ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ФИНАНСИРУЕМых ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА ИССЛЕДОВАНИй или РАЗРАБОТОК
Это изобретение было осуществлено при государственной поддержке по гранту № AI085596, присужденному Национальными институтами здравоохранения. Государство имеет определенные права на изобретение.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Система комплемента является частью врожденного иммунитета, играющей ключевую роль в защите хозяина. Комплемент может быть активирован тремя разными путями, классическим, альтернативным и лектиновым путями. Среди них, альтернативный путь уникален тем, что он не только представляет собой независимый путь, посредством которого комплемент активируется по механизму «холостой» активации, но он также усиливает активацию комплемента, инициированную за счет двух других путей. Для альтернативного пути требуется участие C3, фактора B (fB), фактора D (fD) и пропердина (fP). Все пути сходятся на стадии активации C3, начиная с которой вступает в действие петля усиления альтернативного пути. Независимо от действующего пути активации комплемента активированный комплемент осуществляет эффекторные функции трех типов: опсонизацию мишеней за счет C3b/iC3b/C3d для облегчения фагоцитоза и клиренса, продуцирование провоспалительных медиаторов C3a и C5a, а также прямую атаку клеток эффектором терминальной стадии активации комплемента C5b-9, также известным как мембраноатакующий комплекс (MAC). Посредством активации рецепторов комплемента (CR), таких как CR2 на B-клетках и фолликулярных дендритных клетках, и рецепторов анафилотоксинов - рецептора C3a (C3aR) и рецептора C5a (C5aR) на лейкоцитах, таких как макрофаги и моноциты, комплемент также взаимодействует с, и перекрестно регулирует систему адаптивного иммунитета и, таким образом, играет модулирующую роль в иммунных функциях B- и T-клеток.
Нарушение регуляции комплемента является причиной целого ряда заболеваний человека, приводя к опосредованному комплементом аутологичному повреждению тканей. Нарушение регуляции комплемента может возникать вследствие мутаций, либо соматических, либо в зародышевой линии, в генах регуляторов или генах, связанных с регуляторами, комплемента, в результате чего эти регуляторы перестают нормально функционировать. Примеры из этой категории включают мутации в гемопоэтических стволовых клетках гена PIG-A, который кодирует ключевой фермент в биосинтезе GPI-якоря, и такие мутации приводят к отсутствию экспрессии DAF и CD59 на клетках крови у пациентов, страдающих пароксизмальной ночной гемоглобинурией (PNH). В результате у пациентов с PNH эритроциты и тромбоциты не защищены от атаки комплемента, и у них развивается внутрисосудистый гемолиз и активация тромбоцитов, что приводит к анемии и тромботическим приступам. Вторым примером является мутация в мембранном регуляторе MCP или жидкофазных регуляторах fH или fI, что вызывает избыточную активацию альтернативного пути комплемента в почках, приводя к патогенезу C3-гломерулопатии или атипичного гемолитико-уремического синдрома (aHUS). В дополнение к таким редким и высокопенетрантным мутациям, приводящим к отсутствию экспрессии или дисфункции DAF, CD59, fH, fI и MCP, существуют однонуклеотидные полиморфизмы (SNP) в fH, которые являются более распространенными и менее пенетрантными, но, тем не менее, как установлено, вносят свой вклад в патогенез заболеваний по механизму, опосредованному комплементом. Очень хорошо изученным примером является сильная связь Y420H полиморфизма в fH с возрастной дегенерацией желтого пятна (AMD). Таким образом, дисфункция или изменение последовательности регулятора комплемента может приводить к распространенным, а также редким, заболеваниям человека.
Нарушение регуляции комплемента может возникать не только вследствие мутаций/полиморфизмов регуляторов, но также вследствие мутаций в генах, которые кодируют критические компоненты альтернативного пути, а именно, C3 и fB, а также за счет присутствия аутоантител против регуляторов или белков комплемента, таких как fH, C3 или fB. Теперь понятно, что некоторые мутации в C3 или fB приводят к появлению белков, которые при активации образуют необычно стабильную C3-конвертазу C3bBb альтернативного пути, устойчивую к регуляции регуляторными белками, что, в свою очередь, может приводить к нарушению регуляции и избыточной активации комплемента. В случае аутоантител против регуляторов комплемента, их действие часто имитирует последствия мутаций в генах, кодирующих такие белки, вследствие чего уменьшается функциональная эффективность таких белков в жидкой фазе или на клеточной поверхности. Отдельно, аутоантитела против C3b, называемые C3-нефритогенными факторами (C3nef), способны связывать и стабилизировать C3-конвертазу C3bBb альтернативного пути, таким образом обеспечивая тот же эффект увеличения времени полужизни и активности конвертазы, который создают мутации в генах C3 или fB. В целом, существуют распространенные и редкие заболевания человека, причиной которых является избыточная активация комплемента, возникающая в результате нарушения регуляции каскада активации комплемента. Механизмы, лежащие в основе нарушения регуляции системы комплемента, различны, некоторые обусловлены генными мутациями, а другие аутоантителами, при этом продуктами мутантных генов или мишенями аутоантител могут быть регуляторные белки или компоненты альтернативного пути активации комплемента.
Современные терапевтические подходы направлены на разработку реагентов, таких как мАт, пептиды или другие малые молекулы, которые связывают и блокируют конкретные компоненты альтернативного пути или терминального пути активации комплемента. Клинически валидированным примером является экулизумаб, гуманизированное мАт против компонента C5 комплемента, одобренное для лечения PNH и aHUS. Другие подходы, которые были описаны, включают мАт против fB, fD или fP, а также циклический пептид, который связывает и ингибирует C3. Недостатком этих подходов является то, что они требуют постоянного и причиняющего неудобства в/в введения доз пациентам. Кроме того, поскольку они блокируют альтернативный путь или терминальный путь, они рискуют поставить под угрозу защитную систему хозяев. Действительно, пациентов, получающих лечение экулизумабом, необходимо вакцинировать против бактериальных штаммов, вызывающих смертельно опасный менингит, и эти пациенты также получают профилактическую терапию антибиотиками прежде, чем начать получать лечение одобренным лечебным мАт.
В других подходах были протестированы рекомбинантные регуляторные белки, такие как растворимые DAF, CR1, CRIg и белки, содержащие минимальные домены fH (N-концевой короткий консенсусный повтор (SCR) 1-5 и C-концевой SCR 19-20), или слитые белки из fH и CR2 (TT30). См., например, публикацию патента США № US2013/0296255; публикацию патента США № 2008/0221011. Однако крупномасштабная гетерологичная экспрессия таких белков в качестве терапевтических лекарственных средств требует приложения значительных усилий, и исследования на животных показали, что скорость их выведения после введения in vivo очень высока (Nichols EM, Barbour TD, Pappworth IY, Wong EK, Palmer JM, Sheerin NS, Pickering MC, Marchbank KJ. Kidney Int. 2015 Jul 29. doi: 10.1038/ki.2015.233.; Fridkis-Hareli M, Storek M, Mazsaroff I, Risitano AM, Lundberg AS, Horvath CJ, Holers VM, Blood. 2011 Oct 27; 118(17): 4705-13. doi: 10.1182/blood-2011-06-359646. Epub 2011 Aug 22), это делает такие терапевтические стратегии затруднительными и менее практичными, поскольку потребовалось бы многократное и частое введение подобных белковых лекарственных средств.
В данной области сохраняется потребность в более эффективных композициях продолжительного действия, полезных для лечения заболеваний, опосредованных комплементом.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте изобретение относится к рекомбинантному вектору с упакованной в нем экспрессионной кассетой, содержащей рекомбинантный ген человеческого регулятора комплемента фактора H (fH), функционально связанный с последовательностями контроля экспрессии, которые управляют его экспрессией, при этом указанный ген hfH кодирует растворимый вариант белка hfH, сохраняющий функцию регуляции комплемента, причем указанный вариант fH содержит короткий консенсусный повтор (SCR) 1, 2, 3, 4, 19 и 20, и по меньшей мере один из SCR7, SCR17 и/ SCR18, при этом после введения вектора субъекту и экспрессии поддающиеся обнаружению уровни в плазме варианта hfH присутствуют у субъекта в течение по меньшей мере недели.
В другом аспекте изобретение относится к рекомбинантному AAV вектору с упакованной в нем экспрессионной кассетой, содержащей рекомбинантный ген человеческого регулятора комплемента фактора H (fH), функционально связанный с последовательностями контроля экспрессии, которые управляют его экспрессией, при этом указанный ген hfH кодирует растворимый вариант белка hfH, сохраняющий функцию регуляции комплемента, причем указанный вариант fH содержит короткий консенсусный повтор (SCR) 1, 2, 3, 4, 19 и 20, при этом после введения вектора субъекту и экспрессии поддающиеся обнаружению терапевтически полезные уровни в плазме варианта hfH присутствуют у субъекта в течение по меньшей мере примерно месяца.
В следующем аспекте изобретение относится к фармацевтической композиции, содержащей носитель и/или эксципиент и рекомбинантный вектор, описанный в настоящем документе, который экспрессирует вариант fH.
В другом аспекте изобретение относится к способу лечения связанного с комплементом заболевания путем доставки в организм пациента вектора, описанного в настоящем описании. Связанное с комплементом заболевание может представлять собой, среди прочего, мембранопролиферативный гломерулонефрит, атипичный гемолитико-уремический синдром (aHUS), возрастную дегенерацию желтого пятна (AMD), микроангиопатическую гемолитическую анемию, тромбоцитопению, острую почечную недостаточность, пароксизмальную ночную гемоглобинурию (PNH), шизофрению, ишемический инсульт и/или бактериальные инфекции, вызываемые рекрутингом бактериальных патогенов.
В следующем аспекте изобретение относится к применению рекомбинантного вектора для лечения AMD. В другом аспекте описано применение rAAV вектора для лечения PNH, aHUS или другого связанного с комплементом заболевания.
В другом аспекте изобретение относится к рекомбинантному варианту hfH, содержащему лидерную последовательность и последовательности SCR человеческого рецептора комплемента, состоящие из: (a) SCR1-4, 7 и 19-20; (b) SCR1-4, 6, 7 и 19-20; (c) SCR1-4, 7, 8 и 19-20; (d) SCR1-4, 6, 7, 8 и 19-20; (e) SCR1-4, 17 и 19-20; (f) SCR1-4 и 18-20; (g) SCR1-4 и 17-20. Другие варианты осуществления включают, например, SCR1-4, 7 и 18-20; SCR1-4, 6, 7 и 18-20; SCR1-4, 7, 8 и 18-20; или SCR1-4, 6, 7, 8 и 18-20, SCR1-4, 7 и 17-20; SCR1-4, 6, 7 и 17-20; SCR1-4, 7, 8 и 17-20; или SCR1-4, 6, 7, 8 и 17-20. Необязательно, в по меньшей мере один из SCR введен по меньшей мере один сайт гликозилирования. В другом аспекте один из рекомбинантных вариантов hfH является пегилированным.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей по меньшей мере один вид рекомбинантного варианта hfH, носитель и/или эксципиент. Такую композицию можно использовать отдельно или в сочетании с другой терапией, в частности, например, векторной терапией, описанной в настоящем описании.
Другие аспекты и преимущества изобретения станут очевидны из следующего далее подробного описания изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На ФИГ. 1A приведено схематическое изображение доменной структуры зрелого белка человеческого фактора H.
На ФИГ. 1B-1E приведены нуклеотидные и аминокислотные последовательности лидерного пептида и указано расположение 20 доменов короткого консенсусного повтора (SCR), использованных для создания вариантов fH, описанных в приведенных ниже примерах. SEQ ID NO: 1 соответствует нуклеотидной последовательности; SEQ ID NO: 2 соответствует аминокислотной последовательности сигнального пептида. Аминокислотные последовательности SCR1-20 включают SEQ ID NO: 3 (SCR1), 5 (SCR2), 7 (SCR3), 9 (SCR4), 11 (SCR5), 13 (SCR6), 14 (SCR7), 16 (SCR8), 17 (SCR9), 19 (SCR10), 21 (SCR11), 23 (SCR12), 25 (SCR13), 27 (SCR14), 29 (SCR15), 31 (SCR16), 33 (SCR17), 35 (SCR18), 37 (SCR19) и 38 (SCR20), соответственно. Расположение этих доменов в изоформе 1 fH указано на основании правил, описанных в статье C. Estaller et al., Eur J Immunol. 1991 Mar; 21(3): 799-802. Аминокислотные последовательности между указанными SCR представляют собой последовательности линкеров, придающие гибкость fH (SEQ ID NO: 4, 6, 8, 10, 12, 15, 18, 20, 22, 24, 26, 28, 30, 32, 34 и 36, соответственно). Линкер между SCR19 и SCR20 содержит всего три аминокислоты (Leu-His-Pro) и, таким образом, не приведен в списке последовательностей.
На ФИГ. 2A приведено схематическое изображение доменной структуры варианта человеческого фактора H, содержащего SCR1-4, 6-8 и 19-20.
На ФИГ. 2B-2C приведены нуклеотидная (нт 53-1804 в SEQ ID NO: 41) и аминокислотная последовательности (SEQ ID NO: 42) лидерного пептида и 9 доменов короткого консенсусного повтора (SCR) варианта fH SCR1-4, 6-8 и 19-20.
На ФИГ. 3A-3B приведены полные последовательности кДНК (нт 53-1804 в SEQ ID NO: 41), а также 5'- (нт 1-52 в SEQ ID NO: 41) и 3'-UTR (нт 1805-2068 в SEQ ID NO: 41) конструкта укороченного человеческого фактора H, содержащего лидерный пептид и SCR1-4, 6-8 и 19-20 (hfH1-4.678.19-20).
На ФИГ. 4 приведена аминокислотная последовательность конструкта укороченного фактора H, содержащего лидерный пептид (подчеркнуто) и SCR1-4, 6-8, 19-20 (hfH1-4.678.19-20) (SEQ ID NO: 42).
На ФИГ. 5A и 5B приведены изображения гелей, подтверждающие экспрессию и стабильность белка hfH1-4.678.19-20. Приведенную на ФИГ. 2 последовательность кДНК укороченного варианта человеческого fH, содержащего SCR1-4, 6-8 и 19-20 (SEQ ID NO: 41), клонировали в эукариотические экспрессионные векторы, которые затем использовали для трансфекции клеток HEK. Супернатант клеточных культур использовали в вестерн-блот анализе для обнаружения экспрессии укороченного белка fH. Панель A: дорожка 1, нетрансфицированные клетки HEK; дорожки 2 и 3, клетки HEK, трансфицированные вектором pCMV Sport6, содержащим кДНК укороченного варианта fH; дорожки 4-6, клетки HEK, трансфицированные вектором pCBARBG, содержащим кДНК варианта fH. Вектор pCBARBG содержит те же 5' и 3'-регуляторные элементы, что и конструкт pAAV вектора, приведенный на ФИГ. 4. Панель B: дорожка 1, нетрансфицированные клетки HEK; дорожка 2, клетки HEK, трансфицированные вектором pCBARG, содержащим кДНК укороченного варианта fH в качестве контроля; дорожка 3, клетки HEK, трансфицированные AAV8-плазмидой, содержащей кДНК укороченного варианта fH.
На ФИГ. 6 приведено изображение SDS-геля, показывающее степень очистки рекомбинантного hfH1-4.678.19-20. Анализ методом электрофореза в SDS-геле выполняли путем окрашивания Кумасси голубым укороченного варианта человеческого fH, содержащего SCR1-4, 6-8 и 19-20, который был экспрессирован клетками HEK, трансфицированными вектором pCBARBG. Рекомбинантный укороченный белок fH очищали из супернатанта пропусканием через колонку с аффинной смолой, полученной с использованием мАт против человеческого фактора H (клон OX-23), которое узнает эпитоп в SCR2-3. Размер и расположение белков-маркеров молекулярной массы показано слева.
На ФИГ. 7 приведено изображение геля, показывающее, что рекомбинантный hfH1-4.678.19-20 сохраняет комплемент-регулирующую активность (кофакторную активность). Укороченный вариант человеческого fH, содержащий SCR1-4, 6-8 и 19-20, тестировали на кофакторную активность для опосредованного фактором I расщепления C3b. Для этого анализа человеческий C3b смешивали с фактором I в присутствии (дорожки 1-6) или в отсутствие (дорожка 7) полноразмерного fH (hfH) или укороченного варианта fH (hfH1-4.678.19-20). Реакционную смесь инкубировали и затем анализировали методом SDS-ПААГ и вестерн-блоттинга. На наличие кофакторной активности указывает появление фрагмента α-цепи iC3b.
На ФИГ. 8 приведен линейный график, показывающий, что рекомбинантный hfH1-4.678.19-20 (квадраты, верхняя линия) имеет сильную гепарин-связывающую активность. Укороченный вариант человеческого fH, содержащий SCR1-4, 6-8 и 19-20, сохраняет гепарин-связывающую активность. Его гепарин-связывающая активность зависит от дозы, и при основанном на концентрации (мкг/мл) сравнении с полноразмерным человеческим fH (ромбы, нижняя линия) он имеет более высокую активность. Гепарин-связывающую активность оценивали методом ELISA с использованием планшета, покрытого гепарином, на который наносили раствор полноразмерного или укороченного белка fH, и после промывания обнаруживали связанный fH или укороченный fH при помощи мАт OX-23 (против эпитопа в SCR2-3).
На ФИГ. 9 приведен линейный график, показывающий, что рекомбинантный hfH1-4.678.19-20 (квадраты, верхняя линия) имеет сильную C3b-связывающую активность. Укороченный вариант человеческого fH, содержащий SCR1-4, 6-8 и 19-20, сохраняет C3b-связывающую активность. Его C3b-связывающая активность зависит от дозы, и при основанном на концентрации (мкг/мл) сравнении с полноразмерным человеческим fH (ромбы, нижняя линия) он имеет более высокую активность. C3b-связывающую активность оценивали методом ELISA с использованием планшета, покрытого C3b, на который наносили раствор полноразмерного или укороченного белка fH, и после промывания обнаруживали связанный fH или укороченный fH при помощи мАт OX-23 (против эпитопа в SCR2-3).
На ФИГ. 10 приведен линейный график, показывающий обнаружение методом ELISA hfH1-4.678.19-20 в крови 3 разных мутантных по fH мышей (fHm/m; F1, F2, F20) через одну неделю после AAV8-опосредованной терапии геном fH. Мыши fHm/m представляют собой линию мутантных по fH мышей, которые имеют кодоны преждевременной остановки в начале последовательности SCR19. У этих мышей продуцируются следовые количества укороченного fH (без SCR19-20) и у них имеет место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе и истощение (вторичный дефицит) C3 и fB. Мышей инфицировали в/в введением через ретроорбитальный синус вируса AAV8, содержащего hfH1-4.678.19-20 (3×1011 геномных копий/мышь), и через одну неделю образцы крови собирали и обрабатывали для обнаружения человеческого белка fH. Для анализа ELISA мАт OX-23 (узнающее эпитоп в SCR2-3 человеческого fH) использовали в качестве улавливающего антитела и биотинилированное мАт L20/3 (узнающее SCR19 человеческого fH) использовали в качестве обнаруживающего антитела. Как показано на фигуре, в крови 3 мышей fHm/m (F1, F2, F20) отсутствует hfH1-4.678.19-20 до введения AAV-hfH1-4.678.19-20 (Пре), но hfH1-4.678.19-20 был обнаружен через одну неделю (1Н) после введения.
На ФИГ. 11 показаны результаты вестерн-блот анализа, демонстрирующие, что AAV8-опосредованная терапия геном человеческого fH у мышей fHm/m приводит к ингибированию альтернативного пути активации комплемента; из-за отсутствия достаточной экспрессии эндогенного мышиного fH у не получавших лечение мышей fHm/m имела место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе, и в результате у них в плазме имело место истощение C3 и fB (сравните дорожку 1 для Д/Т с дорожками 2, 4, 6 для трех мышей fHm/m до проведения генной терапии). Через одну неделю после введения AAV8-hfH1-4.678.19-20 мышам fHm/m уровни в плазме C3 и fB значительно возрастали относительно уровней до начала лечения, свидетельствуя о том, что AAV8-опосредованная терапия геном человеческого fH приводила к ингибированию неконтролируемой регуляции альтернативного пути активации комплемента и истощения C3 и fB. Каждая из трех мышей (F1, F2 и F20) получала по 3×1011 геномных копий в/в введением через ретроорбиитальный синус.
На ФИГ. 12A приведено схематическое изображение доменной структуры варианта человеческого фактора H, содержащего SCR1-4, 6-8 и 17-20, на котором местонахождение сайтов N-гликозилирования указано стрелками.
На ФИГ. 12B-12C приведены нуклеотидные и аминокислотные последовательности лидерного пептида и 11 доменов короткого консенсусного повтора (SCR) варианта fH, содержащего SCR1-4, 6-8 и 17-20 (SEQ ID NO: 45 и 46, соответственно).
На ФИГ. 13 приведены полные последовательности кДНК и 5'UTR варианта человеческого фактора H, содержащего лидерный пептид и SCR1-4, 6-8 и 17-20 (hfH1-4.678.17-20) (5'UTR представлена заглавными буквами) (SEQ ID NO: 47).
На ФИГ. 14 приведена аминокислотная последовательность конструкта укороченного фактора H, содержащего лидерный пептид (подчеркнуто) и SCR1-4, 6-8 и 17-20 (hfH1-4.678.17-20) (SEQ ID NO 48).
На ФИГ. 15A-15C показано определение методом ELISA уровня белка hfH1-4.6-8.17-20 в плазме у 3 мутантных по fH мышей, получавших разные дозы AAV8-hfH1-4.678.17-20. Мыши fHm/m представляют собой линию мутантных по fH мышей, которые имеют кодоны преждевременной остановки в начале последовательности SCR19. У этих мышей продуцируются следовые количества укороченного fH (без SCR19-20) и у них имеет место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе и истощение (вторичный дефицит) C3 и fB. Мышей инфицировали в/в введением через ретроорбитальный синус вируса AAV8, содержащего hfH1-4.678.17-20, в трех дозах, 1×1012 геномных копий (гк)/мышь, 3×1011 гк/мышь и 1×1011 гк/мышь, соответственно. Образцы плазмы собирали для анализа ELISA перед введением AAV (Пре) или через одну неделю (Н1), две недели (Н2), один месяц (М1), два месяца (М2) или 3 месяца (М3) после введения AAV. Для анализа ELISA мАт OX-23 (узнающее эпитоп в SCR2-3 человеческого fH) использовали в качестве улавливающего антитела и биотинилированное мАт L20/3 (узнающее SCR19 человеческого fH) использовали в качестве обнаруживающего антитела. Как показано на фигуре, в крови мышей fHm/m отсутствует hfH1-4.678.17-20 до введения AAV-hfH1-4.678.17-20 (Пре), но высокий уровень hfH1-4.678.17-20 был обнаружен после введения AAV, и экспрессия hfH1-4.678.17-20 оставалась стабильной в течение по меньшей мере 3 месяцев.
На ФИГ. 16A-16C показаны результаты вестерн-блот анализа, демонстрирующие, что генная терапия мышей fHm/m с использованием AAV8-hfH1-4.678.17-20 приводит к ингибированию альтернативного пути активации комплемента. Из-за отсутствия достаточной экспрессии эндогенного мышиного fH у не получавших лечение мышей fHm/m имела место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе, и в результате у них в плазме имело место истощение C3 и fB (дорожка 1). У трех мышей fHm/m, получавших 1×1012 геномных копий (гк)/мышь (ФИГ. 16A), 3×1011 геномных копий (гк)/мышь (ФИГ. 16B) и 1×1011 геномных копий (гк)/мышь (ФИГ. 16C), соответственно, в/в введением через ретроорбитальный синус, альтернативный путь активации комплемента был предотвращен, с соответствующим восстановлением в плазме уровней C3 и fB, когда получавших лечение мышей изучали через одну неделю (Н1), один месяц (М1), 2 месяца (М2) и 3 месяца (М3) после генной терапии с использованием AAV8-hfH1-4.678.17-20. При каждой терапевтической дозе и в каждой временной точке (дорожки 2, 3, 4, 5) уровни в плазме C3 и fB были заметно выше после генной терапии с использованием AAV8-hfH1-4.678.17-20, чем до лечения (Пре, дорожка 1).
На ФИГ. 17A-17C приведены нуклеотидные и аминокислотные последовательности лидерного пептида и 20 доменов короткого консенсусного повтора (SCR) у мышей (SEQ ID NO: 79 и 80, соответственно). Аминокислотные последовательности между указанными SCR представляют собой последовательности линкеров, придающие гибкость fH.
На ФИГ. 18A-18B приведены нуклеотидные и аминокислотные последовательности лидерного пептида и 9 доменов короткого консенсусного повтора (SCR) варианта мышиного fH (SEQ ID NO: 81 и 82, соответственно). Аминокислотные последовательности между указанными SCR представляют собой последовательности линкеров, придающие гибкость белку fH. Этот вариант мышиного fH использован в качестве суррогата для тестирования in vivo функции hfH1-4.678.19-20 в последующих исследованиях.
На ФИГ. 19 приведены кодирующая, а также 5'- и 3'-UTR последовательности конструкта укороченного мышиного фактора H, содержащего лидерный пептид (подчеркнуто) и SCR1-4, 6-8, 19-20 (mfH1-4.678.19-20) (SEQ ID NO: 43).
На ФИГ. 20 приведена аминокислотная последовательность конструкта укороченного мышиного фактора H, содержащего лидерный пептид (подчеркнуто) и SCR1-4, 6-8 и 19-20 (mfH1-4.678.19-20) (SEQ ID NO: 44).
На ФИГ. 21 приведено изображение геля, подтверждающее экспрессию и стабильность белков mfH1-4.19-20 и mfH1-4.678.19-20. Последовательность кДНК укороченного варианта мышиного fH, содержащего SCR1-4, 678 и 19-20, или последовательность кДНК другого укороченного варианта fH, содержащего SCR1-4 и 19-20, клонировали в эукариотический экспрессионный вектор pCBARBG, который затем использовали для трансфекции линии клеток печени мыши, клеток Hepa1C1C7. Супернатант клеточной культуры использовали в вестерн-блот анализе для обнаружения экспрессии укороченного белка мышиного fH. M: маркеры молекулярной массы; дорожка 1, нетрансфицированные клетки Hepa1C1C7 (контроль); дорожки 2 и 3, клетки Hepa1C1C7, трансфицированные pCBARBG-mfH1-4.19-20 клона 3 или клона 4; дорожки 5 и 6, клетки Hepa1C1C7, трансфицированные pCBARBG-mfH1-4.678.19-20 клона 1 (смысловая последовательность) или клона 2 (антисмысловая последовательность).
На ФИГ. 22A приведена блок-схема, показывающая, как образцы крови были собраны и обработаны для обнаружения белка fH. Мыши fHm/m представляют собой линию мутантных по fH мышей, которые имеют кодоны преждевременной остановки в начале последовательности SCR19. У этих мышей продуцируются следовые количества укороченного fH (без SCR19-20) и у них имеет место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе и истощение (вторичный дефицит) C3 и fB. Мышей инфицировали в/в введением через ретроорбитальный синус вируса AAV8, содержащего mfH1-4.678.19-20 (3×1012 геномных копий/мышь), и через одну неделю образцы крови собирали, обрабатывали и анализировали, как показано на блок-схеме.
На ФИГ. 22B показано обнаружение методом вестерн-блоттинга mfH1-4.678.19-20 в крови мутантной по fH мыши (fHm/m) через одну неделю после AAV8-опосредованной терапии геном fH. Как показано на фигуре, mfH1-4.678.19-20 (примерно 70 кДа) отсутствует у Д/Т и не получавших лечение мышей fHm/m. У трех инфицированных вирусом мышей fHm/m, M3, F10, F30 (M обозначает самца и F обозначает самку) mfH1-4.678.19-20 был отчетливо виден.
На ФИГ. 23 показаны результаты вестерн-блот анализа, демонстрирующие, что AAV8-опосредованная терапия геном fH у мышей fHm/m предотвращает неконтролируемую регуляцию альтернативного пути активации комплемента. Из-за отсутствия достаточной экспрессии эндогенного fH у не получавших лечение мышей fHm/m имела место неконтролируемая регуляция альтернативного пути активации комплемента в жидкой фазе, и в результате у них в плазме имело место истощение C3 и fB (сравните дорожку 1 для Д/Т с дорожками 2, 5, 8 для мышей fHm/m до начала генной терапии). После введения мышам fHm/m AAV8-mfH1-4.678.19-20 через одну неделю (1Н, дорожки 3, 6, 9) и один месяц (1М, дорожки 4, 7, 10) уровни в плазме C3 и fB восстанавливались до уровней у Д/Т, свидетельствуя о том, что AAV8-опосредованная терапия геном fH предотвращала неконтролируемую регуляцию альтернативного пути активации комплемента, а также истощение C3 и fB, и что терапевтический эффект был очевиден уже через одну неделю и продолжался по меньшей мере в течение одного месяца.
На ФИГ. 24 показаны результаты вестерн-блот анализа, демонстрирующие, что AAV8-опосредованная терапия геном fH предотвращает неконтролируемую регуляцию альтернативного пути активации комплемента в мышиной модели летальной C3-гломерулопатии (C3-G). У мышей fHm/m, также имеющих дефицит пропердина (fHm/mP-/-), имеет место аналогичная неконтролируемая регуляция альтернативного пути активации комплемента с истощением C3 и fB. В отличие от мышей fHm/m, у мышей fHm/mP-/- развивается летальная форма C3-G и они погибают к возрасту 10-12 недель. В данном эксперименте каждой из двух мышей fHm/mP-/- в возрасте примерно 7 недель вводили AAV8-mfH1-4.678.19-20 или пустой вектор AAV8 (pAAV.TBG.rBG) в качестве контроля (контрольный AAV). Через одну неделю после AAV8-опосредованной генной терапии образцы крови собирали и анализировали на уровни C3 и fB методом вестерн-блоттинга. Как показано на панелях, в сравнении с образцами крови, взятыми до введения AAV8 (пре), отсутствовали различия в уровнях интактных C3 или fB через одну неделю (1Н) после введения контрольного AAV8 (дорожки 2-5). Однако уровни в плазме C3 и fB у мышей через одну неделю после введения AAV8-mfH1-4.678.19-20 были значительно повышены (дорожки 6-9), свидетельствуя о том, что неконтролируемая регуляция альтернативного пути активации комплемента была ингибирована за счет генной терапии. Мыши получали AAV8 (3×1012 геномных копий/мышь) в/в инъекцией через ретроорбитальный синус.
На ФИГ. 25 показаны результаты долгосрочного последующего наблюдения мыши fHm/mP-/- (М3 с ФИГ. 24), получавшей генную терапию с использованием AAV8-mfH1-4.678.19-20. Результаты вестерн-блот анализа уровней в плазме C3 и fB до начала генной терапии (Пре) и через 1 неделю (1Н), 1, 2, 3, 4, 5 и 6 месяцев (1М, 2М, 3M, 4M, 5M, 6M) после введения AAV8-mfH1-4.678.19-20 показывают, что уровни C3 и fB после генной терапии были устойчиво повышены до уровней у мышей дикого типа, свидетельствуя о том, что терапевтический эффект был продолжительным.
На ФИГ. 26 показаны результаты долгосрочного последующего наблюдения мыши fHm/mP-/- (M3 с ФИГ. 24), получавшей генную терапию с использованием AAV8-mfH1-4.678.19-20. Результаты анализа методом ELISA уровней в плазме белка mfH1-4.678.19-20 до введения (Пре) и через 1 неделю, 1, 2, 3, 4, 5 и 6 месяцев (M) после введения AAV8-mfH1-4.678.19-20 показывают устойчивую экспрессию mfH1-4.678.19-20 в качестве терапевтического белкового лекарственного средства.
На ФИГ. 27 показана эффективность генной терапии с использованием AAV8-mfH1-4.678.19-20 для предотвращения почечной патологии при C3-гломерулопатии. Мышь fHm/mP-/-, получавшая контрольный вектор AAV8 (мышь M1 с ФИГ. 24), находилась в предсмертном состоянии в пределах 2 недель после введения, и окрашивание ее почки показало сильные гломерулярные отложения C3, как описано ранее для не получавших лечение мышей fHm/mP-/- (левые панели). Напротив, мышь fHm/mP-/-, получавшая вектор AAV8-mfH1-4.678.19-20 (M3 с ФИГ. 23), выжила и все-еще была здорова через 6 месяцев после введения, когда ее умертвили и провели анализ гистологии почек. Никакие гломерулярные отложения C3 не были обнаружены у этой мыши (правые панели), это свидетельствовало о том, что C3-гломерулопатия была предотвращена за счет генной терапии с использованием AAV8-mfH1-4.678.19-20.
На ФИГ. 28A и 28B показано, что генная терапия с использованием AAV8-mfH1-4.678.19-20 предотвращает альтернативный путь активации комплемента, вызываемый нарушением функции мембранного регулятора. В данном эксперименте мышам с дефицитом двух мембранных регуляторов, DAF и Crry, вводили AAV8-mfH1-4.678.19-20 (ретроорбитальный путь введения, в/в, 3×1012 геномных копий/мышь). Образцы плазмы собирали до и через 1 неделю (1Н) после генной терапии для анализа уровней в плазме C3 (A) и fB (B) методом вестерн-блоттинга. Как показывают данные, мыши с двойной мутацией DAF/Crry отличались избыточной активностью альтернативного пути активации комплемента и низкими уровнями C3 и fB (Пре). После введения AAV8-mfH1-4.678.19-20 уровни как C3, так и fB, были восстановлены до уровней у мышей дикого типа, свидетельствуя о том, что введение AAV8-mfH1-4.678.19-20 может корректировать патологии, вызываемые нарушением функции мембранных регуляторов комплемента. Эти данные указывают на то, что введение AAV8-mfH1-4.678.19-20, в целом, было эффективным для лечения комплемент-опосредованных заболеваний, вызываемых неконтролируемой регуляцией альтернативного пути активации комплемента, независимо от лежащего в основе дефекта регуляторного механизма. Мыши с двойной мутацией DAF/Crry, используемые в данном исследовании, представляют собой потомство от скрещивания между мышами с нокаутом DAF и мышами Crryflox/flox-Tie-2Cre+. Поскольку Tie-2-Cre экспрессируется в зародышевых клетках, результатом является делеция в клетках зародышевой линии гена Crry у некоторых потомков, приводя к общему отсутствию Crry.
На ФИГ. 29A-29B приведено сравнение доз AAV8-mfH1-4.678.19-20 в генной терапии с использованием восстановления уровня C3 в качестве показателя. В данном эксперименте разные дозы AAV8-mfH1-4.678.19-20 вводили мышам fHm/m (ретроорбитальный путь введения, в/в). Каждой из двух мышей в группе вводили следующие дозы: 1×1012 геномных копий/мышь (M#1, M#2), 3×1011 геномных копий/мышь (M#3, M#6) и 1×1011 геномных копий/мышь (M#4, M#5). Проводили вестерн-блоттинг для анализа уровней в плазме C3 до (Пре) и через одну неделю (1Н) или 1 месяц (1М) после генной терапии. Как показано, все протестированные дозы были способны приводить к увеличению уровней C3 в плазме при изучении во временных точках 1Н и 1М.
На ФИГ. 30A-30B приведено сравнение доз AAV8-mfH1-4.678.19-20 в генной терапии с использованием восстановления уровня fB в качестве показателя. Вестерн-блоттинг выполняли, в целом, как описано для ФИГ. 29A-B, где C3 использовали в качестве показателя. Как показано, все протестированные дозы были способны приводить к увеличению уровней fB в плазме при изучении во временных точках 1Н и 1М.
На ФИГ. 31 приведена схематическая диаграмма, показывающая стратегию таргетирования генов, используемую для введения мутации замены W на R в SCR20 мышиного fH (положение 1206, соответствующее положению 1183 в человеческом fH).
На ФИГ. 32 приведены кривые выживаемости однопометных мышей дикого типа и мутантных мышей, несущих мутацию W1206R в fH. У мышей с мутантным fH развивались патологии, характерные для aHUS, и половина из них погибала к возрасту 30 недель.
На ФИГ. 33A приведено сравнение количества тромбоцитов у мышей дикого типа, гетерозиготных и гомозиготных мутантных мышей. У гомозиготных мутантных мышей имело место низкое количество тромбоцитов, свидетельствуя о том, что они страдали от хронической тромбоцитопении.
На ФИГ. 33B приведено сравнение уровней гемоглобина у мышей дикого типа, гетерозиготных и гомозиготных мутантных мышей. У гомозиготных мутантных мышей имели место низкие уровни гемоглобина, свидетельствуя о том, что они страдали от хронической гемолитической анемии.
На ФИГ. 34A-34C показаны срезы почки мышей с мутацией W1206R, демонстрирующие патологии, характерные для aHUS. Патологические признаки включали мезангиальную экспансию и сужение просвета капилляров (панель A), тромбы в мелких сосудах, отмеченные стрелками на панели A и панели C. Электронно-микроскопическое исследование показало, что в стенке клубочковых капилляров наблюдалась экспансия субэндотелиального пространства с рыхлым гранулярным электронно-прозрачным материалом и образование двойных контуров и новой клубочковой базальной мембраны.
На ФИГ. 35A-35D показано, что у мышей, несущих мутацию W1206R в fH, также развилось повреждение сетчатки и образование сгустков крови в глазах. В сравнении с нормальными сетчатками мышей дикого типа (ФИГ. 35A) у мышей с мутацией W1206R в fH наблюдалось множество белых пятен, отек сетчатки и расширенные сосуды в сетчатке (ФИГ. 35B). Кроме того, флуоресцентная ангиография показала, что у мутантных мышей сетчатка не была хорошо перфузируемой, поскольку краситель проникал во все кровеносные сосуды в глазах у мышей дикого типа в пределах 30 секунд (ФИГ. 35C), но не достигал большой части области сетчатки у мутантных мышей даже через 4 мин (ФИГ. 35D).
На ФИГ. 36 показано, что белок mfH1-4.678.19-20 был обнаружен методом ELISA в крови у мышей fHW1206R/W1206R через 1 месяц и 2 месяца после введение вектора AAV8-mfH1-4.678.19-20 в дозе 3×1011 гк/mouse, но не в крови у этих мышей до начала генной терапии с использованием AAV.
На ФИГ. 37A и 37B приведены линейные графики, демонстрирующие, что введение мышам fHW1206R/W1206R вектора AAV8-mfH1-4.678.19-20 в дозе 3×1011 гк/мышь привело к нормализации у них количества тромбоцитов. Все 3 мыши fHW1206R/W1206R, получавшие AAV8-mfH1-4.678.19-20, были живы и здоровы. Количество тромбоцитов (ФИГ. 37A) и уровни гемоглобина (Hb, ФИГ. 37B) у них возросли и сохранялись в нормальном диапазоне. Напротив, 1 из 2 мышей fHW1206R/W1206R, получавших контрольный AAV вектор, погибла (через 4 недели после введения) и оставшаяся мышь имела постоянно низкие уровни тромбоцитов и меняющийся уровень гемоглобина, который был ниже уровня у мышей, получавших AAV8-mfH1-4.678.19-20.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем описании описаны новые рекомбинантные гены и варианты белка фактора H (fH). Эти варианты отличаются увеличенным временем полужизни и повышенной эффективностью в лечении состояний, связанных с фактором H, и других заболеваний, связанных с комплементом.
Описана доставка этих вариантов в организм пациентов, которые нуждаются в этом, разными путями доставки и, в частности, путем экспрессии in vivo, опосредованной рекомбинантным вектором, таким как rAAV вектор. Также предложены способы использования этих вариантов в схемах лечения заболеваний, связанных с фактором H. Предпочтительно, композиции, предложенные в настоящем описании, полезны для одновременного таргетирования нескольких путей и/или изменения, или модулирования, неконтролируемой регуляции альтернативного пути активации комплемента, вызываемой различными факторами.
Используемый в настоящем описании термин «лечение заболеваний, связанных с фактором H комплемента» может охватывать облегчение, уменьшение и/или ослабление симптомов, и/или предотвращение развития дополнительных симптомов, заболевания, связанного с фактором H комплемента, которое может проявляться в виде нескольких разных фенотипов, включая бессимптомный, рецидивирующие бактериальные инфекции и почечную недостаточность. При этом, как правило, имеет место снижение сывороточных уровней фактора H, компонента C3 комплемента, а также уменьшение содержания других компонентов терминального пути активации комплемента, что указывает на активацию альтернативного пути активации комплемента. Данное заболевание связано с целым рядом заболеваний почек с различными клиническими проявлениями и прогрессированием, включая C3-гломерулопатию и атипичный гемолитико-уремический синдром. В настоящем описании также предложены композиции и способы для лечения одного или более из: возрастной дегенерации желтого пятна (AMD), атипичного гемолитико-уремического синдрома (включая, например, микроангиопатическую гемолитическую анемию, тромбоцитопению, острую почечную недостаточность), пароксизмальной ночной гемоглобинурии (PNH), шизофрении, ишемического инсульта, и/или предотвращения или лечения бактериальных инфекций, вызываемых рекрутингом бактериальных патогенов (например, Aspergillus spp.; Borrelia burgdorferi; B. duttonii; B. recurrentis; Candida albicans; Francisella tularensis; Haemophilus influenzae; Neisseria meningitidis; Streptococcus pyogenes) или одного из пяти связывающих фактор H белков B. burgdorferi (CRASP-1, CRASP-2, CRASP-3, CRASP-4 или CRASP-5), в числе прочих.
Используемый в настоящем описании термин «лечение заболеваний, связанных с комплементом» включает облегчение, уменьшение и/или ослабление симптомов, как заболеваний, связанных с фактором H комплемента, указанных выше, так и других заболеваний, связанных с неконтролируемой регуляцией альтернативного пути активации комплемента. Более конкретно, данные, представленные в настоящем описании, свидетельствуют о том, что по меньшей мере один введенный с помощью AAV вариант fH, в целом, эффективен для лечения опосредованных комплементом заболеваний, вызываемых неконтролируемой регуляцией альтернативного пути активации комплемента, независимо от лежащего в основе дефекта регуляторного механизма. См., например, ФИГ. 23.
«Заболевания, опосредованные комплементом» могут включать симптомы, связанные с нарушением регуляции комплемента, которые могут проявляться в виде нескольких разных фенотипов, включая бессимптомный, рецидивирующие бактериальные инфекции, а также различные повреждения тканей, включая, но без ограничения, заболевания почек. Если не указано иначе, это определение применимо как к гомозиготным пациентам, так и гетерозиготным пациентам. Причиной нарушения регуляции комплемента, как правило, являются мутации с потерей функции, или аутоантитела против, регуляторных белков комплемента, включая, но без ограничения, fH, фактор I (fI) и мембранный кофакторный белок (MCP), либо мутации с приобретением функции в других белках комплемента, включая, но без ограничения, C3 и фактор B (fB). Нарушение регуляции комплемента, как правило, хотя и не всегда, характеризуется снижением сывороточных уровней фактора H, компонента C3 комплемента, fB, а также снижением уровней других компонентов терминального пути активации комплемента, что указывает на активацию альтернативного и/или терминального пути активации комплемента. Патологии, опосредованные комплементом, которые можно лечить с помощью композиций и способов по настоящему изобретению, включают, но не ограничиваются ими, следующие заболевания с различными клиническими проявлениями и прогрессированием: C3-гломерулопатию (ранее называемую мембранопролиферативным гломерулонефритом II типа или MPGNII), для которой известны две формы - болезнь плотных депозитов (DDD) и C3-гломерулонефрит (C3-GN); тромботическую микроангиопатию (TMA), включая, но без ограничения, атипичный гемолитико-уремический синдром (aHUS), шигаподобный токсин-продуцирующую E. coli HUS (STEC-HUS) и тромботическую тромбоцитопеническую пурпуру (TTP); дегенеративное заболевание сетчатки глаза, включая возрастную дегенерацию желтого пятна (AMD), дегенерацию ПЭС, хориоретинальную дегенерацию, дегенерацию фоторецепторов; пароксизмальную ночную гемоглобинурию (PNH), ишемическое реперфузионное повреждение всех органов и окружающих тканей, ревматоидный артрит, гемодиализ, диабетическую нефропатию, диабетическую васкулопатию, астму, системную красную волчанку (SLE), ишемический инсульт, аневризму брюшной аорты (AAA), опосредованный антинейтрофильными цитоплазматическими антителами (ANCA) васкулит (ANCA-васкулит), ANCA-опосредованное геморрагическое повреждение и заболевание легких, ANCA-гломерулонефрит, реакцию трансплантат против хозяина (GvHD), острое или отсроченное отторжение трансплантата при трансплантации органов, болезнь Крона, псориаз, рассеянный склероз, антифосфолипидный синдром, преэклампсию, атеросклероз, оптиконевромиелит (NMO), аутоиммунную пузырчатку, буллезный пемфигоид (BP), болезнь Альцгеймера (AD), а также бактериальные инфекции, вызываемые рекрутингом бактериальных патогенов (например, Aspergillus spp.; Borrelia burgdorferi; B. duttonii; B. recurrentis; Candida albicans; Francisella tularensis; Haemophilus influenzae; Neisseria meningitidis; Streptococcus pyogenes). Другие примеры таких заболеваний описаны более подробно ниже.
Аминокислотная последовательность зрелого человеческого фактора H комплемента (изоформы 1) «дикого типа» приведена в настоящем описании в виде http://www.uniprot.org/uniprot/P08603 и служит в качестве эталона для нумерации аминокислот изоформы 1 hfH (приведена в SEQ ID NO: 39). Лидерная последовательность соответствует аминокислотам 1-18 фактора H, представленного SEQ ID NO: 39. Аминокислотная последовательность лидерной области приведена в SEQ ID NO: 2. Зрелый (секретируемый) белок hfH соответствует аминокислотам 19-1231 в SEQ ID NO: 39. Существуют альтернативные способы определения расположения 20 коротких консенсусных повторов (SCR). Местоположение доменов, использованных в экспериментах, приведенных ниже, аннотировано на ФИГ. 1 и основано на системе нумерации, используемой в статье C. Estaller et al., Eur J Immunol. 1991 Mar; 21(3): 799-802.
Аминокислотная последовательность человеческого фактора H комплемента дикого типа, изоформы 1, приведена в SEQ ID NO: 39. Выделенный раздел в SEQ ID NO: 1 также иллюстрирует альтернативную систему идентификации начала/конца каждого из 20 доменов коротких консенсусных повторов (SCR). В этой системе последовательности линкеров отсутствуют между всеми SCR.
Необязательно, рекомбинантный вариант hfH, предложенный в настоящем описании, может иметь гетерологичную лидерную последовательность, заменяющую природную лидерную последовательность hfH. Дополнительно, или необязательно, другая изоформа hfH (например, изоформа 2), последовательность которой доступна, например, на сайте http://www.uniprot.org/uniprot/P08603, и/или один из ее природных аминокислотных вариантов, которые не связаны с заболеванием. См. SEQ ID NO: 40. В следующих далее описаниях замены могут быть описаны следующим образом: (первая аминокислота, идентифицированная однобуквенным кодом) - № положения остатка - (вторая аминокислота, идентифицированная однобуквенным кодом), при этом первая аминокислота представляет собой замещенную аминокислоту и вторая аминокислота представляет собой замещающую аминокислоту в указанном положении эталонной изоформы 1; однако за счет общепринятых этапов выравнивания соответствующие аминокислотные остатки, идентифицированные в настоящем описании на основании нумерации изоформы 1, могут быть расположены в изоформе 2 и не вызывающих заболевания природных вариантах SCR изоформы 1 или 2 fH.
В настоящем описании при упоминании SCR №-№№ перечень доменов включает крайние члены, и это равноценно написанию «SCR №,…,SCR №№». В конкретных вариантах осуществления при определении доменов используют цифры, разделенные чертой. Например, SCR1-4 означает «SCR1, SCR2, SCR3 и SCR4» и равноценно написанию «SCR1,2,3,4» или «SCR1.2.3.4.». SCR19-20 означает SCR19 и SCR20, и равноценно написанию «SCR19,20». Например, варианты написания «SCR6-8», «SCR6.7.8» и «SCR6,7,8» означают одни и те же домены.
Используемый в настоящем описании термин «функциональный вариант fH» включает варианты fH, которые характеризуются наличием регулирующей комплемент активности (кофакторной активности), обусловленной SCR1-4, и, необязательно, функциональной C3b-связывающей и GAG-связывающей способности (обусловленной SCR7 и SCR19-20 дикого типа), которые присущи fH дикого типа. В некоторых вариантах осуществления рекомбинантные варианты fH имеют более 100% кофакторной активности и/или GAG-связывающей способности fH дикого типа. См., например, фигуру 8 и примеры ниже, которые показывают, что вариант fH, описанный в настоящем описании, имеет GAG- и C3b-связывающую активность, статистически достоверно превышающую активность полноразмерного человеческого fH, например, превышающую примерно на 10-40%. В другом варианте осуществления рекомбинантный вариант fH имеет менее примерно 95-100% активности функционального fH дикого типа. Например, рекомбинантный вариант fH может иметь по меньшей мере 50% от кофакторной активности, имеющейся у функционального fH дикого типа, и более желательно, по меньшей мере примерно 60%, по меньшей мере примерно 75%, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 95% или по меньшей мере примерно 99%. Альтернативно или дополнительно, в другом варианте осуществления рекомбинантный вариант fH может иметь по меньшей мере 50% от GAG-связывающей активности функционального fH, и более желательно, по меньшей мере примерно 60%, по меньшей мере примерно 75%, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 95% или по меньшей мере примерно 99%. Методы определения кофакторной активности, связывающей активности и/или определения увеличенного времени полужизни в системе циркуляции по сравнению с белками hfH известны в данной области, и по меньшей мере один из таких анализов описан в приведенных ниже примерах.
Примеры функциональных вариантов fH включают варианты, содержащие SCR1-4 и 19-20 белка fH, с одним или более из доменов SCR7, SCR17 или SCR18. Дополнительные варианты включают те, которые содержат один или более из SCR6, SCR8, SCR16, SCR17, SCR18, либо их фрагментов, а также их комбинации. Например, такие варианты могут содержать, например, fH SCR1-4, 6-8, 19-20; fH SCR1-4, 6-8, 18-20; fH SCR1-4, 6-8, 17-20; fH SCR1-4, 6-7, 19-20; fH SCR1-4, 6-7, 18-20; fH SCR1-4, 6-7, 17-20; fH SCR1-4, 7-8, 19-20; fH SCR1-4, 7-8, 18-20; fH SCR1-4, 7-8, 17-20; fH SCR1-4, 7, 19-20; fH SCR1-4, 7, 18-20; fH SCR1-4, 7, 17-20; SCR1-4, 17, 19-20; SCR1-4, 18-20; SCR1-4, 17-20 и/или fH SCR1-4, 7, 16-20, в числе прочих. В конкретных вариантах осуществления вариант hfH, кроме того, содержит дополнительные hfH SCR, например, SCR 6, SCR8, SCR16, или их комбинацию. В предпочтительных вариантах осуществления hfH SCR5 отсутствует. Однако в некоторых вариантах осуществления hfH SCR5 может присутствовать целиком или в виде фрагмента. В конкретных вариантах осуществления hfH SCR9, SCR10, SCR11, SCR12, SCR13, SCR14 и/или SCR15 отсутствуют или по меньшей мере функционально делетированы. Необязательно, один или более из SCR в этих вариантах может представлять собой «функциональный фрагмент» SCR, а не полноразмерный SCR, представленный на ФИГ. 1 или выделенных участках SEQ ID NO: 1. «Функциональный фрагмент» означает аминокислотную последовательность (или кодирующую ее последовательность) меньшего размера, чем полноразмерный SCR, которая характеризуется наличием одной или более из комплемент-ингибирующей активности, способности к связыванию гепарина и/или C3b-связывающей активности.
Эти и другие варианты могут содержать другие последовательности fH. Например, при экспрессии с вирусного вектора кодирующая последовательность варианта fH также содержит лидерную последовательность. Такая лидерная последовательность может представлять собой лидерную последовательность fH. Необязательно, лидерная последовательность может происходить из другого источника, например, представлять собой лидерную последовательность IL-2 (см., например, указатель лидерных последовательностей млекопитающих, представленный на сайте http://www.signalpeptide.de/, включенный в настоящий документ посредством ссылки). В одном варианте осуществления выбранная лидерная последовательность имеет менее примерно 26 аминокислот в длину (например, от примерно 1 до примерно 26 аминокислот), более предпочтительно менее примерно 20 аминокислот (от примерно 1 до примерно 20 аминокислот) и наиболее предпочтительно менее примерно 18 аминокислот в длину (от примерно 1 до примерно 18 аминокислот). Термин «функциональная делеция» относится к аминокислотной последовательности (или кодирующей ее последовательности), которая лишена комплемент-ингибирующей активности, C3b-связывающей активности и, необязательно, также лишена гепарин-связывающей активности.
В случае вариантов, домены могут быть смежными друг с другом (например, карбокси-конец одного домена может непосредственно следовать за амино-концом предыдущего домена). Альтернативно, один или более из доменов SCR могут иметь линкер, содержащий от одной до примерно 12-18 аминокислот, расположенный между ними. Например, вариант может содержать SCR1-(L1)-SCR2-(L2)-SCR3-(L3)-SCR4-(L4)-(SCR6-(L4'))-SCR7-(L5)-(SCR8-(L5'))-(SCR16-(L5"))-(SCR17-(L5"'))-(SCR18-(L5""))-SCR19-(L6)-SCR20, при этом () обозначает необязательный компонент, «L» обозначает линкер и каждый из L1, L2, L3, L4, L4', L5, L5', L5", L5"', L5"" и L6 может отсутствовать или быть независимо выбран из аминокислотной последовательности, содержащей от примерно 1 до примерно 12-18 аминокислот. Иными словами, если вариант содержит несколько линкеров, все линкеры могут иметь одинаковые или разные последовательности. В конкретных вариантах осуществления вариант содержит по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять линкеров, по меньшей мере шесть линкеров. Примеры подходящих линкеров включают природные линкеры, представленные на ФИГ. 1 или ФИГ. 17, SEQ ID NO: 4, 6, 8, 10, 12, 15, 18, 20, 22, 24, 26, 28, 30, 32, 34 и 36, или синтетические линкеры. Каждый из этих линкеров дикого типа может находиться в его естественном положении. Альтернативно, один или более из этих линкеров дикого типа может быть использован в другом положении линкера или в нескольких разных положениях линкеров.
Необязательно, один или более из этих линкеров могут представлять собой последовательности fH и быть независимо выбраны. Альтернативно, один или более из этих линкеров могут быть гетерологичными в отношении fH, например, происходить из иного источника, либо быть искусственными, синтетическими, либо происходить из другого белка, что придает соответствующую гибкость варианту fH. Примеры других подходящих линкеров могут включать, например, поли-Gly линкер и другие линкеры, обеспечивающие соответствующую гибкость (например, http://parts.igem.org/Protein_domains/Linker, электронный адрес включен в настоящий документ посредством ссылки). В конкретных вариантах осуществления линкеры лишены какой-либо функции fH.
Использованный выше термин «аминокислотная замена», и его синонимы, означает модификацию аминокислотной последовательности путем замены аминокислоты другой, замещающей, аминокислотой. Замена может представлять собой консервативную замену. Она также может представлять собой неконсервативную замену. Термин «консервативная» замена, используемый применительно к двум аминокислотам, должен означать, что аминокислоты имеют общее свойство, признаваемое специалистами в данной области. Например, аминокислоты, имеющие гидрофобные некислотные боковые цепи, аминокислоты, имеющие гидрофобные кислотные боковые цепи, аминокислоты, имеющие гидрофильные некислотные боковые цепи, аминокислоты, имеющие гидрофильные кислотные боковые цепи, и аминокислоты, имеющие гидрофильные основные боковые цепи. Обладающими общими свойствами также могут быть аминокислоты, имеющие гидрофобные боковые цепи, аминокислоты, имеющие алифатические гидрофобные боковые цепи, аминокислоты, имеющие ароматические гидрофобные боковые цепи, аминокислоты с полярными нейтральными боковыми цепями, аминокислоты с заряженными боковыми цепями, аминокислоты с заряженными кислотными боковыми цепями и аминокислоты с заряженными основными боковыми цепями. Как природные, так и неприродные аминокислоты известны в данной области и могут быть использованы в качестве замещающих аминокислот в вариантах осуществления. Методы замены аминокислот хорошо известны специалистам в данной области и включают, но не ограничиваются ими, мутации нуклеотидной последовательности, кодирующей аминокислотную последовательность. Выражение «один или более» в настоящем описании должно охватывать, например, 1, 2, 3, 4, 5, 6 или более отдельных вариантов осуществления.
Помимо вариантов белка fH, предложенных в настоящем описании, также предложены нуклеотидные последовательности, кодирующие эти варианты белка fH. Кодирующие последовательности для этих вариантов могут быть из последовательностей дикого типа для лидерной последовательности и/или одного или более SCR изоформы 1, изоформы 2, либо не связанных с заболеваниями вариантов. Альтернативно или дополнительно, можно использовать имеющиеся в сети интернет или коммерчески доступные компьютерные программы, а также компании, предлагающие такого рода сервис, для обратной трансляции аминокислотных последовательностей лидерной последовательности и/или одного или более из SCR в кодирующие нуклеотидные последовательности, включая РНК и/или кДНК. См., например, сайты для обратной трансляции от компаний EMBOSS, http://www.ebi.ac.uk/Tools/st/; Gene Infinity (http://www.geneinfinity.org/sms-/sms_backtranslation.html); ExPasy (http://www.expasy.org/tools/). В одном варианте осуществления кодирующие последовательности РНК и/или кДНК разработаны для оптимальной экспрессии в клетках человека.
Кодон-оптимизацию кодирующих областей можно проектировать различными методами. Эту оптимизацию можно проводить с использованием методов, доступных в сети интернет, опубликованных методов, или компаний, которые предлагают услуги кодон-оптимизации. Один метод кодон-оптимизации описан, например, в WO 2015/012924 A2, который включен в настоящий документ посредством ссылки. Вкратце, нуклеотидную последовательность, кодирующую продукт, модифицируют последовательностями синонимичных кодонов. Предпочтительно, изменяется вся длина открытой рамки считывания (ORF) для продукта. Однако в некоторых вариантах осуществления может быть изменен лишь фрагмент ORF. За счет использования одного из этих методов можно применять принцип частоты встречаемости кодонов к любой конкретной полипептидной последовательности, и получать фрагмент нуклеотидной последовательности кодон-оптимизированной кодирующей области, которая кодирует полипептид.
Термины «процент (%) идентичности», «идентичность последовательностей», «процент идентичности последовательностей» или «процентная идентичность» в контексте нуклеотидных последовательностей относится к основаниям в двух последовательностях, которые являются одинаковыми при выравнивании для сравнения. Сравнение последовательностей для определения идентичности можно выполнять на протяжении всей длины генома, полноразмерной кодирующей последовательности гена или фрагмента из по меньшей мере примерно 500-5000 нуклеотидов, либо в зависимости от обстоятельств. Однако также может быть желательно определять идентичность более коротких фрагментов, например, длиной по меньшей мере примерно девять нуклеотидов, как правило, по меньшей мере примерно 20-24 нуклеотидов, по меньшей мере примерно 28-32 нуклеотидов, по меньшей мере примерно 36 или более нуклеотидов. Многие программы выравнивания последовательностей также доступны для нуклеотидных последовательностей. Примеры таких программ включают «Clustal W», «CAP Sequence Assembly», «BLAST», «MAP» и «MEME», которые доступны в сети интернет. Другие источники для таких программ известны специалистам в данной области. Альтернативно, также можно использовать сервисные программы Vector NTI. Также существует ряд алгоритмов, известных в данной области, которые можно использовать для определения идентичности нуклеотидных последовательностей, включая те, которые содержатся в программах, описанных выше. В качестве другого примера, полинуклеотидные последовательности можно сравнивать с использованием Fasta™, программы в GCG версии 6.1. Fasta™ производит выравнивание и определение процентной идентичности последовательностей в областях наилучшего совпадения между запрашиваемой последовательностью и последовательностью из базы данных. Например, процент идентичности последовательностей между нуклеотидными последовательностями можно определять с использованием программы Fasta™ с ее параметрами по умолчанию (размер слова 6 и фактор NOPAM для матрицы замен), предложенными в GCG версии 6.1, включенной в настоящий документ посредством ссылки.
Термины «процент (%) идентичности», «идентичность последовательностей», «процент идентичности последовательностей» или «процентная идентичность» в контексте аминокислотных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для сравнения. Процент идентичности можно с легкостью определять для аминокислотных последовательностей на протяжении полноразмерного белка, полипептида, примерно 32 аминокислот, примерно 330 аминокислот или пептидного фрагмента, либо для соответствующих нуклеотидных последовательностей, кодирующих эти последовательности. Подходящий аминокислотный фрагмент может иметь по меньшей мере примерно 8 аминокислот в длину, и может содержать до примерно 700 аминокислот. Как правило, когда речь идет об «идентичности», «гомологии» или «сходстве» между двумя разными последовательностями, «идентичность», «гомологию» или «сходство» определяют для «выровненных» последовательностей. Термины «выровненные» последовательности или «выравнивания» относятся к нескольким нуклеотидным последовательностям или белковым (аминокислотным) последовательностям, часто имеющим коррекции для утраченных или дополнительных оснований, либо аминокислот, по сравнению с эталонной последовательностью. Выравнивания производят с использованием любой из множества общедоступных или коммерчески доступных программ множественного выравнивания последовательностей. Программы выравнивания последовательностей доступны для аминокислотных последовательностей, например, программы «Clustal X», «MAP», «PIMA», «MSA», «BLOCKMAKER», «MEME» и «Match-Box». Как правило, любую из этих программ используют с параметрами по умолчанию, хотя специалист в данной области может изменять эти параметры по мере необходимости. Альтернативно, специалист в данной области может использовать другой алгоритм или компьютерную программу, которая обеспечивает по меньшей мере такой же уровень определения идентичности или выравнивания, который обеспечивают эталонные алгоритмы и программы. См., например, J. D. Thomson et al., Nucl. Acids. Res., «A comprehensive comparison of multiple sequence alignments», 27(13): 2682-2690 (1999).
В одном варианте осуществления нуклеотидные последовательности, кодирующие варианты fH (например, вариант гена hfH), описанные в настоящем описании, введены генно-инженерными методами в любой подходящий генетический элемент, например, «голую» ДНК, фаг, транспозон, космиду, молекулу РНК (например, мРНК), эписому и так далее, который переносит находящиеся в нем последовательности hfH в клетку-хозяина, например, для образования наночастиц, несущих ДНК или РНК, вирусных векторов в упаковывающей клетке-хозяине и/или для доставки в клетки-хозяева у пациента. В одном варианте осуществления генетический элемент представляет собой плазмиду. Выбранный генетический элемент может быть доставлен любым подходящим способом, включая трансфекцию, электропорацию, доставку с помощью липосом, методы слияния мембран, введенные с высокой скоростью гранулы с нанесенной на них ДНК, вирусную инфекцию и слияние протопластов. Методы, используемые для получения таких конструктов, известны специалистам, умеющим работать с нуклеиновыми кислотами, и включают генетическую инженерию, рекомбинантную инженерию и методы синтеза. См., например, Green and Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY (2012).
Используемый в настоящем описании термин «экспрессионная кассета» означает молекулу нуклеиновой кислоты, которая содержит кодирующие последовательности варианта hfH, промотор и может содержать другие регуляторные последовательности (например, последовательности 5' и/или 3'-UTR), эту кассету можно вводить в генетический элемент и/или упаковывать в капсид вирусного вектора (например, вирусную частицу). Как правило, такая экспрессионная кассета для создания вирусного вектора содержит последовательности hfH, описанные в настоящем описании, фланкированные сигналами упаковывания вирусного генома и другими последовательностями контроля экспрессии, такими как те, которые описаны в настоящем описании.
Экспрессионная кассета, как правило, содержит последовательность промотора, как часть последовательностей контроля экспрессии. В иллюстративных плазмиде и векторе, описанных в настоящем описании, используют промотор бета-актина кур. Альтернативно, можно выбирать другой конститутивный промотор. В конкретных вариантах осуществления избегать нацеливания на не являющиеся мишенью клетки можно за счет использования соответствующих элементов вектора, например, микроРНК. Дополнительно или альтернативно, выбранный вектор может иметь преимущественное нацеливание на нужную ткань, например, AAV8, AAV9 или AAVrh10 для печени, AAV8, AAV1 или другой AAV для глаза, или тому подобное.
Нацеливание вектора на нужную ткань может быть желательным для максимального увеличения экспрессии белка. И, следовательно, можно выбирать специфичный для печени промотор. Примеры соответствующих промоторов включают промотор для тироксинсвязывающего глобулина (TBG), альфа-1-антитрипсина (A1AT); человеческого альбумина, Miyatake et al., J. Virol., 71: 5124 32 (1997), humAlb; и ядерный промотор вируса гепатита B, Sandig et al., Gene Ther., 3: 1002-9 (1996), минимальный энхансер/промотор TTR, промотор альфа-антитрипсина, LSP (845 нт) 25 (необходим безинтронный скAAV). Альтернативно, можно использовать другие специфичные для печени промоторы (см., например, The Liver Specific Gene Promoter Database, Cold Spring Harbor, http://rulai.schl.edu/LSPD). Альтернативно, если необходимо нацеливание на другую ткань, можно выбирать другие тканеспецифичные промоторы. Промотор может происходить из любых биологических видов. Например, для использования в глазах можно выбирать, например, промотор для белков пигментного эпителия сетчатки (ПЭС) или фоторецепторов. В другом варианте осуществления промотор представляет собой промотор человеческого связанного с G-белками рецептора протеинкиназы 1 (GRK1) (регистрационный номер в Genbank AY327580). В другом варианте осуществления промотор представляет собой 292-нт фрагмент (положения 1793-2087) в промоторе GRK1 (см. также Beltran et al., Gene Therapy 2010 17: 1162-74, статья включена в настоящий документ посредством ссылки). В другом предпочтительном варианте осуществления промотор представляет собой проксимальный промотор человеческого интерфоторецепторного ретиноид-связывающего белка (IRBP). В другом варианте осуществления промотор представляет собой природный промотор гена, который должен быть экспрессирован. В одном варианте осуществления промотор представляет собой проксимальный промотор RPGR (Shu et al., IOVS, May 2012, статья включена в настоящий документ посредством ссылки). Другие промоторы, которые можно использовать по изобретению, включают, без ограничения, промотор опсина палочек, промотор красного/зеленого опсина, промотор синего опсина, промотор цГМФ-β-фосфодиэстеразы, промотор мышиного опсина (Beltran et al. 2010, выше), промотор родопсина (Mussolino et al., Gene Ther, July 2011, 18(7): 637-45); промотор альфа-субъединицы трансдуцина колбочек (Morrissey et al., BMC Dev, Biol, Jan 2011, 11: 3); промотор бета-фосфодиэстеразы (PDE); промотор белка пигментного ретинита 1 (RP1) (Nicord et al., J. Gene Med, Dec 2007, 9(12): 1015-23); промотор NXNL2/NXNL1 (Lambard et al., PLoS One, Oct. 2010, 5(10): e13025), промотор RPE65; промотор гена, обуславливающего медленную дегенерацию сетчатки/периферина 2 (Rds/perph2) (Cai et al., Exp Eye Res. 2010 Aug; 91(2): 186-94), и промотор VMD2 (Kachi et al., Human Gene Therapy, 2009 (20: 31-9)). Примеры промоторов, специфичных для фоторецепторов, включают, без ограничения, промотор опсина палочек, промотор красного/зеленого опсина, промотор синего опсина, промотор интерфоторецепторного ретиноид-связывающего белка (IRBP) и промотор цГМФ-β-фосфодиэстеразы. Альтернативно, другие промоторы, такие как вирусные промоторы, конститутивные промоторы, регулируемые промоторы (см., например, WO 2011/126808 и WO 2013/04943), или промотор, реагирующий на физиологические сигналы, могут быть использованы в векторах, описанных в настоящем описании.
Помимо промотора, экспрессионная кассета и/или вектор может содержать другие соответствующие последовательности инициации и терминации транскрипции, последовательности энхансеров, сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования (полиA); последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (например, консенсусную последовательность Козак); последовательности, повышающие стабильность белка; и, если необходимо, последовательности, обеспечивающие увеличение секреции закодированного продукта. Примеры соответствующих последовательностей сигнала полиA включают, например, последовательности сигнала полиA SV40, бычьего гормона роста (bGH), кроличьего бета-глобулина и TK. Примеры соответствующих энхансеров включают, например, энхансер альфа-фетопротеина, минимальный промотор/энхансер TTR, LSP (промотор TH-связывающего глобулина/энхансер альфа1-микроглобулина/бикунина), в числе прочих.
Эти контрольные последовательности «функционально связаны» с последовательностями генов fH. Используемый в настоящем описании термин «функционально связанные» относится как к последовательностям контроля экспрессии, которые расположены рядом с интересующим геном, так и последовательностям контроля экспрессии, которые действуют со стороны или на расстоянии, контролируя интересующий ген.
Экспрессионную кассету можно вводить в плазмиду, которую используют для доставки лекарственного средства или для получения вирусного вектора. Подходящие вирусные векторы предпочтительно являются репликативно-дефектными и выбраны из векторов, нацеленных на клетки глаза. Вирусные векторы, которые можно использовать, могут включать любой вирус, подходящий для генной терапии, включая, но без ограничения, аденовирус, вирус герпеса, лентивирус, ретровирус, парвовирус и так далее.
Предпочтительно, если создают один из этих векторов, его получают в виде репликативно-дефектного вирусного вектора. Термины «репликативно-дефектный вирус» или «вирусный вектор» относятся к синтетической или рекомбинантной вирусной частице, в которой экспрессионная кассета, содержащая интересующий ген, упакована в вирусный капсид или оболочку, при этом любые вирусные геномные последовательности, также упакованные в вирусный капсид или оболочку, являются репликативно-дефектными; то есть, они не способны к продуцированию вирионов-потомков, но сохраняют способность инфицировать клетки-мишени. В одном варианте осуществления геном вирусного вектора не содержит гены, кодирующие ферменты, необходимые для репликации (рекомбинантный геном может быть «выпотрошенным» - содержащим лишь интересующий трансген, фланкированный сигналами, необходимыми для амплификации и упаковки искусственного генома), но эти гены могут быть предоставлены во время продуцирования. Вследствие этого, такие вирусы считаются безопасными для использования в генной терапии, поскольку репликация и инфицирование вирионами-потомками не может происходить в отсутствие вирусного фермента, необходимого для репликации.
В одном варианте осуществления вирусный вектор представляет собой аденоассоциированный вирус (AAV). Аденоассоциированный вирусный (AAV) вектор представляет собой устойчивую к ДНКазе AAV частицу, имеющую белковый капсид AAV, в котором упакованы нуклеотидные последовательности для доставки в клетки-мишени. Капсид AAV состоит из 60 субъединиц белков капсида, VP1, VP2 и VP3, которые расположены в икосаэдрической симметрии в соотношении примерно от 1:1:10 до 1:1:20, в зависимости от выбранного AAV.
В исследованиях, описанных в настоящем описании, используют AAV8 в качестве иллюстративного вектора. Используемый в настоящем описании термин «капсид AAV8» означает капсид AAV8, имеющий закодированную аминокислотную последовательность с регистрационным номером GenBank: YP_077180, включенным в настоящий документ посредством ссылки. Некоторые вариации данной закодированной последовательности охвачены настоящим изобретением, к ним относятся последовательности, имеющие примерно 99% идентичности с эталонной аминокислотной последовательностью с регистрационным номером GenBank: YP_077180; патенты США 7282199, 7790449, 8319480, 8962330, US 8962332, (то есть, менее примерно 1% изменений по сравнению с эталонной последовательностью). В другом варианте осуществления капсид AAV8 может иметь последовательность VP1 варианта AAV8, описанного в WO 2014/124282, который включен в настоящий документ посредством ссылки. Способы создания капсида, его кодирующие последовательности и способы получения rAAV вирусных векторов известны. См., например, Gao, et al., Proc. Natl. Acad. Sci. U.S.A. 100 (10), 6081-6086 (2003), US 2013/0045186A1 и WO 2014/124282. В конкретных вариантах осуществления выбирают вариант AAV8, демонстрирующий тропизм в отношении нужных клеток-мишеней, например, печени, фоторецепторов, ПЭС или других клеток глаза. Например, капсид AAV8 может иметь мутации Y447F, Y733F и T494V (другое название «AAV8(C&G+T494V)» и «rep2-cap8(Y447F+733F+T494V)»), как описано в статье Kay et al., Targeting Photoreceptors via Intravitreal Delivery Using Novel, Capsid-Mutated AAV Vectors, PLoS One. 2013; 8(4): e62097. Опубликовано онлайн 26 апреля 2013 г., содержание включено в настоящий документ посредством ссылки. См., например, Mowat et al., Tyrosine capsid-mutant AAV vectors for gene delivery to the canine retina from a subretinal or intravitreal approach, Gene Therapy 21, 96-105 (January 2014), содержание включено в настоящий документ посредством ссылки. В другом варианте осуществления капсид AAV представляет собой капсид AAV8bp, который предпочтительно нацелен на биполярные клетки. См. WO 2014/024282, содержание включено в настоящий документ посредством ссылки.
Можно выбирать другие серотипы AAV в качестве источников для капсидов AAV вирусных векторов (устойчивых к ДНКазе вирусных частиц), включая, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV6.2, AAV7, AAV8, AAV9, rh10, AAVrh64R1, AAVrh64R2, rh8 (см., например, публикацию патентной заявки США № 2007-0036760-A1; публикацию патентной заявки США № 2009-0197338-A1; EP 1310571). См. также WO 2003/042397 (AAV7 и другие обезьяньи AAV), патент США 7790449 и патент США 7282199 (AAV8), WO 2005/033321, US 7906111 (AAV9) и WO 2006/110689, а также rh10 (WO 2003/042397), их варианты, или обнаруженные в будущем вирусы, или рекомбинантные AAV на их основе, можно использовать в качестве источников для капсида AAV. В этих документах также описаны другие AAV, которые могут быть выбраны для создания AAV, и они включены в настоящий документ посредством ссылки. В некоторых вариантах осуществления капсид AAV для использования в вирусном векторе может быть получен путем мутагенеза (то есть, путем вставок, делеций или замен) одного из вышеупомянутых капсидов AAV или кодирующих их нуклеиновых кислот. В некоторых вариантах осуществления капсид AAV является химерным, содержащим домены из двух или трех, или четырех, или более из вышеупомянутых капсидных белков AAV. В некоторых вариантах осуществления капсид AAV представляет собой мозаику из мономеров VP1, VP2 и VP3 из двух или трех разных AAV или рекомбинантных AAV. В некоторых вариантах осуществления в состав rAAV входит более одного из вышеупомянутых капсидов.
Для упаковки экспрессионной кассеты в вирионы единственными компонентами AAV, которые должны находиться рядом в том же конструкте, что и ген, являются ITR. В одном варианте осуществления кодирующие последовательности для репликации (rep) и/или капсида (cap) удалены из генома AAV и предоставлены со стороны, или линией клеток-упаковщиков, для создания AAV вектора. Например, как описано выше, псевдотипированный AAV может содержать ITR из источника, отличного от источника капсида AAV. Дополнительно или альтернативно, можно использовать химерный капсид AAV. Могут быть выбраны и другие компоненты AAV. Источники таких последовательностей AAV описаны в настоящем описании, и они также могут быть выделены или получены из академических, коммерческих или общедоступных источников (например, Американской коллекции типовых культур, Manassas, VA). Альтернативно, последовательности AAV могут быть получены путем синтеза или другими подходящими методами на основании опубликованных последовательностей, которые доступны в литературе или в базах данных, таких как, например, GenBank®, PubMed® или тому подобное.
Минимальными последовательностями, необходимыми для упаковки экспрессионной кассеты в AAV вирусную частицу, являются 5' и 3'-ITR AAV, которые могут происходить из того же AAV, что и капсид, или из других AAV (для получения псевдотипа AAV). В одном варианте осуществления последовательности ITR из AAV2, или их делетированный вариант (ΔITR), используют для удобства и для ускорения получения одобрения регулирующими органами. Однако можно выбирать ITR из других источников AAV. Если источником ITR является AAV2, а источником капсида AAV является другой AAV, полученный вектор может быть назван псевдотипированным. Как правило, экспрессионная кассета для AAV вектора содержит 5'-ITR AAV, кодирующие последовательности и любые регуляторные последовательности, а также 3'-ITR AAV. Однако и другие конфигурации из этих элементов могут быть подходящими. Был описан укороченный вариант 5'-ITR, называемый ΔITR, в котором D-последовательность и сайт концевого разрешения (trs) делетированы. В других вариантах осуществления используют полноразмерные 5' и 3'-ITR AAV.
Сокращение «ск» означает самокомплементарные. Термин «самокомплементарные AAV» означает плазмиды или векторы, имеющие экспрессионную кассету, в которой кодирующая область, находящаяся в нуклеотидной последовательности рекомбинантного AAV, была сконструирована для образования внутримолекулярной двухцепочечной ДНК-матрицы. При инфицировании, вместо ожидания опосредованного клеткой синтеза второй цепи, две комплементарные половины скAAV будут связываться, с образованием одной единицы двухцепочечной ДНК (дцДНК), которая готова для немедленной репликации и транскрипции. См., например, D.M. McCarty et al., «Self-complementary recombinant adeno-associated virus (scAAV) vectors promote efficient transduction independently of DNA synthesis», Gene Therapy, (August 2001), Vol. 8, № 16, P. 1248-1254. Самокомплементарные AAV описаны, например, в патентах США №№ 6596535, 7125717 и 7456683, содержание которых включено в настоящий документ посредством ссылки в полном объеме.
Способы получения и выделения AAV вирусных векторов, подходящих для доставки в организм пациента, известны в данной области. См., например, патент США 7790449, патент США 7282199, WO 2003/042397, WO 2005/033321, WO 2006/110689 и US 7588772 B2. В одной системе линию клеток-продуцентов временно трансфицируют конструктом, кодирующим трансген, фланкированный ITR, и конструктом(ами), кодирующим rep и cap. Во второй системе линию клеток-упаковщиков, которые стабильно продуцируют rep и cap, временно трансфицируют конструктом, кодирующим трансген, фланкированный ITR. В каждой из этих систем вирионы AAV продуцируются в ответ на инфицирование хелперным аденовирусом или вирусом герпеса, и требуется отделение rAAV от примесного вируса. Совсем недавно были разработаны системы, для которых не требуется инфицирование хелперным вирусом для получения AAV - необходимые хелперные функции (то есть, E1, E2a, VA и E4 аденовируса или UL5, UL8, UL52 и UL29 вируса герпеса, а также полимераза вируса герпеса) также предоставляются, со стороны, системой. В этих более новых системах хелперные функции могут быть предоставлены за счет временной трансфекции клеток конструктами, кодирующими необходимые хелперные функции, или клетки могут быть сконструированы для постоянного содержания генов, кодирующих хелперные функции, экспрессия которых может контролироваться на транскрипционном или посттранскрипционном уровне. В другой системе трансген, фланкированный ITR и генами rep/cap, вводят в клетки насекомых путем инфицирования векторами на основе бакуловируса. Для обзора таких продуцирующих систем см., в основном, например, Zhang et al., 2009, «Adenovirus-adeno-associated virus hybrid for large-scale recombinant adeno-associated virus production», Human Gene Therapy 20: 922-929, содержание включено в настоящий документ посредством ссылки в полном объеме. Способы получения и использования этих и других систем продуцирования AAV также описаны в следующих патентах США, содержание каждого из которых включено в настоящий документ посредством ссылки в полном объеме: 5139941, 5741683, 6057152, 6204059, 6268213, 6491907, 6660514, 6951753, 7094604, 7172893, 7201898, 7229823 и 7439065. См. в основном, например, Grieger & Samulski, 2005, «Adeno-associated virus as a gene therapy vector: Vector development, production and clinical applications», Adv. Biochem. Engin/Biotechnol. 99: 119-145; Buning et al., 2008, «Recent developments in adeno-associated virus vector technology», J. Gene Med. 10: 717-733, а также ссылки, приведенные ниже, содержание которых включено в настоящий документ посредством ссылки в полном объеме. Методы, используемые для конструирования любого варианта осуществления данного изобретения, известны специалистам, умеющим работать с нуклеиновыми кислотами, и включают генетическую инженерию, рекомбинантную инженерию и методы синтеза. См., например, Green and Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY (2012). Аналогично, методы получения вирионов rAAV хорошо известны, и выбор соответствующего метода не ограничен в настоящем изобретении. См., например, K. Fisher et al., (1993) J. Virol., 70: 520-532 и патент США № 5478745.
Необязательно, гены fH, описанные в настоящем описании, могут быть доставлены при помощи вирусных векторов, отличных от rAAV. Такие другие вирусные векторы могут включать любой вирус, подходящий для использования в генной терапии, в том числе, но без ограничения, аденовирус; вирус герпеса; лентивирус; ретровирус и так далее. Предпочтительно, если создают один из этих векторов, его получают в виде репликативно-дефектного вирусного вектора.
Термины «репликативно-дефектный вирус» или «вирусный вектор» относятся к синтетической или искусственной вирусной частице, в которой экспрессионная кассета, содержащая интересующий ген, упакована в вирусный капсид или оболочку, при этом любые вирусные геномные последовательности, также упакованные в вирусный капсид или оболочку, являются репликативно-дефектными; то есть, они не способны к продуцированию вирионов-потомков, но сохраняют способность инфицировать клетки-мишени. В одном варианте осуществления геном вирусного вектора не содержит гены, кодирующие ферменты, необходимые для репликации (рекомбинантный геном может быть «выпотрошенным» - содержащим лишь интересующий трансген, фланкированный сигналами, необходимыми для амплификации и упаковки искусственного генома), но эти гены могут быть предоставлены во время продуцирования. Вследствие этого, такие вирусы считаются безопасными для использования в генной терапии, поскольку репликация и инфицирование вирионами-потомками не может происходить в отсутствие вирусного фермента, необходимого для репликации.
Фармацевтические композиции, описанные в настоящем описании, спроектированы для доставки в организм пациентов, которые нуждаются в этом, любым подходящим путем введения или сочетанием разных путей введения, например, прямой доставкой в печень (необязательно, внутривенным введением, через печеночную артерию или с помощью трансплантата), пероральным путем, ингаляцией, интраназальным, интратрахеальным, внутриартериальным, внутриглазным, внутривенным, внутримышечным, подкожным, внутрикожным и другими парентеральными путями введения. Вирусные векторы, описанные в настоящем описании, могут быть доставлены в одной композиции или в нескольких композициях. Необязательно, могут быть доставлены два или более разных AAV, либо несколько вирусов (см., например, WO 2011/126808 и WO 2013/049493). В другом варианте осуществления несколько вирусов могут включать разные репликативно-дефектные вирусы (например, AAV и аденовирус).
Репликативно-дефектные вирусы могут быть сформулированы с физиологически приемлемым носителем для использования в переноске генов и генной терапии. В случае AAV вирусных векторов в качестве меры дозы, содержащейся в препарате, может быть использовано количество геномных копий (гк). Любой метод, известный в данной области, можно использовать для определения числа геномных копий (гк) в композициях репликативно-дефектного вируса по изобретению. Одним из методов проведения титрования числа гк AAV является следующий: образцы очищенного AAV вектора сначала обрабатывают ДНКазой для элиминации не инкапсулированной геномной ДНК AAV или примесной плазмидной ДНК, появившейся в процессе получения. Устойчивые к ДНКазе частицы затем подвергают воздействию высоких температур для высвобождения генома из капсида. Вслед за этим высвобожденные геномы количественно определяют методом ПЦР в реальном времени с использованием наборов праймеров/зондов, направленных на определенную область вирусного генома (как правило, сигнал полиA).
Кроме того, композиции репликативно-дефектного вируса могут быть сформулированы в единицах дозирования, содержащих количество репликативно-дефектного вируса, находящееся в диапазоне от примерно 1,0×109 гк до примерно 1,0×1015 гк (для лечения среднего пациента с массой тела 70 кг), и предпочтительно от 1,0×1012 гк до 1,0×1014 гк для пациента-человека. В другом варианте осуществления доза составляет менее примерно 1,5×1011 гк/кг. Например, доза AAV вируса может составлять примерно 1×109 гк, примерно 5×109 гк, примерно 1×1010 гк, примерно 5×1010 гк или примерно 1×1011 гк. В другом примере варианты могут быть доставлены в количестве от примерно 0,001 мг до примерно 10 мг/кг.
Вышеописанные рекомбинантные векторы могут быть доставлены в клетки-хозяева описанными в литературе способами. rAAV, предпочтительно суспендированный в физиологически совместимом носителе, можно вводить человеку или пациенту-млекопитающему, не являющемуся человеком. Соответствующие носители может с легкостью выбирать специалист в данной области с учетом состояния, для которого предписано введение вектора. Например, одним из подходящих носителей является солевой раствор, который может быть сформулирован с различными буферными растворами (например, фосфатно-солевой буфер). Другие иллюстративные носители включают стерильный солевой раствор, лактозу, сахарозу, фосфат кальция, желатин, декстран, агар, пектин, арахисовое масло, кунжутное масло и воду. Выбор носителя не ограничен по настоящему изобретению.
Необязательно, композиции по изобретению могут содержать, в дополнение к rAAV и/или вариантам и носителю(ям), эксципиенты, включая другие неактивные общепринятые фармацевтические ингредиенты, такие как консерванты, химические стабилизаторы, суспендирующие средства и/или сурфактанты. Подходящие иллюстративные консерванты включают хлорбутанол, сорбат калия, сорбиновую кислоту, диоксид серы, пропилгаллат, парабены, этилванилин, глицерин, фенол и парахлорфенол. Подходящие химические стабилизаторы включают желатин и альбумин. Необязательно, для композиций на основе белков или на основе антител могут быть выбраны эксципиенты, подходящие для твердых композиций, включая, например, наполнители, гранулы, объемообразующие средства, дезинтегрирующие средства, вещества, способствующие скольжению, ароматизаторы, красители или другие компоненты.
Вирусные векторы и другие конструкты, описанные в настоящем описании, могут быть использованы в получении лекарственного средства для доставки варианта fH в организм пациента, который нуждается в этом, доставляющего вариант fH с увеличенным временем полужизни в организм пациента, и/или для лечения заболеваний, связанных с комплементом.
Курс лечения может, необязательно, включать повторное введение одного и того же вирусного вектора (например, AAV8 вектора) или разных вирусных векторов (например, AAV8 и AAVrh10). Например, если мишенью является печень, повторное введение может быть желательным на протяжении 18 месяцев, 2 лет или более длительного периода времени из-за ослабления экспрессии, вызванного естественной пролиферацией гепатоцитов. Можно выбирать и другие сочетания вирусного и белкового лечения с использованием вирусных векторов, описанных в настоящем описании. Необязательно, композицию, описанную в настоящем описании, можно комбинировать в схеме лечения, включающей использование других лекарственных средств против комплемента (например, моноклональных антител и так далее) или терапию белковыми препаратами (включая, например, доставку композиции, содержащей один или более вариантов fH, описанных в настоящем описании).
Например, рекомбинантный вариант hfH, описанный в настоящем описании, может быть доставлен в белковой форме. Необязательно, при доставке в организм пациента в белковой форме вариант fH может иметь лидерную последовательность или может быть лишен всей или части лидерной последовательности. Необязательно, терапию белковыми препаратами можно использовать в сочетании с опосредованным вирусом введением варианта hfH. В одном варианте осуществления белок fH может обеспечивать немедленно высвобождающуюся форму hfH в организме пациента, например, поддающиеся обнаружению уровни в плазме в пределах 2 часов после введения; при этом выведение этой формы из организма пациента, как правило, будет начинаться в пределах от примерно 24 часов до примерно 48 часов, или до примерно 72 часов, если будет обнаружено, что существует некоторое время задержки до начала экспрессии с опосредованной вирусом системы доставки. В другом варианте осуществления вариант hfH дополнительно модифицирован для продления его времени полужизни за счет введения в вариант по меньшей мере одного сайта гликозилирования, при этом сайт вводят в по меньшей мере один из SCR, находящихся в варианте, по меньшей мере два из SCR, находящихся в варианте, по меньшей мере три из SCR, находящихся в варианте, или более. Например, сайт гликозилирования может быть введен в один или более из SCR1, SCR2, SCR3, SCR4, SCR19 и/или SCR20. В другом варианте осуществления дополнительно или альтернативно гликозилированы SCR17 и/или SCR18. В следующем варианте осуществления гликозилированы SCR4, 17 и 18. В конкретных вариантах осуществления сайт гликозилирования может быть введен в линкер. Однако в этом случае линкер предпочтительно имеет в длину от по меньшей мере шести аминокислот до примерно 18 аминокислот, например, 8-18, 10-15 или 12 аминокислот. Дополнительно или альтернативно, рекомбинантный белковый вариант hfH может быть пегилирован, то есть, модифицирован фрагментом полиэтиленгликоля с использованием известных методов (см., например, Fee, Conan J.; Van Alstine, James M. (2006). «PEG-proteins: Reaction engineering and separation issues». Chemical Engineering Science 61 (3): 924).
Используемый в настоящем описании термин «сайт гликозилирования» означает точку присоединения олигосахаридов к атому углерода (C-связанное), атому азота (N-связанное) или атому кислорода (O-связанное), или точку гликирования (не ферментативного присоединения восстанавливающих сахаров к атому азота белка (например, атому азота боковой цепи аспарагина (Asn), который является частью последовательности Asn-X-Ser/Thr, где X представляет собой любую аминокислоту, за исключением Pro). В конкретных вариантах осуществления желательными являются сайты N-гликозилирования. Различные методы известны в данной области для создания сайтов N-гликозилирования. См., например, Y. Liu et al., Biotech Prog 2009 Sep-Oct; 25(5): 1468-1475; Sala RJ, Griebenos K. Glycoslylation of therapeutic proteins: an effective strategy to optimize efficacy. BioDrugs. 2010 Feb 1; 24(1): 9-21.
Кроме того, рекомбинантный вариант hfH, предложенный в настоящем описании, может быть сформулирован с соответствующим носителем и/или эксципиентом для доставки в организм пациента любым подходящим путем введения. Помимо общепринятых суспензионных носителей, носителем может быть липосома или наноноситель. Подходящие дозы варианта hfH включают те, которые позволяют достигать уровней в плазме, достаточных для лечения заболевания, связанного с комплементом. Примеры доз вариантов hfH включают, но не ограничиваются ими, эффективное количество в диапазоне доз от примерно 0,01 мкг/кг до примерно 300 мг/кг или в пределах от примерно 0,1 мкг/кг до примерно 40 мг/кг, или в пределах от примерно 1 мкг/кг до примерно 20 мг/кг, или в пределах от примерно 1 мкг/кг до примерно 10 мг/кг. Например, при внутриглазном введении композицию можно вводить в диапазоне низких значений микрограмм, включая, например, примерно 0,1 мкг/кг или менее, примерно 0,05 мкг/кг или менее, или 0,01 мкг/кг или менее. В некоторых вариантах осуществления количество варианта hfH, вводимого индивидууму, составляет от примерно 10 мкг до примерно 500 мг на дозу или от примерно 10 мкг до примерно 50 мкг, от примерно 50 мкг до примерно 100 мкг, от примерно 100 мкг до примерно 200 мкг, от примерно 200 мкг до примерно 300 мкг, от примерно 300 мкг до примерно 500 мкг, от примерно 500 мкг до примерно 1 мг, от примерно 1 мг до примерно 10 мг, от примерно 10 мг до примерно 50 мг, от примерно 50 мг до примерно 100 мг, от примерно 100 мг до примерно 200 мг, от примерно 200 мг до примерно 300 мг, от примерно 300 мг до примерно 400 мг или от примерно 400 мг до примерно 500 мг на дозу.
Фармацевтические композиции можно вводить отдельно. Необязательно, композиции, описанные в настоящем описании, можно вводить в сочетании с другими молекулами, которые, как известно, оказывают полезное действие. Например, полезные кофакторы включают вызывающие облегчение симптомов кофакторы, в том числе антисептики, антибиотики, противовирусные и противогрибковые средства и анальгетики, противовоспалительные средства, анестетики. В другом варианте осуществления, в котором предусмотрено внутриглазное введение, может быть желательным использование молекул, способствующих прикреплению сетчатки или лечению поврежденной ткани сетчатки. Примеры полезных кофакторов включают анти-VEGF средства (такие как антитело против VEGF), основной фактор роста фибробластов (bFGF), цилиарный нейротрофический фактор (CNTF), аксокин (мутеин CNTF), лейкоз-ингибирующий фактор (LIF), нейротрофин-3 (NT-3), нейротрофин-4 (NT-4), фактор роста нервов (NGF), инсулиноподобный фактор роста II, простагландин E2, 30-кДа фактор выживания, таурин и витамин A. Другие подходящие терапевтические средства могут включать антитело против комплемента, например, антитело против регулятора комплемента C3 (например, такое как коммерчески доступное антитело экулизумаб).
Композиции, описанные в настоящем описании (как на основе вектора, так и на основе белка), можно вводить пациенту любым путем введения, включая, но без ограничения, внутривенный (например, с помощью инфузионных насосов), внутрибрюшинный, внутриглазной, внутриартериальный, внутрилегочный, пероральный, ингаляцию, интравезикулярный, внутримышечный, внутритрахеальный, подкожный, внутриглазной (включая интравитреальный и интраретинальный), интратекальный, чрескожный, трансплевральный, внутриартериальный, топический, ингаляционный (например, в виде аэрозолей или спреев), мукозальный (например, через слизистую оболочку носа), подкожный, чрескожный, желудочно-кишечный, внутрисуставной, интрацистернальный, интравентрикулярный, ректальный (то есть, с помощью суппозитория), вагинальный (то есть, с помощью пессария), интракраниальный, внутриуретральный, внутрипеченочный и внутриопухолевый. В некоторых вариантах осуществления композиции вводят системно (например, путем внутривенной инъекции). В некоторых вариантах осуществления композиции вводят локально (например, путем внутриартериальной или внутриглазной инъекции).
Таким образом, следующий аспект относится к применению фармацевтической композиции в лечении заболевания, связанного с комплементом, включая, например, заболевание, связанное с фактором H комплемента, такое, как описано в настоящем описании, и другие заболевания, связанные с комплементом, включая, без ограничения: повреждение тканей вследствие ишемии-реперфузии после острого инфаркта миокарда, аневризму, инсульт, геморрагический шок, повреждение с размозжением тканей, полиорганную недостаточность, ишемию кишечника при гиповолемическом шоке, повреждение спинного мозга и травматическое повреждение головного мозга; воспалительные заболевания, например, ожоги, эндотоксемию и септический шок, респираторный дистресс-синдром взрослых, сердечно-легочный шунт, гемодиализ; анафилактический шок, тяжелую астму, ангионевротический отек, болезнь Крона, серповидноклеточную анемию, постстрептококковый гломерулонефрит, мембранозный нефрит и панкреатит; отторжение трансплантата, например, сверхострое отторжение ксенотрансплантата; заболевания, связанные с беременностью, такие как рецидивирующая потеря плода и преэклампсия; побочные реакции на лекарственное средство, например, аллергию на лекарственное средство, индуцированный IL-2 синдром транссудации и аллергию на рентгеноконтрастное вещество; а также аутоиммунные заболевания, включая, но без ограничения, миастению, болезнь Альцгеймера, рассеянный склероз, эмфизему, ожирение, ревматоидный артрит, системную красную волчанку, рассеянный склероз, миастению, инсулинозависимый сахарный диабет, острый диссеминированный энцефаломиелит, болезнь Аддисона, синдром антифосфолипидных антител, аутоиммунный гепатит, болезнь Крона, синдром Гудпасчера, болезнь Грэйвса, синдром Гийена-Барре, болезнь Хашимото, идиопатическую тромбоцитопеническую пурпуру, пемфигус, синдром Шегрена и артериит Такаясу, осложнения после сердечно-легочного шунтирования; инфаркт миокарда; повреждения вследствие ишемии/реперфузии; инсульт; острый респираторный дистресс-синдром (ARDS); сепсис; ожог; воспаление, связанное с сердечно-легочным шунтированием и гемодиализом; плазмоферез; тромбаферез; лейкоферез; экстракорпоральную мембранную оксигенацию (ECMO); индуцированное гепарином экстракорпоральное осаждение ЛНП (HELP); аллергическую реакцию на рентгеноконтрастное вещество; отторжение трансплантата.
Необходимо отметить, что упомянутое существительное в единственном числе следует понимать, как «один или более». Следовательно, в настоящем описании при упоминании существительного в единственном числе взаимозаменяемыми терминами являются «один или более» и «по меньшей мере один».
Слова «включают», «включает» и «включая» должны толковаться как включающие, а не исключающие. Слова «состоит», «состоящий» и их варианты должны толковаться как исключающие, а не включающие. Хотя различные варианты осуществления в данной спецификации представлены с использованием выражений типа «включая», при других обстоятельствах родственный вариант осуществления также должен быть интерпретирован и описан с использованием выражений типа «состоящий из» или «состоящий по существу из».
Используемый в настоящем описании термин «примерно» означает вариабельность в пределах 10% от указанного значения, если нет иных указаний.
Используемый в настоящем описании термин «регуляция», или его варианты, означает способность композиции ингибировать один или более компонентов биологического пути.
Если в настоящем описании не указано иначе, как гомозиготные пациенты, так и гетерозиготные пациенты, охвачены понятием «пациент, имеющий заболевание, опосредованное комплементом».
«Пациент» представляет собой млекопитающее, например, человека, мышь, крысу, морскую свинку, собаку, кошку, лошадь, корову, свинью или не являющегося человеком примата, такого как обезьяна, шимпанзе, бабуин или горилла.
В настоящем описании термины «заболевание», «нарушение», «нарушение функции» и «состояние» используют взаимозаменяемо для обозначения аномального состояния у пациента, если нет иных указаний.
Если в настоящей спецификации не указано иначе, технические и научные термины, используемые в настоящем описании, имеют то значение, которое им обычно придают специалисты в данной области и которое приведено в литературных источниках, предоставляющих специалисту в данной области общее руководство для понимания терминов, используемых в настоящей заявке.
Следующие далее примеры являются исключительно иллюстративными и не предназначены для ограничения настоящего изобретения.
ПРИМЕРЫ
Конструирование и клонирование укороченного варианта человеческого фактора H (hfH1-4.678.19-20)
Укороченные варианты получали методом обращенной ПЦР с использованием высокоточной ДНК-полимеразы Phusion® (каталожный № M0530S, New England Biolabs) в соответствии с протоколом производителя. кДНК полноразмерного человеческого фактора H комплемента в pCMV Sport6, использованную в качестве матрицы для обращенной ПЦР, получали от Thermo Fisher Scientific (каталожный № MHS6278-202800294, клон № 40148771). Праймеры для ПЦР, используемые для получения hfH1-4.678.19-20, приведены в Таблице 1. После того, как ПЦР-фрагменты разделяли в 0,8% агарозном геле и экстрагировали при помощи набора AccuPrep gel extraction kit (каталожный № K-3035, Bioneer), 50 нг очищенного в геле фрагмента использовали для лигирования при помощи набора Rapid DNA ligation Kit (каталожный № K-4123, Thermo Fisher Scientific) и вводили трансформацией в компетентные клетки DH5α (каталожный № 1825801, Invitrogen). Положительные клоны подтверждали либо расщеплением ферментами рестрикции, либо методом ПЦР-скрининга с использованием специфических праймеров. Затем вставку hfH1-4.678.19-20 из pCMV Sport6 вырезали расщеплением EcoR I и Not I, у очищенного в геле фрагмента затупляли концы при помощи модуля End-repair module (каталожный № E6050S, New England Biolabs), и фрагмент очищали. Данный фрагмент субклонировали в вектор pCBABG (который имел промотор бета-актина кур с энхансером CMV и частичную последовательность интрона того же гена, а также сигнальную последовательность полиаденилирования из гена бета-глобулина кролика) в сайте EcoR V. Положительные клоны отбирали методами расщепления ферментами рестрикции и ПЦР.
Таблица 1:
Конструирование и клонирование укороченного варианта человеческого фактора H hfH1-4.678.17-20
Укороченные варианты получали методом обращенной ПЦР с использованием высокоточной ДНК-полимеразы Phusion® (каталожный № M0530S, New England Biolabs) в соответствии с протоколом производителя. кДНК полноразмерного человеческого фактора H комплемента в pCMV Sport6, использованную в качестве матрицы для обращенной ПЦР, получали от Thermo Fisher Scientific (каталожный № MHS6278-202800294, клон № 40148771). Праймеры для ПЦР, используемые для получения hfH1-4.678.17-20, приведены в Таблице 1. После того, как ПЦР-фрагменты разделяли в 0,8% агарозном геле и экстрагировали при помощи набора AccuPrep gel extraction kit (каталожный № K-3035, Bioneer), 50 нг очищенного в геле фрагмента использовали для лигирования при помощи набора Rapid DNA ligation Kit (каталожный № K-4123, Thermo Fisher Scientific) и вводили трансформацией в компетентные клетки DH5α (каталожный № 1825801, Invitrogen). Положительные клоны подтверждали либо расщеплением ферментами рестрикции, либо методом ПЦР-скрининга с использованием специфических праймеров. Затем рекомбинантный вариант hfH1-4.678.17-20 в pCMV Sport6 субклонировали в вектор pCBABG в сайте EcoRI методом клонирования In-Fusion™ (Clontech каталожный № 638909). Праймеры для получения укороченного белка и клонирования в экспрессионный вектор приведены в Таблице 2.
Таблица 2:
Экспрессия и очистка рекомбинантного белка hfH1-4.678.19-20
Положительные клоны (hfH1-4.678.19-20 в векторе pCBARBG) трансфицировали в клетки HEK для оценки стабильности и функциональной активности белка hfH1-4.678.19-20. Клетки HEK на стадии образования примерно 80% конфлюэнтного слоя в 6-луночном планшете (Falcon, каталожный № 353046) трансфицировали кДНК hfH1-4.678.19-20 в pCBARBG с использованием липофектамина 2000 (каталожный № 11668019, Invitrogen) в соответствии с инструкциями производителя. Экспрессию белка подтверждали методом вестерн-блоттинга с использованием IgG козы против человеческого фактора H (каталожный № A237, Complement tech). Для крупномасштабной экспрессии белка клетки HEK на стадии образования примерно 80% конфлюэнтного слоя в 150-см чашках (Falcon, каталожный № 353025) трансфицировали свободной от эндотоксина кДНК hfH1-4.678.19-20 в плазмиде pCBARBG с PEI (каталожный № 23966, Polysciences) в соответствии с инструкциями производителя. Через два дня после трансфекции супернатант собирали с чашек, фильтровали через 0,2-мкм фильтр и наносили на уравновешенную PBS колонку для аффинной хроматографии с Ox23-сефарозой (мАт мыши против человеческого fH, специфичное для SCR 2/3, каталожный № 10402-1VL, Sigma). После промывания PBS, содержащим 500 мМ NaCl, в объеме, равном 25 объемам колонки, связанный hfH1-4.678.19-20 элюировали 100 мМ глицин-HCl буфером, pH 2,7, и элюированные фракции (2 мл на фракцию) нейтрализовали 200 мкл 1,5 M трис-HCl, pH 8,5. Чистоту элюированного белка проверяли методом SDS-ПААГ, чистые фракции объединяли и диализовали против PBS в течение ночи, 2 раза меняя буфер.
Конструирование и клонирование укороченного варианта мышиного фактора H (mfH1-4.678.19-20)
Укороченные варианты получали методом обращенной ПЦР с использованием высокоточной ДНК-полимеразы Phusion® в соответствии с протоколом производителя. кДНК полноразмерного мышиного фактора H комплемента в pBluescript SK(-), использованная в качестве матрицы для обращенной ПЦР, была любезно предоставлена Dr M. Nonaka (University of Tokyo, Japan, нуклеотиды 110-4361 из NCBI NM 009888.3). Все праймеры для ПЦР, используемые для получения варианта mfH1-4.678.19-20, приведены в Таблице 3. После того, как ПЦР-фрагменты разделяли в 0,8% агарозном геле и экстрагировали при помощи набора AccuPrep gel extraction kit, 50 нг очищенного в геле фрагмента использовали для лигирования при помощи набора Rapid DNA ligation Kit и вводили трансформацией в компетентные клетки DH5α. Положительные клоны подтверждали либо расщеплением ферментами рестрикции, либо методом ПЦР-скрининга с использованием специфических праймеров. Затем вставку mfH1-4,678.19-20 из pBluescript SK(-) вырезали расщеплением Sma I и EcoR V и очищали в геле. Данный фрагмент субклонировали в вектор pCBARBG в сайте EcoR V. Положительные клоны отбирали методами расщепления ферментами рестрикции и ПЦР.
Таблица 3:
Экспрессия рекомбинантного белка mfH1-4.678.19-20
Положительные клоны (mfH1-4.678.19-20 в векторе pCBARBG) трансфицировали в клетки Hepa1C1C7 (линия клеток гепатомы мышей, ATCC® CRL-2026) для оценки стабильности и функциональной активности белка mfH1-4.678.19-20. Клетки на стадии образования примерно 80% конфлюэнтного слоя в 6-луночном планшете трансфицировали кДНК mfH1-4.678.19-20 с использованием липофектамина 2000 в соответствии с инструкциями производителя. Экспрессию белка подтверждали методом вестерн-блоттинга с использованием IgG кролика против мышиного fH (Ref №1). Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus (каталожный № 80196, Thermo Fisher Scientific).
Получение AAV плазмиды и вируса для переноса
Экспрессионную кассету mfH1-4.678.19-20 или hfH1-4.678.19-20 из вектора pCBARBG вырезали расщеплением Hinc II и Pst I, у очищенного в геле фрагмента затупляли концы при помощи модуля End-repair module (каталожный № E6050S, NEB) и лигировали в расщепленный Nhe I и Xho I вектор pAAV TBG.PI.EGFP.WPRE.BGH (каталожный № PL-C-PV0146) с затупленными концами от Головного отдела изучения векторов Университета Пенсильвании http://www.med.upenn.edu/gtp/vectorcore/production.shtml). Скрининг положительных клонов проводили путем расщепления Sma I.
Вектор pCBABG с hfH1-4.678.17-20 модифицировали в AAV-плазмиду для переноса путем вставки ITR (инвертированных концевых повторов) на 5'-конце (SEQ ID NO: 61:
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagg-gagtggccaactcc-atcactaggggttccttgtagttaat, в сайте HincII) и 3'-конце (SEQ ID NO: 62:
attaactacaaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcag, в сайте Pst I) экспрессионной кассеты с использованием метода клонирования In-Fusion™. Праймеры, используемые для амплификации ITR AAV из вектора pENN.AAV.TBG.PI.RBG, используемого в качестве матрицы, приведены в Таблице 4. Вектор pENN.AAV.TBG.PI.RBG был получен от Головного отдела изучения векторов Университета Пенсильвании http://www.med.upenn.edu/gtp/vectorcore/production.shtml) (каталожный № PL-C-PV1015).
Таблица 4
Суперскрученную свободную от эндотоксина AAV-плазмиду получали с использованием набора Endo free plasmid kit (каталожный № 12362, Qiagen) и использовали для получения AAV вируса в Головном отделе изучения векторов Университета Пенсильвании или в Головном отделе изучения векторов Центра генной терапии Университета Массачусетса. Упаковку, очистку и определение титра AAV, кодирующего mfH1-4.678.19-20, hfH1-4.678.19-20 или hfH1-4.678.17-20, выполняли с использованием стандартных методов, как описано (http://www.med.upenn.edu/gtp/vectorcore/production.shtml).
Терапевтическая эффективность AAV hfH1-4.678.19-20 и hfH1-4.678.17-20 у мышей fHm/m
Создание мышей fHm/m, у которых развивается C3-гломерулопатия, описано ранее в статье Lesher et al. (2013) «Combination of factor H mutation and properdin deficiency causes severe C3 glomerulonephritis», J Am Soc Nephrol. 2013 Jan; 24(1): 53-65. Epub 2012 Nov 30. Для тестирования уровней экспрессии, срока действия и терапевтической эффективности hfH1-4.678.19-20 и hfH1-4.678.17-20 при лечении C3-гломерулопатии мышам fHm/m в возрасте 10-12 недель вводили инъекцией 3×1012 геномных копий/мышь (hfH1-4.678.19-20) или 1×1011-1×1012 геномных копий/мышь (hfH1-4.678.17-20) через ретроорбитальный синус. Для отдельных групп мышей в качестве контроля использовали контрольный AAV вектор (pAAV.TBG.NULL.rBG). Из предыдущих исследований известно, что природный человеческий fH функционально активен для ингибирования альтернативного пути активации (AP) комплемента у мышей (Fakhouri, F., et al., Kidney International (2010) 78, 279-286; опубликовано онлайн 5 мая 2010 г.). Кровь собирали из ретроорбитального синуса перед инъекцией и через 1 неделю после инъекции (в случае hfH1-4.678.19-20) или 1, 2 недели, 1, 2, 3 месяца после инъекции (в случае hfH1-4.678.17-20). У мышей fHm/m спонтанно развивается C3-гломерулопатия, характеризующаяся неконтролируемой активацией в плазме AP комплемента, приводящей к истощению C3, фактора B (fB) и C5, и значительным гломерулярным отложениям C3 и C5b-9 (Lesher et al., 2013). Если hfH1-4.678.19-20 или hfH1-4.678.17-20 функционально активны у мышей fHm/m, то можно ожидать уменьшения истощения C3 и fB. Вследствие этого, в качестве показателя терапевтической эффективности hfH1-4.678.19-20 и hfH1-4.678.17-20 авторы изобретения определяли уровни в плазме C3 и fB методом вестерн-блотинга до и после инъекции AAV мышам fHm/m. Плазму мышей (1 мкл) разбавляли буфером для образцов и кипятили перед нанесением на 4-20% градиентные SDS-ПАА гели в восстанавливающих условиях. Затем образцы переносили на PVDF мембраны и проводили обработку соответствующими антителами. Для обнаружения C3 и fB HRP-конъюгированное Ат козы против мышиного C3 (1:4000, MP Biomedicals, каталожный № 0855557) или аффинно очищенное Ат козы против человеческого fB (перекрестно реагирует с мышиным fB; 1:2500, каталожный № A235, Complement Technology) использовали в качестве первичных антител, с последующей обработкой HRP-конъюгированными антителами кролика против IgG козы (1:4000, каталожный № 1721034, Bio-Rad). Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus.
Обнаружение белка hfH1-4.678.19-20 или hfH1-4.678.17-20 в крови мышей
Для обнаружения присутствия hfH1-4.678.19-20 или hfH1-4.678.17-20 в организме получавших AAV мышей fHm/m был разработан и использован метод ELISA. Вкратце, 96-луночные планшеты (MaxiSorp) предварительно покрывали 4 мкг/мл раствором мАт против человеческого фактора H (OX-23) при RT в течение 2 час. Незанятые сайты связывания на планшетах блокировали 1% раствором бычьего сывороточного альбумина (БСА) в PBS при RT в течение 1 часа. Образцы плазмы мышей, серийно разведенные в блокирующем буфере, содержащем 10 мМ ЭДТА, добавляли в лунки и инкубировали при RT в течение 1 ч, затем добавляли 2 мкг/мл раствор меченых биотином мАт против hfH (клон L20/3, специфичное для SCR19 человеческого фактора H, каталожный № 518504, Bio-Legend) и инкубировали при RT в течение 1 часа. После промывания планшеты инкубировали с конъюгатом авидин-HRP (1/1000, каталожный № 554058, BD Biosciences) при RT в течение 1 часа, затем использовали субстрат TMB для развития окраски (каталожный № 51-2606KC и BD каталожный № 51-2607KC, BD Biosciences).
Терапевтическая эффективность mfH1-4.678.19-20, доставленного с помощью AAV в организм мышей fHm/m или fHm/mP-/-
Для тестирования терапевтической эффективности mfH1-4.678.19-20 в качестве суррогата hfH1-4.678.19-20 мышей fHm/m и мышей fHm/mP-/- инфицировали AAV вектором, содержащим кодирующие последовательности для mfH1-4.678.19-20. Как ранее описано в статье Lesher et al. (Lesher et al., 2013, выше), в то время как у мышей fHm/m развивается не летальная C3-гломерулопатия с истощением C3 и fB, у мышей с двойной мутацией fHm/mP-/- (мыши fHm/m с дефицитом пропердина) развивается обостренная и летальная форма C3-гломерулопатии, и они погибают к возрасту 10-12 недель (Lesher et al. 2013). Вследствие этого, использование мышей fHm/mP-/- также позволило авторам изобретения использовать смертность в качестве другого показателя терапевтической эффективности AAV mfH1-4.678.19-20. Мышам fHm/m или fHm/mP-/- в возрасте 7 недель вводили инъекцией либо контрольный AAV (pAAV.TBG.NULL.rBG), либо mfH1-4.678.19-20 AA, в дозе 3×1012 геномных копий/мышь через ретроорбитальный синус. Кровь собирали через ретроорбитальный синус перед инъекцией и в разных временных точках, начиная с 1 недели после инъекции. Для оценки уровней в плазме C3 и fB плазму мышей (1 мкл) разбавляли буфером для образцов и кипятили перед нанесением на 4-20% градиентные SDS-ПАА гели в восстанавливающих условиях. Затем образцы переносили на PVDF мембраны и проводили обработку соответствующими антителами. Для обнаружения C3 и fB HRP-конъюгированное Ат козы против мышиного C3 или аффинно очищенное Ат козы против человеческого fB (перекрестно реагирует с мышиным fB) использовали в качестве первичных антител, с последующей обработкой HRP-конъюгированными антителами кролика против IgG козы. В некоторых случаях мышей, получавших инъекцию, наблюдали в течение 6 или 10 месяцев для контроля эффективности AAV mfH1-4.678.19-20 в предотвращении гибели и/или активации AP комплемента с использованием уровней в плазме C3 и fB в качестве показателей.
Определение дозы AAV mfH1-4.678.19-20 для мышей fHm/m
В экспериментах по титрованию количества копий AAV mfH1-4.678.19-20, необходимого для достижения терапевтической эффективности, мышам fHm/m в возрасте 10-12 недель (Lesher, 2013) вводили инъекцией 1×1012, 3×1011 или 1×1011 геномных копий/мышь AAV через ретроорбитальный синус. Кровь собирали из ретроорбитального синуса перед инъекцией и в указанных временных точках (1 неделя и 1 месяц после инъекции). Плазму мышей (1 мкл) разбавляли буфером для образцов и кипятили перед нанесением на 4-20% градиентные SDS-ПАА гели в восстанавливающих условиях. Затем образцы переносили на PVDF мембраны и проводили обработку соответствующими антителами. Для обнаружения C3 и fB HRP-конъюгированное Ат козы против мышиного C3 (1:4000, MP Biomedicals, каталожный № 0855557) или аффинно очищенное Ат козы против человеческого fB (перекрестно реагирует с мышиным fB; 1:2500, каталожный № A235, Complement Technology) использовали в качестве первичных антител, с последующим обнаружением при помощи HRP-конъюгированных антител кролика против IgG козы (1:4000, каталожный № 1721034, Bio-Rad). Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus.
Обнаружение белка mfH1-4.678.19-20 в крови мышей методом ELISA
Для обнаружения присутствия белка mfH1-4.678.19-20 в крови мышей был разработан и использован метод ELISA. Вкратце, 96-луночные планшеты предварительно покрывали 2 мкг/мл раствором мышиного мАт против SCR19-20 мышиного fH (клон 12, полученный в собственной лаборатории путем иммунизации мышей fHm/m рекомбинантным белком SCR19-20 мышиного fH (Barata, L., et al., J. Immunol 190(6): 2886-95 (2013)) при 37°C в течение 1-2 ч при комнатной температуре. Незанятые сайты связывания на планшетах блокировали 1% раствором БСА в PBS при RT в течение 1 часа. Образцы плазмы мышей, серийно разведенные в блокирующем буфере, содержащем 10 мМ ЭДТА, добавляли в лунки и инкубировали при RT в течение 1 ч, затем добавляли меченые биотином Ат кролика против мышиного fH (Lesher et al., 2013) при RT на 1 час. Планшеты инкубировали с конъюгатом авидин-HRP при RT в течение 1 часа, затем использовали субстрат TMB для развития окраски.
Обнаружение белка mfH1-4.678.19-20 в плазме мышей методом вестерн-блоттинга
Для обнаружения присутствия белка mfH1-4.678.19-20 в крови мышей методом вестерн-блоттинга 10 мкл плазмы мышей разбавляли 90 мкл PBS, содержащего 10 мМ ЭДТА, и инкубировали с гранулами сефарозы®, связанными с мАт против мышиного fH (клон 12), в течение 30 мин при комнатной температуре. После 2-кратного промывания PBS, содержащим 500 мМ NaCl, гранулы сефарозы® кипятили в буфере для образцов для SDS-ПААГ в течение 5 мин и проводили электрофорез образов в SDS-ПААГ. Затем образцы переносили на PVDF мембрану и проводили обнаружение белка mfH1-4.678.19-20 с использованием предварительно абсорбированного на БСА Ат кролика против доменов 19-20 мышиного fH (Lesher et al., 2013). Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus.
Иммунофлуоресцентное окрашивание C3 в почках
Почки мышей fHm/m или fHm/mP-/-, получавших контрольный AAV или AAV mfH1-4.678.19-20, быстро замораживали в среде OCT и хранили при -80°C. Для иммунофлуоресцентного анализа готовили 4-мкм срезы и использовали их для окрашивания. Для окрашивания на C3 использовали конъюгированные с FITC Ат козы против мышиного C3 (1:500, каталожный № 855500, MP Biomedicals), и эксперимент проводили, как описано (Lesher et al. 2013).
Анализ выживаемости мышей
В следующей далее Таблице приведены данные по выживаемости мышей fHm/mP-/-, получавших контрольный AAV8 вектор или AAV8-mfH1-4.678.19-20 вектор. Все 8 мышей fHm/mP-/-, получавших контрольный AAV8 вектор, погибли в течение 2-3 недель после инъекции, в то время как 7 из 9 мышей fHm/mP-/-, получавших AAV8-mfH1-4.678.19-20 вектор, избежали развития летальной C3-гломерулопатии. Всем мышам вводили инъекцией 3×1012 геномных копий/мышь соответствующего AAV вируса в/в через ретроорбитальный синус. Показатели выживаемости получавших контрольный AAV или AAV mfH1-4.678.19-20 мышей fHm/mP-/- регистрировали после введения AAV в течение 10 месяцев. Данные относили к категориям «умерщвление (эвтаназия)» или «естественная смерть» и анализировали с использованием GraphPad Prism (La Jolla, CA).
2 - здоровы через 6 месяцев после генной терапии (умерщвлены через 6 месяцев)
1 - здорова через 5 месяцев после генной терапии (продолжает жить)
1 - предсмертное состояние через 3 месяца после генной терапии
1 - предсмертное состояние через 2 недели после генной терапии
Анализ связывания гепарина
Для тестирования гепарин-связывающей активности белков hfH1-4.678.19-20 и mfH1-4.678.19-20 96-луночные планшеты предварительно покрывали 100 мкг гепарина (Sigma, H3393) в бикарбонатном буфере (pH 9,6) при 37°C в течение 1 часа. Незанятые сайты связывания на планшетах блокировали 1% раствором БСА в PBS при RT в течение 1 часа. Добавляли разные количества белка hfH1-4.678.19-20 или mfH1-4.678.19-20 и инкубировали при RT в течение 1 часа, затем добавляли 2 мкг/мл раствор мышиного мАт против человеческого fH (OX-23) и инкубировали при RT в течение 1 часа. Планшеты инкубировали с HRP-конъюгированными антителами кролика против IgG мыши (1/4000, каталожный №A9044, Sigma) при RT в течение 1 часа, затем использовали субстрат TMB для развития окраски.
Анализ связывания C3b
Для тестирования C3b-связывающей активности белков hfH1-4.678.19-20 и mfH1-4.678.19-20 96-луночные планшеты предварительно покрывали 2 мкг/мл раствором человеческого C3b (каталожный № A114, CompTech) при 37°C в течение 1 часа. Незанятые сайты связывания на планшетах блокировали 1% раствором БСА в PBS при RT в течение 1 часа. Добавляли разные количества белка hfH1-4.678.19-20 или mfH1-4.678.19-20 и инкубировали при RT в течение 1 часа, затем добавляли 2 мкг/мл раствор мышиного мАт против человеческого fH (OX-23) и инкубировали при RT в течение 1 часа. Планшеты инкубировали с HRP-конъюгированными антителами кролика против IgG мыши при RT в течение 1 часа, затем использовали субстрат TMB для развития окраски.
Анализ жидкофазной кофакторной активности белка fH при опосредованном фактором I расщеплении C3b
Для оценки жидкофазной кофакторной активности белков hfH1-4.678.19-20 и mfH1-4.678.19-20 при опосредованном фактором I расщеплении C3b 0,5 или 0,25 мкг очищенного белка hfH1-4.678.19-20 или mfH1-4.678.19-20 смешивали с 2 мкг человеческого C3b в 15 мкл PBS, затем добавляли 1 мкг человеческого фактора I (каталожный № A138, CompTech) и инкубировали при 37°C в течение 15 минут. Реакцию останавливали добавлением 5x восстанавливающего буфера для образцов для SDS-ПААГ. Протеолиз C3b определяли, анализируя расщепление α-цепи и образование фрагментов α41 и α39 методом электрофореза в 4-20% градиентных SDS-ПАА гелях в восстанавливающих условиях, с последующим обнаружением методом вестерн-блоттинга с использованием HRP-конъюгированных IgG козы против человеческого C3 (1/4000, каталожный № 855237, MP biomedicals). Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus.
Оценка терапевтической эффективности mfH1-4.678.19-20 для предотвращения активации AP комплемента, вызываемой дефектами мембранного регулятора комплемента
Для определения того, может ли введение AAV fH1-4.678.19-20 также быть эффективным для предотвращения активации AP комплемента, вызываемой дефектами мембранных регуляторов комплемента, mfH1-4.678.19-20 тестировали на линии мышей с дефицитом двух мембранных регуляторов комплемента DAF и Crry (мыши с двойной мутацией DAF/Crry). Создание мышей с двойной мутацией DAF/Crry (DAF-/--Crryflox/flox-Tie-2Cre+) описано ранее, животные имеют фенотип вторичной недостаточности компонентов комплемента вследствие избыточной активации AP комплемента (Barata et al., 2013). Как и у мышей fHm/m, у мышей с двойной мутацией DAF/Crry имеет место истощение C3 и fB (Barata et al., 2013). Мышам с двойной мутацией DAF/Crry (в возрасте 10 недель) вводили инъекцией AAV mfH1-4.678.19-20 в дозе 3×1012 геномных копий/мышь через ретроорбитальный синус. Кровь собирали через ретроорбитальный синус перед инъекцией и через 1 неделю после инъекции. Терапевтическую эффективность оценивали путем измерения уровней в плазме C3 и fB до и после введения AAV mfH1-4.678.19-20 методом вестерн-блоттинга. Для вестерн-блоттинга плазму мышей (1 мкл) разбавляли буфером для образцов и кипятили перед нанесением на 4-20% градиентные SDS-ПАА гели в восстанавливающих условиях. Затем образцы переносили на PVDF мембраны и проводили обработку соответствующими антителами. Для обнаружения мышиных C3 и fB HRP-конъюгированные Ат козы против мышиного C3 (1:4000, каталожный № 0855557, MP Biomedicals) или аффинно очищенные Ат козы против человеческого fB (каталожный № A235, CompTech, Texas, перекрестно реагируют с мышиным fB) использовали в качестве первичных антител, затем проводили обнаружение с использованием HRP-конъюгированных антител кролика против IgG козы. Блоты визуализировали с использованием субстрата для вестерн-блоттинга Pierce ECL plus.
Создание мышиной модели aHUS
Для создания мышиной модели aHUS для тестирования терапевтической эффективности AAV-опосредованной терапии геном fH получали мутантную линию мышей, несущих точечную мутацию в SCR20 fH, соответствующую мутации W1183R в человеческом fH, которая имеет место у пациентов с aHUS, путем основанного на гомологичной рекомбинации метода таргетирования гена (Lesher et al., 2013; Dunkelberger, et al., J Immunol. 2012 Apr 15; 188(8): 4032-4042; Takashi et al., Blood. 2009 Mar 19; 113(12): 2684-2694; Kimura Y1, et al., Blood. 2008 Jan 15; 111(2): 732-40. Epub 2007 Oct 4; Kimura Y1, et al., J Clin Invest. 2010 Oct; 120(10): 3545-54). Для этого эксперимента фрагменты гена fH амплифицировали из геномной ДНК мышей C57BL/6 с использованием системы ПЦР Expand Long Template PCR system (Roche, Indianapolis, IN) для конструирования таргетирующего ген вектора. Длинное плечо таргетирующего вектора состояло из 6-т.п.н. фрагмента, содержащего 21-й экзон и фланкирующие интронные последовательности гена мышиного fH. Его амплифицировали методом ПЦР с использованием следующих праймеров: SEQ ID NO: 67: 5'-gcggccgccctatccattagtgagtgtgg-3' и SEQ ID NO: 68: 5'-ctcgaggacagcgatgtaagaacaatc-3'. Продукт ПЦР лигировали в вектор PCR2.1 (Invitrogen), затем вставку вырезали из вектора PCR2.1 расщеплением ферментами рестрикции Not I и XhoI, очищали и субклонировали в вектор pND1 выше кассеты NEO. Использование вектора pND1 описано в предыдущих публикациях, посвященных экспериментам по таргетированию генов (Lesher et al., 2013; Dunkelberger et al., 2012; Miwa et al., 2009; Kimura et al., 2008; Kimura et al. 2012): данный вектор содержит кассеты гена, определяющего устойчивость к неомицину (NEO), и гена дифтерийного токсина (DT) для положительной и отрицательной селекции, соответственно (Lesher et al. 2013; Dunkelberger et al., 2012; Miwa et al., 2009; Kimura et al., 2008; Kimura et al., 2012). Вектор pND1 также содержит сайт loxP и два сайта мишени узнавания флиппазы (FRT), фланкирующие кассету NEO для потенциального удаления NEO рекомбиназой FLPe (Rodríguez C.I., et al., Nat Genet. 2000 Jun; 25(2): 139-40).
Последовательность короткого плеча состояла из 3,85-т.п.н. фрагмента, содержащего 22-й экзон, кодирующий SCR20, и фланкирующие интронные последовательности гена мышиного fH. Эта последовательность была ПЦР-амплифицирована с использованием следующих праймеров: SEQ ID NO: 69: 5'-ggtaccaagcttattgaccagctacagacagta-3' и SEQ ID NO: 70: 5'-ggtaccctcactcaggtgtattactc-3'. Продукт ПЦР клонировали в вектор PCR2.1 и затем вносили мутацию замены триптофана (W) на аргинин (R) в положении 1206, соответствующую мутации W1183R в SCR20 человеческого fH, методом сайт-направленного мутагенеза с использованием набора Stratagene QuickChange Site-Directed Mutagenesis kit (Agilent Technologies, CA) со следующими двумя праймерами, SEQ ID NO: 71: 5'-ggAATCACACAATATAATTCTCAAAAggAgACACACTg-3' и SEQ ID NO: 72: 5'-CAgTgTgTCTCCTTTTgAgAATTATATTgTgTgATTCC-3'. После подтверждения мутации замены W на R фрагмент короткого плеча вырезали из PCR2.1 расщеплением Kpn I и субклонировали в вектор pND1 ниже кассеты NEO в том же сайте рестрикции. Таргетирующий вектор затем линеаризовали расщеплением Not I и трансфицировали в эмбриональные стволовые (ЭС) клетки мышей C57BL/6 (EmbroMAX Embryonic stem cell line-strain C57BL/6, каталожный № CMTI-2, Millipore) методом электропорации. Трансфицированные ЭС клетки подвергали селекции при помощи G418, начиная через 48 часов после электропорации. ЭС клетки с гомологичной рекомбинацией повергали скринингу методом саузерн-блоттинга геномной ДНК после расщепления HindIII с 480-п.н. 3′-зондом в реакции амплификации с использованием SEQ ID NO: 73: 5'-ATAGCATGTGCCAGGAGACAC-3' и SEQ ID NO: 83: 5'-AGTGTTGACTCGTGGAGACCA-3' в качестве праймеров. В случае аллеля дикого типа был получен 12,5-т.п.н. фрагмент, в то время как в случае таргетированного аллеля был получен 10,2-т.п.н. фрагмент. В Головном отделе трансгенных исследований Школы медицины Университета Пенсильвании правильно таргетированные ЭС клетки (fHW1201R(Neo-положительные)/+) вводили инъекцией в бластоцисты мышей C57BL/6J через 3,5 дня после оплодотворения для получения химер. У полученных химер имела место трансмиссия зародышевой линии, что оценивали по сочетанию цвета шерсти и результатов ПЦР-скрининга для обнаружения NEO с использованием следующих двух праймеров: праймер для Neo-4: SEQ ID NO: 74: 5-CTTGGGTGGAGAGGCTATTC-3' и SEQ ID NO: 75: праймер для Neo-5: 5'-AGGTGAGATGACAGGAGATC-3'. Кассета гена устойчивости к неомицину (NEO) в тагретирующем векторе была фланкирована 2 сайтами-мишенями рекомбиназы флиппазы (FLP) (FRT), позволяющими в последствии удалять ее с помощью рекомбиназы FLP. Гетерозиготные мыши с таргетированным FH (fHW1206R(Neo-положительные)/+) были скрещены с трансгенными мышами FLPe (экспрессирующими более эффективный вариант FLP, на фоновом генотипе C57BL/6) для удаления NEO из аллеля fH, в результате чего были получены гетерозиготные мутантные по fH мыши без кассеты гена устойчивости к NEO (fHW1206R/+). Мыши fHW1206R/+ были скрещены между собой для получения гомозиготных мышей fHW1206R/W1206R на фоновом генотипе C57BL/6. Для генотипирования были использованы следующие праймеры с целью обнаружения дикого типа и мутантных аллелей fH методом ПЦР: WR1 (FH-специфичный) SEQ ID NO: 76: 5'-GATATGGTCAATTTAGGGAAAGT, SEQ ID NO: 77: Neo7 (NEO-специфичный) 5'-gggTgggATTAgATAAATgCC-3' и SEQ ID NO: 78: WR4 (FH-специфичный) 5'-TACTGTCTGTAGCTGGTCAAT-3'.
В следующей Таблице приведены результаты лечения мышей fHW1206R/W1206R, получавших контрольный AAV вектор или AAV8-mfH1-4.678.19-20 вектор в дозе 3×1011 гк/мышь.
Гомозиготные мыши fHW1206R/W1206R имели плохое общее состояние со значительно сниженной массой тела в возрасте 4-6 недель и с показателем смертности почти 50% к возрасту 30 недель. Все мыши fHW1206R/W1206R демонстрировали один или более из характерных признаков aHUS, то есть, поражение почек (повышенные уровни в крови азота мочевины и/или гистологические признаки тромботической микроангиопатии в клубочках), тромбоцитопению и анемию. Примерно у одной трети мышей fHW1206R/W1206R также развились тяжелые неврологические симптомы, свидетельствующие об инсульте. Помимо тромботической микроангиопатии в почечных клубочках у мышей fHW1206R/W1206R имели место многочисленные тромбы в крупных сосудах в разных органах (печень, легкие, селезенка, почки, головной мозг и глаза).
К моментам времени, указанным выше, все 3 мыши fHW1206R/W1206R, получавшие AAV8-mfH1-4.678.19-20, были живы и здоровы, с нормальными количествами тромбоцитов, в то время как 1 из 2 мышей fHW1206R/W1206R, получавших контрольный AAV вектор, погибла (через 4 недели после инъекции) и оставшаяся мышь проявляла симптомы aHUS, включая тромбоцитопению.
Терапевтическая эффективность mfH1-4.678.19-20, доставляемого с помощью AAV в организм мышей fHW1206R/W1206R
Для тестирования терапевтической эффективности mfH1-4.678.19-20 в качестве суррогата для hfH1-4.678.19-20 авторы изобретения вводили его инъекцией 4-недельным гомозиготным мышам fHW1206R/W1206R в дозе 3×1011 генные копии/мышь через ретроорбитальный синус. Если mfH1-4.678.19-20 является функционально активным у мышей fHW1206R/W1206R, то можно ожидать уменьшения тромбоцитопении и поражения почек. Вследствие этого, в качестве показателей терапевтической эффективности mfH1-4.678.19-20 авторы изобретения подсчитывали количество тромбоцитов и измеряли уровень азота мочевины в сыворотке крови. Поскольку мыши fHW1206R/W1206R имели плохое общее состояние со значительно сниженной массой тела в возрасте 4-6 недель и с показателем смертности почти 50% к возрасту 30 недель, авторы изобретения также имели возможность использовать уровень смертности мышей fHW1206R/W1206R в качестве другого показателя терапевтической эффективности AAV mfH1-4.678.19-20.
Количество тромбоцитов у получавших контрольный AAV и AAV mfH1-4.678.19-20 мышей fHW1206R/W1206R
Для определения количества тромбоцитов у получавших контрольный AAV и AAV mfH1-4.678.19-20 мышей fHW1206R/W1206R кровь собирали в пробирки с ЭДТА (конечная концентрация: 0,02 M) через ретроорбитальный синус до инъекции и в разных временных точках после инъекции, начиная с 1 месяца после инъекции, и анализировали на автоматическом гематологическом анализаторе Sysmex XT-2000iV в Центральной лаборатории трансляционных исследований CTRC Детской больницы Филадельфии (https://ctrc.research.chop.edu/services-facilities/translational-core-laboratory-tcl/hematology).
Измерение азота мочевины в крови (BUN) у получавших контрольный AAV и AAV mfH1-4.678.19-20 мышей fHW1206R/W1206R
Для измерения уровня азота мочевины в сыворотке крови образцы крови собирали из ретроорбитального синуса перед инъекцией и в разных временных точках после инъекции, начиная с 1 месяца после инъекции. Сывороточные уровни BUN измеряли с использованием реагентов для определения азота мочевины (Sigma-Aldrich) в соответствии с инструкциями производителя.
Гистологическое исследование почек и других органов мышей fHW1206R/W1206R
У мышей fHW1206R/W1206R собирали парные почки и другие органы. Один орган из пары фиксировали в растворе формалина в течение ночи и обрабатывали для заливки в парафин, а другой быстро замораживали в среде OCT (Sakura Finetek). Проводили гистологическую оценку почек и других органов на признаки aHUS/тромботической микроангиопатии с использованием светового микроскопа, а также иммуногистохимические исследования, включая иммунофлуоресцентное и иммунопероксидазное окрашивание.
(Бесплатный текст списка последовательностей)
Следующая информация приведена для последовательностей, содержащих бесплатный текст под номером <223>.
(содержит бесплатный текст)
<221> СИГНАЛ
<222> (1)..(18)
<220>
<221> ДОМЕН
<222> (19)..(82)
<223> Sushi 1
<220>
<221> ДОМЕН
<222> (83)..(143)
<223> Sushi 1
<220>
<221> ДОМЕН
<222> (144)..(207)
<223> Sushi 3
<220>
<221> ДОМЕН
<222> (208)..(264)
<223> Sushi 4
<220>
<221> ДОМЕН
<222> (265)..(322)
<223> Sushi 5
<220>
<221> ДОМЕН
<222> (324)..(386)
<223> Sushi 6
<220>
<221> ДОМЕН
<222> (387)..(444)
<223> Sushi 7
<220>
<221> ДОМЕН
<222> (446)..(507)
<223> Sushi 8
<220>
<221> ДОМЕН
<222> (515)..(566)
<223> Sushi 9
<220>
<221> ДОМЕН
<222> (576)..(625)
<223> Sushi 10
<220>
<221> ДОМЕН
<222> (628)..(686)
<223> Sushi 11
<220>
<221> ДОМЕН
<222> (689)..(746)
<223> Sushi 12
<220>
<221> ДОМЕН
<222> (751)..(805)
<223> Sushi 13
<220>
<221> ДОМЕН
<222> (809)..(866)
<223> Sushi 14
<220>
<221> ДОМЕН
<222> (868)..(928)
<223> Sushi 15
<220>
<221> ДОМЕН
<222> (929)..(986)
<223> Sushi 16
<220>
<221> ДОМЕН
<222> (987)..(1045)
<223> Sushi 17
<220>
<221> ДОМЕН
<222> (1046)..(1104)
<223> Sushi 18
<220>
<221> ДОМЕН
<222> (1107)..(1165)
<223> Sushi 19
<220>
<221> ДОМЕН
<222> (1170)..(1230)
<223> Sushi 20
Все публикации, цитированные в настоящем описании, включены в настоящий документ посредством ссылки. Предварительная патентная заявка США № 62/232008, поданная 24 сентября 2015 г., также включена посредством ссылки. Аналогично, SEQ ID NO, упомянутые в настоящем описании и приведенные в прилагаемом Списке последовательностей, включены посредством ссылки. При том, что изобретение описано со ссылкой на конкретные варианты осуществления, следует понимать, что могут быть произведены модификации без отклонения от сущности изобретения. Такие модификации должны входить в объем прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНТРАТЕКАЛЬНОЕ ВВЕДЕНИЕ ВЕКТОРОВ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ГЕННОЙ ТЕРАПИИ | 2016 |
|
RU2775138C2 |
| ЛЕЧЕНИЕ КОМПЛЕМЕНТ-ОПОСРЕДУЕМЫХ РАССТРОЙСТВ | 2017 |
|
RU2768982C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ MPS1 | 2014 |
|
RU2708318C2 |
| ФАКТОР КОМПЛЕМЕНТА I И КОФАКТОР ФАКТОРА КОМПЛЕМЕНТА I, КОДИРУЮЩИЕ ИХ ВЕКТОРЫ И ПРИМЕНЕНИЕ В ТЕРАПЕВТИЧЕСКИХ ЦЕЛЯХ | 2019 |
|
RU2809247C2 |
| ГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ A | 2017 |
|
RU2762257C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА D И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799044C2 |
| ОПТИМИЗИРОВАННЫЕ ГЕНЫ И ЭКСПРЕССИОННЫЕ КАССЕТЫ CLN1, И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2764919C2 |
| КОМПОЗИЦИИ, СПОСОБЫ И ПРИМЕНЕНИЯ ПЕРЕНОСА ГЕНОВ ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2805606C2 |
| АНТИ-C5a-АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2773779C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛЕЗНИ ДАНОНА И ДРУГИХ НАРУШЕНИЙ АУТОФАГИИ | 2017 |
|
RU2777571C2 |



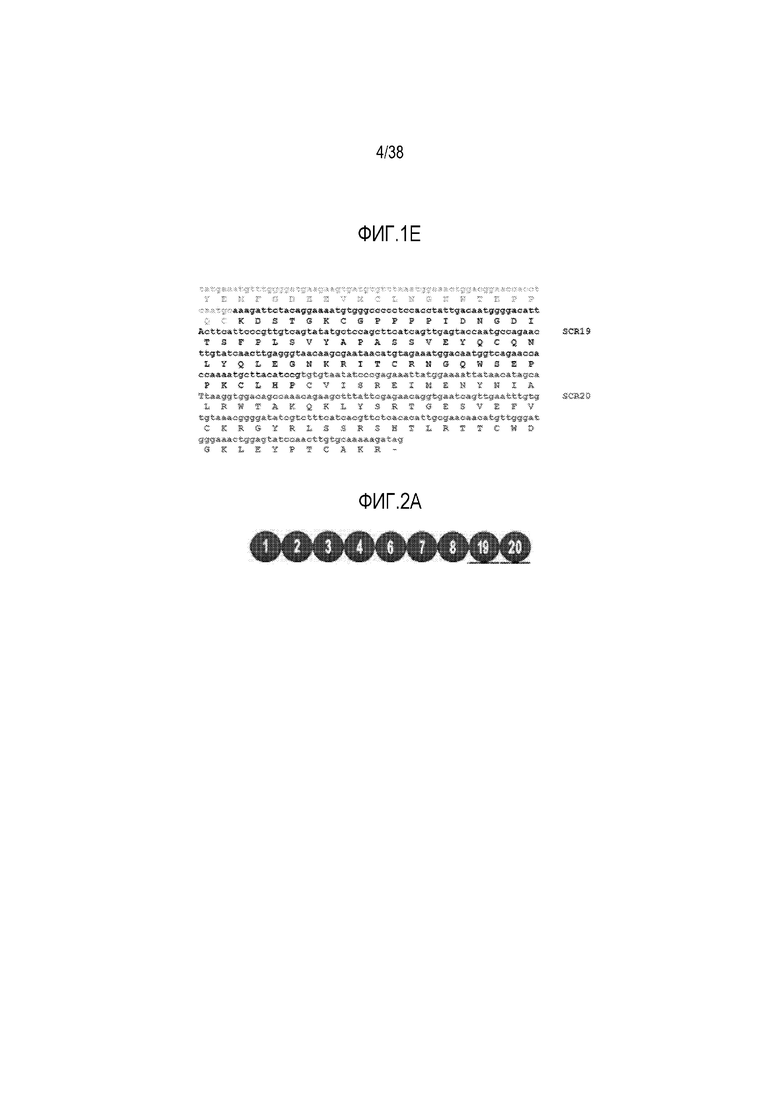




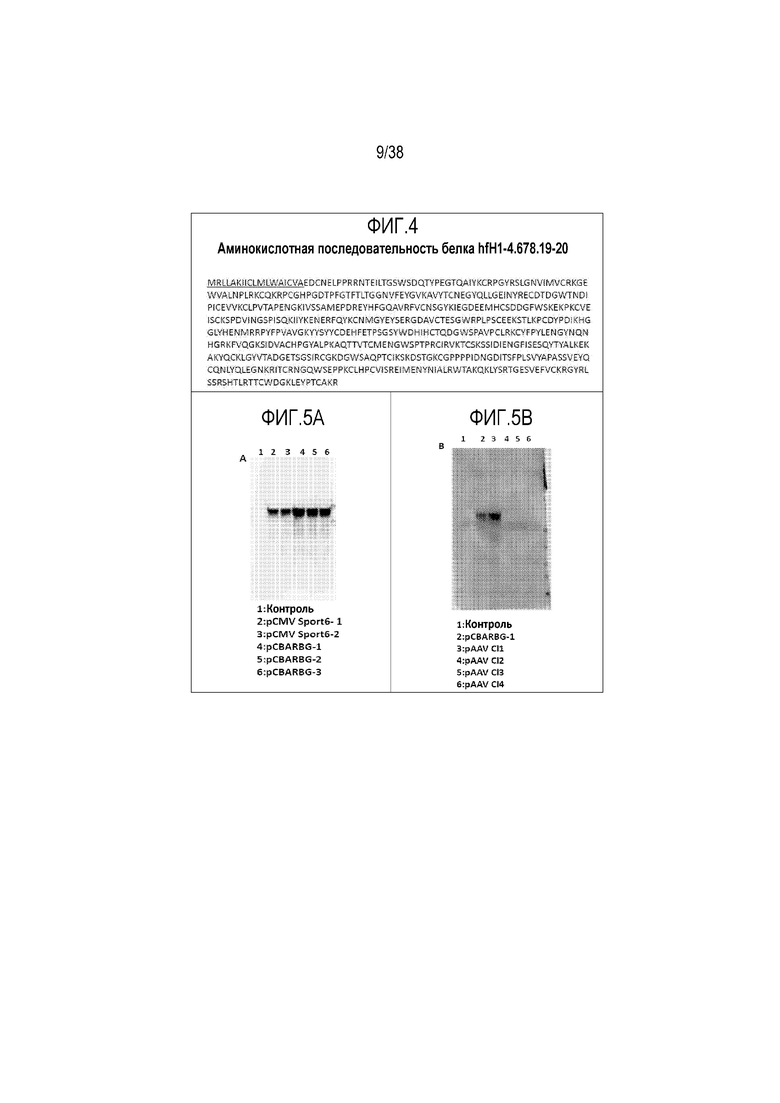
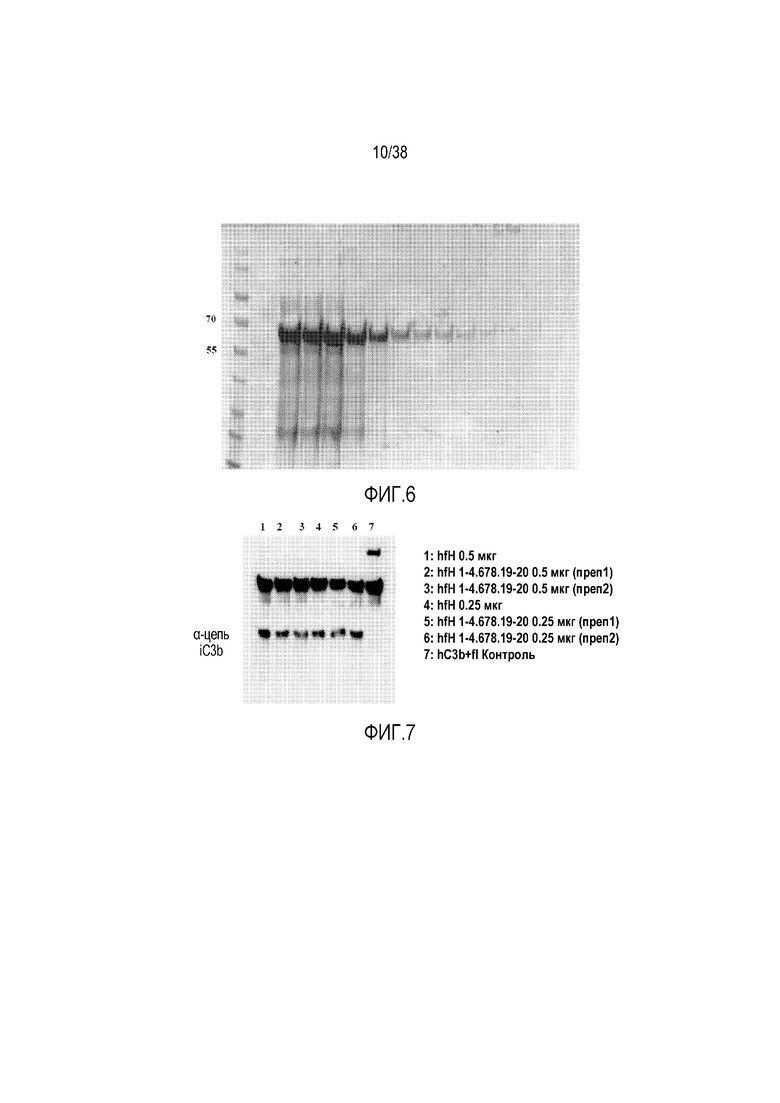
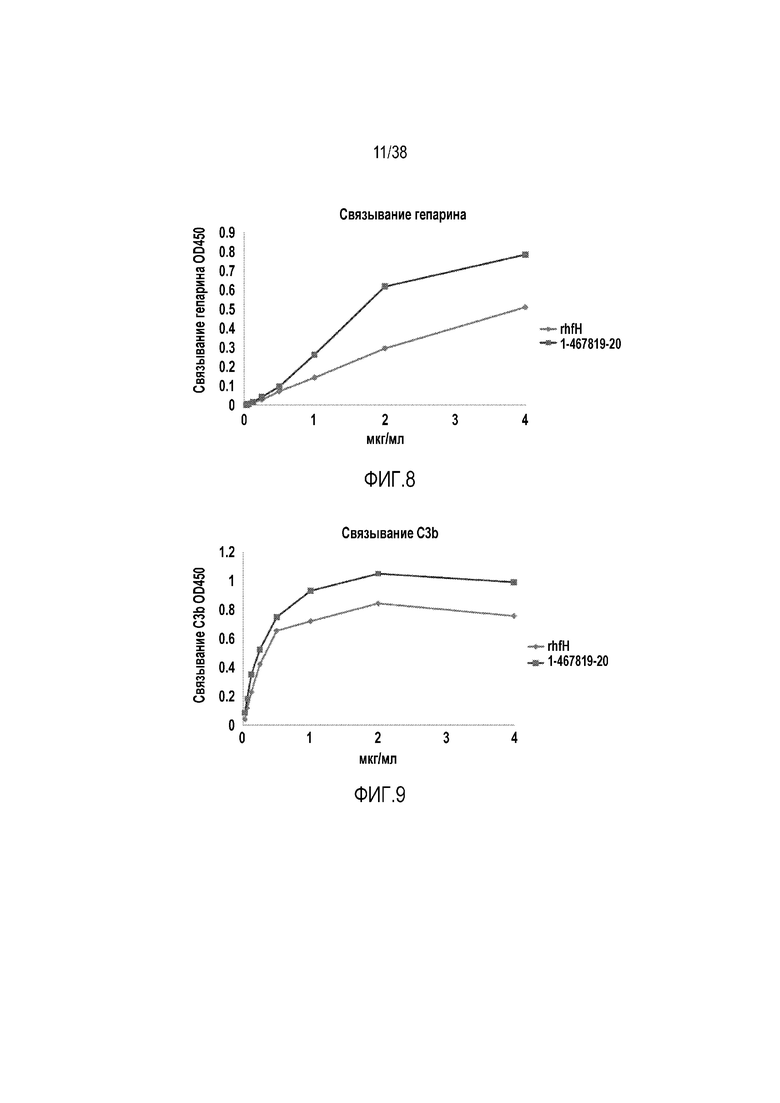
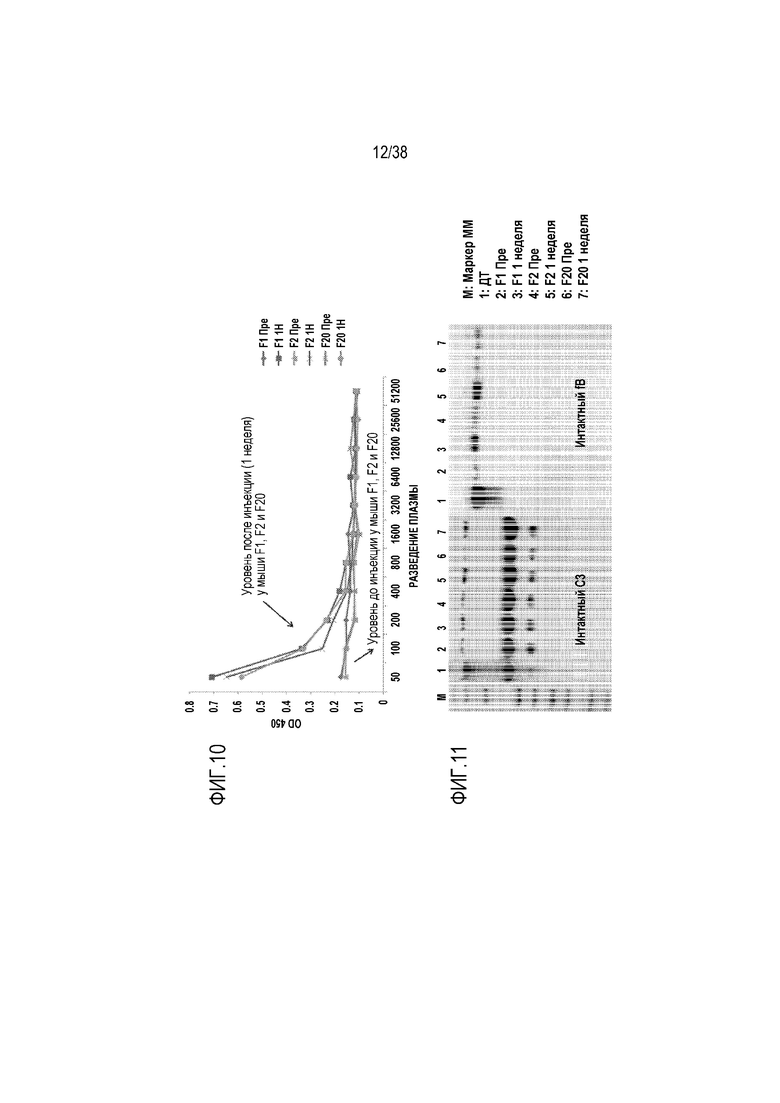





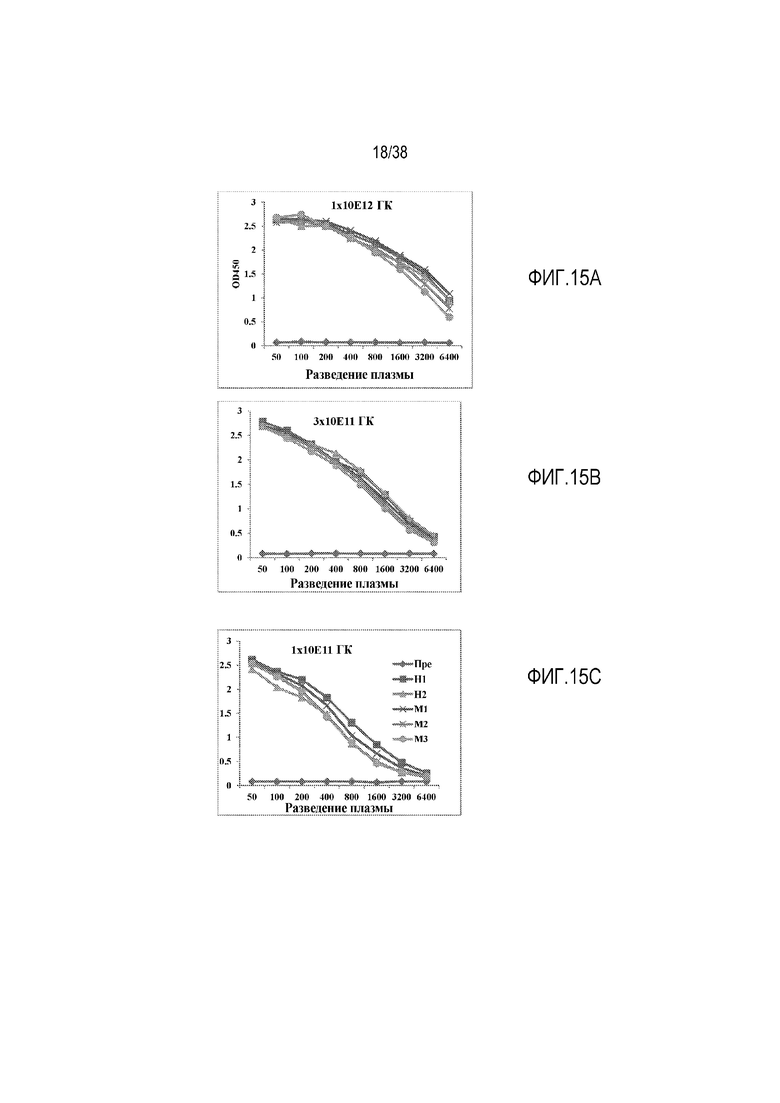
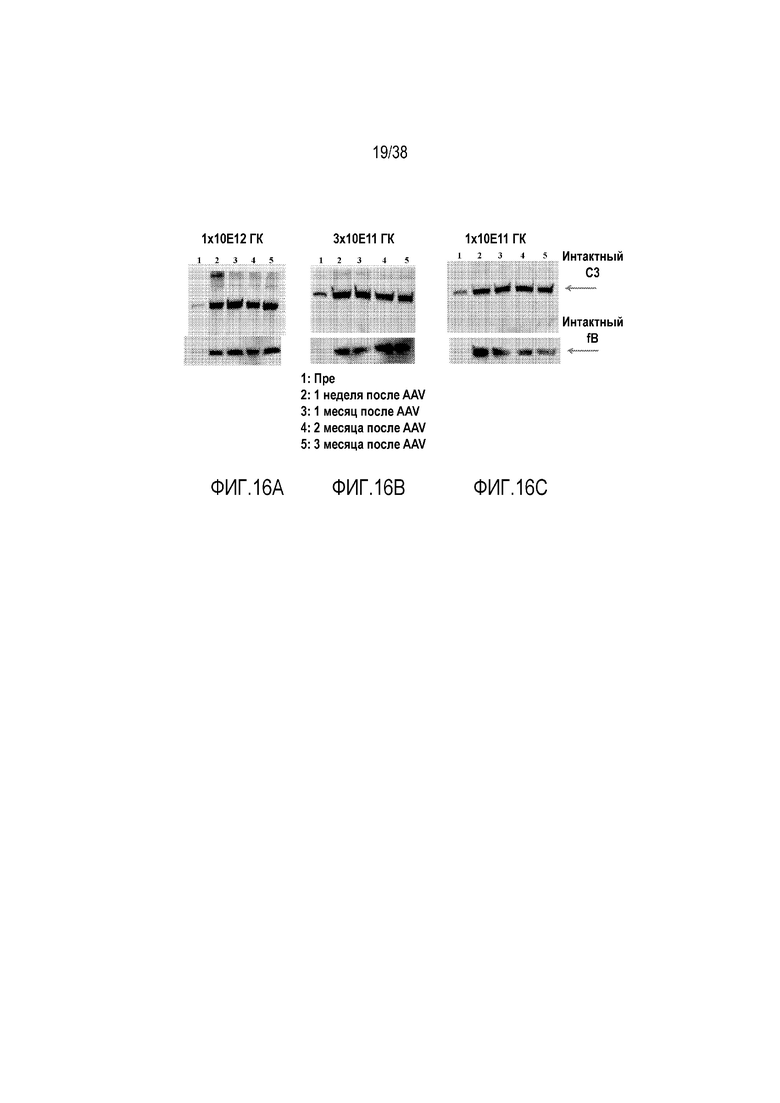







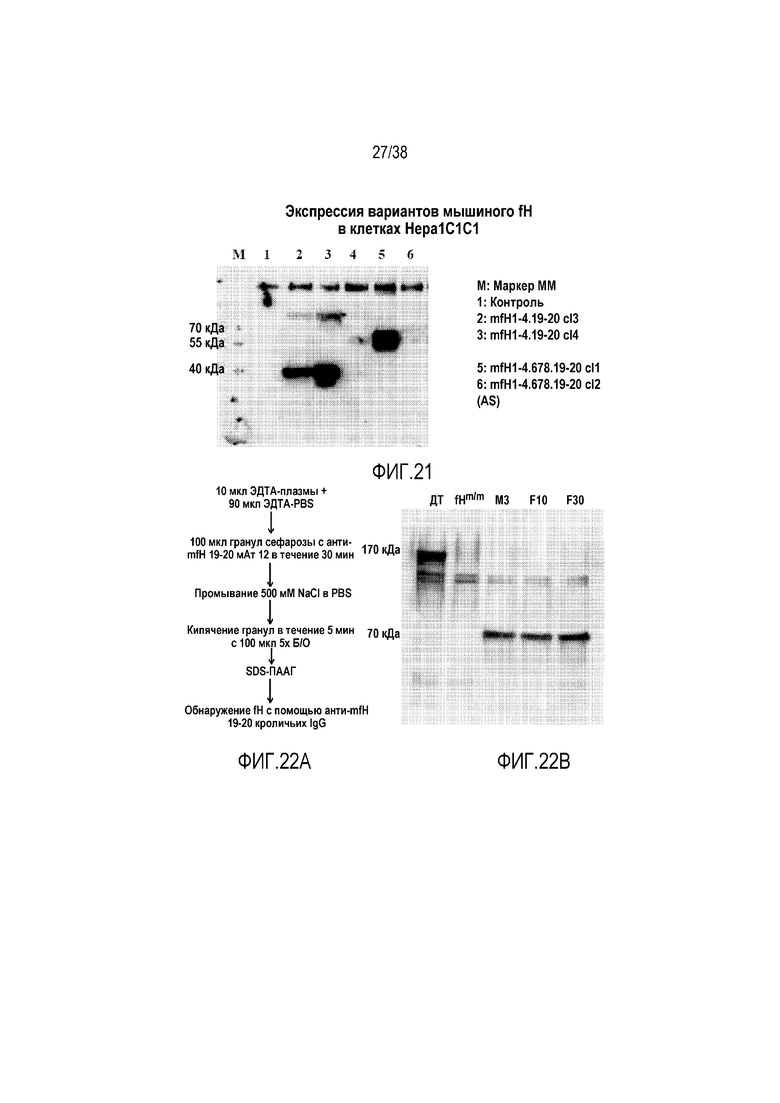

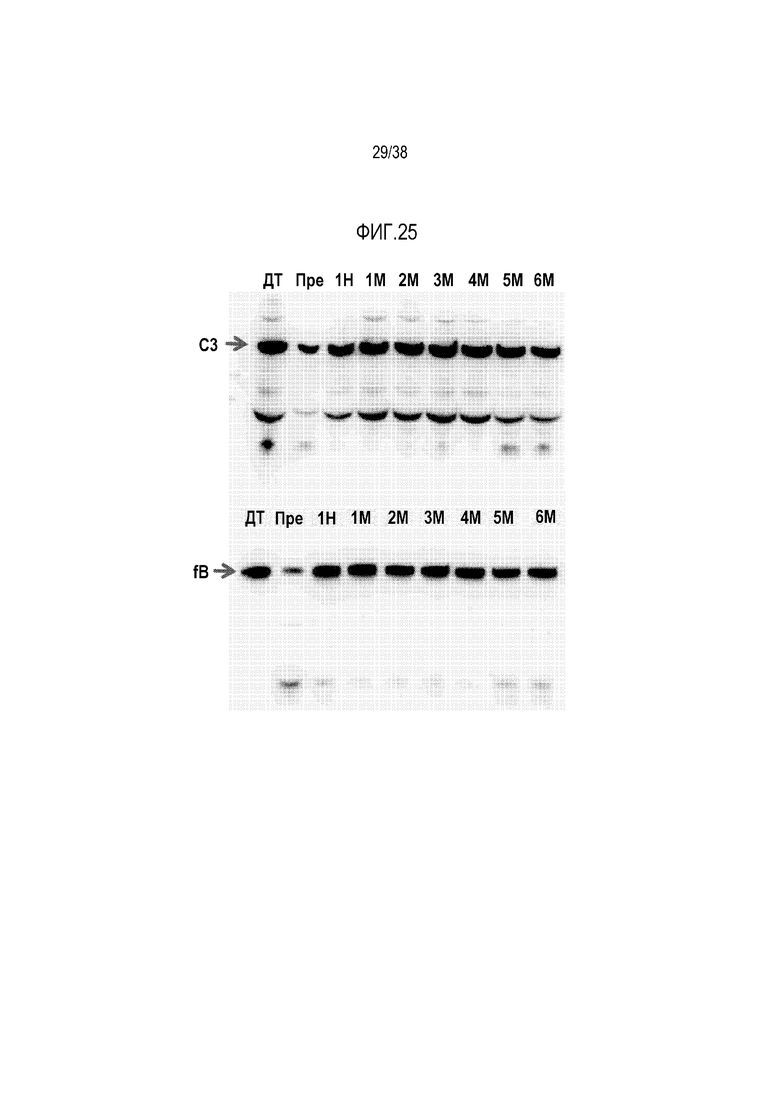

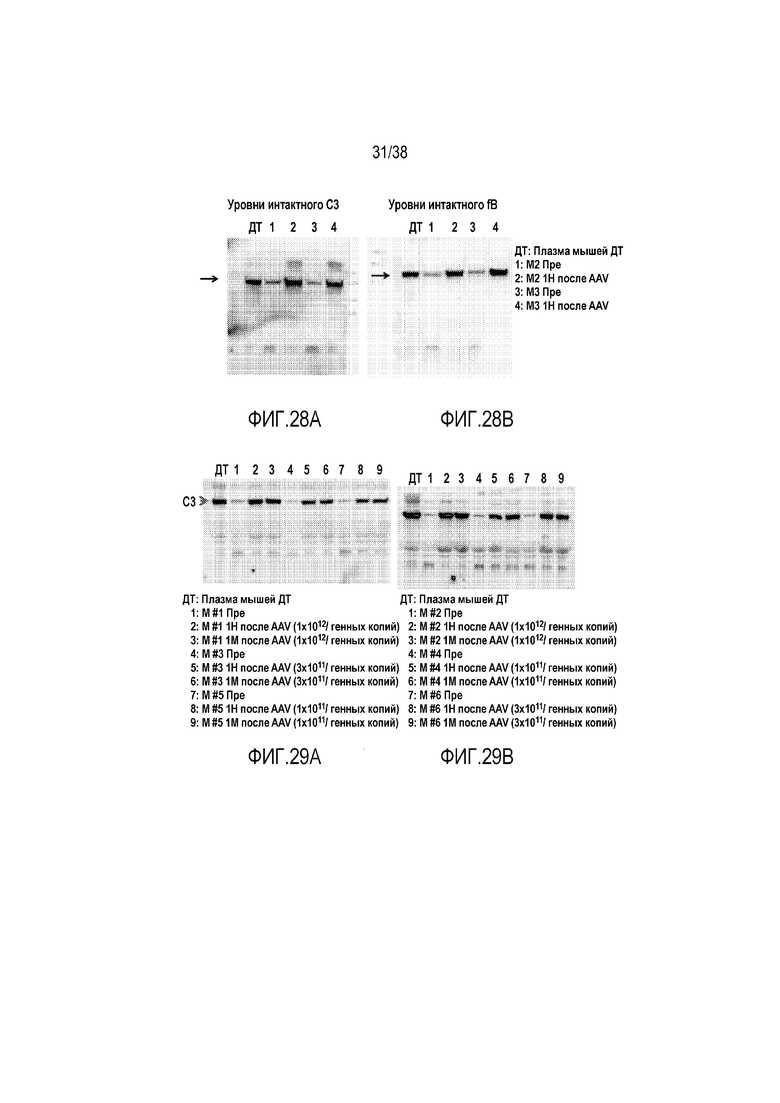

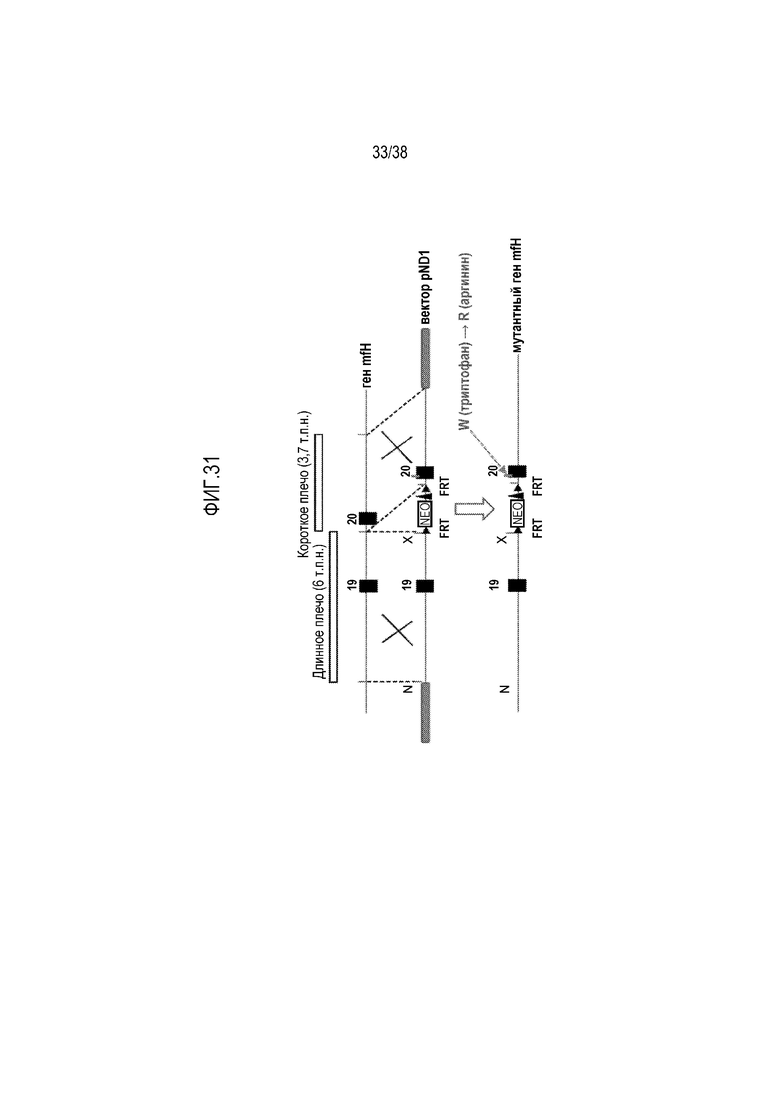
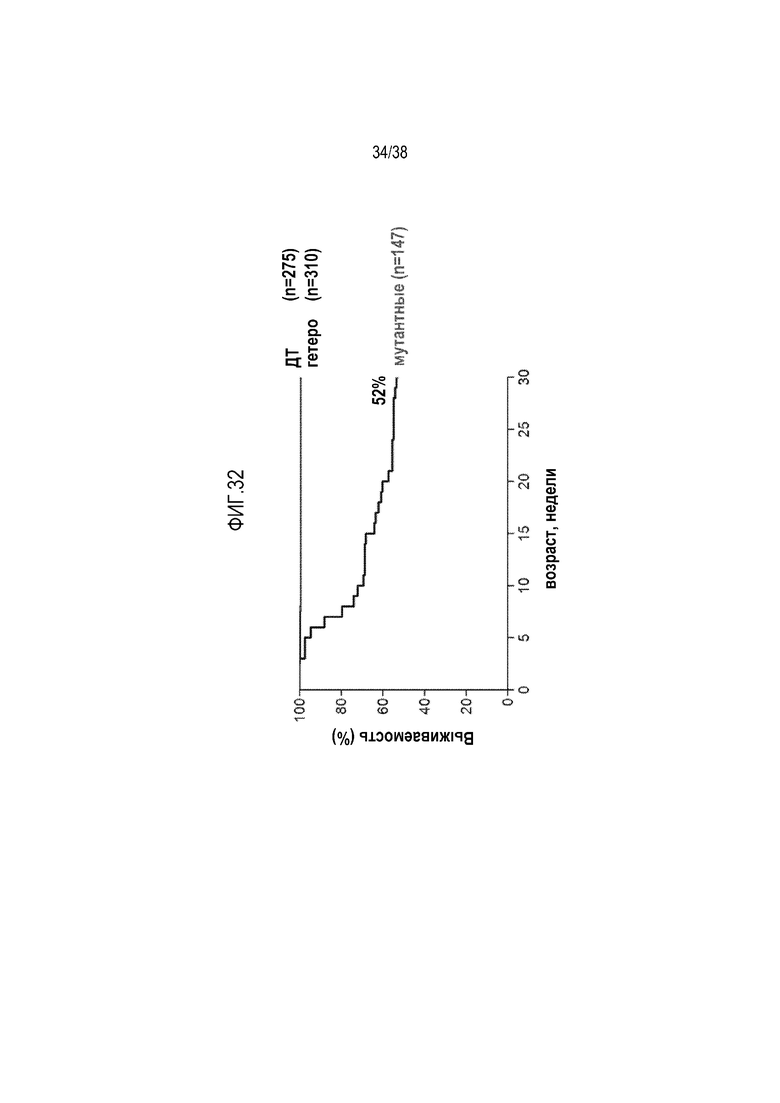
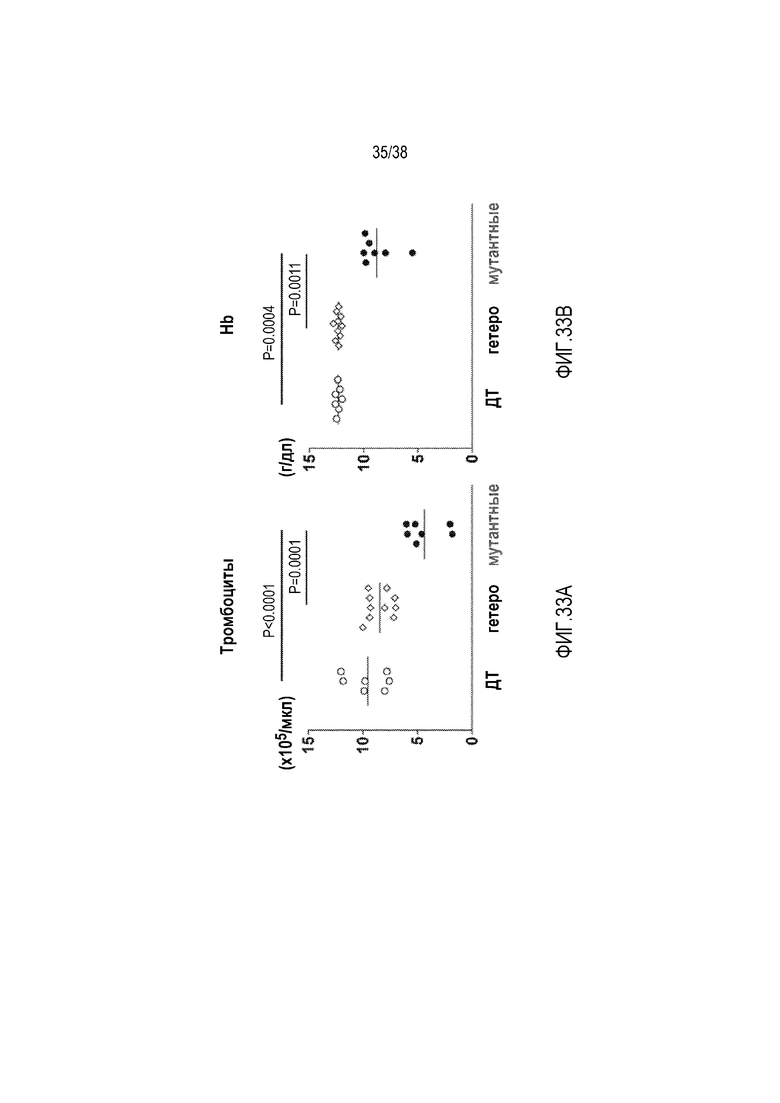
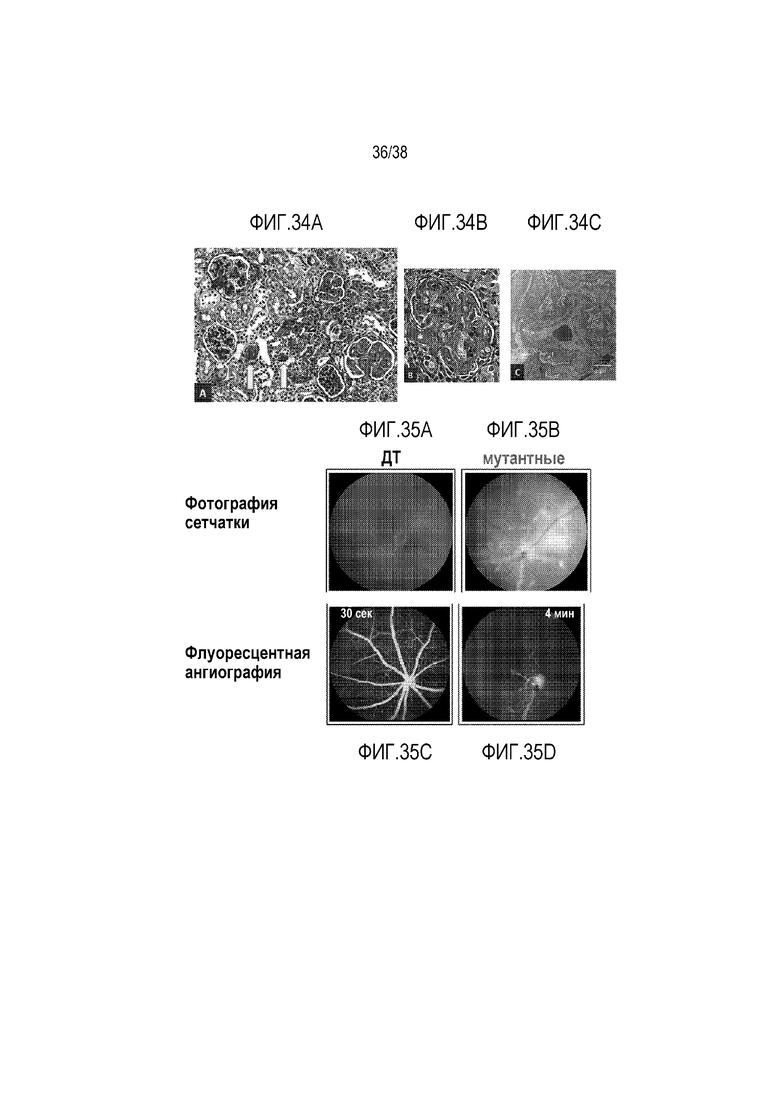
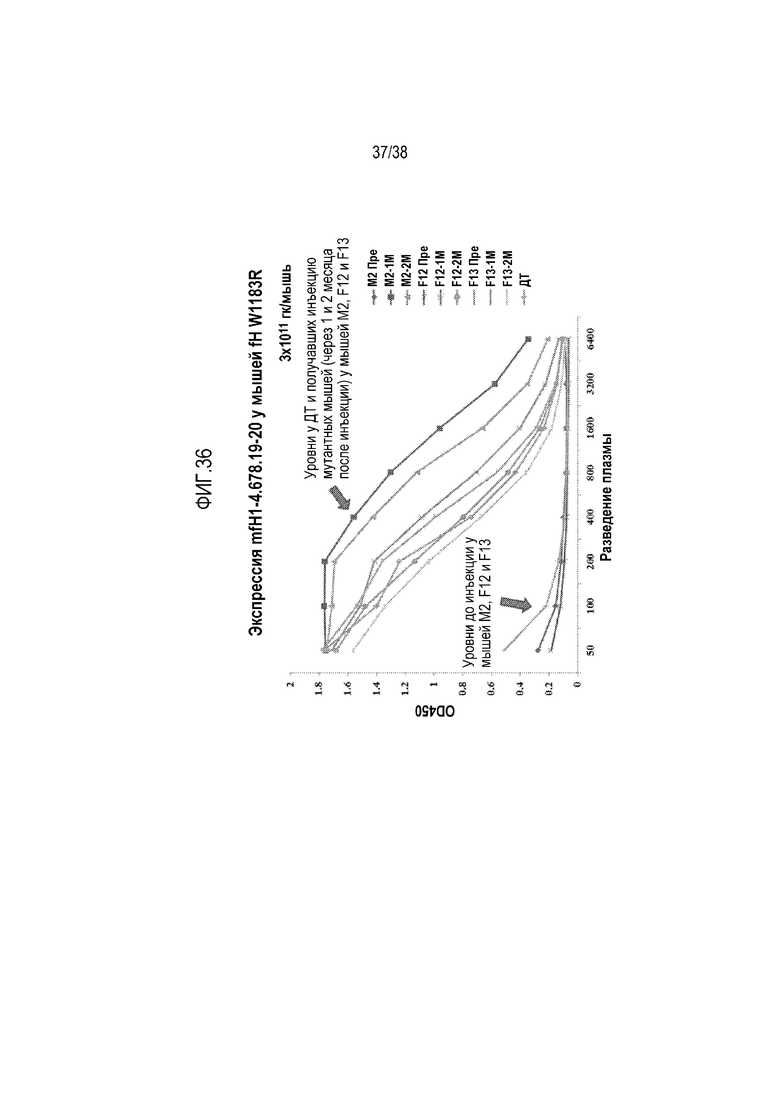
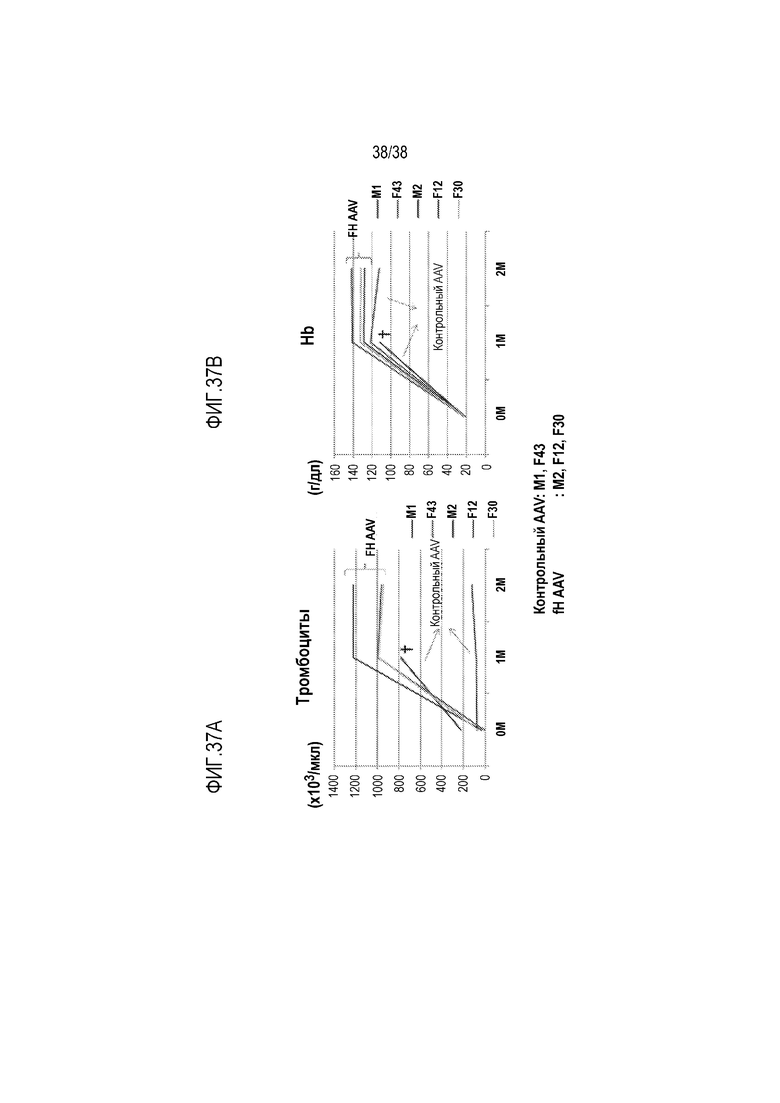
Изобретение относится к области биотехнологии, конкретно к рекомбинантным экспрессирующим AAV векторам, и может быть использовано в медицине в лечении заболевания, связанного с комплементом. Рекомбинантный AAV вектор с упакованной в нем экспрессионной кассетой, содержит нуклеиновую кислоту, кодирующую вариант человеческого регулятора комплемента фактора H (hfH), функционально связанный с последовательностями контроля экспрессии, которые управляют его экспрессией. При этом указанный вариант hfH содержит короткий консенсусный повтор (SCR) 1, 2, 3, 4, 19 и 20, а SCR 9, 10, 11, 12, 13, 14 и/или 15 отсутствуют. Изобретение обеспечивает экспрессию растворимого варианта белка hfH, сохраняющего функцию регуляции комплемента. 5 н. и 12 з.п. ф-лы, 37 ил., 7 табл.
1. Рекомбинантный AAV вектор с упакованной в нем экспрессионной кассетой, содержащий нуклеиновую кислоту, кодирующую вариант человеческого регулятора комплемента фактора H (hfH), функционально связанный с последовательностями контроля экспрессии, которые управляют его экспрессией, при этом указанный вариант hfH представляет собой растворимый вариант белка hfH, сохраняющий функцию регуляции комплемента, причем указанный вариант hfH содержит:
короткий консенсусный повтор (SCR) 1, 2, 3, 4, 19 и 20, и причем SCR 9, 10, 11, 12, 13, 14 и/или 15 отсутствуют.
2. Рекомбинантный вектор по п. 1, в котором вариант hfH содержит комбинацию доменов SCR, выбранную из одной или более из:
(a) SCR1-4, 7 и 19-20;
(b) SCR1-4, 6, 7 и 19-20;
(c) SCR1-4, 7, 8 и 19-20;
(d) SCR1-4, 6, 7, 8 и 19-20;
(e) SCR1-4, 17, 19-20;
(f) SCR1-4, 18-20;
(g) SCR1-4, 17-20;
(h) SCR1-4, 7 и 18-20;
(i) SCR1-4, 6, 7 и 18-20;
(j) SCR1-4, 7, 8 и 18-20;
(k) SCR1-4, 6, 7, 8 и 18-20,
(l) SCR1-4, 7 и 17-20;
(m) SCR1-4, 6, 7 и 17-20;
(n) SCR1-4, 7, 8 и 17-20; или
(o) SCR1-4, 6, 7, 8 и 17-20.
3. Рекомбинантный вектор AAV по п. 1 или 2, в котором вариант hfH содержит SCR1-4, 6, 7, 8 и 19-20.
4. Рекомбинантный вектор AAV по п. 3, в котором экспрессируемый вариант hfH содержит аминокислотную последовательность SEQ ID NO:42.
5. Рекомбинантный вектор AAV по п. 1 или 2, в котором вариант hfH содержит SCR1-4, 6-8 и 17-20.
6. Рекомбинантный вектор AAV по п. 5, в котором экспрессируемый вариант hfH содержит аминокислотную последовательность SEQ ID NO:48.
7. Рекомбинантный вектор по любому из пп. 1-6, причем вектор дополнительно содержит промотор, специфичный для тканей глаза.
8. Рекомбинантный AAV вектор по п. 2, в котором вариант hfH содержит по меньшей мере один линкер, содержащий от 1 до 18 аминокислот, расположенный между одним или более из SCR, причем в случае необходимости вариант hfH содержит SCR1-(L1)-SCR2-(L2)-SCR3-(L3)-SCR4-(L4)-(SCR6-(L4'))-SCR7-(L5)-(SCR8-(L5'))-(SCR16-(L5"))-(SCR17-(L5"'))-(SCR18-(L5""))-SCR19-(L6)-SCR20, при этом () обозначает необязательный компонент, «L» обозначает линкер, и каждый из L1, L2, L3, L4, L4', L5, L5', L5", L5"', L5"" и L6 может отсутствовать или быть независимо выбран из аминокислотной последовательности, содержащей от 1 до 18 аминокислот.
9. Рекомбинантный AAV вектор по пп. 1-8, причем рекомбинантный AAV имеет капсид, выбранный из капсида AAV8, капсида rh64R1, капсида AAV9 или капсида rh10, причем в случае необходимости экспрессионная кассета содержит промотор, специфически управляющий экспрессией варианта hfH в клетках печени.
10. Рекомбинантный AAV вектор по любому из пп. 1-8, сконструированный для экспрессии в глазах, и капсид AAV выбран из AAV1, AAV2 и AAV5.
11. Фармацевтическая композиция, содержащая фармацевтически приемлемый носитель и рекомбинантный вектор по любому из пп. 1-10.
12. Рекомбинантный вектор по любому из пп. 1-10 для применения в лечении заболевания, связанного с комплементом.
13. Рекомбинантный вектор по любому из пп. 1-10 для применения по п. 12, в котором (i) заболевание, связанное с комплементом, представляет собой C3-гломерулопатию, включая болезнь плотных депозитов и C3-гломерулонефрит, атипичный гемолитико-уремический синдром (aHUS), возрастную дегенерацию желтого пятна (AMD), микроангиопатическую гемолитическую анемию, тромботическую тромбоцитопеническую пурпуру (TTP), острую почечную недостаточность, пароксизмальную ночную гемоглобинурию (PNH), шизофрению, ишемический инсульт и/или бактериальные инфекции, вызываемые рекрутингом бактериальных патогенов; (ii) вектор доставляют путем введения, выбранным из внутривенного, внутриглазного, внутримышечного, подкожного пути введения, или их комбинаций; или (iii) терапевтическое средство на основе белка fH доставляют указанному пациенту практически одновременно с введением вектора.
14. Рекомбинантный вектор по любому из пп. 1-10 для применения по п. 12, в котором заболевание, связанное с комплементом, представляет собой возрастную дегенерацию желтого пятна, причем в случае необходимости вектор вводят субретинально.
15. Рекомбинантный вектор по любому из пп. 1-10 для применения по п. 12, в котором заболевание, связанное с комплементом, представляет собой почечную недостаточность.
16. Рекомбинантный вектор по любому из пп. 1-10 для применения в схеме лечения AMD.
17. rAAV вектор по любому из пп. 6-10 для применения в схеме лечения связанной с комплементом почечной недостаточности.
| WO2013142362 A1, 26.09.2013 | |||
| WO2011107591 A1, 09.09.2011 | |||
| WO2011163412 А1, 29.12.2011 | |||
| NICHOLS E.-M.ET AL., An extended mini-complement factor H molecule ameliorates experimental C3 glomerulopathy, KIDNEY INTERNATIONAL, 29.07.2015, v | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| WO2015092335 А2, 25.06.2015 | |||
| ПРИМЕНЕНИЕ ИНГИБИТОРОВ ПУТИ КОМПЛЕМЕНТА ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2006 |
|
RU2417099C2 |

