ОБЛАСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении раскрыты полинуклеотидные последовательности генов, кодирующие Terminal Flower 1 (TFL1) из Nicotiana tabacum, и их варианты, гомологи и фрагменты. Также раскрыты полипептидные последовательности, кодируемые таким образом, и их варианты, гомологи и фрагменты. Также раскрыта модификация у растения экспрессии одного или более генов или активности белка(-ов), кодируемого(-ых) таким образом, с целью обеспечения модуляции периода времени до наступления цветения. В одном варианте осуществления экспрессия одного или более генов или активность белка(-ов), кодируемого(-ых) таким образом, снижена с целью сокращения периода времени до наступления цветения. Растения, растительный материал и т. п. с измененным периодом времени до наступления цветения также описаны.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Период времени до цветения представляет собой строго контролируемый механизм у растений, который имеет непосредственное влияние на выживаемость и размножение. Переход к цветению также непосредственно связан с урожайностью культур. У растений образовались специализированные сигнальные пути, которые приводят к образованию репродуктивных структур вместо листьев. Локус цветения T (FT) и TFL1 являются членами семейства фосфатидилэтаноламинсвязывающих белков (PEBP), которые сходны с PEBP млекопитающих и функционируют в качестве факторов транскрипции. TFL1 действует антагонистически путем задержки вовлечения в цветение. Белок FT взаимодействует с фактором транскрипции bZIP локуса цветения D (FD) в апикальной меристеме побега с обеспечением цветения. Белок TFL1 также связывается с FD с целью подавления генов, расположенных ниже по цепи, таких как LEAFY (LFY) и APETALA1 (AP1). После перехода к цветению TFL1 подвергается повышающей регуляции для уравновешивания активности FT.
Существует общая необходимость в данной области техники в разработке растений, которые характеризуются сокращенным периодом времени до наступления цветения, поскольку это может приводить к ряду преимуществ, особенно связанных с коммерческим получением растений. Например, это может обеспечивать более короткий период времени от посева/посадки до сбора урожая, что может сокращать вегетационный период. Это может давать возможность более быстрого введения новых признаков путем скрещивания. Это может приводить к снижению затрат при коммерческом получении растений. Настоящее изобретение направлено на удовлетворение этой потребности.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Было идентифицировано семь генов TFL-1 в Nicotiana tabacum, называемых TFL1-1S (SEQ ID NO: 1 или 2), TFL1-1T (SEQ ID NO: 4 или 5), TFL1-2S (SEQ ID NO: 7 или 8), TFL1-2T (SEQ ID NO: 10 или 11), TFL1-3T (SEQ ID NO: 13 или 14), TFL1-4S (SEQ ID NO: 16 или 17) и TFL1-4T (SEQ ID NO: 19 или 20). Неожиданно авторы настоящего изобретения обнаружили, что при нарушении (например, снижении) экспрессии каждого из таких генов только TFL1-2S (SEQ ID NO: 7 или 8), и TFL1-2T (SEQ ID NO: 10 или 11), и TFL1-4T (SEQ ID NO: 19 или 20) оказывают влияние на период времени до наступления цветения путем изменения (например, ускорения) развития цветка и, таким образом, изменения (например, сокращения) периода времени до наступления цветения. Неожиданно TFL1-1S (SEQ ID NO: 1 или 2) и TFL1-1T (SEQ ID NO: 4 или 5), TFL1-3T (SEQ ID NO: 13 или 14) и TFL1-4S (SEQ ID NO: 16 или 17) почти не оказывали влияния на период времени до наступления цветения. Были идентифицированы некоторые мотивы в TFL1-2S (SEQ ID NO: 7 или 8), и TFL1-2T (SEQ ID NO: 10 или 11), и TFL1-4T (SEQ ID NO: 19 или 20), на которые можно нацеливаться для разрушения генов (например, нокаута RNAi, мутагенеза и т. п., описанных в данном документе) с целью изменения периода времени до наступления цветения (см., например, пример 4). Такие мотивы можно использовать в качестве целевых областей с целью изменения экспрессии соответствующих им генов с целью разработки стабильных линий, которые цветут раньше или позже. Не желая ограничиваться теорией, полагают, что нарушение экспрессии одного или более генов TFL1, которые ответственны за поддержание вегетативного состояния, будет облегчать взаимодействие генов FT с промоторами генов цветения с обеспечением, таким образом, сокращения периода времени до наступления цветения.
АСПЕКТЫ И ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Аспекты и варианты осуществления настоящего изобретения изложены в прилагаемой формуле изобретения.
В первом аспекте предусмотрено мутантное, не встречающееся в природе или трансгенное растение или его часть, характеризующиеся сниженной экспрессией гена, кодирующего Terminal Flower 1 (TFL1), или сниженной активностью белка, кодируемого TFL1, при этом указанный TFL1 содержит, состоит или по существу состоит из (i) полинуклеотидной последовательности, содержащей, состоящей или по существу состоящей из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептида, кодируемого полинуклеотидом, представленным в (i); или (iii) полипептида, характеризующегося по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; где экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii) или (iii), снижены по сравнению с контрольным растением, у которого экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii) или (iii), не были снижены.
Соответственно, сниженная экспрессия полинуклеотида или сниженная активность полипептида обеспечивают сокращение периода времени до наступления цветения по сравнению с контрольным растением, соответственно, где период времени до наступления цветения сокращен на по меньшей мере 8%, или по меньшей мере 20%, или по меньшей мере 28%, или по меньшей мере 30%.
Соответственно, число листьев уменьшается на по меньшей мере 16% или по меньшей мере 22%.
Соответственно, высота растения уменьшается на по меньшей мере 13%, или по меньшей мере 23%, или остается приблизительно той же.
Соответственно, растение содержит по меньшей мере одну генетическую альтерацию в полинуклеотидной последовательности, кодирующей TFL1.
Соответственно, растение содержит по меньшей мере одну мутацию в полинуклеотидной последовательности, кодирующей TFL1.
Соответственно, по меньшей мере одна мутация выбрана из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
Соответственно, растение содержит по меньшей мере одну мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
Соответственно, растение содержит по меньшей мере одну мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L. Соответственно, растение относится к роду Nicotiana или происходит из него, соответственно, где растение представляет собой Nicotiana tabacum.
В дополнительном аспекте предусмотрен растительный материал, полученный или который можно получить из растения, описанного в данном документе.
В дополнительном аспекте предусмотрен растительный продукт, содержащий по меньшей мере часть растения или растительного материала, описанного в данном документе
В дополнительном аспекте предусмотрен способ сокращения периода времени до наступления цветения у растения, включающий модификацию растения путем снижения экспрессии по меньшей мере одного гена TFL1 или активности по меньшей мере одного белка, кодируемого им, у указанного растения.
Соответственно, способ включает (a) получение растения или его части, содержащих (i) полинуклеотидную последовательность, содержащую, состоящую или по существу состоящую из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептид, кодируемый полинуклеотидом, представленным в (i); или (iii) полипептид, характеризующийся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; и (b) снижение экспрессии гена TFL1 или активности белка TFL1 у растения; и (c) получение растения с сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, у которого экспрессия гена TFL1 или активность белка TFL1 не были снижены.
В дополнительном аспекте предусмотрено использование сниженной экспрессии по меньшей мере одного гена TFL1 или активности по меньшей мере одного белка, кодируемого им, для сокращения периода времени до наступления цветения у растения.
Соответственно, экспрессию TFL1 или активность TFL1 снижают с помощью способа, выбранного из группы, состоящей из a) мутирования гена TFL1 у растения; b) экспрессии экзогенного полинуклеотида или полипептида у растения; и c) удаления гена TFL1 у растения, или комбинации одного или более из них.
Соответственно, по меньшей мере одна мутация выбрана из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
Соответственно, по меньшей мере одна мутация представляет собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
Соответственно, по меньшей мере одна мутация представляет собой мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L. Соответственно, мутация представляет собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S и мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
В дополнительном аспекте предусмотрен способ получения растительного материала с сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, при этом указанный способ включает (a) получение растения или растительного материала, описанных в данном документе; (b) сбор растительного материала от растения; (c) необязательно сушку или высушивание растительного материала в течение периода времени; и(d) получение растительного материала, который характеризуется сокращенным периодом времени до наступления цветения по сравнению с контрольным растением.
В дополнительном аспекте предусмотрен растительный материал, полученный или который можно получить с помощью способа или применения, описанных в данном документе.
В дополнительном аспекте предусмотрена выделенная полинуклеотидная последовательность, содержащая, состоящая или по существу состоящая из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20.
В дополнительном аспекте предусмотрен выделенный полипептид, кодируемый полинуклеотидом по п. 16, или полипептид, характеризующийся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21.
Соответственно, в выделенном полипептиде по меньшей мере одна мутация выбрана из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
Соответственно, по меньшей мере одна мутация представляет собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
Соответственно, по меньшей мере одна мутация представляет собой мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L. Соответственно, мутация представляет собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S и мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
В дополнительном аспекте предусмотрено антитело, которое специфически связывается с выделенным полипептидом, описанным в данном документе.
В дополнительном аспекте предусмотрены конструкция, вектор или вектор экспрессии, содержащие выделенный полинуклеотид, описанный в данном документе.
В дополнительном аспекте предусмотрены растение, или растительный материал, или клетка растения, содержащие конструкцию, вектор или вектор экспрессии, описанные в данном документе.
В дополнительном аспекте предусмотрена клетка растения, полученная или которую можно получить из растения или растительного материала, описанных в данном документе.
В дополнительном аспекте предусмотрен растительный материал, содержащий клетку, описанную в данном документе.
В дополнительном аспекте предусмотрены табачный продукт или курительное изделие, содержащие растительный материал, описанный в данном документе.
В дополнительном аспекте предусмотрена конструкция RNAi для подавления экспрессии гена TFL-1, содержащая последовательность, которая гибридизируется с целевой последовательностью в mRNA гена TFL-1 и обеспечивает подавление экспрессии гена TFL-1 посредством механизма РНК-интерференции, где указанная целевая последовательность выбрана из группы, состоящей из SEQ ID NO: 7, 8, 10, 11, 19 и/или 20.
В дополнительном аспекте предусмотрена двухнитевая РНК, содержащая по меньшей мере две последовательности, которые по меньшей мере частично комплементарны друг другу, и где смысловая нить содержит первую последовательность, и антисмысловая нить содержит вторую последовательность, и где по меньшей мере одна из последовательностей содержит по меньшей мере 10 смежных нуклеотидов из РНК TFL1, соответственно, где по меньшей мере одна из последовательностей содержит 21-23 смежных нуклеотидов из РНК TFL1.
Соответственно, двухнитевая РНК содержит первую последовательность, имеющую по меньшей мере 10 нуклеотидов из TFL1, соответственно, 21-23 нуклеотида из TFL-1; вторую последовательность; и третью последовательность, содержащую последовательность, обратно комплементарную первой последовательности, расположенной в той же ориентации, что и первая последовательность, где вторая последовательность расположена между первой последовательностью и третьей последовательностью, и вторая последовательность функционально связана с первой последовательностью и третьей последовательностью.
Соответственно, первая последовательность выбрана из группы, состоящей из SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20, и/или где третья последовательность представляет собой последовательность, обратно комплементарную последовательности, соответствующей SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20.
Соответственно, первая последовательность содержит или состоит из SEQ ID NO: 22, и третья последовательность содержит или состоит из SEQ ID NO: 23; или первая последовательность содержит или состоит из SEQ ID NO: 25, и третья последовательность содержит или состоит из SEQ ID NO: 26; или первая последовательность содержит или состоит из SEQ ID NO: 27, и третья последовательность содержит или состоит из SEQ ID NO: 28; или первая последовательность содержит или состоит из SEQ ID NO: 29, и третья последовательность содержит или состоит из SEQ ID NO: 30; или первая последовательность содержит или состоит из SEQ ID NO: 32, и третья последовательность содержит или состоит из SEQ ID NO: 33; или первая последовательность содержит или состоит из SEQ ID NO: 34, и третья последовательность содержит или состоит из SEQ ID NO: 35; или первая последовательность содержит или состоит из SEQ ID NO: 36, и третья последовательность содержит или состоит из SEQ ID NO: 37; или первая последовательность содержит или состоит из SEQ ID NO: 39, и третья последовательность содержит или состоит из SEQ ID NO: 40.
Соответственно, двухнитевая РНК содержит или состоит из последовательностей, выбранных из группы, состоящей из SEQ ID NO: 24, SEQ ID NO: 35, SEQ ID NO: 31, SEQ ID NO: 38 и SEQ ID NO: 41.
В дополнительном аспекте предусмотрена выделенная полинуклеотидная последовательность, содержащая, состоящая или по существу состоящая из последовательности, имеющей по меньшей мере 21 смежный нуклеотид из SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39 или SEQ ID NO:40, соответственно, где последовательность содержит, состоит или по существу состоит из от по меньшей мере 21 до 23 смежных нуклеотидов.
В дополнительном аспекте предусмотрен способ идентификации молекулы, которая обеспечивает модуляцию активности или экспрессии полинуклеотида TFL1 или полипептида TFL1, при этом способ включает (a) приведение молекулы в контакт с растением, содержащим полинуклеотид или полипептид, описанные в данном документе, такие как полинуклеотидная последовательность, содержащая, состоящая или по существу состоящая из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20, или полипептид, кодируемый полинуклеотидом, или полипептид, характеризующийся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; (b) отслеживание одного или более из (i) уровня экспрессии полинуклеотида TFL1 у растения; (ii) уровня экспрессии полипептида TFL1 у растения; (iii) модуляции активности полипептида TFL1 у растения; или (iv) модуляции активности полинуклеотида TFL1 у растения; и (c) идентификацию молекулы, которая обеспечивает модуляцию активности или экспрессии полинуклеотида TFL1 или полипептида TFL1.
Также раскрыты комбинации одного или более изложенных вариантов осуществления.
НЕКОТОРЫЕ ПРЕИМУЩЕСТВА
Признак быстрого цветения может обеспечивать селекцию сортов растений с поздним цветением.
Признак быстрого цветения может обеспечивать получение коммерческих сортов растений, приспособленных к климатическим условиям.
Управление периодом времени до цветения может обеспечивать увеличение продуктивности в отношении семян или плодов, а также продуктивность в отношении экстракта из цветков.
Управление периодом времени до цветения может обеспечивать отсутствие необходимости в обработке для созревания.
Признак быстрого цветения может обеспечивать более короткий период времени от посева/посадки до сбора урожая, что может сокращать вегетационный период.
Признак быстрого цветения может обеспечивать более быстрое введение новых признаков путем скрещивания.
Более короткий жизненный цикл растения мог бы приводить к нескольким урожаям растения в год, что может в результате приводить к более устойчивому производству.
Признак быстрого цветения может обеспечивать более раннее срезание цветов, что может в результате приводить к более высокому качеству растительных продуктов.
Преимущественной является разработка подходов на основе не являющихся генетически модифицированными организмов (не относящихся к ГМО) для сокращения периода времени до наступления цветения у растений посредством применения инактивации генов. Из-за трудностей выращивания и получения прибыли с генетически модифицированных сельскохозяйственных культур в некоторых странах, включая европейские, может быть желательным работать с мутантами с наличием однонуклеотидных полиморфизмов, полученных с помощью обработки этилметансульфонатом (EMS) или тому подобным, а не за счет использования методик генной инженерии. Мутанты не рассматриваются как ГМО, даже когда мутации вызваны искусственно. В странах ЕС, например, отсутствуют специальные постановления для растений, полученных вследствие мутационной селекции. До настоящего времени единственным известным решением сокращения периода времени до наступления цветения является сверхэкспрессия генов FT, что может быть неподходящим в среде, не относящейся к ГМО. Осуществление нокаута генов TFL1 с помощью, например, отбора линий EMS/облучения или с применением отбора на основе природных вариантов TFL1 из разных сортов Nicotiana tabacum или интрогрессивной формы из других видов Nicotiana представляет собой подходящее решение, не относящееся к ГМО. В качестве альтернативы, также можно рассматривать любую технологию редактирования генов, но регулирование таких технических подходов все еще неясно. Наличие вариантов TFL1 обеспечивает быструю селекцию с помощью ДНК-тестирования без ожидания цветения и необходимости в утомительных стадиях самоопыления.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Экспрессия TFL1 и FT в листьях цветущих растений N. tabacum (TN90) в поле. Данные по транскрипту получены с помощью считывания фрагментов на тысячу оснований экзона на миллион картированных фрагментов (FPKM) (см. Nat Biotechnol. 2010 28(5):511-5). Экспрессия TFL1 и FT определена с помощью анализов РНК-последовательностей в незрелых цветках, листьях в нижней части стебля, листьях в средней части стебля, листьях в верхней части стебля, лепестках, корнях, чашелистиках и стебле.
Фигура 2. Фенотипические анализы T0 RNAi TFL1-1S/T, TFL1-2S/T, TFL1-3T и TFL1-4T (20) и контрольных (10) растений, выращенных в теплице при контролируемых условиях. Период времени до цветения (дни после пересадки, DAT) четырех трансгенных линий показан на A и D, значения числа листьев показаны на B и E, и высота четырех линий показана на C и F. Показаны средние значения и стандартные отклонения для каждого растения.
Фигура 3. Изображения растений RNAi TFL1-2S/T в теплице. Быстрое цветение линий RNAi TFL1-2S/T по сравнению с контрольными растениями (Coltabaco 23RM), 117 дней после пересадки в горшки (A) и после получения шариков семян, сравнение единичных растений (B).
Фигура 4. Диаграмма, иллюстрирующая мутацию TFL1-2T-P131S. Мутирование кодона CCT > Pro в TCT > Ser.
Фигура 5. Столбчатый график, иллюстрирующий, что мутантные растения TFL1-2T-P131S зацветают на примерно 30% быстрее как у гомозиготных растений (мутантных по обоим аллелям), так и у гетерозиготных растений (мутантных по одному аллелю). Мутантные растения имеют меньше листьев, но не наблюдалось влияния на высоту растений по сравнению с растением табака дикого типа. WT=Nicotiana tabacum; TFL1-2T-P131S wt=внешний сегрегант мутированного растения без мутации и считается дополнительным контрольным растением на том же фенотипическом фоне, что и мутантные растения; TFL1-2T-P131S mut homo=гомозиготное мутантное растение; TFL1-2T-P131S mut hetero=гетерозиготное мутантное растение. N=4.
Фигура 6. Диаграмма, иллюстрирующая мутацию TFL1-4T-P110L. Мутирование кодона CCA > Pro в CTA > Leu.
Фигура 7. Столбчатый график, иллюстрирующий, что мутантные растения TFL1-4T-P110L зацветают на примерно 30% быстрее у гетерозиготных растений (мутантных по одному аллелю). Мутантные растения имеют меньше листьев, но не наблюдалось влияния на высоту растений по сравнению с растением табака дикого типа. WT=Nicotiana tabacum; TFL1-4T-P110L mut hetero=гетерозиготное мутантное растение. N=3.
ОПРЕДЕЛЕНИЯ
Техническим терминам и выражениям, используемым в пределах объема данной заявки, следует придавать значение, которое обычно применяется к ним в данной области биологии растений и молекулярной биологии. Все нижеследующие определения терминов применяют ко всему содержанию данной заявки. Слово «содержащий» не исключает другие элементы или стадии, а формы единственного числа не исключают множественное. Одна стадия может выполнять функции нескольких признаков, изложенных в формуле изобретения. Термины «приблизительно», «по существу» и «примерно» в контексте данного цифрового значения или диапазона относятся к значению или диапазону, которые находятся в пределах 20%, в пределах 10%, или в пределах 5%, 4%, 3%, 2% или 1% от заданного значения или диапазона.
Термин «сокращенный период времени до наступления цветения» или его эквиваленты означают сокращенный период времени от посева до зацветания первых цветков по сравнению с контрольным растением. Период можно сократить на по меньшей мере приблизительно 5%, 6%, 7%, 8%, 9% 10%, 20%, 28%, 30%, 40%, 50%, 60%, 70%, 80%, или 90%, или более по сравнению с контрольным растением.
Термин «продленный период времени до наступления цветения» или его эквиваленты означают более длительный период времени от посева до зацветания первых цветков по сравнению с контрольным растением. Период можно продлить на по меньшей мере приблизительно 5%, 6%, 7%, 8%, 9% 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, или 90%, или более по сравнению с контрольным растением.
Термин «выделенный» относится к любому объекту, который взят из его естественной среды, но этот термин не подразумевает какой-либо степени очистки.
«Вектор экспрессии» представляет собой средство доставки нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения экспрессии нуклеиновой кислоты. Подходящие векторы экспрессии включают эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды из двухнитевой нуклеиновой кислоты; линеаризованные плазмиды из двухнитевой нуклеиновой кислоты и другие функционально эквивалентные векторы экспрессии любого происхождения. Вектор экспрессии содержит по меньшей мере промотор, расположенный выше по цепи и функционально связанный с нуклеиновой кислотой, конструкции нуклеиновых кислот или конъюгат нуклеиновых кислот, как определено ниже.
Термин «конструкция» относится к двухнитевому фрагменту рекомбинантной нуклеиновой кислоты, содержащему один или более полинуклеотидов. Конструкция содержит «матричную нить», основания которой спарены с комплементарной «смысловой или кодирующей нитью». Данная конструкция может быть встроена в вектор в двух возможных ориентациях, либо в той же (или смысловой) ориентации, или в обратной (или антисмысловой) ориентации по отношению к ориентации промотора, расположенного в векторе, таком как вектор экспрессии.
«Вектор» относится к средству доставки нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения переноса нуклеиновой кислоты, конструкций нуклеиновых кислот и конъюгатов нуклеиновых кислот и тому подобного. Подходящие векторы включают эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды из двухнитевой нуклеиновой кислоты; линеаризованные плазмиды из двухнитевой нуклеиновой кислоты и другие векторы любого происхождения.
«Промотор» относится к элементу/последовательности нуклеиновой кислоты, как правило расположенной выше по цепи и функционально связанной с фрагментом двухнитевой ДНК. Промоторы могут быть получены целиком из участков вблизи нативного представляющего интерес гена, или могут состоять из разных элементов, полученных из разных нативных промоторов или сегментов синтетической ДНК.
Термины «гомология, идентичность или сходство» относятся к степени сходства последовательностей между двумя полипептидами или между двумя молекулами нуклеиновых кислот, сравниваемых путем выравнивания последовательностей. Соответственно, термины «гомология, идентичность или сходство» относятся к степени сходства последовательности между полной последовательностью, например, полноразмерной последовательностью, двух полипептидов или между двумя молекулами нуклеиновых кислот. Степень идентичности между двумя отдельными сравниваемыми последовательностями нуклеиновых кислот является функцией количества идентичных или совпадающих нуклеотидов в сопоставимых положениях. Процент идентичности может быть определен путем визуального осмотра и математических расчетов. В качестве альтернативы, процент идентичности двух последовательностей нуклеиновых кислот можно определить путем сравнения информации о последовательностях с использованием компьютерной программы, такой как ClustalW, BLAST, FASTA или Smith-Waterman. Процент идентичности двух последовательностей может принимать разные значения в зависимости от (i) способа, используемого для выравнивания последовательностей, например, ClustalW, BLAST, FASTA, Smith-Waterman (используемых в разных программах), или структурного выравнивания из 3D-сравнения; и (ii) параметров, используемых в способе выравнивания, например, локального по сравнению с глобальным выравниванием, используемой матрицы замен пар (например, BLOSUM62, PAM250, Gonnet и т. д.), и штрафа за гэп, например, функциональной формы и констант. После проведения выравнивания существуют разные способы подсчета процента идентичности между двумя последовательностями. Например, можно разделить число идентичностей на (i) длину самой короткой последовательности; (ii) длину выравнивания; (iii) среднюю длину последовательности; (iv) число положений без пропусков; или (iv) число эквивалентных положений, за исключением выступающих концов. Кроме того, будет понятно, что процент идентичности также сильно зависит от длины. Следовательно, чем короче пара последовательностей, тем выше можно ожидать идентичность последовательности, которая случайным образом будет иметь место. Популярная программа множественного выравнивания ClustalW (Nucleic Acids Research (1994) 22, 4673-4680; Nucleic Acids Research (1997), 24, 4876-4882) представляет собой подходящий способ получения множественных выравниваний полипептидов или полинуклеотидов. Подходящие параметры для ClustalW могут являться следующими. Для выравниваний полинуклеотидов: штраф за открытие гэпа=15,0, штраф за продолжение гэпа=6,66 и матрица=идентичность. Для выравниваний полипептидов: штраф за открытие гэпа=10. o, штраф за продолжение гэпа=0,2 и матрица=Gonnet. Для выравниваний ДНК и белка: ENDGAP = -1 и GAPDIST=4. Специалист в данной области будет знать, что может быть необходимым варьировать эти и другие параметры для оптимального выравнивания последовательностей. Соответственно, затем проводят расчет процента идентичностей на основании такого выравнивания в виде (N/T), где N представляет собой количество положений, в которых в последовательностях присутствует идентичный остаток, и T представляет собой общее количество сравниваемых положений, включая гэпы, но за исключением выступающих концов.
«Вариант» означает практически аналогичную последовательность. Вариант может обладать аналогичной функцией или практически аналогичной функцией относительно последовательности дикого типа. В случае TFL1 аналогичная функция составляет по меньшей мере приблизительно 50%, 60%, 70%, 80% или 90% от функции белка дикого типа при тех же условиях. В случае TFL1 практически аналогичная функция составляет по меньшей мере приблизительно 90%, 95%, 96%, 97%, 98% или 99% от функции белка дикого типа при тех же условиях. Варианты могут иметь одну или более благоприятных мутаций, которые приводят к получению сниженного уровня активности TFL1 по сравнению с полипептидом дикого типа. Варианты могут иметь одну или более благоприятных мутаций, которые приводят к нокауту активности TFL1 (т. е. 100% подавлению и, таким образом, нефункциональному полипептиду).
Термин «растение» относится к любому растению или части растения на любой стадии его жизненного цикла или развития, а также к его потомству. В одном варианте осуществления растение представляет собой «растение табака», которое относится к растению, принадлежащему к роду Nicotiana. Предпочтительные виды растения табака описаны в данном документе. Соответственно, растение представляет собой мутантное, не встречающееся в природе или трансгенное растение, в котором экспрессию одного или более генов или активность одного или более белков модулируют по сравнению с контрольным растением. Соответственно, альтерация, которая делает растение мутантным, не встречающимся в природе или трансгенным растением, приводит к модуляции экспрессии одного или более генов TFL1 или модуляции активности одного или более белков TFL1. В определенных вариантах осуществления альтерация представляет собой генетическую альтерацию или генетическую модификацию. Примеры мутаций, которые можно вводить в растения для сокращения периода времени до наступления цветения, описаны в данном документе.
«Части растения» включают клетки растения, протопласты растений, тканевые культуры клеток растений, из которых можно регенерировать целое растение, каллюсы растений, корневища растений и клетки растений, которые являются интактными в растениях или частях растений, таких как зародыши, пыльца, пыльники, семяпочки, семена, листья, цветки, стебли, ветви, плод, корни, кончики корней и т. п. Потомство, варианты и мутанты регенерированных растений также включены в объем настоящего изобретения при условии, что они содержат введенные полинуклеотиды, описанные в данном документе. Листья растений являются в особенности предпочтительными для применения в настоящем изобретении.
«Клетка растения» относится к структурной и физиологической единице растения. Клетка растения может находиться в виде протопласта без клеточной стенки, выделенной отдельной клетки или культивируемой клетки или может быть представлена как часть более высокоорганизованной единицы, такой как без ограничения растительная ткань, орган растения или целое растение.
Термин «растительный материал» относится к любой твердой, жидкой или газообразной композиции, или их комбинации, получаемой из растения, включая биомассу, листья, стебли, корни, цветки или части цветка, плоды, пыльцу, яйцеклетки, зиготы, семена, черенки, секреты, экстракты, клеточные или тканевые культуры, или любые другие части или продукты растения. В одном варианте осуществления растительный материал содержит или состоит из биомассы, стеблей, семян или листьев. В другом варианте осуществления растительный материал содержит или состоит из листьев.
Термин «разновидность» относится к популяции растений, которые разделяют постоянные характеристики, отделяющие их от других растений того же вида. Имея один или более отличительных признаков, разновидность дополнительно характеризуется очень небольшим общим варьированием между особями в пределах этой разновидности. Разновидность часто продается на коммерческой основе.
Термин «линия» или «селекционная линия», используемый в данном документе, обозначает группу растений, которые используют при селекции растений. Линия отличается от разновидности, поскольку демонстрирует небольшую вариабельность между особями по одному или более представляющим интерес признакам, хотя может присутствовать некоторая вариабельность между особями по другим признакам.
Термин «не встречающийся в природе», используемый в данном документе, описывает объект (например, полинуклеотид, генетическую мутацию, полипептид, растение и клетку растения и растительный материал), который не образован естественным путем или не существует в природе. Такие не встречающиеся в природе объекты или искусственные объекты можно создать, синтезировать, произвести, модифицировать, подвергнуть вмешательству или манипуляции способами, описанными в данном документе, или которые известны в данной области. Такие не встречающиеся в природе объекты или искусственные объекты можно создать, синтезировать, произвести, модифицировать, подвергнуть вмешательству или манипуляции человеком. Таким образом, в качестве примера не встречающееся в природе растение, не встречающуюся в природе клетку растения или не встречающийся в природе растительный материал можно создать с применением методик манипуляции с генами - таких как с использованием антисмысловой РНК, интерферирующей РНК, мегануклеазы и т. п. В качестве дополнительного примера, не встречающееся в природе растение, не встречающуюся в природе клетку растения или не встречающийся в природе растительный материал можно создать с использованием интрогрессии или путем переноса одной или более генетических мутаций (например, одного или более полиморфизмов) от первого растения или клетки растения ко второму растению или клетке растения (которые сами по себе могут быть встречающимися в природе), таким образом, что полученное растение, клетка растения или растительный материал или их потомство содержит генетическую структуру (например, геном, хромосому или ее сегмент), которая не образована естественным путем, или которая не существует в природе. Полученное растение, клетка растения или растительный материал, таким образом, являются искусственными или не встречающимися в природе. Соответственно, искусственные или не встречающиеся в природе растение или клетку растения можно создать путем модификации генетической последовательности в первом встречающемся в природе растении или клетке растения, даже если полученная генетическая последовательность встречается в природе во втором растении или клетке растения, которая содержит отличающийся генетический фон из первого растения или клетки растения.
Термин «модулирование» может относиться к снижению, ингибированию, увеличению или иному влиянию на экспрессию или активность полипептида. Этот термин может также относиться к снижению, ингибированию, увеличению или иному влиянию на активность гена, кодирующего полипептид, которое может включать, помимо прочего, модулирование транскрипционной активности. Термин «модулирование» также может относиться к сокращению или продлению периода времени до наступления цветения.
Термины «снижение», или «сниженный», или «уменьшение», или «уменьшенный», используемые в данном документе, относятся к снижению от приблизительно 10% до приблизительно 99%, или снижению, составляющему по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25% или 28%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, или по меньшей мере 100% или более от количества или активности, такой как без ограничения полипептидная активность, транскрипционная активность и экспрессия белка.
Термины «ингибировать» или «ингибированный», используемые в данном документе, относятся к снижению, составляющему от приблизительно 98% до приблизительно 100%, или снижению, составляющему по меньшей мере 98%, по меньшей мере 99%, но в особенности 100% от количества или активности, такой как, помимо прочего, полипептидная активность, транскрипционная активность и экспрессия белка.
Трансформация клетки может быть стабильной или временной. Термины «временная трансформация» или «временно трансформированный» или варианты указанных терминов относятся к введению одного или более экзогенных полинуклеотидов в клетку при отсутствии встраивания экзогенного полинуклеотида в геном клетки-хозяина. Напротив, термины «стабильная трансформация» или «стабильно трансформированный» относятся к введению и интеграции одного или более экзогенных полинуклеотидов в геном клетки. Термин «стабильный трансформант» относится к клетке, в которую стабильно интегрировали один или более экзогенных полинуклеотидов в геномную ДНК или ДНК органелл. Следует понимать, что организм или его клетка, трансформированные с помощью нуклеиновых кислот, конструкций и/или векторов по настоящему изобретению, могут являться временно, а также стабильно трансформированными. В определенных вариантах осуществления стабильная трансформация является предпочтительной.
Термины «увеличение» или «увеличенный», используемые в данном документе, относятся к увеличению, составляющему от приблизительно 5% до приблизительно 99%, или увеличению, составляющему по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, или по меньшей мере 100% или больше от количества или активности, такой как, помимо прочего, полипептидная активность, транскрипционная активность и экспрессия белка.
Термин «практически», используемый в данном документе и при использовании в контексте количества означает, что количество составляет по меньшей мере приблизительно 10%, по меньшей мере приблизительно 9%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 1% или по меньшей мере приблизительно 0,1% от количества, с которым его сравнивают.
Термин «контроль» в контексте контрольного растения или контрольной клетки растения и т. п. означает растение или клетку растения, в которых экспрессия или активность гена или белка, представляющих интерес, не были модулированы, и поэтому данный контроль может обеспечить сравнение или эталон относительно растения или клетки растения, в которых экспрессия или активность были модифицированы. Таким образом, в контексте настоящего изобретения контроль не будет содержать по меньшей мере одну модификацию или генетическую альтерацию, что снижает экспрессию или активность TFL1. Контрольное растение или клетка растения могут содержать пустой вектор. Контрольное растение или клетка растения могут соответствовать растению дикого типа или клетке растения дикого типа и т. п. Во всех таких случаях исследуемое растение и контрольное растение культивируют и собирают с использованием тех же протоколов для целей сравнения. Изменения уровней, соотношений, активности или распределения генов или полипептидов, описанных в данном документе, или изменений в фенотипе растений можно измерять с помощью сравнения исследуемого растения с контрольным растением, соответственно, где исследуемое растение и контрольное растение культивировали и/или собирали с применением одних и тех же протоколов. Контрольное растение может обеспечивать эталонную точку для измерения изменений в фенотипе исследуемого растения. Показатель изменений в фенотипе можно измерять в любой момент времени для растения, в том числе, во время развития растения, старения или после сушки. Показатель изменений в фенотипе можно измерять для растений, выращенных в любых условиях, в том числе для растений, выращенных в камере для выращивания, теплице или на поле.
ПОДРОБНОЕ ОПИСАНИЕ
В одном варианте осуществления предложен выделенный полинуклеотид, содержащий полинуклеотидную последовательность, состоящий из нее или состоящий по существу из нее, которая характеризуется по меньшей мере 72% идентичностью последовательности с любой из последовательностей, описанных в данном документе, включая любой из полинуклеотидов, показанный в перечне последовательностей. Соответственно, выделенный полинуклеотид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с ним.
В другом варианте осуществления предусмотрен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из полинуклеотидной последовательности, характеризующейся по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20, или любой из SEQ ID NO: 22-41.
В другом варианте осуществления предусмотрен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из полинуклеотидной последовательности, характеризующейся по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из полинуклеотидной последовательности, характеризующейся по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с SEQ ID NO: 7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 19, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрен выделенный полинуклеотид, содержащий, состоящий или по существу состоящий из полинуклеотидной последовательности, характеризующейся по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% идентичностью последовательности с SEQ ID NO:8 или SEQ ID NO: 11, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрены полинуклеотидные варианты, которые характеризуются по меньшей мере приблизительно 72%, 75%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% идентичностью последовательности с последовательностью под SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20, или любой из SEQ ID NO: 22-41.
В другом варианте осуществления предусмотрены полинуклеотидные варианты, которые характеризуются по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% идентичностью последовательности с последовательностью под SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрены полинуклеотидные варианты, которые характеризуются по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% идентичностью последовательности с последовательностью под SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрены фрагменты SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20, или любой из SEQ ID NO: 22-41 с существенной степенью гомологии (то есть, сходства последовательности) или существенной степенью идентичности с ними, которые характеризуются по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с соответствующими фрагментами SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20, или любой из SEQ ID NO: 22-41. В определенных вариантах осуществления длина фрагментов может составлять 21-23 смежных нуклеотида. В определенных вариантах осуществления длина фрагментов может составлять по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов. В определенных вариантах осуществления длина фрагментов может составлять по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или более смежных нуклеотидов.
В другом варианте осуществления предусмотрены фрагменты SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20 с существенной степенью гомологии (то есть, сходства последовательности) или существенной степенью идентичности с ними, которые характеризуются по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с соответствующими фрагментами SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрены фрагменты SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20 с существенной степенью гомологии (то есть, сходства последовательности) или существенной степенью идентичности с ними, которые характеризуются по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с соответствующими фрагментами SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрены полинуклеотиды, предусматривающие достаточную или существенную степень идентичности или сходства с SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20, которые кодируют полипептид, который функционирует как белок Terminal Flower 1.
В другом варианте осуществления предусмотрены полинуклеотиды, предусматривающие достаточную или существенную степень идентичности или сходства с SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20, которые кодируют полипептид, который функционирует как белок Terminal Flower 1.
В другом варианте осуществления предусмотрены полинуклеотиды, предусматривающие достаточную или существенную степень идентичности или сходства с SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20, которые кодируют полипептид, который функционирует как белок Terminal Flower 1.
В другом варианте осуществления предусмотрен(-ы) полинуклеотид(-ы), который(-ые) кодирует(-ют) белок с активностью белка Terminal Flower 1, которая составляет по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% или более от активности белка, представленного под SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20.
В другом варианте осуществления предусмотрен(-ы) полинуклеотид(-ы), который(-ые) кодирует(-ют) белок с активностью белка Terminal Flower 1, которая составляет по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% или более от активности белка, представленного под SEQ ID NO: 3, или SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:15, или SEQ ID NO:18, или SEQ ID NO:21.
В другом варианте осуществления предусмотрен(-ы) полинуклеотид(-ы), описанный(-ые) в данном документе, который(-ые) кодирует(-ют) белок с активностью белка Terminal Flower 1, которая составляет по меньшей мере приблизительно 72%, 73%, 74%, 75%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% или более от активности белка, представленного под SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21.
Для определения того, является ли полипептид функциональным белком Terminal Flower 1, можно использовать анализы BLAST (основное средство поиска локального выравнивания) для обнаружения участков сходства между биологическими последовательностями. Программу можно использовать для сравнения нуклеотидных или белковых последовательностей с базами данных последовательностей и расчета статистической значимости. Активность фактора транскрипции TFL1 можно определять с помощью способности TFL1 обладать повышенной или сниженной функцией связывания - такой как повышенная или сниженная функция связывания с другими белками (например, факторами транскрипции) или повышенное или сниженное связывание с одной или более нуклеиновыми кислотами. Транскрипционную активность TFL1 можно определять либо биохимически путем определения связывающих свойств, либо путем оценки результата активности фактора транскрипции - такого как повышенная или сниженная экспрессия целевого гена, который отвечает на активность фактора транскрипции. Например, у Arabidopsis thaliana TFL1 действует путем подавления активности генов LFY и AP1 (Development (1998) 125: 1609-1615; Development (1999) 126: 1109-1120). Транскрипционную активность TFL1, следовательно, можно определять с помощью измерения активности генов LFY и/или AP1 в присутствии и в отсутствие TFL1. Конечным явлением является уменьшенный период времени до цветения, так что основная биологическая активность TFL1 может быть определена как подавление цветения. Биологическую роль TFL1/FT в развитии цветков можно протестировать с применением приобретения функции и потери функции соответствующих аллелей (см. The Plant Journal (2010) 63: 241-253). Полинуклеотид, описанный в данном документе, может включать полимер из нуклеотидов, который может быть немодифицированной или модифицированной дезоксирибонуклеиновой кислотой (ДНК) или рибонуклеиновой кислотой (РНК). Соответственно, полинуклеотид может быть, без ограничения, геномной ДНК, комплементарной ДНК (cDNA), mRNA или антисмысловой РНК или их фрагментом(-ами). Кроме того, полинуклеотид может представлять собой однонитевую или двухнитевую ДНК, ДНК, которая является смесью однонитевых и двухнитевых участков, гибридной молекулой, содержащей ДНК и РНК, или гибридной молекулой со смесью однонитевых и двухнитевых участков или их фрагментом(-ами). Кроме того, полинуклеотид может быть составлен из трехнитевых участков, содержащих ДНК, РНК или обе, или их фрагмент(-ы). Полинуклеотид может содержать одно или более модифицированных оснований, таких как фосфоротиоаты, и может представлять собой пептидную нуклеиновую кислоту. Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов cDNA, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов, или комбинации вышеперечисленного. Хотя полинуклеотидные последовательности, описанные в данном документе, представлены в виде последовательностей ДНК, последовательности включают их соответствующие последовательности РНК и их комплементарные (например, полностью комплементарные) последовательности ДНК или РНК, в том числе, их обратно комплементарные цепи. Полинуклеотиды, описанные в данном документе, могут содержать одну или более модификаций, представляющих собой замены. Полинуклеотиды, описанные в данном документе, могут содержать одну или более меток.
Полинуклеотид, описанный в данном документе, обычно содержит фосфодиэфирные связи, хотя в некоторых случаях включены полинуклеотидные аналоги, которые могут иметь альтернативные остовы, содержащие, например, фосфороамидатные, фосфоротиоатные, фосфородитиоатные или О-метилфосфороамидитные связи; и пептидные полинуклеотидные остовы и связи. Другие аналоги полинуклеотидов включают полинуклеотиды с положительно заряженными каркасами, неионными каркасами и безрибозными каркасами. Модификации рибозофосфатного остова можно делать по целому ряду причин, например, для повышения стабильности и периода полужизни таких молекул в физиологических средах или в качестве зондов на биочипе. Можно получать смеси природных полинуклеотидов и аналогов; в качестве альтернативы, можно получать смеси разных полинуклеотидных аналогов и смеси природных полинуклеотидов и аналогов.
Известно множество полинуклеотидных аналогов, включая, например, фосфороамидатные, фосфоротиоатные, фосфородитиоатные или О-метилфосфороамидитные связи и пептидные полинуклеотидные каркасы и связи. Другие аналоги полинуклеотидов включают полинуклеотиды с положительно заряженными каркасами, неионными каркасами и безрибозными каркасами. Полинуклеотиды, содержащие один или более карбоциклических сахаров, также включены.
Другие аналоги включают пептидные полинуклеотиды, которые представляют собой пептидные полинуклеотидные аналоги. Эти остовы являются главным образом неионными в нейтральных условиях в отличие от высокозаряженного фосфодиэфирного каркаса встречающихся в природе полинуклеотидов. Это может давать преимущества. Во-первых, пептидный полинуклеотидный остов может характеризоваться улучшенной кинетикой гибридизации. Пептидные полинуклеотиды характеризуются более значительными изменениями температуры плавления для несоответствующих по сравнению с идеально совпадающими парами оснований. ДНК и РНК, как правило, проявляют понижения температуры плавления на 2-4 C из-за внутреннего несовпадения. В случае неионного пептидного полинуклеотидного каркаса падение составляет ближе к 7-9°C. Подобным образом, из-за их неионной природы, гибридизация оснований, присоединенных к этим каркасам, является относительно нечувствительной к концентрации солей. Кроме того, пептидные полинуклеотиды могут не разрушаться или разрушаться в меньшей степени клеточными ферментами, и, таким образом, могут быть более стабильными.
Среди применений раскрытых полинуклеотидов и их фрагментов находится применение фрагментов в качестве зондов в анализах с гибридизацией нуклеиновых кислот или праймеров для применения в анализах с амплификацией нуклеиновых кислот. Такие фрагменты обычно содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов из последовательности ДНК. В других вариантах осуществления фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или больше смежных нуклеотидов из последовательности ДНК. Такие фрагменты, в целом, содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов из SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20. В других вариантах осуществления фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или более смежных нуклеотидов из SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 13, или SEQ ID NO: 14, или SEQ ID NO: 16, или SEQ ID NO: 17, или SEQ ID NO: 19, или SEQ ID NO: 20. Такие фрагменты, в целом, содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов из SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20. В других вариантах осуществления фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или более смежных нуклеотидов из SEQ ID NO:2, или SEQ ID NO:5, или SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 14, или SEQ ID NO: 17, или SEQ ID NO: 20. Такие фрагменты, в целом, содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов из SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20. В других вариантах осуществления фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или более смежных нуклеотидов из SEQ ID NO:8, или SEQ ID NO: 11, или SEQ ID NO: 20.
Основные параметры, влияющие на выбор условий гибридизации в отношении полинуклеотидов, и руководство для разработки подходящих условий описаны в Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). Используя данные о генетическом коде в комбинации с аминокислотными последовательностями, описанными в данном документе, можно получить наборы вырожденных олигонуклеотидов. Такие олигонуклеотиды применимы в качестве праймеров, например, в полимеразных цепных реакциях (ПЦР), с помощью которых выделяют и амплифицируют фрагменты ДНК. В определенных вариантах осуществления вырожденные праймеры можно использовать в качестве зондов для генетических библиотек. Такие библиотеки будут включать без ограничения библиотеки cDNA, геномные библиотеки и даже электронные библиотеки меток экспрессируемых последовательностей или ДНК. Гомологичные последовательности, идентифицированные этим способом, будут затем использованы в качестве зондов для идентификации гомологов последовательностей, указанных в данном документе.
Кроме того, потенциал использования имеют полинуклеотиды и олигонуклеотиды (например, праймеры или зонды), которые гибридизируются в условиях пониженной жесткости, как правило, в условиях средней жесткости, и обычно, в условиях высокой жесткости с полинуклеотидом(-ами), описанным(-и) в данном документе. Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий могут быть легко определены обычными специалистами в данной области на основе, например, длины или состава оснований полинуклеотида. Один из путей достижения умеренно жестких условий включает использование раствора для предварительного отмывания, содержащего 5x стандартного цитрата натрия, 0,5% додецилсульфата натрия, 1,0 мМ этилендиаминтетрауксусной кислоты (pH 8,0), гибридизационного буфера с приблизительно 50% формамида, 6x стандартного цитрата натрия и температуры гибридизации, составляющей приблизительно 55°C (или других подобных гибридизационных растворов, таких как содержащих приблизительно 50% формамида с температурой гибридизации, составляющей приблизительно 42°С) и условий отмывания при приблизительно 60°C в 0,5x стандартного цитрата натрия, 0,1% додецилсульфата натрия. Как правило, условия высокой жесткости определяются как условия гибридизации, как описано выше, но с отмыванием при примерно 68 С, 0,2x стандартным цитратом натрия, 0,1% додецилсульфатом натрия. SSPE (1x SSPE представляет собой 0,15 M хлорида натрия, 10 мМ фосфата натрия, 1,25 мМ этилендиаминтетрауксусной кислоты, рН 7,4) можно заменить стандартным цитратом натрия (1x стандартный цитрат натрия представляет собой 0,15 M хлорида натрия и 15 мМ цитрата натрия) в гибридизационных буферах и отмывочных буферах; отмывания выполняют в течение 15 минут после завершения гибридизации. Следует понимать, что температуру отмывания и концентрацию солей при отмывании можно регулировать по мере необходимости для достижения требуемой степени жесткости с применением основных принципов, которые управляют реакциями гибридизации и стабильностью дуплексов, как это известно специалистам в данной области и дополнительно описано ниже (см., например, Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y). При гибридизации полинуклеотида с целевым полинуклеотидом с неизвестной последовательностью длина гибрида предполагается такой, как у гибридизируемого полинуклеотида. Когда гибридизируют полинуклеотиды с известными последовательностями, длина гибрида может быть определена путем выравнивания последовательностей полинуклеотидов и идентификации участка или участков с оптимальной комплементарностью последовательности. Температура гибридизации для гибридов, которые предположительно составят менее чем 50 пар оснований в длину, должна быть на 5-10 C меньше, чем температура плавления гибрида, где температуру плавления определяют в соответствии со следующими уравнениями. Для гибридов, длина которых составляет менее 18 пар оснований, температура плавления (°С) = 2(количество оснований А+Т)+4(количество оснований G+C). Для гибридов, длина которых составляет более 18 пар оснований, температура плавления (°C) = 81,5+16,6(log10 [Na+])+0,41(% G+C)-(600/N), где N - количество оснований в гибриде и [Na+] - концентрация ионов натрия в буфере для гибридизации ([Na+] для 1x стандартного цитрата натрия=0,165M). Как правило, каждый такой гибридизирующийся полинуклеотид имеет длину, которая составляет по меньшей мере 25% (обычно по меньшей мере 50%, 60% или 70% и наиболее часто по меньшей мере 80%) от длины полинуклеотида, с которым он гибридизируется, и характеризуется по меньшей мере 60% идентичностью последовательности (например, по меньшей мере 70%, 72%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%) с полинуклеотидом, с которым он гибридизируется.
Как будет понятно специалисту в данной области, линейная ДНК имеет две возможные ориентации: направление 5'-3' и направление 3'-5'. Например, если эталонная последовательность расположена в направлении 5'-3', и если вторая последовательность расположена в направлении 5'-3' в той же полинуклеотидной молекуле/нити, то эталонная последовательность и вторая последовательность ориентированы в одном направлении или имеют одинаковую ориентацию. Как правило, последовательность промотора и представляющий интерес ген, находящийся под контролем данного промотора, расположены в одинаковой ориентации. Однако если по отношению к эталонной последовательности, расположенной в направлении 5'-3', вторая последовательность расположена в направлении 3'-5' в той же полинуклеотидной молекуле/нити, тогда эталонная последовательность и вторая последовательность ориентированы в антисмысловом направлении или имеют антисмысловую ориентацию. Две последовательности, имеющие антисмысловые ориентации по отношению друг к другу, могут быть альтернативно описаны как имеющие одинаковую ориентацию, если эталонная последовательность (направление 5'-3') и последовательность, обратно комплементарная эталонной последовательности (эталонной последовательности, расположенной в 5'-3'), расположены в пределах одной полинуклеотидной молекулы/нити. Последовательности, изложенные в данном документе, показаны в направлении 5'-3'.
Рекомбинантные конструкции, предложенные в данном документе, могут быть использованы для трансформации растений или клеток растений для модулирования уровней экспрессии и/или активности белка. Рекомбинантная полинуклеотидная конструкция может содержать полинуклеотид, кодирующий один или более полинуклеотидов, описанных в данном документе, функционально связанных с регуляторным участком, подходящим для экспрессии полипептида. Таким образом, полинуклеотид может содержать кодирующую последовательность, которая кодирует полипептид, описанный в данном документе. Растения или клетки растений, в которых модулируют уровни экспрессии и/или активности белка, могут включать мутантные, не встречающиеся в природе, трансгенные, созданные человеком или полученные с помощью методик генной инженерии растения или клетки растений. Соответственно, растение или клетка растения содержит геном, который был изменен путем стабильной интеграции рекомбинантной ДНК. Рекомбинантная ДНК включает ДНК, полученную с помощью методик генной инженерии и сконструированную вне клетки, и включает ДНК, содержащую встречающуюся в природе ДНК, или cDNA, или синтетическую ДНК. Растение может включать растение, регенерированное из первоначально трансформированной клетки растения, и растения-потомки от более поздних поколений или скрещиваний трансформированного растения. Соответственно, модификация изменяет экспрессию или активность полинуклеотида или полипептида, описанного в данном документе, по сравнению с контрольным растением.
Полипептид, кодируемый рекомбинантным полинуклеотидом, может быть нативным полипептидом или может быть гетерологичным по отношению к клетке. В некоторых случаях рекомбинантная конструкция содержит полинуклеотид, который обеспечивает модуляцию экспрессии, функционально связанный с регуляторным участком. Примеры подходящих регуляторных участков описаны в данном документе.
Также предусмотрены векторы, содержащие рекомбинантные полинуклеотидные конструкции, такие как описанные в данном документе. Подходящие основы для векторов включают, например, те, которые обычно используют в данной области, такие как плазмиды, вирусы, искусственные хромосомы, искусственные хромосомы бактерий, искусственные хромосомы дрожжей или искусственные хромосомы бактериофагов. Подходящие векторы экспрессии включают, без ограничения, плазмиды и вирусные векторы, полученные из, например, бактериофага, бакуловирусов и ретровирусов. Многочисленные векторы и системы экспрессии являются коммерчески доступными. Векторы могут включать, например, точки начала репликации, участки связывания с ядерным матриксом или маркеры. Маркерный ген может придавать клетке растения селектируемый фенотип. Например, маркер может придавать биоцидную устойчивость, такую как устойчивость к антибиотику (например, к канамицину, G418, блеомицину или гигромицину), или к гербициду (например, к глифосату, хлорсульфурону или фосфинотрицину). Кроме того, вектор экспрессии может включать последовательность метки, предназначенную для облегчения манипуляций или выявления (например, очистки или локализации) экспрессированного полипептида. Последовательности меток, такие как последовательности люциферазы, бета-глюкуронидазы, зеленого флуоресцентного белка, глутатион S-трансферазы, полигистидина, c-myc или гемагглютинина, как правило, экспрессируются в виде слияния с кодируемым полипептидом. Такие метки могут быть вставлены в любом месте в пределах полипептида, в том числе на карбоксильном или амино-конце.
Растение или клетку растения можно трансформировать путем интегрирования рекомбинантного полинуклеотида в ее геном с получением стабильной трансформации. Растение или клетка растения, описанные в данном документе, могут быть стабильно трансформированными. Стабильно трансформированные клетки, как правило, сохраняют введенный полинуклеотид с каждым клеточным делением. Растение или клетку растения также можно временно трансформировать таким образом, что рекомбинантный полинуклеотид не интегрируется в их геном. Временно трансформированные клетки, как правило, теряют весь введенный рекомбинантный полинуклеотид или некоторую его часть с каждым клеточным делением, так что введенный рекомбинантный полинуклеотид нельзя выявить в дочерних клетках после достаточного числа клеточных делений. Также в данном документе предусматривается применение редактирования генома.
Существует ряд способов, доступных в данной области для трансформации клетки растения, каждый из которых охвачен в данном документе, в том числе, биолистика, методы с применением генной пушки, Agrobacterium-опосредованная трансформация, опосредованная вирусным вектором трансформация и электропорация. Система Agrobacterium для интеграции чужеродной ДНК в хромосомы растений была тщательно изучена, модифицирована и использована для генной инженерии растений. Молекулы депротеинизированной рекомбинантной ДНК, содержащие последовательности ДНК, соответствующие исследуемому очищенному белку, функционально связанные в смысловой или антисмысловой ориентации с регуляторными последовательностями, соединяют с соответствующими последовательностями Т-ДНК с помощью обычных способов. Их вводят в протопласты с помощью методики с применением полиэтиленгликоля или методики с применением электропорации, обе из которых являются стандартными. В качестве альтернативы, такие векторы, содержащие молекулы рекомбинантной ДНК, кодирующие исследуемый очищенный белок, вводят в живые клетки Agrobacterium, которые затем переносят ДНК в клетки растения. Трансформацию с помощью депротеинизированной ДНК без сопутствующих векторных последовательностей Т-ДНК можно выполнять посредством слияния протопластов с ДНК-содержащими липосомами или посредством электропорации. Депротеинизированную ДНК без сопутствующих векторных последовательностей Т-ДНК также можно использовать для трансформации клеток с помощью инертных, высокоскоростных микрочастиц.
Если клетку или культивируемую ткань используют в качестве реципиентной ткани для трансформации, растения можно регенерировать из трансформированных культур, при необходимости, с помощью методик, известных специалистам в данной области.
Выбор регуляторных участков, которые следует включить в рекомбинантную конструкцию, зависит от нескольких факторов, в том числе без ограничения от эффективности, селектируемости, индуцируемости, требуемого уровня экспрессии и предпочтительной экспрессии в определенных клетках или тканях. Обычным делом для специалиста в данной области является модулирование экспрессии кодирующей последовательности путем соответствующего выбора и размещения регуляторных участков по отношению к кодирующей последовательности. Транскрипцию полинуклеотида можно модулировать подобным образом. Некоторые подходящие регуляторные участки инициируют транскрипцию исключительно или преимущественно в определенных типах клеток. Способы идентификации и установления характеристик регуляторных участков в геномной ДНК растений хорошо известны в данной области.
Примеры промоторов включают тканеспецифичные промоторы, распознаваемые тканеспецифичными факторами, присутствующими в разных типах тканей или клеток (например, специфичные для корней промоторы, специфичные для побегов промоторы, специфичные для ксилем промоторы), или присутствующими на разных стадиях развития, или присутствующими в ответ на разные условия окружающей среды. Примеры промоторов включают конститутивные промоторы, которые могут активироваться в большинстве типов клеток без необходимости в специфических индукторах. Примеры промоторов для управления выработкой полипептида RNAi включают промотор 35S вируса мозаики цветной капусты (CaMV/35S), SSU, OCS, lib4, usp, STLS1, B33, nos или промоторы гена убиквитина или фазеолина. Специалисты в данной области могут создавать множество вариантов рекомбинантных промоторов.
Тканеспецифичные промоторы представляют собой элементы управления транскрипцией, которые активны только в определенных клетках или тканях в определенное время в ходе развития растений, например, в вегетативных тканях или репродуктивных тканях. Тканеспецифичная экспрессия может быть преимущественной, например, когда экспрессия полинуклеотидов в определенных тканях является предпочтительной. Примеры тканеспецифичных промоторов, связанных с развитием, включают промоторы, которые могут инициировать транскрипцию только (или в основном только) в определенных тканях, таких как вегетативные ткани, например, корни или листья, или репродуктивные ткани, такие как плоды, семяпочки, семена, пыльца, тычинки, цветки или любая эмбриональная ткань. Промоторы, специфичные для репродуктивных тканей, могут быть, например, специфичными для пыльника, специфичными для семяпочки, специфичными для зародыша, специфичными для эндосперма, специфичными для интегумента, специфичными для семян и кожуры семян, специфичными для пыльцы, специфичными для лепестка, специфичными для чашелистика или их комбинациями.
Примеры специфичных для листа промоторов включают промотор гена пируватортофосфатдикиназы (PPDK) из C4-растения (маиса), промотор cab-m1Ca+2 из маиса, промотор родственных myb генов из Arabidopsis thaliana (Atmyb5), промоторы генов рибулозобифосфаткарбоксилазы (RBCS) (например, генов томата RBCS 1, RBCS2 и RBCS3A, экспрессирующихся в листьях и выращенных на свету саженцах, RBCS1 и RBCS2, экспрессирующихся в развивающихся плодах томата, или промотор гена рибулозобифосфаткарбоксилазы, экспрессирующегося на высоких уровнях почти исключительно в мезофильных клетках листовых пластинок и листовых пазух).
Примеры специфичных для стареющих тканей промоторов включают промотор из томата, активный во время созревания плодов, старения и опадения листьев, промотор гена, кодирующего цистеиновую протеазу маиса, промотор гена 82E4 и промотор генов SAG.
Специфичные для пыльников промоторы представляют собой дополнительные примеры. Можно выбрать предпочтительные для корней промоторы, известные специалистам в данной области. Предпочтительные для семян промоторы включают как специфичные для семян промоторы (промоторы, активные во время развития семян, такие как промоторы запасных белков семян), так и промоторы прорастающих семян (промоторы, активные во время прорастания семян). Такие предпочтительные для семян промоторы включают без ограничения промотор гена Cim1 (цитокинин-индуцированный сигнал); cZ19B1 (19 кДа зеин кукурузы); milps (миоинозитол-1-фосфатсинтаза); mZE40-2, также известный как Zm-40; nuclc и celA (целлюлозосинтаза). Промотор гена гамма-зеина представляет собой специфичный для эндосперма промотор. Промотор гена Glob-1 представляет собой специфичный для зародыша промотор. Для двудольных растений специфичные для семян промоторы включают без ограничения промотор гена бета-фазеолина фасоли, гена напина, гена β-конглицинина, гена лектина сои, гена круциферина и им подобные. Для однодольных растений специфичные для семян промоторы включают без ограничения промотор гена 15 кДа зеина кукурузы, промотор гена 22 кДа зеина, промотор гена 27 кДа зеина, промотор гена g-зеина, промотор гена 27 кДа гамма-зеина (такой как промотор gzw64A, см. номер доступа S78780 в Genbank), промотор гена waxy, промотор гена shrunken 1, промотор гена shrunken 2, промотор гена глобулина 1 (см. номер доступа L22344 в Genbank), промотор гена Itp2, промотор гена cim1, промоторы генов end1 и end2 кукурузы, промотор гена nuc1, промотор гена Zm40, промотор гена eep1 и eep2; промотор гена lec1, промотор гена тиоредоксина H; промотор гена mlip15, промотор гена PCNA2 и промотор гена shrunken-2.
Примеры индуцибельных промоторов включают промоторы, реагирующие на воздействие патогенов, анаэробные условия, повышенную температуру, свет, засуху, низкую температуру или высокую концентрацию солей. Патоген-индуцибельные промоторы включают промоторы связанных с патогенезом белков (белки PR), которые индуцируются после инфицирования патогеном (например, белки PR, белки SAR, бета-1,3-глюканаза, хитиназа).
В дополнение к растительным промоторам можно получить другие подходящие промоторы бактериального происхождения (например, промотор гена октопинсинтазы, промотор гена нопалинсинтазы и другие промоторы, полученные из Ti-плазмид), или их можно получить из вирусных промоторов (например, 35S и 19S РНК промоторов вируса мозаики цветной капусты (CaMV), конститутивных промоторов вируса табачной мозаики, промоторов 19S и 35S вируса мозаики цветной капусты (CaMV) или промотора 35S вируса мозаики норичника). В определенных вариантах осуществления промотор вируса мозаики мирабилис (MMV) является предпочтительным. В определенных вариантах осуществления промотор 35S является предпочтительным.
В другом аспекте предложен выделенный полипептид, содержащий полипептидную последовательность, состоящий или по существу состоящий из нее, которая характеризуется по меньшей мере 72% идентичностью последовательности с любой из полипептидных последовательностей, описанных в данном документе, включая любые полипептиды, приведенные в перечне последовательностей. Соответственно, выделенный полипептид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5% 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с ним.
В одном варианте осуществления выделенный полипептид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 72%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO: 3, или SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:15, или SEQ ID NO:18, или SEQ ID NO:21.
В другом варианте осуществления выделенный полипептид содержит, состоит или по существу состоит из последовательности, характеризующейся по меньшей мере 72%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21.
В определенных вариантах осуществления активность полипептида, содержащего, состоящего или по существу состоящего из последовательности, кодирующей белок Terminal Flower 1 и характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO: 3, или SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:15, или SEQ ID NO:18, или SEQ ID NO:21, модулировали. В другом варианте осуществления активность полипептида, содержащего, состоящего или по существу состоящего из последовательности, кодирующей белок Terminal Flower 1 и характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21, модулировали.
Полипептид может содержать фрагменты последовательностей, предусматривающих достаточную или существенную степень идентичности или сходства для функционирования в качестве белка Terminal Flower 1. Фрагменты полипептида(-ов), как правило, сохраняют некоторую или всю активность полноразмерной последовательности - такую как по меньшей мере приблизительно 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% активность.
Как рассматривается в данном документе, полипептиды также включают мутантов, полученных путем введения альтераций любого типа (например, одной или более вставок, делеций или замен аминокислот; изменений структуры гликозилирования, изменений, которые влияют на рефолдинг или изомеризацию, трехмерную структуру или состояния самоассоциации) при условии, что они все еще сохраняют свою функцию или активность в качестве белка Terminal Flower 1 в некоторой степени или полностью. Соответственно, функция или активность в качестве белка Terminal Flower 1 модулирована, снижена или подавлена. Соответственно, функция или активность в качестве белка Terminal Flower 1 подавлена таким образом, что активность белка Terminal Flower 1 не является выявляемой. Иллюстративные мутанты описаны в данном документе.
Полипептиды включают варианты, полученные путем введения любого типа альтераций (например, вставок, делеций или замен аминокислот, изменений структуры гликозилирования, изменений, которые влияют на рефолдинг или изомеризацию, трехмерные структуры или состояния самоассоциации), которые можно намеренно сконструировать или легко выделить. Альтерация может представлять собой один или более стоп-кодонов. Вариант может иметь альтерации, которые производят молчащее изменение и приводят в результате к образованию функционально эквивалентного белка. Намеренные аминокислотные замены можно осуществлять, исходя из сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и амфипатической природы остатков при условии, что вторичная активность в отношении связывания вещества сохраняется. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту, положительно заряженные аминокислоты включают лизин и аргинин, и аминокислоты с незаряженными полярными концевыми группами с подобными значениями гидрофильности включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин. Консервативные замены можно осуществлять, например, в соответствии с приведенной ниже таблицей. Аминокислоты в одном и том же блоке во втором столбце и предпочтительно в одной и той же строке в третьем столбце можно заменять одна на другую.
Полипептид может быть зрелым белком или незрелым белком, или белком, полученным из незрелого белка. Полипептиды могут быть в линейной форме или циклизированы с использованием известных способов. Полипептиды, как правило, содержат по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, или по меньшей мере 40, или по меньшей мере 50, или по меньшей мере 100, или по меньшей мере 200, или по меньшей мере 300, или по меньшей мере 400, или по меньшей мере 500, или по меньшей мере 600 смежных аминокислот.
Также раскрыт полипептид, кодируемый SEQ ID NO: 3, или SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:15, или SEQ ID NO:18, или SEQ ID NO:21, который характеризуется 100% идентичностью последовательности с ним, или полипептид, содержащий, состоящий или по существу состоящий из последовательности, представленной под SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:15, или SEQ ID NO:21.
Полипептид можно получить путем культивирования трансформированных или рекомбинантных клеток-хозяев в условиях культивирования, подходящих для экспрессии полипептида. Полученный экспрессированный полипептид затем можно очистить из такой культуры с помощью известных способов очистки. Очистка полипептида может включать использование аффинной колонки, содержащей средства, которые связываются с полипептидом; одну или более стадий с использованием колонки с такими аффинными смолами; одну или более стадий с участием хроматографии с гидрофобным взаимодействием или иммуноаффинную хроматографию. В качестве альтернативы, полипептид может быть экспрессирован в форме, которая будет облегчать очистку. Например, он может быть экспрессирован в виде полипептида слияния, например, с мальтозосвязывающим полипептидом, глутатион-5-трансферазой, his-тагом или тиоредоксином. Наборы для экспрессии и очистки полипептидов слияния являются коммерчески доступными. Полипептид можно метить эпитопом и затем очистить с использованием специфического антитела к такому эпитопу. Одну или более стадий жидкостной хроматографии, такой как обращенно-фазовая высокоэффективная жидкостная хроматография, можно использовать для дополнительной очистки полипептида. Некоторые или все из вышеуказанных стадий очистки в различных комбинациях можно использовать для получения практически гомогенного рекомбинантного полипептида. Очищенный таким образом полипептид может быть практически свободным от других полипептидов и определен в данном документе как «практически очищенный полипептид»; такие очищенные полипептиды включают полипептиды, фрагменты, варианты и т. п. Экспрессию, выделение и очистку полипептидов и фрагментов можно выполнять с помощью любой подходящей методики, включая без ограничения способы, описанные в данном документе.
Кроме того, можно использовать аффинную колонку, например, с моноклональным антителом, созданным против полипептидов, для аффинной очистки экспрессированных полипептидов. Эти полипептиды можно удалять из аффинной колонки с помощью общепринятых методик, например, в элюирующем буфере с высоким содержанием солей с последующим диализом в буфере с низким содержанием солей для использования, или посредством изменения рН или других компонентов в зависимости от используемой аффинной матрицы, или путем конкурентного удаления с помощью естественного субстрата аффинного фрагмента.
Описаны выделенные или практически очищенные полинуклеотиды или композиции белков. «Выделенный» или «очищенный» полинуклеотид или белок, или его биологически активная часть, практически или по существу не содержит компоненты, которые обычно дополняют полинуклеотид или белок или взаимодействуют с ним, обнаруживаемые в его естественном окружении. Таким образом, выделенный или очищенный полинуклеотид или белок практически не содержит другой клеточный материал или культуральную среду при получении с помощью методик рекомбинации, или практически не содержит веществ-предшественников или других веществ при получении путем химического синтеза. В оптимальном случае «выделенный» полинуклеотид не содержит последовательности (оптимально кодирующие белок последовательности), которые в естественных условиях фланкируют полинуклеотид (например, последовательности, расположенные на 5'- и 3'-концах полинуклеотида) в геномной ДНК организма, из которого получен полинуклеотид. Например, в различных вариантах осуществления выделенный полинуклеотид может содержать менее приблизительно 5 т.п.о., 4 т.п.о., 3 т.п.о., 2 т.п.о., 1 т.п.о., 0.5 т.п.о., или 0,1 т.п.о. нуклеотидной последовательности, которая в естественных условиях фланкирует полинуклеотид в геномной ДНК клетки, из которой получен полинуклеотид. Белок, который практически не содержит клеточный материал, включает препараты белка, характеризующегося содержанием контаминирующего белка, составляющим менее приблизительно 30%, 20%, 10%, 5% или 1% (из расчета на сухой вес).
Полипептид можно также получать с помощью известных общепринятых методик химического синтеза. Способы конструирования полипептидов или их фрагментов способами синтеза известны специалистам в данной области. Синтетически сконструированные полипептидные последовательности благодаря общим первичным, вторичным или третичным структурным, или конформационным характеристикам с нативными полипептидами могут обладать общими с ними биологическими свойствами, включая биологическую активность.
Различия в генетическом фоне можно выявить по фенотипическим различиям или с помощью методик молекулярной биологии, известных из уровня техники, таких как секвенирование нуклеиновых кислот, определение наличия или отсутствия генетических маркеров (например, маркеров микросателлитной РНК).
Также предусмотрены антитела, которые являются иммунореактивными по отношению к полипептидам, описанным в данном документе. Полипептиды, фрагменты, варианты, полипептиды слияния и им подобные, изложенные в данном документе, можно использовать в качестве «иммуногенов» в получении антител, иммунореактивных по отношению к ним. Такие антитела могут специфически связываться с полипептидом через антигенсвязывающие сайты антитела. Специфически связывающимися антителами являются такие антитела, которые будут специфически распознавать полипептид, гомологи и варианты и связываться с ними, а не с другими молекулами. В одном варианте осуществления антитела являются специфичными для полипептидов, имеющих аминокислотную последовательность, изложенную в данном документе, и не взаимодействуют перекрестно с другими полипептидами.
Более конкретно, полипептиды, фрагмент, варианты, полипептиды слияния и им подобные содержат антигенные детерминанты или эпитопы, которые вызывают образование антител. Данные антигенные детерминанты или эпитопы могут быть линейными или конформационными (прерывистыми). Линейные эпитопы состоят из одного отрезка аминокислот полипептида, тогда как конформационные или прерывистые эпитопы состоят из аминокислотных частей из разных участков полипептидной цепи, которые были приведены в непосредственную близость при сворачивании полипептида. Эпитопы можно идентифицировать любыми способами, известными в данной области. Кроме того, эпитопы из полипептидов можно использовать в качестве реагентов для исследования, в анализах и для очистки специфически связывающихся антител из веществ, таких как поликлональные сыворотки или надосадочные жидкости из культивируемых гибридом. Такие эпитопы или их варианты можно получать с использованием методик, известных в данной области, например, путем твердофазного синтеза, химического или ферментативного расщепления полипептида, или с помощью технологии рекомбинантной ДНК.
Как поликлональные, так и моноклональные антитела к полипептидам можно получать с помощью общепринятых методик. Клеточные линии гибридом, которые продуцируют моноклональные антитела, специфичные для полипептидов, также рассматриваются в данном документе. Такие гибридомы можно получать и идентифицировать с помощью общеизвестных методик. Для получения антител различных животных-хозяев можно иммунизировать путем введения полипептида, его фрагмента, варианта или мутантов. К примеру, такие животные-хозяева могут включать без ограничения кроликов, мышей и крыс. Различные адъюванты можно использовать для повышения иммунологического ответа. В зависимости от вида хозяина такие адъюванты включают, помимо прочего, адъюванты Фрейнда (полные и неполные), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюрониловые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки, динитрофенол и потенциально пригодные адъюванты человека, такие как BCG (бацилла Кальметта-Герена) и Corynebacterium parvum. Моноклональные антитела можно выделять с помощью общепринятых методик. Такие моноклональные антитела могут быть иммуноглобулинами любого класса, в том числе IgG, IgM, IgE, IgA, IgD, и любого их подкласса.
Антитела можно также использовать в анализах для выявления присутствия полипетидов или фрагментов либо in vitro, либо in vivo. Антитела можно также использовать для очистки полипептидов или фрагментов с помощью иммуноаффинной хроматографии.
Также раскрыты фрагменты полинуклеотидов, описанные в данном документе, и полипептиды, кодируемые таким образом. Фрагменты полинуклеотида могут кодировать фрагменты белка, которые сохраняют биологическую активность нативного белка. В качестве альтернативы, фрагменты полинуклеотида, применимые в качестве зондов для гибридизации или ПЦР-праймеров, как правило, не кодируют фрагмент белка, сохраняющий биологическую активность. Кроме того, фрагменты раскрытых нуклеотидных последовательностей включают такие фрагменты, которые могут быть собраны в рекомбинантные конструкции, как описано в данном документе. Фрагменты полинуклеотидной последовательности могут находиться в диапазоне от по меньшей мере приблизительно 21 нуклеотида, приблизительно 22 нуклеотидов, приблизительно 23 нуклеотидов, приблизительно 25 нуклеотидов, приблизительно 50 нуклеотидов, приблизительно 75 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 150 нуклеотидов, приблизительно 200 нуклеотидов, приблизительно 250 нуклеотидов, приблизительно 300 нуклеотидов, приблизительно 400 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 600 нуклеотидов, приблизительно 700 нуклеотидов, приблизительно 800 нуклеотидов, приблизительно 900 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 1100 нуклеотидов, приблизительно 1200 нуклеотидов, приблизительно 1300 нуклеотидов или приблизительно 1500 нуклеотидов, приблизительно 2000 нуклеотидов, приблизительно 3000 нуклеотидов, приблизительно 4000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 6000 нуклеотидов, приблизительно 7000 нуклеотидов, приблизительно 8000 нуклеотидов, приблизительно 9000 нуклеотидов, приблизительно 10000 нуклеотидов, приблизительно 15000 нуклеотидов, приблизительно 20000 нуклеотидов и до полноразмерного полинуклеотида, кодирующего полипептиды, описанные в данном документе.
Фрагменты полипептидной последовательности могут находится в диапазоне от по меньшей мере приблизительно 25 аминокислот, приблизительно 50 аминокислот, приблизительно 75 аминокислот, приблизительно 100 аминокислот приблизительно 150 аминокислот, приблизительно 200 аминокислот, приблизительно 250 аминокислот, приблизительно 300 аминокислот, приблизительно 400 аминокислот, приблизительно 500 аминокислот, приблизительно 600 аминокислот или до полноразмерного полипептида, описанного в данном документе.
Модулирование экспрессии или активности одного или более белков TFL1 или одной более последовательностей нуклеиновых кислот TFL1 является преимущественным по причинам, описанным в данном документе. Экспрессию TFL1-2S (SEQ ID NO: 7 или 8), или TFL1-2T (SEQ ID NO: 10 или 11), или TFL1-4T (SEQ ID NO: 19 или 20) можно модулировать по отдельности у растения таким образом, что модулируют экспрессию только одного из TFL1-2S (SEQ ID NO: 7 или 8), или TFL1-2T (SEQ ID NO: 10 или 11), или TFL1-4T (SEQ ID NO: 19 или 20). Экспрессию двух или более из TFL1-2S (SEQ ID NO: 7 или 8), или TFL1-2T (SEQ ID NO: 10 или 11), или TFL1-4T (SEQ ID NO: 19 или 20) можно модулировать у растения таким образом, что модулируют экспрессию двух или более из TFL1-2S (SEQ ID NO: 7 или 8), или TFL1-2T (SEQ ID NO: 10 или 11), или TFL1-4T (SEQ ID NO: 19 или 20). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 7 или 8) и TFL1-2T (SEQ ID NO: 10 или 11). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 7 или 8) и TFL1-4T (SEQ ID NO: 19 или 20). Например, можно модулировать экспрессию TFL1-2T (SEQ ID NO: 10 или 11) и TFL1-4T (SEQ ID NO: 19 или 20). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 7 или 8) и TFL1-4T (SEQ ID NO: 19 или 20).
Активность TFL1-2S(SEQ ID NO: 9), или TFL1-2T (SEQ ID NO: 12), или TFL1-4T(SEQ ID NO: 21) можно модулировать по отдельности у растения таким образом, что модулируют экспрессию только одного из TFL1-2S(SEQ ID NO: 9), или TFL1-2T (SEQ ID NO: 12), или TFL1-4T(SEQ ID NO: 21). Экспрессию двух или более из TFL1-2S(SEQ ID NO: 9), или TFL1-2T (SEQ ID NO: 12), или TFL1-4T(SEQ ID NO: 21) можно модулировать у растения таким образом, что модулируют экспрессию двух или более из TFL1-2S(SEQ ID NO: 9), или TFL1-2T (SEQ ID NO: 12), или TFL1-4T(SEQ ID NO: 21). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 9) и TFL1-2T (SEQ ID NO: 12). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 9) и TFL1-4T (SEQ ID NO: 21). Например, можно модулировать экспрессию TFL1-2T (SEQ ID NO: 12) и TFL1-4T (SEQ ID NO: 21). Например, можно модулировать экспрессию TFL1-2S (SEQ ID NO: 21) и TFL1-4T (SEQ ID NO: 21).
В соответствии с определенными вариантами осуществления модулирование (например, снижение) экспрессии белка Terminal Flower 1 можно осуществлять на геномном уровне и/или уровне транскрипта с применением ряда молекул, которые нарушают транскрипцию и/или трансляцию, включая без ограничения антисмысловые молекулы, молекулы siRNA, молекулы рибозима или дезоксирибозима. Также можно применять вставку одной или более мутаций в по меньшей мере один ген, включая делеции, вставки, сайт-специфические мутации, нуклеазы с «цинковыми пальцами» и т. п. В соответствии с другими вариантами осуществления экспрессию можно подавить на уровне белка с применением антагонистов или ферментов, которые расщепляют полипептид, и т. п.
В одном аспекте описано мутантное растение или его часть, содержащие по меньшей мере одну мутацию в (i) полинуклеотидной последовательности, содержащей, состоящей или по существу состоящей из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептиде, кодируемом полинуклеотидом, представленным в (i); или (iii) полипептиде, характеризующемся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; или (iv) конструкции, векторе или векторе экспрессии, содержащих выделенный полинуклеотид, представленный в (i), где по меньшей мере одна мутация обеспечивает снижение экспрессии или активности белка Terminal Flower 1 по сравнению с контрольным растением, которое не содержит по меньшей мере одну мутацию. Растение или клетка растения, следовательно, могут содержать одну или более мутаций в TFL1-2S (SEQ ID NO: 7 или 8), и/или TFL1-2T (SEQ ID NO: 10 или 11), и/или TFL1-4T (SEQ ID NO: 19 или 20), где указанная мутация приводит к сниженной экспрессии или сниженной функции указанного гена или белка, кодируемого им.
Экспрессию или функцию мутанта(-ов) можно модулировать, подавлять или уменьшать. Мутантные растение или клетка растения могут иметь одну или более дополнительных мутаций в одном или более других генах или полипептидах. В определенных вариантах осуществления мутанты могут иметь одну или более дополнительных мутаций в одном или более других генах или полипептидах.
Указанные мутантные растение или клетка растения могут быть гетерозиготными или гомозиготными по мутации(-ям). Указанные мутантные растение или клетка растения могут быть гетерозиготными по меньшей мере по одной мутации и гомозиготными по меньшей мере по одной отличающейся мутации. Соответственно, мутантные растение или клетка растения являются гомозиготными по мутации(-ям).
Иллюстративные мутанты и мутации описаны в данном документе.
В одном варианте осуществления по меньшей мере одна мутация выбрана из группы, состоящей из мутации в положении T143 и/или G129 в SEQ ID NO: 9; или мутации в положении R120, и/или G129, и/или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21 или комбинацию двух или более из них.
В одном варианте осуществления по меньшей мере одна мутация в SEQ ID NO: 9 выбрана из мутаций в положениях {T143,G129} {T143,H84} {G129,H84} {T143,G129,H84}.
В одном варианте осуществления по меньшей мере одна мутация в SEQ ID NO: 12 выбрана из мутаций в положениях {R120,G129} {R120,P131} {G129,P131} {R120,G129,P131} {R120,P131,D142}
В одном варианте осуществления по меньшей мере одна мутация в SEQ ID NO: 12 выбрана из мутаций в положениях {P110,H86}.
В одном варианте осуществления по меньшей мере одна мутация представляет собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
В одном варианте осуществления по меньшей мере одна мутация представляет собой мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
В одном варианте осуществления мутации представляют собой мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S и мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
Раскрыты все возможные комбинации таких мутаций, которые включают любые 2, 3, 4, 5, 6 или 7 мутаций, выбранных из положений T143 и G129 в SEQ ID NO: 9, и положений R120, и/или G129, и/или P131 в SEQ ID NO: 12, и положений P110 или H86 в SEQ ID NO: 21.
В одном варианте осуществления мутация представляет собой T143I в SEQ ID NO: 9. В одном варианте осуществления мутация представляет собой G129R в SEQ ID NO: 9. В одном варианте осуществления мутация представляет собой G129E в SEQ ID NO: 9. В одном варианте осуществления мутация представляет собой H84STOP в SEQ ID NO: 9. В одном варианте осуществления мутация представляет собой R120C в SEQ ID NO: 12. В одном варианте осуществления мутация представляет собой G129E в SEQ ID NO: 12. В одном варианте осуществления мутация представляет собой P131S в SEQ ID NO: 12. В одном варианте осуществления мутация представляет собой P110L в SEQ ID NO: 21. В одном варианте осуществления мутация представляет собой H86STOP в SEQ ID NO: 21.
Раскрыты все возможные комбинации таких мутаций, которые включают любые 2, 3, 4, 5, 6, 7, 8 или 9 мутаций, выбранных из T143I, и/или G129R, и/или G129E, и/или H84STOP в SEQ ID NO: 9, и/или R120C, и/или G129E, и/или P131S в SEQ ID NO: 12, и/или P110L, и/или H86STOP в SEQ ID NO: 21.
В другом аспекте предусмотрен способ сокращения периода времени до наступления цветения у растения или у растительного материала, полученного из растения, при этом указанный способ включает введение в геном указанного растения одной или более мутаций, которые обеспечивают снижение экспрессии по меньшей мере одного гена TFL-1, где указанный по меньшей мере один ген TFL-1 кодирует TFL1-2S (SEQ ID NO: 7 или 8), TFL1-2T (SEQ ID NO: 10 или 11) и TFL1-4T (SEQ ID NO: 19 или 20).
Также предусмотрен способ идентификации растения с сокращенным периодом времени до наступления цветения, при этом указанный способ включает скрининг образца нуклеиновой кислоты из растения, представляющего интерес, в отношении наличия одной или более мутаций в TFL1-2S (SEQ ID NO: 7 или 8), TFL1-2T (SEQ ID NO: 10 или 11) и TFL1-4T (SEQ ID NO: 19 или 20).
Также раскрыты растение или клетка растения, которые являются гетерозиготными или гомозиготными в отношении одной или более мутаций в гене, кодирующем TFL1-2S (SEQ ID NO: 7 или 8), TFL1-2T (SEQ ID NO: 10 или 11) и TFL1-4T (SEQ ID NO: 19 или 20), где указанная(-ые) мутация(-и) приводят к сниженной экспрессии гена или сниженной функции белка, кодируемого им.
В некоторых вариантах осуществления благоприятную(-ые) мутацию(-ии) вводят в растение или клетку растения с использованием подхода, предусматривающего мутагенез, и введенную мутацию идентифицируют или подвергают отбору с применением способов, известных специалисту в данной области, таких как блоттинг Саузерна, секвенирование ДНК, ПЦР-анализ или фенотипический анализ. Мутации, которые влияют на экспрессию генов, или которые нарушают функцию кодируемого белка, можно определять с использованием способов, хорошо известных в данной области. Мутации по типу вставки в экзон гена, как правило, приводят к образованию нефункциональных мутантов. Мутации в консервативных остатках могут быть особенно эффективными для подавления или снижения метаболической функции кодируемого белка.
Также раскрыты способы получения мутантных полинуклеотидов и полипептидов. Любое представляющее интерес растение, в том числе клетку растения или растительный материал, можно генетически модифицировать различными способами, с помощью которых, как известно, индуцируется мутагенез, в том числе сайт-направленный мутагенез, олигонуклеотид-направленный мутагенез, индуцированный химическими соединениями мутагенез, индуцированный ионизирующим излучением мутагенез, мутагенез с использованием модифицированных оснований, мутагенез с использованием содержащей разрывы двухспиральной ДНК, мутагенез с двухнитевыми разрывами, мутагенез с использованием штаммов хозяев с нарушенной репарацией, мутагенез посредством синтеза полного гена, шаффлинг ДНК и другие эквивалентные способы.
Мутантные варианты полипептида можно использовать для создания мутантных, не встречающихся в природе или трансгенных растений (например, мутантных, не встречающихся в природе, трансгенных, созданных человеком или полученных с помощью методик генной инженерии) или клеток растения, содержащих один или более мутантных вариантов полипептида. Соответственно, мутантные варианты полипептида сохраняют активность немутантного полипептида. Активность мутантного варианта полипептида может быть выше, ниже или приблизительно такой же, как немутантного полипептида.
Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут включать внесенные человеком мутации или синтетические мутации, или мутации, созданные с помощью методик генной инженерии. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут быть мутациями, которые получены или которые можно получить посредством способа, который включает стадию манипуляции in vitro или in vivo. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут быть мутациями, которые получены или которые можно получить посредством способа, который включает вмешательство человека. В качестве примера, способ может включать мутагенез с применением экзогенно добавляемых веществ, таких как мутагенные, тератогенные или канцерогенные органические соединения, например этилметансульфонат (EMS), которые приводят к случайным мутациям в генетическом материале. В качестве дополнительного примера способ может включать одну или более стадий, предусматривающих генную инженерию, таких как одну или более стадий, предусматривающих генную инженерию, которые описаны в данном документе, или их комбинации. В качестве дополнительного примера способ может включать одну или более стадий скрещивания растений.
Активность одного или более полипептидов Terminal Flower 1 у растения является сниженной или подавленной согласно настоящему изобретению, если активность превращения является статистически более низкой, чем активность превращения того(-ех) же полипептида(-ов) Terminal Flower 1 у растения, которое не было модифицировано с подавлением активности превращения этого полипептида Terminal Flower 1 и которое было культивировано и собрано с применением тех же протоколов. Активность полипептида Terminal Flower 1 у растения считается устраненной, если данная активность не является выявляемой с помощью аналитических способов, описанных в данном документе. Способы определения активности полипептида Terminal Flower 1 описаны в данном документе.
Кроме продуктов мутагенеза композиции, которые могут модулировать экспрессию или активность одного или более полинуклеотидов или полипептидов, описанных в данном документе, включают без ограничения специфичные в отношении последовательности полинуклеотиды, которые могут нарушать транскрипцию одного или более эндогенных генов; специфичные в отношении последовательности полинуклеотиды, которые могут нарушать трансляцию РНК-транскриптов (например, двухнитевые РНК, siRNA, рибозимы); специфичные в отношении последовательности полинуклеотиды, которые могут нарушать стабильность одного или более белков; специфичные в отношении последовательности полинуклеотиды, которые могут нарушать ферментативную активность одного или более белков или активность связывания одного или более белков в отношении субстратов или регуляторных белков; антитела, которые проявляют специфичность в отношении одного или более белков; низкомолекулярные соединения, которые могут нарушать стабильность одного или более белков или ферментативную активность одного или более белков или активность связывания одного или более белков; белки «цинковые пальцы», которые связывают один или более полинуклеотидов; и мегануклеазы, которые обладают активностью в отношении одного или более полинуклеотидов. Технологии редактирования гена, генетические технологии редактирования и технологии редактирование генома хорошо известны в данной области.
Один из способов редактирования гена предусматривает использование подобных активатору транскрипции эффекторных нуклеаз (TALEN), вызывающих двухнитевые разрывы, на которые клетка может отвечать с привлечением механизмов репарации. Негомологичное соединение концов повторно сцепляет ДНК с обеих сторон двухнитевого разрыва, где имеет место незначительное, или вовсе отсутствует, перекрывание последовательности для комплементарного связывания. Этот механизм репарации индуцирует ошибки в геноме посредством вставки или делеции, либо хромосомной перестройки. Любые такие ошибки могут сделать генные продукты, закодированные в этом месте, не функциональными. Другой способ редактирования гена предусматривает использование бактериальной системы CRISPR/Cas. Бактерии и архебактерии характеризуются хромосомными элементами, называемыми короткими палиндромными повторами, регулярно расположенными группами (CRISPR), которые являются частью адаптивной иммунной системы, защищающей от проникновения вирусной и плазмидной ДНК. В системах CRISPR II типа, CRISPR РНК (crRNA) функционирует с трансактивирующей crRNA (tracrRNA) и CRISPR-ассоциированными (Cas) белками с внесением двухнитевых разрывов в целевую ДНК. Целевое расщепление с помощью Cas9 требует спаривания оснований crRNA и tracrRNA, а также спаривания оснований crRNA и целевой ДНК. Целевое распознавание облегчается при наличии короткого мотива, называемого прилегающим к протоспейсеру мотивом (PAM), который соответствует последовательности NGG. Данную систему можно использовать для редактирования генома. Cas9 обычно программируется двойной РНК, состоящей из crRNA и tracrRNA. Тем не менее, коровые компоненты этих РНК могут быть объединены в единую гибридную «направляющую РНК» для нацеливания Cas9. Использование некодирующей направляющей РНК с нацеливанием ДНК для сайт-специфического расщепления обещает быть значительно более простым, чем существующие технологии, такие как TALEN. Использование стратегии CRISPR/Cas с перенацеливанием нуклеазного комплекса требует только введения новой последовательности РНК и нет необходимости в переконструировании специфичности факторов транскрипции белка.
Антисмысловая технология представляет собой другой известный способ, который можно использовать для модулирования экспрессии полипептида. Полинуклеотид гена, который подлежит репрессии, клонируют и функционально связывают с регуляторным участком и последовательностью терминации транскрипции, так что антисмысловая нить РНК транскрибируется. Затем рекомбинантной конструкцией трансформируют клетку растения и получают антисмысловую нить РНК. Полинуклеотид не обязательно является полной последовательностью гена, который подлежит репрессии, но, как правило, будет практически комплементарным по меньшей мере части смысловой нити гена, который должен быть репрессирован.
Полинуклеотид может быть транскрибирован в рибозим или каталитическую РНК, которая влияет на экспрессию mRNA. Рибозимы можно сконструировать для специфического спаривания практически с любой целевой РНК и расщепления фосфодиэфирного остова в определенном месте, тем самым функционально инактивируя целевую РНК. Гетерологичные полинуклеотиды могут кодировать рибозимы, сконструированные для расщепления конкретных транскриптов mRNA, предотвращая таким образом экспрессию полипептида. Рибозимы типа hammerhead являются применимыми для разрушения конкретных mRNA, хотя можно использовать различные рибозимы, которые расщепляют mRNA в последовательностях сайт-специфического распознавания. Рибозимы типа hammerhead расщепляют mRNA в местах, определяемых фланкирующими участками, которые формируют комплементарные пары оснований с целевой mRNA. Единственным требованием является то, что целевая РНК должна содержать нуклеотидную последовательность 5'-UG-3'. Конструирование и получение рибозимов типа hammerhead известно в данной области. Последовательности рибозимов типа hammerhead можно встроить в стабильную РНК, такую как транспортная РНК (tRNA) для повышения эффективности расщепления in vivo.
В одном варианте осуществления специфичный к последовательности полинуклеотид, который может нарушать трансляцию транскрипта(-ов) РНК, представляет собой интерферирующую РНК. РНК-интерференция или РНК-сайленсинг представляет собой эволюционно консервативный процесс, в котором конкретные mRNA могут быть нацелены для ферментативной деградации. Двухнитевую РНК (двухнитевая РНК) вводят в клетку или получают в клетке (например, двухнитевой РНК-вирус или полинуклеотиды интерферирующей РНК) для инициации пути интерферирующей РНК. Двухнитевую РНК можно преобразовывать в несколько дуплексов малых интерферирующих РНК длиной 21-23 п. о. с помощью РНКаз III, которые представляют собой эндонуклеазы, специфичные к двухнитевой РНК. Малые интерферирующие РНК можно затем распознать РНК-индуцированными комплексами сайленсинга, которые способствуют раскручиванию малых интерферирующих РНК с помощью АТФ-зависимого процесса. Раскрученная антисмысловая нить малой интерферирующей РНК направляет активированные РНК-индуцированные комплексы сайленсинга к нацеленной mRNA, содержащей последовательность, комплементарную антисмысловой нити малой интерферирующей РНК. Нацеленная mRNA и антисмысловая нить могут образовывать А-форму спирали, и большую бороздку А-формы спирали можно распознать активированными РНК-индуцированными комплексами сайленсинга. Целевую mRNA можно расщепить с помощью активированных РНК-индуцированных комплексов сайленсинга в одном сайте, определенном сайтом связывания 5'-конца нити малой интерферирующей РНК. Активированные РНК-индуцированные комплексы сайленсинга могут повторно использоваться для катализа еще одного события расщепления.
Пример целевой последовательности смысловой RNAi для TFL1-1S/T представлен под SEQ ID NO: 22. Пример целевой последовательности антисмысловой RNAi для TFL1-1S/T представлен под SEQ ID NO: 23. Пример конструкции TFL1-1S/T RNAi представлен под SEQ ID NO: 24.
Пример целевой последовательности смысловой RNAi для TFL1-1S представлен под SEQ ID NO: 25. Пример целевой последовательности антисмысловой RNAi для TFL1-1S представлен под SEQ ID NO: 26.
Пример целевой последовательности смысловой RNAi для TFL1-1T представлен под SEQ ID NO: 27. Пример целевой последовательности антисмысловой RNAi для TFL1-1T представлен под SEQ ID NO: 28.
Пример целевой последовательности смысловой RNAi для TFL1-2S/T представлен под SEQ ID NO: 29. Пример целевой последовательности антисмысловой RNAi для TFL1-2S/T представлен под SEQ ID NO: 30. Пример конструкции TFL1-2S/T RNAi представлен под SEQ ID NO: 31.
Пример целевой последовательности смысловой RNAi для TFL1-2S представлен под SEQ ID NO: 32. Пример целевой последовательности антисмысловой RNAi для TFL1-2S представлен под SEQ ID NO: 33.
Пример целевой последовательности смысловой RNAi для TFL1-2T представлен под SEQ ID NO: 34. Пример целевой последовательности антисмысловой RNAi для TFL1-2T представлен под SEQ ID NO: 35.
Пример целевой последовательности смысловой RNAi для TFL1-3T представлен под SEQ ID NO: 36. Пример целевой последовательности антисмысловой RNAi для TFL1-3T представлен под SEQ ID NO: 37. Пример конструкции TFL1-3T RNAi представлен под SEQ ID NO: 38.
Пример целевой последовательности смысловой RNAi для TFL1-4T представлен под SEQ ID NO: 39. Пример целевой последовательности антисмысловой RNAi для TFL1-4T представлен под SEQ ID NO: 40. Пример конструкции TFL1-4T RNAi представлен под SEQ ID NO: 41.
Раскрыты последовательности длиной от приблизительно 21 до 23 нуклеотидов для любого из SEQ ID NO: 22, 23, 25, 26, 27, 28, 29, 30, 32, 33, 34, 35, 36, 37, 39, 40 или 41.
Также рассмотрены способы сайленсинга генов с применением таких кодирующих последовательностей и пути их применения.
Векторы экспрессии интерферирующей РНК могут содержать конструкции интерферирующей РНК, кодирующие полинуклеотиды интерферирующей РНК, которые проявляют активностью в отношении РНК-интерференции путем снижения уровня экспрессии mRNA, пре-mRNA или родственных вариантов РНК. Векторы экспрессии могут содержать промотор, расположенный выше по цепи и функционально связанный с конструкцией интерферирующей РНК, как дополнительно описано в данном документе. Векторы экспрессии интерферирующей РНК могут содержать подходящий минимальный коровый промотор, представляющую интерес конструкцию интерферирующей РНК, расположенный выше по цепи (5') регуляторный участок, расположенный ниже по цепи (3') регуляторный участок, в том числе сигналы терминации транскрипции и полиаденилирования, и другие последовательности, известные специалистам в данной области, такие как различные селективные маркеры.
Примеры конструкций интерферирующей РНК представлены под SEQ ID NO: 24, 31, 38 и 41.
В одном варианте осуществления вектор экспрессии содержит промотор, такой как сильный конститутивный промотор MMV (вируса мозаики мирабилис), расположенный выше по цепи и функционально связанный с конструкцией интерферирующей РНК, и регуляторный участок ниже по цепи (3'), такой как терминаторная 3'-последовательность nos гена нопалинсинтазы Agrobacterium tumefaciens.
Полинуклеотиды можно получить в различных формах, в том числе в виде двухнитевых структур (то есть, двухнитевая молекула РНК, содержащая антисмысловую нить и комплементарную ей смысловую нить), двухнитевых шпилькообразных структур или однонитевых структур (то есть, молекула ssRNA, содержащая только антисмысловую нить). Структуры могут содержать вторичную структуру в виде двойной спирали, асимметричной двойной спирали, шпильки или асимметричной шпильки, имеющую самокомплементарную смысловую и антисмысловую нити. Двухнитевую интерферирующую РНК можно превратить ферментативным путем в двухнитевые малые интерферирующие РНК. Одну из нитей двойной спирали малой интерферирующей РНК можно отжечь с комплементарной последовательностью в целевой mRNA и родственных вариантах РНК. Двойные спирали малой интерферирующей РНК/mRNA распознаются РНК-индуцированными комплексами сайленсинга, которые могут расщеплять РНК в нескольких сайтах зависимым от последовательности образом, приводя в результате к разрушению целевой mRNA и родственных вариантов РНК.
Молекулы двухнитевой РНК могут включать молекулы малой интерферирующей РНК, собранные из одного олигонуклеотида в структуре стебель-петля, где самокомплементарные смысловой и антисмысловой участки молекулы малой интерферирующей РНК соединены с помощью основанного на полинуклеотидах или не основанного на полинуклеотидах линкера(-ов), а также кольцевую однонитевую РНК с двумя или более петлевыми структурами и стеблем, содержащую самокомплементарные смысловую и антисмысловую нити, где кольцевую РНК можно процессировать либо in vivo , либо in vitro с образованием активной молекулы малой интерферирующей РНК, способной опосредовать интерференции РНК.
Двухнитевая РНК может содержать по меньшей мере две последовательности, которые по меньшей мере частично комплементарны друг другу. Смысловая нить может содержать первую последовательность, и антисмысловая нить может содержать вторую последовательность. По меньшей мере одна из последовательностей может содержать по меньшей мере 10 смежных нуклеотидов из РНК TFL1. По меньшей мере одна из последовательностей может содержать приблизительно 21-23 смежных нуклеотида из РНК TFL1.
В одном варианте осуществления двухнитевая РНК имеет первую последовательность, содержащую по меньшей мере приблизительно 10 смежных нуклеотидов из TFL1, соответственно, приблизительно 21-23 смежных нуклеотида из TFL-1. Двухнитевая РНК может иметь вторую последовательность. Двухнитевая РНК может иметь третью последовательность, содержащую последовательность, обратно комплементарную первой последовательности, расположенную в той же ориентации, что и первая последовательность. Вторая последовательность может быть расположена между первой последовательностью и третьей последовательностью. Вторая последовательность может быть функционально связана с первой последовательностью и третьей последовательностью.
Первая последовательность может быть выбрана из группы, состоящей из SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20, и/или где третья последовательность представляет собой последовательность, обратно комплементарную последовательности, соответствующей SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20.
Первая последовательность может содержать или состоять из SEQ ID NO: 22, и третья последовательность может содержать или состоять из SEQ ID NO: 23; или первая последовательность может содержать или состоять из SEQ ID NO: 25, и третья последовательность может содержать или состоять из SEQ ID NO: 26; или первая последовательность может содержать или состоять из SEQ ID NO: 27, и третья последовательность может содержать или состоять из SEQ ID NO: 28; или первая последовательность может содержать или состоять из SEQ ID NO: 29, и третья последовательность может содержать или состоять из SEQ ID NO: 30; или первая последовательность может содержать или состоять из SEQ ID NO: 32, и третья последовательность может содержать или состоять из SEQ ID NO: 33; или первая последовательность может содержать или состоять из SEQ ID NO: 34, и третья последовательность может содержать или состоять из SEQ ID NO: 35; или первая последовательность может содержать или состоять из SEQ ID NO: 36, и третья последовательность может содержать или состоять из SEQ ID NO: 37; или первая последовательность может содержать или состоять из SEQ ID NO: 39, и третья последовательность может содержать или состоять из SEQ ID NO: 40.
Двухнитевая РНК может содержать или состоять из последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, SEQ ID NO: 35, SEQ ID NO: 31, SEQ ID NO: 38 и SEQ ID NO: 41.
Также предполагается использование молекул малой РНК, образующей шпильки. Они содержат специфичную антисмысловую последовательность в дополнение к обратно комплементарной (смысловой) последовательности, как правило, отделенной спейсером или последовательностью петли. Расщепление спейсера или петли обеспечивает молекулу однонитевой РНК и ее обратно комплементарную нить, так что их можно отжечь с образованием двухнитевой молекулы РНК (необязательно, с дополнительными стадиями обработки, которые могут привести в результате к добавлению или удалению одного, двух, трех или более нуклеотидов 3'-конца или 5'-конца одной или обеих нитей). Спейсер может иметь достаточную длину для обеспечения возможности комплементарного связывания антисмысловой и смысловой последовательностей и образования двухнитевой структуры (или стебля) до расщепления спейсера (и, необязательно, последующих стадий обработки, которые могут привести в результате к добавлению или удалению одного, двух, трех, четырех или более нуклеотидов с 3'-конца или с 5'-конца одной или обеих нитей). Спейсерная последовательность представляет собой, как правило, неродственную нуклеотидную последовательность, которая находится между двух комплементарных участков нуклеотидной последовательности, которая при комплементарном связывании в двухнитевой полинуклеотид содержит малую РНК, образующую шпильки. Спейсерная последовательность обычно содержит от приблизительно 3 до приблизительно 100 нуклеотидов.
Любой представляющий интерес полинуклеотид РНК можно получить путем подбора подходящей композиции последовательности, размера петли и длины стебля для получения двойной спирали шпильки. Иллюстративная последовательность ДНК антисмысловых и смысловых целевых последовательностей RNAi и конструкций показаны под SEQ ID No: 22-41. Подходящий диапазон для конструирования длины стебля двойной спирали шпильки включает длину стебля, составляющую по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов, например, приблизительно 14-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов, приблизительно 200-300 нуклеотидов, приблизительно 300-400 нуклеотидов, приблизительно 400-500 нуклеотидов, приблизительно 500-600 нуклеотидов и приблизительно 600-700 нуклеотидов. Подходящий диапазон для конструирования длины петли двойной спирали шпильки включает длину петли, составляющую приблизительно 4-25 нуклеотидов, приблизительно 25-50 нуклеотидов или длиннее, если длина стебля двойной спирали шпильки является значительной. В определенных вариантах осуществления молекула двухнитевой РНК или ssRNA составляет от приблизительно 15 до приблизительно 40 нуклеотидов в длину. В другом варианте осуществления молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 15 до приблизительно 35 нуклеотидов в длину. В другом варианте осуществления молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 17 до приблизительно 30 нуклеотидов в длину. В другом варианте осуществления молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 19 до приблизительно 25 нуклеотидов в длину. В другом варианте осуществления молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 21 до приблизительно 23 нуклеотидов в длину. В определенных вариантах осуществления структуры в виде шпильки с участками двойной спирали длиннее, чем 21 нуклеотид, могут способствовать эффективному сайленсингу, управляемому малой интерферирующей РНК, вне зависимости от последовательности и длины петли. Иллюстративные последовательности, применяемые для РНК-интерференции, изложены под SEQ ID NO: 22-33.
Целевая последовательность mRNA составляет, как правило, от приблизительно 14 до приблизительно 50 нуклеотидов в длину. Целевую mRNA можно, таким образом, проверить на наличие участков от приблизительно 14 до приблизительно 50 нуклеотидов в длину, которые предпочтительно соответствуют одному или более из следующих критериев для целевой последовательности: соотношение A+T/G+С составляет от приблизительно 2:1 до приблизительно 1:2; динуклеотид AA или динуклеотид CA на 5'-конце целевой последовательности; последовательность из по меньшей мере 10 смежных нуклеотидов, уникальных для целевой mRNA (то есть, последовательность отсутствует в других последовательностях mRNA из этого же растения); и нет «полос» из более чем трех смежных гуаниновых (G) нуклеотидов или более чем трех смежных цитозиновых (C) нуклеотидов. Данные критерии можно оценивать с помощью различных методик, известных в данной области, например, компьютерные программы, такие как BLAST, можно использовать для поиска по общедоступным базам данных, чтобы определить, является ли выбранная целевая последовательность уникальной для целевой mRNA. В качестве альтернативы, можно выбрать целевую последовательность (и сконструировать последовательность малой интерферирующей РНК) с помощью коммерчески доступного компьютерного программного обеспечения (например, OligoEngine, Target Finder и Design Tool для малых интерферирующих РНК, которые являются коммерчески доступными).
В одном варианте осуществления выбраны целевые последовательности mRNA, которые составляют от приблизительно 14 до приблизительно 30 нуклеотидов в длину и соответствуют одному или более критериям, указанным выше. В другом варианте осуществления выбраны целевые последовательности, которые составляют от приблизительно 16 до приблизительно 30 нуклеотидов в длину и соответствуют одному или более критериям, указанным выше. В дополнительном варианте осуществления выбраны целевые последовательности, которые составляют от приблизительно 19 до приблизительно 30 нуклеотидов в длину и соответствуют одному или более критериям, указанным выше. В другом варианте осуществления выбраны целевые последовательности, которые составляют от приблизительно 19 до приблизительно 25 нуклеотидов в длину и соответствуют одному или более критериям, указанным выше.
В иллюстративном варианте осуществления молекулы малой интерферирующей РНК содержат специфическую антисмысловую последовательность, которая комплементарна по меньшей мере 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежным нуклеотидам из любой из описанных в данном документе полинуклеотидных последовательностей.
Специфическая антисмысловая последовательность, содержащаяся в молекуле малой интерферирующей РНК, может быть идентичной или практически идентичной последовательности, комплементарной целевой последовательности. В одном варианте осуществления специфическая антисмысловая последовательность, содержащаяся в молекуле малой интерферирующей РНК, является на по меньшей мере приблизительно 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, комплементарной последовательности целевой mRNA. Способы определения идентичности последовательности известны в данной области, и ее можно определить, например, с помощью программы BLASTN программного обеспечения Computer Group (GCG) Университета штата Висконсин, или предоставленной на веб-сайте NCBI.
Специфическая антисмысловая последовательность молекул малой интерферирующей РНК может проявлять изменчивость в отношении различий (например, нуклеотидной замены, в том числе, транзиции или трансверсии) по одному, двум, трем, четырем или более нуклеотидам последовательности целевой mRNA. Если такие нуклеотидные замены находятся в антисмысловой нити молекулы двухнитевой РНК, то комплементарный нуклеотид в смысловой нити, с которым замещенный нуклеотид, как правило, должен образовывать водородные связи при спаривании оснований, может быть или может не быть соответственно замещенным. Молекулы двухнитевой РНК, в которых одна или более нуклеотидных замен происходят в смысловой последовательности, но не в антисмысловой нити, также рассматриваются. Если антисмысловая последовательность молекулы малой интерферирующей РНК содержит одно или более несовпадений между нуклеотидной последовательностью малой интерферирующей РНК и целевой нуклеотидной последовательностью, как описано выше, эти несоответствия могут быть обнаружены на 3'-конце, 5'-конце, или в центральной части антисмысловой последовательности.
В другом варианте осуществления молекулы малой интерферирующей РНК содержат специфическую антисмысловую последовательность, которая способна селективно гибридизироваться в жестких условиях с частью встречающегося в природе целевого гена или целевой mRNA. Как известно специалистам в данной области, вариации жесткости условий гибридизации могут быть достигнуты путем изменения времени, температуры или концентрации растворов, используемых для стадий гибридизации и отмывания. Подходящие условия могут также частично зависеть от конкретных используемых нуклеотидных последовательностей, например, последовательности целевой mRNA или гена.
Один способ индукции сайленсинга двухнитевой РНК у растений представляет собой трансформацию с помощью генной конструкции, продуцирующей РНК, образующую шпильки (см. Smith et al. (2000) Nature, 407, 319-320). Такие конструкции содержат инвертированные участки последовательности целевого гена, отделенные соответствующим спейсером. Вставка функционального интронного участка растения в качестве спейсерного фрагмента дополнительно повышает эффективность индукции сайленсинга гена в связи с выработкой РНК, образующей шпильки, сплайсированной с интроном (Wesley et al. (2001) Plant J., 27, 581-590). Соответственно, длина стебля составляет от приблизительно 50 нуклеотидов до приблизительно 1 тысячи нуклеотидов в длину. Способы получения РНК, образующей шпильки, сплайсированной с интроном, хорошо описаны в данной области (см., например, Bioscience, Biotechnology, and Biochemistry (2008) 72, 2, 615-617).
Молекулы интерферирующей РНК, имеющие двуспиральную или двухнитевую структуру, например, двухнитевую РНК или малую РНК, образующую шпильки, могут иметь тупые концы, или могут иметь липкие 3'- или 5'-концы. Используемый в данном документе термин «липкий конец» относится к неспаренному нуклеотиду или нуклеотидам, которые выступают из двухспиральной структуры, если 3'-конец одной нити РНК выходит за пределы 5'-конца другой нити (липкий 3'-конец), или наоборот (липкий 5'-конец). Нуклеотиды, представляющие собой липкие концы, могут быть рибонуклеотидами, дезоксирибонуклеотидами или их модифицированными версиями. В одном варианте осуществления по меньшей мере одна нить молекулы интерферирующей РНК имеет липкий 3'-конец от приблизительно 1 до приблизительно 6 нуклеотидов в длину. В других вариантах осуществления липкий 3'-конец составляет от приблизительно 1 до приблизительно 5 нуклеотидов, от приблизительно 1 до приблизительно 3 нуклеотидов и от приблизительно 2 до приблизительно 4 нуклеотидов в длину.
Если молекула интерферирующей РНК содержит липкий 3'-конец на одном конце молекулы, другой конец может быть с тупым концом, или также иметь липкий конец (5' или 3'). Если молекула интерферирующей РНК содержит липкий конец с обоих концов молекулы, длина липких концов может быть одинаковой или разной. В одном варианте осуществления молекула интерферирующей РНК содержит липкие 3'-концы от приблизительно 1 до приблизительно 3 нуклеотидов в длину на обоих концах молекулы. В дополнительном варианте осуществления молекула интерферирующей РНК представляет собой двухнитевую РНК, имеющую липкий 3'-конец из 2 нуклеотидов с обоих концов молекулы. В еще одном варианте осуществления нуклеотиды, представляющие собой липкий конец интерферирующей РНК, являются динуклеотидами TT или динуклеотидами UU.
При определении процента идентичности молекулы интерферирующей РНК, содержащей один или более липких концов, с последовательностью целевой mRNA, липкий(-е) конец(-цы) может учитываться или не учитываться. Например, нуклеотиды с липкого 3'-конца и до 2 нуклеотидов с 5'- или 3'-конца двойной нити могут быть модифицированы без существенной потери активности молекулы малой интерферирующей РНК.
Молекулы интерферирующей РНК могут содержать одну или более 5' или 3'-кэп-структур. Молекула интерферирующей РНК может содержать кэп-структуру на 3'-конце смысловой нити, антисмысловой нити или как смысловой, так и антисмысловой нитей; или на 5'-конце смысловой нити, антисмысловой нити или как смысловой, так и антисмысловой нитей молекулы интерферирующей РНК. В качестве альтернативы, молекула интерферирующей РНК может содержать кэп-структуру как на 3'-конце, так и на 5'-конце молекулы интерферирующей РНК. Термин «кэп-структура» относится к химической модификации, включенной с любого конца олигонуклеотида, которая защищает молекулу от разрушения эндонуклеазами и может также облегчать доставку или локализацию внутри клетки.
Другой модификацией, применяемой к молекулам интерферирующей РНК, является химическое связывание с молекулой интерферирующей РНК одного или более фрагментов или конъюгатов, которые повышают активность, клеточное распределение, клеточный захват, биодоступность или стабильность молекулы интерферирующей РНК. Полинуклеотиды можно синтезировать или модифицировать с помощью способов, общепринятых в данной области. Химические модификации могут включать без ограничения 2'-модификации, введение неприродных оснований, ковалентное присоединение лиганда и замещение фосфатных связей тиофосфатными связями. В данном варианте осуществления целостность двухспиральной структуры усиливают с помощью по меньшей мере одной и, как правило, двух химических связей. Химическое связывание может обеспечиваться с помощью любого из множества хорошо известных методик, например, путем введения ковалентных, ионных или водородных связей; гидрофобных взаимодействий, Ван-дер-Ваальсовых или стекинговых взаимодействий; посредством координации ионами металлов или благодаря использованию аналогов пуринов.
Нуклеотиды одной или обеих из двух одинарных нитей можно модифицировать для модулирования активации клеточных ферментов, таких как, например, без ограничения определенные нуклеазы. Методики для снижения или ингибирования активации клеточных ферментов известны в данной области и включают без ограничения 2'-аминомодификации, 2'-фтормодификации, 2'-алкилмодификации, модификации незаряженного каркаса, морфолиновые модификации, 2'-О-метилмодификации и фосфорамидат. Таким образом, по меньшей мере одну 2'-гидроксильную группу нуклеотидов двухнитевой РНК замещают химической группой. Также, по меньшей мере один нуклеотид можно модифицировать с образованием закрытого нуклеотида. Такой закрытый нуклеотид содержит метиленовый или этиленовый мостик, который соединяет 2'-кислород рибозы с 4'-углеродом рибозы. Введение закрытого нуклеотида в олигонуклеотид улучшает аффиность к комплементарным последовательностям и повышает температуру плавления на несколько градусов.
Можно конъюгировать лиганды с молекулой интерферирующей РНК, например, для повышения в отношении нее клеточной абсорбции. В определенных вариантах осуществления гидрофобный лиганд конъюгируют с молекулой для облегчения прямого проникновения через клеточную мембрану. Данные подходы использованы для облегчения проникновения в клетку антисмысловых олигонуклеотидов. В определенных случаях конъюгация катионного лиганда с олигонуклеотидами часто приводит к улучшенной устойчивости к нуклеазам. Типичные примеры катионных лигандов включают пропиламмоний и диметилпропиламоний. Антисмысловые олигонуклеотиды могут сохранять свою высокую аффинность связывания с mRNA, когда катионный лиганд распределен по всему олигонуклеотиду.
Молекулы и полинуклеотиды, описанные в данном документе, можно получить с помощью хорошо известных методик твердофазного синтеза. Любые другие средства для такого синтеза, известные в данной области, можно использовать дополнительно или в качестве альтернативы.
Различные варианты осуществления направлены на векторы экспрессии, содержащие один или более полинуклеотидов или одну или более конструкций интерферирующей РНК, описанных в данном документе. Иллюстративные конструкции показаны на фигуре 21.
Различные варианты осуществления направлены на векторы экспрессии, содержащие один или более полинуклеотидов или одну или более конструкций интерферирующей РНК, кодирующих один или более полинуклеотидов интерферирующей РНК, описанных в данном документе, которые способны к самоотжигу с образованием структуры в виде шпильки, в которых конструкция содержит (a) один или более полинуклеотидов, описанных в данном документе; (b) вторую последовательность, кодирующую спейсерный элемент, который образует петлю структуры в виде шпильки; и (c) третью последовательность, содержащую последовательность, обратно комплементарную первой последовательности, расположенную в той же ориентации, что и первая последовательность, где вторая последовательность расположена между первой последовательностью и третьей последовательностью, и третья последовательность функционально связана с первой последовательностью и третьей последовательностью.
Раскрытые последовательности можно использовать для конструирования различных полинуклеотидов, которые не образуют структуры в виде шпильки. Например, двухнитевая РНК может быть образована посредством (1) транскрибирования первой нити ДНК путем функционального связывания с первым промотором и (2) транскрибирования последовательности, обратно комплементарной последовательности фрагмента ДНК первой нити, путем функционального связывания со вторым промотором. Каждую нить полинуклеотида можно транскрибировать из одного вектора экспрессии, или из разных векторов экспрессии. Двойную спираль РНК, обладающую активностью в отношении РНК-интерференции, можно ферментативно превратить в малые интерферирующие РНК для модулирования уровней РНК.
Таким образом, различные варианты осуществления направлены на векторы экспрессии, содержащие один или более полинуклеотидов или конструкций интерферирующей РНК, описанных в данном документе, кодирующих полинуклеотиды интерферирующей РНК, способные к самоотжигу, в которых конструкция содержит (a) один или более полинуклеотидов, описанных в данном документе; и (b) вторую последовательность, содержащую комплементарную (например, обратно комплементарную) последовательность первой последовательности, расположенную в той же ориентации, что и первая последовательность.
Предусмотрены различные композиции и способы для модулирования уровней эндогенной экспрессии одного или более полипептидов, описанных в данном документе (или любой их комбинации, описанной в данном документе) путем стимулирования косупрессии экспрессии гена. Явление косупрессии возникает как результат введения множества копий трансгена в клетку растения-хозяина. Интеграция множества копий трансгена может приводить к модулированию экспрессии трансгена и нацеленного эндогенного гена. Степень косупрессии зависит от степени идентичности последовательности между трансгеном и нацеленным эндогенным геном. Сайленсинг как эндогенного гена, так и трансгена может происходить из-за обширного метилирования подавляемых локусов (то есть, эндогенного промотора и эндогенного представляющего интерес гена), что может нарушать транскрипцию. В качестве альтернативы, в некоторых случаях косупрессия эндогенного гена и трансгена может происходить посредством пост-транскрипционного сайленсинга гена, при котором транскрипты могут быть получены, но повышенные уровни разрушения препятствуют накоплению транскриптов. Механизм косупрессии посредством пост-транскрипционного сайленсинга генов, как полагают, напоминает РНК-интерференцию в том, что РНК, по-видимому, важна как инициатор и мишень в этих процессах, и может быть опосредован, по меньшей мере частично, тем же молекулярным механизмом, возможно, через РНК-направляемое разрушение mRNA.
Косупрессия нуклеиновых кислот может быть достигнута путем введения множества копий нуклеиновой кислоты или ее фрагментов в качестве трансгенов в геном представляющего интерес растения. Растение-хозяин можно трансформировать вектором экспрессии, содержащим промотор, функционально связанный с такой нуклеиновой кислотой или ее фрагментами. Различные варианты осуществления направлены на векторы экспрессии для стимулирования косупрессии эндогенных генов, содержащих промотор, функционально связанный с полинуклеотидом.
Различные варианты осуществления направлены на способы модулирования уровня экспрессии одного или более полинуклеотидов, описанных в данном документе (или любой их комбинации, как описано в данном документе), посредством интеграции множества копий полинуклеотида(-ов) в геном растения, предусматривающие трансформацию клетки растения-хозяина с помощью вектора экспрессии, который содержит промотор, функционально связанный с полинуклеотидом.
Различные композиции и способы предусмотрены для модулирования уровня экспрессии эндогенного гена путем модулирования трансляции mRNA. Клетку растения-хозяина можно трансформировать с помощью вектора экспрессии, содержащего промотор, функционально связанный с полинуклеотидом, расположенного в антисмысловой ориентации по отношению к промотору, для обеспечения возможности экспрессии полинуклеотидов РНК, характеризующихся последовательностью, комплементарной части mRNA.
Различные векторы экспрессии для модулирования трансляции mRNA могут содержать промотор, функционально связанный с полинуклеотидом, в антисмысловой ориентации по отношению к промотору. Длина полинуклеотидов антисмысловой РНК может варьировать и составлять от приблизительно 15-20 нуклеотидов, приблизительно 20-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-75 нуклеотидов, приблизительно 75-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов и приблизительно 200-300 нуклеотидов.
Также на гены можно целенаправленно воздействовать для инактивации путем введения транспозонов (например, IS-элементов) в геномы представляющих интерес растений. Данные мобильные генетические элементы можно ввести с помощью полового перекрестного опыления и мутантов со вставками можно подвергнуть скринингу в отношении потери активности белка. Разрушенный ген родительского растения можно ввести в другие растения путем скрещивания родительского растения с растением, не подвергнутым индуцированному транспозоном мутагенезу, например, путем полового перекрестного опыления. Можно использовать любые стандартные методики селекции, известные специалистам в данной области. В одном варианте осуществления один или более генов можно инактивировать путем вставки одного или более транспозонов. Мутации могут привести к гомозиготному разрушению одного или более генов, к гетерозиготному разрушению одного или более генов, или к комбинации гомозиготных и гетерозиготных разрушений, если разрушен более чем один ген. Подходящие мобильные элементы включают ретротранспозоны, ретропозоны и SINE-подобные элементы. Такие способы известны специалистам в данной области.
В качестве альтернативы, на гены можно целенаправленно воздействовать для инактивации путем введения рибозимов, полученных из ряда малых кольцевых РНК, которые способны к саморасщеплению и репликации в растениях. Данные РНК могут реплицироваться либо самостоятельно (РНК вироида), либо с участием вируса-помощника (сателлитные РНК). Примеры подходящих РНК включают полученные из вироида солнечной пятнистости авокадо и сателлитные РНК, полученные из вируса кольцевой пятнистости табака, вируса временной полосатости люцерны, вируса бархатной пятнистости табака, вируса пятнистости Solanum nodiflorum и вируса пятнистости клевера подземного. Различные специфичные к целевой РНК рибозимы известны специалистам в данной области.
Как обсуждалось в данном документе, экспрессию одного или более полипептидов можно модулировать нетрансгенными способами, как например, путем создания одной или более мутаций в одном или более генах, как обсуждалось в данном документе. Способы, с помощью которых вводят случайную мутацию в генную последовательность, могут включать химический мутагенез, мутагенез с помощью EMS и радиационный мутагенез. Способы, с помощью которых вводят одну или более целенаправленно воздействующих мутаций в клетку, включают без ограничения технологию редактирования генома, в частности, мутагенез, опосредованный нуклеазой с мотивом цинкового пальца, TILLING (целенаправленные индуцированные локальные повреждения в геноме), гомологичную рекомбинацию, олигонуклеотид-направленный мутагенез и мутагенез, опосредованный мегануклеазой. В одном варианте осуществления используют TILLING. Он представляет собой технологию, которую можно использовать для создания и/или идентификации полинуклеотидов, кодирующих полипептиды с модифицированной экспрессией и/или активностью. TILLING также допускает отбор растений, несущих такие мутации. В TILLING комбинируют мутагенез высокой плотности со способами высокопроизводительного скрининга. Способы для проведения TILLING хорошо известны из уровня техники (см. McCallum et al., (2000) Nat Biotechnol 18: 455-457 и Stemple (2004) Nat Rev Genet 5(2): 145-50).
Некоторые неограничивающие примеры мутаций представляют собой делеции, вставки и миссенс-мутации по меньшей мере одного нуклеотида, однонуклеотидные полиморфизмы и простую повторяющуюся последовательность. После внесения мутации можно провести скрининг для идентификации мутаций, которые создают ранние стоп-кодоны или другим образом нефункциональные гены. После внесения мутации можно провести скрининг для идентификации мутаций, которые создают функциональные гены, способные экспрессироваться на повышенных уровнях. Скрининг мутантов можно выполнить путем секвенирования, или с использованием одного или более зондов или праймеров, специфичных для данного гена или белка. Можно также создать в полинуклеотидах конкретные мутации, которые приводят в результате к модулированию экспрессии гена, модулированию стабильности mRNA или модулированию стабильности белка. Такие растения названы в данном документе как «не встречающиеся в природе» или «мутантные» растения. Как правило, мутантные или не встречающиеся в природе растения будут содержать по меньшей мере часть чужеродной или синтетической или созданной человеком нуклеиновой кислоты (например, ДНК или РНК), которая не представлена в растении до проведения с ним манипуляций. Чужеродная нуклеиновая кислота может представлять собой один нуклеотид, два или более нуклеотидов, два или более смежных нуклеотидов или два или более несмежных нуклеотидов, таких как по меньшей мере 10, 20, 30, 40, 50,100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400 или 1500 или более смежных или несмежных нуклеотидов.
Мутантные или не встречающиеся в природе растения или клетки растения могут иметь любую комбинацию одной или более мутаций в одном или более генах, которая приводит в результате к модулированию уровней белка. Например, мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну мутацию в одном гене; несколько мутаций в одном гене; одну мутацию в двух или более, или трех или более, или четырех или более генах; или несколько мутаций в двух или более, или трех или более, или четырех или более генах. Примеры таких мутаций описаны в данном документе. В качестве дополнительного примера, мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций в конкретной части гена(-ов), например, в участке гена, который кодирует активный сайт белка или его часть. В качестве дополнительного примера, мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций в участке вне одного или более генов, в таком как участок выше по цепи или ниже по цепи гена, который он регулирует, при условии, что они модулируют активность или экспрессию гена(-ов). Элементы, расположенные выше по цепи, могут включать промоторы, энхансеры или факторы транскрипции. Некоторые элементы, такие как энхансеры, могут располагаться выше по цепи или ниже по цепи гена, который они регулируют. Элемент(-ы) не обязательно расположен(-ы) рядом с геном, который он(-и) регулирует(-ют), так как было обнаружено, что некоторые элементы расположены на расстоянии в несколько тысяч пар оснований выше по цепи или ниже по цепи относительно гена, который они регулируют. Мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций, расположенных в первых 100 нуклеотидах гена(-ов), в первых 200 нуклеотидах гена(-ов), в первых 300 нуклеотидах гена(-ов), в первых 400 нуклеотидах гена(-ов), в первых 500 нуклеотидах гена(-ов), в первых 600 нуклеотидах гена(-ов), в первых 700 нуклеотидах гена(-ов), в первых 800 нуклеотидах гена(-ов), в первых 900 нуклеотидах гена(-ов), в первой 1000 нуклеотидов гена(-ов), в первых 1100 нуклеотидах гена(-ов), в первых 1200 нуклеотидах гена(-ов), в первых 1300 нуклеотидах гена(-ов), в первых 1400 нуклеотидах гена(-ов) или в первых 1500 нуклеотидах гена(-ов). Мутантные или не встречающиеся в природе растения или клетки растения могут иметь одну или более мутаций, расположенных в первом, втором, третьем, четвертом, пятом, шестом, седьмом, восьмом, девятом, десятом, одиннадцатом, двенадцатом, тринадцатом, четырнадцатом или пятнадцатом наборе из 100 нуклеотидов гена(-ов) или их комбинации. Раскрыты мутантные или не встречающиеся в природе растения или клетки растения (например, мутантные, не встречающиеся в природе или трансгенные растения или клетки растения и им подобные, как описано в данном документе), содержащие варианты мутантного полипептида.
В одном варианте осуществления семена растений подвергают мутагенезу, и затем выращивают из них мутантные растения первого поколения. Затем обеспечивают возможность самоопыления растений первого поколения и из семян от растений первого поколения выращивают растения второго поколения, которые затем подвергают скринингу на наличие мутаций в их локусах. Не смотря на то, что подвергнутый мутации растительный материал можно проверить на наличие мутаций, преимуществом скрининга растений второго поколения является то, что все соматические мутации соответствуют мутациям в зародышевых линиях. Специалисту в данной области будет понятно, что различный растительный материал, в том числе без ограничения семена, пыльцу, растительные ткани или клетки растения можно подвергнуть мутагенезу для создания мутантных растений. Однако тип растительного материала, подвергнутого мутагенезу, может иметь значение, когда нуклеиновую кислоту растения подвергают скринингу на наличие мутаций. Например, если пыльцу подвергают мутагенезу до проведения опыления не подвергаемого мутагенезу растения, из семян, полученных в результате такого опыления, выращивают растения первого поколения. Каждая клетка растений первого поколения будет содержать мутации, созданные в пыльце; таким образом, эти растения первого поколения можно затем подергать скринингу на наличие мутаций вместо того, чтобы дожидаться второго поколения.
Для создания мутаций можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, трансверсии и/или транзиции, включая химические мутагены и облучение. Мутагены включают без ограничения этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, мономер акриламида, мельфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитро-нитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметил-бенз(a)антрацен, этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и т. п.), 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина дигидрохлорид и формальдегид.
Также предусматриваются спонтанные мутации в локусе, которые могут не быть непосредственно вызванными мутагеном, при условии, что они приводят к требуемому фенотипу. Подходящие мутагенные средства могут также включать, например, ионизирующее излучение, такое как рентгеновское излучение, гамма-излучение, излучение быстрых нейтронов и ультрафиолетовое излучение. Любой способ получения нуклеиновой кислоты растения, известный специалистам в данной области, можно использовать для получения нуклеиновой кислоты растения для скрининга в отношении мутаций.
Полученную нуклеиновую кислоту из отдельных растений, клеток растения или растительного материала можно необязательно объединить для того, чтобы ускорить скрининг в отношении мутаций в популяции растений, происходящих из подвергнутых мутагенезу растительных тканей, клеток или материала. Можно подвергать скринингу одно или более следующих поколений растений, клеток растения или растительного материала. Размер необязательно объединенной группы зависит от чувствительности используемого способа скрининга.
После того, как образцы нуклеиновых кислот необязательно объединили, их можно подвергнуть методикам полинуклеотид-специфичной амплификации, таким как полимеразная цепная реакция. Любой один или более праймеров или зондов, специфичных в отношении гена или последовательностей, непосредственно примыкающих к гену, можно использовать для амплификации последовательностей в пределах необязательно объединенного образца нуклеиновых кислот. Соответственно, один или более праймеров или зондов конструируют для амплификации участков локуса, в которых с наибольшей вероятностью возникают полезные мутации. Наиболее предпочтительно праймер конструируют для выявления мутаций в участках полинуклеотида. Дополнительно, предпочтительным для праймера(-ов) и зонда(-ов) было бы избегать известных полиморфных сайтов для облегчения скрининга точечных мутаций. Для облегчения выявления продуктов амплификации один или более праймеров или зондов можно метить с использованием любого общепринятого способа введения метки. Праймер(-ы) или зонд(-ы) можно сконструировать на основе последовательностей, описанных в данном документе, с применением способов, которые хорошо известны в данной области.
Для облегчения выявления продуктов амплификации праймер(-ы) или зонд(-ы) можно пометить с использованием любого общепринятого способа внесения метки. Их можно сконструировать на основе последовательностей, описанных в данном документе, с применением способов, которые хорошо известны в данной области. Полиморфизмы можно идентифицировать с помощью средств, известных в данной области, и некоторых из описанных в литературе.
В дополнительном аспекте предусмотрен способ получения мутантного растения. Способ включает получение по меньшей мере одной клетки растения, содержащей ген, кодирующий функциональный полинуклеотид, описанный в данном документе (или любую их комбинацию, описанную в данном документе). Далее эту по меньшей мере одну клетку растения обрабатывают в условиях, эффективных для модулирования активности полинуклеотида(-ов), описанного(-ых) в данном документе. По меньшей мере одну мутантную клетку растения затем подвергают размножению в мутантное растение, где мутантное растение характеризуется модулированным уровнем описанного(-ых) полипептида(-ов) (или любой их комбинации, описанной в данном документе) по сравнению с уровнем у контрольного растения. В одном варианте осуществления данного способа получения мутантного растения стадия обработки включает воздействие на по меньшей мере одну клетку химическим мутагенным средством, описанным выше, и в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. В другом варианте осуществления данного способа стадия обработки включает воздействие на по меньшей мере одну клетку источником ионизирующего излучения в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. Термин «мутантное растение» включает мутантные растения, в которых генотип модифицирован по сравнению с контрольным растением, соответственно, с помощью способов, отличных от способов генной инженерии и генетической модификации.
В определенных вариантах осуществления мутантное растение, клетка мутантного растения или мутантный растительный материал может содержать одну или более мутаций, которые встречаются в природе в другом растении, клетке растения или растительном материале и обеспечивают требуемый признак. Данную мутацию можно ввести (например, путем интрогрессии) в другое растение, клетку растения или растительный материал (например, растение, клетку растения или растительный материал с генетическим фоном, отличающимся от такового у растения, из которого получена мутация) для создания мутации, которая не встречается в природе у данного растения и для обеспечения у них данного признака. Таким образом, в качестве примера, мутацию, которая встречается в природе в первом растении, можно ввести во второе растение, такое как второе растение с генетическим фоном, отличающимся от такового у первого растения. Специалист в данной области, таким образом, может осуществлять поиск и идентифицировать растение, несущее в естественных условиях в своем геноме один или более мутантных аллелей генов, описанных в данном документе, которые обеспечивают требуемый признак. В определенных вариантах осуществления одной или более мутаций в одном аллеле достаточно для сокращения периода времени до наступления цветения. Мутантный(-е) аллель(-и), который(-е) встречается(-ются) в природе, можно перенести во второе растение различными способами, включая селекцию, обратное скрещивание и интрогрессию с получением линий, разновидностей или гибридов, которые имеют одну или более мутаций в генах, описанных в данном документе. Растения, демонстрирующие требуемый признак, можно отобрать из пула мутантных растений. Соответственно, отбор осуществляют с использованием данных о нуклеотидных последовательностях, описанных в данном документе. Следовательно, можно осуществлять скрининг в отношении генетического признака по сравнению с контролем. Такой подход, предусматривающий применение скрининга, может включать применение общепринятых методик амплификации и/или гибридизации нуклеиновых кислот, как обсуждается в данном документе. Таким образом, дополнительный аспект относится к способу идентификации мутантного растения с сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, включающему (a) получение образца из растения, подлежащего скринингу; (b) определение того, содержит ли указанный образец одну или более мутаций в одном или более полинуклеотидах, описанных в данном документе; и (c) определение скорости наступления цветения указанного растения.
В другом аспекте предусмотрен способ получения мутантного растения, которое характеризуется сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, включающий стадии (a) получения образца из первого растения; (b) определения того, содержит ли указанный образец одну или более мутаций в одном или более полинуклеотидах, описанных в данном документе, что приводит к сокращению периода времени до наступления цветения; и (c) переноса одной или более мутаций во второе растение. Мутацию(-и) можно перенести во второе растение с применением различных способов, которые известны в данной области, с помощью таких как способы генной инженерии, манипуляции с генами, интрогрессии, селекции растений, обратного скрещивания и им подобных. В одном варианте осуществления первое растение является встречающимся в природе растением. В одном варианте осуществления второе растение имеет генетический фон, отличающийся от такового первого растения.
В другом аспекте предусмотрен способ получения мутантного растения, которое характеризуется сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, включающий стадии (a) получения образца из первого растения; (b) определения того, содержит ли указанный образец одну или более мутаций в одном или более полинуклеотидах, описанных в данном документе, что приводит к сокращению периода времени до наступления цветения; и (c) интрогрессии одной или более мутаций из первого растения во второе растение. В одном варианте осуществления стадия интрогрессии предусматривает селекцию растений, необязательно включая обратное скрещивание и т. п. В одном варианте осуществления первое растение является встречающимся в природе растением. В одном варианте осуществления второе растение имеет генетический фон, отличающийся от такового первого растения. В одном варианте осуществления первое растение не представляет собой сорт или элитный сорт. В одном варианте осуществления второе растение представляет собой сорт или элитный сорт.
Дополнительный аспект относится к мутантному растению (включая мутантное растение сорта или элитного сорта), полученному или которое можно получить с помощью способов, описанных в данном документе. В определенных вариантах осуществления «мутантное растение» может содержать одну или более мутаций, локализованных только в конкретном участке растения, таком как в последовательности одного или более полинуклеотидов, описанных в данном документе. Согласно данному варианту осуществления остальная геномная последовательность мутантного растения будет такой же или практически такой же, как у растения до мутагенеза.
В определенных вариантах осуществления мутантные растения могут иметь одну или более мутаций, локализованных в более чем одном участке растения, таком как в последовательности одного или более полинуклеотидов, описанных в данном документе, и в одном или более дополнительных участках генома. Согласно данному варианту осуществления остальная геномная последовательность мутантного растения не будет такой же или не будет практически такой же, как у растения до мутагенеза. В определенных вариантах осуществления мутантные растения могут не иметь одну или более мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более экзонах полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более интронах полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в промоторе полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в 3'-нетранслируемом участке полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в 5'-нетранслируемом участке полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в кодирующем участке полинуклеотида(-ов), описанного(-ых) в данном документе; или могут не иметь одну или более мутаций в некодирующем участке полинуклеотида(-ов), описанного(-ых) в данном документе; или любую комбинацию двух или более, трех или более, четырех или более, пяти или более, или шести или более из их частей.
В дополнительном аспекте предусмотрен способ идентификации растения, клетки растения или растительного материала, содержащих мутацию в гене, кодирующем полинуклеотид, описанный в данном документе, включающий (a) осуществление мутагенеза растения, клетки растения или растительного материала; (b) получение образца нуклеиновой кислоты из указанного растения, клетки растения или растительного материала или их потомков; и (c) определение последовательности нуклеиновых кислот гена, кодирующего полинуклеотид, описанный в данном документе, или его вариантов или фрагментов, где отличие в указанной последовательности свидетельствует об одной или более мутациях в ней.
Белки «цинковые пальцы» также можно применять для модулирования экспрессии или активности одного или более полинуклеотидов, описанных в данном документе. В различных вариантах осуществления последовательность геномной ДНК, содержащую часть или всю кодирующую последовательность полинуклеотида, модифицируют путем мутагенеза, опосредованного нуклеазой с мотивом цинкового пальца. В последовательности геномной ДНК осуществляют поиск уникального сайта связывания белка «цинковый палец». В качестве альтернативы, в последовательности геномной ДНК осуществляют поиск двух уникальных сайтов для связывания белка «цинковый палец», при этом оба сайта находятся на противоположных нитях и близко друг к другу, например, на расстоянии 1, 2, 3, 4, 5, 6 или более пар оснований друг от друга. Соответственно, предусмотрены белки «цинковые пальцы», которые связываются с полинуклеотидами.
Белок «цинковый палец» можно сконструировать для распознавания выбранного целевого сайта в гене. Белок «цинковый палец» может содержать любую комбинацию мотивов, полученных из природных ДНК-связывающих доменов «цинковых пальцев» и неприродных ДНК-связывающих доменов «цинковых пальцев», полученных за счет усечения или расширения или способа сайт-направленного мутагенеза в сочетании со способом отбора, таким как без ограничения отбор с помощью фагового дисплея, отбор с помощью бактериальной дигибридной системы или отбор с помощью бактериальной одногибридной системы. Термин «неприродный ДНК-связывающий домен цинковый палец» относится к ДНК-связывающему домену «цинковому пальцу», который связывает последовательность из трех пар оснований в целевой нуклеиновой кислоте и который не встречается в клетке или организме, содержащих нуклеиновую кислоту, которая подлежит модификации. Способы конструирования белка «цинковый палец», который связывает специфические нуклеотидные последовательности, которые являются уникальными для целевого гена, известны из уровня техники.
Нуклеазу с мотивом цинкового пальца можно сконструировать путем создания слияния первого полинуклеотида, кодирующего белок «цинковый палец», который связывается с полинуклеотидом, и второго полинуклеотида, кодирующего неспецифическую эндонуклеазу, такую как без ограничения эндонуклеаза типа IIS. Слитый белок между белком «цинковый палец» и нуклеазой может содержать спейсер, состоящий из двух пар оснований или, в качестве альтернативы, спейсер, который может состоять из трех, четырех, пяти, шести, семи или более пар оснований. В различных вариантах осуществления нуклеаза цинкового пальца вносит двухнитевой разрыв в регуляторный участок, кодирующий участок или некодирующий участок последовательности геномной ДНК полинуклеотида и приводит к снижению уровня экспрессии полинуклеотида или снижению активности белка, кодируемого им. Расщепление с помощью нуклеаз с мотивом цинкового пальца часто приводит в результате к делеции ДНК в сайте расщепления с последующей репарацией ДНК путем соединения негомологичных концов.
В других вариантах осуществления белок «цинковый палец» можно выбирать для связывания с регуляторной последовательностью полинуклеотида. Более конкретно, регуляторная последовательность может содержать сайт инициации транскрипции, стартовый кодон, участок экзона, границу раздела экзон-интрон, терминатор или стоп-кодон. Соответственно, настоящее изобретение предусматривает мутантное, не встречающееся в природе или трансгенное растение или клетку растения, полученные с помощью мутагенеза, опосредованного нуклеазой с мотивом «цинкового пальца», вблизи от или в пределах одного или более полинуклеотидов, описанных в данном документе, и способы получения такого растения или клетки растения с помощью мутагенеза, опосредованного нуклеазой с мотивом «цинкового пальца». Способы доставки белка «цинковый палец» и нуклеазы с мотивом «цинкового пальца» в растение подобны описанным ниже для доставки мегануклеазы.
В другом аспекте описаны способы получения мутантных, не встречающихся в природе или трансгенных или другим образом генетически модифицированных растений с использованием мегануклеаз, таких как I-CreI. Встречающиеся в природе мегануклеазы, а также рекомбинантные мегануклеазы можно использовать для того, чтобы специфически вызывать двухнитевый разрыв в одном сайте или в относительно небольшом числе сайтов в геномной ДНК растения с обеспечением разрушения одного или более полинуклеотидов, описанных в данном документе. Мегануклеаза может быть сконструированной мегануклеазой с измененными свойствами распознавания ДНК. Белки мегануклеаз можно доставлять в клетки растения с помощью ряда разных механизмов, известных из уровня техники.
Настоящее изобретение также охватывает использование мегануклеаз для инактивации полинуклеотида(-ов), описанного(-ых) в данном документе (или любой их комбинации, описанной в данном документе), в клетке растения или у растения. В частности, настоящее изобретение предусматривает способ инактивации полинуклеотида у растения с применением мегануклеазы, включающий (a) получение клетки растения, содержащей полинуклеотид, описанный в данном документе; (b) введение мегануклеазы или конструкции, кодирующей мегануклеазу, в указанную клетку растения и (c) обеспечение возможности мегануклеазе практически инактивировать полинуклеотид(-ы)
Мегануклеазы можно использовать для расщепления сайтов распознавания мегануклеазы в кодирующих участках полинуклеотида. Такое расщепление часто приводит в результате к делеции ДНК в сайте распознавания мегануклеазы с последующей репарацией мутагенной ДНК путем соединения негомологичных концов. Такие мутации в кодирующей последовательности гена являются, как правило, достаточными для инактивации гена. Этот способ модификации клетки растения включает, во-первых, доставку кассеты экспрессии мегануклеазы в клетку растения с помощью подходящего способа трансформации. Для достижения максимальной эффективности требуется связать кассету экспрессии мегануклеазы с селектируемым маркером и отобрать успешно трансформированные клетки в присутствии селективного средства. Этот подход приведет к интеграции кассеты экспрессии мегануклеазы в геном, что, однако, может быть нежелательным, если для растения, скорее всего, потребуется официальное разрешение. В таких случаях кассету экспрессии мегануклеазы (и связанный селектируемый маркерный ген) можно выделить из последующих поколений растения с применением традиционных методик селекции. В качестве альтернативы, клетки растения можно изначально трансформировать с помощью кассеты экспрессии мегануклеазы без селектируемого маркера и можно вырастить на средах без селективного средства. В таких условиях часть обработанных клеток приобретет кассету экспрессии мегануклеазы и будет экспрессировать сконструированную мегануклеазу временно без интеграции кассеты экспрессии мегануклеазы в геном. Поскольку это не обеспечивает эффективность трансформации, то для этой последней процедуры трансформации требуется скрининг большего количества обработанных клеток для получения требуемой модификации генома. Описанный выше подход можно также применить для модификации клетки растения при использовании белка «цинковый палец» или нуклеазы с мотивом цинкового пальца.
После доставки кассеты экспрессии мегануклеазы клетки растения выращивают, изначально, в условиях, которые являются типичными для конкретной процедуры трансформации, которую использовали. Это может означать, что трансформированные клетки растут на средах при температуре ниже 26 C, зачастую в темноте. Такие стандартные условия можно использовать в течение периода времени, составляющего предпочтительно 1-4 дня, для обеспечения восстановления клетки растения после процесса трансформации. В любой момент после этого начального периода восстановления температуру роста можно повысить для стимулирования активности сконструированной мегануклеазы в отношении расщепления и мутирования сайта распознавания мегануклеазы.
Для определенных видов применения может требоваться точное удаление полинуклеотида из генома растения. Такие виды применения возможны с применением пары сконструированных мегануклеаз, каждая из которых расщепляет сайт распознавания мегануклеазы по обе стороны от предполагаемой делеции. Также можно использовать эффекторные нуклеазы TAL (TALEN), которые способны распознавать и связываться с геном и вносить двухнитевой разрыв в геном. Таким образом, в другом аспекте предлагаются способы получения мутантных, не встречающихся в природе или трансгенных или другим образом генетически модифицированных растений, описанных в данном документе, с помощью эффекторных нуклеаз TAL.
Растения, подходящие для применения в настоящем изобретении включают без ограничения однодольные и двудольные растения и системы на основе клеток растения, включая виды из одного из следующих семейств: Acanthaceae, Alliaceae, Alstroemeriaceae, Amaryllidaceae, Apocynaceae, Arecaceae, Asteraceae, Berberidaceae, Bixaceae, Brassicaceae, Bromeliaceae, Cannabaceae, Caryophyllaceae, Cephalotaxaceae, Chenopodiaceae, Colchicaceae, Cucurbitaceae, Dioscoreaceae, Ephedraceae, Erythroxylaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Linaceae, Lycopodiaceae, Malvaceae, Melanthiaceae, Musaceae, Myrtaceae, Nyssaceae, Papaveraceae, Pinaceae, Plantaginaceae, Poaceae, Rosaceae, Rubiaceae, Salicaceae, Sapindaceae, Solanaceae, Taxaceae, Theaceae или Vitaceae.
Подходящие виды могут включать представителей рода Abelmoschus, Abies, Acer, Agrostis, Allium, Alstroemeria, Ananas, Andrographis, Andropogon, Artemisia, Arundo, Atropa, Berberis, Beta, Bixa, Brassica, Calendula, Camellia, Camptotheca, Cannabis, Capsicum, Carthamus, Catharanthus, Cephalotaxus, Chrysanthemum, Cinchona, Citrullus, Coffea, Colchicum, Coleus, Cucumis, Cucurbita, Cynodon, Datura, Dianthus, Digitalis, Dioscorea, Elaeis, Ephedra, Erianthus, Erythroxylum, Eucalyptus, Festuca, Fragaria, Galanthus, Glycine, Gossypium, Helianthus, Hevea, Hordeum, Hyoscyamus, Jatropha, Lactuca, Linum, Lolium, Lupinus, Lycopersicon, Lycopodium, Manihot, Medicago, Mentha, Miscanthus, Musa, Nicotiana, Oryza, Panicum, Papaver, Parthenium, Pennisetum, Petunia, Phalaris, Phleum, Pinus, Poa, Poinsettia, Populus, Rauwolfia, Ricinus, Rosa, Saccharum, Salix, Sanguinaria, Scopolia, Secale, Solanum, Sorghum, Spartina, Spinacea, Tanacetum, Taxus, Theobroma, Triticosecale, Triticum, Uniola, Veratrum, Vinca, Vitis и Zea.
Подходящие виды могут включать Panicum spp., Sorghum spp., Miscanthus spp., Saccharum spp., Erianthus spp., Populus spp., Andropogon gerardii (бородач), Pennisetum purpureum (слоновая трава), Phalaris arundinacea (двукисточник тростниковидный), Cynodon dactylon (свинорой пальчатый), Festuca arundinacea (овсяница тростниковая), Spartina pectinata (спартина гребешковая), Medicago sativa (люцерна), Arundo donax (арундо тростниковый), Secale cereale (рожь), Salix spp. (ива), Eucalyptus spp. (эвкалипт), Triticosecale (тритикале), бамбук, Helianthus annuus (подсолнечник), Carthamus tinctorius (сафлор красильный), Jatropha curcas (ятрофа), Ricinus communis (клещевина), Elaeis guineensis (масличная пальма), Linum usitatissimum (лен), Brassica juncea, Beta vulgaris (сахарная свекла), Manihot esculenta (маниок), Lycopersicon esculentum (томат), Lactuca sativa (латук), Musyclise alca (банан), Solanum tuberosum (картофель), Brassica oleracea (брокколи, цветная капуста, брюссельская капуста), Camellia sinensis (чай), Fragaria ananassa (земляника), Theobroma cacao (какао), Coffea ycliseca (кофе), Vitis vinifera (виноград), Ananas comosus (ананас), Capsicum annum (острый и сладкий перец), Allium cepa (лук), Cucumis melo (дыня), Cucumis sativus (огурец), Cucurbita maxima (тыква гигантская), Cucurbita moschata (тыква мускатная), Spinacea oleracea (шпинат), Citrullus lanatus (арбуз), Abelmoschus esculentus (бамия), Solanum melongena (баклажан), Rosa spp. (роза), Dianthus caryophyllus (гвоздика), Petunia spp. (петуния), Poinsettia pulcherrima (пуансеттия), Lupinus albus (люпин), Uniola paniculata (овес), полевица (Agrostis spp.), Populus tremuloides (тополь осинообразный), Pinus spp. (сосна), Abies spp. (пихта), Acer spp. (клен), Hordeum vulgare (ячмень), Poa pratensis (мятлик), Lolium spp. (плевел) и Phleum pratense (тимофеевка), Panicum virgatum (просо), Sorghu52yclise52or (сорго, суданская трава), Miscanthus giganteus (мискантус), Saccharum sp. (сахарный тростник), Populus balsamifera (тополь), Zea mays (кукуруза), Glycine max (соя), Brassica napus (канола), Triticum aestivum (пшеница), Gossypium hirsutum (хлопок), Oryza sativa (рис), Helianthus annuus (подсолнечник), Medicago sativa (люцерна), Beta vulgaris (сахарная свекла) или Pennisetum glaucum (просо жемчужное).
Различные варианты осуществления направлены на мутантные, не встречающиеся в природе трансгенные растения или клетки растения, модифицированные для модулирования уровней экспрессии гена, в результате чего получают растения или клетки растения, такие как растение табака или клетка растения табака, в которых уровень экспрессии полипептида модулирован в тканях, представляющих интерес, по сравнению с контролем. Раскрытые композиции и способы можно применять в отношении любого вида рода Nicotiana, включая N. rustica и N. tabacum (например, LA B21, LN KY171, TI 1406, Basma, Galpao, Perique, Beinhart 1000-1 и Petico). Другие виды включают N. acaulis, N. acuminata, N. africana, N. alata, N. ameghinoi, N. amplexicaulis, N. arentsii, N. attenuata, N. azambujae, N. benavidesii, N. benthamiana, N. bigelovii, N. bonariensis, N. cavicola, N. clevelandii, N. cordifolia, N. corymbosa, N. debneyi, N. excelsior, N. forgetiana, N. fragrans, N. glauca, N. glutinosa, N. goodspeedii, N. gossei, N. hybrid, N. ingulba, N. kawakamii, N. knightiana, N. langsdorffii, N. linearis, N. longiflora, N. maritima, N. megalosiphon, N. miersii, N. noctiflora, N. nudicaulis, N. obtusifolia, N. occidentalis, N. occidentalis subsp. hesperis, N. otophora, N. paniculata, N. pauciflora, N. petunioides, N. plumbaginifolia, N. quadrivalvis, N. raimondii, N. repanda, N. rosulata, N. rosulata subsp. ingulba, N. rotundifolia, N. setchellii, N. simulans, N. solanifolia, N. spegazzinii, N. stocktonii, N. suaveolens, N. sylvestris, N. thyrsiflora, N. tomentosa, N. tomentosiformis, N. trigonophylla, N. umbratica, N. undulata, N. velutina, N. wigandioides и N. x sanderae.
Применение сортов табака и элитных сортов табака также предусмотрено в данном документе. Трансгенное, не встречающееся в природе или мутантное растение, следовательно, может представлять собой разновидность табака или элитный сорт табака, которые содержат один или более трансгенов или одну или более генетических мутаций или их комбинацию. Генетическая(-ие) мутация(-ии) (например, один или более полиморфизмов) может(-гут) представлять собой мутации, которые не существуют в природе в отдельной разновидности табака или сорте табака (например, элитном сорте табака), или может(-гут) представлять собой генетическую(-ие) мутацию(-ии), которая(-ые) существует(-ют) в природе при условии, что мутация не существует в природе в отдельной разновидности табака или сорте табака (например, в элитном сорте табака).
Особенно применимые разновидности Nicotiana tabacum включают табак типа Burley, табак темного типа, табак трубоогневой сушки и табак восточного типа. Неограничивающими примерами разновидностей или сортов являются BD 64, CC 101, CC 200, CC 27, CC 301, CC 400, CC 500, CC 600, CC 700, CC 800, CC 900, Coker 176, Coker 319, Coker 371 Gold, Coker 48, CD 263, DF911, DT 538 LC табак Galpao, GL 26H, GL 350, GL 600, GL 737, GL 939, GL 973, HB 04P, HB 04P LC, HB3307PLC, гибрид 403LC, гибрид 404LC, гибрид 501 LC, K 149, K 326, K 346, K 358, K394, K 399, K 730, KDH 959, KT 200, KT204LC, KY10, KY14, KY 160, KY 17, KY 171, KY 907, KY907LC, KY14xL8 LC, Little Crittenden, McNair 373, McNair 944, msKY 14xL8, Narrow Leaf Madole, Narrow Leaf Madole LC, NBH 98, N-126, N-777LC, N-7371LC, NC 100, NC 102, NC 2000, NC 291, NC 297, NC 299, NC 3, NC 4, NC 5, NC 6, NC7, NC 606, NC 71, NC 72, NC 810, NC BH 129, NC 2002, Neal Smith Madole, OXFORD 207, PD 7302 LC, PD 7309 LC, PD 7312 LC, табак «Перик», PVH03, PVH09, PVH19, PVH50, PVH51, R 610, R 630, R 7-11, R 7-12, RG 17, RG 81, RG H51, RGH 4, RGH 51, RS 1410, Speight 168, Speight 172, Speight 179, Speight 210, Speight 220, Speight 225, Speight 227, Speight 234, Speight G-28, Speight G-70, Speight H-6, Speight H20, Speight NF3, TI 1406, TI 1269, TN 86, TN86LC, TN 90, TN 97, TN97LC, TN D94, TN D950, TR (Tom Rosson) Madole, VA 309, VA359, AA 37-1, B13P, Xanthi (Mitchell-Mor), Bel-W3, 79-615, Samsun Holmes NN, KTRDC номер 2 гибрид 49, Burley 21, KY8959, KY9, MD 609, PG01, PG04, PO1, PO2, PO3, RG11, RG 8, VA509, AS44, Banket A1, Basma Drama B84/31, Basma I Zichna ZP4/B, Basma Xanthi BX 2A, Batek, Besuki Jember, C104, Coker 347, Criollo Misionero, Delcrest, Djebel 81, DVH 405,  Comum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1, LA BU 21, NC 2326, NC 297, PVH 2110, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1, Prilep P12-2/1, Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, TI-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141, Basma xanthi, GR149, GR153, Petit Havana. Также предполагаются подразновидности вышеуказанного с низким уровнем превращения никотина в норникотин, даже если они специально не указаны в данном документе.
Comum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1, LA BU 21, NC 2326, NC 297, PVH 2110, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1, Prilep P12-2/1, Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, TI-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141, Basma xanthi, GR149, GR153, Petit Havana. Также предполагаются подразновидности вышеуказанного с низким уровнем превращения никотина в норникотин, даже если они специально не указаны в данном документе.
Варианты осуществления также направлены на композиции и способы получения мутантных растений, не встречающихся в природе растений, гибридных растений и трансгенных растений, которые были модифицированы для модулирования экспрессии или активности полинуклеотида(-ов), описанного(-ых) в данном документе (или любой их комбинации, как описано в данном документе). Различные фенотипические характеристики, такие как степень зрелости, количество листьев на растении, высота стебля, угол отхождения листьев, размер листьев (ширина и длина), длина междоузлия и соотношение листовая пластина-главная жилка, можно оценивать путем полевых наблюдений.
Один аспект относится к семенам мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения, описанного в данном документе. Предпочтительно, семена представляют собой семена табака. Дополнительный аспект относится к пыльце или семяпочке мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения, описанного в данном документе. Кроме того, предусмотрено мутантное растение, не встречающееся в природе растение, гибридное растение или трансгенное растение, описанное в данном документе, которое дополнительно содержит нуклеиновую кислоту, обеспечивающую мужскую стерильность.
Также предложена тканевая культура регенерируемых клеток мутантного растения, не встречающегося в природе растения, гибридного растения или трансгенного растения или его части, как описано в данном документе, при этом из культуры регенерируют растения, способные экспрессировать все морфологические и физиологические характеристики родителя. Регенерируемые клетки включают без ограничения клетки из листьев, пыльцы, зародышей, семядолей, гипокотилей, корней, кончиков корней, пыльников, цветков и их части, семяпочек, побегов, стеблей, черешков, сердцевины и семенных коробочек или каллюса или протопластов, полученных из них.
Еще один дополнительный аспект относится к высушенному растительному материалу, такому как высушенный лист или высушенный табак, полученному или который можно получить из мутантного, не встречающегося в природе или трансгенного растения или клетки.
Варианты осуществления также направлены на композиции и способы получения мутантных, не встречающихся в природе или трансгенных растений или клеток растения, которые были модифицированы для модулирования экспрессии или активности одного или более полинуклеотидов или полипептидов, описанных в данном документе, что может приводить к получению растений с сокращенным периодом времени до наступления цветения.
В другом аспекте предусмотрен способ сокращения периода времени до наступления цветения у растения, включающий (i) модулирование (например, снижение) экспрессии или активности одного или более полипептидов или полинуклеотидов, описанных в данном документе; (ii) измерение скорости наступления цветения мутантного, не встречающегося в природе или трансгенного растения, полученного на стадии (i); и (iii) идентификацию мутантного, не встречающегося в природе или трансгенного растения, которое характеризуется сокращенным периодом времени до наступления цветения по сравнению с контрольным растением. Соответственно, растение представляет собой растение табака.
Снижение уровня экспрессии по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или снижение может составлять по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%, которое включает снижение транскрипционной активности или уровня экспрессии полинуклеотида или уровня экспрессии полипептида или их комбинации.
Снижение активности по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или снижение может составлять по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%.
Полинуклеотиды и рекомбинантные конструкции, описанные в данном документе, можно использовать для модулирования экспрессии у видов растений, представляющих интерес, соответственно, у табака.
Целый ряд способов, основанных на использовании полинуклеотидов, можно применять, например, для повышения уровня экспрессии генов у растений и в клетках растений. В качестве примера, можно получать конструкцию, вектор или вектор экспрессии, совместимые с подлежащим трансформации растением, которое содержит представляющий интерес ген вместе с расположенным выше по цепи промотором, способным к сверхэкспрессии гена у растения или в клетке растения. Иллюстративные промоторы описаны в данном документе. После трансформации и при условии роста в подходящих условиях промотор может управлять экспрессией. В одном иллюстративном варианте осуществления создают вектор, несущий один или более полинуклеотидов, описанных в данном документе (или любую их комбинацию, как описано в данном документе), для сверхэкспрессии гена у растения или в клетке растения. Вектор несет подходящий промотор, такой как промотор 35S вируса мозаики цветной капусты CaMV, выше по цепи относительно трансгена, управляющего его конститутивной экспрессией во всех тканях растения. Вектор также несет ген устойчивости к антибиотику для обеспечения возможности отбора трансформированных каллюсов и клеточных линий.
Различные варианты осуществления направлены на способы снижения уровня экспрессии одного или более полинуклеотидов, описанных в данном документе, посредством интеграции множества копий полинуклеотида в геном растения, предусматривающие трансформацию клетки растения-хозяина с помощью вектора экспрессии, который содержит промотор, функционально связанный с одним или более полинуклеотидами, описанными в данном документе. Полипептид, кодируемый рекомбинантным полинуклеотидом, может быть нативным полипептидом или может быть гетерологичным по отношению к клетке.
Растение, несущее мутантный аллель одного или более полинуклеотидов, описанных в данном документе (или любой их комбинации, как описано в данном документе), можно применять в программе селекции растений для создания применимых линий, разновидностей и гибридов. В частности, мутантный аллель интрогрессируют в коммерчески важные разновидности, описанные выше. Таким образом, предложены способы селекции растений, которые предусматривают скрещивание мутантного растения, не встречающегося в природе растения или трансгенного растения, как описано в данном документе, с растением, характеризующимся отличающейся генетической идентичностью. Способ может дополнительно предусматривать скрещивание растения-потомка с другим растением и необязательно повторное скрещивание до тех пор, пока не будет получено потомство с необходимыми генетическими признаками или генетическим фоном. Одной целью, для которой служат такие способы селекции, является введение необходимого генетического признака в другие разновидности, селекционные линии, гибриды или сорта, особенно те, которые представляют коммерческий интерес. Другой целью является облегчение накопления генетических модификаций разных генов в отдельных разновидностях, линиях, гибридах или сортах растений. Предусмотрены внутривидовые, а также межвидовые скрещивания. Растения-потомки, которые возникают в результате таких скрещиваний, также называемые селекционными линиями, являются примерами не встречающихся в природе растений по настоящему изобретению.
В одном варианте осуществления предложен способ получения не встречающегося в природе растения, включающий (a) скрещивание мутантного или трансгенного растения со вторым растением с получением семени-потомка табака; (b) выращивание семени-потомка в условиях роста растений с получением не встречающегося в природе растения. Способ может дополнительно включать (c) скрещивание предыдущего поколения не встречающегося в природе растения с самим собою или с другим растением с получением семени-потомка; (d) выращивание семени-потомка из стадии (c) в условиях роста растений с получением дополнительных не встречающихся в природе растений; и (e) повторение стадий скрещивания и выращивания (c) и (d) много раз с получением следующих поколений не встречающихся в природе растений. Способ может необязательно предусматривать перед стадией (a) стадию получения родительского растения, которое характеризуется генетической идентичностью, которая является характерной, и которое не идентично мутантному или трансгенному растению. В некоторых вариантах осуществления в зависимости от программы селекции стадии скрещивания и выращивания повторяют от 0 до 2 раз, от 0 до 3 раз, от 0 до 4 раз, от 0 до 5 раз, от 0 до 6 раз, от 0 до 7 раз, от 0 до 8 раз, от 0 до 9 раз или от 0 до 10 раз для получения поколений не встречающихся в природе растений. Обратное скрещивание является примером такого способа, где потомство скрещивают с одним из его родителей или с другим растением, генетически подобным его родителю, для получения растения-потомка в следующем поколении, которое характеризуется генетической идентичностью, более близкой к одному из родителей. Методики селекции растений, в частности, селекции растений, хорошо известны и могут быть использованы в способах по настоящему изобретению. В настоящем изобретении дополнительно предложены не встречающиеся в природе растения, полученные с помощью данных способов. Определенные варианты осуществления исключают стадию отбора растения.
В некоторых вариантах осуществления способов, описанных в данном документе, линии, полученные в результате селекции и скрининга вариантных генов, оценивают в поле с использованием стандартных полевых процедур. Включены контрольные генотипы, в том числе исходных не подвергнутых мутагенезу родителей, и места посадки растений расположены в поле согласно рандомизированному полноблочному плану или согласно другому соответствующему планированию поля. Для табака используют стандартные агротехнические приемы, например, табак собирают, взвешивают и отбирают образцы для химического и другого общепринятого тестирования до и во время сушки или высушивания. Статистические анализы данных выполняют для подтверждения сходства выбранных линий с родительской линией. Цитогенетические анализы выбранных растений необязательно выполняют для подтверждения взаимосвязей набора хромосом и конъюгации хромосом.
ДНК-фингерпринтинг, однонуклеотидный полиморфизм, микросателлитные маркеры или аналогичные технологии можно применять в программе селекции с отбором с помощью маркера (MAS) для переноса или размножения мутантных аллелей гена в других растениях, как описано в данном документе. Например, селекционер может создать расщепляющиеся популяции при гибридизации генотипа, содержащего мутантный аллель, с требуемым с точки зрения агрономии генотипом. Растения из F2 или обратно скрещенных поколений можно подвергнуть скринингу с использованием маркера, полученного из геномной последовательности или ее фрагмента, с использованием одной из методик, перечисленных в данном документе. Растения, идентифицированные как обладающие мутантным аллелем, можно подвергнуть обратному скрещиванию или самоопылению для создания второй популяции, подлежащей скринингу. В зависимости от предполагаемого характера наследования или используемой технологии MAS может быть необходимым самоопыление отобранных растений перед каждым циклом обратного скрещивания для облегчения идентификации необходимых отдельных растений. Обратное скрещивание или другую процедуру селекции можно повторять до тех пор, пока необходимый фенотип рекуррентного родителя не восстановится.
В программе селекции успешные скрещивания дают растения F1, которые являются фертильными. Отобранные растения F1 можно скрещивать с одним из родителей, и растения первого поколения обратного скрещивания можно самоопылять с получением популяции, которую снова подвергают скринингу в отношении экспрессии вариантного гена (например, нулевой версии гена). Процесс обратного скрещивания, самоопыления и скрининга повторяют, например, по меньшей мере 4 раза до тех пор, пока при конечном скрининге не получат растение, которое является фертильным и в достаточной степени сходным с рекуррентным родителем. Это растение, при необходимости, самоопыляют и затем потомство снова подвергают скринингу, чтобы подтвердить, что растение демонстрирует экспрессию вариантного гена. В некоторых вариантах осуществления популяцию растений в поколении F2 подвергают скринингу на наличие экспрессии вариантного гена, например, растение, которое не экспрессирует полипептид из-за отсутствия гена, идентифицируют согласно стандартным способам, например, с помощью методики ПЦР с использованием праймеров, созданных на основе информации о последовательности нуклеотидов для полинуклеотида(-ов), описанного в данном документе (или любой их комбинации, описанной в данном документе).
Гибридные разновидности можно получить путем предотвращения самоопыления женских родительских растений (то есть материнских родителей) первой разновидности, позволяя пыльце с мужских родительских растений второй разновидности оплодотворить женские родительские растения и обеспечивая образование гибридных семян F1 на женских растениях. Самоопыление женских растений можно предотвратить путем кастрации цветков на ранней стадии развития цветка. В качестве альтернативы, образование пыльцы на женских родительских растениях можно предотвратить с применением вида мужской стерильности. Например, мужскую стерильность можно получить с помощью цитоплазматической мужской стерильности (CMS) или трансгенной мужской стерильности, где трансген подавляет микроспорогенез и/или образование пыльцы, или вызывает самонесовместимость. Женские родительские растения, характеризующиеся CMS, являются особенно применимыми. В вариантах осуществления, в которых женские родительские растения характеризуются CMS, пыльцу собирают с мужских фертильных растений и наносят вручную на рыльца женских родительских растений с CMS и собирают полученные семена F1.
Разновидности и линии, описанные в данном документе, можно использовать для образования простых гибридов F1. В таких вариантах осуществления растения родительских разновидностей можно выращивать в виде практически однородно соединенных популяций для облегчения естественного перекрестного опыления мужскими родительскими растениями женских родительских растений. Семена F1, образовавшиеся на женских родительских растениях, выборочно собирают с помощью обычных средств. Можно также вырастить две разновидности родительских растений в общей массе и собрать смесь гибридных семян F1, образовавшихся на женском родителе, и семян, образовавшихся на мужском родителе в результате самоопыления. В качестве альтернативы, можно осуществить трехлинейное скрещивание, где простой гибрид F1 используют в качестве женского родителя и скрещивают с отличающимся мужским родителем. В качестве другой альтернативы, можно создать гибриды двойного скрещивания, где потомство F1 двух разных простых гибридов скрещивают само с собой.
Популяцию мутантных, не встречающихся в природе или трансгенных растений можно подвергнуть скринингу или отбору в отношении тех представителей популяции, которые имеют требуемый признак или фенотип. Требуемым признаком или фенотипом может быть сокращенный период времени до наступления цветения (например, на по меньшей мере 30% сокращенный период времени до наступления цветения по сравнению с растением дикого типа) и необязательно меньшее число листьев, чем у растения дикого типа, и необязательно та же высота, что у растения дикого типа.
Например, популяцию потомства линии с одной трансформацией можно подвергнуть скринингу в отношении тех растений, которые имеют необходимый уровень экспрессии или активности полипептида(-ов), кодируемого(-ых) с ее помощью. Физические и биохимические способы можно использовать для определения уровней экспрессии или активности. Они включают анализ по Саузерну или ПЦР-амплификацию для выявления полинуклеотида; нозерн-блоттинг, анализ с защитой от РНКазы S1, анализ методом удлинения праймера или ПЦР-амплификацию в режиме реального времени для выявления РНК-транскриптов; ферментные анализы для выявления ферментативной или рибозимной активности полипептидов и полинуклеотидов; и гель-электрофорез белков, вестерн-блоттинг, иммунопреципитацию и иммуноферментные анализы для выявления полипептидов. Другие методики, такие как гибридизация in situ, ферментативное окрашивание и иммуноокрашивание, и ферментативный анализ также можно использовать для выявления присутствия, или экспрессии, или активности полипептидов или полинуклеотидов.
В данном документе описаны мутантные, не встречающиеся в природе или трансгенные клетки растения и растения, содержащие один или более рекомбинантных полинуклеотидов, одну или более полинуклеотидных конструкций, одну или более двухнитевых РНК, один или более конъюгатов или один или более векторов/векторов экспрессии.
Без ограничения растения, описанные в данном документе, можно модифицировать для других целей либо до того, либо после того, как экспрессия или активность была модулирована согласно настоящему изобретению. Одна или более следующих генетических модификаций могут присутствовать в мутантных, не встречающихся в природе или трансгенных растениях. В одном варианте осуществления один или более генов, которые вовлечены в превращение промежуточных продуктов азотного обмена, модифицируют с получением растений (таких как листья), которые в высушенном состоянии дают более низкие уровни по меньшей мере одного специфичного для табака нитрозамина, чем контрольные растения. Неограничивающие примеры генов, которые можно модифицировать, включают гены, кодирующие аспарагинсинтетазу, такие как CYP82E4, CYP82E5 и CYP82E10, которые принимают участие в превращении никотина в норникотин и описаны в публикациях WO2006091194, WO2008070274, WO2009064771 и PCT/US2011/021088, и описанные в данном документе. В другом варианте осуществления один или более генов, которые принимают участие в поглощении тяжелых металлов или транспорте тяжелых металлов, модифицируют с получением в результате растений или частей растений (таких как листья), имеющих более низкое содержание тяжелых металлов, чем у контрольных растений или их частей без модификации(-ий). Неограничивающие примеры включают гены семейства белков, ассоциированных с множественной лекарственной устойчивостью, семейства посредников диффузии катионов (CDF), семейства Zrt-, Irt-подобных белков (ZIP), семейства катионообменников (CAX), семейства транспортеров меди (COPT), семейства АТФаз тяжелых металлов Р-типа (например, HMA, как описано в WO2009074325), семейства гомологов белков макрофагов, ассоциированных с естественной устойчивостью (NRAMP), и семейства транспортеров с АТФ-связывающей кассетой (ABC) (например, MRP, описанных в WO2012/028309, которые участвуют в транспорте тяжелых металлов, таких как кадмий). Термин «тяжелый металл», используемый в данном документе, включает переходные металлы. Примеры других модификаций включают выносливость в отношении гербицидов, например, глифосат является активным ингредиентом многих гербицидов широкого спектра действия. Устойчивые к глифосату трансгенные растения были разработаны путем переноса гена aroA (глифосат EPSP-синтетаза из Salmonella typhimurium и E.coli). Устойчивые к сульфонилмочевине растения были получены путем трансформации мутантного гена ALS (ацетолактатсинтетаза) из Arabidopsis. Белок OB фотосистемы II из мутантного Amaranthus hybridus был перенесен в растения с получением атразин-устойчивых трансгенных растений; и бромоксинил-устойчивые трансгенные растения были получены путем встраивания гена bxn из бактерии Klebsiella pneumoniae. Другие иллюстративные модификации приводят в результате к получению растений, которые устойчивы к насекомым. Токсины Bacillus thuringiensis (Bt) могут обеспечить эффективный путь отсрочки появления Bt-устойчивых вредителей, как недавно показано на брокколи, где гены Btcry1Ac и cry1C в составе «пирамиды» контролировали капустную моль, устойчивую к любому одному белку, и существенно задерживали развитие устойчивых насекомых. Другая иллюстративная модификация приводит к получению растений, которые устойчивы к заболеваниям, вызванным патогенами (например, вирусами, бактериями, грибами). Были разработаны растения, экспрессирующие ген Xa21 (устойчивость к бактериальному некрозу), с растениями, экспрессирующими как ген слияния Bt, так и ген хитиназы (устойчивость к желтой огневке-травянке и выносливость в отношении ризоктониоза). Другая иллюстративная модификация приводит к измененной репродуктивной способности, такой как мужская стерильность. Другая иллюстративная модификация приводит к получению растений, которые выносливы в отношении абиотического стресса (например, засухи, изменения температуры, засоленности), и выносливые трансгенные растения были получены путем переноса фермента глицеролфосфат-ацилтрансферазы из Arabidopsis; генов, кодирующих маннитдегидрогеназу и сорбитдегидрогеназу, которые вовлечены в синтез маннита и сорбита, улучшающих устойчивость к засухе. Другие иллюстративные модификации могут приводить в результате к получению растений с улучшенным накоплением белков и масел, растений с повышенной эффективностью фотосинтеза, растений с длительным сроком хранения, растений с повышенным содержанием углеводов и растений, устойчивых к грибам; растений, кодирующих фермент, участвующий в биосинтезе алкалоидов. Также рассматриваются трансгенные растения, в которых экспрессия S-аденозил-L-метионин (SAM) и/или цистатионин-гамма-синтазы (CGS) была модулирована.
Один или более таких признаков можно интрогрессировать в мутантные, не встречающиеся в природе или трансгенные растения другого сорта или можно непосредственно трансформировать в них. Интрогрессия признака(-ов) в мутантные, не встречающиеся в природе или трансгенные растения может быть достигнута любым способом селекции растений, известным из уровня техники, например, селекцией на основе родословной, обратным скрещиванием, селекцией двойных гаплоидов и т. п. (см. Wernsman, E. A. и Rufty, R. C. 1987. Chapter Seventeen. Tobacco. страницы 669-698 в: Cultivar Development. Crop Species. W. H. Fehr (ed.), MacMillan Publishing Co, Inc., New York, N.Y стр. 761). Методики, основанные на молекулярной биологии, описанные выше, в частности, с применением RFLP и микросателлитных маркеров, можно использовать при таких обратных скрещиваниях для идентификации потомков, характеризующихся самой высокой степенью генетической идентичности с рекуррентным родителем. Это позволяет ускорить получение разновидностей, характеризующихся по меньшей мере 90%, предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 99% генетической идентичностью с рекуррентным родителем, еще более предпочтительно, генетически идентичных рекуррентному родителю и дополнительно характеризующихся признаком(-ами), интрогрессированным(-ми) от родителя-донора. Такое определение генетической идентичности может основываться на молекулярных маркерах, известных из уровня техники.
Последнее поколение обратного скрещивания можно подвергнуть самоопылению с получением потомства чистого разведения для подлежащей переносу нуклеиновой(-ых) кислоты(-т). Полученные растения обычно обладают по существу всеми морфологическими и физиологическими характеристиками мутантных, не встречающихся в природе или трансгенных растений в дополнение к перенесенному(-ым) признаку(-ам) (например, одному или более признакам, определяемым одним геном). Точный протокол обратного скрещивания будет зависеть от изменяемого признака для определения соответствующего протокола испытаний. Хотя способы обратного скрещивания упрощаются, если переносимый признак представляет собой доминантный аллель, рецессивный аллель также можно переносить. В данном случае может быть необходимо применять исследование потомства для определения успешности переноса требуемого признака.
В различных вариантах осуществления предложены мутантные растения, не встречающиеся в природе растения или трансгенные растения, а также биомасса, в которых уровень экспрессии полинуклеотида (или любой их комбинации, как описано в данном документе) снижен для сокращения периода времени до наступления цветения.
В различных вариантах осуществления предложены мутантные растения, не встречающиеся в природе растения или трансгенные растения, а также биомасса, в которых активность полипептида (или любой их комбинации, как описано в данном документе) снижена для сокращения периода времени до наступления цветения.
Части таких растений, в частности растений табака, и более конкретно, листовую пластинку и среднюю жилку растений табака, можно включить в изготовление или использовать в изготовлении различных продуктов потребления, включая без ограничения материалы, образующие аэрозоль, устройства, образующие аэрозоль, курительные изделия, изделия для курения, бездымные продукты и табачные продукты. Примеры материалов, образующих аэрозоль, включают без ограничения табачные композиции, виды табака, табачный экстракт, резаный табак, резаный наполнитель, просушенный или высушенный табак, взорванный табак, гомогенизированный табак, восстановленный табак и виды трубочного табака. Курительные изделия и изделия для курения являются типами устройств, образующих аэрозоль. Примеры курительных изделий и изделий для курения включают без ограничения сигареты, сигариллы и сигары. Примеры бездымных продуктов включают виды жевательного табака и виды нюхательного табака. В определенных устройствах, образующих аэрозоль, вместо сгорания (или сжигания) табачную композицию или другой материал, образующий аэрозоль, подвергают нагреванию, например, с помощью одного или более электрических нагревательных элементов или углеродного источника тепла с получением аэрозоля. Как правило, в таких нагреваемых курительных изделиях аэрозоль образуется в результате передачи тепла от источника тепла на физически отделенный субстрат или материал, образующий аэрозоль, который может быть расположен внутри, вокруг или ниже по потоку относительно источника тепла. Во время курения летучие соединения высвобождаются из субстрата, образующего аэрозоль, в результате передачи тепла от источника тепла и захватываются в воздух, втягиваемый через курительное изделие. По мере охлаждения высвобождаемых соединений, они конденсируются с образованием аэрозоля, который вдыхается пользователем. Такие устройства включают, например, электрически нагреваемые устройства, генерирующее аэрозоль, в которых аэрозоль образуется в результате передачи тепла от устройства, генерирующего аэрозоль, на субстрат, образующий аэрозоль, нагреваемого курительного изделия. Соответственно, во время нагревания субстрата, образующего аэрозоль, не происходит сгорание или сжигание табака. Подходящее образующее аэрозоль изделие описано в WO2013/098405 и содержит субстрат, образующий аэрозоль, для генерирования вдыхаемого аэрозоля при нагревании с помощью внутреннего нагревательного элемента устройства, генерирующего аэрозоль. Оно может содержать электрически нагреваемое устройство, генерирующее аэрозоль, содержащее внутренний нагревательный элемент. Оно может дополнительно содержать в линейном последовательном расположении субстрат, образующий аэрозоль, опорный элемент, размещенный непосредственно ниже по потоку относительно субстрата, образующего аэрозоль, элемент, охлаждающий аэрозоль, размещенный ниже по потоку относительно опорного элемента, и наружную обертку, окружающую субстрат, образующий аэрозоль, опорный элемент и элемент, охлаждающий аэрозоль. Опорный элемент может примыкать к субстрату, образующему аэрозоль. В субстрат, образующий аэрозоль, проникает нагревательный элемент устройства, генерирующего аэрозоль.
Используемый в данном документе термин «сгорание» относится к окислительно-восстановительной химической реакции, где реагирующие молекулы, а именно топливо и окислитель, смешиваются и перегруппировываются, становясь молекулами продукта с одновременным высвобождением тепла. Сгорание можно положительно отметить присутствием соответствующих количеств оксидов азота в газообразных продуктах, не образовавшихся в результате разложения нитратов, присутствующих в исходном реагирующем субстрате, и явным доказательством одновременного общего экзотермического процесса. Образование соответствующих количеств оксидов азота в газообразных продуктах сгорания может быть определено с помощью сравнения общих количеств оксидов азота, образовавшихся в условиях, представляющих интерес (например, в воздухе), и оксидов азота, образовавшихся в тех же условиях, но в отсутствие кислорода (например, в атмосфере чистого азота или гелия). В другом типе нагреваемого устройства, образующего аэрозоль, аэрозоль образуется в результате передачи тепла от горючего топливного элемента или источника тепла к физически отделенному материалу, образующему аэрозоль, который может быть расположен внутри, вокруг или ниже по потоку относительно источника тепла. Бездымные табачные продукты и различные табаксодержащие материалы, образующие аэрозоль, могут содержать табак в любом виде, в том числе в виде высушенных частиц, обрезков, гранул, порошков или суспензии, нанесенный на другие ингредиенты, смешанный с ними, окруженный ими или иным образом объединенный с ними в любом формате, таком как хлопья, пленки, таблетки, пены или шарики. Используемый в данном документе термин «дым» используют для описания типа аэрозоля, получаемого с помощью курительных изделий, таких как сгораемые сигареты, или при сгорании материала, образующего аэрозоль.
В определенных вариантах осуществления нагревание без сгорания или сжигания растительного материала является предпочтительным.
В одном варианте осуществления также предложен сушеный растительный материал из мутантных, трансгенных и не встречающихся в природе растений табака, описанных в данном документе. Способы сушки зеленых табачных листьев известны средним специалистам в данной области и включают без ограничения воздушную сушку, огневую сушку, дымовую сушку и сушку на солнце. Способ сушки зеленых табачных листьев зависит от типа собранного табака. Например, Burley и определенные темные разновидности обычно сушат посредством воздушной сушки, а трубочный табак, жевательный табак и нюхательный табак сушат посредством огневой сушки.
В другом варианте осуществления также предложен высушенный растительный материал из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Способы высушивания листьев известны средним специалистам в данной области и включают без ограничения высушивание на воздухе и высушивание на солнце. Соответствующий способ высушивания листьев зависит от типа растения, которое собирают. Соответственно, растительный материал высушивают после сбора. Таким образом, применение высушенного материала и высушенного после сбора материала рассматривается в данном документе.
В другом варианте осуществления описаны табачные продукты, в том числе табаксодержащие материалы, образующие аэрозоль, содержащие растительный материал, такой как листья, предпочтительно сушеные или высушенные листья, из мутантных растений табака, трансгенных растений табака или не встречающихся в природе растений табака, описанных в данном документе. Табачные продукты, описанные в данном документе, могут представлять собой смешанный табачный продукт, который может дополнительно содержать немодифицированный табак.
В настоящем изобретении также предложены способы получения семян, включающие культивирование мутантного растения, не встречающегося в природе растения или трансгенного растения, описанных в данном документе, и сбор семян культивируемых растений. Семена растений, описанных в данном документе, можно кондиционировать и упаковать в упаковочный материал с помощью средств, известных из уровня техники, с получением промышленного изделия. Упаковочный материал, такой как бумага или ткань, хорошо известен из уровня техники. Упаковка с семенами может иметь этикетку, например, бирку или этикетку, прикрепленную к упаковочному материалу, этикетку, напечатанную на упаковке, которая описывает происхождение содержащихся в ней семян.
Композиции, способы и наборы для генотипирования растений для идентификации, отбора или селекции могут включать средства выявления присутствия полинуклеотида (или любой их комбинации, как описано в данном документе) в образце полинуклеотида. Соответственно, описана композиция, содержащая один или более праймеров для специфической амплификации по меньшей мере части одного или более полинуклеотидов и, необязательно, один или более зондов и, необязательно, один или более реагентов для проведения амплификации или выявления.
Соответственно, раскрыты специфичные в отношении гена олигонуклеотидные праймеры или зонды, содержащие приблизительно 10 или больше смежных полинуклеотидов, соответствующих полинуклеотиду(-ам), описанному(-ым) в данном документе. Указанные праймеры или зонды могут содержать или состоять из приблизительно 15, 20, 25, 30, 40, 45 или 50 или больше смежных полинуклеотидов, которые гибридизируются (например, специфически гибридизируются) с полинуклеотидом(-ами), описанным(-и) в данном документе. В некоторых вариантах осуществления праймеры или зонды могут содержать или состоять из приблизительно 10-50 смежных нуклеотидов, приблизительно 10-40 смежных нуклеотидов, приблизительно 10-30 смежных нуклеотидов или приблизительно 15-30 смежных нуклеотидов, которые можно использовать в зависимых от последовательности способах идентификации гена (например, Саузерн-гибридизация) или выделения (например, гибридизация in situ бактериальных колоний или бляшек бактериофага) или обнаружения гена (например, в виде одного или более амплифицирующих праймеров при амплификации и обнаружении нуклеиновой кислоты). Один или более специфических праймеров или зондов можно спроектировать и использовать для амплификации и выявления части или всего полинуклеотида(-ов). В качестве конкретного примера, два праймера можно использовать в протоколе полимеразной цепной реакции для амплификации фрагмента нуклеиновой кислоты, кодирующего нуклеиновую кислоту, такую как ДНК или РНК. Полимеразную цепную реакцию можно также проводить с применением одного праймера, который получен из последовательности нуклеиновой кислоты, и второго праймера, который гибридизируется с последовательностью выше по цепи или ниже по цепи относительно последовательности нуклеиновой кислоты, такой как промоторная последовательность, 3'-конец mRNA-предшественника или последовательность, полученная из вектора. Примеры термических и изотермических методик, применимых для амплификации полинуклеотидов in vitro, хорошо известны из уровня техники. Образец может представлять собой растение, клетку растения или растительный материал или табачный продукт или его можно получить из них, при этом табачный продукт изготовлен или получен из растения, клетки растения или растительного материала, как описано в данном документе.
В дополнительном аспекте также предложен способ выявления полинуклеотида(-ов), описанного(-ых) в данном документе (или любой их комбинации, описанной в данном документе) в образце, предусматривающий стадию (a) получения образца, содержащего, или предположительно содержащего полинуклеотид; (b) приведения указанного образца в контакт с одним или более праймерами или одним или более зондами для специфического выявления по меньшей мере части полинуклеотида(-ов) и (c) выявления присутствия продукта амплификации, где присутствие продукта амплификации свидетельствует о присутствии полинуклеотида(-ов) в образце. В дополнительном аспекте также предложено использование одного или более праймеров или зондов для специфического выявления по меньшей мере части полинуклеотида(-ов). Также предложены наборы для выявления по меньшей мере части полинуклеотида(-ов), которые содержат один или более праймеров или зондов для специфического выявления по меньшей мере части полинуклеотида(-ов). Набор может содержать реагенты для амплификации полинуклеотидов, такой как ПЦР, или реагенты для технологии выявления с применением гибридизационного зонда, такой как Саузерн-блоттинг, нозерн-блоттинг, гибридизация in situ или микрочип. Набор может содержать реагенты для технологии выявления связывания антител, такой как вестерн-блоттинг, ELISA, SELDI-масс-спектрометрия или тест-полоски. Набор может содержать реагенты для секвенирования ДНК.
В некоторых вариантах осуществления набор может содержать инструкции для одного или более описанных способов. Описанные наборы могут быть применимы для определения генетической идентичности, филогенетических исследований, генотипирования, гаплотипирования, анализа родословной или селекции растений, в частности, с количественной оценкой кодоминантных признаков.
В настоящем изобретении также предложен способ генотипирования растения, клетки растения или растительного материала, содержащих полинуклеотид, описанный в данном документе. Генотипирование обеспечивает средства различения гомологов пары хромосом и может быть использовано для различения сегрегантов в популяции растений. Молекулярные способы с использованием маркеров можно применять для филогенетических исследований, характеризующих генетические связи между разновидностями сельскохозяйственных культур, идентификации гибридов или соматических гибридов, локализации хромосомных сегментов, влияющих на моногенные признаки, клонирования на основе генетических карт и изучения количественного наследования. В определенном способе генотипирования может использоваться любое количество аналитических методик с молекулярным маркером, включая полиморфизмы длины амлифицированного фрагмента (AFLP). AFLP являются продуктом аллельных различий между фрагментами амплификации, вызванных изменчивостью нуклеотидной последовательности. Таким образом, в настоящем изобретении дополнительно предложены способы отслеживания сегрегации одного или более генов или нуклеиновых кислот, а также хромосомных последовательностей, генетически связанных с этими генами или нуклеиновыми кислотами, с применением таких методик, как анализ AFLP.
В одном варианте осуществления также предложен сушеный или высушенный растительный материал из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Например, способы сушки или высушивания табачных листьев известны специалистам в данной области и включают без ограничения воздушную сушку, огневую сушку, дымовую сушку и сушку на солнце. Способ сушки зеленых табачных листьев зависит от типа собранного табака, как описано в данном документе.
В другом варианте осуществления описаны табачные продукты, в том числе табачные продукты, содержащие растительный материал, такой как листья, соответственно, сушеный растительный материал, такой как сушеные или высушенные листья, из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе, или которые получены посредством способов, описанных в данном документе. Табачные продукты, описанные в данном документе, могут дополнительно содержать немодифицированный табак.
В другом варианте осуществления описаны табачные продукты, содержащие растительный материал, предпочтительно листья, такие как сушеные или высушенные листья, из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Например, растительный материал можно добавить внутрь или снаружи табачного продукта, вследствие чего при сгорании высвобождается необходимый аромат. Табачный продукт в соответствии с этим вариантом осуществления может даже представлять собой немодифицированный табак или модифицированный табак. Табачный продукт в соответствии с данным вариантом осуществления может быть даже получен из мутантного, трансгенного или не встречающегося в природе растения, которое имеет модификации в одном или более генах, отличающихся от генов, раскрытых в данном документе.
В дополнительном аспекте предложен способ идентификации молекулы, которая обеспечивает модуляцию активности или экспрессии полинуклеотида TFL1 или полипептида TFL1. Иллюстративные молекулы, которые можно применять в таком способе, описаны в данном документе, в том числе антисмысловые молекулы, siRNA, молекулы рибозима, молекулы дезоксирибозима, TALEN, нуклеазы с «цинковыми пальцами» и т. п. Также рассматривается применение химических соединений.
Также описано мутантное, не встречающееся в природе или трансгенное растение или его часть, обладающие модулированной (например, сниженной или повышенной) экспрессией гена, кодирующего Terminal Flower 1 (TFL1), или модулированной (например, сниженной или повышенной) активностью белка, кодируемого TFL1, при этом указанный TFL1 содержит, состоит или по существу состоит из (i) полинуклеотидной последовательности, содержащей, состоящей или по существу состоящей из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептида, кодируемого полинуклеотидом, представленным в (i); или (iii) полипептида, характеризующегося по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; где экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii) или (iii), модулирована (например, снижена или повышена) по сравнению с контрольным растением, у которого экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii) или (iii), не была модулирована (например, снижена или повышена).
Модулированная экспрессия полинуклеотида или модулированная активность полипептида обеспечивали модулирование периода времени до наступления цветения по сравнению с контрольным растением, соответственно, где период времени, предшествующий цветению, модулирован на по меньшей мере 8% или по меньшей мере 20%.
Повышенная экспрессия полинуклеотида или повышенная активность полипептида обеспечивают продление периода времени до наступления цветения по сравнению с контрольным растением, соответственно, где период времени, предшествующий цветению, продлен на по меньшей мере 8% или по меньшей мере 20%.
Также раскрыт способ модулирования (например, увеличения или уменьшения) периода времени до наступления цветения у растения, включающий модификацию растения путем модулирования экспрессии по меньшей мере одного гена TFL1 или активности по меньшей мере одного белка, кодируемого им, в указанном растении; соответственно, где способ включает: (a) получение растения или его части, содержащих (i) полинуклеотидную последовательность содержащую, состоящую или по существу состоящую из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептид, кодируемый полинуклеотидом, представленным в (i); или (iii) полипептид, характеризующийся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; и (b) модулирование (например, повышение или снижение) экспрессии гена TFL1 или активности белка TFL1 у растения; и (c) получение растения с модулированным периодом времени до наступления цветения по сравнению с контрольным растением, у которого экспрессия гена TFL1 или активность белка TFL1 не были модулированы.
Соответственно, экспрессию TFL1 или активность TFL1 модулируют с помощью способа, выбранного из группы, состоящей из a) мутирования гена TFL1 у растения; b) экспрессии экзогенного полинуклеотида или полипептида у растения; и c) удаления гена TFL1 у растения, или комбинации одного или более из них.
Также раскрыт способ увеличения периода времени до наступления цветения у растения, включающий модификацию растения путем повышения экспрессии по меньшей мере одного гена TFL1 или активности по меньшей мере одного белка, кодируемого им, в указанном растении; соответственно, где способ включает (a) получение растения или его части, содержащих (i) полинуклеотидную последовательность содержащую, состоящую или по существу состоящую из последовательности, характеризующейся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:7, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:11, или SEQ ID NO:19, или SEQ ID NO:20; или (ii) полипептид, кодируемый полинуклеотидом, представленным в (i); или (iii) полипептид, характеризующийся по меньшей мере 72% идентичностью последовательности с SEQ ID NO:9, или SEQ ID NO:12, или SEQ ID NO:21; и (b) повышение экспрессии гена TFL1 или активности белка TFL1 у растения; и (c) получение растения с продленным периодом времени до наступления цветения по сравнению с контрольным растением, у которого экспрессия гена TFL1 или активность белка TFL1 не были модулированы.
Соответственно, экспрессию TFL1 или активность TFL1 повышают с помощью способа, выбранного из группы, состоящей из a) мутирования гена TFL1 у растения; b) экспрессии экзогенного полинуклеотида или полипептида у растения; и c) удаления гена TFL1 у растения, или комбинации одного или более из них.
Также описан способ получения растительного материала с модулированным периодом времени до наступления цветения по сравнению с контрольным растением, при этом указанный способ включает (a) получение растения или растительного материала, как описано в данном документе; (b) сбор растительного материала от растения; (c) необязательно сушку или высушивание растительного материала в течение периода времени; и(d) получение растительного материала, который имеет модулированный период времени до наступления цветения по сравнению с контрольным растением.
Также описан способ получения растительного материала с продленным периодом времени до наступления цветения по сравнению с контрольным растением, при этом указанный способ включает (a) получение растения или растительного материала, как описано в данном документе; (b) сбор растительного материала от растения; (c) необязательно сушку или высушивание растительного материала в течение периода времени; и(d) получение растительного материала, который имеет продленный период времени до наступления цветения по сравнению с контрольным растением.
Настоящее изобретение дополнительно описано в примерах ниже, которые представлены для более подробного описания настоящего изобретения. Эти примеры, в которых изложен предпочтительный режим, предполагаемый в настоящее время для осуществления настоящего изобретения, предназначены для иллюстрации, а не для ограничения настоящего изобретения.
ПРИМЕРЫ
Пример 1 - Идентификация генов TFL1 табака
Идентифицировали семейство генов TFL1 в табаке и определяли идентичность 7 генов TFL1 табака. Они называются TFL1-1S, TFL1-1T, TFL1-2S, TFL1-2T, TFL1-3T, TFL1-4T и TFL1-3S. Последовательности данных генов описаны в данном документе.
Пример 2 - Анализ уровней экспрессии TFL1
Для идентификации активных форм TFL1 в вегетативных тканях листьев и корней растения N. tabacum трех разных разновидностей (Burley-TN90, Virginia-K326 и восточного типа - BX_Basma Xanthi) выращивали в ящиках Magenta с бактоагаром-MS и рассчитывали данные экспрессии (FPKM) после выделения РНК и секвенирования соответствующих библиотек cDNA (Nat Commun. (2014) 5: 3833). Данные показаны в таблице 1. TFL1 главным образом экспрессировался в тканях корней. Основные гены TFL1, экспрессировавшиеся в корнях при этих условиях (вегетативное состояние), представляют собой TFL1-2S, TFL1-2T, TFL1-3T и TFL1-4T. Можно заметить, что транскрипты TFL1-4S не идентифицированы в табаке Virginia (K326), Burley (TN90) или Dark, при этом лишь очень небольшое количество РНК-копий найдено в корнях Basma Xanthi (BX) восточного типа. Причина, по которой не выявлены транскрипты в K326 и TN90, является неясной, но это может происходить из-за разрушения промотора, поскольку 5'-конец 2 т. о. промотора у K326 и TN90 не смогли идентифицировать.
Для определения того, отличаются ли профили экспрессии генов у цветущих растений, N. tabacum (сорт TN90) выращивали в поле и выделяли РНК из незрелых цветков, листьев в нижней части стебля, листьев в средней части стебля, листьев в верхней части стебля, лепестков, корней, чашелистиков и стебля непосредственно после появления первых цветков. Результаты показаны на фигуре 1. В этих растениях уровень экспрессии TFL1-2T и TFL1-4T оставался невысоким в корнях, стебле и незрелых цветках (значения FPKM не превышают 1,1).
Пример 3 - Генетическая манипуляция с TFL1: растений табака с TFL1-RNAi
Были разработаны четыре специфических фрагмента ДНК для подвергания сайленсингу шести экспрессированных копий TFL1 (см. SEQ ID NO: 24, 27, 30 и 33) и клонированы между сильным конститутивным промотором MMV (вирус мозаики мирабилис) и терминаторной 3'-последовательностью nos гена нопалинсинтазы Agrobacterium tumefaciens (Plant Mol Biol. (1999) 40: 771-82): (1) C100S3-MMVp-GW-pDONR221 - TFL1-1 S/T; (2) C100S3-MMVp-GW-pDONR221 - TFL1-2 S/T; (3) C100S3-MMVp-GW-pDONR221 - TFL1-3 T; (4) C100S3-MMVp-GW-pDONR221 - TFL1-4 T.
Разновидность темного табака трансформировали с помощью каждой из четырех конструкций с применением стандартных протоколов опосредованной Agrobacterium трансформации (Cold Spring Harb Symp Quant Biol. (1985) 50: 433-437). В случае каждой конструкции отбирали 20 независимых растений T0 на канамицине и затем переносили в почву в отделение теплицы с установками 9 ч. света (24°C +/- 3°C) и 15 ч. темноты (18°C +/- 3°C) в течение трех первых месяцев, затем 15 ч. света и 9 ч. темноты. Контрольные растения подвергали той же процедуре in vitro (без стадии трансформации) и также одновременно переносили в теплицу. Сперва сеянцы культивировали в лотках и пересаживали в горшки объемом 5 л через 25 дней. Растения поливали каждый день раствором удобрения (Yara, Нидерланды). Каждое растение для четырех конструкций и их соответствующие контроли анализировали фенотипически и регистрировали данные в виде появления цветков после пересадки (DAT), высоты и числа листьев (см. фигуру 2 и таблицу 2). Данные растений RNAi TFL1-1S/T и TFL1-4T и их соответствующих контролей (фигуры 2A,B,C) необходимо было сравнивать отдельно от группы TFL1-2S/T и TFL1-3T (фигуры 2D,E,F), поскольку первую группу растений пересаживали и культивировали в теплице через 3 недели после второй группы растений. Все растения выращивали при аналогичных условиях в теплице. Данные показали, что трансгенные линии RNAi TFL1-2S/T зацветали значительно быстрее, чем контрольные растения, демонстрируя 20% сокращение периода времени до наступления цветения. Кроме того, линия RNAi TFL1-2S/T также продемонстрировала значительное уменьшение высоты и числа листьев, что соотносится с сокращенным периодом времени до наступления цветения (фигуры 2A, B, C). Данные являются статистически значимыми для всех измеренных параметров (P<0,001), и фенотипы были четко видны в теплице (фигура3). Кроме того, другой продукт гена TFL1, TFL1-4T, по видимому, играет роль в регуляции периода времени до цветения. Действительно, наблюдали 8% сокращение периода времени до цветения, но не в значительной степени по сравнению с контролем, при этом два других измеренных параметра, высота (16% сокращение по сравнению с контролем) и число листьев (13% уменьшение по сравнению с контролем) являются статистически значимыми.
Данные, собранные на растениях RNAi, в сочетании с данными экспрессии, дают основания предполагать, что TFL1-1S и TFL1-1T вносят минимальный вклад в регуляцию периода времени до цветения и поддержание вегетативного состояния растений.
В заключение, данные указывают, что наиболее эффективными генами TFL1 в отношении поддержания вегетативного состояния у табака при описанных условиях роста являются TFL1-2S, TFL1-2T > TFL1-4T > TFL1-3T > TFL1-1S, TFL1-1T. Следовательно, подвергание сайленсингу или нокауту TFL1-2S, и/или TFL1-2T, и/или TFL1-4-T, вероятно, является решением для обеспечения сокращения периода времени до наступления цветения.
Пример 4 - мутации TFL1 для сокращения периода времени до наступления цветения
Для селекции табака популяцию обработанного EMS сорта табака подвергали скринингу на наличие мутаций в TFL1. Все аминокислотные замены, идентифицированные в TFL1-2S, TFL1-2T и TFL-4T, анализировали в отношении возможного влияния на функцию белка с применением программы SIFT (Nucleic Acids Res. (2003) 1;31(13):3812-4). Небольшой показатель SIFT (<0,05) подразумевает, что аминокислотный остаток, вероятно, недопустим на функциональном уровне. В TFL1-2S, TFL1-2T и TFL-4T идентифицировали семь мутаций с соотношением показателя SIFT ниже 0,05, как показано ниже:
TFL1-2S_G129E 0,0002
TFL1-2S_G129R 0,0002
TFL1-2S_T143I 0,0011
TFL1-2T_R120C 0
TFL1-2T_P131S 0,0007
TFL1-2T_G129E 0,0002
TFL1-4T_P110L 0
Показатель SIFT используется в качестве инструмента для облегчения отбора мутаций. В эксперименте в теплице тестировали мутации и комбинации мутаций и измеряли скорость наступления цветения.
В TFL1-2S, TFL1-2T и TFL1-4T мотив His88 является консервативным в положениях 84, 88 и 86 соответственно. Мутация (например, миссенс-мутация) в данном аминокислотном положении может обеспечивать разрушение трехмерной структуры фактора транскрипции, влияя таким образом на свойства связывания на промоторе. Соответственно, мутация может представлять собой стоп-мутацию.
Мотив Asp144 является консервативным в TFL1-2S и TFL1-2T в положениях 138 и 142 соответственно. Он заменен на глутамат в TFL1-4T в положении 139. Три мутации, расположенные близко к TFL1-2S (D138), TFL1-2T (D142), соответственно, идентифицировали вблизи данного участка, и они могут влиять на трехмерную структуру фактора транскрипции и, тем самым, изменять связывание с промотором.
Пример 5 - мутация TFL1-2T-P131S
Растение табака, содержащее мутацию TFL1-2T-P131S (как показано на фигуре 4), выращивали в условиях теплицы. Период времени до цветения, число листьев и высоту мутантных растений отслеживали относительно растения дикого типа. Растения табака, содержащие данную мутацию, зацветали на 30% быстрее и имели меньше листьев (6-7), если копия TFL1-2T содержала мутацию P131S как у гомозигот, так и у гетерозигот. Не наблюдалось влияния на высоту растений. Мутантное растение табака имело сокращенный период времени до наступления цветения по сравнению с растением дикого типа.
Пример 6 - мутация TFL1-4T-P110L Растение табака, содержащее мутацию TFL1-4T-P110L (как показано на фигуре 6), выращивали в условиях теплицы. Период времени до цветения, число листьев и высоту мутантных растений отслеживали относительно растения дикого типа. Для этого мутанта не были доступны гомозиготные растения, но, как наблюдалось для TFL1-2T-P131S, растения табака зацветают на приблизительно 30% быстрее (например, на приблизительно 28% быстрее) и имеют меньше листьев (8-9), если TFL1-4T содержит гетерозиготную мутацию P110L. Не наблюдалось влияния на высоту. Мутантное растение табака имеет сокращенный период времени до наступления цветения по сравнению с растением дикого типа. Данные дают основания предполагать, что только одного аллеля достаточно для ускорения наступления цветения в случае обеих мутаций TFL1-2T-P131S и TFL1-4T-P110L.
Любая публикация, цитированная или описанная в данном документе, предоставляет соответствующую информацию, раскрытую до даты подачи настоящей заявки. Заявления, сделанные в данном документе, не должны быть истолкованы как признание того, что авторы настоящего изобретения не имеют оснований для его противопоставления таким раскрытиям как более раннего. Все публикации, упомянутые в вышеприведенном описании, включены в данный документ посредством ссылки. Различные модификации и варианты настоящего изобретения будут очевидны специалистам в данной области без отступления от объема и сути настоящего изобретения. Несмотря на то, что настоящее изобретение описано применительно к конкретным предпочтительным вариантам осуществления, следует понимать, что заявленное изобретение не должно неправомерно ограничиваться такими конкретными вариантами осуществления. В действительности различные модификации описанных способов осуществления изобретения, которые очевидны специалистам в клеточной, молекулярной биологии и биологии растений или в смежных областях, предназначены находиться в пределах объема прилагаемой формулы изобретения.
ТАБЛИЦА 1
Данные экспрессии (FPKM) TFL1 у растений в вегетативном состоянии (нецветущих), выращенных в условиях бактоагара-MS. Не были выявлены транскрипты в TN90 и K326 (нет).
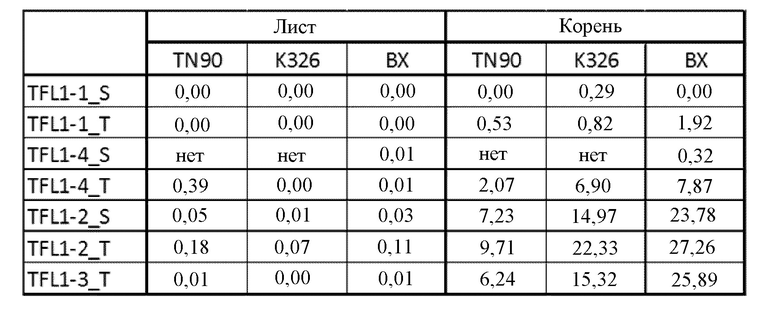
Таблица 2
Статистические анализы данных, представленных на фигуре 2, проводили с помощью модифицированного двухвыборочного критерия Уэлча (Biometrika (1947) 34 (1-2): 28-35; BioMed Central Medical Research Methodology (2012) 12: 78)

ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO: 1
Геномная последовательность гена TFL1-1S N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
tgtttattgtacattttgaactgcatcgagctgcacgattgacagatctagtctgcaacagagtactttggtttctaattatagtaatcctaacacctcagaaatttccaaaattggaattatgggtgcaaatcatctttatatatactctcaaatgacttctcagatcttacggtcactaattgtctgtaaattaagtatggtggagttcactttatatactctgagcggtgtgcccatcctcctccatcagaatatcacttaaaacaattaacgtactttataatctttccctattgtttacacacaatttaggaaagtttctaacaaagataagcccatatttcaagattagtgtcttaggattaccctaaatgaaaaaagcaatagattccctggcaatatgtttacttatattttgaatttttgcaaaaaaaataaataaatgtgtgtcaacttgcttagttgggaccacatgcaaaaactaattcaaggatttcatctgataattttatagtggagaggaaaaggcttggattaatttaagtacttattatgtagggcaaatatcacttttagcctgcggccaccatatttatattcaagccgtaaaagtgtataaaatttgtatttttttatatataatatacggaaatgtgtgtatatatatatatatatatatatatatatatatataagaaatataaaaaaaattggctattatttttcagagcagctatacaatatcattttccatatcaggtaagctgaacaacaaatcatggaccaattcgaagagcaccagtcaggtgtgaaagagaactgacatgatagcataaaatacatatcactaactcctccacatctaagagcatacatttaatttcctatatggaagataagattaattagaaaagtgtttgataaagctaagatatgttagtaccagtttgtagtatccagttgaaattttagtgtcaacttaaggataattttactatgattagaagacagttatacattcacaaagtttctgaaaggaacttataagtcttctttttattgtacttacatttgtcaagtcatatatagtacatatgacccctcctaaaggaaaagaaagtagggaaagtaccaagctagggcatatttaaaggaaaataaaatggcaattttaagatgttagagtaagggcaggggtaggccaaagaagtattggggagaagtgattagacaagacatgccactgcttcacctaaccgcggacataactagcgataggaagatgtggaggtcgagaattaaggttgtgagttgacaggtagttatgagtttactagtaatactagtactattcttgtattcttttattcttagttttttattactttgttatgtcactcgcttccattactagttatctgttgttattgcttgttgctttatttttaccatttttttagccgagggtctatcaaaaacagtctctctgcctttataggataggggtaaggctgcgtacacactaccctttccagaccccacgtatgggattaatccgagtttgttgttgttgttgttgttgatgttgttgttgttgaaaaggaaacgtttataattaagtacatgtattaagatttcattattttcggggaagaatttcgaatgtatttaaaccataaaacgtcattctcctgcaagtatttttggtgatcaagataagtatagttaaaaattgagcaatcttctttgcctacgatgtcctgtataaaactaacaaagaaaaaattcagtttatttttcaggggtagggttactttttttttaaaaaatattctatttgaagttccataaaatcccattttttatttgcaactatattgagtagatgttccgagacagcaactataagtgaggctctatttcactgttcaaccaaaatctctcacaagctaagttccttgcaatccaaaagatctctctctctctctctctctctaatggcaaggaatgtagagcctctagtagtagggagagtagtaggggatgttcttgattcatttagtcccacagttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttcccttctgcagtcactattagacctagggttgaagttcaaggtggtgatatgagaactttcttcacattggtaacttttctaattttccttcaggttattaacttttcattgctatatagacatctttcaaacctgtctagaagattcttttcttgagctgagggttcggagacagcctctctatcccacacaagataaggttaacgtctgcatataccttgccgtcccctgacaccacatatgggattatactgcgtatgttgttgttgttgtattgctatatagacatctttgataaatcactaatcatgcatgatattattcttcttattgtaggtcatcacagatcctgatgtacctggccctagtgatccgtatctgagagaacatgttcactggtatgtactacattcaagaaaatctatggaaaaaaaataactagaagagatgagtccaaaagaaaggagattaattattgtttgatttgtattttttatttgcattagattaagtcatgcaattacctagtaataaattcgttataaggacatacacaaaaggtatgctcaacataattaaagaaagaaaagcataagcattatattcttatatgcaagctcccaatctgattgtggcgaagctaagaatttcttcgggtgtttaaatttaaaagaagtgaaaaaaaatccgataaaaaatttatgtgttatatacctctaaaacttaatattgtacctatatacatattgtaatttttcgacgaatgaccactcttattgcccatgctgccaatgccaatatgtaaaatacttcgccaatagacaaatatgtatcacattaattagttacctaaataggtaattatccataataagtgggactcgtaacttgaaaaatatggttagtaatttgctacaacatgttaagatacagtgagaatgtaaatgtatatatgtcctttcacgccgtcgtaagtttatataagttaaatctggttaacaatgtaggtcatcataacactcacaattgggaataataactcttaatcatacttctttacattcatatttgcaggatagtgactgatattccaggaactacagatgccacctttggtaaattggctattttggtttcttttattaagctggtgtttgacatgctagaattaatgtactttaattttgagcaggaaaagagttggttagctatgagatcccaaggcctaatattggaatacataggtttgtgtttgttctctttaagcagaaatgcagacaatcagtcagcccacctacttcaagggatcatttcaacactcgcaactttgccaacgtaaatgaccttggtccgcctgtcgccgccgtcttcttcaatgcacaacgagagaccgccgccaggaggcgctaa
SEQ ID NO: 2
Геномная последовательность гена TFL1-1S N. tabacum
atggcaaggaatgtagagcctctagtagtagggagagtagtaggggatgttcttgattcatttagtcccacagttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttcccttctgcagtcactattagacctagggttgaagttcaaggtggtgatatgagaactttcttcacattggtaacttttctaattttccttcaggttattaacttttcattgctatatagacatctttcaaacctgtctagaagattcttttcttgagctgagggttcggagacagcctctctatcccacacaagataaggttaacgtctgcatataccttgccgtcccctgacaccacatatgggattatactgcgtatgttgttgttgttgtattgctatatagacatctttgataaatcactaatcatgcatgatattattcttcttattgtaggtcatcacagatcctgatgtacctggccctagtgatccgtatctgagagaacatgttcactggtatgtactacattcaagaaaatctatggaaaaaaaataactagaagagatgagtccaaaagaaaggagattaattattgtttgatttgtattttttatttgcattagattaagtcatgcaattacctagtaataaattcgttataaggacatacacaaaaggtatgctcaacataattaaagaaagaaaagcataagcattatattcttatatgcaagctcccaatctgattgtggcgaagctaagaatttcttcgggtgtttaaatttaaaagaagtgaaaaaaaatccgataaaaaatttatgtgttatatacctctaaaacttaatattgtacctatatacatattgtaatttttcgacgaatgaccactcttattgcccatgctgccaatgccaatatgtaaaatacttcgccaatagacaaatatgtatcacattaattagttacctaaataggtaattatccataataagtgggactcgtaacttgaaaaatatggttagtaatttgctacaacatgttaagatacagtgagaatgtaaatgtatatatgtcctttcacgccgtcgtaagtttatataagttaaatctggttaacaatgtaggtcatcataacactcacaattgggaataataactcttaatcatacttctttacattcatatttgcaggatagtgactgatattccaggaactacagatgccacctttggtaaattggctattttggtttcttttattaagctggtgtttgacatgctagaattaatgtactttaattttgagcaggaaaagagttggttagctatgagatcccaaggcctaatattggaatacataggtttgtgtttgttctctttaagcagaaatgcagacaatcagtcagcccacctacttcaagggatcatttcaacactcgcaactttgccaacgtaaatgaccttggtccgcctgtcgccgccgtcttcttcaatgcacaacgagagaccgccgccaggaggcgctaa
SEQ ID NO: 3
Аминокислотная последовательность гена TFL1-1S N. tabacum, полученная из SEQ ID NO: 1 или SEQ ID NO: 2.
MARNVEPLVVGRVVGDVLDSFSPTVKMTVTYNNKQVCNGQELFPSAVTIRPRVEVQGGDMRTFFTLVITDPDVPGPSDPYLREHVHWIVTDIPGTTDATFGKELVSYEIPRPNIGIHRFVFVLFKQKCRQSVSPPTSRDHFNTRNFANVNDLGPPVAAVFFNAQRETAARRR
SEQ ID NO: 4
Геномная последовательность гена TFL1-1T N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
ttcgagtcgtgataagtgttttgcaaaaatataagataagactgcatgcgtacaatagactcttttggtccggcccttttctggaccctgcgcataacggaagcttagtgcacccggcaaccgtttttcaacgttccagatgcaagaatattataaggaggctttttgacacttttaaatttaatctaatagagtttaagtttcatgcatcgactgtcaaaaaaatatttatataatcactaatacgataattacaagtgaaatgctatactaaacattaagtagtaacctaataaaacggtagctagctaatctactatatatcacgtagtattaaaattacacttatgtaaaaatctttacggtgttaagaataacttaaaagcaatttagtaatacattaatgagtgaaaaaggtgaagaggaaacgtggtgtgttttgaactgcatcgagctgcacgattgacagagctagtctggaacagagtactttggtttctaattatagtaatcctaacatctcagaaatttccataataggaattatgggtgcaaatcatctttatatatactctcaaattaatgacttctcagcaagtactgtcattgtctgtaaattaagtatggtggagttcactttatactctgagcagtgtgcccatcctcctccatcacttaaaacaattaatgtactttataatctttccctattgtttacacacacaatttaggaaagtttttaacaaagacaagcccatgtttcaagatttgtttcttaggattaccctaaatgaaaaaagcaatagattccctggcaatttacttatattttgaatttttgcaaaaacaaaaagaaaaagtgacaacttgcttagttgggaccacatgcaaaaactaatttaaggaattcatctgatatttttatagtggagaggaaaagggctggattaatttaaatattccttatttgccaaaattatttatattcgatagctgtaaaaatatataaaatttgtatatttgtttttgtatagtatacacggaaatgtatatatatacaagaaattaaaaaaaaactattattttcagagcagttatacaatattattttccctatcatgtaagcttagcttatcaacaaatcatggaccaattctaagagctccattcaggtgtgaaagagagctgacatgatagtataaaatacacatcactaactcctccacatttgagagctatagattaatttcctatatggaagataagattaattagaaaagtgtttgaataagctaagatgtgttagtaccagtttgtaatatcaagctaaaattttagtgtcaatttaaggataattttactatgattagaagacaagttatcattcacaaatttctgaaaggaacttataagactttttatttttattttttattgtacttacatttgtcaattaagtacatgtgacccctcctaaaggaaaagaaagtagggaaattaaagtaccaagttagcgcatacttaaagccaaggtaagcaaaatggcaattttaaggtgttgtagaggaaacgtttataattaagtacacgtactaagatttcattattttcgtggaagaaatttagaatgtatttaaaccataaaacgtcattctcgtgcaagtatttttggtgatcaagataagtatagttaacgttgaacaatcttctttgcttactatgtcttgtataaaactaacaaagaaaaattcagtttatttttcaggggtagggttacttttttaaaaaaaaatattctatttgaagttccataagatcccatttttaatttgtaactatattgagtagatattccaagacagcaactataaatgaggatctatttcaccgttcaatcaaaatctctcacaagctaagttcctagcaatccaaaaaagatctctctctctctctctctctctctctctctctctctctctctctctctctctctctctctaatggcaaggaatgtagagcctctagttgtagggagagtagtaggggatgttcttgattcattcagtcccaaagttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttcccttctgcggtcaccattagacctagggttgaggttcaaggtggtgatatgagaactttcttcacattggtaacttttctaaattctccttaaggttattaacttttcatttctatatagacacatcgagggtcaacagagacaactctatctcacacaaggtaggggtaaggtattcgtataccctacccggcccacatgtgagatcacactgaatatgttgttgttgtcgcatttaggggtgtacaaatgaaactgacaaactgcaccaatctgataatccgagtcaaatcgagaaaaaatccgattatggtttggtttgatttggtttggtgatggaaaaaaccccgacatatttggttttgtttggttttaactaaaaaaagtcaaaccgaaaccaaaccaaccagacattatatgtgtagaaattttaaatatatttaatacataaaaatatttatggtagtgtaatttataaatatttcttaagatttttcatagtttatcttttaacgtattatttcaaacttgggcttataatttttggatgctccaataagttttatagtccataaatgttagtaactcaaataaatcctaaaccaaaatcaaatcaatactaatgctaataaaagacattcaattcaattgtactatgaatgaaaatagtgttggatatatatttttatagtttttccacggtttagataaaatgtataacttatttttctttgagtatggttagtcatgtaaataatcttattaatcataattttaaattatgtttatttttattatggcttattaataatatttaattttttgtgcaattttattatctttattgttgaatattttagtacaatgccacgactcatctcatatttatgttattttattgaaaaacacctcatatagttctgcctcattaggattaaaaaaatatttggagcacaaattttactttttgtgttatgaagactttatgaaaaaaaaataaaataaaaacccgaaaacccgaaacctcgagaaaaatcgagattaaaaatccgacttttattggtttggtttggtatttagatttaataacccgatacaattagtttggtttggtaattagaaaatccgaatcaaacccctaaccgtgtacacccctagtcgtattggtatatagacatctttgataaatcattaatcatgcatgatattcttcgatctccttattgtaggtcatgacagaccctgatgttcctggccctagtgatccgtatctgagagaacatcttcactggtatgcactatattcaagaaaagctatggaaaaaaataactagatgagataggtaaaaaagaaaggagtttaatgattgtttgatttgcattaattatattaagtcatgccattatctagtaataaactggttataaggacatacacaaaaggtatggtcaacatataatcaaagaaagaaaagcataagcatccttatgcaagctgccaatgtcatgtaaaatacttcgctaatagacaaatatatattacattagttacctaaaagataggtatataatcagtgggactcctaacttaaaaaataaggttagtaatctgctataacgatacactgagaatcgtcgtcgtcagtttataagttaaaaattaatgtaggtcatcacaacactcacaaagagtgcctcaattgggaagagtatgttatatagttagaatttatgttacatatggaaccacagtactacagaaggataactcttaaacatacttctttaccatcatatttgcaggatagtgactgatattccaggaacaacagatgccacctttggtaagctcactattttggcattttcatttttccttcaatttcttttagtatatagctaggttggtttttgacatgctaattttgagcaggaaaagagttggttagctatgagatcccacggcctaatattggaatacataggtttgtgtttgttctgtttaagcagaaatgcagacagtcagttagtccacatgatgtttccagagatcacttcaacactcgcaactttgccaacgtaaacgatcttggcccgcctgtcgccgccgtcttcttcaatgcacaacgagagaccgccgccaggagacgctaa
SEQ ID NO: 5
Геномная последовательность гена TFL1-1T N. tabacum
atggcaaggaatgtagagcctctagttgtagggagagtagtaggggatgttcttgattcattcagtcccaaagttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttcccttctgcggtcaccattagacctagggttgaggttcaaggtggtgatatgagaactttcttcacattggtaacttttctaaattctccttaaggttattaacttttcatttctatatagacacatcgagggtcaacagagacaactctatctcacacaaggtaggggtaaggtattcgtataccctacccggcccacatgtgagatcacactgaatatgttgttgttgtcgcatttaggggtgtacaaatgaaactgacaaactgcaccaatctgataatccgagtcaaatcgagaaaaaatccgattatggtttggtttgatttggtttggtgatggaaaaaaccccgacatatttggttttgtttggttttaactaaaaaaagtcaaaccgaaaccaaaccaaccagacattatatgtgtagaaattttaaatatatttaatacataaaaatatttatggtagtgtaatttataaatatttcttaagatttttcatagtttatcttttaacgtattatttcaaacttgggcttataatttttggatgctccaataagttttatagtccataaatgttagtaactcaaataaatcctaaaccaaaatcaaatcaatactaatgctaataaaagacattcaattcaattgtactatgaatgaaaatagtgttggatatatatttttatagtttttccacggtttagataaaatgtataacttatttttctttgagtatggttagtcatgtaaataatcttattaatcataattttaaattatgtttatttttattatggcttattaataatatttaattttttgtgcaattttattatctttattgttgaatattttagtacaatgccacgactcatctcatatttatgttattttattgaaaaacacctcatatagttctgcctcattaggattaaaaaaatatttggagcacaaattttactttttgtgttatgaagactttatgaaaaaaaaataaaataaaaacccgaaaacccgaaacctcgagaaaaatcgagattaaaaatccgacttttattggtttggtttggtatttagatttaataacccgatacaattagtttggtttggtaattagaaaatccgaatcaaacccctaaccgtgtacacccctagtcgtattggtatatagacatctttgataaatcattaatcatgcatgatattcttcgatctccttattgtaggtcatgacagaccctgatgttcctggccctagtgatccgtatctgagagaacatcttcactggtatgcactatattcaagaaaagctatggaaaaaaataactagatgagataggtaaaaaagaaaggagtttaatgattgtttgatttgcattaattatattaagtcatgccattatctagtaataaactggttataaggacatacacaaaaggtatggtcaacatataatcaaagaaagaaaagcataagcatccttatgcaagctgccaatgtcatgtaaaatacttcgctaatagacaaatatatattacattagttacctaaaagataggtatataatcagtgggactcctaacttaaaaaataaggttagtaatctgctataacgatacactgagaatcgtcgtcgtcagtttataagttaaaaattaatgtaggtcatcacaacactcacaaagagtgcctcaattgggaagagtatgttatatagttagaatttatgttacatatggaaccacagtactacagaaggataactcttaaacatacttctttaccatcatatttgcaggatagtgactgatattccaggaacaacagatgccacctttggtaagctcactattttggcattttcatttttccttcaatttcttttagtatatagctaggttggtttttgacatgctaattttgagcaggaaaagagttggttagctatgagatcccacggcctaatattggaatacataggtttgtgtttgttctgtttaagcagaaatgcagacagtcagttagtccacatgatgtttccagagatcacttcaacactcgcaactttgccaacgtaaacgatcttggcccgcctgtcgccgccgtcttcttcaatgcacaacgagagaccgccgccaggagacgctaa
SEQ ID NO: 6
Аминокислотная последовательность гена TFL1-1T N. tabacum, полученная из SEQ ID NO: 4 или SEQ ID NO: 5.
MARNVEPLVVGRVVGDVLDSFSPKVKMTVTYNNKQVCNGQELFPSAVTIRPRVEVQGGDMRTFFTLVMTDPDVPGPSDPYLREHLHWIVTDIPGTTDATFGKELVSYEIPRPNIGIHRFVFVLFKQKCRQSVSPHDVSRDHFNTRNFANVNDLGPPVAAVFFNAQRETAARRR
SEQ ID NO: 7
Геномная последовательность гена TFL1-2S N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
catgaccttttagctactcttaactcttctgattgttctgctgtaacttgtccccttgagttaaatgtaaagttaaaggctaaagaaggggatcctctccctaatcctgaaaattatagaggcctcgttggtaagctaaatttcctcactcacactaggcctgacataagttttgttgtgcaacatcttagtcagttcatgcaacagccctgctttcctcacatgaaggcagctttgcacctgttgaggtatctcagagacacttctaattttggcctcttatactcgaattctactgatctctctttgcaggcttattgtgatagtgattggggatcctgccctgataactggagatttgtttctgatttctgtttattctttggtggcagtctcattgggtggaaatctaagaaacatgcagtggtctctttatcttcggctgaagttgagtatagatctatgagcaaggctgtggctgaaattacttgggtgtgtaggcgtctatctgatcttggggtctcttctgcttctcttgttcctctccattgtgacagtatctctgccattcacattgcctacaatcctgtcttctatgagcggaccaaagacattgagttggattgccattttgaacgtaccaagcttgctgaaggtctcatcagtttatctcacatttccagtgcttctcagctcgcgaatgtcttcatcaaacccctgtgtgggccttctcaccatcttcatattcgaaagttgggagttctctcaccctcctacttgagggggggctgttgagataggctgaaatcagtgtggctcagacccaattattatttatttatgtacatcagattaggcccattagttagtctttagttagtcttttatttctttacatatattgggccatgtatacatacatagagacccgattttgtaatagttagatgattcatttttcggttcttaatcaataagaaatatctcgaactttctctctatctctctttaaccctaaattcttcttcgttgaatctacgagaatgatgaacattaacattagaaaatgtagatttgatcaaatcttcttaatcttttgtttatcatcttttctaattgttttgtatctgattgtatattagttaaccaccaaaattgctcaaacaatctggcttcaaatttatctaacgtttgaatatatatatatggttgaaacatgaaaaataaatttttgaagatgagatgaaaaataatttttgaaagttaaaattgtatttgaacacgtattttacttgaaaagaatttgaaattttgtgagcagaaaacttaaaaaattactctaaaacttttttttgagatttgaggattttattttcaaaattttccataaaatggcttaaatctataagcaaaagatatttgaaaataattttttttttaaaaaagctctcaaattttacagccaaacggaagcttaggataaaaaagggggaaatgggggagatgggtgggcaggttgggctgaagagaaatagacaacagtgcattaacatgtcaaatcatctttatccctctttctaaaccttacaaggagtactttttattttcttttttctttttttggccctaataaaaattaaaacacatattctctagctgctaagctataactttaactcattggcaccacgacgagtaggagaataacctttttgggcttttcttttcttttctttggtcccctttttttgaactatcaatattttagtccaaacacacctgactctacagtgatctgatggccactataaatattggctttttgcaactctcttctcaccaaaatacaaatcggttgaactcttcatatataatattcccactactattactcttaactttaaatagatttcttatatatgggttcaaaaatgtctgatccccttgtgattggtagagtgattggggaagttgttgattatttcactccaagtgttaagatgtctgttacttataacagcagcaagcatgtttataatgggcatgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttacaatggtacatactgcttccttcgattttcaatacttttattaggggtggagcttagcggcggagccaagattttaactaaggggagtcaaaatataaataagtaagcacacaaaaaaatcaagggggtcaacgtatagtatatacacataaaattaagaatttaacatatttataccgtgtaattttccagcgaaggggtgtcaattgactctccttgccaatgagtggctccgccactggcggagctagagttctagttacggttcgttgtattgtgttaagaagtccacttatactgtcttttctagaatttagaattcataaattcaaaattatggctctgcccttaaatttatttttatacatttctattatatagtaaatcgtttatattgaccccttatttttcttttttaccttaattgacagatcatgatagacccagatgttcctggtcctagtgatccatatctcagggaacacctacactggtaaagaaataagttttttaattactaactcattcaattttatcgtcccttcttttccttgtttacttggagggaaaataatacgatctcatcgaaaagataaaaattcttcaggcttgttatctaaaaacttgttaaaaaataccgtaatgaaaagacatatgagtttgttattaggtatttgactaaatatgatcgatcatatggtgttcggacaagaaatattttgtgaaaaggtccgcatacttttaaaaaaagaaaatctgccttgactcttgagtttgtgcttctcgggaaaacaatttcttccttctttttttttttttttttggtttattgacctttacatattaaagacaccactgagacacatatctagaaaaattgtatttgggaacgcaaaagcaaagaaaacatgtgttattaatcttatgtcaatgccaccagcagctcaggaaaaatatggtcgatatattgtgatttgcttgcaaaaggagcaaagaagaaatcttttgataatgtttgttatgacgatgtacttaaagcaaataagttagaggtcgtttggtacatgggataaggataataattttgggataaagtttaggattaactttatcttatatttggtttggagtattagctaaccgcgaggtatttttcaaactaaaatagtgggattagctatcccatataaaaagtaggatagctaatcccatgggatatcccaccctatcggatagtaatagtccaataggagacaactctaatttgtacagacataatgtccagtcacaccttgtttttttgtcatgacacatattaagcatgaataataatatttcgacaatcttgtagcatttgattagacttagcaaattataaatatgtccaataattggtcacattgttctaataattacttgtttcccttatcattatatatagtgcttcattcactaaacagaacccaaaaaaaaaaaaaaaaaaactgcaaaatggtcatatcatgtagtaacggaataaaaacgtactcagttttatgataaaatcaaagtgacatatttgtacgctttgatagttgacaaatacctgaaaaaagaatttgaccatctttacaggattgtcacagacattccaggcactacagattgctcgtttggtatgtatctttaacccaaatttcaagcttcgaaatagtaacagcttttgtttttaatattttatttgtcttaaatacatattttccttattataaatttcttcgcctagtggtaacgggatcaggtattgattcgtatttattttttattgatcaacaaaaaaagagtacaaaagaaagaattgtttttctacacttagatttatatatatgcaatgtctagaaattaatgagtttacaaattcattgatgtgtatatctcacaatcaaatccaaaatactgatccaaaaattttgatcagggaaagaaatagttggctatgaaatgccaaggccaaatattggaattcacaggtttgtatttctgctgttcaagcagaagaagaggcaaacagtattgactgcacctctctccagggatcgatttaatacgcgtaaattcgcagaagaaaatgagcttgggtctcctgttgcagcagttttcttcaattgccagagggaaactgctgccagaaggcgttga
SEQ ID NO: 8
Геномная последовательность гена TFL1-2S N. tabacum
atgtctgttacttataacagcagcaagcatgtttataatgggcatgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttacaatggtacatactgcttccttcgattttcaatacttttattaggggtggagcttagcggcggagccaagattttaactaaggggagtcaaaatataaataagtaagcacacaaaaaaatcaagggggtcaacgtatagtatatacacataaaattaagaatttaacatatttataccgtgtaattttccagcgaaggggtgtcaattgactctccttgccaatgagtggctccgccactggcggagctagagttctagttacggttcgttgtattgtgttaagaagtccacttatactgtcttttctagaatttagaattcataaattcaaaattatggctctgcccttaaatttatttttatacatttctattatatagtaaatcgtttatattgaccccttatttttcttttttaccttaattgacagatcatgatagacccagatgttcctggtcctagtgatccatatctcagggaacacctacactggtaaagaaataagttttttaattactaactcattcaattttatcgtcccttcttttccttgtttacttggagggaaaataatacgatctcatcgaaaagataaaaattcttcaggcttgttatctaaaaacttgttaaaaaataccgtaatgaaaagacatatgagtttgttattaggtatttgactaaatatgatcgatcatatggtgttcggacaagaaatattttgtgaaaaggtccgcatacttttaaaaaaagaaaatctgccttgactcttgagtttgtgcttctcgggaaaacaatttcttccttctttttttttttttttttggtttattgacctttacatattaaagacaccactgagacacatatctagaaaaattgtatttgggaacgcaaaagcaaagaaaacatgtgttattaatcttatgtcaatgccaccagcagctcaggaaaaatatggtcgatatattgtgatttgcttgcaaaaggagcaaagaagaaatcttttgataatgtttgttatgacgatgtacttaaagcaaataagttagaggtcgtttggtacatgggataaggataataattttgggataaagtttaggattaactttatcttatatttggtttggagtattagctaaccgcgaggtatttttcaaactaaaatagtgggattagctatcccatataaaaagtaggatagctaatcccatgggatatcccaccctatcggatagtaatagtccaataggagacaactctaatttgtacagacataatgtccagtcacaccttgtttttttgtcatgacacatattaagcatgaataataatatttcgacaatcttgtagcatttgattagacttagcaaattataaatatgtccaataattggtcacattgttctaataattacttgtttcccttatcattatatatagtgcttcattcactaaacagaacccaaaaaaaaaaaaaaaaaaactgcaaaatggtcatatcatgtagtaacggaataaaaacgtactcagttttatgataaaatcaaagtgacatatttgtacgctttgatagttgacaaatacctgaaaaaagaatttgaccatctttacaggattgtcacagacattccaggcactacagattgctcgtttggtatgtatctttaacccaaatttcaagcttcgaaatagtaacagcttttgtttttaatattttatttgtcttaaatacatattttccttattataaatttcttcgcctagtggtaacgggatcaggtattgattcgtatttattttttattgatcaacaaaaaaagagtacaaaagaaagaattgtttttctacacttagatttatatatatgcaatgtctagaaattaatgagtttacaaattcattgatgtgtatatctcacaatcaaatccaaaatactgatccaaaaattttgatcagggaaagaaatagttggctatgaaatgccaaggccaaatattggaattcacaggtttgtatttctgctgttcaagcagaagaagaggcaaacagtattgactgcacctctctccagggatcgatttaatacgcgtaaattcgcagaagaaaatgagcttgggtctcctgttgcagcagttttcttcaattgccagagggaaactgctgccagaaggcgttga
SEQ ID NO: 9: Аминокислотная последовательность гена TFL1-2S N. tabacum, полученная из SEQ ID NO: 7 или SEQ ID NO: 8
MSVTYNSSKHVYNGHELFPSSVTSKPRVEVHGGDLRSFFTMIMIDPDVPGPSDPYLREHLHWIVTDIPGTTDCSFGKEIVGYEMPRPNIGIHRFVFLLFKQKKRQTVLTAPLSRDRFNTRKFAEENELGSPVAAVFFNCQRETAARRR
SEQ ID NO: 10: Геномная последовательность гена TFL1-2T N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
agggcacgaccctaagaccttcttcatagctataaatagtgagctcaggtttcattgtaaatggaacgactattctggcaaacttatacaatattttatacaaaactcaattcaatcttatcttctgatttctagattctttttgtttttgtgcccgaaaaccttgttcctggaattgttgcttctgttgtttcgtccatatcttaaggctaagtgttatataattcttcaattatttatttatttatttcaggttcaaattaattcacttatctaaaaatcatgtataaatttaattgtaccattttacgggtgaacagtttggcgcccatcgtggggcctagataaccgtgtaactaaaggacaaacgtcttttcgggaacttttctattttcaagaactcaaacccgagatttagacctctgagggatctgatcatctcactacatcgctgagtggtagttgattccatatacgattaacctagtttacaactaaattaaattatgtgcattaatccaagcaacttttgatgatcagctgatcaacctaacgtaagaaagcaattaatttagatgcatatattctacaaatggaaattagtaggagcaagcaagttatgcaaaagaaaggaaaagagaaaacattagaagtaggccaaagaaagaagaaggaagaggaagcaatcagccactgttctagaatggaatatggagaaaaataataaattaaattcagatttctataagtagtaatcctcttctttctattaccggttaaagctgcagaaattttctttttcttgacatgacctgaccatagcttccaccattgtttgcaggctggtggtggagtccctttataccctcatctctcctacctaagaaccataggattaggtgattcaagtttttatttttaacaaaaaatgaaaaatttatgaaggaagttcaactttttattaccttaaataaaaaagaccttgatgctttaagtagctccaagacggtagctgcaaattccatctgcttttcctttttaataaaataatgtactacctactatctgaaagtttaacttctatgattctgtaggttttgtaaaacacttgggggtatttatattttataggggattgcaattagaggcagatacaatttggtttagttaaccaccgatattactcaaacaatttggctttaaatctggttagtgtttggatatagatttggttgaaatttgaagaaaaaaaatgagtttttaaaaatgagatgaaaaataatttttgaaagttaaaattgtatttggacatgcattttatttgaaaagaatttgaagttttgtaagttaaaattttcaaaaacttcaaaaagttatttttgagatttgaagattttattttcaaaatttgcattataatctataaacaaatagatactatttgagaacaaaatttaaaaaataaagctttcaaacttatgacgaaagggaagcttaggataaaaaggggggaaatggcctgggagatgggtgggcaggttgggctgaagagaaatagacaacagtgcattaacatgtcaaatcatctttatccctctttaaaaacattattaggagtacttctttttttcttggggtgcaaaagcctaatacaagttaaaacacatattctctagctgctaagctataactttaactcattggtaccacgacgagtaggagaataaactttttgggcttttcttttcttttctttggttcccattttttgaactatcaatattttagtccaaacacacctgactctacagtgatctgatggccactataaatattggctttttgcagctccaaaatacaaatcggtcgaactcttcatatatattactcttaactttaaataaatagatttcttatatatgggttcaaaaatgtctgatccccttgtgattggtagagtgataggggaagttgttgattatttcactccaagtgttaagatgtctgttacttataacagcagcaagcatgtctataatggacatgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttacactggtacatactccttcgattttcactacttttaatttattaggggcgaagctagagttctagctacgggttcgttgtattaattgtgttaagaagtccacttaagctgtcttttttagaatttagaatccataaactcaaaatagtgactttgcttctaaattaatttttatgcatttctcttatatcgtgtatgtgaatattgaccccttattttttcttttttaccttaattgacagatcatgatagacccagatgttcctggtcctagtgatccatatctcagggaacatctacactggtaaagacatacgttttttaattactaactcattcaattttatcgccccttcttttccttgtttacttggagggaaaataatacgatctcgtcaagaagatcaaaaatcttcaggcttgttatttaggaacttgttcaaaaataccgttttgaaaagaacatatgagtttgttattaggtatttgactaaataggaacgatcatatggtgttcggacaagaaaatttttgtgaaaaggtccgcatactttaaaaaaaaaaaaaaaaaaaaaaaatccgccttgactcttgagtttctgcttcttggaaaaaacatttcttctttttttttttgggttttttgacctttatatattaattaaagacaccactgagacacttaattaaaaaattgtatatgggaacgcaaaaagaaaaaaaaacatgtgttattaatcttatgtcaatgccatcagcaactcaggaaaatacggtcgatatactgtgatttgcttgcgaaaggagcaaagaagaaatcttttgataatgtttgttatgacgatgcacttaacctaaaataagttaggggccgtttggtaaatgaaataaggataataatctcggaacaaagtttaggattaactttatcccatatttgatttggagtattagttaattgcgggataactttcaaattaaaatagtaggattagttatctcatatataaagtaaaatacctaatcccaataatataataggagacaactctaatttgcgtagacataatgtccagtctcactttgtatatttgtcatgacgcatattaagcatgaatgataatatttcgacaatcttgtggcatttgattacactcagcaaattataaatatgtccaataattgcattaataattacttgttcctcttatcattatagtgcctcattcactaaaccgaacccaaaagaacactgcaaaatggtcatatcatgtagtaacagaaaaaaaaaacgtactcgattttatgataaaatcaaagtgacatatgtgtcgctttgataattgacaaatacctgaaaaaagaatttgaccatctttacaggattgtcacagacattccaggcactacagattgctcgtttggtatgtatctttaacccaaagttcaagctatgaaatagtaacagcttttctttttaatattttatttgtcttaaatacatattttccttattataaatttattcgcctagtggtaacgggatcaggtattgattcgtatttaatttttattgttcaacaaaaaagagtacaaaaagaaagaattgattttctacacttagatttatatgcaatatctagaaatcagaagatcagcaatgagtttactaattcatcgatgtgtatatcgcacaatcaaatccaattactaataatactgatctaaaaatttcgatcagggagagaaatagttgggtatgaaatgccaaggccaaatattggaatccacaggtttgtatttctgctgttcaagcagaagaagaggcaaacattattgagtgcacctctctccagggatcgatttaatacgcgcaaattctcagaagaaaatgagcttgggtctcctgttgcagcagctttcttcaattgccagagggaaaccgctgccagaaggcgttga
SEQ ID NO: 11: Геномная последовательность гена TFL1-2T N. tabacum
atgtctgttacttataacagcagcaagcatgtctataatggacatgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttacactggtacatactccttcgattttcactacttttaatttattaggggcgaagctagagttctagctacgggttcgttgtattaattgtgttaagaagtccacttaagctgtcttttttagaatttagaatccataaactcaaaatagtgactttgcttctaaattaatttttatgcatttctcttatatcgtgtatgtgaatattgaccccttattttttcttttttaccttaattgacagatcatgatagacccagatgttcctggtcctagtgatccatatctcagggaacatctacactggtaaagacatacgttttttaattactaactcattcaattttatcgccccttcttttccttgtttacttggagggaaaataatacgatctcgtcaagaagatcaaaaatcttcaggcttgttatttaggaacttgttcaaaaataccgttttgaaaagaacatatgagtttgttattaggtatttgactaaataggaacgatcatatggtgttcggacaagaaaatttttgtgaaaaggtccgcatactttaaaaaaaaaaaaaaaaaaaaaaaatccgccttgactcttgagtttctgcttcttggaaaaaacatttcttctttttttttttgggttttttgacctttatatattaattaaagacaccactgagacacttaattaaaaaattgtatatgggaacgcaaaaagaaaaaaaaacatgtgttattaatcttatgtcaatgccatcagcaactcaggaaaatacggtcgatatactgtgatttgcttgcgaaaggagcaaagaagaaatcttttgataatgtttgttatgacgatgcacttaacctaaaataagttaggggccgtttggtaaatgaaataaggataataatctcggaacaaagtttaggattaactttatcccatatttgatttggagtattagttaattgcgggataactttcaaattaaaatagtaggattagttatctcatatataaagtaaaatacctaatcccaataatataataggagacaactctaatttgcgtagacataatgtccagtctcactttgtatatttgtcatgacgcatattaagcatgaatgataatatttcgacaatcttgtggcatttgattacactcagcaaattataaatatgtccaataattgcattaataattacttgttcctcttatcattatagtgcctcattcactaaaccgaacccaaaagaacactgcaaaatggtcatatcatgtagtaacagaaaaaaaaaacgtactcgattttatgataaaatcaaagtgacatatgtgtcgctttgataattgacaaatacctgaaaaaagaatttgaccatctttacaggattgtcacagacattccaggcactacagattgctcgtttggtatgtatctttaacccaaagttcaagctatgaaatagtaacagcttttctttttaatattttatttgtcttaaatacatattttccttattataaatttattcgcctagtggtaacgggatcaggtattgattcgtatttaatttttattgttcaacaaaaaagagtacaaaaagaaagaattgattttctacacttagatttatatgcaatatctagaaatcagaagatcagcaatgagtttactaattcatcgatgtgtatatcgcacaatcaaatccaattactaataatactgatctaaaaatttcgatcagggagagaaatagttgggtatgaaatgccaaggccaaatattggaatccacaggtttgtatttctgctgttcaagcagaagaagaggcaaacattattgagtgcacctctctccagggatcgatttaatacgcgcaaattctcagaagaaaatgagcttgggtctcctgttgcagcagctttcttcaattgccagagggaaaccgctgccagaaggcgttga
SEQ ID NO: 12: Аминокислотная последовательность гена TFL1-2T N. tabacum, полученная из SEQ ID NO: 10 или SEQ ID NO: 11
MSVTYNSSKHVYNGHELFPSSVTSKPRVEVHGGDLRSFFTLIMIDPDVPGPSDPYLREHLHWIVTDIPGTTDCSFGREIVGYEMPRPNIGIHRFVFLLFKQKKRQTLLSAPLSRDRFNTRKFSEENELGSPVAAAFFNCQRETAARRR
SEQ ID NO: 13: Геномная последовательность гена TFL1-3T N. tabacum, содержащая 2 т. о. перед ATG
tgaagttgtgtttggacatgcgttgtatttgagaaaaaattgaagttttgtgagaggaatttttttgaccccaaaactacataatttgaattattatttaaaaaaaatgatcatattacatgaacaaacagtgttttcaatttatttttgaaaaaaacagccaaaatctagccaaatgggagctaagtgtatgatcaagattcatgtcccaaatggaaaagaatattaacaaaaaaaagggcagtaaagaagaggtggctacaataggatcgcgcaaaagaaagatggaaaaaagaggaacaggaggggggaataagcagcacaagaagttattataagtcagctcttccagaaaggaatatggagaaaagttaacctcaggtttctataaataggaatgtccaactttcatttactagtttaagctgcagaaattctctttttcttgacatgaccttttcaccatctttaatttggtgggctttgtggtggagtccctttataccataggctctcctagaggatccataacattagattggtaaggttctaagtggactcacgatatgaaatttgtgatcgaacctataactcgtctgagttactgaatttgtaataaaatatttatacatatttaataaattttctaatataaatacagaatctaaacaaaaactattgagttcatccgtaccgatacctaatactctagctccaaccctgattctaatataatgaaaataaaaccacatctaggaagttcatgacctgttcttaccttaaatgccaaaggccttaaacctttgatagcttgagaatagccaaacaagtatagattccatctactttaattttctttctgattaagatatattgcaactcctgtaaatgcgcaaggagtcagctggttcttcccccatttctatattttttagtatcactttcttttcttaattattccttcttacatttgaatctttttccatcagctagctgttttgatagtagtaaaaatgcgaaggctcttcttactaatatttcaatgaccaatgaatttagatggagaagcaagttctattaaacgttcatgctagaaaataacaagtataatatttcattttcattttatataacgctcttgtcttttcttgtctatttaaacaagaatcaatttaactctcttaatgacatgctctttagtcacagaaaaattataacaaattgaagacattagttattaatgttcttttcgcactaaaagttttttaaaaattttttatcttaaatgtttgtgactaatcaaatatcatcatataaaattaatctgggaatatgacatttttcaatataactaatatggtgcaaattgcatacactacgcaataaatttgtggttagggtcatatcattagcctgtcaaatcatctttatccctctttctctaaacgacttcttttctcccttttttttcgcccctcaaacaaagcaaatagactattctctagctgctaattagctaaacaatgactttaactcgttgtgcccagaggagaataacctttatctctcttctcttttctttgtttccatctttaatttagacttctttttttggttttttatcccatattcggtatttattggagttcgattaaattcaaatttataataggaagtctcacattgagagtacgatgactccatactcaggattcgaatttgagatctttagttaaagatgaaagaatatcattcaaccacaacttttgttggtcccatctttatatctatatgttcttactatattttaatccatttcccacttccaatgatttaaagaagctataggataggtgcatttggaccactataaatataggttttgcagttctatgctccatacaaatatccagcaagaaactaaactatatatttactgagttactactaatagttttcactcaatctatttccactctttctcctcttcattatattatatggctcaaatgacagatccccttgtgattagtagggtggttggagatgttgttgattatttctctccaagtgttaagatgtgtgttatttataaccccagtaagcatgtctataatgggcatgaactctttccatcccttgttacctctaaacctaaggttgaagttcatggaggtgacatgagatccttctttacactggtaattaattcacactacttcaatagttttcttgttcttatattttattatctatctatatatatataataaaggagcggcaaagccaccatataaatgacaaatgtaaacttttaggacaaaactccaaaaaagttggagttttaaaattattttatatataaaataaataaataaataaataaataaactatcaattcaaattggggagtagtttcttactaatatgatagctatatctatatctatatctatctatatatgtaaaacatttatatgatgccaagtggcataaccactgataagatttttaaatttgaatatgaatgaattttaaatgaagttctaacttcttaaaaataaaccctaatataggttactatttttagtaatgattgaaattattattaaaatattttgttgaaaacaacatagagataaaatttgattattaaatttatgtattacaacaataataattattgaaaatattgctaaaattttcatgaaaggattcacccataattattagtataatagaaaactaaaaaattattaagtctaaagttctagatctctatatttataaacgtataaactgttattttattttctgaaaaaagcaaaaatactgaagagaaaaatgataaaaatattttaaaatatgtaagtcatgtgcaaataataaagtgaacaaatgatgtagtagtatactgaataagatatgtttttttgtcataaaataagtatatgcataactcatctcaataatttgctgactccatctgagtcaaaatatcttctaaattcaagcgaagataattatctatcgcattatttttttatcattaatataaggcaagacgaatctatatctcatatgggactttttaaatagatacatctttataaatgaaccactttatgagttttatcacgaattacaagtaagaataacttgaagattgaaagaatttttggatttatttaattataatatatttttattcattttaaattaatttatattttcaaattatttgtagcaacctatataattatgatatttgagtattatcttataagttatttgatgattgtcgtttgatttaattattgaactattactacaggggacataagatgataattataattttgtagaaacatatgatctaatgtgctcaaataaattactatcatactttgatatgactaatattctttaataatttttgcgcatcgggcgggtactaatactagttcttaaaaaaagggtagcgcgatgcacaaagcattccgcattcacacaggatcctaggaattgggtcgcaccccacagtctaccctaatgcaaacattagcgactactttcacggctcgaactcgtcacttatagatcatacagagacaaatttactgttgctccaagttccctttcttattttattattcttataatttctattcttatattgttataaattatttttttctttttgatagatcatgactgaccctgatgttcctggtcctagcgatccatatcttagggagcacttacattggtatgtatcatactatcatcaactttgaaagcttaaaacactgtaaagttgatgattcacaccaaagattttaatcgtcgtcgtgttacttccatataaatcagtatcgagaagtatgtggccatcactccatcaacgacaccaaaatgaaataaagagtccctatatcaatacaatataaattaatcttaaacatgaagttgactttaaattggataaattgtttccactactaagcttagcgtataaattagtcctttgactttcaattttgtataataatgcgaagctttttttcttgtaaatgcaatttttgtccttgagggtttgcaacttcttttttaaggaaaaaaaaaagactaaagttgtgtgacactaaaaccaagagttagcttaatacttcatggacacacgttagcataaaacatataaccgatattcaaaattacaaaaatgatagaatcataatttttgtttctatttaaaaaggaagtaagccaaattactactaacatagtggacttaaagggtattaattttttgttattttaatgatatctgttcatgacttcttgactacttctactcctttatatcaatcaaattataatttacttcgtttgactatctaatttacagggtaattacagacattccaggcactacagattcctcgtttggtatggaataatattgtattccttttttacttttctgcctagcatttctaaatagagtagtccgatacacgaaatatttcactttacgcaggatccagaaataaaggaccataccccaattgggtgtaatataagtagtcatgggcgcatgcagtattttagtgacgggtttaattgcactcataattttggacgcttagcataaagtagtagatatgtatccataacttcaaaaatataataggttcaatgttaaaaatttcaaaagagatgaactcatagagtttaaatcatgatccgcctctgtaggcagtctaccctaatgaaagaatcagtggctgatttcacagttcaaaaccgtaacctatgaatcacataaagccaactttaccatcgctccaagactcgccttcttctgcctaacattactactgctaataaagagaattttaataaaactactaatgctaattattattctttgctaaaatcttcatcaggaaaagaagtggtgggctatgaaatgccaatgcctaacattggaatccataggtttgtgtttctgctcttcaagcagaagaagaggcaaacagtgagcgcaccattatccagggaccgattcaatacgcggaaatacgcagaagaaaatgagcttggctctccagttgctgctgttttcttcaactgccaaagggaaaccgcggccagaaagcgttga
SEQ ID NO: 14: Геномная последовательность гена TFL1-3T N. tabacum
atggctcaaatgacagatccccttgtgattagtagggtggttggagatgttgttgattatttctctccaagtgttaagatgtgtgttatttataaccccagtaagcatgtctataatgggcatgaactctttccatcccttgttacctctaaacctaaggttgaagttcatggaggtgacatgagatccttctttacactggtaattaattcacactacttcaatagttttcttgttcttatattttattatctatctatatatatataataaaggagcggcaaagccaccatataaatgacaaatgtaaacttttaggacaaaactccaaaaaagttggagttttaaaattattttatatataaaataaataaataaataaataaataaactatcaattcaaattggggagtagtttcttactaatatgatagctatatctatatctatatctatctatatatgtaaaacatttatatgatgccaagtggcataaccactgataagatttttaaatttgaatatgaatgaattttaaatgaagttctaacttcttaaaaataaaccctaatataggttactatttttagtaatgattgaaattattattaaaatattttgttgaaaacaacatagagataaaatttgattattaaatttatgtattacaacaataataattattgaaaatattgctaaaattttcatgaaaggattcacccataattattagtataatagaaaactaaaaaattattaagtctaaagttctagatctctatatttataaacgtataaactgttattttattttctgaaaaaagcaaaaatactgaagagaaaaatgataaaaatattttaaaatatgtaagtcatgtgcaaataataaagtgaacaaatgatgtagtagtatactgaataagatatgtttttttgtcataaaataagtatatgcataactcatctcaataatttgctgactccatctgagtcaaaatatcttctaaattcaagcgaagataattatctatcgcattatttttttatcattaatataaggcaagacgaatctatatctcatatgggactttttaaatagatacatctttataaatgaaccactttatgagttttatcacgaattacaagtaagaataacttgaagattgaaagaatttttggatttatttaattataatatatttttattcattttaaattaatttatattttcaaattatttgtagcaacctatataattatgatatttgagtattatcttataagttatttgatgattgtcgtttgatttaattattgaactattactacaggggacataagatgataattataattttgtagaaacatatgatctaatgtgctcaaataaattactatcatactttgatatgactaatattctttaataatttttgcgcatcgggcgggtactaatactagttcttaaaaaaagggtagcgcgatgcacaaagcattccgcattcacacaggatcctaggaattgggtcgcaccccacagtctaccctaatgcaaacattagcgactactttcacggctcgaactcgtcacttatagatcatacagagacaaatttactgttgctccaagttccctttcttattttattattcttataatttctattcttatattgttataaattatttttttctttttgatagatcatgactgaccctgatgttcctggtcctagcgatccatatcttagggagcacttacattggtatgtatcatactatcatcaactttgaaagcttaaaacactgtaaagttgatgattcacaccaaagattttaatcgtcgtcgtgttacttccatataaatcagtatcgagaagtatgtggccatcactccatcaacgacaccaaaatgaaataaagagtccctatatcaatacaatataaattaatcttaaacatgaagttgactttaaattggataaattgtttccactactaagcttagcgtataaattagtcctttgactttcaattttgtataataatgcgaagctttttttcttgtaaatgcaatttttgtccttgagggtttgcaacttcttttttaaggaaaaaaaaaagactaaagttgtgtgacactaaaaccaagagttagcttaatacttcatggacacacgttagcataaaacatataaccgatattcaaaattacaaaaatgatagaatcataatttttgtttctatttaaaaaggaagtaagccaaattactactaacatagtggacttaaagggtattaattttttgttattttaatgatatctgttcatgacttcttgactacttctactcctttatatcaatcaaattataatttacttcgtttgactatctaatttacagggtaattacagacattccaggcactacagattcctcgtttggtatggaataatattgtattccttttttacttttctgcctagcatttctaaatagagtagtccgatacacgaaatatttcactttacgcaggatccagaaataaaggaccataccccaattgggtgtaatataagtagtcatgggcgcatgcagtattttagtgacgggtttaattgcactcataattttggacgcttagcataaagtagtagatatgtatccataacttcaaaaatataataggttcaatgttaaaaatttcaaaagagatgaactcatagagtttaaatcatgatccgcctctgtaggcagtctaccctaatgaaagaatcagtggctgatttcacagttcaaaaccgtaacctatgaatcacataaagccaactttaccatcgctccaagactcgccttcttctgcctaacattactactgctaataaagagaattttaataaaactactaatgctaattattattctttgctaaaatcttcatcaggaaaagaagtggtgggctatgaaatgccaatgcctaacattggaatccataggtttgtgtttctgctcttcaagcagaagaagaggcaaacagtgagcgcaccattatccagggaccgattcaatacgcggaaatacgcagaagaaaatgagcttggctctccagttgctgctgttttcttcaactgccaaagggaaaccgcggccagaaagcgttga
SEQ ID NO: 15: Аминокислотная последовательность гена TFL1-3T N. tabacum, полученная из SEQ ID NO: 13 или SEQ ID NO: 14
MAQMTDPLVISRVVGDVVDYFSPSVKMCVIYNPSKHVYNGHELFPSLVTSKPKVEVHGGDMRSFFTLIMTDPDVPGPSDPYLREHLHWVITDIPGTTDSSFGKEVVGYEMPMPNIGIHRFVFLLFKQKKRQTVSAPLSRDRFNTRKYAEENELGSPVAAVFFNCQRETAARKR
SEQ ID NO: 16: Геномная последовательность гена TFL1-4S N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
ccttatgggttctcagcatttgggcaaaagtgatactttaagcaagtgaggaagttttttaatgttggccggaaagatgcctgtgggtgctgtttgggggaaaaaaacaaaagctcgggtaaattatcaatacccgagttggttcctttcgtttgccactggaccaactcctgattttgcttatatatgggttccaaactaaaaatacttatatatttaataaatttatcaatacaaatacaaggctcgggtaaaagttattaggttctcggaaacccatacccgatactatggatccgcccctgcttatccctaccttgtgtgaggtagaaacgcttttgataaggttatttaaaagtaaaagaataagtttaatgtgacaatttgaatggttgagacaacatgccaaaagctaatttaagggattttatttgacatttatatatgggagagaagaaaaagtattgcccagtatattattttaagctctacaaccaatcaagaatcaattcctagaatccattcaggtgtcaaagaatactgacatgatataataaaatacaatttatatcacatcagtatttgctttttcttgggagattagataaaaagagatcagaaatggagttttatggtactaggagaattcaaggatttactacttttgtggcacaacataagctcccaatttttttaaggaatttataaaagttggttttctaagtacttacaattgtcaaatttacaagtcatttagtacataaaaagaaacccaatgatgaggttcaggaaaaaaaaaaatcctatactgtgatttcctagttggcgttcggacataaaaattatgaaattccgaaaaaaaaaattgttttaagttgaaaatggtatgtgaaaattaaagttatatatggacataaatataatttggagctgtttttgaatttttgtgagtgctttgaagtgaaattttctaaaaacagctttttggagtttttcaaattccggagttcaacttcaagcgaaaaattaaaattttcatgatcaaatgttgattccgaaaaaagtgaaaaaattcgaaaaaaagatttttttttttatggccaaacagacctaactagtttcattttagtcattaagggtagaattgaaagaattttaaattaaagtatttttagatatataaaaataatgtactttttaaaacacacaaaaaaaggagtgccatatattaatttaatataaggatatatagtggatgcattcataactaacattaaccaaaagcatttattgatcctattttgacaccattttattttaatacaattcataaatttcaagaatttgaatacattagcttaatctcacttaaattttgaggtgatgcctgttctctttctagtcacaactttaatgtacattttatatgtcaaattaatacctgaatttgtaacccatcaaatatcgccacataatatgaaacagtgaaaatatcttatattcctgtattttatgactaagacattaagtagctaacaacgatcgaaaaacattcctaataacaagcgaattacaactctgtcggataatcgtctgaaaccctaaaaagctactgaaatgatttcctactagtataattccgatgaaattttgttcgaaaattctataagaaatacacgtatttttagtagtgaaaaaagatttgttgtaatttttttaggtggggtggggtgatttggggagggttggggagtaggacctcaaaaacaaagaattttaatactttggagtttccttaggtcccatgttttatactttcttttattctccttcaccattatagctataacttagtacatatatatatatatatatatatatatatatatatatatnatataaggtgtccatctgatcaagtatccaatacaaaccattcttaagtctttgaaaatttctctttttttccttatctctatctctgtctaattttctttattatggcaagaagtttagagcctctaattgttgggagagtagtaggagatgttcttgattcatttagtcctataatgaaaatgacaatatcatataacaacaaattagtgtgcaatggccatgaactccttccttctgttgtcactgctagacctaaagttgaagttcaagggggagatttgagaactttcttcacattggtatttttttcttgatttctacttaatttccaagatcatcaagttcccattatttctttaaaaaaaaaaaaagcagttcggtgcactaaactcccgctatgcgcggggttcggtgaagcaccgaaccataagggtctattgtacgcaaccttaccctgcatttatgcaagaggcttgctcaccattacaagttatattaatttaacatgttatatataaccacaaaggctgtcgtgggatggtaaatatccttctatccttaatcagaagtttcgggttcaagttatagccctaggaatatagtcgtctttggtagggatcctttacccccaaaactttccgccgtgaatccagattagtaaacctcaaagcgggtatcgggcattggatgacaaaccaaaaaaacttcaacgtgttatagcatgttataacttattacagttaatttagttttccagtcgatactatattaaatagagtgcctgtaatttactttggagtgatttgattgttattttttcgcatcgtcagtacataaaacttatattaattttcgaatatgtaggtcatgacagaccctgatgttcctggccctagtgatccttatctaagagagcatctccactggtatgccctaaactcaatttttttttaaaaaaaaaaaatagaaaatgagaaaaaatatgtaaaaatctacaaatatgagaagatcatgattaattggaactatttttactgactatttgacaggatagtaactgacattccaggtaccactgatgctacttttggtaagttctctgtatcttctgcaaaattacaagcacatgtgaagataaaagaagtttttctattattcacttattttgtctagctagttatatagaataattataagatcaacaattttgtatagtagtgaatgttggacttctaaagtcgaacatgtccacttgatgagtgtcacaaaaatgtagaaactaaacaatcgtttggacataaaaaaaaaagtaagtttttttgagttaaattgaaaaagaaaatatttagaatttgaaattgtggatatacatttaaattgaaaagcattgcagttttgtaaggaaaataaactttcatatacataaaaaagtgattttttggaaactcatcttcaagaatatttttaaaaatttccgtccaatgtataaccaaacattattttgaaaaagattaaaaaaaggaaaaactttaggaacaacgggtcccaagataaatgtgtctagtcatataagattagataaattaggattttattatatttggtagaaggtgcaagaagcatatgtaaataataaattgagaagtcacttaagatattttgatcatgtcccacatcgataacaagaggtaccattctatatatgttaaatcatggtaagttaaagtattatatcacatattaaatggtgatataatagacctaaatcacatgaaacgaaattgtcccgaaaggtctataaatttttgaaattcatgtagacgaagctaaaagtaggatacaataaaaaaaaaattaaagatctatattggcgatactatttagttgggattgcattttagttattctagtacatttactttaatctaatttttgctagctaggagtcttttaatcttattagaaatttacataccaaaaaatttagagaacttgctaggacaattggtatttctttatataatattgtggaagttgtattagagtatgttgtttacattacactctttgagtgcgttccttctccgaactagctaatgcatgaacacgagatgccttctgcaccgtgctaccctattaatatataaaaaaatggtagcccggtgcattaagctcccgctatgcgcgggttccgaaaaaggatcagaccacaagggtctatgtttgcaaccttacttgtatttctgcaagaaactgtttccacggctcgaacccatgatcttctggtcacatgacaataactttaccggttacaccaaggttccccttcacgcgctgcccttttaatattgtctattaatatttcctactagagttatacacccctttgttattactcactcttagggtgattattaacatataatatgtttaatatttatactaaaaacaggacgagaattggttagctatgagattccaatgccaaatattggaatccataggtttgtatttgtacttttcaagcaaaaacgaagacaatcagttagctctcctacttcaagggatcacttcaacactagaaattttgctgaagaaaatgatcttggccaacctgttgctgctgttttcttcaatgcacagcgagaaaccgccgcacgaagacgctaa
SEQ ID NO: 17: Геномная последовательность гена TFL1-4S N. tabacum
atggcaagaagtttagagcctctaattgttgggagagtagtaggagatgttcttgattcatttagtcctataatgaaaatgacaatatcatataacaacaaattagtgtgcaatggccatgaactccttccttctgttgtcactgctagacctaaagttgaagttcaagggggagatttgagaactttcttcacattggtatttttttcttgatttctacttaatttccaagatcatcaagttcccattatttctttaaaaaaaaaaaaagcagttcggtgcactaaactcccgctatgcgcggggttcggtgaagcaccgaaccataagggtctattgtacgcaaccttaccctgcatttatgcaagaggcttgctcaccattacaagttatattaatttaacatgttatatataaccacaaaggctgtcgtgggatggtaaatatccttctatccttaatcagaagtttcgggttcaagttatagccctaggaatatagtcgtctttggtagggatcctttacccccaaaactttccgccgtgaatccagattagtaaacctcaaagcgggtatcgggcattggatgacaaaccaaaaaaacttcaacgtgttatagcatgttataacttattacagttaatttagttttccagtcgatactatattaaatagagtgcctgtaatttactttggagtgatttgattgttattttttcgcatcgtcagtacataaaacttatattaattttcgaatatgtaggtcatgacagaccctgatgttcctggccctagtgatccttatctaagagagcatctccactggtatgccctaaactcaatttttttttaaaaaaaaaaaatagaaaatgagaaaaaatatgtaaaaatctacaaatatgagaagatcatgattaattggaactatttttactgactatttgacaggatagtaactgacattccaggtaccactgatgctacttttggtaagttctctgtatcttctgcaaaattacaagcacatgtgaagataaaagaagtttttctattattcacttattttgtctagctagttatatagaataattataagatcaacaattttgtatagtagtgaatgttggacttctaaagtcgaacatgtccacttgatgagtgtcacaaaaatgtagaaactaaacaatcgtttggacataaaaaaaaaagtaagtttttttgagttaaattgaaaaagaaaatatttagaatttgaaattgtggatatacatttaaattgaaaagcattgcagttttgtaaggaaaataaactttcatatacataaaaaagtgattttttggaaactcatcttcaagaatatttttaaaaatttccgtccaatgtataaccaaacattattttgaaaaagattaaaaaaaggaaaaactttaggaacaacgggtcccaagataaatgtgtctagtcatataagattagataaattaggattttattatatttggtagaaggtgcaagaagcatatgtaaataataaattgagaagtcacttaagatattttgatcatgtcccacatcgataacaagaggtaccattctatatatgttaaatcatggtaagttaaagtattatatcacatattaaatggtgatataatagacctaaatcacatgaaacgaaattgtcccgaaaggtctataaatttttgaaattcatgtagacgaagctaaaagtaggatacaataaaaaaaaaattaaagatctatattggcgatactatttagttgggattgcattttagttattctagtacatttactttaatctaatttttgctagctaggagtcttttaatcttattagaaatttacataccaaaaaatttagagaacttgctaggacaattggtatttctttatataatattgtggaagttgtattagagtatgttgtttacattacactctttgagtgcgttccttctccgaactagctaatgcatgaacacgagatgccttctgcaccgtgctaccctattaatatataaaaaaatggtagcccggtgcattaagctcccgctatgcgcgggttccgaaaaaggatcagaccacaagggtctatgtttgcaaccttacttgtatttctgcaagaaactgtttccacggctcgaacccatgatcttctggtcacatgacaataactttaccggttacaccaaggttccccttcacgcgctgcccttttaatattgtctattaatatttcctactagagttatacacccctttgttattactcactcttagggtgattattaacatataatatgtttaatatttatactaaaaacaggacgagaattggttagctatgagattccaatgccaaatattggaatccataggtttgtatttgtacttttcaagcaaaaacgaagacaatcagttagctctcctacttcaagggatcacttcaacactagaaattttgctgaagaaaatgatcttggccaacctgttgctgctgttttcttcaatgcacagcgagaaaccgccgcacgaagacgctaa
SEQ ID NO: 18: Аминокислотная последовательность гена TFL1-4S N. tabacum, полученная из SEQ ID NO: 16 или SEQ ID NO: 17.
MARSLEPLIVGRVVGDVLDSFSPIMKMTISYNNKLVCNGHELLPSVVTARPKVEVQGGDLRTFFTLVMTDPDVPGPSDPYLREHLHWIVTDIPGTTDATFGRELVSYEIPMPNIGIHRFVFVLFKQKRRQSVSSPTSRDHFNTRNFAEENDLGQPVAAVFFNAQRETAARRR
SEQ ID NO: 19: Геномная последовательность гена TFL1-4T N. tabacum, содержащая 2 т. о. перед ATG (выделено жирным шрифтом)
atccccagaggcggatctaggatttgaaccttatgggttctcagcatttgggcaaaagtgatactttaagcaagtgaggaagttttttaatgttggccggaaagatgcctgtgggtgctgtttgggggaaaaaaacaaaagctcgggtaaattatcaatacccgagttggttcctttcgtttgccactggaccaactcctgattttgcttatatatgggttccaaactaaaaatacttatatatttaataaatttatcaatacaaatacaaggctcgggtaaaagttattaggttctcggaaacccatacccgatactatggatccgcccctgcttatccctaccttgtgtgaggtagaaacgcttttgataaggttatttaaaagtaaaagaataagtttaatgtgacaatttgaatggttgagacaacatgccaaaagctaatttaagggattttatttgacatttatatatgggagagaagaaaaagtattgcccagtatattattttaagctctacaaccaatcaagaatcaattcctagaatccattcaggtgtcaaagaatactgacatgatataataaaatacaatttatatcacatcagtatttgctttttcttgggagattagataaaaagagatcagaaatggagttttatggtactaggagaattcaaggatttactacttttgtggcacaacataagctcccaatttttttaaggaatttataaaagttggttttctaagtacttacaattgtcaaatttacaagtcatttagtacataaaaagaaacccaatgatgaggttcaggaaaaaaaaaaaatcctatactgtgatttcctagttggcgttcggacataaaaattatgaaattccgaaaaaaaaaattgttttaagttgaaaatggtatgtgaaaattaaagttatatatggacataaatataatttggagctgtttttgaatttttgtgagtgctttgaagtgaaattttctaaaaacagctttttggagtttttcaaattccggagttcaacttcaagcgaaaaattaaaattttcatgatcaaatgttgattccgaaaaaagtgaaaaaattcgaaaaaaagatttttttttttatggccaaacagacctaactagtttcattttagtcattaagggtagaattgaaagaattttaaattaaagtatttttagatatataaaaataatgtactttttaaaacacacaaaaaaaggagtgccatatattaatttaatataaggatatatagtggatgcattcataactaacattaaccaaaagcatttattgatcctattttgacaccattttattttaatacaattcataaatttcaagaatttgaatacattagcttaatctcacttaaattttgaggtgatgcctgttctctttctagtcacaactttaatgtacattttatatgtcaaattaatacctgaatttgtaacccatcaaatatcgccacataatatgaaacagtgaaaatatcttatattcctgtattttatgactaagacattaagtagctaacaacgatcgaaaaacattcctaataacaagcgaattacaactctgtcggataatcgtctgaaaccctaaaaagctactgaaatgatttcctactagtataattccgatgaaattttgttcgaaaattctataagaaatacacgtatttttagtagtgaaaaaagatttgttgtaatttttttaggtggggtggggtgatttggggagggttggggagtaggacctcaaaaacaaagaattttaatactttggagtttccttaggtcccatgttttatactttcttttattctccttcaccattatagctataacttagtacatatatatatatggtggccctctgatccaatgtaaaatgcaaaccattcttaagatctttgaaatttctctcttttttttctttatctctatctctgtctaattctctctattatggcaagaagtttggagcctctaatagttgggagagtagtaggagatgttcttgattcatttagtcctatagtgaaaatgacaattacttataacaacaaattagtgtgcaatggtcatgaattctttccttctattgtcacttctagacctaaggttgaagttcaaggaggagatttgagaactttcttcacactggtaatttttcttgattttttccttaattccaagatcatcaagttccatttatttctttacaagttatattaatttaaccctttataatcaccaaaggctggcgtgggttgctaagtaaccttccatcgttaatcagatgtttcgggttcgagctagccctgggattacaatcgttttttgtaggaagcgctttaacccccaaaatttttcagcacgaacccggattagtaaacctcaaaactcgtgccaaatactagatgacaaaccaaaagagttttcaacctgttataacatatgttacttgttacaattattagttttccggtcaatagtatattatgtaattttctttgaagtgacttgattgttattttttcacattatcagtgcataaaacttatactattattttttaatatgtaggtcatgacagaccctgatgttcccggccctagtgatccttatctacgagagcatctccactggtacactctctataatagtttcatttgttccgaattttcttggctgttatataaaaaatatattataacatagcatgaaaattggttccacaaaaacttaatttttatagtgaataattgttatatattgatattgttatagagaggtctgtctatatgccctaaactcaatgaaaaaaaatagaaaatgagaaaaaatatgtaaaatctacaaatatgagaagatcatttttagttgaaactatccttatatactactgaatatttagctggcaaataaaattgacagtgttttactgattgtttgacaggatagtaactgacattccaggtaccactgatgctacttttggtaagttttattagtttcttctgcaagattacaagcacatgtgaagaagatacaagatgtttttccattactcacttattttgtcttgctaataattatatagaacaattgtaagatcaacagtgttatataatagtgaatgttggacttctaaagtcgaacatgtccacatgatgagcgtcacaaaaatgcagatacgagctcgtttggattgacttaaaaaatgtggtttttcagcaaaaataacttttaagccaaaaaacaataagttagggttgtccacctttttgcttttggcttaatttaagcattttaaaatttattttaagcaatttttgacttagccaaacaccgaaaaaagctaaaagaaacttaaaagctgatttgactagcttaaaagtaaatccaaacaccctctaactaagcatttggacataaaaaaaatatgtcatttttgaaaaaagtagttcttttgagttaagtcaaaaaagaatatataaaatttgaaattgtatttagacatgcatttcacttgaaaattattagagttttatgagaaaaatgaacttttagatgaaaaagtggtttttggaaactcatcttcaagaatttttccaaaacttcagtccaatcgtataaccaaacattattttgataaaaacatcgaaaataaaaataaatctatggagaaacgggtcccaagataaatgtgtctagtcatataagattattcaaaattaagaatttatcacatttgtaaaagatgtaagtagcatatgtaaatgataaaatgagaagtcacttgagatgttttgatcatgtcctacgtcgatcttcagaggtaccattccgtatacgtgattggtaagtaaaggtattaaaaagagacataatggacctaaattacgtgaaacgaaattgtcttgaaaagtctttcaaatttttgaaatccatgtagacgaatcgaaaagtagggcacaatgaaatatgatcaaaggtttataatggtgatacaagttagttgggattacgttttagttatgccagtatatttactttaatctaatattttcttggagttttttaatcttattagaaatttacttaccaaaaatttagagaacttgctagaacaatataattgataattcttcatatatattgtcttcgagctgtagaaacagccactaatgtttgcattaggatatgttgtctacatcacacttattgtgtgttgccctcaccggaccctgcatgaacgtatgatgccttatgcaccgcgccccttttaatattatttattaattaatatttcctgctagagttatactcctttgttattactcattcttaggttgatgattaacttataatatgcttaatctttatactaaaaataggaagagaattggttagctatgagattccaaggccaaatattggaatccataggtttgtatttgtacttttcaagcaaagacgaagacaatcagttagccctcctacttcaagggaaaacttcaacactagaaattttgccgaagaaaatgatcttagccaacctgttgctgctgttttcttcaatgcacagcgagaaaccgccgcgcgaagacgctaa
SEQ ID NO: 20: Геномная последовательность гена TFL1-4T N. tabacum
atggcaagaagtttggagcctctaatagttgggagagtagtaggagatgttcttgattcatttagtcctatagtgaaaatgacaattacttataacaacaaattagtgtgcaatggtcatgaattctttccttctattgtcacttctagacctaaggttgaagttcaaggaggagatttgagaactttcttcacactggtaatttttcttgattttttccttaattccaagatcatcaagttccatttatttctttacaagttatattaatttaaccctttataatcaccaaaggctggcgtgggttgctaagtaaccttccatcgttaatcagatgtttcgggttcgagctagccctgggattacaatcgttttttgtaggaagcgctttaacccccaaaatttttcagcacgaacccggattagtaaacctcaaaactcgtgccaaatactagatgacaaaccaaaagagttttcaacctgttataacatatgttacttgttacaattattagttttccggtcaatagtatattatgtaattttctttgaagtgacttgattgttattttttcacattatcagtgcataaaacttatactattattttttaatatgtaggtcatgacagaccctgatgttcccggccctagtgatccttatctacgagagcatctccactggtacactctctataatagtttcatttgttccgaattttcttggctgttatataaaaaatatattataacatagcatgaaaattggttccacaaaaacttaatttttatagtgaataattgttatatattgatattgttatagagaggtctgtctatatgccctaaactcaatgaaaaaaaatagaaaatgagaaaaaatatgtaaaatctacaaatatgagaagatcatttttagttgaaactatccttatatactactgaatatttagctggcaaataaaattgacagtgttttactgattgtttgacaggatagtaactgacattccaggtaccactgatgctacttttggtaagttttattagtttcttctgcaagattacaagcacatgtgaagaagatacaagatgtttttccattactcacttattttgtcttgctaataattatatagaacaattgtaagatcaacagtgttatataatagtgaatgttggacttctaaagtcgaacatgtccacatgatgagcgtcacaaaaatgcagatacgagctcgtttggattgacttaaaaaatgtggtttttcagcaaaaataacttttaagccaaaaaacaataagttagggttgtccacctttttgcttttggcttaatttaagcattttaaaatttattttaagcaatttttgacttagccaaacaccgaaaaaagctaaaagaaacttaaaagctgatttgactagcttaaaagtaaatccaaacaccctctaactaagcatttggacataaaaaaaatatgtcatttttgaaaaaagtagttcttttgagttaagtcaaaaaagaatatataaaatttgaaattgtatttagacatgcatttcacttgaaaattattagagttttatgagaaaaatgaacttttagatgaaaaagtggtttttggaaactcatcttcaagaatttttccaaaacttcagtccaatcgtataaccaaacattattttgataaaaacatcgaaaataaaaataaatctatggagaaacgggtcccaagataaatgtgtctagtcatataagattattcaaaattaagaatttatcacatttgtaaaagatgtaagtagcatatgtaaatgataaaatgagaagtcacttgagatgttttgatcatgtcctacgtcgatcttcagaggtaccattccgtatacgtgattggtaagtaaaggtattaaaaagagacataatggacctaaattacgtgaaacgaaattgtcttgaaaagtctttcaaatttttgaaatccatgtagacgaatcgaaaagtagggcacaatgaaatatgatcaaaggtttataatggtgatacaagttagttgggattacgttttagttatgccagtatatttactttaatctaatattttcttggagttttttaatcttattagaaatttacttaccaaaaatttagagaacttgctagaacaatataattgataattcttcatatatattgtcttcgagctgtagaaacagccactaatgtttgcattaggatatgttgtctacatcacacttattgtgtgttgccctcaccggaccctgcatgaacgtatgatgccttatgcaccgcgccccttttaatattatttattaattaatatttcctgctagagttatactcctttgttattactcattcttaggttgatgattaacttataatatgcttaatctttatactaaaaataggaagagaattggttagctatgagattccaaggccaaatattggaatccataggtttgtatttgtacttttcaagcaaagacgaagacaatcagttagccctcctacttcaagggaaaacttcaacactagaaattttgccgaagaaaatgatcttagccaacctgttgctgctgttttcttcaatgcacagcgagaaaccgccgcgcgaagacgctaa
SEQ ID NO: 21: Аминокислотная последовательность гена TFL1-4T N. tabacum, полученная из SEQ ID NO: 19 или SEQ ID NO: 20.
MARSLEPLIVGRVVGDVLDSFSPIVKMTITYNNKLVCNGHEFFPSIVTSRPKVEVQGGDLRTFFTLVMTDPDVPGPSDPYLREHLHWIVTDIPGTTDATFGRELVSYEIPRPNIGIHRFVFVLFKQRRRQSVSPPTSRENFNTRNFAEENDLSQPVAAVFFNAQRETAARRR
SEQ ID NO: 22: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-1S/T N. tabacum
agttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttc
SEQ ID NO: 23: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-1S/T N. tabacum
Gaagagctcttggccattgcaaacttgtttattgttgtaagtgactgtcattttaact
SEQ ID NO: 24: Последовательность ДНК конструкции RNAi TFL1-1S/T N. tabacum
Ggtaccacaagtttgtacaaaaaagcaggctaagcttgtcgaccatggagttaaaatgacagtcacttacaacaataaacaagtttgcaatggccaagagctcttctggtaacctttaatgtttaaccgttcacatttctaatatttacttatttgtaacatgtcgtcacgtgttagtttcattctttttatgaaccaaacatgcatgcaaagatatttttagatatttggacggcgagtgagatttgaaactaggaccgtttgcctgatacaatattaaaatatgtaaccattttatgtacaagtttaaactgttgatagtagcatattttttacttttatttaagtatactatattccaacaggtaagttaacgaagagctcttggccattgcaaacttgtttattgttgtaagtgactgtcattttaactggcgcgcccgggcaattgacccagctttcttgtacaaagtggtgagctc
SEQ ID NO: 25: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-1S N. tabacum
tgcagtcactattagacctagggttgaagttcaaggtggtgatatgagaactttcttcacattggtcatcacagatcctgatgtacct
SEQ ID NO: 26: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-1S N. tabacum
aggtacatcaggatctgtgatgaccaatgtgaagaaagttctcatatcaccaccttgaacttcaaccctaggtctaatagtgactgca
SEQ ID NO: 27: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-1T N. tabacum
tgcggtcaccattagacctagggttgaggttcaaggtggtgatatgagaactttcttcacattggtcatgacagaccctgatgttcct
SEQ ID NO: 28: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-1T N. tabacum
aggaacatcagggtctgtcatgaccaatgtgaagaaagttctcatatcaccaccttgaacctcaaccctaggtctaatggtgaccgca
SEQ ID NO: 29: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-2S/T N. tabacum
catgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttaca
SEQ ID NO: 30: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-2S/T N. tabacum
tgtaaagaaagatctcaaatcacctccatgaacttcaaccctaggtttagaggtgactgaggaaggaaagagttcatg
SEQ ID NO: 31: Последовательность ДНК конструкции RNAi TFL1-2S/T N. tabacum
ggtaccacaagtttgtacaaaaaagcaggctaagcttgtcgaccatggcatgaactctttccttcctcagtcacctctaaacctagggttgaagttcatggaggtgatttgagatctttctttacatggtaacctttaatgtttaaccgttcacatttctaatatttacttatttgtaacatgtcgtcacgtgttagtttcattctttttatgaaccaaacatgcatgcaaagatatttttagatatttggacggcgagtgagatttgaaactaggaccgtttgcctgatacaatattaaaatatgtaaccattttatgtacaagtttaaactgttgatagtagcatattttttacttttatttaagtatactatattccaacaggtaagttaactgtaaagaaagatctcaaatcacctccatgaacttcaaccctaggtttagaggtgactgaggaaggaaagagttcatgggcgcgcccgggcaattgacccagctttcttgtacaaagtggtgagctc
SEQ ID NO: 32: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-2S N. tabacum
gaaagaaatagttggctatgaaatgccaaggccaaatattggaattcacaggtttgtatttctgctgttcaagcagaagaagaggcaaacagtattgactgcacctctctccagggatcga
SEQ ID NO: 33: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-2S N. tabacum
tcgatccctggagagaggtgcagtcaatactgtttgcctcttcttctgcttgaacagcagaaatacaaacctgtgaattccaatatttggccttggcatttcatagccaactatttctttc
SEQ ID NO: 34: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-2T N. tabacum
gagagaaatagttgggtatgaaatgccaaggccaaatattggaatccacagcagctttcttcaattgccagagggaaaccgctgccagaaggcgttgaagaagatgttta
SEQ ID NO: 35: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-2T N. tabacum
taaacatcttcttcaacgccttctggcagcggtttccctctggcaattgaagaaagctgctgtggattccaatatttggccttggcatttcatacccaactatttctctc
SEQ ID NO: 36: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-3T N. tabacum
atggctcaaatgacagatccccttgtgattagtagggtggttggagatgttgttgattatttctctccaagtgttaagatgtgtgttatttataaccccagtaagcatgtctataatgggcatgaactctttccatcc
SEQ ID NO: 37: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-3T N. tabacum
ggatggaaagagttcatgcccattatagacatgcttactggggttataaataacacacatcttaacacttggagagaaataatcaacaacatctccaaccaccctactaatcacaaggggatctgtcatttgagccat
SEQ ID NO: 38: Последовательность ДНК конструкции RNAi TFL1-3T N. tabacum
ggtaccacaagtttgtacaaaaaagcaggctaagcttgtcgaccatggatggctcaaatgacagatccccttgtgattagtagggtggttggagatgttgttgattatttctctccaagtgttaagatgtgtgttatttataaccccagtaagcatgtctataatgggcatgaactctttccatcctggtaacctttaatgtttaaccgttcacatttctaatatttacttatttgtaacatgtcgtcacgtgttagtttcattctttttatgaaccaaacatgcatgcaaagatatttttagatatttggacggcgagtgagatttgaaactaggaccgtttgcctgatacaatattaaaatatgtaaccattttatgtacaagtttaaactgttgatagtagcatattttttacttttatttaagtatactatattccaacaggtaagttaacggatggaaagagttcatgcccattatagacatgcttactggggttataaataacacacatcttaacacttggagagaaataatcaacaacatctccaaccaccctactaatcacaaggggatctgtcatttgagccatggcgcgcccgggcaattgacccagctttcttgtacaaagtggtgagctc
SEQ ID NO: 39: Последовательность ДНК целевой последовательности смысловой RNAi TFL1-4TN. tabacum
tagtcctatagtgaaaatgacaattacttataacaacaaattagtgtgcaatggtcatgaattctttccttctattgtcacttctagacctaa
SEQ ID NO: 40: Последовательность ДНК целевой последовательности антисмысловой RNAi TFL1-4TN. tabacum
ttaggtctagaagtgacaatagaaggaaagaattcatgaccattgcacactaatttgttgttataagtaattgtcattttcactataggacta
SEQ ID NO: 41: Последовательность ДНК конструкции RNAi TFL1-4T N. tabacum
Ggtaccacaagtttgtacaaaaaagcaggctaagcttgtcgaccatggtagtcctatagtgaaaatgacaattacttataacaacaaattagtgtgcaatggtcatgaattctttccttctattgtcacttctagacctaatggtaacctttaatgtttaaccgttcacatttctaatatttacttatttgtaacatgtcgtcacgtgttagtttcattctttttatgaaccaaacatgcatgcaaagatatttttagatatttggacggcgagtgagatttgaaactaggaccgtttgcctgatacaatattaaaatatgtaaccattttatgtacaagtttaaactgttgatagtagcatattttttacttttatttaagtatactatattccaacaggtaagttaacttaggtctagaagtgacaatagaaggaaagaattcatgaccattgcacactaatttgttgttataagtaattgtcattttcactataggactaggcgcgcccgggcaattgacccagctttcttgtacaaagtggtgagctc
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Филип Моррис Продактс S.A.
<120> Растения с сокращенным периодом времени до наступления цветения
<130> P10400EP
<140> EP16205377.1
<141> 2017-01-05
<160> 41
<170> PatentIn версия 3.5
<210> 1
<211> 3539
<212> ДНК
<213> Nicotiana tabacum
<400> 1
tgtttattgt acattttgaa ctgcatcgag ctgcacgatt gacagatcta gtctgcaaca 60
gagtactttg gtttctaatt atagtaatcc taacacctca gaaatttcca aaattggaat 120
tatgggtgca aatcatcttt atatatactc tcaaatgact tctcagatct tacggtcact 180
aattgtctgt aaattaagta tggtggagtt cactttatat actctgagcg gtgtgcccat 240
cctcctccat cagaatatca cttaaaacaa ttaacgtact ttataatctt tccctattgt 300
ttacacacaa tttaggaaag tttctaacaa agataagccc atatttcaag attagtgtct 360
taggattacc ctaaatgaaa aaagcaatag attccctggc aatatgttta cttatatttt 420
gaatttttgc aaaaaaaata aataaatgtg tgtcaacttg cttagttggg accacatgca 480
aaaactaatt caaggatttc atctgataat tttatagtgg agaggaaaag gcttggatta 540
atttaagtac ttattatgta gggcaaatat cacttttagc ctgcggccac catatttata 600
ttcaagccgt aaaagtgtat aaaatttgta tttttttata tataatatac ggaaatgtgt 660
gtatatatat atatatatat atatatatat atatataaga aatataaaaa aaattggcta 720
ttatttttca gagcagctat acaatatcat tttccatatc aggtaagctg aacaacaaat 780
catggaccaa ttcgaagagc accagtcagg tgtgaaagag aactgacatg atagcataaa 840
atacatatca ctaactcctc cacatctaag agcatacatt taatttccta tatggaagat 900
aagattaatt agaaaagtgt ttgataaagc taagatatgt tagtaccagt ttgtagtatc 960
cagttgaaat tttagtgtca acttaaggat aattttacta tgattagaag acagttatac 1020
attcacaaag tttctgaaag gaacttataa gtcttctttt tattgtactt acatttgtca 1080
agtcatatat agtacatatg acccctccta aaggaaaaga aagtagggaa agtaccaagc 1140
tagggcatat ttaaaggaaa ataaaatggc aattttaaga tgttagagta agggcagggg 1200
taggccaaag aagtattggg gagaagtgat tagacaagac atgccactgc ttcacctaac 1260
cgcggacata actagcgata ggaagatgtg gaggtcgaga attaaggttg tgagttgaca 1320
ggtagttatg agtttactag taatactagt actattcttg tattctttta ttcttagttt 1380
tttattactt tgttatgtca ctcgcttcca ttactagtta tctgttgtta ttgcttgttg 1440
ctttattttt accatttttt tagccgaggg tctatcaaaa acagtctctc tgcctttata 1500
ggataggggt aaggctgcgt acacactacc ctttccagac cccacgtatg ggattaatcc 1560
gagtttgttg ttgttgttgt tgttgatgtt gttgttgttg aaaaggaaac gtttataatt 1620
aagtacatgt attaagattt cattattttc ggggaagaat ttcgaatgta tttaaaccat 1680
aaaacgtcat tctcctgcaa gtatttttgg tgatcaagat aagtatagtt aaaaattgag 1740
caatcttctt tgcctacgat gtcctgtata aaactaacaa agaaaaaatt cagtttattt 1800
ttcaggggta gggttacttt ttttttaaaa aatattctat ttgaagttcc ataaaatccc 1860
attttttatt tgcaactata ttgagtagat gttccgagac agcaactata agtgaggctc 1920
tatttcactg ttcaaccaaa atctctcaca agctaagttc cttgcaatcc aaaagatctc 1980
tctctctctc tctctctcta atggcaagga atgtagagcc tctagtagta gggagagtag 2040
taggggatgt tcttgattca tttagtccca cagttaaaat gacagtcact tacaacaata 2100
aacaagtttg caatggccaa gagctcttcc cttctgcagt cactattaga cctagggttg 2160
aagttcaagg tggtgatatg agaactttct tcacattggt aacttttcta attttccttc 2220
aggttattaa cttttcattg ctatatagac atctttcaaa cctgtctaga agattctttt 2280
cttgagctga gggttcggag acagcctctc tatcccacac aagataaggt taacgtctgc 2340
atataccttg ccgtcccctg acaccacata tgggattata ctgcgtatgt tgttgttgtt 2400
gtattgctat atagacatct ttgataaatc actaatcatg catgatatta ttcttcttat 2460
tgtaggtcat cacagatcct gatgtacctg gccctagtga tccgtatctg agagaacatg 2520
ttcactggta tgtactacat tcaagaaaat ctatggaaaa aaaataacta gaagagatga 2580
gtccaaaaga aaggagatta attattgttt gatttgtatt ttttatttgc attagattaa 2640
gtcatgcaat tacctagtaa taaattcgtt ataaggacat acacaaaagg tatgctcaac 2700
ataattaaag aaagaaaagc ataagcatta tattcttata tgcaagctcc caatctgatt 2760
gtggcgaagc taagaatttc ttcgggtgtt taaatttaaa agaagtgaaa aaaaatccga 2820
taaaaaattt atgtgttata tacctctaaa acttaatatt gtacctatat acatattgta 2880
atttttcgac gaatgaccac tcttattgcc catgctgcca atgccaatat gtaaaatact 2940
tcgccaatag acaaatatgt atcacattaa ttagttacct aaataggtaa ttatccataa 3000
taagtgggac tcgtaacttg aaaaatatgg ttagtaattt gctacaacat gttaagatac 3060
agtgagaatg taaatgtata tatgtccttt cacgccgtcg taagtttata taagttaaat 3120
ctggttaaca atgtaggtca tcataacact cacaattggg aataataact cttaatcata 3180
cttctttaca ttcatatttg caggatagtg actgatattc caggaactac agatgccacc 3240
tttggtaaat tggctatttt ggtttctttt attaagctgg tgtttgacat gctagaatta 3300
atgtacttta attttgagca ggaaaagagt tggttagcta tgagatccca aggcctaata 3360
ttggaataca taggtttgtg tttgttctct ttaagcagaa atgcagacaa tcagtcagcc 3420
cacctacttc aagggatcat ttcaacactc gcaactttgc caacgtaaat gaccttggtc 3480
cgcctgtcgc cgccgtcttc ttcaatgcac aacgagagac cgccgccagg aggcgctaa 3539
<210> 2
<211> 1539
<212> ДНК
<213> Nicotiana tabacum
<400> 2
atggcaagga atgtagagcc tctagtagta gggagagtag taggggatgt tcttgattca 60
tttagtccca cagttaaaat gacagtcact tacaacaata aacaagtttg caatggccaa 120
gagctcttcc cttctgcagt cactattaga cctagggttg aagttcaagg tggtgatatg 180
agaactttct tcacattggt aacttttcta attttccttc aggttattaa cttttcattg 240
ctatatagac atctttcaaa cctgtctaga agattctttt cttgagctga gggttcggag 300
acagcctctc tatcccacac aagataaggt taacgtctgc atataccttg ccgtcccctg 360
acaccacata tgggattata ctgcgtatgt tgttgttgtt gtattgctat atagacatct 420
ttgataaatc actaatcatg catgatatta ttcttcttat tgtaggtcat cacagatcct 480
gatgtacctg gccctagtga tccgtatctg agagaacatg ttcactggta tgtactacat 540
tcaagaaaat ctatggaaaa aaaataacta gaagagatga gtccaaaaga aaggagatta 600
attattgttt gatttgtatt ttttatttgc attagattaa gtcatgcaat tacctagtaa 660
taaattcgtt ataaggacat acacaaaagg tatgctcaac ataattaaag aaagaaaagc 720
ataagcatta tattcttata tgcaagctcc caatctgatt gtggcgaagc taagaatttc 780
ttcgggtgtt taaatttaaa agaagtgaaa aaaaatccga taaaaaattt atgtgttata 840
tacctctaaa acttaatatt gtacctatat acatattgta atttttcgac gaatgaccac 900
tcttattgcc catgctgcca atgccaatat gtaaaatact tcgccaatag acaaatatgt 960
atcacattaa ttagttacct aaataggtaa ttatccataa taagtgggac tcgtaacttg 1020
aaaaatatgg ttagtaattt gctacaacat gttaagatac agtgagaatg taaatgtata 1080
tatgtccttt cacgccgtcg taagtttata taagttaaat ctggttaaca atgtaggtca 1140
tcataacact cacaattggg aataataact cttaatcata cttctttaca ttcatatttg 1200
caggatagtg actgatattc caggaactac agatgccacc tttggtaaat tggctatttt 1260
ggtttctttt attaagctgg tgtttgacat gctagaatta atgtacttta attttgagca 1320
ggaaaagagt tggttagcta tgagatccca aggcctaata ttggaataca taggtttgtg 1380
tttgttctct ttaagcagaa atgcagacaa tcagtcagcc cacctacttc aagggatcat 1440
ttcaacactc gcaactttgc caacgtaaat gaccttggtc cgcctgtcgc cgccgtcttc 1500
ttcaatgcac aacgagagac cgccgccagg aggcgctaa 1539
<210> 3
<211> 172
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 3
Met Ala Arg Asn Val Glu Pro Leu Val Val Gly Arg Val Val Gly Asp
1. 5 10 15
Val Leu Asp Ser Phe Ser Pro Thr Val Lys Met Thr Val Thr Tyr Asn
20 25 30
Asn Lys Gln Val Cys Asn Gly Gln Glu Leu Phe Pro Ser Ala Val Thr
35 40 45
Ile Arg Pro Arg Val Glu Val Gln Gly Gly Asp Met Arg Thr Phe Phe
50 55 60
Thr Leu Val Ile Thr Asp Pro Asp Val Pro Gly Pro Ser Asp Pro Tyr
65 70 75 80
Leu Arg Glu His Val His Trp Ile Val Thr Asp Ile Pro Gly Thr Thr
85 90 95
Asp Ala Thr Phe Gly Lys Glu Leu Val Ser Tyr Glu Ile Pro Arg Pro
100 105 110
Asn Ile Gly Ile His Arg Phe Val Phe Val Leu Phe Lys Gln Lys Cys
115 120 125
Arg Gln Ser Val Ser Pro Pro Thr Ser Arg Asp His Phe Asn Thr Arg
130 135 140
Asn Phe Ala Asn Val Asn Asp Leu Gly Pro Pro Val Ala Ala Val Phe
145 150 155 160
Phe Asn Ala Gln Arg Glu Thr Ala Ala Arg Arg Arg
165 170
<210> 4
<211> 4283
<212> ДНК
<213> Nicotiana tabacum
<400> 4
ttcgagtcgt gataagtgtt ttgcaaaaat ataagataag actgcatgcg tacaatagac 60
tcttttggtc cggccctttt ctggaccctg cgcataacgg aagcttagtg cacccggcaa 120
ccgtttttca acgttccaga tgcaagaata ttataaggag gctttttgac acttttaaat 180
ttaatctaat agagtttaag tttcatgcat cgactgtcaa aaaaatattt atataatcac 240
taatacgata attacaagtg aaatgctata ctaaacatta agtagtaacc taataaaacg 300
gtagctagct aatctactat atatcacgta gtattaaaat tacacttatg taaaaatctt 360
tacggtgtta agaataactt aaaagcaatt tagtaataca ttaatgagtg aaaaaggtga 420
agaggaaacg tggtgtgttt tgaactgcat cgagctgcac gattgacaga gctagtctgg 480
aacagagtac tttggtttct aattatagta atcctaacat ctcagaaatt tccataatag 540
gaattatggg tgcaaatcat ctttatatat actctcaaat taatgacttc tcagcaagta 600
ctgtcattgt ctgtaaatta agtatggtgg agttcacttt atactctgag cagtgtgccc 660
atcctcctcc atcacttaaa acaattaatg tactttataa tctttcccta ttgtttacac 720
acacaattta ggaaagtttt taacaaagac aagcccatgt ttcaagattt gtttcttagg 780
attaccctaa atgaaaaaag caatagattc cctggcaatt tacttatatt ttgaattttt 840
gcaaaaacaa aaagaaaaag tgacaacttg cttagttggg accacatgca aaaactaatt 900
taaggaattc atctgatatt tttatagtgg agaggaaaag ggctggatta atttaaatat 960
tccttatttg ccaaaattat ttatattcga tagctgtaaa aatatataaa atttgtatat 1020
ttgtttttgt atagtataca cggaaatgta tatatataca agaaattaaa aaaaaactat 1080
tattttcaga gcagttatac aatattattt tccctatcat gtaagcttag cttatcaaca 1140
aatcatggac caattctaag agctccattc aggtgtgaaa gagagctgac atgatagtat 1200
aaaatacaca tcactaactc ctccacattt gagagctata gattaatttc ctatatggaa 1260
gataagatta attagaaaag tgtttgaata agctaagatg tgttagtacc agtttgtaat 1320
atcaagctaa aattttagtg tcaatttaag gataatttta ctatgattag aagacaagtt 1380
atcattcaca aatttctgaa aggaacttat aagacttttt atttttattt tttattgtac 1440
ttacatttgt caattaagta catgtgaccc ctcctaaagg aaaagaaagt agggaaatta 1500
aagtaccaag ttagcgcata cttaaagcca aggtaagcaa aatggcaatt ttaaggtgtt 1560
gtagaggaaa cgtttataat taagtacacg tactaagatt tcattatttt cgtggaagaa 1620
atttagaatg tatttaaacc ataaaacgtc attctcgtgc aagtattttt ggtgatcaag 1680
ataagtatag ttaacgttga acaatcttct ttgcttacta tgtcttgtat aaaactaaca 1740
aagaaaaatt cagtttattt ttcaggggta gggttacttt tttaaaaaaa aatattctat 1800
ttgaagttcc ataagatccc atttttaatt tgtaactata ttgagtagat attccaagac 1860
agcaactata aatgaggatc tatttcaccg ttcaatcaaa atctctcaca agctaagttc 1920
ctagcaatcc aaaaaagatc tctctctctc tctctctctc tctctctctc tctctctctc 1980
tctctctctc tctctctcta atggcaagga atgtagagcc tctagttgta gggagagtag 2040
taggggatgt tcttgattca ttcagtccca aagttaaaat gacagtcact tacaacaata 2100
aacaagtttg caatggccaa gagctcttcc cttctgcggt caccattaga cctagggttg 2160
aggttcaagg tggtgatatg agaactttct tcacattggt aacttttcta aattctcctt 2220
aaggttatta acttttcatt tctatataga cacatcgagg gtcaacagag acaactctat 2280
ctcacacaag gtaggggtaa ggtattcgta taccctaccc ggcccacatg tgagatcaca 2340
ctgaatatgt tgttgttgtc gcatttaggg gtgtacaaat gaaactgaca aactgcacca 2400
atctgataat ccgagtcaaa tcgagaaaaa atccgattat ggtttggttt gatttggttt 2460
ggtgatggaa aaaaccccga catatttggt tttgtttggt tttaactaaa aaaagtcaaa 2520
ccgaaaccaa accaaccaga cattatatgt gtagaaattt taaatatatt taatacataa 2580
aaatatttat ggtagtgtaa tttataaata tttcttaaga tttttcatag tttatctttt 2640
aacgtattat ttcaaacttg ggcttataat ttttggatgc tccaataagt tttatagtcc 2700
ataaatgtta gtaactcaaa taaatcctaa accaaaatca aatcaatact aatgctaata 2760
aaagacattc aattcaattg tactatgaat gaaaatagtg ttggatatat atttttatag 2820
tttttccacg gtttagataa aatgtataac ttatttttct ttgagtatgg ttagtcatgt 2880
aaataatctt attaatcata attttaaatt atgtttattt ttattatggc ttattaataa 2940
tatttaattt tttgtgcaat tttattatct ttattgttga atattttagt acaatgccac 3000
gactcatctc atatttatgt tattttattg aaaaacacct catatagttc tgcctcatta 3060
ggattaaaaa aatatttgga gcacaaattt tactttttgt gttatgaaga ctttatgaaa 3120
aaaaaataaa ataaaaaccc gaaaacccga aacctcgaga aaaatcgaga ttaaaaatcc 3180
gacttttatt ggtttggttt ggtatttaga tttaataacc cgatacaatt agtttggttt 3240
ggtaattaga aaatccgaat caaaccccta accgtgtaca cccctagtcg tattggtata 3300
tagacatctt tgataaatca ttaatcatgc atgatattct tcgatctcct tattgtaggt 3360
catgacagac cctgatgttc ctggccctag tgatccgtat ctgagagaac atcttcactg 3420
gtatgcacta tattcaagaa aagctatgga aaaaaataac tagatgagat aggtaaaaaa 3480
gaaaggagtt taatgattgt ttgatttgca ttaattatat taagtcatgc cattatctag 3540
taataaactg gttataagga catacacaaa aggtatggtc aacatataat caaagaaaga 3600
aaagcataag catccttatg caagctgcca atgtcatgta aaatacttcg ctaatagaca 3660
aatatatatt acattagtta cctaaaagat aggtatataa tcagtgggac tcctaactta 3720
aaaaataagg ttagtaatct gctataacga tacactgaga atcgtcgtcg tcagtttata 3780
agttaaaaat taatgtaggt catcacaaca ctcacaaaga gtgcctcaat tgggaagagt 3840
atgttatata gttagaattt atgttacata tggaaccaca gtactacaga aggataactc 3900
ttaaacatac ttctttacca tcatatttgc aggatagtga ctgatattcc aggaacaaca 3960
gatgccacct ttggtaagct cactattttg gcattttcat ttttccttca atttctttta 4020
gtatatagct aggttggttt ttgacatgct aattttgagc aggaaaagag ttggttagct 4080
atgagatccc acggcctaat attggaatac ataggtttgt gtttgttctg tttaagcaga 4140
aatgcagaca gtcagttagt ccacatgatg tttccagaga tcacttcaac actcgcaact 4200
ttgccaacgt aaacgatctt ggcccgcctg tcgccgccgt cttcttcaat gcacaacgag 4260
agaccgccgc caggagacgc taa 4283
<210> 5
<211> 2283
<212> ДНК
<213> Nicotiana tabacum
<400> 5
atggcaagga atgtagagcc tctagttgta gggagagtag taggggatgt tcttgattca 60
ttcagtccca aagttaaaat gacagtcact tacaacaata aacaagtttg caatggccaa 120
gagctcttcc cttctgcggt caccattaga cctagggttg aggttcaagg tggtgatatg 180
agaactttct tcacattggt aacttttcta aattctcctt aaggttatta acttttcatt 240
tctatataga cacatcgagg gtcaacagag acaactctat ctcacacaag gtaggggtaa 300
ggtattcgta taccctaccc ggcccacatg tgagatcaca ctgaatatgt tgttgttgtc 360
gcatttaggg gtgtacaaat gaaactgaca aactgcacca atctgataat ccgagtcaaa 420
tcgagaaaaa atccgattat ggtttggttt gatttggttt ggtgatggaa aaaaccccga 480
catatttggt tttgtttggt tttaactaaa aaaagtcaaa ccgaaaccaa accaaccaga 540
cattatatgt gtagaaattt taaatatatt taatacataa aaatatttat ggtagtgtaa 600
tttataaata tttcttaaga tttttcatag tttatctttt aacgtattat ttcaaacttg 660
ggcttataat ttttggatgc tccaataagt tttatagtcc ataaatgtta gtaactcaaa 720
taaatcctaa accaaaatca aatcaatact aatgctaata aaagacattc aattcaattg 780
tactatgaat gaaaatagtg ttggatatat atttttatag tttttccacg gtttagataa 840
aatgtataac ttatttttct ttgagtatgg ttagtcatgt aaataatctt attaatcata 900
attttaaatt atgtttattt ttattatggc ttattaataa tatttaattt tttgtgcaat 960
tttattatct ttattgttga atattttagt acaatgccac gactcatctc atatttatgt 1020
tattttattg aaaaacacct catatagttc tgcctcatta ggattaaaaa aatatttgga 1080
gcacaaattt tactttttgt gttatgaaga ctttatgaaa aaaaaataaa ataaaaaccc 1140
gaaaacccga aacctcgaga aaaatcgaga ttaaaaatcc gacttttatt ggtttggttt 1200
ggtatttaga tttaataacc cgatacaatt agtttggttt ggtaattaga aaatccgaat 1260
caaaccccta accgtgtaca cccctagtcg tattggtata tagacatctt tgataaatca 1320
ttaatcatgc atgatattct tcgatctcct tattgtaggt catgacagac cctgatgttc 1380
ctggccctag tgatccgtat ctgagagaac atcttcactg gtatgcacta tattcaagaa 1440
aagctatgga aaaaaataac tagatgagat aggtaaaaaa gaaaggagtt taatgattgt 1500
ttgatttgca ttaattatat taagtcatgc cattatctag taataaactg gttataagga 1560
catacacaaa aggtatggtc aacatataat caaagaaaga aaagcataag catccttatg 1620
caagctgcca atgtcatgta aaatacttcg ctaatagaca aatatatatt acattagtta 1680
cctaaaagat aggtatataa tcagtgggac tcctaactta aaaaataagg ttagtaatct 1740
gctataacga tacactgaga atcgtcgtcg tcagtttata agttaaaaat taatgtaggt 1800
catcacaaca ctcacaaaga gtgcctcaat tgggaagagt atgttatata gttagaattt 1860
atgttacata tggaaccaca gtactacaga aggataactc ttaaacatac ttctttacca 1920
tcatatttgc aggatagtga ctgatattcc aggaacaaca gatgccacct ttggtaagct 1980
cactattttg gcattttcat ttttccttca atttctttta gtatatagct aggttggttt 2040
ttgacatgct aattttgagc aggaaaagag ttggttagct atgagatccc acggcctaat 2100
attggaatac ataggtttgt gtttgttctg tttaagcaga aatgcagaca gtcagttagt 2160
ccacatgatg tttccagaga tcacttcaac actcgcaact ttgccaacgt aaacgatctt 2220
ggcccgcctg tcgccgccgt cttcttcaat gcacaacgag agaccgccgc caggagacgc 2280
taa 2283
<210> 6
<211> 173
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 6
Met Ala Arg Asn Val Glu Pro Leu Val Val Gly Arg Val Val Gly Asp
1. 5 10 15
Val Leu Asp Ser Phe Ser Pro Lys Val Lys Met Thr Val Thr Tyr Asn
20 25 30
Asn Lys Gln Val Cys Asn Gly Gln Glu Leu Phe Pro Ser Ala Val Thr
35 40 45
Ile Arg Pro Arg Val Glu Val Gln Gly Gly Asp Met Arg Thr Phe Phe
50 55 60
Thr Leu Val Met Thr Asp Pro Asp Val Pro Gly Pro Ser Asp Pro Tyr
65 70 75 80
Leu Arg Glu His Leu His Trp Ile Val Thr Asp Ile Pro Gly Thr Thr
85 90 95
Asp Ala Thr Phe Gly Lys Glu Leu Val Ser Tyr Glu Ile Pro Arg Pro
100 105 110
Asn Ile Gly Ile His Arg Phe Val Phe Val Leu Phe Lys Gln Lys Cys
115 120 125
Arg Gln Ser Val Ser Pro His Asp Val Ser Arg Asp His Phe Asn Thr
130 135 140
Arg Asn Phe Ala Asn Val Asn Asp Leu Gly Pro Pro Val Ala Ala Val
145 150 155 160
Phe Phe Asn Ala Gln Arg Glu Thr Ala Ala Arg Arg Arg
165 170
<210> 7
<211> 4296
<212> ДНК
<213> Nicotiana tabacum
<400> 7
catgaccttt tagctactct taactcttct gattgttctg ctgtaacttg tccccttgag 60
ttaaatgtaa agttaaaggc taaagaaggg gatcctctcc ctaatcctga aaattataga 120
ggcctcgttg gtaagctaaa tttcctcact cacactaggc ctgacataag ttttgttgtg 180
caacatctta gtcagttcat gcaacagccc tgctttcctc acatgaaggc agctttgcac 240
ctgttgaggt atctcagaga cacttctaat tttggcctct tatactcgaa ttctactgat 300
ctctctttgc aggcttattg tgatagtgat tggggatcct gccctgataa ctggagattt 360
gtttctgatt tctgtttatt ctttggtggc agtctcattg ggtggaaatc taagaaacat 420
gcagtggtct ctttatcttc ggctgaagtt gagtatagat ctatgagcaa ggctgtggct 480
gaaattactt gggtgtgtag gcgtctatct gatcttgggg tctcttctgc ttctcttgtt 540
cctctccatt gtgacagtat ctctgccatt cacattgcct acaatcctgt cttctatgag 600
cggaccaaag acattgagtt ggattgccat tttgaacgta ccaagcttgc tgaaggtctc 660
atcagtttat ctcacatttc cagtgcttct cagctcgcga atgtcttcat caaacccctg 720
tgtgggcctt ctcaccatct tcatattcga aagttgggag ttctctcacc ctcctacttg 780
agggggggct gttgagatag gctgaaatca gtgtggctca gacccaatta ttatttattt 840
atgtacatca gattaggccc attagttagt ctttagttag tcttttattt ctttacatat 900
attgggccat gtatacatac atagagaccc gattttgtaa tagttagatg attcattttt 960
cggttcttaa tcaataagaa atatctcgaa ctttctctct atctctcttt aaccctaaat 1020
tcttcttcgt tgaatctacg agaatgatga acattaacat tagaaaatgt agatttgatc 1080
aaatcttctt aatcttttgt ttatcatctt ttctaattgt tttgtatctg attgtatatt 1140
agttaaccac caaaattgct caaacaatct ggcttcaaat ttatctaacg tttgaatata 1200
tatatatggt tgaaacatga aaaataaatt tttgaagatg agatgaaaaa taatttttga 1260
aagttaaaat tgtatttgaa cacgtatttt acttgaaaag aatttgaaat tttgtgagca 1320
gaaaacttaa aaaattactc taaaactttt ttttgagatt tgaggatttt attttcaaaa 1380
ttttccataa aatggcttaa atctataagc aaaagatatt tgaaaataat ttttttttta 1440
aaaaagctct caaattttac agccaaacgg aagcttagga taaaaaaggg ggaaatgggg 1500
gagatgggtg ggcaggttgg gctgaagaga aatagacaac agtgcattaa catgtcaaat 1560
catctttatc cctctttcta aaccttacaa ggagtacttt ttattttctt ttttcttttt 1620
ttggccctaa taaaaattaa aacacatatt ctctagctgc taagctataa ctttaactca 1680
ttggcaccac gacgagtagg agaataacct ttttgggctt ttcttttctt ttctttggtc 1740
cccttttttt gaactatcaa tattttagtc caaacacacc tgactctaca gtgatctgat 1800
ggccactata aatattggct ttttgcaact ctcttctcac caaaatacaa atcggttgaa 1860
ctcttcatat ataatattcc cactactatt actcttaact ttaaatagat ttcttatata 1920
tgggttcaaa aatgtctgat ccccttgtga ttggtagagt gattggggaa gttgttgatt 1980
atttcactcc aagtgttaag atgtctgtta cttataacag cagcaagcat gtttataatg 2040
ggcatgaact ctttccttcc tcagtcacct ctaaacctag ggttgaagtt catggaggtg 2100
atttgagatc tttctttaca atggtacata ctgcttcctt cgattttcaa tacttttatt 2160
aggggtggag cttagcggcg gagccaagat tttaactaag gggagtcaaa atataaataa 2220
gtaagcacac aaaaaaatca agggggtcaa cgtatagtat atacacataa aattaagaat 2280
ttaacatatt tataccgtgt aattttccag cgaaggggtg tcaattgact ctccttgcca 2340
atgagtggct ccgccactgg cggagctaga gttctagtta cggttcgttg tattgtgtta 2400
agaagtccac ttatactgtc ttttctagaa tttagaattc ataaattcaa aattatggct 2460
ctgcccttaa atttattttt atacatttct attatatagt aaatcgttta tattgacccc 2520
ttatttttct tttttacctt aattgacaga tcatgataga cccagatgtt cctggtccta 2580
gtgatccata tctcagggaa cacctacact ggtaaagaaa taagtttttt aattactaac 2640
tcattcaatt ttatcgtccc ttcttttcct tgtttacttg gagggaaaat aatacgatct 2700
catcgaaaag ataaaaattc ttcaggcttg ttatctaaaa acttgttaaa aaataccgta 2760
atgaaaagac atatgagttt gttattaggt atttgactaa atatgatcga tcatatggtg 2820
ttcggacaag aaatattttg tgaaaaggtc cgcatacttt taaaaaaaga aaatctgcct 2880
tgactcttga gtttgtgctt ctcgggaaaa caatttcttc cttctttttt tttttttttt 2940
tggtttattg acctttacat attaaagaca ccactgagac acatatctag aaaaattgta 3000
tttgggaacg caaaagcaaa gaaaacatgt gttattaatc ttatgtcaat gccaccagca 3060
gctcaggaaa aatatggtcg atatattgtg atttgcttgc aaaaggagca aagaagaaat 3120
cttttgataa tgtttgttat gacgatgtac ttaaagcaaa taagttagag gtcgtttggt 3180
acatgggata aggataataa ttttgggata aagtttagga ttaactttat cttatatttg 3240
gtttggagta ttagctaacc gcgaggtatt tttcaaacta aaatagtggg attagctatc 3300
ccatataaaa agtaggatag ctaatcccat gggatatccc accctatcgg atagtaatag 3360
tccaatagga gacaactcta atttgtacag acataatgtc cagtcacacc ttgttttttt 3420
gtcatgacac atattaagca tgaataataa tatttcgaca atcttgtagc atttgattag 3480
acttagcaaa ttataaatat gtccaataat tggtcacatt gttctaataa ttacttgttt 3540
cccttatcat tatatatagt gcttcattca ctaaacagaa cccaaaaaaa aaaaaaaaaa 3600
aactgcaaaa tggtcatatc atgtagtaac ggaataaaaa cgtactcagt tttatgataa 3660
aatcaaagtg acatatttgt acgctttgat agttgacaaa tacctgaaaa aagaatttga 3720
ccatctttac aggattgtca cagacattcc aggcactaca gattgctcgt ttggtatgta 3780
tctttaaccc aaatttcaag cttcgaaata gtaacagctt ttgtttttaa tattttattt 3840
gtcttaaata catattttcc ttattataaa tttcttcgcc tagtggtaac gggatcaggt 3900
attgattcgt atttattttt tattgatcaa caaaaaaaga gtacaaaaga aagaattgtt 3960
tttctacact tagatttata tatatgcaat gtctagaaat taatgagttt acaaattcat 4020
tgatgtgtat atctcacaat caaatccaaa atactgatcc aaaaattttg atcagggaaa 4080
gaaatagttg gctatgaaat gccaaggcca aatattggaa ttcacaggtt tgtatttctg 4140
ctgttcaagc agaagaagag gcaaacagta ttgactgcac ctctctccag ggatcgattt 4200
aatacgcgta aattcgcaga agaaaatgag cttgggtctc ctgttgcagc agttttcttc 4260
aattgccaga gggaaactgc tgccagaagg cgttga 4296
<210> 8
<211> 2296
<212> ДНК
<213> Nicotiana tabacum
<400> 8
atgtctgtta cttataacag cagcaagcat gtttataatg ggcatgaact ctttccttcc 60
tcagtcacct ctaaacctag ggttgaagtt catggaggtg atttgagatc tttctttaca 120
atggtacata ctgcttcctt cgattttcaa tacttttatt aggggtggag cttagcggcg 180
gagccaagat tttaactaag gggagtcaaa atataaataa gtaagcacac aaaaaaatca 240
agggggtcaa cgtatagtat atacacataa aattaagaat ttaacatatt tataccgtgt 300
aattttccag cgaaggggtg tcaattgact ctccttgcca atgagtggct ccgccactgg 360
cggagctaga gttctagtta cggttcgttg tattgtgtta agaagtccac ttatactgtc 420
ttttctagaa tttagaattc ataaattcaa aattatggct ctgcccttaa atttattttt 480
atacatttct attatatagt aaatcgttta tattgacccc ttatttttct tttttacctt 540
aattgacaga tcatgataga cccagatgtt cctggtccta gtgatccata tctcagggaa 600
cacctacact ggtaaagaaa taagtttttt aattactaac tcattcaatt ttatcgtccc 660
ttcttttcct tgtttacttg gagggaaaat aatacgatct catcgaaaag ataaaaattc 720
ttcaggcttg ttatctaaaa acttgttaaa aaataccgta atgaaaagac atatgagttt 780
gttattaggt atttgactaa atatgatcga tcatatggtg ttcggacaag aaatattttg 840
tgaaaaggtc cgcatacttt taaaaaaaga aaatctgcct tgactcttga gtttgtgctt 900
ctcgggaaaa caatttcttc cttctttttt tttttttttt tggtttattg acctttacat 960
attaaagaca ccactgagac acatatctag aaaaattgta tttgggaacg caaaagcaaa 1020
gaaaacatgt gttattaatc ttatgtcaat gccaccagca gctcaggaaa aatatggtcg 1080
atatattgtg atttgcttgc aaaaggagca aagaagaaat cttttgataa tgtttgttat 1140
gacgatgtac ttaaagcaaa taagttagag gtcgtttggt acatgggata aggataataa 1200
ttttgggata aagtttagga ttaactttat cttatatttg gtttggagta ttagctaacc 1260
gcgaggtatt tttcaaacta aaatagtggg attagctatc ccatataaaa agtaggatag 1320
ctaatcccat gggatatccc accctatcgg atagtaatag tccaatagga gacaactcta 1380
atttgtacag acataatgtc cagtcacacc ttgttttttt gtcatgacac atattaagca 1440
tgaataataa tatttcgaca atcttgtagc atttgattag acttagcaaa ttataaatat 1500
gtccaataat tggtcacatt gttctaataa ttacttgttt cccttatcat tatatatagt 1560
gcttcattca ctaaacagaa cccaaaaaaa aaaaaaaaaa aactgcaaaa tggtcatatc 1620
atgtagtaac ggaataaaaa cgtactcagt tttatgataa aatcaaagtg acatatttgt 1680
acgctttgat agttgacaaa tacctgaaaa aagaatttga ccatctttac aggattgtca 1740
cagacattcc aggcactaca gattgctcgt ttggtatgta tctttaaccc aaatttcaag 1800
cttcgaaata gtaacagctt ttgtttttaa tattttattt gtcttaaata catattttcc 1860
ttattataaa tttcttcgcc tagtggtaac gggatcaggt attgattcgt atttattttt 1920
tattgatcaa caaaaaaaga gtacaaaaga aagaattgtt tttctacact tagatttata 1980
tatatgcaat gtctagaaat taatgagttt acaaattcat tgatgtgtat atctcacaat 2040
caaatccaaa atactgatcc aaaaattttg atcagggaaa gaaatagttg gctatgaaat 2100
gccaaggcca aatattggaa ttcacaggtt tgtatttctg ctgttcaagc agaagaagag 2160
gcaaacagta ttgactgcac ctctctccag ggatcgattt aatacgcgta aattcgcaga 2220
agaaaatgag cttgggtctc ctgttgcagc agttttcttc aattgccaga gggaaactgc 2280
tgccagaagg cgttga 2296
<210> 9
<211> 148
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 9
Met Ser Val Thr Tyr Asn Ser Ser Lys His Val Tyr Asn Gly His Glu
1. 5 10 15
Leu Phe Pro Ser Ser Val Thr Ser Lys Pro Arg Val Glu Val His Gly
20 25 30
Gly Asp Leu Arg Ser Phe Phe Thr Met Ile Met Ile Asp Pro Asp Val
35 40 45
Pro Gly Pro Ser Asp Pro Tyr Leu Arg Glu His Leu His Trp Ile Val
50 55 60
Thr Asp Ile Pro Gly Thr Thr Asp Cys Ser Phe Gly Lys Glu Ile Val
65 70 75 80
Gly Tyr Glu Met Pro Arg Pro Asn Ile Gly Ile His Arg Phe Val Phe
85 90 95
Leu Leu Phe Lys Gln Lys Lys Arg Gln Thr Val Leu Thr Ala Pro Leu
100 105 110
Ser Arg Asp Arg Phe Asn Thr Arg Lys Phe Ala Glu Glu Asn Glu Leu
115 120 125
Gly Ser Pro Val Ala Ala Val Phe Phe Asn Cys Gln Arg Glu Thr Ala
130 135 140
Ala Arg Arg Arg
145
<210> 10
<211> 4080
<212> ДНК
<213> Nicotiana tabacum
<400> 10
agggcacgac cctaagacct tcttcatagc tataaatagt gagctcaggt ttcattgtaa 60
atggaacgac tattctggca aacttataca atattttata caaaactcaa ttcaatctta 120
tcttctgatt tctagattct ttttgttttt gtgcccgaaa accttgttcc tggaattgtt 180
gcttctgttg tttcgtccat atcttaaggc taagtgttat ataattcttc aattatttat 240
ttatttattt caggttcaaa ttaattcact tatctaaaaa tcatgtataa atttaattgt 300
accattttac gggtgaacag tttggcgccc atcgtggggc ctagataacc gtgtaactaa 360
aggacaaacg tcttttcggg aacttttcta ttttcaagaa ctcaaacccg agatttagac 420
ctctgaggga tctgatcatc tcactacatc gctgagtggt agttgattcc atatacgatt 480
aacctagttt acaactaaat taaattatgt gcattaatcc aagcaacttt tgatgatcag 540
ctgatcaacc taacgtaaga aagcaattaa tttagatgca tatattctac aaatggaaat 600
tagtaggagc aagcaagtta tgcaaaagaa aggaaaagag aaaacattag aagtaggcca 660
aagaaagaag aaggaagagg aagcaatcag ccactgttct agaatggaat atggagaaaa 720
ataataaatt aaattcagat ttctataagt agtaatcctc ttctttctat taccggttaa 780
agctgcagaa attttctttt tcttgacatg acctgaccat agcttccacc attgtttgca 840
ggctggtggt ggagtccctt tataccctca tctctcctac ctaagaacca taggattagg 900
tgattcaagt ttttattttt aacaaaaaat gaaaaattta tgaaggaagt tcaacttttt 960
attaccttaa ataaaaaaga ccttgatgct ttaagtagct ccaagacggt agctgcaaat 1020
tccatctgct tttccttttt aataaaataa tgtactacct actatctgaa agtttaactt 1080
ctatgattct gtaggttttg taaaacactt gggggtattt atattttata ggggattgca 1140
attagaggca gatacaattt ggtttagtta accaccgata ttactcaaac aatttggctt 1200
taaatctggt tagtgtttgg atatagattt ggttgaaatt tgaagaaaaa aaatgagttt 1260
ttaaaaatga gatgaaaaat aatttttgaa agttaaaatt gtatttggac atgcatttta 1320
tttgaaaaga atttgaagtt ttgtaagtta aaattttcaa aaacttcaaa aagttatttt 1380
tgagatttga agattttatt ttcaaaattt gcattataat ctataaacaa atagatacta 1440
tttgagaaca aaatttaaaa aataaagctt tcaaacttat gacgaaaggg aagcttagga 1500
taaaaagggg ggaaatggcc tgggagatgg gtgggcaggt tgggctgaag agaaatagac 1560
aacagtgcat taacatgtca aatcatcttt atccctcttt aaaaacatta ttaggagtac 1620
ttcttttttt cttggggtgc aaaagcctaa tacaagttaa aacacatatt ctctagctgc 1680
taagctataa ctttaactca ttggtaccac gacgagtagg agaataaact ttttgggctt 1740
ttcttttctt ttctttggtt cccatttttt gaactatcaa tattttagtc caaacacacc 1800
tgactctaca gtgatctgat ggccactata aatattggct ttttgcagct ccaaaataca 1860
aatcggtcga actcttcata tatattactc ttaactttaa ataaatagat ttcttatata 1920
tgggttcaaa aatgtctgat ccccttgtga ttggtagagt gataggggaa gttgttgatt 1980
atttcactcc aagtgttaag atgtctgtta cttataacag cagcaagcat gtctataatg 2040
gacatgaact ctttccttcc tcagtcacct ctaaacctag ggttgaagtt catggaggtg 2100
atttgagatc tttctttaca ctggtacata ctccttcgat tttcactact tttaatttat 2160
taggggcgaa gctagagttc tagctacggg ttcgttgtat taattgtgtt aagaagtcca 2220
cttaagctgt cttttttaga atttagaatc cataaactca aaatagtgac tttgcttcta 2280
aattaatttt tatgcatttc tcttatatcg tgtatgtgaa tattgacccc ttattttttc 2340
ttttttacct taattgacag atcatgatag acccagatgt tcctggtcct agtgatccat 2400
atctcaggga acatctacac tggtaaagac atacgttttt taattactaa ctcattcaat 2460
tttatcgccc cttcttttcc ttgtttactt ggagggaaaa taatacgatc tcgtcaagaa 2520
gatcaaaaat cttcaggctt gttatttagg aacttgttca aaaataccgt tttgaaaaga 2580
acatatgagt ttgttattag gtatttgact aaataggaac gatcatatgg tgttcggaca 2640
agaaaatttt tgtgaaaagg tccgcatact ttaaaaaaaa aaaaaaaaaa aaaaaatccg 2700
ccttgactct tgagtttctg cttcttggaa aaaacatttc ttcttttttt ttttgggttt 2760
tttgaccttt atatattaat taaagacacc actgagacac ttaattaaaa aattgtatat 2820
gggaacgcaa aaagaaaaaa aaacatgtgt tattaatctt atgtcaatgc catcagcaac 2880
tcaggaaaat acggtcgata tactgtgatt tgcttgcgaa aggagcaaag aagaaatctt 2940
ttgataatgt ttgttatgac gatgcactta acctaaaata agttaggggc cgtttggtaa 3000
atgaaataag gataataatc tcggaacaaa gtttaggatt aactttatcc catatttgat 3060
ttggagtatt agttaattgc gggataactt tcaaattaaa atagtaggat tagttatctc 3120
atatataaag taaaatacct aatcccaata atataatagg agacaactct aatttgcgta 3180
gacataatgt ccagtctcac tttgtatatt tgtcatgacg catattaagc atgaatgata 3240
atatttcgac aatcttgtgg catttgatta cactcagcaa attataaata tgtccaataa 3300
ttgcattaat aattacttgt tcctcttatc attatagtgc ctcattcact aaaccgaacc 3360
caaaagaaca ctgcaaaatg gtcatatcat gtagtaacag aaaaaaaaaa cgtactcgat 3420
tttatgataa aatcaaagtg acatatgtgt cgctttgata attgacaaat acctgaaaaa 3480
agaatttgac catctttaca ggattgtcac agacattcca ggcactacag attgctcgtt 3540
tggtatgtat ctttaaccca aagttcaagc tatgaaatag taacagcttt tctttttaat 3600
attttatttg tcttaaatac atattttcct tattataaat ttattcgcct agtggtaacg 3660
ggatcaggta ttgattcgta tttaattttt attgttcaac aaaaaagagt acaaaaagaa 3720
agaattgatt ttctacactt agatttatat gcaatatcta gaaatcagaa gatcagcaat 3780
gagtttacta attcatcgat gtgtatatcg cacaatcaaa tccaattact aataatactg 3840
atctaaaaat ttcgatcagg gagagaaata gttgggtatg aaatgccaag gccaaatatt 3900
ggaatccaca ggtttgtatt tctgctgttc aagcagaaga agaggcaaac attattgagt 3960
gcacctctct ccagggatcg atttaatacg cgcaaattct cagaagaaaa tgagcttggg 4020
tctcctgttg cagcagcttt cttcaattgc cagagggaaa ccgctgccag aaggcgttga 4080
<210> 11
<211> 2080
<212> ДНК
<213> Nicotiana tabacum
<400> 11
atgtctgtta cttataacag cagcaagcat gtctataatg gacatgaact ctttccttcc 60
tcagtcacct ctaaacctag ggttgaagtt catggaggtg atttgagatc tttctttaca 120
ctggtacata ctccttcgat tttcactact tttaatttat taggggcgaa gctagagttc 180
tagctacggg ttcgttgtat taattgtgtt aagaagtcca cttaagctgt cttttttaga 240
atttagaatc cataaactca aaatagtgac tttgcttcta aattaatttt tatgcatttc 300
tcttatatcg tgtatgtgaa tattgacccc ttattttttc ttttttacct taattgacag 360
atcatgatag acccagatgt tcctggtcct agtgatccat atctcaggga acatctacac 420
tggtaaagac atacgttttt taattactaa ctcattcaat tttatcgccc cttcttttcc 480
ttgtttactt ggagggaaaa taatacgatc tcgtcaagaa gatcaaaaat cttcaggctt 540
gttatttagg aacttgttca aaaataccgt tttgaaaaga acatatgagt ttgttattag 600
gtatttgact aaataggaac gatcatatgg tgttcggaca agaaaatttt tgtgaaaagg 660
tccgcatact ttaaaaaaaa aaaaaaaaaa aaaaaatccg ccttgactct tgagtttctg 720
cttcttggaa aaaacatttc ttcttttttt ttttgggttt tttgaccttt atatattaat 780
taaagacacc actgagacac ttaattaaaa aattgtatat gggaacgcaa aaagaaaaaa 840
aaacatgtgt tattaatctt atgtcaatgc catcagcaac tcaggaaaat acggtcgata 900
tactgtgatt tgcttgcgaa aggagcaaag aagaaatctt ttgataatgt ttgttatgac 960
gatgcactta acctaaaata agttaggggc cgtttggtaa atgaaataag gataataatc 1020
tcggaacaaa gtttaggatt aactttatcc catatttgat ttggagtatt agttaattgc 1080
gggataactt tcaaattaaa atagtaggat tagttatctc atatataaag taaaatacct 1140
aatcccaata atataatagg agacaactct aatttgcgta gacataatgt ccagtctcac 1200
tttgtatatt tgtcatgacg catattaagc atgaatgata atatttcgac aatcttgtgg 1260
catttgatta cactcagcaa attataaata tgtccaataa ttgcattaat aattacttgt 1320
tcctcttatc attatagtgc ctcattcact aaaccgaacc caaaagaaca ctgcaaaatg 1380
gtcatatcat gtagtaacag aaaaaaaaaa cgtactcgat tttatgataa aatcaaagtg 1440
acatatgtgt cgctttgata attgacaaat acctgaaaaa agaatttgac catctttaca 1500
ggattgtcac agacattcca ggcactacag attgctcgtt tggtatgtat ctttaaccca 1560
aagttcaagc tatgaaatag taacagcttt tctttttaat attttatttg tcttaaatac 1620
atattttcct tattataaat ttattcgcct agtggtaacg ggatcaggta ttgattcgta 1680
tttaattttt attgttcaac aaaaaagagt acaaaaagaa agaattgatt ttctacactt 1740
agatttatat gcaatatcta gaaatcagaa gatcagcaat gagtttacta attcatcgat 1800
gtgtatatcg cacaatcaaa tccaattact aataatactg atctaaaaat ttcgatcagg 1860
gagagaaata gttgggtatg aaatgccaag gccaaatatt ggaatccaca ggtttgtatt 1920
tctgctgttc aagcagaaga agaggcaaac attattgagt gcacctctct ccagggatcg 1980
atttaatacg cgcaaattct cagaagaaaa tgagcttggg tctcctgttg cagcagcttt 2040
cttcaattgc cagagggaaa ccgctgccag aaggcgttga 2080
<210> 12
<211> 148
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 12
Met Ser Val Thr Tyr Asn Ser Ser Lys His Val Tyr Asn Gly His Glu
1. 5 10 15
Leu Phe Pro Ser Ser Val Thr Ser Lys Pro Arg Val Glu Val His Gly
20 25 30
Gly Asp Leu Arg Ser Phe Phe Thr Leu Ile Met Ile Asp Pro Asp Val
35 40 45
Pro Gly Pro Ser Asp Pro Tyr Leu Arg Glu His Leu His Trp Ile Val
50 55 60
Thr Asp Ile Pro Gly Thr Thr Asp Cys Ser Phe Gly Arg Glu Ile Val
65 70 75 80
Gly Tyr Glu Met Pro Arg Pro Asn Ile Gly Ile His Arg Phe Val Phe
85 90 95
Leu Leu Phe Lys Gln Lys Lys Arg Gln Thr Leu Leu Ser Ala Pro Leu
100 105 110
Ser Arg Asp Arg Phe Asn Thr Arg Lys Phe Ser Glu Glu Asn Glu Leu
115 120 125
Gly Ser Pro Val Ala Ala Ala Phe Phe Asn Cys Gln Arg Glu Thr Ala
130 135 140
Ala Arg Arg Arg
145
<210> 13
<211> 5167
<212> ДНК
<213> Nicotiana tabacum
<400> 13
tgaagttgtg tttggacatg cgttgtattt gagaaaaaat tgaagttttg tgagaggaat 60
ttttttgacc ccaaaactac ataatttgaa ttattattta aaaaaaatga tcatattaca 120
tgaacaaaca gtgttttcaa tttatttttg aaaaaaacag ccaaaatcta gccaaatggg 180
agctaagtgt atgatcaaga ttcatgtccc aaatggaaaa gaatattaac aaaaaaaagg 240
gcagtaaaga agaggtggct acaataggat cgcgcaaaag aaagatggaa aaaagaggaa 300
caggaggggg gaataagcag cacaagaagt tattataagt cagctcttcc agaaaggaat 360
atggagaaaa gttaacctca ggtttctata aataggaatg tccaactttc atttactagt 420
ttaagctgca gaaattctct ttttcttgac atgacctttt caccatcttt aatttggtgg 480
gctttgtggt ggagtccctt tataccatag gctctcctag aggatccata acattagatt 540
ggtaaggttc taagtggact cacgatatga aatttgtgat cgaacctata actcgtctga 600
gttactgaat ttgtaataaa atatttatac atatttaata aattttctaa tataaataca 660
gaatctaaac aaaaactatt gagttcatcc gtaccgatac ctaatactct agctccaacc 720
ctgattctaa tataatgaaa ataaaaccac atctaggaag ttcatgacct gttcttacct 780
taaatgccaa aggccttaaa cctttgatag cttgagaata gccaaacaag tatagattcc 840
atctacttta attttctttc tgattaagat atattgcaac tcctgtaaat gcgcaaggag 900
tcagctggtt cttcccccat ttctatattt tttagtatca ctttcttttc ttaattattc 960
cttcttacat ttgaatcttt ttccatcagc tagctgtttt gatagtagta aaaatgcgaa 1020
ggctcttctt actaatattt caatgaccaa tgaatttaga tggagaagca agttctatta 1080
aacgttcatg ctagaaaata acaagtataa tatttcattt tcattttata taacgctctt 1140
gtcttttctt gtctatttaa acaagaatca atttaactct cttaatgaca tgctctttag 1200
tcacagaaaa attataacaa attgaagaca ttagttatta atgttctttt cgcactaaaa 1260
gttttttaaa aattttttat cttaaatgtt tgtgactaat caaatatcat catataaaat 1320
taatctggga atatgacatt tttcaatata actaatatgg tgcaaattgc atacactacg 1380
caataaattt gtggttaggg tcatatcatt agcctgtcaa atcatcttta tccctctttc 1440
tctaaacgac ttcttttctc cctttttttt cgcccctcaa acaaagcaaa tagactattc 1500
tctagctgct aattagctaa acaatgactt taactcgttg tgcccagagg agaataacct 1560
ttatctctct tctcttttct ttgtttccat ctttaattta gacttctttt tttggttttt 1620
tatcccatat tcggtattta ttggagttcg attaaattca aatttataat aggaagtctc 1680
acattgagag tacgatgact ccatactcag gattcgaatt tgagatcttt agttaaagat 1740
gaaagaatat cattcaacca caacttttgt tggtcccatc tttatatcta tatgttctta 1800
ctatatttta atccatttcc cacttccaat gatttaaaga agctatagga taggtgcatt 1860
tggaccacta taaatatagg ttttgcagtt ctatgctcca tacaaatatc cagcaagaaa 1920
ctaaactata tatttactga gttactacta atagttttca ctcaatctat ttccactctt 1980
tctcctcttc attatattat atggctcaaa tgacagatcc ccttgtgatt agtagggtgg 2040
ttggagatgt tgttgattat ttctctccaa gtgttaagat gtgtgttatt tataacccca 2100
gtaagcatgt ctataatggg catgaactct ttccatccct tgttacctct aaacctaagg 2160
ttgaagttca tggaggtgac atgagatcct tctttacact ggtaattaat tcacactact 2220
tcaatagttt tcttgttctt atattttatt atctatctat atatatataa taaaggagcg 2280
gcaaagccac catataaatg acaaatgtaa acttttagga caaaactcca aaaaagttgg 2340
agttttaaaa ttattttata tataaaataa ataaataaat aaataaataa actatcaatt 2400
caaattgggg agtagtttct tactaatatg atagctatat ctatatctat atctatctat 2460
atatgtaaaa catttatatg atgccaagtg gcataaccac tgataagatt tttaaatttg 2520
aatatgaatg aattttaaat gaagttctaa cttcttaaaa ataaacccta atataggtta 2580
ctatttttag taatgattga aattattatt aaaatatttt gttgaaaaca acatagagat 2640
aaaatttgat tattaaattt atgtattaca acaataataa ttattgaaaa tattgctaaa 2700
attttcatga aaggattcac ccataattat tagtataata gaaaactaaa aaattattaa 2760
gtctaaagtt ctagatctct atatttataa acgtataaac tgttatttta ttttctgaaa 2820
aaagcaaaaa tactgaagag aaaaatgata aaaatatttt aaaatatgta agtcatgtgc 2880
aaataataaa gtgaacaaat gatgtagtag tatactgaat aagatatgtt tttttgtcat 2940
aaaataagta tatgcataac tcatctcaat aatttgctga ctccatctga gtcaaaatat 3000
cttctaaatt caagcgaaga taattatcta tcgcattatt tttttatcat taatataagg 3060
caagacgaat ctatatctca tatgggactt tttaaataga tacatcttta taaatgaacc 3120
actttatgag ttttatcacg aattacaagt aagaataact tgaagattga aagaattttt 3180
ggatttattt aattataata tatttttatt cattttaaat taatttatat tttcaaatta 3240
tttgtagcaa cctatataat tatgatattt gagtattatc ttataagtta tttgatgatt 3300
gtcgtttgat ttaattattg aactattact acaggggaca taagatgata attataattt 3360
tgtagaaaca tatgatctaa tgtgctcaaa taaattacta tcatactttg atatgactaa 3420
tattctttaa taatttttgc gcatcgggcg ggtactaata ctagttctta aaaaaagggt 3480
agcgcgatgc acaaagcatt ccgcattcac acaggatcct aggaattggg tcgcacccca 3540
cagtctaccc taatgcaaac attagcgact actttcacgg ctcgaactcg tcacttatag 3600
atcatacaga gacaaattta ctgttgctcc aagttccctt tcttatttta ttattcttat 3660
aatttctatt cttatattgt tataaattat ttttttcttt ttgatagatc atgactgacc 3720
ctgatgttcc tggtcctagc gatccatatc ttagggagca cttacattgg tatgtatcat 3780
actatcatca actttgaaag cttaaaacac tgtaaagttg atgattcaca ccaaagattt 3840
taatcgtcgt cgtgttactt ccatataaat cagtatcgag aagtatgtgg ccatcactcc 3900
atcaacgaca ccaaaatgaa ataaagagtc cctatatcaa tacaatataa attaatctta 3960
aacatgaagt tgactttaaa ttggataaat tgtttccact actaagctta gcgtataaat 4020
tagtcctttg actttcaatt ttgtataata atgcgaagct ttttttcttg taaatgcaat 4080
ttttgtcctt gagggtttgc aacttctttt ttaaggaaaa aaaaaagact aaagttgtgt 4140
gacactaaaa ccaagagtta gcttaatact tcatggacac acgttagcat aaaacatata 4200
accgatattc aaaattacaa aaatgataga atcataattt ttgtttctat ttaaaaagga 4260
agtaagccaa attactacta acatagtgga cttaaagggt attaattttt tgttatttta 4320
atgatatctg ttcatgactt cttgactact tctactcctt tatatcaatc aaattataat 4380
ttacttcgtt tgactatcta atttacaggg taattacaga cattccaggc actacagatt 4440
cctcgtttgg tatggaataa tattgtattc cttttttact tttctgccta gcatttctaa 4500
atagagtagt ccgatacacg aaatatttca ctttacgcag gatccagaaa taaaggacca 4560
taccccaatt gggtgtaata taagtagtca tgggcgcatg cagtatttta gtgacgggtt 4620
taattgcact cataattttg gacgcttagc ataaagtagt agatatgtat ccataacttc 4680
aaaaatataa taggttcaat gttaaaaatt tcaaaagaga tgaactcata gagtttaaat 4740
catgatccgc ctctgtaggc agtctaccct aatgaaagaa tcagtggctg atttcacagt 4800
tcaaaaccgt aacctatgaa tcacataaag ccaactttac catcgctcca agactcgcct 4860
tcttctgcct aacattacta ctgctaataa agagaatttt aataaaacta ctaatgctaa 4920
ttattattct ttgctaaaat cttcatcagg aaaagaagtg gtgggctatg aaatgccaat 4980
gcctaacatt ggaatccata ggtttgtgtt tctgctcttc aagcagaaga agaggcaaac 5040
agtgagcgca ccattatcca gggaccgatt caatacgcgg aaatacgcag aagaaaatga 5100
gcttggctct ccagttgctg ctgttttctt caactgccaa agggaaaccg cggccagaaa 5160
gcgttga 5167
<210> 14
<211> 3167
<212> ДНК
<213> Nicotiana tabacum
<400> 14
atggctcaaa tgacagatcc ccttgtgatt agtagggtgg ttggagatgt tgttgattat 60
ttctctccaa gtgttaagat gtgtgttatt tataacccca gtaagcatgt ctataatggg 120
catgaactct ttccatccct tgttacctct aaacctaagg ttgaagttca tggaggtgac 180
atgagatcct tctttacact ggtaattaat tcacactact tcaatagttt tcttgttctt 240
atattttatt atctatctat atatatataa taaaggagcg gcaaagccac catataaatg 300
acaaatgtaa acttttagga caaaactcca aaaaagttgg agttttaaaa ttattttata 360
tataaaataa ataaataaat aaataaataa actatcaatt caaattgggg agtagtttct 420
tactaatatg atagctatat ctatatctat atctatctat atatgtaaaa catttatatg 480
atgccaagtg gcataaccac tgataagatt tttaaatttg aatatgaatg aattttaaat 540
gaagttctaa cttcttaaaa ataaacccta atataggtta ctatttttag taatgattga 600
aattattatt aaaatatttt gttgaaaaca acatagagat aaaatttgat tattaaattt 660
atgtattaca acaataataa ttattgaaaa tattgctaaa attttcatga aaggattcac 720
ccataattat tagtataata gaaaactaaa aaattattaa gtctaaagtt ctagatctct 780
atatttataa acgtataaac tgttatttta ttttctgaaa aaagcaaaaa tactgaagag 840
aaaaatgata aaaatatttt aaaatatgta agtcatgtgc aaataataaa gtgaacaaat 900
gatgtagtag tatactgaat aagatatgtt tttttgtcat aaaataagta tatgcataac 960
tcatctcaat aatttgctga ctccatctga gtcaaaatat cttctaaatt caagcgaaga 1020
taattatcta tcgcattatt tttttatcat taatataagg caagacgaat ctatatctca 1080
tatgggactt tttaaataga tacatcttta taaatgaacc actttatgag ttttatcacg 1140
aattacaagt aagaataact tgaagattga aagaattttt ggatttattt aattataata 1200
tatttttatt cattttaaat taatttatat tttcaaatta tttgtagcaa cctatataat 1260
tatgatattt gagtattatc ttataagtta tttgatgatt gtcgtttgat ttaattattg 1320
aactattact acaggggaca taagatgata attataattt tgtagaaaca tatgatctaa 1380
tgtgctcaaa taaattacta tcatactttg atatgactaa tattctttaa taatttttgc 1440
gcatcgggcg ggtactaata ctagttctta aaaaaagggt agcgcgatgc acaaagcatt 1500
ccgcattcac acaggatcct aggaattggg tcgcacccca cagtctaccc taatgcaaac 1560
attagcgact actttcacgg ctcgaactcg tcacttatag atcatacaga gacaaattta 1620
ctgttgctcc aagttccctt tcttatttta ttattcttat aatttctatt cttatattgt 1680
tataaattat ttttttcttt ttgatagatc atgactgacc ctgatgttcc tggtcctagc 1740
gatccatatc ttagggagca cttacattgg tatgtatcat actatcatca actttgaaag 1800
cttaaaacac tgtaaagttg atgattcaca ccaaagattt taatcgtcgt cgtgttactt 1860
ccatataaat cagtatcgag aagtatgtgg ccatcactcc atcaacgaca ccaaaatgaa 1920
ataaagagtc cctatatcaa tacaatataa attaatctta aacatgaagt tgactttaaa 1980
ttggataaat tgtttccact actaagctta gcgtataaat tagtcctttg actttcaatt 2040
ttgtataata atgcgaagct ttttttcttg taaatgcaat ttttgtcctt gagggtttgc 2100
aacttctttt ttaaggaaaa aaaaaagact aaagttgtgt gacactaaaa ccaagagtta 2160
gcttaatact tcatggacac acgttagcat aaaacatata accgatattc aaaattacaa 2220
aaatgataga atcataattt ttgtttctat ttaaaaagga agtaagccaa attactacta 2280
acatagtgga cttaaagggt attaattttt tgttatttta atgatatctg ttcatgactt 2340
cttgactact tctactcctt tatatcaatc aaattataat ttacttcgtt tgactatcta 2400
atttacaggg taattacaga cattccaggc actacagatt cctcgtttgg tatggaataa 2460
tattgtattc cttttttact tttctgccta gcatttctaa atagagtagt ccgatacacg 2520
aaatatttca ctttacgcag gatccagaaa taaaggacca taccccaatt gggtgtaata 2580
taagtagtca tgggcgcatg cagtatttta gtgacgggtt taattgcact cataattttg 2640
gacgcttagc ataaagtagt agatatgtat ccataacttc aaaaatataa taggttcaat 2700
gttaaaaatt tcaaaagaga tgaactcata gagtttaaat catgatccgc ctctgtaggc 2760
agtctaccct aatgaaagaa tcagtggctg atttcacagt tcaaaaccgt aacctatgaa 2820
tcacataaag ccaactttac catcgctcca agactcgcct tcttctgcct aacattacta 2880
ctgctaataa agagaatttt aataaaacta ctaatgctaa ttattattct ttgctaaaat 2940
cttcatcagg aaaagaagtg gtgggctatg aaatgccaat gcctaacatt ggaatccata 3000
ggtttgtgtt tctgctcttc aagcagaaga agaggcaaac agtgagcgca ccattatcca 3060
gggaccgatt caatacgcgg aaatacgcag aagaaaatga gcttggctct ccagttgctg 3120
ctgttttctt caactgccaa agggaaaccg cggccagaaa gcgttga 3167
<210> 15
<211> 173
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 15
Met Ala Gln Met Thr Asp Pro Leu Val Ile Ser Arg Val Val Gly Asp
1. 5 10 15
Val Val Asp Tyr Phe Ser Pro Ser Val Lys Met Cys Val Ile Tyr Asn
20 25 30
Pro Ser Lys His Val Tyr Asn Gly His Glu Leu Phe Pro Ser Leu Val
35 40 45
Thr Ser Lys Pro Lys Val Glu Val His Gly Gly Asp Met Arg Ser Phe
50 55 60
Phe Thr Leu Ile Met Thr Asp Pro Asp Val Pro Gly Pro Ser Asp Pro
65 70 75 80
Tyr Leu Arg Glu His Leu His Trp Val Ile Thr Asp Ile Pro Gly Thr
85 90 95
Thr Asp Ser Ser Phe Gly Lys Glu Val Val Gly Tyr Glu Met Pro Met
100 105 110
Pro Asn Ile Gly Ile His Arg Phe Val Phe Leu Leu Phe Lys Gln Lys
115 120 125
Lys Arg Gln Thr Val Ser Ala Pro Leu Ser Arg Asp Arg Phe Asn Thr
130 135 140
Arg Lys Tyr Ala Glu Glu Asn Glu Leu Gly Ser Pro Val Ala Ala Val
145 150 155 160
Phe Phe Asn Cys Gln Arg Glu Thr Ala Ala Arg Lys Arg
165 170
<210> 16
<211> 4569
<212> ДНК
<213> Nicotiana tabacum
<220>
<221> прочие признаки
<222> (1896)..(1896)
<223> n представляет собой a, c, g, or t
<400> 16
ccttatgggt tctcagcatt tgggcaaaag tgatacttta agcaagtgag gaagtttttt 60
aatgttggcc ggaaagatgc ctgtgggtgc tgtttggggg aaaaaaacaa aagctcgggt 120
aaattatcaa tacccgagtt ggttcctttc gtttgccact ggaccaactc ctgattttgc 180
ttatatatgg gttccaaact aaaaatactt atatatttaa taaatttatc aatacaaata 240
caaggctcgg gtaaaagtta ttaggttctc ggaaacccat acccgatact atggatccgc 300
ccctgcttat ccctaccttg tgtgaggtag aaacgctttt gataaggtta tttaaaagta 360
aaagaataag tttaatgtga caatttgaat ggttgagaca acatgccaaa agctaattta 420
agggatttta tttgacattt atatatggga gagaagaaaa agtattgccc agtatattat 480
tttaagctct acaaccaatc aagaatcaat tcctagaatc cattcaggtg tcaaagaata 540
ctgacatgat ataataaaat acaatttata tcacatcagt atttgctttt tcttgggaga 600
ttagataaaa agagatcaga aatggagttt tatggtacta ggagaattca aggatttact 660
acttttgtgg cacaacataa gctcccaatt tttttaagga atttataaaa gttggttttc 720
taagtactta caattgtcaa atttacaagt catttagtac ataaaaagaa acccaatgat 780
gaggttcagg aaaaaaaaaa atcctatact gtgatttcct agttggcgtt cggacataaa 840
aattatgaaa ttccgaaaaa aaaaattgtt ttaagttgaa aatggtatgt gaaaattaaa 900
gttatatatg gacataaata taatttggag ctgtttttga atttttgtga gtgctttgaa 960
gtgaaatttt ctaaaaacag ctttttggag tttttcaaat tccggagttc aacttcaagc 1020
gaaaaattaa aattttcatg atcaaatgtt gattccgaaa aaagtgaaaa aattcgaaaa 1080
aaagattttt ttttttatgg ccaaacagac ctaactagtt tcattttagt cattaagggt 1140
agaattgaaa gaattttaaa ttaaagtatt tttagatata taaaaataat gtacttttta 1200
aaacacacaa aaaaaggagt gccatatatt aatttaatat aaggatatat agtggatgca 1260
ttcataacta acattaacca aaagcattta ttgatcctat tttgacacca ttttatttta 1320
atacaattca taaatttcaa gaatttgaat acattagctt aatctcactt aaattttgag 1380
gtgatgcctg ttctctttct agtcacaact ttaatgtaca ttttatatgt caaattaata 1440
cctgaatttg taacccatca aatatcgcca cataatatga aacagtgaaa atatcttata 1500
ttcctgtatt ttatgactaa gacattaagt agctaacaac gatcgaaaaa cattcctaat 1560
aacaagcgaa ttacaactct gtcggataat cgtctgaaac cctaaaaagc tactgaaatg 1620
atttcctact agtataattc cgatgaaatt ttgttcgaaa attctataag aaatacacgt 1680
atttttagta gtgaaaaaag atttgttgta atttttttag gtggggtggg gtgatttggg 1740
gagggttggg gagtaggacc tcaaaaacaa agaattttaa tactttggag tttccttagg 1800
tcccatgttt tatactttct tttattctcc ttcaccatta tagctataac ttagtacata 1860
tatatatata tatatatata tatatatata tatatnatat aaggtgtcca tctgatcaag 1920
tatccaatac aaaccattct taagtctttg aaaatttctc tttttttcct tatctctatc 1980
tctgtctaat tttctttatt atggcaagaa gtttagagcc tctaattgtt gggagagtag 2040
taggagatgt tcttgattca tttagtccta taatgaaaat gacaatatca tataacaaca 2100
aattagtgtg caatggccat gaactccttc cttctgttgt cactgctaga cctaaagttg 2160
aagttcaagg gggagatttg agaactttct tcacattggt atttttttct tgatttctac 2220
ttaatttcca agatcatcaa gttcccatta tttctttaaa aaaaaaaaaa gcagttcggt 2280
gcactaaact cccgctatgc gcggggttcg gtgaagcacc gaaccataag ggtctattgt 2340
acgcaacctt accctgcatt tatgcaagag gcttgctcac cattacaagt tatattaatt 2400
taacatgtta tatataacca caaaggctgt cgtgggatgg taaatatcct tctatcctta 2460
atcagaagtt tcgggttcaa gttatagccc taggaatata gtcgtctttg gtagggatcc 2520
tttaccccca aaactttccg ccgtgaatcc agattagtaa acctcaaagc gggtatcggg 2580
cattggatga caaaccaaaa aaacttcaac gtgttatagc atgttataac ttattacagt 2640
taatttagtt ttccagtcga tactatatta aatagagtgc ctgtaattta ctttggagtg 2700
atttgattgt tattttttcg catcgtcagt acataaaact tatattaatt ttcgaatatg 2760
taggtcatga cagaccctga tgttcctggc cctagtgatc cttatctaag agagcatctc 2820
cactggtatg ccctaaactc aatttttttt taaaaaaaaa aaatagaaaa tgagaaaaaa 2880
tatgtaaaaa tctacaaata tgagaagatc atgattaatt ggaactattt ttactgacta 2940
tttgacagga tagtaactga cattccaggt accactgatg ctacttttgg taagttctct 3000
gtatcttctg caaaattaca agcacatgtg aagataaaag aagtttttct attattcact 3060
tattttgtct agctagttat atagaataat tataagatca acaattttgt atagtagtga 3120
atgttggact tctaaagtcg aacatgtcca cttgatgagt gtcacaaaaa tgtagaaact 3180
aaacaatcgt ttggacataa aaaaaaaagt aagttttttt gagttaaatt gaaaaagaaa 3240
atatttagaa tttgaaattg tggatataca tttaaattga aaagcattgc agttttgtaa 3300
ggaaaataaa ctttcatata cataaaaaag tgattttttg gaaactcatc ttcaagaata 3360
tttttaaaaa tttccgtcca atgtataacc aaacattatt ttgaaaaaga ttaaaaaaag 3420
gaaaaacttt aggaacaacg ggtcccaaga taaatgtgtc tagtcatata agattagata 3480
aattaggatt ttattatatt tggtagaagg tgcaagaagc atatgtaaat aataaattga 3540
gaagtcactt aagatatttt gatcatgtcc cacatcgata acaagaggta ccattctata 3600
tatgttaaat catggtaagt taaagtatta tatcacatat taaatggtga tataatagac 3660
ctaaatcaca tgaaacgaaa ttgtcccgaa aggtctataa atttttgaaa ttcatgtaga 3720
cgaagctaaa agtaggatac aataaaaaaa aaattaaaga tctatattgg cgatactatt 3780
tagttgggat tgcattttag ttattctagt acatttactt taatctaatt tttgctagct 3840
aggagtcttt taatcttatt agaaatttac ataccaaaaa atttagagaa cttgctagga 3900
caattggtat ttctttatat aatattgtgg aagttgtatt agagtatgtt gtttacatta 3960
cactctttga gtgcgttcct tctccgaact agctaatgca tgaacacgag atgccttctg 4020
caccgtgcta ccctattaat atataaaaaa atggtagccc ggtgcattaa gctcccgcta 4080
tgcgcgggtt ccgaaaaagg atcagaccac aagggtctat gtttgcaacc ttacttgtat 4140
ttctgcaaga aactgtttcc acggctcgaa cccatgatct tctggtcaca tgacaataac 4200
tttaccggtt acaccaaggt tccccttcac gcgctgccct tttaatattg tctattaata 4260
tttcctacta gagttataca cccctttgtt attactcact cttagggtga ttattaacat 4320
ataatatgtt taatatttat actaaaaaca ggacgagaat tggttagcta tgagattcca 4380
atgccaaata ttggaatcca taggtttgta tttgtacttt tcaagcaaaa acgaagacaa 4440
tcagttagct ctcctacttc aagggatcac ttcaacacta gaaattttgc tgaagaaaat 4500
gatcttggcc aacctgttgc tgctgttttc ttcaatgcac agcgagaaac cgccgcacga 4560
agacgctaa 4569
<210> 17
<211> 2569
<212> ДНК
<213> Nicotiana tabacum
<400> 17
atggcaagaa gtttagagcc tctaattgtt gggagagtag taggagatgt tcttgattca 60
tttagtccta taatgaaaat gacaatatca tataacaaca aattagtgtg caatggccat 120
gaactccttc cttctgttgt cactgctaga cctaaagttg aagttcaagg gggagatttg 180
agaactttct tcacattggt atttttttct tgatttctac ttaatttcca agatcatcaa 240
gttcccatta tttctttaaa aaaaaaaaaa gcagttcggt gcactaaact cccgctatgc 300
gcggggttcg gtgaagcacc gaaccataag ggtctattgt acgcaacctt accctgcatt 360
tatgcaagag gcttgctcac cattacaagt tatattaatt taacatgtta tatataacca 420
caaaggctgt cgtgggatgg taaatatcct tctatcctta atcagaagtt tcgggttcaa 480
gttatagccc taggaatata gtcgtctttg gtagggatcc tttaccccca aaactttccg 540
ccgtgaatcc agattagtaa acctcaaagc gggtatcggg cattggatga caaaccaaaa 600
aaacttcaac gtgttatagc atgttataac ttattacagt taatttagtt ttccagtcga 660
tactatatta aatagagtgc ctgtaattta ctttggagtg atttgattgt tattttttcg 720
catcgtcagt acataaaact tatattaatt ttcgaatatg taggtcatga cagaccctga 780
tgttcctggc cctagtgatc cttatctaag agagcatctc cactggtatg ccctaaactc 840
aatttttttt taaaaaaaaa aaatagaaaa tgagaaaaaa tatgtaaaaa tctacaaata 900
tgagaagatc atgattaatt ggaactattt ttactgacta tttgacagga tagtaactga 960
cattccaggt accactgatg ctacttttgg taagttctct gtatcttctg caaaattaca 1020
agcacatgtg aagataaaag aagtttttct attattcact tattttgtct agctagttat 1080
atagaataat tataagatca acaattttgt atagtagtga atgttggact tctaaagtcg 1140
aacatgtcca cttgatgagt gtcacaaaaa tgtagaaact aaacaatcgt ttggacataa 1200
aaaaaaaagt aagttttttt gagttaaatt gaaaaagaaa atatttagaa tttgaaattg 1260
tggatataca tttaaattga aaagcattgc agttttgtaa ggaaaataaa ctttcatata 1320
cataaaaaag tgattttttg gaaactcatc ttcaagaata tttttaaaaa tttccgtcca 1380
atgtataacc aaacattatt ttgaaaaaga ttaaaaaaag gaaaaacttt aggaacaacg 1440
ggtcccaaga taaatgtgtc tagtcatata agattagata aattaggatt ttattatatt 1500
tggtagaagg tgcaagaagc atatgtaaat aataaattga gaagtcactt aagatatttt 1560
gatcatgtcc cacatcgata acaagaggta ccattctata tatgttaaat catggtaagt 1620
taaagtatta tatcacatat taaatggtga tataatagac ctaaatcaca tgaaacgaaa 1680
ttgtcccgaa aggtctataa atttttgaaa ttcatgtaga cgaagctaaa agtaggatac 1740
aataaaaaaa aaattaaaga tctatattgg cgatactatt tagttgggat tgcattttag 1800
ttattctagt acatttactt taatctaatt tttgctagct aggagtcttt taatcttatt 1860
agaaatttac ataccaaaaa atttagagaa cttgctagga caattggtat ttctttatat 1920
aatattgtgg aagttgtatt agagtatgtt gtttacatta cactctttga gtgcgttcct 1980
tctccgaact agctaatgca tgaacacgag atgccttctg caccgtgcta ccctattaat 2040
atataaaaaa atggtagccc ggtgcattaa gctcccgcta tgcgcgggtt ccgaaaaagg 2100
atcagaccac aagggtctat gtttgcaacc ttacttgtat ttctgcaaga aactgtttcc 2160
acggctcgaa cccatgatct tctggtcaca tgacaataac tttaccggtt acaccaaggt 2220
tccccttcac gcgctgccct tttaatattg tctattaata tttcctacta gagttataca 2280
cccctttgtt attactcact cttagggtga ttattaacat ataatatgtt taatatttat 2340
actaaaaaca ggacgagaat tggttagcta tgagattcca atgccaaata ttggaatcca 2400
taggtttgta tttgtacttt tcaagcaaaa acgaagacaa tcagttagct ctcctacttc 2460
aagggatcac ttcaacacta gaaattttgc tgaagaaaat gatcttggcc aacctgttgc 2520
tgctgttttc ttcaatgcac agcgagaaac cgccgcacga agacgctaa 2569
<210> 18
<211> 172
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 18
Met Ala Arg Ser Leu Glu Pro Leu Ile Val Gly Arg Val Val Gly Asp
1. 5 10 15
Val Leu Asp Ser Phe Ser Pro Ile Met Lys Met Thr Ile Ser Tyr Asn
20 25 30
Asn Lys Leu Val Cys Asn Gly His Glu Leu Leu Pro Ser Val Val Thr
35 40 45
Ala Arg Pro Lys Val Glu Val Gln Gly Gly Asp Leu Arg Thr Phe Phe
50 55 60
Thr Leu Val Met Thr Asp Pro Asp Val Pro Gly Pro Ser Asp Pro Tyr
65 70 75 80
Leu Arg Glu His Leu His Trp Ile Val Thr Asp Ile Pro Gly Thr Thr
85 90 95
Asp Ala Thr Phe Gly Arg Glu Leu Val Ser Tyr Glu Ile Pro Met Pro
100 105 110
Asn Ile Gly Ile His Arg Phe Val Phe Val Leu Phe Lys Gln Lys Arg
115 120 125
Arg Gln Ser Val Ser Ser Pro Thr Ser Arg Asp His Phe Asn Thr Arg
130 135 140
Asn Phe Ala Glu Glu Asn Asp Leu Gly Gln Pro Val Ala Ala Val Phe
145 150 155 160
Phe Asn Ala Gln Arg Glu Thr Ala Ala Arg Arg Arg
165 170
<210> 19
<211> 4652
<212> ДНК
<213> Nicotiana tabacum
<400> 19
atccccagag gcggatctag gatttgaacc ttatgggttc tcagcatttg ggcaaaagtg 60
atactttaag caagtgagga agttttttaa tgttggccgg aaagatgcct gtgggtgctg 120
tttgggggaa aaaaacaaaa gctcgggtaa attatcaata cccgagttgg ttcctttcgt 180
ttgccactgg accaactcct gattttgctt atatatgggt tccaaactaa aaatacttat 240
atatttaata aatttatcaa tacaaataca aggctcgggt aaaagttatt aggttctcgg 300
aaacccatac ccgatactat ggatccgccc ctgcttatcc ctaccttgtg tgaggtagaa 360
acgcttttga taaggttatt taaaagtaaa agaataagtt taatgtgaca atttgaatgg 420
ttgagacaac atgccaaaag ctaatttaag ggattttatt tgacatttat atatgggaga 480
gaagaaaaag tattgcccag tatattattt taagctctac aaccaatcaa gaatcaattc 540
ctagaatcca ttcaggtgtc aaagaatact gacatgatat aataaaatac aatttatatc 600
acatcagtat ttgctttttc ttgggagatt agataaaaag agatcagaaa tggagtttta 660
tggtactagg agaattcaag gatttactac ttttgtggca caacataagc tcccaatttt 720
tttaaggaat ttataaaagt tggttttcta agtacttaca attgtcaaat ttacaagtca 780
tttagtacat aaaaagaaac ccaatgatga ggttcaggaa aaaaaaaaaa tcctatactg 840
tgatttccta gttggcgttc ggacataaaa attatgaaat tccgaaaaaa aaaattgttt 900
taagttgaaa atggtatgtg aaaattaaag ttatatatgg acataaatat aatttggagc 960
tgtttttgaa tttttgtgag tgctttgaag tgaaattttc taaaaacagc tttttggagt 1020
ttttcaaatt ccggagttca acttcaagcg aaaaattaaa attttcatga tcaaatgttg 1080
attccgaaaa aagtgaaaaa attcgaaaaa aagatttttt tttttatggc caaacagacc 1140
taactagttt cattttagtc attaagggta gaattgaaag aattttaaat taaagtattt 1200
ttagatatat aaaaataatg tactttttaa aacacacaaa aaaaggagtg ccatatatta 1260
atttaatata aggatatata gtggatgcat tcataactaa cattaaccaa aagcatttat 1320
tgatcctatt ttgacaccat tttattttaa tacaattcat aaatttcaag aatttgaata 1380
cattagctta atctcactta aattttgagg tgatgcctgt tctctttcta gtcacaactt 1440
taatgtacat tttatatgtc aaattaatac ctgaatttgt aacccatcaa atatcgccac 1500
ataatatgaa acagtgaaaa tatcttatat tcctgtattt tatgactaag acattaagta 1560
gctaacaacg atcgaaaaac attcctaata acaagcgaat tacaactctg tcggataatc 1620
gtctgaaacc ctaaaaagct actgaaatga tttcctacta gtataattcc gatgaaattt 1680
tgttcgaaaa ttctataaga aatacacgta tttttagtag tgaaaaaaga tttgttgtaa 1740
tttttttagg tggggtgggg tgatttgggg agggttgggg agtaggacct caaaaacaaa 1800
gaattttaat actttggagt ttccttaggt cccatgtttt atactttctt ttattctcct 1860
tcaccattat agctataact tagtacatat atatatatgg tggccctctg atccaatgta 1920
aaatgcaaac cattcttaag atctttgaaa tttctctctt ttttttcttt atctctatct 1980
ctgtctaatt ctctctatta tggcaagaag tttggagcct ctaatagttg ggagagtagt 2040
aggagatgtt cttgattcat ttagtcctat agtgaaaatg acaattactt ataacaacaa 2100
attagtgtgc aatggtcatg aattctttcc ttctattgtc acttctagac ctaaggttga 2160
agttcaagga ggagatttga gaactttctt cacactggta atttttcttg attttttcct 2220
taattccaag atcatcaagt tccatttatt tctttacaag ttatattaat ttaacccttt 2280
ataatcacca aaggctggcg tgggttgcta agtaaccttc catcgttaat cagatgtttc 2340
gggttcgagc tagccctggg attacaatcg ttttttgtag gaagcgcttt aacccccaaa 2400
atttttcagc acgaacccgg attagtaaac ctcaaaactc gtgccaaata ctagatgaca 2460
aaccaaaaga gttttcaacc tgttataaca tatgttactt gttacaatta ttagttttcc 2520
ggtcaatagt atattatgta attttctttg aagtgacttg attgttattt tttcacatta 2580
tcagtgcata aaacttatac tattattttt taatatgtag gtcatgacag accctgatgt 2640
tcccggccct agtgatcctt atctacgaga gcatctccac tggtacactc tctataatag 2700
tttcatttgt tccgaatttt cttggctgtt atataaaaaa tatattataa catagcatga 2760
aaattggttc cacaaaaact taatttttat agtgaataat tgttatatat tgatattgtt 2820
atagagaggt ctgtctatat gccctaaact caatgaaaaa aaatagaaaa tgagaaaaaa 2880
tatgtaaaat ctacaaatat gagaagatca tttttagttg aaactatcct tatatactac 2940
tgaatattta gctggcaaat aaaattgaca gtgttttact gattgtttga caggatagta 3000
actgacattc caggtaccac tgatgctact tttggtaagt tttattagtt tcttctgcaa 3060
gattacaagc acatgtgaag aagatacaag atgtttttcc attactcact tattttgtct 3120
tgctaataat tatatagaac aattgtaaga tcaacagtgt tatataatag tgaatgttgg 3180
acttctaaag tcgaacatgt ccacatgatg agcgtcacaa aaatgcagat acgagctcgt 3240
ttggattgac ttaaaaaatg tggtttttca gcaaaaataa cttttaagcc aaaaaacaat 3300
aagttagggt tgtccacctt tttgcttttg gcttaattta agcattttaa aatttatttt 3360
aagcaatttt tgacttagcc aaacaccgaa aaaagctaaa agaaacttaa aagctgattt 3420
gactagctta aaagtaaatc caaacaccct ctaactaagc atttggacat aaaaaaaata 3480
tgtcattttt gaaaaaagta gttcttttga gttaagtcaa aaaagaatat ataaaatttg 3540
aaattgtatt tagacatgca tttcacttga aaattattag agttttatga gaaaaatgaa 3600
cttttagatg aaaaagtggt ttttggaaac tcatcttcaa gaatttttcc aaaacttcag 3660
tccaatcgta taaccaaaca ttattttgat aaaaacatcg aaaataaaaa taaatctatg 3720
gagaaacggg tcccaagata aatgtgtcta gtcatataag attattcaaa attaagaatt 3780
tatcacattt gtaaaagatg taagtagcat atgtaaatga taaaatgaga agtcacttga 3840
gatgttttga tcatgtccta cgtcgatctt cagaggtacc attccgtata cgtgattggt 3900
aagtaaaggt attaaaaaga gacataatgg acctaaatta cgtgaaacga aattgtcttg 3960
aaaagtcttt caaatttttg aaatccatgt agacgaatcg aaaagtaggg cacaatgaaa 4020
tatgatcaaa ggtttataat ggtgatacaa gttagttggg attacgtttt agttatgcca 4080
gtatatttac tttaatctaa tattttcttg gagtttttta atcttattag aaatttactt 4140
accaaaaatt tagagaactt gctagaacaa tataattgat aattcttcat atatattgtc 4200
ttcgagctgt agaaacagcc actaatgttt gcattaggat atgttgtcta catcacactt 4260
attgtgtgtt gccctcaccg gaccctgcat gaacgtatga tgccttatgc accgcgcccc 4320
ttttaatatt atttattaat taatatttcc tgctagagtt atactccttt gttattactc 4380
attcttaggt tgatgattaa cttataatat gcttaatctt tatactaaaa ataggaagag 4440
aattggttag ctatgagatt ccaaggccaa atattggaat ccataggttt gtatttgtac 4500
ttttcaagca aagacgaaga caatcagtta gccctcctac ttcaagggaa aacttcaaca 4560
ctagaaattt tgccgaagaa aatgatctta gccaacctgt tgctgctgtt ttcttcaatg 4620
cacagcgaga aaccgccgcg cgaagacgct aa 4652
<210> 20
<211> 2653
<212> ДНК
<213> Nicotiana tabacum
<400> 20
atggcaagaa gtttggagcc tctaatagtt gggagagtag taggagatgt tcttgattca 60
tttagtccta tagtgaaaat gacaattact tataacaaca aattagtgtg caatggtcat 120
gaattctttc cttctattgt cacttctaga cctaaggttg aagttcaagg aggagatttg 180
agaactttct tcacactggt aatttttctt gattttttcc ttaattccaa gatcatcaag 240
ttccatttat ttctttacaa gttatattaa tttaaccctt tataatcacc aaaggctggc 300
gtgggttgct aagtaacctt ccatcgttaa tcagatgttt cgggttcgag ctagccctgg 360
gattacaatc gttttttgta ggaagcgctt taacccccaa aatttttcag cacgaacccg 420
gattagtaaa cctcaaaact cgtgccaaat actagatgac aaaccaaaag agttttcaac 480
ctgttataac atatgttact tgttacaatt attagttttc cggtcaatag tatattatgt 540
aattttcttt gaagtgactt gattgttatt ttttcacatt atcagtgcat aaaacttata 600
ctattatttt ttaatatgta ggtcatgaca gaccctgatg ttcccggccc tagtgatcct 660
tatctacgag agcatctcca ctggtacact ctctataata gtttcatttg ttccgaattt 720
tcttggctgt tatataaaaa atatattata acatagcatg aaaattggtt ccacaaaaac 780
ttaattttta tagtgaataa ttgttatata ttgatattgt tatagagagg tctgtctata 840
tgccctaaac tcaatgaaaa aaaatagaaa atgagaaaaa atatgtaaaa tctacaaata 900
tgagaagatc atttttagtt gaaactatcc ttatatacta ctgaatattt agctggcaaa 960
taaaattgac agtgttttac tgattgtttg acaggatagt aactgacatt ccaggtacca 1020
ctgatgctac ttttggtaag ttttattagt ttcttctgca agattacaag cacatgtgaa 1080
gaagatacaa gatgtttttc cattactcac ttattttgtc ttgctaataa ttatatagaa 1140
caattgtaag atcaacagtg ttatataata gtgaatgttg gacttctaaa gtcgaacatg 1200
tccacatgat gagcgtcaca aaaatgcaga tacgagctcg tttggattga cttaaaaaat 1260
gtggtttttc agcaaaaata acttttaagc caaaaaacaa taagttaggg ttgtccacct 1320
ttttgctttt ggcttaattt aagcatttta aaatttattt taagcaattt ttgacttagc 1380
caaacaccga aaaaagctaa aagaaactta aaagctgatt tgactagctt aaaagtaaat 1440
ccaaacaccc tctaactaag catttggaca taaaaaaaat atgtcatttt tgaaaaaagt 1500
agttcttttg agttaagtca aaaaagaata tataaaattt gaaattgtat ttagacatgc 1560
atttcacttg aaaattatta gagttttatg agaaaaatga acttttagat gaaaaagtgg 1620
tttttggaaa ctcatcttca agaatttttc caaaacttca gtccaatcgt ataaccaaac 1680
attattttga taaaaacatc gaaaataaaa ataaatctat ggagaaacgg gtcccaagat 1740
aaatgtgtct agtcatataa gattattcaa aattaagaat ttatcacatt tgtaaaagat 1800
gtaagtagca tatgtaaatg ataaaatgag aagtcacttg agatgttttg atcatgtcct 1860
acgtcgatct tcagaggtac cattccgtat acgtgattgg taagtaaagg tattaaaaag 1920
agacataatg gacctaaatt acgtgaaacg aaattgtctt gaaaagtctt tcaaattttt 1980
gaaatccatg tagacgaatc gaaaagtagg gcacaatgaa atatgatcaa aggtttataa 2040
tggtgataca agttagttgg gattacgttt tagttatgcc agtatattta ctttaatcta 2100
atattttctt ggagtttttt aatcttatta gaaatttact taccaaaaat ttagagaact 2160
tgctagaaca atataattga taattcttca tatatattgt cttcgagctg tagaaacagc 2220
cactaatgtt tgcattagga tatgttgtct acatcacact tattgtgtgt tgccctcacc 2280
ggaccctgca tgaacgtatg atgccttatg caccgcgccc cttttaatat tatttattaa 2340
ttaatatttc ctgctagagt tatactcctt tgttattact cattcttagg ttgatgatta 2400
acttataata tgcttaatct ttatactaaa aataggaaga gaattggtta gctatgagat 2460
tccaaggcca aatattggaa tccataggtt tgtatttgta cttttcaagc aaagacgaag 2520
acaatcagtt agccctccta cttcaaggga aaacttcaac actagaaatt ttgccgaaga 2580
aaatgatctt agccaacctg ttgctgctgt tttcttcaat gcacagcgag aaaccgccgc 2640
gcgaagacgc taa 2653
<210> 21
<211> 172
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 21
Met Ala Arg Ser Leu Glu Pro Leu Ile Val Gly Arg Val Val Gly Asp
1. 5 10 15
Val Leu Asp Ser Phe Ser Pro Ile Val Lys Met Thr Ile Thr Tyr Asn
20 25 30
Asn Lys Leu Val Cys Asn Gly His Glu Phe Phe Pro Ser Ile Val Thr
35 40 45
Ser Arg Pro Lys Val Glu Val Gln Gly Gly Asp Leu Arg Thr Phe Phe
50 55 60
Thr Leu Val Met Thr Asp Pro Asp Val Pro Gly Pro Ser Asp Pro Tyr
65 70 75 80
Leu Arg Glu His Leu His Trp Ile Val Thr Asp Ile Pro Gly Thr Thr
85 90 95
Asp Ala Thr Phe Gly Arg Glu Leu Val Ser Tyr Glu Ile Pro Arg Pro
100 105 110
Asn Ile Gly Ile His Arg Phe Val Phe Val Leu Phe Lys Gln Arg Arg
115 120 125
Arg Gln Ser Val Ser Pro Pro Thr Ser Arg Glu Asn Phe Asn Thr Arg
130 135 140
Asn Phe Ala Glu Glu Asn Asp Leu Ser Gln Pro Val Ala Ala Val Phe
145 150 155 160
Phe Asn Ala Gln Arg Glu Thr Ala Ala Arg Arg Arg
165 170
<210> 22
<211> 58
<212> ДНК
<213> Nicotiana tabacum
<400> 22
agttaaaatg acagtcactt acaacaataa acaagtttgc aatggccaag agctcttc 58
<210> 23
<211> 58
<212> ДНК
<213> Nicotiana tabacum
<400> 23
gaagagctct tggccattgc aaacttgttt attgttgtaa gtgactgtca ttttaact 58
<210> 24
<211> 482
<212> ДНК
<213> Nicotiana tabacum
<400> 24
ggtaccacaa gtttgtacaa aaaagcaggc taagcttgtc gaccatggag ttaaaatgac 60
agtcacttac aacaataaac aagtttgcaa tggccaagag ctcttctggt aacctttaat 120
gtttaaccgt tcacatttct aatatttact tatttgtaac atgtcgtcac gtgttagttt 180
cattcttttt atgaaccaaa catgcatgca aagatatttt tagatatttg gacggcgagt 240
gagatttgaa actaggaccg tttgcctgat acaatattaa aatatgtaac cattttatgt 300
acaagtttaa actgttgata gtagcatatt ttttactttt atttaagtat actatattcc 360
aacaggtaag ttaacgaaga gctcttggcc attgcaaact tgtttattgt tgtaagtgac 420
tgtcatttta actggcgcgc ccgggcaatt gacccagctt tcttgtacaa agtggtgagc 480
tc 482
<210> 25
<211> 88
<212> ДНК
<213> Nicotiana tabacum
<400> 25
tgcagtcact attagaccta gggttgaagt tcaaggtggt gatatgagaa ctttcttcac 60
attggtcatc acagatcctg atgtacct 88
<210> 26
<211> 88
<212> ДНК
<213> Nicotiana tabacum
<400> 26
aggtacatca ggatctgtga tgaccaatgt gaagaaagtt ctcatatcac caccttgaac 60
ttcaacccta ggtctaatag tgactgca 88
<210> 27
<211> 88
<212> ДНК
<213> Nicotiana tabacum
<400> 27
tgcggtcacc attagaccta gggttgaggt tcaaggtggt gatatgagaa ctttcttcac 60
attggtcatg acagaccctg atgttcct 88
<210> 28
<211> 88
<212> ДНК
<213> Nicotiana tabacum
<400> 28
aggaacatca gggtctgtca tgaccaatgt gaagaaagtt ctcatatcac caccttgaac 60
ctcaacccta ggtctaatgg tgaccgca 88
<210> 29
<211> 78
<212> ДНК
<213> Nicotiana tabacum
<400> 29
catgaactct ttccttcctc agtcacctct aaacctaggg ttgaagttca tggaggtgat 60
ttgagatctt tctttaca 78
<210> 30
<211> 78
<212> ДНК
<213> Nicotiana tabacum
<400> 30
tgtaaagaaa gatctcaaat cacctccatg aacttcaacc ctaggtttag aggtgactga 60
ggaaggaaag agttcatg 78
<210> 31
<211> 522
<212> ДНК
<213> Nicotiana tabacum
<400> 31
ggtaccacaa gtttgtacaa aaaagcaggc taagcttgtc gaccatggca tgaactcttt 60
ccttcctcag tcacctctaa acctagggtt gaagttcatg gaggtgattt gagatctttc 120
tttacatggt aacctttaat gtttaaccgt tcacatttct aatatttact tatttgtaac 180
atgtcgtcac gtgttagttt cattcttttt atgaaccaaa catgcatgca aagatatttt 240
tagatatttg gacggcgagt gagatttgaa actaggaccg tttgcctgat acaatattaa 300
aatatgtaac cattttatgt acaagtttaa actgttgata gtagcatatt ttttactttt 360
atttaagtat actatattcc aacaggtaag ttaactgtaa agaaagatct caaatcacct 420
ccatgaactt caaccctagg tttagaggtg actgaggaag gaaagagttc atgggcgcgc 480
ccgggcaatt gacccagctt tcttgtacaa agtggtgagc tc 522
<210> 32
<211> 121
<212> ДНК
<213> Nicotiana tabacum
<400> 32
gaaagaaata gttggctatg aaatgccaag gccaaatatt ggaattcaca ggtttgtatt 60
tctgctgttc aagcagaaga agaggcaaac agtattgact gcacctctct ccagggatcg 120
a 121
<210> 33
<211> 121
<212> ДНК
<213> Nicotiana tabacum
<400> 33
tcgatccctg gagagaggtg cagtcaatac tgtttgcctc ttcttctgct tgaacagcag 60
aaatacaaac ctgtgaattc caatatttgg ccttggcatt tcatagccaa ctatttcttt 120
c 121
<210> 34
<211> 110
<212> ДНК
<213> Nicotiana tabacum
<400> 34
gagagaaata gttgggtatg aaatgccaag gccaaatatt ggaatccaca gcagctttct 60
tcaattgcca gagggaaacc gctgccagaa ggcgttgaag aagatgttta 110
<210> 35
<211> 110
<212> ДНК
<213> Nicotiana tabacum
<400> 35
taaacatctt cttcaacgcc ttctggcagc ggtttccctc tggcaattga agaaagctgc 60
tgtggattcc aatatttggc cttggcattt catacccaac tatttctctc 110
<210> 36
<211> 138
<212> ДНК
<213> Nicotiana tabacum
<400> 36
atggctcaaa tgacagatcc ccttgtgatt agtagggtgg ttggagatgt tgttgattat 60
ttctctccaa gtgttaagat gtgtgttatt tataacccca gtaagcatgt ctataatggg 120
catgaactct ttccatcc 138
<210> 37
<211> 138
<212> ДНК
<213> Nicotiana tabacum
<400> 37
ggatggaaag agttcatgcc cattatagac atgcttactg gggttataaa taacacacat 60
cttaacactt ggagagaaat aatcaacaac atctccaacc accctactaa tcacaagggg 120
atctgtcatt tgagccat 138
<210> 38
<211> 642
<212> ДНК
<213> Nicotiana tabacum
<400> 38
ggtaccacaa gtttgtacaa aaaagcaggc taagcttgtc gaccatggat ggctcaaatg 60
acagatcccc ttgtgattag tagggtggtt ggagatgttg ttgattattt ctctccaagt 120
gttaagatgt gtgttattta taaccccagt aagcatgtct ataatgggca tgaactcttt 180
ccatcctggt aacctttaat gtttaaccgt tcacatttct aatatttact tatttgtaac 240
atgtcgtcac gtgttagttt cattcttttt atgaaccaaa catgcatgca aagatatttt 300
tagatatttg gacggcgagt gagatttgaa actaggaccg tttgcctgat acaatattaa 360
aatatgtaac cattttatgt acaagtttaa actgttgata gtagcatatt ttttactttt 420
atttaagtat actatattcc aacaggtaag ttaacggatg gaaagagttc atgcccatta 480
tagacatgct tactggggtt ataaataaca cacatcttaa cacttggaga gaaataatca 540
acaacatctc caaccaccct actaatcaca aggggatctg tcatttgagc catggcgcgc 600
ccgggcaatt gacccagctt tcttgtacaa agtggtgagc tc 642
<210> 39
<211> 93
<212> ДНК
<213> Nicotiana tabacum
<400> 39
tagtcctata gtgaaaatga caattactta taacaacaaa ttagtgtgca atggtcatga 60
attctttcct tctattgtca cttctagacc taa 93
<210> 40
<211> 93
<212> ДНК
<213> Nicotiana tabacum
<400> 40
ttaggtctag aagtgacaat agaaggaaag aattcatgac cattgcacac taatttgttg 60
ttataagtaa ttgtcatttt cactatagga cta 93
<210> 41
<211> 552
<212> ДНК
<213> Nicotiana tabacum
<400> 41
ggtaccacaa gtttgtacaa aaaagcaggc taagcttgtc gaccatggta gtcctatagt 60
gaaaatgaca attacttata acaacaaatt agtgtgcaat ggtcatgaat tctttccttc 120
tattgtcact tctagaccta atggtaacct ttaatgttta accgttcaca tttctaatat 180
ttacttattt gtaacatgtc gtcacgtgtt agtttcattc tttttatgaa ccaaacatgc 240
atgcaaagat atttttagat atttggacgg cgagtgagat ttgaaactag gaccgtttgc 300
ctgatacaat attaaaatat gtaaccattt tatgtacaag tttaaactgt tgatagtagc 360
atatttttta cttttattta agtatactat attccaacag gtaagttaac ttaggtctag 420
aagtgacaat agaaggaaag aattcatgac cattgcacac taatttgttg ttataagtaa 480
ttgtcatttt cactatagga ctaggcgcgc ccgggcaatt gacccagctt tcttgtacaa 540
agtggtgagc tc 552
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ АМИНОКИСЛОТ В РАСТЕНИИ | 2019 |
|
RU2799785C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ | 2019 |
|
RU2801948C2 |
| РАСТЕНИЯ СО СНИЖЕННЫМ СОДЕРЖАНИЕМ АСПАРАГИНА | 2016 |
|
RU2742725C2 |
| СНИЖЕНИЕ ТАБАК-СПЕЦИФИЧНЫХ НИТРОЗАМИНОВ В РАСТЕНИЯХ | 2013 |
|
RU2735254C2 |
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| МОДУЛИРОВАНИЕ УРОВНЕЙ НИТРАТОВ В РАСТЕНИЯХ ПОСРЕДСТВОМ МУТАЦИИ НИТРАТРЕДУКТАЗЫ | 2019 |
|
RU2815762C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ САХАРОВ И АМИНОКИСЛОТ В РАСТЕНИИ (SULTR3) | 2020 |
|
RU2826107C1 |
| МОДУЛИРОВАНИЕ БЕТА-ДАМАСЦЕНОНА У РАСТЕНИЙ | 2012 |
|
RU2681497C2 |
| ТРЕОНИНСИНТАЗА ИЗ NICOTIANA TABACUM И ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2689719C2 |
| ВЫСОКОАМИЛОЗНАЯ ПШЕНИЦА-III | 2017 |
|
RU2811010C2 |
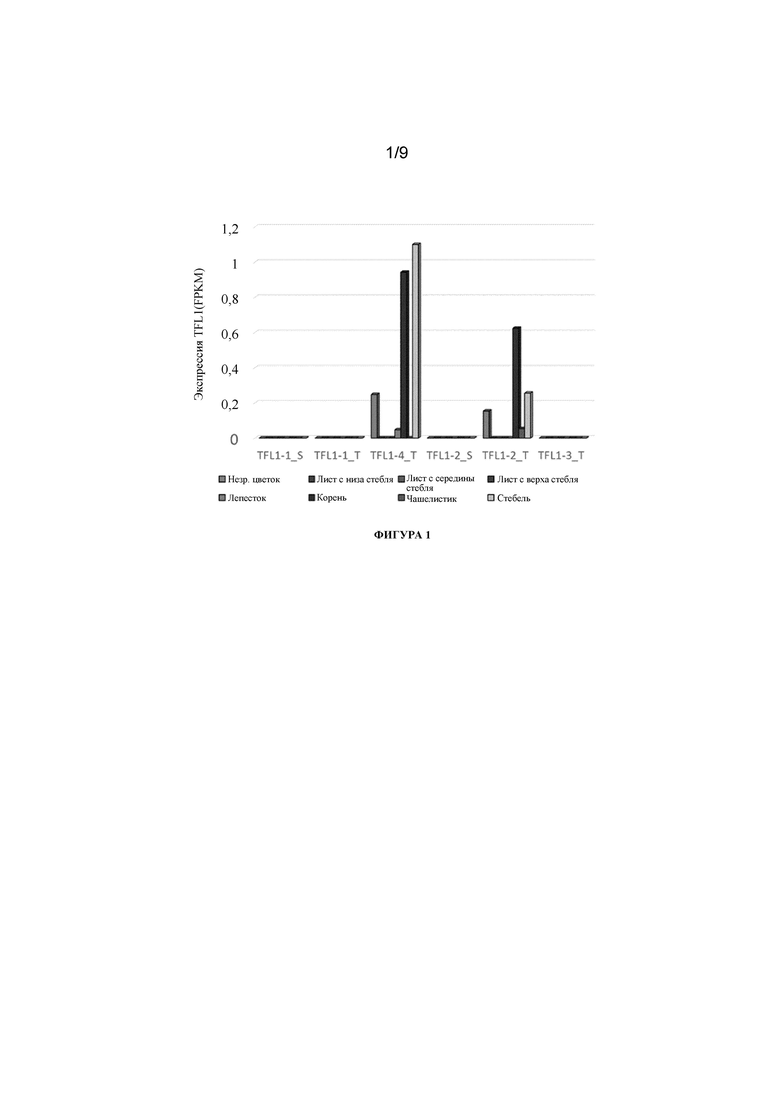







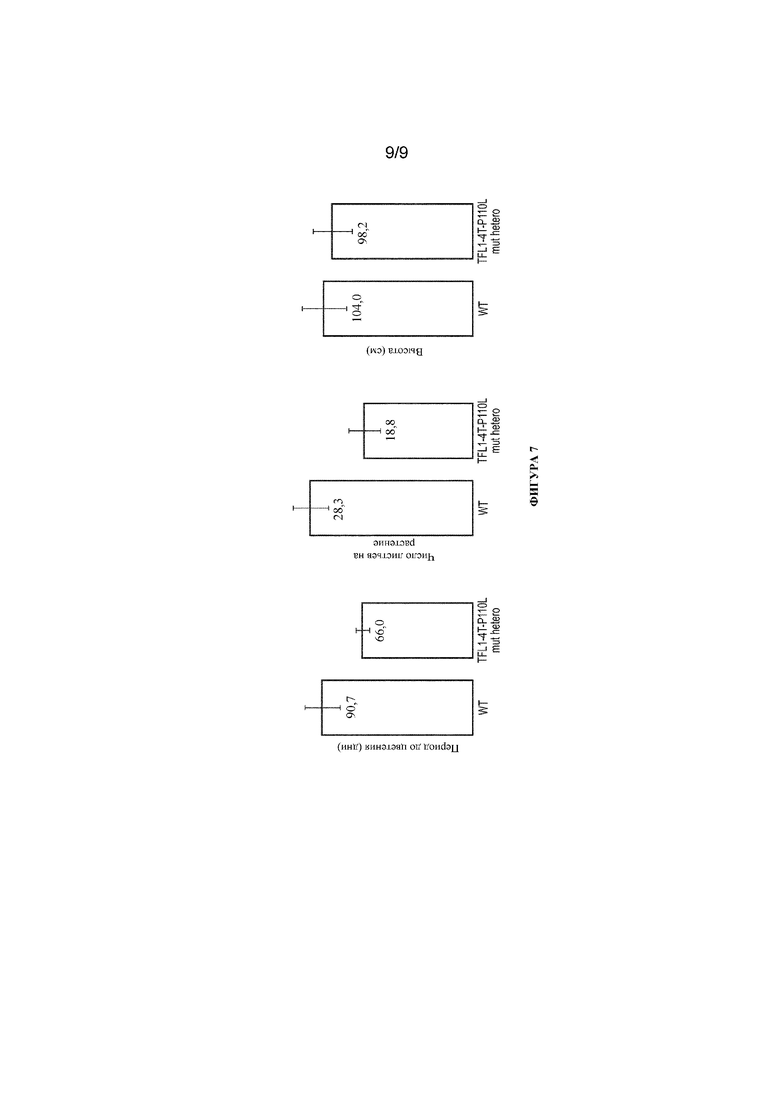
Изобретение относится к области биохимии, в частности к трансгенному растению, имеющему сокращенный период времени до наступления цветения. Раскрыты часть указанного растения; выделенный полинуклеотид или выделенный полипептид, которые содержит указанное растение, конструкция РНКи, двухнитевая РНК, содержащие указанный полинуклеотид. Также раскрыты применение растительного материала в табачном продукте или курительном изделии; применение растительного продукта в табачном продукте или курительном изделии; применение табачного продукта в курительном изделии; способ получения растительного материала из указанного растения; способ идентификации молекулы, которая обеспечивает модуляцию экспрессии полинуклеотида TFL1 или активности полипептида TFL1 в указанном растении. Изобретение позволяет эффективно сокращать период времени до наступления цветения у трансгенного растения. 13 н. и 13 з.п. ф-лы, 7 ил., 2 табл., 6 пр.
1. Трансгенное растение, имеющее сокращенный период времени до наступления цветения по сравнению с контрольным растением, где указанное трансгенное растение характеризуется сниженной экспрессией гена, кодирующего Terminal Flower 1 (TFL1), или сниженной активностью белка, кодируемого TFL1, при этом указанное растение содержит:
(i) полинуклеотидную последовательность, содержащую SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20; или
(ii) полипептид, кодируемый полинуклеотидом, представленным в (i);
где экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii), снижена по сравнению с контрольным растением, у которого экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii), не была снижена.
2. Трансгенное растение по п. 1, где сниженная экспрессия полинуклеотида или сниженная активность полипептида сокращает период времени до наступления цветения по сравнению с контрольным растением, соответственно, где период времени до наступления цветения сокращается по меньшей мере на 8% или по меньшей мере на 20%; и/или где растение содержит по меньшей мере одну генетическую альтерацию в полинуклеотидной последовательности, кодирующей TFL1.
3. Трансгенное растение по п. 1 или 2, где растение содержит по меньшей мере одну мутацию в полинуклеотидной последовательности, кодирующей TFL1; соответственно, где по меньшей мере одна мутация выбрана из группы, состоящей из: мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
4. Трансгенное растение по п. 1 или 2, где растение содержит по меньшей мере одну мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
5. Трансгенное растение по п. 1, или 2, или 4, где растение содержит по меньшей мере одну мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
6. Трансгенное растение по любому одному из предыдущих пунктов, где растение относится к роду Nicotiana или происходит из него, соответственно, где растение представляет собой Nicotiana tabacum.
7. Часть трансгенного растения по п. 1, характеризующаяся сниженной экспрессией гена, кодирующего Terminal Flower 1 (TFL1), или сниженной активностью белка, кодируемого TFL1, где указанная часть содержит:
(i) полинуклеотидную последовательность, содержащую SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20; или
(ii) полипептид, кодируемый полинуклеотидом, представленным в (i);
где экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii), снижена по сравнению с контрольным растением, у которого экспрессия или активность полинуклеотида или полипептида, представленных в (i), (ii), не была снижена.
8. Часть по п. 7, где сниженная экспрессия полинуклеотида или сниженная активность полипептида обеспечивает сокращение периода времени до наступления цветения по сравнению с контрольным растением, соответственно, где период времени до наступления цветения сокращен на по меньшей мере 8% или по меньшей мере 20%; и/или
где растение содержит по меньшей мере одну генетическую альтерацию в полинуклеотидной последовательности, кодирующей TFL1.
9. Часть по п. 7 или 8, где часть содержит по меньшей мере одну мутацию в полинуклеотидной последовательности, кодирующей TFL1; соответственно,
где по меньшей мере одна мутация выбрана из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
10. Часть по п. 7 или 8, где часть содержит по меньшей мере одну мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
11. Часть по п. 7, или 8, или 10, где часть содержит по меньшей мере одну мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
12. Часть по пп. 7-11, где растение относится к роду Nicotiana или происходит из него, соответственно, где растение представляет собой Nicotiana tabacum.
13. Применение растительного материала в табачном продукте или курительном изделии, где указанный растительный материал получен или может быть получен из растения по любому из пп. 1–6; где заявленный растительный материал представляет собой лист.
14. Применение растительного продукта в табачном продукте или курительном изделии, где указанный растительный продукт содержит по меньшей мере растительный материал по п. 13.
15. Применение табачного продукта в курительном изделии, где указанный табачный продукт содержит растительный материал по п. 13.
16. Способ сокращения периода времени до наступления цветения у растения, включающий модификацию растения путем снижения экспрессии по меньшей мере одного гена TFL1 или активности по меньшей мере одного белка, кодируемого им, у указанного растения; соответственно, где способ включает
(a) получение растения, содержащего
(i) полинуклеотидную последовательность SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20; или
(ii) полипептид, кодируемый полинуклеотидом, представленным в (i); и
(b) снижение экспрессии гена TFL1 или активности белка TFL1 у растения; и
(c) получение растения с сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, у которого экспрессия гена TFL1 или активность белка TFL1 не была снижена.
17. Способ по п. 16, где экспрессию TFL1 или активность TFL1 снижают с помощью способа, выбранного из группы, состоящей из a) мутирования гена TFL1 у растения; b) экспрессии экзогенного полинуклеотида или полипептида у растения; и c) удаления гена TFL1 у растения, или комбинации одного или более из них, соответственно,
где растение мутировано в положении, выбранном из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
18. Способ получения растительного материала из растения по п. 1 с сокращенным периодом времени до наступления цветения по сравнению с контрольным растением, при этом указанный способ включает
(a) получение растения по любому из пп. 1–6 или растительного материала по п. 13;
(b) сбор растительного материала от растения;
(c) необязательно сушку или высушивание растительного материала в течение некоторого периода времени; и
(d) получение растительного материала, который характеризуется сокращенным периодом времени до наступления цветения по сравнению с контрольным растением.
19. Применение растительного материала в табачном продукте или курительном изделии, где указанный растительный материал получен или может быть получен с помощью способа по любому из пп. 16–18.
20. Выделенный полинуклеотид для снижения экспрессии или активности определенных полинуклеотидов TFL1, где указанный полинуклеотид содержит SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20.
21. Выделенный полипептид для ингибирования экспрессии TFL-1, где выделенный полипептид содержит по меньшей мере одну мутацию, выбранную из группы, состоящей из мутации в положении T143 или G129 в SEQ ID NO: 9; или мутации в положении R120, или G129, или P131 в SEQ ID NO: 12; или мутации в положении P110 или H86 в SEQ ID NO: 21, или комбинации двух или более из них; соответственно, где мутация представляет собой T143I, или G129R, или G129E, или H84STOP в SEQ ID NO: 9; или где мутация представляет собой R120C, или G129E, или P131S в SEQ ID NO: 12; или где мутация представляет собой P110L или H86STOP в SEQ ID NO: 21 или комбинацию двух или более из них.
22. Выделенный полипептид по п. 21, где выделенный полипептид содержит по меньшей мере одну мутацию в положении P131 в SEQ ID NO: 12, соответственно, где мутация представляет собой P131S.
23. Выделенный полипептид по п. 21 или 22, где растение содержит по меньшей мере одну мутацию в положении P110 в SEQ ID NO: 21, соответственно, где мутация представляет собой P110L.
24. Конструкция РНКи для подавления экспрессии TFL–1, содержащая последовательность, которая гибридизируется с целевой последовательностью в мРНК гена TFL–1 и обеспечивает подавление экспрессии гена TFL–1 посредством механизма РНК–интерференции, где указанная целевая последовательность выбрана из группы, состоящей из SEQ ID NO: 7, 8, 10, 11, 19 и/или 20.
25. Двухнитевая РНК для ингибирования экспрессии TFL-1, где указанная РНК содержит по меньшей мере две последовательности, которые по меньшей мере частично комплементарны друг другу, и где смысловая нить содержит первую последовательность, и антисмысловая нить содержит вторую последовательность, и где по меньшей мере одна из последовательностей содержит по меньшей мере 10 смежных нуклеотидов из РНК TFL1, соответственно, где по меньшей мере одна из последовательностей содержит 21–23 смежных нуклеотида из РНК TFL1; соответственно, где двухнитевая РНК содержит
первую последовательность, содержащую по меньшей мере 10 смежных нуклеотидов из TFL1, соответственно, 21–23 смежных нуклеотида из TFL–1;
вторую последовательность; и
третью последовательность, содержащую последовательность, обратно комплементарную первой последовательности, расположенную в той же ориентации, что и первая последовательность,
где вторая последовательность расположена между первой последовательностью и третьей последовательностью и вторая последовательность функционально связана с первой последовательностью и с третьей последовательностью; соответственно,
где первая последовательность выбрана из группы, состоящей из SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20, и/или где третья последовательность представляет собой последовательность, обратно комплементарную последовательности, соответствующей SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20; соответственно,
где первая последовательность содержит или состоит из SEQ ID NO: 22 и третья последовательность содержит или состоит из SEQ ID NO: 23; или первая последовательность содержит или состоит из SEQ ID NO: 25 и третья последовательность содержит или состоит из SEQ ID NO: 26; или первая последовательность содержит или состоит из SEQ ID NO: 27 и третья последовательность содержит или состоит из SEQ ID NO: 28; или первая последовательность содержит или состоит из SEQ ID NO: 29 и третья последовательность содержит или состоит из SEQ ID NO: 30; или первая последовательность содержит или состоит из SEQ ID NO: 32 и третья последовательность содержит или состоит из SEQ ID NO: 33; или первая последовательность содержит или состоит из SEQ ID NO: 34 и третья последовательность содержит или состоит из SEQ ID NO: 35; или первая последовательность содержит или состоит из SEQ ID NO: 36 и третья последовательность содержит или состоит из SEQ ID NO: 37; или первая последовательность содержит или состоит из SEQ ID NO: 39 и третья последовательность содержит или состоит из SEQ ID NO: 40; соответственно,
где двухнитевая РНК содержит или состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, SEQ ID NO: 35, SEQ ID NO: 31, SEQ ID NO: 38 и SEQ ID NO: 41.
26. Способ идентификации молекулы, которая обеспечивает модуляцию экспрессии полинуклеотида TFL1 или активности полипептида TFL1, при этом способ включает
(a) приведение молекулы в контакт с растением, содержащим полинуклеотидную последовательность, содержащую SEQ ID NO: 7, или SEQ ID NO: 8, или SEQ ID NO: 10, или SEQ ID NO: 11, или SEQ ID NO: 19, или SEQ ID NO: 20, или полипептид, кодируемый полинуклеотидом;
(b) отслеживание одного или более из (i) уровня экспрессии полинуклеотида TFL1 у растения; (ii) уровня экспрессии полипептида TFL1 у растения; (iii) модуляции активности полипептида TFL1 у растения; или (iv) модуляции активности полинуклеотида TFL1 у растения; и
(c) идентификацию молекулы, которая обеспечивает модуляцию экспрессии полинуклеотида TFL1 или активности полипептида TFL1.
| WO 2010000794, 07.01.2010 | |||
| ГЕНЫ CORYNEBACTERIUM GLUTAMICUM, КОДИРУЮЩИЕ БЕЛКИ, УЧАСТВУЮЩИЕ В МЕТАБОЛИЗМЕ УГЛЕРОДА И ПРОДУЦИРОВАНИИ ЭНЕРГИИ | 2005 |
|
RU2321634C2 |
| Способ изготовления электрического изолирующего состава | 1924 |
|
SU4799A1 |
| ПОЛИНУКЛЕОТИДЫ И ПОЛИПЕПТИДЫ ФАГА φ-mru, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2520738C2 |

