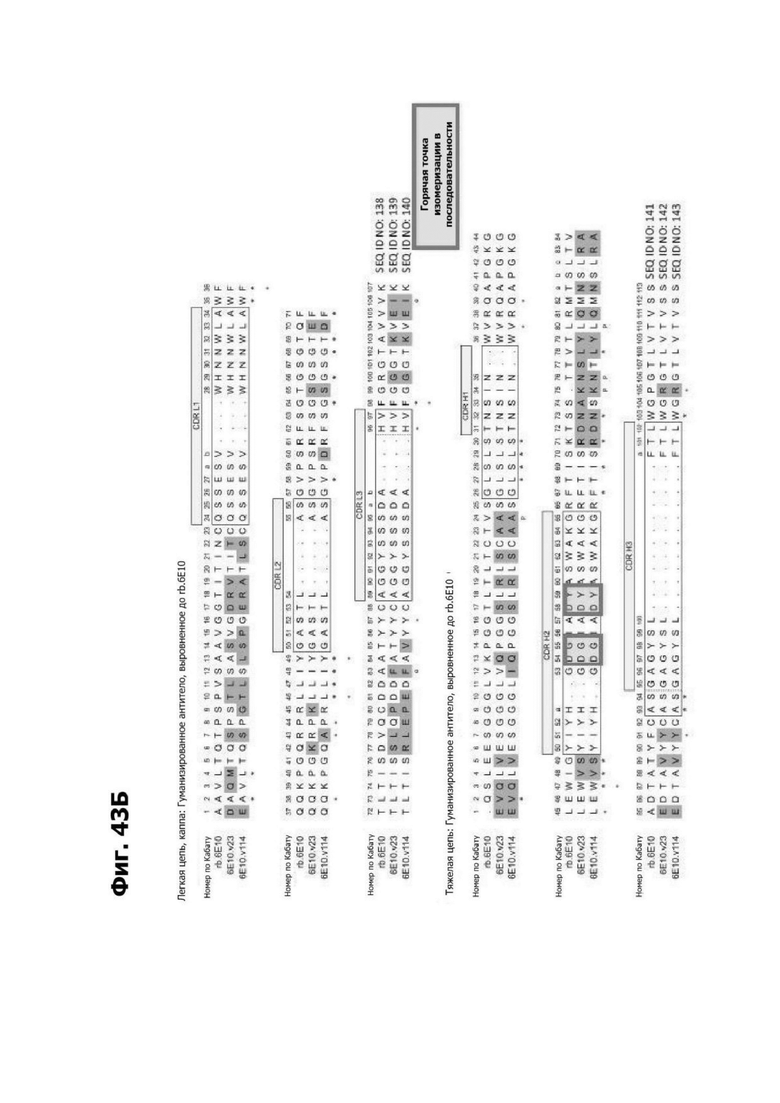
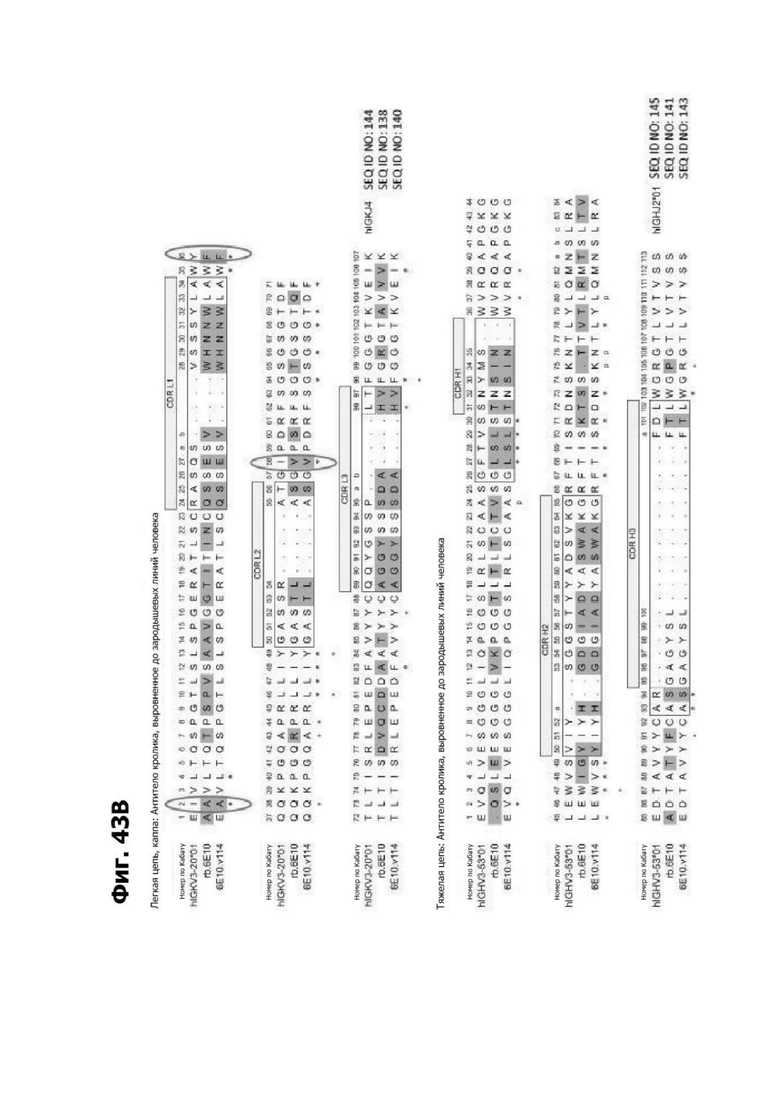
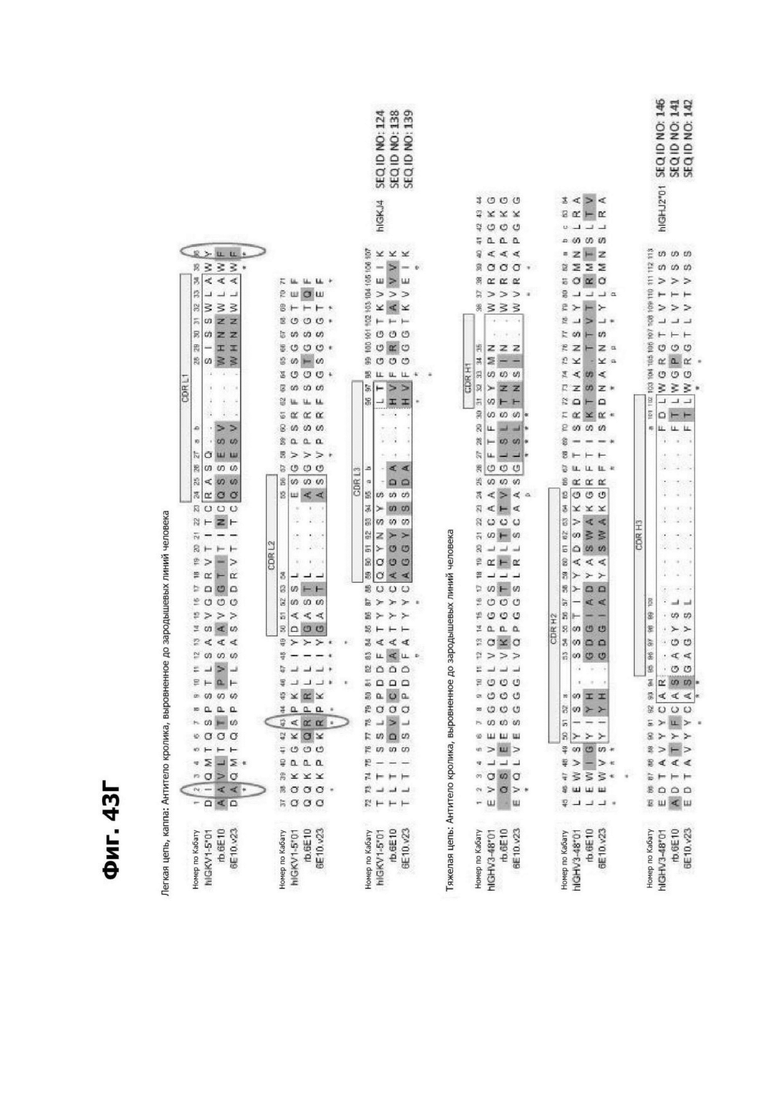
Изобретение относится к области биотехнологии, в частности к биспецифическому антителу, которое связывается с членом G6D семейства лимфоцитарных антигенов 6 (LY6G6D) и кластером дифференциации CD3, к композиции, его содержащей, а также к способу его получения. Также раскрыты выделенная нуклеиновая кислота, кодирующая вышеуказанное антитело, а также клетка и вектор, ее содержащие. Изобретение эффективно для лечения LY6G6D-положительного рака у субъекта. 11 н. и 32 з.п. ф-лы, 43 ил., 11 табл., 9 пр.
1. Биспецифическое антитело, которое связывается с членом G6D семейства лимфоцитарных антигенов 6 (LY6G6D) и кластером дифференциации CD3, где биспецифическое антитело содержит LY6G6D-связывающий домен, содержащий полипептид тяжелой цепи (H1) и полипептид легкой цепи (L1), и CD3-связывающий домен, содержащий полипептид тяжелой цепи (Н2) и полипептид легкой цепи (L2), где каждая H1 и Н2 содержит вариабельный домен тяжелой цепи (VH) и константный домен тяжелой цепи (СН1) и где каждая L1 и L2 содержит вариабельный домен легкой цепи (VL) и константный домен легкой цепи (CL1), где:
(а) LY6G6D-связывающий домен содержит следующие шесть CDRs:
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4;
(ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5;
(iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
(iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 1;
(v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 2; и
(vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 3 и
(б) CD3-связывающий домен содержит следующие шесть CDRs:
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 15;
(ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 16;
(iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 17;
(iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 12;
(v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и
(vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 14.
2. Биспецифическое антитело по п. 1, в котором
(а) СН1 из H1 содержит аминокислотную замену в S183 и CL из L1 содержит аминокислотную замену в положении V133 и/или СН1 из Н2 содержит аминокислотную замену в положении S183 и CL из L2 содержит аминокислотную замену в V133, где замененный остаток в положении S183 в СН1 из H1 формирует зарядную пару с знаменным остатком в положении V133 в CL из L1 и/или замененный остаток в положении S183 в СН1 из Н2 формирует зарядную пару с знаменным остатком в положении V133 в CL из L2, где нумерация положений S183 и V133 соответствует системе нумерации EU;
и
(б) VH из H1 содержит аминокислотную замену в положении Q39 и VL из L1 содержит аминокислотные замены в положении Q38 и/или VH из Н2 содержит аминокислотную замену в положении Q39 и VL из L2 содержит аминокислотную замену в положении Q38, где замененный остаток в положении Q39 в VH из H1 формирует зарядную пару с знаменным остатком в положении Q38 в VL из L1 и/или замененный остаток в положении Q39 в VH из Н2 формирует зарядную пару с знаменным остатком в положении Q38 в VL из L2, где нумерация положений Q39 и Q38 соответствует системе нумерации Кабат.
3. Биспецифическое антитело по п. 2, в котором
(а) СН1 из H1 содержит мутацию S183K; CL из L1 содержит мутацию V133E; СН1 из Н2 содержит мутацию S183E и CL из L2 содержит мутацию V133K,
где нумерация положений S183 и V133 соответствует системе нумерации EU;
(б) VH из H1 содержит мутацию Q39E; VL из L1 содержит мутацию Q38K; VH из Н2 содержит мутацию Q39K; и VL из L2 содержит мутацию Q38E, где нумерация положений Q39 и Q38 соответствует системе нумерации Кабат.
4. Биспецифическое антитело по п. 2, в котором
(а) СН1 из H1 содержит мутацию S183E и CL из L1 содержит мутацию V133K и СН1 из Н2 содержит мутацию S183K и CL из L2 содержит мутацию V133E, где нумерация положений S183 и V133 соответствует системе нумерации EU;
и
(б) VH из H1 содержит мутацию Q39K и VL из L1 содержит мутацию Q38E, VH из Н2 содержит мутацию Q39E и VL из L2 содержит мутацию Q38K, где нумерация положений Q39 и Q38 соответствует системе нумерации Кабат.
5. Биспецифическое антитело по любому из пп. 1-4, в котором аминокислотный остаток в S43 в VL из L2 заменен пролином и где нумерация положения S43 соответствует системе нумерации Кабат.
6. Биспецифическое антитело по любому из пп. 1-3 и 5, в котором
VH1 содержит аминокислотную последовательность SEQ ID NO: 10,
VL1 содержит аминокислотную последовательность SEQ ID NO: 11;
VH2 содержит аминокислотную последовательность SEQ ID NO: 20 и
VL2 содержит аминокислотную последовательность SEQ ID NO: 21.
7. Биспецифическое антитело по любому из пп. 1, 2, 4 и 5, в котором
VH1 содержит аминокислотную последовательность SEQ ID NO: 59,
VL1 содержит аминокислотную последовательность SEQ ID NO: 60;
VH2 содержит аминокислотную последовательность SEQ ID NO: 89 и
VL2 содержит аминокислотную последовательность SEQ ID NO: 90.
8. Биспецифическое антитело по любому из пп. 1-7, причем антитело является полноразмерным антителом.
9. Биспецифическое антитело по любому из пп. 1-8, в котором каждый из первого домена СН3 (СН31) Fc-области H1 и второго домена СН3 (СН32) Fc-области Н2 содержит выступ или впадину, и причем выступ или впадина в СН31 может располагаться во впадине или выступе соответственно в СН32.
10. Биспецифическое антитело по п. 9, в котором СН31 и СН32 сходятся на поверхности между выступом и впадиной.
11. Биспецифическое антитело по п. 9 или 10, в котором СН31 Fc-области H1 содержит выступ и СН32 Fc-области Н2 содержит впадину.
12. Биспецифическое антитело по любому из пп. 9-11, в котором (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотной замены T366W, где нумерация положения Т336 соответствует системе нумерации EU; (б) СН32 Fc-области Н2 содержит впадину, содержащую мутацию аминокислотной замены T366S, L368A или Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU, или их комбинацию; или (в) как (а), так и (б).
13. Биспецифическое антитело по п. 12, в котором (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотой замены T366W, где нумерация положения Т336 соответствует системе нумерации EU; (б) СН32 Fc-области Н2 содержит впадину, содержащую мутации аминокислотной замены T366S, L368A и Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU; или (в) как (а), так и (б).
14. Биспецифическое антитело по п. 13, в котором (а) СН31 Fc-области H1 содержит выступ, содержащий мутацию аминокислотой замены T366W, где нумерация положения Т336 соответствует системе нумерации EU, и (б) СН32 Fc-области Н2 содержит впадину, содержащую мутации аминокислотой замены T366S, L368A и Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU.
15. Биспецифическое антитело по п. 9 или 10, в котором СН31 Fc-области H1 содержит впадину и СН32 Fc-области Н2 содержит выступ.
16. Биспецифическое антитело по любому из пп. 9, 10 или 15, в котором (а) СН31 Fc-области H1 содержит впадину, содержащую мутацию аминокислотой замены T366S, L368A или Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU, или их комбинацию; (б) СН32 Fc-области Н2 содержит выступ, содержащий мутацию аминокислотой замены T366W, где нумерация положения Т336 соответствует системе нумерации EU; или (в) как (а), так и (б).
17. Биспецифическое антитело по п. 16, в котором (а) СН31 Fc-области H1 содержит впадину, содержащую мутации аминокислотой замены T366S, L368A и Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU; (б) СН32 Fc-области Н2 содержит выступ, содержащий мутацию аминокислотой замены T366W, где нумерация положения Т336 соответствует системе нумерации EU; или (в) как (а), так и (б).
18. Биспецифическое антитело по п. 17, в котором (а) СН31 Fc-области H1 содержит впадину, содержащую мутации аминокислотой замены T366S, L368A и Y407V, где нумерация положений Т366, L368 и Y407 соответствует системе нумерации EU, и (б) СН32 Fc-области второго полипептида тяжелой цепи содержит выступ, содержащий мутацию аминокислотой замены T366W, где нумерация положения Т336 соответствует системе нумерации EU.
19. Биспецифическое антитело по любому из пп. 9-18, в котором Fc-области являются областями Fc изотипа IgG человека или их вариантами Fc-области.
20. Биспецифическое антитело по п. 19, в котором Fc-области являются вариантами Fc-области изотипа IgG человека.
21. Биспецифическое антитело по п. 20, в котором каждый из вариантов Fc-области изотипа IgG человека содержит мутацию в аминокислотном остатке N297, что приводит к отсутствию гликозилирования, где нумерация положения N297 соответствует системе нумерации EU.
22. Биспецифическое антитело по п. 21, в котором мутация в аминокислотном остатке N297 представляет собой мутацию по типу замены.
23. Биспецифическое антитело по п. 21 или 22, в котором мутация в аминокислотном остатке N297 снижает эффекторную функцию Fc-области.
24. Биспецифическое антитело по п. 21 или 23, в котором мутация по типу замены представляет собой мутацию N297G или N297A.
25. Биспецифическое антитело по п. 24, в котором каждый из вариантов Fc-области изотипа IgG человека содержит мутацию N297G.
26. Биспецифическое антитело по любому из пп. 1-3, 5, 6, 8-14 и 19-25, где
(а) H1 содержит аминокислотную последовательность SEQ ID NO: 7;
(б) L1 содержит аминокислотную последовательность SEQ ID NO: 9;
(в) Н2 содержит аминокислотную последовательность SEQ ID NO: 18; и
(г) L2 содержит аминокислотную последовательность SEQ ID NO: 19.
27. Биспецифическое антитело по любому из пп. 1, 2, 4, 5, 7-14 и 19-25, где:
(а) H1 содержит аминокислотную последовательность SEQ ID NO: 64;
(б) L1 содержит аминокислотную последовательность SEQ ID NO: 65;
(в) Н2 содержит аминокислотную последовательность SEQ ID NO: 69; и
(г) L2 содержит аминокислотную последовательность SEQ ID NO: 70.
28. Биспецифическое антитело по любому из пп. 1-3, 5, 6, 8-10 и 15-25, где:
(а) H1 содержит аминокислотную последовательность SEQ ID NO: 8;
(б) L1 содержит аминокислотную последовательность SEQ ID NO: 9;
(в) Н2 содержит аминокислотную последовательность SEQ ID NO: 67; и
(г) L2 содержит аминокислотную последовательность SEQ ID NO: 19.
29. Биспецифическое антитело по любому из пп. 1, 2, 4, 5, 7-10 и 15-25, где:
(а) H1 содержит аминокислотную последовательность SEQ ID NO: 66;
(б) L1 содержит аминокислотную последовательность SEQ ID NO: 65;
(в) Н2 содержит аминокислотную последовательность SEQ ID NO: 68; и
(г) L2 содержит аминокислотную последовательность SEQ ID NO: 70.
30. Выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по любому из пп. 1-29.
31. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 30,
где вектор экспрессии предназначен для продуцирования биспецифического антитела по любому из пп. 1-29.
32. Клетка-хозяин, предназначенная для продуцирования биспецифического антитела по любому из пп. 1-29, содержащая вектор по п. 31.
33. Способ получения биспецифического антитела, которое связывает LY6G6D и CD3, причем способ включает культивирование клетки-хозяина по п. 32 в культуральной среде.
34. Композиция, предназначенная для лечения LY6G6D-положительного рака и включающая эффективное количество биспецифического антитела по любому из пп. 1-29, а также фармацевтически приемлемый носитель или разбавитель.
35. Применение биспецифического антитела по любому из пп. 1-29 в качестве лекарственного средства для лечения LY6G6D-положительного рака у субъекта, нуждающегося в этом.
36. Применение биспецифического антитела по любому из пп. 1-29 для производства лекарственного средства для лечения LY6G6D-положительного рака у субъекта, нуждающегося в этом.
37. Применение биспецифического антитела по любому из пп. 1-29 для производства лекарственного средства для замедления прогрессирования LY6G6D-положительного рака у субъекта, нуждающегося в этом.
38. Применение по п. 36 или 37, где LY6G6D-положительный рак у субъекта является LY6G6D-положительным колоректальным раком (CRC).
39. Применение по п. 38, в котором LY6G6D-положительный колоректальный рак имеет статус микросателлитной нестабильности (MSI): микросателлитная стабильность (MSS) или низкий уровень микросателлитной нестабильности (MSI-L).
40. Способ лечения LY6G6D-положительного рака у субъекта, нуждающегося в этом, причем способ включает введение субъекту биспецифического антитела по любому из пп. 1-29.
41. Способ замедления прогрессирования LY6G6G-положительного рака у субъекта, нуждающегося в этом, причем способ включает введение субъекту биспецифического антитела по любому из пп. 1-29.
42. Способ по п. 40 или 41, в котором LY6G6G-положительный рак представляет LY6G6G-положительный CRC.
43. Способ по п. 42, в котором LY6G6D-положительный рак имеет статус MSI: MSS или MSI-L.
| US 2014221244 A1, 07.08.2014 | |||
| WO 2016204966 A1, 22.12.2016 | |||
| GUIDO GIORDANO et al., JAK/Stat5-mediated subtype-specific lymphocyte antigen 6 complex, locus G6D (LY6G6D) expression drives mismatch repair proficient colorectal cancer, J Exp Clin Cancer Res, Published online 2019 Jan 22, Vol | |||
| Способ сужения чугунных изделий | 1922 |
|
SU38A1 |
| БЕЛЯКОВ И.С | |||
| и др., Экзон-интронная структура гена | |||

