Изобретение относится к области сельского хозяйства, в частности, технологии получения семян и плодов сельскохозяйственных растений, обладающих генетически детерминированным барьером самонесовместимости. Может быть использовано для снятия самонесовместимости у плодовых деревьев, размножаемых вегетативно, самнесовместимость делает необходимой закладку многосортовых садов, так как высадка деревьев одного сорта делает такие сады бесплодными (к примеру, вишни, сливы и др.)
Уровень техники изобретения
Процессы опыления и оплодотворения у растений чрезвычайно сложны, они ведут за собой изменения на клеточном, физиологическом и транскриптомном уровнях, путем активации специфических генов. У многих цветущих растений встречается самонесовместимость (СН) - внутривидовой репродуктивный барьер, который является генетическим механизмом, помогающим избежать депрессии инбридинга путем отказа от самоопыления. Это явление способствует изменению внутри видов, но для некоторых растений, таких как слива, вишня, груша и др. СН является скорее помехой, чем преимуществом в сельскохозяйственном производстве. В последние годы накоплено некоторое количество доказательств указывающих на участие каспазо-подобных протеаз в программируемой клеточной гибели (ПКГ) растений (Del Pozo and Lam 1998; Bonneau et al. 2008; Dickman et al. 2017; Zhang et al. 2020). В наших предыдущих исследованиях было показано, что ПГК является фактором самонесовместимости на основе S-РНКазы у Petunia hybrida L. Остановка роста самонесовместимых пыльцевых трубок происходит при повышении уровня активности каспазо-подобных протеаз в первые часы опыления и к моменту остановки роста пыльцевых трубок наблюдаются все признаки ПКС (нарушение целостности плазматической мембраны, деградация/фрагментация ДНК, разрушение внутренней структуры пыльцевой трубки: отсутствие вакуолей, нарушение тургора и отделение клеточной плазматической мембраны от клеточной стенки) (Kovaleva at al. 2020; Zakharova et al. 2021).
До настоящего времени не известно работ и патентных материалов, в которых бы был предложен способ решения проблемы снятия самонесовместимости у растений.
Del Pozo и Lam (Del Pozo О, Lam E (1998) Caspases and programmed cell death in the hypersensitive response of plants to pathogens. Curr Biol 8:1129-1132) выявили, что гиперчувствительный ответ индуцируется определенными патогенами растений и включает запрограммированную гибель клеток для ограничения распространения патогенов из очага инфекции. Авторы в своей работе исследовали участие каспаз в гибели клеток растений при гиперчувствительном ответе. Также показали, что пептидные ингибиторы, специфичные для каспаз, Ac-YVAD-CMK и Ac-DEVD-CHO, могут устранять индуцированный бактериями ПКС растений, но не оказывают значительного влияния на индукцию других аспектов гиперчувствительного ответа. Для нас эта работа интересна тем, что наблюдается эффект снятия ПКС при обработке ингибиторами каспаз, что происходит и в нашем случае, однако это совершенно другая система и в целом область исследования.
В 2008 году вышла обзорная статья по растительным каспазо-подобным протеазам Bonneau с соавторами (Bonneau L, Ge Y, Drury GE, Gallois P (2008) What happened to plant caspases? J Exp Bot 59:491-499), где раскрыты такие аспекты как ПКС и каспазоподобная активность у растений, возникновение метакаспаз, обнаружение саспазы - сериновой протеазы с каспазо-подобной активностью, субклеточная локализация каспазо-подобной активности. В целом обзор очень полезный и интересный, но в нем абсолютно не освещен момент участия каспазо-подобных протеаз в процессах опыления и оплодотворения.
Zhang с соавторами (Zhang QF, Li J, Bi FC, Liu Z, Chang ZY, Wang LY, Huang LQ, Yao N (2020) Ceramide-induced cell death depends on calcium and caspase-like activity in rice. Front Plant Sci 11:145) также изучали ПКС у растений, участие каспазо-подобных протеаз, роль кальция и индукцию ПКС в протопластах риса (Oryza sativa). Исследователь показали, что активность протеазы, подобной каспазе-3, значительно увеличивалась в протопластах, обработанных церамидом С6, а специфический ингибитор каспазы предотвращал гибель клеток, вызванную церамидом Сб. Однако, система культивирования протопластов далека от процессов, происходящих в целом растении.
Наиболее близким к предлагаемому техническому решению является работа Bosch и Franklin-Tong (Bosch М, Franklin-Tong VE (2007) Temporal and spatial activation of caspase-like enzymes induced by self-incompatibility in Papaver pollen. Proc Natl Acad Sci U S A 104:8327-1832). В данном исследовании ученые показали, что у Papaver rhoeas самонесовместимость активирует каспазо-3-подобную протеазу/DEVDase в несовместимой пыльце, которая играет ключевую роль в регулировании ПКС. В работе детально охарактеризована каспазо-подобная активность, индуцированная СН; исследование дает представление о временной и пространственной активации растительных каспазо-подобных ферментов. Авторы показали, что СН также активирует VEIDase и LEVDase и что VEIDase играет роль в индуцированной самонесовместимостью ПКС.DEVDase и VEIDase активируются очень быстро: обнаруживаются в течение 1-2 часов после индукции самонесовместиости; пик активности LEVDase наступает позже. Важно то, что исследователи показывают визуализацию активности DEVDase в клетках высших растений на живых клетках; DEVDase имеет цитозольную и ядерную локализацию. Авторы также демонстрируют, что СН вызывает быстрое и значительное цитозольное подкисление, которое соответствует оптимуму рН in vitro каспазной активности. В нашей работе мы наблюдали подобный эффект - активизирование каспазо-3-подобной протеазы в первые 2-4 часа самонесовместимого опыления и ее локализацию в цитозоле пыльцевой трубки. Но система самонесовмесимости у мака (Papaver rhoeas) отличается от таковой у петунии (Petunia hybrida L.), крое того, все работы авторы выполняют в экспериментальной системе in vitro, где искусственно вызывается реакция самонесвместимости в пыльцевых трубках, ученые не работают in vivo с целыми растениями и не проводили обработку ингибиторами каспаз на предмет снятия самонесовместимости.
Техническим результатом нашей работы является снятие реакции самонесовместимости у Petunia hybrida L. по средством обработки ингибитором каспазы-3 и получение семян от самоопыления самонесовместимых растений.
Технический результат достигается тем, что при обработке ингибитором фермента ПКС каспазы-3, не происходит гибель самонесовместимых пыльцевых трубок, они дорастают до завязи пестика, где происходит оплодотворение и завязывание семян.
Описание чертежей
На Фигуре 1 показаны результаты несовместимого опыления после обработки ингибитором каспазы-7 в концентрации 0,25 mM за 2 часа до опыления. А) Рост пыльцевых трубок в тканях пестика (флуоресцентная микроскопия): Б) Фотография коробочки петунии, полученной после обработки ингибитором каспазы-3 в концентрации 0,25 mM за 2 часа до самонесовместимого опыления.
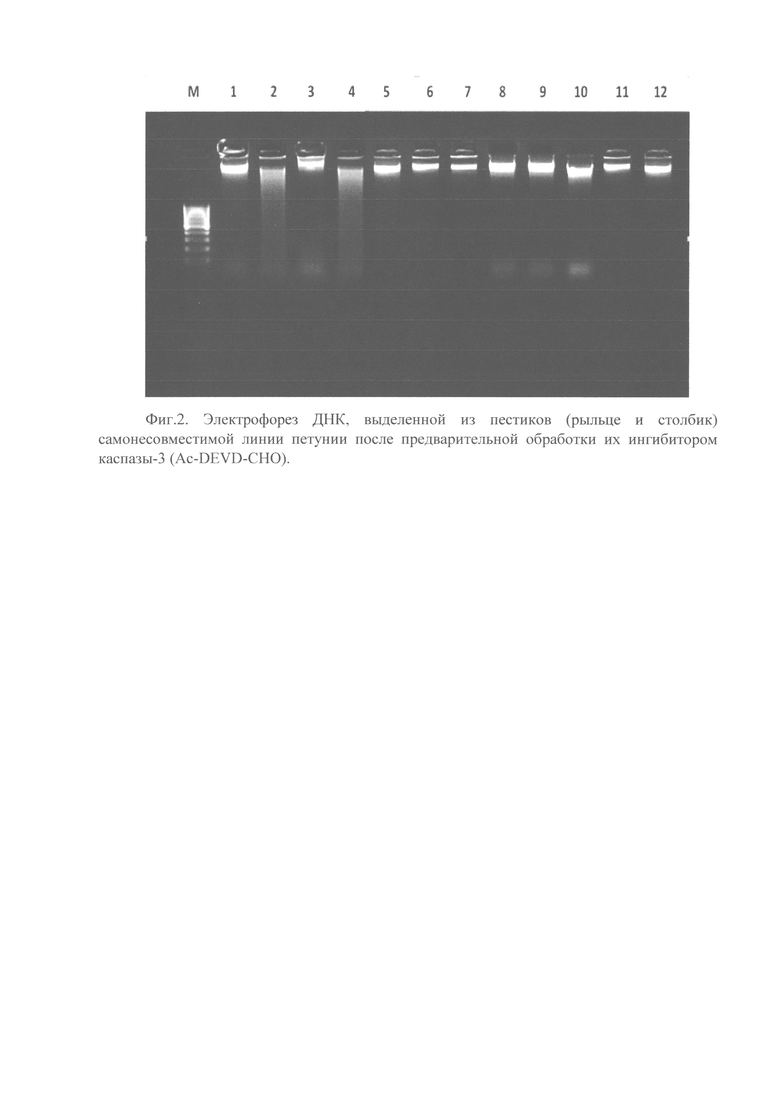
На Фигуре 2 показана электрофореграмма ДНК, выделенной из пестиков (рыльце и столбик) самонесовместимой линии петунии после предварительной обработки их ингибитором каспазы-3 (Ac-DEVD-CHO). М, молекулярный маркер 1 кб ДНК-лестница (Евроген, Россия); 1 - неопыленные пестики из почек с антоцианом самонесовместимой линии петунии (- деградация ДНК); 2 - пыльники из почек без антоциана самонесовместимой линии петунии (+деградация ДНК); 3 - пестики через 6 ч после перекрестно совместимого опыления (- деградация ДНК); 4 - пестики через 6 ч после самонесовместимого опыления (+деградация ДНК); 5 - пестики через 6 ч после перекрестно совместимого опыления + ингибитор 0,25 мм (Ac-DEVD-CHO) за 2 ч до опыления; 6 - пестики через 6 ч после перекрестно совместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) во время опыления; 7 - пестики через 6 ч после перекрестно совместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) через 2 ч после опыления; 8 - пестики через 6 ч после самонесовместимого опыления+0,25 мм ингибитор (Ac-DEVD-CHO) за 2 ч до опыления; 9 - пестики через 6 ч после самонесовместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) во время опыления; 10 - пестики через 6 ч после самонесовместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) через 2 ч после опыления; 11 - пестики через 24 ч после самонесовместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) во время опыления; 12 - пестики через 24 ч после самонесовместимого опыления+ингибитор 0,25 мм (Ac-DEVD-CHO) за 2 ч до опыления.
Сущность изобретения
Сущность нашего изобретения заключается в том, что обработка рылец пестиков петунии ингибитором каспазо-3-подобной протеазы за 2 часа до самонесовместимого опыления в концентрации 0,25 mM ингибирует процесс ПКС в самонесовместимых пыльцевых трубках, что снимает механизм СН, пыльцевые трубки (ПТ) дорастают до завязи, где происходит оплодотворение и завязывание семян. В норме (без обработки) самонесовместимые ПТ у самонесовместимой петунии прорастают на рыльце пестика, врастают в столбик, растут в течение 6-8 часов и останавливают свой рост на расстоянии примерно 8 мм от поверхности рыльца (ПТ гибнут по средствам ПКС) в результате функционирования механизма гаметофитной самонесовместимости S-РНКазного типа.
Примеры
Далее описание настоящего изобретения будет продолжено на примере его предпочтительного варианта осуществления.
В данной работе эксперименты проводились на системе пыльца-пестик петунии in vivo после самонесовместимого, и перекрестного совместимого опылений. Petunia hybrida является классическим объектом при изучении биохимических механизмов регуляции роста мужского гаметофита, в особенности - механизмов несовместимости при самоопылении.
Клонально размноженные растения Petunia hybrida двух линий, самосовместимая и самонесовместимая, были выращены и адаптированы к почвенным условиям при естественном освещении в теплице. Микроклональное размножение производили с использованием среды Мурасиге-Скуга, выращивали микроклоны в климатической камере при температуре 26°С и 16-ти часовом световом дне. Эксперименты проводились на системе пыльца-пестик in vivo после самонесовместимого, и перекрестного совместимого опылений. За день до эксперимента, с распустившихся цветков собирали пыльцу. Чтобы избежать самоопыления, из цветков удалялись пыльники. После кастрации на цветок одевался марлевый изолятор. К опылению приступали на следующий день после кастрации.
Согласно плану обработки на рыльце цветка наносилось 5 мкл ингибирующего раствора с разной концентрацией Ac-DEVD-CHO (Caspase-3 Inhibitor I, N-Ac-Asp-Glu-Val-Asp-CHO) (EMD Biosciences and BIOMOL, USA) 0.25, 0.5, 1 и 1,99 mM за 2 часа, через 2 часа и во время опыления. Контролем в данном опыте служили неопыленные цветки, на рыльца пестиков которых наносили каплю дистиллированной воды. Сбор и фиксацию материала (опыленных пестиков) производили через 2, 4, 6 и 24 часа после опыления.
Помимо контролирования завязываемости семян проводили визуализацию роста ПТ в тканях пестика методом флуоресцентной микроскопии (окрашивание пыльцевых трубок анилиновым голубым). Метод визуализации растущих ПТ с помощью окрашивания анилиновым голубым основан на способности флуорохрома анилинового голубого связываться с коллозой, входящей в состав оболочки пыльцевого зерна и пыльцевых трубок.
В опытах использовали опыленные пестики, зафиксированные в уксусном алкоголе (90% раствор этанола и уксусной кислоты в соотношении 3:1). Мацерацию проводили в 20%-ном спиртовом растворе КОН в течение 20-40 минут, после чего пестики дважды промывали дистиллированной водой и окрашивали 0,01%-ным раствором анилинового голубого в течение 30-40 минут. Окрашенные пестики переносили на предметное стекло в каплю глицерина, смешанного с водой (1:1), накрывали покровным стеклом, слегка раздавливали и исследовали под флуоресцентным микроскопом Zeiss Axioplan (Carl Zeiss, Германия) с длиной волны 365/420 nm. В каждом варианте эксперимента исследовали не менее 200 пыльцевых зерен/пыльцевых трубок.
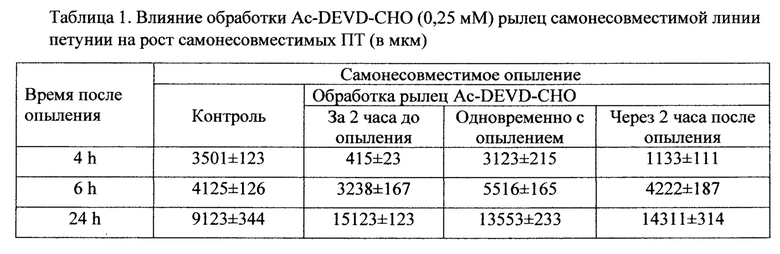
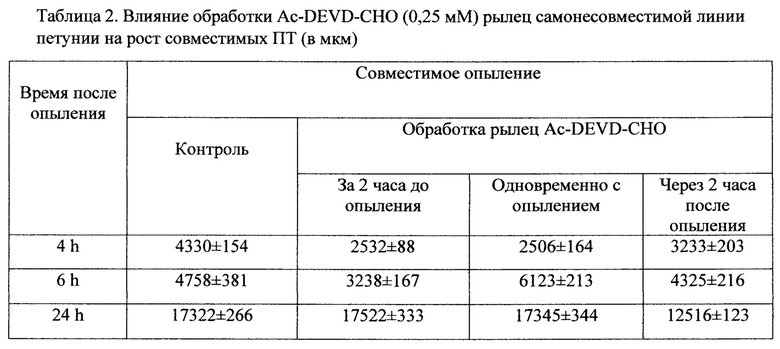
Визуализация растущих в тканях пестика ПТ петунии, с окрашиванием анилиновым голубым и флуоресцентной микроскопией, показала, что после перекрестного совместимого опыления и самонесовместимого опыления почти все пыльцевые зерна прорастают, а ПТ растут в рыльце и тканях столбика. При совместимом опылении длина ПТ после 24 часов опыления равнялась 17322±266 мкм и составляла 80,95% от длины всего пестика, при самонесовместимом опылении ПТ останавливают свой рост через 8 часов после опыления на расстоянии равном примерно 8 мм от поверхности рыльца и остаются на этом уровне и на 24 часах опыления (Таб. 2).
В данном исследовании длина ПТ в контрольном варианте через 24 часа после самонесовместимого опыления равнялась 9123±344 мкм и составила 31,97% от всей длины пестика (Таб. 1).
Обработка ингибитором каспаз Ac-DEVD-CHO в концентрации 1,99 mM оказалась губительной для пестиков петунии, они темнели и рыльце отмирало, пыльца не прорастала. Концентрация 1 гаМ сильно ингибировала рост как совместимых, так и самонесовместимых ПТ, их длина не превышала 7-9% от длины пестика к 24 часам после опыления. Концентрация 0,5 mM была не так драматична для роста ПТ, обработка ингибитором каспаз через 2 часа после опыления замедляла рост совместимых ПТ до 6,85% от длины пестика, самонесовместимых ПТ до 2,07%. Обработка за 2 часа до опыления не влияла на рост совместимых ПТ, их длина была сходна с контролем 17565±413 мкм, при этом рост самонесовместимых ПТ ингибировался до 2333±134 мкм. Вариант обработки одновременно с опылением привел к ингибированию совместимых ПТ до 14344±403 мкм и небольшой стимуляции самонесовместимых ПТ до 10105±630 мкм.
Наиболее интересные результаты получены при обработке ингибитором каспаз Ac-DEVD-CHO в концентрации 0,25 mM. Во всех вариантах обработок (за 2 часа до опыления, одновременно с опылением, через 2 часа до опыления) рост самонесовместимых ПТ увеличивался практически до уровня совместимых (Таб. 1) и почти не оказывала эта концентрация влияния на рост совместимых ПТ (Таб. 2).
Во всех вариантах обработок были оставлены опыленные пестики на завязываемость семян. Обработка ингибитором в концентрации 0,5 mM и 0,25 тМ во всех вариантах совместимых опылений приводила к нормальной завязываемости семян. Только один вариант обработки ингибитором каспаз Ac-DEVD-CHO в концентрации 0,25 mM приводил к 100% завязываемости семян при самонесовместимом опылении (Фиг. 1).
Маркерами присутствия ПКС являются деградация ДНК и активность касазо-подобньгх протеаз в растении.
Собранный материал помещали в жидкий азот. Выделение ДНК из пестиков или их частей (рыльца и столбиков) проводили по стандартной методике. ДНК разделяли в 1,0% агарозном геле 1 × в трис-боратном буфере.
Использование электрофоретического анализа в качестве метода разделения фрагментов ДНК позволил выявить наличие ДНК-деградации в образце через 6 часов после самонесовместимого опыления, в той же точке после самосовместимого опыления ДНК-деградация отсутствует (Фиг. 2). Положительным контролем наличия ДНК-деградации являлись образцы пыльников из бутонов без антоциана самонесовместимого клона, так как широко известно, что при развитии пыльников клетки тапетума гибнут посредством ПГК. Однако, в образцах, подвергнутых обработке ингибитором каспазы-3 Ac-DEVD-CHO в концентрациях 1, 0,5, 0,25 mM ДНК-деградация отсутствует как после самосовместимого, так и самонесовместимого опылений.
Активность каспазо-3 подобной протеазы в экстрактах пестика оценивали по расщеплению флуорогенного субстрата каспазы-3 Ac-DEVD-AMC (Ac-Asp-GluVal Asp-АМС; АМС, 7-амино-4-метилкумарин; EMD Biosciences и BIOMOL, США). Образец (50 мкл) и 400 мкМ Ac-DEVD-AMC (50 мкл) помещали в лунку 96-луночного аналитического планшета (Costar, США) для измерения флуоресценции. Высвобождение флуорофора при расщеплении регистрировали (возбуждение при 360 нм и испускание при 460 нм) на микропланшетном ридере BioTek Synergy HI (США) с многоканальным детектором. Флуоресценцию АМС регистрировали в течение 3 ч с 10-минутными интервалами.
Все эксперименты проводили в трех - пяти биологических проворностях. Достоверность результатов оценивали с помощью t-критерия Стьюдента (Р<0,05). Данные представлены в виде средних значений и их стандартных ошибок.
Образование самонесовместимой ПТ и ее рост в рыльце и столбике в течение 4 ч сопровождались резким повышением активности каспазо-3-подобной протеазы. В этот период активность каспазо-3-подобной протеазы в системе пыльца-пестик увеличилась почти в восемь раз по сравнению с растущими совместимыми пыльцевыми трубками. Резкое снижение активности каспазо-3-подобной протеазы наблюдалось в системе пыльца-пестик в течение последующих 2 часов (Таб. 3). При обработке ингибитором каспаз Ac-DEVD-CHO в концентрации равной 0,25 mM в пестиках после самонесовместимого опыления наблюдается резкое снижение уровня каспазоподобной активности на протяжении всего времени после опыления (Таб. 3).
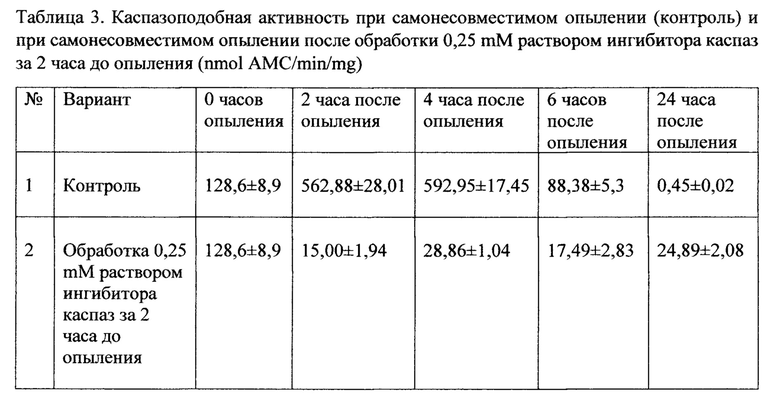
Таким образом, полученные данные показывают, что обработка ингибитором каспазы-3 в концентрации 0.25 mM за 2 часа до самонесовместимого опыления ингибирует активность каспазы-3 (Ac-DEVD-CHO), снимает деградацию ДНК, характерную для самонесовместимого опыления в момент остановки роста ПТ, увеличивает их рост до уровня совместимых ПТ, тем самым снимает явление СН и позволяет получить семена.
В результате, применения заявляемого способа можно предотвратить ПКС в самонесовместимых ПТ, снять самонесовместимсть S-РНКазного типа у и получить семена петунии при самонесовместимом опылении.
Литература
1. Bonneau L, Ge Y, Drury GE, Gallois P (2008) What happened to plant caspases? J Exp Bot 59:491-499
2. Bosch M, Franklin-Tong VE (2007) Temporal and spatial activation of caspase-like enzymes induced by self-incompatibility in Papaver pollen. Proc Natl Acad Sci U S A 104:8327-1832
3. Del Pozo O, Lam E (1998) Caspases and programmed cell death in the hypersensitive response of plants to pathogens. Curr Biol 8:1129-1132
4. Kovaleva LV, Zakharova EV, Timofeeva GV et al (2020) Aminooxyacetic acid (AOA), inhibitor of 1-aminocyclopropane-l-carboxilic acid (ACC) synthesis, suppresses self-incompatibility-induced programmed cell death in self-incompatible Petunia hybrida L. pollen tubes. Protoplasma 257:213-227
5. Zakharova, E. V., Timofeeva, G. V., Fateev, A. D., & Kovaleva, L. V. (2021). Caspase-like proteases and the phytohormone cytokinin as determinants of S-RNAse-based self-incompatibility-induced PCD in Petunia hybrida L. Protoplasma, 258(3), 573-586
6. Zhang QF, Li J, Bi FC, Liu Z, Chang ZY, Wang LY, Huang LQ, Yao N (2020) Ceramide-induced cell death depends on calcium and caspase-like activity in rice. Front Plant Sci 11:145
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПЫЛЕНИЯ ЛИНИЙ ПЕТУНИИ С ГАМЕТОФИТНЫМ ТИПОМ САМОНЕСОВМЕСТИМОСТИ | 2015 |
|
RU2608507C1 |
| Способ получения семян самонесовместимых форм петунии гибридной | 1989 |
|
SU1692402A1 |
| Способ получения семян самонесовместимых форм табака душистого | 1991 |
|
SU1789142A1 |
| ПОЛИНУКЛЕОТИД, СПОСОБЫ ПОЛУЧЕНИЯ РАСТЕНИЙ, ГЕНОМ РАСТЕНИЯ, КЛЕТКИ, ПЛОДЫ, СЕМЕНА, СПОСОБЫ ПОЛУЧЕНИЯ СЕМЯН, ПРИМЕНЕНИЯ ОЛИГОНУКЛЕОТИДА, ПЛАЗМИДА, МИКРООРГАНИЗМ | 1992 |
|
RU2170255C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИБРИДОВ МЕЖДУ КУЛЬТУРНЫМ КАРТОФЕЛЕМ Solanum tuberosum И 1ЕВN ДИПЛОИДНЫМИ ДИКИМИ ВИДАМИ КАРТОФЕЛЯ | 2012 |
|
RU2505957C1 |
| СПОСОБ ТРАНСФОРМАЦИИ РАСТЕНИЙ | 1997 |
|
RU2123781C1 |
| Способ искусственного опыления растений YIcca Spp.L. | 1987 |
|
SU1470245A1 |
| СПОСОБ ПОДБОРА ЛУЧШИХ СОРТОВ ОПЫЛИТЕЛЕЙ ДЛЯ НАСАЖДЕНИЙ ЯБЛОНИ | 2011 |
|
RU2475020C2 |
| Способ получения диплоидных апомиктов табака душистого | 1990 |
|
SU1787385A1 |
| СПОСОБ ТРАНСФОРМАЦИИ РАЗМНОЖАЮЩИХСЯ ПУТЕМ ОПЫЛЕНИЯ РАСТЕНИЙ | 1988 |
|
RU2054482C1 |

Изобретение относится к области биотехнологии и сельского хозяйства. Предложен способ снятия самонесовместимости S-РНКазного типа у Petunia hybrida L., заключающийся в обработке поверхности рыльца пестика 5 мкл раствора ингибитора каспазы-3 Ac-DEVD-CHO в концентрации 0,25 мМ за 2 часа до самонесовместимого опыления. Изобретение позволяет предотвратить программируемую клеточную смерть в самонесовместимых пыльцевых трубках, снять самонесовместимсть S-РНКазного типа у петунии и получить семена петунии при самонесовместимом опылении. 2 ил., 3 табл.
Способ снятия самонесовместимости S-РНКазного типа у Petunia hybrida L., заключающийся в обработке поверхности рыльца пестика 5 мкл раствора ингибитора каспазы-3 Ac-DEVD-CHO в концентрации 0,25 мМ за 2 часа до самонесовместимого опыления.
| ФАТЕЕВ А.Д | |||
| и др | |||
| Влияние аминоуксусной кислоты (АОА), ингибитора синтеза АЦК (этилена), на рост in vivo несовместимых пыльцевых трубок Petunia hybridа L | |||
| Биотехнология в растениеводстве, животноводстве и сельскохозяйственной микробиологии | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |

