Область техники
Настоящее изобретение относится к новому промотору, к вектору, содержащему этот промотор, к микроорганизму, содержащему этот промотор или вектор, и к способу получения целевого продукта с использованием этого микроорганизма.
Предшествующий уровень техники
Предпринимались непрерывные попытки получения целевых продуктов, таких как аминокислоты или полезные вещества, которые можно применять для разных целей, включая корма, фармацевтические лекарственные средства, продукты питания и так далее, с высоким титром, используя микроорганизмы (корейский патент 10-0924065). Одним из таких способов является способ индуцирования сверхэкспрессии целевого гена в микроорганизме, и для этой цели необходима высокоэффективная система экспрессии гена. Так как промоторы являются одним из факторов, которые в значительной мере вовлечены в системы экспрессии генов, необходима разработка подходящего промотора.
В качестве сильного промотора широко известен происходящий из E. coli tac-промотор. В случае коринеформного микроорганизма сильные промоторы были разработаны посредством модификации промоторов собственных генов (Gene, 102, 93-98, 1991; Microbiology, 142, 1297-1309, 1996). Например, в случаях промоторов, происходящих из Corynebacterium ammoniagenesis, раскрыто, что имеет место примерно 10% улучшение по сравнению с tac-промотором, описанном в E.coli (Biotechnol. Lett. 25, 1311-1316, 2003). Кроме того, в качестве сильных промоторов, происходящих из Corynebacterium ammoniagenesis, были разработаны промоторы от Pcj1 до Pcj7 с различной силой, и они обладают сильной промоторной активностью, которая по меньшей мере в 10 раз выше, чем активность tac-промотора (корейский патент 10-0620092). Кроме того, был разработан промотор Po2, который синтезировали в Corynebacterium glutamicum, чтобы иметь сильную промоторную активность (корейский патент 10-1632642). Однако по-прежнему существует потребность в разработке промотора, так как необходима система, которая проявляет высокую эффективность экспрессии в Corynebacterium по сравнению с системой экспрессии генов в E. coli.
В данных обстоятельствах авторами изобретения было предпринято много попыток обнаружения промоторов, которые могут сильно индуцировать экспрессию генов в микроорганизме рода Corynebacterium. В результате они разработали новый синтезированный промотор по настоящему изобретению и подтвердили, что этот промотор имеет более высокую активность экспрессию по сравнению с активностью известных промоторов, завершая таким образом настоящее изобретение.
Описание изобретения
Техническая задача
Целью настоящего изобретения является предложение новой молекулы нуклеиновой кислоты, имеющей промоторную активность; кассеты экспрессии гена, содержащей эту молекулу нуклеиновой кислоты и целевой ген; рекомбинантного вектора, содержащего эту молекулу нуклеиновой кислоты или кассету экспрессии гена; рекомбинантного микроорганизма, содержащего промотор или вектор; и способа получения целевого продукта с использованием этого рекомбинантного микроорганизма.
Техническое решение
Для достижения целей настоящего изобретения, в одном аспекте настоящего изобретения предлагается молекула нуклеиновой кислоты, обладающая промоторной активностью, состоящая из любой нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-3.
При использовании в настоящем изобретении термин “промотор” относится к нетранслируемой нуклеиновокислотной последовательности, расположенной выше кодирующей области, которая включает участок связывания полимеразы и облает активностью инициации транскрипции гена, расположенного ниже промотора в мРНК, то есть домен ДНК, с которым полимераза связывается и инициирует транскрипцию гена. Промотор может быть расположен в 5'-домене домена инициации транскрипции мРНК.
В настоящем изобретении молекулы нуклеиновой кислоты, которые обладают промоторной активностью, состоящие из любой нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-3 (то есть нуклеотидной последовательности SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 3), были обозначены SPL1, SPL7 и SPL13, соответственно. Молекулы нуклеиновой кислоты, обладающие промоторной активностью, также могут называться промоторами, и все термины, описанные выше, могут быть использованы в настоящем описании изобретения.
Промотор по настоящему изобретению обеспечивает возможность экспрессии целевого гена, который функционально связан с молекулой нуклеиновой кислоты, обладающей промоторной активностью, в целевом микроорганизме, и может использоваться в качестве промотора общего применения.
Кроме того, промоторную последовательность по настоящему изобретению можно модифицировать посредством общеизвестного мутагенеза, например прямой эволюции, сайт-направленного мутагенеза и так далее. Таким образом, промотор может включать, без ограничения, любую нуклеотидную последовательность, имеющую гомологию 70% или выше, конкретно 80% или выше, более конкретно 90% или выше, еще более конкретно 95% или выше, даже еще более конкретно 98% или выше и наиболее конкретно 99% или выше, с нуклеотидной последовательностью SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 3, и обладающую аналогичной промоторной активностью. Кроме того, любую нуклеотидную последовательность, имеющую указанную выше гомологию, в которой часть последовательности делегирована, модифицирована, замещена или встроена, следует считать включенной в объем термина молекула нуклеиновой кислоты по настоящему изобретению, пока эта последовательность имеет промоторную активность.
В частности, выражение “состоящая из нуклеотидной последовательности SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 3” не исключает случаи добавления, и/или делении, и/или модификации, и так далее, нуклеотида, что может происходить при связывании ее с целевым геном наряду с использованием фермента рестрикции, когда соответствующий промотор используют посредством связывания с целевым геном.
Конкретно, в дополнение к промотору для осуществления транскрипции гена может быть включена любая последовательность оператора для контроля транскрипцией, последовательность, кодирующая подходящий участок связывания мРНК рибосомы, и последовательность для контроля транскрипции и трансляции. Например, контрольная последовательность, подходящая для прокариотов, может включать любую последовательность оператора или рибосома-связывающий домен, но не ограничена ими. Молекула нуклеиновой кислоты, имеющая промоторную активность по настоящему изобретению, может состоять из последовательности для контроля экспрессии гена, как описано выше, в соответствии с потребностью специалиста обычной квалификации в данной области техники.
Молекула нуклеиновой кислоты, которая состоит из любой нуклеотидной последовательности, выбранной из группы, состоящей из нуклеотидных последовательностей SEQ ID NO: 1-3, имеющих промоторную активность, может включать, без ограничения им, зонд, который может быть получен из известной последовательности гена, например любой нуклеотидной последовательности, которая имеет промоторную активность по настоящему изобретению, посредством гибридизации с комплементарной последовательностью до полной или частичной нуклеотидной последовательности SEQ ID NO: 1-3 по настоящему изобретению в жестких условиях.
При использовании в данном описании изобретения термин “гомология” относится к проценту идентичности между двумя полинуклеотидами или полипептидными группами. Гомология последовательности между одной и другой группой может быть определена с помощью методики, известной в данной области техники. Например, гомология может быть подтверждена с использованием стандартной программы вычисления параметров, таких как счет, идентичность и подобие (конкретно BLAST 2.0), или путем сравнения последовательностей с помощью экспериментов саузерн-гибридизации в определенных жестких условиях, и определенные подходящие условия гибридизации могут быть определены способом, хорошо известным специалисту обычной квалификации в данной области техники в пределах объема соответствующей технологии (например, J. Sambrook et al, Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al, Current Protocols in Molecular Biology, John Wiley & Sons, Inc, New York).
Термин “жесткие условия” относится к условиям, которые обеспечивают возможность специфической гибридизации между полинуклеотидами. Эти условия подробно описаны в ссылках (например, J. Sambrook et al, выше). Например, могут быть включены условия гибридизации, в которых гибридизируются гены с высокой степенью гомологии (например, гены с гомологией 80% или выше, конкретно 90% или выше, более конкретно 95% или выше, еще более конкретно 97% или выше, и еще более конкретно 99% или выше), а гены со степенью гомологии ниже, чем описанная выше, не гибридизируются; или условия промывки, в которых промывку выполняют единоразово, конкретно 2-3 раза, в условиях концентраций солей и температуры, соответствующих обычно используемым условиям промывки для саузерн-гибридизации (т.е. 60°C, 1×SSC (хлорид натрия/цитрат натрия) и 0,1% SDS (додецилсульфат натрия)), конкретно 60°C, 0,1×SSC и 0,1% SDS, более конкретно 68°C, 0,1×SSC и 0,1% SDS. Хотя из-за жесткости гибридизации может происходить ошибочное спаривание нуклеотидов, требуется, чтобы две нуклеиновые кислоты имели комплементарные последовательности. Термин “комплементарный” используют для описания взаимоотношения между нуклеотидными основаниями, которые могут гибридизироваться друг с другом. Например, применительно к нуклеотидным основаниям, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение может включать не только по существу подобные нуклеиновокислотные последовательности, но также выделенные фрагменты нуклеиновых кислот, которые комплементарны полной последовательности. Конкретно, полинуклеотид, имеющий гомологию, может быть обнаружен с использованием условий гибридизации, включающих условия гибридизация при значении Tm (температура плавления) 55°C и условия, описанные выше. Кроме того, значение Tm может представлять собой 60°C, 63°C или 65°C, но без ограничения ими, и может подходящим образом регулироваться специалистом в данной области техники в соответствии с целями. Хорошо известно, что жесткость гибридизации полинуклеотидов зависит от длины и степени комплементарности этих полинуклеотидов и переменные хорошо известны специалисту в данной области техники (см. Sambrook et al, выше, 9.50 - 9.51, 11.7 - 11.8).
Молекула нуклеиновой кислоты по настоящему изобретению, обладающая промоторной активностью, может быть выделена или получена с использованием стандартных методик молекулярной биологии. Например, молекула нуклеиновой кислоты может быть получена с использованием стандартной технологии синтеза, в которой используется автоматический синтезатор ДНК, но получение не ограничено этим.
В другом аспекте настоящего изобретения предлагается кассета экспрессии гена, включающая молекулу нуклеиновой кислоты и целевой ген по настоящему изобретению.
Молекула нуклеиновой кислоты по настоящему изобретению является такой же, как описано выше.
При использовании в данном описании изобретения термин “кассета экспрессии гена” относится к единичной кассете, которая включает промотор и целевой ген и, таким образом, может экспрессировать целевой ген, функционально связанный с промотором ниже него. Такая кассета экспрессии гена может включать различные факторы, которые могут способствовать эффективной экспрессии целевого гена, в пределах или за пределами кассеты. Кассета экспрессии гена обычно может включать сигнал терминации транскрипции, рибосома-связывающий домен и сигнал терминации трансляции в дополнение к промотору, функционально связанному с целевым геном.
При использовании в данном описании изобретения термин “целевой ген” относится к гену, кодирующему белок, который должен экспрессироваться в микроорганизме.
Например, целевой ген может представлять собой ген, вовлеченный в продуцирование продукта, выбранного из группы, состоящей из сахаридов (например, псикозы или тагатозы), L-аминокислот (L-лизина, L-валина и так далее), органических кислот, ферментов и их комбинации, но без ограничения ими. Конкретно, целевой ген может представлять собой ген, кодирующий фермент превращения сахаров, или фермент, ассоциированный с биосинтезом аминокислот, ген, кодирующий фермент, ассоциированный с восстанавливающей способностью, ген, кодирующий фермент, ассоциированный с биосинтезом органических кислот, или ген, кодирующий фермент, ассоциированный с высвобождением целевого продукта, но без ограничения ими. Более конкретно, целевой ген может представлять собой ген, кодирующий псикоза-эпимеразу, ген, кодирующий тагатоза-эпимеразу, или ген, кодирующий тагатуронат-эпимеразу, ген, кодирующий НАДФ-зависимую глицеральдегид-3-фосфатдегидрогеназу, или ген, кодирующий аминотрансферазу аминокислот с разветвленной цепью, но без ограничения ими.
Псикоза-эпимераза может быть указана как ATPE и это относится к псикоза-3-эпимеразе, которая обладает активностью превращения фруктозы в псикозу. Кроме того, тагатуронат-эпимераза или тагатоза-эпимераза (гексуроновая кислота C4-эпимераза; корейский патент 10-1550796) может быть обозначена как UxaE, и это относится к ферменту, который имеет активность превращения фруктуроната в тагатуронат или превращения фруктозы в тагатозу. НАДФ-зависимая глицеральдегид-3-фосфатдегидрогеназа может быть обозначена как GapN, и это относится к ферменту, который обладает активностью превращения в 3-фосфоглицерат, используя глицеральдегид-3-фосфат в качестве субстрата. Аминотрансфераза аминокислот с разветвленной цепью может быть указана как IlvE, и это относится к ферменту последней стадии пути биосинтеза аминокислот с разветвленной цепью. Последовательности генов, кодирующих ATPE, UxaE, GapN и IlvE, могут быть легко получены специалистом обычной квалификации в данной области техники из известной базы данных, такой как GenBank NIH (Национальных институтов здравоохранения) (США). Гены, кодирующие ATPE, UxaE, GapN и IlvE, являются иллюстративными генами-мишенями, которые могут быть функционально связанны с молекулой нуклеиновой кислоты, обладающей промоторной активностью, по настоящему изобретению, и промотор по настоящему изобретению может использовать без ограничения любой ген, который может экспрессироваться в микроорганизме посредством промотора универсального назначения, в качестве целевого гена. При использовании в данном описании изобретения термин “функционально связанный” означает, что последовательность указанных выше генов и последовательность промотора функционально связаны так, что нуклеиновокислотная последовательность, имеющая промоторную активность, по настоящему изобретению, может инициировать и опосредовать транскрипцию целевого гена. Функциональное связывание может быть получено с использованием генетической рекомбинантной технологии, хорошо известной в данной области техники, а сайт-специфичное расщепление и связывание ДНК может быть получено с использованием ферментов расщепления и связывания, и так далее, в данной области техники, но без ограничения ими.
В другом аспекте настоящего изобретения предлагается рекомбинантный вектор, который включает молекулу нуклеиновой кислоты по настоящему изобретению или кассету экспрессии гена по настоящему изобретению.
Молекула нуклеиновой кислоты и кассета экспрессии гена являются такими же, как указано выше.
При использовании в данном описании изобретения термин “вектор” представляет собой искусственную молекулу ДНК, которая имеет генетический материал, обеспечивающий возможность экспрессии целевого гена в подходящей клетке-хозяине и, конкретно, конструкцию ДНК, которая включает нуклеотидную последовательность гена, функционально связанного с подходящей регуляторной последовательностью. Регуляторная последовательность может включать, в дополнение к промотору, способному инициировать транскрипции, любую последовательность оператора для регулирования такой транскрипции, последовательность, кодирующую подходящий связывающий домен мРНК рибосомы, и последовательность для регулирования транскрипции и трансляции, но без ограничения ими.
Вектор, используемый в настоящем изобретении, не может быть конкретным образом ограничен, пока этот вектор способен экспрессироваться в клетке-хозяине, и клетка-хозяин может быть трансформирована с использованием любого вектора, известного в данной области техники. Примеры обычно используемого вектора могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, M13, λBL3, λBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A, Charon21A и так далее; и в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и так далее. Вектор для использования в настоящем изобретении не ограничен конкретным образом, и можно использовать любой известный вектор экспрессии. Кроме того, эндогенный промотор в хромосоме может быть заменен молекулой нуклеиновой кислоты, обладающей промоторной активностью, по настоящему изобретению, при помощи вектора для введения хромосомы в клетку-хозяина. Введение молекулы нуклеиновой кислоты в хромосому может быть выполнено способом, хорошо известным в данной области техники, например посредством гомологичной рекомбинации. Например, можно использовать векторы pECCG117, pDZ, pACYC177, pACYC184, pCL, pUC19, pBR322, pMW118, pCC1BAC, pCES208, pXMJ19 и так далее, но вектор не ограничивается ими. Так как вектор по настоящему изобретению может быть введен в хромосому с помощью гомологичной рекомбинации, дополнительно может быть включен селективный маркер для подтверждения введения в хромосому. Селективный маркер используют для отбора трансформированной клетки, то есть для подтверждения введения целевой молекулы нуклеиновой кислоты, и можно использовать маркеры, способные обеспечить селектируемые фенотипы, такие как устойчивость к лекарственному средству, потребность в питательных веществах, устойчивость к цитотоксическим агентам и экспрессия поверхностных белков. В условиях обработки селективными агентами, только клетки, способные экспрессировать селективные маркеры, могут выживать или экспрессировать другие фенотипические признаки, и таким образом могут быть выбраны трансформированные клетки.
При использовании в данном описании изобретения термин “трансформация” относится к процессу введения вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, обеспечивая тем самым экспрессию полинуклеотида, кодирующего белок, в клетке-хозяине. Для трансформирующего полинуклеотида не имеет значение, введен ли он в хромосому клетки-хозяина и находится в ней, или находится вне хромосомы, пока он может экпрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид можно вводить в любой форме, пока он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую все существенные элементы, необходимые для самоэкспрессии, или в форме вектора, включающего экспрессионную кассету. Экспрессионная кассета или вектор, включающий полинуклеотид, могут представлять собой кассету или вектор, которые включают, например, молекулу нуклеиновой кислоты, состоящую из нуклеотидной последовательности, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 3 по настоящему изобретению, имеющую промоторную активность, и могут представлять собой вектор, с которым целевой ген не связан функционально. Даже в этом случае молекула нуклеиновой кислоты, имеющая промоторную активность, может быть заменена эндогенным промотором в клетке-хозяине (например, микроорганизме рода Corynebacterium) и посредством гомологичной рекомбинации. Как таковой, эндогенный ген в клетке-хозяине может экспрессироваться.
Способ трансформации может включать любой способ, которым можно вводить нуклеиновые кислоты в клетку, и трансформация может быть выполнена посредством выбора подходящего стандартной методики, которая известна в данной области техники, в зависимости от клетки-хозяина. Например, способ может включать электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, способ с использованием полиэтиленгликоля (PEG), способ с DEAE-декстраном, способ с катионными липосомами и способ с ацетат лития-DMSO, и так далее, но без ограничения ими.
В еще одном аспекте настоящего изобретения предлагается рекомбинантный микроорганизм, который включает молекулу нуклеиновой кислоты, обладающую промоторной активностью, по настоящему изобретению, кассету экспрессии гена, или рекомбинантный вектор, включающий кассету экспрессии гена.
Молекула нуклеиновой кислоты, имеющая промоторную активность, кассета экспрессии гена и рекомбинантный вектор являются такими же, как указано выше.
Кассета экспрессии гена и рекомбинантный вектор могут быть введены в микроорганизм путем трансформации.
Кроме того, трансформация является такой же, как указано выше.
При использовании в данном описании изобретения термин “микроорганизм” представляет собой понятие, включающее как микроорганизм дикого типа, так и естественно или искусственно генетически модифицированный микроорганизм, и он может представлять собой микроорганизм, имеющий особый ослабленный или усиленный механизм вследствие введения чужеродного гена, или усиления или ослабления активности эндогенного гена. При использовании в данном описании изобретения микроорганизм может включать, без ограничения, любой микроорганизм, в который молекула нуклеиновой кислоты, имеющая промоторную активность по настоящему изобретению, введена и где она способна функционировать в качестве промотора.
Конкретно, микроорганизм может представлять собой микроорганизм рода Corynebacterium и, более конкретно, Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens и так далее. Еще более конкретно, микроорганизм может представлять собой Corynebacterium glutamicum, но без ограничения ими.
В еще одном аспекте настоящего изобретения предложен способ продуцирования целевого продукта, включающий (а) культивирование рекомбинантного микроорганизма по настоящему изобретению в среде; и (б) извлечение целевого продукта из микроорганизма или среды, в которой культивировали микроорганизм.
При использовании в данном описании изобретения “целевой продукт” может быть выбран из группы, состоящей из сахаридов (например, псикозы или тагатозы), L-аминокислот (например L-лизина, L-валина), органических кислот, ферментов и их комбинации. “Сахарид” относится к углеводу, имеющему сладкий вкус, и может быть выбран из группы, состоящей из, например, глюкозы, фруктозы, галактозы, аллулозы, тагатозы, ксилозы, лактозы, сахарозы и их комбинации, но без ограничения ими.
“Аминокислота” или “L-аминокислота” обычно относится к основной составляющей единице белка, в которой аминогруппа и карбоксильная группа связаны с одним и тем же атомом углерода. Аминокислота может быть выбрана из группы, состоящей из, например, глицина, аланина, валина, лейцина, изолейцина, треонина, серина, цистеина, глутамина, метионина, аспарагиновой кислоты, аспарагина, глутаминовой кислоты, лизина, аргинина, гистидина, фенилаланина, тирозина, триптофана, пролина и их комбинации, но без ограничения ими. Органические кислоты могут представлять собой органические соединения, имеющие кислые свойства, например соединения, в которые включены карбоксигруппа и сульфоновая группа. Конкретные примеры органических кислот могут включать молочную кислоту, уксусную кислоту, янтарную кислоту, масляную кислоту, пальмитиновую кислоту, щавелевую кислоту, винную кислоту, лимонную кислоту, пропионовую кислоту, гексеновую кислоту, каприновую кислоту, каприловую кислоту, валериановую кислоту или лимонную кислоту, но не ограничены ими. “Фермент” относится к белковым катализаторам, которые опосредуют химические взаимодействия, происходящие в живых организмах, и, конкретно, ферменты играют роль катализаторов, снижающих энергию активации, необходимую для взаимодействия, путем образования фермент-субстратного комплекса посредством связывания с субстратами. Например, некоторые ферменты могут быть вовлечены в продуцирование сахаридов (например псикозы или тагатозы) и, более конкретно, эти ферменты могут представлять собой псикоза-эпимеразу, тагатоза-эпимеразу или тагатуронат-эпимеразу, но не ограничены ими. Целевые продукты могут включать любой целевой продукт, который может быть получен путем экспрессии целевого гена, функционально связанного с промотором по настоящему изобретению, но целевые продукты не ограничены ими.
При использовании в данном описании изобретения термин “культивирование” относится к выращиванию микроорганизма в среде с подходящим образом и искусственно регулируемыми окружающими условиями. В настоящем изобретении процесс культивирования может быть осуществлен на основе подходящих культуральных сред и условий культивирования, широко известных в данной области техники. Конкретно, процесс культивирования может быть осуществлен непрерывно в периодическом процессе, в периодическом процессе с подпиткой или в повторяющемся периодическом процессе с подпиткой, но без ограничения ими.
Среды, используемые в культивировании, должны надлежащим образом соответствовать потребностям конкретных штаммов. Описаны культуральные среды для микроорганизмов рода Corynebacterium и рода Escherichia (например Manual of Methods for General Bacteriology. American Society for Bacteriology. Washington D.C, USA, 1981). В качестве источника углерода для использования в средах могут быть включены сахара и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этиловый спирт; и органические кислоты, такие как глюконовая кислота, уксусная кислота и пировиноградная кислота, но без ограничения ими. Эти вещества можно использовать по отдельности или в виде смеси. Для использования в качестве источника азота могут быть включены пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный экстракт, порошок соевого шрота, и мочевина или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония, но источник азота не ограничен ими. Указанный источник азота также можно использовать отдельно или в виде смеси. В качестве источника фосфора можно использовать дигидрофосфат калия или гидрофосфат калия или соответствующие натрий-содержащие соли, но источник фосфора не ограничен ими. Кроме того, культуральные среды могут включать соль металла, такую как сульфат магния или сульфат железа, которая является необходимой для роста. Кроме того, необходимые для роста вещества, такие как аминокислоты и витамины, можно использовать в дополнение к вышеупомянутым веществам. Кроме того, в культуральную среду можно добавлять подходящие предшественники. Конкретно, когда фермент продуцируется в качестве целевого продукта, в средах может содержаться субстрат этого фермента. Например, в среды можно включать фруктозу, которая может служить в качестве субстрата для псикоза-эпимеразы, тагатоза-эпимеразы или тагатуронат-эпимеразы. Описанные выше вещества-источники можно в достаточной мере подавать в культуру периодически или непрерывно во время процесс культивирования. Эти различные процессы культивирования раскрыты, например, в ссылке (“Biochemical Engineering” by James M. Lee, Prentice-Hall International Editions, pp 138-176).
Значение pH культуры можно регулировать при помощи подходящего основного соединения, такого как гидроксид натрия, гидроксид калия или аммиак, или кислого соединения, такого как фосфорная кислота или серная кислота. Кроме того, пенообразование можно регулировать при помощи пеногасителя, такого как сложный эфир жирной кислоты и полигликоля. Для поддержания аэробных условий культивирования можно вводить кислород или кислородсодержащие газовые смеси (например воздух). Температура культивирования обычно может находиться в диапазоне от 20°C до 45°C, конкретно от 25°C до 40°C, но температура не ограничена этими значениями и может меняться в соответствии с условиями культивирования.
Способ получения целевого продукта по настоящему изобретению может включать стадию извлечения целевого продукта из микроорганизма по настоящему изобретению или сред, в которых микроорганизм культивировали. Способ получения целевого продукта из микроорганизма или сред, в которых микроорганизм культивировали, предназначен для выделения или извлечения целевого продукта с использованием подходящего взаимодействия, раскрытого в данной области техники. Например, способы могут включать обработку посредством осаждения белка (метод высаливания), центрифугирование, экстракцию, обработку ультразвуком, ультрафильтрацию, диализ, различные способы хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, аффинная хроматография и так далее, и их комбинацию, но без ограничения ими. Стадия извлечения может включать процесс очистки, и специалист в данной области техники может выбрать процесс из различных процессов очистки и использовать его при необходимости.
Полезные эффекты изобретения
Новый промотор по настоящему изобретению может иметь различную активность в соответствии с микроорганизмами, которые индуцируют экспрессию целевого гена. Таким образом, если требуется регулировать активность целевого гена по необходимости во время получения целевого продукта, целевой продукт можно эффективно получать, используя новый промотор по настоящему изобретению.
Краткое описание графических материалов
На Фиг. 1 показаны результаты анализа с GFP (зеленый флуоресцентный белок), показывающие измеренную силу новых промоторов. На Фиг. 1(A) показаны результаты GFP-анализа новых промоторов на основе Corynebacterium glutamicum ATCC13032, а на Фиг. 1(Б) показаны результаты GFP-анализа новых промоторов на основе Corynebacterium glutamicum ATCC13869.
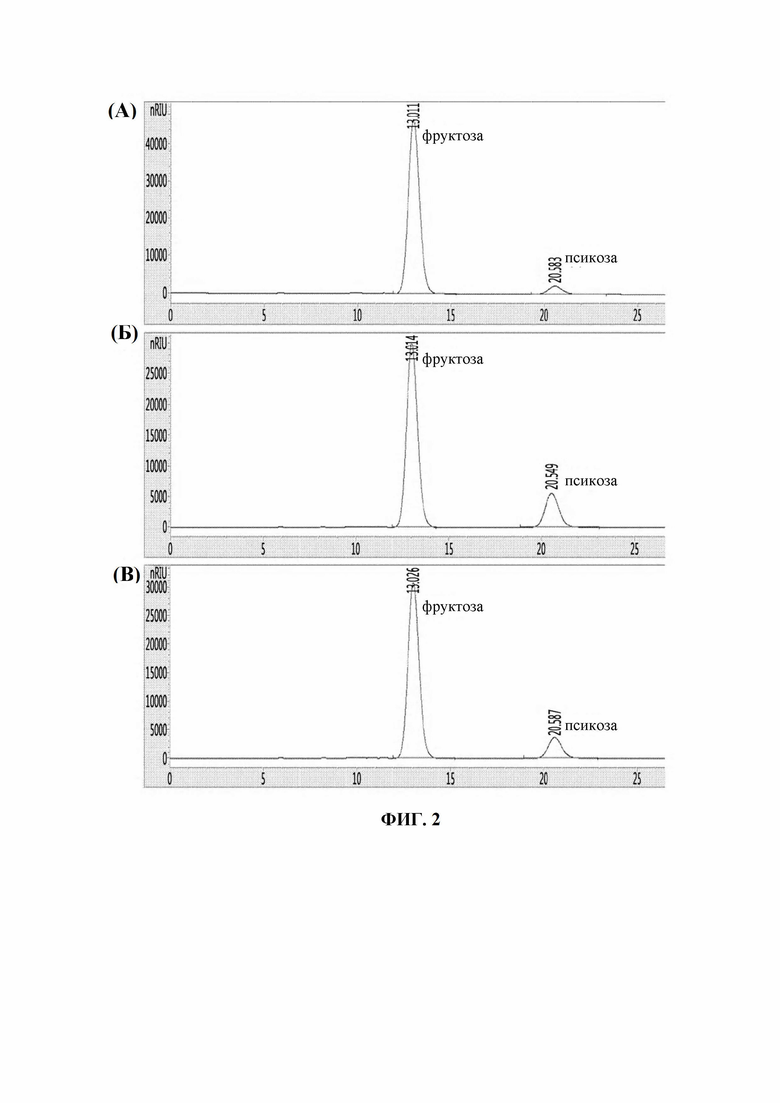
На Фиг. 2 показаны результаты ВЭЖХ, подтверждающие продуцирование псикозы. На Фиг. 2(А) показан результат взаимодействия с фруктозой в качестве субстрата с использованием Corynebacterium glutamicum ATCC13032/CJ4-ATPE-2, на Фиг. 2(Б) показан результат взаимодействия с фруктозой в качестве субстрата с использованием Corynebacterium glutamicum ATCC13032/SPL1-ATPE-2, и на Фиг. 2(В) показан результат взаимодействия с фруктозой в качестве субстрата с использованием Corynebacterium glutamicum ATCC13032/SPL7-ATPE-2.
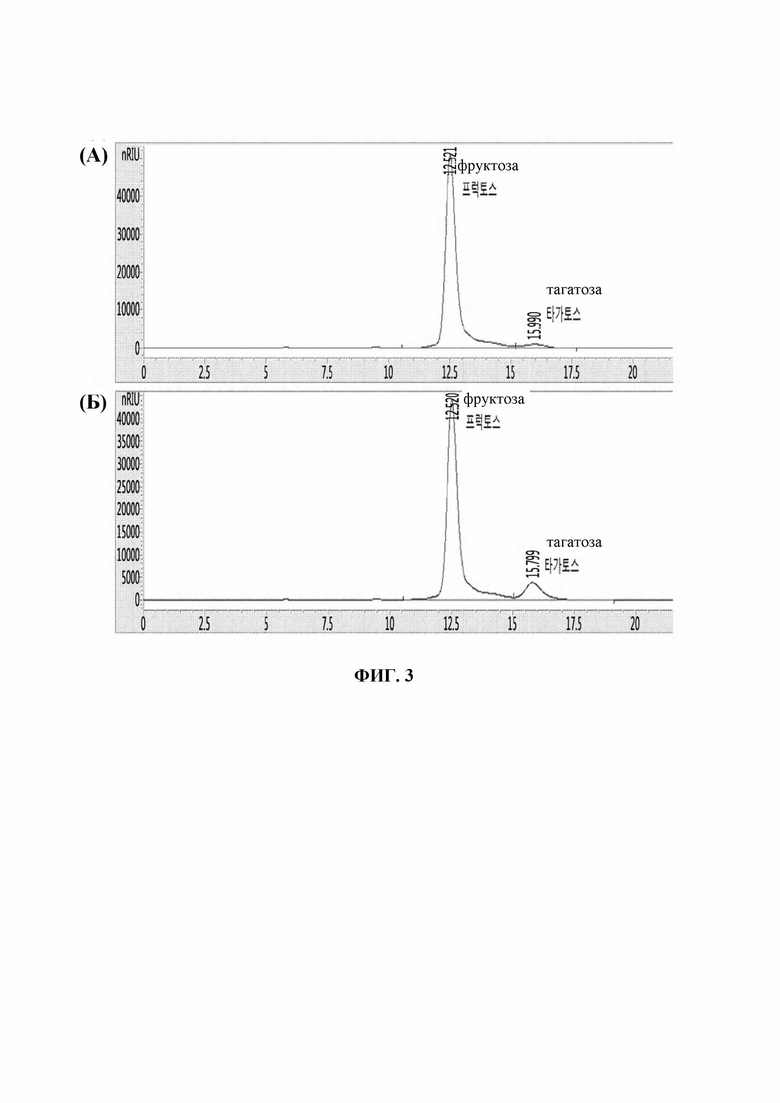
На Фиг. 3 показаны результаты ВЭЖХ, подтверждающие продуцирование тагатозы. На Фиг. 3(А) показан результат взаимодействия с фруктозой в качестве субстрата с использованием Corynebacterium glutamicum ATCC13032/CJ4-TN(m), и на Фиг. 3(Б) показан результат взаимодействия с фруктозой в качестве субстрата с использованием ATCC13032/SPL13-TN(m).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Ниже настоящее изобретение будет описано более подробно со ссылкой на следующие ниже примеры и так далее, чтобы улучшить понимание настоящего изобретения. Однако эти примеры могут быть модифицированы в различные другие формы, и объем настоящего изобретения не следует истолковывать как ограниченный этими примерами. Примеры настоящего изобретения представлены с целью максимально глубокого объяснения для лиц, имеющих средний уровень знаний в данной области техники.
Пример 1: Подтверждение экспрессии целевого гена, индуцированной новым промотором
1-1. Получение рекомбинантных векторов, содержащих новые промоторные последовательности
Для синтеза нового промотора, способного индуцировать экспрессию целевого гена, анализировали различные промоторные последовательности, происходящие из микроорганизма рода Corynebacterium и микроорганизма рода Escherichia. Промоторы, имеющие нуклеотидные последовательности, представленные SEQ ID NO: 1, 2 и 3, синтезировали и обозначали как SPL1, SPL7 и SPL13 соответственно.
На основе промоторов SPL1, SPL7 и SPL13, полученных путем синтеза в качестве матриц, выполняли ПЦР, используя праймеры SEQ ID NO: 4 и SEQ ID NO: 5, которые включают сайты рестрикции KpnI/EcoRV [Sambrook et al, Molecular Cloning, a Laboratory Manual (1989), Cold Spring Harbor Laboratories]. ПЦР выполняли в следующих условиях: денатурация при 94°C в течение 5 мин; 30 циклов денатурации при 94°C в течение 30 секунд, отжиг при 60°C в течение 30 секунд и достройка при 72°C в течение 30 секунд; и достройка при 72°C в течение 7 мин. В результате получали SPL1, SPL7 и SPL13 с размером примерно 300 п.о.
Открытую рамку считывания (ORF) гена GFP получали путем выполнения ПЦР с использованием вектора pGFPuv (Clontech, USA) в качестве матрицы вместе с праймерами SEQ ID NO: 6 и SEQ ID NO: 7, которые включают сайты рестрикции PstI/EcoRV. ПЦР выполняли в следующих условиях: денатурация при 94°C в течение 5 мин; 30 циклов денатурации при 94°C в течение 30 секунд, отжиг при 55°C в течение 30 секунд и достройка при 72°C в течение 1 мин; и достройка при 72°C в течение 7 мин. В результате получали фрагмент гена GFP (SEQ ID NO: 14) примерно 716 п.о.
В сайтах рестрикции PstI и KpnI бифункционального вектора pECCG117 (Biotechnology letters, vol 13, No. 10, p. 721-726 (1991), (корейский патент 10-1992-0007401)), который может экспрессироваться в E. coli и коринеформных микроорганизмах, каждый из SPL1, SPL7 и SPL13, обработанных ферментами рестрикции KpnI и EcoRV, и ORF гена GFP, обработанной PstI и EcoRV, были функционально связаны друг с другом с использованием ДНК-лигазы, и таким образом получали рекомбинантные векторы, в которых каждый из SPL1, SPL7 и SPL13 связан с GFP, и обозначали их как pSPL1-GFP, pSPL7-GFP и pSPL13-GFP соответственно.
1-2. Получение трансформированных штаммов
Вектор pECCG117, рекомбинантные векторы (pSPL1-GFP, pSPL7-GFP и pSPL13-GFP), полученные выше, и p117-CJ4-GFP, которые включают раскрытый ранее промотор pcj4 (корейский патент 10-0620092), трансформировали в Corynebacterium glutamicum ATCC13032 и Corynebacterium glutamicum ATCC13869 электроимпульсным способом (Appl. Microbiol. Biotechnol. (1999) 52: 541-545), соответственно, и трансформированные штаммы получали в чашке с агаром Луриа-Бертани (LB), содержащей канамицин (25 мг/л). Штаммы, полученные на основе ATCC13032, обозначали как Corynebacterium glutamicum ATCC13032/pECCG117, Corynebacterium glutamicum ATCC13032/SPL1-GFP, Corynebacterium glutamicum ATCC13032/SPL7-GFP, Corynebacterium glutamicum ATCC13032/SPL13-GFP и Corynebacterium glutamicum ATCC13032/CJ4-GFP соответственно. Также, штаммы, полученные на основе ATCC13869, обозначали как Corynebacterium glutamicum ATCC13869/pECCG117, Corynebacterium glutamicum ATCC13869/SPL1-GFP, Corynebacterium glutamicum ATCC13869/SPL7-GFP, Corynebacterium glutamicum ATCC13869/SPL13-GFP и Corynebacterium glutamicum ATCC13869/CJ4-GFP, соответственно.
6 видов штаммов, полученных посредством трансформации выше, то есть ATCC13032/SPL7-GFP, ATCC13032/SPL13-GFP, ATCC13032/SPL1-GFP, ATCC13869/SPL7-GFP, ATCC13869/SPL13-GFP и ATCC13869/SPL1-GFP, обозначали как CA01-2301, CA01-2302, CA01-2303, CA01-2304, CA01-2305 и CA01-2306 соответственно, и затем депонировали в Корейском центре культур микроорганизмов (KCCM), международном депозитарном органе согласно Будапештскому договору, 2 июля 2017 года, с учетными номерами KCCM11971P, KCCM11972P, KCCM11973P, KCCM11974P, KCCM11975P и KCCM11976P.
1-3. Подтверждение активности новых промоторов
Для подтверждения активности промоторов SPL1, SPL7 и SPL13, трансформированные штаммы, полученные в Примере 1-2 (то есть Corynebacterium glutamicum ATCC13032/pECCG117, Corynebacterium glutamicum ATCC13032/CJ4-GFP, Corynebacterium glutamicum ATCC13032/SPL1-GFP, Corynebacterium glutamicum ATCC13032/SPL7-GFP, Corynebacterium glutamicum ATCC13032/SPL13-GFP, Corynebacterium glutamicum ATCC13869/pECCG117, Corynebacterium glutamicum ATCC13869/CJ4-GFP, Corynebacterium glutamicum ATCC13869/SPL1-GFP, Corynebacterium glutamicum ATCC13869/SPL7-GFP и Corynebacterium glutamicum ATCC13869/SPL13-GFP), культивировали способом, описанным ниже, и определяли их GFP-активности.
Трансформированные штаммы инокулировали в каждую колбу, содержащую 25 мл культуральной среды (глюкоза (20 г), сульфат аммония ((NH4)2SO4) (5 г), дрожжевой экстракт (5 г), мочевина (1,5 г), KH2PO4 (4 г), K2HPO4 (8 г), MgSO4⋅7H2O (0,5 г), биотин (150 мкг), тиамина HCl соль (1,5 мг), пантотенат кальция (3 мг) и никотинамид (3 мг) (в расчете на 1 л дистиллированной воды), pH 7,2) и культивировали во встряхивателе-инкубаторе при 30°C в течение 20 часов. Бактериальные клетки извлекали центрифугированием (5000 об/мин, 15 мин), дважды промывали буфером 50 мМ трис-HCl (pH 8,0) и суспендировали в том же буфере. К суспензии (1,25 г/1,5 мл) добавляли стеклянные гранулы и разрушали бактериальные клетки, используя шариковую мельницу, в течение 6 минут. Затем полученную суспензию подвергали центрифугированию (15000 об/мин, 20 минут), извлекали из нее супернатант и концентрации белков количественно определяли методом Брэдфорда. Равные количества экстрактов бактериальных клеток облучали возбуждающим светом при 488 нм согласно способу, предложенному Laure Gory et al. (FEMS Microbiology Letters, 194, 127-133, 2001) и определяли излучаемый свет при 511 нм, используя спектрофотометр LS-50B (Perkin-Elmer), таким образом определяя уровень экспрессии гена GFP (Таблица 1).
Таблица 1
Как показано в Таблице 1 выше, все из SPL1, SPL7 и SPL13 демонстрировали свои промоторные активности в двух разных Corynebacterium glutamicum и также показывали более высокую флуоресцентную чувствительность, чем промотор pcj4, который известен как сильный промотор. Исходя из этих результатов было обнаружено, что SPL1, SPL7 и SPL13 представляют собой очень сильные промоторы, которые могут экспрессировать целевые гены в Corynebacterium glutamicum.
Пример 2. Оценка способности продуцировать целевые продукты
2-1. Оценка способности продуцировать псикозу
1) Получение векторов и трансформированных штаммов для экспрессии ATPE, включающих промоторные последовательности SPL1 и SPL7
Векторы для штаммов Corynebacterium с усиленной экспрессией ATPE (псикоза-эпимераза, происходящая из Agrobacterium tumefaciens) получали, используя SPL1 и SPL7. Открытую рамку считывания (ORF) гена ATPE амплифицировали путем выполнения ПЦР (30 циклов взаимодействий по 30 секунд при 94°C, 30 секунд при 55°C, и 1 мин при 72°C), используя вектор pET24-ATPE-2 (SEQ ID NO: 8) в качестве матрицы вместе с праймерами SEQ ID NO: 9 и 10. Амплифицированный ген ATPE и векторы pSPL1-GFP и pSPL7-GFP для штаммов Corynebacterium, полученных в Примере 1, обрабатывали ферментами рестрикции EcoRV и PstI, и ATPE-2, полученную посредством ПЦР, функционально связывали с ними, используя набор BD In-Fusion, и таким образом получали в итоге векторы pSPL1-ATPE-2 и pSPL7-ATPE-2 для штаммов Corynebacterium.
Полученные таким образом векторы pSPL1-ATPE-2 и pSPL7-ATPE-2 интродуцировали в штамм ATCC13032 посредством электропорации, и таким образом получали штаммы SPL1-ATPE-2 и SPL7-ATPE-2.
2) Оценка способности продуцирования псикозы трансформированными штаммами
Штаммы, полученные описанным выше способом, культивировали, используя среды с таким же составом, что и в Примере 1, и определяли их ATPE-активности. Штаммы ATCC13032/pECCG117 и ATCC13032/CJ4-ATPE-2 использовали в качестве контрольных групп.
Штаммы культивировали в течение ночи на твердой среде LB, помещенной в 30°C инкубатор, и ночную культуру каждого штамма инокулировали в 25 мл среды и культивировали во встряхивателе-инкубаторе при 30°C в течение 24 часов. Культуру центрифугировали и супернатант удаляли. Извлеченные бактериальные тельца промывали раствором EPPS ((4-(2-гидроксиэтил)-1-пиперазинпропансульфоновая кислота)) (pH 8,0), и полученный таким образом осадок растворяли в растворе EPPS (pH 8,0). Добавляли в него POESA (полиоксиэтиленстеариламин) (1 мг/мл), оставляли взаимодействовать при комнатной температура в течение 1 часа и центрифугировали. Затем полученный в результате центрифугирования осадок растворяли в растворе EPPS (pH 8,0), добавляли в него раствор фруктозы (350 г/л) в качестве субстрата, оставляли взаимодействовать при 50°C в течение 3 часов и взаимодействие останавливали тепловой обработкой. Затем посредством центрифугирования извлекали супернатант и величину продуцирования псикозы определяли посредством анализа ВЭЖХ (Фиг. 2(А), 2(Б) и 2(В)). Величина продуцирования псикозы после взаимодействия указана в Таблице 2 ниже.
Таблица 2
Как показано в Таблице 2, было подтверждено, что продуктивности по псикозе у Corynebacterium glutamicum ATCC13032/SPL1-ATPE-2 и ATCC13032/SPL7-ATPE-2 улучшались на 321% и 258% соответственно по сравнению с Corynebacterium glutamicum ATCC13032/CJ4-ATPE-2. Исходя из вышеуказанного, было подтверждено, что когда использовали промоторы SPL1 и SPL7 по настоящему изобретению, величина экспрессии гена, кодирующего ATPE, увеличивалась, подтверждая таким образом, что активность ATPE значительно увеличивалась.
2-2. Оценка способности продуцирования тагатозы
1) Получение векторов и трансформированных штаммов для экспрессии UxaE, включающих промоторную последовательность SPL13
Векторы для штаммов Corynebacterium получали клонированием гена тагатоза-эпимеразы (UxaE), происходящего из Thermotoga neapolitana, используя CJ4-GFP, в который введен GFP, и SPL13-GFP, полученный в Примере 1. Открытую рамку считывания (ORF) гена TN(m) амплифицировали путем выполнения ПЦР (30 циклов реакции по 30 секунд при 94°C, 30 секунд при 55°C и 1 мин при 72°C), используя вектор pET28a-TN(m) (SEQ ID NO: 11) в качестве матрицы вместе с праймерами SEQ ID NO: 12 и 13. Амплифицированный ген TN(m) и векторы CJ4-GFP и SPL13-GFP для штаммов Corynebacterium обрабатывали ферментами рестрикции EcoRV и PstI и затем лигировали, и таким образом в итоге получали векторы pCJ4-TN(m) и pSPL13-TN(m) для штаммов Corynebacterium.
Полученные таким образом векторы pCJ4-TN(m) и pSPL13-TN(m) интродуцировали в штамм ATCC13032 посредством электропорации и таким образом получали штаммы ATCC13032/CJ4-TN(m) и SPL13-TN(m).
2) Оценка способности продуцирования тагатозы трансформированными штаммами
Штаммы, полученные описанным выше способом, культивировали и предварительно обрабатывали в таких же средах и условиях культивирования, описанных в Примере 1, и получали штаммы для активирования UxaE. Оценку активности выполняли путем изменения только количества субстрата, температуры взаимодействия и времени таким же образом, как в Примере 2-1 (путем взаимодействия при 60°C в течение 2 часов после добавления раствора фруктозы (100 г/л)). Затем супернатант извлекали путем центрифугирования и величину продуцирования тагатозы определяли посредством анализа ВЭЖХ (Фиг. 3(А) и 3(Б)). Величина продуцирования тагатозы после взаимодействия показана в Таблице 3 ниже.
Таблица 3
Как показано в Таблице 3, возможность продуцирования тагатозы у Corynebacterium glutamicum ATCC13032/SPL13-TN(m) была улучшена на 143% по сравнению с Corynebacterium glutamicum ATCC13032/CJ4-TN(m). Исходя из указанного выше, было подтверждено, что, когда использовали промотор SPL13 по настоящему изобретению, величина экспрессии гена, кодирующего UxaE, увеличивалась, таким образом подтверждая, что активность UxaE была значительно повышенной.
2-3. Оценка способности продуцировать валин
1) Получение вектора pECCG117-SPL7-ilvE и трансформированных штаммов, включающих последовательность промотора SPL7
Для подтверждения способности продуцировать L-валин в качестве примера L-аминокислот, получали векторы pECCG117-CJ7-ilvE и pECCG117-SPL7-ilvE, как описано ниже, для того чтобы усилить ферментативную активность ilvE (NCgl2123), кодирующего аминотрансферазу аминокислот с разветвленной цепью, который представляет собой основной ген биосинтеза валина. Конкретно, в результате выполнения ПЦР (30 циклов реакции по 30 секунд при 94°C, 30 секунд при 55°C и 1 мин при 72°C), используя хромосому ATCC14067 в качестве матрицы вместе с праймерами SEQ ID NO: 14 и 15, амплифицировали ПЦР-фрагмент с размером примерно 1104 п.о, который имеет сайт рестрикции EcoRV на 5'-конце и сайт рестрикции PstI на 3'-конце гена NCgl2123. Полученный таким образом ПЦР-фрагмент очищали и смешивали с pECCG117-CJ7-GFP (корейский патент 10-0620092) и pECCG117-SPL7-GFP, которые обрабатывали ферментами рестрикции EcoRV и PstI, соответственно, и получали векторы, используя набор для клонирования In-fusion. Полученные таким образом векторы были обозначены как pECCG117-CJ7-ilvE и pECCG117-SPL7-ilvE соответственно.
SEQ ID NO: 14 5' GAGATCAAAACAGATATCATGACGTCATTAGAGTTC 3'
SEQ ID NO: 15 5' ATCCCCCGGGCTGCAGTTAGCCAACCAGTGGGTA 3'
Полученные таким образом рекомбинантные векторы pECCG117-CJ7-ilvE и pECCG117-SPL7-ilvE и вектор pECCG117 трансформировали в валин-продуцирующий штамм Corynebacterium glutamicum KCCM11201P (корейский патент 10-1117022) электроимпульсным способом и трансформированные штаммы получали в чашке с агаром LB, содержащей канамицин (25 мг/л). Полученные таким образом штаммы были обозначены как KCCM11201P/pECCG117, KCCM11201P/CJ7-ilvE и KCCM11201P/SPL7-ilvE соответственно.
2) Оценка способности продуцирования валина трансформированными штаммами
Способность продуцирования L-валина 3 разными трансформированными штаммами анализировали посредством культивирования, описанного ниже.
Каждый из штаммов в количестве платиновой петли инокулировали в 250 мл колбу с угловыми перегородками, содержащую 25 мл среды для продуцирования, и культивировали во встряхивателе-инкубаторе (200 об/мин) при 30°C в течение 72 часов. После завершения культивирования концентрацию L-валина в каждой культуре анализировали посредством ВЭЖХ (SHIMADZU LC-20AD).
<Среда для продуцирования (pH 7,2)>
Глюкоза (50 г), (NH4)2SO4 (20 г), сухая кукурузная патока (20 г), KH2PO4 (1 г), MgSO4⋅7H2O (0,5 г), биотин (200 мкг) (в расчете на 1 л дистиллированной воды)
Описанное выше культивирование и анализ выполняли несколько раз и анализируемые концентрации L-валина показаны в Таблице 4 ниже.
Таблица 4
Как показано в Таблице 4, было подтверждено, что способность продуцирования валина штаммом KCCM11201P/SPL7-ilvE, в который введен промотор по настоящему изобретению, была улучшена на 21,8% по сравнению с Corynebacterium glutamicum KCCM11201P/CJ7-ilvE, в который введен известный промотор, и кроме того, она была улучшена на 39,2% по сравнению с контрольной группой, KCCM11201P/pECCG117. Исходя из указанных выше результатов, было подтверждено, что промотор SPL7 усиливал экспрессию гена ilvE, таким образом значительно увеличивая активность фермента, кодируемого соответствующим геном.
2-4. Оценка способности продуцирования лизина
1) Получение вектора pDZTn-SPL13-gapN1 и трансформированных штаммов, включающих последовательность промотора SPL13
Для подтверждения способности продуцировать L-лизин в качестве типичного примера L-аминокислот, получали векторы как описано ниже, чтобы усилить ферментативную активность НАДФ-зависимой глицеральдегид-3-фосфатдегидрогеназы (GapN), которая происходит из известных Streptococcus mutants.
Для введения в «прыгающий» ген NCgl2392 в микроорганизме рода Corynebacterium, выполняли ПЦР (30 циклов реакции по 30 секунд при 94°C, 30 секунд при 55°C и 1 мин при 72°C), используя хромосому дикого типа Corynebacterium glutamicum ATCC13032 в качестве матрицы вместе со следующими праймерами SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 и SEQ ID NO: 19, на основе NIH Genbank из NIH (США), и в результате амплифицировали фрагменты, включающие 5'-конец и 3'-конец гена NCgl2392. В результате выполнения ПЦР (30 циклов реакции по 30 секунд при 94°C, 30 секунд при 55°C и 2 мин при 72°C) с использованием вектора pECCG122-Pcj7-gapN1 (корейский патент 10-1182033) вместе со следующими праймерами SEQ ID NO: 20 и SEQ ID NO: 21, амплифицировали Pcj7-gapN1. В результате выполнения ПЦР (30 циклов реакции по 30 секунд при 94°C, 30 секунд при 55°C и 1 мин при 72°C), используя вектор pECCG122-Pcj7-gapN1 и вектор SPL13-GFP, полученные в Примере 1, вместе со следующими праймерами SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24 и SEQ ID NO: 21, амплифицировали гены SPL13 и gapN соответственно, и эти гены клонировали в вектор pDZ (корейский патент 0924065), который не реплицируется в Corynebacterium glutamicum, вместе с фрагментами гена cNCgl2392, полученными выше, и таким образом получали векторы pDZTn-Pcj7-gapN1 и pDZTn-SPL13-gapN1.
SEQ ID NO: 16 5' ATCCTCTAGAGTCGACCAAATGCTCCAACCGTCCGT 3'
SEQ ID NO: 17 5' CTCGAGGAACTCATTCCTTCTGCTCG 3'
SEQ ID NO: 18 5' TCTAGAACTAGTGGGCCCGACATCTAATAACCGGGCAG 3'
SEQ ID NO: 19 5' ATGCCTGCAGGTCGACGCAGACGCACTCGACTACAC 3'
SEQ ID NO: 20 5' GAATGAGTTCCTCGAGAGAAACATCCCAGCGCTACT 3'
SEQ ID NO: 21 5' GCCCACTAGTTCTAGATTATTTGATATCAAATACGA 3'
SEQ ID NO: 22 5' GAATGAGTTCCTCGAGGGCGCTTCATGTCAACAATC 3'
SEQ ID NO: 23 5' ATTGTTTTGTCATATGTGTTTTGATCTCCTCCAATA 3'
SEQ ID NO: 24 5' CATATGACAAAACAATATAAAAA 3'
Каждый из описанных выше векторов (pDZTn-Pcj7-gapN1 и pDZTn-SPL13-gapN1) трансформировали посредством электроимпульсного способа (Appl. Microbiol. Biotechnol. (1999) 52: 541-545), используя в качестве родительского штамма штамм KCCM11016P с усиленной способностью продуцирования лизина (микроорганизм был раскрыт как KFCC10881, передепонированный в международном органе по депонированию согласно Будапештскому договору и получивший учетный номер KCCM11016P; корейский патент 10-0159812), и трансформированные штаммы получали в селективной среде, содержащей 25 мг/л канамицина. Для отбора колоний, в которых ген gapN1 был введен в геном посредством вторичного рекомбинационного способа (кроссинговер), те колонии, в которые введены гены Pcj7-gapN1 и SPL13-gapN1 соответственно, были получены с использованием пары праймеров SEQ ID NO: 20 и 21, и SEQ ID NO: 21 и 22. Полученные таким образом колонии были обозначены как KCCM11016P/CJ7-gapN1 и KCCM11016P/SPL13-gapN1 соответственно.
2) Оценка способности продуцирования лизина трансформированными штаммами
Способность продуцирования L-лизина 3 разными трансформированными штаммами анализировали путем культивирования, описанного ниже.
Каждый из штаммов инокулировали в 250 мл колбы с угловыми перегородками, содержащие 25 мл среды для посева, и культивировали во встряхивателе-инкубаторе (200 об/мин) при 30°C в течение 20 часов. Затем 1 мл культуры для посева инокулировали в 250 мл колбу с угловыми перегородками, содержащую 24 мл среды для продуцирования и культивировали во встряхивателе-инкубаторе (200 об/мин) при 30°C в течение 72 часов. Концентрацию L-лизина в каждой культуре анализировали посредством ВЭЖХ (SHIMADZU, LC-20AD).
<Среда для посева (pH 7,0)>
Глюкоза (20 г), пептон (10 г), дрожжевой экстракт (5 г), мочевина (1,5 г), KH2PO4 (4 г), K2HPO4 (8 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамина HCl (1000 мкг), пантотенат кальция (2000 мкг), никотинамид (2000 мкг) (в расчете на 1 л дистиллированной воды).
<Среда для продуцирования (pH 7,0)>
Глюкоза (100 г), (NH4)2SO4 (40 г), соевый белок (2,5 г), сухая кукурузная патока (5 г), мочевина (3 г), KH2PO4 (1 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамина гидрохлорид (1000 мкг), пантотенат кальция (2000 мкг), никотинамид (3000 мкг) и CaCO3 (30 г) (в расчете на 1 л дистиллированной воды)
Описанное выше культивирование и анализ выполняли несколько раз и анализируемые концентрации L-лизина показаны в Таблице 5 ниже.
Таблица 5
Как показано в Таблице 5, было подтверждено, что способность продуцировать лизин у штамма KCCM11016P/SPL13-gapN1, в который введен промотор по настоящему изобретению, была улучшена на 7,2% по сравнению с Corynebacterium glutamicum KCCM11016P/CJ7-gapN1, в который введен известный промотор, и кроме того, она была улучшена на 22,7% по сравнению с контрольной группой KCCM11016P. Исходя из описанных выше результатов, было подтверждено, что промотор SPL13 усиливал экспрессию гена gapN1, таким образом значительно увеличивая активность фермента, кодируемого соответствующим геном.
Обобщая описанные выше результаты, промоторы SPL1, SPL7 и SPL13 по настоящему изобретению могут значительно усиливать экспрессию целевого гена в рекомбинантном микроорганизме по сравнению с обычно используемыми известными промоторами. Таким образом, промоторы по настоящему изобретению могут не только обеспечивать эффективную систему экспрессию, но также эффективно применяться в различных отраслях промышленности для продупирования с высоким выходом целевых продуктов, таких как сахариды, функциональные вещества и аминокислоты.
--->
<110> CJ CheilJedang Corporation
<120> Novel promoter and use thereof
<130> OPA17051
<150> KR 10-2016-0111810
<151> 2016-08-31
<160> 14
<170> KopatentIn 2.0
<210> 1
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> SPL1
<400> 1
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagtgagt 240
cctgatacaa ttgaaaacgt gcaaaagcat aaattattgg aggagatcaa aaca 294
<210> 2
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> SPL7
<400> 2
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagcaatt 240
cctgctacaa ttgaaaacgt gcaaaagcat agattattgg aggagatcaa aaca 294
<210> 3
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> SPL13
<400> 3
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagcgggg 240
cctggtataa ttgaaaacgt gcaaaagcat agattattgg aggagatcaa aaca 294
<210> 4
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> SPL праймер-1
<400> 4
ggtaccggcg cttcatgtca acaatc 26
<210> 5
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> SPL праймер-2
<400> 5
gatatctgtt ttgatctcct ccaat 25
<210> 6
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> GFP праймер-1
<400> 6
gatatcatga gtaaaggaga aga 23
<210> 7
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> GFP праймер-2
<400> 7
ctgcagttat ttgtagagct cat 23
<210> 8
<211> 870
<212> DNA
<213> Artificial Sequence
<220>
<223> pET24-ATPE-2
<400> 8
atgaaacacg gcatctatta ttcttactgg gaacatgagt ggagcgccaa gttcggtccc 60
tatatcgaga aggtcgccaa gctcggtttc gacatcctcg aagtcgccgc ccaccatatc 120
aacgaataca gcgacgccga actcgcgacc atcaggaaga gcgcgaagga taacggcatc 180
atcctcaccg ccggcatcgg tccgtcgaaa accaagaacc tgtcgtcgga agatgctgcg 240
gtgcgtgcgg ccggcaaggc gttctttgaa agaacccttt cgaacgtcgc caagctcgat 300
atccacacca tcggcggcgc attgcattcc tattggccaa tcgattattc gcagcccgtc 360
gacaaggcag gcgattatgc gcgcggcgtc gagggtatca acggcattgc cgatttcgcc 420
aatgatctcg gcatcaacct gtgcatcgaa gtcctcaacc gctttgaaaa ccacgtcctc 480
aacacggcgg cggaaggcgt cgcttttgtg aaggatgtcg gcaagaacaa tgtgaaagtc 540
atgctggata ccttccacat gaacatcgag gaagacagtt tcggtgacgc catccgcacg 600
gccggcccgc ttctggggca cttccatacc ggtgaatgca atcgccgcgt accgggcaag 660
ggcagaatgc cgtggcacga aatcggcctt gcgctgcgtg atatcaacta caccggcgcg 720
gtaatcatgg agcctttcgt caagacaggc ggcaccatcg gctcggatat caaggtgtgg 780
cgcgacctga gcggtggcgc cgacatcgcg aaaatggatg aagatgcccg caatgcgctg 840
gcattctccc gcttcgttct tggtggctga 870
<210> 9
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> ATPE праймер-1
<400> 9
atctaggaga ttaagatatc atgaaacacg gcatctatta tt 42
<210> 10
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> ATPE праймер-2
<400> 10
gtggatcccc cgggctgcag tcagccacca agaacg 36
<210> 11
<211> 1473
<212> DNA
<213> Artificial Sequence
<220>
<223> pET28a-TN(m)
<400> 11
atggtcttga aagtgttcaa agatcacttt ggaaggggat acgaagttta cgaaaagtct 60
tatagagaaa aggattctct ctctttcttc ttgacaaagg gagaggaagg aaaaattctg 120
gtagtggctg gagaaaaggc acctgagggt ctgtcgtttt tcaaaaaaca gcgggtggag 180
ggtgtttcgt tctttttctg tgagagaaat catgagaact tggaagttct cagaaaatac 240
tttccagatc tcaaaccagt tcgagcagga ttgagagcgt cttttggaac aggtgacaga 300
ctcggtatca ccacaccggc tcacgtgagg gcgttgaagg attcagggct ttttcccatc 360
tttgcgcagc aggacgtgag ggagaacgaa agaacgggaa ggacctggag agatgtgctg 420
gacgatgcca catggggagt tttccaggag ggatacagtg agggattcgg agcagacgcc 480
gatcacgcga agcggccgga ggatcttgtt tcggctgcaa gggaaggttt caccatgttc 540
acaatcgatc cttcgaatca tgtgaggaat ctttcaaaac tcagtgaaag agaaaagaac 600
gagatgttcg aggaaatact gaaaaaagag cgaatcgaca ggatctatct tgggaaaaaa 660
tacaccgtcc tcggtgaaag actggagttc gacgagaaaa atttgaggga tgctgctctg 720
gtgtactatg atgcgatcgc ccacgtggat atgatgtatc aaattttgaa agacgaaacc 780
ccggatatcg acttcgaagt gtcagttgac gaaacagaaa ctcccacgag tcctctcttc 840
cacattttcg ttgtggaaga actcagacga agaggtgtgg agttcaccaa tcttgccctg 900
agattcatcg gcgaatggga aaagggaata ggttacaagg gggatcttgc acagttcgag 960
agagaaatca aaatgcacgc agaaatcgca aggatgttcg aaggatacaa aatatcactc 1020
cactctggaa gcgacaaatt ttccgtgtat cctgcttttg cttccgcgac aggaggcctt 1080
ttccacgtga agacagccgg aacgagttat cttgaggcgg tgaaggtcat atccatggtc 1140
aacccggagc tcttccggga gatctacagg tgtgctctcg atcactttga ggaagacaga 1200
aagtcctatc acatatctgc ggatctgtcg aaagttccgg aagtagagaa agtgaaagat 1260
gaagatcttc caggtctttt tgaagacatc aacgtgagac agttgatcca tgtcacctat 1320
ggctctgttc tgaaagatgc atctttgaaa gaacggctgt ttaagacgct tgaacaaaat 1380
gaggaactct tctacgagac cgtggcaaaa catataaaaa ggcacgtaga cctgttgaag 1440
gggtgactcg agcaccacca ccaccaccac tga 1473
<210> 12
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> TN(m) праймер-1
<400> 12
gatatcatgg tcttgaaagt gttcaaag 28
<210> 13
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> TN(m) праймер-2
<400> 13
ctgcagtcac cccttcaaca ggtctacgtg 30
<210> 14
<211> 717
<212> DNA
<213> Artificial Sequence
<220>
<223> GFP
<400> 14
atgagtaaag gagaagaact tttcactgga gttgtcccaa ttcttgttga attagatggt 60
gatgttaatg ggcacaaatt ttctgtcagt ggagagggtg aaggtgatgc aacatacgga 120
aaacttaccc ttaaatttat ttgcactact ggaaaactac ctgttccatg gccaacactt 180
gtcactactt tctcttatgg tgttcaatgc ttttcccgtt atccggatca tatgaaacgg 240
catgactttt tcaagagtgc catgcccgaa ggttatgtac aggaacgcac tatatctttc 300
aaagatgacg ggaactacaa gacgcgtgct gaagtcaagt ttgaaggtga tacccttgtt 360
aatcgtatcg agttaaaagg tattgatttt aaagaagatg gaaacattct cggacacaaa 420
ctcgagtaca actataactc acacaatgta tacatcacgg cagacaaaca aaagaatgga 480
atcaaagcta acttcaaaat tcgccacaac attgaagatg gatccgttca actagcagac 540
cattatcaac aaaatactcc aattggcgat ggccctgtcc ttttaccaga caaccattac 600
ctgtcgacac aatctgccct ttcgaaagat cccaacgaaa agcgtgacca catggtcctt 660
cttgagtttg taactgctgc tgggattaca catggcatgg atgagctcta caaataa 717
<---

Изобретение относится к биотехнологии, в частности к новому промотору, к вектору, содержащему этот промотор, к микроорганизму, содержащему этот промотор или вектор, и к способу получения целевого продукта с использованием этого микроорганизма. Изобретение позволяет получать целевой продукт с высокой степенью эффективности. 6 н. и 2 з.п. ф-лы, 5 табл., 2 пр., 3 ил.
1. Молекула нуклеиновой кислоты, имеющая промоторную активность, состоящая из любой нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-3.
2. Кассета экспрессии гена, содержащая молекулу нуклеиновой кислоты по п. 1 и целевой ген.
3. Рекомбинантный вектор для экспрессии, содержащий молекулу нуклеиновой кислоты по п. 1 или кассету экспрессии гена по п. 2.
4. Рекомбинантный вектор для введения, содержащий молекулу нуклеиновой кислоты по п.1 или кассету экспрессии гена по п. 2.
5. Рекомбинантный микроорганизм рода Corynebacterium для получения целевого продукта, содержащий молекулу нуклеиновой кислоты по п. 1 или вектор по п. 3 или 4.
6. Рекомбинантный микроорганизм по п. 4, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum или Corynebacterium ammoniagenesis.
7. Способ получения целевого продукта, включающий:
а) культивирование рекомбинантного микроорганизма по п. 4 в среде и
б) извлечение целевого продукта из микроорганизма или среды.
8. Способ по п. 6, где целевой продукт представляет собой псикозу, тагатозу или аминокислоту.
| MIROSLAV PATEK et al., Corynebacterium glutamicum promoters: a practical approach, Microbial Biotechnology Vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| SUNG SUN YIM et al., Isolation of Fully Synthetic Promoters for High-Level Gene Expression in Corynebacterium glutamicum, Biotechnology and Bioengineering Vol.110, Issue 11, pp | |||
| Курительный мундштук | 1925 |
|
SU2959A1 |
| KR | |||

