ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к D-псикозо-3-эпимеразе и к способу получения D-псикозы с ее использованием.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
D-псикоза (далее именуемая «псикоза») представляет собой моносахарид, известный как редкий сахар, присутствующий в очень маленьком количестве в природе. Он имеет практически ноль калорий при наличии почти 70% сладости сахара и привлек много внимания в качестве нового пищевого ингредиента из-за его функциональных свойств, таких как ингибирование содержания глюкозы в крови, ингибирование синтеза липидов и т.д.
Благодаря указанным свойствам псикоза рассматривается для применения в качестве заменителя сахара в разных пищевых продуктах. Однако имеется возрастающая потребность в способе эффективного получения псикозы, так как она существует в природе в очень маленьком количестве.
Известный способ получения псикозы включает способ применения катализа молибдатными ионами (Bilik, V., Tihlarik, K., 1973, Reaction of Saccharides Catalyzed by Molybdate Ions. IX. Epimerization of Ketohexoses. Chem. Zvesti. 28:106–109), химический способ получения псикозы из D-фруктозы посредством совместного нагревания этанола и триэтиламина (Doner, L.W., 1979, Isomerization of D-Fructose by Base: Liquid-Chromatographic Evaluation and The Isolation of D-Psicose. Carbohydr. Res. 70:209–216) и биологический способ получения псикозы из D-фруктозы с использованием микроорганизма, который продуцирует D-псикозо-3-эпимеразу (выложенная в открытый доступ публикация корейского патента №10-2011-0035805). Получение псикозы химическим способом имеет недостатки в том, что образуется большое количество побочных продуктов, и, таким образом, требуется проводить сложную очистку. Кроме того, биологический способ также имеет недостатки в том, что выход является очень низким, а стоимость получения высокой.
В данных обстоятельствах авторы настоящего изобретения приложили много усилий для разработки способа для увеличения выхода псикозы при получении и, в результате, подтвердили то, что при использовании новой D-псикозо-3-эпимеразы (далее именуемой «псикозоэпимераза») по настоящему изобретению скорость, с которой D-фруктоза превращается в псикозу (далее именуемая скорость превращения псикозы из D-фруктозы), увеличивается, и, посредством этого, смогли заметно увеличить выход псикозы при получении и осуществили настоящее изобретение.
ТЕХНИЧЕСКАЯ ПРОБЛЕМА
В одном воплощении настоящего изобретения предложены новая псикозоэпимераза, полинуклеотид, кодирующий псикозоэпимеразу, рекомбинантный вектор, включающий данный полинуклеотид, и микроорганизм, в который введен данный вектор.
В другом воплощении настоящего изобретения предложены композиция для получения D-псикозы, включающая псикозоэпимеразу, микроорганизм, экспрессирующий данную псикозоэпимеразу, или культуру данного микроорганизма, и способ получения D-псикозы с использованием псикозоэпимеразы.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
Согласно типичному воплощению настоящего изобретения предложена D-псикозо-3-эпимераза, состоящая из аминокислотной последовательности SEQ ID NO: 1.
В типичном воплощении псикозоэпимераза может включать полипептид, имеющий по меньшей мере 80%, 90%, 95%, 97% или 99% гомологии с аминокислотной последовательностью SEQ ID NO: 1. Очевидно то, что аминокислотная последовательность, имеющая активность превращения D-фруктозы в псикозу и вышеописанную гомологию, может включать случай, где часть аминокислотной последовательности SEQ ID NO: 1 заменена, вставлена, модифицирована и/или удалена. Кроме того, полипептиды, имеющие активность псикозоэпимеразы, также могут быть включены, без ограничения, в виде полипептида, кодируемого полинуклеотидом, который гибридизуется в жестких условиях с комплементарной последовательностью ко всей нуклеотидной последовательности или ее части, кодирующей зонд, который можно получать из известной последовательности гена, например, псикозоэпимеразы по настоящему изобретению.
Термин «полинуклеотид» в том виде, в котором он здесь используется, относится к полирибонуклеотиду или полидезоксирибонуклеотиду, в котором нуклеотидный мономер является немодифицированным или модифицированным до полимера из нуклеотидов, вытянутых в длинную цепь посредством ковалентных связей.
Термин «жесткие условия» в том виде, в котором он здесь используется, означает условия, которые обеспечивают специфичную гибридизацию между полинуклеотидами. Данные условия зависят от длины полинуклеотида и степени комплементарности. Их параметры хорошо известны в данной области и конкретно описаны в данном документе (например, J. Sambrook et al., выше). Например, жесткие условия могут описывать условия для гибридизации генов друг с другом, причем каждый имеет высокую гомологию – 80%, 90%, 95%, 97% или 99%, или более, при условиях негибридизации генов друг с другом каждый их которых имеет гомологию меньше, чем эта, или общие условия промывки саузерн-гибридизации, т.е. условия для промывки один раз, в частности, от двух до трех раз при таких концентрации соли и температуре, как 60°С, 1(SSC (раствор хлорида натрия и цитрата натрия), 0,1% SDS (додецилсульфат натрия), в частности, 0°С, 0,1× SSC, 0,1% SDS, и, более конкретно, 68°С, 0,1× SSC, 0,1% SDS. Зонд, используемый в гибридизации, может представлять собой часть комплементарной последовательности оснований. Такой зонд может быть сконструирован посредством ПЦР с использованием фрагмента гена, включающего последовательность оснований в качестве матрицы, посредством применения олигонуклеотида, полученного на основе известной последовательности в качестве праймера. Кроме того, специалисты в данной области могут откорректировать температуру и концентрацию соли промывочного раствора по необходимости, в зависимости от таких факторов, как длина зонда.
Термин «гомология» в том виде, в котором он здесь используется, относится к процентной доле идентичности между двумя полинуклеотидами или полипептидными группировками. Гомология между последовательностями из одной группировки и другой группировки может быть определена известными методиками. Например, гомология может быть определена путем прямого выравнивания параметров информации по последовательности между двумя полинуклеотидными молекулами или двумя полипептидными молекулами, таких как балл, идентичность, сходство и т.д., с использованием компьютерной программы, которая сортирует информацию по последовательности и доступна в настоящее время (например, BLAST 2.0). Кроме того, гомология между полинуклеотидами может быть определена посредством гибридизации полинуклеотидов в условиях образования стабильной двойной цепи между гомологичными областями, с последующим расщеплением нуклеазой, специфичной в отношении одной цепи, для определения размера расщепленного фрагмента.
Кроме того, при условии, что белок имеет активность, соответствующую псикозоэпимеразе, состоящей из аминокислотной последовательности SEQ ID NO: 1 по настоящему изобретению, перед и после аминокислотной последовательности SEQ ID NO: 1 можно добавлять несмысловую последовательность или включать ее встречающуюся в природе мутацию или молчащую мутацию. Белок, включающий аминокислотную последовательность SEQ ID NO: 1, также включен в пределы объема настоящего изобретения.
Кроме того, D-псикозо-3-эпимераза по настоящему изобретению может кодироваться полинуклеотидной последовательностью SEQ ID NO: 2 или полинуклеотидной последовательностью, имеющей по меньшей мере 80%-ную, 90%-ную, 95%-ную, 97%-ную или 99%-ную гомологию с ней, но настоящее изобретение не ограничивается ими. Кроме того, в отношении полинуклеотида, кодирующего D-псикозо-3-эпимеразу по настоящему изобретению, очевидно то, что полинуклеотид, способный к трансляции в белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, или в белок, имеющий гомологию с ним, посредством вырожденности кодонов, также может быть включен в пределы интервала полинуклеотидной последовательности по настоящему изобретению. Специалистам в данной области будет понятно то, что можно получать полинуклеотид, кодирующий фермент, имеющий по существу эквивалентный интервал активности, посредством замены, добавления и/или удаления одной или более чем одной нуклеотидной последовательности SEQ ID NO: 2 с использованием известных методик генной инженерии.
В другом типичном воплощении псикозоэпимераза по настоящему изобретению может иметь происхождение из микроорганизма рода Kaistia. В частности, псикозоэпимераза по настоящему изобретению может иметь происхождение из Kaistia granuli и, более конкретно, может иметь происхождение из Kaistia granuli KCTC 12575.
В другом типичном воплощении псикозоэпимераза по настоящему изобретению может иметь молекулярную массу от 25 кДа до 37 кДа, от 27 кДа до 35 кДа или от 30 кДа до 35 кДа при измерении посредством электрофореза на полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE).
Согласно другому типичному воплощению настоящего изобретения предложен полинуклеотид, кодирующий D-псикозо-3-эпимеразу, состоящую из аминокислотной последовательности SEQ ID NO: 1.
В типичном воплощении предложенный здесь полинуклеотид может представлять собой полинуклеотид, состоящий из последовательности оснований SEQ ID NO: 2 или последовательности, имеющей 80%, 90%, 95%, 97%, 99% или более гомологии с последовательностью оснований SEQ ID NO: 2. Кроме того, очевидно, что в отношении предложеного здесь полинуклеотида, полинуклеотид, способный к трансляции в белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, или в белок, имеющий с ним гомологию посредством вырожденности кодонов, также включается в пределы интервала по настоящему изобретению.
Согласно другому типичному воплощению настоящего изобретения предложен рекомбинантный вектор, включающий полинуклеотид, кодирующий D-псикозо-3-эпимеразу по настоящему изобретению.
Рекомбинантный вектор по настоящему изобретению может иметь форму, в которой полинуклеотид, кодирующий псикозоизомеразу, вставлен в клонирующий вектор или экспрессионный вектор с использованием известных стандартных способов. Термин «клонирующий вектор» в том виде, в котором он здесь используется, относится к вектору, способному переносить фрагмент ДНК в клетку-хозяина и регенерировать его. Данный клонирующий вектор может дополнительно включать сигнал полиаденилирования, последовательность терминации транскрипции и/или сайт множественного клонирования. Здесь сайт множественного клонирования может включать по меньшей мере один сайт эндонуклеазы и рестрикционного фермента. В типичном воплощении полинуклеотид, кодирующий псикозоэпимеразу, может находиться выше сигнала полиаденилирования и последовательности терминации транскрипции. Термин «экспрессионный вектор» в том виде, в котором он здесь используется, относится к последовательности ДНК, необходимой для транскрипции и трансляции клонированной ДНК в подходящем хозяине. Кроме того, термин «экспрессионный вектор» в том виде, в котором он здесь используется, относится к конструкции гена, включающей важный регуляторный элемент, связанный функциональным образом со вставкой таким образом, что данная вставка экспрессируется при наличии в клетке субъекта. Термин «связанный функциональным образом» означает то, что одна функция регулируется другой функцией посредством ассоциации полинуклеотидной последовательности на полинуклеотиде. Экспрессионный вектор может быть получен и очищен с использованием стандартных методик генной инженерии. Данный экспрессионный вектор может включать по меньшей мере один из промотора, кодона инициации, гена, кодирующего цитозинэпимеразу, и кодона терминации.
Согласно другому типичному воплощению настоящего изобретения предложен микроорганизм, в который введен рекомбинантный вектор, как описано выше.
В типичном воплощении микроорганизм, в который введен рекомбинантный вектор по настоящему изобретению, может быть микроорганизмом, который трансформирован рекомбинантным вектором, включающим полинуклеотид, кодирующий псикозоэпимеразу, включающую аминокислотную последовательность SEQ ID NO: 1, или рекомбинантным вектором, включающим полинуклеотид, состоящий из последовательности оснований SEQ ID NO: 2.
Термин «трансформация» в том виде, в котором он здесь используется, означает то, что ген или рекомбинантный вектор, включающий данный ген, введен в клетку-хозяина таким образом, что данный ген способен экспрессироваться в клетке-хозяине. Настоящее изобретение включает любой трансформированный ген, при условии, что данный трансформированный ген способен экспрессироваться в клетке-хозяине, без ограничения, независимо от того, вставлен ли он в хромосому клетки-хозяина или расположен вне хромосомы клетки-хозяина. Способ трансформации по настоящему изобретению включает временную трансформацию, микроинъекцию, трансдукцию, слияние клеток, осаждение фосфатом кальция, трансфекцию, опосредованную липосомами, трансфекцию, опосредованную DEAE декстраном, электроперфорацию (electric perforation), электроинъекцию, химическую обработку и т.д., но настоящее изобретение не ограничивается ими. Клетка-хозяин, способная к трансформации рекомбинантным вектором, может включать прокариотическую клетку, растительную клетку, животную клетку и т.д. Может использоваться клетка-хозяин, имеющая высокую эффективность введения ДНК и высокую скорость экспрессии введенной ДНК. Например, данная клетка-хозяин может представлять собой E. coli, штамм рода Bacillus, штамм рода Corynebacterium, штамм рода Salmonella и т.д., например, может представлять собой такую E. coli, как W3110, BL21, JM109, K-12, LE392, RR1 и DH5α и т.д. Более конкретно микроорганизм по настоящему изобретению может представлять собой E. coli BL21 (DE3) / KGDPE, депонированную с номером депонирования KCCM11918P.
Согласно другому типичному воплощению настоящего изобретения предложена композиция для получения D-псикозы, включающая: D-псикозо-3-эпимеразу, включающую аминокислотную последовательность SEQ ID NO: 1, микроорганизм, экспрессирующий D-псикозо-3-эпимеразу, культуру микроорганизма.
В типичном воплощении микроорганизм по настоящему изобретению сам может представлять собой штамм, его культуру или продукт разрушения микроорганизма. Культура или продукт разрушения по настоящему изобретению может включать D- псикозо-3-эпимеразу по настоящему изобретению. Кроме того, культура микроорганизма по настоящему изобретению может включать или может не включать микроорганизм. Кроме того, продукт разрушения микроорганизма по настоящему изобретению может представлять собой продукт разрушения, полученный разрушением микроорганизма или его культуры, или супернатант, полученный центрифугированием данного продукта разрушения.
В другом типичном воплощении композиция для получения D-псикозы по настоящему изобретению может дополнительно включать D-фруктозу в качестве субстрата псикозоэпимеразы.
В другом типичном воплощении микроорганизм по настоящему изобретению может быть иммобилизирован на носителе, подлежащем применению. Пример носителя, который можно использовать в настоящем изобретении, включает агар, агарозу, k-каррагенан, альгинат или хитозан, но не ограничивается ими.
Кроме того, композиция для получения D-псикозы по настоящему изобретению может дополнительно включать любой компонент, способный поддерживать получение псикозы. В частности, композиция для получения D-псикозы по настоящему изобретению может дополнительно включать металл. Более конкретно, металл по настоящему изобретению может представлять собой по меньшей мере один металл, выбранный из группы, состоящей из марганца, кальция, магния, железа, лития и натрия. Кроме того, металл по настоящему изобретению может представлять собой ион металла или соль металла. Металл по настоящему изобретению может иметь концентрацию от 0,1 мМ до 10 мМ, от 0,1 мМ до 7 мМ, от 0,1 мМ до 4 мМ, от 0,5 мМ до 10 мМ, от 0,5 мМ до 7 мМ, от 0,5 мМ до 4 мМ, от 1 мМ до 10 мМ, от 1 мМ до 7 мМ, от 1 мМ до 4 мМ, от 2 мМ до 10 мМ, от 2 мМ до 7 мМ или от 2 мМ до 4 мМ. Более конкретно, соль металла по настоящему изобретению может представлять собой по меньшей мере одну соль металла, выбранную из группы, состоящей из LiCl, Na2SO4, MgCl2, NaCl, FeSO4, MgSO4, MnCl2, MnSO4 и CaCl2.
Согласно другому типичному воплощению настоящего изобретения предложен способ получения D-псикозы, включающий: приведение D-псикозо-3-эпимеразы, состоящей из аминокислотной последовательности SEQ ID NO: 1, микроорганизма, экспрессирующего D-псикозо-3-эпимеразу, или культуры микроорганизма в контакт с D-фруктозой.
В типичном воплощении способ получения может дополнительно включать добавление металла до, после или одновременно с приведением в контакт с D-фруктозой.
В другом типичном воплощении способ получения может дополнительно включать, после приведения в контакт с D-фруктозой или добавления металла, выделение и/или очистку результата контакта, включающего псикозу. Выделение и/или очистку можно проводить одним или более чем одним известным способом, таким как диализ, осаждение, адсорбция, электрофорез, ионообменная хроматография, фракционная кристаллизация и т.д., но не ограничиваясь ими.
Кроме того, способ получения по настоящему изобретению может дополнительно включать, до или после выделения и/или очистки, проведение обесцвечивания и/или обессоливания. Посредством проведения обесцвечивания и/или обессоливания можно получать более очищенную псикозу без примесей.
В другом типичном воплощении способ получения по настоящему изобретению может дополнительно включать, после приведения в контакт с D-фруктозой, добавления металла, выделения и/или очистки, или проведения обесцвечивания и/или обессоливания, кристаллизацию D-псикозы. Кристаллизацию можно проводить с использованием способа кристаллизации, который традиционно используется. Например, кристаллизацию можно проводить с использованием способа кристаллизации охлаждением.
В другом типичном воплощении способ получения по настоящему изобретению может дополнительно включать, перед кристаллизацией, концентрирование псикозы. Концентрирование может увеличивать эффективность кристаллизации.
В другом воплощении способ получения по настоящему изобретению может дополнительно включать, после выделения и/или очистки, приведение непрореагировавшей D-фруктозы в контакт с псикозоэпимеразой или может дополнительно включать, после кристаллизации, повторное применение маточного раствора, из которого осуществляется выделение кристаллизацией, в выделении и/или очистке, или их комбинацию. Посредством данных дополнительных стадий псикозу можно получать с большим выходом, и может быть уменьшено количество D-фруктозы, подлежащей отбрасыванию, получая, посредством этого, экономическую пользу.
В типичном воплощении приведение в контакт по настоящему изобретению можно проводить при рН от 5,0 до 9,0 при 40-90°С и/или в течение 0,5-48 часов.
В частности, приведение в контакт по настоящему изобретению можно проводить при рН от 6,0 до 8,5, при рН от 6,0 до 8,0 или при рН от 7,0 до 8,0.
Кроме того, приведение в контакт по настоящему изобретению может проводиться при температуре от 40°С до 80°С, от 40°С до 75°С, от 40°С до 65°С, от 50°С до 90°С, от 50°С до 80°С, от 50°С до 75°С, от 50°С до 65°С, от 55°С до 90°С, от 55°С до 80°С, от 55°С до 75°С, от 55°С до 65°С, от 60°С до 90°С, от 60°С до 80°С, от 60°С до 75°С, от 60°С до 65°С, от 65°С до 90°С, от 65°С до 80°С или от 65°С до 75°С.
Кроме того, приведение в контакт по настоящему изобретению может проводиться в течение 0,5 часа или более, 1 часа или более, 3 часов или более, 5 часов или более, или 6 часов или более, и/или 48 часов или менее, 36 часов или менее, 24 часов или менее, 12 часов или менее, 9 часов или менее.
Псикозоэпимераза, металл и носитель, описанные в способе получения D-псикозы по настоящему изобретению, являются такими же, как описано в вышеописанных типичных воплощениях.
Согласно еще одному другому типичному воплощению по настоящему изобретению предложено применение псикозоэпимеразы, микроорганизма, экспрессирующего псикозоэпимеразу, или культуры микроорганизма, как здесь описано, для превращения D-фруктозы при получении псикозы.
ЭФФЕКТ ИЗОБРЕТЕНИЯ
Псикозоэпимераза по настоящему изобретению обладает превосходной активностью превращения D-фруктозы в псикозу, обладает стабильностью при высокой температуре в той степени, в которой она доступна в промышленности, и имеет высокую скорость реакции превращения. Соответственно, при использовании псикозоэпимеразы по настоящему изобретению для получения псикозы можно получать псикозу с высокой эффективностью и высоким выходом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
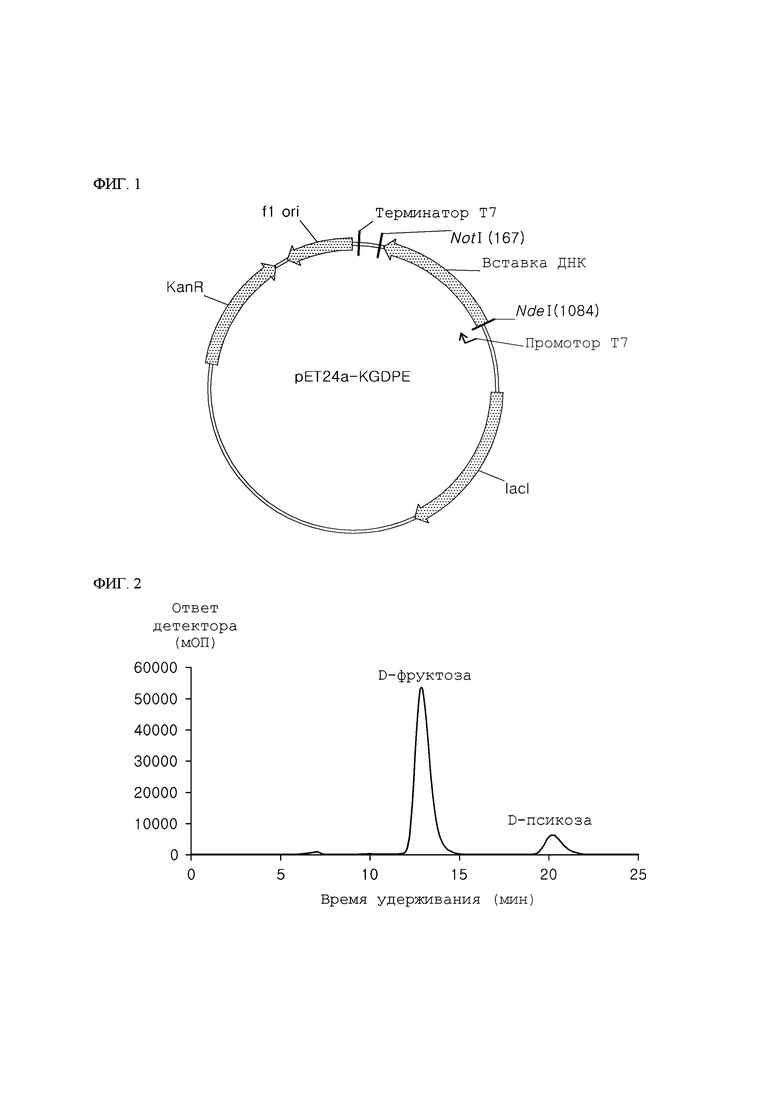
ФИГ. 1 представляет собой диаграмму рекомбинантного вектора для осуществления экспрессии псикозоэпимеразы (KGDPE), состоящей из аминокислотной последовательности SEQ ID NO: 1 по настоящему изобретению.
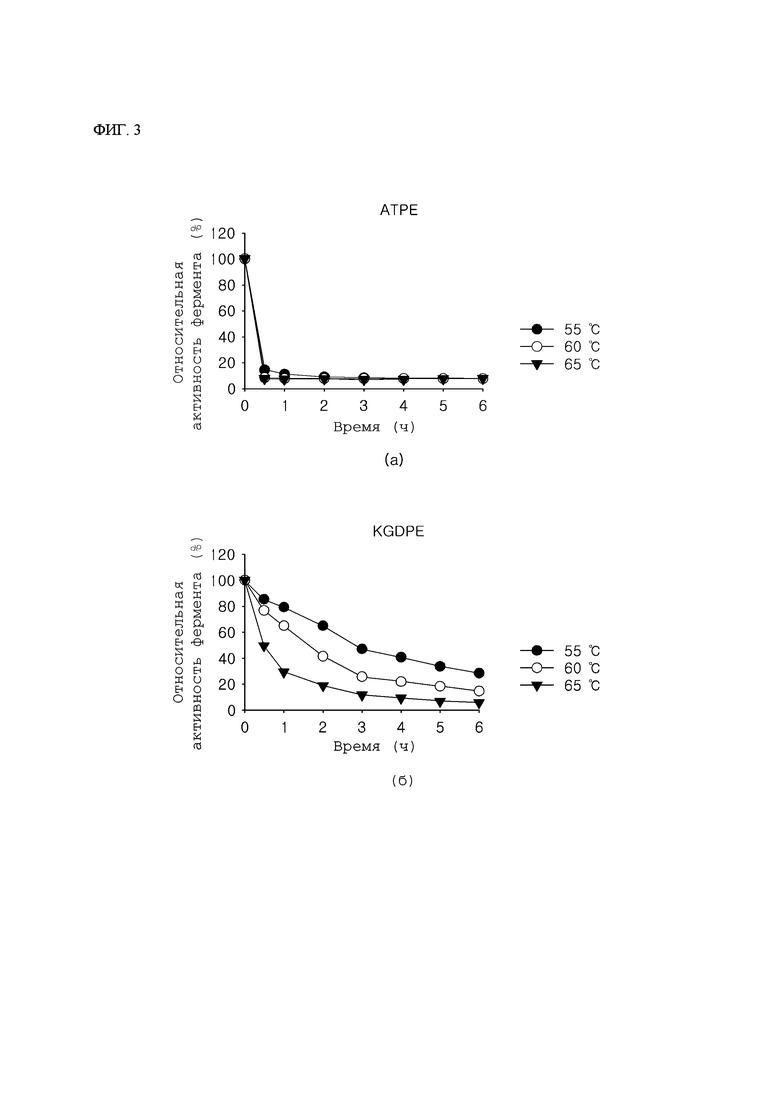
ФИГ. 2 представляет собой данные анализа ВЭЖХ (высокоэффективная жидкостная хроматография) по результатам получения псикозы с использованием D- фруктозы в качестве субстрата, используя KGDPE по настоящему изобретению.
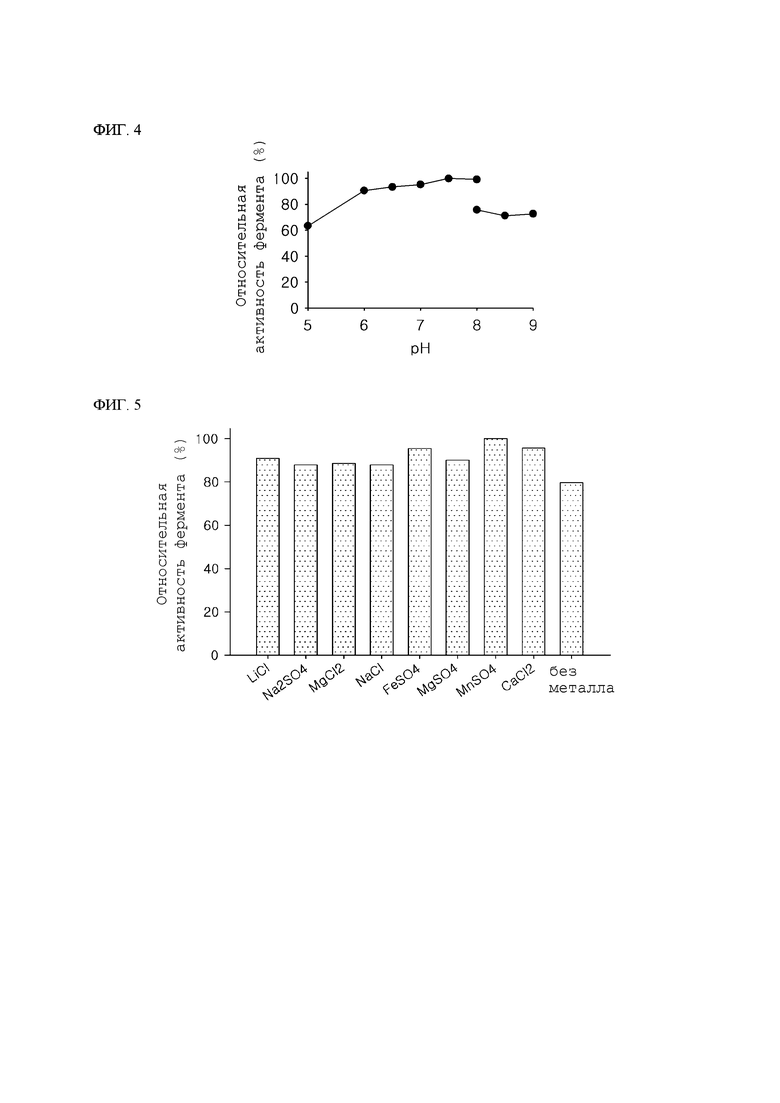
ФИГ. 3А и 3Б представляют собой графики, показывающие относительную ферментативную активность псикозоэпимеразы согласно температуре, где на ФИГ. 3А показана активность D-псикозо-3-эпимеразы (АТРЕ), имеющей происходение из Agrobacterium tumefaciens в предшествующем уровне техники, и на ФИГ. 3Б показана активность KGDPE в настоящем изобретении.
ФИГ. 4 представляет собой график, показывающий относительную ферментативную активность KGDPE по настоящему изобретению согласно изменению рН.
ФИГ. 5 представляет собой график, показывающий относительную ферментативную активность KGDPE по настоящему изобретению согласно добавлению разных металлов.
ПОДРОБНОЕ ОПИСАНИЕ ВОПЛОЩЕНИЙ
Далее настоящее изобретение будет более подробно описано посредством следующих примеров. Однако настоящее изобретение не ограничивается Примерами, приведенными ниже, и следует понимать то, что специалистами в данной области могут быть сделаны разные модификации и изменения в пределах объема и сущности настоящего изобретения.
Во всем описании настоящего изобретения, если не отмечено иное, «%», используемый для обозначения концентрации конкретного вещества, относится к % твердое вещество/твердое вещество (масс./масс.), % твердое вещество/жидкость (масс./об.) и % жидкость/жидкость (об./об.).
ПРИМЕРЫ
Пример 1. Получение трансформированного штамма, для получения псикозоэпимеразы, имеющей происхождение из микроорганизма рода Kaistia.
Выбирали ген из микроорганизма рода Kaistia, который, как ожидалось, имеет активность псикозоэпимеразы, которая превращает D-фруктозу в псикозу, и получали рекомбинантный экспрессионный вектор, включающий данный ген, и трансформированный микроорганизм.
В частности, ген kgdpe Kaistia granuli KCTC 12575, который, как ожидалось, представляет собой псикозоэпимеразу, выбирали из последовательностей гена микроорганизма рода Kaistia, зарегистрированного в Genbank, и конструировали и синтезировали прямой праймер (SEQ ID NO: 3) и обратный праймер (SEQ ID NO: 4) на основе аминокислотной последовательности (SEQ ID NO: 1) и нуклеотидной последовательности (SEQ ID NO: 2) данного гена. Посредством применения синтезированного праймера ген амплифицировали проведением ПЦР (полимеразная цепная реакция) реакции (33 цикла: 1 цикл, включающий 94°С в течение 1 минуты, 58°С в течение 30 секунд и 72°С в течение 1 минуты) с использованием геномной ДНК Kaistia granuli KCTC 12575 в качестве матрицы. Данный амплифицированный ген очищали с использованием набора для очистки ПЦР (Quiagen) и вставляли в pET24a (+) (Novagen, США), используя рестрикционные ферменты NdeI и notI для конструирования рекомбинантного вектора pET24a (+) –KGDPE (ФИГ. 1).
Рекомбинантный вектор трансформировали в Escherichia coli BL21 (DE3) посредством трансформации на основе теплового шока (Sambrook and Russell: Molecular Cloning, 2001), затем хранили в замороженном состоянии в 50%-ном глицерине и использовали. Трансформированный штамм был назван E. coli BL21(DE3)/KGDPE, и был депонирован 20 октября 2016 г. в Корейском центре культур микроорганизмов (KCCM), который представляет собой международный депозиторий, работающий согласно Будапештскому соглашению, и ему был присвоен номер доступа KCCM11918P.
Пример 2. Получение и очистка псикозоэпимеразы
Для получения псикозоэпимеразы из E. coli BL21(DE3)/KGDPE, полученной в Примере 1, E. coli BL21(DE3)/KGDPE инокулировали в 5 мл среды LB-канамицин и подвергали культивированию со встряхиванием при 37°С, 200 об./мин, пока оптическая плотность, измеряемая при 600 нм, не достигала 1,5. Затем жидкость от культивирования со встряхиванием инокулировали в 500 мл среды LB-канамицин, и, когда оптическая плотность при 600 нм составляла 0,7, добавляли 0,5 мМ изопропил-β-D-1-тиогалактопиранозид (IPTG), и осуществляли главное культивирование клеток при 16°С и 150 об./мин в течение 16 часов.
Жидкость от главного культивирования центрифугировали при 8000 об./мин в течение 20 минут для выделения только клеток, и клетки дважды промывали 0,85% (масс./об.) NaCl, затем лизировали в лизисном буфере (50 мМ Tris-HCl, pH 7,0, 300 мМ NaCl) и разрушали при 4°С в течение 20 минут с использованием звукового вибратора. Жидкость от разрушенных клеток центрифугировали при 4°С, 13000 об./мин, в течение 20 минут для выделения супернатанта. Затем супернатант наносили на колонку Ni-NTA (Ni-NTA Superflow, Qiagen), ранее уравновешенную приведенным выше лизисным буфером, и последовательно пропускали буферный раствор (50 мМ Tris-HCl, 300 мМ NaCl, pH 7,0), содержащий 250 мМ имидазол, с получением очищенной псикозоэпимеразы (далее именуемой KGDPE). SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) KGDPE подтвердил то, что размер мономера составлял примерно 32 кДа.
Пример 3. Подтверждение активности KGDPE
3-1. Подтверждение активности превращения из D-фруктозы в псикозу
Чтобы подтвердить, создает ли KGDPE псикозу с использованием D-фруктозы в качестве субстрата, KGDPE (50 мМ Tris-HCl, pH 7,0), полученную в Примере 2, добавляли в 50 мМ буфер Tris-HCl (pH 8,0), содержащий 50% масс. D-фруктозы и 3 мМ MnSO4, и проводили реакцию при 55°С в течение 6 часов. Затем реакцию останавливали нагреванием при 100°С в течение 5 минут, и затем получение псикозы подтверждали анализом ВЭЖХ (высокоэффективная жидкостная хроматография). Анализ ВЭЖХ проводили с использованием ВЭЖХ (Agilent, США) с детектором показателя преломления (Agilent 1260 RID), оснащенного колонкой Aminex HPX-87C (BIO-RAD), где растворителем подвижной фазы была вода, температура составляла 80°С, и скорость тока составляла 0,6 мл/мин.
В результате подтвердили то, что псикоза могла быть получена из D-фруктозы с использованием KGDPE (ФИГ. 2).
3-2. Подтверждение активности превращения из D-фруктозы в псикозу
Чтобы подтвердить, превосходит ли способность KGDPE к получению над способностью обычной псикозоэпимеразы (ATPE, SEQ ID NO: 5, выложенная в открытый доступ публикация корейского патента №10-2011-0035805), используемой в получении псикозы, подтвердили скорость превращения из D-фруктозы в псикозу.
В частности, E. coli BL21 (DE3), трансформированную рекомбинантным экспрессионным вектором pET24a-ATPE, инокулировали в среду LB, содержащую канамицин, имеющий концентрацию 10 мкг/мл, фермент затем экспрессировался, и его очищали таким же способом, что и в Примере 2. Полученный фермент добавляли в 50 мМ буфер Tris-HCl (pH 8,0), содержащий 50% масс. D-фруктозы и 3 мМ MnSO4, и проводили взаимодействие при 55°С в течение 6 часов. Затем реакцию останавливали нагреванием при 100°С в течение 5 минут, и затем получение псикозы подтверждали анализом ВЭЖХ. Анализ ВЭЖХ проводили при тех же самых условиях, что и в Примере 3-1. Скорость превращения в псикозу рассчитывали как количество (мг/мин) псикозы, полученной ферментом за минуту, и скорость реакции KGDPE была показана в виде относительного значения, где значение скорости реакции АТРЕ было принято за 100%.
В результате подтвердили то, что количество псикозы, полученное за минуту при использовании KGDPE, составляло 117,6% по сравнению с применением АТРЕ, и, таким образом, скорость превращения из D-фруктозы в псикозу была заметно увеличена при использовании KGDPE (Таблица 1).
Таблица 1
Пример 4. Анализ характеристик KGDPE
4-1. Анализ активности фермента в соответствии с температурой
KGDPE и субстрат – D-фруктозу – подвергали взаимодействию в течение 2 часов при условиях разной температуры (40°С, 45°С, 50°С, 55°С, 60°С, 65°С, 70°С и 75°С), и сравнивали активности фермента в соответствии с температурами. Приведенные выше реакции проводили таким же способом, что и в Примере 3-1, за исключением температуры и времени реакции, и активности фермента измеряли как скорость превращения от D-фруктозы в псикозу. Скорость превращения рассчитывали как процентную долю массы псикозы, полученной после реакции, относительно массы субстрата (D-фруктозы) до реакции.
В результате KGDPE демонстрировала высокую активность превращения 25% или более во всех интервалах температуры измерения, и подтвердили то, что по мере увеличения температуры возрастала активность, и максимальную скорость превращения наблюдали при максимальной температуре 75°С (Таблица 2).
Таблица 2
4-2. Анализ термостабильности фермента
Для сравнения термостабильности KGDPE с термостабильностью обычного фермента – АТРЕ – соответствующие ферменты обрабатывали нагреванием при разных температурах (55°С, 60°С и 65°С), и растворы ферментативной обработки отбирали для каждого времени тепловой обработки (0,5 часа, 1 час, 2 часа, 3 часа, 4 часа, 5 часов и 6 часов) для определения остаточной активности каждого фермента. Данную реакцию проводили в течение 30 минут, изменяя время реакции только таким же способом, как и в Примере 3-1, и остаточную активность фермента измеряли по скорости превращения от D-фруктозы в псикозы.
В результате, уменьшение времени полужизни KGDPE согласно увеличению температуры было примечательно меньше, чем время полужизни АТРЕ, и, таким образом, было подтверждено то, что KGDPE имела высокую термостабильность (ФИГ. 3А и 3Б).
4-3. Анализ ферментативной активности в соответствии с рН
Для определения ферментативной активности в соответствии с рН, субстрат - D-фруктозу – подвергали взаимодействию с KGDPE при разных рН. В это время реакцию проводили таким же способом, что и в Примере 3-1, за исключением времени реакции и рН.
В частности, ферментативную реакцию проводили при 55°С в течение 30 минут посредством применения 50 мМ фосфата калия при рН 5,0, рН 6,0, рН 6,5, рН 7,0, рН 7,5 и рН 8,0, и используя 50 мМ буфер Tris-HCl при рН 8,0, рН 8,5 и рН 9,0. Затем ферментативную активность измеряли как скорость превращения из D-фруктозы в псикозу.
В результате подтвердили то, что KGDPE демонстрировала активность 70% или больше по сравнению с максимальной активностью при рН 6 – рН 8,5 и демонстрировала наивысшую активность при рН 8,0 (Таблица 3, ФИГ. 4).
Таблица 3
4-4. Анализ активности фермента в соответствии с добавлением металла
Чтобы подтвердить активность KGDPE в соответствии с добавлением металла при тех же самых условиях реакции, что и в Примере 3-1, MnSO4 заменяли разными солями металлов (LiCl, Na2SO4, MgCl2, NaCl, FeSO4 и CaCl2) и добавляли в конечной концентрации 3 мМ. Затем измеряли ферментативную активность. Контрольную группу не обрабатывали солями металлов.
В результате подтвердили то, что добавление Li, Na, Mg, Fe и Ca, а также Mn увеличивало активность KGDPE по сравнению с контрольной группой, и среди них могло быть подтверждено то, что Mn больше всего увеличивал ферментативную активность (Таблица 4 и ФИГ. 5).
Таблица 4
Из приведенного выше описания специалистам в данной области будет понятно то, что настоящее изобретение может быть выполнено в других конкретных формах без модификации его технической идеи или существенных характеристик. В данном отношении следует понимать то, что описанные выше воплощения являются во всех аспектах иллюстративными и не ограничивающими. Объем настоящего изобретения следует интерпретировать как охватывающий все модификации или вариации, происходящие из значения и объема приложенной формулы изобретения и ее эквивалентов, а не подробного описания.
--->
<110> CJ CHEILJEDANG CORPORATION
<120> A Novel D-Psicose 3-Epimerase and Methods for Preparing D-Psicose
Using The Same
<130> OPA18334
<150> KR 10-2016-0152947
<151> 2016-11-16
<160> 5
<170> KopatentIn 2.0
<210> 1
<211> 297
<212> PRT
<213> Amino acid sequences of KGDPE
<400> 1
Met Lys Asn Lys Leu Gly Val His Ala Gln Val Trp Val Gly Gly Trp
1 5 10 15
Ser His Ala Glu Ala Glu Arg Ala Ile Ala Ser Thr Ala Ser Leu Gly
20 25 30
Tyr Asp Tyr Ile Glu Ala Pro Ala Leu Asp Pro Ser Leu Ile Asp Ile
35 40 45
Asp Phe Thr Arg Lys Ala Leu Glu Lys His Gly Leu Gly Ile Thr Thr
50 55 60
Ser Leu Gly Leu Asp Asp Ser Cys Asp Ile Ser Ser Gly Asp Pro Asp
65 70 75 80
Lys Lys Ala Arg Gly Gln Ala His Leu Met Lys Val Val Ser Thr Thr
85 90 95
Arg Asp Leu Gly Gly Thr His Ile Thr Gly Ile Leu Tyr Ser Gly Phe
100 105 110
Gln Lys Tyr Phe Thr Pro Ala Thr Pro Glu Gly Val Ala Gly Ala Val
115 120 125
Glu Val Leu His His Val Ala Glu Glu Ala Ala Lys Ser Asn Ile Thr
130 135 140
Leu Gly Leu Glu Val Val Asn Arg Tyr Glu Thr Asn Val Ile Asn Thr
145 150 155 160
Ala Ala Gln Gly Val Glu Leu Cys Lys Arg Val Gly Met Pro Asn Val
165 170 175
Lys Val His Leu Asp Cys Tyr His Met Asn Ile Glu Glu Ala Asp Ala
180 185 190
Glu Arg Ala Ile Ile Asp Thr Gly Asp Tyr Leu Gly Tyr Phe His Thr
195 200 205
Gly Glu Ser His Arg Gly Tyr Leu Gly Thr Gly Ser Ile Asp Phe Thr
210 215 220
Arg Ile Phe Arg Gly Leu Val Lys Ala Asn Tyr Gln Gly Pro Ile Cys
225 230 235 240
Phe Glu Ser Phe Ser Ser Ala Val Ala Gly Glu Pro Leu Ser Gly Ile
245 250 255
Leu Gly Ile Trp Arg Asn Leu Trp Thr Asp Ser Thr Asp Leu Cys Arg
260 265 270
His Ala Met Gln Phe Thr Gln Ala Gln Met Gln Ala Ala Glu Gln Ala
275 280 285
Gln Ser Ile Arg Thr Gly Ala Asp Trp
290 295
<210> 2
<211> 894
<212> DNA
<213> Nucleotide sequences of KGDPE
<400> 2
atgaagaaca agctgggtgt gcacgcacag gtctgggtcg gcggctggag ccatgcggag 60
gcggagcgcg ccatcgccag caccgcctcg ctcggctacg actatatcga ggcgccggcg 120
ctcgacccgt cgctgatcga catcgacttc acccgcaaag cgctggaaaa gcatggtctc 180
ggcatcacga cgtcgctcgg cctcgacgac agctgcgaca tctcctcggg cgatcccgac 240
aagaaggcgc gcggccaggc gcacctgatg aaggtggtct ccaccacccg tgatctcggc 300
ggcacccaca tcaccggtat cctctattcc ggcttccaga aatacttcac gcccgcaacg 360
ccggagggcg tcgccggcgc cgtcgaggta ttgcaccacg tcgccgagga agcggcgaag 420
agcaacatca cgctcggcct cgaggtggtg aaccgctacg agaccaacgt gatcaacacc 480
gccgcccagg gcgtcgagct ctgcaagcgg gtcggcatgc cgaacgtcaa ggtgcacctc 540
gactgctacc acatgaacat cgaggaagcc gacgccgagc gcgccatcat cgataccggc 600
gactatctgg gttatttcca taccggtgaa tcgcatcgcg gctatctcgg caccggctcg 660
atcgacttca cccgcatctt ccgcggcctg gtgaaggcca actaccaggg tccgatctgc 720
ttcgaatcct tctcgtccgc cgtcgccggc gagccgctct ccggcattct cggcatctgg 780
cgcaatctct ggacggattc gaccgatctc tgccgccacg ccatgcagtt cacgcaggca 840
cagatgcagg cggccgagca ggcccagtcg atccgcaccg gcgcggactg gtag 894
<210> 3
<211> 22
<212> DNA
<213> A forward primer of KGDPE
<400> 3
atgaagaaca agctgggtgt gc 22
<210> 4
<211> 20
<212> DNA
<213> A reverse primer of KGDPE
<400> 4
tcaccagtcc gcgccggtgc 20
<210> 5
<211> 289
<212> PRT
<213> Amino acid sequences of ATPE
<400> 5
Met Lys His Gly Ile Tyr Tyr Ser Tyr Trp Glu His Glu Trp Ser Ala
1 5 10 15
Lys Phe Gly Pro Tyr Ile Glu Lys Val Ala Lys Leu Gly Phe Asp Ile
20 25 30
Leu Glu Val Ala Ala His His Ile Asn Glu Tyr Ser Asp Ala Glu Leu
35 40 45
Ala Thr Ile Arg Lys Ser Ala Lys Asp Asn Gly Ile Ile Leu Thr Ala
50 55 60
Gly Ile Gly Pro Ser Lys Thr Lys Asn Leu Ser Ser Glu Asp Ala Ala
65 70 75 80
Val Arg Ala Ala Gly Lys Ala Phe Phe Glu Arg Thr Leu Ser Asn Val
85 90 95
Ala Lys Leu Asp Ile His Thr Ile Gly Gly Ala Leu His Ser Tyr Trp
100 105 110
Pro Ile Asp Tyr Ser Gln Pro Val Asp Lys Ala Gly Asp Tyr Ala Arg
115 120 125
Gly Val Glu Gly Ile Asn Gly Ile Ala Asp Phe Ala Asn Asp Leu Gly
130 135 140
Ile Asn Leu Cys Ile Glu Val Leu Asn Arg Phe Glu Asn His Val Leu
145 150 155 160
Asn Thr Ala Ala Glu Gly Val Ala Phe Val Lys Asp Val Gly Lys Asn
165 170 175
Asn Val Lys Val Met Leu Asp Thr Phe His Met Asn Ile Glu Glu Asp
180 185 190
Ser Phe Gly Asp Ala Ile Arg Thr Ala Gly Pro Leu Leu Gly His Phe
195 200 205
His Thr Gly Glu Cys Asn Arg Arg Val Pro Gly Lys Gly Arg Met Pro
210 215 220
Trp His Glu Ile Gly Leu Ala Leu Arg Asp Ile Asn Tyr Thr Gly Ala
225 230 235 240
Val Ile Met Glu Pro Phe Val Lys Thr Gly Gly Thr Ile Gly Ser Asp
245 250 255
Ile Lys Val Trp Arg Asp Leu Ser Gly Gly Ala Asp Ile Ala Lys Met
260 265 270
Asp Glu Asp Ala Arg Asn Ala Leu Ala Phe Ser Arg Phe Val Leu Gly
275 280 285
Gly
<---
Изобретение относится к биотехнологии. Описано применение фермента D-псикозо-3-эпимеразы для получения D-псикозы из D-фруктозы, а также способ получения последней с использованием этого фермента или микроорганизма, его продуцирующего. Изобретение позволяет получить моносахарид D-псикозу из D-фруктозы с использованием указанного фермента. 6 н. и 3 з.п. ф-лы, 5 ил., 4 табл., 4 пр.
1. Применение D-псикозо-3-эпимеразы, состоящей из аминокислотной последовательности SEQ ID NO: 1, для получения D-псикозы из D-фруктозы.
2. Применение по п. 1, где D-псикозо-3-эпимераза кодируется полинуклеотидной последовательностью SEQ ID NO: 2.
3. Применение полинуклеотида, кодирующего D-псикозо-3-эпимеразу по п. 1, для получения D-псикозы из D-фруктозы.
4. Применение рекомбинантного вектора, содержащего полинуклеотид по п. 3, для получения D-псикозы из D-фруктозы.
5. Применение микроорганизма, экспрессирующего D-псикозо-3-эпимеразу, состоящую из аминокислотной последовательности SEQ ID NO: 1, для получения D-псикозы из D-фруктозы.
6. Композиция для получения D-псикозы, содержащая: D-фруктозу и D-псикозо-3-эпимеразу, состоящую из аминокислотной последовательности SEQ ID NO: 1, или микроорганизм, экспрессирующий D-псикозо-3-эпимеразу.
7. Способ получения D-псикозы, включающий: приведение D-псикозо-3-эпимеразы, состоящей из аминокислотной последовательности SEQ ID NO: 1, или микроорганизма, экспрессирующего D-псикозо-3-эпимеразу, в контакт с D-фруктозой.
8. Способ по п. 7, в котором приведение в контакт проводят при рН от 5,0 до 9,0, при температуре от 40 °С до 90 °С или в течение 0,5-48 часов.
9. Способ по п. 8, дополнительно включающий: до, после или одновременно с приведением в контакт с D-фруктозой, приведение D-псикозо-3-эпимеразы, состоящей из аминокислотной последовательности SEQ ID NO: 1, или микроорганизма, экспрессирующего D-псикозо-3-эпимеразу, в контакт с металлом.
| NCBI, GenBank ac.no | |||
| WP_018181373.1, 28.06.2013 | |||
| EP 2990483 A1, 02.03.2016 | |||
| US 9217166 B2, 22.12.2015 | |||
| WO 2015099256 A1, 02.07.2015 | |||
| KR 101455624 B1, 28.10.2014 | |||
| МИКРООРГАНИЗМ, ЭКСПРЕССИРУЮЩИЙ КСИЛОЗОИЗОМЕРАЗУ | 2009 |
|
RU2553537C2 |

