Область изобретения
Настоящее изобретение относится к антителам, которые имеют нейтрализующую активность широкого спектра действия против вируса гриппа B, и путям применения таких антител.
Предпосылки изобретения
Вирусы гриппа вызывают ежегодные эпидемии гриппа и периодические пандемии, которые представляют значительную угрозу для здоровья населения во всем мире. Сезонная инфекция, вызванная вирусом гриппа, каждый год сопряжена с 200000-500000 смертей, особенно у маленьких детей, пациентов с ослабленным иммунитетом и у лиц пожилого возраста. Уровень смертности обычно возрастает дополнительно в периоды со вспышками пандемического гриппа. Сохраняется значительная неудовлетворенная медицинская потребность в эффективных противовирусных терапевтических средствах для предупреждения и лечения инфекций, вызванных вирусом гриппа, в частности, у малообеспеченного населения.
Существует три типа вирусов гриппа: типы A, B и C. Большинство заболеваний гриппом вызывают вирусы гриппа A и B (Thompson et al. (2004) JAMA. 292:1333-1340; и Zhou et al. (2012) Clin Infect. Dis. 54:1427-1436). Общая структура вирусов гриппа A, B и C является подобной и включает вирусную оболочку, которая окружает центральную часть. Вирусная оболочка включает в себя два поверхностных гликопротеина, гемагглютинин (HA) и нейраминидазу (NA); при этом HA опосредует связывание вируса с целевыми клеткам и его проникновение в целевые клетки, тогда как NA вовлечена в высвобождение вирусного потомства из инфицированных клеток.
Белок HA является тримерным по структуре и включает в себя три идентичных копии одного полипептида-предшественника HA0, который в ходе протеолитического созревания расщепляется на pH-зависимое метастабильное промежуточное соединение, содержащее глобулярную головку (HA1) и стволовой участок (HA2) (Wilson et al. (1981) Nature. 289:366-373). Дистальная по отношению к мембране глобулярная головка составляет большую часть структуры HA1 и содержит связывающий карман для сиаловой кислоты для вхождения вируса и основные антигенные домены.
Вирусы гриппа A можно разделить на подтипы на основании генетических вариаций генов гемагглютинина (HA) и нейраминидазы (NA). В настоящее время, в период сезонных эпидемий, подтипы H1 и H3 HA вируса гриппа A главным образом связаны с заболеванием у людей, тогда как вирусы, кодирующие H5, H7, H9 и H10, связаны со спорадическими вспышками заболевания у людей, вследствие прямой передачи от животных.
В отличие от вирусов гриппа A вирусы гриппа B не разделяются на подтипы на основании двух поверхностных гликопротеинов, и вплоть до 1970-х их классифицировали как одну однородную группу. На протяжении 1970-х вирусы гриппа B начали дивергировать на две линии, отличающиеся по антигенам, которые назвали линиями Victoria и Yamagata по названию их первых представителей, B/Victoria/2/87 и B/Yamagata/16/88 соответственно. (Biere et al. (2010) J Clin Microbiol. 48(4):1425-7; doi: 10.1128/JCM.02116-09. Epub 2010 Jan 27). Вирусы гриппа B ограничены способностью вызывать инфекции только у людей, при этом обе линии вносят вклад в ежегодные эпидемии. Несмотря на то, что частота заболеваний, вызываемых вирусами гриппа B, более низкая, чем частота заболеваний, связанных с вирусом гриппа A H3N2, она выше, чем частота заболеваний, связанных с вирусом гриппа A H1N1 (Zhou et al. (2012) Clin Infect. Dis. 54:1427-1436).
Нейтрализующие антитела, активированные инфекцией, вызванной вирусом гриппа, в норме направлены на вариабельную глобулярную головку HA1 для предупреждения связывания рецептора вируса и обычно штаммоспецифичны. Перекрестно реагирующие антитела широкого спектра действия, которые нейтрализуют один или несколько подтипов или линий, являются большой редкостью. В последнее время было разработано небольшое число человеческих антител, которые могут нейтрализовать несколько подтипов вирусов гриппа B из обеих линий (Dreyfus et al. (2012) Science. 337(6100):1343-8; и Yasugi et al. (2013) PLoS Path. 9(2):e1003150). Несмотря на то, что данные антитела распознают многие вирусы гриппа B, они характеризуются ограниченным спектром действия и эффективностью, а также не нейтрализуют никакие штаммы вируса гриппа A. К настоящему времени отсутствуют антитела широкого спектра действия, которые полностью нейтрализуют или ингибируют инфекции, вызванные всеми вирусами гриппа B, или ослабляют симптомы заболеваний, вызванных вирусом гриппа B. Таким образом, существует потребность в идентификации новых антител, обеспечивающих защиту от ряда вирусов гриппа.
Краткое описание изобретения
Настоящее изобретение, описанное в данном документе, предусматривает выделенное антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином (HA) вируса гриппа B и нейтрализации вирусов гриппа B двух филогенетически разных линий. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент способны к связыванию с гемагглютинином вируса гриппа B и нейтрализации вируса гриппа B обеих линий Yamagata и Victoria. Линия Yamagata включает без ограничения B/AA/94 (ca B/Ann Arbor/2/94 (yamagata)); B/YSI/98 (ca B/Yamanashi/166/98 (yamagata)); B/JHB/99 (ca B/Johannesburg/5/99 (yamagata)); B/SC/99 (B/Sichuan/379/99 (yamagata)); B/FL/06 (B/Florida/4/2006 (yamagata)). Линия Victoria включает без ограничения B/BJ/97 (ca B/Beijing/243/97 (victoria)), B/HK/01 (B/Hong Kong/330/2001 (victoria)); B/MY/04 (B/Malaysia/2506/2004 (victoria)); B/BNE/08 (ca B/Brisbane/60/2008 (victoria)).
В другом варианте осуществления настоящее изобретение предусматривает выделенное антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином вируса гриппа B и нейтрализации вируса гриппа B предивергентного штамма. Как используется в данном документе, выражение "предивергентный" относится к штаммам вируса гриппа B, которые были идентифицированы до дивергенции вируса гриппа B на линии Yamagata и Victoria. Предивергентные штаммы вируса гриппа B включают без ограничения B/Lee/40 (B/Lee/40); B/AA/66 (ca B/Ann Arbor/1/66) и B/HK/72 (B/Hong Kong/5/72).
В одном варианте осуществления антитело или антигенсвязывающий фрагмент связывают вирус гриппа B при EC50 в диапазоне от приблизительно 1 мкг/мл до приблизительно 50 мкг/мл антитела. В другом варианте осуществления антитело или антигенсвязывающий фрагмент характеризуются эффективностью нейтрализации, выраженной в виде 50% ингибирующей концентрации (IC50, мкг/мл), в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 5 мкг/мл антитела для нейтрализации вируса гриппа B в анализе микронейтрализации, описанном в примере 3. В примере 1 также описаны и другие анализы микронейтрализации.
В одном варианте осуществления антитело способно к связыванию с гемагглютинином вируса гриппа A. Гемагглютинин вируса гриппа A включает гемагглютинин подтипа 1 и подтипа 2. Подтипы группы 1 вируса гриппа A включают H1, H2, H5, H6, H8, H9, H11, H12, H13, H16 и их варианты. Подтипы группы 2 вируса гриппа A включают H3, H4, H7, H10, H14 и H15 и их вариантов. В одном варианте осуществления антитело способно к связыванию с одним или несколькими подтипами группы 1 вируса гриппа A. В другом варианте осуществления антитело способно к связыванию с одним или несколькими подтипами группы 2 вируса гриппа A. В одном варианте осуществления антитело способно к связыванию с подтипом H9 группы 1 вируса гриппа A. В одном варианте осуществления настоящее изобретение предусматривает выделенное антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином (HA) вируса гриппа B и гемагглютинином (HA) вируса гриппа A и нейтрализации по меньшей мере одного вируса гриппа B линии Yamagata; по меньшей мере одного вируса гриппа B линии Victoria; по меньшей мере одного подтипа вируса гриппа A или их комбинаций. В одном варианте осуществления настоящее изобретение предусматривает выделенное антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином (HA) вируса гриппа B и одним или несколькими гемагглютининами (HA) подтипа 1 вируса гриппа A и нейтрализации по меньшей мере одного вируса гриппа B линии Yamagata или по меньшей мере одного вируса гриппа B линии Victoria и по меньшей мере одного подтипа 1 вируса гриппа A. В одном варианте осуществления настоящее изобретение предусматривает выделенное антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином (HA) вируса гриппа B и гемагглютинином (HA) подтипа H9 вируса гриппа A и нейтрализации по меньшей мере одного вируса гриппа B линии Yamagata; по меньшей мере одного вируса гриппа B линии Victoria; подтипа H9 вируса гриппа A или их комбинаций.
В одном варианте осуществления антитело или антигенсвязывающий фрагмент связывают HA вируса гриппа A при EC50 в диапазоне от приблизительно 1 мкг/мл до приблизительно 50 мкг/мл антитела. В другом варианте осуществления антитело или антигенсвязывающий фрагмент характеризуются IC50 в диапазоне от приблизительно 0,01 мкг/мл до приблизительно 5 мкг/мл антитела для нейтрализации вируса гриппа A в анализе микронейтрализации.
В одном варианте осуществления антитело или его фрагмент по настоящему изобретению связываются с участком HA, представляющим собой глобулярную головку, и нейтрализуют инфекцию, вызванную вирусами гриппа B двух филогенетически разных линий. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с участком HA, представляющим собой глобулярную головку, и нейтрализуют инфекцию, вызванную вирусом гриппа B обеих линий Yamagata и Victoria. Антитела по настоящему изобретению, которые представляют собой антитела к вирусу гриппа B, связывающиеся с глобулярной головкой HA, характеризуются более широким спектром действия или лучшей нейтрализующей активностью в отношении вирусов гриппа B по сравнению с известными антителами к вирусам гриппа B.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент включают в себя совокупность из шести CDR: HCDR-1, HCDR-2, HCDR-3, LCDR-1, LCDR-2, LCDR-3, при этом совокупность из шести CDR выбрана из
(a) HCDR-1 с SEQ ID NO: 3, HCDR-2 с SEQ ID NO: 4, HCDR-3 с SEQ ID NO: 5, LCDR-1 с SEQ ID NO: 8, LCDR-2 с SEQ ID NO: 9 и LCDR-3 с SEQ ID NO: 10;
(b) HCDR-1 с SEQ ID NO: 13, HCDR-2 с SEQ ID NO: 14, HCDR-3 с SEQ ID NO: 15, LCDR-1 с SEQ ID NO: 18, LCDR-2 с SEQ ID NO: 19, LCDR-3 с SEQ ID NO: 20;
(c) HCDR-1 с SEQ ID NO: 23, HCDR-2 с SEQ ID NO: 24, HCDR-3 с SEQ ID NO: 25, LCDR-1 с SEQ ID NO: 28, LCDR-2 с SEQ ID NO: 29 и LCDR-3 с SEQ ID NO: 30;
(d) HCDR-1 с SEQ ID NO: 33, HCDR-2 с SEQ ID NO: 34, HCDR-3 с SEQ ID NO: 35, LCDR-1 с SEQ ID NO: 38, LCDR-2 с SEQ ID NO: 39 и LCDR-3 с SEQ ID NO: 40;
(e) HCDR-1 с SEQ ID NO: 43, HCDR-2 с SEQ ID NO: 44, HCDR-3 с SEQ ID NO: 45, LCDR-1 с SEQ ID NO: 48, LCDR-2 с SEQ ID NO: 49 и LCDR-3 с SEQ ID NO: 50;
(f) HCDR-1 с SEQ ID NO: 53, HCDR-2 с SEQ ID NO: 54, HCDR-3 с SEQ ID NO: 55, LCDR-1 с SEQ ID NO: 58, LCDR-2 с SEQ ID NO: 59 и LCDR-3 с SEQ ID NO: 60;
(g) HCDR-1 с SEQ ID NO: 63, HCDR-2 с SEQ ID NO: 64, HCDR-3 с SEQ ID NO: 65, LCDR-1 с SEQ ID NO: 68, LCDR-2 с SEQ ID NO: 69 и LCDR-3 с SEQ ID NO: 70;
(h) HCDR-1 с SEQ ID NO: 75, HCDR-2 с SEQ ID NO: 76, HCDR-3 с SEQ ID NO: 77, LCDR-1 с SEQ ID NO: 83, LCDR-2 с SEQ ID NO: 84 и LCDR-3 с SEQ ID NO: 85;
(i) HCDR-1 с SEQ ID NO: 91, HCDR-2 с SEQ ID NO: 92, HCDR-3 с SEQ ID NO: 93, LCDR-1 с SEQ ID NO: 99, LCDR-2 с SEQ ID NO: 100 и LCDR-3 с SEQ ID NO: 101;
(j) HCDR-1 с SEQ ID NO: 107, HCDR-2 с SEQ ID NO: 108, HCDR-3 с SEQ ID NO: 109, LCDR-1 с SEQ ID NO: 115, LCDR-2 с SEQ ID NO: 116 и LCDR-3 с SEQ ID NO: 117;
(k) HCDR-1 с SEQ ID NO: 121, HCDR-2 с SEQ ID NO: 122, HCDR-3 с SEQ ID NO: 123, LCDR-1 с SEQ ID NO: 124, LCDR-2 с SEQ ID NO: 125 и LCDR-3 с SEQ ID NO: 126;
(l) HCDR-1 с SEQ ID NO: 127, HCDR-2 с SEQ ID NO: 128, HCDR-3 с SEQ ID NO: 129, LCDR-1 с SEQ ID NO: 130, LCDR-2 с SEQ ID NO: 131 и LCDR-3 с SEQ ID NO: 132;
(m) HCDR-1 с SEQ ID NO: 133, HCDR-2 с SEQ ID NO: 134, HCDR-3 с SEQ ID NO: 135, LCDR-1 с SEQ ID NO: 136, LCDR-2 с SEQ ID NO: 137 и LCDR-3 с SEQ ID NO: 138;
(n) HCDR-1 с SEQ ID NO: 139, HCDR-2 с SEQ ID NO: 140, HCDR-3 с SEQ ID NO: 141, LCDR-1 с SEQ ID NO: 142, LCDR-2 с SEQ ID NO: 143 и LCDR-3 с SEQ ID NO: 144;
(o) HCDR-1 с SEQ ID NO: 145, HCDR-2 с SEQ ID NO: 146, HCDR-3 с SEQ ID NO: 147, LCDR-1 с SEQ ID NO: 148, LCDR-2 с SEQ ID NO: 149 и LCDR-3 с SEQ ID NO: 150;
(p) HCDR-1 с SEQ ID NO: 78, HCDR-2 с SEQ ID NO: 79, HCDR-3 с SEQ ID NO: 80, LCDR-1 с SEQ ID NO: 86, LCDR-2 с SEQ ID NO: 87 и LCDR-3 с SEQ ID NO: 88;
(q) HCDR-1 с SEQ ID NO: 94, HCDR-2 с SEQ ID NO: 95, HCDR-3 с SEQ ID NO: 96, LCDR-1 с SEQ ID NO: 102, LCDR-2 с SEQ ID NO: 103 и LCDR-3 с SEQ ID NO: 104;
(r) HCDR-1 с SEQ ID NO: 110, HCDR-2 с SEQ ID NO: 111, HCDR-3 с SEQ ID NO: 112, LCDR-1 с SEQ ID NO: 118, LCDR-2 с SEQ ID NO: 119 и LCDR-3 с SEQ ID NO: 120; и
(s) совокупности из шести CDR в соответствии с любым из (a) -(r), включающих в себя одну или несколько замен, делеций или вставок аминокислот.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержат VH по меньшей мере c 75%, 80%, 85%, 90%, 95% или 100% идентичностью и/или VL по меньшей мере c 75%, 80%, 85%, 90%, 95% или 100% идентичностью с VH и/или VL соответственно, выбранными из
(a) VH с SEQ ID NO.: 2 и VL с SEQ ID NO: 7,
(b) VH с SEQ ID NO.: 12 и VL с SEQ ID NO: 17,
(c) VH с SEQ ID NO.: 22 и VL с SEQ ID NO: 27,
(d) VH с SEQ ID NO.: 32 и VL с SEQ ID NO: 37,
(e) VH с SEQ ID NO.: 42 и VL с SEQ ID NO: 47,
(f) VH с SEQ ID NO.: 52 и VL с SEQ ID NO: 57,
(g) VH с SEQ ID NO.: 62 и VL с SEQ ID NO: 67,
(h) VH с SEQ ID NO.: 74 и VL с SEQ ID NO: 82,
(i) VH с SEQ ID NO.: 90 и VL с SEQ ID NO: 98 и
(j) VH с SEQ ID NO.: 106 и VL с SEQ ID NO: 114.
В более конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент включают в себя VH и VL, выбранные из
(a) VH с SEQ ID NO.: 2 и VL с SEQ ID NO: 7,
(b) VH с SEQ ID NO.: 12 и VL с SEQ ID NO: 17,
(c) VH с SEQ ID NO.: 22 и VL с SEQ ID NO: 27,
(d) VH с SEQ ID NO.: 32 и VL с SEQ ID NO: 37,
(e) VH с SEQ ID NO.: 42 и VL с SEQ ID NO: 47,
(f) VH с SEQ ID NO.: 52 и VL с SEQ ID NO: 57,
(g) VH с SEQ ID NO.: 62 и VL с SEQ ID NO: 67,
(h) VH с SEQ ID NO.: 74 и VL с SEQ ID NO: 82,
(i) VH с SEQ ID NO.: 90 и VL с SEQ ID NO: 98 и
(j) VH с SEQ ID NO.: 106 и VL с SEQ ID NO: 114.
В одном варианте осуществления настоящее изобретение предусматривает антитело или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином (HA) вируса гриппа B и нейтрализации вирусов гриппа B двух филогенетически разных линий, где антитело имеет аминокислотную последовательность VH с SEQ ID NO:71, где Xaa1 в SEQ ID NO:71 представляет собой Val или Glu; Xaa2 в SEQ ID NO:71 представляет собой Leu или Phe; Xaa3 в SEQ ID NO:71 представляет собой Ser или Thr; Xaa4 в SEQ ID NO:71 представляет собой Leu или Ser; Xaa5 в SEQ ID NO:71 представляет собой Ser или Thr; Xaa6 в SEQ ID NO:71 представляет собой Met или Thr; Xaa7 в SEQ ID NO:71 представляет собой Phe или Tyr; Xaa8 в SEQ ID NO:71 представляет собой His или Gln; Xaa9 в SEQ ID NO:71 представляет собой Ser или Asn; Xaa10 в SEQ ID NO:71 представляет собой Arg или Lys и Xaa11 в SEQ ID NO:71 представляет собой Ala или Thr; и аминокислотную последовательность VL с SEQ ID NO:72, где Xaa1 в SEQ ID NO:72 представляет собой Phe или Tyr. В одном варианте осуществления Xaa9 в SEQ ID NO:71 представляет собой Ser. В другом варианте осуществления Xaa4 в SEQ ID NO:71 представляет собой Leu. В еще одном варианте осуществления Xaa1 в SEQ ID NO:71 представляет собой Glu; Xaa5 в SEQ ID NO:71 представляет собой Thr; Xaa6 в SEQ ID NO:71 представляет собой Thr; Xaa7 в SEQ ID NO:71 представляет собой Tyr; Xaa8 в SEQ ID NO:71 представляет собой Gln; Xaa10 в SEQ ID NO:71 представляет собой Lys; Xaa11 в SEQ ID NO:71 представляет собой Thr или их комбинации. В другом варианте осуществления Xaa1 в SEQ ID NO:71 представляет собой Glu; Xaa5 в SEQ ID NO:71 представляет собой Thr; Xaa6 в SEQ ID NO:71 представляет собой Thr; Xaa7 в SEQ ID NO:71 представляет собой Tyr; Xaa8 в SEQ ID NO:71 представляет собой Gln; Xaa9 в SEQ ID NO:71 представляет собой Ser; Xaa10 в SEQ ID NO:71 представляет собой Lys и Xaa11 в SEQ ID NO:71 представляет собой Thr.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент выбраны из молекулы иммуноглобулина, моноклонального антитела, химерного антитела, антитела с привитыми CDR, гуманизированного антитела, Fab, Fab', F(ab')2, Fv, Fv с дисульфидной связью, scFv, однодоменного антитела, диатела, полиспецифического антитела, антитела с двойной специфичностью и биспецифического антитела. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент включают в себя Fc-участок. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент представляют собой IgG1, IgG2 или IgG4 или их фрагмент.
В одном варианте осуществления настоящее изобретение предусматривает антитело к вирусу гриппа B или его антигенсвязывающий фрагмент, которые способны к связыванию с вирусом гриппа B и нейтрализации по меньшей мере одного вируса гриппа B линии Yamagata и по меньшей мере одного вируса гриппа B линии Victoria, где антитело или его антигенсвязывающий фрагмент связывают эпитоп, являющийся консервативным среди по меньшей мере одного вируса гриппа B линии Yamagata и по меньшей мере одного вируса гриппа B линии Victoria. В одном варианте осуществления один или несколько контактирующих остатков эпитопа расположены в участке HA, представляющем собой головку, вируса гриппа B. В одном варианте осуществления эпитоп включает в себя одну или несколько аминокислот, выбранных из 128-й, 141-й, 150-й и 235-й из последовательности участка HA, представляющего собой головку, в качестве контактирующих остатков (Wang et al. (2008) J. Virol. 82(6):3011-20).
В другом варианте осуществления настоящее изобретение предусматривает антитело к вирусу гриппа B или его антигенсвязывающий фрагмент, которые способны к связыванию с гемагглютинином вируса гриппа B и нейтрализации вирусов гриппа B двух филогенетически разных линий, которые связываются с тем же эпитопом или конкурируют за связывание с гемагглютинином вируса гриппа B с антителом по настоящему изобретению. В одном варианте осуществления антитело или антигенсвязывающий фрагмент связываются с тем же эпитопом или конкурируют за связывание с гемагглютинином вируса гриппа A с антителом, имеющим аминокислотную последовательность, выбранную из
(a) VH с SEQ ID NO.: 2 и VL с SEQ ID NO: 7,
(b) VH с SEQ ID NO.: 12 и VL с SEQ ID NO: 17,
(c) VH с SEQ ID NO.: 22 и VL с SEQ ID NO: 27,
(d) VH с SEQ ID NO.: 32 и VL с SEQ ID NO: 37,
(e) VH с SEQ ID NO.: 42 и VL с SEQ ID NO: 47,
(f) VH с SEQ ID NO.: 52 и VL с SEQ ID NO: 57,
(g) VH с SEQ ID NO.: 62 и VL с SEQ ID NO: 67,
(h) VH с SEQ ID NO.: 74 и VL с SEQ ID NO: 82,
(i) VH с SEQ ID NO.: 90 и VL с SEQ ID NO: 98 и
(j) VH с SEQ ID NO.: 106 и VL с SEQ ID NO: 114.
Настоящее изобретение также предусматривает выделенную нуклеиновую кислоту, кодирующую антитело или его антигенсвязывающий фрагмент по настоящему изобретению, а также вектор, который включает в себя такую выделенную нуклеиновую кислоту, и клетку-хозяина, которая включает в себя такую нуклеиновую кислоту или вектор. В одном варианте осуществления вектор представляет собой вектор экспрессии. В другом варианте осуществления вектор представляет собой не встречающийся в природе рекомбинантный вектор. В одном варианте осуществления вектор представляет собой плазмиду. В одном варианте осуществления вектор или плазмида включают в себя нуклеотидную последовательность, кодирующую молекулу антитела по настоящему изобретению или его антигенсвязывающего фрагмента, тяжелую или легкую цепь молекулы антитела по настоящему изобретению, вариабельный домен тяжелой или легкой цепей антитела по настоящему изобретению или его часть, или CDR тяжелой или легкой цепей, функционально связанный с одним или несколькими элементами контроля экспрессии (например, промотором, энхансером, терминаторами транскрипции, сайтами-полиаденилирования и т.д.), ген селектируемого маркера или их комбинации. В одном варианте осуществления вектор или плазмида включают в себя по меньшей мере один гетерологичный элемент контроля экспрессии, селектируемый маркер или их комбинации.
В одном варианте осуществления настоящее изобретение предусматривает способ получения антитела или его антигенсвязывающего фрагмента путем культивирования клетки-хозяина, описанной в данном документе, в условиях, подходящих для экспрессии данного антитела или его фрагмента. В одном варианте осуществления способ включает выделение антитела или его антигенсвязывающего фрагмента из культуры клеток-хозяев. В одном варианте осуществления клетка-хозяин выделена из тканей, в которых данная клетка встречается в природных условиях. Например, клетку-хозяин можно выделить из организма и поддерживать ex vivo в культуре клеток.
Настоящее изобретение также предусматривает композицию, которая включает в себя антитело или его антигенсвязывающий фрагмент по настоящему изобретению и фармацевтически приемлемый носитель. В одном варианте осуществления композиция включает в себя антитело или его антигенсвязывающий фрагмент по настоящему изобретению, и 25 мМ His, и 0,15 М NaCl при pH 6,0.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению применяются в профилактике или лечении инфекции, вызванной вирусом гриппа B, у субъекта. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент применяются в профилактике или лечении инфекции, вызванной вирусом гриппа А и вирусом гриппа B, у субъекта. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению применяются в получении лекарственного препарата, предназначенного для профилактики или лечения инфекции, вызванной вирусом гриппа B, у субъекта. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению применяются в получении лекарственного препарата, предназначенного для профилактики или лечения инфекции, вызванной вирусом гриппа А и вирусом гриппа B, у субъекта.
В одном варианте осуществления настоящее изобретение предусматривает способ профилактики или лечения инфекции, вызванной вирусом гриппа B, у субъекта, включающий введение субъекту эффективного количества антитела или его антигенсвязывающего фрагмента по настоящему изобретению. В другом варианте осуществления настоящее изобретение предусматривает способ профилактики или лечения инфекции, вызванной вирусом гриппа А и вирусом гриппа B, у субъекта, включающий введение субъекту эффективного количества антитела или его антигенсвязывающего фрагмента по настоящему изобретению.
В одном варианте осуществления антитело или его фрагмент по настоящему изобретению применяются для in vitro диагностики инфекции, вызванной вирусом гриппа B, у субъекта. В другом варианте осуществления антитело или его фрагмент по настоящему изобретению применяются для in vitro диагностики инфекции, вызванной вирусом гриппа А, у субъекта. В еще одном варианте осуществления антитело или его фрагмент по настоящему изобретению применяются для in vitro диагностики инфекции, вызванной вирусом гриппа А, и инфекции, вызванной вирусом гриппа B, у субъекта.
Короткое описание фигур
На фигуре 1 показана антителозависимая клеточная цитотоксичность (ADCC), измеренная по активации NK-клеток после инкубации с B/Hong Kong/330/2001 (линия Victoria) и серийным разведением антител к HA, FBD-94 и FBC-39, а также вариантов, не обладающих эффекторной функцией Fc (FBD-94 LALA и FBC-39 LALA).
На фигуре 2 показан процент выживших животных, которым вводили разные концентрации FBC-39 (A и C), FBD-94 (B и D) и нерелевантного контрольного антитела за 4 часа до инфицирования смертельной дозой вируса гриппа B/Sichuan/379/99 (Yamagata) (A и B) или вируса гриппа B/Hong Kong/330/2001 (Victoria) (C и D).
На фигуре 3 показан процент выживших животных, которых инфицировали смертельной дозой B/Sichuan/379/99 (Yamagata) (A и B) или B/Hong Kong/330/2001 (Victoria) (C и D) и через 2 дня после инфицирования обрабатывали разными дозами FBC-39 (A и C), или FBD-94 (B и D), или нерелевантного контрольного антитела.
На фигуре 4 показан процент выживших животных, которых инфицировали смертельной дозой B/Hong Kong/330/2001 (Victoria) и через 1, 2, 3 или 4 дня после инфицирования обрабатывали с помощью 3 мкг/кг FBC-39 (A), или FBD-94 (B), или нерелевантного контрольного антитела.
На фигуре 5 показаны CDR (заключенные в рамки) в последовательностях VH и VL антител к вирусу гриппа B, FBD-56, FBD-94, FBC-39, FBC-39 LSL, FBC-39 FSL, FBC-39 LTL, FBC-39 FTL, FBC-39 FSS, FBC-39 LTS и FBC-39 FTS, определенные с помощью систем нумерации по Kabat и IMGT. Модифицированные аминокислоты в последовательностях VH и VL выделены жирным и подчеркнуты.
На фигуре 6 показана аминокислотная последовательность тяжелой цепи обобщенного антитела к вирусу гриппа B (SEQ ID NO:71) на основе аминокислотной последовательности тяжелой цепи FBC-39 (SEQ ID NO:22), где Xaa1 может представлять собой Val или Glu; Xaa2 может представлять собой Leu или Phe; Xaa3 может представлять собой Thr или Ser; Xaa4 может представлять собой Ser или Leu; Xaa5 может представлять собой Thr или Ser; Xaa6 может представлять собой Thr или Met; Xaa7 может представлять собой Tyr или Phe; Xaa8 может представлять собой Gln или His; Xaa9 может представлять собой Asn или Ser; Xaa10 может представлять собой Lys или Arg и Xaa11 может представлять собой Thr или Ala.
На фигуре 7 показана аминокислотная последовательность легкой цепи обобщенного антитела к вирусу гриппа B (SEQ ID NO:72) на основе аминокислотной последовательности легкой цепи FBC-39 (SEQ ID NO:27), где Xaa1 может представлять собой Phe или Tyr.
На фигуре 8 показано выравнивание белков HA1 вирусов, применяемых для выделения устойчивых к моноклональным антителам мутантов (MARM). Аминокислотные положения, которые, как обнаружено посредством отбора MARM, являются контактирующими остатками, заключены в рамку.
Подробное описание
Введение
Настоящее изобретение предусматривает антитела, в том числе человеческие формы, а также их антигенсвязывающие фрагменты, производные/конъюгаты и композиции, которые связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирусы гриппа B двух филогенетически разных линий, описанных в данном документе. В одном варианте осуществления антитела или их антигенсвязывающие фрагменты связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирус гриппа B обеих линий Yamagata и Victoria, описанных в данном документе; при этом такие антитела к вирусу гриппа B и их фрагменты называются в данном документе антителами по настоящему изобретению. В другом варианте осуществления антитела или их антигенсвязывающие фрагменты связывают гемагглютинин (HA) вируса гриппа B и гемагглютинин (HA) вируса гриппа A и нейтрализуют по меньшей мере один вирус гриппа B линии Yamagata; по меньшей мере один вирус гриппа B линии Victoria; по меньшей мере один подтип вируса гриппа A или их комбинации. Такие антитела к вирусу гриппа B и их фрагменты также называют в данном документе антителами по настоящему изобретению.
Как используется в данном документе, выражение "нейтрализовать" относится к способности антитела или его антигенсвязывающего фрагмента связываться с возбудителем инфекции, таким как вирус гриппа A и/или вирус гриппа B, и снижать биологическую активность, например, вирулентность, возбудителя инфекции. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению иммуноспецифично связывают по меньшей мере один определенный эпитоп или антигенную детерминанту вируса гриппа А, вируса гриппа B или их комбинаций. В более конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению иммуноспецифично связывают по меньшей мере один определенный эпитоп или антигенную детерминанту гемагглютинина (HA) вируса гриппа B. В другом более конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению иммуноспецифично связывают по меньшей мере один определенный эпитоп или антигенную детерминанту глобулярной головки HA вируса гриппа B.
Антитело может нейтрализовать активность возбудителя инфекции, такого как вирус гриппа А и/или вирус гриппа B, в различные моменты в течение жизненного цикла вируса. Например, антитело может нарушать прикрепление вируса к клетке-мишени, препятствуя взаимодействию вируса и одного или нескольких рецепторов клеточной поверхности. Альтернативно, антитело может нарушать один или несколько видов взаимодействия вируса со своими рецепторами после прикрепления, например, препятствуя интернализации вируса в результате эндоцитоза, опосредованного рецепторами.
Как используется в данном документе, выражения "антитело" и "антитела", также известные как иммуноглобулины, охватывают моноклональные антитела (в том числе моноклональные антитела полной длины), человеческие антитела, гуманизированные антитела, верблюжьи антитела, химерные антитела, одноцепочечные Fv (scFv), одноцепочечные антитела, однодоменные антитела, доменные антитела, Fab-фрагменты, F(аb')2-фрагменты, фрагменты антител, которые проявляют необходимую биологическую активность (например, антигенсвязывающую часть), Fv c дисульфидными связями (dsFv) и антиидиотипические (анти-Id) антитела (включая, например, анти-Id антитела к антителам по настоящему изобретению), внутриклеточные антитела и эпитопсвязывающие фрагменты любого из вышеупомянутого. В частности, антитела включают молекулы иммуноглобулинов и иммунологически активные фрагменты молекул иммуноглобулинов, т.е. молекулы, которые содержат по меньшей мере один антигенсвязывающий участок. Молекулы иммуноглобулинов могут быть любого изотипа (например, IgG, IgE, IgM, IgD, IgA и IgY), субизотипа (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или аллотипа (например, Gm, например, G1m(f, z, a или x), G2m(n), G3m(g, b или c), Am, Em и Km(1, 2 или 3)).
Человеческие антитела, как правило, представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 дальтон, которые включают в себя две идентичные легкие (L) цепи и две идентичные тяжелые (H) цепи. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, при этом среди тяжелых цепей разных изотипов иммуноглобулина количество дисульфидных связей различается. Каждая тяжелая и легкая цепь также имеет расположенные с равными интервалами дисульфидные мостики между цепями. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым расположены несколько константных доменов (CH). Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен (CL) на своем другом конце; причем константный домен легкой цепи расположен на одной линии с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи расположен на одной линии с вариабельным доменом тяжелой цепи. Легкие цепи относят к лямбда-цепям либо каппа-цепям на основании аминокислотной последовательности константного участка легкой цепи. Вариабельный домен легкой каппа-цепи также может обозначаться в данном документе как VK.
Антитела по настоящему изобретению включают антитело полной длины или интактное антитело, фрагменты антител, в том числе антигенсвязывающие фрагменты, антитело с нативной последовательностью или аминокислотными вариантами, человеческие, гуманизированные, подвергнутые посттрансляционной модификации, химерные или гибридные антитела, иммуноконъюгаты и их функциональные фрагменты. Антитела могут быть модифицированы в Fc-участке для обеспечения желаемых эффекторных функций или периода полувыведения из сыворотки. Как обсуждается более подробно в следующих далее разделах в отношении соответствующих Fc-участков, "голое" антитело, связанное на клеточной поверхности, может индуцировать цитотоксичность, например, посредством антителозависимой клеточной цитотоксичности (ADCC), или с помощью привлечения компонентов системы комплемента в комплементзависимой цитотоксичности (CDC), или с помощью привлечения неспецифических цитотоксических клеток, которые экспрессируют один или несколько эффекторных лигандов, распознающих антитело, связанное с вирусом гриппа А и/или вирусом гриппа В, и впоследствии вызвать фагоцитоз клетки по типу антителозависимого клеточноопосредованного фагоцитоза (ADCP) или какого-либо другого механизма. Альтернативно, если необходимо исключить или ослабить эффекторную функцию для того, чтобы минимизировать побочные эффекты или терапевтические осложнения, можно применять определенные другие Fc-участки. Способы повышения, а также снижения или устранения эффекторной функции Fc описаны в данном документе. Кроме того, Fc-участок антител по настоящему изобретению может быть модифицирован для увеличения аффинности связывания FcRn и, таким образом, увеличения периода полувыведения из сыворотки. Альтернативно, Fc-участок может быть конъюгирован с PEG или альбумином для увеличения периода полувыведения из сыворотки или подвергнут какому-либо другому конъюгированию, которое приводит к желаемому эффекту.
В одном варианте осуществления антитела являются пригодными для диагностики, предупреждения, лечения и/или облегчения одного или нескольких симптомов инфекции, вызванной вирусом гриппа B, у млекопитающего. В другом варианте осуществления антитела являются пригодными для диагностики, предупреждения, лечения и/или облегчения одного или нескольких симптомов инфекции, вызванной вирусом гриппа A и вирусом гриппа B, у животного. Как используется в данном документе, выражение "животное" относится к млекопитающим, в том числе без ограничения людям, отличным от человека приматам, собакам, кошкам, лошадям, кроликам, мышам и крысам; и не относящимся к млекопитающим видам, в том числе без ограничения видам птиц, таким как куры, индейки, утки и перепела.
Настоящее изобретение предусматривает композицию, которая включает в себя антитело по настоящему изобретению и носитель. С целью предупреждения или лечения инфекции, вызванной вирусом гриппа B, композиции можно вводить пациенту, нуждающемуся в таком лечении. В одном варианте осуществления композицию можно вводить пациенту для предупреждения или лечения инфекции, вызванной вирусом гриппа A; инфекции, вызванной вирусом гриппа B; и их комбинаций. Настоящее изобретение также предусматривает составы, которые включают в себя антитело по настоящему изобретению и носитель. В одном варианте осуществления состав представляет собой терапевтический состав, который включает в себя фармацевтически приемлемый носитель.
В определенных вариантах осуществления настоящее изобретение предусматривает способы, пригодные для предупреждения или лечения инфекции, вызванной вирусом гриппа B, у млекопитающего, включающие введение млекопитающему терапевтически эффективного количества антитела. В других вариантах осуществления настоящее изобретение предусматривает способы, пригодные для предупреждения или лечения инфекции, вызванной вирусом гриппа A, инфекции, вызванной вирусом гриппа B, и их комбинаций у млекопитающего, включающие введение млекопитающему терапевтически эффективного количества антитела. Терапевтические композиции антитела можно вводить кратковременно (однократно), длительно или периодически, как указывается врачом.
В определенных вариантах осуществления настоящее изобретение также предусматривает готовые изделия, которые включают в себя по меньшей мере антитело по настоящему изобретению, такие как стерильные лекарственные формы и наборы. Могут быть представлены наборы, которые содержат антитела для выявления и количественного определения вируса гриппа in vitro, например, в ELISA или вестерн-блоттинге. Такое антитело, пригодное для выявления, можно обеспечить меткой, такой как флуоресцентная или радиоизотопная метка.
Терминология
Прежде чем подробно описывать настоящее изобретение, следует понимать, что настоящее изобретение не ограничивается конкретными композициями или этапами способов, поскольку таковые могут изменяться. Следует отметить, что используемые в данном описании и прилагаемой формуле изобретения формы единственного числа включают определяемые объекты во множественном числе, если из контекста явно не следует иное.
Если не определено иное, все технические и научные выражения, используемые в данном документе, имеют то же значение, которое обычно понимает специалист в данной области техники, к которой относится настоящее изобретение. Например, The Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show (2002), 2nd ed., CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed. (1999), Academic Press и Oxford Dictionary Of Biochemistry And Molecular Biology, Revised (2000), Oxford University Press, обеспечивают специалиста общим словарем многих выражений, используемых в настоящем изобретении.
Аминокислоты могут называться в данном документе по их общеизвестным трехбуквенным символам либо по однобуквенным символам, рекомендованным комиссией по биохимической номенклатуре IUPAC-IUB. Аналогично, нуклеотиды могут называться по их общепринятым однобуквенным кодам.
Антитела к вирусу гриппа B
В определенных вариантах осуществления антитела являются выделенными, и/или очищенными, и/или апирогенными антителами. Выражение "очищенный", как используется в данном документе, относится к другим молекулам, например, молекуле полипептида, нуклеиновой кислоты, которая была идентифицирована и отделена и/или извлечена из компонента своего естественного окружения. Таким образом, в одном варианте осуществления антитела по настоящему изобретению представляют собой очищенные антитела, при этом они были отделены от одного или нескольких компонентов их естественного окружения. Выражение "выделенное антитело", как используется в данном документе, относится к антителу, которое фактически не содержит других молекул антител, имеющих отличающуюся антигенную специфичность (например, выделенное антитело, специфично связывающееся с вирусом гриппа B, которое фактически не содержит антител, которые специфично связывают антигены, отличные от связываемых антителом к HA вируса гриппа В). Таким образом, в одном варианте осуществления антитела по настоящему изобретению являются выделенными антителами, которые были отделены от антител с отличающейся специфичностью. Обычно выделенное антитело представляет собой моноклональное антитело. Кроме того, выделенное антитело по настоящему изобретению может фактически не содержать одного или нескольких других материалов и/или химических веществ клетки и называется в данном документе выделенным и очищенным антителом. В одном варианте осуществления настоящего изобретения комбинация "выделенных" моноклональных антител относится к антителам, имеющим отличающиеся специфичности и объединенным в четко определенную композицию. Способы получения и очистки/выделения антител более подробно описаны ниже.
Выделенные антитела по настоящему изобретению включают в себя аминокислотные последовательности антител, раскрытые в данном документе, кодируемые любым подходящим полинуклеотидом, или любое выделенное или составленное антитело.
Антитела по настоящему изобретению иммуноспецифично связывают по меньшей мере один определенный эпитоп, специфичный для белка HA вируса гриппа B. Выражение "эпитоп", как используется в данном документе, относится к детерминанте белка, способной связываться с антителом. Эпитопы обычно включают химически активные поверхностные группы молекул, таких как аминокислоты или боковые цепи сахаров, и обычно обладают специфическими характеристиками трехмерной структуры, а также специфическими характеристиками заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первыми, но не с последними утрачивается в присутствии денатурирующих растворителей.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим на вирусах гриппа B по меньшей мере двух филогенетически разных линий. В более конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим по меньшей мере в одном штамме вируса гриппа B линии Yamagata и по меньшей мере в одном штамме вируса гриппа B линии Victoria. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим в вирусе гриппа B как линии Yamagata, так и линии Victoria. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, который является консервативным среди вирусов гриппа B как линии Yamagata, так и линии Victoria.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются по меньшей мере с одним вирусом гриппа B штамма Yamagata и по меньшей мере с одним вирусом гриппа B штамма Victoria при полумаксимальной эффективной концентрации (EC50) от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл, 100 нг/мл, 50 нг/мл, 40 нг/мл, 30 нг/мл, 20 нг/мл или 15 мкг/мл. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с вирусом гриппа B линий Yamagata и Victoria при EC50 от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл, 100 нг/мл, 50 нг/мл, 40 нг/мл, 30 нг/мл, 20 нг/мл или 15 мкг/мл. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим у вируса гриппа B как линии Yamagata, так и линии Victoria, при EC50 от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл, 100 нг/мл, 50 нг/мл, 40 нг/мл, 30 нг/мл, 20 нг/мл или 15 мкг/мл.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим на вирусе гриппа B линии Yamagata, при EC50 от приблизительно 1 нг/мл до приблизительно 100 нг/мл, от 1 нг/мл до приблизительно 50 нг/мл, или от приблизительно 1 нг/мл до приблизительно 25 нг/мл, или менее чем приблизительно 50 нг/мл или 25 нг/мл; и эпитопом, присутствующим на вирусе гриппа B линии Victoria, при EC50 от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл, 100 нг/мл или 50 нг/мл.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим на вирусе гриппа B линии Yamagata, при EC50 от приблизительно 1 нг/мл до приблизительно 100 нг/мл, от 1 нг/мл до приблизительно 50 нг/мл, или от приблизительно 1 нг/мл до приблизительно 25 нг/мл, или менее чем приблизительно 50 нг/мл или 25 нг/мл; эпитопом, присутствующим на вирусе гриппа B линии Victoria, при EC50 от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл или 100 нг/мл; и эпитопом, присутствующим на HA вируса гриппа A, при EC50 от приблизительно 1 мкг/мл до приблизительно 50 мкг/мл, или менее чем приблизительно 50 мкг/мл, 25 мкг/мл, 15 мкг/мл или 10 мкг/мл. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с эпитопом, присутствующим на вирусе гриппа B линии Yamagata, при EC50 от приблизительно 1 нг/мл до приблизительно 100 нг/мл, от 1 нг/мл до приблизительно 50 нг/мл, или от приблизительно 1 нг/мл до приблизительно 25 нг/мл, или менее чем приблизительно 50 нг/мл или 25 нг/мл; эпитопом, присутствующим на вирусе гриппа B линии Victoria, при EC50 от приблизительно 1 нг/мл до приблизительно 500 нг/мл, или от приблизительно 1 нг/мл до приблизительно 250 нг/мл, или от приблизительно 1 нг/мл до приблизительно 50 нг/мл, или менее чем приблизительно 500 нг/мл, 250 нг/мл или 100 нг/мл; и эпитопом, присутствующим на HA H9 вируса гриппа A, при EC50 от приблизительно 1 мкг/мл до приблизительно 50 мкг/мл, или менее чем приблизительно 50 мкг/мл, 25 мкг/мл, 15 мкг/мл или 10 мкг/мл.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент распознают эпитоп, который является линейным эпитопом или непрерывным эпитопом. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент распознают нелинейный или конформационный эпитоп. В одном варианте осуществления эпитоп расположен на гликопротеине гемагглютинина (HA) вируса гриппа B. В более конкретном варианте осуществления эпитоп расположен в участке гликопротеина HA, представляющем собой головку, вируса гриппа B. В одном варианте осуществления эпитоп включает в себя одну или несколько аминокислот в положениях 128, 141, 150 или 235 участка HA, представляющего собой головку, вируса гриппа B в качестве контактирующих остатков, которые пронумерованы в соответствии с системой нумерации H3, как описано в Wang et al. (2008) J. Virol. 82(6):3011-20. В одном варианте осуществления эпитоп включает в себя аминокислоту 128 из последовательности участка HA, представляющего собой головку, вируса гриппа B в качестве контактирующего остатка. В другом варианте осуществления эпитоп включает в себя аминокислоты 141, 150 и 235 из последовательности участка HA, представляющего собой головку, вируса гриппа B в качестве контактирующих остатков.
Эпитоп или эпитопы, распознаваемые антителом или его антигенсвязывающим фрагментом по настоящему изобретению, могут иметь множество путей применения. Например, эпитоп в очищенной или синтетической форме можно применять для усиления иммунных ответов (т.е. в качестве вакцины, или для выработки антител для других вариантов применения) или для скрининга сывороток на антитела, иммунореактивных в отношении эпитопа. В одном варианте осуществления эпитоп, распознаваемый антителом или его антигенсвязывающим фрагментом по настоящему изобретению, или антиген, имеющий такой эпитоп, можно применять в качестве вакцины для индуцирования иммунного ответа. В другом варианте осуществления антитела и антигенсвязывающие фрагменты по настоящему изобретению можно применять для отслеживания качества вакцин, например, с помощью определения того, содержит ли антиген в вакцине надлежащий иммуногенный эпитоп в надлежащей конформации.
Вариабельные участки
Как используется в данном документе, выражение ʺисходное антителоʺ относится к антителу, которое кодируется аминокислотной последовательностью, применяемой для получения варианта или производного, определенных в данном документе. Исходный полипептид может включать в себя нативную последовательность антитела (т.е. встречающуюся в природе, в том числе встречающийся в природе аллельный вариант) или последовательность антитела с ранее существовавшими модификациями аминокислотной последовательности (такими как другие вставки, делеции и/или замены) во встречающейся в природе последовательности. Исходное антитело может быть гуманизированным антителом или человеческим антителом. В одном варианте осуществления антитела по настоящему изобретению являются вариантами исходного антитела. Как используется в данном документе, выражение "вариант" относится к антителу, которое отличается по аминокислотной последовательности от аминокислотной последовательности "исходного" антитела вследствие добавления, делеции и/или замены одного или нескольких аминокислотных остатков в последовательности исходного антитела.
Антигенсвязывающая часть антитела включает в себя один или несколько фрагментов антитела, которые сохраняют способность специфично связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может осуществляться с помощью фрагментов антитела полной длины. Примеры антигенсвязывающих фрагментов, охватываемые выражением "антигенсвязывающая часть" антитела, включают (i) Fab-фрагмент, моновалентный фрагмент, включающий в себя VL-, VH-, CL- и CH1-домены; (ii) F(ab')2-фрагмент, бивалентный фрагмент, включающий в себя два Fab-фрагмента, связанные дисульфидным мостиком в шарнирном участке; (iii) Fd-фрагмент, включающий в себя VH- и CH1-домены; (iv) Fv-фрагмент, включающий в себя VL- и VH-домены одного плеча антитела, (v) dAb-фрагмент (Ward et al. (1989) Nature. 341:544-546), включающий в себя VH-домен; и (vi) выделенный участок, определяющий комплементарность (CDR). Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их можно соединить с применением рекомбинантных способов с помощью синтетического линкера, который обеспечивает возможность получения из них одной белковой цепи, в которой VL- и VH-участки спариваются с образованием моновалентной молекулы (известной как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science. 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Подразумевается, что такие одноцепочечные антитела также охвачены выражением "антигенсвязывающая часть" антитела. Такие фрагменты антител получают с помощью традиционных методик, известных специалистам в данной области, и фрагменты проверяют на пригодность таким же образом, как и интактные антитела. Антигенсвязывающие части можно получить с помощью методик рекомбинантной ДНК или с помощью ферментативного или химического расщепления интактных иммуноглобулинов.
Антитела по настоящему изобретению включают в себя по меньшей мере один антигенсвязывающий домен, включающий VH- и VL-домены, описанные в данном документе. Иллюстративные VH- и VL-домены показаны в таблице 1 ниже.
Таблица 1. VH- и VL-домены
В определенных вариантах осуществления очищенные антитела включают в себя VH и/или VL, которые характеризуются определенным процентом идентичности по меньшей мере с одной из последовательностей VH и/или VL, раскрытых в данном документе. Как используется в данном документе, выражение "процент (%) идентичности последовательности", также включающее "гомологию", определено как процентная доля аминокислотных остатков или нуклеотидов в последовательности-кандидате, идентичных аминокислотным остаткам или нуклеотидам в эталонных последовательностях, таких как последовательность исходного антитела, после выравнивания последовательностей и введения гэпов, если необходимо, для достижения максимального процента идентичности последовательности, и не учитывает какие-либо консервативные замены как часть идентичности последовательности. Оптимальное выравнивание последовательностей для сравнения можно выполнить, кроме как ручным способом, с помощью алгоритма поиска локальной гомологии Смита-Уотермана (1981) Ads App. Math. 2: 482, с помощью алгоритма поиска локальной гомологии Нидлмана-Вунша (1970), J. Mol. Biol. 48: 443, с помощью способа поиска подобия Липмана-Пирсона (1988), Proc. Natl Acad. Sci. USA 85: 2444, или с помощью компьютерных программ, в которых применяются эти алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Drive, Мэдисон, Висконсин).
Антитела по настоящему изобретению могут включать в себя аминокислотную последовательность VH по меньшей мере c 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичностью с аминокислотными последовательностями VH, описанными в данном документе. В другом варианте осуществления антитела по настоящему изобретению могут иметь аминокислотную последовательность VH по меньшей мере c 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с аминокислотными последовательностями VH, описанными в данном документе.
Антитела по настоящему изобретению могут включать в себя аминокислотную последовательность VL по меньшей мере c 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичностью с аминокислотными последовательностями VL, описанными в данном документе. В другом варианте осуществления антитела по настоящему изобретению могут иметь аминокислотную последовательность VL по меньшей мере c 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с аминокислотными последовательностями VL, описанными в данном документе.
Участки, определяющие комплементарность (CDR)
Несмотря на то, что вариабельный домен (VH и VL) включает в себя антигенсвязывающий участок, вариабельность распределяется по вариабельным доменам антител неравномерно. Она сосредоточена в сегментах, называемых участками, определяющими комплементарность (CDR), в вариабельных доменах как легкой цепи (VL или VK), так и тяжелой цепи (VH). Более высококонсервативные части вариабельных доменов называются каркасными областями (FR). Каждый из вариабельных доменов нативной тяжелой и легкой цепей включает в себя четыре FR, по большей части принимающих конфигурацию β-листа, соединенных тремя CDR, которые образуют петли, объединяющие структуру β-листа и в некоторых случаях образующие ее часть. В каждой цепи CDR удерживаются в непосредственной близости с помощью FR и вместе с CDR из другой цепи участвуют в образовании антиген-связывающего участка антител. Три CDR тяжелой цепи обозначают HCDR-1, HCDR-2 и HCDR-3, а три CDR легкой цепи обозначают LCDR-1, LCDR-2 и LCDR-3.
В одном варианте осуществления аминокислоты в вариабельном домене, участках, определяющих комплементарность (CDR), и каркасных участках (FR) антитела можно идентифицировать в соответствии с Kabat et al. (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. При применении этой системы нумерации фактическая линейная аминокислотная последовательность может содержать меньшее количество аминокислот или дополнительные аминокислоты, соответствующие укорочению FR или CDR вариабельного домена или вставке в них. Например, вариабельный домен тяжелой цепи может содержать вставку одной аминокислоты (остаток 52a согласно Kabat) после остатка 52 в H2 и вставленные остатки (например, остатки 82a, 82b и 82c и т.д., согласно Kabat) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Kabat можно определить для данного антитела путем выравнивания участков антитела с гомологией последовательности антитела со "стандартной" последовательностью с нумерацией по Kabat. Максимальное выравнивание остатков каркасных участков часто требует вставки "спейсерных" остатков в систему нумерации. Кроме того, идентичность определенных отдельных остатков в любом указанном участке с нумерацией по Kabat может изменяться от цепи антитела к цепи антитела вследствие межвидовой или аллельной дивергенции.
Согласно системе нумерации по Kabat et al. HCDR-1 начинается примерно с аминокислоты 31 (т.е. примерно через 9 остатков от первого цистеинового остатка), включает в себя примерно 5-7 аминокислот и заканчивается на следующем тирозиновом остатке. HCDR-2 начинается с пятнадцатого остатка от конца CDR-H1, включает в себя примерно 16-19 аминокислот и заканчивается на следующем аргининовом или лизиновом остатке. HCDR-3 начинается примерно с тридцать третьего аминокислотного остатка от конца HCDR-2; включает в себя 3-25 аминокислот и заканчивается на последовательности W-G-X-G, где X представляет собой любую аминокислоту. LCDR-1 начинается примерно с остатка 24 (т.е. после цистеинового остатка), включает в себя примерно 10-17 остатков и заканчивается на следующем тирозиновом остатке. LCDR-2 начинается примерно с шестнадцатого остатка от конца LCDR-1 и включает в себя примерно 7 остатков. LCDR-3 начинается примерно с тридцать третьего остатка от конца LCDR-2, включает в себя примерно 7-11 остатков и заканчивается на последовательности F-G-X-G, где X представляет собой любую аминокислоту. Следует отметить, что CDR значительно изменяются от антитела к антителу (и по определению не будут проявлять гомологию с консенсусными последовательностями по Kabat). Последовательности CDR тяжелой цепи и легкой цепи антител по настоящему изобретению, пронумерованные с помощью системы Kabat, показаны в таблицах 2 и 3 ниже.
Таблица 2. HCDR-1-3, определенные по Kabat et al.
SEQ ID NO:
SEQ ID NO:
SEQ ID NO:
Таблица 3. LCDR-1-3, определенные по Kabat et al.
SEQ ID NO:
SEQ ID NO:
SEQ ID NO:
Несмотря на широкое применение схемы нумерации по Kabat, она имеет некоторые недостатки. Во-первых, поскольку схема нумерации была разработана на основании данных о последовательностях, в отсутствие информации о структуре, положение, в котором вставки находятся в LCDR-1 и HCDR-1, не всегда совпадает с положением вставки с точки зрения структуры. Таким образом, топологически эквивалентным остаткам в этих петлях может быть не присвоен один и тот же номер. Во-вторых, система нумерации не является гибкой, позволяя вносить только ограниченное количество вставок. Если имеется больше остатков, чем предназначенное в рамках системы нумерации для вставок, то не существует стандартного способа их нумерации.
В другом варианте осуществления аминокислоты в вариабельном домене, участках, определяющих комплементарность (CDR), и каркасных участках (FR) антитела можно идентифицировать с помощью базы данных Immunogenetics (IMGT) (http://imgt.cines.fr). Lefranc et al. (2003) Dev Comp Immunol. 27(1):55-77. База данных IMGT была разработана с применением информации о последовательностях иммуноглобулинов (IgG), T-клеточных рецепторов (TcR) и молекул главного комплекса гистосовместимости (MHC), и она унифицирует нумерацию в легких цепях лямбда и каппа, тяжелых цепях антител и цепях T-клеточных рецепторов, а также позволяет избежать применения кодов вставки для всех, за исключением чрезвычайно длинных вставок. IMGT также учитывает и объединяет в себе определение каркасных участков (FR) и участков, определяющих комплементарность (CDR), согласно Kabat et al., определение характеристик гипервариабельных петель согласно Chothia et al., а также данные о структуре из рентгеноструктурных исследований. Последовательности CDR тяжелой цепи и легкой цепи антител по настоящему изобретению, пронумерованные с помощью системы IMGT, показаны в таблицах 4 и 5 ниже. На фигуре 5 представлено выравнивание последовательностей FBD-56, FBD-94 и FBC-39 с указанием последовательностей CDR, идентифицированных по Kabat и IMGT.
Таблица 4. HCDR-1-3, определенные по IMGT
SEQ ID NO:
SEQ ID NO:
SEQ ID NO:
Таблица 5. LCDR-1-3, определенные по IMGT
SEQ ID NO:
SEQ ID NO:
SEQ ID NO:
Настоящее изобретение охватывает нейтрализующие антитела к вирусу гриппа B, включающие в себя аминокислоты в последовательности, которая по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентична аминокислотной последовательности VH с SEQ ID NO: 2; SEQ ID NO: 12; SEQ ID NO: 22; SEQ ID NO: 32; SEQ ID NO: 42; SEQ ID NO: 52; SEQ ID NO: 62; SEQ ID NO: 74; SEQ ID NO: 90 или SEQ ID NO: 106; и/или по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентична аминокислотной последовательности VL с SEQ ID NO: 7; SEQ ID NO: 17; SEQ ID NO: 27; SEQ ID NO: 37; SEQ ID NO: 47; SEQ ID NO: 57; SEQ ID NO: 67; SEQ ID NO: 82; SEQ ID NO: 98 или SEQ ID NO: 114. В другом варианте осуществления настоящее изобретение охватывает нейтрализующие антитела к вирусу гриппа B, включающие в себя аминокислоты в последовательности, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности VH с SEQ ID NO: 2; SEQ ID NO: 12; SEQ ID NO: 22; SEQ ID NO: 32; SEQ ID NO: 42; SEQ ID NO: 52; SEQ ID NO: 62; SEQ ID NO: 74; SEQ ID NO: 90 или SEQ ID NO: 106; и/или по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности VL с SEQ ID NO: 7; SEQ ID NO: 17; SEQ ID NO: 27; SEQ ID NO: 37; SEQ ID NO: 47; SEQ ID NO: 57; SEQ ID NO: 67; SEQ ID NO: 82; SEQ ID NO: 98 или SEQ ID NO: 114.
В другом варианте осуществления настоящее изобретение предусматривает антитела и их антигенсвязывающие фрагменты, которые включают в себя совокупность из шести CDR: HCDR-1, HCDR-2, HCDR-3, LCDR-1, LCDR-2, LCDR-3, где CDR выбраны из HCDR и LCDR, приведенных в таблицах 2-5. В другом варианте осуществления настоящее изобретение предусматривает антитела и их антигенсвязывающие фрагменты, которые включают в себя совокупность из шести CDR: HCDR-1, HCDR-2, HCDR-3, LCDR-1, LCDR-2, LCDR-3, где CDR включают в себя аминокислоты в последовательности, которая по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентична аминокислотной последовательности HCDR и LCDR, приведенных в таблицах 2-5. В другом варианте осуществления настоящее изобретение предусматривает антитела и их антигенсвязывающие фрагменты, которые включают в себя совокупность из шести CDR: HCDR-1, HCDR-2, HCDR-3, LCDR-1, LCDR-2, LCDR-3, где CDR включают в себя аминокислоты в последовательности, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности HCDR и LCDR, приведенных в таблицах 2-5.
Каркасные области
Каждый из вариабельных доменов тяжелой и легкой цепей включает в себя четыре каркасные участки (FR1, FR2, FR3, FR4), которые являются наиболее высоко консервативными частями вариабельных доменов. Четыре FR тяжелой цепи обозначают FR-H1, FR-H2, FR-H3 и FR-H4, а четыре FR легкой цепи обозначают FR-L1, FR-L2, FR-L3 и FR-L4.
В одном варианте осуществления для идентификации каркасных участков можно применять систему нумерации по Kabat. Согласно Kabat et al. FR-H1 начинается в положении 1 и заканчивается примерно на аминокислоте 30, FR-H2 расположен примерно от аминокислоты 36 до 49, FR-H3 расположен примерно от аминокислоты 66 до 94 и FR-H4 расположен примерно от аминокислоты 103 до 113. FR-L1 начинается с аминокислоты 1 и заканчивается примерно на аминокислоте 23, FR-L2 расположен примерно от аминокислоты 35 до 49, FR-L3 расположен примерно от аминокислоты 57 до 88, и FR-L4 расположен примерно от аминокислоты 98 до 107. В определенных вариантах осуществления каркасные области могут содержать замены в соответствии с системой нумерации по Kabat, например, вставку в 106A в FR-L1. Кроме встречающихся в природе замен, в антитело по настоящему изобретению также можно ввести одно или несколько изменений (например, замен) остатков FR, при условии, что оно сохранит нейтрализующую способность. В определенных вариантах осуществления они приводят к улучшению или оптимизации аффинности связывания антитела с HA вируса гриппа B. Примеры подлежащих модифицированию остатков каркасного участка включают остатки, которые непосредственно не ковалентно связывают антиген (Amit et al. (1986) Science. 233:747-753); взаимодействуют с CDR/влияют на его конформацию (Chothia et al. (1987) J. Mol. Biol. 196:901-917); и/или участвуют в образовании области контакта VL-VH (патент США № 5225539). В других вариантах осуществления каркасные участки могут быть идентифицированы с помощью системы нумерации IMGT.
В другом варианте осуществления FR может включать в себя одно или несколько аминокислотных изменений с целью "приведения в соответствие с зародышевым типом". Например, аминокислотные последовательности тяжелой и легкой цепей выбранного антитела можно сравнить с аминокислотными последовательностями тяжелой и легкой цепей зародышевого типа; при этом если определенные каркасные остатки выбранных VL и/или VH цепей отличаются от конфигурации зародышевого типа (например, в результате соматической мутации иммуноглобулина), может быть желательно "мутировать к первоначальному виду" измененные каркасные остатки выбранных антител до конфигурации зародышевого типа (т.е. изменить аминокислотные последовательности каркасных участков выбранных антител так, чтобы они были такими же, как аминокислотные последовательности каркасных участков зародышевого типа), например, для снижения риска иммуногенности. Такую "обратную мутацию" (или "приведение в соответствие с зародышевым типом") остатков каркасных областей можно осуществить с помощью стандартных способов молекулярной биологии для внедрения специфических мутаций (например, сайт-направленного мутагенеза; ПЦР-опосредованного мутагенеза и т.п.).
На фигуре 6 показана аминокислотная последовательность VH-домена обобщенного антитела к вирусу гриппа B (SEQ ID NO:71), в которой несвойственные зародышевому типу остатки FBC-39 (SEQ ID NO: 22) обозначены как Xaa1-11. В одном варианте осуществления один или несколько несвойственных зародышевому типу (Xaa1-11) остатков "подвергают мутации к первоначальному виду" с получением остатков, свойственных зародышевому типу. В одном варианте осуществления Xaa1 в SEQ ID NO: 71 представляет собой Val или Glu; Xaa2 в SEQ ID NO: 71 представляет собой Leu или Phe; Xaa3 в SEQ ID NO: 71 представляет собой Ser или Thr; Xaa4 в SEQ ID NO: 71 представляет собой Leu или Ser; Xaa5 в SEQ ID NO: 71 представляет собой Ser или Thr; Xaa6 в SEQ ID NO: 71 представляет собой Met или Thr; Xaa7 в SEQ ID NO: 71 представляет собой Phe или Tyr; Xaa8 в SEQ ID NO: 71 представляет собой His или Gln; Xaa9 в SEQ ID NO: 71 представляет собой Ser или Asn; Xaa10 в SEQ ID NO: 71 представляет собой Arg или Lys и Xaa11 в SEQ ID NO: 71 представляет собой Ala или Thr. В другом варианте осуществления Xaa1 в SEQ ID NO:71 представляет собой Glu; Xaa5 в SEQ ID NO:71 представляет собой Thr; Xaa6 в SEQ ID NO:71 представляет собой Thr; Xaa7 в SEQ ID NO:71 представляет собой Tyr; Xaa8 в SEQ ID NO:71 представляет собой Gln; Xaa10 в SEQ ID NO:71 представляет собой Lys; Xaa11 в SEQ ID NO:71 представляет собой Thr или их комбинации. В более конкретном варианте осуществления Xaa9 в SEQ ID NO:71 представляет собой Ser. В еще одном варианте осуществления Xaa4 в SEQ ID NO:71 представляет собой Leu.
На фигуре 7 показана аминокислотная последовательность VL-домена обобщенного антитела к вирусу гриппа B (SEQ ID NO:72), в которой несвойственные зародышевому типу остатки FBC-39 (SEQ ID NO: 27) представлены Xaa1. В одном варианте осуществления несвойственный зародышевому типу остаток (Xaa1) "подвергают мутации к первоначальному виду" с получением остатков, свойственных зародышевому типу. В одном варианте осуществления Xaa1 в SEQ ID NO:72 представляет собой Phe или Tyr. В более конкретном варианте осуществления Xaa1 в SEQ ID NO:72 представляет собой Tyr.
Нуклеотидные последовательности, кодирующие антитела по настоящему изобретению
Кроме аминокислотных последовательностей, описанных выше, настоящее изобретение дополнительно предусматривает нуклеотидные последовательности, соответствующие аминокислотным последовательностям и кодирующие человеческие антитела по настоящему изобретению. В одном варианте осуществления настоящее изобретение предусматривает полинуклеотиды, которые включают в себя нуклеотидную последовательность, кодирующую антитело, описанное в данном документе, или его фрагменты. Они включают, без ограничения, нуклеотидные последовательности, которые кодируют упомянутые выше аминокислотные последовательности. Таким образом, настоящее изобретение также предусматривает нуклеотидные последовательности, кодирующие каркасные области VH и VL, в том числе CDR и FR антител, описанных в данном документе, а также векторы экспрессии для их эффективной экспрессии в клетках (например, клетках млекопитающих). Способы получения антител с применением полинуклеотидов более подробно описаны ниже.
Настоящее изобретение также охватывает полинуклеотиды, которые гибридизируются в жестких условиях или условиях низкой жесткости гибридизации, например, как определено в данном документе, с полинуклеотидами, кодирующими антитело по настоящему изобретению, описанное в данном документе. Выражение "жесткость", как используется в данном документе, относится к экспериментальным условиям (например, температуре и концентрации солей) эксперимента по гибридизации для обозначения степени гомологии между зондом и связанной на фильтре нуклеиновой кислотой; чем выше жесткость, тем выше процент гомологии между зондом и связанной на фильтре нуклеиновой кислотой.
Жесткие условия гибридизации включают без ограничения гибридизацию со связанной на фильтре ДНК в 6X хлориде натрия/цитрате натрия (SSC) при приблизительно 45°C, за которой следует одна или несколько промывок в 0,2X SSC/0,1% SDS при приблизительно 50-65°C, условия высокой жесткости, такие как гибридизация со связанной на фильтре ДНК в 6X SSC при приблизительно 45°C, за которой следует одна или несколько промывок в 0,1X SSC/0,2% SDS при приблизительно 65°C, или любые другие жесткие условия гибридизации, известные специалисту в данной области (см., например, Ausubel et al., eds. (1989) Current Protocols in Molecular Biology, vol. 1, Green Publishing Associates, Inc. and John Wiley and Sons, Inc., NY на страницах 6.3.1-6.3.6 и 2.10.3).
Фактически идентичные последовательности могут быть полиморфными последовательностями, т.е. альтернативными последовательностями или аллелями в популяции. Различие между аллелями может составлять до одной пары оснований. Фактически идентичные последовательности могут также включать подвергнутые мутагенезу последовательности, в том числе последовательности, содержащие молчащие мутации. Мутация может включать изменения одного или нескольких остатков, делецию одного или нескольких остатков или вставку одного или нескольких дополнительных остатков.
С помощью любого способа, известного из уровня техники, можно получить полинуклеотиды и можно определить нуклеотидную последовательность полинуклеотидов. Например, если известна нуклеотидная последовательность антитела, то полинуклеотид, кодирующий антитело, можно собрать из химически синтезированных олигонуклеотидов (например, как описано в Kutmeier et al. (1994) BioTechniques. 17:242), что, вкратце, включает синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей антитело, отжиг и лигирование этих олигонуклеотидов, а затем амплификацию лигированных олигонуклеотидов с помощью ПЦР.
Полинуклеотид, кодирующий антитело, можно также получить из нуклеиновой кислоты из подходящего источника. Если клон, содержащий нуклеиновую кислоту, кодирующую конкретное антитело, недоступен, а последовательность молекулы антитела известна, то нуклеиновую кислоту, кодирующую иммуноглобулин, можно химически синтезировать или получить из подходящего источника (например, библиотеки кДНК антител, или библиотеки кДНК, полученной из, или нуклеиновой кислоты, в одном варианте осуществления поли(A)+РНК, выделенной из любой ткани или клеток, экспрессирующих антитело, таких как клетки гибридомы, выбранные для экспрессии антитела) с помощью ПЦР-амплификации с применением синтетических праймеров, гибридизируемых с 3'- и 5'-концами последовательности, или с помощью клонирования с применением олигонуклеотидного зонда, специфичного в отношении конкретной последовательности гена, подлежащей идентификации, например, клона кДНК из библиотеки кДНК, кодирующего антитело. Амплифицированные нуклеиновые кислоты, полученные с помощью ПЦР, можно затем клонировать в реплицируемые клонирующие векторы с помощью любого способа, хорошо известного из уровня техники.
Когда нуклеотидная последовательность и соответствующая аминокислотная последовательность антитела определены, с этой нуклеотидной последовательностью для антитела можно производить манипуляции с помощью хорошо известных из уровня техники способов манипуляции с нуклеотидными последовательностями, например, методик рекомбинантных ДНК, сайт-направленного мутагенеза, ПЦР и т.д. (см., например, методики, описанные в Sambrook et al. (1990) Molecular Cloning, A Laboratory Manual, 2d Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. и Ausubel et al. eds. (1998), Current Protocols in Molecular Biology, John Wiley & Sons, NY) с образованием антител, имеющих отличающуюся аминокислотную последовательность, например, чтобы произвести замены, делеции и/или вставки аминокислот.
Характеристики связывания
Как описано выше антитела или антигенсвязывающие фрагменты по настоящему изобретению иммуноспецифично связывают по меньшей мере один определенный эпитоп или антигенную детерминанту белка, пептида, субъединицы, фрагмента, части HA вируса гриппа B или любой их комбинации исключительно или преимущественно по отношению к другим полипептидам. В конкретном варианте осуществления эпитоп или антигенная детерминанта белка HA вируса гриппа B представляют собой глобулярную головку. Выражение "эпитоп" или "антигенная детерминанта", как используется в данном документе, относится к белковой детерминанте, способной связываться с антителом. В одном варианте осуществления выражение "связывание" в данном документе относится к специфичному связыванию. Эти белковые детерминанты или эпитопы обычно включают в себя химически активные поверхностные группы молекул, таких как аминокислоты или боковые цепи сахаров, и обычно обладают специфическими характеристиками трехмерной структуры, а также специфическими характеристиками заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первыми, но не с последними утрачивается в присутствии денатурирующих растворителей. Выражение "прерывистый эпитоп", как используется в данном документе, относится к конформационному эпитопу на белковом антигене, который образован по меньшей мере из двух разделенных участков в первичной последовательности белка.
Взаимодействия между антигенами и антителами являются такими же, как и другие нековалентные белок-белковые взаимодействия. В целом, между антигенами и антителами существуют четыре типа связывающих взаимодействий: (i) водородные связи, (ii) дисперсионные силы, (iii) электростатические силы между кислотами Льюиса и основаниями Льюиса и (iv) гидрофобные взаимодействия. Гидрофобные взаимодействия являются основной движущей силой для взаимодействия антитела и антигена и основаны на отталкивании воды неполярными группами, а не на притяжении молекул (Tanford, (1978) Science. 200:1012-8). Однако, определенные физические силы также способствуют связыванию антигена и антитела, например, совместимость или комплементарность форм эпитопов и различных связывающих участков антител. Кроме того, другие материалы и антигены могут перекрестно реагировать с антителом, конкурируя, таким образом, за доступное свободное антитело.
Измерение константы аффинности и специфичности связывания между антигеном и антителом может облегчить определение эффективности профилактических, терапевтических, диагностических и исследовательских способов, в которых применяются антитела по настоящему изобретению. "Аффинность связывания" в целом относится к силе суммы всех нековалентных взаимодействий между одним связывающим участком молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иное, как используется в данном документе, "аффинность связывания" относится к присущей аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y в целом можно представить равновесной константой диссоциации (Kd), которую рассчитывают в виде соотношения koff/kon. См., например, Chen et al. (1999) J. Mol Biol. 293:865-881. Антитела с низкой аффинностью, как правило, связывают антиген медленно и проявляют тенденцию к легкой диссоциации, тогда как антитела с высокой аффинностью, как правило, связывают антиген быстрее и проявляют тенденцию к тому, чтобы оставаться связанными дольше. Из уровня техники известен ряд способов измерения аффинности связывания, любой из которых можно применять для целей настоящего изобретения.
Один способ определения аффинности связывания включает измерение константы диссоциации ʺKdʺ c помощью анализа связывания меченного радиоактивным изотопом антигена (RIA),проводимого с Fab-вариантом антитела, представляющего интерес, и его антигеном, как описано в Chen et al. (1999) J. Mol Biol. 293:865-881. Альтернативно, значение Kd можно измерить с помощью анализов поверхностного плазмонного резонанса с применением BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Пискатауэй, Нью-Джерси). Если на основании анализа поверхностного плазмонного резонанса скорость прямой реакции превышает 106 М-1 с-1, то скорость прямой реакции можно определить путем применения методики гашения флуоресценции, в которой измеряют повышение или снижение интенсивности излучения флуоресценции в присутствии возрастающих концентраций антигена. "Скорость прямой реакции", или "скорость ассоциации", или "kon" можно также определить с помощью такой же методики поверхностного плазмонного резонанса, описанной выше.
Способы и реактивы, подходящие для определения характеристик связывания антитела по настоящему изобретению или его измененного/мутантного производного, известны из уровня техники и/или коммерчески доступны (патенты США №№ 6849425; 6632926; 6294391; 6143574). Кроме того, коммерчески доступно оборудование и программное обеспечение, предназначенное для таких кинетических анализов (например, приборы Biacore® A100 и Biacore® 2000; Biacore International AB, Уппсала, Швеция).
В одном варианте осуществления антитела по настоящему изобретению, в том числе их антигенсвязывающие фрагменты или варианты, также могут быть описаны или определены с точки зрения их аффинности связывания с полипептидами вируса гриппа A; полипептидами вируса гриппа B или их комбинацией. Обычно антитела с высокой аффинностью имеют Kd менее 10-7 М. В одном варианте осуществления антитела или их антигенсвязывающие фрагменты связывают полипептиды вируса гриппа A; полипептиды вируса гриппа B; их фрагменты или варианты или их комбинацию при константе диссоциации или Kd, меньшей или равной 5×10-7 М, 10-7 M, 5×10-8 M, 10-8 M, 5×10-9 M, 10-9 M, 5×10-10 M, 10-10 M, 5×10-11 M, 10-11 M, 5×10-12 M, 10-12 M, 5×10-13 M, 10-13 M, 5×10-14 M, 10-14 M, 5×10-15 M или 10-15 M. В более конкретном варианте осуществления антитела или их антигенсвязывающие фрагменты связывают полипептиды вируса гриппа A; полипептиды вируса гриппа B, их фрагменты или варианты или их комбинации при константе диссоциации или Kd, меньшей или равной 5×10-10 М, 10-10 М, 5×10-11 М, 10-11 М, 5×10-12 М или 10-12 М. Настоящее изобретение охватывает антитела, которые связывают полипептиды вируса гриппа A; полипептиды вируса гриппа В или их комбинацию при константе диссоциации или Kd, которая находится в диапазоне между любыми из отдельных упомянутых значений.
В другом варианте осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению связывают полипептиды вируса гриппа A, полипептиды вируса гриппа В; их фрагменты или варианты, или их комбинации со скоростью обратной реакции (koff), меньшей или равной 5×10-2 с-1, 10-2 с-1, 5×10-3 с-1 или 10-3 с-1, 5×10-4 с-1, 10-4 с-1, 5×10-5 с-1 или 10-5 с-1, 5×10-6 с-1, 10-6 с-1, 5×10-7 с-1 или 10-7 с-1. В более конкретном варианте осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению связывают полипептиды вируса гриппа A или их фрагменты или варианты со скоростью обратной реакции (koff), меньшей или равной 5×10-4 с-1, 10-4 с-1, 5×10-5 с-1 или 10-5 c-1, 5×10-6 c-1, 10-6 c-1, 5×10-7 c-1 или 10-7 с-1. Настоящее изобретение также охватывает антитела, которые связывают полипептиды вируса гриппа A, полипептиды вируса гриппа В или их комбинации со скоростью обратной реакции (koff), которая находится в диапазоне между любыми из отдельных упомянутых значений.
В другом варианте осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению связывают полипептиды вируса гриппа A, полипептиды вируса гриппа B, их фрагменты или варианты или их комбинации со скоростью прямой реакции (kon), большей или равной 103 M-1 с-1, 5×103 M-1 с-1, 104 M-1 с-1, 5×104 M-1 с-1, 105 M-1 с-1, 5×105 M-1 с-1, 106 M-1 с-1, 5×106 M-1 с-1, 107 M-1 с-1 или 5×107 M-1 с-1. В более конкретном варианте осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению связывают полипептиды вируса гриппа A; полипептиды вируса гриппа В; их фрагменты или варианты или их комбинации со скоростью прямой реакции (kon), большей или равной 105 M-1 с-1, 5×105 M-1 с-1, 106 M-1 с-1, 5×106 M-1 с-1, 107 M-1 с-1 или 5×107 M-1 с-1. Настоящее изобретение охватывает антитела, которые связывают полипептиды вируса гриппа A, полипептиды вируса гриппа В или их комбинации со скоростью прямой реакции (kon), которая находится в диапазоне между любыми из отдельных упомянутых значений.
В одном варианте осуществления анализ связывания можно проводить или в виде анализа прямого связывания, или в виде анализа конкурентного связывания. Связывание можно выявить с применением стандартных анализов ELISA или стандартных анализов по методу проточной цитометрии. В анализе прямого связывания антитело-кандидат исследуют в отношении связывания с его когнатным антигеном. В анализе конкурентного связывания, с другой стороны, оценивают способность антитела-кандидата конкурировать с известным антителом или другим соединением, которое связывается с конкретным антигеном, например, HA вируса гриппа В. В целом, любой способ, который позволяет выявить связывание антитела с HA вируса гриппа B, включен в объем настоящего изобретения для выявления и измерения характеристик связывания антител. Специалисту в данной области понятны эти хорошо известные способы, и по этой причине они не представлены подробно в данном документе. Эти способы также используются для скрининга панели антител с выявлением обеспечивающих желаемые характеристики.
В одном варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается с HA вируса гриппа B и способно к нейтрализации инфекции, вызванной вирусом гриппа B. В одном варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается по меньшей мере c одним вирусом гриппа B линии Yamagata и по меньшей мере с одним вирусом гриппа B линии Victoria. В другом варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается c вирусом гриппа B линии Yamagata и линии Victoria.
В другом варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается c HA вируса гриппа B и HA вируса гриппа A и способно к нейтрализации инфекции, вызванной вирусом гриппа B и вирусом гриппа A. В одном варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается по меньшей мере c одним вирусом гриппа B линии Yamagata; по меньшей мере с одним вирусом гриппа B линии Victoria и по меньшей мере с одним подтипом вируса гриппа А.
Подтипы гемагглютинина вирусов гриппа A разделяются на две основные филогенетические группы, идентифицируемые как группа 1, которая включает подтипы H1, H2, H5, H6, H8, H9, H11, H12, H13, H16 и H17, и группа 2, которая включает подтипы H3, H4, H7, H10, H14 и H15. В одном варианте осуществления антитело или антигенсвязывающий фрагмент в соответствии с настоящим изобретением способны к связыванию и/или нейтрализации одного или нескольких подтипов группы 1 вируса гриппа A, выбранных из H8, H9, H11, H12, H13, H16 и H17 и их вариантов. В другом варианте осуществления антитело или антигенсвязывающий фрагмент в соответствии с настоящим изобретением способны к связыванию и/или нейтрализации одного или нескольких подтипов группы 2 вируса гриппа A, выбранных из H4, H10, H14 и H15 и их вариантов. В одном варианте осуществления антитело по настоящему изобретению связывается с подтипом H9 группы 1 вируса гриппа A. В одном варианте осуществления антитело по настоящему изобретению связывается с подтипом H9 группы 1 вируса гриппа A и нейтрализует его.
В одном варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается с HA вируса гриппа B и способно к нейтрализации инфекции, вызванной вирусом гриппа B. В другом варианте осуществления антитело по настоящему изобретению иммуноспецифически связывается c HA вируса гриппа A и вируса гриппа B и способно к нейтрализации инфекции, вызванной вирусом гриппа B и вирусом гриппа A. Анализы нейтрализации можно проводить, как описано в данном документе в разделе "Примеры", или с помощью других способов, известных из уровня техники. Выражение ʺ50% ингибирующая концентрацияʺ (сокращенно "IC50") представляет концентрацию ингибитора (например, антитела по настоящему изобретению), которая требуется для 50% нейтрализации вируса гриппа A и/или вируса гриппа B. Специалисту в данной области будет понятно, что более низкое значение IC50 соответствует более эффективному ингибитору.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент в соответствии с настоящим изобретением характеризуются IC50 для нейтрализации вируса гриппа B в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 5 мкг/мл, или в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 1 мкг/мл антитела, или менее 5 мкг/мл, менее 2 мкг/мл, менее 1 мкг/мл, менее 0,5 мкг/мл, менее 0,1 мкг/мл, менее 0,05 мкг/мл или менее 0,01 мкг/мл в анализе микронейтрализации.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент в соответствии с настоящим изобретением характеризуются IC50 для нейтрализации вируса гриппа B в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 5 мкг/мл, или в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 1 мкг/мл антитела, или менее 5 мкг/мл, менее 2 мкг/мл, менее 1 мкг/мл, менее 0,5 мкг/мл, менее 0,1 мкг/мл, менее 0,05 мкг/мл или менее 0,01 мкг/мл в анализе микронейтрализации; и IC50 для нейтрализации вируса гриппа A в диапазоне от приблизительно 0,1 мкг/мл до приблизительно 5 мкг/мл, или в диапазоне от приблизительно 0,1 мкг/мл до приблизительно 2 мкг/мл антитела, или менее 5 мкг/мл, менее 2 мкг/мл, менее 1 мкг/мл или менее 0,5 мкг/мл для нейтрализации вируса гриппа A в анализе микронейтрализации.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент в соответствии с настоящим изобретением характеризуются IC50 для нейтрализации вируса гриппа B в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 50 мкг/мл, или в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 5 мкг/мл антитела, или в диапазоне от приблизительно 0,001 мкг/мл до приблизительно 1 мкг/мл антитела, или менее 10 мкг/мл, менее 5 мкг/мл, менее 1 мкг/мл, менее 0,5 мкг/мл, менее 0,1 мкг/мл, менее 0,05 мкг/мл или менее 0,01 мкг/мл в анализе микронейтрализации; и IC50 для нейтрализации вируса гриппа A в диапазоне от приблизительно 0,01 мкг/мл до приблизительно 50 мкг/мл, или в диапазоне от приблизительно 0,05 мкг/мл до приблизительно 5 мкг/мл антитела, или в диапазоне от приблизительно 0,1 мкг/мл до приблизительно 2 мкг/мл антитела, или менее 50 мкг/мл, менее 25 мкг/мл, менее 10 мкг/мл, менее 5 мкг/мл или менее 2 мкг/мл для нейтрализации вируса гриппа A в анализе микронейтрализации.
В определенных вариантах осуществления антитела по настоящему изобретению могут индуцировать гибель клеток. Антитело, которое "индуцирует гибель клеток", приводит к тому, что жизнеспособная клетка становится нежизнеспособной. Гибель клеток in vitro может быть определена в отсутствие системы комплемента и иммунных эффекторных клеток, чтобы различить гибель клеток, индуцированную антителозависимой клеточноопосредованной цитотоксичностью (ADCC) или комплементзависимой цитотоксичностью (CDC). Таким образом, анализ гибели клеток можно выполнить с применением термоинактивированной сыворотки (т.е. в отсутствие системы комплемента) и в отсутствие иммунных эффекторных клеток. Чтобы определить, способно ли антитело индуцировать гибель клеток, можно оценить потерю целостности мембран, определяемую с помощью поглощения йодида пропидия (PI), трипанового синего (см., Moore et al. (1995) Cytotechnology 17:1-11), 7AAD или других способов, хорошо известных из уровня техники, в сравнении с необработанными клетками.
В конкретных вариантах осуществления антитела по настоящему изобретению могут индуцировать гибель клеток путем апоптоза. Антитело, которое "индуцирует апоптоз", индуцирует программируемую гибель клеток, определяемую по связыванию аннексина V, фрагментации ДНК, сжатию клеток, расширению эндоплазматического ретикулума, фрагментации клеток и/или образованию мембранных пузырьков (называемых апоптозными тельцами). Для оценки клеточных событий, связанных с апоптозом, доступны различные способы. Например, транслокацию фосфатидилсерина (PS) можно измерить по связыванию аннексина; фрагментацию ДНК можно оценить с помощью электрофоретической картины расщепления ДНК; и конденсацию ядерного хроматина наряду с фрагментацией ДНК можно оценить по какому-либо увеличению количества гиподиплоидных клеток. В одном варианте осуществления антитело, которое индуцирует апоптоз, представляет собой антитело, которое приводит к приблизительно 2-50-кратной, в одном варианте осуществления приблизительно 5-50-кратной и в одном варианте осуществления приблизительно 10-50-кратной индукции связывания аннексина по сравнению с необработанной клеткой в анализе связывания аннексина.
В другом конкретном варианте осуществления антитела по настоящему изобретению могут индуцировать гибель клеток посредством антителозависимой клеточной цитотоксичности (ADCC), и/или комплементзависимой клеточноопосредованной цитотоксичности (CDC), и/или антителозависимого клеточноопосредованного фагоцитоза (ADCP). Проявление ADCC-активности и CDC-активности человеческого антитела подкласса IgG1 обычно включает связывание Fc-участка антитела с рецептором для антитела (далее называемого ʺFcγRʺ), присутствующим на поверхности эффекторных клеток, таких как клетки-киллеры, естественные клетки-киллеры или активированные макрофаги. Могут быть связанными различные компоненты системы комплемента. В отношении связывания предполагалось, что важными являются несколько аминокислотных остатков в шарнирном участке и втором домене C-участка (далее называемом "Cγ2-домен") антитела (Greenwood et al. (1993) Eur. J. Immunol. 23(5):1098-104; Morgan et al. (1995) Immunology. 86(2):319-324; Clark, M. (1997) Chemical Immunology. 65:88-110) и что также важной является сахарная цепь в Cγ2-домене (Clark, M. (1997) Chemical Immunology. 65:88-110).
Для оценки ADCC-активности антитела, представляющего интерес, можно использовать анализ ADCC in vitro, такой как описанный в патенте США № 5500362. Анализ также можно проводить с применением коммерчески доступного набора, например, CytoTox 96 ® (Promega). Подходящие эффекторные клетки для таких анализов включают, без ограничения, мононуклеарные клетки периферической крови (PBMC), естественные клетки-киллеры (NK) и линии клеток NK. Линии клеток NK, экспрессирующие трансгенный Fc-рецептор (например, CD16) и соответствующий сигнальный полипептид (например, FCεRI-γ), могут также выполнять роль эффекторных клеток (WO 2006/023148). В одном варианте осуществления линия NK-клеток включает CD16 и содержит ген люциферазы под контролем промотора NFAT, и это можно использовать для измерения активации NK-клеток, вместо лизиса клеток или гибели клеток. Подобную технологию реализует Promega, в ней применяются клетки Jurkat вместо NK-клеток (биологический анализ ADCC с репортером №G7010 от Promega). Например, можно определить способность любого конкретного антитела опосредовать лизис с помощью активации системы комплемента и/или ADCC. Клетки, представляющие интерес, выращивают и метят in vitro; антитело добавляют к культуре клеток в комбинации с иммунными клетками, которые могут быть активированы комплексами антиген-антитело, т.е., эффекторными клетками, участвующими в ADCC-ответе. Антитело также можно исследовать в отношении активации системы комплемента. В любом случае цитолиз выявляют по высвобождению метки из лизированных клеток. Степень лизиса клеток может быть также определена путем выявления высвобождения цитоплазматических белков (например, LDH) в надосадочную жидкость. Фактически, антитела можно подвергать скринингу с применением собственной сыворотки пациента в качестве источника системы комплемента и/или иммунных клеток. Антитела, которые способны опосредовать ADCC у человека в тесте in vitro, затем можно применять с терапевтической целью у этого конкретного пациента. Может быть также проведена оценка ADCC-активности молекулы, представляющей интерес, in vivo, например, в животной модели, такой как раскрытая в Clynes et al. (1998) Proc. Natl. Acad. Sci. USA 95:652-656. Кроме того, методики для модулирования (т.е. увеличения или уменьшения) уровня ADCC, и, необязательно, CDC-активности антитела хорошо известны из уровня техники (например, патенты США №№ 5624821; 6194551; 7317091). Антитела по настоящему изобретению могут быть способны или могли быть модифицированы с получением способности к индукции ADCC и/или CDC. Анализы для определения ADCC-функции можно выполнять с применением человеческих эффекторных клеток для оценки ADCC-функции у человека. Такие анализы могут также включать те, которые направлены на скрининг антител, которые индуцируют, опосредуют, усиливают, блокируют гибель клеток с помощью механизмов некроза и/или апоптоза. Такие способы, включающие анализы, в которых используются прижизненные красители, способы выявления и анализа каспаз, а также анализы, в которых измеряют разрывы ДНК, можно применять для оценки апоптотической активности клеток, культивируемых in vitro с антителом, представляющим интерес.
Получение антител
Далее описаны иллюстративные методики получения антител, пригодных в настоящем изобретении.
Моноклональные антитела
Моноклональные антитела можно получать при помощи широкого разнообразия методик, известных из уровня техники, включающих применение гибридом (Kohler et al. (1975) Nature. 256:495; Harlow et al. (1988) Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed.); Hammerling et al. (1981) в Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y.), рекомбинантной методики и методики фагового дисплея или их комбинации. Выражение ʺмоноклональное антителоʺ, как используется в данном документе, относится к антителу, полученному из популяции фактически однородных или выделенных антител, например, отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, будучи направленными на один антигенный участок. Более того, в отличие от препаратов поликлональных антител, которые включают разные антитела, направленные на разные детерминанты (эпитопы), каждое моноклональное антитело направлено на одну и ту же детерминанту на антигене. Кроме своей специфичности, моноклональные антитела являются преимущественными в том, что их можно синтезировать незагрязненными другими антителами. Модификатор ʺмоноклональноеʺ не следует понимать как нуждающееся в получении каким-либо конкретным способом антитело. Далее следует описание иллюстративных способов получения моноклональных антител, которое не предназначено быть ограничивающим и может быть использовано для получения, например, моноклональных антител млекопитающих, химерных, гуманизированных, человеческих, доменных антител, диател, вакцинных антител, линейных и полиспецифических антител.
Гибридомные методики
Способы получения и скрининга в отношении специфических антител с использованием гибридомной технологии являются обычными и хорошо известными из уровня техники. В гибридомном способе мышей или других подходящих животных-хозяев, таких как хомяк, иммунизируют, как описано выше, чтобы получить лимфоциты, которые вырабатывают или способны вырабатывать антитела, которые будут специфично связываться с антигеном, применяемым для иммунизации. Альтернативно, лимфоциты можно иммунизировать in vitro. После иммунизации лимфоциты выделяют и затем сливают с линией миеломных клеток с применением подходящего фактора, вызывающего слияние клеток, или партнера по слиянию, такого как полиэтиленгликоль, с образованием клетки гибридомы (Goding (1986) Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press)). В определенных вариантах осуществления выбранные миеломные клетки сливаются эффективно, поддерживают стабильный высокий уровень выработки антитела выбранными антителообразующими клетками и являются чувствительными к селективной среде, с помощью которой проводят отбор, направленный против неслившихся исходных клеток. В одном аспекте линиями миеломных клеток являются мышиные миеломные линии, как, например, происходящие от мышиных опухолей MOPC-21 и MPC-11, доступные от Центра распределения клеток при Институте Солка, Сан-Диего, Калифорния, США, а также SP-2 и происходящие от нее линии, например, клетки X63-Ag8-653, доступные от Американской коллекции типовых культур, Роквилл, Мэриленд, США. Линии клеток миеломы человека и гетеромиеломные линии клеток мыши и человека также были описаны для получения человеческих моноклональных антител (Kozbor (1984) J. Immunol. 133:3001; и Brodeur et al. (1987) Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York).
Непосредственно после идентификации клеток гибридомы, которые вырабатывают антитела с желаемой специфичностью, аффинностью и/или активностью, клоны можно пересевать с помощью процедур предельного разведения и выращивать с помощью стандартных способов (Goding, выше). Подходящие культуральные среды для этой цели включают, например, среду D-MEM или RPMI-1640. Кроме того, клетки гибридомы можно выращивать in vivo в виде асцитных опухолей у животных, например, путем i.p. инъекции клеток мышам.
Моноклональные антитела, секретируемые очередным пассажем, подходящим образом отделяют от культуральной среды, асцитической жидкости или сыворотки с помощью стандартных процедур очистки антител, таких как, например, аффинная хроматография (например, с применением белка A или белка G-сефарозы) или ионообменная хроматография, аффинное мечение, хроматография на гидроксиапатите, гель-электрофорез, диализ и т.д. Иллюстративные способы очистки более подробно описаны ниже.
Методики рекомбинантных ДНК
Способы получения и скрининга в отношении специфических антител с применением технологии рекомбинантных ДНК являются обычными и хорошо известными из уровня техники (например, патент США № 4816567). ДНК, кодирующую моноклональные антитела, можно легко выделить и/или секвенировать с применением стандартных процедур (например, путем применения олигонуклеотидных зондов, которые способны к специфическому связыванию с генами, кодирующими тяжелую и легкую цепи мышиных антител). Непосредственно после выделения ДНК можно поместить в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьяны, клетки яичника китайского хомячка (CHO) или миеломные клетки, которые в других случаях не вырабатывают белковое антитело, для достижения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии у бактерий ДНК, кодирующей антитело, включают Skerra et al. (1993) Curr. Opinion in Immunol. 5:256-262 и Pluckthun (1992) Immunol. Revs. 130:151-188. Как описано ниже для антител, получаемых с помощью фагового дисплея и гуманизации антител, ДНК или генетический материал для рекомбинантных антител можно получить из отличного(отличных) от гибридом источника(источников) с тем, чтобы получить антитела по настоящему изобретению.
Рекомбинантная экспрессия антитела или его варианта, как правило, требует конструирования вектора экспрессии, содержащего полинуклеотид, который кодирует антитело. Настоящее изобретение, таким образом, предусматривает реплицируемые векторы, которые включают в себя нуклеотидную последовательность, кодирующую молекулу антитела, тяжелую или легкую цепи антитела, вариабельный домен тяжелой или легкой цепей антитела, или его часть, или CDR тяжелой или легкой цепей, функционально связанную с промотором. Такие векторы могут содержать нуклеотидную последовательность, кодирующую константный участок молекулы антитела (см., например, патенты США №№ 5981216; 5591639; 5658759 и 5122464), а вариабельный домен антитела можно клонировать в такой вектор для экспрессии полной тяжелой, полной легкой цепи или полных как тяжелой, так и легкой цепей.
Непосредственно после переноса вектора экспрессии в клетку-хозяина с помощью традиционных методик трансфицированные клетки затем культивируют с помощью традиционных методик с получением антитела. Таким образом, настоящее изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий антитело по настоящему изобретению или его фрагменты, или его тяжелую или легкую цепи или их часть, или одноцепочечное антитело по настоящему изобретению, функционально связанный с гетерологичным промотором. В определенных вариантах осуществления для экспрессии антител с двумя цепями векторы, кодирующие как тяжелую, так и легкую цепи, можно совместно экспрессировать в клетке-хозяине для экспрессии целой молекулы иммуноглобулина, как подробно описано ниже.
Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии рекомбинантных антител, хорошо известны из уровня техники и включают многие иммортализованные линии клеток, доступные от Американской коллекции типовых культур (ATCC), в том числе, без ограничения, клетки яичника китайского хомячка (CHO), клетки HeLa, клетки почки новорожденного хомячка (BHK), клетки почки обезьяны (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), клетки почечного эпителия человека 293 и ряд других линий клеток. Разные клетки-хозяева имеют характерные и конкретные механизмы посттрансляционного процессинга и модификации белков и продуктов генов. Подходящие линии клеток или системы-хозяева можно выбрать для того, чтобы обеспечить правильную модификацию и процессинг экспрессируемого антитела или его части. В связи с этим можно применять эукариотические клетки-хозяева, которые обладают клеточным аппаратом для надлежащего процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена. Такие клетки-хозяева млекопитающих включают, без ограничения, клетки CHO, VERY, BHK, HeLa, COS, MDCK, 293, 3T3, W138, BT483, Hs578T, HTB2, BT2O и T47D, NS0 (линия клеток мышиной миеломы, которая не вырабатывает эндогенно каких-либо функциональных цепей иммуноглобулинов), SP20, CRL7O3O и HsS78Bst. Линии клеток человека, разработанные путем иммортализации человеческих лимфоцитов, можно применять для рекомбинантного получения моноклональных антител. Линию клеток человека PER.C6® (Crucell, Нидерланды) можно применять для рекомбинантного получения моноклональных антител.
Дополнительные линии клеток, которые можно применять в качестве хозяев для экспрессии рекомбинантных антител, включают без ограничения клетки насекомых (например, Sf21/Sf9, Trichoplusia ni Bti-Tn5b1-4) или клетки дрожжей (например, S. cerevisiae, Pichia, US7326681 и т.д.), растительные клетки (US20080066200) и куриные клетки (WO2008142124).
В определенных вариантах осуществления антитела по настоящему изобретению экспрессируются в линии клеток со стабильной экспрессией антитела. Стабильную экспрессию можно применять для длительного высокопродуктивного получения рекомбинантных белков. Например, можно создать линии клеток, стабильно экспрессирующих молекулу антитела. Клетки-хозяева можно трансформировать подходящим образом сконструированным вектором, который включает в себя элементы контроля экспрессии (например, промотор, энхансер, терминаторы транскрипции, сайты полиаденилирования и т.д.) и ген селектируемого маркера. После внедрения чужеродной ДНК клеткам можно позволить расти в течение 1-2 суток на обогащенной среде, и затем их переводят на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к отбору и позволяет клеткам со стабильно интегрированной плазмидой в их хромосомах расти и образовывать фокусы, которые, в свою очередь, можно клонировать и наращивать с получением линий клеток. Способы получения стабильных линий клеток с высокой продуктивностью хорошо известны из уровня техники, и реактивы, как правило, являются коммерчески доступными.
В определенных вариантах осуществления антитела по настоящему изобретению экспрессируются в линии клеток с транзиентной экспрессией антитела. Транзиентная трансфекция представляет собой процесс, при котором нуклеиновая кислота, внедряемая в клетку, не интегрируется в геномную или хромосомную ДНК такой клетки. Она фактически сохраняется в виде внехромосомного элемента, например, в виде эписомы, в клетке. Процессы транскрипции нуклеиновой кислоты эписомы не затрагиваются, и вырабатывается белок, кодируемый нуклеиновой кислотой эписомы.
Линию клеток, подвергнутую стабильной или транзиентной трансфекции, поддерживают в среде для культивирования клеток и условиях, хорошо известных из уровня техники, что приводит к экспрессии и выработке моноклональных антител. В определенных вариантах осуществления в основе среды для культивирования клеток млекопитающих лежат коммерчески доступные составы сред, в том числе, например, DMEM или среды Хэма F12. В других вариантах осуществления среду для культивирования клеток модифицируют для поддержки как роста клеток, так и биологической экспрессии белка. Как используется в данном документе, выражения "среда для культивирования клеток", "культуральная среда" и "состав среды" относятся к питательному раствору для поддержания, роста, размножения или наращивания клеток в искусственном окружении in vitro вне многоклеточного организма или ткани. Среду для культивирования клеток можно оптимизировать для применения в конкретной культуре клеток, в том числе, например, ростовую среду для культивирования клеток, которая составлена для стимуляции клеточного роста, или продуктивную среду для культивирования клеток, которая составлена для стимуляции выработки рекомбинантного белка. Выражения "питательное вещество", "ингредиент" и "компонент", используемые в данном документе взаимозаменяемо, относятся к составляющим, которые образуют среду для культивирования клеток.
В одном варианте осуществления линии клеток поддерживают с применением способа периодического культивирования с подпиткой. Как используется в данном документе, ʺспособ периодического культивирования с подпиткойʺ относится к способу, при котором культуру клеток подпитывают дополнительными питательными веществами после изначального инкубирования с базовой средой. Например, способ периодического культивирования с подпиткой может включать добавление среды с добавками в соответствии с определенным режимом подпитки в течение определенного периода времени. Таким образом, "подпитываемая культура клеток" относится к культуре клеток, где в сосуд для культивирования изначально добавляют клетки, обычно млекопитающего, и культуральную среду, и дополнительные питательные вещества для культуры подаются непрерывно или пошагово в культуру во время культивирования с периодическим сбором или без периодического сбора клеток и/или продукта до завершения культивирования.
Применяемая среда для культивирования клеток и питательные вещества, содержащиеся в ней, известны специалисту в данной области. В одном варианте осуществления среда для культивирования клеток включает базовую среду и по меньшей мере один гидролизат, например, гидролизат на основе сои, гидролизат на основе дрожжей или комбинацию двух типов гидролизатов, что приводит в результате к модифицированной базовой среде. В другом варианте осуществления дополнительные питательные вещества могут включать только базовую среду, такую как концентрированная базовая среда, или могут включать только гидролизаты или концентрированные гидролизаты. Подходящие базовые среды включают, без ограничения, среду Игла в модификации Дульбекко (DMEM), DME/F12, минимальную питательную среду (MEM), базовую среду Игла (BME), RPMI 1640, F-10, F-12, минимальную питательную среду α (α-MEM), минимальную питательную среду Глазго (G-MEM), PF CHO (см., например, не содержащую белков среду для CHO (Sigma) или не содержащую белков бессывороточную среду для клеток CHO EX-CELL™ 325 PF CHO (SAFC Bioscience)) и среду Дульбекко в модификации Искова. Другие примеры базовых сред, которые можно применять в настоящем изобретении, включают базовую среду BME (Gibco-Invitrogen; см. также Eagle, H (1965) Proc. Soc. Exp. Biol. Med. 89, 36); среду Игла в модификации Дульбекко (DMEM, порошок) (Gibco-Invitrogen (№ 31600); см. также Dulbecco and Freeman (1959) Virology. 8:396; Smith et al. (1960) Virology. 12:185. Tissue Culture Standards Committee, In Vitro 6:2, 93); среду CMRL 1066 (Gibco-Invitrogen (№ 11530); см. также Parker et al. (1957) Special Publications, N.Y. Academy of Sciences, 5:303).
Базовая среда может быть бессывороточной, что означает, что среда не содержит сыворотки (например, фетальной бычьей сыворотки (FBS), лошадиной сыворотки, козьей сыворотки или сыворотки, полученной от любого другого животного, известной специалисту в данной области), или средой, не содержащей животных белков, или средой с определенным химическим составом.
Базовую среду можно модифицировать с целью удаления определенных компонентов, не являющихся питательными веществами, находящихся в стандартной базовой среде, таких как разнообразные неорганические и органические буферы, поверхностно-активное(поверхностно-активные) вещество(вещества) и хлорид натрия. Удаление таких компонентов из базовой среды для клеток позволяет повысить концентрацию остальных питательных компонентов и может улучшить общий рост клеток и экспрессию белка. Кроме того, изъятые компоненты можно добавить обратно в среду для культивирования клеток, содержащую модифицированную базовую среду для клеток, в соответствии с требованиями условий культивирования клеток. В определенных вариантах осуществления среда для культивирования клеток содержит модифицированную базовую среду для клеток и по меньшей мере одно из следующих питательных веществ: источника железа, рекомбинантного фактора роста; буфера; поверхностно-активного вещества; регулятора осмолярности; источника энергии и гидролизатов неживотного происхождения. Кроме того, модифицированная базовая среда для клеток может необязательно содержать аминокислоты, витамины или комбинацию как аминокислот, так и витаминов. В другом варианте осуществления модифицированная базовая среда дополнительно содержит глутамин, например, L-глутамин, и/или метотрексат.
Получение антител может проводиться в большом количестве с помощью процессов в биореакторе с использованием способов периодического культивирования с подпиткой, периодического культивирования в биореакторе, культивирования в перфузионном биореакторе или биореакторе с непрерывной подачей, известных из уровня техники. Крупномасштабные биореакторы имеют емкость, составляющую по меньшей мере 1000 литров, в одном варианте осуществления емкость, составляющую приблизительно 1000-100000 литров. В этих биореакторах могут применяться лопастные мешалки для распределения кислорода и питательных веществ. Биореакторы небольшого масштаба относятся, как правило, к культивированию клеток в рабочем объеме, составляющем не более чем примерно 100 литров, который может варьировать от приблизительно 1 литра до приблизительно 100 литров. Альтернативно, биореакторы одноразового применения (SUB) можно использовать для крупномасштабного либо мелкомасштабного культивирования.
Температура, pH, перемешивание, аэрация и плотность посева будут изменяться в зависимости от применяемых клеток-хозяев и рекомбинантных белков, которые нужно экспрессировать. Например, культуру клеток с рекомбинантным белком можно поддерживать при температуре от 30°C до 45°C. pH культуральной среды можно отслеживать в ходе процесса культивирования так, чтобы pH оставался на оптимальном уровне, который для определенных клеток-хозяев может находиться в пределах диапазона pH от 6,0 до 8,0. Осуществляемое лопастной мешалкой смешивание можно применять для таких способов культивирования для перемешивания. Скорость вращения лопастной мешалки может быть окружной скоростью концов лопастей, составляющей примерно 50-200 см/с, но можно применять и другие аэролифтные или другие системы перемешивания/аэрации, известные из уровня техники, в зависимости от типа культивируемых клеток-хозяев. Достаточная аэрация обеспечивается для поддержания концентрации растворенного кислорода при насыщении культуры воздухом примерно от 20% до 80%, опять-таки в зависимости от выбранного типа культивируемых клеток-хозяев. Альтернативно, биореактор может барботировать воздух или кислород непосредственно в культуральную среду. Существуют другие способы подачи кислорода, включающие системы беспузырьковой аэрации, в которых используются аэраторы с мембраной из полых волокон.
Методики фагового дисплея
Моноклональные антитела или фрагменты антител могут быть выделены из фаговых библиотек антител, полученных с помощью методик, описанных в McCafferty et al. (1990) Nature. 348:552-554; Clackson et al. (1991) Nature. 352:624-628 и Marks et al. (1991) J. Mol. Biol. 222:581-597. В таких способах антитела могут быть выделены путем скрининга рекомбинантной комбинаторной библиотеки антител. В одном варианте осуществления библиотеку фагового дисплея scFv получали с использованием кДНК человеческих VL и VH, полученных из мРНК, происходящей из человеческих лимфоцитов. Методы получения и скрининга таких библиотек известны из уровня техники. Кроме коммерчески доступных наборов для получения фаг-дисплейных библиотек (например, системы фаговых рекомбинантных антител Pharmacia, номер по каталогу 27-9400-01; и набора для фагового дисплея SurfZAPTM от Stratagene, номер по каталогу 240612), примеры способов и реактивов, особенно пригодных для применения в получении и скрининге дисплейных библиотек антител, можно найти, например, в патентах США №№ 6248516; US 6545142; 6291158; 6291159; 6291160; 6291161; 6680192; 5969108; 6172197; 6806079; 5885793; 6521404; 6544731; 6555313; 6593081; 6582915; 7195866. Таким образом, эти методики являются эффективными альтернативами традиционным гибридомным методикам получения моноклональных антител для получения и выделения моноклональных антител.
В способах фагового дисплея функциональные домены антител представлены на поверхности фаговых частиц, которые несут последовательности полинуклеотидов, кодирующие их. В конкретном варианте осуществления такой фаг можно использовать для представления антиген-связывающих доменов, экспрессируемых в репертуаре или комбинаторной библиотеке антител (например, человеческих или мышиных). Фаг, экспрессирующий антигенсвязывающий домен, который связывается с антигеном, представляющим интерес, можно отобрать или идентифицировать с помощью антигена, например, с помощью меченого антигена или антигена, связанного с твердой поверхностью или гранулой или зафиксированного на них. Фаг, применяемый в этих способах, обычно является нитчатым фагом, содержащим связывающие домены fd и M13, экспрессируемые в фаге с доменами антитела Fab, Fv или Fv, стабилизированными дисульфидными связями, слитыми рекомбинантным путем с белком гена III или гена VIII фага.
Как описано в указанных выше источниках, после отбора фага участки, кодирующие антитело, из фага можно выделять и применять для получения целых антител, в том числе человеческих антител, гуманизированных антител, или любого другого предпочтительного антигенсвязывающего фрагмента и экспрессировать у любого желаемого хозяина, в том числе в клетках млекопитающих, клетках насекомых, растительных клетках, у дрожжей и бактерий, например, как подробно описано ниже. Например, также можно использовать методики рекомбинантного получения фрагментов Fab, Fab' и F(ab')2 с применением способов, известных из уровня техники, таких как раскрытые в PCT публикации WO 92/22324; Mullinax et al. (1992) BioTechniques. 12(6):864-869; и Better et al. (1988) Science. 240:1041-1043.
Примеры методик, которые можно применять для получения одноцепочечных Fv и антител, включают описанные в патентах США №№ 4946778 и 5258498. Таким образом, методики, описанные выше, и таковые, хорошо известные из уровня техники, можно применять для получения рекомбинантных антител, где связывающий домен, например, scFv, был выделен из фаг-дисплейной библиотеки.
Очистка и выделение антител
Непосредственно после получения молекулы антитела путем рекомбинантной или гибридомной экспрессии ее можно очистить любым способом, известным из уровня техники, для очистки молекулы иммуноглобулина, например, с помощью хроматографии (например, ионообменной, аффинной, в частности, по аффинности к специфическим антигенам белку A или белку G, и колоночной хроматографией размеров), центрифугирования, дифференциальной растворимости или любой другой стандартной методики очистки белков. Кроме того, антитела по настоящему изобретению или их фрагменты могут быть слиты с гетерологичными полипептидными последовательностями (называемыми в данном документе ʺметкамиʺ) для облегчения очистки.
При применении рекомбинантных методик антитело может быть получено внутриклеточно, в периплазматическом пространстве или непосредственно секретировано в среду. Если антитело получено внутриклеточно, в качестве первого этапа удаляют частицы дебриса, клетки-хозяева или лизированные фрагменты, например, путем центрифугирования или ультрафильтрации. В Carter et al. (1992) Bio/Technology. 10:163-167 описана процедура выделения антител, которые секретируются в периплазматическое пространство E. coli. Если антитело секретируется в среду, надосадочные жидкости из таких систем экспрессии, как правило, вначале концентрируют с помощью коммерчески доступного фильтра для концентрирования белка, например, ультрафильтрационного блока Amicon или Millipore Pellicon. Ингибитор протеаз, такой как PMSF, может быть включен в любой из последующих этапов для ингибирования протеолиза, и антибиотики могут быть включены для предупреждения роста случайных загрязнителей.
Композицию антитела, полученную из клеток, можно очистить с помощью, например, хроматографии на гидроксиапатите, хроматографии гидрофобных взаимодействий, ионообменной хроматографии, гель-электрофореза, диализа и/или аффинной хроматографии в отдельности или в комбинации с другими этапами очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого домена Fc иммуноглобулина, который присутствует в антителе, и будет понятна специалисту в данной области. Матрицей, к которой прикрепляется аффинный лиганд, чаще всего является агароза, но доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и меньшее время обработки, чем можно получить с агарозой. Если антитело включает в себя CH3-домен, то для очистки полезна смола Bakerbond ABX (J.T. Baker, Филипсбург, Нью-Джерси). Также доступны другие методики для очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая HPLC, хроматография на силикагеле, хроматография на гепарине, хроматография с использованием сефарозы на анионо- или катионообменной смоле (как, например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония, в зависимости от антитела, которое подлежит извлечению.
После любой (любых) стадии (стадий) предварительной очистки смесь, включающую в себя представляющее интерес антитело и примеси, можно подвергнуть хроматографии гидрофобных взаимодействий при низком pH с применением элюирующего буфера при pH приблизительно 2,5-4,5, проводимой при низких концентрациях солей (например, приблизительно 0-0,25 М соли).
Таким образом, в определенных вариантах осуществления представлены антитела по настоящему изобретению, которые являются фактически очищенными/выделенными. В одном варианте осуществления эти выделенные/очищенные рекомбинантно экспрессируемые антитела можно вводить пациенту с тем, чтобы опосредовать профилактический или терапевтический эффект. Профилактика представляет собой использование лекарственных препаратов или лечение, разработанное и применяемое для предупреждения возникновения заболевания, нарушения или инфекции. Терапевтическое средство связано определенным образом с лечением конкретного заболевания, нарушения или инфекции. Терапевтическая доза представляет собой количество, необходимое для лечения конкретного заболевания, нарушения или инфекции. В другом варианте осуществления эти выделенные/очищенные антитела можно применять для диагностики инфекции, вызванной вирусом гриппа, например, инфекции, вызванной вирусом гриппа B, или в других вариантах осуществления инфекции, вызванной вирусом гриппа A и вирусом гриппа B.
Человеческие антитела
Человеческие антитела могут быть получены с помощью способов, хорошо известных из уровня техники. Человеческие антитела лишены некоторых проблем, связанных с антителами, которые содержат мышиные или крысиные вариабельные и/или константные участки. Присутствие таких полученных от мыши или крысы белков может приводить к быстрому выведению антител или может приводить к формированию иммунного ответа на антитело у пациента.
Человеческие антитела могут быть получены с помощью способов in vitro. Подходящие примеры включают, без ограничения, фаговый дисплей (MedImmune (ранее CAT), Morphosys, Dyax, Biosite/Medarex, Xoma, Symphogen, Alexion (ранее Proliferon), Affimed), рибосомный дисплей (MedImmune (ранее CAT)), дрожжевой дисплей и т.п. Технология фагового дисплея (см., например, патент США № 5969108) может применяться для получения человеческих антител и фрагментов антител in vitro из репертуаров генов вариабельных (V) доменов иммуноглобулинов от неиммунизированных доноров. В соответствии с этой методикой, гены V-доменов антител клонируют внутрирамочно в ген основного либо минорного оболочечного белка нитчатого бактериофага, такого как M13 или fd, и представляют в качестве функциональных фрагментов антител на поверхности фаговой частицы. Поскольку нитевидная частица содержит копию одноцепочечной ДНК фагового генома, отборы на основе функциональных свойств антитела также приводят к отбору гена, кодирующего антитело, проявляющее эти свойства. Таким образом, фаг имитирует некоторые из свойств B-клетки. Фаговый дисплей может быть выполнен во множестве форматов, описанных в обзорах, например, в Johnson and Chiswell (1993) Current Opinion in Structural Biology. 3:564-571. Для фагового дисплея можно применять некоторые источники V-сегментов генов. Clackson et al. (1991), Nature. 352:624-628 (1991), выделили неоднородную совокупность антител к оксазолону из небольшой произвольной комбинаторной библиотеки V-генов, полученных из селезенок иммунизированных мышей. Может быть сконструирован репертуар V-генов от неиммунизированных доноров-людей, и антитела к неоднородной совокупности антигенов (в том числе собственных антигенов) могут быть выделены преимущественно в соответствии со следующими методиками, описанными Marks et al. (1991) J. Mol. Biol. 222:581-597 или Griffith et al. (1993) EMBO J. 12:725-734. См. также патенты США №№ 5565332 и 5573905.
Как обсуждается выше, человеческие антитела также могут вырабатываться активированными in vitro B-клетками (см. патенты США №№ 5567610 и 5229275).
Гены иммуноглобулинов претерпевают различные модификации при развитии иммунного ответа, в том числе рекомбинацию между сегментами генов V, D и J, переключение изотипов и гипермутацию в вариабельных участках. Рекомбинация и соматическая гипермутация являются основанием для формирования разнообразия антител и созревания аффинности, но они также могут приводить к формированию факторов неустойчивости последовательностей, которые могут осложнить коммерческое получение таких иммуноглобулинов в качестве терапевтических средств или повысить риск иммуногенности антитела. Как правило, мутации в CDR-участках, вероятно, способствуют улучшению аффинности и функции, тогда как мутации в каркасных областях могут повышать риск иммуногенности. Этот риск может быть снижен путем возвращения мутантных участков каркасных областей к зародышевому типу при обеспечении того, что активность антитела не подвергается неблагоприятному влиянию. Благодаря процессам внесения разнообразия также могут формироваться некоторые факторы структурной неустойчивости, или эти факторы структурной неустойчивости могут существовать в последовательностях зародышевого типа, участвующих в образовании вариабельных доменов тяжелой и легкой цепи. Независимо от источника, может быть желательным устранение потенциальных факторов структурной неустойчивости, которые могут приводить к нестабильности, агрегации, неоднородности продукта или повышению иммуногенности. Примеры нежелательных факторов неустойчивости включают неспаренные цистеиновые остатки (которые могут обуславливать перестановку дисульфидных связей или образование неодинаковых сульфгидрильных аддуктов), сайты N-сцепленного гликозилирования (обуславливающие неоднородность структуры и активности), а также сайты дезамидирования (например, NG, NS), изомеризации (DG), окисления (доступный метионин) и гидролиза (DP).
Следовательно, для снижения риска иммуногенности и улучшения фармацевтических свойств антител может быть желательным возвращение последовательности каркасной области к зародышевому типу, возвращение CDR к зародышевому типу и/или устранение структурной неустойчивости.
Таким образом, в одном варианте осуществления, если конкретное антитело отличается от его соответствующей последовательности зародышевого типа на аминокислотном уровне, последовательность антитела можно подвергнуть обратной мутации в последовательность зародышевого типа. Такие корректирующие мутации могут происходить в одном, двух, трех или более положениях или в комбинации каких-либо из мутантных положений с применением стандартных методик молекулярной биологии.
Фрагменты антител
В определенных вариантах осуществления антитела по настоящему изобретению являются фрагментами антител или антителами, включающими в себя такие фрагменты. Фрагмент антитела включает в себя часть антитела полной длины, которая, как правило, является его антигенсвязывающим или вариабельным участком. Примеры фрагментов антител включают Fab-, Fab'-, F(ab')2-, Fd- и Fv-фрагменты, диатела, линейные антитела (патент США № 5641870) и одноцепочечные молекулы антител.
Традиционно, эти фрагменты получали путем протеолитического расщепления интактных антител с применением методик, хорошо известных из уровня техники. Однако, эти фрагменты теперь могут быть получены в данном документе непосредственно с помощью рекомбинантных клеток-хозяев. Все Fab-, Fv- и scFv-фрагменты антител могут экспрессироваться и секретироваться E. coli, что, таким образом, обеспечивает легкое получение больших количеств этих фрагментов. В одном варианте осуществления фрагменты антител могут быть выделены из обсуждаемых выше фаговых библиотек антител. Альтернативно, Fab'-SH-фрагменты также могут быть непосредственно извлечены из E. coli и химически соединены с образованием F(ab')2-фрагментов (Carter et al. (1992) Bio/Technology. 10:163-167). В соответствии с другим подходом F(ab')2-фрагменты могут быть выделены непосредственно из культуры рекомбинантных клеток-хозяев. Другие методики получения фрагментов антител будут очевидны практикующему специалисту. В других вариантах осуществления выбранное антитело представляет собой одноцепочечный Fv-фрагмент (scFv). В определенных вариантах осуществления антителом не является Fab-фрагмент. Fv и scFv являются единственными молекулами с интактными антигенсвязывающими активными центрами, которые лишены константных участков; таким образом, они являются подходящими для ослабленного неспецифического связывания во время применения in vivo. Гибридные белки scFv могут быть сконструированы с получением слияния эффекторного белка с амино- или карбоксильным концом scFv.
В определенных вариантах осуществления антитела в соответствии с настоящим изобретением являются доменными антителами, например, антителами, содержащими небольшие функциональные связывающие единицы антител, соответствующие вариабельным участкам тяжелой (VH) или легкой (VL) цепей человеческих антител. Примеры доменных антител включают, без ограничения, антитела Domantis (см., например, WO04/058821; WO04/081026; WO04/003019; WO03/002609; патенты США №№ 6291158; 6582915; 6696245 и 6593081).
В определенных вариантах осуществления настоящего изобретения антителами по настоящему изобретению являются линейные антитела. Линейные антитела включают в себя пару тандемных сегментов Fd (VH-CH1-VH-CH1), которые образуют пару антигенсвязывающих участков. См., Zapata et al. (1995) Protein Eng. 8(10):1057-1062.
Другие модификации аминокислотных последовательностей
Кроме описанных выше человеческих, гуманизированных и/или химерных антител, настоящее изобретение также охватывает дополнительные модификации, а также варианты и фрагменты антител по настоящему изобретению, включающие одну или несколько замен, добавлений и/или делеций аминокислотных остатков и/или полипептидов в вариабельном домене легкой цепи (VL), и/или вариабельном домене тяжелой цепи (VH), и/или Fc-участке и посттрансляционных модификаций. Такие модификации включают конъюгаты антител, где антитело ковалентно присоединено к компоненту. Компоненты, подходящие для присоединения к антителам, включают, без ограничения, белки, пептиды, лекарственные средства, метки и цитотоксины. Эти изменения антител могут быть выполнены для изменения или точной регулировки характеристик (биохимических характеристик, характеристик связывания и/или функциональных характеристик) антител как подходящих для лечения и/или диагностики инфекции, вызванной вирусом гриппа. Способы образования конъюгатов, осуществления аминокислотных и/или полипептидных изменений и посттрансляционных модификаций хорошо известны из уровня техники, причем некоторые из них подробно описаны ниже.
Аминокислотные изменения антител неизбежно приводят к образованию последовательностей, менее чем на 100% идентичным определенным выше последовательностям антител или последовательности исходного антитела. В определенных вариантах осуществления в данном контексте антитела могут характеризоваться идентичностью последовательности с аминокислотной последовательностью вариабельного домена тяжелой либо легкой цепи антитела, описанного в данном документе, составляющей от приблизительно 25% до приблизительно 95%. Таким образом, в одном варианте осуществления модифицированное антитело может иметь аминокислотную последовательность по меньшей мере с 25%, 35%, 45%, 55%, 65%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью или подобием аминокислотной последовательности с аминокислотной последовательностью вариабельного домена тяжелой либо легкой цепи антитела, описанного в данном документе. В другом варианте осуществления измененное антитело может иметь аминокислотную последовательность по меньшей мере с 25%, 35%, 45%, 55%, 65%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью или сходством аминокислотной последовательности с аминокислотной последовательностью CDR-1, CDR-2 или CDR-3 тяжелой или легкой цепи антитела, описанного в данном документе. В другом варианте осуществления измененное антитело может иметь аминокислотную последовательность по меньшей мере с 25%, 35%, 45%, 55%, 65%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью или подобием аминокислотной последовательности с аминокислотной последовательностью FR1, FR2, FR3 или FR4 тяжелой либо легкой цепи антитела, описанного в данном документе.
В определенных вариантах осуществления измененные антитела получают с помощью одного или нескольких аминокислотных изменений (например, замен, делеций и/или добавлений), введенных в один или несколько вариабельных участков антитела. В другом варианте осуществления аминокислотные изменения вводят в каркасные области. Одно или несколько изменений остатков каркасных областей могут приводить к улучшению аффинности связывания антитела по отношению к антигену. Это особенно может быть справедливым, если такие изменения выполняют для гуманизированных антител, где каркасная область и CDR-участки могут происходить от разных видов. Примеры подлежащих модифицированию остатков каркасного участка включают остатки, которые непосредственно не ковалентно связывают антиген (Amit et al. (1986) Science. 233:747-753); взаимодействуют с CDR/влияют на его конформацию (Chothia et al. (1987) J. Mol. Biol. 196:901-917) и/или участвуют в образовании области контакта VL-VH (патенты США №№ 5225539 и 6548640). В одном варианте осуществления могут быть изменены от приблизительно одного до приблизительно пяти остатков каркасных областей. Иногда этого может быть достаточно для получения мутантного антитела, подходящего для применения в доклинических испытаниях, даже если не был изменен ни один остаток гипервариабельного участка. Обычно, однако, измененное антитело будет включать дополнительное (дополнительные) изменение (изменения) гипервариабельного участка.
Одна подходящая процедура для получения измененных антител называется "аланин-сканирующий мутагенез" (Cunningham and Wells (1989) Science. 244:1081-1085). В этом способе один или несколько остатков гипервариабельного участка замещаются аланиновым(аланиновыми) или полиаланиновым(полиаланиновыми) остатком(остатками) для изменения взаимодействия аминокислот с целевым антигеном. Этот (эти) остаток(остатки) гипервариабельного участка, демонстрирующий(демонстрирующие) функциональную чувствительность к заменам, затем улучшают путем введения дополнительных или других мутаций в сайтах или вместо сайтов замены. Таким образом, хотя сайт для введения варианта аминокислотной последовательности устанавливается предварительно, нет необходимости в предварительном определении природы мутации как таковой. Ala-мутанты, получаемые таким путем, подвергают скринингу в отношении их биологической активности, как описано в данном документе.
В определенных вариантах осуществления вариант с заменой предусматривает замену одного или нескольких остатков гипревариабельного участка исходного антитела (например, гуманизированного или человеческого антитела). Как правило, полученный (полученные) вариант (варианты), выбранный (выбранные) для дополнительной разработки, будет (будут) обладать улучшенными биологическими свойствами относительно исходного антитела, из которого они получены. Удобный способ для получения таких вариантов с заменами включает созревание аффинности с применением фагового дисплея (Hawkins et al. (1992) J. Mol. Biol. 254:889-896 и Lowman et al. (1991) Biochemistry. 30(45):10832-10837)). Вкратце, несколько сайтов гипервариабельного участка (например, 6-7 сайтов) подвергают мутации с образованием всех возможных замен аминокислот в каждом сайте. Мутантные антитела, полученные таким образом, представляют в одновалентной форме на частицах нитевидного фага в виде гибридов с продуктом гена III M13, упакованных в каждой частице. Фаг-дисплейных мутантов затем подвергают скринингу в отношении их биологической активности (например, аффинности связывания), как раскрыто в данном документе.
Мутации в последовательностях антител могут включать замены, делеции, в том числе внутренние делеции, добавления, в том числе добавления, обуславливающие получение гибридных белков, или консервативные замены аминокислотных остатков в аминокислотной последовательности и/или рядом с ней, которые, однако, приводят к "молчащему" изменению в том смысле, что изменение дает функционально эквивалентное антитело. Консервативные замены аминокислот могут быть выполнены на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы включенных остатков. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; а отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту. Кроме того, глицин и пролин являются остатками, которые могут влиять на ориентацию цепи. Неконсервативные замены повлекут за собой замену члена одного из этих классов членом другого класса. Более того, если желательно, неклассические аминокислоты или химические аналоги аминокислот могут быть введены в последовательность антитела в виде замены или добавления. Неклассические аминокислоты включают, без ограничения, D-изомеры стандартных аминокислот, α-аминоизомасляную кислоту, 4-аминомасляную кислоту, Abu, 2-аминомасляную кислоту, γ-Abu, ε-Ahx, 6-аминогексановую кислоту, Aib, 2-аминоизомасляную кислоту, 3-аминопропионовую кислоту, орнитин, норлейцин, норвалин, гидроксипролин, саркозин, цитруллин, цистеиновую кислоту, трет-бутилглицин, трет-бутилаланин, фенилглицин, циклогексилаланин, β-аланин, фторсодержащие аминокислоты, искусственные аминокислоты, такие как β-метиламинокислоты, Cα-метиламинокислоты, Nα-метиламинокислоты и аналоги аминокислот, как правило.
В другом варианте осуществления любой цистеиновый остаток, не участвующий в поддержании надлежащей конформации антитела, также может быть замещен, как правило, серином, для улучшения устойчивости к окислению молекулы и предупреждения аномального сшивания. Наоборот, цистеиновая(цистеиновые) связь(связи) может(могут) быть добавлена(добавлены) к антителу для улучшения его стабильности (особенно если антитело представляет собой фрагмент антитела, такой как Fv-фрагмент).
Варианты Fc-участков
Известно, что варианты Fc-участка (например, замены, и/или добавления, и/или делеции аминокислот) усиливают или ослабляют эффекторную функцию антитела (см., например, патенты США №№ 5624821; 5885573; 6538124; 7317091; 5648260; 6538124; WO 03/074679; WO 04/029207; WO 04/099249; WO 99/58572; публикации заявок на патенты США №№ 2006/0134105; 2004/0132101; 2006/0008883) и могут изменять фармакокинетические свойства (например, период полувыведения) антитела (см. патенты США 6277375 и 7083784). Таким образом, в определенных вариантах осуществления антитела по настоящему изобретению включают в себя измененный Fc-участок (также называемый в данном документе ʺвариантом Fc-участкаʺ), в котором одно или несколько изменений были осуществлены в Fc-участке для изменения функциональных и/или фармакокинетических свойств антител. Подобные изменения могут приводить к ослаблению или усилению связывания Clq и комплементзависимой цитотоксичности (CDC) или связывания FcγR для IgG и антителозависимой клеточной цитотоксичности (ADCC) или антителозависимого клеточноопосредованного фагоцитоза (ADCP). Настоящее изобретение охватывает антитела, описанные в данном документе, с вариантами Fc-участков, где изменения были осуществлены для точной регулировки эффекторной функции, усиления или ослабления, и обеспечения необходимой эффекторной функции. Соответственно, антитела по настоящему изобретению включают в себя вариант Fc-участка (т.е. Fc-участки, которые были изменены, как обсуждается ниже). Антитела по настоящему изобретению, содержащие вариант Fc-участка, также называются в данном документе "антителами с вариантом Fc". Как используется в данном документе, нативный относится к немодифицированной исходной последовательности, а антитело, содержащее нативный Fc-участок, называется в данном документе "антителом с нативным Fc". Антитела с вариантами Fc могут быть получены при помощи многочисленных способов, хорошо известных специалисту в данной области. Неограничивающие примеры включают выделение участков, кодирующих антитело (например, из гибридомы), и осуществление одной или нескольких необходимых замен в Fc-участке выделенного участка, кодирующего антитело. Альтернативно, антигенсвязывающую часть (например, вариабельные участки) антитела можно субклонировать в вектор, кодирующий вариант Fc-участка. В одном варианте осуществления вариант Fc-участка характеризуется сходным уровнем индукции эффекторной функции по сравнению с нативным Fc-участком. В другом варианте осуществления вариант Fc-участка характеризуется более высокой индукцией эффекторной функции по сравнению с нативным Fc. Некоторые конкретные варианты осуществления вариантов Fc-участков подробно описаны ниже. Способы измерения эффекторной функции хорошо известны из уровня техники.
Эффекторную функцию антитела модифицируют путем изменений в Fc-участке, в том числе, без ограничения, замен аминокислот, добавлений аминокислот, делеций аминокислот и изменений по типу посттрансляционных модификаций аминокислот в Fc (например, гликозилирования). Способы, описанные ниже, можно применять для точной регулировки эффекторной функции антитела по настоящему изобретению, соотношения свойств связывания Fc-участка для FcR (например, аффинности и специфичности), что приводит к образованию терапевтического антитела с необходимыми свойствами.
Следует понимать, что Fc-участок, как используется в данном документе, включает в себя полипептиды, составляющие константный участок антитела за исключением первого домена константного участка иммуноглобулина. Таким образом, Fc относится к последним двум доменам константной области иммуноглобулинов IgA, IgD и IgG и последним трем доменам константной области иммуноглобулинов IgE и IgM, а также гибкой шарнирной области на N-конце этих доменов. В случае IgA и IgM Fc может включать в себя J-цепь. В случае IgG Fc включает в себя домены иммуноглобулина C-гамма-2 и C-гамма-3 (Cγ2 и Cγ3) и шарнирный участок между C-гамма-1 (Cγ1) и C-гамма-2 (Cγ2).
Несмотря на то, что границы Fc-участка могут варьировать, Fc-участок тяжелой цепи человеческого IgG можно определить как включающий в себя от остатков C226 или P230 до его карбоксильного конца, где нумерация приведена согласно EU-индексу, как изложено у Kabat. Fc может относиться к этому выделенному участку или этому участку в контексте антитела, фрагмента антитела или гибридного белка на основе Fc. Наблюдались полиморфизмы в ряде различных положений Fc, в том числе, без ограничения, в положениях 270, 272, 312, 315, 356 и 358, пронумерованным согласно EU-индексу, и, следовательно, могут существовать незначительные различия между представленной последовательностью и последовательностями из уровня техники.
В одном варианте осуществления антитела с вариантами Fc проявляют измененную аффинность связывания в отношении одного или нескольких Fc-рецепторов, в том числе, без ограничения, FcRn, FcγRI (CD64), в том числе изоформ FcγRIA, FcγRIB и FcγRIC; FcγRII (CD32, в том числе изоформ FcγRIIA, FcγRIIB и FcγRIIC) и FcγRIII (CD16, в том числе изоформ FcγRIIIA и FcγRIIIB), по сравнению с антителом с нативным Fc.
В одном варианте осуществления антитело с вариантом Fc характеризуется повышенным связыванием с одним или нескольким лигандами Fc по сравнению с антителом c нативным Fc. В другом варианте осуществления антитело с вариантом Fc характеризуется повышенной или пониженной аффинностью в отношении лиганда Fc, при этом она по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, и не более чем в 25 раз, или не более чем в 50 раз, или не более чем в 75 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше или ниже, чем таковая у антитела c нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении лиганда Fc, по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 40%, по меньшей мере на 30%, по меньшей мере на 20%, по меньшей мере на 10% или по меньшей мере на 5% большими или меньшими, чем у антитела c нативным Fc. В определенных вариантах осуществления антитело с вариантом Fc характеризуется повышенной аффинностью в отношении лиганда Fc. В других вариантах осуществления антитело с вариантом Fc характеризуется пониженной аффинностью в отношении лиганда Fc.
В конкретном варианте осуществления антитело с вариантом Fc характеризуется повышенным связыванием с Fc-рецептором FcγRIIIA. В другом конкретном варианте осуществления антитело с вариантом Fc характеризуется повышенным связыванием с Fc-рецептором FcγRIIB. В дополнительном конкретном варианте осуществления антитело с вариантом Fc характеризуется повышенным связыванием с Fc-рецепторами как FcγRIIIA, так и FcγRIIB. В определенных вариантах осуществления антитела c вариантами Fc, характеризующиеся повышенным связыванием с FcγRIIIA, не характеризуются при этом повышенным связыванием с рецептором FcγRIIB по сравнению с антителом c нативным Fc. В конкретном варианте осуществления антитело с вариантом Fc характеризуется пониженным связыванием с Fc-рецептором FcγRIIIA. В дополнительном конкретном варианте осуществления антитело с вариантом Fc характеризуется пониженным связыванием с Fc-рецептором FcγRIIB. В еще одном конкретном варианте осуществления антитело с вариантом Fc, проявляющее измененную аффинность в отношении FcγRIIIA и/или FcγRIIB, характеризуется повышенным связыванием с Fc-рецептором FcRn. В еще одном конкретном варианте осуществления антитело с вариантом Fc, проявляющее измененную аффинность в отношении FcγRIIIA и/или FcγRIIB, характеризуется измененным связыванием с C1q по сравнению с антителом c нативным Fc.
В одном варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении рецептора FcγRIIIA, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 60 раз, или не более чем в 70 раз, или не более чем в 80 раз, или не более чем в 90 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше или ниже, чем таковые у антитела c нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении FcγRIIIA, по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 40%, по меньшей мере на 30%, по меньшей мере на 20%, по меньшей мере на 10% или по меньшей мере на 5% большими или меньшими, чем у антитела c нативным Fc.
В одном варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении рецептора FcγRIIB, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 60 раз, или не более чем в 70 раз, или не более чем в 80 раз, или не более чем в 90 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше или ниже, чем таковые у антитела c нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении FcγRIIB, по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 40%, по меньшей мере на 30%, по меньшей мере на 20%, по меньшей мере на 10% или по меньшей мере на 5% большими или меньшими, чем у антитела c нативным Fc.
В одном варианте осуществления антитела c вариантами Fc характеризуются повышенными или пониженными значениями аффинности в отношении C1q по сравнению с антителом с нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении рецептора C1q, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 7 раз, или по меньшей мере в 10 раз, или по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или по меньшей мере в 40 раз, или по меньшей мере в 50 раз, или по меньшей мере в 60 раз, или по меньшей мере в 70 раз, или по меньшей мере в 80 раз, или по меньшей мере в 90 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 60 раз, или не более чем в 70 раз, или не более чем в 80 раз, или не более чем в 90 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше или ниже, чем таковые у антитела c нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями аффинности в отношении C1q, по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 40%, по меньшей мере на 30%, по меньшей мере на 20%, по меньшей мере на 10% или по меньшей мере на 5% большими или меньшими, чем у антитела c нативным Fc. В еще одном конкретном варианте осуществления антитело с вариантом Fc, проявляющее измененную аффинность в отношении C1q, характеризуется повышенным связыванием с Fc-рецептором FcRn. В еще одном конкретном варианте осуществления антитело с вариантом Fc, проявляющее измененную аффинность в отношении C1q, характеризуется измененным связыванием с FcγRIIIA и/или FcγRIIB по сравнению с антителом c нативным Fc.
Хорошо известно из уровня техники, что антитела способны к управлению атакой и разрушением посредством нескольких процессов, в совокупности известных из уровня техники как эффекторные функции антител. Один из этих процессов, известный как "антителозависимая клеточноопосредованная цитотоксичность" или "ADCC", относится к форме цитотоксичности, при которой секретируемый Ig, связываемый с Fc-рецепторами (FcR), представленными на определенных цитотоксических клетках (например, естественных клетках-киллерах (NK), нейтрофилах и макрофагах), дает возможность данным цитотоксическим эффекторным клеткам специфично связываться с клетками, несущими антиген, и затем уничтожать клетки с помощью цитотоксинов. Специфические высокоаффинные антитела IgG, направленные на поверхность клеток, "активизируют" цитотоксические клетки и требуются для такого уничтожения. Лизис клетки является внеклеточным, требует непосредственного межклеточного контакта и не вовлекает систему комплемента.
Другим процессом, охватываемым выражением "эффекторная функция", является комплементзависимая цитотоксичность (далее в данном документе называемая "CDC"), которая относится к биохимическому явлению разрушения клетки с помощью системы комплемента. Система комплемента представляет собой сложную систему белков, обнаруживаемых в нормальной плазме крови, которая в сочетании с антителами разрушает патогенные бактерии и другие чужеродные клетки.
Еще одним процессом, охватываемым выражением "эффекторная функция", является антителозависимый клеточноопосредованный фагоцитоз (ADCP), который относится к клеточноопосредованной реакции, где неспецифические цитотоксические клетки, которые экспрессируют один или несколько эффекторных лигандов, распознают связанное антитело на клетке и затем вызывают фагоцитоз клетки.
Предполагается, что характеристики антител с вариантами Fc получают при помощи функциональных анализов in vitro для определения одной или нескольких эффекторных клеточных функций, опосредованных FcγR. В определенных вариантах осуществления антитела c вариантами Fc в моделях in vivo (таких как описанные и раскрытые в данном документе) характеризуются свойствами связывания и эффекторными клеточными функциями, сходными с таковыми в анализах in vitro. Однако, настоящее изобретение не исключает антитела c вариантами Fc, которые не проявляют желаемый фенотип в анализах in vitro, но проявляют желаемый фенотип in vivo.
В определенных вариантах осуществления антитело, имеющее вариант Fc, обеспечивает повышенную цитотоксичность или фагоцитарную активность (например, ADCC, CDC и ADCP) по сравнению с антителом с нативным Fc-участком. В конкретном варианте осуществления антитело с вариантом Fc обеспечивает цитотоксичность или фагоцитарную активность, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 75 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше, чем антитело c нативным Fc. Альтернативно, антитело с вариантом Fc характеризуется пониженной цитотоксичностью или фагоцитарной активностью по сравнению с антителом c нативным Fc. В конкретном варианте осуществления антитело с вариантом Fc обеспечивает цитотоксичность или фагоцитарную активность, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 75 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз ниже, чем антитело c нативным Fc.
В определенных вариантах осуществления антитела c вариантами Fc характеризуются пониженными значениями ADCC-активности по сравнению с антителом c нативным Fc. В другом варианте осуществления антитела с вариантами Fc проявляют значения ADCC-активности, которые по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 75 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз ниже, чем таковые у антитела c нативным Fc. В еще одном варианте осуществления антитела c вариантами Fc характеризуются значениями ADCC-активности, сниженными по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 200%, или по меньшей мере на 300%, или по меньшей мере на 400%, или по меньшей мере на 500% по сравнению с антителом c нативным Fc. В определенных вариантах осуществления антитела c вариантами Fc не характеризуются выявляемой ADCC-активностью. В конкретных вариантах осуществления снижение и/или устранение ADCC-активности может быть объяснено сниженной аффинностью, которую антитела c вариантами Fc проявляют по отношению к лигандам и/или рецепторам Fc.
В альтернативном варианте осуществления антитела c вариантами Fc характеризуются повышенными значениями ADCC-активности по сравнению с антителом c нативным Fc. В другом варианте осуществления антитела c вариантами Fc характеризуются значениями ADCC-активности, по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз большими, чем у антитела c нативным Fc. В еще одном варианте осуществления антитела c вариантами Fc характеризуются значениями ADCC-активности, повышенными по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 200%, или по меньшей мере на 300%, или по меньшей мере на 400%, или по меньшей мере на 500% по сравнению с антителом c нативным Fc. В конкретных вариантах осуществления повышение ADCC-активности может быть объяснено повышенной аффинностью, которую антитела c вариантами Fc проявляют по отношению к лигандам и/или рецепторам Fc.
В конкретном варианте осуществления антитело с вариантом Fc характеризуется повышенным связыванием с Fc-рецептором FcγRIIIA и характеризуется повышенной ADCC-активностью по сравнению с антителом c нативным Fc. В других вариантах осуществления антитело с вариантом Fc характеризуется как повышенной ADCC-активностью, так и увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc. В другом конкретном варианте осуществления антитело с вариантом Fc характеризуется пониженным связыванием с Fc-рецептором FcγRIIIA и характеризуется пониженной ADCC-активностью по сравнению с антителом c нативным Fc. В других вариантах осуществления антитело с вариантом Fc характеризуется как пониженной ADCC-активностью, так и увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc.
В определенных вариантах осуществления цитотоксичность опосредована CDC, при этом антитело с вариантом Fc характеризуется повышенной либо пониженной CDC-активностью по сравнению с антителом c нативным Fc. Путь активации системы комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой, например, антителом, образующим комплекс с когнатным антигеном. Для оценки активации системы комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al. (1996) J. Immunol. Methods, 202:163.
В одном варианте осуществления антитела по настоящему изобретению характеризуются повышенной CDC-активностью по сравнению с антителом c нативным Fc. В другом варианте осуществления антитела с вариантами Fc проявляют CDC-активность, которая по меньшей мере в 2 раза, или по меньшей мере в 3 раза, или по меньшей мере в 5 раз, или по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз, или не более чем в 50 раз, или не более чем в 75 раз, или не более чем в 100 раз, или не более чем в 200 раз, или в 2-10 раз, или в 5-50 раз, или в 25-100 раз, или в 75-200 раз, или в 100-200 раз выше, чем таковая у антитела c нативным Fc. В еще одном варианте осуществления антитела c вариантами Fc характеризуются CDC-активностью, повышенной по меньшей мере на 10%, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100%, или по меньшей мере на 200%, или по меньшей мере на 300%, или по меньшей мере на 400%, или по меньшей мере на 500% по сравнению с антителом c нативным Fc. В конкретных вариантах осуществления повышение CDC-активности может быть объяснено повышенной аффинностью, которую антитела c вариантами Fc проявляют по отношению к C1q.
Антитела по настоящему изобретению могут характеризоваться повышенной CDC-активностью по сравнению с антителом c нативным Fc благодаря технологии COMPLEGENT® (Kyowa Hakko Kirin Co., Ltd.), которая усиливает один из основных механизмов действия антитела, CDC. С использованием подхода, называемого изотипическим химеризмом, в котором части IgG3, изотипа антитела, вводят в соответствующие участки IgG1, стандартного изотипа для терапевтических антител, технология COMPLEGENT® значительно повышает CDC-активность с превышением таковой как у IgG1, так и у IgG3, при этом сохраняя желаемые характеристики IgG1, такие как ADCC, PK-профиль и связывание белка А. Кроме того, ее можно применять вместе с технологией POTELLIGENT®, создавая терапевтическое mAb еще более высокого качества (ACCRETAMAB®) с повышенными значениями ADCC- и CDC-активности.
Антитело с вариантом Fc по настоящему изобретению может характеризоваться повышенной ADCC-активностью и увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc.
Антитело с вариантом Fc по настоящему изобретению может характеризоваться повышенной CDC-активностью и увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc.
Антитело с вариантом Fc по настоящему изобретению может характеризоваться повышенной ADCC-активностью, повышенной CDC-активностью и увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc.
Период полувыведения из сыворотки у белков, имеющих Fc-участки, можно увеличить путем повышения аффинности связывания Fc-участка с FcRn. Выражение ʺвремя полужизни антителаʺ, используемое в данном документе, означает фармакокинетическое свойство антитела, которое представляет собой меру среднего времени жизни молекул антитела после их введения. Время полужизни антитела можно выразить как время, необходимое для выведения 50 процентов известного количества иммуноглобулина из организма пациента (или другого млекопитающего) или его определенной части, например, измеренное в сыворотке крови, т.е. время полужизни в кровотоке, или других тканях. Период полувыведения может различаться среди разных иммуноглобулинов или классов иммуноглобулинов. Как правило, увеличение периода полувыведения антитела приводит к увеличению среднего времени удержания (MRT) введенного антитела в кровотоке.
Увеличение периода полувыведения позволяет уменьшить количество лекарственного средства, даваемого пациенту, а также снизить частоту введения. Для увеличения периода полувыведения антитела из сыворотки в состав антитела (в частности, фрагмента антитела) может быть введен эпитоп, связывающийся с рецептором реутилизации, как описано, например, в патенте США № 5739277. Как используется в данном документе, выражение "эпитоп, связывающийся с рецептором реутилизации" относится к эпитопу Fc-участка молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за увеличение периода полувыведения из сыворотки молекулы IgG in vivo.
Альтернативно, антитела по настоящему изобретению с увеличенным периодом полувыведения можно получить с помощью модификации аминокислотных остатков, идентифицированных как вовлеченные во взаимодействие между Fc и FcRn-рецептором (см., например, патенты США №№ 6821505 и 7083784; а также WO 09/058492). Кроме того, период полувыведения антител по настоящему изобретению можно увеличить путем конъюгирования с PEG или альбумином при помощи методик, широко используемых в данной области техники. В некоторых вариантах осуществления антитела, имеющие варианты Fc-участков, по настоящему изобретению характеризуются периодом полувыведения, увеличенным на приблизительно 5%, на приблизительно 10%, на приблизительно 15%, на приблизительно 20%, на приблизительно 25%, на приблизительно 30%, на приблизительно 35%, на приблизительно 40%, на приблизительно 45%, на приблизительно 50%, на приблизительно 60%, на приблизительно 65%, на приблизительно 70%, на приблизительно 80%, на приблизительно 85%, на приблизительно 90%, на приблизительно 95%, на приблизительно 100%, на приблизительно 125%, на приблизительно 150% или более по сравнению с антителом, имеющим нативный Fc-участок. В некоторых вариантах осуществления антитела, имеющие варианты Fc-участков, характеризуются периодом полувыведения, увеличенным в приблизительно 2 раза, в приблизительно 3 раза, в приблизительно 4 раза, в приблизительно 5 раз, в приблизительно 10 раз, в приблизительно 20 раз, в приблизительно 50 раз или более, или не более чем в приблизительно 10 раз, в приблизительно 20 раз или в приблизительно 50 раз, или в 2-10 раз, или в 5-25 раз, или в 15-50 раз по сравнению с антителом с нативным Fc-участком.
В одном варианте осуществления настоящее изобретение предусматривает варианты Fc, где Fc-участок включает в себя модификацию (например, замены аминокислот, вставки аминокислот, делеции аминокислот) в одном или нескольких положениях, выбранных из 221, 225, 228, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 247, 250, 251, 252, 254, 255, 256, 257, 262, 263, 264, 265, 266, 267, 268, 269, 279, 280, 284, 292, 296, 297, 298, 299, 305, 308, 313, 316, 318, 320, 322, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 339, 341, 343, 370, 373, 378, 392, 416, 419, 421, 428, 433, 434, 435, 436, 440, и 443 согласно нумерации с помощью EU-индекса, как изложено у Kabat. Необязательно, Fc-участок может включать в себя модификацию в дополнительных и/или альтернативных положениях, известных специалисту в данной области (см., например, патенты США 5624821; 6277375; 6737056; 7083784; 7317091; 7217797; 7276585; 7355008; 2002/0147311; 2004/0002587; 2005/0215768; 2007/0135620; 2007/0224188; 2008/0089892; WO 94/29351 и WO 99/58572). Дополнительные подходящие положения и конкретные замены аминокислот приведены в качестве примеров в таблицах 2 и 6-10 US 6737056; таблицах, представленных на фигуре 41 US 2006/024298; таблицах, представленных на фигурах 5, 12 и 15 US 2006/235208; таблицах, представленных на фигурах 8, 9 и 10 US 2006/0173170, и таблицах, представленных на фигурах 8-10, 13 и 14 WO 09/058492.
В конкретном варианте осуществления настоящее изобретение предусматривает вариант Fc, где Fc-участок включает в себя по меньшей мере одну замену, выбранную из 221K, 221Y, 225E, 225K, 225W, 228P, 234D, 234E, 234N, 234Q, 234T, 234H, 234Y, 234I, 234V, 234F, 235A, 235D, 235R, 235W, 235P, 235S, 235N, 235Q, 235T, 235H, 235Y, 235I, 235V, 235E, 235F, 236E, 237L, 237M, 237P, 239D, 239E, 239N, 239Q, 239F, 239T, 239H, 239Y, 240I, 240A, 240T, 240M, 241W, 241L, 241Y, 241E, 241R. 243W, 243L 243Y, 243R, 243Q, 244H, 245A, 247L, 247V, 247G, 250E, 250Q, 251F, 252L, 252Y, 254S, 254T, 255L, 256E, 256F, 256M, 257C, 257M, 257N, 262I, 262A, 262T, 262E, 263I, 263A, 263T, 263M, 264L, 264I, 264W, 264T, 264R, 264F, 264M, 264Y, 264E, 265A, 265G, 265N, 265Q, 265Y, 265F, 265V, 265I, 265L, 265H, 265T, 266I, 266A, 266T, 266M, 267Q, 267L, 268E, 269H, 269Y, 269F, 269R, 270E, 280A, 284M, 292P, 292L, 296E, 296Q, 296D, 296N, 296S, 296T, 296L, 296I, 296H, 296G, 297S, 297D, 297E, 298A, 298H, 298I, 298T, 298F, 299I, 299L, 299A, 299S, 299V, 299H, 299F, 299E, 305I, 308F, 313F, 316D, 318A, 318S, 320A, 320S, 322A, 322S, 325Q, 325L, 325I, 325D, 325E, 325A, 325T, 325V, 325H, 326A, 326D, 326E, 326G, 326M, 326V, 327G, 327W, 327N, 327L, 328S, 328M, 328D, 328E, 328N, 328Q, 328F, 328I, 328V, 328T, 328H, 328A, 329F, 329H, 329Q, 330K, 330G, 330T, 330C, 330L, 330Y, 330V, 330I, 330F, 330R, 330H, 331G, 331A, 331L, 331M, 331F, 331W, 331K, 331Q, 331E, 331S, 331V, 331I, 331C, 331Y, 331H, 331R, 331N, 331D, 331T, 332D, 332S, 332W, 332F, 332E, 332N, 332Q, 332T, 332H, 332Y, 332A, 333A, 333D, 333G, 333Q, 333S, 333V, 334A, 334E, 334H, 334L, 334M, 334Q, 334V, 334Y, 339T, 370E, 370N, 378D, 392T, 396L, 416G, 419H, 421K, 428L, 428F, 433K, 433L, 434A, 424F, 434W, 434Y, 436H, 440Y и 443W согласно нумерации с помощью EU-индекса, как изложено у Kabat. Необязательно, Fc-участок может включать в себя дополнительные и/или альтернативные замены аминокислот, известные специалисту в данной области, в том числе без ограничения приведенные в качестве примеров в таблицах 2 и 6-10 патента США 6737056; таблицах, представленных на фигуре 41 US 2006/024298; таблицах, представленных на фигурах 5, 12 и 15 US 2006/235208; таблицах, представленных на фигурах 8, 9 и 10 US 2006/0173170, и таблицах, представленных на фигурах 8, 9 и 10 WO 09/058492.
В конкретном варианте осуществления настоящее изобретение предусматривает антитело с вариантом Fc, где Fc-участок включает в себя по меньшей мере одну модификацию (например, замены аминокислот, вставки аминокислот, делеции аминокислот) в одном или нескольких положениях, выбранных из 228, 234, 235 и 331 согласно нумерации с помощью EU-индекса, как изложено у Kabat. В одном варианте осуществления модификация представляет собой по меньшей мере одну замену, выбранную из 228P, 234F, 235E, 235F, 235Y и 331S согласно нумерации с помощью EU-индекса, как изложено у Kabat.
В другом конкретном варианте осуществления настоящее изобретение предусматривает антитело с вариантом Fc, где Fc-участок представляет собой Fc-участок IgG4 и включает в себя по меньшей мере одну модификацию в одном или нескольких положениях, выбранных из 228 и 235 согласно нумерации с помощью EU-индекса, как изложено у Kabat. В еще одном конкретном варианте осуществления Fc-участок представляет собой Fc-участок IgG4, а не встречающиеся в природе аминокислоты выбраны из 228P, 235E и 235Y согласно нумерации с помощью EU-индекса, как изложено у Kabat.
В другом конкретном варианте осуществления настоящее изобретение предусматривает вариант Fc, где Fc-участок включает в себя по меньшей мере одну не встречающуюся в природе аминокислоту в одном или нескольких положениях, выбранных из 239, 330 и 332 согласно нумерации с помощью EU-индекса, как изложено у Kabat. В одном варианте осуществления модификация представляет собой по меньшей мере одну замену, выбранную из 239D, 330L, 330Y и 332E согласно нумерации с помощью EU-индекса, как изложено у Kabat.
В конкретном варианте осуществления настоящее изобретение предусматривает антитело с вариантом Fc, где Fc-участок включает в себя по меньшей мере одну не встречающуюся в природе аминокислоту в одном или нескольких положениях, выбранных из 252, 254 и 256 согласно нумерации с помощью EU-индекса, как изложено у Kabat. В одном варианте осуществления модификация представляет собой по меньшей мере одну замену, выбранную из 252Y, 254T и 256E согласно нумерации с помощью EU-индекса, как изложено у Kabat. В особо предпочтительных антителах по настоящему изобретению модификация представляет собой три замены 252Y, 254T и 256E согласно нумерации с помощью EU-индекса, как изложено у Kabat (известные как "YTE"), см. U.S. 7083784.
В определенных вариантах осуществления эффекторные функции, проявляемые IgG-антителами, в значительной степени зависят от углеводного фрагмента, соединенного с Fc-участком белка (Ferrara et al. (2006) Biotechnology and Bioengineering. 93:851-861). Таким образом, гликозилирование Fc-участка можно модифицировать для усиления или ослабления эффекторной функции (см., например, Umana et al. (1999) Nat. Biotechnol. 17:176-180; Davies et al. (2001) Biotechnol Bioeng. 74:288-294; Shields et al. (2002) J Biol Chem. 277:26733-26740; Shinkawa et al. (2003) J Biol Chem. 278:3466-3473; патенты США №№ 6602684; 6946292; 7064191; 7214775; 7393683; 7425446; 7504256; публикации США №№ 2003/0157108; 2003/0003097; 2009/0010921; технологию Potelligent™ (Biowa, Inc. Принстон, Нью-Джерси); инженерную технологию гликозилирования GlycoMAb™ (GLYCART Biotechnology AG, Цюрих, Швейцария)). Соответственно, в одном варианте осуществления Fc-участки антител по настоящему изобретению включают в себя измененное гликозилирование аминокислотных остатков. В другом варианте осуществления измененное гликозилирование аминокислотных остатков приводит к ослаблению эффекторной функции. В другом варианте осуществления измененное гликозилирование аминокислотных остатков приводит к усилению эффекторной функции. В конкретном варианте осуществления Fc-участок характеризуется пониженным фукозилированием. В другом варианте осуществления Fc-участок является нефукозилированным (см., например, публикацию заявки на патент США № 2005/0226867). В одном аспекте эти антитела с усиленной эффекторной функцией, в частности ADCC, полученные в клетках-хозяевах (например, клетках CHO, Lemna minor), сконструированных для выработки антитела с высокой степенью дефукозилирования, обеспечивающего более чем в 100 раз повышенную ADCC по сравнению с антителом, вырабатываемым исходными клетками (Mori et al. (2004) Biotechnol Bioeng. 88:901-908; Cox et al. (2006) Nat Biotechnol. 24:1591-7).
Добавление сиаловой кислоты к олигосахаридам на молекулах IgG может усиливать их противовоспалительную активность и изменяет их влияние на цитотоксичность (Keneko et al. (2006) Science. 313:670-673; Scallon et al. (2007) Mol. Immuno. 44(7):1524-34). Исследования, упомянутые выше, показывают, что молекулы IgG с повышенным сиалированием обладают противовоспалительными свойствами, тогда как молекулы IgG с пониженным сиалированием обладают усиленными иммуностимулирующими свойствами (например, повышают ADCC-активность). Таким образом, антитело можно модифицировать с помощью подходящего профиля сиалирования для конкретного терапевтического применения (публикация США № 2009/0004179 и международная публикация № WO 2007/005786).
В одном варианте осуществления Fc-участки антител по настоящему изобретению имеют измененный профиль сиалирования по сравнению с нативным Fc-участком. В одном варианте осуществления Fc-участки антител по настоящему изобретению имеют профиль повышенного сиалирования по сравнению с нативным Fc-участком. В другом варианте осуществления Fc-участки антител по настоящему изобретению имеют профиль пониженного сиалирования по сравнению с нативным Fc-участком.
В одном варианте осуществления варианты Fc по настоящему изобретению можно комбинировать с другими известными вариантами Fc, такими как раскрытые в Ghetie et al. (1997) Nat Biotech. 15:637-40; Duncan et al. (1988) Nature. 332:563-564; Lund et al. (1991) J. Immunol. 147:2657-2662; Lund et al. (1992) Mol Immunol. 29:53-59; Alegre et al. (1994) Transplantation. 57:1537-1543; Hutchins et al. (1995) Proc Natl. Acad. Sci. USA. 92:11980-11984; Jefferis et al. (1995) Immunol Lett. 44:111-117; Lund et al. (1995) Faseb. J. 9:115-119; Jefferis et al. (1996) Immunol. Lett. 54:101-104; Lund et al. (1996) J. Immunol. 157:4963-4969; Armour et al. (1999) Eur. J. Immunol. 29:2613-2624; Idusogie et al. (2000) J. Immunol. 164:4178-4184; Reddy et al. (2000) J. Immunol. 164:1925-1933; Xu et al. (2000) Cell. Immunol. 200:16-26; Idusogie et al. (2001) J. Immunol. 166:2571-2575; Shields et al. (2001) J. Biol. Chem. 276:6591-6604; Jefferis et al. (2002) Immunol. Lett. 82:57-65; Presta et al. (2002) Biochem. Soc. Trans. 30:487-490); патентах США №№ 5,624,821; 5,885,573; 5,677,425; 6,165,745; 6,277,375; 5,869,046; 6,121,022; 5,624,821; 5,648,260; 6,528,624; 6,194,551; 6,737,056; 7,122,637; 7,183,387; 7,332,581; 7,335,742; 7,371,826; 6,821,505; 6,180,377; 7,317,091; 7,355,008; 2004/0002587; и WO 99/58572. Другие модификации, и/или замены, и/или добавления, и/или делеции в Fc-домене будут абсолютно очевидны специалисту в данной области.
Гликозилирование
Кроме способности гликозилирования изменять эффекторную функцию антитела, модифицированное гликозилирование в вариабельном участке может изменять аффинность антитела к антигену. В одном варианте осуществления паттерн гликозилирования в вариабельном участке антител по настоящему изобретению является модифицированным. Например, можно создать негликозилированное антитело (т.е. антитело, у которого отсутствует гликозилирование). Гликозилирование можно изменить, например, с тем, чтобы повысить аффинность антитела к антигену. Таких углеводных модификаций можно достичь при помощи, например, изменения одного или нескольких сайтов гликозилирования в пределах последовательности антитела. Например, можно провести одну либо несколько замен аминокислот, которые приводят к удалению одного или нескольких сайтов гликозилирования в каркасной области вариабельного участка с устранением, таким образом, гликозилирования в этом сайте. Такое гликозилирование может повышать аффинность антитела к антигену. Такой подход более подробно описан в патентах США №№ 5714350 и 6350861. Также могут быть получены одна или несколько замен аминокислот, результатом чего является удаление сайта гликозилирования, представленного в Fc-участке (например, аспарагина 297 IgG). Более того, негликозилированные антитела можно получать в бактериальных клетках, которые не имеют необходимого аппарата гликозилирования.
Конъюгаты антител
В определенных вариантах осуществления антитела по настоящему изобретению являются конъюгированными или ковалентно связанными с веществом с использованием способов, хорошо известных из уровня техники. В одном варианте осуществления присоединенное вещество представляет собой терапевтическое средство, детектируемую метку (также называемую в данном документе репортерной молекулой) или твердую подложку. Подходящие вещества для присоединения к антителам включают, без ограничения, аминокислоту, пептид, белок, полисахарид, нуклеозид, нуклеотид, олигонуклеотид, нуклеиновую кислоту, гаптен, лекарственное средство, гормон, липид, ансамбль липидов, синтетический полимер, полимерную микрочастицу, биологическую клетку, вирус, флуорофор, хромофор, краситель, токсин, гаптен, фермент, антитело, фрагмент антитела, радиоактивный изотоп, твердые матрицы, полутвердые матрицы и их комбинации. Способы конъюгирования или ковалентного присоединения другого вещества к антителу хорошо известны из уровня техники.
В определенных вариантах осуществления антитела по настоящему изобретению конъюгированы с твердой подложкой. Антитела могут быть конъюгированы с твердой подложкой в качестве части способа скрининга, и/или очистки, и/или производства. Альтернативно, антитела могут быть конъюгированы с твердой подложкой в качестве части способа или композиции для диагностики. Твердая подложка, подходящая для применения в настоящем изобретении, обычно фактически нерастворима в жидких фазах. Специалисту в данной области доступно и известно большое число подложек. Таким образом, твердые подложки включают твердые и полутвердые матрицы, такие как аэрогели и гидрогели, смолы, гранулы, биочипы (в том числе биочипы с тонкопленочным покрытием), микрожидкостный чип, кремниевый чип, многолуночные планшеты (также называемые титрационными микропланшетами или микропланшетами), мембраны, проводящие и непроводящие металлы, стекло (в том числе предметные стекла) и намагниченные подложки. Более конкретные примеры твердых подложек включают силикагели, полимерные мембраны, частицы, дериватизированные полимерные пленки, стеклянные бусы, хлопок, полимерные гранулы, алюмогели, полисахариды, такие как сефароза, поли(акрилат), полистирол, поли(акриламид), полиол, агарозу, агар, целлюлозу, декстран, крахмал, фиколл, гепарин, гликоген, амилопектин, маннан, инулин, нитроцеллюлозу, диазоцеллюлозу, поливинилхлорид, полипропилен, полиэтилен (в том числе поли(этиленгликоль)), нейлон, латексную гранулу, магнитную гранулу, парамагнитную гранулу, суперпарамагнитную гранулу, крахмал и т.п.
В некоторых вариантах осуществления твердая подложка может содержать реакционноспособную функциональную группу, в том числе, без ограничения, гидроксил, карбоксил, амино, тиол, альдегид, галоген, нитро, циано, амидо, мочевину, карбонат, карбамат, изоцианат, сульфон, сульфонат, сульфонамид, сульфоксид и т.д., для присоединения антител по настоящему изобретению.
Подходящая твердофазная подложка может быть выбрана, исходя из желаемого конечного применения и пригодности для различных протоколов синтеза. Например, если образование амидной связи желательно для присоединения антител по настоящему изобретению к твердой подложке, можно использовать смолы, обычно применимые в синтезе пептидов, такие как полистирол (например, PAM-смола, получаемая из Bachem Inc., Peninsula Laboratories и др.), смола POLYHIPE™ (получаемая из Aminotech, Канада), полиамидная смола (получаемая из Peninsula Laboratories), полистирольная смола с привитым полиэтиленгликолем (TentaGel™, Rapp Polymere, Тюбинген, Германия), полидиметилакриламидная смола (доступная от Milligen/Biosearch, Калифорния) или гранулы PEGA (получаемые из Polymer Laboratories).
В определенных вариантах осуществления антитела по настоящему изобретению конъюгируют с метками с целью диагностики и других анализов, где антитело и/или его соответствующий лиганд могут быть выявлены. Метка, конъюгированная с антителом и применяемая в способах и композициях по настоящему изобретению, описанных в данном документе, представляет собой любой химический компонент, органический или неорганический, который характеризуется максимумом поглощения при длинах волн более 280 нм и сохраняет свои спектральные свойства при ковалентном присоединении к антителу. Метки включают, без ограничения, хромофор, флуорофор, флуоресцентный белок, фосфоресцентный краситель, тандемный краситель, частицу, гаптен, фермент и радиоактивный изотоп.
В определенных вариантах осуществления антитела конъюгированы с флуорофором. Соответственно, флуорофоры, применяемые для мечения антител по настоящему изобретению, включают, без ограничения, пирен (в том числе любое из соответствующих производных соединений, раскрытых в патенте США 5132432), антрацен, нафталин, акридин, стильбен, индол или бензиндол, оксазол или бензоксазол, тиазол или бензотиазол, 4-амино-7-нитробенз-2-окса-1,3-диазол (NBD), цианин (в том числе любые соответствующие соединения из патентов США №№ 6977305 и 6974873), карбоцианин (в том числе любые соответствующие соединения из заявки на патент США с регистрационным номером 09/557275; патентов США №№ 4981977; 5268486; 5569587; 5569766; 5486616; 5627027; 5808044; 5877310; 6002003; 6004536; 6008373; 6043025; 6127134; 6130094; 6133445 и публикаций WO 02/26891, WO 97/40104, WO 99/51702, WO 01/21624; EP 1065250 A1), карбостирил, порфирин, салицилат, антранилат, азулен, перилен, пиридин, хинолин, бораполиазаиндацен (в том числе любые соответствующие соединения, раскрытые в патентах США №№ 4774339; 5187288; 5248782; 5274113 и 5433896), ксантен (в том числе любые соответствующие соединения, раскрытые в патентах США №№ 6162931; 6130101; 6229055; 6339392; 5451343; 5227487; 5442045; 5798276; 5846737; 4945171; заявках на патенты США с регистрационным номером 09/129015 и 09/922333), оксазин (в том числе любые соответствующие соединения, раскрытые в патенте США № 4714763) или бензоксазин, карбазин (в том числе любые соответствующие соединения, раскрытые в патенте США №4810636), феналенон, кумарин (в том числе любые соответствующие соединения, раскрытые в патентах №№ 5696157; 5459276; 5501980 и 5830912), бензофуран (в том числе любые соответствующие соединения, раскрытые в патентах США №№ 4603209 и 4849362) и бензфеналенон (в том числе любые соответствующие соединения, раскрытые в патенте США № 4812409) и их производные. Как используется в данном документе, оксазины включают резоруфины (в том числе любые соответствующие соединения, раскрытые в 5242805), аминооксазиноны, диаминооксазины и их бензозамещенные аналоги.
В конкретном варианте осуществления флуорофоры, конъюгированные с антителами, описанными в данном документе, включают ксантен (родол, родамин, флуоресцеин и их производные), кумарин, цианин, пирен, оксазин и бораполиазаиндацен. В других вариантах осуществления такие флуорофоры представляют собой сульфированные ксантены, фторированные ксантены, сульфированные кумарины, фторированные кумарины и сульфированные цианины. Также включены красители, продаваемые под торговыми марками, и, как правило, известные как ALEXA FLUOR®, DyLight, красители CY®, BODIPY®, OREGON GREEN®, PACIFIC BLUE™, IRDYE®, FAM, FITC и ROX™.
Выбор флуорофора, присоединяемого к антителу, будет определять свойства поглощения и испускания флуоресценции конъюгированного антитела. Физические свойства флуорофорной метки, которую можно применять для антитела и связываемых антителом лигандов, включают, без ограничения, спектральные характеристики (поглощение, испускание и стоксов сдвиг), интенсивность, длительность, поляризацию флуоресценции и скорость обесцвечивания или их комбинацию. Все эти физические свойства можно применять для того, чтобы отличить один флуорофор от другого, и, таким образом, способствовать мультиплексному анализу. В определенных вариантах осуществления флуорофор имеет максимум поглощения при длинах волн более 480 нм. В других вариантах осуществления флуорофор поглощает при 488 нм - 514 нм или вблизи этих значений (что особенно подходит для возбуждения излучением аргонового ионного лазерного источника возбуждения) или вблизи 546 нм (что особенно подходит для возбуждения ртутной дуговой лампой). В другом варианте осуществления флуорофор может испускать в NIR (ближней инфракрасной области) для применения в тканях или во всем организме. Другие желаемые свойства флуоресцентной метки могут включать клеточную проницаемость и низкую токсичность, например, если мечение антитела необходимо проводить в клетке или организме (например, живом животном).
В определенных вариантах осуществления фермент представляет собой метку и конъюгирован с антителом, как описано в данном документе. Ферменты являются желательными метками, поскольку можно получить усиление детектируемого сигнала, приводящее к повышению чувствительности анализов. Сам фермент не вызывает детектируемый ответ, а действует путем разрушения субстрата при контакте с подходящим субстратом, так что превращаемый субстрат испускает флуоресцентный, колориметрический или люминесцентный сигнал. Ферменты усиливают детектируемый сигнал, поскольку один фермент в метящем реактиве может приводить к превращению нескольких субстратов в детектируемый сигнал. Субстрат для фермента выбирают так, чтобы получить предпочтительный измеримый продукт, например, колориметрический, флуоресцентный или хемилюминесцентный. Такие субстраты широко применяются в данной области техники и хорошо известны специалисту в данной области.
В одном варианте осуществления в комбинации колориметрического или флуорогенного субстрата и фермента применяются оксидоредуктазы, такие как пероксидаза хрена, и субстрат, такой как 3,3'-диаминобензидин (DAB) и 3-амино-9-этилкарбазол (AEC), которые дают отличительный цвет (коричневый и красный, соответственно). Другие колориметрические субстраты оксидоредуктаз, которые дают детектируемые продукты, включают, без ограничения: 2,2-азино-бис(3-этилбензотиазолин-6-сульфоновую кислоту) (ABTS), o-фенилендиамин (OPD), 3,3',5,5'-тетраметилбензидин (TMB), o-дианизидин, 5-аминосалициловую кислоту, 4-хлор-1-нафтол. Флуорогенные субстраты включают, без ограничения, гомованилиновую кислоту или 4-гидрокси-3-метоксифенилуксусную кислоту, восстановленные феноксазины и восстановленные бензотиазины, в том числе, реактив Amplex® Red и его варианты (патент США № 4384042) и восстановленные дигидроксантены, в том числе дигидрофлуоресцеины (патент США № 6162931) и дигидрородамины, в том числе дигидрородамин 123. Субстраты пероксидазы, которыми являются тирамиды (патенты США №№ 5196306; 5583001 и 5731158) представляют уникальный класс субстратов пероксидазы, поскольку по своей природе они могут быть выявлены до действия фермента, но под действием пероксидазы "фиксируются на месте" в процессе, описанном как тирамидное усиление сигнала (TSA). Эти субстраты широко используются для мечения антигена в образцах, которыми являются клетки, ткани или монослои, для их последующего выявления с помощью микроскопии, проточной цитометрии, оптического сканирования и флуорометрии.
В другом варианте осуществления в комбинации колориметрического (и в некоторых случаях флуорогенного) субстрата и фермента применяется фермент фосфатаза, такой как кислая фосфатаза, щелочная фосфатаза или рекомбинантный вариант такой фосфатазы, в комбинации с колориметрическим субстратом, таким как 5-бром-6-хлор-3-индолилфосфат (BCIP), 6-хлор-3-индолилфосфат, 5-бром-6-хлор-3-индолилфосфат, п-нитрофенилфосфат или o-нитрофенилфосфат, или с флуорогенным субстратом, таким как 4-метилумбеллиферилфосфат, 6,8-дифтор-7-гидрокси-4-метилкумаринилфосфат (DiFMUP, патент США № 5830912), флуоресцеиндифосфат, 3-O-метилфлуоресцеинфосфат, резоруфинфосфат, 9H-(1,3-дихлор-9,9-диметилакридин-2-он-7-ил)фосфат (DDAO-фосфат) или ELF 97, ELF 39 или родственные фосфаты (патенты США №№ 5316906 и 5443986).
Гликозидазы, в частности, бета-галактозидаза, бета-глюкуронидаза и бета-глюкозидаза, являются дополнительными подходящими ферментами. Подходящие колориметрические субстраты включают, без ограничения, 5-бром-4-хлор-3-индолил-бета-D-галактопиранозид (X-gal) и подобные индолилгалактозиды, глюкозиды и глюкурониды, o-нитрофенил-бета-D-галактопиранозид (ONPG) и п-нитрофенил-бета-D-галактопиранозид. В одном варианте осуществления флуорогенные субстраты включают резоруфин-бета-D-галактопиранозид, флуоресцеиндигалактозид (FDG), флуоресцеиндиглюкуронид и их структурные варианты (патенты США №№ 5208148; 5242805; 5362628; 5576424 и 5773236), 4-метилумбеллиферил-бета-D-галактопиранозид, карбоксиумбеллиферил-бета-D-галактопиранозид и фторированные кумарин-бета-D-галактопиранозиды (патент США № 5830912).
Дополнительные ферменты включают, без ограничения, гидролазы, такие как холинэстеразы и пептидазы, оксидазы, такие как глюкозооксидаза и цитохромоксидазы, и редуктазы, для которых известны подходящие субстраты.
Ферменты и их соответствующие субстраты, которые дают хемилюминесценцию, являются предпочтительными для некоторых анализов. Они включают, без ограничения, природные и рекомбинантные формы люцифераз и экворинов. Дополнительно подходят дающие хемилюминесценцию субстраты фосфатаз, гликозидаз и оксидаз, как, например, содержащие стабильные диоксетаны, люминол, изолюминол и сложные эфиры акридиния.
В другом варианте осуществления гаптены, такие как биотин, также используются в качестве меток. Биотин является подходящим, поскольку он может функционировать в ферментной системе для дополнительного усиления детектируемого сигнала, и он может функционировать в качестве метки для применения в аффинной хроматографии с целью выделения. В целях выявления применяют конъюгат фермента, который характеризуется аффинностью к биотину, такой как авидин-HRP. Затем добавляют субстрат пероксидазы для получения детектируемого сигнала.
Гаптены также включают гормоны, встречающиеся в природе и синтетические лекарственные средства, загрязнители, аллергены, эффекторные молекулы, факторы роста, хемокины, цитокины, лимфокины, аминокислоты, пептиды, промежуточные химические соединения, нуклеотиды и т.п.
В определенных вариантах осуществления флуоресцентные белки могут быть конъюгированы с антителами в качестве метки. Примеры флуоресцентных белков включают зеленый флуоресцентный белок (GFP) и фикобилипротеины и их производные. Флуоресцентные белки, в частности, фикобилипротеин, особенно полезны для получения метящих реактивов на основе тандемных красителей. Эти тандемные красители включают в себя флуоресцентный белок и флуорофор с целью получения большего стоксового сдвига, где спектр испускания дальше смещен от длины волны спектра поглощения флуоресцентного белка. Это является особенно преимущественным для выявления небольшого количества антигена в образце, где испускаемый флуоресцентный свет максимально оптимизирован, другими словами, испускаемый свет повторно поглощается флуоресцентным белком в незначительном количестве или совсем не поглощается. С этой целью флуоресцентный белок и флуорофор функционируют в качестве пары для переноса энергии, при этом флуоресцентный белок испускает при длине волны, при которой флуорофор поглощает, а затем флуорофор испускает при длине волны, смещенной относительно флуоресцентных белков дальше, чем можно было бы получить с помощью только флуоресцентного белка. Особенно подходящей комбинацией является фикобилипротеины, раскрытые в патентах США №№ 4520110; 4859582; 5055556, и сульфородаминовые флуорофоры, раскрытые в патенте США № 5798276, или сульфированные цианиновые флуорофоры, раскрытые в патентах США №№ 6977305 и 6974873; или сульфированные производные ксантена, раскрытые в патенте США № 6130101, и их комбинации, раскрытые в патенте США № 4542104. Альтернативно, флуорофор функционирует в качестве донора энергии, а флуоресцентный белок является акцептором энергии.
В определенных вариантах осуществления меткой является радиоактивный изотоп. Примеры подходящих радиоактивных материалов включают, без ограничения, йод (121I, 123I, 125I, 131I), углерод (14C), серу (35S), тритий (3H), индий (111In, 112In, 113mIn, 115mIn), технеций (99Tc, 99mTc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (135Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh и 97Ru.
Способы медицинского лечения и пути применения
Антитела и их антигенсвязывающие фрагменты по настоящему изобретению, а также их варианты можно применять для лечения инфекции, вызванной вирусом гриппа B, для предупреждения инфекции, вызванной вирусом гриппа B, для выявления, диагностики и/или прогнозирования течения инфекции, вызванной вирусом гриппа B, или их комбинаций. В одном варианте осуществления антитела и их антигенсвязывающие фрагменты по настоящему изобретению, а также их варианты можно применять для лечения инфекции, вызванной вирусом гриппа А и вирусом гриппа B, для предупреждения инфекции, вызванной вирусом гриппа А и вирусом гриппа B, для выявления, диагностики и/или прогнозирования течения инфекции, вызванной вирусом гриппа А и вирусом гриппа B, или их комбинаций.
Способы диагностики могут включать приведение антитела или фрагмента антитела в контакт с образцом. Такими образцами могут быть образцы тканей, полученные, например, из носовых ходов, околоносовых пазух, слюнных желез, легкого, печени, поджелудочной железы, почки, уха, глаза, плаценты, пищеварительного канала, сердца, яичников, гипофиза, надпочечников, щитовидной железы, головного мозга или кожи. Способы выявления, диагностики и/или прогнозирования могут также включать выявление комплекса антиген/антитело.
В одном варианте осуществления настоящее изобретение предусматривает способ лечения субъекта путем введения субъекту эффективного количества антитела или его антигенсвязывающего фрагмента в соответствии с настоящим изобретением или фармацевтической композиции, которая включает в себя антитело или его антигенсвязывающий фрагмент. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент являются фактически очищенными (т.е. фактически не содержащими соединений, которые ограничивают их эффект или вызывают нежелательные побочные эффекты). В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят после воздействия, или после того, как субъект претерпел воздействие вируса гриппа B или был инфицирован вирусом гриппа B. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят после воздействия, или после того, как субъект претерпел воздействие вируса гриппа B линий Yamagata и/или Victoria или был инфицирован вирусом гриппа B линий Yamagata и/или Victoria. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят после воздействия, или после того, как субъект претерпел воздействие по меньшей мере одного подтипа вируса гриппа A; вируса гриппа B линии Yamagata; вируса гриппа B линии Victoria или их комбинаций или был инфицирован по меньшей мере одним подтипом вируса гриппа A и/или вирусом гриппа B линий Yamagata и/или Victoria.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят до воздействия, или субъекту, который еще не претерпел воздействие вируса гриппа B или еще не был инфицирован вирусом гриппа B. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят до воздействия, или субъекту, который еще не претерпел воздействие вируса гриппа B линий Yamagata и/или Victoria или еще не был инфицирован вирусом гриппа B линий Yamagata и/или Victoria. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят до воздействия, или субъекту, который еще не претерпел воздействие вируса гриппа А; вируса гриппа B линии Yamagata; вируса гриппа B линии Victoria или их комбинаций или еще не был инфицирован вирусом гриппа А; вирусом гриппа B линии Yamagata; вирусом гриппа В линии Victoria или их комбинациями.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серонегативным в отношении одного или нескольких вирусов гриппа B. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серонегативным в отношении одной или нескольких линий вируса гриппа B. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серонегативным в отношении одного или нескольких подтипов вируса гриппа А и/или одного или нескольких вирусов гриппа В.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серопозитивным в отношении одного или нескольких вирусов гриппа B. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серопозитивным в отношении одной или нескольких линий вируса гриппа B. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту, который является серопозитивным в отношении одного или нескольких подтипов вируса гриппа A и/или одного или нескольких вирусов гриппа B. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту в рамках периода 1, 2, 3, 4, 5 дней после появления инфекции или симптома. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению можно вводить субъекту через 1, 2, 3, 4, 5, 6 или 7 дней, или в рамках периода 2, 3, 4, 5, 6, 7, 8, 9 или 10 дней после появления инфекции или симптома.
В одном варианте осуществления с помощью способа ослабляют инфекцию, вызванную вирусом гриппа B, у субъекта. В другом варианте осуществления с помощью способа ослабляют инфекцию, вызванную вирусом гриппа А, и/или инфекцию, вызванную вирусом гриппа B, у субъекта. В другом варианте осуществления с помощью способа предупреждают инфекцию, вызванную вирусом гриппа B, снижают риск ее развития или замедляют ее развитие у субъекта. В другом варианте осуществления с помощью способа предупреждают инфекцию, вызванную вирусом гриппа A и/или вирусом гриппа B, снижают риск ее развития или замедляют ее развитие у субъекта. В одном варианте осуществления субъектом является животное. В одном варианте осуществления субъектом является млекопитающее. В более конкретном варианте осуществления субъектом является человек. В одном варианте осуществления субъект включает без ограничения субъекта, который особенно подвержен риску развития инфекции, вызванной вирусом гриппа А и/или вирусом гриппа В, или восприимчив к ней, в том числе, например, субъекта с ослабленным иммунитетом.
Лечение может проходить по схеме введения однократной дозы или по схеме введения многократных доз, и антитело или его антигенсвязывающий фрагмент по настоящему изобретению можно применять при пассивной иммунизации или активной вакцинации.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту в комбинации с одним или несколькими противовирусными лекарственными препаратами. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению вводят субъекту в комбинации с одним или несколькими низкомолекулярными противовирусными лекарственными препаратами. Низкомолекулярные противовирусные лекарственные препараты включают ингибиторы нейраминидазы, такие как озельтамивир (TAMIFLU®), занамивир (RELENZA®), и адамантаны, такие как амантадин и римантадин.
В другом варианте осуществления настоящее изобретение предусматривает композицию для применения в качестве лекарственного препарата для предупреждения или лечения инфекции, вызванной вирусом гриппа А и/или вирусом гриппа B. В другом варианте осуществления настоящее изобретение предусматривает применение антитела или его антигенсвязывающего фрагмента по настоящему изобретению и/или белка, включающего в себя эпитоп, с которым связываются антитело или его антигенсвязывающий фрагмент по настоящему изобретению, в получении лекарственного препарата для лечения субъекта и/или диагностики у субъекта.
Антитела и их фрагменты, как описано в настоящем изобретении, можно также применять в наборе для диагностики инфекции, вызванной вирусом гриппа А; инфекции, вызванной вирусом гриппа В, или их комбинаций. Кроме того, эпитопы, способные к связыванию антитела по настоящему изобретению, можно применять в наборе для отслеживания эффективности процедур вакцинации путем выявления наличия защитных антител к вирусу гриппа A и/или вирусу гриппа В. Антитела, фрагменты антител или их варианты и производные, как описано в настоящем изобретении, можно также применять в наборе для отслеживания производства вакцин с желаемой иммуногенностью.
Настоящее изобретение предусматривает способ получения фармацевтической композиции, который включает этап смешивания моноклонального антитела с одним или несколькими фармацевтически приемлемыми носителями, при этом антитело представляет собой моноклональное антитело в соответствии с настоящим изобретением, описанное в данном документе.
Для введения антитела или его антигенсвязывающего фрагмента по настоящему изобретению известны и могут применяться различные системы доставки, в том числе без ограничения инкапсулирование в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные к экспрессии антитела или фрагмента антитела, опосредованный рецепторами эндоцитоз, конструирование нуклеиновой кислоты как части ретровирусного или другого вектора и т.д. Способы введения включают без ограничения внутрикожные, внутримышечные, внутрибрюшинные, внутривенные, подкожные, интраназальные, эпидуральные и пероральные пути. В другом варианте осуществления вакцину можно вводить в виде ДНК-вакцины, например, с использованием технологии электропорации, в том числе без ограничения in vivo электропорации. Композиции можно вводить любым удобным путем, например, путем инфузии или болюсной инъекции, путем всасывания через эпителиальные или кожно-слизистые покровы (например, слизистую оболочку ротовой полости, слизистую оболочку прямой кишки и кишечника и т.д.), и их можно вводить совместно с другими биологически активными средствами, в том числе без ограничения низкомолекулярными противовирусными композициями. Введение может быть системным или местным. Также можно использовать легочное введение, например, путем применения ингалятора или распылителя и состава с аэрозольным средством. В еще одном варианте осуществления композиция может доставляться в системе с контролируемым высвобождением.
Настоящее изобретение также предусматривает фармацевтические композиции. Такие композиции включают в себя терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента по настоящему изобретению и фармацевтически приемлемый носитель. Выражение "фармацевтически приемлемый", как используется в данном документе, означает одобренный регулирующим ведомством федерального правительства или правительства штата или указанный в Фармакопее США или другой общепризнанной фармакопее как применяемый у животных и более конкретно у людей. Выражение "носитель" относится к разбавителю, вспомогательному средству, наполнителю или основе, с которыми вводят терапевтическое средство. Такие фармацевтические носители могут быть стерильными жидкостями, такими как вода и масла, в том числе нефтяного, животного, растительного или синтетического происхождения, такими как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем, если фармацевтическую композицию вводят внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также можно использовать в качестве жидких носителей, в частности, для растворов для инъекций. Подходящие фармацевтические наполнители включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и т.п. Композиция при желании также может содержать незначительные количества смачивающих или эмульгирующих веществ или рН-буферных средств. Эти композиции могут иметь форму растворов, суспензий, эмульсии, таблеток, пилюль, капсул, порошков, составов с замедленным высвобождением и т.п. Композицию также можно составить в виде суппозитория с традиционными связывающими средствами и носителями, такими как триглицериды. Состав для перорального применения может включать в себя стандартные носители, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния фармацевтической степени чистоты и т.д. В одном варианте осуществления фармацевтическая композиция содержит терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента, предпочтительно в очищенной форме, вместе с подходящим количеством носителя, чтобы обеспечить придание формы для надлежащего введения пациенту. Состав должен соответствовать способу введения.
Обычно для терапевтических средств на основе антител доза, вводимая пациенту, составляет от приблизительно 0,1 мг/кг до 100 мг/кг веса тела пациента.
Примеры
Пример 1. Конструирование и оптимизация человеческих моноклональных антител, выделенных из B-клеток памяти
CD22+ IgG+ B-клетки отсортировывали из криоконсервированных мононуклеарных клеток периферической крови (PBMC) донора, выбранного по высоким титрам нейтрализующих антител как к B/Florida/4/2006 линии Yamagata (B/FL/06), так и к B/Brisbane/60/2008 линии Victoria (B/BNE/08), и иммортализировали по 3 клетки/лунка с использованием вируса Эпштейна-Барр (EBV), олигодезоксинуклеотида CpG 2006 и питающих клеток. Надосадочные жидкости культур, содержащие антитела, собирали через 14 дней и подвергали скринингу с помощью анализа микронейтрализации (MNA), который представлял собой модификации ранее описанного анализа ускоренного ингибирования вируса с применением активности нейраминидазы (NA) в качестве регистрируемого показателя (Hassantoufighi et al. (2010) Vaccine. 28:790-7), для идентификации клонов антител, которые могли бы нейтрализовать вирусы B как линии Yamagata, так и линии Victoria.
Вкратце, 10 мкл надосадочной жидкости культуры инкубировали с 400 TCID50 вируса гриппа B/BNE/08 (линия Victoria) или B/FL/06 (линия Yamagata) в течение одного часа при 37°C. В планшеты добавляли клетки Мадин-Дарби почек собак (MDCK) (20000 клеток на лунку), инкубировали в течение 4 часов, дважды промывали средой, содержащей TPCK-трипсин, а затем инкубировали в течение 2 дней при 37°C. После инкубации активность NA измеряли путем добавления флуоресцентно-меченого субстрата, метилумбеллиферил-N-ацетилнейраминовой кислоты (MU-NANA) (Sigma), из расчета 25 мкл/лунка (10 мкМ) и планшеты считывали с помощью флуориметра.
Было обнаружено, что три клона B-клеток (FBC-39, FBD-56 и FBD-94) обладали нейтрализующей активностью в отношении вируса гриппа B как линии Victoria, так и линии Yamagata. Гены VH и VL этих клонов секвенировали и клонировали в векторы экспрессии IgG1. Рекомбинантные антитела получали с помощью транзиентной трансфекции линий клеток млекопитающих, происходящих из эмбриональных клеток почки человека (HEK) или клеток яичника китайского хомячка (CHO). Надосадочные жидкости от трансфицированных клеток собирали спустя 7-10 дней культивирования и антитела (IgG) подвергали аффинной очистке с помощью хроматографии с белком А и диализу против фосфатно-солевого буферного раствора (PBS).
Алгоритм BLAST для Ig применяли для выравнивания последовательностей mAb относительно последовательностей человеческих антител зародышевого типа из базы данных. Наиболее близкие матрицы зародышевого типа идентифицировали для каждого из участков генов VH и VL (таблица 6). Несвойственные зародышевому типу аминокислоты идентифицировали путем выравнивания относительно этих эталонных последовательностей.
Таблица 6 Идентификация наиболее близких человеческих генов, содержащих VH и VL
Конструирование варианта FBC-39
В VL антитела FBC-39 присутствовал только один несвойственный зародышевому типу остаток каркасного участка, F в положении 87 легкой цепи, при этом аминокислота, свойственная зародышевому типу, представляет собой Y (или L87F(Y)), согласно нумерации по Kabat (положение 103 при нумерации IMGT). Последовательность зародышевого типа в данном документе называют FBC-39-L87Y. Несвойственную зародышевому типу последовательность называют FBC-39-L87F.
В VH антитела FBC-39 присутствовали 11 несвойственных зародышевому типу остатков каркасного участка, как определено по Kabat: H6V(E), H27L(F), H28S(T), H30L(S), H68S(T), H77M(T), H79F(Y), H81H(Q), H82aS(N), H83R(K) и H93A(T) согласно нумерации по Kabat. При применении IMGT для определения остатков каркасного участка присутствовало 7 несвойственных зародышевому типу остатков: H6V(E), H77S(T), H86M(T), H88F(Y), H90H(Q), H92S(N) и H95R(K).
Конструировали вариант, в котором все 12 несвойственных зародышевому типу остатков каркасного участка, как определено по Kabat, обращали в свойственные зародышевому типу аминокислоты. Эта конструкция антитела проявляла значительное снижение нейтрализующей активности и спектра действия в отношении вирусов гриппа B линии Victoria, что свидетельствует о том, что один или несколько несвойственных зародышевому типу остатков являются важными для активности.
Три несвойственные зародышевому типу остатка каркасного участка, расположенные в положениях H27, H28 и H30, находились в области, определенной как каркасный участок 1 VH согласно системе Kabat, однако согласно системе IMGT они считаются частью HCDR-1. Варианты антитела получали путем обращения всех несвойственных зародышевому типу аминокислот каркасного участка, как определено по Kabat, на их соответствующие свойственные зародышевому типу остатки, за исключением остатков в следующих трех положениях: H27L(F), H28S(T), H30L(S), которые "видоизменяли" от свойственных зародышевому типу остатков до несвойственных зародышевому типу остатков с получением семи вариантов тяжелой цепи: FBC-39 LSL, FBC-39 FSL; FBC-39 LTL; FBC-39 FTL; FBC-39-FSS; FBC-39-LTS; FBC-39-FTS, в которых свойственные зародышевому типу остатки подчеркнуты, так что FBC-39 FTS содержит три свойственные зародышевому типу аминокислоты, а FBC-39 LSL содержит три остатка дикого типа в положениях H27, H28 и H30.
Кроме того, свойственный зародышевому типу остаток N в H82 (H92 по IMGT) создавал потенциальный сайт дезамидирования (NS) в VH. Следовательно, его заменяли на S дикого типа из FBC-39 во всех семи вариантах: FBC-39 LSL, FBC-39 FSL, FBC-39 LTL, FBC-39 FTL, FBC-39-FSS, FBC-39-LTS и FBC-39-FTS. Кроме того, все семь вариантов FBC-39 имели одинаковую последовательность легкой цепи (FBC-39-L87Y), которая отличалась от легкой цепи FBC-39 одной аминокислотой.
Полученные в результате варианты антитела экспрессировали и очищали, как описано выше, и подвергали дальнейшему определению характеристик.
Пример 2. Выделенные антитела к HA связывались с HA вируса гриппа В обеих линий
Анализ ELISA в отношении связывания с HA проводили для определения способности связывания и перекрестной реактивности выделенных антител. 384-луночный планшет для ELISA Maxisorb (Nunc) покрывали на протяжении ночи при 4°C с помощью 0,5 мкг/мл рекомбинантного HA, полученного из штамма B/FL/06 линии Yamagata или штамма B/BNE/08 линии Victoria, в PBS. Планшет промывали с помощью PBS, содержащего 0,1% объем/объем Tween-20, для удаления несвязавшегося белка, и добавляли блокирующий раствор, содержащий 1% (вес/объем) казеина (Thermo Scientific), на 1 час при комнатной температуре. Блокирующий раствор удаляли, и добавляли 3-кратные серийные разведения в PBS каждого из антител к HA (FBC-39, FBD-56 и FBD-94), и инкубировали в течение 1 часа при комнатной температуре. Планшет промывали три раза, и связанные антитела выявляли с помощью конъюгированного с пероксидазой мышиного антитела к человеческому IgG (Jackson). Активность связывания антитела рассчитывали либо путем измерения хемилюминесцентного сигнала после добавления субстрата Supersignal Pico (Thermo Scientific), либо путем измерения изменения цвета при 450 нм после инкубирования с однокомпонентным субстратом тетраметилбензидином (TMB) (доступен от Kirkegaard and Perry Laboratories, Inc. (KPL) Гейтерсберг, Мэриленд) с последующим добавлением 2 н. серной кислоты для остановки реакции.
Таблица 7

В таблице 7 показаны средние значения EC50 на основании трех независимых экспериментов. Все три IgG к HA (FBC-39, FBD-56 и FBD-94) связывали рекомбинантный HA вирусов гриппа B обеих линий. Подобные значения EC50 наблюдали у всех трех антител против HA Yamagata (B/FL/06). Более низкое значение EC50 наблюдали в отношении Victoria (B/BNE/08) в случае FBD-94, а не в случае как FBD-56, так и FBC-39.
Семь свойственных зародышевому типу вариантов антитела FBC-39 испытывали в отношении активности связывания с помощью ELISA. В таблице 8 показаны результаты связывания неочищенных вариантов IgG FBC-39 к HA, где FBC-39 FTS содержит свойственные зародышевому типу аминокислоты каркасного участка, как определено по Kabat, а FBC-39 LSL содержит свойственные зародышевому типу аминокислоты каркасного участка, как определено по Kabat, за исключением остатков дикого типа в положениях H27, H28 и H30 (нумерация по Kabat). Эти результаты показывают, что все варианты связывались с белком HA линии Yamagata (B/FL/06), но варианты, содержащие остаток S в положении H30, теряли аффинность связывания с белком HA линии Victoria (B/BNE/08). Четыре из вариантов, FBC-39 LSL, FSL, LTL и FTL, продемонстрировали равную или лучшую аффинность связывания с белками HA обеих линий, чем FBC-39.
Таблица 8

Пример 3. In vitro нейтрализующая активность перекрестно реагирующих IgG к HA вируса гриппа B в отношении вирусов двух разных линий
Для тестирования активности очищенных mAb применяли анализ микронейтрализации, подобный описанному в примере 1. Вкратце, клетки MDCK культивировали в среде MEM (Invitrogen), дополненной антибиотиками, глутамином (полная среда MEM) и 10% (объем/объем) фетальной бычьей сыворотки. 60 TCID50 (50% инфицирующие дозы для культуры тканей) вируса добавляли в 384-луночном планшете к трехкратных разведениям антитела в полной среде MEM, содержащей 0,75 мкг/мл обработанного TPCK трипсина (Worthington), в дублирующих лунках, после инкубирования в течение 30 минут при комнатной температуре в планшет добавляли 2×104 клеток/лунка. После инкубирования при 33°C в инкубаторе с 5% CO2 в течение примерно 40 часов активность NA измеряли путем добавления в каждую лунку флуоресцентно-меченного субстрата, метилумбеллиферил-N-ацетилнейраминовой кислоты (MU-NANA) (Sigma), и инкубирования при 37°C в течение 1 часа. Репликацию вируса, представленную активностью NA, количественно определяли путем считывания данных о флуоресценции с помощью флуориметра Envision (PerkinElmer) с применением следующих установочных параметров: длина волны возбуждения 355 нм, длина волны испускания 460 нм; 10 вспышек на лунку. Титр нейтрализации (50% ингибирующую концентрацию [IC50]) выражали в виде конечной концентрации антитела, которая ослабляла сигнал флуоресценции на 50% по сравнению с лунками с контрольными клетками. Штаммы вируса гриппа B, используемые в таблице 9, перечислены ниже: B/Lee/40 (B/Lee/40); B/AA/66 (ca B/Ann Arbor/1/66); B/HK/72 (B/Hong Kong/5/72); B/BJ/97 (ca B/Beijing/243/97 (victoria)), B/HK/01 (B/Hong Kong/330/2001 (victoria)); B/MY/04 (B/Malaysia/2506/2004 (victoria)); B/BNE/08 (ca B/Brisbane/60/2008 (victoria)); B/AA/94 (ca B/Ann Arbor/2/94 (yamagata)); B/YSI/98 (ca B/Yamanashi/166/98 (yamagata)); B/JHB/99 (ca B/Johannesburg/5/99 (yamagata)); B/SC/99 (B/Sichuan/379/99 (yamagata)); B/FL/06 (B/Florida/4/2006 (yamagata)).
Таблица 9
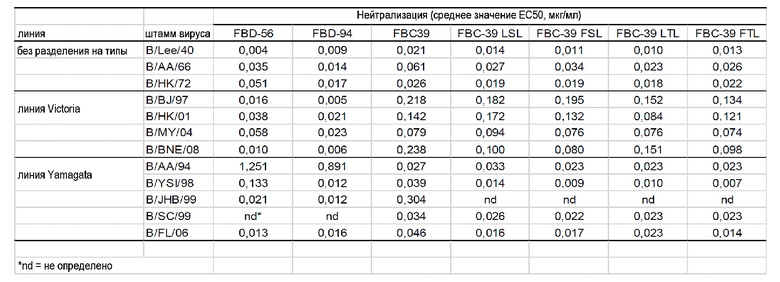
В таблице 9 показаны средние значения IC50 на основании двух независимых экспериментов. Антитела к HA нейтрализовали все испытываемые вирусы гриппа B. FBD-56 и FBD-94 были более эффективными, чем FBC-39, но продемонстрировали несколько сниженную активность в отношении штамма B/AA/94. Варианты FBC-39, LSL, FSL, LTL и FTL, нейтрализовали все вирусы при значениях IC50, подобных или более низких, чем FBC-39.
Пример 4. Связывание и нейтрализация штаммов H9 вируса гриппа A
Анализы ELISA в отношении связывания HA проводили с применением подобной методики, как в примере 2, за исключением того, что 384-луночные планшеты покрывали в течение 2 часов при комнатной температуре с помощью 3 мкг/мл рекомбинантного HA, полученного из подтипа H9 вируса гриппа A (A/chicken/HK/G9/97(H9N2)), в PBS. Результаты показывают, что FBC-39 и свойственные зародышевому типу варианты LTL связывали HA H9 при подобных значениях EC50, составляющих 6,2 и 6,3 мкг/мл соответственно, а FBC-39 LSL и FTL связывали при более высоких значениях EC50, составляющих 41,7 и 46,1 мкг/мл соответственно (таблица 10). В тоже время при наивысшей испытываемой дозе 50 мкг/мл FBC-39 FSL связывался лишь слабо, и никакого связывания не наблюдали в случае FBD-56 и FBD-94.
Таблица 10
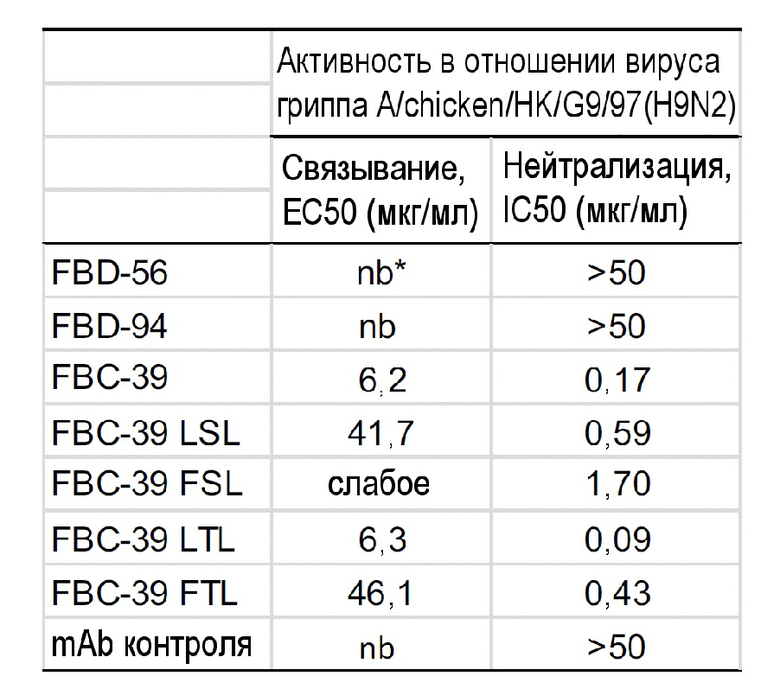
*nb=отсутствие связывания
Подтверждение связывания белка HA H9 вируса гриппа A было важным с точки зрения функциональности, при этом анализ микронейтрализации проводили с применением методики, подобной описанной в примере 3. Для этого анализа с помощью способов обратной генетики получали холодоадаптированный (ca) живой аттенуированный вакцинный вирус гриппа, содержащий вирусные гены HA и NA от вируса A/chicken/Hong Kong/G9/97 (H9N2) в сочетании с шестью генами внутренних белков вируса ca A/Ann Arbor/6/60 (H2N2), с применением методики, подобной описанной Jin et al. (2003) Virology. 306:18-24). Результаты анализа микронейтрализации показаны в таблице 10. В соответствии с профилем связывания FBC-39 и варианты эффективно нейтрализовали вирус H9N2 при биологически значимых значениях IC50. FBC-39 и FBC-39 LTL имели наиболее высокую активность со значениями IC50, составлявшими 0,17 и 0,09 мкг/мл соответственно. Как и ожидалось, FBD-56, FBD-94 и контрольные антитела не демонстрировали нейтрализующей активности при наиболее высокой испытываемой концентрации (50 мкг/мл).
Пример 5. Идентификация эпитопов с помощью отбора мутантов, устойчивых к моноклональным антителам (MARM)
Вирус гриппа B линии Yamagata (B/Florida/4/2006; B/FL/06) и вирус гриппа B линии Victoria (B/Malaysia/2506/2004; B/MY/04) инкубировали с FBC-39, FBD-56 и FBD-94 при высоких концентрациях (125xIC50) за 1 час до поглощения смеси вируса и антитела клетками MDCK из расчета 30000 TCID50 на лунку в 10×96-луночных планшетах и культивирования в присутствии FBC-39, FBD-56 и FBD-94 (10xIC50). Выделяли предполагаемые MARM, проявляющие цитопатический эффект (CPE) в отношении инфицированных клеток в период не более 3 дней после инфицирования. Ген HA амплифицировали посредством RT-PCR и далее секвенировали, а затем подтверждали устойчивость выделенного вируса с помощью анализа микронейтрализации. Ни одного MARM не выделяли из вируса B/FLA/09 при культивировании в присутствии FBC-39, FBD-56 или FBD-94. При применении вируса линии Victoria (B/MY/04) выделяли MARM в присутствии FBD-56 и FBD-94, но не в присутствии FBC-39. С помощью анализа последовательности выявили, что два MARM относительно FBD-56 содержали замену одной аминокислоты в положении 128, замену глутаминовой кислоты (E) на лизин (K) или валин (V) (таблица 11). MARM относительно FBD-94 содержал замену одной аминокислоты в положении 128, замену E на K (таблица 11). Вариант B/Florida/4/2006 линии Yamagata, содержащий замену одной аминокислоты в положении 141, замену глицина (G) на E (B/FL/06 G141E), обеспечивал 8-кратное снижение нейтрализации с помощью FBC-39 по сравнению с вирусом дикого типа (B/FL/06). С применением этого варианта B/FLA/09 G141E и вируса B/Jiangsu/10/2003 (B/JIN/03) линии Yamagata, циркулирующего в природе вируса, который содержит R в положении 141 (G141R), выделение MARM повторяли только с mAb FBC-39. Один MARM-вирус выделяли с применением вируса B/FLA/09 G141E с изменением одной аминокислоты, заменой G на аргинин (R) в положении 235 (таблица 11). Идентифицировали два "ускользнувших" мутанта вируса B/JIN/03 с заменами одной аминокислоты, заменой серина (S) на изолейцин (I) в положении 150 или E на лейцин (L) в положении 235 (таблица 11) соответственно. Замены аминокислот, идентифицированные в этих MARM-вирусах гриппа B, расположены в участке HA, представляющем собой головку (Wang et al. (2008) J. Virol. 82(6):3011-20), что позволяет предположить, что FBC-39, FBD-56 и FBD-94 распознают эпитопы на головке HA вируса гриппа B, при этом у FBD-56 и FBD-94 ключевой контакт находится в положении 128 и они имеют перекрывающийся эпитоп, а у FBC-39 имеется конформационный эпитоп с важными контактирующими остатками в положениях 141, 150 и 235.
Таблица 11
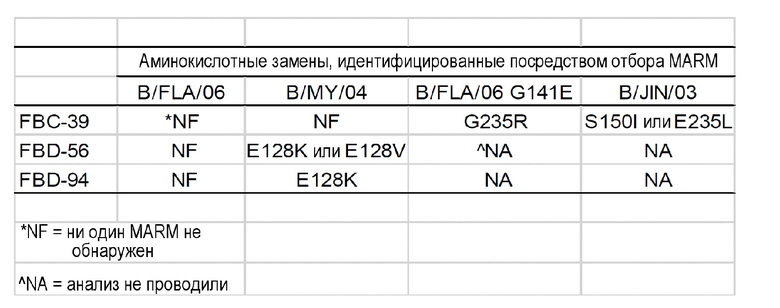
Пример 6. Антитела к HA вируса гриппа B проявляют эффекторную функцию Fc
Антитела обладают потенциалом к устранению инфицированных вирусом клеток посредством эффекторной функции Fc, такой как антителозависимая клеточная цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз и комплементзависимый цитолиз. Для подтверждения того, что антитела к HA проявляли ADCC-активность, исследовали их способность к активации NK-клеток в присутствии вируса гриппа B с помощью биологического анализа ADCC. В этом анализе применяли линию человеческих NK-клеток (NK92), которая была стабильно трансфицирована человеческим высокоаффинным рецептором FcgIIIA и трансгеном люциферазы под контролем промотора NFAT, для измерения активации эффекторного механизма Fc. 96-луночные планшеты покрывали с помощью 5,0×104 TCID50/лунка исходного раствора вируса B/Hong Kong/330/2001 (Victoria). Серийные разведения FBD-94, FBC-39, а также вариантов, не обладающих эффекторной функцией Fc, которые содержали две замены в участке Fc, L234A и L235A (FBD-94 LALA и FBC-39 LALA) (Hezareh et al. (2001) J. Virol. 75(24):12161) вносили к вирусу, а затем добавляли NK-клетки виз расчета 5,0×104 клеток/лунка и инкубировали при 37°C в течение 4 часов. Люциферазу выявляли путем добавления реактива Steady-Glo (Promega) и измеряли с помощью планшет-ридера Envision. На фигуре 1 показано, что как FBD-94, так и FBC-39 проявляли дозозависимую ADCC-активность в отношении вируса гриппа B, тогда как варианты LALA не продемонстрировали никакой активности при таких же концентрациях.
Пример 7. In vivo профилактический и терапевтический эффект IgG к вирусу гриппа B на мышиной модели инфекции, вызванной смертельной дозой вируса гриппа
Эффективность защиты нейтрализующих моноклональных антител к вирусу гриппа B оценивали на мышиной модели инфекции, вызванной смертельной дозой вируса гриппа B.
Профилактическая активность (фигуры 2A-D). Для исследования профилактической эффективности мышам BALB/c (Harlan Laboratories) возрастом шесть-восемь недель в группах по восемь особей вводили одно из антител FBC-39 или FBD-94 в виде однократной внутрибрюшинной (IP) инъекции в дозах 3, 1, 0,3 или 0,1 мг/кг в объеме 100 мкл. Для каждого исследования группу контрольных животных обрабатывали IP нерелевантным человеческим IgG изотипического контроля из расчета 3 мг/кг в объеме 100 мкл. Через четыре часа после введения дозы мышам интраназально инокулировали 15-кратную пятидесятипроцентную смертельную дозу для мыши (15 MLD50) B/Sichuan/379/99 (Yamagata) (B/Sic/99) или 10 MLD50 B/Hong Kong/330/2001 (Victoria) (B/HK/01) в объеме 50 мкл. Мышей взвешивали в день заражения вирусом и в течение 14 дней ежедневно отслеживали потерю веса и выживание (мышей с потерей веса тела ≥ 25% подвергали эвтаназии). Оба mAb FBC-39 и FBD-94 обеспечивали защиту дозозависимым образом. Введение FBC-39 и FBD-94 из расчета 0,3 мг/кг или больше обеспечивало 90% - 100% защиту для животных, зараженных B/Sic/99 (фигуры 2A и B) и B/HK/01 (фигуры 2 C и D). Введение FBC-39 и FBD94 при более низкой дозе 0,1 мг/кг также обладало высокой степенью защиты от B/HK/01, при этом уровень выживания составлял 90% и 80% соответственно. Как и ожидалось, ни одна из мышей, которые получали mAb изотипического контроля из расчета 3 или 30 мг/кг, не выжила после заражения B/Sic/99 или B/HK/01 соответственно.
Терапевтическая активность (фигуры 3A-D и фигуры 4A и B). Для оценки терапевтической эффективности антител мышам инокулировали 10 MLD50 B/Sic/99 (Yamagata) или 5 MLD50 B/HK/01 (Victoria) и через два дня после инфицирования (pi) инъецировали 10, 3, 1 или 0,3 мг/кг FBC-39 или FBD-94. FBC-39 и FBD-94 обеспечивали полную защиту для животных, зараженных B/Sic/99, при введении в количестве 1 мг/кг или больше (фигуры 3A и B). В отношении инфекции, вызванной B/HK/01, введение FBC-39 и FBD-94 при дозах 0,3 мг/кг и больше обеспечивало полную защиту (фигуры 3C и D). Как и ожидалось, mAb изотипического контроля, введенное из расчета 10 или 30 мг/кг, не смогло обеспечить защиту мышей, при этом уровень выживания составлял 10% или 20% в случае инфекций, вызванных B/Sic/99 и B/HK/01 соответственно.
Для исследования способности антител к вирусу гриппа B обеспечивать защиту с течением времени мышам инокулировали 5 MLD50 B/HK/01 и инъецировали IP 3 мг/кг FBC-39 или FBD-94, начиная в 1, 2, 3 или 4 дни pi. FBC-39 обеспечивало защиту 100% мышей при введении в день 1 pi, а также 80% и 70% в дни 2 и 3 pi соответственно (фигура 4A). FBD-94 обеспечивало защиту 100% мышей при введении в день 1 и день 2 pi, а также 80% и 60% в день 3 и день 4 pi соответственно (фигура 4В). Как и ожидалось, обработка мышей такой же дозой нерелевантного антитела изотипического контроля не смогла обеспечить защиту мышей, при этом уровень выживания составлял 10%.
Пример 8. Активность в отношении ингибирования гемагглютинации
Для определения возможного механизма действия антител к вирусу гриппа B, функциональной способности антител по настоящему изобретению, проводили анализы ингибирования гемагглютинации (HAI) с применением разных групп штаммов вируса гриппа B. С помощью HAI-анализа выявляли антитела, которые блокируют связывание с вирусными рецепторами, представляющими собой экспрессируемую на поверхности клеток сиаловую кислоту, путем измерения ингибирования агглютинации эритроцитов, опосредованной вирусами. Концентрацию вирусов гриппа B (сокращения описаны в таблице 12 ниже) доводили до 4 HA-единиц, что определяли путем инкубирования с 0,05% эритроцитов индюка (Lampire Biological Laboratories) в отсутствие антитела. В 96-луночном планшете с U-образным дном серийно разводили IgG FBD-94 и FBC-39 с двукратным шагом и добавляли в лунки разведенный вирус. Спустя 30-60 мин. инкубирования добавляли 50 мкл 0,05% эритроцитов индюка. Планшеты инкубировали в течение дополнительных 30-60 минут и исследовали на предмет агглютинации. Титр для HAI определяли как минимально эффективную концентрацию (нМ) антитела, при которой происходило полное ингибирование агглютинации. В таблице 12 показано, что FBD-94 и FBC39 обладали HAI-активностью в отношении всех испытываемых штаммов вируса гриппа B, что доказывает связывание с глобулярной головкой HA вируса гриппа B. Другие антитела по настоящему изобретению характеризовались подобной активностью в HAI-анализах.
Таблица 12
Титр для ингибирования гемагглютинации (нМ)
B/Lee/40 (B/Lee/40); B/AA/66 (ca B/Ann Arbor/1/66); B/HK/72 (B/Hong Kong/5/72); B/BJ/97 (ca B/Beijing/243/97 (victoria)), B/HK/01 (B/Hong Kong/330/2001 (victoria)); B/Mal/04 (B/Malaysia/2506/2004 (victoria)); B/OH/05 (B/Ohio/1/2005 (victoria)); B/BNE/08 (ca B/Brisbane/60/2008 (victoria)); B/Yam/88 (B/Yamagata/16/88 (yamagata)); B/AA/94 (ca B/Ann Arbor/2/94 (yamagata)); B/geo/98 (ca B/Georgia/02/98 (yamagata)); B/Ysh/98 (ca B/Yamanashi/166/98 (yamagata)); B/Joh/99 (ca B/Johannesburg/5/99 (yamagata)); B/Sic/99 (B/Sichuan/379/99 (yamagata)); B/Vic/00 (ca B/Victoria/504/2000 (yamagata)); B/Shg/02 (B/Shanghai/361/02 (yamagata)) и B/FL/06 (B/Florida/4/06 (yamagata)).
Все публикации, патенты и заявки на патенты, упоминаемые в настоящем описании, включены в данный документ посредством ссылки в настоящем описании в такой же степени, как если бы каждая отдельная публикация, патент или заявка на патент были конкретно и отдельно указаны как включенные в данный документ посредством ссылки.
Информация о последовательностях
SEQ ID NO:1 (FBD-56 VH DNA)
GAAGTGCAGCTGGTGGAGTCTGGGGGACACTTGGTGCAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGAGGATTATGCCATGAATTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGTCATTAGTTGGGACAGTGGTAGGATAGGCTATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCTCGTATCTGCAAATGAA CAGTCTGAGACCTGAGGACACTGCCTTGTATTATTGTGTAAGAGATATGTTGGCTTATTATTCTGACAATAGTGGCAAAAAATACAACGTCTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAG
SEQ ID NO:2 (VH FBD-56, белок)
EVQLVESGGHLVQPGRSLRLSCAASGFTFEDYAMNWVRQAPGKGLEWVSVISWDSGRIGYADSVKGRFTISRDNAKNSSYLQMNSLRPEDTALYYCVRDMLAYYSDNSGKKYNVYGMDVWGQGTTVTVSS
SEQ ID NO:3 (FBD-56 HCDR-1- Kabat): DYAMN
SEQ ID NO:4 (FBD-56 HCDR-2 - Kabat): VISWDSGRIGYADSVKG
SEQ ID NO:5 (FBD-56 HCDR-3 - Kabat): DMLAYYSDNSGKKYNVYGMDV
SEQ ID NO:6 (FBD-56 VL DNA)
GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCAC CCTCTCCTGCAGGGCCAGTCAGAGTGTTTCCACCTTCTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATGTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAACCTGAAGATTTTGC AATTTACTACTGTCAGCAGCGTAGCCACTGGCCTCCTATCTTCGGCCAAGGGACACGACTGGAGATTAAAC
SEQ ID NO:7 (VL FBD-56, белок)
EIVLTQSPATLSLSPGERATLSCRASQSVSTFLAWYQQKPGQAPRLLMYDASNRATGIP ARFSGSGSGTDFTLTISSLEPEDFAIYYCQQRSHWPPIFGQGTRLEIK
SEQ ID NO:8 (FBD-56 LCDR-1 - Kabat): ASQSVSTFLA
SEQ ID NO:9 (FBD-56 LCDR-2 - Kabat): DASNRAT
SEQ ID NO:10 (FBD-56 LCDR-3 - Kabat): QQRSHWPPI
SEQ ID NO:11 (FBD-94 VH DNA)
GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTGCAACCTGGCAGGTCCCTGAGACT CTCCTGTGCAGTTTCTGGATTCATCTTTGAAGATTATGCCATAAACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAATTATTAGTTGGGACAGTGGTAGGATAGGCTACGCGGACT CTGTGAGGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCTCGTTTCTGCAAATGAACAGTCTGAGACCCGAAGACACGGCCGTGTATTATTGTGTAAAAGATATGTTGGCGTATTATTATGATGGTAGCGGCATCAGGTACAACCTCTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAG
SEQ ID NO:12 (VH FBD-94, белок)
EVQLVESGGGLVQPGRSLRLSCAVSGFIFEDYAINWVRQAPGKGLEWVSIISWDSGRIGYADSVRGRFTISRDNAKNSSFLQMNSLRPEDTAVYYCVKDMLAYYYDGSGIRYNLYGMDVWGQGTTVTVSS
SEQ ID NO:13 (FBD-94 HCDR-1 - Kabat): DYAIN
SEQ ID NO:14 (FBD-94 HCDR-2 - Kabat): IISWDSGRIGYADSVRG
SEQ ID NO:15 (FBD-94 HCDR-3 - Kabat): DMLAYYYDGSGIRYNLYGMDV
SEQ ID NO:16 (FBD-94 VL DNA)
GAAATTGTGTTGACACAGTCTCCAGCCACTCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCGGAGTATTACCACCTTCTTAGCCTGGTACCAACAAAAACCTGGCCAGGCTCCCAGGCTCCTCATCTACGATGCATCCAACAGGGCCACTGGCGTCCCAGCCAGGTTCA GTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAACAGCCTAGAGCCTGACGATTTTGCAATTTATTACTGTCAGCAGCGTGACCACTGGCCTCCGATCTTCGGCCAAGGGACACGACTGGAGATTAAAC
SEQ ID NO:17 (VL FBD-94, белок)
EIVLTQSPATLSLSPGERATLSCRASRSITTFLAWYQQKPGQAPRLLIYDASNRATGVP ARFSGSGSGTDFTLTINSLEPDDFAIYYCQQRDHWPPIFGQGTRLEIK
SEQ ID NO:18 (FBD-94 LCDR-1 - Kabat): RASRSITTFLA
SEQ ID NO:19 (FBD-94 LCDR-2 - Kabat): DASNRAT
SEQ ID NO:20 (FBD-94 LCDR-3 - Kabat): QQRDHWPPI
SEQ ID NO:21 (FBC-39 VH DNA)
GAGGTGCAGCTGGTGGTGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGACTCAGTTTCCTTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCAGCATCTCAAGAGACGATTCAAAGAACATGCTGTTTCTGCATATGAGCAGCCTGAGAACCGAGGACACAGCCGTCTATTACTGCGCCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAG
SEQ ID NO:22 (VH FBC-39, белок)
EVQLVVSGGGLVKPGGSLRLSCAASGLSFLNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFSISRDDSKNMLFLHMSSLRTEDTAVYYCATDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO:23 (FBC-39 HCDR-1 - Kabat): NAWMS
SEQ ID NO:24 (FBC-39 HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO:25 (FBC-39 HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:26 (FBC-39 VL DNA)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTTTTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO:27 (VL FBC-39, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYFCQQANSFPPTFGQGTKLEIK
SEQ ID NO:28 (FBC-39 LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO:29 (FBC-39 LCDR-2 - Kabat): AASSLQS
SEQ ID NO:30 (FBC-39 LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO: 31 (FBC-39 LSL VH DNA)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGACTCTCTTTCCTTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO:32 (VH FBC-39 LSL, белок)
EVQLVESGGGLVKPGGSLRLSCAASGLSFLNAWMSWVRQAPGKGLEWVGRIKSNTDGGT TDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO:33 (FBC-39 LSL HCDR-1 - Kabat): NAWMS
SEQ ID NO:34 (FBC-39 LSL HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO:35 (FBC-39 LSL HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:36 (VL FBC-39 LSL, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO:37 (VL FBC-39 LSL, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO:38 (FBC-39 LSL LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO:39 (FBC-39 LSL LCDR-2 - Kabat): AASSLQS
SEQ ID NO:40 (FBC-39 LSL LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO:41 (VH FBC-39 FSL, ДНК)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGATTCTCTTTCCTTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO:42 (VH FBC-39 FSL, белок)
EVQLVESGGGLVKPGGSLRLSCAASGFSFLNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO:43 (FBC-39 FSL HCDR-1 - Kabat): NAWMS
SEQ ID NO:44 (FBC-39 FSL HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO:45 (FBC-39 FSL HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:46 (VL FBC-39 FSL, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO:47 (VL FBC-39 FSL, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO:48 (FBC-39 FSL LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO:49 (FBC-39 FSL LCDR-2 - Kabat): AASSLQS
SEQ ID NO:50 (FBC-39 FSL LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO:51 (VH FBC-39 LTL, ДНК)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGACTCACTTTCCTTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO:52 (VH FBC-39 LTL, белок)
EVQLVESGGGLVKPGGSLRLSCAASGLTFLNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO:53 (FBC-39 LTL HCDR-1 - Kabat): NAWMS
SEQ ID NO:54 (FBC-39 LTL HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO:55 (FBC-39 LTL HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:56 (VL FBC-39 LTL, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO:57 (VL FBC-39 LTL, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO:58 (FBC-39 LTL LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO:59 (FBC-39 LTL LCDR-2 - Kabat): AASSLQS
SEQ ID NO:60 (FBC-39 LTL LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO:61 (VH FBC-39 FTL, ДНК)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGATTCACTTTCCTTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO:62 (VH FBC-39 FTL, белок)
EVQLVESGGGLVKPGGSLRLSCAASGFTFLNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO:63 (FBC-39 FTL HCDR-1 - Kabat): NAWMS
SEQ ID NO:64 (FBC-39 FTL HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO:65 (FBC-39 FTL HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:66 (VL FBC-39 FTL, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO:67 (VL FBC-39 FTL, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO:68 (FBC-39 FTL LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO:69 (FBC-39 FTL LCDR-2 - Kabat): AASSLQS
SEQ ID NO:70 (FBC-39 FTL LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO:71 (VH FBC-39, белок с варьирующим составом аминокислот)
(См. фигуру 6)
SEQ ID NO:72 (VL FBC-39, белок с варьирующим составом аминокислот)
(См. фигуру 7)
SEQ ID NO: 73 (VH FBC-39 FSS, ДНК)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGATTCTCTTTCAGTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO: 74 (VH FBC-39 FSS, белок)
EVQLVESGGGLVKPGGSLRLSCAASGFSFSNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO: 75 (FBC-39 FSS HCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO: 76 (FBC-39 FSS HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO: 77 (FBC-39 FSS HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 78 (FBC-39 FSS HCDR-1 - IMGT): GFSFSNAW
SEQ ID NO: 79 (FBC-39 FSS HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO: 80 (FBC-39 FSS HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 81 (VL FBC-39 FSS, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO: 82 (VL FBC-39 FSS, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO: 83 (FBC-39 FSS LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO: 84 (FBC-39 FSS LCDR-2 - Kabat): AASSLQS
SEQ ID NO: 85 (FBC-39 FSS LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO: 86 (FBC-39 FSS LCDR-1 - IMGT): QDISTW
SEQ ID NO: 87 (FBC-39 FSS LCDR-2 - IMGT): AAS
SEQ ID NO: 88 (FBC-39 FSS LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO: 89 (VH FBC-39 LTS, ДНК):
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGACTCACTTTCAGTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO: 90 (VH FBC-39 LTS, белок):
EVQLVESGGGLVKPGGSLRLSCAASGLTFSNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO: 91 (FBC-39 LTS HCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO: 92 (FBC-39 LTS HCDR-2 - Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO: 93 (FBC-39 LTS HCDR-3 - Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 94 (FBC-39 LTS HCDR-1 - IMGT): GLTFSNAW
SEQ ID NO: 95 (FBC-39 LTS HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO: 96 (FBC-39 LTS HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 97 (VL FBC-39 LTS, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO: 98 (VL FBC-39 LTS, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO: 99 (FBC-39 LTS LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO: 100 (FBC-39 LTS LCDR-2 - Kabat): AASSLQS
SEQ ID NO: 101 (FBC-39 LTS LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO: 102 (FBC-39 LTS LCDR-1 - IMGT): QDISTW
SEQ ID NO: 103 (FBC-39 LTS LCDR-2 - IMGT): AAS
SEQ ID NO: 104 (FBC-39 LTS LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO: 105 (VH FBC-39 FTS, ДНК):
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGCCTGGAGTGGGTTGGCCGTATTAAAAGTAATACTGATGGTGGGACAACAGACTACGCCGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGACGATTCAAAGAACACGCTGTATCTGCAAATGAGCAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGCACCACAGATGGACCTTACTCTGACGATTTTAGAAGTGGTTATGCCGCACGCTACCGTTATTTCGGAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA
SEQ ID NO: 106 (VH FBC-39 FTS, белок):
EVQLVESGGGLVKPGGSLRLSCAASGFTFSNAWMSWVRQAPGKGLEWVGRIKSNTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMSSLKTEDTAVYYCTTDGPYSDDFRSGYAARYRYFGMDVWGQGTTVTVSS
SEQ ID NO: 107 (FBC-39 FTS HCDR-1- Kabat): RASQDISTWLA
SEQ ID NO: 108 (FBC-39 FTS HCDR-2- Kabat): RIKSNTDGGTTDYAAPVKG
SEQ ID NO: 109 (FBC-39 FTS HCDR-3- Kabat): DGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 110 (FBC-39 FTS HCDR-1 - IMGT): GFTFSNAW
SEQ ID NO: 111 (FBC-39 FTS HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO: 112 (FBC-39 FTS HCDR-3 - IMGT): TDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO: 113 (VL FBC-39 FTS, ДНК)
GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGATATTAGCACCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGATTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAGCAGGCTAACAGTTTCCCTCCGACTTTTGGCCAGGGGACCAAGCTGGAGATCAAAC
SEQ ID NO: 114 (VL FBC-39 FTS, белок)
DIQMTQSPSSVSASVGDRVTITCRASQDISTWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPPTFGQGTKLEIK
SEQ ID NO: 115 (FBC-39 FTS LCDR-1 - Kabat): RASQDISTWLA
SEQ ID NO: 116 (FBC-39 FTS LCDR-2 - Kabat): AASSLQS
SEQ ID NO: 117 (FBC-39 FTS LCDR-3 - Kabat): QQANSFPPT
SEQ ID NO: 118 (FBC-39 FTS LCDR-1 - IMGT): QDISTW
SEQ ID NO: 119 (FBC-39 FTS LCDR-2 - IMGT): AAS
SEQ ID NO: 120 (FBC-39 FTS LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO:121 (FBC-39 HCDR-1 - IMGT): GLSFLNAW
SEQ ID NO:122 (FBC-39 HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO:123 (FBC-39 HCDR-3 - IMGT): TDGPYSDDFRSGYAARYRYFGMDVW
SEQ ID NO:124 (FBC-39 LCDR-1 - IMGT): QDISTW
SEQ ID NO:125 (FBC-39 LCDR-2 - IMGT): AAS
SEQ ID NO:126 (FBC-39 LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO:127 (FBC-39 LSL HCDR-1 - IMGT): GLSFLNAW
SEQ ID NO:128 (FBC-39 LSL HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO:129 (FBC-39 LSL HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:130 (FBC-39 LSL LCDR-1 - IMGT): QDISTW
SEQ ID NO:131 (FBC-39 LSL LCDR-2 - IMGT): AAS
SEQ ID NO:132 (FBC-39 LSL LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO:133 (FBC-39 FSL HCDR-1 - IMGT): GFSFLNAW
SEQ ID NO:134 (FBC-39 FSL HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO:135 (FBC-39 LSL HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:136 (FBC-39 FSL LCDR-1 - IMGT): QDISTW
SEQ ID NO:137 (FBC-39 FSL LCDR-2 - IMGT): AAS
SEQ ID NO:138 (FBC-39 FSL LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO:139 (FBC-39 LTL HCDR-1 - IMGT): GLTFLNAW
SEQ ID NO:140 (FBC-39 LTL HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO:141 (FBC-39 LTL HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:142 (FBC-39 LTL LCDR-1 - IMGT): QDISTW
SEQ ID NO:143 (FBC-39 LTL LCDR-2 - IMGT): AAS
SEQ ID NO:144 (FBC-39 LTL LCDR-3 - IMGT): QQANSFPPT
SEQ ID NO:145 (FBC-39 FTL HCDR-1 - IMGT): GFTFLNAW
SEQ ID NO:146 (FBC-39 FTL HCDR-2 - IMGT): IKSNTDGGTT
SEQ ID NO:147 (FBC-39 FTL HCDR-3 - IMGT): TTDGPYSDDFRSGYAARYRYFGMDV
SEQ ID NO:148 (FBC-39 FTL LCDR-1 - IMGT): QDISTW
SEQ ID NO:149 (FBC-39 FTL LCDR-2 - IMGT): AAS
SEQ ID NO:150 (FBC-39 FTL LCDR-3 - IMGT): QQANSFPPT
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> МЕДИММЬЮН, ЭЛ.ЭЛ.СИ
<120> НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА B И ПУТИ ИХ ПРИМЕНЕНИЯ
<130> FLUB-100WO1
<140> PCT/US2015/040385
<141> 2015-07-14
<150> 62/024,804
<151> 2014-07-15
<160> 154
<170> PatentIn version 3.5
<210> 1
<211> 391
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 1
gaagtgcagc tggtggagtc tgggggacac ttggtgcagc ctggcaggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttgag gattatgcca tgaattgggt ccggcaagct 120
ccagggaagg gcctggagtg ggtctcagtc attagttggg acagtggtag gataggctat 180
gcggactctg tgaagggccg attcaccatc tccagagaca acgccaagaa ctcctcgtat 240
ctgcaaatga acagtctgag acctgaggac actgccttgt attattgtgt aagagatatg 300
ttggcttatt attctgacaa tagtggcaaa aaatacaacg tctacggtat ggacgtctgg 360
ggccaaggga ccacggtcac cgtctcctca g 391
<210> 2
<211> 130
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Gly His Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Glu Asp Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Trp Asp Ser Gly Arg Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Ser Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Asp Met Leu Ala Tyr Tyr Ser Asp Asn Ser Gly Lys Lys Tyr
100 105 110
Asn Val Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val
115 120 125
Ser Ser
130
<210> 3
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 3
Asp Tyr Ala Met Asn
1 5
<210> 4
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 4
Val Ile Ser Trp Asp Ser Gly Arg Ile Gly Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 5
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 5
Asp Met Leu Ala Tyr Tyr Ser Asp Asn Ser Gly Lys Lys Tyr Asn Val
1 5 10 15
Tyr Gly Met Asp Val
20
<210> 6
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 6
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgtttcc accttcttag cctggtacca acagaaacct 120
ggccaggctc ccaggctcct catgtatgat gcatccaaca gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagaacct 240
gaagattttg caatttacta ctgtcagcag cgtagccact ggcctcctat cttcggccaa 300
gggacacgac tggagattaa ac 322
<210> 7
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 7
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Thr Phe
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Met
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Ile Tyr Tyr Cys Gln Gln Arg Ser His Trp Pro Pro
85 90 95
Ile Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 8
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 8
Ala Ser Gln Ser Val Ser Thr Phe Leu Ala
1 5 10
<210> 9
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 9
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 10
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 10
Gln Gln Arg Ser His Trp Pro Pro Ile
1 5
<210> 11
<211> 391
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 11
gaagtgcagc tggtggagtc tgggggaggc ttggtgcaac ctggcaggtc cctgagactc 60
tcctgtgcag tttctggatt catctttgaa gattatgcca taaactgggt ccggcaagct 120
ccagggaagg gcctggagtg ggtctcaatt attagttggg acagtggtag gataggctac 180
gcggactctg tgaggggccg attcaccatc tccagagaca acgccaagaa ctcctcgttt 240
ctgcaaatga acagtctgag acccgaagac acggccgtgt attattgtgt aaaagatatg 300
ttggcgtatt attatgatgg tagcggcatc aggtacaacc tctacggtat ggacgtctgg 360
ggccaaggga ccacggtcac cgtctcctca g 391
<210> 12
<211> 130
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Ile Phe Glu Asp Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ser Trp Asp Ser Gly Arg Ile Gly Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Ser Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Lys Asp Met Leu Ala Tyr Tyr Tyr Asp Gly Ser Gly Ile Arg Tyr
100 105 110
Asn Leu Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val
115 120 125
Ser Ser
130
<210> 13
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 13
Asp Tyr Ala Ile Asn
1 5
<210> 14
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 14
Ile Ile Ser Trp Asp Ser Gly Arg Ile Gly Tyr Ala Asp Ser Val Arg
1 5 10 15
Gly
<210> 15
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 15
Asp Met Leu Ala Tyr Tyr Tyr Asp Gly Ser Gly Ile Arg Tyr Asn Leu
1 5 10 15
Tyr Gly Met Asp Val
20
<210> 16
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 16
gaaattgtgt tgacacagtc tccagccact ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtcg gagtattacc accttcttag cctggtacca acaaaaacct 120
ggccaggctc ccaggctcct catctacgat gcatccaaca gggccactgg cgtcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcaacag cctagagcct 240
gacgattttg caatttatta ctgtcagcag cgtgaccact ggcctccgat cttcggccaa 300
gggacacgac tggagattaa ac 322
<210> 17
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 17
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Ile Thr Thr Phe
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Pro
65 70 75 80
Asp Asp Phe Ala Ile Tyr Tyr Cys Gln Gln Arg Asp His Trp Pro Pro
85 90 95
Ile Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 18
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 18
Arg Ala Ser Arg Ser Ile Thr Thr Phe Leu Ala
1 5 10
<210> 19
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 19
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 20
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 20
Gln Gln Arg Asp His Trp Pro Pro Ile
1 5
<210> 21
<211> 403
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 21
gaggtgcagc tggtggtgtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggact cagtttcctt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc agcatctcaa gagacgattc aaagaacatg 240
ctgtttctgc atatgagcag cctgagaacc gaggacacag ccgtctatta ctgcgccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct cag 403
<210> 22
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 22
Glu Val Gln Leu Val Val Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Ser Phe Leu Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Ser Ile Ser Arg Asp Asp Ser Lys Asn Met
65 70 75 80
Leu Phe Leu His Met Ser Ser Leu Arg Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 23
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 23
Asn Ala Trp Met Ser
1 5
<210> 24
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 24
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 25
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 25
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 26
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 26
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttactt ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 27
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 28
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 28
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 29
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 29
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 30
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 30
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 31
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 31
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggact ctctttcctt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 32
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Ser Phe Leu Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 33
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 33
Asn Ala Trp Met Ser
1 5
<210> 34
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 34
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 35
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 35
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 36
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 36
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 37
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 37
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 38
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 38
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 39
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 39
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 40
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 40
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 41
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 41
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt ctctttcctt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 42
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Leu Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 43
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 43
Asn Ala Trp Met Ser
1 5
<210> 44
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 44
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 45
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 45
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 46
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 46
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 47
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 48
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 48
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 49
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 49
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 50
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 50
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 51
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 51
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggact cactttcctt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 52
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Leu Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 53
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 53
Asn Ala Trp Met Ser
1 5
<210> 54
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 54
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 55
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 55
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 56
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 56
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 57
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 57
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 58
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 58
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 59
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 59
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 60
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 60
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 61
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 61
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt cactttcctt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 62
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Leu Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 63
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 63
Asn Ala Trp Met Ser
1 5
<210> 64
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 64
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 65
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 65
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 66
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 66
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 67
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 68
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 68
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 69
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 69
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 70
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 70
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 71
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<220>
<221> МОДИФ_ОСТ
<222> (6)..(6)
<223> Val или Glu
<220>
<221> МОДИФ_ОСТ
<222> (27)..(27)
<223> Leu или Phe
<220>
<221> МОДИФ_ОСТ
<222> (28)..(28)
<223> Thr или Ser
<220>
<221> МОДИФ_ОСТ
<222> (30)..(30)
<223> Ser или Leu
<220>
<221> МОДИФ_ОСТ
<222> (71)..(71)
<223> Thr или Ser
<220>
<221> МОДИФ_ОСТ
<222> (80)..(80)
<223> Thr или Met
<220>
<221> МОДИФ_ОСТ
<222> (82)..(82)
<223> Tyr или Phe
<220>
<221> МОДИФ_ОСТ
<222> (84)..(84)
<223> Gln или His
<220>
<221> МОДИФ_ОСТ
<222> (86)..(86)
<223> Asn или Ser
<220>
<221> МОДИФ_ОСТ
<222> (89)..(89)
<223> Lys или Arg
<220>
<221> МОДИФ_ОСТ
<222> (99)..(99)
<223> Thr или Ala
<400> 71
Glu Val Gln Leu Val Xaa Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Xaa Xaa Phe Xaa Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Xaa Ile Ser Arg Asp Asp Ser Lys Asn Xaa
65 70 75 80
Leu Xaa Leu Xaa Met Xaa Ser Leu Xaa Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Xaa Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 72
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<220>
<221> МОДИФ_ОСТ
<222> (87)..(87)
<223> Phe или Tyr
<400> 72
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Xaa Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 73
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 73
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt ctctttcagt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 74
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 75
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 75
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 76
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 76
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 77
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 77
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 78
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 78
Gly Phe Ser Phe Ser Asn Ala Trp
1 5
<210> 79
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 79
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 80
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 80
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 81
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 81
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 82
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 82
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 83
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 83
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 84
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 84
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 85
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 85
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 86
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 86
Gln Asp Ile Ser Thr Trp
1 5
<210> 87
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 87
Ala Ala Ser
1
<210> 88
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 88
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 89
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 89
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggact cactttcagt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 90
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 90
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 91
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 91
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 92
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 92
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 93
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 93
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 94
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 94
Gly Leu Thr Phe Ser Asn Ala Trp
1 5
<210> 95
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 95
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 96
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 96
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 97
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 97
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 98
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 98
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 99
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 99
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 100
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 100
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 101
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 101
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 102
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 102
Gln Asp Ile Ser Thr Trp
1 5
<210> 103
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 103
Ala Ala Ser
1
<210> 104
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 104
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 105
<211> 402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 105
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt cactttcagt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg gcctggagtg ggttggccgt attaaaagta atactgatgg tgggacaaca 180
gactacgccg cacccgtgaa aggcagattc accatctcaa gagacgattc aaagaacacg 240
ctgtatctgc aaatgagcag cctgaaaacc gaggacacag ccgtctatta ctgcaccaca 300
gatggacctt actctgacga ttttagaagt ggttatgccg cacgctaccg ttatttcgga 360
atggacgtct ggggccaagg gaccacggtc accgtctcct ca 402
<210> 106
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr
100 105 110
Ala Ala Arg Tyr Arg Tyr Phe Gly Met Asp Val Trp Gly Gln Gly Thr
115 120 125
Thr Val Thr Val Ser Ser
130
<210> 107
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 107
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 108
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 108
Arg Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 109
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 109
Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg Tyr
1 5 10 15
Arg Tyr Phe Gly Met Asp Val
20
<210> 110
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 110
Gly Phe Thr Phe Ser Asn Ala Trp
1 5
<210> 111
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 111
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 112
<211> 24
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 112
Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg
1 5 10 15
Tyr Arg Tyr Phe Gly Met Asp Val
20
<210> 113
<211> 322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 113
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtgggaga cagagtcacc 60
atcacttgtc gggcgagtca ggatattagc acctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
agattcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcagcag gctaacagtt tccctccgac ttttggccag 300
gggaccaagc tggagatcaa ac 322
<210> 114
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 114
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 115
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 115
Arg Ala Ser Gln Asp Ile Ser Thr Trp Leu Ala
1 5 10
<210> 116
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 116
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 117
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 117
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 118
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 118
Gln Asp Ile Ser Thr Trp
1 5
<210> 119
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 119
Ala Ala Ser
1
<210> 120
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 120
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 121
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 121
Gly Leu Ser Phe Leu Asn Ala Trp
1 5
<210> 122
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 122
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 123
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 123
Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala Arg
1 5 10 15
Tyr Arg Tyr Phe Gly Met Asp Val Trp
20 25
<210> 124
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 124
Gln Asp Ile Ser Thr Trp
1 5
<210> 125
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 125
Ala Ala Ser
1
<210> 126
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 126
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 127
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 127
Gly Leu Ser Phe Leu Asn Ala Trp
1 5
<210> 128
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 128
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 129
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 129
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 130
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 130
Gln Asp Ile Ser Thr Trp
1 5
<210> 131
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 131
Ala Ala Ser
1
<210> 132
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 132
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 133
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 133
Gly Phe Ser Phe Leu Asn Ala Trp
1 5
<210> 134
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 134
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 135
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 135
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 136
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 136
Gln Asp Ile Ser Thr Trp
1 5
<210> 137
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 137
Ala Ala Ser
1
<210> 138
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 138
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 139
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 139
Gly Leu Thr Phe Leu Asn Ala Trp
1 5
<210> 140
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 140
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 141
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 141
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 142
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 142
Gln Asp Ile Ser Thr Trp
1 5
<210> 143
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 143
Ala Ala Ser
1
<210> 144
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 144
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 145
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 145
Gly Phe Thr Phe Leu Asn Ala Trp
1 5
<210> 146
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 146
Ile Lys Ser Asn Thr Asp Gly Gly Thr Thr
1 5 10
<210> 147
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 147
Thr Thr Asp Gly Pro Tyr Ser Asp Asp Phe Arg Ser Gly Tyr Ala Ala
1 5 10 15
Arg Tyr Arg Tyr Phe Gly Met Asp Val
20 25
<210> 148
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 148
Gln Asp Ile Ser Thr Trp
1 5
<210> 149
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 149
Ala Ala Ser
1
<210> 150
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 150
Gln Gln Ala Asn Ser Phe Pro Pro Thr
1 5
<210> 151
<211> 347
<212> БЕЛОК
<213> Вирус гриппа B
<400> 151
Asp Arg Ile Cys Thr Gly Ile Thr Ser Ser Asn Ser Pro His Val Val
1 5 10 15
Lys Thr Ala Thr Gln Gly Glu Val Asn Val Thr Gly Val Ile Pro Leu
20 25 30
Thr Thr Thr Pro Thr Lys Ser His Phe Ala Asn Leu Lys Gly Thr Glu
35 40 45
Thr Arg Gly Lys Leu Cys Pro Lys Cys Leu Asn Cys Thr Asp Leu Asp
50 55 60
Val Ala Leu Gly Arg Pro Lys Cys Thr Gly Asn Ile Pro Ser Ala Arg
65 70 75 80
Val Ser Ile Leu His Glu Val Arg Pro Val Thr Ser Gly Cys Phe Pro
85 90 95
Ile Met His Asp Arg Thr Lys Ile Arg Gln Leu Pro Asn Leu Leu Arg
100 105 110
Gly Tyr Glu His Ile Arg Leu Ser Thr His Asn Val Ile Asn Ala Glu
115 120 125
Asn Ala Pro Gly Gly Pro Tyr Lys Ile Gly Thr Ser Gly Ser Cys Pro
130 135 140
Asn Val Thr Asn Gly Asn Gly Phe Phe Ala Thr Met Ala Trp Ala Val
145 150 155 160
Pro Lys Asn Asp Asn Asn Lys Thr Ala Thr Asn Ser Leu Thr Ile Glu
165 170 175
Val Pro Tyr Ile Cys Thr Glu Gly Glu Asp Gln Ile Thr Val Trp Gly
180 185 190
Phe His Ser Asp Asn Glu Ile Gln Met Ala Lys Leu Tyr Gly Asp Ser
195 200 205
Lys Pro Gln Lys Phe Thr Ser Ser Ala Asn Gly Val Thr Thr His Tyr
210 215 220
Val Ser Gln Ile Gly Gly Phe Pro Asn Gln Thr Glu Asp Gly Gly Leu
225 230 235 240
Pro Gln Ser Gly Arg Ile Val Val Asp Tyr Met Val Gln Lys Ser Gly
245 250 255
Lys Thr Gly Thr Ile Thr Tyr Gln Arg Gly Ile Leu Leu Pro Gln Lys
260 265 270
Val Trp Cys Ala Ser Gly Arg Ser Lys Val Ile Lys Gly Ser Leu Pro
275 280 285
Leu Ile Gly Glu Ala Asp Cys Leu His Glu Lys Tyr Gly Gly Leu Asn
290 295 300
Lys Ser Lys Pro Tyr Tyr Thr Gly Glu His Ala Lys Ala Ile Gly Asn
305 310 315 320
Cys Pro Ile Trp Val Lys Thr Pro Leu Lys Leu Ala Asn Gly Thr Lys
325 330 335
Tyr Arg Pro Pro Ala Lys Leu Leu Lys Glu Arg
340 345
<210> 152
<211> 346
<212> БЕЛОК
<213> Вирус гриппа B
<400> 152
Asp Arg Ile Cys Thr Gly Ile Thr Ser Ser Asn Ser Pro His Val Val
1 5 10 15
Lys Thr Ala Thr Gln Gly Glu Val Asn Val Thr Gly Val Ile Pro Leu
20 25 30
Thr Thr Thr Pro Thr Lys Ser Tyr Phe Ala Asn Leu Lys Gly Thr Arg
35 40 45
Thr Arg Gly Lys Leu Cys Pro Asp Cys Leu Asn Cys Thr Asp Leu Asp
50 55 60
Val Ala Leu Gly Arg Pro Met Cys Val Gly Thr Thr Pro Ser Ala Lys
65 70 75 80
Ala Ser Ile Leu His Glu Val Lys Pro Val Thr Ser Gly Cys Phe Pro
85 90 95
Ile Met His Asp Arg Thr Lys Ile Arg Gln Leu Pro Asn Leu Leu Arg
100 105 110
Gly Tyr Glu Asn Ile Arg Leu Ser Thr Gln Asn Val Ile Asp Ala Glu
115 120 125
Lys Ala Pro Gly Gly Pro Tyr Arg Leu Gly Thr Ser Gly Ser Cys Pro
130 135 140
Asn Ala Thr Ser Lys Ser Gly Phe Phe Ala Thr Met Ala Trp Ala Val
145 150 155 160
Pro Lys Asp Asn Asn Lys Asn Ala Thr Asn Pro Leu Thr Val Glu Val
165 170 175
Pro Tyr Ile Cys Thr Glu Gly Glu Asp Gln Ile Thr Val Trp Gly Phe
180 185 190
His Ser Asp Asp Lys Thr Gln Met Lys Asn Leu Tyr Gly Asp Ser Asn
195 200 205
Pro Gln Lys Phe Thr Ser Ser Ala Asn Gly Val Thr Thr His Tyr Val
210 215 220
Ser Gln Ile Gly Ser Phe Pro Asp Gln Thr Glu Asp Gly Gly Leu Pro
225 230 235 240
Gln Ser Gly Arg Ile Val Val Asp Tyr Met Met Gln Lys Pro Gly Lys
245 250 255
Thr Gly Thr Ile Val Tyr Gln Arg Gly Val Leu Leu Pro Gln Lys Val
260 265 270
Trp Cys Ala Ser Gly Arg Ser Lys Val Ile Lys Gly Ser Leu Pro Leu
275 280 285
Ile Gly Glu Ala Asp Cys Leu His Glu Lys Tyr Gly Gly Leu Asn Lys
290 295 300
Ser Lys Pro Tyr Tyr Thr Gly Glu His Ala Lys Ala Ile Gly Asn Cys
305 310 315 320
Pro Ile Trp Val Lys Thr Pro Leu Lys Leu Ala Asn Gly Thr Lys Tyr
325 330 335
Arg Pro Pro Ala Lys Leu Leu Lys Glu Arg
340 345
<210> 153
<211> 346
<212> БЕЛОК
<213> Вирус гриппа B
<400> 153
Asp Arg Ile Cys Thr Gly Ile Thr Ser Ser Asn Ser Pro His Val Val
1 5 10 15
Lys Thr Ala Thr Gln Gly Glu Val Asn Val Thr Gly Val Ile Pro Leu
20 25 30
Thr Thr Thr Pro Thr Lys Ser Tyr Phe Ala Asn Leu Lys Gly Thr Arg
35 40 45
Thr Arg Gly Lys Leu Cys Pro Asp Cys Leu Asn Cys Thr Asp Leu Asp
50 55 60
Val Ala Leu Gly Arg Pro Met Cys Val Gly Thr Thr Pro Ser Ala Lys
65 70 75 80
Ala Ser Ile Leu His Glu Val Lys Pro Val Thr Ser Gly Cys Phe Pro
85 90 95
Ile Met His Asp Arg Thr Lys Ile Arg Gln Leu Pro Asn Leu Leu Arg
100 105 110
Gly Tyr Glu Asn Ile Arg Leu Ser Thr Gln Asn Val Ile Asp Ala Glu
115 120 125
Lys Ala Pro Gly Gly Pro Tyr Arg Leu Gly Thr Ser Glu Ser Cys Pro
130 135 140
Asn Ala Thr Ser Lys Ser Gly Phe Phe Ala Thr Met Ala Trp Ala Val
145 150 155 160
Pro Lys Asp Asn Asn Lys Asn Ala Thr Asn Pro Leu Thr Val Glu Val
165 170 175
Pro Tyr Ile Cys Thr Glu Gly Glu Asp Gln Ile Thr Val Trp Gly Phe
180 185 190
His Ser Asp Asp Lys Thr Gln Met Lys Asn Leu Tyr Gly Asp Ser Asn
195 200 205
Pro Gln Lys Phe Thr Ser Ser Ala Asn Gly Val Thr Thr His Tyr Val
210 215 220
Ser Gln Ile Gly Ser Phe Pro Asp Gln Thr Glu Asp Gly Gly Leu Pro
225 230 235 240
Gln Ser Gly Arg Ile Val Val Asp Tyr Met Met Gln Lys Pro Gly Lys
245 250 255
Thr Gly Thr Ile Val Tyr Gln Arg Gly Val Leu Leu Pro Gln Lys Val
260 265 270
Trp Cys Ala Ser Gly Arg Ser Lys Val Ile Lys Gly Ser Leu Pro Leu
275 280 285
Ile Gly Glu Ala Asp Cys Leu His Glu Lys Tyr Gly Gly Leu Asn Lys
290 295 300
Ser Lys Pro Tyr Tyr Thr Gly Glu His Ala Lys Ala Ile Gly Asn Cys
305 310 315 320
Pro Ile Trp Val Lys Thr Pro Leu Lys Leu Ala Asn Gly Thr Lys Tyr
325 330 335
Arg Pro Pro Ala Lys Leu Leu Lys Glu Arg
340 345
<210> 154
<211> 346
<212> БЕЛОК
<213> Вирус гриппа B
<400> 154
Asp Arg Ile Cys Thr Gly Ile Thr Ser Ser Asn Ser Pro His Val Val
1 5 10 15
Lys Thr Ala Thr Gln Gly Glu Val Asn Val Thr Gly Val Ile Pro Leu
20 25 30
Thr Thr Thr Pro Thr Lys Ser Tyr Phe Ala Asn Leu Lys Gly Thr Arg
35 40 45
Thr Arg Gly Lys Leu Cys Pro Asp Cys Leu Asn Cys Thr Asp Leu Asp
50 55 60
Val Ala Leu Gly Arg Pro Met Cys Val Gly Thr Thr Pro Ser Ala Lys
65 70 75 80
Ala Ser Ile Leu His Glu Val Arg Pro Val Thr Ser Gly Cys Phe Pro
85 90 95
Ile Met His Asp Arg Thr Lys Ile Arg Gln Leu Pro Asn Leu Leu Arg
100 105 110
Gly Tyr Glu Asn Ile Arg Leu Ser Thr Gln Asn Val Ile Asp Ala Glu
115 120 125
Asn Ala Pro Gly Gly Pro Tyr Arg Leu Gly Thr Ser Arg Ser Cys Pro
130 135 140
Asn Ala Thr Ser Lys Ser Gly Phe Phe Ala Thr Met Ala Trp Ala Val
145 150 155 160
Pro Lys Asp Asn Asn Lys Asn Ala Thr Asn Pro Leu Thr Val Glu Val
165 170 175
Pro Tyr Val Cys Thr Glu Gly Glu Asp Gln Ile Thr Val Trp Gly Phe
180 185 190
His Ser Asp Asn Lys Thr Gln Met Lys Asn Leu Tyr Gly Asp Ser Asn
195 200 205
Pro Gln Lys Phe Thr Ser Ser Ala Asn Gly Val Thr Thr His Tyr Val
210 215 220
Ser Gln Ile Gly Gly Phe Pro Ala Gln Thr Glu Asp Glu Gly Leu Pro
225 230 235 240
Gln Ser Gly Arg Ile Val Val Asp Tyr Met Val Gln Lys Pro Arg Lys
245 250 255
Thr Gly Thr Ile Val Tyr Gln Arg Gly Val Leu Leu Pro Gln Lys Val
260 265 270
Trp Cys Ala Ser Gly Arg Ser Lys Val Ile Lys Gly Ser Leu Pro Leu
275 280 285
Ile Gly Glu Ala Asp Cys Leu His Glu Lys Tyr Gly Gly Leu Asn Lys
290 295 300
Ser Lys Pro Tyr Tyr Thr Gly Glu His Ala Lys Ala Ile Gly Asn Cys
305 310 315 320
Pro Ile Trp Val Lys Thr Pro Leu Lys Leu Ala Asn Gly Thr Lys Tyr
325 330 335
Arg Pro Pro Ala Lys Leu Leu Lys Glu Arg
340 345
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА B И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2739952C2 |
| НОВЫЕ АНТИТЕЛА К BSSL | 2020 |
|
RU2826992C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| АНТИ-GDF15 АНТИТЕЛА, КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2795457C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛО К ЦИТОМЕГАЛОВИРУСУ ЧЕЛОВЕКА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2817217C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С HLA-A2/WT1 | 2018 |
|
RU2815176C2 |
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА, И ИХ КОМПОЗИЦИИ, КОМБИНАЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2720282C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
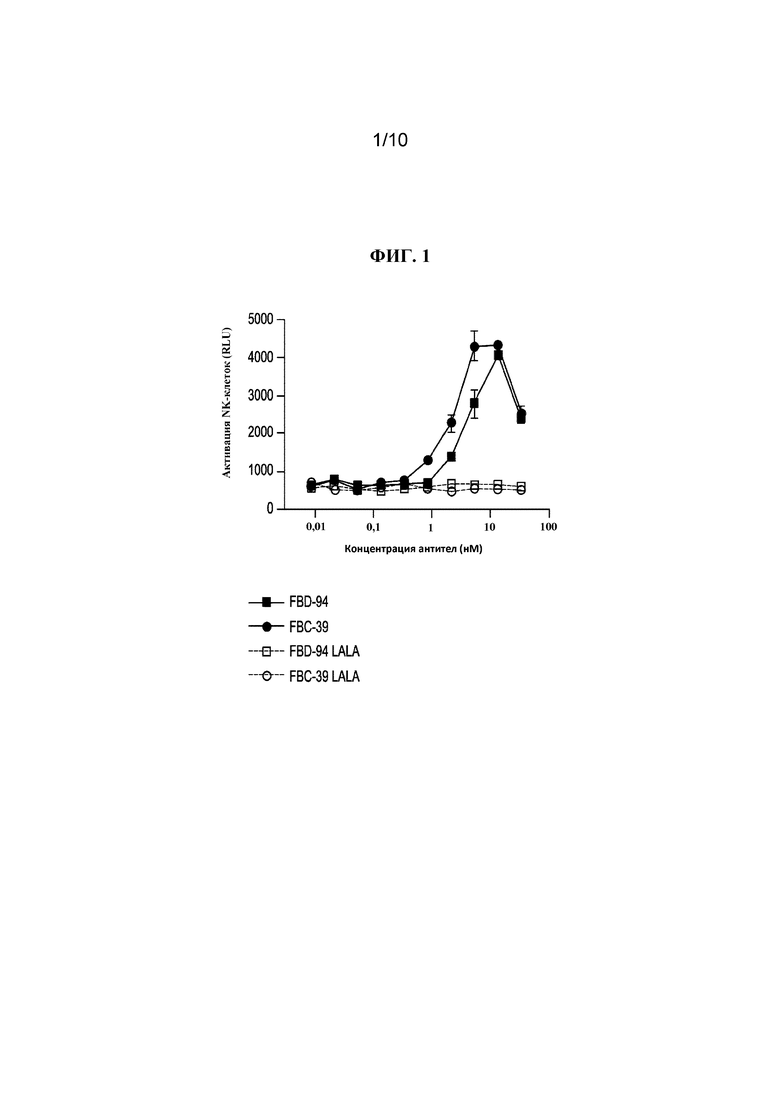
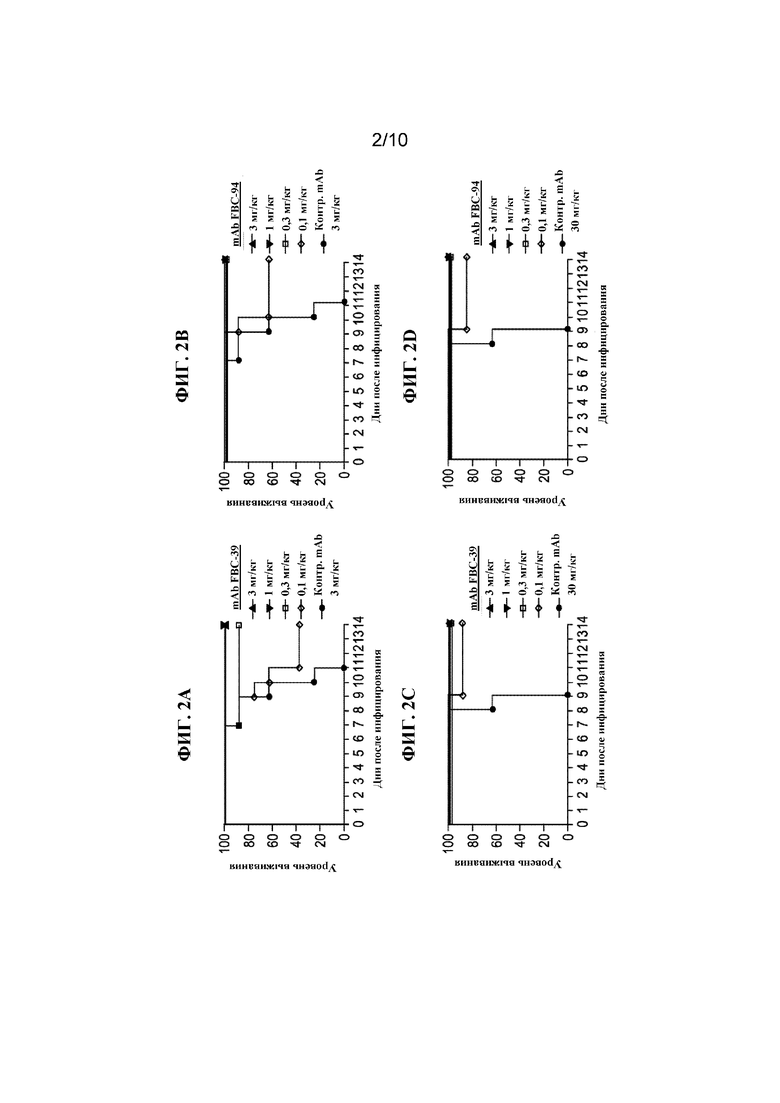
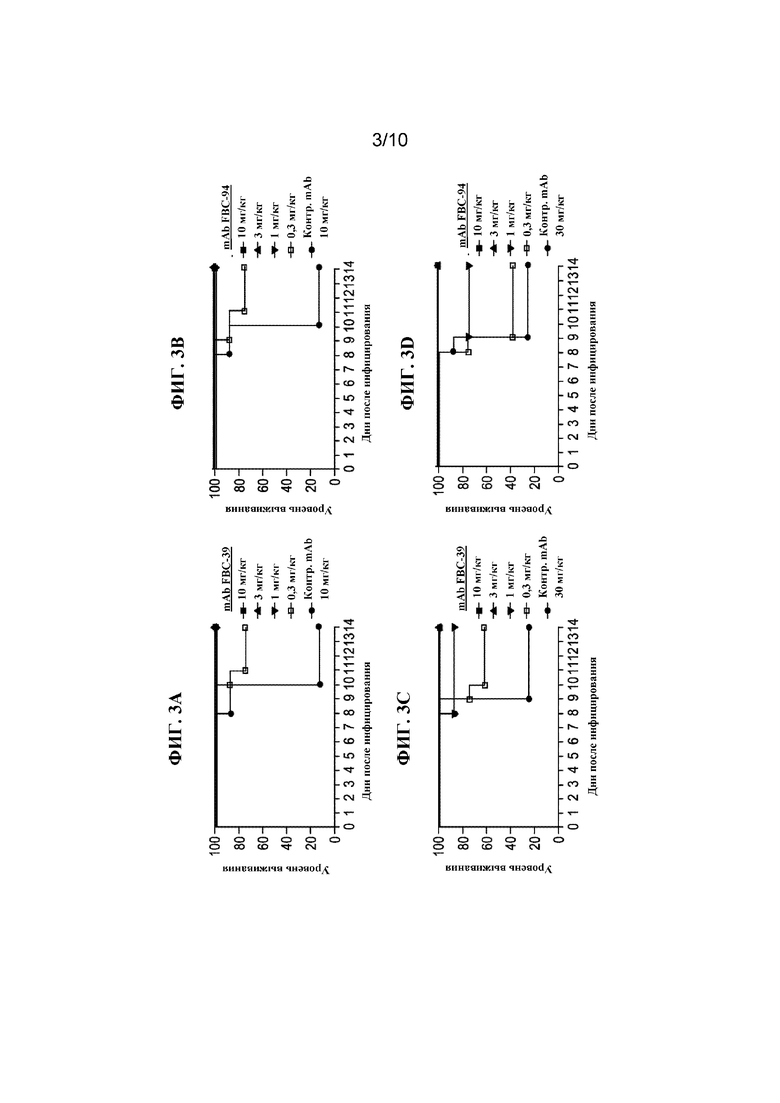
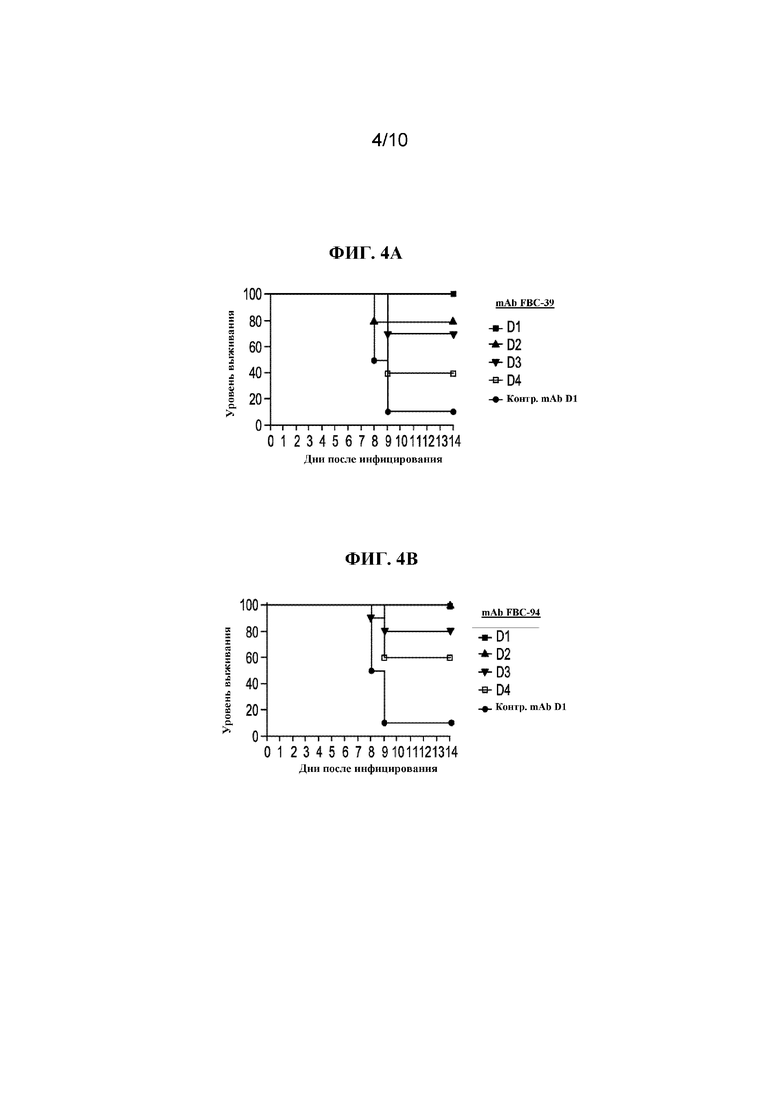
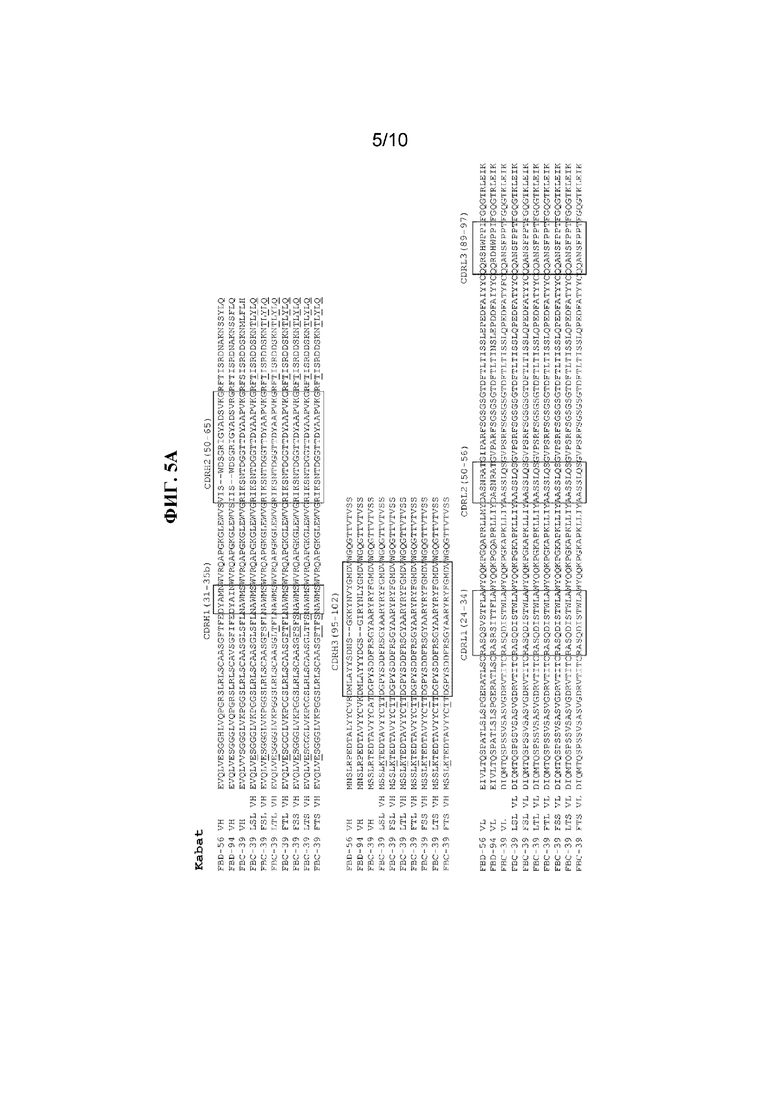

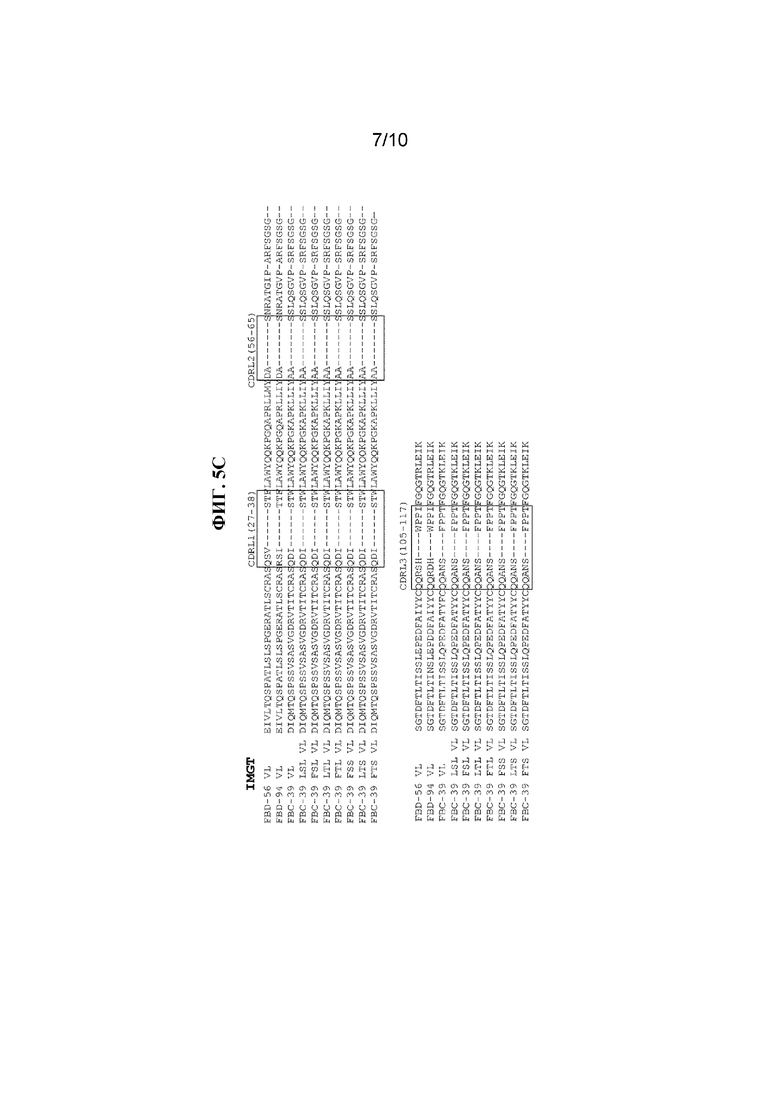


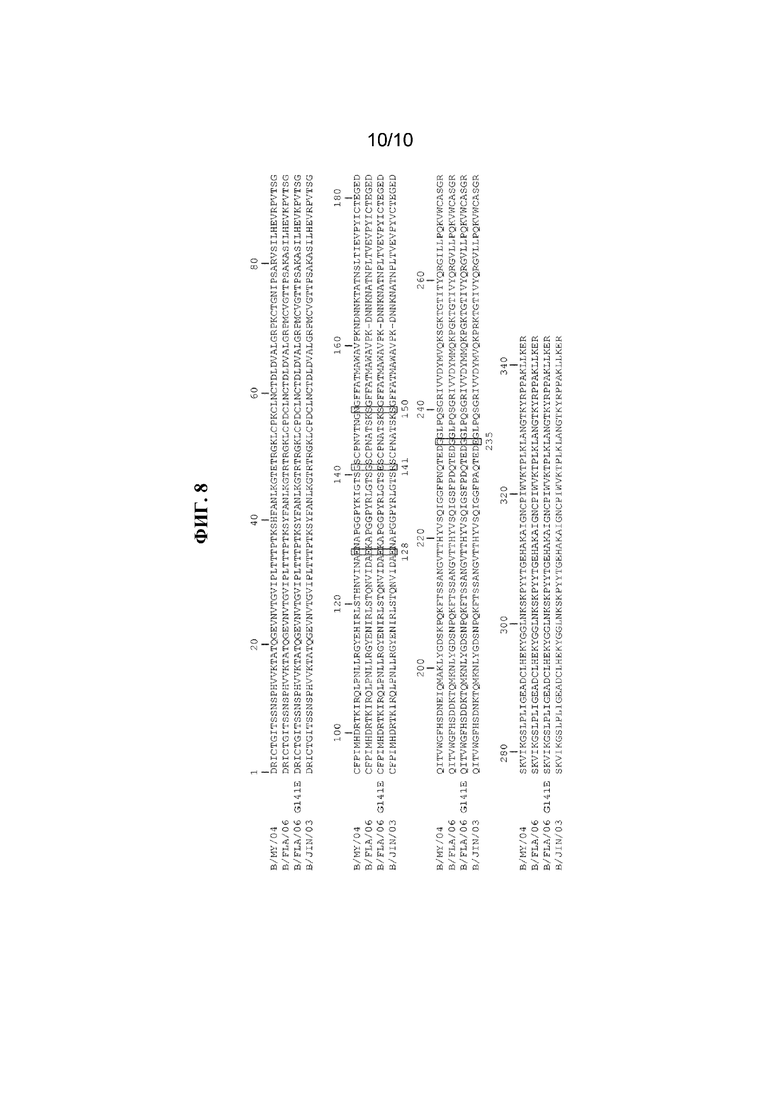
Изобретения относятся к области биотехнологии. Описана группа изобретений, включающая выделенное антитело или его антигенсвязывающий фрагмент, обладающие вариантом Fc-участка, где антитело или его связывающий фрагмент связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирус гриппа B в двух филогенетически разных линиях и обладают увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc, выделенную нуклеиновую кислоту, кодирующую вышеуказанное антитело или его антигенсвязывающий фрагмент, вектор для получения вышеуказанного выделенного антитела или антигенсвязывающего фрагмента, клетку-хозяина, экспрессирующую выделенное антитело или антигенсвязывающий фрагмент, способ получения антитела или его антигенсвязывающего фрагмента и композицию для лечения инфекции, вызванной вирусом гриппа B (варианты). Изобретения расширяют арсенал средств, связывающихся с гемагглютинином (HA) вируса гриппа B и нейтрализующих вирус гриппа B в двух филогенетически разных линиях. 7 н. и 17 з.п. ф-лы, 8 ил., 12 табл., 8 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, обладающие вариантом Fc-участка, где антитело или его связывающий фрагмент связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирус гриппа B в двух филогенетически разных линиях и обладают увеличенным периодом полувыведения из сыворотки по сравнению с антителом c нативным Fc, где антитело или его связывающий фрагмент включает совокупность из шести CDR, где набор из шести CDR включает:
HCDR-1 SEQ ID NO: 145, HCDR-2 SEQ ID NO: 146, HCDR-3 SEQ ID NO: 147, LCDR-1 SEQ ID NO: 148, LCDR-2 SEQ ID NO: 149 и LCDR-3 SEQ ID NO: 150.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1, где антитело связывается с гемагглютинином вируса гриппа В и нейтрализует вирус гриппа B как в Yamagata, так и в Victoria линиях.
3. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1, где антитело или его антигенсвязывающий фрагмент связываются с вирусом гриппа B линии Yamagata, выбранным из: B/AA/94; B/YSI/98; B/JHB/99; B/SC/99; B/FL/06; вирусом гриппа B линии Victoria, выбранным из B/BJ/97, B/HK/01; B/MY/04; B/BNE/08; предивергентными штаммами вируса гриппа B, выбранными из B/Lee/40; B/AA/66; B/HK/72; и их комбинациями.
4. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, содержащее VH SEQ ID NO: 62 и VL SEQ ID NO: 67.
5. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-4, где вариант Fc-участка включает модификацию в одном или нескольких положениях, выбранных из 221, 225, 228, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 247, 250, 251, 252, 254, 255, 256, 257, 262, 263, 264, 265, 266, 267, 268, 269, 279, 280, 284, 292, 296, 297, 298, 299, 305, 308, 313, 316, 318, 320, 322, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 339, 341, 343, 370, 373, 378, 392, 416, 419, 421, 428, 433, 434, 435, 436, 440 и 443 согласно нумерации с помощью EU-индекса, как изложено у Kabat.
6. Выделенное антитело или его связывающий фрагмент по п. 5, где модификация выбрана из замены, вставки и делеции.
7. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-участка включает по меньшей мере одну замену, выбранную из 221K, 221Y, 225E, 225K, 225W, 228P, 234D, 234E, 234N, 234Q, 234T, 234H, 234Y, 234I, 234V, 234F, 235A, 235D, 235R, 235W, 235P, 235S, 235N, 235Q, 235T, 235H, 235Y, 235I, 235V, 235E, 235F, 236E, 237L, 237M, 237P, 239D, 239E, 239N, 239Q, 239F, 239T, 239H, 239Y, 240I, 240A, 240T, 240M, 241W, 241L, 241Y, 241E, 241R. 243W, 243L 243Y, 243R, 243Q, 244H, 245A, 247L, 247V, 247G, 250E, 250Q, 251F, 252L, 252Y, 254S, 254T, 255L, 256E, 256F, 256M, 257C, 257M, 257N, 262I, 262A, 262T, 262E, 263I, 263A, 263T, 263M, 264L, 264I, 264W, 264T, 264R, 264F, 264M, 264Y, 264E, 265A, 265G, 265N, 265Q, 265Y, 265F, 265V, 265I, 265L, 265H, 265T, 266I, 266A, 266T, 266M, 267Q, 267L, 268E, 269H, 269Y, 269F, 269R, 270E, 280A, 284M, 292P, 292L, 296E, 296Q, 296D, 296N, 296S, 296T, 296L, 296I, 296H, 296G, 297S, 297D, 297E, 298A, 298H, 298I, 298T, 298F, 299I, 299L, 299A, 299S, 299V, 299H, 299F, 299E, 305I, 308F, 313F, 316D, 318A, 318S, 320A, 320S, 322A, 322S, 325Q, 325L, 325I, 325D, 325E, 325A, 325T, 325V, 325H, 326A, 326D, 326E, 326G, 326M, 326V, 327G, 327W, 327N, 327L, 328S, 328M, 328D, 328E, 328N, 328Q, 328F, 328I, 328V, 328T, 328H, 328A, 329F, 329H, 329Q, 330K, 330G, 330T, 330C, 330L, 330Y, 330V, 330I, 330F, 330R, 330H, 331G, 331A, 331L, 331M, 331F, 331W, 331K, 331Q, 331E, 331S, 331V, 331I, 331C, 331Y, 331H, 331R, 331N, 331D, 331T, 332D, 332S, 332W, 332F, 332E, 332N, 332Q, 332T, 332H, 332Y, 332A, 333A, 333D, 333G, 333Q, 333S, 333V, 334A, 334E, 334H, 334L, 334M, 334Q, 334V, 334Y, 339T, 370E, 370N, 378D, 392T, 396L, 416G, 419H, 421K, 428L, 428F, 433K, 433L, 434A, 434W, 434Y, 436H, 440Y и 443W согласно нумерации с помощью EU-индекса, как изложено у Kabat.
8. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-участка включает по меньшей мере одну модификацию в одном или более положениях, выбранных из 228, 234, 235 и 331 согласно нумерации с помощью EU-индекса, как изложено у Kabat.
9. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-участка включает одну или более замен, выбранных из 228P, 234F, 235E, 235F, 235Y и 331S согласно нумерации с помощью EU-индекса, как изложено у Kabat.
10. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-участка включает Fc-участок IgG4.
11. Выделенное антитело или его связывающий фрагмент по п. 10, где вариант Fc-области содержит по меньшей мере одну модификацию в одном или более положениях, выбранных из 228 и 235 согласно нумерации с помощью EU-индекса, как изложено у Kabat.
12. Выделенное антитело или его связывающий фрагмент по п. 10, где вариант Fc-области содержит одну или несколько модификаций, выбранных из 228P, 235E и 235Y согласно нумерации с помощью EU-индекса, как изложено у Kabat.
13. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-области содержит по меньшей мере одну модификацию в одном или нескольких положениях, выбранных из 239, 330 и 332 согласно нумерации с помощью EU-индекса, как изложено у Kabat.
14. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-области содержит по меньшей мере одну замену, выбранную из 239D, 330L, 330Y и 332E согласно нумерации с помощью EU-индекса, как изложено у Kabat.
15. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-области содержит по меньшей мере одну модификацию в одном или нескольких положениях, выбранных из 252, 254 и 256 согласно нумерации с помощью EU-индекса, как изложено у Kabat.
16. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-области содержит по меньшей мере одну замену, выбранную из 252Y, 254T и 256E согласно нумерации с помощью EU-индекса, как изложено у Kabat.
17. Выделенное антитело или его связывающий фрагмент по любому из пп. 1-6, где вариант Fc-области содержит три замены: 252Y, 254T и 256E согласно нумерации с помощью EU-индекса, как изложено у Kabat.
18. Выделенная нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17.
19. Вектор для получения выделенного антитела или антигенсвязывающего фрагмента, которые связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирус гриппа B в двух филогенетически разных линиях, где вектор содержит выделенную нуклеиновую кислоту по п. 18.
20. Клетка-хозяин, экспрессирующая выделенное антитело или антигенсвязывающий фрагмент, которые связываются с гемагглютинином (HA) вируса гриппа B и нейтрализуют вирус гриппа B в двух филогенетически разных линиях, где клетка-хозяин содержит нуклеиновую кислоту по п. 18 или вектор по п. 19.
21. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-17, включающий культивирование клетки-хозяина по п. 20 в условиях, подходящих для экспрессии антитела или его фрагмента.
22. Способ по п. 21, дополнительно включающий выделение антитела или его антигенсвязывающего фрагмента из культуры клеток-хозяев.
23. Композиция для лечения инфекции, вызванной вирусом гриппа B, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17 и фармацевтически приемлемый носитель.
24. Композиция для лечения инфекции, вызванной вирусом гриппа B, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17, и 25 мM His, и 0,15 M NaCl при pH 6,0.
| Привод | 1982 |
|
SU1167382A1 |
| WO 2013132007 A1, 12.09.2013 | |||
| EP 2919813 B1, 24.10.2018 | |||
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ПОВЕРХНОСТНЫМ АНТИГЕНОМ ВИРУСА ГЕПАТИТА B, FAB-ФРАГМЕНТ И СПОСОБ СНИЖЕНИЯ УРОВНЯ ЦИРКУЛИРУЮЩЕГО ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B У ПАЦИЕНТА | 1992 |
|
RU2146706C1 |
| Тарантул В.З | |||
| Толковый биотехнологический словарь | |||
| Русско-английский | |||
| - М., Языки славянских культур, 2009 | |||
| PAKULA A.A., SAUER R.T., Genetic analysis of protein stability and function, Annual review of genetics, 1989, vol.23, pp | |||

