Изобретение относится к онкологии, а именно к экспериментальной онкологии, и может быть использовано для изучения противоопухолевого воздействия на меланому в эксперименте.
Митохондрии - это важнейшие органеллы, отвечающие за выживание клеток и апоптоз. Здоровые митохондрии необходимы для поддержания нормального функционирования клеток. В то же время известно, что митохондрии опухолей претерпевают адаптивные изменения для еще большего ускорения быстрого размножения опухолевых клеток в кислой и гипоксической микросреде (см. Jing X, Yang F, Shao C, Wei K, Xie M, Shen H. et al. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol Cancer. 2019; 18: 157. https://doi.org/10.1186/s12943-019-1089-9).
Все больше накапливается данных свидетельствующих о том, что метаболизм митохондрий, функции митохондрий незаменимы при онкогенезе и прогрессировании рака, что выделяет митохондрии и митохондриальные функции как перспективные мишени для противоопухолевой терапии (см. Dong, L., Gopalan, V., Holland, O., & Neuzil, J. Mitocans Revisited: Mitochondrial Targeting as Efficient Anti-Cancer Therapy. International journal of molecular sciences. 2020; 21(21): 7941. https://doi.org/10.3390/ijms21217941).
Генетические и/или метаболические изменения в митохондриях способствуют возникновению многих заболеваний человека, включая рак (см. Singh B., Modica-Napolitano J.S., Singh K.K. Defining the Momiome: Promiscuous Information Transfer by Mobile Mitochondria and Mitochondrial Genome. Semin. Cancer Biol. 2017; 47: 1-17. https://doi.org/10.1016/j.semcancer.2017.05.004). Хотя гликолиз традиционно считался основным источником энергии в злокачественных клетках, в соответствии с так называемым «эффектом Варбурга», впервые предложенным почти столетие назад, имея в виду повышенное поглощение глюкозы, которое характерно для большинства видов рака, митохондриальная функция известная как окислительное фосфорилирование (OXPHOS), недавно признана ключевым фактором в онкогенезе (см. Porporato P.E., Filigheddu N., Pedro J.M.B., Kroemer G., Galluzzi L. Mitochondrial metabolism and cancer. Cell Res. 2018;28:265-280. https://doi.org/10.1038/cr.2017.155; Dong L.F., Jiri Neuzil J. Targeting mitochondria as an anticancer strategy. Cancer Commun. 2019;39:63. https://doi.org/10.1186/s40880-019-0412-6).
Кроме того, опухолевые клетки уникальным образом перепрограммируют свою клеточную активность, чтобы поддерживать собственное быстрое распространение и миграцию, а также противодействовать метаболическому и генотоксическому стрессу во время прогрессирования злокачественного процесса (см. Wang Y., Xia Y., Lu Z. Metabolic features of cancer cells. Cancer Commun. 2018;38:65. https://doi.org/10.1186/s40880-018-0335-7).
Митохондрии злокачественных клеток обладают способностью гибко переключаться между гликолизом и OXPHOS для повышения выживаемости (см. Dong L.F., Jiri Neuzil J. Targeting mitochondria as an anticancer strategy. Cancer Commun. 2019;39:63. https://doi.org/10.1186/s40880-019-0412-6). Группой исследователей была подтверждена важность митохондрий в межклеточной коммуникации, так оказалось, что мтДНК внутри целых митохондрий подвижна и может подвергаться горизонтальному переносу между клетками. Обнаружено, что злокачественные клетки, лишенные своей мтДНК и, следовательно, утрачивающие канцерогенный потенциал, могут повторно получить это свойство, приобретая здоровую мтДНК из стромальных клеток хозяина посредством переноса целых митохондрий, что приводит к восстановлению митохондриального дыхания (см. Tan A.S., Baty J.W., Dong L.F., Bezawork-Geleta A., Endaya B., Yan B., Goodwin J., Vondrusova M., Bajzikova M., Peterka M., et al. Mitochondrial genome acquisition restores respiratory function and tumorigenic potential of cancer cells without mitochondrial DNA. Cell Metab. 2015;21:81-94. https://doi.org/10.1016/j.cmet.2014.12.003; Dong L.F., Kovarova J., Bajzikova M., Bezawork-Geleta A., Svec D., Endaya B., Schaphibulkij K., Coelho A., Sebkova N., Ruzickova A., et al. Horizontal transfer of whole mitochondria restores tumorigenic potential in mitochondrial DNA-deficient cancer cells. Elife. 2017;6:e22187. https://doi.org/10.7554/eLife.22187).
Недавно был предложен термин «митоканы», аббревиатуру, образованную от терминов митохондрии и рак, группы соединений с противораковой активностью, проявляющейся через их молекулярные мишени в митохондриях, причем некоторые митоканы являются селективными для злокачественных тканей (см. Neuzil J., Dong L.F., Rohlena J., Truksa J., Ralph S.J. Classification of mitocans, anti-cancer drugs acting on mitochondria. Mitochondrion.2013;13:199208.https://doi.org/10.1016/j.mito.2012.07.112; Mani S., Swargiary G., Singh K.K. Natural Agents Targeting Mitochondria in Cancer. Int. J. Mol. Sci. 2020; 21: E6992. https://doi.org/10.3390/ijms2119699). Эти различные агенты, нацеленные на митохондрии и их всевозможные функции, вносят вклад в новые противоопухолевые программы с высоким терапевтическим потенциалом. Новые программы включают агенты, которые нацелены на ETC и OXPHOS, гликолиз, цикл трикарбоновой кислоты (TCA), пути апоптоза, гомеостаз активных форм кислорода (ROS), комплекс пор с переходом проницаемости, мтДНК, а также синтез пиримидина (см. Cui Q., Wen S., Huang P. Targeting cancer cell mitochondria as a therapeutic approach: Recent updates. Future Med Chem. 2017;9:929-949. https://doi.org/10.4155/fmc-2017-0011). Все большее количество исследований сосредоточено на доставке противоопухолевых препаратов в митохондрии для лечения злокачественных опухолей, и на данное инновационное направление возлагаются большие надежды в сфере разработки новых эффективных противоопухолевых терапевтических средств (см. Du X., Zhang P., Fu H., Ahsan H.M., Gao J., Chen Q. Smart mitochondrial targeted cancer therapy: Subcellular distribution, selective TrxR2 inhibition accompany with declined antioxidant capacity. Int. J. Pharm. 2019;555:346-355. doi: 10.1016/j.ijpharm.2018.11.057; Noh I., Lee D.Y., Kim H., Jeong C.U., Lee Y., Ahn J.O., Hyun H., Park J.H., Kim Y.C. Enhanced Photodynamic Cancer Treatment by Mitochondria-Targeting and Brominated Near-Infrared Fluorophores. Adv.Sci. 2018;5:1700481. https://doi.org/10.1002/advs.20170048).
Таким образом, введение здоровых митохондрий в опухолевые клетки, как предполагается, имеет высокую эффективность в предотвращении роста опухоли (см.Singh B, Modica-Napolitano JS, Singh KK. Defining the momiome: promiscuous information transfer by mobile mitochondria and the mitochondrial genome. Semin Cancer Biol Semin Cancer Biol. 2017;47:1-17. https://doi.org/10.1016/j.semcancer.2017.05.004; Roushandeh AM, Kuwahara Y, Roudkenar MH. Mitochondrial transplantation as a potential and novel master key for treatment of various incurable diseases. Cytotechnology. 2019;71:647-663. https://doi.org/ 10.1007/s10616-019-00302-9).
В настоящее время экзогенные здоровые митохондрии были использованы при лечении нескольких видов рака, включая рак молочной железы, рак поджелудочной железы, глиомы и в результате была показана превосходная противоопухолевая активность здоровых митохондрий (см. Fu A, Hou Y, Yu Z, Zhao Z, Liu X. Healthy mitochondria inhibit the metastatic melanoma in lungs. Int J Biol Sci. 2019;15:2707-2718 https://doi.org/10.7150/ijbs.38104; Chang JC, Chang HS, Wu YC, Cheng WL, Lin TT, Chang HJ. et al. Mitochondrial transplantation regulates antitumour activity, chemoresistance and mitochondrial dynamics in breast cancer. J Exp Clin Cancer Res. 2019;38:30. https://doi.org/10.1186/s13046-019-1028-z; см. Sun C, Liu X, Wang B, Wang Z, Liu Y, Di C. et al. Endocytosis-mediated mitochondrial transplantation: Transferring normal human astrocytic mitochondria into glioma cells rescues aerobic respiration and enhances radiosensitivity. Theranostics. 2019;9:3595-3607. https://doi.org/10.7150/thno.33100). Хотя биохимические процессы в здоровых митохондриях после трансплантации несколько изменяются в сторону снижения способности к окислительному фосфорилированию (OXPHOS), тем не менее, сохраняется возможность запуска апоптоза в опухолевых клетках. Несмотря на выявленные факты, механизм молекулярного сигнала для митохондриальной трансплантационной терапии до сих пор неясен.
Митохондрии выполняют центральную роль в передаче сигнала опухолевых клеток (см Qureshi MA, Haynes CM, Pellegrino MW. The mitochondrial unfolded protein response: Signaling from the powerhouse. J Biol Chem. 2017;292:13500-13506. https://doi.org/10.1074/jbc.R117.791061). Разрастание опухолей в значительной степени зависит от функционального состояния митохондрий, которое может быстро и эффективно влиять на передачу молекулярных сигналов в клетках (Yoshida GJ. Metabolic reprogramming: the emerging concept and associated therapeutic strategies. J Exp Clin Cancer Res. 2015;34:111. https://doi.org/10.1186/s13046-015-0221-y; Vyas S, Zaganjor E, Haigis MC. Mitochondria and cancer. 2016; 166: 555-566. https://doi.org/ 10.1016/j.cell.2016.07.002). Из-за неограниченной быстрой пролиферации опухолевые клетки находятся в бескислородной и кислой микросреде и в процессе адаптации к этой враждебной среде функция нативных митохондрий опухолевых клеток способна подстраиваться под новые условия и происходить это может как с усилением, так и, напротив, с замедлением ряда метаболических процессов (см. Avnet S, Baldini N, Brisson L, De Milito A, Otto AM, Pastoreková S. et al. Annual meeting of the international society of cancer metabolism (ISCaM): cancer metabolism. Front Oncol. 2018;8:329. https://doi.org/10.3389/fonc.2018.00329).
В настоящее время меланома стала основной причиной смерти от кожных заболеваний. Было приложено много усилий, чтобы найти способы вылечить агрессивную болезнь. В меланоме часто встречаются делеция и мутации митохондриальной ДНК (мтДНК). При исследовании 67 пациентов с меланомой в тканях меланомы, индуцированных солнечным излучением, были обнаружены делеции 4977 п.н. и 5128 п.н. в мтДНК (см. Hubbard K, Steinberg ML, Hill H, Orlow I. Mitochondrial DNA deletions in skin from melanoma patients. Ethn Dis. 2008;18(S2): 38-43.). Кроме того, мутации мтДНК вносят большой вклад в независимый от ультрафиолетового излучения канцерогенез меланомы (см. Mitra D, Luo X, Morgan A, Wang J, Hoang MP, Lo J. et al. An ultraviolet-radiation-independent pathway to melanoma carcinogenesis in the red hair/fair skin background. Nature. 2012;491:449-453. https://doi.org/10.1038/nature11624).
Более того, дисфункциональные митохондрии накапливаются посредством неселективного слияния из-за активации передачи сигналов WNT / β-catenin в меланоме, что ведет к ремоделированию митохондрий и затем к быстрой проферации и инвазии клеток меланомы (см. Brown K, Yang P, Salvador D, Kulikauskas R, Ruohola-Baker H, Robitaille AM. et al. WNT/β-catenin signaling regulates mitochondrial activity to alter the oncogenic potential of melanoma in a PTEN-dependent manner. Oncogene. 2017;36:3119-3136. https://doi.org/10.1038/onc.2016.450).
Однако микросреда может вызвать дисфункцию здоровых митохондрий, а затем привести к митохондриальному стрессу, когда здоровые митохондрии попадут в опухолевые клетки.
Ретроградная транскрипционная передача сигналов митохондрий играет жизненно важную роль в качестве сенсоров стресса для адаптации своего окружения (см. Vyas S, Zaganjor E, Haigis MC. Mitochondria and Cancer. Cell. 2016;166:555-566. https://doi.org/10.1016/j.cell.2016.07.002). Ретроградные сигналы между митохондриями и ядром, в том числе АТФ, ROS, 2-hydroxyglutarate, повышение митохондриальной проницаемости через открытие переходной поры, могут регулировать экспрессию генов в ядре (см. Zhao X, Ma X, Guo J, Mi M, Wang K, Zhang C. et al. Circular RNA circEZH2 suppresses transmissible gastroenteritis coronavirus-induced opening of mitochondrial permeability transition pore via targeting miR-22 in IPEC-J2. Int J Biol Sci. 2019;15:2051-2064. https://doi.org/10.7150/ijbs.36532; см Zhou Q, Zhu L, Qiu W, Liu Y, Yang F, Chen W, Xu R. Nicotinamide riboside enhances mitochondrial proteostasis and adult neurogenesis through activation of mitochondrial unfolded protein response signaling in the brain of ALS SOD1G93A mice. Int J Biol Sci. 2020; 16: 284-297. https://doi.org/ 10.7150/ijbs.38487).
Недавно была описана вероятная связь между митохондриальным стрессом и широко распространенными изменениями в структуре хроматина (см. Merkwirth C, Jovaisaite V, Durieux J, Matilainen O, Jordan SD, Quiros PM. et al. Two conserved histone demethylases regulate mitochondrial stress-induced longevity. Cell. 2016;165:1209-1223. https://doi.org/ 10.1016/j.cell.2016.04.012). Митохондриальный стрессорный ответ активирует реорганизацию хроматина посредством диметилирования гистона - H3K9, которое связано с подавлением генов (см. Tian Y, Garcia G, Bian Q, Steffen KK, Joe L, Wolff S. et al. Mitochondrial Stress Induces Chromatin Reorganization to Promote Longevity and UPR(mt) Cell. 2016;165:1197-1208. https://doi.org/ 10.1016/j.cell.2016.04.011). Изменения эпигенома, вызванные митохондриальным стрессом, обычно обширны и продолжительны, что приводит к замедлению пролиферации опухолевых клеток (см. Tatar M, Sedivy JM. Mitochondria: masters of epigenetics. Cell. 2016;165:1052-1054. https://doi.org/ 10.1016/j.cell.2016.05.021; Mayorga L, Salassa BN, Marzese DM, Loos MA, Eiroa HD, Lubieniecki F. et al. Mitochondrial stress triggers a pro-survival response through epigenetic modifications of nuclear DNA. Cell Mol Life Sci. 2019;76:1397-1417. https://doi.org/ 10.1007/s00018-019-03008-5). Тем не менее, части хроматина могут открываться, чтобы модулировать клеточные ответы на стресс, даже если хроматин глобально находится в покоящемся состоянии.
Среди сигналов, опосредованных митохондриальным стрессом, ответная реакция митохондриального развернутого белка (UPR mt) является потенциальным митохондриально-ядерным ретроградным путем, который оказывает существенное влияние на функцию клеток посредством регуляции экспрессии ядерных генов (см. Kenny TC, Germain D. mtDNA, mtastasis, and the mtochondrial unfolded protein response (UPRmt) Front Cell Dev Biol. 2017; 5: 1-6. https://doi.org/10.3389/fcell.2017.00037). Активированный UPR mt может индуцировать повышенную регуляцию метаболизма глутатиона, аутофагию и опухолевую супрессию. Аутофагия в бескислородной среде и экспрессия опухолевого супрессора могут вызвать ингибирование роста опухоли (см. Munch C, Harper JW. Mitochondrial unfolded protein response controls matrix pre-RNA processing and translation. Nature. 2016;534:710-713. https://doi.org/10.1038/nature18302).
Более того, митохондрии являются ключевыми игроками в процессе апоптоза клеток (см. Holley AK, Dhar SK, Clair D. Manganese superoxide dismutase versus p53: the mitochondrial center. Ann N Y Acad. Sci. 2010;201:72-78. https://doi.org/ 10.1111/j.1749-6632.2010.05612.x). Здоровые митохондрии обычно запускают апоптоз клеток в условиях патологической гипоксии для устранения поврежденных клеток. Однако опухолевые клетки подавляют апоптотическую передачу сигналов митохондрий, особенно активацию антиапоптотических систем, что позволяет клеткам ускользать от апоптоза, а это ведет к неконтролируемой пролиферации в микроокружении опухоли. Выявлено, что при трансплантации здоровых митохондрий транскрипция гена антиапоптотических белков, как правило, блокировалась, это могло быть связано с подавлением хромосом, после чего возможна активация процесса апоптоза для уничтожения опухолевых клеток (см. Mohammada RM, Muqbila I, Lowec L, Yedjoud C, Hsue HY, Linf LT. et al. Broad targeting of resistance to apoptosis in cancer. Semin Cancer Biol. 2015;35:S78-S103.https://doi.org/ 10.1016/j.semcancer.2015.03.001).
Исследования показали, что экзогенные митохондрии могут напрямую проникать в культивируемые ксеногенные клетки и улучшать их митохондриальную функцию. Например, митохондрии, выделенные из мезенхимальных клеток (EMC), происходящих из эндометриальных желез матки человека, попадают в кардиомиобласты (H9c2) путем простой совместной инкубации и восстанавливают дыхательную функцию митохондрий клеток (см. Kitani T., Kami D., Matoba S., Gojo S. Internalization of isolated functional mitochondria: involvement of macropinocytosis. J. Cell. Mol. Med. 2014; 18: 1694-1703. https://doi.org/10.1111/jcmm.12316). Другой пример, (см. Katrangi E., D’Souza G., Boddapati S. V., Kulawiec M., Singh K. K., Bigger B., et al. Xenogenic transfer of isolated murine mitochondria into human rho0 cells can improve respiratory function. Rejuvenation Res. 2007; 10: 561-570. https://doi.org/10.1089/rej.2007.0575) исследователи переносят митохондрии мышей в клетки rho0 человека и определяют, что митохондрии могут улучшать дыхательную функцию. Существуют исследования с использованием генетически меченных GFP митохондрий, которые были выделены из клеток, трансфицированных лентивирусным вектором. Авторы определили in vivo распределение и функцию экзогенных митохондрий после системной инъекции. Результаты показали, что все мыши, с трансплантированными митохондриями, выжили и никаких аномалий не появилось, а митохондрии проникли в клетки различных тканей, включая печень, легкие, мозг, мышцы и почки (см. Fu, A., Shi, X., Zhang, H., & Fu, B. Mitotherapy for Fatty Liver by Intravenous Administration of Exogenous Mitochondria in Male Mice. Frontiers in pharmacology. 2017;8: 241. https://doi.org/10.3389/fphar.2017.00241).
Основываясь на экспериментах по контролируемому переносу митохондрий, системный подход к трансплантации митохондрий также использовался для определения ключевых характеристик, окружающих микросреду опухоли и ее развитие.
В линии клеток метастатической меланомы мыши B16 ρ0 с дефицитом митохондрий и линии клеток рака молочной железы мыши 4T1 ρ0 опухолевые клетки приобретали мтДНК из клеток в микроокружении опухоли, что приводило к восстановлению их митохондриального дыхания (см. Tan A.S., Baty J.W., Dong L.F., Bezawork-Geleta A., Endaya B., Goodwin J., Bajzikova M., Kovarova J., Peterka M., Yan B., et al. Mitochondrial genome acquisition restores respiratory function and tumorigenic potential of cancer cells without mitochondrial DNA. Cell Metab. 2015;21:81-94. https://doi.org/10.1016/j.cmet.2014.12.003). Этот эффект лучше развивался в циркулирующих раковых клетках по сравнению с первичными опухолями, что свидетельствовало об адаптивном ответе на постепенное приобретение мтДНК из микроокружения опухоли как часть инвазивности раковых клеток. Еще одним фактом, подтверждающим участие митохондрий в метастазировании является последнее исследование, где отмечается, что перенос интактных митохондрий от ассоциированных с раком фибробластов к клеткам рака предстательной железы человека PC3 указывают на то, что митохондрии играют ключевую роль в метастазировании рака (см. Ippolito L., Morandi A., Taddei M.L., Parri M., Comito G., Iscaro A., Raspollini M.R., Magherini F., Rapizzi E., Masquelier J., et al. Cancer-associated fibroblasts promote prostate cancer malignancy via metabolic rewiring and mitochondrial transfer. Oncogene. 2019;38:5339-5355. https://doi.org/10.1038/s41388-019-0805-7).
Ishikawa K. et al. (2008) использовали цитоплазматическую гибридную (цибридную) технологию для замены эндогенной мтДНК в линии опухолевых клеток мыши, которая была со слабым метастатическим потенциалом, на мтДНК из линии клеток, которая была высокометастатической, и наоборот.
В ходе мониторинга по наличию метастазирования у мышей, было обнаружено, что опухолевые клетки-реципиенты приобрели метастатический потенциал перенесенной мтДНК. МтДНК, обеспечивающая высокий метастатический потенциал, содержала мутации G13997A и 13885insC в гене, кодирующем NADH (восстановленная форма никотинамидадениндинуклеотида) субъединицы 6 дегидрогеназы (ND6). Эти мутации вызывали подавление активности респираторного комплекса I, что было связано с избыточной продукцией активных форм кислорода (АФК).
Предварительная обработка высокометастатических опухолевых клеток акцепторами АФК подавляла их метастатический потенциал у мышей. Эти результаты показывают, что мутации мтДНК могут способствовать прогрессированию опухоли за счет увеличения метастатического потенциала опухолевых клеток (см. Ishikawa K, Takenaga K, Akimoto M, Koshikawa N, Yamaguchi A, Imanishi H, Nakada K, Honma Y, Hayashi J. ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis. Science. 2008 May 2;320(5876):661-4. https://doi.org/ 10.1126/science.1156906. Epub 2008 Apr 3. PMID: 18388260).
Техническим результатом настоящего изобретения является создание способа экспериментальной противоопухолевой биотерапии, основанной на трансплантации митохондрий, выделенных из печени интактных животных.
Технический результат достигается тем, что самкам и самцам мышей линии BALB/c Nude подкожно под лопатку слева вводят суспензию опухолевых клеток мышиной меланомы В16/F10 в 0,5 мл физиологического раствора в разведении 1:20, через 24 часа после перевивки меланомы В16/F10 мышам внутрибрюшинно вводят свежевыделенные митохондрии из печени крыс из расчёта 3,3 мг белка на 1 животное в 0,3 мл физиологического раствора, далее митохондриальную терапию проводят по схеме на 3, 5, 9, 13, 16, 19, 21 сутки. Животным перевивают меланому В16/F10 и, начиная с 1 суток после перевивки, 2 раза в неделю внутрибрюшинно вводят свежевыделенные митохондрии из печени крыс обоего пола.
Изобретение «Способ экспериментальной биотерапии меланомы В16/F10» является новым, так как оно неизвестно в области экспериментальных исследований в онкологии в модификации роста меланомы В16/F10 у мышей линии BALB/c Nude обоего пола с помощью внутрибрюшинного введения митохондрий, выделенных их печени крыс обоего пола.
Новизна изобретения заключается в использовании мышей обоего пола линии BALB/c Nude, развитии у них злокачественного процесса - меланомы В16/F10 и проведении экспериментальной биотерапии митохондриями, выделенными из печени крыс. Проведение экспериментальной биотерапии способствовало удлинению латентного периода выхода опухоли, замедлению ее роста и выраженным онколитическим процессом, доказанным при морфологическом исследовании.
Изобретение «Способ экспериментальной биотерапии меланомы В16/F10» является промышленно применимым, так как может быть использовано в научно-исследовательских учреждениях онкологического профиля для воспроизведения экспериментальной биотерапии, изучения ее воздействия на первичную опухоль, особенностей метастазирования, механизмов ее противоопухолевого действия.
Способ иллюстрируется фигурами.
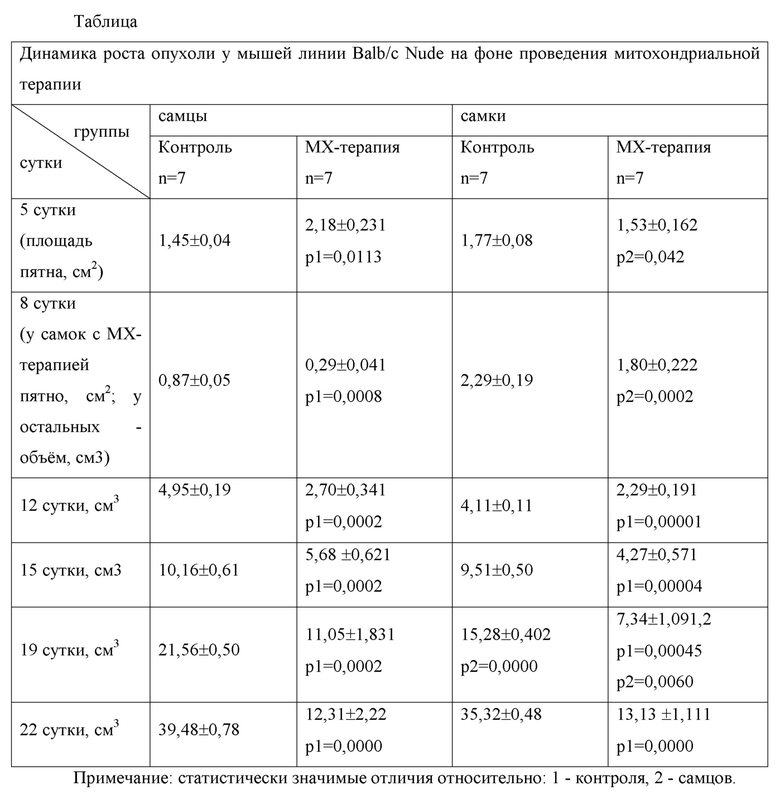
Фигура 1.Подкожный рост перевивной меланомы В-16 у крыс-самцов (а) и самок (б) в контроле (без проведения МХТ); в) фрагмент сплошного опухолевого поля с инфильтрацией опухолевыми клетками подкожной клетчатки. Окр. гематоксилин-эозином. Ув.х 200; г) клетки меланомы неправильной угольчатой формы с отростчатой цитоплазмой и эксцентричным расположением ядер. Окр. гематоксилин-эозином. Ув.х 400.
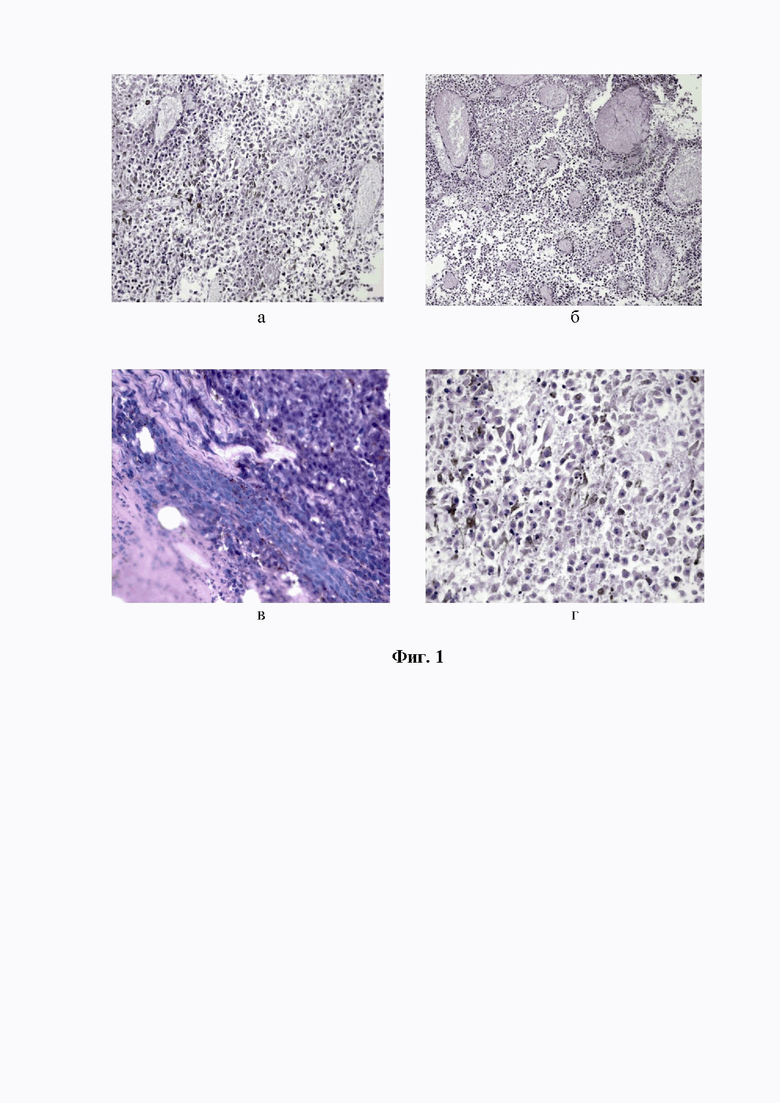
Фигура 2. Фрагмент меланомы В16/F10 у мышей BALB/c Nude-самцов после проведения МХ-терапии. Признаки распада опухолевой ткани с выраженным гиалинозом матрикса (а, б), некроза и ишемии опухолевых клеток (в, г), образованием крупных полостей, заполненных детритом ( г, д, е). Окр. гематоксилин-эозином. Ув.х 400.
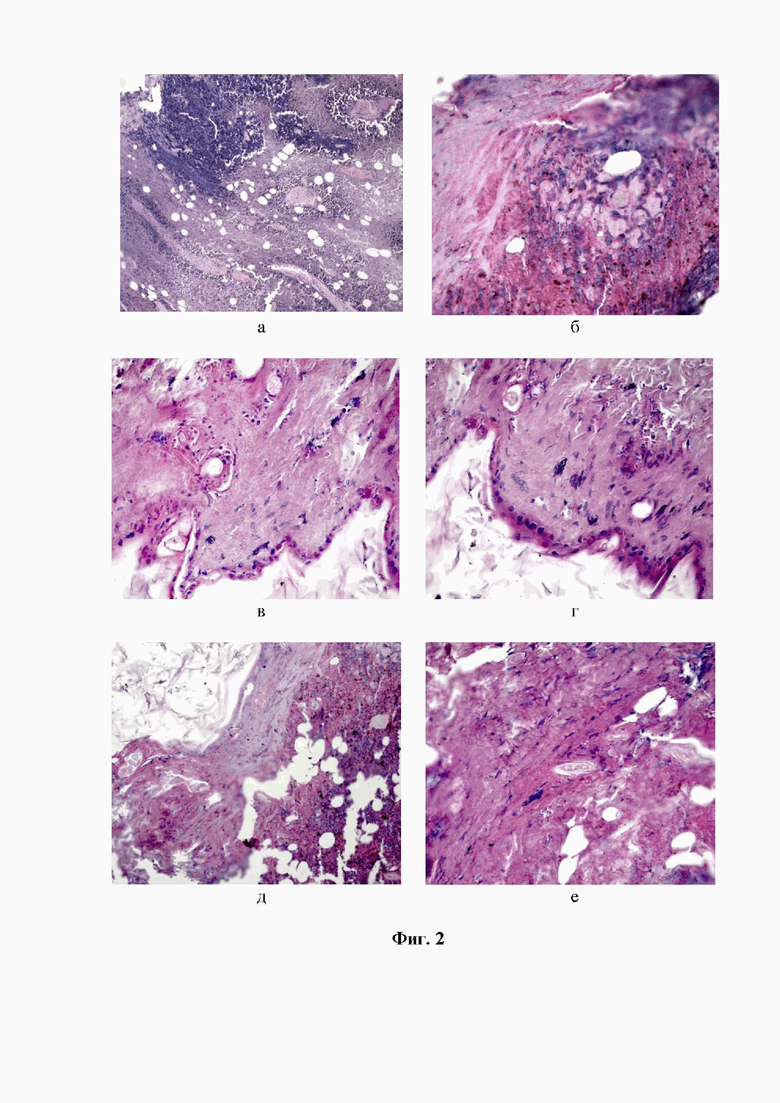
Фигура 3. Фрагмент меланомы В16/F10 у мышей BALB/c Nude -самок после проведения МХТ. Регрессивные изменения опухолевых клеток, включающие фрагментацию ядра и цитоплазмы, образование апоптотических телец, высвобождение пигмента меланина (а, б), скопление опустошенных аутофагоцитирующих клеток и гранулоцитов (в), заполнение крупных опухолевых сосудов зернами меланина (г), полную деградацию опухоли со слабым проявлением теней клеток и глыбок меланина (д). Окр. Гематоксилин-эозином. Ув. х 200,х400.
«Способ экспериментальной биотерапии меланомы В16/F10» выполняется следующим образом.
Воспроизведение меланомы В16/F10. Самцам и самкам мышей линии BALB/c Nude под кожу спины ниже угла левой лопатки вводят 0,5 мл взвеси опухолевых клеток мышиной меланомы В16/F10 в физиологическом растворе в разведении 1:20. Для этого, соблюдая все условия асептики, ассистент фиксирует мышь спиной кверху, предварительно обработав кожу 5% спиртовым раствором йода.
Экспериментатор рукой в стерильной перчатке захватывает кожную складку, в центре которой прокалывает кожу и вводит опухолевую взвесь В16/F10. После извлечения иглы места введения плотно прижимает ватным тампоном, смоченным в 70% спирте с небольшим добавлением йода, на 1 минуту, чтобы исключить вытекание опухолевого материала.
Выделение митохондрий. Крысу умерщвляют с помощью гильотины. Печень перфузируют ледяным стерильным 0,9% раствором KCl. Митохондрии выделяют с применением дифференциального центрифугирования на высокоскоростной рефрижераторной центрифуге Avanti J-E, BECMAN COULTER, USA по методу Егоровой М.В. и Афанасьева С.А. (см. Егорова М.В., Афанасьев С.А. Выделение митохондрий из клеток и тканей животных и человека: Современные методические приемы. Сибирский медицинский журнал. 2011;26(1-1):22-28).
Для разрушения межклеточных связей, клеточной стенки и плазматических мембран применяют механическую обработку тканей с измельчением ножницами и гомогенизацией в стеклянном гомогенизаторе с тефлоновым пестиком (гомогенизатор Поттера-Эльвегейма). На каждый грамм ткани добавляют по 10 мл стерильной среды выделения (0,22 М маннитол, 0,3 М сахароза, 1мМ ЭДТА, 2 мМ TRIS-HCL, 10мМ HEPES, pH 7,4).
Ткани гомогенизируют и центрифугируют первый раз 10 мин. при скорости 1000 g, температура 0-2°С, второе и третье центрифугирование осуществляется при 20000 g, 20 мин., температура 0-2°С. Между центрифугированием проводят процедуру ресуспендирования осадка митохондрий в среде выделения. Митохондрии дополнительно очищают от лизосом, пероксисом, меланосом и т.п., центрифугируя в 23% градиенте Перколла.
Суспензию субклеточных структур наслаивают на градиент Перколла, центрифугируют 15 мин. при 21000 g, после этого наблюдается разделение на 3 фазы, оставляют нижний слой митохондрий и ресуспендируют средой выделения.
Следующую промывку митохондрий осуществляют путем центрифугирования в течение 10 мин. при 15000 g, температура 0-2°С. Митохондриальные образцы разводят 0,9% раствором NaCl до концентрация белка 3,3 мг белка в 0,3 мл физиологического раствора.
Проведение биотерапии митохондриями. Через 24 часа после перевивки меланомы В16/F10 мышам внутрибрюшинно вводили свежевыделенные митохондрии (3,3 мг белка на 1 животное в 0,3 мл физиологического раствора).
Далее митохондриальную терапию (МХ-терапия) проводили по схеме на 3, 5, 9, 13, 16, 19, 21 сутки.
Контролем служат мыши-самцы линии BALB/c Nude с перевивкой меланомы В16/F10, которым 2 раза в неделю внутрибрюшинно вводят 0,3 мл физиологического раствора.
Особенности динамики роста опухолей при самостоятельном росте меланомы В16/F10 у мышей линии BALB/c Nude при проведении биотератии митохондриями представлены в таблице.
Подкожная опухоль у мышей обоего пола стала определяться на 5-е сутки с момента перевивки опухоли, а регрессивный эффект от проведения МХ-терапии начали фиксировать у самцов с 8 суток роста опухоли. Средний объем опухоли в группе самцов с МХ-терапией на 8 сутки был в 3,0 раза меньше, чем в соответствующей контрольной группе. На 12 и 15 сутки эксперимента регрессивная разница в объеме опухолевой массы между группами составила 1,8 раза (p<0,05) (МХ-терапия). На 19 сутки фиксировали замедление роста опухоли в группе с МХ-терапией, где объем опухоли был ниже в 2 раза по сравнению с контрольными объемами. В конце эксперимента на 22 сутки разница средних объемов опухолевого узла составила 3,2 раза, т.е. в группе самцов с МХ-терапией определялось существенное торможение роста опухоли.
У всех самцов некрозы появлялись на 12 сутки эксперимента. Однако они отличались по количеству и характеристикам у мышей, получавших МХ-терапию и без лечения. Так, у самцов контрольной группы некрозы были сухие и имели вид тёмной корочки. Тогда как у мышей основной группы некрозы, практически с первого дня появления, были множественными, у ряда мышей несколько очагов сливались в один большой некроз, занимающий всю поверхность кожи над опухолью. Эти некрозы были влажными, сочились.
У самок на 5 сутки роста опухоли различий в объеме опухолевого очага между группами контрольная и МХ-терапия не фиксировали, однако выявлена статистически значимая разница при межполовом сравнении групп животных с МХ-терапией. Определено, что у самок с МХ-терапией площадь опухолевого пятна в этот срок (5 суток) была в 1,4 раза (p<0,05) меньше, чем в соответствующей группе самцов. На 8 сутки у самок с МХ-терапией опухоль еще не сконцентрировалось в объемное образование, а оставалось в виде опухолевого плоского образования, лишь к 12 суткам опухоль из плоской формы сформировалось в объемное образование. На 12 сутки роста опухоли у самок с МХ-терапией отмечали меньшие объемы опухолевых узлов, и разница с соответствующими контрольными значениями составила 1,8 раза (p<0,05). В срок эксперимента - 15 сутки объем опухолевой массы в группе самок с МХ-терапией снижался в 2,2 раза по сравнению с контрольными величинами. На 19 сутки были выявлены как межполовые различия в развитии опухоли, так и различия в росте опухоли между контрольной группой и группой с МХ-терапией. В контрольных группах животных установлено, что по сравнению с самцами на 19 сутках эксперимента у самок объем опухолевой массы был в 1,4 раза (p<0,05) меньше. Кроме того в группах с МХ-терапией у самок объем опухолевого очага был меньше в 1,5 раза (p<0,05), чем у самцов, а по сравнению с соответствующей контрольной группой самок меньше в 2,1 раза. В результате к концу эксперимента - 22 сутки меньшие объемы опухолевых узлов сохранились у группы самок, получавших МХ-терапию, и разница с контрольной группой составила 2,7 раза (p<0,05).
У всех самок некрозы появлялись на 15 сутки эксперимента, так же, как и у самцов в контрольной группе. Некрозы были представлены 1 пятном. У самок основной группы некрозы были сухими.
Рост меланомы В16/F10 у мышей BALB/c Nude - самцов и самок контрольной группы (без проведения митохондриальной терапии) характеризовался однотипной морфологической картиной.
По всей поверхности препарата меланомы В16/F10 отмечалось плотное и местами разрозненное заселение крупными опухолевыми клетками эпителиоподобного типа. Форма клеток была неправильная, в основном, пластинчато-округлая или угольчатая с отростчатой цитоплазмой и гиперхромными ядрами с эксцентричным расположением. В ядрах обнаруживались фигуры патологического митоза. Отмечалась обильная васкуляризация опухоли с образованием «муфт» из опухолевой ткани вокруг множественных сосудов с малым и широким просветом, заполненных кровью. Количество пигментсодержащих клеток было умеренным, а сам меланин во внеклеточном пространстве не агрегировался. На отдельных участках можно было видеть мелкие очаги некроза (см. фиг. 1а-г).
Структурные изменения меланомы В16/F10 у мышей BALB/c Nude - самцов под влиянием митохондриальной терапии.
Изучение морфологических изменений меланомы В-16 у крыс-самцов после проведения МХ-терапии выявило массовую гибель опухолевых клеток на фоне гиалиноза матрикса, выраженного отёка подкожной клетчатки с сегрегацией и некротизацией жировых скоплений. Оставшиеся единичные опухолевые клетки выявлялись в виде теней с признаками деградации ядерного материала. При этом, в микрокатине опухоли можно было обнаружить участки, указывающие на апоптотическую гибель клеток меланомы В16/F10, однако значительно преобладали признаки тотального некроза. Дегенеративные изменения опухолевых клеток, по-видимому, были связаны с процессами аутолиза и аутоокисления как последствия острой энергетической гипоксии. На признаки тотального некроза и ишемии опухоли указывало образование крупной полости, по краю которой формировалась фиброзная капсула с включениями лейкоцитов и распадающихся опухолевых клеток, продукты деградации которых заполняли полость. Регрессия опухоли сопровождалась исчезновением опухолевых «муфт» вокруг кровеносных сосудов, которые были тромбированы и склерозированы. На некоторых участках отмечалось скопление гиперхромных гранул, указывающих на активацию гранулоцитарных и фагоцитарных элементов в механизме регрессии опухоли под влиянием МХ-терапии (см. фиг. 2 а-е).
Структурные изменения меланомы В16/F10 у мышей BALB/c Nude - самок под влиянием митохондриальной терапии.
На микропрепарате меланомы В-16/F10 у мышей-самок, прошедших курс МХТ, определяются обширные поля гибели опухолевых клеток. Однако, на участках с оставшимися опухолевыми клетками четко прослеживается структурная деградация иного, чем у мышей-самцов, характера. Наряду с множественными мелкими участками некротических изменений, наблюдается выраженная фрагментация ядер и цитоплазмы с образованием апоптотических телец. На доминирование процессов апоптоза указывало и то, что на отдельных участках можно было обнаружить слегка сохранившие свои очертания клетки, подвергшиеся аутофагоцитозу, по контуру которых располагались мелкие агрегации коричневого цвета - пигмента меланина. По всей поверхности гистологического среза были рассеяны крупные конгломерации и мелкие гранулы высвобожденного меланина, который скапливался в расширенном русле сосудов опухоли, которые прекращали доставку в опухоль жизненно необходимых продуктов, образуя своеобразные хранилища пигмента. Процесс регрессии меланомы подтверждался значительным увеличением свободных фиброзированных зон, где сохранялись лишь слабые тени клеток и присущего ей меланина (см. фиг. 3а-д).
Приведенный иллюстративный материал убедительно демонстрирует возможность ингибирования роста меланомы В16/F10 у мышей BALB/c Nude - самцов и самок под влиянием митохондриальной терапии. Обращает внимание, что исходное состояние перевитой под кожу опухоли деградирует под влиянием МХ-терапии у мышей BALB/c Nude различного пола по-разному. Именно гендерные, по сути, гормональные различия обуславливают доминирование различных путей гибели клеток - некроза (у самцов) и апоптоза (у самок). Однако, несмотря на преимущество того или иного пути, четко проявляется доступность митохондриальной терапии к процессам предотвращения опухолевого роста.
Технико-экономическая эффективность «Способа экспериментальной биотерапии меланомы В16/F10» заключается в том, что применение митохондриальной терапии у мышей линии BALB/c Nude с меланомой В16/F10 замедляет рост опухоли у мышей обоего пола, по сравнению с ростом меланомы В16/F10 в контрольной группе, получавших инъекции физиологического раствора. Это дает возможность изучать патогенез злокачественного роста под влиянием введения суспензии митохондрий и механизм митохондриального воздействия, что важно для клиники.
Способ экономичен, доступен.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ предотвращения развития инфаркта миокарда мышей с меланомой, развившейся на фоне хронической нейрогенной боли | 2022 |
|
RU2786322C1 |
| Способ образования опухолевых узлов меланомы в организме экспериментальных животных | 2022 |
|
RU2796892C1 |
| СПОСОБ ВЫБОРА МИТОХОНДРИЙ ДЛЯ ЭКСПЕРИМЕНТАЛЬНОЙ ТЕРАПИИ МЕЛАНОМЫ В16-F10 | 2022 |
|
RU2794762C1 |
| Способ формирования опухолевого роста в легких экспериментальных животных | 2023 |
|
RU2810431C1 |
| Способ индуцирования роста аденокарциномы ободочной кишки человека в органах экспериментальных животных | 2023 |
|
RU2799251C1 |
| Способ моделирования первично-множественного роста злокачественных опухолей с подавлением одной опухоли другой в условиях первичного иммунодефицита | 2021 |
|
RU2750127C1 |
| Способ усиления роста меланомы В16/F10 по сравнению с ростом меланомы В16/F10 при самостоятельной перевивке и замедления роста LLC (карциномы Льюиса) по сравнению с ростом LLC при самостоятельной перевивке при первично-множественных злокачественных опухолях на фоне первичного иммунодефицита | 2021 |
|
RU2759487C1 |
| Способ создания полинеоплазии со стимуляцией опухолевого роста в условиях первичного иммунодефицита в эксперименте | 2021 |
|
RU2751930C1 |
| Способ обнаружения свойств митохондрий глиобластомы человека индуцировать экстракраниальный диффузный опухолевый рост в органах экспериментальных животных | 2023 |
|
RU2794641C1 |
| Способ отмены генетически детерминированного ингибирования роста злокачественной опухоли в эксперименте | 2019 |
|
RU2718671C1 |
Изобретение относится к онкологии, а именно к экспериментальной онкологии, и может быть использовано для изучения противоопухолевого воздействия на меланому в эксперименте. Для этого самкам и самцам мышей линии BALB/c Nude подкожно под лопатку слева вводят суспензию опухолевых клеток мышиной меланомы В16/F10 в 0,5 мл физиологического раствора в разведении 1:20. Через 24 часа после перевивки меланомы В16/F10 мышам внутрибрюшинно вводят свежевыделенные митохондрии из печени крыс из расчёта 3,3 мг белка на 1 животное в 0,3 мл физиологического раствора. Далее митохондриальную терапию проводят по схеме на 3, 5, 9, 13, 16, 19, 21 сутки. Изобретение позволяет замедлить рост опухоли у мышей обоего пола, изучить патогенез злокачественного роста под влиянием введения суспензии митохондрий и механизм митохондриального воздействия. 1 табл., 3 ил.
Способ экспериментальной биотерапии меланомы В16/F10, заключающийся в том, что самкам и самцам мышей линии BALB/c Nude подкожно под лопатку слева вводят суспензию опухолевых клеток мышиной меланомы В16/F10 в 0,5 мл физиологического раствора в разведении 1:20, через 24 часа после перевивки меланомы В16/F10 мышам внутрибрюшинно вводят свежевыделенные митохондрии из печени крыс из расчёта 3,3 мг белка на 1 животное в 0,3 мл физиологического раствора, далее митохондриальную терапию проводят по схеме на 3, 5, 9, 13, 16, 19, 21 сутки.
| СПОСОБ МОДИФИКАЦИИ ХРОНИЧЕСКОЙ БОЛЬЮ ЗЛОКАЧЕСТВЕННОГО РОСТА МЕЛАНОМЫ В У МЫШЕЙ | 2017 |
|
RU2650587C1 |
| Способ моделирования лимфогенного и гематогенного метастазирования мышиной меланомы В у белых нелинейных крыс | 2016 |
|
RU2615908C1 |
| ФРАНЦИЯНЦ Е.М | |||
| и др | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| LEO L.D | |||
| et al | |||
| The Complex Role of Autophagy in | |||

