Изобретение относится к биотехнологии и может быть использовано в исследовательских и диагностических целях для эффективного получения генетически- модифицированных иммортализованных первичных Т-лимфоцитов.
Лабораторно-клиническая практика получения аутологичных или гетерологичных пациент-специфичных лимфоидных линий требует разработки и создания инструментов, обеспечивающих возможность эффективной иммортализации и генетической модификации первичных культур. Эффективным методом иммортализации первичных Т-лимфоцитов является их заражение Гамма-2 герпесвирусом беличьих обезьян Herpesvirus saimiri (HVS), обеспечивающим антигеннезависимую пролиферацию инфицированных культур при сохранении ими функциональной активности и зрелого фенотипа. Использование рекомбинантного HVS также возможно для доставки в клетки протяженных экспрессионных конструкций, благодаря наличию в геноме вируса участков, использование которых в качестве сайтов интеграции целевых генетических последовательностей не оказывает влияния на жизнеспособность вируса. Сочетание этих особенностей делает Herpesvirus Saimiri перспективным инструментом, позволяющим совместить трансдукцию первичных Т- лимфоцитов с их иммортализацией и добиться получения пролиферирующих популяций Т-клеток, экспрессирующих химерные антигенные рецепторы против заданных антигенов.
Распознавание единичного антигена и последующая полная активация цитотоксического действия иммунной клетки целесообразна, если в качестве мишени для нацеливания клеточного иммунотерапевтического препарата использован опухолевый неоантиген, не экспрессирующийся на поверхности нормальных клеток организма. Многие новообразования характеризуются отсутствием уникальных антигенов, однако обладают отличительным паттерном экспрессии нескольких редких рецепторов, каждый из которых по отдельности может встречаться на поверхности нормальных клеток. В этом случае более эффективным оказывается таргетирование клеточных препаратов на сочетание двух антигенов, при условии, что связывание каждого из них в отдельности не должно приводить к полной активации цитотоксического действия во избежание неспецифической активности препарата и побочных эффектов.
Перспективной мишенью для нацеливания клеточных иммунотерапевтических препаратов служит комбинация поверхностных гликопротеинов CD44 и CD133. Экспрессия CD 133 характерна для популяций раковых стволовых клеток опухолей головного мозга; содержание CD133(+) клеток при глиомах отражает степень анаплазии новообразования и служит прогностическим фактором, обратно коррелирующим с продолжительностью жизни больных. CD44 - другой распространенный маркер раковых стволовых клеток, служит рецептором гиалуроновой кислоты, повышенный уровень которой оказывается характерен для микроокружения стволовых клеток и новообразований, в частности, первичных опухолей головного мозга. Использование парных химерных рецепторов против опухолевых антигенов CD44 и CD 133 должно обеспечивать эффективное нацеливание Т-лимфоцитов на наиболее злокачественные клетки глиобластом при минимизации побочных эффектов со стороны организма.
Возможным решением проблемы является техническое решение, предлагаемое в заявке /US5670352A, C12N15/65, опубл.23.09.1997/, описывающее использование штаммов Herpesvirus saimiri с целью трансформации и генетической модификации Т- лимфоцитов, а также с целью обеспечения антигеннезависимой пролиферации инфицированных культур при сохранении ими функциональной активности и зрелого фенотипа. В заявке не оговаривается возможность использования изобретения для получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов CD44 и CD 133; более того, предметом заявки является методика, а не рекомбинантный штамм Herpesvirus Saimiri.
Известен генетически модифицированный штамм Herpesvirus Saimiri, обеспечивающий возможность доставки в клетки гетерологичной ДНК /международная заявка WO1998010083A1, C12N15/86, опубл. 12.03.1998 г./. В заявке отсутствует описание возможности использования изобретения с целью одновременной трансдукции и иммортализации первичных Т-лимфоцитов, в частности получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов CD44 и CD133.
Техническое решение, приведенное в заявке /US20200032219A1, C12N7/00, опубл. 30.01.2020 г./, описывает генетически модифицированный, в частности в ходе гомологичной рекомбинации, штаммам вируса герпеса, используемый для противоопухолевой терапии. Заявленный терапевтический эффект достигается за счет прямого цитотоксического воздействия вируса на клетки опухоли in vivo; в заявке не оговаривается возможность использования изобретения с целью одновременной трансдукции и иммортализации первичных Т-лимфоцитов.
Другие аналоги, в частности, техническое решение /JPH11513565A, C12N15/86, опубл. 24.11.1999г./ раскрывают возможности использования рекомбинантных генетических векторов на основе вируса герпеса, в частности, Herpesvirus saimiri (HVS), с целью трансдукции и генетической модификации широкого спектра культур клеток человека и животных, в частности, клеток лимфоидных линий. Предметом данной заявки служит методика использования генетических векторов на основе герпесвирусов, не включающая описание рекомбинантных штаммов Herpesvirus Saimiri.
Известна генетическая конструкция, кодирующая структуру Т-клеточного химерного рецептора на основе одноцепочечного VHH-антитела, специфичного к опухолевому рецептору CD47, содержащая в качестве антиген-распознающей части последовательность моноклонального одноцепочечного VHH-антитела, а также последовательности, кодирующие сигнальную последовательность рецептора CD8, последовательность шарнирного домена IgG4 длиной 12 аминокислот, последовательность трансмембранного домена рецептора CD28, и последовательности, кодирующие сигнальные домены рецепторов CD28, CD137 и CD3Z /патент 2707535, заявка 2017146641, опубл. 28.12.2017/, которая позволяет осуществлять экспрессию Т-клеточного химерного рецептора в Т-лимфоцитах путем трансдукции их лентивирусным вектором.
Таким образом, проведенные исследования патентной литературы позволяют заключить, что заявленное техническое решение характеризуется научной новизной, не имеет полных аналогов и обладает конкурентными преимуществами для получения рекомбинантных герпесвирусов, обеспечивающих возможность одновременной трансдукции и иммортализации первичных Т-лимфоцитов.
Технической проблемой, на решение которой направлено заявляемое изобретение, является создание инструмента, обеспечивающего возможность одновременной иммортализации и генетической модификации первичных Т-лимфоцитов, а именно: получение рекомбинантного штамма Herpesvirus Saimiri, способного вызывать иммортализацию человеческих периферических Т-лимфоцитов и содержащего в своем геноме генетическую последовательность для экспрессии парного химерного антигенного рецептора против опухолевых антигенов CD44 и CD133, предназначенного для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов для терапии злокачественных новообразований, в частности, глиобластом.
Заявленная техническая проблема решается за счет создания рекомбинантного штамма Herpesvirus Saimiri, предназначенного для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов, характеризующегося следующими параметрами:
Гетерологичная ДНК, предназначенная для введения в геном вируса "дикого" штамма Herpesvirus Saimiri, включает в себя генетические последовательности, кодирующие парные химерные рецепторы против опухолевых антигенов CD44 (с последовательностью антиген-распознающей части химерного рецептора на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD44 SEQ ID NO.1) и CD133 (с последовательностью антиген-распознающей часть химерного рецептора на основе моноклонального одноцепочечного VHH- антитела, специфичного к опухолевому антигену CD 133 SEQ ID NO.2).
В составе указанной ДНК, последовательность, кодирующая первый активирующий химерный рецептор, включает в себя N-концевую сигнальную последовательность рецептора CD8, а также участки, кодирующие антиген-связывающий домен на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD44, спейсерную область, представленную 12 аминокислотной последовательностью шарнирного домена иммуноглобулина G4, трансмембранный домен CD28 и сигнальный домен CD3zeta; последовательность, кодирующая второй костимуляторный химерный рецептор включает в себя N-концевую сигнальную последовательность рецептора CD8, а также участки, кодирующие антиген- связывающий домен на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD133, шарнирную область длиной 228 аминокислот, трансмембранный и сигнальный домены рецептора CD28, а также сигнальный домен рецептора CD137.
В составе указанной ДНК, последовательность, предназначенная для введения в геном вируса "дикого" штамма Herpesvirus Saimiri, фланкирована участками протяженностью 300-900 пар нуклеотидных оснований, гомологичных тому участку генома Herpesvirus Saimiri, в который производится интеграция целевого фрагмента посредством гомологичной рекомбинации (SEQ ID NO.3).
В составе указанной ДНК, в качестве последовательности протяженностью 300900 пар нуклеотидных оснований, гомологичной участку генома Herpesvirus Saimiri, используют последовательность вирусного гена vCD59 (SEQ ID NO.4).
В составе указанной ДНК, в состав целевого участка, предназначенного для введения в геном "дикого" штамма вируса Herpesvirus Saimiri, входит последовательность, кодирующая флуоресцентный белок (SEQ ID NO.5).
Предлагается рекомбинантный штамм Herpesvirus Saimiri, предназначенный для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов CD44 и CD133, содержащий в своем геноме последовательность ДНК, указанную выше.
Получение рекомбинантного штамма Herpesvirus Saimiri, способного вызывать иммортализацию человеческих периферических Т-лимфоцитов и содержащего в своем геноме генетическую последовательность для экспрессии парного химерного антигенного рецептора против опухолевых антигенов CD44 и CD133.
Пермиссивная к литической инфекции HVS культура клеток OMK была инфицирована "диким" штаммом вируса HVS-C488 (VR-1414) ATCC (США).
Использованные генетические конструкции:
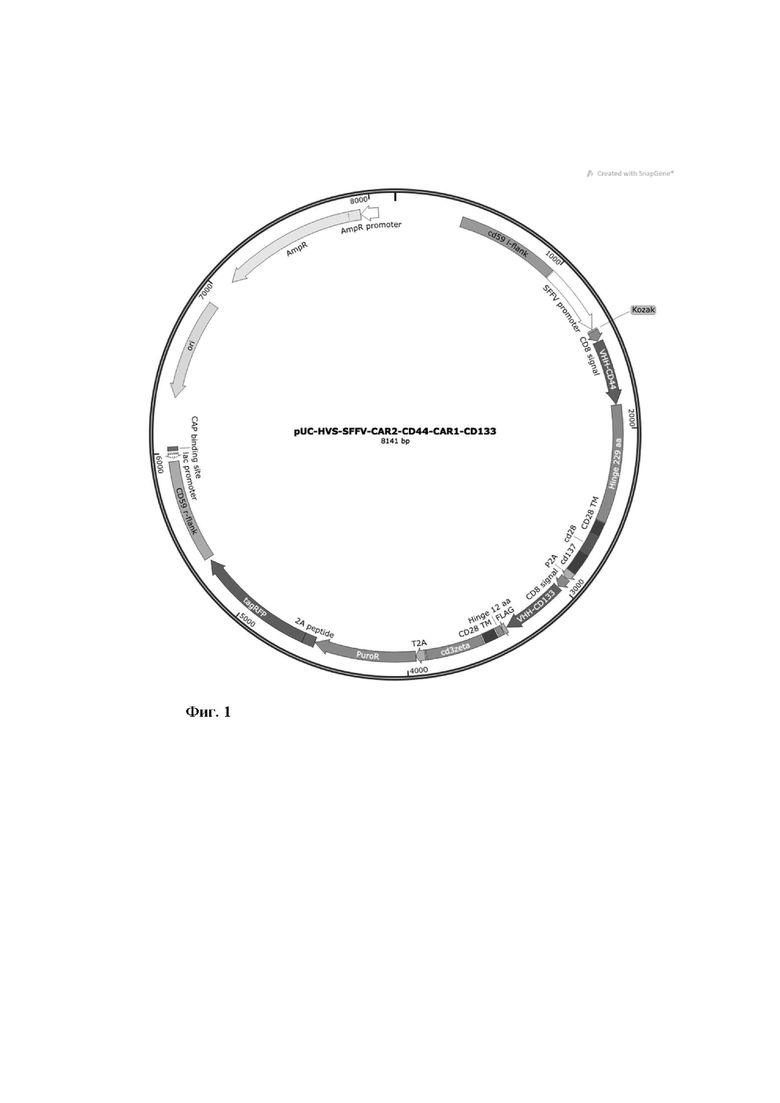
экспрессионная плазмида pUC-HVS-SFFV-CAR2@CD44- CAR1@CD133, включающая в себя ген устойчивости к пуромицину (pac), эпитоп FLAG (DYKDDDDK) для осуществления иммуномагнитной сепарации трансдуцированных клеток и генетическую последовательность, кодирующую парные химерные рецепторы против опухолевых антигенов CD44 и CD 133, фланкированные участками (600 п.н.), гомологичными области vCD59 генома HVS (Фиг. 1);
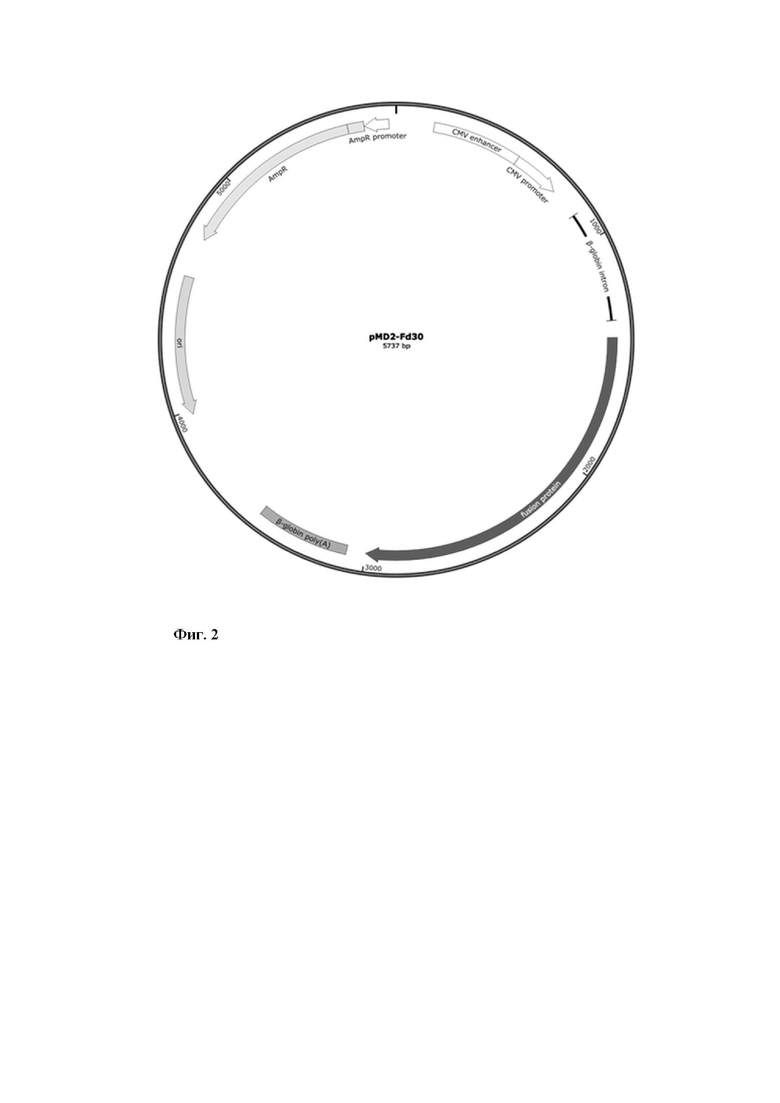
экспрессионная плазмида pMD2-FA30, кодирующая поверхностный гликопротеин F вакцинного штамма вируса кори (Фиг. 2);
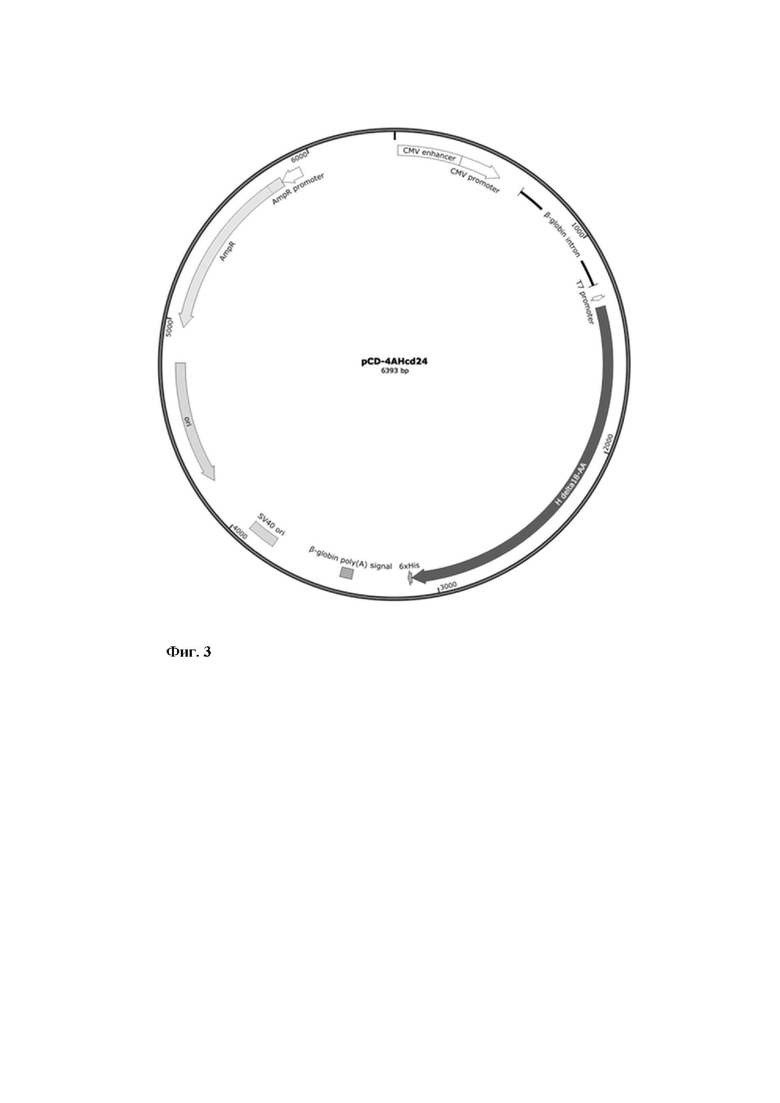
экспрессионная плазмида pCG-4AHcA24, кодирующая поверхностный гликопротеин H вакцинного штамма вируса кори (Фиг. 3).
Перечисленными экспрессионными плазмидами (2a, 2b и 2c) с использованием полиэтиленимина 25 кДа (PEI-25, Polysciences, США) были котрансфицированы клетки линии HEK-293T; плазмиды смешивались в соотношении 8 : 7 : 1, соответственно.
Культуры инфицированных клеток OMK и трансфицированных клеток HEK- 293T были подвергнуты совместному культивированию в соотношении 1:7, соответственно; для пассирования клеток была использована ростовая среда Eagle's Minimum Essential Medium, Catalog No. 30-2003. В ходе микроскопического исследования совместно культивируемых линий было зафиксировано формирование многоядерных синцитиев из клеток обеих культур.
Совместно культивируемые клетки, сформировавшие многоядерные синцитии, были использованы для многократного отбора ростовой среды, содержащей рекомбинантные вирионы. Вируссодержащая среда была подвергнута центрифугированию (5000 g; 15 минут) для удаления клеточного дебриса с последующим определением вирусного титра методом Рида-Менча (1.28E+04).
Вируссодержащим супернатантом были заражены культуры ОМК; по прошествии семи дней в конфлюэнтной клеточной культуре было зафиксировано формирование бляшек.
Для клонирования рекомбинантного HVS, на лунки 96-луночного планшета, в которые за 24 часа до этого были рассеяны свежие клетки линии ОМК в субкофлюэнтном количестве, была добавлена культуральная среда, содержащая отсортированные в ходе иммуномагнитной сепарации инфицированные клетки ОМК из расчета 0.1 инфицированная клетка на лунку. Через 10 дней после инфекции в образцах, подвергшихся заражению, были зафиксированы единичные бляшки, образованные индивидуальными рекомбинировавшими вирусными частицами. Вируссодержащая среда с лунок, в которых наблюдалось наиболее выраженное развитие литического процесса, была использована для наработки препаративных количеств рекомбинантного вирусного препарата и его анализа.
Наработанный образец вируса был проанализирован в ходе секвенирования, показавшего успешную интеграцию целевой экспрессионной кассеты в области vCD59 генома.
Анализ эффективности использования рекомбинантного штамма Herpesvirus Saimiri для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов CD44 и CD133, для терапии глиобластом.
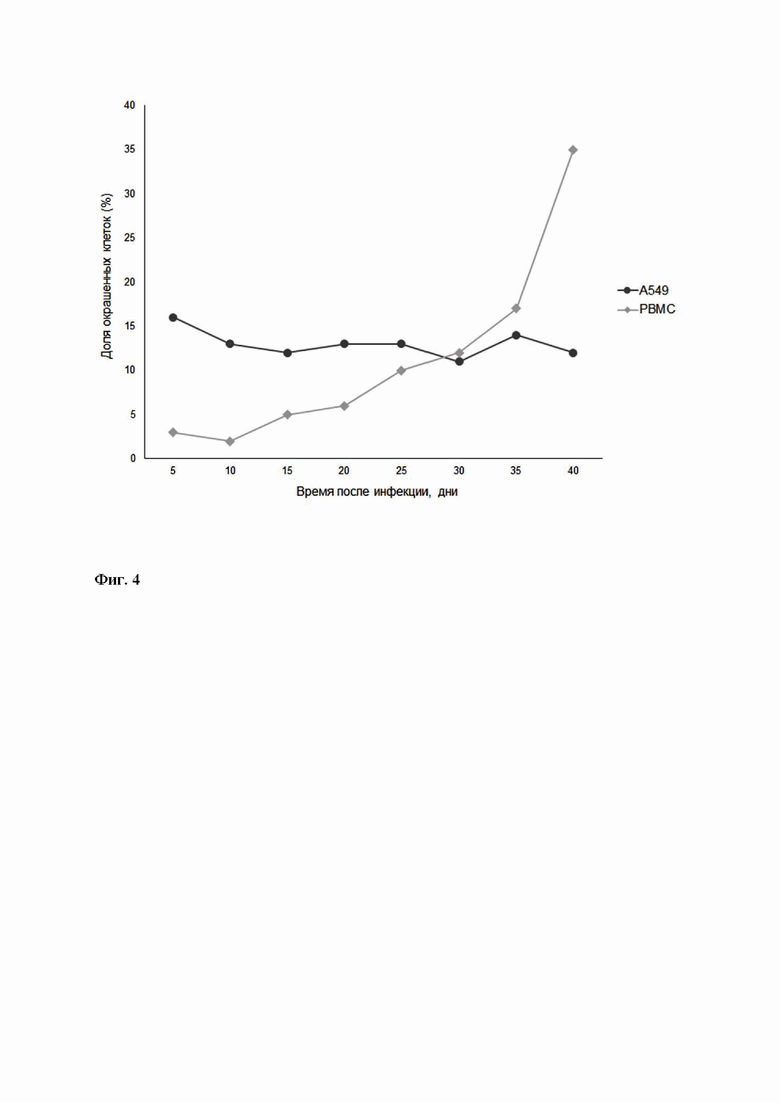
В ходе оценки возможности использования заявленного рекомбинантного вируса для одновременной иммортализации и генетической модификации первичных Т- лимфоцитов полученный препарат был использован для трансдукции периферических мононуклеарных клеток (PBMC), а также линии А549, использованной в качестве контроля. Трансдукция клеток была проведена в течение 8 часов в бессывороточной питательной среде AIM-V, после чего клетки были осаждены центрифугированием и рассажены на свежую порцию питательной среды AIM-V. Для верификации эффективности введения в клетки экспрессионной кассеты инфецированные клетки были подвергнуты окрашиванию антителами к эпитопу FLAG с последующим анализом в ходе проточной цитометрии. Проведенный анализ показал успешное появление фракций флуоресцирующих клеток: для линии А549 их доля сохранялась постоянной в течение 5 недель после инфекции, а для PBMC - постоянно увеличивалась (Фиг.4), свидетельствуя об эффективности вирус-опосредованной иммортализации периферических мононуклеарных клеток и успешности введения в клетки целевой экспрессионной кассеты.
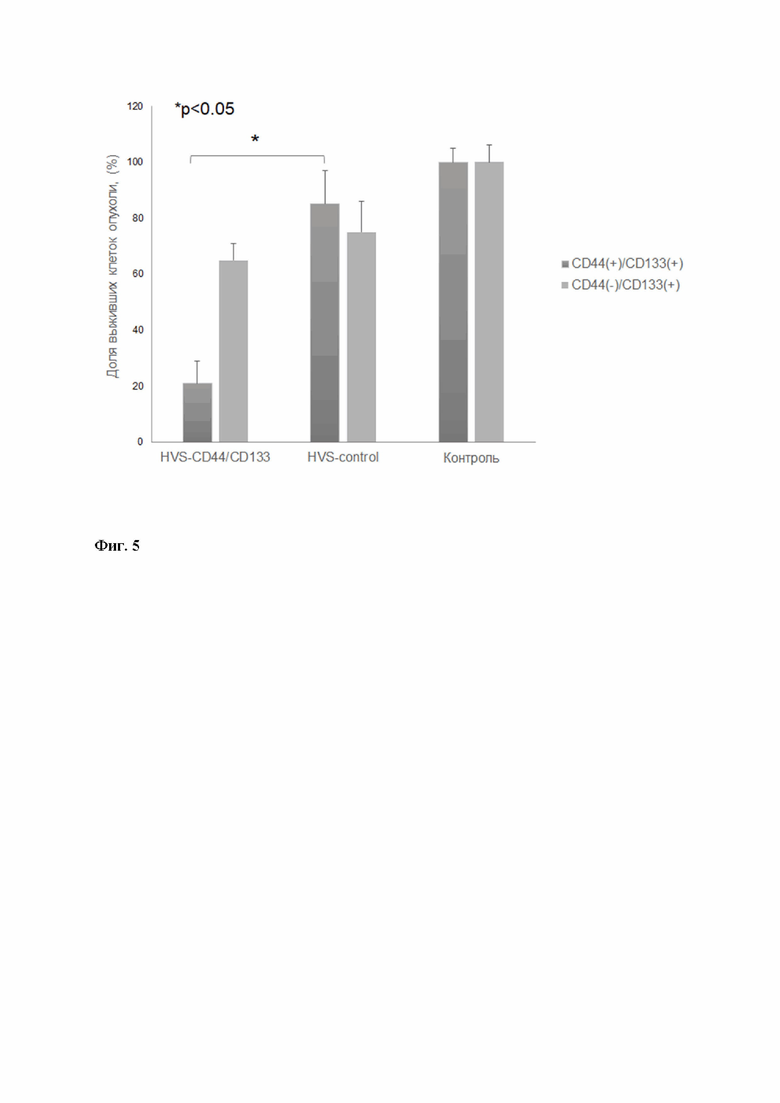
В ходе анализа эффективности вирус-опосредованного нацеливания Т-клеток на опухолевые антигены CD44 и CD 133 было проведено культивирование клеток линии LN-18 (глиобластома, IV стадия; ATCC® CRL-2610™), характеризующейся CD44 (+)/CD133(+) фенотипом, и предварительно отсортированной CD44(-)/CD133(+) фракции LN-18 в присутствии PBMC, полученных на предыдущем этапе; в качестве контроля использовали Т-лимфоциты, трансдуцированные контрольным препаратом HVS, не кодирующим последовательность парных химерных рецепторов. Совместное культивирование клеток было проведено в течение 30 часов в соотношении 1:1; по завершении эксперимента была проведена оценка выживаемости клеток линии LN-18 с помощью набора CellTiter-Glo® (Promega, USA), согласно протоколу, рекомендованному производителем (Фиг.5).
Результаты анализа свидетельствуют о эффективности элиминирования CD44(+)/CD133(+) раковых клеток линии LN-18 клетками PBMC, инфицированным тестируемым препаратом HVS по сравнению с контрольными неинфицированными PBMC (значение точного критерия Фишера составило 0.02809, р < 0,05), что подтверждает эффективность использования тестируемого рекомбинантного вируса для таргетирования клеток лимфоидных линий. Важно отметить значительно более низкую цитотоксичность PBMC в отношении CD44(-)/CD133(+) фракции клеток LN-18, свидетельствующей о том, что связывание каждого из парных антигенных рецепторов в отдельности не приводит к полной активации цитотоксического действия, минимизируя неспецифическую активности препарата и вероятность потенциальных побочных эффектов.
Возможность использования изобретения для решения существующих задач в сочетании с наблюдаемой терапевтической эффективностью подтверждают конкурентное преимущество заявленного технического решения и позволяют рекомендовать его для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов для терапии злокачественных новообразований, в частности, глиобластом.
Изобретение иллюстрировано следующими материалами:
Фиг. 1 - Схематическое изображение генетической конструкции pUC-HVS-SFFV- CAR2@CD44-CAR1@CD 133.
Фиг. 2 - Схематическое изображение генетической конструкции pMD2-Fd30.
Фиг. 3 - Схематическое изображение генетической конструкции pCD-4AHcd24.
Фиг. 4 - Динамика изменения численности флуоресцентных популяций клеток после инфекции рекомбинантным HVS.
Фиг. 5 - Результаты анализа эффективности элиминирования CD44/CD133(+) и CD44/CD133(-) раковых клеток линии LN-18 PBMC, инфицированным тестируемым или контрольным препаратами HVS; высота столбцов соответствует относительному уровню люминесценции, отражающему количество живых клеток линии LN-18 в культуре; каждое значение получено в трех повторностях, планки погрешностей соответствуют стандартной ошибке среднего.
SEQ ID NO.1 Нуклеотидная последовательность, кодирующая антиген-распознающую часть химерного рецептора на основе моноклонального одноцепочечного VHH- антитела, специфичного к опухолевому антигену CD44.
SEQ ID NO.2 Нуклеотидная последовательность, кодирующая антиген-распознающую часть химерного рецептора на основе моноклонального одноцепочечного VHH- антитела, специфичного к опухолевому антигену CD133.
SEQ ID NO.3 Нуклеотидная последовательность участка генома Herpesvirus Saimiri, в который производится интеграция целевого фрагмента.
SEQ ID NO.4 Последовательность вирусного гена vCD59.
SEQ ID NO.5 Последовательность, кодирующая флуоресцентный белок.
Последовательности
Последовательность 1: "Нуклеотидная последовательность, кодирующая антиген-распознающую часть химерного рецептора на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену СD44 "
Характеристики
--->
Последовательность
atgcaagttc aactggtaga aagtggtggc ggtctcgtcc aaccgggtgg gtcccttcgc 60
ttgagttgtg aagcgagcgg tttcaatttg gggtattacg gtatctgttg gtttcggcga 120
gctgccggga aagagcggga gggcgtcgcg tgttctgcca cacctgatga ttcaacttac 180
tacgctgatt ccgtcaaggg ccggtttacg atttcaaaag acaaggcaaa gaacatgatc 240
tttttgcaaa tgaatgactt gaaaccagaa gacactgccg tctattattg cgtcacagac 300
cggaaaccca aatacggcaa acagtggttg gagttttccg agtacgacta ctggggacag 360
ggaacccaag tcatagtgtc cagcgagcct aaaacgccga agccacaacc a 411
<---
Последовательность 2: "Нуклеотидная последовательность, кодирующая антиген-распознающую часть химерного рецептора на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD133"
Характеристики
--->
Последовательность
atgcaggtgc aggtgcagga gtcgggggga ggctgggtgc agcctggggg gtctctgacg 60
ctctcctgtg cagcccccag atcaatcttc agtgtccttg acataagctg gtaccgccag 120
gctacaggga accagcgcga gttggtcgca gcaattacta tcggtggtgg cacaagctat 180
acagactccg tgaagggccg attcaccatc tccagagacg acgccaagaa gtcggcgtat 240
ctccaaatga gcagcctgaa acctgaggac acggccgtct atgtctgtaa tgcccgatgg 300
cgtgcaggtg ggcagaacta ctggggccag gggacccggg tcaccgtctc ctcagcgcac 360
cacagcgaag accccagctc c 381
<---
Последовательность 3: "Нуклеотидная последовательность участка генома Herpesvirus Saimiri, в который производится интеграция целевого фрагмента"
Характеристики
--->
Последовательность
acaggctgct cttcaggagc accagaagaa ggtcgaattg cgttccttct caaaagagga 60
ggcatgtttg caaatgaaat gagaatctgg tgagataaga tgtatttgca tgaagcttct 120
atttatacta cattagaggc atttttcaga agcaaaaatg cctctaatta tatacactgt 180
actatttacc tctattacac attttctatt ttaagtctga aagtgattaa tcaagaaaga 240
agtttgtggt tccctggaga ttagttcaca agctgtctga ggttaaaggt ggttctttag 300
cactgacaca caagttgcta taagaattga agcttgcttt ataaaaagtt acttgtgttt 360
aattactatt aaaataaaga gaatgtatat tttgtttaag ttgatgcttg cttctgtttt 420
ttgcaagtca agctacagct tgcaatgcta caactgttct cactcaacta tgcagtgtac 480
aacatctact agttgtacat ctaatcttga ttcttgtctc attgctaaag ctgggtcaaa 540
agtatattac aggtgctgga agtttgatga ctgtagcttc aaacgcatct caaatcaatt 600
gtctgaaaca cagttaaagt atcattgttg taagaaaaac ttgtgtaatg ttaacaaagt 660
gattgaaaat ggaaaaagaa caatatcaga taaggctctt ttactattag cattgttttt 720
agtaactgct tagaaccttt ctctttaaaa gtgaacaaca tacctatatt gtaacattta 780
tttttgcgta gcttattcgt attgctatta caagttaaaa tattgtgttt ttaactataa 840
tttttaaaaa gataaaatga gatgtagtat attacccata gtcaaaatta aagtgctaga 900
tattattagc atttttatca acaacgcaaa taaaagttaa gatacattat ttttttgata 960
tttggattat tgtgtgcttt ttatcatatg ttaaaagttt tatgtcattt tattcttaca 1020
tatataaagc taaattttaa agcaacttat cagtagcatc ttagcttctg atctgtacag 1080
acttatataa tatgggttta tccttaagaa aaaacaaaga agaaaaaaat aacacagtgc 1140
caaacttgcc agttaattac atgttaagaa atcctgttta taaaaagaaa tatctgccag 1200
<---
Перечень остатков был усечен, поскольку он превышает заданный предел: максимум 1200 остатка(-ов) для печати
Последовательность 4: "Последовательность вирусного гена vCD59"
Характеристики
--->
Последовательность
atgtatattt tgtttaagtt gatgcttgct tctgtttttt gcaagtcaag ctacagcttg 60
caatgctaca actgttctca ctcaactatg cagtgtacaa catctactag ttgtacatct 120
aatcttgatt cttgtctcat tgctaaagct gggtcaaaag tatattacag gtgctggaag 180
tttgatgact gtagcttcaa acgcatctca aatcaattgt ctgaaacaca gttaaagtat 240
cattgttgta agaaaaactt gtgtaatgtt aacaaagtga ttgaaaatgg aaaaagaaca 300
atatcagata aggctctttt actattagca ttgtttttag taactgctta g 351
<---
Последовательность 5: "Последовательность, кодирующая флуоресцентный белок"
Характеристики
--->
Последовательность
atggtgtcta agggcgaaga gctgattaag gagaacatgc acatgaagct gtacatggag 60
ggcaccgtga acaaccacca cttcaagtgc acatccgagg gcgaaggcaa gccctacgag 120
ggcacccaga ccatgagaat caaggtggtc gagggcggcc ctctcccctt cgccttcgac 180
atcctggcta ccagcttcat gtacggcagc agaaccttca tcaaccacac ccagggcatc 240
cccgacttct ttaagcagtc cttccctgag ggcttcacat gggagagagt caccacatac 300
gaagacgggg gcgtgctgac cgctacccag gacaccagcc tccaggacgg ctgcctcatc 360
tacaacgtca agatcagagg ggtgaacttc ccatccaacg gccctgtgat gcagaagaaa 420
acactcggct gggaggccaa caccgagatg ctgtaccccg ctgacggcgg cctggaaggc 480
agaagcgaca tggccctgaa gctcgtgggc gggggccacc tgatctgcaa cttcaagacc 540
acatacagat ccaagaaacc cgctaagaac ctcaagatgc ccggcgtcta ctatgtggac 600
cacagactgg aaagaatcaa ggaggccgac aaagagacct acgtcgagca gcacgaggtg 660
gctgtggcca gatactgcga cctccctagc aaactggggc acaaacttaa ttaa 714
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения рекомбинантных герпесвирусов для одновременной трансдукции и иммортализации первичных Т-лимфоцитов и НК-клеток | 2019 |
|
RU2751482C2 |
| Набор олигонуклеотидных праймеров и зондов для выявления Herpesvirus Saimiri в клеточных культурах методом ПЦР в реальном времени | 2020 |
|
RU2753767C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ, КОДИРУЮЩАЯ СТРУКТУРУ Т-КЛЕТОЧНОГО ХИМЕРНОГО РЕЦЕПТОРА НА ОСНОВЕ ОДНОЦЕПОЧЕЧНЫХ VHH-АНТИТЕЛ, СПЕЦИФИЧНЫХ ОПУХОЛЕВОМУ РЕЦЕПТОРУ CD47, ДЛЯ НАЦЕЛЕННОЙ ИММУНОТЕРАПИИ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2017 |
|
RU2707535C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЦИТОТОКСИЧЕСКИХ Т-ЛИМФОЦИТОВ, ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2714380C1 |
| Генетический вектор Ad6/3-hTERT-GMCSF, содержащий геномные последовательности рекомбинантного аденовируса 6 серотипа, промотор теломеразы человека, ген гранулоцитарно-макрофагального колониестимулирующего фактора человека, а также ген белка файбер со встройкой домена fiber knob аденовируса 3 серотипа, обладающий повышенной трансдукцией в опухолевые клетки | 2023 |
|
RU2814581C1 |
| СИСТЕМА ДЛЯ СТАБИЛЬНОЙ ЭКСПРЕССИИ ОПУХОЛЬ-АССОЦИИРОВАННЫХ АНТИГЕНОВ НА ОСНОВЕ ЛЕНТИВИРУСНОГО ВЕКТОРА | 2018 |
|
RU2725493C2 |
| Вирусоподобные химерные частицы для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2, содержащие белки коронавируса и ротавируса | 2022 |
|
RU2779810C1 |
| СИСТЕМЫ ЭКСПРЕССИИ РНК РЕКОМБИНАНТНОГО ВИРУСА НЬЮКАСЛСКОЙ БОЛЕЗНИ И ВАКЦИНЫ | 1999 |
|
RU2270864C2 |
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |
| Искусственный ген EctoS_SC2, кодирующий эктодомен гликопротеина S коронавируса SARS-CoV-2 с C-концевым тримеризующим доменом, рекомбинантная плазмида pStem-rVSV-EctoS_SC2, обеспечивающая экспрессию искусственного гена, и рекомбинантный штамм вируса везикулярного стоматита rVSV-EctoS_SC2, используемый для создания вакцины против коронавируса SARS-CoV-2 | 2020 |
|
RU2733834C1 |
Группа изобретений относится к биотехнологии. Предложена гетерологичная ДНК, предназначенная для введения в геном вируса “дикого” штамма Herpesvirus saimiri, включает в себя генетические последовательности, кодирующие парные химерные рецепторы против опухолевых антигенов CD44 и CD133, фланкированная участками протяженностью 300-900 пар нуклеотидных оснований, гомологичных тому участку генома Herpesvirus saimiri, в который производится интеграция целевого фрагмента посредством гомологичной рекомбинации. Также предложен рекомбинантный штамм Herpesvirus saimiri, содержащий в своем геноме последовательность указанной ДНК, предназначенный для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против антигенов CD44 и CD133. Изобретения могут быть использованы в исследовательских и диагностических целях для эффективного получения генетически-модифицированных иммортализованных первичных Т-лимфоцитов для терапии злокачественных новообразований, в частности глиобластом. 2 н. и 3 з.п. ф-лы, 5 ил.
1. Гетерологичная ДНК, предназначенная для введения в геном вируса "дикого" штамма Herpesvirus saimiri, включающая в себя генетические последовательности, кодирующие парные химерные рецепторы: против опухолевого антигена CD44, который включает в себя N-концевую сигнальную последовательность рецептора CD8, нуклеотидную последовательность антиген-распознающей части на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD44, с SEQ ID NO.1, спейсерную область, представленную 12 аминокислотной последовательностью шарнирного домена иммуноглобулина G4, трансмембранный домен CD28 и сигнальный домен CD3zeta; и против опухолевого антигена CD133, который включает в себя N-концевую сигнальную последовательность рецептора CD8, нуклеотидную последовательность антиген-распознающей части на основе моноклонального одноцепочечного VHH-антитела, специфичного к опухолевому антигену CD133, с SEQ ID NO.2, шарнирную область длиной 228 аминокислот, трансмембранный и сигнальный домены рецептора CD28, а также сигнальный домен рецептора CD137.
2. ДНК по п. 1, в которой участок гетерологичной ДНК, предназначенный для введения в геном вируса "дикого" штамма Herpesvirus saimiri, фланкирован участками протяженностью 300-900 пар нуклеотидных оснований, гомологичных участку генома Herpesvirus saimiri с нуклеотидной последовательностью SEQ ID NO.3, в который производится интеграция целевого фрагмента посредством гомологичной рекомбинации.
3. ДНК по п. 2, в которой в качестве последовательности протяженностью 300-900 пар нуклеотидных оснований, гомологичной участку генома Herpesvirus saimiri, используется последовательность вирусного гена vCD59, имеющая последовательность SEQ ID NO.4.
4. ДНК по п. 1, в которой в состав участка гетерологичной ДНК, предназначенного для введения в геном "дикого" штамма вируса Herpesvirus saimiri, входит последовательность SEQ ID NO.5, кодирующая флуоресцентный белок.
5. Рекомбинантный штамм Herpesvirus saimiri, предназначенный для быстрого получения пролиферирующих популяций аутологичных Т-лимфоцитов, экспрессирующих парные химерные антигенные рецепторы против опухолевых антигенов CD44 и CD133, содержащий в своем геноме последовательность ДНК по пп. 1-4.
| US 6379967 B1, 30.04.2002 | |||
| US 5670352 A, 23.09.1997 | |||
| MITTRÜCKER H.-W | |||
| et al | |||
| Herpes virus saimiri-transformed human T lymphocytes: normal functional phenotype and preserved T cell receptor signalling | |||
| International Immunology, 1993, 5(8), p.985-990 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| КРАВЧЕНКО Ю | |||
| Е | |||
| и др | |||
| Проведение рекомбинации Herpesvirus saimiri | |||

