ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
1. Область изобретения
Настоящее изобретение относится к рибулозофосфат-3-эпимеразе, в частности к рибулозофосфат-3-эпимеразе с низкой 3-эпимеризационной активностью в отношении псикозы, к композиции для получения псикозо-6-фосфата или псикозы, содержащей указанную рибулозофосфат-3-эпимеразу, и к способу получения получения псикозо-6-фосфата или псикозы с использованием указанной рибулозофосфат-3-эпимеразы или указанной композиции.
2. Описание предшествующего уровня техники
Псикоза (аллюлоза), являющаяся эпимером фруктозы по положению С3, представляет собой моносахарид, известный как редкий сахар, обнаруживаемый в природе в крайне малых количествах. Псикоза обладает приблизительно 70% сладости сахарозы при почти нулевой энергетической ценностью и привлекла значительное внимание в качестве нового пищевого сырья, используемого в функциональных продуктах питания вследствие таких ее функций, как ингибирование повышения глюкозы в крови, синтеза жиров и так далее.
Ввиду этих свойств псикозы рассматривается возможность ее применения в различных продуктах питания в качестве заменяющего сахар подсластителя. Тем не менее, поскольку в природе она существует в очень малых количествах, присутствувет постоянная потребность в способе, обеспечивающем эффективное получение псикозы.
Один из известных способов получения псикозы представляет собой способ получения псикозо-6-фосфата посредством превращения в глюкозу или глюкозо-1-фосфат, глюкозо-6-фосфат и фруктозо-6-фосфат (публикация патента Кореи №10-2018-0004023), но существует все большая потребность в разработке технологии более эффективного и экономичного получения псикозы.
Псикозо-3-эпимераза (D-псикозо-3-эпимераза, ЕС 5.1.3.30) известна как фермент, позволяющий получать аллюлозу посредством 3-эпимеризации (С3-эпимеризации) фруктозы (D-фруктозы). При получении аллюлозы из фруктозы посредством одной ферментативной реакции с использованием указанного выше фермента существует определенный уровень равновесия реакции между субстратом фруктозой и продуктом аллюлозой (продукт/субстрат приблизительно от 20% до 35%). Таким образом, в случае получения аллюлозы с высокой степенью чистоты с использованием одной ферментативной реакции необходим дополнительный процесс очистки с отделением и удалением фруктозы в высокой концентрации из полученной реакционной смеси.
Кроме того, поскольку известные ранее псикозо-6-фосфат-3-эпимеразы обладают 3-эпимеризационной активностью в отношении псикозы, их нельзя назвать ферментами, специфичными в отношении 3-эпимеризации псикозо-6-фосфата, и они не являются подходящими для практического получения псикозы (WO2018/129275, WO2018/112139).
Исследуя последовательности конкретных мотивов, которые могут влиять на 3-эпимеризационную активность в отношении псикозы, авторы настоящего изобретения обнаружили, что определенные мотивы являются особенно критически важными для 3-эпимеризации псикозо-6-фосфата.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Одна задача настоящего изобретения состоит в том, чтобы предложить рибулозофосфат-3-эпимеразу.
Другая задача настоящего изобретения состоит в том, чтобы предложить нуклеиновую кислоту, кодирующую указанную рибулозофосфат-3-эпимеразу.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить трансформанта, содержащего указанную нуклеиновую кислоту, кодирующую указанную рибулозофосфат-3-эпимеразу.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить композицию для получения псикозо-6-фосфата, содержащую указанную рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить способ получения псикозо-6-фосфата, включающий стадию приведения фруктозо-6-фосфата в контакт с указанной рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить композицию для получения псикозы, содержащую указанную рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить способ получения псикозы, включающий стадию приведения фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
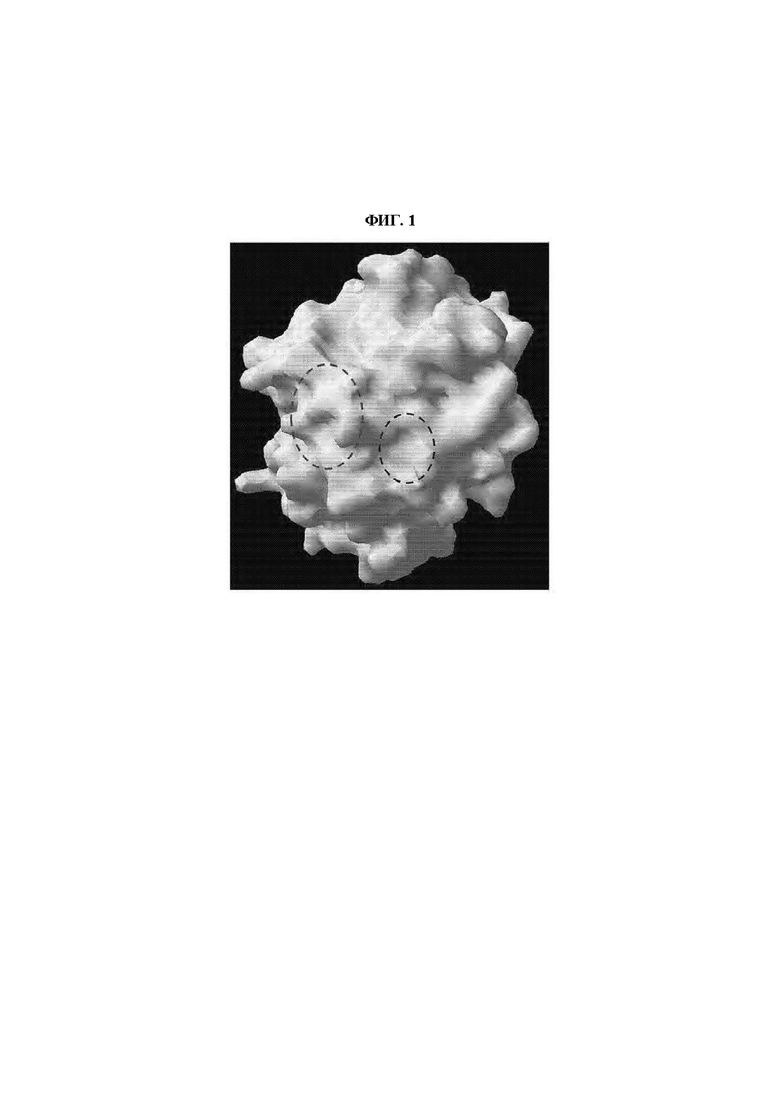
На ФИГ. 1 показана структура белка, спрогнозированная по аминокислотной последовательности рибулозофосфат-3-эпимеразы (SEQ ID NO:9: KPL22606); и
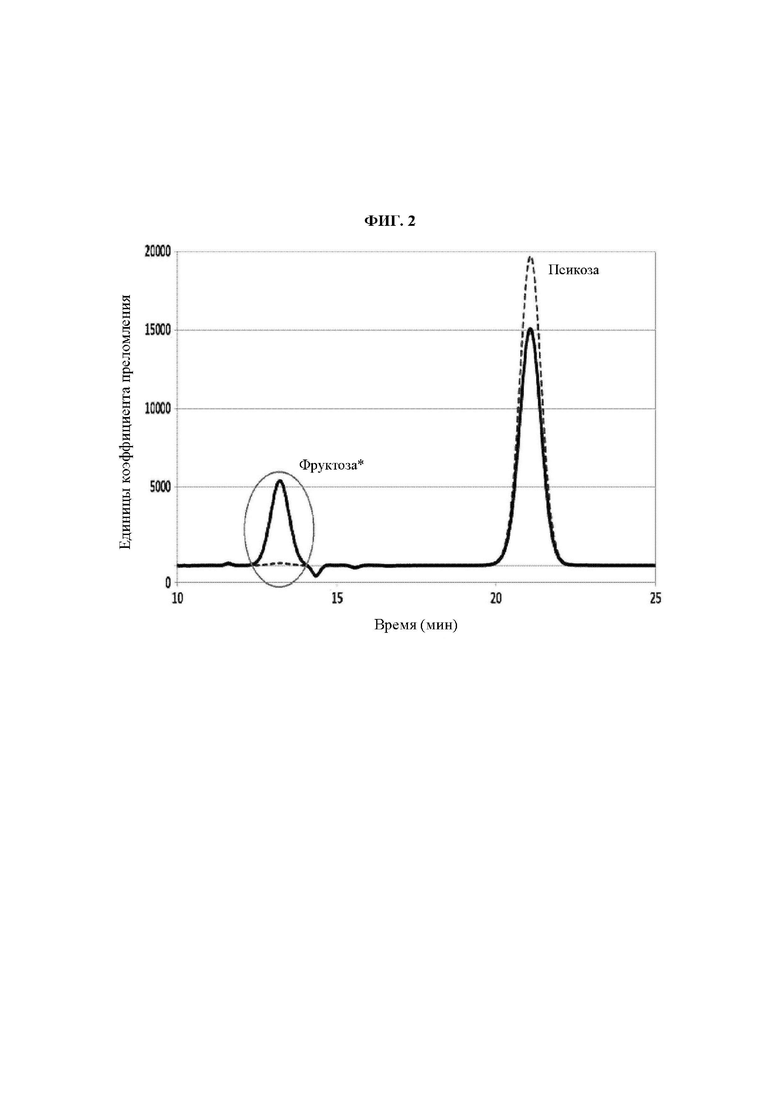
на ФИГ. 2 показана HPLC(высокоэффективная жидкостная хроматография)-гистограмма, на которой показаны известная ранее псикозо-6-фосфат-3-эпимераза (ADL69228; сплошная линия) и фермент по настоящему изобретению (SEQ ID NO:20; прерывистая линия).
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
Далее настоящее изобретение будет описано подробно, как изложено ниже. В то же время, каждое из описаний и воплощений, раскрытых в данном изобретении, может также быть применено к другим описаниям и воплощениям. То есть объем настоящего изобретения включает все комбинации различных элементов, раскрытых в данном изобретении. Кроме того, объем настоящего изобретения не ограничен конкретным описанием, приведенным ниже.
Для решения указанных выше задач согласно одному аспекту настоящего изобретения предложена рибулозофосфат-3-эпимераза.
Конкретно, рибулозофосфат-3-эпимераза по настоящему изобретению может представлять собой рибулозофосфат-3-эпимеразу, содержащую мотив I, состоящий из аминокислотной последовательности SEQ ID NO:1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO:3, и обладающую высокой активностью и устойчивостью к нагреванию, и более конкретно не содержащую мотив II, состоящий из аминокислотной последовательности SEQ ID NO:2, но без ограничения этим.
При использовании здесь термин «рибулозофосфат-3-эпимераза» относится к ферменту, рибулозофосфат-3-эпимеразная активность которого известна, или который обладает рибулозофосфат-3-эпимеразной активностью, и, в частности, к ферменту, который может действовать как фруктозо-6-фосфат-3-эпимераза или псикозо-6-фосфат-3-эпимераза. Когда рибулозофосфат-3-эпимераза обладает фруктозо-6-фосфат-3-эпимеразной активностью или псикозо-6-фосфат-3-эпимеразной активностью, она может содержать аминокислотную последовательность, имеющую делецию, модификацию, замену, консервативную замену или добавление некоторых последовательностей.
Конкретно, фермент по настоящему изобретению представляет собой фермент, обладающий обратимой конверсионной активностью, заключающейся в обратимом превращении псикозо-6-фосфата во фруктозо-6-фосфат или фруктозо-6-фосфат в псикозо-6-фосфат, и в настоящем изобретении термин «рибулозофосфат-3-эпимераза» может быть использован взаимозаменяемо с терминами «псикозо-6-фосфат-3-эпимераза» или «фермент».
Фермент по настоящему изобретению может представлять собой фермент, превращающий глюкозо-1-фосфат (D-глюкозо-1-фосфат), глюкозо-6-фосфат (D-глюкозо-6-фосфат) или фруктозо-6-фосфат (D-фруктозо-6-фосфат) в псикозо-6-фосфат при их смешивании друг с другом. Например, фермент по настоящему изобретению может демонстрировать показатель превращения в псикозо-6-фосфат, при смешивании равного количества псикозо-6-фосфата, глюкозо-1-фосфата, глюкозо-6-фосфата и фруктозо-6-фосфата, составляющий 1% или более, 10% или более или 30% или более. Как описано, благодаря избирательной активности фермента по настоящему изобретению он может демонстрировать высокий показатель превращения в псикозу при ферментативном превращении в одной емкости, где используют множество ферментов и субстратов одновременно.
При использовании здесь «мотив» относится к части (области) последовательности фермента, имеющей определенную последовательность, и может относиться к последовательности, обладающей определенной белковой функцией или активностью, и может представлять собой последовательность, консервативную у разных видов микроорганизмов, но без ограничения этим. Рибулозофосфат-3-эпимераза по настоящему изобретению может содержать мотив I, состоящий из аминокислотной последовательности SEQ ID NO:1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO:3. Кроме того, рибулозофосфат-3-эпимераза может характеризоваться тем, что она не содержит мотив II, состоящий из аминокислотной последовательности SEQ ID NO:2. Поскольку указанный фермент не содержит мотив II, он может характеризоваться низкой псикозо-3-эпимеразной активностью, но без ограничения этим.
Конкретно, указанный фермент может обладать 5% или менее, 4% или менее, 3% или менее, 2% или менее или 1% или менее активности по превращению псикозы во фруктозу или не обладать такой активностью по сравнению с ферментом без мотива I и мотива III, но без ограничения этим.
Кроме того, фермент по настоящему изобретению может дополнительно содержать мотив, состоящий из аминокислотной последовательности SEQ ID NO:4 или 5.
Фермент по настоящему изобретению характеризуется тем, что он содержит или по существу содержит определенный мотив, а также не содержит определенный мотив. Конкретно, мотивы I и III по настоящему изобретению могут быть включены в сайт (сайт связывания) фермента, который частично или полностью связывается с субстратом и/или ионом металла (например, Mg, Mn, Zn и так далее) и взаимодействует с ними, и могут сохранять собственную ферментативную активность при снижении побочной реакционной способности. Более конкретно, мотивы I и III могут быть включены в структуру TIM-бочки в сайте связывания. Фермент, «содержащий» определенный мотив, может дополнительно содержать или может не содержать другой мотив, домен, аминокислотную последовательность, фрагмент и так далее, в дополнение к соответствующему мотиву, и фермент, «по существу содержащий» определенный мотив, может по существу содержать соответствующий мотив с получением желаемого свойства или характеристики и может также содержать или не содержать другой мотив, домен, аминокислотную последовательность, фрагмент и так далее, в дополнение к соответствующему мотиву, но без ограничения этим. Фермент, «не содержащий» определенный мотив, не может содержать последовательность, соответствующую соответствующему мотиву, и может содержать вставку, замену или делецию с другой аминокислотной последовательностью в месте соответствующего мотива или их комбинацию, но без ограничения этим.
Кроме того, каждый из мотивов, включенных или не включенных в фермент по настоящему изобретению, может быть включен или может не быть включен независимым образом, и они не ограничены определенным порядком расположения или положением.
Например, фермент по настоящему изобретению может представлять собой фермент, содержащий только мотив I с SEQ ID NO:1, фермент, содержащий мотив I с SEQ ID NO:1 и мотив III с SEQ ID NO:3 без мотива II с SEQ ID NO:2, или фермент, содержащий мотив I с SEQ ID NO:1, мотив III с SEQ ID NO:3 и мотивы с SEQ ID NO:4 и 5 без мотива II с SEQ ID NO:2, но без ограничения этим.
При использовании здесь «мотив I» может состоять из аминокислотной последовательности следующей SEQ ID NO:1. Очевидно, что последовательность, содержащая вставку, замену, делецию и так далее незначащих аминокислотных остатков, может также быть включена в мотив I по настоящему изобретению, при условии, что это не влияет на активность аминокислотной последовательности SEQ ID NO:1.
Мотив I (SEQ ID NO:1): V-D-G.
Мотив I может быть включен в сайт связывания, взаимодействующий с субстратом рибулозофосфат-3-эпимеразы и ионом металла, но без ограничения этим. Конкретно, указанный мотив может быть расположен в аминокислотных положениях 173-184 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Иными словами, валин (V), являющийся первым аминокислотным остатком мотива I, может быть расположен между положениями 173-182, и глицин (G), являющийся последним остатком мотива I, может быть расположен между положениями 175-184 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим.
В домене, где связаны первая бета-складчатая структура - спиральная структура - альфа-спиральная структура - вторая бета-складчатая структура, первый аминокислотный остаток мотива I может начинаться на С-конце относительно второй бета-складчатости, и домен может быть включен в сайт связывания или может иметь структуру, в которой некоторые области перекрываются (ФИГ. 1).
При использовании здесь «мотив II» может состоять из аминокислотной последовательности следующей SEQ ID NO:2. Очевидно, что последовательность, содержащая вставку, замену, делецию и так далее незначащих аминокислотных остатков, может также быть включена в мотив II по настоящему изобретению, при условии, что это не влияет на активность аминокислотной последовательности SEQ ID NO:2.
Мотив II (SEQ ID NO:2): M-X-X-D-P-G (X представляет собой любую аминокислоту).
Мотив II может быть включен в N-концевую область рибулозофосфат-3-эпимеразы, но без ограничения этим. Конкретно, указанный мотив может быть расположен в аминокислотных положениях 136-150 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Иными словами, метионин (М), являющийся первым аминокислотным остатком мотива II, может быть расположен между положениями 136-145, и глицин (G), являющийся последним остатком мотива II, может быть расположен между положениями 141-150 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. В домене, где связаны первая бета-складчатая структура спиральная структура альфа-спиральная структура вторая бета-складчатая структура, первый аминокислотный остаток мотива II может начинаться на С-конце первой бета-складчатости, и домен может быть включен в сайт связывания или может иметь структуру, в которой некоторые области перекрываются, и домен может быть идентичен домену, содержащему мотив I (ФИГ. 1).
Конкретно, X может включать любую аминокислоту без ограничения и, конкретно, треонин (Т) или валин (V), но без ограничения этим. Более конкретно, мотив II может содержать последовательность M-(T/A/M/L)-(V/N/I)-D-P-G, но без ограничения этим.
При использовании здесь «мотив III» может состоять из аминокислотной последовательности следующей SEQ ID NO:3. Очевидно, что последовательность, содержащая вставку, замену, делецию и так далее незначащих аминокислотных остатков, может также быть включена в мотив III по настоящему изобретению, при условии, что это не влияет на активность аминокислотной последовательности SEQ ID NO:3.
Мотив III (SEQ ID NO:3): M-X-X-X'-P-G (X представляет собой любую аминокислоту).
Мотив III может быть включен в N-концевую область рибулозофосфат-3-эпимеразы, но без ограничения этим. Конкретно, указанный мотив может быть расположен в аминокислотных положениях 136-150 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Иными словами, метионин (M), являющийся первым аминокислотным остатком мотива III, может быть расположен между положениями 136-145, и глицин (G), являющийся последним остатком мотива III, может быть расположен между положениями 141-150 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. В домене, где связаны первая бета-складчатая структура спиральная структура альфа-спиральная структура вторая бета-складчатая структура, первый аминокислотный остаток мотива III может начинаться на С-конце первой бета-складчатости, и домен может быть включен в сайт связывания или может иметь структуру, в которой некоторые области перекрываются, и домен может быть идентичен домену, содержащему мотив I (ФИГ. 1).
Более конкретно, мотивы II и III могут представлять собой мотивы, включенные в одну и ту же область при их выравнивании в ферменте по настоящему изобретению, без ограничения этим.
Конкретно, X может включать любую аминокислоту без ограничения и, конкретно, треонин (Т), аланин (А), метионин (М), лейцин (L), валин (V), аспарагин (N) или изолейцин (I), но без ограничения этим.
Кроме того, X' может включать любую аминокислоту, за исключением аспарагиновой кислоты (D), без ограничения и, конкретно, незаряженную аминокислоту или положительно заряженную аминокислоту. Конкретно, незаряженная аминокислота может включать все полярные аминокислоты и неполярные аминокислоты и может представлять собой любую из серина, треонина, цистеина, аспарагина, глутамина, глицина, аланина, пролина, валина, лейцина, изолейцина и метионина. Кроме того, положительно заряженная аминокислота может представлять собой любое из лизина, аргинина и гистидина. Например, положительно заряженная аминокислота может включать аспарагин (N) и лизин (K). Более конкретно, мотив III может содержать последовательность M-(T/A/M/L)-(V/N/I)-N-P-G, но без ограничения этим.
Фермент по настоящему изобретению может дополнительно содержать мотив, состоящий из аминокислотной последовательности следующей SEQ ID NO:4. Очевидно, что последовательность, содержащая вставку, замену, делецию и так далее незначащих аминокислотных остатков, может также быть включена в фермент по настоящему изобретению, при условии, что это не влияет на активность аминокислотной последовательности SEQ ID NO:4.
SEQ ID NO:4: S-X-M/I-C (X представляет собой любую аминокислоту).
Мотив, имеющий аминокислотную последовательность SEQ ID NO:4, может быть включен в N-концевую область рибулозофосфат-3-эпимеразы, но без ограничения этим. Конкретно, указанный мотив может быть расположен в аминокислотных положениях 5-20 и более конкретно в аминокислотных положениях 7-19 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Иными словами, серии (S), являющийся первым аминокислотным остатком мотива, имеющего аминокислотную последовательность SEQ ID NO:4, может быть расположен между положениями 7-16, и цистеин (С), являющийся последним остатком мотива, имеющего аминокислотную последовательность SEQ ID NO:4, может быть расположен между положениями 10-19 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Кроме того, мотив SEQ ID NO:4 может быть образован после бета-складчатой структуры, и, конкретно, часть последовательности мотива SEQ ID NO:4 может быть включена в альфа-спиральную структуру, образованную после бета-складчатой структуры.
Конкретно, X может включать любую аминокислоту без ограничения, конкретно, метионин, изолейцин, лейцин или валин, но без ограничения этим. Более конкретно, мотив, состоящий из аминокислотной последовательности SEQ ID NO:4, может содержать последовательность SIMC (SEQ ID NO:27), SMMC (SEQ ID NO:28), SLMC (SEQ ID NO:29) или SVMC (SEQ ID NO:30), но без ограничения этим.
Фермент по настоящему изобретению может дополнительно содержать мотив, состоящий из аминокислотной последовательности следующей SEQ ID NO:5. Очевидно, что последовательность, содержащая вставку, замену, делецию и так далее незначащих аминокислотных остатков, может также быть включена в фермент по настоящему изобретению, при условии, что это не влияет на активность аминокислотной последовательности SEQ ID NO:5.
SEQ ID NO:5: G-X-X-X-X-F/L (X представляет собой любую аминокислоту).
Мотив, состоящий из аминокислотной последовательности SEQ ID NO:5, может быть включен в С-концевую область рибулозофосфат-3-эпимеразы, но без ограничения этим. Конкретно, указанный мотив может быть расположен в аминокислотных положениях 190-210 и, более конкретно, в аминокислотных положениях 196-210 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Иными словами, глицин (G), являющийся первым аминокислотным остатком мотива, состоящего из аминокислотной последовательности SEQ ID NO:5, может быть расположен между положениями 196-205, и фенилаланин (F), являющийся последним остатком мотива, состоящего из аминокислотной последовательности SEQ ID NO:5, может быть расположен между положениями 201-210 от первой аминокислоты N-конца рибулозофосфат-3-эпимеразы, но без ограничения этим. Кроме того, мотив SEQ ID NO:5 может быть образован после бета-складчатой структуры, и, конкретно, часть последовательности мотива SEQ ID NO:5 может быть включена в альфа-спиральную структуру, образованную после бета-складчатой структуры.
Конкретно, каждый X мотива, состоящего из аминокислотной последовательности SEQ ID NO:5, может независимо представлять собой треонин, серии, глицин, лейцин, цистеин, изолейцин, аспарагин, лизин, аланин, валин или глутамин, но без ограничения этим. Более конкретно, мотив, состоящий из аминокислотной последовательности SEQ ID NO:5, может содержать аминокислотную последовательность GNSGLF (SEQ ID NO:31), GSSGLFGSSSLF (SEQ ID NO:32), GSTSLF (SEQ ID NO:33), GTAGLF (SEQ ID NO:34), GTKGLF (SEQ ID NO:35), GTQSLF (SEQ IDNO:36), GTSCLF (SEQ ID NO:37), GTSGLF (SEQ ID NO:38), GTSSIF (SEQ ID NO:39), GTSGIF (SEQ ID NO:40), GTSSLF (SEQ ID NO:41) или GTSSVF (SEQ ID NO:42), но без ограничения этим. Фермент по настоящему изобретению характеризуется тем, что он обладает высокой активностью по превращению фруктозо-6-фосфата в псикозо-6-фосфат благодаря тому, что он содержит мотив(ы) SEQ ID NO:4 и/или 5.
Соответствующие аминокислотные остатки, включенные в мотив по настоящему изобретению, можно независимо комбинировать друг с другом с получением мотива.
Рибулозофосфат-3-эпимераза по настоящему изобретению может не содержать определенный мотив, то есть мотив II, вследствие чего она демонстрирует очень низкую 3-эпимеризационную активность в отношении псикозы, что указывает на возможность получения конечного продукта псикозы с высоким выходом за счет уменьшения побочного взаимодействия во время процесса получения псикозо-6-фосфата из фруктозо-6-фосфата. Кроме того, рибулозофосфат-3-эпимеразу можно использовать в комбинации с другими ферментами (например, псикозо-6-фосфатфосфатазой) для эффективного применения для получения псикозы.
Альфа-спиральную (α-спиральную) структуру, бета-складчатую (β-складчатую) структуру и спиральную структуру по настоящему изобретению можно понимать в соответствии с общепринятым определением, приведенным в Kwangsoo Kim et al. (Crystal Structure of d-Psicose 3-epimerase from Agrobacterium tumefaciens and its Complex with True Substrate d-Fructose, Volume 361, Issue 5, 1 September 2006, Pages 920-931) и так далее, и альфа-спираль может представлять собой правостороннюю альфа-спираль. Структуру можно получить путем прямого прогнозирования с применением общеизвестного метода, такого как NMR (ядерно-магнитный резонанс), рентгеновская кристаллография и так далее, или с использованием алгоритмов Rosetta или интернет-сервера (I-TASSER, ROBETTA и так далее), исходя из аминокислотной последовательности.
Кроме того, в настоящем изобретении домен, в котором собраны структуры, может быть экспрессирован как структура 1 структура 2.
Кроме того, рибулозофосфат-3-эпимераза по настоящему изобретению может иметь происхождение из любого, выбранного из группы, состоящей из Chthonomonas, Geobacillus, Mahella, Thermoanaerobacterium, Tepidanaerobacter, Ardenticatenia, Firmicutes, Aeribacillus, Epulopiscium и Thermoflavimicrobium, и более конкретно может иметь происхождение из любого, выбранного из группы, состоящей из Chthonomonas calidirosea Т49, Geobacillus sp. 8, Geobacillus thermocatenulatus, Mahella australiensis 50-1 BON, Thermoanaerobacterium sp. PSU-2, Thermoanaerobacterium thermosaccharolyticum, Tepidanaerobacter syntrophicus, бактерии Ardenticatenia, бактерии Firmicutes HGW-Firmicutes-5, Aeribacillus pallidus, Epulopiscium sp. SCG-B05WGA-EpuloA1 и Thermoflavimicrobium dichotomicum, но без ограничения этим.
Кроме того, рибулозофосфат-3-эпимераза по настоящему изобретению может содержать любую последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:15-26, или может состоять из любой последовательности, выбранной из группы, состоящей из аминокислотных последовательностей SEQ ID NO:15-26, но без ограничения этим. Более конкретно, фермент может состоять из аминокислотной последовательности SEQ ID NO:19, 20 или 22, но без ограничения этим.
Присутствие или отсутствие определенного мотива в рибулозофосфат-3-эпимеразе по настоящему изобретению оказывает важное влияние на ферментативную активность, и поэтому последовательности ферментов, исключающие область мотива, могут иметь низкую идентичность. Конкретно, фермент может состоять из любой последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, имеющих 26%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% или более идентичности последовательности с областью, исключающей мотивы I и III в аминокислотной последовательности, и аминокислотных последовательностей, имеющих 24%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% или более идентичности последовательности с областью, исключающей мотивы I и III, и областями SEQ ID NO:4 и 5, но без ограничения этим.
Кроме того, фермент может содержать любую из аминокислотных последовательностей SEQ ID NO:15-26 или аминокислотную последовательность, имеющую 70% или более гомологии или идентичности с такой последовательностью, но без ограничения этим. Конкретно, аминокислотная последовательность может содержать последовательности SEQ ID NO:15-26 и аминокислотные последовательности, имеющие по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99%, или более гомологии или идентичности с указанными последовательностями. Кроме того, очевидно, что белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или добавлением части последовательности может также быть включен в объем настоящего изобретения при условии, что его аминокислотная последовательность имеет такую гомологию или идентичность и демонстрирует эффективность, соответствующую рассматриваемому белку.
Иными словами, несмотря на то, что в настоящем изобретении раскрыт «белок, имеющий аминокислотную последовательность, обозначенную определенным номером последовательности», при условии, что белок обладает активностью, идентичной или эквивалентной активности белка, состоящего из аминокислотной последовательности с соответствующим номером последовательности, очевидно, что объем настоящего изобретения включает также белки, имеющие аминокислотную последовательность с частичной делецией, модификацией, заменой, консервативной заменой или добавлением. Например, когда белок обладает активностью, идентичной или эквивалентной активности фермента, очевидно, что не исключены добавление последовательности, не меняющей функцию белка, до и после аминокислотной последовательности, ее естественная мутация, молчащая мутация или консервативная замена, и объем настоящего изобретения включает добавление или мутацию последовательности.
При использовании здесь термин «гомология» или «идентичность» относится к степени соответствия двух заданных аминокислотных последовательностей или нуклеотидных последовательностей, и она может быть выражена в процентах.
Термины «гомология» и «идентичность» могут часто быть использованы взаимозаменяемо друг с другом.
Гомология или идентичность последовательности консервативных полинуклеотидов или полипептидов может быть определена с помощью стандартных алгоритмов выравнивания и может быть использована вместе с штрафом за разрыв, установленным в используемой программе по умолчанию. По существу, гомологичные или идентичные последовательности будут предположительно гибридизоваться по всей длине или на протяжении приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, или приблизительно 90% или более от полной длины последовательностей в условиях умеренной или высокой жесткости. Также рассматриваются полинуклеотиды, содержащие вырожденные кодоны вместо кодонов гибридизующихся полинуклеотидов.
Присутствие гомологии, сходства или идентичности любых двух полинуклеотидных или полипептидных последовательностей можно определить с использованием известного компьютерного алгоритма, такого как программа «FASTA», как описано в Pearson et al. (1988) Proc. Natl. Acad. Sci. USA 85:2444, с использованием параметров по умолчанию. Альтернативно, его можно определить с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-153) с использованием программы Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) (версии 5.0.0 или более поздней), пакета программ GCG (Devereux, J. et al., Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F. et al., J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, и CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, гомологию, сходство или идентичность можно определить с использованием BLAST или Clusta1W Национального центра биотехнологической информации.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определить путем сравнения информации о последовательностях с применением, например, компьютерной GAP-программы, такой как Needleman et al. (1970), J Mol Biol. 48:443, как раскрыто в Smith and Waterman, Adv. Appl. Math (1981) 2:482. Кратко, GAP-программа определяет гомологию, сходство или идентичность как значение, полученное делением числа сходно выровненных символов (то есть нуклеотидов или аминокислот) на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для GAP-программы могут включать: (1) одинарную матрицу сравнения (включающую значение 1 для идентичных положений и 0 для неидентичных) и взвешенную матрицу сравнения по Gribskov et al. (1986) Nucl. Acids Res. 14:6745, как описано в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979), (альтернативно, матрицу замен EDNAFULL (EMBOSS-версия NCBI NUC4.4)); (2) штраф 3,0 за каждый разрыв и штраф 0,10 для каждого символа каждого разрыва (или штраф за начало разрыва 10 и штраф за продолжение разрыва 0,5); и (3) отсутствие штрафов за окончание разрывов. Соответственно, при использовании здесь термин «гомология» или «идентичность» относится к сходству последовательностей.
В то же время, рибулозофосфат-3-эпимераза по настоящему изобретению может обладать устойчивостью к нагреванию, но без ограничения этим.
При использовании здесь термин «устойчивость к нагреванию» обозначает способность демонстрировать исходную активность фермента без потери его активности даже в условиях высокой температуры, и устойчивость фермента к нагреванию имеет множество преимуществ в процессе получения желаемого продукта. Конкретно, рибулозофосфат-3-эпимераза по настоящему изобретению может обладать 3-эпимеризационной активностью в отношении псикозо-6-фосфата при 40°С или более, более конкретно при 50°С или более, и намного более конкретно при 60°С или более, но без ограничения этим. Еще намного более конкретно, фермент по настоящему изобретению может обладать 3-эпимеризационной активностью в отношении псикозо-6-фосфата в условиях рН от 5,0 до 10,0 и от 50°С до 90°С в течение от 1 минуты до 24 часов, но без ограничения этим.
Рибулозофосфат-3-эпимераза по настоящему изобретению может быть получена путем трансформации штамма самим ферментом или ДНК, экспрессирующей фермент, культивированием штамма с получением культуры, разрушением культуры и последующей очисткой продукта с использованием колонки и так далее. Штамм для трансформации может представлять собой Escherichia coli, Corynebacterium glutamicum, Aspergillus oryzae, Saccharomyces cerevisiae, Yarrowia lipolytica, Pichia pastoris или Bacillus subtilis, но без ограничения этим, и в будущем может потенциально быть трансформирован в GRAS-штамм (Generally Recognized as Safe, общепризнанно безопасный).
Конкретных ограничений относительно метода очистки рибулозофосфат-3-эпимеразы по настоящему изобретению нет, и может быть применен метод, обычно применяемый в области, к которой относится настоящее изобретение. Их неограничивающие примеры могут включать хроматографию, обработку нагреванием, адсорбцию, фильтрацию, ионную очистку и так далее. Может быть применен только один из методов очистки, и могут быть применены два или более чем два метода в комбинации.
Согласно другому аспекту настоящего изобретения предложены нуклеиновая кислота, кодирующая рибулозофосфат-3-эпимеразу, или вектор, содержащий указанную нуклеиновую кислоту.
При использовании здесь «нуклеиновая кислота» имеет значение, включающее как молекулы ДНК, так и молекулы РНК. Нуклеотиды, являющиеся основными структурными единицами нуклеиновых кислот, могут включать не только естественные нуклеотиды, но также их модифицированные аналоги, в которых модифицированы сахарные или основные группировки.
Нуклеиновая кислота по настоящему изобретению может представлять собой последовательность ДНК или РНК, где нуклеотидные единицы связаны друг с другом ковалентными связями, и, конкретно, она может представлять собой любую из всех нуклеотидных последовательностей, возможных при преобразовании аминокислотных последовательностей SEQ ID NO:15-26 в ДНК (с заменой аминокислот на 61 кодон), и, более конкретно, она может включать нуклеиновую кислоту, имеющую на 90% или более, 95% или более, 97% или более, 99% или более или 100% или более гомологии, сходства или идентичности с каждым нуклеотидом, который может быть транслирован в любую аминокислотную последовательность из аминокислотных последовательностей SEQ ID NO:15-26 по настоящему изобретению, в то же время способную демонстрировать желаемую ферментативную активность после трансляции. Также очевидно, что полинуклеотид, который, в силу вырожденности генетического кода, может быть транслирован с получением белка, обладающего такой же активностью или имеющего такую же аминокислотную последовательность после трансляции, конкретно, состоящего из любой аминокислотной последовательности из SEQ ID NO:15-26, или белка, имеющего гомологию, сходство или идентичность с ним, может также быть включен в объем настоящего изобретения. Более конкретно, последовательность нуклеиновой кислоты по настоящему изобретению отдельно не указана, и нуклеиновая кислота может, без ограничения, состоять из любого числа кодонов ДНК, которые могут быть транслированы с получением аминокислотных последовательностей SEQ ID NO:15-26.
Альтернативно, может быть включен, без ограничения, зонд, который может быть получен из известной генной последовательности, например любой последовательности, которая гибридизуется в жестких условиях с последовательностью, комплементарной всей или части нуклеотидной последовательности, кодирующей фермент по настоящему изобретению.
«Жесткие условия» относятся к условиям, позволяющим осуществлять специфичную гибридизацию полинуклеотидов. Такие условия подробно раскрыты в литературе (например, J. Sambrook et al., указанная ниже). Например, жесткие условия могут включать, условия при которых гены, имеющие высокую степень гомологии или идентичности, гены, имеющие гомологию или идентичность 80% или более или 85% или более, конкретно 90% или более, более конкретно 95% или более, еще более конкретно 97% или более и, особенно конкретно 99% или более, гибридизуются друг с другом, в то время как гены, имеющие более низкую гомологию или идентичность, чем указано выше, не гибридизуются друг с другом, или могут включать обычные условия промывки при Саузерн-гибридизации, то есть однократную промывку, конкретно двукратную или трехкратную промывку, при концентрации соли и температуре, соответствующих 60°С, 1xSSC (физиологический раствор - цитрат натрия), 0,1% SDS (додецилсульфат натрия), конкретно 60°С, 0,1xSSC, 0,1% SDS, и более конкретно 68°С, 0,1xSSC, 0,1% SDS.
Для гибридизации необходимо, чтобы два полинуклеотида имели комплементарные последовательности, однако, в зависимости от жесткости гибридизации, возможны несовпадения оснований. Термин «комплементарный» использован для описания отношения между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, применительно к ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом настоящее изобретение может также включать выделенный фрагмент нуклеиновой кислоты, комплементарный полной последовательности, а также последовательность нуклеиновой кислоты, по существу сходную с ним.
Конкретно, полинуклеотид, имеющий гомологию или идентичность, можно определить с использованием условий гибридизации, включающих стадию гибридизации при значении Tm 55°С в условиях, описанных выше. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, но без ограничения этим, и специалисты в данной области могут надлежащим образом его контролировать в зависимости от поставленной задачи.
Подходящая строгость гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов, и эти параметры хорошо известны в данной области.
При использовании здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность нуклеиновой кислоты, кодирующей фермент по настоящему изобретению, функционально связанную с подходящей последовательностью регуляции экспрессии для экспрессии желаемого варианта белка в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции такой транскрипции, последовательность, кодирующую подходящий сайт связывания рибосом на мРНК (матричная РНК), и последовательность для регулирования терминации транскрипции и трансляции. Вектором может быть трансформирована подходящая клетка-хозяин, и затем он может реплицироваться или функционировать независимо от генома хозяина или может быть интегрирован в сам геном.
Конкретных ограничений относительно вектора, используемого в настоящем изобретении, нет при условии, что вектор способен к репликации в клетке-хозяине, и может быть использован любой вектор, известный в данной области. Примеры обычно используемых векторов могут включать нативные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее, а в качестве плазмидного вектора могут быть использованы плазмиды на основе pBR, на основе pUC, на основе pBluescriptII, на основе pGEM, на основе pTZ, на основе pCL, на основе рЕТ и так далее. Конкретно, могут быть использованы векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
Согласно еще одному аспекту настоящего изобретения предложен трансформант, содержащий нуклеиновую кислоту, кодирующую фермент по настоящему изобретению, или вектор, содержащий нуклеиновую кислоту, кодирующую фермент по настоящему изобретению.
При использовании здесь «трансформант, содержащий нуклеиновую кислоту, кодирующую фермент», или «трансформант, содержащий вектор, содержащий нуклеиновую кислоту, кодирующую фермент», может относиться к микроорганизму, рекомбинированному для экспрессии рибулозофосфат-3-эпимеразы по настоящему изобретению, и, например, относится к клетке-хозяину или микроорганизму, которые могут содержать нуклеиновую кислоту, кодирующую рибулозофосфат-3-эпимеразу, или могут быть трансформированы вектором, содержащим нуклеиновую кислоту, кодирующую рибулозофосфат-3-эпимеразу, для экспрессии рибулозофосфат-3-эпимеразы. Применительно к задачам настоящего изобретения, рибулозофосфат-3-эпимераза, экспрессированная трансформантом, может состоять из любой аминокислотной последовательности SEQ ID NO:15-26, но без ограничения этим.
При использовании здесь «трансформация» относится к введению вектора, содержащего нуклеиновую кислоту, кодирующую рибулозофосфат-3-эпимеразу по настоящему изобретению, в клетку-хозяина, таким образом, что происходит экспрессия белка, кодируемого нуклеиновой кислотой, в клетке-хозяине. При условии, что нуклеиновая кислота, используемая для трансформации, может быть экспрессирована в клетке-хозяине, она может включать любые нуклеиновые кислоты, используемые для трансформации, независимо от того, введена и расположена нуклеиновая кислота, используемая для трансформации, в хромосоме клетки-хозяина или расположена вне хромосомы. Кроме того, нуклеиновая кислота включает ДНК и РНК, кодирующие нуклеиновую кислоту, кодирующую рибулозофосфат-3-эпимеразу по настоящему изобретению. Нуклеиновая кислота может быть введена в любой форме, при условии, что нуклеиновая кислота может быть введена в клетку-хозяина для экспрессии. Например, нуклеиновая кислота может быть введена в клетку-хозяина в форме экспрессионной кассеты, представляющей собой геномную структуру, содержащую все элементы, необходимые для автономной экспрессии. Обычно экспрессионная кассета может содержать промотор, функционально связанный с нуклеиновой кислотой, сигнал терминации транскрипции, сайт связывания рибосом и сигнал терминации трансляции. Экспрессионная кассета может представлять собой самореплицируемый вектор экспрессии. Кроме того, нуклеиновая кислота может также быть введена в клетку-хозяина сама по себе и может быть функционально связана с последовательностью, необходимой для экспрессии в клетке-хозяине, но без ограничения этим.
Кроме того, термин «функционально связана» означает, что генная последовательность функционально связана с промоторной последовательностью, инициирующей и опосредующей транскрипцию нуклеиновой кислоты, кодирующей псикозо-6-фосфатфосфатазу по настоящему изобретению.
Введение нуклеиновой кислоты или вектора в хромосому можно осуществить любым методом, известным в данной области, например путем гомологичной рекомбинации, но без ограничения этим. Может быть дополнительно включен селекционный маркер для подтверждения введения в хромосому. Селекционный маркер предназначен для отбора клеток, трансформированных вектором, то есть для подтверждения введения целевой молекулы нуклеиновой кислоты. Могут быть использованы маркеры, обеспечивающие селективные фенотипы, такие как устойчивость к лекарственным средствам, ауксотрофность, устойчивость к цитотоксическим лекарственным средствам или экспрессия модифицированных поверхностных белков. Отбор трансформированных клеток можно проводить в среде, обработанной селективным агентом, поскольку только клетки, экспрессирующие селекционный маркер, выживают или демонстрируют другие фенотипы.
Метод трансформации вектором по настоящему изобретению включает любой метод введения нуклеиновой кислоты в клетку-хозяина и может быть осуществлен путем выбора подходящей стандартной методики, известной в данной области, в зависимости от клетки-хозяина. Примеры метода могут включать электропорацию, преципитацию с фосфатом кальция (СаРО4), преципитацию с хлоридом кальция (CaCl2), ретровирусную инфекцию, микроинъекцию, методику с полиэтиленгликолем (PEG), методику с DEAE-декстраном, методику с катионными липосомами, методику с ацетатом лития и DMSO (диметилсульфоксид) и так далее, но без ограничения этим.
В качестве клетки-хозяина предпочтительно использовать хозяина с высокой эффективностью введения ДНК в клетку-хозяина и высокой эффективностью экспрессии введенной ДНК. Например, он может представлять собой микроорганизм Corynebacterium sp., микроорганизм Escherichia sp., микроорганизм Serratia sp., микроорганизм Bacillus sp., микроорганизм Saccharomyces cerevisiae sp.или микроорганизм Pichia sp., и, конкретно, E. coli, но без ограничения этим. Могут быть использованы все GRAS-штаммы.
Более конкретно, трансформант по настоящему изобретению может представлять собой Е. coli BL21(DE3)/pET-CJ-ef7, E. coli BL21(DE3)/pET-CJ-ef12 или E. coli BL21(DE3)/pET-CJ-ef15, но без ограничения этим.
Согласно еще одному аспекту настоящего изобретения предложена композиция для получения псикозо-6-фосфата, содержащая рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма.
Композиция для получения псикозо-6-фосфата по настоящему изобретению может содержать рибулозофосфат-3-эпимеразу, демонстрирующую активность по превращению фруктозо-6-фосфата в псикозо-6-фосфат, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма, и, таким образом, при приведении указанной композиции в контакт (взаимодействие) с фруктозо-6-фосфатом из фруктозо-6-фосфата может быть получен псикозо-6-фосфат.
Конкретно, композиция может дополнительно содержать фруктозо-6-фосфат в качестве субстрата, но без ограничения этим.
Композиция для получения псикозо-6-фосфата по настоящему изобретению может дополнительно содержать любой подходящий эксципиент, обычно используемый в композиции для получения псикозо-6-фосфата. Эксципиент может включать, например, консервант, смачивающий агент, диспергирующий агент, суспендирующий агент, буфер, стабилизирующий агент, изотонический агент и так далее, но без ограничения этим.
Композиция для получения псикозо-6-фосфата по настоящему изобретению может дополнительно содержать ион металла или соль металла. В одном воплощении ион металла может представлять собой двухвалентный катион и, конкретно, ионы одного или более чем одного металла, выбранного из группы, состоящей из Ni, Mg, Ni, Со, Mn, Fe и Zn. Более конкретно, композиция для получения псикозо-6-фосфата по настоящему изобретению может дополнительно содержать соль металла. Еще более конкретно, соль металла может представлять собой одно или более, выбранное из группы, состоящей из NiSO4, MgSO4, MgCl2, NiCl2, COSO4, COCl2, MnCl2, MnSO4, FeSO4 и ZnSO4.
Согласно еще одному аспекту настоящего изобретения предложен способ получения псикозо-6-фосфата, включающий стадию приведения фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма.
Рибулозофосфат-3-эпимераза по настоящему изобретению может демонстрировать активность по превращению фруктозо-6-фосфата в псикозо-6-фосфат, и, таким образом, при приведении рибулозофосфат-3-эпимеразы, микроорганизма, экспрессирующего указанную рибулозофосфат-3-эпимеразу, или культуры указанного микроорганизма в контакт с фруктозо-6-фосфатом из фруктозо-6-фосфата может быть получен псикозо-6-фосфат.
Условия приведения в контакт и взаимодействия с фруктозо-6-фосфатом и псикозо-6-фосфатом могут быть надлежащим образом выбраны специалистом в данной области с учетом субстрата, фермента и так далее.
Конкретно, стадию получения псикозо-6-фосфата посредством приведения фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой можно осуществлять при рН от 5,0 до 9,0 и температуре от 40°С до 80°С и/или в течение 2-24 часов, более конкретно при рН от 6,0 до 8,0 и температуре от 40°С до 60°С и/или в течение 20-24 часов, и более конкретно при рН 7,0 и температуре 50°С в течение 24 часов, но без ограничения этим.
Способ получения псикозо-6-фосфата по настоящему изобретению может дополнительно включать стадию выделения и/или очистки полученного псикозо-6-фосфата, но без ограничения этим.
Стадию выделения и/или очистки псикозо-6-фосфата можно осуществлять с использованием метода, известного в данной области, но без ограничения каким-либо конкретным методом.
Согласно еще одному аспекту настоящего изобретения предложена композиция для получения псикозы, содержащая рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма. Конкретно, композиция может дополнительно содержать фруктозо-6-фосфат в качестве субстрата, но без ограничения этим.
Композиция для получения псикозы по настоящему изобретению может обеспечивать получение псикозы, поскольку дополнительно содержит фермент, необходимый для получения псикозы, например псикозо-6-фосфатфосфатазу, обеспечивающую получение псикозы посредством дефосфорилирования псикозо-6-фосфата, в дополнение к рибулозофосфат-3-эпимеразе, обеспечивающей получение псикозо-6-фосфата из фруктозо-6-фосфата.
Конкретно, композиция для получения псикозы по настоящему изобретению может дополнительно содержать один или более чем один фермент, выбранный из группы, состоящей из глюкозо-6-фосфатизомеразы, фосфоглюкомутазы, полифосфатглюкокиназы, α-глюканфосфорилазы, крахмалфосфорилазы, мальтодекстринфосфорилазы или сахарозофосфорилазы, α-амилазы, пуллуланазы, изоамилазы, а-глюкантрансферазы, глюкоамилазы, сахаразы и псикозо-6-фосфатфосфатазы, микроорганизм, экспрессирующий указанный фермент, или культуру указанного микроорганизма и, конкретно, может дополнительно содержать псикозо-6-фосфатфосфатазу, но без ограничения этим.
Более конкретно, композиция для получения псикозы по настоящему изобретению может дополнительно содержать: (а) (1) крахмал, мальтодекстрин, сахарозу или их комбинацию, глюкозу, глюкозо-1-фосфат, глюкозо-6-фосфат или фруктозо-6-фосфат; (2) фосфат; (3) псикозо-6-фосфатфосфатазу; (4) глюкозо-6-фосфатизомеразу; (5) фосфоглюкомутазу или глюкокиназу; и/или (6) α-глюканфосфорилазу, крахмалфосфорилазу, мальтодекстринфосфорилазу, сахарозофосфорилазу, α-амилазу, пуллуланазу, изоамилазу, глюкоамилазу или сахаразу; или
(б) микроорганизм, экспрессирующий фермент по (а), или культуру микроорганизма, экспрессирующего фермент по (а), но без ограничения этим.
Тем не менее, это приведено лишь в целях иллюстрации, и, при условии, что псикоза может быть получена с использованием рибулозофосфат-3-эпимеразы по настоящему изобретению, ограничений относительно ферментов, включенных в композицию для получения псикозы по настоящему изобретению, и субстрата, используемого для получения псикозы, нет.
Конкретно, крахмал/мальтодекстринфосфорилаза (ЕС 2.4.1.1) и α-глюканфосфорилаза по настоящему изобретению могут включать любой белок, при условии, что он представляет собой белок, обладающий активностью по образованию глюкозо-1-фосфата из крахмала или мальтодекстрина посредством фосфорильного переноса фосфата на глюкозу. Крахмал/мальтодекстринфосфорилаза (ЕС 2.4.1.1) и α-глюканфосфорилаза могут включать любой белок, при условии, что он представляет собой белок, обладающий активностью по образованию глюкозо-1-фосфата из крахмала или мальтодекстрина посредством фосфорильного переноса фосфата на глюкозу. Сахарозофосфорилаза (ЕС 2.4.1.7) может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по образованию глюкозо-1-фосфата из сахарозы посредством фосфорильного переноса фосфата на глюкозу. α-амилаза (ЕС 3.2.1.1), пуллуланаза (ЕС 3.2.1.41), изоамилаза (ЕС 3.2.1.68), 4-α-глюкантрансфераза (ЕС 2.4.1.25) и глюкоамилаза (ЕС 3.2.1.3), являющиеся ферментами, расщепляющими крахмал, могут включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению крахмала или мальтодекстрина в неразветвленный мальтоолигосахарид или глюкозу. Сахараза (ЕС 3.2.1.26) может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению сахарозы в глюкозу. Фосфоглюкомутаза (ЕС 5.4.2.2) по настоящему изобретению может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению глюкозо-1-фосфата в глюкозо-6-фосфат. Полифосфатглюкокиназа (ЕС 2.7.1.63) может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению глюкозы в глюкозо-6-фосфат посредством переноса фосфата от полифосфата на глюкозу. Глюкозо-6-фосфатизомераза по настоящему изобретению может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению глюкозо-6-фосфата во фруктозо-6-фосфат. Псикозо-6-фосфатфосфатаза по настоящему изобретению может включать любой белок, при условии, что он представляет собой белок, обладающий активностью по превращению псикозо-6-фосфата в псикозу. Более конкретно, псикозо-6-фосфатфосфатаза может представлять собой белок, обладающий активностью по необратимому превращению псикозо-6-фосфата в псикозу.
Ферменты, включенные в композицию для получения псикозы по настоящему изобретению, могут демонстрировать высокий показатель превращения в псикозу при ферментативном превращении в одной емкости, в которой используют множество ферментов и субстратов одновременно.
Композиция для получения псикозы по настоящему изобретению может дополнительно содержать любой подходящий эксципиент, обычно используемый в композиции для получения псикозы. Эксципиент может включать, например, консервант, смачивающий агент, диспергирующий агент, суспендирующий агент, буфер, стабилизирующий агент, изотонический агент и так далее, но без ограничения этим.
Композиция для получения псикозы по настоящему изобретению может дополнительно содержать ион металла или соль металла. В одном воплощении ион металла может представлять собой двухвалентный катион и, конкретно, ионы одного или более чем одного металла, выбранного из группы, состоящей из Ni, Mg, Ni, Со, Mn, Fe и Zn. Более конкретно, композиция для получения псикозы по настоящему изобретению может дополнительно содержать соль металла, и, еще более конкретно, соль металла может представлять собой одно или более, выбранное из группы, состоящей из NiSO4, MgSO4, MgCl2, NiCl2, CoSO4, CoCl2, MnCl2, MnSO4, FeSO4 и ZnSO4.
Согласно еще одному аспекту настоящего изобретения предложен способ получения псикозы, включающий стадию приведения фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма.
Конкретно, способ получения псикозы по настоящему изобретению может последовательно включать стадии:
приведение фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой микроорганизма, экспрессирующего указанную рибулозофосфат-3-эпимеразу, с превращением фруктозо-6-фосфата в псикозо-6-фосфат;
приведение псикозо-6-фосфата в контакт с псикозо-6-фосфатфосфатазой, микроорганизмом, экспрессирующим указанную псикозо-6-фосфатфосфатазу, или культурой указанного микроорганизма с превращением псикозо-6-фосфата в псикозу; но без ограничения этим.
Кроме того, способ получения псикозы по настоящему изобретению может дополнительно включать стадию приведения глюкозо-6-фосфата в контакт с глюкозо-6-фосфатизомеразой, микроорганизмом, экспрессирующим указанную глюкозо-6-фосфатизомеразу, или культурой микроорганизма, экспрессирующего указанную глюкозо-6-фосфатизомеразу, с превращением глюкозо-6-фосфата во фруктозо-6-фосфат перед стадией превращения фруктозо-6-фосфата в псикозо-6-фосфат.
Способ получения псикозы по настоящему изобретению может дополнительно включать стадию приведения глюкозо-1-фосфата в контакт с фосфоглюкомутазой, микроорганизмом, экспрессирующим указанную фосфоглюкомутазу, или культурой микроорганизма, экспрессирующего указанную фосфоглюкомутазу, с превращением глюкозо-1-фосфата в глюкозо-6-фосфат перед стадией превращения глюкозо-6-фосфата во фруктозо-6-фосфат.
Способ получения псикозы по настоящему изобретению может дополнительно включать стадию приведения глюкозы в контакт с полифосфатглюкокиназой, микроорганизмом, экспрессирующим указанную полифосфатглюкокиназу, или культурой микроорганизма, экспрессирующего указанную полифосфатглюкокиназу, и полифосфатом с превращением глюкозы в глюкозо-6-фосфат перед стадией превращения глюкозо-6-фосфата во фруктозо-6-фосфат.
Способ получения псикозы по настоящему изобретению может дополнительно включать стадию приведения крахмала, мальтодекстрина, сахарозы или их комбинации в контакт с α-глюканфосфорилазой, крахмалфосфорилазой, мальтодекстринфосфорилазой или сахарозофосфорилазой, микроорганизмом, экспрессирующим указанную фосфорилазу, или культурой микроорганизма, экспрессирующего указанную фосфорилазу, и фосфатом с превращением крахмала, мальтодекстрина, сахарозы или их комбинации в глюкозо-1-фосфат перед стадией превращения глюкозо-1-фосфата в глюкозо-6-фосфат.
Способ получения псикозы по настоящему изобретению может дополнительно включать стадию приведения крахмала, мальтодекстрина, сахарозы или их комбинации в контакт с α-амилазой, пуллуланазой, изоамилазой, глюкоамилазой или сахаразой, микроорганизмом, экспрессирующим указанную α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, или культурой микроорганизма, экспрессирующего указанную α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, с превращением крахмала, мальтодекстрина, сахарозы или их комбинации в глюкозу перед стадией превращения крахмала, мальтодекстрина, сахарозы или их комбинации в глюкозо-1-фосфат.
Рибулозофосфат-3-эпимераза, псикозо-6-фосфатфосфатаза, α-глюканфосфорилаза, фосфоглюкомутаза (или фосфоманномутаза), глюкозо-6-фосфатизомераза, псикозо-6-фосфат-3-эпимераза (или рибулозо-5-фосфат-3-эпимераза), пуллуланаза (или изоамилаза), 4-α-глюкантрансфераза, полифосфатглюкокиназа и так далее, используемые в способе получения псикозы по настоящему изобретению, могут не обладать побочной реакционной способностью или обладать меньшей побочной реакционной способностью в отношении конечного продукта псикозы.
Способ получения псикозы по настоящему изобретению разлагает высокую концентрацию крахмала с получением оптимального/максимального количества псикозы в сложной комбинации с глюкозофосфат-превращающими ферментами. Для обеспечения максимальной продуктивности в отношении псикозы возможно использование до восьми видов ферментов в комбинации.
Во-первых, глюканфосфорилаза (гликогенфосфорилаза, ЕС 2.4.1.1), представляющая собой фермент, расщепляющий крахмал с получением глюкозо-1-фосфата, специфична в отношении α-1,4-связанного крахмала с получением глюкозо-1-фосфата. Во-вторых, в промежуточной комплексной ферментативной реакции используют фосфоглюкомутазу (ЕС 2.7.5.1) или фосфоманномутазу (ЕС 5.4.2.8), превращающую полученный таким образом глюкозо-1-фосфат в глюкозо-6-фосфат. В-третьих, используют глюкозо-6-фосфатизомеразу (ЕС 5.3.1.9), представляющую собой фермент, превращающий глюкозо-6-фосфат во фруктозо-6-фосфат. В-четвертых, фруктозо-6-фосфат-3-эпимеразу, представляющую собой фермент, превращающий фруктозо-6-фосфат в псикозо-6-фосфат, используют для получения псикозо-6-фосфата в обратимой реакции.
Кроме того, для повышения скорости утилизации крахмала также используют фермент пуллуланазу (ЕС 3.2.1.41) или изоамилазу (ЕС 3.2.1.68) для расщепления разветвленных α-1,6-связей в дополнение к α-1,4-связям амилопектина. Кроме того, для повышения утилизации крахмала глюканфосфорилазой может быть использована глюкантрансфераза (4-альфа-глюкантрансфераза, ЕС 2.4.1.25). Скорость утилизации сегментированных крахмальных субстратов может быть повышена за счет связывания олигосахаридов в α-1,4-связанной форме с мальтозой или другими олигосахаридами, представляющими собой субстраты с относительно низкой активностью. Кроме того, возможно получение дополнительного количества псикозы с использованием полифосфатглюкокиназы (полифосфатглюкозофосфотрансферазы, ЕС 2.7.1.63) посредством комплексной ферментативной реакции с расщепленной глюкозой, полученной при утилизации крахмала.
Кроме того, в способе получения псикозы по настоящему изобретению приведение в контакт по настоящему изобретению можно осуществлять при рН от 5,0 до 9,0, конкретно при рН от 6,0 до 8,0.
В способе получения псикозы по настоящему изобретению приведение в контакт по настоящему изобретению можно осуществлять при температуре от 40°С до 80°С, конкретно при температуре от 40°С до 60°С или от 50°С до 60°С.
В способе получения псикозы по настоящему изобретению приведение в контакт по настоящему изобретению можно осуществлять в течение от 2 до 24 часов, конкретно от 6 часов до 24 часов.
В способе получения псикозы по настоящему изобретению приведение в контакт по настоящему изобретению можно осуществлять при рН от 5,0 до 9,0 и температуре от 40°С до 80°С и/или в течение 2-24 часов. Конкретно, приведение в контакт можно осуществлять при рН от 6,0 до 8,0 и температуре от 40°С до 60°С или от 50°С до 60°С и/или в течение 6-24 часов.
Способ получения псикозы по настоящему изобретению может дополнительно включать стадию очистки псикозы. Конкретных ограничений относительно очистки по настоящему изобретению нет, и ее можно осуществлять с применением метода, обычно применяемого в области, к которой относится настоящее изобретение. Его неограничивающие примеры могут включать хроматографию, фракционную кристаллизацию, ионную очистку и так далее. Эти методы очистки можно осуществлять по отдельности, или можно осуществлять комбинацию двух или более чем двух из этих методов. Например, полученную псикоза можно очищать посредством хроматографии, а выделение Сахаров посредством хроматографии можно осуществлять с использованием различия сил слабого связывания между выделяемыми сахарами и ионами металла, связанными с ионной смолой.
Кроме того, настоящее изобретение может дополнительно включать осуществление обесцвечивания, обессоливания или как обесцвечивания, так и обессоливания до или после стадии очистки по настоящему изобретению. Осуществление обесцвечивания и/или обессоливания позволяет получить более очищенный продукт псикозу без примесей.
Далее настоящее изобретение будет описано более подробно со ссылкой на типичные воплощения. Тем не менее, эти типичные воплощения приведены только для лучшего понимания настоящего изобретения, и объем настоящего изобретения не следует ограничивать ими.
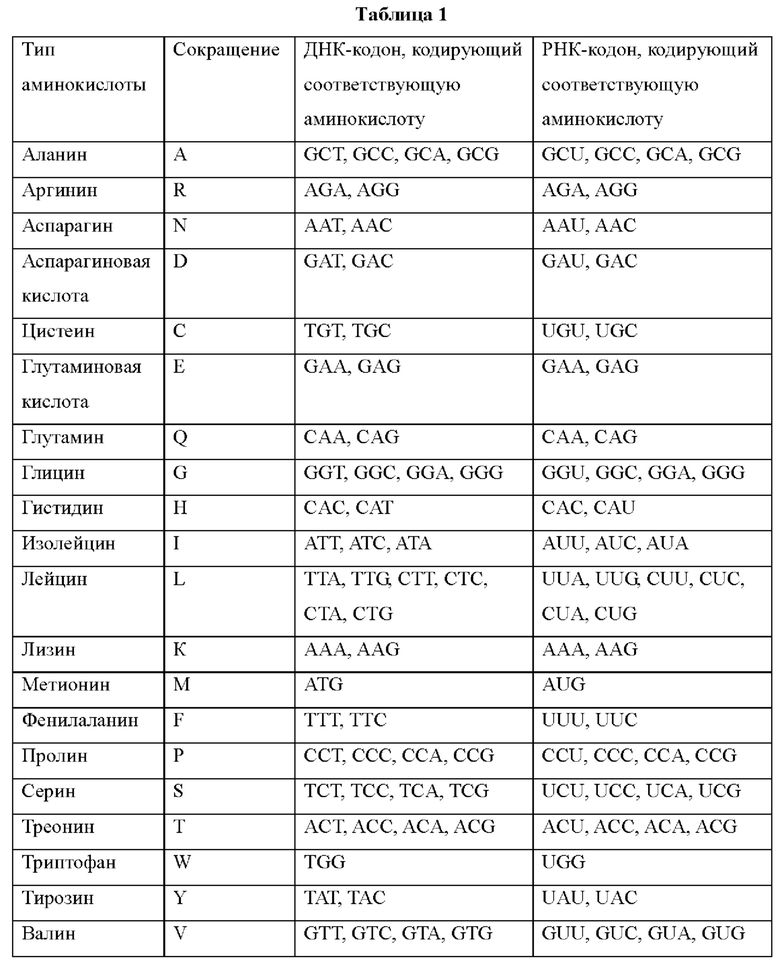
В настоящем изобретении аминокислоты могут быть представлены следующими сокращениями или названиями аминокислот.
Пример 1: Получение рекомбинантного вектора экспрессии каждого фермента и трансформированного микроорганизма
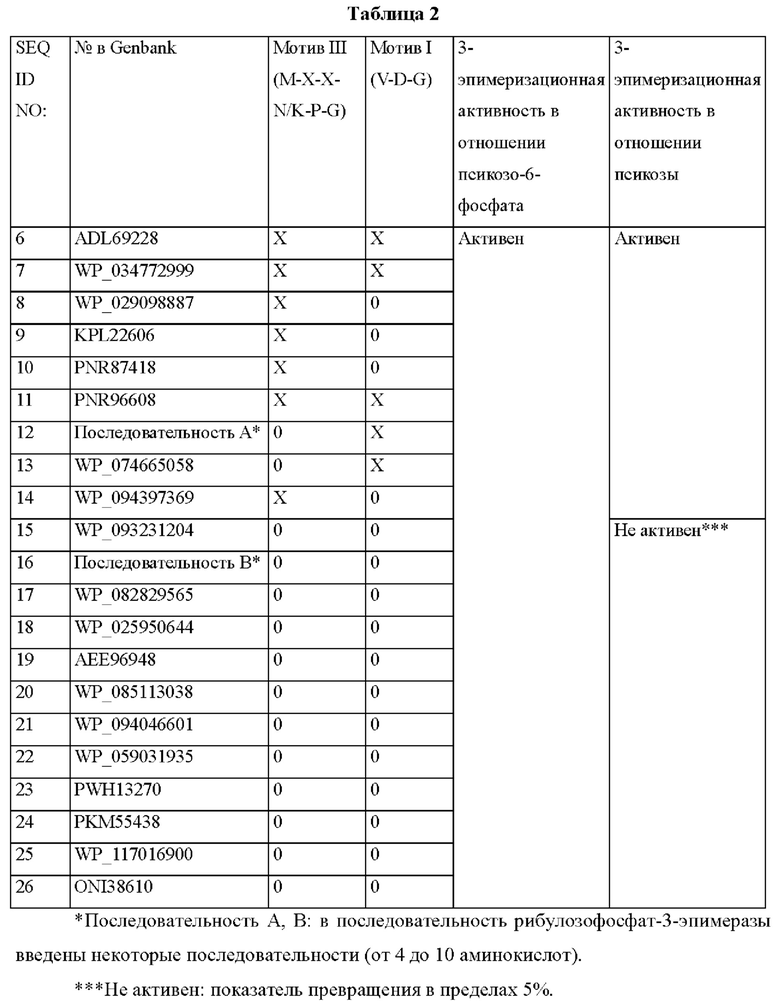
Для обеспечения соответствующих ферментов, необходимых для путей получения псикозы по настоящему изобретению, отбирали гены ферментов, устойчивых к нагреванию. Кроме того, для высокой эпимеризационной активности отбирали гены ферментов, имеющих аминокислотную последовательность S-X-M-C (SEQ ID NO:4) или G-X-X-X-X-F (SEQ ID NO:5) (Таблица 2).
Отобранные гены аминокислот амплифицировали посредством синтеза генов или полимеразной цепной реакции (PCR) в отношении каждого гена из геномной ДНК каждого штамма, и каждую амплифицированную ДНК вводили в плазмидный вектор рЕТ21а (Novagen) для экспрессии в Е. coli с использванием методов сборки ДНК, получая в каждом случае рекомбинантный вектор экспрессии. Этим вектором экспрессии трансформировали штамм E.coli BL21(DE3) в соответствии с обычным методом трансформации (Sambrook et al. 1989), получая в каждом случае трансформированный микроорганизм.
Конкретно, штамм Е. coli BL21(DE3) трансформировали псикозо-6-фосфат-3-эпимеразами с SEQ ID NO:6-26, соответственно. Из них микроорганизмы, трансформированные описанным выше методом получения с использованием SEQ ID NO:19, 20 и 22, были депонированы в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM), являющемся международным органом по депонированию, 16 апреля 2019 г. под регистрационными номерами KCCM12494P (Е. coli BL21(DE3)/pET-CJ-fep19), KCCM12495P (E. coli BL21(DE3)/pET-CJ-fep20) и KCCM12496P (E. coli BL21(DE3)/pET-CJ-fep22), соответственно.
Пример 2: Получение рекомбинантных ферментов
Для получения рекомбинантных ферментов каждый трансформированный микроорганизм, полученный в Примере 1, вносили в культуральную пробирку, содержавшую 5 мл жидкой среды LB, и посевную культуру культивировали в инкубаторе с покачиванием при 37°С до достижения оптической плотности при 600 им 2,0. Культуральную среду посевной культуры вносили в культуральную колбу, содержавшую жидкую среду LB, для осуществления основного культивирования. При достижении оптической плотности при 600 им 2,0 вносили 1 мМ IPTG (Изопропил-β-D-тиогалактозид) для индуцирования экспрессии и получения рекомбинантного фермента. Во время культивирования скорость перемешивания составляла 180 об/мин, и поддерживали температуру культуры 37°С. Культуральную среду центрифугировали при 8000 g и 4°С в течение 20 минут для получения клеточного осадка. Полученный клеточный осадок промывали буфером с 50 мМ трис-HCl (рН 8,0), суспендировали в том же буфере и разрушали клетки ультразвуком. Клеточный лизат центрифугировали при 13000 g и 4°С в течение 20 минут, отбирая только супернатант. Рекомбинантный фермент из супернатанта очищали с использованием аффинной хроматографии с His-метками, подвергали его диализу против буфера с 50 мМ трис-HCl (рН 8,0) и затем использовали в реакциях.
Пример 3: Моделирование псикозо-6-фосфат-3-эпимеразы, анализ 3-эпимеризационной активности в отношении псикозы и сравнение их последовательностей
Аминокислотную последовательность SEQ ID NO:9, раскрытую как рибулозофосфат-3-эпимераза, вводили на сервере I-TASSER, Phyre2, Galaxyweb для анализа структуры белка.
В результате, было подтверждено, что фермент имел структуру TIM-бочки, где в центре расположена β-складчатость, которую окружает α-спираль, и в структуре (структуре TIM-бочки), были выбраны мотивы I (V-D-G) и III (М-Х-Х-X'-P-G), предположительно являющиеся сайтом связывания псикозы или псикозо-6-фосфата (ФИГ. 1, синяя пунктирная линия (мотив I), красная пунктирная линия (мотив III), пример структурной модели (SEQ ID NO:9)).
В то же время, поскольку известные псикозо-6-фосфат-3-эпимеразы (ADL69228, WP_034772999, WP_029098887; WO2018/129275, WO2018/112139) проявляют активность по образованию фруктозы посредством эпимеризации псикозы, выход получения псикозы снижается.
Соответственно, авторы настоящего изобретения предположили, что аспарагиновая кислота (D) в структуре (M-X-X-D-P-G), имеющая отрицательный заряд, может влиять на 3-эпимеризационную активность в отношении псикозы, и, исходя из этого, были выбраны ферменты, предположительно не влияющие на 3-эпимеризационную активность в отношении псикозы (WP_085113038, PKM55438, WP 117016900), где X' мотива III (M-X-X-X'-P-G) представляет собой N/K, заряд которого отличается от аспарагиновой кислоты.
В результате, было подтверждено, что 3-эпимеризационная активность в отношении псикозы может варьировать в зависимости от типа определенного мотива в ферменте.
Пример 4: Анализ активности рибулозофосфат-3-эпимеразы по превращению в псикозо-6-фосфат
Для анализа активности рибулозофосфат-3-эпимеразы по настоящему изобретению, являющейся ферментом, осуществляющим превращение в псикозо-6-фосфат, 50 мМ фруктозо-6-фосфата или 20 мМ глюкозо-1-фосфата суспендировали в буфере с 50 мМ трис-HCl (рН 7,0), или 50 мМ фосфата натрия (рН 6-7), или 50 мМ фосфата калия (рН 6-7), добавляли фосфоглюкомутазу или фосфоманномутазу, и глюкозо-6-фосфатизомеразу, и псикозо-6-фосфатфосфатазу, и рекомбинантную рибулозофосфат-3-эпимеразу, полученную в Примере 2, каждую в количестве 0,1 единицы/мл, и давали им взаимодействовать при 45-70°С в течение 1-24 часов.
Для анализа активности псикозо-3-эпимеразы псикозу суспендировали в концентрации 1% (масс./об.) в буфере с 50 мМ трис-HCl (рН 7,0), или 50 мМ фосфата натрия (рН 6-7), или 50 мМ фосфата калия (рН 6-7), добавляли рибулозофосфат-3-эпимеразу, 0,1 единицы/мл, и давали им взаимодействовать при 45-70°С в течение 1-24 часов. Образование глюкозы, фруктозы или псикозы анализировали посредством HPLC. HPLC-анализ осуществляли с использованием колонки SP_0810 (Shodex) и колонки Aminex НРХ-87С (Bio-RAD) при 80°С с использованием подвижной фазы со скоростью потока 0,6 мл/мин и выявление осуществляли с использованием детектора коэффициента преломления (RID). Осуществляли качественный и количественный анализ глюкозы, фруктозы и псикозы, являющихся основными сахарами, каждый из которых получен при смешивании с указанными выше ферментами. Для количественной оценки допустимое отклонение по скорости превращения во фруктозу относительно исходной концентрации псикозы было установлено как 5% с учетом спонтанной ошибки эксперимента в соответствии с чувствительностью LC (жидкостная хроматография), высокой скорости превращения субстрата и концентрации субстрата.
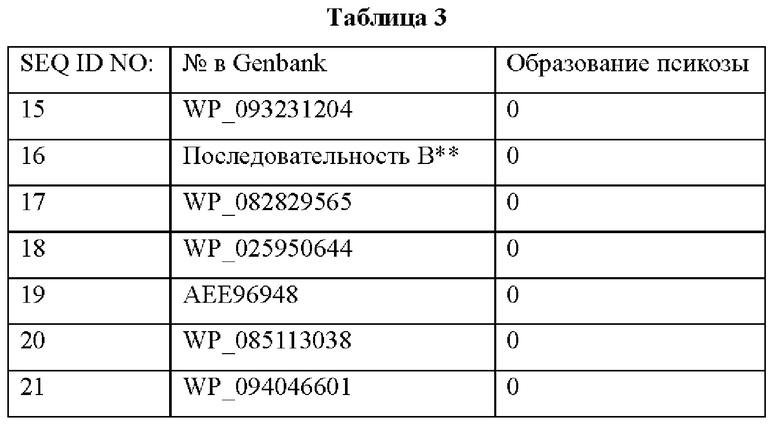
В Таблице 2 среди нескольких рибулозофосфат-3-эпимераз, демонстрирующих 3-эпимеризационную активность в отношении псикозо-6-фосфата, ферменты, не обладающие 3-эпимеризационной активностью в отношении псикозы, были разделены, исходя из присутствия или отсутствия мотивов M-X-X-N/K-P-G и V-D-G. Было подтверждено, что из этих ферментов фермент, содержащий определенный мотив, конкретно мотив с SEQ ID NO:1 без мотива с SEQ ID NO:2, и содержащий мотив с SEQ ID NO:3 в том же месте вместо мотива с SEQ ID NO:2, обладал высокой эпимеризационной активностью, специфичной в отношении псикозо-6-фосфата. Кроме того, как подтверждено в случае последовательностей А и В, когда фермент содержит мотивы I и III, он обладает высокой эпимеризационной активностью, специфичной в отношении псикозо-6-фосфата.
Кроме того, было подтверждено, что фермент, имеющий аминокислотную последовательность SEQ ID NO:20, по настоящему изобретению демонстрировал значительно меньшее образование фруктозы по сравнению с известной псикозо-6-фосфат-3-эпимеразой (ФИГ. 2).
Это показывает, что присутствие или отсутствие мотива оказывает существенное влияние на выход получения аллюлозы.
Кроме того, было подтверждено, что последовательность, содержащая SEQ ID NO:4 и/или SEQ ID NO:5, демонстрировала высокую 3-эпимеризационную активность в отношении псикозо-6-фосфата.
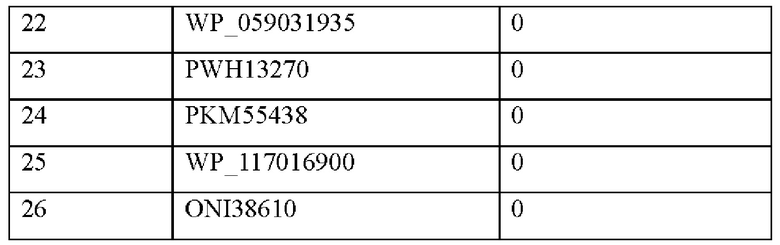
Пример 4: Анализ активности по образованию псикозы в комплексной ферментативной реакции (с множеством ферментов)
Для получения псикозы из мальтодекстрина осуществляли реакцию с глюканфосфорилазой, пуллуланазой, 4-альфа-глюкантрансферазой, фосфоглюкомутазой, глюкозо-6-фосфатизомеразой, псикозо-6-фосфатфосфатазой и фруктозо-6-фосфат-3-эпимеразой по настоящему изобретению в одной емкости.
Конкретно, каждый из семи типов ферментов, 0,1 единицы/мл, добавляли в раствор, где 5% (масс/об.) мальтодекстрин добавляли к 1-5 мМ MgCl2, 10-50 мМ фосфату натрия (рН 7,0), и давали им взаимодействовать при температуре 50°С в течение 12 часов.
По завершении реакции псикозу, присутствовавшую в продуктах реакции, анализировали посредством HPLC. HPLC-анализ осуществляли с использованием колонки Aminex НРХ-87С (Bio-Rad) при 80°С с использованием подвижной фазы со скоростью потока 0,6 мл/мин и выявление осуществляли с использованием детектора коэффициента преломления. В результате было подтверждено, что псикоза была получена из мальтодекстрина посредством комплексной ферментативной реакции.
На основании приведенного выше описания специалистам в данной области будет ясно, что настоящее изобретение может быть осуществлено в различных конкретных формах без изменения его технической идеи или существенных признаков. В этой связи, следует понимать, что во всех аспектах описанные выше воплощения приведены для наглядности и не являются ограничивающими. Объем изобретения определен в приложенной формуле изобретения, а не в предшествующем описании, и поэтому подразумевают, что формула изобретения включает все изменения и модификации, не выходящие за рамки границ и пределов формулы изобретения или эквивалентов таких границ и пределов.
Эффект изобретения
Поскольку рибулозофосфат-3-эпимераза по настоящему изобретению не содержит определенный мотив, она обладает низкой 3-эпимеризационной активностью в отношении псикозы и обладает устойчивостью к нагреванию и, таким образом, обладает преимуществами при ее промышленном применении, таком как получение псикозы и так далее.
Изобретение относится к биотехнологии и представляет собой применение рибулозофосфат-3-эпимеразы для получения псикозо-6-фосфата или псикозы. Изобретение относится также к применению микроорганизма, продуцирующего рибулозофосфат-3-эпимеразу для получения псикозо-6-фосфата или псикозы. Изобретение позволяет эффективно получать псикозо-6-фосфат или псикозу. 7 н. и 10 з.п. ф-лы, 2 ил., 3 табл., 4 пр.
1. Применение рибулозофосфат-3-эпимеразы, для продуцирования псикозо-6-фосфата, содержащей мотив I, состоящий из аминокислотной последовательности SEQ ID NO: 1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO: 3, при этом рибулозофосфат-3-эпимераза не содержит мотив II, состоящий из аминокислотной последовательности SEQ ID NO: 2.
2. Применение по п. 1, где рибулозофосфат-3-эпимераза не обладает активностью по превращению псикозы во фруктозу или обладает 5% или менее указанной активности.
3. Применение по п. 1, где рибулозофосфат-3-эпимераза дополнительно содержит мотив, состоящий из аминокислотной последовательности SEQ ID NO: 4 или 5.
4. Применение по п. 1, где рибулозофосфат-3-эпимераза содержит мотив I в положениях 173-184 от N-концевой аминокислоты.
5. Применение по п. 1, где рибулозофосфат-3-эпимераза содержит мотив III в положениях 136-150 от N-концевой аминокислоты.
6. Применение по п. 1, где рибулозофосфат-3-эпимераза обладает 3-эпимеризационной активностью в отношении псикозо-6-фосфата при температуре от 50 до 90°C.
7. Применение по п. 1, где рибулозофосфат-3-эпимераза имеет происхождение из любого, выбранного из группы, состоящей из Chthonomonas, Geobacillus, Mahella, Thermoanaerobacterium, Tepidanaerobacter, Ardenticatenia, Firmicutes, Aeribacillus, Epulopiscium и Thermoflavimicrobium.
8. Применение по п. 1, где рибулозофосфат-3-эпимераза имеет происхождение из любого, выбранного из группы, состоящей из Chthonomonas calidirosea T49, Geobacillus sp. 8, Geobacillus thermocatenulatus, Mahella australiensis 50-1 BON, Thermoanaerobacterium sp. PSU-2, Thermoanaerobacterium thermosaccharolyticum, Tepidanaerobacter syntrophicus, бактерии Ardenticatenia, бактерии Firmicutes HGW-Firmicutes-5, Aeribacillus pallidus, Epulopiscium sp. SCG-B05WGA-EpuloA1 и Thermoflavimicrobium dichotomicum.
9. Применение по п. 1, где рибулозофосфат-3-эпимераза состоит из любой последовательности, выбранной из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 15-26 и аминокислотных последовательностей, имеющих по меньшей мере 26% идентичности с областью, исключающей мотивы I и III в аминокислотных последовательностях.
10. Применение нуклеиновой кислоты, кодирующей рибулозофосфат-3-эпимеразу, как она определена в пп. 1-9, для продуцирования псикозо-6-фосфата.
11. Применение микроорганизма, содержащего нуклеиновую кислоту, как она определена в п. 10, для продуцирования псикозо-6-фосфата.
12. Композиция для получения псикозо-6-фосфата, содержащая рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма, где рибулозофосфат-3-эпимераза содержит мотив I, состоящий из аминокислотной последовательности SEQ ID NO: 1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO: 3, где композиция дополнительно содержит фруктозо-6-фосфат, при этом рибулозофосфат-3-эпимераза не содержит мотив II аминокислотной последовательности SEQ ID NO: 2.
13. Способ получения псикозо-6-фосфата, включающий стадию приведения фруктозо-6-фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма, где рибулозофосфат-3-эпимераза содержит мотив I, состоящий из аминокислотной последовательности SEQ ID NO: 1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO: 3, при этом рибулозофосфат-3-эпимераза не содержит мотив II аминокислотной последовательности SEQ ID NO: 2.
14. Композиция для получения псикозы, содержащая рибулозофосфат-3-эпимеразу, микроорганизм, экспрессирующий указанную рибулозофосфат-3-эпимеразу, или культуру указанного микроорганизма, и псикозо-6-фосфатфосфатазу, микроорганизм, экспрессирующий указанную псикозо-6-фосфатфосфатазу, или культуру указанного микроорганизма, где рибулозофосфат-3-эпимераза содержит мотив I, состоящий из аминокислотной последовательности SEQ ID NO: 1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO: 3, при этом рибулозофосфат-3-эпимераза не содержит мотив II аминокислотной последовательности SEQ ID NO: 2.
15. Композиция по п. 14, дополнительно содержащая один или более чем один фермент, выбранный из группы, состоящей из глюкозо-6-фосфатизомеразы, фосфоглюкомутазы, полифосфатглюкокиназы, α-глюканфосфорилазы, крахмалфосфорилазы, мальтодекстринфосфорилазы или сахарозофосфорилазы, α-амилазы, пуллуланазы, изоамилазы, α-глюкантрансферазы, глюкоамилазы, сахаразы и псикозо-6-фосфатфосфатазы, микроорганизм, экспрессирующий указанный фермент, или культуру указанного микроорганизма.
16. Способ получения псикозы, включающий стадии приведения фруктозо-6- фосфата в контакт с рибулозофосфат-3-эпимеразой, микроорганизмом, экспрессирующим указанную рибулозофосфат-3-эпимеразу, или культурой указанного микроорганизма и приведения псикозо-6-фосфата, полученного из фруктозо-6-фосфата, в контакт с псикозо-6-фосфатфосфатазой, микроорганизмом, экспрессирующим указанную псикозо-6-фосфатфосфатазу, или культурой указанного микроорганизма, где рибулозофосфат-3-эпимераза содержит мотив I, состоящий из аминокислотной последовательности SEQ ID NO: 1, и мотив III, состоящий из аминокислотной последовательности SEQ ID NO: 3, при этом рибулозофосфат-3-эпимераза не содержит мотив II аминокислотной последовательности SEQ ID NO: 2.
17. Способ по п. 16, дополнительно включающий стадию выделения псикозы, полученной из псикозо-6-фосфата.
| WO 2004042043 A2, 21.05.2004 | |||
| US 3970522 A1, 20.07.1976 | |||
| WO 2003084986 A2, 16.10.2003 | |||
| D-ПСИКОЗА В ЗАМОРОЖЕННЫХ НАПИТКАХ НУЛЕВОЙ ИЛИ НИЗКОЙ КАЛОРИЙНОСТИ | 2014 |
|
RU2664311C2 |

