Область техники.
Группа изобретений относится к биотехнологии, иммунологии и вирусологии. Созданы однодоменные антитела, а также их модификации, специфически связывающиеся со стеблевым доменом гемагглютинина вируса гриппа А и обладающие вируснейтрализующей активностью, предложено применение таких антител для экстренной профилактики заболеваний, вызываемых вирусом гриппа А.
Предшествующий уровень техники.
Вирус гриппа А остается серьезной проблемой для мирового здравоохранения, несмотря на широкую доступность ежегодно обновляющихся средств его терапии и профилактики По данным ВОЗ, ежегодно, на долю гриппа и ОРВИ приходится до 90% случаев регистрируемой инфекционной патологии, а количество смертей в результате респираторных осложнений достигает 650000 ежегодно [https://www.who.int/influenza/surveillance_monitoring/bod/en/]. Помимо сезонных вспышек эпидемий, вирусы гриппа А, способны вызывать пандемии, благодаря высокой скорости изменчивости в РНК-геноме, образовывать реассортанты и возможности преодолевать межвидовой барьер. Также потенциальную опасность представляют высокопатогенные вирусы гриппа А птиц субтипов H5N1 и H7N9, летальность которых среди инфицированных людей достигает 60% и 47% соответственно [Li, F. C. K., Choi, B. C. K., Sly, T., & Pak, A. W. P. (2008). Finding the real case-fatality rate of H5N1 avian influenza. Journal of Epidemiology & Community Health, 62(6), 555-559. doi:10.1136/jech.2007.064030; Wang X, Jiang H, Wu P, Uyeki TM, Feng L, Lai S, et al. Epidemiology of avian influenza A H7N9 virus in human beings across five epidemics in mainland China, 2013-17: an epidemiological study of laboratory-confirmed case series. Lancet Infect Dis 2017;17:822- 832. doi: 10.1016/s1473-3099(17)30323-7.]. Пока случаев передачи таких субтипов от человека к человеку не зафиксировано, однако такую возможность нельзя отрицать, учитывая быструю антигенную изменчивость вируса гриппа А.
На сегодняшний день, основным способом контроля распространения гриппозной инфекции является вакцинопрофилактика, которую дополняют противовирусные препараты, действующие как на сезонные, так и на пандемические штаммы вируса гриппа А. Используемые на сегодняшний день противогриппозные лекарственные средства делятся на три группы: блокаторы М2-каналов, ингибиторы NA и ингибиторы вирусной полимеразы. Блокаторы М2-каналов утратили свою эффективность из-за повсеместно распространенного высокого уровня резистентности вирусов гриппа [Zaraket, H., Saito, R., Suzuki, Y., Suzuki, Y., Caperig-Dapat, I., Dapat, C., … Suzuki, H. (2010). Genomic events contributing to the high prevalence of amantadine-resistant influenza A/H3N2. Antiviral Therapy, 15(3), 307-319. doi:10.3851/imp1538]. Подавляющее большинство терапевтических противогриппозных средств на рынке представлено ингибиторами NA (осельтамивир, занамивир), в 2020 году в России одобрили применение балоксавира марбоксила - ингибитора кэп-зависимой эндонуклеазы. Однако, устойчивость к таким типам противогриппозной терапии неуклонно растет с каждым годом [Whitley, R. J., Boucher, C. A., Lina, B., Nguyen-Van-Tam, J. S., Osterhaus, A., Schutten, M., & Monto, A. S. (2013). Global Assessment of Resistance to Neuraminidase Inhibitors, 2008-2011: The Influenza Resistance Information Study (IRIS). Clinical Infectious Diseases, 56(9), 1197-1205. doi:10.1093/cid/cis1220; Uehara T, Hayden FG, Kawaguchi K, Omoto S, Hurt AC, De Jong MD, Hirotsu N, Sugaya N, Lee N, Baba K, Shishido T, Tsuchiya K, Portsmouth S, Kida H. Treatment-Emergent Influenza Variant Viruses With Reduced Baloxavir Susceptibility: Impact on Clinical and Virologic Outcomes in Uncomplicated Influenza. J Infect Dis. 2020 Jan 14;221(3):346-355. doi: 10.1093/infdis/jiz244. PMID: 31309975.]. Более того, блокаторы М2-каналов малоэффективны при применении у пожилых и детей, а действие ингибиторов NA напрямую зависит от времени начала их приема после возникновения симптомов [Yen, H.-L. (2016). Current and novel antiviral strategies for influenza infection. Current Opinion in Virology, 18, 126-134. doi:10.1016/j.coviro.2016.05.004 ]. Таким образом, разработка новых средств терапии инфекции вызванной вирусом гриппа А, является на сегодняшний день весьма актуальной задачей.
Перспективным направление в создании противовирусных средств является получение моноклональных антител, специфичных к высоко консервативным участкам вирусных белков. Гемагглютинин (НА) - один из мажорных белков вириона вируса гриппа, представленный его на поверхности в качестве тримера, мономеры которого состоят из двух субъединиц: НА1 и НА2. В пространственной конфигурации НА можно разделить на апикальный глобулярный домен, включающий в себя центральную часть НА1 субъединицы, и дистальный стеблевой домен, который сформирован НА2 субъединицей и N- и C-концевыми участками субъединицы НА1. Подавляющее большинство антител в организме вырабатываются против высоко иммуногенного участка вокруг рецептор-связывающего сайта, расположенного в глобулярном домене, тем самым обеспечивая нейтрализацию вируса и в тоже время оказывая селективное давление, приводящее к возникновению эскейп-мутантов. Такие антитела почти всегда штаммо- или субтип-специфичны. Стеблевой домен, напротив, менее иммуногенен, однако антитела к нему зачастую распознают несколько субтипов НА, за счет наличия высоко консервативных эпитопов. Таким образом, разработка антител к стеблевому домену является перспективным направлением в создании новых противовирусных препаратов.
Известные на данный момент моноклональные антитела специфичные к стеблевому домену обладают различным репертуаром кросс реактивности: от связывания с субтипами НА внутри одной филогенетической группы [Ekiert DC, Friesen RH, Bhabha G, Kwaks T, Jongeneelen M, Yu W, Ophorst C, Cox F, Korse HJ, Brandenburg B, Vogels R, Brakenhoff JP, Kompier R, Koldijk MH, Cornelissen LA, Poon LL, Peiris M, Koudstaal W, Wilson IA, Goudsmit J. A highly conserved neutralizing epitope on group 2 influenza A viruses. Science. 2011 Aug 12;333(6044):843-50. doi: 10.1126/science.1204839. Epub 2011 Jul 7. PMID: 21737702; PMCID: PMC3210727.; Throsby M, van den Brink E, Jongeneelen M, Poon LLM, Alard P, Cornelissen L, et al. (2008) Heterosubtypic Neutralizing Monoclonal Antibodies Cross-Protective against H5N1 and H1N1 Recovered from Human IgM+ Memory B Cells. PLoS ONE 3(12): e3942.; Sui, J., Hwang, W. C., Perez, S., Wei, G., Aird, D., Chen, L., … Marasco, W. A. (2009). Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nature Structural & Molecular Biology, 16(3), 265-273. doi:10.1038/nsmb.1566 ; Wyrzucki, A., Dreyfus, C., Kohler, I., Steck, M., Wilson, I.A., Hangartner, L., 2014. Alternative recognition of the conserved stem epitope in influenza A virus hemagglutinin by a VH3-30-encoded heterosubtypic antibody. J. Virol. 88, 7083-7092. ; Corti, D., Suguitan Jr., A.L., Pinna, D., Silacci, C., Fernandez-Rodriguez, B.M., Vanzetta, F., Santos, C., Luke, C.J., Torres-Velez, F.J., Temperton, N.J., et al., 2010. Heterosubtypic neutralizing antibodies are produced by individuals immunized with a seasonal in- fluenza vaccine. J. Clin. Invest. 120, 1663-1673.; De Marco, Donata et al. “A non-VH1-69 heterosubtypic neutralizing human monoclonal antibody protects mice against H1N1 and H5N1 viruses.” PloS one vol. 7,4 (2012): e34415. doi:10.1371/journal.pone.0034415; Kashyap AK, Steel J, Rubrum A, Estelles A, Briante R, Ilyushina NA, Xu L, Swale RE, Faynboym AM, Foreman PK, Horowitz M, Horowitz L, Webby R, Palese P, Lerner RA, Bhatt RR. Protection from the 2009 H1N1 pandemic influenza by an antibody from combinatorial survivor-based libraries. PLoS Pathog. 2010 Jul 8;6(7):e1000990. doi: 10.1371/journal.ppat.1000990. PMID: 20628565; PMCID: PMC2900296.; Ekiert DC, Friesen RH, Bhabha G, Kwaks T, Jongeneelen M, Yu W, Ophorst C, Cox F, Korse HJ, Brandenburg B, Vogels R, Brakenhoff JP, Kompier R, Koldijk MH, Cornelissen LA, Poon LL, Peiris M, Koudstaal W, Wilson IA, Goudsmit J. A highly conserved neutralizing epitope on group 2 influenza A viruses. Science. 2011 Aug 12;333(6044):843-50. doi: 10.1126/science.1204839. Epub 2011 Jul 7. PMID: 21737702; PMCID: PMC3210727.; Friesen RH, Lee PS, Stoop EJ, Hoffman RM, Ekiert DC, Bhabha G, Yu W, Juraszek J, Koudstaal W, Jongeneelen M, Korse HJ, Ophorst C, Brinkman-van der Linden EC, Throsby M, Kwakkenbos MJ, Bakker AQ, Beaumont T, Spits H, Kwaks T, Vogels R, Ward AB, Goudsmit J, Wilson IA. A common solution to group 2 influenza virus neutralization. Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):445-50. doi: 10.1073/pnas.1319058110. Epub 2013 Dec 11. PMID: 24335589; PMCID: PMC3890827.], до распознавания НА среди обеих групп [Corti, D., Voss, J., Gamblin, S. J., Codoni, G., Macagno, A., Jarrossay, D., … Lanzavecchia, A. (2011). A Neutralizing Antibody Selected from Plasma Cells That Binds to Group 1 and Group 2 Influenza A Hemagglutinins. Science, 333(6044), 850-856. doi:10.1126/science.1205669; Nakamura, G., Chai, N., Park, S., Chiang, N., Lin, Z., Chiu, H., … Swem, L. R. (2013). An In Vivo Human-Plasmablast Enrichment Technique Allows Rapid Identification of Therapeutic Influenza A Antibodies. Cell Host & Microbe, 14(1), 93-103. doi:10.1016/j.chom.2013.06.004; Wu, Ying et al. “A potent broad-spectrum protective human monoclonal antibody crosslinking two haemagglutinin monomers of influenza A virus.” Nature communications vol. 6 7708. 21 Jul. 2015, doi:10.1038/ncomms8708; Tharakaraman, Kannan et al. “A broadly neutralizing human monoclonal antibody is effective against H7N9.” Proceedings of the National Academy of Sciences of the United States of America vol. 112,35 (2015): 10890-5. doi:10.1073/pnas.1502374112; Wyrzucki, A., Bianchi, M., Kohler, I., Steck, M., Hangartner, L., 2015. Heterosubtypic anti- bodies to influenza A virus have limited activity against cell-bound virus but are not impaired by strain-specific serum antibodies. J. Virol. 89, 3136-3144.; Hu, W., Chen, A., Miao, Y., Xia, S., Ling, Z., Xu, K., Wang, T., Xu, Y., Cui, J., Wu, H., et al., 2013. Fully human broadly neutralizing monoclonal antibodies against influenza A viruses generated from the memory B cells of a 2009 pandemic H1N1 influenza vaccine re- cipient. Virology 435, 320-328.; Li, G.M., Chiu, C., Wrammert, J., McCausland, M., Andrews, S.F., Zheng, N.Y., Lee, J.H., Huang, M., Qu, X., Edupuganti, S., et al., 2012. Pandemic H1N1 influenza vaccine in- duces a recall response in humans that favors broadly cross-reactive memory B cells. Proc. Natl. Acad. Sci. U. S. A. 109, 9047-9052.; Henry Dunand, C.J., Leon, P.E., Kaur, K., Tan, G.S., Zheng, N.Y., Andrews, S., Huang, M., Qu, X., Huang, Y., Salgado-Ferrer, M., et al., 2015. Preexisting human antibodies neutralize recently emerged H7N9 influenza strains. J. Clin. Invest. 125, 1255-1268.; Clementi, N., De Marco, D., Mancini, N., Solforosi, L., Moreno, G.J., Gubareva, L.V., Mishin, V., Di Pietro, A., Vicenzi, E., Siccardi, A.G., et al., 2011. A human monoclonal antibody with neutralizing activity against highly divergent influenza subtypes. PLoS One 6, e28001.; Kallewaard, N.L., Corti, D., Collins, P.J., Neu, U., McAuliffe, J.M., Benjamin, E., Wachter- Rosati, L., Palmer-Hill, F.J., Yuan, A.Q., Walker, P.A., et al., 2016. Structure and function analysis of an antibody recognizing all influenza A subtypes. Cell 166, 596-608.; Joyce, M.G., Wheatley, A.K., Thomas, P.V., Chuang, G.Y., Soto, C., Bailer, R.T., Druz, A., Georgiev, I.S., Gillespie, R.A., Kanekiyo, M., et al., 2016. Vaccine-induced antibodies that neutralize group 1 and group 2 influenza A viruses. Cell 166, 609-623.], и даже перекрестной активности между НА типа А и В [Dreyfus, Cyrille et al. “Highly conserved protective epitopes on influenza B viruses.” Science (New York, N.Y.) vol. 337,6100 (2012): 1343-8. doi:10.1126/science.1222908]. Большинство этих моноклональных антител узнают конформационные консервативные эпитопы в стеблевом домене.
Однако моноклональные антитела имеют ряд недостатков, к которым относятся:
- трудоемкость генно-инженерных манипуляций;
- трудности, связанные с узнаванием некоторых «скрытых» эпитопов;
- необходимость внутривенного введения, что влечет за собой дополнительную
нагрузку на систему здравоохранения;
- дорогостоящее производство антител.
Решением проблемы может стать использование однодоменных антител (наноантител), которые в природе можно найти у представителей семейства верблюдовые. [Hamers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hammers, C., Songa, E. B., … Hammers, R. (1993). Naturally occurring antibodies devoid of light chains. Nature, 363(6428), 446-448. doi:10.1038/363446a0]. Благодаря относительно небольшому размеру однодоменные антитела обладают благоприятными биофизическими свойствами и дешевле в производстве, чем стандартные моноклональные антитела. Они могут быть получены с использованием прокариотических или эукариотических систем экспрессии. Несмотря на свой небольшой размер (12-15 кДа), Однодоменные антитела не уступают в аффиности и специфичности каноническим антителам. Благодаря уникальной стабильности в широком диапазоне температур, в присутствии различных детергентов, а также устойчивости к протеолитическому расщеплению, становится возможной доставка однодоменных антител в организм перорально или при помощи ингаляции [Harmsen, M.M., van Solt, C.B., van Zijderveld-van Bemmel, A.M., Niewold, T.A., van Zijderveld, F.G., 2006. Selection and optimization of proteolytically stable llama single-domain antibody fragments for oral immunotherapy. Appl. Microbiol. Biotechnol. 72, 544-551.; Van Heeke, G., Allosery, K., De Brabandere, V., De Smedt, T., Detalle, L., & de Fougerolles, A. (2017). Nanobodies® † †Nanobody is a registered trademark of Ablynx NV. as inhaled biotherapeutics for lung diseases. Pharmacology & Therapeutics, 169, 47-56. doi:10.1016/j.pharmthera.2016.06.012]. Однодоменные антитела нашли применение в терапии онкологических, гематологических, инфекционных и аутоиммунных заболеваний, некоторые из препаратов в данный момент проходят клинические испытания или одобрены для применения в европейских странах и США [Arbabi-Ghahroudi M. Camelid Single-Domain Antibodies: Historical Perspective and Future Outlook. Front Immunol. 2017 Nov 20;8:1589. doi: 10.3389/fimmu.2017.01589. PMID: 29209322; PMCID: PMC5701970; Jovčevska I, Muyldermans S. The Therapeutic Potential of Nanobodies. BioDrugs. 2020 Feb;34(1):11-26. doi: 10.1007/s40259-019-00392-z. PMID: 31686399; PMCID: PMC6985073].
Существует ряд работ по получению однодоменных антител к стеблевому домену НА [Laursen, N. S., Friesen, R. H. E., Zhu, X., Jongeneelen, M., Blokland, S., Vermond, J., … Wilson, I. A. (2018). Universal protection against influenza infection by a multidomain antibody to influenza hemagglutinin. Science, 362(6414), 598-602. doi:10.1126/science.aaq0620; Gaiotto T, Hufton SE. Cross-Neutralising Nanobodies Bind to a Conserved Pocket in the Hemagglutinin Stem Region Identified Using Yeast Display and Deep Mutational Scanning. PLoS One. 2016 Oct 14;11(10):e0164296. doi: 10.1371/journal.pone.0164296. PMID: 27741319; PMCID: PMC5065140.]. Все описанные наноантитела обладают кросс-реактивностью и нейтрализуют вирусы внутри одной филогенетической группы НА или, в результате создания мультиспецифичных конструкций, приобретают способность нейтрализовать вирусы как обеих групп НА типа А, так и типа В.
Известно решение (RU 2683498 C2, 28.03.2019), в котором описано выделенное антитело или его связывающий фрагмент, способные связываться с гемагглютинином вируса гриппа А и нейтрализовать по меньшей мере один подтип группы 1 и по меньшей мере 1 подтип группы 2 вируса гриппа A. Также раскрывается, что антитело или его связывающий фрагмент могут содержать Fc-участок, предпочтительно, антитело представляет собой IgG1, IgG2 или IgG4 или их связывающий фрагмент. Кроме того, антитело или его фрагмент применяют в профилактике или лечении инфекции, вызванной вирусом гриппа A.
Также известно решение (EA 201791735 A1, 30.11.2017), в котором описано однодоменное антитело, способное специфично связываться с гемагглютинином (НА) по меньшей мере двух штаммов вируса гриппа А. При этом однодоменное антитело, связывается с эпитопом в стеблевой области НА. Также однодоменное антитело имеет Fc-концевую часть, которая представляет собой Fc-концевую часть IgG человека.
В работе doi:10.1126/science.aaq0620 были получены рекомбинантные антитела, представляющие собой однодоменные антитела слитые с Fc- фрагментом IgG1 человека и показана возможность их применения для экстренной профилактики гриппозной инфекции. Данное решение, как наиболее близкое к заявляемому, было выбрано авторами патента за прототип. Однако у существующего решения есть недостатки, связанные с возможным явлением антител-зависимого усиления гриппозной инфекции (ADE), связанного с Fc-опосредованными функциями моноклональных антител и в частности связывания с Fc-рецепторами на поверхности иммунокомпетентных клеток (J Med Virol Infection enhancement of influenza A NWS virus in primary murine macrophages by anti-hemagglutinin monoclonal antibody 1992 Mar;36(3):217-21. doi: 10.1002/jmv.1890360312; J Med Virol 1990 Apr;30(4):258-65. Infection enhancement of influenza A H1 subtype viruses in macrophage-like P388D1 cells by cross-reactive antibodies,doi: 10.1002/jmv.1890300406).
Таким образом, в уровне техники существует потребность в создании нового антитела, специфически связывающегося стеблевой домена гемагглютинина типа А, для экстренной профилактики инфекции, вызываемой вирусом гриппа типа А.
Раскрытие сущности изобретения
Технической задачей заявленной группы изобретений является расширение арсенала антител для экстренной профилактики заболеваний, вызываемых вирусом гриппа типа А.
Технический результат заключается в создании однодоменного антитела и его модификаций, которые эффективно связывают стеблевой домен гемагглютинина вируса гриппа типа А, нейтрализуют вирус гриппа типа А и могут быть использованы для экстренной профилактики заболеваний, вызываемых вирусом гриппа типа А. Кроме того, технический результат заключается в том, что разработано однодоменное антитело, слитое с Fc-фрагментом IgG2 человека, который обладает менее выраженной аффинностью к Fc-рецептору человека по сравнению с IgG1 и, таким образом, обеспечивают меньшую вероятность возникновения антител-зависимого усиления инфекции.
Указанный технический результат достигается тем, что создано однодоменное антитело для нейтрализации вирусов, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, и имеющее аминокислотную последовательность SEQ ID NO:1, или SEQ ID NO:2.
Также технический результат достигается тем, что создано олигомеризованное (димеризованное) однодоменное антитело для нейтрализации вирусов, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, содержащее в качестве мономерного блока любой вариант созданного однодоменного антитела, и имеющее конечную аминокислотную последовательность SEQ ID NO:3 или SEQ ID NO:4.
Кроме того, создано рекомбинантное антитело для нейтрализации вирусов, представляющее собой любой вариант созданного однодоменноого антитела, слитоого с Fc-фрагментом иммуноглобулина G2 человека, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, имеющее конечную аминокислотную последовательность SEQ ID NO:5, или SEQ ID NO:6.
А также разработан способ экстренной профилактики заболеваний, вызываемых вирусом гриппа А, заключающийся во введении в организм млекопитающих в эффективном количестве любого созданного антитела.
Краткое описание чертежей
На фиг. 1 представлена электрореграмма анализа препарата стеблевого домена гемагглютинина вируса гриппа А.
1 - образец препарата стеблевого домена гемагглютинина вируса гриппа А в не восстанавливающих условиях;
2 - образец препарата стеблевого домена гемагглютинина вируса гриппа А в восстанавливающих условиях;
М - маркер молекулярного веса.
На фиг. 2 представлены данные по определению титра специфических антител к стеблевому домену гемагглютинина вируса грипп А, в сыворотке крови альпака после иммунизации препаратом рекомбинантного стеблевого домена гемагглютинина вируса гриппа А.
Ось ординат - оптическая плотность раствора при длине волны 450 нм
Ось абсцисс - разведения сыворотки крови альпака
 - Уровень сигнала сыворотки альпака на BSA (отрицательный контроль)
- Уровень сигнала сыворотки альпака на BSA (отрицательный контроль)
 - Уровень специфического сигнала сыворотки альпака на рекомбинантный стеблевой домен гемагглютинина вируса гриппа А
- Уровень специфического сигнала сыворотки альпака на рекомбинантный стеблевой домен гемагглютинина вируса гриппа А
На фиг. 3 представлена электрофореграмма анализа результатов очистки однодоменных антител аффинной хроматографией,
где
1 - препарат однодоменных антитела клон 1;
2 - препарат однодоменных антитела клон 4;
3 - препарат однодоменных антитела клон 5;
4 - препарат однодоменных антитела клон 8;
5 - препарат однодоменных антитела клон 12;
6 - препарат однодоменных антитела клон 17;
7 - препарат однодоменных антитела клон 22;
8 - препарат однодоменных антитела клон 23.
На фиг. 4 представлено схематическое изображение аминокислотной последовательности однодоменных антител,
где
1 - N-конец;
2 - последовательность однодоменного антитела;
3 - His-tag (гистидиновая метка);
4 - C-конец.
На фиг. 5 представлены результаты измерения специфической активности клонов однодоменных антител методом непрямого ИФА.
Ось ординат - оптическая плотность раствора при длине волны 450 нм
Ось абсцисс - разведения клонов
 - Уровень сигнала однодоменных антител на BSA (отрицательный контроль)
- Уровень сигнала однодоменных антител на BSA (отрицательный контроль)
- Уровень специфического сигнала однодоменных антител на стеблевой домен гемагглютинина вируса гриппа А (stem).
Клон 1 - препарат однодоменного антитела клон 1;
Клон 2 - препарат однодоменного антитела клон 4;
Клон 3 - препарат однодоменного антитела клон 5;
Клон 4 - препарат однодоменного антитела клон 8;
Клон 5 - препарат однодоменного антитела клон 12;
Клон 6 - препарат однодоменного антитела клон 17;
Клон 7 - препарат однодоменного антитела клон 22;
Клон 8 - препарат однодоменного антитела клон 23.
На фиг. 6 представлены результаты измерения специфической активности однодоменного антитела (клон 12) на различные подтипы гемагглютинина вируса гриппа А, методом непрямого ИФА.
Ось ординат - оптическая плотность раствора при длине волны 450 нм
Ось абсцисс - клоны однодоменных антител
 - Уровень сигнала однодоменного антитела клон 12 на BSA (отрицательный контроль);
- Уровень сигнала однодоменного антитела клон 12 на BSA (отрицательный контроль);
 - Уровень специфического сигнала однодоменного антитела клон 12 на рекомбинантный гемагглютинин H1N1 (A/California/04/2009);
- Уровень специфического сигнала однодоменного антитела клон 12 на рекомбинантный гемагглютинин H1N1 (A/California/04/2009);
- Уровень специфического сигнала однодоменного антитела клон 12 на рекомбинантный гемагглютинин H5N1 (A/Vietnam/1203/2004)
1 - концентрация однодоменного антитела 1000 нг/мл;
2 - концентрация однодоменного антитела 500 нг/мл;
3 - концентрация однодоменного антитела 250 нг/мл;
4 - концентрация однодоменного антитела 125 нг/мл;
5 - концентрация однодоменного антитела 62,5 нг/мл;
6 - концентрация однодоменного антитела 31,25 нг/мл;
7 - концентрация однодоменного антитела 15,625 нг/мл;
8 - концентрация однодоменного антитела 7,8 нг/мл;
9 - концентрация однодоменного антитела 3,9 нг/мл;
10 - концентрация однодоменного антитела 2 нг/мл;
11 - концентрация однодоменного антитела 1 нг/мл.
На фиг. 7 представлены результаты измерения специфической активности однодоменного антитела (клон 23) на различные подтипы гемагглютинина вируса гриппа А, методом непрямого ИФА.
Ось ординат - оптическая плотность раствора при длине волны 450 нм
Ось абсцисс - клоны однодоменных антител
- Уровень сигнала однодоменного антитела клон 23 на BSA (отрицательный контроль);
- Уровень специфического сигнала однодоменного антитела клон 23 на рекомбинантный гемагглютинин H1N1 (A/California/04/2009);
- Уровень специфического сигнала однодоменного антитела клон 23 на рекомбинантный гемагглютинин H5N1 (A/Vietnam/1203/2004)
1 - концентрация однодоменного антитела 1000 нг/мл;
2 - концентрация однодоменного антитела 500 нг/мл;
3 - концентрация однодоменного антитела 250 нг/мл;
4 - концентрация однодоменного антитела 125 нг/мл;
5 - концентрация однодоменного антитела 62,5 нг/мл;
6 - концентрация однодоменного антитела 31,25 нг/мл;
7 - концентрация однодоменного антитела 15,625 нг/мл;
8 - концентрация однодоменного антитела 7,8 нг/мл;
9 - концентрация однодоменного антитела 3,9 нг/мл;
10 - концентрация однодоменного антитела 2 нг/мл;
11 - концентрация однодоменного антитела 1 нг/мл.
На фиг. 8 представлено схематическое изображение аминокислотной последовательности олигомеризованных однодоменных антител,
где
1 - N-конец;
2 - последовательность однодоменного антитела;
3 - глицин-сериновый линкер;
4 - последовательность однодоменного антитела;
5 - His-tag (гистидиновая метка);
6 - C-конец.
На фиг. 9 представлено схематическое изображение аминокислотной последовательности олигомеризованных однодоменных антител, модифицированных Fc-фрагментом IgG2 человека,
где
1 - глицин-сериновый линкер;
2 - димеризованное через серин-глициновый линкер однодоменное антитело;
2 - шарнирный регион;
3 - Fc-фрагмент IgG2 человека.
На фиг. 10 представлены результаты эксперимента по изучению протективной активности однодоменных антител и их модификаций против летальной дозы вируса гриппа А подтипа H1N1.
Ось ординат - процент выживаемости мышей
Ось абсцисс - дни после введения вируса и антител
 животные, получившие однодоменное антитело клон 12;
животные, получившие однодоменное антитело клон 12;
 животные, получившие однодоменное антитело клон 23;
животные, получившие однодоменное антитело клон 23;
 животные, получившие димеризованное однодоменное антитело 12-dim;
животные, получившие димеризованное однодоменное антитело 12-dim;
 животные, получившие димеризованное однодоменное антитело 23-dim;
животные, получившие димеризованное однодоменное антитело 23-dim;
 животные, получившие рекомбинантное антитело 12-Fc;
животные, получившие рекомбинантное антитело 12-Fc;
 животные, получившие рекомбинантное антитело 23-Fc;
животные, получившие рекомбинантное антитело 23-Fc;
 животные, получившие фосфатно-солевой буфер (отрицательный контроль)
животные, получившие фосфатно-солевой буфер (отрицательный контроль)
На фиг. 11 представлены результаты эксперимента по изучению протективной активности однодоменных антител и их модификаций против летальной дозы вируса гриппа А подтипа H5N2.
Ось ординат - процент выживаемости мышей
Ось абсцисс - дни после введения вируса и антител
животные, получившие однодоменное антитело клон 12;
животные, получившие однодоменное антитело клон 23;
животные, получившие димеризованное однодоменное антитело 12-dim;
животные, получившие димеризованное однодоменное антитело 23-dim;
животные, получившие рекомбинантное антитело 12-Fc;
животные, получившие рекомбинантное антитело 23-Fc;
животные, получившие фосфатно-солевой буфер (отрицательный контроль)
Осуществление изобретения
Вариантом данного изобретения является однодоменное антитело, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, обладающее вируснейтрализующей активностью и имеющее аминокислотную последовательность SEQ ID NO:1, или SEQ ID NO:2.
Для получения данного антитела осуществляли иммунизацию альпака рекомбинантным стеблевым доменом гемагглютинина вируса гриппа А, полученным в клеточной линии СНО и очищенным металл-аффинной и эксклюзионной хроматографией. Эффективность иммунизации подтверждали оценкой титра антител, специфических к стеблевому доменом гемагглютинина вируса гриппа А, в сыворотке крови животного. Далее у животных выделяли мононуклеарные клетки периферической крови, из которых получали РНК. На основе РНК синтезировали кДНК. На матрице, полученной кДНК ставили гнездовую ПЦР, позволяющую амплифицировать последовательности однодоменных антител. Таким образом была получена библиотека ампликонов последовательностей однодоменных антител.
Специалистам в данной области известно, что в целях осуществления настоящего изобретения, возможно использование не только мононуклеарных клеток, а также цельной крови, лимфатических узлов, селезенки, тимуса или других клеток и органов иммунизированного животного, экспрессирующих однодоменные антитела. Также специалистам в данной области известно, что в целях осуществления настоящего изобретения возможно использование синтетических библиотек последовательностей однодоменных антител.
Селекцию антител проводили методом фагового дисплея с использованием паннинга на антигене (рекомбинантный стеблевой домен гемагглютинина вируса гриппа А). Специалистам в данной области известно, что в целях осуществления настоящего изобретения, возможно использование других подходов селекции, таких как паннинг на вирусных частицах, паннинг в растворе биотинилированного антигена, паннинг с использованием конкурентного связывания и элюции. Также специалистам в данной области известно, что в целях осуществления настоящего изобретения возможно использование других методов селекции, таких как рибосомный дисплей, дрожжевой дисплей. В результате селекции были отобраны однодоменные антитела, специфически связывающиеся со стеблевым доменом гемагглютинина вируса гриппа А (SEQ ID NO:1, SEQ ID NO:2). В последующих экспериментах было показано, что данные антитела обладают вируснейтрализующей активностью.
Специалистам в данной области известно, что в целях осуществления настоящего изобретения последовательности однодоменных антител, со стеблевым доменом гемагглютинина вируса гриппа А, могут содержать в себе аминокислотные замены, которые не сказываются на вторичной и третичной структуре однодоменных антител и не влияют на их способность связывать со стеблевым доменом гемагглютинина вируса гриппа А и нейтрализовать вирус гриппа А. Более того, поскольку в связывании с антигеном участвуют только антигенсвязывающие петли (CDR домены) однодоменных антител, то, как известно специалистам в данной области, любой иммуноглобулин или его аналог, содержащий такие же аминокислотные последовательности CDR доменов, попадает под объем настоящего изобретения. Более того, последовательности CDR доменов могут содержать замены/делеции/вставки аминокислот, которые при парном выравнивании аминокислотных последовательностей обеспечивают гомологичность не менее 70% и не оказывают качественного влияния на способность связываться с антигеном. Таким образом, под объем настоящего изобретения попадают все иммуноглобулины или их аналоги, содержащие последовательности CDR доменов, гомология которых составляет не менее 70% с предлагаемыми последовательностями.
Специалистам в данной области известно, что в целях осуществления настоящего изобретения последовательности однодоменных антител, специфических к стеблевому домену гемагглютинина вируса гриппа А, могут быть модифицированы путем присоединения различных белков, белковых доменов и тэгов, таких, например, как НА-тэг, Flag-тэг, GST-тэг, GFP белок, RFP белок, биотин и другие.
Нуклеотидную последовательность однодоменных антител определяли методом секвенирования. Специалистам в данной области известно, что в целях осуществления настоящего изобретения нуклеотидные последовательности, кодирующие однодоменные антитела, могут отличаться, основываясь на принципе вырожденности генетического кода. Таким образом, под объем настоящего изобретения попадают все последовательности нуклеотидов, кодирующие аминокислотные последовательности однодоменных антител SEQ ID NO:1, SEQ ID NO:2. Например, нуклеотидные последовательности SEQ ID NO:7, SEQ ID NO:8.
С помощью методов генной инженерии была получена клетка-продуцент, содержащая нуклеотидную последовательность однодоменного антитела и экспрессирующая однодоменное антитело SEQ ID NO:1, или SEQ ID NO:2. Таким образом, например, были получены: клетка-продуцент на основе Escherichia coli (E.coli), экспрессирующая однодоменное антитело SEQ ID NO:1; клетка-продуцент на основе Escherichia coli (E.coli), экспрессирующая однодоменное антитело SEQ ID NO:2. Специалистам в данной области известно, что в целях осуществления настоящего изобретения в качестве продуцента можно использовать другие про- и эукариотические системы экспрессии, такие как лактобактерии, бациллы, растения, дрожжи, культуры клеток и другие. При необходимости способ получения дополнительно включает в себя выделение и очистку любым способом, известным в данной области.
Другим вариантом данного изобретения является олигомеризованное однодоменное антитело специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, обладающее вируснейтрализующей активностью, содержащее в качестве мономерного блока любой вариант однодоменного антитела, выбранный из SEQ ID NO:1, SEQ ID NO:2. Частными случаями данного варианта изобретения являются: димеризованное однодоменное антитело, в котором мономеры соединены глицин-сериновым линкером (SEQ ID NO 3), димеризованное однодоменное антитело, в котором мономеры соединены глицин-сериновым линкером (SEQ ID NO 4), рекомбинантное антитело, представляющее собой однодоменное антитело, слитое с Fc-фрагментом иммуноглобулина G2 человека (SEQ ID NO 5) и рекомбинантное антитело, представляющее собой однодоменное антитело, слитое с Fc-фрагментом иммуноглобулина G2 человека (SEQ ID NO 6). Специалистам в данной области известно, что в целях осуществления настоящего изобретения, олигомеризацию однодоменных антител можно осуществлять при помощи других линкеров. Олигомеризованное однодоменное антитело было получено путем экспрессии соответствующей нуклеотидной последовательности в клетках-продуцентах. В последующих экспериментах было показано, что данные антитела обладают вируснейтрализующей активностью.
Нуклеотидная последовательность олигомеризованного однодоменного антитела была получена генно-инженерным методом. Специалистам в данной области известно, что в целях осуществления настоящего изобретения нуклеотидные последовательности, кодирующие однодоменные антитела, могут отличаться, основываясь на принципе вырожденности генетического кода. Таким образом, под объем настоящего изобретения попадают все последовательности нуклеотидов, кодирующие аминокислотные последовательности олигомеризованных однодоменных антител, в которых мономерами являются однодоменные антитела (SEQ ID NO:1, или SEQ ID NO:2). Например, нуклеотидные последовательности SEQ ID NO:7, SEQ ID NO:8.
С помощью методов генной инженерии была получена клетка-продуцент, содержащая нуклеотидную последовательность олигомеризованного однодоменного антитела, в котором мономерами являются однодоменные антитела (SEQ ID NO:1, или SEQ ID NO:2) и экспрессирующая олигомеризованное однодоменное антитело, в котором мономерами являются однодоменные антитела (SEQ ID NO:1, или SEQ ID NO:2). Таким образом, например, были получены: клетка-продуцент на основе Escherichia coli (E.coli), экспрессирующая олигомеризованное однодоменное антитело, в котором мономерами являются однодоменные антитела SEQ ID NO:1; клетка-продуцент на основе Escherichia coli (E.coli), экспрессирующая олигомеризованное однодоменное антитело, в котором мономерами являются однодоменные антитела SEQ ID NO:2. Специалистам в данной области известно, что в целях осуществления настоящего изобретения в качестве продуцента можно использовать другие про- и эукариотические системы экспрессии, такие как лактобактерии, бациллы, растения, дрожжи, культуры клеток и другие. При необходимости способ получения дополнительно включает в себя выделение и очистку любым способом, известным в данной области.
Кроме того, вариантом изобретения является рекомбинантное антитело, представляющее собой любой вариант из созданного однодоменное антитело, слитое с Fc-фрагментом иммуноглобуллина G2 человека, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, обладающее вируснейтрализующей активностью, имеющее конечную аминокислотную последовательность SEQ ID NO:5, или SEQ ID NO:6. Данное антитело было получено путем экспрессии нуклеотидной последовательности рекомбинантного антитела в клетках-продуцентах.
Нуклеотидные последовательности рекомбинантных антител были созданы генно-инженерным путем на основе нуклеотидной последовательности однодоменных антител и нуклеотидной последовательности Fc-фрагмента иммуноглобуллина G2 человека. Специалистам в данной области известно, что в целях осуществления настоящего изобретения нуклеотидные последовательности, кодирующие рекомбинантные антитела, могут отличаться, основываясь на принципе вырожденности генетического кода. Таким образом, под объем настоящего изобретения попадают все последовательности нуклеотидов, кодирующие аминокислотные последовательности рекомбинантных антител (SEQ ID NO:5, SEQ ID NO:6). Например, нуклеотидные последовательности SEQ ID NO:11, SEQ ID NO:12.
С помощью методов генной инженерии была получена клетка-продуцент, содержащая нуклеотидную последовательность рекомбинантного антитела и экспрессирующая рекомбинатное антитело SEQ ID NO:5, или SEQ ID NO:6. Таким образом, например, были получены: клетка-продуцент на основе клеточной линии СНО, экспрессирующая рекомбинатное антитело SEQ ID NO:5; клетка-продуцент на основе клеточной линии СНО, экспрессирующая рекомбинатное антитело SEQ ID NO:6. Специалистам в данной области известно, что в целях осуществления настоящего изобретения в качестве продуцента можно использовать другие эукариотические системы экспрессии. При необходимости способ получения дополнительно включает в себя выделение и очистку любым способом, известным в данной области.
Изучена фармакокинетика однодоменных антител и их модификаций. Продемонстрировано что модификация однодоменных антител и их олигомеров Fc-фрагментом IgG2 человека, позволяет увеличить время их циркуляции в организме с нескольких часов до нескольких недель.
Изучена аффинность рекомбнинатных антител, слитых с Fc-фрагментом IgG2 человека с рекомбинантными Fc-рецепторами. Продемонстрировано, что рекомбинантные антитела, слитые с Fc-фрагментом IgG2 человека, обладают меньшей аффинностью к рекомбинантным Fc-рецепторам по сравнению IgG1 человека. Таким образом, модификация однодоменных антител Fc-фрагментом IgG2 человека, позволяет снизить риск возникновения антител-зависимого усиления инфекции.
Таким образом, авторами патента были получены варианты антител, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, обладающее вируснейтрализующей активностью, обладающие улучшенной фармакокинетикой по сравнению с однодоменными антителами, а также обладающими пониженной аффинностью к Fc-рецепторам, что обуславливает снижения риска возникновения антител-зависимого усиления инфекции.
Кроме того, авторами патента разработан способ экстренной профилактики заболеваний, вызываемых вирусом гриппа А, заключающийся во введении в организм млекопитающих в эффективном количестве любого из разработанных вариантов антител.
Осуществление изобретения подтверждается следующими примерами.
Пример 1. Получение стеблевого домена гемагглютинина вируса гриппа А
Большинство моноклональных антител, способных нейтрализовать вирусы гриппа А разных подтипов, узнают консервативные конформационные эпитопы в стеблевом домене гемагглютинина, однако, в процессе гриппозной инфекции или иммунизации полноразмерным гемагглютинином, они трудно доступны, в следствии экспонирования преимущественно вариабельных эпитопов глобулярного домена гемагглютинина. Таким образом, для иммунизации и отбора антител широкого спектра действия, необходимо получить стеблевой домен в оптимальной стабилизированной конформации с сохранением эпитопов нейтрализующих антител.
Аминокислотную последовательность тримеризующегося стеблевого домена вируса гриппа А штамма H1N1 (A/Brisbane/59/2007), модифицированную гистидиновой меткой синтезированную в ЗАО «Евроген» (Россия) и клонировали в плазмиду pShuttle-CMV (Stratagene, США) и получали, таким образом, плазмиду pShuttle-CMV-HAstem. Далее, культуру клеток CHO-S (Thermo Fisher Scientific, США) транзиентно трансфецировали плазмидой pShuttle-CMV- HAstem с использованием системы CHO Gro (Mirus Bio, США) в соответствии с протоколом производителя. Клетки культивировали в колбах Эрленмейера при 125 об/мин, 5% СО2, 80% влажности, 37°C, затем, спустя 24 часа, снижали температуру до 32°C и продолжали культивирование в течение 10 дней. Начиная с 3-го дня, добавляли подпитки Cell boosts 7a (2%), 7b (0.2%) (HyClone, США), и 0,5% CHO Bioreactor Feed (Sigma, США) один раз в день. По истечении 10 дней культивирования, культуральную жидкость осветляли центрифугированием при 5000g. Рекомбинантный стеблевой домен очищали металл-аффинной хроматографией на системе AKTA start (Cytiva, Швеция), используя колонки HisTrap 1 мл (Cytiva, Швеция) в соответствии с протоколом производителя. Дополнительную очистку и замену буфера на 20 мМ фосфат натрия, 150 мМ натрия хлорид проводили на колонке ХК 26/100 (Cytiva, Швеция), упакованной сорбентом Superdex 200 pg (Cytiva, Швеция). Полученный препарат анализировали при помощи белкового электрофореза в восстанавливающих и не восстанавливающих условиях. Результаты анализа представлены на фигуре 1. На электрофореграмме видно, что в не восстанавливающих условиях образец имеет молекулярную массу свыше 75 кДа, что соответствует тримереризованной форме. В восстанавливающих условиях образец имеет молекулярную массу около 30 кДа, что соответствует мономеру.
Таким образом, был получен препарат стеблевого домена гемагглютинина вируса гриппа А. В результате анализа препарата методом электрофореза в полиакриламидном геле, подтверждена его подлинность и способность тримеризоваться.
Пример 2. Получение иммунной библиотеки кДНК однодоменных антител альпака.
На следующем этапе работы альпака (Lama pacos - представитель семейства Cameliedae) иммунизировали полученным рекомбинантным стеблевым доменом гемагглютинина вируса гриппа А (100мкг на иммунизацию) 5 раз с промежутками в 10-14 дней. Через неделю после последней иммунизации определили титр антител в сыворотке крови альпака, который составил 1/12500 (фигура 2).
Через 7 дней после последней иммунизации у альпака отбирали 50,0 мл периферической крови и выделяли мононуклеарные клетки на градиенте фиколла 1,077 (Панэко, Россия). Изолированные мононуклеарные клетки использовали для выделения тотальной РНК реагентом Trizol (Invitrogene, США) согласно протоколу фирмы-производителя. Выделенную РНК использовали в качестве матрицы для синтеза кДНК со случайными праймерами и ревертазой SuperScriptIII (Invitrogene, США). На матрице, полученной кДНК, ставили гнездовую ПЦР, позволяющую амплифицировать последовательности однодоменных антител. Для этой процедуры использовали высокоточную полимеразу Q5 (NEB, Великобритания) и специфические праймеры, содержащие на концах сайты рестрикции.
Таким образом, в результате проведенной работы были получены ампликоны, которые являются иммунной библиотекой кДНК однодоменных антител альпака.
Пример 3. Получение библиотеки рекомбинантных бактерифагов.
Ампликоны, полученные в примере 2 гидролизовали рестриктазами и клонировали в фагмидный вектор, гидролизованный по тем же сайтам. Затем трансформировали клетки E.coli (штамм TG1) фагмидными векторами, полученными в результате клонирования.
Суспензию бактериальных клонов-трансформантов клеток E.coli (штамм TG1) использовали для продукции рекомбинантных бактериофагов с использованием бактериофага-помощника M13KO7 (NEB, Великобритания) согласно протоколу фирмы-производителя. Рекомбинантные бактериофаги осаждали путем преципитации полиэтиленгликолем (PEG/NaCl).
Таким образом, получена библиотека рекомбинантных бактерифагов с титром 1012 трансдуцирующих единиц/мл суспензии.
Пример 4. Селекция и определение последовательности специфических однодоменных антител.
Селекцию специфических рекомбинантных бактериофагов осуществляли путем паннинга на антигене исходной библиотеки бактериофагов. Для этого рекомбинантный стеблевой домен гемагглютинина вируса гриппа А (пример 1) иммобилизовали в лунке иммунологического планшета в 50,0 мМ карбонатно-бикарбонатном буфере в течение ночи. Не связавшийся белок удаляли, а лунку блокировали 5% раствором сухого молока в фосфатном буфере в течение 1 часа при комнатной температуре. Затем добавляли суспензию 1011 рекомбинантных бактерифагов и инкубировали 1 час при комнатной температуре. Не связавшиеся бактерифаги отмывали коллоидным 0,1% раствором детергента Твин-20 в фосфатном буфере. Связавшиеся бактериофаги элюировали при помощи раствора трипсина (0,1 мг/мл). Элюированные бактериофаги использовали для трансдукции клеток E.coli TG1, которые затем высевали на агаризованные чашки в разведении, позволяющем изолировать индивидуальные колонии. Полученные колонии использовали для наращивания бактериальной массы и выделения плазмидной ДНК. Плазмиды секвенировали для определения нуклеотидных последовательностей, кодирующих однодоменные антитела. Было определено 8 индивидуальных последовательностей.
Таким образом, в результате было получено 8 индивидуальных последовательностей однодоменных антител специфичных к стеблевому домену гемагглютинина вируса гриппа А.
Пример 5. Продукция и очистка однодоменных антител.
Для продукции однодоменных антител к интенсивно делящимся клеткам E.coli TG1, полученных из колоний на предыдущем этапе, добавляли 1,0 мкМ изопропил тиогалактопиранозид (IPTG) (Хеликон, Россия), спустя 4 часа бактериальную массу использовали для выделения рекомбинантных белков на колонке HisTrap FF 5 мл («GE Healthcare Life Sciences», США) с использованием хроматографической системы AKTA start («GE Healthcare Life Sciences», США), согласно протоколу фирмы-производителя. Результаты хроматографической очистки однодоменных антител представлены на фигуре 3. Данный пример демонстрирует простоту и эффективность продукции и очистки однодоменных антител в бактериальной системе экспрессии.
Таким образом были получены очищенные препараты однодоменных антител клоны 1, 4, 5, 8, 12, 17, 22, 23. Была подтверждена их чистота и подлинность методом вертикального электрофореза в полиакриламидном геле. Схематичное изображение аминокислотных последовательностей однодоменных антител представлено на фигуре 4.
Пример 6. Определение специфической активности очищенных клонов однодоменных антител в непрямом ИФА.
Для определения специфической активности 8 очищенных клонов однодоменных антител применяли метод непрямого ИФА. Для этого рекомбинантный стеблевой домен гемагглютинина иммобилизовали в лунке иммунологического планшета в 50,0 мМ карбонатно-бикарбонатном буфере в течение ночи. Не связавшийся белок удаляли, а лунку блокировали 5% раствором сухого молока в фосфатном буфере в течение 1 часа при комнатной температуре. Затем лунки промывали 0,1% раствором детергента Твин-20 в фосфатном буфере и добавляли раствор 5% раствором сухого молока в фосфатном буфере, содержащим очищенные препараты отобранных антител 1 мкг/мл. Каждый препарат антител вносили в 3-х повторах, инкубировали 1 час при 37°С при слабом перемешивании. Затем проводили 3-х кратную отмывку 0,1% раствором детергента Твин-20 в фосфатном буфере. После отмывки вносили раствор конъюгата, меченного перексидазой и инкубировали 1 час при 37°С при слабом перемешивании. Далее проводили 5-х кратную отмывку 0,1% раствором детергента Твин-20 в фосфатном буфере. Далее вносили раствор субстрата ТМБ и инкубировали 10 мин при комнатной температуре без перемешивания, после чего реакцию останавливали добавлением раствора серной кислоты и проводили измерение на спектрофотометре. В результате было продемонстрировано, что 3 клона из 8 обладают наиболее выраженной специфической активностью в непрямом ИФА (фигура 4).
Таким образом, 3 наиболее специфичных в ИФА клона однодоменных антител (клон 12, 22 и 23) были отобраны в дальнейшее исследование.
Пример 7. Определение констант взаимодействия однодоменных антител.
Константы взаимодействия (KD) однодоменных антител клоны 12, 22 и 23 определяли путем детекции изменения показателей поверхностного плазмонного резонанса на приборе Biacore3000 (General Electric, Швеция). Для этого рекомбинантный стеблевой домен гемагглютинина ковалентно иммобилизировали на поверхности декстранового матрикса чипа СМ5 (General Electric, Швеция), а затем пропускали над поверхностью чипа различные концентрации полученных однодоменных антител. Обработку данных и вычисление констант проводили в автоматическом режиме при помощи программы Bioevaluation (General Electric, Швеция). Полученные данные представлены в таблице 1
Полученные данные демонстрируют высокие значения констант равновесных диссоциаций отобранных однодоменных антител с рекомбинантным стеблевым доменом гемагглютинина вируса гриппа А.
Таким образом, полученные данные свидетельствуют, что наибольшей аффинностью к рекомбинантному стеблевому домену гемагглютинина вируса гриппа А обладают клоны 12 и 23, которые были взяты в дальнейшее исследование.
Пример 8. Изучение кросс-реактивности отобранных клонов однодоменных антител в непрямом ИФА.
Способность отобранных клонов однодоменных антител взаимодействовать с гемагглютининами различных подтипов вируса гриппа А изучали в непрямом ИФА.
Для этого рекомбинантные полноразмерные гемагглютинины штаммов вируса гриппа H1N1 (A/California/04/2009), H5N1 (A/Vietnam/1203/2004) иммобилизовали в лунке иммунологического планшета в 50,0 мМ карбонатно-бикарбонатном буфере в течение ночи. Не связавшийся белок удаляли, а лунку блокировали 5% раствором сухого молока в фосфатном буфере в течение 1 часа при комнатной температуре. Затем лунки промывали 0,1% раствором детергента Твин-20 в фосфатном буфере и добавляли раствор 5% раствором сухого молока в фосфатном буфере, содержащим очищенные препараты отобранных антител 1 мкг/мл. Каждый препарат антител вносили в различных разведениях, инкубировали 1 час при 37°С при слабом перемешивании. Затем проводили 3-х кратную отмывку 0,1% раствором детергента Твин-20 в фосфатном буфере. После отмывки вносили раствор конъюгата, меченного перексидазой и инкубировали 1 час при 37°С при слабом перемешивании. Далее проводили 5-х кратную отмывку 0,1% раствором детергента Твин-20 в фосфатном буфере. Далее вносили раствор субстрата ТМБ и инкубировали 10 мин при комнатной температуре без перемешивания, после чего реакцию останавливали добавлением раствора серной кислоты и проводили измерение на спектрофотометре. Результаты непрямого ИФА представлены на фигурах 6 и 7. В результате было продемонстрировано, что оба препарата способны взаимодействовать с гемагглютининами вируса гриппа А подтипов H1N1 и H5N1.
Таким образом, продемонстрировано, что отобранные клоны антител 12 и 23 обладают кросс-реактивностью к различным подтипам вируса гриппа А.
Пример 9. Получение, продукция и очистка олигомеризованных однодоменных антител.
Данный пример иллюстрирует получение олигомеризованных однодоменных антител, в частности димеризованных однодоменных антител 12-dimer (SEQ ID NO:3), и 23-dimer (SEQ ID NO:4). На первом этапе разработали дизайн нуклеотидной последовательности димеризованных антител, которые представляют собой две аминокислотные последовательности однодоменного антитела (SEQ ID NO:1 или SEQ ID NO:2, соединенные через глицин-сериновый линкер (GGGGSGGGGSGGGGSGGGGS). Нуклеотидные последовательности димеризованных антител были синтезированы компанией ЗАО «Евроген». Полученные нуклеотидные последовательности клонировали в экспрессионный вектор pET30a, получая таким образом плазмидные векторы pET30a-12-dimer, и pET30a-23-dimer. Полученными экспрессионными векторами были трансформированы клетки E.coli штамма BL21, экспрессия была индуцирована путем добавления 1,0 мкМ IPTG (Хеликон, Россия). Спустя 4 часа из бактериальной массы выделяли димеризованные однодоменные антитела на колонке HisTrap FF 5 мл («GE Healthcare Life Sciences», США) с использованием хроматографической системы AKTA start («GE Healthcare Life Sciences», США), согласно протоколу фирмы-производителя. Так были полученны препараты димеризованных однодоменных антител 12-dimer (SEQ ID NO:3) и 23-dimer (SEQ ID NO:4). Схематичное изображение аминокислотных последовательностей димеризованных однодоменных антител представлено на фигуре 8.
Таким образом, в результате проведенной работы были получены олигомеризованные однодоменные антитела, мономерами которых являются разработанные однодоменные антитела (SEQ ID NO:1 или SEQ ID NO:2).
Пример 10. Получение, продукция и очистка рекомбинантных антител.
Рекомбинатное антитело представляет собой однодоменное антитело, модифицированное Fc-фрагментом иммуноглобуллина человека. Данная модификация однодоменных антител позволит улучшить их фармакокинетические свойства за счет добавления Fc-фрагмента IgG2 человека и его правильного гликозилирования в эукариотических клетках-продуцентах. Кроме того, модификация Fc-фрагментом иммуноглобуллина человека IgG2, позволит обеспечить низкую аффинность к Fc-рецепторам и, таким образом, снизить вероятность возникновения антител-зависимого усиления инфекции.
На первом этапе работы разработали дизайн аминокислотной последовательности рекомбинатного антитела (SEQ ID NO:5, или SEQ ID NO:6), которое представляет собой однодоменное антитело (SEQ ID NO:1, или SEQ ID NO:2), модифицированное Fc-фрагментом иммуноглобулина G2 человека. На основе аминокислотной последовательности были получены нуклеотидные рекомбинатных антител, которые были синтезированы в компании ЗАО «Евроген». Полученные нуклеотидные последовательности клонировали в вектор для экспрессии в эукариотических клетках. Далее проводили трансфекцию клеток линии СНО полученными экспрессионными векторами с использованием системы CHO Gro (Mirus Bio, США) в соответствии с протоколом производителя. Клетки культивировали в колбах Эрленмейера в течение 10 дней. После чего культуральную жидкость осветляли центрифугированием при 5000g. Антитело очищали аффинной хроматографией на системе AKTA start (Cytiva, Швеция), используя колонки MAbSelect SuRe 1 мл (Cytiva, Швеция) в соответствии с протоколом производителя. Дополнительную очистку и замену буфера проводили на колонке ХК 26/100 (Cytiva, Швеция), упакованной сорбентом Superdex 200 pg (Cytiva, Швеция). Чистоту полученных рекомбинатных антител 12-Fc (SEQ ID NO:5), 23-Fc (SEQ ID NO:6), определяли методом вертикального электрофореза в полиакриламидном геле в не денатурирующих условиях и денатурирующих условиях. Схематичное изображение аминокислотной последовательности рекомбинантных антител представлено на фигуре. 9.
Таким образом, в результате проведенной работы было получено рекомбинатное антитело, представляющее собой разработанное однодоменное антитело, модифицированное Fc-фрагментом иммуноглобулина G2 человека, имеющее конечную аминокислотную последовательность SEQ ID NO:5, или SEQ ID NO:6.
Пример 11. Определение аффинности рекомбинатных антител к рекомбинантным Fc-рецепторам.
Целью данного эксперимента являлось изучение аффинности полученных рекомбинатных антител к рекомбинантным Fc-рецепторам, для оценки потенциальной возможности антител-зависимого усиления инфекции. Эксперимент проводили при помощи определения констант равновесных диссоциацийрекомбинатных антител 12-Fc и 23-Fc c рекомбинантными Fc-рецепторами FcRn, CD64A, CD16a, CD32a, CD32b, используя систему оценки молекулярных взаимодействий Octet Red 96 (ForteBio) и биосенсоры Ni-NTA (ForteBio). Анализ проводили в 96-луночных черных плоскодонных полипропиленовых микропланшетах (Greiner Bio-One part no. 655209). Белки FcRn, CD64A, CD16a, CD32a, CD32b (10 мкг/мл) загружали на сенсоры в 1х кинетическом буфере (ФСБ с 0,002% Tween-20 и 1 мг/мл BSA). Связывание 12-Fc и 23-Fc и контрольного антитела (антитело человека IgG1) в концентрациях 100, 50, 25 и 0 мкг/мл осуществляли также осуществляли в 1X кинетическом буфере. Для каждого белка и для исследуемых и контрольного образца антител проводили отдельный анализ. Программное обеспечение Data Analysis 10.0 было использовано для оценки констант диссоциации антител и белков. Схема анализа приведена в таблице 2.
Данные по результатам анализа констант равновесных диссоциации (KD) для рекомбинатных антител 12-Fc и 23-Fc и контрольного антитела человека с изотипом IgG1 с белками Fc-рецепторов человека представлены в таблице 3.
В результате была продемонстрирована высокая аффинность Fc-фрагмента антитела 12-Fc и 23-Fc к рецептору FcRn человека и отсутствие взаимодействия с белками Fc-рецепторов CD64A, CD16a, CD32a, CD32b, в то время как контрольное антитело человека с изотипом IgG1 облает достаточно высокой аффинностью ко всем перечисленным белкам.
Таким образом, была продемонстрирована высокая аффинность рекомбинантных антител к неонатальному рецептору FcRn человека и отсутствие взаимодействия с рецепторами CD64A, CD16a, CD32a, CD32b, что означает низкую вероятность взаимодействия рекомбинантных антител 12-Fc и 23-Fc с иммунокомпетентными клетками и как следствие низкую вероятность возникновения антител-зависимого усиления инфекции. Кроме того, взаимодействие антител с FcRn, обуславливает их защиту от деградации и обеспечивает продолжительный период их циркуляции в организме (Roopenian, D., Akilesh, S. FcRn: the neonatal Fc receptor comes of age. Nat Rev Immunol 7,715-725 (2007). https://doi.org/10.1038/nri2155).
Пример 12. Способ экстренной профилактики заболеваний, вызываемых вирусом гриппа А.
Целью данной работы являлась разработка способа экстренной профилактики заболеваний, вызываемый вирусом гриппа А.
С этой целью использовали модель летальной инфекции мешей адаптированными вирусами гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018) и H5N2 (A/Mallard duck/Pennsylvania/10218/84). Животных случайным образом распределяли по 5 особей в каждую группу (опытные и контрольные) и заражали интраназально предварительно инкубированными 1 ч при 37°С смесью антител (200 мкг) и вируса (15LD50) в объёме 50 мкл/мышь (в случае опытных групп) или вирусом в дозе 15LD50 в ФСБ (для групп контроля). За мышами наблюдали в течение 14 дней после инфицирования, ежедневно осматривали и взвешивали. Таким образом, в экспериментах участвовали следующие группы животных:
1) однодоменное антитело 12 + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
2) однодоменное антитело 23 + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
3) димеризованное однодоменное антитело 12-dim + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
4) димеризованное однодоменное антитело 23-dim + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
5) рекомбинатное антитело 12-Fc + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
6) рекомбинатное антитело 23-Fc + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
7) отрицательный контроль, ФСБ + вирус гриппа H1N1 (A/Duck: mallard/Moscow/4970/2018);
8) однодоменное антитело 12 + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
9) однодоменное антитело 23 + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
10) димеризованное однодоменное антитело 12-dim + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
11) димеризованное однодоменное антитело 23-dim + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
12) рекомбинатное антитело 12-Fc + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
13) рекомбинатное антитело 23-Fc + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
14) отрицательный контроль, ФСБ + вирус гриппа H5N2 (A/Mallard duck/Pennsylvania/10218/84);
Полученные данные представлены на фигурах 10 и 11.
В результате эксперимента в опытных группах наблюдалась 100% выживаемость и 100% гибель животных в контрольных группах, что говорит об эффективной нейтрализации вирусов H1N1 и H5N2 однодоменными антитела и их модификациями.
Полученные результаты свидетельствуют о том, что введение препаратов однодоменных антител и их модификаций позволяет защитить млекопитающих от клинических проявлений инфекции, вызванной вирусом гриппа А.
Таким образом, в результате проведенных экспериментов был разработан способ экстренной профилактики заболеваний, вызываемых вирусом гриппа А, заключающийся во введении в организм млекопитающих в эффективном количестве любого созданного однодоменного антитела или его модификаций.
Исходя из экспериментальных данных и данных других исследований по применению однодоменных и моноклональных антител для терапии инфекционных заболеваний в том числе вируса гриппа А (https://science.sciencemag.org/content/sci/suppl/2018/10/31/362.6414.598.DC1/aaq0620-Laursen-SM.pdf, ), антитела могут вводиться человеку в концентрации от 1 до 100 мг/мл.
Промышленная применимость
Все приведенные примеры подтверждают эффективность созданных однодоменных антител и их модификаций, обладающих протективными свойствами против вируса гриппа А и их промышленную применимость.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1.
<110> Федеральное Государственное Бюджетное Учреждение
«Национальный исследовательский центр эпидемиологии и
микробиологии имени почетного академика Н.Ф. Гамалеи»
Министерства здравоохранения Российской Федерации
<120>Однодоменное антитело для нейтрализации вирусов
и его модификации, и способ их применения для экстренной
профилактики заболеваний, вызываемых вирусом гриппа А.
<160> 1
<170> BiSSAP 1.3.6
<210> 1
<211> 136
<212> PRT
<213> Vicugnapacos
<220>
<223> аминокислотная последовательность однодоменного антитела
клон 12
<400> 1
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Gly Phe Ser Val Phe
20 25 30
Ala Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Gly Ile Ser Ser Asp Phe Ser Thr Ser Tyr Thr Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Lys
85 90 95
Gln Ser Val Asp Gly Lys Ser Leu Asn Ile Gly Ser Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser Ala Ala Ala Glu Gln Lys Leu Ile Ser
115 120 125
Glu Glu Asp Leu Asn Gly Ala Ala
130 135 140
SEQ ID NO:2.
<210> 2
<211> 141
<212> PRT
<213> Vicugna pacos
<220>
<223> аминокислотная последовательность однодоменного антитела клон
23
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Ile Phe Arg Thr Tyr
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Arg Ile Ser Trp Ser Gly Gly Ser Thr Asn Tyr Ala Asp Phe Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Asn Ala Asp Pro Ser Ser Val Val Gly Ala Ile Gly Ala Gly Tyr Val
100 105 110
Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Ala Ala Glu
115 120 125
Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Gly Ala Ala
130 135 140
SEQ ID NO:3.
<210> 3
<211> 275
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность олигомеризованного
однодоменного антитела клон 12 (12-dim)
<400> 1
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Gly Phe Ser Val Phe
20 25 30
Ala Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Gly Ile Ser Ser Asp Phe Ser Thr Ser Tyr Thr Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Lys
85 90 95
Gln Ser Val Asp Gly Lys Ser Leu Asn Ile Gly Ser Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val
130 135 140
Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Ser Gly Phe Ser Val Phe Ala Met Gly Trp Tyr
165 170 175
Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val Ala Gly Ile Ser Ser
180 185 190
Asp Phe Ser Thr Ser Tyr Thr Asp Ser Val Lys Gly Arg Phe Thr Ile
195 200 205
Ser Arg Asp Asn Ala Arg Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
210 215 220
Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Lys Gln Ser Val Asp Gly
225 230 235 240
Lys Ser Leu Asn Ile Gly Ser Trp Gly Gln Gly Thr Gln Val Thr Val
245 250 255
Ser Ser Ala Ala Ala Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn
260 265 270
Gly Ala Ala
275 280 285
SEQ ID NO:4.
<210> 4
<211> 285
<212> PRT
<213> ArtificialSequence
<220>
<223> аминокислотная последовательность олигомеризованного
однодоменного антитела клон 23 (23-dim)
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Ile Phe Arg Thr Tyr
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Arg Ile Ser Trp Ser Gly Gly Ser Thr Asn Tyr Ala Asp Phe Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Asn Ala Asp Pro Ser Ser Val Val Gly Ala Ile Gly Ala Gly Tyr Val
100 105 110
Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
145 150 155 160
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Ile Phe Arg Thr Tyr
165 170 175
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
180 185 190
Ala Arg Ile Ser Trp Ser Gly Gly Ser Thr Asn Tyr Ala Asp Phe Val
195 200 205
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Thr Val Tyr
210 215 220
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Phe Cys
225 230 235 240
Asn Ala Asp Pro Ser Ser Val Val Gly Ala Ile Gly Ala Gly Tyr Val
245 250 255
Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Ala Ala Glu
260 265 270
Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Gly Ala Ala
275 280 285
SEQ ID NO:5.
<210> 5
<211> 347
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность вьюжно антитела 12-Fc,
представляющего собой однодоменное антитело клон 12,
модифицированное Fc-фрагментом IgG2 человека
<400> 1
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Gly Phe Ser Val Phe
20 25 30
Ala Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Gly Ile Ser Ser Asp Phe Ser Thr Ser Tyr Thr Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Lys
85 90 95
Gln Ser Val Asp Gly Lys Ser Leu Asn Ile Gly Ser Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser Glu Arg Lys Cys Cys Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
SEQ ID NO:6.
<210> 6
<211> 352
<212> PRT
<213> ArtificialSequence
<220>
<223> аминокислотная последовательность фьюжн антитела 23-Fc,
представляющего собой однодоменное антитело клон 23,
модифицированное Fc-фрагментом IgG2 человека
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Ile Phe Arg Thr Tyr
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Arg Ile Ser Trp Ser Gly Gly Ser Thr Asn Tyr Ala Asp Phe Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Asn Ala Asp Pro Ser Ser Val Val Gly Ala Ile Gly Ala Gly Tyr Val
100 105 110
Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Glu Arg Lys Cys
115 120 125
Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser
130 135 140
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
145 150 155 160
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
165 170 175
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
180 185 190
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val
195 200 205
Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
210 215 220
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr
225 230 235 240
Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
245 250 255
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
260 265 270
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser
275 280 285
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp
290 295 300
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
305 310 315 320
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
325 330 335
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
SEQ ID NO:7.
<400> 1
<210> 7
<211> 408
<212> DNA
<213> Vicugnapacos
<220>
<223> нуклеотидная последовательность однодоменного антитела
клон 12
<400> 1
CAGGTGCAGC TGGTGCAGTC TGGGGGAGGC TTGGTGCAGC CTGGGGGGTC TCTGAGACTC 60
TCCTGTGCAG CCTCTGGAAG CGGGTTCAGT GTCTTTGCCA TGGGCTGGTA CCGCCAGGCT 120
CCAGGGAAGC AGCGCGAGTT GGTCGCAGGT ATTTCTAGTG ATTTTAGCAC AAGTTATACA 180
GACTCCGTGA AGGGCCGATT CACCATCTCC AGAGACAACG CCCGGAACAC GGTGTATCTG 240
CAAATGAACA GCCTGAAACC TGAGGATACG GCCGTATATT ACTGTAAACA ATCGGTAGAT 300
GGCAAGTCTC TAAATATTGG TTCCTGGGGC CAGGGGACCC AGGTCACTGT CTCCTCAGCG 360
GCCGCAGAAC AAAAACTCAT CTCAGAAGAG GATCTGAATG GGGCCGCA 408
SEQ ID NO:8.
<210> 8
<211> 423
<212> DNA
<213> Vicugnapacos
<220>
<223> нуклеотидная последовательность однодоменного антитела
клон 23
GAGGTGCAGCTGGTGGAGTCGGGGGGAGGATTGGTGCAGGCTGGGGGCTCTCTGAGACTC 60
TCCTGTGCAGCCTCTGGACGCATTTTCCGAACGTATGACATGGGCTGGTACCGCCAGGCT 120
CCAGGGAAGGAGCGTGAGTTTGTCGCACGAATTAGTTGGAGTGGTGGTAGCACAAACTAT 180
GCAGACTTCGTGAAGGGCCGGTTCACCATCTCCAAAGACAGCGCCAAGAACACGGTGTAT 240
CTGCAAATGAACAGCCTGAAACCTGAGGACACGGCCATGTATTTCTGTAATGCAGATCCC 300
TCCAGCGTCGTTGGGGCAATCGGCGCGGGATATGTCTACTGGGGCCAGGGGACCCAGGTC 360
ACCGTCTCCTCAGCGGCCGCAGAACAAAAACTCATCTCAGAAGAGGATCTGAATGGGGCC 420
GCA 423
SEQ ID NO:9.
<210> 9
<211> 825
<212> DNA
<213> Vicugnapacos
<220>
<223> нуклеотидная последовательность олигомеризованного
однодоменного антитела 12-dim
CAGGTTCAGC TGGTTCAGTC TGGTGGTGGT CTGGTTCAGC CGGGTGGTTC TCTGCGTCTG 60
TCTTGCGCGG CGTCTGGTTC TGGTTTCTCT GTTTTCGCGA TGGGTTGGTA CCGTCAGGCG 120
CCGGGTAAAC AGCGTGAACT GGTTGCGGGT ATCTCTTCTG ACTTCTCTAC CTCTTACACC 180
GACTCTGTTA AAGGTCGTTT CACCATCTCT CGTGACAACG CGCGTAACAC CGTTTACCTG 240
CAGATGAACT CTCTGAAACC GGAAGACACC GCGGTTTACT ACTGCAAACA GTCTGTTGAC 300
GGTAAATCTC TGAACATCGG TTCTTGGGGT CAGGGTACCC AGGTTACCGT TTCTTCTGGT 360
GGTGGTGGTT CTGGTGGTGG TGGTTCTGGT GGTGGTGGTT CTGGTGGTGG TGGTTCTCAG 420
GTTCAGCTGG TTCAGTCTGG TGGTGGTCTG GTTCAGCCGG GTGGTTCTCT GCGTCTGTCT 480
TGCGCGGCGT CTGGTTCTGG TTTCTCTGTT TTCGCGATGG GTTGGTACCG TCAGGCGCCG 540
GGTAAACAGC GTGAACTGGT TGCGGGTATC TCTTCTGACT TCTCTACCTC TTACACCGAC 600
TCTGTTAAAG GTCGTTTCAC CATCTCTCGT GACAACGCGC GTAACACCGT TTACCTGCAG 660
ATGAACTCTC TGAAACCGGA AGACACCGCG GTTTACTACT GCAAACAGTC TGTTGACGGT 720
AAATCTCTGA ACATCGGTTC TTGGGGTCAG GGTACCCAGG TTACCGTTTC TTCTGCGGCG 780
GCGGAACAGA AACTGATCTC TGAAGAAGAC CTGAACGGTG CGGCG 825
SEQ ID NO:10.
<210> 10
<211> 855
<212> DNA
<213> ArtificialSequence
<220>
<223> нуклеотидная последовательность олигомеризованного
однодоменного антитела 23-dim
<400>
GAAGTTCAGC TGGTTGAATC TGGTGGTGGT CTGGTTCAGG CGGGTGGTTC TCTGCGTCTG 60
TCTTGCGCGG CGTCTGGTCG TATCTTCCGT ACCTACGACA TGGGTTGGTA CCGTCAGGCG 120
CCGGGTAAAG AACGTGAATT CGTTGCGCGT ATCTCTTGGT CTGGTGGTTC TACCAACTAC 180
GCGGACTTCG TTAAAGGTCG TTTCACCATC TCTAAAGACT CTGCGAAAAA CACCGTTTAC 240
CTGCAGATGA ACTCTCTGAA ACCGGAAGAC ACCGCGATGT ACTTCTGCAA CGCGGACCCG 300
TCTTCTGTTG TTGGTGCGAT CGGTGCGGGT TACGTTTACT GGGGTCAGGG TACCCAGGTT 360
ACCGTTTCTT CTGGTGGTGG TGGTTCTGGT GGTGGTGGTT CTGGTGGTGG TGGTTCTGGT 420
GGTGGTGGTT CTGAAGTTCA GCTGGTTGAA TCTGGTGGTG GTCTGGTTCA GGCGGGTGGT 480
TCTCTGCGTC TGTCTTGCGC GGCGTCTGGT CGTATCTTCC GTACCTACGA CATGGGTTGG 540
TACCGTCAGG CGCCGGGTAA AGAACGTGAA TTCGTTGCGC GTATCTCTTG GTCTGGTGGT 600
TCTACCAACT ACGCGGACTT CGTTAAAGGT CGTTTCACCA TCTCTAAAGA CTCTGCGAAA 660
AACACCGTTT ACCTGCAGAT GAACTCTCTG AAACCGGAAG ACACCGCGAT GTACTTCTGC 720
AACGCGGACC CGTCTTCTGT TGTTGGTGCG ATCGGTGCGG GTTACGTTTA CTGGGGTCAG 780
GGTACCCAGG TTACCGTTTC TTCTGCGGCG GCGGAACAGA AACTGATCTC TGAAGAAGAC 840
CTGAACGGTG CGGCG 855
SEQ ID NO:11.
<210> 11
<211> 1041
<212> DNA
<213> ArtificialSequence
<220>
<223> нуклеотидная последовательность фьюжн антитела 12-Fc,
представляющего собой однодоменное антитело клон 12,
модифицированное Fc-фрагментом IgG2 человека
<400>
CAGGTGCAGC TGGTGCAGTC CGGCGGCGGC CTGGTGCAGC CCGGCGGCTC CCTGCGCCTG 60
TCCTGCGCCG CCTCCGGCTC CGGCTTCTCC GTGTTCGCCA TGGGCTGGTA CCGCCAGGCC 120
CCCGGCAAGC AGCGCGAGCT GGTGGCCGGC ATCTCCTCCG ACTTCTCCAC CTCCTACACC 180
GACTCCGTGA AGGGCCGCTT CACCATCTCC CGCGACAACG CCCGCAACAC CGTGTACCTG 240
CAGATGAACT CCCTGAAGCC CGAGGACACC GCCGTGTACT ACTGCAAGCA GTCCGTGGAC 300
GGCAAGTCCC TGAACATCGG CTCCTGGGGC CAGGGCACCC AGGTGACCGT GTCCTCCGAG 360
CGCAAGTGCT GCGTGGAGTG CCCCCCCTGC CCCGCCCCCC CCGTGGCCGG CCCCTCCGTG 420
TTCCTGTTCC CCCCCAAGCC CAAGGACACC CTGATGATCT CCCGCACCCC CGAGGTGACC 480
TGCGTGGTGG TGGACGTGTC CCACGAGGAC CCCGAGGTGC AGTTCAACTG GTACGTGGAC 540
GGCGTGGAGG TGCACAACGC CAAGACCAAG CCCCGCGAGG AGCAGTTCAA CTCCACCTTC 600
CGCGTGGTGT CCGTGCTGAC CGTGGTGCAC CAGGACTGGC TGAACGGCAA GGAGTACAAG 660
TGCAAGGTGT CCAACAAGGG CCTGCCCGCC CCCATCGAGA AGACCATCTC CAAGACCAAG 720
GGCCAGCCCC GCGAGCCCCA GGTGTACACC CTGCCCCCCT CCCGCGAGGA GATGACCAAG 780
AACCAGGTGT CCCTGACCTG CCTGGTGAAG GGCTTCTACC CCTCCGACAT CTCCGTGGAG 840
TGGGAGTCCA ACGGCCAGCC CGAGAACAAC TACAAGACCA CCCCCCCCAT GCTGGACTCC 900
GACGGCTCCT TCTTCCTGTA CTCCAAGCTG ACCGTGGACA AGTCCCGCTG GCAGCAGGGC 960
AACGTGTTCT CCTGCTCCGT GATGCACGAG GCCCTGCACA ACCACTACAC CCAGAAGTCC 1020
CTGTCCCTGT CCCCCGGCAA G 1041
SEQ ID NO:12.
<210> 12
<211> 1056
<212> DNA
<213> ArtificialSequence
<220>
<223> нуклеотидная последовательность фьюжн антитела 23-Fc,
представляющего собой однодоменное антитело клон 23,
модифицированное Fc-фрагментом IgG2 человека
<400>
GAGGTGCAGC TGGTGGAGTC CGGCGGCGGC CTGGTGCAGG CCGGCGGCTC CCTGCGCCTG 60
TCCTGCGCCG CCTCCGGCCG CATCTTCCGC ACCTACGACA TGGGCTGGTA CCGCCAGGCC 120
CCCGGCAAGG AGCGCGAGTT CGTGGCCCGC ATCTCCTGGT CCGGCGGCTC CACCAACTAC 180
GCCGACTTCG TGAAGGGCCG CTTCACCATC TCCAAGGACT CCGCCAAGAA CACCGTGTAC 240
CTGCAGATGA ACTCCCTGAA GCCCGAGGAC ACCGCCATGT ACTTCTGCAA CGCCGACCCC 300
TCCTCCGTGG TGGGCGCCAT CGGCGCCGGC TACGTGTACT GGGGCCAGGG CACCCAGGTG 360
ACCGTGTCCT CCGAGCGCAA GTGCTGCGTG GAGTGCCCCC CCTGCCCCGC CCCCCCCGTG 420
GCCGGCCCCT CCGTGTTCCT GTTCCCCCCC AAGCCCAAGG ACACCCTGAT GATCTCCCGC 480
ACCCCCGAGG TGACCTGCGT GGTGGTGGAC GTGTCCCACG AGGACCCCGA GGTGCAGTTC 540
AACTGGTACG TGGACGGCGT GGAGGTGCAC AACGCCAAGA CCAAGCCCCG CGAGGAGCAG 600
TTCAACTCCA CCTTCCGCGT GGTGTCCGTG CTGACCGTGG TGCACCAGGA CTGGCTGAAC 660
GGCAAGGAGT ACAAGTGCAA GGTGTCCAAC AAGGGCCTGC CCGCCCCCAT CGAGAAGACC 720
ATCTCCAAGA CCAAGGGCCA GCCCCGCGAG CCCCAGGTGT ACACCCTGCC CCCCTCCCGC 780
GAGGAGATGA CCAAGAACCA GGTGTCCCTG ACCTGCCTGG TGAAGGGCTT CTACCCCTCC 840
GACATCTCCG TGGAGTGGGA GTCCAACGGC CAGCCCGAGA ACAACTACAA GACCACCCCC 900
CCCATGCTGG ACTCCGACGG CTCCTTCTTC CTGTACTCCA AGCTGACCGT GGACAAGTCC 960
CGCTGGCAGC AGGGCAACGT GTTCTCCTGC TCCGTGATGC ACGAGGCCCT GCACAACCAC 1020
TACACCCAGA AGTCCCTGTC CCTGTCCCCC GGCAAG 1056
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ ВАКЦИНЫ ПРОТИВ ГРИППА | 2021 |
|
RU2834969C1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА B И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2739952C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА B И ПУТИ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2784915C2 |
| Однодоменное антитело и его модификации, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, и способ их применения для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2763001C1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2683498C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2803850C2 |
| Однодоменное антитело и его модификации, специфически связывающиеся с ботулиническим нейротоксином типа А, и способ их применения для терапии или экстренной профилактики интоксикации, вызванной ботулиническим нейротоксином типа А. | 2021 |
|
RU2766348C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Выделенный рекомбинантный вирус на основе вируса гриппа для индукции специфического иммунитета к вирусу гриппа и/или профилактики заболеваний, вызванных вирусом гриппа | 2021 |
|
RU2813150C2 |
| СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ БЕЛКОВОГО/ПЕПТИДНОГО АНТИГЕНА | 2021 |
|
RU2830627C1 |
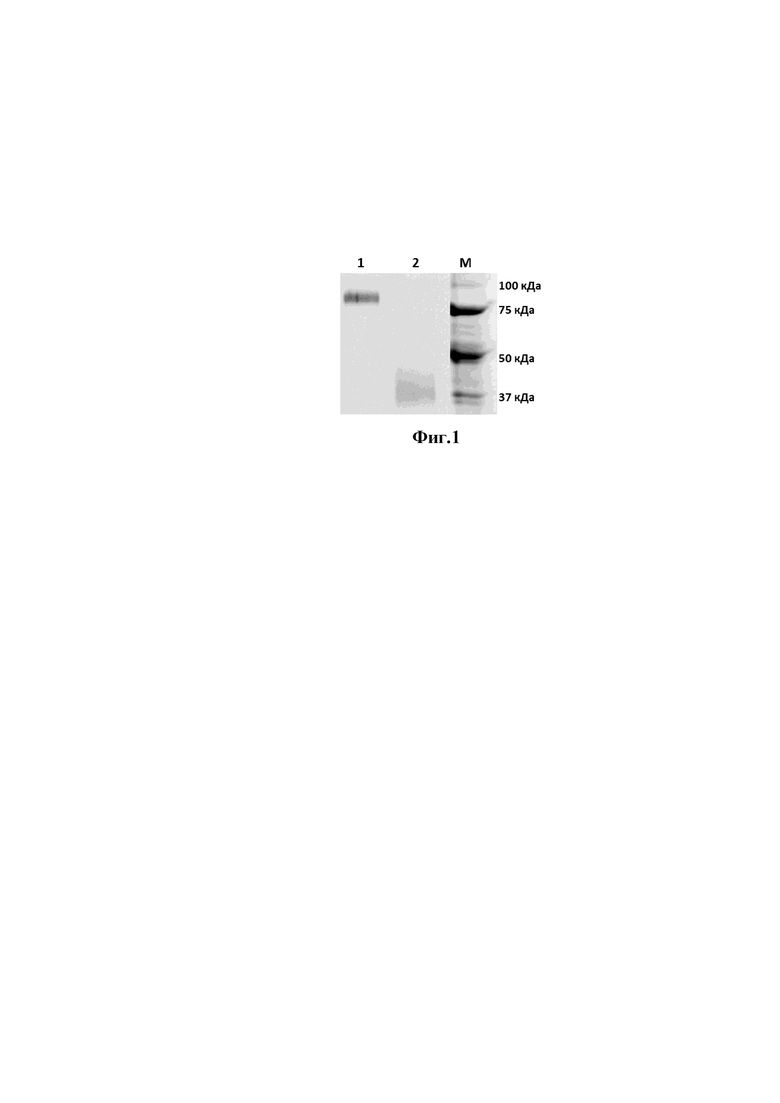
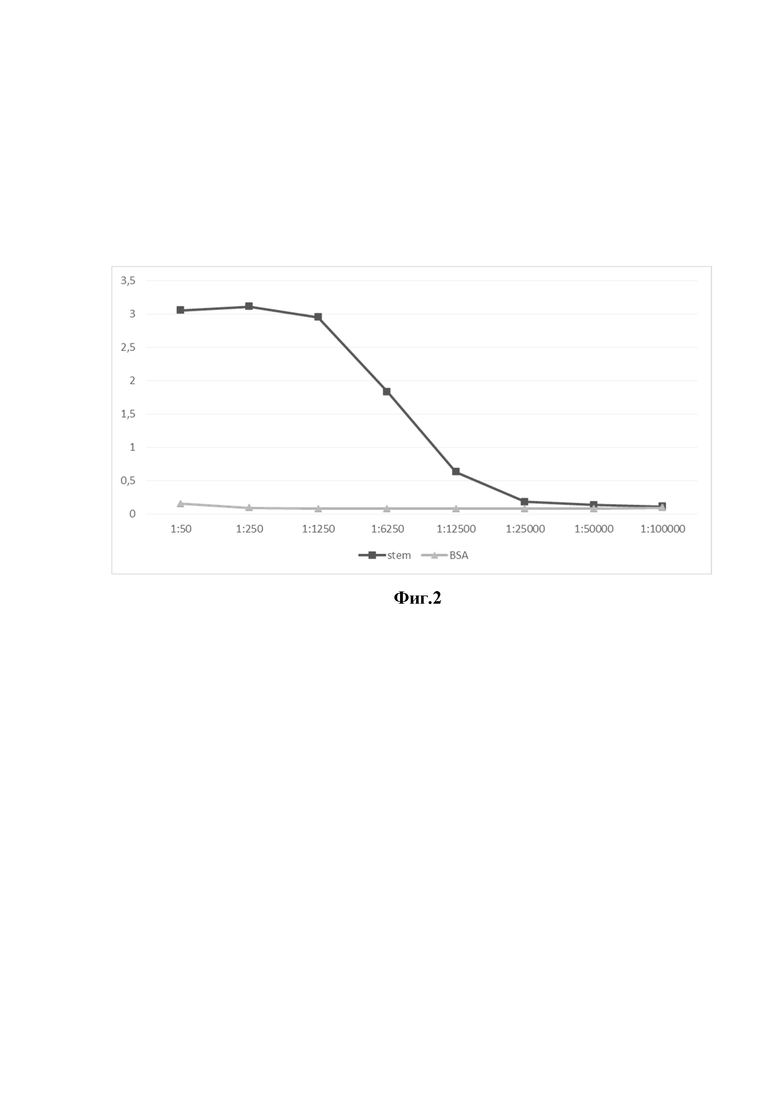
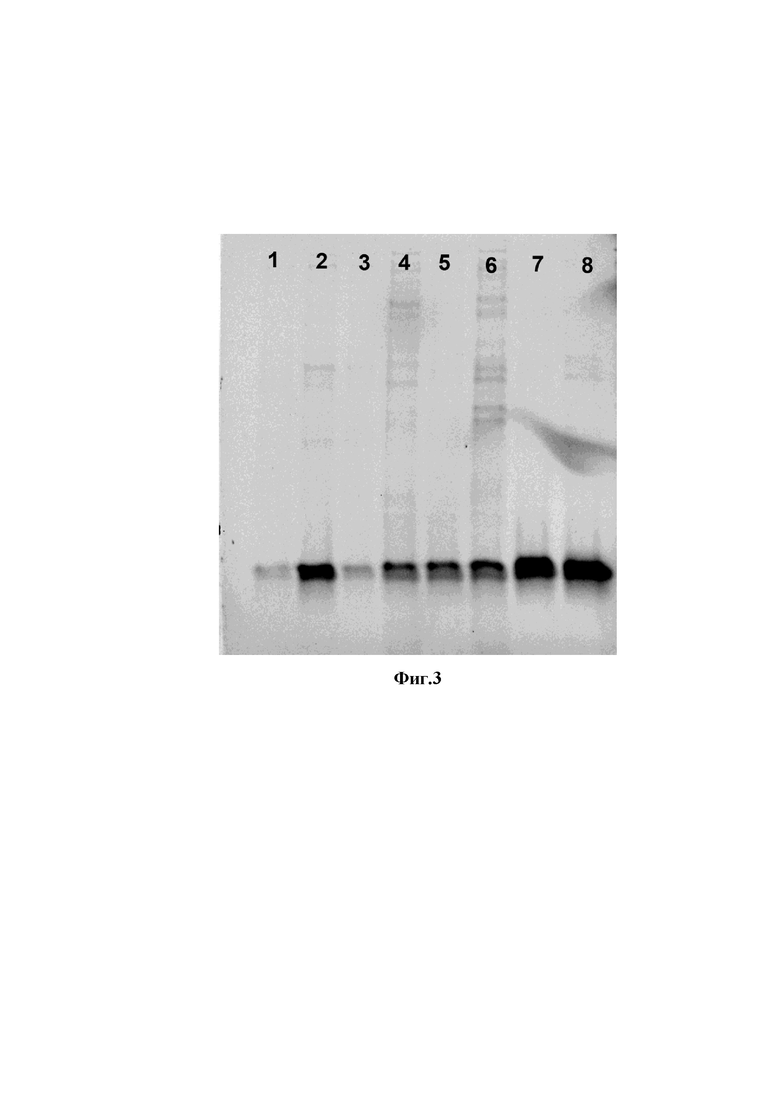
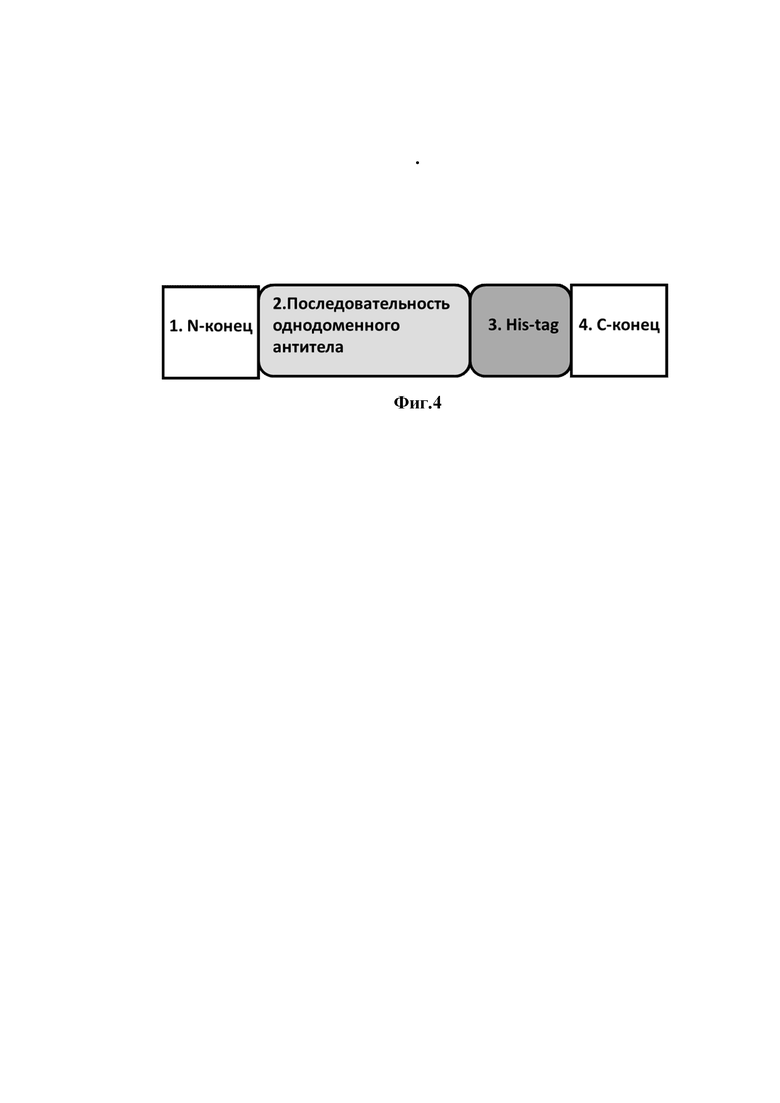

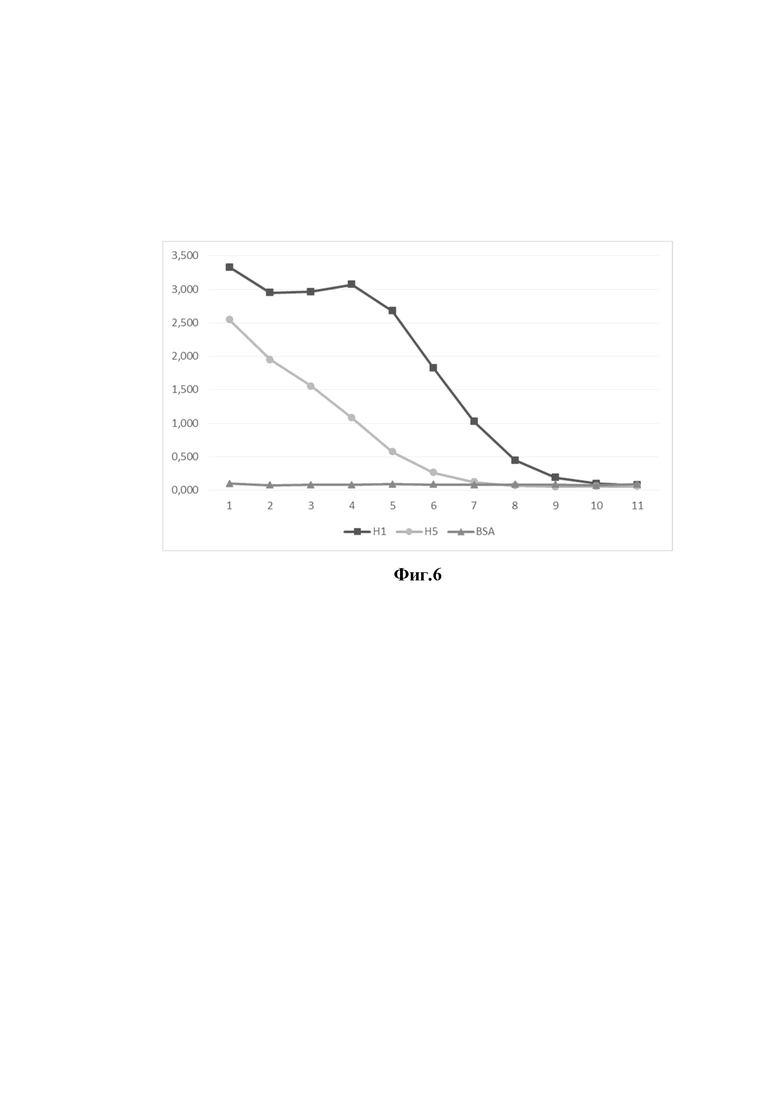
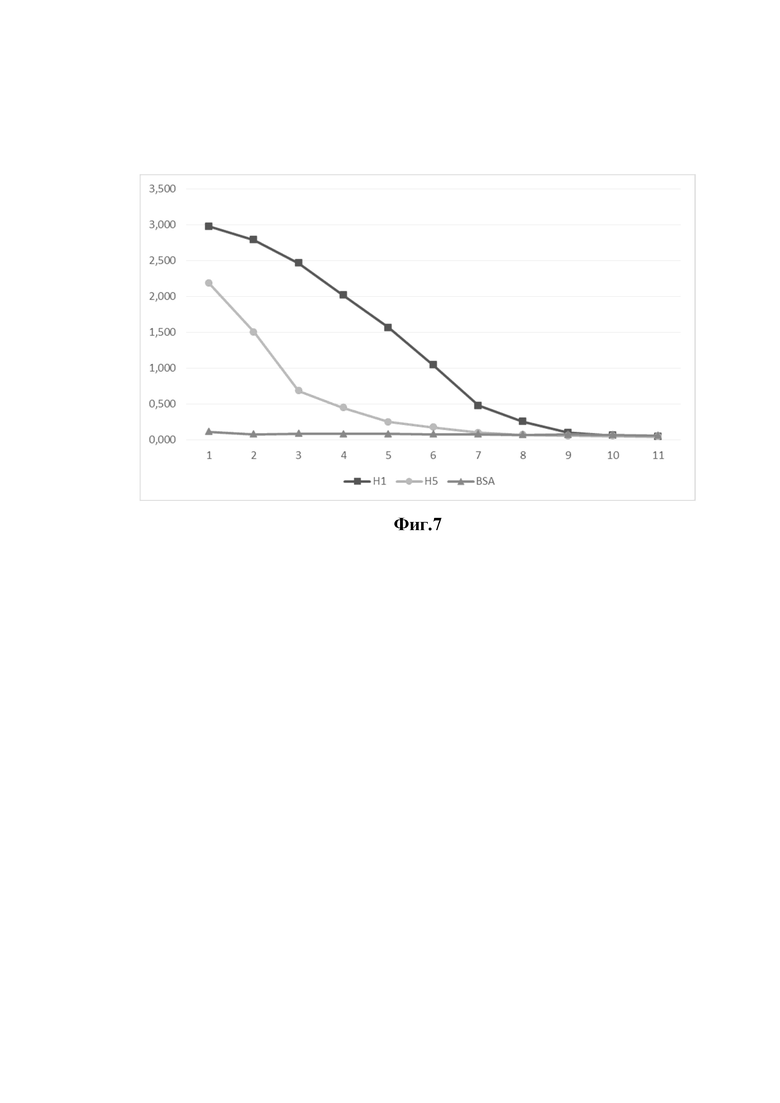

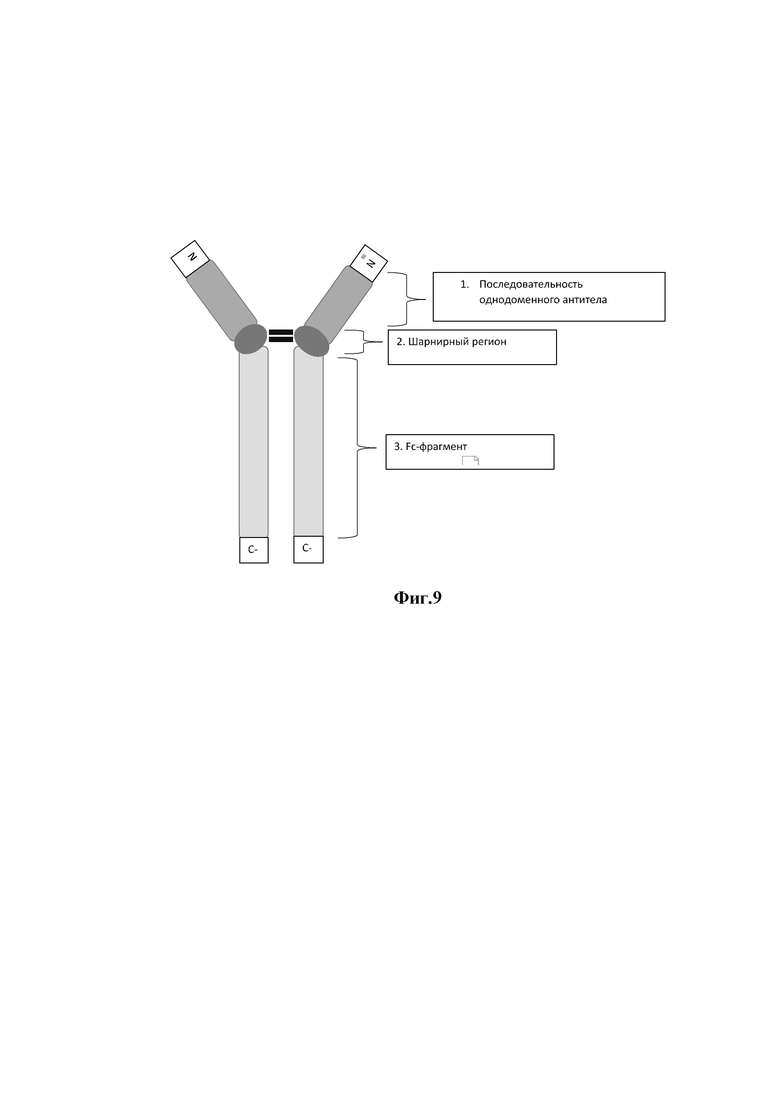
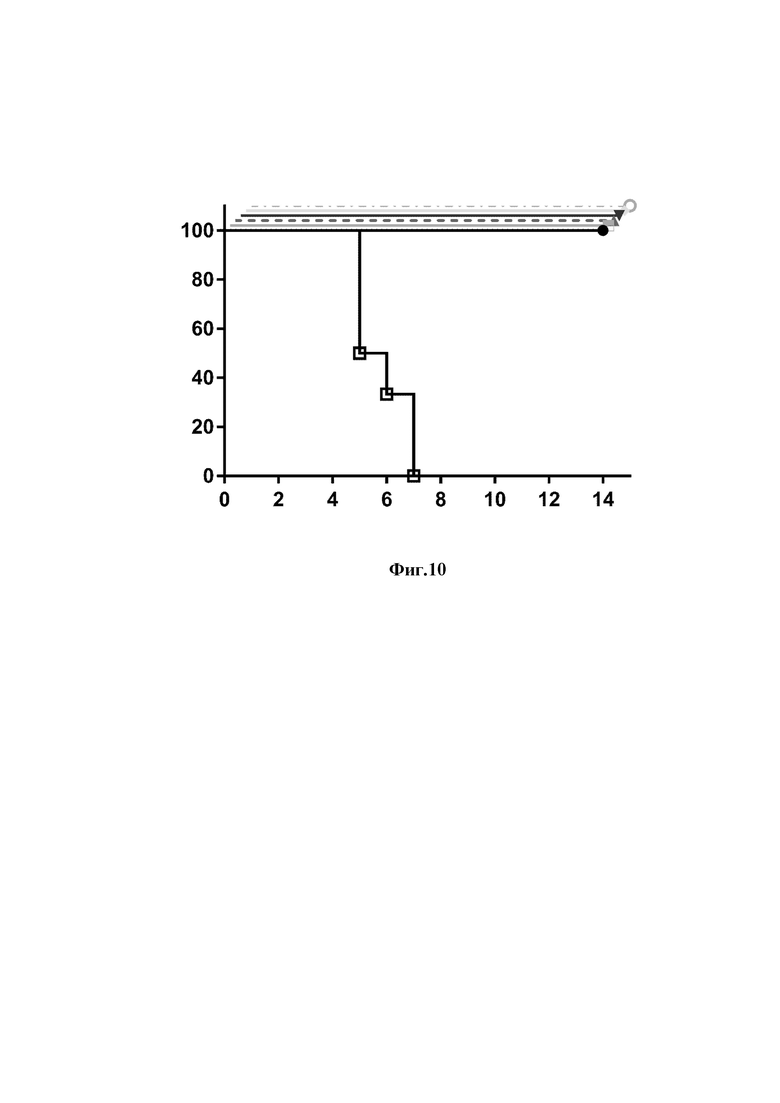
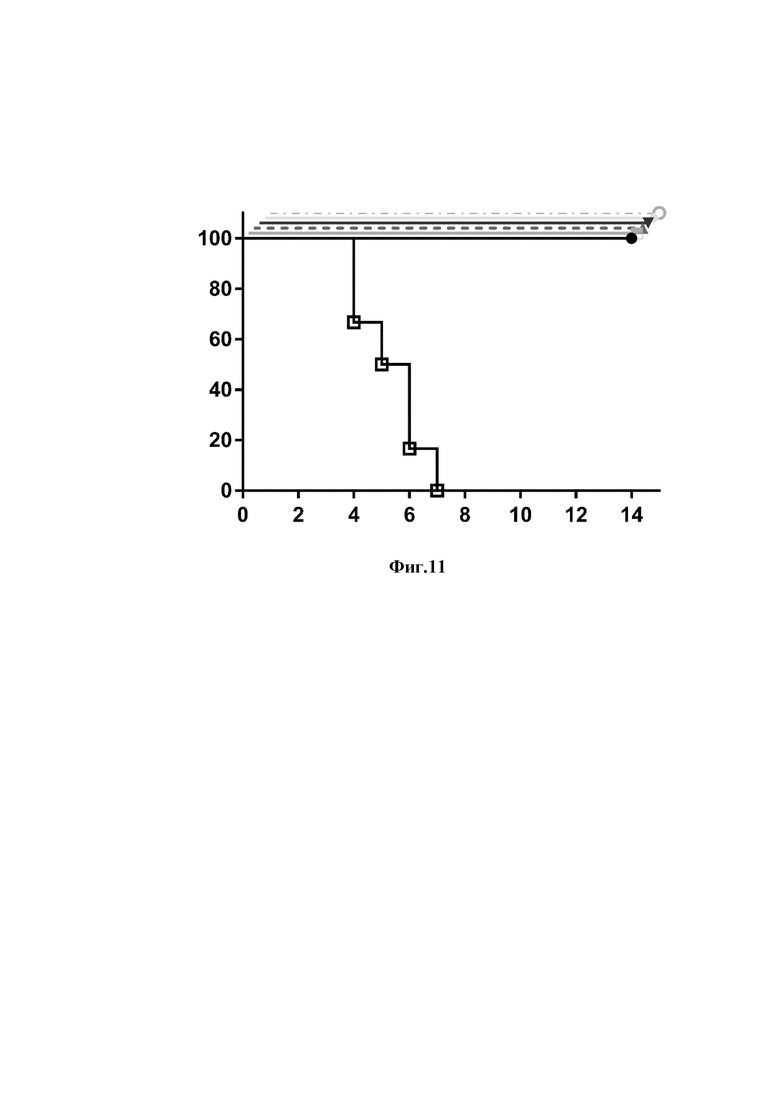
Группа изобретений относится к области биотехнологии, иммунологии и микробиологии и касается нейтрализации вируса гриппа А. Предложено однодоменное антитело, которое специфически связывается со стеблевым доменом гемагглютинина вируса гриппа А и имеет аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2. Также предложено олигомеризованное однодоменное антитело, которое специфически связывается со стеблевым доменом гемагглютинина вируса гриппа А, содержащее в качестве мономерного блока однодоменное антитело и имеющее конечную аминокислотную последовательность SEQ ID NO: 3 или SEQ ID NO: 4. Также создано рекомбинантное антитело, представляющее собой однодоменное антитело, слитое с Fc-фрагментом иммуноглобулина G2 человека, которое связывается со стеблевым доменом гемагглютинина вируса гриппа А и имеет конечную аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 6. Также раскрыт способ экстренной профилактики заболевания, вызванного вирусом гриппа А, заключающийся во введении в организм млекопитающих в эффективном количестве созданного антитела. Группа изобретений позволяет создать однодоменные антитела и его модификации, которые эффективно связывают стеблевой домен гемагглютинина вируса гриппа типа А, нейтрализуют вирус гриппа типа А и могут быть использованы для экстренной профилактики заболеваний, вызываемых вирусом гриппа типа А, обладают менее выраженной аффинностью к Fc-рецептору человека по сравнению с IgG1. 4 н.п. ф-лы, 11 ил., 3 табл., 14 пр.
1. Однодоменное антитело для нейтрализации вирусов, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А и имеющее аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2.
2. Олигомеризованное однодоменное антитело для нейтрализации вирусов, специфически связывающееся со стеблевым доменом гемагглютинина вируса гриппа А, содержащее в качестве мономерного блока однодоменное антитело по п.1 и имеющее аминокислотную последовательность SEQ ID NO: 3 или SEQ ID NO: 4.
3. Рекомбинантное антитело для нейтрализации вирусов, представляющее собой однодоменное антитело по п.1, слитое с Fc-фрагментом иммуноглобулина G2 человека, связывающееся со стеблевым доменом гемагглютинина вируса гриппа А и имеющее аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 6.
4. Способ экстренной профилактики заболевания, вызванного вирусом гриппа А, заключающийся во введении в организм млекопитающих в эффективном количестве антитела по любому из пп.1-3.
| Д | |||
| Н | |||
| ЩЕРБИНИН, С | |||
| В | |||
| АЛЕКСЕЕВА и др | |||
| Анализ В-клеточных эпитопов гемагглютинина вирусов гриппа, ACTA NATURAE, ТОМ 8, номер 1 (28), 2016, стр.14-22 | |||
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА B И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2739952C2 |
| WO 2016196470 A1, 08.12.2016 | |||
| WO 2015051010 A1, 09.04.2015. | |||

