Область техники, к которой относится изобретение
Настоящее изобретение относится к области биотехнологии для применения в сельском хозяйстве, в частности селекции и семеноводстве, а также генетике растений и может быть использовано для идентификации генотипов льна со сниженным содержанием линоленовой кислоты в масле.
Уровень техники
Лен обыкновенный (Linum usitatissimum L.) является одним из древнейших растений, возделываемых человеком, семена которого традиционно используют для получения пищевого масла [Weiss Е., Zohary D., Hopf М. 2012. Domestication of Plants in the Old World - The Origin and Spread of Domesticated Plants in South-west Asia, Europe, and the Mediterranean Basin. Oxford University Press, New York]. Семена льна являются одним из богатейших растительных источников полезных для здоровья ω-3 жирных кислот, а также содержат ряд других полезных для человека соединений (прежде всего лигнаны), поэтому употребление продуктов из семян льна препятствует возникновению и развитию сердечно-сосудистых, онкологических и других заболеваний [Muir A.D., Westcott N.D. 2003. Flax: The genus Linum. CRC Press, London; Goyal A., Sharma V., Upadhyay N., et al. 2014. Flax and flaxseed oil: an ancient medicine & modern functional food. J Food Sci Technol. 51, 1633-1653; Imran M., Ahmad N., Anjum F.M., et al. 2015. Potential protective properties of flax lignan secoisolariciresinol diglucoside. Nutr J. 14, 71; Abdelhamid A.S., Martin N., Bridges C., et al. 2018. Polyunsaturated fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst Rev. 7, CD012345; Kezimana P., Dmitriev A.A., Kudryavtseva A.V., et al. 2018. Secoisolariciresinol Diglucoside of Flaxseed and Its Metabolites: Biosynthesis and Potential for Nutraceuticals. Front Genet. 9, 641; Locke A., Schneiderhan J., Zick S.M. 2018. Diets for Health: Goals and Guidelines. Am Fam Physician. 97, 721-728; Parikh M., Netticadan Т., Pierce G.N. 2018. Flaxseed: its bioactive components and their cardiovascular benefits. Am J Physiol Heart Circ Physiol. 314, Н146-Н159; Campos J.R., Severino P., Ferreira C.S., et al. 2019. Linseed Essential Oil - Source of Lipids as Active Ingredients for Pharmaceuticals and Nutraceuticals. Curr Med Chem. 26, 4537-4558; Cullis C.A. 2019. Genetics and Genomics of Linum. Springer, Cham; Mali A.V., Padhye S.B., Anant S., et al. 2019. Anticancer and antimetastatic potential of enterolactone: Clinical, preclinical and mechanistic perspectives. Eur J Pharmacol. 852, 107-124]. Семена масличного льна применяются в производстве экологически безопасных красок, линолеума, композитных материалов, пищевых продуктов, биологически активных добавок, фармацевтических препаратов, а также функциональных кормов для животных [Kouba М., Mourot J. 2011. A review of nutritional effects on fat composition of animal products with special emphasis on n-3 polyunsaturated fatty acids. Biochimie. 93, 13-17; Singh K.K., Mridula D., Rehal J., et al. 2011. Flaxseed: a potential source of food, feed and fiber. Crit Rev Food Sci Nutr. 51, 210-222; Corino C., Rossi R., Cannata S., et al. 2014. Effect of dietary linseed on the nutritional value and quality of pork and pork products: systematic review and meta-analysis. Meat Sci. 98,679-688; Goyal A., Sharma V., Upadhyay N., et al. 2014. Flax and flaxseed oil: an ancient medicine & modern functional food. J Food Sci Technol. 51, 1633-1653; Campos J.R., Severino P., Ferreira C.S., et al. 2019. Linseed Essential Oil - Source of Lipids as Active Ingredients for Pharmaceuticals and Nutraceuticals. Curr Med Chem. 26, 4537-4558; Fombuena V., Petrucci R., Dominici F., et al. 2019. Maleinized Linseed Oil as Epoxy Resin Hardener for Composites with High Bio Content Obtained from Linen Byproducts. Polymers (Basel). 11]. Основными жирными кислотами льняного масла являются ненасыщенные линоленовая (ω-3, в среднем около 50%), линолевая (ω-6,15%), олеиновая (ω-9, 24%), а также насыщенные стеариновая (4%) и пальмитиновая (6%). Жирнокислотный состав масла значительно варьируется у различных сортов, например, разница в содержании линоленовой кислоты может достигать 10 и более раз, и именно содержание той или иной жирной кислоты определяет направление использования масла [Muir A.D., Westcott N.D. 2003. Flax: The genus Linum. CRC Press, London; Cloutier S., Ragupathy R., Niu Z., et al. 2011. SSR-based linkage map of flax (Linum usitatissimum L.) and mapping of QTLs underlying fatty acid composition traits. Molecular Breeding. 28, 437-451; Dmitriev A.A., Kezimana P., Rozhmina T.A., et al. 2020. Genetic diversity of SAD and FAD genes responsible for the fatty acid composition in flax cultivars and lines. BMC Plant Biol. 20, 301]. Масло с высоким содержанием линоленовой кислоты (более 50%) находит применение в фармацевтической, лакокрасочной и полимерной промышленности, однако быстрое «прогоркание» ограничивает его использование в пищевой промышленности, где более перспективным является использование семян со сниженным содержанием линоленовой кислоты [Muir A.D., Westcott N.D. 2003. Flax: The genus Linum. CRC Press, London; Goyal A., Sharma V., Upadhyay N., et al. 2014. Flax and flaxseed oil: an ancient medicine & modern functional food. J Food Sci Technol. 51, 1633-1653; Chen Y., Zhou X.R., Zhang Z.J., et al. 2015. Development of high oleic oil crop platform in flax through RNAi-mediated multiple FAD2 gene silencing. Plant Cell Rep. 34, 643-653; Брач Н.Б., Пороховинова E.A., Шеленга Т.В. 2016. Перспективы создания сортов масличного льна специализированного назначения Аграрный вестник Юго-Востока 1-2, 50-52; Cullis С.А. 2019. Genetics and Genomics of Linum. Springer, Cham; Fombuena V., Petrucci R., Dominici F., et al. 2019. Maleinized Linseed Oil as Epoxy Resin Hardener for Composites with High Bio Content Obtained from Linen Byproducts. Polymers (Basel). 11].
Известно, что ключевые гены, участвующие в синтезе жирных кислот льна, кодируют десатуразы, вносящие двойные связи в углеводородную цепь. FAD3A и FAD3B (десатуразы жирных кислот) катализируют превращение линолевой кислоты в линоленовую [Vrinten Р., Hu Z., Munchinsky М.А., et al. 2005. Two FAD3 desaturase genes control the level of linolenic acid in flax seed. Plant Physiol. 139, 79-87; Banik M., Duguid S., Cloutier S. 2011. Transcript profiling and gene characterization of three fatty acid desaturase genes in high, moderate, and low linolenic acid genotypes of flax (Linum usitatissimum L.) and their role in linolenic acid accumulation. Genome. 54, 471-483]. Длина паралогов FAD3A и FAD3B составляет около 3 тысяч нуклеотидов, они имеют 6 экзонов [Thambugala D., Duguid S., Loewen E., et al. 2013. Genetic variation of six desaturase genes in flax and their impact on fatty acid composition. Theor Appl Genet. 126, 2627-2641; Rajwade A.V., Kadoo N.Y., Borikar S.P., et al. 2014. Differential transcriptional activity of SAD, FAD2 and FAD3 desaturase genes in developing seeds of linseed contributes to varietal variation in alpha-linolenic acid content. Phytochemistry. 98, 41-53]. Показано, что полиморфизмы генов FAD3A и FAD3B ассоциированы с содержанием линолевой и линоленовой кислот в льняном масле, например, мутация в первом экзоне гена FAD3A (замена G на А в сайте СР027631.1:16092348 в сборке генома льна GCA_000224295.2/ASM22429v2 из базы NCBI, https://www.ncbi.nlm.nih.gov/), которая приводит к замене триптофана на стоп-кодон (результат замены триплета TGG на TGA), определяет сниженное содержание линоленовой кислоты в масле [Vrinten Р., Hu Z., Munchinsky М.А., et al. 2005. Two FAD3 desaturase genes control the level of linolenic acid in flax seed. Plant Physiol. 139, 79-87; Banik M., Duguid S., Cloutier S. 2011. Transcript profiling and gene characterization of three fatty acid desaturase genes in high, moderate, and low linolenic acid genotypes of flax (Linum usitatissimum L.) and their role in linolenic acid accumulation. Genome. 54, 471-483; Thambugala D., Duguid S., Loewen E., et al. 2013. Genetic variation of six desaturase genes in flax and their impact on fatty acid composition. Theor Appl Genet. 126, 2627-2641; Raj wade A.V., Kadoo N.Y., Borikar S.P., et al. 2014. Differential transcriptional activity of SAD, FAD2 and F AD3 desaturase genes in developing seeds of linseed contributes to varietal variation in alpha-linolenic acid content. Phytochemistry. 98, 41-53; You F.M., Xiao J., Li P., et al. 2018. Genome-Wide Association Study and Selection Signatures Detect Genomic Regions Associated with Seed Yield and Oil Quality in Flax. Int J Mol Sci. 19; Porokhovinova E.A., Shelenga T.V., Matveeva T.V., et al. 2019. Polymorphism of genes controlling low level of linolenic acid in lines from VIR flax genetic collection. Ecological Genetics. 17, 5-19; Dmitriev A.A., Kezimana P., Rozhmina T.A., et al. 2020. Genetic diversity of SAD and FAD genes responsible for the fatty acid composition in flax cultivars and lines. BMC Plant Biol. 20, 301].
Оценка жирнокислотного состава льняного масла, как правило, проводится хроматографическим методом [Ермаков А.И., Арасимович В.В., Ярош Н.П. 1987. Методы биохимического исследования растений. Л.: Агропромиздат, Ленинград; Bjelkova М., Nozkova J., Fatrcova-Sramkova K., et al. 2012. Comparison of linseed (Linum usitatissimum L.) genotypes with respect to the content of polyunsaturated fatty acids. Chemical Papers. 66, 972-976] и имеет ряд недостатков: не позволяет идентифицировать образцы с детерминирующими содержание жирных кислот мутациями в гетерозиготном состоянии; требует значительного числа семян для анализа; анализ пула семян, часть из которых имеет высокое, а часть низкое содержание линоленовой кислоты, приводит к получению неверного результата о том, что данный генотип характеризуется средним содержанием линоленовой кислоты в масле. В связи с вышеперечисленными особенностями оценка жирнокислотного состава часто выполняется лишь на поздних этапах селекционного процесса, что затрудняет эффективное создание сортов с необходимым содержанием линоленовой кислоты. Использование ДНК-маркеров решит обозначенные проблемы - позволит проводить анализ отдельных растений, используя для выделения ДНК лишь несколько листьев, давая возможность выполнять другие исследования этого генотипа и получить с него семена, а также различать искомые мутации в гетерозиготном и гомозиготном состояниях, что существенно повышает точность анализа и позволяет выполнять отбор ценных генотипов, начиная с ранних этапов селекционного процесса (например, с поколения F2 гибридов).
Таким образом, разработка надежной и удобной в применении ДНК-тест-системы для идентификация мутации в первом экзоне гена FAD3A, приводящей к снижению содержания линоленовой кислоты, откроет новые возможности для маркер-ориентированной селекции льна с целью создания сортов пищевого, фармацевтического или промышленного назначения, а также контроля сортовой чистоты при ведении семеноводства.
Раскрытие сущности изобретения
Целью создания изобретения являлась разработка ДНК-тест-системы, которая позволяет идентифицировать мутацию в первом экзоне гена FAD3A (замена G на А в сайте СР027631.1-.16092348 в сборке генома льна GCA_000224295.2/ASM22429v2 из базы NCBI), приводящую к сниженному содержанию линоленовой кислоты в масле, и делает возможным отбор растений льна с наличием искомой мутации в гомозиготном состоянии или ее отсутствием и, соответственно, необходимым жирнокислотным составом масла даже на ранних этапах селекционного процесса. Для идентификации мутации в гене FAD3А, приводящей к снижению содержания линоленовой кислоты, авторами был выполнен анализ полиморфизма гена FAD3A на основе данных глубокого секвенирования для более чем 400 сортов, линий и гибридов льна, различающихся по содержанию линоленовой кислоты в масле, и установлено, что мутация в первом экзоне этого гена, приводящая к замене триптофана на стоп-кодон, определяет сниженное содержание линоленовой кислоты, а также выявлены другие ДНК-полиморфизмы в гене FAD3A, информация о которых необходима для успешного подбора праймеров для создания ДНК-тест-системы. На основе полученных данных разработаны специфичные праймеры для амплификации участка гена FAD3A, содержащего сайт искомой мутации и при этом не содержащего сайты других ДНК-полиморфизмов, для последующей оценки полученных ампликонов методом анализа кривых плавления высокого разрешения (HRM, High Resolution Melting), а также два стандарта, представляющие собой участки гена FAD3A, один из которых несет мутацию, а в другом она отсутствует.
Настоящее изобретение относится к новому способу идентификации генотипов льна со сниженным содержанием линоленовой кислоты в масле, который представляет собой анализ кривых плавления высокого разрешения участка первого экзона гена FAD3A. Сдвиг кривой плавления при анализе участка гена FAD3A, несущего мутацию в первом экзоне, по сравнению с тем же участком гена без мутации служит маркером сниженного содержания линоленовой кислоты в льняном масле.
Еще одним аспектом настоящего изобретения является возможность использования кривой плавления участка первого экзона гена FAD3A в качестве маркера для определения содержания линоленовой кислоты в масле различных образцов льна.
Способ определения мутации включает следующие стадии:
а) получение материала растений льна (например, несколько листьев);
б) выделение ДНК;
в) проведение реакции амплификации участка гена FAD3A с использованием пары генспецифичных праймеров и матрицы (ДНК, полученной на стадии б), или стандартов, представляющих собой участки гена FAD3A, один из которых несет мутацию, а в другом она отсутствует);
г) проведение анализа кривых плавления высокого разрешения амплифицированного фрагмента гена FAD3A для исследуемого образца и двух стандартов;
д) сравнение полученной кривой плавления для изучаемого генотипа льна с кривыми плавления двух стандартов; сходство кривой плавления анализируемого образца и стандарта, несущего мутацию, свидетельствует о присутствии мутации в изучаемом генотипе в гомозиготном состоянии; сходство кривой плавления анализируемого образца и стандарта, не имеющего мутации, свидетельствует об отсутствии мутации в изучаемом генотипе; форма и положение кривой плавления анализируемого образца отличные от двух стандартов свидетельствуют о наличии мутации в изучаемом генотипе в гетерозиготном состоянии.
В отдельном предпочтительном воплощении данного изобретения последовательности праймеров представлены SEQ ID NO: 1 и 2 (Перечень последовательностей).
Еще одним аспектом настоящего изобретения является набор праймеров для осуществления полимеразной цепной реакции (ПНР) с целью последующего определения наличия мутации в первом экзоне гена FAD3A, имеющих последовательности SEQ ID NO: 1 и 2 (Перечень последовательностей).
В еще одном воплощении способа определения мутации на стадии в) реакция амплификации участка гена FAD3A представляет собой ПЦР.
В следующем воплощении заявленного способа на стадии д) в качестве стандартов используют последовательности ДНК SEQ ID NO: 3 и 4 (Перечень последовательностей).
Краткое описание фигур и таблиц
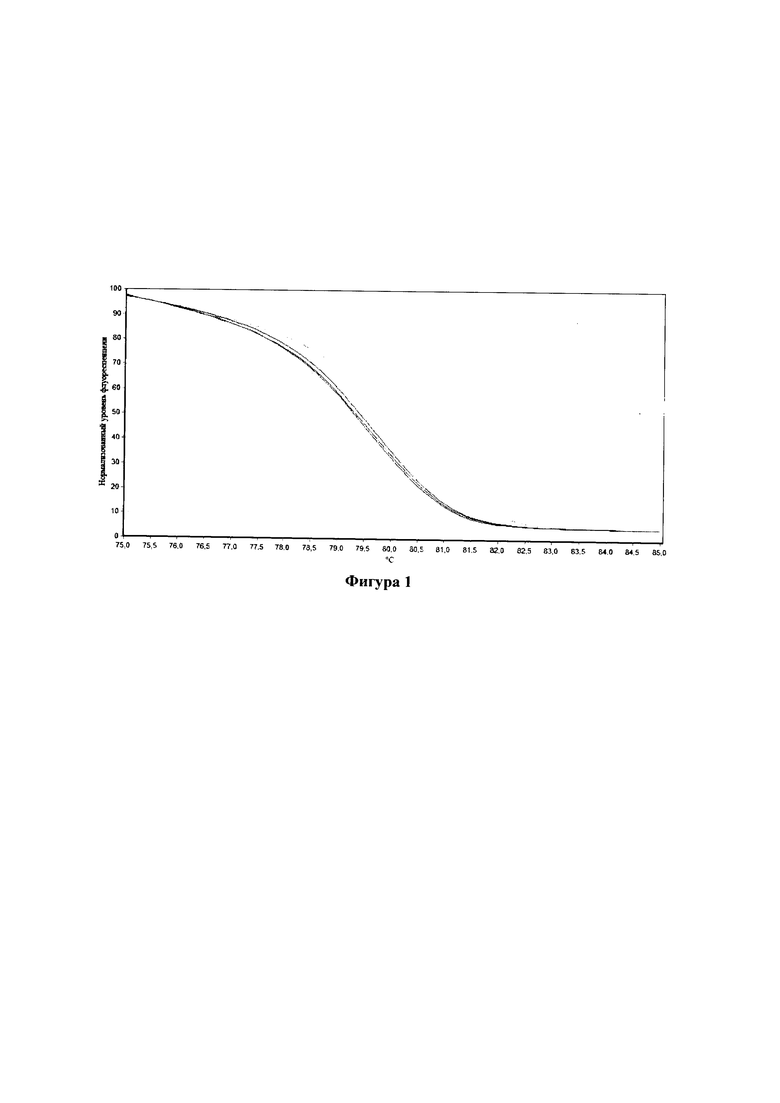
Фигура 1. Кривые плавления высокого разрешения ампликонов двух стандартов в трех технических повторностях: референсного FAD3A_E1_ref (без мутации, справа) и альтернативного FAD3A_E1_alt (с мутацией, слева)
Осуществление изобретения
Данное изобретение предлагает способ идентификации генотипов льна с определенным содержанием линоленовой кислоты в масле, основанный на анализе кривых плавления высокого разрешения для ампликона участка гена FADS А, несущего мутацию, которая приводит к замене триптофана на стоп-кодон.
Образцы тканей для анализа
В качестве материала для анализа могут быть использованы различные ткани растения льна, однако предпочтительным вариантом является использование нескольких листьев, что дает возможность выполнить другие исследования этого генотипа и получить с растения семена.
Выделение ДНК
Способы выделения ДНК из тканей растений хорошо известны и, как правило, включают следующие стадии: измельчение растительного материала, лизис клеток, выделение ДНК и ее очистку, проверку качества ДНК электрофорезом в 1%-ном агарозном геле в присутствии красителя бромистого этидия или его аналогов, а также спектрофотометрическое определение количества ДНК. Гомогенизацию материала можно проводить вручную с использованием пестиков или с помощью гомогенизаторов, например, MagNA Lyser (Roche, Швейцария) или TissueLyser II (Qiagen, Германия) в пробирках с керамическими или стальными шариками. Для выделения ДНК могут быть использованы различные протоколы или наборы реактивов. В классических методах выделения ДНК из растений в качестве лизирующего агента используется буфер, содержащий гуанидин хлорид или ЦТАБ (цетилтриметиламмоний бромид) с последующей экстракцией хлороформом. Для выделения ДНК из растений также доступны коммерческие наборы, например, ДНК-Экстран-3 (Синтол, Россия) и DNeasy Plant Mini Kit (Qiagen, Германия).
Выбор специфичных праймеров
Выбор специфичных праймеров осуществлялся с использованием полученных авторами и другими исследователями данных о полиморфизмах гена FAD3A, а также программ NCBI Primer-BLAST [Ye J., Coulouris G., Zaretskaya I., et al. 2012. Primer-BLAST: a tool to design target-specific primers for polymerase chain reaction. BMC Bioinformatics. 13, 134] и MethyMer [Krasnov G.S., Melnikova N.V., Lakunina V.A., et al. 2018. MethyMer: Design of combinations of specific primers for bisulfite sequencing of complete CpG islands. J Bioinform Comput Biol. 16, 1840004]. Подбор праймеров для амплификации участка гена FAD3A, в котором локализована мутация, приводящая к замене триптофана на стоп-кодон, с целью дальнейшего анализа кривых плавления высокого разрешения проводился в соответствии со стандартными рекомендациями и требованиями метода. Кроме того, праймеры подбирались таким образом, чтобы в ампликоне отсутствовали сайты ДНК-полиморфизмов помимо сайта искомой мутации. В предпочтительном воплощении используются праймеры SEQ ID NO: 1 и 2 (Перечень последовательностей).
Определение мутации в первом экзоне гена FAD3A. приводящей к замене триптофана на стоп-кодон
Метод анализа кривых плавления высокого разрешения (HRM, High Resolution Melting) широко применяется для идентификации ДНК-полиморфизмов и генотипирования образцов [Montgomery J.L., Sanford L.N., Wittwer С.Т. 2010. High-resolution DNA melting analysis in clinical research and diagnostics. Expert Rev Mol Diagn. 10, 219-240; Druml В., Cichna-Markl M. 2014. High resolution melting (HRM) analysis of DNA-its role and potential in food analysis. Food Chem. 158, 245-254; Simko I. 2016. High-Resolution DNA Melting Analysis in Plant Research. Trends Plant Sci. 21, 528-537; Tamburro M., Ripabelli G. 2017. High Resolution Melting as a rapid, reliable, accurate and cost-effective emerging tool for genotyping pathogenic bacteria and enhancing molecular epidemiological surveillance: a comprehensive review of the literature. Ann Ig. 29, 293-316]. В настоящем изобретении для образцов растений льна используется анализ кривых плавления высокого разрешения ампликона размером 59 п. н., полученного с использованием праймеров FAD3A_E1_F (SEQ ID NO: 1) и FAD3A_E1_R (SEQ ID NO: 2) (Перечень последовательностей) посредством полимеразной цепной реакции (ПЦР).
Выбор образцов сравнения
Для сравнения результатов анализа кривых плавления высокого разрешения, полученных для образцов льна, с целью идентификации мутации в первом экзоне гена FAD3A, приводящей к замене триптофана на стоп-кодон, используются два стандарта, первый из которых представляет собой вариант последовательности ДНК, в которой отсутствует искомая мутация, а второй - вариант последовательности ДНК, в которой искомая мутация присутствует. В предпочтительном воплощении используются стандарты FAD3A_E1_ref (SEQ ID NO: 3) и FAD3A_E1_alt (SEQ ID NO: 4) (Перечень последовательностей).
Далее настоящее изобретение детально проиллюстрировано с использованием конкретных примеров, представляющих предпочтительные воплощения изобретения. В то же время, изобретение не ограничивается описанными воплощениями, а включает любые альтернативные варианты и модификации, применение которых возможно с учетом сущности изобретения.
Пример 1. Подготовка образцов
Семена сортов и линий льна обрабатывали 3% раствором гипохлорита натрия, промывали дистиллированной водой и проращивали в чашках Петри на фильтровальной бумаге, смоченной водой. Для выделения ДНК использовали листья 7-дневных проростков.
Пример 2. Выделение ДНК
Листья измельчали на гомогенизаторе MagNA Lyser (Roche, Швейцария), после чего проводили экстракцию ДНК набором ДНК-Экстран-3 (Синтол, Россия) согласно протоколу производителя. Качество ДНК оценивали методом электрофореза в 1% агарозном геле. Концентрацию ДНК определяли на флуориметре Qubit 2.0 (Life Technologies, США) или спектрофотометре NanoDrop ND-1000 (Thermo Fisher Scientific, США).
Пример 3. Амплификация участка гена FAD3A, в котором локализована мутация, приводящая к замене триптофана на стоп-кодон, и HRM-анализ
1. Готовили реакционную смесь для амплификации, смешивая все компоненты реакции, за исключением ДНК, из расчета 1 реакция объемом 20 мкл для каждого образца и стандарта, плюс 10%. Состав реакционной смеси для амплификации участка гена FAD3A, в котором локализована мутация, приводящая к замене триптофана на стоп-кодон, был следующим: 1x Taq Turbo буфер (Евроген, Россия), 250 нМ дезоксинуклеозидтрифосфатов (Евроген), 350 нМ праймера FAD3A_E1_F (SEQ ID NO: 1), 350 нМ праймера FAD3A_E1_R (SEQ ID NO: 2), 1x краситель EvaGreen (Biotium, США), 2 ед. HS Taq ДНК-полимеразы (Евроген).
2. В оптически прозрачные пробирки для амплификации вносили по 15 мкл приготовленной реакционной смеси без ДНК.
3. Вносили в пробирки по 5 мкл ДНК (концентрация 5 нг/мкл для образцов и 5 фг/мкл для стандартов), плотно закрывали пробирки.
4. Помещали пробирки в приборное отделение амплификатора, задавали названия ячеек и программу: +95°С - 10 мин - первичная денатурация ДНК; 50 циклов амплификации: +95°С - 15 сек - денатурация, +60°С - 1 мин - отжиг праймеров и элонгация цепи ДНК (детекция на канале Green); HRM-анализ (+65°С - +90°С, шаг 0,1°С, детекция на канале Green). Затем запускали амплификатор Rotor-Gene Q (Qiagen, Германия).
5. Проводили обработку данных с использованием программного обеспечения амплификатора (Rotor-Gene Q Series Software) и представляли результаты в графическом и/или табличном виде. В качестве двух стандартов (с референсным и альтернативным аллелем) использовали ампликоны, полученные с последовательностей FAD3A_E1_ref (SEQ ID NO: 3) и FAD3A_E1_alt (SEQ ID NO: 4) соответственно (Фиг. 1).
Приведенное выше подробное описание изобретения и его конкретных воплощений, представленных в примерах, предназначено исключительно для более полного уяснения сущности заявленного изобретения, но не для его ограничения. Могут быть внесены изменения, которые, однако, будут соответствовать сущности настоящего изобретения, определяющейся прилагаемой формулой изобретения.
--->
Перечень последовательностей
<210> 1
<211> 19
<212> FAD3A_E1_F
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 1
AAG CTG GAC AGC TGG ACT G (SEQ ID NO: 1)
<210> 2
<211> 23
<212> FAD3A_E1_R
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 2
CAG AAC ATG GTT CCT TGA GCA AT (SEQ ID NO: 2)
<210> 3
<211> 59
<212> FAD3A_E1_ref
<213> Искусственная последовательность
<220>
<223> ДНК-последовательность стандарта для референсного аллеля
<400> 3
AAG CTG GAC AGC TGG ACT GTC TGG CCT CTC TAC TGG ATT GCT CAA GGA
ACC ATG TTC TG (SEQ ID NO: 3)
<210> 4
<211> 59
<212> FAD3A_E1_alt
<213> Искусственная последовательность
<220>
<223> ДНК-последовательность стандарта для альтернативного аллеля
<400> 4
AAG CTG GAC AGC TGG ACT GTC TGA CCT CTC TAC TGG ATT GCT CAA GGA
ACC ATG TTC TG (SEQ ID NO: 4)
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ идентификации мутации в пятом экзоне гена FAD3A, приводящей к снижению содержания линоленовой кислоты в льняном масле | 2022 |
|
RU2806563C1 |
| Способ идентификации мутации во втором экзоне гена FAD3B, приводящей к снижению содержания линоленовой кислоты в льняном масле | 2022 |
|
RU2806561C1 |
| Способ мультиплексной идентификации 32 генетических маркеров льна | 2022 |
|
RU2804939C1 |
| Способ мультиплексной идентификации 52 генетических маркеров хозяйственно ценных признаков льна | 2023 |
|
RU2826718C1 |
| Способ генотипирования крупного рогатого скота по аллелям 878 СТ гена scd1 (rs41255693) методом ПЦР в режиме реального времени | 2020 |
|
RU2744174C1 |
| SNP-панель для генотипирования и геномной селекции подсолнечника по содержанию жирных кислот в масле семян | 2018 |
|
RU2717642C1 |
| Brassica juncea КАЧЕСТВА ОМЕГА-9 | 2009 |
|
RU2557316C2 |
| Тест-система и способ выявления A, B, D мутаций гена NPM1 для количественного определения минимальной остаточной болезни | 2023 |
|
RU2830545C1 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| СПОСОБ АНАЛИЗА СОМАТИЧЕСКИХ МУТАЦИЙ В ГЕНЕ PI3K С ИСПОЛЬЗОВАНИЕМ LNA-БЛОКИРУЮЩЕЙ МУЛЬТИПЛЕКСНОЙ ПЦР И ПОСЛЕДУЮЩЕЙ ГИБРИДИЗАЦИЕЙ С ОЛИГОНУКЛЕОТИДНЫМ БИОЛОГИЧЕСКИМ МИКРОЧИПОМ (БИОЧИПОМ) | 2013 |
|
RU2549682C1 |
Изобретение относится к области биотехнологии. Предложен способ идентификации мутации в первом экзоне гена FAD3A - замена триплета TGG на TGA и, как следствие, замена триптофана на стоп-кодон, приводящей к снижению содержания линоленовой кислоты в льняном масле. Способ основан на амплификации фрагмента гена FAD3A исследуемого образца и двух стандартов, с последующим анализом кривых плавления высокого разрешения. Изобретение позволяет, начиная с ранних этапов селекционного процесса, с высокой точностью отбирать растения льна с искомой мутацией гена FAD3A в гомозиготном состоянии или без нее, которые будут иметь определенное содержание линоленовой кислоты в масле. 1 ил., 1 пр.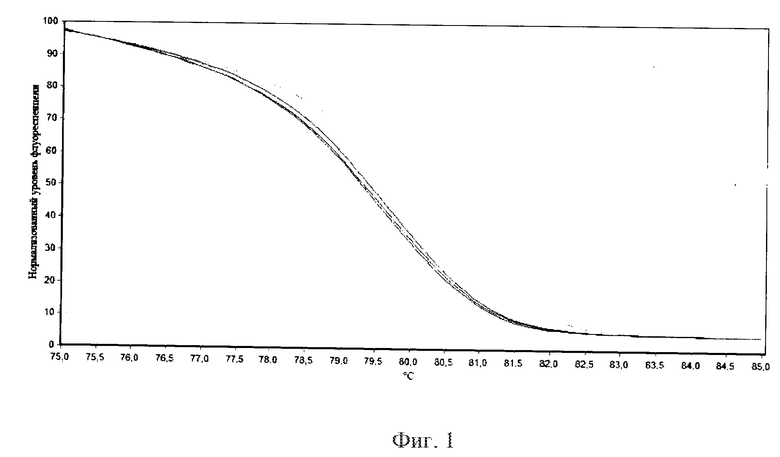
Способ идентификации мутации в первом экзоне гена FAD3A, приводящей к снижению содержания линоленовой кислоты в льняном масле, путем проведения амплификации участка первого экзона гена FAD3A, несущего сайт СР027631.1:16092348 в сборке генома льна GCA_000224295.2/ASM22429v2, замена G на А в котором в результате изменения триплета TGG на TGA приводит к замене триптофана на стоп-кодон, с использованием праймеров, представленных последовательностями SEQ ID NO: 1 и SEQ ID NO: 2, с последующим анализом кривых плавления высокого разрешения для исследуемых образцов льна в сравнении с ампликонами, полученными с двух стандартов, в качестве которых использовали последовательности с референсным аллелем SEQ ID NO: 3 и альтернативным аллелем SEQ ID NO: 4.
| ПОРОХОВИНОВА Е.А | |||
| "Генетическая коллекция льна (LINUM USITATISSIMUM L.): создание, анализ и перспективы использования", Диссертация: ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ НАУЧНОЕ УЧРЕЖДЕНИЕ "ФЕДЕРАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ ЦЕНТР ВСЕРОССИЙСКИЙ ИНСТИТУТ ГЕНЕТИЧЕСКИХ РЕСУРСОВ РАСТЕНИЙ ИМЕНИ Н | |||
| И | |||
| ВАВИЛОВА" (ВИР), Санкт-Петербург 2019 г., с.307-311 | |||
| RU |

