Область техники, к которой относится изобретение
Изобретение относится к экспериментальной биологии, а именно к экспериментальной биохимии и молекулярной физиологии, и может быть использовано для выявления профиля активности пептидов, имитирующих N-концевой фрагмент рецепторов, активируемых протеазами, 1 типа, на основе зависимости их эффектов от активности β-аррестина-2 в экспериментальных условиях, моделирующих ишемические.
Уровень техники
В настоящее время показано, что рецепторы, активируемые протеазами, 1 типа (ПАР1) регулируют множество физиологических процессов. В частности, установлено, что ПАР1 экспрессируются различными клетками нервной ткани: нейронами, астроцитами, а также эндотелием сосудов мозга и регулируют их функционирование как в условиях нормы, так и в условиях патологии (Ossovskaya V.S., Bunnett N.W., 2004. Protease-activated receptors: contribution to physiology and disease // Physiol. Rev. V. 84. P. 579–621; Traynelis S.F., Trejo J., 2007. Protease-activated receptor signaling: new roles and regulatory mechanisms // Curr. Opin. Hematol. V. 14. P. 230–235).
С этой точки зрения представляется особенно важным изучение роли активации ПАР1 в условиях развивающегося ишемического инсульта, поскольку данная патология по данным Всемирной Организации Здравоохранения является лидирующей причиной смертности и инвалидизации населения, уступая лишь ишемической болезни сердца. Вместе с тем, несмотря на распространенность ишемии головного мозга, набор лекарственных препаратов, улучшающих исход и течение инсульта, крайне ограничен.
ПАР1 относятся к семейству семидоменных трансмембранных рецепторов, сопряженных с G-белками (GPCRs), а в качестве их основных эндогенных активаторов рассматриваются сериновые протеазы гемостаза, тромбин и активированный протеин С (АПС). Внеклеточная N-концевая последовательность ПАР1 содержит пептидную связь «активационного сайта», которая расщепляется сериновой протеазой. При этом различные протеазы характеризуются своим уникальным сайтом расщепления. Вновь образованный N-конец, так называемый «привязанный лиганд», взаимодействуя со второй внеклеточной петлей рецептора, приводит к изменению конформации молекулы ПАР1, активации рецептора и запуску внутриклеточного сигнала (Ossovskaya V.S., Bunnett N.W., 2004. Protease-activated receptors: contribution to physiology and disease // Physiol. Rev. V. 84. P. 579–621).
Ряд исследований указывают на важнейшую роль ПАР1 в развитии ишемического инсульта головного мозга. При этом описанный механизм протеолитической активации ПАР1 предопределяет запуск различных внутриклеточных путей. В случае расщепления ПАР1 тромбином (между Арг41 и Сер42) происходит активация Gq- или G12/13-белка, следующие за этим мобилизация внутриклеточных ионов кальция, повышение активности протеинкиназы С и ГТФазы RhoA. Описанный внутриклеточный каскад ведет к гибели нейронов, активации астроцитов и микроглии, а также к нарушению целостности эндотелиальных барьеров, что усугубляет тяжесть экспериментальной ишемии (Griffin J.H., Zlokovic B. V., Mosnier L.O., 2015. Activated protein C: Biased for translation // Blood. V. 125. P. 2898–2907). При этом белок β-аррестин-2, связываясь с ПАР1, после фосфорилирования его внутриклеточных петель по специфическим сайтам киназами GPCRs, препятствует дальнейшей активации G-белков путем инициации десенситизации и интернализации рецептора (Ossovskaya V.S., Bunnett N.W., 2004. Protease-activated receptors: contribution to physiology and disease // Physiol. Rev. V. 84. P. 579–621; Traynelis S.F., Trejo J., 2007. Protease-activated receptor signaling: new roles and regulatory mechanisms // Curr. Opin. Hematol. V. 14. P. 230–235).
После расщепления ПАР1 при помощи АПС (Арг46-Асн47), напротив, наблюдается реализация цитопротекторных сигнальных путей. Так, АПС посредством активации ПАР1 способствует выживанию нейронов, снижению уровня активации астроцитов, а также поддержанию целостности эндотелиальных барьеров в условиях, имитирующих ишемические. В этом случае β-аррестин-2 после АПС-опосредованной активации ПАР1 рекрутирует ряд эффекторных белков, активирующих ГТФазу Rac1, которая, в свою очередь, вовлечена в регуляцию множества протекторных внутриклеточных каскадов. В этом случае для активации ПАР1 с помощью АПС необходима колокализация рецептора с эндотелиальным рецептором протеина С и тромбомодулином в кавеоле, т.е. требуется особое «молекулярное окружение» ПАР1 (Griffin J.H., Zlokovic B. V., Mosnier L.O., 2015. Activated protein C: Biased for translation // Blood. V. 125. P. 2898–2907; Горбачева Л.Р., Киселева Е.В., Савинкова И.Г., Струкова С.М., 2017. Новая концепция действия протеаз гемостаза на процессы воспаления, нейротоксичности и регенерации тканей // Биохимия. V. 82. P. 1018–1032).
Таким образом, активация ПАР1 может оказывать на развитие ишемии головного мозга прямо противоположное действие: в случае активации рецептора тромбином наблюдается ухудшение тяжести патологии, а для АПС-зависимой активации – напротив, облегчение. При этом в первом случае β-аррестин-2 препятствует поддержанию активного состояния ПАР1, а во втором – сам вовлечен в запуск внутриклеточного сигналинга. Однако использование сериновых протеаз гемостаза в качестве терапевтических средств представляется затруднительным ввиду их влияния на систему свертывания крови. В частности, исследования на животных, проводимые с целью тестирования АПС как средства для терапии ишемического инсульта, показали, что он обладает выраженным антикоагулянтным эффектом, что сопряжено с риском развития геморрагий (Williams P.D., Zlokovic B.., Griffin J.H., Pryor K.E., Davis T.P., 2012. Preclinical safety and pharmacokinetic profile of 3K3A-APC, a novel, modified activated protein C for ischemic stroke // Curr. Pharm. Des. V. 18. P. 4215–4222).
Вместе с тем описанный механизм активации ПАР делает возможным использовать пептиды, имитирующие «привязанный лиганд», в качестве активаторов рецепторов. В настоящее время предложено множество таких пептидов (Ramachandran R., Hollenberg M., 2008. Proteinases and signalling: pathophysiological and therapeutic implications via PARs and more // Br. J. Pharmacol. V. 153. P. 263–282). При этом показано, что некоторые пептиды, например, TRAP (группа пептидов, имитирующих N-конец ПАР1 после его расщепления тромбином) проявляют «тромбиновую» активность, приводя к нарушению целостности эндотелиальных барьеров (Bae J.S., Rezaie A.R., 2008. Protease activated receptor 1 (PAR-1) activation by thrombin is protective in human pulmonary artery endothelial cells if endothelial protein C receptor is occupied by its natural ligand // Thromb. Haemost. V. 100. P. 101–109) и активации астроцитов (Debeir T., Vigé X., Benavides J., 1997. Pharmacological characterization of protease-activated receptor (PAR-1) in rat astrocytes // Eur. J. Pharmacol. V. 323. P. 111–117). Некоторые пептиды, напротив, проявляют профиль активности схожий с таковым для АПС. Так, пептид TR47, имитирующий фрагмент «привязанного лиганда», высвобождаемого АПС, способствует поддержанию целостности эндотелиальных барьеров in vitro и in vivo (Mosnier L.O., Sinha R.K., Burnier L., Bouwens E.A., Griffin J.H., 2013. Biased agonism of protease-activated receptor 1 by activated protein C caused by noncanonical cleavage at Arg46 // Blood. V. 120. P. 5237–5246).
Таким образом, именно пептиды-агонисты могут рассматриваться как наиболее перспективные агенты для терапии ишемического инсульта, основанной на активации ПАР1 на различных клеточных компонентах нервной ткани. Однако в этом случае критически важным является определение профиля их активности, т.е. характера сигнального пути, по которому пептиды запускают активацию рецептора (тромбин- или АПС-подобный путь), в экспериментальных условиях. Именно это можно рассматривать как определяющую характеристику того или иного пептида, позволяющую считать его положительным или негативным регулятором развивающихся вследствие ишемии нарушений.
В настоящее время определение функциональной активности пептидов-агонистов ПАР1 осуществляется с помощью нескольких уже известных подходов. Наиболее распространенным методом служит определение активности пептидов на основе их аминокислотной последовательности. Принято считать, что пептиды, имитирующие последовательность «привязанного лиганда», высвобождаемого тромбином, запускают сигнальные каскады, характерные для тромбина (с участием протеинкиназы С, RhoA и внутриклеточных ионов кальция). Если же пептид является аналогом N-конца ПАР1, формирующегося после его АПС-зависимого расщепления, то тогда представляется наиболее вероятным демонстрация им профиля активности данной протеазы с вовлечением Rac1. Вместе с тем такой подход не всегда полностью применим.
Как уже было отмечено выше, запуск того или иного внутриклеточного сигнала после активации ПАР1 определяется не только видом «привязанного лиганда», но и тем молекулярным окружением, т.е. ансамблем сопряженных с рецептором белков, в котором он пребывает. Имеющиеся экспериментальные данные указывают на то, что именно такое окружение оказывается решающим в определении молекулярного каскада, реализующегося после активации ПАР1.
В частности, на культуре эндотелиальных клеток было показано, что тромбин через ПАР1 запускает цитопротекторные пути, аналогичные таковым, описанным для АПС. Это проявляется в стабилизации в 5 раз по сравнению с контрольными значениями проницаемости клеточного пласта, чьи барьерные свойства были нарушены добавлением липополисахарида в культуральную среду, после действия на него тромбина (Roy R.V., Ardeshirylajimi A., Dinarvand P., Yang L., Rezaie A.R., 2016. Occupancy of human EPCR by protein C induces β-arrestin-2 biased PAR1 signaling by both APC and thrombin // Blood. V. 128. P. 1884–1893). Кроме того, пептид из группы TRAP, как было продемонстрировано ранее, оказывал на нейроны протекторное, а не ожидаемое цитотоксическое действие (выживаемость клеток повышалась от 108% до 124% после действия пептида по сравнению с контрольным значением, принятым за 100%) (García P.S., Ciavatta V.T., Fidler J.A., Woodbury A., Levy J.H., Tyor W.R., 2015. Concentration-dependent dual role of thrombin in protection of cultured rat cortical neurons // Neurochem. Res. V. 40. P. 2220–2229).
Таким образом, определение профиля активности пептидов на основании сравнения их структуры с последовательностью «привязанного лиганда» ПАР1, высвобождаемого той или иной протеазой, представляет большую сложность, т.к. запускаемые молекулярные каскады зависят не только от самой структуры агониста, но и от наличия пространственной и сигнальной сопряженности ПАР1 с другими белками (например, с эндотелиальным рецептором протеина С и тромбомодулином, как было описано выше). Интерпретация полученных данных трудна, поскольку от исследователя требуется владение большим объемом специальных знаний о внутриклеточных сигнальных путях.
Помимо этого, в настоящее время разработаны различные способы модификации пептидов, изменяющие их способность проходить через различные тканевые барьеры, а также регулирующие их биодоступность (Bickel U., Yoshikawa T., Pardridge W.M., 2001. Delivery of peptides and proteins through the blood-brain barrier // Adv. Drug Deliv. V. 46. P. 247–279). С этой точки зрения, реализация описанного метода представляется затруднительной ввиду неполного структурного соответствия немодифицированных пептидов (полные аналоги фрагментов «привязанного лиганда») и молекул, подвергшихся химической модификации.
Еще один подход для определения характера эффектов пептидов, имитирующих N-концевой фрагмент ПАР1, заключается в качественном сравнении их наблюдаемых эффектов с эффектами, которые в аналогичных экспериментальных условиях демонстрируют АПС и тромбин. Данный подход позволяет учесть тот «молекулярный ансамбль», в котором находится ПАР1 на мембранах используемых в эксперименте клеток. Однако основным недостатком такого метода является большое количество экспериментальных групп, которые должны быть использованы в опыте. Так, чтобы выявить профиль активности пептида, имитирующего фрагмент «привязанного лиганда» ПАР1, высвобождаемого АПС, в первичных культурах кератиноцитов потребовалось формирование 8 экспериментальных групп: помимо группы, предполагавшей воздействие самого пептида, были сформированы группы, в которых на клеточные культуры воздействовали АПС в различных концентрациях (Киселева Е.В., Сидорова М.В., Горбачева Л.Р., Струкова С.М., 2014. Пептид-агонист рецептора, активируемого протеазами, стимулирует пролиферацию кератиноцитов и заживление раны эпителиального пласта подобно активированному протеину C // Биомедицинская химия. V. 60. P. 702–706). Очевидно, что для более полного тестирования изучаемых пептидов, необходимо включение в опыт и дополнительных групп, на которые будет осуществляться воздействие тромбином. Это приведет к использованию 10-12 групп, тогда как настоящее изобретение может быть осуществлено с использованием только 4-х экспериментальных групп.
Такое усложнение дизайна эксперимента увеличивает его продолжительность до 40-55 дней. Помимо этого, при росте числа экспериментальных групп возрастает в 2,5-3 раза и расход соответствующих реактивов (растворы АПС, тромбина и пептида, среда для культивирования, раствор фотосенсибилизатора и т.д.) и биоматериала (новорожденные крысы или мыши). Становится ясно, что при установлении профиля активности большого количества пептидов в режиме скрининга существующий подход ввиду его сложности и продолжительности является крайне неудобным.
Помимо этого, иногда используется метод блокады ПАР1 блокаторами или специфическими антителами. Такой подход, действительно, позволяет установить, являются ли эффекты изучаемых пептидов ПАР1-зависимыми. Так, с помощью специфического блокатора ПАР1 SCH 79797 было установлено, что его применение отменяет цитопротекторные эффекты АП9 в культурах нейронов при их кокультивировании с активированными тучными клетками (при добавлении АП9 относительная гибель нейронов составляла около 0,8, при совместно добавлении АП9 и SCH 79797 она возрастала до 1,2) (Babkina I.I., Strukova S.M., Pinelis V.G., Reiser G., Gorbacheva L.R., 2016. New synthetic peptide protects neurons from death induced by toxic influence of activated mast cells via protease-activated receptor // Biochem. V. 10. P. 126–134). Однако такой подход не позволяет установить характер тех каскадов, которые запускают изучаемые пептиды. Также стоит учитывать, что подавление активности ПАР1 само по себе оказывает протекторные эффекты в условиях ишемии. В частности, нокаут гена данного рецептора приводил к снижению размеров очага поражения мозга у мышей после окклюзии средней мозговой артерии (данный показатель составил 83,8±11,2 мм3 у контрольных животных и 27,1±6,1 мм3 у животных с нокаутом целевого гена) (Junge C.E., Sugawara T., Mannaioni G., Alagarsamy S., Conn P.J., Brat D.J., Chan P.H., Traynelis S.F., 2003. The contribution of protease-activated receptor 1 to neuronal damage caused by transient focal cerebral ischemia // PNAS. V. 100. P. 13019–13024). Все это делает получаемые данные недостаточными для однозначного определения профиля функциональной активности тестируемого пептида.
Таким образом, существует потребность в разработке простого, удобного, не требующего проведения большого количества экспериментов и надежного метода для определения характера каскадов, запускаемых пептидами, имитирующими «привязанный лиганд» ПАР1, в условиях, моделирующих ишемические. Разработка такого подхода значительно упростит поиск и создание таких пептидов, которые можно рассматривать в качестве перспективных нейропротекторных агентов.
Раскрытие изобретения
Задачей изобретения является разработка способа определения функциональной активности пептидов, имитирующих фрагменты «привязанного лиганда» рецепторов, активируемых протеазами, 1 типа (ПАР1), на основе зависимости их эффектов от наличия β-аррестина-2 в экспериментальных условиях, моделирующих ишемические.
Техническим результатом является тест-система, позволяющая определять функциональную активность пептидов за короткий промежуток времени (15-25 дней) с использованием минимального количества экспериментальных групп, реактивов и биоматериалов (в 2,5-3 раза меньшими по сравнению с известными способами) и характеризующаяся легкостью в интерпретации результатов, что не требует от исследователя наличия специальных знаний в области молекулярной биологии. В настоящем изобретении для определения характера запускаемых пептидами-агонистами ПАР1 внутриклеточных сигнальных путей используют подход, основанный на определении сохранности или отмены действия пептидов при использовании в качестве тест-системы первичных культур клеток нервной ткани или животных, у которых подавлена активность β аррестина-2. Т.е. в основе предлагаемого метода лежит регистрация зависимости эффектов пептидов от активности β-аррестина-2, вовлеченного во все сигнальные каскады ПАР1.
Если тот или иной пептид в условиях экспериментальной ишемии демонстрирует цитопротекторные свойства, которые отменяет подавление активности β-аррестина-2 (например, за счет нокаута гена данного белка), то следует считать, что данный пептид проявляет АПС-подобный профиль активности и запускает сигнальные пути с вовлечением Rac1. Это связано с тем, что в случае активации ПАР1 с помощью АПС, как было отмечено выше, β-аррестин-2 необходим для запуска его сигнальных каскадов. Если же пептид, напротив, усугубляет тяжесть экспериментальной ишемии, и этот эффект только усиливается при отсутствии β-аррестина-2, то пептид запускает тромбин подобные сигнальные пути, т.к. в этом случае β-аррестин-2 нужен для десенситизации и интернализации ПАР1.
В настоящем изобретении определение функциональной активности пептидов может быть проведено как в условиях in vitro, так и в условиях in vivo. Выбор подхода диктуется наличием доступных материалов, а таже исследовательской целью дальнейшего использования тестируемого пептида. В случае если функциональный профиль пептида важен для изучения его эффектов на культуры нервной ткани, подвергнутые ишемическим условиям, то необходимо производить тестирование с использованием культивируемых нейронов. Если в дальнейшем предполагается изучение эффектов пептида на уровне всего организма, то качестве тест-системы необходимо использовать модель экспериментального инсульта на животных. Однако тестирование того или иного пептида-агониста ПАР1, проведенное как с использованием культивируемых нейронов, так и опытных животных позволит получить более надежные результаты.
Подавление активности β-аррестина-2 в первичных нейрональных культурах может быть осуществлено за счет нокдауна его гена с использованием малых интерферирующих РНК (siRNA). Для этого используют siRNA с последовательностью 5’-AAGGACCGCAAAGUGUUUGUG-3’, проводя трансфекцию клеток по ранее описанному протоколу (Wang W.Y., Wu X.M., Jia L.J., Zhang H.H., Cai F., Mao H., Xu W.C., Chen L., Zhang J., Hu S.F., 2016. Beta-arrestin-1 and 2 differently modulate metabotropic glutamate receptor 7 signaling in rat developmental sevoflurane-induced neuronal apoptosis // Neuroscience. V. 313. P. 199–212). Помимо этого, возможно использование аптамеров (Kotula J.W., Sun J., Li M., Pratico E.D., Fereshteh M.P., Ahrens D.P., Sullenger B.A., Kovacs J.J., 2014. Targeted disruption of β-arrestin-2-mediated signaling pathways by aptamer chimeras leads to inhibition of leukemic cell growth // PLoS One. V. 9. P. 1–11) или системы CRISPR/Cas9 (Luttrell L.M. и др., 2018. Manifold roles of β-arrestins in GPCR signaling elucidated with siRNA and CRISPR/Cas9 // Sci. Signal. V. 11. P. 1–22) для блокады активности β-аррестина-2. Кроме того, нейроны могут быть выделены из мозга мышей, имеющих нокаут гена β-аррестина-2, выполненный методом гомологичной рекомбинации по ранее описанной методике (Bohn L.M., Lefkowitz R.J., Gainetdinov R.R., Peppel K., Caron M.G., Lin F.-T., 1999. Enhanced morphine analgesia in mice lacking β-arrestin-2 // Science. V. 286. P. 2495–2498). Данную линию мышей, т.е. животных с нокаутом целевого гена, следует использовать и при моделировании ишемии головного мозга in vivo.
Для определения функциональной активности исследуемого пептида в условиях in vitro в данном изобретении необходимо использовать в качестве тест-системы первичную культуру нейронов коры больших полушарий новорожденных мышей или крыс (0-3 дня от рождения). Культура клеток готовится и поддерживается по стандартному протоколу, который включает в себя получение суспензии кортикальных нейронов (концентрация клеток около 106 клеток/мл) и их дальнейшее культивирование при 370С, 5% СО2 (Gorbacheva L.R., Storozhevykh T.P., Pinelis V. G., Davydova O.N., Ishiwata S., Strukova S.M., 2008. Activated protein C via PAR1 receptor regulates survival of neurons under conditions of glutamate excitotoxicity // Biochem. V. 73. P. 717–724). Дальнейшая работа производится на 9-14-дневных культурах.
Далее клетки подвергают условиям, имитирующим условия ишемического инсульта. Такие условия могут быть созданы с использованием кислородно-глюкозной депривации, которая предполагает пребывание нейрональных культур в течение 2-4 ч в газовом инкубаторе без доступа кислорода в атмосфере N2/CO2 (95%/5%) в культуральной среде, лишенной глюкозы (Wang Q., Gong Q., Wu Q., Shi J., 2010. Neuroprotective effects of Dendrobium alkaloids on rat cortical neurons injured by oxygen-glucose deprivation and reperfusion // Phytomedicine. V. 17. P. 108–115). Помимо этого, возможно использование модели глутаматной эксайтотоксичности, которая рассматривается как один из основных негативных факторов развивающегося ишемического инсульта. Данная модель основана на инкубации культивируемых нейронов в течение 30-40 мин в среде с высоким содержанием глутамата (100 мкМ) (Gorbacheva L.R., Storozhevykh T.P., Pinelis V. G., Davydova O.N., Ishiwata S., Strukova S.M., 2008. Activated protein C via PAR1 receptor regulates survival of neurons under conditions of glutamate excitotoxicity // Biochem. V. 73. P. 717–724). Выживаемость клеток, определенная описанным ниже методом, при ишемических условиях должна составлять 40-60% по сравнению с клетками, которые не находились в данных условиях. Необходимо заранее подобрать оптимальные параметры: длительность кислородно-глюкозной депривации или концентрацию глутамата в среде, чтобы достичь приведенных значений. Далее подобранные значения используются для всех экспериментальных групп.
Инкубация с исследуемым пептидом должна осуществляться на протяжении 15-30 мин непосредственно до создания условий кислородно-глюкозной депривации или добавления глутамата в среду, при этом концентрация 10-30 мкМ в культуральной среде, пептид растворяют или непосредственно в культуральной среде до нужной концентрации, а далее помещают клетки в данную среду, или его предварительно растворяют в дистиллированной воде или в водном буферном растворе до такой концентрации, чтобы вносимый объем его раствора составил 3-5% от общего объема среды, в которой пребывают клетки.
Далее настоящее изобретение предполагает оценку клеточной гибели через 24-36 ч после воздействия тестируемого пептида на клетки на фоне кислородно-глюкозной депривации или высоких концентраций глутамата. Клеточная гибель может быть оценена с помощью реагента МТТ (3-(4,5-диметилтиазол-2-ил)-2,5-дифенил-тетразолиум бромид) с использованием спектрофотометра по стандартному протоколу (Gorbacheva L.R., Storozhevykh T.P., Pinelis V. G., Davydova O.N., Ishiwata S., Strukova S.M., 2008. Activated protein C via PAR1 receptor regulates survival of neurons under conditions of glutamate excitotoxicity // Biochem. V. 73. P. 717–724) (МТТ может быть заменен на водорастворимые тетразолиевые соли, например, WST-1 (Galkov M.D., Ivanova A.E., Gulyaev M.V., Kiseleva E.V., Savinkova I.G., Gorbacheva L.R., 2020. The influence of β-arrestin-2 gene knockout in mice on survival of cultured astrocytes exposed to thrombin and on the cerebral thrombosis aftereffects in vivo // Biochem. V. 14. P. 17–23). MTT и WST-1, будучи тетразолиевыми солями, восстанавливаются до яркоокрашенных формазанов дегидрогеназами живых клеток. Следовательно, чем выше оптическая плотность, тем больше живых клеток находится в среде. Полученное значение оптической плотности (длины волн для измерения: 550-600 нм для MTT и 450 нм для WST-1) нормируют к таковому, измеренному в контрольной группе (принимается за 100%), тем самым рассчитывая относительный уровень клеточной гибели.
Для определения функциональной активности пептида в условиях in vivo в данном изобретении необходимо использовать в качестве тест-системы группу животных с нокаутом гена β-аррестина-2, проведенным в ходе гомологичной рекомбинации (Bohn L.M., Lefkowitz R.J., Gainetdinov R.R., Peppel K., Caron M.G., Lin F.-T., 1999. Enhanced morphine analgesia in mice lacking β-arrestin-2 // Science. V. 286. P. 2495–2498), и группу, где активность гена целевого белка не подавлена. Моделирование ишемического инсульта может быть проведено с помощью окклюзии средней мозговой артерии в течение 40-60 мин по стандартному протоколу (Rousselet E., Kriz J., Seidah N.G., 2012. Mouse model of intraluminal MCAO: cerebral infarct evaluation by cresyl violet staining // J. Vis. Exp. V. 69. P. 1–5) или с использованием фототромбоза, в ходе которого сенсомоторная область коры больших полушарий животного после внутривенного введения фотосенсибилизатора в дозе 10-15 мг/кг подвергается действию лазерного излучения (освещенность у поверхности черепа 55-65 лк) (Galkov M.D., Ivanova A.E., Gulyaev M. V., Kiseleva E. V., Savinkova I.G., Gorbacheva L.R., 2020. The influence of β-arrestin-2 gene knockout in mice on survival of cultured astrocytes exposed to thrombin and on the cerebral thrombosis aftereffects in vivo // Biochem. V. 14. P. 17–23). Две инъекции тестируемого пептида, растворенного в физиологическом растворе, в дозе от 10 до 30 мг/кг (в такой концентрации, чтобы объем вводимого за 1 раз раствора не превышал 1 мл на 100 г массы животных) необходимо производить внутривенно (через катетер, вживленный в яремную или бедренную вену, или посредством инъекции в хвостовую вену) за 10-15 мин до индукции ишемического поражения и через 40-60 мин после нее.
Далее через 24 ч после индукции ишемии необходимо оценить объем ишемического поражения, который непосредственно отражает тяжесть экспериментальной ишемии. Данный показатель может быть определен прижизненно с помощью метода магнитно-резонансной томографии или путем окрашивания секций мозга 0,5-1% раствором хлористого 2,3,5-трифенилтетразолия (ТТС) по описанным протоколам (Galkov M.D., Ivanova A.E., Gulyaev M. V., Kiseleva E. V., Savinkova I.G., Gorbacheva L.R., 2020. The influence of β-arrestin-2 gene knockout in mice on survival of cultured astrocytes exposed to thrombin and on the cerebral thrombosis aftereffects in vivo // Biochem. V. 14. P. 17–23). Объем подсчитывается на основе площади повреждения, измеренной на каждом МРТ-снимке или срезе, окрашенном ТТС. Далее нормируют полученное значение на таковое, рассчитанное в контрольной группе, где оно принимается за 100%.
Таким образом, настоящая тест-система основана на оценке эффекта тестируемого пептида-агониста ПАР1 в зависимости от активности β-аррестина-2 в первичной культуре нервной ткани или у животных в условиях развития ишемии. В экспериментах должно быть использовано не менее четырех экспериментальных групп:
1. культивируемые нейроны/животные без подавления активности β-аррестина-2, подвергнутые действию ишемических условий, но не подвергнутые действию тестируемого пептида;
2. культивируемые нейроны/животные без подавления активности β-аррестина-2, подвергнутые действию ишемии и действию тестируемого пептида;
3. культивируемые нейроны/животные с подавленной активностью β-аррестина-2, подвергнутые действию ишемических условий, но не подвергнутые действию тестируемого пептида;
4. культивируемые нейроны/животные с подавленной активностью β-аррестина-2, подвергнутые действию ишемии и действию тестируемого пептида.
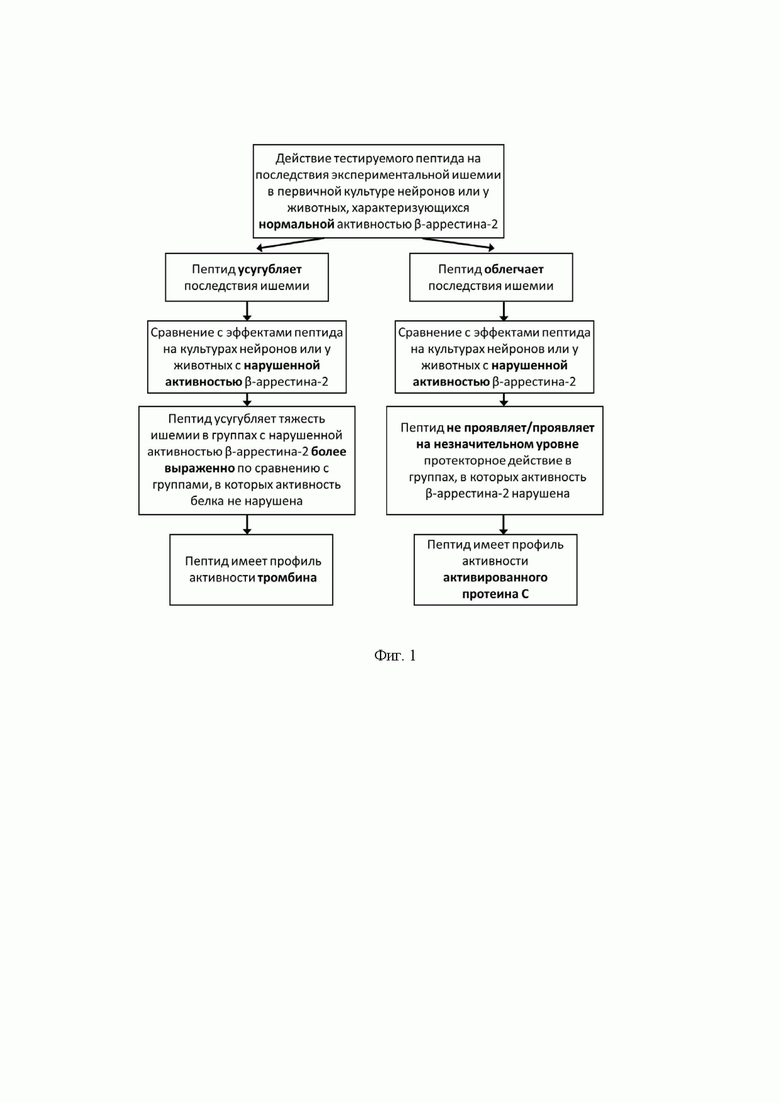
Для интерпретации данных рассчитывают уровень клеточной гибели или объем повреждения в группе 2 относительно группы 1 (контрольная группа) и в группе 4 относительно группы 3 (контрольная группа). В случае, если тестируемый пептид, действительно, является агонистом ПАР1, то возможно получение двух принципиально различающихся эффектов. Так, если пептид проявляет функциональную активность тромбина, то он будет усугублять последствия ишемических условий у первичных культур или у животных. Т.е. уровень гибели клеток или объем ишемического повреждения будут на 15-40% выше в группах 2 и 4 по сравнению с группами 1 и 3, соответственно. При этом в группе, где подавлена активность β-аррестина-2 (группа 4) такое усугубление тяжести экспериментальной ишемии будет на 10-20% выше, чем в группе, характеризующейся нормальной активностью целевого белка (группа 2) (фиг.1).
Наблюдаемый эффект связан с тем, что тромбин, активируя ПАР1, приводит к запуску зависимых от Gq-белка каскадов. В данные сигнальные пути вовлечена ГТФаза RhoA и мобилизация внутриклеточных ионов кальция. Это, в свою очередь, может приводить к гибели нейронов вследствие запуска апоптотических программ, активации глиальных клеток, развития нейровоспаления и повреждения эндотелиальных барьеров. В этом случае β-аррестин-2 необходим для десенситизации и интернализации ПАР1, т.е. для ингибирования активированного рецептора, что объясняет усугубление тяжести ишемии пептидами с функциональной активностью тромбина в группах с подавленной активностью β-аррестина-2.
Если при использовании тестируемого пептида наблюдается облегчение тяжести ишемических условий в экспериментальной группе 2 по сравнению с группой 1 (группы с нормальной активностью β-аррестина-2), выражающееся в уменьшении гибели клеток или объема ишемического повреждения на 15-40%, и отсутствие таких эффектов/их незначительное проявление (уменьшение менее, чем на 15%) в группах с нарушенной активностью белка (группы 3 и 4), то данный пептид характеризуется АПС-подобным профилем (фиг.1). Это связано с запуском пептидом цитопротекторных внутриклеточных каскадов, включающих в себя сборку молекулярной платформы с помощью β-аррестина-2 и рекрутинг эффекторных белков в группе, где β-аррестин-2 экспрессируется на нормальном уровне, но не в группе, где данный белок отсутствует или неактивен.
Если в ходе тестирования пептида при сравнении уровня клеточной гибели или объема ишемического поражения между группами получают значения, отличные от описанных, то пептид не является специфическим агонистом ПАР1 и должен быть исключен из дальнейших исследований.
Дальнейшая реализация протекторного сигнала в этом случае сходна с таковой, описанной для АПС. В частности, возможно инициируемое пептидом подавление экспрессии генов, индуцируемых NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells, фактор транскрипции), модуляция путей апоптоза, включая активацию эндотелиального гомолога Bcl-2 (B-cell lymphoma 2, белок-регулятор апоптоза), NO-синтазы и ингибиторов апоптоза семейства IAP (inhibitors of apoptosis). Кроме того, возможна активация пептидом с АПС-подобными эффектами оси ангиопоэтин-1/Tie-2 (angiopoietin receptor-2), увеличение синтеза сфингозин-1-фосфата и отмена транслокации в ядро нейронов субъединицы p65 транскрипционного фактора NF-κB. Все описанные внутриклеточные сигнальные пути зависимы от β-аррестина-2 и могут вносить свой вклад в реализацию цитопротекторных эффектов тестируемых пептидов с профилем функциональной активности АПС.
Таким образом, предлагаемое изобретение позволяет установить характер активности пептидов-аналогов «привязанного лиганда» ПАР1 на основе зависимости их эффектов от активности β-аррестина-2. Если цитопротекторные эффекты пептида зависят от уровня экспрессии данного белка, то тогда запускаемые данным пептидом каскады сходны с таковыми, описанными для АПС. В этом случае такой пептид можно рассматривать как потенциальный нейропротекторный агент. Если пептид усугубляет тяжесть ишемии, и такое ухудшение более выражено в экспериментальных группах, характеризующихся нарушенной активностью β-аррестина-2, то пептид имеет «тромбиновый» профиль эффектов и запускаемых сигнальных путей. Данное изобретение может использоваться как простой, быстрый и легкий в интерпретации результатов тест для скрининга различных пептидов-миметиков N-конца ПАР1 с целью установления характера запускаемых ими внутриклеточных путей (АПС- или тромбин-подобных).
Краткое описание чертежей
Фиг. 1. Схема, отражающая дизайн проведения исследования с использованием настоящего изобретения и основные принципы интерпретации результатов.
Фиг. 2. Подтверждение структуры и гомогенности пептида NPNDKYEPF-NH2 (АП9): А – профиль ВЭЖХ пептида, Б – масс-спектр пептида.
Фиг. 3. Процесс индукции фототромбоза как модели ишемического поражения.
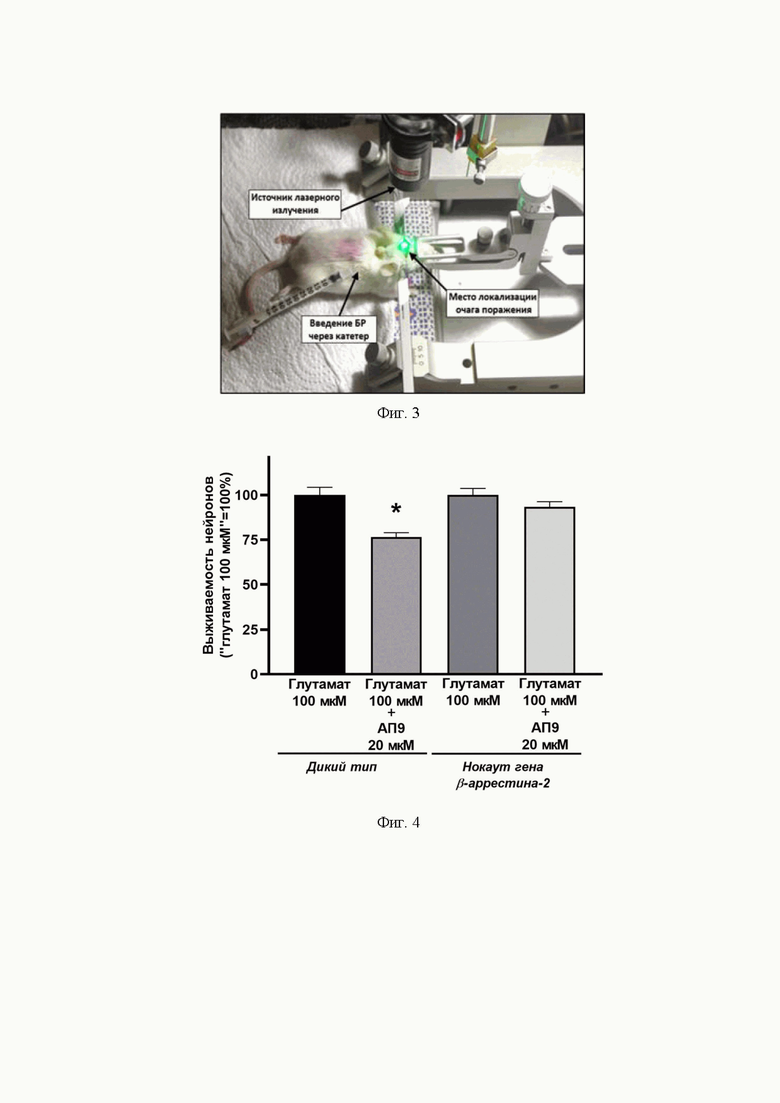
Фиг. 4. Эффект АП9 (20 мкМ) на выживаемость культивируемых нейронов, полученных от мышей с полноценной экспрессией β-aррестина-2 и с нокаутом его гена, в условиях глутаматной эксайтотоксичности (100 мкМ) (тест Краскала-Уоллиса, 6 независимых первичных культур для каждой экспериментальной группы, * – по сравнению с группой с нормальной экспрессией β-аррестина-2, подвергшейся действию глутамата, но не пептида, p<0,05; дикий тип – группа с полноценной экспрессией β-aррестина-2).
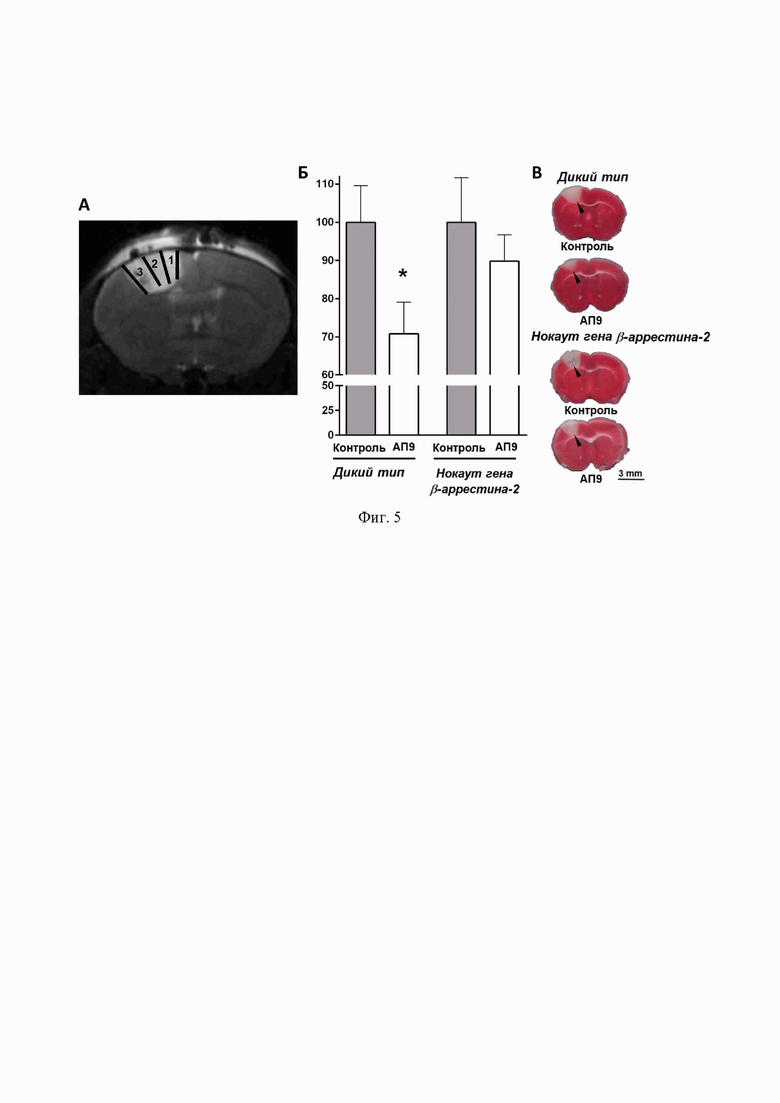
Фиг. 5. Эффект АП9 (две инъекции в дозе 20 мг/кг: за 10 мин до и через 1 ч после индукции ишемии) на объем повреждения мозга у животных с полноценной экспрессией β-aррестина-2 (дикий тип) и мышей с нокаутом его гена через 24 ч после фототромбоза. А – прижизненная визуализация мозга методом МРТ, подтверждающая локализацию фотоиндуцированного поражения в сенсомоторной коре больших полушарий (1 – первичная моторная кора, 2 и 3 – соматосенсорная кора задних и передних конечностей, соответственно), Б – объем поражения мозга у животных разных групп, выявленный методом окрашивания ТТС (тест Краскала-Уоллиса, 6 животных в каждой группе, * – по сравнению с контрольной группой, p<0,05), В – серия репрезентативных секций мозга, окрашенных ТТС (треугольником отмечена область поражения).
Осуществление изобретения
В настоящем изобретении в качестве частного варианта его реализации показана эффективность данного подхода на основе выявления зависимости эффектов пептида АП9, состоящего из девяти аминокислот, от наличия экспрессии β-аррестина-2 в условиях, имитирующих ишемические. Ранее была установлена структура АП9 (NPNDKYEPF-NH2), которая совпадает со структурой N-конца ПАР1, расщепляемого АПС. Также было установлено, что АП9 подобно АПС снижает гибель нейронов в условиях глутаматной эксайтотоксичности (Savinkova I.G., Gorbacheva L.R., Bespalova Z.D., Pinelis V.G., Strukova S.M., 2014. Peptides analogous to tethered ligands liberated by activated protein C exert neuroprotective effects in glutamate induced excitotoxicity // Biochem. V. 8. P. 116–120) или при действии на них активированных тучных клеток (Babkina I.I., Strukova S.M., Pinelis V.G., Reiser G., Gorbacheva L.R., 2016. New synthetic peptide protects neurons from death induced by toxic influence of activated mast cells via protease-activated receptor // Biochem. V. 10. P. 126–134) in vitro. Помимо этого, действие АП9, схожее с эффектами АПС, было показано и в модели фотоиндуцированной ишемии у мышей (Galkov M., Gorbacheva L., Gulyaev M., Sidorova M., 2018. Neuroprotective effects of protease-activated receptor 1 peptide-agonist in a murine model of focal ischemic stroke // Книга тезисов Объединенного международного конгресса «Congress Open Issues in Thrombosis and Hemostasis совместно с 9-ой Всероссийской конференцией по клинической гемостазиологии и гемореологии». P. 76–77; Galkov M., Sidorova M., Kiseleva E., Gorbacheva L., 2019. New peptide-agonist of PAR1 demonstrates neuroprotective effect at the photothrombosis-induced damage of mouse brain // Res. Pract. Thromb. Haemost. (сборник тезисов конференции). P. 7–7).
Однако в данных исследованиях вывод об АПС-подобном действии АП9 был сделан лишь на основании сравнения их функциональных эффектов (в том числе описанных ранее в других исследованиях), сопоставления структуры пептида и «привязанного лиганда» ПАР1 или с использованием блокаторов рецептора. Данные подходы, как было отмечено выше, имеют ряд недостатков, а именно, большое количество формируемых экспериментальных групп, сложность интерпретации полученных результатов и невозможность принять во внимание «молекулярный ансамбль», в котором пребывает рецептор.
Предложенный же в данном изобретении подход, подтвердивший АПС-подобный профиль активности АП9, основывается на анализе зависимости действия пептидов от активности белка β-аррестина-2.
Этап 1. Получение культуры нейронов из коры головного мозга мышей.
Весь процесс выделения клеток проводили на льду, центрифугу предварительно охлаждали до 4°С. Мышат (от 0 до 3 дней от рождения) с полноценной экспрессией β-аррестина-2 или ранее описанной линии с нокаутом его гена (Bohn L.M., Lefkowitz R.J., Gainetdinov R.R., Peppel K., Caron M.G., Lin F.-T., 1999. Enhanced morphine analgesia in mice lacking β-arrestin-2 // Science. V. 286. P. 2495–2498) декапитировали, после чего выделяли и очищали полушария в буфере без магния и кальция. При этом наличие нокаута гена β-аррестина-2, выполненного методом гомологичной рекомбинации, ранее было нами подтверждено с помощью методов Вестерн-блоттинга и полимеразной цепной реакции (Galkov M.D., Ivanova A.E., Gulyaev M. V., Kiseleva E. V., Savinkova I.G., Gorbacheva L.R., 2020. The influence of β-arrestin-2 gene knockout in mice on survival of cultured astrocytes exposed to thrombin and on the cerebral thrombosis aftereffects in vivo // Biochem. V. 14. P. 17–23).
Далее инкубировали измельченную кору больших полушарий в растворе трипсин-ЭДТА 15 мин при 37°С. Затем клетки переносили в пробирку с бычьей фетальной сывороткой (Invitrogen, США) и инкубировали 3-5 мин при комнатной температуре. Клетки отмывали от сыворотки 3 раза, заливали средой, содержащей 10% бычьей фетальной сыворотки и диспергировали до получения однородной клеточной суспензии, затем центрифугировали 7 мин при 1000 об/мин. Клеточный осадок диспергировали в среде культивирования.
Полученную суспензию клеток переносили на покровные стекла, покрытые поли-D-лизином (10 мг/мл). Клеткам давали сесть на стекло в течение 2 ч при 37°С, 5% СО2, затем отбирали неприкрепившиеся клетки и добавляли 1 мл культуральной среды (нейробазальная среда, содержащая 2% Supplement B-27 и 0,5 мМ L-глютамина, Gibco, США). Арабинозид добавляли на 3-4 день на 24 ч для подавления роста глиальных клеток, затем меняли среду. Далее через каждые 4-5 дней меняли 1/3 объема среды на свежую. Культивирование клеток осуществляли в течение 14 дней.
Этап 2. Осуществление воздействий на культивируемые нейроны и оценка уровня их гибели.
Гибель нейронов определяли через 24 ч после воздействия на их культуры высокой концентрации глутамата (100 мкМ) в течение 40 мин (модель глутаматной эксайтотоксичности). В экспериментальных группах инкубацию с АП9 (20 мкМ) проводили в течение 15 мин в 48-луночных планшетах (60*103 клеток/лунка, 250 мкл среды в каждой лунке, 10 мкл раствора АП9 в концентрации 561,5 мкг пептида на 1 мл дистиллированной воды добавляли в одну лунку) непосредственно перед добавлением глутамата. Пептид-агонист ПАР1 – АП9 (NPNDKYEPF-NH2), высвобождаемый АПС при расщеплении ПАР1, был синтезирован в Национальном медицинском исследовательском центре кардиологии Минздрава России твердофазным методом с использованием Fmoc-технологии М.В. Сидоровой (Savinkova I.G., Gorbacheva L.R., Bespalova Z.D., Pinelis V.G., Strukova S.M., 2014. Peptides analogous to tethered ligands liberated by activated protein C exert neuroprotective effects in glutamate induced excitotoxicity // Biochem. V. 8. P. 116–120). Гомогенность АП9 и соответствие его структуры, заявленной были подтверждены данными аналитической ВЭЖХ и масс-спектрометрии, соответственно (фиг. 2).
Выживаемость клеток оценивали биохимическим методом с помощью тетразолиевого красителя МТТ. МТТ (Sigma, США) добавляли в культуральную среду до конечной концентрации 1 мг/мл и инкубировали клетки 3 ч при 37°С. Затем среду отбирали и добавляли диметилсульфоксид для растворения образовавшихся формазанов. Оптическую плотность измеряли на иммуноферментном анализаторе «Униплан» АИФР-01 (ЗАО «Пикон», Россия) при 590 нм. Оценивали выживаемость клеток в процентах внутри групп с нормальной экспрессией β-арресина-2 (данный параметр был принят в группе, подвергшейся действию глутамата, но не пептида, за 100%) и внутри групп с нокаутом целевого гена (выживаемость также была принята в группе, подвергшейся действию глутамата, но не пептида, за 100%).
Этап 3. Моделирование ишемического инсульта и схема фармакологического воздействия in vivo.
Настоящее изобретение было разработано с использованием самцов мышей массой 25-30 г и возрастом 2,5-3 месяца с полноценной экспрессией β-аррестина-2 или с нокаутом его гена. Животные содержались в стандартных условиях (температура 22°С, световой день – 12/12 ч) без ограничения доступа к воде и корму. Все эксперименты проводились в полном соответствии с Директивой 2010/63/EU Европейского Парламента и Совета Европейского Союза.
В качестве анестезии при выполнении исследований использовали смесь коммерческих препаратов «Золетил100» (Virbac Sante Animale, Франция) и «Ксила» (Interchemie werken «De Adelaar» B.V., Нидерланды) в дозах 35 и 3,5 мг/кг (внутрибрюшинно), соответственно. Моделирование ишемического инсульта осуществляли посредством фототромбоза (фиг. 3). Наркотизированных животных фиксировали в стереотаксической установке на термоподдерживающей панели, после чего рассекали кожу головы вдоль сагиттального шва черепа и удаляли надкостницу с его поверхности при помощи 3% раствора Н2О2. Через катетер, предварительно вживленный животным в яремную вену за три дня до индукции тромбоза, вводили фотосенсибилизатор бенгальский розовый в дозе 10 мг/кг, растворенный в физиологическом растворе в концентрации 0,15%. По истечении 3 мин с использованием лазерной установки (λ=532 нм, освещенность у поверхности черепа – 50 лк) облучали локализованную с помощью светонепроницаемой мембраны область черепа диаметром 3 мм над сенсомоторной корой больших полушарий (латеральнее на 2,2 мм от точки bregma) в течение 5 мин. Рану зашивали прерывистым швом и обрабатывали 5% спиртовым раствором йода. После восстановления двигательных функций животных помещали в домашние клетки. Введение АП9, растворенного в физиологическом растворе (6 мг/мл), осуществлялось внутривенно посредством катетера в дозе 20 мг/кг дважды – за 10 мин до и через 1 ч после тромбоза, т.к. ранее данные доза и схема введения пептида показали протекторное действие в аналогичной модели инсульта (Galkov M., Gorbacheva L., Gulyaev M., Sidorova M., 2018. Neuroprotective effects of protease-activated receptor 1 peptide-agonist in a murine model of focal ischemic stroke // Книга тезисов Объединенного международного конгресса «Congress Open Issues in Thrombosis and Hemostasis совместно с 9-ой Всероссийской конференцией по клинической гемостазиологии и гемореологии». P. 76–77; Galkov M., Sidorova M., Kiseleva E., Gorbacheva L., 2019. New peptide-agonist of PAR1 demonstrates neuroprotective effect at the photothrombosis-induced damage of mouse brain // Res. Pract. Thromb. Haemost. (сборник тезисов конференции). P. 7–7).
Этап 4. Прижизненная визуализация ишемического повреждения головного мозга и измерение его объема после экспериментальной ишемии.
Прижизненная визуализация повреждения головного мозга проводилась методом магнитно-резонансной томографии (МРТ). Визуализацию проводили у наркотизированных животных, размещенных на термоподдерживающем столике томографа BioSpec 70/30 USR (Bruker, Германия) с индукцией магнитного поля 7,05 Tл, на основе Т2-взвешенных изображений, используя 2D RARE метод (градиентная система – 105 мТл/м, TR/TEeff = 3000/50 мс). Число срезов – 16, толщина – 0,4 мм, rare-factor – 4; in-plane resolution – 100 на 100 мкм. Общее время сканирования для каждого животного – 9 мин.
Для измерения объема поражения мозга с помощью окрашивания его секций хлористым 2,3,5-трифенилтетразолием (ТТС) животных умерщвляли парами эфира, декапитировали и быстро извлекали мозг из черепной коробки. Далее мозг при помощи тонкого лезвия рассекали на секции толщиной 1 мм, которые переносили в 1% раствор ТТС (Sigma, США) на фосфатно-солевом буфере. Инкубацию срезов производили в течение 15 мин при 37°С, после чего останавливали реакцию путем замены рабочего раствора на 4% раствор формалина (Panreac, Испания). Далее секции мозга фотографировали и проводили анализ изображений в программе ImageJ (Национальный институт здоровья, США). Объем ишемического очага определяли на основе измеренной площади поражения на полученных фотографиях и принимали в контрольных группах (животные, которым не вводили пептид) за 100%.
Примеры реализации изобретения
В качестве примера реализации данного изобретения был определен профиль функциональной активности пептид-агониста ПАР1, АП9, который по результатам ранее проведенных многочисленных исследований проявляет АПС-подобную активность. Настоящее изобретение также позволило определить его профиль активности, который, как и ожидалось, был аналогичен АПС. Все необходимые эксперименты были проведены как в условиях in vitro за 16 дней, так и в условиях in vivo за 18 дней.
Влияние нокаута гена β-аррестина-2 на выживаемость культивируемых нейронов в условиях глутаматной эксайтотоксичности при действии на них АП9. В ходе реализации изобретения было показано, что цитопротекторный эффект АП9 зависит от экспрессии β-аррестина-2: в случае нокаута гена данного белка эффект пептида не проявляется. В частности, АП9 при концентрации в культуральной среде 20 мкМ снижал гибель нейронов, полученных от мышей с полноценной экспрессией β-аррестина-2, в условиях глутаматной эксайтотоксичности (концентрация глутамата в среде – 100 мкМ). В контрольной группе, т.е. группе, находящейся в среде с высокой концентрацией глутамата, но без добавления АП9, выживаемость клеток была равна 100,0±4,3%. В группе, подвергшейся действию АП9, данный показатель статистически значимо снизился до 76,5±2,4% (p<0,05) (фиг. 4).
Вместе с тем у нейронов, не экспрессирующих β-аррестин-2, проявления защитных эффектов АП9 выявить не удалось (уровень выживаемости клеток составил 93,3±2,9% и статистически значимо не отличался от такового, измеренного в группе, подвергнутой только действию глутамата – 100,0±3,7%) (фиг. 4). Результаты проведенных экспериментов подтвердили, что АП9 демонстрирует АПС-подобную функциональную активность в условиях in vitro, имитирующих ишемические.
Влияние нокаута гена β-аррестина-2 на объем поражения головного мозга после фототромбоза и внутривенного введения АП9. Для выявления эффективности предложенного в настоящем изобретении метода определения профиля активности пептидов, имитирующих «привязанный лиганд» ПАР1, в условиях in vivo была выбрана модель фотоиндуцированной ишемии. Сначала методом МРТ была доказана локализация поражения головного мозга исключительно в сенсомоторной области коры больших полушарий, что позволило считать выбранную модель адекватной и воспроизводимой.
Далее было установлено, что две внутривенные инъекции АП9 в дозе 20 мг/кг (за 10 мин до индукции ишемии и через 1 ч после) статистически значимо снижают объем повреждения мозга у мышей с полноценной экспрессией белка β-аррестина-2. Данный показатель составил 100,00±9,56% у контрольных животных (введение физиологического раствора вместо раствора АП9) и 70,80±8,28% у группы, подвергнутой инъекциям пептида. У мышей с нокаутом целевого гена АП9, напротив, не вызывал значимого снижения объема поврежденной области (100,00±11,63% – контрольная группа и 91,82±6,41% – животные, которым инъецировали АП9) (фиг. 5).
Результаты проведенных экспериментов подтвердили, что описанный в данном изобретении подход для определения характера запускаемых каскадов пептидов-агонистов ПАР1 проявляет эффективность в экспериментальной модели ишемии in vivo.
Предлагаемый способ прост в исполнении, легок для интерпретации результатов, а также учитывает то молекулярное окружение ПАР1, от которого зависит эффекты тестируемого пептида.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ КОМПЛЕКСНОЙ ЗАЩИТЫ КЛЕТОК ПРИ НЕЙРОВОСПАЛЕНИИ НА МОДЕЛИ IN VITRO ПРИ ВОЗДЕЙСТВИИ АГОНИСТОМ ПАР1 | 2022 |
|
RU2796524C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ НЕЙРОПРОТЕКТОРНЫМИ СВОЙСТВАМИ В ЭКСПЕРИМЕНТЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2015 |
|
RU2614694C1 |
| СПОСОБ НЕЙРОПРОТЕКЦИИ В ЭКСПЕРИМЕНТЕ | 2015 |
|
RU2616509C1 |
| Способ применения тромбина и активированного протеина С в качестве антивоспалительных средств | 2020 |
|
RU2780490C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНТИАПОПТОТИЧЕСКОЙ АКТИВНОСТЬЮ | 2018 |
|
RU2685428C1 |
| ПРИМЕНЕНИЕ ПРЕПАРАТА "СЕМАКС" В КАЧЕСТВЕ СРЕДСТВА АКТИВАЦИИ АДАПТИВНОГО ИММУНИТЕТА | 2015 |
|
RU2584234C1 |
| БИЦИКЛИЧЕСКИЕ ПИРИМИДИНЫ ИЛИ ИХ ФАРМАЦЕВТИЧЕСКИ ПРИЕМЛЕМЫЕ СОЛИ-АКТИВАТОРЫ АНТИОКСИДАНТНОЙ ПРОГРАММЫ И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ЦИТОПРОТЕКТОРОВ | 2014 |
|
RU2545758C1 |
| СОЕДИНЕНИЯ И ФАРМАЦЕВТИЧЕСКИЕ КОМБИНАЦИИ ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ И ИШЕМИЧЕСКИХ БОЛЕЗНЕЙ МОЗГА | 2012 |
|
RU2569302C2 |
| Средство, обладающее нейропротекторной активностью | 2019 |
|
RU2800955C2 |
| КОМБИНАЦИИ ЭДАРАВОНА ДЛЯ ЛЕЧЕНИЯ ИШЕМИЧЕСКИХ ПОВРЕЖДЕНИЙ МОЗГА | 2017 |
|
RU2693627C2 |

Изобретение относится к области биохимии, а именно к способу определения профиля функциональной активности пептидов-аналогов N-концевого фрагмента рецепторов, активируемых протеазами 1 типа на основе зависимости их эффектов от активности β-аррестина-2. Способ включает определение профиля функциональной активности пептидов-аналогов N-концевого фрагмента рецепторов, активируемых протеазами 1 типа в экспериментальных условиях, имитирующих ишемические, in vitro и/или in vivo не менее чем на 4 экспериментальных группах, из которых формируют по меньшей мере две подгруппы, которые включают по меньшей мере группы 1 и 2 с нормальной активностью β-аррестина-2 и по меньшей мере группы 3 и 4 с подавленной активностью β-аррестина-2, при этом по меньшей мере группы 1 и 3 подвергают воздействию только ишемических условий, а группы 2 и 4 подвергают воздействию и ишемических условий, и тестируемого пептида, далее проводят определение профиля активности пептида путем сравнения степени тяжести ишемических поражений между группами, и если исследуемый пептид облегчает последствия экспериментальной ишемии в группе без подавления активности β-аррестина-2 по сравнению с группой без подавления активности β-аррестина-2 и без воздействия пептида и этот эффект не выражен или менее выражен в группе с подавленной активностью β-аррестина-2 по сравнению с группой с подавленной активностью β-аррестина-2 и без воздействия пептида, то делают вывод, что пептид проявляет профиль активности активированного протеина С, если пептид усугубляет тяжесть экспериментальной ишемии в группе без подавления активности β-аррестина-2 по сравнению с группой без подавления активности β-аррестина-2 и без воздействия пептида и этот эффект более выражен в группе с подавленной активностью β-аррестина-2 по сравнению с группой с подавленной активностью β-аррестина-2 и без воздействия пептида, то делают вывод, что пептид проявляет функциональную активность тромбина. Изобретение обеспечивает создание тест-системы, позволяющей определять функциональную активность пептидов за короткий промежуток времени с использованием минимального количества экспериментальных групп, реактивов и биоматериалов, характеризующейся легкостью в интерпретации результатов. 14 з.п. ф-лы, 5 ил., 6 пр.
1. Способ определения профиля функциональной активности пептидов-аналогов N-концевого фрагмента рецепторов, активируемых протеазами, 1 типа на основе зависимости их эффектов от активности β-аррестина-2 в экспериментальных условиях, имитирующих ишемические, характеризующийся тем, что определение проводят в условиях in vitro и/или in vivo не менее чем на 4 экспериментальных группах, из которых формируют по меньшей мере две подгруппы, которые включают по меньшей мере группы 1 и 2 с нормальной активностью β-аррестина-2 и по меньшей мере группы 3 и 4 с подавленной активностью β-аррестина-2, при этом по меньшей мере группы 1 и 3 подвергают воздействию только ишемических условий, а группы 2 и 4 подвергают воздействию и ишемических условий, и тестируемого пептида, далее проводят определение профиля активности пептида путем сравнения степени тяжести ишемических поражений между группами, и если исследуемый пептид облегчает последствия экспериментальной ишемии в группе без подавления активности β-аррестина-2 по сравнению с группой без подавления активности β-аррестина-2 и без воздействия пептида и этот эффект не выражен или менее выражен в группе с подавленной активностью β-аррестина-2 по сравнению с группой с подавленной активностью β-аррестина-2 и без воздействия пептида, то делают вывод, что пептид проявляет профиль активности активированного протеина С (АПС), если пептид усугубляет тяжесть экспериментальной ишемии в группе без подавления активности β-аррестина-2 по сравнению с группой без подавления активности β-аррестина-2 и без воздействия пептида и этот эффект более выражен в группе с подавленной активностью β-аррестина-2 по сравнению с группой с подавленной активностью β-аррестина-2 и без воздействия пептида, то делают вывод, что пептид проявляет функциональную активность тромбина.
2. Способ по п. 1, характеризующийся тем, что для определения функциональной активности пептидов в условиях in vitro используют первичную культуру нейронов коры больших полушарий новорожденных мышей или крыс, а для определения функциональной активности пептидов в условиях in vivo используют экспериментальных мышей или крыс.
3. Способ по п. 1, характеризующийся тем, что для моделирования используют не менее четырех экспериментальных групп:
- культивируемые нейроны/животные без подавления активности β-аррестина-2, подвергнутые действию ишемических условий, но не подвергнутые действию тестируемого пептида, - группа 1;
- культивируемые нейроны/животные без подавления активности β-аррестина-2, подвергнутые действию ишемии и действию тестируемого пептида, - группа 2;
- культивируемые нейроны/животные с подавленной активностью β-аррестина-2, подвергнутые действию ишемических условий, но не подвергнутые действию тестируемого пептида, - группа 3;
- культивируемые нейроны/животные с подавленной активностью β-аррестина-2, подвергнутые действию ишемии и действию тестируемого пептида, - группа 4.
4. Способ по п. 1, характеризующийся тем, что если гибель клеток или объем ишемического повреждения на 15-40% выше в группах 2 и 4 по сравнению с группами 1 и 3, а в группе 4 такое усугубление тяжести ишемии будет на 10-20% выше, чем в группе 2, то делают вывод о том, что исследуемый пептид проявляет функциональную активность тромбина, если гибель клеток или объем ишемического повреждения на 15-40% меньше в группе 2 по сравнению с группой 1 и менее чем на 15% меньше в группе 4 по сравнению с группой 3, то делают вывод о том, что пептид обладает АПС-подобной функциональной активностью.
5. Способ по п. 1, характеризующийся тем, что для воздействия тестируемого пептида в условиях in vitro исследуемый пептид или растворяют непосредственно в культуральной среде до концентрации 10-30 мкМ, далее помещают клетки в данную среду, или его предварительно растворяют в дистиллированной воде или в водном буферном растворе до такой концентрации, чтобы после внесения раствора в культуральную среду концентрация пептида в ней составила 10-30 мкМ и вносимый объем его раствора составлял 3-5% от общего объема культуральной среды, далее осуществляют инкубацию нейронов с пептидом в течение 15-30 мин и начинают моделирование условий ишемии, а оценку клеточной гибели проводят через 24-36 ч после воздействия исследуемого пептида.
6. Способ по п. 2, характеризующийся тем, что для моделирования условий ишемического инсульта используют 9-14-дневную культуру нейронов.
7. Способ по п. 2, характеризующийся тем, что подавление активности β-аррестина-2 в первичных нейрональных культурах осуществляют за счет нокдауна его гена с использованием малых интерферирующих РНК (siRNA), или аптамеров, или системы CRISPR/Cas9 или выделения нейронов из мозга животных, имеющих нокаут гена β-аррестина-2, выполненный методом гомологичной рекомбинации.
8. Способ по п. 7, характеризующийся тем, что для нокдауна используют siRNA с последовательностью 5’-AAGGACCGCAAAGUGUUUGUG-3’.
9. Способ по п. 2, характеризующийся тем, что для моделирования условий ишемического инсульта используют кислородно-глюкозную депривацию или модель глутаматной эксайтотоксичности.
10. Способ по п. 5, характеризующийся тем, что для определения функциональной активности пептидов используют клетки, выживаемость которых составляет 40-60% по сравнению с клетками, которые не подвергались моделированию ишемического инсульта.
11. Способ по п. 5, характеризующийся тем, что оценку клеточной гибели проводят с помощью реагента МТТ или WST-1.
12. Способ по п. 2, характеризующийся тем, что для определения функциональной активности пептидов в условиях in vivo используют экспериментальных животных с нокаутом гена β-аррестина-2, проведенным в ходе гомологичной рекомбинации, со смоделированным ишемическим инсультом, при этом введение исследуемого пептида проводят внутривенно в дозе от 10 до 30 мг/кг за 10-15 мин до моделирования ишемического поражения и через 40-60 мин после него, а оценку объема ишемического поражения проводят через 24 ч после индукции ишемии.
13. Способ по п. 12, характеризующийся тем, что моделирование ишемического инсульта проводят с помощью окклюзии средней мозговой артерии или с использованием фототромбоза.
14. Способ по п. 12, характеризующийся тем, что внутривенное введение осуществляют через катетер, вживленный в яремную или бедренную вену, или посредством инъекции в хвостовую вену экспериментального животного.
15. Способ по п. 12, характеризующийся тем, что объем ишемического поражения оценивают на основе подсчета площади повреждения, измеренной на каждом МРТ-снимке или срезе, окрашенном ТТС.
| КИСЕЛЕВА Е | |||
| В | |||
| и др | |||
| Пептид-агонист рецептора, активируемого протеазами, стимулирует пролиферацию кератиноцитов и заживление раны эпителиального пласта подобно активированному протеину C //Биомедицинская химия | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| - Т | |||
| Способ получения молочной кислоты | 1922 |
|
SU60A1 |
| - N | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| - С | |||
| 702-706 | |||
| ГАЛКОВ М | |||
| Д | |||
| и др | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

