Изобретение относится к биотехнологии и может быть использовано в исследовательских и диагностических целях для эффективного получения генетически-модифицированных иммортализованных первичных Т-лимфоцитов и НК-клеток.
Лабораторно-клиническая практика создания аутологичных или гетерологичных пациент-специфичных лимфоидных линий требует разработки и оптимизации протоколов, обеспечивающих возможность эффективной иммортализации и генетической модификации первичных культур. Эффективным методом иммортализации первичных Т-лимфоцитов и НК-клеток является их заражение Гамма-2 герпесвирусом беличьих обезьян Herpesvirus saimiri (HVS), обеспечивающим антигеннезависимую пролиферацию инфицированных культур при сохранении ими функциональной активности и зрелого фенотипа. Использование рекомбинантного HVS также возможно для доставки в клетки протяженных экспрессионных конструкций, благодаря наличию в геноме вируса участков, использование которых в качестве сайтов интеграции целевых рекомбинантных последовательностей не оказывает влияния на жизнеспособность вируса. Сочетание этих особенностей Herpesvirus Saimiri позволяет рассматривать получение рекомбинантных герпесвирусов в качестве перспективного подхода по совмещению методик трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток.
В настоящее время разработаны различные алгоритмы использования герпесвирусов для трансдукции первичных Т-лимфоцитов и НК-клеток; наиболее близкими к заявленному изобретению по технической сущности и результатам, достигаемым аналогами, являются технические решения, описанные в заявках: US5670352А (C12N15/65, опубл. 23.09.1997), JPH02171190A (C12N15/86, опубл. 14.08.1991) и WO2004078911A3 (C12N15/65, опубл. 16.12.2004).
Применение существующих методик сопряжено с рядом ограничений, обусловленных, главным образом, особенностями линий клеток, служащих для получения рекомбинантного вируса. Используемые культуры клеток должны эффективно подвергаться трансфекции экспрессионными плазмидами, кодирующими генетические последовательности, предназначенные для введения в геном “дикого” штамма вируса, а также должны быть пермиссивны к литической инфекции Herpesvirus Saimiri. Необходимость сочетания этих свойств является серьезным ограничением, значительно сужающим спектр клеточных линий, подходящих для наработки рекомбинантного вируса.
Возможным решением проблемы являются технические решения, приведенные в заявках US5670352A и JPH02171190A, описывающие использование Herpesvirus saimiri для трансформации и генетической модификации Т-лимфоцитов. Для наработки вируса авторами изобретений предложено использование клеточной линии OMK (ATCC® CRL-1556™), пермиссивной к литической инфекции Herpesvirus Saimiri. Вместе с тем, удобство использования клеток этой линии для введения в них высокомолекулярной рекомбинантной ДНК вызывает сомнения, значительно уступая культурам, традиционно использующимся в лабораторной практике, таким как как HEK-293 (CRL-1573) или CHO (CRL-11965).
Заявка на изобретение JPH11513565A раскрывает возможности использования рекомбинантных генетических векторов на основе вируса герпеса, в частности, Herpesvirus saimiri (HVS), с целью трансдукции и генетической модификации широкого спектра культур клеток человека и животных, в частности, клеток лимфоидных линий. В заявке не раскрываются способы преодоления трудностей введения генетических конструкций в вирус-пермиссивные линии, а также не оговаривается возможность использования изобретения с целью одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток.
Другие аналоги, в частности, техническое решение WO2004078911A3, раскрывающее способ трансформации Т-лимфоцитов штаммом Herpesvirus saimiri, модифицированным с целью обеспечения их стабильной интерлейкин-независимой пролиферации, не предусматривают доставку генетических конструкций в клетки инфицируемых линий, что не позволяет использовать данный метод для выполнения задач, решаемых заявленным изобретением, в частности, для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток.
Таким образом, проведенные исследования патентной литературы позволяют заключить, что заявленное техническое решение характеризуется научной новизной, не имеет полных аналогов и обладает конкурентными преимуществами для получения рекомбинантных герпесвирусов, обеспечивающих возможность одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток.
Техническая проблема, решаемая настоящим изобретением - совмещение производительной наработки вируса в клетках линий, пермиссивных к литической инфекции HVS, с высокой эффективностью трансфекции клеток экспрессионными плазмидами, кодирующими генетические последовательности, предназначенные для введения в геном “дикого” штамма вируса, для получения рекомбинантных герпесвирусов.
Техническим результатом изобретения является оптимизация процесса получения иммортализованных генетически модифицированных первичных Т-лимфоцитов и НК-клеток, имеющих потенциал использования в терапевтических и исследовательских целях, за счет усовершенствования методики эффективного получения рекомбинантных герпесвирусов.
Техническая проблема решается за счет того, что пермиссивную к литической инфекции вирусом Herpesvirus Saimiri культуру клеток заражают штаммом вируса Herpesvirus Saimiri HVS-C488, а другую перевиваемую линию клеток трансфицируют смесью экспрессионных плазмид, одна из которых содержит целевой участок, предназначенный для введения в геном вируса штамма Herpesvirus Saimiri HVS-C488 и фланкированный последовательностями протяженностью 300-900 пар нуклеотидных оснований, гомологичных тому участку генома Herpesvirus Saimiri, в который производится введение целевого участка, а две другие экспрессионные плазмиды кодируют поверхностные гликопротеины F и H вакцинного штамма вируса кори, после чего инфицированную Herpesvirus Saimiri пермиссивную клеточную культуру и трансфицированную экспрессионными плазмидами клеточную культуру смешивают и подвергают ко-культивации, в результате чего происходит образование многоядерных синцитиев из клеток обеих культур, что обеспечивает прохождение рекомбинации гомологичных участков, внедрение целевого гена в геном Herpesvirus Saimiri и выход во внеклеточную жидкость рекомбинантных герпесвирусов; в качестве пермиссивной к литической инфекции Herpesvirus Saimiri культуры клеток может быть использована культура клеток ОМК (owl monkey kidney cells, почечный эпителий ночных обезьян); в качестве перевиваемой клеточной линии, которую трансфицируют смесью экспрессионных плазмид, может быть использована клеточная линия HEK-293, либо ее производные; в качестве последовательности протяженностью 300-900 пар нуклеотидных оснований, гомологичной участку генома Herpesvirus Saimiri, может быть использована последовательность вирусного гена vCD59; в состав целевого участка, предназначенного для введения в геном штамма вируса Herpesvirus Saimiri HVS-C488, может входить последовательность, кодирующая флуоресцентный белок; в состав целевого участка, предназначенного для введения в геном штамма вируса Herpesvirus Saimiri HVS-C488, может входить последовательность, кодирующая химерный антигенный рецептор.
Разработанная методика получения рекомбинантных герпесвирусов для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и
НК-клеток
1. Заражение пермиссивной к литической HVS инфекции культуры клеток №1 штаммом вируса Herpesvirus Saimiri HVS-C488.
2. Подготовка генетических конструкций:
a. экспрессионная плазмида, содержащая целевой участок, предназначенный для введения в геном вируса штамма Herpesvirus Saimiri HVS-C488, фланкированный последовательностями протяженностью 300-900 пар нуклеотидных оснований, гомологичных тому участку генома Herpesvirus Saimiri, в который производится введение целевого участка;
b. экспрессионная плазмида, кодирующая поверхностный гликопротеин F вакцинного штамма вируса кори;
c. экспрессионная плазмида, кодирующая поверхностный гликопротеин H вакцинного штамма вируса кори.
3. Трансфекция перевиваемой линии клеток №2 смесью экспрессионных плазмид, перечисленных в п. 2 методики.
4. Совместная культивация культур №1 и 2; визуальный контроль формирования многоядерных синцитиев из клеток обеих культур.
5. Отбор и центрифугирование ростовой среды совместно культивируемых культур.
6. Заражение вируссодержащим супернатантом пермиссивной культуры, сортировка GFP-позитивных клеток.
7. Клонирование и экспансия клонов, наработка и выделение рекомбинантного вируса, определение титра.
8. Секвенирование участка генома рекомбинантного вируса, содержащего сайт интеграции целевых рекомбинантных последовательностей.
9. Трансдукция первичных культур Т-клеток/NK-клеток наработанным рекомбинантным герпесвирусом.
10. Оценка пролиферативной активности трансдуцированных клеток.
11. Верификация экспрессии целевых трансгенов в трансдуцированных клетках.
Осуществление изобретения
1. Пермиссивная к литической инфекции HVS культура клеток OMK была инфицирована “диким” штаммом вируса HVS-C488 (VR-1414) ATCC (США).
2. Использованные генетические конструкции:
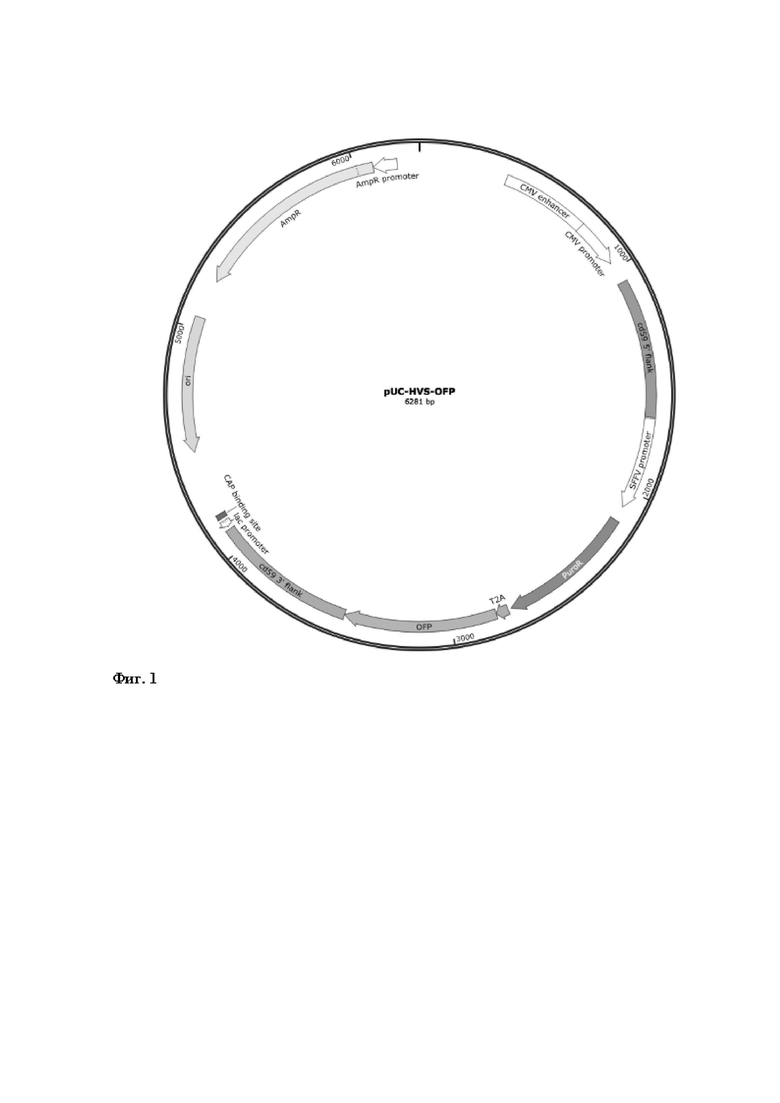
a. Рекомбинантная плазмидная ДНК pUC-HVS-OFP (Фиг.1), состоящая из следующих ключевых генетических элементов:
1. гена устойчивости к пуромицину (pac);
2. гена оранжевого флуоресцентного белка (OFP);
3. фланкирующими участками (600 п.н.), гомологичными области vCD59 генома HVS (SEQ ID NO : 1 и SEQ ID NO : 2);
4. гена устойчивости к антибиотику ампициллину (AmpR) и бактериального промотора гена устойчивости к ампициллину (AmpR promoter);
5. ориджина репликации бактериофага f1 (ori);
6. промотора цитомегаловируса (CMV).
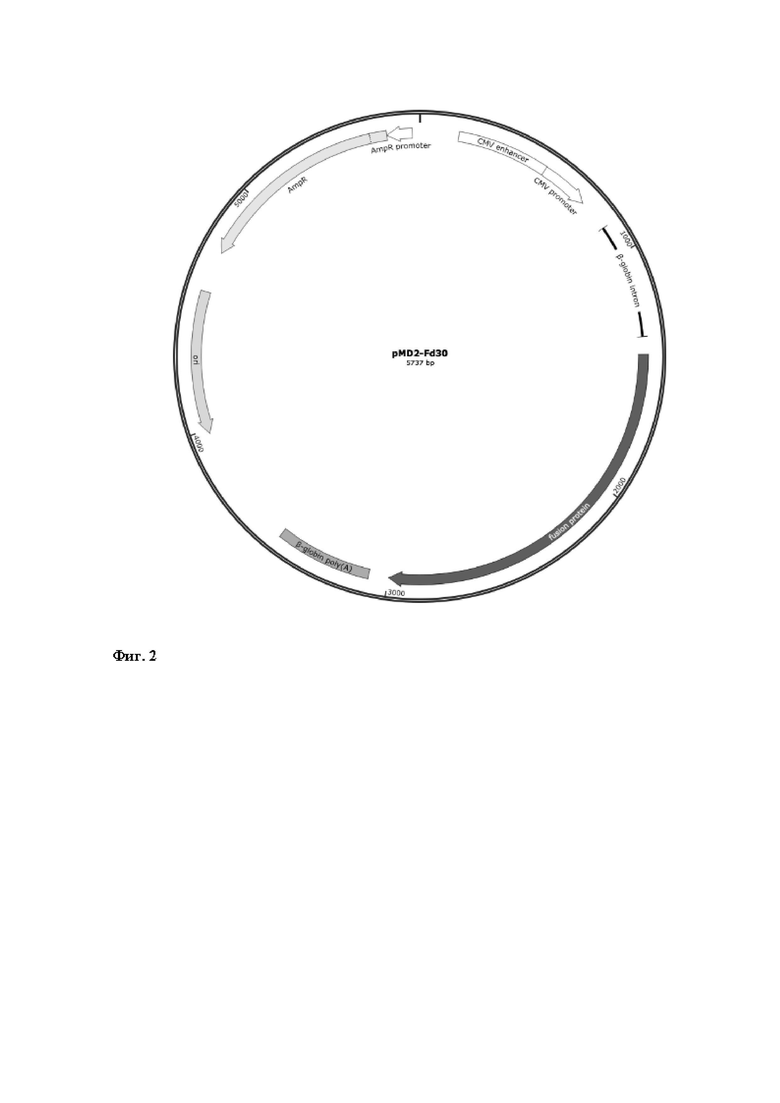
b. Рекомбинантная плазмидная ДНК pMD2-F∆30 (Фиг. 2) состоящая из следующих ключевых генетических элементов:
1. ориджина репликации ColE1/pMB1/pBR322/pUC;
2. гена устойчивости к антибиотику ампициллину (AmpR) и бактериального промотора гена устойчивости к ампициллину (AmpR promoter);
3. промотора цитомегаловируса (CMV);
4. последовательности, кодирующей поверхностный гликопротеин F вакцинного штамма вируса кори с укороченным на 30 аминокислот С-концевым цитоплазматическим доменом (F∆30).
c. Плазмидная ДНК pCG-4AHc∆24 (Journal of Virology Jul 2002, 76 (14) 7174-7186), кодирующая поверхностный гликопротеин H вакцинного штамма вируса кори (Фиг. 3).
3. Перечисленными плазмидными ДНК (Фиг. 1, 2 и 3) с использованием полиэтиленимина 25 кДА (PEI-25, Polysciences, США) были котрансфицированы клетки линии HEK-293T; плазмиды смешивались в соотношении 8 : 7 : 1, соответственно.
4. Культуры инфицированных клеток OMK и трансфицированных клеток HEK-293 были подвергнуты совместному культивированию в соотношении 1:7, соответственно; для пассирования клеток была использована ростовая среда Eagle's Minimum Essential Medium, Catalog No. 30-2003. В ходе микроскопического исследования совместно культивируемых линий было зафиксировано формирование многоядерных синцитиев из клеток обеих культур.
5. Совместно культивируемые клетки, сформировавшие многоядерные синцитии, были использованы для многократного отбора ростовой среды, содержащей рекомбинантные вирионы. Вируссодержащая среда была подвергнута центрифугированию (5000 g; 15 минут) для удаления клеточного дебриса с последующим определением вирусного титра методом Рида-Менча (1.28E+04).
6. Вируссодержащим супернатантом были заражены культуры ОМК; по прошествии семи дней в конфлюэнтной клеточной культуре было зафиксировано формирование бляшек, сформированных преимущественно флуоресцентными клетками.
7. Для клонирования рекомбинантного HVS-OFP, на лунки 96-луночного планшета, в которые за 24 часа до этого были рассеяны свежие клетки линии ОМК в субконфлюэнтном количестве, была добавлена культуральная среда, содержащая отсортированные инфицированные и флуоресцирующие клетки ОМК из расчета 0.1 инфицированная клетка на лунку. Через 10 дней после инфекции в образцах, подвергшихся заражению, были зафиксированы единичные флуоресцентные бляшки, образованные индивидуальными рекомбинировавшими вирусными частицами. Вируссодержащая среда с лунок, в которых наблюдалось наиболее выраженное развитие литического процесса, была использована для наработки препаративных количеств рекомбинантного вирусного препарата и его анализа.
8. Наработанный образец вируса был проанализирован в ходе секвенирования, показавшего успешную интеграцию целевой экспрессионной кассеты в области vCD59 генома (SEQ ID NO : 3).
9. Заражение полученным препаратом рекомбинантного герпесвируса Т-лимфоцитов и НК-клеток было проведено из расчета 10 вирусных частиц на 1 клетку. Процесс заражения был проведен в течение 8 часов в бессывороточной питательной среде AIM-V, после чего клетки были осаждены центрифугированием и были рассажены на свежую порцию питательной среды AIM-V.
10. Пролиферативная активность инфицированных культур была оценена в ходе стандартизированных протоколов систематического подсчета клеток в камере Горяева.
11. Верификация эффективности введения в клетки экспрессионной кассеты была проведена с помощью флуоресцентной микроскопии, в ходе которой был определен процент OFP-позитивных лимфоцитов к общему числу инфицированных клеток.
Анализ эффективности разработанной методики получения рекомбинантных герпесвирусов для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток
Конкурентное преимущество предложенной методики базируется на использовании системы из двух линий клеток: линии, пермиссивной к литической инфекции вирусом Herpesvirus Saimiri, и вспомогательной линии, характеризующейся высокой эффективностью трансфекции. Верификация ценности изобретения была построена на сопоставлении предложенной методики с традиционными способами наработки рекомбинантных герпесвирусов, подразумевающими использование одной линии клеток.
В рамках тестирования традиционной методики вирус-пермиссивные клетки линии ОМК были трансфицированы экспрессионной плазмидой pUC-HVS-OFP, несущей ген устойчивости к пуромицину (pac), ген оранжевого флуоресцентного белка (OFP), фланкированные участками (600 п.н.), гомологичными области vCD59 генома HVS; процедура была проведена с использованием катионного полимера PEI-25 в заранее подобранных условиях (соотношение PEI:ДНК 6:1, 10 мкг ДНК на 2 млн клеток). Ростовая среда, собранная с трансфицированных и зараженных вирусом ОМК клеток, была использована для реинфекции клеток. Эффективность получения рекомбинантного вируса была оценена по процентному соотношению флуоресцентных клеток в реинфицированной популяции ОМК; полученные значения не превышали 3% (Фиг. 4 А).
Тестирование эффективности изобретения было проведено согласно пунктам 1-6 разработанной методики получения рекомбинантных герпесвирусов, также подробно описанным в разделе “Осуществление изобретения”. Эффективность получения рекомбинантного вируса была оценена по процентному соотношению флуоресцентных клеток в инфицированной популяции ОМК; полученные результаты (Фиг. 4 В) достоверно превышали значения, полученные при использовании традиционного протокола.
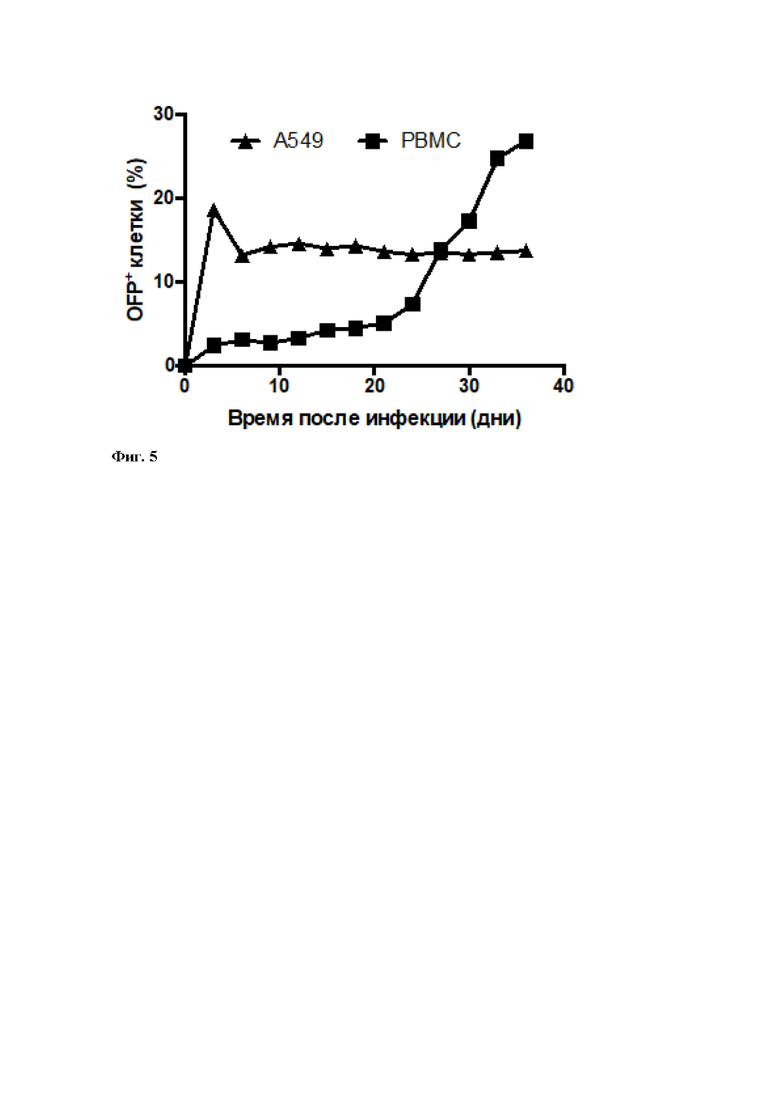
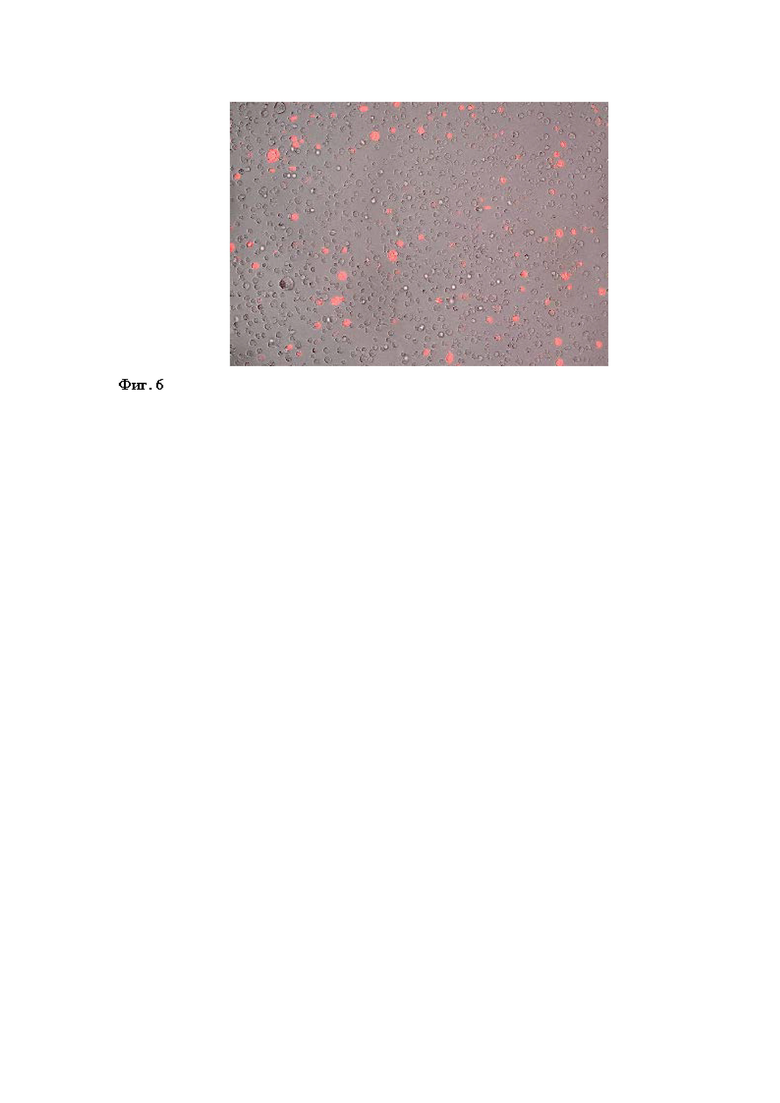
Для оценки возможности использования вируса для одновременной трансдукции и иммортализации лимфоцитов, полученный препарат был использован для трансдукции периферических мононуклеарных клеток, а также линии А549, использованной в качестве контроля. Анализ популяций клеток в ходе проточной цитометрии показал образование фракций, флуоресцирующих OFP-позитивных клеток: их доля для линии А549 сохранялась постоянной в течение 5 недель после инфекции, а для PBMC - постоянно увеличивалась (Фиг. 5, 6). Полученные данные позволяют заключить, что вирус вызвал иммортализацию человеческих периферических мононуклеарных клеток, в то время как введенная в геном вируса экспрессионная кассета сохранила работоспособность как в процессе литической инфекции, так и в латентной форме существования герпесвируса.
Возможность использования изобретения для решения заявленных задач в сочетании с эффективностью, превосходящей эффективность существующих протоколов, подтверждают конкурентное преимущество предложенной методики и позволяют рекомендовать ее для получения рекомбинантных герпесвирусов для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток.
Изобретение иллюстрировано следующими материалами:
Фиг. 1 – Схематичное изображение генетической конструкции pUC-HVS-OFP.
Фиг. 2 – Схематическое изображение генетической конструкции pMD2-Fd30.
Фиг. 3 – Схематическое изображение генетической конструкции pCD-4AHcd24.
Фиг. 4 – Доля OFP-позитивных клеток в общей популяции ОМК, инфицированных препаратами рекомбинантного герпесвируса, наработанным согласно традиционному протоколу (А) и разработанной методики (В).
Фиг. 5 – Динамика изменения численности флуоресцентных популяций после инфекции HVS-OFP.
Фиг. 6 - Доля OFP-позитивных клеток в общей популяции PBMC, инфицированных препаратом рекомбинантного герпесвируса HVS-OFP.
Перечень последовательностей
SEQ ID NO: 1 – 5'-концевой фланкирующий участок, гомологичный области vCD59 генома HVS ACAGGCTGCTCTTCAGGAGCACCAGAAGAAGGTCGAATTGCGTTCCTTCTCAAAAGAGGAGGCATGTTTGCAAATGAAATGAGAATCTGGTGAGATAAGATGTATTTGCATGAAGCTTCTATTTATACTACATTAGAGGCATTTTTCAGAAGCAAAAATGCCTCTAATTATATACACTGTACTATTTACCTCTATTACACATTTTCTATTTTAAGTCTGAAAGTGATTAATCAAGAAAGAAGTTTGTGGTTCCCTGGAGATTAGTTCACAAGCTGTCTGAGGTTAAAGGTGGTTCTTTAGCACTGACACACAAGTTGCTATAAGAATTGAAGCTTGCTTTATAAAAAGTTACTTGTGTTTAATTACTATTAAAATAAAGAGAATGTATATTTTGTTTAAGTTGATGCTTGCTTCTGTTTTTTGCAAGTCAAGCTACAGCTTGCAATGCTACAACTGTTCTCACTCAACTATGCAGTGTACAACATCTACTAGTTGTACATCTAATCTTGATTCTTGTCTCATTGCTAAAGCTGGGTCAAAAGTATATTACAGGTGCTGGAAGTTTGATGACTGTAGCTTCAAACGCATCTCAAATCAATTG
SEQ ID NO: 2 – 3'-концевой фланкирующий участок, гомологичный области vCD59 генома HVS
TCTGAAACACAGTTAAAGTATCATTGTTGTAAGAAAAACTTGTGTAATGTTAACAAAGTGATTGAAAATGGAAAAAGAACAATATCAGATAAGGCTCTTTTACTATTAGCATTGTTTTTAGTAACTGCTTAGAACCTTTCTCTTTAAAAGTGAACAACATACCTATATTGTAACATTTATTTTTGCGTAGCTTATTCGTATTGCTATTACAAGTTAAAATATTGTGTTTTTAACTATAATTTTTAAAAAGATAAAATGAGATGTAGTATATTACCCATAGTCAAAATTAAAGTGCTAGATATTATTAGCATTTTTATCAACAACGCAAATAAAAGTTAAGATACATTATTTTTTTGATATTTGGATTATTGTGTGCTTTTTATCATATGTTAAAAGTTTTATGTCATTTTATTCTTACATATATAAAGCTAAATTTTAAAGCAACTTATCAGTAGCATCTTAGCTTCTGATCTGTACAGACTTATATAATATGGGTTTATCCTTAAGAAAAAACAAAGAAGAAAAAAATAACACAGTGCCAAACTTGCCAGTTAATTACATGTTAAGAAATCCTGTTTATAAAAAGAAATATCTGCCAGC
SEQ ID NO : 3 – нуклеотидная последовательность участка генома рекомбинантного вируса HVS-OFP с интегрированной кассетой, расположенная между последовательностями вирусных генов IE14-C и vBcl-2.
CTGGTGAGATAAGATGTATTTGCATGAAGCTTCTATTTATACTACATTAGAGGCATTTTTCAGAAGCAAAAATGCCTCTAATTATATACACTGTACTATTTACCTCTATTACACATTTTCTATTTTAAGTCTGAAAGTGATTAATCAAGAAAGAAGTTTGTGGTTCCCTGGAGATTAGTTCACAAGCTGTCTGAGGTTAAAGGTGGTTCTTTAGCACTGACACACAAGTTGCTATAAGAATTGAAGCTTGCTTTATAAAAAGTTACTTGTGTTTAATTACTATTAAAATAAAGAGAATGTATATTTTGTTTAAGTTGATGCTTGCTTCTGTTTTTTGCAAGTCAAGCTACAGCTTGCAATGCTACAACTGTTCTCACTCAACTATGCAGTGTACAACATCTACTAGTTGTACATCTAATCTTGATTCTTGTCTCATTGCTAAAGCTGGGTCAAAAGTATATTACAGGTGCTGGAAGTTTGATGACTGTAGCTTCAAACGCATCTCAAATCAATTGCTCGAGGTAACGCCATTTTGCAAGGCATGGAAAAATACCAAACCAAGAATAGAGAAGTTCAGATCAAGGGCGGGTACATGAAAATAGCTAACGTTGGGCCAAACAGGATATCTGCGGTGAGCAGTTTCGGCCCCGGCCCGGGGCCAAGAACAGATGGTCACCGCAGTTTCGGCCCCGGCCCGAGGCCAAGAACAGATGGTCCCCAGATATGGCCCAACCCTCAGCAGTTTCTTAAGACCCATCAGATGTTTCCAGGCTCCCCCAAGGACCTGAAATGACCCTGCGCCTTATTTGAATTAACCAATCAGCCTGCTTCTCGCTTCTGTTCGCGCGCTTCTGCTTCCCGAGCTCTATAAAAGAGCTCACAACCCCTCACTCGGCGCGCCAGTCCTCCGACAGACTGAGTCGGAATTCGCAACAAACTTCTCTCTGCTGAAACAAGCCGGAGATGTCGAAGAGAATCCTGGACCGATGACCGAGTACAAGCCCACGGTGCGCCTCGCCACCCGCGACGACGTCCCCAGGGCCGTACGCACCCTCGCCGCCGCGTTCGCCGACTACCCCGCCACGCGCCACACCGTCGATCCGGACCGCCACATCGAGCGGGTCACCGAGCTGCAAGAACTCTTCCTCACGCGCGTCGGGCTCGACATCGGCAAGGTGTGGGTCGCGGACGACGGCGCCGCGGTGGCGGTCTGGACCACGCCGGAGAGCGTCGAAGCGGGGGCGGTGTTCGCCGAGATCGGCCCGCGCATGGCCGAGTTGAGCGGTTCCCGGCTGGCCGCGCAGCAACAGATGGAAGGCCTCCTGGCGCCGCACCGGCCCAAGGAGCCCGCGTGGTTCCTGGCCACCGTCGGCGTCTCGCCCGACCACCAGGGCAAGGGTCTGGGCAGCGCCGTCGTGCTCCCCGGAGTGGAGGCGGCCGAGCGCGCCGGGGTGCCCGCCTTCCTGGAGACCTCCGCGCCCCGCAACCTCCCCTTCTACGAGCGGCTCGGCTTCACCGTCACCGCCGACGTCGAGTGCCCGAAGGACCGCGCGACCTGGTGCATGACCCGCAAGCCCGGTGCCTCTAGAGGCAGTGGAGAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGCCCAATGAACCTGAGCAAAAACGTGAGCGTGAGCGTGTATATGAAGGGGAACGTCAACAATCATGAGTTTGAGTACGACGGGGAAGGTGGTGGTGATCCTTATACAGGTAAATATTCCATGAAGATGACGCTACGTGGTCAAAATTCCCTACCCTTTTCCTATGATATCATTACCACGGCATTTCAGTATGGTTTCCGCGTATTTACAAAATACCCTGAGGGAATTGTTGACTATTTTAAGGACTCGCTTCCCGACGCATTCCAGTGGAACAGACGAATTGTGTTTGAAGATGGTGGAGTACTAAACATGAGCAGTGATATCACATATAAAGATAATGTTCTGCATGGTGACGTCAAGGCTGAGGGAGTGAACTTCCCGCCGAATGGGCCAGTGATGAAGAATGAAATTGTGATGGAGGAACCGACTGAAGAAACATTTACTCCAAAAAACGGGGTTCTTGTTGGCTTTTGTCCCAAAGCGTACTTACTTAAAGACGGTTCCTATTACTATGGAAATATGACAACATTTTACAGATCCAAGAAATCTGGCCAGGCACCTCCTGGGTATCACTTTGTTAAGCATCGTCTCGTCAAGACCAATGTGGGACATGGATTTAAGACGGTTGAGCAGACTGAATATGCCACTGCTCATGTCAGTGATCTTCCCAAGTTCGAAGCTTGACCTGCAGGTCTGAAACACAGTTAAAGTATCATTGTTGTAAGAAAAACTTGTGTAATGTTAACAAAGTGATTGAAAATGGAAAAAGAACAATATCAGATAAGGCTCTTTTACTATTAGCATTGTTTTTAGTAACTGCTTAGAACCTTTCTCTTTAAAAGTGAACAACATACCTATATTGTAACATTTATTTTTGCGTAGCTTATTCGTATTGCTATTACAAGTTAAAATATTGTGTTTTTAACTATAATTTTTAAAAAGATAAAATGAGATGTAGTATATTACCCATAGTCAAAATTAAAGTGCTAGATATTATTAGCATTTTTATCAACAACGCAAATAAAAGTTAAGATACATTATTTTTTTGATATTTGGATTATTGTGTGCTTTTTATCATATGTTAAAAGTTTTATGTCATTTTATTCTTACATATATAAAGCTAAATTTTAAAGCAACTTATCAGTAGCATCTTAGCTTCTGATCTGTACAGACTTATATAATATGGGTTTATCCTTAAGAAAAAACAAAGAAGAAAAAAATAACACAGTGCCAAACTTGCCAGTTAATTACATGTTAAGAAATCCTGTTTATAAAAAGAAATATCTGCCAGCAAATATTTGGGAATCTAGAAATTCTGTTTTTTGGAACAGTGTCTATGTTGTTAATTCAAAAGCTAAATAAAAGCTTATTTATAGCATGTTGTTATTTAGAAGAGATAATTTTATGAAAACTATAAATTTGAAAGTAAGAAATCTAATGCTTATTAAAAGTGTGTTATAAAAAGCAAGAAGCTACATTATGTACTTAAAAATAAAATTAGTATATGAATTTATGTTTAACAAGCATATTCATAACAGCAGCTGAGTTACCACATCTAAGAAAAT
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный штамм Herpesvirus saimiri для получения популяций иммортализованных аутологичных Т-лимфоцитов, экспрессирующих парные химерные рецепторы против опухолевых антигенов CD44 и CD133 | 2020 |
|
RU2771081C2 |
| Набор олигонуклеотидных праймеров и зондов для выявления Herpesvirus Saimiri в клеточных культурах методом ПЦР в реальном времени | 2020 |
|
RU2753767C1 |
| РЕКОМБИНАНТНЫЙ ГЕРПЕСВИРУС КОИ (KHV) И ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЯ, ВЫЗЫВАЕМОГО KHV | 2012 |
|
RU2662768C2 |
| ПОЛУЧЕНИЕ ВИРУСНОГО ВЕКТОРА | 2014 |
|
RU2658487C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЦИТОТОКСИЧЕСКИХ Т-ЛИМФОЦИТОВ, ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2714380C1 |
| СПОСОБ СОЗДАНИЯ РЕКОМБИНАНТНОГО АДЕНОВИРУСА ПТИЦ ДЛЯ ВАКЦИНАЦИИ И ГЕННОЙ ТЕРАПИИ | 2007 |
|
RU2326942C1 |
| СПОСОБ СОЗДАНИЯ РЕКОМБИНАНТНОГО АДЕНОВИРУСА ПТИЦ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИРУСА ГРИППА ПТИЦ Н5N1 | 2006 |
|
RU2326943C1 |
| ХИМЕРНЫЕ ВАКЦИННЫЕ АНТИГЕНЫ ПРОТИВ ВИРУСА КЛАССИЧЕСКОЙ ЧУМЫ СВИНЕЙ | 2007 |
|
RU2406534C2 |
| ХИМЕРНЫЙ ГЕН CR3 И КОДИРУЕМЫЙ ИМ ХИМЕРНЫЙ БЕЛОК CR3 (ВАРИАНТЫ), ИНДУЦИРУЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ-1 | 2002 |
|
RU2302461C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |

Изобретение относится к биотехнологии, а именно к способу получения рекомбинантных герпесвирусов для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток. Рекомбинантная плазмидная ДНК pUC-HVS-OFP состоит из следующих ключевых генетических элементов: гена устойчивости к пуромицину (рас); гена оранжевого флуоресцентного белка (OFP); фланкирующих участков (600 п.н.), гомологичных области vCD59 генома HVS; гена устойчивости к антибиотику ампициллину (AmpR) и бактериального промотора гена устойчивости к ампициллину (AmpR promoter); ориджина репликации бактериофага f1 (ori); промотора цитомегаловируса (CMV). Пермиссивную к литической инфекции вирусом Herpesvirus Saimiri культуру клеток заражают штаммом вируса Herpesvirus Saimiri HVS-C488. Другую перевиваемую линию клеток трансфицируют смесью из экспрессионных плазмид и плазмиды pUC-HVS-OFP. Затем инфицированную Herpesvirus Saimiri пермиссивную клеточную культуру и трансфицированную экспрессионными плазмидами клеточную культуру смешивают и подвергают ко-культивации. Совмещение производительной наработки вируса в клетках линий, пермиссивных к литической инфекции HVS, с высокой эффективностью трансфекции клеток экспрессионными плазмидами, кодирующими генетические последовательности, предназначенные для введения в геном «дикого» штамма вируса, позволяет получить рекомбинантные герпесвирусы с повышенной эффективностью наработки. 2 н. и 4 з.п. ф-лы, 6 ил.
1. Рекомбинантная плазмидная ДНК pUC-HVS-OFP, предназначенная для введения генетических последовательностей в область vCD59 генома Herpesvirus Saimiri, состоящая из следующих ключевых генетических элементов:
а) гена устойчивости к пуромицину (pac);
б) гена оранжевого флуоресцентного белка (OFP);
в) фланкирующих участков (600 п.н.), гомологичных области vCD59 генома HVS, имеющих последовательности SEQ ID NO: 1 и SEQ ID NO: 2;
г) гена устойчивости к антибиотику ампициллину (AmpR) и бактериального промотора гена устойчивости к ампициллину (AmpR promoter);
д) ориджина репликации бактериофага f1 (ori);
е) промотора цитомегаловируса (CMV);
как представлено на Фиг.1.
2. Способ получения рекомбинантных герпесвирусов, характеризующийся следующими параметрами:
пермиссивную к литической инфекции вирусом Herpesvirus Saimiri культуру клеток заражают штаммом вируса Herpesvirus Saimiri HVS-C488, а другую перевиваемую линию клеток трансфицируют смесью из экспрессионных плазмид и плазмиды по п.1, которая содержит целевой участок, предназначенный для введения в геном вируса штамма Herpesvirus Saimiri HVS-C488, фланкированный последовательностями протяженностью 300-900 пар нуклеотидных оснований, гомологичных тому участку генома Herpesvirus Saimiri, в который производится введение целевого участка, а две другие экспрессионные плазмиды кодируют поверхностные гликопротеины F и H вакцинного штамма вируса кори, после чего инфицированную Herpesvirus Saimiri пермиссивную клеточную культуру и трансфицированную экспрессионными плазмидами клеточную культуру смешивают и подвергают ко-культивации, в результате чего происходит образование многоядерных синцитиев из клеток обеих культур, что обеспечивает прохождение рекомбинации гомологичных участков, внедрение целевого гена в геном Herpesvirus Saimiri и выход во внеклеточную жидкость рекомбинантных герпесвирусов.
3. Способ по п. 2, отличающийся тем, что в качестве пермиссивной к литической инфекции Herpesvirus Saimiri культуры клеток используют культуру клеток ОМК (owl monkey kidney cells, почечный эпителий ночных обезьян).
4. Способ по п. 2, отличающийся тем, что в качестве перевиваемой клеточной линии, которую трансфицируют смесью экспрессионных плазмид и плазмиды по п. 1, используют клеточную линию HEK-293 либо ее производные.
5. Способ по п. 2, отличающийся тем, что в качестве последовательности протяженностью 300-900 пар нуклеотидных оснований, гомологичной участку генома Herpesvirus Saimiri, используют последовательность вирусного гена vCD59.
6. Способ по п. 2, отличающийся тем, что в состав целевого участка, предназначенного для введения в геном штамма вируса Herpesvirus Saimiri HVS-C488, входят последовательности, кодирующие флуоресцентный белок и химерный антигенный рецептор.
| WO 2004078911 A, 16.09.2004 | |||
| US 5670352 A, 23.09.1997 | |||
| ХАМАД А | |||
| и др | |||
| Получение рекомбинантного штамма Herpesvirus saimiri путем совместной культивации трансфицированной и пермиссивной клеточной культур, Вестник РГМУ, 6, 2019, с | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |

