Перекрестная ссылка на родственные заявки
Настоящая заявка претендует на приоритет по предварительной заявке на патент США с серийным номером 63/022328, поданной 8 мая 2020 г., которая полностью включена в настоящий документ посредством ссылки для всех целей.
Перечень последовательностей
Рассматриваемая заявка содержит Перечень последовательностей, который был подан в электронном виде в формате ASCII и полностью включен в настоящий документ посредством ссылки. Указанная копия ASCII, созданная 6 мая 2021 года, называется G4590-08600PCT_SeqListing.txt и имеет размер 28 килобайт.
Область техники, к которой относится изобретение
Настоящее изобретение относится к химерному полипептиду гемагглютинина (HA) вируса гриппа, содержащей его иммуногенной/вакцинной композиции и ее применению.
Предпосылки создания изобретения
Традиционный способ получения вакцин против гриппа заключается в культивировании вируса в яйцах кур с эмбрионами, свободными от специфических патогенов (SPF), и этот процесс часто требует более шести месяцев для массового производства. Однако некоторые штаммы вакцинных вирусов плохо растут в яйцах, и у людей с аллергией на куриные яйца могут возникнуть проблемы, связанные с безопасностью. Для замены метода на основе яиц были разработаны новые подходы, основанные на культивировании вирусов в культурах клеток; но метод культивирования в культурах клеток по-прежнему сопряжен с риском продукции потенциально опасных вирусов. Чтобы преодолеть эти проблемы, изучение альтернативных стратегий показало, что рекомбинантные вакцины на основе HA могут индуцировать нейтрализующие антитела против инфекции, вызванной вирусом гриппа. Однако антитела, индуцированные конкретным подтипом вируса гриппа, обычно не могли эффективно нейтрализовать другие подтипы гриппа. Кроме того, вакцину приходится ежегодно обновлять из-за постоянной мутации вируса.
Таким образом, по-прежнему существует потребность в разработке универсальной вакцины против широкого спектра штаммов вируса гриппа.
Краткое описание изобретения
В одном аспекте настоящее изобретение обеспечивает химерный полипептид гемагглютинина (HA) вируса гриппа, содержащий одну или несколько последовательностей стеблевого домена, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью стеблевого домена HA подтипа H1 (H1 HA) и/или HA подтипа H5 (H5 HA), слитых с одной или несколькими последовательностями глобулярного головного домена, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью глобулярного головного домена HA подтипа H1 (H1 HA) или HA подтипа H5 (H5 HA).
В некоторых вариантах осуществления HA представляет собой HA гриппа A, HA гриппа B или HA гриппа C.
В некоторых вариантах осуществления гомология составляет по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%.
В некоторых вариантах осуществления последовательность стеблевого домена представляет собой N-концевой стеблевой сегмент H1 HA или С-концевой стеблевой сегмент H1 HA; N-концевой стеблевой сегмент H1 HA или C-концевой стеблевой сегмент последовательностей H1+H5 HA; или N-концевой стеблевой сегмент H5 НА или С-концевой стеблевой сегмент последовательностей H1+H5 HA.
В некоторых вариантах осуществления консенсусная последовательность стеблевого домена H1 HA и/или H5 HA содержит аминокислотную последовательность SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 9 или SEQ ID NO: 10.
В некоторых вариантах осуществления консенсусная последовательность глобулярного головного домена H1 HA или H5 HA содержит аминокислотную последовательность SEQ ID NO: 3, SEQ ID NO: 7 или SEQ ID NO: 11.
В одном варианте осуществления химерный полипептид НА вируса гриппа содержит аминокислотную последовательность SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12.
В некоторых вариантах осуществления один или несколько гликосайтов на HA являются моногликозилированными. В другом варианте осуществления моногликозилированный НА имеет только N-ацетилглюкозамин (GlcNAc) на каждом гликосайте.
В одном варианте осуществления химерный полипептид НА вируса гриппа используют в качестве иммуногена.
В другом аспекте настоящее изобретение обеспечивает иммуногенную композицию, содержащую химерный полипептид НА вируса гриппа и адъювант. В одном варианте осуществления адъювант представляет собой гликолипидный адъювант.
В другом аспекте настоящее изобретение обеспечивает рекомбинантный полинуклеотид, содержащий последовательность нуклеиновой кислоты, кодирующую полипептид по настоящему изобретению, и необязательно последовательность нуклеиновой кислоты, кодирующую сигнальный пептид. В некоторых вариантах осуществления сигнальный пептид содержит последовательность SEQ ID NO: 13 или SEQ ID NO: 14.
В другом аспекте настоящее изобретение обеспечивает вектор, содержащий рекомбинантный полинуклеотид по настоящему изобретению. Также обеспечена клетка-хозяин, которая содержит вектор по настоящему изобретению.
В другом аспекте настоящее изобретение обеспечивает способ иммунизации субъекта против вируса гриппа, включающий введение субъекту эффективного количества химерного полипептида гемагглютинина (HA) вируса гриппа или иммуногенной композиции по настоящему изобретению.
В другом аспекте настоящее изобретение обеспечивает способ предупреждения заболевания, вызванного вирусом гриппа, у субъекта, включающий введение субъекту эффективного количества химерного полипептида гемагглютинина (HA) вируса гриппа или иммуногенной композиции по настоящему изобретению.
В одном варианте осуществления способы, описанные в настоящем документе, вызывают CD4+ и/или CD8+ Т-клеточные иммунные ответы.
В одном варианте осуществления способы, описанные в настоящем документе, индуцируют стебель-специфические антитела с более высокой антителозависимой клеточной цитотоксичностью (ADCC), лучшей нейтрализующей и более сильной кросс-протективной активностями против штаммов и подтипов H1, H3, H5 и H7.
В одном варианте осуществления способы, описанные в настоящем документе, повышают эффективность вакцины за счет продукции большего количества IFN-гамма, IL-4 и CD8+ Т-клеток памяти.
Краткое описание чертежей
Фиг. 1 (А)-(I). Химерная конструкция H5/1 с консенсусом глобулярной головки H5 и консенсусом стебля H1 (cHA), и широко кросс-протективные, стебель-специфические антитела, вызванные вакцинацией иммуногенами cHAmg. (A) Конструкции замены H1/5 (глобулярная головка H1 и ствол H1+H5[HA2]), замены H5/1 (глобулярная головка H5 и ствол H5+H1[HA2]) и химерного H5/1 (cHA: глобулярная головка H5 и ствол H1). (B) Нейтрализующая активность против вирусов H1N1 California/07/2009 и H5N1 Vietnam/1194/2004. (C) Количество продуцирующих гранзим B (GrzB) CD8+ T-клеток в спленоцитах, стимулированных HA (черный столбец) или контролем PBS (белый столбец) в течение 2 дней у мышей, вакцинированных PBS (контроль), HA+Alu или HA+С34, оценивали с помощью анализа методом проточной цитометрии. (D-I) Титры антител от мышей, вакцинированных cHAfg и cHAmg с адъювантом Al(OH)3 по сравнению с cHAfg и cHAmg с адъювантом C34, измеряли на 42-й день с помощью ELISA с белком HA A/California/07/2009 H1N1 (D), белком НА A/Brisbane/59/2007 H1N1 (E), белком НА A/Brisbane/10/2007 H3N2 (F), белком НА A/Vietnam/1194/2004 H5N1 (G), белком HA A/Shanghai/2/2013 H7N9 (H) и белком стебля HA (no. 4900) A/Brisbane/59/2007 (Bris/07) (I) в качестве покрывающего антигена. Конечный титр антител определялся как последнее разведение антисыворотки, при котором оптическая плотность в 2,5 раза превышала оптическую плотность отрицательного контроля (неиммунная сыворотка). Данные анализировали с использованием t-критерия Стьюдента и двухфакторного дисперсионного анализа ANOVA от Prism; различия считались статистически значимыми при *P <0,05; **Р <0,01. Данные представляют среднее значение ± SEM.
Фиг. 2 (А)-(С). Репортерный анализ ADCC антисыворотки от cHA-вакцинированных мышей против клеток-мишеней, экспрессирующих HA вирусов H1N1, H3N2 или H5N1 и их подтипов. Антисыворотки, собранные у мышей, иммунизированных белками cHAfg или cHAmg с адъювантом гидроксидом алюминия или C34, инкубировали в течение 30 мин с клетками MDCK, инфицированными (A) вирусом H1N1, (B) вирусом H5N1 или (C) вирусом H3N2. Затем репортерный анализ ADCC проводили с использованием эффекторных клеток Jurkat, экспрессирующих мышиный FcγRIII, и измеряли относительные единицы люминесценции (RLU), и значения представляли собой средние значения ± SEM. ***Р <0,001. Значение P рассчитывали с помощью программного обеспечения Prism с использованием двухфакторного дисперсионного анализа ANOVA.
Фиг. 3 (А)-(Е). Было вызвано больше CD4+ и CD8+ Т-клеточных ответов и широко нейтрализующих антител, обеспечив более широкую кросс-протекцию с помощью cHAmg с адъювантом C34. Мышей BALB/c иммунизировали cHAfg и cHAmg с адъювантом Al(OH)3 или C34; клетки из селезенок иммунизированных мышей получали после трех иммунизаций, и клетки, секретирующие IFN-гамма (A), IL-4 (B) и GzB (C), определяли с помощью анализа ELISpot с использованием специфических пептидов. Количество клеток, образующих пятна (SFC), выражали как среднее значение ± SEM. Нейтрализующие активности антисыворотки от мышей, вакцинированных cHAfg и cHAmg, анализировали против (D) вируса H1N1 и (E) вируса H5N1. Данные представляли в виде среднего значения ± SEM. Результаты рассчитывали с помощью программного обеспечения Prism с использованием t-критерия Стьюдента и двухфакторного дисперсионного анализа ANOVA; значимые различия отмечены как *P <0,05; **Р <0,01; ***Р <0,001.
Фиг. 4 (А)-(F). Кросс-протективная эффективность у мышей, зараженных летальными дозами вирусов H1N1 и H5N1. Мышей BALB/c иммунизировали тремя дозами cHAfg и cHAmg с адъювантом Al(OH)3 или C34 с 2-недельными интервалами. Иммунизированных мышей заражали вирусами H1N1 A/California/07/2009 (A), H1N1 A/New Caledonia/1999 (B), H1N1 A/WSN/1933 (C), H1N1 A/Solomon Islands/03/2006 (D), H5N1 A/Vietnam/1194/2004/NIBRG14 (E) или H5N1 A/Turkey/1/2005/NIBRG23 (F), и эффективность оценивали путем регистрации уровня выживаемости в течение 14 дней после инфицирования. **Р <0,01. Значимые различия в уровне выживаемости анализировали с помощью логарифмического рангового критерия (Мантела-Кокса).
Фиг. 5 (А)-(Е). Дизайн и получение химерных белков НА. (A) Разработанные последовательности НА вируса гриппа конструировали с использованием консенсусной последовательности H1N1 и консенсусной последовательности H5N1 pCHA5-II для создания химерного НА. Домен глобулярной головки состоит из аминокислотной последовательности между остатками C52 и C277 (нумерация H3). Стеблевая область состоит из частей субъединиц НА1 и НА2. Белковые структуры загружали из базы данных белков Protein Data Bank с ID кодом 2IBX (VN1194 H5 HA) и 3LZG (A/California/04/2009). Конечные изображения генерировали с помощью PyMol. Поскольку нет опубликованной структуры консенсуса HA, для конструкции химерного HA использовали изображение домена головки от птичьего гриппа H5 (Vietnam/1194/2004) и стеблевой области пандемического H1N1 (California/07/2009). (B-D) Очистка химерного белка HA и анализ методом гель-фильтрационной хроматографии. (B) Очищенные белки HA анализировали с помощью SDS/PAGE. М: маркер молекулярной массы. Слева: cHAfg, полностью гликозилированный cHA, непосредственно выделенный из клеток HEK293T; (C) cHAmg, моногликозилированный cHA, выделенный из клеток HEK293S и расщепленный эндогликозидазой H. (D) Гель-фильтрационный анализ очищенных секретируемых белков HA. Полностью гликозилированный сНА из клеток HEK293T и моногликозилированный сНА существовали в виде тримера (>200 кДа), как показано на хроматограмме. На фиг. представлены наложенные друг на друга профили элюирования белков cHA, экспрессируемых клетками HEK293T, перекрывающиеся с калибровочными стандартами (пунктирная линия). (E) Схематическая фиг. для обозначения основных гликанов на гликосайтах cHAfg и cHAmg, определенных с помощью LC-MS/MS. Соблюдались общие символы гликанов.
Фиг. 6 (А) и (В). Конструкции секретируемого НА и очистка. (A) Последовательность, кодирующую эктодомен HA, изготавливали в векторе экспрессии pcDNA и трансфицировали в клетки HEK293T. Белок конструировали таким образом, чтобы он содержал сигнал стабилизации/тримеризации, фолдон, а также С-концевую (His)6-метку для очистки. (B) Очищенные белки HA анализировали с помощью SDS/PAGE. М: маркер молекулярной массы. Дорожка 1: белок НА H1N1 (A/Brisbane/59/2007); дорожка 2: белок НА H1N1 (A/California/07/2009); дорожка 3: белок НА H3N2 (Brisbane/10/2007); дорожка 4: белок НА H5N1 (Vietnam/1194/2004); дорожка 5: белок НА H7N9 (A/Shanghai/2/2013).
Фиг. 7 (А)-(F). Связывающие НА активности антисыворотки от мышей, вакцинированных cHAfg и cHAmg. Мышей BALB/c (n=10 на группу) иммунизировали с двухнедельными интервалами cHAfg или cHAmg с адъювантом Al(OH)3 или C34. Титры антител от мышей, вакцинированных cHAfg и cHAmg с адъювантом Al(OH)3 по сравнению с cHAfg и cHAmg с адъювантом C34, измеряли на 28-й день с помощью ELISA с белком HA H1N1 A/California/07/2009 (A), белком HA H1N1 A/Brisbane/59/2007 (B), белком HA H3N2 A/Brisbane/10/2007 (C), белком HA H5N1 A/Vietnam/1194/2004 (D), белком HA H7N9 A/Shanghai/2/2013 (E) и белком ствола HA (#4900) A/Brisbane/59/2007 (Bris/07) (F) в качестве покрывающего антигена. Конечный титр антител определялся как самое высокое разведение сыворотки, при котором оптическая плотность в 2,5 раза превышала оптическую плотность (OD) отрицательного контроля (неиммунная сыворотка). Данные анализировали с использованием двухфакторного дисперсионного анализа ANOVA от Prism; различия считались статистически значимыми при **P <0,01; ***Р <0,001. Данные представляют собой среднее значение ± SEM.
Фиг. 8 (А) и (В). Связывание ствол-реактивных антител (F10 IgG) с рекомбинантными H1, H5 и cHA. (A) Очищенный F10 анализировали с помощью SDS/PAGE. М: маркер молекулярной массы. Дорожка 1: антитело F10. (B) Аффинности связывания F10 IgG и различных HA измеряли с помощью ELISA. По оси х отложена концентрация различных белков НА, а по оси у показано значение поглощения при OD405 нм.
Фиг. 9 (А)-(D). Дозозависимые эффекты C34 на титры антител. Мышам BALB/c (n=10 в группе) с двухнедельными интервалами вводили 20 мкг cHA с 0,5 мкг, 2 мкг или 10 мкг адъюванта C34. Мышиную сыворотку собирали через две недели после второй (Д28) и третьей (Д42) иммунизаций. Титры антител измеряли с помощью ELISA с белками НА H1N1 A/California/07/2009 (A и C) и H5N1 Vietnam/1194/2004 (B и D). Значение P титров антител рассчитывали с использованием двухфакторного дисперсионного анализа ANOVA от Prism; различия считались статистически значимыми при *P <0,05; **Р <0,01. Данные представляют собой среднее значение ± SEM.
Фиг. 10 (А)-(С). Дозозависимые эффекты C34 на клетки, секретирующие антиген-специфические цитокины. Мышам BALB/c (n=5 в группе) с двухнедельными интервалами вводили 20 мкг очищенного cHA с добавленными в качестве адъюванта тремя различными дозами C34, составляющими 0,5, 2 и 10 мкг. Спленоциты мышей, иммунизированных сНА, получали после второй (Д28) и третьей (Д42) иммунизаций. Клетки, секретирующие (A) IFN-гамма и (B) IL4, оценивали с помощью анализа Elispot. (С) Количество продуцирующих гранзим В CD8+ Т-клеток в спленоцитах определяли с помощью анализа Elispot с использованием специфических пептидов. ***Р <0,001. Значение P рассчитывали с помощью программного обеспечения Prism с использованием двухфакторного дисперсионного анализа ANOVA.
Фиг. 11 (А)-(F). Масса тела мышей, вакцинированных cHAfg или cHAmg, зараженных вирусами H1N1 и H5N1 в летальной дозе. Изменения массы тела иммунизированных мышей, зараженных вирусами H1N1 A/California/07/2009 (A), H1N1 A/New Caledonia/1999 (B), H1N1 A/WSN/1933 (C), H1N1 A/Solomon Islands/03/2006 (D), H5N1 A/Vietnam/1194/2004 (E) или H5N1 A/Turkey/1/2005 (F), наблюдали в течение 14 дней после инфицирования. Изменение массы тела представлено как среднее значение ± SEM.
Подробное описание изобретения
На практике в настоящем изобретении будут использованы, если не указано иное, общепринятые методы молекулярной биологии, микробиологии, рекомбинантной ДНК и иммунологии, которые находятся в пределах компетенции специалистов в данной области. Такие методы подробно описаны в литературе. См., например, Molecular Cloning A Laboratory Manual, 2nd Ed., ed. By Sambrook, Fritsch and Maniatis (Cold Spring Harbor Laboratory Press, 1989); DNA Cloning, Volumes I and II (D. N. Glover ed., 1985); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); the treatise, Methods in Enzymology (Academic Press, Inc., N.Y.); Gene Transfer Vectors For Mammalian Cells (J. H. Miller and M. P. Calos eds., 1987, Cold Spring Harbor Laboratory); Methods In Enzymology, Vols. 154 and 155 (Wu et al. eds.), Immunochemical Methods in Cell and Molecular Biology (Mayer and Walker, eds., Academic Press, London, 1987); Antibodies: A Laboratory Manual, by Harlow and Lane s (Cold Spring Harbor Laboratory Press, 1988); и Handbook of Experimental Immunology, Volumes I-IV (D. M. Weir and C. C. Blackwell, eds., 1986).
Определения
Используемые в описании и формуле изобретения формы единственного числа включают ссылки во множественном числе, если из контекста явно не следует иное. Например, термин «химерный трансмембранный рецептор» включает множество химерных трансмембранных рецепторов.
Используемые в настоящем документе термины «гемагглютинин» и «НА» относятся к любому гемагглютинину, известному специалистам в данной области. В некоторых вариантах осуществления гемагглютинин представляет собой гемагглютинин гриппа, такой как гемагглютинин гриппа А, гемагглютинин гриппа В или гемагглютинин гриппа С. Типичный гемагглютинин содержит домены, известные специалистам в данной области, включая сигнальный пептид, стеблевой домен, домен глобулярный головки, люминальный домен, трансмембранный домен и цитоплазматический домен.
Используемые в настоящем документе термины «полипептид стеблевого домена», «стеблевой домен HA», «полипептид стеблевого домена гемагглютинина вируса гриппа» и «стволовой домен HA» относятся к полипептиду, содержащему или состоящему из одной или нескольких полипептидных цепей, которые составляют стеблевой домен гемагглютинина вируса гриппа. Полипептид стеблевого домена может представлять собой одну полипептидную цепь, две полипептидные цепи или несколько полипептидных цепей.
Используемые в настоящем документе термины «полипептид домена головки гемагглютинина вируса гриппа», «домен головки гемагглютинина вируса гриппа», «домен глобулярной головки HA» и «домен головки HA» относятся к домену глобулярной головки полипептида гемагглютинина вируса гриппа.
Используемый в настоящем документе термин «антиген» определяется как любое вещество, способное вызывать иммунный ответ.
Используемый в настоящем документе термин «иммуногенность» относится к способности иммуногена, антигена или вакцины стимулировать иммунный ответ.
Используемый в настоящем документе термин «эпитоп» определяется как части молекулы антигена, которые контактируют с антигенсвязывающим сайтом антитела или Т-клеточного рецептора.
Используемый в настоящем документе термин «вакцина» относится к препарату, который содержит антиген, состоящий из целых болезнетворных организмов (убитых или ослабленных) или компонентов таких организмов, таких как белки, гликопротеины, пептиды, гликопептиды, гликолипиды, полисахариды, или любой их комбинации, которая используется для наделения иммунитетом против заболевания, вызываемого организмами. Вакцинные препараты могут быть природными, синтетическими или полученными с помощью технологии рекомбинантной ДНК.
Используемый в настоящем документе термин «антигенспецифический» относится к свойству клеточной популяции, такому как поставка конкретного антигена или фрагмента антигена, что приводит к специфической клеточной пролиферации.
«Эффективное количество» относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата.
«Терапевтически эффективное количество» вещества/молекулы по изобретению может варьироваться в зависимости от таких факторов, как состояние заболевания, возраст, пол и масса тела индивидуума, а также способности вещества/молекулы вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество также представляет собой такое количество, при котором любые токсические или вредные эффекты вещества/молекулы перевешиваются терапевтически полезными эффектами. «Профилактически эффективное количество» относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого профилактического результата. Как правило, но не обязательно, поскольку профилактическая доза используется у субъектов до появления или на ранней стадии заболевания, профилактически эффективное количество будет меньше, чем терапевтически эффективное количество.
Консенсусную последовательность ДНК птичьего гриппа H5 (pCHA5-II) использовали в качестве вакцины для введения мышам, и результаты показали наличие широкой защиты от различных подтипов H5 (Chen, M.W. et al. Broadly neutralizing DNA vaccine with specific mutation alters the antigenicity and sugar-binding activities of influenza hemagglutinin. Proc.Natl Acad. Sci. USA 108, 3510-3515 (2011)). В настоящем изобретении представлен дизайн и оценка различных химерных вакцин на основе наиболее распространенных последовательностей птичьего гриппа H5 и человеческого гриппа H1. Среди этих конструкций лучшей оказалась вакцина на основе химерного HA (cHA) с консенсусом H5 в качестве глобулярной головки и консенсусом H1 в качестве стебля, и было показано, что она вызывает сильные CD4+ и CD8+ Т-клеточные иммунные ответы. Удивительно, что вакцина на основе моногликозилированного cHA (cHAmg), содержащая только GlcNAc на каждом гликосайте, индуцировала больше стебель-специфических антител с более высокой антителозависимой клеточной цитотоксичностью (ADCC), лучшей нейтрализующей и более сильной кросс-протективной активностями против штаммов и подтипов H1, H3, H5 и H7. Кроме того, вакцина на основе cHAmg в сочетании с гликолипидным адъювантом, разработанным для переключения классов, дополнительно повышала эффективность вакцины за счет продукции большего количества IFN-гамма, IL-4 и CD8+ T-клеток памяти.
Химерный полипептид гемагглютинина (HA) вируса гриппа
Настоящее изобретение обеспечивает химерный полипептид гемагглютинина (HA) вируса гриппа, используемый в качестве иммуногена или вакцины для вызывания CD4+ и CD8+ T-клеточных иммунных ответов. Соответственно, химерный полипептид НА вируса гриппа может предупреждать заболевание, вызванное вирусом гриппа, у субъекта.
Химерный полипептид гемагглютинина (HA) вируса гриппа по настоящему изобретению содержит одну или несколько последовательностей стеблевого домена, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью стеблевого домена HA подтипа H1 (H1 HA) и/или HA подтипа H5 (H5 HA), слитых с одной или несколькими последовательностями домена глобулярной головки, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью HA подтипа H1 (H1 HA) или HA подтипа H5 (H5 HA).
Используемый в настоящем документе термин «гомология» относится к общему родству между полимерными молекулами, например, между молекулами нуклеиновых кислот (например, молекулами ДНК и/или молекулами РНК) и/или между молекулами полипептидов. Полимерные молекулы (например, молекулы нуклеиновых кислот (например, молекулы ДНК и/или молекулы РНК) и/или молекулы полипептидов), которые имеют общий пороговый уровень сходства или идентичности, определяемый выравниванием совпадающих остатков, называются гомологичными. Гомология представляет собой качественный термин, который описывает взаимоотношения между молекулами и может быть основан на количественном сходстве или идентичности. Сходство и идентичность представляют собой количественные термины, которые определяют степень совпадения последовательностей между двумя сравниваемыми последовательностями. В некоторых вариантах осуществления полимерные молекулы считаются «гомологичными» друг другу, если их последовательности являются идентичными или схожими по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99%.
В некоторых вариантах осуществления полипептид в соответствии с настоящим изобретением может содержать одну или несколько последовательностей, имеющих по меньшей мере 60% гомологию с консенсусной последовательностью H1 HA или H5 HA по сравнению с известными штаммами вируса гриппа человека и птичьего гриппа. В некоторых вариантах осуществления гомология составляет по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%. В некоторых вариантах осуществления последовательность стеблевого домена представляет собой N-концевой стеблевой сегмент H1 HA или С-концевой стеблевой сегмент H1 HA; N-концевой стеблевой сегмент H1 HA или C-концевой стеблевой сегмент H1+H5 HA последовательностей; или N-концевой стеблевой сегмент H5 НА или С-концевой стеблевой сегмент H1+H5 HA последовательностей.
В некоторых вариантах осуществления консенсусная последовательность стеблевого домена H1 HA и/или H5 HA содержит аминокислотную последовательность SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 9 или SEQ ID NO: 10.
SEQ ID NO: 1 (H1 STEM)
DTLCIGYHANNSTDTVDTVLEKNVTVTHSVNLLEDKHNGKL
SEQ ID NO: 2 (H1 STEM)
NTTCQTPKGAINTSLPFQNIHPITIGKCPKYVKSTKLRLATGLRNVPSIQSRGLFGAIAGFIEGGWTGMVDGWYGYHHQNEQGSGYAADLKSTQNAIDKITNKVNSVIEKMNTQFTAVGKEFNHLEKRIENLNKKVDDGFLDIWTYNAELLVLLENERTLDYHDSNVKNLYEKVRNQLKNNAKEIGNGCFEFYHKCDNTCMESVKNGTYDYPKYSEEAKLNREEIDGVKLESTRIYQ
SEQ ID NO: 5 (H1 STEM)
DTLCIGYHANNSTDTVDTVLEKNVTVTHSVNLLEDKHNGKL
SEQ ID NO: 6 (H1+H5 STEM)
NTTCQTPKGAINTSLPFQNIHPITIGKCPKYVKSTKLRLATGLRNVPSIQSRGLFGAIAGFIEGGWQGMVDGWYGYHHSNEQGSGYAADKESTQKAIDGVTNKVNSIIDKMNTQFEAVGREFNNLERRIENLNKKMEDGFLDVWTYNAELLVLMENERTLDFHDSNVKNLYDKVRLQLRDNAKELGNGCFEFYHKCDNECMESVRNGTYDYPQYSEEARLKREEISGV
SEQ ID NO: 9 (H5 STEM)
DQICIGYHANNSTEQVDTIMEKNVTVTHAQDILEKTHNGKL
SEQ ID NO: 10 (H5+H1 STEM)
NTKCQTPMGAINSSMPFHNIHPLTIGECPKYVKSNRLVLATGLRNSPQRERRRKKR GLFGAIAGFIEGGWTGMVDGWYGYHHQNEQGSGYAADLKSTQNAIDKITNKVNSVIEKMNTQFTAVGKEFNHLEKRIENLNKKVDDGFLDIWTYNAELLVLLENERTLDYHDSNVKNLYEKVRNQLKNNAKEIGNGCFEFYHKCDNTCMESVKNGTYDYPKYSEEAKLNREEIDGV
В одном варианте осуществления консенсусная последовательность домена глобулярной головки H1 HA или H5 HA содержит аминокислотную последовательность SEQ ID NO:3, SEQ ID NO:7 или SEQ ID NO:11.
SEQ ID NO: 3 (H5 GLOBULAR HEAD)
CDLDGVKPLILRDCSVAGWLLGNPMCDEFINVPEWSYIVEKANPANDLCYPGNFNDYEELKHLLSRINHFEKIQIIPKSSWSDHEASSGVSSACPYQGKSSFFRNVVWLIKKNSTYPTIKRSYNNTNQEDLLVLWGIHHPNDAAEQTRLYQNPTTYISVGTSTLNQRLVPKIATRSKVNGQSGRMEFFWTILKPNDAINFESNGNFIAPEYAYKIVKKGDSTIMKSELEYGNC
SEQ ID NO: 7 (H1 GLOBULAR HEAD)
CKLRGVAPLHLGKCNIAGWILGNPECESLSTASSWSYIVETSSSDNGTCYPGDFIDYEELREQLSSVSSFERFEIFPKTSSWPNHDSNKGVTAACPHAGAKSFYKNLIWLVKKGNSYPKLSKSYINDKGKEVLVLWGIHHPSTTADQQSLYQNADAYVFVGTSRYSKKFKPEIAIRPKVRDQEGRMNYYWTLVEPGDKITFEATGNLVVPRYAFAMERNAGSGIIISDTPVHDC
SEQ ID NO: 11 (H5 GLOBULAR HEAD)
CDLDGVKPLILRDCSVAGWLLGNPMCDEFINVPEWSYIVEKANPANDLCYPGNFNDYEELKHLLSRINHFEKIQIIPKSSWSDHEASSGVSSACPYQGKSSFFRNVVWLIKKNSTYPTIKRSYNNTNQEDLLVLWGIHHPNDAAEQTRLYQNPTTYISVGTSTLNQRLVPKIATRSKVNGQSGRMEFFWTILKPNDAINFESNGNFIAPEYAYKIVKKGDSTIMKSELEYGNC
В одном варианте осуществления химерный полипептид НА вируса гриппа содержит аминокислотную последовательность SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12.
SEQ ID NO: 4 (Химерный H5/1)
DTLCIGYHANNSTDTVDTVLEKNVTVTHSVNLLEDKHNGKLCDLDGVKPLILRDCSVAGWLLGNPMCDEFINVPEWSYIVEKANPANDLCYPGNFNDYEELKHLLSRINHFEKIQIIPKSSWSDHEASSGVSSACPYQGKSSFFRNVVWLIKKNSTYPTIKRSYNNTNQEDLLVLWGIHHPNDAAEQTRLYQNPTTYISVGTSTLNQRLVPKIATRSKVNGQSGRMEFFWTILKPNDAINFESNGNFIAPEYAYKIVKKGDSTIMKSELEYGNCNTTCQTPKGAINTSLPFQNIHPITIGKCPKYVKSTKLRLATGLRNVPSIQSRGLFGAIAGFIEGGWTGMVDGWYGYHHQNEQGSGYAADLKSTQNAIDKITNKVNSVIEKMNTQFTAVGKEFNHLEKRIENLNKKVDDGFLDIWTYNAELLVLLENERTLDYHDSNVKNLYEKVRNQLKNNAKEIGNGCFEFYHKCDNTCMESVKNGTYDYPKYSEEAKLNREEIDGVKLESTRIYQ
SEQ ID NO: 8 (Замена H1/5)
DTLCIGYHANNSTDTVDTVLEKNVTVTHSVNLLEDKHNGKLCKLRGVAPLHLGKCNIAGWILGNPECESLSTASSWSYIVETSSSDNGTCYPGDFIDYEELREQLSSVSSFERFEIFPKTSSWPNHDSNKGVTAACPHAGAKSFYKNLIWLVKKGNSYPKLSKSYINDKGKEVLVLWGIHHPSTTADQQSLYQNADAYVFVGTSRYSKKFKPEIAIRPKVRDQEGRMNYYWTLVEPGDKITFEATGNLVVPRYAFAMERNAGSGIIISDTPVHDCNTTCQTPKGAINTSLPFQNIHPITIGKCPKYVKSTKLRLATGLRNVPSIQSRGLFGAIAGFIEGGWQGMVDGWYGYHHSNEQGSGYAADKESTQKAIDGVTNKVNSIIDKMNTQFEAVGREFNNLERRIENLNKKMEDGFLDVWTYNAELLVLMENERTLDFHDSNVKNLYDKVRLQLRDNAKELGNGCFEFYHKCDNECMESVRNGTYDYPQYSEEARLKREEISGV
SEQ ID NO: 12 (Замена H5/1)
DQICIGYHANNSTEQVDTIMEKNVTVTHAQDILEKTHNGKLCDLDGVKPLILRDCSVAGWLLGNPMCDEFINVPEWSYIVEKANPANDLCYPGNFNDYEELKHLLSRINHFEKIQIIPKSSWSDHEASSGVSSACPYQGKSSFFRNVVWLIKKNSTYPTIKRSYNNTNQEDLLVLWGIHHPNDAAEQTRLYQNPTTYISVGTSTLNQRLVPKIATRSKVNGQSGRMEFFWTILKPNDAINFESNGNFIAPEYAYKIVKKGDSTIMKSELEYGNCNTKCQTPMGAINSSMPFHNIHPLTIGECPKYVKSNRLVLATGLRNSPQRERRRKKRGLFGAIAGFIEGGWTGMVDGWYGYHHQNEQGSGYAADLKSTQNAIDKITNKVNSVIEKMNTQFTAVGKEFNHLEKRIENLNKKVDDGFLDIWTYNAELLVLLENERTLDYHDSNVKNLYEKVRNQLKNNAKEIGNGCFEFYHKCDNTCMESVKNGTYDYPKYSEEAKLNREEIDGV
В некоторых вариантах осуществления для усиления иммуногенности один или несколько гликосайтов на HA являются моногликозилированными. Предпочтительно моногликозилированный НА имеет только N-ацетилглюкозамин (GlcNAc) на каждом гликосайте.
Химерный полипептид НА вируса гриппа может быть получен с помощью любого подходящего способа, многие из которых известны специалистам в данной области. Например, белки могут быть химически синтезированы или получены с использованием технологии рекомбинантной ДНК (например, в бактериальных клетках, в клеточной культуре (клетки млекопитающих, дрожжей или насекомых), в растениях или растительных клетках, или с помощью бесклеточных прокариотических или эукариотических систем экспрессии, с помощью других систем in vitro, и т.д.). Соответственно, настоящее изобретение обеспечивает рекомбинантный полинуклеотид, содержащий последовательность нуклеиновой кислоты, кодирующую полипептид по настоящему изобретению, и необязательно последовательность нуклеиновой кислоты, кодирующую сигнальный пептид. Настоящее изобретение обеспечивает вектор, содержащий рекомбинантный полинуклеотид по настоящему изобретению. Варианты осуществления полипептида по настоящему изобретению описаны в настоящем документе. В одном варианте осуществления сигнальный пептид содержит последовательность SEQ ID NO: 13 (MEKIVLLLAIVSLVKS) или SEQ ID NO: 14 (MKAILVVLLYTFATANA). Также обеспечена клетка-хозяин, содержащая вектор по настоящему изобретению.
Иммуногенные композиции
Иммуногенная композиция предпочтительно содержит по меньшей мере один фармацевтически приемлемый носитель и/или адъювант. В одном варианте осуществления адъювант представляет собой гликолипидный адъювант. Примеры адъюванта включают, но без ограничения, Al(OH)3, AlPO4, C34, сквален и QS21.
Химерный полипептид НА вируса гриппа по настоящему изобретению может быть составлен или введен в комбинации с одним или несколькими фармацевтически приемлемыми вспомогательными веществами. Иммуногенные/вакцинные композиции могут быть стерильными, апирогенными или одновременно стерильными и апирогенными. Общие соображения, касающиеся составления и/или изготовления фармацевтических агентов, таких как вакцинные композиции, можно найти, например, в Remington: The Science and Practice of Pharmacy 21st ed., Lippincott Williams & Wilkins, 2005 (включен в настоящий документ посредством ссылки в полном объеме).
Иммуногенную композицию вводят способом, совместимым с дозированной лекарственной формой, и в количестве, которое является терапевтически эффективным, протективным и иммуногенным. Вводимое количество зависит от субъекта, подвергаемого лечению, включая, например, способность иммунной системы индивидуума синтезировать антитела и, при необходимости, продуцировать клеточно-опосредованный иммунный ответ. Точные количества активного ингредиента, подлежащие введению, зависят от суждения практикующего врача. Однако подходящие диапазоны доз могут быть легко определены специалистом в данной области. Подходящие режимы для начального введения и бустерных доз также варьируются, но могут включать начальное введение, за которым следуют последующие введения. Дозировка вакцины также может зависеть от пути введения и варьируется в зависимости от размера хозяина.
Составы вакцинных композиций, описанных в настоящем документе, могут быть изготовлены с помощью любого способа, известного или впоследствии разработанного в области фармакологии. Как правило, такие подготовительные способы включают стадию приведения в ассоциацию активного ингредиента со вспомогательным веществом и/или одним или несколькими другими дополнительными ингредиентами, а затем, если это необходимо и/или желательно, разделение, формование и/или упаковку продукта в желаемую одно- или многодозовую единицу.
Применения
Некоторое время было известно, что цитотоксические Т-лимфоциты (CTL) могут обеспечивать иммунный ответ против штаммов вируса гриппа. Недавние исследования показали, что ответ CTL у человека может быть направлен на несколько эпитопов.
В настоящем документе обеспечены способы предупреждения заболевания, вызванного вирусом гриппа, у людей и других млекопитающих. Также обеспечен способ вызывания иммунного ответа у субъекта против вируса гриппа. Способ включает введение субъекту эффективного количества химерного полипептида НА вируса гриппа или иммуногенной композиции/вакцины по настоящему изобретению, тем самым вызывая у субъекта иммунный ответ, специфический к штаммам вируса гриппа (таким как штаммы и подтипы H1, H3, H5 и H7). Предпочтительно способы индуцируют CD4+ и CD8+ T-клеточные иммунные ответы. Более предпочтительно, способы индуцируют стебель-специфические антитела с более высокой антителозависимой клеточной цитотоксичностью (ADCC), лучшей нейтрализующей и более сильной кросс-протективной активностями против штаммов и подтипов H1, H3, H5 и H7. Эти способы также повышают эффективность вакцины за счет увеличения продукции IFN-гамма, IL-4 и CD8+ T-клеток памяти.
Титр антител у субъекта повышается после вакцинации. В иллюстративных аспектах иммунные композиции или вакцины по настоящему изобретению применяют для обеспечения профилактической защиты от гриппа. Профилактическая защита от гриппа может быть достигнута после введения вакцины или комбинированной вакцины по настоящему изобретению. Вакцины (включая комбинированные вакцины) можно вводить один, два, три, четыре или более раз, но вероятно будет достаточно однократного введения вакцины (необязательно с последующим однократным бустером). Возможно, потребуется скорректировать соответствующим образом дозирование.
Профилактически эффективная доза представляет собой терапевтически эффективную дозу, которая защищает от вируса гриппа на клинически приемлемом уровне. В некоторых вариантах осуществления терапевтически эффективная доза представляет собой дозу, указанную в листке-вкладыше для вакцины.
Химерный полипептид НА вируса гриппа или иммуногенную композицию/вакцину по настоящему изобретению можно вводить любым путем, который приводит к терапевтически эффективному результату. Указанные пути введения включают, но без ограничения, внутрикожное, внутримышечное и/или подкожное введение. В некоторых вариантах осуществления химерный полипептид НА вируса гриппа или иммуногенную композицию/вакцину по настоящему изобретению можно вводить внутримышечно или внутрикожно аналогично введению инактивированных вакцин, известных в данной области.
Данное изобретение не ограничивается в своем применении деталями конструкции и расположением компонентов, изложенных в следующем описании или проиллюстрированных на чертежах. Изобретение допускает другие варианты осуществления и может быть осуществлено на практике или реализовано различными способами.
Примеры
Способы
Вакцина и конструирование плазмиды. Все 102 полноразмерные последовательности HA из вирусов H1N1, доступные в начале 2009~2013 гг., загружали из базы данных NCBI и выравнивали с помощью алгоритма ClustalW из программы BioEdit. Наиболее консервативную аминокислоту в каждом положении выбирали для создания консенсусной последовательности H1. Консенсусную последовательность гемагглютинина H5 (pCHA5-II) генерировали, как описано ранее. Нуклеотидные последовательности консенсуса гемагглютинина H5 (pCHA5-II) и консенсуса H1 клонировали в вектор экспрессии pcDNA, и полученные плазмиды использовали в качестве матриц для замены и конструирования химерного HA. Замена H1/5 состоит из H1 в виде HA1 (аминокислота 1~327 из SEQ ID NO: 8) и H5 в виде HA2 (аминокислота 328~503 из SEQ ID NO: 8), что дает H1 в виде глобулярной головки и стебля H1+H5 (HA2). Замена H5/1 состоит из H5 в виде HA1 (аминокислота 1~330 из SEQ ID NO: 12) и H1 в виде HA2 (аминокислота 331~506 из SEQ ID NO: 12), что дает H5 в виде глобулярной головки и стебля H5+H1 (HA2). Для химерной конструкции H5/1 глобулярный головной домен состоит из аминокислотной последовательности между остатками C42 и C274 из SEQ ID NO: 4 (нумерация H3), и стеблевой участок состоит из частей субъединиц HA1 и HA2 (аминокислоты 1~41 из SEQ ID NO: 4 и 275~511 из SEQ ID NO: 4). Трансмембранный домен был заменен дополнительными остатками из последовательности тримеризации фибритина бактериофага Т4, сайта расщепления тромбином и (His)6-метки на С-конце НА. Обе последовательности ДНК консенсуса НА были оптимизированы для экспрессии путем использования кодонов, предпочтительных для человека, и различные области были амплифицированы с помощью PCR и впоследствии клонированы в вектор pcDNA для экспрессии. Кроме того, ген HA сезонного вируса гриппа H1N1 Brisbane/59/2007, пандемического H1N1 California/07/2009, H3N2 Brisbane/10/2007, H7N9 A/Shanghai/2/2013 и птичьего гриппа H5N1 Vietnam/1194/2004 также был оптимизирован, синтезирован и клонирован в вектор экспрессии pcDNA. Последовательности были подтверждены секвенированием ДНК и подготовлены в высоком качестве для экспрессии и очистки белка.
Экспрессия рекомбинантного секретируемого НА из экспрессируемых клеток. Эпителиальные клетки почек человека (HEK) 293T и HEK293S обычно поддерживали в среде DMEM (Gibco) с добавлением 10% фетальной бычьей сыворотки (Gibco). Для транзиентной трансфекции клетки 293T или 293S высевали в чашку диаметром 10 см (Nunc, Roskilde, Denmark) и все процедуры выполняли в соответствии с протоколом производителя. Вкратце, клетки 293T или 293S при 80% конфлюэнтности трансфицировали с помощью реагента для трансфекции Mirus TransIT®-LT1 (Mirus Bio) с использованием соотношения 3:1 реагента к плазмидной ДНК. Реагенты TransIT®-LT1 разбавляли Opti-MEM (Gibco) и смесь инкубировали в течение 5-20 минут при комнатной температуре. В раствор добавляли плазмидную ДНК и полностью смешивали с последующей инкубацией в течение 15-30 минут. Перед трансфекцией клеткам заменяли среду на свежую среду DMEM (Gibco) с добавлением 10% эмбриональной бычьей сыворотки. К клеткам добавляли комплекс реагента TransIT®-LT1/ДНК и инкубировали в течение 48 ч при 37°С. Экспрессию гемагглютинина подтверждали с помощью иммуноблотов с использованием антител против (his)6 (Qiagen) или специфических антител против гемагглютинина и вторичных антител, конъюгированных с пероксидазой хрена (HRP) (PerkinElmer).
Очистка рекомбинантных секретируемых гемагглютининов. Для экспрессии в клетках 293T человека получали pcDNA высокого качества, несущую представляющий интерес ген, и трансфицировали в клетки с помощью Mirus TransIT®-LT1 (Mirus Bio). Через 48 ч после трансфекции среду собирали и клетки очищали центрифугированием при 1000×g в течение 10 мин. Супернатант очищали на колонке для аффинной хроматографии Ni-NTA (никель-нитрилотриуксусная кислота) (GE Healthcare). Супернатанты загружали на колонку для аффинной хроматографии Ni-NTA, предварительно уравновешенную в 20 мМ Tris-HCl, pH 8,0, и 300 мМ NaCl. Несвязанные белки вымывали градиентом имидазола от 25 до 50 мМ в 20 мМ Tris-HCl pH 8,0 и 300 мМ NaCl (буфер А). Затем белок HA элюировали градиентом имидазола от 100 до 300 мМ в буфере А. Очищенные белки HA концентрировали на установке ультрафильтрации Amicon Ultrafiltration Unit (отсечка MW30K) (Millipore) в PBS, pH 7,4. Чистоту контролировали с помощью SDS-PAGE, и наличие белков подтверждали методом Вестерн-блоттинга с помощью анти-(his)6 антител (Qiagen) или специфических антител против гемагглютинина и вторичных антител, конъюгированных с пероксидазой хрена (PerkinElmer). Наконец, тримерную форму белков HA получали с использованием эксклюзионной колонки, колонки для гель-фильтрации Superdex 200 Increase 10/300 GL (GE Healthcare).
Получение моногликозилированных белков НА. Клетки HEK293S, дефицитные по N-ацетилглюкозаминилтрансферазе I, использовали для получения HA с высоким содержанием маннозы гликанов. Очищенный белок HA из клеток HEK293S обрабатывали Endo H (NEB) при 20°С в течение ночи с получением моногликозилированного HAmg. Соотношение белков к Endo H составляло 3 к 1 (масса/объем) для HA. Затем Endo H и моногликозилированный белок HA разделяли на колонке для гель-фильтрации Superdex 200 Increase 10/300 GL (GE Healthcare). Белки HAmg концентрировали на установке ультрафильтрации Amicon Ultrafiltration Unit (отсечка MW30K) (Millipore) в PBS, pH 7,4, и подтверждали с помощью SDS-PAGE и анализа LC-MS/MS.
Идентификация N-связанного гликозилирования на белках НА. Десять микрограмм белка анализировали на SDS-PAGE и готовили для расщепления в геле. Желаемые полосы белков иссекали острым скальпелем, разрезали кубиками на кусочки размером 1 мм и помещали в пробирки Эппендорфа объемом 1,3 мл. После двукратной промывки с помощью 500 мкл 25 мМ бикарбоната аммония в 50% ACN (ацетонитрил) в течение 3 мин кусочки геля высушивали с использованием испарителя SpeedVac (Thermo). Высушенные образцы восстанавливали добавлением 100 мкл 50 мМ дитиотреитола (DTT) в 25 мМ бикарбоната аммония (рН 8,5) при 37°С в течение 1 ч с последующим центрифугированием при 10000 g в течение 1 мин. Раствор удаляли, и образцы геля подвергали стадии алкилирования путем добавления 100 мкл 100 мМ йодацетамида (IAA) в 25 мМ бикарбоната аммония (pH 8,5) и инкубировали в темноте при комнатной температуре в течение 1 ч. После промывки 500 мкл 50% ацетонитрила в 25 мМ бикарбоната аммония (pH 8,5) и 500 мкл 100% ацетонитрила образцы центрифугировали при 10000 g в течение 1 мин и полностью удаляли надосадочную жидкость. Образцы геля высушивали на испарителе SpeedVac и повторно растворяли в 200 мкл 25 мМ бикарбоната аммония (pH 8,5). Затем образцы геля обрабатывали 0,5 мкг трипсина (Promega, Madison, WI, USA) и 1 мкг химотрипсина (Promega, Madison, WI, USA) в течение ночи. После разложения в течение ночи к образцам добавляли 100 мкл 50% ацетонитрила в 5% TFA. Образцы обрабатывали ультразвуком в течение 10 секунд, а затем останавливали на 10 секунд. Процессы повторяли 10 раз. Супернатант, содержащий смеси пептидов, удаляли из пробирок с образцами и переносили в новые пробирки. Процедуру повторяли дважды. Объединенные супернатанты высушивали в концентраторе SpeedVac и обрабатывали для анализа LC-MS/MS.
Измерение эндотоксина. Уровни эндотоксина определяли с использованием набора Pierce® LAL Chromogenic Endotoxin Quantitation Kit (Thermo Scientific). Образцы белка разводили в 10, 20, 100 и 1000 раз, а стандарты эндотоксина готовили в концентрациях 10, 5, 2,5, 1,25, 0,63, 0,31, 0,15 и 0 нг/мл. После уравновешивания микропланшета в нагревательном блоке в течение 10 минут при 37°С образцы белка или стандарты смешивали с ЛАЛ-реактивом (Limulus Amebocye Lasate, LAL) (конечный объем 100 мкл) (1:1) в лунках, свободных от эндотоксинов, при 37°С в течение 10 минут. В каждую лунку добавляли по 100 мкл раствора субстрата и планшеты инкубировали при 37°С в течение 6 мин. Реакцию останавливали добавлением 50 мкл стоп-реагента (25% уксусная кислота). Поглощение лунок измеряли при 405 нм с использованием SpectraMax M5 (Molecular Devices, Sunnyvale, CA, USA). Стандартную кривую получали путем построения графика зависимости поглощения от соответствующих концентраций стандартов. Стандартную кривую использовали для определения концентрации эндотоксина в образцах. Значения эндотоксина всех очищенных белков составили <0,5 нг/мл.
Вакцинация мышей. Адъювант С34 химически синтезировали, как описано, и растворяли в DMSO. Самок мышей BALB/c в возрасте от 6 до 8 недель (n=10 на группу) иммунизировали внутримышечно 20 мкг очищенных химерных белков HAfg или HAmg в PBS, pH 7,4, и смешанных с 50 мкг гидроксида алюминия (Alum; Sigma) или 2 мкг C34. Контрольным мышам вводили фосфатно-солевой буферный раствор (PBS). Делали три прививки с двухнедельными интервалами. Кровь собирали через 14 дней после второй или третьей иммунизации. Кровь инкубировали при 37°С в течение 30 минут и центрифугировали при 12000 об/мин в течение 10 минут для сбора сыворотки. HA-специфические антитела в сыворотке, собранной от вакцинированных мышей, оценивали с помощью твердофазного иммуноферментного анализа (ELISA) и анализа нейтрализации.
Определение HA-специфических антител с помощью ELISA. Титры НА-специфических антител определяли с помощью ELISA с использованием белков НА H1N1 A/Brisbane/59/2007, H1N1 A/California/07/2009, H3N2 Brisbane/10/2007, H7N9 A/Shanghai/2/2013 и H5N1 Vietnam/1194/2004 в качестве субстратов. Планшет для ELISA на 96 лунок (Greiner bio-one, Frickenhausen, Germany) покрывали 100 мкл белка, разведенного в покрывающем буфере для ELISA, 100 мМ бикарбоната натрия (pH 8,8) в концентрации 5 мкг/мл на лунку, и накрывали герметично пластиковой крышкой при 4°С на ночь. После блокирования планшетов с помощью 1% BSA в TBST (137 мМ NaCl, 20 мМ Tris-основание, 0,05% Tween 20, pH 7,4) при 37°С в течение 1 ч и промывки 3 раза TBST, планшеты инкубировали с 200 мкл мышиной сыворотки в 2-кратных серийных разведениях при 37°С в течение 2 ч. После переноса сыворотки и промывки 6 раз планшета, измеряли HA-специфический IgG с использованием 200 мкл вторичного HRP-меченого антимышиного антитела (1:8000) (PerkinElmer, Waltham, MA, USA). После 1 ч инкубации при 37°С планшеты 6 раз промывали TBST и проявляли с помощью 100 мкл субстрата Super Aquablue ELISA (eBioscience, San Diego, CA, USA) в течение 1 мин. Реакцию останавливали добавлением 100 мкл 0,625 М щавелевой кислоты. Поглощение лунок измеряли при 405 нм с использованием SpectraMax M5 (Molecular Devices, Sunnyvale, CA, USA). Конечный титр антител определялся как самое высокое разведение сыворотки, при котором оптическая плотность в 2,5 раза превышала оптическую плотность (OD) отрицательного контроля (неиммунная сыворотка). Титр фоновых антител в конечной точке был определен как менее 1:50.
Сбор дендритных клеток костного мозга. Дендритные клетки костного мозга (BMDC), культивированные в присутствии GM-CSF, получали, как описано ранее. Вкратце, суспензии одиночных клеток костного мозга подвергали лизису RBC для удаления эритроцитов (RBC). Остальные клетки культивировали в 10 мл RPMI 1640 с добавлением 20 нг/мл мышиного GM-CSF (eBioscience), 10% FBS (BenchMark), 50 мкМ 2-ME, 100 единиц/мл пенициллина и 100 мкг/мл стрептомицина. Клетки высевали в каждую чашку Петри для достижения конечной плотности клеток 2 × 106 клеток/чашку Петри. Культуру пополняли, добавляя 10 мл свежей культуральной среды, содержащей 20 нг/мл мышиного GM-CSF, на 3-й день, и на 6-й день пополняли половиной объема полной культуральной среды, как описано выше. На 8-й день незрелые BMDC собирали путем сбора неприлипших клеток путем осторожного пипетирования, и повторно высевали клетки при плотности 106/мл. Для анализа CD8+ T-клеток незрелые BMDC совместно культивировали с CD8+ T-клетками и химерными белками HA (0,1 мг/лунку в 100 мкл) в течение 48 часов. Количество продуцирующих гранзим В CD8+ Т-клеток определяли с помощью анализа методом проточной цитометрии после промывки.
Иммуноферментный анализ пятен (ELISpot). Планшеты ELISPOT покрывали антимышиным IFN-гамма, IL-4 (Mabtech AB, Stockholm, Sweden) или гранзимом B (R&D Systems) в соответствии с инструкциями производителя. Планшеты промывали четыре раза и инкубировали в течение 30 мин с RPMI-1640 с добавлением 10% фетальной бычьей сыворотки (Gibco). Для обнаружения клеток, секретирующих IFN-гамма, IL-4 и гранзим B, у мышей, иммунизированных химерой, собирали спленоциты и культивировали при плотности 5×105 на лунку при 37°С в 5% CO2 в течение 24 ч со специфическими пептидами из HA для повторной стимуляции. Клетки удаляли и инкубировали с биотинилированным специфическим антителом против мышиного IFN-гамма, IL-4 (Mabtech AB) или гранзима B (R&D Systems). Планшеты промывали пять раз перед добавлением конъюгата стрептавидин-ALP и проявляли готовым к использованию субстратом BCIP/NPT. После высушивания количество образовавшихся пятен анализировали с помощью считывателя Immune Spot Reader (Cellular Technology Ltd.). Данные получали из лунок в трех повторностях.
Анализ нейтрализации. Супернатант клеточной культуры, содержащий 100 TCID50 вируса, смешивали с равным объемом двукратно серийно разбавленной сыворотки и инкубировали при 37°С в течение 1 ч. Затем смеси добавляли к клеткам MDCK в каждую лунку 96-луночного планшета и инкубировали при 37°С в течение 3 дней. К клеткам добавляли 30 мкл CellTiter-Glo (Promega) для определения количества жизнеспособных клеток на основе количественного определения присутствующего ATP. Нейтрализующую активность сыворотки определяли как максимальную кратность разведения, которая значительно защищала клетки от гибели, вызванной вирусом.
Анализ микронейтрализации. Инфекционную среду (DMEM с добавлением 0,3% BSA, 2 мкг/мл TPCK-трипсина), содержащую вирус при 100 TCID50, смешивали в равном объеме с двукратными серийными разведениями сыворотки и инкубировали при 37°С в течение 1 ч. Затем смесь добавляли к клеткам MDCK (1,5×104 клеток на лунку) в каждую лунку 96-луночного планшета и инкубировали при 37°С в течение 16-20 ч. Клетки промывали PBS, фиксировали в растворе ацетон/метанол (об./об. 1:1) и блокировали 5% обезжиренным молоком. Через 1 ч инкубации при 37°С лунки 6 раз промывали PBST, и титр вируса определяли с использованием 100 мкл mAb к белку NP вируса гриппа А (1:2500). После 1 ч инкубации при 37°С лунки 6 раз промывали PBST и добавляли 100 мкл вторичного HRP-меченого антикроличьего антитела (1:5000) (PerkinElmer, Waltham, MA, USA). После 1 ч инкубации при 37°С лунки снова промывали 6 раз PBST и проявляли с помощью 50 мкл субстрата 1-Step Ultra TMB (Thermo) в течение 1 мин. Реакцию останавливали добавлением 50 мкл 1 М H2SO4. Поглощение лунок измеряли при 450 нм на приборе SpectraMax M5 (Molecular Devices, Sunnyvale, CA, USA).
Репортерный анализ антителозависимой клеточно-опосредованной цитотоксичности. Клетки MDCK (1×104 клеток на лунку) в каждой лунке 96-луночного плоскодонного планшета инкубировали при 37°С в течение 24 ч. На следующий день 1×104 клеток MDCK инфицировали вирусами гриппа при множественности заражения (MOI) 1 в течение 24 ч. Затем эту среду заменяли средой Roswell Park Memorial Institute (RPMI) 1640, дополненной 4% сывороткой с низкой концентрацией IgG, с последующим добавлением серийных разведений антисыворотки от мышей, вакцинированных химерным белком HA, и инкубировали при 37°С в течение 30 минут. Эффекторные клетки Jurkat, экспрессирующие мышиный FcγRIII (Promega), суспендировали в среде RPMI 1640, содержащей 4% FBS с низкой концентрацией IgG, и к инфицированным клеткам MDCK добавляли клетки-мишени и эффекторные клетки в соотношении 1:5. После инкубации при 37°С в течение 6 ч аналитические планшеты удаляли из инкубатора при 37°С и уравновешивали в течение 15 минут при температуре окружающей среды, после чего добавляли буфер Bio-Glo™ Luciferase Assay Buffer (Promega) в соотношении 1:1. Люминесценцию измеряли на планшет-ридере CLARIOstar.
Эксперименты с заражением вирусом. Через две недели после трех вакцинаций с двухнедельными интервалами иммунизированных мышей интраназально заражали 10 LD50 (дозы вируса, приводящие к 50% гибели мышей) H1N1 California/07/2009, H1N1 A/New Caledonia/1999, H1N1 A/WSN/1933, H1N1 A/Solomon Islands/03/2006 и реассортантный вирус H5N1 A/Vietnam/1194/2004/NIBRG14 и H5N1 A/Turkey/1/2005/NIBRG23. После инфицирования мышей наблюдали ежедневно в течение 14 дней, и регистрировали выживаемость и массу тела. Процент от массы тела рассчитывали для каждого отдельного животного в группе путем сравнения ежедневной массы с массой до заражения, и мышей, потерявших более 25% от своей исходной массы, умерщвляли и оценивали как мертвых. Исследования на мышах были одобрены Институциональным комитетом по уходу и использованию животных (Institutional Animal Care and Use Committee of Academia Sinica). Все эксперименты на животных проводились в условиях уровня биобезопасности 3.
Экспрессия и очистка рекомбинантных антител F10. Плазмиду, кодирующую антитело F10, трансфицировали в клетки линии 293F, адаптированные к бессывороточной среде FreeStyle™, с использованием полиэтиленимина и культивировали в экспрессионной среде FreeStyleTM 293 (Gibco) в стерильных колбах Эрленмейера объемом 125 мл, вращающихся со скоростью 135 об/мин на платформе орбитального шейкера. Супернатант собирали через 72 ч после трансфекции и клетки осветляли центрифугированием при 1000×g в течение 10 мин. Супернатанты загружали на колонку Protein-A (GE Healthcare), предварительно уравновешенную 5 объемами колонки (CV) промывочного буфера, забуференного фосфатом (PBS) (pH 7,0), с последующими 5 CV промывочного буфера. Антитело F10 элюировали 0,2 М глициновым буфером (рН 2,5) и фракции собирали в пробирки, содержащие 0,5 мл 1 М Tris-HCl, рН 9,0, для нейтрализации. Чистоту контролировали с помощью SDS-PAGE.
Статистический анализ. Эксперименты на животных, используемые для оценки иммунных ответов, повторяли по меньшей мере три раза (n = 5 на группу), и исследования с заражением вирусом проводили по меньшей мере дважды (n = 10 на группу). Ответ каждой мыши подсчитывали как отдельную точку данных для статистического анализа. Данные, полученные в исследованиях на животных, анализировали с использованием двухфакторного дисперсионного анализа ANOVA от Prism; данные представляли как среднее значение ± SEM, а различия считались значимыми при *P <0,05; **Р <0,01; ***Р <0,001.
Пример 1. Получение и характеристика моногликозилированного химерного НА
Для разработки универсальной вакцины сначала цель состояла в получении вакцины с широкой защитой от вируса гриппа А группы 1 (H1 и H5 являются основными подтипами, тогда как H2, H6 и H9 являются второстепенными). Таким образом, последовательности НА из вирусов H1N1, доступные с начала 2009 по 2013 год, использовали для создания консенсусной последовательности H1. Консенсус H5 и консенсус H1 затем использовали в качестве матриц для разработки вакцины. При репликации вируса гриппа предшественник НА (НА0) протеолитически расщепляется на две субъединицы, НА1 и НА2; субъединица НА1 несет сайт связывания с 5-N-ацетилнейраминовой кислотой (сиаловой кислотой), и субъединица НА2 отвечает за слияние вируса с клеточной мембраной хозяина (фиг. 5А). С другой стороны, HA можно разделить на два структурных домена, глобулярную головку и стебель, на основе трехмерной (3D) структуры. Стеблевая область содержит домен HA2, 36~50 N-концевых остатков и короткий участок C-конца домена HA1. Таким образом, были разработаны вакцины на основе различных комбинаций доменов из H1 и H5. Сначала создавали замену H1/5 (глобулярная головка H1 и [стебель H1+H5(HA2)]), замену H5/1 (глобулярная головка H5 и [стебель H5+H1(HA2)]) и химерный H5/1 (глобулярная головка H5 и стебель H1) для сравнения (фиг. 1A и фиг. 5A). Результат показал, что иммунизация с консенсусом H1N1 и заменой H1/5 не индуцировала кросс-протективные активности, а замена H5/1 и химерный H5/1 вызывали кросс-нейтрализующую активность против вирусов H1N1 и H5N1 (фиг. 1B). Далее исследовали, была ли эта кросс-протекция обусловлена ответом CD8+ T-клеток, и обнаружили, что гранзим B больше секретировался у мышей, иммунизированных химерным H5/1, на основании чего можно предположить, что химерная вакцина H5/1 индуцировала более сильный ответ CD8+ Т-клеток по сравнению с вакциной на основе замены H5/1 (фиг. 1C).
Пример 2. Эффект гликозилирования на иммунный ответ химерного H5/1 (cHA)
Для изучения иммуногенности вакцины на основе химерного H5/1 (cHA) с различными состояниями гликозилирования, сравнивали вакцину на основе моногликозилированного cHA (cHAmg) и полностью гликозилированного cHA (cHAfg) (фиг. 5). Известно, что эндогликозидаза (Endo-H) является специфической для гликанов с высоким содержанием маннозы, но не для гликанов сложного типа. Гликопротеин HA, экспрессируемый в клетках HEK293S, дефицитных по N-ацетилглюкозаминилтрансферазе I и продуцирующих гликопротеины с N-гликанами с высоким содержанием маннозы, обрабатывали Endo-H для расщепления N-гликанов до одного остатка GlcNAc. Для создания cHAmg, cHA получали из клеток человека (HEK293S), и очищенный cHA с гликанами с высоким содержанием маннозы обрабатывали Endo-H для удаления внешних частей N-гликанов с получением HA только с одним N-ацетилглюкозамином (GlcNAc), связанным с остатком аспарагина каждого гликосайта. После обработки Endo-H смесь подвергали гель-фильтрации для отделения Endo-H от тримерного cHAmg. После концентрирования белки cHAmg подвергали электрофорезу в полиакриламидном геле с додецилсульфатом натрия (SDS/PAGE) и жидкостной хроматографии-тандемной масс-спектрометрии (LC-MS/MS) для обеспечения чистоты и композиции гликанов (фиг. 5C). Поскольку НА вируса гриппа существует в виде тримера на поверхности вируса, выполняли гель-фильтрацию, чтобы подтвердить, что cHAfg и cHAmg существуют в виде тримера (> 200 кДа) (фиг. 5D). Авторы изобретения также создали другой полностью гликозилированный cHAfg из клеток человека (HEK293T) для сравнения (фиг. 5B), и культура клеток показала содержание cHAfg ~6 мг/л.
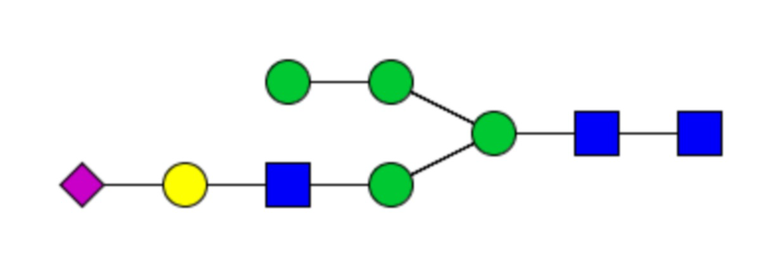
Сайты N-связанного гликозилирования и гликановый профиль рекомбинантных cHAfg и cHAmg анализировали с помощью LC-MS/MS, и было показано семь сайтов гликозилирования (N28, N40, N171, N182, N292, N303 и N497); N-гликаны cHAfg в основном были сложного типа, и cHAmg можно было получить в ~99% в виде одной гликоформы только с GlcNAc в каждом из его сайтов N-гликозилирования (фиг. 5E и таблица 1).
Таблица 1. N-связанные гликановые структуры cHA в полностью гликозилированных и моногликозилированных белках, проанализированные с помощью LC–MS/MS

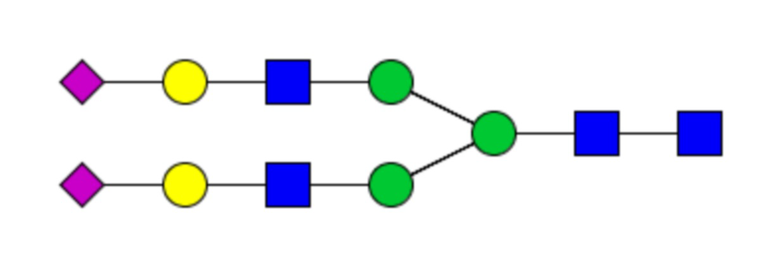
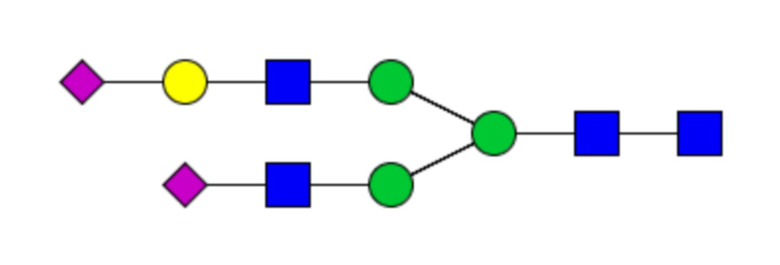
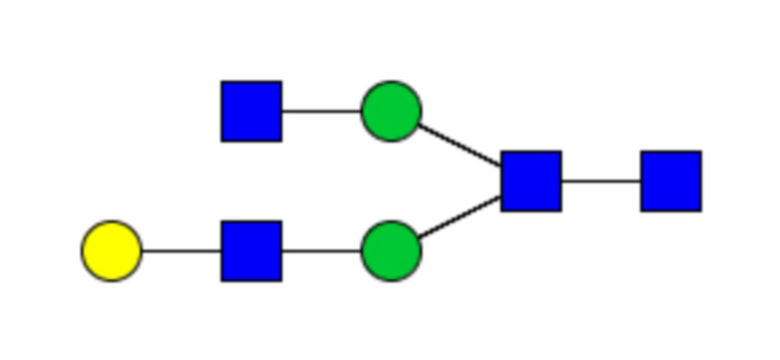
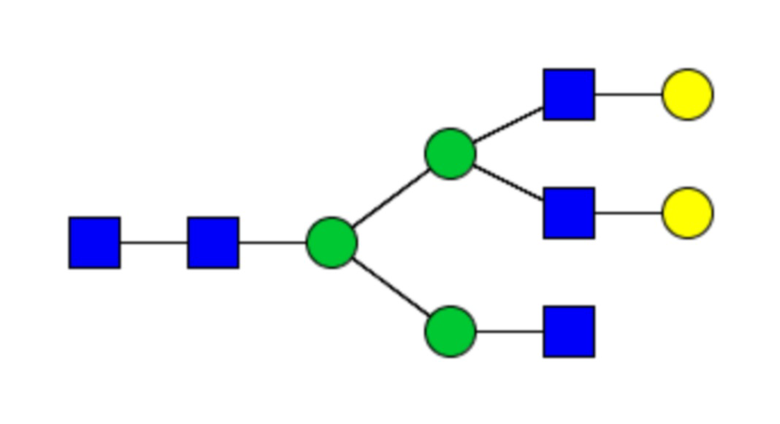
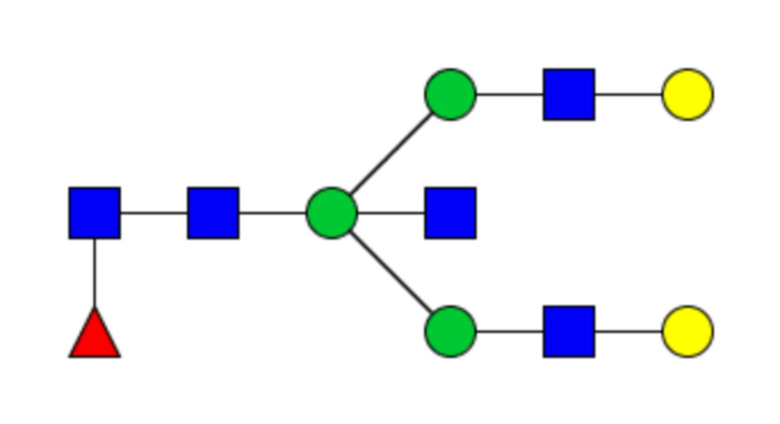
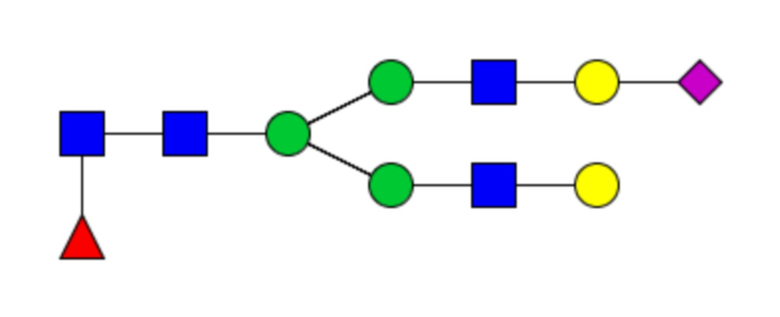
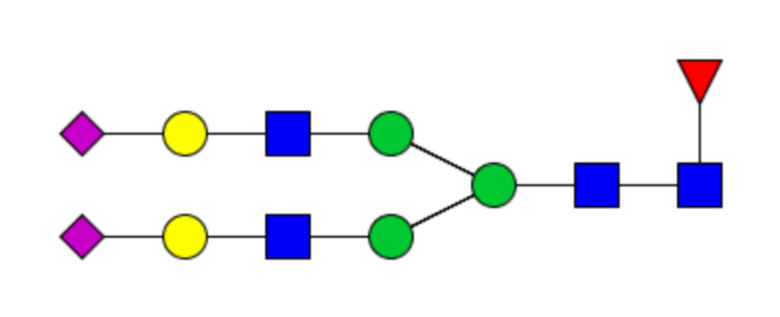
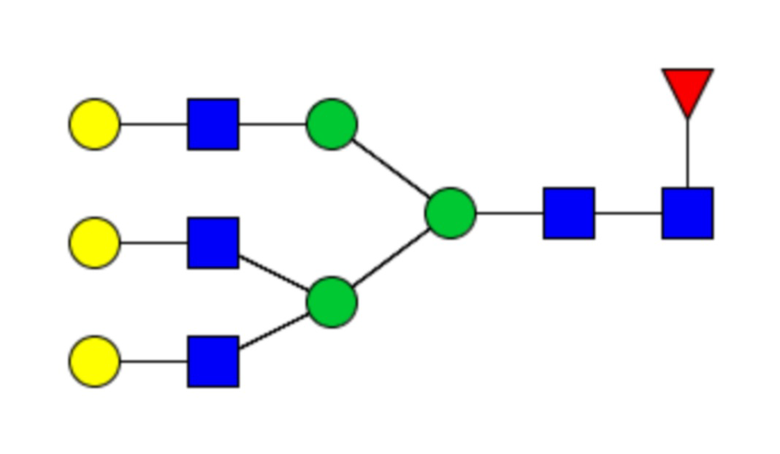
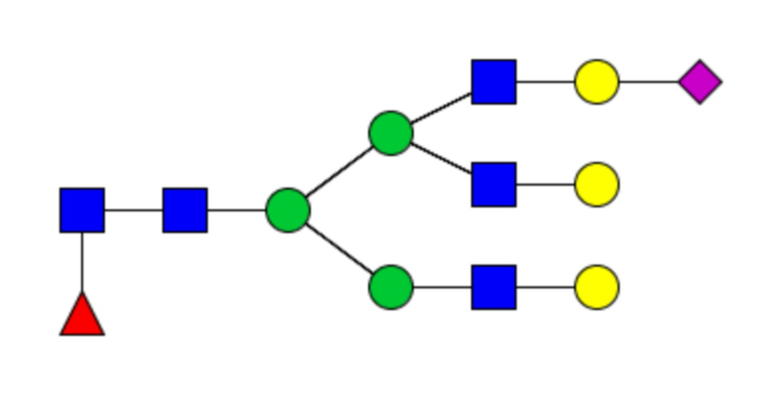
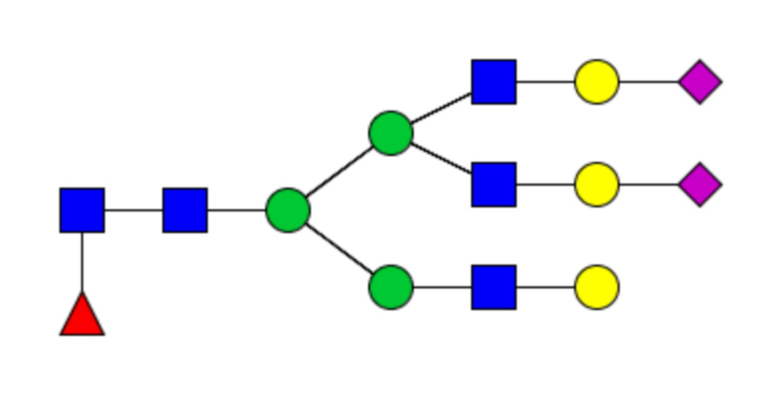
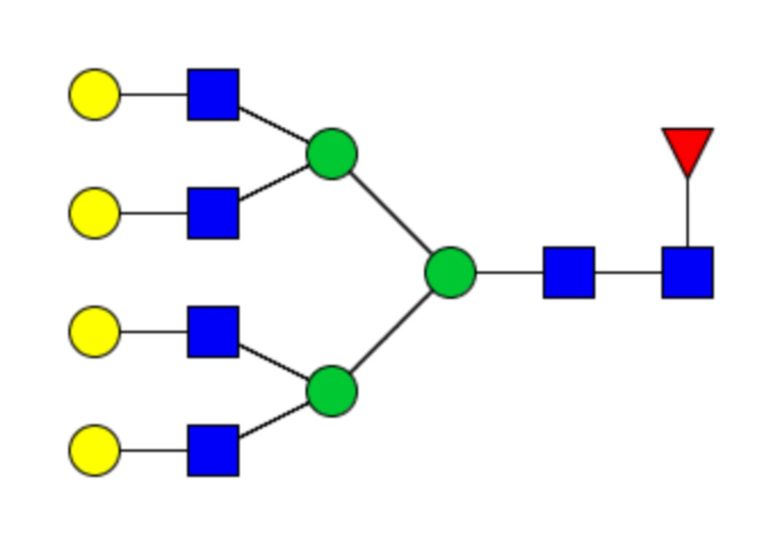
Пример 3. Кросс-реактивность антисывороток от мышей, иммунизированных полностью гликозилированным химерным H5/1 (cHAfg) и моногликозилированным химерным H5/1 (cHAmg)
Для оценки связывающей активности антитела, вызванной конструкциями cHA, мышей BALB/c внутримышечно иммунизировали 20 мкг белка cHAfg или cHAmg с адъювантом Al(OH)3 или C34, аналогом альфа-галактозилцерамида (альфа-GalCer). Мышей иммунизировали на 0, 2 и 4 неделе, и HA-индуцированную сыворотку получали на 28-й и 42-й день, и измеряли с помощью твердофазного иммуноферментного анализа (ELISA) с различными рекомбинантными HA (фиг. 6). По сравнению с максимальным разведением антисыворотки после двух иммунизаций, три иммунизации действительно продуцировали антисыворотку с более высокими титрами HA-специфических антител (фиг. 1 D-I и фиг. 7), и вакцинация cHAmg индуцировала лучший ответ антител по сравнению с cHAfg (фиг. 1 D, E и G). Кроме того, антисыворотка от cHAmg показала немного лучшее связывание с белками H3 и H7 HA (фиг. 1 F и H), и не наблюдалось существенных различий между адъювантами Al(OH)3 и C34. Эти данные указывают на то, что вакцина cHA может вызывать кросс-реактивные антитела, распознающие HA из штаммов H1N1, H3N2, H5N1, а также H7N9.
F10 представляет собой широко нейтрализующее IgG-антитело, которое, как известно, нацеливается на стеблевую область НА, которая является высококонсервативной среди различных подтипов вирусов гриппа. Чтобы сравнить связывание F10 с рекомбинантными H1, H5 и cHA, измеряли авидности связывания F10 с различными HA, и результаты показали, что F10 может связываться с белками H1, H5 и cHA (фиг. 8). Для исследования того, были ли F10-подобные антитела вызваны вакцинацией cHA, связывание cHA-индуцированной сыворотки со стеблем HA no. 4900 измеряли с помощью ELISA. Результаты показали, что вакцина cHAmg может индуцировать более высокий титр стебель-специфических антител, чем cHAfg (фиг. 1I и фиг. 7F), и лучшие результаты наблюдались при использовании вакцины cHA с адъювантом C34 (фиг. 1I), которая индуцировала большее количество стебель-специфических антител.
Пример 4. Вакцинация мышей cHAmg и адъювантом С34 вызывает сильные CD4+ и CD8+ Т-клеточные ответы и антителозависимые эффекторные функции, и нейтрализующие активности против вирусов H1, H3, H5, а также их подтипов
Помимо антитело-опосредованной нейтрализации, Fc-опосредованные эффекторные функции также играют важную роль в защите от инфекции, вызванной вирусом гриппа. Поэтому проводили исследование, будут ли антитела вызывать иммунный ответ, опосредованный Fc-рецептором. Анализ ADCC, адаптированный для мышей, проводили с использованием эффекторных клеток Jurkat, экспрессирующих FcγRIII, для оценки активностей ADCC сывороток от мышей, иммунизированных cHAfg и cHAmg (фиг. 2). Как и ожидалось, сыворотка от мышей, вакцинированных cHAfg или cHAmg, индуцировала сопоставимые уровни активностей ADCC против вирусов H5N1 NIBRG14 (A/Vietnam/1194/2004), NIBRG23 (A/Turkey/1/2005), RG5 (A/Anhui/1/2005) или RG2 (A/Indonesia/5/2005). Интересно, что лучшие активности ADCC наблюдались в группе cHAmg с адъювантом Al(OH)3 (фиг. 2B), и аналогичные результаты наблюдались в экспериментах против вирусов H1N1 A/California/07/2009, A/Brisbane/59/2007, A/Solomon Islands/3/2006, A/New Caledonia/20/1999 (фиг. 2A), H3N2 A/Wisconsin/67/2005 и A/Victoria/361/2011 (фиг. 2C).
Для оценки роли антиген-специфических цитокин-секретирующих клеток у cHA-иммунизированных мышей, спленоциты собирали после двух и трех иммунизаций, и клетки, секретирующие IFN-гамма, IL-4 и гранзим B (GzB), оценивали с помощью иммуноферментного анализа пятен (ELISpot) со специфическими пептидами из НА для стимуляции. Как показано на фиг. 3, вакцины cHAfg и cHAmg с адъювантом Al(OH)3 продуцировали сходные уровни цитокин-секретирующих клеток. Однако большее количество CD4+/IFN-гамма+ Th1 клеток (фиг. 3A), CD4+/IL-4+ Th2 (фиг. 3B) и CD8+ GzB-секретирующих клеток (фиг. 3C) было вызвано при вакцинации cHAmg с адъювантом C34, чем с Al(OH)3. Эти результаты подтвердили, что cHAmg с адъювантом C34 может стимулировать больший ответ CD4+ T-хелперов и более сильные цитотоксические эффекты CD8+ по сравнению с cHAfg.
Для оценки зависимости дозы С34 от титров антител и клеточно-опосредованного иммунитета мышей иммунизировали внутримышечно cHAfg с адъювантом С34 в трех различных дозах, равных 0,5, 2 и 10 мкг. Результат показал, что cHAfg с 2 мкг адъюванта C34 индуцировал более высокие титры, чем с 0,5 и 10 мкг C34 после двух или трех иммунизаций (фиг. 9). Кроме того, вакцина cHAfg с 2 мкг адъюванта C34 индуцировала больше IFN-гамма, чем с 0,5 и 10 мкг C34 (фиг. 10А), и 2 и 10 мкг C34 индуцировали больше IL-4, чем с 0,5 мкг C34 после трех иммунизаций (фиг. 10В). С другой стороны, не было различий в отношении увеличения CD8+ GzB-секретирующих клеток, когда вакцину cHAfg дополняли 0,5, 2 или 10 мкг C34 в качестве адъюванта после двух и трех иммунизаций (фиг. 10C). На основании этих наблюдений на протяжении всех экспериментов использовали 2 мкг C34.
Далее исследовали нейтрализующие активности cHA-индуцированных антисывороток. Было показано, что антисыворотки после вакцинации cHAmg обладали лучшими нейтрализующими активностями против гомологичных вирусов H1N1 A/California/07/2009 (фиг. 3D) и гетерологичных вирусов H5N1 NIBRG14 (A/Vietnam/1194/2004), NIBRG23 (A/Turkey/1/2005), RG5 (A/Anhui/1/2005) или RG2 (A/Indonesia/5/2005) (фиг. 3E). Кроме того, антисыворотки от мышей, вакцинированных cHAmg, проявили значительные нейтрализующие активности против гетерологичных вирусов H1N1 A/Brisbane/59/2007, A/New Caledonia/20/1999 и A/Solomon Islands/3/2006 (фиг. 3D). Антисыворотки от cHA-иммунизированных мышей явно были способны блокировать инфицирование вирусами H1N1 и H5N1, и нейтрализующая активность cHAmg в целом была лучше, чем cHAfg, особенно против гетерологичных вирусов.
Пример 5. Вакцинация мышей cHAmg/C34 обеспечивает кросс-протекцию от H1N1 и H5N1, а также их подтипов в исследовании с заражением.
Чтобы оценить, обеспечивает ли вакцинация cHAmg широкий кросс-протективный иммунитет против различных вирусов H1N1 и H5N1, вакцинированных мышей заражали путем интраназальной инокуляции летальными дозами нескольких вирусов H1N1 и H5N1, и протективную эффективность вакцины оценивали в течение 14 дней путем регистрации выживаемости и изменения массы тела (фиг. 4 и фиг. 11). Для мышей, зараженных вирусами H1N1 A/California/07/2009, все вакцины cHA обеспечивали 100% защиту (фиг. 4A). Кроме того, мыши, иммунизированные cHAmg с адъювантом C34, показали минимальные степени потери массы тела по сравнению с cHAfg (фиг. 11A). Мыши, иммунизированные cHAfg с адъювантом C34, дали только 30% защиту от заражений A/New Caledonia/1999; однако вакцина cHAmg с адъювантом C34 обеспечила 90% защиту от разных штаммов вирусов A/New Caledonia/1999, и аналогичные результаты наблюдались при вакцинации cHA с адъювантом Al(OH)3 (фиг. 4B). Для мышей, зараженных разными штаммами вирусов A/WSN/1933, все мыши, иммунизированные cHA с адъювантом Al(OH)3, выжили; однако мыши, иммунизированные CHAfg с адъювантом C34, дали только 80% защиту (фиг. 4C). Заражения летальными дозами также выполняли с использованием A/Solomon Islands/03/2006. Все мыши, иммунизированные сНА с адъювантом Al(OH)3, показали более низкую защиту; однако мыши, иммунизированные cHAmg с адъювантом С34, показали лучшую защиту от разных штаммов вирусов A/Solomon Islands/03/2006 (фиг. 4D). Что касается мышей, зараженных H5N1 NIBRG14 (A/Vietnam/1194/2004) и NIBRG23 (A/Turkey/1/2005), то все иммунизированные мыши выжили (фиг. 4 E и F). Также оценивали изменения массы тела после вирусных заражений (фиг. 11). Данные показали, что cHA эффективно вызывал значительный защитный иммунитет против различных вирусов H1N1 и H5N1, а cHAmg обеспечивал более широкую кросс-протективную способность по сравнению с cHAfg.
В настоящее время представляет интерес разработка универсальной вакцины против гриппа для обеспечения защиты от нескольких штаммов и подтипов вирусов гриппа, и эпитопы, используемые для разработки универсальной вакцины, включают высококонсервативный эктодомен M2, содержащий 24 негликозилированные аминокислоты, нуклеопротеин NP, и различные конструкции НА, которые, как было показано, индуцируют более высокие титры широко нейтрализующих антител для нацеливания на стеблевую область НА или блокирования проникновения вируса. Например, было показано, что вакцина на основе растворимого тримерного HA (мини-HA) с субъединицей повторно выровненного стебля полностью защищает мышей от летального заражения гетерологичными и гетеросубтипическими вирусами, и прайм-бустерная вакцинация «DNA prime-protein boost» химерным HA и подвергание той же стеблевой области и дивергентным экзотическим головным доменам вызывает широко протективные стебель-специфические антитела. Однако результат показал, что CD8+ Т-клетки не играют ключевой роли в кросс-протективных активностях. Хотя ДНК-вакцины являются многообещающими, они все еще находятся на ранней стадии разработки. В этом исследовании конструкции cHA, которые экспрессируют консенсус H5 глобулярной головки и консенсус H1 стеблевой области, были разработаны для имитации реального статуса передачи вируса гриппа от птичьего вируса к человеку. Для сравнения были подготовлены как полностью гликозилированный cHAfg, так и моногликозилированный cHAmg, и результат показал, что вакцина cHAmg вызывает более высокие титры кросс-реактивных антител против подтипов H1, H3, H5 и H7 (фиг. 1 D–H) через CD4+ и CD8+ Т-клеточные ответы (фиг. 3 A-C).
Было показано, что гликозилирование HA играет важную роль в фолдинге и стабильности белка, а также в модулировании его биологических активностей, включая защиту антигенных сайтов от нейтрализующих антител для снижения иммуногенности. Кроме того, гипергликозилированный HA был разработан для маскировки антигенных сайтов в высоко вариабельном головном домене и, таким образом, иммунный ответ был перенаправлен на консервативную стеблевую область. Согласно полученным результатам, нейтрализующие активности антисыворотки cHAmg значительно превосходили cHAfg-индуцированную антисыворотку, особенно против гетерологичного H1N1 A/Brisbane/59/2007, A/Solomon Islands/03/2006 и A/New Caledonia/20/1999 (фиг. 3D). Более широкие нейтрализующие активности вакцины cHAmg, вероятно, связаны с индукцией большего количества вариантов антител, как сообщалось ранее. IgG является преобладающим антителом, присутствующим у мышей, и является важным подтипом HA-специфических антител с высокой авидностью к рецептору FcγRIII на иммунных клетках для индукции ADCC. Было показано, что иммунизация cHAmg индуцировала более высокую ADCC и больше стебель-специфических антител с лучшей протективной активностью (фиг. 1I и 2), что согласуется с исследованиями, показывающими, что ADCC необходима для защиты от гриппа in vivo. Известно, что гидроксид алюминия (Alum) стимулирует ответ Th2 и был одобрен FDA для использования в качестве адъюванта вакцины; однако его механизм действия недостаточно изучен. Гликолипид C34 является лигандом и презентируется CD1d на дендритных клетках для взаимодействия с рецептором на инвариантных естественных клетках-киллерах T (iNKT), что приводит к стимуляции iNKT-клеток для продукции цитокинов Th1 (например, IFN-гамма) с адъювантным эффектом и цитокинов Th2 (например, IL-4) с активностью переключения классов. В полученных результатах количество секретирующих IFN-гамма (цитокин Th1), IL-4 (цитокин Th2) клеток и продуцирующих гранзим B CD8+ Т-клеток было значительно увеличено путем иммунизации cHAmg с адъювантом C34 по сравнению с Al(OH)3 (фиг. 3 А-С).
Таким образом, в настоящее время представляет интерес разработка вакцин против гриппа следующего поколения с широко-протективными иммунными ответами, и сообщалось о некоторых многообещающих результатах, что делает разработку универсальной вакцины вполне достижимой. Стремясь к достижению этой цели, в этом исследовании было успешно продемонстрировано доказательство того, что вакцина на основе моногликозилированного cHA с консенсусом головки H5 и консенсусом стебля H1 является эффективной вакциной против гриппа, проявляющей широкую протективную активность против гетерологичных вирусов гриппа, включая вирусы и подтипы H1, H3, H5 и H7 в исследовании нейтрализации, и H1N1, H5N1 и подтипы в исследовании с заражением. В связи с успехом в разработке широко протективной вакцины против различных штаммов и подтипов вируса гриппа А, нашей целью является применение стратегии, разработанной в данном исследовании, для разработки более широкой универсальной вакцины против вирусов гриппа А и В.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ACADEMIA SINICA
WONG, CHI-HUEY
<120> CHIMERIC INFLUENZA VACCINES
<130> 63/022,328
<160> 14
<170> PatentIn version 3.5
<210> 1
<211> 41
<212> PRT
<213> Influenza A virus
<400> 1
Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr Val
1 5 10 15
Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn Leu
20 25 30
Leu Glu Asp Lys His Asn Gly Lys Leu
35 40
<210> 2
<211> 237
<212> PRT
<213> Influenza A virus
<400> 2
Asn Thr Thr Cys Gln Thr Pro Lys Gly Ala Ile Asn Thr Ser Leu Pro
1 5 10 15
Phe Gln Asn Ile His Pro Ile Thr Ile Gly Lys Cys Pro Lys Tyr Val
20 25 30
Lys Ser Thr Lys Leu Arg Leu Ala Thr Gly Leu Arg Asn Val Pro Ser
35 40 45
Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu Gly
50 55 60
Gly Trp Thr Gly Met Val Asp Gly Trp Tyr Gly Tyr His His Gln Asn
65 70 75 80
Glu Gln Gly Ser Gly Tyr Ala Ala Asp Leu Lys Ser Thr Gln Asn Ala
85 90 95
Ile Asp Lys Ile Thr Asn Lys Val Asn Ser Val Ile Glu Lys Met Asn
100 105 110
Thr Gln Phe Thr Ala Val Gly Lys Glu Phe Asn His Leu Glu Lys Arg
115 120 125
Ile Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe Leu Asp Ile Trp
130 135 140
Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu Glu Asn Glu Arg Thr Leu
145 150 155 160
Asp Tyr His Asp Ser Asn Val Lys Asn Leu Tyr Glu Lys Val Arg Asn
165 170 175
Gln Leu Lys Asn Asn Ala Lys Glu Ile Gly Asn Gly Cys Phe Glu Phe
180 185 190
Tyr His Lys Cys Asp Asn Thr Cys Met Glu Ser Val Lys Asn Gly Thr
195 200 205
Tyr Asp Tyr Pro Lys Tyr Ser Glu Glu Ala Lys Leu Asn Arg Glu Glu
210 215 220
Ile Asp Gly Val Lys Leu Glu Ser Thr Arg Ile Tyr Gln
225 230 235
<210> 3
<211> 233
<212> PRT
<213> Influenza A virus
<400> 3
Cys Asp Leu Asp Gly Val Lys Pro Leu Ile Leu Arg Asp Cys Ser Val
1 5 10 15
Ala Gly Trp Leu Leu Gly Asn Pro Met Cys Asp Glu Phe Ile Asn Val
20 25 30
Pro Glu Trp Ser Tyr Ile Val Glu Lys Ala Asn Pro Ala Asn Asp Leu
35 40 45
Cys Tyr Pro Gly Asn Phe Asn Asp Tyr Glu Glu Leu Lys His Leu Leu
50 55 60
Ser Arg Ile Asn His Phe Glu Lys Ile Gln Ile Ile Pro Lys Ser Ser
65 70 75 80
Trp Ser Asp His Glu Ala Ser Ser Gly Val Ser Ser Ala Cys Pro Tyr
85 90 95
Gln Gly Lys Ser Ser Phe Phe Arg Asn Val Val Trp Leu Ile Lys Lys
100 105 110
Asn Ser Thr Tyr Pro Thr Ile Lys Arg Ser Tyr Asn Asn Thr Asn Gln
115 120 125
Glu Asp Leu Leu Val Leu Trp Gly Ile His His Pro Asn Asp Ala Ala
130 135 140
Glu Gln Thr Arg Leu Tyr Gln Asn Pro Thr Thr Tyr Ile Ser Val Gly
145 150 155 160
Thr Ser Thr Leu Asn Gln Arg Leu Val Pro Lys Ile Ala Thr Arg Ser
165 170 175
Lys Val Asn Gly Gln Ser Gly Arg Met Glu Phe Phe Trp Thr Ile Leu
180 185 190
Lys Pro Asn Asp Ala Ile Asn Phe Glu Ser Asn Gly Asn Phe Ile Ala
195 200 205
Pro Glu Tyr Ala Tyr Lys Ile Val Lys Lys Gly Asp Ser Thr Ile Met
210 215 220
Lys Ser Glu Leu Glu Tyr Gly Asn Cys
225 230
<210> 4
<211> 511
<212> PRT
<213> Artificial Sequence
<220>
<223> chimeric influenza virus HA polypeptide
<400> 4
Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr Val
1 5 10 15
Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn Leu
20 25 30
Leu Glu Asp Lys His Asn Gly Lys Leu Cys Asp Leu Asp Gly Val Lys
35 40 45
Pro Leu Ile Leu Arg Asp Cys Ser Val Ala Gly Trp Leu Leu Gly Asn
50 55 60
Pro Met Cys Asp Glu Phe Ile Asn Val Pro Glu Trp Ser Tyr Ile Val
65 70 75 80
Glu Lys Ala Asn Pro Ala Asn Asp Leu Cys Tyr Pro Gly Asn Phe Asn
85 90 95
Asp Tyr Glu Glu Leu Lys His Leu Leu Ser Arg Ile Asn His Phe Glu
100 105 110
Lys Ile Gln Ile Ile Pro Lys Ser Ser Trp Ser Asp His Glu Ala Ser
115 120 125
Ser Gly Val Ser Ser Ala Cys Pro Tyr Gln Gly Lys Ser Ser Phe Phe
130 135 140
Arg Asn Val Val Trp Leu Ile Lys Lys Asn Ser Thr Tyr Pro Thr Ile
145 150 155 160
Lys Arg Ser Tyr Asn Asn Thr Asn Gln Glu Asp Leu Leu Val Leu Trp
165 170 175
Gly Ile His His Pro Asn Asp Ala Ala Glu Gln Thr Arg Leu Tyr Gln
180 185 190
Asn Pro Thr Thr Tyr Ile Ser Val Gly Thr Ser Thr Leu Asn Gln Arg
195 200 205
Leu Val Pro Lys Ile Ala Thr Arg Ser Lys Val Asn Gly Gln Ser Gly
210 215 220
Arg Met Glu Phe Phe Trp Thr Ile Leu Lys Pro Asn Asp Ala Ile Asn
225 230 235 240
Phe Glu Ser Asn Gly Asn Phe Ile Ala Pro Glu Tyr Ala Tyr Lys Ile
245 250 255
Val Lys Lys Gly Asp Ser Thr Ile Met Lys Ser Glu Leu Glu Tyr Gly
260 265 270
Asn Cys Asn Thr Thr Cys Gln Thr Pro Lys Gly Ala Ile Asn Thr Ser
275 280 285
Leu Pro Phe Gln Asn Ile His Pro Ile Thr Ile Gly Lys Cys Pro Lys
290 295 300
Tyr Val Lys Ser Thr Lys Leu Arg Leu Ala Thr Gly Leu Arg Asn Val
305 310 315 320
Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile
325 330 335
Glu Gly Gly Trp Thr Gly Met Val Asp Gly Trp Tyr Gly Tyr His His
340 345 350
Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Leu Lys Ser Thr Gln
355 360 365
Asn Ala Ile Asp Lys Ile Thr Asn Lys Val Asn Ser Val Ile Glu Lys
370 375 380
Met Asn Thr Gln Phe Thr Ala Val Gly Lys Glu Phe Asn His Leu Glu
385 390 395 400
Lys Arg Ile Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe Leu Asp
405 410 415
Ile Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu Glu Asn Glu Arg
420 425 430
Thr Leu Asp Tyr His Asp Ser Asn Val Lys Asn Leu Tyr Glu Lys Val
435 440 445
Arg Asn Gln Leu Lys Asn Asn Ala Lys Glu Ile Gly Asn Gly Cys Phe
450 455 460
Glu Phe Tyr His Lys Cys Asp Asn Thr Cys Met Glu Ser Val Lys Asn
465 470 475 480
Gly Thr Tyr Asp Tyr Pro Lys Tyr Ser Glu Glu Ala Lys Leu Asn Arg
485 490 495
Glu Glu Ile Asp Gly Val Lys Leu Glu Ser Thr Arg Ile Tyr Gln
500 505 510
<210> 5
<211> 41
<212> PRT
<213> Influenza A virus
<400> 5
Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr Val
1 5 10 15
Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn Leu
20 25 30
Leu Glu Asp Lys His Asn Gly Lys Leu
35 40
<210> 6
<211> 228
<212> PRT
<213> Artificial Sequence
<220>
<223> stem domain consensus sequence of H1 HA and H5 HA
<400> 6
Asn Thr Thr Cys Gln Thr Pro Lys Gly Ala Ile Asn Thr Ser Leu Pro
1 5 10 15
Phe Gln Asn Ile His Pro Ile Thr Ile Gly Lys Cys Pro Lys Tyr Val
20 25 30
Lys Ser Thr Lys Leu Arg Leu Ala Thr Gly Leu Arg Asn Val Pro Ser
35 40 45
Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu Gly
50 55 60
Gly Trp Gln Gly Met Val Asp Gly Trp Tyr Gly Tyr His His Ser Asn
65 70 75 80
Glu Gln Gly Ser Gly Tyr Ala Ala Asp Lys Glu Ser Thr Gln Lys Ala
85 90 95
Ile Asp Gly Val Thr Asn Lys Val Asn Ser Ile Ile Asp Lys Met Asn
100 105 110
Thr Gln Phe Glu Ala Val Gly Arg Glu Phe Asn Asn Leu Glu Arg Arg
115 120 125
Ile Glu Asn Leu Asn Lys Lys Met Glu Asp Gly Phe Leu Asp Val Trp
130 135 140
Thr Tyr Asn Ala Glu Leu Leu Val Leu Met Glu Asn Glu Arg Thr Leu
145 150 155 160
Asp Phe His Asp Ser Asn Val Lys Asn Leu Tyr Asp Lys Val Arg Leu
165 170 175
Gln Leu Arg Asp Asn Ala Lys Glu Leu Gly Asn Gly Cys Phe Glu Phe
180 185 190
Tyr His Lys Cys Asp Asn Glu Cys Met Glu Ser Val Arg Asn Gly Thr
195 200 205
Tyr Asp Tyr Pro Gln Tyr Ser Glu Glu Ala Arg Leu Lys Arg Glu Glu
210 215 220
Ile Ser Gly Val
225
<210> 7
<211> 234
<212> PRT
<213> Influenza A virus
<400> 7
Cys Lys Leu Arg Gly Val Ala Pro Leu His Leu Gly Lys Cys Asn Ile
1 5 10 15
Ala Gly Trp Ile Leu Gly Asn Pro Glu Cys Glu Ser Leu Ser Thr Ala
20 25 30
Ser Ser Trp Ser Tyr Ile Val Glu Thr Ser Ser Ser Asp Asn Gly Thr
35 40 45
Cys Tyr Pro Gly Asp Phe Ile Asp Tyr Glu Glu Leu Arg Glu Gln Leu
50 55 60
Ser Ser Val Ser Ser Phe Glu Arg Phe Glu Ile Phe Pro Lys Thr Ser
65 70 75 80
Ser Trp Pro Asn His Asp Ser Asn Lys Gly Val Thr Ala Ala Cys Pro
85 90 95
His Ala Gly Ala Lys Ser Phe Tyr Lys Asn Leu Ile Trp Leu Val Lys
100 105 110
Lys Gly Asn Ser Tyr Pro Lys Leu Ser Lys Ser Tyr Ile Asn Asp Lys
115 120 125
Gly Lys Glu Val Leu Val Leu Trp Gly Ile His His Pro Ser Thr Thr
130 135 140
Ala Asp Gln Gln Ser Leu Tyr Gln Asn Ala Asp Ala Tyr Val Phe Val
145 150 155 160
Gly Thr Ser Arg Tyr Ser Lys Lys Phe Lys Pro Glu Ile Ala Ile Arg
165 170 175
Pro Lys Val Arg Asp Gln Glu Gly Arg Met Asn Tyr Tyr Trp Thr Leu
180 185 190
Val Glu Pro Gly Asp Lys Ile Thr Phe Glu Ala Thr Gly Asn Leu Val
195 200 205
Val Pro Arg Tyr Ala Phe Ala Met Glu Arg Asn Ala Gly Ser Gly Ile
210 215 220
Ile Ile Ser Asp Thr Pro Val His Asp Cys
225 230
<210> 8
<211> 503
<212> PRT
<213> Artificial Sequence
<220>
<223> chimeric influenza virus HA polypeptide
<400> 8
Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr Val
1 5 10 15
Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Val Asn Leu
20 25 30
Leu Glu Asp Lys His Asn Gly Lys Leu Cys Lys Leu Arg Gly Val Ala
35 40 45
Pro Leu His Leu Gly Lys Cys Asn Ile Ala Gly Trp Ile Leu Gly Asn
50 55 60
Pro Glu Cys Glu Ser Leu Ser Thr Ala Ser Ser Trp Ser Tyr Ile Val
65 70 75 80
Glu Thr Ser Ser Ser Asp Asn Gly Thr Cys Tyr Pro Gly Asp Phe Ile
85 90 95
Asp Tyr Glu Glu Leu Arg Glu Gln Leu Ser Ser Val Ser Ser Phe Glu
100 105 110
Arg Phe Glu Ile Phe Pro Lys Thr Ser Ser Trp Pro Asn His Asp Ser
115 120 125
Asn Lys Gly Val Thr Ala Ala Cys Pro His Ala Gly Ala Lys Ser Phe
130 135 140
Tyr Lys Asn Leu Ile Trp Leu Val Lys Lys Gly Asn Ser Tyr Pro Lys
145 150 155 160
Leu Ser Lys Ser Tyr Ile Asn Asp Lys Gly Lys Glu Val Leu Val Leu
165 170 175
Trp Gly Ile His His Pro Ser Thr Thr Ala Asp Gln Gln Ser Leu Tyr
180 185 190
Gln Asn Ala Asp Ala Tyr Val Phe Val Gly Thr Ser Arg Tyr Ser Lys
195 200 205
Lys Phe Lys Pro Glu Ile Ala Ile Arg Pro Lys Val Arg Asp Gln Glu
210 215 220
Gly Arg Met Asn Tyr Tyr Trp Thr Leu Val Glu Pro Gly Asp Lys Ile
225 230 235 240
Thr Phe Glu Ala Thr Gly Asn Leu Val Val Pro Arg Tyr Ala Phe Ala
245 250 255
Met Glu Arg Asn Ala Gly Ser Gly Ile Ile Ile Ser Asp Thr Pro Val
260 265 270
His Asp Cys Asn Thr Thr Cys Gln Thr Pro Lys Gly Ala Ile Asn Thr
275 280 285
Ser Leu Pro Phe Gln Asn Ile His Pro Ile Thr Ile Gly Lys Cys Pro
290 295 300
Lys Tyr Val Lys Ser Thr Lys Leu Arg Leu Ala Thr Gly Leu Arg Asn
305 310 315 320
Val Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe
325 330 335
Ile Glu Gly Gly Trp Gln Gly Met Val Asp Gly Trp Tyr Gly Tyr His
340 345 350
His Ser Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Lys Glu Ser Thr
355 360 365
Gln Lys Ala Ile Asp Gly Val Thr Asn Lys Val Asn Ser Ile Ile Asp
370 375 380
Lys Met Asn Thr Gln Phe Glu Ala Val Gly Arg Glu Phe Asn Asn Leu
385 390 395 400
Glu Arg Arg Ile Glu Asn Leu Asn Lys Lys Met Glu Asp Gly Phe Leu
405 410 415
Asp Val Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Met Glu Asn Glu
420 425 430
Arg Thr Leu Asp Phe His Asp Ser Asn Val Lys Asn Leu Tyr Asp Lys
435 440 445
Val Arg Leu Gln Leu Arg Asp Asn Ala Lys Glu Leu Gly Asn Gly Cys
450 455 460
Phe Glu Phe Tyr His Lys Cys Asp Asn Glu Cys Met Glu Ser Val Arg
465 470 475 480
Asn Gly Thr Tyr Asp Tyr Pro Gln Tyr Ser Glu Glu Ala Arg Leu Lys
485 490 495
Arg Glu Glu Ile Ser Gly Val
500
<210> 9
<211> 41
<212> PRT
<213> Influenza A virus
<400> 9
Asp Gln Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Glu Gln Val
1 5 10 15
Asp Thr Ile Met Glu Lys Asn Val Thr Val Thr His Ala Gln Asp Ile
20 25 30
Leu Glu Lys Thr His Asn Gly Lys Leu
35 40
<210> 10
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> stem domain consensus sequence of H1 HA and H5 HA
<400> 10
Asn Thr Lys Cys Gln Thr Pro Met Gly Ala Ile Asn Ser Ser Met Pro
1 5 10 15
Phe His Asn Ile His Pro Leu Thr Ile Gly Glu Cys Pro Lys Tyr Val
20 25 30
Lys Ser Asn Arg Leu Val Leu Ala Thr Gly Leu Arg Asn Ser Pro Gln
35 40 45
Arg Glu Arg Arg Arg Lys Lys Arg Gly Leu Phe Gly Ala Ile Ala Gly
50 55 60
Phe Ile Glu Gly Gly Trp Thr Gly Met Val Asp Gly Trp Tyr Gly Tyr
65 70 75 80
His His Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Leu Lys Ser
85 90 95
Thr Gln Asn Ala Ile Asp Lys Ile Thr Asn Lys Val Asn Ser Val Ile
100 105 110
Glu Lys Met Asn Thr Gln Phe Thr Ala Val Gly Lys Glu Phe Asn His
115 120 125
Leu Glu Lys Arg Ile Glu Asn Leu Asn Lys Lys Val Asp Asp Gly Phe
130 135 140
Leu Asp Ile Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu Glu Asn
145 150 155 160
Glu Arg Thr Leu Asp Tyr His Asp Ser Asn Val Lys Asn Leu Tyr Glu
165 170 175
Lys Val Arg Asn Gln Leu Lys Asn Asn Ala Lys Glu Ile Gly Asn Gly
180 185 190
Cys Phe Glu Phe Tyr His Lys Cys Asp Asn Thr Cys Met Glu Ser Val
195 200 205
Lys Asn Gly Thr Tyr Asp Tyr Pro Lys Tyr Ser Glu Glu Ala Lys Leu
210 215 220
Asn Arg Glu Glu Ile Asp Gly Val
225 230
<210> 11
<211> 233
<212> PRT
<213> Influenza A virus
<400> 11
Cys Asp Leu Asp Gly Val Lys Pro Leu Ile Leu Arg Asp Cys Ser Val
1 5 10 15
Ala Gly Trp Leu Leu Gly Asn Pro Met Cys Asp Glu Phe Ile Asn Val
20 25 30
Pro Glu Trp Ser Tyr Ile Val Glu Lys Ala Asn Pro Ala Asn Asp Leu
35 40 45
Cys Tyr Pro Gly Asn Phe Asn Asp Tyr Glu Glu Leu Lys His Leu Leu
50 55 60
Ser Arg Ile Asn His Phe Glu Lys Ile Gln Ile Ile Pro Lys Ser Ser
65 70 75 80
Trp Ser Asp His Glu Ala Ser Ser Gly Val Ser Ser Ala Cys Pro Tyr
85 90 95
Gln Gly Lys Ser Ser Phe Phe Arg Asn Val Val Trp Leu Ile Lys Lys
100 105 110
Asn Ser Thr Tyr Pro Thr Ile Lys Arg Ser Tyr Asn Asn Thr Asn Gln
115 120 125
Glu Asp Leu Leu Val Leu Trp Gly Ile His His Pro Asn Asp Ala Ala
130 135 140
Glu Gln Thr Arg Leu Tyr Gln Asn Pro Thr Thr Tyr Ile Ser Val Gly
145 150 155 160
Thr Ser Thr Leu Asn Gln Arg Leu Val Pro Lys Ile Ala Thr Arg Ser
165 170 175
Lys Val Asn Gly Gln Ser Gly Arg Met Glu Phe Phe Trp Thr Ile Leu
180 185 190
Lys Pro Asn Asp Ala Ile Asn Phe Glu Ser Asn Gly Asn Phe Ile Ala
195 200 205
Pro Glu Tyr Ala Tyr Lys Ile Val Lys Lys Gly Asp Ser Thr Ile Met
210 215 220
Lys Ser Glu Leu Glu Tyr Gly Asn Cys
225 230
<210> 12
<211> 506
<212> PRT
<213> Artificial Sequence
<220>
<223> chimeric influenza virus HA polypeptide
<400> 12
Asp Gln Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Glu Gln Val
1 5 10 15
Asp Thr Ile Met Glu Lys Asn Val Thr Val Thr His Ala Gln Asp Ile
20 25 30
Leu Glu Lys Thr His Asn Gly Lys Leu Cys Asp Leu Asp Gly Val Lys
35 40 45
Pro Leu Ile Leu Arg Asp Cys Ser Val Ala Gly Trp Leu Leu Gly Asn
50 55 60
Pro Met Cys Asp Glu Phe Ile Asn Val Pro Glu Trp Ser Tyr Ile Val
65 70 75 80
Glu Lys Ala Asn Pro Ala Asn Asp Leu Cys Tyr Pro Gly Asn Phe Asn
85 90 95
Asp Tyr Glu Glu Leu Lys His Leu Leu Ser Arg Ile Asn His Phe Glu
100 105 110
Lys Ile Gln Ile Ile Pro Lys Ser Ser Trp Ser Asp His Glu Ala Ser
115 120 125
Ser Gly Val Ser Ser Ala Cys Pro Tyr Gln Gly Lys Ser Ser Phe Phe
130 135 140
Arg Asn Val Val Trp Leu Ile Lys Lys Asn Ser Thr Tyr Pro Thr Ile
145 150 155 160
Lys Arg Ser Tyr Asn Asn Thr Asn Gln Glu Asp Leu Leu Val Leu Trp
165 170 175
Gly Ile His His Pro Asn Asp Ala Ala Glu Gln Thr Arg Leu Tyr Gln
180 185 190
Asn Pro Thr Thr Tyr Ile Ser Val Gly Thr Ser Thr Leu Asn Gln Arg
195 200 205
Leu Val Pro Lys Ile Ala Thr Arg Ser Lys Val Asn Gly Gln Ser Gly
210 215 220
Arg Met Glu Phe Phe Trp Thr Ile Leu Lys Pro Asn Asp Ala Ile Asn
225 230 235 240
Phe Glu Ser Asn Gly Asn Phe Ile Ala Pro Glu Tyr Ala Tyr Lys Ile
245 250 255
Val Lys Lys Gly Asp Ser Thr Ile Met Lys Ser Glu Leu Glu Tyr Gly
260 265 270
Asn Cys Asn Thr Lys Cys Gln Thr Pro Met Gly Ala Ile Asn Ser Ser
275 280 285
Met Pro Phe His Asn Ile His Pro Leu Thr Ile Gly Glu Cys Pro Lys
290 295 300
Tyr Val Lys Ser Asn Arg Leu Val Leu Ala Thr Gly Leu Arg Asn Ser
305 310 315 320
Pro Gln Arg Glu Arg Arg Arg Lys Lys Arg Gly Leu Phe Gly Ala Ile
325 330 335
Ala Gly Phe Ile Glu Gly Gly Trp Thr Gly Met Val Asp Gly Trp Tyr
340 345 350
Gly Tyr His His Gln Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Leu
355 360 365
Lys Ser Thr Gln Asn Ala Ile Asp Lys Ile Thr Asn Lys Val Asn Ser
370 375 380
Val Ile Glu Lys Met Asn Thr Gln Phe Thr Ala Val Gly Lys Glu Phe
385 390 395 400
Asn His Leu Glu Lys Arg Ile Glu Asn Leu Asn Lys Lys Val Asp Asp
405 410 415
Gly Phe Leu Asp Ile Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Leu
420 425 430
Glu Asn Glu Arg Thr Leu Asp Tyr His Asp Ser Asn Val Lys Asn Leu
435 440 445
Tyr Glu Lys Val Arg Asn Gln Leu Lys Asn Asn Ala Lys Glu Ile Gly
450 455 460
Asn Gly Cys Phe Glu Phe Tyr His Lys Cys Asp Asn Thr Cys Met Glu
465 470 475 480
Ser Val Lys Asn Gly Thr Tyr Asp Tyr Pro Lys Tyr Ser Glu Glu Ala
485 490 495
Lys Leu Asn Arg Glu Glu Ile Asp Gly Val
500 505
<210> 13
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> signal peptide
<400> 13
Met Glu Lys Ile Val Leu Leu Leu Ala Ile Val Ser Leu Val Lys Ser
1 5 10 15
<210> 14
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> signal peptide
<400> 14
Met Lys Ala Ile Leu Val Val Leu Leu Tyr Thr Phe Ala Thr Ala Asn
1 5 10 15
Ala
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ ГЕМАГГЛЮТИНИНА ГРИППА С ГЕТЕРОЛОГИЧНЫМИ ЭПИТОПАМИ И/ИЛИ ИЗМЕНЕННЫМИ САЙТАМИ РАСЩЕПЛЕНИЯ ПРИ СОЗРЕВАНИИ | 2017 |
|
RU2769406C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА СВИНОГО ГРИППА, СОДЕРЖАЩАЯ КОНСТРУКЦИЮ НУКЛЕИНОВОЙ КИСЛОТЫ, ИМЕЮЩУЮ ОПРЕДЕЛЕННЫЙ ПОРЯДОК РАСПОЛОЖЕНИЯ ГЕНОВ | 2021 |
|
RU2833693C1 |
| ВАКЦИНА ПРОТИВ ВИРУСА СВИНОГО ГРИППА А, СОДЕРЖАЩАЯ ДВЕ РАЗНЫЕ ЧАСТИЦЫ РНК-РЕПЛИКОНА | 2021 |
|
RU2839757C1 |
| ВАКЦИНА ПРОТИВ ВИРУСА СВИНОГО ГРИППА А, СОДЕРЖАЩАЯ КОНСТРУКЦИЮ НУКЛЕИНОВОЙ КИСЛОТЫ, СОДЕРЖАЩУЮ ПЕРВУЮ, ВТОРУЮ И ТРЕТЬЮ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВЫХ КИСЛОТ, КОДИРУЮЩИЕ РАЗНЫЕ АНТИГЕНЫ, НЕЙРАМИНИДАЗЫ, ВИРУСА | 2021 |
|
RU2841382C1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2683498C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2803850C2 |
| ВАКЦИНА К ВИРУСУ СВИНОГО ГРИППА A | 2018 |
|
RU2787596C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ВИРУСА ПТИЧЬЕГО ГРИППА, И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2761869C2 |
| ВАКЦИННЫЕ КОМПОЗИЦИИ, ХАРАКТЕРИЗУЮЩИЕСЯ УЛУЧШЕННОЙ СТАБИЛЬНОСТЬЮ И ИММУНОГЕННОСТЬЮ | 2016 |
|
RU2730625C2 |
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА, И ИХ КОМПОЗИЦИИ, КОМБИНАЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2720282C1 |
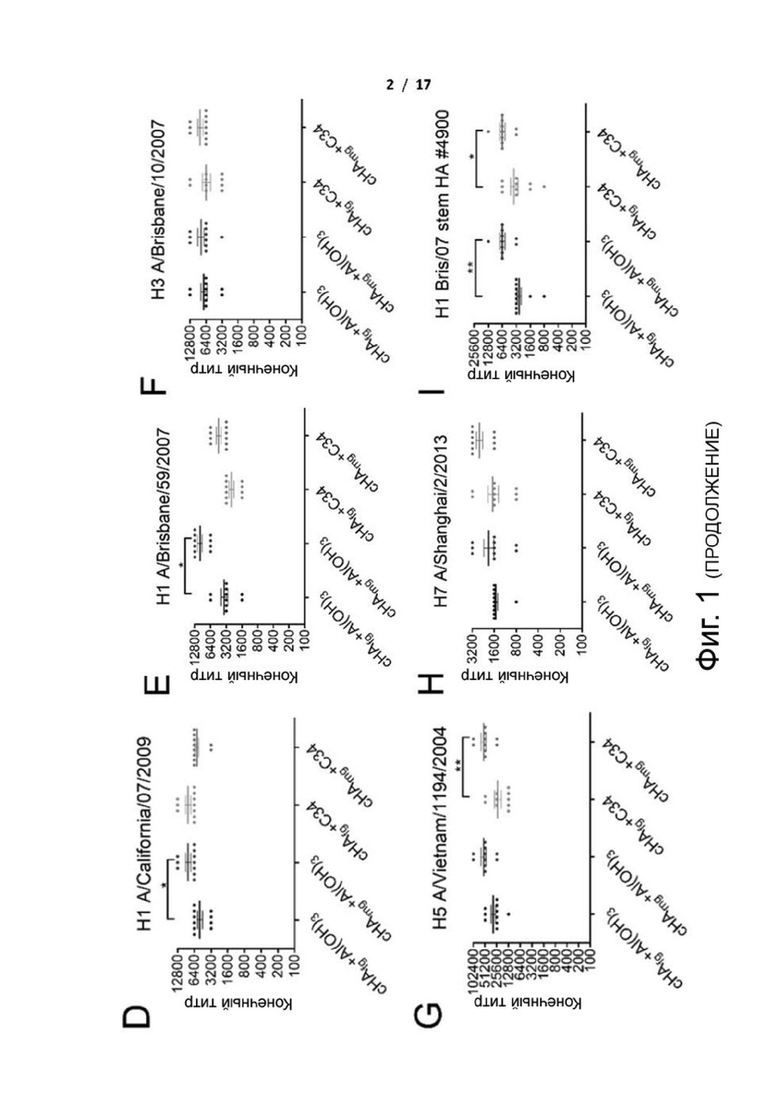
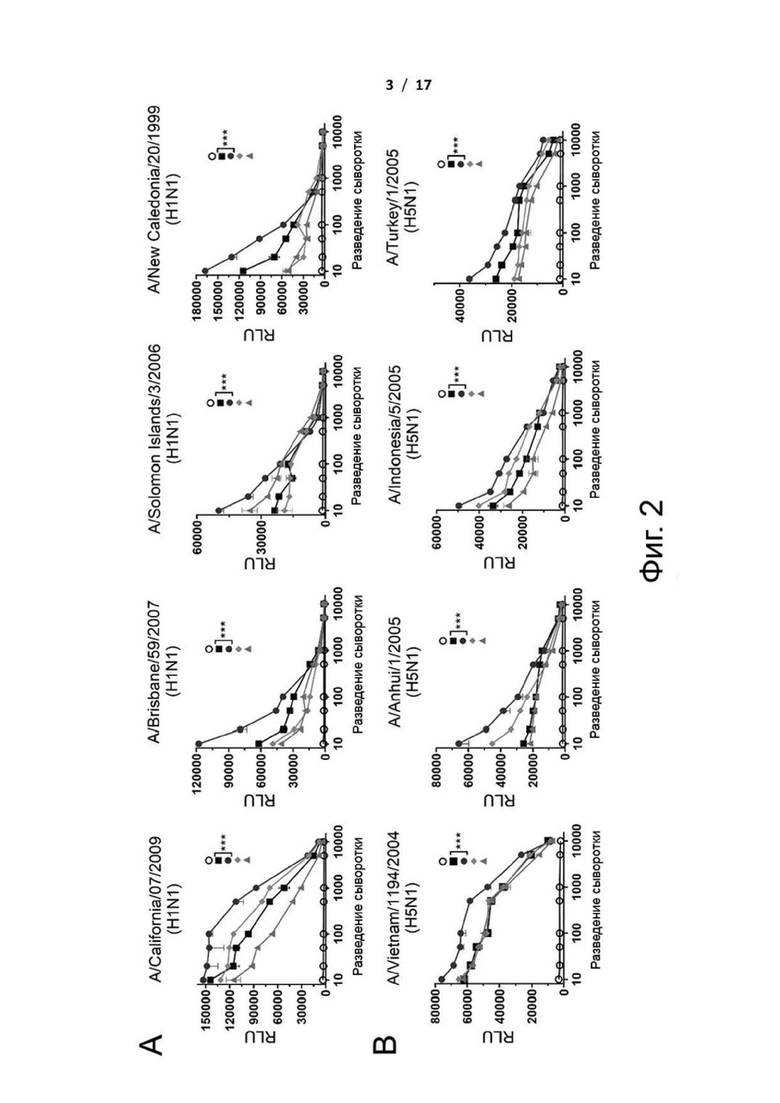
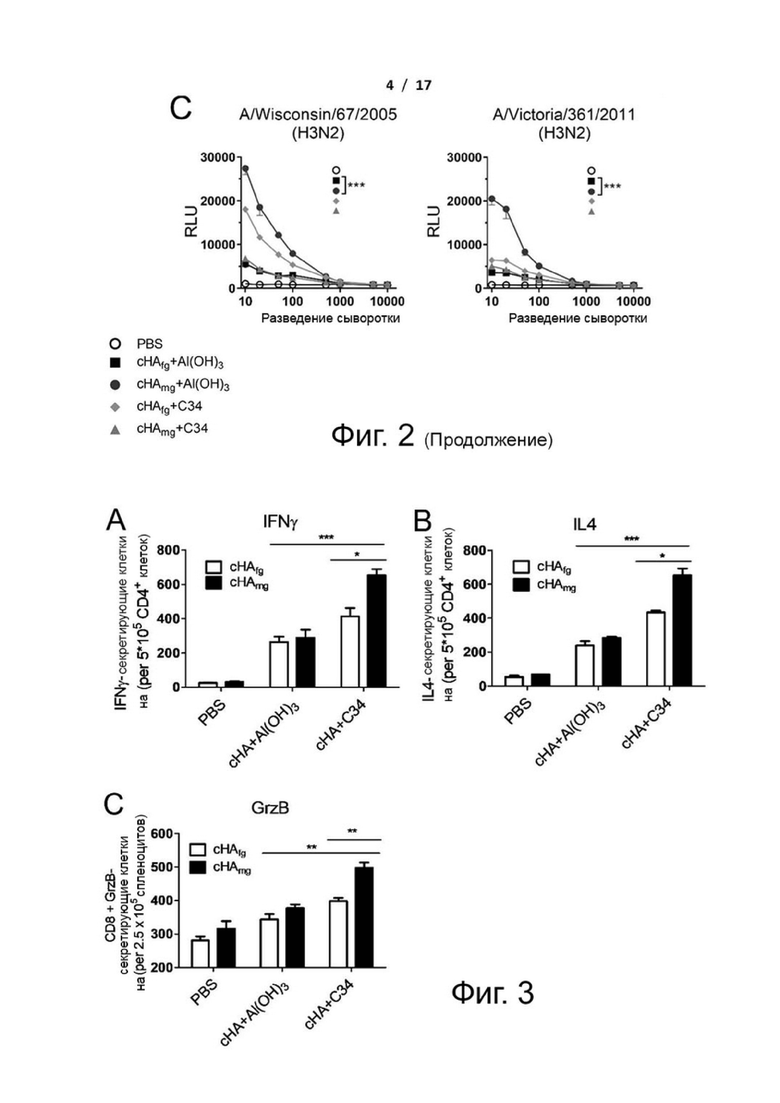
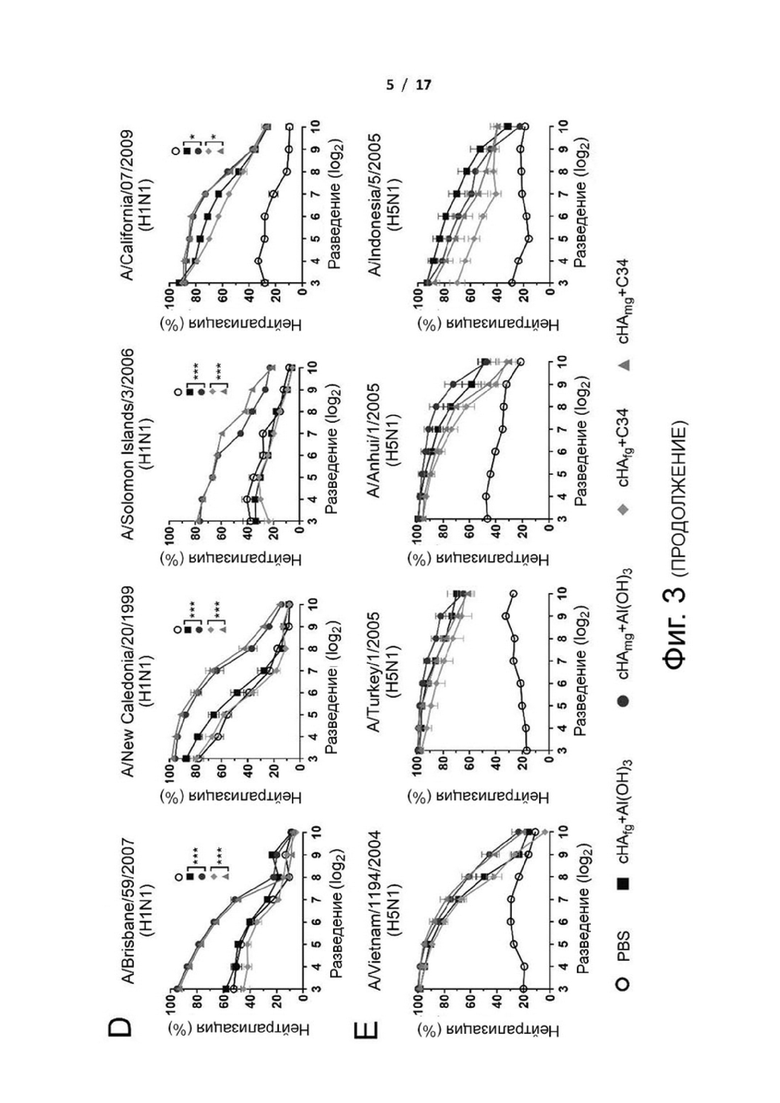
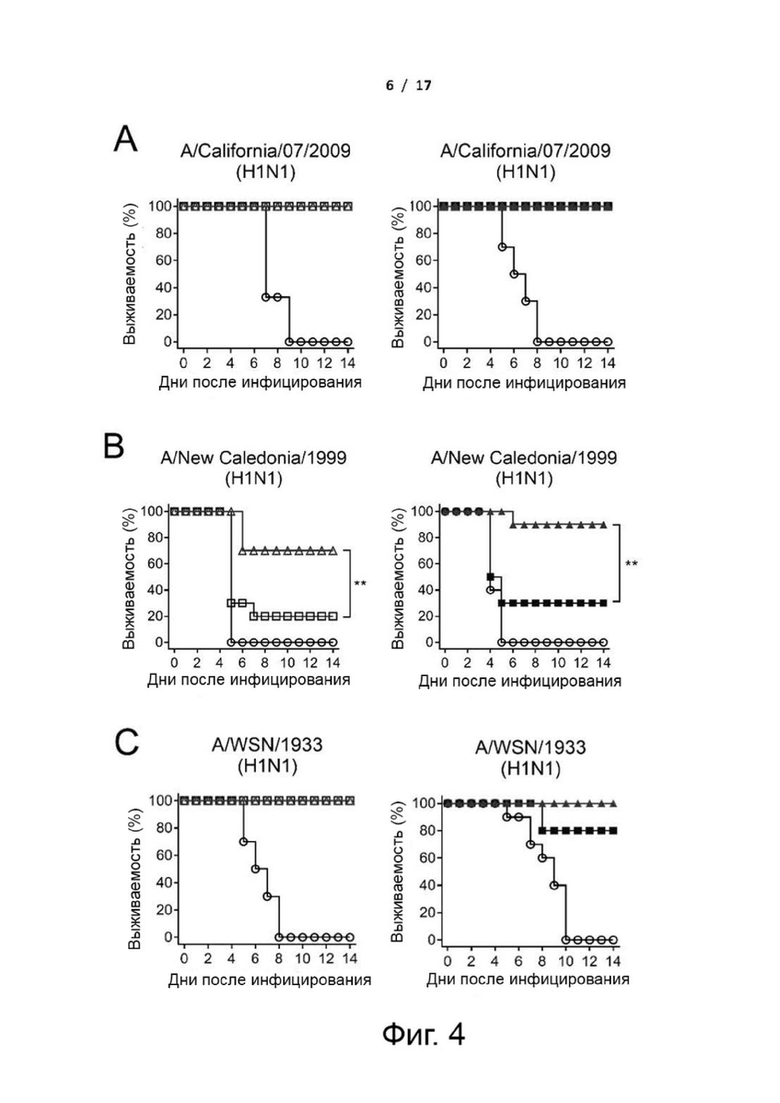
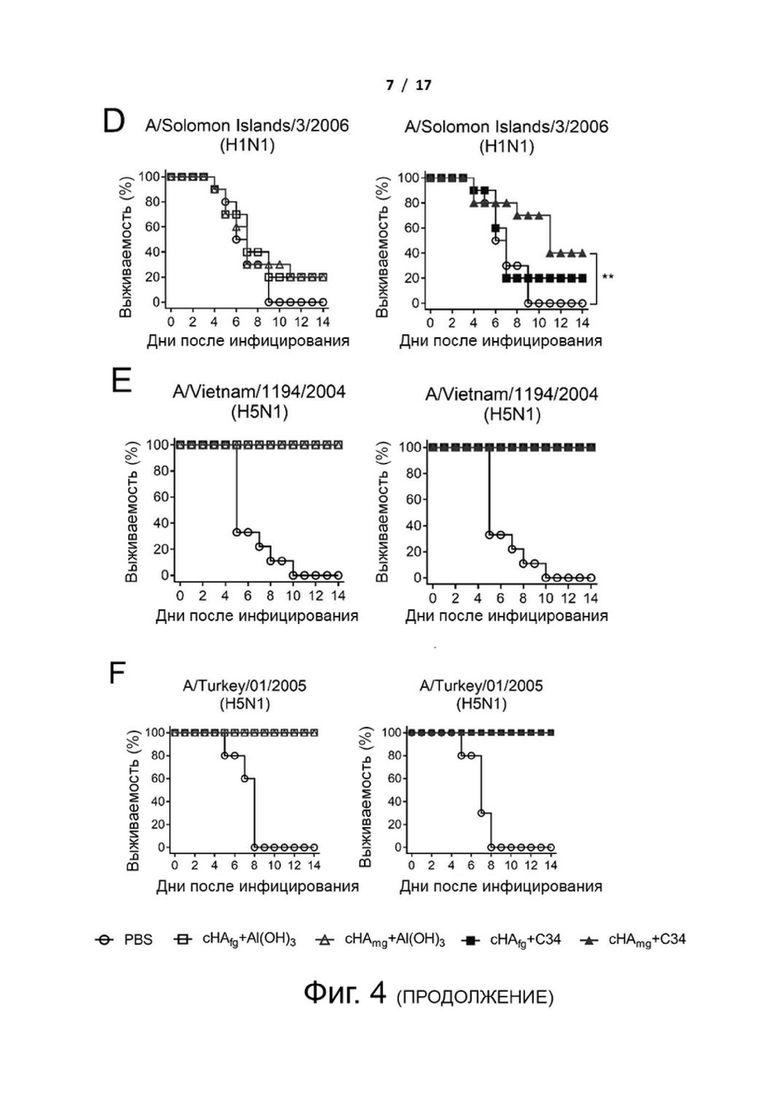
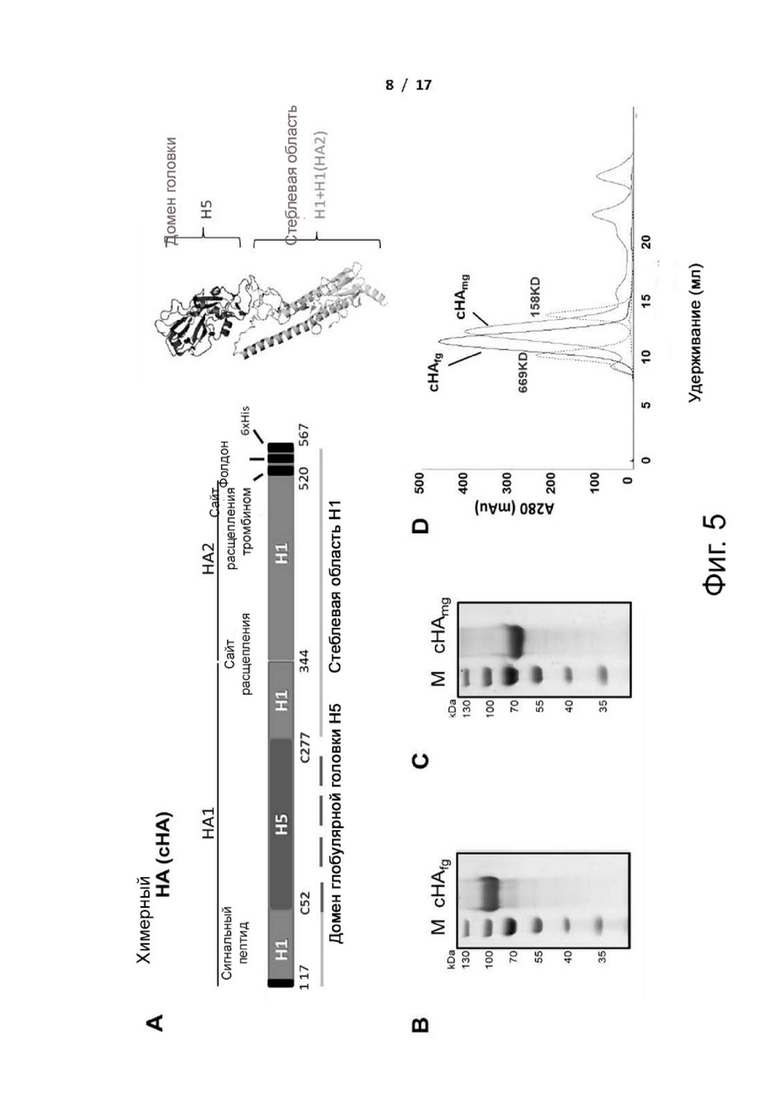
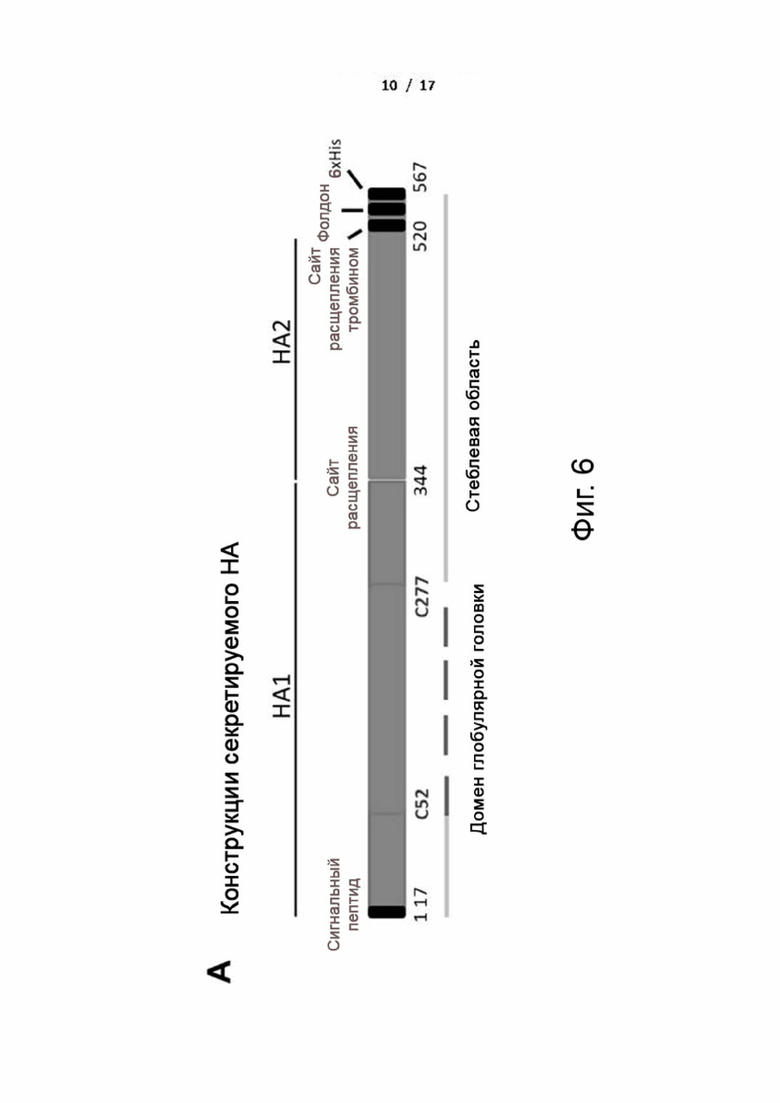
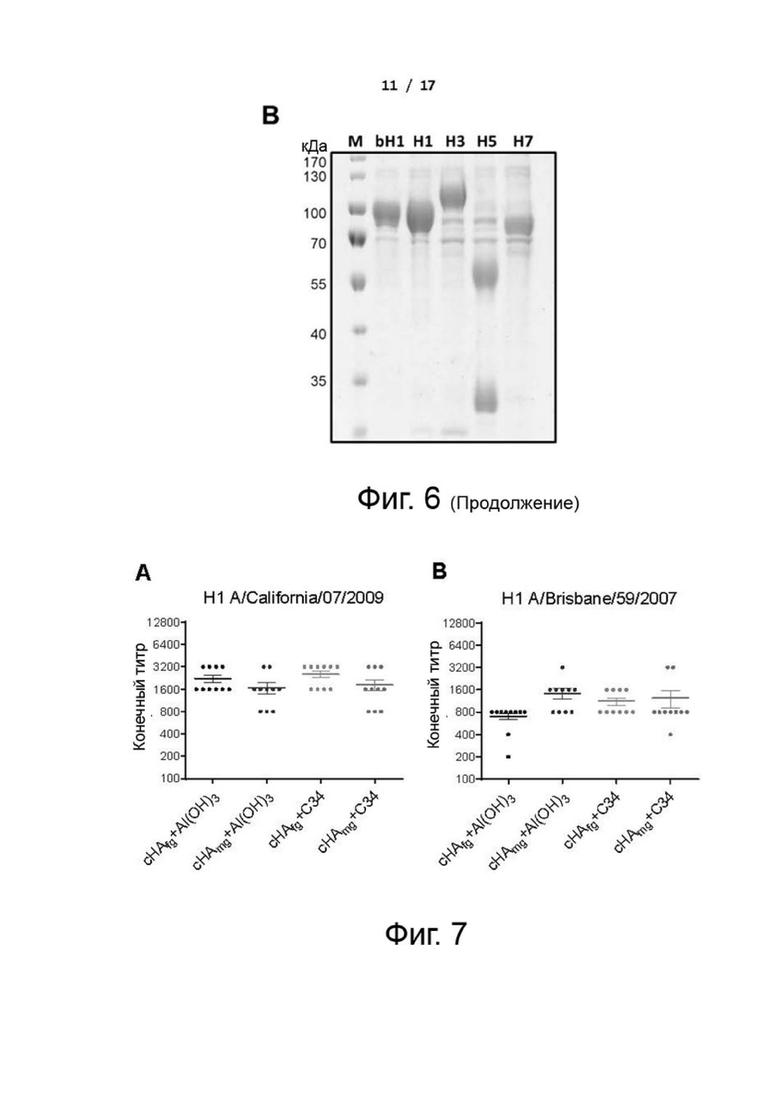
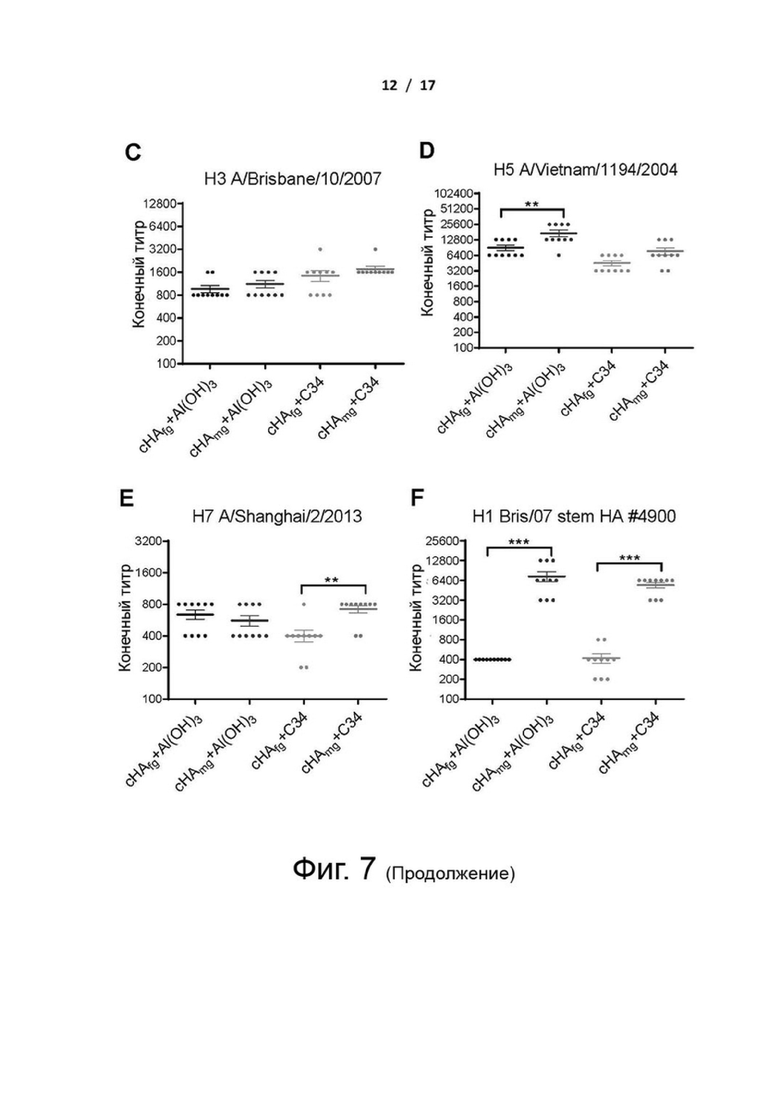
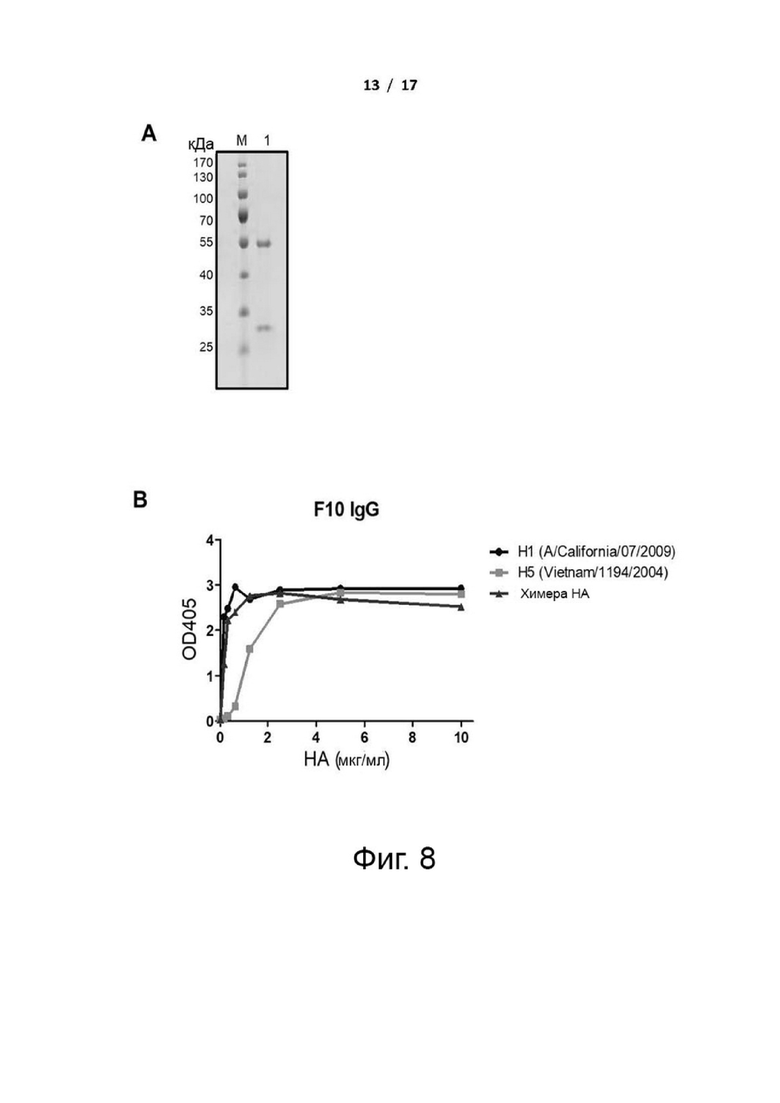
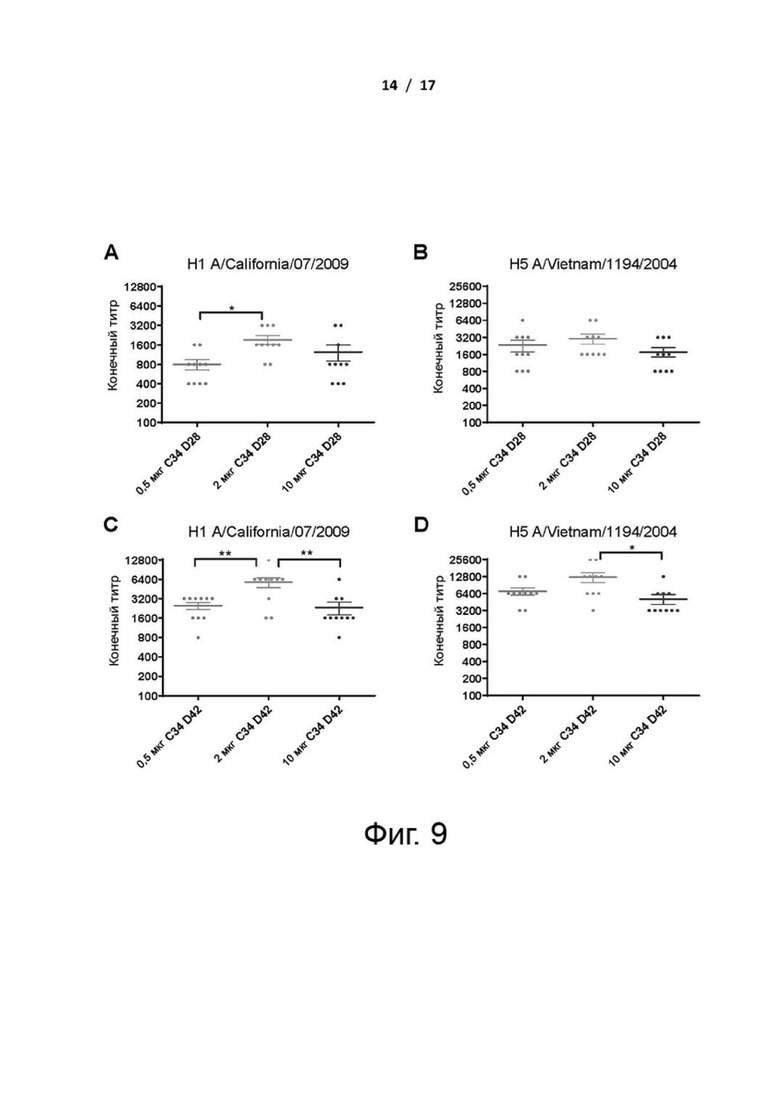
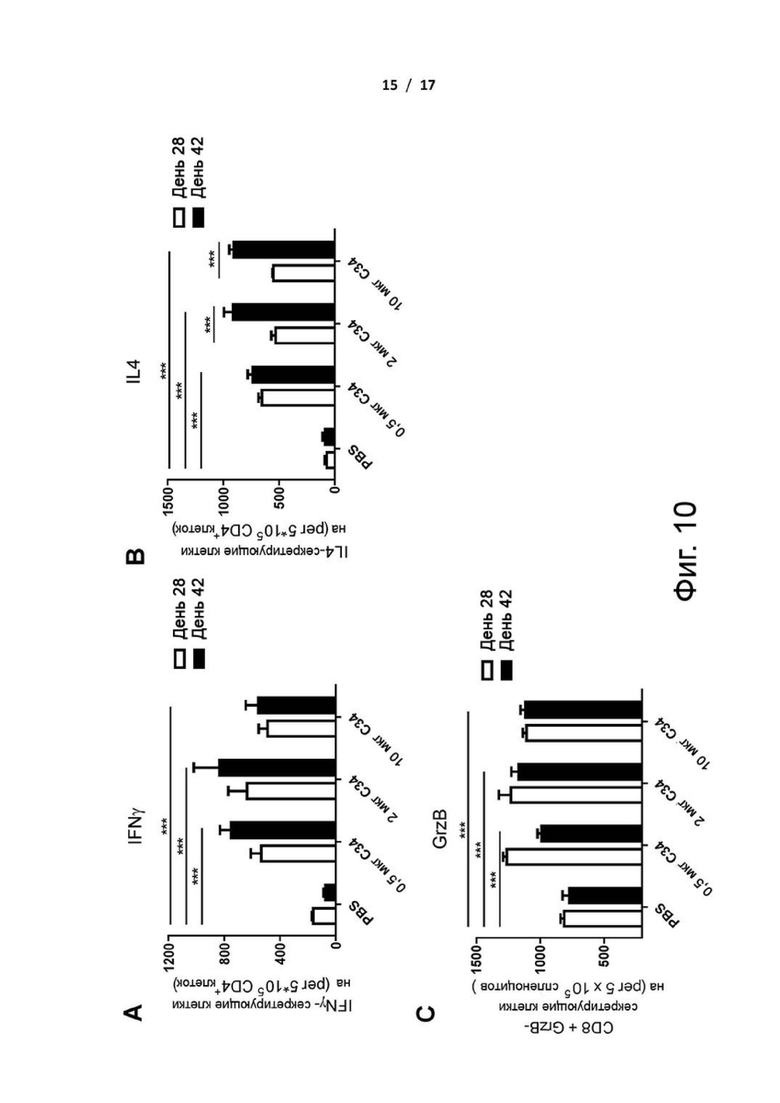
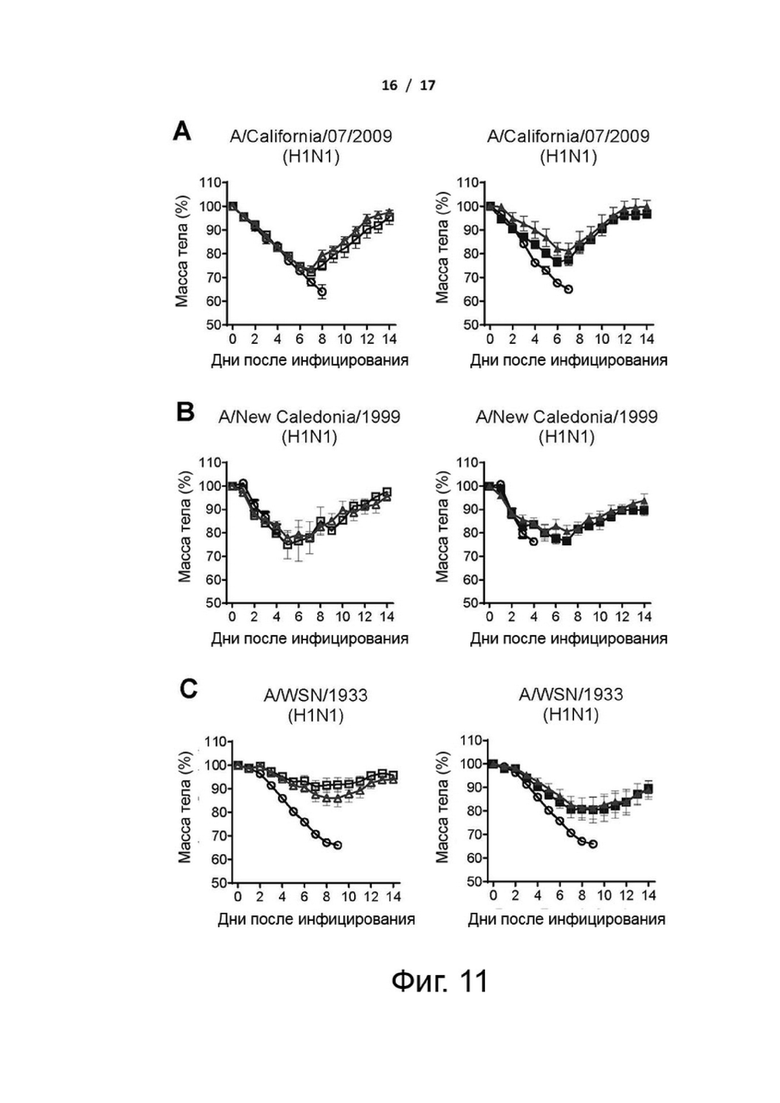
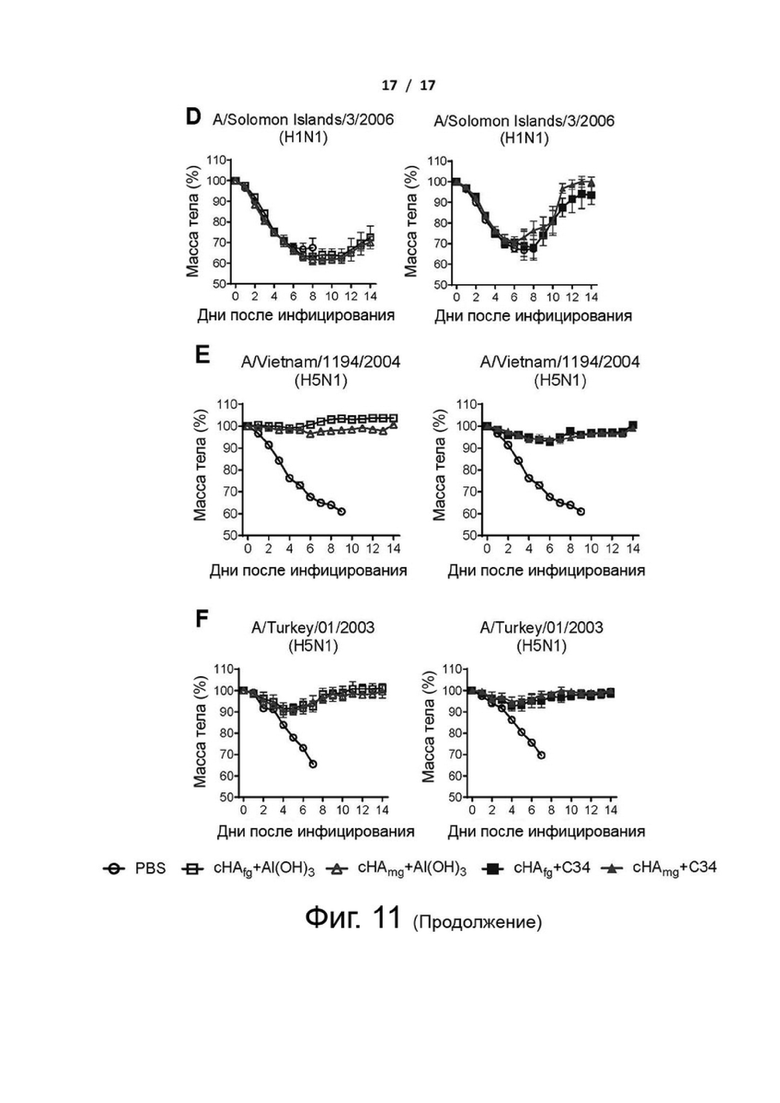
Изобретение относится к биотехнологии. Описан химерный полипептид гемагглютинина (HA) вируса гриппа, используемый в качестве иммуногена, содержащий одну или несколько последовательностей стеблевого домена, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью стеблевого домена HA подтипа H1 (H1 HA) и/или HA подтипа H5 (H5 HA), слитых с одной или несколькими последовательностями домена глобулярной головки, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью домена глобулярной головки HA подтипа H1 (H1 HA) или HA подтипа H5 (H5 HA), где химерный полипептид гемагглютинина (HA) вируса гриппа содержит аминокислотную последовательность SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12. Также описана иммуногенная композиция, содержащая химерный полипептид НА вируса гриппа. Раскрыт способ иммунизации субъекта против вируса гриппа или предупреждения заболевания, вызванного вирусом гриппа, включающий введение субъекту эффективного количества химерного полипептида гемагглютинина (HA) вируса гриппа по любому из пп. 1, 2 или иммуногенной композиции по п. 3, где способ увеличивает продукцию IFN-гамма, IL-4 и CD8+ T-клеток памяти. Также раскрыт рекомбинантный полинуклеотид, кодирующий полипептид. Представлен вектор экспрессии, содержащий рекомбинантный полинуклеотид, и клетка-хозяин, содержащая вектор. 6 н. и 5 з.п. ф-лы, 11 ил., 1 табл., 5 пр.
1. Химерный полипептид гемагглютинина (HA) вируса гриппа, используемый в качестве иммуногена, содержащий одну или несколько последовательностей стеблевого домена, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью стеблевого домена HA подтипа H1 (H1 HA) и/или HA подтипа H5 (H5 HA), слитых с одной или несколькими последовательностями домена глобулярной головки, каждая из которых имеет по меньшей мере 60% гомологию с консенсусной последовательностью домена глобулярной головки HA подтипа H1 (H1 HA) или HA подтипа H5 (H5 HA), где химерный полипептид гемагглютинина (HA) вируса гриппа содержит аминокислотную последовательность SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12.
2. Полипептид по п. 1, где один или несколько гликосайтов на HA являются моногликозилированными, при этом моногликозилированный HA содержит только N-ацетилглюкозамин (GlcNAc) на каждом гликосайте.
3. Иммуногенная композиция, содержащая химерный полипептид НА вируса гриппа по любому из пп. 1, 2.
4. Иммуногенная композиция по п. 3, где иммуногенная композиция дополнительно содержит гликолипидный адъювант.
5. Способ иммунизации субъекта против вируса гриппа или предупреждения заболевания, вызванного вирусом гриппа, включающий введение субъекту эффективного количества химерного полипептида гемагглютинина (HA) вируса гриппа по любому из пп. 1, 2 или иммуногенной композиции по п. 3, где способ увеличивает продукцию IFN-гамма, IL-4 и CD8+ T-клеток памяти.
6. Способ по п. 5, где способ вызывает CD4+ и CD8+ T-клеточные иммунные ответы.
7. Способ по п. 5, который индуцирует стебель-специфические антитела, которые индуцируют антителозависимую клеточную цитотоксичность (ADCC) и имеют нейтрализующую и кросс-протективную активности против штаммов и подтипов H1, H3, H5 и H7.
8. Рекомбинантный полинуклеотид, кодирующий полипептид по любому из пп. 1, 2, содержащий последовательность нуклеиновой кислоты, кодирующую полипептид по любому из пп. 1, 2.
9. Рекомбинантный полинуклеотид по п. 8, где рекомбинантный полинуклеотид также содержит последовательность нуклеиновой кислоты, кодирующую сигнальный полипептид, и где сигнальный пептид содержит последовательность SEQ ID NO: 13 или SEQ ID NO: 14.
10. Вектор экспрессии, содержащий рекомбинантный полинуклеотид по п. 9.
11. Клетка-хозяин, содержащая вектор по п. 10, где клетка-хозяин предназначена для продукции рекомбинантного полипептида по любому из пп. 1, 2.
| WO 2019246363 A1, 26.12.2019 | |||
| Высоковольтный кабель | 1931 |
|
SU34639A1 |
| RU 2017141447 A, 13.02.2019. | |||

