Область техники, к которой относится изобретение
Настоящая заявка относится к новой псикозо-6-фосфат фосфатазе, композиции для получения D-псикозы, содержащей фермент, и способам получения D-псикозы с использованием фермента.
Предшествующий уровень техники
D-псикозо-3-эпимераза (EC 5.1.3.30) и D-тагатозо-3-эпимераза (EC 5.1.3.31) известны в качестве ферментов, которые катализируют 3-эпимеризацию D-фруктозы с получением D-псикозы. Когда D-псикозу получают посредством единственной ферментативной реакции с использованием фермента, равновесие реакции между субстратом (то есть D-фруктозой) и продуктом (то есть D-псикозой) находится на постоянном уровне (продукт/субстрат = ~20-35%). Таким образом, получение сверхчистой D-псикозы требует дополнительного процесса разделения и удаления относительно высокой концентрации D-фруктозы из продукта ферментативной реакции.
С другой стороны, Chan et al. (2008. Biochemistry. 47:9608-9617) сообщил о D-рибулозо-5-фосфат-3-эпимеразе (EC 5.1.3.1), полученной из Streptococcus pyogenes, и о D-псикозо-6-фосфат-3-эпимеразе (EC 5.1.3.-), полученной из E. coli, способной катализировать 3-эпимеризацию D-фруктозо-6-фосфата и D-псикозо-6-фосфата. Однако, эти ферменты не применимы в промышленности из-за их слабой термоустойчивости.
В этих обстоятельствах авторы настоящего изобретения провели серьезное исследование для разработки способа увеличения скорости преобразования D-псикозы в промышленном масштабе экономичным способом. В результате авторы настоящего изобретения обнаружили, что после преобразования сахарозы или крахмала (например, мальтодекстрина) в качестве недорогого исходного материала в D-псикозо-6-фосфат, использование псикозо-6-фосфат фосфатазы, специфичной к D-псикозо-6-фосфату и участвующей в необратимом пути реакции, обеспечивает получение D-псикозы посредством однореакторного ферментативного преобразования с двумя или более ферментами, вовлеченными в пути получения D-псикозы, и может значительно увеличить скорость преобразования в D-псикозу. Настоящая заявка была выполнена на основе этого открытия.
Сущность изобретения
Техническая задача
Одной целью настоящей заявки является предоставление новой псикозо-6-фосфат фосфатазы, содержащей мотив A и мотив B.
Другой целью настоящей заявки является предоставление нуклеиновой кислоты, кодирующей псикозо-6-фосфат фосфатазу, описанную в данном документе, и трансформанта, содержащего нуклеиновую кислоту.
Дополнительной целью настоящей заявки является предоставление композиции для получения D-псикозы, содержащей инозитол-монофосфатазу, микроорганизм, экспрессирующий инозитол-монофосфатазу или культуру микроорганизмов.
Еще одной целью настоящей заявки является предоставление способа получения D-псикозы, включающего введение в контакт инозитол-монофосфатазы, микроорганизма, экспрессирующего инозитол-монофосфатазу, или культуры микроорганизмов с D-псикозо-6-фосфатом для преобразования D-псикозо-6-фосфата в D-псикозу.
Техническое решение
Далее настоящая заявка будет подробно описана. Между тем, объяснения аспектов и вариантов осуществления, раскрытых в настоящей заявке, также можно применить к объяснениям других аспектов и вариантов осуществления. Кроме того, все комбинации разных элементов, раскрытых в настоящей заявке, попадают в объем настоящей заявки. Кроме того, не следует считать, что объем настоящей заявки ограничен следующим подробным описанием.
Для достижения указанных выше и других целей настоящей заявки в одном аспекте настоящей заявки представлена псикозо-6-фосфат фосфатаза, содержащая мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или гэп, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
Конкретно, в мотиве A Xa1 может быть W или F, Xa2 может быть I или V, Xa3 может быть V или I, а Xa4 может быть T. Конкретно, в мотиве B Ya1 может быть W, Ya2 может быть V или I, Wa1 может быть AAG, Ya3 может быть W, I или V, Wa2 может быть LLV, LIV, LII или LLI, Ya4 может быть E, R или S, а Wa3 может быть EAGG или EGGG.
Между мотивом A и мотивом B может находиться одна или более аминокислот, на одном конце мотива A или на одном конце мотива B. Из последовательностей известных инозитол-монофосфатаз можно образовать аминокислотные последовательности, не являющиеся мотивом A и мотивом B (например, последовательности, не являющиеся мотивом A и мотивом B в последовательностях, приведенных в SEQ ID NO: 1-20).
Мотив A и/или мотив B представляет активный сайт в последовательности инозитол-монофосфатазы. Мотив A и/или мотив B также известен как сайт связывания инозитолфосфата в качестве субстрата фермента (см. Federation of European Biochemical Societies, Volume 294, number 1,2, 16-18, December 1991). Авторы настоящего изобретения обнаружили, что инозитол-монофосфатаза проявляет активность псикозо-6-фосфат фосфатазы. Авторы настоящего изобретения также обнаружили, что мотив A и/или мотив B инозитол-монофосфатазы может представлять собой сайт связывания субстрата фосфатазы.
Псикозо-6-фосфат фосфатаза может содержать, например, любую одну из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20.
Псикозо-6-фосфат фосфатаза согласно настоящей заявке может представлять собой фермент, который имеет функции известных инозитол-монофосфатаз.
Псикозо-6-фосфат фосфатаза согласно настоящей заявке более избирательно катализирует дефосфорилирование D-псикозо-6-фосфата и может быть неспецифической для D-глюкозо-1-фосфата, D-глюкозо-6-фосфата или D-фруктозо-6-фосфата.
Псикозо-6-фосфат фосфатазу согласно настоящей заявке можно получать путем трансформации штамма ферментом как он есть или ДНК, экспрессирующей фермент (например, SEQ ID NO: 21-40), культивирования трансформированного штамма, разрушения культуры с последующей очисткой. Очистку можно выполнять путем колоночной хроматографии. Штаммом может быть, например, Escherichia coli, Corynebacterium glutamicum, Aspergillus oryzae или Bacillus subtilis.
Согласно одному варианту осуществления настоящей заявки псикозо-6-фосфат фосфатаза согласно настоящей заявке может представлять собой фермент, который имеет гомологию, сходство или идентичность по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% с последовательностью мотива A и/или мотива B и проявляет активность псикозо-6-фосфат фосфатазы. Альтернативно, псикозо-6-фосфат фосфатаза согласно настоящей заявке может представлять собой фермент, который имеет гомологию, сходство или идентичность по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% с последовательностями, не являющимися мотивом A и/или мотивом B. В одном варианте осуществления псикозо-6-фосфат фосфатаза согласно настоящей заявке может содержать белок, состоящий из последовательности, которая имеет гомологию, сходство или идентичность по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% с последовательностью мотива A и/или мотива B в последовательностях, приведенных в SEQ ID NO: 1-20, и имеет гомологию, сходство или идентичность по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% с аминокислотной последовательностью, не являющейся мотивом A и/или мотивом B в последовательностях, приведенных в SEQ ID NO: 1-20, или гомологию, сходство или идентичность в диапазоне, образованном любыми двумя из значений выше. Например, в объеме настоящей заявки также находится любая аминокислотная последовательность, которая имеет гомологию, сходство или идентичность, определенные выше, и проявляет эффективность, соответствующую эффективности белка псикозо-6-фосфат фосфатаза, например, белка, состоящего из любой одной из последовательностей, приведенных в SEQ ID NO: 1-20, и имеет частичную делецию, модификацию, замещение или добавление.
Белки, в которых нерелевантные последовательности добавлены впереди и позади последовательностей мотива A и мотива B, находятся в объеме настоящей заявки, и существующие в природе мутации или их молчащие мутации не исключены из объема настоящей заявки при условии, что они имеют активность, соответствующую активности псикозо-6-фосфат фосфатазы, содержащей мотив A и мотив B. В частности, белки, в которых нерелевантные последовательности добавлены впереди и позади любой одной из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20, существующие в природе мутации и их молчащие мутации не исключены из объема настоящей заявки при условии, что они имеют активность, соответствующую активности псикозо-6-фосфат фосфатазы, содержащей мотив A и мотив B. Белки, содержащие мотив A и мотив B, или белки, содержащие аминокислотные последовательности, приведенные в SEQ ID NO: 1-20, также находятся в объеме настоящей заявки при условии, что они имеют активность, соответствующую активности псикозо-6-фосфат фосфатазы, содержащей мотив A и мотив B.
В другом аспекте настоящей заявки представлена нуклеиновая кислота, кодирующая псикозо-6-фосфат фосфатазу.
В рамках настоящего изобретения термин «нуклеиновая кислота» охватывает молекулы ДНК и РНК, и нуклеотид в качестве основной единицы нуклеиновой кислоты включает в себя натуральный нуклеотид, а также аналог с модифицированным сахаром или основанием (см. Scheit, Nucleotide Analogs, John Wiley, New York(1980); Uhlman and Peyman, Chemical Reviews, 90:543-584(1990)).
Конкретно, нуклеиновая кислота, кодирующая псикозо-6-фосфат фосфатазу, может содержать последовательности, содержащие нуклеотиды, которые можно транслировать в мотив A и мотив B. В одном варианте осуществления нуклеиновая кислота согласно настоящей заявке может состоять из любой одной из нуклеотидных последовательностей, приведенных в SEQ ID NO: 21-40. Более конкретно, нуклеиновая кислота согласно настоящей заявке может содержать нуклеиновую кислоту, которая имеет гомологию, сходство или идентичность по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% с нуклеотидами, которые можно транслировать в мотив A и мотив B, и может проявлять требуемую ферментативную активность после трансляции. Конкретно, нуклеиновая кислота согласно настоящей заявке может содержать нуклеиновую кислоту, которая имеет гомологию, сходство или идентичность по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% с нуклеотидами, которые можно транслировать в мотив A и мотив B в последовательностях, приведенных в SEQ ID NO: 21-40. Альтернативно, нуклеиновая кислота согласно настоящей заявке может представлять собой нуклеиновую кислоту, которая имеет гомологию, сходство или идентичность по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% с нуклеотидами, которые можно транслировать в мотивы, не являющиеся мотивом A и мотивом B в последовательностях, приведенных в SEQ ID NO: 21-40. Белки, которые можно транслировать в белки, содержащие мотив A и мотив B, из-за вырожденности кодона, конкретно, белки, состоящие из любой одной из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20, или полинуклеотиды, которые можно транслировать в белки имеющие гомологию, сходство или идентичность с белками, состоящими из любой одной из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20, также находятся в объеме настоящей заявки.
Фермент, содержащий мотив A и мотив B согласно настоящей заявке, можно получить из термоустойчивого или теплолюбивого штамма. Конкретно, ферментом, состоящим из любой одной из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20, может быть фермент, полученный из Rhodothermus marinus, Thermotoga lettingae, Meiothermus ruber, Dictyoglomus turgidum, Pyrobaculum ferrireducens, Thermoanaerobacter wiegelii, Thermus thermophilus, Thermococcus litoralis, Geobacillus stearothermophilus, Anaerolinea thermophila, Sulfolobus acidocaldarius, Thermosulfidibacter takaii, Pyrococcus furiosus, Archaeoglobus fulgidus, Alicyclobacillus acidocaldarius, Meiothermus silvanus, Meiothermus rufus, Meiothermus taiwanensis, Meiothermus chliarophilus или Meiothermus cerbereus.
В рамках настоящего изобретения термин «гомология» или «идентичность» указывает степень родства между двумя заданными аминокислотными последовательностями или нуклеотидными последовательностями, которую можно выразить в процентах.
Термины «гомология» и «идентичность» часто используют взаимозаменяемо.
Гомологию или идентичность последовательностей консервативных полинуклеотидов или полипептидов определяют с помощью стандартных алгоритмов выравнивания, и их можно использовать со штрафами за гэпы по умолчанию, устанавливаемыми с помощью используемых программ. По существу, гомологичная или идентичная последовательность обычно будет гибридизироваться со средней жесткостью или с высокой жесткостью на всем протяжении по меньшей мере приблизительно 50%, 60%, 70%, 80% или 90% полноразмерного интересующего полинуклеотида или полипептида. Также предусмотрены полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов в гибридизирующихся полинуклеотидах.
Являются ли любые две полинуклеотидные или полипептидные последовательности гомологичными, сходными или идентичными, можно определить с использованием известных компьютерных алгоритмов, таких как программа «FASTA», с использованием например, параметров по умолчанию, как в Pearson et al. (1988), Proc. Natl. Acad. Sci. USA 85: 2444. Альтернативно, гомологию, сходство или идентичность последовательностей можно определить с использованием алгоритма Needleman-Wunsch (Needleman и Wunsch, 1970, J. Mol. Biol. 48: 443-453), который реализован в программе Needle пакета EMBOSS (EMBOSS: European Molecular Biology Open Software Suite, Rice et al. 2000, Trends Genet. 16: 276-277), версия 5.0.0 или более поздняя. Другие программы включают пакет программ GCG (Devereux, J., et al, Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, and [CARILLO et al.] (1988) SIAM J Applied Math 48: 1073). Например, для определения гомологии, сходства или идентичности можно использовать инструмент BLAST базы данных NCBI. Также можно использовать другие коммерческие или общедоступные программы, такие как ClustalW.
Процент гомологии, сходства или идентичности полинуклеотидов или полипептидов можно определить, например, путем сравнения информации о последовательности с использованием компьютерной программы GAP (например, Needleman et al. (1970) J. Mol. Biol. 48:443, пересмотренной Smith и Waterman, Adv. Appl. Math (1981) 2:482). Кратко, программа GAP определяет сходство, как количество выровненных символов (то есть нуклеотидов или аминокислот), которые являются сходными, разделенных на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) унарную матрицу сравнения (содержащую значение 1 для тождеств и 0 для не тождеств) и взвешенную матрицу сравнения Gribskov et al. (1986) Nucl. Acids Res.14:6745, которая описана в Schwartz и Dayhoff, eds. Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979); (2) штраф 3,0 за каждый гэп и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф 10,0 за открытие гэпа, штраф 0,5 за продление гэпа); и (3) отсутствие штрафа за концевые гэпы. Следовательно, термины «гомология» или «идентичность», которые использованы в данном документе, означают сравнение между полипептидами или полинуклеотидами.
В дополнительном аспекте настоящей заявки представлен вектор, содержащий нуклеиновую кислоту, кодирующую псикозо-6-фосфат фосфатазу, описанную в данном документе, или трансформант, содержащий нуклеиновую кислоту, кодирующую псикозо-6-фосфат фосфатазу, или вектор, содержащий нуклеиновую кислоту, кодирующую псикозо-6-фосфат фосфатазу.
В рамках настоящего изобретения термин «вектор» относится к любому носителю для клонирования и/или переноса оснований нуклеиновой кислоты в клетку-хозяин. Вектор может представлять собой репликон, к которому может быть присоединен другой сегмент ДНК, чтобы вызвать репликацию присоединенного сегмента. «Репликон» относится к любому генетическому элементу (например, плазмиде, фагу, космиде, хромосоме или вирусу), который функционирует в качестве автономной единицы репликации ДНК in vivo, то есть способен к репликации под своим собственным управлением. Термин «вектор» может включать как вирусные, так и невирусные носители для введения нуклеиновой кислоты в клетку-хозяин in vitro, ex vivo или in vivo. Термин «вектор» может также включать миникольцевые ДНК. Конкретно, вектором, содержащим нуклеиновую кислоту, кодирующую псикозо-6-фосфат фосфатазу согласно настоящей заявке, может быть pET21a-CJ_Rma, pET21a-CJ_Tle, pET21a-CJ_Mrub, pET21a-CJ_Dtu, pET21a-CJ_Msi, pET21a-CJ_Mruf, pET21a-CJ_Mta, pET21a-CJ_Mch, pET21a-CJ_Mce, pBT7-C-His-CJ_Pfe, pBT7-C-His-CJ_Twi, pBT7-C-His-CJ_Tth, pBT7-C-His-CJ_Tli, pBT7-C-His-CJ_Gst, pBT7-C-His-CJ_Ath, pBT7-C-His-CJ_Sac, pBT7-C-His-CJ_Tta, pBT7-C-His-CJ_Pfu, pBT7-C-His-CJ_Afu или pBT7-C-His-CJ_Aac.
В рамках настоящего изобретения термин «трансформация» относится к введению вектора, содержащего нуклеиновую кислоту, кодирующую белок-мишень, в клетку-хозяин для экспрессии белка, кодируемого нуклеиновой кислотой в клетке-хозяине. Трансформированная нуклеиновая кислота либо может быть вставлена и локализована в хромосоме клетки-хозяина, либо может существовать вне хромосом при условии, что она может экспрессироваться в клетке-хозяине. Нуклеиновая кислота включает ДНК и РНК, кодирующие белок-мишень. Нуклеиновую кислоту можно вводить в любой форме при условии, что она может быть введена и экспрессироваться в клетке-хозяине. Например, нуклеиновую кислоту можно вводить в клетку-хозяин в форме кассеты экспрессии, которая представляет собой генную конструкцию, содержащую все элементы, необходимые для ее автономной экспрессии, но ее форма этим не ограничена. Обычно экспрессионная кассета содержит промотор, функционально связанный с нуклеиновой кислотой, сигнал терминации транскрипции, домен, связывающий рибосому, и сигнал терминации трансляции. Кассета экспрессии может быть в форме самореплицируемого вектора экспрессии. Нуклеиновую кислоту можно вводить в клетку-хозяин в том виде, как она есть, и функционально связывать с последовательностью, необходимой для экспрессии в клетке-хозяине.
В рамках настоящего изобретения термин «функционально связанный» относится к функциональной связи между промоторной последовательностью, инициирующей и опосредующей транскрипцию нуклеиновой кислоты, кодирующей белок-мишень согласно настоящей заявке, и последовательностью генов.
Для аминокислот, упомянутых в настоящей заявке, используют следующие сокращения и названия.
[Таблица 1]
Для введения нуклеиновой кислоты в клетку можно использовать любой метод трансформации. Способ трансформации можно осуществлять с помощью подходящего стандартного метода, известного в данной области техники, в зависимости от типа клетки-хозяина. Примеры таких способов трансформации включают, но без ограничения, электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), ретровирусную инфекцию, микроинъекцию, полиэтиленгликолевый метод (PEG), диэтиламиноэтилдекстрановый метод, катионный липосомный метод и метод ацетата лития диметилсульфоксида.
Клетками-хозяевами предпочтительно являются клетки-хозяева, в которые с высокой эффективностью вводят ДНК и в которых введенная ДНК экспрессируется на высоком уровне. Примеры клеток-хозяев включают, но без ограничения, клетки микроорганизмов, относящихся к родам Corynebacterium, Escherichia и Serratia. Конкретно, клетками-хозяевами могут быть клетки E. coli.
Трансформантом согласно настоящей заявке может быть E. coli BL21(DE3)/pET21a-CJ_Rma(E. coli_P1_CJ_Rma, KCCM12057P), E. coli BL21(DE3)/pET21a-CJ_Tle(E. coli_P2_CJ_Tle, KCCM12058P), E. coli BL21(DE3)/pET21a-CJ_Mrub(E. coli_P3_CJ_Mrub, KCCM12059P), E. coli BL21(DE3)/pET21a-CJ_Dtu(E. coli_P4_CJ_Dtu, KCCM12060P), E. coli BL21(DE3)/pBT7-C-His-CJ_Pfe(E. coli_P5_CJ_Pfe, KCCM12061P), E. coli BL21(DE3)/pBT7-C-His-CJ_Twi(E. coli_P6_CJ_Twi, KCCM12062P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tth(E. coli_P7_CJ_Tth, KCCM12063P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tli(E. coli_P8_CJ_Tli, KCCM12064P), E. coli BL21(DE3)/pBT7-C-His-CJ_Gst(E. coli_P9_CJ_Gst, KCCM12065P), E. coli BL21(DE3)/pBT7-C-His-CJ_Ath(E. coli_P10_CJ_Ath, KCCM12066P), E. coli BL21(DE3)/pBT7-C-His-CJ_Sac(E. coli_P11_CJ_Sac, KCCM12067P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tta(E. coli_P12_CJ_Tta, KCCM12068P), E. coli BL21(DE3)/pBT7-C-His-CJ_Pfu(E. coli_P13_CJ_Pfu, KCCM12069P), E. coli BL21(DE3)/pBT7-C-His-CJ_Afu(E. coli_P14_CJ_Afu, KCCM12070P), E. coli BL21(DE3)/pBT7-C-His-CJ_Aac(E. coli_P15_CJ_Aac, KCCM12071P), E. coli BL21(DE3)/pET21a-CJ_Msi(E. coli_P16_CJ_Msi, KCCM12072P), E. coli BL21(DE3)/pET21a-CJ_Mruf(E. coli_P17_CJ_Mruf, KCCM12073P), E. coli BL21(DE3)/pET21a-CJ_Mta(E. coli_P18_CJ_Mta, KCCM12074P), E. coli BL21(DE3)/pET21a-CJ_Mch(E. coli_P19_CJ_Mch, KCCM12075P) или E. coli BL21(DE3)/pET21a-CJ_Mce(E. coli_P20_CJ_Mce, KCCM12076P).
В другом аспекте настоящей заявки представлена композиция для получения D-псикозы, содержащая инозитол-монофосфатазу, микроорганизм, экспрессирующий инозитол-монофосфатазу, или культуру микроорганизмов, экспрессирующих инозитол-монофосфатазу.
Инозитол-монофосфатаза согласно настоящей заявке может содержать мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или гэп, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV. То есть, инозитол-монофосфатазу можно использовать взаимозаменяемо с псикозо-6-фосфат фосфатазой. Таким образом, объяснения псикозо-6-фосфат фосфатазы можно применять к инозитол-монофосфатазе. В одном варианте осуществления инозитол-монофосфатазой может быть фермент, состоящий из любой одной из аминокислотных последовательностей, приведенных в SEQ ID NO: 1-20.
Композиция согласно настоящей заявке может дополнительно содержать фермент и/или субстрат, вовлеченный в путь получения D-псикозы (см. ФИГ. 1) [(i) крахмал, мальтодекстрин, сахарозу или их комбинацию; (ii) фосфат; (iii) D-фруктозо-6-фосфат-3-эпимеразу; (iv) D-глюкозо-6-фосфат-изомеразу; (v) фосфоглюкомутазу или глюкокиназу; и/или (vi) α-глюканфосфорилазу, фосфорилазу крахмала, фосфорилазу мальтодекстрина, фосфорилазу сахарозы, α-амилазу, пуллуланазу, изоамилазу, глюкоамилазу или сахаразу]; микроорганизм, экспрессирующий фермент, вовлеченный в путь получения D-псикозы; или культуру микроорганизмов, экспрессирующих фермент, вовлеченный в путь получения D-псикозы. Дополнительный фермент и субстрат являются всего лишь иллюстративными и не являются ограничением при условии, что D-псикозу можно получать с использованием псикозо-6-фосфат фосфатазы согласно настоящей заявке.
Фосфорилаза крахмала/мальтодекстрина (EC 2.4.1.1) и α-глюканфосфорилаза может включать в себя любой белок, активный при переносе фосфорила в глюкозу с получением D-глюкозо-1-фосфата из крахмала или мальтодекстрина. Конкретно, белок активный при получении D-глюкозо-1-фосфата из крахмала или мальтодекстрина, может состоять из аминокислотной последовательности, приведенной в SEQ ID NO: 59. Белок, активный при получении D-глюкозо-1-фосфата из крахмала или мальтодекстрина, может кодироваться последовательностью, приведенной в SEQ ID NO: 60.
Фосфорилаза сахарозы (EC 2.4.1.7) может содержать любой белок, активный при переносе фосфорила в глюкозу с получением D-глюкозо-1-фосфата из сахарозы.
α-амилаза (EC 3.2.1.1), пуллуланаза (EC 3.2.1.41), глюкоамилаза (EC 3.2.1.3) и изоамилаза представляют собой разжижающие крахмал ферменты и могут содержать любой белок, активный для преобразования крахмала или мальтодекстрина в глюкозу.
Сахараза (EC 3.2.1.26) может содержать любой белок, активный для преобразования сахарозы в глюкозу.
Фосфоглюкомутаза (EC 5.4.2.2) может содержать любой белок, активный для преобразования D-глюкозо-1-фосфата в D-глюкозо-6-фосфат. Конкретно, белок, активный для преобразования D-глюкозо-1-фосфата в D-глюкозо-6-фосфат, может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 61. Белок, активный для преобразования D-глюкозо-1-фосфата в D-глюкозо-6-фосфат, может кодироваться последовательностью, приведенной в SEQ ID NO: 62.
Глюкокиназа может содержать любой белок, активный при переносе фосфорила в глюкозу для преобразования глюкозы в D-глюкозо-6-фосфат. Конкретно, глюкокиназа может представлять собой полифосфат-зависимую глюкокиназу. Более конкретно, глюкокиназа может представлять собой белок, состоящий из аминокислотной последовательности, приведенной в SEQ ID NO: 77 или 78. D-глюкозо-6-фосфат-изомераза может содержать любой белок, активный для преобразования D-глюкозо-6-фосфата в D-фруктозо-6-фосфат. Конкретно, D-глюкозо-6-фосфат-изомераза может представлять собой белок, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 63. D-глюкозо-6-фосфат-изомераза может кодироваться последовательностью, приведенной в SEQ ID NO: 64.
D-фруктозо-6-фосфат-3-эпимераза может содержать любой белок, активный для преобразования D-фруктозо-6-фосфата в D-псикозо-6-фосфат. Конкретно, D-фруктозо-6-фосфат-3-эпимераза может представлять собой белок, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 65. D-фруктозо-6-фосфат-3-эпимераза может кодироваться последовательностью, приведенной в SEQ ID NO: 66.
Композиция для получения D-псикозы согласно настоящей заявке может дополнительно содержать белок, активный для преобразования глюкозы в крахмал, мальтодекстрин или сахарозу. Белком может быть, например, 4-α-глюканотрансфераза. Белком, активным для преобразования глюкозы в крахмал, мальтодекстрин или сахарозу, может быть фермент, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 67. Конкретно, белок, активный для преобразования глюкозы в крахмал, мальтодекстрин или сахарозу, может кодироваться нуклеотидной последовательностью, приведенной в SEQ ID NO: 68. Композиция для получения D-псикозы согласно настоящей заявке может дополнительно содержать любое подходящее вспомогательное вещество, известное в данной области. Примеры таких вспомогательных веществ включают, но без ограничения, консерванты, смачивающие средства, диспергаторы, суспендирующие средства, буферы, стабилизаторы изотонические средства.
Композиция для получения D-псикозы согласно настоящей заявке может дополнительно содержать ион металла или соль металла. В одном варианте осуществления ион металла может представлять собой катион двухвалентного металла. Конкретно, ион металла можно выбирать из группы, состоящей из ионов Ni, Mg, Co, Mn, Fe и Zn. Более конкретно, композиция для получения D-псикозы согласно настоящей заявке может дополнительно содержать соль металла. Еще более конкретно, соль металла можно выбирать из группы, состоящей из NiSO4, MgSO4, MgCl2, NiCl2, CoSO4, CoCl2, MnCl2, FeSO4, ZnSO4 и их смесей.
В другом аспекте настоящей заявки представлен способ получения D-псикозы, включающий введение в контакт инозитол-монофосфатазы, микроорганизма, экспрессирующего фермент, или культуры микроорганизмов с D-псикозо-6-фосфатом для преобразования D-псикозо-6-фосфата в D-псикозу.
Инозитол-монофосфатаза может содержать мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3. В данном документе инозитол-монофосфатазу можно использовать взаимозаменяемо с псикозо-6-фосфат фосфатазой.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием D-псикозо-6-фосфата в D-псикозу введение D-фруктозо-6-фосфат-3-эпимеразы, микроорганизма, экспрессирующего D-фруктозо-6-фосфат-3-эпимеразу, или культуры микроорганизмов, экспрессирующих D-фруктозо-6-фосфат-3-эпимеразу, в контакт с D-фруктозо-6-фосфатом для преобразования D-фруктозо-6-фосфата в D-псикозо-6-фосфат.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием D-фруктозо-6-фосфата в D-псикозо-6-фосфат введение D-глюкозо-6-фосфат-изомеразы, микроорганизма, экспрессирующего D-глюкозо-6-фосфат-изомеразу, или культуры микроорганизмов, экспрессирующих D-глюкозо-6-фосфат-изомеразу, в контакт с D-глюкозо-6-фосфатом для преобразования D-глюкозо-6-фосфата в D-фруктозо-6-фосфат.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием D-глюкозо-6-фосфата в D-фруктозо-6-фосфат введение фосфоглюкомутазы, микроорганизма, экспрессирующего фосфоглюкомутазу, или культуры микроорганизмов, экспрессирующих фосфоглюкомутазу, в контакт с D-глюкозо-1-фосфатом для преобразования D-глюкозо-1-фосфата в D-глюкозо-6-фосфат.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием D-глюкозо-6-фосфата в D-фруктозо-6-фосфат введение глюкокиназы, микроорганизма, экспрессирующего глюкокиназу, или культуры микроорганизмов, экспрессирующих глюкокиназу, и фосфата в контакт с глюкозой для преобразования глюкозы в D-глюкозо-6-фосфат.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием D-глюкозо-1-фосфата в D-глюкозо-6-фосфат введение α-глюканфосфорилазы, фосфорилазы крахмала, фосфорилазы мальтодекстрина или фосфорилазы сахарозы, микроорганизма, экспрессирующего фосфорилазу, или культуры микроорганизмов, экспрессирующих фосфорилазу, и фосфата в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией для преобразования крахмала, мальтодекстрина, сахарозы или их комбинации в D-глюкозо-1-фосфат.
Способ согласно настоящей заявке может дополнительно включать перед преобразованием глюкозы в D-глюкозо-6-фосфат введение α-амилазы, пуллуланазы, глюкоамилазы, сахаразы или изоамилазы, микроорганизма, экспрессирующего α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, или культуры микроорганизмов, экспрессирующих α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией для преобразования крахмала, мальтодекстрина, сахарозы или их комбинации в глюкозу.
Еще в одном аспекте настоящей заявки представлен способ получения D-псикозы включающий введение (a) инозитол-монофосфатазы, D-фруктозо-6-фосфат-3-эпимеразы, D-глюкозо-6-фосфат-изомеразы, фосфоглюкомутазы или глюкокиназы и α-глюканфосфорилазы, фосфорилазы крахмала, фосфорилазы мальтодекстрина, фосфорилазы сахарозы, α-амилазы, пуллуланазы, изоамилазы, глюкоамилазы или сахаразы или (b) микроорганизма, экспрессирующего ферменты (a), или культуры микроорганизмов в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией и фосфатом.
В способе согласно настоящей заявке контактную реакцию можно проводить при pH 5,0-9,0, конкретно pH 6,0-8,0.
В способе согласно настоящей заявке контактную реакцию можно проводить при температуре 40°C-80°C, конкретно при температуре 40°C-60°C или 50°C-60°C.
В способе согласно настоящей заявке контактную реакцию можно проводить в течение от 2 часов до 24 часов, конкретно 6-24 часов.
В способе согласно настоящей заявке контактную реакцию можно проводить при pH 5,0-9,0, при температуре 40°C-80°C и/или в течение от 2 часов до 24 часов. Конкретно, контактную реакцию можно проводить при pH 6,0-8,0, при температуре 40°C-60°C или 50°C-60°C и/или в течение от 6 часов до 24 часов.
Способ согласно настоящей заявке может дополнительно включать очистку продукта реакции D-псикозы. Нет особого ограничения способа очистки D-псикозы. D-псикозу можно очищать любым подходящим способом, известным в данной области. Неограничивающие примеры таких способов очистки включают хроматографию, фракционную кристаллизацию и ионную очистку, которые можно проводить отдельно или в комбинации. Например, D-псикозу можно очищать путем хроматографии. В этом случае сахарид-мишень можно отделять на основе небольших отличий в силе связывания между сахаридами и ионами металлов, присоединенными к ионной смоле.
Каждый из способов согласно настоящей заявке может дополнительно включать отбеливание и/или дименирализацию перед или после стадии очистки. Отбеливание и/или дименирализация делает D-псикозу более чистой без примесей.
Предпочтительные результаты
Новая псикозо-6-фосфат фосфатаза согласно настоящей заявке является термоустойчивой инозитол-монофосфатазой. Из-за своей термоустойчивости фермент согласно настоящей заявке может участвовать в пути преобразования D-псикозо-6-фосфата в D-псикозу, обеспечивая получение D-псикозы в промышленном масштабе. Использование фермента согласно настоящей заявке обеспечивает прохождение пути синтеза D-псикозы из глюкозы или крахмала (например, мальтодекстрина) в качестве недорогого исходного материала. Кроме того, при использовании фермента согласно настоящей заявке D-псикозу можно получать посредством необратимого дефосфорилирования D-псикозо-6-фосфата. Вследствие этого, использование фермента согласно настоящей заявке значительно увеличивает скорость преобразования в D-псикозу.
Кроме того, способы согласно настоящей заявке на основе использование инозитол-монофосфатазы обеспечивают получение высокой концентрации D-псикозы при высокой скорости преобразования, упрощая или исключая разделение и очистку продукта реакции. Вследствие этого, способы согласно настоящей заявке можно проводить простым способом, и они является предпочтительными с экономической точки зрения.
Описание чертежей
На ФИГ. 1 схематично представлены пути ферментативной реакции для получения D-псикозы из крахмала (например, мальтодекстрина) или глюкозы.
На ФИГ. 2a, 2b и 2c представлены изображения SDS-PAGE маркеров размеров (M), экспрессируемых коферментов (S) и очищенных рекомбинантных ферментов (E) (pET21a-CJ_Rma, pET21a-CJ_Tle, pET21a-CJ_Mrub, pET21a-CJ_Dtu, pBT7-C-His-CJ_Pfe, pBT7-C-His-CJ_Twi, pBT7-C-His-CJ_Tth, pBT7-C-His-CJ_Tli, pBT7-C-His-CJ_Gst, pBT7-C-His-CJ_Ath, pBT7-C-His-CJ_Sac, pBT7-C-His-CJ_Tta, pBT7-C-His-CJ_Pfu, pBT7-C-His-CJ_Afu, pBT7-C-His-CJ_Aac, pET21a-CJ_Msi, pET21a-CJ_Mruf, pET21a-CJ_Mta, pET21a-CJ_Mch, pET21a-CJ_Mce) для определения их молекулярных масс, которые получали после электрофореза белка.
На ФИГ. 3a представлена относительная активность инозитол-монофосфатаз (pET21a-CJ_Rma, pET21a-CJ_Tle, pET21a-CJ_Mrub, pET21a-CJ_Dtu, pBT7-C-His-CJ_Pfe, pBT7-C-His-CJ_Twi, pBT7-C-His-CJ_Tth, pBT7-C-His-CJ_Tli, pBT7-C-His-CJ_Gst, pBT7-C-His-CJ_Ath, pBT7-C-His-CJ_Sac, pBT7-C-His-CJ_Tta, pBT7-C-His-CJ_Pfu, pBT7-C-His-CJ_Afu, pBT7-C-His-CJ_Aac) для дефосфорилирования (%, левая ось Y, гистограмма) и избирательные скорости дефосфорилирования с помощью инозитол-монофосфатаз в смеси дефосфорилирования, содержащей D-глюкозо-6-фосфат, D-глюкозо-1-фосфат, D-фруктозо-6-фосфат и D-псикозо-6-фосфат (%, правая ось Y, квадратные точки).
На ФИГ. 3b представлено сравнение относительной активности (%) инозитол-монофосфатаз (pET21a-CJ_Mrub, pET21a-CJ_Msi, pET21a-CJ_Mruf, pET21a-CJ_Mta, pET21a-CJ_Mch, pET21a-CJ_Mce) для дефосфорилирования.
На ФИГ. 4a представлены HPLC хроматограммы, подтверждающие получение D-псикозы из мальтодекстрина посредством множества ферментативных реакций с α-глюканфосфорилазой, фосфоглюкомутазой, D-глюкозо-6-фосфат-изомеразой, 4-α-глюканотрансферазой, D-фруктозо-6-фосфат-3-эпимеразой и псикозо-6-фосфат фосфатазой.
На ФИГ. 4b представлены HPLC хроматограммы, подтверждающие получение D-псикозы посредством избирательного дефосфорилирования D-псикозо-6-фосфата в реакционном растворе, содержащем D-глюкозо-1-фосфат, D-глюкозо-6-фосфат, D-фруктозо-6-фосфат и D-псикозо-6-фосфат в присутствии инозитол-монофосфатазы согласно настоящей заявке.
Способ осуществления изобретения
Настоящая заявка будет подробно объяснена со ссылкой на следующие примеры. Однако, эти примеры представлены, чтобы помочь понять настоящую заявку, а не ограничить объем настоящей заявки.
Примеры
Пример 1: Получение рекомбинантных векторов экспрессии инозитол-монофосфатаз и трансформированных микроорганизмов
Для получения псикозо-6-фосфат фосфатазы, необходимой для пути получения D-псикозы, скринировали гены термоустойчивой инозитол-монофосфатазы. Конкретно, гены инозитол-монофосфатазы (Rma, Tle, Mrub, Dtu, Msi, Mruf, Mta, Mch, и Mce) скринировали из геномных последовательностей Rhodothermus marinus, Thermotoga lettingae, Meiothermus ruber, Dictyoglomus turgidum, Pyrobaculum ferrireducens, Thermoanaerobacter wiegelii, Thermus thermophilus, Thermococcus litoralis, Geobacillus stearothermophilus, Anaerolinea thermophila, Sulfolobus acidocaldarius, Thermosulfidibacter takaii, Pyrococcus furiosus, Archaeoglobus fulgidus, Alicyclobacillus acidocaldarius, Meiothermus silvanus, Meiothermus rufus, Meiothermus taiwanensis, Meiothermus chliarophilus и Meiothermus cerbereus, зарегистрированных в GenBank.
На основе информации о нуклеотидных последовательностях (SEQ ID NO: 21, 22, 23, 24, 36, 37, 38, 39 и 40 в порядке генов) и аминокислотных последовательностях (SEQ ID NO: 1, 2, 3, 4, 16, 17, 18, 19 и 20 в порядке генов) скринированных генов, конструировали прямые праймеры (SEQ ID NO: 41, 43, 45, 47, 49, 51, 53, 55 и 57) и обратные праймеры (SEQ ID NO: 42, 44, 46, 48, 50, 52, 54, 56 и 58). Гены амплифицировали из геномных ДНК Rhodothermus marinus, Thermotoga lettingae, Meiothermus ruber, Dictyoglomus turgidum, Meiothermus silvanus, Meiothermus rufus, Meiothermus taiwanensis, Meiothermus chliarophilus и Meiothermus cerbereus посредством полимеразной цепной реакции (ПЦР) с использованием синтезированных праймеров. Амплифицированные гены инозитол-монофосфатазы вставляли в плазмидный вектор pET21a (Novagen) для экспрессии E. coli с использованием рестрикционных ферментов NdeI и XhoI или Sa1I для конструирования рекомбинантных векторов экспрессии, которые назвали pET21a-CJ_Rma(Nde I/Xho I), pET21a-CJ_Tle(Nde I/Xho I), pET21a-CJ_Mrub(Nde I/Xho I), pET21a-CJ_Dtu(Nde I/Xho I), pET21a-CJ_Msi(Nde I/Sal I), pET21a-CJ_Mruf(Nde I/Sal I), pET21a-CJ_Mta(Nde I/Sal I), pET21a-CJ_Mch(Nde I/Sal I) и pET21a-CJ_Mce(Nde I/Sal I).
Кроме того, скринировали гены инозитол-монофосфатазы (Pfe, Twi, Tth, Tli, Gst, Ath, Sac, Tta, Pfu, Afu и Aac), полученные из Pyrobaculum ferrireducens, Thermoanaerobacter wiegelii, Thermus thermophilus, Thermococcus litoralis, Geobacillus stearothermophilus, Anaerolinea thermophila, Sulfolobus acidocaldarius, Thermosulfidibacter takaii, Pyrococcus furiosus, Archaeoglobus fulgidus и Alicyclobacillus acidocaldarius. На основе информации о нуклеотидных последовательностях (SEQ ID NO: 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 в порядке генов) и аминокислотных последовательностях (SEQ ID NO: 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 и 15 в порядке генов) скринированных генов запросили синтез ДНК в Bioneer (Корея). ДНК вставляли в вектор pBT7-C-His (Bioneer) для конструирования рекомбинантных векторов экспрессии, которые назвали pBT7-C-His-CJ_Pfe, pBT7-C-His-CJ_Twi, pBT7-C-His-CJ_Tth, pBT7-C-His-CJ_Tli, pBT7-C-His-CJ_Gst, pBT7-C-His-CJ_Ath, pBT7-C-His-CJ_Sac, pBT7-C-His-CJ_Tta, pBT7-C-His-CJ_Pfu, pBT7-C-His-CJ_Afu и pBT7-C-His-CJ_Aac.
Векторы экспрессии трансформировали в штамм E. coli BL21(DE3) с помощью обычной методики трансформации (см. Sambrook et al. 1989) с получением трансформированных микроорганизмов, которые назвали E. coli BL21(DE3)/pET21a-CJ_Rma (E. coli_P1_CJ_Rma, KCCM12057P), E. coli BL21(DE3)/pET21a-CJ_Tle (E. coli_P2_CJ_Tle, KCCM12058P), E. coli BL21(DE3)/pET21a-CJ_Mrub (E. coli_P3_CJ_Mrub, KCCM12059P), E. coli BL21(DE3)/pET21a-CJ_Dtu (E. coli_P4_CJ_Dtu, KCCM12060P), E. coli BL21(DE3)/pBT7-C-His-CJ_Pfe (E. coli_P5_CJ_Pfe, KCCM12061P), E. coli BL21(DE3)/pBT7-C-His-CJ_Twi (E. coli_P6_CJ_Twi, KCCM12062P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tth (E. coli_P7_CJ_Tth, KCCM12063P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tli (E. coli_P8_CJ_Tli, KCCM12064P), E. coli BL21(DE3)/pBT7-C-His-CJ_Gst (E. coli_P9_CJ_Gst, KCCM12065P), E. coli BL21(DE3)/pBT7-C-His-CJ_Ath (E. coli_P10_CJ_Ath, KCCM12066P), E. coli BL21(DE3)/pBT7-C-His-CJ_Sac (E. coli_P11_CJ_Sac, KCCM12067P), E. coli BL21(DE3)/pBT7-C-His-CJ_Tta (E. coli_P12_CJ_Tta, KCCM12068P), E. coli BL21(DE3)/pBT7-C-His-CJ_Pfu (E. coli_P13_CJ_Pfu, KCCM12069P), E. coli BL21(DE3)/pBT7-C-His-CJ_Afu (E. coli_P14_CJ_Afu, KCCM12070P), E. coli BL21(DE3)/pBT7-C-His-CJ_Aac (E. coli_P15_CJ_Aac, KCCM12071P), E. coli BL21(DE3)/pET21a-CJ_Msi (E. coli_P16_CJ_Msi, KCCM12072P), E. coli BL21(DE3)/pET21a-CJ_Mruf (E. coli_P17_CJ_Mruf, KCCM12073P), E. coli BL21(DE3)/pET21a-CJ_Mta (E. coli_P18_CJ_Mta, KCCM12074P), E. coli BL21(DE3)/pET21a-CJ_Mch (E. coli_P19_CJ_Mch, KCCM12075P) и E. coli BL21(DE3)/pET21a-CJ_Mce (E. coli_P20_CJ_Mce, KCCM12076P).
Трансформированные штаммы депонировали в центре культур микроорганизмов Кореи (KCCM) 10 июля 2017 года по Будапештскому договору (регистрационный номер: KCCM12057P - KCCM12076P).
Пример 2: Получение ферментов, необходимых для пути получения D-псикозы
Для получения α-глюканфосфорилазы, фосфоглюкомутазы, D-глюкозо-6-фосфат-изомеразы и D-фруктозо-6-фосфат-3-эпимеразы, полученных из Thermotoga neapolitana в качестве термоустойчивых ферментов, необходимых для путей получения D-псикозы, скринировали гены, соответствующие ферментам (ct1, ct2, tn1 и fp3e в порядке ферментов).
На основе нуклеотидных последовательностей (SEQ ID NO: 60, 62, 64 и 66 в порядке ферментов) и аминокислотных последовательностей (SEQ ID NO: 59, 61, 63 и 65 в порядке ферментов) скринированных генов конструировали прямые праймеры (SEQ ID NO: 69, 71, 73 и 75) и обратные праймеры (SEQ ID NO: 70, 72, 74 и 76). Гены ферментов амплифицировали из геномной ДНК Thermotoga neapolitana в качестве матрицы посредством полимеразной цепной реакции (ПЦР) с использованием праймеров. ПЦР проводили в течение в общей сложности 25 циклов с использованием следующих условий: денатурирование при 95°C в течение 30 сек, отжигание при 55°C в течение 30 сек и полимеризация при 68°C в течение 2 мин. Амплифицированные гены ферментов вставляли в плазмидный вектор pET21a (Novagen) для экспрессии E. coli с использованием рестрикционных ферментов NdeI и XhoI для конструирования рекомбинантных векторов экспрессии, которые назвали pET21a-CJ_ct1, pET21a-CJ_ct2, pET21a-CJ_ tn1 и pET21a-CJ_ fp3e. Рекомбинантные векторы экспрессии трансформировали в штамм E. coli BL21(DE3) посредством обычной методики трансформации (см. Sambrook et al. 1989) с получением трансформированных микроорганизмов, которые назвали E. coli BL21(DE3)/pET21a-CJ_ct1 (KCCM11990P), E. coli BL21(DE3)/pET21a-CJ_ct2 (KCCM11991P), E. coli BL21(DE3)/pET21a-CJ_tn1 (KCCM11992P), и E. coli BL21(DE3)/CJ_tn_fp3e (KCCM11848P). Штаммы депонировали в центре культур микроорганизмов Кореи (KCCM) 23 июня 2016 года по Будапештскому договору.
Пример 3: Получение рекомбинантных ферментов
В этом примере получали рекомбинантные ферменты. Сначала культуральную пробирку, содержащую 5 мл жидкой среды LB, инокулировали каждым из трансформированных микроорганизмов, полученных в примерах 1 и 2. Инокулят культивировали в шейкере-инкубаторе при 37°С до достижения поглощения 2,0 при 600 нм. Культуральный бульон добавляли в жидкую среду LB в культуральной колбе с последующей основной культурой. Когда поглощение культуры при 600 нм достигло 2,0, добавляли 1 мМ IPTG для индукции экспрессии и получения рекомбинантного фермента. Температуру культуры поддерживали при 37°С при перемешивании со скоростью 180 об/мин. Культуральный бульон центрифугировали при 8000 × g и 4°С в течение 20 минут для сбора бактериальных клеток. Собранные бактериальные клетки дважды промывали 50 мМ трис-HCl-буфером (рН 8,0) и суспендировали в том же буфере. Затем клетки разрушали с помощью ультразвукового гомогенизатора. Клеточный лизат центрифугировали при 13000 × g и 4°С в течение 20 минут. Рекомбинантный фермент очищали от супернатанта с помощью аффинной хроматографии His-tag. Очищенный рекомбинантный фермент подвергали диализу против 50 мМ буфера Tris-HCl (рН 8,0) и затем использовали для последующей реакции. Молекулярную массу очищенного рекомбинантного фермента определяли с помощью SDS-PAGE.
Далее следуют названия и молекулярные массы очищенных ферментов, полученных с использованием трансформированных микроорганизмов (ФИГ. 2a, 2b и 2c):
30,3 кДа для фермента (RMA), получаемого из E. coli BL21(DE3)/pET21a-CJ_Rma (E. coli_P1_CJ_Rma);
28,5 кДа для фермента (TLE), получаемого из E. coli BL21(DE3)/pET21a-CJ_Tle (E. coli_P2_CJ_Tle);
28 кДа для фермента (MRUB), получаемого из E. coli BL21(DE3)/pET21a-CJ_Mrub (E. coli_P3_CJ_Mrub);
30,2 кДа для фермента (DTU), получаемого из E. coli BL21(DE3)/pET21a-CJ_Dtu (E. coli_P4_CJ_Dtu);
27,2 кДа для фермента (PFE), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Pfe (E. coli_P5_CJ_Pfe);
28,8 кДа для фермента (TWI), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Twi (E. coli_P6_CJ_Twi);
28,6 кДа для фермента (TTH), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Tth (E. coli_P7_CJ_Tth);
28 кДа для фермента (TLI), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Tli (E. coli_P8_CJ_Tli);
29,6 кДа для фермента (GST), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Gst (E. coli_P9_CJ_Gst);
28,7 кДа для фермента (ATH), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Ath (E. coli_P10_CJ_Ath);
29,9 кДа для фермента (SAC), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Sac (E. coli_P11_CJ_Sac);
28,6 кДа для фермента (TTA), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Tta (E. coli_P12_CJ_Tta);
27,9 кДа для фермента (PFU), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Pfu (E. coli_P13_CJ_Pfu);
28 кДа для фермента (AFU), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Afu (E. coli_P14_CJ_Afu);
29 кДа для фермента (AAC), получаемого из E. coli BL21(DE3)/pBT7-C-His-CJ_Aac (E. coli_P15_CJ_Aac);
28,1 кДа для фермента (MSI), получаемого из E. coli BL21(DE3)/pET21a-CJ_Msi (E. coli_P16_CJ_Msi);
28 кДа для фермента (MRUF), получаемого из E. coli BL21(DE3)/pET21a-CJ_Mruf (E. coli_P17_CJ_Mruf);
28,1 кДа для фермента (MTA), получаемого из E. coli BL21(DE3)/pET21a-CJ_Mta (E. coli_P18_CJ_Mta);
28,4 кДа для фермента (MCH), получаемого из E. coli BL21(DE3)/pET21a-CJ_Mch (E. coli_P19_CJ_Mch);
28,1 кДа для фермента (MCE), получаемого из E. coli BL21(DE3)/pET21a-CJ_Mce (E. coli_P20_CJ_Mce);
Фермент (CT1), получаемый из E. coli BL21(DE3)/pET21a-CJ_ct1 (KCCM11990P);
Фермент (CT2), получаемый из E. coli BL21(DE3)/pET21a-CJ_ct2 (KCCM11991P);
Фермент (TN1), получаемый из E. coli BL21(DE3)/pET21a-CJ_tn1 (KCCM11992P); и
Фермент (FP3E), получаемый из E. coli BL21(DE3)/CJ_tn_fp3e (KCCM11848P).
Пример 4: Анализ активности инозитол-монофосфатаз
4-1. Анализ активности псикозо-6-фосфат фосфатаз
Псикозо-6-фосфат трудно приобрести. Поэтому авторы изобретения сами получали D-псикозо-6-фосфат из D-фруктозо-6-фосфат и исследовали активность инозитол-монофосфатаз для получения D-псикозы.
В частности, 50 мМ D-фруктозо-6-фосфат суспендировали в 50 мМ Tris-HCl (рН 7,0), а затем туда добавляли D-фруктозо-6-фосфат-3-эпимеразу (FP3E), полученную в примере 3, и 0,1 ед/мл каждой из 20 инозитол-монофосфатаз. Смеси давали реагировать при 70°С в течение 1 часа. Получение D-псикозы подтверждали с помощью HPLC (колонка SP_0810 (Shodex), колонка Aminex HPX-87C (Bio-RAD), 80°С, скорость потока подвижной фазы 0,6 мл/мин, детектор показателя преломления).
Исследовали эффективность дефосфорилирования всех 20 инозитол-монофосфатаз для D-псикозо-6-фосфата (фиг. 3a и 3b).
4-2. Анализ активности инозитол-монофосфатаз для специфического дефосфорилирования D-псикозо-6-фосфата
Измеряли удельные скорости дефосфорилирования D-псикозо-6-фосфата в смеси, содержащей D-глюкозо-6-фосфат, D-глюкозо-1-фосфат, D-фруктозо-6-фосфат и D-псикозо-6-фосфат в присутствии инозитол-монофосфатаз.
Конкретно, 0,1 ед/мл каждой из инозитол-монофосфатаз и 5 мМ MgCl2 добавляли к смеси 1% (м/о) D-глюкозо-6-фосфата, D-глюкозо-1-фосфата, D-фруктозо-6-фосфата и D-псикозо-6-фосфата. Обеспечивали возможность протекания реакции при 50°С в течение 12 часов. Продукты реакции анализировали с помощью HPLC (колонка Aminex HPX-87C (Bio-RAD), 80°C, скорость потока подвижной фазы 0,6 мл/мин). Для обнаружения выработки D-псикозы и других сахаридов (фруктозы и глюкозы) использовали детектор показателя преломления.
В результате фермент MRUB показал наивысшую удельную скорость дефосфорилирования D-псикозо-6-фосфата (фиг. 3a).
Пример 5: Анализ активности ферментов посредством множества ферментативных реакций
Для получения D-псикозы из мальтодекстрина обеспечивали возможность одновременной реакции ферментов CT1, CT2, TN1, FP3E и MRUB с мальтодекстрином. 5% (м/о) мальтодекстрина добавляли к 0,1 ед/мл каждого фермента, 5 мМ MgCl2 и 20 мМ фосфата натрия (рН 7,0). Обеспечивали возможность реакции смеси при температуре 50°С в течение 12 часов. Продукты реакции анализировали с помощью HPLC (колонка Aminex HPX-87C (Bio-RAD), 80°C, скорость потока подвижной фазы 0,6 мл/мин, детектор показателя преломления).
В результате было подтверждено получение D-псикозы из мальтодекстрина посредством множества ферментативных реакций (фиг. 4а).
Хотя вариант осуществления настоящей заявки был подробно описан, специалистам в данной области техники будет понятно, что заявка может быть реализована в других конкретных формах без изменения сущности или существенных особенностей заявки. Следовательно, следует отметить, что вышеприведенные варианты осуществления являются просто иллюстративными во всех аспектах, и их не следует рассматривать в качестве ограничения заявки. Объем заявки ограничен прилагаемой формулой изобретения, а не подробным описанием заявки. Все изменения или модификации или их эквиваленты, сделанные в рамках значений и объема формулы изобретения, следует рассматривать в границах объема заявки.
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_________________ , строка ______3__________________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12057P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______4______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12058P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______5______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12059P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______6______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12060P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_________________ , строка ______7_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12061P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______8______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12062P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______9______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12063P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _______10_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12064P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _______11_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12065P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _______12_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12066P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _______13_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12067P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка _______14____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12068P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _______15_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12069P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка _______16____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12070P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка _______17____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12071P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ______18_____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12072P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка _____19______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12073P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9_______________ , строка _____20_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12074P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ____21_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12075P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __9________________ , строка ____22_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
10 июля 2017 г.
KCCM12076P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __18________________ , строка ____2_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
20 марта 2017 г.
KCCM11990P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __18_________________ , строка ______3____________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
20 марта 2017 г.
KCCM11991P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __18________________ , строка ____4_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
20 марта 2017 г.
KCCM11992P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
УКАЗАНИЯ, ОТНОСЯЩИЕСЯ К ДЕПОНИРОВАННОМУ МИКРООРГАНИЗМУ ИЛИ ДРУГОМУ БИОЛОГИЧЕСКОМУ МАТЕРИАЛУ
(PCT Правило 13bis)
на странице __18_______________ , строка _____5_______________ .
Корейский центр культур микроорганизмов (KCCM)
Юрим Билдинг, Хондженэ-2га-гил 45, Содэмун-Гу, Сеул, 120-091, Республика Корея
23 июня 2016 г.
KCCM11848P
□ Данный лист получен с международной заявкой
□ Данный лист получен Международным Бюро:
Форма PCT/RO/134 (Июль 1998 г.; репринт январь 2004 г.)
--->
<110> CJ CHEILJEDANG CORPORATION
<120> НОВАЯ ПСИКОЗО-6-ФОСФАТ ФОСФАТАЗА, КОМПОЗИЦИЯ ДЛЯ ПОЛУЧЕНИЯ ПСИКОЗЫ,
СОДЕРЖАЩАЯ УКАЗАННЫЙ ФЕРМЕНТ, СПОСОБ ПОЛУЧЕНИЯ ПСИКОЗЫ С ИСПОЛЬЗОВАНИЕМ
УКАЗАННОГО ФЕРМЕНТА
<130> P18-6064
<150> KR 2017/0097334
<151> 2017-07-31
<160> 78
<170> KoPatentIn 3.0
<210> 1
<211> 272
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности RMA
<400> 1
Met Met Asp Ala Tyr Thr Leu Tyr Glu Glu Ala Arg Glu Val Ala Ala
1 5 10 15
Arg Leu Ala Arg Asp Ala Gly Gln Ile Ala Arg Tyr Tyr Ala Gly Arg
20 25 30
Val Thr Val Arg Glu Lys Gly Tyr Asn Glu Leu Val Thr Gln Ala Asp
35 40 45
Glu Glu Val Gln Arg Phe Leu Ile Glu Gln Ile His Arg His Phe Pro
50 55 60
Glu His Ala Ile Leu Ala Glu Glu Asn Leu Ser Asp Met Gln Asp Gly
65 70 75 80
Arg Glu Gly Ala Ser Phe Arg Trp Ile Ile Asp Pro Ile Asp Gly Thr
85 90 95
Thr Asn Phe Thr His Gly Val Pro Pro Tyr Gly Ile Ser Leu Ala Leu
100 105 110
Gln His Glu Gly Arg Thr Val Val Gly Val Val Tyr Asp Val Pro His
115 120 125
Asp Glu Leu Phe Thr Ala Val Arg Gly Gly Gly Leu Tyr Val Asn Gly
130 135 140
Val Arg Ala Arg Val Ser Gln Thr Glu Thr Leu Arg Glu Ala Leu Ile
145 150 155 160
Thr Thr Gly Phe Pro Tyr Arg Glu Val Val His Leu Glu Glu Tyr Leu
165 170 175
Glu Ala Leu Gly Arg Val Ile Arg Ala Thr Arg Gly Val Arg Arg Pro
180 185 190
Gly Ala Ala Ser Val Asp Leu Ala Trp Val Ala Cys Gly Arg Phe Asp
195 200 205
Gly Phe Phe Glu Thr Gly Leu Ser Pro Trp Asp Val Ala Ala Gly Ile
210 215 220
Leu Leu Val Glu Glu Gly Gly Gly Arg Val Thr Asp Phe His Gly Arg
225 230 235 240
Pro Asp Pro Ile Phe Ala Arg Gln Met Leu Ala Thr Asn Gly Arg Ile
245 250 255
His Glu Ala Leu Cys Glu Leu Val Ala Pro Leu His His Val Tyr Ala
260 265 270
<210> 2
<211> 255
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TLE
<400> 2
Met Asp Arg Met Asp Phe Ser Ile Lys Ile Ala Arg Lys Val Gly Leu
1 5 10 15
Tyr Leu Met Glu His Trp Gly Asn Ala Glu Asn Val Arg Gln Lys Ser
20 25 30
Ser Phe Gln Asp Leu Val Ser Asp Cys Asp Lys Gln Ala Gln Lys Met
35 40 45
Ile Val Gln Lys Ile Lys Asp His Phe Pro Asp Asp Ala Ile Leu Ala
50 55 60
Glu Glu Gly Leu Phe Glu Lys Gly Asp Arg Met Trp Ile Ile Asp Pro
65 70 75 80
Ile Asp Gly Thr Met Asn Tyr Val His Gly Leu Pro Ser Phe Ala Ile
85 90 95
Gly Ile Ala Tyr Val Glu Lys Glu Gln Val Ile Leu Gly Val Ala His
100 105 110
Asp Pro Val Leu Asn Glu Thr Tyr Tyr Ala Ile Lys Gly Gln Gly Ala
115 120 125
Tyr Lys Asn Gly Glu Arg Ile Asn Val Ser Glu Asn Ser Leu Leu Lys
130 135 140
Asp Ser Ile Gly Asn Thr Gly Phe Tyr Thr Asp Phe Thr Gly Ile Phe
145 150 155 160
Ile Ser Ala Ile Glu Lys Lys Val Arg Arg Val Arg Met Thr Gly Ser
165 170 175
Ala Ile Leu Ala Gly Ala Tyr Val Ala Cys Gly Arg Phe Asp Phe Phe
180 185 190
Ile Ala Lys Arg Ala Asn Ser Trp Asp Val Ala Pro Leu Phe Val Leu
195 200 205
Val Pro Glu Ala Gly Gly Ile Val Thr Asp Leu Ser Gly Asn Gln Ala
210 215 220
His Leu Asn Thr Gly Asn Phe Leu Phe Ser Asn Gly Leu Leu His Asp
225 230 235 240
Gln Val Leu Glu Val Ile Arg Glu Val Asn Lys Lys Val Arg Lys
245 250 255
<210> 3
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MRUB
<400> 3
Met Asp Leu Arg Ala Tyr Leu Gln Thr Ala Leu Asp Ala Ala Tyr Leu
1 5 10 15
Ala Lys Gly Ile His Gln Tyr Tyr Gln Glu Lys Gly Phe Thr Gln Ser
20 25 30
Thr Lys Ser Thr Pro Thr Asp Leu Val Thr Gln Ala Asp His Glu Ser
35 40 45
Glu Ala Ala Ile Arg Glu Leu Ile Ala Ser Arg His Pro Asp His Val
50 55 60
Val Leu Gly Glu Glu Gln Gly Gln Asp Lys Glu Gly Ala Phe Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Ala Val Ser Ile Gly Leu Glu Ala His Gly Glu Val Val Leu
100 105 110
Gly Val Val Leu Asp Thr Ala Arg Gly Glu Leu Phe Thr Ala Thr Lys
115 120 125
Gly Gly Gly Ala Tyr Leu Asn Gly Arg Pro Ile Arg Val Ser Thr Arg
130 135 140
Ser Thr Leu Val Gly Ser Leu Leu Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ser Lys Asp Thr Glu Asn Leu Val Tyr Phe Gln Arg Ala Leu Thr Lys
165 170 175
Gly Leu Met Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Tyr
180 185 190
Val Ala Ala Gly Arg Leu Asp Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Ile Val Ser Glu Ala Gly Gly Arg
210 215 220
Ile Thr Gly Leu Gln Gly Glu Asp Tyr Arg Leu Gly Asn Arg Tyr Leu
225 230 235 240
Val Ala Ser Asn Gly Leu Ile His Gly Pro Leu Leu Asp Thr Ile His
245 250 255
Gly Arg
<210> 4
<211> 265
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности DTU
<400> 4
Met Lys Arg Ile Leu Glu Val Ala Ile Lys Thr Ile Lys Glu Ser Gly
1 5 10 15
Asn Ile Leu Leu Asn Tyr Ile Gly Glu Glu Lys Glu Ile Glu Leu Lys
20 25 30
Gly Ile Ser Asn Leu Val Thr Gln Val Asp Lys Leu Ser Glu Arg His
35 40 45
Ile Leu Lys Ser Ile Glu Glu Asn Phe Pro Asp His Ser Ile Leu Thr
50 55 60
Glu Glu Thr Gly Phe Ile Asn Lys Asn Ser Glu Tyr Thr Trp Ile Val
65 70 75 80
Asp Pro Leu Asp Gly Thr Thr Asn Tyr Ala His Asn Phe Pro Phe Phe
85 90 95
Gly Ile Ser Ile Ala Leu Ile Lys Asn Lys Glu Ile Ile Leu Gly Leu
100 105 110
Ile Tyr Asp Pro Ile Arg Asp Glu Leu Phe Tyr Ala Ile Lys Asn Glu
115 120 125
Gly Ala Tyr Leu Asn Asp Arg Arg Ile Glu Val Ser Lys Thr Glu Ser
130 135 140
Leu Glu Asn Ser Leu Ile Ser Phe Ala Phe Pro Tyr Glu Leu Ser Leu
145 150 155 160
Glu Glu Lys Asn Phe Ile Pro Phe Ile Asn Phe Ser Ser Arg Thr His
165 170 175
Gly Ile Arg Arg Thr Gly Ser Ala Ala Ile Glu Ile Ala Tyr Val Gly
180 185 190
Cys Gly Arg Leu Asp Gly Phe Trp Ala Lys Lys Leu Lys Pro Trp Asp
195 200 205
Ile Ser Ala Gly Ile Leu Ile Val Glu Glu Ala Lys Gly Lys Val Thr
210 215 220
Asp Phe Ser Gly Asn Asn Ile Asp Ile His Thr Asp Asn Ile Leu Phe
225 230 235 240
Ser Asn Gly Lys Ile His Gln Glu Met Ile Lys Ile Leu Asn Leu Gly
245 250 255
Lys Ile Phe Ile Arg Asn Glu Lys Phe
260 265
<210> 5
<211> 251
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности PFE
<400> 5
Met Leu Gly Val Leu Glu Ala Val Ala Val Arg Ala Ser His Phe Leu
1 5 10 15
Met Glu Tyr Phe Arg Ala Gly Arg Gly Val Asp Val Val Ser Arg Lys
20 25 30
Glu Asp Asp Val Thr Arg Glu Val Asp Ile Ala Ala Glu Thr Leu Ile
35 40 45
Tyr Lys Met Leu Arg Glu Ala Phe Lys Glu Gly Gly Val Leu Tyr Ala
50 55 60
Glu Glu Gly Gly Ile Tyr Arg Trp Gly Asp Glu Arg Tyr Ile Phe Val
65 70 75 80
Leu Asp Pro Leu Asp Gly Ser Leu Asn Tyr Ala Val Gly Val Pro Phe
85 90 95
Phe Ala Val Ser Ile Ala Ala Gly Lys His Arg Glu Gly Thr Leu Ala
100 105 110
Asp Leu Glu Tyr Ala Val Val Ala Ile Pro Pro Thr Gly Asp Val Tyr
115 120 125
Thr Ala Ala Pro Gly Val Gly Ala Arg Lys Asn Gly Lys Pro Leu Arg
130 135 140
Arg Thr Pro Arg Ser Asn Ile Val Phe Val Ala Val Ser Asn Ser Phe
145 150 155 160
Pro Pro Lys Thr Cys Glu Val Val Arg Arg Leu Gly Leu Arg Gly Arg
165 170 175
Ser Leu Gly Ser Ser Ala Ala Glu Leu Ala Tyr Thr Val Glu Gly Ile
180 185 190
Ala Arg Gly Phe Leu Asp Leu Arg Gly Lys Leu Arg Leu Leu Asp Val
195 200 205
Ala Gly Ala Leu Thr Ile Gly Lys Tyr Val Asp Gly Phe Arg Tyr Val
210 215 220
Val Met Gly Asp Thr Lys Pro His Ser Lys Val Ser Leu Val Ala Gly
225 230 235 240
Asp Val Asp Phe Val Asn Ala Ala Thr Thr Asp
245 250
<210> 6
<211> 257
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TWI
<400> 6
Met Lys Asp Glu Lys Gly Ile Val Val Asp Ile Ile Lys Lys Ala Gly
1 5 10 15
Glu Ile Leu Gln Asp Gly Trp Asn Lys Lys Asn Phe Lys Ile Tyr Arg
20 25 30
Lys Gly Thr Ile Asn Leu Val Thr Glu Ile Asp Lys Lys Ile Glu Phe
35 40 45
Leu Ile Ile Gln Leu Leu Lys Gln Tyr Phe Pro Asp Tyr Gly Ile Leu
50 55 60
Thr Glu Glu Ser Lys Glu Ile Asn Ser Lys Ala Asn Val Arg Trp Ile
65 70 75 80
Ile Asp Pro Leu Asp Gly Thr Thr Asn Tyr Ile Lys Gln Tyr Pro Phe
85 90 95
Val Ala Ile Ser Ile Ala Leu Glu Lys Glu Gly Glu Leu Ile Leu Gly
100 105 110
Val Val Tyr Asn Pro Ile Leu Asn Glu Met Phe Ile Ala Gln Lys Gly
115 120 125
Cys Gly Ala Thr Tyr Asn Gly Lys Ser Ile His Val Ser Lys Ile Lys
130 135 140
Glu Leu Gly Ser Ala Val Leu Ala Ser Gly Phe Pro Tyr Asp Ala Trp
145 150 155 160
Glu Asn Pro Asp Asn Asn Ala Lys Gln Trp Arg Gln Phe Leu Thr Arg
165 170 175
Ser Leu Ser Leu Arg Cys Asp Gly Ser Ala Ala Leu Asp Leu Cys Arg
180 185 190
Val Ala Cys Gly Gln Leu Asp Gly Tyr Trp Glu Lys Gly Ile Ser Pro
195 200 205
Trp Asp Val Ala Ala Gly Ile Val Ile Leu Arg Glu Ala Gly Gly Ile
210 215 220
Ile Thr Asp Tyr Leu Gly Glu Glu Asn Phe Phe Lys Arg Gly Glu Val
225 230 235 240
Val Ala Ala Asn Pro Val Leu His Ala Gln Met Leu Lys Val Leu Asn
245 250 255
Asn
<210> 7
<211> 264
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TTH
<400> 7
Met Ile Gly Arg Arg His Pro Tyr Tyr Pro Tyr Leu Glu Ala Ala Leu
1 5 10 15
Glu Ala Ala Ser Leu Ala Arg Gly Ile His Leu Tyr Tyr Leu Glu Lys
20 25 30
Gly Phe Thr Glu Gly Thr Lys Ser Gly Pro Thr Asp Leu Val Thr Gln
35 40 45
Ala Asp Arg Glu Ala Glu Glu Ala Val Lys Gly Leu Leu Leu Ser Arg
50 55 60
Phe Pro Glu Ala Gly Phe Leu Gly Glu Glu Gly Gly Ser Glu Gly Gly
65 70 75 80
Lys Ala Leu Arg Phe Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr
85 90 95
Ala His Gly Phe Pro Phe Phe Ala Val Ser Ile Ala Leu Glu Ala Glu
100 105 110
Gly Ala Ile Gln Met Gly Val Val Met Asp Thr Ala Arg Gly Glu Val
115 120 125
Phe Tyr Ala Leu Arg Gly Glu Gly Ala Tyr Leu Asn Gly Arg Pro Ile
130 135 140
Arg Val Thr Gly Arg Glu Ser Leu Val Gly Ser Leu Leu Ala Thr Gly
145 150 155 160
Phe Pro Tyr Asp Val Ala Lys Asp Pro Glu Asn Leu Thr Tyr Phe Glu
165 170 175
Arg Ala Leu Gly Lys Gly Leu Leu Val Arg Arg Pro Gly Ala Ala Ala
180 185 190
Leu Asp Leu Ala Tyr Val Ala Ala Gly Arg Leu Glu Gly Phe Trp Glu
195 200 205
Val Lys Leu Asn Pro Trp Asp Val Ala Ala Gly Trp Leu Leu Val Glu
210 215 220
Glu Ala Gly Gly Arg Val Thr Asp Leu Glu Gly Asn Pro Tyr Arg Leu
225 230 235 240
Gly Ser Arg Tyr Ile Leu Ala Thr Asn Gly Arg Val His Glu Ala Leu
245 250 255
Arg Arg Thr Leu Leu Gly Leu Asp
260
<210> 8
<211> 251
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TLI
<400> 8
Met Tyr Glu Trp Asn Glu Ile Ala Leu Asn Leu Ala Lys Asp Ile Glu
1 5 10 15
Arg Glu Val Met Pro Leu Phe Gly Thr Lys Lys Ala Gly Glu Phe Ile
20 25 30
Gly Phe Ser Pro Ser Gly Asp Lys Thr Lys Leu Val Asp Lys Val Ala
35 40 45
Glu Asp Val Val Leu Glu Tyr Leu Arg Pro Leu Gly Val Asn Val Val
50 55 60
Ser Glu Glu Ile Gly Asn Ile Glu Ala Gly Ser Glu Tyr Thr Ile Val
65 70 75 80
Val Asp Pro Ile Asp Gly Ser Phe Asn Phe Ile Gln Gly Ile Pro Ile
85 90 95
Phe Gly Phe Ser Phe Ala Val Phe Lys Asn Glu Lys Pro Val Tyr Ala
100 105 110
Met Ile Tyr Glu Phe Ile Thr Lys Asn Val Tyr Glu Gly Ile Pro Gly
115 120 125
Glu Gly Ala Tyr Leu Asn Gly Glu Arg Ile Arg Val Arg His Leu Asn
130 135 140
Glu Lys Ser Ile Ser Ile Ser Phe Tyr Thr Arg Gly Arg Gly Ala Arg
145 150 155 160
Leu Val Glu Lys Val Lys Arg Thr Arg Val Leu Gly Ala Ile Ala Val
165 170 175
Glu Leu Ala Tyr Leu Ala Arg Gly Ser Leu Asp Gly Val Ile Asp Ile
180 185 190
Arg Asn Tyr Val Arg Pro Thr Asp Ile Ala Ala Gly Tyr Ile Ile Ala
195 200 205
Lys Glu Ala Gly Ala Ile Ile Thr Asp Asp Ser Gly Glu Glu Ile Lys
210 215 220
Phe Arg Leu Asp Ala Arg Glu Lys Leu Asn Ile Ile Ala Val Asn Asp
225 230 235 240
Lys Arg Leu Leu Lys Leu Ile Leu Glu Val Ile
245 250
<210> 9
<211> 264
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности GST
<400> 9
Met Ala Glu Lys Trp Glu Glu Ile Asp Arg Tyr Ala Arg Gln Trp Ile
1 5 10 15
Asp Glu Ala Gly Lys Arg Ile Arg Ala Ser Phe Ala Lys Gln Leu Thr
20 25 30
Val Glu Ala Lys Glu Asn Pro Asn Asp Leu Val Thr Asn Val Asp Arg
35 40 45
Ala Ile Glu Gln Phe Phe Ala Glu His Ile Arg Arg Gln Phe Pro Ser
50 55 60
His Arg Leu Leu Gly Glu Glu Gly Phe Gly Asp Arg Ile Asp Ala Leu
65 70 75 80
Asp Gly Val Val Trp Val Ile Asp Pro Ile Asp Gly Thr Met Asn Phe
85 90 95
Val His Gln Arg Arg His Phe Ala Val Ser Ile Gly Ile Phe Glu Asp
100 105 110
Gly Ile Gly Gln Leu Gly Tyr Val Tyr Asp Val Val Phe Asp Glu Leu
115 120 125
Tyr Ala Ala Gln Lys Gly Arg Gly Val Phe Leu Asn Gly Glu Pro Leu
130 135 140
Gly Leu Leu Gln Pro Ala Pro Val Ala Glu Ser Ile Ile Ala Ile Asn
145 150 155 160
Gly Thr Trp Leu Met Glu Asn Lys Arg Leu Asp His Arg Pro Leu Met
165 170 175
Arg Leu Ala Lys Glu Ala Arg Gly Thr Arg Ser Tyr Gly Ser Ala Ala
180 185 190
Leu Glu Leu Ala Tyr Val Ala Ala Gly Arg Leu Asp Ala Tyr Ile Ser
195 200 205
Pro Arg Leu Ser Pro Trp Asp Phe Ala Gly Gly Met Ile Leu Ile Glu
210 215 220
Glu Ala Gly Gly Met Val Thr Thr Leu Asp Gly Lys Pro Leu Asp Leu
225 230 235 240
Leu Gly Arg Asn Ser Val Leu Ala Ala Lys Pro Gly Val His Glu Glu
245 250 255
Ile Leu Arg Arg Tyr Leu His Asp
260
<210> 10
<211> 259
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности ATH
<400> 10
Met Thr Thr Ser Leu Arg Asp Leu Leu Asp Phe Ala Val Glu Thr Ala
1 5 10 15
Tyr Leu Ala Gly Arg Thr Thr Leu Ala Tyr Phe Gln Thr Gly Val Gln
20 25 30
Ala Glu Phe Lys Ala Asp Ser Ser Pro Val Thr Leu Ala Asp Arg Ala
35 40 45
Ala Glu Gln Leu Ile Arg Gln Arg Ile Glu Lys Arg Phe Pro His His
50 55 60
Ala Ile Val Gly Glu Glu Phe Gly Val Gln Gly Ser Ala Asp Ala Thr
65 70 75 80
His Arg Trp Phe Ile Asp Pro Ile Asp Gly Thr Lys Ser Phe Leu Arg
85 90 95
Gly Ile Pro Leu Tyr Ala Val Leu Leu Gly Leu Glu Ile Glu Gly Arg
100 105 110
Val Gln Val Gly Val Ala Tyr Tyr Pro Ala Met Asp Glu Met Leu Ser
115 120 125
Ala Ala Asp Gly Glu Gly Cys Trp Trp Asn Gly Arg Arg Ala Arg Val
130 135 140
Ser Thr Ala Ser Arg Leu Ala Glu Ala Trp Val Thr Ser Thr Asp Pro
145 150 155 160
Tyr Asn Phe Gln Lys Thr Gly Lys Asp Ala Ala Trp Gln Arg Ile Gln
165 170 175
Ala Val Ser Tyr His Arg Gly Gly Trp Gly Asp Ala Tyr Gly Tyr Leu
180 185 190
Leu Val Ala Thr Gly Arg Ala Glu Val Met Leu Asp Pro Ile Met Asn
195 200 205
Glu Trp Asp Cys Ala Pro Phe Pro Pro Ile Phe Arg Glu Ala Gly Gly
210 215 220
Phe Phe Gly Asp Trp Gln Gly Asn Glu Thr Ile Tyr Gly Gly Glu Ala
225 230 235 240
Leu Ala Thr Thr Gln Val Leu Leu Pro Glu Val Leu Glu Cys Leu His
245 250 255
Ser Ser Leu
<210> 11
<211> 267
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности SAC
<400> 11
Met Lys Arg Glu Asp Val Glu Lys Val Ala Ser Glu Ala Ser Lys Tyr
1 5 10 15
Ile Tyr Glu Glu Arg Glu Asn Lys Asp Val Asp Arg Val Ile Asn Val
20 25 30
His Gly Asn Asp Val Thr Arg Ile Ile Asp Lys Arg Ser Glu Asp Phe
35 40 45
Ile Val Asp Arg Leu Lys Ser Leu Gly Tyr Asn Ile Leu Ile Val Thr
50 55 60
Glu Glu Ser Gly Val Ile Asp Ser Tyr Gly Lys Asn Tyr Asp Tyr Ile
65 70 75 80
Ala Ile Val Asp Pro Leu Asp Gly Ser Thr Asn Phe Val Ser Gly Ile
85 90 95
Pro Trp Ser Ser Val Ser Ile Ala Ile Tyr Asn Arg Asp Glu Glu Asp
100 105 110
Ile Leu Ser Ser Asn Val Gly Ala Val Ser Ser Ile Phe Thr Pro Tyr
115 120 125
Thr Phe Ser Tyr Asp Glu Gly Ser Ala Tyr Val Asn Gly Val Lys Ile
130 135 140
Ala Glu Ile Lys Lys Pro Glu Lys Ile Leu Leu Leu Ala Tyr Phe Ser
145 150 155 160
Arg Ser Lys Leu Pro Asn Leu Lys Leu Phe Phe Glu Lys Ile Gly Gln
165 170 175
Gly Tyr Lys Ile Arg Ser Leu Gly Ser Ala Ser Leu Asp Met Ile Leu
180 185 190
Val Cys Thr Gly Arg Ala Thr Met Phe Phe Asp Ile Arg Gly Lys Leu
195 200 205
Arg Asn Val Asp Ile Ala Ala Ser Ser Asn Phe Cys Ser Arg Leu Gly
210 215 220
Val Ile Pro Tyr Asp Ile Gly Leu Arg Lys Ile Lys Ser Ser Leu Thr
225 230 235 240
Glu Val Ser Val Val Lys Asp Leu Val Ile Ser Leu Asp Glu Ser Leu
245 250 255
Leu Arg Ser Phe Ser Leu Ala Leu Gln Thr Val
260 265
<210> 12
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TTA
<400> 12
Met Leu Glu Leu Leu Lys Asp Thr Ala Lys Lys Ala Gly Arg Leu Leu
1 5 10 15
Lys Ile Arg Phe Leu Glu Leu Leu Asn Gly Met Glu Thr Glu Val Arg
20 25 30
Glu Lys Gly Lys Ser Asp Phe Val Thr Arg Val Asp Val Glu Val Glu
35 40 45
Asn Tyr Ile Lys Glu Leu Leu Ala Asn Thr Asn Ile Ser Val Val Gly
50 55 60
Glu Glu Ser Phe Lys Gly Glu Ile Pro Gly Thr Cys Ile Phe Ile Asp
65 70 75 80
Pro Ile Asp Gly Thr Arg Asn Phe Met Arg Lys Asn Pro His Phe Ala
85 90 95
Ile Asn Leu Ala Tyr Gln Glu Lys Gly Lys Leu Leu Ala Gly Val Thr
100 105 110
Tyr Asp Pro Met Lys Asn Glu Met Phe Ser Ala Ala Phe Ser Lys Gly
115 120 125
Ala Phe Leu Asn Gly Glu Arg Ile Tyr Ala Ser Thr Asn Lys Asp Ile
130 135 140
Gly Lys Ala Ile Ile Ala Ile Gly Leu Pro Tyr Arg Gly Arg Glu Leu
145 150 155 160
Ile Asp Ile Gln Thr Asn Leu Tyr Arg Asn Ile Phe Leu Asn Gly Ala
165 170 175
Ala Thr Arg His Thr Gly Ser Ala Ala Leu Asp Leu Ala Tyr Ile Ser
180 185 190
Cys Gly Arg Tyr Asp Ala Ala Ile Tyr Phe Tyr Leu Ser Pro Trp Asp
195 200 205
Val Ala Pro Gly Ile Leu Leu Val Glu Glu Ala Gly Gly Glu Val Glu
210 215 220
Gly Thr Met Gly Arg Glu Pro Ile Gln Gly Trp Ile Ile Ala Ser Asn
225 230 235 240
Lys Val Ile His Pro Glu Val Lys Asp Ile Leu Glu Gly Ser Leu Lys
245 250 255
Ala Val
<210> 13
<211> 254
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности PFU
<400> 13
Met Lys Leu Lys Phe Trp Arg Glu Val Ala Ile Asp Ile Ile Ser Asp
1 5 10 15
Phe Glu Thr Thr Ile Met Pro Phe Phe Gly Asn Pro Asp Gly Gly Lys
20 25 30
Leu Val Lys Ile Ser Pro Ser Gly Asp Glu Thr Lys Leu Val Asp Lys
35 40 45
Leu Ala Glu Asp Leu Ile Leu Ser Arg Ile Thr Glu Leu Gly Val Asn
50 55 60
Val Val Ser Glu Glu Val Gly Val Ile Asp Asn Glu Ser Glu Tyr Thr
65 70 75 80
Val Ile Val Asp Pro Leu Asp Gly Ser Tyr Asn Phe Ile Ala Gly Ile
85 90 95
Pro Phe Phe Ala Leu Ser Leu Ala Val Phe Lys Lys Asp Lys Pro Ile
100 105 110
Tyr Ala Ile Ile Tyr Glu Pro Met Thr Glu Arg Phe Phe Glu Gly Ile
115 120 125
Pro Gly Glu Gly Ala Phe Leu Asn Gly Lys Arg Ile Lys Val Arg Lys
130 135 140
Thr Pro Asp Glu Lys Pro Ser Ile Ser Phe Tyr Ser Arg Gly Lys Gly
145 150 155 160
His Glu Ile Val Lys His Val Lys Arg Thr Arg Thr Leu Gly Ala Ile
165 170 175
Ala Leu Glu Leu Ala Tyr Leu Ala Met Gly Ala Leu Asp Gly Val Val
180 185 190
Asp Val Arg Lys Tyr Val Arg Pro Thr Asp Ile Ala Ala Gly Thr Ile
195 200 205
Ile Ala Lys Glu Ala Gly Ala Leu Ile Lys Asp Ser Ala Gly Lys Asp
210 215 220
Ile Asp Ile Ser Phe Asn Ala Thr Asp Arg Leu Asp Val Ile Ala Val
225 230 235 240
Asn Ser Glu Glu Leu Leu Lys Thr Ile Leu Ser Leu Leu Glu
245 250
<210> 14
<211> 252
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности AFU
<400> 14
Met Asp Glu Arg Asp Ala Leu Arg Ile Ser Arg Glu Ile Ala Gly Glu
1 5 10 15
Val Arg Lys Ala Ile Ala Ser Met Pro Leu Arg Glu Arg Val Lys Asp
20 25 30
Val Gly Met Gly Lys Asp Gly Thr Pro Thr Lys Ala Ala Asp Arg Val
35 40 45
Ala Glu Asp Ala Ala Leu Glu Ile Leu Arg Lys Glu Arg Val Thr Val
50 55 60
Val Thr Glu Glu Ser Gly Val Leu Gly Glu Gly Asp Val Phe Val Ala
65 70 75 80
Leu Asp Pro Leu Asp Gly Thr Phe Asn Ala Thr Arg Gly Ile Pro Val
85 90 95
Tyr Ser Val Ser Leu Cys Phe Ser Tyr Ser Asp Lys Leu Lys Asp Ala
100 105 110
Phe Phe Gly Tyr Val Tyr Asn Leu Ala Thr Gly Asp Glu Tyr Tyr Ala
115 120 125
Asp Ser Ser Gly Ala Tyr Arg Asn Gly Glu Arg Ile Glu Val Ser Asp
130 135 140
Ala Glu Glu Leu Tyr Cys Asn Ala Ile Ile Tyr Tyr Pro Asp Arg Lys
145 150 155 160
Phe Pro Phe Lys Arg Met Arg Ile Phe Gly Ser Ala Ala Thr Glu Leu
165 170 175
Cys Phe Phe Ala Asp Gly Ser Phe Asp Cys Phe Leu Asp Ile Arg Pro
180 185 190
Gly Lys Met Leu Arg Ile Tyr Asp Ala Ala Ala Gly Val Phe Ile Ala
195 200 205
Glu Lys Ala Gly Gly Lys Val Thr Glu Leu Asp Gly Glu Ser Leu Gly
210 215 220
Asn Lys Lys Phe Asp Met Gln Glu Arg Leu Asn Ile Val Ala Ala Asn
225 230 235 240
Glu Lys Leu His Pro Lys Leu Leu Glu Leu Ile Lys
245 250
<210> 15
<211> 265
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности AAC
<400> 15
Met Asp Lys Ala Trp Leu Asp Gly Leu Ala Glu Val Val Arg Glu Ala
1 5 10 15
Gly Arg Leu Val Glu Glu Ile Ala Arg Gln Gly Phe Asp Thr Gln Phe
20 25 30
Lys His Pro Glu Glu Arg Arg Asp Pro Val Thr Thr Ala Asp Leu Ala
35 40 45
Cys Asp Ala Phe Leu Lys Glu Arg Leu Leu Thr Leu Leu Pro Glu Ala
50 55 60
Gly Trp Leu Ser Glu Glu Thr Lys Asp Arg Pro Asp Arg Leu Glu Lys
65 70 75 80
Arg Trp Val Trp Ile Val Asp Pro Ile Asp Gly Thr Arg Glu Phe Val
85 90 95
Arg Arg Ile Pro Glu Tyr Ala Ile Ser Val Ala Leu Ala Arg Asp Gly
100 105 110
Glu Pro Val Ala Gly Ala Val Val Asn Pro Ala Thr Gly Asp Leu Phe
115 120 125
Leu Gly Ala Val Gly Val Gly Ala Trp Arg Asn Gly Thr Pro Met Val
130 135 140
Cys Ser Arg Ile Arg Gly Glu Arg Leu Thr Ile Leu Gly Ser Arg Ser
145 150 155 160
Glu Met Asn Arg Gly Glu Phe Glu Pro Phe Ala Gly Ile Leu Glu Val
165 170 175
Arg Ala Val Gly Ser Ile Ala Tyr Lys Leu Ala Leu Val Ala Ala Gly
180 185 190
Glu Ala Asp Gly Thr Phe Ser Leu Gly Pro Lys His Glu Trp Asp Ile
195 200 205
Ala Ala Gly Val Ala Leu Val Leu Ala Ala Gly Gly Arg Val His Asp
210 215 220
Gly Ala Gly Arg Pro Phe Arg Phe Asn Gln Pro His Thr Leu Thr Arg
225 230 235 240
Gly Ile Val Ala Ala Thr Arg Glu Ala Tyr Gly Asp Leu Ala Leu Leu
245 250 255
Ile Glu Arg His Ala Pro Arg Arg Ala
260 265
<210> 16
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MSI
<400> 16
Met Asp Leu Gln Arg Tyr Leu Glu Ala Ala Leu Asp Ala Ala Tyr Leu
1 5 10 15
Ala Arg Gly Ile His Leu Tyr Tyr Arg Glu Lys Gly Phe Ala Leu Glu
20 25 30
Ser Lys Thr Ser Pro Thr Asp Leu Val Thr Gln Ala Asp Arg Glu Ser
35 40 45
Glu Glu Ala Ile Arg Ser Leu Leu Leu Glu Arg Phe Pro Asp His Val
50 55 60
Val Leu Gly Glu Glu Gly Gly Gln Glu Gly His Ser Asp Phe Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Ala Val Ser Ile Ala Leu Glu Val Gly Gly Glu Val Val Val
100 105 110
Gly Ala Val Leu Asp Thr Ala Arg Gly Glu Leu Phe Thr Ala Leu Lys
115 120 125
Gly Glu Gly Ala Phe Gln Asn Gly Arg Pro Ile Arg Val Ser Ser Thr
130 135 140
Ala Thr Leu Ile Gly Ser Leu Leu Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ala Lys Asp Ala Glu Asn Leu Thr Tyr Phe Gln Arg Ala Leu Ala Lys
165 170 175
Gly Leu Thr Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Tyr
180 185 190
Val Ala Ala Gly Arg Leu Glu Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Leu Ile Thr Glu Ala Gly Gly Thr
210 215 220
Val Ser Gly Ile Gln Gly Glu Pro Tyr Arg Leu Gly Asn Arg Tyr Leu
225 230 235 240
Val Ala Ser Asn Gly Lys Ile His Glu Gln Leu Leu Asp Thr Leu His
245 250 255
Gly Arg
<210> 17
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MRUF
<400> 17
Met Asp Leu Arg Pro Tyr Leu Glu Ala Ala Leu Asp Ala Ala Tyr Leu
1 5 10 15
Ala Lys Gly Ile His Gln Tyr Tyr Gln Glu Lys Gly Phe Thr His Ser
20 25 30
Thr Lys Ser Thr Pro Thr Asp Val Val Thr Gln Ala Asp His Glu Ala
35 40 45
Glu Ala Ala Ile Arg Ala Leu Ile Ala Glu Arg Phe Pro Asp His Val
50 55 60
Val Leu Gly Glu Glu Gln Gly Gln Glu Gly Glu Gly Glu Phe Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Gly Val Ser Ile Gly Leu Glu Val Arg Gly Glu Val Val Leu
100 105 110
Gly Val Val Leu Asp Thr Ala Arg Gly Asp Leu Phe Thr Ala Thr Lys
115 120 125
Gly Gly Gly Ala Tyr Leu Asn Gly Arg Pro Leu Arg Val Ser Gln Arg
130 135 140
Thr Thr Leu Val Gly Ser Leu Leu Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ala Arg Asp Pro Glu Asn Leu Thr Tyr Phe Gln Arg Ala Leu Ala Lys
165 170 175
Gly Leu Thr Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Asn
180 185 190
Val Ala Ala Gly Arg Leu Asp Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Leu Ile Thr Glu Ala Gly Gly Gln
210 215 220
Val Thr Gly Phe Gln Gly Glu Ala Tyr Arg Leu Gly Asn Arg Tyr Leu
225 230 235 240
Val Ala Ser Asn Gly Arg Ile His Gln Ala Leu Leu Glu Ala Leu Gln
245 250 255
Gly Arg
<210> 18
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MTA
<400> 18
Met Asp Leu Arg Ala Tyr Leu Gln Thr Ala Leu Asp Ala Ala Tyr Leu
1 5 10 15
Ala Lys Gly Ile His Gln Tyr Tyr Gln Glu Lys Gly Phe Thr Gln Ser
20 25 30
Ser Lys Ser Thr Pro Thr Asp Leu Val Thr Gln Ala Asp His Glu Ser
35 40 45
Glu Ala Ala Ile Arg Glu Leu Ile Ala Ala Arg His Pro Asp His Val
50 55 60
Val Leu Gly Glu Glu Gln Gly Gln Asp Lys Glu Gly Ala Phe Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Ala Val Ser Ile Gly Leu Glu Ala His Gly Glu Val Val Leu
100 105 110
Gly Val Val Leu Asp Thr Ala Arg Gly Glu Leu Phe Thr Ala Thr Lys
115 120 125
Gly Gly Gly Ala Tyr Leu Asn Gly Arg Pro Ile Arg Val Ser Thr Arg
130 135 140
Ser Thr Leu Val Gly Ser Leu Leu Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ser Lys Asp Thr Glu Asn Leu Thr Tyr Phe Gln Arg Ala Leu Thr Lys
165 170 175
Gly Leu Met Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Tyr
180 185 190
Val Ala Ala Gly Arg Leu Asp Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Ile Val Ser Glu Ala Gly Gly Arg
210 215 220
Val Ser Gly Leu Gln Gly Glu Asp Tyr Arg Leu Gly Asn Arg Tyr Leu
225 230 235 240
Val Ala Ser Asn Gly Leu Ile His Gln Pro Leu Leu Asp Thr Leu His
245 250 255
Gly Arg
<210> 19
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MCH
<400> 19
Met Asp Leu Arg Pro Phe Leu Glu Thr Ala Leu Ala Ala Ala Tyr Val
1 5 10 15
Ala Arg Gly Ile His Gln Leu Tyr Gln Asp Lys Gly Phe Thr Gln Ser
20 25 30
Thr Lys Ser Thr Pro Thr Asp Val Val Thr Gln Ala Asp Lys Glu Ala
35 40 45
Glu Ala Ala Ile Arg Ala Leu Ile Glu Gln Arg His Pro Gly His Val
50 55 60
Val Leu Gly Glu Glu Glu Gly Gln Gln Gly Glu Gly Glu Tyr Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Cys Val Ser Ile Gly Leu Glu Val Arg Gly Glu Val Met Val
100 105 110
Gly Val Val Leu Asp Thr Ala Arg Gly Asp Leu Phe Thr Ala Thr Arg
115 120 125
Gly Gly Gly Ala Tyr Phe Asn Gly Arg Pro Met Arg Val Ser Arg Ser
130 135 140
Pro Lys Leu Leu Gly Ser Leu Ile Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ala Arg Asp Arg Glu Asn Leu Thr Tyr Leu Glu Arg Val Leu Phe Lys
165 170 175
Gly Ile Thr Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Tyr
180 185 190
Val Ala Cys Gly Arg Leu Asp Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Leu Val Glu Glu Ala Gly Gly Arg
210 215 220
Ile Thr Gly Ile His Gly Glu Pro Tyr Arg Met Gly Asn Arg Tyr Leu
225 230 235 240
Met Ala Ser Asn Gly His Ile His Glu Glu Leu Leu Ala Thr Ile His
245 250 255
Gly Arg
<210> 20
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности MCE
<400> 20
Met Asp Leu Arg Val Tyr Leu Gln Thr Ala Leu Asp Ala Ala Tyr Leu
1 5 10 15
Ala Lys Gly Ile His Gln Tyr Tyr Gln Glu Lys Gly Phe Thr Leu Ser
20 25 30
Ser Lys Ser Thr Pro Thr Asp Leu Val Thr Gln Ala Asp His Glu Ser
35 40 45
Glu Ala Ala Ile Arg Glu Leu Ile Ala Ala Arg His Pro Asp His Val
50 55 60
Val Leu Gly Glu Glu Gln Gly Gln Asp Arg Glu Gly Ala Phe Arg Trp
65 70 75 80
Ile Val Asp Pro Leu Asp Gly Thr Val Asn Tyr Ala His Gly Phe Pro
85 90 95
Phe Tyr Ala Val Ser Ile Gly Leu Glu Ala His Gly Glu Val Val Leu
100 105 110
Gly Val Ile Leu Asp Thr Ala Arg Gly Asp Leu Phe Thr Ala Thr Lys
115 120 125
Gly Gly Gly Ala Phe Met Asn Gly Arg Pro Ile Arg Val Ser Pro Arg
130 135 140
Ala Thr Leu Val Gly Ser Leu Leu Ala Thr Gly Phe Pro Tyr Asp Val
145 150 155 160
Ser Lys Asp Thr Glu Asn Leu Val Tyr Phe Gln Arg Ala Leu Thr Arg
165 170 175
Gly Leu Met Val Arg Arg Pro Gly Ala Ala Ala Leu Asp Leu Ala Tyr
180 185 190
Val Ala Ala Gly Arg Leu Asp Gly Phe Trp Glu Val Lys Leu Asn Pro
195 200 205
Trp Asp Val Ala Ala Gly Trp Leu Ile Ile Arg Glu Ala Gly Gly Thr
210 215 220
Val Thr Gly Met Glu Gly Glu Ala Tyr Arg Leu Gly Asn Arg Tyr Leu
225 230 235 240
Val Ala Ser Asn Gly Leu Ile His Gln Pro Leu Leu Asp Thr Ile His
245 250 255
Gly Arg
<210> 21
<211> 819
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности RMA
<400> 21
atgatggatg cctacacgct ttacgaagaa gcccgtgagg tggcggcccg gctggcccgc 60
gatgccggac agatcgcccg ctactatgcc ggtcgggtga ccgtccgcga aaaaggctat 120
aacgagctgg tcacgcaggc cgacgaagaa gtacagcgct tcctgatcga gcagatccac 180
cggcatttcc ccgagcacgc gattctggcc gaggagaacc tgtccgacat gcaggacggc 240
agggaagggg cgtcgtttcg atggatcatc gatccgatcg acggcacgac gaacttcacg 300
cacggtgtgc cgccctatgg catcagtctg gcactccagc atgaaggccg gacggtcgtg 360
ggggtcgtct acgacgtgcc ccacgacgag ctgttcaccg cagtgcgggg cggcgggctg 420
tacgtcaacg gggtgcgcgc ccgggttagc cagaccgaaa cgctccggga ggcgctcatc 480
acgaccggct tcccctaccg ggaagtcgtg catctggaag aatatctgga ggcgctcggg 540
cgtgtgattc gagcgacgcg aggggtgcgt cggccgggcg cggcttcggt cgatctggcc 600
tgggtggcct gcggacgctt cgacggattc ttcgagacgg gcctgagtcc ctgggatgta 660
gcggccggca tcctgctggt cgaagaaggc ggaggacggg tgacggactt tcacgggcgg 720
cccgatccga tctttgcccg ccagatgctg gccacgaacg ggcgcatcca cgaggcgctc 780
tgcgagctgg tcgcgccgct gcaccacgtc tacgcctga 819
<210> 22
<211> 768
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TLE
<400> 22
atggacagaa tggatttttc aataaagatc gccagaaaag tcggtctcta cctgatggaa 60
cactggggca atgcagaaaa tgttcggcaa aaaagctctt ttcaggacct ggtaagtgac 120
tgcgacaagc aagcacagaa aatgattgtt cagaagataa aagaccattt tccagatgac 180
gctattttag ccgaggaagg gctattcgaa aaaggagaca gaatgtggat aattgaccca 240
atagatggca caatgaacta cgtccatggt ctaccttctt ttgcgattgg aatcgcctac 300
gttgaaaaag agcaagttat ccttggcgta gcacacgatc cagttctcaa cgagacatac 360
tatgcaatca aagggcaggg tgcgtataaa aatggggaaa gaataaatgt ttcagaaaat 420
tcgcttttga aagactctat tggaaatact ggcttttata cagattttac agggatcttc 480
atcagcgcaa tcgaaaagaa agtcaggcgg gtgagaatga caggcagtgc tatacttgct 540
ggcgcttatg ttgcctgtgg aaggtttgat ttctttatag caaaaagagc caattcttgg 600
gatgtagctc ctttattcgt cctcgtccca gaagctggtg gaatcgtaac ggacctgtcc 660
ggaaaccagg cacatctcaa caccggtaat tttcttttca gtaacggtct attacacgat 720
caggtcttag aagtcataag agaggtgaat aaaaaagtaa gaaaatga 768
<210> 23
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MRUB
<400> 23
atggacttgc gtgcctatct gcaaaccgcc ctcgacgccg cctatttagc caaaggcatc 60
caccagtact accaggaaaa aggctttacc cagagcacca agtccacccc caccgatcta 120
gtcacccagg ccgaccacga gtcggaggcg gccatccgag aactcattgc ttcgcgccac 180
cccgaccacg tggtgctggg cgaggaacaa ggccaggaca aagagggggc cttccgctgg 240
attgtagacc ccctggacgg caccgtaaac tacgcccatg gcttcccttt ttatgcggtg 300
agcatcggcc tggaagccca cggcgaggtg gtgctggggg tggttctgga taccgcccgg 360
ggcgagctat ttaccgctac caaggggggc ggggcctacc tgaatggtcg tcccattcgg 420
gtatccaccc gttcaaccct ggtgggcagc cttctagcca ccggattccc ctacgatgtg 480
agcaaggaca ccgaaaacct ggtctacttt cagcgggccc tgaccaaggg gctcatggtg 540
cggcgaccgg gcgcggcagc cctcgacctg gcttatgtgg ccgctgggcg cctggacggc 600
ttctgggaag tgaagctcaa cccctgggat gtggccgcgg gctggctcat cgttagcgag 660
gccggagggc ggataaccgg ccttcaaggg gaggattacc ggctgggcaa ccgctaccta 720
gtggcctcca acggcctgat ccacgggccg ctgctggaca ccatccacgg gcgatag 777
<210> 24
<211> 798
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности DTU
<400> 24
atgaagagaa tacttgaagt agctataaaa acaattaaag aatctggtaa tatacttttg 60
aactacatag gcgaggaaaa agaaatagaa cttaaaggaa tttccaattt agtcactcaa 120
gtagataaac tctcagaaag acatatacta aaatccatag aagaaaattt tccggatcat 180
tcaatcttga cagaagaaac aggctttata aataaaaatt ctgaatatac ctggattgta 240
gatcccttag atggaaccac aaattatgcc cataatttcc ctttttttgg aattagtatt 300
gccctcataa aaaataaaga aataatactt ggattaatct atgatcccat aagagacgag 360
ttattttatg ccataaaaaa tgaaggtgca tacctaaatg ataggagaat agaggtatca 420
aaaacagaaa gtcttgaaaa ttctctcatt agttttgcct ttccttacga attaagtctt 480
gaggagaaaa attttattcc ttttataaat ttttcctcta gaactcatgg tataagaagg 540
acaggttcgg cagcaataga aatagcctat gtagggtgtg gaagacttga tggattttgg 600
gcaaagaaac taaaaccatg ggatattagt gcaggcattc taatagtgga agaggcaaaa 660
ggtaaagtta cagattttag tggaaacaat attgatattc atacagacaa tattttgttt 720
tcaaacggta aaatacacca agagatgata aaaatcttaa atttaggtaa aatattcatt 780
agaaatgaaa aattttga 798
<210> 25
<211> 756
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности PFE
<400> 25
atgctcggcg tcttggaggc cgtggcggtc cgcgcctcgc attttttaat ggagtacttc 60
agagccggca gaggcgtaga cgtcgtgtcg cgtaaagagg acgacgtcac cagggaggtg 120
gacatcgccg cggagacgct catatacaaa atgctcagag aggcctttaa agaaggcggc 180
gtcctatacg cagaggaagg cggcatctac aggtggggag acgagcgcta catcttcgtc 240
ctagacccac tcgacgggtc gctgaactac gccgtgggcg tgcccttctt cgccgtatct 300
atcgcggcag ggaaacatag agagggcacc ctggccgatt tagaatacgc cgtcgtggcg 360
atacccccca cgggagacgt ctacacggcg gcccccggcg tcggcgcccg caaaaacgga 420
aagccgttga ggaggacccc gaggagcaat atagtattcg tagcggtgag caacagcttc 480
cccccaaaga cctgcgaagt cgtgaggcgg ctggggctca gggggaggag cctcggcagc 540
tccgccgccg agctggcgta cacggtggaa ggcatagccc gcggcttcct agacctccgg 600
ggcaagctca gactgctcga cgtggcgggg gcattgacga taggcaaata cgtagacggc 660
ttccggtacg tggtgatggg cgacaccaag ccgcactcca aggtgtctct agtggcgggc 720
gacgtagatt tcgtaaatgc cgcaaccaca gattga 756
<210> 26
<211> 774
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TWI
<400> 26
atgaaagatg agaaaggaat tgtagtagat attataaaaa aagcaggaga gattcttcaa 60
gatgggtgga ataaaaaaaa ttttaaaatt tatcggaaag gaacaattaa tttagttaca 120
gaaatagata agaaaattga atttcttata atacaacttt taaaacaata ttttcctgat 180
tatggtattc ttacagaaga aagcaaagaa attaacagca aagcaaacgt gcgatggatt 240
atagatcccc tagatggtac tactaattat ataaaacaat atccgtttgt agccatatca 300
attgctttag agaaagaagg agaattaatt ttgggagttg tttataatcc gattttaaat 360
gaaatgttta tagctcaaaa aggatgtggt gcgacttata atgggaagtc aattcatgtt 420
tctaaaataa aagaattggg atcggcagtt ctagcatcgg gctttcctta tgatgcgtgg 480
gaaaatcctg ataataatgc taaacaatgg aggcaatttc ttactcgtag tttatctctt 540
cgatgcgatg gatcagcagc tctcgatctt tgtagagtag cttgtggaca attggatggt 600
tattgggaaa agggaattag cccgtgggat gttgcagctg gtattgtaat attgcgcgaa 660
gcaggaggaa taattactga ttatttgggt gaggaaaatt tctttaaacg tggagaagtt 720
gttgctgcaa atcctgttct ccacgcacaa atgttgaaag ttttaaataa ttag 774
<210> 27
<211> 795
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TTH
<400> 27
atgatcggga ggcgccaccc ctactacccg tacctggagg cggccctgga ggcggcgagc 60
ctcgcccggg gcatccacct ctactacctg gaaaagggct tcaccgaggg gaccaagtcc 120
ggccccaccg acctggtgac ccaggcggac cgggaggcgg aagaggcggt gaagggcctc 180
ctcctctccc gcttccccga ggcggggttt ctgggggagg aagggggaag cgagggcggg 240
aaggccctcc gcttcatcgt ggacccgctg gacgggacgg tgaactacgc ccacggcttc 300
cccttcttcg ccgtatccat cgccctcgag gccgagggcg ccatccagat gggcgtggtg 360
atggacaccg cccgggggga ggtcttctac gccctgaggg gggaaggggc ctacctcaac 420
ggccgcccca tccgggtcac ggggcgggag agcctcgtgg ggagcctcct cgccacgggc 480
ttcccctacg acgtggccaa ggacccggaa aacctcacct actttgaacg ggccctgggc 540
aagggcctgc tcgtccgcag gcctggggcg gcggccctgg acctggccta cgtggccgcg 600
gggcggctgg agggcttctg ggaggtgaag ctgaacccct gggacgtggc ggcggggtgg 660
ctcctcgtgg aagaggcggg aggaagggtg acggacctcg aggggaaccc ctaccgcctg 720
ggcagccgct acatcctcgc caccaacggg cgggtccacg aggccctgcg ccggaccctc 780
ctgggcctgg actga 795
<210> 28
<211> 756
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TLI
<400> 28
atgtatgaat ggaatgaaat agccctaaat ctcgcaaagg atattgaaag agaagtaatg 60
cctctctttg gaacaaagaa ggctggagaa ttcatcgggt ttagcccgag cggggacaaa 120
actaaactcg tagataaagt tgcagaagac gttgttctgg agtatctcag accccttggc 180
gtaaacgtag taagtgagga gattgggaac atagaggctg gaagtgaata cactattgtt 240
gtcgacccca tcgatggttc tttcaacttt atccaaggca tccctatctt tggattcagc 300
tttgcagttt tcaaaaatga aaaacccgtc tatgctatga tttacgagtt catcacaaag 360
aacgtttatg aaggaattcc cggtgaggga gcctatctaa acggggagag aatacgagtt 420
aggcatttaa atgaaaagtc catctcaata agcttctaca caagaggtag gggagcgagg 480
cttgttgaaa aagttaagag aacgagggtt ttaggtgcta tagcagttga gcttgcttat 540
ctcgctagag gttctctaga cggtgtaata gacataagga actatgtaag gccgacagac 600
atagcagcag gttacataat cgcaaaagaa gctggagcta ttataaccga cgatagcgga 660
gaggaaataa agtttagatt agatgcaagg gaaaagctca acataatagc ggtaaatgat 720
aagagactcc ttaagctcat acttgaagtt atttaa 756
<210> 29
<211> 795
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности GST
<400> 29
atggcagaaa aatgggaaga aatcgaccga tacgcccggc aatggatcga cgaagcagga 60
aagcgcatcc gcgcctcgtt tgccaaacag ctgacggtgg aagcgaaaga gaacccgaac 120
gacttagtga cgaatgttga ccgcgccatt gagcagtttt tcgccgagca catccgtcgc 180
cagttcccta gccaccgcct gttgggggaa gaaggatttg gcgaccggat cgatgccttg 240
gacggcgttg tttgggtgat tgacccgatc gacggcacga tgaatttcgt tcatcagcgg 300
cgccattttg ctgtatcgat cggcattttt gaagatggca tcgggcagct cggctacgtg 360
tatgacgtcg tgtttgacga actgtatgcg gcgcaaaaag ggcggggggt gtttttgaac 420
ggggagccgc tcggcctcct gcagcccgcg ccagtggcgg agtcgatcat cgccatcaac 480
gggacatggc tcatggaaaa caagcgcctc gaccatcgcc cgctcatgag gctggcgaaa 540
gaggcgcgcg gcacgcgctc gtatggttca gcggcgcttg agctcgcgta tgttgccgcc 600
ggccgtttgg acgcctacat ttcgccgcgc ctgtcgccgt gggatttcgc cggcgggatg 660
attttgattg aagaagcggg cggaatggtg acgacccttg acgggaagcc gctggatctg 720
cttggccgca attcggtgct tgccgcgaaa cctggggtgc acgaagaaat tttgcggcgc 780
tatcttcacg attga 795
<210> 30
<211> 780
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности ATH
<400> 30
atgaccactt cccttcgcga tttgctcgat tttgctgtgg aaacggcgta tctggcagga 60
cgaaccaccc tggcgtattt tcagacgggt gttcaggcgg agttcaaagc cgattcttcc 120
cccgtgactc ttgccgaccg cgccgccgag caattgattc gtcaacgcat tgaaaaacgc 180
ttccctcatc atgccattgt gggtgaggaa tttggtgttc agggaagcgc tgatgctacc 240
caccgctggt tcattgaccc gattgacggc accaaatcct tcctgcgcgg cattcccctg 300
tacgctgtgc tgttgggctt ggagattgaa ggcagggtgc aggtgggagt ggcttattac 360
cccgctatgg acgagatgct gtctgccgcc gatggcgagg gatgctggtg gaacgggcgg 420
cgggcgcgtg tctccacggc gagccgcctg gcagaagcct gggtgaccag caccgacccc 480
tacaatttcc agaaaaccgg caaagatgcc gcctggcagc gcattcaggc ggtgtcttat 540
caccgcggcg gctgggggga tgcctacggc tatctgctgg ttgctactgg gagagccgaa 600
gtgatgctcg atcccatcat gaacgagtgg gattgcgcgc cgttcccgcc catcttccgc 660
gaagccggtg gttttttcgg agactggcag ggtaacgaaa ccatctatgg cggcgaggcg 720
ctggcaacaa ctcaggttct tctccctgag gtgctggagt gtttacattc ttccctttga 780
780
<210> 31
<211> 804
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности SAC
<400> 31
atgaaaagag aagatgtaga gaaggtagct agcgaagctt ctaaatatat atatgaagag 60
agagaaaaca aggacgtcga cagagtaatt aatgtacatg gtaatgacgt gactagaatt 120
atagataaga gatctgaaga ctttattgtg gacaggttaa aaagtctagg atataatata 180
ctcatagtta cagaagaatc aggtgtaatt gacagttatg ggaaaaatta cgattacatt 240
gcaattgtag atccactgga tggaagtaca aattttgtat caggaatacc atggtcttca 300
gtatctattg ctatatacaa tagggacgaa gaagatatac tcagttcaaa tgtaggtgct 360
gtaagcagta ttttcactcc ttacacattt tcctacgatg aaggaagcgc ttacgtaaat 420
ggcgtaaaga tagcagagat aaaaaagcct gaaaaaatac tattattagc ctatttctcc 480
aggtcaaagc tacctaatct gaaattattt tttgagaaga ttggtcaggg ttataaaata 540
agaagtttag gaagtgcatc cttagatatg atattagtgt gtacaggcag ggcaacaatg 600
ttttttgata taaggggaaa actgagaaat gtggatattg cagcctcatc taacttctgt 660
tccagattag gggtaattcc atacgacata ggtttaagaa aaattaaaag tagtttgacg 720
gaagtaagtg ttgtaaaaga tttagttata agtcttgacg aaagtcttct aagatctttc 780
tctcttgctt tacagacggt ttaa 804
<210> 32
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TTA
<400> 32
atgctagaac tgctaaagga tacagcaaaa aaagcaggac gactgctaaa aataagattc 60
ttggaacttt tgaacggcat ggaaaccgag gtaagagaaa agggaaaaag cgattttgta 120
acaagggtag atgttgaagt ggaaaactac ataaaggaac tgcttgctaa cacaaatata 180
tcggtggtgg gagaagaaag ctttaaaggc gagataccgg gcacctgcat cttcatagac 240
cccatagatg gcacaagaaa cttcatgaga aagaaccccc acttcgccat taaccttgct 300
tatcaggaga agggaaagct tttggctgga gtgacctatg accctatgaa gaatgaaatg 360
ttttctgctg catttagcaa aggtgcattt ttaaacggag agcgcatcta cgcttccaca 420
aacaaggata ttggcaaagc catcatcgct ataggacttc catacagagg aagagaactc 480
atagatattc aaaccaacct ttacaggaac atcttcctaa acggcgctgc tacccgccac 540
acaggctcag cagcccttga tctggcctac atatcctgcg gaagatacga tgctgctatc 600
tatttttatc tttccccctg ggacgtagct ccgggcatac tgctggtaga agaggcaggc 660
ggagaagttg aaggcaccat gggacgagaa cccatccagg ggtggataat agcatctaac 720
aaagtcatcc accccgaagt taaagacatc cttgaaggct ctttaaaagc ggtataa 777
<210> 33
<211> 765
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности PFU
<400> 33
atgaagctta agttctggag ggaagtagcc attgacataa tctcagattt cgaaactaca 60
ataatgcctt tctttggaaa ccctgatggg ggaaagctcg taaagataag ccccagtgga 120
gatgagacaa agctggttga taagctcgcc gaggatttaa tactttcaag aatcacagag 180
cttggagtca acgttgtgag cgaggaagtt ggtgttattg ataacgagag tgaatacacc 240
gttatagtag acccactaga tggttcatat aacttcattg ctggaattcc tttttttgct 300
ttgagtctag cagtatttaa aaaagataag ccaatatatg caattatcta tgaacctatg 360
acagagagat tctttgaggg aatcccagga gagggagctt ttttaaatgg aaaaagaata 420
aaggtaagaa aaaccccgga cgagaagcca tcaataagct tttattctcg aggtaaaggc 480
catgaaatcg ttaaacatgt aaagagaact agaaccttag gggcaatcgc tctagaattg 540
gcatacctag ctatgggtgc attagatgga gtagttgatg tgaggaaata cgtaaggcca 600
acggacatag ctgctggaac gataattgca aaagaggcag gagcccttat taaggactcc 660
gcgggaaagg atatagatat ttcatttaat gcaactgata ggcttgatgt gatagccgtg 720
aacagcgaag agttgctaaa aacaatttta agcttactgg agtag 765
<210> 34
<211> 759
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности AFU
<400> 34
atggatgaaa gggatgcgct gagaatttcg agggagattg caggagaggt cagaaaggcc 60
attgcgtcaa tgcccttgag ggagagagta aaggacgtgg gaatggggaa ggatggcact 120
ccaaccaagg cagccgacag agttgcggaa gatgctgcgc ttgaaatttt gagaaaggag 180
agggttacgg tggttacaga ggagtcggga gttttggggg agggagatgt ctttgttgcc 240
ctcgaccccc tcgatggaac cttcaatgct acgagaggga ttccagttta ttcagtcagc 300
ctctgcttct cctattcgga taaactgaaa gacgccttct ttggctacgt ttacaacctc 360
gcaacagggg atgaatacta cgcggactcc agcggggctt acagaaacgg ggagaggatt 420
gaggtgagcg atgctgagga gctttactgc aacgccataa tctactatcc cgacaggaag 480
tttcccttta agaggatgag gatttttgga agtgctgcaa cggagctttg cttctttgct 540
gacggctcct ttgactgctt cctcgacatc cgccccggaa agatgcttag aatctacgat 600
gccgctgcgg gtgtttttat tgcagaaaag gcaggaggaa aggttaccga acttgatgga 660
gaaagcttgg ggaataaaaa atttgatatg caggaaaggc tcaatatcgt cgccgcaaat 720
gaaaaactcc atccaaagct gctggagctg attaaatga 759
<210> 35
<211> 798
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности AAC
<400> 35
atggacaagg catggctcga tggcctggcg gaggtcgtgc gggaggctgg ccgcctggtg 60
gaggagatcg ccaggcaggg cttcgatacg caattcaagc atccggaaga gaggcgagat 120
ccggtcacga cagccgatct cgcctgtgac gcgtttctga aagagcggct gctcacgctg 180
cttccggagg cgggctggct gtccgaggag acgaaggatc ggccggatcg attggagaaa 240
aggtgggtgt ggatcgtcga tcccatcgac ggcacgcggg agttcgtccg ccgcattccc 300
gagtacgcca tctcggtggc gctcgcgcgg gacggcgagc cggtggcggg cgcggtggtg 360
aatcccgcga cgggggatct gtttctcggc gccgtgggcg tcggcgcatg gcggaatgga 420
acgcccatgg tttgttcccg catccgcggc gagcggctca cgattctcgg cagccggtcg 480
gagatgaacc gcggcgagtt cgagccgttt gccggcattc tcgaggtccg ggcggtgggc 540
tccatcgcgt ataagctcgc gctcgtcgcg gcgggcgagg cggatggcac gttcagcctc 600
ggcccgaaac acgagtggga tatcgcggcc ggcgtcgcgc tcgtcctggc ggcgggcggc 660
cgggtacacg atggcgccgg gcgcccgttt cgcttcaacc agccgcacac gctcacccgt 720
ggcatcgtcg cggccacgcg cgaggcgtat ggcgatctcg ccctgctcat cgagcggcac 780
gccccacggc gcgcctag 798
<210> 36
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MSI
<400> 36
atggacttgc aacgctactt ggaagccgca ctcgatgctg cgtacttggc tcgaggcatc 60
cacctctact accgcgaaaa aggcttcgcc ctggagagca aaaccagtcc caccgacttg 120
gtcacccaag ccgaccggga gagcgaagaa gctatccgga gtttactgct cgagcgcttc 180
cccgatcatg tggtgctggg tgaagaaggt gggcaggaag gccacagcga ttttcgctgg 240
atcgtggacc cgctcgacgg cacggtcaac tacgcccacg gcttcccctt ctacgcggta 300
agcatcgccc tcgaggtggg gggcgaggtc gtggtggggg cagtgctgga caccgcacgg 360
ggcgagctgt ttaccgccct caaaggagag ggagcattcc aaaatggacg ccccattcgc 420
gtttcgagca ccgcgaccct catcggaagc ctgctggcaa ccggcttccc ctacgatgtg 480
gccaaggacg ccgagaacct gacctacttc cagcgggctc ttgccaaagg ccttaccgtg 540
cgccgccccg gagccgccgc gctggatttg gcttacgtgg cggcagggcg gctggagggt 600
ttctgggagg tcaagctcaa cccctgggac gtggccgcgg gctggctcct catcaccgag 660
gcgggcggaa ccgtgagcgg catccagggc gagccttacc ggctaggaaa ccgctacttg 720
gtagccagca acgggaagat tcatgagcag ctcctcgaca cgctccacgg gcgatga 777
<210> 37
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MRUF
<400> 37
atggatctgc gtccctacct cgaggccgct ctcgatgcgg cctacctagc caaaggcatt 60
caccagtact accaggaaaa aggcttcacc cacagcacca aatctacccc taccgatgtg 120
gtcacccagg ccgaccacga ggccgaggca gccatccggg ctcttatcgc cgagcgcttc 180
cccgaccacg tggtgttagg cgaggagcag ggccaggaag gggagggcga gtttcgctgg 240
attgtagacc ccttggatgg cacggtgaac tacgcccacg gctttccctt ctatggggtg 300
agcatcgggc tggaggtacg aggagaggtg gtgttggggg tggtgctgga taccgcccgg 360
ggcgatctct tcaccgctac caaaggcggc ggggcctacc tgaatggccg cccgctgcgg 420
gtttcccagc gaaccacttt ggtgggcagt ctgctggcta ccggttttcc ctacgatgtg 480
gcacgagacc cggagaacct cacctacttc cagcgggctt tggccaaggg gctgaccgtg 540
cggcggcccg gggcagcggc cctcgacctg gccaatgtgg ccgcgggccg gctggacggt 600
ttctgggagg ttaagctcaa cccctgggat gtggctgcgg gctggctcct catcaccgag 660
gcaggcggcc aggtcaccgg gttccagggc gaggcctacc gcctgggtaa ccgctattta 720
gtggcctcca atgggcggat tcaccaggcg ctgctcgagg cccttcaggg ccgatag 777
<210> 38
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MTA
<400> 38
atggacttgc gtgcctacct gcaaaccgcc ctcgacgccg cctatttagc caaaggcatc 60
caccagtact accaggaaaa aggctttacc cagagcagca agtccacccc caccgatctg 120
gtcacccagg ccgaccacga gtcggaggcg gccatccgag aactcatcgc cgcgcgccac 180
cccgaccacg tggtgctggg cgaggagcag ggccaggaca aagaaggggc cttccgctgg 240
attgtagacc ccctggacgg caccgtaaac tacgcccatg gttttccttt ttatgcggtg 300
agcattggcc tggaagccca cggcgaggtg gtgctggggg tggttctgga taccgctcga 360
ggcgagctgt ttaccgctac caaagggggt ggggcttacc tgaacggtcg tcccatccgg 420
gtttccactc gttcaaccct ggtgggcagc ctcctggcca ccggatttcc ctacgacgta 480
agcaaggata ccgaaaacct gacctacttc cagcgggcat tgaccaaagg gctcatggta 540
cggcgccccg gcgcggcggc cctcgacctg gcctatgtgg ccgccgggcg cctggacggc 600
ttctgggagg tgaagctgaa cccctgggac gtggccgcgg gctggctcat cgtcagtgag 660
gcgggggggc gggtaagcgg ccttcagggc gaggattacc ggctgggcaa ccgctacctg 720
gtggcctcca acggcctaat tcatcagccc ttgctggaca ccctccacgg gcgatag 777
<210> 39
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MCH
<400> 39
atggacctgc gccccttcct cgagaccgcg ctcgccgcgg cttacgtggc tcgaggcatc 60
caccagctct accaggacaa aggcttcacc cagagcacca agtccacccc caccgacgtc 120
gtgacccaag ccgacaagga ggccgaggcc gccatccgcg ccctcatcga gcagcggcac 180
cccggccacg tggtgctcgg cgaggaggag ggccagcagg gcgagggcga gtaccgctgg 240
atcgtggacc cgctcgacgg caccgtgaat tacgcccacg gcttcccctt ctactgcgtg 300
agcatcgggc tcgaggtgcg cggcgaggtg atggtgggcg tggtgctcga caccgcccgc 360
ggcgacctct tcaccgcgac ccgcggcggc ggggcctact tcaacgggcg gcccatgcgg 420
gtctcgcggt cgcccaagct gctgggcagc ctcatcgcca ccggcttccc ctacgacgtc 480
gcccgcgacc gcgaaaacct gacctacctc gagcgcgtcc tcttcaaggg catcaccgtg 540
cgccgccccg gcgccgccgc cctcgacctc gcctacgtgg cctgcgggcg gctcgacggc 600
ttctgggagg tcaagctcaa cccctgggac gtggcggcgg gctggctgct ggtcgaggag 660
gcgggcggca ggatcaccgg catccacggc gagccctacc gcatgggcaa ccgctacctc 720
atggcctcca acggccacat ccacgaggaa ttgctggcga cgatccacgg ccggtag 777
<210> 40
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности MCE
<400> 40
atggacttgc gtgtatacct gcagaccgcc ctcgatgcag cctatctggc caaaggcatt 60
caccagtact accaggagaa gggttttacc ctaagcagca agtccacccc cacagacctg 120
gttactcagg ccgaccacga gtcggaagcg gccatccgag aactcattgc cgcgcgccac 180
cccgaccacg tggtgctggg agaggagcag ggccaggaca gagaaggggc ttttcgctgg 240
attgtagacc ccctggatgg caccgtaaac tacgcccacg gttttccctt ttatgcggtg 300
agtatcggcc tcgaggccca cggcgaggtg gtgctgggag tgattctcga taccgcccgg 360
ggcgacctat tcaccgctac caaggggggc ggggccttca tgaatggccg ccccatccgg 420
gtttcccctc gagccaccct ggtgggaagc ctgctggcca ccggcttccc ctatgacgtt 480
agcaaagaca ccgaaaacct ggtctacttc cagcgggccc tgaccagagg gcttatggtg 540
cggcgtcccg gcgctgccgc cctcgacctg gcctacgtgg ccgccgggcg tctggatggt 600
ttctgggagg tgaagctcaa cccctgggat gtggccgcag gctggctcat catccgcgag 660
gccgggggta cggttacggg catggagggg gaggcctacc ggctgggcaa ccgctacctg 720
gtagcctcca acggcctgat tcaccagcct ctgctggaca ccatacacgg gcgatag 777
<210> 41
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Rma
<400> 41
ctccgccata tgatggatgc ctacac 26
<210> 42
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Rma
<400> 42
gccacctcga gggcgtagac gtggt 25
<210> 43
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Tle
<400> 43
gtgcaaatca tatggacaga atggattttt caa 33
<210> 44
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Tle
<400> 44
gatattctcg agttttctta cttttttatt cacc 34
<210> 45
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Mrub
<400> 45
tgaccccata tggacttgcg tgcctat 27
<210> 46
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Mrub
<400> 46
ttctgcctcg agtcgcccgt ggatg 25
<210> 47
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Dtu
<400> 47
taagctcata tgaagagaat acttgaagta gcta 34
<210> 48
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Dtu
<400> 48
acaaaagact cgagaaattt ttcatttcta atga 34
<210> 49
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Msi
<400> 49
atatacatat ggacttgcaa cgctacttgg aagc 34
<210> 50
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Msi
<400> 50
tggtggtcga ctcgcccgtg gagcgtgtcg a 31
<210> 51
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Mruf
<400> 51
atatacatat ggatctgcgt ccctacctcg aggc 34
<210> 52
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Mruf
<400> 52
tggtggtcga ctcggccctg aagggcctcg 30
<210> 53
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Mta
<400> 53
atatacatat ggacttgcgt gcctacctgc aaac 34
<210> 54
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Mta
<400> 54
tggtggtcga ctcgcccgtg gagggtgtcc a 31
<210> 55
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Mch
<400> 55
atatacatat ggacctgcgc cccttcctcg agac 34
<210> 56
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Mch
<400> 56
tggtggtcga cccggccgtg gatcgtcgcc a 31
<210> 57
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера Mce
<400> 57
atatacatat ggacttgcgt gtatacctgc agac 34
<210> 58
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера Mce
<400> 58
tggtggtcga ctcgcccgtg tatggtgtcc a 31
<210> 59
<211> 823
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CT1
<400> 59
Met Leu Lys Lys Leu Pro Glu Asn Leu Glu His Leu Glu Glu Leu Ala
1 5 10 15
Tyr Asn Leu Trp Trp Ser Trp Ser Arg Pro Ala Gln Arg Leu Trp Arg
20 25 30
Lys Ile Asp Pro Glu Gly Trp Glu Glu His Arg Asn Pro Val Lys Ile
35 40 45
Leu Lys Glu Val Ser Asp Glu Arg Leu Glu Glu Leu Ser Lys Asp Asp
50 55 60
Asp Phe Ile Ser Leu Tyr Glu Leu Thr Ile Glu Arg Phe Lys Asp Tyr
65 70 75 80
Met Glu Lys Glu Asp Thr Trp Phe Asn Val Asn Tyr Pro Glu Trp Asp
85 90 95
Glu Lys Ile Val Tyr Met Cys Met Glu Tyr Gly Leu Thr Lys Ala Leu
100 105 110
Pro Ile Tyr Ser Gly Gly Leu Gly Ile Leu Ala Gly Asp His Leu Lys
115 120 125
Ser Ala Ser Asp Leu Gly Leu Pro Leu Ile Ala Ile Gly Leu Leu Tyr
130 135 140
Lys His Gly Tyr Phe Thr Gln Gln Ile Asp Arg Asp Gly Lys Gln Ile
145 150 155 160
Glu Ile Phe Pro Asp Tyr Asn Pro Glu Asp Leu Pro Met Lys Pro Leu
165 170 175
Lys Asp Glu Lys Gly Asn Gln Val Ile Val Glu Val Pro Leu Asp Ser
180 185 190
Thr Val Val Lys Ala Arg Val Phe Glu Val Lys Val Gly Arg Val Ser
195 200 205
Leu Tyr Leu Leu Asp Pro Asp Ile Glu Glu Asn Glu Glu Arg Tyr Arg
210 215 220
Lys Ile Cys Asn Tyr Leu Tyr Asn Pro Glu Pro Asp Val Arg Val Ser
225 230 235 240
Gln Glu Ile Leu Leu Gly Ile Gly Gly Met Lys Leu Leu Arg Ala Leu
245 250 255
Asn Leu Lys Pro Gly Val Ile His Leu Asn Glu Gly His Pro Ala Phe
260 265 270
Ser Ser Leu Glu Arg Ile Lys Asn Tyr Met Glu Glu Gly Tyr Ser Phe
275 280 285
Thr Glu Ala Leu Glu Ile Val Arg Gln Thr Ser Val Phe Thr Thr His
290 295 300
Thr Pro Val Pro Ala Gly His Asp Arg Phe Pro Phe Asp Leu Val Glu
305 310 315 320
Lys Lys Leu Ser Lys Phe Phe Glu Gly Phe Glu Lys Arg Asn Leu Leu
325 330 335
Met Asp Leu Gly Lys Asp Glu Thr Gly Ser Phe Asn Met Thr Tyr Leu
340 345 350
Ala Leu Arg Thr Ser Ser Phe Ile Asn Gly Val Ser Lys Leu His Ala
355 360 365
Glu Val Ser Arg Arg Met Phe Lys Asn Val Trp Gln Gly Val Pro Val
370 375 380
Glu Glu Ile Pro Ile Glu Gly Ile Thr Asn Gly Val His Met Gly Thr
385 390 395 400
Trp Ile Asn Arg Glu Met Arg Lys Leu Tyr Asp Arg Tyr Leu Gly Arg
405 410 415
Val Trp Arg Asp His Thr Asp Leu Glu Gly Ile Trp Tyr Gly Val Asp
420 425 430
Arg Ile Pro Asp Glu Glu Leu Trp Gln Ala His Leu Arg Ala Lys Lys
435 440 445
Arg Phe Ile Glu Tyr Ile Lys Glu Ser Val Arg Arg Arg Asn Glu Arg
450 455 460
Leu Gly Ile Asp Glu Asp Val Pro Asn Ile Asp Glu Asn Ser Leu Ile
465 470 475 480
Ile Gly Phe Ala Arg Arg Phe Ala Thr Tyr Lys Arg Ala Val Leu Leu
485 490 495
Leu Ser Asp Leu Glu Arg Leu Lys Lys Ile Leu Asn Asp Pro Glu Arg
500 505 510
Pro Val Tyr Val Val Tyr Ala Gly Lys Ala His Pro Arg Asp Asp Ala
515 520 525
Gly Lys Glu Phe Leu Lys Arg Ile Tyr Glu Val Ser Gln Met Pro Glu
530 535 540
Phe Lys Asn Arg Ile Ile Val Leu Glu Asn Tyr Asp Ile Gly Met Ala
545 550 555 560
Arg Leu Met Val Ser Gly Val Asp Val Trp Leu Asn Asn Pro Arg Arg
565 570 575
Pro Met Glu Ala Ser Gly Thr Ser Gly Met Lys Ala Ala Ala Asn Gly
580 585 590
Val Leu Asn Ala Ser Val Tyr Asp Gly Trp Trp Val Glu Gly Tyr Asn
595 600 605
Gly Arg Asn Gly Trp Val Ile Gly Asp Glu Ser Val Leu Pro Glu Thr
610 615 620
Glu Val Asp Asp Pro Arg Asp Ala Glu Ala Leu Tyr Asp Leu Leu Glu
625 630 635 640
Asn Glu Ile Ile Pro Thr Tyr Tyr Glu Asn Lys Glu Lys Trp Ile Phe
645 650 655
Met Met Lys Glu Ser Ile Lys Ser Val Ala Pro Arg Phe Ser Thr Thr
660 665 670
Arg Met Leu Lys Glu Tyr Thr Glu Lys Phe Tyr Ile Lys Gly Leu Val
675 680 685
Asn Lys Glu Trp Leu Glu Arg Lys Glu Asn Ala Glu Arg Phe Gly Ala
690 695 700
Trp Lys Glu Arg Ile Leu Arg Asn Trp Ser Ser Val Ser Ile Glu Arg
705 710 715 720
Ile Val Leu Glu Asp Thr Arg Ser Val Glu Val Thr Val Lys Leu Gly
725 730 735
Asp Leu Ser Pro Asp Asp Val Leu Val Glu Leu Leu Ile Gly Arg Gly
740 745 750
Glu Ser Met Glu Asp Leu Glu Ile Trp Lys Val Ile Gln Ile Arg Lys
755 760 765
His Arg Arg Glu Gly Asp Leu Phe Ile Tyr Ser Tyr Val Asn Gly Ala
770 775 780
Leu Gly His Leu Gly Ser Pro Gly Trp Phe Tyr Ala Val Arg Val Leu
785 790 795 800
Pro Tyr His Pro Lys Leu Pro Thr Arg Phe Leu Pro Glu Ile Pro Val
805 810 815
Val Trp Lys Lys Val Leu Gly
820
<210> 60
<211> 2472
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности CT1
<400> 60
atgctgaaga aactcccgga gaatctggag catctggaag aactcgccta caacctctgg 60
tggagctggt ctaggcccgc tcagagactc tggagaaaga tagatccgga aggctgggag 120
gaacacagaa accccgttaa aatactgaaa gaagtttctg atgaaaggct cgaagaactt 180
tcaaaagatg atgatttcat atccctctac gaactcacca ttgaaaggtt caaggattac 240
atggagaaag aagacacctg gttcaacgtg aactaccccg aatgggacga gaagatcgtc 300
tacatgtgta tggagtacgg tttgaccaaa gcccttccga tctactccgg tggtcttgga 360
atcctcgcgg gagaccatct caaatccgca agcgatcttg gacttcctct catagcgatc 420
ggacttctct acaaacatgg atatttcacc cagcagatcg acagagatgg aaaacagata 480
gagattttcc ctgattacaa cccagaggac ttacccatga agcccctgaa ggatgaaaag 540
ggaaaccagg tgatcgtgga ggttcctctc gacagtaccg tggtgaaggc acgtgttttt 600
gaagtgaagg taggaagggt gagtctgtac ctgctcgatc cggacatcga ggaaaacgag 660
gaacgataca gaaagatctg caactacctt tacaacccgg aacccgatgt gagggtctcc 720
caggagatac tcctcggaat tgggggaatg aagcttctca gggctctgaa cctgaaacca 780
ggagtcatcc atctgaacga aggacatccg gcgttctctt ccctcgaaag gataaagaac 840
tacatggaag aaggatattc cttcacagag gcccttgaga tcgtgagaca gacgagtgtg 900
tttacaaccc acacacccgt tcccgctgga cacgacagat ttccctttga cctcgtggaa 960
aagaaacttt cgaaattctt cgaaggattc gaaaagagaa atcttctcat ggatcttggg 1020
aaagatgaaa caggcagttt caacatgacg tatcttgccc tgagaacgtc ctctttcata 1080
aacggcgtga gcaaactgca tgcggaagtt tccagaagga tgttcaaaaa cgtgtggcag 1140
ggtgttcccg tggaggaaat accgatcgaa gggataacga acggcgttca catgggaacc 1200
tggatcaacc gtgagatgag aaaactgtac gacagatatc tcggaagggt atggagagat 1260
cacaccgacc ttgagggtat ctggtacggt gttgacagga ttccagatga agaactctgg 1320
caggctcacc tgagggcaaa gaagagattc atcgagtaca taaaagaatc ggtaagaaga 1380
agaaacgaga gactgggaat cgacgaagat gtgccgaaca tcgatgaaaa ttcgctcatc 1440
ataggttttg caagaaggtt tgccacttac aagagggcag ttctcctgct cagcgatctg 1500
gagagactca agaagatcct caacgatcca gaaagacccg tttacgtggt ctatgcgggg 1560
aaggcccatc caagggacga tgcggggaag gaatttttga aacgcatcta cgaagtctcg 1620
cagatgcctg agttcaaaaa caggatcatc gtactggaaa actacgacat tggaatggca 1680
cggctcatgg tgtcgggagt ggatgtgtgg ctgaacaacc cgagaagacc catggaagca 1740
agtggaacaa gcggaatgaa ggcagcagcc aacggagttc ttaacgcgag tgtttacgat 1800
ggatggtggg ttgaagggta caacggcaga aacggctggg tcataggcga tgaaagcgtt 1860
cttccagaga cggaagtgga cgatcccagg gacgcagaag cactctacga tctcctcgaa 1920
aacgaaatca tcccaaccta ctacgaaaac aaagaaaagt ggatcttcat gatgaaagag 1980
agcataaaga gtgttgctcc aagattcagc accaccagaa tgctcaaaga atacacggag 2040
aagttctaca taaagggact tgtgaacaaa gaatggcttg aaagaaaaga aaacgccgaa 2100
aggtttggtg catggaagga aaggatcctc agaaactgga gcagcgtttc catagaaaga 2160
atcgtccttg aggacacaag gagtgttgag gtgacggtga aactgggaga cctttcacct 2220
gatgatgtac tggttgaact tttgattgga agaggagaaa gcatggaaga tctggagatc 2280
tggaaggtga tacagataag aaagcacaga agggaagggg atctgttcat ctacagttat 2340
gtcaacggtg ccctcggtca tcttggctct ccgggatggt tctacgcggt gagggtgcta 2400
ccttatcatc cgaaacttcc caccagattc ttgccggaga tacctgtggt gtggaaaaag 2460
gttctcgggt ga 2472
<210> 61
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CT2
<400> 61
Met Ile Gly Tyr Gln Ile Tyr Val Arg Ser Phe Arg Asp Gly Asn Phe
1 5 10 15
Asp Gly Val Gly Asp Phe Lys Gly Leu Lys Gly Ala Ile Ser Tyr Leu
20 25 30
Lys Glu Leu Gly Val Asp Phe Val Trp Leu Met Pro Val Phe Ser Ser
35 40 45
Ile Ser Phe His Gly Tyr Asp Val Val Asp Phe Tyr Ser Phe Lys Ala
50 55 60
Glu Tyr Gly Asp Glu Lys Asp Phe Arg Glu Met Ile Glu Ala Phe His
65 70 75 80
Asp Asn Gly Ile Lys Val Val Leu Asp Leu Pro Ile His His Thr Gly
85 90 95
Phe Leu His Val Trp Phe Gln Lys Ala Leu Lys Gly Asp Pro His Tyr
100 105 110
Arg Asp Tyr Tyr Val Trp Ala Ser Glu Lys Thr Asp Leu Asp Glu Arg
115 120 125
Arg Glu Trp Asp Asn Glu Arg Ile Trp His Pro Leu Glu Asp Gly Arg
130 135 140
Phe Tyr Arg Gly Leu Phe Gly Pro Leu Ser Pro Asp Leu Asn Tyr Asp
145 150 155 160
Asn Pro Gln Val Phe Glu Glu Met Lys Lys Val Val Tyr His Leu Leu
165 170 175
Glu Met Gly Val Asp Gly Phe Arg Phe Asp Ala Ala Lys His Met Arg
180 185 190
Asp Thr Leu Glu Gln Asn Val Arg Phe Trp Arg Tyr Phe Leu Ser Asp
195 200 205
Ile Glu Gly Ile Phe Leu Ala Glu Ile Trp Ala Glu Ser Lys Val Val
210 215 220
Asp Glu His Gly Arg Ile Phe Gly Tyr Met Leu Asn Phe Asp Thr Ser
225 230 235 240
His Cys Ile Lys Glu Ala Val Trp Lys Glu Asn Phe Lys Val Leu Ile
245 250 255
Glu Ser Ile Glu Arg Ala Leu Val Gly Lys Asp Tyr Leu Pro Val Asn
260 265 270
Phe Thr Ser Asn His Asp Met Ser Arg Leu Ala Ser Phe Glu Gly Gly
275 280 285
Leu Ser Glu Glu Lys Val Lys Leu Ser Leu Ser Ile Leu Phe Thr Leu
290 295 300
Pro Gly Val Pro Leu Ile Phe Tyr Gly Asp Glu Leu Gly Met Lys Gly
305 310 315 320
Ile Tyr Arg Lys Pro Asn Thr Glu Val Val Leu Asp Pro Phe Pro Trp
325 330 335
Ser Glu Asn Met Cys Val Glu Gly Gln Thr Phe Trp Lys Trp Pro Ala
340 345 350
Tyr Asn Asp Pro Phe Ser Gly Val Ser Val Glu Tyr Gln Arg Arg Asn
355 360 365
Arg Asp Ser Ile Leu Ser His Thr Met Arg Trp Ala Gly Phe Arg Gly
370 375 380
Glu Asn His Trp Leu Asp Arg Ala Asn Ile Glu Phe Leu Cys Lys Glu
385 390 395 400
Glu Lys Leu Leu Val Tyr Arg Leu Val Asp Glu Gly Arg Ser Leu Lys
405 410 415
Val Ile His Asn Leu Ser Asn Gly Glu Met Val Phe Glu Gly Val Arg
420 425 430
Val Gln Pro Tyr Ser Thr Glu Val Val
435 440
<210> 62
<211> 1326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности CT2
<400> 62
atgataggct accagatcta cgtgagatca ttcagggatg gaaacttcga tggtgtgggg 60
gatttcaaag gattgaaagg tgcgatttcc tacctgaaag aactgggtgt tgattttgtc 120
tggctcatgc ccgtcttttc ctccatttcc ttccacgggt atgacgtggt ggatttttat 180
tctttcaaag ccgagtacgg agacgagaaa gactttagag agatgatcga ggcgttccac 240
gacaacggta taaaagtcgt tctcgatctt cccatccatc atactggttt cctccatgtg 300
tggtttcaga aagccctgaa aggagatcca cactacaggg attattacgt atgggcgagt 360
gaaaaaacgg atctggacga aagaagagag tgggacaacg aaaggatctg gcatcctctg 420
gaggacggaa ggttctacag aggacttttc ggtcccctct cacccgatct gaactacgat 480
aacccgcagg tttttgaaga gatgaagaag gtggtttatc accttcttga aatgggagtg 540
gacggattca gattcgacgc agcaaagcac atgagagata ctctggaaca gaacgttcgc 600
ttttggaggt atttcctctc cgatattgag ggaatattcc ttgcggaaat ctgggcagaa 660
tccaaagttg tggatgaaca cggcaggata ttcggctaca tgctaaattt cgatacctca 720
cactgtatta aggaagcggt gtggaaggaa aacttcaaag tgttgatcga gtcgatcgaa 780
agggccctgg ttggaaaaga ttatctgccg gtgaacttca catcgaacca tgatatgtca 840
aggcttgcga gtttcgaagg agggttgagt gaagagaagg tgaaactctc actttccatt 900
ctgttcacgc ttcccggggt tcctctcata ttctacggag acgaactggg aatgaaagga 960
atctatcgaa aaccgaacac ggaagtcgtg ctggatccgt tcccctggag cgaaaacatg 1020
tgtgttgaag gccagacatt ttggaaatgg cccgcgtata acgatccatt ctccggtgtt 1080
tctgttgagt atcagaggag aaatcgtgat tcgattctct cacacacgat gaggtgggca 1140
ggattcagag gggaaaatca ctggctggac agggcaaaca tcgaatttct gtgcaaagaa 1200
gaaaaactgc tcgtgtacag actggtcgat gaagggcgtt ctctgaaagt gatacacaac 1260
ctgtcgaatg gtgaaatggt gtttgaggga gtgcgcgtac aaccctacag cacggaggtg 1320
gtttga 1326
<210> 63
<211> 451
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TN1
<400> 63
Met Lys Lys Met Ala Leu Lys Phe Asp Phe Ser Asn Leu Phe Glu Pro
1 5 10 15
Asn Ile Ser Gly Gly Leu Arg Glu Glu Asp Leu Glu Ser Thr Lys Glu
20 25 30
Lys Val Ile Glu Ala Ile Lys Asn Phe Thr Glu Asn Thr Pro Asp Phe
35 40 45
Ala Arg Leu Asp Arg Lys Trp Ile Asp Ser Val Lys Glu Leu Glu Glu
50 55 60
Trp Val Val Asn Phe Asp Thr Val Val Val Leu Gly Ile Gly Gly Ser
65 70 75 80
Gly Leu Gly Asn Leu Ala Leu His Tyr Ser Leu Arg Pro Leu Asn Trp
85 90 95
Asn Glu Met Ser Arg Glu Glu Arg Asn Gly Tyr Ala Arg Val Phe Val
100 105 110
Val Asp Asn Val Asp Pro Asp Leu Met Ala Ser Val Leu Asp Arg Ile
115 120 125
Asp Leu Lys Thr Thr Leu Phe Asn Val Ile Ser Lys Ser Gly Ser Thr
130 135 140
Ala Glu Val Met Ala Asn Tyr Ser Ile Ala Arg Gly Ile Leu Glu Ala
145 150 155 160
Asn Gly Leu Asp Pro Lys Glu His Ile Leu Ile Thr Thr Asp Pro Glu
165 170 175
Lys Gly Phe Leu Arg Lys Val Val Lys Glu Glu Gly Phe Arg Ser Leu
180 185 190
Glu Val Pro Pro Gly Val Gly Gly Arg Phe Ser Val Leu Thr Pro Val
195 200 205
Gly Leu Phe Ser Ala Met Ala Glu Gly Ile Asp Ile Glu Glu Leu His
210 215 220
Asp Gly Ala Arg Asp Ala Phe Glu Arg Cys Lys Lys Glu Asp Leu Phe
225 230 235 240
Glu Asn Pro Ala Ala Met Ile Ala Leu Thr His Tyr Leu Tyr Leu Lys
245 250 255
Arg Gly Lys Ser Ile Ser Val Met Met Ala Tyr Ser Asn Arg Met Thr
260 265 270
Tyr Leu Val Asp Trp Tyr Arg Gln Leu Trp Ala Glu Ser Leu Gly Lys
275 280 285
Arg Tyr Asn Leu Lys Gly Glu Glu Val Phe Thr Gly Gln Thr Pro Val
290 295 300
Lys Ala Ile Gly Ala Thr Asp Gln His Ser Gln Ile Gln Leu Tyr Asn
305 310 315 320
Glu Gly Pro Asn Asp Lys Val Ile Thr Phe Leu Arg Leu Glu Asn Phe
325 330 335
Asp Arg Glu Ile Ile Ile Pro Asp Thr Gly Arg Glu Glu Leu Lys Tyr
340 345 350
Leu Ala Arg Lys Arg Leu Ser Glu Leu Leu Leu Ala Glu Gln Thr Gly
355 360 365
Thr Glu Glu Ala Leu Arg Lys Asn Asp Arg Pro Asn Met Lys Val Ile
370 375 380
Phe Asp Arg Leu Thr Ser Tyr Asn Val Gly Gln Phe Phe Ala Tyr Tyr
385 390 395 400
Glu Ala Ala Thr Ala Phe Met Gly Tyr Leu Leu Glu Ile Asn Pro Phe
405 410 415
Asp Gln Pro Gly Val Glu Leu Gly Lys Lys Ile Thr Phe Ala Leu Met
420 425 430
Gly Arg Glu Gly Tyr Glu Tyr Glu Ile Lys Asp Arg Thr Lys Lys Val
435 440 445
Ile Ile Glu
450
<210> 64
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TN1
<400> 64
atgaaaaaga tggctttgaa atttgatttt tcaaatcttt ttgaaccgaa catctccggt 60
ggactgagag aggaagatct ggaaagcaca aaagaaaagg tgatagaggc gataaagaat 120
ttcactgaga acacaccgga ttttgccaga ctggacagaa aatggatcga ttcggtgaag 180
gaactcgagg agtgggtggt gaacttcgac acggtggtcg ttctgggaat tgggggatcc 240
ggtcttggaa accttgccct tcattattcg ttgagaccac tgaactggaa cgagatgtcg 300
agagaggaaa gaaacggtta tgcgagagtc ttcgtggtgg acaacgtaga tcccgatctc 360
atggcctccg tccttgatag gatagatctg aagacaacgc tgttcaacgt gatctcaaaa 420
tctggatcca cggctgaggt tatggcgaat tactcgatcg caaggggaat cctggaggct 480
aatggtctgg acccgaaaga acacatcctc atcacaacag atccagagaa gggctttttg 540
agaaaagtag tgaaagaaga gggcttcaga agtcttgagg tccctcccgg cgttggagga 600
aggttcagcg tgctgacgcc cgttggcctc ttctctgcca tggcggaggg tatcgacata 660
gaagaactcc acgacggtgc ccgggatgcg ttcgagagat gcaagaagga agacctgttc 720
gaaaatccag cggcgatgat cgccctcaca cactatctct atctgaagag aggaaagagc 780
atctccgtca tgatggccta ctccaacagg atgacctacc tcgtggactg gtacagacag 840
ctgtgggcag aaagtctggg aaagagatac aacctgaaag gagaggaggt cttcacgggt 900
cagaccccgg tgaaggcaat aggagccacc gatcagcact ctcagataca gctttacaac 960
gagggcccaa acgacaaagt gataacgttt ttgcggttgg aaaacttcga tagagagatc 1020
ataataccgg acaccggaag agaagagctc aaataccttg caagaaaaag actctctgaa 1080
cttctccttg cagaacagac aggaacagag gaagccctaa ggaaaaacga cagaccgaac 1140
atgaaggtga tcttcgacag actcacctct tacaatgtgg gccagttctt cgcttattat 1200
gaagccgcaa ctgctttcat ggggtatctc ctcgagatca acccgtttga tcagccgggt 1260
gtggaacttg gaaagaagat cacgtttgcc ctcatgggaa gggaaggtta cgaatacgaa 1320
ataaaagatc gcaccaagaa ggtgatcata gaatga 1356
<210> 65
<211> 221
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности FP3E
<400> 65
Met Met Val Lys Ile Ala Ala Ser Ile Leu Ala Cys Asp Leu Ala Arg
1 5 10 15
Leu Ala Asp Glu Val Lys Arg Val Glu Glu His Ile Asp Met Val His
20 25 30
Phe Asp Val Met Asp Gly His Phe Val Pro Asn Ile Ser Phe Gly Leu
35 40 45
Pro Val Leu Lys Ala Leu Arg Lys Glu Thr Ser Leu Pro Ile Ser Val
50 55 60
His Leu Met Ile Thr Asn Pro Glu Asp Tyr Val Asp Arg Phe Val Glu
65 70 75 80
Glu Gly Ala Asp Met Val Ala Val His Tyr Glu Thr Thr Pro His Leu
85 90 95
His Arg Ile Val His Arg Ile Lys Asp Leu Gly Ala Lys Ala Phe Val
100 105 110
Ala Leu Asn Pro His Thr Pro Val Phe Leu Leu Ser Glu Ile Ile Thr
115 120 125
Asp Val Asp Gly Val Leu Val Met Ser Val Asn Pro Gly Phe Ser Gly
130 135 140
Gln Arg Phe Ile Ala Arg Ser Leu Glu Lys Ile Arg Ser Leu Lys Lys
145 150 155 160
Met Ile Arg Asp Leu Gly Leu Glu Thr Glu Ile Met Val Asp Gly Gly
165 170 175
Val Asn Glu Glu Asn Ala Ser Ile Leu Ile Lys Asn Gly Ala Thr Ile
180 185 190
Leu Val Met Gly Tyr Gly Ile Phe Lys Asn Glu Asn Tyr Val Glu Leu
195 200 205
Val Arg Ser Ile Lys Gln Glu Arg Gly Glu Ser Ala Gly
210 215 220
<210> 66
<211> 666
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности FP3E
<400> 66
atgatggtaa agatcgccgc ttcaatcctt gcgtgtgatc ttgcaagact cgccgatgag 60
gtaaaaaggg tggaagaaca catagacatg gttcacttcg atgtcatgga tggacacttc 120
gttccgaaca tctcgttcgg attgcccgtt ctcaaagccc tgagaaaaga aaccagcctt 180
cctataagtg ttcatctgat gatcacaaat ccagaggact atgtggaccg tttcgtggaa 240
gagggagcgg acatggtggc ggtccactac gagacaacgc cgcaccttca caggatagtg 300
cacaggataa aggatctcgg ggcgaaggcg ttcgtcgccc tcaacccaca cacaccggtt 360
tttctcctgt ctgagatcat aacggatgtg gatggcgtac tcgtgatgag tgtgaacccg 420
ggcttttctg gtcagagatt cattgcaagg agtctggaaa aaataaggag tctgaagaag 480
atgataaggg atctgggact cgaaacggag atcatggtcg atggtggtgt caacgaagaa 540
aacgcttcta tcttaataaa gaacggtgcg acgatccttg taatggggta cggtatcttc 600
aaaaacgaaa actatgtgga actggtgaga tccatcaagc aggaaagagg ggaatctgct 660
ggctga 666
<210> 67
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности TN2
<400> 67
Met Ile Gly Tyr Gln Ile Tyr Val Arg Ser Phe Arg Asp Gly Asn Phe
1 5 10 15
Asp Gly Val Gly Asp Phe Lys Gly Leu Lys Gly Ala Ile Ser Tyr Leu
20 25 30
Lys Glu Leu Gly Val Asp Phe Val Trp Leu Met Pro Val Phe Ser Ser
35 40 45
Ile Ser Phe His Gly Tyr Asp Val Val Asp Phe Tyr Ser Phe Lys Ala
50 55 60
Glu Tyr Gly Asp Glu Lys Asp Phe Arg Glu Met Ile Glu Ala Phe His
65 70 75 80
Asp Asn Gly Ile Lys Val Val Leu Asp Leu Pro Ile His His Thr Gly
85 90 95
Phe Leu His Val Trp Phe Gln Lys Ala Leu Lys Gly Asp Pro His Tyr
100 105 110
Arg Asp Tyr Tyr Val Trp Ala Ser Glu Lys Thr Asp Leu Asp Glu Arg
115 120 125
Arg Glu Trp Asp Asn Glu Arg Ile Trp His Pro Leu Glu Asp Gly Arg
130 135 140
Phe Tyr Arg Gly Leu Phe Gly Pro Leu Ser Pro Asp Leu Asn Tyr Asp
145 150 155 160
Asn Pro Gln Val Phe Glu Glu Met Lys Lys Val Val Tyr His Leu Leu
165 170 175
Glu Met Gly Val Asp Gly Phe Arg Phe Asp Ala Ala Lys His Met Arg
180 185 190
Asp Thr Leu Glu Gln Asn Val Arg Phe Trp Arg Tyr Phe Leu Ser Asp
195 200 205
Ile Glu Gly Ile Phe Leu Ala Glu Ile Trp Ala Glu Ser Lys Val Val
210 215 220
Asp Glu His Gly Arg Ile Phe Gly Tyr Met Leu Asn Phe Asp Thr Ser
225 230 235 240
His Cys Ile Lys Glu Ala Val Trp Lys Glu Asn Phe Lys Val Leu Ile
245 250 255
Glu Ser Ile Glu Arg Ala Leu Val Gly Lys Asp Tyr Leu Pro Val Asn
260 265 270
Phe Thr Ser Asn His Asp Met Ser Arg Leu Ala Ser Phe Glu Gly Gly
275 280 285
Leu Ser Glu Glu Lys Val Lys Leu Ser Leu Ser Ile Leu Phe Thr Leu
290 295 300
Pro Gly Val Pro Leu Ile Phe Tyr Gly Asp Glu Leu Gly Met Lys Gly
305 310 315 320
Ile Tyr Arg Lys Pro Asn Thr Glu Val Val Leu Asp Pro Phe Pro Trp
325 330 335
Ser Glu Asn Met Cys Val Glu Gly Gln Thr Phe Trp Lys Trp Pro Ala
340 345 350
Tyr Asn Asp Pro Phe Ser Gly Val Ser Val Glu Tyr Gln Arg Arg Asn
355 360 365
Arg Asp Ser Ile Leu Ser His Thr Met Arg Trp Ala Gly Phe Arg Gly
370 375 380
Glu Asn His Trp Leu Asp Arg Ala Asn Ile Glu Phe Leu Cys Lys Glu
385 390 395 400
Glu Lys Leu Leu Val Tyr Arg Leu Val Asp Glu Gly Arg Ser Leu Lys
405 410 415
Val Ile His Asn Leu Ser Asn Gly Glu Met Val Phe Glu Gly Val Arg
420 425 430
Val Gln Pro Tyr Ser Thr Glu Val Val
435 440
<210> 68
<211> 1326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидные последовательности TN2
<400> 68
atgataggct accagatcta cgtgagatca ttcagggatg gaaacttcga tggtgtgggg 60
gatttcaaag gattgaaagg tgcgatttcc tacctgaaag aactgggtgt tgattttgtc 120
tggctcatgc ccgtcttttc ctccatttcc ttccacgggt atgacgtggt ggatttttat 180
tctttcaaag ccgagtacgg agacgagaaa gactttagag agatgatcga ggcgttccac 240
gacaacggta taaaagtcgt tctcgatctt cccatccatc atactggttt cctccatgtg 300
tggtttcaga aagccctgaa aggagatcca cactacaggg attattacgt atgggcgagt 360
gaaaaaacgg atctggacga aagaagagag tgggacaacg aaaggatctg gcatcctctg 420
gaggacggaa ggttctacag aggacttttc ggtcccctct cacccgatct gaactacgat 480
aacccgcagg tttttgaaga gatgaagaag gtggtttatc accttcttga aatgggagtg 540
gacggattca gattcgacgc agcaaagcac atgagagata ctctggaaca gaacgttcgc 600
ttttggaggt atttcctctc cgatattgag ggaatattcc ttgcggaaat ctgggcagaa 660
tccaaagttg tggatgaaca cggcaggata ttcggctaca tgctaaattt cgatacctca 720
cactgtatta aggaagcggt gtggaaggaa aacttcaaag tgttgatcga gtcgatcgaa 780
agggccctgg ttggaaaaga ttatctgccg gtgaacttca catcgaacca tgatatgtca 840
aggcttgcga gtttcgaagg agggttgagt gaagagaagg tgaaactctc actttccatt 900
ctgttcacgc ttcccggggt tcctctcata ttctacggag acgaactggg aatgaaagga 960
atctatcgaa aaccgaacac ggaagtcgtg ctggatccgt tcccctggag cgaaaacatg 1020
tgtgttgaag gccagacatt ttggaaatgg cccgcgtata acgatccatt ctccggtgtt 1080
tctgttgagt atcagaggag aaatcgtgat tcgattctct cacacacgat gaggtgggca 1140
ggattcagag gggaaaatca ctggctggac agggcaaaca tcgaatttct gtgcaaagaa 1200
gaaaaactgc tcgtgtacag actggtcgat gaagggcgtt ctctgaaagt gatacacaac 1260
ctgtcgaatg gtgaaatggt gtttgaggga gtgcgcgtac aaccctacag cacggaggtg 1320
gtttga 1326
<210> 69
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера ct1
<400> 69
aggagaaact catatgctga agaaactccc ggag 34
<210> 70
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера ct1
<400> 70
agccccctcg agcccgagaa c 21
<210> 71
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера ct2
<400> 71
aaagggcata tgatcctgtt tggaac 26
<210> 72
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера ct2
<400> 72
ataccagtct cgagcagttt caggatc 27
<210> 73
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера tn1
<400> 73
ttactgaggg catatgaaaa agatggcttt gaaa 34
<210> 74
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера tn1
<400> 74
aagacgcgtc gacttctatg atcaccttct 30
<210> 75
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК прямого праймера fp3e
<400> 75
ggaacatatg atggtaaaga tcgccgcttc aatc 34
<210> 76
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК обратного праймера fp3e
<400> 76
catactcgag cttcccctct cctatct 27
<210> 77
<211> 270
<212> PRT
<213> Искусственная последовательность
<220>
<223> Полифосфат-зависимая глюкокиназа, полученная из Deinococcus geothermalis
<400> 77
Met Leu Ala Ala Ser Asp Ser Ser Gln His Gly Gly Lys Ala Val Thr
1 5 10 15
Leu Ser Pro Met Ser Val Ile Leu Gly Ile Asp Ile Gly Gly Ser Gly
20 25 30
Ile Lys Gly Ala Pro Val Asp Thr Ala Thr Gly Lys Leu Val Ala Glu
35 40 45
Arg His Arg Ile Pro Thr Pro Glu Gly Ala His Pro Asp Ala Val Lys
50 55 60
Asp Val Val Val Glu Leu Val Arg His Phe Gly His Ala Gly Pro Val
65 70 75 80
Gly Ile Thr Phe Pro Gly Ile Val Gln His Gly His Thr Leu Ser Ala
85 90 95
Ala Asn Val Asp Lys Ala Trp Ile Gly Leu Asp Ala Asp Thr Leu Phe
100 105 110
Thr Glu Ala Thr Gly Arg Asp Val Thr Val Ile Asn Asp Ala Asp Ala
115 120 125
Ala Gly Leu Ala Glu Ala Arg Phe Gly Ala Gly Ala Gly Val Pro Gly
130 135 140
Glu Val Leu Leu Leu Thr Phe Gly Thr Gly Ile Gly Ser Ala Leu Ile
145 150 155 160
Tyr Asn Gly Val Leu Val Pro Asn Thr Glu Phe Gly His Leu Tyr Leu
165 170 175
Lys Gly Asp Lys His Ala Glu Thr Trp Ala Ser Asp Arg Ala Arg Glu
180 185 190
Gln Gly Asp Leu Asn Trp Lys Gln Trp Ala Lys Arg Val Ser Arg Tyr
195 200 205
Leu Gln Tyr Leu Glu Gly Leu Phe Ser Pro Asp Leu Phe Ile Ile Gly
210 215 220
Gly Gly Val Ser Lys Lys Ala Asp Lys Trp Gln Pro His Val Ala Thr
225 230 235 240
Thr Arg Thr Arg Leu Val Pro Ala Ala Leu Gln Asn Glu Ala Gly Ile
245 250 255
Val Gly Ala Ala Met Val Ala Ala Gln Arg Ser Gln Gly Asp
260 265 270
<210> 78
<211> 253
<212> PRT
<213> Искусственная последовательность
<220>
<223> Полифосфат-зависимая глюкокиназа, полученная из Anaerolinea thermophila
<400> 78
Met Gly Arg Gln Gly Met Glu Ile Leu Gly Ile Asp Ile Gly Gly Ser
1 5 10 15
Gly Ile Lys Gly Ala Pro Val Asp Val Glu Thr Gly Gln Leu Thr Ala
20 25 30
Glu Arg Tyr Arg Leu Pro Thr Pro Glu Asn Ala Leu Pro Glu Glu Val
35 40 45
Ala Leu Val Val Ala Gln Ile Val Glu His Phe Gln Trp Lys Gly Arg
50 55 60
Val Gly Ala Gly Phe Pro Ala Ala Ile Lys His Gly Val Ala Gln Thr
65 70 75 80
Ala Ala Asn Ile His Pro Thr Trp Ile Gly Leu His Ala Gly Asn Leu
85 90 95
Phe Ser Glu Lys Cys Gly Cys Pro Val Ser Val Leu Asn Asp Ala Asp
100 105 110
Ala Ala Gly Leu Ala Glu Met Ile Phe Gly Ala Gly Lys Gly Gln Lys
115 120 125
Gly Val Val Leu Met Ile Thr Ile Gly Thr Gly Ile Gly Thr Ala Leu
130 135 140
Phe Thr Asp Gly Ile Leu Val Pro Asn Thr Glu Leu Gly His Ile Glu
145 150 155 160
Ile Arg Gly Lys Asp Ala Glu Gln Arg Ser Ser Glu Ala Ala Arg Gln
165 170 175
Arg Lys Asp Trp Thr Trp Gln Gln Trp Ala Lys Arg Leu Asn Glu His
180 185 190
Leu Glu Arg Leu Glu Ala Leu Phe Trp Pro Asp Leu Phe Ile Leu Gly
195 200 205
Gly Gly Ala Val Lys Asn His Glu Lys Phe Phe Pro Tyr Leu Lys Leu
210 215 220
Arg Thr Pro Phe Val Ala Ala Lys Leu Gly Asn Leu Ala Gly Ile Val
225 230 235 240
Gly Ala Ala Trp Tyr Ala His Thr Gln Glu Thr Gln Ala
245 250
<---
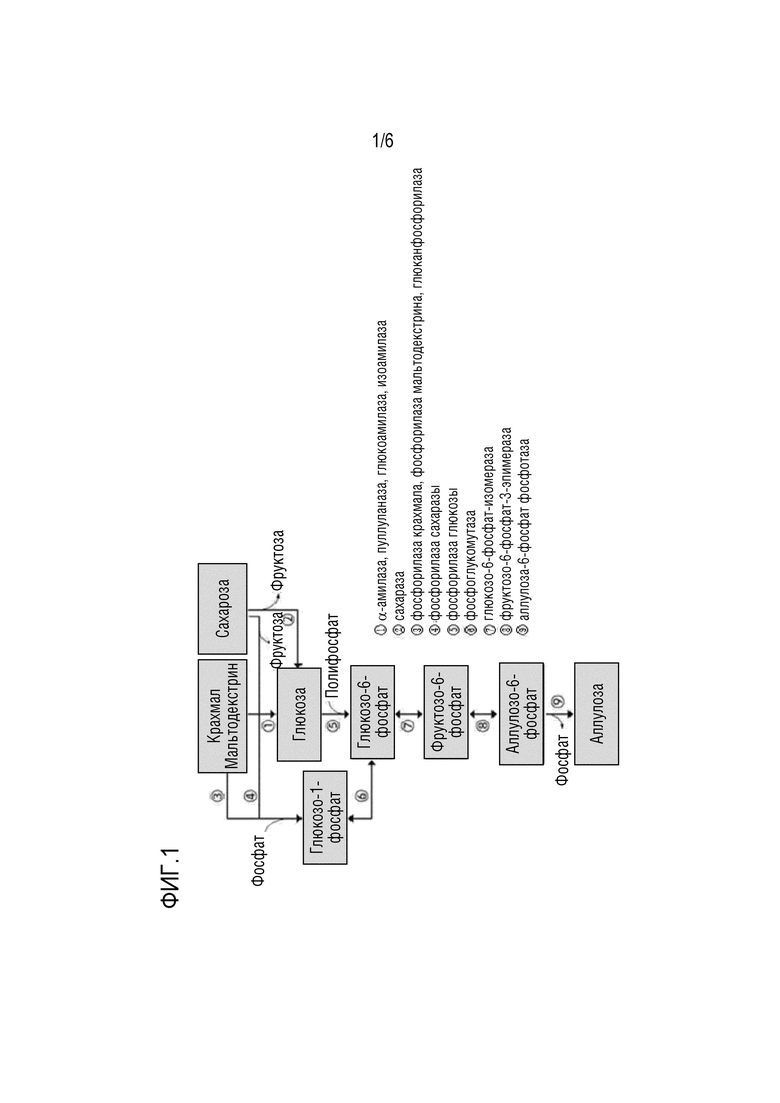
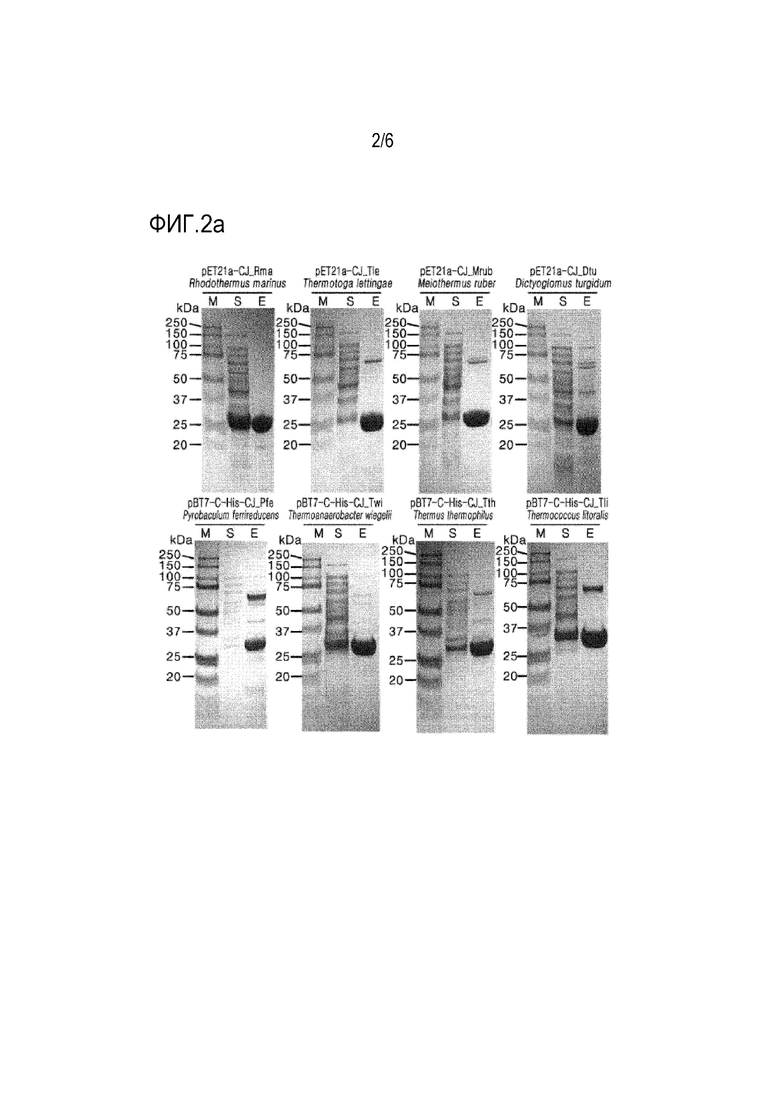

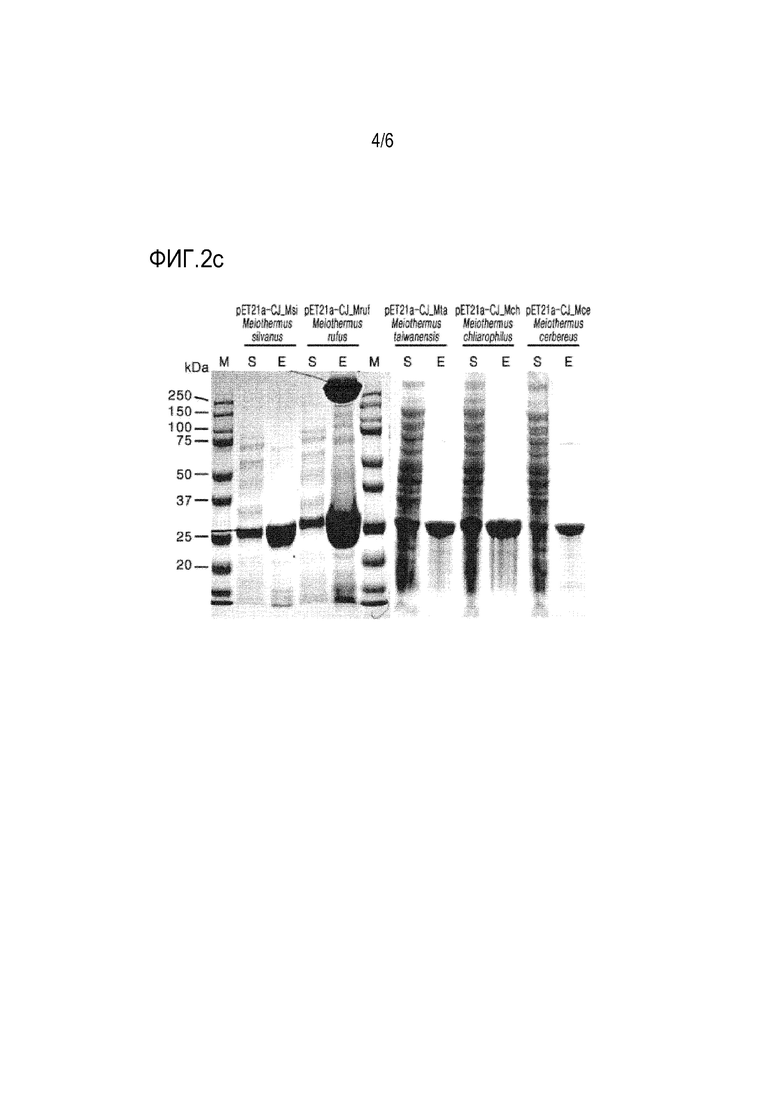

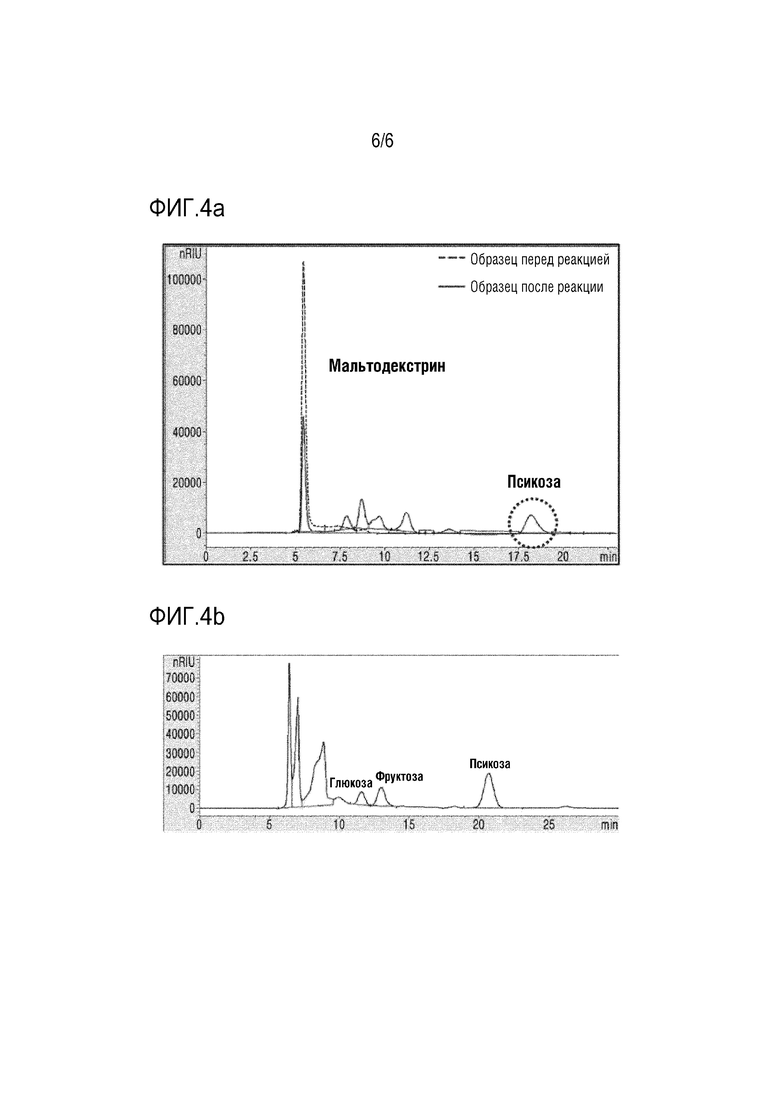
Изобретение относится к биотехнологии и представляет собой применение псикозо-6-фосфат фосфатазы для получения D-псикозы, где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV. Изобретение позволяет повысить выход D-псикозы. 5 н. и 11 з.п. ф-лы, 4 ил., 1 табл., 5 пр.
1. Применение псикозо-6-фосфат фосфатазы для получения D-псикозы, где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
2. Применение по п. 1, причем в мотиве A Xa1 представляет W или F, Xa2 представляет I или V, Xa3 представляет V или I, а Xa4 представляет T; и в мотиве B Ya1 представляет W, Ya2 представляет V или I, Wa1 представляет AAG, Ya3 представляет W, I или V, Wa2 представляет LLV, LIV, LII или LLI, Ya4 представляет E, R или S, а Wa3 представляет EАGG или EGGG.
3. Применение по п. 1, причем псикозо-6-фосфат фосфатаза содержит любую одну из аминокислотных последовательностей, приведенных в SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 и 20, или последовательность, имеющую идентичность по меньшей мере 85% с аминокислотной последовательностью, не являющейся мотивом A и мотивом B в последовательностях, приведенных в SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 и 20.
4. Применение по п. 3, причем псикозо-6-фосфат фосфатаза состоит из аминокислотной последовательности, приведенной в SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, кодируемой нуклеотидной последовательностью, приведенной в SEQ ID NO: 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 соответственно.
5. Применение нуклеиновой кислоты для получения D-псикозы, где нуклеиновя кислота кодирует псикозо-6-фосфат фосфатазу, где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
6. Комбинация для получения D-псикозы, содержащая (а) псикозо-6-фосфат фосфатазу, микроорганизм, экспрессирующий псикозо-6-фосфат фосфатазу, или культуру микроорганизма, экспессирующего псикозо-6-фосфат фосфатазу; и (b) D-псикозо-6-фосфат; где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
7. Комбинация по п. 6, дополнительно содержащая:
(a) (i) крахмал, мальтодекстрин, сахарозу или их комбинацию, (ii) фосфат, и
(b) (i) D-фруктозо-6-фосфат-3-эпимеразу, (ii) D-глюкозо-6-фосфат-изомеразу, (iii) фосфоглюкомутазу или глюкокиназу и (iv) α-глюканфосфорилазу, фосфорилазу крахмала, фосфорилазу мальтодекстрина, фосфорилазу сахарозы, α-амилазу, пуллуланазу, изоамилазу, глюкоамилазу или сахаразу; или
(a) (i) крахмал, мальтодекстрин, сахарозу или их комбинацию, (ii) фосфат; и
(с) микроорганизм, экспрессирующий ферменты, перечисленные в (b), или культуру микроорганизма, экспрессирующего ферменты, перечисленные в (b).
8. Способ получения D-псикозы, включающий введение в контакт псикозо-6-фосфат фосфатазы, микроорганизма, экспрессирующего псикозо-6-фосфат фосфатазу, или культуры микроорганизма, экспрессирующего псикозо-6-фосфат фосфатазу, с D-псикозо-6-фосфатом для преобразования D-псикозо-6-фосфата в D-псикозу, где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
9. Способ по п. 8, дополнительно включающий перед преобразованием D-псикозо-6-фосфата в D-псикозу введение D-фруктозо-6-фосфат-3-эпимеразы, микроорганизма, экспрессирующего D-фруктозо-6-фосфат-3-эпимеразу, или культуры микроорганизма, экспрессирующего D-фруктозо-6-фосфат-3-эпимеразу, в контакт с D-фруктозо-6-фосфатом для преобразования D-фруктозо-6-фосфата в D-псикозо-6-фосфат.
10. Способ по п. 9, дополнительно включающий перед преобразованием D-фруктозо-6-фосфата в D-псикозо-6-фосфат введение D-глюкозо-6-фосфат-изомеразы, микроорганизма, экспрессирующего D-глюкозо-6-фосфат-изомеразу, или культуры микроорганизма, экспрессирующего D-глюкозо-6-фосфат-изомеразу, в контакт с D-глюкозо-6-фосфатом для преобразования D-глюкозо-6-фосфата в D-фруктозо-6-фосфат.
11. Способ по п. 10, дополнительно включающий перед преобразованием D-глюкозо-6-фосфата в D-фруктозо-6-фосфат введение фосфоглюкомутазы, микроорганизма, экспрессирующего фосфоглюкомутазу, или культуры микроорганизма, экспрессирующего фосфоглюкомутазу, в контакт с D-глюкозо-1-фосфатом для преобразования D-глюкозо-1-фосфата в D-глюкозо-6-фосфат.
12. Способ по п. 10, дополнительно включающий перед преобразованием D-глюкозо-6-фосфата в D-фруктозо-6-фосфат введение глюкокиназы, микроорганизма, экспрессирующего глюкокиназу, или культуры микроорганизма, экспрессирующего глюкокиназу, и фосфата в контакт с глюкозой для преобразования глюкозы в D-глюкозо-6-фосфат.
13. Способ по п. 11, дополнительно включающий перед преобразованием D-глюкозо-1-фосфата в D-глюкозо-6-фосфат введение
(а) α-глюканфосфорилазы, фосфорилазы крахмала, фосфорилазы мальтодекстрина или фосфорилазы сахарозы;
(b) микроорганизма, экспрессирующего α-глюканфосфорилазу, фосфорилазу крахмала, фосфорилазу мальтодекстрина или фосфорилазу сахарозы; или
(с) культуры микроорганизма, экспрессирующего α-глюканфосфорилазу, фосфорилазу крахмала, фосфорилазу мальтодекстрина или фосфорилазу сахарозы, и фосфата в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией для преобразования крахмала, мальтодекстрина, сахарозы или их комбинации в D-глюкозо-1-фосфат.
14. Способ по п. 12, дополнительно включающий перед преобразованием глюкозы в D-глюкозо-6-фосфат введение
(а) α-амилазы, пуллуланазы, глюкоамилазы, сахаразы или изоамилазы;
(b) микроорганизма, экспрессирующего α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, или
(с) культуры микроорганизма, экспрессирующего α-амилазу, пуллуланазу, глюкоамилазу, сахаразу или изоамилазу, в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией для преобразования крахмала, мальтодекстрина, сахарозы или их комбинации в глюкозу.
15. Способ получения D-псикозы, включающий введение
(a) ферментов
(i) псикозо-6-фосфат фосфатазы,
(ii) D-фруктозо-6-фосфат-3-эпимеразы,
(iii) D-глюкозо-6-фосфат-изомеразы,
(iv) фосфоглюкомутазы или глюкокиназы и
(iv) α-глюканфосфорилазы, фосфорилазы крахмала, фосфорилазы мальтодекстрина, фосфорилазы сахарозы, α-амилазы, пуллуланазы, изоамилазы, глюкоамилазы или сахаразы,
(b) микроорганизмов, экспрессирующих ферменты, перечисленные в (a), или
(с) культуры микроорганизмов, экспрессирующих ферменты, перечисленные в (а), в контакт с крахмалом, мальтодекстрином, сахарозой или их комбинацией и фосфатом;
где псикозо-6-фосфат фосфатаза содержит мотив A, представленный Xa1-Xa2-Xa3-DPLDG-Xa4 или Xa1-Xa2-Xa3-DPIDG-Xa4, причем Xa1 представляет W, F, V, I или A, Xa2 представляет I, F, V, A или отсутствует, Xa3 представляет V, I или L, а Xa4 представляет T или S, и мотив B, представленный Ya1-D-Ya2-Wa1-Ya3-Wa2-Ya4-Wa3, причем Ya1 представляет W, Y, T, L или V, Ya2 представляет V, I, C, F или A, Wa1 представляет AAG, AAS, SAG, APG, APF, AGG, APL или AGA, Ya3 представляет W, I, P, M, V, Y, F, R, L, T или S, Wa2 представляет LLV, LIV, LLI, LII, ILI, FIA, ALV, IIA, VLV, VIL, TIG, NFC или PIF, Ya4 представляет E, R, S, T, L, K или P, а Wa3 представляет EАGG, EGGG, EAKG, KAGG, AAGG, YVDG, EAGA или RLGV.
16. Способ по любому из пп. 8-15, в котором контактную реакцию проводят при pH 5,0-9,0, при температуре 40°C-80°C и/или в течение от 2 часов до 24 часов.
| NCBIReferenceSequence: WP_013012427.1, 19/10/2015, https://www.ncbi.nlm.nih.gov/protein/WP_013012427.1?report=genbank&log$=protalign&blast_rank=1&RID=SER6864D016 | |||
| WO 2015099256 A1, 02.07.2015 | |||
| US 9701953 B2, 11.07.2017 | |||
| EP 3135762 A1, 01.03.2017 | |||
| D-ПСИКОЗА В ЗАМОРОЖЕННЫХ НАПИТКАХ НУЛЕВОЙ ИЛИ НИЗКОЙ КАЛОРИЙНОСТИ | 2014 |
|
RU2664311C2 |

